Область техники
Настоящее изобретение относится к варианту белка-рецептора цАМФ (циклического аденозинмонофосфата), микроорганизму, включающему этот вариант, и способу получения L-аминокислоты с использованием этого варианта.
Предшествующий уровень техники
CRP (белок-рецептор циклического АМФ), также называемый САР (белком-активатором катаболитных оперонов), является наиболее хорошо известным регулятором транскрипции у Е. coli. CRP характеризуется наличием регулирующего механизма, зависимого от источника углерода, который представлен «катаболитной репрессией». Это действие инициируется посредством внутриклеточной концентрации циклического АМФ (в дальнейшем в этом документе называется «цАМФ»). В присутствии предпочтительного источника углерода, такого как глюкоза, активность аденилатциклазы ингибируется с понижением цАМФ, и этот сигнал ингибирует экспрессию катаболических генов. В обратном случае, активность аденилатциклазы повышается, и в результате подавляются репрессоры и инициируется экспрессия катаболических генов. Кроме того, известно, что CRP играет различные роли, такие как внутриклеточная передача сигнала посредством цАМФ, осморегуляция, ответы на экстренные состояния клеток, образование биопленки, фиксация азота, транспорт железа и так далее.
По имеющимся данным известно, что 418 генов E. coli регулируются посредством CRP, но соответствующие механизмы еще не были полностью выявлены (J Biol Eng. (2009) 24; 3:13). При таком широком диапазоне регуляторных способностей, CRP обладает потенциалом для демонстрации большого количества фенотипов в результате мутаций. Благодаря его эффектам CRP был изучен в качестве мишени, подходящей для перестройки штаммов на клеточном уровне, которые являются применимыми к различным средам. В последнее время были проведены различные экспериментальные работы, такие как способ изменения экспрессии генов, подлежащих регулированию, посредством изменения степени связывания ДНК аминокислотной вариацией CRP, выбранной с помощью биоинформатики (Nucleic Acids Research, (2009) 37: 2493-2503), способ селекции E. coli, устойчивой к нагреванию, осмосу и низкой температуре, с использованием искусственного фактора транскрипции (ATF), полученного посредством слияния ДНК-связывающего сайта в виде мотива «цинковые пальцы» и CRP ((Nucleic Acids Research, (2008) 36: e102), и так далее. Другими словами, так как изменения экспрессии CRP стимулируют широкий диапазон изменений экспрессии подконтрольных генов, CRP, вероятно, будет хорошим средством для получения микроорганизмов с полезными признаками.
Техническая задача
Авторы настоящего изобретения разработали новый вариант белка, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, и они обнаружили, что этот вариант белка может увеличивать продуктивность L-аминокислоты, тем самым дополняя настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить вариант белка-рецептора цАМФ.
Другая задача настоящего изобретения заключается в том, чтобы предложить полинуклеотид, кодирующий вариант белка-рецептора цАМФ.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить вектор, включающий полинуклеотид.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить микроорганизм рода Escherichia, включающий вариант.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить способ получения L-аминокислоты, включающий культивирование микроорганизма рода Escherichia в среде.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить применение варианта или микроорганизма рода Escherichia, включающего вариант, в продуцировании L-аминокислоты.
Полезные эффекты изобретения
При культивировании микроорганизма рода Escherichia, продуцирующего L-аминокислоту, включающего вариант белка-рецептора цАМФ по настоящему изобретению, можно получать L-аминокислоту с высоким выходом. Соответственно, в промышленных аспектах, наряду с удобством производства, можно ожидать снижение стоимости производства.
Лучший вариант осуществления изобретения
Настоящее изобретение будет описано подробно, как изложено ниже. При этом каждое описание и воплощение, раскрытые в этом описании изобретения, также можно применять к другим описаниям и воплощениям. То есть, все комбинации различных элементов, раскрытых в этом описании изобретения, попадают в объем настоящего изобретения. Кроме того, объем настоящего изобретения не ограничивается частичным описанием, описанным ниже.
Для достижения указанных выше задач, в одном аспекте настоящего изобретения, предложен вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1. Более точно, в настоящем изобретении предложен вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, где аминокислотная замена включает замену аминокислоты в позиции 35 от N-конца на аланин. Более конкретно, в настоящем изобретении предложен вариант рецепторного белка цАМФ, включающий замену аминокислоты в позиции 35 на аланин в аминокислотной последовательности SEQ ID NO: 1.
При использовании в данном описании изобретения, термин «белок-рецептор цАМФ (CRP)» представляет собой наиболее хорошо известный регулятор транскрипции у E. coli, и CRP также называется «двойным регулятором», гак как CRP как таковой имеет одновременно функции активатора и ингибитора. CRP обычно связывается с симметричной последовательностью ДНК, имеющей 22 основания, против хода транскрипции от структурного гена для индукции изгибания ДНК, и CRP действует в качестве активатора, обеспечивая возможность первому активному сайту в С-конце и второму активному сайту в N-конце взаимодействовать с РНК-полимеразой, отвечающей за транскрипцию, и он действует в качестве ингибитора путем захвата ранее других позиции, чтобы уберечь активный белок от связывания с активным сайтом, или путем связывания с активным белком с превращением структуры в структуру, которая не связывается с активным сайтом. Белок-рецептор цАМФ представляет собой белок-рецептор цАМФ, кодируемый геном crp.
«Белок-рецептор цАМФ (белок-рецептор циклического АМФ (аденозинмонофосфата), CRP)» по настоящему изобретению можно применять взаимозаменяемо с белком-активатором катаболитных оперонов (САР), белком CRP, белком САР и так далее.
В настоящем изобретении последовательность CRP может быть получена из известной базы данных GenBank в NCBI (Национальном центре биотехнологической информации). Например, CRP может представлять собой CRP, полученный из рода Escherichia (Escherichia sp.), и более конкретно, полипептид/белок, включающий аминокислотную последовательность, представленную SEQ ID NO: 1, но без ограничения этим. Кроме того, последовательность, имеющая такую же активность, как указанная выше аминокислотная последовательность, может быть включена без ограничения. Кроме того, может быть включена аминокислотная последовательность SEQ ID NO: 1 или аминокислотная последовательность, имеющая с ней 80% или более гомологии или идентичности, но без ограничения этим. Более точно, аминокислота может включать аминокислоту из SEQ ID NO: 1 и аминокислоту, имеющую гомологию или идентичность с SEQ ID NO: 1 по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более. Кроме того понятно, что белок, имеющий аминокислотную последовательность, часть которой делетирована, модифицирована, заменена или добавлена, может являться частью объема настоящего изобретения, до тех пор, пока аминокислотная последовательность имеет указанную выше гомологию или идентичность и проявляет эффективность, соответствующую указанному выше белку.
