Область техники
Настоящее изобретение относится к биотехнологии, в частности к бактерии рода Bacillus, в частности, бактерии Bacillus subtilis, продуцирующей гиалуроновую кислоту, а также к способу микробиологического синтеза гиалуроновой кислоты путем культивирования такой бактерии.
Предшествующий уровень техники
Раскрытие функций и механизмов биологического действия ряда биополимеров способствует созданию все новых продуктов и препаратов на их основе. Одним из таких биополимеров животного происхождения является гиалуроновая кислота. Гиалуроновая кислота, гиалуронат, или гиалуронан - (C14H21NO11)n - органическое соединение, относящееся к группе несульфатированных гликозоаминогликанов. Наличие многочисленных сульфатированных групп у родственных гликозоаминогликанов является причиной существования многочисленных разнообразных изомеров, чего не наблюдается у гиалуроновой кислоты, которая всегда химически идентична, вне зависимости от методов и источников получения. Молекула гиалуроновой кислоты построена из повторяющихся фрагментов D-глюкуроновой кислоты и N-ацетил-Dглюкозоамина, соединенных β-(1-3) гликозидной связью. Основы фрагментов сахаров - это глюкопиранозное кольцо с различными заместителями (ацетамидная группа, гидроксильные и карбоксильные функциональные группы). Для молекулы гиалуроновой кислоты характерно образование большого количества водородных связей как внутри молекулы, так и между соседними углеводными остатками, находящимися на значительном расстоянии друг от друга, а в водном растворе даже между соседними молекулами через карбоксил и ацетамидную группу. Имеет кислую реакцию среды ввиду наличия непротонированной карбоксильной группы. Кислотные свойства гиалуроновой кислоты позволяют получать растворимые в воде соли с щелочными металлами. В организме гиалуроновая кислота присутствует в виде натриевой соли – гиалуроната. Гиалуроновая кислота – это анионный линейный полисахарид с различной молекулярной массой. Молекулярная масса зависит от способа получения, причем, ввиду отсутствия изомерии, получаемый гиалуронат всегда химически идентичен стандартному.
Определены многочисленные функции гиалуроната в организме. Гиалуронат присутствует в гиалиновом хряще, синовиальной суставной жидкости и кожной ткани как в дерме, так и в эпидермисе. Гиалуронат также участвует во многих физиологических функциях, таких как адгезия, развитие, клеточная подвижность, канцерогенез, ангиогенез и заживление ран. Вследствие уникальных физических и биологических свойств гиалуронат используется в глазной и суставной хирургии и разрабатывается для применения в других медицинских процедурах. Препараты гиалуроната также разработаны для применения в ортопедии, ревматологии и дерматологии. В качестве активного ингредиента гиалуронат входит в состав многих косметических композиций.
Все известные способы получения гиалуроновой кислоты можно разделить на две группы: физико-химический метод, который заключается в экстрагировании гиалуроната из тканей животного сырья млекопитающих и птиц, например, из стекловидного тела глаза крупного рогатого скота, гребней кур или пупочных канатиков новорожденных, и микробный метод получения гиалуроната на основе бактерийпродуцентов.
Ранее наибольшее распространение, в силу доступности сырья и высокого содержания биополимера, получил метод выделения гиалуроновой кислоты из петушиных гребней. Экстракция производится смесью ацетона с хлороформом (удаление белка), водой, либо водноспиртовой смесью (пропионовый, трет-бутиловый спирты) с последующей сорбцией на активированном угле, посредством электрофореза или на ионообменной смоле. При анализе животных источников гиалуроната становится очевидным, что сырьевая база промышленного получения данного полисахарида весьма ограничена и не может полностью удовлетворить постоянно растущий спрос на гиалуронат. Кроме того, выделение гиалуроната из животного сырья часто осложняется тем, что в тканях и органах млекопитающих и птиц биополимер находится в комплексе с белками; присутствие родственных гликозаминогликанов также затрудняет выделение гиалуроната. Еще одним потенциальным риском использования препаратов животного происхождения является их контаминация нуклеиновыми кислотами, прионами и вирусами.
Этих недостатков лишен биотехнологический способ получения гиалуроната, основанный на культивировании микроорганизмовпродуцентов гиалуроната. Сырьем для получения полисахарида микробиологическим синтезом являются доступные соединения и компоненты: глюкоза, дрожжевой экстракт, минеральные соли и т. п. Производство гиалуроната на основе микробиологического синтеза не зависит от поставок животного сырья, а также легко масштабируется и перепрофилируется в случае изменения конъюнктуры рынка. В международных публикациях WO 99/51265 и WO 00/27437 описаны способы ферментации для получения гиалуроновой кислоты с использованием штамма Pasteurella multocida. В патенте US 4801539 и в международной публикации WO 99/23227 описаны способы ферментации для получения гиалуроновой кислоты с использованием штаммов Streptococcus zooepidemicus и Streptococcus equisimilis с приведенными выходами, составляющими примерно 3,6 г гиалуроновой кислоты на литр. В Европейских патентах EP 0694616 и EP 2046969 описаны способы ферментации, использующие улучшенный штамм Streptococcus zooepidemicus с приведенными выходами, составляющими примерно 3,5 г гиалуроновой кислоты на литр.
Микроорганизмы, используемые для получения гиалуроновой кислоты путем ферментации, представляют собой штаммы патогенных бактерий, основные из которых представляют собой различные виды Streptococcus. Стрептококки группы A и группы C окружают себя неантигенной капсулой, состоящей из гиалуроната, который по композиции идентичен тому, который обнаружен в соединительной ткани и суставах
Ключевым и последним ферментом на пути биосинтеза гиалуроновой кислоты является гиалуронансинтаза. Описаны гиалуронансинтазы позвоночных, бактериальных патогенов и вирусов водорослей (DeAngelis, P. L., 1999, Cell. Mol. Life Sci, 56: 670682). В международной публикации WO 99/23227 описана гиалуронатсинтаза группы I Streptococcus equisimilis. В международных публикациях WO 99/51265 и WO 00/27437 описана гиалуронатсинтаза группы II Pasteurella multocida. Описан оперон гиалуронансинтазы Streptococcus pyogenes, который состоит из трех генов hasA, hasB и hasC, кодирующие гиалуронатсинтазу, UDPглюкозодегидрогеназу и UDP-глюкозопирофосфорилазу, соответственно (Proc. Natl. Acad. Sci. USA. 98, 4658-4663,2001). В международной публикации WO 99/51265 описан сегмент нуклеиновой кислоты, содержащий кодирующую область гиалуронатсинтазы Streptococcus equisimilis.
После идентификации генов, вовлеченных в биосинтез гиалуроната, оказалось возможным проводить генетическую модификацию большой группы бактерий (Bacillus, Agrobacterium, E. coli and Lactococcus) с целью их использования в качестве продуцентов гиалуроната (WO 2012/032153, Appl. Microbiol. Biotechnol. 84:63–69, 2009).
Перспективные для продукции гиалуроната результаты получены при создании рекомбинантных штаммов бацилл B. subtilis (публикация заявки US 2014/0038235 и патент RU 2346049 компании Novozymes). Экспрессионная система таких штаммов содержит гетерологичный ген hasA, клонированный из S. equisimilis и входящий в единый оперон с одним или несколькими природными генами B. subtilis (гомологами генов hasB, hasC и hasD Streptococcus). Преимуществами этих продуцентов является то обстоятельство, что получаемый продукт не содержит экзотоксинов и эндотоксинов стрептококков.
Хотя продукция гиалуроната была существенно улучшена путем получения вышеуказанных штаммов микроорганизмов или улучшения процесса его получения, необходимо разрабатывать более эффективные процессы получения гиалуроната для того, чтобы удовлетворить растущие потребности в этом соединении для нужд медицины, ветеринарии и косметологии.
