Данное изобретение относится к иммуногенным композициям с компонентом(компонентами) вируса гепатита В (HBV), которые могут быть основаны на нуклеиновой кислоте или полипептиде. Указанные иммуногенные композиции могут быть использованы для стимуляции или повышения иммунного ответа на HBV с целью обеспечения защитного или терапевтического эффекта против HBV-инфекции и любого состояния или заболевания, вызванного или связанного с инфекцией HBV. Данное изобретение также относится к экспрессионному вектору для экспрессии такого HBV-компонента(ов) и его терапевтического или профилактического применения. Изобретение представляет особый интерес в области иммунотерапии и, в частности, для лечения пациентов, инфицированных HBV, особенно хронически инфицированных.
Уровень техники
Гепатит В является одной из основных проблем общественного здравоохранения с более чем 350 миллионами хронически инфицированных людей во всем мире, при этом от 20 до 40% имеют риск развития хронического заболевания печени, цирроза печени и гепатоцеллюлярной карциномы. Несмотря на наличие эффективных профилактических вакцин, инфекция вирусом гепатита В (HBV) по-прежнему процветает в многих странах, даже в развитых, с оценкой 4,5 миллионов новых случаев заражения в год по всему миру. В отличие от рекомендации ВОЗ, по которой должна осуществляться всеобщая вакцинация, охват полным курсом профилактической вакцинации колеблется от 25% в Азии до 75-90% в Европе. В настоящее время гепатит В является десятой по счету причиной смертности (примерно 1 миллион смертей/год) и HBV-связанной карциномы печени, пятым среди наиболее распространенных раков. Географическое распространение HBV-инфекции неравномерно с распространенностью менее 1% в западных странах и более 10% в юго-восточных странах, большей части Африки и Экваториальной Южной Америки. В областях с высокой распространенностью хронических носителей HBV вертикальная передача от инфицированной матери новорожденному является наиболее частым способом заражения и почти всегда приводит к хроническому гепатиту (90% случаев). Этот показатель может быть снижен до 15% путем профилактической вакцинации инфицированных детей сразу после рождения. В западных странах заражение происходит чаще всего во взрослом возрасте путем горизонтальной передачи через жидкости организма, например через кровь, сперму, слюну, что приводит к острой и самостоятельно восстанавливающейся инфекции у 85% пациентов, но, тем не менее, к хронической инфекции в 15% случаев.
Вирус гепатита В (HBV) является членом Hepadnaviridae и в первую очередь поражает печень, реплицируясь в гепатоцитах. Инфекционные частицы являются так называемыми частицами Дейна размером 42-45 нм, которые состоят из внешней липопротеиновой оболочки, которая содержит три различных поверхностных белка (HBs), и внутреннего нуклеокапсида, основным структурным белком которого является сердцевинный белок (HBcAg). В нуклеокапсиде имеется одна копия генома HBV, связанная с вирусным полимеразным белком (Р). В дополнение к вирионам размером 42-45 нм кровь HBV-инфицированных пациентов содержит сферы размером 20 нм из HBsAg и липидов хозяина, которые выделяются из инфицированных клеток. Эти сферы численно превосходят вирионы с коэффициентом 104-106.
HBV-геном является релаксированной (открытой) кольцевой, частично двухцепочечной ДНК примерно с 3200 нуклеотидов, состоящей из минус-нити полной длины и более короткой плюс-нити. Он содержит четыре перекрывающиеся открытые рамки считывания (ORF), С, S, Р и X. ORF С кодирует сердцевинный белок (или HBcAg), белок длиной 183 аминокислоты, который составляет нуклеокапсид HBV, и второй белок, содержащийся в сыворотке крови больных во время репликации вируса, известный как HBcAg, который содержит М-концевое удлинение в области precore и часть HBcAg. С-конец сердцевинного белка очень прост и состоит из четырех Arg-богатых областей, которые, по прогнозам, связываются с нуклеиновыми кислотами, а также многочисленных сайтов фосфорилирования (состояние фосфорилирования сердцевины связано с конформационными изменениями в капсидной частице, как описано в Yu and Sommers, 1994, J. Virol. 68:29650. ORF S кодирует три поверхностных белка, которые имеют одинаковый С-конец, но различаются по N-концу в -связи с наличием трех старт-кодонов ATG в рамке считывания, которые разделяют ORF S на три области, S (226 аминокислот), preS2 (55 аминокислот) и preS1 (108 аминокислот), соответственно. Большой антигенный белок поверхности (L) продуцируется после инициации трансляции на первом старт-кодоне ATG и включает 389 аминокислотных остатков (preS1-preS2-S). Средний антигенный белок поверхности (М) получается при трансляции области S и области pre-S2 со второго старт-кодона ATG, в то время как малый антигенный белок поверхности из 226 аминокислот (S, также обозначаемый как HBsAg) получается при трансляции области S с третьего старт-кодона ATG. Поверхностные белки HBV являются гликопротеинами с углеводными боковыми цепями (гликанами), которые присоединяются путем N-гликозидных связей. ORF Р кодирует вирусную полимеразу, а ORF Х содержит белок, известный как белок X, который считается активатором транскрипции.
После входа вирионов в гепатоциты через неизвестные пока рецепторы нуклеокапсиды переносят геномную ДНК HBV в ядро, где релаксированная кольцевая ДНК превращается в ковалентно замкнутую кольцевую ДНК (кзкДНК, cccDNA). кзкДНК функционирует в качестве матрицы для транскрипции четырех вирусных РНК, которые переносятся в цитоплазму и используются как мРНК для трансляции HBV-белков. Самая длинная (прегеномная) РНК также функционирует как матрица для репликации HBV, которая происходит в нуклеокапсидах в цитоплазме. Некоторые из ДНК HBV и содержащие полимеразу капсиды затем переносятся в ядро, где они высвобождают вновь образованную релаксированную кольцевую ДНК для формирования дополнительной кзкДНК. Со временем полужизни больше, чем у гепатоцитов, кзкDNA отвечает за сохранение HBV. Другие капсиды окружаются оболочкой путем почкования в эндоплазматический ретикулум и секретируются после прохождения через комплекс Гольджи.
Большое число докпинических и клинических исследований подчеркивают важность CD4+ и CD8+ Т-клеточных иммунных реакций для эффективного противовирусного ответа (Ferrari et al., 1990, J ImmuL 145:3442; Penna et al., 1996, J Clin Invest, 98:1185; Penna et al., 1997, Hepatology, 25:1022). Т.е. у пациентов, естественным образом поправившихся после гепатита В, установлены полиспецифичные и замедленные ответы, опосредованные Т-хелперами (TH) и цитотоксическими Т-лимфоцитами (CTL), которые легко обнаруживаются в периферической крови. При распознавании вирусных пептидов CTL приобретают способность или вылечить HBV-инфицированные клетки через нецитопатическое цитокин-опосредованное ингибирование репликации HBV и/или убить их через пути, опосредованные перфорин-Fas-лигандом и ФНОα. И эффекторные функции, наблюдаемые при разрешении острого гепатита В, и этот ответ Т-клеток 1 типа (Th1) сохраняюся и после клинического восстановления. Оно часто совпадает с повышением уровней сывороточной аланинаминотрансферазы (АЛТ) и с появлением HBcAg-специфических IgM и IgG. Антитела анти-НВе и анти-HBs появляются позже и указывают на благоприятный исход инфекции. HBsAg-специфические антитела являются нейтрализующими, опосредуют иммунитет и сохраняются после клинического выздоровления на всю жизнь. Тем не менее, хроническая HBV-инфекция редко разрешается иммунной системой. Когда это происходит, вирусный клиренс связан с повышенной активностью CTL и повышенными уровнями АЛТ, вызванными разрушением инфицированных гепатоцитов иммунной системой. Тем не менее, подавляющее большинство хронически инфицированных пациентов демонстрируют слабый и временный CD4+ и CD8+ Т-клеточный иммунный ответ, который-антигенно ограничен и неэффективен в плане выведения вирусной инфекции, хотя отдельные HBV-специфичные Т-клеточные клоны были выделены из биоптата печени. Причина этого изменения эффекторных функций клеточного иммунного ответа при хроническом гепатите В в настоящее время не известна. Тем не менее, было показано, что функциональные Т-клеточные ответы могут быть частично восстановлены у некоторых пациентов при вирусной нагрузке ниже порога в 106 МЕ/мл (Webster et al., 2004, J. Virol. 78:5707). Эти данные явно обнадеживают и подчеркивают необходимость иммуномодуляторных стратегий, способных индуцировать эффективный Т-клеточный ответ.
В идеале, лечение хронического вирусного гепатита В прежде всего должно позволить ингибировать репликацию HBV до необратимого повреждения печени, таким образом, чтобы избавиться от вируса, предотвратить прогрессирование заболевания в цирроз или рак печени и улучшить выживаемость пациентов. Стандартное лечение хронического гепатита В включает пегилированный интерферон-альфа (IFNα) и нуклеозидные/нуклеотидные аналоги (NUC), такие как ламивудин, а в последнее время энтекавир, телбивудин, адефовир и тенофовир (EASL Clinical Practice Guidelines: management of chronic hepatitis B, 2009). IFNα является мощной противовирусной молекулой, приводящей к ингибированию репликации вируса, которая, тем не менее, вызывает серьезные побочные эффекты у 25-30% пациентов. NUC выступают в качестве конкурентных ингибиторов полимеразы HBV, направленных на подавление обратной транскрипции прегеномной РНК в минус-нить ДНК, а затем в двухцепочечную вирусную ДНК. Они ограничивают формирование новых вирионов, но неэффективны в устранении суперскрученной кзкДНК, скрытой в ядрах инфицированных гепатоцитов, которая является источником новых потомств вируса. Этим можно объяснить, почему эффективность NUC носит временный характер, и вирусная нагрузка возникает сразу после прекращения лечения, требуя оставлять пациентов на пожизненном лечении. Кроме того, долгосрочная эффективность также ограничена в связи с появлением устойчивых мутантов HBV (более 24% после одного года и примерно 66% после четырех лет лечения ламивудином, как описано в Leung et al., 2001, Hepatology 33:1527), хотя новые NUC (энтекавир, телбивудин и тенофовир) демонстрируют гораздо меньше случаев лекарственно-устойчивых мутантов HBV при увеличении подавления ДНК HBV. Тем не менее, данные по длительному лечению этими новыми лекарствами ограничены, и эта более высокая эффективность не коррелирует с существенно более высоким уровнем HBs-сероконверсии.
Помимо противовирусной терапии в настоящее время прикладываются усилия по разработке дополнительной терапии, направленной на улучшение иммунного ответа хозяина, особенно того, который опосредован цитотоксическими Т-лимфоцитами и хелперными Т-лимфоцитами. Подавляющее большинство существующих подходов иммунотерапии сосредоточено на применении поверхностного белка(белков) HBV, S, рге81 и/или preS2 (Smith et al., 1983, Nature 302:490; Luoeck et al., 1989, Proc. NatI Acad, Sci. USA 86:б7б3; Adkins et al., 1998, BioDrugs 10:137; Loirat et al., 2000, J. Immunol. 165:4748; Funuy-Ren et al. 2003, J. Med. Viral. 71:376; Kasaks et al., 2004, J. Gen, Viral. 85:2665; Xiangming Li et al., 2005, Intern. Irnrnunol. 17:1293; Mancini-Bourguine et al., 2006, Vaccine 24:4482; Vandepapeliere et al., 2007, Vaccine 25:8585). Обнадеживающие результаты были получены по меньшей мере по отношению к стимуляции иммунных ответов. Например, Mancini-Bourguine et al. (2006, Vaccine24:4482) сообщили об индукции и/или возвращении Т-клеточного ответа у хронически HBV-инфицированных пациентов, которым вводили ДНК-вакцину, кодирующую preS2-S, что является хорошим признаком того, что иммунная система все еще действует у этих пациентов.
HBcAg также используется как иммуноген (Yi-Ping Xing et al., 2005, World J, Gastro, 11:4583), также как и химерные HBcAg-капсиды, несущие чужеродные эпитопы на своей поверхности (WO 92/11368; WOOO/32625; Koletzki et al., 1997, J. Gen, Virol. 78:2049). Наиболее перспективной локализацией для вставки эпитопов с точки зрения иммуногенности, по-видимому, является место внешней петли, которое, по прогнозам, находится на поверхности HBcAg вблизи позиции 80 (Argos et al, 1988, 10 EMBO J. 7:819). Schodel et al. (1992, J. Virol. 66:106) и Borisova et al. (1993, J. Virol, 67:3696) смогли вставить эпитопы preSI и HBsAg в эту область и сообщили об успешной иммунизации химерными частицами.
Также были исследованы претенденты на многовалентные вакцины, нацеленные одновременно на несколько HBV-антигенов. Примечательно, было показано, что полиэпитопная ДНК-вакцина, кодирующая гибридный полипептид множества эпитопов цитотоксических Т-лимфоцитов (CTL) и хелперных Т-лимфоцитов (HTL), присутствующих в оболочечном, сердцевинном и полимеразном белках, вызывает многочисленные CTL- и HTL-ответы на доклинических мышиных моделях (Depla et al, 2008, J. Virol. 82:435). Некоторые вакцинные составы на основе смеси ДНК-плазмид, кодирующих HBsAg, HBcAg и HBV-полимеразу, были разработаны (WO 2005/056051; WO 2008/020656) и показали специфический анти-HBV клеточный и гуморальный ответ на трансгенной мышиной модели хронического гепатита В (Спае Young Kirn et al., 2008, Exp.Mol. Medicine 40:669). Фаза I клинических испытаний была начата в Южной Корее на носителях HBV в комбинации с лечением ламивудином (Yang et al., 2006, Gene Ther. 13:1110).
Соответственно, все еще существует необходимость в альтернативных иммунотерапевтических подходах для индукции иммунного ответа более мощным и эффективным образом, особенно клеточно-опосредованного иммунного ответа, у лиц, нуждающихся в этом, например у пациентов, хронически инфицированных HBV. Кроме того, существует необходимость обеспечения композицией на основе вектора, способного экспрессировать HBV-антиген стабильным и устойчивым образом.
Эта техническая задача решается путем предоставления воплощений, определенных в формуле изобретения.
Другие и дополнительные аспекты, особенности и преимущества данного изобретения станут понятны из последующего описания предпочтительных воплощений изобретения. Эти воплощения приведены для раскрытия изобретения.
Сущность изобретения
Соответственно, в первом аспекте данное изобретение предусматривает иммуногенную композицию, включающую по меньшей мере один полипептид или нуклеиновокислотную молекулу, кодирующую указанный по меньшей мере один полипептид, где указанный по меньшей мере один полипептид выбран из группы, состоящей из:
(i) полимеразной части, включающей по меньшей мере 450 аминокислотных остатков полимеразного белка, полученного из первого вируса HBV;
(ii) сердцевинной части, включающей по меньшей мере 100 аминокислотных остатков сердцевинного белка, полученного из второго вируса HBV; и
(iii) оболочечной части, включающей один или более иммуногенный домен от 15 до 100 последовательных аминокислотных остатков белка HBsAg, полученного из третьего вируса HBV; или
любой комбинации указанных полимеразной части, сердцевинной части, оболочечной части, указанной нуклеиновокислотной молекулы, кодирующей указанную полимеразную часть, указанной нуклеиновокислотной молекулы, кодирующей указанную сердцевинную часть, и/или указанной нуклеиновокислотной молекулы, кодирующей указанную оболочечную часть.
Определения
Используемые в данное заявке термины в единственном числе применяются в значении «по меньшей мере один», «по меньшей мере первый», «один или более» или «множество» упоминаемых соединений или этапов, если контекст ясно не указывает иное. Например, термин «клетка» включает множество клеток, содержащихся в их смеси.
Термин «и/или», используемый в данном документе, включает значение «и», «или» или «все или любые комбинации элементов, связанных с указанным термином».
Термины «примерно» или «приблизительно», используемые в данном документе, означают в пределах 10%, предпочтительно в пределах 8% и более предпочтительно в пределах 5% данного значения.
Используемые в данной заявке для обозначения продуктов, композиций и способов термины «содержащий» (и его любые формы, в т.ч. «содержит» или «содержат»), «имеющий» (и его любые формы, в т.ч. «имеет» или «имеют»), «включающий» (и его любые формы, в т.ч. «включает» и «включают») или «состоящий из» (и его любые формы, в т.ч. «состоит из» или «состоят из») не являются ограничивающими и не исключают дополнительные, не упоминаемые элементы или этапы способа. Выражение «содержащий по существу» будет означать исключение других компонентов или этапов любого существенного значения. Таким образом, композиция, состоящая преимущественно из упоминаемых компонентов, не будет исключать следы контаминантов и фармацевтически приемлемых носителей. Выражение «состоящий из» будет означать исключение более чем следовых элементов других компонентов или этапов. Например, полипептид «состоит из» аминокислотной последовательности, когда полипептид не содержит никаких аминокислот не упоминаемой аминокислотной последовательности. Полипептид «состоит по существу» из аминокислотной последовательности, когда такая аминокислотная последовательность присутствует в конечном итоге только с несколькими дополнительными аминокислотными остатками. Полипептид «содержит» аминокислотную последовательность, когда аминокислотная последовательность является по меньшей мере частью конечной аминокислотной последовательности полипептида. Такой полипептид может иметь от нескольких до нескольких сотен дополнительных аминокислотных остатков.
Термины «аминокислоты», «остатки» и «аминокислотные остатки» являются синонимами и охватывают природные аминокислоты, а также аналоги аминокислот (например, неприродные, синтетические и модифицированные аминокислоты, в том числе D- или L-оптические изомеры).
Термины «полипептид», «пептид» и «белок» используются в данном документе взаимозаменяемо для обозначения полимеров аминокислотных остатков, которые состоят из девяти или более аминокислот, связанных посредством пептидных связей. Полимер может быть линейным, разветвленным или циклическим, может включать природные аминокислоты и/или аминокислотные аналоги и может быть прерван не аминокислотами. В качестве общего указания, если аминокислотный полимер длинный (например, более 50 аминокислотных остатков), то предпочтительно его называть полипептидом или белком, а если его длина 50 аминокислот или меньше, он называется «пептидом».
В контексте данного изобретения термины «нуклеиновая кислота», «нуклеиновокислотная молекула», «полинуклеотид» и «нуклеотидная последовательность» используются как синонимы и обозначают полимер любой длины или полидезоксирибонуклеотидной (ДНК) (например, кДНК, геномной ДНК, плазмид, векторов, вирусных геномов, выделенных ДНК, зондов, праймеров и любой их смеси) или полирибонуклеотидной (РНК) молекул (например, мРНК антисмысловой РНК) или смешанной полирибо-полидезоксирибонуклеотидной молекулы. Они охватывают одно- или двухцепочечные, линейные или кольцевые, природные или синтетические полинуклеотиды. Кроме того, полинуклеотид может содержать неприродные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги (см. US 5525711, US 4711955 или ЕРА 302175 в качестве примеров модификаций), и может быть прерван ненуклеотидными компонентами. При наличии изменений в нуклеотиде они могут быть сделаны до или после полимеризации.
Используемый в данном документе термин «иммуногенная композиция» относится к составу, который содержит 1, 2, 3, 4 или более компонентов, описанных далее (например, полимеразную часть, сердцевинную часть, оболочечную часть, нуклеиновокислотную молекулу, кодирующую полимеразную часть, нуклеиновокислотную молекулу, кодирующую сердцевинную часть, и/или нуклеиновокислотную молекулу, кодирующую оболочечную часть), и, возможно, другие компоненты (например, адъювант, носитель, разбавитель и т.д.). Иммуногенная композиция данного изобретения, как правило, находится в форме, которая может быть введена в организм хозяина и индуцирует защитный или терапевтический иммунный ответ, достаточный, чтобы индуцировать или стимулировать анти-HBV-иммунитет, давая терапевтические преимущества, такие как профилактика HBV-инфекции, сокращение и/или улучшение по меньшей мере одного состояния, вызванного или связанного с HBV-инфекцией (например, снижение вирусной нагрузки, уменьшение или отдаление риска поражения печени, такого как цирроз или рак печени, улучшение качества жизни и т.д.), и/или уменьшение сывороточных уровней HBcAg, HBsAg или обоих, и/или индукция сероконверсии HBe, сероконверсии HBs или обеих, и/или повышение эффективности других подходов к лечению и профилактике HBV. После введения в организм хозяина иммуногенная композиция данного изобретения может вызвать иммунный ответ, в том числе, но не ограничиваясь этим, продукцию антител и/или цитокинов и/или активацию цитотоксических Т-клеток, В-, Т-лимфоцитов, антиген-представляющих клеток, хелперных Т-клеток, дендритных клеток, NK-клеток, что приводит к развитию врожденного иммунного ответа и/или специфического гуморального и/или клеточного иммунного ответа по меньшей мере против одного антигена/эпитопа HBV.
Понятие «иммуногенный домен» относится к структурной части белка HBV, которая может быть связана антителом или Т-клеточным рецептором. Как правило, такой иммуногенный домен содержит один или более В- и/или Т-эпитоп(ы), в частности. CTL или TH или оба эпитопа участвуют в распознавании конкретным антителом, или, в контексте МНС (главный комплекс гистосовместимости), Т-клеточным рецептором. Понятие «эпитоп» соответствует минимальному пептидному мотиву (как правило, набору из 9-11 аминокислотных остатков), которые вместе образуют сайт, распознаваемый антителом, Т-клеточным рецептором или HLA-молекулой. Такие остатки могут быть последовательными (линейный эпитоп) или непоследовательными (конформационный эпитоп, который включает остатки, не расположенные в непосредственной близости друг к другу). Распознавание Т-клеточного эпитопа Т-клеткой, как правило, происходит через механизм, в котором Т-клетки распознают пептидные фрагменты антигенов, которые связаны с молекулами МНС 1 класса или II класса, экспрессированными на антиген-представляющих клетках.
Используемые в данном документе понятия «HBV» и «вирус гепатита В» являются взаимозаменяемыми и относятся к любому члену семейства Hepadnaviridae (см., например, Ganem and Schneider in Hepadnaviridae (2001) "The viruses and their replication" (pp2923-2969), Knipe DM et al., eds. Fields Virology, 4th ed. 5 Philadelphia, Lippineott Williams & Wilkins, или последующие издания). Обширные филогенетические анализы привели к классификации вирусов гепатита В на 8 основных генотипов (от А до Н), которые демонстрируют расхождения в последовательности по меньшей мере 8%. Различные генотипы HBV демонстрируют различное географическое распределение и могут отображать симптомы и/или клинические результаты гетерогенных заболеваний. Разнообразные HBV были классифицированы на девять различных подтипов (ayw1, ayw2, ayw3, ayw4, ayr, adw2, adw4, adrq+ и adqr-) в связи с HBsAg-связанной серологией (см. обзор Marrsum-AI Mahtab et. al., 2008, Hepatobiliary Panerease Dis Int 5:457; Schaeffer, 2007, World Gastroenterol. 7:14; Norder et al., 1993, J. Gen Virol.74:1341). Каждый генотип и серотип охватывает различные штаммы и изоляты HBV. Изолят соответствует конкретному вирусу, выделенному из определенного источника HBV (например, из образца от пациента или из другого биологического резервуара HBV), в то время как штамм охватывает различные изоляты, которые очень близки друг к другу с точки зрения геномных последовательностей.
В данной области, особенно в Genbank, было описано большое число HBV, подходящих для применения в контексте данного изобретения. Типичные HBV генотипа А включают, но не ограничиваясь ими, изолят HB-J1444AF и штамм НВ-JI444A (регистрационный номер АР 007263). Типичные HBV генотипа В включают, но не ограничиваясь ими, клон pJDW233 (регистрационный номер D00329), изолят HBV/14611 (регистрационный номер AF 121243), HBV-B1, выявленный в 2001 году Нои et al. (регистрационный номер GenBank AF282917.1), штамм HBV Whutj-37 (регистрационный номер GenBank AY 2933309.1), выявленный Zhang et al. (2005, Arch. Virol. 150, 721-741), китайский штамм HBV GDH1, выявленный Не et al. (регистрационный номер GenBank AY 766463.1), и изолят HBV 57-1 подтипа adw, выявленный Jiang et al. (регистрационный номер GenBank AYS 18556.1). Типичные HBV генотипа С включают, но не ограничиваясь ими, изолят AH-1-ON980424 (регистрационный номер АВ 113879), штамм НСС-3-ТТ (регистрационный номер АВ 113877), изолят HBV SWT3.3, выявленный Fang et al. (регистрационный номер GenBank EU 916241.1), изолят HBV H85, выявленный Zhu et al. (регистрационный номер GenBank AY 306136.1), штамм HBV C1248, выявленный Tu et al. (регистрационный номер GenBank DQ 975272.1), изолят HBV CHN-H155 (регистрационный номер GenBank DQ 478901.1), выявленный Wang et al. (2007, j. Viral Hepat 14, 426-434) и изолят HBV GZ28-1, выявленный Zhou et al. (регистрационный номер GenBank EF 688062). Типичные HBV генотипа D включают, но не ограничиваясь ими, изолят КАМСНАТКА27 (регистрационный номер АВ 188243), ALTAY36 (регистрационный номер АВ 188245) и Y07587, описанный Stoll-Becker et al. (1997, J. ViroL 71:5399) и доступный в Genbank под регистрационным номером Y07587, а также изолят HBV, описанный под регистрационным номером АВ 267090. Типичные HBV генотипа Е включают, но не ограничиваясь ими, изолят HB-JI411F и штамм HB-JI411 (регистрационный номер АР 007262). Типичные HBV генотипа F включают, но не ограничиваясь ими, изоляты HBV-BL597 (регистрационный номер АВ 214516) и HBV-BL592 (регистрационный номер АВ 166850). Типичные HBV генотипа G включают, но не ограничиваясь ими, изолят HB-JI444GF и штамм HB-J1444G (регистрационный номер АР 007264). Типичные HBV генотипа Н включают, но не ограничиваясь ими, изолят HBV ST0404 (регистрационный номер АВ 298362) и изолят HB-JI260F и штамм HB-JI260 (регистрационный номер АР 007261). Тем не менее, данное изобретение не ограничивается этими типичными HBV. Действительно, нуклеотидные и аминокислотные последовательности могут варьировать в различных изолятах и генотипах HBV, и эта природная генетическая изменчивость входит в рамки изобретения, как и неприродные модификации, такие как описаны ниже.
Используемое в данном документе понятие «нативный белок HBV» относится к белку, полипептиду или пептиду (например, полимеразному белку, сердцевинному белку или HBsAg, и т.д.), которые можно найти, выделить, получить из источника HBV в природе, в отличие от искусственно модифицированных или измененных человеком в лабораторных условиях. Таким образом, этот термин будет включать природные белки, полипептиды или пептиды HBV, если не указано иное. Такими природными источниками являются биологические образцы (например, крови, плазмы, сыворотки крови, спермы, слюны, срезов тканей, биопсии и т.д.), взятые у субъекта, инфицированного или подвергшегося воздействию HBV, культивируемые клетки (например, HepG2.2.15, HuH6-C15 (Sureau et al., 1986, Cell 47:37; Sells et al., 1987, Proc. Nail. Acad. Sci. 84(4): 1005); HuH7.TA61 или HuH7.TA62 (Sun et al., 2006, J Hepatol. 45(5):636), культуры тканей, а также рекомбинантные материалы. Рекомбинантные материалы включают, но не ограничиваясь ими, изоляты HBV (например, имеющиеся в депозитных учреждениях), геном HBV, библиотеки геномной РНК или кДНК, плазмиды, содержащие геном HBV или его фрагмент(ы), или любой вектор, используемый в данной области для включения таких элементов.