При использовании в данном описании изобретения термин «вариант» относится к полипептиду, одна или более аминокислот которого отличается от описанной последовательности по консервативным заменам и/или модификациям, но он сохраняет функции или свойства белка. Вариантные полипептиды отличаются от идентифицированной последовательности по замене, делеции или добавлению нескольких аминокислот. Такие варианты обычно могут быть идентифицированы путем модификации одной из указанных выше полипептидных последовательностей и оценки свойств модифицированного полипептида. Другими словами, потенциал варианта может быть увеличен, неизменен или уменьшен по сравнению с таковым у нативного белка. Такие варианты обычно могут быть идентифицированы путем модификации одной из указанных выше полипептидных последовательностей и оценки реакционной способности модифицированного полипептида. Кроме того, некоторые варианты могут включать полипептиды, в которых были удалены одна или более частей, таких как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, в которых была удалена часть из N- и/или С-конца зрелого белка.
При использовании в данном описании изобретения термин «консервативная замена» означает замену одной аминокислоты другой аминокислотой, которая имеет подобные структурные и/или химические свойства. Вариант может иметь, например, одну или более консервативных замен, сохраняя при этом одну или более биологических активностей. Обычно такие аминокислотные замены могут быть произведены исходя из подобия полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; отрицательно заряженные (кислые) аминокислоты включают глутаминовую кислоту и аспарагиновую кислоту; ароматические аминокислоты включают фенилаланин, триптофан и тирозин; и гидрофобные аминокислоты включают аланин, валин, изолейцин, лейцин, метионин, фенилаланин, пролин, глицин и триптофан.
Кроме того, варианты могут включать делецию или добавление аминокислот, которые имеют минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид можно конъюгировать с сигнальной (или лидерной) последовательностью на N-конце белка, который котрансляционно или поcттрансляционно направляет перенос белка. Полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
При использовании в данном описании изобретения термин «вариант белка-рецептора цАМФ» представляет собой вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности полипептида, имеющего активность белка-рецептора цАМФ, где аминокислотные замены включают замену аминокислоты в позиции 35 с N-конца другой аминокислотой. Более точно, вариант может включать вариант белка, в котором другая аминокислота заменяет аминокислоту в позиции 35 в аминокислотной последовательности полипептида, имеющего активность белка-рецептора АМФ. Например, вариант белка может включать вариант белка, в котором имеет место вариация в позиции 35 с N-конца аминокислотной последовательности SEQ ID NO: 1. Более конкретно, вариант белка может представлять собой белок, в котором другая аминокислота заменяет аминокислоту в позиции 35 аминокислотной последовательности SEQ ID NO: 1. «Другая аминокислота» не ограничена до тех пор, пока она представляет собой аминокислоту, отличную от L-глутаминовой кислоты, которая представляет собой аминокислоту в позиции 35. Более точно, вариант может представлять собой белок, в котором гидрофобная аминокислота заменяет аминокислоту в позиции 35 в аминокислотной последовательности SEQ ID NO: 1. Гидрофобная аминокислота может представлять собой одну из L-аланина, L-глицина, L-валина, L-изолейцина, L-лейцина, L-метионина, L-пролина, L-фенилаланина и L-триптофана. Более конкретно, вариант может представлять собой белок, в котором аланин заменяет аминокислоту в позиции 35 в аминокислотной последовательности SEQ ID NO: 1, но без ограничения этим.
Кроме того, вариант означает вариант, имеющий вариацию аминокислоты в позиции 35 с N-конца в описанной выше аминокислотной последовательности SEQ ID NO: 1 и/или в аминокислотной последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% или более гомологии или идентичности с SEQ ID NO: 1.
При использовании в данном описании изобретения термин «вариант белка-рецептора цАМФ» можно использовать взаимозаменяемо с вариантным белком CRP, вариантом CRP, вариантом рецепторного белка цАМФ, вариантным белком САР, вариантом САР, вариантом белка-активатора катаболитных оперонов, вариантным белком-активатором катаболитных оперонов и так далее.
Что касается задач настоящего изобретения, микроорганизм, включающий вариант белка-рецептора цАМФ, отличается наличием высокой продуктивности по L-аминокислоте по сравнению с микроорганизмом, не включающим вариант белка-рецептора цАМФ. Вариант CRP отличается наличием регуляторной активности гена для увеличения продуктивности по L-аминокислоте по сравнению с нативным белком дикого типа или не вариантом белка-рецептора цАМФ. Это является значимым, так как продуктивность по L-аминокислоте может быть увеличена посредством введения варианта CRP по настоящему изобретению в микроорганизм. Более точно, L-аминокислота может представлять собой L-треонин или L-триптофан. Однако можно включать без ограничения любую L-аминокислоту до тех пор, пока она может быть продуцирована в результате введения или включения варианта белка-рецептора цАМФ.
Вариант белка-рецептора цАМФ может представлять собой, например, вариант, включающий аминокислотную последовательность, в которой другая аминокислота заменяет аминокислоту в позиции 35 в аминокислотной последовательности, представленной SEQ ID NO: 1, вариант, состоящий из SEQ ID NO: 3. Вариант, в котором аланин заменяет аминокислоту в позиции 35 в аминокислотной последовательности, представленной SEQ ID NO: 1, может состоять из SEQ ID NO: 3, но без ограничения этим. Кроме того, вариант CRP может включать аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, имеющую 80% или более гомологии или идентичности с ней, но без ограничения этим. Более точно, вариант CRP по настоящему изобретению может включать белок, имеющий SEQ ID NO: 3, и белок, имеющий по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% или более гомологии или идентичности с ней. Кроме того понятно, что белок, имеющий аминокислотную последовательность, часть которой делетирована, модифицирована, заменена или добавлена в дополнение к аминокислотной последовательности в позиции 35, может являться частью объема настоящего изобретения до тех пор, пока аминокислотная последовательность имеет указанную выше гомологию или идентичность и проявляет эффективность, соответствующую указанному выше белку.
Другими словами, даже если «белок, имеющий аминокислотную последовательность конкретной SEQ ID NO», описан в данном описании изобретения, понятно, что белок, имеющий аминокислотную последовательность, часть которой делетирована, модифицирована, заменена, консервативно заменена или добавлена, можно применять в настоящем изобретении до тех пор, пока он имеет активность, идентичную или соответствующую таковой у белка, состоящего из аминокислотной последовательности соответствующей SEQ ID NO. Например, до тех пор, пока белок имеет активность, идентичную или соответствующую таковой вариантного белка, добавление последовательности, которая не изменяет функцию белка до и после аминокислотной последовательности, его природные мутации, молчащие мутации или консервативные замены не исключаются. Понятно, что даже если белок имеет такое добавление последовательности или мутацию, он попадает в объем настоящего изобретения.
При использовании в данном описании изобретения термин «гомология» или «идентичность» обозначает степень соответствия между двумя данными аминокислотными последовательностями или нуклеотидными последовательностями и может быть выражена в процентах.
Термины «гомология» и «идентичность» часто можно использовать взаимозаменяемо.
Гомология или идентичность последовательностей консервативного полинуклеотида или полипептида можно определять с помощью стандартных алгоритмов выравнивания и можно использовать со штрафами за гэп, установленными по умолчанию в используемой программе. По существу, гомологичные или идентичные последовательности могут гибридизоваться в умеренно или очень жестких условиях, таких, чтобы могла гибридизоваться полная длина последовательности или по меньшей мере примерно 50%, 60%, 70%, 80% или 90% или более полной длины. Также рассматриваются полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов в гибридизации.
