Область техники
Изобретение относится к области биоинформатики и иммунологии, а именно, к in silico способу идентификации и отбора высоко консервативных эпитопов белков вируса гриппа, а также к рекомбинантному вектору экспрессии, кодирующему один или несколько эпитопов, идентифицированных указанным способом, к вакцинной композиции, предназначенной для иммунизации позвоночного, например, млекопитающего, содержащей рекомбинантный вектор экспрессии, кодирующий набор эпитопов белков вируса гриппа, идентифицированных указанным способом, совместно с иммунологически приемлемым носителем, а также к способу вакцинации позвоночного, например, млекопитающего, иммунологически эффективным количеством указанной вакцинной композиции.
Уровень техники
Инфекционные заболевания, в том числе вирусные, согласно данным Всемирной организации здравоохранения (далее - ВОЗ), являются одной из основных причин смерти или инвалидизации людей [Health statistics and information systems. Disease burden and mortality estimates. - URL: https://www.who.int/healthinfo/global_burden_disease/estimates/en]. Среди всех патогенных для людей вирусов необходимо отметить РНК-содержащие вирусы как группу, имеющую важнейшее клиническое значение, представляющую собой существенную угрозу для человека.
Генетический материал РНК-содержащих вирусов представлен рибонуклеиновой кислотой (РНК), как правило, одноцепочечной или, реже, двухцепочечной [Patton J.T. (editor). Segmented Double-stranded RNA Viruses: Structure and Molecular Biology. - Caister Academic Press, 2008].
Группа РНК-содержащих вирусов включает, без ограничения, коронавирусы, в частности, открытый в конце 2019 г. вирус SARS Cov-2, ротавирусы, вирусы гриппа, вирусы геморрагических лихорадок, вирус полиомиелита, вирус гепатита С, флавивирусы, в том числе, вирус Эбола, ретровирусы, в том числе, вирус иммунодефицита человека, а также вирус паротита (свинки), вирус кори и др. Особо следует отметить арбовирусы, которые переносятся членистоногими, например, клещами или москитами. К числу арбовирусов, имеющих важное клиническое значение, относятся вирус клещевого энцефалита, вирус Денге, вирус Зика и вирус желтой лихорадки.
Также очень важное клиническое значение имеют вирусы гриппа: по оценкам ВОЗ, во всем мире ежегодные эпидемии гриппа приводят к примерно 3-5 миллионам случаев тяжелой болезни и примерно от 290 000 до 650 000 смертей [Influenza. Burden of disease. // Электронный ресурс; URL: https://www.who.int/influenza/surveillance_monitoring/bod/en/]. Уровень смертности варьирует среди разных возрастных групп и достигает максимума у детей до 5 лет, пожилых людей и лиц с хроническими заболеваниями органов дыхания и сердечно-сосудистой системы [Nair Н., et al., The Lancet, 2011. - Vol. 378. - No. 9807. - P. 1917-1930]. Расчеты экспертов в США показали, что за последние годы эпидемии гриппа наносят ежегодно ущерб этой стране, составляющий от 71 до 167 миллиардов долларов. В России на грипп и острые респираторные вирусные инфекции (ОРВИ) ежегодно приходится до 90% от всей регистрируемой инфекционной заболеваемости [Организация и практическая реализация системы эпидемического надзора за гриппом и острыми респираторными заболеваниями (ОРЗ) в России. - Грипп и другие респираторные вирусные инфекции: эпидемиология, профилактика, диагностика и терапия / И.Г. Маринич, В.А. Кондратьев, Д.Ф. Житенев / Под ред. О.И. Киселева, И.Г. Маринича, А.А. Сомининой. СПб., 2003. - Гл. 6. - С. 147-156]. По данным Роспотребнадзора, в 2018 г. экономические потери от гриппа и ОРВИ составили более 80% от всего ущерба, наносимого инфекционными болезнями [О состоянии санитарно-эпидемиологического благополучия населения в Российской Федерации в 2018 году: Государственный доклад. - М.: Федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека, 2019. - 254 с.].
В уровне техники в работах различных авторов неоднократно предпринимались попытки предложить такие способы отбора высоко консервативных эпитопов для различных белков вирусов позвоночных (в частности, птиц или млекопитающих и, особенно, человека), имеющих важное клиническое значение, которые позволяли бы подобрать высоко консервативные эпитопы вирусных белков, которые затем можно было бы использовать для создания вакцины против соответствующего вируса. При отборе таких эпитопов необходимо помнить, что эффективная вакцинация против патогена подразумевает хорошую индукцию двух веток приобретенного иммунного ответа (гуморального и клеточного) против него. Известно, что В-клеточные эпитопы, а также CD4+ Т-клеточные и CD8+ Т-клеточные эпитопы играют ключевую роль в стимуляции этих иммунных реакций. Также известно, что существует положительная взаимосвязь между дозой антигена и силой иммунного ответа [A. Jegerlehner, Т. Storni, G. Lipowsky et al., European Journal of Immunology (2002), vol. 32, pp. 3305-3314].
Один из современных подходов к созданию вакцин против патогенов, в том числе вирусных патогенов, заключается в использовании для создания вакцин эпитопов, соответствующих консервативным иммуногенным последовательностям вирусных белков, - так называемых эпитопных вакцин [Hans D, Young Р, Fairlie D., Med Chem. (2006); vol. 2, issue 6, pp. 627-46] (так называемый эпитопный подход).
Эпитопный подход использует Т-клеточные эпитопы - короткие пептиды длиной от 8 до 10 аминокислотных остатков, активирующие Т-клетки, и В-клеточные эпитопы - длинные пептиды длиной до 20 аминокислотных остатков [Rötzschke О, Falk K, Stevanović S, Jung G, Walden P, Rammensee H., Eur J Immunol. (1991); 21(11):2891-4]. В уровне техники неоднократно, как на животных моделях, так и в клинических исследованиях, была продемонстрирована эффективность таких вакцин против различных инфекционных заболеваний человека, в т.ч., против вирусных заболеваний [Engler О, Dai W, Sette A, Hunziker I, Reichen J, Pichler W, Cemy A., Mol Immunol. (2001); 38(6):457-65] или ВИЧ [Gahery H, Daniel N, Charmeteau B, Ourth L, Jackson A, Andrieu M, Choppin J, Salmon D, Pialoux G, Guillet J., AIDS Res Hum Retroviruses (2006); 22(7):684-94; Pinto L, Berzofsky J, Fowke K, et al., AIDS (1999); 13(15): 2003-12].
Из работы Molero-Abraham М., Lafuente Е.М. et al., Clin Dev Immunol. 2013; 2013: 601943 известен способ отбора высоко консервативных эпитопов РНК-содержащего вируса, имеющего важнейшее клиническое значение, - вируса гепатита С, включающий в себя:
(i) отбор в специализированных базах данных, таких как EPIMHC ["EPIMHC: A Database of Naturally Processed МНС-restricted Peptide Ligands and Epitopes for Customized Computational Vaccinology" // Электронный ресурс; URL: http://bio.med.ucm.es/epimhc/; P. A. Reche, H. Zhang, J. Glutting, and E. L. Reinherz, Bioinformatics, vol. 21, no. 9, pp. 2140-2141, 2005] и IEDB ["The Immune Epitope Database". // Электронный ресурс; URL: www.iedb.org; В. Peters, J. Sidney, P. Bourne et al., PLoS Biology, vol. 3, no. 3, p. e91, 2005], HCV-специфических эпитопов CD8 Т-лимфоцитов;
(ii) определение согласно методике, предложенной в работе С.Е. Shannon "The mathematical theory of communication" - The Bell System Technical Journal, vol. 27, pp. 379-423; 623-656, 1948, вариабельности каждого эпитопа с использованием энтропии Шеннона (Н) в качестве меры вариабельности эпитопов, где энтропию Шеннона определяют по формуле
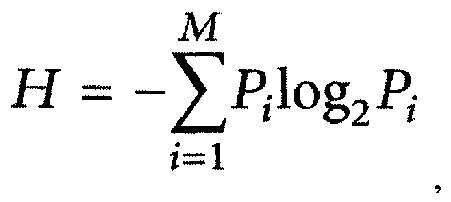
где Pi - доля остатков аминокислоты типа i, а М - общее число протеиногенных аминокислот, равное 20, с последующим отбором тех эпитопов, для которых значение Н не превышает 0,5;
(iii) прогнозирование способности эпитопов, отобранных на этапе (ii), связываться с молекулами HLA I класса, причем указанное прогнозирование осуществляют с использованием специальных весовых матриц согласно работам Р. А. Reche, J.-P. Glutting, Н. Zhang, and Е. L. Reinherz, Immunogenetics, vol. 56, no. 6, pp. 405-419, 2004, P. A. Reche, J. P. Glutting, and E. L. Reinherz, "Prediction of MHC class I binding peptides using profile motifs" - Human Immunology, vol. 63, no. 9, pp. 701-709, 2002 и P. A. Reche and E. L. Reinherz, Methods in Molecular Biology, vol. 409, pp. 185-200, 2007;
(iv) выявление оптимальных комбинаций эпитопов с заданным покрытием популяционного иммунитета, причем оптимальные комбинации эпитопов с заданным покрытием популяционного иммунитета в работе Molero-Abraham М., Lafuente Е.М. et al., 2013 выявляли, используя исходный набор CD8 Т-клеточных эпитопов и строя профили связывания указанных эпитопов с молекулами HLA I, что позволило авторам данной работы вычислить кумулятивную частоту фенотипа и покрытие популяционного иммунитета для конкретного эпитопа. Зная профили связывания эпитопов HLA I, Molero-Abraham М., Lafuente Е.М. et al., начиная с одного эпитопа и итеративно увеличивая на каждом шаге число рассматриваемых эпитопов, отыскивали в исходном наборе такие комбинации эпитопов, которые обеспечивали бы достижение заданного значения РРС, при этом на каждом шаге такой итерации Molero-Abraham М., Lafuente Е.М. et al. вычисляли максимально возможное для заданного числа эпитопов значение РРС, принимая также во внимание, что в разных этнических группах различные аллели гена HLA I встречаются с разной частотой.
Высоко консервативные эпитопы белков вируса гепатита С, отобранные согласно Molero-Abraham М., Lafuente Е.М. et al., затем могут быть использованы для создания вакцин против вируса гепатита С, обладающих широким спектром действия.
Однако в самой работе Molero-Abraham М., Lafuente Е.М. et al. протективная активность отобранных высоко консервативных эпитопов белков вируса гепатита С, т.е. способность указанных эпитопов обеспечивать реципиенту, иммунитет к вирусу гепатита С, не была исследована. Таким образом, работа Molero-Abraham М., Lafuente Е.М. et al. ограничилась in silico отбором высоко консервативных эпитопов белков вируса гепатита С без последующей практической проверки протективной активности этих эпитопов на адекватной экспериментальной модели, и не привела к созданию вакцины против вируса гепатита С на основе отобранных эпитопов.
Кроме того, способ отбора высоко консервативных эпитопов белков вируса гепатита С, предложенный Molero-Abraham М., Lafuente Е.М. et al., использует для выравнивания последовательностей программное обеспечение "Muscle", которое предполагает ограничение на число выравниваемых последовательностей.
Также из уровня техники из работы Chong Li Chuin и Khan Asif M. Identification of highly conserved, serotype-specific dengue virus sequences: implications for vaccine design. - BMC Genomics 2019, 20 (Suppl 9): 921, известен способ отбора высоко консервативных эпитопов другого РНК-содержащего вируса, имеющего важное клиническое значение, -вируса денге, возбудителя лихорадки денге. Способ, предложенный Chong Li Chuin и Khan Asif M., схематически представленный на фигуре 1 указанной работы, включает в себя:
(а) сбор данных, включающий получение выборки последовательностей белков вируса денге из базы данных "NCBI Entrez Protein database" (URL: https://www.ncbi.nlm.nih.gov/protein/);
(б) обработку данных, включающую построение на выборки последовательностей, полученной на этапе (а), базы данных с возможностью поиска по ней;
(в) поиск с помощью алгоритма "Blastp" [URL: https://blast.ncbi.nlm.nih.gov/Blast.cgi?], описанного в работе S.F. Altschul, W. Gish, W. Miller, et al., J Mol Biol. 1990 Oct 5; 215(3): 403 - 10, с целью извлечения из базы данных, построенной на этапе (б), каждого серотипа белка вируса денге, с последующим исключением из отобранного набора дублирующих и нерелевантных последовательностей;
(г) построение для каждого полученного массива данных множественного выравнивания последовательностей с последующей проверкой вручную построенного выравнивания и, при необходимости, исключением ошибочного выравнивания;
(д) анализ собранной на этапах (а) - (г) информации, включающий расчет энтропии Шеннона и анализ взаимной информации (согласно работам Shannon СЕ., Bell Syst Tech J 1948; 27: 379-423, 623-56, Miotto О, Heiny AT, Tan TW, August JT, Brusic V., BMC Bioinform. 2008; 9 (Suppl 1):S18 и Miotto O, Heiny AT, Albrecht R, García-Sastre A, Tan TW, August JT, et al., PLoS One 2010; 5(2):e9025). Энтропию на данном этапе рассчитывали для перекрывающихся нонамерных фрагментов белков вируса денге, при этом нонамерные фрагменты (нонамерные пептиды) согласно работе Chong Li Chuin и Khan Asif M. были выбраны из-за их соответствия по длине эпитопам HLA I класса, а также из-за их соответствия по длине "ядра" эпитопам HLA II класса, причем энтропию нонамерного фрагмента белка вируса денге Н(х) в заданном положении выравнивания х вычисляли по формуле 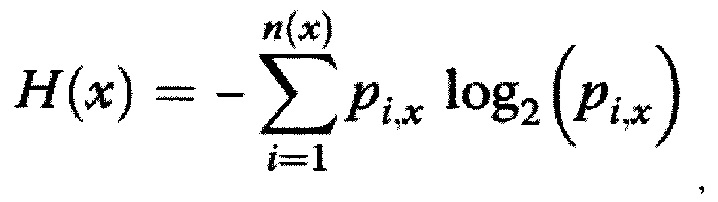
где p(i,x) - вероятность [наличия в последовательности] конкретного нонамерного пептида i с начальной аминокислотой х. Высоко консервативные позиции (высоко консервативные аминокислотные остатки) будут давать низкие значения энтропии, определяемой согласно указанной формуле, а нулевое значение энтропии будет соответствовать полностью консервативным позициям (полностью консервативным аминокислотным остаткам).
