Настоящее изобретение описывает неиммуногенную РНК. Эта РНК служит основой для разработки терапевтических агентов для индукции толерантности к аутоантигену и, следовательно, для лечения аутоиммунных заболеваний.
Предшествующий уровень техники
Эволюция иммунной системы привела позвоночных к высокоэффективной сети, основанной на двух типах защиты: врожденном и адаптивном иммунитете. В отличие от эволюционной древней врожденной иммунной системы, которая опирается на инвариантные рецепторы, распознающие общие молекулярные паттерны, связанные с патогенами, адоптивный иммунитет основан на высокоспецифичных рецепторах антигена на В-клетках (В-лимфоциты) и Т-клетках (Т-лимфоциты) и клональном отборе. В то время как В-клетки вызывают гуморальные иммунные ответы путем секреции антител, Т-клетки опосредуют клеточные иммунные ответы, ведущие к разрушению распознаваемых клеток, и играют центральную роль в клеточно-опосредованном иммунитете у людей и животных.
Зрелые Т-клетки распознают антиген и отвечают на него через свои антигенспецифические рецепторы (TCR), которые взаимодействуют с иммуногенными пептидами (эпитопами), связанными с молекулами основного комплекса гистосовместимости (MHC), и представлены на поверхности клеток-мишеней. Например, цитотоксические Т-клетки реагируют на антиген, который представлен в ассоциации с белками MHC-I. Хелперные Т-клетки распознают антиген, представленный на белках MHC-II. Самым непосредственным следствием активации TCR является инициация сигнальных путей, приводящих к клональной экспансии Т-клеток, активация маркеров активации на клеточной поверхности, дифференцировке в эффекторные клетки, индукции цитотоксичности или секреции цитокинов и индукция апоптоза. TCR является частью сложного сигнального механизма, который включает гетеродимерный комплекс α- и β-цепей TCR, корецептор CD4 или CD8 и модуль передачи сигнала CD3. В то время как цепи CD3 передают сигнал активации внутри клетки, гетеродимер TCR α/β отвечает исключительно за распознавание антигена.
В дополнение к критической роли, которую в иммунном ответе играют Т-клетки, также важны дендритные клетки (DC). DC являются профессиональными антигенпрезентирующими клетками, играющими ключевую регулирующую роль в поддержании толерантности к аутоантигенам и в активации врожденного и адаптивного иммунитета против чужеродных антигенов.
Иммунная система также может вызывать нежелательные эффекты. Например, аутоиммунные расстройства характеризуются потерей толерантности к аутоантигенам (собственным антигенам), активацией лимфоцитов, реагирующих на аутоантигены, и патологическим повреждением в органах-мишенях.
Современные методы лечения аутоиммунных расстройств в основном сосредоточены на симптоматическом ответе и ослаблении иммунной системы в целом. Антигенспецифическая терапия недавно стала потенциальной терапией для аутоиммунных расстройств, но выявление соответствующих иммунных реакций оказалось трудным, и эти методы лечения имели ограниченный успех.
Таким образом, существует необходимость в эффективной терапии аутоиммунных заболеваний.
Целью настоящего изобретения было создание агентов для терапии аутоиммунных заболеваний. Эта цель достигается согласно изобретению субъектом формулы изобретения.
Настоящее изобретение охватывает композиции, способы и применения для индукции толерантности к аутоантигену. Согласно изобретению аутоантигены, против которых иммунный ответ характерен для аутоиммунных заболеваний, вводят в форме неиммуногенной РНК, кодирующей пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента. РНК, которую вводят в одном предпочтительном воплощении, делают неиммуногенной за счет включения в РНК модифицированных нуклеотидов, подавляющих РНК-опосредованную активацию врожденных иммунных рецепторов и удаляющих двухцепочечную РНК (дцРНК). Согласно изобретению показано, что иммунизация мышей неиммуногенной РНК полностью блокирует любые признаки аутоиммунного заболевания. Также продемонстрировано, что введение одного эпитопа, вызывающего заболевание, может быть достаточным.
Один аспект изобретения относится к способу лечения аутоиммунного заболевания у субъекта, включающему введение субъекту неиммуногенной РНК, кодирующей пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента.
В одном воплощении аутоиммунное заболевание представляет собой аутоиммунное заболевание, опосредованное Т-клетками.
Один аспект изобретения относится к способу индукции толерантности к аутореактивным Т-клеткам у субъекта, включающему введение субъекту неиммуногенной РНК, кодирующей пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента.
В одном воплощении субъект имеет аутоиммунное заболевание. В одном воплощении аутоиммунное заболевание представляет собой опосредованное Т-клетками аутоиммунное заболевание. В одном воплощении Т-клетки являются аутореактивными с аутоантигеном или клетками, экспрессирующими и предпочтительно презентирующими аутоантиген.
В одном воплощении способов по изобретению аутоиммунное заболевание представляет собой аутоиммунное заболевание ЦНС. В одном воплощении способов по изобретению аутоиммунное заболевание представляет собой рассеянный склероз.
В одном воплощении способов по изобретению неиммуногенная РНК при введении не приводит к активации дендритных клеток, активации Т-клеток и/или секреции IFN-альфа.
В одном воплощении способов по изобретению неиммуногенная РНК становится неиммуногенной за счет включения модифицированных нуклеотидов и удаления дцРНК. В одном воплощении модифицированные нуклеотиды подавляют РНК-опосредованную активацию врожденных иммунных рецепторов. В одном воплощении модифицированные нуклеотиды содержат замену одного или нескольких уридинов нуклеозидом, содержащим модифицированное нуклеиновое основание. В одном воплощении модифицированная нуклеиновое основание представляет собой модифицированный урацил. В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, выбран из группы, состоящей из 3-метил-уридина (m3U), 5-метоксиуридина (mo5U), 5-азауридина, 6-азауридина, 2-тио -5-азауридина, 2-тиоуридина (s2U), 4-тиоуридина (s4U), 4-тио-псевдоуридина, 2-тио-псевдоуридина, 5-гидроксиуридина (ho5U), 5-аминоаллил- уридина, 5-галоген-уридина (например, 5-йод-уридина или 5-бром-уридин), уридин-5-оксиуксусной кислоты (cmo5U), метилового эфира уридин-5-оксиуксусной кислоты (mcmo5U), 5-карбоксиметил-уридина (cm5U), 1-карбоксиметил-псевдоуридина, 5-карбоксигидроксиметил-уридина (chm5U), 5-карбоксигидроксиметил-уридина, метилового эфира (mchm5U), 5-метоксикарбонилметил-уридина (mcm5U), 5-метоксикарбонилметил-2-тио-уридина (mcm5s2U), 5-аминометил-2-тио-уридина (nm5s2U), 5-метиламинометил-уридина (mnm5U), 1-этил-псевдоуридина, 5-метиламинометил-2-тио-уридина (mnm5s2U), 5-метиламинометил-2-селеноурид (mnm5se2U), 5-карбамоилметил-уридина (ncm5U), 5-карбоксиметиламинометил-уридина (cmn m5U), 5-карбоксиметиламинометил-2-тиоуридина (cmnm5s2U), 5-пропинил-уридина, 1-пропинил-псевдоуридина, 5-тауринометил-уридина (τm5U), 1-тауринометил-псевдоуридина, 5-тауроина, 5-тауроин -уридина (τm5s2U), 1-тауринометил-4-тио-псевдоуридина, 5-метил-2-тио-уридина (m5s2U), 1-метил-4-тио-псевдоуридина (m1s4ψ), 4-тио-1-метил- псевдоуридина, 3-метил-псевдоуридина (m3ψ), 2-тио-1-метил-псевдоуридина, 1-метил-1-деаза-псевдоуридина, 2-тио-1-метил-1-деаза-псевдоуридина, дигидроуридина (D), дигидропсевдуридина, 5,6-дигидроуридина, 5-метил-дигидроуридина (m5D), 2-тио-дигидроуридина, 2-тио-дигидропсевдуридина, 2-метоксиуридина, 2-метокси-4-тиоуридина, 4-метокси-псевдоуридина, 4-метокси-2-тио-псевдоуридина, N1-метил-псевдоуридина, 3- (3-амино-3-карбоксипропил)уридина (acp3U), 1-метил-3- (3-амино-3-карбоксипропил) псевдоуридина (acp3 ψ), 5- (изопентениламинометил)уридина (inm5U), 5-(изопентениламинометил)-2-тиоуридина (inm5s2U), α-тиоуридина, 2'-O-метил-уридина (Um), 5,2'-O-диметилуридина (m5Um), 2'-O-метил-псевдоуридина (ψm), 2-тио-2'-O-метил-уридина (s2Um), 5-метоксикарбонилметил-2'-O-метилуридина (mcm5Um), 5-карбамоилметил-2'-O-метилуридина (ncm5Um), 5-карбоксиметиламинометил-2'-O-метилуридина (cmmm5Um), 3,2'-O-диметилуридина (m3Um), 5- (изопентениламинометил) -2'-O-метилуридина (inm5Um), 1-тиоуридина, дезокситимидина, 2'-F-арауридина, 2'-F-уридина, 2'-ОН-арауридина, 5-(2-карбометоксивинил) уридина и 5-[3-(1-E-пропениламино)уридина. В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, представляет собой псевдоуридин (ψ), N1-метил-псевдоуридин (m1ψ) или 5-метил-уридин (m5U). В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, представляет собой 1-метил-псевдоуридин.
В одном воплощении способов по изобретению неиммуногенная РНК представляет собой мРНК. В одном воплощении способов по изобретению неиммуногенная РНК представляет собой in vitro транскрибированную РНК.
В одном воплощении способов по изобретению аутоантиген связан с аутоиммунным заболеванием. В одном воплощении способов по изобретению аутоантиген представляет собой Т-клеточный антиген. В одном воплощении способов по изобретению аутоантиген происходит из ЦНС. В одном воплощении способов по изобретению аутоантиген представляет собой миелиновый антиген. В одном воплощении способов по изобретению аутоантиген представляет собой миелин-олигодендроцитарный гликопротеин (MOG). В одном воплощении способов по изобретению пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, содержит аминокислоты 35-55 миелин-олигодендроцитарного гликопротеина (MOG).
В одном воплощении способов по изобретению неиммуногенная РНК транзиторно экспрессируется в клетках субъекта.
В одном воплощении способов по изобретению неиммуногенная РНК доставляется в дендритные клетки. В одном воплощении дендритные клетки являются незрелыми дендритными клетками.
В одном воплощении способов по изобретению неиммуногенную РНК включают в состав с носителем для доставки. В одном воплощении средство доставки содержит частицы. В одном воплощении средство доставки содержит липид. В одном воплощении липид содержит катионный липид. В одном воплощении липид образует комплекс с и/или инкапсулирует неиммуногенную РНК. В одном воплощении способов согласно изобретению неиммуногенные РНК включаются в состав с липосомами.
Один аспект изобретения относится к фармацевтической композиции, содержащей неиммуногенную РНК, кодирующую пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента.
В одном воплощении неиммуногенная РНК при введении не приводит к активации дендритных клеток, активации Т-клеток и/или секреции IFN-альфа.
В одном воплощении неиммуногенная РНК становится неиммуногенной за счет включения модифицированных нуклеотидов и удаления дцРНК. В одном воплощении модифицированные нуклеотиды подавляют РНК-опосредованную активацию врожденных иммунных рецепторов. В одном воплощении модифицированные нуклеотиды содержат замену одного или нескольких уридинов нуклеозидом, содержащим модифицированное нуклеиновое основание. В одном воплощении модифицированное нуклеиновое основание представляет собой модифицированный урацил. В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, выбран из группы, состоящей из 3-метил-уридина (m3U), 5-метоксиуридина (mo5U), 5-азауридина, 6-азауридина, 2-тио -5-азауридина, 2-тиоуридина (s2U), 4-тиоуридина (s4U), 4-тио-псевдоуридина, 2-тио-псевдоуридина, 5-гидроксиуридина (ho5U), 5-аминоаллил- уридина, 5-галоген-уридина (например, 5-йод-уридина или 5-бром-уридин), уридин-5-оксиуксусной кислоты (cmo5U), метилового эфира уридин-5-оксиуксусной кислоты (mcmo5U), 5-карбоксиметил-уридина (cm5U), 1-карбоксиметил-псевдоуридина, 5-карбоксигидроксиметил-уридина (chm5U), 5-карбоксигидроксиметил-уридина, метилового эфира (mchm5U), 5-метоксикарбонилметил-уридина (mcm5U), 5-метоксикарбонилметил-2-тио-уридина (mcm5s2U), 5-аминометил-2-тио-уридина (nm5s2U), 5-метиламинометил-уридина (mnm5U), 1-этил-псевдоуридина, 5-метиламинометил-2-тио-уридина (mnm5s2U), 5-метиламинометил-2-селеноурид (mnm5se2U), 5-карбамоилметил-уридина (ncm5U), 5-карбоксиметиламинометил-уридина (cmn m5U), 5-карбоксиметиламинометил-2-тиоуридина (cmnm5s2U), 5-пропинил-уридина, 1-пропинил-псевдоуридина, 5-тауринометил-уридина (τm5U), 1-тауринометил-псевдоуридина, 5-тауроина, 5-тауроин -уридина (τm5s2U), 1-тауринометил-4-тио-псевдоуридина, 5-метил-2-тио-уридина (m5s2U), 1-метил-4-тио-псевдоуридина (m1s4ψ), 4-тио-1-метил- псевдоуридина, 3-метил-псевдоуридина (m3ψ), 2-тио-1-метил-псевдоуридина, 1-метил-1-деаза-псевдоуридина, 2-тио-1-метил-1-деаза-псевдоуридина, дигидроуридина (D), дигидропсевдуридина, 5,6-дигидроуридина, 5-метил-дигидроуридина (m5D), 2-тио-дигидроуридина, 2-тио-дигидропсевдуридина, 2-метоксиуридина, 2-метокси-4-тиоуридина, 4-метокси-псевдоуридина, 4-метокси-2-тио-псевдоуридина, N1-метил-псевдоуридина, 3- (3-амино-3-карбоксипропил)уридина (acp3U), 1-метил-3- (3-амино-3-карбоксипропил) псевдоуридина (acp3 ψ), 5- (изопентениламинометил)уридина (inm5U), 5-(изопентениламинометил)-2-тиоуридина (inm5s2U), α-тиоуридина, 2'-O-метил-уридина (Um), 5,2'-O-диметилуридина (m5Um), 2'-O-метил-псевдоуридина (ψm), 2-тио-2'-O-метил-уридина (s2Um), 5-метоксикарбонилметил-2'-O-метилуридина (mcm5Um), 5-карбамоилметил-2'-O-метилуридина (ncm5Um), 5-карбоксиметиламинометил-2'-O-метилуридина (cmmm5Um), 3,2'-O-диметилуридина (m3Um), 5- (изопентениламинометил) -2'-O-метилуридина (inm5Um), 1-тиоуридина, дезокситимидина, 2'-F-арауридина, 2'-F-уридина, 2'-ОН-арауридина, 5-(2-карбометоксивинил) уридина и 5- [3- (1-E-пропениламино) уридина. В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, представляет собой псевдоуридин (ψ), N1-метил-псевдоуридин (m1ψ) или 5-метил-уридин (m5U). В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, представляет собой 1-метил-псевдоуридин.
В одном воплощении фармацевтической композиции по изобретению неиммуногенная РНК представляет собой мРНК. В одном воплощении фармацевтической композиции по изобретению неиммуногенная РНК представляет собой in vitro транскрибированную РНК.
В одном воплощении фармацевтической композиции по изобретению аутоантиген связан с аутоиммунным заболеванием. В одном воплощении фармацевтической композиции по изобретению аутоантиген представляет собой Т-клеточный антиген. В одном воплощении фармацевтической композиции по изобретению аутоантиген происходит из ЦНС. В одном воплощении фармацевтической композиции по изобретению аутоантиген представляет собой миелиновый антиген. В одном воплощении фармацевтической композиции по изобретению аутоантиген представляет собой миелин-олигодендроцитарный гликопротеин (MOG). В одном воплощении фармацевтической композиции по изобретению пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, содержит аминокислоты 35-55 миелин-олигодендроцитарного гликопротеина (MOG).
В одном воплощении фармацевтической композиции по изобретению неиммуногенная РНК транзиторно экспрессируется в клетках субъекта, которому вводят фармацевтическую композицию.
В одном воплощении фармацевтической композиции по изобретению неиммуногенная РНК доставляется в дендритные клетки субъекта, которому вводят фармацевтическую композицию. В одном воплощении дендритные клетки являются незрелыми дендритными клетками.
В одном воплощении фармацевтической композиции по изобретению неиммуногенная РНК включается в состав с носителем для доставки. В одном воплощении средство доставки содержит частицы. В одном воплощении средство доставки содержит липид. В одном воплощении липид содержит катионный липид. В одном воплощении липид образует комплекс с и/или инкапсулирует неиммуногенную РНК. В одном воплощении фармацевтической композиции по настоящему изобретению, неиммуногенная РНК включается в состав с липосомами.
Один аспект изобретения относится к фармацевтической композиции по изобретению для применения в способах по изобретению.
Другие признаки и преимущества настоящего изобретения будут очевидны из следующего подробного описания и формулы изобретения.
Подробное описание изобретения
Хотя настоящее изобретение подробно описано ниже, следует понимать, что это изобретение не ограничено конкретными методологиями, протоколами и реагентами, описанными в данном документе, поскольку они могут различаться. Кроме того, следует понимать, что терминология, используемая в настоящем документе, предназначена для целей описания только конкретных воплощений, и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения. Если не указано иное, все технические и научные термины, используемые в данном документе, имеют те же значения, которые обычно понимаются специалистом в данной области техники.
Ниже будут описаны элементы настоящего изобретения. Эти элементы перечислены в конкретных воплощениях, однако следует понимать, что они могут быть объединены любым способом и любым числом для создания дополнительных воплощений. Различные описанные примеры и предпочтительные воплощения не должны толковаться как ограничивающие настоящее изобретение только явно описанными воплощениями. Это описание следует понимать как поддерживающее и охватывающее воплощения, которые объединяют явно описанные воплощения с любым количеством раскрытых и/или предпочтительных элементов. Кроме того, любые перестановки и комбинации всех описанных элементов в этом приложении должны рассматриваться раскрытыми описанием настоящей заявки, если контекст не указывает иное.
Предпочтительно термины, используемые в данном документе, определяются в соответствии с описанием «Многоязычного глоссария биотехнологических терминов: (Рекомендации IUPAC)», HGW Leuenberger, B. Nagel и H. Kölbl, Eds., Helvetica Chimica Acta, CH-4010 Базель, Швейцария (1995).
При практическом осуществлении настоящего изобретения будут использоваться, если не указано иное, обычные методы биохимии, клеточной биологии, иммунологии и методики рекомбинантной ДНК, которые описаны в литературе в данной области (см., например, Molecular Cloning:A Laboratory Manual, 2nd Edition, J. Sambrook et al. eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1989).
Всюду по этому описанию и последующей формуле изобретения, если контекст не требует иного, слово «включает», и варианты, такие как «включают» и «включающий», будут подразумеваться как включение заявленного элемента, целого или стадии или группы элементов, целых или стадий, но не исключение любого элемента, целого или стадии или группы элементов, целых или стадий, хотя в некоторых воплощениях может быть исключен такой другой элемент, целое или стадия или группа элементов, целые или стадии, т.е. субъект изобретения заключается во включении указанного элемента, целого или стадии или группы элементов, целых или стадий. Термины «a» и «an» и «the» и аналогичные ссылки, используемые в контексте описания изобретения (особенно в контексте формулы изобретения), должны толковаться как охватывающие как единственное, так и множественное число, если иное не указано в данном документе или нет явного противоречия контексту. Изложение диапазонов значений в данном документе просто предназначено для применения в качестве сокращенного способа обращения индивидуально к каждому отдельному значению, входящему в диапазон. Если не указано иное, каждое индивидуальное значение включается в описание, как если бы оно было индивидуально описано в данном документе.
Все описанные в данном документе способы могут быть выполнены в любом подходящем порядке, если в данном документе не указано иное или иное явно противоречит контексту. Использование любых и всех примеров или примерной формулировки (например, «такой как»), приведенных в данном документе, предназначено просто для лучшей иллюстрации изобретения и не представляет собой ограничение объема изобретения, заявленного иным образом. Ни одна формулировка в описании не должна быть истолкована как указание на любой незаявленный элемент как существенный при практическом осуществлении изобретения.
В тексте этого описания приводятся несколько документов. Каждый из документов, процитированных в данном документе (включая все патенты, заявки на патент, научные публикации, спецификации производителей, инструкции и т.д.), независимо от того, находятся они выше или ниже, полностью включены в настоящее описание ссылкой. Никакую часть настоящего документа не следует считать признанием того, что изобретение не может претендовать на более раннюю дату настоящего описания в силу предшествующего изобретения.
Настоящее изобретение предусматривает лечение или профилактику аутоиммунных заболеваний путем индукции толерантности иммунной системы к аутоантигену, связанному с аутоиммунным заболеванием. Толерантность к аутоантигену индуцируется введением неиммуногенной РНК. Неиммуногенная РНК содержит последовательности, кодирующие пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, который подавляет иммунный ответ на указанный аутоантиген при введении субъекту.
В различных воплощениях неиммуногенная РНК, описанная в настоящем документе, имеет длину от 200 до 20000 нуклеотидов, от 500 до 5000 нуклеотидов, от 500 до 2500 нуклеотидов, в частности от 600 до 2500 нуклеотидов или от 800 до 2000 нуклеотидов.
В соответствии с изобретением предпочтительно вводить неиммуногенную РНК, описанную в данном документе, включенную в состав с носителями или средствами доставки, такими как состав с наночастицами, в частности состав с липоплексом. Соответственно, неиммуногенные молекулы РНК, описанные в настоящем документе, могут присутствовать в препаратах в носителях или средствах доставки, таких как композиции с наночастицами или композиции с наночастицами, в частности состав с липоплексом, как описано в данном документе.
В одном воплощении можно использовать носители для доставки, которые доставляют неиммуногенные молекулы РНК к антигенпрезентирующим клеткам, таким как дендритные клетки (DC) в селезенке, после системного введения. Например, композиции РНК с наночастицами с определенным размером частиц, в которых суммарный заряд частиц близок к нулю или отрицательный, такие как электронейтральные или отрицательно заряженные липоплексы из РНК и липосом, например, липоплексы, содержащие DOTMA и DOPE или DOTMA и холестерин, приводят к существенной доставке РНК в DC селезенки после системного введения. В соответствии с изобретением особенно предпочтительным является препарат РНК из наночастиц, в котором соотношение зарядов положительных зарядов к отрицательным зарядам в наночастицах составляет 1,4:1 или менее, и/или дзета-потенциал наночастиц составляет 0 или менее. В одном воплощении отношение зарядов положительных зарядов к отрицательным зарядам в наночастицах составляет от 1,4:1 до 1:8, предпочтительно от 1,2:1 до 1:4, например от 1:1 до 1:3, например, от 1:1,2 и 1:2, 1:1,2 и 1:1,8, 1:1,3 и 1:1,7, в частности, от 1:1,4 до 1:1,6, например, около 1:1,5. В одном воплощении дзета-потенциал наночастиц составляет -5 или менее, -10 или менее, -15 или менее, -20 или менее или -25 или менее. В различных воплощениях дзета-потенциал наночастиц составляет -35 или более, -30 или более или -25 или более. В одном воплощении наночастицы имеют дзета-потенциал от 0 мВ до -50 мВ, предпочтительно от 0 мВ до -40 мВ или от -10 мВ до -30 мВ. В одном воплощении положительные заряды вносятся, по меньшей мере, одним катионным липидом, присутствующим в наночастицах, а отрицательные заряды вносятся РНК. В одном воплощении наночастицы содержат, по меньшей мере, один вспомогательный липид. Вспомогательный липид может быть нейтральным или анионным липидом.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTMA и DOPE в молярном соотношении от 10:0 до 1:9, предпочтительно от 8:2 до 3:7 и более предпочтительно от 7:3 до 5:5 и в котором соотношение зарядов положительных зарядов в DOTMA к отрицательным зарядам в РНК составляет от 1,8:2 до 0,8:2, более предпочтительно от 1,6:2 до 1:2, еще более предпочтительно от 1,4:2 до 1,1:2 и еще более предпочтительно около 1,2:2.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTMA и холестерин в молярном соотношении от 10:0 до 1:9, предпочтительно от 8:2 до 3:7 и более предпочтительно от 7:3 до 5:5, причем соотношение заряда от положительных зарядов в DOTMA до отрицательных зарядов в РНК составляет от 1,8:2 до 0,8:2, более предпочтительно от 1,6:2 до 1:2, еще более предпочтительно от 1,4:2 до 1,1:2 и еще более предпочтительно около 1,2:2.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTAP и DOPE в молярном соотношении от 10:0 до 1:9, предпочтительно от 8:2 до 3:7 и более предпочтительно от 7:3 до 5:5, причем соотношение заряда от положительных зарядов в DOTMA до отрицательных зарядов в РНК составляет от 1,8:2 до 0,8:2, более предпочтительно от 1,6:2 до 1:2, еще более предпочтительно от 1,4:2 до 1,1:2 и еще более предпочтительно около 1,2:2.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTMA и DOPE в молярном соотношении от 2:1 до 1:2, предпочтительно от 2:1 до 1:1, и в котором соотношение зарядов положительных зарядов в DOTMA к отрицательным зарядам в РНК составляет 1,4:1 или меньше.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTMA и холестерин в молярном соотношении от 2:1 до 1:2, предпочтительно от 2:1 до 1:1, и где соотношение зарядов положительных зарядов в DOTMA к отрицательным зарядам в РНК составляет 1,4:1 или меньше.
В одном воплощении наночастицы представляют собой липоплексы, содержащие DOTAP и DOPE в молярном соотношении от 2:1 до 1:2, предпочтительно от 2:1 до 1:1, и где соотношение зарядов положительных зарядов в DOTAP к отрицательным зарядам в РНК составляет 1,4:1 или меньше.
В одном воплощении неиммуногенная РНК по изобретению составлена в липосомы F12 или F5, предпочтительно липосомы F12.
Согласно изобретению термин «F12» обозначает липосомы, содержащие DOTMA и DOPE в молярном соотношении 2:1, и липоплексы с РНК, которые образуются с использованием таких липосом.
Согласно изобретению термин «F5» обозначает липосомы, содержащие DOTMA и холестерин в молярном соотношении 1:1, и липоплексы с РНК, которые образуются с использованием таких липосом.
Используемый в данном документе термин «наночастица» относится к любой частице, имеющей диаметр, делающий частицу пригодной для системного, в частности парентерального, введения, в частности, нуклеиновых кислот, обычно диаметром менее 1000 нанометров (нм). В некоторых воплощениях наночастица имеет диаметр менее 600 нм. В некоторых воплощениях наночастица имеет диаметр менее 400 нм. В некоторых воплощениях наночастица имеет средний диаметр в диапазоне от около 50 нм до около 1000 нм, предпочтительно от около 50 нм до около 400 нм, предпочтительно от около 100 нм до около 300 нм, например от около 150 нм до около 200 нм. В некоторых воплощениях наночастица имеет диаметр в диапазоне от около 200 до около 700 нм, от около 200 до около 600 нм, предпочтительно от около 250 до около 550 нм, в частности от около 300 до около 500 нм или от около 200 до около 400 нм.
Используемый в данном документе термин «композиция из наночастиц» или подобные термины относятся к любому веществу, которое содержит, по меньшей мере, одну наночастицу. В некоторых воплощениях композиция наночастиц представляет собой однородную совокупность наночастиц. В некоторых воплощениях композиции из наночастиц представляют собой дисперсии или эмульсии. В общем, дисперсия или эмульсия образуются, когда объединяются, по меньшей мере, два несмешивающихся материала.
Термин «липоплекс» или «липоплекс нуклеиновой кислоты», в частности «РНК-липоплекс», относится к комплексу липидов и нуклеиновых кислот, в частности РНК. Липоплексы образуются спонтанно, когда катионные липосомы, которые часто также включают нейтральный «хелперный» липид, смешиваются с нуклеиновыми кислотами.
Если настоящее изобретение относится к заряду, такому как положительный заряд, отрицательный заряд или нейтральный заряд, или к катионному соединению, отрицательному соединению или нейтральному соединению, это обычно означает, что упомянутый заряд присутствует при выбранном рН, таком как физиологический рН. Например, термин «катионный липид» означает липид, имеющий чистый положительный заряд при выбранном рН, таком как физиологический рН. Термин «нейтральный липид» означает липид, не имеющий чистого положительного или отрицательного заряда, и может присутствовать в виде не заряженного или нейтрального амфотерного иона при выбранном рН, таком как физиологический рН. Под «физиологическим pH» в данном документе подразумевается pH около 7,5.
Носители с наночастицами, такие как липидные носители, предполагаемые для применения в настоящем изобретении, включают любые вещества или носители, с которыми может быть связана нуклеиновая кислота, такая как РНК, например, путем образования комплексов с нуклеиновой кислотой или образования везикул, в которых нуклеиновая кислота заключена или инкапсулирована. Это может привести к повышенной стабильности нуклеиновой кислоты по сравнению с «голой» нуклеиновой кислотой. В частности, стабильность нуклеиновой кислоты в крови может быть увеличена.
Катионные липиды, катионные полимеры и другие вещества с положительными зарядами могут образовывать комплексы с отрицательно заряженными нуклеиновыми кислотами. Эти катионные молекулы могут быть использованы для комплексирования нуклеиновых кислот, таким образом, образуя, например, так называемые липоплексы или полиплексы соответственно, и было показано, что эти комплексы доставляют нуклеиновые кислоты в клетки.
Составы нуклеиновых кислот с наночастицами для применения в настоящем изобретении могут быть получены различными протоколами и из различных комплексообразующих соединений нуклеиновых кислот. Липиды, полимеры, олигомеры или амфифилы являются типичными комплексообразующими агентами. В одном воплощении комплексообразующее соединение включает, по меньшей мере, один агент, выбранный из группы, состоящей из протамина, полиэтиленимина, поли-L-лизина, поли-L-аргинина или гистона.
Согласно изобретению протамин полезен в качестве катионного агента-носителя. Термин «протамин» относится к любому из различных сильноосновных белков с относительно низкой молекулярной массой, которые богаты аргинином и обнаруживают, что они связаны, в частности, с ДНК вместо соматических гистонов в сперматозоидах различных животных (например, рыб). В частности, термин «протамин» относится к белкам, обнаруженным в сперме рыб, которые являются сильно основными, растворимыми в воде, не коагулируют при нагревании и дают в основном аргинин при гидролизе. В очищенном виде они используются в составе инсулина длительного действия и для нейтрализации антикоагулянтного действия гепарина. В соответствии с изобретением термин «протамин», используемый в данном документе, означает любую аминокислотную последовательность протамина, полученную или полученную из природных или биологических источников, включая ее фрагменты и мультимерные формы указанной аминокислотной последовательности или ее фрагмента. Кроме того, термин охватывает (синтезированные) полипептиды, которые являются искусственными и специально предназначены для конкретных целей и не могут быть выделены из природных или биологических источников. Протамин, используемый согласно настоящему изобретению, может представлять собой сульфатированный протамин или гидрохлорид протамина. В предпочтительном воплощении источником протамина, используемым для получения описанных в данном документе наночастиц, является протамин 5000, который содержит протамин в количестве более 10 мг/мл (5000 единиц, нейтрализующих гепарин на мл) в изотоническом солевом растворе.
Липосомы представляют собой микроскопические липидные везикулы, часто имеющие один или несколько бислоев липидообразующего липида, такого как фосфолипид, и способны инкапсулировать лекарственное средство. Различные типы липосом могут быть использованы в контексте настоящего изобретения, включая, помимо прочего, мультиламеллярные везикулы (MLV), небольшие однослойные везикулы (SUV), большие однослойные везикулы (LUV), стерически стабилизированные липосомы (SSL), мультивезикулярные везикулы (MV) и большие мультивезикулярные везикулы (LMV), а также другие двухслойные формы, известные в данной области. Размер и ламеллярность липосомы будут зависеть от способа приготовления, а выбор типа используемых везикул будет зависеть от предпочтительного способа введения. Существует несколько других форм супрамолекулярной организации, в которых липиды могут присутствовать в водной среде, включая пластинчатые фазы, гексагональные и обратные гексагональные фазы, кубические фазы, мицеллы, обратные мицеллы, состоящие из монослоев. Эти фазы также могут быть получены в комбинации с ДНК или РНК, и взаимодействие с РНК и ДНК может существенно влиять на фазовое состояние. Описанные фазы могут присутствовать в композициях нуклеиновых кислот с наночастицами по настоящему изобретению.
