ОБЛАСТЬ ТЕХНИКИ
Данное описание относится к варианту белка-рецептора цАМФ, к микроорганизму, содержащему его, и к способу получения L-аминокислоты с их применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
CRP (белок-рецептор циклического АМФ), также называемый САР (белок-активатор катаболизма), является наиболее хорошо известным регулятором транскрипции у Е. coli. CRP характеризуется наличием зависимого от источника углерода регуляторного механизма, представленного "подавлением катаболитами". Это действие инициируется внутриклеточной концентрацией циклического АМФ (ниже обозначаемого как "цАМФ"). В присутствии предпочтительного источника углерода, такого как глюкоза, активность аденилатциклазы ингибируется с понижением уровня цАМФ, и этот сигнал подавляет экспрессию катаболических генов. В противном случае активность аденилатциклазы возрастает, в результате чего репрессоры подавляются и инициируется экспрессия катаболических генов. Кроме того, известно, что CRP играет различные роли, такие как внутриклеточная передача сигнала, опосредованная цАМФ, осмотическая регуляция, ответы клеток на стрессовые ситуации, образование биопленок, фиксация азота, транспорт железа и т.д.
Согласно сообщениям, CRP осуществляет регуляцию 418 генов Е. coli, однако соответствующие механизмы еще полностью не изучены (J Biol Eng. (2009) 24;3:13). При таком широком спектре регуляторных способностей, мутации CRP могут проявляться различными фенотипами. Благодаря его преимуществам, CRP изучали в качестве мишени, подходящей для реконструкции штаммов на клеточном уровне, которые можно применять в различных условиях. Недавно были проведены различные эксперименты, такие как способ изменения экспрессии регулируемых генов путем изменения степени связывания ДНК посредством аминокислотных вариаций CRP, выбранных с использованием биоинформатики (Nucleic Acids Research, (2009) 37: 2493-2503), способ отбора Е. coli с устойчивостью к нагреванию, осмосу и низкой температуре с использованием искусственного транскрипционного фактора (ATF), полученного путем слияния сайта связывания ДНК "цинковый палец" и CRP ((Nucleic Acids Research, (2008) 36: e102) и т.д. Другими словами, поскольку изменения экспрессии CRP вызывают широкий спектр изменений экспрессии генов, расположенных в направлении 3', CRP вероятно является хорошим инструментом для получения микроорганизмов с полезными характеристиками.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Техническая задача
Авторы данного изобретения разработали новый вариант белка, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, и обнаружили, что данный белок может повышать продуцирование L-аминокислот, тем самым завершив данное изобретение.
Техническое решение
Задачей данного изобретения является предложение варианта белка-рецептора цАМФ.
Другой задачей данного изобретения является предложение полинуклеотида, кодирующего этот вариант белка-рецептора цАМФ.
Еще одной задачей данного изобретения является предложение вектора, включающего в себя этот полинуклеотид.
Еще одной задачей данного изобретения является предложение микроорганизма рода Escherichia, включающего этот вариант.
Еще одной задачей данного изобретения является предложение способа получения L-аминокислоты, включающего культивирование микроорганизма рода Escherichia в среде.
Еще одной задачей данного изобретения является предложение применения варианта микроорганизма рода Escherichia, включающего в себя указанный вариант, в получении L-аминокислоты.
Полезные эффекты
При культивировании микроорганизма рода Escherichia, продуцирующего L-аминокислоту, включающего вариант белка-рецептора цАМФ по данному изобретению, возможно получение L-аминокислоты с высоким выходом. Соответственно, в промышленных аспектах можно ожидать снижения себестоимости продукции наряду с удобством получения.
ПРЕДПОЧТИТЕЛЬНЫЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее данное изобретение будет описано более подробно следующим образом. При этом каждое из описаний и воплощений, изложенных в данной заявке, могут быть также применены к другим описаниям и воплощениям. Таким образом, все комбинации различных элементов, раскрытых в данном описании, входят в объем данного изобретения. Кроме того, объем данного изобретения не ограничен конкретным описанием, приведенным ниже.
Для достижения вышеуказанных задач в одном аспекте данного изобретения предложен вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1. В частности, в данном изобретении предложен вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, где аминокислотные замены включают замену аминокислоты в положении 196 от N-конца на глутамин. Более конкретно, в данном изобретении предложен вариант белка-рецептора цАМФ, включающий замену аминокислоты в положении 196 в аминокислотной последовательности SEQ ID NO: 1 на глутамин.
В данном описании изобретения термин "белок-рецептор цАМФ (CRP)" представляет собой наиболее хорошо известный регулятор транскрипции у Е. coli, и CRP также называют "двойным регулятором", поскольку сам CRP обладает функциями как активатора, так и ингибитора. CRP обычно связывается с симметричной последовательностью ДНК, имеющей 22 основания в направлении 5' от структурного гена, что индуцирует сгибание ДНК, и CRP действует как активатор, обеспечивая возможность взаимодействия первого активного сайта на С-конце и второго активного сайта на N-конце с РНК-полимеразой, отвечающей за транскрипцию, и он действует как ингибитор, занимая данное положение и предотвращая связывание активного белка с активным сайтом или связываясь с активным белком с превращением его структуры в структуру, которая не связывается с активным сайтом. Белок-рецептор цАМФ представляет собой белок-рецептор цАМФ, кодируемый геном стр.
Термин "белок-рецептор цАМФ (белок-рецептор циклического АМФ, CRP)" по данному изобретению может быть использован взаимозаменяемо с термином "белок-активатор катаболизма (САР)", "белок CRP", "белок САР" и т.д.
В данном описании изобретения последовательность CRP может быть получена из известной базы данных GenBank NCBI. Например, CRP может представлять собой CRP, происходящий из рода Escherichia (Escherichia sp.), и более конкретно, полипептид/белок, включающий аминокислотную последовательность, представленную в SEQ ID NO: 1, но не ограничивается ими. Кроме того, он может включать последовательность, обладающую такой же активностью, как указанная выше аминокислотная последовательность, без ограничения. Кроме того, он может включать аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, обладающую гомологией или идентичностью с ней 80% или более, без ограничения ими. В частности, аминокислотная последовательность может включать аминокислотную последовательность SEQ ID NO: 1 и аминокислотную последовательность, обладающую гомологией или идентичностью по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более с SEQ ID NO: 1. Кроме того, очевидно, что белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или добавлением части последовательности, также входит в объем данного изобретения, при условии, что аминокислотная последовательность имеет указанную выше гомологию или идентичность и демонстрирует эффективность, соответствующую эффективности указанного выше белка.
В данном описании изобретения термин "вариант" относится к полипептиду, в котором одна или более аминокислот отличаются от приведенной последовательности консервативными заменами и/или модификациями, но он сохраняет функции или свойства этого белка. Варианты полипептидов отличаются от установленной последовательности заменой, делецией или добавлением нескольких аминокислот. Такие варианты обычно могут быть идентифицированы путем модификации одной из вышеупомянутых полипептидных последовательностей и оценки свойств модифицированного полипептида. Другими словами, способность варианта может быть повышена, оставлена без изменений или ослаблена по сравнению с нативным белком. Такие варианты обычно могут быть идентифицированы путем модификации одной из вышеупомянутых полипептидных последовательностей и оценки реакционной способности модифицированного полипептида. Кроме того, некоторые варианты могут включать варианты, в которых были удалены один или более участков, таких как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, в которых был удален участок с N- и/или С-конца зрелого белка.
