ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Высокоагрессивные Т-клетки часто являются причиной нежелательных иммунологических реакций, которые в свою очередь вызывают различные нарушения, например аутоиммунные заболевания, отторжение трансплантатов, аллергические заболевания и Т-клеточные виды рака. Вследствие этого сокращение количества агрессивных Т-клеток является важным фактором при лечении таких нарушений. Активность данных клеток может ограничиваться за счет иммунодепрессии либо индуцирования иммунологической толерантности. Альтернативным решением является индуцирование апоптоза, который, по-видимому, участвует в удалении нежелательных клеток, включая высокоагрессивные Т-клетки. См., например, Kabelitz и др. (1993) hnmunol Today 14, 338-340; and Raff (1992) Nature 356, 397-399.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение относится к антителам и их производным, индуцирующим апоптоз за счет присоединения к гликопротеиновому лиганду-1 Р-селектина (PSGL-1) на активированных Т-клетках.
С одной стороны, изобретение представляет собой иммуноглобулиновую цепь, имеющую три последовательности, которые: (1) содержат, соответственно, RSSQSIVHNDGNTYFE, KVSNRFS и FQGSYVPLT (SEQ ID №№1-3); (2) содержат, соответственно, SFGMH, YINGGSSTIFYANAVKG и YASYGGGAMDY (SEQ ID №№4-6); (3) содержат, соответственно, RASSTVNSTYLH, GSSNLAS и QQYSGYPLT (SEQ ID №№7-9); (4) содержат, соответственно, AYYIH, VNPNTGGTSYNPKFKG и SGSPYYRYDD (SEQ ID №№10-12); (5) содержат, соответственно, RSSQSIVNSNGNTYLE, KVSNRFS и FQGSHVPWT (SEQ ID №№13-15) или (6) содержат, соответственно, TNAMNWVRQAPGKGLE, TYYADSVKD и GGSYWYFDV (SEQ ID №№16-18).
Каждая из указанных шести групп последовательностей соответствует трем гипервариабельным участкам (hv-участкам) легкой или тяжелой цепи антитела, которое связывается с PSGL-1, например участкам трех антител мыши 15А7, 43В6 и 9F9, описанных ниже в примерах. Ниже показаны вариабельные области (V) легких и тяжелых цепей трех указанных антител (SEQ ID №№ 19-26, hv-участки подчеркнуты и выделены):
SEQ ID № 19 (V область легкой цепи антитела мыши 15А7):

SEQ ID № 21 (V область легкой цепи антитела мыши 43В6):

SEQ ID №23 (V область легкой цепи антитела мыши 9F9):

Так как антигенсвязывающая специфичность антител определяется hv-участками легкой и тяжелой цепи, описанные выше hv-участки позволяют генерировать производные антител, сохраняющие антигенсвязывающую специфичность. В качестве примеров производных антител можно привести химерные антитела, гуманизированные антитела, а также их функциональные эквиваленты. Ниже показаны V области легкой цепи (SEQ ID № 25) и тяжелой цепи (SEQ ID № 26) гуманизированного антитела 15А7, которые соответственно включают SEQ ID №№ 1-3 и SEQ ID №№ 4-6:

К области данного изобретения относится также выделенная нуклеиновая кислота, которая включает последовательность, кодирующую одну из вышеуказанных иммуноглобулиновых цепей. Термин «антитело» либо «иммуноглобулиновая цепь» относится к изолированному полипептиду, т.е. полипептиду, в значительной степени отделенному от прочих белков, липидов и нуклеиновых кислот, с которыми он связан в природе. Полипептид может составлять не менее чем 50, 70 либо 95% сухой массы очищенного препарата. «Выделенная нуклеиновая кислота» относится к нуклеиновой кислоте, структура которой отличается от структуры любой природной нуклеиновой кислоты или любого фрагмента природной геномной нуклеиновой кислоты. Таким образом, термин относится, например, к (а) ДНК, включающей последовательность части природной геномной молекулы ДНК, но не фланкированной кодирующими последовательностями, которые фланкируют указанную часть молекулы в геноме организма, где она находится в природе; (б) нуклеиновой кислоте, включенной в вектор либо в геномную ДНК прокариота или эукариота таким образом, что образующаяся молекула не идентична ни одному природному вектору или геномной ДНК; (в) отдельной молекуле, такой как кДНК, геномному фрагменту, фрагменту, образованному за счет полимеразной цепной реакции (ПЦР), либо фрагменту рестрикции; (д) рекомбинантной нуклеотидной последовательности, являющейся частью гибридного гена, т.е. гена, кодирующего слитый белок. Нуклеиновая кислота в соответствии с данным изобретением пригодна для экспрессирования полипептида в соответствии с данным изобретением. С этой целью нуклеиновая кислота может быть связана с соответствующей регуляторной последовательностью для генерирования экспрессирующего вектора.
Вектор представляет собой молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой он связан, а также способную к независимой репликации или включению в хозяйскую ДНК. В качестве примеров можно назвать плазмидные, космидные и вирусные векторы. Вектор в соответствии с данным изобретением включает нуклеиновую кислоту в форме, пригодной для экспрессии нуклеиновой кислоты в клетке-хозяине. Предпочтительно, вектор включает одну (или более) регуляторную последовательность, оперативно связанную с последовательностью нуклеиновой кислоты, подлежащей экспрессии. В качестве примеров регуляторных последовательностей можно привести промоторы, энхансеры и другие регуляторы экспрессии (например, сигналы полиаденилирования). К регуляторным последовательностям относятся также последовательности, которые регулируют конституционную экспрессию нуклеотидной последовательности, а также тканеспецифические регуляторные и/или индуцированные последовательности. Разработка такого экспрессирующего вектора основана на исследованиях, включающих подбор клетки-хозяина и требуемого уровня экспрессии. Экспрессирующий вектор можно ввести в клетку-хозяин для получения полипептида - предмета данного изобретения. Изобретение включает также клетку-хозяин, содержащую вышеописанную нуклеиновую кислоту. Клетка-хозяин - клетка, содержащая экзогенную кодирующую последовательность либо некодирующую последовательность. Экзогенную последовательность можно вводить в клетку методом трансфекции с фосфатом кальция, с DEAE декстраном либо методом электропорации. К таким клеткам-хозяевам относятся бактериальные клетки (например, Е. coli. Bacillus subtilis и Salmonella typhimurium), дрожжевые клетки (пивные дрожжи - Saccharomyces cerevisiae и Schizosaccharomyces pombe), клетки растений (табак - Nicotiana tabacum и хлопчатник - Gossypium hirsutum), a также клетки млекопитающих (гибридомы мышей, яичника китайского хомячка и фибробласты 3Т3).
Для получения иммуноглобулиновой цепи в соответствии с данным изобретением клетку-хозяин культивируют в условиях, позволяющих экспрессию полипептида, кодируемого описанной выше нуклеиновой кислотой, и затем полипептид выделяют из среды культивации. Вместо этого, возможна транскрипция и трансляция нуклеиновой кислоты - предмета данного изобретения in vitro, например, с помощью регуляторной последовательности промотора Т7 и Т7-полимеразы.
Данное изобретение охватывает также антитело. Его образует первая иммуноглобулиновая цепь и вторая иммуноглобулиновая цепь, которые соответственно содержат hv-участки легкой цепи и hv-участки тяжелой цепи антител 15А7, 43В6 и 9F9 мышей, упомянутых выше. Предпочтительно, данное антитело образовано легкими и тяжелыми цепями 15А7. Помимо этого, к предмету изобретения относится другое антитело, которое (1) специфически связывается с гликопротеиновым лигандом-1 Р-селектина, не нарушая связывания гликопротеинового лиганда-1 Р-селектина и Р-селектина, (2) при связывании с гликопротеиновым лигандом-1 Р-селектина (PSGL-1) на активированных Т-клетках индуцирует гибель Т-клеток. В одном из вариантов данное антитело специфически связывается с гликопротеиновым лигандом-1 Р-селектина человека.