Имеют или нет любые две полинуклеотидные или полипептидные последовательности гомологию, подобие или идентичность, можно определять с использованием известных компьютерных алгоритмов, таких как программа «FASTA», используя, например, параметры по умолчанию, такие как B Pearson et al. (1988) [Proc. Natl. Acad. Sci. USA 85]: 2444, или определять с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), который задействован в программе Нидлмана пакета программ EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или более поздняя) (включая пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, и [CARILLO ETA/.](1988) SIAM J Applied Math 48: 1073). Например, BLAST из базы данных Национального центра биотехнологической информации или ClustalW можно использовать для определения гомологии, подобия или идентичности.
Гомологию, подобие или идентичность полинуклеотидов или полипептидов можно определять, например, посредством сравнения информации о последовательностях с использованием компьютерной программы GAP, такой как Needleman et al. (1970), J Mol Biol. 48: 443, которая раскрыта в Smith and Waterman, Adv. Appl. Math (1981) 2: 482. Кратко, программа GAP определяет подобие в виде количества выровненных символов (то есть нуклеотидов или аминокислот), которые являются подобными, разделенного на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) унарную матрицу сравнения (содержащую значение 1 для идентичностей и 0 для неидентичностей) и взвешенную матрицу сравнения из Gribskov et al (1986) Nucl. Acids Res. 14: 6745, как раскрыто в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или матрица замен EDNAFULL (EMBOSS версия NCBI NUC4.4)); (2) штраф 3,0 за каждый гэп и дополнительный штраф 0,10 за каждый символ в каждом гэпе (или штраф за открытие гэпа 10, штраф за продление гэпа 0,5); и (3) отсутствие штрафа для концевых гэпов. Таким образом, при использовании в данном описании изобретения термин «гомология» или «идентичность» представляет собой соответствие между последовательностями.
В другом аспекте настоящего изобретения предложен полинуклеотид, кодирующий вариант CRP, или вектор, включающий полинуклеотид.
При использовании в данном описании изобретения термин «полинуклеотид» относится к нити ДНК или РНК, имеющей заранее установленную длину или длиннее, которая представляет собой длинноцепочечный полимер нуклеотидов, образованный путем соединения нуклеотидных мономеров посредством ковалентных связей. Более конкретно, полинуклеотид относится к фрагменту полинуклеотида, кодирующему вариантный белок.
Полинуклеотид, кодирующий вариант CRP по настоящему изобретению, может включать любую полинуклеотидную последовательность без ограничения до тех пор, пока она представляет собой полинуклеотидную последовательность, кодирующую вариант белка-рецептора цАМФ по настоящему изобретению. Полинуклеотид, кодирующий вариант CRP, может включать любую последовательность без ограничения до тех пор, пока она представляет собой последовательность, кодирующую вариантный белок, в котором другая аминокислота замещает аминокислоту в позиции 35 в аминокислотной последовательности SEQ ID NO: 1. Более точно, полинуклеотид может представлять собой полинуклеотидную последовательность, кодирующую вариант, в котором аланин заменяет аминокислоту в позиции 35 в аминокислотной последовательности SEQ ID NO: 1. Например, полинуклеотид, кодирующий вариант CRP по настоящему изобретению может представлять собой полинуклеотидную последовательность, кодирующий аминокислотную последовательность SEQ ID NO: 3, но без ограничения этим. Более конкретно, полинуклеотид может состоять из полинуклеотидной последовательности SEQ ID NO: 4, но без ограничения этим. В полинуклеотиде в кодирующей области могут быть выполнены различные модификации при условии, что они не изменяют аминокислотную последовательность белка, из-за вырождения кодона или с учетом кодонов, предпочитаемых организмом, в котором должен экспрессироваться белок. Таким образом, понятно, что, из-за вырождения кодона, также может быть включен полинуклеотид, который может быть транслирован в полипептид, состоящий из аминокислотной последовательности SEQ ID NO: 3, или полипептид, имеющий гомологию или идентичность с ней.
Кроме того, также может быть включен, без ограничения, зонд, который может быть получен из известной нуклеотидной последовательности, например, последовательности, которая гибридизуется с комплементарной последовательностью до полной или части нуклеотидной последовательности в жестких условиях, чтобы кодировать вариант CRP, в котором другая аминокислота заменяет аминокислоту в позиции 35 в аминокислотной последовательности SEQ ID NO: 1.
Термин «жесткие условия» обозначают условия, при которых обеспечивается конкретная гибридизация между полинуклеотидами. Такие условия подробно описаны в литературе (например J. Sambrook et al., выше). Например, жесткие условия могут включать, например, условия, при которых гены, имеющие высокую гомологию или идентичность, гомологию или идентичность 80% или выше, 85% или выше, более точно 90% или выше, более конкретно 95% или выше, гораздо более конкретно 97% или выше, особенно конкретно 99% или выше, гибридизуются друг с другом, и гены, имеющие гомологию или идентичность ниже, чем указанные выше гомология или идентичность, не гибридизуются друг с другом, или стандартные условия отмывки Саузерн-гибридизации, то есть отмывки один раз, конкретно, два или три раза при концентрации солей и температуре, соответствующим 60°С, 1×SSC, 0,1% SDS, более точно, 60°С, 0,1×SSC, 0,1% SDS и более конкретно 68°С, 0,1×SSC, 0,1% SDS.
Хотя может иметь место ошибка спаривания между нуклеотидами вследствие жесткости гибридизации, требуется, чтобы две нуклеиновые кислоты имели комплементарные последовательности. Термин «комплементарный» используют для описания взаимоотношения между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину, и цитозин комплементарен гуанину. Соответственно, настоящее изобретение может включать не только по существу подобные последовательности нуклеиновых кислот, но также выделенные фрагменты нуклеиновых кислот, которые комплементарны целой последовательности.
Более точно, полинуклеотид, имеющий гомологию или идентичность, может быть обнаружен с использованием условий гибридизации, включающих гибридизацию при значении Тпл. 55°С и условий, описанных выше. Дополнительно, значение Тпл. может представлять собой 60°С, 63°С или 65°С, но без ограничения этим, и может подходящим образом регулироваться специалистом в данной области техники согласно целям.
Подходящая жесткость для гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов, и параметры хорошо известны в данной области техники (см. Sambrook et al., выше, 9.50-9.51, 11.7-11.8).
При использовании в данном описании изобретения термин «вектор» относится к ДНК-конструкции, которая включает нуклеотидную последовательность полинуклеотида, кодирующего целевой вариантный белок, функционально связанный с подходящей регуляторной последовательностью для обеспечения экспрессии целевого белка в подходящей клетке-хозяине. Регуляторная последовательность может включать промотор, способный к инициации транскрипции, любую операторную последовательность для регуляции такой транскрипции, последовательность, кодирующую подходящий домен связывания рибосомы мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После того как вектор трансформирован в подходящую клетку-хозяина, он может реплицироваться или действовать независимо от генома хозяина и может сам интегрироваться в геном.