Нуклеотидные последовательности, кодирующие различные белки HBV, можно найти в специализированных банках данных (например, упомянутых выше) и в литературе (см. например, Valenzuela et al., 1980, The nucleotide sequence of the hepatitis В viral genome and the identification of the major viral genes (pp57~7G) в "Animal Vims Genetics"; eds B. Fields, et al.; Academic Press Inc., New-York and Vaudin et al., 1988, J. Gen, ViroL 69:1383). Типичные примеры природных полимеразного, сердцевинного и HBsAg-полипептидов приведены в SEQ ID №1-3, соответственно (SEQ ID №1 предусматривает аминокислотную последовательность нативного полимеразного белка изолята HBV Y07587, SEQ ID №2 предусматривает аминокислотную последовательность нативного сердцевинного белка изолята HBV Y07587, a SEQ ID №3 предусматривает аминокислотную последовательность нативного оболочечного белка (HBsAg) изолята HBV Y07587). Нуклеотидные последовательности, кодирующие нативный полимеразный, сердцевинный и HBsAg из HBV Y07587, показаны с иллюстративной целью в SEQ ID №4, 5 и 6, соответственно. Тем не менее, как уже говорилось выше, HBV-белки не ограничиваются этими типичными последовательностями, и генетические вариации входят в рамки изобретения.
Используемый в данном документе термин «часть» (например полимеразная, сердцевинная и/или оболочечная части) относится к белку, полипептиду или пептиду, который получен из нативного белка, полипептида или пептида HBV после того, как он был искусственно модифицирован или изменен человеком в лаборатории, как описано в данном документе. Термин «модифицированный» включает удаление, замену или добавление одного или более нуклеотидного/аминокислотного остатка(ов), любую комбинацию этих возможностей (например, дегенерацию нативной нуклеотидной последовательности для уменьшения гомологии между последовательностями HBV, кодируемыми композицией изобретения, введение соответствующих сайтов рестрикции), а также неприродные механизмы (например, слияние двух или более белков, полипептидов и пептидов HBV или части/частей). Когда предусмотрено несколько модификаций, они могут касаться последовательных остатков и/или непоследовательных остатков. Модификация (модификации) может быть получена разными способами, известными специалистам в данной области, такими как сайт-направленный мутагенез (например, с помощью системы мутагенеза in vitro Sculptor™ от Amersharn, Лес-Улис, Франция), ПЦР-мутагенез, перемешивание ДНК и синтетические химические методики (например, дающие синтетические нуклеиновокислотные молекулы). Модификация(и), предусмотренная данным изобретением, включает молчащие модификации, которые не изменяют аминокислотную последовательность закодированных полипептидов HBV, а также модификации, которые будут транслироваться в закодированный полипептид, образуя модифицированную аминокислотную последовательность по сравнению с соответствующей нативной. Термин «образует» (или образующий») используется для определения первоначального источника молекулы, но не предназначен для ограничения способа, посредством которого делают молекулу, который может быть, например, химическим синтезом или рекомбинантным средством.
Предпочтительно каждая из частей HBV, используемых в данном изобретении, сохраняет высокую степень идентичности аминокислотной последовательности с соответствующим нативным белком HBV, с белком полной длины или с его частью (частями). Процент идентичности двух полипептидов зависит от числа одинаковых позиций, общих для последовательностей, с учетом ряда пробелов, которые должны быть введены для оптимального выравнивания, и длины каждого пробела. Для определения процента идентичности аминокислотных последовательностей в данной области доступны различные компьютерные программы и математические алгоритмы, такие как, например, программа Blast (например, Altschul et al., 1997, Nucleic Acid Res. 25:3389; Altschul et al., 2005, FEES J, 5 272:5101), доступная в NCBI. То же самое может быть применено для нуклеотидных последовательностей. Программы для определения гомологии нуклеотидных последовательностей также доступны в специализированной базе данных (Genbank или Wisconsin Sequence Analysis Package), например программы BESTFIT, FASTA и GAP.
Например, в дополнение к модификациям, описанным ниже (например, сниженной ферментативной активности и т.д.), любая или все части HBV, содержащиеся или кодирующиеся композицией изобретения, могут быть модифицированы таким образом, чтобы представлять конкретный генотип, и, следовательно, включают аминокислотную последовательность, соответствующую консенсусной или неконсенсусной последовательности, которая обычно определяется после выравнивания последовательностей различных полипептидов HBV определенного генотипа.
Термин «комбинация», используемый в данном документе, относится к любой комбинации по меньшей мере двух компонентов, содержащихся или кодирующихся иммуногенной композицией изобретения и любой возможной компоновкой различных компонентов с определенным предпочтением 2 или 3 из указанных компонентов. Он охватывает смесь двух или более полипептидов, смесь двух или более нуклеиновокислотных молекул/векторов, смесь одного или более полипептида(ов) и одной или более нуклеиновокислотной молекулы/вектора, а также слияние двух или более нуклеиновокислотных молекул, так чтобы получить единую полипептидную цепь, несущую две или более части HBV (например, неприродная компоновка).
В предпочтительном воплощении иммуногенная композиция изобретения содержит комбинацию по меньшей мере двух из указанных полимеразной части, сердцевинной части, оболочечной части или по меньшей мере двух из указанных нуклеиновокислотной молекулы, кодирующей указанную полимеразную часть, нуклеиновокислотной молекулы, кодирующей указанную сердцевинную часть, и/или нуклеиновокислотной молекулы, кодирующей указанную оболочечную часть. Особенно предпочтительная композиция изобретения выбрана из группы, состоящей из (i) композиции, содержащей комбинацию полимеразной части и сердцевинной части, определенных в данном документе, или комбинацию нуклеиновокислотных молекул, кодирующих указанную полимеразную часть и указанную сердцевинную часть; (ii) композиции, содержащей комбинацию сердцевинной части и оболочечной части, определенных в данном документе, или комбинацию нуклеиновокислотных молекул, кодирующих указанную сердцевинную часть и указанную оболочечную часть; (iii) композиции, содержащей комбинацию полимеразной части, сердцевинной части и оболочечной части, определенных в данном документе, или комбинацию нуклеиновокислотных молекул, кодирующих указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть.
Термины «слияние» или «гибридный белок», используемые в данном документе, относятся к комбинации друг с другом по меньшей мере двух полипептидов (или их фрагментов) в одну полипептидную цепь. Предпочтительно слияние различных полипептидов осуществляется с помощью генетических средств, т.е. путем слияния в рамке считывания нуклеотидных последовательностей, кодирующих каждый из указанных полипептидов. Под выражением «слиты в рамке считывания» подразумевается, что экспрессия слитых кодирующих последовательностей дает один белок без какого-либо терминатора трансляции между каждым из слитых полипептидов. Слияние может быть прямым (т.е. без каких-либо дополнительных аминокислотных остатков между ними), либо через линкер. Наличие линкера может способствовать правильному формированию, сворачиванию и/или функционированию гибридного белка. Данное изобретение не ограничивается формой, размером или количеством используемых линкерных последовательностей, и на стыке между слитыми полипептидами может быть вставлено множество копий линкерной последовательности. Подходящие линкеры в соответствии с изобретением имеют от 3 до 30 аминокислот в длину и состоят из повторов аминокислотных остатков, таких как глицин, серин, треонин, аспарагин, аланин и/или пролин (см., например, Wiederrecht et al., 1988, Cell 54, 841; Aumailly et al., 1990 FEBS Lett. 262, 82; и Dekker et al., 1993, Nature 362, 852), например, линкер Ser-Gly-Ser или Gly-Ser-Gly-Ser-GIy.
Используемый в данном документе термин «гетерологичная гидрофобная последовательность» относится к пептиду гидрофобной природы (который содержит большое количество остатков гидрофобных аминокислот, таких как остатки Val, Leu, lie, Met, Phe, Tyr и Trp). Понятие «гетерологичный» относится к последовательности, чужеродной для нативного белка, полипептида или пептида HBV, из которой получают выбранную часть HBV. Это может быть пептид, чужеродный вирусу HBV (например, пептид из вируса кори или вируса бешенства), или пептид из HBV-вируса, но в позиции, в которой он обычно не встречается в вирусном геноме. Гетерологичная гидрофобная последовательность может быть слита в рамке считывания на N-конце, на С-конце или в пределах части HBV, и может играть роль, среди прочего, в переносе полипептида, облегчении продукции или очистки полипептида, удлинении периода полувыведения. Подходящие гетерологичные гидрофобные последовательности в соответствии с изобретением имеют от 15 до 100 аминокислот в длину и содержат высоко гидрофобный домен.
Термин «вектор», используемый в данном документе, относится и к экспрессионным, и к неэкспрессионным векторам, и включает вирусные, а также невирусные векторы, в том числе внехромосомные векторы (например, многокопийные плазмиды) и интегрирующие векторы, предназначенные для включения их в принимающую хромосому(ы). Особенно важными в контексте данного изобретения являются векторы для переноса нуклеиновокислотных(ой) молекул(ы) в вирусный геном (так называемые векторы для переноса), векторы для применения в иммунотерапии (т.е. которые способны доставлять нуклеиновокислотные молекулы в организм хозяина), а также экспрессионные векторы для применения в различных экспрессионных системах или в организме хозяина.
Используемый в данном документе термин «вирусный вектор» охватывает векторную ДНК, а также вирусные частицы, образованные ею. Вирусные векторы могут быть репликативно-компетентными, либо могут быть генетически неполноценными, т.е. могут быть репликативно-дефектными или репликативно-ослабленными. Термин «репликативно-компетентный», используемый в данном документе, включает селективно-репликативные и условно-репликативные вирусные векторы, которые спроектированы для лучшей или селективной репликации в конкретных клетках-хозяевах (например, опухолевых клетках).
Используемый в данном документе термин «регуляторная последовательность» относится к любой последовательности, которая позволяет, способствует или модулирует экспрессию нуклеиновокислотной молекулы в данной клетке-хозяине или организме, включая репликацию, дупликацию, транскрипцию, сплайсинг, трансляцию, стабильность и/или транспорт нуклеиновой кислоты или одного из ее производных (например, мРНК) в клетку-хозяина или организм.
Используемый в данном документе термин «клетка-хозяин» следует понимать в широком смысле без всяких ограничений, касающихся конкретной организации ткани, органа или выделенных клеток. Такие клетки могут быть уникальным типом клеток или группой различных типов клеток и включают бактерии, низшие и высшие эукариотические клетки, а также культивируемые клеточные линии, первичные клетки и пролиферативные клетки. Этот термин включает клетки, которые могут быть или были реципиентами композиции(й), нуклеиновокислотных молекул(ы), вектора(ов) или инфекционных вирусных частиц(ы) изобретения, а также потомство таких клеток.
Термин «организм-хозяин» относится к позвоночным, в частности, к представителям видов млекопитающих, и особенно к домашним животным, сельскохозяйственным животным, спортивным животным и приматам, включая человека. Предпочтительно организм-хозяин является пациентом, страдающим от хронической HBV-инфекции. Инфицирующий HBV может быть того же генотипа или серотипа, что и по меньшей мере один из первого, второго или третьего HBV, используемых в данном изобретении.
Используемый в данном документе термин «выделенный» относится к белку, полипептиду, пептиду, нуклеиновокислотной молекуле, клетке-хозяину или вирусу, который был выделен из своей природной среды (например, отделен по меньшей мере от одного компонента(ов), с которым он природно связан).
Используемое в данном документе понятие «терапевтически эффективное количество» является дозой, достаточной для облегчения одного или более симптома, обычно связанного с HBV-инфекцией или заболеванием или состоянием, вызванным или связанным с HBV-инфекцией. При профилактическом применении этот термин означает дозу, достаточную для предотвращения или отсрочки появления HBV-инфекции. «Терапевтические» композиции разработаны для введения в организм хозяина, уже инфицированного HBV, с целью сокращения или смягчения по меньшей мере одного заболевания или состояния, вызванного или связанного с указанной HBV-инфекцией, в конечном счете в сочетании с одним или более традиционным терапевтическим подходом, описанным выше (например, лечением нуклеозидными или нуклеотидными аналогами). Например, терапевтически эффективное количество для индукции иммунного ответа может быть таким количеством, которое необходимо для активации иммунной системы (например, приводящей к развитию анти-HBV-реакции). Термин «рак» охватывает любые раковые состояния, включая диффузные или локализованные опухоли, метастазы, раковые полипы, а также предраковые поражения (например, цирроз печени).
Используемый в данном документе термин «фармацевтически приемлемый носитель» предназначен для включения любого и всех носителей, растворителей, разбавителей, эксципиентов, адъювантов, дисперсных сред, покрытий, антибактериальных и противогрибковых агентов, а также агентов, задерживающих абсорбцию, и т.п., совместимых с фармацевтическим введением.
В соответствии с данным изобретением указанные полимеразная, сердцевинная и/или оболочечная части, содержащиеся или кодируемые композицией изобретения, могут происходить независимо из любого генотипа, штамма и изолята HBV, определенного в настоящее время, такого как описанные выше в связи с термином «HBV». Кроме того, каждая из полимеразной, сердцевинной и/или оболочечной частей может происходить из нативного соответствующего белка HBV или из модифицированного полипептида HBV (т.е. измененного таким образом, чтобы быть представителем конкретного генотипа). Таким образом, первый, второй и третий HBV-вирусы, из которых происходят полимеразная, сердцевинная и/или оболочечная части, содержащиеся или кодируемые композицией данного изобретения, могут быть независимо из одного и того же или из разных генотипов, серотипов и/или изолятов. Например, с помощью HBV-частей, полученных из двух или трех различных генотипов HBV, можно обеспечить защиту против широкого диапазона генотипов HBV.
В одном воплощении может быть интересно адаптировать иммуногенную композицию изобретения к конкретной географической области, используя по меньшей мере одну часть HBV из генотипа(ов) HBV, эндемичного для этой области. Для наглядности, генотипы А и С являются наиболее распространенными в США, в то время как пациенты из стран Западной Европы заражаются, главным образом, генотипами А и D, а в бассейне Средиземного моря генотипом D. Ограниченные данные из Индии подтверждают, что генотипы А и D являются наиболее распространенными в Индии. С другой стороны, генотипы В и С являются наиболее распространенными в Китае. Например, композиция изобретения, предназначенная для европейских стран, может содержать полимеразную часть, которая произошла из генотипа А, и сердцевинную и оболочечную части, которые произошли из генотипа D, или наоборот. Кроме того, полимеразная и сердцевинная части могут быть из генотипа А, а оболочечная часть из генотипа D. В качестве другого примера, композиция изобретения, предназначенная для европейских стран и США, может включать полимеразную, сердцевинную и оболочечную части, которые независимо произошли из генотипов А, С и D. С другой стороны, композиция изобретения, предназначенная для Китая, может включать или кодировать полимеразную, сердцевинную и оболочечную части, которые независимо друг от друга произошли из генотипа В и/или С.
Можно также адаптировать иммуногенную композицию изобретения к популяции пациентов, подлежащих лечению. Например, генотип А чаще встречается среди американских белых и афро-американцев и получивших HBV-инфекцию половым путем, тогда как генотипы В и С, с другой стороны, распространены среди американцев азиатского происхождения, пациентов, родившихся в Азии, и получивших HBV-инфекцию путем передачи от матери ребенку. HBV-генотипы также связаны с различными клиническими исходами (Schaeffer et ai, 2005, J, Viral, Hepatitis 12:111), при этом генотипы D и F связаны с более серьезным прогрессированием заболевания и худшим прогнозом, чем генотип A (Sanchez-Tapias et al., 2002, Gastroenterology 123:1848). Специалисты в данной области смогут адаптировать композицию изобретения в соответствии с рассматриваемым населением и/или географической областью путем выбора соответствующих генотипов, серотипов, штаммов и/или изолятов HBV.
В соответствии с предпочтительным воплощением по меньшей мере два из первого, второго и третьего HBV, а предпочтительно все, имеют один и тот же HBV-генотип, и в частности генотип D. Независимо друг от друга они могут происходить из одного и того же изолята, при этом с особым предпочтением первый, второй и третий вирусы HBV взяты из HBV-изолята Y07587. Предпочтительно, полимеразная часть, используемая в данном изобретении, включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №1, или с ее частью(частями), включающей не менее 450 аминокислотных остатков. Альтернативно или в сочетании, сердцевинная часть, используемая в данном изобретении, включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №2, или ее частью (частями), включающей не менее 100 аминокислотных остатков. Альтернативно или в сочетании одна или более иммуногенная область оболочечной части, используемая в данном изобретении, включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с частью(частями) длиной в 15-100 аминокислотных остатков из аминокислотной последовательности, приведенной в SEQ ID №3.
Полимеразная часть
В соответствии с одним воплощением полимеразная часть, содержащаяся или кодируемая композицией изобретения, модифицирована по сравнению с соответствующей нативной HBV-полимеразой.
Соответствующая модификация представляет собой отсечение по меньшей мере 20 аминокислотных остатков и максимум 335 аминокислотных остатков, обычно присутствующих на М-конце нативной HBV-полимеразы. Эта модификация особенно важна для композиции изобретения, которая также включает второй полипептид для уменьшения или удаления перекрывающихся частей между полимеразной и сердцевинной частями. Специалист в данной области сможет адаптировать усечение для композиции изобретения в диапазоне от по меньшей мере 20 аминокислотных остатков до максимум 335 аминокислотных остатков. Что касается нативной HBV-полимеразы, полимеразная часть, используемая в контексте данного изобретения, является предпочтительно усеченной по меньшей мере на 30 аминокислотных остатков и не более чем на 200 аминокислотных остатков, предпочтительно по меньшей мере на 35 аминокислот и не более чем на 100 аминокислотных остатков, предпочтительно по меньшей мере на 40 аминокислотных остатков и не более чем на 60 аминокислотных остатков, и более предпочтительно по меньшей мере на 45 и не более чем на 50 аминокислотных остатков, при этом особое предпочтение имеет усечение, включающее первые 47 или 46 аминокислотных остатков после инициаторного остатка Met, расположенного на М-конце нативной HBV-полимеразы. Предпочтительно усечение простирается от позиции 1 (инициатор Met) или 2 до позиции 47 в SEQ ID №1.
Предпочтительное воплощение направлено на полимеразную часть, включающую аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с частью аминокислотной последовательности, приведенной в SEQ ID №1, простираясь от приблизительно 48 позиции до приблизительно 832 позиции; еще более предпочтительно полимеразная часть включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с частью аминокислотной последовательности, приведенной в SEQ ID №7.
Альтернативно или в сочетании, полимеразная часть, используемая в изобретении, модифицирована таким образом, чтобы демонстрировать сниженную обратно-транскриптазную (ОТазную) ферментативную активность по сравнению с нативной HBV-полимеразой. Преимущественно указанное снижение ОТазной активности обеспечивается одной или более мутацией (мутациями) в области, ответственной за ОТазную ферментативную активность.
Структурная и функциональная организация HBV-полимераз была исследована почти 20 лет назад (см., например, Radziwill et al., 1990, J. Virol. 64:613). Они представляют собой многофункциональные белки с тремя функциональными областями, которые катализируют основные этапы репликации HBV (прайминг, синтез ДНК и удаление РНК-матриц), и не основными разделителями, которые расположены в следующем порядке:
- первая область, которая продолжается от позиции 1 приблизительно до позиции 177, является ответственной за концевую активность белка HBV,
- разделитель, расположенный приблизительно от 178 позиции приблизительно до 335 позиции,
- ДНК-полимеразная область, которая продолжается приблизительно от позиции 336 приблизительно до позиции 679, несет ответственность за ОТазную активность, и
- область РНКазы Н приблизительно от позиции 680 до С-конца (приблизительно позиция 832) участвует в активности РНКазы Н.
Четыре остатка вовлечены в ОТазную активность, формируя мотив «YMDD» (для остатков Туг, Met, Asp и Asp), обычно присутствующий приблизительно от позиции 538 приблизительно до позиции 541 нативной HBV-полимеразы (например, соответствующий позициям 538, 539, 540 и 541 в SEQ ID №1 и позициям 492, 493, 494 и 495 в SEQ ID №7), и данное изобретение охватывает любые мутации в этом мотиве или в другом месте ОТазной области, которые коррелируют со значительным сокращением (т.е. сокращением по меньшей мере в 10 раз) или удалением ОТазной активности при сохранении иммуногенных свойств. Типичные примеры подходящих ОТаза-дефицитных полимеразных мутантов описаны в литературе, например, в Radziwill et al. (1990, J. Virol. 64:613), в Bartenschlager et al. (1990, J. Virol, 64:5324) и в Jeong et al. (1996, Biochem Bioph Res Commun. 223(2):264). Предпочтительно полимеразная часть, используемая в данном изобретении, включает замену первого остатка Asp из мотива YMDD (который соответствует позиции 540 в SEQ ID №1 и позиции 494 в SEQ ID №7) или аминокислотного остатка, который находится в эквивалентной позиции в нативной HBV-полимеразе, на любой аминокислотный остаток, кроме Asp, при этом особо предпочтительна замена на остаток His (мутация D540H). Уменьшение или удаление ОТазной активности можно выявить с помощью анализов, хорошо известных в данной области (например, эндогенных полимеразных анализов, описанных в Radziwill et al., 1990, J Virol. 64:613).
Альтернативно или в сочетании, полимеразная часть, используемая в изобретении, модифицирована таким образом, чтобы демонстрировать сниженную ферментативную активность РНКазы Н по сравнению с нативной HBV-полимеразой. Преимущественно указанное снижение активности РНКазы Н обеспечивается одной или более мутацией (мутациями) в области, ответственной за ферментативную активность РНКазы Н. Как уже говорилось выше, функциональная область, вовлеченная в активность РНКазы Н, была спроецирована на С-концевую часть HBV-полимеразы, более конкретно с позиции 680 до С-концевой позиции 832, и данное изобретение охватывает любые мутации в этой области, которые коррелируют со значительным сокращением (т.е. сокращением по меньшей мере в 10 раз) или удалением активности РНКазы Н при сохранении иммуногенных свойств. Типичные примеры подходящих РНКаза Н-дефицитных полимеразных мутантов описаны в литературе, например, в Radziwill et al. (1990, J. Virol. 64:613), в Bartenschlager et al. (1990, J. Virol, 64:5324). Предпочтительно полимеразная часть, используемая в данном изобретении, включает замену остатка Glu, соответствующего позиции 718 в SEQ ID №1 и позиции 672 в SEQ ID №7, или аминокислотного остатка, который находится в эквивалентной позиции в нативной HBV-полимеразе, на любой аминокислотный остаток, кроме Glu, при этом особо предпочтительна замена на остаток His (мутация Е718Н). Уменьшение или удаление активности РНКазы Н может выявить с помощью анализов, хорошо известных в данной области (например, анализов активности РНКазы Н in vitro или анализа тандемной молекулы ДНК-РНК, описанного в Radziwill et al., 1990, J Virol, 64:613 или в Lee et al., 1997, Bioehem. Bioph. Res, Commun. 233(2):401).
Предпочтительно полимеразная часть, используемая в изобретении, мутирована таким образом, чтобы уменьшить или удалить и ОТазную, и РНКазную активность, и включает модификации, описанные выше в связи с этими ферментативными функциями, с особым предпочтением мутаций D540H и Е718Н.
Предпочтительное воплощение данного изобретения направлено на полимеразную часть, включающую, альтернативно состоящую по существу или же альтернативно состоящую из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №8, или с аминокислотной последовательностью, приведенной в SEQ ID №7, с заменой остатка Asp в позиции 494 на остаток His и заменой остатка Glu в позиции 672 на остаток His.
В другом и предпочтительном воплощении полимеразная часть, используемая в данном изобретении, слита в рамке считывания с гетерологичной гидрофобной последовательностью (последовательностями) для улучшения синтеза и/или стабильности и/или презентации на поверхности экспрессирующей клетки-хозяина и/или презентации антигена хозяйским МНС класса 1 и/или МНС класса II. Подходящие гетерологичные гидрофобные последовательности включают последовательности, такие как сигнальные и/или трансмембранные пептиды, которые позволяют направлять полимеразную часть на секреторный путь. Такие пептиды известны в данной области. Вкратце, сигнальные пептиды, как правило, присутствуют на М-конце представленных на мембране или секретированных полипептидов, и инициируют их переход в эндоплазматический ретикулум (ER). Они включают от 15 до 35 существенно гидрофобных аминокислот, которые затем удаляются определенной ER-расположенной эндопептидазой, образуя зрелый полипептид. Трансмембранные пептиды в природе обычно являются высоко гидрофобными и служат для закрепления полипептидов в клеточной мембране (см., например, Brandon and Tooze, 1991, в Introduction to Protein Structure p.202-214, NY Garland; WO 99/03885). Выбор трансмембранного и/или сигнального пептидов, которые могут быть использованы в контексте данного изобретения, огромен. Они могут быть получены из любого заякоренного в мембране и/или секретированного полипептида (например, клеточного или вирусного полипептида), такого как иммуноглобулины, тканевой активатор плазминогена, инсулин, гликопротеин вируса бешенства, оболочечный гликопротеин ВИЧ или F-белок вируса кори, или могут быть синтетическими. Предпочтительным местом вставки сигнального пептида является М-конец после кодона инициации трансляции, а трансмембранного пептида - С-конец, например, непосредственно перед стоп-кодоном. Кроме того, пептидный линкер может быть использован для связывания сигнального и/или трансмембранного пептидов с полимеразной частью.
В контексте изобретения может быть использована другая гидрофобная последовательность(и), такая как те, которые обычно присутствуют в белках, связанных с оболочкой или мембраной, в том числе HBsAg. Особый интерес в контексте данного изобретения представляет слияние полимеразной части с одной или более иммуногенной областью, описанной в данном документе (env1, env2, env3 и/или env4), которая имеет гидрофобную природу. Одна или более гидрофобная область может быть слита в рамке считывания на М-конце, на С-конце или в полимеразной части.
В контексте изобретения предпочтительно полимеразная часть, используемая в изобретении, слита в рамке считывания с сигнальным и трансмембранным пептидами гликопротеина вируса бешенства. Как показано в прилагаемом разделе с примерами, указанная сигнальная последовательность из вируса бешенства слита в рамке считывания с N-концом, а указанная трансмембранная последовательность из вируса бешенства слита в рамке считывания с С-концом указанной полимеразной части. Наиболее предпочтительное воплощение направлено на полимеразную часть, которая включает, альтернативно состоит по существу или же альтернативно состоит из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №9.
Сердцевинная часть
Альтернативно или в сочетании с композицией, описанной выше, данное изобретение также предусматривает композицию, включающую сердцевинную часть, происходящую из второго HBV. Как описано в данном документе в связи с вирусом HBV, сердцевинная часть, используемая в изобретении, происходит из HBV, который является таким же или отличается от вируса HBV, из которого берет начало полимеразная часть. Предпочтительно, сердцевинная и полимеразная части происходят из HBV генотипа D, при этом особое предпочтение отдается HBV-изоляту Y07587 или, альтернативно, из генотипа HBV, распространенного в Китае (например, генотипа В или С).