Кроме того, данное изобретение включает еще одно антитело, которое специфически связывается с аминокислотными остатками 115-126 зрелого гликопротеинового лиганда-1 Р-селектина человека. Предпочтительно, антитело специфически связывается с аминокислотными остатками 117-123. Более предпочтительно, оно специфически связывается с аминокислотными остатками 119-121 - обобщающей типичной последовательностью по всем исследованным детерминантам. Действительно, мутация одного или более из этих аминокислотных остатков прекращает связывание антител. В одном примере данные антитела при связывании с гликопротеиновым лигандом-1 Р-селектина на активированной Т-клетке вызывают гибель таких клеток.
В одном из вариантов настоящего изобретения, одно из двух указанных антител образовано легкой цепью и тяжелой цепью, которые соответственно содержат SEQ ID №№1-3 и SEQ ID №№4-6 (например, SEQ ID №№ 19 и 20 либо SEQ ID №№ 25 и 26). Далее, изобретение включает способ стимулирования гибели активированных Т-клеток. Способ основан на контактировании одного из трех описанных выше антител с активированной Т-клеткой, при этом связывание антитела с активированной Т-клеткой приводит к гибели клетки.
Данное изобретение включает также способ модулирования Т-клеточного иммунного ответа в организме пациента. Способ включает (1) выбор пациента, находящегося в состоянии или имеющего риск развития состояния, связанного с избыточным Т-клеточным иммунным ответом, и (2) введение эффективного количества одного из трех описанных выше антител. «Избыточный Т-клеточный иммунный ответ» означает иммунную реакцию, вызванную избыточным уровнем активированных Т-клеток. «Избыточный уровень» означает (1) уровень выше нормального и (2) уровень выше, чем требуется в организме в данном случае, даже если он не превышает нормальный уровень. В качестве примеров таких состояний можно назвать воспалительные заболевания, аутоиммунные заболевания, аллергические заболевания и Т-клеточные виды рака, а также ситуации наличия или планирования аллогенной или ксеногенной трансплантации.
Ниже приведено более подробное описание одного или нескольких вариантов воплощения изобретения. В процессе описания приведены прочие признаки, объекты воздействия и преимущества данного изобретения.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение основано, по крайней мере частично, на неожиданном открытии возможности вызвать апоптоз активированных Т-клеток, которые удаляются за счет связывания антител или их производных с лигандом PSGL-1 на активированных Т-клетках. Антитела и их производные могут использоваться для лечения состояний, вызванных избыточным или нежелательным Т-клеточным иммунным ответом или пролиферацией Т-клеток.
Соответственно, изобретение относится к полипептидам, содержащим hv-участки легких и тяжелых иммуноглобулиновых цепей антител анти-PSGL-1, а также к нуклеиновым кислотам, их кодирующим. Как цепи иммуноглобулина, так и нуклеиновые кислоты позволяют получать антитела и производные.
Иммуноглобулиновая цепь в соответствии с данным изобретением может быть получена как синтетический полипептид или рекомбинантный полипептид. Для получения рекомбинантного полипептида кодирующую его нуклеиновую кислоту присоединяют к другой нуклеиновой кислоте, кодирующей партнера слияния, например глютатион-8-трансферазу (GST), эпитоп 6х-His, белок М13 Gene 3 либо С-область тяжелой цепи иммуноглобулина. Полученную таким образом слитую нуклеиновую кислоту вводят в клетку для экспрессии белка. Слитый белок отделяют от клетки-хозяина известными в данной области методами. Выделенный слитый белок можно далее обрабатывать, например, под действием ферментов для удаления партнера слияния с получением целевого рекомбинантного полипептида. Помимо этого, иммуноглобулиновая цепь может быть получена из соответствующей клетки-хозяина путем активации эндогенной экспрессии нуклеиновой кислоты, кодирующей данную цепь.
Аминокислотный состав иммуноглобулиновой цепи в соответствии с изобретением может варьировать без нарушения способности к образованию антител для связывания с лигандом PSGL-1. Например, продукт может содержать одну или более консервативных замен аминокислот.«Консервативной заменой аминокислот» считается замена, при которой аминокислотный остаток замещается на другой аминокислотный остаток с аналогичной боковой группой. Известны семейства аминокислотных остатков с аналогичными боковыми группами. Семейства представляют собой аминокислоты с боковыми группами, имеющими свойства оснований (например, лизин, аргинин, гистидин), кислотными боковыми группами (например, аспарагиновая кислота, глютаминовая кислота), незаряженными полярными боковыми группами (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми группами (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), боковыми группами с бета-разветвлением (например, треонин, валин, изолейцин) и ароматическими боковыми группами (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом заданный остаток заменимой аминокислоты в полипептиде предпочтительно заменяют на другой аминокислотный остаток, принадлежащий к тому же семейству по своей боковой группе. Помимо этого, мутации могут быть введены случайным образом по всей или части полипептида в соответствии с изобретением, например, методом насыщающего мутагенеза, и полученные мутанты могут быть исследованы на образование антител, способных к связыванию с лигандом PSGL-1 с целью выявления возможных вариаций данного изобретения, как описано ниже в приведенных примерах. Так, выражение «иммуноглобулиновая цепь, содержащая SEQ ID №19» включает цепи иммуноглобулинов, содержащие различные вариации SEQ ID №19.
Описанные выше иммуноглобулиновые цепи и их вариации позволяют получать антитело в соответствии с данным изобретением и его производные. Понятие «антитело» включает как целые молекулы, так и их фрагменты, такие как Fab, F(ab')2, Fv, scFv (одноцепочечное антитело) и dAb (доменное антитело; Ward и др. (1989) Nature, 341, 544). Производное антитело означает белок или белковый комплекс, где полипептид представляет собой вариацию данного изобретения. Антитела и производные в соответствии с данным изобретением можно получить методом коэкспрессии соответствующих полипептидов, содержащих hv-участки легких и тяжелых цепей в подходящей клетке-хозяине, как показано в приведенных ниже примерах. Помимо этого, они могут быть получены известными в данной области методами получения моноклональных и поликлональных антител и фрагментов. См., например, Harlow and Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York.
Для приготовления антител в соответствии с данным изобретением лиганд PSGL-1 или его антигенный фрагмент присоединяют к белку-носителю, например KLH, смешанному с адъювантом, и вводят в организм животного. Антитела, которые вырабатываются в организме этого животного, очищают с помощью аффинной хроматографии пептидов. В качестве таких животных как правило используют кроликов, мышей, морских свинок и крыс. Выбор адъюванта для усиления иммунного ответа зависит от вида животного, это может быть адъювант Фрейнда (полный и неполный), неорганический гель, такой как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюрониловые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки и динитрофенол. Полезными адъювантами, пригодными для людей, являются вакцина БЦЖ (бацилла Кальметта-Герена) и микроорганизмы рода Corynebacterium parvum.
Поликлональные антитела, гетерогенные популяции антител, присутствуют в сыворотках иммунизированных объектов. Моноклональные антитела, гомогенные популяции антител к определенному антигену, можно приготовить по стандартной технологии с помощью гибридов. См., например, Kohler et al. (1975) Nature 256, 495; Kohler et al. (1976) Eur. J. Immunol. 6, 511; Kohler et al. (1976) Eur. J. Immunol. 6, 292; and Hammerling et al. (1981) Monoclonal antibodies and Т Cell Hybridomas, Elsevier, N.Y. В частности, моноклональные антитела могут быть получены любым методом, который обеспечивает производство молекул антител стабильным клеточным штаммом в культуре, такой как описана в Патенте США №4376110; The human B-cell hybridoma technique (Kosbor et al. (1983) Immunol Today 4, 72; Cole et al. (1983) Proc. Natl. Acad. Sci. USA 80, 2026) и The EBV-hybridoma technique (Cole et al. (1983) Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77-96). Такие антитела могут принадлежать к любому классу иммуноглобулинов, в т.ч. IgG, IgM, IgE, IgA, IgD, включая любой подкласс данных классов. Гибридомы, продуцирующие моноклональные антитела, относящиеся к данному изобретению, можно культивировать in vitro или in vivo. Благодаря способности продуцировать моноклональные антитела с высоким титром in vivo, такой способ получения является эффективным.
