Область техники, к которой относится изобретение
Изобретение касается создания и применения антител, которые могут эффективно связываться с доменом СεmX на мембраносвязанном IgE (mIgE), экспрессированном на поверхности человеческих В лимфоцитов. Обнаружено, что только моноклональные антитела, специфические для определенных сегментов СεmX, такие как GLAGGSAQSQRAPDRVL и HSGQQQGLPRAAGGSVPHPR, могут эффективно связываться с mIgE на человеческих В клетках и, следовательно, пригодны для направленного воздействия на эти В клетки для лечения заболеваний, опосредованных IgE.
Уровень техники
IgE играет центральную роль в опосредовании реакций гиперчувствительности типа I, ответственных за возникновение аллергических заболеваний, включающих аллергическую астму, аллергические риниты, атопические дерматиты и другие. Аллергические реакции - это реакции иммунной системы на безвредные вещества внешней среды, такие как пылевые клещи, пыльца деревьев и травянистых растений, некоторые пищевые продукты и лекарства и укусы пчел и огненных муравьев. В таких реакциях связывание аллергена с IgE на поверхности базофилов и тучных клеток вызывает образование поперечных связей в IgE и агрегацию соответствующих IgE.Fc, рецепторов типа I IgE.Fc или FcεRI. Эта агрегация рецепторов затем активирует сигнальный путь, приводящий к экзоцитозу гранул и высвобождению фармакологических медиаторов, таких как гистамин, лейкотриены, триптаза, цитокины и хемокины. Высвобождение этих медиаторов из тучных клеток и базофилов вызывает различные патологические проявления аллергии.
Анти-IgE антитела, которые связываются со свободным IgE в крови и в интерстициальной жидкости и с mIgE на В клетках, но не с IgE, связанным с FcεRI на базофилах и тучных клетках, были разработаны для лечения опосредованных IgE аллергических заболеваний. Лечение гуманизированным анти-IgE антителом omalizumab (коммерческое название Xolair) продемонстрировало множество фармакологических эффектов по ослаблению гиперчувствительности типа I при различных аллергических проявлениях. Антитело связывается с IgE с высоким сродством в сайте СН3 домена Fc, который перекрывается со связывающим сайтом FcεRI. Следовательно, терапия основана на связывании антитела со свободным IgE и с mIgE на В лимфобластах и на В клетках памяти, что приводит к снижению общего уровня свободного IgE в крови и в интерстициальной жидкости.
Связывание анти-IgE со свободным IgE предотвращает связывание IgE с FcεRI на поверхности базофилов и тучных клеток. Поскольку FcεRI, не занятый IgE, не стабилен и впоследствии интернализуется и деградирует, уменьшение свободного IgE при связывании анти-IgE также постепенно снижает содержание FcεRI на базофилах и тучных клетках. Были найдены свидетельства других эффектов терапии антителами, включая нейтрализацию цитокинэргической активности, ослабление общей воспалительной активности и, возможно, удаление аллергенов путем накопления IgE-анти-IgE иммунных комплексов.
Один из авторов (T.W.Chang) данного изобретения обнаружил, что, кроме антигенного сайта на СН3 IgE, с которым связывается omalizumab, на mIgE человека имеется другой антигенный сайт, обозначаемый как СεmX, пригодный для направленного действия на В лимфоциты, экспрессирующие mIgE. CεmX - это сегмент из 52 аминокислот, локализованный между доменом СН4 и С-терминальным мембраносвязанным сегментом связанной с мембраной ε цепи человека (mε). Было показано, что у большинства исследованных лиц mε без CεmX (mεs) присутствует в минимальных количествах, в то время как цепь mε с CεmX (mεL) экспрессируется доминантно. Все m РНК для ε цепи свободного секретируемого IgE и для mεs и mεL цепей mIgE возникают в результате альтернативного сплайсинга ε РНК транскрипта. Аминокислотная и нуклеотидная последовательности CεmX уникальны среди всего разнообразия белков и ДНК. Таким образом, CεmX образует уникальный антигенный сайт для направленного воздействия на mIgE и mIgE-экспрессирующие В клетки.
Исследовательская группа Chang ранее сообщила о нескольких СεmX-специфичных мышиных моноклональных антителах, включающих а20, которые могут связываться с рекомбинантными белками, содержащими сегмент CεmX, и с клетками клеточной линии SKO-007, которые являются производными миеломы человека, экспрессирующими человеческий mIgE, и с клетками клеточной линии СНО, которые трансфектированы геном, соответствующим сегменту домена СН2 через цитоплазматический конец mεL (mεL(CH2-CM); СМ; цитоплазма). Показано, что моноклональное антитело а20 и все разработанные ранее антитела связываются с пептидным сегментом 8-а, RADWPGPP, остатками #45-52 на С-терминальном конце 52 а.а. домена CεmX
Раскрытие изобретения
Данное изобретение относится к разработке и идентификации антител, специфичных к домену СεmX человеческого mIgE, способных связываться с mIgE на человеческих В лимфоцитах. Оно также относится к использованию этих антител для лечения аллергических и других заболеваний, опосредованных IgE.
