Заявляемая группа изобретений относится к биотехнологии и представляет собой бактерию Bacillus subtilis, продуцирующую АИКАР и представленную рядом сконструированных штаммов, а также способ микробиологического синтеза АИКАР путем культивирования такой бактерии.
Пуриновый нуклеозид АИКАР является природным аналогом аденозина, который накапливается в клетках любых организмов при ограничении биосинтеза пуриновых нуклеотидов на этапе образования инозин-5-монофосфата (ИМФ). Превращение АИКАР в ИМФ в бактериях контролируется геном purH, кодирующим белок, который последовательно осуществляет формилирование АИКАР-Ф и циклизацию пуринового гетероцикла.
При введении препаратов АИКАР в организм животных и человека происходит его фосфорилирование с образованием нуклеотида АИКАР-Ф. Будучи аналогом АМФ, АИКАР-Ф способен активировать фермент АМФ-зависимую протеинкиназу (АМФК) - регулятор метаболизма углеводов и липидов у эукариот (Eur. J. Biochem., 229, 558-565 (1995)). Благодаря способности активировать АМФК, препараты АИКАР имеют широкий терапевтический потенциал, нормализуя углеводный и липидный метаболизм и ограничивая пролиферацию клеток. В опытах на животных показано, АИКАР эффективно влияет на составляющие метаболического синдрома (Trends Pharmacol. Sci., 26, 69-76 (2005)). АИКАР ослабляет последствия гипоксии, снижает риск инфаркта миокарда (US 20040072138), снижает синтез липидов и провоцирует их окисление, понижает резистентность к инсулину (US 20030212034, Diabetes, 51, 2199-2206 (2002)). Показана эффективность АИКАР в предупреждении диабета 2 типа (Diabetes, 54, 928-934 (2005)). АИКАР специфически индуцирует апоптоз и эффективен как противораковый препарат при лечении хронических и острых лейкозов (US 20050233987, Molecular Cancer 6,:46, 1-12, (2007), J. Biol. Chem., 280, 39582-39593 (2005)). В основе большинства терапевтических эффектов АИКАР лежит коррекция энергетического метаболизма всего организма.
Описаны методы микробиологического синтеза АИКАР путем ферментации бактерий Arthrobacter albidus (US 3173848) и Brevibacterium ammoniagenes (US 3748232), накапливающих от 2 до 11 г/л АИКАР в культуральной жидкости за 96 часов ферментации. Продуцировать АИКАР также способны бактерии Bacillus subtilis - штамм АТСС №15116, и Bacillus megaterium - штамм АТСС №15117, накапливащие до 10 г/л АИКАР за 80 часов ферментации (US 323810). Все указанные выше штаммы получены путем традиционного мутагенеза и являются условными ауксотрофами по пуринам, что затрудняет процесс ферментации и требует сложных по составу сред для культивации.
Известен рекомбинантный штамм Bacillus subtilis АМ25-41 (RU 2405833), накапливаний за 48 часов ферментации на среде с глюкозой до 3.5 г/л АИКАР. Этот продуцент получен на основе родительского штамма В.subtilis ВНИИгенетика-304 (ВКПМ В-2116) путем генетического конструирования: содержит мутацию purH::EmR, приводящую к нарушению экспрессии гена purH, мутацию в гене purR, кодирующем репрессор биосинтеза пуриновых нуклеотидов, делецию терминатора транскрипции в лидерной области pur-оперона и дополнительную копию гена prs, обеспечивающего синтез фосфорибозилпирофосфата (ФРПФ) под контролем промотора гена rpsF рибосомного белка S6. В ходе последующей работы в геном штамма В.subtilis АМ25-41 интегрированы модифицированные гены prs и purF E.coli, кодирующие ключевые ферменты синтеза предшественников пуринов, под контролем сильного промотора PrpsF, обеспечивающего высокий уровень их экспрессии в клетках В.subtilis. В итоге работы был получен штамм-продуцент Bacillus subtilis АМ815, накапливающий 11-13 г/л АИКАР в культуральной жидкости за 72 часов ферментации. Штамм Bacillus subtilis АМ815 (генотипа leu met purR::neo ΔΤ-purE ∆purH атуЕ::[PrpsF-prsE-purFE]), описанный в статье Лобанова с соавт. «Реконструкция пуринового метаболизма у Bacillus subtilis с целью получения штамма-продуцента AICAR - нового препарата широкого терапевтического применения» (2011) Acta Naturae, т.3, №2 (9), стр. 83-93 и зарегистрированный во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ В-11156, который рассматривается в качестве ближайшего аналога заявляемой бактерии. В качестве ближайшего аналога заявляемого способа рассматривается описанный в патенте RU 2405833 способ микробиологического синтеза АИКАР путем культивирования штамма Bacillus subtilis В-11156 на среде следующего состава (мас. %):
Уровень синтеза АИКАР, достигаемый при использовании в способе ближайшего аналога, недостаточен для удовлетворения растущих потребностей экспериментальной и практической медицины в этом нуклеозиде.
Задача настоящей группы изобретений - разработать более продуктивный способ микробиологического синтеза АИКАР.