В соответствии с одним воплощением сердцевинная часть, используемая в данном изобретении, может быть нативной сердцевинной частью HBV (например, приведенной в SEQ ID №2) или сердцевинной частью, модифицированной по сравнению с соответствующей нативной сердцевинной частью HBV, которая сохраняет по меньшей мере 100 аминокислотных остатков сердцевинного белка, при этом особое предпочтение отдается сердцевинной части, содержащей от 120 до 180 аминокислотных остатков, предпочтительно от 125 до 148 аминокислотных остатков, и предпочтительно от 130 до 143 аминокислотных остатков (например, приблизительно 140 аминокислотных остатков).
Соответствующей модификацией является усечение. Предпочтительно усечение охватывает по меньшей мере 10 аминокислотных остатков и не более 40 аминокислотных остатков, обычно присутствующих на С-конце нативной сердцевинной части HBV или в С-концевой части (т.е. части, охватывающей последние 40 аминокислотных остатков). Эта модификация особенно важна для композиций изобретения, которые также включают полимеразную часть, для уменьшения или удаления перекрывающейся части между полимеразной и сердцевинной частями. Она также может иметь значение для удаления NLS (сигнала ядерной локализации) в этой сердцевинной области и/или для подавления взаимодействия с HBV-полимеразой. Специалисты смогут адаптировать усечение для композиции изобретения в упоминаемом диапазоне по меньшей мере от 10 аминокислотных остатков и до 40 аминокислотных остатков. Соответствующие усечения включают 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 последовательных аминокислотных остатков, обычно присутствующих на С-конце нативной сердцевины HBV или в С-концевой части. Что касается нативной сердцевины HBV, сердцевинная часть, используемая в контексте данного изобретения, предпочтительно усечена по меньшей мере на 20 аминокислотных остатков и не более чем на 40 аминокислотных остатков, предпочтительно по меньшей мере на 30 аминокислотных остатков и не более чем на 38 аминокислотных остатков, и более предпочтительно по меньшей мере на 34 аминокислотных остатка и не более чем на 37 аминокислотных остатков, расположенных на С-конце нативного HBV или в С-концевой части, при этом особым предпочтением пользуется усечение, включающее последние 35 аминокислотных остатков нативной сердцевины HBV (в других терминах, усечение простирается приблизительно от позиции 149 до С-конца сердцевинного полипептида).
Особый интерес представляет сердцевинная часть, включающая аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность, и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №2, простирающейся с позиции 1 предпочтительно до позиции 148, и даже более предпочтительной является сердцевинная часть, включающая аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность, и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №10.
Альтернативно или в сочетании, сердцевинная часть модифицирована таким образом, чтобы демонстрировать сниженное распознавание и/или взаимодействие с белком оболочки HBV по сравнению с нативной HBV-сердцевиной. Преимущественно указанное снижение распознавания/взаимодействия обеспечивается одной или более мутацией (мутациями) в области, расположенной вблизи от остатка 80, которая, по-видимому, формирует внешнюю петлю, доступную на поверхности сердцевинных частиц (Argos et al., 1988, EMBO J. 7:819). Уменьшение или удаление распознавания или взаимодействия с оболочечным белком можно выявить с помощью анализов, хорошо известных в данной области (например, электронной микроскопии, анализа формирования нуклеокапсидов в клетках, секретированных вирионов после временной трансфекции в HuH7, как описано в Seitz et al., 2007, EMBO J., 26:416 или в Ponsel et al, 2003, J. Virol.77(1):416).
Предпочтительная модификация включает удаление одного или более аминокислотного остатка в области сердцевины, продолжающееся приблизительно от позиции 75 приблизительно до позиции 85 (соответствующее остаткам 75-85 из SEQ ID №2 и SEQ ID №10), с особо предпочтительным удалением части сердцевины, продолжающимся от приблизительно 77 позиции до приблизительно 84 позиции (соответствующим остаткам 77-84 SEQ ID №2 и SEQ ID №10). Предпочтительно сердцевинная часть включает, альтернативно состоит по существу или же альтернативно состоит из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №11, или аминокислотной последовательностью, указанной в SEQ ID №2, в которой отсутствуют остатки 77-84.
Оболочечная часть
Альтернативно или в сочетании с композицией, описанной выше, данное изобретение также предусматривает композицию, содержащую оболочечную часть. Указанная оболочечная часть состоит из одной или более иммуногенной области (областей) по меньшей мере из 15 последовательных аминокислотных остатков, присутствующих в белке HBs, полученном из третьего вируса гепатита. Предпочтительно, каждая из указанных иммуногенных областей соответствует части по меньшей мере из 20 аминокислот и максимум 100 аминокислот, присутствующих в белке HBsAg в нативной или модифицированной форме (например, модифицированной таким образом, чтобы представлять конкретный генотип). Все иммуногенные области могут происходить из одного и того же или разных вирусов HBV, которые могут быть такими же или могут отличаться от вирусов HBV, из которых получены сердцевинная и полимеразная части. Предпочтительно, все иммуногенные области оболочечной части получены из генотипа D HBV, и особенно из изолята вируса гепатита Y07587 или, альтернативно, из генотипа HBV, распространенного в Китае (например, генотипа В или С).
Преимущественно каждая из одной или более иммуногенных областей включает Т-клеточные эпитопы, специфичные для Т-хелперных клеток (TH) и/или цитотоксических Т-клеток (CTL), которые могут быть ограничены различными антигенами МНС класса I и/или класса II (например, А2, А24, DR, DP и т.д.). Такие эпитопы были описаны в данной области (W093/03764; W094/19011; Desombere et al., 2000, Clin. Exp.Immunol 122:390; Loirat et al., 2000, J. Immunol, 165:4748; Schirmbeck et ah, 2002, J, Immunol 168:6253; Depla et al., 2008, J. Virol 82:435), и специалист сможет разработать подходящую иммуногенную область (области), описанную выше, в диапазоне от 15 до 100 аминокислотных остатков, включающую В-, TH- и/или CTL-эпитопы из нативного оболочечного белка (например, приведенного в SEQ ID №3). Предпочтительно, оболочечная часть, содержащаяся или кодируемая композицией изобретения, не содержит иммуногенную область (области), происходящую из областей preS1 и preS2.
Данное изобретение предусматривает оболочечную часть, содержащую одну иммуногенную область, а также содержащую две, три или более.
Предпочтительно, одна или более иммуногенная область (области), используемая в данном изобретении, выбрана из группы, состоящей из:
- части оболочечного белка (HBsAg), продолжающейся от позиции 14 до 51 (область env1);
- части оболочечного белка (HBsAg), продолжающейся от позиции 165 до 194 (область env2);
- части оболочечного белка (HBsAg), продолжающейся от позиции 81 до 106 (область env3);
- части оболочечного белка (HBsAg), продолжающейся от позиции 202 до 226 (область env4); а также
- их любых комбинаций.
Особенно подходящая иммуногенная область (области), используемая в данном изобретении, включает, альтернативно по существу состоит из или же состоит из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность, и даже более предпочтительно 100% идентичность с любой из аминокислотных последовательностей, приведенных в SEQ ID №12-15.
В контексте изобретения сочетание иммуногенных областей может быть в форме смеси отдельных иммуногенных областей в композиции изобретения или в форме слияния в рамке считывания двух или более иммуногенных областей в любом возможном порядке (например, слияние или смесь env1-env2, env2-env1, env1-env3, env3-env1, env1-env4. env4-env1, env2-env3, env3-env2, env2-env4, env4-env2, env3-env4, env4-env3, env1-env2-env3, env2-env1-env3, env1-env2-env4 и т.д.). Кроме того, сочетание может включать одну копию или несколько копий (например, env1-env-2-env1, env1-env4-env1 и т.д.). Слияние всех иммуногенных областей может быть прямым или через линкер.
Оболочечные части, представляющие особый интерес в контексте изобретения, включают слияние двух или трех иммуногенных областей, приведенных в SEQ ID №12-15, с особым предпочтением слияния env1-env2, включающего, альтернативно по существу состоящего из или же состоящего из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №16, или слияния env1-env2-env4, включающего, альтернативно по существу состоящего из или же состоящего из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №17.
В соответствии с предпочтительным воплощением полимеразная, сердцевинная и/или оболочечная части, содержащиеся или кодируемые композицией изобретения, могут быть слиты в рамке считывания попарно или все вместе. Например, может быть предусмотрено слияние полимеразной и оболочечной части в единую полипептидную цепь. Альтернативно, сердцевинная и оболочечная части могут быть слиты в рамке считывания в единую полипептидную цепь. Опять же, кодирующие нуклеиновокислотные последовательности могут быть слиты либо непосредственно, либо через линкер. Предпочтительно, оболочечная часть слита в рамке считывания с С-концом сердцевинной или полимеразной части.
Предпочтительные примеры гибридных полипептидов сердцевинной части с оболочечной частью выбраны из группы, состоящей из:
- полипептида, включающего, альтернативно по существу состоящего или же состоящего из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №18 (core*t-env1);
- полипептида, включающего, альтернативно по существу состоящего или же состоящего из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №19 (core*t-env1-env2); и
- полипептида, включающего, альтернативно по существу состоящего или же состоящего, из аминокислотной последовательности, которая демонстрирует по меньшей мере 80% идентичность, предпочтительно по меньшей мере 85% идентичность, предпочтительно по меньшей мере 90% идентичность, более предпочтительно по меньшей мере 95% идентичность и даже более предпочтительно 100% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №20 (core-env1-env2-env4), или частью аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251 (core-env1-env2), или частью аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221 (core-env1); или с его делецированной версией, в которой отсутствуют остатки 77-84 в сердцевинной части.
В контексте изобретения полимеразная, сердцевинная и/или оболочечная части, содержащиеся или кодируемые композицией изобретения, могут также содержать дополнительные модификации. Подходящими модификациями являются те, которые положительно влияют на синтез, процессинг, стабильность и растворимость полученного полипептида (например, те, которые направлены на изменение потенциальных сайтов расщепления, потенциальных сайтов гликозилирования и/или мембранного заякоривания, как описано выше), а также те, которые являются выгодными для иммуногенности полученной композиции (например, включение или слияние с одним или более соединением, способным повышать иммуногенные свойства). Такие соединения, способные повышать иммуногенные свойства, были описаны в литературе и включают, но не ограниваясь ими, калретикулин (Cheng et al., 2001, J. Clin. Invest. 108:669), белок теплового шока 70 (HSP70) Mycobacterium tuberculosis (Chen et al., 2000, Cancer Res, 60:1035), убиквитин (Rodriguez et al., 1997, J. Virol. 71:8497), бактериальный токсин, такой как транслоцированная область экзотоксина A (ETA(dIII)) Pseudomonas aeruginosa (Hung et al., 2001 Cancer Res. 61:3698), а также Т-хелперный эпитоп(ы), такой как паптид Pan-Dr (Sidney et al., 1994, Immunity 1:751), эпитоп pstS1 GCG (Vordermeier et al., 1992, Eur, J, Immunol. 22:2631), дифтерийно-столбнячные пептиды Р2ТТ (Panina-Bordignon et al., 1989, Eur. J. Immunol, 19:2237) и РЗОТТ (Demote et al., 1993, Eur, J, Immunol. 23:425), эпитоп вируса гриппа (Lamb et al., 1982, Nature 300:66) и гемагглютининовый эпитоп (Rothbard et al., 1989, Int. Immunol. 1:479).
Нуклеиновокислотная молекула
Данное изобретение также предусматривает выделенные нуклеиновокислотные молекулы, кодирующие независимо или в сочетании полимеразную, сердцевинную и/или оболочечную части, используемые в данном изобретении, а также композиции, содержащие такие нуклеиновокислотные молекулы. Особый интерес представляют:
- нуклеиновокислотные молекулы, которые кодируют полимеразые части, описанные выше, с особым предпочтением тех, которые содержат любую из аминокислотных последовательностей, приведенных в SEQ ID №7, 8, 9, или аминокислотную последовательность, приведенную в SEQ ID №7, с заменой остатка Asp в позиции 494 на остаток His и заменой остатка Glu в позиции 672 на остаток His;
- нуклеиновокислотные молекулы, которые кодируют сердцевинные части, описанные в данном документе, с особым предпочтением тех, которые содержат аминокислотную последовательность, приведенную в SEQ ID №10 или 11;
- нуклеиновокислотные молекулы, которые кодируют оболочечные части, описанные в данном документе, с особым предпочтением тех, которые содержат аминокислотную последовательность, приведенную в SEQ ID №12-17; и
- нуклеиновокислотные молекулы, которые кодируют слитые сердцевинную и оболочечную части, описанные в данном документе, с особым предпочтением тех, которые содержат аминокислотную последовательность, приведенную в любой из SEQ ID №18, 19, 20, или часть аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251 (core-env1-env2), или часть аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221 (core-env1); или их депонированные версии, в которых отсутствуют сердцевинные остатки 77-84 из SEQ ID №20.
Предпочтительно, нуклеиновокислотные молекулы изобретения могут быть оптимизированы для обеспечения высокого уровня экспрессии в конкретной клетке-хозяине или организме, например, млекопитающего, дрожжей (например, Saccharomyces cerevisiae, Saccharornyces pombe или Pichia pastoris) или бактерий (например, E.coll. Bacillus subtilis или Listeria). В самом деле, было отмечено, что когда более чем один кодон доступен для кодирования данной аминокислоты, модели частоты использования кодонов у организмов очень неслучайны (см., например, Wad a et al., 1992, Nucleic Acids Res. 20:2111), и использование кодонов может существенно отличаться у различных хозяев (см., например, Nakamura et al., 1996, Nucleic Acids Res. 24:214). Так как нуклеотидные последовательности, используемые в изобретении, имеют в основном вирусное происхождение (HBV), они могут иметь неподходящие модели частоты использования кодонов для эффективной экспрессии в клетках хозяина, таких как бактериальные, высшие или низшие эукариотические клетки. Как правило, оптимизация кодонов осуществляется путем замены одного или более «нативного» (например, HBV) кодона, соответствующего кодону, редко используемому в интересующей клетке-хозяине, на один или более кодон, кодирующий ту же аминокислоту, который используется чаще. Не обязательно заменять все нативные кодоны, соответствующие нечасто используемым кодонам, так как повышенная экспрессия может быть достигнута даже с частичной заменой. Более того, некоторые отклонения от строгого соблюдения оптимизированного использования кодонов можно сделать для того, чтобы ввести сайт(ы) рестрикции в полученную нуклеиновокислотную молекулу.
В дополнение к оптимизации кодонов, экспрессия в клетке-хозяине или организме может также быть улучшена за счет дополнительных изменений в нуклеотидной последовательности. Например, нуклеиновокислотная молекула изобретения может быть модифицирована таким образом, чтобы предотвратить кластеризацию редких, неоптимальных кодонов, присутствующих в концентрированных областях, и/или подавить или изменить, по меньшей мере частично, отрицательные элементы последовательности, которые, по-видимому, отрицательно влияют на уровни экспрессии. Такие отрицательные элементы последовательности включают, но не ограничиваясь ими, области с очень высоким (>80%) или очень низким (<30%) содержанием GC; АТ-богатые или GC-богатые участки последовательности; нестабильные прямые или инвертированные повторные последовательности; вторичные структуры РНК и/или внутренние скрытые регуляторные элементы, такие как внутренние ТАТА-боксы, chi-сайты, сайты посадки рибосомы и/или донорные/акцепторные сайты сплайсинга. Другое воплощение изобретения относится к фрагментам нуклеиновокислотной молекулы изобретения, например, к фрагментам, образуемым с помощью рестриктаз и ПЦР. Такие фрагменты могут быть использованы в качестве зондов, праймеров или фрагментов, кодирующих иммуногенную часть первого и/или второго полипептида.
Предпочтительная нуклеиновокислотная молекула в соответствии с изобретением выбрана из группы, состоящей из:
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №21 (кодирующей усеченную Pol SEQ ID №7);
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №22 (кодирующей усеченную Pol SEQ ID №8);
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №21, с заменой нуклеотида G в позиции 1480 на С, нуклеотида G в позиции 2014 на С и нуклеотида А в позиции 2016 на Т (кодирующей усеченную Pol SEQ ID №7 с мутациями D540H и Е718Н);
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №23 (кодирующей усеченную и мутированную Pol-TMR SEQ ID №9);
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №24 (кодирующей core*t-env1 SEQ ID №18);
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №25 (кодирующей core*t-env1-env2 SEQ ID №19); и
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №26 (кодирующей core*t-env1-env2-env4 SEQ ID №20), или с частью нуклеотидной последовательности, приведенной в SEQ ID №26, начиная с нуклеотида 1 и заканчивая нуклеотидом 753 (кодирующей core*t-env1-env2), или с частью нуклеотидной последовательности, приведенной в SEQ ID №26, начиная с нуклеотида 1 и заканчивая нуклеотидом 663 (кодирующей core*t-env1); или с их депонированными версиями, в которых отсутствует часть от G в позиции 229 до А в позиции 252 SEQ ID №26 (что соответствует удалению остатков 77-84 в сердцевинной части).
Нуклеиновокислотные молекулы данного изобретения могут быть созданы с помощью данных последовательности, доступных в данной области, и информации о последовательности, приведенной в данном документе. Последовательность ДНК, кодирующая каждый из полипептидов HBV, может быть выделена непосредственно из HBV-содержащих клеток, библиотек кДНК и геномных библиотек, вирусных геномов или любых имеющихся векторов, о которых известно, что они включают ее, с помощью обычных методик молекулярной биологии или ПЦР, и может быть изменена (например, как описано в данном документе). Кроме того, нуклеиновокислотная молекула изобретения также может быть получена путем химического синтеза в автоматизированном процессе (например, собрана из перекрывающихся синтетических олигонуклеотидов, как описано, например, в Edge, 1981, Nature 292, 756; Nambair et al., 1984, Science 223:1299; Jay et al., 1984, J. Biol, Chem. 259:6311).
Кроме того, данным изобретением предусмотрены векторы, содержащие одну или более нуклеиновокислотную молекулу изобретения, а также композиции, содержащие такой вектор(ы).
В контексте данного изобретения могут быть использованы различные системы векторов-хозяев, в том числе бактериофаговые, плазмидные или космидные векторы, адаптированные к экспрессии в прокариотическом организме-хозяине, таком как бактерии (например, £. coil, Bacillus subtilis или Listeria); векторы, адаптированные к экспрессии в дрожжах (например, Saccharomyces cerevisiae, Saccharomyces pombe, Pichia pastoris); вирусные экспрессионные векторы (например, бакуловирусные), адаптированные к экспрессии в клеточных системах насекомых (например, клетках Sf 9); вирусные или плазмидные экспрессионные векторы (например, Ti-плазмида, вирус мозаики цветной капусты CaMV; вирус табачной мозаики TMV), адаптированные к экспрессии в системах растительных клеток; а также плазмидные и вирусные векторы, адаптированные к экспрессии в клетках или организмах высших эукариотов. Такие векторы в основном описаны в литературе и коммерчески доступны (например, от Stratagene, Amersham Bioscienees, Promega и т.д.). Типичные примеры подходящих плазмидных векторов включают, но не ограничиваясь ими, pREP4, pCEP4 (Invitrogene), PCI (Promega), pCDM8 (Seed, 1987, Nature 329, 840) и рМТ2РС (Kaufman et al., 1987, EMBO J. 6:187), pVAX и pgWiz (Gene Therapy System Inc; Himoudi et al., 2002, J. Virol. 76:12735). В контексте данного изобретения также может быть использовано большое количество вирусных векторов, полученных из различных вирусов (например, ретровируса, аденовируса, AAV, поксвируса, вируса герпеса, вируса кори, пенистого вируса, альфавируса, вируса везикулярного стоматита и т.п.).
Особый интерес представляют аденовирусные векторы, которые обладают целым рядом хорошо задокументированных преимуществ для генного переноса или рекомбинантной продукции (см. обзор в Adenoviral vectors for gene therapy, 2002, Ed D, Curiel and J. Douglas, Academic Press). Аденовирусные векторы для применения в соответствии с данным изобретением могут быть получены из разнообразных источников человеческого или животного происхождения (например, аденовирус собак, овец, обезьян и т.д.). Может быть использован любой серотип с особым предпочтением человеческих аденовирусов и особенно с предпочтением аденовирусов подрода С, таких как Ad2 (Ad2), 5 (Ad5), 6 (Ad6), подрода В, таких как 11 (Ad11), 34 (Ad34) и 35 (Ad35), и подрода D, таких как 19 (Ad 19), 24 (Ad24), 48 (Ad48) и 49 (Ad49). Также может быть выгодным применение Ad животных с особым предпочтением Ad шимпанзе, таким как Ad3 шимпанзе (Peruzzi et al., 2009, Vaccine 2.7:1293) и Ad63 шимпанзе (Dudareva et al., 2009, vaccine 27:3501). Упомянутые аденовирусы доступны из Американской коллекции типовых культур (АТСС, Роквилл, штат Мэриленд) или были объектом многочисленных публикаций, описывающих их последовательность, организацию и способы производства, что позволяет производителям их применять (см., например, US 6133028; US 6110735; WO 02/40665; WO 00/50573; EP 1016711; Vogels et al., 2003, J. Viral. 77:8263; WO 00/70071; WO 02/40665; WO 2004/001032; WO 2004/083418; WO 2004/097016; WO 2005/010149).
В одном воплощении аденовирусной вектор данного изобретения является репликативно-дефектным. Предпочтительными репликативно-дефектными аденовирусными векторами являются El-дефектные (см., например, US 6136594 и US 6013638) с El-делецией, которая простирается от приблизительно позиции 459 до 3328 или от приблизительно позиции 459 до 3510 (со ссылкой на последовательность человеческого аденовируса типа 5, раскрытую в GeneBank под номером М 73260 и в Chroboczek et al., 1992, Virol. 186:280). Мощность клонирования также может быть повышена путем удаления дополнительной части (частей) аденовирусного генома (полностью или часть несущественной области ЕЗ или других важных областей Е2, Е4, как описано в WO 94/28152; Lusky et al., 1998, J, Virol 72:2022).
Нуклеиновокислотная молекула(ы) данного изобретения может быть вставлена в любое место аденовирусного генома, с особым предпочтением для вставки El-области. Она/они могут быть расположены в смысловой или антисмысловой ориентации по отношению к природному транскрипционному направлению обсуждаемой области.
Другие подходящие вирусные векторы в контексте изобретения получены из поксвирусов (см., например, Сох et al., in "Viruses in Human Gene Therapy" Ed J.М.Bos, Carolina Academic Press). В контексте данного изобретения поксвирусный вектор может быть получен из любого члена Poxviridae, в частности вируса оспы канареек, вируса оспы кур и вируса осповакцины, причем последняя предпочтительнее. Подходящие вирусы осповакцины включают, но не ограничиваясь ими, штамм Copenhagen (Goebel et al., 1990, Virol. 179:247 and 517;
Johnson et al., 1993, Virol. 196:381), штамм Wyeth и модифицированный штамм Анкара (MVA) (Antoine et al., 1998, Virol. 244:365). Общие условия для построения рекомбинантного поксивируса хорошо известны в данной области (см., например, EP 206 920; Mayr et al., 1975, Infection 3:6; Sutter and Moss, 1992, Proe. Natl. Acad. Sci. USA 89:10847; US 6440422). Нуклеиновокислотная молекула данного изобретения предпочтительно вставляется в поксвирусный геном в несущественных локусах. Тимидинкиназный ген является особенно подходящим для включения в векторы на основе вируса осповакцины Copenhagen (Hruby et al., 1983, Proc. Natl. Acad. Sci USA 80:3411; Weir et ah, 1983, J. Virol. 46:530), и делеция II или III для включения в вектор MVA (Meyer et al., 1991, J.Gen, Virol. 72:1031; Sutler et al., 1994, Vaccine 12:1032).
Данное изобретение также предусматривает векторы (например, плазмидные ДНК) в комплексе с липидами или полимерами для формирования структурных частиц, таких как липосомы, липоплексы или наночастицы. Такие технологии имеются в данной области (см., например, Arangoa et al., 2003, Gene Ther. 10:5; Eliaz et al., 2002, Gene Ther. 9:1230 и Betageri et al., 1993, "Liposome drug delivery 5 systems", Technornic Publishing Company, Inc).
В соответствии с предпочтительным воплощением векторы изобретения содержат нуклеиновокислотную молекулу (молекулы) изобретения в форме, пригодной для экспрессии в клетке-хозяине или организме, что означает, что Нуклеиновокислотная молекула(ы) находится под контролем одной или более регуляторной последовательности, соответствующей вектору и/или клетке-хозяину. Специалисты в этой области примут во внимание, что выбор регуляторных последовательностей может зависеть от таких факторов, как клетка-хозяин, необходимый уровень экспрессии и т.д.
Промотор имеет особое значение, и подходящие промоторы, используемые в контексте данного изобретения, включают конститутивные промоторы, которые направляют экспрессию нуклеиновокислотной молекулы (молекул) во многих типах клеток-хозяев, и те, которые направляют экспрессию нуклеиновокислотной молекулы (молекул) в определенных клетках-хозяевах (например, специфичные для печени регуляторные последовательности) или в ответ на определенные события или экзогенные факторы (например, температуру, питательные добавки, гормоны или другие лиганды).
Промоторы, подходящие для конститутивной экспрессии в клетках млекопитающих, включают, но не ограничиваясь ими, цитомегаловирусный (CMV) немедленный ранний промотор (Boshart et al., 1985, Cell 41:521), промотор RSV, аденовирусный основной поздний промотор, фосфоглицерокиназный (PGK) промотор (Adra et al., 1987, Gene 60:65) и тимидинкиназный (ТК) промотор вируса простого герпеса (HSV)-1. Промоторы вируса осповакцины особенно приспособлены для экспрессии поксвирусных векторов. Типичые примеры включают, но не ограничиваясь ими, промоторы вируса осповакцины 7.5К, H5R, 11К7.5 (Erbs et al., 2008, Cancer Gene Ther. 15:18), TK, p28, p11 и K1L, а также синтетические промоторы, такие как описаны в Chakrabarti et al. (1997, Biotechniques 23:1094, в связи с промотором pSE/L), Hammond et al. (1997, J. Virological Methods 66:135) и Kumar and Boyle (1990, Virology 179:151), а также ранний/поздний химерные промоторы. Специфичные для печени промоторы включают, но не ограничиваясь ими, промоторы генов ГМГ-КоА-редуктазы (Luskey, 1987, MoL Cell. Biol 7:1881); стирол-регулирующего элемента 1 (SRE-1, Smith et al., 1990, J.Biol. Chem. 265:2306); альбумина (Pinkert et al., 1987, Genes Dev. 1:268);
фосфоенолпируваткарбоксикиназы (PEPCK) (Eisenberger et al., 1992, Mol. Cell Biol. 12:1396); человеческого С-реактивного белка (CRP) (Li et al., 1990, J. Biol. Chem. 265:4136); человеческой глюкокиназы (Tanizawa et al., 1992, Mol. Endocrinology 6:1070); холестерин-7-альфа-гидролазы (CYP-7) (Lee et al., 1994, J. Biol. Chem. 269:14681), альфа-1-антитрипсина (Ciliberto et al., 1985, Cell 41:531), инсулиноподобного ростового фактора (IGFBP-1) (Babajko et al., 1993, Biochem Biophys. Res. Comm. 196:480); человеческого трансферрина (Mendelzon et al., 1990, Nucleic Acids Res. 18:5717); коллагена типа I (Houglum et al., 1994, J. Clin. Invest. 94:808) и FIX (US 5814716).