В свою очередь, анализ взаимной информации, использованный в работе Chong Li Chuin и Khan Asif M., является мерой, определяющей зависимость между двумя переменными. Для таких двух переменных величин (А и В) такая мера (MI) в общем случае определяется по формуле:
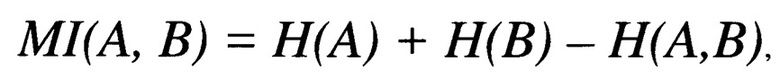
где Н(А) - энтропия переменной А, Н(В) - энтропия переменной В, а Н(А,В) - общая энтропия А и В, вычисляемая согласно вышеприведенной формуле путем замены i на (А, В), где (А, В) - набор всех возможных пар значений А и В. Высокая степень различий между двумя массивами данных (А и В) даст высокое значение MI (максимально возможное значение будет равно 1), тогда как низкое значение MI (вплоть до 0) будет указывать на сходное распределение аминокислот в двух наборах.
Согласно работе Chong Li Chuin и Khan Asif M., эпитопы белков вируса денге отбирали из открытых баз данных иммуногенных эпитопов вирусных белков, таких как база иммуногенных эпитопов "IEDB" (Vita R., Overton J.A., Greenbaum J.A., Ponomarenko J., Clark J.D., Cantrell J.R., et al., Nucleic Acids Res. 2015; 43 (Database issue): D405-12).
Однако работа Chong Li Chuin и Khan Asif M. также ограничилась in silico отбором высоко консервативных эпитопов белков вируса денге, без последующей практической проверки протективной активности этих эпитопов на адекватной экспериментальной модели, и не привела к созданию вакцины против вируса денге на основе отобранных эпитопов.
Кроме того, работа Chong Li Chuin и Khan Asif M. использует программное обеспечение, предполагающее ограничение на число выравниваемых последовательностей. Другим минусом этой работы может быть использование программы blastp для подбора выборки, которую потом авторы использовали для оценки консервативности. Алгоритм blast заведомо ищет наиболее похожие последовательности, тем самым завышая в выборке общий уровень консервативности.
Также из уровня техники из работы Sheikh Q.M., Gatherer D., et al., Bioinformatics 2016 Nov 1; 32(21): 3233-3239 известен способ отбора иммуногенных эпитопов еще одного РНК-содержащего вируса, имеющего важное клиническое значение, - вируса гриппа А. Способ, предложенный Sheikh Q.M., Gatherer D., et al., включал в себя отбор CD4+ и CD8+ Т-клеточных эпитопов вируса гриппа А из онлайн-базы IEDB (URL: www.iedb.org; см. также: Peters В., Sidney J., Bourne P. et al., PLoS Biology, vol. 3, no. 3, p. e91, 2005), построение множественного выравнивания с использованием полипротеина вируса гриппа А, полученного из Ресурса вируса гриппа при Национальном центре биотехнологической информации [Bao, Y., Bolotov, P., Dernovoy, D., Kiryutin, В., Zaslavsky, L., Tatusova, Т. et al., Journal of Virology (2008), 82, 596-601] и из базы данных исследований гриппа [Squires, R. В., Noronha, J., Hunt, V., Garcia-Sastre, A., Macken, C., Baumgarth, N. et al., Influenza and Other Respiratory Viruses (2012), 6,404-416], исключение идентичных последовательностей, которые могут привести к сдвигу при выявлении консервативных областей, и неполных последовательностей, и обработку наборов данных, относящиеся к 12 различным белкам вируса гриппа А, с помощью двух различных веб-серверов кластеризации, включая "BLASTclust" [Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D.J., J Mol Biol (1990), 215, 403-410; URL: http://toolkit.tuebingen.mpg.de/blastclust] и "CD-HIT" [Fu, L. M., Niu, B. F., Zhu, Z. W., Wu, S. T. & Li, W. Z., Bioinformatics (2012), 28, 3150-3152; URL: http://weizhong-lab.ucsd.edu/cdhit_suite/cgibin/index.cgi?cmd=ed-hit].
Для каждого из анализируемых белков вируса гриппа А в работе Sheikh Q.M., Gatherer D., et al. выполняли анализ вариабельности последовательностей, используя веб-сервис "Protein Variability Server" (PVS) (Garcia-Boronat, M. et al., Nucleic Acids Research (2008), 36, W35-W41), используя в качестве меры вариабельности последовательностей энтропию Шеннона (согласно методике, предложенной в работе Stewart J.J., Lee C.Y., Ibrahim S., Watts P., Shlomchik M., Weigert M., Litwin S., Mol Immunol. (1997) 34:1067-1082); порог вариабельности принимали равным 0,5. Согласно Sheikh Q.M., Gatherer D., et al., энтропия Шеннона находилась в диапазоне от 4,3 (при данной величине энтропии все аминокислоты представлены с равной частотой в данной позиции) до 0,0 (инвариантная позиция, в которой может находиться только какой-то один определенный аминокислотный остаток). На первом этапе работы Sheikh Q.M., Gatherer D., et al. получали консенсусные последовательности, в которых вариабельные позиции, для которых Н имело положительные значения, экранировали, затем отбирали те CD4+ и CD8+ Т-клеточные эпитопы, которые точно совпадали по всей длине с генерированными консенсусными последовательностями; таким образом, были отобраны только те эпитопы, в которых не было аминокислотных остатков со значением Н, превышающим 0,5.
Покрытие популяционного иммунитета, соответствующее суммарной фенотипической частоте HLA-аллелей, ограничивающих Т-клеточные эпитопы, в работе Sheikh Q.M., Gatherer D., et al. вычисляли, используя данные, касающиеся генетических частот релевантных аллелей HLA в популяции, с помощью сервисов "EPISOPT" (версия 1) (URL: http://imed.med.ucm.es/EPISOPT.html) и средств прогнозирования покрытия популяционного иммунитета, предусмотренных базой данных "IEDB" (URL: http://tools.immuneepitope.org/tools/population/iedb_input), согласно методике, предложенной в работе Сао, K., Hollenbach, J., Shi, X., et al., Human Immunology (2001), 62, 1009-1030. Использование Sheikh Q.M., Gatherer D., et al. референсного набора аллелей HLA I класса, представляющего собой аллели, которые часто встречаются в популяции, позволило авторам данной работы идентифицировать в качестве наиболее консервативных 6 CD8+ Т-клеточных эпитопа (GILGFVFTL, FMYSDFHFI, GMDPRMCSL, SVKEKDMTK, FYIQMCTEL и DTVNRTHQY) и 3 CD4+ Т-клеточных эпитопа (KGILGFVFTLTVPSE, EYIMKGVYINTALLN, ILGFVFTLTVPSERG). Было показано, что указанные наборы CD8+ и CD4+ Т-клеточных эпитопов позволяют охватить соответственно 96% и 95% подтипов вируса гриппа А, циркулирующих в США, и 97% и 88% подтипов вируса гриппа А, циркулирующих в мире.
Однако работа Sheikh Q.M., Gatherer D., et al., как и названные выше работы, также ограничилась in silico отбором высоко консервативных Т-клеточных эпитопов белков вируса гриппа А, без поиска и отбора В-клеточных эпитопов и без последующей практической проверки протективной активности отобранных Т-клеточных эпитопов на адекватной экспериментальной модели, и не привела к созданию вакцины против вируса гриппа А на основе отобранных эпитопов.
Наконец, как и работы, указанные выше, работа Sheikh Q.M., Gatherer D., et al. использует для построения множественного выравнивания последовательностей программное обеспечение, содержащее ограничение на число выравниваемых последовательностей.
В уровне техники в принадлежащем компании BIONDVAX PHARMACEUTICALS LTD [IL] патенте US 9353159 В2, 31.05.2016 и в работе Atsmon J., Kate-Ilovitz Е., Shaikevich D., et al., Journal of Clinical Immunology (2012), vol. 32, No. 3, pp.595-603 описана предлагаемая в качестве ближайшего аналога вакцины, заявляемой в настоящем изобретении, эпитопная вакцина "Multimeric-001" против гриппа, содержащая 9 консервативных линейных эпитопов белков вируса гриппа, включая 5 эпитопов из гена гемагглютинина (НА), в том числе четыре В-клеточных эпитопа и один CD4+ Т-клеточный эпитоп, а также 3 эпитопа из гена нуклеопротеина (NP), включая два CD8+ Т-клеточных эпитопа и один CD4+ Т-клеточный эпитоп, и 1 эпитоп из гена матричного белка (M1), содержащий признаки как В-клеточного, так и CD8+ Т-клеточного эпитопа, экспрессируемые в клетке-хозяине в виде единого полипротеина. В то же время, ни в работе Atsmon J., Kate-Ilovitz Е., Shaikevich D., et al., Journal of Clinical Immunology (2012), vol. 32, No. 3, pp. 595-603, ни в патенте US 9353159 B2 не раскрывается сам алгоритм отбора указанных линейных эпитопов белков вируса гриппа.
При этом вакцина "Multimeric-001" является пептидной вакциной, т.е. содержит сами вышеуказанные В-клеточные и Т-клеточные эпитопы белков вируса гриппа в виде отдельных пептидов. Такие пептидные вакцины, как хорошо известно специалистам в данной области, часто бывают мало иммуногенными и требуют дополнительного использования адъювантов для усиления их иммуногенности [Purcell A.W., McCluskey J., Rossjohn J., Nat. Rev. Drug Discov. 2007; 6: 404 - 414; Aguilar J.C., Rodriguez E.G., Vaccine 2007; 25: 3752 - 3762].
Ввиду вышеуказанных недостатков известных из уровня техники способов отбора эпитопов белков РНК-содержащих вирусов и эпитопных вакцин, содержащих эпитопы белков патогенных вирусов, отобранные такими способами, по-прежнему остается актуальной задача разработки способа, который позволял бы идентифицировать и отбирать высоко консервативные и иммуногенные эпитопы вирусных белков с целью дальнейшего создания вакцин широкого спектра действия против соответствующих вирусов, содержащих такой набор эпитопов. Такой способ должен давать возможность сократить сроки создания новых вакцин, а также, при появлении новых вариантов патогенных вирусов, отличающихся от известных вариантов данных вирусов в области выбранных эпитопов, должен давать возможность в кратчайшие сроки добавлять дополнительные эпитопы или целые антигены-, в конечном счете сокращая, таким образом, сроки создания вакцин против новых вариантов известных патогенных вирусов.
При осуществлении такого способа должны достигаться следующие технические результаты:
(i) обеспечение способа идентификации и отбора высоко консервативных и высоко иммуногенных эпитопов вирусных белков;
(ii) обеспечение отбора эпитопов вирусных белков, пригодных для создания вакцин широкого спектра действия;
Также остается актуальной задача, заключающаяся в создании на основе отобранных эпитопов вакцины, обеспечивающей иммунитет реципиентам -позвоночным животным, например, таким как птицы или млекопитающие, против РНК-содержащих вирусов и при этом лишенной недостатков таких известных из уровня техники вакцин, как "Multimeric-001".
Одним из способов преодолеть указанные недостатки является использование векторных вакцин, т.е. вакцин, содержащих не сами полипептиды эпитопов вирусных белков, а полинуклеотиды, кодирующие такие полипептиды. Преимущества векторных вакцин, как хорошо известно специалистам в данной области, заключаются в их высокой иммуногенности, в их способности индуцировать устойчивый иммунный ответ и усиливать клеточный иммунный ответ, а также в их способности обеспечивать высокоспецифичную доставку таких полинуклеотидов в целевые клетки [Ura Т., Okuda K., Shimada М., Vaccines (Basel) 2014 Sep; 2(3): 624 - 641].
На сегодняшний день, из уровня техники неизвестны векторные вакцины против РНК-содержащих вирусов, которые были бы получены с использованием эпитопного подхода. В связи с этим, одна из задач предлагаемого изобретения заключается в создании такой вакцины против вируса гриппа.
В рамках настоящего изобретения указанные задачи решаются, а технические результаты - достигаются за счет предлагаемого в настоящем изобретении способа in silico идентификации и отбора высоко консервативных белков вируса гриппа, согласно которому:
а) указанные линейные эпитопы вирусных белков отбирают таким образом, чтобы они стимулировали обе ветви приобретенного иммунитета человека, включая гуморальный иммунитет и клеточный иммунитет,
при этом, если на этапе (а) такие эпитопы отобраны,
б) среди эпитопов вирусных белков, отобранных на этапе (а), отбирают высоко консервативные CD8+ и CD4+ Т-клеточные и высоко консервативные В-клеточные эпитопы, причем отбор высоко консервативных CD8+ и CD4+ Т-клеточных и высоко консервативные В-клеточных эпитопов включает в себя:
б1) составление множественного выравнивания последовательностей аминокислотных остатков эпитопов вирусных белков, отобранных на этапе (а);
б2) оценку консервативности указанных эпитопов вирусных белков, причем, оценку консервативности указанных эпитопов на этапе (б2) осуществляют, определяя количество типов аминокислотных остатков k*, встречающихся в данной колонке выравнивания, причем k* вычисляют по формуле (1)
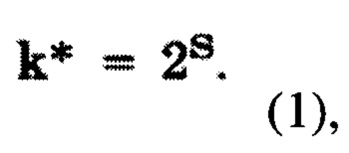
где S - информационная энтропия соответствующей колонки, определяемая по формуле (2)
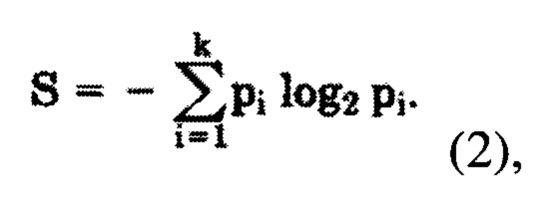
где pi - частота встречаемости i-того аминокислотного остатка в исследуемой колонке выравнивания,
k - количество всех типов остатков, встречающихся в исследуемой колонке выравнивания,
при этом 1-й аминокислотный остаток в колонке выравнивания считают консервативным, если для него выполняется условие 1≤ ki*<1,5,
а затем
в) картируют высоко консервативные В-клеточные эпитопы, отобранные на этапе (б), на данные рентгеноструктурного анализа антигена для проверки условия их доступности для антител,
а если на этапе (а) такие эпитопы не отобраны,
г) выполняют биоинформатическое предсказание для CD8+ и CD4+ Т-клеточных и для В-клеточных эпитопов, получая таким образом набор эпитопов, характеризующихся заданным баллом связывания,
после чего
д) выполняют в отношении набора эпитопов, полученного на этапе (г), этапы (б) - (в), получая таким образом итоговый набор иммуногенных высоко консервативных линейных эпитопов вирусных белков.