Раскрытие изобретения
Технической задачей настоящей группы изобретений являлась разработка более продуктивного способа микробиологического синтеза гиалуроновой кислоты.
Указанная задача была решена путем конструирования бактерии рода Bacillus, в частности B. subtilis, продуцирующей гиалуроновую кислоту, которая содержит в составе хромосомы слияние гетерологичного гена hasA из S. equisimilis с геном tuaD B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB и которая дополнительно необязательно содержит в составе хромосомы :
- ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB; и/или
- оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
Указанная задача также была решена путем разработки способа микробиологического синтеза гиалуроновой кислоты путем культивирования указанной бактерии в питательной среде с последующим выделением гиалуроновой кислоты из культуральной жидкости.
Настоящее изобретение относится к бактерии рода Bacillus, в частности B. subtilis, продуцирующей гиалуроновую кислоту, которая содержит в составе хромосомы слияние гетерологичного гена hasA из S. equisimilis с геном tuaD из B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB и дополнительно необязательно содержит в составе хромосомы :
- ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB; и/или
- оперон gcaD-prs под контролем гибридного тандемного промотора
PrpsF-gsiB.
Также настоящее изобретение относится к описанной выше бактерии, содержащей в хромосоме ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB.
Также настоящее изобретение относится к описанной выше бактерии, содержащей в хромосоме оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
Также настоящее изобретение относится к описанной выше бактерии, содержащей в хромосоме ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB и оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
Также настоящее изобретение относится к описанной выше бактерии, которая представляет собой B. subtilis.
Также настоящее изобретение относится к способу получения гиалуроновой кислоты, включающему:
а) культивирование описанной выше бактерии в питательной среде,
пригодной для продукции гиалуроновой кислоты, и
б) выделение гиалуроновой кислоты из культуральной жидкости.
Также настоящее изобретение относится к способу получения гиалуроновой кислоты, в котором используют питательную среду следующего состава, мас. %:
Сахароза – 8,0;
Мальтоза – 4,0;
Дрожжевой экстракт – 0,5;
Мочевина – 0,6;
KH2PO4 – 0,6;
K2HPO4 – 1,4;
(NH4)2S04 – 0,3;
Na-цитрат – 0,2;
Казаминовые кислоты – 0.04;
MgSO4 – 0.3.
Краткое описание чертежей
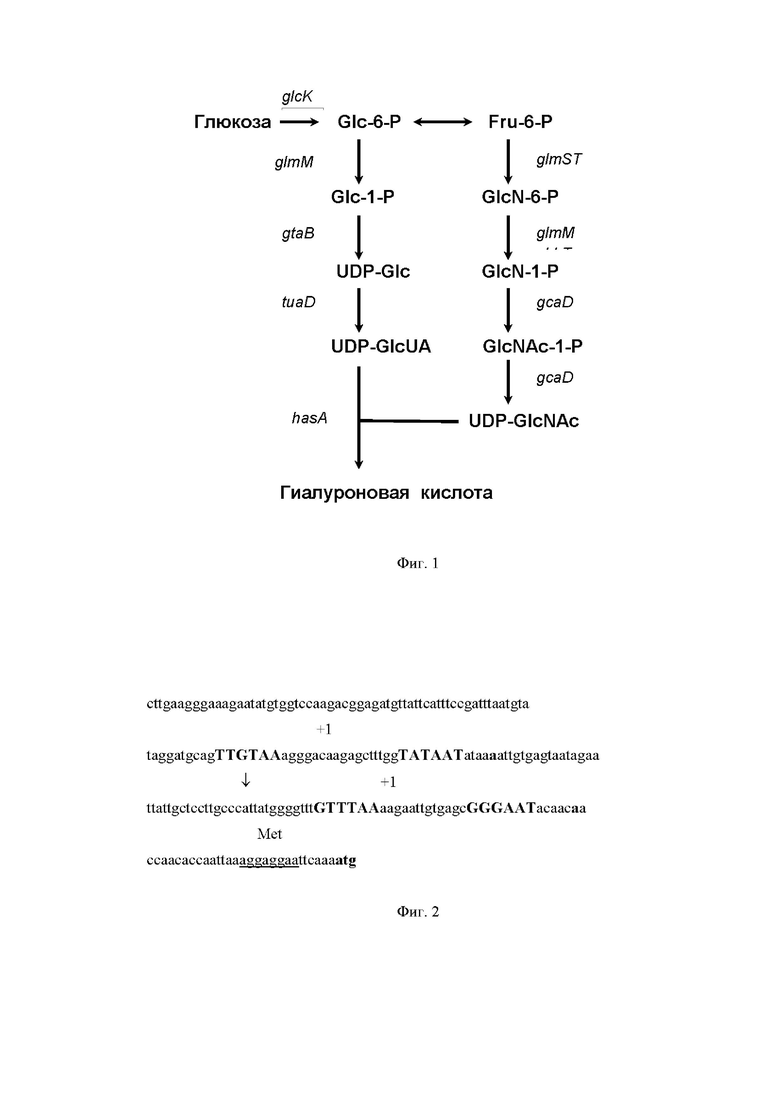
На Фиг. 1 показана схема биосинтеза гиалуроновой кислоты. Указаны гены и контролируемые ими реакции биосинтеза гиалуроновой кислоты.
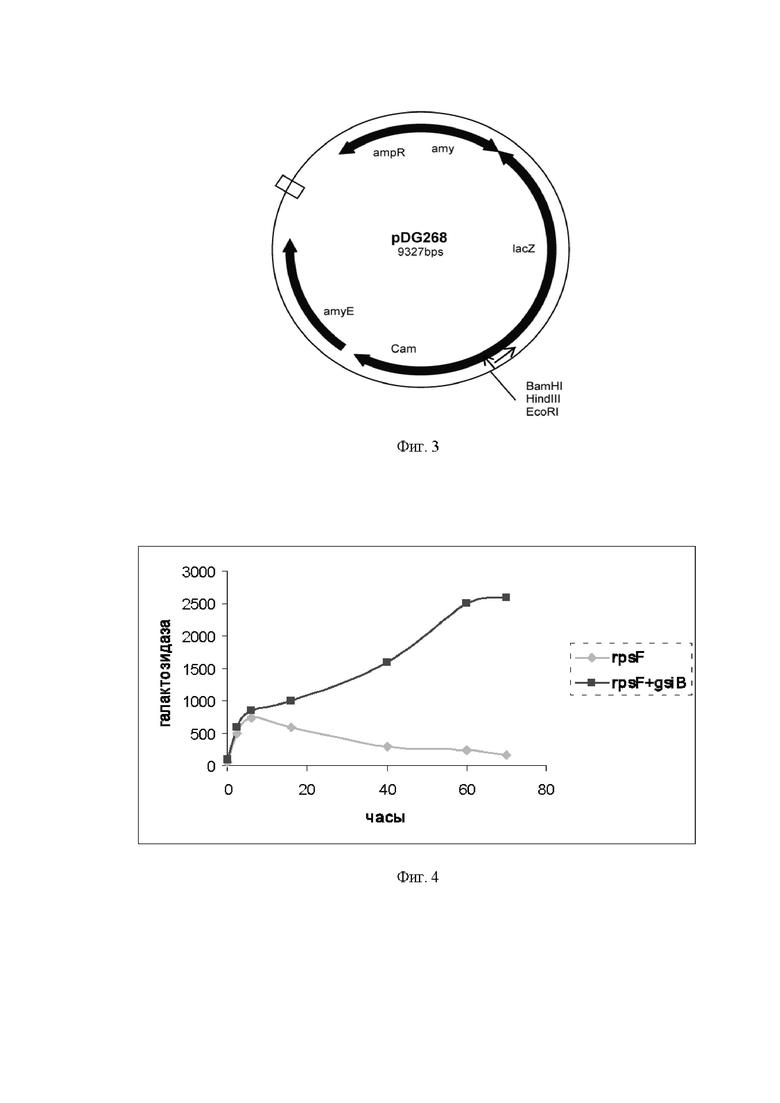
На Фиг. 2 показана нуклеотидная последовательность гибридного промотора PrpsF-gsiB. Заглавными жирными буквами обозначены -35 и -10 участки промоторов, подчеркнут сайт связывания рибосомы. Старты транскрипции обозначены как +1. Место стыка промоторов PrpsF и PgsiB обозначено стрелкой.
