Область техники
Настоящее изобретение относится к микроорганизму, способному продуцировать L-треонин или L-триптофан и к способу получения L-треонина или L-триптофана с применением этого микроорганизма.
Предшествующий уровень техники
Известно, что микроорганизмы, которые продуцируют полезные продукты посредством брожения, при интенсифицикации пути биосинтеза требуют очень больших количеств источников энергии, таких как АТФ.
Как известно в данной области техники, очень важно, чтобы соблюдался внутриклеточный баланс между никотинамидадениндинуклеотидом (НАД-H), синтезируемом в катаболических реакциях и никотинамидадениндинуклеотидфосфатом НАДФ-H, который используется в анаболических реакциях в микробиальных метаболических процессах. НАД-H является промежуточным соединением в катаболических реакциях, которые образуют АТФ путем окисления пищи и служит источником энергии. А НАДФ-H играет роль в обеспечении восстанавливающих соединений в метаболических процессах in vivo, предоставляя высокоэнергетические электроны, необходимые для синтеза молекул в результате реакции с ферментом, который в основном катализирует анаболические реакции. Баланс между ними регулируется или фосфорилированием НАД, как показано в приведенном ниже уравнении 1) или дефосфорилированием НАДФ, как показано в приведенном ниже уравнении 2).
Уравнение 1.
NAD+ + ATP ⇔ NADP+ + ADP
Уравнение 2
NADP+ ⇔ NAD+ + фосфат
Таким образом, с целью эффективного получения восстанавливающих соединений, таких как НАДФ-H, одновременно с ними должен быть увеличено количество источника фосфата, такого как АТФ.
АТФ (аденозин-5′-трифосфат) имеет высокоэнергетическую фосфатную связь, и вырабатывает энергию при гидролизе с образованием АДФ и фосфата. АТФ получают в основном хемиосмотическим фосфорилированием через систему переноса электронов в микроорганизмах или субстратным фосфорилированием. Полученный АТФ разлагается для выделения энергии, необходимой клеткам и повторно используется посредством регенерации через путь гликолиза или окислительное фосфорилирование.
Основываясь на этом факте, были проведены исследования применения в массовом производстве полезных продуктов процесса регенерации энергии АТФ бактерий с целью способствовать энергоснабжению (Biosci Biotechnol Biochem., (1997) 61: 840-845). В исследованиях регенерации АТФ у E. coli выявлено, что уровень АТФ в микроорганизме приблизительно на 150% выше, чем в родительском штамме, когда были инактивированы несколько генов, включая ysaA (NCBI Gene ID: 948085), ydaS (NCBI Gene ID: 945923) и ybiX (NCBI Gene ID: 947502), соответственно, и это открытие применили к получению глутатиона (FEMS Microbiol Lett., (2009) 297: 217-224). Однако не было прямого сообщения, определенно объясняющего прирост в образовании аминокислоты, вызванного ослаблением активностей белков, кодируемых этими генами.
Описание
Технические задачи
Авторы настоящего изобретения обнаружили, что повышение внутриклеточного уровня АТФ, который применяют как наиболее богатый источник энергии в клетках, продуцирующих L-аминокислоту, эффективно для увеличения синтеза L-треонина или L-триптофана, таким образом завершая настоящее изобретение.
Целью настоящего изобретения является получение рекомбинантного штамма E. coli, продуцирующего L-треонин или L-триптофан, повышенную путем увеличения синтеза АТФ.
Другой целью настоящего изобретения является создание способа получения L-треонина или L-триптофана с использованием рекомбинантного штамма E. coli.
Техническое решение
Для достижения вышеуказанных целей, вариант осуществления настоящего изобретения предлагает рекомбинантый штамм E. coli, продуцирующий L-треонин или L-триптофан, где штамм модифицирован для понижения активности по меньшей мере одного белка, выбранного из группы, состоящей из белка YsaA, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 2, белка YdaS, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 4, и белка YbiX, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 6.
Вариант осуществления настоящего изобретения также относится к способу получения L-треонина или L-триптофана, который включает культивирование рекомбинантного штамма E. coli.
Полезные эффекты
Настоящее изобретение относится к рекомбинантному микроорганизму, у которого синтез L-треонина или L-триптофана улучшен путем повышения внутриклеточного уровня АТФ в микроорганизме, продуцирующем L-треонин или L-триптофан. По настоящему изобретению предложен способ увеличить продуцирование L-треонина или L-триптофана путем восстановления баланса энергетического обмена для увеличения клеточной активности и уменьшения времени культивирования.
Описание фигур
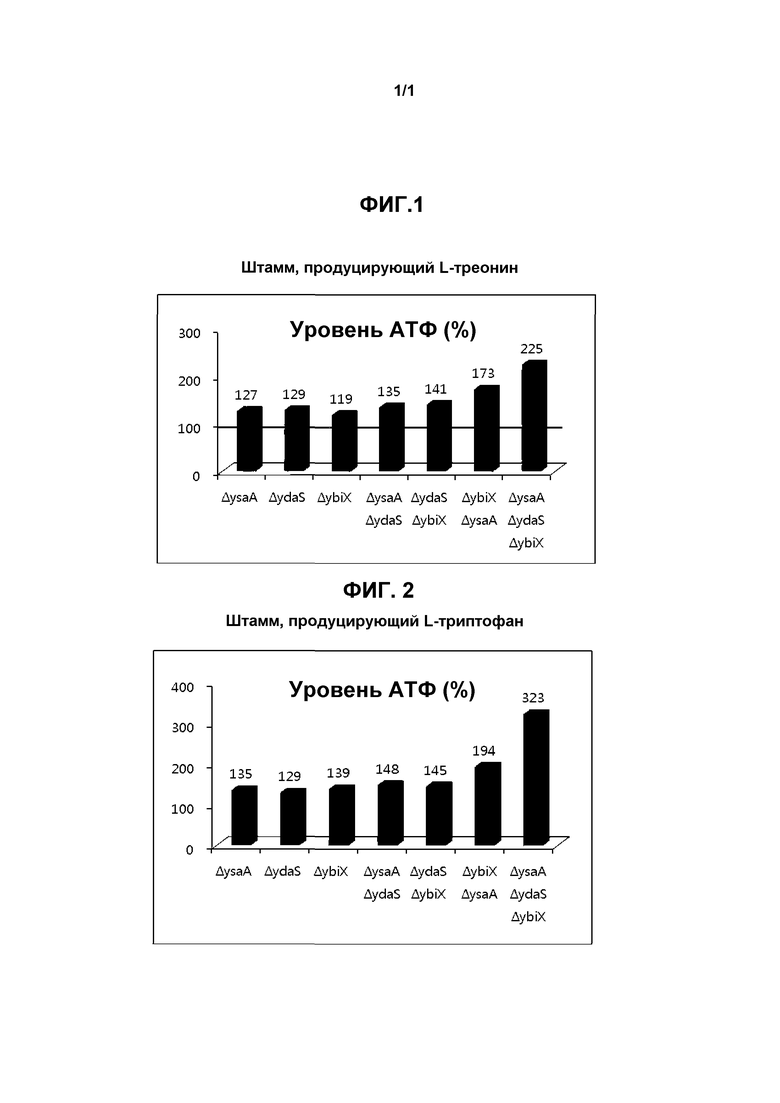
Фиг. 1 показывает относительный уровень АТФ (%) в продуцирующем L-треонин штамме относительно его родительского штамма.
Фиг. 2 показывает относительный уровень АТФ (%) в продуцирующем L-триптофан штамме относительно его родительского штамма.
Лучший вариант осуществления изобретения
Далее настоящее изобретение будет описано подробно.
Вариант осуществления настоящего изобретения относится к продуцирующему L-треонин или L-триптофан рекомбинантному штамму E. coli, где штамм модифицирован для понижения активности по меньшей мере одного белка, выбранного из группы, состоящей из белка YsaA, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 2, белка YdaS, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 4, и белка YbiX, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 6.
Микроорганизм, продуцирующий L-треонин или L-триптофан, который можно использовать в настоящем изобретении, может быть любым микроорганизмом, способным продуцировать L-треонин или L-триптофан, таким как бактерия рода Escherichia, E. coli, коринеформная бактерия, бактерия рода Serratia, бактерия рода Providencia, или т.п. В частности, можно использовать микроорганизм, принадлежащий к роду Escherichia.
В конкретном варианте осуществления настоящего изобретения применяют продуцирующий L-триптофан рекомбинантный штамм E. coli CJ600 (KCCM 10812P) (Корейский патент, регистрационный №10-0792095), полученный путем генетической инженерии рекомбинантного штамма E. coli (KFCC 10066), продуцирующего L-фенилаланин, с тем, чтобы десенсибилизировать ауксотрофию по триптофану, блокировать биосинтез L-фенилаланина и стимулировать гены, связанные с биосинтезом триптофана.
В другом конкретном варианте осуществления настоящего изобретения применяют рекомбинантный штамм E. coli FTR2533 (KCCM-10541) (Корейский патент, регистрационный №10-057 6342) продуцирующий L-треонин, полученный путем генетической инженерии мутантного штамма E. coli (KFCC 10718), продуцирующего L-треонин, с тем, чтобы блокировать активность гена дикого типа galR.
YsaA, белок, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 2, является предполагаемой гидрогеназой компонента 4Fe-4S ферредоксинового типа, но его точные функции еще не установлены.
YdaS, белок, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 4, является предполагаемым ДНК-связывающим регулятором транскрипции, но его точные функции еще не установлены.
YbiX, белок, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 6, является одним из суперсемейства Fe(II)-зависимых оксигеназ, функции которого подобны оксидоредуктазе, которая окисляет ее субстрат, используя кислород.
Полипептиды YsaA, YdaS и YbiX настоящего изобретения имеют аминокислотные последовательности, приведенные в SEQ ID NOS: 2, 4 и 6, соответственно, но не ограничиваются ими, так как аминокислотные последовательности белков могут зависеть от видов или штаммов микроорганизмов.
Другими словами, белки настоящего изобретения могут быть мутантными или искусственным вариантами, кодирующими белок, который имеет аминокислотную последовательность, включающую замену, делецию, вставку, добавление или перестановку одной или нескольких аминокислот в одной или нескольких положениях аминокислотной последовательности, приведенных в SEQ ID NO 2, 4 или 6, при условии, что мутантные или искусственные варианты могут использоваться для увеличения продукции аминокислоты путем понижения активностей, описанных в настоящем изобретении. Здесь количество "несколько" аминокислот различается в зависимости от положения или типа аминокислотных остатков в трехмерной структуре белка, но составляет в частности 2-20, конкретно 2-10, и более конкретно 2-5. Кроме того, эта замена, делеция, вставка, добавление или перестановка аминокислот также включает эти модификации, вызванные природной мутацией или искусственным изменением, основанным на различии между индивидуумами или видами микроорганизмов, имеющими активность полипептидов.
Термин "понижение" применяют в настоящем описании, чтобы обозначить, что активность белка ослабляют или удалением части или всех генов, кодирующих белок, или модифицированием регулирующей экспрессию последовательности для понижения экспрессии гена, или модификацией хромосомной последовательности генов для понижения активности белка или их сочетания.
В настоящем изобретении снижение активности может быть достигнуто способом, выбранным из группы, состоящей из: 1) удаления части или всех полинуклеотидов, кодирующих белок; 2) модификации регулирующей экспрессию последовательности для уменьшения экспрессии полинуклеотида; 3) модификации хромосомной полинуклеотидной последовательности для понижения активности белка; и 4) их сочетания.
Способ делеции части или всех полинуклеотидов, кодирующих белок, можно осуществить заменой полинуклеотида, который кодирует эндогенный белок-мишень в хромосоме, или полинуклеотидом, в котором удаляют часть нуклеотидной последовательности, или маркерным геном с помощью хромосомного инсерционного вектора.
В настоящем описании термин "часть" нуклеотидной последовательности означает в зависимости от типа гена, но не зависит от его положения, и составляет конкретно 1-200, более конкретно 1-100, и еще более конкретно 1-50.
Также способ модификации регулирующей экспрессию последовательности для понижения экспрессии полинуклеотида можно осуществлять или вызывая мутацию в регулирующей экспрессию последовательности делецией, вставкой, неконсервативной или консервативной заменой, или их сочетанием одного или нескольких нуклеотидов для снижения активности регулирующей экспрессию последовательности, или заменой регулирующей экспрессию последовательностью с пониженной активностью. Регулирующая экспрессию последовательность включает промотор, последовательность операторов, последовательность, кодирующую участок связывания рибосомы, последовательность, регулирующую терминацию транскрипции и трансляции.
Кроме того, способ модификации хромосомной полинуклеотидной последовательности, кодирующей белок по настоящему изобретению можно осуществлять или вызывая мутацию в последовательности делецией, вставкой, неконсервативной или консервативной заменой, или их сочетанием одного или нескольких нуклеотидов для снижения активности последовательности, или заменой последовательности модифицированной нуклеотидной последовательностью, имеющей пониженную активность.
Полинуклеотид, кодирующий белок по настоящему изобретению, может быть внесен в клетку-хозяин и может быть заменен кодоном, с затрудненной экспрессией в хозяине. Кроме того, их N-конец или C-конец могут быть расширены или удалены, и стартовый кодон может быть модифицирован для регуляции уровня экспрессии.
Каждый из полинуклеотидов по настоящему изобретению может иметь полинуклеотидную последовательность, кодирующую белок, обладающий гомологией по меньшей мере 80%, конкретно по меньшей мере 90%, более конкретно по меньшей мере 95%, и еще более конкретно по меньшей мере 97% с каждой аминокислотной последовательностью, приведенной в SEQ ID NOs: 2, 4 и 6, при условии, что полинуклеотид может снижать активность измененного белка. Более конкретно, полинуклеотиды имеют полинуклеотидную последовательность, показанную в SEQ ID NOs: 1, 3 и 5, соответственно.
В настоящем документе термин "гомология" применяют относительно к идентичности между двумя аминокислотными последовательностями. Гомологию можно определять, используя хорошо известные способы, например, компьютерную программу BLAST 2.0, которая вычисляет такие параметры как оценка сходства, идентичность и подобие.
Также полинуклеотидные последовательности по настоящему изобретению могут быть гибридизированы с полинуклеотидными последовательностями, приведенными в SEQ ID NOs: 1, 3 и 5 и зондами, полученными из вышеописанных нуклеотидных последовательностей в жестких условиях, и могут быть модифицированными последовательностями, кодирующими нормально функционирующие белки.
В настоящем документе, термин "жесткие условия" применяют относительно к условиям, которые позволяют специфичную гибридизацию между полинуклеотидами. Альтернативно, термин относится к полипептидам или белкам, включая их производные (Molecular Cloning, A Laboratory Manual, J. Sambrook et al.r Editors, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; or Current Protocols in Molecular Biology, F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York).
В частности, "жесткие условия" относятся к гибридизации при 65°C в гибридизационном буфере (3,5SSC, 0,02% Фиколл, 0,02% поливинилпирролидон, 0,02% бычий сывороточный альбумин, 2,5 мМ NaH2PO4 (pH 7), 0,5% SDS, 2 мМ EDTA). SSC представляет собой 0,15M хлорид натрия/0,15M цитрат натрия с pH 7. После гибридизации мембрану, на которую была перенесена ДНК, отмывают в 2SSC при комнатной температуре, а затем в 0,1-0,5×SSC/0,1×SDS при 68°C.
В настоящем документе, термин "вектор" применяют по отношению к ДНК конструкции, содержащей нуклеотидную последовательность гена, кодирующего белок-мишень, функционально связанную с подходящей регуляторной последовательностью, способной экспрессировать целевой ген в подходящей клетке-хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любой оператор для регулирования этой транскрипции, последовательность, кодирующую подходящий участок связывания рибосомы мРНК, и последовательность для регулирования терминации транскрипции и трансляции. После трансформации подходящего хозяина, вектор может реплицироваться или функционировать независимо от генома хозяина, или может, в некоторых случаях, сам встраиваться в геном.
Вектор, который применяют в настоящем изобретении, не ограничен конкретно и может быть любым вектором, известным в данной области техники, при условии, что он может реплицироваться в хозяине. Примеры широко используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, фаговый вектор или космидные векторы включают pWE15, M13, λBL3, λBL4, λXII, λSHII, λPII, λ10, λ11, Charon4A, и Charon21A, и плазмидные векторы включают pBR, pUC, pBluescriptII, pGEM, pTZ, pCL1920 и плазмиды типа pET. Векторы, которые можно использовать, не ограничены конкретно, и можно использовать любые известные экспрессирующие векторы. В частности, можно использовать векторы pACYC177, pACYC184, pCL1920, pECCG117, pUC19, pBR322, pMW118 или pCClBAC. Более конкретно, можно использовать векторы pACYC177, pCL1920 и pCClBAC.
Кроме того, вектор, который применяют в настоящем изобретении, является вектором, способным изменять клетки-хозяева, для вставки полинуклеотида, кодирующего белок-мишень, в хромосому клетки-хозяина. Конкретные примеры вектора в качестве неограничивающих примеров включают челночный вектор pECCG112, который может самореплицироваться и в E. coli, и в бактериях типа коринебактерии (Kap-Soo, Noh, Kor. Jour. Microbiol. July 1991, pl49-154).
Также полинуклеотид, кодирующий эндогенный белок-мишень в хромосоме, может быть заменен новым полинуклеотидным вектором для вставки в бактериальную хромосому. Вставку полинуклеотида в хромосому можно проводить любым известным в данной области способом, например, гомологичной рекомбинацией. Так как вектор по настоящему изобретению может быть вставлен в хромосому путем гомологичной рекомбинации, он может дополнительно содержать селективный маркер для подтверждения его вставки в хромосому. Селективный маркер применяют, чтобы выбрать клетку, измененную вектором, то есть, подтвердить вставку целевого полинуклеотида. Селективный маркер, который применяют в настоящем изобретении, можно выбирать из маркеров, которые обеспечивают селектируемые фенотипы, такие как устойчивость к лекарственному средству, ауксотрофия, устойчивость к цитотоксическим средствам, или экспрессию поверхностного белка. Только клетки, экспрессирующие селективный маркер, способны выживать или показывать различные фенотипы в среде, обработанной селективным агентом, и таким образом можно выбирать трансформированные клетки.
В настоящем документе термин "трансформация" применяют для обозначения внесения вектора, содержащего полинуклеотид, кодирующий белок-мишень, в клетку-хозяина, с тем, чтобы экспрессировать кодируемый полинуклеотидом белок в клетке-хозяине. Трансформированные полинуклеотиды включают все гены, вставленные в хромосому клетки-хозяина или расположенные снаружи хромосомы, при условии, что они могут быть экспрессированы в клетке-хозяине. Кроме того, полинуклеотиды включают ДНК и РНК, которые кодируют белок-мишень. Ген можно вносить в любой форме при условии, что полинуклеотид может быть внесен в клетку-хозяина и экспрессироваться в нем.
Например, полинуклеотид может быть внесен в клетку-хозяина в форме экспрессионной кассеты, которая является полинуклеотидной конструкцией, включающей все элементы для экспрессии гена. Экспрессионная кассета включает промотор, который функционально связан с геном, сигнал терминации транскрипции, участок связывания рибосомы, и сигнал терминации трансляции. Экспрессионная кассета может быть в форме экспрессионного вектора, способного к саморепликации. Полинуклеотид также может быть внесен в клетку-хозяина сам по себе, и быть функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине.
В частности, снижение активности белка, кодируемого ysaA, ydaS или ybiX геном, может быть достигнуто делецией гена. В частности, мутация в гене может быть вызвана с использованием химических реагентов или света, такого как ультрафиолетовый свет, тем самым получая вариант, имеющий удаленный ген. Альтернативно, вариант, у которого отсутствует активность белка, можно получать заменой хромосомного гена на нуклеотид, у которого отсутствует активность, методом рекомбинации генов, способом замены гена путем гомологичной рекомбинации.
Также вариант осуществления настоящего изобретения также относится к способу получения L-треонина или L-триптофана, включающему культивирование продуцирующего L-треонин или L-триптофан рекомбинантного штамма E. coli, где штамм модифицирован для понижения активности по меньшей мере одного белка, выбранного из группы, состоящей из белка YsaA, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 2, белка YdaS, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 4, и белка YbiX, имеющего аминокислотную последовательность, приведенную в SEQ ID NO 6.
Процесс культивирования по настоящему изобретению можно проводить в подходящей среде и условиях культивирования, известных в данной области техники. Процесс культивирования может быть легко модифицирован любым специалистом в данной области в зависимости от типа выбранного штамма. Примеры процесса культивирования в качестве неограничивающих примеров включают периодическую культуру, непрерывную культуру, и подпитываемую культуру.
Среда и условия культивирования, которые применяют для выращивания микроорганизма по настоящему изобретению, могут быть такими, которые применяют для культивирования микроорганизмов, принадлежащих к роду Escherichia, но они должны полностью удовлетворять требованиям микроорганизма по настоящему изобретению.
В конкретном варианте осуществления настоящего изобретения микроорганизм можно культивировать в общепринятой среде, содержащей подходящие источники углерода, источники азота, аминокислоты, витамины и т.п. при аэробных условиях при регулируемых температуре, pH и т.п.
Источники углерода, которые можно использовать в настоящем изобретении, включают углеводы, такие как глюкоза, фруктоза, сахароза, мальтоза, маннит, сорбит; спирты, такие как сахарный спирт, глицерин, пировиноградная кислота, молочная кислота и лимонная кислота; и аминокислоты, такие как органическая кислота, глутаминовая кислота, метионин и лизин. Кроме того, можно использовать природные органические источники питательных веществ, такие как гидролизаты крахмала, меласса, сырая меласса, рисовые отруби, маниока, багасса и жидкий кукурузный экстракт. В частности, органические источники питательных веществ включают глюкозу и стерильную предварительно обработанную мелассу (т.е., мелассу преобразованную в восстанавливающие сахара), и подходящие количества источников углерода можно использовать без ограничения.
Источники азота, которые можно использовать в настоящем изобретении, включают неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; аминокислоты, такие как глутаминовая кислота, метионин и глутамин; и органические источники азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыбная мука или продукты ее расщепления, обезжиренный соевый жмых или продукты его расщепления, и т.д. Эти источники азота можно использовать по одному или в сочетании. Среда может содержать в качестве источников фосфора дигидрофосфат калия, гидрофосфат калия и соответствующие натрийсодержащие соли.
Неорганические соединения, которые можно использовать в настоящем изобретении, включают хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца и карбонат кальция. Кроме того, среда может содержать аминокислоты, витамины и подходящие предшественники. Эти среды или предшественники можно добавлять в среду порционно или непрерывно.
Для регулирования pH среды для культивирования в процессе культивирования можно надлежащим образом добавлять в среду соединения, такие как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота.
Кроме того, для подавления образования пузырьков в процессе культивирования можно использовать противопенную добавку, такую как сложный эфир жирной кислоты с полигликолем. Также, с целью поддерживать среду для культивирования в аэробном состоянии, можно вводить в среду для культивирования кислород или кислородсодержащий газ. Кроме того, с целью поддерживать среду для культивирования в анаэробном или неаэробном состоянии, не вводится газ, или можно вводить газы азот, водород или диоксид углерода в среду для культивирования. Как правило, среду для культивирования поддерживают при температуре в диапазоне от 27°C до 37°C, и в частности от 30°C до 35°C. Культивирование микроорганизма может продолжаться до достижения желаемого уровня полезного вещества. Конкретно, время культивирования составляет от 10 до 100 часов.
Способ по настоящему изобретению может дополнительно содержать средство, очищающее или извлекающее L-аминокислоту, полученную на этапе культивирования. Процесс очистки или извлечения можно проводить очищением или извлечения желаемой L-аминокислоты из среды для культивирования с использованием подходящего способа, выбранного в зависимости от способа, использованного для культивирования микроорганизма, например, способ периодической культуры, непрерывной культуры, и подпитываемой культуры.
Далее в настоящем описании настоящее изобретение будет описано в подробных деталях со ссылкой на примеры. Однако следует понимать, что эти примеры приведены для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения.
Примеры
Пример 1: Конструирование продуцирующих L-треонин и L-триптофан штаммов, обладающих пониженной активностью белка, кодируемого генами ysaA, ydaiS или ybiX
В этом Примере каждый из генов ysaA, ydaS и ybiX в продуцирующем L-триптофан штамме KCCM10812P (Корейский патент, регистрационный №10-0792095) и продуцирующем L-треонин штамме KCCM10541 (Корейский патент, регистрационный № 10-0576342) был удален путем гомологичной рекомбинации.
Продуцирующий L-триптофан родительский штамм KCCM10812P является штаммом, полученным из варианта E. coli KFCC 10066, продуцирующего L-фенилаланин. Это рекомбинантный штамм E. coli, продуцирующий L-триптофан, характеризуется тем, что хромосомная ауксотрофия по триптофану была десенсибилизирована, гены pHeA, trpR, mtr и tnaAB были аттенуированны и гены aroG и trpE были модифицированы.
Также продуцирующий L-треонин родительский штамм KCCM10541 является штаммом, полученным из E. coli KFCC10718 (корейская выложенная патентная публикация №1992-0008365). Он имеет устойчивость к аналогу L-метионина, ауксотрофный фенотип по метионину, устойчивость к аналогу L-треонина, текучий ауксотрофный фенотип по изолейцину, устойчивость к аналогу L-лизина, и устойчивость к α-аминомасляной кислоте, и способен продуцировать L-треонин.
Удаляемые гены ysaA, ydaS и ybiX имеют полинуклеотидные последовательности, приведенные в SEQ ID NOs: 1, 2 и 3, соответственно.
Для этой цели применяли одноступенчатый способ инактивации (разработанный Datsenko KA et al.), который представляет собой метод мутагенеза с использованием лямбда Red рекомбиназы (Proc Natl Acad Sci USA., (2000) 97: 6640-6645). В качестве маркера для подтверждения вставки в гены применяли ген устойчивости к хлорамфениколу pUCprmfmloxC (корейская выложенная патентная публикация №2009-0075549).
Фрагменты генов размером приблизительно 1200 п.н. амплифицировали полимеразной цепной реакцией (далее в настоящем документе обозначаемой как ПЦР) с использованием pUCprmfmloxC в качестве матрицы и пары праймеров 1 и 2, пары праймеров 7 и 8 и пары праймеров 13 и 14, которые имели часть каждого из трех генов и часть гена устойчивости к хлорамфениколу pUCprmfmloxC. ПЦР проводили в 30 циклов, каждый состоящий из денатурации при 94°C в течение 30 сек, отжига при 55°C в течение 30 сек и элонгации при 72°C в течение 1 мин.
GCAAAGGAGTGATCAGGTGACACTATAGAACGCG-3′
CGCCCGGAAAACGTTTAGTGGATCTGATGGGTACC-3′
TGGATCAAAGCAGTAGGGACGCGCTCTCTGGC-3′
TACGCCGCATCTGGCATAAACAAAGCGCACTG-3′
TGGAGTTCATTCGGAGGTGACACTATAGAACGCG-3′
CCTCTTGATGTTTAGTGGATCTGATGGGTACC-3′
CAGCCTATTGACAATTGCGTTAGGCGTCGCCTA-3′
CAAACAACACTGTTCGATGTGCTCATGCTTG-3′
TGGAGAAATTGTAGGTGACACTATAGAACGCG-3′
GGGCAGTTCAAGCATAGTGGATCTGATGGGTACC-3′
TCACAGCCAATATGCATTTCTGATTCAGATGTGG-3′
CCATATCAATACTTGTACAGTTAAGTGTAGCTA-3′
Также фрагменты ДНК, полученные ПЦР амплификацией, подвергали электрофорезу в 0,8% агарозном геле, а затем элюировали и использовали в качестве матриц во вторичной ПЦР. Вторичную ПЦР проводили так, чтобы 5′ и 3′ концевые области первичных фрагментов ДНК имели 20 пар комплементарных нуклеотид оснований. Фрагменты генов размером приблизительно 1300-п.н. амплифицировали ПЦР используя элюированные продукты первичной ПЦР в качестве матриц и пару праймеров 3 и 4, пару праймеров 9 и 10 и пару праймеров 15 и 16, которые включали 5′ и 3′ области генов. ПЦР проводили в 30 циклов, каждый состоящий из денатурации при 94°C в течение 30 сек, отжига при 55°C в течение 30 сек и элонгации при 72°C в течение 1 мин. Фрагменты ДНК, полученные ПЦР амплификация подвергали электрофорезу в 0,8% агарозном геле, а затем элюировали и использовали в рекомбинации.
Согласно способу, разработанному Datsenko KA et al. (Proc Natl Acad Sci USA., (2000) 97:6640 6645), штамм E. coli, трансформированный pKD4 6 делали компетентным, а затем трансформировали фрагментами гена размером 1300 п.н., полученными ПЦР. Полученные в результате штаммы, имеющие устойчивость к хлорамфениколу, выбирали в среде Лурия-Бертани. ПЦР проводили, используя пару праймеров 5 и 6, пару праймеров 11 и 12 и пару праймеров 17 и 18, и продукты амплификации имели размеры 1450, 1530 и 1640 п.н., соответственно, и было подтверждено, что гены были удалены.
Из первичных рекомбинантных штаммов, имеющих устойчивость к хлорамфениколу, удаляли pKD4 6, а затем в штаммы вносили вектор pJW168, и удаляли маркерный ген устойчивости к хлорамфениколу из бактериальных клеток (Gene, (2000) 247, 255-264). Полученные в результате бактериальные клетки были продуктами амплификации размером приблизительно 400 п.н., 500 п.н. и 600 п.н., полученными с помощью пары праймеров 5 и 6, пары праймеров 11 и 12 и пары праймеров 17 и 18, и подтверждали, что делеция желаемого гена успешно выполнена.
Согласно вышеописанному способу конструировали продуцирующие L-треонин штаммы KCCM10541 ΔysaA, KCCM10541 ΔydaS и KCCM10541 ΔybiX. Кроме того, конструировали продуцирующие L-триптофан штаммы KCCM10812P ΔysaA, KCCM10812P ΔydaS и KCCM10812P ΔybiX.
Пример 2: Конструирование рекомбинантных продуцирующих L-треонин и L-триптофан штаммов с делецией двух или более из генов ysaA, ydaS и ybiX.
Согласно способу, описанному в примере, конструировали рекомбинантные штаммы с делецией двух или более генов.
Вектор pKD4 6 для использования лямбда Red рекомбиназы вносили в штаммы с делецией любого из генов, а затем штаммы делали компетентным. Также, фрагменты гена, амплифицированные ПЦР для включения части трех генов и гена устойчивости к хлорамфениколу pUCprmfmloxC трансформировали в различные штаммы с делецией одного из генов. Полученные штаммы, имеющие устойчивость к хлорамфениколу, отбирали в среде Лурия-Бертани, и делецию сочетания генов подтверждали, используя пары праймеров, описанные в примере 1.
Согласно вышеописанному способу конструировали продуцирующие L-треонин штаммы KCCM10541 ΔysaA ΔydaS, KCCM10541 ΔydaS ΔybiX, KCCM10541 ΔybiX ΔysaA и KCCM10541 ΔysaA ΔydaS ΔybiX. Также конструировали продуцирующие L-триптофан штаммы KCCM10812P ΔysaA ΔydaS, KCCM10812P ΔydaS ΔybiX, KCCM10812P ΔybiX ΔysaA и KCCM10812P ΔysaA ΔydaS ΔybiX.
Среди рекомбинантных штаммов, полученных как описано выше, KCCM10541 ΔysaA ΔydaS ΔybiX и KCCM10812P ΔysaA ΔydaS ΔybiX были названы "E. coli CA03-4257P" и E. coli CA04-2002", соответственно, и депонированы в Корейском Центре Культур Микроорганизмов (361-221, Hongje 1-dong, Seodaemun-gu, Seoul, Korea), международный депозитарий, 29 декабря 2011 года под регистрационными номерами KCCM11243P и KCCM11245P, соответственно.
Пример 3: Измерение уровней АТФ в сконструированных продуцирующих L-треонин штаммах и продуцирующих L-триптофан штаммах.
В этом Примере были количественно измерены уровни АТФ в штаммах, сконструированных в Примерах 1 и 2.
Для этой цели применяли способ, разработанный Kiyotaka Y. Hara et al., использующий люциферазу, ("An Efficient Method for Quantitative determination of Cellular ATP Synthetic Activity", J Biom Sere, (2006) VII: № 3: PP310-17).
Конкретно, штаммы, имеющие различные генетические признаки, культивировали в течение ночи в жидкой среде Лурия-Бертани, содержащей глюкозу. Надосадочную жидкость удаляли центрифугированием, бактериальные клетки промывали 100 мМ Tris-Cl (pH 7,5), а затем обрабатывали PB буфером (проницаемый буфер: 40% [об./об.] глюкоза, 0,8% [об./об.] Triton X-100) в течение 30 минут для высвобождения внутриклеточного АТФ. Затем надосадочную жидкость удаляли посредством центрифугирования, и добавляли в клетки люциферин в качестве субстрата для люциферазы. Клетки выдерживали в течение 10 минут, а затем активность люциферазы в клетках измеряли люминометром для количественного определения уровня АТФ. Результаты измерений показаны на фиг. 1 и 2. Все результаты были зафиксированы как среднее по трем повторным экспериментам.
Как можно видеть на фиг. 1 и 2, уровни АТФ в штаммах, сконструированных из продуцирующего L-треонин штамма и продуцирующего L-триптофан штамма в Примерах 1 и 2, увеличились. Кроме того, уровень АТФ был увеличен в штаммах с делецией комбинации генов, по сравнению со штаммами с делецией одного гена.
Пример 4: Изучение титра продуцирующего L-треонин штамма, имеющего сниженную активность одного или комбинации ферментов, кодируемых генами ysaA, ydaS и ybiX в глюкозосодержащей среде.
Согласно способам, описанным в примерах 1 и 2, из продуцирующего L-треонин штамма KCCM10541 (Корейский патент, регистрационный № 10-0576342) был удален один ген или комбинация генов ysaA, ydaS и ybiX для повышения внутриклеточного уровня АТФ. Титры полученных штаммов оценивали, используя в качестве источника углерода глюкозу.
Конкретно, штаммы, имеющие различные генетические признаки, культивировали в течение ночи в твердой среде Лурия-Бертани в инкубаторе при 33°C и высевали платиновой петлей в 25 мл глюкозосодержащей среды, имеющей состав, показанный в таблице 2 ниже. Затем штаммы выращивали в инкубаторе при 33°C и 200 об./мин в течение 50 часов. Результаты показаны в таблице 3 ниже. Все результаты были зафиксированы как средний результат по трем колбам.
** измерено через 50 часов
Как видно из таблицы 3 выше, было показано, что использование глюкозы рекомбинантными продуцирующими L-треонин штаммами E. coli, сконструированными по настоящему изобретению, увечилось на приблизительно до 22% по сравнению с родительским штаммом, и продуцирование треонина рекомбинантными штаммами увечилось на приблизительно до 7% по сравнению с родительским штаммом. Принимая во внимание уровень АТФ, показанный на фиг. 1, эти результаты показывают, что скорость расхода глюкозы или продуктивность по аминокислоте рекомбинантных штаммов увеличилась в соответствии с повышенным уровнем АТФ.
Пример 5: Изучение титра продуцирующего L-треонин штамма, имеющего сниженную активность одного или сочетания ферментов, кодируемых генами ysaA, ydaS и ybiX в сахарозосодержащей среде.
Согласно способам, описанным в примерах 1 и 2, из продуцирующего L-треонин штамма KCCM10541 (Korean Патент Registration № 10-057 6342) был удален один ген или сочетание генов ysaA, ydaS и ybiX для повышения внутриклеточного уровня АТФ. Титры полученных штаммов оценивали, используя в качестве источника углерода сахарозу.
Конкретно, штаммы, имеющие различные генетические признаки, культивировали в течение ночи в твердой среде Лурия-Бертани в инкубаторе при 33°C и высевали платиновой петлей в 25 мл глюкозосодержащей среды, имеющей состав, показанный в таблице 4 ниже. Затем штаммы выращивали в инкубаторе при 33°C и 200 об./мин в течение 48 часов. Результаты показаны в таблице 5 ниже. Все результаты были зафиксированы как средний результат по трем колбам.
** измерено через 48 часов
Как можно видеть в таблице 5 выше, авторы показали, что использование сахарозы рекомбинантными продуцирующими L-треонин штаммами E. Coli, сконструированными по настоящему изобретению, увечилось на приблизительно до 10% по сравнению с родительским штаммом, и продуцирование треонина рекомбинантыми штаммами увечилось на приблизительно до 5% по сравнению с родительским штаммом. Принимая во внимание уровень АТФ, показанный на фиг. 1, эти результаты показывают, что активность и скорость расхода сахарозы или продуктивность по аминокислоте рекомбинантных штаммов увеличилась в соответствии с повышенным уровнем АТФ.
Пример 6: Изучение титра продуцирующего L-триптофан штамма, имеющего сниженную активность одного или сочетания ферментов, кодируемых генами ysaA, ydaS и ybiX в глюкозосодержащей среде.
Согласно способам, описанным в примерах 1 и 2, из продуцирующего L-триптофан штамма KCCM10812P (Korean Патент Registration № 10 0792095) был удален один ген или комбинация генов ysaA, ydaS и ybiX для повышения внутриклеточного уровня АТФ. Титры полученных штаммов оценивали, используя в качестве источника углерода глюкозу.
С целью изучения титра, штаммы высевали платиновой петлей в твердую среду Лурия-Бертани, а затем культивировали в течение ночи в инкубаторе и высевали платиновой петлей в колбу с 25 мл среды титрования, имеющей состав, показанный в таблице 6 ниже. Затем штаммы выращивали в инкубаторе при 37°C и 200 об./мин в течение 48 часов. Результаты показаны в таблице 7 ниже. Все результаты были зафиксированы как средний результат по трем колбам.
** измерено через 48 часов
Тогда как настоящее изобретение было описано со ссылкой на частные иллюстративные варианты осуществления, специалисты в данной области техники, к которой относится настоящее изобретение, могут понять, что настоящее изобретение можно осуществлять в других конкретных формах без отступления от технической сущности или существенных характеристик по настоящему изобретению. Таким образом, варианты осуществления, описанные выше, рассматривают как иллюстративные во всех отношениях и не ограничивающие. Кроме того, объем настоящего изобретения определяется приложенной формулой изобретения, а не подробным описанием, и следует понимать, что все модификации или изменения, полученные из смысла и объема настоящего изобретения, и их эквиваленты включены в объем приложенной формулы изобретения.
Регистрационный номер
Депозитарий: Корейский Центр Культур Микроорганизмов (Международный)
Регистрационный номер: KCCM11243P
Дата депонирования: 29 декабря 2011
Депозитарий: Корейский Центр Культур Микроорганизмов (Международный)
Регистрационный номер: KCCM11245P
Дата депонирования: 29 декабря 2011








Изобретение относится к биотехнологии и представляет собой продуцирующую L-треонин или L-триптофан рекомбинантную клетку-хозяин Е. coli, где в клетке-хозяине делетирован по меньшей мере один ген, выбранный из группы, состоящей из генов, кодирующих белок YsaA, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 2, белок YdaS, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 4, и белок YbiX, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 6. Изобретение относится также к способу получения L-треонина или L-триптофана путем культивирования указанной продуцирующей L-треонин или L-триптофан рекомбинантной клетки-хозяина Е. сoli. 2 н. и 5 з.п. ф-лы, 2 ил., 7 табл., 6 пр.
1. Продуцирующая L-треонин или L-триптофан рекомбинантная клетка-хозяин Е. coli, где в клетке-хозяине делетирован по меньшей мере один ген, выбранный из группы, состоящей из генов, кодирующих белок YsaA, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 2, белок YdaS, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 4, и белок YbiX, имеющий аминокислотную последовательность, приведенную в SEQ ID NO 6.
2. Продуцирующая L-треонин или L-триптофан рекомбинантная клетка-хозяин Е. coli по п. 1, где белок YsaA кодирован полинуклеотидной последовательностью, приведенной в SEQ ID NO 1, белок YdaS кодирован полинуклеотидной последовательностью, приведенной в SEQ ID NO 3, и белок YbiX кодирован полинуклеотидной последовательностью, приведенной в SEQ ID NO 5.
3. Продуцирующая L-треонин или L-триптофан рекомбинантная клетка-хозяин Е. coli по п. 1, где рекомбинантная клетка-хозяин Е. coli является продуцирующим L-треонин Escherichia coli СА03-4257Р (KCCM11243P).
4. Продуцирующая L-треонин или L-триптофан рекомбинантная клетка-хозяин Е. coli по п. 1, где рекомбинантная клетка-хозяин Е. coli является продуцирующим L-триптофан Escherichia coli СА04-2002 (KCCM11245P).
5. Способ получения L-треонина или L-триптофана, содержащий культивирование продуцирующей L-треонин или L-триптофан рекомбинантной клетки-хозяина Е. Coli по п. 1.
6. Способ по п. 5, где рекомбинантная клетка-хозяин Е. coli является продуцирующим L-треонин Escherichia coli СА03-4257Р (KCCM11243P).
7. Способ по п. 5, где рекомбинантная клетка-хозяин Е. coli является продуцирующим L-триптофан Escherichia coli СА04-2002 (KCCM11245P).
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОДИН ИЛИ НЕСКОЛЬКО ГЕНОВ, КОДИРУЮЩИХ МАЛЫЕ РНК | 2007 |
|
RU2395567C2 |
| HARA KY et al | |||
| Glutathione production by efficient ATP-regenerating Escherichia coli mutants, FEMS Microbiol Lett | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ РАЗРУШЕН ПУТЬ БИОСИНТЕЗА ГЛИКОГЕНА | 2006 |
|
RU2315809C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН lrhA | 2006 |
|
RU2337956C2 |