Специалистам в данной области будет понятно, что регуляторные элементы, контролирующие экспрессию нуклеиновокислотной молекулы (молекул) изобретения, могут также содержать дополнительные элементы для надлежащей инициации, регуляции и/или терминации транскрипции (например, полиА-последовательности для терминации транскрипции), транспорта мРНК (например, сигнальные последовательности ядерной локализации), процессинга (например, сигналы сплайсинга), а также стабильности (например, интроны и некодирующие 5'- и 3'-последовательности), трансляции (например, инициатор Met, трехчастные лидерные последовательности, рибосом-связывающие сайты, последовательности Шайна-Дальгарно и т.д.) в клетке-хозяине или организме и на этапах очистки (например, метки).
В соответствии с данным изобретением нуклеиновокислотные молекулы данного изобретения, кодирующие указанную полимеразную часть, указанную сердцевинную часть и/или указанную оболочечную часть, могут быть осуществлены одним и тем же вектором или по меньшей мере двумя векторами (например, двумя или тремя независимыми векторами). Таким образом, данное изобретение предусматривает вектор, который несет нуклеиновокислотные молекулы, кодирующие указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть, а также независимые векторы, каждый из которых несет только одну или две нуклеиновокислотные молекулы, кодирующие указанную полимеразную часть указанную сердцевинную часть и/или указанную оболочечную часть. Такой вектор(ы) также предусмотрен данным изобретением, как и композиции, содержащие такой вектор(ы). При использовании различных векторов они могут иметь различное или одно и то же происхождение. Например, можно представить экспрессию одной HBV-части из дефектного поксвируса (например, MVA) и экспрессию двух других частей из другого поксвирусного вектора (например, вектора Copenhagen). В качестве другого примера можно представить композицию, включающую аденовирусный вектор, кодирующий полимеразную часть, и аденовирусный вектор, кодирующий сердцевинную часть и оболочечную часть. Кроме того, экспрессия из различных вирусных векторов (например, экспрессия полимеразной части из аденовирусного вектора и экспрессия сердцевинной и/или оболочечной части из MVA, или наоборот) также входит в контекст изобретения, как и экспрессия HBV-частей из плазмидных и вирусных векторов.
Предпочтительные воплощения изобретения направлены на векторы, выбранные из группы, состоящей из:
(i) вектора MVA, включающего нуклеиновокислотную молекулу, находящуюся под контролем промотора вируса осповакцины, такого как промотор 7.5К, и кодирующую полимеразную часть, содержащую аминокислотную последовательность, приведенную в SEQ ID №7, 8 или 9 или в SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и с заменой остатка Glu в позиции 672 на остаток His. Предпочтительно, указанная нуклеиновокислотная молекула вставлена в делецию III генома MVA.
(ii) вектора MVA, включающего нуклеиновокислотную молекулу, находящуюся под контролем промотора вируса осповакцины, такого как промотор рН5 г, и кодирующую сердцевинную часть и оболочечную часть, включая аминокислотную последовательность, приведенную в SEQ ID №18 или 19. Предпочтительно, указанная нуклеиновокислотная молекула вставлена в делецию III генома MVA.
(iii) E1-дефектный Ad-вектор, содержащий вставленную вместо области Е1 нуклеиновокислотную молекулу, которая находится под контролем промотора CMV и кодирует полимеразную часть, включая аминокислотную последовательность, приведенную в SEQ ID №7, 8 или 9 или в SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и заменой остатка Glu в позиции 672 на остаток His.
(iv) E1-дефектный Ad-вектор, содержащий вставленную вместо области Е1 нуклеиновокислотную молекулу, которая находится под контролем промотора CMV и кодирует сердцевинную часть и оболочечную часть, включая аминокислотную последовательность, приведенную в SEQ ID №18, 19, 20, или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251 (core-env1-env2), или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221 (core-env1),
а также к композиции, содержащей векторы (i) и (ii) или (iii) и (iv).
При необходимости вектор или композиция изобретения может также содержать один или более трансген, например интересующий ген, который будет экспрессирован вместе с нуклеиновокислотной молекулой(ами) изобретения в клетке-хозяине или организме-хозяине. Предпочтительно, экспрессия трансгена имеет терапевтическую или защитную активность по отношению к HBV-инфекции или заболеванию или состоянию, вызванному или связанному с HBV-инфекцией, или способна повышать иммуногенность композиции изобретения. Подходящие трансгены включают, но не ограничиваясь ими, один или более дополнительный HBV-полипептид/пептид или кодирующую нуклеиновокислотную молекулу(ы), например белок Х или его фрагмент, иммуномодуляторы, такие как цитокины (например, IL-2, IL-7, IL-12, IL-15, IL-18, IL-21, IFNγ), гибридные цитокины (такие как описаны в WO 2005/14642) и кодирующие их нуклеиновокислотные молекулы, а также продукты суицидальных генов или кодирующих нуклеиновокислотных молекул, особенно используемых в контексте лечения карциномы печени (например, цитозиндезаминазу (CDase), урацилфосфорибозилтрансферазу (UPRTase), продукт гена FCU-1 (описан в WO 99/54481) и его производные (описаны в WO 2006/048768), которые используют с предшественником 5-фторцитозином (5-FC). Если используется трансген, он может быть экспрессирован в форме, слитой с любой нуклеиновокислотной молекулой изобретения, или может быть экспрессирован самостоятельно под контролем соответствующих регуляторных элементов. Кроме того, он может быть вставлен в любое место в векторе изобретения или в независимый вектор, который используется в комбинации с вектором(ами) или композицией изобретения.
В другом аспекте данное изобретение предусматривает инфекционные вирусные частицы, содержащие нуклеиновокислотные молекулы или векторы данного изобретения, а также композиции, содержащие такие инфекционные вирусные частицы.
Как правило, такие вирусные частицы получают в процессе, включающем следующие этапы:
(а) введение вирусного вектора изобретения в подходящую клеточную линию,
(б) культивирование указанной клеточной линии в условиях, подходящих для продукции указанной инфекционной вирусной частицы,
(в) восстановление полученной инфекционной вирусной частицы из культуры указанной клеточной линии, и
(г) возможно, очистка указанной восстановленной инфекционной вирусной частицы.
Когда вирусный вектор является дефектным, инфекционные частицы, как правило, производятся в комплементационной клеточной линии или через применение вируса-помощника, который поставляет in trans нефункциональные вирусные гены. Например, подходящие клеточные линии для дополнения аденовирусного вектора с Е1-делецией включают клетки 293 (Graham et al., 1997, J. Gen. Virol. 36, 59-72), а также клетки HER-96 и PER-C6 (например, Fallaux et al., 1998, Human Gene Ther. 9, 1909-1917; WO 97/OQ326). Клетки, подходящие для размножения поксвирусных векторов, являются птичьими клетками, и наиболее предпочтительны первичные фибробласты куриного эмбриона (CEF), изготовленные из куриных эмбрионов, полученных из оплодотворенных яиц.
Инфекционные вирусные частицы могут быть восстановлены из культурального супернатанта или из клеток после лизиса. Они могут быть также очищены в соответствии со стандартными методиками (хроматография, ультрацентрифугирование в градиенте хлорида цезия, как описано, например, в WO 96/27677, WO 98/00524, WO 98/22588, WO 98/26048, WO 00/40702, EP 1016700 и WO 00/50573).
Данное изобретение также предусматривает векторы или вирусные частицы, которые были модифицированы для предпочтительного направления в конкретные клетки-мишени (см., например Wickam et al., 1997, J. Virol. 71, 8221-8229; Arnberg et al., 1997, Virol. 227, 239-2.44; Michael et al., 1995, Gene Therapy 2, 660-668; WO 94/10323; WO 02/96939 и EP 1146125). Характерной чертой целевых векторов и вирусных частиц изобретения является наличие на их поверхности лигандов, способных распознавать и связывать клеточные и поверхностно-экспонированные компоненты, такие как клеточный специфический маркер (например, на HBV-инфицированных клетках), тканеспецифический маркер (например, печеночный специфический маркер), а также вирусные (например, HBV) антигены. Примеры подходящих лигандов включают антитела или их фрагменты, направленные на антигенный домен HBV. Нацеливание клеток может быть осуществлено путем генетической вставки лиганда в полипептид, представленный на поверхности вируса (например, аденовирусные волокна, пентон, plX или продукт гена р14 вируса осповакцины).
Данное изобретение также предусматривает клетки-хозяева, которые содержат нуклеиновокислотные молекулы, векторы или инфекционные вирусные частицы изобретения, а также композиции, содержащие такие клетки-хозяева. В контексте изобретения клетки-хозяева включают прокариотические клетки, низшие эукариотические клетки, такие как дрожжи, и другие эукариотические клетки, такие как клетки насекомых, растений и млекопитающих (например, человеческие или нечеловеческие), а также дополняющие клетки, способные дополнять по меньшей мере одну дефектную функцию репликативно-дефектного вектора изобретения (например, аденовирусного вектора), такие как клетки 293 и PERC.6.
В соответствии с конкретным воплощением изобретения клетки-хозяева также могут быть инкапсулированы. Технологии клеточной инкапсуляции были описаны ранее (Tresco et al., 1992, ASAIO J. 38, 17-23; Aebischer et al., 1996, Human GeneTher.7.851-860).
Еще одним аспектом данного изобретения является способ получения рекомбинантной полимеразной, сердцевинной и/или оболочечной части с использованием векторов, инфекционных вирусных частиц и/или клеток-хозяев изобретения. Способ данного изобретения включает (а) введение вектора или инфекционной вирусной частицы изобретения в подходящую клетку-хозяина для получения трансфицированной или инфицированной клетки-хозяина, (б) культивирование in vitro указанной трансфицированной или инфицированной клетки-хозяина в условиях, подходящих для роста клетки-хозяина, (в) восстановление полимеразной, сердцевинной и/или оболочечной части из культуры клеток, и (г) возможно, очистку восстановленного полипептида(ов).
Ожидается, что специалисты в данной области хорошо осведомлены о многочисленных доступных экспрессионных системах для продукции HBV-части (частей) в соответствующих клетках-хозяевах и о способах введения вектора или инфекционной вирусной частицы в клетку-хозяина. Такие способы включают, но не ограничиваясь ими, микроинъекцию (Capechi et al., 1980, Cell 22:479), CaPO4-опосредованную трансфекцию (Chen and Qkayama, 1987, Mol. Cell Biol. 7:2745), DEAE-декстран-опосредованную трансфекцию, электропорацию (Chu et al., 1987, Nucleic Acid Res. 15:1311), липофекцию/липосомное слияние (Feigner et al., 1987, Proc. Natl. Acad. Sci. USA 84:7413), бомбардировку частицами (Yang et al., 1990, Proc, Natl. Acad. Sci. USA 87:9568), генные пушки, трансдукцию, вирусное инфицирование, а также прямое введение в организм-хозяин с помощью различных средств.
Векторы изобретения для облегчения введения вектора в клетку-хозяина могут быть использованы в сочетании с реагентами для трансфекции, такими как поликатионные полимеры (например, хитозан, полиметакрилат, PEI (полиэтиленимин) и т.д.) и катионные липиды (например, DC-Chol/DOPE, липофектамин для трансфекции Transfectam, теперь доступный от Promega). Кроме того, как уже говорилось выше, технологии рекомбинантной ДНК могут быть использованы для повышения экспрессии нуклеиновокислотной молекулы (молекул) в клетке-хозяине или организме-хозяине, например, с помощью высококопийных векторов, замены или изменения одной или более транскрипционной регуляторной последовательности (например, промотора, энхансера и т.п.), оптимизации частоты использования кодонов в нуклеиновокислотной молекуле (молекулах) в клетке-хозяине, и супрессии негативных последовательностей, которые могут дестабилизировать траснкрипт.
Клетки-хозяева данного изобретения можно культивировать в обычных ферментативных биореакторах, флаконах и, чашках Пети. Культивирование может осуществляться при температуре, рН и содержании кислорода, подходящем для данной клетки-хозяина. В данном документе не будет дано подробного описания различных способов, известных для продукции белков в прокариотических и эукариотических клетках.
HBV-часть (части) может быть очищена с помощью хорошо известных способов очистки, включая осаждение сульфатом аммония, экстракцию кислотой, гель-электрофорез; фильтрацию и хроматографические способы (например, обратно-фазовую, гель-проникающую, ионнообменную, аффинную, фосфоцеллюлозную, гидрофобных взаимодействий, гидроксиапатитную или высокоэффективную жидкостную хроматографию). Используемые условия и технология будут зависеть от таких факторов, как результирующий заряд, молекулярная масса, гидрофобность, гидрофильность, и будут очевидны специалистам в данной области. Кроме того, уровень очистки будет зависеть от предполагаемого применения.
В другом аспекте данное изобретение предусматривает композицию, содержащую по меньшей мере одну из полимеразной, сердцевинной и/или оболочечной частей, кодирующие нуклеиновокислотные молекулы, вектор(ы), инфекционные вирусные частицы или клетку-хозяина изобретения (также называемую в данном документе «активным агентом») или любую их комбинацию (например, комбинацию полипептидов или векторов/вирусных частиц, кодирующих различные HBV-части, описанные в данном документе, или комбинацию различных генотипов).
Предпочтительно, композиция представляет собой фармацевтическую композицию, которая содержит фармацевтически приемлемый носитель в дополнение к эффективному количеству активного агента(ов).
Соответственно, композиция изобретения включает разбавитель, подходящий для применения у человека или животного. Предпочтительно он является изотоническим, гипотоническим или слабо гипертоническим и имеет относительно низкую ионную силу. Типичные примеры включают стерильную воду, физиологический раствор (например, хлорид натрия), раствор Рингера, растворы глюкозы, трегалозы или сахарозы, раствор Хэнка и другие водные физиологически сбалансированные солевые растворы (см., например, последнее издание Remington: The Science and Practice of Pharmacy, A. Gennaro, Lippincott, Williams&Wilkins). Композиция изобретения соответствующим образом буферизована, чтобы подходить для применения у человека, до физиологического или слабощелочного рН (например, от приблизительно рН 7 до приблизительно рН 9). Подходящие буферы включают, но не ограничиваясь ими, фосфатный буфер (например, PBS), бикарбонатный буфер и/или Трис-буфер.
Композиция также может содержать другие фармацевтически приемлемые эксципиенты для обеспечения предпочтительных фармацевтических или фармакодинамических свойств, включая, например, изменение или поддержание рН, осмолярности, вязкости, прозрачности, цвета, стерильности, стабильности, скорости растворения состава, изменение или поддержание высвобождения или абсорбции в организм человека или животного, помощь в транспорте через барьер или в проникновении в тот или иной орган (например, печень). Подходящие эксципиенты включают аминокислоты.
Фармацевтически приемлемые носители, включенные в композицию изобретения, должны также позволять сохранять стабильность в условиях производства и длительного хранения (например, по меньшей мере в течение одного месяца) при замораживании (например, при -70ºС, -20ºС), охлаждении (например, при 4ºС) или при температуре окружающей среды. В связи с этим составы, которые особенно адаптированы к композиции изобретения, включают:
- 1М сахарозу, 150 мМ NaCl, IMM MgCl2, 54 мг/л Твина 80, 10 мМ Трис с рН 8,5 (особенно если активным агентом является аденовирусный вектор), и
- 10 мг/мл маннитола, 1 мг/мл HSA, 20 мМ Трис с рН 7,2 и 150 мМ NaCl,
- физиологический раствор.
Кроме того, композиция изобретения может включать один или более адъювант, подходящий для применения у человека системно или на слизистой. Предпочтительно адъювант способен стимулировать иммунитет к композиции изобретения, особенно Т-клеточный иммунитет, например через Toll-подобные рецепторы (TLR), такие как TLR-7, TLR-8 и TLR-9. Типичные примеры используемых адъювантов включают, но не ограничиваясь ими, алюминий, эмульсии минеральных масел, такие как полный и неполный адъювант Фрейнда (IFA), липополисахарид или его производное (Ribi et al., 1986, Immunology and Immunopharmacology of Bacterial Endotoxins, Plenum Publ. Corp., NY, p407-419), сапонины, такие как QS21 (Sumino et al., 1998, J.Virol. 72:4931; WO 98/56415), имидазо-хинолиновые соединения, такие как Imiquimod (Suader, 2000, J. Am Acad Dermatol. 43:S6), S-27609 (Smorlesi, 2005, Gene Ther. 12:1324) и родственные соединения, такие как описанные в WO 2007/147529, цитозин-фосфат-гуанозин олигодезоксинуклеотиды, такие как CpG (Chu et al., 1997, J. Exp.Med. 186:1623; Tritel et al., 2003, J. Immunol. 171:2358), и катионные пептиды, такие как 1С-31 (Kritsch et al., 2005, J. Chromatogr Anal. Technol Biomed Life Sci 822:263).
Композиция данного изобретения подходит для различных режимов введения, в том числе системного, местного и локализованного введения. Инъекция может быть выполнена с помощью любого средства, например путем подкожной, внутрикожной, внутримышечной, внутривенной, внутрибрюшинной, внутриопухолевой, внутрисосудистой, внутриартериальной инъекции или путем прямого введения в артерию (например, путем инфузии в печеночную артерию) или печеночную вену (например, путем инъекции в воротную вену). Инъекции могут быть выполнены с помощью обычных шприцев и игл, или с помощью любых других соответствующих устройств, доступных в данной области. Кроме того, композиция данного изобретения может быть введена через слизистую, например перорально/алиментарно, интраназально, интратрахеально, внутрилегочно, интравагинально или ректально. Введение в дыхательные пути может осуществляться посредством распыления или аэрозолизации капель, спрея или сухих порошковых композиций с помощью напорных емкостей (например, с подходящим пропеллентом, таким как дихлордифторметан, пропан, азот и т.п.) или ненапорных дозаторов. Местное введение также может быть выполнено с использованием трансдермальных средств (например, пластырей и т.п.). В контексте изобретения предпочтительная композиция составлена для внутримышечного и подкожного путей.
Композиция изобретения может быть в различных формах, например твердой, жидкой или замороженной. Твердые (например, сухие порошковые или лиофилизированные) композиции могут быть получены в процессе, включающем вакуумную сушку и сублимационную сушку. Для введения через слизистую композиции могут быть составлены как гастрорезистентные капсулы и гранулы для перорального приема, суппозитории для ректального или вагинального введения, наконец, в сочетании с усилителями абсорбции, использующимися для увеличения размера пор слизистых оболочек. Такие усилители абсорбции, как правило, представляют собой вещества, имеющие структурное сходство с фосфолипидными областями слизистых оболочек, например дезоксихолат натрия, гликохолат натрия, диметил-бета-циклодекстрин, лаурил-1-лизофосфатидилхолин).
Подходящая дозировка может быть адаптирована в зависимости от различных параметров, в частности, от способа введения; используемой композиции; возраста, состояния здоровья и веса организма-хозяина; характера и степени проявления симптомов; вида одновременного лечения; частоты лечения; и/или необходимости профилактики или лечения. Дальнейшее уточнение расчетов, необходимых для определения соответствующей дозировки для лечения, проводится стандартным образом врачом с учетом соответствующих обстоятельств. В качестве общего руководства, подходящая доза композиции, содержащей вирус, варьирует от примерно 105 до примерно 1013 vp (вирусных частиц), iu (инфекционных единиц) или pfu (бляшкообразующих единиц) в зависимости от используемого вектора и методик количественной оценки. Методики, доступные для оценки количества vp, iu и pfu, присутствующих в образце, являются стандартными в этой области. Например, количество аденовирусных частиц (vp) обычно определяют путем измерения абсорбции при А260, титры iu оценивают путем количественной иммунофлуоресценции DBP, а pfu путем подсчета количества бляшек после инфицирования пермиссивных (чувствительных) клеток. Предпочтительно, соотношение vp/iu составляет менее 100 в соответствии с принципами FDA. Дозы от примерно 5×105 до примерно 109 pfu являются предпочтительными для основанной на MVA композиции, с особым предпочтением доз примерно 107, примерно 5×107, примерно 108 или примерно 5×108 pfu. Что касается композиций на основе Ad, предпочтительные дозы включают от 106 до 1012 vp, с особым предпочтением доз примерно 109, примерно 5×109, примерно 1010, примерно 5×1010 или примерно 1011 vp. Композиция на основе векторных плазмид может быть введена в дозах от 10 мкг до 20 мг, предпочтительно от 100 мкг до 2 мг. Белковая композиция может быть введена в одной или более дозах от 10 нг до 20 мг, с особым предпочтением дозировки от примерно 0,1 мкг до примерно 2 мг терапевтического белка на кг массы тела. Введение можно проводить в виде разовой дозы или дозы, повторяющейся один или несколько раз через определенный промежуток времени.
Композиция изобретения может быть использована в способах лечения различных заболеваний и патологических состояний, особенно тех, которые вызваны или связаны с HBV-инфекцией. Используемый в данном документе термин «лечение» включает профилактику и/или терапию. Особенно он используется для лечения хронической HBV-инфекции и/или поражений печени у HBV-инфицированных пациентов, включая цирроз и рак печени. Предпочтительно, при введении в организм хозяина в соответствии с порядком, изложенным в данном документе, композиция изобретения обеспечивает терапевтическое улучшение для подвергающегося лечению хозяина по сравнению с состоянием до лечения. Терапевтическое улучшение может быть доказано несколькими способами, например, снижением вирусной нагрузки HBV, выявленной в крови, плазме, сыворотке или печени инфицированного субъекта, и/или выявлением анти-HBV-иммунного ответа (например, продукции анти-HBV-антител и/или Т-клеточного иммунитета), или замедлением развития симптомов, связанных с HBV-инфекцией (например, задержкой развития цирроза печени или рака), или уменьшением состояния воспаления/стеатоза/фиброза печени, обычно связанного с HBV-инфекцией, или улучшением реакции индивидуума на стандартные способы лечения.
Соответственно, данное изобретение также предусматривает применение по меньшей мере одной из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев или композиций изобретения для изготовления препарата, предназначенного для лечения или профилактики HBV-инфекции, HBV-ассоциированных заболеваний и патологических состояний в соответствии с порядком, изложенным в данном документе.
Данное изобретение также предусматривает способ лечения или профилактики HBV-инфекции, в частности хронической HBV-инфекции, HBV-ассоциированных заболеваний и патологических состояний, который включает введение в организм человека или животного, нуждающегося в этом, терапевтически эффективного количества по меньшей мере одной из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев или композиций изобретения.
Способ или применение изобретения включает одно или более введение (1, 2, 3, 4, 5, 6, 7, 8, 9, 10 и т.д.) терапевтически эффективного количества указанного активного агента (агентов), при этом указанные введения отделены друг от друга соответствующим промежутком времени и осуществляются одним и тем же или различными путями введения (например, внутримышечным и подкожным путем), в одно и то же или в разные места. Три введения, отделенные друг от друга промежутком времени от 3 до 10 дней (например, три еженедельных введения), особенно подходят для композиций и вектора (векторов) на основе MVA. За этой первой серией введений может следовать одно или более последующих введений с применением тех же активных агентов (агента), которые могут осуществляться в течение одного или нескольких месяцев, чтобы вызвать анти-HBV-иммунный ответ, начатый тремя последовательными введениями. Что касается композиций и вектора(ов) на основе Ad, предпочтительный способ или применение включает одно введение, за которым в конце концов следует одно или два последующих введения через один месяц или шесть месяцев.
При необходимости способ или применение изобретения может быть осуществлено в сочетании с одним или более традиционным терапевтическим способом (например, радиацией, химиотерапией и/или хирургическим вмешательством). Применение множественных терапевтических подходов предусматривает пациента с более широким вмешательством. В одном воплощении способ изобретения может следовать до или после хирургического вмешательства. В другом воплощении он может следовать до или после лучевой терапии (например, гамма-излучения). Специалисты в данной области смогут легко разработать соответствующие протоколы лучевой терапии и используемые параметры (см., например, Perez and Brady, 1992, Principles and Practice of Radiation Oncology, 2nd Ed, JB Lippincott Co; с использованием соответствующих адаптации и изменений, которые будут очевидны специалистам в данной области).
В еще одном воплощении способ или применение изобретения связаны с химиотерапией одним или более HBV-препаратами, которые обычно используются для лечения или профилактики HBV-инфекций, HBV-ассоциированных заболеваний и патологических состояний. Их введение может предшествовать, сопутствовать или следовать за введением активного агента, используемого в изобретении. Типичные примеры HBV-препаратов включают, но не ограничиваясь ими, ингибиторы полимеразы, ингибиторы РНКазы Н, нуклеозидные аналоги, нуклеотидные аналоги, TLR-агонисты, ингибиторы N-гликозилирования, миРНК, антисмысловые олигонуклеотиды, анти-HBV-антитела, иммуномодуляторы, терапевтические вакцины и противоопухолевые агенты, которые обычно используются в лечении HBV-ассоциированных раков печени (например, адриамицин, адриамицин с липиодолом или сорафенибом). Примеры подходящих терапевтических вакцин включают, но не ограничиваясь ими, рекомбинантные антигены, вирус-подобные частицы, векторы или синтетические пептиды, основанные на HBV-белках или кодирующие их (core, preS1, preS2, S и/или полимераза), которые особенно подходят для запуска анти-HBV гуморального иммунного ответа. Такие HBV-препараты могут быть предоставлены в виде разовой дозы или, наоборот, в множественных дозах в соответствии со стандартными протоколами, дозировками и схемами на несколько часов, дней и/или недель. Особенно подходящий способ или применение изобретения используется в сочетании со стандартной медицинской помощью, которая может следовать раньше, параллельно или последовательно со способом или применением изобретения. Хотя такой стандарт лечения может изменяться от пациента к пациенту, он обычно включает лечение цитокинами (например, IFNα, пегилированным IFNα2) и/или нуклеотидными или нуклеозидными аналогами, такими как ламивудин, энтекавир, телбивудин, адефовир, дипивоксил или тенофовир.
В другом воплощении способ или применение изобретения осуществляется в соответствии с терапевтическим способом первого бустирования, который включает последовательное введение одной или более начальной композиции и одной или более бустирующей композиции. Как правило, начальные и бустирующие композиции используют различные носители, которые содержат или кодируют по меньшей мере антигенный домен в целом. Кроме того, начальные и бустирующие композиции могут быть введены в одно и то же место или в альтернативные места одним и тем же или различными путями. Например, композиции на основе полипептида можно вводить через слизистую, в то время как композиции на основе векторов предпочтительно вводить, например, путем подкожных инъекций для вектора МВА, внутримышечных инъекций для ДНК-плазмид и подкожных или внутримышечных инъекций для аденовирусного вектора.
Данное изобретение также предусматривает способ индукции или стимуляции иммунного ответа против HBV в организме хозяина, включающий введение в указанный организм по меньшей мере одной из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев или композиций изобретения, таким образом индуцируя или стимулируя указанный иммунный ответ. Иммунный ответ может быть специфическим и/или неспецифическим, гуморальным и/или клеточным, и в этом контексте он может быть опосредован CD4+ или CD8+, или обоими. Иммунный ответ предпочтительно является Т-клеточным ответом, направленным против HBV-антигена.