Уровень техники может без ограничения включать в себя патенты, опубликованные патентные заявки, а также доступные онлайн базы данных и опубликованные в научной периодике и в непериодической печати монографии, статьи, диссертации на соискание ученой степени и иные источники информации.
Далее, указанные задачи в настоящем изобретении решаются, а технические результаты - достигаются также за счет предлагаемого в настоящем изобретении рекомбинантного вектора экспрессии, кодирующего полипептид, составленный из эпитопов, идентифицированных указанным способом.
Далее, указанные задачи в настоящем изобретении решаются, а технические результаты - достигаются также за счет предлагаемой в настоящем изобретении вакцины (вакцинной композиции) для иммунизации позвоночных против вирусов гриппа, содержащая рекомбинантный вектор экспрессии, кодирующий полипептид, составленный из эпитопов, идентифицированных указанным способом, и иммунологически приемлемый носитель.
Наконец, указанные задачи в настоящем изобретении решаются, а технические результаты достигаются за счет предлагаемого в настоящем изобретении способа вакцинации позвоночного против вируса гриппа, где указанный способ включает введение указанному позвоночному иммунологически эффективного количества вакцинной композиции.
Предпочтительные варианты осуществления изобретения
Ниже в настоящем описании приведены предпочтительные варианты осуществления настоящего изобретения. При этом специалисту понятно, что данные варианты являются неограничивающими и что представленные в настоящем описании предпочтительные варианты осуществления изобретения служат лишь целям иллюстрации изобретения, но не целям ограничения объема притязаний. В предпочтительных вариантах осуществления изобретения, относящегося к способу идентификации и отбора высоко консервативных эпитопов вирусных белков, но без ограничения этими вариантами, отбор линейных эпитопов вирусных белков на этапе (а) осуществляют из уровня техники, включая, без ограничения, базы данных вирусных белков, доступных посредством сети Интернет, или, также без ограничения, статьи или патенты. В более предпочтительных неограничивающих вариантах осуществления изобретения, отбор линейных эпитопов вирусных белков на этапе (а) осуществляют из онлайн-баз данных вирусных белков, доступных посредством сети Интернет, например (без ограничения), из таких, как база данных IEDB [URL: www.iedb.org], описанная в работе Peters В., Sidney J., Bourne P. et al., PLoS Biology, vol. 3, no. 3, p. e91, 2005, и база данных EPIMHC [URL: bio.med.ucm.es/epimhc], описанная в работе Reche P.A., Zhang H., Glutting J., and Reinherz E.L., Bioinformatics, vol. 21, no. 9, pp. 2140-2141, 2005.
В предпочтительных вариантах осуществления изобретения, но без ограничения ими, отбор иммуногенных эпитопов вирусных белков на этапе (а) осуществляют на основании упоминания соответствующего эпитопа в качестве иммуногенного в непатентной и патентной литературе, являющейся частью предшествующего уровня техники, при этом непатентную и патентную литературу, являющуюся частью предшествующего уровня техники.
Предпочтительно, хотя и без ограничения данным сроком, уровень техники изучают с ретроспективной глубиной не менее 5, более предпочтительно, не менее 10, еще более предпочтительно, не менее 15, еще более предпочтительно, не менее 20, наиболее предпочтительно, не менее 25 лет.
Также в предпочтительных вариантах осуществления изобретения, но без ограничения ими, составление множественного выравнивания последовательностей аминокислотных остатков эпитопов вирусных белков, отобранных на этапе (а), на этапе (б1) осуществляют с использованием программного обеспечения MAFFT версии 7 или выше [Kazutaka Katoh and Daron M. Standley, Molecular Biology and Evolution, vol. 30, Issue 4, April 2013, Pages 772-780] с последующим редактированием выравнивания вручную в программе Jalview [Waterhouse, A.M., Procter, J.B., Martin, D.M.A, Clamp, M. and Barton, G. J. (2009) "Jalview Version 2 - a multiple sequence alignment editor and analysis workbench" - Bioinformatics, 25 (9) 1189-1191, doi: 10.1093/bioinformatics/btp033], при этом в ходе такого редактирования удаляются последовательности с выравниванием меньше 40% относительно других, подстроки последовательности размером меньше 5, расположенные внутри гэпов, сдвигаются к ближайшему выравненному блоку, столбцы, содержащие только гэпы или аминокислотные остатки не больше 5, удаляются.
В отличие от программных средств, используемых для построения множественного выравнивания в уровне техники, например, в работах Molero-Abraham М., Lafuente Е.М. et al., 2013, Chong Li Chuin и Khan Asif M., 2019 и Sheikh Q.M., Gatherer D., et al., 2016, программное обеспечение, используемое в настоящем изобретении, не содержит ограничений на число выравниваемых последовательностей.
Также в предпочтительных вариантах осуществления изобретения, но без ограничения ими, оценку консервативности указанных эпитопов на этапе (б2) осуществляют с помощью скрипта, написанного на языке программирования Python [https://github.com/lioj/bioinformatics/blob/master/py/MSA_stat.py, опубликовано: 20.07.2020 г.]. На входе скрипту подаются множественное выравнивание последовательностей и идентификатор референсной последовательности, присутствующей в множественном выравнивании. Для каждого столбца множественного выравнивания вычисляются частоты аминокислотных остатков или гэпов (пробелов), которые есть в этом столбце и консервативность с помощью формул (1) и (2) этапа (б2). На выходе скрипт выдает массив данных в виде таблицы, в которой нумерация осуществляется по референсной последовательности и для каждой позиции имеется информация по частотному составу аминокислотных остатков или гэпов и оценки консервативности.
Также в предпочтительных вариантах осуществления изобретения, но без ограничения ими, биоинформатическое предсказание на этапе (г) выполняют с помощью вычислительных алгоритмов, позволяющих предсказать положение линейных В-клеточных эпитопов [Jespersen МС, Peters В, Nielsen М, Marcatili P. BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes. - Nucleic acids research 45 2017], а также с помощью вычислительных алгоритмов, позволяющих предсказать положение Т-клеточных эпитопов, способных связываться с определенной молекулой главного комплекса гистосовместимости (МНС) [Rammensee Н, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: database for MHC ligands and peptide motifs. - Immunogenetics 50 1999]. В более предпочтительных, также неограничивающих, вариантах осуществления изобретения биоинформатическое предсказание на этапе (г) выполняют с помощью программно-вычислительных средств, доступных посредством сети Интернет и обладающих пользовательским интерфейсом, в которых реализованы такие вычислительные алгоритмы. В наиболее предпочтительных вариантах осуществления изобретения, но без ограничения ими, биоинформатическое предсказание для В-клеточных и для Т-клеточных эпитопов на этапе (г) выполняют с помощью доступных онлайн-посредством сети Интернет сервисов, таких как "BepiPred-2.0" [BepiPred-2.0: Sequential B-Cell Epitope Predictor. - URL: http://www.cbs.dtu.dk/services/BepiPred/], описанный в работах Pernille Haste Andersen, Morten Nielsen, and Ole Lund, Protein Sci. 2006 Nov; 15(11): 2558-2567; Jens Erik Pontoppidan Larsen, Ole Lund, and Morten Nielsen, Immunome Res. 2006; 2: 2, и "Syfpeithi" [Epitope prediction. - URL: http://www.svfpeithi.de/bin/MHCServer.dll/EpitopePrediction.htm], описанный в работе H. - G. Rammensee, J. Bachmann, N. P. N. Emmerich, O. A. Bachor, and S. Stevanovic, Immuno genetics (1999), vol. 50, pp. 213-219, соответственно. Онлайн-сервисы "BepiPred-2.0" и "Syfpeithi" были разработаны для предсказания положения и структуры В-клеточных и Т-клеточных эпитопов соответственно. Указанные онлайн-сервисы позволяют определить для каждого эпитопа предсказанный балл связывания (англ. predicted binding score).
На сегодняшний день BepiPred 2.0 является единственной программой, которая предсказывает линейные В-клеточные эпитопы, основываясь на алгоритме, обученном на эпитопах, аннотированных из белковых структур антиген-антитело. Данные о таких эпитопах считаются более качественными, что позволяет улучшить предсказание по сравнению с другими инструментами (НММ, LBTope) [Jespersen МС, Peters В, Nielsen М, Marcatili P. BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic acids research. 45 2017; Larsen JE, Lund O, Nielsen M. Improved method for predicting linear B-cell epitopes. Immunome research. 2: (2). 2006; Singh H, Ansari HR, Raghava GP. Improved method for linear B-cell epitope prediction using antigen's primary sequence. PloS one 8(5):e62216 2013]. На выходе программа выдает таблицу, в которой для каждой позиции последовательности выдается определенная оценка антигенности. Порог 0.5 является оптимальным. При этом большее значение указывает на большую антигенность.
Программа SYFPEITHI предсказывает эпитопы МНС класса I и II, используя подход на основе мотивов. Вероятностная модель строится на основе данных по экспериментально подтвержденным эпитопам. Для алеллей МНС строятся матрицы мотивов (вероятностный образ эпитопа для определенного аллеля МНС). На вход программа принимает последовательность, а также список аллелей МНС и предполагаемый размер эпитопа. На выходе получается список эпитопов, сортированный по оценке, полученной после применения матрицы мотива для конкретного аллеля в соответствии с алгоритмом, предложенным разработчиками программы [Rammensee Н, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: database for MHC ligands and peptide motifs. - Immuno genetics 50, 1999]. Также программа указывает жирным шрифтом вероятностные якорные аминокислотные остатки, через которые осуществляется основное взаимодействие с молекулой МНС.По статистике, для 2% первых эпитопов подтверждается иммуногенность в 90% случаев.
В альтернативных вариантах осуществления изобретения биоинформатическое предсказание для В-клеточных и для Т-клеточных эпитопов на этапе (г) может быть выполнено с помощью иных сервисов, аналогичных, соответственно, "BepiPred-2.0" и "Syfpeithi", реализующих вышеуказанные алгоритмы. Можно привести следующие неограничивающие примеры таких сервисов: программа LBTope [Singh Н, Ansari HR, Raghava GP. Improved method for linear B-cell epitope prediction using antigen's primary sequence. PloS one 8(5):e62216, 2013] для предсказания В клеточных эпитопов и программы на основе алгоритмов машинного обучения (консенсусный метод [Wang Р, Sidney J, Dow С, Mothe В, Sette A, Peters В. A systematic assessment of МНС class II peptide binding predictions and evaluation of a consensus approach. PLoS computational biology. 4:4(el000048). 2008] и метод на основе искусственной нейронной сети [Jensen KK, Andreatta М, Marcatili Р, Buus S, Greenbaum JA, Yan Z, Sette A, Peters B, Nielsen M. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology. 154:3(394-406). 2018] для эпитопов класса MHCII и метод на основе искусственной нейронной сети NetMHC-3.0 [Lundegaard С, Lamberth K, Harndahl М, Buus S, Lund О, Nielsen M. NetMHC-3.0: accurate web accessible predictions of human, mouse and monkey MHC class I affinities for peptides of length 8-11. Nucleic acids research 36(Web Server issue):W509-12 2008])
При этом специалисту в данной области понятно, что в уровне техники могут существовать и другие сервисы аналогичного назначения, использующие аналогичные алгоритмы, что эти сервисы могут использоваться согласно предлагаемому способу в качестве альтернативы "BepiPred-2.0" и/или "Syfpeithi", и что способы, использующие такие сервисы вместо "BepiPred-2.0" и "Syfpeithi", также являются частью настоящего изобретения.
Также в предпочтительных вариантах осуществления изобретения, но без ограничения ими, картирование В-клеточных эпитопов на этапе (в) осуществляют методами, известными специалистам в данной области, включая, без ограничения, методы, описанные в "Epitope Mapping Protocols" (in: Methods in Molecular Biology, Vol.66, edited by G. E. Morris, 1996), а также путем определения кристаллической структуры комплекса "антитело - антиген", анализом экспрессии фрагмента гена, анализом на основе синтетических пептидов, а также другими методами, описанными Harlow и Lane в монографии "Using Antibodies: A Laboratory Manual" (New York, "Cold Spring Harbor Laboratory Press", 1998). Также без ограничения, для определения аминокислотной последовательности эпитопа можно использовать эпитопное картирование. В отдельных предпочтительных неограничивающих вариантах осуществления изобретения такое эпитопное картирование может быть выполнено при помощи коммерчески доступной системы "Pepscan Systems" (www.pepscan.com/epitope-mapping; см. также: Carter J.M., Methods Mol Biol 1994; 36:207-23; Loomis-Price L.D., Methods Mol Med. 1999; 17:293-307; John Mark Carter, Larry Loomis-Price, Curr Protoc Immunol. 2004 May; Chapter 9: Unit 9.4) либо с помощью аналогичных систем, известных специалисту в данной области.
Определяемый эпитоп может, без ограничения, представлять собой линейный эпитоп.Иные способы картирования эпитопов, которые могут использоваться на этапе (в) предлагаемого в настоящем изобретении способа, включают, без ограничения, систематический скрининг с использованием перекрывающихся пептидов, либо с использованием библиотек последовательностей случайных пептидов, а также такие экспериментальные и расчетные способы картирования эпитопов, как мутагенез, рентгеноструктурный анализ, спектроскопию, ядерно-магнитный резонанс, масс-спектрометрию на основе обмена водорода на дейтерий. В альтернативных вариантах осуществления способа, предлагаемого в настоящем изобретении, для картирования эпитопа также могут использоваться разные способы конкурентного связывания, хорошо известные специалистам в данной области. В предпочтительных вариантах осуществления способа согласно настоящему изобретению, но без ограничения ими, картирование В-клеточных эпитопов на этапе (в) осуществляют с использованием методик рентгеноструктурного анализа или ядерного магнитного резонанса, доступных специалистам в данной области техники.
В предпочтительных вариантах осуществления изобретения, относящегося к рекомбинантному вектору экспрессии, указанный вектор представляет собой молекулу ДНК или вирус, содержащие последовательности нуклеиновой кислоты, необходимые для экспрессии эпитопов вирусных белков, отобранных предлагаемым в настоящем изобретении способом, в клетке-хозяине. Вектор кодирует эпитопы вирусных белков, отобранные способом по изобретению, а также функционально связанные с ним регуляторные элементы, такие как промоторы, энхансеры и терминаторы.
