Перекрестная ссылка на родственные патенты и патентные заявки
В настоящей заявке испрашивается приоритет предварительной заявки на патент США No. 61/189357, поданной 18 августа 2008, которая во всей своей полноте включена в настоящее описание посредством ссылки.
Соглашение о совместном проведении исследований
Представленные здесь описание и формула изобретения являются результатом работы, предпринятой в рамках соглашения о совместных исследованиях сотрудниками компании Pfizer Inc. и Abgenix Inc., которые были проведены до подачи заявки на рассматриваемый предмет изобретения.
Предшествующий уровень техники
Очевидно, что инфильтрация лейкоцитов в участки воспаления регулируется белками размером 8-10 кДа, известными как хемокины. Эти хемокины подразделяются на четыре группы, в зависимости от пространственной локализации их N-концевых цистеиновых остатков, обозначаемых CC, CXC, XC и CX3C. Хемокины могут опосредовать ряд провоспалительных функций лейкоцитов, таких как запуск хемотаксиса, дегрануляция, синтез липидных медиаторов и активация интегринов (Oppenheim, J. J. et al., Annu. Rev. Immunol., 9:617-648 (1991); Baggiolini, M., et al., Adv. Imunol., 55:97-179 (1994); Miller, M. D. & Krangel, M. S., Crit. Rev. Immunol., 12:17-46 (1992)).
Один из хемокинов, а именно, моноцитарный хемотаксический фактор 1 (MCP-1), также известный как CCL2, действует на моноциты, лимфоциты и дендритные клетки и тем самым индуцирует хемотаксис, высвобождение гранул, респираторную «вспышку» и высвобождение цитокинов. Результаты исследований дают основание предположить, что MCP-1 участвует в патогенезе заболеваний, таких как ревматоидный артрит, атеросклероз, гранулематозные заболевания, хроническая обструктивная болезнь легких (ХОБЛ), ожирение/диабет, невропатическая боль, рак и рассеянный склероз (Koch, J. Clin. Invest. 90:772-79 (1992); Hosaka et al., Clin. Exp. Immunol. 97:451-457 (1994); Schwartz et al., Am. J. Cardiol. 71 (6):9B-14B (1993); Schimmer ef al., J. Immunol. 160:1466-1471 (1998); Flory et al., Lab. Invest. 69:396-404 (1993); Gong et al., J. Exp. Med. 186:131-137 (1997); Salcedo et al. Blood 96(1) 34-40 (2000); Bracke et al., Inflammation & Allergy - Drug Targets 6: 75-79 (2007); Chung Current Drug Targets - Inflammation & Allergy 4: 619-625 (2005).
CCR2 представляет собой связанный с G-белком хемотаксический рецептор, который состоит из семи трансмембранных доменов и связывается с MCP-1, а также с другими хемокинами, включая CCL8 (MCP-2), CCL7 (MCP-3) и CCL13 (MCP-4) (Charo, I. F., et al., Proc. Natl. Acad. Sci. USA 91 :2752-2756 (1994); Myers, S. J., et al., J. Biol. Chem. 270:5786-5792 (1995); Gong et al., J. Biol Chem 272:11682-11685 (1997); Garcia-Zepeda et al., J. Immunol. 157:5613-5626 (1996)). CCR2 также известен как CMKBR2 и CKR2. Были клонированы две альтернативно сплайсированные формы CCR2, CCR2A и CCR2B, которые отличаются друг от друга по своим С-концам (Wong et al (1997) J. Biol. Chem. 272:1038-1045). В исследованиях по передаче сигнала, CCR2A и CCR2B опосредуют зависимую от агониста мобилизацию кальция и ингибирование аденилилциклазы. CCR2 экспрессируется на моноцитах, Т-клетках и дендритных клетках, и взаимодействует с хемокинами, секретируемыми эндотелиальными клетками, моноцитами и синовиальными фибробластами.
Биологическая роль CCR2 была исследована на CCR2-дефицитных мышах (Boring et al., J Clin Invest. 100(10):2552-61 (1997); Boring et al., Nature 394(6696): 894-7 (1998); De Paolo et al., J. Immunol. 171(7):3560-7 (2003); Gaupp et al., Am. J. Pathol. 162(1): 139-50(2003)). У CCR2-/--мышей наблюдается в значительной степени недостаточная гиперчувствительность замедленного типа и недостаточное продуцирование цитокинов типа Th1, и такие мыши обычно менее восприимчивы к развитию экспериментального аутоиммунного энцефаломиелита (ЭАЭ). CCR2, помимо своей способности модулировать иммунные ответы, является также ко-рецептором для ВИЧ (Connor et al., J. Exp. Med. 185:621-628 (1997); Frade et al., J Clin Invest. 100(3):497-502(1997)).
Благодаря участию MCP-1 и его рецептора CCR2 в нежелательных иммунных ответах, антагонисты CCR2 могут рассматриваться как перспективные терапевтические средства. Однако в литературе описаны лишь немногие антагонисты CCR2 (см. Ogilvie et al., Blood 97(7): 1920-4 (2001)). Таким образом, необходимо разработать новые и улучшенные композиции, которые были бы способны связываться с CCR2 и блокировать передачу CCR2-сигнала, опосредуемого его лигандом.
Описание сущности изобретения
Настоящее изобретение относится к выделенным антителам или их антигенсвязывающим частям, которые специфически связываются с CCR2, а в частности, с человеческим CCR2, и могут действовать как антагонист CCR2, а также к композициям, содержащим указанные антитела или их части. В настоящем изобретении также описаны антитела или их антигенсвязывающие части, которые связываются с CCR2 на эпитопе, отличающемся от эпитопа N-концевой части или третьей петли CCR2. Такие антитела могут связываться с первой и/или второй внеклеточными петлями CCR2.
Настоящее изобретение относится к композициям, содержащим (i) тяжелую и/или легкую цепь, вариабельные домены или антигенсвязывающие части указанного анти-CCR2 антитела, или молекулы нуклеиновой кислоты, кодирующие такие антитела, и (ii) фармацевтически приемлемый носитель. Эти композиции могут также включать другой компонент, такой как терапевтическое средство или диагностическое средство.
Настоящее изобретение также относится к способам диагностики и терапии. Аналогичным образом, настоящее изобретение также относится к анти-CCR2 антителам и их частям, используемым в целях приготовления лекарственных препаратов для лечения воспалительных и невоспалительных расстройств.
Настоящее изобретение относится к векторам и к клеткам-хозяевам, содержащим молекулы нуклеиновой кислоты, а также к способам рекомбинантного продуцирования полипептидов, кодируемых молекулами нуклеиновой кислоты. Настоящее изобретение также относится к выделенным клеточным линиям, продуцирующим анти-CCR2 антитело или его антигенсвязывающую часть.
Краткое описание графического материала
На фигурах 1A, 1B и 1C представлены графики, иллюстрирующие связывание анти-CCR2 антитела с клетками, как было проанализировано с помощью FACS-анализа. На фигуре 1A представлен график, иллюстрирующий связывание AF-488 (ALEXA FLUOR® 488, Invitrogen)-конъюгированного анти-CCR2 антитела 4.40 A68G S230P с моноцитами цельной человеческой крови, по сравнению с контрольным антителом против гемоцианина лимфы улитки (KLH), как было проанализировано с помощью FACS-анализа. На фигуре 1B представлен график, иллюстрирующий связывание AF-488-конъюгированного анти-CCR2 антитела 4.40 A68G S230P с клетками 300-19, экспрессирующими человеческий CCR2, как было проанализировано с помощью FACS-анализа. На фигуре 1C представлен график, иллюстрирующий связывание антитела 4.40 A68G S230P в различных концентрациях с CCR2-трансфецированными клетками 300-19, детектируемое с использованием античеловеческого антитела, конъюгированного с ФЭ.
На фигуре 2 проиллюстрировано дозозависимое связывание антитела 4.40 A68G S230P с CCR2-трансфецированными клетками 300-19 в анализе на насыщенное связывание.
На фигуре 3 проиллюстрирована способность антитела 4.40 A68G S230P ингибировать хемотаксис THP-1-клеток в ответ на действие CCR2-лиганда MCP-1, но не в ответ на действие CCR1/CCR5-лиганда MIP-1a.
На фигуре 4 проиллюстрирована способность антитела 4.40 A68G S230P ингибировать хемотаксис первичных человеческих моноцитов в ответ на действие MCP-1.
На фигуре 5 показана плазмидная карта ретровирусного вектора для экспрессии химер CCR1/CCR2.
На фигурах 6A и 6B проиллюстрировано связывание антитела 4.40 A68G S230P с клетками 300-19, экспрессирующими химерный рецептор, состоящий только из внеклеточных 1-й и 2-й петель CCR2 и N-конца и третьей петли CCR1. На фигуре 6C проиллюстрирован анализ на насыщенное связывание 4.40 A68G S230P с клетками 300-19, трансфецированными химерным рецептором, как было определено с помощью FACS-анализа.
На фигуре 7 проиллюстрирован анализ на насыщенное связывание (кривая насыщения, достигаемого за 3 часа) антитела 4.40 A68G S230P с (A) контрольными клетками 300-19; (B) трансфецированными клетками 300-19, экспрессирующими полноразмерный CCR2; (C) с трансфецированными клетками 300-19, экспрессирующими flag-меченную (M1) химеру MRRR [flag-меченый N-конец CCR2 (M), для гарантии экспрессии рецептора, и области петли CCR1 (R)]; (D) с трансфецированными клетками 300-19, экспрессирующими flag-меченную (M1) химеру RRRM [flag-меченый N-конец и 1-я и 2-я петли CCR1 (R), для гарантии экспрессии рецептора, и 3-я петля CCR2 (M)]; и (E) с трансфецированными клетками 300-19, экспрессирующими flag-меченную (M1) химеру RMMR [flag-меченый N-конец N и 3-я петля CCR1 (R), для гарантии экспрессии рецептора, и 1-я и 2-я петли CCR2 (M)].
На фигуре 8 проиллюстрировано связывание антитела 4.40 A68G S230P с пептидными областями либо петли 2, либо петли 3 CCR2, как было определено с помощью ELISA с захватом.
На фигуре 9 представлены два графика, иллюстрирующие FACS-иммуноокрашивание клеток 300-19, экспрессирующих рекомбинантный CCR5, либо AF-488-конъюгированным антителом 4.40.3 A68G S230P (панель A), либо анти-CCR5 антителом (панель B).
На фигуре 10 проиллюстрировано ингибирование MCP-1-индуцированной активности под действием антитела 4.40 A68G S230P, как было определено по мобилизации кальция на CCR2-трансфецированных клетках 300-19.
На фигуре 11 проиллюстрировано ингибирование MCP-3-индуцированного хемотаксиса CCR2-трансфецированных клеток 300-19 под действием антитела 4.40 A68G S230P.
На фигуре 12 проиллюстрировано ингибирование полимеризации актина человеческих моноцитов в ответ на MCP-1 в цельной крови под действием антитела 4.40 A68G S230P.
На фигуре 13 проиллюстрировано ингибирование полимеризации актина моноцитов у самок собакоподобных обезьян в ответ на MCP-1 в цельной крови под действием антитела 4.40 A68G S230P.
На фигуре 14 проиллюстрировано дозозависимое ингибирование синтеза мРНК коллагена 1 в клеточной линии hHSC, LI90, под действием антитела 4.40 A68G S230P.
На фигуре 15 проиллюстрировано дозозависимое ингибирование фосфорилирования pERK в человеческой цельной крови в концентрации 10 нМ MCP-1 под действием антитела 4.40 A68G S230P.
На фигуре 16 проиллюстрировано снижение активности аланин-трансаминазы (ALT) и аспартат-аминотрансферазы (AST) в плазме у мышей, дефицитных по человеческому CCR2, под действием антитела 4.40 A68G S230P через 24 часа после одной инъекции ConA.
На фигурах 17A-17D проиллюстрировано выравнивание аминокислотных последовательностей вариабельных областей тяжелой и легкой цепи антител зародышевой линии с соответствующими последовательностями вариабельных областей тяжелой и легкой цепи антител 4.22.3, 4.40.2, 4.39.3 и 4.9.2 (для антител 4.22.3, 4.40.2, 4.39.3 и 4.9.2 показаны только несоответствия). CDR подчеркнуты, а несоответствующией(ие) остаток(тки) (пробелы) показан(ы) знаком фунта (#).
Подробное описание изобретения
Определения и общие методы
Если это не оговорено особо, то используемые в настоящем описании научные и технические термины имеют общепринятые значения, известные среднему специалисту в данной области. Кроме того, если это не очевидно из контекста описания, то существительные, употребляемые в единственном числе, могут означать и существительные во множественном числе, а существительные, употребляемые во множественном единственном числе, могут означать существительные в единственном числе. Все упомянутые здесь публикации и другие работы во всей своей полноте вводятся в настоящее описание посредством ссылки. В общих чертах, описанные здесь номенклатура и способы культивирования клеток и тканей, а также методы, применяемые в молекулярной биологии, иммунологии, микробиологии, генетике, химии белков и нуклеиновых кислот, и методы их гибридизации, хорошо известны специалистам и широко применяются в данной области. Если при определении любого термина в данном описании возникают противоречия с другими определениями, то они могут быть разрешены в соответствии с описанием, представленным ниже.
Осуществляемые методы и технологии обычно являются стандартными методами, хорошо известными специалистам, и описаны в различных общей и специальной литературе, цитируемой и обсуждаемой в настоящем описании, если это не оговорено особо. См., например, руководства Sambrook et al., Molecular Cloning: A Laboratory Manual, second ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates (1992); и Harlow and Lane, Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1990), каждое из которых включено в настоящее описание посредством ссылки. Ферментативные реакции и методы очистки осуществляют в соответствии с инструкциями производителей, в основном, как описано в литературе или в настоящей заявке. Используемая в данном описании номенклатура, лабораторные процедуры и методы аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, описанные в настоящей заявке, хорошо известны специалистам и широко используются в данной области. Химический синтез, химические анализы, приготовление фармацевтических препаратов, составление композиций, их доставку и лечение пациентов осуществляют стандартными методами.
В описании настоящего изобретения используются указанные термины, которые, если это не оговорено особо, имеют значения, определенные ниже.
Термин «полипептид» охватывает нативные или искусственные белки, фрагменты белка и полипептидные аналоги последовательности белка. Полипептид может быть мономерным или полимерным.
Используемые в данном описании термины «выделенный белок», «выделенный полипептид» или «выделенное антитело» означают белок, полипептид или антитело, которые, по своей природе или источнику происхождения, (1) не ассоциируются с природными компонентами, с которыми они ассоциируются в природе, (2) изолированы от других белков, происходящих от того же источника, (3) экспрессируются клетками различных типов или (4) не встречаются в природе. Таким образом, полипептид, который был синтезирован химически или был синтезирован в клеточной системе, отличающейся от клеток, в которых он синтезируется в природе, может быть «отделен» от его природных компонентов. Белок может быть также, по существу, отделен от его природных компонентов путем выделения методами очистки белков, хорошо известными специалистам.
Примерами выделенных антител являются анти-CCR2 антитела, которые были подвергнуты аффинной очистке с использованием CCR2 или его части; анти-CCR2 антитела, которые были синтезированы гибридомой или другой клеточной линией in vitro; и человеческие анти-CCR2 антитела, происходящие от трансгенных мышей.
Белок или полипептид рассматривается как «в основном, чистый», «в основном, гомогенный» или «в основном, очищенный», если по меньшей мере, примерно 60-75% образца содержит полипептиды одного типа. Полипептид или белок могут быть мономерными или мультимерными. В основном, чистый полипептид или белок обычно составляет примерно 50%, 60%, 70%, 80% или 90% по массе образца белка, а обычно его чистота составляет примерно 95%, но может составлять более 99%. Чистота и гомогенность белка могут быть оценены различными методами, хорошо известными специалистам, такими как электрофорез образца белка в полиакриламидном геле с последующей визуализацией одной полипептидной полосы после окрашивания геля красителем, хорошо известным специалистам. Для некоторых целей, более высокое разрешение может быть достигнуто с помощью ВЭЖХ или другими средствами, хорошо известными специалистам в области очистки.
Используемый в данном описании термин «аналог антитела» означает антитело, содержащее сегмент, который, по существу, идентичен части аминокислотной последовательности и который обладает по меньшей мере одним из нижеследующих свойств, а именно (1) обладает способностью специфически связываться с CCR2 в подходящих условиях связывания, (2) обладает способностью ингибировать по меньшей мере одну биологическую активность CCR2. Обычно, аналоги антитела, в отличие от природной последовательности, содержат консервативную аминокислотную замену (инсерцию или делецию). Аналоги обычно имеют длину по меньшей мере в 20 или 25 аминокислот, а также по меньшей мере в 50, 60, 70, 80, 90, 100, 150 или 200 аминокислот или более, а в большинстве случаев, их длина равна длине полноразмерных тяжелых или легких цепей антител. В некоторых случаях, такими аналогами являются аналоги, в аминокислотной последовательности которых присутствуют 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 замен, по сравнению с аминокислотной последовательностью зародышевой линии.
В некоторых случаях, аминокислотными заменами в анти-CCR2 антителе или в его антигенсвязывающей части являются замены, которые приводят: (1) к снижению чувствительности к протеолизу, (2) к снижению чувствительности к окислению, (3) к изменению аффинности связывания для образования белковых комплексов, (4) к добавлению или удалению сайтов гликозилирования и (5) к сообщению или модификации других физико-химических или функциональных свойств таких аналогов, но которые, при этом, сохраняют специфичность связывания с CCR2. Указанными аналогами могут быть различные мутеины с последовательностью, отличающейся от природной пептидной последовательности. Так, например, в природную последовательность той части полипептида, которая расположена за пределами домен-образующих межмолекулярных контактов, могут быть внесены одна или множество аминокислотных замен, таких как консервативные аминокислотные замены. Консервативная аминокислотная замена не должна вносить значительные изменения в структурные свойства родительской последовательности, например, аминокислотная замена не должна приводить к изменению антипараллельной β-складки, которая составляет связывающий домен иммуноглобулина, присутствующий в родительской последовательности, или к разрушению вторичной структуры других типов, характеризующих родительскую последовательность. В основном, глицин и пролин не встречаются в антипараллельной β-складке. Примеры известных вторичных и третичных структур полипептида описаны в работах Protein, Structures and Molecular Principles (Creighton ed., W.H. Freeman and Company, New York 1984); Introduction to Protein Structure (C. Branden & J. Tooze eds., Garland Publishing, New York, N,Y. (1991)) и в работе Thornton et al., Nature 354:105 (1991), которые включена в настоящее описание посредством ссылки.
При употреблении термина «антитело» также подразумевается, что этот термин может также включать антигенсвязывающую часть антитела. Антигенсвязывающая часть конкурирует с интактным антителом за специфическое связывание. В общих чертах, см. издание Fundamental Immunology. Ch. 7 (Paul, W., ed., second ed. Raven Press, N.Y. (1989)) (которое во всей своей полноте включено в настоящее описание посредством ссылки). Антигенсвязывающие части могут быть продуцированы методами рекомбинантных ДНК или методами ферментативного или химического расщепления интактного антитела. В некоторых случаях, антигенсвязывающими частями являются Fab, Fab'-, F(ab')2-, Fd-, Fv-, dAb-фрагменты, и их гипервариабельные области (комплементарность-определяющие области (CDR)), одноцепочечные антитела (например, scFv), химерные антитела, диантитела и полипептиды, которые содержат по меньшей мере часть антитела, которая является достаточной для сообщения специфичности связывания антигена с полипептидами.
Вариабельные домены зрелых тяжелых и легких цепей антитела, от N-конца до C-конца, включают области FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. В настоящем описании, аминокислоты приписываются к каждому домену в соответствии с определениями по Кэбату, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 and 1991)); Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987); или по Чотия, Chothia et al., Nature 342:878-883 (1989).
Используемое в данном описании антитело обозначается номером, аналогичным номеру моноклонального антитела, полученного из гибридомы, обозначаемой тем же самым номером. Так, например, моноклональное антитело 4.40 представляет собой антитело, аналогичное антителу, полученному из гибридомы 4.40, или его субклон. Следующие друг за другом субклоны обозначаются, например, 4.40.1, 4.40.2 и 4.40.3, и имеют, в основном, те же самые последовательности и функциональные свойства.
Используемый в данном описании Fd-фрагмент означает фрагмент антитела, состоящий из доменов VH и CH1; Fv-фрагмент означает фрагмент антитела, состоящий из доменов VL и VH одной ветви антитела, а dAb-фрагмент (Ward et al., Nature 341: 544-546 (1989)) означает фрагмент антитела, состоящий из VH-домена.
В некоторых случаях, антителом является одноцепочечное антитело (например, scFv), в котором домены VL и VH спариваются с образованием одновалентных молекул посредством синтетического линкера, который позволяет им образовывать одну полипептидную цепь (см., например, Bird et al., Science 242:423-426 (1988) and Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988). В некоторых случаях, антителами являются диантитела, то есть, двухвалентные антитела, в которых домены VH и VL экспрессируются на одной полипептидной цепи, но посредством линкера, который является слишком коротким для спаривания двух доменов на одной и той же цепи, а поэтому эти домены вынуждены спариваться с комплементарными доменами другой цепи и образовывать два антигенсвязывающих сайта (см., например (см. например, Holliger P. et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993), и Poljak R. J. et al., Structure 2:1121-1123 (1994).) В некоторых случаях, одна или более CDR от описанного в данном описании антитела могут быть включены в молекулу посредством ковалентного или нековалентного связывания с образованием иммуноадгезина, который специфически связывается с CCR2. В таких случаях, последовательность(и) CDR может(могут) быть включена(ы) как часть более крупной полипептидной цепи, либо она(они) может(могут) быть ковалентно присоединена(ы) к другой полипептидной цепи, либо она(они) может(могут) быть включена(ы) посредством нековалентного связывания. В случае присутствия одного или нескольких сайтов связывания, такие сайты связывания могут быть идентичными, либо они могут отличаться друг от друга.
Используемый в данном описании термин «человеческое антитело» означает любое антитело, в котором последовательности вариабельного и константного доменов представляют собой человеческие последовательности. Этот термин охватывает антитела, последовательности которых происходят от генов человека, но которые были модифицированы, например, для снижения возможной иммуногенности, повышения аффинности, удаления цистеинов, которые могут образовывать нежелательную укладку и т.п. Этот термин также охватывает антитела, которые были продуцированы рекомбинантными методами в не-человеческих клетках, и которые могут сообщать характер гликозилирования, нетипичный для человеческих клеток. Эти антитела могут быть получены различными методами, описанными в настоящей заявке.
Используемый в данном описании термин «химерное антитело» означает антитело, содержащее области, происходящие от двух или более различных антител. В одном случае, одна или более CDR химерного антитела происходят от CDR человеческого анти-CCR2 антитела. В другом случае, все CDR происходят от CDR человеческого анти-CCR2 антитела. В другом варианте, CDR от более чем одного человеческого анти-CCR2 антитела объединяют друг с другом с получением химерного антитела. Так, например, химерное антитело может содержать CDR1 легкой цепи первого человеческого анти-CCR2 антитела, CDR2 легкой цепи второго человеческого анти-CCR2 антитела, и CDR3 легкой цепи третьего человеческого анти-CCR2 антитела, а CDR тяжелой цепи могут происходить от CDR одного или нескольких других анти-CCR2 антител. Кроме того, каркасные области могут происходить от одного из анти-CCR2 антител, от которого происходят одна или более CDR, или от одного или нескольких других человеческих антител.
В некоторых случаях, химерным антителом является гуманизованное анти-CCR2 антитело. Гуманизованное анти-CCR2 антитело содержит аминокислотную последовательность одной или нескольких каркасных областей и/или аминокислотную последовательность по меньшей мере части константной области одного или нескольких человеческих анти-CCR2 антител и CDR, происходящие от нечеловеческого анти-CCR2 антитела.
Фрагменты или аналоги антител или молекул иммуноглобулинов могут быть легко получены средним специалистом в данной области исходя из описания, приведенного в настоящей заявке. Предпочтительные амино- и карбокси-концы фрагментов или аналогов расположены почти на границе функциональных доменов.
Используемый в данном описании термин «поверхностный плазмонный резонанс» означает оптическое явление, позволяющее осуществлять анализ биоспецифических взаимодействий в реальном времени посредством детектирования изменений концентраций белка в биосенсорной матрице, например, с использованием системы BIACORE™ (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, N.J.). Более подробное описание этого метода можно найти у Jonsson U. et al., Ann. Biol. Clin. 51:19-26 (1993); Jonsson U. et al., Biotechniques 11:620-627 (1991); Jonsson B. et al., J. Mol. Recognit. 8:125-131 (1995); и Johnsson B. et al., Anal. Biochem. 198:268-277 (1991).
Термин «КD» означает константу равновесной диссоциации конкретного взаимодействия «антитело-антиген». Считается, что антитело специфически связывается с антигеном, если константа диссоциации составляет ≤1 мМ, ≤100 нМ или ≤10 нМ. В некоторых случаях, КD составляет от 1 пМ до 500 пМ. В других случаях, КD составляет от 500 пМ до 1 мкМ, от 1 мкМ до 100 нМ или от 100 мМ до 10 нМ.
Термин «эпитоп» включает любую белковую детерминанту, обладающую способностью специфически связываться с иммуноглобулином или с Т-клеточным рецептором или взаимодействовать с молекулой каким-либо другим образом. Эпитопные детерминанты обычно состоят их химически активных поверхностных групп молекул, таких как аминокислоты или углеводные или сахарные боковые цепи, и обычно обладают специфическими свойствами трехмерной структуры, а также специфическими зарядовыми свойствами. Эпитоп может быть «линейным» или «конформационным». В линейном эпитопе, все сайты взаимодействия белка и взаимодействующей молекулы (такой как антитело) расположены в один ряд на первичной аминокислотной последовательности белка. В конформационном эпитопе, сайты взаимодействия, присутствующие на белке, состоят из аминокислотных остатков, которые отделены друг от друга на цепи, но стерически близки друг к другу. После определения нужного эпитопа на антигене могут быть получены антитела против этого эпитопа, например, методами, описанными в настоящей заявке. Альтернативно, в процессе идентификации, продуцирование и характеризация антител позволяет получить данные о желаемых эпитопах. Исходя из этих данных, можно осуществлять скрининг на конкурентное связывание антител с одним и тем же эпитопом. Для достижения этого могут быть проведены исследования на конкурентное и перекрестное конкурентное связывание с получением антител, которые будут конкурировать или перекрестно конкурировать друг с другом за связывание с CCR2, например, антител, конкурентно связывающихся с антигеном. Высокоэффективный способ «картирования» антител, основанный на их перекрестной конкуренции за связывание, описан в Международной патентной заявке No. WO 03/48731.
Используемые в данном описании двадцать незаменимых аминокислот и их обозначения определены в настоящем описании в соответствии со стандартной практикой. См. руководство Immunology - A Synthesis (second Edition, E.S. Golub and D. R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991)), которое вводится в настоящее описание посредством ссылки.
Термин «полинуклеотид» означает полимерную форму нуклеотидной последовательности, состоящей по меньшей мере, 10 нуклеотидов, либо рибонуклеотидной или дезоксинуклеотидной последовательности, либо модифицированную форму, состоящую из нуклеотидов любого типа. Этот термин включает одноцепочечные и двухцепочечные формы ДНК.
Используемый в данном описании термин «полинуклеотид» означает полинуклеотид, происходящий от геномной ДНК, кДНК или синтетической ДНК или некоторых их комбинаций, где «выделенный полинуклеотид», по своей природе (1) не ассоциируется с полноразмерными полинуклеотидами или их частями, с которыми данный «выделенный полинуклеотид» встречается в природе, (2) функционально присоединен к полинуклеотиду, с которым он не связан в природе или (3) не встречается в природе как часть более крупной последовательности.
Используемый в данном описании термин «природные нуклеотиды» означает дезоксирибонуклеотиды и рибонуклеотиды. Используемый в данном описании термин «модифицированные нуклеотиды» означает нуклеотиды с модифицированными или с замещенными сахарными группами и т.п. Используемый в данном описании термин «олигонуклеотидные связи» включает такие олигонуклеотидные связи, как фосфортиоат, фосфордитиоат, фосфорселеноат, фосфордиселеноат, фосфоранилотиоат, фосфоранилидат, фосфорамидат и т.п. См., например, публикации LaPlanche et al., Nucl. Acids Res. 14:9081 (1986); Stec et al., J. Am. Chem. Soc. 106:6077 (1984); Stein et al., Nucl. Acids Res. 16:3209 (1988); Zon et al., Anti-Cancer Drug Design 6:539 (1991); Zon et al. Oligonucleotides and Analogues: A Practical Approach, pp. 87-108 (F. Eckstein, Ed., Oxford University Press, Oxford England (1991)); патент США No. 5151510; Uhlmann and Peyman, Chemical Reviews 90:543 (1990), которые включены в настоящее описание посредством ссылки. Если это необходимо, то олигонуклеотид может включать метку для детектирования.
«Функционально присоединенные» последовательности включают последовательности регуляции экспрессии, которые являются смежными с представляющим интерес геном, и последовательности регуляции экспрессии, которые действуют in trans или на расстоянии от представляющего интерес гена, находящегося под контролем данных последовательностей. Используемый в данном описании термин «последовательность регуляции экспрессии» означает полинуклеотидные последовательности, необходимые для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они связаны. Последовательностями регуляции экспрессии являются соответствующие последовательности инициации и терминации транскрипции, промоторные и энхансерные последовательности; сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (то есть, консенсусная последовательность Козака); последовательности, повышающие стабильность белка, и, если это необходимо, последовательности, усиливающие секрецию белка. Природа таких регуляторных последовательностей варьируется в зависимости от организма-хозяина; причем, в прокариотах, такие регуляторные последовательности обычно включают промотор, сайт связывания с рибосомой и последовательность терминации транскрипции; а в эукариотах, такими регуляторными последовательностями обычно являются промоторы и последовательности терминации транскрипции. Термин “регуляторные последовательности" включает, как минимум, все компоненты, присутствие которых является необходимым для осуществления экспрессии и процессинга, а также он может включать дополнительные компоненты, присутствие которых является предпочтительным, например, лидерные последовательности и последовательности, являющиеся партнерами для гибридных последовательностей.
Используемый в данном описании термин «вектор» означает молекулу нуклеиновой кислоты, способную осуществлять транспорт другой нуклеиновой кислоты, с которой она связана. В некоторых случаях, вектором является плазмида, то есть, кольцевая двухцепочечная молекула ДНК, с которой могут быть лигированы дополнительные сегменты ДНК. В некоторых случаях, вектором является вирусный вектор, в вирусный геном которого могут быть лигированы дополнительные сегменты ДНК. В некоторых случаях, векторы обладают способностью автономно реплицироваться в клетке-хозяине, в которую они были введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации и эписомные векторы млекопитающих). В других случаях, векторы (например, не-эписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина после введения в данную клетку-хозяина, и таким образом, могут реплицироваться вместе с геномом хозяина. Кроме того, некоторые векторы обладают способностью направлять экспрессию генов, к которым они функционально присоединены. Такие векторы называются в данном описании «рекомбинантными экспрессионными векторами» (или просто «экспрессионными векторами»).
Используемый в данном описании термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») означает клетку, в которую был введен рекомбинантный экспрессионный вектор. Следует отметить, что термины «рекомбинантная клетка-хозяин» и «клетка-хозяин» относятся не только к клетке конкретного индивидуума, но также и к потомству такой клетки. Поскольку в последующих генерациях могут присутствовать некоторые модификации, обусловленные либо мутациями, либо воздействием окружающей среды, то это потомство, фактически, не может быть идентичным родительской клетке, однако, такое потомство все же входит в объем используемого в данном описании термина «клетка-хозяин».
Термин «процент идентичности последовательностей», используемый по отношению к нуклеотидным последовательностям, означает остатки в двух последовательностях, которые являются идентичными при их выравнивании на максимальное соответствие. Для определения идентичности нуклеотидных последовательностей существует ряд различных алгоритмов, известных специалистам. Так, например, полинуклеотидные последовательности могут быть подвергнуты сравнению с использованием компьютерных программ FASTA, Gap или Bestfit, которые входят в пакет программ Wisconsin Package Version 10.0, Genetics Computer Group (GCG), Madison, Wisconsin. Программа FASTA, которая включает, например, программы FASTA2 и FASTA3, позволяет осуществлять выравнивание и определять процент идентичности последовательностей в областях наибольшего перекрывания между запрашиваемой последовательностью и исследуемыми последовательностями (Pearson, Methods Enzymol. 183:63-98 (1990); Pearson, Methods Mol. Biol. 132:185-219 (2000); Pearson, Methods Enzymol. 266:227-258 (1996); Pearson, J. Mol. Biol. 276:71-84 (1998); эти работы вводятся в настоящее описание посредством ссылки). Если это не оговорено особо, то в конкретных программах и алгоритмах использовались параметры по умолчанию. Так, например, процент идентичности между нуклеотидными последовательностями может быть определен с использованием программы FASTA с параметрами по умолчанию (размер «слова» из 6 «букв» и фактор NOPAM для оценочной матрицы) или с использованием программы Gap с параметрами по умолчанию, указанными в пакете программ GCG Version 6.1, которые включены в настоящее описание посредством ссылки.
Термин «нуклеотидная последовательность» включает ее комплемент, если это не оговорено особо. Таким образом, термин «нуклеиновая кислота, имеющая конкретную последовательность», должен включать ее комплементарную цепь вместе с ее комплементарной последовательностью.
Используемые в данном описании термины «процент идентичности последовательностей» и «процент гомологии последовательностей» являются синонимами.
Термины «по существу, идентичный» или «по существу, идентичные последовательности», если они употребляются по отношению к нуклеиновой кислоте или к ее фрагменту, означают что при оптимальном выравнивании их последовательностей, имеющих соответствующие инсерции или делеции нуклеотидов, с последовательностью другой нуклеиновой кислоты (или с ее комплементарной цепи), идентичность нуклеотидных оснований в таких нуклеотидных последовательностях составляет по меньшей мере примерно 85%, по меньшей мере примерно 90%, и по меньшей мере примерно 95%, 96%, 97%, 98% или 99%, как можно определить с помощью любого хорошо известного алгоритма идентичности последовательностей, такого как FASTA, BLAST или Gap, обсуждаемых выше.
Термин «по существу, идентичный», если он употребляется по отношению к полипептидам, означает, что две пептидных последовательности, при их оптимальном выравнивании, например, с помощью программ GAP или BESTFIT с использованием “весов” пробелов по умолчанию, имеют, по меньшей мере, 70%, 75% или 80%-ную идентичность последовательностей, по меньшей мере, 90%-ную или 95%-ную идентичность последовательностей, и по меньшей мере 97%, 98% или 99%-ную идентичность последовательностей. В некоторых случаях, положения остатков, которые не являются идентичными, отличаются в результате консервативных аминокислотных замен. Термин «консервативная аминокислотная замена» означает замену, при которой аминокислотный остаток заменен другим аминокислотным остатком, имеющим группу R боковой цепи с аналогичными химическими свойствами (например, с зарядом или гидрофобностью). В общих чертах, консервативная аминокислотная замена, по существу, не оказывает влияния на функциональные свойства белка. В случае если две или более аминокислотных последовательностей отличаются друг от друга консервативными заменами, то процент идентичности последовательностей может быть скорректирован в расчете на консервативную природу такой замены. Методы осуществления такой коррекции хорошо известны специалистам. См., например, Pearson, Methods Mol. Biol. 243:307-31 (1994). Примерами групп аминокислот, имеющих боковые цепи с аналогичными химическими свойствами, являются аминокислоты, имеющие (1) алифатические боковые цепи: глицин, аланин, валин, лейцин и изолейцин; (2) алифатические гидроксильные боковые цепи: серин и треонин; (3) амид-содержащие боковые цепи: аспарагин и глутамин; (4) ароматические боковые цепи: фенилаланин, тирозин и триптофан; (5) основные боковые цепи: лизин, аргинин и гистидин; (6) кислотные боковые цепи: аспарагиновая кислота и глютаминовая кислота; и (7) серосодержащие боковые цепи: цистеин и метионин. Консервативными аминокислотными заменами являются замены в пределах таких групп, как валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутамат-аспартат и аспарагин-глутамин.