Задача решена путем:
- конструирования бактерии Bacillus subtilis, продуцирующей АИКАР и содержащей дерегулированный pur-оперон на фоне инактивированного гена purH, модифицированные гетерологичные гены prs и purF E.coli под контролем сильного промотора PrpsF в составе хромосомы и обладающей по крайней мере одной из следующих характеристик:
- содержит ген zwf под контролем сильного промотора PrpsF,
- содержит гетерологичный ген udhA под контролем сильного промотора PrpsF,
- содержит делецию гена sacB,
- разработки способа микробиологического синтеза АИКАР путем культивирования сконструированной бактерии в среде следующего состава, мас. %:
Термин «бактерия Bacillus subtilis» означает бактерию, которую относят к виду Bacillus subtilis в соответствии с классификацией, известной для специалиста в области микробиологии.
Термин «pur-оперон» означает фрагмент хромосомы Bacillus subtilis, содержащий гены, кодирующие белки, существенные для продукции АИКАР. В бактериях, принадлежащих к роду Bacillus, pur-оперон содержит следующие гены: purP-L, кодирующий регуляторный элемент, ген purE, кодирующий N5-карбоксиаминоимидазолрибонуклеотидмутазу (КФ 4.1.1.21), ген purK, кодирующий N5-карбоксиаминоимидазолрибонуклеотидсинтазу (КФ 4.1.1.21), ген purB, кодирующий аденилсукцинатлиазу (КФ 4.3.2.2), ген purC, кодирующий фосфорибозиламиноимидазолсукцинокарбоксамидсинтазу (КФ 6.3.2.6), ген purS, кодирующий гипотетический белковый фактор, необходимый для функционирования фосфорибозилформилглицинамидинсинтетазы (КФ 6.3.5.3), ген purQ, кодирующий фософорибозилформилглицинамидинсинтазу 1 (КФ 6.3.5.3), ген purL, кодирующий фософорибозилформилглицинамидинсинтазу 2 (КФ 6.3.5.3), ген purF, кодирующий амидофосфорибозилтрансферазу (КФ 2.4.2.14), ген purM, кодирующий фософорибозилформилглицинамидинциклолигазу (КФ 6.3.3.1), ген purN, кодирующий фософорибозилглицинамидформилтрансферазу (КФ 2.1.2.2), ген purH, кодирующий бифункциональный фермент фосфорибозиламиноимидазолкарбоксиформилформилтрансферазу/инозинмонофосфатциклогидролазу (КФ 2.1.2.3 и КФ 3.5.4.10), и ген purD, кодирующий фосфорибозиламинглицинлигазу (КФ 6.3.4.13). Нуклеотидные последовательности описанных выше генов purP-L-purD представлены в базе данных «GenBank» под номерами GeneID: 8302912, 939481, 936039, 936048, 939389, 936041, 938760, 939233, 936046, 936044, 936045, 936053, 938741, соответственно (нуклеотиды с 698369 по 698471, с 698538 по 699172, с 698921 по 700405, с 700037 по 701720, с 701601 по 702326, с 702319 по 702573, с 702570 по 703253, с 703237 по 705465, с 705441 по 706871, с 706973 по 708013, с 708010 по 708597, с 708594 по 710132, с 710148 по 711416 в последовательности NC_000964.3).
Термин «дерегулирован» означает, что уровень продукции пуринов, включая АИКАР, выше, чем уровень продукции, наблюдаемый у бактерий с природной регуляторной системой синтеза АИКАР, а именно у бактерий, встречающихся в природе.
Дерегуляцию контроля синтеза АИКАР осуществляют путем изменения регуляторного участка pur-оперона, например, делецией терминатора транскрипции, расположенного в этом участке, замены указанного регуляторного участка другим сильным участком с конститутивным уровнем экспрессии, инактивации гена purR, кодирующего репрессор, получения мутации в белке-репрессоре и т.д. Указанную дерегуляцию осуществляют традиционными методами, такими как мутагенная обработка с использованием УФ-излучения или обработкой нитрозогуанидином (Ν-метил-Ν′-нитро-Ν-нитрозогуанидин), сайт-направленным мутагенезом, разрушением гена с помощью гомологичной рекомбинации и/или инсерционно-делеционным мутагенезом. Нуклеотидная последовательность гена purR B.subtilis представлена в базе данных «GenBank» под номером GeneID: 937000 (нуклеотиды с 54441 по 55298 в последовательности NC_000964.3).
Термин «на фоне инактивированного гена purH» означает, что ферментативная активность белка АИКАР-фосфат трансформилазы - ИМФ циклогидролазы, кодируемого геном purH, полностью отсутствует или понижена в результате генетической модификации гена purH (делеции всего гена или его части, сдвига рамки считывания гена, введения миссенс/нонсенс мутаций или модификации примыкающих к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промоторы, энхансеры, аттенуаторы, сайты связывания рибосом и т.д.).
Термин «гетерологичный ген» означает, что указанный ген выделен из хромосомы организма, отличного от В.subtilis. Бактерию, содержащую гетерологичный ген в хромосоме, получают стандартными методами рекомбинантных ДНК, трансформацией, трансфекцией.
Термин «модифицированные гетерологичные гены prs и purF Ε.coli» означает, что ген prs E.coli, кодирующий фермент фосфорибозилпирофосфат (PRPP) синтазы (КФ 2.7.6.1), содержит специфическую мутацию, приводящую к аминокислотной замене Asp128→Ala в белке PRPP-синтазы, которая обусловливает снятие ретроингибирования фермента пуриновыми нуклеотидами, а ген purF Ε.coli, кодирующий первый фермент пуринового биосинтеза - амидофосфорибозилпирофосфат трансферазы (КФ 2.4.2.14), содержит специфические мутации, приводящие к аминокислотным заменам Lys326→Gln и Pro410→Trp в белке глютамин-PRPP-аминотрансферазы, которые обусловливают снятие ретроингибирования фермента аденозинмонофосфатом и гуанозинмонофосфатом. Оба модифицированных гена интегрированы в составе хромосомы В.subtilis под контролем сильного промотора PrpsF. Нуклеотидные последовательности генов prs и purF E.coli представлены в базе данных «GenBank» под номерами GeneID: 945772 и 946794 соответственно (нуклеотиды с 1260151 по 1261098 и с 2426743 по 2428260 в последовательности NC_000913.2).