При изучении анти-СεmX моноклонального антитела а20, которое было разработано исследовательской группой Chang, было обнаружено, что а20 хорошо связывается с клеточными линиями, трансфектированными геном mεL(CH2-CM), такими как клеточная линия СНО или клеточная линия NS0, которые не экспрессируют Igα (CD79a), Igβ (CD79b), CD21, CD19, CD81 и другие белки, ассоциированные с рецепторами В клеток (BCR). Однако было обнаружено, что а20 плохо связывается с клеточными линиями, трансфектированными геном mεL(CH2-CM), которые экспрессируют Igα, Igβ и другие ассоциированные с BCR белки, такими как клеточная линия Ramos. Авторами было сделано предположение, что антигенный эпитоп на СεmX, распознаваемый а20, может быть блокирован определенным(и) белком(ами), ассоциированным(и) с BCR. Поэтому моноклональное антитело а20 и его гибридные или гуманизированные версии не пригодны для использования у больных людей in vivo с целью направленного действия на В лимфобласты и клетки памяти, экспрессирующие mIgE.
Если пептидный эпитоп RADWPGPP - единственный эпитоп для индуцирования ответа антител, все моноклональные антитела, генерируемые с помощью гибридомной техники с использованием мышей, иммунизированных человеческими CεmX-содержащими белками, будут специфичны к этому пептидному сегменту. Однако если этот эпитоп - доминантный эпитоп, но не единственный иммуногенный эпитоп, могут образоваться дополнительные моноклональные антитела, специфичные к другим антигенным эпитопам на СεmX. Возможно, что на СεmX имеется эпитоп(ы), связывание которого с антителом не блокируется ассоциированными с BCR белками. Если это так, то может быть разработано антитело, которое связывается с IgE на В клетках и которое может быть использовано для направленного воздействия на эти В клетки.
В приведенных далее примерах авторы успешно показали, что хотя RADWPGPP - доминантный эпитоп, он не является единственным иммуногенным и антигенным эпитопом на СεmX. Кроме того, были обнаружены моноклональные антитела 4В12 и 26Н2, которые связываются с CεmX на антигенных эпитопах, не локализованных в сегменте RADWPGPP. Эти моноклональные антитела не конкурируют с антителом а20 при связывании с CεmX. Они связываются с mIgE на В клетках значительно сильнее, чем а20, и значительно эффективнее, чем а20, вызывают зависящий от антитела цитолиз и апоптоз mIgE-экспрессирующих клеток.
Эти примеры показывают, что моноклональные антитела, такие как 4В12 и 26Н2, могут связываться с mIgE на В лимфоцитах человека и пригодны для использования их с целью направленного воздействия на экспрессирующие mIgE В лимфобласты и В клетки памяти с целью подавления синтеза IgE. Антитела в гибридной или гуманизированной форме будут полезны для применения больными с опосредованными IgE аллергическими заболеваниями, такими как аллергическая астма, аллергические риниты и атопические дерматиты. Поскольку показано, что нейтрализация IgE с помощью анти-IgE эффективно лечит холодовую крапивницу, хроническую крапивницу, холинэргическую крапивницу, хронические риносинуситы, системный мастоцитоз, кожный мастоцитоз, аллергический бронхолегочный аспергиллез, рекуррентную идиопатическую ангиоэдему и интерстициальный цистит или связанные с эозинофилами желудочно-кишечные нарушения, такие антитела, как 4В12 и 26Н2, тоже могут быть использованы для лечения этих разнообразных заболеваний.
Приведенные далее примеры позволяют предположить существование потенциально полезных свойств пептидов, распознаваемых 4В12 и 26Н2, для индуцирования иммунного ответа против CεmX и, следовательно, против экспрессирующих mIgE В клеток. Пептиды и их аналоги со сходными антигенными свойствами, то есть со способностью связываться с антителами анти-CεmX, такими как 4В12 и 26Н2, могут быть использованы индивидуально или в комбинации с молекулярными конструктами, также содержащими компоненты, способные индуцировать Т-хелперы. Такие конструкты могут вызывать активную иммунизацию против mIgE-экспрессирующих В клеток и достигать, таким образом, эффектов подавления общего синтеза IgE.
Пример 1: Новые анти-CεmX моноклональные антитела, связывающие антигенные сайты, иные чем RADWPGPP
Чтобы вызвать анти-CεmX иммунный ответ, мышей BALB/c дважды иммунизировали подкожным введением 50 мкг солюбилизированных в n-андецил-β-d-мальтопиранозиде (UDM; Anatrance) рекомбинантных белков IgE.FcL, которые были эмульсифицированы в TiterMax Gold адьюванте (Sigma-Aldrich), с двухнедельными интервалами в соответствии с рекомендациями производителя. Не следуя протоколу гипер-иммунизации, авторы получали мышей, продуцировавших антитела не только к доминантному эпитопу RADWPGPP. Последнее стимулирование осуществляли внутрибрюшинно с помощью 0,1 мг солюбилизированных в UDM рекомбинантных белков IgE.FcL без адьюванта. За день до слияния NS0 клетки пересевали в свежую среду DMEM (Invitrogen) с добавлением 10% инактивированной - нагреванием телячьей сыворотки (FBS; Invitrogen) и 1% пенициллин-стрептомициновой смеси (100×Pen-Strep раствор; Invitrogen) при плотности клеток 5×105 клеток/мл. Через три дня после последнего стимулирования собирали клетки селезенки иммунизированных мышей и дважды промывали их средой DMEM без сыворотки. После промывания клетки селезенки и клетки NS0 подвергали слиянию, добавляя 1 мл предварительно нагретого 50% полиэтиленгликоля 1500 (PEG 1500, Roche Applied Science) при постоянном мягком перемешивании клеток кончиком пипетки в течение 1 мин, добавляя 2 мл предварительно нагретой не содержащей сыворотки DMEM в течение 2 мин и, наконец, добавляя 8 мл не содержащей сыворотки DMEM в течение 2 мин. После центрифугирования при 200×g в течение 10 мин слившиеся клетки ресуспендировали в 600 мл HAT среды [среды DMEM с добавлением 2% гипоксантин-амидоптерин-тим идиновой смеси (50×HAT раствор; Invitrogen), 10% BM-Condimed H1 (Roche Applied Science), 10% инактивированной нагреванием FBS и 1% пенициллин-стрептомициновой смеси] и размещали в 30 96-ячеечных культуральных планшетах по 200 мкл/ячейку. На третий день в каждую ячейку добавляли по 100 мкл среды HAT. На 7 и 10 день среду обновляли, отсасывая половину объема из каждой ячейки и замещая ее средой HAT. На 14 день использовали супернатанты гибридомы для отбора анти-СεmX mAbc, связывающихся с UDM-солюбилизированными белками mIgE.FcL или mIgE.Fcs с помощью фермент-зависимого иммуносорбентного анализа (ELISA).