Альтернативно, консервативной заменой является любая замена, имеющая положительную величину в матрице логарифмического правдоподобия PAM250, описанной в публикации Gonnet et al., Science 256:1443-45 (1992), которая включена в настоящее описание посредством ссылки. «Умеренно консервативной заменой» является любая замена, имеющая неотрицательную величину в матрице логарифмического правдоподобия PAM250.
Идентичность последовательностей полипептидов обычно определяют с использованием компьютерной программы для анализа последовательностей. Компьютерная программа для анализа позволяет проводить сравнение последовательностей с использованием параметров сходства, присвоенных различным заменам, делециям и другим модификациям, включая консервативные аминокислотные замены. Так, например, пакет программ GCG содержит программы, такие как «Gap» и «Bestfit», которые могут быть использованы с параметрами по умолчанию, установленными в этих программах для определения гомологии последовательностей или идентичности последовательностей между близкородственными полипептидами, такие как гомологичные полипептиды, происходящие от различных организмов, или последовательностей белков дикого типа и их мутеинов. См., например, GCG Version 6.1 (University of Wisconsin, Wl). Полипептидные последовательности могут быть также подвергнуты сравнению с использованием программы FASTA с параметрами по умолчанию или с рекомендованными параметрами, см., например, GCG Version 6.1. Программа FASTA (например, FASTA2 и FASTA3) позволяет осуществлять выравнивание и определять процент идентичности последовательностей в областях наибольшего перекрывания между запрашиваемой последовательностью и исследуемыми последовательностями (Pearson, Methods Enzymol. 183:63-98 (1990); Pearson, Methods Mol. Biol. 132:185-219 (2000)). Другим предпочтительным алгоритмом, используемым для сравнения последовательности с последовательностями, имеющимися в базе данных, содержащей большое число последовательностей от других организмов, является компьютерная программа BLAST, а в частности, blastp или tblastn, в которых используются параметры по умолчанию, установленные в данной программе. См., например, Altschul et al., J. Mol. Biol. 215:403-410 (1990); Altschul et al., Nucleic Acids Res. 25:3389-402 (1997).
CCR2 представляет собой белок из семи трансмембранных доменов, и в соответствии с этим имеет 6 петель. Петли 1, 3 и 5, если считать от внеклеточного N-конца, представляют собой внутриклеточные петли, а петли 2, 4 и 6 являются внеклеточными. Первая, вторая и третья внеклеточные петли CCR2 обозначаются петлями 2, 4 и 6, соответственно.
В настоящем описании, при употреблении слова «содержать» или его вариантов, таких как «содержит» или «содержащий», подразумевается, что данный объект включает определенное целое число элементов или группу элементов, но не исключает присутствия любого другого элемента или группы элементов.
Человеческие анти-CCR2 антитела и их характеризация
В некоторых вариантах, настоящее изобретение относится к человеческим анти-CCR2 антителам. В некоторых вариантах, человеческие анти-CCR2 антитела продуцируют путем иммунизации не-человеческого трансгенного животного, например, кролика, геном которого включает гены человеческого иммуноглобулина, так, чтобы данное трансгенное животное продуцировало человеческие антитела. В некоторых вариантах, анти-CCR2 антителами или антигенсвязывающими частями являются, но не ограничиваются ими, антитела или антигенсвязывающие части, которые (i) связываются с первой или второй внеклеточной петлей CCR2, или с обеими петлями; (ii) не связываются с N-концом или с третьей внеклеточной петлей CCR2 или с тем и с другим; или (iii) осуществляют функции, указанные в (i) и (ii). В другом своем варианте, настоящее изобретение относится к человеческим анти-CCR2 антителам, которые связываются с полипептидом, содержащим аминокислотную последовательность SEQ ID NO:128 или SEQ ID NO: 129. В другом варианте изобретения, человеческие анти-CCR2 антитела связываются с полипептидом, содержащим аминокислотную последовательность, которая на 80, 85, 90, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 128 или SEQ ID NO: 129. В другом варианте, анти-CCR2 антителами и их антигенсвязывающими частями являются, но не ограничиваются ими, антитела или их антигенсвязывающие части, связывающиеся с третьей внеклеточной петлей CCR2.
Гены VH, VК, Vλ подразделяются на семейства исходя из их гомологии последовательностей. Два гена VH, VК или Vλ принадлежат к одному и тому же семейству, если их нуклеотидные последовательности идентичны в более чем 80% положений. Анти-CCR2 антитело может содержать человеческую легкую цепь каппа (VК) или человеческую легкую цепь лямбда (Vλ) или происходящую от них аминокислотную последовательность. В некоторых вариантах антител, содержащих легкую цепь лямбда, в вариабельном домене легкой цепи (VL) используется человеческий ген семейства Vλ1, Vλ2, Vλ3, Vλ4, Vλ5, Vλ6, Vλ7, Vλ8, Vλ9 или Vλ10 (Williams S.C. et al. J. Mol. Bio. 264:220-232 (1996)).
В некоторых вариантах антител, содержащих легкую цепь каппа, в вариабельном домене легкой цепи (VL) используется человеческий ген семейства VKI, VKII, VKIII, VKIV, VKV или VKVI (Cox J. P. L., et al., Eur. J. Immunol 24:827-836 (1994)), предпочтительно, VKI, VKII, VKIV или VKVI, а более предпочтительно, ген семейства VKI или VKVI. В некоторых вариантах, последовательность зародышевой линии легкой цепи выбрана из человеческих последовательностей VK, включая, но не ограничиваясь ими, A1, A10, A11, A14, A17, A18, A19, A2, A20, A23, A26, A27, A3, A30, A5, A7, B2, B3, L1, L10, L11, L12, L14, L15, L16, L18, L19, L2, L20, L22, L23, L24, L25, L4/18a, L5, L6, L8, L9, О1, О11, О12, О14, О18, О2, О4 и О8. В некоторых случаях, этот ген легкой цепи человеческой зародышевой линии выбран из V1-11, V1-13, V1-16, V1-17, V1-18, V1-19, V1-2, V1-20, V1-22, V1-3, V1-4, V1-5, V1-7, V1-9, V2-1, V2-11, V2-13, V2-14, V2-15, V2-17, V2-19, V2-6, V2-7, V2-8, V3-2, V3-3, V3-4, V4-1, V4-2, V4-3, V4-4, V4-6, V5-1, V5-2, V5-4 и V5-6. В некоторых случаях, в легкой цепи используется ген человеческой зародышевой линии VKI О12, VKII A1, VKIV B3 или VKVI A26.
Анти-CCR2 антитело может содержать вариабельный домен тяжелой цепи (VH), в котором используется человеческий ген семейства VH1, VH2, VH3, VH4, VH5, VH6 или VH7. В конкретных примерах, этот ген тяжелой цепи человеческой зародышевой линии выбран из VH1-18, VH1-2, VH1-24, VH1-3, VH1-45, VH1-46, VH1-58, VH1-69, VH1-8, VH2-26, VH2-5, VH2-70, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-7, VH3-72, VH3-73, VH3-74, VH3-9, VH4-28, VH4-31, VH4-34, VH4-39, VH4-4, VH4-59, VH4-61, VH5-51, VH6-1 и VH7-81. В некоторых случаях, в тяжелой цепи используется человеческий ген VHI 1-46 или VH III 3-30.
В конкретных случаях, вариабельная область легкой цепи и/или вариабельная область тяжелой цепи содержит каркасную область или по меньшей мере часть каркасной области (например, содержащую 2 или 3 подобласти, такие как FR2 и FR3). В некоторых случаях, по меньшей мере FRL1, FRL2, FRL3 или FRL4 являются полностью человеческими. В других примерах, по меньшей мере FRH1, FRH2, FRH3 или FRH4 являются полностью человеческими. В некоторых случаях, по меньшей мере FRL1, FRL2, FRL3 или FRL4 представляют собой последовательности зародышевой линии (например, человеческой зародышевой линии) или содержат человеческие консенсусные последовательности для конкретной каркасной области (легко доступные из источников известных человеческих последовательностей Ig, описанных в настоящей заявке). В других примерах, по меньшей мере FRH1, FRH2, FRH3 или FRH4 представляют собой последовательности зародышевой линии (например, человеческой зародышевой линии) или содержат человеческие консенсусные последовательности для конкретной каркасной области.
В некоторых случаях, VL анти-CCR2 антитела содержит одну или более аминокислотных замен, делеций и/или инсерций по сравнению с аминокислотной последовательностью зародышевой линии человеческого гена. В некоторых случаях, VL анти-CCR2 антитела содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных замен по сравнению с аминокислотной последовательностью зародышевой линии. В некоторых случаях, в CDR легкой цепи имеется одна или более таких замен, делеций и/или инсерций. В некоторых случаях, аминокислотные замены, делеции и/или инсерции, по сравнению с зародышевой линией, находятся в одном или нескольких положениях, идентичных положениям с заменами, делециями и/или инсерциями по сравнению с зародышевой линией в любой одной и/или нескольких VL антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P. Так, например, VL анти-CCR2 антитела может содержать одну или более аминокислотных замен, делеций и/или инсерций по сравнению с аминокислотами зародышевой линии, присутствующими в VL антитела 4.40. В некоторых случаях, аминокислотные модификации присутствуют в одном или нескольких одинаковых положениях, но представляют собой другие модификации, а именно другие замены, делеции и/или инсерции по сравнению с аминокислотами зародышевой линии. В некоторых случаях, замена может представлять собой консервативные аминокислотные замены в указанном(ых) положении(ях) по сравнению с положениями аминокислот в сравниваемом антителе. Так, например, если в конкретном положении в одном из этих антител имеется замена по сравнению с положением в зародышевой линии, и такой заменой является замена на глутамат, то в этом положении может быть осуществлена замена на аспартат. Аналогичным образом, если аминокислотной заменой по сравнению с аминокислотной последовательностью зародышевой линии, является серин, то в этом положении может быть осуществлена консервативная замена серина треонином.
В некоторых случаях, легкая цепь человеческого анти-CCR2 антитела содержит аминокислотную последовательность вариабельного домена (VL) антитела 4.40 (SEQ ID NO: 101); 4.9 (SEQ ID NO:29); 4.22 (SEQ ID NO:65); 4.39 (SEQ ID NO:194); или 4.40 A68G S230P (SEQ ID NO:113); либо указанную аминокислотную последовательность, содержащую до 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 консервативных аминокислотных замен и/или всего до 3 неконсервативных аминокислотных замен. В некоторых случаях, легкая цепь может содержать CDR1, CDR2 и CDR3, независимо выбранные из CDR1, CDR2 и CDR3 легкой цепи, соответственно, антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P, или CDR, каждая из которых имеет менее 4 или менее 3 консервативных аминокислотных замен и/или всего три или менее неконсервативных аминокислотных замен. В некоторых случаях, легкая цепь анти-CCR2 антитела содержит CDR1, CDR2 и CDR3 легкой цепи, каждая из которых независимо выбрана из областей CDR1, CDR2 и CDR3 легкой цепи моноклонального антитела 4.40 (SEQ ID NO:100); 4.9 (SEQ ID NO:28); 4.22 (SEQ ID NO:64); 4.39 (SEQ ID NO:193); или 4.40 A68G S230P (SEQ ID NO:112). В некоторых случаях, легкая цепь анти-CCR2 антитела включает CDR1, CDR2 и CDR3 легкой цепи антитела, содержащего аминокислотную последовательность области VL антитела, выбранного из антител 4.40 (SEQ ID NO: 101); 4.9 (SEQ ID NO:29); 4.22 (SEQ ID NO:65); 4.39 (SEQ ID NO:194); или 4.40 A68G S230P (SEQ ID NO:113); либо указанные CDR, каждая из которых содержит менее 4 или менее 3 консервативных аминокислотных замен и/или всего три или менее неконсервативных аминокислотных замен. Идентификационные номера последовательностей для CDR некоторых антител представлены в таблице 8.
Что касается тяжелой цепи, то в некоторых случаях, в вариабельном домене (VH) используется последовательность человеческих генов VH 3-30 или VH 1-46. В некоторых случаях, последовательность VH анти-CCR2 антитела содержит одну или более аминокислотных замен, делеций и/или инсерций (добавлений) по сравнению с аминокислотной последовательностью зародышевой линии. В некоторых случаях, вариабельный домен тяжелой цепи содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 замен, делеций и/или инсерций по сравнению с аминокислотной последовательностью зародышевой линии. В некоторых случаях, указанной заменой, в отличие от аминокислотной последовательности зародышевой линии, является неконсервативная замена. В некоторых случаях, в CDR тяжелой цепи имеются замены, делеции и/или инсерции. В некоторых случаях, аминокислотная замена, делеция и/или инсерция присутствуют в одном или нескольких одинаковых положениях в качестве мутаций зародышевой линии в любом одном или нескольких VH антител 4.40, 4.22, 4.39 или 4.9. В других случаях, аминокислотные замены, делеции и/или инсерции присутствуют в одном или нескольких одинаковых положениях, но представляют собой другие замены, делеции и/или инсерции по сравнению с эталонным антителом. В некоторых случаях, антитело содержит CDR3 тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO:202 или SEQ ID NO:203.
В некоторых случаях, тяжелая цепь содержит аминокислотную последовательность VH антитела 4.40 (SEQ ID NO:83); 4.22 (SEQ ID NO:47); 4.9 (SEQ ID NO:11); 4.39 (SEQ ID NO:176); либо указанную аминокислотную последовательность VH, имеющую до 1, 2, 3, 4, 6, 8 или 10 консервативных аминокислотных замен и/или до 3 неконсервативных аминокислотных замен. В некоторых случаях, тяжелая цепь содержит аминокислотную последовательность, начинающуюся от CDR1 и заканчивающуюся CDR3 любого одного из вышеуказанных антител.
В некоторых случаях, тяжелая цепь содержит CDR1, CDR2 и CDR3 антитела 4.40, 4.22, 4.39 или 4.9, либо указанные CDR, каждая из которых имеет менее 8, менее 6, менее 4 или менее 3 консервативных аминокислотных замен и/или всего три или менее неконсервативных аминокислотных замен.
В некоторых случаях, CDR тяжелой цепи независимо выбраны из CDR антител 4.40, 4.22, 4.39 или 4.9. В другом случае, тяжелая цепь содержит CDR, независимо выбранные из двух или более областей VH, выбранных из областей антител 4.40 (SEQ ID NO:83); 4.22 (SEQ ID NO:47); 4.39 (SEQ ID NO:176) или 4.9 (SEQ ID NO:11). В другом случае, антитело содержит легкую цепь, описанную выше, и тяжелую цепь, описанную выше. В другом случае, CDR легкой цепи и CDR тяжелой цепи происходят от одного и того же антитела.
Аминокислотной заменой одного типа, которая может быть введена, является замена одного или нескольких цистеинов в антителе, которые могут быть химически реакционноспособными, другим остатком, таким как, но не ограничивающимся ими, аланин или серин. В одном из вариантов, такой заменой является замена не-канонического цистеина. Такая замена может быть сделана в CDR или в каркасной области вариабельного домена или в константном домене антитела. В некоторых случаях, указанный цистеин является каноническим.
Аминокислотная замена другого типа, которая может быть сделана, может представлять собой замену любого из потенциальных протеолитических сайтов в антителе. Такие сайты могут присутствовать в CDR или в каркасной области вариабельного домена или в константном домене антитела. Замена цистеиновых остатков и удаление протеолитических сайтов может снизить риск возникновения любой гетерогенности в продукте антитела, и тем самым повышать его гомогенность. Аминокислотная замена другого типа может быть сделана в целях элиминации аспарагин-глициновой пары, которая может образовывать потенциальные сайты дезамидирования, и может быть введена путем замены одного или обоих этих остатков. В некоторых случаях, аминокислотную замену осуществляют для введения или удаления сайта гликозилирования. В некоторых случаях, C-концевой лизин тяжелой цепи анти-CCR2 антитела может быть протеолитически или генетически удален. В различных случаях, тяжелая и легкая цепи анти-CCR2 антител могут включать, но необязательно, сигнальную последовательность.
В одном из аспектов изобретения, антитела получают гибридомным методом.
В таблице 1 приводятся идентификационные номера последовательностей (SEQ ID NOs) нуклеиновых кислот, кодирующих полноразмерные антитела и их части, содержащие вариабельный домен тяжелой и легкой цепей, и соответствующие выведенные аминокислотные последовательности репрезентативных антител.
Настоящее изобретение также относится к вариантам тяжелой цепи и/или легкой цепи некоторых вышеперечисленных человеческих анти-CCR2 антител, содержащих одну или более аминокислотных замен. При обозначении этих вариантов, первой буквой является однобуквенный символ аминокислоты цепи природного антитела, цифра обозначает положение аминокислоты (где положение 1 означает N-концевую аминокислоту), а второй буквой является однобуквенный символ аминокислотного варианта. В некоторых своих вариантах, настоящее изобретение относится к вариантам тяжелой цепи. Одним из таких вариантов тяжелой цепи является шарнирная область, стабилизирующая мутацию для снижения уровня образования полумономера (Angal, S. et al. Molecular Immunology 30: 105-108 (1993)). В одной из таких шарнирных областей, стабилизирующих мутацию варианта тяжелой цепи антитела 4.40, имеется замена серина пролином в положении 230 SEQ ID NO:82. Последовательность ДНК, кодирующая вариант S230P, имеет кодон CCA, начинающийся в положении 688 SEQ ID NO:115.
Настоящее изобретение также относится к вариантам легких цепей моноклонального антитела 4.40. A68G представляет собой вариант легкой цепи антитела 4.40, представленный последовательностью SEQ ID NO:113, где остаток в положении 68 представляет собой глициновый остаток. В последовательности ДНК, A68G-вариант 4.40 кодируется последовательностью SEQ ID NO:109, где кодоном, начинающимся с положения 252, является кодон GGG.
В других случаях могут быть получены антитела, содержащие комбинации аминокислотных вариантов. Примером комбинации вариантов является анти-CCR2 антитело 4.40 A68G S230P, содержащее легкую цепь с заменой A68G и тяжелую цепь с заменой S230P в последовательности антитела 4.40. Были включены и другие комбинации вариантов тяжелой цепи и легкой цепи антитела 4.40.
В одном из вариантов, анти-CCR2 антителом являются антитела 4.40, 4.22, 4.39, 4.40 A68G S230P или 4.9. В других вариантах, настоящее изобретение относится к антителам, содержащим вариабельный домен, аминокислотная последовательность которого более, чем на 80%, более, чем на 85%, более, чем на 90%, более, чем на 95%, более, чем на 96%, более, чем на 97%, более, чем на 98% или более, чем на 99% идентична аминокислотной последовательности вариабельного домена любого из вышеупомянутых человеческих анти-CCR2 антител.
Классы и подклассы анти-CCR2 антител
Классы и подклассы анти-CCR2 антител могут быть определены любым методом, известным специалистам. В основном, классы и подклассы антитела могут быть определены с использованием антител, которые являются специфичными для конкретного класса и подкласса антитела. Такие антитела являются коммерчески доступными. Классы и подклассы могут быть определены с помощью ELISA или вестерн-блот-анализа, а также другими методами. Альтернативно, классы и подклассы антитела могут быть определены путем секвенирования полноразмерных константных доменов тяжелой и/или легкой цепи антител или их частей; сравнения их аминокислотных последовательностей с известными аминокислотными последовательностями иммуноглобулинов различных классов и подклассов, и определения класса и подкласса антител.
В некоторых случаях, анти-CCR2 антителом является моноклональное антитело. Анти-CCR2 антителом может быть молекула IgG, IgM, IgE, IgA или IgD. В одном из вариантов, анти-CCR2 антителом является IgG, принадлежащий, например, к подклассу IgG1, IgG2, IgG3 или IgG4. В другом случае, анти-CCR2 антителом является IgG4. В другом случае, указанным антителом является изоаллотип IgG4 (Ellison J. and Hood L., PNAS 79:1984-1988 (1982); & Brusco A. et al., Eur J. Immunogenticsl 25:349- 355 (1998)).
Аффинность связывания анти-CCR2 антител с CCR2
В некоторых случаях, анти-CCR2 антитела связываются с CCR2 с высокой аффинностью.
В некоторых случаях, анти-CCR2 антитела обладают высокой аффинностью связывания с первой внеклеточной петлей CCR2, со второй внеклеточной петлей CCR2, или с эпитопом, образованным 1-й и 2-й внеклеточными петлями.
В родственном варианте, анти-CCR2 антитела связываются с полипептидом, состоящим из аминокислотной последовательности, представленной в SEQ ID NO: 128 или в SEQ ID NO:129.
В некоторых случаях, анти-CCR2 антитела не связываются с третьей внеклеточной петлей CCR2 или с N-концевым доменом CCR2.
В другом случае, анти-CCR2 антитела не связываются с пептидом, состоящим из последовательности, представленной в SEQ ID NO:127 или в SEQ ID NO:130. В другом случае, анти-CCR2 антитело связывается с первой и/или второй внеклеточными петлями CCR2, с KD примерно 2×10-7 М или менее, KD примерно 2×10-8 M или менее, KD примерно 2×10-9 M или менее, KD примерно 1×10-9 M или менее, KD примерно 9×10-10 M или менее, KD примерно 8×10-10 M или менее, KD примерно 7×10-10 M или менее, KD примерно 6×10-10 M или менее, KD примерно 5×10-10 M или менее, KD примерно 4×10-10 M или менее, KD примерно 3×10-10 M или менее, или KD примерно 2×10-10 M или менее. В некоторых случаях, указанное антитело связывается с CCR2, или с первой и/или второй внеклеточными петлями CCR2, в основном, при КD, аналогичном КD антитела, выбранного из антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P. В другом случае, указанное антитело связывается с CCR2, или с первой и/или второй внеклеточными петлями CCR2, в основном, при КD, аналогичном КD антитела, которое содержит вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность области VH, присутствующей в SEQ ID NO:83, SEQ ID NO:11, SEQ ID NO: 176 или SEQ ID NO:47. В еще одном варианте, указанное антитело связывается с CCR2, в основном, при КD, аналогичном КD антитела, которое содержит CDR вариабельного домена легкой цепи, имеющего последовательность области VL, присутствующей в SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:29, SEQ ID NO:194 или SEQ ID NO:65, или которое содержит CDR вариабельного домена тяжелой цепи, имеющего аминокислотную последовательность области VH, присутствующей в SEQ ID NO:83, SEQ ID NO:11, SEQ ID NO:176 или SEQ ID NO:47.
В некоторых случаях, анти-CCR2 антитело может иметь низкую константу скорости диссоциации (koff). В некоторых случаях, анти-CCR2 антитело может связываться с CCR2, или более предпочтительно, с первой и/или второй внеклеточными петлями CCR2, с koff 1,0×10-3 с-1 или менее, koff 5,0×10-4 с-1 или менее, или с koff 2×10-4 с-1 или менее. В некоторых случаях, koff может быть, в основном, аналогичен koff описанного здесь антитела, включая антитело, выбранное из антител 4.40, 4.9, 4.22, 4.39 и 4.40 A68G S230P. В некоторых случаях, антитело может связываться с CCR2 или с первой и/или второй внеклеточными петлями CCR2, с koff, в основном, аналогичным koff антитела, которое содержит CDR тяжелой цепи или CDR легкой цепи и которое выбрано из антител 4.40, 4.9 и 4.40 A68G S230P. В некоторых случаях, указанное антитело может связываться с CCR2 или с первой и/или второй внеклеточными петлями CCR2, с koff, в основном, аналогичным koff антитела, которое содержит (i) вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность области VH, присутствующей в SEQ ID NO:83 или SEQ ID NO:11, (ii) вариабельный домен легкой цепи, имеющий аминокислотную последовательность области VL, присутствующей в SEQ ID NO: 101, SEQ ID NO: 113, SEQ ID NO:29, или (iii) вариабельные домены (i) и (ii). В другом случае, указанное антитело может связываться с CCR2, или с первой и/или второй внеклеточными петлями CCR2, с koff, в основном, аналогичным koff антитела, которое содержит CDR вариабельного домена легкой цепи, имеющего аминокислотную последовательность области VL, присутствующей в SEQ ID NO:101, SEQ ID NO:113 или SEQ ID NO:29; и CDR вариабельного домена тяжелой цепи, имеющего аминокислотную последовательность области VH, присутствующей в SEQ ID NO:83 или SEQ ID NO:11.
Аффинность связывания анти-CCR2 антитела с CCR2 и скорость его диссоциации могут быть определены методами, известными специалистам. Аффинность связывания может быть измерена с помощью ELISA, РИА, проточной цитометрии и поверхностного плазмонного резонанса, такого как BIACORE™. Скорость диссоциации может быть измерена методом поверхностного плазмонного резонанса. С применением известных методов можно определить, имеет ли антитело, в основном, такой же КD, как и анти-CCR2 антитело. В примере 5 описан метод определения констант аффинности для моноклональных анти-CCR2 антител.
Идентификация эпитопов CCR2, распознаваемых анти-CCR2 антителами
Настоящее изобретение относится к человеческим моноклональным анти-CCR2 антителам, которые связываются с CCR2 и могут конкурировать или перекрестно конкурировать за связывание и/или связываться с таким же эпитопом, как и эпитоп для (a) антитела, выбранного из m 4.40, 4.9, 4.22, 4.39 и 4.40 A68G S230P; (b) антитела, содержащего вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность вариабельного домена, присутствующего в SEQ ID NO:83, SEQ ID NO:11, SEQ ID NO:176 или SEQ ID NO:47, (c) антитела, содержащего вариабельный домен легкой цепи, имеющий аминокислотную последовательность вариабельного домена, присутствующего в SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:29, SEQ ID NO:194 или SEQ ID NO:65, или (d) антитела, содержащего вариабельный домен тяжелой цепи, определенный в (b) и вариабельный домен легкой цепи, определенный в (c). Если два антитела взаимно конкурируют друг с другом за связывание с CCR2, то говорят, что эти антитела перекрестно конкурируют.
С применением известных методов можно определить, связывается ли антитело с тем же эпитопом, и конкурирует ли оно или перекрестно конкурирует с описанным здесь анти-CCR2 антителом за связывание. В одном из вариантов обеспечивают связывание рассматриваемого в данном описании анти-CCR2 антитела с CCR2 в условиях насыщения, а затем определяют способность этого тестируемого антитела связываться с CCR2. Если тестируемое антитело обладает способностью связываться с CCR2 одновременно с описанным здесь анти-CCR2 антителом, то такое тестируемое антитело связывается с эпитопом, отличающимся от эпитопа, с которым связывается анти-CCR2 антитело. Однако если тестируемое антитело неспособно одновременно связываться с CCR2, то такое тестируемое антитело связывается с тем же эпитопом, с перекрывающимся эпитопом или с эпитопом, расположенным в непосредственной близости к эпитопу, с которым связывается описанное здесь человеческое анти-CCR2 антитело. Для того чтобы определить, может ли тестируемое антитело перекрестно конкурировать с эталонным антителом, был проведен эксперимент по связыванию антител в обратном порядке, то есть, сначала обеспечивали связывание тестируемого антитела с CCR2, а затем определяли способность рассматриваемого в данном описании анти-CCR2 антитела связываться с CCR2. Эти эксперименты могут быть проведены с помощью ELISA, РИА, BIACORE™ или проточной цитометрии (FACS).
Ингибирование активности CCR2 под действием анти-CCR2 антитела
В некоторых случаях, настоящее изобретение относится к анти-CCR2 антителам, которые ингибируют CCR2-опосредуемую передачу сигнала. В других своих вариантах, настоящее изобретение относится к анти-CCR2 антителам, которые ингибируют MCP-1-, MCP-2-, MCP-3-, и/или MCP-4-опосредуемую передачу сигнала посредством CCR2. В других своих вариантах, настоящее изобретение относится к анти-CCR2 антителам, которые ингибируют связывание MCP-1 MCP-2, MCP-3 и/или MCP-4 с CCR2. В одном из вариантов, CCR2 представляет собой человеческий CCR2. В некоторых случаях, CCR2 представляет собой человеческий CCR2A, CCR2B или т.п. В другом случае, анти-CCR2 антителом является человеческое антитело.
IC50 анти-CCR2 антитела может быть измерена в анализах на связывание с лигандом, таких как ELISA, РИА или в родственных анализах, а также в клеточных анализах, таких как анализы на хемотаксис клеток, экспрессирующих CCR2. В различных вариантах, антитело или его антигенсвязывающая часть ингибируют связывание лиганда MCP-1 с CCR2 при IC50 не более чем 5 мкг/мл, не более, чем 1 мкг/мл, не более, чем 0,5 мкг/мл или не более, чем 0,20 мкг/мл, как было определено с помощью ELISA.
В другом своем варианте, настоящее изобретение относится к анти-CCR2 антителу, которое подавляет активацию CCR2 в присутствии лигандов CCR2, таких как MCP-1 (CCL2), MCP-2, MCP-3, и/или MCP-4. В одном из вариантов изобретения, анти-CCR2 антитело может ингибировать индуцируемую лигандом CCR2 (i) активацию G-белка, (ii) активацию аденилатциклазы, (iii) активацию протеинкиназы (MAPK) митогеном, (iv) мобилизацию кальция в цитозоле, (v) фосфорилирование ERK, (vi) хемотаксис, или (vii) полимеризацию актина. Для того чтобы определить, может ли анти-CCR2 антитело предупреждать, ингибировать или подавлять активацию CCR2 в присутствии MCP-1, вычисляют отношение GTP/GDP для G-белков в клетках, меченных радиоактивно меченным GTP, измеряют уровень включения GTPgS, измеряют приток кальция в цитозоль с использованием кальций-содержащих хромофоров, или измеряют уровень фосфорилирования MAPK в клетке. Анализы для детектирования активации CCR2 и/или связывания MCP-1 с CCR2 описаны, например, Gabrilin et al., Biochem. Biophys. Res. Commun. 327(2):533-40 (2005), и Jimenez-Sainz et al., Mol. Pharmacol. 64(3):773-82 (2003).
В одном из вариантов, уровни активации CCR2 определяют с помощью анализа на хемотаксис. В некоторых случаях, IC50, измеренная с помощью анализа на хемотаксис, составляет не более, чем 5 мкг/мл, не более, чем 1 мкг/мл, не более, чем 0,5 мкг/мл, или не более, чем 0,20 мкг/мл. В примере 10 проиллюстрирован один тип анализа, который позволяет измерять уровень ингибирования CCR2 под действием анти-CCR2 антитела путем мониторинга мобилизации кальция.
В другом аспекте изобретения, контактирование клеток с антителом может приводить к ингибированию экспрессии CCR2 на клеточной поверхности после его инкубирования с указанным антителом. В некоторых случаях, инкубирование может быть проведено в течение короткого периода времени (например, 4 часов) или более длительного периода времени (например, 24 часов). Ингибирование экспрессии CCR2 на клеточной поверхности может быть измерено с помощью вестерн-блот-анализа, ELISA или FACS. В конкретных случаях, контактирование клеток с антителом может приводить по меньшей мере к 6% снижению, по меньшей мере к 10% снижению, по меньшей мере к 20% снижению, по меньшей мере к 30% снижению, или по меньшей мере к 50% снижению уровня экспрессии CCR2 на клеточной поверхности, как было определено с помощью вестерн-блот-анализа или ELISA.
В другом аспекте изобретения, указанное антитело снижает уровень MCP-1-индуцированного фосфорилирования pERK. Ингибирование MCP-1-индуцированного фосфорилирования pERK может быть измерено с помощью вестерн-блот-анализа, ELISA или FACS-анализа. В конкретных случаях, указанное антитело обеспечивает по меньшей мере 6%-ное снижение, по меньшей мере 10%-ное снижение, по меньшей мере 20%-ное снижение, по меньшей мере 30%-ное снижение или по меньшей мере 50%-ное снижение MCP-1-индуцированного фосфорилирования pERK, как было определено с помощью FACS-анализа.
Ингибирование хемотаксиса in vivo под действием анти-CCR2 антител
В соответствии с некоторыми своими вариантами, настоящее изобретение относится к анти-CCR2 антителам, которые ингибируют хемотаксис иммунных клеток in vivo. Иммунными клетками, хемотаксис которых подвергают ингибированию, являются мононуклеарные клетки периферической крови, клетки THP, моноциты, T-лимфоциты памяти, дендритные клетки, базофилы, природные киллеры и адаптивно трансфецированные CCR2+-клетки. В одном из вариантов, анти-CCR2 антитело ингибирует хемотаксис иммунных клеток в ответ на один или более MCP-1, MCP-2, MCP-3 и MCP-4. В одном из вариантов, хемокином является MCP-1. В другом варианте, хемокином является MCP-3. Анти-CCR2 антитело может ингибировать хемотаксис в участки воспаления или повреждения.
В соответствии с некоторыми своими вариантами, настоящее изобретение также включает анти-CCR2 антитела, которые ингибируют хемотаксис неиммунных клеток, включая, но не ограничиваясь ими, фибробласт-подобные синовиоциты (FLS) (см. Garcia-Vicuna et al., Arthritis Rheum. 50(12):3866-77 (2004)), стволовые клетки нервной системы взрослого человека (см. Widera et al., Eur. J. Cell Biol. 83(8):381-7 (2004)) и астроциты человеческого плода (см. Andjelkovic et al., J. Neurosci. Res. 70(2):219-31 (2002)).
В одном из вариантов, ингибирование антителом хемотаксиса клеток сравнивают с ингибированием хемотаксиса клеток у необработанных животных. В другом варианте, анти-CCR2 антитело подавляет хемотаксис по меньшей мере на 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%. В одном из вариантов, ингибирование хемотаксиса определяют по меньшей мере через 1 час после начала обработки животных антителом. В другом варианте, ингибирование хемотаксиса определяют по меньшей мере через 7 дней после начала обработки животных антителом. В другом случае, анти-CCR2 антитело ингибирует хемотаксис по меньшей мере на 10%-100%.
Видовая и молекулярная селективность
В другом аспекте изобретения было продемонстрировано, что анти-CCR2 антитела обладают видовой и молекулярной селективностью. В некоторых случаях, анти-CCR2 антитело связывается с CCR2 человека и собакоподобных обезьян. Анти-CCR2 антитело может связываться с дополнительным CCR2 приматов, не являющихся человеком. В некоторых случаях, анти-CCR2 антитело не связывается с мышиным или крысиным CCR2. В соответствии с описанием, приведенным в данном описании, можно определить видовую селективность для анти-CCR2 антитела методами, хорошо известными специалистам. Так, например, видовая селективность может быть определена с помощью вестерн-блот-анализа, проточной цитометрии, ELISA, иммунопреципитации или РИА. В одном из вариантов, видовая селективность может быть определена с помощью проточной цитометрии. В другом варианте, видоспецифичность может быть определена путем оценки способности антитела ингибировать функциональные ответы MCP-1 с использованием клеток, происходящих от этих видов. Таким ответом может быть хемотаксис, полимеризация актина, мобилизация кальция и т.п.
В другом случае, анти-CCR2 антитело может обладать селективностью по отношению к полипептиду, состоящему из аминокислотной последовательности, представленной в SEQ ID NO:126 (человеческого CCR2B), по сравнению с селективностью к полипептиду, состоящему из аминокислотной последовательности, представленной в SEQ ID NO: 131 (человеческого CCR5). В другом варианте, анти-CCR2 антитело обладает селективностью по отношению к CCR2, которая по меньшей мере в 2 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 25 раз, по меньшей мере в 50 раз или по меньшей мере в 100 раз превышает селективность по отношению к CCR5. В другом варианте, человеческие анти-CCR2 антитела связываются с полипептидом, содержащим аминокислотную последовательность, которая на 80, 85, 90, 95, 96, 97, 98 или 99% идентична последовательности SEQ ID NO: 126.
В другом случае, анти-CCR2 антитело может обладать селективностью по отношению к полипептиду, состоящему из аминокислотной последовательности, представленной в SEQ ID NO: 125 (человеческого CCR2A), где указанная селективность по меньшей мере в 2 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 25 раз, по меньшей мере в 50 раз или по меньшей мере в 100 раз превышает селективность по отношению к полипептиду, состоящему из аминокислотной последовательности, представленной в SEQ ID NO:131 (человеческого CCR5). В другом варианте, человеческие анти-CCR2 антитела связываются с полипептидом, содержащим аминокислотную последовательность, которая на 80, 85, 90, 95, 96, 97, 98 или 99% идентична последовательности SEQ ID NO: 125.