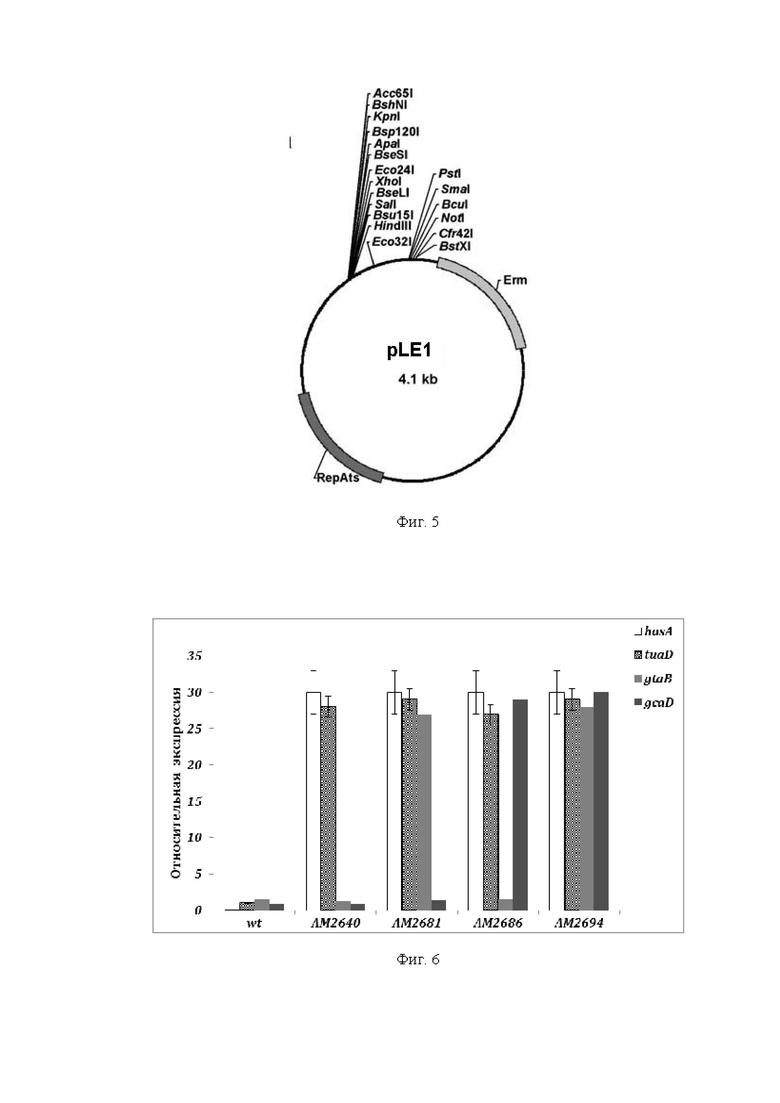
На Фиг. 3 показана структура плазмиды pDG268.
На Фиг. 4 показано сравнение эффективности функционирования гибридного промотора PrpsF-gsiB и одиночного промотора PrpsF в процессе ферментации в штамме B. subtilis на примере репортерного гена lacZ, кодирующего β-галактозидазу.
На Фиг. 5 показана структура вектора pLE1.
На Фиг. 6 показаны относительные уровни транскрипции генов hasA, tuaD, gtaB и gcaD в штаммах АM2640, AM2681, AM2686 и АМ2694.
Осуществление изобретения
Термин «бактерия рода Bacillus» означает бактерию, которую относят к роду Bacillus в соответствии с классификацией, известной для специалиста в области микробиологии. В частности, к бактерии рода Bacillus относится бактерия, выбранная из группы, включающей Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis и Bacillus thuringiensis.
Известно, что род Bacillus включает в себя больше 200 видов бактерий, наиболее практически важные из которых, кроме общих морфологических, физиологических культуральных свойств, отличаются определенным сходством в организации генома и механизмах экспрессии генов. В прямых экспериментах показано, что, например, промотор гена внеклеточной протеиназы aprE B. subtilis способен инициировать синтез репортерного белка в клетках бацилл различного вида, иногда даже в большей степени, чем в клетках B. subtilis (Genome Biology, 5, 77, 2004).
Термин «бактерия Bacillus subtilis» означает бактерию, которую относят к виду Bacillus subtilis в соответствии с классификацией, известной для специалиста в области микробиологии.
Термин «гиалуроновая кислота» определяется здесь как несульфатированный глюкозаминогликан, состоящий из повторяющихся дисахаридных единиц N-ацетилглюкозамина (GlcNAc) и глюкуроновой кислоты (GlcUA), связанных вместе перемежающимися бета-1,4- и бета1,3-гликозидными связями. Натриевая соль гиалуроновой кислоты также известна как гиалуронан, или гиалуронат.
Термин «гиалуронансинтаза» определяется здесь как синтаза (КФ 2.4.1.212), которая катализирует удлинение цепи гиалуронана путем добавления сахарных предшественников GlcUA и GlcNAc. Для продукции гиалуроновой кислоты в клетках рода Bacillus, в частности, B. subtilis в настоящем изобретении используется слияние гетерологичного гена hasA из S. equisimilis с геном tuaD из B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB.
Гены, участвующие в биосинтезе предшественников сахара для получения гиалуроновой кислоты, включают ген tuaD B. subtilis, кодирующий UDP-глюкозо-6-дегидрогеназу (КФ1.1.1.22), ген gtaB B. subtilis, кодируюший UDP-глюкозопирофосфорилазу (КФ 2.7.7.9), ген gcaD B. subtilis, кодирующий UDP-N-ацетилгюкозамин-пирофосфорилазу (КФ 2.7.7.23), и ген prs, кодирующий фосфорибозилпирофосфат синтазу (КФ 2.7.6.1) (см. Фиг. 1).
Термин «гетерологичный ген», означает, что указанный ген выделен из хромосомы организма, отличного от бактерии рода Bacillus, в частности, B. subtilis. Бактерию, содержащую гетерологичный ген hasA из S. equisimilis в хромосоме, получают стандартными методами рекомбинантных ДНК, в частности трансформацией, трансфекцией.
