Настоящее изобретение относится к иммуногенным композициям, которые применяются для того, чтобы вызывать иммунные ответы на патогенные организмы, и, в частности, к иммуногенным композициям, способным вызывать защитные иммунные ответы.
Neisseria meningitidis (менингококк) представляет собой инкапсулированную грамотрицательную бактерию диплококк, которая обитает в носоглотке человека. Показатели носительства для популяции в целом составляют, как правило, около 10%. Сложное взаимоотношение хозяин-патоген, как правило, имеет комменсальную природу. Иногда, однако, ношение менингококка может привести к инвазивному заболеванию. Это явление, как правило, связано со штаммами, принадлежащими небольшому ряду гипервирулентных клональных линий (Caugant, D. A. et al. (1987) J Bacteriol 169:2781-2792). N. meningitidis является причиной пиогенного менингита, распространенного во всем мире, и единственной бактерией, способной вызывать вспышки менингита и сепсиса. Показатели пораженности варьируются от 1 до 3 на 105 популяции в зависимости от эндемической или эпидемической распространенности заболевания в любой географической области. Поэтому существует необходимость в разработке профилактических и терапевтических стратегий уменьшения заболеваемости, болезненности и смертности, связанных с инвазивным менингококковым заболеванием.
Организм Neisseria meningitidis чувствителен к некоторым антибиотикам первой линии, однако, несмотря на это, значительная часть пациентов, у которых диагностирована менингококковая инфекция, умирают от неизлечимого заболевания или страдают от серьезных осложнений. Коэффициент смертности составляет от около 2-3% в случае менингита без осложнений до 50% или более в случае септического шока (Cartwright, K. A. and D. A. Ala'Aldeen (1997) J Infect 34:15-19).
Современные лицензированные вакцины основываются на полисахаридной капсуле, которая окружает внешнюю бактериальную мембрану, и поэтому обеспечивают защиту только против подгруппы штаммов, которые экспрессируют соответствующую капсулу. Кроме того, не существует вполне эффективных вакцин против Neisseria meningitidis серогруппы В, которая является наиболее частой причиной менингококкового заболевания в развитых странах. Причиной этого является то, что капсула, в случае серогруппы В, состоит из полимера сиаловой кислоты, который структурно связан с модификацией адгезивных молекул нервных клеток человека, экспрессированных в грудном возрасте и в ходе эмбрионального развития. Существует концепция, что иммунизация этим антигеном будет вызывать аутоиммунные ответы.
Настоящее изобретение относится к новым композициям и, в частности, к новым иммуногенным композициям, содержащим модифицированный белок, связывающий фактор Н, и к применению этих композиций для того, чтобы вызывать иммунный ответ против Neisseria meningitidis.
Задачей настоящего изобретения является обеспечение одной или более композиций, которые могут применяться для того, чтобы вызывать иммунный ответ против Neisseria meningitidis.
Согласно первому объекту настоящее изобретение обеспечивает иммуногенную композицию, содержащую, по меньшей мере, один модифицированный белок, связывающий фактор Н, где композиция способна вызывать иммунный ответ, при введении человеку или не принадлежащему к человеческому роду животному.
Предпочтительно, упоминаемый в настоящем изобретении белок, связывающий фактор Н, относится к белку, связывающему фактор Н, из Neisseria meningitidis или Neisseria gonorrhoeae.
Neisseria meningitidis приводит к нарушению иммунного ответа организма хозяина, путем подражания хозяину. Neisseria meningitidis использует белок в форме белка, связывающего фактор Н, вместо углеводов для привлечения регулятора системы комплемента хозяина, фактора Н.
У здоровых субъектов активация комплемента строго контролируется мембраносвязанными белками и регуляторными растворимыми белками плазмы, включая фактор Н (fH). Фактор Н представляет собой белок массой 155 кДа, состоящий из двадцати доменов (терминальные повторы комплемент-контролирующих белков или CCPs). Некоторые патогены адаптировались и могут избегать комплемент-опосредованной гибели посредством связывания фактора Н с их поверхностью.
Как обсуждалось выше, Neisseria meningitidis представляет собой адптировавшийся к организму человека патоген, который имеет большое значение, так как является причиной бактериального менингита и септического шока (Stephens et al. (2007) Lancet 369, 2196-210). Благодаря изменчивости штамма Neisserial имеющиеся в настоящее время вакцины, пригодные для применения против менингококкового заболевания, не обеспечивают широкий спектр защиты и поэтому их применение ограничено. Один из подходов состоит в применении белка, связывающего фактор Н (известного как fHbp, GNA1870 или R2086) в качестве антигена в составе вакцины. Белок, связывающий фактор Н, представляет собой поверхностный липопротеин массой 27 кДа, присутствующий на всей поверхности N. Meningitidis, который вызывает защитные бактериальные антитела (Fletcher et al. (2004) Infect Immun 72, 2088-100; Masignani et al. (2003) J Exp Med 197, 789-99). Белок, связывающий фактор Н, служит для привлечения отрицательного регулятора комплемента, фактора Н, к бактериальной поверхности и вносит свой вклад в способность менингококка избегать врожденных иммунных ответов путем ингибирования комплемент-опосредованного лизиса в плазме человека (Madico et al. (2006) J Immunol 177, 501-10; Schneider et al. (2006) J Immunol 176, 7566-75).