Термин «под контролем сильного промотора PrpsF» означает, что природные промоторы указанных генов заменены на промотор гена rpsF, кодирующего синтез рибосомного белка S6 у бацилл. Известно, что промоторы генов рибосомных белков относятся к наиболее сильным промоторам в составе хромосомы В.subtilis, их экспрессия координирована со скоростью роста бактерий и достигает максимума на логарифмической стадии роста бактерий. Нуклеотидная последовательность гена rpsF В.subtilis представлена в базе данных «GenBank» под номером GeneID: 937919 (нуклеотиды с 4199445 по 4199732 в последовательности NC_000964.3).
Термин «ген zwf под контролем сильного промотора» означает, что структурный ген zwf В.subtilis, контролирующий синтез фермента глюкозо-6-фосфат 1-дегидрогеназы (КФ 1.1.1.49), подставлен под контроль сильного бациллярного промотра PrpsF (см. выше). Нуклеотидная последовательность гена zwf В.subtilis представлена в базе данных «GenBank» под номером GeneID: 938690 (нуклеотиды с 2479156 по 2480625 в последовательности NC_000964.3). Известно, что фермент глюкозо-6-фосфат 1-дегидрогеназа играет ключевую роль в перераспределении потока глюкозы в направлении пентозофосфатного пути, ответственного за окислительное фосфорилирование пентоз. Усиление этого пути должно приводить к существенному увеличению пула фосфорилированных производных рибозы, которые являются непосредственными предшественниками пуриновых соединений, в том числе и АИКАР. Поскольку природный ген zwf под контролем собственного промотора подвержен довольно сложной негативной регуляции, решено усилить его экспрессию путем подстановки под контроль сильного промотора PrpsF.
Термин «гетерологичный ген udhA E.coli под контролем сильного промотора» означает, что структурный ген udhA E.coli, контролирующий синтез растворимой NADPH/NADH трансгидрогеназы (КФ 1.6.1.1), клонированный из E.coli, подставлен под контроль сильного бациллярного промотора PrpsF. Нуклеотидная последовательность гена udhA E.coli представлена в базе данных «GenBank» под номером GeneID: 948461 (нуклеотиды с 4157413 по 4158813 в последовательности NC_000913.2). Этот фермент играет важную роль в установлении внутриклеточного баланса между пулами NADPH/NADH в клетках E.coli (Sauer U. et al. The soluble and membrane-bound transhydrogenases UdhA and PntAB have divergent functions in NADPH metabolism of Escherichia coli // JBiol. Chem. 2004. 279 (8) 6613-6619). В то же время, ортолог гена udhA (и соответствующий фермент) у В.subtilis до настоящего времени не идентифицирован. На основании вышеизложенного сделано предположение, что клонирование гетерологичного гена udhA E.coli и его последующая интеграция в хромосому В.subtilis под контролем сильного бациллярного промотора PrpsF приведет к усилению реакции реокисления пула NADPH, накапливаемого в процессе биосинтеза пуринов, в том числе и АИКАР.
Термин «содержит делецию гена sacB» означает, что структурный ген sacB B.subtilis, кодирующий синтез фермента левансахаразы (КФ 2.4.1.10), инактивирован путем внесения прецизионной делеции, полностью перекрывающей рамку считывания гена sacB. Нуклеотидная последовательность гена sacB B.subtilis представлена в базе данных «GenBank» под номером GeneID: 936413 (нуклеотиды с 3536012 по 3537433 в последовательности NC_000964.3). Известно, что левансахараза катализирует образование высокомолекулярного фруктана леванного типа путем полимеризации фруктозильных остатков сахарозы. В результате этого в процессе культивирования на более дешевом по сравнению с глюкозой источнике углерода - сахарозе, вязкость бактериальной культуры значительно возрастает, что существенным образом затрудняет высвобождение АИКАР из культуральной жидкости и приводит к большим потерям в ходе его выделения и очистки.
В геноме полученных штаммов Bacillus subtilis АМ891, АМ887, АМ893 и АМ895 - продуцентов АИКАР - ген sacB инактивирован, что позволяет существенно упростить процедуру выделения и очистки целевого продукта.
Характеристика сконструированных штаммов
Сконструированные штаммы Bacillus subtilis АМ878, Bacillus subtilis AM883, Bacillus subtilis AM891, Bacillus subtilis AM887, Bacillus subtilis AM893, а также Bacillus subtilis AM890 и Bacillus subtilis AM895 продуцируют АИКАР и имеют следующие свойства.
Культурально-морфологические признаки
Спорообразующая грамположительная палочка с закругленными концами и краевым расположением спор. На LB-агаре (Миллер Дж. 1979. Эксперименты в молекулярной генетике. Мир) при 37°C на вторые сутки образует колонии с неровным краем диаметром 3-4 мм, а на минимальной среде Спицайзена (J.Bacteriol. 81:741-746, 1961) такие же колонии диаметром 2-3 мм.