Для отбора гибридом, секретирующих анти-СεmX mAbc, с помощью ELISA, очищенные UDM-солюбилизированные белки mIgE.Fcs или mIgE.Fcs наносили на 96-ячеечные планшеты MaxiSorp (Nunc) в количестве 50 нг/ячейку в 0,1 М NaCO3 при 4°С и оставляли на ночь. Покрытые белком ячейки блокировали 200 мкл 1% BSA в PBS при комнатной температуре в течение 1 часа. Планшеты промывали три раза, добавляя по 200 мкл/ячейку PBS с 0,05% Твин-20 и добавляя затем в ячейки по 100 мкл супернатантов гибридомы. Инкубацию выполняли при комнатной температуре в течение 2 часов. Все ячейки аспирировали и промывали 6 раз, добавляя по 200 мкл/ячейку PBS с 0,05% Твин-20. Планшеты инкубировали с разбавленными 1:10000 конъюгированными с пероксидазой хрена (HRP) анти-мышиными IgG антителами козы (Chemicon) в течение 1 часа (100 мкл/ячейку). Затем все ячейки аспирировали и промывали 6 раз, добавляя по 200 мкл/ячейку PBS с 0,05% Твин-20. Наконец, ячейки проявляли с помощью 50 мкл/ячейку раствора субстрата тетраметил бензидина (ТМВ) (Sure Blue™, KPL) и реакцию останавливали добавлением 50 мкл/ячейку 1N HCl. Измеряли поглощение при OD450 на ридере ELISA. Из >4000 гибридомных клонов, выделенных из двух слияний, 17 клонов продемонстрировали по показаниям ELISA специфичность к mIgE.FcL, а не к mIgE.Fcs.
Чтобы исследовать специфичность анти-СεmX mAbc к СεmX, различные СεmX-специфические клоны тестировали затем на способность реагировать с 3 синтетическими пептидами, представляющими 3 последовательных сегмента СεmX, разделенных С остатком, локализованным в позиции #18, и сегментом СНС, локализованным в #39-41. Конкретно, пептид Р1 содержит последние 4 аминокислотных остатка СН4 цепи mε и первые 17 аминокислотных остатков (#1-17), а именно GLAGGSAQSQRAPDRVL из СεmX; пептид Р2 содержит 20 аминокислотных остатков #19-38, а именно HSGQQQGLPRAAGGSVPHPR, из СεmX; пептид Р3 содержит терминальные 11 аминокислотных остатков (#42-52), а именно GAGRADWPGPP из СεmX, и первые 4 аминокислотных остатка из следующих migis областей, а именно N-терминальную внеклеточную область мембраносвязывающего пептида mε цепи. Все пептиды синтезировали в Исследовательском центре геномики (Genomics Reseqarch Center, Academica Sinica (Taipei, Taiwan)). Пептиды растворяли в PBS при концентрации 10 мг/мл. Все пептиды наносили на 96-ячеечные MaxiSorp планшеты по 500 нг/ячейку в 0,1 М NaCO3 (pH 9,6) при 4°С и оставляли на ночь. Покрытые белком ячейки блокировали, добавляя по 200 мкл 1% BSA в PBS при комнатной температуре в течение 1 часа. Планшеты промывали три раза, добавляя по 200 мкл/ячейку PBS с 0,05% Твин-20 и добавляя затем к ячейкам по 100 мкл 1 мкг/мл анти-CεmX mAbs. Инкубацию выполняли при комнатной температуре в течение 2 часов. Все ячейки аспирировали и промывали 6 раз, добавляя по 200 мкл/ячейку PBS с 0,05% Твин-20. Планшеты инкубировали с разбавленными 1:10000 конъюгированными с пероксидазой хрена (HRP) анти-мышиными IgG антителами козы (Chemicon) в течение 1 часа. После 6-кратного промывания PBS с 0,05% Твин-20 добавлением по 200 мкл/ячейку в каждую ячейку добавляли по 50 мкл/ячейку раствора ТМВ субстрата. Реакцию останавливали добавлением 50 мкл/ячейку 1N HCl. Измеряли поглощение при OD450 на ридере ELISA. Из многих CεmX - специфических моноклональных антител, приготовленных в наших экспериментах, только 4В12 и 26Н2 не реагировали с Р3 пептидом, содержащим RADWPGPP. 4B12 реагировал с пептидом Р1, а 26Н2 с пептидом Р2. Все остальные CεmX-специфические моноклональные антитела реагировали с Р3 (Фигура 1). Таким образом, инкубировали с разбавленными 1:10000 конъюгированными с пероксидазой хрена (HRP) анти-мышиными IgG антителами козы (Chemicon) в течение 1 часа.