В отдельных частных вариантах осуществления изобретения, относящегося к рекомбинантному вектору экспрессии, указанный вектор представляет собой, без ограничения, рекомбинантный вектор для экспрессии in vivo, например, полинуклеотидный вектор или плазмиду (например, согласно ЕР 1001025 А2, 17.05.2000 или согласно Chaudhuri P., Res. Vet. Sci. 2001, 70: 255-256) или вирусный вектор, например, вектор на основе аденовируса, вектор на основе аденоассоциированного вируса, вектор на основе ретровируса, вектор на основе лентивируса, вектор на основе цитомегаловируса, вектор на основе альфавируса или вектор на основе поксвируса.
В альтернативных вариантах осуществления изобретения вектор может представлять собой иной известный специалисту в данной области вирусный вектор, пригодный для экспрессии рекомбинантных генов в клетках позвоночных. Предпочтительно, хотя и без ограничения данным вариантом, используемый вирусный вектор пригоден для экспрессии рекомбинантных генов в клетках птиц или млекопитающих. Наиболее предпочтительно, хотя и без ограничения данным вариантом, используемый вирусный вектор пригоден для экспрессии рекомбинантных генов в клетках человека.
В предпочтительных вариантах осуществления изобретения, но без ограничения ими, вектор представляет собой вектор на основе поксвируса (поксвирусный вектор), например, поксвирусный вектор MVA Ankara, полученный из вируса коровьей оспы (CVA) штамма Анкара на фибробластах куриного эмбриона (Mayr, A., et al., Infection 1975, 3:6-14), модифицированный для применения его в качестве вирусного вектора для экспрессии рекомбинантных генов или в качестве рекомбинантной вакцины согласно работе Sutter, G. et al., Vaccine 1994, 12:1032-40. Поксвирусные векторы, безопасность и эффективность которых были показаны в уровне техники [Rerks-Ngarm S., Pitisuttithum P., Nitayaphan S., Kaewkungwal J., Chiu J., Paris R., Premsri N., Namwat C, de Souza M., Adams E., et al., N. Engl. J. Med. 2009; 361:2209 - 2220], считаются одними из наиболее перспективных среди вирусных векторов, используемых для создания векторных вакцин. К преимуществам поксвирусных векторов относятся высокая эффективность трансдукции, емкость и способность заражать большинство типов клеток [Davidson B.L., Breakefield Х.О., Nat Rev Neurosci 2003; 4(5): 353 - 364].
Поксвирусные векторы во многих работах зарекомендовали себя как перспективные векторы для создания векторных вакцин против широкого спектра патогенных для человека вирусов, таких как вирус иммунодефицита человека [Fidler S., Stohr W., et al., The Lancet 2020 Mar 14, 395(10227): 888 - 898], вирус ближневосточного острого респираторного синдрома (MERS) [Koch Т., Dahlke С, et al., The Lancet Infectious Diseases 2020 Jul 01, 20(7): 827 - 838], вирус Эбола [Wagstaffe H.R., Clutterbuck E.A., et al., J Clin Invest. 2020 Jul 1; 130(7): 3936 - 3946], вирус гриппа [Puksuriwong S., Ahmed M.S. et al., J Infect Dis. 2020 Sep 1; 222(5): 807 - 819], цитомегаловирус [Aldoss I., La Rosa C, et al., Annals of Internal Medicine 2020 Mar 3; 172(5): 306 - 316] и респираторный синцитиальный вирус [Jordan, Е., Lawrence, S. J., Meyer, Т. P. H., et al., J Infect Dis. 2021 Mar 29; 223(6): 1062-1072], а также векторных вакцин против значимых для человека невирусных патогенов, например, против малярии [Alfred В. Tiono, Issa Nébié, et al., PLoS One. 2018; 13(12): e0208328].
В других неограничивающих вариантах осуществления изобретения вектор может представлять собой вирусный вектор MVA согласно патентам US 6761893, 13.07.2004 и US 6193752, 27.02.2001, имеющий улучшенные профили безопасности и предназначенный для разработки более безопасных продуктов, таких как вакцины или лекарственные средства.
Вектор MVA, далее в настоящем описании также указываемый как вирус осповакцины, обладает рядом преимуществ, которые делают его подходящим кандидатом для использования в векторных вакцинах. Вирус осповакцины имеет линейный двухцепочечный ДНК-геном длиной примерно 190 тысяч пар нуклеотидов (т.п.н.), из-за чего способен вмещать приблизительно 25 т.п.н. чужеродной ДНК без потери инфекционности. Вакцины на основе вектора MVA могут вызывать устойчивый иммунный ответ против чужеродных антигенов из-за высокой экспрессии трансгена [Gómez С.Е., Nájera J.L., et al., Curr Gene Ther. 2011 Jun; 11(3): 189-217]. Кроме того, вектор MVA активирует сильный врожденный иммунный ответ, опосредованный Toll-подобными рецепторами (TLR) и инфламмасомой, что приводит к адъювантному эффекту [Zhu J., Martinez J., et al., Blood 2007 Jan 15; 109(2): 619-625; Philip J R Price 1, Lino E Torres-Dommguez, et al., Vaccine 2013 Sep 6; 31(39): 4231-4]. Отсутствие геномной интеграции из-за репликации в цитоплазме, ограниченный тропизм в клетках млекопитающих [Sutter G., Moss В., PNAS USA 1992 Nov 15; 89(22): 10847-51], точная и контролируемая экспрессия рекомбинантной ДНК, регулируемая сильным поксвирусным промотором, делают MVA безопасным вектором для создания вакцин.
Специалисту в данной области должно быть понятно, что в рамках настоящего изобретения могут также использоваться и иные векторы, известные в данной области в качестве векторов для экспрессии рекомбинантных генов, в т.ч., векторы, известные в качестве векторов для экспрессии рекомбинантных генов в клетках млекопитающих. Неограничивающими примерами таких векторов являются аденовирусы [Mercado NB, Zahn R, et al. et al., Nature 586(7830):583-588 2020; Logunov DY, Dolzhikova IV, et al. et al., - Lancet (London, England), 397(10275):671-681. 2021], вирус везикулярного стоматита [Yahalom-Ronen Y, Tamir H, et al. et al. - Nature communications, 11(1):6402. 2020].
В наиболее предпочтительных (неограничивающих) вариантах осуществления изобретения вектор может быть получен, наработан и очищен с использованием протоколов получения, наработки и очистки вирусных векторов, известных специалисту в данной области.
В предпочтительных (неограничивающих) вариантах осуществления изобретения, относящегося к вакцинной композиции для иммунизации позвоночного, например (без ограничения), млекопитающего, против вирусов гриппа, указанная вакцинная композиция содержит иммунологически эффективное количество рекомбинантного вектора, кодирующий указанные высоко консервативные эпитопы вирусных белков, совместно с иммунологически приемлемым носителем.
В некоторых наиболее предпочтительных вариантах осуществления изобретения, относящегося к вакцинной композиции, указанная вакцинная композиция может дополнительно содержать один или несколько адъювантов из числа фармацевтически приемлемых адъювантов, известных специалисту и используемых в данной области техники при создании вакцинных композиций. Без ограничения перечисленными вариантами, адъювант может быть выбран из группы, включающей монтанид®, квасцы, мурамилпептид, гелвакас®, микрочастицы хитина, микрочастицы хитозана, субъединицу В холерного токсина, интралипида® и липофундина®; гидрохлорид алюминия.
В других вариантах осуществления изобретения, относящегося к вакцинной композиции, указанная вакцинная композиция может не содержать адъювант.
Также в некоторых наиболее предпочтительных вариантах осуществления изобретения, относящегося к вакцинной композиции, указанная вакцинная композиция может быть выполнена в форме, пригодной для внутримышечного, интраназального, перорального, внутрибрюшинного, подкожного, местного, внутрикожного или чрескожного введения пациенту. Специалисту в данной области понятно также, что вакцинная композиция по изобретению может быть выполнена и в формах, пригодных для введения пациенту иными способами.
Изобретение иллюстрируется приведенными ниже примерами, которые, однако, приводятся лишь в целях иллюстрации предлагаемого изобретения и не могут использоваться в целях ограничения объема притязаний. Также изобретение поясняется фигурами 1-4.
На фигуре 1 приведена общая схема генетических конструкций k1 - k2 и k5. Обозначения: ORF - открытая рамка считывания; Kozak - консенсусная последовательность  .
.
На фигуре 2 приведена карта вектора pShuttle.
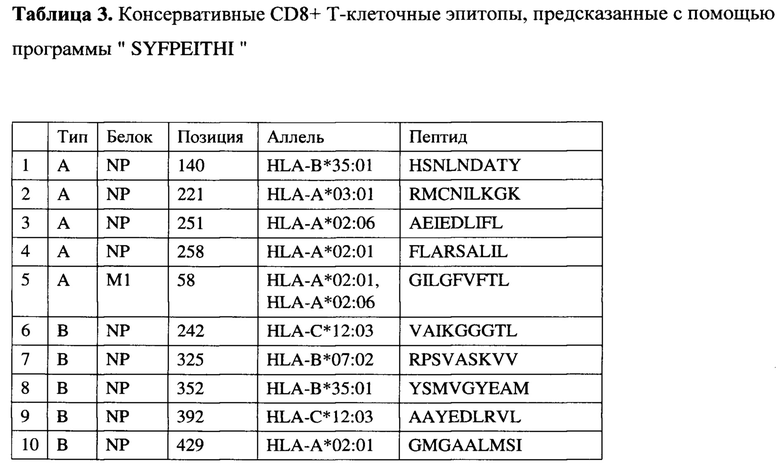
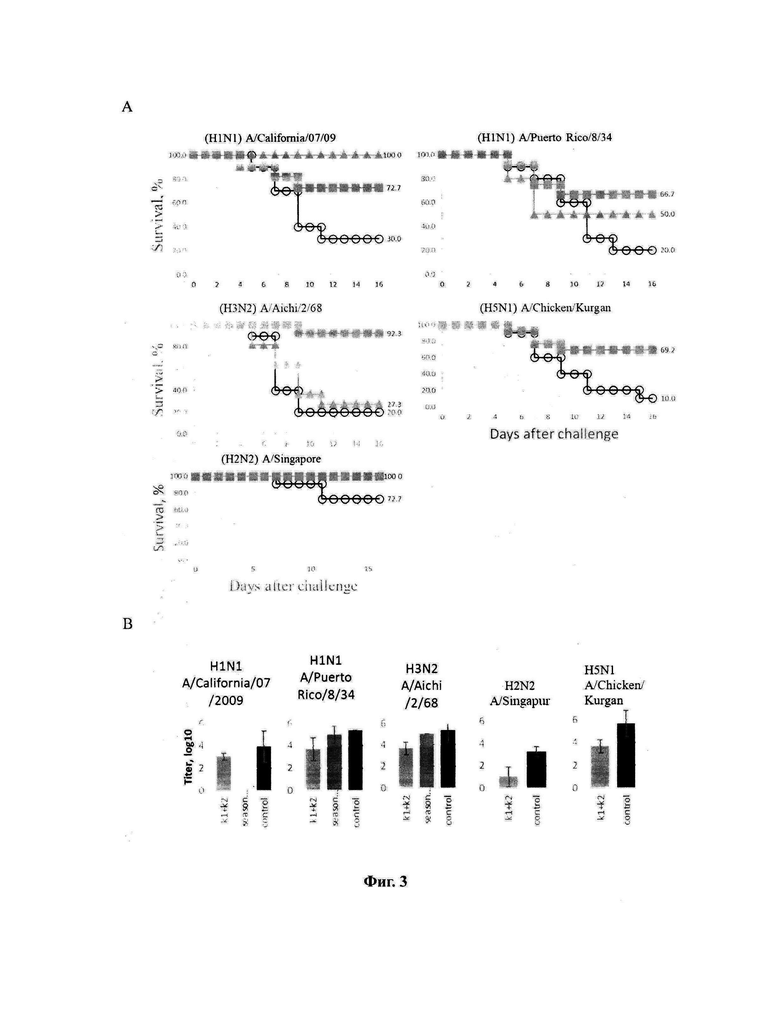
На фигуре 3 приведены сравнительные данные по выживаемости животных в различных группах, включая группу животных, иммунизированных конструктом MVA-k1+k2 (зеленый квадрат), группу животных, иммунизированных сезонной вакциной Flu-М (синий треугольник) и группу невакцинированных животных (панель А), а также сравнительные титры вируса в легких животных в указанных группах (панель Б).