Селективность анти-CCR2 антитела по отношению к CCR2 может быть определена методами, хорошо известными специалистам в соответствии с описанием, приведенным в настоящей заявке. Так, например, селективность может быть определена с помощью вестерн-блот-анализа, проточной цитометрии, ELISA, иммунопреципитации или РИА, и/или функциональных анализов, таких как анализ на хемотаксис, мобилизацию кальция или полимеризацию актина.
Методы получения антител и антитело-продуцирующих клеточных линий
Иммунизация
В некоторых случаях, человеческие антитела получают путем иммунизации трансгенного животного, не являющегося человеком и содержащего в своем геноме некоторые или все локусы тяжелой и легкой цепи человеческого иммуноглобулина, антигеном CCR2. В одном из вариантов, животным, не являющимся человеком, является животное XENOMOUSE™. (Amgen Fremont, Inc. (прежнее название Abgenix, Inc.), Fremont, CA).
Мышь XENOMOUSE™ была создана методом генной инженерии из мышиных штаммов, которые содержат крупные фрагменты локусов тяжелой и легкой цепи человеческого иммуноглобулина, и которые являются дефицитными по продуцированию мышиных антител. См., например, Green et al., Nature Genetics 7:13-21 (1994) и патенты США №№ 5916771, 5939598, 5985615, 5998209, 6075181, 6091001, 6114598, 6130364, 6162963 и 6150584. См. также WO 91/10741, WO 94/02602, WO 96/34096, WO 96/33735, WO 98/16654, WO 98/24893, WO 98/50433, WO 99/45031, WO 99/53049, WO 00/09560 и WO 00/037504.
В другом своем аспекте, настоящее изобретение относится к способу продуцирования анти-CCR2 антител у животных, не являющихся человеком и мышью, путем иммунизации трансгенных животных, которые не являются человеком, и которые содержат локус человеческого иммуноглобулина, антигеном CCR2. Такие животные могут быть созданы методами, описанными в цитируемых выше документах. Методы, описанные в этих документах, могут быть модифицированы как описано в патенте США 5994619, который включен в данное описание посредством ссылки. В патенте США 5994619 описаны методы продуцирования новых клеток и клеточных линий из культивированной внутренней клеточной массы (CICM), полученной от свиней и коров с последующим введением в эти трансгенные клетки CICM гетерологичной ДНК. Трансгенные клетки CICM могут быть использованы для продуцирования клонированных трансгенных эмбрионов, плода и потомства. В патенте '619 также описаны методы создания трансгенных животных, способных передавать гетерологичную ДНК своему потомству. В предпочтительных вариантах изобретения, животные, не являющиеся человеком, представляют собой млекопитающие, а в частности, крысы, овцы, свиньи, козы, крупный рогатый скот или лошади.
Мыши XENOMOUSE™ продуцируют репертуар полностью человеческих антител, подобных антителам взрослого человека, и антигенспецифические человеческие антитела. В некоторых вариантах, мыши XENOMOUSE™ содержат приблизительно 80% репертуара гена V человеческих антител, что обусловлено введением фрагментов локусов человеческой тяжелой цепи и локусов легкой цепи каппа, имеющих конфигурацию зародышевой линии. В других вариантах изобретения, мыши XENOMOUSE™ также содержат приблизительно все локусы человеческой легкой цепи лямбда. См. публикацию Mendez et al., Nature Genetics 15:146-156 (1997), Green and Jakobovits, J. Exp. Med. 188:483-495 (1998), и заявку WO 98/24893, описание которых вводится в настоящее описание посредством ссылки.
В другом случае, антитела получают с применением технологии VELOCIMOUSE™ (Regeneron Pharmaceuticals, Tarrytown, NY.) для быстрого создания генетически модифицированных мышей непосредственно из модифицированных эмбриональных стволовых (ES) клеток (Poueymirou W.T., et al., Nature Biotechnology 25:91-99 (2007)).
В некоторых случаях, животные, не являющиеся человеком и содержащие гены человеческого иммуноглобулина, представляют собой животные, имеющие «минилокус» человеческого иммуноглобулина. В методе с применением минилокуса, экзогенный локус Ig имитируют путем включения отдельных генов от локуса Ig. Так, например, из одного или нескольких генов VH, из одного или нескольких генов DH, из одного или нескольких генов JH, из константного домена «мю» и второго константного домена (предпочтительно, константного домена гамма) создают конструкцию для введения животному. Такой способ описан, inter alia, в патентах США №№ 5545807, 5545806, 5569825, 5625126, 5633425, 5661016, 5770429, 5789650, 5814318, 5591669, 5612205, 5721367, 5789215 и 5643763, которые вводятся в настоящее описание посредством ссылки.
В другом своем аспекте, настоящее изобретение относится к способу получения гуманизованных анти-CCR2 антител. В некоторых случаях, животных, не являющихся человеком, иммунизируют описанным здесь антигеном CCR2 в условиях, обеспечивающих продуцирование антитела. Антитело-продуцирующие клетки выделяют у животных, после чего, из этих клеток выделяют нуклеиновые кислоты, кодирующие тяжелую и легкую цепи представляющего интерес анти-CCR2 антитела. Затем, эти нуклеиновые кислоты конструируют методами, известными специалистам и более подробно описанными ниже, в целях уменьшения количества не-человеческих последовательностей, то есть, в целях гуманизации антитела для снижения иммунного ответа у человека.
В некоторых случаях, антигеном CCR2 является выделенный и/или очищенный CCR2. В одном из вариантов, антигеном CCR2 является человеческий CCR2. В некоторых случаях, антигеном CCR2 является фрагмент CCR2. В некоторых случаях, фрагментом CCR2 является внеклеточная петля, N-концевой домен или С-конец CCR2. В некоторых случаях, фрагмент CCR2 содержит первую или вторую внеклеточную петлю CCR2. В других случаях, фрагмент CCR2 содержит аминокислотную последовательность, представленную в SEQ ID NO:128 или SEQ ID NO:129.
В других случаях, фрагмент CCR2 не содержит третью внеклеточную петлю или N-концевой домен CCR2.
В других случаях, фрагмент CCR2 не содержит аминокислотную последовательность, представленную в SEQ ID NO: 127 или SEQ ID NO: 130. В некоторых случаях, фрагмент CCR2 содержит по меньшей мере один эпитоп CCR2. В других случаях, антигеном CCR2 является клетка, которая экспрессирует или сверхэкспрессирует на своей поверхности CCR2 или его иммуногенный фрагмент. В некоторых случаях, антигеном CCR2 является гибридный белок CCR2. В некоторых случаях, CCR2 представляет собой синтетический пептидный иммуноген.
Иммунизация животных может быть осуществлена любым методом, известным специалистам. См., например, Harlow and Lane, Antibodies: A Laboratory Manual. New York: Cold Spring Harbor Press, 1990. Методы иммунизации животных, не являющихся человеком, таких как мыши, крысы, овцы, козы, свиньи, крупный рогатый скот и лошади, хорошо известны специалистам. См., например, Harlow and Lane, supra, и патент США № 5994619. В одном из вариантов, антиген CCR2 вводят вместе с адъювантом для стимуляции иммунного ответа.
Примерами адъювантов являются полный или неполный адъювант Фрейнда, RIBI (мурамил-дипептиды) или ISCOM (иммуностимулирующие комплексы). Такие адъюванты могут защищать полипептид от быстрого диспергирования посредством его секвестрации в локальном участке, либо они могут содержать вещества, которые стимулируют у хозяина секрецию факторов, обеспечивающих хемотаксис макрофагов и других компонентов иммунной системы. При введении полипептида, схема иммунизации включает два или более введений полипептида с интервалом времени в несколько недель. В примере 1 проиллюстрирован метод продуцирования моноклональных анти-CCR2 антител у мышей XENOMOUSE™.
Получение антител и антитело-продуцирующих клеточных линий
После иммунизации животного антигеном CCR2, от такого животного могут быть получены антитела и/или антитело-продуцирующие клетки. В некоторых случаях, сыворотку, содержащую анти-CCR2 антитело, получают от животного путем забора крови или его умерщвления. Эта сыворотка может быть использована сразу после ее взятия у животного, либо из этой сыворотки может быть выделена фракция иммуноглобулина, либо из этой сыворотки могут быть выделены анти-CCR2 антитела.
В некоторых случаях, антитело-продуцирующие иммортализованные клеточные линии получают из клеток, выделенных у иммунизованного животного. После иммунизации, животное умерщвляют, и клетки периферической крови, клетки лимфоузлов и/или В-клетки селезенки подвергают иммортализации любыми методами, известными специалистам. Методы иммортализации клеток включают, но не ограничиваются ими, трансфекцию онкогенами, инфицирование онкогенным вирусом, и культивирование в условиях, благоприятствующих отбору иммортализованных клеток с последующей их обработкой канцерогенными соединениями или соединениями, вызывающими мутацию, слиянием этих клеток с иммортализованными клетками, например, с миеломными клетками, и инактивацией геном-супрессором опухоли. См., например, Harlow and Lane, supra. Если применяется слияние с миеломными клетками, то миеломные клетки, предпочтительно, не должны секретировать полипептиды иммуноглобулинов (не-секреторная клеточная линия). Иммортализованные клетки скринируют с использованием CCR2 и его части, или с использованием клеток, экспрессирующих CCR2. В одном из вариантов, часть CCR2 (i) содержит первую и/или вторую внеклеточные петли CCR2; (ii) содержит аминокислотную последовательность, представленную в SEQ ID NO: 128 и/или SEQ ID NO: 129; (iii) не содержит третью внеклеточную петлю и/или N-концевой домен CCR2; (iv) не содержит аминокислотную последовательность, представленную в SEQ ID NO:127 и/или SEQ ID NO:130; или (v) имеет комбинированную функцию. В одном из вариантов, начальный скрининг осуществляют с помощью твердофазного иммуноферментного анализа (ELISA) или радиоиммуноанализа. Пример ELISA-скрининга описан в заявке WO 00/37504, которая вводится в настоящее описание посредством ссылки.
Клетки, продуцирующие анти-CCR2 антитела, например, гибридомы, отбирают, клонируют, а затем подвергают скринингу на желаемые свойства, включая устойчивый рост, высокий уровень продуцирования антител и желательные свойства антител, более подробно обсуждаемые ниже. Гибридомы могут быть размножены in vivo у сингенных животных, у которых отсутствует иммунная система, например, у «голых» мышей, или в клеточной культуре in vitro. Методы отбора, клонирования и размножения гибридом хорошо известны среднему специалисту в данной области.
В одном из аспектов, иммунизованным животным является животное, не являющееся человеком, у которого экспрессируются гены человеческого иммуноглобулина, и у которого В-клетки селезенки слиты с миеломной клеточной линией, происходящей от животного, принадлежащего к тому же виду, к которому принадлежит животное, не являющееся человеком. В репрезентативном случае, иммунизованным животным является мышь XENOMOUSE™, а миеломной клеточной линией является не-секреторная мышиная миелома. В одном из вариантов, миеломной клеточной линией является P3-X63-Ag8.653 (Американская коллекция типовых культур). См., например, пример 1.
Скрининг иммортализованных антитело-продуцирующих клеток для идентификации антитела против первой и/или второй внеклеточных петель CCR2 может быть достигнут путем тестирования продуцируемых клетками антител на связывание с пептидом, содержащим аминокислотную последовательность первой или второй внеклеточных петель CCR2. Альтернативно, или в комбинации, антитела, продуцируемые этими клетками, могут быть протестированы на связывание с химерными рецепторами химокинов, которые содержат, главным образом, последовательность другого рецептора хемокина, но не последовательности первой и/или второй внеклеточных петель CCR2. См. пример 8, где продемонстрировано применение химер в целях картирования эпитопа для анти-CCR2 антител. В дополнительном варианте, антитела, которые продуцируются клетками, и которые, как известно, связываются с CCR2, могут быть протестированы на связывание с химерными рецепторами хемокинов, которые содержат, главным образом, последовательность CCR2, но в которых отсутствуют первая и/или вторая внеклеточные петли CCR2 дикого типа, например, которые, в одной или обеих этих внеклеточных петлях имеют первую и/или вторую внеклеточные петли от рецептора другого цитокина, или которые содержат мутации в указанных петлях.
В другом своем аспекте, настоящее изобретение относится к клеткам и к клеточным линиям (включая гибридомы), которые продуцируют человеческое анти-CCR2 антитело. В одном из вариантов, человеческим анти-CCR2 антителом, продуцируемым клеткой, клеточной линией или гибридомой, является антагонист CCR2. В другом варианте, человеческое анти-CCR2 антитело (i) связывается с первой и/или второй внеклеточными петлями CCR2; (ii) связывается с аминокислотной последовательностью, представленной в SEQ ID NO: 128 и/или SEQ ID NO: 129; (iii) не связывается с третьей внеклеточной петлей и/или N-концевым доменом CCR2; (iv) не связывается с аминокислотной последовательностью, представленной в SEQ ID NO:127 и/или SEQ ID NO:130; или (v) обладает комбинацией указанных функций. В другом варианте, человеческое анти-CCR2 антитело, продуцируемое клеткой, клеточной линией или гибридомой, не связывается с третьей внеклеточной петлей с высокой аффинностью, не связывается с N-концевым доменом CCR2 с высокой аффинностью, или не связывается ни с одним из этих компонентов с высокой аффинностью.
В еще одном аспекте изобретения, трансгенное животное иммунизируют CCR2, и первичные клетки, например, клетки селезенки или периферической крови выделяют у иммунизованного трансгенного животного, а затем идентифицируют отдельные клетки, продуцирующие антитела, специфичные к нужному антигену. Так, например, из каждой отдельной клетки выделяют полиаденилированную мРНК и осуществляют полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР) с использованием смысловых праймеров, которые гибридизуются с последовательностями вариабельной области, например, вырожденных праймеров, которые распознают большую часть генов областей FR1 вариабельной области человеческой тяжелой и легкой цепи или все эти гены, и антисмысловых праймеров, которые гибридизуются с последовательностями константной области или области стыка (J). Затем, кДНК вариабельных доменов тяжелой и легкой цепи клонируют и экспрессируют в любой подходящей клетке-хозяине, например, в миеломной клетке, в виде химерных антител, содержащих соответствующие константные области иммуноглобулина, такие как константные домены тяжелой цепи и цепи κ или λ. См. публикацию Babcook, J. S. et al., Proc. Natl. Acad. Sci. USA 93: 7843-48 (1996), которая вводится в настоящее описание посредством ссылки. Анти-CCR2 антитела могут быть затем идентифицированы и выделены как описано в настоящей заявке.
В другом аспекте изобретения, для получения библиотек, содержащих репертуар антител с различными аффинностями по отношению к CCR2, могут быть применены методы фагового представления. В качестве источника ДНК могут быть использованы непосредственно первичные В-клетки. Смесь кДНК, полученную из В-клеток, например, происходящих от селезенки, используют для получения экспрессионной библиотеки, например, библиотеки фагового представления, трансфецированной в E. coli. Полученные клетки тестируют на иммунореактивность с CCR2. Методы идентификации высокоаффинных человеческих антител из таких библиотек описаны в публикациях Griffiths et al., EMBO J. 13:3245-3260 (1994); Nissim et al., ibid, pp. 692-698 и Griffiths et al., ibid, 12:725-734, которые вводятся в настоящее описание посредством ссылки. В конечном счете, идентифицируют клоны библиотеки, которые имеют нужную величину аффинности связывания с антигеном, а затем ДНК, кодирующую продукт, ответственный за такое связывание, выделяют и подвергают модификации для проведения стандартной рекомбинантной экспрессии. Библиотеки фагового представления могут быть также сконструированы с использованием ранее модифицированных нуклеотидных последовательностей и скринированы аналогичным образом. Вообще говоря, для получения Fv-аналогов в целях создания фаговой библиотеки, кДНК, кодирующие тяжелую и легкую цепи, используются независимо или в связанной форме.
Затем фаговую библиотеку скринируют на антитела с наивысшей аффинностью по отношению к CCR2, и из соответствующего клона выделяют генетический материал. Последующие раунды скрининга могут повышать аффинность исходного выделенного антитела.
Нуклеиновые кислоты, векторы, клетки-хозяева и рекомбинантные методы получения антител
Нуклеиновые кислоты
Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующим анти-CCR2 антитела или их антигенсвязывающие части. В некоторых случаях, различные молекулы нуклеиновой кислоты кодируют тяжелую цепь и легкую цепь анти-CCR2 иммуноглобулина. В других случаях, одна и та же молекула нуклеиновой кислоты кодирует тяжелую цепь и легкую цепь анти-CCR2 иммуноглобулина. В одном из вариантов изобретения, нуклеиновая кислота кодирует анти-CCR2 антитело или его антигенсвязывающую часть.
В некоторых вариантах изобретения, молекула нуклеиновой кислоты, кодирующая вариабельный домен легкой цепи (VL), содержит сегмент, принадлежащий к семейству человеческих генов Vκ1, Vκ2, Vκ3, Vκ4, Vκ5 или Vκ6, и сегмент генов Jκ1, Jκ2, Jκ4, или Jκ5, содержащий или не содержащий мутации зародышевой линии.
В некоторых случаях, молекула нуклеиновой кислоты, кодирующая легкую цепь, кодирует аминокислотную последовательность, содержащую 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 замен, внесенных в аминокислотную(ые) последовательность(и) зародышевой линии. В некоторых случаях, молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность VL, содержащую 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 консервативных аминокислотных замен и/или 1, 2 или 3 не-консервативных замены по сравнению с последовательностями Vκ, и Jκ зародышевой линии. Замены могут быть сделаны в CDR, в каркасных областях или в константном домене.
В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность VL, содержащую один или более вариантов, по сравнению с последовательностью зародышевой линии, где указанные варианты идентичны вариантам, имеющимся в VL одного из антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P. В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность VL, содержащую мутацию в одном или нескольких положениях, аналогичных положениям мутаций в описанном здесь анти-CCR2 антителе зародышевой линии, но, при этом, содержащую другую замену, а в некоторых случаях, консервативную замену, по сравнению с заменой в указанном антителе.
В некоторых случаях, молекула нуклеиновой кислоты кодирует по меньшей мере три аминокислотных замены по сравнению с последовательностью зародышевой линии, присутствующей в VL одного из антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P.
В некоторых случаях, молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность VL моноклонального антитела 4.40 (SEQ ID NO:101), 4.9 (SEQ ID NO:29), 4.22 (SEQ ID NO:65), 4.39 (SEQ ID NO: 194) или 4.40 A68G S230P (SEQ ID NO: 113), или его варианта или части. В некоторых случаях, нуклеиновая кислота кодирует аминокислотную последовательность, содержащую CDR легкой цепи одного из указанных вышеперечисленных антител. В некоторых случаях, указанной частью является часть из смежных последовательностей, содержащая CDR1-CDR3.
В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность VL, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98% или 99% идентична аминокислотной последовательности VL любой одной из областей VL антител 4.40, 4.9, 4.22. 4.39 или 4.40 A68G S230P, или аминокислотной последовательности области VL любой одной из SEQ ID NO:101 , SEQ ID NO:113, SEQ ID NO:29, SEQ ID NO:194 или SEQ ID NO:65. Молекулы нуклеиновой кислоты включают нуклеиновые кислоты, кодирующие область VL, имеющуюся в SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:29 или SEQ ID NO:65.
В другом случае, нуклеиновая кислота кодирует полноразмерную легкую цепь антитела, выбранного из антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P, или легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:100, SEQ ID NO:112, SEQ ID NO:64, SEQ ID NO:193 или SEQ ID NO:28, или указанную последовательность легкой цепи, содержащую мутацию, такую как мутация, описанная в настоящей заявке.
В другом случае, молекула нуклеиновой кислоты кодирует вариабельный домен тяжелой цепи (VH), содержащий последовательность VH 3-30 или 1-46 человеческого гена или происходящую от нее последовательность. В различных случаях, молекула нуклеиновой кислоты содержит последовательность 3-30 VH человеческого гена, последовательность D1-7 человеческого гена и последовательность JH3B человеческого гена; последовательность 1-46 VH человеческого гена, последовательность D1-7 человеческого гена и последовательность JH3B человеческого гена; или последовательность, происходящую от указанных человеческих генов.
В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность, содержащую 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 , 12, 13, 14, 15, 16, 17 или 18 мутаций по сравнению с аминокислотной последовательностью человеческих генов V, D или J зародышевой линии. В некоторых случаях, указанные мутации присутствуют в области VH. В некоторых случаях, указанные мутации присутствуют в CDR.
В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность, содержащую одну или более аминокислотных мутаций по сравнению с последовательностью зародышевой линии, которые идентичны аминокислотным мутациям, имеющимся в VH моноклонального антитела 4.40, 4.22, 4.39 или 4.9. В некоторых случаях, молекула нуклеиновой кислоты кодирует по меньшей мере три аминокислотных мутации, по сравнению с последовательностями зародышевой линии, которые идентичны по меньшей мере трем аминокислотным мутациям, присутствующим в одном из вышеперечисленных моноклональных антител.
В некоторых случаях, молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует по меньшей мере часть аминокислотной последовательности VH моноклонального антитела, выбранного из 4.40 (SEQ ID NO:83), 4.22 (SEQ ID NO:47), 4.39 (SEQ ID NO:176) или 4.9 (SEQ ID NO:11) и его варианта, либо указанная последовательность имеет консервативные аминокислотные мутации и/или все три или менее неконсервативных аминокислотных замен. В различных случаях, указанная последовательность кодирует одну или более CDR, область CDR3, все три CDR, непрерывную часть, включающую CDR1-CDR3, или полноразмерную область VH, в присутствии или в отсутствии сигнальной последовательности.
В некоторых случаях, молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность одной из SEQ ID NO:82, SEQ ID NO:46, SEQ ID NO:10, SEQ ID NO:175 или SEQ ID NO:116, или указанную последовательность, имеющую сигнальную последовательность. В некоторых предпочтительных вариантах, молекула нуклеиновой кислоты содержит по меньшей мере часть нуклеотидной последовательности SEQ ID NO:73 или SEQ ID NO:115, или указанную последовательность, имеющую сигнальную последовательность. В некоторых случаях, указанная часть кодирует область VH (содержащую или не содержащую сигнальную последовательность), область CDR3, все три CDR или непрерывную область, включающую CDR1-CDR3.
В некоторых случаях, молекула нуклеиновой кислоты кодирует аминокислотную последовательность VH, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98% или 99% идентична аминокислотным последовательностям VH любой одной из областей VH антител 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P, или аминокислотную последовательность области VH любой одной из SEQ ID NO:83, SEQ ID NO:47, SEQ ID NO:176 или SEQ ID NO:11. Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим аминокислотную последовательность 4.40 (SEQ ID NO:83), 4.22 (SEQ ID NO:47), 4.39 (SEQ ID NO:176) или 4.9 (SEQ ID NO:11), или их область VH, или содержащим нуклеотидную последовательность SEQ ID NO:74, SEQ ID NO:38, SEQ ID NO:167, SEQ ID NO:2 или последовательность, кодирующую область VH.
В другом случае, нуклеиновая кислота кодирует полноразмерную тяжелую цепь антитела, выбранного из антител 4.40, 4.22, 4.9, 4.39, или 4.40 A68G S230P, или тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 10, SEQ ID NO:46, SEQ ID NO:82, SEQ ID NO:175 или SEQ ID NO:116, в присутствии или в отсутствии сигнальной последовательности, или тяжелую цепь, содержащую мутацию, такую как один из обсуждаемых здесь вариантов. Кроме того, нуклеиновая кислота может содержать нуклеотидную последовательность SEQ ID NO:1, SEQ ID NO:37, SEQ ID NO:73, SEQ ID NO:166 или SEQ ID NO:115, или молекулу нуклеиновой кислоты, кодирующую тяжелую цепь, содержащую мутацию, такую как один из обсуждаемых здесь вариантов мутаций.
Молекула нуклеиновой кислоты, кодирующая тяжелую или легкую цепь анти-CCR2 антитела или его части, может быть выделена из любого источника, продуцирующего такое антитело. В различных вариантах, молекулы нуклеиновой кислоты выделяют из В-клеток, выделенных у животного, иммунизованного CCR2, или из иммортализованной клетки, происходящей от таких В-клеток, экспрессирующих анти-CCR2 антитело. Методы выделения мРНК, кодирующей антитело, хорошо известны специалистам. См., например, Sambrook et al., мРНК может быть использована для продуцирования кДНК, применяемой в полимеразной цепной реакции (ПЦР), или для клонирования кДНК генов антител. В одном из вариантов, молекулу нуклеиновой кислоты выделяют из гибридомы, которая в качестве одного из партнеров по слиянию содержит клетку, продуцирующую человеческий иммуноглобулин и происходящую от трансгенного животного, не являющегося человеком. В других случаях, клетку, продуцирующую человеческий иммуноглобулин, выделяют у животного XENOMOUSE™. В другом случае, клетку, продуцирующую человеческий иммуноглобулин, выделяют у описанного здесь трансгенного животного, не являющегося человеком и мышью. В другом случае, нуклеиновую кислоту выделяют у не-трансгенного животного, не являющегося человеком. Молекулы нуклеиновой кислоты, выделенные у не-трансгенного животного, не являющегося человеком, могут быть использованы, например, для получения гуманизованных антител.
В некоторых случаях, нуклеиновая кислота, кодирующая тяжелую цепь анти-CCR2 антитела, содержит нуклеотидную последовательность, кодирующую домен VH, присоединенный, с сохранением рамки считывания, к нуклеотидной последовательности, кодирующей константный домен тяжелой цепи от любого источника. Аналогичным образом, молекула нуклеиновой кислоты, кодирующая легкую цепь анти-CCR2 антитела, может содержать нуклеотидную последовательность, кодирующую домен VL, присоединенный, с сохранением рамки считывания, к нуклеотидной последовательности, кодирующей константный домен легкой цепи от любого источника.
В другом аспекте изобретения, молекулы нуклеиновой кислоты, кодирующие вариабельный домен тяжелой (VH) и/или легкой (VL) цепей, «превращают» в полноразмерные гены антител. В одном из вариантов, молекулы нуклеиновой кислоты, кодирующие домены VH или VL, «превращают» в полноразмерные гены антител путем встраивания в экспрессионный вектор, уже кодирующий константный домен тяжелой цепи (CH) или константный домен легкой цепи (CL), соответственно, так, чтобы сегмент VH был функционально присоединен к сегменту(ам) CH в указанном векторе и/или так, чтобы сегмент VL был функционально присоединен к сегменту CL в указанном векторе. В другом случае, молекулы нуклеиновой кислоты, кодирующие домены VH и/или VL, превращают в полноразмерные гены антител путем присоединения, например, лигирования молекулы нуклеиновой кислоты, кодирующей домены VH и/или VL, с молекулой нуклеиновой кислоты, кодирующей домен CH и/или CL, стандартными методами, применяемыми в молекулярной биологии. Нуклеотидные последовательности генов константных доменов тяжелой и легкой цепи человеческого иммуноглобулина известны специалистам. См., например, Kabat et al., Sequences of Proteins of Immunological Interest. 5th Ed., NIH Publ. No. 91-3242, 1991. Затем, молекулы нуклеиновой кислоты, кодирующие полноразмерные тяжелые и/или легкие цепи, могут экспрессироваться в клетках, в которые они были введены, после чего может быть выделено анти-CCR2 антитело.
Молекулы нуклеиновой кислоты могут быть использованы для рекомбинантной экспрессии анти-CCR2 антител. Молекулы нуклеиновой кислоты могут быть также использованы для продуцирования химерных антител, биспецифических антител, одноцепочечных антител, иммуноадгезинов, диантител, мутантных антител и производных антител, которые более подробно описаны ниже. Если молекулы нуклеиновой кислоты происходят от не-трансгенного животного, не являющегося человеком, то такие молекулы нуклеиновой кислоты могут быть использованы для гуманизации антител, также описанной в настоящей заявке.
В некоторых своих вариантах, настоящее изобретение относится к нуклеиновым кислотам, кодирующим полноразмерную тяжелую цепь или полноразмерную легкую цепь, где указанная нуклеотидная последовательность, кодирующая аминокислотную последовательность константной области, в отличие от нуклеотидной последовательности, кодирующей последовательность константной области человеческой зародышевой линии, содержит одну или более мутаций. Так, например, нуклеиновая кислота может кодировать аминокислотную замену, которая улучшает свойства антитела, в результате добавления или удаления сайта гликозилирования, или замены, которые повышают стабильность или увеличивают время полужизни антитела. Нуклеиновая кислота может также содержать «молчащие» мутации, вводимые для добавления или удаления сайта рестриктирующего фермента, например, для облегчения клонирования нуклеиновой кислоты в конкретном экспрессионном векторе.
Векторы
Настоящее изобретение относится к векторам, включающим молекулы нуклеиновой кислоты, кодирующие тяжелую цепь и/или легкую цепь анти-CCR2 антитела или его антигенсвязывающих частей. Настоящее изобретение также относится к векторам, включающим молекулы нуклеиновой кислоты, кодирующие гибридные белки, модифицированные антитела, фрагменты антител и их зонды.
В некоторых случаях, анти-CCR2 антитела или их антигенсвязывающие части экспрессируют путем встраивания ДНК, кодирующих неполные или полноразмерные легкие и/или тяжелые цепи, полученные как описано в настоящей заявке, в экспрессионные векторы, так, чтобы гены были функционально присоединены к необходимым последовательностям регуляции экспрессии, таким как последовательности регуляции транскрипции и трансляции. Экспрессионными векторами являются плазмиды, ретровирусы, аденовирусы, аденоассоциированные вирусы (AAV), растительные вирусы, такие как вирус мозаики цветной капусты, вирус мозаики табака; космиды, YAC, эписомы, происходящие от вируса EBV, и т.п. В некоторых случаях, ген антитела лигируют в вектор так, чтобы последовательности регуляции транскрипции и трансляции, присутствующие в этом векторе, выполняли функцию регуляции транскрипции и трансляции гена антитела. Экспрессионный вектор и последовательности регуляции экспрессии выбирают так, чтобы они были совместимы с клетками-хозяевами, используемыми для экспрессии. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть встроены в отдельные векторы или в один и тот же экспрессионный вектор. Гены антител встраивают в экспрессионный вектор стандартными методами (например, путем лигирования комплементарных рестрикционных сайтов на фрагменте и векторе генов антитела, или лигирования затупленного конца, если такие рестрикционные сайты отсутствуют).
Стандартным вектором является вектор, кодирующий функционально полноразмерную последовательность CH или CL человеческого иммуноглобулина, где соответствующие рестрикционные сайты конструируют так, чтобы любая последовательность VH или VL могла быть встроена и могла экспрессироваться как описано в настоящей заявке. В таких векторах, сплайсинг обычно происходит между донорным сайтом сплайсинга во встроенной J-области, и акцепторным сайтом сплайсинга, расположенным за человеческим доменом С, а также в областях сплайсинга, которые присутствуют в человеческих экзонах CH. Полиаденилирование и терминация транскрипции происходят в нативных хромосомных сайтах, расположенных ниже кодирующих областей. Рекомбинантный экспрессионный вектор может также кодировать сигнальный пептид, который облегчает секрецию цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор так, чтобы этот сигнальный пептид был присоединен к амино-концу иммуноглобулиновой цепи с сохранением рамки считывания. Сигнальным пептидом может быть сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть, сигнальный пептид, происходящий от не-иммуноглобулинового белка).
Помимо генов цепи антитела, рекомбинантные экспрессионные векторы могут нести регуляторные последовательности, которые регулируют экспрессию генов цепи антитела в клетках-хозяевах. Для специалиста в данной области очевидно, что конструирование экспрессионного вектора, включающего набор регуляторных последовательностей, может зависеть от таких факторов, как выбор трансформируемой клетки-хозяина, уровень экспрессии нужного белка и т.п. Предпочтительными регуляторными последовательностями для экспрессии в клетках-хозяевах млекопитающих являются вирусные элементы, которые определяют высокие уровни экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, происходящие от LTR ретровирусов, цитомегаловируса (CMV) (такие как промотор/энхансер CMV), обезьяньего вируса 40 (SV40) (такие как промотор/энхансер SV40) и аденовируса (например, главный поздний промотор аденовируса (AdMLP)); промоторы полиомы и сильные промоторы млекопитающих, такое как нативные промоторы иммуноглобулина и актина. Более подробное описание вирусных регуляторных элементов и их последовательностей можно найти, например, в патенте США № 5168062, в патенте США № 4510245 и в патенте США № 4968615. Методы экспрессии антител в растениях, включая описание промоторов и векторов, а также методы трансформации растений известны специалистам. См., например, патент США № 6517529, который вводится в настоящее описание посредством ссылки. Методы экспрессии полипептидов в бактериальных клетках или в клетках грибов, например, в дрожжевых клетках, также хорошо известны специалистам.
Рекомбинантные экспрессионные векторы, помимо генов цепи антител и регуляторных последовательностей, могут содержать и другие последовательности, такие как последовательности, регулирующие репликацию вектора в клетках-хозяевах (например, ориджины репликации), и селективные маркерные гены. Селективный маркерный ген облегчает отбор клеток-хозяев, в которые был введен указанный вектор (см., например, патенты США №№ 4399216, 4634665 и 5179017, которые вводится в настоящее описание посредством ссылки). Так, например, обычно, селективный маркерный ген сообщает резистентность к лекарственным средствам, таким как G418, гигромицин или метотрексат, на клетках-хозяевах, в которые был введен данный вектор. Предпочтительными селективными маркерными генами являются ген дигидрофолатредуктазы (DHFR) (ген dhfr, применяемый в клетках-хозяевах для отбора на метотрексат/амплификации), ген neo (для отбора на G418), и ген глутамат-синтетазы (GS).
Не-гибридомные клетки-хозяева и методы рекомбинантного продуцирования белка
Молекулы нуклеиновой кислоты, кодирующие анти-CCR2 антитела, и векторы, содержащие такие молекулы нуклеиновой кислоты, могут быть использованы для трансфекции подходящих клеток-хозяев млекопитающих, растений, бактерий, насекомых или дрожжей. Трансформация может быть осуществлена любым известным методом введения полинуклеотидов в клетку-хозяина. Методы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны специалистам, и такими методами являются опосредуемая декстраном трансфекция, преципитация фосфатом кальция, трансфекция, опосредуемая полибреном, слияние протопластов, электропорация, инкапсуляция полинуклеотида(ов) в липосомы и прямая микроинжекция ДНК в ядро. Кроме того, молекулы нуклеиновой кислоты могут быть введены в клетки млекопитающих посредством вирусных векторов. Методы трансформации клеток хорошо известны специалистам. См., например, патенты США №№ 4399216, 4912040, 4740461 и 4959455, которые включены в настоящее описание посредством ссылки. Методы трансформации клеток растений хорошо известны специалистам, включая, например, трансформацию, опосредуемую Agrobacterium, биобаллистическую трансформацию, прямую инжекцию, электропорацию и вирусную трансформацию. Методы трансформации клеток бактерий, клеток насекомых и дрожжевых клеток также хорошо известны специалистам.
Клеточные линии млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны специалистам, и такими клетками являются многие иммортализованные клеточные линии, имеющиеся в Американской коллекции типовых культур (ATCC). Такими клетками являются, inter alia, клетки яичника китайского хомячка (CHO), клетки NS0, клетки NSO, клетки SP2, клетки HEK-293T, клетки NIH-3T3, клетки HeLa, клетки почек детенышей хомячка (BHK), клетки почек африканской зеленой мартышки (COS), клетки человеческой гепатоцеллюлярной карциномы (например, Hep G2), клетки A549, и ряд других клеточных линий. Особенно предпочтительные клеточные линии выбирают путем выявления клеточных линий, в которых наблюдаются высокие уровни экспрессии. Другими клеточными линиями, которые могут быть использованы, являются клеточные линии насекомых, такие как клетки Sf9 или Sf21. При введении рекомбинантных экспрессионных векторов, кодирующих гены антител, в клетки-хозяева млекопитающих, антитела получают путем культивирования клеток-хозяев в течение периода времени, достаточного для экспрессии антитела в клетках-хозяевах или секреции антитела в культуральную среду, в которой были выращены такие клетки-хозяева. Антитела могут быть выделены из культуральной среды стандартными методами очистки белка. Растительными клетками-хозяевами являются, например, клетки Nicotiana, клетки Arabidopsis, клетки ряски, кукурузы, пшеницы, картофеля и т.п. Бактериальными клетками-хозяевами являются клетки E. coli и клетки вида Streptomyces. Дрожжевыми клетками-хозяевами являются клетки Schizosaccharomyces pombe, Saccharomyces cerevisiae и Pichia pastoris.