Физиолого-биохимические признаки
Сбраживает сахариды, такие как глюкоза, лактоза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза и гидролизат крахмала; спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая, фумаровая, лимонная и янтарная кислоты, и подобные соединения.
Хранение
В лиофилизированном виде или в парах жидкого азота.
Стабильность
Полученные штаммы стабильны и не теряют способности синтезировать АИКАР после 10 пересевов на полноценной среде.
Особенности конструирования:
- Штамм Bacillus subtilis АМ878 получен путем введения в хромосому штамма-реципиента Bacillus subtilis В-11156 (В-11156) гена zwf под контролем сильного промотора PrpsF.
- Штамм Bacillus subtilis АМ883 получен путем введения в хромосому штамма-реципиента Bacillus subtilis В-11156 (В-11156) гетерологичного гена udhA E.coli под контролем сильного промотора PrpsF.
- Штамм Bacillus subtilis АМ891 получен путем введения в хромосому штамма-реципиента Bacillus subtilis B-11156 (В-11156) делеции гена sacB.
- Штамм Bacillus subtilis АМ890 получен путем введения в хромосому штамма-реципиента Bacillus subtilis АМ878 гетерологичного гена udhA E.coli под контролем сильного промотора PrpsF.
- Штамм Bacillus subtilis АМ887 получен путем введения в хромосому штамма-реципиента Bacillus subtilis АМ878 делеции гена sacB.
- Штамм Bacillus subtilis АМ893 получен путем введения в хромосому штамма-реципиента Bacillus subtilis АМ883 делеции гена sacB.
- Штамм Bacillus subtilis АМ895 получен путем введения в хромосому штамма-реципиента Bacillus subtilis АМ890 делеции гена sacB.
Способ в общем виде
Посевную культуру штамма заявляемой бактерии Bacillus subtilis, продуцирующей АИКАР, получают путем выращивания при встряхивании (200-220 об/мин) в течение 18 часов при 37°C на посевной среде следующего состава, мас. %:
Полученной посевной культурой в количестве 5-20% засевают среду для культивирования следующего состава, мас. %:
Культивирование осуществляют в аэробных условиях (900 об/мин) при температуре 20-40°C, предпочтительно в пределах 30-38°C. pH среды поддерживают в пределах 5-9, предпочтительно 6.5-7.2. pH среды регулируют аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Культивирование в течение от 1-3 дней приводит к накоплению целевого продукта - АИКАР в культуральной среде. Способ позволяет накапливать АИКАР как в процессе роста культуры, так и после наращивания биомассы.
Концентрацию АИКАР в освобожденной от клеток культуральной жидкости определяют методом количественной тонкослойной хроматографии на пластинках «Сорбфил».
В качестве альтернативы используют количественный метод ВЭЖХ на хроматографе системы «ALLIANCE» (Separations Module Waters 2695, Photodiode Array detector Waters 2996). Заявляемый способ позволяет синтезировать АИКАР в количестве до 20 г/л.
Пример 1. Конструирование штамма B.subtilis АМ878 путем введения в хромосому штамма-реципиента Bacillus subtilis В-11156 гена zwf под контролем сильного промотора PrpsF
Ген zwf в геноме штамма B.subtilis В-11156 подставляют под контроль сильного промотора PrpsF для усиления экспрессии. Для этого на первом этапе проводят клонирование промоторной области гена rpsF, кодирующего синтез рибосомного белка S6, в экспрессионный вектор pNZT1, способный к репликации как в клетках B.subtilis, так и E.coli и содержащий ген устойчивости к эритромицину (Лобанов и др. Реконструкция пуринового метаболизма у Bacillus subtilis с целью получения штамма-продуцента AICAR - нового препарата широкого терапевтического применения // Acta Naturae Т.3. №2 (9). С. 83-93. 2011). С этой целью с хромосомной ДНК штамма B.subtilis168 trpC2 (American Journal of Botany. 34:345-348, 1947) проводят ПЦР амплификацию с участием фланкирующих промотор гена rpsF праймеров R3 (5′-cccctgcagttgcttatgaggatcttctt-3′), содержащего сайт рестрикции PstI, и R4 (5′-cgcaagcttataatgggcaaggagcaat-3′), содержащего сайт рестрикции HindIII, проводят ПЦР амплификацию промотора гена rpsF. Полученный ПЦР-продукт размером 192 п.н. очищают в агарозном геле, обрабатывают рестриктазами PstI и HindIII и клонируют в векторе pNZT1 по сайтам PstI и HindIII.
На следующем этапе с хромосомной ДНК штамма B.subtilis168 trpC2 проводят ПЦР амплификацию промотор-проксимального участка гена zwf с помощью праймеров Z1(5′-cgcccgcggctccccagaagcccc-3′), содержащего сайт SacII, и Z2 (5′-cgcctgcagtacaccccgtatatttcctt-3′), содержащего сайт PstI. Полученный ПЦР-продукт размером 427 п.н. очищают в агарозном геле, обрабатывают рестриктазами SacII и PstI и клонируют в векторе pNZT1, содержащем промотор rpsF, по сайтам SacII и PstI.