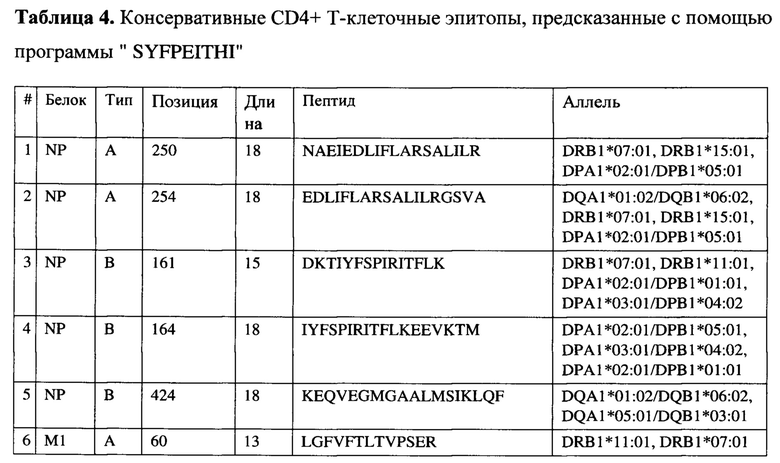
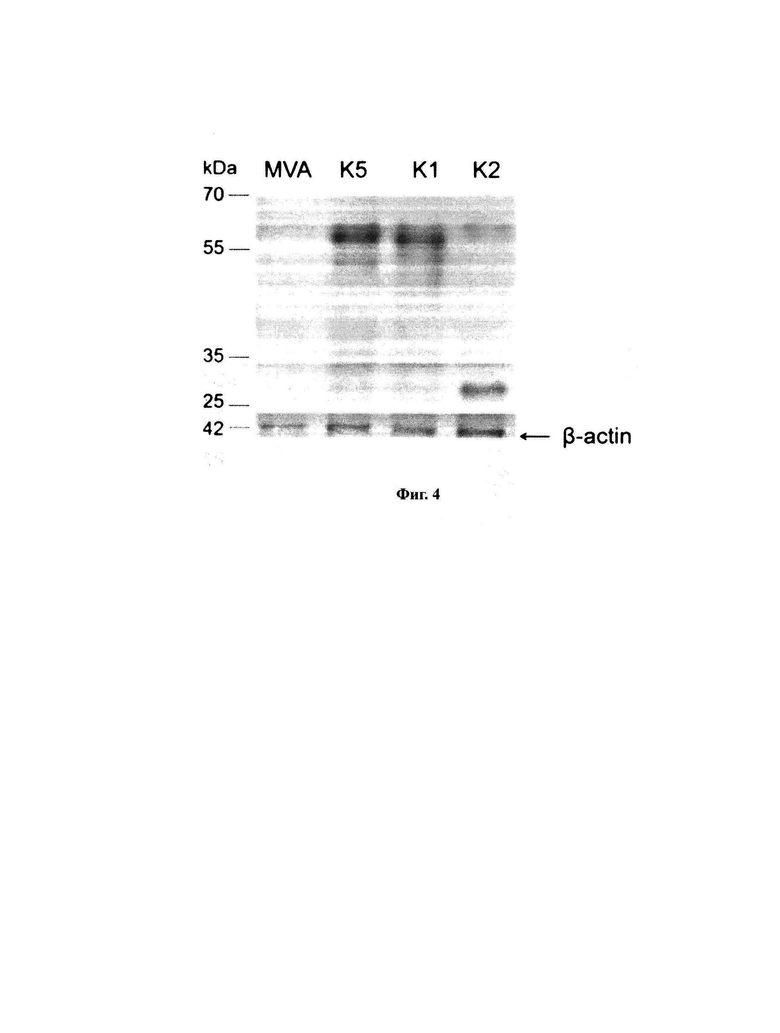
На фигуре 4 приведены результаты вестерн-блот анализа экспрессии антигенов в клетках ВНК-21, инфицированных вирусами MVA-k5, MVA-k1, MVA-k2 и MVA дикого типа.
Ниже приведены примеры осуществления предлагаемых изобретений. Настоящие примеры служат целям иллюстрации изобретения и не могут использоваться для ограничения объема притязаний в соответствии с настоящим изобретением.
Пример 1. Идентификация и отбор иммуногенных эпитопов вируса гриппа А и В.
Используя базу данных "IEDB" ("The Immune Epitope Database", URL: www.iedb.org), осуществляли поиск по линейным эпитопам белков вирусов гриппа А и В, включая линейные эпитопы из белков гемагглютинина (НА), нуклеопротеина (NP) и матриксного белка M1 вируса гриппа, принимая во внимание, что в специальной литературе именно для этих белков вируса гриппа показано наличие иммуногенных эпитопов. Число линейных эпитопов для вируса гриппа А в базе данных по иммуногенным эпитопам (IEDB) составляет 3678, для гриппа В - 51.
Для составления множественного выравнивания использовали белковые последовательности из базы данных по вирусу гриппа ("Influenza Research Database", URL: www.fludb.org/brc/home.spg?decorator=influenza). Число записей последовательностей составило: для гемагглютинина типа А подтипов H1 - 13066, Н2 - 486, Н3 - 11028, Н5 - 3834, Н7 - 1457, Н9 - 2713, для гемагглютинина типа В - 2929, для нуклеопротеина типа А - 10955, для типа В - 1316, для матриксного белка M1 типа А - 3551.
Данные рентгеноструктурного анализа белка гемагглютинина, использовавшиеся для картирования В-клеточных эпитопов гемагглютинина, получали из базы данных "PDB" ("Protein Database", http://www.rcsb.org).
Для построения множественного выравнивания последовательностей использовали программу "MAFFT" на базе алгоритма FFT-NS-2 (Kazutaka Katoh and Daron M. Standley, Molecular Biology and Evolution, vol. 30, Issue 4, April 2013, Pages 772-780; Multiple alignment program for amino acid or nucleotide sequences. - URL: mafft.cbrc.jp/alignment/software). Улучшения множественного выравнивания выполняли с помощью программы, написанной на языке С++, а также вручную в программе "Jalview" (URL: www.jalview.org). В ходе такого улучшения удаляли последовательности с выравниванием меньше 40% относительно других, а также сдвигали подстроки последовательностей размером меньше 5, расположенных внутри гэпов, к ближайшему выравненному блоку, и удаляли столбцы, содержащие только гэпы или не больше 5 аминокислотных остатков. Анализ консервативности эпитопов осуществляли с использованием с помощью скриптов, написанных на языке "Python" [https://github.com/lioj/bioinformatics/blob/master/py/MSA_stat.py, дата публикации: 20.07.2020 г.], определяя количество типов аминокислотных остатков k*, встречающихся в данной колонке выравнивания, где k* вычисляли по формуле (1)
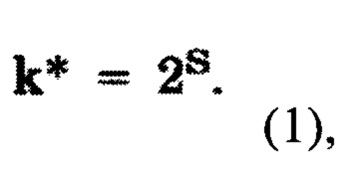
где S - информационная энтропия соответствующей колонки, определяемая по формуле (2)
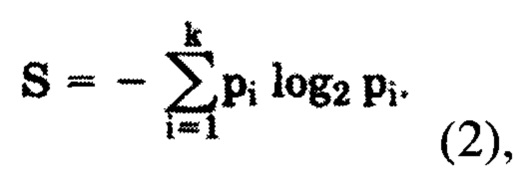
где pi - частота встречаемости i-того аминокислотного остатка в исследуемой колонке выравнивания,
k - количество всех типов остатков, встречающихся в исследуемой колонке выравнивания,
считая 1-й аминокислотный остаток в колонке выравнивания консервативным, если для него выполняется условие 1≤ ki*<1,5.
Визуализацию расположения В-клеточных эпитопов на глобуле белков вируса гриппа (в частности, на глобуле белка гемагглютинина) выполняли в программе "PyMol" [URL: pymol.org/2/].
Для предсказания В-клеточных эпитопов использовали сервис "BepiPred-2.0" [URL: http://www.cbs.dtu.dk/services/BepiPred/]. Для предсказания Т-клеточных эпитопов использовали сервис "SYFPEITHI" [URL: http://www.syfpeithi.de/bin/MHCServer.dll/EpitopePrediction.htm].
Используя предложенный в настоящем изобретении алгоритм, по пунктам "а" - "в" отбирали из уровня техники 9 эпитопов для гуморального иммунного ответа (В-клеточные эпитопы), 8 эпитопов для Т-клеточного иммунного ответа, среди которых 4 эпитопа для CD4+ Т-лимфоцитов (CD4+ Т-клеточные эпитопы) и 4 - для цитотоксических Т-лимфоцитов (CD8+ Т-клеточные эпитопы). В сумме число эпитопов составляло 15, поскольку в двух случаях эпитопы совпадали. Полный список эпитопов, отобранных из литературы с использованием предлагаемого способа, приведен ниже в таблице 1. Номер эпитопа, приведенный в каждой из строк, соответствует номеру последовательности (SEQ ID NO:…), например, эпитоп №1 - SEQ ID NO: 1, эпитоп №2 - SEQ ID NO: 2, и т.д.
В представленном иллюстративном и неограничивающем примере использовали В-клеточные и Т-клеточные эпитопы, описанные в уровне техники. В более общем случае такие эпитопы могут быть неизвестны из уровня техники. В этом случае выполняют биоинформатическое предсказание для CD8+ и CD4+ Т-клеточных и для В-клеточных эпитопов, получая таким образом набор эпитопов, характеризующихся заданным баллом связывания, а затем выполняют в отношении набора предсказанных таким образом эпитопов этапы (б) и (в) предлагаемого способа. Предпочтительно, биоинформатическое предсказание выполняют с помощью используемого в данной области программного обеспечения, неограничивающим (иллюстративным) примером которого являются программы "BepiPred", в частности, "BepiPred-2.0", и "Syfpeithi".
Частные (неограничивающие) варианты предсказания В-клеточных и Т-клеточных эпитопов вируса гриппа с помощью биоинформатических подходов настоящего изобретения, соответствующих этапам (г) и (д) предлагаемого способа, приведенные исключительно в целях иллюстрации заявляемого способа, становятся ясны из Таблиц 2-4, приведенных ниже.
Для проверки иммуногенных свойств, отобранных из уровня техники эпитопов указанные эпитопы объединяли в полипептиды k1, k2 и k5,
где полипептид k1 содержал все отобранные из уровня техники В-клеточные и все CD4+Т-клеточные эпитопы, т.е. эпитопы №1 - №8, №9 - №11 и №13,
полипептид k2 содержал отобранные из уровня техники эпитопы для CD4+ Т-клеточного ответа и все отобранные из уровня техники эпитопы для CD8+ Т-клеточного ответа, т.е. эпитопы №9 - №11 и №13, а также эпитопы №14 и №15,
а полипептид к5 содержал все отобранные из уровня техники В-клеточные эпитопы, т.е. эпитопы №1 - №8, но в ином порядке, нежели в полипептиде k1, а также эпитопы для CD4+ Т-клеточного ответа, т.е. эпитопы №9 - №11 и №13, как представлено ниже в таблице 5, отражающей последовательность расположения эпитопов в полипептидах k1 - k2 и k5, и на фигуре 1, отражающей общую схему конструкций k1 - k2 и k5.
Полипептид k1: Эпитопы расположены в последовательности: эпитоп №8 - эпитоп №1 - эпитоп №10 - эпитоп №7 - эпитоп №2 - эпитоп №9 - эпитоп №4 - эпитоп №5, т.е. так, чтобы в конце находились гидрофильные остатки.
Полипептид k2: Эпитопы расположены в последовательности: эпитоп №11 - эпитоп №12 - эпитоп №13 - эпитоп №14 - эпитоп №15 (собраны эпитопы для CD4+ и эпитопы для CD8 клеток).
Полипептид k5, включающий эпитопы №8, №1, №9, №10, №5, №7, №4 и №2, получают из полипептида k1 с помощью перестановки эпитопов, причем условно соблюдается чередование гидрофобного и гидрофильного эпитопа.
Разделение на варианты k1 и k5 было сделано для того, чтобы увеличить вероятность получения оптимальной конструкции, где В-клеточные эпитопы наилучшим образом будут представлены на поверхности молекулы для взаимодействия с В-клетками. Кроме того, в одной молекуле с В-клеточными эпитопами должны быть CD4+ Т-клеточные эпитопы для того, чтобы при созревании клонов В-клеток было возможно стимулирование пролиферации соответствующими клонами CD4+ Т-клеток.
Из уровня техники известно, что наилучший вариант структуры полипептида выглядит следующим образом: (Е1)3-(Е2)3-…-(En)3, где Е - некоторый эпитоп, который повторяется 3 раза, и линкером между каждым эпитопом служит аминокислотный остаток пролин (Р), который характеризуется минимальными областями разрешенных значений двугранных углов фи и пси на карте Рамачандрана [Ramachandran, G.N.; Ramakrishnan, С; Sasisekharan, V., J Mol Biol. 1963 Jul; 7:95-9], т.е. приводит к резкому повороту полипептидной цепи в пространстве (патент US 9353159 В2, 31.05.2016, A61K 39/00, A61K 39/12, A61K 39/145, A61K 39/295, С07Н 21/04, C07K 14/005).
Известно также, что секретируемые белки более иммуногенны, чем белки, экспрессируемые внутри клетки. Поэтому в начало конструкции было решено вставить последовательность для направления полипептида по секреторному пути. Для этого был выбран лидерный пептид из Ig kappa мышей. Для контроля экспрессии полипептидов к С-концу были присоединены тэги для детекции белков антителами с помощью Вестерн блота: НА tag (эпитоп из НА, на который имеются коммерчески доступные моноклональные антитела) и Мус tag (эпитоп из с-Мус гена, на который имеются коммерчески-доступные моноклональные антитела), а также 6xHisTag (последовательность из 6 аминокислотных остатков гистидина) для выделения белков с помощью ионообменной никелевой смолы.
Аминокислотный состав разных полипептидов, включающих кодируемые конструктами последовательности эпитопов, лидерный пептид и тэги для контроля экспрессии, представленный последовательностями SEQ ID NO: 16 - SEQ ID NO: 17 и SEQ ID NO: 18, приведен в таблице 6.
Аминокислотные последовательности полипептидов, включающих эпитопы, переводили в полинуклеотидные последовательности, используя таблицу стандартного генетического кода и программы Snapgene. При создании полинуклеотидов программа учитывала частоту встречаемости разных кодонов у человека.
Пример 2. Получение рекомбинантного вектора MVA. Оценка экспрессии целевых генов методом вестерн-блот анализа.
Модифицированный вирус осповакцины штамма Анкара (MVA) получали из Американской коллекции клеточных культур (АТСС® VR-1566). Для репликации вируса MVA использовали клеточную линию фибробластов сирийского хомячка BHK-21, которую культивировали в стандартной среде Игла, модифицированной по способу Дульбекко, содержащей 10% эмбриональной сыворотки крупного рогатого скота, 4 мМ L-глютамин 1 мМ пируват натрия, 10 мМ HEPES при температуре 37°С и атмосфере 5% CO2.
Рекомбинантный вирус MVA (далее также указывается как рекомбинант) получали, очищали и нарабатывали согласно протоколу, предложенному Kremer, М., Volz A. et al., Methods in Molecular Biology (2012), vol. 890, pp. 59-92. Подсчет титра вируса проводили согласно протоколу, также предложенному Kremer М., Volz A. et al., 2012, кратко изложенному ниже. Рекомбинанты получали на клетках линии ВНК-21. Клетки заражали диким вирусом MVA, через 90 мин трансфицировали шаттл-вектором и собирали смесь, содержащую рекомбинантные вирусы через 48 часов. Рекомбинантную смесь рассевали до получения отдельных бляшек, которые анализировали на наличие вставки с помощью ПЦР. После получения из бляшки вируса, не содержащего дикого MVA, его анализировали и нарабатывали в несколько этапов. Озвучивание проводили на приборе "Qsonica ql25-220" (США) согласно рекомендациям производителя.