Кроме того, экспрессия антител из клеточных линий-продуцентов может быть усилена с применением ряда известных методов. Так, например, стандартным методом повышения уровня экспрессии в определенных условиях является применение системы экспрессии гена глутамин-синтетазы (системы GS). Система GS подробно или частично обсуждается в описании Европейских патентов №№ 216846, 256055, 323997 и 338841.
Очевидно, что антитела, экспрессируемые различными клеточными линиями или в организме трансгенных животных, имеют различный характер гликозилирования. Однако, все антитела, кодируемые описанными здесь молекулами нуклеиновой кислоты, или содержащие описанные здесь аминокислотные последовательности, являются частью настоящего изобретения, независимо от характера гликозилирования таких антител.
Трансгенные животные и растения
Анти-CCR2 антитела могут быть также получены рекомбинантными методами путем создания животного или растения, которые являются трансгенными по представляющим интерес последовательностям тяжелой и легкой цепи иммуноглобулина, и продуцируют антитела в форме, которую можно выделить из организма этих животных или растений. Что касается трансгенных млекопитающих, то анти-CCR2 антитела могут продуцироваться в молоке коз, коров или других млекопитающих, и могут быть выделены из этого молока. См., например, патенты США №№ 5827690, 5756687, 5750172 и 5741957, которые включены в настоящее описание посредством ссылки. В некоторых случаях, трансгенных животных, которые не являются человеком, и которые содержат локусы человеческого иммуноглобулина, иммунизируют CCR2 или его иммуногенной частью, как описано в настоящей заявке. Методы получения антител в растениях описаны, например, в патентах США №№ 6046037 и 5959177, которые включены в настоящее описание посредством ссылки.
В некоторых случаях, трансгенные животные, не являющиеся человеком, или трансгенные растения получают путем введения одной или нескольких молекул нуклеиновой кислоты, кодирующих анти-CCR2 антитело, животному или в растение стандартными методами трансгенной технологии. См. Hogan и патент США № 6417429, см. выше. Трансгенными клетками, используемыми для создания трансгенного животного, могут быть эмбриональные стволовые клетки или соматические клетки или оплодотворенная яйцеклетка. Трансгенными не-человеческими организмами могут быть химерные и не-химерные гетерозиготы, а также не-химерные гомозиготы. См., например, Hogan et al., Manipulating the Mouse Embryo: A Laboratory Manual second ed., Cold Spring Harbor Press (1999); Jackson et al., Mouse Genetics and Transgenics: A Practical Approach. Oxford University Press (2000); и Pinkert, Transgenic Animal Technology: A Laboratory Handbook. Academic Press (1999), и все эти публикации включены в настоящее описание посредством ссылки. В некоторых случаях, трансгенные животные, не являющиеся человеком, имеют нужную дизрупцию и замену, полученные путем введения конструкции, которая кодирует представляющую интерес тяжелую цепь и/или легкую цепь. В одном из вариантов, трансгенные животные содержат и экспрессируют молекулы нуклеиновой кислоты, кодирующие тяжелую и легкую цепи, которые специфически связываются с CCR2, а предпочтительно, (i) связываются с первой и/или второй внеклеточными петлями CCR2; (ii) не связываются с N-концом или с третьей внеклеточной петлей CCR2 или с тем и другим; или (iii) обладают комбинацией указанных функций. В одном из вариантов, трансгенные животные содержат и экспрессируют молекулы нуклеиновой кислоты, кодирующие тяжелую и легкую цепи, которые специфически связываются с человеческим CCR2. В некоторых случаях, трансгенные животные содержат молекулы нуклеиновой кислоты, кодирующие модифицированное антитело, такое как одноцепочечное антитело, химерное антитело или гуманизованное антитело. Анти-CCR2 антитела могут быть продуцированы у любого трансгенного животного. В одном из вариантов, животными, не относящимися к человеку, являются мыши, крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. У трансгенных животных, не являющихся человеком, указанные кодируемые полипептиды экспрессируются в крови, молоке, моче, слюне, слезах, слизи и в других физиологических жидкостях.
Библиотеки фагового представления
Настоящее изобретение относится к способу продуцирования анти-CCR2 антитела или его антигенсвязывающей части, где указанный способ включает стадии синтеза библиотеки человеческих антител на фаге, скрининга библиотеки с использованием CCR2 или его части, выделения фага, который связывается с CCR2, и получения антитела из этого фага. Так, например, один из способов получения библиотеки антител, используемой в методах фагового представления, включает стадии иммунизации животного, не являющегося человеком и включающего локусы человеческого иммуноглобулина, антигеном CCR2 или его антигенной частью, для вырабатывания иммунного ответа; экстракции антитело-продуцирующих клеток от иммунизованного животного; выделения РНК, кодирующей тяжелые и легкие цепи антител, из экстрагированных клеток; реакции обратной транскрипции РНК с образованием кДНК; амплификации кДНК с использованием праймеров; и встраивания кДНК в вектор фагового представления так, чтобы указанные антитела экспрессировались на этом фаге. Таким образом, могут быть получены рекомбинантные анти-CCR2 антитела.
Рекомбинантные человеческие анти-CCR2 антитела могут быть выделены путем скрининга рекомбинантной комбинаторной библиотеки антител. Такой библиотекой может быть библиотека фагового представления scFv, созданная с использованием кДНК человеческих VL и VH, полученных из мРНК, выделенной из В-клеток. Методы создания и скрининга таких библиотек известны специалистам. Наборы для создания библиотек фагового представления являются коммерчески доступными (например, рекомбинантная система фаговых антител Pharmacia Recombinant Phage Antibody System, номер по каталогу 27-9400-01; и набор для фагового представления Stratagene SurfZAP™, номер по каталогу 240612). Существуют также и другие методы и реагенты, которые могут быть использованы для создания и скрининга библиотека представления антител (см., например, патент США № 5223409; публикации заявок РСТ №№ WO 92/18619, WO 91/17271, WO 92/20791, WO 92/15679, WO 93/01288, WO 92/01047, WO 92/09690; Fuchs et al., Bio/Technology 9:1370-1372 (1991); Hay et al., Hum. Antibod. Hybridomas 3:81-85 (1992); Huse et al., Science 246:1275-1281 (1989); McCafferty et al., Nature 348:552-554 (1990); Griffiths et al., EMBO J. 12:725-734 (1993); Hawkins et al., J. Mol. Biol. 226:889-896 (1992); Clackson et al., Nature 352:624-628 (1991); Gram et al., Proc. Natl. Acad. Sci. USA 89:3576-3580 (1992); Garrad et al., Bio/Technology 9:1373-1377 (1991); Hoogenboom et al., Nuc. Acid Res. 19:4133-4137 (1991); и Barbas et al., Proc. Natl. Acad. Sci. USA 88:7978-7982 (1991), и все эти документы вводятся в настоящее описание посредством ссылки.
В одном из вариантов, для выделения и продуцирования человеческих анти-CCR2 антител с нужными свойствами сначала используют описанное здесь человеческое анти-CCR2 антитело в целях отбора последовательностей тяжелой и легкой цепи, обладающих аналогичной активностью связывания с CCR2, с применением методов импринтинга эпитопов, описанных в публикации заявки РСТ No. WO 93/06213, которая включена в настоящее описание посредством ссылки. Библиотеками антител, используемыми в этом методе, могут быть библиотеки scFv, полученные и скринированные как описано в публикации заявки PCT No. WO 92/01047, McCafferty et al., Nature 348:552-554 (1990); и в работе Griffiths et al., EMBO J. 12:725-734 (1993), которые вводятся в настоящее описание посредством ссылки. Библиотеки антител scFv могут быть скринированы с использованием человеческого CCR2 в качестве антигена.
После предварительного отбора доменов VL и VH проводили эксперименты по определению «точного соответствия», в которых различные пары предварительно отобранных VL- и VH-сегментов скринировали на связывание с CCR2 в целях отбора предпочтительных комбинаций пар VL/VH. Кроме того, для дальнейшего улучшения качества антитела, VL- и VH-сегменты предпочтительной(ых) пары(пар) VL/VH могут быть подвергнуты неспецифической мутации, предпочтительно, в области CDR3 VH и/или VL, по механизму, аналогичном процессу соматической мутации in vivo, ответственному за созревание аффинности антител при вырабатывании природного иммунного ответа. Такое созревание аффинности in vitro может быть осуществлено путем амплификации доменов VH и/или VL с использованием ПЦР-праймеров, комплементарных VH CDR3 или VL CDR3, соответственно, где указанные праймеры в некоторых положениях были «нагружены» рандомизированной смесью из четырех нуклеотидных оснований для получения конечных ПЦР-продуктов, кодирующих сегменты VH и VL, в которых неспецифические мутации были введены в области CDR3 VH и/или VL. Эти сегменты VH и VL с неспецифическими мутациями могут быть подвергнуты повторному скринингу на связывание с CCR2.
После скрининга и выделения анти-CCR2 антитела из библиотеки представления рекомбинантного иммуноглобулина, нуклеиновые кислоты, кодирующие выбранное антитело, могут быть выделены из упаковки для представления ДНК (например, из фагового генома) и субклонированы в другие экспрессионные векторы стандартными методами рекомбинантных ДНК. При необходимости, нуклеиновая кислота может быть также подвергнута модификации для создания других форм антител, описанных в настоящей заявке. Для экспрессии рекомбинантного человеческого антитела, выделенного путем скрининга комбинаторной библиотеки, ДНК, кодирующую антитело, клонируют в рекомбинантный экспрессионный вектор и вводят в клетки-хозяева млекопитающих, описанной в настоящей заявке.
Переключение классов
В другом своем аспекте, настоящее изобретение относится к способу переключения класса или подкласса анти-CCR2 антитела на другой класс или подкласс. В некоторых случаях, молекулу нуклеиновой кислоты, кодирующую VL или VH, и не включающую последовательности, кодирующие CL или CH, выделяют методами, хорошо известными специалистам. Затем молекулу нуклеиновой кислоты функционально присоединяют к нуклеотидной последовательности, кодирующей CL или CH нужного иммуноглобулина определенного класса или подкласса. Это может быть достигнуто с использованием вектора или молекулы нуклеиновой кислоты, содержащих цепи CL или CH, как описано в настоящей заявке. Так, например, анти-CCR2 антитело, которое по своей природе принадлежит к классу IgM, может быть переключено на класс IgG. Затем, переключение класса может быть применено для переключения IgG одного подкласса на другой, например, с IgG1 или IgG2 на IgG4. Другой способ продуцирования антитела, содержащего нужный изотип, включает стадии выделения нуклеиновой кислоты, кодирующей тяжелую цепь анти-CCR2 антитела, и нуклеиновой кислоты, кодирующей легкую цепь анти-CCR2 антитела; выделения последовательности, кодирующей область VH, лигирования последовательности VH с последовательностью, кодирующей константный домен тяжелой цепи нужного изотипа, экспрессии гена легкой цепи и конструкции тяжелой цепи в клетке, и сбора анти-CCR2 антитела с нужным изотипом.
Деиммунизованные антитела
В другом аспекте изобретения, антитело может быть подвергнуто деиммунизации в целях снижения его иммуногенности методами, описанным, например, в публикациях заявок РСТ №№ WO98/52976 и WO00/34317 (которые вводятся в настоящее описание посредством ссылки).
Мутированные антитела
В другом аспекте изобретения, молекулы нуклеиновой кислоты, векторы и клетки-хозяева могут быть использованы для получения мутированных анти-CCR2 антител. Антитела могут быть мутированы в вариабельных доменах тяжелой и/или легкой цепей, например, для модификации связывающих свойств антитела. Так, например, мутация может быть введена в одну или более CDR для увеличения или снижения КD антитела против CCR2, для увеличения или снижения koff или для изменения специфичности связывания антитела. Методы сайт-направленного мутагенеза хорошо известны специалистам. См., например Sambrook et al. and Ausubel et al., supra. В другом варианте, одну или более мутаций вводят в положение аминокислотного остатка, который, как известно, должен быть модифицирован по сравнению с остатком в последовательности моноклонального антитела 4.40, 4.9, 4.22, 4.39 или 4.40 A68G S230P зародышевой линии. Эти мутации могут быть введены в CDR или в каркасную область вариабельного домена или в константный домен. В одном из вариантов, одну или более мутаций вводят в положение аминокислотного остатка, который, как известно, должен быть модифицирован по сравнению с остатком последовательности CDR или каркасной области вариабельного домена аминокислотной последовательности зародышевой линии, выбранной из последовательностей SEQ ID NO:11, SEQ ID NO:47, SEQ ID NO:83, SEQ ID NO:176, SEQ ID NO:29, SEQ ID NO:65, SEQ ID NO:101, SEQ ID NO:194 или SEQ ID NO:113.
В другом аспекте, каркасную область мутируют так, чтобы полученная(ые) каркасная(ые) область(и) имела(ли) аминокислотную последовательность соответствующего гена зародышевой линии. Мутация может быть введена в каркасную область или в константный домен для увеличения времени полужизни анти-CCR2 антитела. См., например, публикацию заявки PCT No. WO 00/09560, которая вводится в настоящее описание посредством ссылки. Мутация в каркасной области или в константном домене может быть также осуществлена для модификации или снижения иммуногенности антитела, для получения сайта, в котором происходит ковалентное или нековалентное связывание с другой молекулой, для добавления или удаления одного или нескольких сайтов гликозилирования или для изменения таких свойств, как фиксация комплемента, связывание с FcR и антитело-зависимая клеточно-опосредуемая цитотоксичность (ADCC). Одно антитело может иметь мутацию в любом одном или нескольких CDR или в каркасных областях вариабельного домена, или в константном домене.
В некоторых случаях, в любом из доменов VH или VL мутированного анти-CCR2 антитела, по сравнению с анти-CCR2 антитела перед введением мутации, имеются от 1 до 8 аминокислотных мутаций, включая любое число в данном интервале. В любом из вышеуказанных антител, мутации могут присутствовать в одной или нескольких CDR. Кроме того, любые из таких мутаций могут быть консервативными аминокислотными заменами. В некоторых случаях, в константные домены могут быть введены не более чем 5, 4, 3, 2 или 1 аминокислотных замен.
Модифицированные антитела
В другом аспекте изобретения может быть получено гибридное антитело или иммуноадгезин, которые содержат полноразмерное анти-CCR2 антитело или его часть, связанные с другим полипептидом. В одном из вариантов, с этим полипептидом связаны только вариабельные домены анти-CCR2 антитела. В другом варианте, домен VH анти-CCR2 антитела связан с первым полипептидом, а домен VL анти-CCR2 антитела связан со вторым полипептидом, который, в свою очередь, связан с первым полипептидом так, что домены VH и VL могут взаимодействовать друг с другом с образованием антигенсвязывающего сайта. В другом варианте, домен VH отделен от домена VL линкером, что позволяет доменами VH и VL взаимодействовать друг с другом (см. ниже раздел «Одноцепочечные антитела»). Затем последовательность VH-линкер-VL антитела присоединяют к представляющему интерес полипептиду. Гибридное антитело может быть использовано для доставки полипептида в CCR2-экспрессирующую клетку или ткань. Полипептидом может быть терапевтическое средство, такое как токсин, хемокин или другой регуляторный белок, либо им может быть диагностическое средство, такое как фермент, который может быть легко визуализирован, например, пероксидаза хрена. Кроме того, могут быть созданы гибридные антитела, в которых два (или более) одноцепочечных антитела связаны друг с другом. Это может быть осуществлено в том случае, если желательно получить двухвалентное или поливалентное антитело на одной полипептидной цепи, или если желательно получить биспецифическое антитело.
Для получения одноцепочечного антитела (scFv), VH- и VL-кодирующие фрагменты ДНК функционально присоединяют к другому фрагменту, кодирующему гибкий линкер, например, кодирующему аминокислотную последовательность (Gly4 -Ser)3, так, чтобы последовательности VH и VL могли экспрессироваться в виде непрерывного одноцепочечного белка, а домены VL и VH были связаны посредством гибкого линкера. См., например, Bird et al., Science 242:423-426 (1988); Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988); McCafferty et al., Nature 348:552-554 (1990). Одноцепочечное антитело может быть одновалентным, если используются отдельно взятые VH и VL; двухвалентным, если используются два VH и VL, или поливалентным, если используются более, чем два VH и VL. Могут быть получены биспецифические или поливалентные антитела, которые специфически связываются с CCR2 и с другой молекулой.
В другом аспекте изобретения, другие модифицированные антитела могут быть получены с использованием молекул нуклеиновой кислоты, кодирующих анти-CCR2 антитело. Так, например, «антитела каппа» Ill et al., Protein Eng. 10: 949-57 (1997)), «миниантитела» (Martin et al., EMBO J. 13:5303-9 (1994)), «диантитела» (Holliger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993)), или «янусины» (Traunecker et al., EMBO J. 10:3655-3659 (1991) и Traunecker et al., Int. J. Cancer (Suppl.) 7:51-52 (1992)) могут быть получены стандартными методами молекулярной биологии в соответствии с описанием в настоящей заявке.
Биспецифические антитела или их антигенсвязывающие фрагменты могут быть получены различными методами, включая слияние гибридом или связывание Fab'-фрагментов. См., например, Songsivilai & Lachmann, Clin. Exp. Immunol. 79: 315-321 (1990), Kostelny et al., J. Immunol. 148:1547-1553 (1992). Кроме того, биспецифические антитела могут быть получены в виде «диантител» или «янусинов». В некоторых случаях, биспецифическое антитело связывается с двумя различными эпитиопами CCR2. В некоторых случаях, биспецифическое антитело имеет первую тяжелую цепь и первую легкую цепь от моноклонального антитела 4.40, 4.9 или 4.40 A68G S230P и тяжелую и легкую цепь другого антитела. В некоторых случаях, дополнительная легкая и тяжелая цепи также происходят от идентифицированных выше моноклональных антител, но отличаются от первой тяжелой и легкой цепей.
В некоторых случаях, описанные здесь модифицированные антитела получают с использованием одного или нескольких вариабельных доменов или CDR, происходящих от описанного здесь человеческого моноклонального анти-CCR2 антитела.
Дериватизированные и меченые антитела
Анти-CCR2 антитело или его антигенсвязывающая часть могут быть дериватизированы другой молекулой или связаны с другой молекулой (например, с другим пептидом или белком). В общих чертах, антитела или их части дериватизируют так, чтобы такая дериватизация или мечение не оказывали негативного влияния на связывание с CCR2. В соответствии с этим, антитела и их части получают так, чтобы они включали интактные и модифицированные формы описанных здесь человеческих анти-CCR2 антител. Так, например, антитело или его часть могут быть функционально присоединены (посредством химического связывания, сцепления генов, нековалентного связывания или каким-либо другим способом) с одной или несколькими другими молекулами, такими как другое антитело (например, биспецифическое антитело или диантитело), детектирующий агент, цитотоксическое средство, фармацевтическое средство и/или белок или пептид, которые могут опосредовать взаимодействие антитела или его части с другой молекулой (такой как коровая область стрептавидина или полигистидиновая метка).
Дериватизированное антитело одного из типов получают путем перекрестного связывания двух или более антител (одного и того же типа или различных типов, например, для получения биспецифических антител). Подходящими перекрестно-сшивающими агентами являются агенты, которые представляют собой гетеробифункциональные молекулы, имеющие две различных реакционноспособных группы, разделенные соответствующим спейсером (например, м-малеимидобензоил-N-гидроксисукцинимидоэфиром) или гомобифункциональные молекулы (например, дисукцинимидилсуберат). Такие линкеры могут быть закуплены у компании Pierce Chemical Company, Rockford, II.
Дериватизированным антителом другого типа является меченое антитело. Подходящими детектирующими агентами, с использованием которых может быть дериватизировано антитело или его антигенсвязывающая часть, являются флуоресцентные соединения, включая флуоресцеин, флуоресцеин-изотиоцианат, родамин, фикоэритрин, 5-диметиламин-1-нафталинсульфонилхлорид, флуоресцирующие соединения лантанидной группы и т.п. Антитело может быть также помечено ферментами, которые могут быть использованы для детектирования, такими как пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза, глюкозо-оксидаза и т.п. Если антитело метят детектируемым ферментом, то оно может быть детектировано путем добавления дополнительных реагентов, используемых ферментами для образования реакционного продукта, который может быть идентифицирован. Так, например, если присутствует такой фермент, как пероксидаза хрена, то добавление пероксида водорода и диаминобензидина приводит к образованию окрашенного реакционного продукта, который является детектируемым. Антитело может быть также помечено биотином и детектировано посредством непрямого измерения уровня связывания авидина или стрептавидина. Антитело может быть также помечено предварительно определенным полипептидным эпитопом, распознаваемым вторичным репортером (например, последовательностями пар «лейциновых молний», сайтами связывания со «вторыми» антителами, металл-связывающими доменами, эпитопами и метками). В некоторых случаях, метки присоединяют посредством спейсерных ножек различной длины для снижения вероятности возникновения стерического затруднения.
Анти-CCR2 антитело может быть также помечено радиоактивно меченной аминокислотой. Радиоактивная метка может быть использована как в диагностике, так и в терапии. Так, например, радиоактивная метка может быть использована для детектирования CCR2-экспрессирующих опухолей с применением рентгенографии или других методов диагностики. Кроме того, радиоактивная метка может быть использована в терапевтических целях в качестве токсина для раковых клеток или опухолей. Примерами меток для полипептидов являются, но не ограничиваются ими, нижеследующие радиоизотопы или радионуклиды 3H, 14C,15N, 35S, 90Y, 99Tc, 111In, 125I или 131I.
Анти-CCR2 антитело может быть также дериватизировано химической группой, такой как полиэтиленгликоль (ПЭГ), метильная или этильная группы, или углеводная группа. Эти группы могут быть использованы для улучшения биологических свойств антитела, например, для увеличения времени полужизни в сыворотке или для повышения уровня связывания с тканью.
В некоторых вариантах, анти-CCR2 антитело может быть помечено парамагнитными, радиоактивными или флуоресцирующими ионами или молекулами, которые могут быть детектированы после визуализации.
В некоторых вариантах, парамагнитным ионом является ион хрома (III), марганца (II), железа (III), железа (II), кобальта (II), никеля (II), меди (II), неодима (III), самария (III), иттербия (III), гадолиния (III), ванадия (II), тербия (III), диспрозия (III), гольмия (III) или эрбия (III). В других вариантах, радиоактивным ионом является иод-123, технеций-99, индий-111, рений-188, рений-186, медь-67, иод-131, иттрий-90, иод-125, астат-211, и галлий-67. В других вариантах, анти-CCR2 антитело метят рентгеновским визуализирующим агентом, таким как лантан (III), золото (III), свинец (II) и висмут (III).
Фармацевтические композиции и наборы
Настоящее изобретение относится к композициям, содержащим человеческое анти-CCR2 антитело с антагоничтическими свойствами. Такие композиции могут быть использованы для лечения состояний, в патогенезе которых определенную роль играет CCR2, включая, но не ограничиваясь ими, такие состояния, как фиброз печени, фиброз почек, фиброз легких, псориаз, воспалительные расстройства, аллергии, аутоиммунные заболевания, расстройства, связанные с отторжением трансплантата, атеросклероз, ожирение, ВИЧ-инфекции, невропатические боли, воспаления, ассоциированные с ишемией, стеноз и рестеноз, рак, сепсис, склеродермия и диабет. В некоторых случаях, такие композиции могут быть использованы для лечения фиброза печени, опосредуемого вирусом гепатита С HCV), вирусом гепатита B (HBV), не-алкогольным стеатогепатитом (NASH), и/или алкогольным стеатогепатитом (ASH). В некоторых случаях, индивидуум нуждается в снижении уровня инфильтрации лейкоцитов в ткани, такие как ткани, в которых вырабатываются воспалительные ответы. В некоторых случаях, индивидуумом, подвергаемым лечению, является человек. В других вариантах, таким индивидуумом является животное. Примерами тканей, нуждающихся в снижении уровня воспаления или инфильтрации лейкоцитов, являются, но не ограничиваются ими, соединительная ткань, хрящ, печень, легкие, почки, нервная ткань, включая нервную ткань головного мозга, спинного мозга и периферическую нервную ткань, сердце, кровеносные сосуды, пищевод, желудок, тонкий кишечник, толстый кишечник, толстая кишка, предстательная железа, поджелудочная железа, мочевые пути, яичник, молочная железа, матка, яички, пенис, кость, мышцы, щитовидная железа, надпочечник, гипофиз, жировая ткань, костный мозг, кровь, тимус, селезенка, лимфоузлы, кожа, глаза, уши или нос. В одном из вариантов, указанными тканями являются ткани, имеющие слизистую оболочку.
Используемый в данном описании термин «фармацевтически приемлемый носитель» означает любые и все растворители, дисперсионные среды, материалы для покрытий, антибактериальные и противогрибковые средства, агенты, придающие изотоничность и замедляющие абсорбцию, и т.п., которые являются физиологически совместимыми. Некоторыми примерами фармацевтически приемлемых носителей, приведенными лишь в целях иллюстрации, являются вода, физиологический раствор, забуференный фосфатом физиологический раствор, декстроза, глицерин, этанол и т.п., а также их комбинации. Во многих случаях, в композицию предпочтительно, включать агенты, придающие изотоничность, например, сахара, многоатомные спирты, такие как маннит и сорбит, или хлорид натрия. Дополнительными примерами фармацевтически приемлемых веществ являются смачивающие агенты или небольшие количества добавок, таких как смачивающие или эмульгирующие агенты, консерванты или буферы, которые увеличивают срок хранения или повышают эффективность антитела.
Композиции могут быть получены в различных формах, например, в виде жидких, полутвердых и твердых лекарственных форм, таких как жидкие растворы (например, растворы для инъекций и вливаний), дисперсии или суспензии, таблетки, драже, порошки, липосомы и суппозитории. Такая форма зависит от выбранного способа введения и целей терапевтического применения. Обычно, композиции приготавливают в форме растворов для инъекции или инфузии, такие как композиции, аналогичные композициям, используемым для пассивной иммунизации человека. Предпочтительным способом введения является парентеральное введение (например, внутривенное введение, подкожное введение, внутрибрюшинное введение и внутримышечное введение). В одном из вариантов, указанное антитело вводят путем внутривенного вливания или внутривенной инъекции. В другом варианте, антитело вводят путем внутримышечной или подкожной инъекции.
Обычно, терапевтические композиции должны быть стерильными и стабильными в условиях их получения и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для введения лекарственного средства в высокой концентрации. Стерильные растворы для инъекций могут быть получены путем включения анти-CCR2 антитела в необходимом количестве в соответствующий растворитель с одним из перечисленных выше ингредиентов или с их комбинациями, если это необходимо, с последующей стерилизацией на фильтре. Обычно, дисперсии получают путем включения активного соединения в стерильный носитель, содержащий основную дисперсионную среду, и других необходимых ингредиентов из числа ингредиентов, перечисленных выше. В случае использования стерильных порошков для получения стерильных растворов для инъекций, предпочтительными методами их получения являются вакуумная сушка и лиофилизация, в результате чего из раствора, предварительно подвергнутого стерильной фильтрации, получают порошок, содержащий активный ингредиент плюс любой дополнительный нужный ингредиент. Собственная текучесть раствора может поддерживаться, например, путем нанесения покрытия, такого как, например, лицитин, путем сохранения требуемого размера частиц в случае дисперсии и использования поверхностно-активных веществ. Пролонгированная абсорбция композиций для инъекций может достигаться путем включения в данную композицию агента, замедляющего абсорбцию, например, моностеаратных солей и желатина.
Антитела могут быть введены различными методами, известными специалистам, хотя для многих терапевтических целей, предпочтительным способом/предпочтительной схемой введения являются подкожное, внутримышечное или внутривенное вливание. Как известно специалистам в данной области, такой способ или такая схема введения могут варьироваться в зависимости от желаемых результатов. Другими способами введения являются внутрибрюшинное введение, внутрибронхиальное введение, введение через слизистую, интраспинальное введение, введение вовнутрь синовиальной жидкости, введение вовнутрь аорты, интраназальное введение, офтальмологическое введение, внутриушное введение, местное введение, трансбуккальное введение и введение вовнутрь опухоли.
В некоторых случаях, активное соединение композиций антитела может быть получено с использованием носителя, который защищает антитело от быстрого высвобождения, например, в таких композициях, как препарат пролонгированного высвобождения, включая имплантаты, чрезкожные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биологически разлагаемые и биологически совместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие методы получения таких препаратов запатентованы или, в основном, известны специалисту в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems (J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978).
В композиции могут быть также введены дополнительные активные соединения. В некоторых случаях, ингибирующее анти-CCR2 антитело приготавливают и/или вводят вместе с одним или более дополнительными терапевтическими, диагностическими или профилактическими средствами. Терапевтическими средствами являются, но не ограничиваются ими, анти-CCR2 антитело с другой узкой специфичностью, антитела, которые связываются с другими мишенями, светочувствительные агенты, андроген, эстроген, нестероидные противовоспалительные средства, антигипертензивные средства, аналгетики, антидепрессанты, антибиотики, противораковые средства, анестетики, противорвотные средства, противоинфекционные средства, контрацептивы, средства против диабета, стероиды, средства против аллергии, химиотерапевтические средства, средства против мигрени, средства для прекращения курения, противовирусные средства, иммунодепрессанты, тромболитические средства, средства, снижающие уровень холестерина и средства против ожирения.
Терапевтическими средствами также являются пептидные аналоги, которые ингибируют CCR2, антитела или другие молекулы, которые связываются с MCP-1, MCP-2, MCP-3 или MCP-4 и предупреждают их связывание с CCR2, и агенты, ингибирующие экспрессию CCR2. В одном из вариантов, дополнительные агенты, которые ингибируют экспрессию CCR2, включают антисмысловую нуклеиновую кислоту, способную гибридизоваться с мРНК CCR2, такой как «шпилечная» РНК или киРНК. Последовательность-специфические нуклеиновые кислоты, способные ингибировать функцию генов посредством интерференции РНК, хорошо известны специалистам. Такая комбинированная терапия может потребовать введения более низких доз ингибирующего анти-CCR2 антитела, а также совместного введения агентов, что позволяет избежать токсического действия или развития осложнений, ассоциированных с различными монотерапиями.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) приготавливают и/или вводят вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой противомикробное(ые) средство(а). Противомикробными средствами являются антибиотики (например, антибактериальное средство), противовирусные средства, противогрибковые средства и средства против простейших. Неограничивающими примерами противомикробных средств являются сульфонамиды, триметоприм-сульфаметоксазол, хинолоны, пенициллины и цефалоспорины.
В некоторых конкретных случаях, терапевтическое(ие) средство(а) приготавливают и/или вводят вместе с ингибирующим анти-CCR2 антителом и антагонистом хемокина, антагонистом CCR2, антагонистом MCP-1 или антагонистом CCR5. Антагонистом CCR2 или антагонистом MCP-1 являются, но не ограничиваются ими, антитела против MCP-1 (патент США 7202343, US2005/0025768, US2006/0039913, US2006/0246069, и US2004/004/0047860); тетрагидропиранилциклопентилбензиламидные соединения (US2006/0116421); гетероарилпиперидиновые соединения (US2005/0250781); пиперидинилциклопентиларилбензиламидные соединения (US2006/0173013); соединения циклического амина (патент США 6140349 и патент США 6476054); циклопентильные соединения (US2002/0049222 и патент США 6545023); тетрагидропиранилциклопентилтетрагидропиридопиридиновые соединения (US2004/0167156, патент США 6812234 и патент США 7230008); аминоциклопентил-конденсированные гетеротрициклоамидные соединения (US2007/0004714); пиперидинил-альфа-аминоамидные соединения (US2005/0250814); замещенные пиразоловые соединения (WO06/88813); гетероциклические тетрагидропиранилциклопентиламидные соединения (US2006/0178363); 7- и 8-членные гетероциклические циклопентилбензиламидные соединения (US2006/0183731); гетероциклические бензоксазинил-амидоциклопентильные соединения (US2006/0069088); 3-аминоциклопентанкарбоксамидные соединения (US2007/0149532); азот-содержащие гетероциклические соединения (US2007/0155713); триазолфенилбензолсульфонамидные соединения (WO08/10934); замещенные пирролидиновые производные (US2004/0186140); замещенные бензамидные и замещенные фенилкарбаматные производные (патент США 7087604); замещенные циклоалкиламиновые соединения (US2005/0054626); замещенные бициклоалкиламиновые соединения (US2005/0227960); пирролидиноновые и пирролидинтионовые соединения (US2003/0149081, патент США 6727275 и патент США 6936633); меркаптоимидазоловые соединения (US2007/0244138); замещенные дипиперидиновые соединения (US2007/0197590); фениламино-замещенные соединения четвертичных солей (US2006/0293379); бициклические и мостиковые азот-содержащие гетероциклы (US2006/0074121); 3-аминоциклопентанкарбоксамидные соединения (US2006/0020133); 3-(4-гетероарилциклогексиламино)циклопентанкарбоксамидные соединения (US2005/0267146); триазоло-соединения (патент США 6492364); гетероарилсульфонамидные соединения (US2006/0173019); арилсульфонамиды (патент США 6939885); бис-арилсульфонамиды (патент США 7227035 и US2004/0167113); замещенные бензамидные соединения (патент США 6821964); замещенные соединения диазепама (US2007/0249589); производные триазаспиро[5.5]ундекана (US2005/267114); замещенные пиперидинкарбоксамидные соединения (US2003/0114443 и US6562978); производные бензазепина (US2004/0235822 и патент США 7262185).