Кроме того, возможно применение методов, разработанных для получения химерных антител. См., например, Morrison et al. (1984) Proc. Natl. Acad. Sci. USA 81, 6851; Neuberger et al. (1984) Nature 312, 604 и Takeda et al. (1984) Nature 314, 452. Химерное антитело представляет собой молекулу, различные части которой получены с помощью разных видов животных, например такие, где V область представляет собой моноклональное антитело мыши, а С область - иммуноглобулин человека. Помимо этого, методы, описанные для производства одноцепочечных антител (Патенты США №4946778 и 4704692), можно модифицировать для получения библиотеки фагов одноцепочечных Fv антител. Одноцепочечные антитела образуются в результате связывания фрагментов тяжелых и легких цепей участка Fv через аминокислотный мостик. Более того, фрагменты антител могут быть сформированы известными методами. Например, такие фрагменты могут представлять собой (но не исключительно) фрагменты F(ab')2, которые могут быть получены пепсиновым гидролизом молекул антител, и фрагменты Fab, которые создаются при восстановлении дисульфидных мостиков фрагментов F(ab')2. Антитела можно также гуманизировать методами, описанными ниже в примерах, либо другими методами, известными в данной области. Например, возможна гуманизация моноклональных антител с требуемой специфичностью связывания в промышленных масштабах (Scotgene, Scotland и Oxford Molecular, Palo Alto, Calif). В объеме данного изобретения применимы полностью гуманизированные антитела, например, экспрессированные в организме генетически измененных животных (см., например, Green et al. (1994) Nature Genetics 7, 13 и Патенты США №5545806 и 5569825).
К тому же, предмет изобретения включает способ стимулирования гибели активированных Т-клеток, например, путем внесения активированных Т-клеток in vitro в контакт с антителом, относящимся к данному изобретению, и введения эффективного количества антител в организм пациента, в этом нуждающегося. Такое воздействие может применяться, если в организме возникло или может возникнуть состояние, связанное с избыточным или нежелательным Т-клеточным иммунным ответом, например, если пациент страдает аутоиммунным заболеванием, в случае отторжения трансплантатов, аллергических заболеваний и Т-клеточных видов рака. Указанный способ может применяться отдельно либо в сочетании с другими лекарствами и видами лечения.
Термин «воздействие» относится к введению композиции пациенту с целью излечения, снятия или облегчения, восстановления, предотвращения либо снижения интенсивности нарушения или симптомов нарушения, вторичного болезненного состояния, вызванного нарушением, либо предрасположенности к появлению нарушения. «Эффективным количеством» считается количество композиции, способное вызвать желаемый медицинский эффект в организме пациента.
Примеры заболеваний для указанного воздействия: сахарный диабет, артрит (в том числе ревматический полиартрит, ювенильный ревматоидный артрит, остеоартрит, а также псориатический артрит), рассеянный склероз, энцефаломиелит, миастения гравис, системная красная волчанка, аутоиммунный тироидит, дерматиты (в том числе атопический дерматит и экзематозный дерматит), псориаз, синдром Шегрена, болезнь Крона, афтозная язва, воспаление радужной оболочки глаза, конъюнктивит, кератоконъюнктивит, диабет первого типа, воспалительные болезни кишечника, язвенный колит, астма, аллергическая астма, кожная красная волчанка, склеродермия, вагинит, проктит, медикаментозный дерматит, эритематозная проказа узелков, аутоиммунный увеит, аллергический энцефаломиелит, острая некротизирующая геморрагическая энцефалопатия, идиопатическая двухсторонняя прогрессирующая нейросенсорная тугоухость, апластическая анемия, истинная эритроцитарная анемия, идиопатическая тромбоцитопения, полихондрия, синдром Вегенера, хронический активный гепатит, злокачественная экссудативная эритема, идиопатические трофические афты, красный плоский лишай, диффузный токсический зоб, доброкачественный гранулематоз, билиарный первичный цирроз печени, задний увеит, интерстициальный легочный фиброз, реакция «трансплантат против хозяина», случаи трансплантации (в т.ч. трансплантация аллогенной или ксеногенной ткани), такие как пересадка костного мозга, печени, либо трансплантация любого органа или ткани, аллергия, например атопическая аллергия, СПИД, и Т клеточные неоплазмы, такие как лейкемия или лимфома.
Согласно одному из методов in vivo, лечебную композицию (например, композицию, содержащую антитела в соответствии с данным изобретением) вводят в организм пациента. В общем случае, антитела суспендируют в фармацевтическом носителе (например, в физиологическом растворе) и вводят орально либо в виде внутривенной, подкожной, внутримышечной, интратекальной или внутрибрюшинной инъекции; интраректально, интравагинально, интраназально, внутрижелудочно, внутритрахеально либо внутрилегочно.
Дозировка препарата зависит от выбранного способа введения; природы лекарственной формы; конкретного заболевания; размера, веса, площади поверхности, возраста и пола пациента; других препаратов, принимаемых одновременно; а также от назначения лечащего врача. Дозы лежат в интервале 0.01-100.0 мг/кг. Значительные изменения дозировки могут быть связаны с разнообразием существующих композиций и разной эффективностью различных путей введения лекарства. Например, дозы при пероральном приеме должны быть выше, чем при внутривенном вливании. Изменения в уровне дозировки могут быть установлены путем стандартных эмпирических процедур оптимизации, хорошо известных в данной области. Капсулирование препарата в соответствующий носитель (например, микрочастицы полимера либо вживляемые приспособления) может повысить эффективность введения, особенно в случае перорального приема.
В дополнение к этому, данное изобретение включает лекарственный препарат, содержащий фармацевтически приемлемый носитель и эффективное количество антител в соответствии с изобретением. Лекарственный препарат пригоден для лечения перечисленных выше заболеваний. Фармацевтически приемлемый носитель может представлять собой растворитель, дисперсную среду, покрытие, антибактериальное и антигрибковое средство, изотоническое средство или средство, замедляющее всасывание.
Лекарственное средство в соответствии с данным изобретением может быть изготовлено в виде различных лекарственных форм для конкретных способов введения с помощью стандартных методов. Например, для перорального приема препарат может быть заключен в капсулу, гель или таблетку. Капсулы могут содержать любой из обычных фармацевтических материалов, например желатин или целлюлозу. Таблетки могут быть сформированы в соответствии с общепринятой практикой путем прессования смеси препарата с твердым наполнителем и смазочным веществом. Твердый наполнитель может представлять собой крахмал и бентонит с добавлением сахара. Также возможно введение препарата в форме таблеток с твердой оболочкой либо капсул со связующим, например лактозой или маннитом, обычным наполнителем и таблетирующей добавкой. Возможно введение лекарственного средства парентеральным способом. Примерами форм для парентерального введения являются водные растворы, изотонический солевой раствор, 5% раствор глюкозы либо другой инертный наполнитель, известный в фармацевтической промышленности. Например, для введения лекарственного препарата пригодны циклодекстрины либо иные солюбилизирующие компоненты, хорошо известные в данной области.
Эффективность лекарственного средства в соответствии с данным изобретением можно оценивать как in vitro, так и in vivo (см. приведенные ниже примеры). Можно провести испытание препарата на способность вызывать гибель активированных Т-клеток in vitro. Для исследования препарата in vivo его вводят в организм животных (например, мышей) и затем измеряют терапевтический эффект. На основании полученных результатов определяют соответствующий интервал дозировок и способ приема.
Конкретные примеры, приведенные ниже, даны с целью иллюстрации, они не ограничивают каким-либо образом описание данного изобретения. Предполагается, что специалист, имеющий квалификацию в данной области, получит возможность полностью применять данное изобретение на основе этого описания без дальнейшей проработки. Все перечисленные здесь публикации полностью включены в виде ссылок.