В настоящем изобретении, по меньшей мере, один модифицированный белок, связывающий Фактор Н, применяется в качестве антигена. Белок модифицирован таким образом, чтобы связывание фактора Н с белком, связывающим фактор Н, предотвращалось или уменьшалось. Связывание фактора Н с белком, связывающим фактор Н, после введения субъекту белка, связывающего фактор Н, в составе вакцины, как полагают, приводит к понижению эффективности вакцины. Во-первых, присутствие фактора Н на белке, связывающем фактор Н, может ограничивать распознавание значимых эпитопов на белке, связывающем фактор Н, иммунной системой хозяина. Антитела, образовывающиеся против этих эпитопов, могут быть как бактерицидными, так и могут ингибировать связывание фактора Н с бактериями, обеспечивая их чувствительность к комплемент-опосредованному лизису. Во-вторых, иммунные ответы против белка, связывающего фактор Н, могут также вызывать ответы против связанного фактора Н подобно гаптену. Это может приводить к потенциальному неожиданному аутоиммунному ответу. Наконец, активация комплемента участвует в иммунных ответах, так как белки комплемента имеют адъювантную активность. Понижение активации комплемента в сайте иммунизации путем привлечения фактора Н могло бы понизить уровень иммунности, образующейся после вакцинации.
Предпочтительно модифицированный белок, связывающий фактор Н, для применения по изобретению модифицирован при одном или более положениях в белке.
Предпочтительно модифицированный белок, связывающий фактор Н, имеет, по меньшей мере, одну аминокислоту, которая была изменена по сравнению с аминокислотой в белке дикого типа.
Предпочтительно одна и более изменяемых аминокислот представляют собой аминокислотные остатки, которые менее выставлены на поверхности, когда белок, связывающий фактор Н, находится в комплексе с фактором Н по сравнению с выделенным белком, связывающим фактор Н.
Одна или более аминокислот, которые меняются в белке, связывающем фактор Н, могут выбираться из группы, состоящей из аминокислотных остатков в положении номер 103, 106, 107, 108, 109, 145, 147, 149, 150, 154, 156, 157, 180, 181, 182, 183, 184, 185, 191, 193, 194, 195, 196, 199, 262, 264, 266, 267, 268, 272, 274, 283, 285, 286, 288, 289, 302, 304 306, 311 и 313, как показано на Фиг.6 (SEQ ID NO:1).
Одна или более аминокислот, которые меняются в белке, связывающем фактор Н, могут представлять собой одну или более аминокислот, которые в белке дикого типа образуют водородные связи с фактором Н.
Одна или более аминокислот, которые меняются в белке, связывающем фактор Н, могут выбираться из группы, содержать номера аминокислот 103, 106, 107, 108, 180, 181, 183, 184, 185, 191, 193, 195, 262, 264, 266, 272, 274, 283, 286, 304 и 306, как показано на Фиг.6 (SEQ ID NO:1).
Две или более, три или более, четыре или более или пять или более из вышеупомянутых аминокислот могут быть изменены в модифицированном белке, связывающим фактор Н, для применения по настоящему изобретению. Предпочтительно, путем изменения одной или более аминокислот в белке, связывающем фактор Н, связывание с фактором Н может предотвращаться или значительно уменьшаться.
В одном варианте выполнения настоящего изобретения, по меньшей мере, одна из аминокислот номер 283 и 204 мутирует/изменяется с образованием аланина вместо глутаминовой кислоты. Предпочтительно эта мутация приводит к почти полному исключению связывания фактора Н.
Предпочтительно, связывание фактора Н с модифицированным белком, связывающим фактор Н, является, по меньшей мере, в пять раз меньше, предпочтительно, по меньшей мере, в десять раз меньше, предпочтительно, по меньшей мере, на два порядка меньше, чем связывание фактора Н с белком дикого типа, связывающим фактор Н. Предпочтительно, уменьшение связывания измеряется путем применения аналита при концентрации около 50 нМ. Уменьшение связывания такого порядка рассматривается как значительное уменьшение.
В дополнение к модификациям, способствующим предотвращению или ингибированию связывания белка, связывающего фактор Н, с фактором Н, белок может также включать другие мутации, которые не влияют на способность белка действовать в качестве иммуногена против инфекции, вызванной N. meningitidis. Например, в белке могут быть сделаны другие изменения консервативных аминокислот. Подобным образом, в белке могут быть сделаны вставки или делеции, которые не влияют на иммуногенность белка.
Предпочтительно, модифицированный белок, связывающий фактор Н, имеет последовательность, на, по меньшей мере, 60%, 70%, 80%, 85%, 90%, 95% или более процентов идентичную последовательности, показанной на Фиг.6, но он модифицирован таким образом, что предпочтительно имеет отличную аминокислоту в, по меньшей мере, одном положении, что приводит к отсутствию или значительному уменьшению связывания фактора Н.
Процент идентичности последовательности определяют как процент аминокислот в последовательности, которые идентичны аминокислотам в контрольной последовательности после выравнивания последовательностей и введения гэпов, если это необходимо для достижения максимального процента идентичности последовательностей. Выравнивание с целью определения процента идентичности последовательностей может осуществляться различными путями, которые хорошо известны специалистам в данной области техники и включают, например, применение BLAST (поисковый механизм (программа) логического сравнения аминокислотных и нуклеотидных последовательностей национального центра биотехнологической информации).