Способность способа изобретения индуцировать или стимулировать анти-HBV иммунный ответ при введении в организм животного или человека может быть оценена in vitro или in vivo с помощью различных анализов, стандартных в данной области. Для общего описания доступных методик оценки возникновения и активации иммунного ответа, см., например, Coligan et al. (1992 and 1994, Current Protocols in Immunology; ed J Wiley & Sons Inc, National Institute of Health). Оценка клеточного иммунитета может быть выполнена путем оценки профилей цитокинов, секретируемых активированными клетками-эффекторами, в том числе произошедшими из CD4+ и CD8+ Т-клеток (т.е. путем количественной оценки IL-10-или IFNy-продуцирующих клеток путем ELISpot), путем определения статуса активации иммунных клеток-эффекторов (например, в анализе пролиферации Т-клеток классическим поглощением [3[H]-тимидина), путем анализа на антиген-специфические Т-лимфоциты у сенсибилизированного субъекта (например, пептид-специфический лизис в анализе цитотоксичности). Способность стимулировать гуморальный ответ можно определить путем связывания антитела и/или конкуренции в связывании (см., например, Harlow, 1989, Antibodies, Cold Spring Harbor Press). Способ изобретения также может быть дополнительно подтвержден на животных моделях с соответствующими инфекционными или вызывающими опухоли агентами (например, вирусом осповакцины или бактерией Listeria monocytogenes, экспрессирующей генные продукты HBV), чтобы определить нейтрализацию инфекционного и вызывающего опухоль агента и, в конечном счете, частичную устойчивость к сопутствующим симптомам, отражающим индукцию или увеличение анти-HBV иммунного ответа. Тестирование и проверка композиций изобретения также показаны в прилагаемом разделе Пример.
В другом аспекте изобретение предусматривает набор для применения в лечении и профилактике HBV-инфекций, в том числе хронической HBV-инфекции, HBV-ассоциированных заболеваний и патологических состояний, в соответствии с порядком, изложенным в данном документе, и, в частности, для индукции или развития иммунного ответа у субъекта, инфицированного HBV, где указанный набор включает множество активных агентов, выбранных из группы, состоящей из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев или композиций, описанных в данном документе.
Предпочтительно, указанное множество активных агентов содержится в форме отдельных полипептидов или отдельных векторов, и введение каждого из активных агентов может происходить одновременно (в одно и то же время) или по отдельности (один после другого через определенный промежуток времени), одним и тем же или различными путями введения, в одно и то же место (или в непосредственной близости) или в различные места, и с использованием одних и тех же или различных доз.
Особый интерес в данном изобретении представляет собой набор, который включает первый вектор, содержащий нуклеиновокислотную молекулу, кодирующую полимеразную часть, определенную в данном документе, и второй вектор, содержащий нуклеиновокислотную молекулу, кодирующую сердцевинную и/или оболочечную части, определенные в данном документе.
В соответствии с предпочтительным воплощением указанный первый вектор является MVA-вектором, содержащим нуклеиновокислотную молекулу, которая находится под контролем промотора вируса осповакцины, такого как промотор 7.5К, и кодирует полимеразную часть, включающую аминокислотную последовательность, приведенную в SEQ ID №7, 8 или 9 или в SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и заменой остатка Glu в позиции 672 на остаток His; а указанный второй вектор является MVA-вектором, содержащим нуклеиновокислотную молекулу, которая находится под контролем промотора вируса осповакцины, такого как промотор рН5 г, и кодирует сердцевинную и/или оболочечную части, включающие аминокислотную последовательность, приведенную в SEQ ID №18 или 19.
В соответствии с другим предпочтительным воплощением указанный первый вектор является аденовирусным вектором, содержащим нуклеиновокислотную молекулу, которая находится под контролем подходящего промотора, такого как CMV-промотор, и кодирует полимеразную часть, включающую аминокислотную последовательность, приведенную в SEQ ID №7 или SEQ ID №8 или SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и заменой остатка Glu в позиции 672 на остаток His; а указанный второй вектор является аденовирусным вектором, содержащим нуклеиновокислотную молекулу, которая находится под контролем подходящего промотора, такого как CMV-промотор, и кодирует сердцевинную и/или оболочечную части, включающие аминокислотную последовательность, приведенную в SEQ ID №18, 19, 20 или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251 (core-env1-env2), или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221 (core-env1).
Набор данного изобретения может также содержать третий вектор, экспрессирующий иммуномодулятор, как указано выше. Для наглядности, предпочтительные дозы каждого активного ингредиента, входящего в набор, являются дозами того же порядка, что и описанные выше в связи с композицией изобретения, с особым предпочтением дозы от 5×105 до 109 pfu для каждого поксвирусного или MVA-вектора и от 106 до 1012 vp для каждого аденовирусного вектора.
Изобретение также предусматривает антитела, которые селективно связываются с HBV-частями, используемыми в данном изобретении, или их пептидными фрагментами. Используемое в данном документе антитело селективно связывает целевой пептид, при этом он связывает целевой пептид и по существу не связывает неродственные белки. В некоторых случаях будет понятно, что антитело, связывающееся с пептидом, по-прежнему селективно, несмотря на некоторую степень перекрестной реактивности. Тем не менее, предпочтительно, чтобы антитело изобретения не связывалось с высокой аффинностью и высокой селективностью с нативным белком HBV.
Используемое в данном документе антитело определяется в соответствии с условиями, признанными в данной области. Антитела данного изобретения включают поликлональные антитела и моноклональные антитела, а также фрагменты таких антител, в том числе, но не ограничиваясь ими, Fab или F(ab)'.sub.2, и Fv-фрагменты. Антитела данного изобретения могут быть получены с использованием стандартных в данной области методик, например после введения животному эффективного количества любой из HBV-частей, описанных в данном документе, и/или их пептидного фрагмента. Антитела предпочтительно получают из областей или дискретных фрагментов HBV-частей, включающих уникальные последовательности, такие как описанные в данном документе направленные на изменения и введенные в нативные белки HBV.
Антитела данного изобретения имеют множество потенциальных применений, которые входят в сферу данного изобретения. Например, такие антитела могут быть использованы (а) в качестве реагентов в анализах для обнаружения первого или второго полипептидов данного изобретения, (б) в качестве реагентов в анализах для обнаружения присутствия HBV в биологическом образце, и/или (в) в качестве инструмента для восстановления рекомбинантно-полученных HBV-частей из смеси белков и других загрязняющих веществ (например, проводя очистку путем афинной хроматографии или иммунопреципитации из культивированных клеток-хозяев).
Данное изобретение также относится к способу обнаружения и/или количественной оценки вируса HBV или анти-HBV-антитела в биологическом образце (например, плазме, сыворотке, ткани), взятом от индивидуума, чувствительного к заражению указанным вирусом HBV, с использованием по меньшей мере одной из HBV-частей нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев, композиций или антител изобретения.
В одном воплощении способ особенно подходит для обнаружения и/или количественной оценки вируса HBV в биологическом образце и включает по меньшей мере этап контактирования указанного биологического образца по меньшей мере с одним антителом изобретения в условиях, позволяющих формировать комплекс между вирусом и антителом, и этап обнаружения и/или количественной оценки образования указанного комплекса любым подходящим способом.
В другом воплощении этот способ особенно подходит обнаружения и/или количественной оценки анти-HBV-антитела в биологическом образце и включает по меньшей мере этап контактирования указанного биологического образца по меньшей мере с одной из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев, композиций изобретения в условиях, позволяющих формировать комплекс между анти-HBV-антителом и HBV-частью, нуклеиновокислотной молекулой, вектором, инфекционной вирусной частицей, клеткой-хозяином, композицией, и этап обнаружения и/или количественной оценки образования указанного комплекса любым подходящим способом.
Специалисту в данной области легко будет определить количество антитела, HBV-части, нуклеиновокислотной молекулы, вектора, инфекционной вирусной частицы, клетки-хозяина, композиции, которое будет использоваться в способах изобретения. Средства обнаружения и/или количественной оценки вируса являются стандартными и хорошо известны специалистам в данной области. В качестве примера можно отметить блоты, ИФА, так называемые сэндвич-методики, конкурентные методики и ПЦР-методики, в частности так называемые методики «в реальном времени». Применение антитела, HBV-части, нуклеиновокислотной молекулы, вектора, инфекционной вирусной частицы, клетки-хозяина или композиции данного изобретения в качестве реагента может быть облегчено путем связывания (например, физического соединения) с обнаруживаемым веществом. Примеры обнаруживаемых веществ включают различные ферменты (например, пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу и ацетилхолинэстеразу), простерические группы (например, стрептавидин/биотин или авидин/биотин), флуоресцентные материалы (например, умбеллиферон, флуоресцеин или производные флуоресцеина), люминесцентные материалы, биолюминесцентные материалы (например, люциферазу, люциферин или экворин) и радиоактивные материалы (например,125I, 131I, 35S или 3H).
Наконец, изобретение относится к применению по меньшей мере одной из HBV-частей, нуклеиновокислотных молекул, векторов, инфекционных вирусных частиц, клеток-хозяев, композиций или антител изобретения для диагностики HBV-инфекции в биологических образцах in vitro.
Изобретение было описано в иллюстративной манере, и необходимо понимать, что используемая терминология обозначает характер описываемого, а не ограничивает его. Очевидно, что в свете вышеизложенных учений возможны многие модификации и вариации данного изобретения. Поэтому следует понимать, что в рамках формулы изобретения изобретение может быть осуществлено другим путем, отличающимся от того, который конкретно описан в данном документе.
Все приведенные выше раскрытия патентов, публикаций и позиции в базе данных конкретно включены в данный документ посредством ссылки во всей их полноте в той же степени, как если бы о каждом таком отдельном патенте, публикации или позиции было конкретно и индивидуально сказано, что они включены посредством ссылки.
Графические материалы
На фиг.1 показана экспрессия HBV-полипептида клетками, инфицированными аденовирусом и MVA. Клетки А549 или фибробласты куриного эмбриона инфицировали аденовирусом с MOI 10 или 50 или MVA с MOI 0,2 или 1, и через 48 часов после инфицирования клетки лизировали. Затем проводили вестерн-блоттинг с клеточными лизатами, полученными из клеток, инфицированных различными конструкциями Ad (фиг.1А) и MVA (фиг.1 В) для обнаружения специфических HBV-белков. Полипептиды, содержащие сердцевину, обнаруживали с помощью анти-Core-антитела (С1-5 или 13А9, разведение 1/200), а полипептиды, содержащие полимеразу, с помощью анти-Ро1-антитела (8D5, разведение 1/200) в качестве первичных антител, а вторичные антитела были связаны с HRP. Ожидаемый размер белков, экспрессируемых Ad TG17909 и Ad TG17910, составлял соответственно 31,6 кДа и 88,5 кДа. Ожидаемые размеры белков, экспрессируемых MVA TG17971, MVA TG17972, MVA TG17993 и MVA TG17994, составляли соответственно 20,2 кДа, 15,8 кДа, 20 кДа и 23,5 кДа. Ожидаемые размеры белков, экспрессируемых MVA TG17842 и MVA TG17843, составляли соответственно 88,5 кДа и 98,2 кДа.
На фиг.2 показана иммуногенность HBV-полипептидов, кодируемых аденовирусом, в анализе Elispots IFNγ. Пять отдельных мышей (HLA-A2-трансгенных мышей) иммунизировали один раз либо только Ad TG17909 (черные столбцы), либо только Ad TG17910 (белые столбцы), либо сочетанием (Ad TG17909 + Ad TG17910) (серые столбцы). На фиг.2А показаны специфические Т-клеточные реакции, направленные на полимеразный белок с использованием HLA-А2-рестриктированного пептида SLY (SEQ ID №55) или нерелевантного пептида (не показан). На фиг.2В показаны специфические Т-клеточные реакции, направленные на сердцевинный белок, с помощью Н1-А-А2-рестриктированных пептидов FLP (SEQ ID №56) или ILC (SEQ ID №57). На фиг.2С показаны специфические Т-клеточные реакции, направленные на оболочечные области, с помощью HLA-А2-рестриктированных пептидов VLQ (SEQ ID №58), FLG (SEQ ID №59) или GLS (SEQ ID №60). Каждый столбец представляет собой отдельную вакцинированную мышь, а заштрихованные столбцы представляют собой среднее значение для каждой группы. Результаты представлены как среднее значение числа пятен, наблюдаемых на 106 клеток селезенки, полученных из трех повторов лунок. Ответ считался положительным, если число пятен было больше 50 на 106 клеток (эта отсечка представлена как толстая черная линия).
На фиг.3 показана иммуногенность HBV-полипептидов, кодируемых аденовирусным вектором, в анализе внутриклеточного окрашивания цитокинов. Пять отдельных мышей (HLA-А2-трансгенных мышей) иммунизировали один раз либо только Ad TG17909 (фиг.3В), либо только Ad TG17910 (фиг.3А), либо сочетанием Ad TG17909 + Ad TG17910 (фиг.3С). Спленоциты культивировали в течение 5 часов с Golgi-Plug и в присутствии каждого из HLA-A2-рестриктированных пептидов (SLY для Pol, FLP, ILC для Core, VLQ, FLG и GLS для Env) или нерелевантного пептида. Процент CD8+ клеток, продуцирующих цитокины (IFNγ и/или TNFα), специфичные для каждого HLA-А2-рестриктированного эпитопа, оценивали в анализе ICS. Каждый столбец представляет собой отдельную вакцинированную мышь, при этом клетки, продуцирующие IFNγ, показаны черным столбцом, клетки, продуцирующие TNFα, показаны белым столбцом, а клетки, продуцирующие и IFNγ, и TNFα, показаны заштрихованным столбцом, и все эти клеточные популяции сложены для каждой мыши.
На фиг.4 показана способность HBV-полипептидов, кодируемых аденовирусным вектором, индуцировать CD8+ и CD4+ Т-клеточные ответы, выявляемые в анализе внутриклеточного окрашивания цитокинов. Пять отдельных мышей (HLA-А2-трансгенных мышей) иммунизировали один раз смесью Ad TG17909 и Ad TG17910. Спленоциты культивировали в течение 5 часов с Golgi-Plug и в присутствии каждого из HLA-А2-рестриктированных пептидов (SLY для Pol, FLP, ILC для Core, VLQ, FLG и GLS для Env) или пула перекрывающихся пептидов (15 аминокислотных, перекрывающихся на 11 аминокислот, 2 пула пептидов для Core и 2 пула пептидов для Env), охватывающих все области антигена, или нерелевантного пептида. Индуцированные специфические CD8+ Т-клетки, продуцирующие IFNγ и/или TNFα (фиг.4А), и индуцированные специфические CD4+ Т-клетки, продуцирующие IFNγ и/или TNFα (фиг.4В) или продуцирующие IFNγ и/или IL2 (фиг.4С), наблюдали в анализе ICS. Каждый столбец представляет собой отдельную вакцинированную мышь, при этом клетки, продуцирующие IFNγ, показаны серым столбцом, клетки, продуцирующие TNFα или IL2, показаны белым столбцом, а клетки, продуцирующие IFNγ+TNFα или IFNγ+IL2, показаны заштрихованным столбцом, и все эти клеточные популяции сложены для каждой мыши. Также показано среднее значение для каждой группы.
На фиг.5 показана способность аденовирусного вектора, кодирующего HBV-полипептиды, индуцировать in vivo функциональный цитолиз против клеток-мишеней, нагруженных HBV HLA-А2-рестриктированными эпитопами. Трех отдельных мышей (Н1А-А2-трансгенных мышей) иммунизировали один раз сочетанием Ad TG17909 и Ad TG17910 (М1-М3), а одну мышь иммунизировали один раз пустым аденовирусным вектором в качестве отрицательного контроля (МО). CFSE-меченые спленоциты от сингенных мышей, нагруженные HBV HLA-A2-эпитопами или без них (отрицательный контроль), вводили внутривенно вакцинированным мышам. Лизис меченых клеток in vivo оценивали для каждой мыши через 24 часа путем проточной цитометрии и рассчитывали, как указано в разделе Материалы и методы. Среднее значение специфического лизиса, наблюдаемого для каждого пептида на трех мышах, вакцинированных AdHBV, рассчитано и показано (среднее М1-М3).
На фиг.6 показана иммуногенность HBV-полипептидов, кодируемых MVA-вектором, определяемая в анализе Elispots IFNγ. Отдельных мышей (HLA-А2-трансгенных мышей) иммунизировали три раза с недельным интервалом либо MVA TG17842, либо MVA TG17843 (фиг.6А), либо MVA TG17971 (фиг.6В), либо MVATGI7972, либо отрицательным контролем MVA TGN33.1 (данные не представлены). На фиг.6А показаны специфические Т-клеточные реакции против полимеразного белка после иммунизации MVA TGI7842 (темно-серые столбцы) или MVA TG17843 (светло-серые столбцы), с использованием HLA-A2-рестриктированного пептида SLY (SEQ ID №55), пула из 8 пептидов, покрывающих С-концевую часть полимеразного белка (25 пептидов из 15 аминокислот с перекрытием на 11 аминокислот/пул), нерелевантного пептида или среды (отрицательные контроли). На фиг.6В показаны специфические Т-клеточные реакции против сердцевинного белка после иммунизации MVA TG17971, с использованием HLA-А2-рестриктированного пептида FLP (SEQ ID №56), ILC (SEQ ID №57), пептидных пулов «core 1 и core 2» (21-22 пептида из 15 аминокислот с перекрытием на 11 аминокислот/пул), нерелевантного пептида или среды (отрицательные контроли). Каждый столбец представляет собой отдельную вакцинированную мышь, а заштрихованные столбцы представляют собой среднее значение для каждой группы. Результаты представлены как среднее значение числа пятен, наблюдаемых на 106 клеток селезенки, полученных из трех повторов лунок. Ответ считался положительным, если число пятен было больше 50 на 106 клеток (эта отсечка представлена как пунктирная черная линия).
На фиг.7 показана иммуногенность HBV-полипептидов, кодируемых MVA-векторами, одновременно введенными мыши, которая определена в анализах Elispots IFNγ. Отдельных мышей (HLA-А2-трансгенных мышей) иммунизировали три раза с недельным интервалом смесью MVA TG17843 и либо MVA TG 17972 (фиг.7А), либо MVA TG17993 (фиг.7В), либо MVATG 17994 (фиг.7С), либо только MVA TG N33.1 в качестве отрицательного контроля (данные не представлены). Специфические Т-клеточные реакции против полимеразного белка выявляли с помощью HLA-А2-рестриктированного пептида SLY (SEQ ID №55), а специфические Т-клеточные реакции против оболочечных частей с помощью HLA-А2-рестриктированного пептида GLS (SEQ ID №60) или пула пептидов, охватывающих область env2 (пул из 15-аминокислотных пептидов с перекрытием на 11 аминокислот). Нерелевантный пептид и среда использовались в качестве отрицательных контролей. Каждый столбец представляет собой отдельную вакцинированную мышь, а заштрихованные столбцы представляют собой среднее значение для каждой группы. Результаты представлены как среднее значение числа пятен, наблюдаемых на 106 клеток селезенки, полученных из трех повторов лунок. Ответ считался положительным, если число пятен было больше 92 на 106 клеток (эта отсечка представлена как пунктирная черная линия).
Примеры 1. Материалы и методы
Конструкции, описанные ниже, осуществляются в соответствии с общими методиками генетической инженерии и молекулярного клонирования, которые подробно описаны в Maniatis et al. (1989, Laboratory Manual, Cold 5 Spring Harbor Laboratory Press, Cold Spring Harbor NY), или в соответствии с рекомендациями изготовителя, если используются коммерческие наборы. Методики ПЦР-амплификации известны специалистам в данной области (см., например, PCR. protocols -A guide to methods and applications, 1990, опубликованный Inniis, Gelfand. Sninsky and White, Academic Press). Рекомбинантные плазмиды, несущие ген устойчивости к ампициллину, реплицируются в Е.coli С600 (Stratagene) на агаре или в жидкой среде, дополненной 100 мкг/мл антибиотика. Конструкции рекомбинантных вирусов осповакцины осуществляются в соответствии со стандартной в данной области технологией, описанной в приведенных выше документах, а также в Mackett et al. (1982, Proc. Natl. Acad. Sci. USA 79, 7415-7419) и Mackett et al. (1984, J. Virol. 49, 857-864). Ген селекции gpt (ксантин-гуанин-фосфорибозилтрансферазы) Е.coli (Falkner and Moss, 35 1988, J. Virol. 62, 1849-185) используется для облегчения отбора рекомбинантных вирусов осповакцины.
1.1. Векторные конструкции и продукция
1.1.1. Выбранные антигены и последовательность штамма HBV Векторы, приведенные далее в качестве примера, были разработаны для экспрессии полимеразы, сердцевинных полипептидов и иммуногенных областей оболочечного белка. Все они происходят из штамма HBV Y07587, последовательность которого описана в международных базах данных (Genbank Y07587) и в различных публикациях. Это вирус генотипа D серотипа ayw.
Сердцевинный полипептид является либо полипептидом дикого типа (АК 1-183), либо сердцевинным полипептидом с удалением аминокислот с 77 по 84 (т.е. сердцевинным белком, содержащим аминокислоты с 1 по 76 и с 85 по 183 и обозначенным Core*), либо усеченным с С-конца полипептидом (1-148) или усеченным с С-конца сердцевинным белком (1-148), в котором также удалены аминокислоты с 77 по 84 (т.е. сердцевинным белком, содержащим аминокислоты с 1 по 76 и с 85 по 148 и обозначенным core*t).
Полимеразный полипептид является либо полипептидом дикого типа, либо усеченным с М-конца полипептидом, в котором отсутствуют первые 47 аминокислот (48-832), либо усеченной с М-конца полимеразой (48-832), также мутированной в позиции 540 (D на Н) и 718 (Е на Н) (позиции 450 и 718 даны по отношению к полимеразе дикого типа), либо усеченной (48-832) и мутированной полимеразой (D540H и Е718Н), которая слита с сигнальным пептидом и трансмембранной областью из гликопротеина вируса бешенства (Pol*TMR).
Выбранными областями оболочечного белка являются: область из аминокислот с 14 по 51 S-белка (env1), область из аминокислот со 165 по 194 HBs-белка (env2) и область из аминокислот с 202 по 226 HBs-белка (env4).
1.1.2. Конструирование и продукция MVA TG 17842, экспрессирующего усеченную и мутированную HBV-полимеразу (Pol*)
Нуклеотидные последовательности, кодирующие модифицированный полипептид HBV-полимеразу, синтезированы компанией Geneart с помощью синтетических олигонуклеотидов и ПЦР-продуктов. Модифицированная HBV-полимераза соответствует полимеразному белку HBV Y07587 (SEQ ID №1), мутированному в позициях 540 (D в Н) и 718 (Е в Н) для устранения активностей РНКазы Н и ОТазы, которые демонстрирует нативная HBV-полимераза (полученная аминокислотная последовательность показана в SEQ ID №8, а нуклеотидная последовательность показана в SEQ ID №22). Заново собранную последовательность Pol клонировали в плазмидный вектор, получив PGA15-Pol (SEQ ID №27). Усеченную версию с удалением первых 47 аминокислот, присутствующих на М-конце нативной HBV-полимеразы, амплифицировали с помощью ПЦР из плазмиды pGA15-Pol с использованием следующих праймеров: OTG19037 (GAGCGATATCCACCATGAATGTTAGTATTCCTTGGAC) (SEQ ID №28) и OTG 19038 (GATCGCTAGCTCACGGTGGTCTCCATGCGAC) (SEQ ID №29). Полученный фрагмент вставляли в MVA-плазмиду для переноса по сайтам рестрикции Nhel и EcoRV после промотора р7.5К (Cochran et al., 1985, J. Virol. 54:30), получая pTG 17842. Мутированная и усеченная полимераза обозначается в дальнейшем как pol*.
MVA-плазмида для переноса предназначена для обеспечения вставки нуклеотидной последовательности, которую нужно перенести путем гомологичной рекомбинации в делецию III генома MVA. Она получена из плазмиды pTG1E (описанной в Braun et al., 2000, Gene Ther. 7:1447), в которую клонировали фланкирующие последовательности (BRG3 и BRD3) по сторонам от делеции III MVA (Sutler and Moss, 1992, Proc. Natl. Acad. Sci. USA 89:10847). Плазмида для переноса содержит также гибрид улучшенного зеленого флуоресцентного белка (гена eGFP, выделенного из pEGP-C1, Clontech) Aequorea victoria и гена ксантин-гуанин-фосфорибозилтрансферазы (гена gpt) под контролем синтетического раннего/позднего промотора вируса осповакцины р11К7.5 (любезно предоставленного R. Wittek, Лозаннский университет). Синтез ксантин-гуанин-фосфорибозилтрансферазы позволяет СРТ+-рекомбинантному MVA формировать бляшки на селективной среде, содержащей микофеноловую кислоту, ксантин и гипоксантин (Falkner et al., 1988, J. Virol. 62, 1849-54), а ген eGFP позволяет визуализировать бляшки с рекомбинантным MVA. Селектируемый маркер eGFP-GPT находится между двумя гомологичными последовательностями в одинаковой ориентации. Когда клональная селекция проведена, селектируемый маркер легко устраняют путем нескольких пассажей без селекции, позволяя расти eGFP-GPT-рекомбинантному MVA.
Создание вируса MVA TG 17842 выполняли путем гомологичной рекомбинации в первичных фибробластах куриных эмбрионов (СЕР), инфицированных MVA и трансфицированных pTG 17842 (в соответствии со стандартным осаждением ДНК фосфатом кальция). Вирусную селекцию выполняли, проводя три цикла клонирования методом бляшкообразования в селективной среде, содержащей микофеноловую кислоту, ксантин и гипоксантин. Как уже упоминалось выше, селектируемый маркер затем устраняли путем пассажа в неселективной среде. Отсутствие контаминации родительским MVA проверяли с помощью ПЦР.
Анализ экспрессии HBV-полимеразы проводили путем вестерн-блоттинга. Клетки А549 (АТСС CCL-185) инфицировали MVATG 17842 (Pol*) с MOI 1 в присутствии протеосомного ингибитора MG-132 (10 мкМ), добавленного в ростовую среду, или в его отсутствие. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Pol-антитела Нер В Pol (8D5, Санта-Крус, №sc-81591).
1.1.3. Конструирование и продукция MVA TG17843, экспрессирующего усеченную и мутированную HBV-полимеразу, слитую с заякоренной в мембране областью гликопротеина оболочки вируса бешенства (Pol*TMR)
Затем последовательность HBV Pol* модифицировали путем слияния его N-конца с сигнальным пептидом (SS) и его С-конца с заякоренной в мембране последовательностью (TMR), полученными из гликопротеина вируса бешенства (изолят ERA; описан в Genbank №М38452). Последовательности SS и TMR амплифицировали из плазмиды pTG8042 (описанной в WO 99/03885) путем ПЦР с использованием, соответственно, пар праймеров OTG19045 (SEQ ID №30) (GAGTGATATCCACCATGGTTCCTCAGGCTCTCCTG)/OTG 19047 (SEQ ID №31) (GTCCAAGGAATACTAACATTAATAGGGAATTTCCCAAAACACAATG) и OTG 19049 (SEQ ID №32) (GTCGCATGGAGACCACCGTATGTATTACTGAGTGCAGGG/OTG 19050 (SEQ ID №33) (GAGTGCTAGCTCACAGTCTGGTCTCACCC). Последовательность Pol* амплифицировали из плазмиды pGA15-Pol путем ПЦР с использованием пары праймеров OTG19046 (SEQ ID №34) (GTTTTGGGAAATTCCCTATTAATGTTAGTATTCCTTGGACTC)/OTG 19048 (SEQ ID №35) (CTGCACTCAGTAATACATACGGTGGTCTCCATGCGACGTGC). Затем последовательность SS-Pol*-TMR собирали посредством тройной ПЦР с использованием следующих праймеров: OTG19045 (SEQ ID №30) и OTG19050 (SEQ ID №33). Полученный фрагмент вставляли в плазмиду для переноса, полученную из вируса осповакцины, по сайтам рестрикции Nhel и EcoRV после промотора р7.5К (Cochran et al., 1985, J. Virol. 54:30), получая pTG17843.