Для образования липоплексов нуклеиновых кислот из нуклеиновой кислоты и липосом может быть использован любой подходящий способ образования липосом, если он обеспечивает предусмотренные липоплексы нуклеиновых кислот. Липосомы могут быть получены с использованием стандартных способов, таких как способ обратного испарения (REV), способ введения этанола, способ дегидратации-регидратации (DRV), обработка ультразвуком или другие подходящие способы.
После образования липосомы могут быть подвергнуты отбору по размеру для получения популяции липосом, имеющих по существу однородный диапазон размеров.
Двухслойные липиды обычно имеют две углеводородные цепи, в частности, ацильные цепи, и головную группу, полярную или неполярную. Двухслойные липиды состоят либо из встречающихся в природе липидов, либо синтетического происхождения, включая фосфолипиды, такие как фосфатидилхолин, фосфатидилэтаноламин, фосфатидная кислота, фосфатидилинозитол и сфингомиелин, где две углеводородные цепи обычно содержат примерно 14-22 атомов углерода в длину и имеют различные степени ненасыщенности. Другие подходящие липиды для применения в композиции по настоящему изобретению включают гликолипиды и стерины, такие как холестерин и его различные аналоги, которые также можно использовать в липосомах.
Катионные липиды обычно имеют липофильную часть, такую как стерол, ацильная или диацильная цепь, и имеют общий чистый положительный заряд. Головная группа липида обычно несет положительный заряд. Катионный липид предпочтительно имеет положительный заряд от 1 до 10 валентностей, более предпочтительно положительный заряд от 1 до 3 валентностей и более предпочтительно положительный заряд от 1 валентности. Примеры катионных липидов включают, без ограничения указанным, 1,2-ди-O-октадеценил-3-триметиламмонийпропан (DOTMA); диметилдиоктадециламмоний (DDAB); 1,2-диолеоил-3-триметиламмонийпропан (DOTAP); 1,2-диолеоил-3-диметиламмонийпропан (DODAP); 1,2-диацилокси-3-диметиламмонийпропан; 1,2-диалкилокси-3-диметиламмонийпропаны; диоктадецилдиметиламмонийхлорид (DODAC), 1,2-димиристоилоксипропил-1,3-диметилгидроксиэтиламмоний (DMRIE) и 2,3-диолеоилокси-N-[2(спермин карбоксамид)этил]-N,N-диметил-1-пропанамий трифторацетат (DOSPA). Предпочтительными являются DOTMA, DOTAP, DODAC и DOSPA. Наиболее предпочтительным является DOTMA.
Кроме того, описанные в данном документе наночастицы предпочтительно дополнительно включают нейтральный липид с точки зрения структурной стабильности и тому подобного. Нейтральный липид может быть соответствующим образом выбран с учетом эффективности доставки комплекса нуклеиновая кислота-липид. Примеры нейтральных липидов включают, без ограничения указанным, 1,2-ди- (9Z-октадеценоил) -sn-глицеро-3-фосфоэтаноламин (DOPE), 1,2- диолеоил-sn-глицеро-3-фосфохолин (DOPC), диацилфосфатидилхолин, диацилфосфатидилэтаноламин, церамид, сфингомиелин, цефалин, стерол и цереброзид. Предпочтительным является DOPE и/или DOPC. Наиболее предпочтительным является допинг. В случае, когда катионная липосома включает как катионный липид, так и нейтральный липид, молярное отношение катионного липида к нейтральному липиду может быть соответствующим образом определено с точки зрения стабильности липосомы и тому подобного.
В соответствии с одним воплощением описанные в данном документе наночастицы могут включать фосфолипиды. Фосфолипиды могут представлять собой глицерофосфолипид. Примеры глицерофосфолипидов включают, без ограничения указанным, три типа липидов: (i) цвиттерионные фосфолипиды, которые включают, например, фосфатидилхолин (PC), фосфатидилхолин яичного желтка, PC, полученный из соевых бобов, в природной, частично гидрированной или полностью гидрогенизированной форме, димиристоилфосфатидилхолин (DMPC) сфингомиелин (SM); (ii) отрицательно заряженные фосфолипиды: которые включают, например, фосфатидилсерин (PS), фосфатидилинозитол (PI), фосфатидную кислоту (PA), фосфатидилглицерин (PG), дипалмипоил PG, димиристоил фосфатидилглицерин (DMPG); синтетические производные, в которых конъюгат придает цвиттерионному фосфолипиду отрицательно заряженный заряд, например, метоксиполиэтилен, гликольдистеароилфосфатидилэтаноламин (mPEG-DSPE); и (iii) катионные фосфолипиды, которые включают, например, фосфатидилхолин или сфингомиелин, из которых фосфомоноэфир был O-метилирован с образованием катионных липидов.
Ассоциация нуклеиновой кислоты с липидным носителем может происходить, например, за счет заполнения нуклеиновой кислотой межузельных пространств носителя, так что носитель физически захватывает нуклеиновую кислоту, или за счет ковалентной, ионной или водородной связи или посредством адсорбции неспецифическими связями.
Термин «иммунный ответ» относится к реакции иммунной системы, предпочтительно на антиген, и предпочтительно относится к клеточному иммунному ответу, гуморальному иммунному ответу или обоим. Согласно изобретению термин «иммунный ответ» или «иммунный ответ против» в отношении мишени, такой как антиген, клетка или ткань, относится к иммунному ответу, направленному против мишени.
Иммунная система делится на две части, врожденную и адаптивную системы. Адаптивный иммунный ответ зависит от В- и Т-лимфоцитов, которые специфичны к определенным антигенам. Врожденная иммунная система реагирует на общие структуры, разделяемые подавляющим большинством угроз. Эти общие структуры называются патоген-ассоциированными молекулярными структурами, или PAMP, и распознаются толл-подобными рецепторами или TLR. В дополнение к клеточным TLR, важной частью врожденной иммунной системы является гуморальная система комплемента, которая опсонизирует и убивает патогены с помощью механизма распознавания PAMP. Эти высококонсервативные растворимые и мембраносвязанные белки в совокупности называются рецепторами распознавания образов (PRR), и именно взаимодействие PAMP/PRR запускает врожденную иммунную систему.
TLR представляют собой трансмембранные белки, экспрессируемые клетками врожденной иммунной системы, которые распознают вторгающиеся микробы и активируют сигнальные пути, которые запускают иммунные и воспалительные реакции для уничтожения захватчиков. Различные TLR служат в качестве рецепторов для различных лигандов, включая компоненты бактериальной клеточной стенки, вирусную двухцепочечную РНК и низкомолекулярные противовирусные или иммуномодулирующие соединения. У людей TLR3, 7, 8 и 9 реагируют, главным образом, на PAMP на основе нуклеиновых кислот от вирусов и бактерий.
Термины «клеточный иммунный ответ», «клеточный ответ», «клеточный иммунитет» или подобные термины предназначены для включения клеточного ответа, направленного на клетки, характеризующиеся экспрессией антигена и/или презентацией антигена MHC класса I или класса II. Клеточный ответ относится к клеткам, называемым Т-клетками или Т-лимфоцитами, которые действуют как «помощники» или «убийцы». Хелперные Т-клетки (также называемые CD4+ Т-клетками) играют центральную роль, регулируя иммунный ответ, а клетки-киллеры (также называемые цитотоксическими Т-клетками, цитолитическими Т-клетками, CD8+ Т-клетками или CTL) убивают клетки, такие как больные клетки.
Термин «гуморальный иммунный ответ» относится к процессу в живых организмах, в котором антитела продуцируются в ответ на агенты и организмы, которые они в конечном итоге нейтрализуют и/или устраняют. Специфичность ответа антител опосредуется Т и/или В-клетками через мембранно-ассоциированные рецепторы, которые связывают антиген одной специфичности. После связывания соответствующего антигена и получения различных других активирующих сигналов делятся В-лимфоциты, которые продуцируют В-клетки памяти, а также клоны плазматических клеток, секретирующих антитела, каждый из которых продуцирует антитела, которые распознают идентичный антигенный эпитоп, который был распознан его антигенным рецептором. В-лимфоциты памяти остаются бездействующими до тех пор, пока они впоследствии не будут активированы их специфическим антигеном. Эти лимфоциты обеспечивают клеточную основу памяти и результирующую эскалацию ответа антител при повторном воздействии специфического антигена.
Используемый в данном документе термин «антитело» относится к молекуле иммуноглобулина, которая способна специфически связываться с эпитопом на антигене. В частности, термин «антитело» относится к гликопротеину, содержащему, по меньшей мере, две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Термин «антитело» включает моноклональные антитела, рекомбинантные антитела, антитела человека, гуманизированные антитела, химерные антитела и комбинации любых из вышеперечисленных. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи (CH). Каждая легкая цепь состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи (CL). Вариабельные области и константные области также упоминаются в данном документе как вариабельные домены и константные домены соответственно. Области VH и VL можно далее подразделить на области гипервариабельности, которые называются областями определения комплементарности (CDR), чередующимися с областями, которые более консервативны, и называются каркасными областями (FR). Каждый VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. CDR VH называются HCDR1, HCDR2 и HCDR3, CDR VL называются LCDR1, LCDR2 и LCDR3. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антитела включают константную область тяжелой цепи (CH) и константную область легкой цепи (CL), где CH может быть дополнительно подразделен на константный домен CH1, шарнирный участок и константные домены CH2 и CH3 (расположенные от аминоконца к карбоксиконцу в следующем порядке: СН1, СН2, СН3). Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента. Антитела могут быть интактными иммуноглобулинами, полученными из природных источников или из рекомбинантных источников, и могут быть иммуноактивными частями интактных иммуноглобулинов. Антитела обычно представляют собой тетрамеры молекул иммуноглобулина. Антитела могут существовать в различных формах, включая, например, поликлональные антитела, моноклональные антитела, Fv, Fab и F(ab)2, а также одноцепочечные антитела и гуманизированные антитела.
Антитела, описанные в данном документе, включают антитела IgA, такие как IgA1 или IgA2, IgG1, IgG2, IgG3, IgG4, IgE, IgM и IgD. В различных воплощениях изобретения антитело представляет собой антитело IgG1, более конкретно изотип IgG1-каппа или IgG1-лямбда (то есть IgG1, κ, λ), антитело IgG2a (например, IgG2a, κ, λ), антитело IgG2b (например, IgG2b, κ, λ), антитело IgG3 (например, IgG3, κ, λ) или антитело IgG4 (например, IgG4, κ, λ).
Термин «иммуноглобулин» относится к белкам суперсемейства иммуноглобулинов, предпочтительно к антигенным рецепторам, таким как антитела или B-клеточный рецептор (BCR). Иммуноглобулины характеризуются структурным доменом, то есть доменом иммуноглобулина, имеющим характерную иммуноглобулиновую складку (Ig). Термин охватывает мембраносвязанные иммуноглобулины, и растворимые иммуноглобулины. Связанные с мембраной иммуноглобулины также называют поверхностными иммуноглобулинами или мембранными иммуноглобулинами, которые обычно являются частью BCR. Растворимые иммуноглобулины обычно называют антителами. Иммуноглобулины обычно содержат несколько цепей, обычно две идентичные тяжелые цепи и две идентичные легкие цепи, которые связаны через дисульфидные связи. Эти цепи в основном состоят из доменов иммуноглобулина, таких как домен VL (вариабельная область легкой цепи), домен CL (константная область легкой цепи), домен VH (вариабельная область тяжелой цепи) и домены CH (константная область тяжелой цепи) CH1, CH2, CH3 и CH4. Существует пять типов тяжелых цепей иммуноглобулина млекопитающих, то есть α, δ, ε, γ и μ, которые обуславливают различные классы антител, т.е. IgA, IgD, IgE, IgG и IgM. В отличие от тяжелых цепей растворимых иммуноглобулинов, тяжелые цепи мембранных или поверхностных иммуноглобулинов содержат трансмембранный домен и короткий цитоплазматический домен на своем карбоксиконце. У млекопитающих есть два типа легких цепей, то есть лямбда и каппа. Цепи иммуноглобулина содержат вариабельную область и константную область. Константная область, по существу, консервативна в различных изотипах иммуноглобулинов, где вариабельная часть сильно отличается и учитывает распознавание антигена.
Согласно изобретению термин «антиген» или «иммуноген» охватывает любое вещество, предпочтительно пептид или белок, которое является мишенью для иммунного ответа и/или которое будет вызывать иммунный ответ. В частности, «антиген» относится к любому веществу, которое специфически реагирует с антителами или Т-лимфоцитами (Т-клетками). Согласно настоящему изобретению термин «антиген» включает любую молекулу, которая включает, по меньшей мере, один эпитоп, такой как В-клеточный или Т-клеточный эпитоп. Предпочтительно, антиген в контексте настоящего изобретения представляет собой молекулу, которая, необязательно после обработки, индуцирует иммунную реакцию, которая предпочтительно является специфической для антигена или клеток, экспрессирующих антиген. Антигены могут включать или могут быть получены из аллергенов, вирусов, бактерий, грибов, паразитов и других инфекционных агентов и патогенов, или антиген также может быть опухолевым антигеном.
«Собственный антиген» или «аутоантиген» представляет собой антиген, полученный из организма, который в нормальных условиях не распознается иммунной системой этого организма, но который может стать целью иммунной атаки, что приводит к аутоиммунному заболеванию.
В предпочтительном воплощении аутоантиген связан с аутоиммунным заболеванием. Термин «аутоантиген, связанный с аутоиммунным заболеванием» относится к аутоантигену, который имеет патологическое значение для аутоиммунного заболевания. В одном воплощении аутоантиген, связанный с аутоиммунным заболеванием, представляет собой молекулу, которая содержит, по меньшей мере, один эпитоп, против которого направлена иммунная реакция у пациента, имеющего аутоиммунное заболевание.
Пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, которое предусмотрено в соответствии с настоящим изобретением субъекту путем введения неиммуногенной РНК, кодирующую пептид или полипептид должен приводить к снижению иммунного ответа на аутоантиген. Таким образом, пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента предлагаемые согласно изобретению, могут соответствовать или включать аутоантиген, связанный с аутоиммунным заболеванием, или его вариантом (в том числе фрагментами аутоантигенов и их вариантами). В одном воплощении такие фрагменты или варианты являются иммунологически эквивалентными аутоантигену, связанному с аутоиммунным заболеванием в том, что - по аналогии с аутоантигеном - их предоставление приводит к толерантности к аутореактивным Т - клеткам, нацеленным на аутоантиген или клеткам, экспрессирующим аутоантиген и, необязательно, презентирующим аутоантиген в контексте молекул МНС. Таким образом, пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, представленный в соответствии с настоящим изобретением может быть идентичен аутоантигеном, связанным с аутоиммунным заболеванием, может включать аутоантиген, связанный с аутоиммунным заболеванием или его часть или фрагмент или может включать антиген, который гомологичен аутоантигену, связанному с аутоиммунным заболеванием, или его частью или фрагментом. Если пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, представленный в соответствии с настоящим изобретением включает часть или фрагмент аутоантигена, связанного с аутоиммунным заболеванием или часть или фрагмент антигена, который является гомологичным в отношении аутоантигена, ассоциированного с аутоиммунным заболеванием, указанная часть или фрагмент может включать эпитоп аутоантигена, ассоциированного с аутоиммунным заболеванием, в частности эпитоп аутоантигена, ассоциированного с аутоиммунным заболеванием, на который нацелены аутореактивные Т-клетки. Таким образом, в соответствии с изобретением, пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, может включать иммуногенный фрагмент аутоантигена, связанный с аутоиммунным заболеванием, таким как пептидный фрагмент аутоантигена, связанного с аутоиммунным заболеванием. «Иммуногенный фрагмент аутоантигена» по изобретению предпочтительно относится к части или фрагменту аутоантигена, который способен стимулировать ответ Т-клеток. Указанная часть или фрагмент, если они предоставляются согласно изобретению, также могут быть способны индуцировать толерантность к аутореактивным Т-клеткам. Пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, представленный в соответствии с настоящим изобретением может представлять собой рекомбинантный пептид или полипептид, и/или неиммуногенную РНК, кодирующую их в соответствии с изобретением, которая может быть рекомбинантной РНК.
Термин «иммунологически эквивалентный» означает, что иммунологически эквивалентная молекула, такая как иммунологически эквивалентная аминокислотная последовательность, демонстрирует одинаковые или по существу одинаковые иммунологические свойства и/или оказывает одинаковые или по существу одинаковые иммунологические эффекты, например, в отношении типа иммунологического эффекта. В контексте настоящего изобретения термин «иммунологически эквивалентный» предпочтительно используется в отношении иммунологических эффектов или свойств антигенов или вариантов антигенов.
В одном воплощении изобретения пептид или полипептид, предоставленный в соответствии с изобретением путем введения неиммуногенной РНК, содержит, по меньшей мере, один эпитоп из аутоантигена, который предпочтительно представляет собой аутоантиген, распознаваемый при аутоиммунном заболевании, таком как описанное аутоиммунное заболевание в данном описании. В одном воплощении пептид или полипептид, предоставленный в соответствии с изобретением, содержит эпитоп, к которому субъект самореактивен.
Термин «эпитоп» относится к антигенной детерминанте в молекуле, такой как антиген, то есть к части или фрагменту молекулы, которая распознается, то есть связана иммунной системой, например, которая распознается антителом или Т-клеточным рецептором. Например, эпитопы представляют собой дискретные трехмерные участки на антигене, которые распознаются иммунной системой. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или сахарные боковые цепи, и обычно имеют специфические трехмерные структурные характеристики, и специфические характеристики заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первым, но не с последним, теряется в присутствии денатурирующих растворителей. Предпочтительно, этот термин относится к иммуногенной части антигена, содержащей эпитоп. Эпитоп белка предпочтительно включает непрерывную или прерывистую часть указанного белка и предпочтительно составляет от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот в длину, например, эпитоп может быть предпочтительно 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот в длину. Предпочтительно, чтобы эпитоп в контексте настоящего изобретения представлял собой Т-клеточный эпитоп.
Используемый в данном документе термин «Т-клеточный эпитоп» относится к пептиду, который связывается с молекулой МНС в конфигурации, распознаваемой рецептором Т-клеток. Обычно Т-клеточные эпитопы представлены на поверхности антигенпрезентирующей клетки. Предпочтительно Т-клеточные эпитопы представляют собой пептиды, представленные МНС класса I и/или класса II. Предпочтительно, Т-клеточные эпитопы содержат аминокислотную последовательность, по существу соответствующую аминокислотной последовательности фрагмента антигена. Предпочтительно указанный фрагмент антигена представляет собой представленный пептид MHC класса I и/или класса II. Пептид, который подходит для связывания с молекулой МНС, в частности с молекулой МНС класса I, предпочтительно имеет длину 7-20 аминокислот, более предпочтительно 7-12 аминокислот, более предпочтительно 8-11 аминокислот, в частности 9 или 10 аминокислот в длину. В одном воплощении Т-клеточный эпитоп, когда он представлен в контексте МНС, такого как МНС антигенпрезентирующих клеток, распознается Т-клеточным рецептором. Т-клеточный эпитоп, если он распознается рецептором Т-клеток, может быть способен индуцировать в присутствии соответствующих костимулирующих сигналов клональную экспансию Т-клеток, несущих рецептор Т-клеток, специфически распознающих Т-клеточный эпитоп.
В соответствии с изобретением эпитоп, такой как Т-клеточный эпитоп аутоантигена, может присутствовать в пептиде или полипептиде, предоставленном в соответствии с изобретением, путем введения неиммуногенной РНК как части более крупного субъекта, такого как последовательность, включающая больше аутоантигена, чем просто эпитоп и/или полипептид, содержащий более одного эпитопа из одного или нескольких аутоантигенов. Представленный пептид или Т-клеточный эпитоп получают после подходящей обработки. Кроме того, эпитопы Т-клеток могут быть модифицированы по одному или нескольким остаткам, которые не являются необходимыми для распознавания TCR или для связывания с МНС. Такие модифицированные Т-клеточные эпитопы можно считать иммунологически эквивалентными.
Неиммуногенная РНК, описанная в данном документе, может быть адаптирована для различных заболеваний, требующих иммунной толерантности, таких как аутоиммунные заболевания, а также может быть адаптирована для отдельных пациентов. Существуют различные иммуноанализы, чтобы определить, развивают ли иммунные клетки, циркулирующие в крови у данного пациента, иммунный ответ на определенные тестируемые пептиды. В ином случае, выбранные антигены или эпитопы могут основываться на наиболее распространенной реактивности, наблюдаемой в классе пациентов.
«Клеточная поверхность» используется в соответствии с ее нормальным значением в данной области техники и, таким образом, включает внешнюю поверхность клетки, которая доступна для связывания белками и другими молекулами. Антиген экспрессируется на поверхности клеток, если он расположен на поверхности указанных клеток и доступен для связывания с антигенспецифическими антителами, добавленными к клеткам. В одном воплощении антиген, экспрессируемый на поверхности клеток, представляет собой цельный мембранный белок, имеющий внеклеточную часть.
Термин «внеклеточная часть» или «экзодомен» в контексте настоящего изобретения относится к части молекулы, такой как белок, которая обращена к внеклеточному пространству клетки и предпочтительно доступна снаружи указанной клетки, например, связывая молекулы, такие как антитела, расположенные вне клетки. Предпочтительно, термин относится к одной или нескольким внеклеточным петлям или доменам или их фрагменту.
Термины «участок» или «часть» используются в данном документе взаимозаменяемо и относятся к непрерывному или прерывистому элементу структуры, такой как аминокислотная последовательность. Термин «фрагмент» относится к непрерывному элементу структуры, такой как аминокислотная последовательность. Сегмент, часть или фрагмент структуры предпочтительно содержит одно или несколько функциональных свойств, например, антигенных, иммунологических и/или связывающих свойств указанной структуры. Сегмент или часть последовательности белка предпочтительно содержит, по меньшей мере, 6, в частности, по меньшей мере, 8, по меньшей мере, 12, по меньшей мере, 15, по меньшей мере, 20, по меньшей мере, 30, по меньшей мере, 50 или, по меньшей мере, 100 последовательных и/или непоследовательных последовательных аминокислот последовательности белка. Фрагмент последовательности белка предпочтительно содержит, по меньшей мере, 6, в частности, по меньшей мере, 8, по меньшей мере, 12, по меньшей мере, 15, по меньшей мере, 20, по меньшей мере, 30, по меньшей мере, 50 или, по меньшей мере, 100 последовательных аминокислот последовательности белка.
Термин «иммуногенность» относится к относительной эффективности антигена в отношении индукции иммунной реакции.
Термин «иммуностимулирующий» используется в данном документе для обозначения увеличения общего иммунного ответа.
«Иммунная толерантность», «иммунологическая толерантность» или просто «толерантность» описывает состояние невосприимчивости иммунной системы к веществам или тканям, которые способны вызывать иммунный ответ. Иммунная толерантность, которая препятствует развитию иммуногенного ответа при самораспознавании, опосредуется несколькими механизмами, в первую очередь включающими презентацию «собственных» пептидов CD4 + или CD8 + Т-клеткам таким образом, который приводит к элиминации, ингибированию или конверсии аутореактивных Т-клеток, которые в противном случае могли бы атаковать клетки и ткани, которые являются источником аутоантигенов и/или поддерживают выработку В-клетками антител, которые реагируют на эти аутоантигены. Механизмы, с помощью которых устанавливается толерантность, различны, но конечный эффект аналогичен. Недостаточная индукция толерантности к аутоантигенам из определенных тканей является основной причиной тканеспецифических аутоиммунных заболеваний. В нормальных условиях тканеспецифичные аутоантигены представлены индуцирующими толерантность (толерогенными) клетками, которые программируют любые реактивные Т-клетки на гибель, нереспонсивность или превращение в тип Treg. При аутоиммунных заболеваниях эти же аутоантигены либо недостаточно презентированы, что ограничивает вовлечение аутореактивных Т-клеток для обучения, либо неправильно презентированы, обучая специфические Т-клетки активировать иммунный ответ вместо того, чтобы переносить антиген как таковой. Антигенспецифическая терапия, которая доставляет эти аутоантигены в мощные толерогенные клетки, систематически или через слизистую оболочку, частично способна восстановить толерантность частично посредством генерации Т-клеток-супрессоров, способных противодействовать неправильно активированным патогенным Т-клеткам.
Термин «мишень» означает агент, такой как клетка, который является мишенью для иммунного ответа. Мишени включают клетки, которые презентируют антиген или эпитоп антигена, то есть пептидный фрагмент, полученный из антигена посредством процессинга антигена.
«Процессинг антигена» относится к деградации антигена в продукты процессирования, которые представляют собой фрагменты указанного антигена (например, деградация белка в пептиды) и ассоциации одного или нескольких из этих фрагментов (например, посредством связывания) с молекулами МНС для презентации клетками, предпочтительно антигенпрезентирующими клетками, специфическим Т-клеткам.
Антигенпрезентирующая клетка (APC) представляет собой клетку, которая презентирует антиген в контексте главного комплекса гистосовместимости (MHC) на его поверхности. Т-клетки могут распознавать этот комплекс, используя их Т-клеточный рецептор (TCR). Антигенпрезентирующие клетки обрабатывают антигены и презентируют их Т-клеткам. Антигенпрезентирующая клетка включает, без ограничения указанным, моноциты/макрофаги, В-клетки и дендритные клетки (DC). Согласно изобретению термин «антигенпрезентирующая клетка» включает профессиональные антигенпрезентирующие клетки и непрофессиональные антигенпрезентирующие клетки.
Профессиональные антигенпрезентирующие клетки очень эффективны при интернализации антигена либо фагоцитозом, либо рецептор-опосредованным эндоцитозом, а затем отображением фрагмента антигена, связанного с молекулой МНС класса II, на своей мембране. Т-клетка распознает и взаимодействует с комплексом антиген-молекула МНС класса II на мембране антигенпрезентирующей клетки. Затем антигенпрезентирующая клетка генерирует дополнительный костимулирующий сигнал, что приводит к активации Т-клетки. Экспрессия костимулирующих молекул является определяющей характеристикой профессиональных антигенпрезентирующих клеток.
Основными типами профессиональных антигенпрезентирующих клеток являются дендритные клетки, которые имеют самый широкий диапазон представления антигена и, вероятно, являются наиболее важными антигенпрезентирующими клетками, макрофагами, B-клетками и некоторыми активированными эпителиальными клетками.
Непрофессиональные антигенпрезентирующие клетки не экспрессируют конститутивно белки МНС класса II, необходимые для взаимодействия с наивными Т-клетками; они экспрессируются только при стимуляции непрофессиональных антигенпрезентирующих клеток некоторыми цитокинами, такими как IFNγ.
Дендритные клетки (DC) представляют собой популяции лейкоцитов, которые представляют антигены, захваченные в периферических тканях, Т-клеткам посредством путей представления антигена как МНС класса II, так и I. Хорошо известно, что дендритные клетки являются мощными индукторами иммунных ответов, и активация этих клеток является критическим шагом для индукции иммунитета.
Дендритные клетки удобно классифицировать как «незрелые» и «зрелые» клетки, которые можно использовать как простой способ различения двух хорошо охарактеризованных фенотипов. Однако эту номенклатуру не следует истолковывать как исключающую все возможные промежуточные стадии дифференцировки.
Незрелые дендритные клетки характеризуются как антигенпрезентирующие клетки с высокой способностью к поглощению и процессингу антигена, что коррелирует с высокой экспрессией рецептора Fcγ и рецептора маннозы. Зрелый фенотип обычно характеризуется более низкой экспрессией этих маркеров, но высокой экспрессией молекул клеточной поверхности, ответственных за активацию Т-клеток, таких как МНС класса I и класса II, молекул адгезии (например, CD54 и CD11) и костимулирующих молекул (например, CD40, CD80, CD86 и 4-1 BB).
Созревание дендритных клеток называется состоянием активации дендритных клеток, при котором такие антигенпрезентирующие дендритные клетки приводят к праймированию Т-клеток, в то время как презентация незрелыми дендритными клетками приводит к толерантности. Созревание дендритных клеток в основном обусловлено биомолекулами с микробными признаками, обнаруживаемыми врожденными рецепторами (бактериальная ДНК, вирусная РНК, эндотоксин и т.д.), провоспалительными цитокинами (TNF, IL-1, IFN), лигированием CD40 на поверхности дендритных клеток с помощью CD40L и веществами, высвобождаемыми из клеток, подвергающихся стрессовой гибели клеток. Дендритные клетки могут быть получены путем культивирования клеток костного мозга in vitro с цитокинами, такими как гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) и альфа-фактор некроза опухоли.
Под «клеткой, характеризуемой презентацией антигена» или «клеткой, презентирующей антиген» или подобными выражениями, подразумевается клетка, такая как антигенпрезентирующая клетка, презентирующая антиген или фрагмент, полученный из указанного антигена, например, путем процессинга антигена в контексте молекул МНС, в частности молекул МНС класса I. Аналогичным образом, термин «заболевание, характеризуемое проявлением антигена» обозначает заболевание, в которое вовлечены клетки, характеризующиеся проявлением антигена, в частности с МНС класса I.
Термин «иммунореактивная клетка» или «эффекторная клетка» в контексте настоящего изобретения относится к клетке, которая выполняет эффекторные функции во время иммунной реакции. «Иммунореактивная клетка» предпочтительно способна связывать антиген или клетку, характеризующуюся экспрессией и/или презентацией антигена или эпитопа, и опосредовать иммунный ответ. Например, такие клетки секретируют цитокины и/или хемокины, убивают микробы, секретируют антитела, распознают инфицированные или злокачественные опухолевые клетки и, необязательно, удаляют такие клетки. Например, иммунореактивные клетки включают Т-клетки (цитотоксические Т-клетки, хелперные Т-клетки, опухолевые инфильтрирующие Т-клетки), В-клетки, природные клетки-киллеры, нейтрофилы, макрофаги и дендритные клетки.
Предпочтительно, «иммунореактивная клетка» распознает антиген или эпитоп с некоторой степенью специфичности, в частности, если она представлена в контексте молекул МНС, таких как на поверхности антигенпрезентирующих клеток. Предпочтительно, указанное распознавание позволяет клетке, которая распознает антиген или эпитоп, быть отзывчивым или реактивным. Если клетка представляет собой хелперную T-клетку (CD4+ T-клетку), несущую рецепторы, которые распознают антиген или эпитоп в контексте молекул MHC класса II, такая реакция или реактивность может включать высвобождение цитокинов и/или активацию лимфоцитов CD8+ (CTL) и/или B-клеток. Если клетка представляет собой CTL, такая респонсивность или реактивность может включать элиминацию клеток, представленных в контексте молекул МНС класса I, то есть клеток, характеризующихся презентацией антигена с МНС класса I, например, посредством апоптоза или лизиса клеток, опосредованного перфорином. В соответствии с изобретением, респонсивность CTL может включать устойчивый поток кальция, деление клеток, образование цитокинов, таких как IFN-γ и TNF-α, положительная регуляция маркеров активации, таких как CD44 и CD69, а также специфическое уничтожение антигенэкспрессирующих клеток-мишеней. Реакция CTL также может быть определена с использованием искусственного репортера, который точно указывает на респонсивность CTL. Такие CTL, которые распознают антиген или эпитоп и являются чувствительными или реактивными, также называют в данном документе «антигенчувствительными CTL». Если клетка представляет собой В-клетку, такая реакция может включать высвобождение иммуноглобулинов.
Термин «Т-клетка» или «Т-лимфоцит» относится к тимусным клеткам, которые участвуют в различных клеточных иммунных реакциях, и включает Т-хелперные клетки (CD4+ Т-клетки) и цитотоксические Т-клетки (CTL, CD8+ Т-клетки), которые содержат цитолитические Т-клетки.
Т-клетки относятся к группе лейкоцитов, известных как лимфоциты, и играют центральную роль в клеточном иммунитете. Их можно отличить от других типов лимфоцитов, таких как В-клетки и естественные клетки-киллеры, по наличию специального рецептора на их клеточной поверхности, называемого Т-клеточным рецептором (TCR). Тимус является основным органом, ответственным за созревание Т-клеток. Было обнаружено несколько различных подмножеств Т-клеток, каждая из которых имела свою функцию.
Т-хелперы помогают другим лейкоцитам в иммунологических процессах, включая, среди прочего, созревание В-клеток в плазматических клетках и активацию цитотоксических Т-клеток и макрофагов. Эти клетки также известны как CD4+ T-клетки, потому что они экспрессируют белок CD4 на своей поверхности. Хелперные Т-клетки становятся активированными, когда молекулы МНС класса II, которые экспрессируются на поверхности антигенпрезентирующих клеток (АРС), презентируют им пептидные антигены. После активации они быстро делятся и секретируют небольшие белки, называемые цитокинами, которые регулируют или помогают в активном иммунном ответе.