Затем с хромосомной ДНК штамма B.subtilis168 trpC2 проводят ПЦР амплификацию фрагмента рамки считывания гена zwf с помощью праймеров Z3 (5′-cgcaagcttaaatgaaaacaaaccaacaac-caaaag-3′), содержащего сайт HindIII, и Z4 (5′-cgcggtaccctctgattttaatgtttttgc-3′), содержащего сайт KpnI. Полученный ПЦР-продукт размером 403 п.н. очищают в агарозном геле, обрабатывают рестриктазами HindIII и KpnI и клонируют по тем же сайтам в полученный на предыдущем этапе вектор pNZT1, содержащий промотор rpsF и промотор-проксимальную область гена zwf. У полученной в результате проведенных манипуляций плазмидной конструкции pLE8 PrpsF-zwf природный промотор гена zwf замещен на промотор PrpsF.
Сконструированной таким образом плазмидой pLE8 проводят трансформацию штамма E.coli TG1 с отбором трансформантов EmR на среде с эритромицином (300 мкг/мл) при 30°C. Полученные клоны проверяют с помощью ПЦР на присутствие в их клетках плазмиды pNZT1 с клонированным в ее составе фрагментом zwf′-PrpsF-′zwf. Выделенными из проверенных клонов плазмидами pLE8 (zwf′-PrpsF-′zwf) проводят трансформацию в родительский штамм В-11156 с отбором трансформантов EmR на среде с эритромицином (3 мкг/мл). Несколько полученных клонов EmR культивируют на жидкой среде LB с эритромицином (3 мкг/мл) при 37°C в течение ночи, а затем рассевают на чашки со средой LB с эритромицином (3 мкг/мл) и инкубируют в течение 24 ч при 37°C. Полученные рекомбинанты EmR формируются в результате интеграции плазмиды pLE8 - (zwf′-PrpsF-′zwf) в соответствующий хромосомный локус zwf, что подтверждают наработкой ПЦР-фрагмента размером 1002 п.н. при использовании праймеров Z1 и Z4. Несколько полученных клонов EmR засевают в жидкую LB среду без антибиотика и инкубируют на качалке при 30°C в течение 48 часов, а затем рассевают на чашки со средой LB без антибиотика и инкубируют 24 часа при 30°C.
Наконец, на последнем этапе работы выросшие клоны проверяют на выщепление интегрированной плазмиды pLE8, о чем судят по появлению эритромицинчувствительных клонов. Выщепление плазмиды происходит либо с сохранением в составе хромосомы промотора дикого типа гена zwf, либо заменой промотора дикого типа на промотор PrpsF. Интеграцию промотора PrpsF перед геном zwf в хромосому штамма В-11156 подтверждают наработкой ПЦР-фрагмента размером 595 п.н. с участием праймеров R1 и Z4. Один из вариантов штамма В-11156, включивший ген zwf под контролем промотора PrpsF, получивший наименование АМ878, накапливает до 12 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 16 г/л на среде по заявляемому способу (Таблица 2).
Пример 2. Конструирование штамма B.subtilis АМ883 путем введения в геном штамма B.subtilis В-11156 гетерологичного гена udhA E.coli под контролем промотора PrpsF
Для клонирования структурной части гена udhA E.coli под контролем промотора PrpsF используют полученную ранее плазмиду pDG268, содержащую в своем составе модифицированные гетерологичные гены prs и purF E.coli под контролем промотора PrpsF (Лобанов и др. Реконструкция пуринового метаболизма у Bacillus subtilis с целью получения штамма-продуцента AICAR - нового препарата широкого терапевтического применения // Acta Naturae. Т.3. №2(9). С. 83-93. 2011). На первом этапе работы с хромосомной ДНК E.coli (штамм MG1655) проводят ПЦР амплификацию структурной области гена udhA с участием праймеров U1 (5′-cgcggatccaggaggtgcaaacagatgggcctggttaagcaaggt-3′), содержащего сайт рестрикции BamHI, и U2 (5′-cccgcggccgcttaaaacaggcggtttaaaccgt-3′), содержащего сайт рестрикции NotI. Полученный ПЦР-продукт размером 1344 п.н. очищают в агарозном геле, обрабатывают рестриктазами BamHI и NotI и клонируют по тем же сайтам в плазмиду pLE4, содержащую в своем составе модифицированные гетерологичные гены prs и purF E.coli под контролем промотора PrpsF. Важно подчеркнуть, что в 5′-область праймера U1 включена оптимизированная для экспрессии в бациллах нуклеотидная последовательность сайта связывания рибосомы (SD). В результате получают плазмиду pLE5, содержащую ген udhA под контролем промотора PrpsF.
На заключительном этапе работы проводят интеграцию полученной плазмиды pLE5 в хромосомный локус amyE штамма-продуцента В-11156 (см. пример 1). Полученный штамм АМ883 накапливает до 13.3 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 16.9 г/л на среде по заявляемому способу (Таблица 2).
Пример 3. Конструирование штамма B.subtilis АМ891 путем введения в геном штамма B.subtilis В-11156 делеции гена sacB
Для получения делеции гена sacB использовали вектор pNZT1, описанный в примере 1. На первом этапе работы проводят клонирование фрагмента хромосомы, расположенного перед рамкой считывания гена sacB. С этой целью с хромосомной ДНК штамма В.subtilis168 trpC2 проводят ПЦР амплификацию соответствующего фрагмента с участием праймера S4 (5′-cgcccgcggcaactttatgcccatgcaa-3′), содержащего сайт рестрикции SacII, и праймера S5 (5′-cgcctgcagtgcggtagtaaaggttaat-3′), содержащего сайт рестрикции PstI. Полученный ПЦР-продукт размером 430 п.н. очищают в агарозном геле, обрабатывают рестриктазами SacII и PstI и клонируют в векторе pNZT1 по сайтам Pst1 и HindIII.