В данном описании изобретения термин "консервативная замена" означает замену одной аминокислоты другой аминокислотой, имеющей аналогичные структурные и/или химические свойства. Например, вариант может иметь одну или более консервативных замен, сохраняя при этом одну или более биологических активностей. Такие аминокислотные замены, как правило, можно осуществлять на основании сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; отрицательно заряженные (кислые) аминокислоты включают глутаминовую кислоту и аспарагиновую кислоту; ароматические аминокислоты включают фенилаланин, триптофан и тирозин; и гидрофобные аминокислоты включают аланин, валин, изолейцин, лейцин, метионин, фенилаланин, пролин, глицин и триптофан.
Кроме того, варианты могут включать делецию или добавление аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью на N-конце белка, которая котрансляционно или посттрансляционно обеспечивает транспорт белка. Полипептид может также быть конъюгирован с другой последовательностью или линкером для обнаружения, очистки или синтеза полипептида.
В данном описании изобретения термин "вариант белка-рецептора цАМФ" представляет собой вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности полипептида, обладающего активностью белка-рецептора цАМФ, где аминокислотные замены включают замену аминокислоты в положении 196 от N-конца на другую аминокислоту. В частности, вариант может включать вариант белка, в котором аминокислота в положении 196 в аминокислотной последовательности полипептида, обладающего активностью белка-рецептора цАМФ, заменена на другую аминокислоту. Например, вариант белка может включать вариант белка, в котором вариация затрагивает положение 196 от N-конца аминокислотной последовательности SEQ ID NO: 1. Более конкретно, вариант белка может представлять собой белок, в котором аминокислота в положении 196 в аминокислотной последовательности SEQ ID NO: 1 заменена на другую аминокислоту. Термин "другая аминокислота" не ограничен, при условии, что другая аминокислота представляет собой аминокислоту отличную от L-лейцина, который является аминокислотой в положении 196. В частности, вариант может представлять собой белок, в котором аминокислота в положении 196 в аминокислотной последовательности SEQ ID NO: 1 заменена на гидрофильную аминокислоту. Гидрофильная аминокислота может представлять собой одну из L-глутамина, L-серина, L-треонина, L-тирозина, L-цистеина и L-аспарагина. Более конкретно, вариант может представлять собой белок, в котором аминокислота в положении 196 в аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин, но не ограничен им.
Кроме того, вариант означает вариант, имеющий аминокислотную вариацию в положении 196 от N-конца в вышеописанной аминокислотной последовательности SEQ ID NO: 1 и/или в аминокислотной последовательности, обладающей по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более гомологией или идентичностью с SEQ ID NO: 1.
В данном описании изобретения термин "вариант белка-рецептора цАМФ" может использоваться взаимозаменяемо с терминами "вариант белка CRP", "вариант CRP", "вариант белка-рецептора цАМФ", "вариант белка САР", "вариант САР", "вариант белка-активатора катаболизма", "белок-вариант активатора катаболизма" и т.д.
Применительно к задачам данного изобретения, микроорганизм, включающий вариант белка-рецептора цАМФ, характеризуется высоким продуцированием L-аминокислот по сравнению с микроорганизмом, не включающим вариант белка-рецептора цАМФ. Вариант CRP характеризуется наличием регулирующей ген активности для увеличения продуцирования L-аминокислот по сравнению с нативным белком-рецептором цАМФ дикого типа или инвариантным белком-рецептором цАМФ. Это имеет значение в том плане, что у микроорганизма с введенным вариантом CRP по данному изобретению может быть увеличено продуцирование L-аминокислот. В частности, L-аминокислота может представлять собой L-треонин или L-триптофан. Однако может быть включена любая L-аминокислота, без ограничения, при условии, что она может быть получена путем введения или включения варианта белка-рецептора цАМФ.
Белок-рецептор цАМФ может представлять собой, например, вариант, включающий аминокислотную последовательность, в которой аминокислота в положении 196 аминокислотной последовательности, представленной в SEQ ID NO: 1, заменена на другую аминокислоту, где вариант состоит из SEQ ID NO: 3. Вариант, в котором аминокислота в положении 196 в аминокислотной последовательности, представленной в SEQ ID NO: 1, заменена на глутамин, может состоять из SEQ ID NO: 3, но не ограничен таковым. Кроме того, вариант CRP может включать аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, обладающую 80% или более гомологией или идентичностью с ней, без ограничения ими. В частности, вариант CRP по данному изобретению может включать белок, имеющий SEQ ID NO: 3, и белок, обладающий гомологией или идентичностью с ним по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более. Кроме того, очевидно, что белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или добавлением части последовательности, дополнительно к положению 196 аминокислотной последовательности, также может входить в объем данного изобретения, при условии, что аминокислотная последовательность обладает указанной выше гомологией или идентичностью и демонстрирует эффективность, соответствующую эффективности указанного выше белка.
Иными словами, даже если в данном описании изобретения описан "белок, имеющий аминокислотную последовательность конкретной SEQ ID NO", очевидно, что белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой, консервативной заменой или добавлением ее части, можно использовать в данном изобретении, при условии, что он обладает активностью идентичной или соответствующей активности белка, состоящего из аминокислотной последовательности соответствующей SEQ ID NO. Например, при условии, что белок обладает активностью идентичной или соответствующей активности варианта белка, не исключено добавление в него последовательности, не меняющей функцию белка, до или после этой аминокислотной последовательности, мутаций естественного происхождения, молчащих мутаций или консервативных замен. Очевидно, что даже если белок имеет такое добавление или мутацию последовательности, он входит в объем данного изобретения.
В данном описании изобретения термины "гомология" или "идентичность" означают степень соответствия двух заданных аминокислотных последовательностей или нуклеотидных последовательностей и могут быть выражены в процентах.
Термины "гомология" и "идентичность" часто могут использоваться взаимозаменяемо.
Гомологию последовательности или идентичность консервативного полинуклеотида или полипептида можно определить при помощи стандартных алгоритмов выравнивания и можно использовать со значениями штрафов за делеции, установленными в используемой программе по умолчанию. По существу, гомологичные или идентичные последовательности могут гибридизоваться в условиях умеренной или высокой жесткости, так что может гибридизоваться вся длина последовательности или по меньшей мере примерно 50%, 60%, 70%, 80% или 90% всей длины. Также рассматриваются полинуклеотиды, которые содержат вырожденные кодоны на месте гибридизируемых кодонов.
Установить, обладают или нет две любые полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, можно, например, при помощи известных компьютерных алгоритмов, таких как программа FASTA, например, с использованием параметров по умолчанию, как описано в Pearson et al. (1988) [Proc. Natl. Acad. Sci. USA 85]: 2444, или с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), используемого в программе Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или позже) (включая пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, and [CARILLO ETA/.](1988) SIAM J Applied Math 48: 1073). Например, для определения гомологии, сходства или идентичности можно использовать BLAST базы данных Национального Центра Биотехнологической Информации или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определить, например, путем сравнения информации о последовательностях с использованием компьютерной программы GAP, такой как в Needleman et al. (1970), J Mol Biol. 48: 443, как описано в Smith and Waterman, Adv. Appl. Math (1981) 2:482. В нескольких словах, программа GAP определяет сходство как количество выровненных символов (т.е. нуклеотидов или аминокислот), которые являются одинаковыми, деленное на общее количество символов в более короткой из этих двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) унарную матрицу сравнения (содержащую значение 1 для идентичных и 0 для неидентичных символов) и взвешенную матрицу сравнения, предложенную в Gribskov et al (1986) Nucl. Acids Res. 14: 6745, как раскрыто в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или подстановочную матрицу EDNAFULL (EMBOSS версия NCBI NUC4.4)); (2) штраф 3,0 для каждого гэпа и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф за открытие гэпа 10, штраф за продолжение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы. Таким образом, при использовании в данном описании изобретения, термины "гомология" или "идентичность" описывают соответствие между последовательностями.