Вектор, используемый в настоящем изобретении, конкретно не ограничивается до тех пор, пока он способен реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры обычно используемых векторов могут включать естественную или рекомбинантную плазмиду, космиду, вирус и бактериофаг. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon 4А, Charon 21А и так далее. В качестве плазмидного вектора можно использовать тип pBR, тип pUC, тип pBluescriptII, тип pGEM, тип pTZ, тип pCL, тип рЕТ и так далее. Более конкретно, можно использовать вектор pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
Например, полинуклеотид, кодирующий целевой вариантный белок в хромосоме, может быть заменен мутировавшим полинуклеотидом, используя вектор для внутриклеточной хромосомной вставки. Хромосомная вставка полинуклеотида может быть выполнена посредством любого способа, известного в данной области техники, например, посредством гомологичной рекомбинации, но без ограничения этим. Кроме того, для подтверждения хромосомной вставки может быть включен селективный маркер. Селективный маркер предназначен для отбора клеток, трансформированных вектором, то есть для подтверждения вставки желаемого полинуклеотида, и селективный маркер может включать маркеры, обеспечивающие селектируемые фенотипы, такие как устойчивость к лекарственным препаратам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия белков с модифицированной поверхностью. Так как только клетки, экспрессирующие селективный маркер, способны выживать или демонстрировать различные фенотипы в условиях окружающей среды, обработанной селективным агентом, можно отбирать трансформированные клетки. В качестве еще одного аспекта настоящего изобретения, в настоящем изобретении предложен микроорганизм, продуцирующий L-аминокислоту, включающий вариантный белок или полинуклеотид, кодирующий вариантный белок. Более точно, микроорганизм, включающий вариантный белок, и/или полинуклеотид, кодирующий вариантный белок, может представлять собой микроорганизм, полученный путем трансформации с помощью вектора, включающего полинуклеотид, кодирующий вариантный белок, но без ограничения этим.
При использовании в данном описании изобретения, термин «трансформация» обозначает введение вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина таким образом, чтобы белок, кодируемый полинуклеотидом, экспрессировался в клетке-хозяине. При условии, что трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, он может интегрироваться и располагаться в хромосоме клетки-хозяина, или он может находиться вне хромосомы или независимо от нее. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме до тех пор, пока он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме кассеты экспрессии, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для ее автономной экспрессии. Обычно кассета экспрессии может включать промотор, функционально связанный с полинуклеотидом, сигналы терминации транскрипции, сайты связывания рибосом и сигналы терминации трансляции. Кассета экспрессии может быть в форме самореплицирующегося вектора экспрессии. Дополнительно, полинуклеотид как таковой может быть введен в клетку-хозяина и функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине, но без ограничения этим.
При использовании в данном описании изобретения, термин «функционально связанный» означает функциональную связь между полинуклеотидной последовательностью, кодирующей целевой вариантный белок по настоящему изобретению, и последовательностью промотора, которая инициирует и опосредует транскрипцию последовательности полинуклеотида.
В еще одном аспекте настоящего изобретения предложен микроорганизм рода Escherichia (Escherichia sp.), включающий вариант белка-рецептора цАМФ.
При использовании в данном описании изобретения термин «микроорганизм, включающий вариант CRP» может относиться к рекомбинантному микроорганизму для экспрессии варианта CRP по настоящему изобретению. Например, микроорганизм относится к клетке-хозяину или микроорганизму, который способен экспрессировать вариант посредством включения полинуклеотида, кодирующего вариант CRP, или путем трансформации с помощью вектора, включающего полинуклеотид, кодирующий вариант CRP. Что касается задач настоящего изобретения, микроорганизм представляет собой микроорганизм, экспрессирующий вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, и микроорганизм может представлять собой микроорганизм, экспрессирующий вариантный белок, имеющий активность белка-рецептора цАМФ, где аминокислотная замена представляет собой замену аминокислоты в позиции 35 с N-конца аланином, но без ограничения этим.
Микроорганизм, включающий вариант CRP, может представлять собой любой микроорганизм до тех пор, пока он включает вариант CRP для экспрессии L-аминокислоты, например, L-треонина или L-триптофана, но без ограничения этим. Например, микроорганизм, включающий вариант CRP, может представлять собой рекомбинантный микроорганизм, имеющий повышенную продуктивность по L-аминокислоте, которую получают посредством экспрессии варианта CRP в природном микроорганизме дикого типа или в микроорганизме, продуцирующем L-аминокислоту. Рекомбинантный микроорганизм, имеющий повышенную продуктивность по L-аминокислоте, может представлять собой микроорганизм, имеющий повышенную продуктивность по L-аминокислоте по сравнению с природным микроорганизмом дикого типа или немодифицированным микроорганизмом, где L-аминокислота может представлять собой L-треонин или L-триптофан, но без ограничения этим.
При использовании в данном описании изобретения термин «микроорганизм, продуцирующий L-аминокислоту» включает микроорганизм дикого типа или микроорганизм, в котором имеет место природная или искусственная генетическая модификация, и он может представлять собой микроорганизм, имеющий особый ослабленный или усиленный механизм в результате вставки чужеродного гена или в результате усиления или инактивации активности эндогенного гена, у которого имеет место генетическая вариация или усиливается активность для получения целевой L-аминокислоты. Что касается задач настоящего изобретения, микроорганизм, продуцирующий L-аминокислоту, может включать вариантный белок, чтобы иметь повышенную продуктивность по целевой L-аминокислоте. Более точно, микроорганизм, продуцирующий L-аминокислоту, или микроорганизм, имеющий продуктивность по L-аминокислоте по настоящему изобретению, может представлять собой микроорганизм, в котором часть генов, вовлеченных в путь биосинтеза L-аминокислоты, усилена или ослаблена, или часть генов, вовлеченных в путь деградации L-аминокислоты, усилена или ослаблена.
«Немодифицированный микроорганизм» относится к природному штамму как таковому, или микроорганизму, не включающему вариант CRP, или микроорганизму, который не трансформирован с помощью вектора, включающего полинуклеотид, кодирующий вариант CRP. «Микроорганизм» может включать любой из прокариотических микроорганизмов и эукариотических микроорганизмов, до тех пор, пока он способен продуцировать L-аминокислоту. Например, микроорганизм может включать микроорганизмы рода Escherichia, рода Erwinia, рода Serratia, рода Providencia, рода Corynebacterium и рода Brevibacterium. Более точно, микроорганизм может представлять собой микроорганизм рода Escherichia, и более конкретно Е. coli, но без ограничения этим.
В еще одном аспекте настоящего изобретения предлагается способ получения L-аминокислоты, где способ включает культивирование микроорганизма рода Escherichia в среде, микроорганизма, продуцирующего L-аминокислоту вследствие включения варианта белка-рецептора цАМФ.
Термины «вариант белка-рецептора цАМФ» и «L-аминокислота» являются такими же, как описано выше.