Для количественной оценки экспрессии целевых генов использовали Вестерн-блотт анализ. Клетки BHK-21 инфицировали вирусами или трансфицировали плазмидами. Через 24 часа получали лизаты клеток, которые разделяли с помощью 12% SDS-PAGE в восстанавливающих условиях, переносили на нитроцеллюлозную мембрану и проводили гибридизацию с антителами к 6xHIS и антителами, специфичными к бета-актину. kDa - маркер молекулярной массы белков. В качестве отрицательного контроля экспрессии для рекомбинантных вирусов использовали MVA дикого типа. Веса белков (кДа): k1 - 45.3, k2 - 31.1, k5 - 45.3.
Эпитопные тэги для детекции на Вестерн-блотте распределяли по конструктам k1 - k2 и k5 таким образом, чтобы можно было дифференцировать их друг от друга в случае одновременного наличия двух целевых генов в одном векторе MVA. Для детекции вариантов MVA с одним целевым геном использовали НА tag или Мус tag. Один или оба этих тэга присутствуют в С-концевой части полипептида в таких векторах. Результаты вестерн-блот анализа приведены на Фигуре 4.
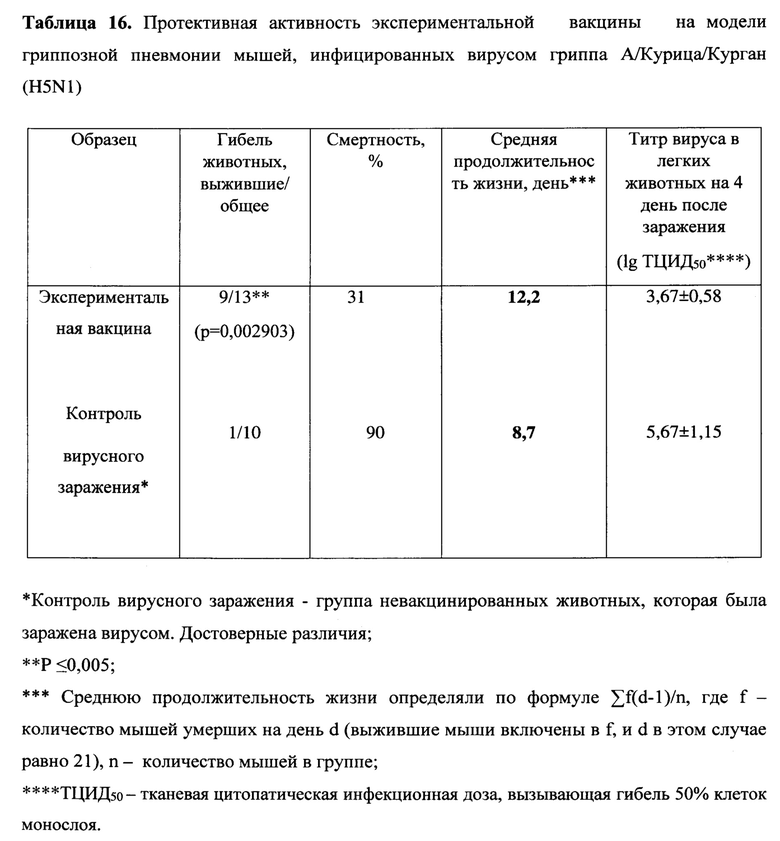
Пример 3. Изучение вариантов вакцин MVA-k1-k2 и MVA-k5-k2 в отношении четырех штаммов вируса гриппа типа А и В на животной модели
Рекомбинантные MVA были использованы для вакцинации мышей. Было сформировано 2 экспериментальные группы мышей, а также контрольная группа чистых (mock) мышей и группа контрольных мышей, которые иммунизировались пустым MVA (MVA mock) (Таблица 7). Мышам из группы 1 дважды вводилась смесь rMVA, содержавших эпитопные варианты k1 и к2 в пропорции 1:1. Группе 2 дважды вводилась смесь rMVA, содержавших эпитопные варианты k5 и k2 (1:1).
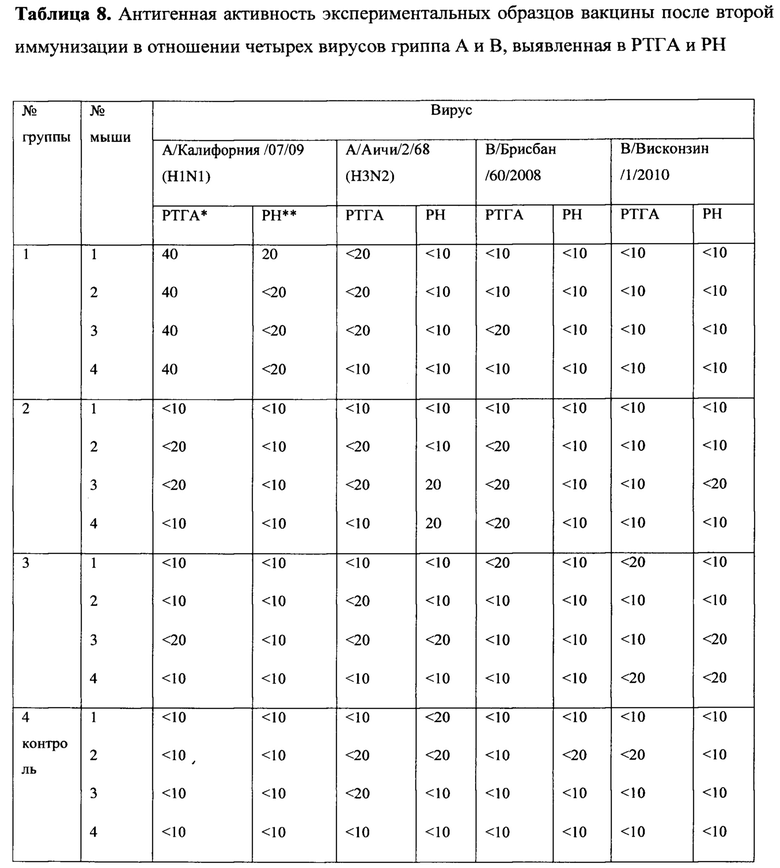
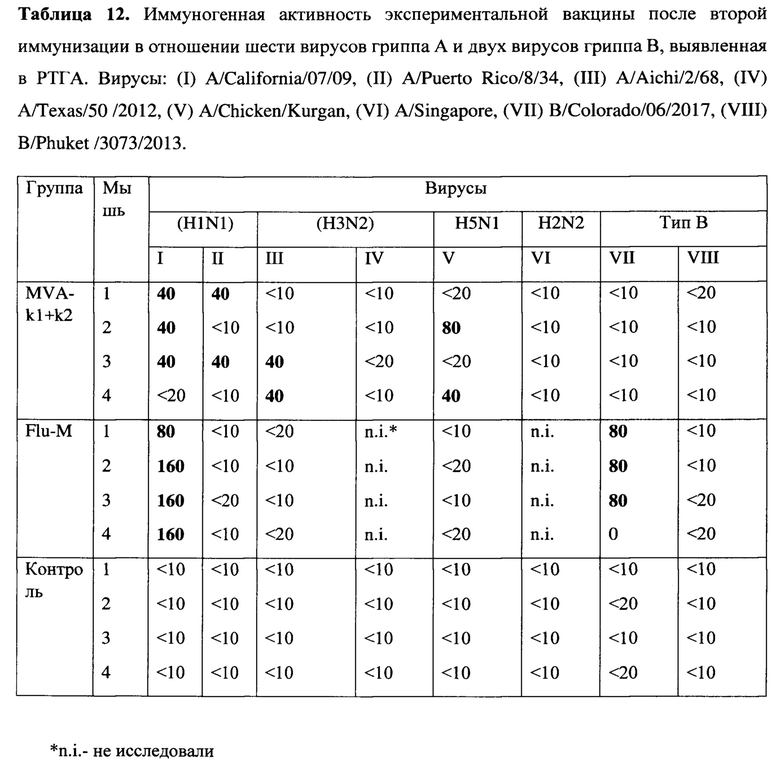
После первой иммунизации во всех группах животных не было выявлено образования антител ни в реакции торможения гемагглютинации (далее - РТГА), ни в реакции нейтрализации (далее - РН) ко всем четырем исследованным вирусам гриппа А и В. Однако, после второй иммунизации были обнаружены антитела у животных в 1 группе, (MVA-k1+k2), к штамму подтипа H1 в постановке РТГА (Таблица 8). У животных остальных групп антител обнаружено не было.
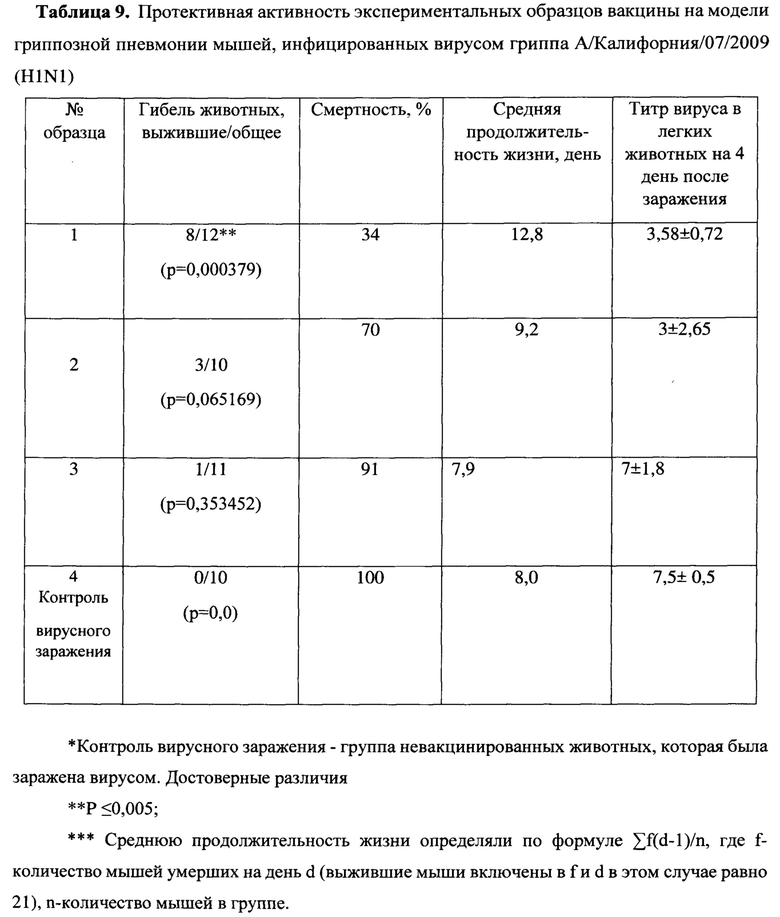
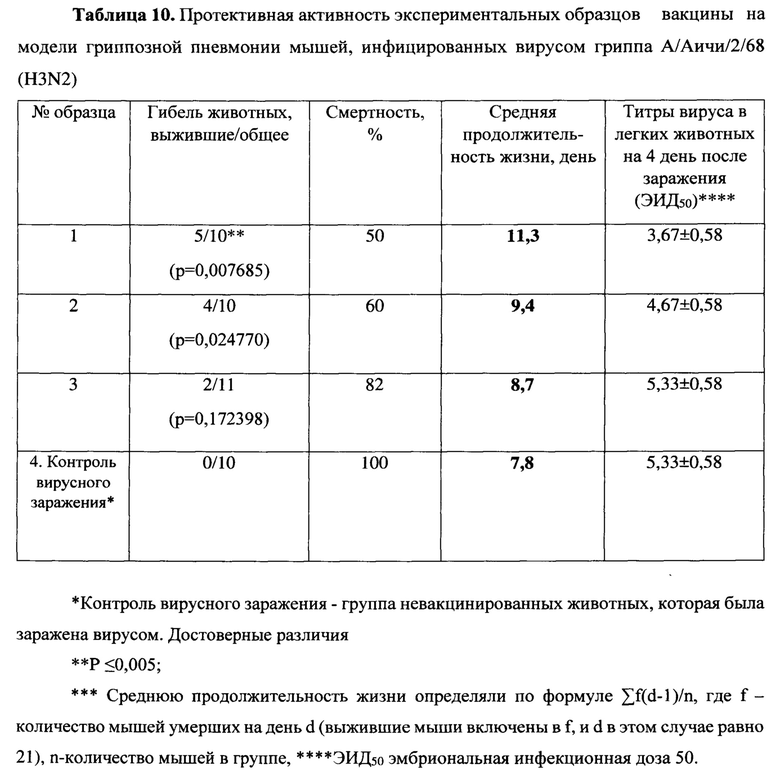
Были изучены защитные свойства созданных нами эпитопных вариантов вакцин на модели гриппозной пневмонии мышей. На 14 день после второй иммунизации мыши каждой группы были разделены на 2 подгруппы, одну из которых заражали вирусом гриппа А подтипа H1N1, другую - H3N2. В течение 14 дней после заражения проводились наблюдения за мышами (Таблицы 9-10).
При заражении вирусами A/California/04/09 (H1N1) и A/Aichi/2/68 (H3N2) в не вакцинированной группе гибель мышей наблюдалась с 6 по 11 и с 4 по 13 дни, соответственно, и достигала 100%. При этом средняя продолжительность жизни составила 8 и 7.8 дней, соответственно. Сходная гибель (91% для H1N1 и 82% для H3N2) и средняя продолжительность жизни (7.9 дня и 8.7) наблюдались в группе 3, вакцинированной пустым вектором MVA, не содержащем антигены вируса гриппа. Среди экспериментальных вариантов самой слабой защитной активностью обладала смесь MVA-k5 и MVA-k2. Падеж мышей в этой группе составлял 70% для H1N1 и 60% для H3N2, а средняя продолжительность жизни была близка к продолжительности жизни животных в контрольных группах (9.2 для H1N1 и 9.4 для H3N2). Статистически значимое увеличение выживаемости относительно контролей (р<0.05) наблюдалось в группе животных 1, дважды вакцинированных смесью MVA-k1+k2 (смертность 34% для H1N1 и 50% для H3N2). Также, в этой группе продолжительность жизни увеличилась в 1.5 раза по сравнению с группами отрицательных контролей.
Титр вируса в легких мышей определяли на 4 день после заражения. В контрольных группах невакцинированных и вакцинированных пустым вектором животных титр вируса в легких (log10) был наибольшим и составлял 7,5±0,5 и 7±1,8 для вируса гриппа A/California/04/09 (H1N1) и 5,33±0,58 и 5,33±0,58 для A/Aichi/2/68 (H3N2), соответственно.