В другом аспекте данного изобретения предложен полинуклеотид, кодирующий вариант CRP, или вектор, включающий этот полинуклеотид.
В данном описании изобретения термин "полинуклеотид" относится к цепи ДНК или РНК, имеющей предварительно определенную длину или более, представляющей собой длинноцепочечный полимер из нуклеотидов, образованный в результате соединения нуклеотидных мономеров ковалентными связями. Более конкретно, полинуклеотид относится к полинуклеотидному фрагменту, кодирующему вариант белка.
Полинуклеотид, кодирующий вариант CRP по данному изобретению, может включать любую полинуклеотидную последовательность без ограничения, при условии, что она представляет собой полинуклеотидную последовательность, кодирующую вариант белка-рецептора цАМФ по данному изобретению. Полинуклеотид, кодирующий вариант CRP, может включать любую последовательность без ограничения, при условии, что он представляет собой последовательность, кодирующую вариант белка, в котором аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена другой аминокислотой. В частности, полинуклеотид может представлять собой полинуклеотидную последовательность, кодирующую вариант, в котором аминокислота в положении 196 в аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин. Например, полинуклеотид, кодирующий вариант CRP по данному изобретению, может представлять собой полинуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 3, но не ограничен ею. Более конкретно, полинуклеотид может состоять из полинуклеотидной последовательности SEQ ID NO: 4, но не ограничен им. В полинуклеотиде могут быть осуществлены различные модификации в кодирующей области, при условии, что они не меняют аминокислотную последовательность белка, вследствие вырожденности кодонов или с учетом кодонов, предпочитаемых организмом, в котором будет экспрессироваться белок. Таким образом, очевидно, что вследствие вырожденности кодонов, изобретение также может охватывать полинуклеотид, который может транслироваться в полипептид, состоящий из аминокислотной последовательности SEQ ID NO: 3, или полипептид, гомологичный или идентичный ему.
Кроме того, в объем изобретения, без ограничения, также может входить зонд, который может быть получен из известной нуклеотидной последовательности, например последовательности, которая в жестких условиях гибридизуется с комплементарной последовательностью по всей или части нуклеотидной последовательности, кодирующей вариант CRP, где аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена на другую аминокислоту.
Термин "жесткие условия" означает условия, в которых обеспечивается специфическая гибридизация полинуклеотидов. Такие условия подробно описаны в литературе (например, в J. Sambrook et al., см. выше). Например, жесткие условия могут включать условия, в которых гены, обладающие высокой гомологией или идентичностью, гомологией или идентичностью 80% или более, 85% или более, в частности 90% или более, более конкретно 95% или более, еще более конкретно 97% или более, особенно 99% или более, гибридизуются друг с другом, а гены, обладающие гомологией или идентичностью ниже указанных, не гибридизуются друг с другом, или стандартные условия отмывки при гибридизации по Саузерну, т.е., в частности, однократную, двукратную или трехкратную отмывку с концентрацией солей и при температуре, соответствующих 60°С, 1X SSC (раствор хлорида и цитрата натрия), 0,1% SDS (додецилсульфат натрия), в частности 60°С, 0,1Х SSC, 0,1% SDS, и более конкретно 68°С, 0,1XSSC, 0,1% SDS.
Хотя возможны несовпадения между нуклеотидами из-за строгости гибридизации, требуется, чтобы две нуклеиновые кислоты имели комплементарную последовательность. Термин "комплементарность" используют для описания соответствия между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в случае ДНК, аденозин комплементарен тимину, и цитозин комплементарен гуанину. Соответственно, данное изобретение может включать не только по существу похожие последовательности нуклеиновых кислот, но также и выделенные фрагменты нуклеиновых кислот, комплементарные полной последовательности.
В частности, полинуклеотид, обладающий гомологией или идентичностью, можно обнаружить с использованием условий гибридизации, включающих гибридизацию при значении Tm 55°С и вышеописанные условия. Кроме того, значение Tm может быть 60°С, 63°С или 65°С, без ограничения ими, и может надлежащим образом контролироваться специалистом в области техники в соответствии с задачами.
Подходящая жесткость гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов и возможные значения хорошо известны в области техники (см. Sambrooket al., выше, 9.50-9.51, 11.7-11.8).
В данном описании изобретения термин "вектор" относится к ДНК-конструкции, которая включает нуклеотидную последовательность полинуклеотида, кодирующего целевой вариант белка, функционально связанную с соответствующей регуляторной последовательностью для обеспечения экспрессии целевого варианта белка в соответствующей клетке-хозяине. Регуляторная последовательность может включать промотор, способный инициировать транскрипцию, любую последовательность оператора для регуляции такой транскрипции, последовательность, кодирующую соответствующий мРНК-связывающий домен рибосомы, и последовательность, регулирующую окончание транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектором, он может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в сам геном.
Вектор, используемый в данном изобретении, не ограничен конкретными векторами, при условии, что он способен реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в области техники. Примеры обычно используемых векторов могут включать природную или рекомбинантную плазмиду, космиду, вирус и бактериофаг. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д. В качестве плазмидного вектора использовать вектор типов pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.д. В частности, можно использовать вектор pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д.
Например, полинуклеотид, кодирующий целевой вариант белка в хромосоме, может быть замещен мутированным полинуклеотидом с использованием вектора для внутриклеточного встраивания в хромосому. Встраивание полинуклеотида в хромосому можно осуществлять любым известным в области техники способом, например посредством гомологичной рекомбинации, без ограничения им. Дополнительно можно включать селективный маркер для подтверждения хромосомного встраивания. Селективный маркер предназначен для отбора клеток, трансформированных вектором, т.е. для подтверждения встраивания необходимого полинуклеотида, и селективный маркер может включать маркеры, придающие селектируемый фенотип, такой как резистентность к лекарственным средствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия белков с модифицированной поверхностью. Поскольку только клетки, экспрессирующие селективный маркер, могут выжить или проявить разные фенотипы в среде, обработанной селективным агентом, трансформированные клетки могут быть отобраны. Еще в одном аспекте данного изобретения предложен микроорганизм, продуцирующий L-аминокислоту, включающий вариант белка или полинуклеотид, кодирующий этот вариант белка. В частности, микроорганизм, включающий вариант белка и/или полинуклеотид, кодирующий вариант белка, может представлять собой микроорганизм, полученный путем трансформации вектором, включающим полинуклеотид, кодирующий вариант белка, но без ограничения им.
В данном описании изобретения термин "трансформация" означает введение вектора, который включает полинуклеотид, кодирующий целевой белок, в клетку-хозяина, так что белок, кодируемый этим полинуклеотидом, экспрессируется в клетке-хозяине. Пока трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, он может быть интегрирован и помещен в хромосому клетки-хозяина, или он может находиться за пределами хромосомы, или безотносительно к ней. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме, при условии, что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую в себя все элементы, необходимые для ее автономной экспрессии. Как правило, экспрессионная кассета включает промотор, функционально связанный с полинуклеотидом, сигналы окончания транскрипции, сайты связывания рибосомы и сигналы окончания трансляции. Экспрессионная кассета может находиться в форме самореплицирующегося экспрессионного вектора. Кроме того, полинуклеотид как таковой может быть введен в клетку-хозяина и функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине, но без ограничения им.
В данном описании изобретения термин "функционально связан" означает функциональную связь между полинуклеотидной последовательностью, кодирующей целевой вариант белка по данному изобретению, и промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотидной последовательности.