Помимо гетерологичного гена hasA Streptococcus equisimilis клетка-хозяин рода Bacillus, в частности, Bacillus subtilis может дополнительно содержать одну или более вторых конструкций нуклеиновой кислоты, содержащих один или более генов, кодирующих ферменты биосинтеза предшественника сахара под контролем гибридного тандемного промотора PrpsF-gsiB
Термины «тандемный промотор PrpsF-gsiB функционально связан» с целевым геном или «ген находится под контролем гибридного тандемного промотора PrpsF-gsiB» означает, что природные промоторы указанных генов заменены на промотор PrpsF-gsiB. Тандемный промотор PrpsF-gsiB представляет из себя слияние промотора гена rpsF, кодирующего синтез рибосомного белка S6, и промотора гена gsiB, ответственного за белок общего клеточного стресса (РgsiB). Промотор PrpsF содержит близкую к консенсусной последовательности область -35 (TTGTAA), консенсусную последовательность области -10 (TATAAT). Промотор PrpsF относится к наиболее сильным промоторам в геноме B. subtilis и обеспечивает максимально высокий уровень транскрипции на логарифмической стадии роста бактерий. Инициация транскрипции этого промотора осуществляется с участием вегетативной сигма-субъединицы РНКполимеразы (ϭA). Нуклеотидная последовательность гена rpsF В. subtilis представлена в базе данных «GenBank» под номером GeneID: 937919 (нуклеотиды с 4199445 по 4199732 в последовательности NC_000964.3). Инициация транскрипции гена gsiB инициируется с участием альтернативной сигма-субъединицы ϭB и достигает максимума на стационарной фазе роста бактерий. Нуклеотидная последовательность гена gsiB В. subtilis представлена в базе данных «GenBank» под номером GeneID: 938235 (нуклеотиды с 494506 по 494877 в последовательности NC_000964.3).
Нуклеотидная последовательность тандемного промотора PrpsF-gsiB, используемого в настоящем изобретении, показана на Фиг. 2 и приведена в Перечне последовательностей под номером SEQ ID NO: 1.
Указанный тандемный промотор PrpsF-gsiB может быть получен любым способом, известным специалисту в данной области техники, в частности путём химического синтеза, методом ПЦР с использованием перекрывающихся праймеров, слияния PrpsF и РgsiB и подобными методами.
Авторами настоящего изобретения показано, что использование гибридного промотора PrpsF-gsiB позволяет получать оптимальный профиль экспрессии целевого гена в ходе культивирования бактерии и представляется весьма перспективным для усиления экспрессии целевых генов, используемых для продукции целевых продуктов, в частности, гиалуроновой кислоты. Так, активность гибридного промотора PrpsF-gsiB в процессе ферментации нарастает и достигает максимума на поздних стадиях стационарного роста. Указанный профиль активности гибридного промотора PrpsF-gsiB, а также сравнение эффективности функционирования гибридного промотора PrpsF-gsiB и одиночного промотора PrpsF в процессе ферментации штаммов B. subtilis показаны на Фиг. 4 на примере репортерного гена lacZ, кодирующего β-галактозидазу.
В рамках настоящего изобретения на основе исходного штамма B. subtilis 168 был получен ряд конструкций, а именно, AM2640, AM2681, AM2686, AM2694, продуцирующих гиалуроновую кислоту. Более детальное описание генетических конструкций и способы их получения подробно описаны в последующих примерах.
Характеристика генетических конструкций
Сконструированные генетические конструкции на основе Bacillus subtilis являются суперпродуцентами гиалуроновой кислоты и имеют следующие свойства.
Культурально-морфологические признаки.
Спорообразующая грамположительная палочка с закругленными концами и краевым расположением спор. При рассеве на LB-агаре (Миллер Дж. 1979. Эксперименты в молекулярной генетике. Мир) и культивировании при 37°C на вторые сутки образует колонии с неровным краем диаметром 3-4 мм, а на минимальной среде Спицайзена (J.
Bacteriol. 81:741-746, 1961) такие же колонии диаметром 2-3 мм.
Физиолого-биохимические признаки.
Сбраживает сахариды, такие как глюкоза, лактоза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза и гидролизат крахмала; спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая, фумаровая, лимонная и янтарная кислоты, и подобные соединения.
Хранение.
В лиофилизированном виде или в парах жидкого азота. Стабильность.
Полученные генетические конструкции на основе B. subtilis стабильны и не теряют способности синтезировать гиалуроновую кислоту после 10 пересевов на полноценной среде.
Способ согласно настоящему изобретению.
К способам согласно настоящему изобретению относится способ получения гиалуроновой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления целевого продукта в указанной питательной среде, и выделения целевого продукта из культуральной жидкости.
Согласно настоящему изобретению выращивание, сбор и очистка целевого продукта из культуральной или подобной ей жидкости могут быть осуществлены способом, подобным традиционным способам культивирования, в которых целевой продукт продуцируется микроорганизмом. Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, которые требуются микроорганизму для роста. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, различные органические кислоты или такие спирты, как этанол и глицерин. В качестве источника азота могут использоваться различные соли аммония, такие как сульфат аммония, другие соединения азота, такие как аммиак, амины, природные источники азота, такие как пептон и ферментолизаты микроорганизмов. В качестве минеральных добавок используются монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные соединения.
Культивирование осуществляют в аэробных условиях с перемешиванием (например, путем встряхивания, с использованием мешалки или барботирования и т.д.) при температуре в пределах 3038°C, предпочтительно при 37°C. pH среды поддерживают в пределах 59, предпочтительно 6.5-7.2. pH среды регулируют буферными растворами или добавлением аммиака, карбоната кальция, различных кислот, оснований. Культивирование в течение от 1 до 3 суток приводит к накоплению целевого продукта – гиалуроновой кислоты в культуральной среде. Способ позволяет накапливать гиалуроновую кислоту как в процессе роста культуры, так и после наращивания биомассы в стационарной фазе роста.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем гиалуроновая кислота может быть собрана и очищена путем ступенчатого осаждения цетилпиридинием и изопропиловым спиртом с последующей очисткой методом колоночной хроматографии на адсорбционных и ионообменных носителях. На последних стадиях выделения также может применяться ультрафильтрация и лиофилизация.
Концентрацию гиалуроновой кислоты в освобожденной от клеток культуральной жидкости определяют карбазольным методом (T. Bitter, H.M. Muir. Analytical Biochemistry, v.4, p. 330-334, 1962).
Настоящее изобретение более детально описано со ссылкой на примеры.
Последующие примеры приведены для целей разъяснения сущности настоящего изобретения и не ограничивают каким-либо образом объём правовой охраны, определяемый формулой настоящего изобретения.
Примеры
________
Пример 1. Конструирование плазмиды pDG268-P, содержащей гибридный промотор PrpsF-gsiB.
В настоящем изобретении используется сконструированный тандемный промотор, функционирующий в бактерии рода Bacillus, в частности, в B. subtilis, содержащий последовательности нуклеотидов промотора гена rpsF и промотора гена gsiB, причем промотор rpsF расположен выше по ходу транскрипции относительно промотора gsiB. Получение указанного промотора описано в находящейся на рассмотрении заявке, озаглавленной «ТАНДЕМНЫЙ ПРОМОТОР, ФУНКЦИОНИРУЮЩИЙ В БАКТЕРИИ РОДА Bacillus, БАКТЕРИЯ РОДА Bacillus - ПРОДУЦЕНТ ПОЛЕЗНОГО МЕТАБОЛИТА, СОДЕРЖАЩАЯ УКАЗАННЫЙ ПРОМОТОР, И СПОСОБ ПОЛУЧЕНИЯ ЦЕЛЕВОГО ПРОДУКТА С ИСПОЛЬЗОВАНИЕМ УКАЗАННОЙ БАКТЕРИИ». Промотор PrpsF относится к наиболее сильным промоторам в геноме B. subtilis и обеспечивает максимально высокий уровень транскрипции на логарифмической стадии роста бактерий. Инициация транскрипции этого промотора осуществляется с участием вегетативной сигма-субъединицы РНК-полимеразы (ϭA). Инициация транскрипции гена gsiB, ответственного за белок общего клеточного стресса, инициируется с участием альтернативной сигма-субъединицы ϭB и достигает максимума на стационарной фазе роста бактерий. Тандемный промотор содержит -35 и -10 участки и старт транскрипции промотора гена rpsF, -35 и -10 участки и старт транскрипции промотора гена gsiB, и сайт связывания рибосомы (Фиг. 2).