На втором этапе с хромосомной ДНК штамма B.subtilis168 trpC2 проводят ПЦР амплификацию фрагмента, расположенного сразу за рамкой считывания гена sacB с участием праймера S6 (5′-cgcctgcagcatccttgaacaaggacaa-3′), содержащего сайт рестрикции PstI, и праймера S7 (5′-cgcctcgagacggcatggcgccatt-3′), содержащего сайт рестрикции XhoI. Полученный ПЦР-продукт размером 387 п.н. очищают в агарозном геле, обрабатывают рестриктазами PstI и XhoI и клонируют по сайтам PstI и XhoI в полученный на предыдущем этапе вектор pNZT1, содержащий вставку фрагмента S4-S5.
Сконструированной таким образом плазмидой pLE6 проводят трансформацию в штамм E.coli TG1 с отбором трансформантов EmR на среде с эритромицином (300 мкг/мл) при 30°C. Полученные клоны проверяют с помощью ПЦР на присутствие в клетках плазмиды pLE6 с клонированными в ее составе фрагментами S4-S5 и S6-S7. Выделенными из проверенных клонов плазмидами проводят трансформацию в штамм В.subtilis ВКПМ В-11156 с отбором трансформантов EmR на среде с эритромицином (3 мкг/мл). Несколько полученных клонов EmR культивируют на жидкой среде LB с эритромицином (3 мкг/мл) при 37°C в течение ночи, а затем рассевают на чашки со средой LB с эритромицином (3 мкг/мл) и инкубируют в течение 24 ч при 37°C. Полученные рекомбинанты EmR сформированы в результате гомологичной рекомбинации между фланкирующими ген sacB последовательностями S4-S5 и S6-S7 и хромосомой. Несколько полученных клонов EmR засевают в жидкую LB среду без антибиотика и инкубируют на качалке при 30°C в течение 48 часов, а затем рассевают на чашки со средой LB без антибиотика и инкубируют 24 часа при 30°C.
Выросшие клоны на последнем этапе работы проверяют на выщепление интегрированной плазмиды pLE6, о чем судят по появлению эритромицинчувствительных клонов. Выщепление плазмиды происходит либо с сохранением в составе хромосомы дикого аллеля гена sacB, либо с включением делеции этого гена. Интеграцию делеции гена sacB в хромосому штамма В.subtilis ВКПМ В-11156 подтверждают наработкой ПЦР-фрагмента размером 817 п.н. с участием праймеров S4 и S7, тогда как в случае штамма дикого типа sacB+ получают ПЦР-фрагмент размером 1776 п.н. Один из вариантов штамма B.subtilis ВКПМ В-11156, включающий делецию гена sacB и получивший наименование АМ891, накапливает до 14,6 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 18,2 г/л на среде по заявляемому способу (Таблица 2).
Пример 4. Конструирование штамма B.subtilis АМ890 путем введения в геном штамма B.subtilis АМ878 гетерологичного гена udhA E.coli под контролем промотора
Для получения штамма B.subtilis, характеризующегося наличием гена zwf под контролем промотора PrpsF и одновременно содержащего гетерологичный ген udhA E.coli под контролем промотора PrpsF, осуществляют необходимые действия аналогично примеру 2.
На заключительном этапе работы проводят интеграцию полученной плазмиды pLE5 в хромосомный локус amyE штамма-продуцента АМ878 (см. пример 2). Полученный штамм АМ890 накапливает до 13.0 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 18.0 г/л на среде по заявляемому способу (Таблица 2).
Пример 5. Конструирование штамма B.subtilis АМ887 путем введения в геном штамма B.subtilis АМ878 делеции гена sacB
Для получения штамма B.subtilis, характеризующегося наличием гена zwf под контролем промотора PrpsF и одновременно содержащего делецию гена sacB, осуществляют необходимые действия аналогично примеру 3.
Сначала проводят трансформацию штамма АМ878 плазмидой pLE6 с отбором трансформантов EmR на среде с эритромицином (3 мкг/мл) при 30°C. Затем осуществляют выщепление интегрированной плазмиды pLE6, сопровождающееся сохранением делеции гена sacB в составе хромосомы (см. пример 3). Один из вариантов штамма АМ878, включивший делецию гена sacB, получивший наименование АМ887, накапливает до 14,9 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 19,3 г/л на среде по заявляемому способу (Таблица 2).
Пример 6. Конструирование штамма B.subtilis АМ893 путем введения в геном штамма B.subtilis АМ883 делеции гена sacB
Для получения штамма B.subtilis, характеризующегося наличием гетерологичного гена udhA E.coli под контролем промотора PrpsF и одновременно содержащего делецию гена sacB, осуществляют необходимые действия аналогично примеру 3.
После трансформации штамма АМ883 плазмидой pLE6 и ее последующего выщепления проводят проверку на включение делеции гена sacB в хромосому (см. пример 3). Один из вариантов штамма АМ883, включивший делецию гена sacB, получивший наименование АМ893, накапливает до 13,9 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 17,9 г/л на среде по заявляемому способу (Таблица 2).
Пример 7. Конструирование штамма B.subtilis АМ895 путем введения в геном штамма B.subtilis АМ890 делеции гена sacB
Для получения штамма B.subtilis, характеризующегося наличием гена zwf и гетерологичного гена udhA E.coli под контролем промотора PrpsF, а также делецией собственного гена sacB, осуществляют необходимые действия аналогично примеру 3. Однако в отличие от примера 3, в качестве реципиентного штамма для трансформации плазмидой pLE6 используют штамм АМ890, получение которого описано в примере 4.