Создание вируса MVA TG 17843 выполняли в CEF путем гомологичной рекомбинации, как описано выше.
Анализ экспрессии Pol*-TMR проводили путем вестерн-блоттинга. Клетки А549 инфицировали MVA TG17843 с MOI 1 в присутствии протеосомного ингибитора MG-132 (10 мкМ), добавленного в ростовую среду, или в его отсутствие. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Pol-антитела Нер В Pol (8D5, Санта-Крус, №sc-81591).
1.1.4 Конструирование и продукция MVA TG17971, экспрессирующего сердцевинный полипептид с делецией (Core*)
Core* соответствует последовательности Core от HBV Y07587 (SEQ ID №2) с удалением аминокислот с 77 по 84.
Последовательности, кодирующие Core*, создавали путем двойной ПЦР на плазмиде pGA4-Core. Эта плазмида была сделана компанией Geneart. Она содержала кодирующую последовательность полной длины модифицированного гена HBV Core, который был собран из синтетических олигонуклеотидов и/или ПЦР-продуктов. Последние два кодона САА TGT кодирующей последовательности были модифицированы таким образом, чтобы избежать гомологии последовательности с Pol (SEQ ID №36).
Последовательность Core от позиции 1 до 76 амплифицировали путем ПЦР с использованием следующих праймеров: OTG19290 (SEQ ID №37) (GACTGTTAACCACCATGGACATTGATCCTTATAAAGAATTTG) и OTG 19292 (SEQ ID №38) (GTTGACATAACTGACTACCAAATTACCACCCACCCAGGTAG). Последовательность Core с позиции 85 по 183 амплифицировали путем ПЦР с использованием следующих праймеров: OTG19291 (SEQ ID №39) (GTGGGTGGTAATTTGGTAGTCAGTTATGTCAACACTAATATG) и OTG 19080 (SEQ ID №61) (GACTCTCGAGTTAGCACTGAGATTCCCGAGATTG). Двойную ПЦР проводили с использованием OTG19290 (SEQ ID №37) и OTG19080 (SEQ ID №61) и обоих полученных ампликонов. Полученный фрагмент вставляли в плазмиду для переноса, полученную из вируса осповакцины, по сайтам рестрикции Xhol и Hpal после промотора pH5R (Rosel et al., 1986, J Virol. 60:436), получая pTG17971.
Создание вируса MVA TG 17971 выполняли в CEF путем гомологичной рекомбинации, как описано выше.
Анализ экспрессии Core* проводили путем вестерн-блоттинга. Фибробласты куриного эмбриона инфицировали MVA TG17971 с MOI 0,2. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Соге-антитела Мер BcAg (13А9) (Санта-Крус, #sc-23946).
1.1.5. Конструирование и продукция MVA TG 17972, экспрессирующего усеченный сердцевинный полипептид с делецией (Core*t)
Core*t соответствует последовательности Core от HBV Y07587 (SEQ Ю№2), усеченной после аминокислоты 148 и с делецией аминокислот с 77 по 84.
Последовательности, кодирующие Core*t, создавали путем двойной ПЦР из плазмиды pGA4-Core, которая содержала последовательность, кодирующую ген HBV Core полной длины, который был собран из синтетических олигонуклеотидов и/или ПЦР-продуктов, за исключением последних двух кодонов САА TGT кодирующей последовательности, которые были модифицированы таким образом, чтобы избежать гомологии последовательности с Pol (SEQ ID №36).
Последовательность Core от позиции 1 до 76 амплифицировали путем ПЦР с использованием следующих праймеров: OTG19290 (SEQ ID №37) (GACTGTTAACCACCATGGACATTGATCCTTATAAAGAATTTG) и OTG19292 (SEQ ID №38) (GTTGACATAACTGACTACCAAATTACCACCCACCCAGGTAG). Последовательность Core с позиции 85 по 183 амплифицировали путем ПЦР с pGA4-Core с использованием следующих праймеров: OTG19291 (SEQ ID №39) (GTGGGTGGTAATTTGGTAGTCAGTTATGTCAACACTAATATG) и OTG19299 (SEQ ID №40) (GACTCTCGAGTTAAACAGTAGTCTCCGGAAGTG). Двойную ПЦР проводили с использованием OTG19290 (SEQ ID №37) и OTG19299 (SEQ ID №40). Полученный фрагмент вставляли в плазмиду для переноса, полученную из вируса осповакцины, по сайтам рестрикции Xhol и Hpal после промотора pH5R (Rosel et al., 1986, J. Virol. 60:436), получая pTG17972.
Создание вируса MVA TG17972 выполняли в CEF путем гомологичной рекомбинации, как описано выше.
Анализ экспрессии Core*t проводили путем вестерн-блоттинга. Фибробласты куриного эмбриона инфицировали MVA TG17972 с MOI 0,2. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Core-антитела Нер BcAg (13А9) (Санта-Крус, №sc-23946).
1.1.6. Конструирование и продукция MVA TG17993, экспрессирующего усеченный сердцевинный полипептид с делецией, слитый с иммуногенной областью env1 (Core*t-env1)
Часть Core*t сливали с областью env1, продолжающейся с аминокислоты 14 по 51 HBs-белка.
Последовательность Core*t-env1 создавали путем двойной ПЦР. Последовательность Core*t амплифицировали путем ПЦР на pTG17972 с использованием следующих праймеров: OTG19317 (SEQ ID №41) (GACGGGATCCACCATGGACATTGATCCTTATAAAGAATTTGG) и OTG19319 (SEQ ID №42) (GCCTGCTTGCAGGACAACAGTAGTCTCCGGAAGTGTTG). Последовательность env1 амплифицировали путем ПЦР на плазмиде рМК-С/Е (SEQ ID №43) с использованием следующих праймеров: OTG19318 (SEQ ID №44) (CCGGAGACTACTGTTGTCCTGCAAGCAGGCTTCTTC) и OTG19320 (SEQ ID №45) (GAGTCATTCTCGACTTGCGGCCGCTTACTGACCCAGGCAAACCGTGG). Двойную ПЦР проводили с использованием OTG19317 (SEQ ID №41) и OTG19320 (SEQ ID №45). Полученный фрагмент вставляли в плазмиду для переноса, полученную из вируса осповакцины, по сайтам рестрикции BamHI и NotI после промотора pH5R промотора (Rosel et al., 1986, J Virol. 60:436), получая pTG17993.
В качестве иллюстративного примера, плазмида рМК-С/Е была сделана компанией Geneart и содержала химерную последовательность, содержащую вставку из трех последовательностей оболочечной области HBV в сердцевинной последовательности (SEQ ID №43). Нативные сердцевинная и оболочечная нуклеотидные последовательности были вырожденными, чтобы избежать гомологии последовательности с последовательностью Pol HBV, а также нестабильности последовательности в связи с участками полиТ или полиGC. Кроме того, из сердцевинной последовательности были удалены аминокислоты с 77 по 84, и она была усечена с АК 148. Выбранными оболочечными областями являются: область с аминокислоты 14 по 51 из белка S (env1), область с аминокислоты 165 по 194 из белка S (env2) и область с аминокислоты 202 по 226 из белка S (env4). Три области вставляли соответственно в позиции нуклеотидов 127, 222 и 416 сердцевинной последовательности. Следует отметить, что вставка этой последовательности в вектор MVA приводит к цитотоксичности в экспрессирующих клетках, подчеркивая тот факт, что разработка гибрида env-core является не прямой.
Создание вируса MVA TG17993 выполняли в CEF путем гомологичной рекомбинации, как описано выше.
Анализ экспрессии Core*t-env1 проводили путем вестерн-блоттинга. Фибробласты куриного эмбриона инфицировали MVA TG17993 с MOI 0,2. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Соге-антитела Нер BcAg (13А9) (Санта-Крус, №sc-23946).
1.1.7. Конструирование и продукция MVA TG17994, экспрессирующего усеченный сердцевинный полипептид с делецией, слитый с иммуногенными областями env1 и env2 (Core*t-env1-env2)
Полипептид Core*t-env1-env2, описанный в 1.1.5, затем сливали с двумя иммуногенными областями, продолжающимися с аминокислоты 14 по 51 (env1) и с аминокислоты 165 по 194 (env2) из HBs-белка.
Последовательность, кодирующую Core*t-env1-env2, собирали путем тройной ПЦР. Последовательность Core*t амплифицировали путем ПЦР с pTG17972 с использованием следующих праймеров: OTG19317 (SEQ ID №41) и OTG19319 (SEQ ID №42). Env1 амплифицировали с плазмиды рМК-С/Е с использованием следующих праймеров: OTG19318 (SEQ ID №44) и OTG19322 (SEQ ID №46) (GCGTGCGCTTGCCCACTGACCCAGGCAAACCGTGG). Env2 амплифицировали с плазмиды рМК-С/Е с использованием следующих праймеров: SEQ ID №47 (CGGTTTGCCTGGGTCAGTGGGCAAGCGCACGCTTTAGC) и OTG19323 (SEQ ID №48) (GAGTCATTCTCGACTTGCGGCCGCTTACACGCTCAGCCACACGGTTGG). Тройную ПЦР проводили с использованием OTG19317 (SEQ ID №41) и OTG19323 (SEQ ID №48). Полученный фрагмент вставляли в плазмиду для переноса, полученную из вируса осповакцины, по сайтам рестрикции BamHI и A/of/ после промотора pH5R (Rosel et al., 1986, J Virol. 60:436), получая pTG17994.
Создание вируса MVA TG 17994 выполняли в CEF путем гомологичной рекомбинации, как описано выше.
Анализ экспрессии Core*t-env1-env2 проводили путем вестерн-блоттинга. Фибробласты куриного эмбриона инфицировали MVA TG 17994 с MOI 0,2. Через 24 часа клетки собирали. Анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Core-антитела Нер BcAg (13А9) (Санта-Крус, №sc-23946).
1.1.8. Конструирование и продукция аденовирусного вектора Ad TG 17909, экспрессирующего Core*1-env1-env2-env4
Синтетический ген (831 нуклеотид), кодирующий гибрид Core*t-env1-env2-env4, создавали путем двойной ПЦР. Core амплифицировали путем ПЦР с pGA4-Core (описан в 1.1.4.) с использованием следующих праймеров: OTG19152 (SEQ ID №49) (GGGGGGCTAGCAAGCTTCCACCATGGACATTGATCCTTATAAAGAATTTG) и OTG19154 (SEQ ID №50) (GAAAGAATCCAGCTTGCAGGACGCACTGAGATTCCCGAGATTGAG). Env1 -env2-env4 амплифицировали путем ПЦР с pGA4-env с использованием следующих праймеров: OTG19153 (SEQ ID №51) (CTCAATCTCGGGAATCTCAGTGCGTCCTGCAAGCTGGATTCTTTC) и OTG19159 (SEQ ID №52) (GAGTCATTCTCGACTTGCGGCCGCTTAGATATAAACCCACAAGC). Двойную ПЦР проводили с использованием OTG19152 (SEQ ID №49) и OTG19159 (SEQ ID №52). Полученный фрагмент вставляли по сайтам рестрикции Nhel и Notl в аденовирусную шаттл-плазмиду, содержащую CMV-управляемую экспрессионную кассету, окруженную аденовирусными последовательностями (аденовирусные нуклеотиды 1-454 и нуклеотиды 3513-5781, соответственно), образуя поколение векторного генома посредством гомологичной рекомбинации (Chartier et al., 1996, J. Virol. 70:4805). Полученный аденовирусный вектор pTG17909 имел делеции Е3 (нуклеотиды 28593-30464) и Е1 (нуклеотиды 455-3512), при этом область Е1 заменена экспрессионной кассетой, содержащей, от 5' к 3', немедленный ранний энхансер/промотор CMV, химерный человеческий β-глобин/интрон IgG (такой как в векторе pCl, доступном от Promega), последовательность, кодирующую Core*t-env1-env2-env4, и поздний сигнал полиаденилирования SV40. Рекомбинантный аденовирус был создан путем трансфекции вирусных геномов, линеаризованных Рас/, в Е1-комплементационную клеточную линию. Размножение, очистка и титрование вируса были проведены в соответствии с описанием выше (Erbs et al., 2000, Cancer Res. 60:3813).
Экспрессию гибридного белка оценивали путем вестерн-блоттинга. 106 клеток А549 (АТСС CCL-185) инфицировали в течение 48 часов Ad TG17909 с MOI 10 или 50 или пустым аденовирусом в качестве отрицательного контроля. Клеточный осадок собирали и исследовали с помощью мышиного моноклонального анти-Core-антитела (С1-5, sc-23945, Санта-Крус).
1.1.9. Конструирование и продукция аденовирусного вектора Ad TG17910, экспрессирующего Pol*
Ген, кодирующий Pol*, полимеразный белок, усеченный на первые 47 аминокислот за исключением инициатора Met (с 48 по 832) и мутированный в позициях 540 (D на Н) и 713 (Е на Н) (по сравнению с полимеразой дикого типа), вставляли в аденовирусный вектор. Ген Pol (2364 нуклеотида) амплифицировали с помощью ПЦР с pGA15-Pol (описан в 1.1.2) с использованием праймеров OTG19155 (SEQ ID №53) (GGGGGGCTAGCAAGCTTCCACCATGAATGTTAGTATTCCTTGGACTCATAAG) и OTG19156 (SEQ ID №54) (GAGTCATTCTCGACTTGCGGCCGCTCACGGTGGTCTCCATGCGACGTGC).
Полученный фрагмент вставляли по сайтам рестрикции Nhel и Notl в аденовирусную шаттл-плазмиду, содержащую CMV-управляемую экспрессионную кассету, окруженную аденовирусными последовательностями (аденовирусные нуклеотиды 1-454 и нуклеотиды 3513-5781, соответственно), образуя поколение векторного генома путем гомологичной рекомбинации (Chartier et al., 1996, J. Virol. 70:4805). Полученный аденовирусный вектор pTG17910 имел делеции Е3 (нуклеотиды 28593-30464) и Е1 (нуклеотиды 455-3512), при этом область Е1 заменена экспрессионной кассетой, содержащей, от 5' к 3', немедленный ранний энхансер/промотор CMV, химерный человеческий β-глобин/интрон IgG (такой как в векторе pCl, доступном от Promega), последовательность, кодирующую усеченный и мутированный Pol, и поздний сигнал полиаденилирования SV40. Рекомбинантный аденовирус был создан путем трансфекции вирусных геномов, линеаризованных Pad, в Е1-комплементационную клеточную линию. Размножение, очистка и титрование вируса были проведены в соответствии с описанием выше (Erbs et al., 2000, Cancer Res. 60:3813).
Экспрессию гибридного белка оценивали в инфицированных аденовирусом клетках в анализе вестерн-блоттинг. Клетки А549 (106 клеток) (АТСС CCL-185) инфицировали в течение 48 часов Ad TG17910 с MOI 10 или 50, а также пустым аденовирусом в качестве отрицательного контроля. Клеточный осадок собирали и исследовали с помощью мышиного моноклонального анти-Ро1-антитела (8D5, sc-81591, Санта-Крус).
7.2. Оценка иммуногенности антигена
Иммуногенность антигена оценивали in vivo в анализе Elispot IFNγ и путем внутриклеточного окрашивания цитокинов (ICS) после иммунизации трансгенных мышей.
1.2.1. Мышиная модель
HLA-A2.1-трансгенные мыши, используемые в исследовании, были описаны Pascolo et al. (1997. J. Exp.Med. 185:2043). Эти мыши были нокаутными по гену Н-2Db и гену мышиного (3-микроглобулина и экспрессировали трансгенную моноцепь молекулы гистосовместимости I класса (HHD-молекулы), в которой С-конец человеческого р2 т ковалентно связан с М-концом химерной тяжелой цепи (HLA-А*0201 α1-α2, H-2Db α3 трансмембранная и интрацитоплазматическая области). Иммунизировали мышей в возрасте от семи до десяти недель (самцов и самок). Средний вес мышей составлял примерно 25-30 г.
1.2.2. Протоколы иммунизации
Мышей разделяли на 4 группы; группу 1 иммунизировали Ad TG 17909 (кодирующим сердцевину HBV, слитую с иммуногенными областями env1, env2 и env4), группу 2 иммунизировали Ad TG17910 (кодирующим усеченный и мутированный Pol*), группу 3 иммунизировали обоими векторами, а группу 4 иммунизировали пустым аденовирусом (Ad TG15149) в качестве отрицательного контроля. Всех животных иммунизировали путем подкожных инъекций в основание хвоста, первая и вторая группы животных получали одну подкожную инъекцию 108 IU каждого аденовируса (TG 17909 или TG17910), третья группа одну подкожную инъекцию смеси, содержащей 2×108 IU каждого аденовируса (всего 2×108 IU: 108 IU Ad TG17 909+108 IU Ad TG17910), а отрицательные контроли получали одну подкожную инъекцию 2×108 IU AdTG15149. Клеточный иммунный ответ оценивали по Elispot IFNγ и по анализу внутриклеточного окрашивания цитокинов (ICS) через две недели после иммунизации.
1.2.3. Пептиды
Пептиды, используемые для стимуляции клеток in vitro, представляли собой либо короткие пептиды длиной от 9 до 10 аминокислот, которые описаны или предсказаны как HLA-А2-рестриктированные эпитопы, либо длинные пептиды длиной 15 аминокислот, включенные в пептидные библиотеки, охватывающие все интересующие антигены.
Короткие пептиды, соответствующие описанным или прогнозируемым HLA-А2-рестриктированным эпитопам полимеразного белка, сердцевинного белка или оболочечных областей, были синтезированы Eurogentec (Бельгия) и растворены в 100% DMSO (Sigma, D2650) в концентрации 10 мМ.
Пептидные библиотеки, охватывающие целиком полимеразный, сердцевинный и оболочечный белки, были синтезированы Prolmmune (Оксфорд, Великобритания). Библиотеки Pol, Core и env состоят из 15-мерных пептидов с перекрытием на 11 аминокислот.Каждый исходный пептид растворяли в 100% DMSO (Sigma, D2650) в концентрации 50 мг/мл. Для каждой библиотеки пептиды объединяли в концентрации по 2 мг/мл на пептид:
Белок HBV Pol покрывается 8 пулами из 24-25 пептидов из библиотеки Pol (пул 1: 24 пептида, охватывающие остатки с 45 по 151; пул 2: 24 пептида, охватывающие остатки со 140 по 251; пул 3: 24 пептида, охватывающие остатки с 241 по 347; пул 4: 24 пептида, охватывающие остатки с 337 по 447; пул 5: 24 пептида, охватывающие остатки с 437 по 543; пул 6: 24 пептида, охватывающие остатки с 533 до 639; пул 7: 24 пептида, охватывающие остатки с 629 по 735; пул 8: 25 пептидов, охватывающих остатки с 725 по 832);
Белок HBV Core покрывается 2 пулами из 21-22 пептидов из библиотеки Core (пул 1: 22 пептида, охватывающие остатки с 1 по 100; пул 2: 21 пептид, охватывающий остатки с 89 по 183);
Белок HBV env покрывается 3 пулами из 6-10 пептидов из библиотеки Env (пул 1: 10 пептидов, охватывающих HBs-остатки с 9 по 59; пул 2: 9 пептидов, охватывающих HBs-остатки со 157 по 194; пул 4: 6 пептидов, охватывающих HBs-остатки со 193 по 226).
1.2.4. Анализы ELISpot IFNγ
Собирали спленоциты от иммунизированных мышей и лизировали эритроциты (Sigma, R7757). 2×105 клеток на лунку культивировали в трех повторах в течение 40 ч в планшетах Multiscreen (Millipore, MSHA S4510), покрытых моноклональным антителом против мышиного IFNy (BD Biosciences; 10 мкг/мл, 551216) в культуральной среде MEM (Gibco, 22571) с добавлением 10% FCS (Sigma, F7524, или JRH, 12003-100M), 80 ЕД/мл пенициллина/80 мкг/мл стрептомицина (PAN, P06-07-100), 2 мМ L-глутамина (Gibco, 25030), 1Х заменимых аминокислот (Gibco, 11140), 10 мМ Hepes (Gibco, 15630), 1 мМ пирувата натрия (Gibco, 31350) и 50 мкМ β-меркаптоэтанола (Gibco, 31350) и в присутствии только 10 ЕД/мл рекомбинантного мышиного IL2 (Peprotech, 212-12) в качестве отрицательного контроля, или с:
- 10 мкМ одного HLA-А2-рестриктированного пептида, присутствующего в HBV-антигенах, кодируемых Ad-векторами (SLY для Pol, FLP, ILC для Core, VLQ, FLG и GLS для env), или нерелевантного пептида;
- пулом пептидов в конечной концентрации 5 мкг/мл каждого пептида;
- 5 мкг/мл конканавалина A (Sigma, C5275) для положительного контроля. IFNγ-продуцирующие Т-клетки количественно оценивали с помощью анализа ELISpot (цитокин-специфического иммуноферментного спот-анализа), как описано выше (Himoudi et al., 2002, J. Virol. 76:12735). Число пятен (соответствующих IFNγ-продуцирующим Т-клеткам) в лунках с отрицательным контролем вычитали из числа пятен, обнаруженных в экспериментальных лунках, содержащих HBV-пептиды. Результаты представлены как среднее значение, полученное в трех лунках с повторами. Экспериментальный порог положительных наблюдаемых реакций (или отсекатель) определяли путем расчета порогового значения, которое соответствует среднему значению пятен, наблюдаемых только для среднего +2 стандартных отклонения для 106 клеток. Техническое отсечение, связанное с CTL ELISpot-ридером, также определяли как 50 пятен/106 клеток (это значение, выше которого CV ридера систематически составляло менее 20%). Наибольшее значение между техническим пороговым значением и экспериментальным порогом, рассчитанным для каждого эксперимента, учитывали для определения порогового значения в каждом эксперименте. Статистический анализ ответов ELISpot проводили с помощью теста Манна-Уитни. Значение р, равное или ниже 0,05, считалось значимым.
1.2.5. Анализы внутриклеточного окрашивания цитокинов (ICS)
ICS выполняли на спленоцитах от каждого животного в каждой группе. После лизиса эритроцитов с помощью литического буфера (Sigma, R7757) 2×106 клеток на лунку в плоскодонном 96-луночном планшете инкубировали в полной культуральной среде альфа-МЕМ (Gibco BRL, 22571) или в присутствии только 10 ЕД/мл мышиного рекомбинантного IL-2 (Peprotech, 212-12) в качестве отрицательного контроля, или с 10 мкМ специфического HBV-пептида, или с пулом пептидов в конечной концентрации 5 мкг/мл каждого пептида, или с 10 мкМ нерелевантного пептида. GolgiPlug (BD Biosciences, 555029) немедленно добавляли в конечной концентрации 1 мкл/мл на 5 часов. Затем клетки собирали в 96-луночные планшеты с V-образным дном лунок и промывали 1% FCS-PBS. Окрашивание проводили с использованием моноклональных антител против CD3 (хомячковое МКА анти-CD3e-РЕ, разведение 1/200), CD8 (крысиное МКА анти-CD8a-APC, разведение 1/600) и CD4 (крысиное МКА анти-CD4-PerCP, разведение 1/600) (все от BD Biosciences, 553063, 553035 и 553052 соответственно) в 50 мкл 1% FCS-PBS в течение 15 мин при комнатной температуре. После промывания клетки фиксировали и пермеабилизировали с помощью Cytofix/Cytoperm и промывали раствором Perm/Wash (BD Biosciences, 554714). Затем антимышиные IFNγ-PE-антитела (BD Bioseiences, 554412557724) и антимышиные ФНОα-А1еха488 антитела (BD Biosciences, 557719) или антимышиные IFNγ-PE-антитела (BD Biosciences 554412557724) и антимышиные IL-2-Alexa488 антитела (BD Bioseiences, 557719) добавляли на 15 мин при комнатной температуре, и после промывания Perm/Wash клетки ресуспендировали в 1% FCS-PBS и анализировали с помощью проточной цитометрии с использованием FACSCalibur (Becton Dickinson), Клетки CD3e+, CD8a+ или клетки CD3e+, CD4+ гейтировали для определения процентов IFNγ+ CD8+ или IFNγ+ CD4+ Т, или TNFα+ CD8+, или TNFα+ CD4+ Т, или IL2+ CD8+, или IL2 CD4+ Т, или IFNγ+ TNFα+ CD8+, или IFNγ+ TNFα+ CD4+, или IFNγ+ IL2+ CD8+, или IFNγ+ IL2+ CD4+ Т-клеточной популяции. Процент, полученный только со средой, считали фоном.
1.2.6. Анализы CTL in vivo
CTL-анализы in vivo проводили, как было описано (Fournillier et al., 2007). Суспензии спленоцитов получали из селезенок сингенных мышей и доводили до 20×106 клеток/мл после лизиса эритроцитов. Треть клеток инкубировали с одним из HBV-специфических пептидов, вторую треть клеток инкубировали с другим HBV-пептидом, все с конечной концентрацией 10 мкМ, в течение 1 часа при температуре 37ºС, а последнюю часть оставляли без воздействия. 5(6)-карбоксифлуоресцеина диацетат сукцинимидил эфир (CFSE) (Molecular probes, С1157) на 10 мин добавляли в концентрации 16 мкМ (CFSE высокий) к клеткам без воздействия, 4 мкМ (CFSE средний) к клеткам, на которые воздействовали пептидом ILC или VLQ, и 1 мкМ (CFSE низкий) к клеткам, на которые воздействовали пептидом SLY или FLP. После промывания PBS три популяции (клетки без воздействия, клетки, на которые воздействовали пептидом ILC и SLY или без воздействия, клетки, на которые воздействовали пептидом FLP и ILC) смешивали, и общее количество клеток 30×106 вводили мышам под наркозом в ретроорбитальные вены (с использованием смеси кетамина-ксилазина-PBS (Ketamine Virbac, Centravet KET204, конечная концентрация 25 мг/мл; Xylazine hydrochloride Rompun Bayer, Centravet, конечная концентрация 5 мг/мл)). Таким образом, популяции с низким и средним CFSE представляли конкретные цели, которые должны быть лизированы цитотоксическими Т-клетками, а популяция с высоким CFSE была внутренним контролем, позволяющим нормировать анализ. Спленоциты от реципиентных мышей анализировали через 24 часа с помощью проточной цитометрии для определения CFSE-меченых клеток. После гейтирования на лимфоцитах (SSC/FSC) второе гейтирование выполняли на основе количества событий/CFSE-флуоресценции (FL1), которое демонстрирует три пика, первый из которых соответствует клеткам с низким CFSE, второй соответствует клеткам со средним CFSE, а третий соответствует клеткам с высоким CFSE. Для каждого животного рассчитывали соотношение CFSE+целей с обработкой пептидом и CFSE + целей без обработки пептидом (R = число клеток с низким CFSE /число клеток с высоким CFSE). Двух наивных мышей использовали для определения контроля R. Процент специфического лизиса для каждого животного определяли по следующей формуле: % лизиса=(1-Rмыши/Rконтрольный)×100. Ответ считался положительным, если процент специфического лизиса был выше 10% (отсечение).