Гибридный промотор PrpsF-gsiB, показанный на Фиг. 2, нуклеотидная последовательность которого приведена в Перечне последовательностей под номером SEQ ID NO: 1, обрабатывали рестриктазами EcoRI и BamHI и клонировали в соответствующие сайты плазмиды pDG268 (Cell 111: 747756, 2002), которая содержит кассету, включающую полилинкер для клонирования, репортерный ген lacZ без собственного промотора и ген устойчивости к хлорамфениколу (CmR) (см. Фиг. 3). Кассета фланкирована фрагментами гена amyE, что позволяет проводить интеграцию вектора с клонированным фрагментом в локус amyE на хромосоме бактерии рода Bacillus, в частности, B. subtilis. Полученную плазмиду, получившую наименование pDG268-P, трансформировали в компетентные клетки штамма E. coli TG1. Трансформанты отбирали на LB-среде, cодержащей 100 мкг/мл ампициллина. Плазмидную ДНК очищали в агарозном геле и очищали с помощью колонок (GFX PCR DNA and Gel Band Purification Kit фирмы Illustra) или шариков DNA Gel Extractionkit. Нуклеотидную последовательность ДНК вставки подтверждали путем секвенирования ДНК с использованием праймера 1 (SEQ ID NO: 2) (прямой) и праймера 2 (SEQ ID NO: 3) (обратный).
Полученная плазмида, получившая наименование pDG268-P, содержащая гибридный промотор PrpsF-gsiB, может быть интегрирована в хромосомный локус amyE бактерии рода Bacillus, в частности, B. subtilis. Так, перед трансформацией в клетки B. subtilis плазмидную ДНК pDG268-P линеаризовали с помощью рестриктазы XbaI для обеспечения интеграции в хромосому путем двойного кроссинговера вместо простого кроссинговера. Компетентные клетки штамма B. subtilis 168 трансформировали обработанной рестриктазой XbaI ДНК плазмиды pDG268-P и проводили отбор трансформантов на среде LB, содержащей 10 мкг/мл хлорамфеникола. Для подтверждения интеграции плазмиды в хромосомный локус amyE использовали индикаторную среду, содержащую амилопектин в концентрации 0,2 %.
Таким образом, в результате проведенных манипуляций была сконструирована плазмида pDG268-Р, содержащая гибридный промотор
PrpsF-gsiB и примыкающий к нему репортерный ген lacZ, что позволяет провести тестирование активности этого гибридного промотора в условиях ферментации.
Как следует из Фиг. 2, гибридный промотор содержит по два -35 и -10 участка и два старта транскрипции, которые, как полагают авторы настоящего изобретения без намерения быть ограниченными какой-либо теорией, могут быть попеременно использованы на различных стадиях роста культуры и обеспечивать оптимальную экспрессию генов, находящихся под контролем указанного промотора (Фиг. 4).
Пример 2. Конструирование генетической конструкции АМ2640 путем введения в хромосому штамма-реципиента Bacillus subtilis 168 слияния гена hasA S. equisimilis с геном tuaD B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB.
Конструирование слияния гена hasA S. equisimilis с геном tuaD
B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB проводили в несколько этапов. Сначала ген hasA синтезировали химически, взяв за основу нуклеотидную последовательность гена hasA S. equisimilis, и клонировали в вектор pUC57. С полученной плазмидной ДНК провели ПЦР-амплификацию гена hasA 1250 п.н. с использованием праймера 3 (SEQ ID NO: 4), содержащего сайт узнавания рестриктазы BamHI, и праймера 4 (SEQ ID NO: 5), содержащего на 5'конце последовательность, комплементарную участку N-конца гена tuaD из B. subtilis.
ПЦР-амплификацию проводили в 50 мкл инкубационной смеси, содержащей 3 нг плазмидной ДНК pUC57 в качестве матрицы, по 20 пмоль праймеров 3 (SEQ ID NO: 4) и 4 (SEQ ID NO: 5), 1x буфер для ПЦР, смесь dNTP (каждый в концентрации 0.5 мМ) и 2 ед. акт. термостабильной Taq-полимеразы. Реакции проводили в термоциклере My Cycler BioRad, запрограммированном на 1 цикл при 95оС в течение 5 мин; 30 циклов, каждый при 95оС в течение 30 сек, при 58оС в течение
30 сек, при 72оС в течение 2 мин, и 1 цикл при 72оС в течение 5 мин. Продукты анализировали гель-электрофорезом в 1.2% агарозе в 1х ТАE.
Ожидаемый фрагмент составлял примерно 1270 п.н.
На следующем этапе c хромосомной ДНК B. subtilis провели ПЦРамплификацию гена tuaD с использованием праймера 5 (SEQ ID NO: 6), содержащего на 5'-конце последовательность, комплементарную С-концу гена hasA S. equisimilis, и праймера 6 (SEQ ID NO: 7), содержащего сайт узнавания рестриктазы NotI.
ПЦР-амплификацию проводили в 50 мкл инкубационной смеси, содержащей 5 нг хромосомной ДНК B. subtilis в качестве матрицы, по 20 пмоль праймеров 5 (SEQ ID NO: 6) и 6 (SEQ ID NO: 7), 1x буфер для ПЦР, смесь dNTP (каждый в концентрации 0.5 мМ) и 2 ед. акт. термостабильной Taq-полимеразы. Реакции проводили в термоциклере My Cycler BioRad, запрограммированном на 1 цикл при 95оС в течение 5 мин; 30 циклов, каждый при 95оС в течение 30 сек, при 58оС в течение
30 сек, при 72оС в течение 2 мин, и 1 цикл при 72оС в течение 5 мин. Продукты анализировали гель-электрофорезом в 1.2% агарозе в 1х ТАE. Ожидаемый фрагмент составлял примерно 1450 п.н.
Затем с помощью ПЦР проводили состыковку фрагментов, полученных с использованием пар праймеров 3-4 и 5-6, путем их отжига друг на друга с использованием следующего режима ПЦР: ПЦР-амплификацию проводили в 50 мкл инкубационной смеси, содержащей по 5 нг ДНК фрагментов, полученных с использованием пар праймеров 3-4 и 5-6, 1x буфер для ПЦР, смесь dNTP (каждый в концентрации 0.5 мМ) и 2 ед. акт. термостабильной Taq-полимеразы. Реакции проводили в термоциклере My Cycler BioRad, запрограммированном на 1 цикл при 95оС в течение 5 мин; 10 циклов, каждый при 95оС в течение 30 сек, при
52оС в течение 30 сек, при 72оС в течение 2 мин, и 1 цикл при 72оС в течение 5 мин. После этого в реакционную смесь добавляли праймер 3 (SEQ ID NO: 4), содержащий сайт узнавания рестриктазы BamHI, и праймер 6 (SEQ ID NO: 7), содержащий сайт узнавания рестриктазы NotI, и проводили ПЦР в режиме: 25 циклов, каждый при 95оС в течение 30 сек, при 56оС в течение 30 сек, при 72оС в течение 2.5 мин, и 1 цикл при 72оС в течение 5 мин. Продукты анализировали гельэлектрофорезом в 1.2% агарозе в 1х ТАE. Ожидаемый фрагмент составлял примерно 2700 п.н.
Полученный ПЦР фрагмент размером 2700 п.н. обрабатывали рестриктазами BamHI и NotI, и клонировали в соответствующие сайты полученной ранее плазмиды pDG268-Р, содержащей гибридный тандемный промотор PrpsF-gsiB. Полученную плазмиду, получившую наименование pDG268-PHT, трансформировали в компетентные клетки штамма E. coli TG1. Трансформанты отбирали на LB-среде, cодержащей 100 мкг/мл ампициллина. Плазмидную ДНК очищали в агарозном геле и элюировали с помощью колонок (GFX PCR DNA and Gel Band Purification Kit фирмы Illustra) или шариков DNA Gel Extractionkit (компания «Fermentas»). Нуклеотидную последовательность ДНК вставки подтверждали путем секвенирования ДНК с использованием праймеров 1 (SEQ ID NO: 2) (прямой) и 2 (SEQ ID NO: 3) (обратный).