Пример 2: 4B12 и 26Н2 связываются с mIgE на mIgE-экспрессирующих В клетках
Авторы далее тестировали способность разных CεmX-специфических моноклональных антител связываться с клеточными линиями СНО и Ramos, которые были трансфектированы рекомбинантной ДНК, кодирующей mεL(CH2-CM) или mεs(CH2-CM). Обе трансфектированные клеточные линии СНО соответственно продуцировали mεL(CH2-CM) или mεs(CH2-CM), ни один из которых не образует полного В клеточного рецептора с корецепторами Igα и Igβ, поскольку клетки СНО не экспрессируют эти белки. Обе трансфектированные клеточные линии Ramos, соответственно, продуцировали mIgE.FcL или mIgE.Fcs, образующие комплексы со своими нативными корецепторами. Чтобы исследовать связывание анти-СεmX mAbc с нативным СεmX, клетки СНО или Ramos, экспрессирующие mIgE.FcL или mIgE.Fcs, ресуспендировали в FACS буфере [PBS, 1% FBS, 0,1% азида натрия и 2 мМ ЭДТА (рН 8,0)] при концентрации 107 клеток/мл. Затем 106 клеток инкубировали 30 мин на льду с 100 мкл супернатантов гибридомы, после чего их отмывали FACS буфером. Связанные антитела регистрировали инкубацией в течение 30 мин на льду с меченным FITC кроличьим фрагментом F(ab')2, специфичным для мышиного IgG (AbD Secrotec), и последующим двукратным отмыванием FACS буфером до начала анализа. Эксперименты с использованием проточной цитометрии выполняли на FACSCanto II проточном цитометре (DD Bioscience) и анализировали с помощью программного обеспечения FCSExpress (De Novo software). Было показано, что все CεmX-специфические моноклональные антитела не связываются с СНО и Ramos клетками, экспрессирующими mIgE.Fcs. Было показано, что все CεmX-специфические моноклональные антитела связываются с СНО клетками, экспрессирующими mIgE.FcL. Однако только 4В12 и 26Н2 могли связываться с Ramos клетками, экспрессирующими mIgE.FcL, в то время как все другие CεmX-специфические моноклональные антитела не могли связываться с Ramos клетками, экспрессирующими mIgE.FcL (фигура 2).
Пример 3: 4В12 и 26Н2 индуцируют зависящую от антител клеточную цитотоксичность против В клеток, экспрессирующих mIgE
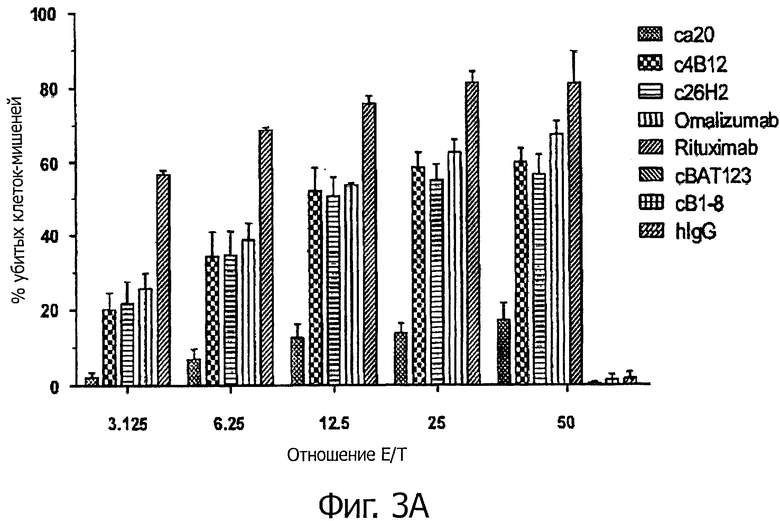
Для исследования активности зависящей от антител клеточной цитотоксичности (ADCC) гибридных анти-CεmX mAbs авторы использовали мононуклеарные клетки периферической крови (PBMCs) в качестве эффекторных клеток для направленного воздействия на Ramos клетки, экспрессирующие mIgE.FcL. PBMCs выделяли из лейкоцитарной пленки здоровых доноров (Taiwan Blood Service Foundation) центрифугированием в градиенте плотности Ficoll-Paque Plus (GE Healthcare) и замораживали в 90% FBS/10% ДМСО (Hybri-Max™; Sigma-Aldrich). Перед использованием PBMCs оттаивали и культивировали в течение ночи при концентрации 2×106 клеток/мл в среде IMDM (Invitrogen) с добавлением 10% инактивированной нагреванием FBS и 1% пенициллин-стрептомициновой смеси. Чтобы идентифицировать клетки-мишени в совместной с PBMCs культуре, Ramos клетки, экспрессирующие mIgE.FcL, метили 2,5 мкМ сукцинимидиловым эфиром диацетата 5-(и 6)-карбоксифлуоресцеина (CFDA, SE; Invitrogen) в 0,1% BSA/PBS в течение 10 мин при 37°С. После трех отмываний холодной средой RPMI (Invitrogen), содержащей 10% FBS, концентрацию клеток доводили до 105 клеток/мл. Для титрования отношений эффектор/мишень (Е/Т) 20000 меченых клеток в 200 мкл полной среды RPMI обрабатывали антителами в концентрации 1 мкг/мл в течение 30 мин при 37°С и затем соединяли с равным объемом