ПРИМЕР 1: Мышиные моноклональные антитела 15А7, 43В6 и 9F9
Получение антител анти-PSGL-1
Мышиные моноклональные антитела, специфически связывающиеся с PSGL-1 человека (hCD162), получали по стандартной методике. Более конкретно, мышей иммунизировали мембранной фракцией человеческих Т-клеток, активированных фитогемагглютинином, и умерщвляли для получения гибридомных клеточных линий. Супернатанты из полученных гибридомных клеточных линий были подвергнуты скринингу на связывание с клетками СНО (яичник китайского хомячка), которые стабильно экспрессировали hCD162. Те линии, которые продуцировали антитела, связывающиеся с клетками СНО, экспрессирующими hCD162, но не с родительскими клетками СНО, идентифицировали, субклонировали и далее анализировали, как описано ниже.
Среди идентифицированных линий были: m152-15А7, m166-43В6 и m128-9F9. Они продуцировали антитела для IgG1: соответственно 15А7, 43В6 и 9F9. Анализ методом иммуноблоттинга показал, что эти три антитела связывают содержащийся в лизате активированных Т-клеток белок, который можно обнаружить при помощи антитела анти-hCD162 (kpl-1, PharMingen, Сан-Диего, Калифорния).
Указанные три антитела были испытаны на способность вызывать апоптоз активированных Т-клеток. Супернатанты культуры, содержащие моноклональные антитела, секретированные тремя гибридомными клеточными линиями, были соответственно инкубированы либо с неактивированными человеческими Т-клетками (День 0), либо с человеческими Т-клетками, активированными in vitro (День 7) в течение 6 часов. Затем клетки были окрашены аннексином V и подвергнуты анализу методом FACS (клеточная сортировка с возбуждением флуоресценции). CD3-положительные клетки были селектированы с целью подсчета либо человеческих Т-клеток, активированных in vitro, либо дремлющих человеческих Т-клеток. Апоптозные клетки проявляли положительную реакцию на окрашивание аннексином V. В Таблице 1 приведены доли апоптозных Т-клеток среди всех исследованных Т-клеток.
Доля апоптозных Т-клеток
Эти результаты показывают, что антитела мыши 15А7, 43В6 и 9F9 (1) являются специфичными к hCD162 и (2) могут связываться с активированными Т-клетками человека и индуцировать апоптоз активированных человеческих Т-клеток, но не дремлющих Т-клеток.
Апоптозный анализ также проведен в отношении мононуклеарных клеток периферической крови человека (РВМС), активированных фитогемагглютинином. Обнаружено, что антитела индуцировали апоптоз только в активированных Т-клетках, но не в дремлющих Т-клетках, В-клетках и нейтрофилах.
Известно, что антитела, разрушающие Т-клетки, такие как анти-CD3, способны вызывать продуцирование растворимых факторов. Терапия с применением таких антител обычно приводит к вредному синдрому цитокинеза. Чтобы проверить, вызывают ли антитела анти-PSGL-1 аналогичные связанные с цитокинезом побочные эффекты, свежеизолированные мононуклеарные клетки периферической крови человека (РВМС) выращивали вместе с 15А7 в течение 24, 48 или 72 часов. Затем определяли содержание цитокинов в супернатанте. В клетках РВМС, активированных фитогемагглютинином, вырабатывались значительные количества IL-2, TNF-a и IFN-y (положительный контроль), тогда как содержание этих цитокинов в клетках, обработанных 15А7, не поддавалось обнаружению. Эти результаты подтвердили, что анти-PSGL-1 не влияет или слабо влияет на дремлющие периферийные клетки крови в отношении как возбуждения апоптоза, так и активации клеток.
Поскольку вышеописанные антитела селективно вызывают апоптоз активированных Т-клеток без вредного воздействия на дремлющие Т-клетки или другие иммунные клетки, маловероятно, чтобы их введение в организм привело к лимфопении или широкому иммунодефициту, как в случае анти-CD3 или иммунодепрессантов.
Картирование эпитопов антител анти-CD162
С целью картирования связывающих эпитопов мыши 15А7, 43В6 и 9F9 по отношению к человеческому гену CD162 экспрессировали и очищали ряд слитых белков, соответствующих разным участкам человеческого гена CD162. Взаимодействия между слитыми белками и данными моноклональными антителами изучены методом сэндвич-иммуноферментного анализа (ELISA).
Вкратце, фрагменты, соответствующие разным участкам человеческого гена CD162, были экспрессированы как слитые белки с С областью тяжелой цепи гамма 1 иммуноглобулина человека в Е. coli. кДНК, кодирующая константную область тяжелой цепи гамма 1 иммуноглобулина человека, была амплифицирована методом ПЦР с праймерами, имеющими сайт BglII и сайт BamHI. Продукт ПЦР был разрезан рестриктазами BglII и BamHI и субклонирован в вектор рЕТ-32а (производитель - Novagen), который был расщеплен теми же ферментами. Затем кДНК, кодирующие разные участки hCD162, были амплифицированы методом ПЦР с праймерами, содержащими сайт NdeI на конце 5' и сайт BglII - на конце 3'. Продукты ПЦР были расщеплены соответствующими ферментами и слиты без сдвига рамки считывания с последовательностью, кодирующей константную область тяжелой цепи гамма 1 иммуноглобулина человека в векторе рЕТ-32а. Праймеры, использованные в каждой из конструкций, приведены в Таблице 2, а последовательности праймеров приведены в Таблице 3.
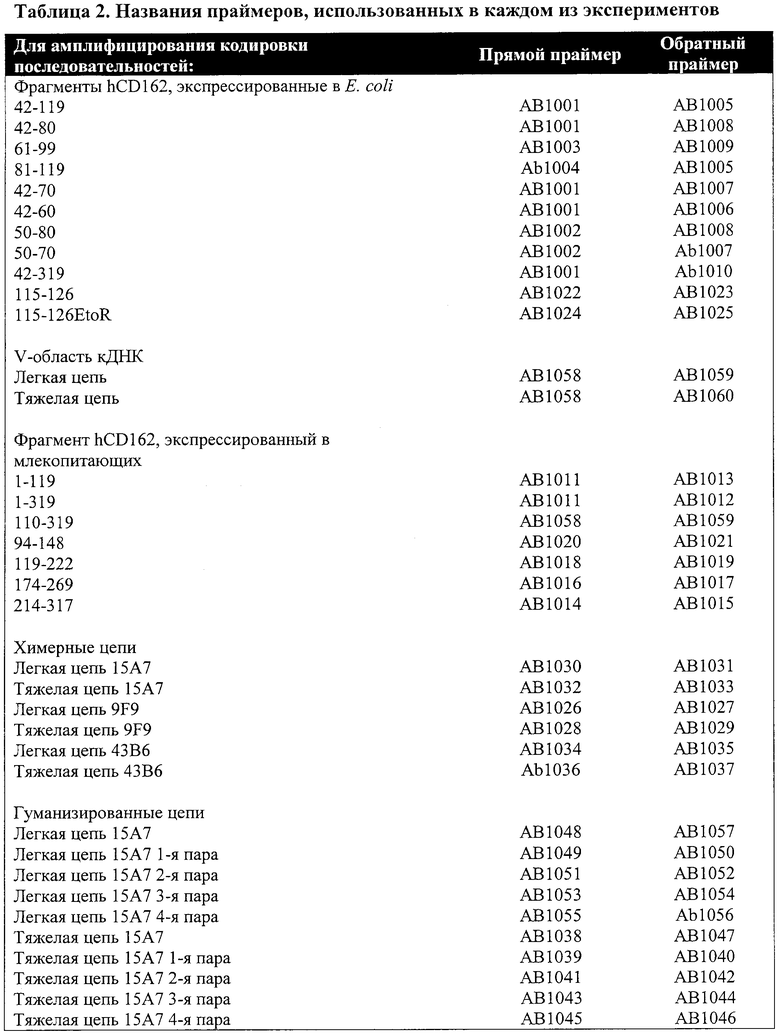


Вышеописанные экспрессионные конструкции были введены в штамм BL21 Escherichia coli (DE3). Трансформированные клетки были собраны через 6 часов после введения ИПТГ (2 мМ) и ресуспендированы в растворе PBS (физиологический раствор с фосфатным буфером). После разрушения клеток ультразвуком и центрифугирования при 14000 g в течение 10 минут полученные супернатанты были собраны для очистки слитых белков. Более точно, супернатанты были сначала инкубированы с гранулами белка G или белка А в течение 3 часов при 4°С. Затем гранулы осаждали центрифугированием при 3000 g и промывали по 5 раз промывочным буфером I (0.05% Тритон Х-100, 50 мМ Трис-HCl, pH 8.5, 400 мМ NaCl, 1 мМ CaCl2 и 1 мг/мл яичного альбумина) и промывочным буфером II (0.05% Тритон Х-100, 50 мМ Трис-HCl, pH 8.5 и 150 мМ NaCl). Затем связанные белки элюировали буфером для элюирования, содержащим 0.1 М соли глицин-HCl с pH 2.7, нейтрализованным 1 М Трис-HCl, pH 8.6. Все очищенные слитые белки количественно определялись при помощи набора для анализа белков фирмы Bio-Rad (Bio-Rad Laboratories, № по каталогу 500-0006) и проверялись методом ПААГ-электрофореза в присутствии додецилсульфата натрия (метод SDS-PAGE).