На заключительном этапе проводили интеграцию полученной плазмиды pDG268-PHT в хромосомный локус amyE B. subtilis. С этой целью перед трансформацией в клетки B. subtilis плазмидную ДНК pDG268-PHT линеаризовали с помощью рестриктазы XbaI для обеспечения интеграции в хромосому путем двойного кроссинговера вместо простого кроссинговера. Компетентные клетки штамма B. subtilis 168 трансформировали обработанной рестриктазой XbaI ДНК плазмиды pDG268P и проводили отбор трансформантов на среде LB, содержащей 10 мкг/мл хлорамфеникола. Для подтверждения интеграции плазмиды в хромосомный локус amyE использовали индикаторную среду, содержащую амилопектин в концентрации 0,2 %.
Таким образом, в результате проведенных манипуляций была получена генетическая конструкция на основе B. subtilis, содержащей в хромосоме слияние гена hasA S. equisimilis с геном tuaD B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB.
Пример 3. Конструирование генетической конструкции АМ2681 путем введения в хромосому реципиента Bacillus subtilis АМ2640 гена gtaB под контролем гибридного тандемного промотора PrpsF-gsiB.
Ген gtaB в геноме полученной выше генетической конструкции B. subtilis АМ2640 подставляли под контроль гибридного тандемного промотора PrpsF-gsiB для усиления его экспрессии. Для этого на первом этапе проводили клонирование промоторной области PrpsF-gsiB в экспрессионный вектор pLE1, способный к репликации как в клетках бактерий рода Bacillus, в частности, B. subtilis, так и E. coli,и содержащий ген устойчивости к эритромицину (Фиг. 5). Вектор pLE1 является делеционным вариантом плазмиды pKS1 (FEMS Microbiol. Letters, 245: 315-319, 2005), у которой делетирован маркер устойчивости к канамицину.
С этой целью с ДНК плазмиды pDG268-Р, содержащей гибридный промотор PrpsF-gsiB, проводили ПЦР амплификацию с участием фланкирующих тандемный промотор праймера 7 (SEQ ID NO: 8), содержащего сайт рестрикции PstI, и праймера 8 (SEQ ID NO: 9), содержащего сайт рестрикции HindIII.
Полученный ПЦР-продукт размером 232 п.н. очищали в агарозном геле, обрабатывали рестриктазами PstI и HindIII и клонировали в векторе pLE1 по сайтам PstI и HindIII. Полученная плазмида, содержащая гибридный тандемный промотор PrpsF-gsiB, получила наименование pLE3.
На следующем этапе с хромосомной ДНК штамма B. subtilis168 trpC2 проводили ПЦР амплификацию промотор-проксимального участка гена gtaB с помощью праймера 9 (SEQ ID NO: 10), содержащего сайт SacII, и праймера 10 (SEQ ID NO: 11), содержащего сайт PstI.
Полученный ПЦР-продукт размером 394 п.н. очищали в агарозном геле, обрабатывали рестриктазами SacII и PstI и клонировали в векторе pLE3, содержащем промотор PrpsF-gsiB, по сайтам SacII и PstI. Затем с хромосомной ДНК штамма B. subtilis168 trpC2 проводят ПЦР амплификацию фрагмента рамки считывания гена gtaB с помощью праймера 11 (SEQ ID NO: 12), содержащего сайт НindIII, и праймера 12 (SEQ ID NO: 13), содержащего сайт KpnI.
Полученный ПЦР-продукт размером 397 п.н. очищали в агарозном геле, обрабатывали рестриктазами НindIII и KpnI и клонировали по тем же сайтам в полученный на предыдущем этапе вектор pLE3, содержащий промотор PrpsF-gsiB и промотор-проксимальную область гена gtaB. У полученной в результате проведенных манипуляций плазмидной конструкции pLE5 gtaB’-PrpsF-gsiB-‘gtaB природный промотор гена gtaB замещен на промотор PrpsF-gsiB.
Сконструированной таким образом плазмидой pLE5 проводили трансформацию штамма E. coli TG1 c отбором трансформантов EmR на среде с эритромицином (300 мкг/мл) при 30оС. Полученные клоны проверяли с помощью ПЦР на присутствие в их клетках плазмиды pLE5 с клонированным в ее составе фрагментом gtaB’-PrpsF-gsiB-‘gtaB. Выделенными из проверенных клонов плазмидами pLE5 (gtaB’-PrpsF-gsiB‘gtaB) проводили трансформацию в родительский штамм B. subtilis АМ2640 c отбором трансформантов EmR на среде с эритромицином (3 мкг/мл). Несколько полученных клонов EmR культивировали на жидкой среде LB с эритромицином (3 мкг/мл) при 37оС в течение ночи, а затем рассевали на чашки со средой LB с эритромицином (3 мкг/мл) и инкубировали в течение 24 ч при 37оС. Полученные рекомбинанты EmR сформировались в результате интеграции плазмиды pLE5 (gtaB’-PrpsF-gsiB‘gtaB) в соответствующий хромосомный локус gtaB, что подтверждали наработкой ПЦР-фрагмента размером 901 п.н. при использовании праймеров 9 (SEQ ID NO: 10) и 12 (SEQ ID NO: 13). Несколько полученных клонов EmR засевали в жидкую LB среду без антибиотика и инкубировали на качалке при 30оС в течение 48 часов, а затем рассевали на чашки со средой LB без антибиотика и инкубировали 24 часа при 30оС.
Наконец, на последнем этапе работы выросшие клоны проверяли на выщепление интегрированной плазмиды pLE5, о чем судили по появлению эритромицин-чувствительных клонов. Выщепление плазмиды происходит либо с сохранением в составе хромосомы промотора дикого типа гена gtaB, либо с заменой промотора дикого типа на промотор PrpsF-gsiB. Интеграцию промотора PrpsF-gsiB перед геном gtaB в хромосому штамма B. subtilis подтверждали наработкой ПЦР-фрагмента размером 791 п.н. с участием праймеров 1 (SEQ ID NO: 2) и 13 (SEQ ID NO: 14) и его последующим секвенированием. Один из вариантов штамма B. subtilis
АМ2640, включивший ген gtaB под контролем промотора PrpsF-gsiB, получивший наименование АМ2681, накапливал до 2,21 г/л гиалуроновой кислоты при ферментации в течение 48 часов (Таблица 1).
Пример 4. Конструирование генетических конструкций АМ2686 и
АМ2694 путем введения в хромосому реципиентов Bacillus subtilis АМ2640 и АМ2681, соответственно, оперона gсaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
Оперон gcaD-prs в геноме полученной выше генетической конструкции B. subtilis АМ2640 подставляли под контроль гибридного тандемного промотора PrpsF-gsiB для усиления его экспрессии. Для этого на первом этапе с хромосомной ДНК штамма B. subtilis168 trpC2 проводили ПЦР амплификацию промотор-проксимального участка оперона gсaD-prs с помощью праймера 14 (SEQ ID NO: 15), содержащего сайт SacII, и праймера 15 (SEQ ID NO: 16), содержащего сайт PstI.
Полученный ПЦР-продукт размером 419 п.н. очищали в агарозном геле, обрабатывали рестриктазами SacII и PstI и клонировали в полученный на предыдущем этапе вектор pLE3, содержащий промотор PrpsFgsiB, по сайтам SacII и PstI.