2. Результаты 2.1 Экспрессия антигена вирусными векторами
2.1.1 Экспрессия антигенов из аденовирусных конструкций Ad TG 17909 и Ad TG17910
Экспрессию гибридного белка core-env1-env2-env4 оценивали в анализе вестерн-блоттинг.Клетки А549 (106 клеток) инфицировали в течение 48 часов Ad TG17909 с MOI 10 или 50 или пустым аденовирусом в качестве отрицательного контроля. Клеточные осадки собирали и исследовали с помощью анти-core мышиного моноклонального антитела (С1-5, sc-23 945, Санта-Крус). Как показано на фиг.1А., главную полосу с ожидаемой молекулярной массой (31,6 кДа) обнаружили в образце клеток, инфицированных Ad TG17909.
Экспрессию полипептида Pol* оценивали в вестерн-блоттинге после инфицирования Ad TG17910 А549-клеток. Затем клеточные осадки собирали и исследовали с анти-Pol мышиным моноклональным антителом (8D5, sc-81591, Санта-Крус). Как показано на фиг.1А, полосу с ожидаемой молекулярной массой (88,5 кДа) обнаружили в образце клеток, инфицированных Ad TG17910, вместе с некоторыми субпродуктами (частичными полимеразными белками).
2.1.2 Экспрессия антигенов из MVA-конструкций
Анализ экспрессии Pol*, Pol*-TMR, Core*t, Core*t-env1, Core*t-env1-env2 выполняли в анализе вестерн-блоттинг. Клетки А549 или СЕР инфицировали с MOI 1 или 0,2 MVA TG17842, MVA TG17843, MVA TG17971, MVA TG17972, MVA TG17993 и MVA TG17994, соответственно, в присутствии или в отсутствие протеосомного ингибитора MG-132 (10 мкМ), добавленного в ростовую среду для MVA TG17842 и MVA TG17843. Через 24 часа клетки собирали.
Для MVA TG17842 анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Pol-антитела Нер В Pol (8D5, Санта-Крус, №sc-81591). Как показано на фиг.1В, экспрессию белка с кажущейся молекулярной массой 88,5 кДа обнаружили только в присутствии MG-132. Эта полоса имела ожидаемую молекулярную массу белка Pol*.
Для MVA TG17843 анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Pol-антитела Нер В Pol (8D5, Санта-Крус, №sc-81591). Как показано на фиг.1В, экспрессию белка с кажущейся молекулярной массой 98,2 кДа обнаружили только в присутствии MG-132. Эта полоса имела ожидаемую молекулярную массу белка Pol*-TMR. Следует отметить, что в присутствии MG132 было обнаружено больше продукта и дополнительный продукт с высокой молекулярной массой более 200 кДа.
Для MVA TG17971 анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Core-антитела Нер В cAg (13А9) (Санта-Крус, №sc-23946). Как показано на фиг.1В, была обнаружена экспрессия белка Core* с молекулярной массой 21 кДа, что соответствовало ожидаемой молекулярной массе.
Для MVA TG17972 анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Core-антитела Нер В cAg (13А9) (Санта-Крус, №sc-23946). Как показано на фиг.1В, была обнаружена экспрессия белка Core*t с молекулярной массой15,8 кДа, что соответствовало ожидаемой молекулярной массе.
Для MVATG 17993 и 17994 MVATG анализ вестерн-блоттинг проводили с использованием коммерческого моноклонального анти-Core-антитела Нер В cAg (13А9) (Санта-Крус, №sc-23946). Как показано на фиг.1В, была обнаружена экспрессия белков с молекулярными массами 19,9 и 23,4 кДа, соответственно. Эти полосыа имели ожидаемую молекулярную массу белков Core*t-env1 и Core*t-env1-env2.
2.2. Иммуногенность антигенов, экспрессированных из аденовирусных векторов Ad TG17909 и Ad TG17910
Иммуногенность HBV-полипептидов, экспрессированных аденовирусными векторами, оценивали на HLA-А2-трансгенных мышах, иммунизированных либо Ad TG17909 или Ad TG17910 в одиночку, либо смесью двух аденовирусов. Специфичные Т-клеточные реакции, индуцированные после одной подкожной инъекции, оценивали с помощью ELISpot IFNγ, ICS и анализов цитолиза in vivo с использованием известных (описанных как мишени специфических Т-клеточных реакций у пациентов) HLA-А2-эпитопов, присутствующих в полимеразной, сердцевинной или оболочечной областях и/или в пулах перекрывающихся пептидов, покрывающих HBV-антигены интереса.
2.2.1 Оценка HBV-специфичных IFNY-продуцирующих клеток в анализах ELISpot
Анализы ELISpot IFNγ показали, что Ad TG17910 может индуцировать IFNγ-продуцирующие клетки, специфичные по HLA-А2-рестриктированному эпитопу (SLYADSPSV) (SEQ ID №55, расположенному в HBV-полимеразе в позициях 816-824) (фиг.2А). Иммунизация Ad TG17909 также приводила к высокой частоте индукции IFNγ-продуцирующих клеток, специфичных для двух сердцевинных HLA-А2-рестриктированных эпитопов (FLPSDFFPSV в позициях 18-27 (SEQ ID №56) и ILCWGELMTL в позициях 99-108 (SEQ ID №57), а также для трех оболочечных HLA-А2-рестриктированных эпитопов (VLQAGFFLL (SEQ ID №58) в позициях 14-22 и FLGGTTVCL (SEQ ID №59) в позициях 41-49 (оба присутствуют в env1) и GLSPTVWLSV (SEQ ID №60) в позициях 185-194 (присутствует в env2) (фиг.2В и С). Иммунизация смесью Ad TG17909 и Ad TG17910 также индуцировала сопоставимый уровень специфических IFNγ-продуцирующих клеток, нацеленных на одни и те же эпитопы в трех антигенах, т.е. на эпитоп SLY, присутствующий в Pol, эпитопы FLP и ILC в сердцевинном белке и три эпитопа оболочечных областей (VLQ, FLG и GLS) (фиг.2А, В и С). Частота Т-клеточных реакций, выявляемых после иммунизации одним Ad или смесью двух, была сопоставима, и это показывает, что среди трех антигенов, экспрессированных из описанных векторов, нет ни одного крупного иммунодоминирующего.
2.2.2. Оценка HBV-специфичных IFNγ/TNFα-продуцирующих клеток в анализах внутриклеточного окрашивания
Количество CD8+ Т-клеток, способных продуцировать либо только IFNγ, либо IFNγ+TNFα, нацеленных на HLA-А2-рестриктированные эпитопы, присутствующие в полимеразной (SLY), сердцевинной (FLP and ILC) и оболочечной областях (VLQ, FLG и GLS), оценивали с помощью анализа ICS. Все эти эпитопы были мишенями двойных и простых секретирующих клеток. Полученные результаты представлены на фиг.3. Животные, иммунизированные Ad TG17909 отдельно или в сочетании с Ad TG17910, примерно соответствовали Core- и env-специфическим CD8+ Т-клеточным реакциям (те же проценты специфических CD8+ Т-клеток, продуцирующих IFNγ или IFNγ+TNFα после повторной стимуляции пептидами FLP, ILC, VLQ, FLG и GLS, как показано на фиг.3В и 3С). С другой стороны, что касается полимеразного специфического CD8+ Т-клеточного ответа (SLY-эпитоп), очень высокий процент CD8+ клеток, продуцирующих IFNγ или IFNγ + TNFα, был обнаружен у мышей, получавших Ad TG1710, экспрессирующий Pol* (фиг.3А), а также у мышей, иммунизированных смесью Ad TG17910 и Ad TG17909, хотя на более низком уровне (фиг.3С).
2.2.3. Оценка HBV-специфичных IFNγ/TNFα-продуцирующих CD8+ и CD4+ Т-клеток после иммунизации смесью аденовирусных векторов путем анализов внутриклеточного окрашивания
Проценты CD8+ и CD4+ Т-клеток, способных продуцировать либо только IFNγ, либо IFNγ+TNFα, либо IFNγ+IL2, нацеленных на HLA-А2-рестриктированные эпитопы, присутствующие в полимеразной (SLY), сердцевинной (FLP и ILC) и оболочечной областях (VLQ, FLG и GLS) или в пулах перекрывающихся пептидов, покрывающих сердцевинную и оболочечную области, оценивали с помощью анализа ICS. Все анализируемые HLA-A2-рестриктированные эпитопы были мишенью клеток, секретирующих один и два цитокина (IFNγ и IFNγ+TNFα), и некоторые пулы перекрывающихся пептидов также были мишенью клеток, секретирующих один и два цитокина (IFNγ и IFNγ+TNF и IFNγ+IL2). Результаты показаны на фиг.4. Пять HLA-А2-трансгенных мышей иммунизировали смесью Ad TG17909 и Ad TG17910, а трех HLA-A2-трансгенных мышей иммунизировали Ad TG15149 (отрицательный контроль).
Животные, иммунизированные Ad TG15149, не демонстрировали HBV-специфичных Т-клеточных реакций (данные не представлены). Животные, иммунизированные Ad TG17909 в сочетании с Ad TG17910, демонстрировали сильный CD8+ Т-клеточный ответ, специфичный к целевым антигенам HBV (фиг.4А), с высоким процентом клеток, продуцирующих один (IFNγ) и два цитокина (IFNγ+TNFα), специфичных к HLA-А2-эпитопам, находящимся в полимеразной, сердцевинной и оболочечной областях, и специфичных к пулу пептидов «Core 1» и к пулу пептидов, покрывающих области env1 и env2. Как показано на фиг.4В и 4С, эти вакцинированные мыши также демонстрировали CD4+ Т-клеточные ответы, специфичные к HBV-антигенам, в частности к клеткам, секретирующим один (IFNγ) и два цитокина (IFNγ+TNFα и IFNγ+IL2), специфичным к пулу пептидов «Core 2» и к пулу пептидов, покрывающих env2.
2.2.4. Индукция цитолиза in vivo, измеряемая в CTL-анализах in vivo Способность аденовирусных векторов Ad TG17909 и Ad TG17910 индуцировать in vivo цитолиз клеток, несущих HLA-А2-эпитопы, оценивали в CTL-анализах in vivo. Было протестировано четыре HLA-А2-эпитопа, соответственно SLY (Pol), FLP и ILC (Core) и VLQ (область env 1). Шесть животных иммунизировали смесью Ad TG17909 + Ad TG17910, и двух животных иммунизировали Ad TG15149 (отрицательный контроль). Половину каждой группы (трех мышей, иммунизированных Ad TG17909 + Ad TG17910, и одну мышь, иммунизированную Ad TG15149) анализировали на способность лизировать in vivo клетки, на которые воздействовали SLY-пептидом, и клетки, на которые воздействовали ILC-пептидом. Другую половину анализировали на способность вакцинированных животных лизировать in vivo клетки, на которые воздействовали FLP-пептидом, и клетки, на которые воздействовали VLQ-пептидом. Полученные результаты представлены на фиг.5. Как и ожидалось, у мышей, иммунизированных Ad TG15149, было обнаружено отсутствие HBV-специфического цитолиза in vivo (данные не представлены). In vivo цитолиз против двух сердцевинных эпитопов FLP и ILC был слабым у мышей, иммунизированных Ad TG17909 + Ad TG17910. Тем не менее, напротив, животные, иммунизированные смесью Ad TG17909 и Ad TG17910, демонстрировали сильный in vivo цитолиз против полимеразного эпитопа SLY (фиг.5А) и envl-эпитопа VLQ (фиг.5В), достигая более 50% специфического лизиса в обоих случаях.
Интересно, что сочетание Ad-векторов, экспрессирующих части pol, core и env, позволяет индуцировать специфические Т-клеточные реакции, нацеленные на три HBV-антигена при совместном введении. Индуцированные Т-клетки способны продуцировать один или два цитокина и лизировать in vivo клетки, нагруженные некоторыми HBV-пептидами. Все вместе эти данные свидетельствуют об иммуногенной активности описанных композиций и об их способности индуцировать CD8+ и CD4+ Т-клеточные реакции, когда они векторизованы Ad.
2.3. Иммуногенность антигенов, экспрессированных из MVA-векторов MVA TG17842, MVA TG17843, MVA TG17971, MVA TG17972, MVATG 17993 и MVA TG17994
Иммуногенную активность композиций на основе MVA оценивали на HLA-А2-трансгенных мышах, иммунизированных одним из MVA-векторов, описанных в примерах с 1.1.2 по 1.1.7 (MVA TG17842, MVA 7843, MVA TG17971 или MVA TG17972 поодиночке), либо смесью двух MVA (MVA TG17843 + MVA TG17972, MVA TG17843 + MVA TG17993 или MVA TG17843 + MVA TG17994). Мышей иммунизировали путем трех подкожных инъекций с интервалом в одну неделю, и оценивали специфические Т-клеточные реакции в ELISpot IFNγ и ICS с использованием описанных выше HLA-А2-эпитопов, присутствующих в полимеразной, сердцевинной или оболочечной областях и/или в пулах перекрывающихся пептидов, покрывающих HBV-антигены интереса.
2.3.1. Оценка HBV-специфических IFNγ-продуцирующих клеток в анализах ELISpot после иммунизации MVA, экспрессирующим полимеразу.
Трех мышей иммунизировали либо MVA TG17842, экспрессирующим усеченный и мутированный полимеразный антиген, либо MVA TG17843, экспрессирующим нацеленную на мембрану версию той же усеченной и мутированной полимеразы, или MVA N33.1 (отрицательный контроль). Полимеразо-специфический Т-клеточный ответ оценивали в анализах IFNγ ELISpot с использованием HLA-А2-ограниченного эпитопа SLY и пула пептидов, покрывающих полимеразу. У мышей, иммунизированных MVA N33.1 и MVA TG17842, не было обнаружено HBV-специфического Т-кпеточного ответа (данные не представлены). Тем не менее, как показано на фиг.6А, IFNY-продуцирующие клетки были индуцированы после иммунизации MVA TG17843, который является специфическими для HLA-А2-ограниченного эпитопа SLY и пептидного пула 8, покрывающего С-концевую часть полимеразы (отсутствие специфического ответа против других пептидных пулов 1-7 может быть обнаружено в экспериментальных условиях испытания). Эти данные подчеркивают преимущество экспрессии полимеразы в качестве связанного с мембраной антигена, по меньшей мере в композициях на основе MVA.
2.3.2. Оценка HBV-специфических IFNY-продуцирующих клеток в анализах ELISpot после иммунизации MVA, экспрессирующим сердцевинный белок.
Восьмерых мышей иммунизировали либо MVA TG17971, экспрессирующим сердцевинную часть с удаленными остатками 77-84, либо MVA TG 17972, экспрессирующим ее усеченный вариант (С-концевое усечение с остатка 149) или MVA N33.1 (отрицательный контроль). Специфические Т-клеточные ответы к сердцевинному белку определяли в анализах ELISpot IFNγ с использованием HLA-А2-ограниченных эпитопов (пептиды FLP и ILC) и описанных выше пептидных пулов Core 1 и Core 2. Как показано на фиг.6В, иммунизация MVA TG17971 может вызвать спорадические специфические Т-клеточные реакции против HLA-A2-ограниченных пептидов FLP и ILC и двух пептидных пулов, охватывающих сердцевинный антиген (фиг.6В). Отсутствие специфических Т-клеточных реакций против сердцевинного белка может быть обнаружено у мышей, иммунизированных MVA TG17972 с использованием тестируемых пептидов и в тестируемых экспериментальных условиях (данные не представлены).
2.3.3. Оценка HBV-специфических IFNY-продуцирующих клеток в анализах ELISpot после иммунизации комбинацией векторов MVA
Трех мышей, иммунизированных смесью MVA TG17843 и либо MVA TG17972, либо MVA TG17993, либо MVA TG17994, и HBV-специфические Т-клеточные реакции оценивали способом ELISpot IFNγ с использованием описанных выше пептидов.
- специфические Т-клеточные реакции против полимеразы были обнаружены у подавляющего большинства животных, вакцинированных сочетанием MVA TG17843 + MVA TG17972 (положительные реакции у 2/3 животных, как показано на фиг.7), MVA TG17843 + MVA TG17993 (положительные реакции у 3/3 животных, как показано на фиг.7В) и MVA TG17843 + MVA TG17994 (положительные реакции у 2/3 животных, как показано на фиг.7С). Частота IFNγ-продуцирующих клеток кажется сравнимой с той, которая наблюдается при введении MVA TG17843 в одиночку (фиг.6А), что показывает, что комбинация векторов не приносит вреда иммунному ответу.
- Отсутствие специфической реакций против сердцевинной части может быть обнаружено в тестируемых экспериментальных условиях у любого из вакцинированных животных с использованием HLA-A2 или пула пептидов (данные не представлены).
- Отсутствие специфической реакций против оболочечной части может быть обнаружено в тестируемых экспериментальных условиях у любого из животных, вакцинированных MVA TG17993-содержащей комбинацией, с использованием env1 HLA-A2 или пептидных пулов (данные не представлены).
- У 2 из 3 животных, иммунизированных MVA TG17994-содержащей комбинацией, как показано на фиг.7С, были обнаружены специфические Т-клеточные реакции против env2, которые были направлены как против HLA-A2-ограниченного эпитопа GLS, так и против env2-охватывающего пептидного пула (отсутствие Т-клеточной реакции против env1-области можно было наблюдать в этих экспериментальных условиях; данные не приведены).
ICS-анализы, проведенные в тех же условиях, подтвердили результаты, наблюдаемые в анализах ELISpot (данные не представлены).
Интересно, что комбинация MVA-векторов, экспрессирующих части pol, core и env, позволяет индуцировать специфические Т-клеточные реакции, направленные на антигены pol и env2 при совместном введении.
Все вместе, эти данные свидетельствуют об иммуногенной активности описанных композиций и об их способности индуцировать Т-клеточные реакции против основных HBV-антигенов.
| название | год | авторы | номер документа |
|---|---|---|---|
| Мутантные полимеразы вируса гепатита В | 2012 |
|
RU2625021C2 |
| ВЕКТОРЫ ДЛЯ МНОЖЕСТВЕННОЙ ГЕННОЙ ЭКСПРЕССИИ | 2008 |
|
RU2462513C2 |
| ГИБРИДИЗАЦИЯ ГЕТЕРООЛИГОМЕРНЫХ МИКОБАКТЕРИАЛЬНЫХ АНТИГЕНОВ | 2015 |
|
RU2695462C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| ПОЛИПЕПТИДЫ Е2 ПАПИЛЛОМАВИРУСА, ПРИМЕНЯЕМЫЕ ДЛЯ ВАКЦИНАЦИИ | 2008 |
|
RU2482189C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| Набор консервативных эпитопов вирусных белков, способ их идентификации и отбора, вакцинная композиция и способ вакцинации | 2022 |
|
RU2814020C2 |
| АНТИГЕННЫЕ TAU-ПЕПТИДЫ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2518291C2 |
| ПРОСТАТОАССОЦИИРОВАННЫЕ АНТИГЕНЫ И ИММУНОТЕРАПЕВТИЧЕСКИЕ СХЕМЫ НА ОСНОВЕ ВАКЦИН | 2013 |
|
RU2737765C2 |
| ПРИМЕНЕНИЕ MVA (МОДИФИЦИРОВАННЫЙ ВИРУС КОРОВЬЕЙ ОСПЫ АНКАРА) ДЛЯ ЛЕЧЕНИЯ РАКА ПРОСТАТЫ | 2008 |
|
RU2499606C2 |









































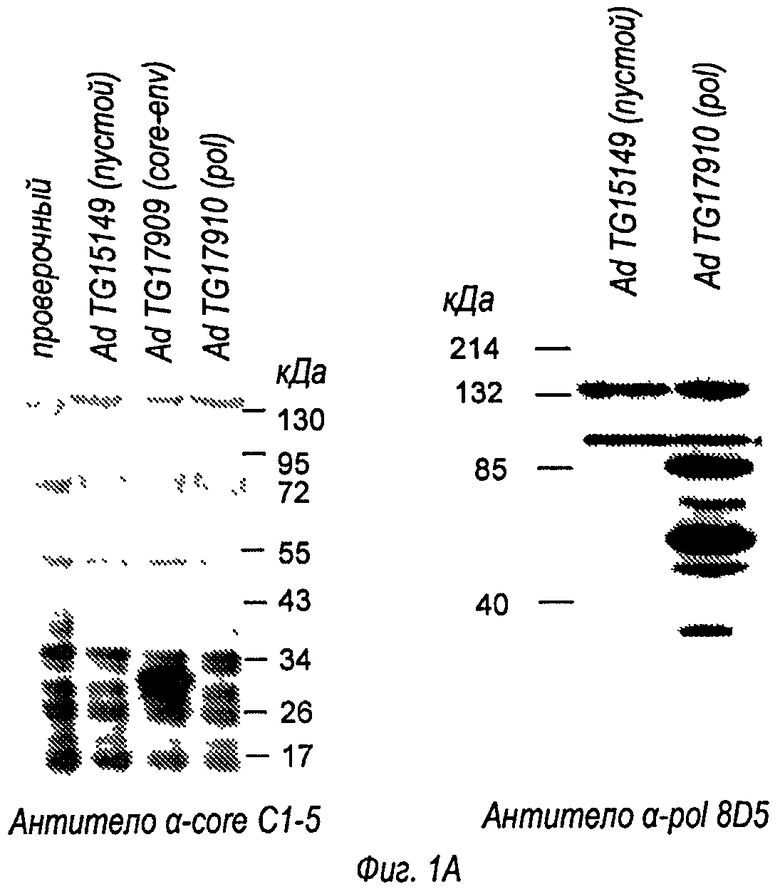
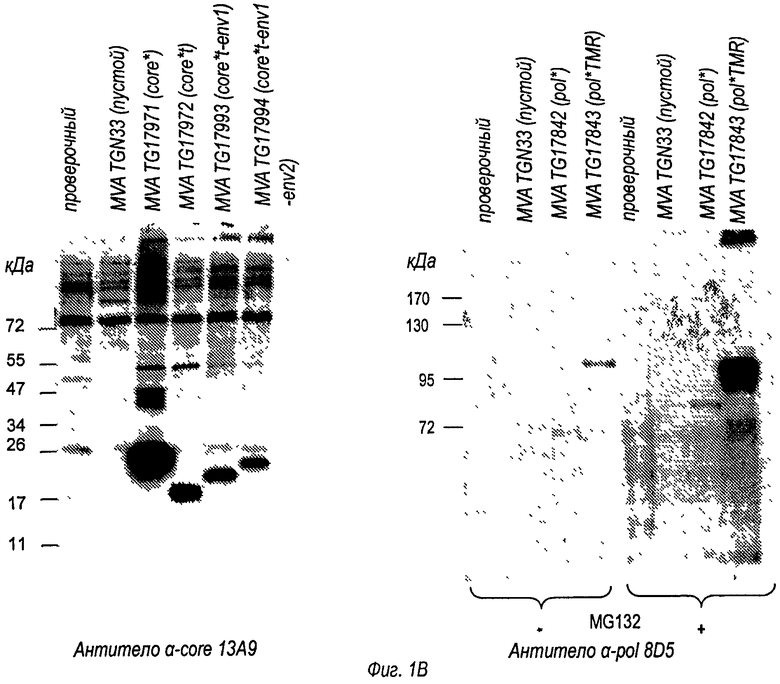
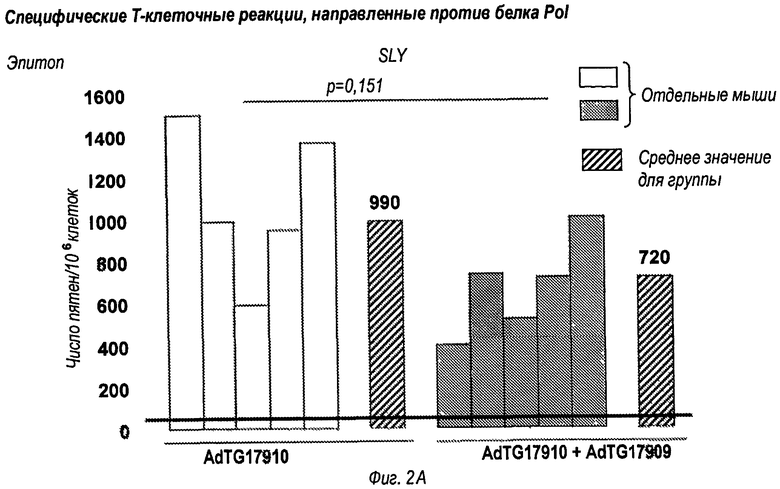

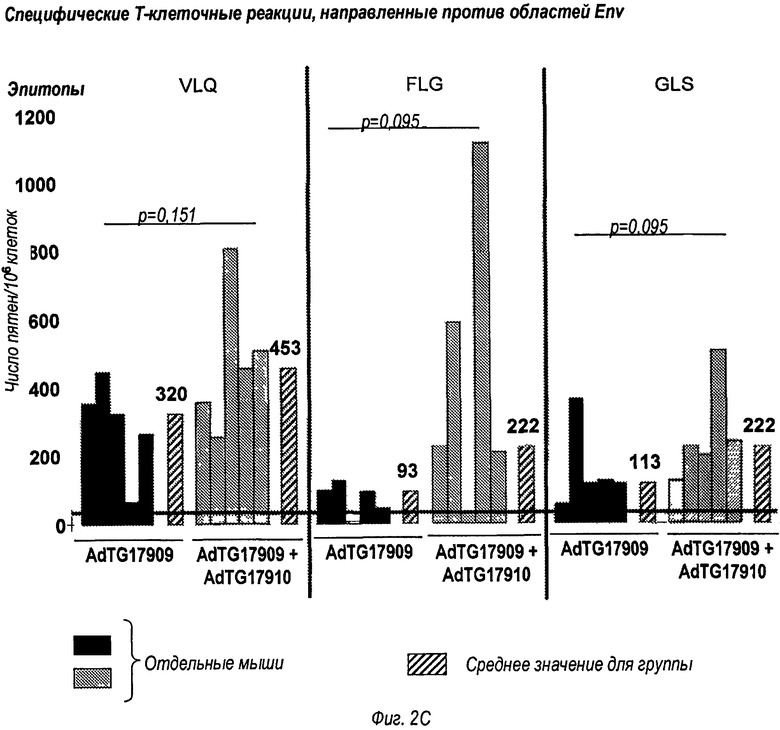
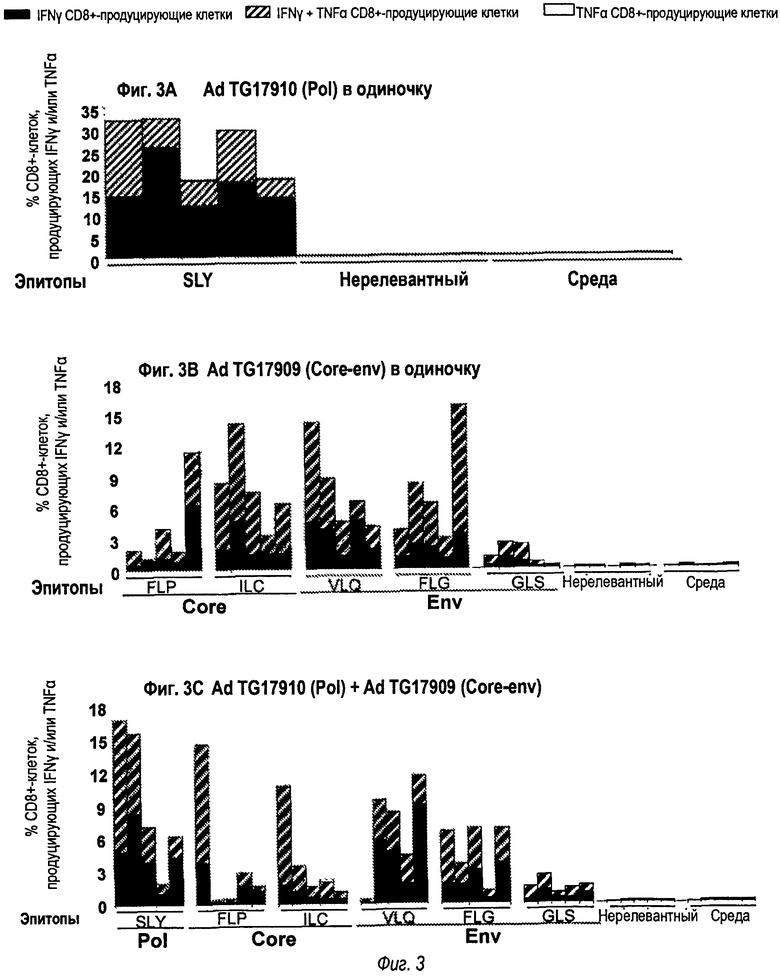
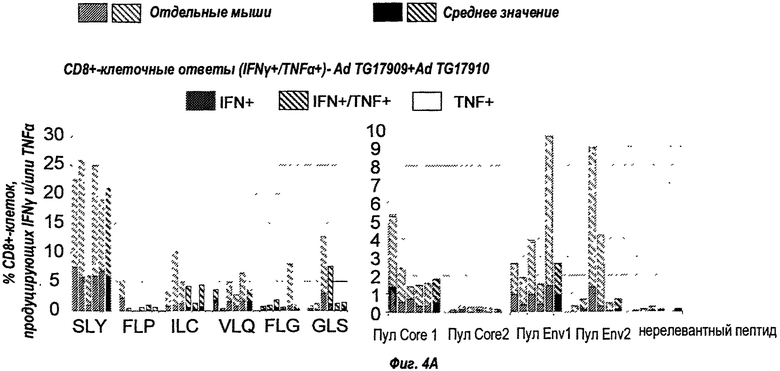
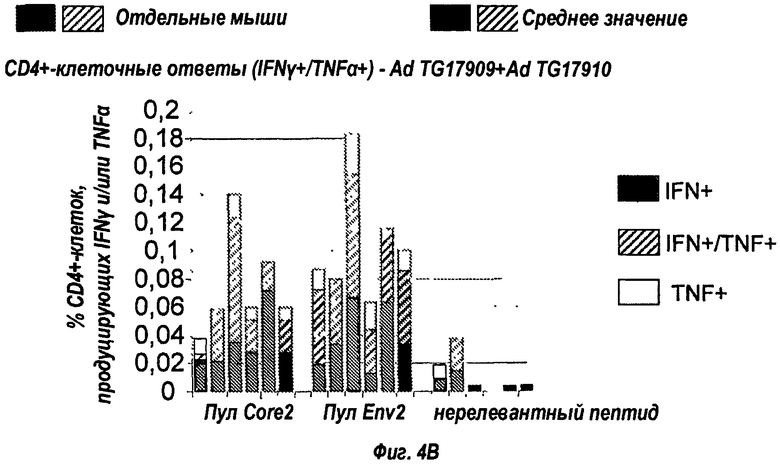