Цитотоксические Т-клетки разрушают инфицированные вирусом клетки и опухолевые клетки, а также участвуют в отторжении трансплантата. Эти клетки также известны как CD8 + T-клетки, поскольку они экспрессируют гликопротеин CD8 на своей поверхности. Эти клетки распознают свои мишени, связываясь с антигеном, связанным с МНС класса I, который присутствует на поверхности почти каждой клетки организма.
Большинство T-клеток имеют T-клеточный рецептор (TCR), существующий в виде комплекса из нескольких белков. Фактический Т-клеточный рецептор состоит из двух отдельных пептидных цепей, которые вырабатываются из независимых альфа- и бета-генов Т-клеточного рецептора (TCRα и TCRβ) и называются α- и β-TCR-цепями. γδ T-клетки (гамма-дельта-T-клетки) представляют собой небольшое подмножество T-клеток, которые обладают определенным рецептором T-клеток (TCR) на своей поверхности. Однако в γδ T-клетках TCR состоит из одной γ-цепи и одной δ-цепи. Эта группа Т-клеток встречается гораздо реже (2% от общего числа Т-клеток), чем αβ Т-клетки.
Структура рецептора Т-клеток очень похожа на фрагменты Fab иммуноглобулина, которые представляют собой области, определяемые как объединенная легкая и тяжелая цепь плеча антитела. Каждая цепь TCR является членом суперсемейства иммуноглобулинов и имеет один N-концевой иммуноглобулиновый (Ig) вариабельный (V) домен, один Ig-константный (C) домен, область трансмембранной/клеточной мембраны и короткую область цитоплазматического хвоста на С-конце. Вариабельный домен как α-цепи, так и β-цепи TCR имеет три гипервариабельных или определяющих комплементарность области (CDR), тогда как вариабельный участок β-цепи имеет дополнительную область гипервариабельности (HV4), которая обычно не связывается с антигеном и поэтому не считается CDR. CDR3 является основным CDR, ответственным за распознавание обработанного антигена, хотя было также показано, что CDR1 α-цепи взаимодействует с N-концевой частью антигенного пептида, тогда как CDR1 β-цепи взаимодействует с C-концевой частью пептида. Считается, что CDR2 распознает MHC. Считается, что CDR4 β-цепи не участвует в распознавании антигена, но было показано, что он взаимодействует с суперантигенами. Константный домен домена TCR состоит из коротких соединительных последовательностей, в которых остаток цистеина образует дисульфидные связи, которые образуют связь между двумя цепями.
Термин «В-клетка» или «В-лимфоцит» относится к типу лейкоцитов подтипа лимфоцитов, которые функционируют в гуморальном иммунитете путем секреции антител. Кроме того, В-клетки представляют антиген и классифицируются как профессиональные антигенпрезентирующие клетки (APC) и секретируют цитокины. В-клетки экспрессируют В-клеточные рецепторы (BCR) на своей клеточной мембране. BCR позволяют B-клетке связывать специфический антиген, против которого она инициирует ответ антител. В-клеточный рецептор состоит из двух частей, мембраносвязанной молекулы иммуноглобулина одного изотипа (IgD, IgM, IgA, IgG или IgE), которые, за исключением присутствия интегрального мембранного домена, идентичны их секретируемым формам и группам сигнальной трансдукции: гетеродимер, называемый Ig-α/Ig-β (CD79), связанный вместе дисульфидными мостиками. Каждый элемент димера охватывает плазматическую мембрану и имеет цитоплазматический хвост, несущий иммунорецепторный мотив активации на основе тирозина (ITAM).
Активация В-клеток происходит во вторичных лимфоидных органах, таких как селезенка и лимфатические узлы. После созревания В-клеток в костном мозге они мигрируют через кровь к вторичным лимфоидным органам, которые получают постоянный запас антигена через циркулирующую лимфу. Активация B-клеток начинается, когда B-клетка связывается с антигеном через BCR. Различные подмножества В-клеток подвергаются Т-клеточно-зависимой активации или Т-клеточно-независимой активации.
Термин «главный комплекс гистосовместимости» и аббревиатура «MHC» включают молекулы MHC класса I и MHC класса II и относятся к комплексу генов, который встречается у всех позвоночных. Белки или молекулы МНС важны для передачи сигналов между лимфоцитами и антигенпрезентирующими клетками или больными клетками в иммунных реакциях, где белки или молекулы МНС связывают пептиды и представляют их для распознавания Т-клеточными рецепторами. Белки, кодируемые MHC, экспрессируются на поверхности клеток и презентируют Т-клетке как собственные антигены (пептидные фрагменты из самой клетки), так и несобственные антигены (например, фрагменты вторгающихся микроорганизмов).
Регион MHC разделен на три подгруппы: класс I, класс II и класс III. Белки MHC класса I содержат α-цепь и β2-микроглобулин (не является частью MHC, кодируемой хромосомой 15). Они представляют фрагменты антигена цитотоксическим Т-клеткам. На большинстве клеток иммунной системы, в частности на антигенпрезентирующих клетках, белки MHC класса II содержат α- и β-цепи и презентируют фрагменты антигена Т-хелперам. Область МНС класса III кодирует другие иммунные компоненты, такие как компоненты комплемента и некоторые, которые кодируют цитокины.
У людей гены в области МНС, которые кодируют антигенпрезентирующие белки на поверхности клетки, называют генами человеческого лейкоцитарного антигена (HLA). Однако аббревиатура MHC часто используется для обозначения продуктов гена HLA. Гены HLA включают девять так называемых классических генов MHC: HLA-A, HLA-B, HLA-C, HLA-DPA1, HLA-DPB1, HLA-DQA1, HLA-DQB1, HLA-DRA и HLA-DRB1.
В одном предпочтительном воплощении всех аспектов изобретения молекула MHC представляет собой молекулу HLA.
Термин «иммунные эффекторные функции» или «эффекторные функции» в контексте настоящего изобретения включает любые функции, опосредованные компонентами иммунной системы, которые приводят, например, к уничтожению клеток. Предпочтительно иммунные эффекторные функции в контексте настоящего изобретения представляют собой опосредованные Т-клетками эффекторные функции. Такие функции включают в случае хелперной Т-клетки (CD4 + Т-клетки) распознавание антигена или пептида антигена, полученного из антигена в контексте молекул МНС класса II Т-клеточными рецепторами, высвобождение цитокинов и/или активация CD8+ лимфоцитов (CTL) и/или B-клеток и, в случае CTL, распознавание антигена или пептида антигена, полученного из антигена в контексте молекул МНС класса I, Т-клеточными рецепторами, устранение клеток, презентирующих в контексте молекул МНС класса I Т-клеточными рецепторами, устранение клеток, презентирующих в контексте молекул МНС класса I, т.е. клетки, характеризующиеся презентацией антигена с МНС класса I, например, через апоптоз или перфорин-опосредованный лизис клеток, продуцирование цитокинов, таких как IFN-γ и TNF-альфа и специфическое цитолитическое уничтожение антигенэкспрессирующих клеток-мишеней.
Термин «толл-подобный рецептор» или «TLR» относится к классу белков, которые играют ключевую роль во врожденной иммунной системе. Они представляют собой единичные некаталитические трансмембранные рецепторы, которые обычно экспрессируются в дозорных клетках, таких как макрофаги и дендритные клетки, которые распознают структурно консервативные молекулы, полученные из микробов. Как только эти микробы преодолевают физические барьеры, такие как слизистая оболочка кожи или кишечного тракта, они распознаются TLR, которые активируют иммунные клеточные ответы.
В одном воплощении изобретения неиммуногенную РНК, которая кодирует пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, вводят субъекту. Продукт трансляции РНК может образовываться в клетках субъекта, и продукт может быть презентирован иммунной системе для индукции толерантности к аутореактивным Т-клеткам, нацеленным на аутоантиген.
В ином случае, настоящее изобретение предусматривает воплощения, в которых неиммуногенная РНК, экспрессирующая пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или фрагмента, вводится в клетки, такие как антигенпрезентирующие клетки ex vivo, например антигенпрезентирующие клетки, взятые у пациента, и клетки, необязательно клонально размноженные ex vivo, трансплантируют обратно тому же пациенту. Трансфицированные клетки могут быть повторно введены пациенту с использованием любых средств, известных в данной области, предпочтительно в стерильной форме путем внутривенного, внутриполостного или внутрибрюшинного введения. Подходящие клетки включают антигенпрезентирующие клетки. Антигенпрезентирующая клетка предпочтительно представляют собой дендритную клетку, макрофаг, В-клетку, мезенхимальную стромальную клетку, эпителиальную клетку, эндотелиальную клетку и фибробластную клетку, и наиболее предпочтительно представляют собой дендритную клетку. Таким образом, изобретение также включает способ лечения аутоиммунного заболевания, включающий введение субъекту, нуждающемуся в этом, выделенных антигенпрезентирующих клеток, которые экспрессируют неиммуногенную РНК, описанную в данном документе. Клетки могут быть аутологичными, аллогенными, сингенными или гетерологичными для субъекта.
Термин «нуклеиновая кислота», при использовании в данном документе, предназначен для включения дезоксирибонуклеиновой кислоты (ДНК) или рибонуклеиновой кислоты (РНК), такой как кДНК, мРНК, рекомбинантно продуцируемых и химически синтезированных молекул. Нуклеиновая кислота может быть одноцепочечной или двухцепочечной. Согласно изобретению РНК включает транскрибированную РНК (РНК IVT) in vitro или синтетическую РНК. Согласно изобретению нуклеиновая кислота предпочтительно представляет собой выделенную нуклеиновую кислоту. Кроме того, описанные в данном документе нуклеиновые кислоты могут быть рекомбинантными молекулами.
Термин «выделенная нуклеиновая кислота» означает, согласно изобретению, что нуклеиновая кислота (i) была амплифицирована in vitro, например, с помощью полимеразной цепной реакции (ПЦР), (ii) была получена рекомбинантным путем клонирования, (iii) была очищена, например, путем расщепления и разделения гель-электрофорезом, или (iv) синтезирована, например, химическим синтезом. Нуклеиновая может быть использована для введения, т.е. трансфекции клеток, например, в форме РНК, которая может быть получена путем транскрипции in vitro матрицы ДНК. Кроме того, РНК может быть модифицирована перед применением путем стабилизации последовательностей, кэппинга и полиаденилирования.
В контексте настоящего изобретения термин «ДНК» относится к молекуле, которая содержит дезоксирибонуклеотидные остатки и предпочтительно полностью или по существу состоит из дезоксирибонуклеотидных остатков. «Дезоксирибонуклеотид» относится к нуклеотиду, в котором отсутствует гидроксильная группа в положении 2'-β-D-рибофуранозильной группы. Термин «ДНК» включает выделенную ДНК, такую как частично или полностью очищенная ДНК, по существу чистая ДНК, синтетическая ДНК и рекомбинантно генерируемая ДНК, и включает модифицированную ДНК, которая отличается от встречающейся в природе ДНК добавлением, делецией, заменой и/или изменением одного или больше нуклеотидов. Такие изменения могут включать добавление ненуклеотидного материала, например, к концу(ам) ДНК или внутри, например, к одному или нескольким нуклеотидам ДНК. Нуклеотиды в молекулах ДНК также могут включать нестандартные нуклеотиды, такие как не встречающиеся в природе нуклеотиды или химически синтезированные нуклеотиды. Эти измененные ДНК могут называться аналогами или аналогами природной ДНК.
В контексте настоящего изобретения термин «РНК» относится к молекуле, которая включает рибонуклеотидные остатки и предпочтительно полностью или по существу состоит из рибонуклеотидных остатков. «Рибонуклеотид» относится к нуклеотиду с гидроксильной группой в положении 2'-β-D-рибофуранозильной группы. Термин включает двухцепочечную РНК, одноцепочечную РНК, выделенную РНК, такую как частично очищенная РНК, по существу очищенную РНК, синтетическую РНК, рекомбинантно продуцируемую РНК, и модифицированную РНК, которая отличается от естественной РНК добавлением, делецией, замещением и/или заменой одного или нескольких нуклеотидов. Такие изменения могут включать добавление ненуклеотидного материала, например, к концу(концам) РНК или внутрь, например, в одном или нескольких нуклеотидах РНК. Нуклеотиды в молекулах РНК могут также включать нестандартные нуклеотиды, такие как неприродные нуклеотиды или химически синтезированные нуклеотиды или дезоксинуклеотиды. Эти измененные РНК могут упоминаться как аналоги или аналоги встречающейся в природе РНК. Согласно настоящему изобретению термин «РНК» включает и предпочтительно относится к «мРНК», что означает «РНК-мессенджер» и относится к транскрипту, который может быть получен с использованием ДНК в качестве матрицы и кодирует пептид или белок. мРНК обычно содержит 5'-нетранслируемую область (5'-UTR), кодирующую область белка или пептида и 3'-нетранслируемую область (3'-UTR). мРНК имеет ограниченный период полувыведения в клетках и in vitro. Предпочтительно, мРНК получают путем транскрипции in vitro с использованием матрицы ДНК. В одном воплощении изобретения РНК получают путем транскрипции in vitro или химического синтеза. Методология транскрипции in vitro известна специалисту. Например, существует множество коммерчески доступных наборов транскрипции in vitro.
Согласно изобретению стабильность и эффективность трансляции РНК могут быть изменены по мере необходимости. Например, РНК может быть стабилизирована, и ее трансляция может быть увеличена за счет одной или нескольких модификаций, обладающих стабилизирующим эффектом и/или повышающих эффективность трансляции РНК. Чтобы увеличить экспрессию РНК, используемой согласно настоящему изобретению, ее можно модифицировать в пределах кодирующей области, то есть последовательности, кодирующей экспрессированный пептид или белок, предпочтительно без изменения последовательности экспрессируемого пептида или белка, чтобы увеличить содержание GC для повышения стабильности мРНК и для оптимизации кодонов и, таким образом, для повышения трансляции в клетках.
Термин «модификация» в контексте РНК, используемой в настоящем изобретении, включает любую модификацию РНК, которая естественно не присутствует в указанной РНК.
В одном воплощении изобретения РНК, используемая согласно изобретению, не имеет некэпированных 5'-трифосфатов. Удаление таких некэпированных 5'-трифосфатов может быть достигнуто путем обработки РНК фосфатазой.
РНК по изобретению может иметь модифицированные рибонуклеотиды для повышения ее стабильности и/или снижения цитотоксичности. Например, в одном воплощении в РНК, используемой согласно изобретению, 5-метилцитидин замещен частично или полностью, предпочтительно полностью, на цитидин. В ином случае или дополнительно в одном воплощении в РНК, используемой согласно изобретению, псевдоуридин замещен частично или полностью, предпочтительно полностью, на уридин.
В одном воплощении термин «модификация» относится к обеспечению РНК 5'-кэп структурой или аналогом 5'-кэп структуры. Термин «5'-кэп» относится к кэп-структуре, обнаруженной на 5'-конце молекулы мРНК, и обычно состоит из нуклеотида гуанозина, связанного с мРНК, посредством необычной 5'-5'-трифосфатной связи. В одном воплощении этот гуанозин метилирован в 7-положении. Термин «традиционный 5'-кэп» относится к природной 5'-кэп РНК, предпочтительно к 7-метилгуанозиновому кэпу (m7G). В контексте настоящего изобретения термин «5'-кэп» включает аналог 5'-кэпа, который напоминает структуру кэпа РНК и модифицируется, чтобы обладать способностью стабилизировать РНК и/или усиливать трансляцию РНК, если он присоединен к нему, предпочтительно in vivo и/или в клетке.
РНК может включать дополнительные модификации. Например, дальнейшая модификация РНК, используемой в настоящем изобретении, может представлять собой удлинение или усечение встречающегося в природе поли (А) хвоста или изменение 5'- или 3'-нетранслируемых областей (UTR), такое как введение UTR, который не связан с кодирующей областью указанной РНК, например, замена существующего 3'-UTR или вставка одной или нескольких, предпочтительно двух копий 3'-UTR, полученных из гена глобина, такой как альфа-2-глобин, альфа-1-глобин, бета-глобин, предпочтительно бета-глобин, более предпочтительно человеческий бета-глобин.
РНК, имеющая немаскированную последовательность поли-А, транслируется более эффективно, чем РНК, имеющая маскированную последовательность поли-А. Термин «поли (А)-хвост» или «последовательность поли-А» относится к последовательности остатков аденила (А), которая обычно расположена на 3'-конце молекулы РНК, и «немаскированная последовательность поли-А» означает, что последовательность поли-А на 3'-конце молекулы РНК заканчивается А из последовательности поли-А, и за ней не следуют нуклеотиды, отличные от А, расположенные на 3'-конце, то есть ниже от последовательности поли-А. Кроме того, длинная поли-A-последовательность из примерно 120 пар оснований обеспечивает оптимальную стабильность транскрипта и эффективность трансляции РНК.
Следовательно, чтобы повысить стабильность и/или экспрессию РНК, используемой согласно настоящему изобретению, ее можно модифицировать так, чтобы она присутствовала в сочетании с последовательностью поли-А, предпочтительно имеющей длину от 10 до 500, более предпочтительно от 30 до 300, еще более предпочтительно от 65 до 200 и особенно от 100 до 150 остатков аденозина. В особенно предпочтительном воплощении последовательность поли-А имеет длину приблизительно 120 остатков аденозина. Для дальнейшего увеличения стабильности и/или экспрессии РНК, используемой в соответствии с изобретением, последовательность поли-А может быть немаскирована.
Термин «стабильность» РНК относится к «периоду полужизни» РНК. «Период полужизни» относится к периоду времени, необходимому для устранения половины активности, количества или числа молекул. В контексте настоящего изобретения период полужизни РНК является показателем стабильности указанной РНК. Период полужизни РНК может влиять на «продолжительность экспрессии» РНК. Можно ожидать, что РНК, имеющая длительный период полужизни, будет экспрессироваться в течение длительного периода времени.
Конечно, если в соответствии с настоящим изобретением желательно уменьшить стабильность и/или эффективность трансляции РНК, можно модифицировать РНК так, чтобы она мешала функционированию элементов, описанных выше, которые увеличивают стабильность и/или эффективность трансляции РНК.
РНК для введения по изобретению не является иммуногенной. Используемый в данном документе термин «неиммуногенная РНК» относится к РНК, которая не вызывает ответ иммунной системы при введении, например, млекопитающему, или индуцирует более слабый ответ, чем тот, который был бы вызван той же РНК, которая отличается только тем, что она не подвергалась модификациям и обработкам, которые делают неиммуногенную РНК неиммуногенной. В одном предпочтительном воплощении неиммуногенная РНК становится неиммуногенной путем включения модифицированных нуклеотидов, подавляющих РНК-опосредованную активацию врожденных иммунных рецепторов, в РНК и удаления двухцепочечной РНК (дцРНК).
Для придания неиммуногенной РНК неиммуногенности путем включения модифицированных нуклеотидов можно использовать любой модифицированный нуклеотид, если он снижает или подавляет иммуногенность РНК. Особенно предпочтительными являются модифицированные нуклеотиды, которые подавляют РНК-опосредованную активацию врожденных иммунных рецепторов. В одном воплощении модифицированные нуклеотиды включают замену одного или нескольких уридинов нуклеозидом, содержащим модифицированную нуклеиновое основание. В одном воплощении модифицированное нуклеиновое основание представляет собой модифицированный урацил. В одном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, выбран из группы, состоящей из 3-метил-уридина (m3U), 5-метоксиуридина (mo5U), 5-азауридина, 6-азауридина, 2-тио -5-азауридина, 2-тиоуридина (s2U), 4-тиоуридина (s4U), 4-тио-псевдоуридина, 2-тио-псевдоуридина, 5-гидроксиуридина (ho5U), 5-аминоаллил- уридина, 5-галоген-уридина (например, 5-йод-уридина или 5-бром-уридин), уридин-5-оксиуксусной кислоты (cmo5U), метилового эфира уридин-5-оксиуксусной кислоты (mcmo5U), 5-карбоксиметил-уридина (cm5U), 1-карбоксиметил-псевдоуридина, 5-карбоксигидроксиметил-уридина (chm5U), 5-карбоксигидроксиметил-уридина, метилового эфира (mchm5U), 5-метоксикарбонилметил-уридина (mcm5U), 5-метоксикарбонилметил-2-тио-уридина (mcm5s2U), 5-аминометил-2-тио-уридина (nm5s2U), 5-метиламинометил-уридина (mnm5U), 1-этил-псевдоуридина, 5-метиламинометил-2-тио-уридина (mnm5s2U), 5-метиламинометил-2-селеноурид (mnm5se2U), 5-карбамоилметил-уридина (ncm5U), 5-карбоксиметиламинометил-уридина (cmn m5U), 5-карбоксиметиламинометил-2-тиоуридина (cmnm5s2U), 5-пропинил-уридина, 1-пропинил-псевдоуридина, 5-тауринометил-уридина (τm5U), 1-тауринометил-псевдоуридина, 5-тауроина, 5-тауроин -уридина (τm5s2U), 1-тауринометил-4-тио-псевдоуридина, 5-метил-2-тио-уридина (m5s2U), 1-метил-4-тио-псевдоуридина (m1s4ψ), 4-тио-1-метил- псевдоуридина, 3-метил-псевдоуридина (m3ψ), 2-тио-1-метил-псевдоуридина, 1-метил-1-деаза-псевдоуридина, 2-тио-1-метил-1-деаза-псевдоуридина, дигидроуридина (D), дигидропсевдуридина, 5,6-дигидроуридина, 5-метил-дигидроуридина (m5D), 2-тио-дигидроуридина, 2-тио-дигидропсевдуридина, 2-метоксиуридина, 2-метокси-4-тиоуридина, 4-метокси-псевдоуридина, 4-метокси-2-тио-псевдоуридина, N1-метил-псевдоуридина, 3- (3-амино-3-карбоксипропил)уридина (acp3U), 1-метил-3- (3-амино-3-карбоксипропил) псевдоуридина (acp3 ψ), 5- (изопентениламинометил)уридина (inm5U), 5-(изопентениламинометил)-2-тиоуридина (inm5s2U), α-тиоуридина, 2'-O-метил-уридина (Um), 5,2'-O-диметилуридина (m5Um), 2'-O-метил-псевдоуридина (ψm), 2-тио-2'-O-метил-уридина (s2Um), 5-метоксикарбонилметил-2'-O-метилуридина (mcm5Um), 5-карбамоилметил-2'-O-метилуридина (ncm5Um), 5-карбоксиметиламинометил-2'-O-метилуридина (cmmm5Um), 3,2'-O- диметилуридина (m3Um), 5- (изопентениламинометил) -2'-O-метилуридина (inm5Um), 1-тиоуридина, дезокситимидина, 2'-F-арауридина, 2'-F-уридина, 2'-ОН-арауридина, 5-(2-карбометоксивинил)уридина и 5- [3- (1-E-пропениламино)уридина. В одном особенно предпочтительном воплощении нуклеозид, содержащий модифицированное нуклеиновое основание, представляет собой псевдоуридин (ψ), N1-метил-псевдоуридин (m1ψ) или 5-метил-уридин (m5U), в частности, 1-метил-псевдоуридин.
Структура типичного нуклеозида, содержащего модифицированное нуклеиновое основание, представляет собой 1-метилпсевдоуридин m1ψ:
 .
.
В одном воплощении замена одного или нескольких уридинов нуклеозидом, содержащим модифицированное нуклеиновое основание, включает замену, по меньшей мере, 1%, по меньшей мере, 2%, по меньшей мере, 3%, по меньшей мере, 4%, по меньшей мере, 5%, по меньшей мере, 10 %, по меньшей мере, 25%, по меньшей мере, 50%, по меньшей мере, 75%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, 99% или 100% уридинов.
Во время синтеза мРНК путем транскрипции in vitro (IVT) с использованием РНК-полимеразы Т7 образуются значительные количества аберрантных продуктов, в том числе двухцепочечной РНК (дцРНК), вследствие нетрадиционной активности фермента. дцРНК индуцирует воспалительные цитокины и активирует эффекторные ферменты, что приводит к ингибированию синтеза белка. дцРНК может быть удалена из РНК, такой как РНК IVT, например, с помощью ВЭЖХ с обращенной фазой с ионной парой, используя непористую или пористую матрицу С-18 полистирол-дивинилбензол (PS-DVB). В ином случае, может быть использован ферментативный способ с использованием РНКазы III E.coli, которая специфически гидролизует дцРНК, но не оцРНК, тем самым удаляя контаминанты дцРНК из препаратов РНК IVT. Кроме того, дцРНК может быть отделена от оцРНК с помощью целлюлозного материала. В одном воплощении препарат РНК приводят в контакт с целлюлозным материалом, а оцРНК отделяют от целлюлозного материала в условиях, которые позволяют связывать дцРНК с целлюлозным материалом и не позволяют связывать оцРНК с целлюлозным материалом.
Используемый в данном документе термин «удалить» или «удаление» относится к характеристике популяции первых веществ, таких как неиммуногенная РНК, отделенных от близости к популяции вторых веществ, таких как дцРНК, где популяция первых веществ не обязательно лишена второго вещества, а популяция вторых веществ не обязательно лишена первого вещества. Однако популяция первых веществ, характеризующаяся удалением популяции вторых веществ, имеет заметно более низкое содержание вторых веществ по сравнению с неразделенной смесью первого и второго веществ.
Нуклеиновые кислоты могут быть перенесены в клетку-хозяина физическими, химическими или биологическими средствами.
Физические способы введения нуклеиновой кислоты в клетку-хозяина включают осаждение фосфата кальция, липофекцию, бомбардировку частицами, микроинъекцию, электропорацию и т.п.
Биологические способы введения интересующей нуклеиновой кислоты в клетку-хозяина включают использование векторов ДНК и РНК. Вирусные векторы, и особенно ретровирусные векторы, стали наиболее широко используемым способом для вставки генов в млекопитающих, например, в клетки человека. Другие вирусные векторы могут быть получены из лентивируса, поксвирусов, вируса простого герпеса I, аденовирусов и аденоассоциированных вирусов и т.п.
Химические средства для введения нуклеиновой кислоты в клетку-хозяина включают коллоидные дисперсионные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, шарики и системы на основе липидов, включая эмульсии масло-в-воде, мицеллы, смешанные мицеллы и липосомы. Предпочтительной коллоидной системой для применения в качестве средства доставки in vitro и in vivo является липосома (то есть искусственная мембрана везикулы). Подготовка и использование таких систем хорошо известны в данной области.
«Кодирование» относится к врожденному свойству конкретных последовательностей нуклеотидов в нуклеиновой кислоте служить в качестве матриц для синтеза других полимеров и макромолекул в биологических процессах, имеющих либо определенную последовательность нуклеотидов, либо определенную последовательность аминокислот. Таким образом, нуклеиновая кислота кодирует белок, если экспрессия (трансляция и, необязательно, транскрипция) нуклеиновой кислоты продуцирует белок в клетке или другой биологической системе.
Термин «экспрессия» используется в соответствии с изобретением в его наиболее общем значении и включает продуцирование РНК и/или пептидов или полипептидов, например, путем транскрипции и/или трансляции. Что касается РНК, то термин «экспрессия» или «трансляция» относится, в частности, к образованию пептидов или белков. Он также включает частичную экспрессию нуклеиновых кислот. Более того, экспрессия может быть транзиторной или стабильной.
В контексте настоящего изобретения термин «транскрипция» относится к процессу, в котором генетический код в последовательности ДНК транскрибируется в РНК. Впоследствии РНК может быть транслирована в белок. Согласно настоящему изобретению термин «транскрипция» включает «транскрипцию in vitro», где термин «транскрипция in vitro» относится к способу, в котором РНК, конкретно, мРНК, синтезируется in vitro в бесклеточной системе, предпочтительно, с использованием подходящих клеточных экстрактов. Предпочтительно векторы клонирования применяются для генерации транскриптов. Эти векторы клонирования обычно обозначаются как векторы транскрипции и согласно настоящему изобретению охватываются термином «вектор». Согласно настоящему изобретению РНК, используемая в настоящем изобретении, предпочтительно представляет собой транскрибированную РНК in vitro (IVT-РНК) и может быть получена путем транскрипции in vitro соответствующей матрицы ДНК. Промотор для контроля транскрипции может быть любым промотором для любой РНК-полимеразы. Конкретными примерами РНК-полимераз являются РНК-полимеразы Т7, Т3 и SP6. Предпочтительно транскрипция in vitro по изобретению контролируется промотором T7 или SP6. ДНК-матрица для транскрипции in vitro может быть получена путем клонирования нуклеиновой кислоты, в частности кДНК, и введения ее в подходящий вектор для транскрипции in vitro. кДНК может быть получена путем обратной транскрипции РНК.
Термин «трансляция» согласно изобретению относится к процессу в рибосомах клетки, посредством которого нить мессенджерной РНК направляет сборку последовательности аминокислот для получения пептида или полипептида.
В соответствии с изобретением предпочтительно, чтобы нуклеиновая кислота, такая как РНК, кодирующая пептид или белок, один раз поглощенная или введенная, то есть трансфицированная или трансдуцированная в клетку, которая может присутствовать in vitro или у субъекта, приводит к экспрессии указанного пептида или белка. Клетка может экспрессировать кодированный пептид или белок внутриклеточно (например, в цитоплазме и/или в ядре), может секретировать кодированный пептид или белок или может экспрессировать его на поверхности.
Согласно изобретению такие термины, как «экспрессия нуклеиновой кислоты» и «кодирование нуклеиновой кислоты» или подобные термины, используются в данном документе взаимозаменяемо и в отношении конкретного пептида или полипептида означают, что нуклеиновая кислота, если она присутствует в соответствующей среде, предпочтительно в пределах клетки, может быть экспрессирована для продуцирования указанного пептида или полипептида.
Термины, такие как «перенос», «введение», «трансфекция» или «трансдукция», используются в данном документе взаимозаменяемо и относятся к введению нуклеиновых кислот, в частности экзогенных или гетерологичных нуклеиновых кислот, таких как РНК, в клетку. Согласно настоящему изобретению клетка может присутствовать in vitro или in vivo, например, клетка может образовывать часть органа, ткани и/или организма. Согласно изобретению трансфекция может быть транзиторной или стабильной. Для некоторых применений трансфекции достаточно, чтобы трансфецированный генетический материал был экспрессирован лишь транзиторно. Так как нуклеиновая кислота, вводимая в процесс трансфекции, обычно не интегрируется в ядерный геном, то чужеродная нуклеиновая кислота будет разведена через митоз или деградирована. Клеточные линии, допускающие эписомную амплификацию нуклеиновых кислот, значительно снижают степень разведения. Если желательно, чтобы трансфецированная нуклеиновая кислота фактически оставалась в геноме клетки и в ее дочерних клетках, то должна иметь место стабильная трансфекция. РНК может быть трансфецирована в клетки для транзиторной экспрессии кодируемого белка.
Согласно настоящему изобретению термин «пептид» относится к веществам, включающим две или более, предпочтительно 3 или более, предпочтительно 4 или более, предпочтительно 6 или более, предпочтительно 8 или более, предпочтительно 10 или более, предпочтительно 13 или более, предпочтительно 16 или более, предпочтительно 21 или более и вплоть до предпочтительно 8, 10, 20, 30, 40 или 50, в частности 100 аминокислот, ковалентно связанных пептидными связями.
Термин «белок» относится к крупным пептидам, то есть полипептидам, предпочтительно к пептидам с более чем со 100 аминокислотными остатками, но в целом термины «пептид», «полипептид» и «белок» являются синонимами и используются в данном документе взаимозаменяемо.
Настоящее изобретение также включает «варианты» пептидов, белков или аминокислотных последовательностей, таких как аутоантигены, описанные в данном документе.
Для целей настоящего изобретения «варианты» аминокислотной последовательности включают варианты с аминокислотными вставками, варианты с добавлением аминокислот, варианты с делецией аминокислот и/или варианты с заменой аминокислот.
Варианты аминокислотных вставок включают вставки одной или двух или более аминокислот в конкретной аминокислотной последовательности. В случае вариантов аминокислотной последовательности, имеющих вставку, один или несколько аминокислотных остатков вводят в конкретный участок в аминокислотной последовательности, хотя также возможна случайная вставка с соответствующим скринированием полученного продукта.
Варианты добавления аминокислот включают амино- и/или карбоксиконцевые слияния одной или нескольких аминокислот, как например, 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот.