Еще в одном аспекте данного изобретения предложен микроорганизм рода Escherichia (Escherichia sp.), включающий вариант белка-рецептора цАМФ.
В данном описании изобретения термин "микроорганизм, включающий вариант CRP" может относиться к рекомбинантному микроорганизму для экспрессии варианта CRP по данному изобретению. Например, этот микроорганизм относится к клетке-хозяину или к микроорганизму, который способен экспрессировать вариант в результате включения полинуклеотида, кодирующего вариант CRP, или в результате трансформации вектором, включающим полинуклеотид, кодирующий вариант CRP. Применительно к задачам данного изобретения, микроорганизм представляет собой микроорганизм, экспрессирующий вариант белка-рецептора цАМФ, включающий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, и микроорганизм может представлять собой микроорганизм, экспрессирующий вариант белка, обладающий активностью белка-рецептора цАМФ, где аминокислотная замена представляет собой замену аминокислоты в положении 196 от N-конца на глутамин, но не ограничен ею.
Микроорганизм, включающий вариант CRP, может быть любым микроорганизмом при условии, что он включает вариант CRP для экспрессии L-аминокислоты, например L-треонина или L-триптофана, не ограничен ими. Например, микроорганизм, включающий вариант CRP, может представлять собой рекомбинантный микроорганизм с повышенным продуцированием L-аминокислот, который получен путем экспрессии варианта CRP в природном микроорганизме дикого типа или в микроорганизме, продуцирующем L-аминокислоту. Рекомбинантный микроорганизм с повышенным продуцированием L-аминокислот может представлять собой микроорганизм, у которого продуцирование L-аминокислот повышено по сравнению с природным микроорганизмом дикого типа или немодифицированным микроорганизмом, где L-аминокислота может представлять собой L-треонин или L-триптофан, без ограничения ими.
В данном описании изобретения термин "микроорганизм, продуцирующий L-аминокислоту" включает микроорганизм дикого типа или микроорганизм, в котором происходит естественная или искусственная генетическая модификация, и он может представлять собой микроорганизм, в котором конкретный механизм ослаблен или усилен в результате встраивания чужеродного гена или в результате усиления или инактивации активности эндогенного гена, где генетическая вариация происходит или активность повышена для продуцирования целевой L-аминокислоты. Применительно к задачам данного изобретения микроорганизм, продуцирующий L-аминокислоту, может включать вариант белка для повышенного продуцирования целевой L-аминокислоты. В частности, в данном изобретении микроорганизм, продуцирующий L-аминокислоту, или микроорганизм, обладающий свойством продуцирования L-аминокислоты, может представлять собой микроорганизм, в котором часть генов, вовлеченных в путь биосинтеза L-аминокислоты, усилена или ослаблена, или часть генов, вовлеченных в путь деградации L-аминокислоты, усилена или ослаблена.
Термин "немодифицированный микроорганизм" относится к природному штамму как таковому или к микроорганизму, который не включает вариант CRP, или к микроорганизму, который не трансформирован вектором, включающим полинуклеотид, кодирующий вариант CRP. "Микроорганизм" может включать любой прокариотический микроорганизм и эукариотический микроорганизм при условии, что он способен продуцировать L-аминокислоту. Например, микроорганизм может включать микроорганизмы рода Escherichia, рода Erwinia, рода Serratia, рода Providencia, рода Cory neb acterium и рода Brevibacterium. В частности, микроорганизм может представлять собой микроорганизм рода Escherichia, и более конкретной, coli, но не ограничен ими.
В еще одном аспекте данного изобретения предложен способ получения L-аминокислоты, включающий культивирование микроорганизма рода Escherichia в среде, где микроорганизм продуцирует L-аминокислоту и включает вариант белка-рецептора цАМФ.
Термины "вариант белка-рецептора цАМФ" и "L-аминокислота" являются такими, как описано выше.
В данном способе культивирование микроорганизма может быть осуществлено посредством известного периодического культивирования, непрерывного культивирования, культивирования с подпиткой и т.д., но не ограничено ими. При этом условия культивирования не ограничены какими-либо конкретными, но можно поддерживать оптимальное значение рН (например, рН 5-9, в частности рН 6-8, и более конкретно рН 6,8), используя щелочное соединение (например, гидроксид натрия, гидроксид калия или аммоний) или кислое соединение (например, фосфорную кислоту или серную кислоту). Кроме того, можно поддерживать аэробные условия путем подачи кислорода или кислородсодержащей газовой смеси в культуру клеток. Температуру культивирования можно поддерживать от 20°С до 45°С, и в частности от 25°С до 40°С. Культивирование можно осуществлять в течение от примерно 10 часов до примерно 160 часов, без конкретного ограничения ими. Продуцируемая посредством описанного выше культивирования L-аминокислота может секретироваться в культуральную среду или может оставаться внутри клеток.
Кроме того, используемая культуральная среда может содержать, в качестве источников углерода, сахара и углеводы (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масло и жир (например, соевое масло, масло семян подсолнечника, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например, глицерин и этанол) и органические кислоты (например, уксусную кислоту) по отдельности или в комбинации, но без ограничения ими. В качестве источников азота можно использовать азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) по отдельности или в комбинации, но без ограничения ими. В качестве источников фосфора можно использовать двузамещенный фосфат калия, однозамещенный фосфат калия и соответствующие натриевые соли по отдельности или в комбинации, но без ограничения ими. Кроме того, среда также может включать незаменимые стимулирующие рост вещества, включающие соли других металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Способ может дополнительно включать сбор L-аминокислоты из микроорганизма или из среды.
Способ сбора L-аминокислоты, полученной при культивировании согласно данному изобретению, может обеспечить сбор целевой L-аминокислоты из культуральной жидкости с использованием подходящего способа, известного в данной области техники, в соответствии со способом культивирования. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию, ВЭЖХ и т.д., и можно собирать целевую L-аминокислоту из среды или микроорганизма с использованием подходящего способа, известного в данной области техники.
Кроме того, сбор может включать процесс очистки и может быть осуществлен с использованием подходящего способа, известного в данной области техники. Таким образом, L-аминокислота для сбора может представлять собой очищенную форму или ферментативный бульон микроорганизма, содержащую(ий) L-аминокислоту (Introduction to Biotechnology and Genetic Engineering, A. J. Nair., 2008).
В еще одном аспекте данного изобретения предложено применение варианта белка-рецептора цАМФ в получении L-аминокислоты, где вариант белка-рецептора цАМФ включает одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1.
В еще одном аспекте данного изобретения предложено применение микроорганизма рода Escherichia в получении L-аминокислоты, где микроорганизм рода Escherichia включает вариант рецептора цАМФ.
Термин "вариант белка-рецептора цАМФ" и "L-аминокислота" являются такими, как описано выше.
ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже данное изобретение будет описано более подробно на примерах. Однако специалистам в данной области техники очевидно, что данные примеры приведены исключительно в иллюстративных целях и не ограничивают объем данного изобретения.
Пример 1. Получение рекомбинантного вектора рСС1ВАС-crp 1-1.
Получение фрагмента гена crp
Для получения примерно 0,96 т.п.н. фрагмента ДНК последовательности SEQ ID NO: 5, включающего ген crp и область регуляции экспрессии, экстрагировали геномную ДНК (гДНК).Е.coli W3110 дикого типа с использованием системы Genomic-tip компании Qiagen и проводили полимеразную цепную реакцию (ПНР) с использованием гДНК в качестве матрицы и набора PCR HL premix kit (производства BIONEER Co., то же применимо ниже). ПЦР для амплификации фрагмента гена crp проводили с использованием праймеров с последовательностями SEQ ID NO: 6 и 7 в течение 27 циклов, состоящих из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 2 мин.