Для изучения взаимодействия между фрагментами hCD162 и каждым из антител 15А7, 9F9 и 43В6 проводили сэндвич-иммуноферментный анализ (ELISA). В лунки 96-луночных микротитровальных планшетов были сорбированы козьи античеловеческие антитела IgG (Southern Biotechnology, № по каталогу 2040-01) (2 мкг/мл, 50 мкл/ячейку) в течение ночи при 4°С. Планшеты блокировали инкубированием с 0.25% БСА в растворе PBS (150 мкл/ячейку) в течение 1 часа при 37°С. Затем блокированные планшеты инкубировали в течение 2 часов при комнатной температуре со слитыми белками, содержащими разные фрагменты человеческого гена CD162 (2 мкг/мл). После 4-кратной промывки раствором PBS, содержащим 0.05% Tween 20 (раствор PBST), планшеты были инкубированы с тестовыми антителами (2 мкг/мл) в течение 1.5 часов при комнатной температуре. После инкубирования планшеты 4 раза промывали раствором PBST. Затем к каждой ячейке добавляли 50 мкл разбавленного в 1-3000 раз козьего антимышиного IgG, конъюгированного с щелочной фосфатазой (Southern Biotechnology, № по каталогу 1031-04), и планшеты инкубировали в течение 1 часа при 37°С. Ферментативная реакция проводилась добавлением 50 мкл раствора субстрата щелочной фосфатазы (1 таблетка субстрата щелочной фосфатазы, растворенная в 5 мл субстратного буфера, содержащего 0.012 М Na2CO3, 0.16 М NaHCO3 и 1 мМ MgCl2, pH 8.6) с измерением поглощения при 405 нм.
Было обнаружено, что антитела 43В6 и 9F9 способны взаимодействовать со всеми слитыми белками, содержащими остатки с 50 по 60 зрелого человеческого гена CD162, что показывает, что эпитопы антител 43В6 и 9F9 расположены в диапазоне остатков с 50 по 60. В отличие от 9F9 и 43В6, антитела 15А7 связывались только со слитыми белками, охватывающими остатки с 42 по 319, но не со слитым белком, охватывающим остатки 42-119, что показывает, что эпитоп 15А7 расположен между остатками с 119 по 319. Затем местоположение эпитопа 15А7 было уточнено до диапазона остатков от 115 до 126. Замена одной аминокислоты в положении 120 (Glu→Arg) ослабила взаимодействие между антителами 15А7 и слитым белком, что показывает, что первичная область контакта 15А7 с человеческим геном CD162 расположена в положении 120 или вблизи него и что остаток Glu является существенным для этого взаимодействия.
Слитые белки, охватывающие разные участки человеческого гена CD162, также были экспрессированы в клетки млекопитающих и испытаны на взаимодействие с 15А7. Фрагменты, охватывающие эти участки, были экспрессированы в виде слитых белков с константной областью тяжелой цепи гамма 1 иммуноглобулина человека в клетках млекопитающих. Прежде всего, кДНК, кодирующая константную область тяжелой цепи гамма 1 иммуноглобулина человека, была введена в вектор pcDNA3 (производитель - Invitrogen). Затем кДНК, кодирующие разные участки hCD162, были амплифицированы методом ПЦР с праймерами, содержащими сайт BamHI на конце 5' и сайт XhoI на конце 3'. Продукты ПЦР были расщеплены соответствующими ферментами и субклонированы в вектор рЕТ-32а, содержащий ген константной области тяжелой цепи гамма 1 иммуноглобулина человека. Названия и последовательности для всех праймеров приведены выше в Таблицах 2 и 3.
Указанные векторы для экспрессии в клетках млекопитающих были трансфицированы в клетки COS-7 с помощью реагента Lipofectamine 2000 (производитель - hivitrogen, № по каталогу 11668-027) по методике, описанной в руководстве производителя. Трансфицированные клетки были выращены в среде со сверхнизким содержанием иммуноглобулина (производитель - Invitrogen, № по каталогу 16250-078). Экспрессированные белки были очищены и подвергнуты сэндвич-иммуноферментному анализу (ELISA), как описано выше.
Результаты анализа методом ELISA показывают, что только слитые белки, содержащие остатки с 94 по 148, могут взаимодействовать с 15А7. Эти результаты согласуются с мыслью, что эпитоп в 15А7 расположен между остатками 115 и 126.
Все описанные выше результаты показывают, что эпитопы антител 9F9, 43В6 и 15А7 являются зависящими от белка, но не зависящими от модификации углевода, поскольку все эти три антитела связывают экспрессированные в бактериях слитые белки. Они также показывают, что хотя антитела 15А7, 9F9 и 43В6 обладают аналогичными свойствами в отношении специфичности связывания и функции индуцирования апоптоза в активированных Т-клетках, они действуют через разные области человеческого гена CD162 и ведут себя различным образом.
ПРИМЕР 2: Химерные антитела 15А7, 43В6 и 9F9
Клонирование вариабельных (V) областей легких и тяжелых цепей антитела анти-СР162
кДНК, кодирующие V области легких и тяжелых цепей (VL и VH) антител 15А7, 43В6 и 9F9, амплифицировали методом ПЦР. 3'-праймеры были гибридизованы к С-участкам, а 5'-праймеры гибридизованы к G-цепи, присоединенной к кДНК с помощью концевой дезокситрансферазы. ПЦР-фрагменты были клонированы в вектор ПЦР-II (производитель - Invitrogen). Для каждой цепи секвенсировали и сравнивали несколько клонов. Была выбрана последовательность, представленная большей частью независимых клонов. Затем транслированную аминокислотную последовательность анализировали, чтобы убедиться, что выбранная последовательность проявляет свойства типичной V области легких и тяжелых цепей мышиных антител и принадлежит к выбранному субтипу. После этого были определены гипервариабельные участки (hv-участки) путем сравнения транслированной аминокислотной последовательности с обобщающей типичной последовательностью для каждого субтипа. Наименование и последовательность для каждого из использованных праймеров приведены выше в Таблицах 2 и 3. Выявленные последовательности аминокислот для V областей легких и тяжелых цепей 15А7, 43В6 и 9F9 (SEQ ID № 19-24) приведены в Реферате.