Варианты аминокислотных делеций характеризуются удалением одной или нескольких аминокислот из последовательности, как например, удаление 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот. Делеции могут быть в любом положении белка. Варианты делеции аминокислот, которые включают делецию на N-конце и/или С-конце белка, также называются N-концевыми и/или С-концевыми укороченными вариантами.
Варианты аминокислот характеризуются, по меньшей мере, одним остатком в удаляемой последовательности и другим остатком, вставленным на его место. Предпочтение отдается модификациям, которые находятся в положениях аминокислотной последовательности, которые не консервативны между гомологичными белками или пептидами и/или заменам аминокислот на другие, обладающие сходными свойствами. Предпочтительно, аминокислотные замены в вариантах белка представляют собой консервативные аминокислотные замены, то есть замены одинаково заряженных или незаряженных аминокислот. Консервативная аминокислотная замена включает замену на одну из аминокислот семейства, которые родственны по своим боковым цепям. Природные аминокислоты обычно делятся на четыре семейства: кислотные (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан) и незаряженные полярные (глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин) аминокислоты. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты.
Предпочтительно, степень сходства, предпочтительно идентичности между данной аминокислотной последовательностью и аминокислотной последовательностью, которая является вариантом указанной аминокислотной последовательности, будет составлять, по меньшей мере, около 60%, 65%, 70%, 80%, 81%, 82 %, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичности дается предпочтительно для аминокислотной области, которая составляет, по меньшей мере, около 10%, по меньшей мере, около 20%, по меньшей мере, около 30%, по меньшей мере, около 40%, по меньшей мере, около 50%, по меньшей мере, около 60% %, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90% или около 100% всей длины эталонной аминокислотной последовательности. Например, если эталонная аминокислотная последовательность состоит из 200 аминокислот, степень сходства или идентичности дается предпочтительно для, по меньшей мере, около 20, по меньшей мере, для около 40, по меньшей мере, для около 60, по меньшей мере, для около 80, по меньшей мере, для около 100, по меньшей мере, для около 120, по меньшей мере, для около 140, по меньшей мере, для около 160, по меньшей мере, для около 180 или для около 200 аминокислот, предпочтительно непрерывных аминокислот. Степень сходства или идентичности дана предпочтительно для сегмента, по меньшей мере, 80, по меньшей мере, 100, по меньшей мере, 120, по меньшей мере, 150, по меньшей мере, 180, по меньшей мере, 250 или, по меньшей мере, 250 аминокислот. В предпочтительных воплощениях степень сходства или идентичности дается для всей длины эталонной аминокислотной последовательности. Выравнивание для определения сходства последовательностей, предпочтительно идентичности последовательности, может быть выполнено с помощью известных инструментов, предпочтительно с использованием наилучшего выравнивания последовательностей, например, с использованием Align, с использованием стандартных настроек, предпочтительно EMBOSS::needle, Матрица: Blosum62, штраф за разрыв 10,0, штраф за продолжение разрыва 0,5.
«Сходство последовательностей» указывает на процент аминокислот, которые либо идентичны, либо представляют консервативные аминокислотные замены. «Идентичность последовательности» между двумя аминокислотными последовательностями указывает процент аминокислот, которые являются идентичными между последовательностями.
Термин «% идентичности» предназначен для обозначения, в частности, процента аминокислотных остатков, которые идентичны в оптимальном выравнивании между двумя сравниваемыми последовательностями, причем указанный процент является чисто статистическим, и различия между этими двумя последовательностями могут быть случайным образом распределены по всей длине последовательности, и сравниваемая последовательность может включать добавления или удаления по сравнению с эталонной последовательностью, чтобы получить оптимальное выравнивание между двумя последовательностями. Сравнение двух последовательностей обычно выполняется путем сравнения указанных последовательностей после оптимального выравнивания относительно сегмента или «окна сравнения», чтобы идентифицировать локальные области соответствующих последовательностей. Оптимальное выравнивание для сравнения может быть выполнено вручную или с помощью алгоритма локальной гомологии Smith and Waterman, 1981, Ads App. Math. 2, 482, с помощью алгоритма локальной гомологии Neddleman и Wunsch, 1970, J. Mol. Biol. 48, 443, и с помощью алгоритма поиска сходства Pearson and Lipman, 1988, Proc. Natl Acad. Sci. США 85, 2444 или с помощью компьютерных программ, использующих указанные алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA в программном комплексе Wisconsin Genetics, Genetics Computer Group, 575 Science Drive, Мэдисон, Висконсин).
Процентную идентичность получают путем определения количества идентичных положений, в которых соответствуют сравниваемые последовательности, деления этого числа на количество сравниваемых положений и умножения этого результата на 100.
Гомологичные аминокислотные последовательности показывают согласно изобретению, по меньшей мере, 40%, в частности, по меньшей мере, 50%, по меньшей мере, 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90% и предпочтительно, по меньшей мере, 95%, по меньшей мере, 98% или, по меньшей мере, 99% идентичности аминокислотных остатков.
Согласно изобретению вариант, фрагмент, часть или участок аминокислотной последовательности, пептида или белка предпочтительно обладает функциональным свойством аминокислотной последовательности, пептида или белка, соответственно, из которой он был получен, т.е. он является функционально эквивалентным. В одном воплощении вариант, фрагмент, часть или участок аминокислотной последовательности, пептида или белка является иммунологически эквивалентным аминокислотной последовательности, пептиду или белку, соответственно, из которой он был получен. В одном воплощении функциональное свойство представляет собой иммунологическое свойство.
Изобретение включает производные пептидов или белков, описанных в данном документе, которые включают термины «пептид» и «белок». Согласно изобретению «производные» белков и пептидов представляют собой модифицированные формы белков и пептидов. Такие модификации включают любую химическую модификацию и включают однократные или множественные замены, делеции и/или добавления любых молекул, связанных с белком или пептидом, таких как углеводы, липиды и/или белки или пептиды. В одном воплощении «производные» белков или пептидов включают такие модифицированные аналоги, которые являются результатом гликозилирования, ацетилирования, фосфорилирования, амидирования, пальмитоилирования, миристоилирования, изопренилирования, липидирования, алкилирования, дериватизации, введения защитных/блокирующих групп, протеолитического расщепления или связывания с антитело или другой клеточный лиганд. Термин «производное» также распространяется на все функциональные химические эквиваленты указанных белков и пептидов. Предпочтительно, модифицированный пептид обладает повышенной стабильностью и/или повышенной иммуногенностью.
Термин «производный» означает в соответствии с изобретением, что конкретный субъект, в частности конкретная последовательность, присутствует в субъекте, из которого он получен, в частности организм или молекула. В случае аминокислотных последовательностей или последовательностей нуклеиновых кислот, особенно определенных областей последовательности, «производное», в частности, означает, что соответствующая аминокислотная последовательность или последовательность нуклеиновой кислоты получена из аминокислотной последовательности или последовательности нуклеиновой кислоты, в которой она присутствует.
Термин «клетка» или «клетка-хозяин» предпочтительно представляет собой интактную клетку, то есть клетку с неповрежденной мембраной, которая не высвободила свои нормальные внутриклеточные компоненты, такие как ферменты, органеллы или генетический материал. Интактная клетка предпочтительно представляет собой жизнеспособную клетку, то есть живую клетку, способную выполнять свои нормальные метаболические функции. Предпочтительно указанный термин относится согласно изобретению к любой клетке, которая может быть трансформирована или трансфицирована экзогенной нуклеиновой кислотой. Термин «клетка» включает согласно изобретению ионные прокариотические клетки (например, E.coli) или эукариотические клетки (например, дендритные клетки, B-клетки, клетки CHO, клетки COS, клетки K562, клетки HEK293, клетки HELA, клетки дрожжей, и клетки насекомых). Экзогенная нуклеиновая кислота может быть обнаружена внутри клетки (i) свободно диспергирована как таковая, (ii) включена в рекомбинантный вектор или (iii) интегрирована в геном клетки-хозяина или митохондриальную ДНК. Особенно предпочтительными являются клетки млекопитающих, такие как клетки человека, мышей, хомяков, свиней, коз и приматов. Клетки могут быть получены из большого количества типов тканей и включают первичные клетки и клеточные линии.
Клетка, которая содержит нуклеиновую кислоту, например, которая была трансфицирована нуклеиновой кислотой, предпочтительно экспрессирует пептид или белок, кодируемый нуклеиновой кислотой.
Термин «экспансия» относится к процессу, в котором конкретный субъект умножается. В одном воплощении настоящего изобретения этот термин используется в контексте иммунологического ответа, при котором лимфоциты стимулируются антигеном, пролиферируют и амплифицируется специфический лимфоцит, распознающий указанный антиген. Предпочтительно, клональная экспансия приводит к дифференцировке лимфоцитов.
Термин «выделенный», используемый в данном документе, предназначен для обозначения молекулы, которая по существу не содержит других молекул, таких как другой клеточный материал.
Термин «рекомбинантный» в контексте настоящего изобретения означает «полученный посредством генной инженерии». Предпочтительно, «рекомбинантный субъект», такой как рекомбинантная нуклеиновая кислота в контексте настоящего изобретения, не встречается в природе.
Используемый в данном документе термин «встречающийся в природе» относится к тому факту, что субъект можно найти в природе. Например, пептид или нуклеиновая кислота, которые присутствуют в организме (включая вирусы) и могут быть выделены из источника в природе и которые не были преднамеренно изменены человеком в лаборатории, встречаются в природе.
Такие термины, как «уменьшение», «ингибирование» или «понижение» относятся к способности вызывать общее снижение, предпочтительно на 5% или более, на 10% или более, на 20% или более, более предпочтительно на 50% или более, и наиболее предпочтительно 75% или более, относительно уровня. Эти термины включают полное или по существу полное ингибирование, то есть снижение до нуля или по существу до нуля.
Такие термины, как «увеличение», «усиление», «промотирование», «стимулирование» или «индуцирование», относятся к способности вызывать общее увеличение, предпочтительно на 5% или более, 10% или более, 20% или более, 50% или более, 75% или более, 100% или более, 200% или более или 500% или более, относительно уровня. Эти термины могут относиться к увеличению, усилению, промотированию, стимулированию или индуцированию от нуля или неизмеримого или не обнаруживаемого уровня до уровня больше нуля или уровня, который можно измерить или обнаружить. В ином случае, эти термины могут также означать, что до увеличения, усиления, промотирования, стимуляции или индукции был определенный уровень, а после увеличения, усиления, промотирования, стимуляции или индукции уровень стал выше.
Агенты и композиции, описанные в данном документе, могут быть использованы в способах супрессии иммунного ответа у млекопитающего, для лечения или профилактики аутоиммунного заболевания. В частности, агенты и композиции, описанные в данном документе, могут быть использованы для лечения субъекта с аутоиммунным заболеванием, характеризующимся наличием иммунной реакции против аутоантигена. Согласно изобретению неиммуногенная РНК, кодирующая пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента, вводится субъекту.
Термин «заболевание» относится к аномальному состоянию, которое влияет на организм человека. Заболевание часто истолковывается как медицинское состояние, связанное со специфическими симптомами и симптомами. Заболевание может быть вызвана факторами, происходящими из внешнего источника, такими как инфекционное заболевание, или может быть вызвано внутренними дисфункциями, такими как аутоиммунные заболевания. У людей «заболевание» часто используется более широко, чтобы ссылаться на любое состояние, которое вызывает боль, дисфункцию, дистресс, социальные проблемы или смерть для отдельных страдающих или подобных проблем для тех, кто находится в контакте с человеком. В этом более широком смысле заболевание иногда включает травмы, инвалидности, расстройства, синдромы, инфекции, изолированные симптомы, девиантное поведение и атипичные вариации структуры и функции, тогда как в других контекстах и для других целей они могут рассматриваться как отличимые категории. Заболевания обычно затрагивают людей не только физически, но и эмоционально, поскольку приобретение и жизнь со многими заболеваниями могут изменить точку зрения на жизнь и личность индивидуума. Согласно изобретению термин «заболевание» включает аутоиммунные заболевания.
Заболевание, которое лечат согласно изобретению, предпочтительно представляет собой заболевание, включающее аутоантиген или ассоциированное с аутоантигеном.
Термин «аутоиммунное заболевание» или «аутоиммунное расстройство» относится к любому заболеванию/расстройству, при котором организм производит иммуногенную (т.е. иммунную систему) реакцию на некоторую составляющую своей собственной ткани. Другими словами, иммунная система теряет способность распознавать какую-то ткань или систему внутри тела как собственную и целится и атакует ее, как если бы она была чужеродной. Аутоиммунные заболевания можно классифицировать на те, у которых поражен преимущественно один орган (например, гемолитическая анемия и антииммунный тиреоидит), и на те, у которых процесс аутоиммунного заболевания распространен во многих тканях (например, системная красная волчанка). Например, считается, что рассеянный склероз вызван Т-клетками, поражающими оболочки, окружающие нервные волокна головного и спинного мозга. Это приводит к потере координации, слабости и помутнения зрения. Примеры аутоиммунных заболеваний включают, без ограничения указанным, болезнь Аддисона, очаговую алопецию, анкилозирующий спондилит, аутоиммунный гепатит, аутоиммунный паротит, болезнь Крона, диабет (тип I), дистрофический эпидермолизный буллезный синдром, эпидидимит, гломерулонефрит, гломерулонефрит, базедову болезнь, синдром Гийена-Барре, болезнь Хашимото, гемолитическую анемию, системную красную волчанку, рассеянный склероз, тяжелую миастению, пузырчатку обыкновенную, псориаз, ревматизм, ревматоидный артрит, саркоидоз, склеродермию, синдром Шегрена, спондилоартропатии, тиреоидит, васкулит, витилиго, микседему, пернициозную анемию, язвенный колит и сахарный диабет I типа, среди прочего.
Ниже приведен список антигенов, которые, как известно, связаны с определенными аутоиммунными заболеваниями. Эпитопы из этих антигенов могут быть включены в пептид или полипептид, содержащий аутоантиген или его фрагмент, или вариант аутоантигена или его фрагмента по изобретению.
Рассеянный склероз (MS)/(+ животная модель EAE (экспериментального аутоиммунного энцефаломиелита))
Возможные кандидаты аутоантигенов при РС включают миелиновые антигены, нейрональные антигены и антигены, полученные из астроцитов. Белковые компоненты белков миелина, такие как основной белок миелина (MBP) (например, MBP84-102, MBP83-99, MBP13-32, MBP144-163, MBP143-168, MBP151-170), протеолипидный белок (PLP) (например, PLP139-151), миелин-олигодендроцитарный гликопротеин (MOG), гликопротеин, ассоциированный с миелином (MAG), основной белок олигодендроцитов, ассоциированный с миелином (MOBP), CNPаза (2',3'-циклический нуклеотид 3'-фосфодиэстераза), белок S100β и Трансальдолаза Н представляют особый интерес для применения согласно изобретению.
Ревматоидный артрит (RA)
Возможные кандидаты аутоантигенов при RA включают коллаген типа II, гликопротеин-39 хряща человека и аггрекан G1.
Диабет I типа
Возможные потенциальные аутоантигены при диабете типа 1 включают инсулин, проинсулин, декарбоксилазу глутаминовой кислоты 65 (GAD65), белок 2, ассоциированный с инсулиномой (IA-2), белок 2 бета, связанный с инсулиномой (IA-2beta), аутоантиген островковых клеток (HSP), глима 38, аутоантиген островковых клеток (ICA69), p52, транспортер цинка 8 (ZnT8), специфичный для островков белок, связанный с каталитической субъединицей глюкозо-6-фосфатазы (IGRP) и хромогранин А.
Тяжелая миастения
Возможные кандидаты аутоантигенов при тяжелой миастении включают nAChR, MuSK и LRP4.
Другие аутоантигены при таких заболеваниях, как MS, RA, диабет 1 типа, тяжелая миастения, IBD и псориаз, некоторые из которых перечислены в данном документе, а другие также известны в данной области, предполагаются для применения с изобретением. Число известных аутоантигенов и аутоиммунных эпитопов растет. Любые такие антигены и эпитопы, которые станут известны в будущем, также предполагаются для применения с изобретением.
Термин «заболевание, связанное с аутоантигеном» или «заболевание, включающее аутоантиген», относится к любому заболеванию, которое подразумевает аутоантиген, например заболеванию, которое характеризуется наличием иммунной реакции против аутоантигена или клеток, экспрессирующих аутоантиген.
Термин «лечение» или «терапевтическое лечение» относится к любому лечению, которое улучшает состояние здоровья и/или продлевает (увеличивает) продолжительность жизни индивидуума. Указанное лечение может устранить заболевание у индивидуума, остановить или замедлить развитие заболевания у индивидуума, ингибировать или замедлить развитие заболевания у индивидуума, уменьшить частоту или тяжесть симптомов у индивидуума и/или уменьшить рецидив у человека, который в настоящее время имеет или который ранее имел заболевание.
Термины «профилактическое лечение» или «превентивное лечение» относятся к любому лечению, которое предназначено для предотвращения возникновения заболевания у индивидуума, в частности у индивидуума, подверженного риску заболевания. Термины «профилактическое лечение» или «превентивное лечение» используются в данном документе взаимозаменяемо.
Под «риском» подразумевается субъект, то есть пациент, который идентифицирован как имеющий более высокий, чем обычно, шанс развития заболевания по сравнению с общей популяцией. Кроме того, субъект, который имел или у которого в настоящее время есть заболевание, в частности злокачественное новообразование, представляет собой субъект, у которого повышен риск развития заболевания, поскольку у такого субъекта может продолжить развиваться заболевание.
Термин «in vivo» относится к ситуации у субъекта.
Термин «индивидуум» или «субъект» относится к позвоночным животным, особенно млекопитающим. Например, млекопитающими в контексте настоящего изобретения являются люди, приматы, отличные от человека, домашние млекопитающие, такие как собаки, кошки, овцы, крупный рогатый скот, козы, свиньи, лошади и т.д., лабораторные животные, такие как мыши, крысы, кролики, морские свинки. свиньи и т.д., а также животные в неволе, такие как животные зоопарков. Термин «субъект» также относится к позвоночным животным, не относящимся к млекопитающим, таким как птицы (особенно одомашненные птицы, такие как курица, утки, гуси, индейки) и рыбы (особенно культивируемые рыбы, например, лосось или сом). Используемый в данном документе термин «животное» также включает людей. Предпочтительно термин «пациент» относится к больному человеку.
Термин «аутологичный» используется для описания всего, что происходит от одного и того же субъекта. Например, «аутологичная трансплантация» относится к трансплантации ткани или органов, полученных от одного и того же субъекта. Такие процедуры являются выгодными, поскольку они преодолевают иммунологический барьер, который в противном случае приводит к отторжению.
Термин «аллогенный» используется для описания всего, что происходит от разных особей одного и того же вида. Говорят, что два или более индивидуумов являются аллогенными по отношению друг к другу, когда гены в одном или нескольких локусах не идентичны.
Термин «сингенный» используется для описания всего, что происходит от индивидуумов или тканей, имеющих идентичные генотипы, т.е. идентичных близнецов или животных одного и того же инбредного штамма или их тканей.
Термин «гетерологичный» используется для описания того, что состоит из нескольких разных элементов. В качестве примера перенос костного мозга одного человека другому человеку представляет собой гетерологичную трансплантацию. Гетерологичный ген представляет собой ген, полученный из источника, отличного от субъекта.
Агенты, описанные в данном документе, могут быть введены в форме любой подходящей фармацевтической композиции. Термин «фармацевтическая композиция» относится к препарату, содержащему терапевтически эффективный агент или его соль, предпочтительно вместе с фармацевтическими наполнителями, такими как буферы, консерванты и модификаторы тоничности. Указанная фармацевтическая композиция полезна для лечения или профилактики заболевания или расстройства путем введения указанной фармацевтической композиции индивидууму. Фармацевтическая композиция также известна в данной области техники как фармацевтическая композиция. Фармацевтическая композиция может вводиться локально или системно.
Термин «системное введение» относится к введению терапевтически эффективного агента, так что агент становится широко распространенным в организме человека в значительных количествах и развивает биологический эффект. Согласно настоящему изобретению предпочтительно, чтобы введение осуществлялось парентеральным введением.
Термин «парентеральное введение» относится к введению терапевтически эффективного агента, так что агент не проходит через кишечник. Термин «парентеральное введение» включает внутривенное введение, подкожное введение, внутрикожное введение или внутриартериальное введение, без ограничения указанным.
Способы по изобретению также предпочтительно включают дополнительное обеспечение субъекта иммуноингибирующим соединением. Если иммуноингибирующее соединение представляет собой пептид или полипептид, оно может быть предоставлено субъекту путем введения иммуноингибирующего соединения или нуклеиновой кислоты, такой как РНК, в частности неиммуногенной РНК, такой как РНК, описанная в данном документе, кодирующей иммуноингибирующее соединение. Иммуноингибирующее соединение может быть выбрано из группы, состоящей из трансформирующего фактора роста бета (TGF-β), интерлейкина 10 (IL-10), антагониста рецептора интерлейкина 1 (IL-1RA), интерлейкина 4 (IL-4), интерлейкина 27 (IL-27), интерлейкина 35 (IL-35), запрограммированного лиганда смерти 1 (PD- L1), индуцибельного лиганда костимуляции Т-клеток (ICOSL), B7-H4, CD39, CD73, FAS, FAS-IL, индоламин 2,3-диоксигеназы 1 (IDO1), индоламин 2,3-диоксигеназы 2 (IDO2), ацетальдегиддегидрогеназы 1 (ALDH1)/ретинальдегиддегидрогеназы (RALDH), аргиназы 1 (ARG1), аргиназы 2 (ARG2), синтазы оксида азота (NOS2), галектина-1, галектина-9, семафорина 4А и любой их комбинации. Кроме того, композиции, описанные в настоящем документе, могут включать такое иммуноингибирующее соединение или нуклеиновую кислоту, как РНК, кодирующая такое иммуноингибирующее соединение.
Фармацевтическая композиция по настоящему изобретению обычно применяется в «фармацевтически эффективном количестве» и в «фармацевтически приемлемом составе».
Термин «фармацевтически эффективное количество» относится к количеству, которое достигает искомой реакции или искомого эффекта отдельно или вместе с дополнительными дозами. В случае лечения конкретного заболевания искомая реакция предпочтительно относится к ингибированию течения заболевания. Это включает замедление прогресса заболевания и, в частности, прекращение или отмену прогресса заболевания. Искомой реакцией при лечении заболевания также может быть задержка начала или предотвращение начала указанного заболевания или указанного состояния. Эффективное количество агента или композиции, описанных в данном документе, будет зависеть от состояния, подлежащего лечению, тяжести заболевания, индивидуальных параметров пациента, включая возраст, физиологическое состояние, размер и массу, продолжительность лечения, типа сопутствующей терапии (если имеется), конкретного пути введения и аналогичных факторов. Соответственно, дозы, вводимые композициями, описанными в данном документе, могут зависеть от различных таких параметров. В случае, если реакция у пациента недостаточна при использовании начальной дозы, могут использоваться более высокие дозы (или эффективные более высокие дозы, достигаемые другим, более локализованным способом введения).
Термин «фармацевтически приемлемый» относится к нетоксичности материала, который не взаимодействует с действием активного компонента фармацевтической композиции.
Фармацевтические композиции по настоящему изобретению могут включать соли, буферы, консерванты, носители и, необязательно, другие терапевтические агенты. Предпочтительно, фармацевтические композиции по настоящему изобретению содержат один или несколько фармацевтически приемлемых носителей, разбавителей и/или наполнителей.
Термин «наполнитель» предназначен для обозначения всех веществ в фармацевтической композиции, которые не являются активными ингредиентами, таких как связующие вещества, смазывающие вещества, загустители, поверхностно-активные вещества, консерванты, эмульгаторы, буферы, ароматизаторы или красители.
Термин «разбавитель» относится к разбавителю и/или разжижителю. Более того, термин «разбавитель» включает любую одну или несколько жидких, жидких или твердых суспензий и/или смешанных сред.
Термин «носитель» относится к одному или нескольким совместимым твердым или жидким наполнителям или разбавителям, которые подходят для введения человеку. Термин «носитель» относится к природному или синтетическому органическому или неорганическому компоненту, который объединяют с активным компонентом, чтобы облегчить нанесение активного компонента. Предпочтительно, компоненты носителя представляют собой стерильные жидкости, такие как вода или масла, включая те, которые получены из минерального масла, животных или растений, такие как арахисовое масло, масло соевых бобов, кунжутное масло, подсолнечное масло и т.д. Солевые растворы и водная декстроза и растворы глицерина также могут быть использованы в качестве водных соединений-носителей.
Фармацевтически приемлемые носители или разбавители для терапевтического применения хорошо известны в области фармацевтики и описаны, например, в Remington's Pharmaceutical Sciences, Mack Publishing Co. (A. R Gennaro edit. 1985). Примеры подходящих носителей включают, например, карбонат магния, стеарат магния, тальк, сахар, лактозу, пектин, декстрин, крахмал, желатин, трагакант, метилцеллюлозу, натрий-карбоксиметилцеллюлозу, легкоплавкий воск, масло какао и тому подобное. Примеры подходящих разбавителей включают этанол, глицерин и воду.
Фармацевтические носители, наполнители или разбавители могут быть выбраны с учетом предполагаемого пути введения и стандартной фармацевтической практики. Фармацевтические композиции по настоящему изобретению могут включать в качестве или в дополнение к носителю(-ям), эксципиенту(-ам) или разбавителю(-ям) любое подходящее связующее(ие), смазывающее(ие) вещество(а), суспендирующий(ие) агент(ы), глазировочный(ые) агент(ы) и/или солюбилизирующий(ие) агент(ы). Примеры подходящих связующих включают крахмал, желатин, натуральные сахара, такие как глюкоза, безводная лактоза, сыпучая лактоза, бета-лактоза, кукурузные подсластители, натуральные и синтетические камеди, такие как акациевая, трагакантовая или альгинат натрия, карбоксиметилцеллюлоза и полиэтиленгликоль. Примеры подходящих смазывающих веществ включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия и т.п. Консерванты, стабилизаторы, красители и даже ароматизаторы могут быть включены в фармацевтическую композицию. Примеры консервантов включают бензоат натрия, сорбиновую кислоту и сложные эфиры п-гидроксибензойной кислоты. Также могут быть использованы антиоксиданты и суспендирующие агенты.
В одном воплощении композиция представляет собой водную композицию. Водная композиция может необязательно включать растворенные вещества, например соли. В одном воплощении композиция находится в форме лиофилизированной композиции. Лиофилизированная композиция может быть получена лиофилизацией соответствующей водной композиции.
Настоящее изобретение дополнительно иллюстрируется следующими фигурами и примерами, которые не следует истолковывать как ограничивающие объем изобретения.
Чертежи
Фиг. 1 - активация иммунных клеток селезенки, высвобождение цитокинов и экспрессия белка после введения люциферазы-РНК в комплексе с липосомами F12. Мышам BALB/c (n = 7) внутривенно вводили в ретроорбитальное сплетение 10 мкг неиммуногенной мРНК-LPX LUC и исследовали через 24 часа (A) на предмет состояния созревания дендритных клеток (n = 3) (выявлено с помощью положительной регуляции CD86), Т-клетки, В-клетки и NK-клетки (выявленные положительной регуляцией CD69). Через 6 часов после инъекции мРНК-LPX мышам BALB/c (B) оценивали сывороточную концентрацию IFNα. (C) Определение активности люциферазы in vivo через 6, 24, 48 и 72 ч после введения мРНК-LPX мышам BALB/c (n = 4) люциферазы-РНК с помощью визуализации биолюминесценции in vivo. Данные представлены как среднее значение ± стандартное отклонение для n = 3 мышей/группу. LLOD = нижний предел обнаружения.
Фиг. 2 - точечный анализ неиммуногенной мРНК MOG35-55, очищенной либо обработкой целлюлозой, либо очисткой ВЭЖХ. Время выдержки 20 сек.
Фиг. 3 - презентация антигена MOG35-55 в DC индуцирует толерантность к EAE. EAE активно индуцировали иммунизацией MOG35-55-CFA (день 0) и токсином коклюша (дни 0 и 2) у мышей C57BL/6. Неиммуногенную MOG35-55-кодирующую мРНК, очищенную методом целлюлозы или ВЭЖХ, внутривенно вводили в ретроорбитальное сплетение на 7 и 10 день после индукции EAE. Контрольные мыши получали 150 мМ NaCl. Данные изображены как средние. n = 6 мышей/группу.
Фиг. 4 - презентация антигена MOG35-55 APC индуцирует антигенспецифическую пролиферацию Т-клеток. Наивные, меченные «Cell Trace Violet» (CTV) Thy1.1+ 2D2+ CD4+ T-клетки были адоптивно перенесены в контрольных мышей Thy1.2+ C57BL/6. На следующий день мышей иммунизировали 10, 20 или 40 мкг неиммуногенной MOG35-55-кодирующей мРНК. Контрольных мышей обрабатывали 20 мкг неиммуногенной нерелевантной контрольной мРНК. Кроме того, вторая контрольная группа получила 150 мМ NaCl. Через 4 дня мышей умерщвляли и селезенку анализировали на пролиферирующие клетки Thy1.1+. Данные представлены как среднее значение ± стандартное отклонение для n = 3 мышей/группу.
Фиг. 5 - подавление пролиферации Т-клеток MOG35-55 in vitro индуцированными антигенспецифическими клетками Treg. (A, B) 2D2 мышей Foxp3-eGFP иммунизировали 4 раза (d0, d3, d6, d9) неиммуногенной MOG35-55-кодирующей мРНК. Контрольных мышей обрабатывали иммуногенной MOG35-55-кодирующей мРНК или неиммуногенной нерелевантной контрольной мРНК. Третья контрольная группа получила дополнительно 150 мМ NaCl. Через 4 дня после последней обработки мРНК мышей умерщвляли и MOG35-55 специфические CD4+ клетки анализировали на экспрессию Foxp3-eGFP (C). Клетки каждой группы обработки (супрессоры) совместно культивировали с меченными CTV 2D2 CD4+ T-клетками необработанных наивных мышей (клетки-респондеры) и повторно стимулировали in vitro с помощью BMDC, нагруженными пептидом MOG35-55, в течение 72 часов. Пролиферацию клеток-респондеров определяли разведением CTV с помощью FACS (отбор на клетках CTV+). Ингибирующую способность супрессорных клеток измеряли при 4 различных соотношениях с клетками-респондерами. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ частоты CD4+ Foxp3+ клеток оценивали с помощью одностороннего анализа ANOVA и множественного сравнения Тьюки, *** p ≤ 0,001.
Фиг. 6 - презентация антигена MOG35-55 в DC индуцирует толерантность к EAE. EAE активно индуцировали иммунизацией MOG35-55-CFA (день 0) и токсином коклюша (дни 0 и 2) у мышей C57BL/6. Неиммуногенную мРНК, кодирующую MOG35-55, а также неиммуногенную нерелевантную мРНК вводили внутривенно в ретроорбитальное сплетение на 7 и 10 день после индукции EAE. Контрольные мыши получали 150 мМ NaCl. Данные изображены как средние. n = 6-8 мышей/группу. Статистический анализ максимальной оценки EAE для каждой мыши в группе определяли с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки. Площадь под кривой (AUC) была использована для определения статистической значимости с помощью одностороннего анализа ANOVA и множественного сравнения Тьюки кривых развития различных заболеваний EAE.
Фиг. 7 - презентация антигена MOG35-55 в DC индуцирует толерантность к EAE в реальных терапевтических условиях. EAE активно индуцировали иммунизацией MOG35- 55-CFA (день 0) и токсином коклюша (дни 0 и 2) у мышей C57BL/6. Неиммуногенное MOG35-55 кодирования мРНК, а также неиммуногенные нерелевантные мРНК вводили внутривенно в ретроорбитальное сплетение после того, как мыши достигали балла EAE 1-2. Контрольные мыши получали 150 мМ NaCI. Данные изображены как средние. n = 6-9 мышей/группу. Статистический анализ максимальной оценки EAE для каждой мыши в каждой группе оценивали с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки. Площадь под кривой (AUC) была использована для определения статистической значимости с помощью одностороннего анализа ANOVA и множественного сравнения Тьюки кривых развития различных заболеваний EAE.