Продукт ПЦР расщепляли с помощью EcoR I, проводили электрофорез в 0,8% агарозном геле и элюировали с получением фрагмента ДНК 0,96 т.п.н. (далее обозначаемого "фрагмент crp").

1-2. Получение рекомбинантного вектора рСС1ВАС-crp
Вектор Copycontrol pCC1BAC (EPICENTRE, USA) обрабатывали EcoR I, проводили электрофорез в 0,8% агарозном геле и элюировали с получением продукта, который затем лигировали с фрагментом crp, полученным в Примере 1-1, таким образом получая плазмиду рСС1ВАС-crp.
Пример 2. Получение библиотеки вариантов рекомбинантного вектора pCC1BACrp
2-1. Получение фрагмента мутантного crp с помощью ПЦР пониженной точности
Выполняли ПЦР с использованием геномной ДНК Е. coli W3110 дикого типа в качестве матрицы и Diversify PCR random mutagenesis kit (набор для увеличения разнообразия случайного мутагенеза при ПЦР) от Clonetech (кат. номер K1830-1, Таблица III, реакции мутагенеза 4). Более конкретно, ПЦР проводили с использованием праймеров с последовательностями SEQ ID NO: 6 и 7, которые использовали в Примере 1-1, в течение 27 циклов, состоящих из денатурации при 94°С в течение 30 сек и элонгации при 68°С в течение 1 мин.
Продукт ПЦР расщепляли с помощью EcoR I, проводили электрофорез в 0,8% агарозном геле и элюировали с получением мутантного фрагмента crp 0,96 т.п.н. (ниже обозначаемого как "фрагмент crpm").
2-2. Получение библиотеки вариантов рекомбинантного вектора рСС1 ВАС-crp
Вектор pCC1BAC обрабатывали рестрикционным ферментом EcoR I, а затем щелочной фосфатазой (NEB). Полученный вектор лигировали с фрагментом crpm, полученным в Примере 2-1, и продуктом лигирования трансформировали электрокомпетентные клетки Е. coli TransforMax EPI300 (EPICENTRE, USA) посредством электрофореза. Трансформированный штамм культивировали на твердой среде LB, содержащей хлорамфеникол (15 мкг/мл), для отбора колоний. Полученные таким образом колонии собирали и осуществляли получение плазмид, таким образом получая библиотеку рСС1ВАС-crpm
Пример 3. Введение библиотеки вариантов crp в штамм, продуцирующий треонин, и отбор штамма с улучшенным ростом
3-1. Введение библиотеки рСС1ВАС-crpm в штамм, продуцирующий треонин Библиотекой рСС1ВАС-crpm, полученной в Примере 2, посредством электропорации трансформировали электрокомпетентные клетки KCCM10541, представляющие собой треонин-продуцирующий микроорганизм. Е. coli KCCM 10541 (патент Кореи №10-0576342), использованный в данном Примере, представляет собой Е. coli, полученный путем инактивации гена galR у L-треонин-продуцирующих Е. coli KFCC10718 (патент Кореи №10-0058286).
В качестве контрольной группы для pCC1BAC-crpm микроорганизма, KCCM10541 трансформировали pCClBAC-стр таким же способом, как описано выше, с получением KCCM10541/рСС1ВАС-crp (WT).
3-2. Сравнение скорости роста рекомбинантного микроорганизма
Минимальную среду М9, содержащую 1% глюкозы и 0,2 г/л дрожжевого экстракта, распределяли в микропланшет с глубокими лунками и затем высевали в них трансформированный и контрольный штаммы, полученные в Примере 3-1, соответственно. Штаммы культивировали в микроинкубаторе-шейкере, поддерживающем постоянную температуру (TAITEC, Japan), в условиях 37°С и 200 об/мин посредством метода HTS (высокопроизводительный скрининг) в течение 20 часов, и отбирали штаммы с улучшенным ростом. Из них окончательно отобрали один тип штамма (Таблица 2).
Штамм KCCM10541 с введенным геном crp дикого типа демонстрировал небольшое увеличение показателя OD (оптическая плотность) благодаря дополнительному введению crp, тогда как трансформант с улучшенным ростом демонстрировал высокое значение OD при таком же времени культивирования по сравнению со штаммом с введенным crp дикого типа. Затем для выбранного варианта crp выполняли мини-получение плазмиды с последующим секвенированием. Результаты обобщены в Таблице 2.

3-3. Сравнение титра треонина у рекомбинантного микроорганизма
Для определения титра треонина у рекомбинантного микроорганизма, выбранного в Примере 3-2, рекомбинантный микроорганизм культивировали в среде для титрования треонина, приготовленной с составом, приведенным в Таблице 3 ниже, для оценки улучшения продуктивности по L-треонину.
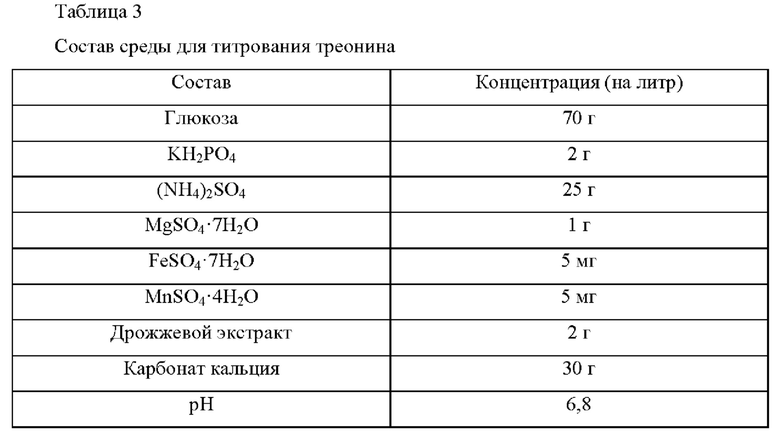
Более конкретно, каждую из платиновой петли Е. coli KCCM10541/рСС1ВАС-crp (WT) и Е. coli KCCM10541/рСС1ВАС-crpTM9, культивированных в течение ночи на твердой среде LB в инкубаторе при 33°С, инокулировали в 25 мл среды для титрования, приведенной в Таблице 3, соответственно и затем культивировали в инкубаторе при 33°С и 200 об/мин в течение 48 часов для сравнения скоростей потребления сахара и концентраций треонина.
В результате, как показано в Таблице 4 ниже, штамм KCCM 10541/рСС1ВАС-crp (WT) в качестве контрольной группы демонстрировал потребление сахара 26,1 г/л в течение 24 ч, тогда как мутантный штамм с введенным crpTM9 демонстрировал улучшение скорости потребления сахара по сравнению с материнским штаммом и штаммом с введенным crp дикого типа примерно на 16% и 11%, соответственно.
Кроме того, при культивировании в течение 48 часов штамм с введенным crp дикого типа продуцировал 29,0 г/л L-треонина, тогда как продуцирование L-треонина мутантным штаммом, полученным выше, повышалось до 30,8 г/л даже при увеличении скорости культивирования, демонстрируя примерно 7% и 6% улучшение концентрации по сравнению с родительским штаммом и со штаммом с введенным crp дикого типа соответственно.
Поскольку введение варианта crp увеличивало выход и потребление штаммом сахара, это, по-видимому, является признаком хорошего варианта, который может вносить существенный вклад в улучшение эффективности продуцирования во время ферментации.