Один из вариантов штамма АМ890, включивший делецию гена sacB, получивший наименование АМ895, накапливает до 15,1 г/л AICAR в культуральной жидкости при ферментации на контрольной среде (RU 2405833) (Таблица 1) и до 20,1 г/л на среде по заявляемому способу (Таблица 2).
Пример 8. Уровень синтеза АИКАР у полученных штаммов при культивировании на контрольной среде, известной из патента RU 2405833
Полученные штаммы с дерегулированным pur-опероном на фоне инактивированного гена purH, содержащие в составе хромосомы модифицированные гетерологичные гены prs и purF E.coli под контролем сильного промотора ΡrpsF и отличающиеся тем, что включают одну или комбинации заявленных характеристик, проверяют на способность синтезировать АИКАР в культуральной жидкости при культивировании в ферментерах Marubishi с использованием контрольной среды. Условия выращивания посевного материала и состав сред приведены выше. Результаты представлены в таблице 1.

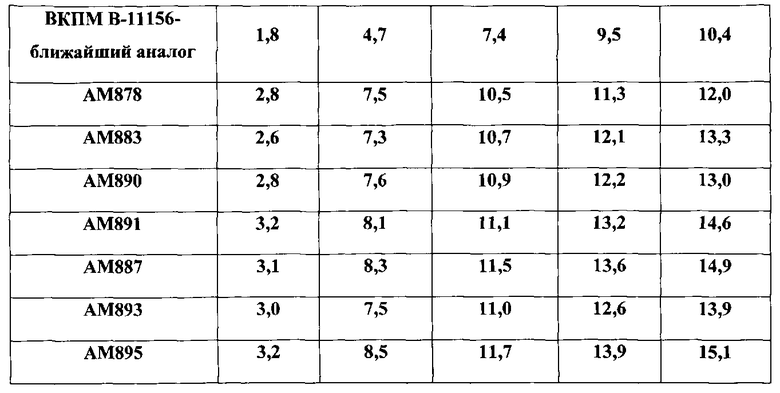
Как следует из данных таблицы 1, штаммы, содержащие любую из заявленных характеристик, обнаруживают более высокий уровень накопления АИКАР в культуральной жидкости по сравнению с контрольным штаммом ближайшего аналога B.subtilis ВКПМ В-11156. Среди штаммов, содержащих комбинацию из двух заявленных характеристик, максимальный уровень синтеза АИКАР (до 14,9 г/л) наблюдается у штамма АМ887, в геноме которого присутствует ген zwf под контролем промотора PrpsF и делеция гена sacB. По продуктивности его превосходит штамм АМ895, накапливающий 15,1 г/л АИКАР, в котором сочетаются все три заявленные характеристики.
Пример 9. Уровень синтеза АИКАР у полученных штаммов при культивировании на среде по заявляемому способу
Уровень синтеза АИКАР у штаммов изучен в тех же условиях, что приведены в примере 8, но с использованием среды по заявляемому способу. Результаты приведены в таблице 2.

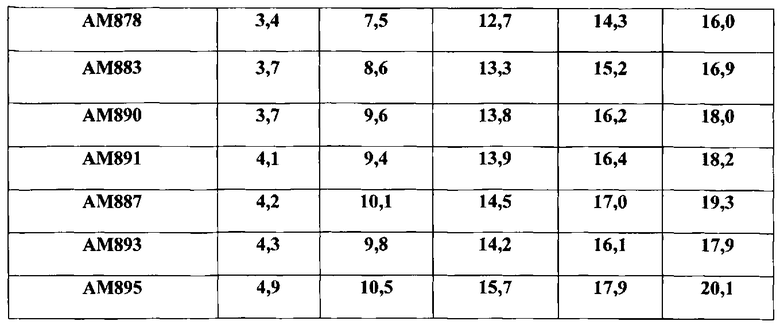
Данные, представленные в таблице 2, свидетельствуют о том, что использование среды по заявляемому способу приводит к заметному повышению уровня синтеза АИКАР как у контрольного штамма ближайшего аналога, так и у всех сконструированных штаммов. Максимальный уровень синтеза АИКАР, как и в случае использования контрольной среды, обнаруживают штаммы АМ887 и АМ895, уровень синтеза АИКАР у которых составляет соответственно 19,3 г/л и 20,1 г/л.
Таким образом, путем поэтапного генетического конструирования, направленного на достижение максимальной экспрессии генов пуринового оперона и усиление метаболического потока предшественников АИКАР от глюкозы до рибозы и фосфорибозилпирофосфата, сконструированы штаммы B.subtilis, превосходящие штамм-ближайший аналог по уровню синтеза АИКАР, а также разработан способ культивирования таких штаммов, позволяющий достичь уровня синтеза АИКАР, равного 19,3 г/л и 20,1 г/л.