Фиг. 8 - представление антигена DC индуцирует толерантность к EAE. EAE активно индуцировали путем иммунизации MOG35-55-CFA у мышей C57BL/6, и мышей обрабатывали на 7 и 10 день после индукции EAE 20 мкг неиммуногенной MOG35-55-кодирующей мРНК или 150 мМ NaCl. На 16-й день после индукции заболевания у мышей, пораженных EAE и мРНК, отбирали органы и культивировали в присутствии пептида MOG35-55 в течение 6 часов в присутствии Брефелдина А. Экспрессию CD40L исследовали путем внутриклеточного окрашивания цитокинов CD4+ CD44+ T-клеток. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ CD4+ Teff-клеток оценивали с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки, * p ≤ 0,05.
Фиг. 9 - количество MOG35-55 пептид-специфичных клеток Th1 и Th17 значительно снижено у мышей, получавших терапевтическую обработку (день 7/день 10). EAE активно индуцировали путем иммунизации MOG35-55-CFA у мышей C57BL/6, и мышей обрабатывали на 7 и 10 день после индукции EAE 20 мкг неиммуногенной мРНК, кодирующей MOG35- 55, или 150 мМ NaCl. Мозг и спинной мозг собирали на 16 день после индукции EAE, и клетки повторно стимулировали in vitro в присутствии пептида MOG35-55 в течение 6 часов в присутствии Брефелдина А. Затем CD4+ Т-клетки анализировали на экспрессию IFN и IL-17А путем внутриклеточного окрашивания цитокинов и рассчитывали процентное содержание IFNγ+ и IL-17A+ CD4+ Т-клеток. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ оценивали односторонним ANOVA и множественным сравнительным тестом Тьюки, * p ≤ 0,05.
Фиг. 10 - активация иммунных клеток селезенки и высвобождение цитокинов после введения различных смесей MOG35-55-мРНК неиммуногенной (m1Y) и иммуногенной (U) мРНК в комплексе с липосомами F12. Мышам C57BL/6 внутривенно вводили в ретроорбитальное сплетение 20 мкг общего мРНК-LPX MOG35- 55 и через 24 часа исследовали на предмет состояния созревания дендритных клеток (выявленных с помощью положительной регуляции CD86), Т-клеток и NK-клеток (выявленных с помощью активации CD69). Через 6 часов после инъекции мРНК-LPX мышам C57BL/6 оценивали сывороточную концентрацию IFNα. Данные представлены как среднее значение ± стандартное отклонение для n = 3 мышей/группу.
Фиг. 11 - дот-блот-анализ различных смесей MOG35-55-мРНК неиммуногенной и иммуногенной мРНК. Время экспонирования 20 сек. Пунктирная линия принята как 100% интенсивности точек для 320 пг дцРНК.
Фиг. 12 - неиммуногенная MOG35-55-мРНК индуцирует полную толерантность к EAE. EAE активно индуцировали иммунизацией MOG35-55-CFA (день 0) и токсином коклюша (дни 0 и 2) у мышей C57BL/6. Различные смеси неиммуногенных (m1Y) и иммуногенных (U) MOG35-55-кодирующих мРНК вводили внутривенно в ретроорбитальное сплетение на 7 и 10 день после индукции EAE. Контрольные мыши получали 150 мМ NaCl. Данные изображены как средние. n = 6-8 мышей/группу. Статистический анализ максимальной оценки EAE для каждой мыши в каждой группе оценивали с помощью одностороннего ANOVA и критерия множественного сравнения Тьюки, ** p ≤ 0,01.
Фиг. 13 - модификация нуклеозида сама по себе не вызывает потери распознавания. Активация иммунных клеток селезенки и высвобождение цитокинов после введения различных нуклеозид-модифицированных мРНК в комплексе с липосомами F12. Мышам C57BL/6 внутривенно инъецировали в ретроорбитальное сплетение 10 мкг общего мРНК-LPX и через 24 часа исследовали на предмет состояния созревания дендритных клеток (выявленных с помощью положительной регуляции CD86), Т-клеток и NK-клеток (выявленных с помощью положительной регуляции CD69). Через 6 часов после инъекции мРНК-LPX мышам C57BL/6 оценивали сывороточную концентрацию IFNα. Данные представлены как среднее значение ± стандартное отклонение для n = 3 мышей/группу.
Фиг. 14 - неиммуногенная обработка MOG35-55-кодирующая мРНК-LPX, не приводит к размножению антигенспецифичных CD4+ эффекторных клеток. Наивных мышей C57BL/6 обрабатывали в 0, 3, 7 и 10 день 20 мкг неиммуногенной MOG35-55-кодирующей мРНК. Контрольных мышей обрабатывали 20 мкг неиммуногенной нерелевантной контрольной мРНК. Вторая контрольная группа получила 150 мМ NaCl. На 13-й день после первой обработки мРНК мышей умерщвляли и анализировали спленоциты на секрецию про- и противовоспалительных цитокинов с помощью IFNγ ELISpot (A) и детектировали цитокины с помощью Multiplex ELISA в надосадочной жидкости клеток, стимулированных пептидом MOG35-55. (В и С). Данные представлены как среднее значение ± стандартное отклонение для n = 4-5 мышей/группу.
Фиг. 15 - соингибирующие молекулы активируются при обработке неиммуногенной MOG35-55-кодирующей мРНК-LPX. Наивных мышей C57BL/6 обрабатывали в 0, 3, 7 и 10 день 20 мкг неиммуногенной MOG35-55-кодирующей мРНК. Контрольных мышей обрабатывали 20 мкг неиммуногенной нерелевантной контрольной мРНК. Кроме того, вторая контрольная группа получила 150 мМ NaCl. На 13-й день после первой обработки мРНК мышей умерщвляли, и CD4+ Т- клетки MOG35-55 из селезенки анализировали с помощью проточной цитометрии на повышенную регуляцию коингибирующих молекул. Данные представлены как среднее значение ± стандартное отклонение для n = 4-5 мышей/группу. Статистический анализ оценивали с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки.
Фиг. 16 - терапевтическая вакцинация неиммуногенной MOG35-55-кодирующей мРНК-LPX, уменьшает инфильтрацию лимфоцитов в ЦНС. Наивные Thy1.1+ 2D2 CD4+ T-клетки были перенесены адоптивно мышам Thy1.2+ C57BL/6 в день d-1. EAE активно индуцировали на следующий день после иммунизации MOG35-55-CFA (день 0) и токсином коклюша (день 0 и 2), и мышей обрабатывали в день 7 и 10 после индукции EAE 20 мкг неиммуногенной MOG35-55-кодирующей мРНК, 20 мкг неиммуногенной нерелевантной мРНК или 150 мМ NaCl. На 16-й день после индукции заболевания брали органы у мышей, получавших различную обработку. Была исследована инфильтрация специфичных для Thy1.1+ CD4+ клеток MOG35-55, и общее количество инфильтрирующих клеток было определено с помощью проточной цитометрии с использованием пробирок Trucount™. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ инфильтрованных клеток CD4 + Teff оценивали с помощью одностороннего анализа ANOVA и множественного сравнения Тьюки.
Фиг. 17 - MOG35-55 специфичные клетки Th1 и Th17 значительно снижены у мышей, получавших терапевтическую обработку (день 7/день 10). EAE активно индуцировали иммунизацией MOG35-55-CFA (день 0) и токсином коклюша (день 0 и 2) у мышей C57BL/6, а мышей обрабатывали на день 7 и день 10 после индукции EAE 20 мкг неиммуногенной MOG35-55-кодирующей мРНК, 20 мкг неиммуногенной нерелевантной мРНК или 150 мМ NaCl. Мозг и спинной мозг собирали на 16 день после индукции EAE, и клетки повторно стимулировали in vitro в присутствии пептида MOG35-55 в течение 6 часов в присутствии Брефелдина А. Затем CD4+ Т-клетки анализировали с помощью проточной цитометрии на предмет их экспрессии. IFNγ и IL-17A путем внутриклеточного окрашивания цитокинов и процентные доли IFNγ+ и IL-17A+ CD4+ T-клеток рассчитывали. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ оценивали односторонним ANOVA и множественным сравнительным тестом Тьюки.
Фиг. 18 - соингибирующие молекулы активируются на MOG35-55-специфичных CD4 + T-клетках после успешной неиммуногенной MOG35-55-специфической обработки мРНК-LPX. EAE активно индуцировали иммунизацией MOG35-55-CFA (день 0) и токсином коклюша (день 0 и 2) у мышей C57BL/6, а мышей обрабатывали на день 7 и день 10 после индукции EAE 20 мкг неиммуногенной MOG35-55-кодирующей мРНК, 20 мкг неиммуногенной нерелевантной мРНК или 150 мМ NaCl. На 16-й день после индукции заболевания иссекали селезенки из групп с разной обработкой. MOG35-55-специфичные CD4 + Т-клетки выделяли с помощью тетрамер-специфического MACS и фенотипически анализировали с помощью проточной цитометрии с помощью окрашивания тетрамера. Данные представлены как среднее значение ± стандартное отклонение для n = 4 мышей/группу. Статистический анализ оценивали односторонним ANOVA и множественным сравнительным тестом Тьюки.
Фиг. 19 - PD-1 и CTLA-4 необходимы для поддержания неиммуногенной мРНК-индуцированной антигенспецифической толерантности. EAE активно индуцировали путем иммунизации MOG35-55-CFA (день 0) и токсином коклюша (день 0 и 2) у мышей C57BL/6, и мышей обрабатывали на 7 день и 10 день после индукции EAE 20 мкг неиммуногенной MOG35-55-кодирующей мРНК или 20 мкг неиммуногенной нерелевантной мРНК (n = 8 мышей/группу). Мышей, обработанных мРНК, обрабатывали одновременно анти-PD-1 или анти-CTLA-4-блокирующими антителами или соответствующими контрольными IgG. Развитие EAE оценивали путем ежедневного мониторинга состояния здоровья. Площадь EAE под кривой (AUC) использовали для определения статистической значимости с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки кривых развития различных заболеваний.
Фиг. 20 - антигенспецифическая индукция толерантности при лечении неиммуногенной антигенспецифической мРНК не зависит от модели заболевания EAE. (A) Терапевтическая вакцинация неиммуногенной PLP139-151, кодирующей мРНК-LPX, индуцирует толерантность к возвратно-ремиттирующему EAE. EAE активно индуцировали иммунизацией PLP139-151-CFA (день 0) и токсином коклюша (день 0 и 2) у мышей SJL. 20 мкг неиммуногенной PLP139-151 мРНК, а также неиммуногенной нерелевантной мРНК вводили внутривенно в ретроорбитальное сплетение 2 раза в неделю, начиная с 7-го и 10-го дня после индукции EAE. (B) Терапевтическая вакцинация неиммуногенной мультиэпитопной кодирующей мРНК-LPX индуцирует толерантность в комплексе EAE. EAE активно индуцировали иммунизацией MOG35-55, PLP139-151, PLP178-191, MBP84-104 и MOBP15-36-CFA (день 0) и токсином коклюша (день 0 и 2) у гибридов F1 у мышей C57BL/6 и SJL/JRJ. 40 мкг неиммуногенной мРНК, кодирующей несколько эпитопов, 20 мкг неиммуногенной мРНК MOG35-55 и 20 мкг неиммуногенной нерелевантной мРНК вводили внутривенно в ретроорбитальное сплетение 2 раза в неделю, начиная с 7 дня и 10 дня после индукции EAE. Развитие EAE оценивали путем ежедневного мониторинга состояния здоровья. Данные изображены как средние. n = 6 - 8 мышей/группу. Площадь EAE под кривой (AUC) использовали для определения статистической значимости с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки кривых развития различных заболеваний.
Фиг. 21 - совместная доставка цитокинов неиммуногенной мРНК улучшает толерогенную активность неиммуногенной антигенспецифической обработки мРНК-LPX в модели EAE. EAE активно индуцировали путем иммунизации MOG35-55-CFA (день 0) и токсином коклюша (день 0 и 2) у мышей C57BL/6, а мышей обрабатывали на 7 день и 10 день после индукции EAE 5 мкг неиммуногенной MOG35-55 мРНК вместе с 15 мкг неиммуногенной мРНК mIL-10 (A) или mIL-27 (B), соответственно. Контрольные мыши получали 5 мкг неиммуногенной MOG35-55 мРНК вместе с 15 мкг неиммуногенной нерелевантной мРНК, 15 мкг неиммуногенной мРНК, кодирующей цитокин, или 20 мкг неиммуногенной нерелевантной мРНК. Развитие EAE оценивали путем ежедневного мониторинга состояния здоровья. Данные изображены как средние. n = 6-8 мышей/группу. Площадь EAE под кривой (AUC) использовали для определения статистической значимости с помощью одностороннего ANOVA и множественного сравнительного теста Тьюки кривых развития различных заболеваний.
Примеры
Пример 1. Материалы и методы
Животные
Мыши дикого типа C57BL/6, BALB/c, SJL/JRj и мыши Hibrid F1 из C57BL/6 и SJL/JRj были приобретены у ENVIGO RMS gmbH, Нидерланды и Janvier Laboratories, Франция, соответственно. Thy1.1+ 2D2 TCR MOG трансгенные мыши C57BL/6, экспрессирующие Т-клеточный рецептор, распознающий MOG35-55 в контексте MHC класса II (IAb), а также Thy1.1+ 2D2 Foxp3-eGFP TCR MOG трансгенные мыши C57BL/6, которые дополнительно включают встраиваемый Foxp3-eGFP, были любезно предоставлены лабораторией профессора д-ра Ari Waisman. В экспериментах использовались животные соответствующего возраста и пола, и мышей содержали в особых условиях отсутствия патогенов (SPF) в помещении для животных BioNTech AG Mainz.
РНК-конструкции и транскрипция in vitro
In vitro транскрипция мРНК LUC была основана на плазмидной цепи pST1-hAg-CDS-FI-A30LA70, которая содержит 5'- UTR человеческого альфа-глобина (hAg), 3'-FI элемент и поли (A) хвост из 100 нуклеотидов, с линкером после 30 нуклеотидов. LUC-конструкция содержит ген люциферазы светлячка. Конструкции MOG35-55, PLP139-151, PLP178-191 и MBP84-104 были сконструированы на основе одной и той же плазмидной цепи (pST1-hAg-CDS-FI-A30LA70) и дополнительно содержали последовательности mmsec (opt) и mmMITD (opt) до и после кодирующих последовательностей MOG35-55, PLP139-151, PLP178-191 и MBP84-104 (pST1-hAg-mmsec (opt) -CDS-mmMITD (opt) -A30LA70), соответственно. Добавление лидерного пептида и сигнала переноса MHC (MITD) значительно улучшает представление антигена APC (Kreiter, S. et al., 2007, J. Immunol. 180, 309-318). Все последовательности антигена содержат дополнительно семь фланкирующих аминокислот до и после эпитопа, соответствующего заболеванию, для облегчения презентации антигена. Антигенная последовательность миелин-олигодендроцитарного гликопротеина (MOG) имеет пептидную последовательность: MOG27-63:SPGKNATGMEVGWYRSPFSRVVHLYRNGKDQDAEAQP. Для пептида мышиного миелинового протеолипидного белка PLP139-151 также были добавлены семь фланкирующих аминокислот для лучшей презентации пептида (PLP131-159:AHSLERVCHCLGKWLGHPDKFVGITYALT). Протеолипидный белок PLP178-191 включает аминокислоты PLP170-199:AVPVYIYFNTWTTCQSIAFPSKTSASIGSL, а основной белок миелина (MBP) 84-104 включает аминокислоты MBP76-112:RTQDENPVVHFFKNIVTPRTPPPSQGKGRGLSLSRFS. Для контрольной иммунизации мРНК использовали пустой вектор, кодирующий неспецифическую антигенную последовательность (pST2-hAg-mmsec (opt) -emtpy-mmMITD (opt) -2hBg-A30LA70). Цитокиновые кодирующие мРНК для мышиного интерлейкина 10 (mIL-10) и мышиного интерлейкина 27 (mIL-27) также были синтезированы на основе плазмидно-остова pST1-hAg-CDS-FI-A30LA70. РНК получали транскрипцией in vitro, как описано Kreiter et al., 2007 (Cancer Immunol. Immunother. 56, 1577-87). Очищенную мРНК элюировали в H2O и хранили при -80°C до дальнейшего применения. In vitro транскрипцию всех описанных конструкций мРНК осуществляли в BioNTech RNA Pharmaceuticals GmbH.
Получение неиммуногенной мРНК
Для получения неиммуногенной мРНК использовали 1-метил-псевдоуридин во время транскрипции in vitro вместо нормального нуклеозида уридина, и мРНК дополнительно очищали с помощью ВЭЖХ или обработки целлюлозой. Для очистки ВЭЖХ был адаптирован протокол Weissman et al., 2013 (Methods Mol Bio. 969, 43-43), и элюирование мРНК проводили с градиентом буфера B 38% -70%. На всех полученных мРНК осуществляли контроль качества (Bioanalyser и дот-блот анализ), чтобы гарантировать чистоту и целостность мРНК.
Для создания иммуногенной мРНК использовали нормальный нуклеозид уридин для транскрипции in vitro, и мРНК не очищали далее с помощью ВЭЖХ или обработки целлюлозой.
Для исследования влияния очистки ВЭЖХ или обработки целлюлозой на активацию спленоцитов использовали мРНК, модифицированную псевдоуридином (pU-мРНК), которая либо получала дополнительную очистку ВЭЖХ (мРНК pU-ВЭЖХ), либо нет. Соответствующую мРНК получали, как уже описано.
Дот-блот-анализ для контроля качества мРНК IVT
In vitro транскрибированную, модифицированную и очищенную ВЭЖХ мРНК анализировали на двухцепочечную мРНК (дцРНК) с помощью дот-блоттинга. 1 мкг различных конструкций мРНК загружали на мембрану NYTRAN SPC (GE Healthcare), блокировали и затем инкубировали с антителом J2 (SCICONS English and Scientific Consulting) для обнаружения дцРНК. В качестве вторичного антитела использовали антитело против мышиного HRP (Jackson ImmunoResearch) и мембраны анализировали с помощью BioRad ChemiDoc.
Подготовка и инъекция РНК-липоплексов
РНК-липоплексы (РНК-LPX) получали в стерильных условиях в отсутствие РНКаз. HEPES (10 мМ)/EDTA (0,1 мМ) использовали для разбавления РНК до концентрации 1 мг/мл. 1,5 М NaCl (5 М NaCl Stock; Ambion) добавляли к определенному объему РНК для получения конечной концентрации 150 мМ NaCl. После кратковременного встряхивания добавляли указанный объем липосом F12, и смесь РНК-липосомы (РНК-LPX) снова кратковременно встряхивали и, наконец, инкубировали в течение 10 мин при комнатной температуре для обеспечения образования РНК-LPX (Kranz et al. 2016, Nature. 16, 396-401). 200 мкл раствора РНК-LPX вводили на мышь внутривенно в ретроорбитальное сплетение.
Проточный цитометрический анализ
Поверхностные и внутриклеточные антитела для сортировки флуоресцентно-активированных клеток (FACS) были приобретены у eBioscience или BD Pharmingen и использовались в соответствии с протоколом производителя. Были использованы следующие антитела: CD11b, CD11c, CD19, CD25, CD4, CD44, CD40L, CD49b, CD69, CD8, CD86, CD90.1, Foxp3, ICOS, IFNγ, IL-17A, Lag-3, PD-1, TIGIT и Tim-3.
Суспензии отдельных клеток из разных органов окрашивали в течение 30 мин при 4°C для внеклеточных маркеров. Для внутриклеточного окрашивания цитокинов IFNγ, IL-17A и CD40L клетки выделяли, как описано, и дополнительно стимулировали в культуральной среде, содержащей пептид MOG35-55 (конц. 15 мкг/мл), монензин (GolgiStop, BD-Bioscience) и Брефельдин А при 37°C, 5% СО2 в течение 6 часов. После проведения окрашивания на живые-мертвые клетки (краситель для оценки жизнеспособности клеток eFluor® 506, eBioscience) и окрашивания маркеров клеточной поверхности, клетки фиксировали и пермеабилизовали с использованием буферов Cytofix/Cytoperm и Perm/Wash от BD Biosciences в соответствии с протоколом производителя. Клетки инкубировали в течение 30 мин при 4°C с внутриклеточными антителами и дважды промывали в Perm/Wash перед анализом FACS. Образцы собирали на BD LSR Fortessa или BD FACS Canto II и анализировали с использованием программного обеспечения FlowJo 7.6.5 или FlowJo 10.4 (Tree Star).
Для внутриклеточного окрашивания Foxp3 использовали буфер для окрашивания фактора транскрипции Foxp3 (Thermo Fisher Scientific) для фиксации и пермеабилизации. Все стадии выполнялись в соответствии с рекомендациями производителя после внеклеточного окрашивания.
MOG35-55-специфические CD4+ Т-клетки детектировали окрашиванием тетрамера. MOG35-55-специфичный APC-конъюгированный pMHC тетрамер класса II был получен из профильного центра тетрамеров Национальных институтов здравоохранения. Суспензии отдельных клеток мышиных спленоцитов инкубировали с APG-конъюгированным тетрамером MOG35-55 в течение 1 часа при комнатной температуре в полной среде DC (состав среды см. в разделе «Получение DC костного мозга (BMDC)»). Клетки промывали и окрашивали анти-АРС и проводили последующие MACS. Клетки инкубировали с внеклеточными или внутриклеточными антителами, как описано выше.
Для определения абсолютного количества CD4+ MOG35-55 (Thy1.1+ 2D2 CD4+ Т- клеток) в селезенке, головном и спинном мозге использовали пробирки Trucount™ (BD Biosciences). Суспензии отдельных клеток окрашивали, как описано выше, затем переносили в пробирки Trucount™ и абсолютное количество клеток определяли проточной цитометрией.
Биолюминесцентная визуализация in vivo (BLI)
Эффективность трансляции неиммуногенной мРНК в клетках селезенки исследовали с использованием системы визуализации Xenogen IVIS Spectrum in vivo (Caliper Life Sciences). Через 6, 24, 48 и 72 ч после РНК-LPX-иммунизации неиммуногенной LUC мРНК мышам вводили внутрибрюшинно водный раствор D-люциферина (250 мкл, 1,6 мг в PBS) (BD Biosciences). Через 5 минут интенсивность сигнала от селезенки определяли и измеряли биолюминесценцией in vivo в областях интереса (ROI) и определяли количественно как общий поток (фотонов/сек) с использованием программного обеспечения IVIS Living Image 4.0. Время сбора составляло 1 мин при биннинге 4. Во время биолюминесцентной визуализации живых мышей анестезировали дозой 2,5% смеси изофлуран/кислород. Интенсивность сигнала LUC испускаемых фотонов живых животных была изображена в виде изображения в градациях серого, где черный цвет является наименее интенсивным, а белый - наиболее интенсивным сигналом биолюминесценции. Изображения мышей анализировали с использованием программного обеспечения IVIS Living Image.
Магнитно-активированная сортировка клеток (MACS)
Thy1.1+ 2D2 CD4+ T-клетки Ty1.1 + 2D2 TCR MOG трансгенных мышей C57BL/6 или Thy1.1+ 2D2 Foxp3-eGFP TCR MOG трансгенных C57BL/6 мышей были выделены из селезенки и лимфатического узла путем положительного отбора с использованием MACS (Miltenyi Biotec) согласно инструкции производителя. CD4+ Т-клетки очищали с использованием CD4 (L3T4) микрогранул.
CD4+ MOG35-55-специфичные Т-клетки были обогащены MACS после окрашивания MOG35-55 специфичным APC-конъюгированным pMHC класса II с использованием анти-APC микрогранул. Обогащение производилось по протоколу производителя.
Получение DC костного мозга (BMDC)
Клетки костного мозга (BM) извлекали из бедренной и большеберцовой костей мышей C57BL/6 и культивировали в колбах для тканевых культур при 37°C в среде RPMI 1640 + GlutaMAX-I (Gibco) с добавлением 10% FBS (Biochrom), 1% 100x пирувата натрия (Gibco), 100x 1% MEM NEAA (Gibco), 0,5% пенициллина-стрептомицина (Gibco), 50 мкМ 2-меркаптоэтанола (Life Technologies) и 1000 Ед./мл фактора дифференцировки дендритных клеток (DC) гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) (Peprotech). В день 6 2×106 DC/мл предварительно инкубировали в течение 3 ч при 37°C с пептидом MOG35-55 (15 мкг/мл) и затем совместно культивировали с Т-клетками в общем объеме 200 мкл среды в 96-луночном планшете.
In vitro анализ супрессии Treg
Для исследований подавления Treg in vitro, CTV-меченные (набор клеточной пролиферации CellTrace Violet, Thermo Fisher Scientific) наивные CD4+ 2D2 трансгенные (Thy1.1+) Т-клетки (3 × 104 клеток на лунку) совместно культивировали с выделенными и CFSE-меченными (5 мкМ) CD4+ 2D2-Foxp3-eGFP трансгенными клетками, повторно иммунизированными мРНК, в различных соотношениях супрессоров и респондеров 2D2 Т-клеток (1:1, 2:1, 4:1, 8:1). Мышам 2D2-Foxp3-eGFP внутривенно вводили в ретроорбитальное сплетение в день 0, день 3, день 6 и день 9 неиммуногенную и иммуногенную MOG35-55 мРНК, а также неиммуногенную нерелевантную мРНК и физиологический раствор. Пролиферацию CTV-меченных респондерных 2D2 Т-клеток анализировали с помощью проточной цитометрии 72 ч после повторной стимуляции с помощью BMDC, нагруженных пептидом MOG35-55 (конечные концентрацией 15 мкг/мл пептида MOG35-55).
Адоптивный перенос Т-клеток и анализ пролиферации in vivo
Для исследований пролиферации обогащенные CD4+ 2D2 трансгенные (Thy1.1+) T-клетки метили CTV (набор пролиферации клеток CellTrace Violet, Thermo Fisher Scientific) в соответствии с инструкцией производителя, подсчитывали и 7 × 106 клеток инъецировали в 200 мкл PBS внутривенно в ретроорбитальное сплетение мышей-реципиентов C57BL/6 (Thy1.2+) под наркозом. На следующий день мышей C57BL/6 иммунизировали 10, 20 или 40 мкг неиммуногенной мРНК, кодирующей эпитоп MOG35-55. Контрольные мыши получали либо 20 мкг неиммуногенной нерелевантной мРНК, либо физиологический раствор. Через 4 дня после переноса Т-клеток мышей умерщвляли и клетки анализировали на пролиферацию с помощью проточной цитометрии.
Для определения абсолютного количества CD4+ MOG35-55-специфических Т-клеток в селезенке, головном и спинном мозге, а также для фенотипического анализа этих клеток после успешной обработки мРНК на 7-й и 10-й день у мышей с EAE, трансгенные CD4+ 2D2 (Thy1.1+) Т-клетки выделяли у трансгенных мышей Th571+ 2D2 TCR MOG C57BL/6, обогащали по CD4-клеткам с помощью MACS, и 7-10 × 106 клеток в 200 мкл PBS инъецировали внутривенно в ретроорбитальное сплетение наивных мышей-реципиентов C57BL/6 (Thy1.2+) под наркозом с использованием испарителя кислород-изофлуран (2,5% изофлуран/кислород). На следующий день у мышей C57BL/6 активно индуцировали EAE.
ELISA
Детекцию мышиного IFN-α проводили с помощью ELISA (PBL) через 6 часов после РНК-иммунизации в мышиной сыворотке с использованием стандартного анализа ELISA в соответствии с инструкциями производителя.
Цитокины Th1/Th2/Th9/Th17/Th22 и Treg, присутствующие в надосадочных жидкостях клеточных культур рестимулированных спленоцитов мышей, повторной подвергнутых обработке мРНК, количественно определяли с использованием 17-плексного мышиного набора для анализа ELISA (Thermo Fisher Scientific). Суспензии отдельных клеток спленоцитов (5 × 105 клеток), выделенных от мышей, стимулировали в течение 72 ч с 0, 5, 10, 20 или 100 мкг/мл пептида MOG35-55, и надосадочные жидкости анализировали на содержание цитокинов.
Твердофазный иммуноанализ IFNγ в пятне (ELISpot)
Частоты отвечающих на MOG35-55 Т-клеток исследовали с помощью ELISpot для интерферона γ (IFNγ). Мышей обрабатывали в 0, 3, 7 и 10 день неиммуногенной мРНК MOG35-55, неиммуногенной нерелевантной мРНК или физиологическим раствором. На 13 день после первой обработки выделяли селезенки разных групп обработки и готовили суспензии отдельных клеток. CD4+ Т-клетки очищали с помощью MACS с использованием микрогранул CD4 (L3T4). Количественную оценку антигенспецифических Т-клеток, отвечающих на IFNγ, проводили после повторной стимуляции пептидом MOG35-55 с использованием набора ELISpot. 96-луночные планшеты ELISpot (MultiScreenHTS IP Filter Plate, Merck Millipore) в течение ночи покрывали IFNγ-специфическим захватывающим антителом (антитело против IFNγ мыши, Mabtech). После блокирования полной средой планшеты промывали и 5 × 105 CD4+ T-клеток высевали вместе с 1 × 105 BMDC, нагруженными пептидом MOG35-55 (концентрация пептида 15 мкг/мл). В качестве контроля CD4+ Т-клетки инкубировали с BMDC, нагруженными нерелевантным пептидом. Все клетки культивировали в течение 16 ч при 37°C, 5% CO2 увлажненного инкубатора. Клетки промывали и инкубировали в течение 2 ч при 37°C и 5% СО2 с биотинилированным антителом, детектирующим анти-IFNγ (биотинилированное антитело против IFNγ мыши, Mabtech). Это вторичное антитело визуализировали, добавляя ExtrAvidin-щелочную фосфатазу (Sigma), и после инкубации в течение 1 часа добавляли жидкий субстрат BCIP/NBT (Sigma), и реакцию останавливали через 1-3 минуты путем промывания планшета под проточной водой. Количественную оценку пятен выполняли на ELISpot Reader (S6 Macro Analyzer; CTL).
Индукция EAE и лечение мышей мРНК
EAE активно индуцировали у самок мышей C57BL/6 в возрасте от восьми до десяти недель с помощью 50 мкг пептида MOG35-55 (аминокислотная последовательность: MEVGWYRSPFSRVVHLYRNGK), эмульгированного в CFA (Difco Laboratories), с добавлением 10 мг/мл инактивированной нагреванием Mycobacterium tuberculosis H37RA (Difco Laboratories) подкожно (п.к.) у основания хвоста. Мышам вводили 150 нг токсина коклюша (List Biological Laboratories, INC., Кэмпбелл, Калифорния) внутрибрюшинно (в.б.) в день иммунизации и через 2 дня. Мышей взвешивали и оценивали ежедневно, начиная с 10-го дня после иммунизации, в соответствии со следующими критериями: 0 - нет заболевания; 1, снижение тонуса хвоста; 2 - нарушение рефлекса восстановления; 3 - частичный паралич задних конечностей; 4 - полный паралич задних конечностей; 5 - паралич задних конечностей с частичным параличом передних конечностей; и 6 - умирающий или мертвый.
Кроме того, EAE активно индуцировали у самок мышей SJL/JRj (8-10 недель) 200 мкг пептида PLP139-151 (аминокислотная последовательность: HSLGKWLGHPDKF), эмульгированного в CFA, дополненном 10 мг/мл инактивированной нагреванием Mycobacterium tuberculosis H37RA п.к. в основание хвоста. Мыши получали 200 нг PTX в.б. в день иммунизации и 2 днями позже. Мышей взвешивали и оценивали ежедневно, начиная с 8-го дня после иммунизации, в соответствии с уже описанными критериями оценки.
Комплекс EAE активно индуцировали у самок гибридных мышей F1 из C57BL/6 и SJL/JRj (8-10 недель) с 250 мкг пептидных смесей (MOG35-55, PLP139-151, PLP178-191 (аминокислотная последовательность: NTWTTCQSIAFPSK), MBP84-104 (аминокислотная последовательность: VHFFKNIVTPRTPPPSQGKGR), MOBP15-36 (аминокислотная последовательность: QKFSEHFSIHCCPPFTFLNSKR)). Из каждого пептида использовали 50 мкг. Пептидную смесь эмульгировали в CFA с добавлением 10 мг/мл инактивированных нагреванием Mycobacterium tuberculosis H37RA и инъецировали подкожно в основание хвоста. Мышам вводили 200 нг PTX в.б. в день иммунизации и через 2 дня. Мышей взвешивали и оценивали ежедневно, начиная с 8-го дня после иммунизации, в соответствии с уже описанными критериями оценки.
Для определения защитного иммунитета у мышей C57BL/6 мышей обрабатывали на 7-й и 10-й день или при балле EAE 1-2 после индукции заболевания 20 мкг неиммуногенной MOG35-55-кодирующей мРНК, 20 мкг неиммуногенной нерелевантной контрольной мРНК или физиологическим раствором.