Различия в проценте идентичности могут быть обусловлены, например, замещениями, вставками или делениями аминокислот. Аминокислотные замещения могут быть консервативными по своей природе, тогда замещенные аминокислоты имеют подобные структурные и/или химические свойства, например, замещение лейцина на изолейцин является консервативным замещением.
Иммуногенная композиция представляет собой композицию, которая способна вызывать иммунный ответ на, по меньшей мере, один модифицированный белок, связывающий фактор Н, когда композиция вводится субъекту. Предпочтительно, субъектом является человек или не принадлежащее к человеческому роду животное, более предпочтительно человек или не принадлежащее к человеческому роду млекопитающее.
Предпочтительно, иммунный ответ, вызываемый композицией по изобретению, влияет на способность N. meningitidis инфицировать иммунизированного человека. Предпочтительно, способность N. meningitidis инфицировать человека, иммунизированного композицией по изобретению, уменьшается или исключается. Это может достигаться несколькими путями. Вызываемый иммунный ответ может приводить к распознаванию и уничтожению N. meningitidis. Альтернативно или дополнительно, вызываемый иммунный ответ может затруднять или предотвращать репликацию N. meningitidis. Альтернативно или дополнительно, вызываемый иммунный ответ может затруднять или предотвращать возникновение заболевания, вызванного N. meningitides, у человека или не принадлежащего к человеческому роду животного.
Модифицированный белок, связывающий фактор Н, может продуцироваться рекомбинантно (например, с помощью сконструированной методами генной инженерии экспрессионной системы) или может быть синтетическим продуктом, например, может продуцироваться путем in vitro пептидного синтеза или in vitro трансляции.
Композиция по настоящему изобретению может, кроме того, содержать один или более антигенов, в дополнение к одному или более модифицированным белкам, связывающим фактор Н. Дополнительные антигены могут также быть получены из N. meningitidis и могут обладать способностью вызывать иммунный ответ, направленный против N. meningitidis.
Композиция может применяться для вызывания/создания защитного иммунного ответа при введении субъекту. Защитный иммунный ответ может вызывать гибель N. meningitidis после инфицирования субъекта или может предотвращать или ингибировать репликацию N. meningitides и/или предотвращать возникновение заболевания.
Композиция может применяться в качестве профилактической или терапевтической вакцины, направленной против N. meningitidis.
Согласно следующему объекту настоящего изобретения настоящее изобретение обеспечивает фармацевтическую композицию, содержащую, по меньшей мере, один модифицированный белок, связывающий фактор Н, и фармацевтически приемлемый носитель или эксципиент.
Предпочтительно, фармацевтическая композиция содержит композицию согласно первому объекту настоящего изобретения.
Предпочтительно, фармацевтическая композиция способна создавать защитный иммунный ответ против N. meningitidis.
Фраза "создание защитного иммунного ответа", как применяется в настоящем изобретении, означает, что композиция способна создавать защитный ответ в организме хозяина, такого как человек или не принадлежащее к человеческому роду млекопитающее, которому она вводится. Предпочтительно, защитный иммунный ответ защищает против возникающей инфекции, вызванной N. meningitidis. Защитный иммунный ответ может исключать или уменьшать уровень инфекции путем уменьшения репликации N. meningitidis или путем воздействия на образ действия N. meningitidis с уменьшением заболевания.
Подходящие приемлемые эксципиенты и носители хорошо известны специалистам в данной области техники. Они могут включать твердые и жидкие носители. Подходящие жидкие носители включают воду и физиологический раствор. Белки могут находиться в композиции в виде эмульсии или в виде биоразлагаемых микросфер или липосом.
Композиция может, кроме того, содержать адъювант. Подходящие адъюванты хорошо известны специалистам в данной области техники и могут включать неполный адъювант Фрейнда (для применения на животных) и соли металлов, такие как соли алюминия или кальция.
Композиция может также содержать полимеры или другие агенты, для того чтобы контролировать консистенцию композиции и/или контролировать высвобождение антигена/секретированного белка из композиции.
Композиция может также содержать другие агенты, такие как разбавители, которые могут включать воду, физиологический раствор, глицерин или другие подходящие спирты и т.д.; смачивающие или эмульгирующие средства; буферизующие средства; загустители, например целлюлозу или производные целлюлозы; консерванты, детергенты, противомикробные средства и тому подобное.
Предпочтительно, активные ингредиенты в композиции составляют более 50% от чистой массы, обычно более 80% от чистой массы, часто более 90% от чистой массы и более предпочтительно более 95%, 98% или 99% от чистой массы. Активные ингредиенты могут достигать 100% от чистой массы, например около 99.5% от чистой массы или около 99.9% от чистой массы, что применяется довольно часто.
Композиция по настоящему изобретению может применяться в качестве вакцины против инфекций, вызываемых N. meningitidis. Композиция может применяться в качестве вакцины, направленной против менингита или других инвазивных менингококковых заболеваний, включая сепсис или септический шок. Вакцина может вводиться в профилактических целях пациентам, которые могут подвергнуться N. meningitidis, и/или в терапевтических целях субъектам, которые уже подверглись действию N. meningitidis.