В этом способе культивирование микроорганизма может быть выполнено, но без конкретного ограничения, посредством известного периодического культивирования, непрерывного культивирования, периодического культивирования с подпиткой и так далее. В этом отношении, условия культивирования конкретно не ограничиваются, но можно поддерживать оптимальный рН (например рН от 5 до 9, более точно рН от 6 до 8 и наиболее точно рН 6,8) посредством использования основного соединения (например гидроксида натрия, гидроксида калия или нашатырного спирта) или кислого соединения (например фосфорной кислоты или серной кислоты). Кроме того, аэробные условия можно поддерживать посредством введения кислорода или кислородсодержащей смеси газов в клеточную культуру. Температуру культивирования можно поддерживать от 20°С до 45°С и, более точно, от 25°С до 40°С. Культивирование можно выполнять в течение от примерно 10 часов до примерно 160 часов, но без ограничения этим. L-аминокислота, полученная посредством вышеуказанного культивирования, может быть выделена в среду или оставаться в клетках.
Кроме того, подлежащая использованию культуральная среда может включать, по отдельности или в комбинации, в качестве источников углерода сахара и углеводы (например глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масла и жиры (например соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например глицерин и этиловый спирт) и органические кислоты (например уксусную кислоту), но без ограничения этим. В качестве источников азота можно использовать, по отдельности или в комбинации, азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), но без ограничения этим. В качестве источников фосфора можно использовать, по отдельности или в комбинации, дигидрофосфат калия, гидрофосфат калия и соответствующие им натрийсодержащие соли, но без ограничения этим. Кроме того, среда может включать другие необходимые стимулирующие рост вещества, включающие соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Кроме того, способ может включать получение L-аминокислоты из микроорганизма или среды.
Способ сбора L-аминокислоты, полученной в ходе культивирования по настоящему изобретению может включать сбор целевой L-аминокислоты из культуральной жидкости с использованием подходящего способа, известного в данной области техники, согласно способу культивирования. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию, ВЭЖХ (высокоэффективную жидкостную хроматографию) и так далее, и целевая L-аминокислота может быть собрана из среды или микроорганизма с использованием подходящего способа, известного в данной области техники.
Кроме того, получение может включать процесс очистки и может быть выполнено, используя подходящий способ, известный в данной области техники. Таким образом, L-аминокислота, подлежащая сбору, может представлять собой очищенную форму или ферментационный бульон микроорганизма, включающего L-аминокислоту (Introduction to Biotechnology and Genetic Engineering, A. J. Nair., 2008).
В еще одном аспекте настоящего изобретения предлагается применение варианта белка-рецептора цАМФ в получении L-аминокислоты, где вариант белка-рецептора цАМФ включает одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1.
В еще одном аспекте настоящего изобретения предлагается применение микроорганизма рода Escherichia в получении L-аминокислоты, где микроорганизм рода Escherichia включает вариант белка-рецептора цАМФ.
Термин «вариант белка-рецептора цАМФ» и «L-аминокислота» являются такими же, как описано выше.
Принцип изобретения
В дальнейшем в этом документе настоящее изобретение будет описано более подробно со ссылкой на Примеры. Однако специалисту в данной области техники понятно, что эти Примеры имеют место только для иллюстративных целей и объем настоящего изобретения не предназначен для ограничения посредством этих Примеров.
Пример 1. Получение рекомбинантного вектора рСС1ВАС- crp
1-1. Получение фрагмента гена crp
Для получения примерно 0,96 т.п.н (тысяч пар нуклеотидов) фрагмента ДНК SEQ ID NO: 5, включающего ген crp и регуляторную область экспрессии, извлекали геномную ДНК (гДНК) W3110 Е. coli дикого типа, используя систему Genomic-tip от Qiagen (компания), и выполняли ПЦР (полимеразную цепную реакцию) с использованием гДНК в качестве матрицы и набор для ПЦР HL premix (изготовлен в BIONEER Co., такой же используют в дальнейшем в этом документе). ПЦР для амплификации фрагмента гена crp выполняли, используя праймеры SEQ ID NOS: 6 и 7 для 27 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 56°С в течение 30 секунд и элонгации при 72°С в течение 2 мин.
Продукт ПЦР расщепляли с помощью EcoR I и выполняли электрофорез на 0,8% агарозном геле и элюирование с получением фрагмента ДНК 0,96 т.п.н (в дальнейшем в этом документе обозначен как «фрагмент crp»).
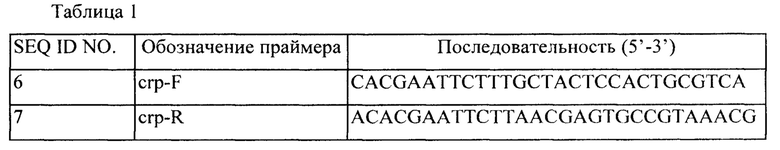
1-2. Получение рекомбинантного вектора рСС1ВАС-crp
Вектор CopyControl pCC1BAC (EPICENTRE, USA) обрабатывали с помощью EcoR I и выполняли электрофорез на 0,8% агарозном геле и элюирование с получением продукта, который затем лигировали с помощью фрагмента crp, полученного в Примере 1-1, посредством этого получая плазмиду рСС1ВАС-crp.
Пример 2. Получение библиотеки вариантов рекомбинантного вектора рСС1ВАС- crp
2-1. Получение мутантного фрагмента crp посредством допускающей ошибки ПЦР
ПЦР выполняли с использованием геномной ДНК Е. coli W3110 дикого типа в качестве матрицы и набор для случайного мутагенеза Diversify PCR (№ в каталоге: К1830-1, Таблица III, реакции мутагенеза 4) от Clonetech. Подробно, ПЦР выполняли с использованием праймеров SEQ ID NOS: 6 и 7, которые использовали в Примере 1-1, для 27 циклов, состоящих из денатурации при 94°С в течение 30 секунд и элонгации при 68°С в течение 1 мин.
Продукт ПЦР расщепляли с помощью EcoR I и выполняли электрофорез на 0,8% агарозном геле и элюирование с получением мутированного фрагмента crp 0,96 т.п.н (в дальнейшем в этом документе обозначен как «фрагмент crpm»).
2-2. Получение библиотеки вариантов рекомбинантного вектора pCC1BAC-crp
Вектор pCC1BAC обрабатывали с помощью фермента рестрикции EcoR I и затем обрабатывали щелочной фосфатазой (NEB). Полученный вектор лигировали с фрагментом crpm, полученного в Примере 2-1, и продукт лигирования трансформировали в электрокомпетентную Е. coli TransforMax EPI300 (EPICENTRE, USA) посредством электрофореза. Трансформированный штамм культивировали на твердой среде LB (Лурия-Бертани) (15 нг/мл), содержащей хлорамфеникол, для селекции колоний. Полученные таким образом колонии собирали и подвергали обработке plasmid prep (набор для выделения и очистки плазмид), посредством этого получая библиотеку pCC1BAC-crpm.