В отдельных случаях, антагонист хемокина, приготавливаемый или вводимый вместе с анти-CCR2 антителами, представляет собой SELZENTRY™ (Maraviroc), которые имеет химическое название 4,4-дифтор-N-{(1S)-3-[экзо-3-(3-изопропил-5-метил-4H-1,2,4-триазол-4-ил)-8-азабицикло[3.2.1]окт-8-ил]-1-фенилпропил}циклогексанкарбоксамид:
В некоторых конкретных вариантах, терапевтическое(ие) средство(а), которое(ые) готовят и/или вводят вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой антагонист рецептора CB-1. Используемый в данном описании термин «рецептор CB-1» означает канабиноидный рецептор типа 1, сопряженный с G-белком. Термин «антагонист» включает полные антагонисты и неполные антагонисты, а также обратные агонисты. Антагонист рецептора CB-1 может быть селективным по отношению к рецептору CB-1. Термин «селективный по отношению к рецептору CB-1» означает, что данное соединение обладает незначительной или вообще не обладает антагонистической активностью по отношению к рецептору канабиноида-2 (CB-2). Антагонист CB-1 может обладать селективностью по отношению к рецептору СВ-1, которая по меньшей мере примерно в 10 раз превышает селективность по отношению к рецептору CB-2. Так, например, ингибирующая концентрация (IC50) антагониста по отношению к рецептору CB-1 примерно в 10 или более раз ниже IC50 антагониста по отношению к рецептору CB-2. Подходящими антагонистами рецептора CB-1 являются соединения, описанные в патентах США №№ 5462960; 5596106; 5624941; 5747524; 6017919; 6028084; 6432984; 6476060; 6479479; 6518264; и 6566356; в публикациях заявок на патент США №№ 2003/0114495; 2004/0077650; 2004/0092520; 2004/0122074; 2004/0157838; 2004/0157839; 2004/0214837; 2004/0214838; 2004/0214855; 2004/0214856; 2004/0058820: 2004/0235926; 2004/0259887; 2005/0080087; 2005/0026983 и 2005/0101592; в публикациях патентных заявок PCT WO 03/075660; WO 02/076949; WO 01/029007; WO 04/048317; WO 04/058145; WO 04/029204; WO 04/012671; WO 03/087037; WO 03/086288; WO 03/082191; WO 03/082190; WO 03/063781; WO 04/012671; WO 04/013120; WO 05/020988; WO 05/039550; WO 05/044785; WO 05/044822; и WO 05/049615: в патентной заявке PCT рег.№ PCT/IB2004/004050, поданной 6 декабря 2004; в PCT/IB2004/004017, поданной 6 декабря, 2004; в PCT/IB2004/004023, поданной 6 декабря, 2004; и PCT/IB2004/004019, поданной 6 декабря, 2004; и в предварительной заявке на патент США № 60/523937, поданной 21 ноября, 2003; 60/529908, поданной 16 декабря, 2003; 60/529909, поданной 16 декабря, 2003; 60/529910, поданной 16 декабря, 2003; 60/530012, поданной 16 декабря, 2003; и 60/564648, поданной 21 апреля, 2004. Все вышеупомянутые патенты и патентные заявки вводятся в настоящее описание посредством ссылки. Предпочтительными антагонистами рецептора CB-1, используемыми в этих способах, являются: римонабант (SR141716A, также известный как средство, выпускаемое под торговым знаком Acomplia™), который может быть закуплен у Sanofi-Synthelabo или может быть получен как описано в патенте США No. 5624941; N-(пиперидин-1-ил)-1-(2,4-дихлорфенил)-5-(4-иодфенил)-4-метил-1H-пиразол-3-карбоксамид (AM251), поставляемый Tocris™, Ellisville, MO; [5-(4-бромфенил)-1-(2,4-дихлорфенил)-4-этил-N- (1-пиперидинил)-1H-пиразол-3-карбоксамид] (SR147778), который может быть получен как описано в патенте США No. 6645985; N-(пиперидин-1-ил)-4,5-дифенил-1-метилимидазол-2-карбоксамид, N-(пиперидин-1-ил)-4-(2,4-дихлорфенил)-5-(4-хлорфенил)-1-метилимидазол-2-карбоксамид, N-(пиперидин-1-ил)-4,5-ди-(4-метилфенил)-1-метилимидазол-2-карбоксамид, N-циклогексил-4,5-ди-(4-метилфенил)-1-метилимидазол-2-карбоксамид, N-(циклогексил)-4-(2,4-дихлорфенил)-5-(4-хлорфенил)-1-метилимидазол-2-карбоксамид и N-(фенил)-4-(2,4-дихлорфенил)-5-(4-хлорфенил)-1-метилимидазол-2-карбоксамид, которые могут быть получены как описано в публикации патентной заявки PCT No. WO 03/075660; гидрохлоридная, мезилатная и безилатная соли 1-[9-(4-хлор-фенил)-8-(2-хлорфенил)-9Н-пурин-6-ил]-4-этиламинопиперидин-4-карбоновой кислоты, которые могут быть получены как описано в публикации заявки на патент США No. 2004/0092520; амид 1-[7-(2-хлорфенил)-8-(4-хлорфенил)-2-метил-пиразоло[1,5-a][1,3,5]триазин-4-ил]-3-этиламиноазетидин-3-карбоновой кислоты и амид 1-[7-(2-хлорфенил)-8-(4-хлорфенил)-2-метил-пиразоло[1,5-a][1,3,5]триазин-4-ил]-3-метиламиноазетидин-3-карбоновой кислоты, которые могут быть получены как описано в публикации заявки на патент США No. 2004/0157839; 3-(4-хлорфенил)-2-(2-хлорфенил)-6-(2,2-дифторпропил)-2,4,5,6-тетрагидропиразоло[3,4-c]пиридин-7-он, 2-(2-хлорфенил)-3-(4-этилфенил)-5-(2,2,2-трифторэтил)-4,5-дигидро-2H-пирроло[3,4-c]пиразол-6-он и 2-(2-хлорфенил)-3-(4-изопропилфенил)-5-(2,2,2-трифторэтил)-4,5-дигидро-2H-пирроло[3,4-c]пиразол-6-он, которые могут быть получены как описано в публикации заявки на патент США No. 2004/0214855; 3-(4-хлорфенил)-2-(2-хлорфенил)-7-(2,2-дифтор-пропил)-6,7-дигидро-2H,5H-4-окса-1,2,7-триаза-азулен-8-one, который может быть получен как описано в публикации заявки на патент США No. 2005/0101592; 2-(2-хлорфенил)-6-(2,2,2-трифтор-этил)-3-(4-трифторметил-фенил)-2,6-дигидро-пиразоло[4,3-d]пиримидин-7-он, который может быть получен как описано в публикации заявки на патент США No. 2004/0214838; (S)-4-хлор-N-{[3-(4-хлорфенил)-4-фенил-4,5-дигидро-пиразол-1-ил]-метиламино-метилен}бензолсульфонамид (SLV-319) и (S)-N-{[3-(4-хлорфенил)-4-фенил-4,5-дигидро-пиразол-1-ил]метиламино-метилен}-4-трифторметил-бензолсульфонамид (SLV-326), который может быть получен как описано в публикации патентной заявки PCT No. WO 02/076949; N-пиперидино-5-(4-бромфенил)-1-(2,4-дихлорфенил)-4-этилпиразол-3-карбоксамид, который может быть получен как описано в публикации заявки на патент США No. 6432984; 1-[бис-(4-хлорфенил)метил]-3-[(3,5-дифторфенил)метансульфонил-метилен]азетидин, который может быть получен как описано в публикации заявки на патент США No. 6518264; 2-(5-(трифторметил)пиридин-2-илокси)-N-(4-(4-хлорфенил)-3-(3-цианофенил)бутан-2-ил)-2-метилпропанамид, который может быть получен как описано в публикации патентной заявки PCT No. WO 04/048317; 4-{[6-метокси-2-(4-метоксифенил)-1-бензофуран-3-ил]карбонил}бензонитрил (LY-320135), который может быть получен как описано в публикации заявки на патент США No. 5747524; 1-[2-(2,4-дихлорфенил)-2-(4-фторфенил)бензо[1,3]диоксол-5-сульфонил]пиперидин, который может быть получен как описано в заявке WO 04/013120; и [3-амино-5-(4-хлорфенил)-6-(2,4-дихлорфенил)фуро[2,3-b]пиридин-2-ил]-фенилметанон, который может быть получен как описано в заявке WO 04/012671.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой ангиогенный фактор. Ангиогенными факторами являются, но не ограничиваются ими, основный фактор роста фибробластов, кислотный фактор фибробластов, васкулярный эндотелиальный фактор роста, ангиогенин, трансформирующий фактор роста, фактор некроза опухоли α и β, ангиопоэтин, тромбоцитарный фактор роста, плацентарный фактор роста, фактор роста гепатоцитов и пролиферин.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой тромболитическое средство. Тромболитическими средствами являются, но не ограничиваются ими, урокиназный активатор плазминогена, урокиназа, стрептакиназа, ингибиторы ингибитора α2-плазмина, и ингибиторы ингибитора активатора плазминогена-1, ингибиторы ангиотензин-конвертирующего фермента (ACE), спиронолактон, тканевый активатор плазминогена (tPA), ингибитор интерлейкин 1β- конвертирующего фермента, антитело против тромбина III и т.п.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство против ожирения. Средствами против ожирения являются, но не ограничиваются ими, ингибитор апо-B/MTP, ингибитор 11β-гидроксистероид-дегидрогеназы-1, пептид YY3-36 или его аналог, агонист MCR-4, агонист CCK-A, ингибитор поглощения моноамина, симпатомиметик, агонист β3-адренергического рецептора, агонист допамина, аналог рецептора меланоцит-стимулирующего гормона, агонист рецептора 5-HT2c, антагонист меланин-концентрирующего гормона, лептин, аналог лептина, агонист рецептора лептина, антагонист галанина, ингибитор липазы, агонист бомбезина, антагонист рецептора нейропептида-Y, тиромиметик, дегидроэпиандростерон или его аналог, антагонист глюкокортикоидного рецептора, антагонист рецептора орексина, агонист рецептора глюкагон-подобного пептида-1, цилиарный нейротропный фактор, антагонист белка, родственного белку человека и агути, антагонист рецептора грелина, антагонист или обратный агонист рецептора гистамина 3 и агонист рецептора нейромедина U.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство, которое ингибирует рекрутинг и/или адгезию нейтрофилов и/или мононуклеарных клеток на участке повреждения сосудов. Такие терапевтические средства могут, например, ингибировать активность (например, активность связывания, активность передачи сигнала) молекулы клеточной поверхности, которая опосредует адгезию, хемотаксис и/или хоминг. Так, например, антагонисты молекул клеточной адгезии (например, интегрины (например, интегрины β1, β2, β3, β4, β5, β6, β7, β8), селектины (например, E-селектин, P-селектин, L-селектин), кадгерины (например, E-, P-, N-кадгерины) и адгезивные молекулы, принадлежащие к суперсемейству иммуноглобулинов (например, LFA-2, LFA-3, CD44)) и антагонисты рецепторов цитокинов (например, антагонисты функции рецептора хемокина) могут быть получены и/или введены вместе с ингибирующим анти-CCR2 антителом. Кроме того, агенты, которые связываются с лигандами молекул клеточной адгезии или цитокинами или хемокинами, и ингибируют связывание лиганда с рецепторами, экспрессирующимися на нейтрофилах и/или мононуклеарных клетках, могут быть также получены и/или введены вместе с ингибирующим анти-CCR2 антителом.
В некоторых конкретных случаях, терапевтическое(ие) средство(а), которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство для лечения болезней сердца. Репрезентативными терапевтическими средствами, подходящими для лечения заболеваний сердца, являются, но не ограничиваются ими, факторы роста, ангиогенные агенты, блокаторы кальциевых каналов, антигипертензивные средства, инотропные средства, антиатерогенные средства, антикоагулянты, β-блокаторы, средства против аритмии, сердечные гликозиды, противовоспалительные средства, антибиотики, противовирусные средства, противогрибковые средства и средства, которые ингибируют инфицирование простейшими, а также противоопухолевые средства.
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой блокатор кальциевых каналов. Блокаторами кальциевых каналов являются, но не ограничиваются ими, дигидропиридины, такие как нифедипин, никардипин, нимодипин и т.п.; бензотиазепины, такие как дилитазем; фенилалкиламины, такие как верапамил; эфиры диариламинопропиламина, такие как бепридил; и бензимидол-замещенные тетралины, такие как мибефрадил.
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой антигипертензивное средство. Антигипертензивными средствами являются, но не ограничиваются ими, диуретики, включая тиазиды, такие как гидрохлортиазид, фуросемид, спиронолактон, триамтерен и амилорид; антиадренергические средства, включая клонидин, гуанабенз, гуанфацин, метилдопу, триметафан, резерпин, гуанетидин, гуанадрел, фентоламин, феноксибензамид, празозин, теразозин, доксазозин, пропанолол, метопролол, надолол, атенолол, тимолол, бетаксолол, картеолол, пиндолол, ацебутолол, лабеталол; вазодилататоры, включая гидрализин, миноксидил, диазоксид, нитропруссид; и ингибиторы ангиотензин-конвертирующего фермента, включая каптоприл, беназеприл, эналаприл, эналаприлат, фозиноприл, лизиноприл, хинаприл, рамиприл; антагонисты рецептора ангиотензина, такие как лозартан; и антагонисты кальциевых каналов, включая нифедин, амлодипин, фелодипин XL, изадипин, никардипин, бензотиазепины (например, дилтиазем) и фенилалкиламины (например, верапамил).
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой антикоагулянт. Антикоагулянтами являются, но не ограничиваются ими, гепарин, варфарин, гирудин, пептид-ангикоагулянт клещей, низкомолекулярные гепарины, такие как эноксапарин, дальтепарин и ардепарин, тиклопидин, данапароид, аргатробан, абциксимаб и тирофибан.
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство против аритмии. Средствами против аритмии являются, но не ограничиваются ими, блокаторы натриевых каналов (например, лидокаин, прокаиномид, энкаинид, флеканид и т.п.), бета-адренергические блокаторы (например, пропранолол), средства, увеличивающие продолжительность действия лекарственного средства (например, амиодарон) и блокаторы кальциевых каналов (например, верпамил, дитиазем, хлорид никеля и т.п.). Интерес также представляет доставка препаратов, вызывающих кардиодепрессивное действие (например, лидокаина), кардиотонических средств (например, изопротеренола, допамина, норэпинефрина и т.п.), и комбинаций множества средств для лечения болезней сердца (например, дигоксина/хинидина для устранения фибрилляции предсердий).
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство для лечения застойной сердечной недостаточности. Средствами для лечения застойной сердечной недостаточности являются, но не ограничиваются ими, сердечный гликозид, инотропные средства, диуретик на основе внеклеточной петли, тиазидный диуретик, диуретик, элиминирующий ионы калия, ингибитор ангиотензин-конвертирующего фермента, антагонист рецептора ангиотензина, нитровазодилататор, ингибитор фосфодиэстеразы, вазодилататор прямого действия, антагонист α1- адренергического рецептора, блокатор кальциевых каналов и симпатомиметик.
В некоторых конкретных случаях, терапевтическое(ие) средство(а) для лечения болезней сердца, которое(ые) может(могут) быть приготовлено(ы) и/или введено(ы) вместе с ингибирующим анти-CCR2 антителом, представляет(ют) собой средство, подходящее для лечения кардиомиопатии, такое как, но не ограничивающееся ими, допамин, эпинефрин, норэпинефрин и фенилэфрин.
Настоящее изобретение также включает композиции для ингибирования вирусных инфекций, а в частности, ВИЧ-инфекций, у млекопитающих, содержащие определенное количество антитела в комбинации с определенным количеством противовирусного средства, где указанные количества анти-CCR2 антитела и противовирусного средства, взятые вместе, являются эффективными в отношении ингибирования репликации вирусов, вирусной инфекции новых клеток или вирусной нагрузки. Многие противовирусные средства известны специалистам, включая нуклеозидные аналоги (например, AZT, 3TC и ddl), ингибиторы протеазы и антагонисты хемокиновых рецепторов, которые могут ингибировать ВИЧ-инфекцию, но не другие вирусные инфекции.
В другом аспекте изобретения, анти-CCR2 антитело или его фрагмент могут быть введены вместе с другими терапевтическими средствами, такими как противовоспалительные лекарственные средства или молекулы, пациенту, страдающему воспалительным расстройством, таким как артрит, аторосклероз или рассеянный склероз. В одном из своих аспектов, настоящее изобретение относится к способам лечения воспалительного расстройства у млекопитающего, где указанные способы включают введение указанному млекопитающему терапевтически эффективного количества соединения в комбинации с противовоспалительным средством. Противовоспалительными средствами являются, но не ограничиваются ими, любое известное нестероидное противовоспалительное средство, такое как производные салициловой кислоты (аспирин), производные пара-аминофенола (ацетаминофен), индол- и инден-уксусные кислоты (индометацин), гетероарилуксусные кислоты (кеторолак), арилпропионовые кислоты (ибупрофен), антраниловые кислоты (мефенаминовая кислота), еноловые кислоты (оксикамы) и алканоны (набуметон) и любое известное стероидное противовоспалительное средство, которое включает кортикостероиды и биологически активные синтетические аналоги, обладающие относительной глюкокортикоидной (метаболической) активностью и минералокортикоидной (регулирующей водно-солевой обмен) активностью. В другом случае, анти-CCR2 антитело вводят в комбинации с нестероидным противовоспалительным средством, таким как аспирин (Bayer, Bufferin), ибупрофен (Motrin, Advil), напроксен-натрий (Aleve), кетопрофен (Orudis KT), индометацин (Indocin), этодолак (Lodine), диклофенак-натрий (Voltaren), рофекоксиб (Vioxx), целекоксиб (Celebrex), набуметон (Relafen), или в комбинации со стероидом, таким как преднизон, преднизолон, дексаметазон, беклометазон, будезонид, флутиказон или триамцинолон.
Кроме того, другими лекарственными средствами, используемыми для лечения воспаления, являются, но не ограничиваются ими, антагонисты, такие как все антагонисты гистаминовых и брадикининовых рецепторов, антагонисты рецепторов лейкотриена и простагландина, и антагонисты рецептора фактора активации тромбоцитов. В другом варианте, лечение антителом или комбинированную терапию осуществляют в комбинации с лучевой терапией, химиотерапией, фотодинамической терапией, хирургическим вмешательством или с другой иммунотерапией, то есть, терапией, направленной на иммунную систему. В еще одном варианте изобретения, антитело вводят вместе с другим антителом. Так, например, анти-CCR2 антитело может быть введено вместе с другим антителом или с другим средством, которые, как известно, ослабляют воспаление, например, с антителом или средством, которые ингибируют рецептор альфа-4-интегрина (US2004/0009169) или IL-8 (US2004/0037830). Дополнительные антитела, которые могут быть введены вместе с анти-CCR2 антителами, описаны в патентах США №№ 6696550; 6406865; 6352832; и 6084075, содержание которых включено в настоящее описание посредством ссылки.
Композиции могут включать «терапевтически эффективное количество» или «профилактически эффективное количество» антитела или антигенсвязывающей части. Термин «терапевтически эффективное количество» означает количество, которое, при его введении в определенных дозах и в течение необходимого периода времени, является эффективным для достижения желаемого терапевтического эффекта. Терапевтически эффективное количество антитела или его части может варьироваться в зависимости от таких факторов, как патологическое состояние, возраст, пол и масса тела индивидуума, и способность антитела или его части вырабатывать у индивидуума нужный ответ. Терапевтически эффективным количеством также является количество, при котором терапевтически благоприятное действие значительно превосходит любое токсическое или какое-либо другое нежелательное действие антитела или его части. Термин «профилактически эффективное количество» означает количество, которое, при его введении в определенных дозах и в течение необходимого периода времени, является эффективным для достижения желаемого терапевтического эффекта. Вообще говоря, поскольку профилактическую дозу вводят индивидуумам до развития заболевания или на ранней стадии его развития, то профилактически эффективное количество может быть меньше терапевтически эффективного количества.
Схемы введения доз могут быть скорректированы в целях достижения оптимального желаемого ответа (например, терапевтического или профилактического ответа). Так, например, может быть введена одна ударная доза, либо могут быть введены несколько дробных доз в течение определенного периода времени, либо вводимая доза может быть пропорциональна снижена или увеличена в зависимости от терапевтической ситуации. Для облегчения введения и для достижения однородности лекарственной формы, особенно предпочтительно, чтобы такая лекарственная унифицированная форма была получена в виде композиций для парентерального введения. Используемый в данном описании термин «унифицированная лекарственная форма» означает физически дискретные единицы, подходящие для их введения млекопитающему в виде унитарных лекарственных форм, где каждая унифицированная лекарственная форма, содержащая предварительно определенное количество активного соединения, была вычислена так, чтобы она давала желаемый терапевтический эффект в комбинации с необходимым фармацевтическим носителем. Спецификация унифицированных лекарственных форм определяется нижеуказанными свойствами и непосредственно зависит: (a) от уникальных свойств анти-CCR2 антитела или его части и конкретного достигаемого терапевтического или профилактического эффекта и (b) от ограничений, известных специалистам в области составления рецептуры лекарственных препаратов, таких как антитело, используемое для лечения индивидуумов, восприимчивых к такому лечению.
Репрезентативный и неограничивающий интервал терапевтически или профилактически эффективных количеств антитела или его части составляет 0,025-50 мг/кг, 0,1-50 мг/кг, 0,1-25, 0,1-10 или 0,1-3 мг/кг. В одном из вариантов, антитело вводят в препарате в виде стерильного водного раствора, имеющего рН примерно от 5,0 до 6,5, и содержащего примерно от 1 мг/мл до 200 мг/мл антитела; примерно от 1 миллимоль до 100 миллимоль гистидинового буфера; примерно от 0,01 мг/мл до 10 мг/мл полисорбата 80 или полисорбата 20; примерно от 100 миллимоль до 400 миллимоль невосстанавливающего сахара, выбранного, из группы сахаров, но не ограничивающейся ими, таких как трегалоза или сахароза; примерно от 0,01 миллимоль дигидрата динатрий-EDTA; и необязательно содержащего фармацевтически приемлемый антиоксидант, а также хелатообразующий агент. Подходящими антиоксидантами являются, но не ограничиваются ими, метионин, тиосульфат натрия, каталаза и платина. Так, например, указанная композиция может содержать метионин в концентрации в пределах от 1 мМ до примерно 100 мМ, а в частности, примерно 27 мМ. В некоторых случаях, указанная композиция содержит 5 мг/мл антитела в буфере, состоящем из 20 мМ цитрата натрия, pH 5,5, 140 мМ NaCl, и 0,2 мг/мл полисорбата 80. Следует отметить, что величины доз могут варьироваться в зависимости от типа и тяжести состояния, подвергаемого лечению. Также очевидно, что для любого конкретного индивидуума, конкретная схема введения доз должна быть скорректирована в процессе проведения лечения в соответствии с потребностями индивидуума и в соответствии с заключением врача-специалиста, осуществляющего введение или наблюдение за введением указанных композиций, и что представленные здесь интервалы доз приводятся лишь в целях иллюстрации и не ограничивают объема или практического применения заявленной композиции.
В другом своем аспекте, настоящее изобретение относится к наборам, содержащим анти-CCR2 антитело или его антигенсвязывающую часть, или композицию, содержащую такое антитело или антигенсвязывающий фрагмент. Набор может включать, помимо такого антитела или такой композиции, диагностические или терапевтические средства. Набор может также включать инструкции по применению методов диагностики или терапии, а также упаковочный материал, такой как, но не ограничивающийся ими, лед, сухой лед, STYROFOAM™, пена, пластик, целлофан, усадочная упаковочная пленка, блистерная упаковка, картон и крахмал из арахисового масла. В одном из вариантов, указанный набор включает антитело или его содержащую композицию и диагностический агент, который может быть использован в описанном здесь методе. В другом варианте, указанный набор включает антитело или его содержащую композицию и терапевтический агент, который может быть использован в описанном здесь методе.
Настоящее изобретение относится к композициям и наборам для лечения рака у млекопитающего, где указанные композиции и наборы включают определенное количество антитела в комбинации с определенным количеством химиотерапевтического агента, где указанные количества соединения, соли, сольвата и пролекарства и количества химиотерапевтического агента, взятые вместе, являются эффективными для ингибирования роста аномальных клеток. Многие химиотерапевтические средства известны специалистам. В некоторых случаях, химиотерапевтическое средство выбирают из группы, состоящей из ингибиторов митоза, алкилирующих агентов, антиметаболитов, интеркалирующих антибиотиков, ингибиторов хемокинов, ингибиторов клеточного цикла, ферментов, ингибиторов топоизомераз, модификаторов биологического ответа, антигормональных средств, например, антиандрогенов и средств против ангиогенеза.
Методы, применяемые в диагностике
В другом своем аспекте, настоящее изобретение относится к методом диагностики. Анти-CCR2 антитела могут быть использованы для детектирования CCR2 в биологическом образце in vitro или in vivo. В одном из своих вариантов, настоящее изобретение относится к способу детектирования присутствия или к способу определения локализации CCR2-экспрессирующих клеток у индивидуума, нуждающегося в этом, где указанный способ включает стадии введения антитела индивидууму, определения экспрессии CCR2 у индивидуума путем определения области локализации связанного антитела, сравнения уровня экспрессии у данного индивидуума с уровнем экспрессии у здорового индивидуума или со стандартным уровнем, и детектирования присутствия или участка локализации данных клеток. Анти-CCR2 антитела могут быть также использованы в качестве маркера воспаления и/или инфильтрации иммунных клеток, таких как моноциты, в ткани.
Анти-CCR2 антитела могут быть использованы в стандартном иммуноанализе, включая, но не ограничиваясь ими, ELISA, РИА, проточная цитометрия, иммуногистохимический анализ ткани, вестерн-блот-анализ или иммунопреципитация. Анти-CCR2 антитела могут быть использованы для детектирования CCR2 у человека. В другом варианте, анти-CCR2 антитела могут быть использованы для детектирования CCR2 у собакоподобных обезьян или у макак-резусов. В другом варианте, анти-CCR2 антитела могут быть использованы для детектирования CCR2 у грызунов, таких как мыши и крысы.
Настоящее изобретение также относится к способу детектирования CCR2 в биологическом образце, включающему контактирование биологического образца с анти-CCR2 антителом и детектирование связанного антитела. В одном из вариантов, анти- CCR2 антитело непосредственно метят детектируемой меткой. В другом варианте, анти-CCR2 антитело (первое антитело) является немеченным, а «второе» антитело или другая молекула, которые могут связываться с анти-CCR2 антителом, являются меченными. Как хорошо известно специалистам в данной области, «второе» антитело выбирают так, чтобы оно обладало способностью специфически связываться с «первым» антителом конкретного вида и класса. Так, например, если анти-CCR2 антителом является человеческий IgG, то «вторым» антителом может быть антитело против человеческого IgG. Другими молекулами, которые могут связываться с антителами, являются, но не ограничиваются ими, белок А и G-белок, которые являются коммерчески доступными, например, могут быть закуплены у Pierce Chemical Co.
Подходящие метки для антитела или «второго» антитела описаны выше, и такими метками являются различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества и радиоактивные вещества. Примерами подходящих ферментов являются пероксидаза хрена, щелочная фосфатаза, β-галактозидаза или ацетилхолинэстераза; примерами подходящих комплексов, содержащих простетическую группу, являются стрептавидин/биотин и авидин/биотин; примерами подходящих флуоресцентных веществ являются умбеллиферон, флуоресцеин, флуоресцеин-изотиоцианат, родамин, дихлортриазениламин-флуоресцеин, данзилхлорид или фикоэритрин; примером люминесцентного вещества является люминол, а примерами подходящих радиоактивных веществ являются 125I, 131I, 35S или 3H.
В других вариантах, CCR2 может быть проанализирован в биологическом образце путем проведения иммуноанализа на конкурентное связывание, в котором используется CCR2-стандарты, меченные детектируемым веществом, и немеченое анти-CCR2 антитело. В этом анализе, биологический образец, меченные CCR2-стандартны и анти-CCR2 антитела объединяют, а затем определяют количество меченного CCR2-стандарта, связанного с немеченным антителом. Количество CCR2 в биологическом образце обратно пропорционально количеству меченного CCR2-стандарта, связанного с анти-CCR2 антителом.
Иммуноанализы, описанные выше, могут применяться в различных целях. Так, например, анти-CCR2 антитела могут быть использованы для детектирования CCR2 в культивированных клетках. В одном из вариантов, анти-CCR2 антитела используются для определения количества CCR2 на поверхности клеток, которые были обработаны различными соединениями. Такой метод может быть использован для идентификации соединений, модулирующих уровни белка CCR2. В соответствии с этим методом, один образец клеток обрабатывают тестируемым соединением в течение определенного периода времени, а другой образец оставляют необработанным. При измерении общего уровня экспрессии CCR2, клетки подвергают лизису, и общий уровень экспрессии CCR2 измеряют с помощью одного из описанных здесь иммуноанализов. Общий уровень экспрессии в обработанном образце сравнивают с уровнем экспрессии в необработанных клетках для определения эффекта тестируемого соединения.
Предпочтительным иммуноанализом для измерения общего уровня экспрессии CCR2 является проточная цитометрия или иммуногистохимический анализ. При измерении уровня экспрессии CCR2 на клеточной поверхности, клетки не подвергают лизису, а уровни CCR2 на клеточной поверхности измеряют с помощью одного из описанных здесь иммуноанализов. Предпочтительный иммуноанализ для определения уровней CCR2 на клеточной поверхности включает стадии мечения белков клеточной поверхности детектируемой меткой, такой как биотин или 125I; иммунопреципитации CCR2 анти-CCR2 антителом, и последующего детектирования меченного CCR2.
Другой иммуноанализ для определения локализации CCR2, например, уровней CCR2 на клеточной поверхности, осуществляют иммуногистохимическим методом. Иммуноанализ для детектирования уровней CCR2 на клеточной поверхности включает связывание анти-CCR2 антитела, меченного соответствующий флуорофором, таким как флуоресцеин или фикоэритрин, и детектирование «первого» антитела с помощью проточной цитометрии. В другом варианте, анти-CCR2 антитело является немеченным, а «второе» антитело или другая молекула, которая может связываться с анти-CCR2 антителом, являются меченными. Такие методы, как ELISA, РИА, проточная цитометрия, вестерн-блот-анализ, иммуногистохимический анализ, мечение клеточной поверхности интегральными мембранными белками и иммунопреципитация хорошо известны специалистам. См., например, Harlow and Lane, supra. Кроме того, иммуноанализы могут быть масштабированы в целях проведения высокоэффективного скрининга для тестирования большого числа соединений на активацию или ингибирование CCR2.
Анти-CCR2 антитела могут быть также использованы для определения уровней CCR2 в ткани или в клетках, происходящих от этой ткани. В некоторых случаях, такой тканью является пораженная ткань. В некоторых случаях, такой тканью является тканевый биоптат. В некоторых вариантах указанного способа, ткань или биоптат берут у пациента. Затем ткань или биоптат используют в иммуноанализе для определения, например, общего уровня экспрессии CCR2, уровней CCR2 на клеточной поверхности или локализации CCR2 методами, обсуждаемыми выше. Такие методы могут быть использованы для того, чтобы определить, способна ли данная ткань экспрессировать высокие уровни CCR2, которые будут служить показателем того, что эта ткань является мишенью для анти-CCR2 антитела, используемого в терапии.
Антитела могут быть также использованы in vivo для идентификации тканей и органов, экспрессирующих CCR2. В некоторых случаях, анти-CCR2 антитела используют для идентификации CCR2-экспрессирующих клеток.
Этот способ включает стадии введения детектируемо меченного анти-CCR2 антитела или композиции, содержащей такое антитело, пациенту, нуждающемуся в диагностическом анализе и визуализирующем анализе, проводимых в целях определения локализации CCR2-экспрессирующих тканей у данного пациента. Визуализирующий анализ хорошо известен специалистам-медикам, и таким анализом является, но не ограничивается ими, рентгеноструктурный анализ, визуализация методом магнитного резонанса (MRI) или компьютерная томография (КТ). Антитело может быть помечено любым агентом, подходящим для визуализации in vivo, например, контрастным веществом, таким как барий, который может быть использован для рентгеноструктурного анализа, или магнитным контрастным веществом, таким как хелатный комплекс гадолиния, который может быть использован для MRI или КТ. Другими агентами для мечения являются, но не ограничиваются ими, радиоизотопы, такие как 99Tc. В другом случае, анти-CCR2 антитело может быть немеченным и может быть визуализировано путем введения «второго» антитела или другой молекулы, которые могут быть детектированы, и которые могут связываться с анти-CCR2 антителом. В другом случае, у пациента берут биоптат для того, чтобы определить, может ли представляющая интерес ткань экспрессировать CCR2.
В некоторых случаях, детектируемо меченное анти-CCR2 антитело содержит флуорофор. В некоторых случаях, флуорофор выбран из группы, состоящей из флуоресцентного красителя, излучающего в ближнем инфракрасном диапазоне, динитрофенила, флуоресцеина и их производных, родамина, производных родамина, фикоэритрина, фикоцианина, аллофикоцианина, o-фтальдегида и флуоресцамина, техасского красного (TEXAS RED™), родаминового зеленого (RHODAMINE GREEN™), зеленого орегона (OREGON GREEN™), CASCADE BLUE™, фикоэритрина, CY3™, CY5™, CY2™, CY7™, кумарина, инфракрасного 40, MR 200, IRD 40, ALEXA FLUOR™, тетраметилродамина, PACIFIC BLUE™, SYBR™ и BODIPY™. В другом случае, флуорофор включает одно из нижеследующих соединений, где их максимальные величины излучения указаны в нм в скобках, CY2™ (506), белок, флуоресцирующий в зеленом диапазоне спектра (GFP) (со сдвигом в красную область) (507), YO-PRO®-1 (509), YOYO®EM (509), кальцеин (517), ФИТЦ (518), FLUORX® (519), ALEXA® (520), родамин 110 (520), 5-FAM (522), OREGON GREEN® 500 (522), OREGON GREEN © 488 (524), RIBOGREEN® (525), родаминовый зеленый (RHODAMINE GREEN®) (527), родамин 123 (529), магниевый зеленый (MAGNESIUM GREEN®) (531), кальциевый зеленый (CALCIUM GREEN®) (533), TO-PRO®-1 (533), TOTO®-1 (533), JOE (548), BODI PY® 530/550 (550), Dil (565), BODIPY® (568), BODIPY® 558/568 (568), BODIPY® 564/570 (570), CY3® (570), ALEXA® 546 (570), TRITC (572), магниевый оранжевый (MAGNESIUM ORANGE®) (575), фикоэритрин R&B (575), родамин-фаллоидин (575), кальциевый оранжевый (CALCIUM ORANGE®)(576), пиронин Y (580), родамин B (580), TAMRA (582), родаминовый красный (RHODAMINE RED®) (590), CY3.5® (596), ROX (608), кальциевый малиновый (CALCIUM CRIMSON™) (615), ALEXA® 594 (615), техасский красный (TEXAS RED™) (615), нильский красный (628), YO-PRO®-3 (631), YOYO®-3 (631), R-фикоцианин (642), C-фикоцианин (648), TO-PRO®-3 (660), TOTO®-3 (660), DiD DilC(5) (665), CY5™ (670), тиадикарбоцианин (671) и Cy5.5™ (694).
Способы, используемые в терапии
В другом своем аспекте, настоящее изобретение относится к способам ингибирования активности CCR2 путем введения анти-CCR2 антитела пациенту, нуждающемуся в этом. В терапии может быть использовано любое из описанных в данном описании антител. В различных вариантах, анти-CCR2 антителом является человеческое, химерное или гуманизованное антитело. В некоторых случаях, антитело или его антигенсвязывающая часть связывается с первой и/или второй внеклеточными петлями CCR2. Предпочтительно, антитело или его антигенсвязывающая часть не должны связываться с третьей внеклеточной петлей или N-концевым доменом CCR2.
В другом варианте, CCR2 является человеческим, а пациентом является человек. Альтернативно, пациентом может быть млекопитающее, экспрессирующее CCR2, с которым перекрестно реагирует анти-CCR2 антитело. Это антитело может быть использовано в ветеринарии и может быть введено млекопитающему, не являющемуся человеком и экспрессирующему CCR2, с которым перекрестно реагирует указанное антитело, а также животному с моделью человеческого заболевания. Такие животные-модели могут быть использованы для оценки терапевтической эффективности антител.
В одном из своих аспектов, настоящее изобретение относится к способам лечения, к способам вспомогательного лечения, а также к способам предупреждения или вспомогательного предупреждения CCR2-опосредуемого заболевания у индивидуума, где указанные способы включают введение данному индивидууму терапевтически эффективного количества анти-CCR2 антитела. Используемый в данном описании термин «CCR2-опосредуемое заболевание» включает заболевание и другие расстройства, при которых наблюдается присутствие высоких уровней экспрессии или активности CCR2 у индивидуума, страдающего расстройством, которое было диагностировано или подозревается у данного индивидуума, где указанные высокие уровни CCR2, либо ответственны за патофизиологию данного расстройства, либо являются фактором, приводящим к ухудшению данного расстройства. Такие расстройства могут быть диагностированы, например, по увеличению уровней CCR2 на клеточной поверхности пораженных клеток или тканей индивидуума, страдающего таким расстройством, или по увеличению CCR2-опосредуемой активности в клетках, таких как базофилы, моноциты или лимфоциты, которые вносят свой вклад в развитие патологии данного расстройства, или которые вызывают ухудшение такого расстройства, или по увеличению уровней лигандов CCR2, таких как MCP-1, на участке воспаления. Повышение уровня экспрессии CCR2 может быть детектировано, например, с использованием анти-CCR2 антитела. Повышение активности CCR2 может быть детектировано по повышению уровня активации G-белка, полимеризации F-актина или по увеличению уровня хемотаксиса CCR2-экспрессирующих клеток, такого как хемотаксис, индуцированный действием MCP-1 или других лигандов CCR2.