Предпочтительно, если композиция применяется в качестве вакцины, то композиция содержит иммунологически эффективное количество антигена, где композиция содержит, по меньшей мере, один модифицированный белок, связывающий фактор Н. "Иммунологически эффективное количество" представляет собой количество антигена, которое при введении субъекту либо в виде однократной дозы, либо в виде серии доз эффективно для лечения или предотвращения инфекции, вызванной N. meningitidis. Это количество в значительной степени зависит от здоровья и физического состояния субъекта, который подлежит лечению, и от антигена. Определение эффективного количества иммуногенной композиции или композиции вакцины для введения в организм субъекта лежит в области компетенции специалиста в данной области техники.
Композиция по настоящему изобретению может вводиться перорально, системно, парентерально, местно, мукозально, внутримышечно, внутривенно, внутрибрюшинно, интрадермально, подкожно, интраназально, интравагинально, интраректально, трансдермально, сублингвально, путем ингаляции или в виде аэрозоля.
Композиция может вводиться либо в виде однократной дозы, либо согласно схеме введения многократных доз. Многократные дозы могут вводиться в виде первичной иммунизации с последующими одной или более активными иммунизациями. Подходящий промежуток времени между первичной иммунизацией и активными иммунизациями может быть определен согласно общепринятому порядку.
Композиция по настоящему изобретению может применяться сама по себе или может быть объединена с одной или более другими иммуногенными композициями или композициями вакцин и/или с одним или более другими терапевтическими режимами.
Композиции по изобретению могут обладать способностью индуцировать бактерицидные ответы сывороточных антител и активировать антитела, которые опосредуют опсонофагоцитоз после их введения субъекту. Эти ответы обычно измеряют на мышах и результаты являются стандартным индикатором эффективности вакцины.
Композиции по настоящему изобретению могут также, или альтернативно, вызывать иммунный ответ, который нейтрализует бактериальные белки или другие молекулы, таким образом мешая им выполнять их нормальные функции и предупреждая или уменьшая развитие заболевания без неизбежного разрушения патогенного организма/бактерии, этот случай относится к N. meningitidis.
Согласно следующему объекту настоящего изобретения настоящее изобретение обеспечивает применение одного или более модифицированных белков, связывающих фактор Н, для получения лекарственного средства, которое способно вызывать иммунный ответ. Лекарственное средство может применяться для профилактической или терапевтической вакцинации субъектов против N. meningitidis. Лекарственное средство может представлять собой профилактическую или терапевтическую вакцину. Эта вакцина может применяться против менингита, сепсиса и/или септического шока, вызванного N. meningitidis.
Согласно еще одному объекту настоящего изобретения настоящее изобретение обеспечивает композицию, содержащую один или более модифицированных белков, связывающих фактор Н, для применения в целях получения иммунного ответа на N. meningitidis. Иммунный ответ может быть профилактическим или терапевтическим. Композиция может применяться в качестве вакцины.
Согласно следующему объекту настоящее изобретение обеспечивает способ защиты человека или не принадлежащего к человеческому роду животного от воздействия инфекции, вызванной N. meningitides, содержащий введение человеку или не принадлежащему к человеческому роду животному композиции согласно любому другому объекту настоящего изобретения. Композиция может представлять собой вакцину.
Согласно другому объекту настоящее изобретение обеспечивает способ повышения иммунного ответа у человека или не принадлежащего к человеческому роду животного, содержащий введение фармацевтической композиции по изобретению человеку или не принадлежащему к человеческому роду животному. Иммунный ответ предпочтительно является защитным. Способ может приводить к повышению бустерного ответа у пациента, который уже подвергался лечению. Иммунный ответ может быть профилактическим или терапевтическим.
Для того чтобы проверить эффективность терапевтического лечения, содержащего введение композиции по изобретению, контролируют инфекцию, вызываемую N. meningitides, после введения композиции. Один способ проверки эффективности терапевтического лечения, содержащего введение композиции по изобретению, включает контроль за иммунными ответами на Neisseria meningitidis после введения композиции.
Согласно другому объекту настоящее изобретение обеспечивает применение одного или более модифицированных белков, связывающих фактор Н, для получения лекарственного средства, применяемого для иммунизации человека или не относящегося к человеческому роду млекопитающего против инфекции, вызываемой N. meningitidis.
Согласно другому объекту настоящее изобретение обеспечивает набор, применяемый для индукции иммунного ответа в организме, содержащий иммуногенную композицию или композицию вакцины по изобретению и инструкции по введению.
В дополнение к их потенциальному применению в качестве вакцины композиции по настоящему изобретению могут быть полезны в качестве диагностических реагентов и для измерения иммунной компетентности вакцины.
Несмотря на то что все приведенные выше утверждения и предпочтительные признаки относятся к to N. meningitidis, специалисту в данной области техники будет очевидно, что они равным образом относятся и к N. gonorrhoeae.
Специалисты в данной области техники могут оценить по достоинству, что любые предпочтительные признаки, обсуждаемые выше, могут применяться к любому объекту настоящего изобретения.
Предпочтительные варианты выполнения настоящего изобретения описываются далее с помощью примеров и ссылок на приведенные чертежи и примеры.