Пример 3. Введение библиотеки вариантов crp в треонин-продуцирующий штамм и селекция штамма с улучшенным ростом
3-1. Введение библиотеки pCClBAC-crpm в треонин-продуцирующий штамм
Библиотеку рСС1ВАС-crpm, полученную в Примере 2, трансформировали в электрокомпетентные клетки KCCM10541, который представляет собой треонин-продуцирующий микроорганизм, посредством электропорации. KCCM10541 Е. coli (Патент Кореи №10-0576342), используемый в этом Примере, представляет собой Е. coli, полученную путем инактивирования гена galR в L-треонин-продуцирующем KFCC10718 Е. coli (Патент Кореи No. 10-0058286).
В качестве контрольной группы библиотеки pCClBAC-crpm, введенной в микроорганизм, рСС1ВАС-crp трансформировали в KCCM 10541 таким же образом, как указано выше, с получением KCCM10541/1CCXBAC-crp(WT).
3-2. Сравнение скорости роста рекомбинантного микроорганизма
Минимальную среду М9, содержащую 1% глюкозу и 0,2 г/л дрожжевого экстракта, распределяли в микропланшет с глубокими лунками и затем засевали туда трансформант и контрольный штамм, полученный в Примере 3-1, соответственно. Штаммы культивировали с использованием микроразмерного шейкер-инкубатора с постоянной температурой (TAITEC, Japan) в условиях 37°С и 200 об/мин, посредством способа HTS (скрининга с высокой пропускной способностью) в течение 20 часов и отбирали штаммы с улучшенным ростом. Из них в конечном счете выбирали один вид штамма (Таблица 2).
Штамм KCCM10541 с введенным геном crp дикого типа демонстрировал незначительное увеличение значения OD вследствие дополнительного введения crp, тогда как трансформант с улучшенным ростом демонстрировал высокое значение OD после одинакового времени культивирования по сравнению со штаммом дикого типа с введенным crp. Кроме того, выбранный вариант crp подвергали обработке plasmid miniprep (набор для быстрого выделения плазмидной ДНК) с последующим анализом секвенирования. Результаты обобщены в Таблице 2.

3-3. Сравнение титра треонина рекомбинантного микроорганизма
Для измерения титра треонина рекомбинантного микроорганизма, отобранного в Примере 3-2, рекомбинантный микроорганизм культивировали в среде для титрования треонина, приготовленной, например, в составе, указанном в следующей Таблице 3, для изучения улучшения продуктивности по L-треонину.

Подробно, каждую платиновую петлю Е. coli KCCM10541/pCC1BAC-crp(WT) и Е. coli KCCM10541/рСС1ВАС-crpTM4, культивированных в течение ночи на твердой среде LB в инкубаторе при 33°С, инокулировали в 25 мл среды для титрования, указанной в Таблице 3, соответственно, и затем культивировали в инкубаторе при 33°С и 200 об/мин в течение 48 часов для сравнения скорости потребления сахара и концентраций треонина.
В результате, как описано в следующей Таблице 4, штамм KCCM10541/рСС1ВАС-crp(WT), в качестве контрольной группы, демонстрировал потребление сахара 26,1 г/л за 24 часа, тогда как мутантный штамм с введенным crpTM4 демонстрировал улучшение скорости потребления сахара примерно на 16% и 11% по сравнению с материнским штаммом и штаммом с введенным crp дикого типа, соответственно.
Кроме того, при культивировании в течение 48 часов штамм с введенным crp дикого типа демонстрировал продуцирование по L-треонину 29,0 г/л, тогда как продуцирование по L-треонину мутантного штамма, полученного выше, увеличивалось вплоть до 31,0 г/л, даже если скорость культивирования была увеличена, демонстрируя улучшение концентрации примерно на 8% и 7% по сравнению с материнским штаммом и штаммом с введенным crp дикого типа, соответственно.
Так как введение варианта crp увеличивало выход и потребление сахара у штамма, очевидно, это может быть хорошим признаком варианта, который может в значительной степени способствовать улучшению эффективности продуцирования во время ферментации.

Пример 4. Введение варианта pCClBAC- crp TM4 в триптофан-продуцирующий штамм
4-1. Введение рСС1ВАС-crpTM4 в штамм для скрининга
рСС1ВАС-crpTM4, полученный в Примере 3, трансформировали электрокомпетентные клетки триптофан-продуцирующего штамма KCCM11166P посредством электропорации. KCCM11166P, используемый в этом Примере, представляет собой L-триптофан-продуцирующий Е. coli, в котором был делетирован ген tehB, а активность НАД-киназы усилена (Патент Кореи No. 10-1261147).
В качестве контрольной группы микроорганизма с введенным рСС1ВАС-crpTM4, pCC1BAC-crp (WT) трансформировали в KCCM11166P таким же образом, как указано выше, с получением KCCM11166P/pCC1BAC-crp(WT).
4-2. Сравнение скорости роста рекомбинантного микроорганизма
Минимальную среду М9, содержащую 1% глюкозу и 0,2 г/л дрожжевого экстракта, распределяли в микропланшет с глубокими лунками и затем засевали туда трансформант и контрольный штамм, полученный как указано в Примере 4-1, соответственно. Штаммы культивировали, используя микроразмерный шейкер-инкубатор с постоянной температурой (TAITEC, Japan), в условиях 37°С и 200 об/мин посредством способа HTS (скрининга с высокой пропускной способностью) в течение 16 часов для подтверждения улучшения роста трансформанта KCCM11166P/рСС1ВАС-crpTM4 (Таблица 5).
Штамм KCCM11166P с введенным геном crp дикого типа демонстрировал эквивалентный уровень OD следствие дополнительного введения crp после одинакового времени культивирования, тогда как трансформант с улучшенным ростом демонстрировал высокое значение OD по сравнению с crp дикого типа.

4-3. Сравнение титра триптофана рекомбинантного микроорганизма
Для измерения титра триптофана рекомбинантного микроорганизма, полученного в Примере 4-2, рекомбинантный микроорганизм культивировали в среде для титрования триптофана, приготовленной, например, в составе, указанном в следующей Таблице 6, для изучения улучшения продуктивности по L-триптофану.

Подробно, каждую платиновую петлю KCCM11166/рСС1ВАС-crpWT) Е. coli и KCCM11166P/рСС1ВАС-crpTM4 Е. coli, культивированных в течение ночи на твердой среде LB в инкубаторе при 37°С, инокулировали в 25 мл среды для титрования, указанной в Таблице 6, соответственно, и затем культивировали в инкубаторе при 37°С и 200 об/мин в течение 48 часов для сравнения скоростей потребления сахара и концентраций триптофана.
В результате, как описано в следующей Таблице 7, штамм KCCM11166/pCC1BAC-crp(WT) в качестве контрольной группы демонстрировал потребление сахара 30,2 г/л за 22 часа, тогда как мутантный штамм с введенным crpTM4 демонстрировал улучшение скорости потребления сахара примерно на 146% и 9% по сравнению с материнским штаммом и штаммом дикого типа с введенным crp, соответственно.