Пример 4. Введение варианта pCC1BAC-crpTM3 в триптофан-продуцирующий штамм
4-1. Введение рСС1ВАС-crpTM3 в штамм, отобранный при скрининге
Посредством электропорации электрокомпетентные клетки триптофан-продуцирующего штамма KCCM11166P трансформировали рСС1ВАС-crpTM9, полученным в Примере 3. KCCM11166P, использованный в данном Примере, представляет собой L-триптофан-продуцирующий Е. coli, в котором ген tehB делетирован и активность NAD-киназы усилена (патент Кореи №10-1261147).
В качестве контрольной группы для микроорганизма с введенным pCC1BAC-crpTM9, KCCM11166P трансформировали при помощи рСС1ВАС-crp (WT) таким же образом, как описано выше, с получением KCCM11166P/рСС1ВАС-crp (WT).
4-2. Сравнение скорости роста рекомбинантного микроорганизма
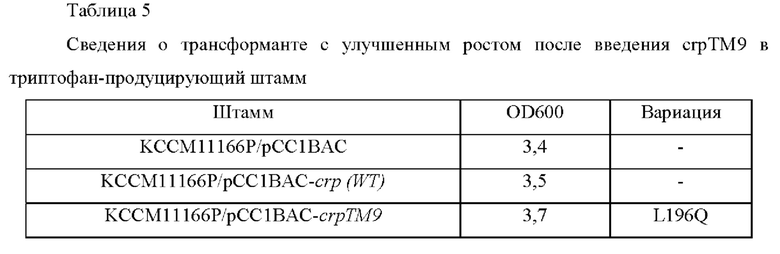
Минимальную среду М9, содержащую 1% глюкозы и 0,2 г/л дрожжевого экстракта распределяли в микропланшет с глубокими лунками и затем высевали в них трансформированный и контрольный штаммы, полученные согласно Примеру 4-1, соответственно. Штаммы культивировали в микроинкубаторе-шейкере, поддерживающем постоянную температуру (TAITEC, Japan), в условиях 37°С и 200 об/мин посредством метода HTS (высокопроизводительный скрининг) в течение 16 часов для подтверждения улучшения роста трансформанта KCCM11166P/рСС1ВАС-crpTM9 (Таблица 5).
Штамм KCCM11166P с введенным геном crp дикого типа демонстрировал эквивалентный уровень OD, благодаря дополнительному введению crp, при таком же времени культивирования, тогда как трансформант с улучшенным ростом демонстрировал высокое значение OD по сравнению со штаммом с введенным crp дикого типа.
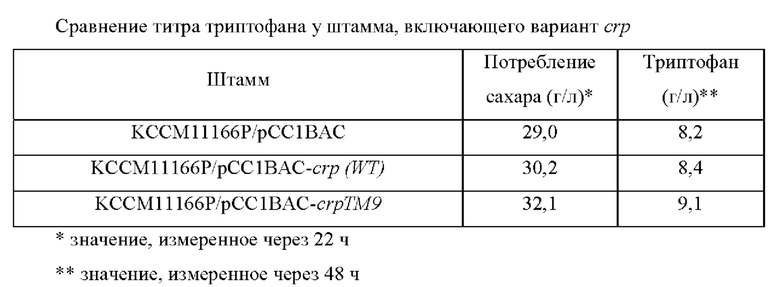
4-3. Сравнение титра триптофана у рекомбинантного микроорганизма
Для определения титра триптофана у рекомбинантного микроорганизма, полученного в Примере 4-2, рекомбинантный микроорганизм культивировали в среде для титрования триптофана, приготовленной с составом, приведенным в Таблице 6, для оценки улучшения продуктивности L-триптофана.

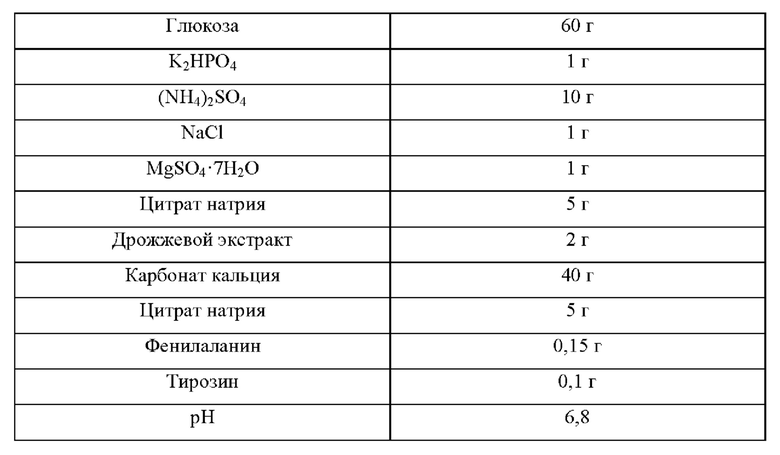
Более конкретно, каждую из платиновой петли Е. coli KCCM10541/рСС1ВАС-crp (WT) и Е. coli KCCM10541/рСС1ВАС-crpTM9, культивированных в течение ночи на твердой среде LB в инкубаторе при 37°С, инокулировали в 25 мл среды для титрования, приведенной в Таблице 6, и затем культивировали в инкубаторе при 37°С и 200 об/мин в течение 48 часов для сравнения скорости потребления сахара и концентраций триптофана.
В результате, как показано в Таблице 7 ниже, штамм KCCM11166P/рСС1ВАС-crp (WT), в качестве контрольной группы, демонстрировал потребление сахара 30,2 г/л в течение 22 ч, тогда как мутантный штамм с введенным crpTM9 демонстрировал примерно 7% и 6% улучшение скорости потребления сахара по сравнению с родительским штаммом и со штаммом с введенным crp дикого типа соответственно.
При культивировании в течение 48 часов штамм с введенным crp дикого типа продуцировал 8,4 г/л L-триптофана, тогда как продуцирование L-триптофана мутантным штаммом, полученным выше, увеличивалось до 9,1 г/л даже при увеличении скорости культивирования, демонстрируя улучшение концентрации примерно на 8% и 10% по сравнению с родительским штаммом и штаммом с введенным crp дикого типа соответственно.
Поскольку введение варианта crp увеличивало потребление сахара штаммом и выход, это, по-видимому, является признаком хорошего варианта, который может вносить существенный вклад в улучшение эффективности продуцирования во время ферментации.
Пример 5. Введение эндогенного вектора с эффективным вариантом crp в Е. coli дикого типа
5-1. Введение эффективного варианта pCC1BAC-crp в треонин-продуцирующий штамм, происходящий из штамма дикого типа
Чтобы установить, демонстрирует ли вектор, включающий вариант crp, отобранный при скрининге в Примере 3, эквивалентные эффекты в штамме дикого типа, штамм, происходящий из штамма дикого типа, способный продуцировать треонин, трансформировали посредством электропорации вектором рСС1ВАС-crp (WT) или рСС1ВАС-crpTM9 соответственно. Кроме того, получали штамм с введенным рСС1ВАС-crp (WT) в качестве контрольной группы.
Штамм, происходящий из штамма дикого типа, способный продуцировать треонин, использованный в данном Примере, представляет собой W3110::PcysK-ppc/pACYC184-thrABC. W3110::PcysK-ppc/pACYC184-thrABC представляет собой штамм, в котором нативный промотор гена ррс, кодирующего в хромосоме фосфоенолпируваткарбоксилазу, заменен промотором гена cysK, и гена оперона биосинтеза треонина введен в форме вектора для увеличения количества копий, таким образом повышая продуцирование треонина. Более подробно, штамм W3110::PcycK-ppc получали, используя pUCpcycKmloxP таким же образом, как описано в патенте Кореи №10-0966324, и трансформировали штамм с помощью pACYC184-thrABC (патент Кореи №10-1865998) посредством электропорации.