Высокий уровень гомологии генетического аппарата и универсальность биохимических процессов биосинтеза пуринов у различных представителей рода Bacillus являются основанием для экстраполяции разработанной схемы получения штаммов-продуцентов АИКАР и на другие виды этого рода микроорганизмов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ПУРИНОВОГО НУКЛЕОЗИДА 5'-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИДА (АИКАР) И ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ АИКАР | 2008 |
|
RU2405833C2 |
| БАКТЕРИЯ РОДА BACILLUS, ПРОДУЦИРУЮЩАЯ ГИАЛУРОНОВУЮ КИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ ГИАЛУРОНОВОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ УКАЗАННОЙ БАКТЕРИИ | 2019 |
|
RU2719140C1 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia,- ПРОДУЦЕНТ L-ГИСТИДИНА И СПОСОБ ПОЛУЧЕНИЯ L-ГИСТИДИНА | 2003 |
|
RU2276688C2 |
| БЕЛОК YdhL ИЗ Bacillus amyloliquefaciens, ФРАГМЕНТ ДНК, БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia ИЛИ Bacillus, - ПРОДУЦЕНТ ПУРИНОВЫХ НУКЛЕОЗИДОВ, СПОСОБ ПОЛУЧЕНИЯ ПУРИНОВЫХ НУКЛЕОЗИДОВ И НУКЛЕОТИДОВ | 2005 |
|
RU2294962C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОЗИДОВ И НУКЛЕОТИДОВ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ Escherichia ИЛИ Bacillus | 2006 |
|
RU2365622C2 |
| СПОСОБ ПОЛУЧЕНИЯ РИБОФЛАВИНА, ШТАММ BACILLUS SUBTILIS - ПРОДУЦЕНТ РИБОФЛАВИНА (ВАРИАНТЫ) | 2002 |
|
RU2261273C2 |
| Фрагмент ДНК ALV 7, кодирующий синтез альфа-амилазы, способ его конструирования и штамм бактерий BacILLUS SUвтILIS - продуцент альфа-амилазы | 1990 |
|
SU1717633A1 |
| Плазмида для редактирования генома бактерий рода Bacillus и способ внесения модификаций в геном бактерий рода Bacillus | 2022 |
|
RU2815835C1 |
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |
Группа изобретений относится к биотехнологии. Предложена бактерия Bacillus subtilis, продуцирующая 5′-аминоимидазол-4-карбоксамидрибозид (АИКАР). Указанная бактерия содержит дерегулированный pur-оперон на фоне инактивированного гена purH, модифицированные гетерологичные гены prs и purF E.coli под контролем сильного промотора PrpsF в составе хромосомы. При этом она обладает по крайней мере одной из следующих характеристик: содержит ген zwf под контролем сильного промотора PrpsF, содержит гетерологичный ген udhA под контролем сильного промотора PrpsF, содержит делецию гена sacB. Предложен также способ синтеза АИКАР путем культивирования в соответствующих условиях указанной бактерии. При этом культивирование осуществляют на среде следующего состава, мас. %: кормовые дрожжи 0,5-1,0, сахароза 10-13, изолят сои 2,5-3,5, кукурузный экстракт 3,0-5,0, мочевина 0,6-0,8, (ΝΗ4)2ΗΡO4 0,8-1,6, пропинол 0,4-0,5, вода - остальное. Группа изобретений обеспечивает высокий уровень синтеза АИКАР до 20 г/л. 2 н. и 3 з.п. ф-лы, 2 табл., 9 пр.
1. Бактерия Bacillus subtilis, продуцирующая 5′-аминоимидазол-4-карбоксамидрибозид, содержащая дерегулированный pur-оперон на фоне инактивированного гена purH, модифицированные гетерологичные гены prs и purF E.coli под контролем сильного промотора PrpsF в составе хромосомы, и отличающаяся по крайней мере одной из следующих характеристик:
- содержит ген zwf под контролем сильного промотора PrpsF,
- содержит гетерологичный ген udhA под контролем сильного промотора PrpsF,
- содержит делецию гена sacB.
2. Бактерия по п. 1, представляющая собой штамм Bacillus subtilis АМ878, продуцирующий 5′-аминоимидазол-4-карбоксамидрибозид и полученный путем введения в хромосому штамма-реципиента Bacillus subtilis ВКПМ В-11156 гена zwf под контролем сильного промотора PrpsF.
3. Бактерия по п. 1, представляющая собой штамм Bacillus subtilis АМ890, продуцирующий 5′-аминоимидазол-4-карбоксамидрибозид и полученный путем введения в хромосому штамма-реципиента Bacillus subtilis АМ878 гетерологичного гена udhA E.coli под контролем сильного промотора PrpsF.
4. Бактерия по п. 1, представляющая собой штамм Bacillus subtilis АМ895, продуцирующий 5′-аминоимидазол-4-карбоксамидрибозид и полученный путем введения в хромосому штамма-реципиента Bacillus subtilis АМ890 делеции гена sacB.
5. Способ микробиологического синтеза пуринового нуклеозида 5′-аминоимидазол-4-карбоксамидрибозида путем культивирования в соответствующих условиях бактерии по п. 1 на среде следующего состава, мас. %:
| ЛОБАНОВ К.В | |||
| И ДР | |||
| Реконструкция пуринового метаболизма у Bacillus subtilis с целью получения штамма-продуцента AICAR - нового препарата широкого терапевтического применения // ACTA NATURAE, 2011, т | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Пуговица | 0 |
|
SU83A1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ПУРИНОВОГО НУКЛЕОЗИДА 5'-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИДА (АИКАР) И ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ АИКАР | 2008 |
|
RU2405833C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПУРИНОВЫХ РИБОНУКЛЕОЗИДОВ И РИБОНУКЛЕОТИДОВ | 2008 |
|
RU2422510C2 |
| US 20040259212 А1, 23.12.2004 | |||