При культивировании в течение 48 часов штамм с введенным crp дикого типа демонстрировал продуцирование L-триптофана 8,4 г/л, тогда как продуцирование L-триптофана мутантным штаммом, полученным выше, увеличивалось вплоть до 9,7 г/л, даже если скорость культивирования была увеличена, демонстрируя примерно 18% и 15% улучшения концентрации, по сравнению с материнским штаммом и штаммом с введенным crp дикого типа, соответственно.
Так как введение варианта crp повышало потребление сахара у штамма и выход, очевидно, это может быть хорошим признаком варианта, который может в значительной степени способствовать улучшению эффективности продуцирования во время ферментации.

Пример 5. Введение эндогенного вектора с эффективным вариантом crp в Е. coli дикого типа
5-1. Введение эффективного варианта pCC1BAC-crp в треонин-продуцирующий штамм, полученный из дикого тина
Для того чтобы изучить, демонстрировал ли вектор, включающий вариант crp, исследованный в Примере 3, также эквивалентные эффекты у штамма дикого типа, вектор pCC1BAC-crp(WT) или рСС1ВАС-crpTM4, соответственно, трансформировали посредством электропорации в полученный из дикого типа штамм, способный продуцировать треонин. Кроме того, в качестве контрольной группы получали штамм с введенным pCC1BAC-crp(WT).
Используемый в этом Примере штамм, полученный из штамма дикого типа, способный продуцировать треонин, представляет собой W3110::PcysK-ppc/pACYC184-thrABC. W3110::PcysK-ppc/pACYC184-thrABC представляет собой штамм, в котором нативный промотор гена ррс, кодирующего фосфоенолпируваткарбоксилазу на хромосоме, был заменен промотором гена cysK, и ген оперона биосинтеза треонина был введен в форме вектора для увеличения количества копий, повышая посредством этого продуктивность по треонину. Подробно, штамм W3110::PcycK-ррс получали с использованием pUCpcycKmloxP таким же образом, как описано в Патенте Кореи №10-0966324, и pACYC184-thrABC (Патент Кореи No. 10-1865998) трансформировали в штамм путем электропорации.
Полученные штаммы культивировали в среде тестирования треонина, полученной с составом, указанным в следующей Таблице 8, и сравнивали скорости роста и продуктивность по L-треонину.

Подробно, каждую платиновую петлю W3110 и соответствующих штаммов, культивированных в течение ночи на твердой среде LB в инкубаторе при 33°С, инокулировали в 25 мл среды для титрования, указанной в Таблице 8, соответственно, и затем культивировали в инкубаторе при 33°С и 200 об/мин в течение 48 часов. Результаты культивирования показаны в следующей Таблице 9. Как видно в следующих результатах, вариантный белок, выбранный в настоящем изобретении, также способен эффективно продуцировать треонин в штамме дикого типа с высоким выходом.

5-2. Введение эффективного варианта рСС1ВАС-crp в триптофан-продуцирующий штамм, полученный из дикого типа
Для того чтобы изучить, демонстрировал ли вектор, включающий вариант crp, исследованный в Примере 4, также эквивалентные эффекты в штамме дикого типа, вектор pCC1BAC-crp(WT) или pCC1BAC-crpTM4, соответственно, трансформировали в штамм, полученный из дикого типа, способный продуцировать триптофан.
Используемый в этом Примере штамм, полученный из дикого типа, способный продуцировать триптофан, представляет собой W3110 trpΔ2/pCL-Dtrp_att-trpEDCBA. W3110 trpΔ2/pCL-Dtrp_att-trpEDCBA представляет собой штамм с введенным вектором, в котором регуляторный механизм регуляторного участка оперона триптофана был ослаблен и экспрессия оперона триптофана была усилена для сверхэкспрессии триптофана (Патент Кореи №10-1532129). Штаммы с введенным вектором культивировали в среде для тестирования триптофана, полученной с составом, указанным в следующей Таблице 10, и сравнивали их продуктивность по L-триптофану.

Подробно, каждую платиновую петлю штаммов, культивированных в течение ночи на твердой среде LB в инкубаторе при 37°С, инокулировали в 25 мл среды для тестирования, указанной в Таблице 9, соответственно, и затем культивировали в инкубаторе при 37°С и 200 об/мин в течение 48 часов. Значения OD и концентрации триптофана сравнивали и представили в Таблице 11. Как видно в следующих результатах, вариантный белок, отобранный в настоящем изобретении, также способен эффективно продуцировать триптофан с высоким выходом в штамме дикого типа.

Авторы настоящего изобретения обозначили штамм с введенным pCC1BAC-crpTM4 на основе KCCM11166P, имеющий улучшенную продуктивность по триптофану и скорость потребления сахара (KCCM11166P/рСС1АС-crpTM4), как «СА04-2809», и затем депонировали штамм в корейском центре культур микроорганизмов (KCCM), который представляет собой международный депозитарный орган в соответствии с Будапештским договором, 7 ноября 2018 с номером доступа KCCM12375P.
Эти результаты показывают, что скорость потребления сахара была улучшена и продуктивность по L-аминокислоте была увеличена у микроорганизма рода Escherichia по настоящему изобретению с введенным вариантом crp и, следовательно, продуктивность по L-аминокислоте была повышена по сравнению с немодифицированным штаммом.