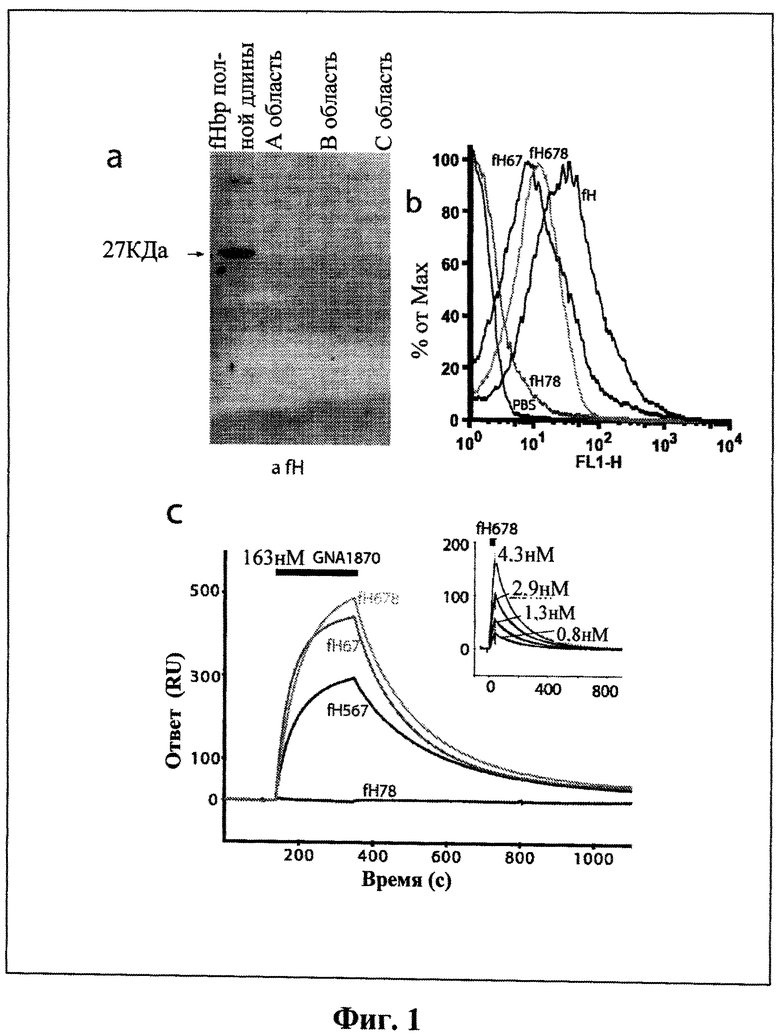
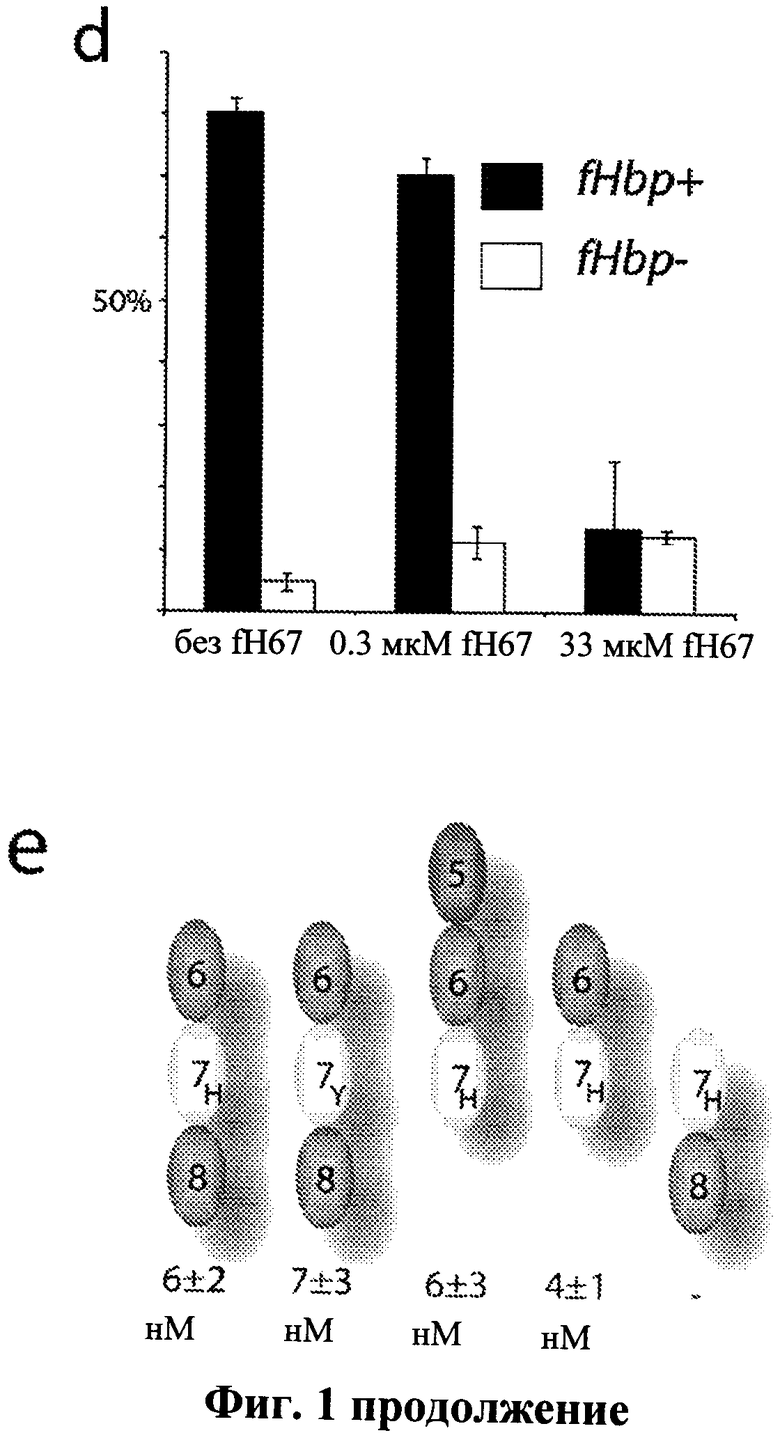
Фиг.1а-1f - Фиг.1а-1f демонстрирует, что сайт связывания с белком, связывающим фактор Н, ограничен до ССР6 в факторе Н и нуждается во всей внеклеточной части белка, связывающего фактор Н. На Фиг.1а показан Фар-Вестерн анализ связывания фактора Н с интактным белком, связывающим фактор Н, и его усеченными вариантами (как показано). Мембраны инкубировали с очищенным фактором Н, который определили с помощью α-fH pAbs. Связывание наблюдалось только для интактного белка, связывающего фактор Н, массой 27 кДа (показано стрелкой). На Фиг.1b приведены результаты FACS анализа с применением α-fH pAbs, которые показали связывание между N. meningitidis и фактором Н. На Фиг.1 с результаты SPR приводятся для того, чтобы показать, что белок, связывающий фактор Н, способен связываться только с конструкциями фактора Н, содержащими ССР6. На вставке показана модель Ленгмюра 1:1, подходящая для серийных разбавлений fH67, инъецируемых по поверхности белка, связывающего фактор Н, для определения кинетических параметров. На Фиг.1d приведены результаты конкурентного FACS анализа (применяя α-fH mAb, направленные против ССР5, и поэтому не способные распознавать конструкцию fH67), которые показывают, что короткая конструкция fH67 (от ~0.3 до 30 мкМ) может конкурировать с фактором Н полной