При заражении H1N1 титр вируса в легких животных был достоверно меньше во всех экспериментальных группах относительно группы чистых (Mock) мышей (Таблицы 9 - 10). Наибольшее различие относительно Mock наблюдалось в группе, вакцинированной смесью MVA-k1+k2: титр был ниже на 3.92 порядков величины (р-value<0.001). Меньшее различие (3 порядка величины) относительно группы Mock наблюдалось в группе MVA-k5-k2 (p-value<0.05). Помимо этого, при заражении H1N1 наблюдалось статистически значимое (p-value<0.01) снижение титра в легких на 3.4 порядков величины в группе, вакцинированной MVA-k1+k2, относительно группы, вакцинированной пустым MVA. Этот факт позволяет утверждать, что достоверное понижение титра вируса H1N1 было вызвано иммунизацией антигенами гриппа k1+k2.
В случае заражения H3N2 титр вируса у мышей, вакцинированных антигенами гриппа, также снижался во всех группах. Стоит отметить, что при отсутствии достоверного снижения титра наблюдалось достоверное (p-value<0.05) повышение выживаемости в группе, вакцинированной MVA-k1+k2, относительно группы mock. (Таблица 10).
Таким образом, в процессе создания вакцины было проверено 2 варианта антигена для выработки гуморального иммунного ответа: k1 и k5, в которых состав В-клеточных и CD4+ Т-клеточных эпитопов был одинаков, но различался порядок. По нашему представлению это могло быть важным, т.к. необходимо было создать и выбрать антиген с оптимальной презентацией В-клеточных эпитопов на поверхности белковой молекулы. Сравнение защитного эффекта вариантов вакцин MVA-k1+k2 и MVA-k5+k2 показало, что и по выживаемости, и по титру вируса при заражении H1N1 и H3N2 штаммами вируса гриппа вариант MVA-k1+k2 был лучше, чем MVA-k5+k2, хотя различия не достигали статистической достоверности. Полученные данные позволяют утверждать, что в случае антигена k1 представление эпитопов было более удачным, соответственно используемый принцип группировки на N-конце гидрофобных эпитопов, а на С-конце гидрофильных оказался лучше, чем их чередование по свойствам гидрофильности и гидрофобности.
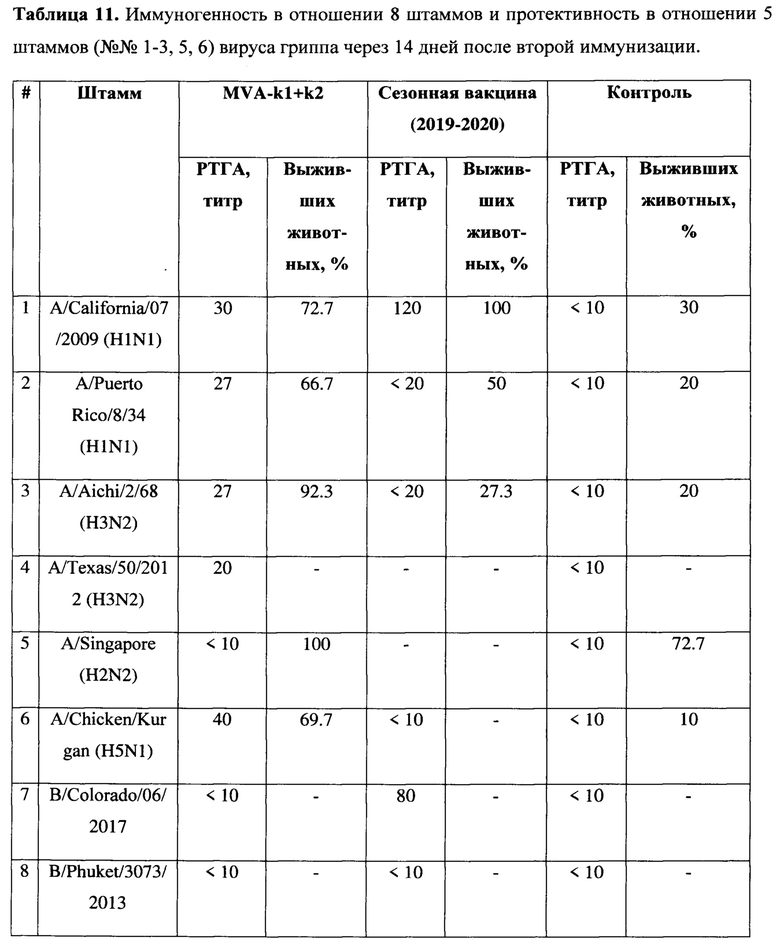
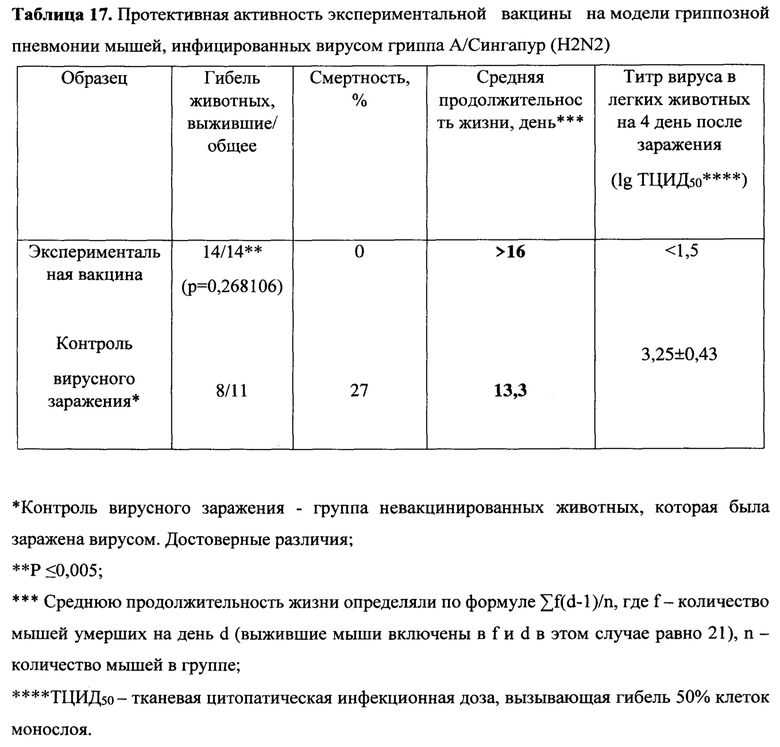
Пример 4. Оценка широты иммуногенности и защиты, обеспечиваемой вариантом вакцины MVA-k1+k2, на животной модели
Иммуногенные характеристики и защита варианта вакцины MVA-k1+k2 были исследованы в отношении 8 штаммов разных подтипов вирусов гриппа типа А и В (Таблица 11). Для сравнения использовалась сезонная вакцина Flu-M (сезон 2019-2020), в состав которой вошли штаммы подтипов H1N1, H3N2 и типа В, в состав которой входили НА A/Brisbane/02/2018 H1N1 pdm09-like, А/Kansas/14/2017 (H3N2)-like, B/Colorado/06/2017-like, далее указываемая как сезонная вакцина. Было сформировано 2 экспериментальные группы мышей (группа MVA-k1+k2 и группа Flu-M), а также 1 контрольная группа чистых (mock) мышей.
Иммунизация проводилась по следующей схеме. Все животные иммунизировались образцами по 0,2 мл внутримышечно (4x10E6 инфекционных частиц MVA, либо гриппозная инактивированная расщепленная вакцина Flu-M производства СПбНИИВС ФМБА России сезона 2019-2020 в состав которой входили НА A/Brisbane/02/2018 H1N1 pdm09-like, А/Kansas/14/2017 (H3N2)-like, B/Colorado/06/2017-like, половина человеческой дозы). Животным из mock группы вместо препаратов вакцины в соответствующие дни вводили по 0,2 мл стерильного ФСБ внутримышечно. Иммунизация животных проводилась дважды: (1) на 0 день проводилась прайм вакцинация, (2) а на 21 день - буст вакцинация. На 21 день после первой иммунизации и на 14 день после второй иммунизации у 4 мышей из каждой группы отбирались образцы крови для проведения РТГА и РН. Также, на 14 день после второй иммунизации оставшиеся мыши заражались вирусами гриппа для оценки защитной эффективности в течение 2х недель. На 3 день после заражения у 3-х мышей из каждой группы определяли титр вируса в легких.
Прочерк означает, что данное исследование не проводилось в отношении этого штамма.
Оценка гуморального иммунитета в РТГА с использованием 8 штаммов вирусов показала, что после первой иммунизации антитела были обнаружены только у мышей, вакцинированных сезонной вакциной, у которых были выявлены антитела к вирусу (H1N1) A/California/07/09 в титрах 40, 80, 80 и 80. После второй иммунизации в группе, вакцинированной MVA-k1+k2, обнаружено образование антител в титрах (от 1:20 до 1:40) к 4 вирусам гриппа A: A/California/07/2009 (H1N1), A/Puerto Rico/8/34 (H1N1), A/Aichi/2/68 (H3N2), A/Chicken/Kurgan (H5N1). В группе, вакцинированной сезонной вакциной, был выявлен титр антител более 1:40 к двум вирусам гриппа: A/California/07/2009 (H1N1) и В/ Colorado /06/2017, в то время как в этой же группе не было выявлено образование антител к четырем другим исследованным вирусам. (Таблица 12).
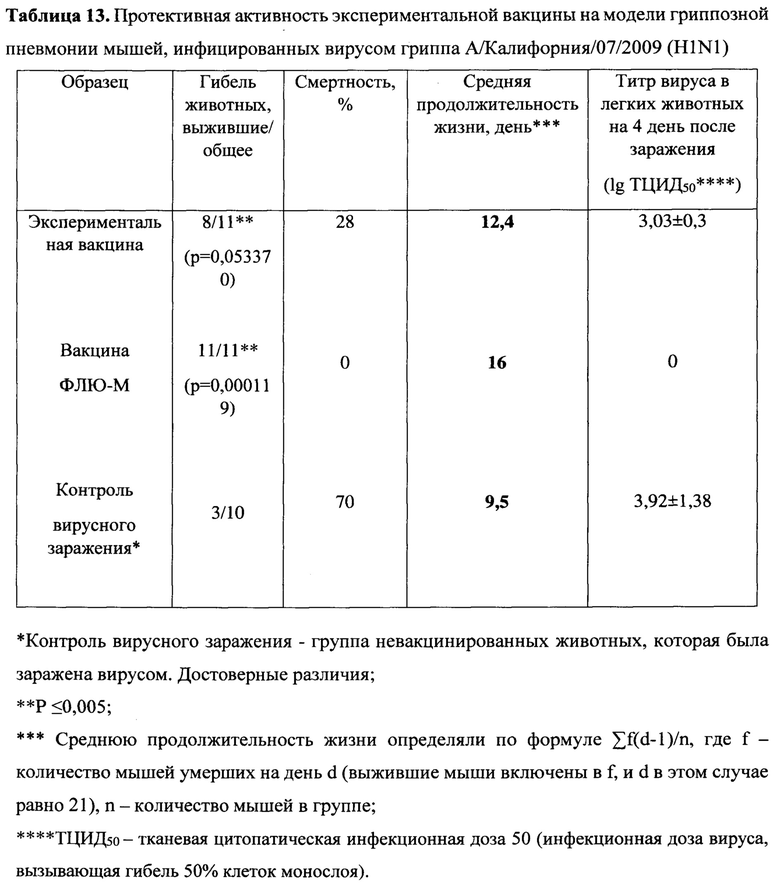
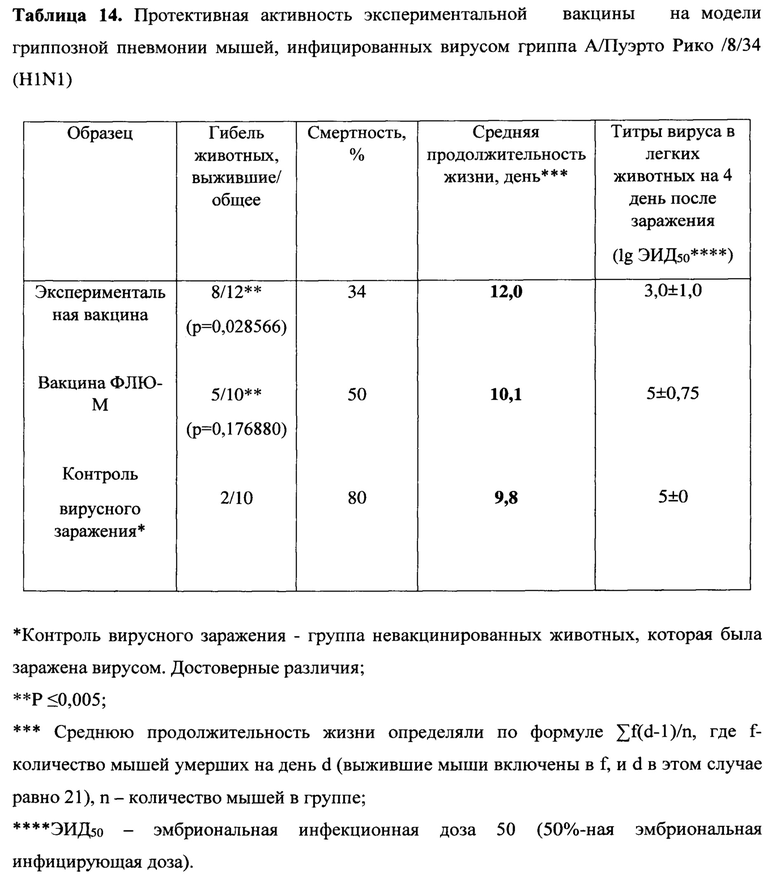
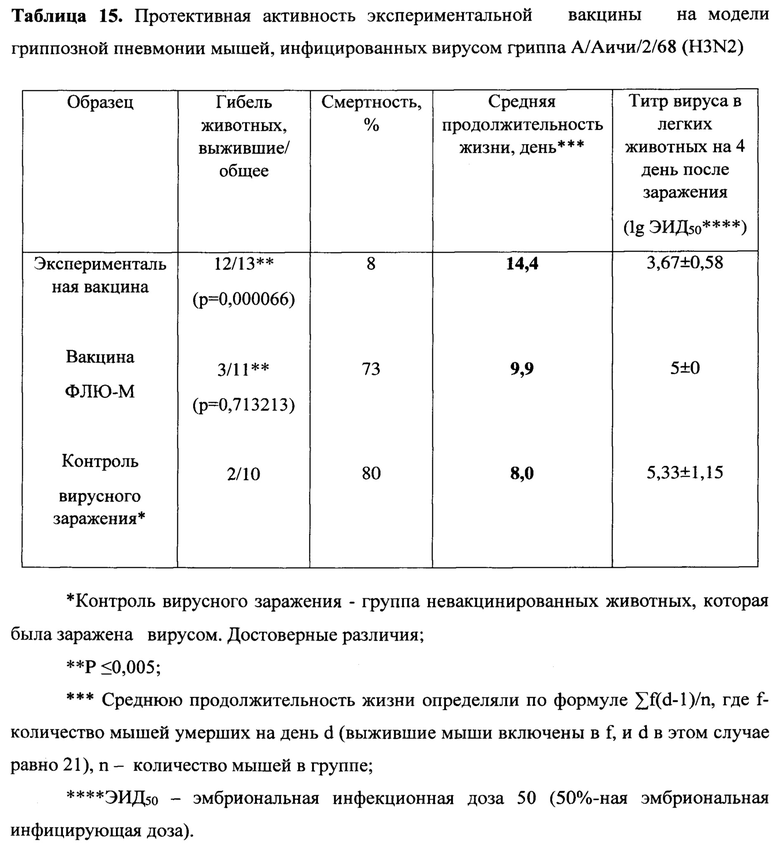
Протективность варианта вакцины MVA-k1+k2 в отношении 5 штаммов гриппа А и сезонной вакцины в отношении 3 штаммов оценивались по выживаемости мышей. Результаты представлены на Фигуре 3А и в таблицах 13-17.