Полученные штаммы культивировали в среде для анализа треонина, приготовленной с составом, представленным в Таблице 8 ниже, и сравнивали их скорости роста и продуцирование L-треонина.
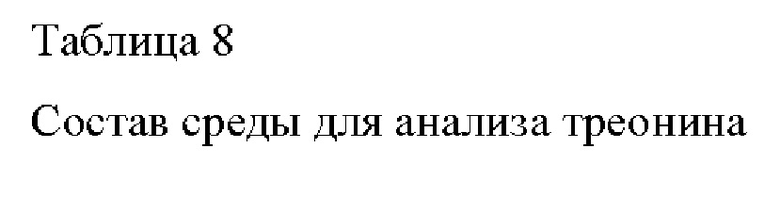
Более подробно, каждую из платиновой петли W3110 и соответствующих штаммов, культивированных в течение ночи на твердой среде LB в инкубаторе при 33°С, инокулировали в 25 мл среды для титрования, приведенной в Таблице 8, соответственно, и затем культивировали в инкубаторе при 33°С и 200 об/мин в течение 48 ч. Результаты представлены в Таблице 9, приведенной ниже. Как свидетельствуют представленные ниже результаты, выбранный вариант белка по настоящему изобретению также способен эффективно продуцировать треонин с высоким выходом в штамме дикого типа.

5-2. Введение эффективного варианта рСС1ВАС-crp в триптофан-продуцирующий штамм, происходящий из штамма дикого типа
Чтобы установить, оказывает ли вектор, включающий вариант crp, отобранный при скрининге в Примере 4, эквивалентные эффекты в штамме дикого типа, штамм, происходящий из штамма дикого типа, способный продуцировать триптофан, трансформировали вектором рСС1ВАС-crp (WT) или рСС1ВАС-crpTM9, соответственно.
Штамм, происходящий из штамма дикого типа, способный продуцировать триптофан, использованный в данном Примере, представляет собой W3110 trpA2/pCL-Dtrp att-trpEDCBA. W3110 trpA2/pCL-rJtrp att-trpEDCBA представляет собой штамм с введенным вектором, в котором отключен регуляторный механизм регуляторной области триптофанового оперона и экспрессия триптофанового оперона усилена для сверхэкспрессии триптофана (патент Кореи №10-1532129). Штаммы с введенным вектором культивировали в среде для анализа триптофана, приготовленной с составом, которой приведен в Таблице 10 ниже, и сравнивали их продуктивности по L-триптофану
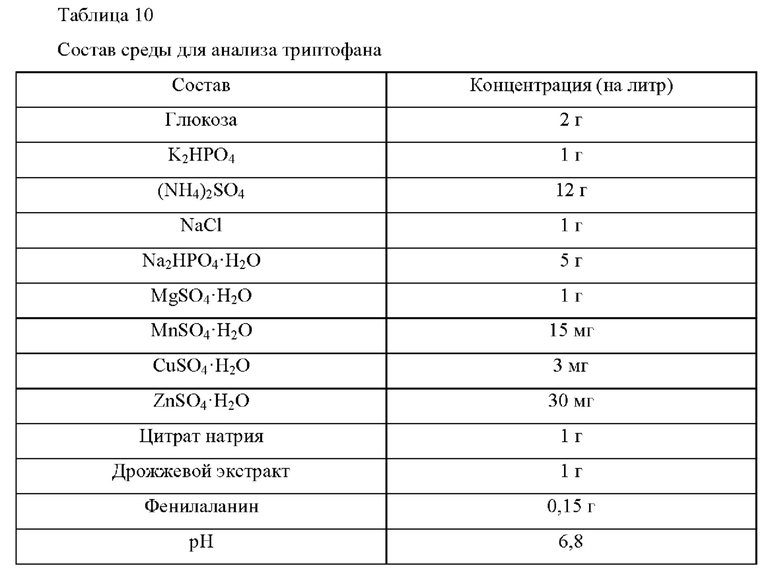
Более подробно, каждую из платиновой петли штаммов, культивированных в течение ночи на твердой среде LB в инкубаторе при 37°С, инокулировали в 25 мл среды для анализа, приведенной в Таблице 10, соответственно, и затем культивировали в инкубаторе при 37°С и 200 об/мин в течение 48 ч. Сравнивали значения OD и концентрации триптофана, они представлены в Таблице 11. Как свидетельствуют представленные ниже результаты, отобранный вариант белка по данному изобретению также способен эффективно продуцировать триптофана с высоким выходом в штамме дикого типа.

Авторы изобретения обозначили штамм на основе KCCM11166P с введенным pCC1BAC-crpTM9, обладающий повышенным продуцированием триптофана и скоростью потребления сахара (KCCM11166P/рСС1ВАС-crpTM9), как "СА04-2808" и затем депонировали этот штамм в Корейском центре культур микроорганизмов (KCCM), являющимся международным депозитарием в соответствии с Будапештским договором, 07 ноября 2018 года, под регистрационным номером KCCM12374P.
Приведенные результаты свидетельствуют о том, что у микроорганизма рода Escherichia по настоящему изобретению с введенным вариантом crp были повышены скорость потребления сахара и продуцирование L-аминокислот, и, следовательно, продуцирование L-аминокислот было повышено по сравнению с немодифицированным штаммом.
На основании приведенного выше описания специалист в данной области техники должен понимать, что настоящее изобретение может быть реализовано в другой конкретной форме, без изменения технической сути или существенных признаков изобретения. Таким образом, следует понимать, что приведенное выше воплощение является не ограничивающим, а только иллюстративным во всех аспектах. Объем данного изобретения определяется прилагаемой формулой изобретения, а не предшествующим описанием, и, следовательно, все изменения и модификации, подпадающие под объем формулы изобретения или их эквиваленты, считаются входящими в объем формулы изобретения.