Терапевтическую обработку возвратно-ремиттирующего EAE у мышей SJL проводили с использованием 20 мкг неиммуногенной мРНК PLP139-151 2 раза в неделю, начиная с 7 дня и 10 дня после индукции EAE. Контрольные мыши получали 20 мкг неиммуногенной нерелевантной контрольной мРНК.
В экспериментах, сравнивающих неиммуногенную мРНК с одним эпитопом с неиммуногенной мультиэпитопной обработкой, мышей с комплексным EAE обрабатывали 40 мкг неиммуногенной мРНК, кодирующей несколько эпитопов (10 мкг каждой неиммуногенной мРНК, кодирующей эпитоп (MOG35) -55, PLP139-151, PLP178-191, MBP84-104), 20 мкг неиммуногенной мРНК MOG35-55 или 20 мкг неиммуногенной нерелевантной контрольной мРНК. мРНК вводили 2 раза в неделю, начиная с 7-го дня и 10-го дня после индукции EAE.
Для тестирования комбинаторных эффектов обработки мРНК, кодирующей цитокины, и обработки антигенспецифической мРНК, 5 мкг неиммуногенной мРНК, кодирующей MOG35-55, инъецировали вместе с 15 мкг неиммуногенной мРНК, кодирующей mIL-10 или mIL-27, на 7 и 10 день после индукции EAE. Контрольные мыши получали 5 мкг неиммуногенной мРНК MOG35-55 вместе с 15 мкг неиммуногенной нерелевантной мРНК, 15 мкг неиммуногенной мРНК, кодирующей цитокин, или 20 мкг неиммуногенной нерелевантной мРНК.
Для всех экспериментов мРНК вводили внутривенно в ретроорбитальное сплетение под наркозом, используя испаритель кислород-изофлуран (2,5% изофлуран/кислород).
Обработка антителами
Мышей C57BL/6 обрабатывали PD-1 (первая обработка 500 мкг, 250 мкг после обработки, клон RMP1-14, BioXcell) или CTLA-4 (первая обработка 500 мкг, 250 мкг после обработки, клон 9H10, BioXcell) блокирующим антителом или изотипными контрольными антителами (IgG2a крысы и IgG сирийского хомяка, BioXCell) на 7, 10, 14 и 17 день после индукции EAE. Антитела разводили в PBS и вводили в.б.
Выделение инфильтратов селезенки, лимфатических узлов и ЦНС
Все клеточные анализы проводили на одноклеточной суспензии селезенки, лимфатических узлов и органов ЦНС. Вкратце, селезенки и лимфоузлы инкубировали с коллагеназой D 1 мг/мл и ДНКазой I 0,1 мг/мл в течение 15 минут и затем протирали через 70 мкм сито для клеток при промывании PBS. Лизис эритроцитов (буфер для гипотонического лизиса:1 г KHCO3 и 8,25 г NH4Cl, растворенного в 1 л H2O и 200 мкл 0,5 М EDTA) проводили на одноклеточной суспензии спленоцитов. После дополнительной стадии промывания PBS клетки подсчитывали с помощью счетчика клеток Vi-Cell (Beckman Coulter).
Для выделения инфильтратов ЦНС из головного и спинного мозга мышей анестезировали Кетамином-Ромпун и перфузировали, используя 0,9% NaCl через левый желудочек сердца. Головной и спинной мозг удаляли вручную, разрезали на мелкие кусочки и гидролизовали в PBS ++ (PBS с кальцием и магнием, Gibco), содержащим коллагеназу D (1 мг/мл) и ДНКазу I (0,1 мг/мл) в течение 20 мин при 37°C. Затем расщепленную ткань гомогенизировали вручную, всасывая кусочки ткани в шприц и снова выдавливали на стенку 15-миллилитровой пробирки Falcon. Это было выполнено вплоть до 10 раз, пока перестали быть видны какие-либо фрагменты. Суспензию отдельных клеток ресуспендировали в 70% перколле и наслаивали в градиенте перколла 30:37. Конечный градиент перколла составлял 30:37:70% и его центрифугировали при 300 g в течение 40 минут при комнатной температуре. Слой мононуклеарных клеток, лежащий на межфазном интервале от 70% до 37% перколла, промывали 2% FCS/PBS перед дальнейшими анализами.
Статистический анализ
Статистическую значимость оценивали с использованием программного обеспечения GraphPad Prism 7 (Graphpad Software, Inc.) с использованием одностороннего ANOVA, скорректированного с помощью критерия сравнения Тьюки, при сравнении более двух групп. Кривые развития EAE сравнивали путем расчета площади под кривой развития заболевания (AUC) для одиночных мышей. Значения p ≤ 0,05 считались статистически значимыми; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001. Используемые тесты упоминаются в соответствующих легендах к фигурам.
Пример 2. Исследование неиммуногенной мРНК в отношении активации спленоцитов
Для анализа активации спленоцитов и эффективности трансляции использованной неиммуногенной мРНК мышам BALB/c внутривенно вводили в ретроорбитальное сплетение мРНК Люцеферазы (10 мкг) в комплексе с липосомами F12. Активность люциферазы оценивали с помощью визуализации in vivo через 6, 24, 48 и 72 часа после инъекции РНК-LPX, и типичные мыши показаны на фиг. 1С. Иммунизация мышей неиммуногенной мРНК приводила к длительной высокой трансляции LUC-мРНК. Следовательно, экспрессия белка LUC может быть обнаружена вплоть до 72 часов после иммунизации мРНК.
Кроме того, как видно на фиг. 1А, иммунизация мышей неиммуногенной мРНК не приводила к усилению активности маркеров активации CD86 на DC и CD69 на лимфоцитах, как у необработанных контрольных мышей. У мышей, иммунизированных неиммуногенной LUC-мРНК, также не был обнаружен IFNα в крови через 6 часов после иммунизации мРНК (фиг. 1B).
Пример 3. Характеризация неиммуногенной мРНК
Неиммуногенная мРНК используется для доставки специфичных для заболевания антигенов в дендритные клетки для обеспечения презентации антигена без иммунной активации в терапевтических применениях. Само собой разумеется, что включение 1-метилпсевдоуридина в мРНК усиливает экспрессию белка в клетках и снижает иммуногенность мРНК в клеточных линиях млекопитающих, а также in vivo в мышах (Andries et al., 2015, J Control Release. 217, 337-344). Этот эффект зависит, скорее всего, от увеличенной способности мРНК, избегать активацию эндосомных толл-подобных рецепторов и нижележащего врожденной иммунной сигнализации (Andries et al., 2015). В дополнение к применению 1-метилпсевдоуридина вместо нуклеозида уридина для включения в мРНК во время транскрипции in vitro, очистка синтетической мРНК методом ВЭЖХ устраняет, кроме того, иммунную активацию и улучшает трансляцию нуклеозид-модифицированной, белок-кодирующей мРНК (Kariko et al., 2011, Nucleic Acids Res. 39, e142). Путем очистки ВЭЖХ оставшиеся двухцепочечные контаминанты мРНК удаляются после транскрипции мРНК in vitro, в результате чего мРНК не индуцирует передачу сигналов интерферона и воспалительные цитокины (Kariko et al., 2011). Альтернативным способом очистки нуклеозид-очищенной мРНК является очистка целлюлозы (PCT/EP2016/059056).
Чтобы выяснить, действительно ли мРНК, используемая для терапевтического применения, является неиммуногенной, были выполнены анализы Bioanalyzer и дот-блоттингом для подтверждения целостности и чистоты мРНК. На фиг. 2 показан дот-блот-анализ неиммуногенной мРНК MOG35-55, очищенной либо обработкой целлюлозой, либо очисткой ВЭЖХ. Антитело J2, специфичное к дцРНК, не показало сигнала после очистки целлюлозой или ВЭЖХ анализируемой MOG35-55 мРНК в диапазоне от 40 нг до 1 мкг используемой мРНК. Этот контроль качества обеспечивает неиммуногенные характеристики мРНК в отношении обеих партий мРНК, при обработке EAE могут быть достигнуты положительные терапевтические эффекты. EAE активно индуцировали у мышей C57BL/6, и обрабатывали мышей на 7 и 10 день после индукции заболевания антигенспецифической MOG35-55-кодирующей неиммуногенной мРНК, очищенной на целлюлозе или методом ВЭЖХ. Обе группы были полностью устойчивы к индукции EAE (фиг. 3). Более подробное описание терапевтической вакцинации неиммуногенной антигенспецифической мРНК будет объяснено в следующих примерах.
Пример 4. Эффекты иммунизации неиммуногенной MOG35-55 мРНК на пролиферацию антигенспецифических Т-клеток
Чтобы изучить, приводит ли иммунизация неиммуногенной мРНК-LPX MOG35-55 к презентации антигена MOG35-55 в DC для T-клеток, исследовали пролиферацию антигенспецифических CD4+ T-клеток (CD4+ 2D2 T-клеток), которые несут миелин-олигодендроцитарный гликопротеин (MOG) -специфический Т-клеточный рецептор. «Cell-Trace-Violet» (CTV)-меченные Т-клетки, Thy1.1+ MOG35-55-специфичные CD4+ 2D2 Т-клетки переносили мышам Thy1.2+ C57BL/6. Через 24 часа мышей-реципиентов иммунизировали различными концентрациями (10, 20 или 40 мкг мРНК) неиммуногенной мРНК, кодирующей эпитоп MOG35-55, и еще через 4 дня MOG35-55-специфичные CD4+ 2D2 T-клетки были проанализированы на предмет размножения. Контрольные мыши получали либо 20 мкг неиммуногенной нерелевантной мРНК, либо физиологический раствор. 2D2 Т-клетки пролиферировали, когда мышей обрабатывали неиммуногенными мРНК MOG35-55 по сравнению с контрольными мышами (фиг. 4), что указывает на то, что антигенпрезентирующие клетки (АРС) транслируют и обрабатывают антигенкодирующую неиммуногенную мРНК правильным образом и презентируют пептид MOG35-55. Кроме того, иммунизация мышей неиммуногенной мРНК MOG35-55 с увеличением дозы от 10 до 40 мкг также показала увеличение антигенспецифической популяции Т-клеток.
Пример 5. Иммунизация антигенспецифической неиммуногенной мРНК индуцирует развитие регуляторных Т-клеток у наивных мышей
Treg являются фундаментальными в контроле различных иммунных реакций, и они важны для индукции и поддержания толерантности. Доклинические исследования на животных моделях показывают, что адоптивный перенос Treg может предотвратить или вылечить несколько заболеваний, опосредованных Т-клетками, включая аутоиммунные заболевания, путем восстановления иммунной толерантности к собственным антигенам. Было описано несколько CD4+ регуляторных T-клеток (Shevach, EM, 2006, Immunity 25, 195-201) и подразделяются на две основные подгруппы: природные и индуцибельные Treg, которые генерируются на периферии в различных толерогенных условиях. Клетки Treg регулируют примирование аутореактивных Т-клеток, ограничивая их экспансию и дифференцировку (Sakaguchi, S. et al., 2008, Cell 133, 775-87).
Для дальнейшего изучения влияния иммунизации антигенспецифической неиммуногенной мРНК на соответствующий фенотип Т-клеток, слитый белок 2D2-Foxp3-eGFP трансфицировали в Т-клетки Th1.1-позитивных мышей. Эти мыши были повторно иммунизированы (d0, d3, d6 и d9) неиммуногенной и иммуногенной мРНК MOG35-55, а также неиммуногенной нерелевантной мРНК или физиологическим раствором. Иммунизация мышей неиммуногенной мРНК MOG35-55 привела к развитию у мышей клеток Foxp3+, проанализированных через 4 дня после последней обработки мРНК (d13). Мыши, обработанные иммуногенной антигенспецифической мРНК или нерелевантной неиммуногенной мРНК, не показали увеличения популяции клеток Foxp3 и имели частоту Foxp3-положительных клеток, таких как контрольные мыши, обработанные физиологическим раствором (фиг. 5А и В).
Действуют ли индуцированные Foxp3+ клетки супрессивно, оценивали с помощью анализа супрессии in vitro (фиг. 5С). Следовательно, суммарные CD4+ клетки иммунизированных мышей метили CFSE и совместно культивировали в течение 72 часов в различных соотношениях супрессор-респондер с исходными MOG35-55-специфичными CD4+ 2D2 T-клетками, которые дополнительно метили CTV. Важно, что индуцированные регуляторные клетки от мышей, иммунизированных неиммуногенной мРНК MOG35-55, опосредовали дозозависимое подавление пролиферации эффекторных Т-клеток 2D2 в присутствии BMDC, нагруженных пептидом MOG35-55. Напротив, все остальные группы не оказывали супрессирующего действия.
В совокупности, эти результаты показывают, что APC-специфическая презентация антигена в невоспалительных условиях опосредовала развитие функциональных регуляторных Т-клеток.
Пример 6. Влияние терапевтической вакцинации неиммуногенной MOG35-55 мРНК на модель заболевания EAE
Чтобы выяснить толерогенный эффект иммунизации мРНК антигенспецифической неиммуногенной мРНК на модели заболевания, у мышей C57BL/6 активно индуцировали EAE, и мышей обрабатывали на 7 и 10 день после индукции заболевания антигенспецифической MOG35-55 кодирующей неиммуногенной мРНК, а также неиммуногенной нерелевантной мРНК. Обработка была начата в момент времени, когда сильные сигналы активации и созревания постоянного тока, а также первое праймирование Т-клеток уже имели место. РНК-иммунизация неиммуногенной антигенспецифической мРНК полностью блокировала любые признаки EAE (фиг. 6), и ни у одной мыши из 6 иммунизированных мРНК мышей заболевание не развивалось.
В то же время обработка мышей неиммуногенной мРНК, кодирующей нерелевантный эпитоп, не защищало мышей от заболевания, и у 5/6 мышей развился EAE (фиг. 6). Эти результаты показывают, что презентация MOG незрелыми DC может подавлять EAE, и только животные, получавшие неиммуногенную антигенспецифичную мРНК, оказались полностью устойчивыми к индукции EAE.
Кроме того было исследовано, можно ли лечить EAE во время более поздних моментов прогрессирования заболевания, когда симптомы заболевания являются видимыми, и мыши достигли балла заболевания 1-2. Примечательно, что можно показать, что после РНК-иммунизации неиммуногенной MOG35-55-кодирующей мРНК у мышей с EAE не развивалось тяжелое EAE, и симптомы можно было уменьшить при обработке при балле 1 (фиг. 7). Даже когда мышей обрабатывали при 2 баллах, быстрое развитие болезни можно было остановить. Максимальный балл заболевания у мышей составил только 2-3, в то время как почти у всех необработанных контрольных мышей развилась полная EAE с максимальным баллом 4. По сравнению с мышами EAE, получавшими неиммуногенную MOG35-55 кодирующую мРНК, терапевтическое лечение неиммуногенной нерелевантной мРНК не оказывало положительного влияния на прогрессирование заболевания, и у животных развивался EAE, как у необработанных контрольных мышей.
Для изучения молекулярного механизма толерантности, вызванной обработкой мРНК, и определения того, влияет ли мРНК-иммунизация неиммуногенной антигенспецифической мРНК и, следовательно, презентация MOG35-55 на DC, непосредственно на эффекторные T-клетки (Teff), EAE снова индуцировали у мышей C57BL/6, затем на 7 и 10 день после индукции заболевания животных обрабатывали неиммуногенной MOG35-55-кодирующей мРНК, и через 6 дней после последней обработки мРНК выделяли селезенку, LN, головной и спинной мозг. Клетки из разных органов реактивировали in vitro пептидом MOG35-55 в течение 6 часов и затем отбирали Teff-клетки по пороговому сигналу на CD40 лиганд (CD40L). CD40L быстро экспрессировался на Т-клетках после активации и поэтому использовался в качестве маркера для MOG-специфических клеток Teff. Как видно на фиг. 8, введение неиммуногенного MOG35-55 мРНК приводит к значительному снижению частоты антигенспецифических CD4+ Т-клеток в группах, получавших неиммуногенную антигенспецифическую мРНК, в головном и спинном мозге. На функциональном уровне можно показать, что CD4+ T-клетки секретируют значительно меньше интерферона-γ (IFNγ) в головном мозге и интерлейкина-17A (IL-17A) в спинном мозге у мышей, получавших неиммуногенную MOG35-55-кодирующую мРНК по сравнению с необработанными контрольными мышами (фиг. 9). Эти данные указывают на то, что обработка мРНК неиммуногенной MOG35-55 мРНК может инактивировать T- и Teff-клетки памяти, что приводит к уменьшению этих клеток.
Пример 7. Влияние чистоты мРНК на эффективность обработки EAE
Эксперименты с комбинациями иммуногенной мРНК, содержащей уридин (U), и неиммуногенной мРНК, содержащей 1-метилпсевдоуридин и очищаемой с помощью целлюлозы (m1Y), показали, что активация спленоцитов (исследована путем активации регулятора активации CD86 на DC и CD69 на лимфоцитах) секреция IFNα коррелирует с количеством используемой иммуногенной мРНК. Активация почти не была достигнута с неиммуногенной мРНК (20 мкг mOG35-55 мРНК), и активация постоянно увеличивалась при добавлении иммуногенной мРНК (U MOG35-55 мРНК) (фиг. 10). 5 мкг мРНК U + 15 мкг мРНК m1Y показали лишь немного меньшую активацию, чем 20 мкг мРНК U, тогда как 20 мкг мРНК m1Y снова вообще не активировали иммунную систему. Соответствующий дот-блот-анализ различных смесей мРНК также показал, что только 100% неиммуногенной (m1Y) мРНК не показывают сигнала для дцРНК (фиг. 11).
Для дальнейшего исследования влияния чистоты использованной мРНК на эффективность обработки EAE были использованы те же самые смеси мРНК для обработки продолжающегося EAE. EAE опять активно индуцировали у мышей C57BL/6, и мышей обрабатывали на 7 и 10 день после индукции заболевания антигенспецифической мРНК. В этом эксперименте были использованы те же смеси мРНК неиммуногенной и иммуногенной MOG35-55 мРНК, что и в предыдущем эксперименте «активации». В отличие от 100% неиммуногенной (m1Y) мРНК, смесь неиммуногенной и иммуногенной мРНК (2,5 мкг U + 17,5 мкг m1Y MOG35-55 мРНК, а также 5 мкг U + 15 мкг m1Y MOG35-55 мРНК) не может предотвратить заболевание развитие полностью, и у некоторых мышей развивается заболевание, подобное заболеванию мышей, получавших 100% иммуногенную мРНК (фиг. 12). Эти результаты показывают, что для эффективного лечения EAE необходима чистая неиммуногенная (m1Y) мРНК.
Было проведено дополнительное сравнение с точки зрения активации селезенки немодифицированной и модифицированной мРНК, которая была очищена или не очищена с помощью ВЭЖХ. Как показано на фиг. 13, модификации нуклеозидов, такие как исключительно псевдоуридин (pU), не вызывают потерю распознавания мРНК иммунной системой, но процесс очистки отличается. Неочищенная pU-мРНК все еще является иммуногенной и работает как обычная мРНК (U-мРНК) в отношении активации селезенки (сильная активация CD86 на DC, активация CD69 на B-клетках, T-клетках и NK-клетках) и секреция IFNα. Напротив, pU-мРНК, которая была дополнительно очищена с помощью ВЭЖХ, чтобы избавиться от дцРНК (мРНК pU- HPLC), предотвращает активацию DC, B-клеток, T-клеток и NK-клеток и, следовательно, секрецию IFNα, как у необработанных контрольных мышей. Эти результаты подчеркивают, что исключительно комбинация уридин-модификации и дополнительной очистки мРНК IVT сводит к минимуму иммуногенность мРНК, как это наблюдалось у наивных мышей, уровень, на котором можно индуцировать толерантность. Таким образом, неиммуногенная мРНК (уридин-модифицированная в сочетании с очисткой мРНК) является оптимальной мРНК с целью индукции толерантности.
Пример 8. Обработка неиммуногенной MOG35-55-специфичной мРНК не вызывает de novo праймирования антигенспецифических CD4+ Т-клеток, но приводит к размножению анергических Т-клеток у наивных мышей
Поддержание и индукция периферической толерантности зависит от представления самоантигена APC, которые экспрессируют низкие уровни костимулирующих молекул, таких как CD86 или CD40, на их поверхности. Отсутствие экспрессии костимулирующей молекулы и продукции цитокинов APC при обработке неиммуногенной мРНК было показано в примере 2, на фиг. 1. Следовательно, при обработке мРНК, АРС процессируют и презентируют эпитоп, кодируемый неиммуногенной мРНК, в отсутствие активирующих стимулов. Вовлечение TCR в отсутствие костимулирующих сигналов не приводит к размножению эффекторных Т-клеток, скорее к апоптозу или анергии Т-клеток (Mueller, DL, 2010, Nat. Immunol. 11, 21-7). Напротив, во время праймирования эффекторных Т-клеток DC высвобождают специфические цитокины, которые вместе с презентацией антигена и экспрессией костимулирующих молекул поляризуют эффекторные иммунные ответы, управляя дифференцировкой CD4+ Т-клеток в клетки Th1, Th2 или Th17. Действительно, получение эффекторных Т-клеток при неиммуногенной антигенспецифической обработке мРНК не предназначена для аутоиммунного заболевания, поскольку большее количество эффекторных Т-клеток может даже привести к более тяжелому течению заболевания вместо лечебной обработки.
Чтобы исследовать получение антигеннаправленных Т-клеточных ответов, наивных мышей C57BL/6 периодически обрабатывали (день 0, 3, 7 и 10) 20 мкг неиммуногенной кодирующей MOG35-55 мРНК, 20 мкг неиммуногенной нерелевантной мРНК или физиологическим раствором. После четырех иммунизаций мРНК CD4+ Т-клетки выделяли на 13 день из селезенки и проводили ELISpot IFNγ. На фиг. 14А показано, что у мышей, получавших неиммуногенную мРНК, кодирующую MOG35-55, не развивались IFNγ+ CD4+ эффекторные Т-клетки, отвечающие на распознавание антигена при обработке антигенспецифической мРНК.
В соответствии с этими результатами также рестимуляция CD4+ T-клеток селезенки с различными концентрациями пептида MOG35-55 (0, 5, 10, 20 и 100 мкг) в течение 48 ч не приводила к секреции других воспалительных цитокинов, таких как TNFα и GM-CSF, как измерено в надосадочных жидкостях с помощью ELISA (фиг. 14 B). Только секрет противовоспалительного цитокина IL-10 был обнаружен у мышей, получавших неиммуногенную антигенспецифичную мРНК, но не с неиммуногенной нерелевантной мРНК (фиг. 14C). Кроме того, только в этой группе, секрецию цитокинов Th2, таких как IL-4 и IL-5 может быть обнаружено (данные не показаны). Продуцирование IL-10 в ответ на неиммуногенную антигенспецифическую мРНК свидетельствует о том, что антигенспецифические регуляторные Т-клетки (Treg) активируются в ответ на обработку. Treg являются основополагающими в контроле различных иммунных реакций. Сообщалось о множестве различных подмножеств индуцибельных Treg, а продуцирование высоких уровней IL-10 особенно характерно для клеток CD25+ Foxp3+ и Tr1. Эти регуляторные Т-клетки регулируют прайминг аутореактивных Т-клеток, ограничивая их экспансию и дифференцировку.
Чтобы исследовать фенотип CD4 после неиммуногенной антигенспецифической обработки мРНК, мышей C57BL/6 периодически иммунизировали 20 мкг неиммуногенной антигенспецифической MOG35-55 мРНК, 20 мкг неиммуногенной нерелевантной мРНК или физиологическим раствором в день 0, 3, 7 и 10. Экспрессию TIGIT, Tim-3, PD-1 и Lag-3 на клетках CD4+ MOG35-55 тетрамер+ мышей, обработанных неиммуногенной кодирующей MOG35-55мРНК, сравнивали с общими CD4+ клетками неиммуногенных, нерелевантных и необработанных контрольных мышей. Повторная иммунизация неиммуногенной антигенспецифической мРНК приводит к усилению регуляции TIGIT, Tim-3, PD-1 и Lag-3 по сравнению с мышами, получавшими неиммуногенную нерелевантную мРНК (фиг. 15).
Повышенная регуляция этих маркеров коррелирует с анергическим фенотипом Т-клеток. Анергия является приобретенным состоянием функциональной нереспонсивности и следствием включения рецептора Т-клеток (TCR) без костимуляции через CD28 (Jenkins, MK et al., 1987, Proc. Natl. Acad. Sci. U. S. A. 84, 5409-13; Quill, H. & Schwartz, R.H., 1987, J. Immunol. 138, 3704-3712; Jenkins, M.K. et al., 1990, J. Immunol. 144, 16-22). Кроме того, она представляет собой одно из средств навязывания периферической толерантности. Анергические Т-клетки функционально неактивны и неспособны инициировать продуктивный иммунный ответ, даже когда антиген встречается в присутствии полной костимуляции. Анергические CD4+ T-клетки с различными фенотипическими и ген-экспрессирующими программами могут даже превращаться в регуляторные T-клетки, которые, в свою очередь, могут стимулировать анергию патогенных CD4 + T-клеток и ингибировать аутоиммунитет (Kalekar, LA et al., 2016, Nat. Immunol., 17, 304-14). Lag-3, Tim-3, ICOS, а также TIGIT опосредуют коингибирующие функции на эффекторных Т-клетках, а избыточная экспрессия этих молекул связана с истощением Т-клеток (Anderson, AC et al., 2016, Immunity 44, 989-1004). Накопление множества коингибирующих рецепторов на поверхности Т-клеток даже связано с повышенной дисфункцией (Blackburn, SD et al., 2009, Nat. Immunol. 10, 29-37). Сам TIGIT действует как коингибирующая молекула, непосредственно подавляя пролиферацию Т-клеток, но также предотвращая созревание DC, снижая секрецию IL-12 и индуцируя выработку иммуносупрессивного цитокина IL-10 (Yu, X. et al., 2009, Nat. Immunol. 10, 48-57; Joller, N. et al., 2011, J. Immunol. 186, 1338-1342). Lag-3 играет роль в модуляции гомеостаза Т-клеток и эффекторных ответов Т-клеток (Workman, CJ et al., 2002, J. Immunol. 169, 5392-5395; Workman, C.J. & Vignali, D.A.A., 2003, Eur. J. Immunol. 33, 970-979; Workman, C.J. & Vignali, D.A.A., 2005, J. Immunol. 174, 688-695).
Кроме того, молекулы иммунной контрольной точки, такие как запрограммированный рецептор смерти 1 (PD-1) и цитотоксический белок 4, ассоциированный с Т-лимфоцитами (CTLA-4), являются коингибирующими молекулами, участвующими в отрицательной регуляции иммунных реакций и, следовательно, в поддержании периферической аутотолерантности. Коингибирующие молекулы в целом активируются при активации Т-клеток и сдерживают эффекторный ответ посредством ингибирования по обратной связи. PD- 1 и его лиганды, например, PD-L1 и PD-L2, доставляют ингибирующие сигналы, которые регулируют баланс между активацией и толерантностью Т-клеток (Keir, ME et al., 2008, Annu. Rev. Immunol. 26, 677-704). Иммунные ответы на аутоантигены требуют специфических и сбалансированных ответов для поддержания толерантности, которая опосредована различными коингибирующими рецепторами и их лигандом. При активизации PD-1 эффекторные ответы Т-клеток будут отключены, и ткани, в свою очередь, защищены от повреждения, вызванного иммунными клетками.
Все эти коингибирующие рецепторы играют центральную роль в регуляции аутоиммунных заболеваний, а дефицит некоторых из этих молекул даже приводит к аутоиммунному заболеванию.
Пример 9. Более подробная характеристика MOG35-55-специфических CD4+ Т-клеток при обработке неиммуногенной антигенспецифической мРНК в модели заболевания EAE
В соответствии с результатами, показанными в примере 6, на фиг. 8 и 9 эксперименты по адоптивному переносу клеток MOG35-55-специфичных к CD4+ T-клеток показали те же результаты. Специфичные для MOG35-55 Thy1.1+ CD4+ T-клетки 2D2 TCR-трансгенных мышей были перенесены в Thy1.2+ C57BL/6 мышей, и через 24 ч индуцировали EAE у мышей-реципиентов. На 7 и 10 день после индукции заболевания мышей обрабатывали 20 мкг неиммуногенной MOG35-55-кодирующей мРНК, 20 мкг неиммуногенной нерелевантной мРНК или физиологическим раствором. Через шесть дней после последней обработки мРНК выделяли мозг и спинной мозг. Клетки из этих тканей повторно активировали in vitro пептидом MOG35-55 в течение 6 часов, а затем отбирали по сигналу выше порогового значения на Thy1.1+ CD4+ Т-клетках. Как показано на фиг. 16, введение неиммуногенного кодирования MOG35-55 мРНК привела к значительному снижению частоты MOG35-55-специфических CD4+ Т- клеток в головном и спинном мозге. На функциональном уровне CD4+ Т-клетки секретируют значительно меньше IFNγ и интерлейкина-17A (IL-17A) в головном и спинном мозге мышей, получавших неиммуногенный MOG35-55 кодирующую мРНК по сравнению с контрольными мышами (фиг. 17). Эти данные снова указывают на то, что обработка неиммуногенной MOG35-55 мРНК может инактивировать клетки Teff, приводя к уменьшенной инфильтрации этих клеток в головной и спинной мозг.
Чтобы исследовать антигенспецифический фенотип CD4 Т-клеток после успешной обработки, EAE активно индуцировали и на 7 и 10 день после индукции заболевания животных обрабатывали 20 мкг неиммуногенной MOG35-55-кодирующей мРНК и 20 мкг неиммуногенной нерелевантной мРНК или физиологическим раствором. На пике заболевания (16-й день после индукции заболевания) фенотип CD4 Т-клеток в селезенке исследовали окрашиванием тетрамером MOG35-55. В соответствии с экспериментами, показанными в примерах 5 и 8, также в условиях заболевания EAE, антигенспецифическая толерантность, индуцированная неиммуногенной антигенспецифической мРНК, опосредуется активацией коингибирующих молекул Lag-3, ICOS, TIGIT и Tim-3 на MOG35-55-специфических CD4+ T-клетках в селезенке (фиг. 18A). Помимо внутреннего клеточного механизма, опосредующего иммунную толерантность, такого как анергия и негативная костимуляция, эффект обработки неиммуногенной антигенспецифической мРНК также могут опосредовать внешние клеточные механизмы (толерантность Treg-клеток). Как уже обсуждалось, Treg играют решающую роль в антигенспецифической толерантности (Sakaguchi, S. et al., 1985, J. Exp. Med. 161, 72-87). FACS-анализ MOG35-55-специфических CD4+ T-клеток, кроме того, показал значительное увеличение популяций Treg (фиг. 18B). Кроме того, Treg в селезенке мышей, получавших антигенспецифическую неиммуногенную мРНК, экспрессируют высокие уровни CD69, указывая на то, что они высокоактивированы.
Чтобы определить, необходима ли экспрессия отрицательных костимулирующих молекул для индукции и поддержания толерантности, PD-1 или CTLA-4 блокировались в ходе обработки EAE с помощью мРНК. Обработка EAE на 7 и 10 день после индукции заболевания неиммуногенной MOG35-55-кодирующей мРНК в сочетании с антителом против PD-1 или против CTLA-4 приводило к изменению защиты от EAE по сравнению с мышами, получавшими иммуногенную MOG35-55-кодирующую мРНК в сочетании с контролями IgG (фиг. 19). Это демонстрирует, что PD-1 и CTLA-4 имеют решающее значение для долгосрочного поддержания антигенспецифической толерантности, индуцированной неиммуногенной антигенспецифической мРНК, поскольку блокирование антител PD-1 и CTLA-4 параллельно с обработкой мРНК приводило к заметному снижению эффективности индукции толерантности, опосредованной неиммуногенной антигенспецифической обработкой мРНК.
В совокупности представленные эксперименты исследовали механизм действия этого нового терапевтического подхода к индукции антигенспецифической толерантности путем обработки антигенспецифической неиммуногенной мРНК. Результаты показывают, что обработка антигенспецифической неиммуногенной мРНК является эффективным способом изменения вредных иммунных ответов при аутоиммунности для придания защиты. Эксперименты, показанные на наивных, а также на пораженных болезнью мышах, демонстрируют, что обработка неиммуногенной антигенспецифической мРНК приводит к индукции анергических и регуляторных Т-клеток, которые опосредуют толерантность на периферии, что приводит к успешной обработке продолжающегося аутоиммунного заболевания. Было показано, что терапевтический эффект основан на уже хорошо описанных механизмах индукции толерантности, представленных в литературе, обсуждаемой в каждом примере. Эти механизмы важны и фундаментальны для индукции толерантности, независимо от типа аутоиммунного заболевания. Механизм толерантности, опосредованный обработкой неиммуногенной антигенспецифической мРНК, таким образом, может быть перенесен на любой другой тип аутоиммунного заболевания.