PBMCs при разных отношениях Е/Т от 50 до 3,125. Для титрования антител 20000 меченых клеток в 200 мкл полной среды RPMI опсонизировали разными концентрациями антитела (1000-0.01 нг/мл) в течение 30 мин при 37°С, а затем соединяли с PBMCs при отношении Е/Т 25:1. Чтобы измерить не зависящий от антитела лизис клеток, меченые клетки-мишени также инкубировали с PBMCs без антител при данных отношениях Е/Т. К концу 24-часовой инкубации мертвые клетки окрашивали с помощью 2,5 мкг/мл 7-амино актиномицина (7-AAD; Invitrogen) в течение 15 мин на льду. Клетки анализировали на Becton Dickinson FACSCanto II проточном цитометре. Живые клетки-мишени определяли как процент CFSE-положительных/7-AAD-отрицательных клеток при анализе точечной диаграммы. Процентную долю клеток, лизированных при данном отношении Е/Т, вычисляли по следующей формуле: 100 × [(% живых клеток-мишеней в не зависящем от антител контроле - % живых клеток-мишеней в образце)/% живых клеток-мишеней в не зависящем от антител контроле]. Наблюдали активность ADCC с4В12, с26Н2 и omalizumab при различных отношениях Е/Т. При отношении Е/Т 50 с4В12, с26Н2 и omalizumab вызывали до 60% специфического лизиса; напротив, са20 был менее активен и вызывал только 10-20% специфического лизиса (фигура 3А). Кроме того, значительная ADCC наблюдалась, если концентрация с4В12 и с26Н2 была выше чем 0,01 мкг/мл. При максимальной дозе 10 мкг/мл специфический лизис клеток-мишеней антителами с4В12 и с26Н2 находился в интервале от 80% до 90%, в то время как са20 вызывал до 50% специфического лизиса (фигура 3В). Положительный контроль rituximab, направленный против CD20, и omalizumab эффективно индуцировали ADCC дозозависимым образом при различных отношениях Е/Т. Таким образом, авторы сделали вывод, что с4В12 и с26Н2 - более мощные анти-CεmX mAbs, чем са20, при опосредовании ADCC, и могут эффективно мобилизовать эффекторные клетки для направленного воздействия на mIgE-экспрессирующие В клетки in vivo.
Пример 4: Гибридные антитела анти-CεmX mAbs индуцируют апоптоз Ramos клеток, экспрессирующих связанные с мембраной IgE.FcL.
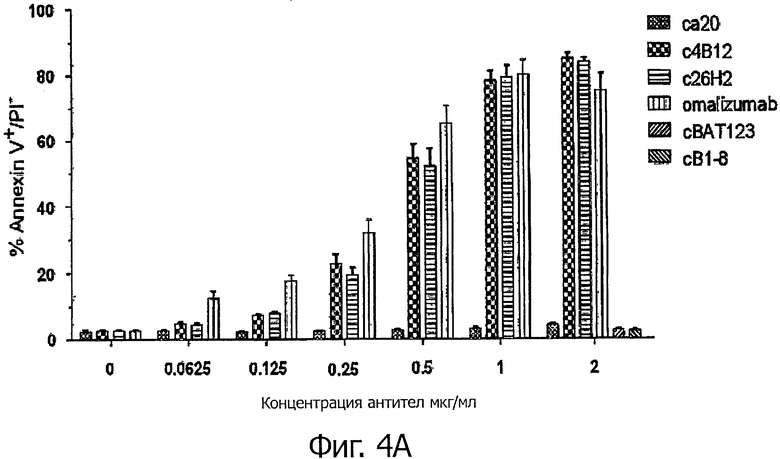
Для регистрации экспозиции фосфатидилсерина (PS) Ramos клетки (5×105 клеток/мл), экспрессирующие mIgE.FcL., инкубировали с гибридными антителами анти-CεmX mAbs, omalizumab или контрольными антителами с указанными концентрациями в полной культуральной среде в течение 1 часа при 37°С. Затем клетки обрабатывали козьим фрагментом F(ab')2, специфическим для Fc фрагмента человеческого IgG (Jackson ImmunoResearch Laboratories Inc.) при концентрации 10 мкг/мл и далее инкубировали 24 часа при 37°С. Регистрацию экспозиции фосфатидилсерина (PS) осуществляли, окрашивая клетки в 200 мкл аннексинового буфера [10 мМ HEPES/NaOH (pH 7,4), 140 мМ NaCl, 5 мМ CaCl2], содержащего Annexin V, меченный флуоресцеин изотиоцианатом (FITC) - (BioVision), разбавленного 1/200, и 2,5 мкг/мл пропидиум иодида (propidium iodide) (PI, Sigma-Aldrich), в течение 15 мин в темноте при комнатной температуре. Клетки анализировали на проточном цитометре FACSCanto II. Апоптотические клетки определяли как процентное отношение Аннексин-положительные/PI отрицательные клетки при анализе точечной диаграммы. Приблизительно 80% Ramos клеток, экспрессирующих mIgE.FcL, были мертвы, поскольку при увеличении концентраций с4В12, с26Н2 или omalizumab, но не са20, наблюдался апоптоз с максимальной индукцией при 1 мкг/мл (фигура 4А).