В одном из своих аспектов, CCR2-опосредуемое расстройство характеризуется фиброзом. Используемый в данном описании термин «фиброз» означает патологическое состояние, характеризующееся чрезмерным отложением и метаболизмом фиброзного материала (например, внутриклеточного матрикса) в ответ на повреждение ткани. Во многих случаях, фиброз представляет собой нормальный процесс репарации ткани (то есть, заживление ран), происходящий с некоторыми отклонениями, обусловленными хроническим или чрезмерным повреждением ткани, которое приводит к активации и пролиферации фибробластов или звездчатых клеток и аккумуляции коллагена. Фиброзными состояниями являются фибропролиферативные расстройства, которые ассоциируются с васкулярными заболеваниями, такими как болезнь сердца, заболевание коры головного мозга и заболевания периферических сосудов, а также всех главных систем тканей и органов, таких как глаза, кожа, почки, легкие, кишечник и печень (Wynn, Nature Reviews 4:583-594 (2004); Bataller, R. and Brenner, D., J. Clin. Invest. 115:209-218 (2005)). Другими причинами, вызывающими фиброз, являются химиотерапевтические средства, облучение, а также травмы и ожоги. Хотя фиброзные состояния охватывают широкую группу патологий, однако, очевидно, что в большинстве случаев, эти заболевания имеют общие механизмы, приводящие к аккумуляции фиброзной ткани и имеющие много общих элементов. Такие состояния часто вызываются реакцией на приток воспалительных клеток и непрерывно поддерживаются последующими путями передачи цитокиновых сигналов между инфильтрирующими клетками (например, макрофагами, Т-клетками) и клетками-резидентами в ткани (например, звездчатыми клетками, миофибробластами или клетками Купфера). Так, например, было показано, что MCP-1 играет определенную роль в развитии некоторых болезней легких (Rose C.E., Jr, Sung S.S., Fu S.M. Microcirculation 10:273-288 (2003)), и было показано, что CCR2-дефицитные мыши защищены от развития фиброза легких (Moore B.B., et al. Protection from Pulmonary Fibrosis in the Absence of CCR2 Signaling. J. Immunol. 167: 4368-4377 (2001)), что дает основание предположить, что этот рецептор играет важную роль в легких. Аналогичным образом, перициты представляют собой ключевые фиброгенные клетки, участвующие в развитии склеродермии, и было показано, что ингибиторы тирозинкиназного рецептора PDGF (RTKI) ослабляют пролиферацию перицитов и снижают уровень кожных поражений у пациентов с таким прогрессирующим заболеванием. В почках, инфильтрация лейкоцитов играет главную роль в опосредовании воспаления интерстициальной ткани трубочек и в индуцировании фиброза при хроническом заболевании почек. Vielhauer и коллеги установили, что у мышей с моделью обструктивной нефропатии, экспрессия CCR2 и CCR5 на аккумулирующихся макрофагах и CD3+-лимфоцитах коррелирует с прогрессирующим фиброзом на участках поражения ткани (Vielhauer V, et al. J. Am. Soc. Nephrol. 12:1173-1187 (2001)).
Используемый в данном описании термин «фиброз» также является синонимом термина «аккумуляция фибробластов и отложение коллагена». Фибробласты представляют собой клетки соединительной ткани, которые распространены в соединительной ткани всего организма. Фибробласты секретируют неригидный внеклеточный матрикс, содержащий коллаген типа I и/или типа III. В ответ на повреждение ткани, находящиеся поблизости фибробласты или звездчатые клетки мигрируют в рану, пролиферируют и продуцируют большое количество коллагенового внеклеточного матрикса. Коллаген представляет собой фиброзный белок, богатый глицином и пролином, которые являются главными компонентами внеклеточного матрикса, а также соединительной ткани, хряща и кости. Молекулы колагена представляют собой трехцепочечные спиральные структуры, называемые α-цепями, которые скручены друг с другом в спираль наподобие каната. Коллаген имеет несколько форм и типов, и наиболее распространенными из них являются коллаген типа I, который присутствует в коже, сухожилии и кости; и коллаген типа III, который присутствует в коже, сосудах и внутренних органах. Примерами фиброзных состояний являются, но не ограничиваются ими, ассоциированные с фиброзом болезни легких, например, идиопатический фиброз легких, фиброз, вызываемый облучением, хроническая обструктивная болезнь легких (ХОБЛ), склеродермия, фиброз легких, вызываемый блеомицином, хроническая астма, силикоз, фиброз легких, индуцированный асбестом, острое поражение легких и острый респираторный дистресс-синдром (включая острый респираторный дистресс-синдром, вызываемый бактериальной пневмонией, травмой, вирусной пневмонией, вентиляцией, не-легочным сепсисом и аспирацией); хронические нефропатии, ассоциированные с повреждением ткани/фиброзом (фиброзом почек), например, с волчанкой, диабетом, склеродермией, гломерулярным нефритом, очаговым сегментарным гломерулярным склерозом, IgA-нефропатией, гипертензией, аллотрансплантацией, волчанкой и болезнью Альпорта; фиброз кишечника, например, склеродермия и фиброз кишечника, вызываемый облучением; фиброз печени, например, цирроз, фиброз печени, вызываемый алкоголем, не-алкогольный стеатогепатит (NASH), повреждение желчных протоков, первичный биллиарный цирроз, фиброз печени, вызываемый инфекцией или вирусом (например, хронической HCV-инфекцией) и аутоиммунный гепатит; фиброз головы и шеи, например, фиброз головы и шеи, индуцированный облучением; образование рубцов на роговице, например, вызываемое LASIX™, трансплантацией роговицы и трабекулэктомией; гипертрофические рубцы и келоидные рубцы, вызываемые, например, ожогами и хирургической операцией; и другие фиброзные заболевания, например, саркоидоз, склеродермия, повреждение спинного мозга/фиброз спинного мозга, миелофиброз, рестеноз сосудов, аторосклероз, гранулематоз Вегенера, смешанное заболевание соединительных тканей и болезнь Пейрони.
В одном из аспектов изобретения, CCR2-опосредуемое расстройство характеризуется патологическим воспалением. Используемый в данном описании термин «патологическое воспаление» означает аномальное и/или хроническое воспаление, ассоциированное с расстройствами, включая, но не ограничиваясь ими, астму, атеросклероз, деменцию при СПИДе, диабет, воспалительное заболевание кишечника, ревматоидный артрит, отторжение трансплантата, реакцию «трансплантат против хозяина», рассеянный склероз (который, в частности, приводит к последующей демиелинизации), опухоли, опухолевые метастазы, нефрит, атопический дерматит, псориаз, ишемию миокарда, хронический простатит, ожирение и метаболический синдром. Такое воспаление характеризуется повышенным ответом воспалительных клеток, включая инфильтрирующие лейкоциты. В течение определенного периода времени, такое патологическое воспаление часто приводит к повреждению ткани в области аномального воспаления. В соответствии с этим, настоящее изобретение относится к способам лечения индивидуума, страдающего патологическим воспалением, где указанные способы включают введение указанному индивидууму терапевтически эффективного количества антитела или его антигенсвязывающей части, которые связываются с CCR2 и ингибируют CCR2.
Антитело или его антигенсвязывающие фрагменты могут быть также использованы для лечения расстройств, развитие которых вызывает активация CCR2 посредством связывания с хемокинами, включая MCP-1, MCP-2, MCP-3 и MCP-4.
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения воспалительных или аллергических заболеваний и состояний, включая респираторные аллергические заболевания, такие как астма, аллергический ринит, заболевания, ассоциированные с гиперчувствительностью легких, гиперчувствительный пневмонит, заболевание интерстициальной ткани легких (ILD) (например, идиопатический фиброз легких или ILD, ассоциированные с ревматоидным артритом, системной красной волчанкой, анкилозирующим спондилитом, системным склерозом, синдромом Сьегрена, полимиозитом или дерматомиозитом), хроническую обструктивную болезнь легких, анафилактический шок или реакции гиперчувствительности, аллергии, вызванные лекарственными средствами (например, пенициллином, цефалоспорином), аллергии от укусов насекомых, воспалительные заболевания кишечника, такие как болезнь Крона и язвенный колит, спондилоартропатии, склеродермия, псориаз и воспалительные дерматозы, такие как дерматит, экзема, атопический дерматит, аллергический контактный дерматит, крапивница, васкулиты (например, некрозирующий васкулит, кожный васкулит и васкулит, ассоциированный с гиперчувствительностью).
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения аутоиммунных заболеваний. Примерами аутоиммунных заболеваний являются, но не ограничиваются ими, болезнь Аддиссона, гемолитическая анемия, антифосфолипидный синдром, ревматоидный артрит, дерматит, аллергический энцефаломиелит, гломерулонефрит, синдром Гудпасчера, болезнь Грейвса, рассеянный склероз, тяжелая миастения, неврит, офтальмологическое заболевание, буллезный пемфигоид, пузырчатка, полиэндокринопатии, пурпура, болезнь Рейтера, синдром негнущегося человека, аутоиммунный тиреоидит, системная красная волчанка, аутоиммунное воспаление легких, синдром Гийена-Барре, инсулинзависимый сахарный диабет, и аутоиммунное воспалительное заболевание глаз.
В некоторых случаях, аутоиммунным заболеванием, подвергаемым лечению антагонистическими анти-CCR2 антителами, является ревматоидный артрит (РА). TH1-индуцированным расстройством, РА, является наиболее распространенное аутоиммунное заболевание у человека, которому подвержено примерно 1% населения индоевропейской расы (Harris, B. J. et al., 1997, In Textbook of Rheumatology 898-932), и которым в настоящее время страдают 2,5 миллиона американцев. РА характеризуется хроническим воспалением синовиальных соединений в суставах и инфильтрацией активированных Т-клеток, макрофагов и клеток плазмы, которое приводит к прогрессирующей деструкции суставного хряща. Это является наиболее тяжелой формой заболевания суставов.
В другом аспекте изобретения, аутоиммунным заболеванием, подвергаемым лечению анти-CCR2 антителами-антагонистами, является рассеянный склероз (РС). РС, который также представляет собой TH1-расстройство, является наиболее распространенным демиелинизирующим заболеванием центральной нервной системы (ЦНС), которым страдает 350000 (0,1%) человек в Северной Америке и 1,1 миллиона человек во всем мире. Вообще говоря, считается, что РС представляет собой аутоиммунное заболевание, частично опосредуемое провоспалительными CD4-Т (Th1)-клетками и моноцитами, которые специфически распознают миелиновые полипептиды в ассоциации с молекулами MHC класса II, экспрессируемыми на антиген (Ag)-презентирующих клетках (АПК).
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения сахарного диабета типа I или инсулинзависимого сахарного диабета (ИЗСД) у человека, то есть, заболевания, характеризующегося аутоиммунной деструкцией бета-клеток в панкреатических островках Лангерганса. Истощение бета-клеток приводит к нарушению регуляции уровней глюкозы в крови. У человека, началу развития диабета предшествует длительный преклинический период. За это период происходит постепенная потеря функций панкреатических бета-клеток. Развитие заболевания осложняется присутствием аутоантител против инсулина, (глутаминовая кислота)-декарбоксилазы и тирозинфосфатазы IA2 (IA2).
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения невропатической боли. Было показано, что у мышей с дефицитом CCR2 наблюдается ослабление невропатической боли (Abbadie et al., Proc. Natl. Acad. Sci. USA. 100(13): 7947-52 (2003)). Используемый в данном описании термин «невропатическая боль» означает боль, возникающую в результате повреждения нервной ткани. Невропатическую боль следует дифференцировать от ноцицептивной боли, которая вызывается сильным повреждением ткани, включая малые кожные нервы или кожные нервы в мышечной или соединительной ткани. Боль, вызываемая ноцицептивным механизмом, обычно ограничивается продолжительностью периода репарации ткани и ослабляется под действием доступных аналгетиков или опиоидов, описанных Myers, Regional Anesthesia 20:173-184 (1995). Невропатическая боль, обычно является продолжительной или хронической и часто длится в течение нескольких дней или месяцев после первого сильного повреждения ткани. Неврологическая боль может быть персистентной, спонтанной, а также может представлять собой аллодинию, являющуюся болевой реакцией на раздражитель, который, по своей природе, не вызывает боль. Невропатическая боль может также проявляться гипералгезией, при которой наблюдается явно выраженный ответ обычно на легкий болевой раздражитель, такой как укол иглой.
Анти-CCR2 антитела-антагонисты могут быть использованы для ослабления невропатической боли, возникающей при заболеваниях периферической нервной системы, спинномозговых узлов, спинного мозга, ствола головного мозга, таламуса или коры головного мозга. Такой способ может быть применен для ослабления невропатической боли независимо от этиологии ее возникновения. Так, например, такой способ может быть применен для ослабления невропатической боли, возникающей в результате заболеваний периферической нервной системы, таких как неврома, сдавливание нерва, разрушение нерва, натяжение нерва или неполный поперечный разрыв нерва; мононевропатия или полиневропатия. Этот способ может быть использован для ослабления невропатической боли, возникающей в результате таких расстройств, как сдавливание спинномозговых узлов; воспаление спинного мозга; контузия; опухоли; поражение одной половины спинного мозга; опухоли ствола головного мозга, таламуса и коры головного мозга; или травма ствола головного мозга, таламуса и коры головного мозга.
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения атеросклероза. Атеросклеротические бляшки развиваются в течение нескольких десятилетий и вызывают инфильтрацию воспалительных клеток, пролиферацию клеток гладких мышц, аккумуляцию внеклеточного матрикса, образование фиброзной чашечки и ангиогенез (Bayes-Genis et al. Circ. Res. 86:125-130 (2000)). Хемотаксис участвует в развитии атеросклероза на ранней стадии. Клеточные популяции мигрируют во внутреннюю часть стенки сосудов и образуют новую внутреннюю оболочку, что приводит к образованию атеросклеротических бляшек. Так, например, хемотаксис моноцитов индуцируется белком-хемоаттрактантом моноцитов 1 (MCP-1), который экспрессируется на ранней стадии развития атеросклероза в пораженных стенках артерий (Furukawa et al. Circ. Res. 84:306-314 (1999); Han et al., J. Lipid Res. 40:1053 (1999)). Трансплантация костного мозга CCR2-/--мышей, но не CCR2+/+-мышей, мышам ApoE3-Leiden, то есть, мышам, восприимчивым к атеросклерозу, индуцированному кормом, приводит к снижению атерогенеза, что позволяет предположить, что развитие атеросклероза вызывается передачей MCP-1-сигнала посредством CCR2 (Guo et al. (2003) Arterioscler Thromb. Vasc. Biol.; 23(3):447-53). В соответствии с этим, антитела-антагонисты против CCR2, а в частности, антитела-антагонисты, которые связываются с первой и/или второй внеклеточной петлей CCR2, могут быть введены индивидууму в целях снижения вероятности развития у него атеросклероза или в целях лечения или вспомогательного лечения атеросклероза.
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения ожирения. Было продемонстрировано, что при ожирении, жировая ткань содержит большое число моноцитов. Эти моноциты могут способствовать либо накоплению жира, либо развитию различных осложнений, обычно ассоциированных с ожирением, которое обычно называют метаболическим синдромом. Метаболический синдром включает такие осложнения, как развитие диабета.
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для лечения стеноза или рестеноза сосудистой системы, а в частности, артритов, например, болезни коронарной артерии, таких как стеноз или рестеноз, которые возникают в результате повреждения сосудов (например, в результате хирургического, терапевтического или механического вмешательства), а также гиперплазии неоинтимы. Так, например, рестеноз, который обычно вызывает сужение просвета сосуда, может быть результатом повреждения сосуда, включая, но не ограничиваясь ими, повреждения, продуцируемые трансплантацией, ангиопластикой, включая ангиопластику, осуществляемую с помощью баллонного катетера, атерэектомии, лазерной техники или другим подходящим методом (например, путем чрезкожной внутрипросветной коронарной ангиопластики (PTCA)), установки стента (например, установки механического или биологического эндоваскулярного стента), сосудистое шунтирование или их комбинаций, а также другие операции, применяемые для лечения стеноза и закупорки сосудов.
Анти-CCR2 антитела или их антигенсвязывающие фрагменты (например, антитела 4.40 и/или 4.9 или их антигенсвязывающие фрагменты), обладающие антагонистическим действием, могут быть использованы для предотвращения отторжения трансплантата (например, при трансплантации), включая отторжение аллотрансплантата или реакцию «трансплантат против хозяина» и артериосклероз, ассоциированный с трансплантацией органов.
Антитела и их антигенсвязывающие фрагменты, которые являются антагонистами CCR2, могут быть использованы в качестве терапевтических средств для лечения ВИЧ-инфекций. ВИЧ-1 и ВИЧ-2 представляют собой этиологические факторы, вызывающие синдром приобретенного иммунодефицита (СПИД) у человека. СПИД возникает отчасти в результате истощения CD4+-Т-лимфоцитов у ВИЧ-инфицированных индивидуумов. ВИЧ-1 инфицирует, главным образом, T-лимфоциты, моноциты/макрофаги, дендритные клетки, а в центральной нервной системе, этот вирус инфицирует микроглиальные клетки. Все эти клетки экспрессируют гликопротеин CD4, который служит в качестве рецептора для ВИЧ-1 и ВИЧ-2. Эффективное проникновение ВИЧ в клетки-мишени зависит от уровня связывания гликопротеина внешней оболочки вируса, gp120, с амино-концевым доменом CD4.
Кроме того, было показано, что CCR2 действует как ко-рецептор для ВИЧ-1 (Frade et al. (1997) J. Clin. Invest.; 100(3):497-502). После связывания с вирусом, гликопротеины оболочки ВИЧ-1 опосредуют слияние вируса и мембран клеток-хозяев, и этим завершается процесс проникновения вируса в клетку. Слияние с мембраной, регулируемой гликопротеинами оболочки ВИЧ-1, экспрессируемым на поверхности инфицированных клеток, приводит к слиянию клеток и образованию синцития.
Антитела и их антигенсвязывающие фрагменты, которые являются антагонистами CCR2, могут быть использованы в качестве терапевтических средств для лечения различных глазных болезней, включая возрастную дегенерацию желтого пятна (AMD), увеит и инфекционное заболевание роговицы.
Возрастная дегенерация желтого пятна (AMD)
Было показано, что CCR2 играет определенную роль в рекрутинге макрофагов с последующим развитием хороидальной неоваскуляризации (CNV), которая наблюдается у пациентов с возрастной дегенерацией желтого пятна, а также у пациентов с ангиоидными полосами сетчатки, сильной близорукостью и глазным гистоплазмозом (Tsutsumi, C. et al. Journal of Leukocyte Biology 74:25-32, 2003).
Увеит
Было показано, что у пациентов, страдающих увеитом, острый идиопатический передний увеит (Yeo, TK, et al. Cytokine 35:29-35, 2006) и увеит заднего сегмента, опосредуемый идиопатическим иммунным заболеванием, ассоциируется со связыванием между CCR2, имеющего полиморфизм в один нуклеотид, и его лигандом (MCP-1) (Ahad, MA, et al. Mol. Vis 13:388-396 2007.
Инфекции роговицы (бактериальные):
Было показано, что CCL2 играет определенную роль в регуляции рекрутинга полиморфонуклеарных нейтрофилов (PMN) в процессе инфицирования роговицы (Xue, M. L, et al., Immunology and Cell Biology 85:525-531 2007). Хотя PMN играют важную роль в элиминации бактерий и в стимуляции заживления ран роговицы, однако, постоянное присутствие этих клеток может приводить к развитию хронического воспалительного заболевания.
Антитела и их антигенсвязывающие фрагменты, которые являются антагонистами CCR2, могут быть использованы в качестве терапевтических средств для лечения различных солидных опухолей и раковых заболеваний, включая хронический лимфоцитарный лейкоз, хронический миелоцитарный лейкоз, множественную миелому, злокачественную миелому, болезнь Ходжкина и карциному предстательной железы, мочевого пузыря, молочной железы, шейки матки, толстой кишки, легких, печени или желудка, солидные опухоли и раковые заболевания.
Рак предстательной железы
Было показано, что МСР-1 действует как паракринный и аутокринный фактор, вызывающий рост и инвазию раковых клеток предстательной железы (Lu, Y. et al. The Prostate 66:1311-1318, 2006), а также было показано, что экспрессия CCR2 коррелирует с прогрессированием рака предстательной железы (Lu, Y. et al. Journal of cellular biochemistry 101: 676-685, 2007). Было показано, что системное введение нейтрализующих анти-MCP-1 антител индуцирует регрессию раковой опухоли предстательной железы у мышей (Loberg, R. D., Cancer Res 67:9417- 9424 2007).
Рак молочной железы
Считается, что опухолеассоциированные макрофаги играют важную роль в «иммунном надзоре» и развитии опухоли, а активация и рекрутинг лимфоцитов регулируется хемокинами, включая MCP-1. При раке молочной железы была продемонстрирована его явная взаимосвязь с полиморфизмом CCR2 (Zafiropoulos, A., N. et al., Journal of medical genetics 41:e59 2009). Было также показано, что путь CCL2/CCR2 играет ведущую роль в рекрутинге миелоид-ингибирующих клеток, приводящем к раковым заболеваниям, включая рак молочной железы, яичника и желудка, где указанные клетки стимулируют прогрессирование опухоли, ангиогенез и васкулоангиогенез (Huang, B., et al., Cancer letters 252:86-92, 2007).
Меланома
Было также показано, что блокирование функции MCP-1 приводит к ингибированию рекрутинга опухолеассоциированных макрофагов и к предупреждению ангиогенеза опухоли и роста злокачественной меланомы у мышей (Koga, M. et al., Biochemical and biophysical research communications 365:279-284, 2008).
Рак печени
Было показано, что блокирование CCR2 приводит к ослаблению транспорта печеночных звездчатых клеток, то есть, главного источника металлопротеиназы матрикса 2, который облегчает неоваскуляризацию в процессе образования опухоли печени (Yang, X. et al. International journal of cancer 118:335-345, 2006).
Рак шейки матки
Было показано, что CCR2 играет определенную роль в рекрутинге макрофагов, приводящем к ангиогенезу опухоли и к развитию неоплазии шейки матки в результате поражений внутри плоскоклеточного эпителия (Coelho, A., et al. Gynecologic oncology 96:760-764, 2005; Coelho, A., et al. Gynecologic and obstetric investigation 64:208-212, 2007).
Рак яичника
Было также показано, что путь CCL2/CCR2 также играет важную роль в рекрутинге миелоидных клеток-супрессоров, приводящем к раковым заболеваниям, включая рак молочной железы, яичника и желудка, где указанные клетки стимулируют прогрессирование опухоли, ангиогенез и васкулоангиогенез (Huang, B., et al., Cancer letters 252:86-92, 2007).
Антитело может быть введено один раз или более раз. Антитело может быть введено от четырех раз в день до одного раза через каждые шесть месяцев или более. Введение может быть осуществлено по такой схеме: введение как три раза в день, два раза в день, один раз в день, один раз через каждые два дня, один раз через каждые три дня, один раз в неделю, один раз в каждые две недели, один раз в месяц, один раз в каждые два месяца, один раз в каждые три месяца и один раз в каждые шесть месяцев. Антитело может быть также введено непрерывно с помощью мининасоса. Антитело может быть введено через слизистую, трансбуккально, интраназально, путем ингаляции, внутривенно, подкожно, внутримышечно, парентерально, вовнутрь опухоли или местно. Антитело может быть введено в отдельный участок или системно.
Антитело может быть введено один раз по меньшей мере два раза или по меньшей мере в течение периода времени, достаточного для лечения, а именно, для паллиативного лечения или полного излечения данного заболевания. Обычно, антитело вводят как часть фармацевтической композиции, описанной в настоящей заявке. Доза антитела обычно составляет в пределах 0,1-100 мг/кг, 0,5-50 мг/кг, 1-20 мг/кг или 1-10 мг/кг. Концентрация антитела в сыворотке может быть измерена любым методом, известным специалистам.
Нижеследующие примеры приводятся лишь в целях иллюстрации и никоим образом не должны рассматриваться как ограничение объема изобретения.
Пример 1
Получение гибридом, продуцирующих анти-CCR2 антитело
8-10 недельных мышей XENOMOUSE™, у которых продуцируются человеческие антитела IgG2 и IgG4, иммунизировали путем инъекции им CCR2B-трансфецированных клеток 300-19 в подушечку задних лап (Genbank MN000648, SEQ ID NO:204) (107 клеток/дозу/мышь в адъюванте TITERMAX™ Gold, Sigma, номер по каталогу #T2684, партия #K1599; в объеме 50/50). Этим мышам вводили 5-9 бустер-инъекций в адъюванте, таком как гель на основе фосфата алюминия (номер по каталогу # 452-250, партия #8937, HCI Biosector (5 мкл/мышь/бустер-дозу)) и qCpG (мышиный адъювант IMMUNEASY™) (номер по каталогу #303101; партия #11551249; Qiagen (15 мкл/мышь/бустер-дозу)) в течение 3-8 недель. За четыре дня до процедуры слияния клеток, мышам вводили конечную инъекцию в PBS. Лимфоциты селезенки и лимфоузлов, взятые у иммунизованных мышей, собирали и подвергали слиянию с несекреторной миеломной клеточной линией P3-X63-Ag8.653. Слитые клетки подвергали HAT-отбору, как описано ранее (Galfre and Milstein, Methods Enzymol. 73:3-46 (1981)). Затем выделяли панель гибридом, каждая из которых секретировала CCR2-специфические человеческие антитела. Ряд антител идентифицировали на связывание с CCR2, как было определено с помощью FACS-анализа. Гибридомы отбирали для последующего исследования, и некоторые из них представлены в таблице 6.
Гибридомы, указанные в таблице 2, были депонированы в Американской коллекции типовых культур (ATCC), 10801 University Blvd., Manassas, VA 20110-2209, в соответствии с Будапештским договором. Этим гибридомам были присвоены нижеследующие регистрационные номера:
Пример 2
Секвенирование анти-CCR2 антител
Для анализа структуры продуцируемых антител, нуклеиновые кислоты, кодирующие фрагменты, содержащие вариабельный домен тяжелой и легкой цепи, клонировали из гибридом, продуцирующих моноклональные анти-CCR2 антитела. Затем клонировали легкие и тяжелые цепи анти-CCR2 антител, и последовательность, подтверждали как описано для антитела 4.40.2, следующим образом. Poly(A)+-мРНК выделяли с использованием мининабора RNEASY™ MINI KIT (Qiagen), и кДНК синтезировали из мРНК с помощью набора для ОТ-ПЦР ADVANTAGE (BD Biosciences) с использованием oligo(dT)-праймеров. Oligo(dT)-праймированную кДНК амплифицировали для клона 4.40.2 с использованием праймеров, перечисленных в таблице 3. Амплификацию осуществляли с использованием полимеразы PFUULTRA HIGH-FIDELITY (Stratagene) на устройстве PTC-200 DNA Engine (MJ Research) с проведением следующих циклов: 2 мин. при 95°C; 25 × (20 сек. при 95°C, 30 сек. при 55°C, 30 сек. при 72°C); 10 минут при 72°C. ПЦР-ампликоны клонировали в экспрессионные векторы тяжелой и легкой цепи. Затем векторы трансформировали в химически компетентные клетки DH5α MAX EFFICIENCY (Invitrogen) в соответствии со стандартным протоколом. Последовательности клонов подтверждали с использованием химических реагентов Grills 16th BDTv3.1/dGTP (Applied Biosystems Inc) на анализаторе ДНК 3730×1 (Applied Biosystems Inc). Исходя из нуклеотидной последовательности и предсказанной аминокислотной последовательности антител определяли частоту встречаемости генов для каждой цепи антитела. Выравнивание с «эталонной последовательностью V-оснований» (MRC Centre for Protein Engineering, Cambridge, UK; Tomlinson, et al., J. Mol. Biol., 227, 776-798 (1992); Hum. Mol. Genet., 3, 853-860 (1994); EMBO J., 14, 4628-638 (1995)) осуществляли с помощью компьютерных программ MACVECTOR™ и GENEWORKS™ (Oxford Molecular Group, Campbell, CA, USA).
Праймеры вариабельного домена, сконструированные для амплификации 4.40.2 (5'-3')
(SEQ ID NO: 147)
(SEQ ID NO: 148)
(SEQ ID NO: 149)
(SEQ ID N0:150)
В таблице 4 представлена встречаемость генов в выбранных клонах антитело-продуцирующих гибридом.
Встречаемость генов тяжелых и легких цепей
Гибридомные клоны получали у мышей с IgG4, и после получения последовательностей, вариабельные домены клонировали в экспрессионные векторы в формате IgG1, IgG2 и IgG4 для сравнения.
Пример 3
Мутагенез анти-CCR2 антител
Восстановление последовательности вариабельной области легкой цепи зародышевой линии
Аланиновый остаток, находящийся в положении 68 легкой цепи анти-CCR2 антитела 4.40.2, снова заменяли глициновым остатком зародышевой линии посредством сайт-направленного мутагенеза как описано ниже. Точковую мутацию в кодоне аминокислотного остатка 68 заменяли с использованием праймеров, представленных в таблице 5. Сайт-направленный мутагенез осуществляли с использованием набора для сайт-направленного мутагенеза QUIKCHANGE™ Il (Stratagene) в соответствии со стандартным протоколом проведения циклов реакций с нижеследующими модификациями: 2 минуты при 96°C; 16X (50 сек. при 96°C, 10 сек. при 68°C); 10 сек. при 68°C. Вектор, подвергнутый мутагенезу, переносили в сверхкомпетентные клетки XL1-Blue. Для подтверждения успешно проведенного мутагенеза, клоны секвенировали с использованием химических реагентов Grills 16th BDTv3.1/dGTP (Applied Biosystems Inc.) на анализаторе ДНК 3730xI (Applied Biosystems Inc.).
Праймеры для мутагенеза (5'-3')
(SEQ ID NO: 151)
(SEQ ID NO: 152)
Полученное антитело, имеющее глицин в положении 68 вариабельной области легкой цепи, обозначали 4.40 A68G (SEQ ID NO:112).
Получение константной области тяжелой цепи IgG4 с мутацией, стабилизирующей шарнирную область
Константные области тяжелой цепи IgG4 выделяли из экспрессионного вектора IgG4, который был получен от pCON-G4(pro) (Lonza, Basel, Switzerland) и, по сравнению с зародышевой линией, имел мутацию, стабилизирующую шарнирную область и введенную путем замены аминокислоты серина на пролин в положении 230 (Angal, S. et al., Molecular Immunology 30:105-108 (1993)) в последовательности аллотипа L309 IgG4 (Brusco A. et al., Eur J. Immunogenticsl 25:349-355 (1998)).
Молчащую мутацию (Lys, AAG→AAA) вводили посредством ПЦР-мутагенеза в 11-й нуклеотидный остаток экзона G4 CH1 вектора pCON-G4(pro) для удаления сайта распознавания Apal в целях облегчения субклонирования с использованием нижеследующих праймеров для амплификации:
G4_прямой ttatgctgggcccagctctgtcccacaccgcggtcacatggcaccacctctcttgcaGCTTCCACC AAAGGCCCATCCGTCTTCCCCC (SEQ ID NO: 153)
(Гибридизующиеся основания показаны заглавными буквами (CAPS) (мутация в остатке #11 CH 1 показана жирным шрифтом); не-гибридизующиеся основания показаны строчными буквами)
G4_обратный:
tcatattctctagaTCATTTACCCAGAGACAGGGAGAGG (SEQ ID NO: 154) (Гибридизующиеся основания показаны заглавными буквами (CAPS). не-гибридизующиеся основания с Xbal-сайтом показаны строчными буквами).
Амплификацию осуществляли с использованием полимеразы Expand Hi-Fi (Roche) на оборудовании PTC-200 DNA ENGINE™ (MJ Research), и реакции проводили в термоячейке в следующих условиях: 3 мин. при 95°C; 22X (20 сек. при 95°C, 30 сек при 58°C, 2 мин. 10 сек при 72°C); 10 мин. при 72°C.
Другую молчащую мутацию (Thr, ACC→ACA) вводили посредством ПЦР-мутагенеза в нуклеотидный остаток 209 экзона G4 CH1 вектора pCON-G4(pro) для удаления BstEII-сайта в целях облегчения использования этого фермента для субклонирования вариабельных областей в экспрессионный вектор с использованием нижеследующих праймеров:
G4delBst_F
CAGCGTGGTGACAGTGCCCTCCAGCAG (SEQ ID NO: 155)
G4delBst_R
CTGCTGGAGGGCACTGTCACCACGCTG (SEQ ID NO: 156).
Амплификацию осуществляли с использованием набора для сайт-направленного мутагенеза QUIKCHANGE™ (Stratagene) на оборудовании PTC-200 DNA ENGINE™ (MJ Research), и реакции проводили в термоячейке в следующих условиях: 1 мин. при 96°C; 14X (50 сек. при 96°C, 14 мин 48 сек. при 68°C), 10 мин. при 72°C.
Затем полученную константную область IgG4, содержащую мутацию в шарнирной области и две молчащие мутации, обозначаемые S230P, субклонировали в виде Apal/Xbal-фрагмента в Apal/Xbal-сайты экспрессионного вектора, кодирующего DHFR- антитело. Затем вариабельную область тяжелой цепи антитела 4.40.2 клонировали в экспрессионный вектор, содержащий константную область G4 S230P с получением полноразмерной конструкции тяжелой цепи. Антитело, происходящее от 4.40 и имеющее замену A68G в вариабельной области тяжелой цепи зародышевой линии и замену S230P в шарнирной области тяжелой цепи, называли антителом 4.40 A68G S230P, имеющим аминокислотную последовательность легкой цепи SEQ ID NO:112 и аминокислотную последовательность тяжелой цепи SEQ ID NO:116.
Супернатанты клеток, трансфецированных экспрессионными векторами, собирали и очищали стандартной аффинной хроматографией на белке А для выделения рекомбинантных иммуноглобулинов. Затем эти белки охарактеризовывали с помощью электрофореза в ДСН-ПААГ, методом рассеяния света и спектрофотометрии.
Пример 4
Связывание In vitro
На фигуре 1 показано in vitro связывание AF-488-конъюгированного антитела 4.40 A68G S230P с человеческими моноцитами в цельной крови (фигура 1А), а именно с CCR2- трансфецированными клетками 300-19 (фигура 1B), и связывание антитела 4.40 A68G S230P в различных концентрациях с CCR2- трансфецированными клетками 300-19, как было детектировано с использованием ФЭ-конъюгированного античеловеческого антитела (фигура 1C). Вкратце, 1 миллион клеток в объеме 100 мкл окрашивали тестируемым антителом с использованием забуференного фосфатом физиологического раствора Дульбекко, содержащего 2% термоинактивированную фетальную бычью сыворотку и 0,002% азид натрия. Через 15 минут добавляли «второе» детектирующее антитело. Через 30 минут, клетки промывали в буфере, ресуспендировали в 500 мкл, и окрашивание оценивали на приборе FACS CALIBUR. Было оценено всего 10000 событий связывания на пробирку. Среднюю интенсивность флуоресценции канала проточной кюветы оценивали для каждой концентрации антитела как показатель величины окрашивания.
Пример 5
Определение констант аффинности (К D ) моноклональных анти-CCR2 антител
Для определения КD антител против CCR2, связывание антитела с CCR2-экспрессирующими клетками 300-19 оценивали с помощью FACS-анализа, проводимого в течение 3 часов. Вкратце, с использованием буфера, состоящего из забуференного фосфатом физиологического раствора Дульбекко, содержащего 2% термоинактивированную фетальную бычью сыворотку, 0,002% азид натрия и 0,005 мг/мл цитохалазина-В (Sigma 6762), 1 миллион клеток в объеме 100 мкл окрашивали тестируемым антителом. Через пятнадцать минут добавляли «второе» детектирующее антитело, то есть, конъюгированный с R-фикоэритрином аффинно очищенный F(ab')-фрагмент ослиного антитела против человеческих IgG H+L (Jackson 709-116-149). Пробирки слегка встряхивали в течение 3 часов при комнатной температуре. После 2 промывок в буфере, клетки ресуспендировали в 500 мкл буфера и оценивали на приборе FACS CALIBUR™. Было оценено всего 10000 событий связывания на пробирку. Среднюю интенсивность флуоресценции канала проточной кюветы оценивали для каждой концентрации антитела как показатель величины окрашивания. Эти исследования по связыванию показали, что антитело 4.40 A68G S230P связывается с человеческим CCR2 на трансфецированных клетках с КD=0,085 мкг/мл (0,58 нМ) (фигура 2). После оценки насыщенного связывания на клетках, экспрессирующих химерный рецептор CCR2/CCR5, имеющий первую и вторую внеклеточные петли CCR2 (см. пример 7), строили кривую для соответствующих концентраций и КD (КD=0,023 мкг/мл (0,16 нМ)), как показано на фигуре 6C.