Затем с хромосомной ДНК штамма B. subtilis 168 trpC2 проводили ПЦР амплификацию фрагмента рамки считывания гена gcaD с помощью праймера 16 (SEQ ID NO: 17), содержащего сайт СlaI, и праймера 17 (SEQ ID NO: 18), содержащего сайт KpnI.
Полученный ПЦР-продукт размером 394 п.н. очищали в агарозном геле, обрабатывали рестриктазами СlaI и KpnI и клонировали по тем же сайтам в полученный на предыдущем этапе вектор pLE3, содержащий промотор PrpsF-gsiB и промотор-проксимальную область оперона gcaD-prs. У полученной в результате проведенных манипуляций плазмидной конструкции pLE8 gcaD’-PrpsF-gsiB-‘gcaD природный промотор оперона gcaD-prs замещен на промотор PrpsF-gsiB.
Сконструированной таким образом плазмидой pLE8 проводили трансформацию штамма E. coli TG1 c отбором трансформантов EmR на среде с эритромицином (300 мкг/мл) при 30оС. Полученные клоны проверяли с помощью ПЦР на присутствие в их клетках плазмиды pLE8 с клонированным в ее составе фрагментом gcaD’-PrpsF-gsiB-‘gcaD.
Выделенными из проверенных клонов плазмидами pLE8 (gcaD’-PrpsF-gsiB‘gcaD) проводили трансформацию в сконструированные ранее генетические конструкции B. subtilis АМ2640 и АМ2681 c отбором трансформантов EmR на среде с эритромицином (3 мкг/мл). Несколько полученных клонов EmR, полученных при трансформации генетических конструкций АМ2640 и АМ2681, культивировали на жидкой среде LB с эритромицином (3 мкг/мл) при 37оС в течение ночи, а затем рассевали на чашки со средой LB с эритромицином (3 мкг/мл) и инкубировали в течение 24 ч при 37оС. Полученные рекомбинанты EmR сформировались в результате интеграции плазмиды pLE8 (gcaD’-PrpsF-gsiB-‘gcaD) в соответствующий хромосомный локус gcaD, что подтверждали наработкой ПЦР-фрагмента размером 891 п.н. при использовании праймера 14 (SEQ ID NO: 15) и праймера 17 (SEQ ID NO: 18). Несколько полученных клонов EmR засевали в жидкую LB среду без антибиотика и инкубировали на качалке при 30оС в течение 48 часов, а затем рассевали на чашки со средой LB без антибиотика и инкубировали 24 часа при 30оС.
Наконец, на последнем этапе работы выросшие клоны проверяли на выщепление интегрированной плазмиды pLE8, о чем судили по появлению эритромицин-чувствительных клонов. Выщепление плазмиды происходит либо с сохранением в составе хромосомы промотора дикого типа оперона gcaD-prs, либо с заменой промотора дикого типа на промотор PrpsF-gsiB. Интеграцию промотора PrpsF-gsiB перед опероном gcaD-prs в хромосому штамма B. subtilis подтверждали наработкой ПЦР-фрагмента размером 772 п.н. с участием праймера 1 (SEQ ID NO: 2) и праймера 18 (SEQ ID NO: 19) и его последующим секвенированием. По одному варианту генетические конструкции B. subtilis АМ2640 и АМ2681, включившие оперон gcaD-prs под контролем промотора PrpsF-gsiB, получившие наименование АМ2686 и АМ2694, накапливали до 2,83 и 3,21 г/л гиалуроновой кислоты, соответственно, при ферментации в течение 48 часов (Таблица 1).
Пример 5. Определение уровня транскрипции целевых генов в полученных генетических конструкциях методом ПЦР в реальном времени.
Уровень транскрипции генов hasA, tuaD, gtaB и gcaD определяли методом ПЦР в реальном времени. С этой целью клеточные культуры анализируемых генетических конструкций растили до ОП600 = 1.4-1.6 и выделяли тотальную РНК. Очистку РНК проводили с использованием набора RNeasy Mini Kit (Qiagen) в соответствии с инструкцией фирмыпроизводителя. Полученные препараты РНК обрабатывали ДНКазой I. Качество полученной тотальной РНК анализировали в 1% агарозном геле с добавлением формамида. Количество определяли спектрофотометрически по величине поглощения при длине волны 260 нм. Перед проведением реакции обратной транскрипции образцы РНК обрабатывали ДНКазой I (Thermo). Реакцию обратной транскрипции осуществляли в присутствии специфичных для исследуемых генов олигонуклеотидов при помощи набора “SuperScript III First-Strand Synthesis Kit for RT-PCR” (Invitrogen). Далее 1 мкл от объема всей реакции обратной транскрипции использовали в качестве матрицы для ПЦР с детекцией в реальном времени. Уровни экспрессии генов rrnA и gyrB использовали для нормализации.
Анализ проводили с использованием набора реагентов для ПЦР в реальном времени производства компании Синтол, Россия. Амплификацию проводили на приборе DTlite (ДНК-технология). Продукты реакции анализировали электрофорезом в 2% агарозном геле для подтверждения того, что полученные продукты имеют ожидаемый размер. Каждая реакция была поставлена трижды, в качестве результата бралось среднее значение. Уровень транскрипции определяли по значениям порогового цикла с учетом того, что концентрация исходных специфических фрагментов ДНК увеличивается приблизительно как 2N, где N - количество циклов. Результаты определения уровня транскрипции генов hasA, tuaD, gtaB и gcaD в штаммах АM2640, AM2681, AM2686 и АМ2694 представлены на Фиг. 6.
Как следует из данных на Фиг. 6, подстановка генов hasA, tuaD, gtaB и gcaD под контроль тандемного гибридного промотора PrpsF-gsiB обеспечивает примерно одинаковое, почти 30-кратное увеличение их уровня транскрипции.
Пример 6. Получение гиалуроновой кислоты с помощью полученных генетических конструкций.
Посевную культуру штамма заявляемой генетической конструкции Bacillus subtilis, продуцирующей гиалуроновую кислоту, получают путем выращивания при встряхивании (200-220 об/мин) в течение 18 часов при 37°C на среде LB следующего состава, мас. %:
Пептон казеиновый – 1,0;
Дрожжевой экстракт – 0,5;
NaCl – 0,5;
Глюкоза – 1,0;
MgSO4 – 0,02.
Полученной посевной культурой засевают ферментационную среду следующего состава, мас.%:
Сахароза – 8,0;
Мальтоза – 4,0;
Дрожжевой экстракт – 0,5;
Мочевина – 0,6;
KH2PO4 – 0,6;
K2HPO4 – 1,4;
(NH4)2S04 – 0,3;
Na-цитрат – 0,2;
Казаминовые кислоты – 0,04, MgSO4 – 0,3.
Ферментацию штамма-продуцента проводили в течение 48 час. при температуре 37оС. Выделение гиалуроновой кислоты из культуральной жидкости включает в себя следующие стадии:
Удаление клеточной массы из культуральной жидкости, в которой выращивались описанные выше генетические конструкции, центрифугированием при 8000 об/мин. Осадок отбрасывают, а супернатант используют далее.
Прецепитирование ГК раствором цетилпиридиния и изопропанола.
К супернатанту добавляют 10% водный раствор цетилпиридиния до конечной концентрации 1%. Раствор оставляют на 1,5 часа при постоянном помешивании на 37°С. После окончания инкубации раствор центрифугируют при 8000 об/мин 30 мин.
Осадок растворяют в 0,5M NaCl (объем в 2-2,5 раза меньший начального объема супернатанта), при помешивании и 37°С, при полном растворении, наливают равный объем изопропанола (1:1), и оставляют при -20°С на ночь. Сформировавшийся осадок ГК центрифугируют при8000 об/мин 30 мин и далее работают с ним.