Пример 10. Индукция антигенспецифической толерантности путем обработки неиммуногенной антигенспецифической мРНК не зависит от модели заболевания EAE
Все ранее описанные эксперименты с EAE проводили на мышах C57BL/6, индуцированных пептидом MOG35-55, что приводило к прогрессированию монофазного заболевания. Другая модель EAE характеризуется рецидивирующим заболеванием из-за другого релевантного эпитопа (PLP139-151), приводящего к течению возвратно-ремиттирующего заболевания у мышей SJL. Эта модель была использована для демонстрации индукции толерантности в более клинически значимых условиях заболевания. На фиг. 20А показано, что обработка мРНК 2 раза в неделю, начиная с 7-го и 10-го дня после индукции EAE с помощью неиммуногенной PLP139-151-кодирующей мРНК в модели рецидивирующего заболевания, оказывает положительное влияние на развитие EAE, поскольку эти мыши демонстрируют значительно более слабую реакцию прогрессирование заболевания, чем мыши, получавших неиммуногенную нерелевантную контрольную мРНК.
Чтобы еще лучше имитировать ситуацию при MS, был использован комплекс EAE, связанный с множеством патогенных аутореактивных клонов Т-клеток. Поскольку при определенном диагнозе MS, заболевания, вероятно уже ассоциированного с комплексной антимиелиновой аутореактивностью, из-за «распространения эпитопа» была исследована эффективность подхода с многоэпитопной обработкой. Как показано на фиг. 20B, комплекс EAE был успешно обработан смесью четырех различных неиммуногенных мРНК, кодирующих эпитопы заболеваний MOG35-55, PLP139-151, PLP178-191 и MBP84-104. Использовали по 10 мкг для каждой неиммуногенной, кодирующей эпитоп заболевания мРНК, что дало 40 мкг эффективной обработки. Ни одна из обработок отдельных эпитопов не дала аналогичные результаты обработки, что показано для 20 мкг неиммуногенной MOG35-55-кодирующей мРНК.
Эти результаты демонстрируют, что антигенспецифическая индукция толерантности, наблюдаемая в разных экспериментах, не зависит от конкретного эпитопа и может быть опосредована различными эпитопами, относящимися к различным заболеваниям. Кроме того, данные подчеркивают, что даже сложное продолжающееся заболевание со сложным патогенным аутоиммунным процессом можно успешно лечить путем введения множества неиммуногенных миелин-специфических конструкций мРНК.
Пример 11. Совместная доставка иммуномодулирующих цитокинов улучшает толерогенный эффект обработки неиммуногенной антигенспецифической мРНК-LPX
Несколько иммунорегуляторных молекул, таких как IL-10, могут поддерживать функцию Treg. Кроме того, известно, что противовоспалительный цитокин IL-10 индуцирует толерогенные DC (Torres-Aguilar, H. et al., 2010, Autoimmun. Rev. 10, 8-17; Torres-Aguilar, H. et al., 2010, J. Immunol. 184, 1765-1775; Steinbrink K. et al., 1997, J. Immunol. 159, 4772-4780) и подавляет пролиферацию Т-клеток и ответы цитокинов (Gu, Y. et al., 2008, Eur. J. Immunol. 38, 1807-1813; Guo, B., 2016, J. Clin. Cell Immunol. 7, 1-16). Также IL-27 является цитокином, который, как известно, обладает противовоспалительными свойствами, и в исследованиях EAE было показано, что этот цитокин снижает тяжесть заболевания (Mascanfroni, ID et al., 2013, Nat. Immunol. 14, 1054-63; Thom, R. et al., 2017, Front. Immunol. 8, 1-14). В ряде экспериментов с EAE следует исследовать совместную доставку неиммуногенной антигенспецифической мРНК с иммуномодулирующими цитокинами, такими как IL-10 и IL-27, кодируемыми неиммуногенной мРНК, с целью модулирования иммунного ответа и повышения эффективности индукция толерантности.
Способность совместной доставки иммуномодулирующих цитокинов усиливать толерогенность обработки одним антигенспецифическим мРНК-LPX оценивали с использованием «субтерапевтической» обработки мышей с EAE. Неиммуногенное антигенспецифическое лечение мРНК-LPX продемонстрировало устойчивую индукцию толерантности в предыдущих экспериментах, поэтому комбинаторный подход дополнительной неиммуногенной обработки мРНК, кодирующей цитокины, был исследован в модели EAE в субтерапевтических дозах для выявления терапевтического вклада.
Чтобы исследовать положительный эффект обработки неиммуногенной цитокин-кодирующей мРНК на неиммуногенную антигенспецифическую мРНК в модели заболевания, у мышей C57BL/6 активно индуцировали EAE, и обрабатывали мышей на 7 и 10 день после индукции заболевания неиммуногенной антигенспецифичной MOG35-55-кодирующей мРНК, неиммуногенной MOG35-55-кодирующей мРНК в сочетании с неиммуногенной mIL-10-кодирующей (фиг. 21A) или mIL-27-кодирующей (фиг. 21B) мРНК, неиммуногенной цитокин-кодирующей мРНК, а также неиммуногенной нерелевантной мРНК.
Результаты, представленные на фиг. 21, позволяют предположить, что совместная доставка иммуномодулирующих цитокинов, кодируемых неиммуногенной мРНК, повышает толерантность в антигенспецифическом контексте. Кроме того, они в целом не подавляют иммунный ответ, поскольку обработка мРНК, кодирующей один цитокин, не показала изменений в прогрессии EAE по сравнению с контрольными мышами.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> БИОНТЕХ РНА ФАРМАСЬЮТИКАЛС ГМБХ И ДР.
<120> РНК для лечения аутоиммунных заболеваний
<130> 674-192PCT2
<150> PCT/EP2017/058651
<151> 2017-04-11
<160> 9
<170> PatentIn version 3.5
<210> 1
<211> 37
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 1
Ser Pro Gly Lys Asn Ala Thr Gly Met Glu Val Gly Trp Tyr Arg Ser
1 5 10 15
Pro Phe Ser Arg Val Val His Leu Tyr Arg Asn Gly Lys Asp Gln Asp
20 25 30
Ala Glu Ala Gln Pro
35
<210> 2
<211> 21
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 2
Met Glu Val Gly Trp Tyr Arg Ser Pro Phe Ser Arg Val Val His Leu
1 5 10 15
Tyr Arg Asn Gly Lys
20
<210> 3
<211> 29
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 3