Пример 6
Ингибирование MCP-1-индуцированного хемотаксиса
Человеческие анти-CCR2 антитела оценивали на их способность ингибировать хемотаксис моноцитов THP-1 (ATCC # TIB 202) в ответ на действие лиганда CCR2, MCP-1 (CCL2). Хемотаксис осуществляли в 96-луночных камерах для хемотаксиса, закупленных у NeuroProbe, Inc. (Gaithersburg, MD), как было описано в литературе (см., Gladue RP et al., J. Biol. Chem. 278:40473-40480 (2003)). Вкратце, CCL2 разводили в среде RPMI 1640 (Roswell Park Memorial Institute), содержащей 0,1% BSA, а затем наносили на дно лунок камеры. Фильтр с 5 мкм порами (Neuroprobe) помещали между верхними и нижними лунками камеры. Затем клетки THP-1 добавляли в верхнюю часть камеры (8×105) в присутствии или в отсутствии различных концентраций тестируемых антител. Устройство инкубировали в течение 3 часов в 5% CO2-инкубаторе с повышенной влажностью при 37°C. После инкубирования, не-мигрировавшие клетки удаляли из верхней части камеры, а верхнюю часть фильтра оборачивали пленкой. Затем в верхние лунки добавляли 2 мМ холодного EDTA, и содержимое камеры для хемотаксиса инкубировали при 4°C в течение 20 минут. Затем EDTA удаляли, и 96-луночный микротитрационный планшет центрифугировали при 800×g в течение 10 минут. После этого, фильтр удаляли, культуральную среду отбрасывали, и в планшеты добавляли 0,2% FDA. Затем планшеты инкубировали в течение 1,5 часа при 37°С до появления желтой окраски. Число мигрировавших клеток количественно оценивали путем считывания интенсивности окраски на ридере для микротитрационного планшета на 490 нм. Как показано в таблице 6, было идентифицировано несколько антител, которые в различной степени ингибировали хемотаксис ТНР-1.
Ингибирование MCP-1-индуцированного хемотаксиса THP-1
Аналогичным образом, антитело 4.40 A68G S230P ингибировало хемотаксис моноцитов THP-1 в ответ на действие MCP-1 (IC50=0,148 мкг/мл), но не в ответ на действие CCR1/CCR5-лиганда MIP-1, как показано на фигуре 3.
Хемотаксис первичных моноцитов
Гепаринизированную цельную человеческую кровь наносили слоями на поверхность пробирок ACCUSPIN HISTOPAQUE 1077 (Sigma; St. Louis, MO) и центрифугировали. Фракцию мононуклеарных клеток собирали, 3 раза промывали PBS, после чего эритроциты (RBC) подвергали лизису водой и клетки ресуспендировали при плотности 4×106/мл в среде RPMI (Gibco; Grand Isle, NY), содержащей 0,1% BSA (Sigma) и 10 мМ HEPES (Gibco). Разведения антител или контроль KLH добавляли к 0,25 нМ MCP-1 (Peprotech; Rocky Hill, NJ), и 30 мкл помещали на дно 48-луночной камеры Бойдена (Neuroprobe; Gaithersburg, MD); а в качестве негативного контроля использовали только среду. Сверху помещали 5 мкм фильтр, не содержащий ПВП (Neuroprobe), и камеру герметично закрывали. Выделенные мононуклеарные клетки инкубировали с разведениями антител или KLH-контролем в течение 30 минут при комнатной температуре. Затем в верхние лунки добавляли 50 мкл. Камеру инкубировали в течение 90 минут при 37°С в 5% CO2-инкубаторе с повышенной влажностью. После инкубирования, клетки удаляли путем аспирации из верхних лунок, верхнюю часть фильтра оборачивали пленкой, сушили воздухом, окрашивали красителем DIFF-QUIK™ (Dade-Behring; Newark, DE), и подсчитывали число мигрировавших клеток в 6 полях. Ингибирование хемотаксиса выделенных первичных человеческих моноцитов антителом 4.40 A68G S230P в ответ на действие MCP-1 показано на фигуре 4.
Кроме того, как было показано для антитела 4.40, какого-либо ингибирования клеток THP-1 CCR1/CCR5-лигандом MIP-1a не наблюдалось (фигура 3).
Пример 7
Конструирование химеры CCR2/CCR1
Антитела, ингибирующие MCP-1-индуцированный хемотаксис, тестировали на связывание с химерами CCR2 для картирования их эпитопов. Это было осуществлено путем оценки связывания антител с клетками 300-19, экспрессирующими различные химерные рецепторы, состоящие из внеклеточной петли (1-й, 2-й и/или 3-й) и N-концевых замен CCR2 с частями CCR1 (M=CCR2, R = CCR1).
Конструирование химерного рецептора MRRR
Химерный рецептор MRRR (N-конец CCR2 (M), 1-ю, 2-ю и 3-ю внеклеточные петли CCR1 (R)) конструировали с использованием набора для сайт-направленного мутагенеза QUIKCHANGE® от Stratagene и нижеследующих праймеров для мутагенеза в целях введения мутации в первый рестрикционный ApaL-сайт человеческого CCR1 дикого типа, а затем клонировали в экспрессионный вектор pcDNA3 (Invitrogen, Carlsbad, CA) в BamHI-сайт с использованием следующих праймеров:
смысловой праймер 40A:
CGAGAGGGCCTTTGGGATCCAACTGCTGCC (SEQ ID NO: 132)
антисмысловой праймер 40B:
GGCAGCAGTTGGATCCCAAAGGCCCTCTCG (SEQ ID NO: 133).
Химерный рецептор, содержащий FLAG-меченную химеру MMRR (N-конец и 1-ю внеклеточную петлю CCR2, 2-ю и 3-ю внеклеточную петли CCR1), клонированную в экспрессионный вектор, полученный из плазмиды pCMV-1 (Pharmacia; Piscataway, NJ), был предоставлен Израэлем Чаро из Института Дэвида Гладстона. Конструирование химеры MMRR (обозначенной авторами «2211») описано Monteclaro F. S. et al. (Methods in Enzymology 228:70-84 (1997)).
Первый ApaL-сайт в этой химере заменяли BamH1-сайтом с использованием нижеследующих праймеров для мутагенеза:
смысловой праймер: 41A:
GCAAATTGGGATCCAACTCCTGCC (SEQ ID NO: 134)
антисмысловой праймер: 41B:
GGCAGGAGTTGGATCCCAATTTGC (SEQ ID NO: 135).
Каждый полученный мутированный клон разрезали ферментами Ndel и BamHI. Фрагмент, содержащий вектор BamHI-мутанта CCR1/pcDNA3 и Ndel/BamHI-фрагмента, состоящего из 632 пар оснований и содержащего лишь N-конец CCR2 от мутированной плазмиды MMRR/pCMV1, выделяли на геле и лигировали друг с другом с получением химеры MRRR/pcDNA3.
BamHI-сайт в химере MRRR снова заменяли на ApaL-сайт с использованием нижеследующих праймеров для мутагенеза:
смысловой праймер 41C:
GCAAATTGGGGCCCAACTGCTGCC (SEQ ID NO: 136)
антисмысловой праймер 41D:
GGCAGCAGTTGGGCCCCAATTTGC (SEQ ID NO: 137).
Полноразмерный фрагмент MRRR подвергали ПЦР-амплификации с использованием смыслового праймера 66C (SEQ ID NO: 138), который был гомологичен области FLAG-метки указанного вектора, и антисмыслового праймера 66D (SEQ ID NO: 139), который был гомологичен 3'-концу CCR1 и содержал HindIII-сайт.
Смысловой праймер 66C
CTCTTGTGCCAGGGTGTGGTCTCCGA (SEQ ID NO: 138)
Антисмысловой праймер 66D
GATCGAAGCTTTCAGAACCCAGCAGAGAGTTCATG (SEQ ID NO: 139).
Затем этот фрагмент субклонировали в PfIMI- и HindIII-сайты ретровирусной конструкции, полученной из плазмиды pMIG (Van Parijs, L et al., Immunity 11:281-188 (1999)), (фигура 5), в которой перед FLAG-меткой была сделана инсерция гена (химера «мышь/человек»), а за ними были расположены Sal1-сайт, и внутренний сайт связывания с рибосомой - сайт активированного белка, флуоресценирующего в зеленом диапазоне спектра (IRESEGFP).
Конструирование химерных рецепторов RRRM и RMMR
Химерные рецепторы RRRM (N-конец, 1-ю и 2-ю внеклеточные петли CCR1, и 3-я внеклеточную петлю CCR2) и RMMR (N-конец и 3-ю внеклеточную петлю CCR1, и 2-ю и 3-ю внеклеточные петли CCR2) получали с применением двухстадийного ПЦР-метода.
Для получения химеры RRRM, третью петлю человеческого CCR2 дикого типа сначала подвергали ПЦР-амплификации с использованием нижеследующих праймеров:
Смысловой химерный праймер 89A:
GACTATACTTATTTCTGTTTTCATTGTCATTCTCCTGAACACC (SEQ ID NO: 140) Антисмысловой праймер 81 B:
CGCCAAGCTTCATTATAAACCAGCCGAGA (SEQ ID NO: 141).
Человеческий CCR2 дикого типа клонировали в ретровирусный экспрессионный вектор pMIG в качестве матрицы. N-концевую область - вторую внеклеточную петлю подвергали ПЦР-амплификации с использованием нижеследующих праймеров:
Смысловой праймер 89C:
ACGCGTCGACGAAACTCCAAACACCACAGAG (SEQ ID NO: 142)
Антисмысловой химерный праймер 89B:
GTTCAGGAGAATGACAATGAAAACAGAAATAAGTATAGTC (SEQ ID NO: 143),
и CCR1 дикого типа клонировали в экспрессионный вектор pcDNA3 в качестве матрицы.
Эти фрагменты очищали на геле, объединяли и подвергали ПЦР-амплификации с использованием 5'- и 3'-концевых праймеров: 89C (SEQ ID NO: 142) и 81B (SEQ ID NO:141). Полученный фрагмент лигировали в SaII/HindII-сайты ретровирусного вектора pMIG, который содержал N-концевую FLAG-метку, за которой был расположен сайт IRESEGFP.
Конструкция химерного рецептора RMMR
Последовательность от третьей внеклеточной петли CCR1 до цитоалазматического хвоста ПЦР-амплифицировали с использованием CCR1 дикого типа, клонированного в экспрессионный вектор pcDNA3 в качестве матрицы, и с использованием нижеследующих праймеров:
Смысловой химерный праймер 85A:
TCATTCTCCTGAACACCTTCCAAGACTTCCTGTTCACCCA (SEQ ID NO: 144)
Антисмысловой праймер 78D:
GCCAAGCTTCCAGTGTGATGGATATCTGA (SEQ ID NO: 145),
которые гибридизовались с областью вектора, расположенной со стороны 3'-конца от вставки в плазмиде CCR1/pcDNA3 дикого типа.
Второй фрагмент, содержащий N-конец CCR1, и область, содержащую первую и вторую внеклеточные петли CCR2, подвергали ПЦР-амплификации с использованием в качестве матрицы FLAG-меченной плазмиды RMMM/pcDNA3, предоставленной Израэлем Чаро из Института Дэвида Гладстона. Конструирование химерной конструкции RMMM (обозначенной авторами «2111») описано Monteclaro, F. S. et al. (Methods in Enzymology 228:70-84 (1997)). Используемыми праймерами являются смысловой праймер 66C (SEQ ID NO: 138), который гибридизовался с FLAG-меткой, и антисмысловой химерный праймер 85B:
TGGGTGAACAGGAAGTCTTGGAAGGTGTTCAGGAGAATGA (SEQ ID NO: 146).
Эти два фрагмента очищали на геле, объединяли и ПЦР-амплифицировали с использованием 5'- и 3'-концевых праймеров: 66C (SEQ ID NO:138) и 78D (SEQ ID NO:144). Этот конечный фрагмент гидролизовали ферментами PfIMI и HindIII и лигировали в ретровирусный вектор pMIG, который содержал N-концевую FLAG-метку, а за ней находился сайт IRESEGFP.
Экспрессия химеры
Из каждой химерной конструкции получали ретровирус, и этот ретровирус использовали для трансдукции клеток 300-19. Экспрессию оценивали с помощью FACS-анализа с использованием антитела против FLAG-эпитопа M1 (Sigma; St. Louis, MO, catalog # F3040).
Пример 8
Картирование эпитопов
Связывание антител с химерными рецепторами CCR2/CCR1 оценивали с FACS-анализа. Все рецепторы были помечены у N-конца меткой FLAG, а также окрашены анти-FLAG антителом M1 для подтверждения экспрессии рецептора. На фигурах 6A и B проиллюстрировано FACS-окрашивание FLAG на N-конце антителом М1 (фигура 6A) по сравнению с окрашиванием антителом 4.40 (фигура 6B). На фигуре 6C показана кривая насыщенного связывания антитела 4.40 A68G S230P на клетках 300-19, трансфецированных химерой RMMR. Насыщенное связывание антитела 4.40 A68G S230P с клетками 300-19, не трансфецированными CCR2, не наблюдалось в концентрациях до 10 мкг/мл (фигура 7A), а клетки 300-19, трансфецированные полностью человеческим CCR2, обнаруживали дозозависимое связывание в концентрациях, превышающих 0,01 мкг/мл (фигура 7B). Кроме того, насыщенное связывание антитела 4.40 A68G S230P с химерными рецепторами, содержащими только N-конец CCR2 (MRRR) вместе с 3 областями петли CCR1 (фигура 7C) или с 3-й внеклеточной петлей CCR2 с N-концом и с областями 1-й и 2-й петли CCR1 (RRRM), почти не обнаруживалось в концентрациях до 10 мкг/мл (фигура 7D). В противоположность этому, наблюдалось значительное дозозависимое насыщенное связывание антитела 4.40 A68G S230P с химерными рецепторами, содержащими N-конец и область 3-й петли CCR1 и области 1-й и 2-й петли CCR2 (RMMR), в концентрациях, превышающих 0,001 мкг/мл (фигура 7E). Экспрессию рецептора во всех указанных трансфектантах рецептора подтверждали путем оценки окрашивания их FLAG-метки у N-конца антителом M1 (Sigma #F3040).
Картирование эпитопа
ELISA пептида
Кроме того, связывание эпитопа с первой и/или второй петлей CCR2 также оценивали с помощью ELISA пептида. ELISA-планшет с покрытием Reacti-Bind NEUTRAVIDIN™, используемый для анализа на высокий уровень связывания (Pierce; Rockford, IL), 3 раза промывали PBS/0,05% твином 20, а затем покрывали 6 мкг/мл биотинилированного пептида петли 2 (SEQ ID NO: 129) или петли 3 (SEQ ID NO: 130) CCR2 (AnaSpec; San Jose, CA) в объеме 100 мкл/лунку. Планшеты инкубировали в течение 1 часа, а затем 3 раза промывали. Затем в планшет добавляли серийно разведенное антитело-антагонист 4.40.3 A68G S230P против CCR2 или другое «первое» антитело против CCR2 в PBS/0,1% BSA/0,05% твин-20, и инкубировали в течение 1 часа, при этом, также использовали другой контроль. Планшет 3 раза промывали, и во все лунки добавляли 100 мкл/лунку ПХ-конъюгированного мышиного «второго» антитела против человеческих IgG4 (Zymed; So. San Francisco, CA) при разведении 1:5000, и инкубировали в течение 1 часа. Затем планшет 3 раза промывали, после чего во все лунки добавляли субстрат TMB и инкубировали ~30 минут. Цветовую реакцию завершали добавлением 2M H2SO4, и определяли оптическую плотность на планшет-ридере на 450 нм (фигура 8). Были идентифицированы антитела, включая антитела 4.9 и 4.40, которые связывались с пептидом второй внеклеточной петли рецептора и не связывались с третьей петлей.
Пример 9
Селективность связывания
Селективность связывания антитела 4.40 A68G S230P с человеческим CCR2 подтверждали по отсутствию связывания с близкородственным хемокиновым рецептором CCR5, как показано на фигуре 9, и по отсутствию ингибирования хемотаксиса, индуцированного CCR5/CCR1-лигандом MIP-1a (фигура 3). Анти-CCR5 антитело (Pharmingen, catalog #555992) связывалось с клетками, экспрессирующими CCR5 (фигура 9b), а анти-CCR2 антитело 4.40.3 A68G S230P не связывалось с этими клетками (фигура 9a). В этих исследованиях, клетки 300.19, сконструированные для экспрессии CCR5, инкубировали либо с CCR2-специфическим антителом, либо с антителом 4.40 A68G S230P, либо с CCR5-специфическим антителом в течение 30 минут. Затем клетки промывали в FACS-буфере (0,02% азид натрия, 2% термоинактивированная фетальная бычья сыворотка в PBS) и анализировали на экспрессию на клеточной поверхности с помощью проточного цитометра стандартными методами.
Пример 10
Мобилизация кальция
Для того чтобы определить, действует ли антитело 4.40 как функциональный антагонист на CCR2 и не обладает ли он какими-либо значимыми агонистическими свойствами, действие антитела 4.40.3 на мобилизацию кальция в CCR2-трансфецированных клетках 300-19 тестировали как описано ранее (Gladue RP et al., J. Biol. Chem. 278:40473-40480 (2003)). Человеческие CCR2-трансфецированные клетки 300-19 центрифугировали и ресуспендировали при плотности 2×106 клеток/мл в буфере PTI (HBSS (Gibco, Grand Island, NY), содержащем 10 мМ Hepes (Gibco) и 4,0 мМ CaCl2 (Sigma; St. Louis, MO)). В эти клетки загружали 2 мкл индо-1 AM (Molecular Probes; Eugene, OR) на мл (конечная концентрация 2 мкМ) и инкубировали в течение 25 минут при 37°C. Затем клетки два раза промывали буфером PTI и суспендировали при плотности 1×107/мл. Затем в клетки добавляли несколько концентраций антитела или соответствующего контроля, и инкубировали в течение 30 минут при комнатной температуре. В квадратную 1 мм-кювету (Sarstedt; Germany) добавляли 1,8 мл предварительно нагретого буфера PTI вместе с 200 мкл клеточной суспензии. Клетки подвергали воздействию флуоресцентного излучения и интенсивность флуоресценции измеряли на устройстве от Photon Technology Corporation International (PTI; Lawrenceville, NJ). Затем работу устройства останавливали и добавляли 20 мкл 100 нМ MCP-1 (Peprotech; Rocky Hill, NJ). После получения ответа добавляли нижеследующие реагенты в указанном порядке для высвобождения и образования хелатного комплекса общего кальция: 20 мкл 18% тритона X-100; 20 мкл 3M триса, pH 8,5; 20 мкл 0,5M EGTA, pH 8,5 (все от Sigma). Антитело 4.40 подавляло способность MCP-1 индуцировать мобилизацию кальция в зависимости от дозы (фигура 10). Аналогичные результаты были получены с использованием антитела 4.40 A68G S230P.
Пример 11
Ингибирование хемотаксиса в сторону CCL2 и CCL7
Для того чтобы определить, является ли ингибирование хемотаксиса под действием антитела 4.40 A68G S230P специфическим по отношению к MCP-1, либо оно может быть специфическим и по отношению к другим лигандам CCR2, хемотаксис клеток THP-1 по отношению к MCP-1 (CCL2) сравнивали с хемотаксисом этих клеток по отношению к CCL3 (MIP-1a), как показано на фигуре 3. Помимо миграции CCR2-трансфецированных клеток 300-19 в сторону другого лиганда CCR2 оценивали также миграцию CCL7 (MCP-3) в присутствии антитела 4.40 A68G S230P с помощью аналогичного анализа на хемотаксис, описанного в примере 6. Антитело 4.40 A68G S230P ингибировало хемотаксис по отношению к обоим известным лигандам CCR2, CCL2 (фигура 3) и CCL7 (фигура 11), но, как показано на фигуре 3, это антитело не блокировало хемотаксис по отношению к лиганду CCR1/CCR5, CCL3 (MIP-1a).
Пример 12
Полимеризация актина в моноцитах
Полимеризация актина в цельной человеческой крови
Моноклональное антитело 4.40 A68G S230P также тестировали на его способность ингибировать полимеризацию актина в цельной человеческой крови как показано на фигуре 12. Кровь собирали в пробирки Vacutainer с EDTA (VWR; Boston, MA), а затем инкубировали с разведениями антитела или KLH-контроля в течение 30 минут при комнатной температуре. Затем в 48-луночный планшет Corning добавляли буфер-контроль или 10 мкл 100 нМ MCP-1 (Peprotech; Rocky Hill, NJ)(VWR). После этого добавляли 100 мкл крови, слегка размешивая, и инкубировали в течение 40 секунд, а затем реакцию прекращали добавлением 0,8 мл реагента для завершения реакции/лизиса (10% FACS-раствора для лизиса (Becton Dickinson; San Jose, CA), 16%-ный параформальдегид (20%) (Electron Microscopy Sciences; Ft. Washington, PA) и H2O (70%)). Планшет инкубировали в течение 10 минут при комнатной температуре и центрифугировали, а затем клетки переносили в 96-луночный круглодонный полипропиленовый планшет, и два раза промывали PBS. После этого добавляли 100 мкл PBS на лунку. Затем добавляли 50 мкл окрашивающего реагента (10% 5 мг/мл лизофосфотидилхолина (Sigma) в 10×HBSS, 16%-ный параформальдегид (80%) и 6,6 мкМ NBD-фаллацидина (10%) (Molecular Probes; Eugene, OR)). Клетки окрашивали в течение одного часа при комнатной температуре в темноте. После инкубирования, клетки 2 раза промывали PBS, содержащим 2% фетальную бычью сыворотку (Hyclone, Logan UT), и флуоресценцию количественно оценивали на устройстве FACSCAN (Becton Dickinson) с установкой дискриминационного окна для моноцитов. Как показано на фигуре 12, антитело 4.40 A68G S230P ингибировало полимеризацию актина с IC50=0,168 мкг/мл.
Полимеризация актина в цельной крови собакоподобных обезьян
Кровь собирали в пробирки Vacutainer с EDTA (VWR; Boston, MA), а затем инкубировали с разведениями антитела или его контрольного изотипа в течение 30 минут при комнатной температуре. В течение этого периода времени, в 48-луночный планшет Corning (VWR) добавляли 10 мкл 100 нМ MCP-1 (Peprotech; Rocky Hill, NJ; конечная концентрация 10 нМ) или буфер-контроль (HBSS без Ca+2 или Mg+2 (Gibco; Grand Island, NY) и 0,2% BSA (Sigma; St. Louis, MO)). После этого добавляли 100 мкл крови, слегка размешивая, и инкубировали в течение 40 секунд. Затем реакцию прекращали добавлением 0,8 мл реагента для завершения реакции/лизиса (10% FACS-раствора для лизиса (Becton Dickinson; San Jose, CA), 16%-ный параформальдегид (20%) (Electron Microscopy Sciences; Ft. Washington, PA) и H2O (70%)). Планшет инкубировали в течение 10 минут, центрифугировали, и два раза промывали PBS. Затем клетки окрашивали в течение одного часа при комнатной температуре в темноте с использованием окрашивающего реагента (10% 5 мг/мл лизофосфотидилхолина (Sigma) в 10×HBSS, 16%-ный параформальдегид (80%) и 6,6 мкМ NBD-фаллацидина (10%) (Molecular Probes; Eugene, OR)). После инкубирования, клетки 2 раза промывали PBS, содержащим 2% фетальную бычью сыворотку (Hyclone, Logan UT), и флуоресценцию количественно оценивали на устройстве FACSCAN (Becton Dickinson) с установкой дискриминационного окна для активированных моноцитов. Как показано на фигуре 13, антитело 4.40 A68G S230P ингибировало полимеризацию актина в цельной крови собакоподобных обезьян с IC50=0,49 мкг/мл.
Пример 13
Количественный анализ мРНК коллагена I
Клеточную линию звездчатых клеток человеческой печени, LI-90 (JCRB Cellbank, Japan), культивировали в колбах, содержащих среду DME, в которую было добавлено 10% дезактивированной FBS, 100 ед/мл пенициллина/100 мкг/мл стрептомицина и 2 мМ L-Gln (Invitrogen) при 37°C в 5% CO2-инубаторе с повышенной влажностью. Клетки выращивали в 96-луночных планшетах для культивирования тканей при плотности 20000 клеток/лунку в течение 3 дней и обрабатывали 1000 нМ MCP-1 (PeproTech) и различными концентрациями антитела 4.40 A68G S230P в течение 48 часов при 37°C. Затем культуральную среду удаляли, и клетки подвергали лизису путем добавления 100 мкл буфера для лизиса, поставляемого в наборе для экспрессии QUANTIGENE™ (Panomics). Анализ разветвленной ДНК осуществляли в соответствии с инструкциями производителей. Вкратце, образцы полноразмерной РНК загружали на 96-луночный микротитрационный планшет, содержащий буфер дли гибридизации и 50 фмоль/мкл набора зондов Col1A1/GAPDH. Иммобилизованные мРНК гибридизовали с разветвленными молекулами ДНК, содержащими молекулы щелочной фосфатазы, путем инкубирования при 46°C в течение 60 минут. После дополнительного инкубирования с хемилюминесцентным субстратом при 46°C в течение 30 минут, люминесценцию количественно оценивали на люминометре (ARVOsx, Perkin Elmer; Massachusetts, USA). Затем вычисляли отношение люминесценции Col1A1/GAPDH, и полученные данные анализировали с помощью компьютерной программы Prism 4.0 для определения величин IC50 (GraphPad). Как показано на фигуре 14, антитело 4.40 A68G S230P ингибировало синтез мРНК коллагена IAI в клетках LI90, индуцированный MCP-1, с IC50=0,89 мкг/мл (6,2 нМ).
Пример 14
Фосфорилирование pERK
Свежую цельную человеческую кровь брали у здоровых добровольцев. Через 6 минут после добавления МСР-1, FACS-анализ указывал на высокие уровни фосфорилирования киназы, регулируемой внеклеточными сигналами (pERK) в CD14+-моноцитах. Антитело 4.40 A68G S230P ингибировало MCP-1-индуцированное фосфорилирование pERK в цельной крови в зависимости от дозы (фигура 15). Фосфорилирование pERK в моноцитах, после их ex vivo стимуляции молекулой MCP-1, может быть использовано в качестве биомаркера данного механизма, что тем самым облегчает определение ФК/ФД-свойств и осуществление медикаментозного лечения, основанного на механизмах трансляции. Величины IC50 и IC90, полученные для антитела 4.40 A68G S230P в данном анализе цельной крови, составляли 0,44 мкг/мл (2,9 нМ) и 0,89 мкг/мл (6,1 нМ), соответственно.
Пример 15
Мыши с моделью острого гепатита
Эффективность антитела 4.40 A68G S230P также оценивали с использованием мышей ConA с моделью острого гепатита. В этом исследовании были использованы трансгенные мыши, в которых мышиный CCR2 был заменен человеческим CCR2, поскольку антитело 4.40 A68G S230P не распознает CCR2 грызунов. Этим животным была введена одна внутрибрюшинная инъекция mAb в дозе 0,1, 0,3 или 1,0 мг/кг в физиологическом растворе. Мышам отдельной группы вводили антитело IgG4 контрольного изотипа. Через тридцать минут, животным в хвостовую вену вводили i.v. инъекцию (0,1 мл) 15 мг/кг ConA в апирогенном физиологическом растворе. Животным контрольной группы вводили физиологический раствор без ConA. Через 24 часа брали пробы крови и проводили анализ на ферменты плазмы печени. Как показано на фигуре 16, уровни ALT и AST значительно увеличивались у группы, которой вводили контрольное антитело, и эти величины составляли примерно 25000 ед./л. В противоположность этому, у мышей, обработанных 0,5 и 1,0 мг/кг антитела 4.40 A68G S230P, наблюдалось значимое снижение уровней этих ферментов в плазме печени приблизительно на 50% и 80%, соответственно.
Пример 17
Терапевтические эффекты у животных с моделью заболевания
Воспаление
Терапевтические эффекты человеческих анти-CCR2 антител или их антигенсвязывающих фрагментов, включая антитела 4.40 и 4.9, тестировали с использованием моделей млекопитающих in vivo с воспалением. После внутрикожной инъекции хемокина, такого как МСР-1 и антитела или его фрагмента, реагирующего с CCR2 млекопитающих, такого как антитело 4.40, подходящим животным, таким как кролики, мыши, крысы, морские свинки или макак-резусы, проводили мониторинг инфильтрации лейкоцитов (см., например, Van Damme, J. et al., J. Exp. Med. 176:59-65 (1992); Zachariae, C. O. C. et al., J. Exp. Med. 171 :2177-2182 (1990); Jose, P. J. et al., J. Exp. Med. 179: 881-887 (1994)). Биоптаты кожи оценивали с помощью гистологического анализа на инфильтрацию лейкоцитов (например, эозинофилов и гранулоцитов). Как и предполагалось, анти-CCR2 антитела-антагонисты обладали способностью снижать уровень инфильтрации лейкоцитов по сравнению с уровнями у контрольных животных.
Рассеянный склероз
Терапевтические эффекты антител или их фрагментов, включая антитела 4.40 и 4.9, тестировали на млекопитающих с моделью in vivo рассеянного склероза. Экспериментальный аутоиммунный энцефаломиелит (ЭАЭ) представляет собой T-клеточно-опосредуемое воспалительное заболевание центральной нервной системы (ЦНС), которое служит у животного в качестве модели рассеянного склероза (РС) (см. Steinman L, Neuron 24:511-514 (1999)). На день 0 и на день 7 после иммунизации (p.i.) мышей C57/J129 и/или C57BL/6 энцефалитогенным пептидом MOG35-55, этих мышей сенсибилизировали для развития у них острого ЭАЭ путем подкожной (s.c.) инъекции (в два участка, в боковую заднюю часть) всего 600 мкг энцефалитогенного пептида MOG35-55, эмульгированного в неполном адъюванте Фрейнда (IFA; Difco, Detroit, MI), содержащем 70 мкг Mycobacterium tuberculosis, H37Ra (Difco). На день 0 и на день 2 p.i., каждой мыши внутривенно (i.v.) в хвостовую вену также вводили 500 нг коклюшного токсина (PTX; List Biological Laboratories, Campbell, CA). Некоторым животным также вводили возрастающие дозы анти-CCR2 антител-антагонистов, таких как антитело 4.40. Животных ежедневно обследовали на клинические признаки и оценивали по следующей шкале оценок: балл 0, патология отсутствует; балл 1, повисание хвоста; балл 2, повисание хвоста до земли и слабость в задних конечностях; балл 3, парапарез задних конечностей; балл 4, тетраплегия; и балл 5, предсмертное состояние или смерть.
Кроме того, наблюдения под оптическом микроскопом проводили на образцах тканей, фиксированных глутаральдегидом/осмием и взятых из зрительного нерва, коры головного мозга, мозжечка и спинного мозга. Образцы тканей подвергали дегидратации и заливали в эпоксидную смолу, из которой делали 1 мкм-срезы, которые затем окрашивали толуидиновым синим. Воспаление, демиелинизацию, уоллеровскую дегенерацию (WD) и ремиелинизацию оценивали по шкале от 0 до 5 баллов, как описано в литературе (Cannella et al., Proc. Natl. Acad. Sci. USA 95:10100-10105 (1998)). Как и предполагалось, у мышей, обработанных анти-CCR2 антителами-антагонистами, наблюдалось ослабление признаков клинической патологии, воспаления, демиелинизаци и WD по сравнению с контрольными животными. Кроме того, как и предполагалось, такое ослабление зависит от дозы.
Невропатическая боль
Терапевтические эффекты антител или их фрагментов, включая антитела 4.40 и 4.9, тестировали на млекопитающих in vivo с моделью невропатической боли. Мышей C57BL разделяли на две группы. Животным экспериментальной группы ежедневно вводили анти-CCR2 антитела-антагонисты, а именно, антитело 4.40, путем инъекции в хвостовую вену, а животным другой группы антитело не вводили. Животных двух групп тестировали через определенные интервалы времени после начала обработки путем проведения нижеследующих анализов:
Вращающийся стержень . Сначала, мышей обучали с использованием стержня, вращающегося в течение 3 минут со скоростью 10 об/мин. Для проведения теста, скорость устанавливали на 10 об/мин на 60 секунд, а затем скорость увеличивали до 600 об/мин. Затем регистрировали время до падения мышей после начала увеличения скорости.
Горячая пластина . Мышей приучали к горячей пластине при температуре, установленной на 45°C, 2 мин. После этого, каждую мышь помещали на горячую пластину и температуру постепенно доводили до 52,5°C и 55,5°C (предельное время 30 секунд), а затем до 58,5°C (предельное время 20 секунд). Время, когда мыши начинали облизывать лапы или подпрыгивать, регистрировали.
Формалиновый тест . За 4 дня до проведения теста, мышей каждый день на 2 часа помещали на тест-платформу для акклиматизации. В день проведения эксперимента, мышей на 1 час помещали на тест-платформу, а затем, в подушечку левой лапы вводили 10 мкл 2% формалина. Время, когда мыши начинали облизывать или поднимать лапы, в которые была сделана инъекция, регистрировали в течение 2-минутного периода времени с 5-минутными интервалами в течение 50 минут.
Тепловая и механическая стимуляция . Термочувствительность оценивали путем измерения времени запаздывания отдергивания лапы в ответ на раздражитель, такой как теплота, вырабатываемая при лучеиспускании (Hargreaves et al., Pain 32:77-88 (1988)). Механическую чувствительность определяли с использованием прокалиброванных волосков фон Фрея в соответствии с парадигмой движения «вверх и вниз» (Chaplan et al., J. Neurosci. Methods 53, 55-63 (1994)).
Повреждение нервной ткани . Мышей анестезировали смесью кетамина (50 мг/кг, i.m.) и медетомидина (1 мг/кг, i.m.). Разрез делали непосредственно ниже кости бедра параллельно седалищному нерву. Затем нерв обнажали, и из этого нерва удаляли любую связующую ткань. После этого, накладывали жесткую лигатуру шелковой нитью для наложения швов 6-0 диаметром, составляющим примерно от 1/3 до 1/2 от диаметра седалищного нерва. Мышцы закрывали нитью для наложения швов, а раны - зажимами для ран. Реакцию мышей на механический раздражитель анализировали до повреждения нерва и через 15 дней после его повреждения.
Как и предполагалось, у мышей, обработанных анти-CCR2 антителами-антагонистами, наблюдалось ослабление признаков невропатической боли по сравнению с контрольными мышами.