длины в отношении связывания, демонстрируя, что эта конструкция содержит полный сайт связывания с белком, связывающим фактор Н. Показанные значения представляют собой среднее значение интенсивности флуоресценции, полученное по трем экспериментам, ± стандартное отклонение. Фиг.1е показывает, что количественное определение с помощью SPR подтверждает, что присутствие ССР6 является необходимым и достаточным для высокой аффинности связывания с белком, связывающим фактор Н, и что общий полиморфизм фактора Н в ССР7 (402His/Tyr) незначительно изменяет аффинность fHbp связывания. На Фиг.1f приведена таблица, иллюстрирующая константы связывания, полученные с применением поверхностного плазменного резонанса, где белок, связывающий фактор Н, находится на поверхности чипа, а конструкции фактора Н в жидкой фазе. Подборы осуществляли с помощью программного обеспечения BiaEvaluation 2.0 и модели Ленгмюра 1:1. Показанные значения представляют собой среднее значение ± стандартное отклонение, Chi2 относится к эффективности подобранных индикаторов, соответствующих программному обеспечению BiaEvaluation 2.0.
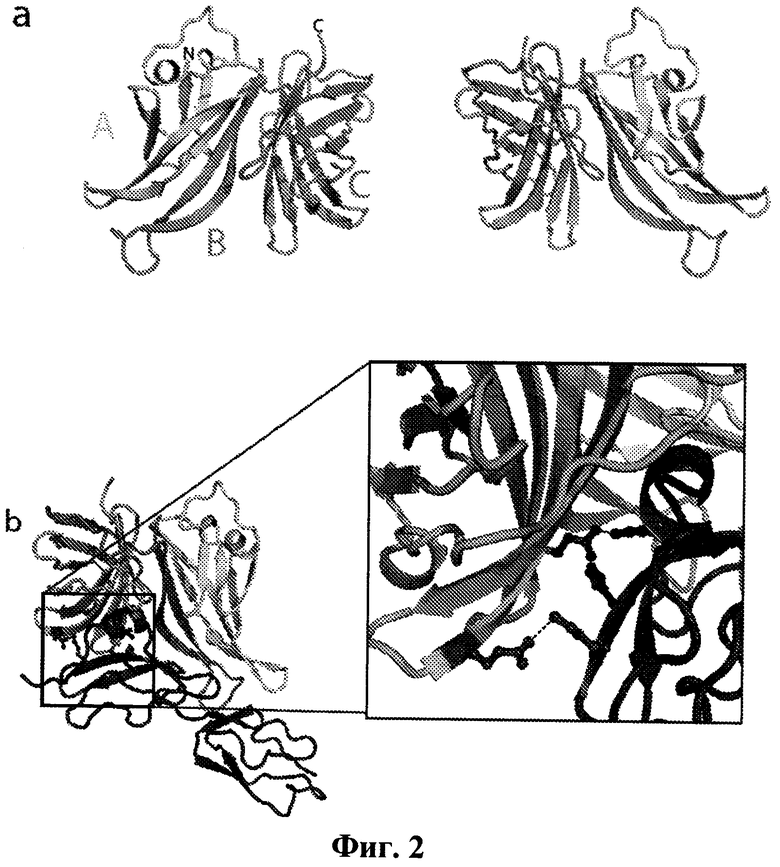
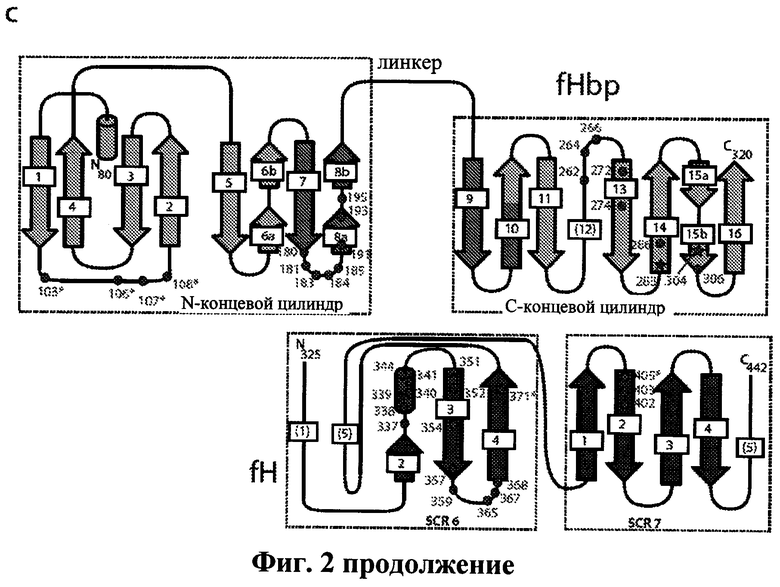