При заражении животных из контрольной группы вирусами гриппа А смертность достигала 70-90%, за исключением заражения вирусом A/Singapore (H2N2), которое приводило к смертности не более 30%. В группах, вакцинированных MVA-k1+k2, выживаемость при заражении различными штаммами составила 67%-100%. Сходная выживаемость в отношение штаммов разных подтипов указывает на универсальные свойства вакцины. Для сезонной вакцины 100% защита наблюдалась только против штамма A/California/07/09, для более старого штамма A/Puerto Rico/8/34 этого же подтипа защита упала до 50%, а при заражении A/Aichi/2/68 подтипа H3N2 выживаемость составила 27% при 20% выживаемости в контрольной группе.
Также в каждой группе у 3 мышей проводили определение титра вируса в легких. У животных, получивших сезонную вакцину, вирус не обнаруживался в легких только при заражении A/California/07/09, в других случаях титр вируса был на уровне контроля. В то же время титр вируса в легких мышей, вакцинированных MVA-k1+k2, снижался на 0.9-2.25 порядка величины по сравнению с контролем, но достоверно отличался (p-value<0.05) только для штаммов A/Puerto Rico/8/34 (H1N1), A/Chicken/Kurgan (H5N1), A/Singapore (H2N2) (Фигура 3 В).
Таким образом, в случае совпадения штамма в составе сезонной вакцины с вирусом гриппа A/California/07/2009 (H1N1), используемым для заражения, как и ожидалось, наблюдалась 100% защита. При заражении штаммами, более отдаленными по антигенным характеристикам, защита сезонной вакцины значительно снижалась и составила 50% выживаемости мышей при заражении (H1N1) A/Puerto Rico/8/34 и лишь 27% выживаемости в случае заражения штаммом A/Aichi/2/68 подтипа (H3N2) (при 20% выживаемости в контрольной группе). В то же время, вакцина MVA-k1+k2 обеспечивала защиту от 67% до 92% животных в случае заражения вирусами подтипов H1N1 и H3N2. При заражении штаммом A/Singapore подтипа H2N2 наблюдалась 100% защита, однако модель в данном случае не является летальной. Тем не менее, наблюдалось снижение титра вируса в легких. Кроме того, важно отметить, что вакцинация MVA-k1+k2 защищала 70% мышей от летального течения птичьего гриппа штамма A/Chicken/Kurgan подтипа H5N1. Защита против этих штаммов чрезвычайно важна, так как сезонные вакцины не формируют защиту против них, однако несмотря на то что штаммы подтипов H2N2 и H5N1 в данный момент не циркулируют в популяции людей, они считаются потенциально пандемическими. [Viboud С, Simonsen L, Fuentes R, Flores J, Miller MA, Chowell G. Global Mortality Impact of the 1957-1959 Influenza Pandemic. - The Journal of Infectious Diseases 213(5):738-45 2016; Kilbourne ED. Influenza pandemics of the 20th century. Emerging infectious diseases 12(1):9-14 2006; Flahault A, Vergu E, Boelle PY. Potential for a global dynamic of Influenza A (H1N1). - BMC infectious diseases. 9: (129). 2009; Krammer F. Emerging influenza viruses and the prospect of a universal influenza virus vaccine. - Biotechnology journal 10:5(690-701). 2015].
Полученные данные показывают, что предлагаемая группа изобретений, в частности, предлагаемый способ обеспечивает возможность идентификации и отбора иммуногенных эпитопов вирусных белков, эффективных для иммунизации реципиента, такого как позвоночное, например, млекопитающее или птица, против широкого спектра патогенных для такого позвоночного РНК-содержащих вирусов, что позволяет сократить сроки разработки новых вакцин, в т.ч., вакцин против новых вариантов известных патогенных вирусов, и обеспечивает возможность включения вновь выявленных вариантов эпитопов вирусных белков в уже разработанные вакцины.



| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов H1, H2, H5 для создания противогриппозных иммуногенных препаратов, способ получения гена | 2018 |
|
RU2713722C1 |
| ВАКЦИНЫ НА ОСНОВЕ НУКЛЕОПРОТЕИНА ВИРУСА ГРИППА | 2014 |
|
RU2662667C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА "СВИНОГО" ГРИППА H1N1 И СПСОБ ЕЕ ПОЛУЧЕНИЯ | 2010 |
|
RU2451027C2 |
| КРОСС-РЕАКТИВНАЯ РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА А ЧЕЛОВЕКА | 2020 |
|
RU2742336C1 |
| СРЕДСТВА И СПОСОБЫ ЛЕЧЕНИЯ ВИРУСА ГЕПАТИТА B | 2017 |
|
RU2740802C2 |
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| Мутантные полимеразы вируса гепатита В | 2012 |
|
RU2625021C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ВИРУСОМ ГЕПАТИТА В | 2010 |
|
RU2555346C2 |
Изобретение относится к области биоинформатики и иммунологии. Описан способ in silico идентификации и отбора иммуногенных высоко консервативных линейных эпитопов белков вируса гриппа, при этом а) указанные линейные эпитопы вирусных белков отбирают таким образом, чтобы они стимулировали обе ветви приобретенного иммунитета человека, включая гуморальный иммунитет и клеточный иммунитет, при этом, если на этапе (а) такие эпитопы отобраны, б) среди эпитопов вирусных белков, отобранных на этапе (а), отбирают высоко консервативные CD8+ и CD4+ Т-клеточные и высоко консервативные В-клеточные эпитопы, причем отбор высоко консервативных CD8+ и CD4+ Т-клеточных и высоко консервативных В-клеточных эпитопов включает в себя: б1) составление множественного выравнивания последовательностей аминокислотных остатков эпитопов вирусных белков, отобранных на этапе (а); б2) оценку консервативности указанных эпитопов вирусных белков, причем оценку консервативности указанных эпитопов на этапе (б2) осуществляют, определяя количество типов аминокислотных остатков k*, встречающихся в данной колонке выравнивания, причем k* вычисляют по формуле k* = 2s (1), где S - информационная энтропия соответствующей колонки, определяемая по формуле
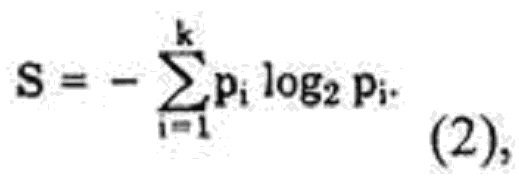
где pi - частота встречаемости i-того аминокислотного остатка в исследуемой колонке выравнивания, k - количество всех типов остатков, встречающихся в исследуемой колонке выравнивания, при этом i-й аминокислотный остаток в колонке выравнивания считают консервативным, если для него выполняется условие 1< ki* < 1,5, а затем в) картируют высоко консервативные В-клеточные эпитопы, отобранные на этапе (б), на данные рентгеноструктурного анализа антигена для проверки условия их доступности для антител, а если на этапе (а) такие эпитопы не найдены или не отобраны, г) выполняют биоинформатическое предсказание для CD8+ и CD4+ Т-клеточных и для В-клеточных эпитопов, получая таким образом набор эпитопов, характеризующихся заданным баллом связывания, после чего д) выполняют в отношении набора эпитопов, полученного на этапе (г), этапы (б) - (в), получая таким образом итоговый набор иммуногенных высоко консервативных линейных эпитопов вирусных белков. Описан рекомбинантный вектор экспрессии, кодирующий полипептид, составленный из эпитопов, идентифицированных описанным способом, при этом указанный полипептид, имеет SEQ ID NO: 16, SEQ ID NO: 17 или SEQ ID NO: 20. Представлена вакцинная композиция для иммунизации позвоночных против вирусов гриппа, содержащая рекомбинантный вектор экспрессии и иммунологически приемлемый носитель. Раскрыт способ вакцинации позвоночного против вирусов гриппа, где указанный способ включает введение указанному позвоночному иммунологически эффективного количества вакцинной композиции. Изобретение обеспечивает отбор эпитопов вирусных белков, пригодных для создания вакцин широкого спектра действия. 4 н. и 5 з.п. ф-лы, 4 ил., 17 табл., 4 пр.
1. Способ in silico идентификации и отбора иммуногенных высоко консервативных линейных эпитопов белков вируса гриппа, при этом
а) указанные линейные эпитопы вирусных белков отбирают таким образом, чтобы они стимулировали обе ветви приобретенного иммунитета человека, включая гуморальный иммунитет и клеточный иммунитет, при этом, если на этапе (а) такие эпитопы отобраны,
б) среди эпитопов вирусных белков, отобранных на этапе (а), отбирают высоко консервативные CD8+ и CD4+ Т-клеточные и высоко консервативные В-клеточные эпитопы, причем отбор высоко консервативных CD8+ и CD4+ Т-клеточных и высоко консервативных В-клеточных эпитопов включает в себя:
б1) составление множественного выравнивания последовательностей аминокислотных остатков эпитопов вирусных белков, отобранных на этапе (а);
б2) оценку консервативности указанных эпитопов вирусных белков, причем оценку консервативности указанных эпитопов на этапе (б2) осуществляют, определяя количество типов аминокислотных остатков k*, встречающихся в данной колонке выравнивания, причем k* вычисляют по формуле (1)
k*= 2s (1),
где S - информационная энтропия соответствующей колонки, определяемая по формуле (2)
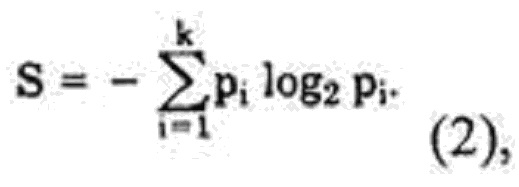
где pi - частота встречаемости i-того аминокислотного остатка в исследуемой колонке выравнивания,
k - количество всех типов остатков, встречающихся в исследуемой колонке выравнивания, при этом i-й аминокислотный остаток в колонке выравнивания считают консервативным, если для него выполняется условие 1 ≤ ki* < 1,5, а затем
в) картируют высоко консервативные В-клеточные эпитопы, отобранные на этапе (б), на данные рентгеноструктурного анализа антигена для проверки условия их доступности для антител, а если на этапе (а) такие эпитопы не найдены или не отобраны,
г) выполняют биоинформатическое предсказание для CD8+ и CD4+ Т-клеточных и для В-клеточных эпитопов, получая таким образом набор эпитопов, характеризующихся заданным баллом связывания, после чего
д) выполняют в отношении набора эпитопов, полученного на этапе (г), этапы (б) - (в), получая таким образом итоговый набор иммуногенных высоко консервативных линейных эпитопов вирусных белков.
2. Способ по п. 1, отличающийся тем, что указанное множественное выравнивание последовательностей аминокислотных остатков эпитопов на этапе (б1) осуществляют с использованием программы "MAFFT" с последующим редактированием вручную.
3. Способ по п. 1, отличающийся тем, что картирование В-клеточных эпитопов на данные рентгеноструктурного анализа на этапе (г) и проверку выполнения условия доступности указанных эпитопов для антител осуществляют с помощью программ молекулярной визуализации и скриптов на данные рентгеноструктурного анализа.
4. Способ по п. 1, отличающийся тем, что сопоставление выбранных В-клеточных и Т-клеточных эпитопов с данными биоинформатического предсказания для В-клеточных и Т-клеточных эпитопов вирусных белков осуществляются для ранжирования списка выбранных эпитопов, в котором первые по списку должны обладать потенциально наиболее иммуногенными свойствами.
5. Рекомбинантный вектор экспрессии, кодирующий полипептид, составленный из эпитопов, идентифицированных способом по любому из пп. 1-4, при этом указанный полипептид, имеет SEQ ID NO: 16, SEQ ID NO: 17 или SEQ ID NO: 20. 6. Вакцинная композиция для иммунизации позвоночных против вирусов гриппа, содержащая рекомбинантный вектор экспрессии по п. 5 и иммунологически приемлемый носитель.
7. Вакцинная композиция по п. 6, где позвоночное животное представляет собой млекопитающее.
8. Способ вакцинации позвоночного против вирусов гриппа, где указанный способ включает введение указанному позвоночному иммунологически эффективного количества вакцинной композиции по любому из пп. 6,7.
9. Способ вакцинации по п. 8, где позвоночное животное представляет собой млекопитающее.
| Christopher S Eickhoff, Frances E Terry | |||
| et al., Highly conserved influenza T cell epitopes induce broadly protective immunity Vaccine | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |
| WO 2021214204 А, 28.10.2021 | |||
| WO 2014108515 А, 17.07.2014 | |||
| Мима К.А | |||
| и | |||