Химерные антитела
Для генерирования векторов для экспрессии химерных антител сДНК, кодирующие VL и VH участки 15А7, 43В6 и 9F9, были амплифицированы методом ПЦР с использованием праймеров с включением последовательности 5'-сигнального пептида и 3' донора сплайсированного фрагмента. Праймеры вводили сайты XbaI по обоим концам ПЦР-продуктов, которые затем были разрезаны ферментом XbaI и лигированы с вектором pVK, pVg1, pVg2, or pVg4, разложенным ферментом XbaI. Более конкретно, VL-участки кДНК 15А7, 43В6 и 9F9 были субклонированы в плазмидную pVk. Указанная плазмида содержала CMV промотор и последовательность, кодирующую человеческую константную область легкой цепи. Области VH кДНК 15А7, 43В6 и 9F9 были субклонированы в плазмиды pVg1, pVg2, or pVg4. Каждая из трех плазмид содержала CMV промотор. Они также содержали соответственно С-области тяжелой цепи IgG1, IgG2 и IgG4 человека.
Каждая из трех описанных выше плазмид, кодирующих легкую цепь, была сотрансфицирована с плазмидой, кодирующей тяжелую цепь в клетки COS-7. Собирали супернатанты трансфицированных клеток. Химерные антитела в супернатантах анализировали на наличие способности к связыванию человеческого гена CD162 и к индуцированию апоптоза активированных Т-клеток.
Было обнаружено, что все химерные антитела, полученные из антител 15А7, 43В6 и 9F9, связанных трансфицированными клетками Sp2/0, но не родительскими клетками Sp2/0, устойчиво экспрессировали человеческий ген CD162, что свидетельствует о сохранении специфичности связывания человеческого гена CD162. Более того, было обнаружено, что химерные антитела вызывали апоптоз в Т-клетках, активированных в течение 7 дней, что указывает на сохранение данной функции в случае их мышиных аналогов.
Гуманизированные антитела
Для получения гуманизированных антител применялись мышиные антитела 15А7, hv-участки которых прививали к человеческому каркасу. Для сохранения связывающей способности и специфичности важно не нарушить конформацию V области при пересадке hv-участков на человеческий каркас. Для выбора донора человеческого каркаса аминокислотную последовательность V областей легкой и тяжелой цепи 15А7 сравнивали с таковыми у 50 мышиных антител, которые были гуманизированы.
Обнаружено, что мышиное антитело mDREG-55 имеет высокою гомологию последовательностей с V областью мышиного антитела 15А7 как в легкой, так и в тяжелой цепи. Ниже приведено сравнение последовательностей мышиного антитела 15А7 с указанным антителом mDREG-55 (hv-участки выделены):
cравнение легких цепей:

cравнение тяжелых цепей:

Мышиное антитело DREG-55 - моноклональное антитело IgG1 против L-селектина. Последовательности VL и VH областей мышиного антитела 15А7 были гомологичны соответственно на 64.3% (только для каркаса: 73.8%) и на 70% (только для каркаса: 81.6%) последовательностям мышиного антитела DREG 55. Гуманизированное антитело DREG-55 (HuDREG-55) конструировали, используя каркасные последовательности VL и VH областей человеческого антитела Gal. Для гуманизации мышиного антитела 15А7 каркасную последовательность легкой и тяжелой цепи человеческого антитела Gal использовали для замены аналогичных участков мышиного антитела 15А7.
Гуманизированные легкие и тяжелые вариабельные области 15А7 были скомпонованы каждый 4-мя парами синтетических олигонуклеотидов (длиной ~80 оснований). Олигонуклеотиды в каждой паре были перекрыты приблизительно на 20 нуклеотидов. Последовательности нуклеотидов были выбраны и синтезированы для кодирования белковой последовательности гуманизированных вариабельных областей, в том числе сигнальных пептидов. Компоновку и амплификацию генов проводили в четыре стадии: (1) четыре пары комплементарных олигонуклеотидов гибридизовали и наращивали фрагментом Кленова в результате 4 отдельных реакций; (2) полученные 4 фрагмента дцДНК попарно смешивали, денатурировали, регибридизовали и наращивали в двух отдельных реакциях; (3) полученные 2 фрагмента дцЦНК смешивали, денатурировали, регибридизовали и наращивали с образованием конечной полной дцЦНК и (4) полученную ДНК амплифицировали методом ПЦР с праймерами с целью введения сайта XbaI с обоих концов. Фрагмент ПЦР затем удаляли с помощью XbaI и вводили в векторы pVk и pVg4, разложенные XbaI. Затем в тех положениях, где взаимодействия между hv-участком и каркасом считали важными, остатки антитела Gal снова заменяли на остатки мышиного антитела 15А7 (т.е. I62V и D74H). Ниже показано сравнение мышиного антитела 15А7 и гуманизированного 15А7 (Нu15А7) с mDREG-55, в котором участки V62 и Н74 подчеркнуты.

Полученные при этом плазмиды кодируют тяжелую и легкую цепь гуманизированного 15А7. Указанные плазмиды были затем сотрансфицированы с клетками COS-7. Собирали супернатанты с культивированных клеток. Гуманизированные 15А7 в супернатантах испытывали на способность связываться с трансфицированными клетками СНО, экспрессирующими hCD162, и вызывать апоптоз в Т-клетках, активированных в течение 7 дней. Показано, что данные способности сохранены.
Получение химерных и гуманизированных антител
Генерировали клетки, продуцирующие гуманизированные и химерные антитела. Более конкретно, клетки Sp2/0 (Sp2/0-Agl4; ATCC CRL 1581) были трансфицированы с подходящими плазмидами путем электропорации с помощью прибора Gene Pulser (Bio-Rad Laboratories) при 360 В и 25 мкФ в соответствии с методикой производителя. Перед трансфекцией плазмиды были линеаризованы с ферментом BamHI. Все трансфекции осуществлялись с 107 клеток в растворе PBS и по 20 мкг плазмидных ДНК. Клетки после каждой трансфекции помещали в два 96-луночных планшета для культивирования. Через 48 часов наносили элективную среду (DMEM - модифицированная по способу Дульбекко среда Игла - 10% FBS/ гипоксантин/тимидин) и 1 мкг/мл микофенольной кислоты. Антителообразующие клетки были отделены, наличие антител в супернатанте культуры было определено методом сэндвич-иммуноферментного анализа (ELISA).
Отделенные клетки культивировали в бессывороточной среде (с низким содержанием Ig), собирая супернатант культуры. Антитела очищали, пропуская через колонку с protein A-Sepharose CL-4B, и промывали по 5 раз буфером I (0.05% Тритон X-100, 50 мМ Трис-HCl, pH 8.5, 400 мМ NaCl, 1 мМ CaCl2 и 1 мг/мл яичного альбумина) и промывочным буфером II (0.05% Тритон Х-100, 50 мМ Трис-HCl, pH 8.5 и 150 мМ NaCl). После этого связанные антитела элюировали буфером для элюирования, содержащим 0.1 М соли глицин-HCl с pH 2.7, нейтрализованным 1 М Трис-HCl, pH 8.6.
Изменения аффинности
Связывающую способность вышеописанных мышиного, химерного и гуманизированного антител 15А7 определяли методом конкурентного связывания.
Мышиные антитела 15А7 биотинилировали действием системы EZ-Link сульфо-NHS-биотин (производитель - Pierce Biotechnology, № по каталогу 21217). Кратко, 0.5 мг (3.3×10-6 нмоль) мышиных антител 15А7 растворяли в 187 мкл PBS и смешивали с 6.8×10-5 нмоль системы сульфо-NНS-биотин. Смесь инкубировали на льду в течение 2 часов, после чего свободный биотин удаляли диализом при 4°С против PBS, оставив на ночь. Биотинилированные мышиные антитела 15А7 хранили при 4°С до использования.