Фиг.2а-2с иллюстрируют структуру белка, связывающего фактор Н, и его комплекс с fH67. Фиг.2а показывает два вида изображения белка, связывающего фактор Н (остатки 80-320). Указаны области А, В и С. Фиг.2b показывает комплекс fHbp:fH67 между белком, связывающим фактор Н, и ССР 6 и 7 фактора Н. Боковые цепи обоих белков, участвующие в формировании солевых мостиков по поверхности взаимодействия, показаны в виде шаростержневой модели (увеличены и перенесены в рамку). Фиг.2 с иллюстрирует топологию белка, связывающего фактор Н, и fh67, а также показаны остатки, участвующие во взаимодействиях между белками, либо в виде Н-связей, либо в виде солевых мостиков. Закрашенные кружки показывают остатки, которые образуют водородную связь с их партнерами. Закрашенные стрелки показывают остатки, которые образуют солевой мостик с их партнерами. Звездочка показывает взаимодействие, которое видно только в некоторых независимых копиях сложной структуры.
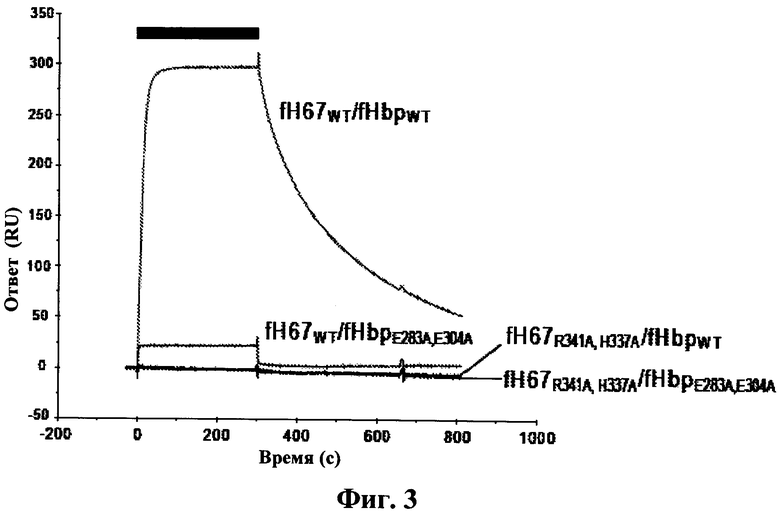
Фиг.3 относится к взаимодействию между белком, связывающим фактор Н, и фактором Н, и демонстрирует, что сайт-направленный мутагенез, связанный с замещением заряженных боковых цепей аланином, как в факторе Н, так и в белке, связывающем фактор Н, вызывает прекращение связывания каждого с формой дикого типа их партнера при концентрациях около KD (константы диссоциации) дикого типа. Черная полоса показывает период времени, в течение которого аналиты фактора Н инъецируются (при 50 нМ, 40 мкл·мин-1) по поверхности белка, связывающего фактор Н.