Последовательности CDR (SEQ ID NO:)
















































































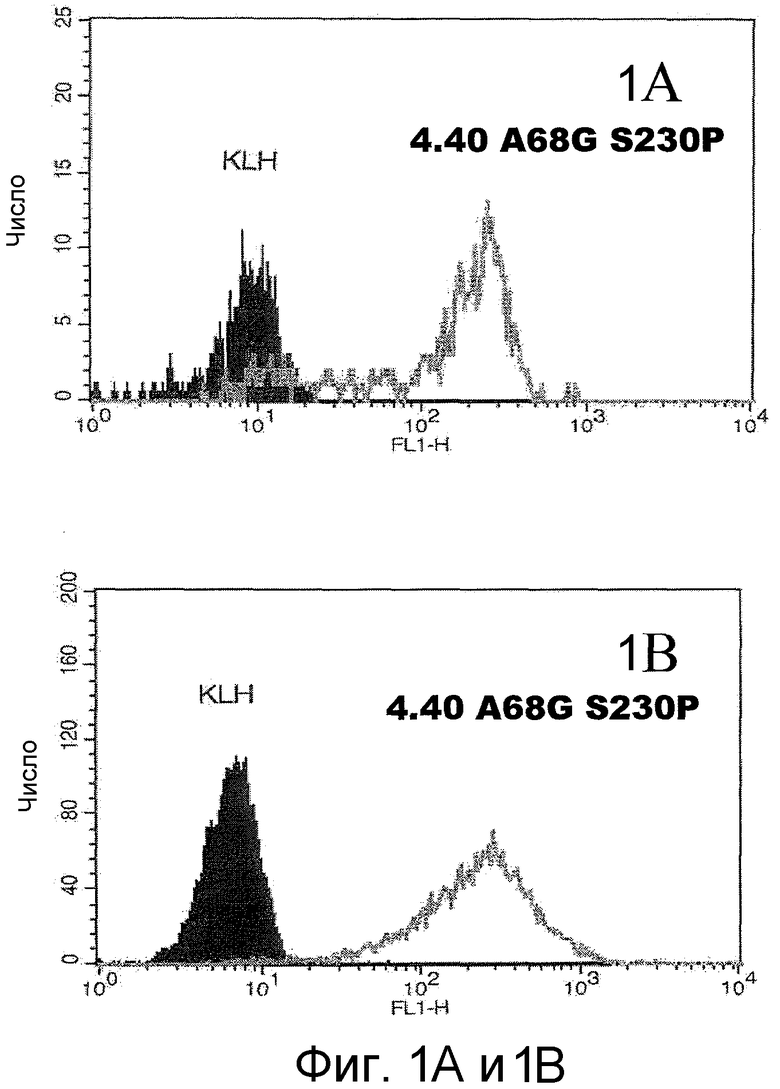
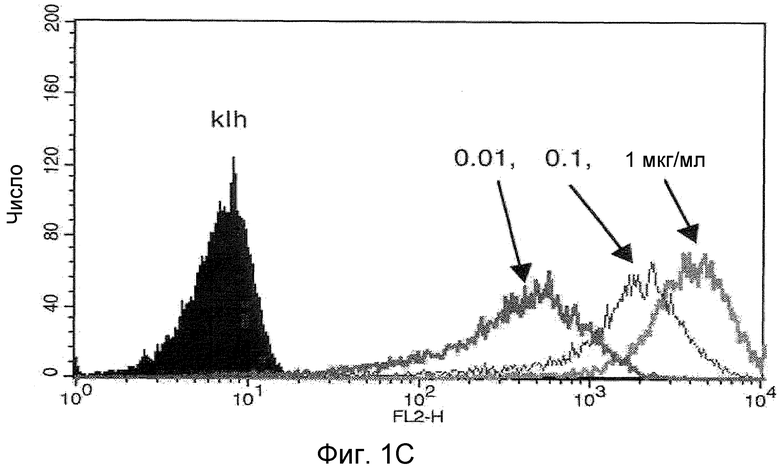
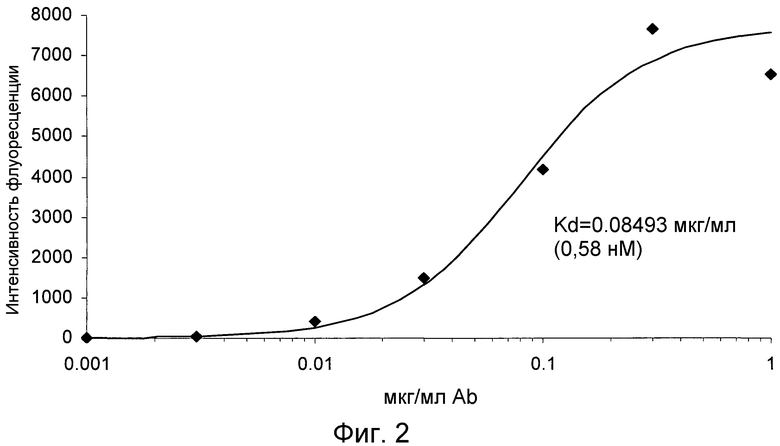
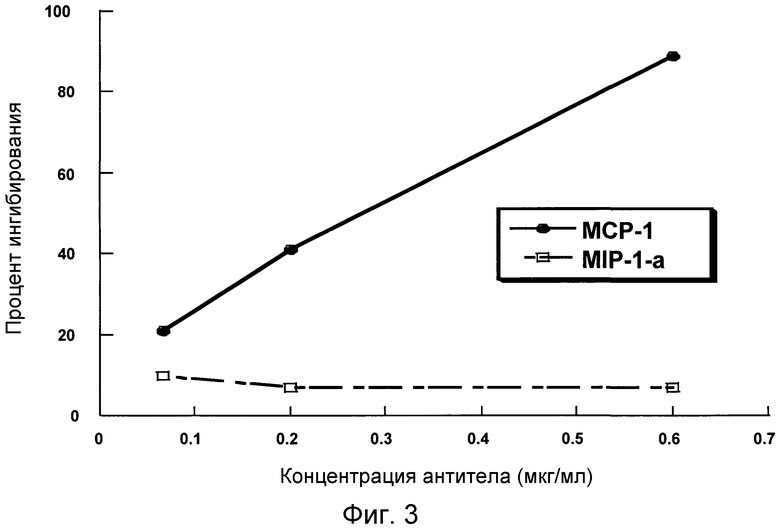
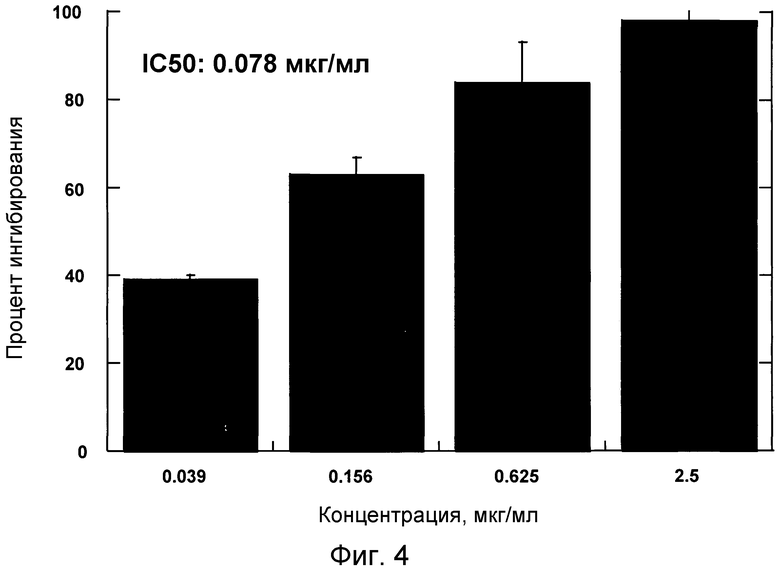
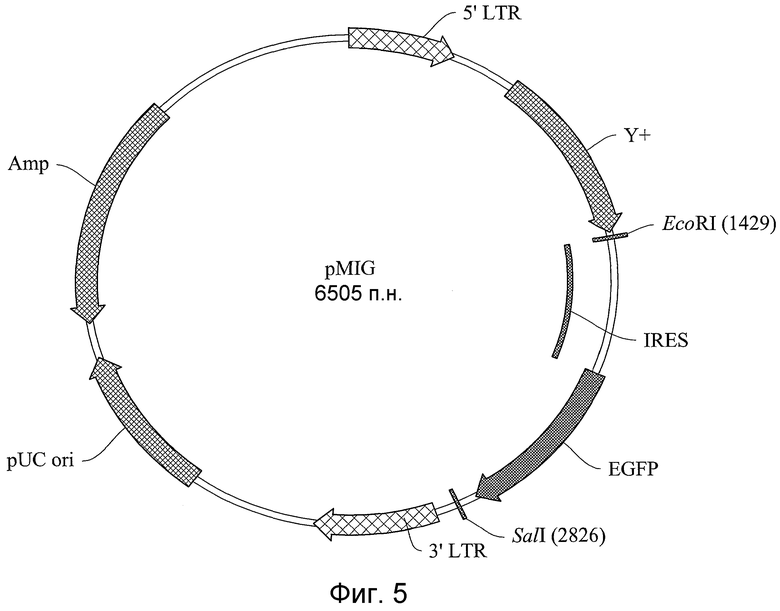
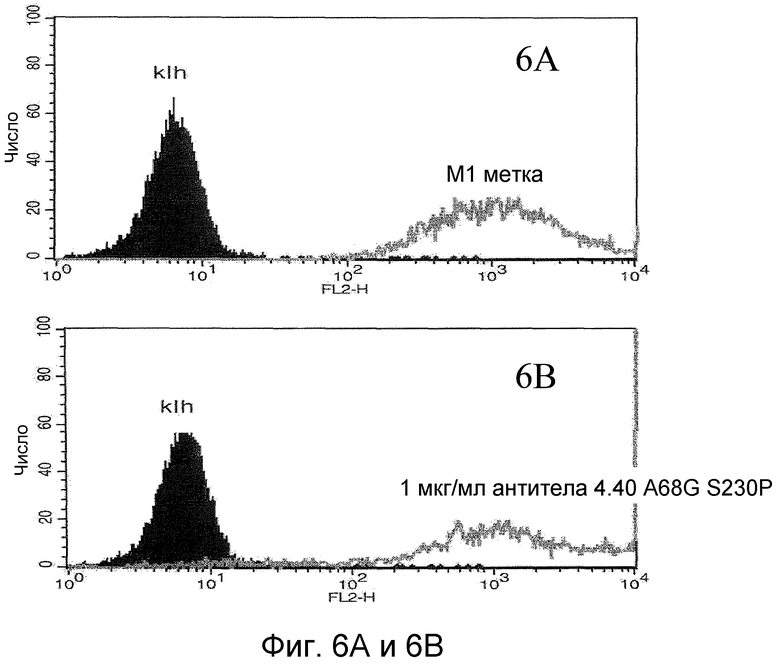
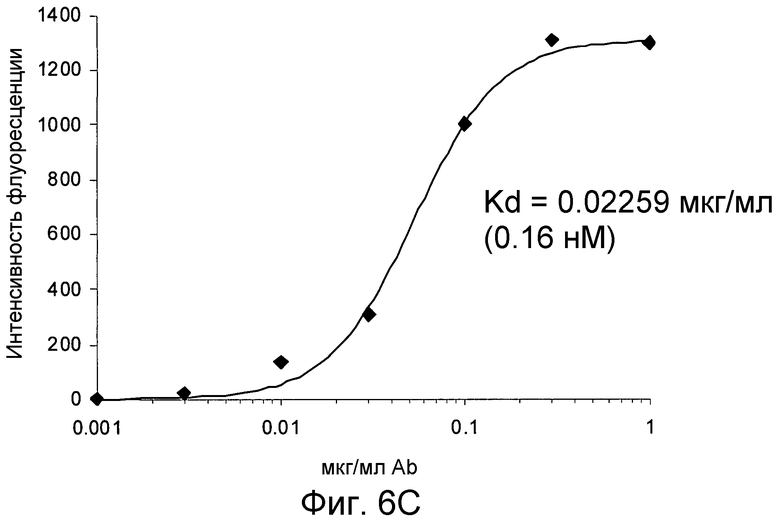
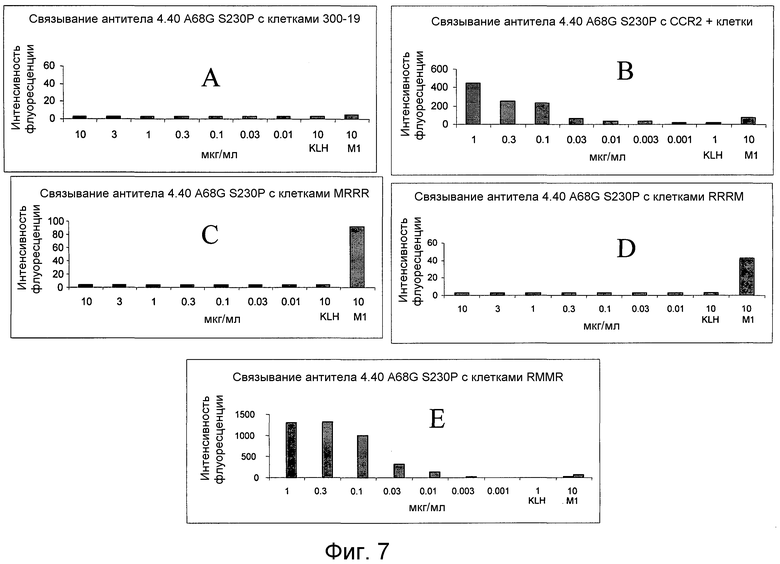
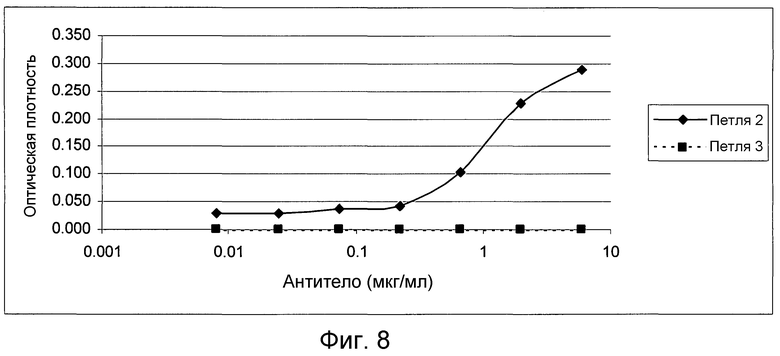
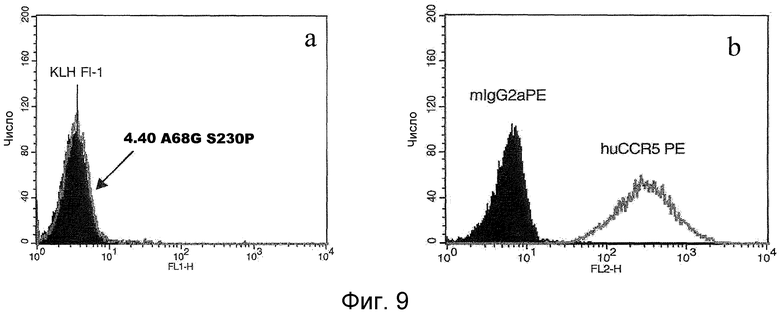
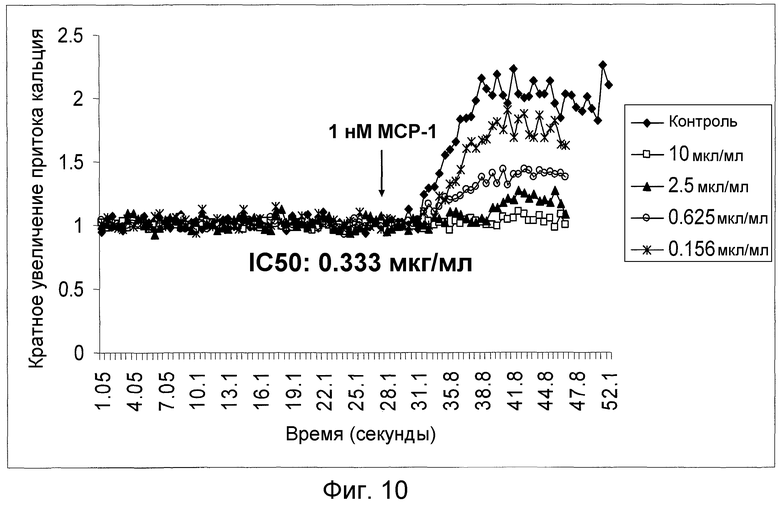
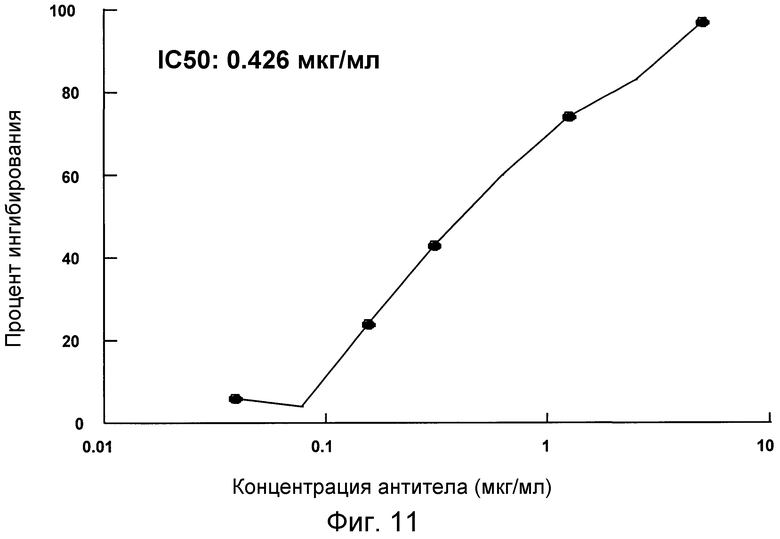
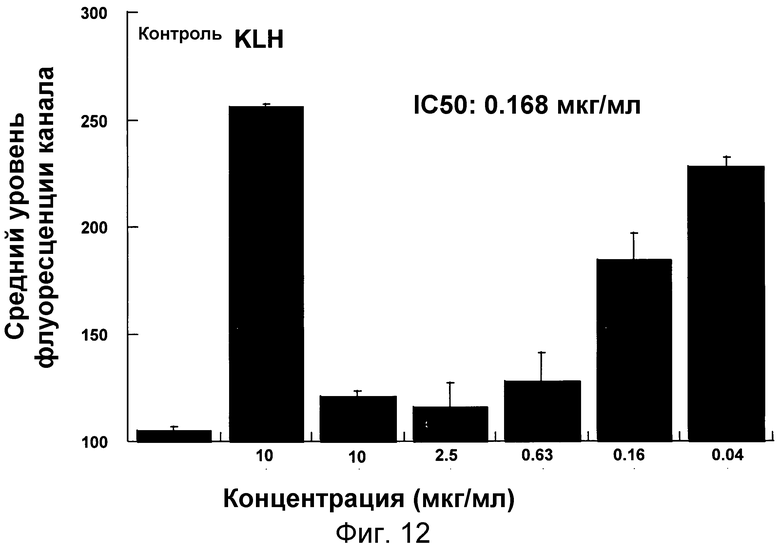
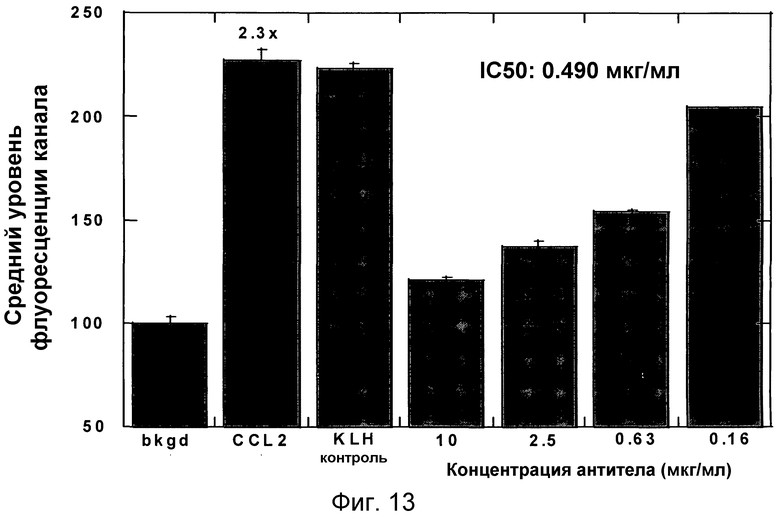
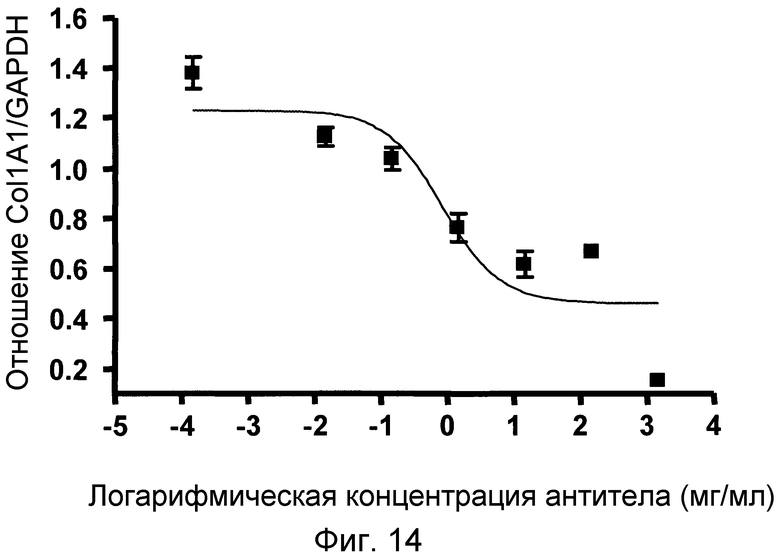
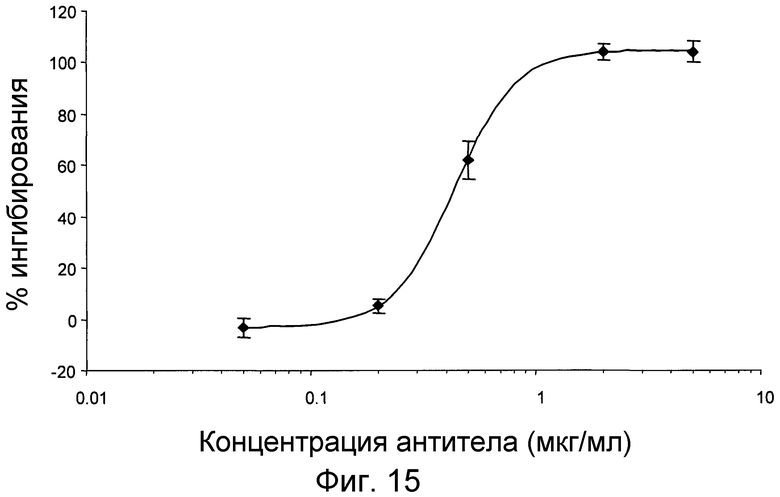
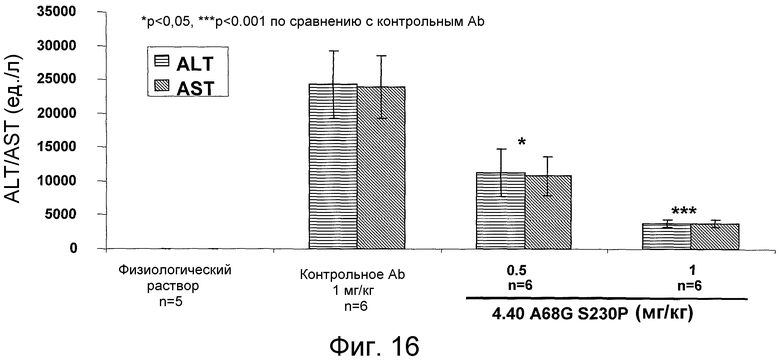


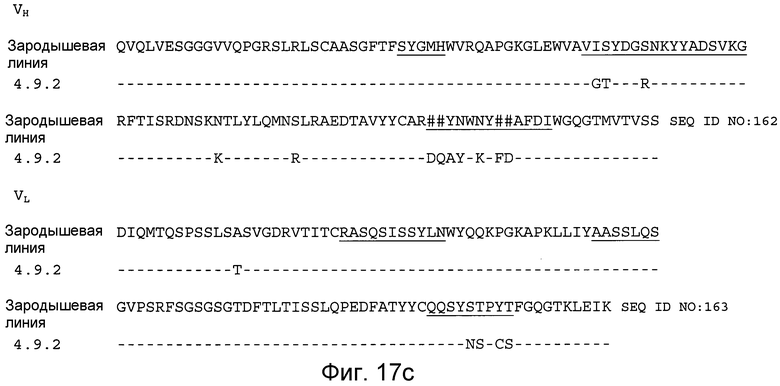

Настоящее изобретение относится к антителам, включая человеческие антитела и их антигенсвязывающие части, которые специфически связываются с CCR2, в частности с человеческим CCR2, и могут действовать как ингибиторы CCR2. Анти-CCR2 антителами являются антитела, которые связываются с первой и/или второй внеклеточными петлями CCR2. Настоящее изобретение также относится к человеческим анти-CCR2 антителам и к их антигенсвязывающим частям. Настоящее изобретение относится к выделенным тяжелой и легкой цепям иммуноглобулина, происходящим от человеческих анти-CCR2 антител, и к молекулам нуклеиновой кислоты, кодирующим такие иммуноглобулины. Настоящее изобретение также относится к способам получения человеческих анти-CCR2 антител или их антигенсвязывающих частей, к композициям, содержащим такие антитела или их антигенсвязывающие части, и к способам применения антител и их антигенсвязывающих частей и композиций для диагностики и лечения. Настоящее изобретение также относится к способам генотерапии с применением молекул нуклеиновой кислоты, кодирующих молекулы тяжелой и/или легкой цепи иммуноглобулина, где указанные молекулы включают человеческие анти-CCR2 антитела или их антигенсвязывающие части. 10 н. и 15 з.п. ф-лы, 24 ил., 8 табл., 17 пр.
1. Выделенное человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с человеческим CCR2, включающие аминокислотную последовательность вариабельного домена (VH) тяжелой цепи, содержащей CDR1, CDR2 и CDR3, и аминокислотную последовательность вариабельного (VL) домена легкой цепи, содержащей CDR1, CDR2 и CDR3, где CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL, CDR3 VL выбраны из группы, состоящей из:
a) CDR1 VH, представленной в SEQ ID NO:84, CDR2 VH, представленной в SEQ ID NO:85, CDR3 VH, представленной в SEQ ID NO:86, CDR1 VL, представленной в SEQ ID NO:102, CDR2 VL, представленной в SEQ ID NO:103, и CDR3 VL, представленной в SEQ ID NO:104;
b) CDR1 VH, представленной в SEQ ID NO:12, CDR2 VH, представленной в SEQ ID NO:13, CDR3 VH, представленной в SEQ ID NO:14, CDR1 VL, представленной в SEQ ID NO:30, CDR2 VL, представленной в SEQ ID NO:31, и CDR3 VL, представленной в SEQ ID NO:32;
c) CDR1 VH, представленной в SEQ ID NO:48, CDR2 VH, представленной в SEQ ID NO:49, CDR3 VH, представленной в SEQ ID NO:50, CDR1 VL, представленной в SEQ ID NO:66, CDR2 VL, представленной в SEQ ID NO:67, и CDR3 VL, представленной в SEQ ID NO:68; и
d) CDR1 VH, представленной в SEQ ID NO:177, CDR2 VH, представленной в SEQ ID NO:178, CDR3 VH, представленной в SEQ ID NO:179, CDR1 VL, представленной в SEQ ID NO:195, CDR2 VL, представленной в SEQ ID NO:196, и CDR3 VL, представленной в SEQ ID NO:197.
2. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.1, содержащие: CDR1 VH, представленную в SEQ ID NO:84, CDR2 VH, представленную в SEQ ID NO:85, CDR3 VH, представленную в SEQ ID NO:86, CDR1 VL, представленную в SEQ ID NO:102, CDR2 VL, представленную в SEQ ID NO:103, и CDR3 VL, представленную в SEQ ID NO:104.
3. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.1, содержащие аминокислотную последовательность домена VH, выбранную из группы, состоящей из
SEQ ID NO:11, SEQ ID NO:47, SEQ ID NO:83, SEQ ID NO:176,
SEQ ID NO:11, имеющей консервативную аминокислотную замену; SEQ ID NO:47, имеющей консервативную аминокислотную замену; SEQ ID NO:83, имеющей консервативную аминокислотную замену; SEQ ID NO:176, имеющей консервативную аминокислотную замену;
SEQ ID NO:11, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:162; SEQ ID NO:47, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:158; SEQ ID NO:83, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:160; SEQ ID NO:176, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:164;
аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:11; аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:47; аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:83; и аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:176;
и
аминокислотную последовательность домена VL, выбранную из группы, состоящей из
SEQ ID NO:29, SEQ ID NO:65, SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:194;
SEQ ID NO:29, имеющей консервативную аминокислотную замену; SEQ ID NO:65, имеющей консервативную аминокислотную замену; SEQ ID NO:101, имеющей консервативную аминокислотную замену; SEQ ID NO:113 имеющей консервативную аминокислотную замену; SEQ ID NO:194, имеющей консервативную аминокислотную замену;
SEQ ID NO:29, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:163; SEQ ID NO:65, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:15 9; SEQ ID NO:101, имеющей консервативную аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:161; SEQ ID NO:113, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:161; SEQ ID NO:194, имеющей аминокислотную замену в зародышевой линии, приведенной в SEQ ID NO:165;
аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:29; аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:65; аминокислотной последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO:101; аминокислотной последовательности, которая по меньшей мере на 90% идентична SEQ ID NO:113; и аминокислотной последовательности, которая по меньшей мере на 90% идентична SEQ ID NO:194.
4. Выделенное человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с человеческим CCR2, включающие
аминокислотную последовательность вариабельного домена (VH) тяжелой цепи, содержащей CDR1, CDR2 и CDR3 из аминокислотной последовательности домена VH, выбранной из группы, состоящей из SEQ ID NO:11, SEQ ID NO:47, SEQ ID NO:83, SEQ ID NO:176, и
аминокислотную последовательность вариабельного (VL) домена легкой цепи, содержащей CDR1, CDR2 и CDR3 из аминокислотной последовательности домена VL, выбранной из группы, состоящей из SEQ ID NO:29, SEQ ID NO:65, SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:194.
5. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.1, где указанное антитело содержит
тяжелую цепь, имеющую аминокислотную последовательность, выбранную из группы, состоящей из
последовательностей SEQ ID NO:10, SEQ ID NO:46, SEQ ID NO:82, SEQ ID NO:116, SEQ ID NO:175;
SEQ ID NO:10, имеющей консервативную аминокислотную замену; SEQ ID NO:46, имеющей консервативную аминокислотную замену; SEQ ID NO:82, имеющей консервативную аминокислотную замену; SEQ ID NO:116, имеющей консервативную аминокислотную замену; SEQ ID NO:175, имеющей консервативную аминокислотную замену;
SEQ ID NO:10, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:162; SEQ ID NO:46, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:158; SEQ ID NO:82, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:160; SEQ ID NO:116, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:160; и SEQ ID NO:175, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:164;
и
легкую цепь, имеющую аминокислотную последовательность, выбранную из группы, состоящей из
последовательностей SEQ ID NO:28, SEQ ID NO:64, SEQ ID NO:100, SEQ ID NO:112, SEQ ID NO:193;
SEQ ID NO:28, имеющей консервативную аминокислотную замену; SEQ ID NO:64, имеющей консервативную аминокислотную замену; SEQ ID NO:100, имеющей консервативную аминокислотную замену; SEQ ID NO:112, имеющей консервативную аминокислотную замену; SEQ ID NO:193, имеющей консервативную аминокислотную замену;
SEQ ID NO:28, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:163; SEQ ID NO:64, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:159; SEQ ID NO:100, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:161; SEQ ID NO:112, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:161; и SEQ ID NO:193, имеющей аминокислотную замену на аминокислоту, которая отлична от соответствующей аминокислотной последовательности зародышевой линии, приведенной в SEQ ID NO:165.
6. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.5, где указанное антитело имеет тяжелую цепь и легкую цепь, выбранные из группы, состоящей из:
a) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:116, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:112,
b) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:10, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:28,
c) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:46, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:64,
d) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:82, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:100,
e) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:82, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:112, и
f) тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO:175, и легкой цепи, имеющей аминокислотную последовательность SEQ ID NO:193.
7. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.6, где аминокислотная последовательность тяжелой цепи представляет собой SEQ ID NO:116, а аминокислотная последовательность легкой цепи представляет собой SEQ ID NO:112.
8. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.1, которые дополнительно содержат одну или более аминокислотных последовательностей FR1, FR2, FR3 или FR4 VH из аминокислотной последовательности вариабельного домена (VH) тяжелой цепи, выбранной из группы, состоящей из SEQ ID NO:11, SEQ ID NO:47, SEQ ID NO:83, SEQ ID NO:176; и/или FR1, FR2, FR3 или FR4 из аминокислотной последовательности вариабельного домена (VL) легкой цепи, выбранной из группы, состоящей из SEQ ID NO:29, SEQ ID NO:66, SEQ ID NO:101, SEQ ID NO:113, SEQ ID NO:194.
9. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.8, которые дополнительно содержат константный домен тяжелой цепи.
10. Выделенное человеческое моноклональное антитело или антигенсвязывающий фрагмент по п.9, которые дополнительно содержат константный домен легкой цепи.
11. Фармацевтическая композиция для лечения, предотвращения или облегчения симптомов опосредованного CCR2 расстройства, содержащая эффективное количество антитела или антигенсвязывающего фрагмента по п.1 и фармацевтически приемлемый носитель.
12. Выделенная клеточная линия, продуцирующая антитело или антигенсвязывающий фрагмент по п.1.
13. Выделенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую антитело или антигенсвязывающий фрагмент по п.1.
14. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п.13, где указанный вектор необязательно содержит последовательность регуляции экспрессии, функционально связанную с указанной молекулой нуклеиновой кислоты.
15. Способ лечения, предупреждения или ослабления симптомов опосредуемого CCR2 расстройства у индивидуума, нуждающегося в этом, где указанный способ включает стадию введения указанному индивидууму терапевтически эффективного количества антитела или антигенсвязывающего фрагмента по п.1.
16. Способ по п.15, где указанный индивидуум страдает одним или более состояниями или нарушениями, выбранными из группы, состоящей из воспалительного расстройства, аллергического расстройства, аутоиммунного расстройства, расстройства, ассоциированного с отторжением трансплантата, атеросклероза, ожирения, ВИЧ-инфекции, невропатической боли, стеноза, рестеноза, рассеянного склероза, фиброзного состояния, возрастной дегенерации желтого пятна, увеита, инфекционного заболевания роговицы и рака.
17. Способ по п.16, где указанное фиброзное состояние представляет собой фиброз печени.
18. Способ по п.17, где указанный фиброз печени вызван инфекцией вирусом гепатита С (HCV), инфекцией вирусом гепатита В (HBV), алкогольным стеатогепатитом (ASH) или не-алкогольным стеатогепатитом (NASH).
19. Применение антитела или антигенсвязывающего фрагмента по п.1 для производства лекарственного средства для лечения, предупреждения или ослабления симптомов опосредуемого CCR2 расстройства у индивидуума, нуждающегося в этом.
20. Применение по п.19, где указанный индивидуум страдает одним или более состояниями или расстройствами, выбранными из группы, состоящей из воспалительного расстройства, аллергического расстройства, аутоиммунного расстройства, расстройства, ассоциированного с отторжением трансплантата, атеросклероза, ожирения, ВИЧ-инфекции, невропатической боли, стеноза, рестеноза, рассеянного склероза, фиброзного состояния, возрастной дегенерации желтого пятна, увеита, инфекционного заболевания роговицы и рака.
21. Применение по п.20, где указанное фиброзное состояние представляет собой фиброз печени.
22. Применение по п.21, где указанный фиброз печени вызван инфекцией вирусом гепатита C (HCV), инфекцией вирусом гепатита B (HBV), алкогольным стеатогепатитом (ASH) или не-алкогольным стеатогепатитом (NASH).
23. Выделенное антитело или антигенсвязывающий фрагмент по п.1, содержащие аминокислотную последовательность вариабельного домена (VH) тяжелой цепи SEQ ID NO:83 и аминокислотную последовательность вариабельного (VL) домена легкой цепи SEQ ID NO:113.
24. Выделенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую антитело или антигенсвязывающий фрагмент по п.7.
25. Выделенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую антитело или антигенсвязывающий фрагмент по п.23.
| HAN K.H | |||
| et al., "Role of the first extracellular loop in the functional activation of CCR2 | |||
| The first extracellular loop contains distinct domains necessary for both agonist binding and transmembrane signaling", J Biol Chem | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| FRADE J.M | |||
| et al., "The amino-terminal domain of the CCR2 chemokine receptor acts as coreceptor | |||