Трансфицированные клетки Sp2/0, устойчиво экспрессирующие человеческий ген CD162, были использованы как источник человеческого антигена CD162. Биотилированные мышиные антитела 15А7 применяли в качестве меченого препарата. Возрастающие количества конкурентных антител (мышиного, химерного или гуманизированного 15А7) смешивали с 35 нг биотилированных мышиных антител 15А7 и инкубировали с 1×10-5 клеток Sp2/0, экспрессирующих CD162, в течение 1.5 ч при 4°С при постоянном встряхивании. После промывки к смеси добавляли вторичные антитела стрептавидин-пероксидаза (производитель - Becton Dickinson Immunocytometry System Inc., № по каталогу 349023). После инкубирования в течение 45 минут при 4°С клетки снова промывали и ресуспендировали в 300 мкл PBS-1% FBS, после чего анализировали методом FACS (клеточная сортировка с возбуждением флуоресценции).
Было обнаружено, что полумаксимальная конкурентная концентрация мышиных антител 15А7 составляет 3.72 мкг/мг, а для химерных и гуманизированных антител - соответственно около 5.71 мкг/мл и 4.51 мкг/мл. Полученные результаты показывают, что аффинности мышиных, химерных и гуманизированных антител 15А7 сравнимы между собой, другими словами, связывающая способность (Kа) для мышиных 15А7 составляет 4.03×107 М-1, а для химерных и гуманизированных антител 15А7 - соответственно 2.62×107 М-1 and 3.33×107 М-1.
Конкурентный анализ
Методом конкурентного анализа изучали взаимодействия трех перечисленных выше видов мышиных антител, PSGL-1 и Р-селектина.
Р-селектин представляет собой основной высокоаффинный лиганд для PSGL-1 на большей части лейкоцитов. Для выяснения, способны ли три указанных вида антител предотвращать связывание Р-селектина с PSGL-1, измеряли степень связывания Р-селектина человека с активированными Т-клетками в присутствии трех видов антител. В качестве положительного контроля применяли антитела KPL-1, которые блокируют взаимодействие Р-селектина с PSGL-1.
Мононуклеарные клетки периферической крови человека (РВМС) активировали действием 1% фитогемагглютинина в течение 2 суток и выдерживали в среде, содержащей IL-2, в течение 3 суток. Клетки инкубировали с титрованными антителами 9F9, 15А7, 43В6, KPL-1 (антагонист PSGL-1) либо с контрольными антителами (9Е10) в течение 30 мин, а затем добавляли рекомбинантный Р-селектин человека (1.25 мкг/мл). Связывание Р-селектина с активированными Т-клетками измеряли с помощью FACS анализа анти-Р-селектин-флуоресцеинизотиоцианата.
В соответствии с предыдущими отчетами, антитела KPL-1 в низкой концентрации (0.31 мкг/мл) почти полностью блокировали связывание Р-селектина с активированными Т-клетками. Антитела 43В6 препятствовали связыванию Р-селектина с активированными Т-клетками столь же эффективно, как KPL-1, в то время как в случае 9F9 для достижения равного эффекта требуется более высокая концентрация антител. Действительно, 0.08 мкг/мл KPL или 43В6 снижает связывание на 50%, а для 9F9 эта величина составляет 5 мкг/мл. Антитела 15А7 не оказывали ингибирующего действия на связывание Р-селектина даже в концентрации 20 мкг/мл. Более того, оказалось, что эти антитела способствуют связыванию Р-селектина с PSGL-1. Таким образом, можно заключить, что механизмы связывания антител 15А7 и Р-селектина с PSGL-1 на активированных Т-клетках различны.
Отсутствие конкурентного связывания антител 15А7 и Р-селектина с PSGL-1 позволяет заключить, что введение 15А7 in vivo, по-видимому, не будет влиять на естественный иммунитет за счет ингибирования Р-селектинзависимого обновления лейкоцитов.
Сообщалось об экспрессии PSGL-1 на низком уровне тромбоцитами. Исследовалось влияние антител 15А7 на тромбоциты. Обнаружено, что антитела не усиливают и не снижают агрегацию тромбоцитов в организме человека.
ПРИМЕР 3: Моноклональные антитела хомячка ТАВ4 против мышиного PSGL-1
Моноклональные антитела ТАВ4 к мышиному PSGL-1 были получены аналогично методу, описанному в Примере 1. Они вызывали апоптоз Т-клеток in vitro и элиминировали Т-клетки in vivo. Для выяснения, ингибируют ли данные антитела связывание между мышиным PSGL-1 и мышиным Р-селектином, проводили конкурентный анализ по методу, аналогичному описанному в Примере 2. Было обнаружено, что ТАВ4 не ингибирует связывание мышиного Р-селектина с мышиным PSGL-1, даже при концентрации до 20 мкг/мл.
ПРИМЕР 4: Моноклональные антитела мыши 4В7, 5С4, 12Е7, 14В3, 17Е5 и 18D12
Охарактеризованы дополнительные моноклональные антитела к PSGL-1 человека: 4В7, 5С4, 12Е7, 14В3, 17Е5 и 18D12. При связывании с активированными Т-клетками все они вызывали гибель активированных Т-клеток. Для выяснения, блокируют ли они взаимодействие между PSGL-1 и Р-селектином, проводили конкурентный анализ, как описано в Примере 2. Обнаружено, что эти антитела оказывают слабое ингибирующее действие на связывание человеческого Р-селектина с человеческим PSGL-1 или вообще не оказывают такого воздействия даже при максимальной из исследованных концентраций (5 мкг/мл).
ВАРИАНТЫ ВОПЛОЩЕНИЯ
Признаки, заявленные в данном описании, могут применяться в любых сочетаниях. Каждый из заявленных признаков может быть заменен альтернативным признаком, осуществляющим ту же, эквивалентную или аналогичную функцию. Таким образом, если не указано иначе, каждый из заявленных признаков является только примером из ряда эквивалентных или аналогичных признаков.
Приведенное описание позволяет специалисту легко определить существенные характеристики данного изобретения и осуществлять различные изменения и модификации изобретения, не выходя за рамки его смысла и объема, для адаптации изобретения к различным условиям использования. Таким образом, варианты реализации также относятся к предмету данного изобретения
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К Р-СЕЛЕКТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ И ИДЕНТИФИКАЦИИ | 2011 |
|
RU2603097C2 |
| АНТИ-PSGL-1-АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2650817C2 |
| МОДУЛЯТОРЫ Р-СЕЛЕКТИН ГЛИКОПРОТЕИН ЛИГАНДА 1 | 2004 |
|
RU2407539C2 |
| ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИТЕЛА К PSGL-1 И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766000C2 |
| РЕКОМБИНАНТНЫЕ IL4-АНТИТЕЛА, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ НАРУШЕНИЙ, СВЯЗАННЫХ С ДЕЙСТВИЕМ IL4 | 1994 |
|
RU2162711C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CTLA4 | 2012 |
|
RU2629768C2 |
| Антитела к интерлейкину-1альфа и способы применения | 2013 |
|
RU2666915C2 |
| ОДНОДОМЕННОЕ АНТИТЕЛО И ЕГО ПРОИЗВОДНЫЕ БЕЛКИ К ЛИГАНДУ-1 БЕЛКА ПРОГРАММИРУЕМОЙ СМЕРТИ КЛЕТОК (PDLI) | 2016 |
|
RU2715595C2 |
| ЭПИТОПЫ, ИНДУЦИРУЮЩИЕ ГИБЕЛЬ Т-КЛЕТОК | 2005 |
|
RU2412945C2 |
| АНТИТЕЛО АНТИ-PVRIG ПРОТЕИН ИЛИ ФРАГМЕНТ АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2022 |
|
RU2827062C1 |























Настоящее изобретение относится к области иммунологии. Предложены антитела, которые связываются с гликопротеиновым лигандом-1 Р-селектина (PSGL-1), а также способы стимулирования гибели активированных Т-клеток и моделирования Т клеточного иммунного ответа у пациента посредством антител по изобретению и фармацевтические композиции, содержащие антитела по изобретению. Также раскрываются нуклеиновые кислоты, векторы экспрессии и клетки-хозяева для получения антител по изобретению. Настоящее изобретение может найти дальнейшее применение в терапии заболеваний, ассоциированных с PSGL-1. 17 н. и 18 з.п. ф-лы, 4 пр., 3 табл.