Осадок ГК растворяют в дистиллированной воде до полного растворения. Полученную вязкую желтоватую жидкость обесцвечивают на колонке с активированным углем. Если жидкость мутная, то перед нанесением на колонку фильтруют через фильтр с диаметром пор 0,45 мкм. Элюцию с колонки ведут дистиллированной водой. Цвет выходящей жидкости должен быть бесцветный или слабо-кремовый. Если цвет более интенсивный, препарат наносят на уголь еще раз.
К образцу ГК добавляют до концентрации 0,1 % 10% суспензию бентонита с 0,5% Na2CO3, предварительно подкислив образец ГК 100% уксусной кислотой до pH=3,5. Инкубируют в течение 2 часов при помешивании 150 об/мин. После инкубации центрифугируют раствор при 8000 об/мин или фильтруют раствор через фильтр с размером пор 0,45 мкм.
Ультрафильтрация через мембрану Pellicon 2 – 300 Кда. Пропущенный через 0,45 мкм препарат ГК фильтруют от остатков цетилпиридиния и соли против 10-20 кратного объема дистиллированной воды. Проводимость препарата ГК не должна превышать 2 мСм/см. Готовый препарат ГК оставляют при температуре +4оС на 1-2 суток. Если через сутки препарат помутнеет, фильтруют его через фильтр с диаметром пор 0,45 мкм или 0,22 мкм.
Пример 7. Уровень синтеза гиалуроновой кислоты при культивировании полученных генетических конструкций.
Полученные генетические конструкции, содержащие в составе хромосомы слияние гетерологичного гена hasA S. equisimilis с геном tuaD B. subtilis под контролем гибридного тандемного промотора PrpsFgsiB ,а также дополнительно содержащие в составе хромосомы ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB и/или оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB, проверяли на способность синтезировать гиалуроновую кислоту при культивировании в лабораторном ферментере объемом 3 л с использованием ферментационной среды, как описано в Примере 7.
Результаты представлены в Таблице 1.
Таблица 1. Динамика накопления гиалуроновой кислоты в культуральной жидкости полученных генетических конструкций – продуцентов гиалуроновой кислоты.
Генетич еская конструкция
subtilis 168 (исходный штамм)
hasA-tuaD
PrpsF-gsiB-gtaB
hasA-tuaD
PrpsF-gsiB-gcaD-prs
hasA-tuaD
PrpsF-gsiB-gtaB PrpsFgsiB-gcaD-prs
Как следует из данных Таблицы 1, все полученные генетические конструкции демонстрировали более высокий уровень накопления гиалуроновой кислоты по сравнению с исходным штаммом B. subtilis 168. Наиболее высокий уровень накопления целевого продукта демонстрировала генетическая конструкция АМ2694, в которой наряду с amyE::PrpsF-gsiB-hasA-tuaD в составе хромосомы присутствовали ген gtaB и оперон gсaD-prs под контролем гибридного тандемного промотора PrpsFgsiB. Указанная генетическая конструкция накапливала 3.21 г/л гиалуроновой кислоты.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТАНДЕМНЫЙ ПРОМОТОР, ФУНКЦИОНИРУЮЩИЙ В БАКТЕРИИ РОДА Bacillus, ТРАНСФОРМИРОВАННАЯ БАКТЕРИЯ РОДА Bacillus - ПРОДУЦЕНТ ЦЕЛЕВОГО ПРОДУКТА, СОДЕРЖАЩАЯ УКАЗАННЫЙ ПРОМОТОР, И СПОСОБ ПОЛУЧЕНИЯ ЦЕЛЕВОГО ПРОДУКТА С ИСПОЛЬЗОВАНИЕМ УКАЗАННОЙ БАКТЕРИИ | 2019 |
|
RU2723722C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГИАЛУРОНАНА В РЕКОМБИНАНТНОЙ КЛЕТКЕ-ХОЗЯИНЕ | 2002 |
|
RU2346049C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ПУРИНОВОГО НУКЛЕОЗИДА 5'-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИДА (АИКАР) И ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ АИКАР | 2008 |
|
RU2405833C2 |
| ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2015 |
|
RU2603004C1 |
| МУТАНТНАЯ ФОСФОРИБОЗИЛПИРОФОСФАТСИНТЕТАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, СПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОЗИДОВ И CПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОТИДОВ | 2008 |
|
RU2403286C2 |
| БАКТЕРИЯ Escherichia coli - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ | 2010 |
|
RU2466186C2 |
| БАКТЕРИЯ Bacillus subtilis, ПРОДУЦИРУЮЩАЯ 5`-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИД (АИКАР), И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА АИКАР ПУТЕМ КУЛЬТИВИРОВАНИЯ ТАКОЙ БАКТЕРИИ | 2013 |
|
RU2542387C1 |
| РЕКОМБИНАНТНАЯ CORYNEBACTERIUM GLUTAMICUM ДЛЯ ЭФФЕКТИВНОГО СИНТЕЗА ВЫСОКООЧИЩЕННОЙ ГИАЛУРОНОВОЙ КИСЛОТЫ И ЕЕ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2823305C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2012 |
|
RU2528056C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
Изобретение относится к биотехнологии. Описана трансформированная клетка бактерии рода Bacillus, продуцирующая гиалуроновую кислоту и содержащая в составе хромосомы слияние гетерологичного гена hasA из S. equisimilis с геном tuaD из B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB; причем указанная клетка дополнительно характеризуется тем, что необязательно содержит в составе хромосомы ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB и/или оперон gcaDprs под контролем гибридного тандемного промотора PrpsF-gsiB. Также представлен способ получения гиалуроновой кислоты с использованием указанной трансформированной клетки. Изобретение позволяет более продуктивно получать гиалуроновую кислоту, до 3.21 г/л гиалуроновой кислоты. 2 н. и 5 з.п. ф-лы, 6 ил., 1 табл., 7 пр.
1. Трансформированная клетка бактерии рода Bacillus, продуцирующая гиалуроновую кислоту и содержащая в составе хромосомы слияние гетерологичного гена hasA из S. equisimilis с геном tuaD из B. subtilis под контролем гибридного тандемного промотора PrpsF-gsiB; причем указанная клетка дополнительно характеризуется тем, что необязательно содержит в составе хромосомы ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB и/или оперон gcaDprs под контролем гибридного тандемного промотора PrpsF-gsiB.
2. Трансформированная клетка по п. 1, содержащая в хромосоме ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB.
3. Трансформированная клетка по п. 1, содержащая в хромосоме оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
4. Трансформированная клетка по п. 1, содержащая в хромосоме ген gtaB под контролем гибридного тандемного промотора PrpsF-gsiB и оперон gcaD-prs под контролем гибридного тандемного промотора PrpsF-gsiB.
5. Трансформированная клетка по любому из пп. 1-4, где указанная бактерия рода Bacillus является бактерией B. subtilis.
6. Способ получения гиалуроновой кислоты, включающий:
а) культивирование трансформированной клетки по любому из пп. 1-5 в питательной среде, пригодной для продукции гиалуроновой кислоты, и
б) выделение гиалуроновой кислоты из культуральной жидкости.
7. Способ по п. 6, характеризующийся тем, что используют жидкую питательную среду следующего состава, мас. %:
| WO 2012032154 A1, 15.03.2012 | |||
| БАКТЕРИЯ Bacillus subtilis, ПРОДУЦИРУЮЩАЯ 5`-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИД (АИКАР), И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА АИКАР ПУТЕМ КУЛЬТИВИРОВАНИЯ ТАКОЙ БАКТЕРИИ | 2013 |
|
RU2542387C1 |
| US 20070259004 A1, 08.11.2007 | |||
| Асинхронный двигатель с короткозамкнутым ротором | 1930 |
|
SU32045A1 |