Ala His Ser Leu Glu Arg Val Cys His Cys Leu Gly Lys Trp Leu Gly
1 5 10 15
His Pro Asp Lys Phe Val Gly Ile Thr Tyr Ala Leu Thr
20 25
<210> 4
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 4
Ala Val Pro Val Tyr Ile Tyr Phe Asn Thr Trp Thr Thr Cys Gln Ser
1 5 10 15
Ile Ala Phe Pro Ser Lys Thr Ser Ala Ser Ile Gly Ser Leu
20 25 30
<210> 5
<211> 37
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 5
Arg Thr Gln Asp Glu Asn Pro Val Val His Phe Phe Lys Asn Ile Val
1 5 10 15
Thr Pro Arg Thr Pro Pro Pro Ser Gln Gly Lys Gly Arg Gly Leu Ser
20 25 30
Leu Ser Arg Phe Ser
35
<210> 6
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 6
His Ser Leu Gly Lys Trp Leu Gly His Pro Asp Lys Phe
1 5 10
<210> 7
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 7
Asn Thr Trp Thr Thr Cys Gln Ser Ile Ala Phe Pro Ser Lys
1 5 10
<210> 8
<211> 21
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 8
Val His Phe Phe Lys Asn Ile Val Thr Pro Arg Thr Pro Pro Pro Ser
1 5 10 15
Gln Gly Lys Gly Arg
20
<210> 9
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope
<400> 9
Gln Lys Phe Ser Glu His Phe Ser Ile His Cys Cys Pro Pro Phe Thr
1 5 10 15
Phe Leu Asn Ser Lys Arg
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКАЯ РНК ДЛЯ РАКА ЯИЧНИКА | 2020 |
|
RU2832172C2 |
| ТЕРАПЕВТИЧЕСКАЯ РНК ДЛЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2020 |
|
RU2838161C2 |
| АНТИГЕННЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2792042C2 |
| ИНДИВИДУАЛИЗИРОВАННЫЕ ПРОТИВООПУХОЛЕВЫЕ ВАКЦИНЫ | 2012 |
|
RU2670745C9 |
| ИНДИВИДУАЛИЗИРОВАННЫЕ ПРОТИВООПУХОЛЕВЫЕ ВАКЦИНЫ | 2012 |
|
RU2779946C2 |
| 5'-КЭП-ТРИНУКЛЕОТИДНЫЕ СОЕДИНЕНИЯ ИЛИ 5'-КЭП-СОЕДИНЕНИЯ С БОЛЬШИМ КОЛИЧЕСТВОМ НУКЛЕОТИДОВ И ИХ ПРИМЕНЕНИЕ ДЛЯ СТАБИЛИЗАЦИИ РНК, ЭКСПРЕССИИ БЕЛКОВ И В ТЕРАПИИ | 2019 |
|
RU2811940C2 |
| РЕЦЕПТУРА ДЛЯ ВВЕДЕНИЯ РНК | 2019 |
|
RU2797147C2 |
| ИММУНОГЕННЫЕ ПЕПТИДЫ ДЛЯ ПРИМЕНЕНИЯ В ПРОФИЛАКТИКЕ И/ИЛИ ЛЕЧЕНИИ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ, АУТОИММУННЫХ ЗАБОЛЕВАНИЙ, ИММУННЫХ ОТВЕТОВ НА АЛЛОГЕННЫЕ ФАКТОРЫ, АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ, ОПУХОЛЕЙ, ОТТОРЖЕНИЯ ТРАНСПЛАНТАТА И ИММУННЫХ ОТВЕТОВ ПРОТИВ ВИРУСНЫХ ВЕКТОРОВ. ИСПОЛЬЗУЕМЫХ ДЛЯ ГЕННОЙ ТЕРАПИИ ИЛИ ГЕННОЙ ВАКЦИНАЦИИ | 2011 |
|
RU2615460C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УСТОЙЧИВОЙ ИММУНОТЕРАПИИ | 2014 |
|
RU2696876C2 |
| АНТИГЕННЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2794945C2 |
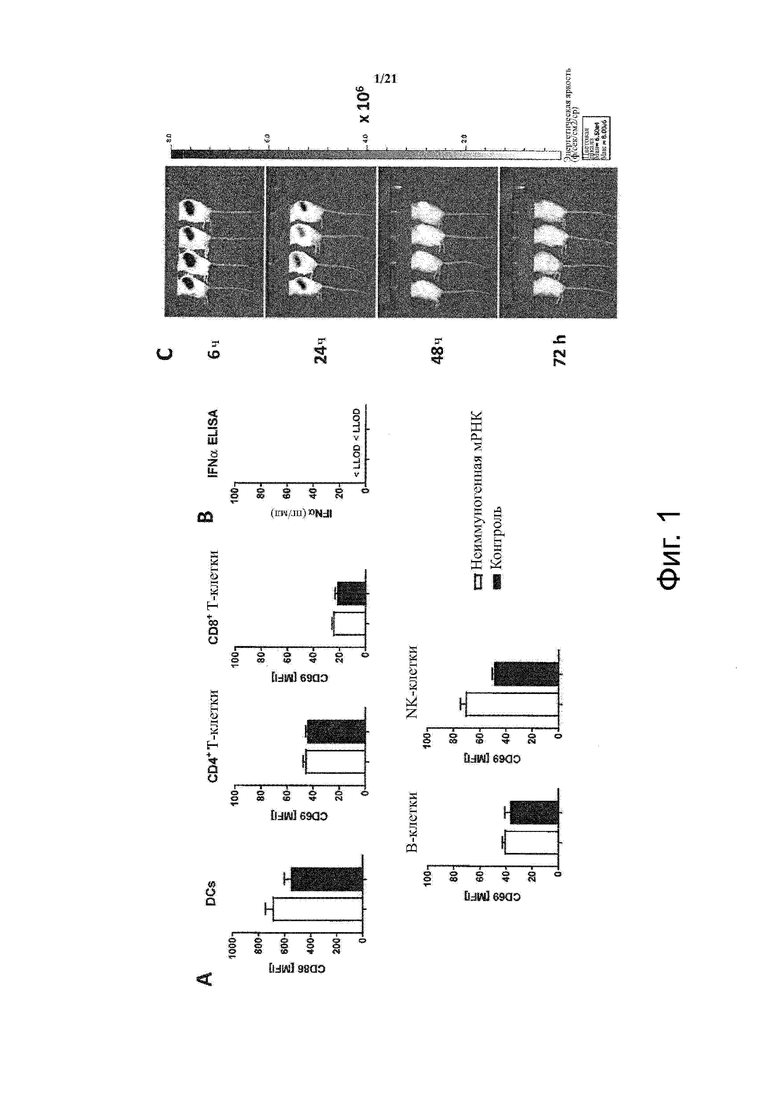

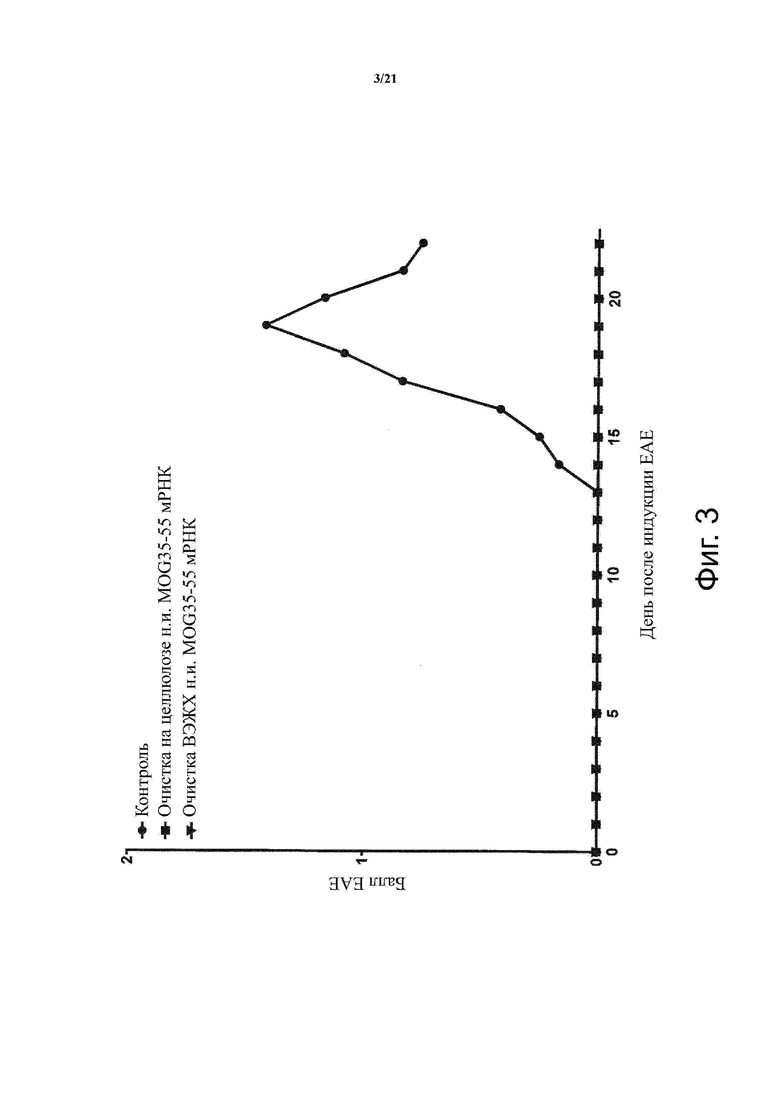
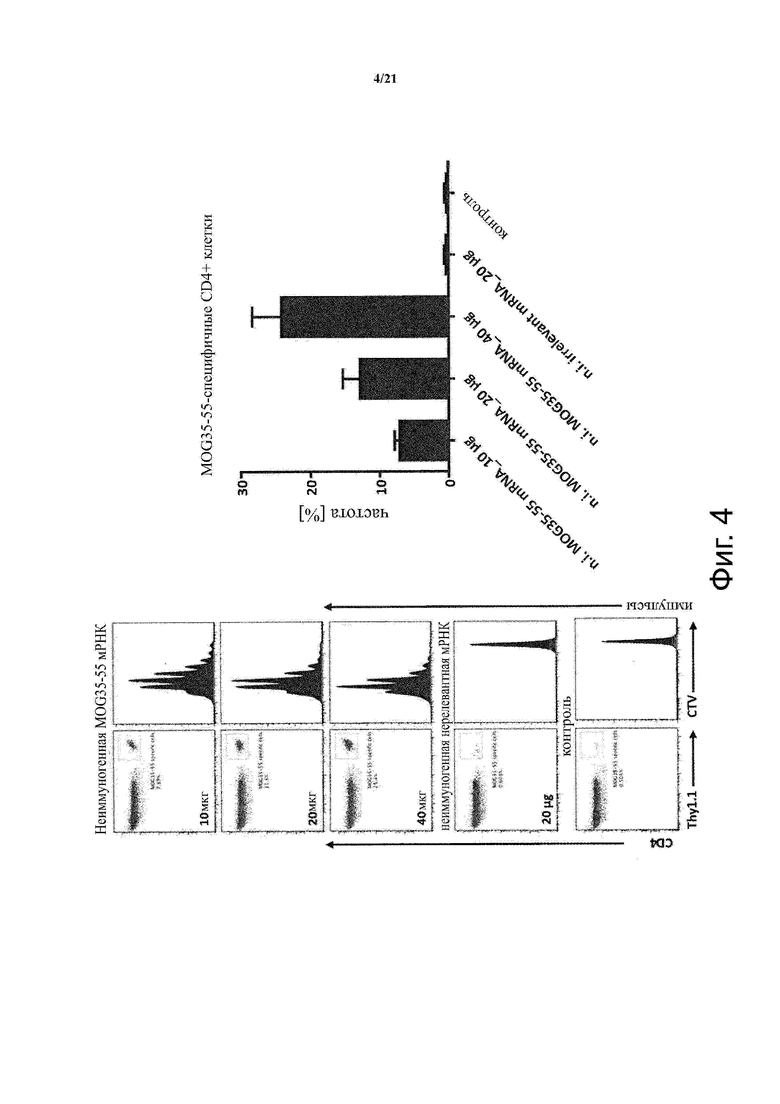
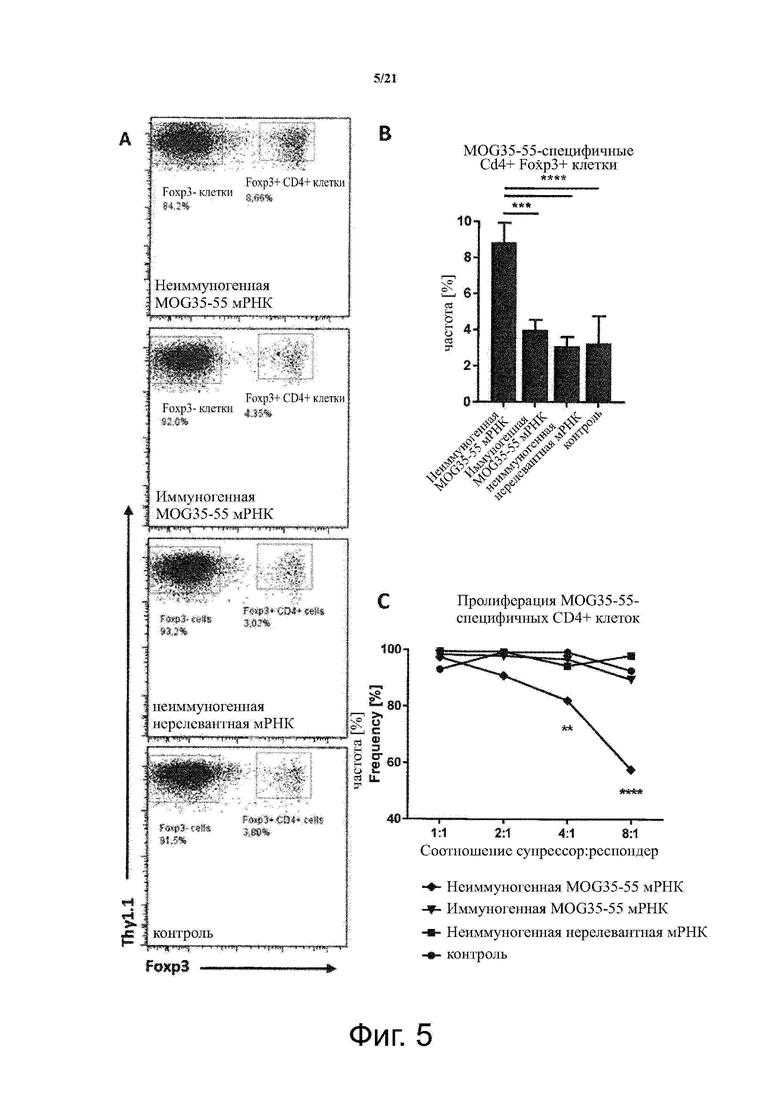
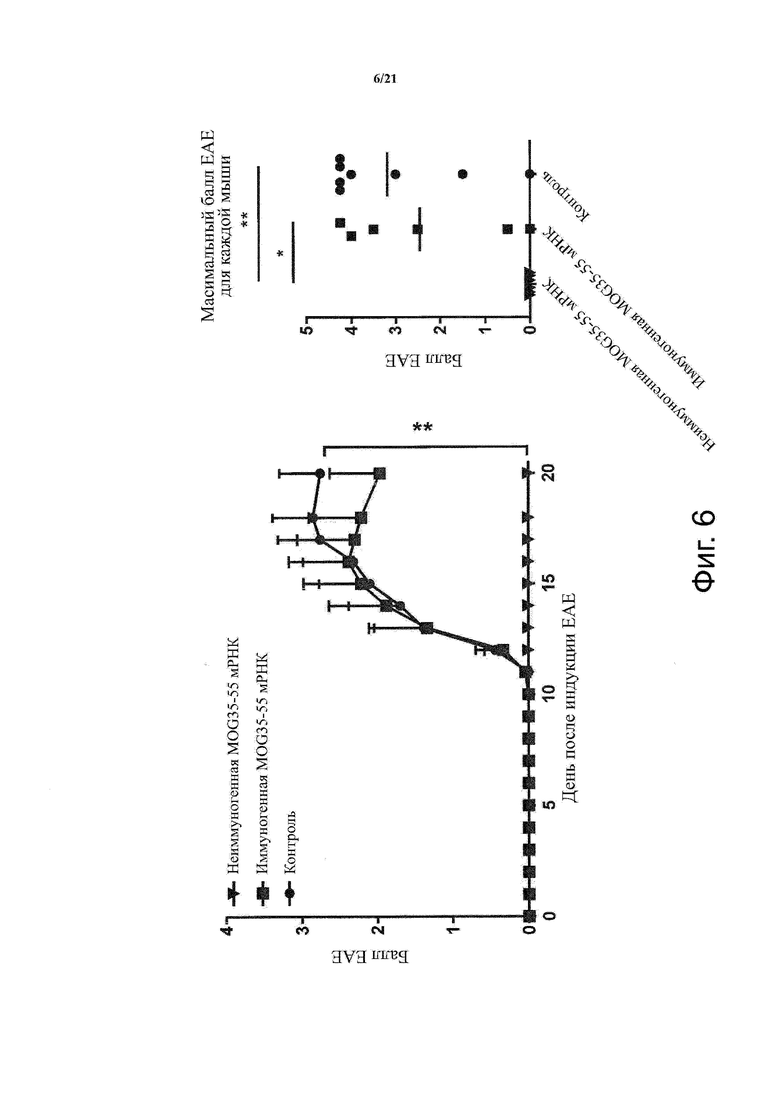
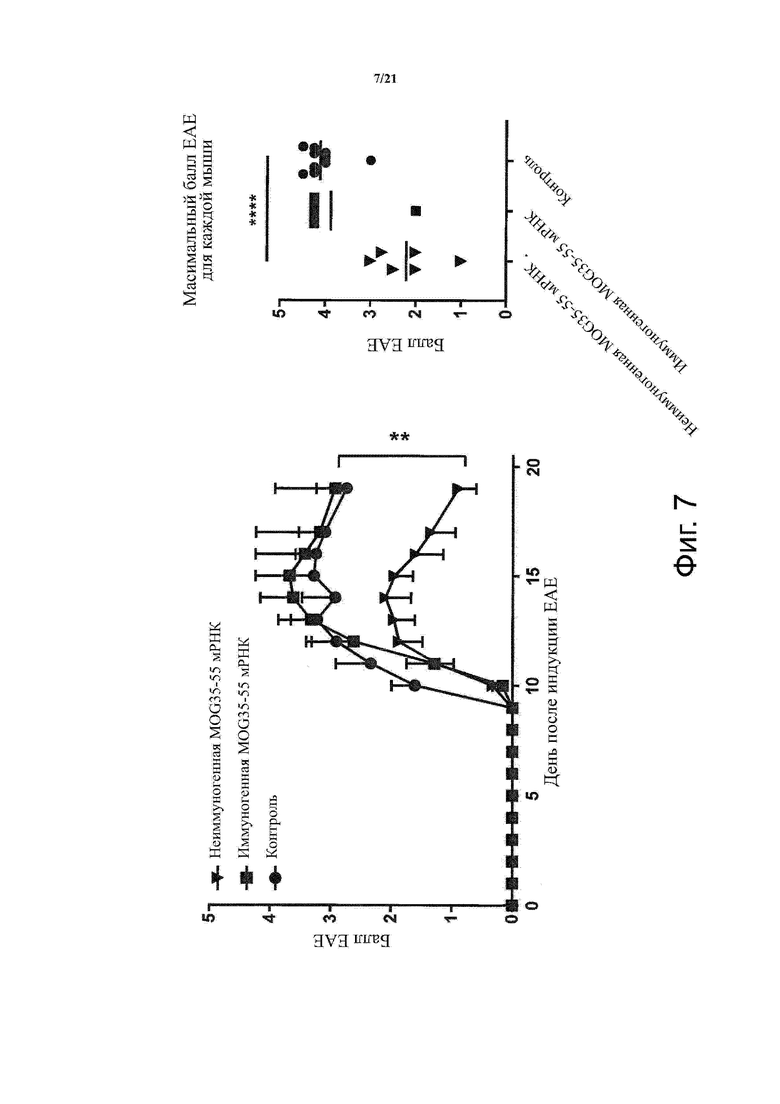
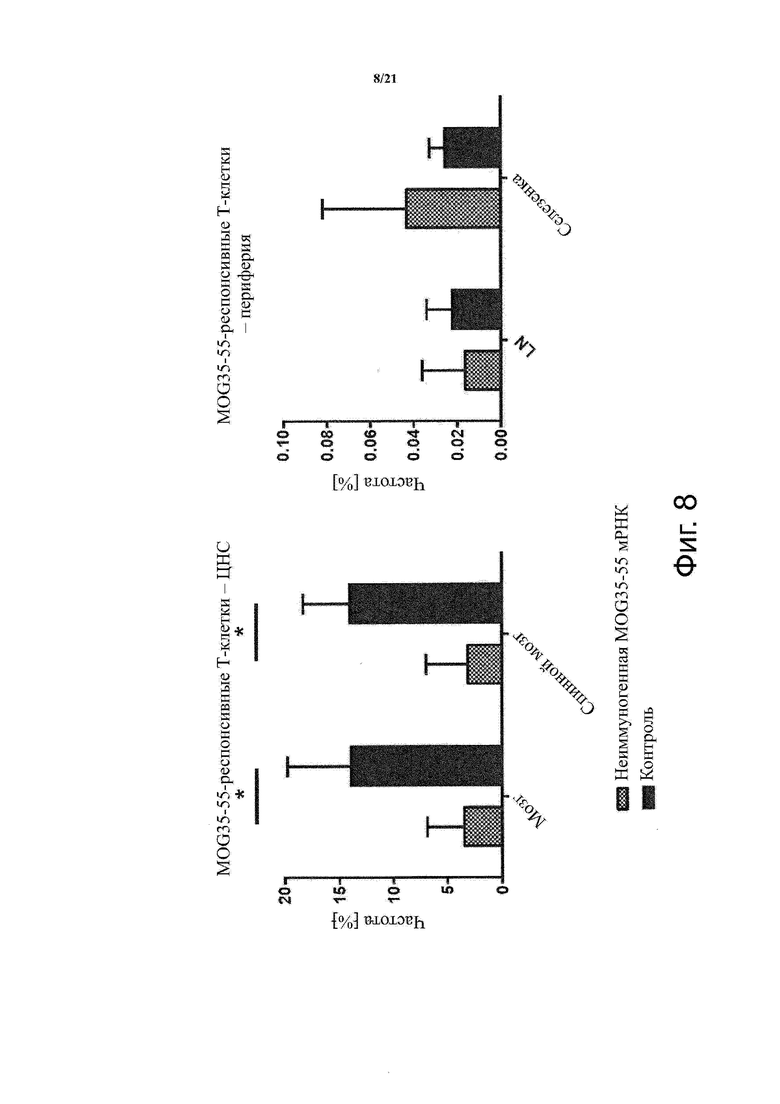
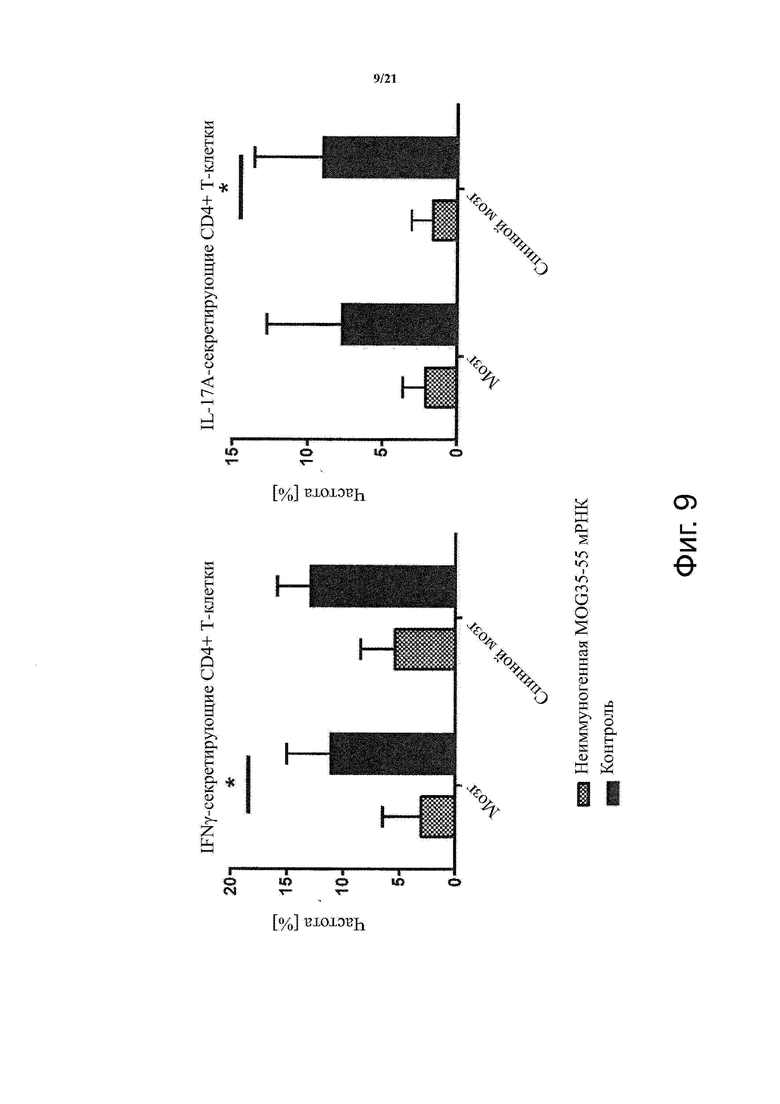

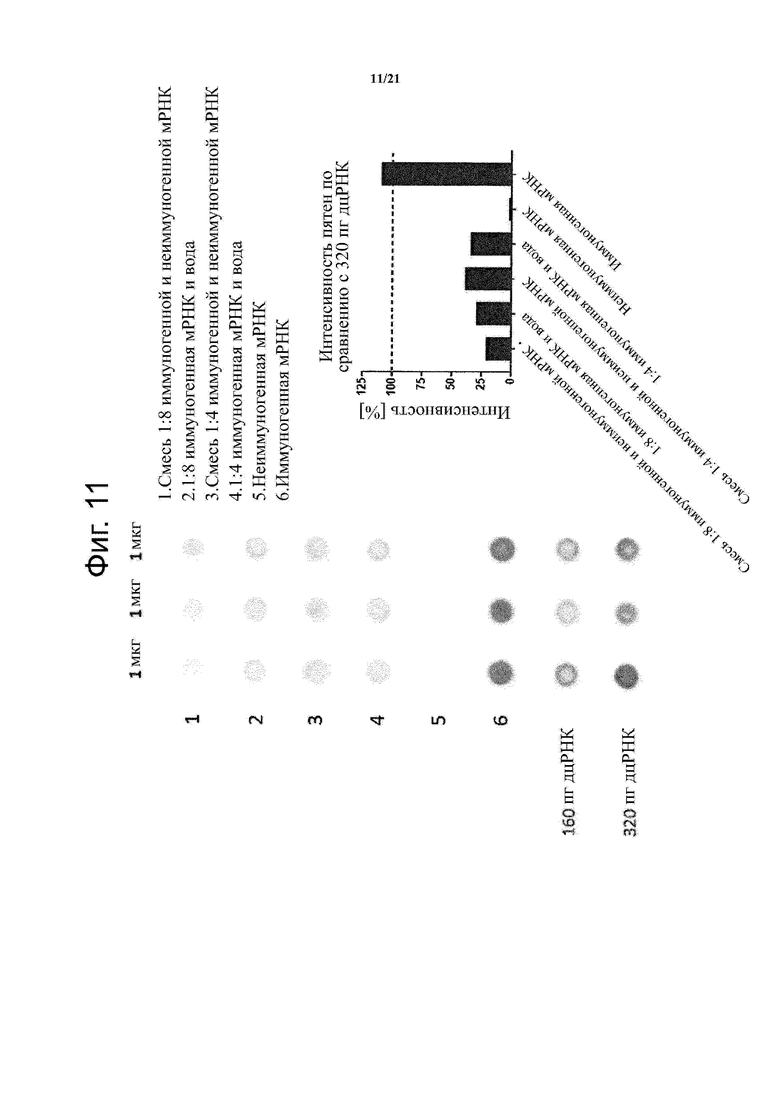
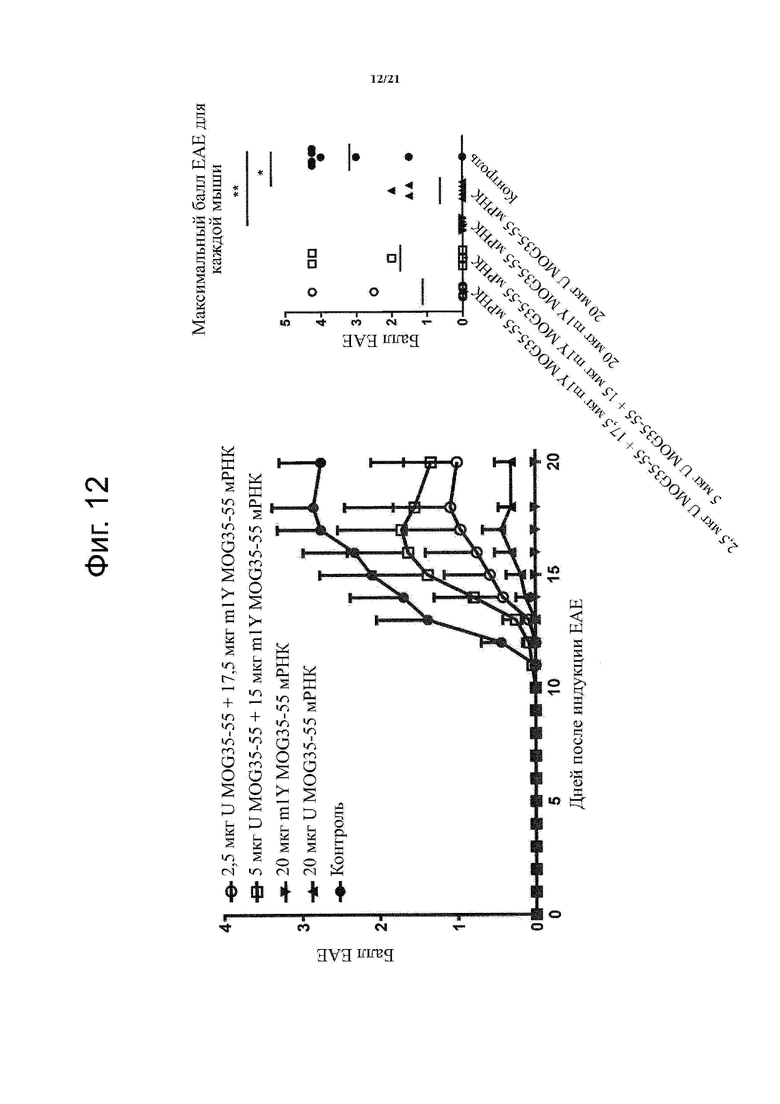
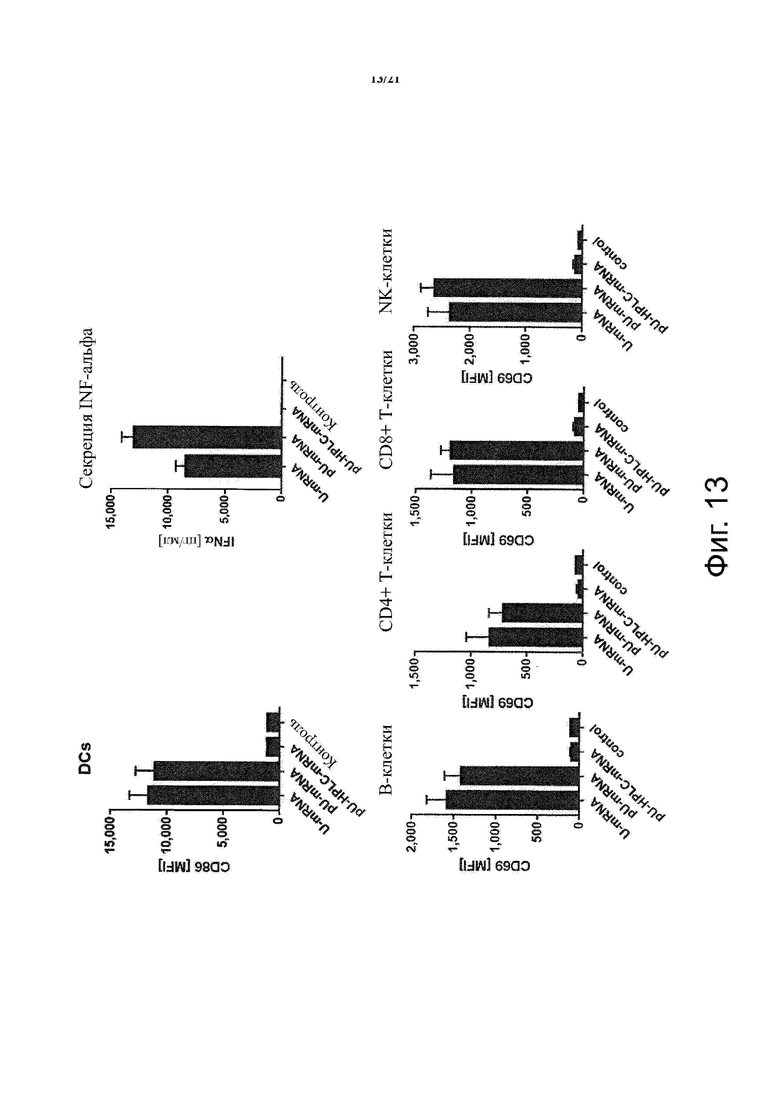
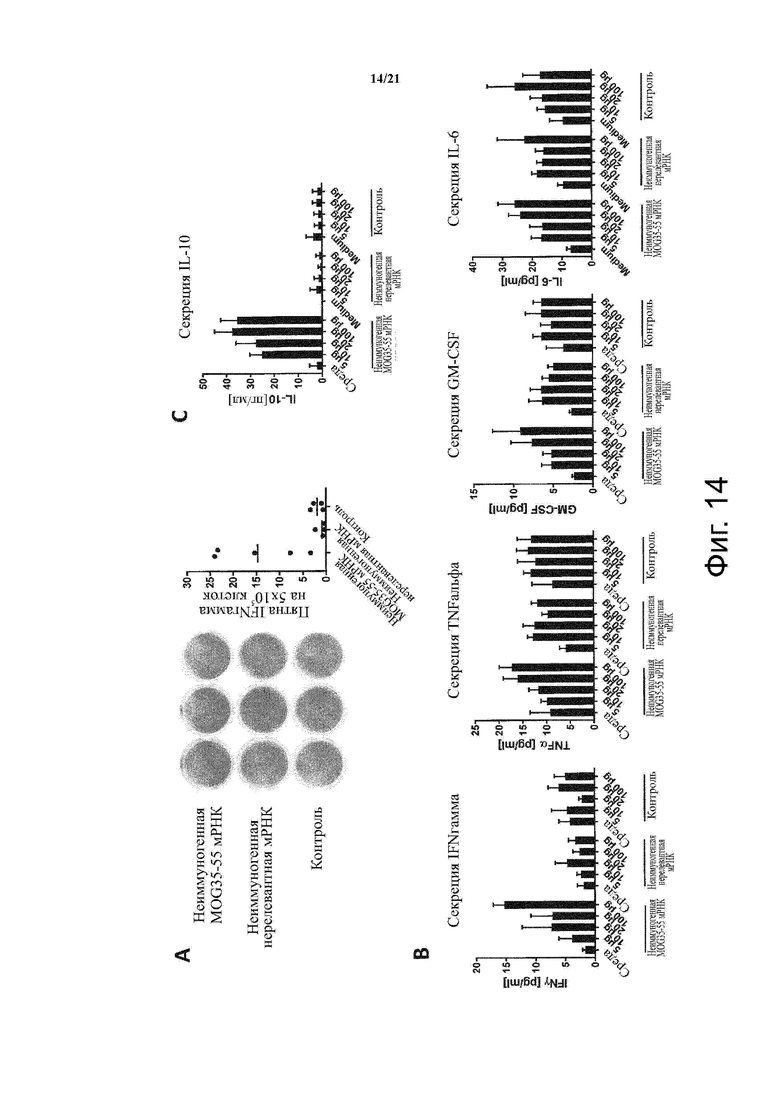
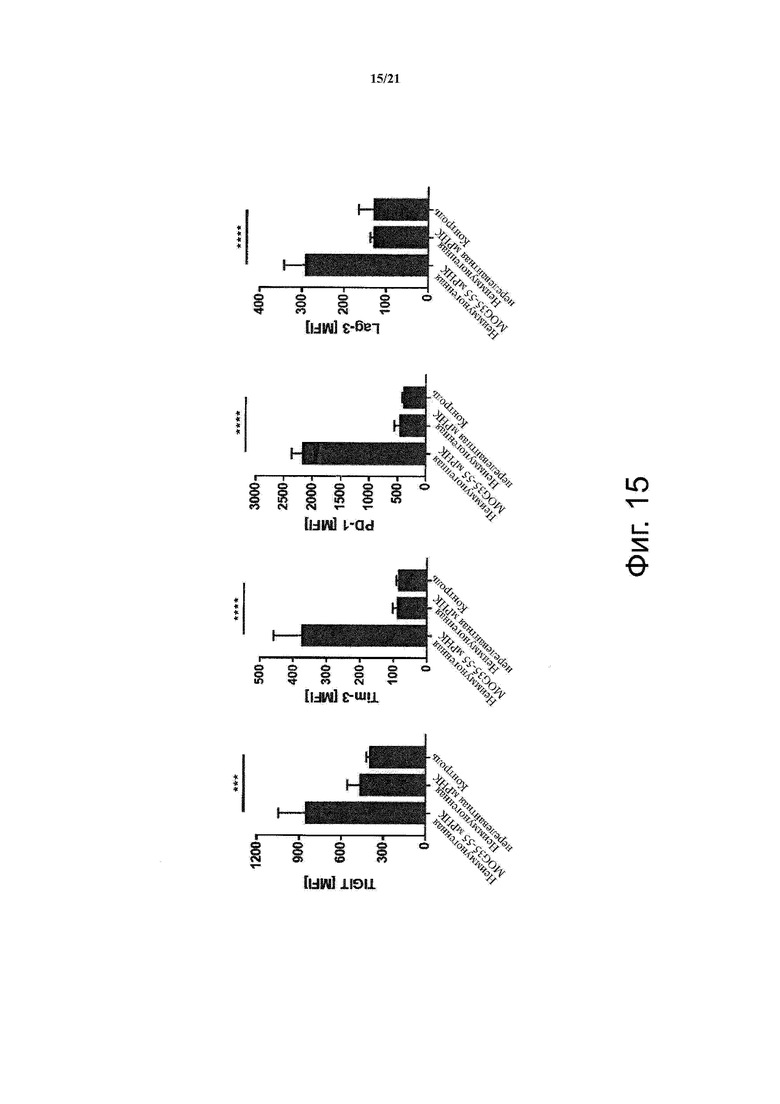
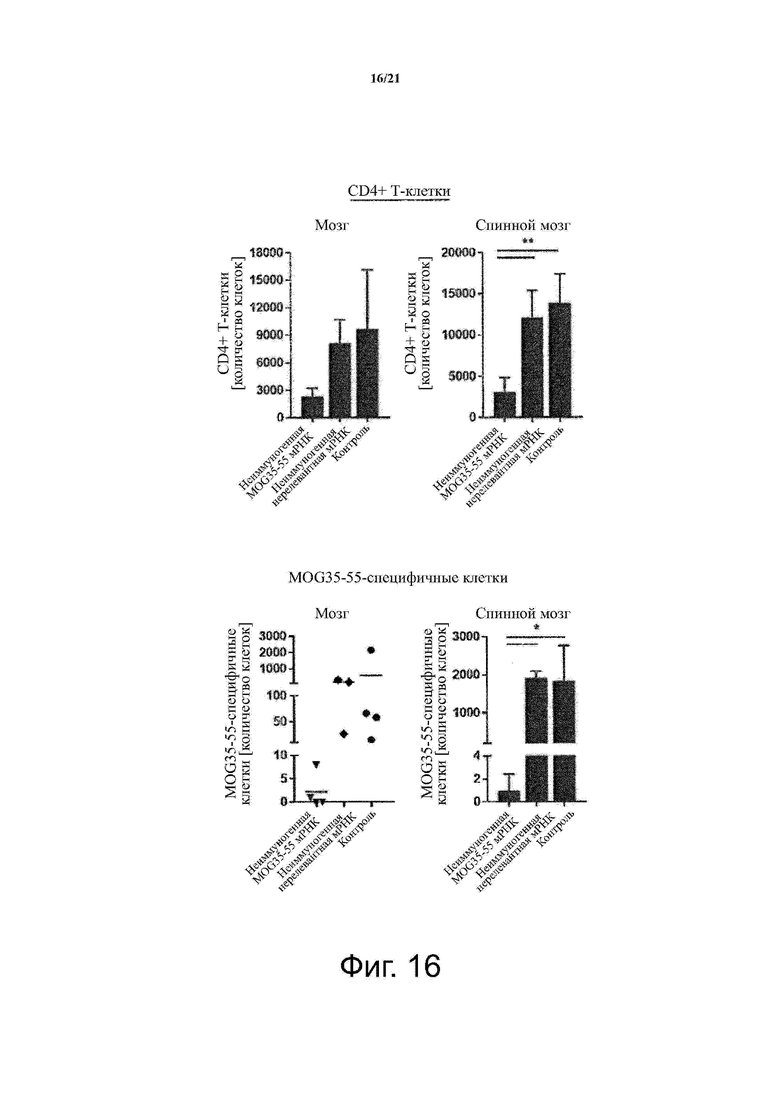
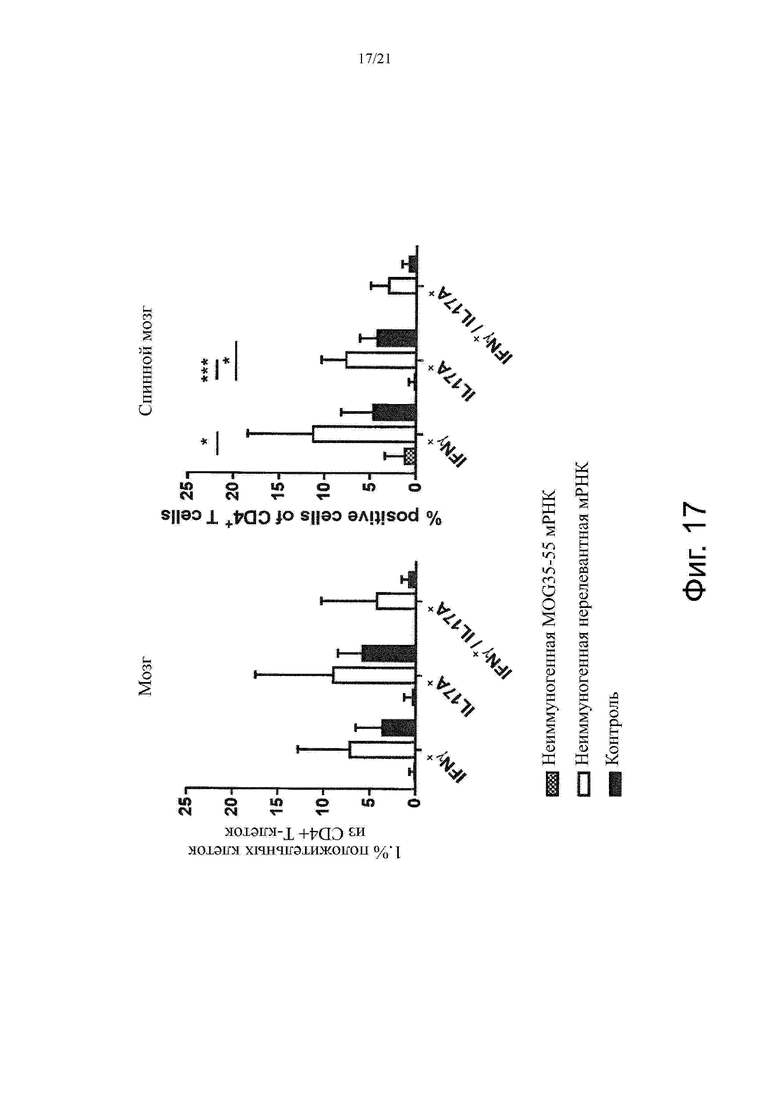
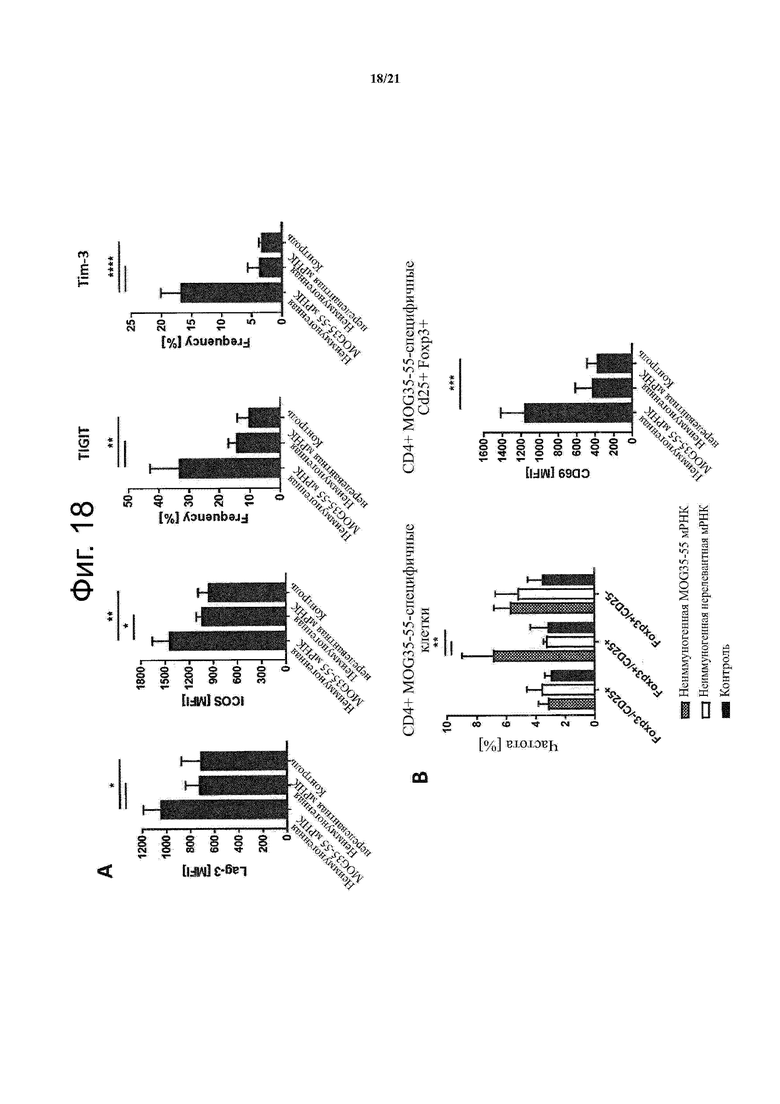
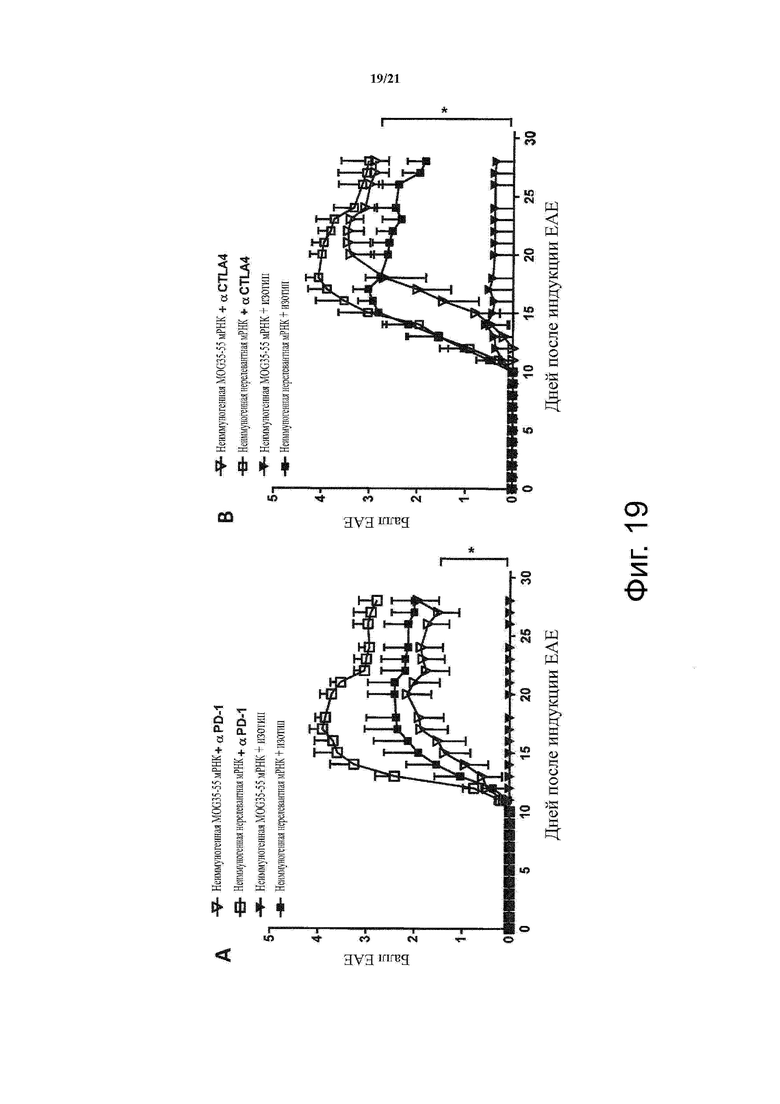
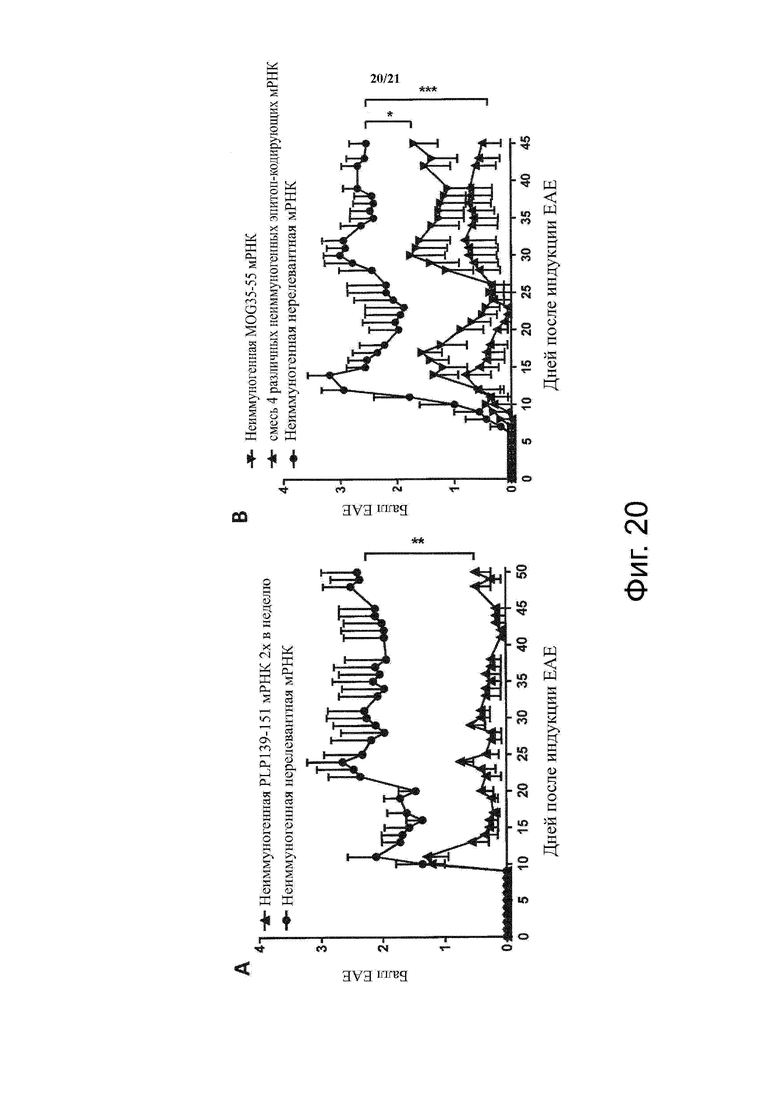
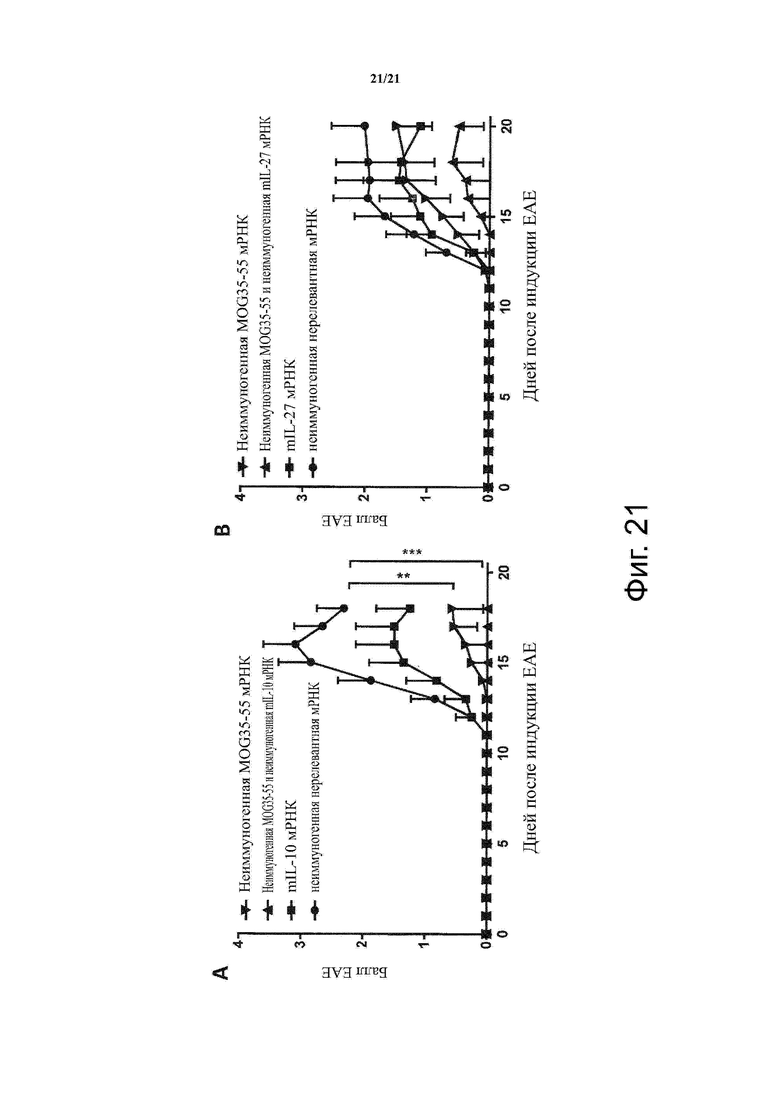
Изобретение относится к области биотехнологии, конкретно к фармацевтической композиции для лечения или предотвращения рассеянного склероза. Фармацевтическая композиция включает неиммуногенную РНК, кодирующую пептид или полипептид, содержащий аутоантиген. Неиммуногенная РНК становится неиммуногенной за счет включения 1-метилпсевдоуридинов и удаления дцРНК. Пептид или полипептид, кодируемый неимуногенной РНК, содержит аминокислотную последовательность миелин-олигодендроцитарного гликопротеина (MOG). Фармацевтическая композиция индуцирует толерантность к аутоантигену и может быть использована в медицине для лечения или предотвращения рассеянного склероза. 2 з.п. ф-лы, 21 ил., 11 пр.
1. Фармацевтическая композиция для лечения или предотвращения рассеянного склероза, которая включает неиммуногенную РНК, кодирующую пептид или полипептид, содержащий аутоантиген, где неиммуногенная РНК становится неиммуногенной за счет включения 1-метилпсевдоуридинов и удаления дцРНК и где пептид или полипептид содержит аминокислотную последовательность SEQ ID NO: 2 миелин-олигодендроцитарного гликопротеина (MOG).
2. Фармацевтическая композиция по п. 1, в которой неиммуногенная РНК представляет собой транскрибируемую in vitro РНК.
3. Фармацевтическая композиция по п. 1 или 2, которая дополнительно включает иммуноингибирующее соединение или неиммуногенную РНК, кодирующую указанное иммуноингибирующее соединение, где указанное иммуноингибирующее соединение выбирают из группы, состоящей из трансформирующего фактора роста бета (TGF-β), интерлейкина 10 (IL10), антагониста рецептора интерлейкина 1 (IL-1RA), интерлейкина 4 (IL-4), интерлейкина 27 (IL-27), интерлейкина 35 (IL-35), лиганда 1 белка программируемой смерти клеток 1 (PDL1), лиганда индуцируемого Т-клеточного ко-стимулятора (ICOSL), B7-H4, CD39, CD73, FAS, FAS-IL, индоламин-2,3-диоксигеназы 1 (IDO1), индоламин-2,3-диоксигеназы 2 (IDO2), ацетальдегиддегидрогеназы 1 (ALDH1)/ретинальдегиддегидрогеназы (RALDH), аргиназы 1 (ARG1), аргиназы 2 (ARG2), синтазы оксида азота (NOS2), галектина-1, галектина-9, семафорина 4A.
| CREUSOT R.J | |||
| et al., "A short pulse of IL-4 delivered by DCs electroporated with modified mRNA can both prevent and treat autoimmune diabetes in NOD mice", Molecular therapy, 2010, V | |||
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Подогреватель нефти для двигателей | 1924 |
|
SU2112A1 |
| LOBELL A | |||
| et al., "Suppressive DNA vaccination in myelin oligodendrocyte glycoprotein peptide-induced experimental autoimmune | |||