1. Антитело, которое специфически связывается с гликопротеиновым лигандом-1 Р-селектина (PSGL-1), где антитело при связывании с PGSL-1 на активированной Т-клетке вызывает гибель активированной Т-клетки, и где антитело включает легкую цепь и тяжелую цепь, содержащие соответственно SEQ ID NO: 7-9 и SEQ ID NO: 10-12.
2. Антитело по п.1, отличающееся тем, что легкая и тяжелая цепь содержат соответственно SEQ ID NO: 21 и 22.
3. Антитело, которое специфически связывается с гликопротеиновым лигандом-1 Р-селектина (PSGL-1), где антитело при связывании с PGSL-1 на активированной Т-клетке вызывает гибель активированной Т-клетки, и где антитело включает легкую цепь и тяжелую цепь, содержащие соответственно SEQ ID NO: 7-9 и SEQ ID NO: 10-12.
4. Антитело по п.3, отличающееся тем, что легкая и тяжелая цепь содержат соответственно SEQ ID NO: 23 и 24.
5. Антитело, которое специфически связывается с гликопротеиновым лигандом-1 Р-селектина (PSGL-1), не препятствуя связыванию PSGL-1 с Р-селектином, и где антитело при связывании с PGSL-1 на активированной Т-клетке вызывает гибель активированной Т-клетки, и при этом антитело включает легкую цепь и тяжелую цепь, содержащие соответственно SEQ ID NO: 1-3 и SEQ ID NO: 4-6.
6. Антитело, которое специфически связывается с гликопротеиновым лигандом-1 Р-селектина (PSGL-1), где антитело включает легкую и тяжелую цепь, содержащие соответственно SEQ ID NO: 1-3 и SEQ ID NO: 4-6, и где антитело при связывании с PSGL-1 на активированной Т-клетке вызывает гибель активированной Т-клетки.
7. Антитело по п.6, отличающееся тем, что легкая цепь и тяжелая цепь содержат соответственно SEQ ID NO: 25 и 26.
8. Антитело, которое специфически связывается с аминокислотными остатками 115-126 гликопротеинового лиганда-1 Р-селектина человека (PSGL-1), где антитело при связывании с PSGL-1 на активированной Т-клетке вызывает гибель активированной Т-клетки, и при этом антитело включает легкую цепь и тяжелую цепь, содержащие соответственно SEQ ID NO: 1-3 и SEQ ID NO: 4-6.
9. Антитело по п.8, отличающееся тем, что оно специфически связывается с аминокислотными остатками 117-123.
10. Антитело по п.9, отличающееся тем, что оно специфически связывается с аминокислотными остатками 119-121.
11. Способ стимулирования гибели активированной Т-клетки, включающий контактирование антитела по любому из пп.5-10 с активированной Т-клеткой, при этом связывание антитела с активированной Т-клеткой вызывает гибель активированной Т-клетки.
12. Способ модулирования Т клеточного иммунного ответа в организме пациента, включающий:
выбор пациента, находящегося в состоянии или имеющего риск развития состояния, связанного с избыточным Т-клеточным иммунным ответом и введение в организм пациента эффективного количества антитела по любому из пп.5-10.
13. Способ по п.12, отличающийся тем, что указанное состояние представляет собой воспалительное заболевание, аутоиммунное заболевание и аллергическое заболевание.
14. Способ по п.12, отличающийся тем, что указанное состояние представляет собой Т-клеточный рак.
15. Способ по п.12, отличающийся тем, что указанное состояние представляет собой отторжение аллогенного или ксеногенного трансплантата.
16. Клетка-хозяин, экспрессирующая антитело по любому из пп.5-10, содержащая нуклеиновую кислоту, кодирующую аминокислотную последовательность, приведенную в SEQ ID NO: 19, 20, 25 или 26.
17. Клетка-хозяин по п.16, отличающаяся тем, что она является бактериальной клеткой, дрожжевой клеткой, растительной клеткой, клеткой насекомого или клеткой млекопитающего.
18. Клетка-хозяин по п.17, отличающаяся тем, что клетка млекопитающего представляет собой клетку гибридомы.
19. Клетка-хозяин, экспрессирующая антитело по любому из пп.1 и 2, содержащая нуклеиновую кислоту, кодирующую аминокислотную последовательность, приведенную в SEQ ID NO: 21 или 22.
20. Клетка-хозяин по п.19, отличающаяся тем, что она является бактериальной клеткой, дрожжевой клеткой, растительной клеткой, клеткой насекомого или клеткой млекопитающего.
21. Клетка-хозяин по п.20, отличающаяся тем, что клетка млекопитающего представляет собой клетку гибридомы.
22. Клетка-хозяин, экспрессирующая антитело по любому из пп.3 и 4, содержащая нуклеиновую кислоту, кодирующую аминокислотную последовательность, приведенную в SEQ ID NO: 23 или 24.
23. Клетка-хозяин по п.22, отличающаяся тем, что она является бактериальной клеткой, дрожжевой клеткой, растительной клеткой, клеткой насекомого или клеткой млекопитающего.
24. Клетка-хозяин по п.23, отличающаяся тем, что клетка млекопитающего представляет собой клетку гибридомы.
25. Антитело по п.6, отличающееся тем, что легкая цепь и тяжелая цепь содержат соответственно SEQ ID NO: 19 и 20.
26. Выделенная нуклеиновая кислота, кодирующая антитело по любому из пп.1-10, содержащая нуклеотидную последовательность, соответствующую аминокислотной последовательности, представленной в SEQ ID NO: 19, 20, 21, 22, 23, 24, 25 или 26.
27. Вектор экспрессии, содержащий нуклеиновую кислоту по п.26.
28. Выделенная нуклеиновая кислота, кодирующая вариабельный домен антитела по любому из пп.1-10, содержащий SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 или 18.
29. Вектор экспрессии, содержащий нуклеиновую кислоту по п.28.
30. Клетка-хозяин, экспрессирующая антитело по любому из пп.1-10, содержащая нуклеиновую кислоту, кодирующую аминокислотную последовательность, приведенную в SEQ ID NO: 1-3, SEQ ID NO: 4-6, SEQ ID NO: 7-9, SEQ ID NO: 10-12, SEQ ID NO: 13-15 или SEQ ID NO: 16-18.
31. Клетка-хозяин по п.30, отличающаяся тем, что она является бактериальной клеткой, дрожжевой клеткой, растительной клеткой, клеткой насекомого или клеткой млекопитающего.
32. Клетка-хозяин по п.31, отличающаяся тем, что клетка млекопитающего представляет собой клетку гибридомы.
33. Выделенное антитело, которое специфически связывается с гликопротеиновым лигандом-1 Р-селектина, причем антитело содержит (i) легкую цепь, включающую вариабельную область с аминокислотной последовательностью SEQ ID NO: 19, SEQ ID NO: 25, SEQ ID NO: 21 или SEQ ID NO: 23, связанную с константной областью легкой цепи каппа иммуноглобулина человека, и (ii) тяжелую цепь, включающую вариабельную область с аминокислотной последовательностью SEQ ID NO: 20, SEQ ID NO: 26, SEQ ID NO: 22 или SEQ ID NO: 24.
34. Антитело по п.33, отличающееся тем, что содержит константную область тяжелой цепи IgG1, IgG2 или IgG4 человека.
35. Фармацевтическая композиция для лечения состояния, связанного с избыточным или нежелательным Т-клеточным иммунным ответом, содержащая эффективное количество антитела по п.33 или 34.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| NORMAN K.E | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Blood | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| ГУМАНИЗИРОВАННЫЙ ИММУНОГЛОБУЛИН, СПЕЦИФИЧНЫЙ ДЛЯ БЕЛКА L-СЕЛЕКТИНА ЧЕЛОВЕКА | 1993 |
|
RU2151612C1 |