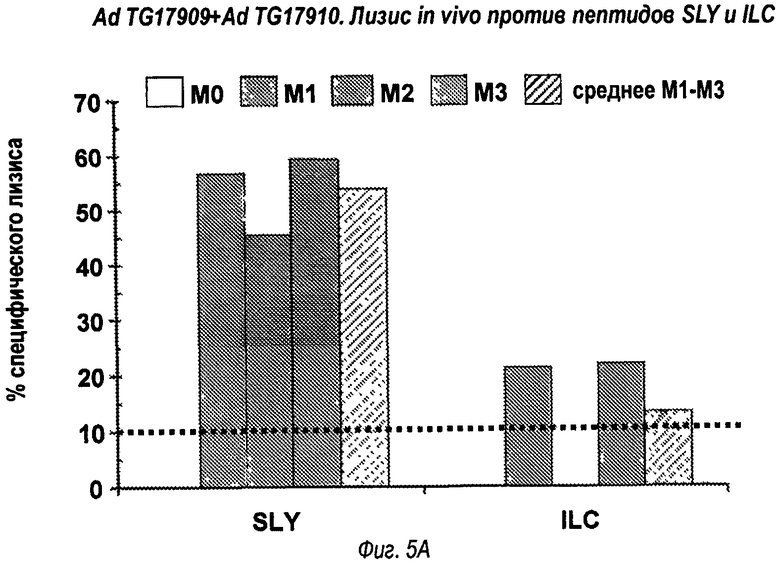

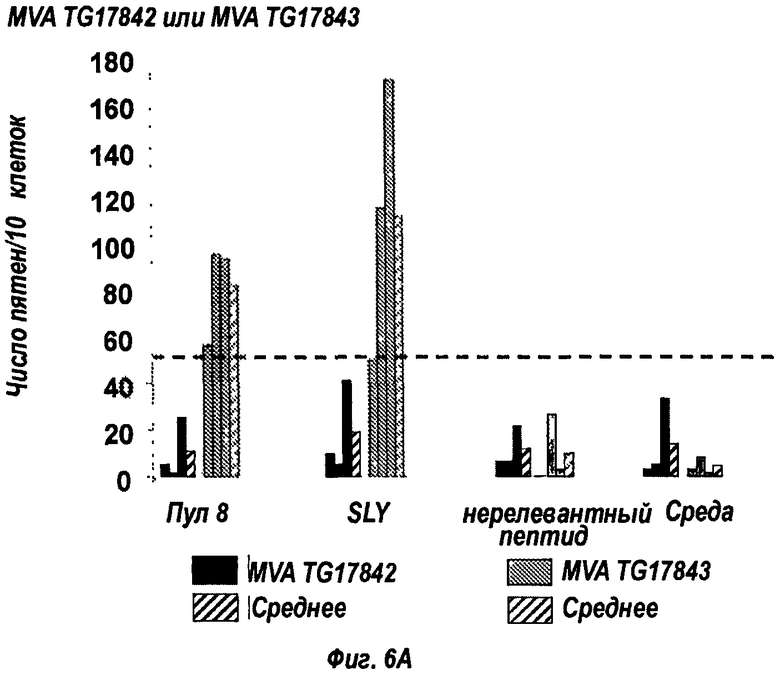
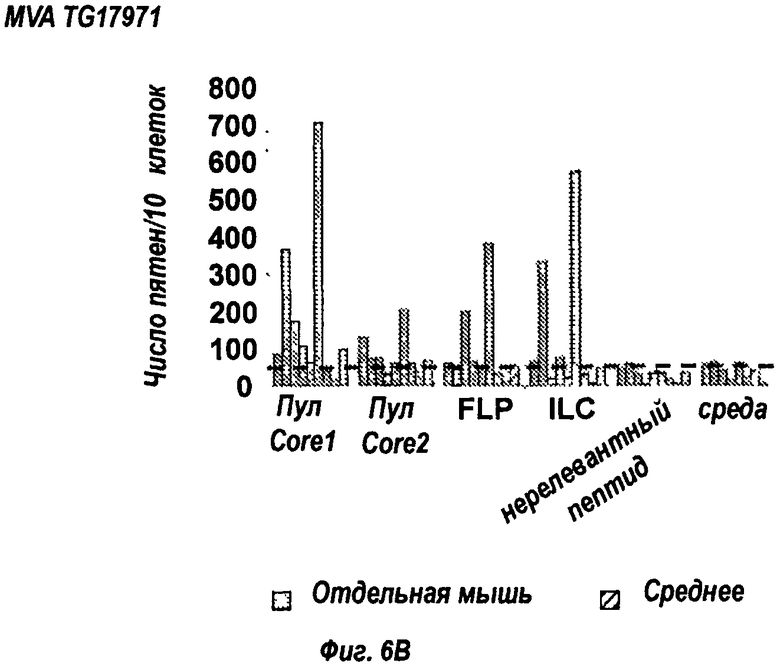
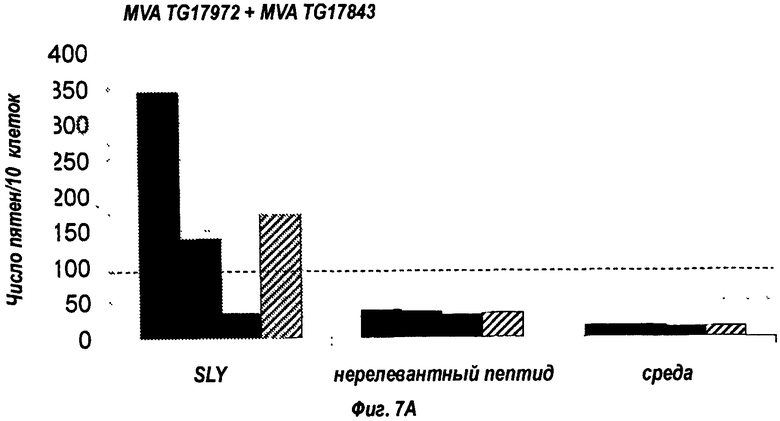

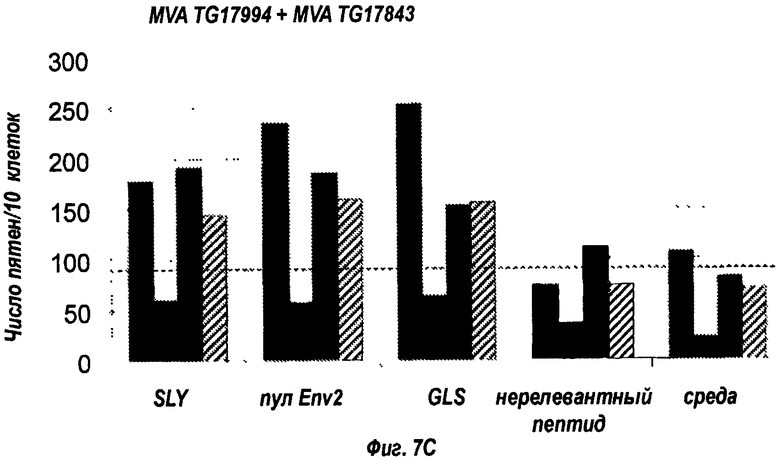
Группа изобретений относится к медицине и касается иммуногенной композиции, которая содержит комбинацию следующих частей: (i) полимеразной части, содержащей по меньшей мере 450 аминокислотных остатков полимеразного белка, полученного из первого HBV-вируса; (ii) сердцевинной части, содержащей по меньшей мере 100 аминокислотных остатков сердцевинного белка, полученного из второго HBV-вируса; и (iii) оболочечной части, содержащей одну или более иммуногенную область из 20-100 последовательных аминокислотных остатков белка HBsAg, полученного из третьего HBV-вируса; или комбинацию нуклеиновокислотных молекул, кодирующих указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть. Группа изобретений также касается нуклеиновокислотной молекулы для стимуляции или повышения иммунного ответа на HBV; вектора для стимуляции или повышения иммунного ответа на HBV, содержащего одну или более нуклеиновокислотную молекулу; инфекционной вирусной частицы для стимуляции или повышения иммунного ответа на HBV; способа лечения или профилактики HBV-инфекции или HBV-связанного заболевания или патологического состояния; набора для применения в лечении HBV-инфекции. Группа изобретений обеспечивает стабильность и сохранение высокой иммуногенности векторов, используемых в композиции. 10 н. и 41 з.п. ф-лы, 7 ил., 2 пр.
1. Иммуногенная композиция, которая содержит комбинацию следующих частей:
(i) полимеразной части, содержащей по меньшей мере 450 аминокислотных остатков полимеразного белка, полученного из первого HBV-вируса;
(ii) сердцевинной части, содержащей по меньшей мере 100 аминокислотных остатков сердцевинного белка, полученного из второго HBV-вируса; и
(iii) оболочечной части, содержащей одну или более иммуногенную область из 20-100 последовательных аминокислотных остатков белка HBsAg, полученного из третьего HBV-вируса;
или комбинацию нуклеиновокислотных молекул, кодирующих указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть.
2. Иммуногенная композиция по п. 1, где указанные первый, второй и третий HBV-вирусы имеют разные генотипы.
3. Иммуногенная композиция по п. 1, где по меньшей мере два из указанных первого, второго и третьего HBV-вирусов имеют один и тот же HBV-генотип и предпочтительно указанные первый, второй и третий HBV-вирусы имеют один и тот же генотип.
4. Иммуногенная композиция по п. 3, где первый, второй и третий HBV-вирусы имеют генотип D и предпочтительно представляют собой HBV-изолят Y07587.
5. Иммуногенная композиция по п. 3, где указанные первый, второй и третий HBV-вирусы имеют генотип В или С.
6. Иммуногенная композиция по. 1, где указанная полимеразная часть модифицирована путем усечения по меньшей мере 20 аминокислотных остатков и не более 335 аминокислотных остатков, обычно присутствующих на N-конце нативной HBV-полимеразы, и предпочтительно первых 47 или 46 аминокислотных остатков после инициаторного остатка Met, расположенного на N-конце нативной HBV-полимеразы.
7. Иммуногенная композиция по п. 6, где указанная полимеразная часть включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №7.
8. Иммуногенная композиция по п. 1, где указанная полимеразная часть модифицирована таким образом, что демонстрирует сниженную обратнотранскриптазную (ОТазную) ферментативную активность по сравнению с нативной HBV-полимеразой, и предпочтительно включает замену остатка Asp, соответствующего позиции 540 в SEQ ID №1 и позиции 494 в SEQ ID №7, на любой аминокислотный остаток, отличный от Asp, и предпочтительно на остаток His.
9. Иммуногенная композиция по п. 1, где указанная полимеразная часть модифицирована таким образом, что демонстрирует сниженную ферментативную активность РНКазы Н по сравнению с нативной HBV-полимеразой, и предпочтительно включает замену остатка Glu, соответствующего позиции 718 в SEQ ID №1 и позиции 672 в SEQ ID №7, на любой аминокислотный остаток, отличный от Glu, и предпочтительно на остаток His.
10. Иммуногенная композиция по п. 9, где указанная полимеразная часть включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №8, или с аминокислотной последовательностью, приведенной в SEQ ID №7, с заменой остатка Asp в позиции 494 на остаток His и с заменой остатка Glu в позиции 672 на остаток His.
11. Иммуногенная композиция по п. 1, где указанная полимеразная часть слита в рамке считывания с гетерологичной гидрофобной последовательностью и предпочтительно в рамке считывания с сигнальным и трансмембранным пептидами гликопротеина вируса бешенства, и где указанная сигнальная последовательность вируса бешенства слита в рамке считывания с N-концом, а указанная трансмембранная последовательность вируса бешенства слита в рамке считывания с С-концом указанной полимеразной части.
12. Иммуногенная композиция по п. 11, где указанная полимеразная часть содержит аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №9.
13. Иммуногенная композиция по п. 1, где указанная сердцевинная часть модифицирована путем усечения по меньшей мере 10 аминокислотных остатков и не более 40 аминокислотных остатков, которые обычно присутствуют на С-конце нативной сердцевины HBV, и предпочтительно 35 аминокислотных остатков нативной сердцевины HBV.
14. Иммуногенная композиция по п. 13 где указанная сердцевинная часть включает аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №10.
15. Иммуногенная композиция по п. 1, где указанная сердцевинная часть модифицирована таким образом, что демонстрирует сниженное распознавание и/или взаимодействие с белком оболочки HBV по сравнению с нативной сердцевиной HBV, и предпочтительно содержит аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №11.
16. Иммуногенная композиция по п. 1, где указанная одна или более иммуногенная область, используемая в данном изобретении, выбрана из группы, состоящей из:
- части оболочечного белка (HBsAg), которая продолжается от позиции 14 до 51 (область env1),
- части оболочечного белка (HBsAg), которая продолжается от позиции 165 до 194 (область env2);
- части оболочечного белка (HBsAg), которая продолжается от позиции 81 до 106 (область env3);
- части оболочечного белка (HBsAg), которая продолжается от позиции 202 до 226 (область env4); а также
- любой их комбинации.
17. Иммуногенная композиция по п. 16, где указанная иммуногенная область(области) содержит аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с любой из аминокислотных последовательностей, приведенных в SEQ ID №12-15.
18. Иммуногенная композиция по п. 17, где указанная оболочечная часть содержит аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №16 или SEQ ID №17.
19. Иммуногенная композиция по п. 1, где указанная полимеразная, сердцевинная и/или оболочечная части слиты в рамке считывания в единую полипептидную цепь попарно или все вместе.
20. Иммуногенная композиция по п. 19, где указанная оболочечная часть слита в рамке считывания с С-концом сердцевинной части.
21. Иммуногенная композиция по п. 20, где указанный гибридный полипептид с сердцевинной и оболочечной частями выбран из группы, состоящей из:
- полипептида, содержащего аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №18;
- полипептида, содержащего аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №19; и
- полипептида, содержащего аминокислотную последовательность, которая демонстрирует по меньшей мере 80% идентичность с аминокислотной последовательностью, приведенной в SEQ ID №20, или с частью аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251, или, альтернативно, с частью аминокислотной последовательности, приведенной в SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221; или с их делецированными версиями, в которых отсутствуют остатки 77-84 в SEQ ID №20.
22. Нуклеиновокислотная молекула для стимуляции или повышения иммунного ответа на HBV, кодирующая комбинацию полимеразной части, определенной в любом из пп. 1 и 6-12, сердцевинной части, определенной в любом из пп. 1 и 13-15, и оболочечной части, определенной в любом из пп. 1 и 16-21.
23. Нуклеиновокислотная молекула по п. 22, где указанная нуклеиновокислотная молекула содержит нуклеиновокислотную молекулу, выбранную из группы, состоящей из:
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №21;
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №22;
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №21, с заменой нуклеотида G в позиции 1480 на С, нуклеотида G в позиции 2014 на С и нуклеотида А в позиции 2016 на Т;
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №23;
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №24;
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №25; и
- нуклеиновокислотной молекулы, содержащей нуклеотидную последовательность, которая демонстрирует по меньшей мере 80% идентичность с нуклеотидной последовательностью, приведенной в SEQ ID №26, или с частью нуклеотидной последовательности, приведенной в SEQ ID №26, начиная с 1 нуклеотида и заканчивая 753 нуклеотидом, или с частью нуклеотидной последовательности, приведенной в SEQ ID №26, начиная с 1 нуклеотида и заканчивая 663 нуклеотидом, или с их делецированными версиями, в которых отсутствует часть, продолжающаяся от G в позиции 229 до А в позиции 252 SEQ ID №26.
24. Вектор для стимуляции или повышения иммунного ответа на HBV, содержащий одну или более нуклеиновокислотную молекулу по п. 22 или 23.
25. Вектор по п. 24, который представляет собой плазмидный вектор.
26. Вектор по п. 24, который представляет собой вирусный вектор.
27. Вектор по п. 26, где указанный вирусный вектор является репликативно-дефектным аденовирусным вектором и предпочтительно аденовирусным вектором, который содержит указанную нуклеиновокислотную молекулу, вставленную вместо Е1-области.
28. Вектор по п. 26, где указанный вирусный вектор является поксвирусным вектором и, в частности, получен из вируса оспы канареек, вируса оспы кур или вируса осповакцины, при этом указанный вирус осповакцины предпочтительно является модифицированным штаммом Анкара (MVA).
29. Вектор по любому из пп. 24-28, который несет нуклеиновокислотные молекулы, кодирующие указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть.
30. Вектор по любому из пп. 24-28, который несет только одну или две нуклеиновокислотные молекулы, кодирующие указанную полимеразную часть, указанную сердцевинную часть и указанную оболочечную часть.
31. Вектор по п. 30, который выбран из группы, состоящей из:
(i) MVA-вектора, содержащего нуклеиновокислотную молекулу, находящуюся под контролем промотора вируса осповакцины, такого как промотор 7.5 К, и кодирующую полимеразную часть, содержащую аминокислотную последовательность, указанную в SEQ ID №7, 8 или 9 или в SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и с заменой остатка Glu в позиции 672 на остаток His;
(ii) MVA-вектора, содержащего нуклеиновокислотную молекулу, находящуюся под контролем промотора вируса осповакцины, такого как промотор pH5r, и кодирующую сердцевинную часть и оболочечную часть, которые содержат аминокислотную последовательность, указанную в SEQ ID №18 или 19;
(iii) Е1-дефектного Ad-вектора, содержащего вставленную вместо Е1-области нуклеиновокислотную молекулу, находящуюся под контролем CMV-промотора и кодирующую полимеразную часть, содержащую аминокислотную последовательность, указанную в SEQ ID №7, 8 или 9 или в SEQ ID №7 с заменой остатка Asp в позиции 494 на остаток His и с заменой остатка Glu в позиции 672 на остаток His; и
(iv) Е1-дефектного Ad-вектора, содержащего вставленную вместо Е1-области нуклеиновокислотную молекулу, находящуюся под контролем CMV-промотора и кодирующую сердцевинную часть и оболочечную часть, которые содержат аминокислотную последовательность, указанную в SEQ ID №18, 19 или 20, или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 251, или часть SEQ ID №20, начиная с остатка 1 и заканчивая остатком 221.
32. Инфекционная вирусная частица для стимуляции или повышения иммунного ответа на HBV, содержащая нуклеиновокислотную молекулу по любому из пп. 22-23 или вектор по любому из пп. 24-31.
33. Инфекционная вирусная частица по п. 32, которая получена в процессе, включающем следующие этапы:
(а) введение вирусного вектора изобретения в подходящую клеточную линию;
(б) культивирование указанной клеточной линии в соответствующих условиях для получения указанной инфекционной вирусной частицы,
(в) восстановление полученной инфекционной вирусной частицы из культуры указанной клеточной линии, и
(г) возможно, очистка указанной восстановленной инфекционной вирусной частицы.
34. Композиция для стимуляции или повышения иммунного ответа на HBV, содержащая нуклеиновокислотную молекулу по любому из пп. 22-23, вектор по любому из пп. 24-31 или инфекционную вирусную частицу по п. 32 или 33 и фармацевтически приемлемый носитель.
35. Композиция по п. 34, которая также содержит один или более адъювант, подходящий для системного применения или применения на слизистой у людей.
36. Композиция по п. 34 или 35, которая разработана для внутримышечного или подкожного введения.
37. Композиция по п. 34 или 35, которая содержит от примерно 105 до примерно 1013 инфекционных единиц вирусного вектора или инфекционной вирусной частицы.
38. Иммуногенная композиция для стимуляции или повышения иммунного ответа на HBV, содержащая комбинацию по меньшей мере двух независимых векторов, вместе кодирующих полимеразную часть, сердцевинную часть и оболочечную часть, определенных в пп. 1 и 2-7.
39. Применение по меньшей мере одной иммуногенной композиции по любому из пп. 1-21, нуклеиновокислотной молекулы по любому из пп. 22-23, вектора по любому из пп. 24-31, инфекционной вирусной частицы по п. 32 или 33 или композиции по любому из пп. 34-38 для изготовления лекарственного препарата, предназначенного для лечения или профилактики HBV-инфекции или HBV-связанного заболевания или патологического состояния.
40. Применение по п. 39, где лекарственный препарат предназначен для лечения пациента с хронической инфекцией вирусом гепатита В.
41. Способ лечения или профилактики HBV-инфекции или HBV-связанного заболевания или патологического состояния, который включает введение в организм человека или животного, который нуждается в этом, эффективного количества по меньшей мере одной иммуногенной композиции по любому из пп. 1-21, нуклеиновокислотной молекулы по любому из пп. 22-23, вектора по любому из пп. 24-31, инфекционной вирусной частицы по п. 32 или 33 или композиции по любому из пп. 34-38.
42. Способ лечения по п. 41, где HBV-инфекция представляет собой хроническую инфекцию вируса гепатита В.
43. Способ или применение по любому из пп. 39-42, которое проводится в комбинации со стандартной медицинской помощью.
44. Способ или применение по п. 43, где стандартная медицинская помощь включает цитокины и/или нуклеотидные или нуклеозидные аналоги, выбранные из группы, состоящей из ламивудина, энтекавира, телбивудина, адефовира, дипивоксила и тенофовира.
45. Способ индукции или стимуляции иммунного ответа против HBV в организме хозяина, включающий введение в указанный организм по меньшей мере одной иммуногенной композиции по любому из пп. 1-21, нуклеиновокислотной молекулы по любому из пп. 22-23, вектора по любому из пп. 24-31, инфекционной вирусной частицы по п. 32 или 33 или композиции по любому из пп. 34-38.
46. Способ индукции или стимуляции иммунного ответа по п. 45, где иммунный ответ представляет собой CD4+ и/или CD8+ - опосредованный ответ.
47. Набор для применения в лечении HBV-инфекции, который включает множество активных агентов, выбранных из группы, состоящей из иммуногенной композиции по любому из пп. 1-21, нуклеиновокислотной молекулы по любому из пп. 22-23, вектора по любому из пп. 24-31, инфекционной вирусной частицы по п. 32 или 33 или композиции по любому из пп. 34-38.
48. Набор по п. 47, который включает первый вектор, содержащий нуклеиновокислотную молекулу, кодирующую полимеразу часть, которая определена в любом из пп. 1 и 6-12, и второй вектор, содержащий нуклеиновокислотную молекулу, кодирующую сердцевинную и оболочечную часть, которые определены в любом из пп. 1 и 13-21.
49. Набор по п. 48, где первый указанный вектор является MVA-вектором, который определен в п. 31(i), а указанный второй вектор является MVA-вектором, который определен в п. 31(ii).
50. Набор по п. 48, где указанный первый вектор является Ad-вектором, который определен в п. 31(iii), а указанный второй вектор является Ad-вектором, который определен в п. 31(iv).
51. Набор по любому из пп. 48-50, также включающий третий вектор, экспрессирующий иммуномодулятор.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| DEPLA E., et al., Rational design of a multiepitope vaccine encoding T-lymphocyte epitopes for treatment of chronic hepatitis B virus infections | |||
| J.Virol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| BARTENSCHLAGER R., et al., The P gene product of hepatitis B | |||