На основании описания выше, специалисту в данной области техники будет понятно, что настоящее изобретение моет быть осуществлено в другой конкретной форме без изменения его технической сущности или его существенных характеристик. Таким образом, следует понимать, что указанное выше воплощение является не ограничивающим, а иллюстративным во всех аспектах. Объем изобретения определяется прилагаемой формулой изобретения, а не описанием, предшествующим ей, и таким образом все изменения и модификации, которые входят в границы и пределы формулы изобретения, или эквиваленты таких границ и пределов, таким образом, рассматривают как охваченные формулой изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> cAMP receptor protein variant and method for producing L-amino
acid using the same
<130> OPA19120-PCT
<150> KR 10-2018-0151043
<151> 2018-11-29
<160> 7
<170> KoPatentIn 3.0
<210> 1
<211> 210
<212> PRT
<213> Unknown
<220>
<223> CRP
<400> 1
Met Val Leu Gly Lys Pro Gln Thr Asp Pro Thr Leu Glu Trp Phe Leu
1 5 10 15
Ser His Cys His Ile His Lys Tyr Pro Ser Lys Ser Lys Leu Ile His
20 25 30
Gln Gly Glu Lys Ala Glu Thr Leu Tyr Tyr Ile Val Lys Gly Ser Val
35 40 45
Ala Val Leu Ile Lys Asp Glu Glu Gly Lys Glu Met Ile Leu Ser Tyr
50 55 60
Leu Asn Gln Gly Asp Phe Ile Gly Glu Leu Gly Leu Phe Glu Glu Gly
65 70 75 80
Gln Glu Arg Ser Ala Trp Val Arg Ala Lys Thr Ala Cys Glu Val Ala
85 90 95
Glu Ile Ser Tyr Lys Lys Phe Arg Gln Leu Ile Gln Val Asn Pro Asp
100 105 110
Ile Leu Met Arg Leu Ser Ala Gln Met Ala Arg Arg Leu Gln Val Thr
115 120 125
Ser Glu Lys Val Gly Asn Leu Ala Phe Leu Asp Val Thr Gly Arg Ile
130 135 140
Ala Gln Thr Leu Leu Asn Leu Ala Lys Gln Pro Asp Ala Met Thr His
145 150 155 160
Pro Asp Gly Met Gln Ile Lys Ile Thr Arg Gln Glu Ile Gly Gln Ile
165 170 175
Val Gly Cys Ser Arg Glu Thr Val Gly Arg Ile Leu Lys Met Leu Glu
180 185 190
Asp Gln Asn Leu Ile Ser Ala His Gly Lys Thr Ile Val Val Tyr Gly
195 200 205
Thr Arg
210
<210> 2
<211> 633
<212> DNA
<213> Unknown
<220>
<223> crp
<400> 2
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 60
attcataagt acccatccaa gagcaagctt attcaccagg gtgaaaaagc ggaaacgctg 120
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 180
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 240
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 300
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 360
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 420
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 480
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 540
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaacctgat ctccgcacac 600
ggtaaaacca tcgtcgttta cggcactcgt taa 633
<210> 3
<211> 210
<212> PRT
<213> Unknown
<220>
<223> modified CRP
<400> 3
Met Val Leu Gly Lys Pro Gln Thr Asp Pro Thr Leu Glu Trp Phe Leu
1 5 10 15
Ser His Cys His Ile His Lys Tyr Pro Ser Lys Ser Lys Leu Ile His
20 25 30
Gln Gly Ala Lys Ala Glu Thr Leu Tyr Tyr Ile Val Lys Gly Ser Val
35 40 45
Ala Val Leu Ile Lys Asp Glu Glu Gly Lys Glu Met Ile Leu Ser Tyr
50 55 60
Leu Asn Gln Gly Asp Phe Ile Gly Glu Leu Gly Leu Phe Glu Glu Gly
65 70 75 80
Gln Glu Arg Ser Ala Trp Val Arg Ala Lys Thr Ala Cys Glu Val Ala
85 90 95
Glu Ile Ser Tyr Lys Lys Phe Arg Gln Leu Ile Gln Val Asn Pro Asp
100 105 110
Ile Leu Met Arg Leu Ser Ala Gln Met Ala Arg Arg Leu Gln Val Thr
115 120 125
Ser Glu Lys Val Gly Asn Leu Ala Phe Leu Asp Val Thr Gly Arg Ile
130 135 140
Ala Gln Thr Leu Leu Asn Leu Ala Lys Gln Pro Asp Ala Met Thr His
145 150 155 160
Pro Asp Gly Met Gln Ile Lys Ile Thr Arg Gln Glu Ile Gly Gln Ile
165 170 175
Val Gly Cys Ser Arg Glu Thr Val Gly Arg Ile Leu Lys Met Leu Glu
180 185 190
Asp Gln Asn Leu Ile Ser Ala His Gly Lys Thr Ile Val Val Tyr Gly
195 200 205
Thr Arg
210
<210> 4
<211> 633
<212> DNA
<213> Unknown
<220>
<223> modified crp
<400> 4
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 60
attcataagt acccatccaa gagcaagctt attcaccagg gtgcaaaagc ggaaacgctg 120
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 180
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 240
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 300
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 360
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 420
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 480
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 540
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaacctgat ctccgcacac 600
ggtaaaacca tcgtcgttta cggcactcgt taa 633
<210> 5
<211> 933
<212> DNA
<213> Unknown
<220>
<223> crp and expression control region
<400> 5
tttgctactc cactgcgtca attttcctga cagagtacgc gtactaacca aatcgcgcaa 60
cggaaggcga cctgggtcat gctgaagcga gacaccagga gacacaaagc gaaagctatg 120
ctaaaacagt caggatgcta cagtaataca ttgatgtact gcatgtatgc aaaggacgtc 180
acattaccgt gcagtacagt tgatagcccc ttcccaggta gcgggaagca tatttcggca 240
atccagagac agcggcgtta tctggctctg gagaaagctt ataacagagg ataaccgcgc 300
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 360
attcataagt acccatccaa gagcaagctt attcaccagg gtgaaaaagc ggaaacgctg 420
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 480
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 540
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 600
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 660
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 720
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 780
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 840
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaacctgat ctccgcacac 900
ggtaaaacca tcgtcgttta cggcactcgt taa 933
<210> 6
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> crp-F
<400> 6
cacgaattct ttgctactcc actgcgtca 29
<210> 7
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> crp-R
<400> 7
acacgaattc ttaacgagtg ccgtaaacg 29
<---
Группа изобретений относится к белку-рецептору цАМФ и его использованию для получения L-аминокислоты. Предложен белок-рецептор цАМФ, который является регулятором транскрипции и в котором аминокислота в позиции 35 в аминокислотной последовательности SEQ ID NO: 1 заменена на аланин. Также предложены микроорганизм рода Escherichia для продуцирования L-аминокислоты, содержащий указанный белок, и способ получения L-аминокислоты с использованием указанного микроорганизма. Также предложены полинуклеотид, кодирующий указанный белок, и экспрессионный вектор, содержащий полинуклеотид, кодирующий указанный белок. Группа изобретений обеспечивает получение L-аминокислоты с высоким выходом. 6 н. и 3 з.п. ф-лы, 11 табл., 5 пр.
1. Белок-рецептор цАМФ, который является регулятором транскрипции и в котором аминокислота в позиции 35 в аминокислотной последовательности SEQ ID NO: 1 заменена на аланин.
2. Полинуклеотид, кодирующий белок-рецептор цАМФ по п. 1.
3. Экспрессионный вектор, содержащий полинуклеотид, кодирующий белок-рецептор цАМФ по п. 1.
4. Микроорганизм рода Escherichia для продуцирования L-аминокислоты, содержащий белок-рецептор цАМФ, в котором аминокислота в позиции 35 в аминокислотной последовательности SEQ ID NO: 1 заменена на аланин.
5. Микроорганизм рода Escherichia по п. 4, представляющий собой E. coli.
6. Микроорганизм рода Escherichia по п. 4, где L-аминокислота представляет собой L-треонин или L-триптофан.
7. Способ получения L-аминокислоты, включающий:
культивирование микроорганизма рода Escherichia в среде, где микроорганизм содержит белок-рецептор цАМФ, в котором аминокислота в позиции 35 в аминокислотной последовательности SEQIDNO: 1 заменена на аланин; и
выделение L-аминокислоты из микроорганизма или среды.
8. Способ по п. 7, где L-аминокислота представляет собой L-треонин или L-триптофан.
9. Применение белка-рецептора цАМФ, в котором аминокислота в позиции 35 в аминокислотной последовательности SEQ ID NO: 1 заменена на аланин, или микроорганизма рода Escherichia, включающего этот белок, в получении L-аминокислоты.
| CN 103114069 B, 03.12.2014 | |||
| US 10125178 B2, 13.11.2018 | |||
| МИКРООРГАНИЗМ, СПОСОБНЫЙ ПРОДУЦИРОВАТЬ L- АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2600875C2 |
| US 8889425 B2, 18.11.2014. | |||