Регистрационный номер
Наименование депозитария: Корейский центр культур микроорганизмов (международный депозитарий)
Номер депонирования: KCCM12374P
Дата депонирования: 7 ноября 2018
--->
<110> CJ CheilJedang Corporation
<120> cAMP receptor protein variant and method for producing L-amino
acid using the same
<130> OPA19121-PCT
<150> KR 10-2018-0151042
<151> 2018-11-29
<160> 7
<170> KoPatentIn 3.0
<210> 1
<211> 210
<212> PRT
<213> Unknown
<220>
<223> CRP (белок-рецептор цАМФ)
<400> 1
Met Val Leu Gly Lys Pro Gln Thr Asp Pro Thr Leu Glu Trp Phe Leu
1 5 10 15
Ser His Cys His Ile His Lys Tyr Pro Ser Lys Ser Lys Leu Ile His
20 25 30
Gln Gly Glu Lys Ala Glu Thr Leu Tyr Tyr Ile Val Lys Gly Ser Val
35 40 45
Ala Val Leu Ile Lys Asp Glu Glu Gly Lys Glu Met Ile Leu Ser Tyr
50 55 60
Leu Asn Gln Gly Asp Phe Ile Gly Glu Leu Gly Leu Phe Glu Glu Gly
65 70 75 80
Gln Glu Arg Ser Ala Trp Val Arg Ala Lys Thr Ala Cys Glu Val Ala
85 90 95
Glu Ile Ser Tyr Lys Lys Phe Arg Gln Leu Ile Gln Val Asn Pro Asp
100 105 110
Ile Leu Met Arg Leu Ser Ala Gln Met Ala Arg Arg Leu Gln Val Thr
115 120 125
Ser Glu Lys Val Gly Asn Leu Ala Phe Leu Asp Val Thr Gly Arg Ile
130 135 140
Ala Gln Thr Leu Leu Asn Leu Ala Lys Gln Pro Asp Ala Met Thr His
145 150 155 160
Pro Asp Gly Met Gln Ile Lys Ile Thr Arg Gln Glu Ile Gly Gln Ile
165 170 175
Val Gly Cys Ser Arg Glu Thr Val Gly Arg Ile Leu Lys Met Leu Glu
180 185 190
Asp Gln Asn Leu Ile Ser Ala His Gly Lys Thr Ile Val Val Tyr Gly
195 200 205
Thr Arg
210
<210> 2
<211> 633
<212> DNA
<213> Unknown
<220>
<223> crp
<400> 2
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 60
attcataagt acccatccaa gagcaagctt attcaccagg gtgaaaaagc ggaaacgctg 120
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 180
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 240
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 300
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 360
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 420
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 480
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 540
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaacctgat ctccgcacac 600
ggtaaaacca tcgtcgttta cggcactcgt taa 633
<210> 3
<211> 210
<212> PRT
<213> Unknown
<220>
<223> модифицированный CRP
<400> 3
Met Val Leu Gly Lys Pro Gln Thr Asp Pro Thr Leu Glu Trp Phe Leu
1 5 10 15
Ser His Cys His Ile His Lys Tyr Pro Ser Lys Ser Lys Leu Ile His
20 25 30
Gln Gly Glu Lys Ala Glu Thr Leu Tyr Tyr Ile Val Lys Gly Ser Val
35 40 45
Ala Val Leu Ile Lys Asp Glu Glu Gly Lys Glu Met Ile Leu Ser Tyr
50 55 60
Leu Asn Gln Gly Asp Phe Ile Gly Glu Leu Gly Leu Phe Glu Glu Gly
65 70 75 80
Gln Glu Arg Ser Ala Trp Val Arg Ala Lys Thr Ala Cys Glu Val Ala
85 90 95
Glu Ile Ser Tyr Lys Lys Phe Arg Gln Leu Ile Gln Val Asn Pro Asp
100 105 110
Ile Leu Met Arg Leu Ser Ala Gln Met Ala Arg Arg Leu Gln Val Thr
115 120 125
Ser Glu Lys Val Gly Asn Leu Ala Phe Leu Asp Val Thr Gly Arg Ile
130 135 140
Ala Gln Thr Leu Leu Asn Leu Ala Lys Gln Pro Asp Ala Met Thr His
145 150 155 160
Pro Asp Gly Met Gln Ile Lys Ile Thr Arg Gln Glu Ile Gly Gln Ile
165 170 175
Val Gly Cys Ser Arg Glu Thr Val Gly Arg Ile Leu Lys Met Leu Glu
180 185 190
Asp Gln Asn Gln Ile Ser Ala His Gly Lys Thr Ile Val Val Tyr Gly
195 200 205
Thr Arg
210
<210> 4
<211> 633
<212> DNA
<213> Unknown
<220>
<223> модифицированный crp
<400> 4
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 60
attcataagt acccatccaa gagcaagctt attcaccagg gtgaaaaagc ggaaacgctg 120
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 180
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 240
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 300
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 360
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 420
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 480
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 540
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaaccagat ctccgcacac 600
ggtaaaacca tcgtcgttta cggcactcgt taa 633
<210> 5
<211> 933
<212> DNA
<213> Unknown
<220>
<223> crp и область регуляции экспрессии
<400> 5
tttgctactc cactgcgtca attttcctga cagagtacgc gtactaacca aatcgcgcaa 60
cggaaggcga cctgggtcat gctgaagcga gacaccagga gacacaaagc gaaagctatg 120
ctaaaacagt caggatgcta cagtaataca ttgatgtact gcatgtatgc aaaggacgtc 180
acattaccgt gcagtacagt tgatagcccc ttcccaggta gcgggaagca tatttcggca 240
atccagagac agcggcgtta tctggctctg gagaaagctt ataacagagg ataaccgcgc 300
atggtgcttg gcaaaccgca aacagacccg actctcgaat ggttcttgtc tcattgccac 360
attcataagt acccatccaa gagcaagctt attcaccagg gtgaaaaagc ggaaacgctg 420
tactacatcg ttaaaggctc tgtggcagtg ctgatcaaag acgaagaggg taaagaaatg 480
atcctctcct atctgaatca gggtgatttt attggcgaac tgggcctgtt tgaagagggc 540
caggaacgta gcgcatgggt acgtgcgaaa accgcctgtg aagtggctga aatttcgtac 600
aaaaaatttc gccaattgat tcaggtaaac ccggacattc tgatgcgttt gtctgcacag 660
atggcgcgtc gtctgcaagt cacttcagag aaagtgggca acctggcgtt cctcgacgtg 720
acgggccgca ttgcacagac tctgctgaat ctggcaaaac aaccagacgc tatgactcac 780
ccggacggta tgcaaatcaa aattacccgt caggaaattg gtcagattgt cggctgttct 840
cgtgaaaccg tgggacgcat tctgaagatg ctggaagatc agaacctgat ctccgcacac 900
ggtaaaacca tcgtcgttta cggcactcgt taa 933
<210> 6
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> crp-F
<400> 6
cacgaattct ttgctactcc actgcgtca 29
<210> 7
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> crp-R
<400> 7
acacgaattc ttaacgagtg ccgtaaacg 29
<---
Предложен вариант белка рецептора цАМФ, микроорганизм, включающий его, и способ получения L-аминокислоты с их применением. Данный белок позволяет получать L-аминокислоты, например, такие как L-треонин или L-триптофан, с высоким выходом. 6 н. и 4 з.п. ф-лы, 11 табл., 5 пр.
1. Вариант белка рецептора цАМФ, представляющего собой регулятор транскрипции, в котором аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин.
2. Полинуклеотид, кодирующий вариант белка рецептора цАМФ по п. 1.
3. Экспрессионный вектор, содержащий полинуклеотид, кодирующий вариант белка рецептора цАМФ по п. 1.
4. Микроорганизм рода Escherichia (Escherichia sp.) для получения L-аминокислоты, содержащий вариант белка рецептора цАМФ, в котором аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин.
5. Микроорганизм рода Escherichia по п. 4, представляющий собой Е. coli.
6. Микроорганизм рода Escherichia по п. 4, где L-аминокислота представляет собой L-треонин или L-триптофан.
7. Способ получения L-аминокислоты, включающий:
культивирование микроорганизма рода Escherichia в среде, где микроорганизм включает вариант белка рецептора цАМФ, в котором аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин.
8. Способ по п. 7, дополнительно включающий сбор L-аминокислоты из микроорганизма или из среды.
9. Способ по п. 7, где L-аминокислота представляет собой L-треонин или L-триптофан.
10. Применение варианта белка рецептора цАМФ, в котором аминокислота в положении 196 аминокислотной последовательности SEQ ID NO: 1 заменена на глутамин, или микроорганизма рода Escherichia, включающего этот вариант, в получении L-аминокислоты.
| CN 103114069 B, 03.12.2014 | |||
| US 20160362456 A1, 15.12.2016 | |||
| RU 2017142878 A, 17.06.2019 | |||
| Микроорганизм рода Escherichia, продуцирующий L-триптофан, и способ получения L-триптофана с его использованием | 2016 |
|
RU2692645C2 |
| МИКРООРГАНИЗМ С ПОВЫШЕННОЙ ПРОДУКЦИЕЙ L-АМИНОКИСЛОТ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ЕГО ПРИМЕНЕНИЕМ | 2012 |
|
RU2549689C2 |
| US 8889425 B2, 18.11.2014 | |||
| EP 3144385 A1, 22.03.2017 | |||
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |