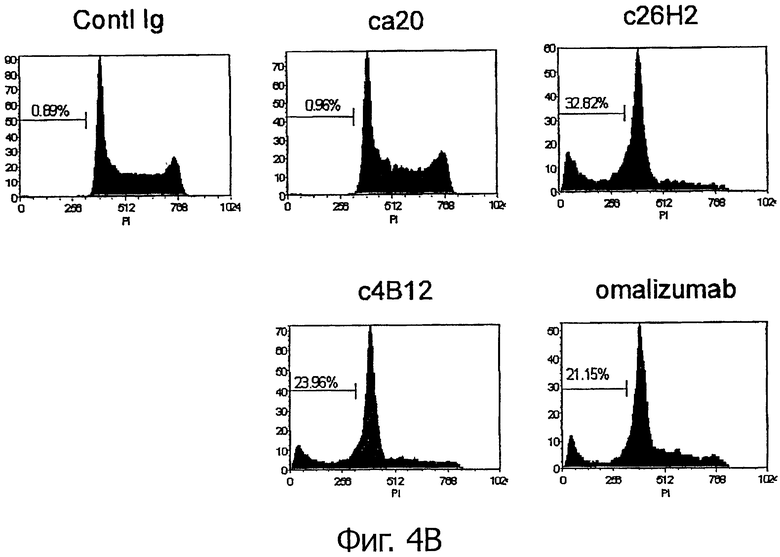
Для регистрации апоптотических ядер Ramos клетки, экспрессирующие mIgE.FcL (5×105 клеток/мл), инкубировали с гибридными антителами анти-CεmX mAbs, omalizumab или контрольными антителами с концентрацией 1 мкг/мл в полной культуральной среде в течение 1 часа при 37°С. Затем клетки обрабатывали козьим фрагментом F(ab')2, специфическим для Fc фрагмента человеческого IgG, при конечной концентрации 10 мкг/мл и далее инкубировали 48 часов при 37°С. 5×105 клеток инкубировали в 0,5 мл пропидиум иодида (PI)/в растворе Тритона (0,1% цитрата натрия, 0,1% Тритона Х-100, 15 мкг/мл PI и 100 мкг/мл РНК-азы А в PBS; все от Sigma-Aldrich) в течение 1 часа в темноте на льду. Флуоресценцию PI определяли на проточном цитометре FACSCanto II. Содержание ДНК в интактных ядрах записывали на линейной шкале. Апоптотические ядра, содержащие гиподиплоидную ДНК, излучающую флуоресценцию в каналах ниже пика G0/G1, определяли в процентах к полной популяции. Значительное увеличение клеточной популяции с гиподиплоидной ДНК наблюдалось в экспрессирующих mIgE.FcL Ramos клетках, обработанных с4В12, с26Н2 или omalizumab (фигура 4В).
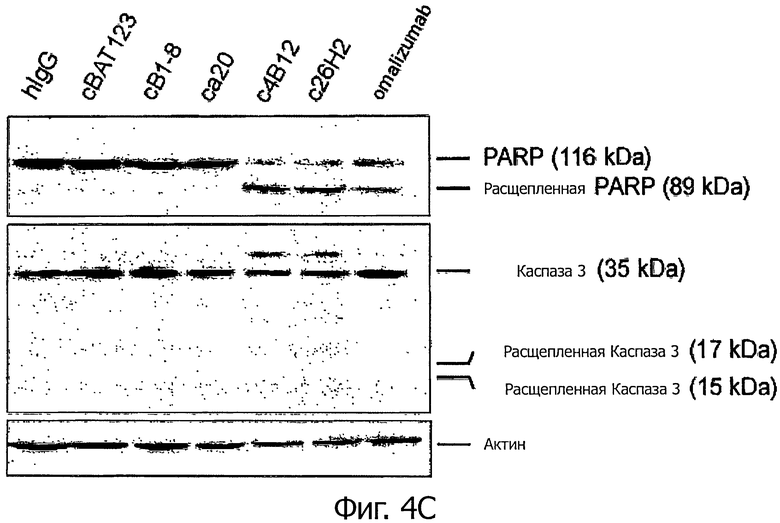
Для регистрации каспазы 3 и расщепления полимеразы поли(АДФ-рибозы) (PARP) Ramos клетки, экспрессирующие mIgE.FcL (5×105 клеток/мл), инкубировали с гибридными антителами анти-CεmX mAbs, omalizumab или контрольными антителами с концентрацией 1 мкг/мл в полной культуральной среде в течение 1 часа при 37°С. Затем клетки обрабатывали козьим фрагментом F(ab')2, специфическим для Fc фрагмента человеческого IgG, при концентрации 10 мкг/мл и далее инкубировали 24 часа при 37°С. 5×106 клеток отмывали охлажденным до 0°С PBS и ресуспендировали в 100 мкл охлажденного до 0°С RIPA буфера [20 мМ Триса (рН 7,4), 150 мМ NaCl, 1% Тритона-Х 100, 0,5% дезоксихолата, 0,1% додецилсульфата натрия (SDS), 5 мМ ЭДТА и ингибитор протеаз (Sigma-Aldrich). Лизаты инкубировали 20 мин на льду. Пробы центрифугировали в течение 20 мин при 16000×g и 4°С. Супернатанты переносили в новые 1,5 мл пробирки и хранили при -80°С. Количество белка в каждом осветленном лизате определяли способом Protein DC (Bio-Rad Laboratories) в соответствии с рекомендациями изготовителя. Каждую пробу нормировали на полное содержание белка и подвергали SDS-электрофорезу в полиакриламидном геле (SDS-PAGE), после чего добавляли к PVDF мембранам (GE Healthcare). Кроличьи поликлональные антитела к каспазе-3 и PARP получали от Cell Signaling Technology и применяли в разведениях 1:500. Конъюгированные с HRP козьи антикроличьи вторичные IgG антитела (Sigma-Aldrich) применяли в разведениях 1:10000. Мембраны готовили с EGL-реагентом (Immobilon™ Western; Millipore). Эквивалентную белковую загрузку проверяли по пятну, соответствующему антителу к β-актину (Sigma-Aldrich). Через 24 часа после обработки экспрессирующих mIgE.FcL Ramos клеток с4В12, с26Н2 или omalizumab, но не са20, наблюдали расщепление каспазы на Mr фрагменты по 19 и 17 - Kda. Кроме того, в обработанных с4В12, с26Н2 или omalizumab экспрессирующих mIgE.FcL Ramos клетках наблюдали расщепление PARP с помощью антитела, различающего Mr 116 кДа интактной PARP и Mr 89 кДа продукта деградации (фигура 4С).
Краткое описание чертежей
Фигура 1 демонстрирует три синтетических пептида, представляющих последовательные сегменты CεmX и способность реагировать различных анти-CεmX mAbc с этими пептидами. Аминокислотные остатки домена CεmX показаны жирным шрифтом.
Фигура 2 демонстрирует связывание различных анти-hu4B12 mAbc с клеточными линиями СНО или Ramos, экспрессирующими белки mIgE.FcL или mIgE.Fcs.
Фигура 3А демонстрирует, что гибридные антитела с4В12 и с26Н2 индуцируют ADCC против экспрессирующих mIgE.FcL Ramos клеток при различных отношениях Е/Т. Фигура 3В демонстрирует, что гибридные с4В12 и с26Н2 индуцируют ADCC против экспрессирующих mIgE.FcL Ramos клеток дозозависимым образом.
Фигура 4А демонстрирует, что экспозиция PS, индуцированная гибридными с4В12 и с26Н2 на экспрессирующих mIgE.FcL Ramos клетках, дозозависима. Фигура 4В демонстрирует, что в обработанных гибридными антителами с4В12 и с26Н2 Ramos клетках, экспрессирующих mIgE.FcL, наблюдаются апоптотические ядра. Фигура 4С демонстрирует, что в обработанных гибридными антителами с4В12 и с26Н2 Ramos клетках, экспрессирующих mIgE.FcL, наблюдается расщепление каспазы 3 и PARP.
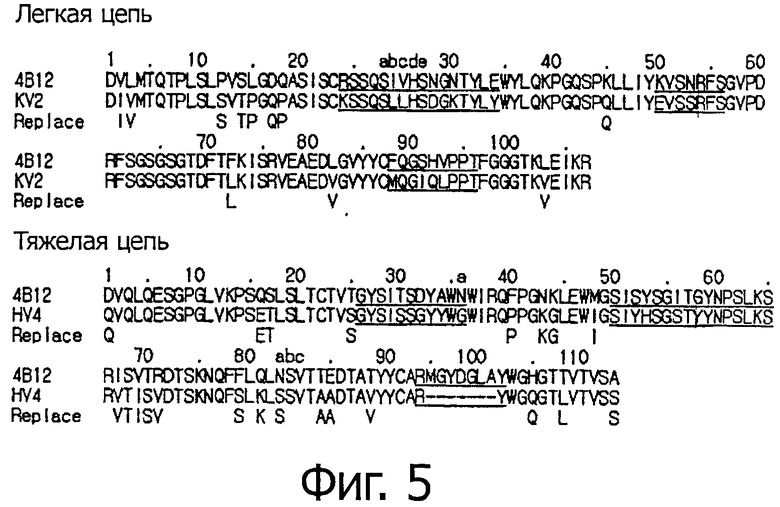
Фигура 5 демонстрирует линейное расположение аминокислотных последовательностей VL и VH родительского мышиного антитела 4В12, выбранных матриц зародышевых клеток человека KV2 и HV4 для VL и VH, соответственно, и гуманизированного антитела 4В12 (hu4B12), обозначенного в последовательностях как «Replace». Это антитело hu4B12 имеет такое же сродство к рекомбинантным белкам СεmX и к Ramos клеткам, экспрессирующим mIgE.FcL, как химерные 4В12 (с4В12).
Литература
Родственные патентные документы
US 5091313 2/1992, Chang
US 5254671 10/1993, Chang
US 5260416 11/1993, Chang
US 5274075 12/1993, Chang
US 5292867 3/1994, Chang
US 5342924 8/1994, Chang
US 2009/0010924 A1, Wu
Другие ссылки
Davis FM, Gossett LA, Chang TW (1991) An epitope on membrane-bound but not secreted IgE: implications in isotype-specific regulation. Bio/Technology 9: 53-56.
Peng С, Davis FM, Sun LK, Liou RS, Kim YW, Chang TW (1992) A new isoform of human membrane-bound IgE. J Immunol 148: 129-136.
Chen, H.Y., Liu, F.T., Hou, C.M.H., Huang J.S.W., Sharma, B.B. and Chang, T.W. (2002) Monoclonal antibodies against CεmX domain in human membrane-bound IgE and their potential on targeting IgE-expressing В cells. Int. Archives Allergy & Immunol. 128, 315-324.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕПТИДНЫЕ ИММУНОГЕНЫ И СОДЕРЖАЩИЕ ИХ СОСТАВЫ, НАЦЕЛЕННЫЕ НА МЕМБРАНОСВЯЗАННЫЕ IgE ДЛЯ ЛЕЧЕНИЯ ОПОСРЕДОВАННЫХ IgE АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2769983C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ B-ЛИМФОЦИТОВ, ПРОДУЦИРУЮЩИХ IGE | 1988 |
|
RU2047177C1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИХСЯ С Ig E | 1988 |
|
RU2082428C1 |
| НОВЫЕ АНТИ-IL 13 АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2004 |
|
RU2487887C2 |
| ИЗОЛИРОВАННОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, СПЕЦИФИЧЕСКОЕ(ИЙ) ДЛЯ ЧЕЛОВЕЧЕСКОГО ИММУНОГЛОБУЛИНА Е (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КЛЕТКА-ХОЗЯИН, СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПРИМЕНЕНИЕ ЕЕ ДЛЯ ЛЕЧЕНИЯ РАССТРОЙСТВ, АССОЦИИРОВАННЫХ С IgE | 2008 |
|
RU2466140C2 |
| АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА FcγRIIB И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2404991C2 |
| АНТИТЕЛА | 2005 |
|
RU2482131C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| АПОПТОТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IgE | 2008 |
|
RU2500686C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2535992C2 |








Изобретение относится к области иммунологии. Рассмотрено применение пептида с аминокислотной последовательностью GLAGGSAQSQRAPDRVL для отбора СεmX-специфических антител и антигенсвязывающих фрагментов таких антител. Предложено СεmX-специфическое антитело, а также способ, предусматривающий отбор антител, специфически связывающих пептид по изобретению, и способ лечения заболеваний, опосредованных IgE, включающий введение субъекту антитела по изобретению. Обнаружено, что моноклональные антитела, специфически связывающие сегмент СεmX GLAGGSAQSQRAPDRVL, могут эффективно связываться с mIgE на человеческих В клетках и пригодны для направленного воздействия на эти В клетки для лечения заболеваний, опосредованных IgE. 4 н. и 10 з.п. ф-лы, 5 ил., 4 пр.
1. Применение пептида GLAGGSAQSQRAPDRVL (SEQ ID NO: 2) для отбора CεmХ-специфического антитела или его антигенсвязывающего фрагмента.
2. Изолированное CεmХ-специфическое антитело или его антигенсвязывающий фрагмент, содержащие одинаковые с антителом 4В12 гипервариабельные области, которые представляют собой 24-39 в SEQ ID NO: 5; 55-61 в SEQ ID NO: 5; 94-102 в SEQ ID NO: 5, 26-36 в SEQ ID NO: 8; 51-66 в SEQ ID NO: 8; 98-106 в SEQ ID NO: 8.
3. Антитело или его антигенсвязывающий фрагмент по п.2, где антитело является гибридным, содержащим вариабельные области мышиного моноклонального антитела и константные области человеческого антитела.
4. Антитело или его антигенсвязывающий фрагмент по п.2, где антитело является гуманизированным моноклональным, содержащим в основном гипервариабельные области мышиного моноклонального антитела и скелетные области и константные области человеческих антител.
5. Антитело или его антигенсвязывающий фрагмент по п.2, где антигенсвязывающий фрагмент представляет собой Fab, F(ab)′2 или одноцепочечный Fv.
6. Антитело по п.2 или его антигенсвязывающий фрагмент, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 8, и вариабельную область легкой цепи, которая содержит SEQ ID NO: 5.
7. Антитело по п.2 или его антигенсвязывающий фрагмент, представляющее собой гуманизированное антитело, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи, которая содержит SEQ ID NO: 7.
8. Способ получения СεmX-специфического антитела, включающий стадии иммунизации субъекта белком IgE и отбор антител, специфически связывающих GLAGGSAQSQRAPDRVL (SEQ ID NO:2), или их антигенсвязывающих фрагментов.
9. Способ лечения заболеваний, опосредованных IgE, включающий введение субъекту, нуждающемуся в этом, эффективного количества антитела по п.2 или его антигенсвязывающего фрагмента.
10. Способ по п.9, в котором заболевание, опосредованное IgE, представляет собой аллергическую астму, аллергические риниты или атопические дерматиты.
11. Способ по п.9, в котором заболевание, опосредованное IgE, представляет собой холодовую крапивницу, хроническую крапивницу, холинэргическую крапивницу, хронические риносинуситы, системный мастоцитоз, кожный мастоцитоз, аллергический бронхолегочный аспергиллиоз, рекуррентную идиопатическую ангиоэдему и интерстициальный цистит или связанные с эозинофилами желудочно-кишечные нарушения.
12. Способ по п.9, в котором антитело вводятся субъекту в количестве, эффективном для индукции антителозависимой клеточной цитотоксичности в IgE экспрессирующих В-клетках.
13. Способ по п.9, в котором антитело или его антигенсвязывающий фрагмент вводят в количестве, эффективном для индукции апоптоза IgE экспрессирующих В-клеток.
14. Способ по п.9, в котором антитело или его антигенсвязывающий фрагмент вводят в количестве, эффективном для уменьшения IgE экспрессирующих В-клеток.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |
| CHEN H.Y | |||
| et al., "Monoclonal antibodies against the C(epsilon)mX domain of human membrane-bound IgE and their potential use for targeting IgE-expressing B cells." Int Arch Allergy Immunol, 2002; 128(4):315-324 | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| POGGIANELLA M | |||
| et al., | |||