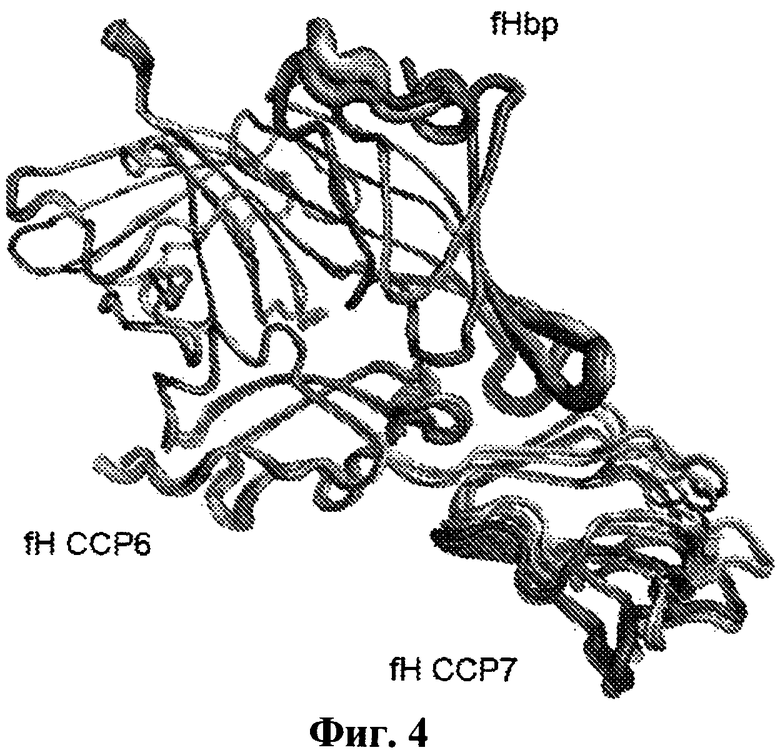
На Фиг.4 рассматривается гибкость комплекса fH67/fHbp, при этом применяется наложение семи кристаллографически независимых копий комплекса fH67/fHbp из двух кристаллических форм (четыре копии комплекса в Р1 форме и 3 копии в С2 форме). Высокая степень подобия, несмотря на различные условия упаковки, указывает на то, что комплекс является биологически релевантным. Перегруппировка белков внутри комплекса ограничена небольшой возможностью движения в положении ССР7 fH67 относительно ССР6 и белка, связывающего фактор Н. Петли белка, связывающего фактор Н, которые упаковываются наиболее близко к ССР7, также относительно подвижны, на что указывают их более высокие температурные факторы (отображенные здесь в виде "более плотного" закрашивания).
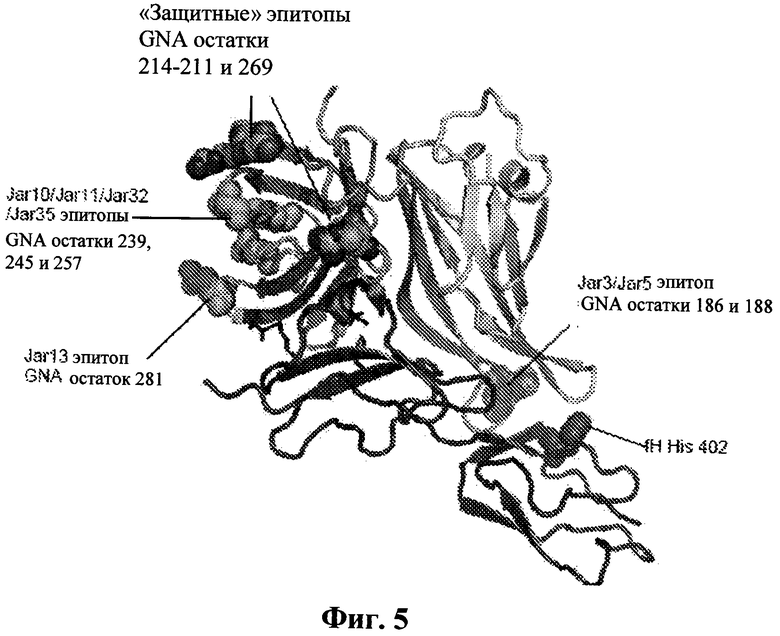
Фиг.5 показывает расположение эпитопов антител, обозначенных на поверхности белка, связывающего фактор Н. Остатки в защитных эпитопах, лишенных ССР в 12, 20, 21, показаны в виде CPK изображения (Glu 211, Arg 214 и Arg 269) с окрашенными элементами (С - зеленый, N - синий, О - красный). Эпитопы для антител, которые блокируют связывание фактора Н, показаны как СРК окрашенные согласно окрашиванию белка, связывающего фактор Н (область "А" - желтая, область "В" - зеленая, область "С" - синия). Несмотря на то что ни один из этих эпитопов не лежит непосредственно в сайте распознавания фактора Н, все они лежат достаточно близко, так что связывание большого антитела будет, вероятно, затруднять распознавание фактора Н. Гистидиновая боковая цепь при полиморфном остатке 402 фактора Н также показана на CPK изображении. Хотя этот остаток лежит близко для дополнительного контакта с белком, связывающим фактор Н (около остатков 103-108, смотрите Фиг.2), очевидность, исходя из SPR, двух полиморфных форм предполагает, что этот контакт не имеет важного значения. Это также поддерживается тем фактом, что эта область комплекса (как GNA петли, так и домен фактора Н) является, как оказалось, наиболее подвижной областью со значительным числом вариантов контакта двух белков друг с другом, как видно в области между кристаллографическими независимыми копиями комплекса (Фиг.4).
На Фиг.6 приведена аминокислотная последовательность белка, связывающего фактор Н (SEQ ID NO:6). Этот белок имеет номер доступа в генетическом банке №: AAF42204.
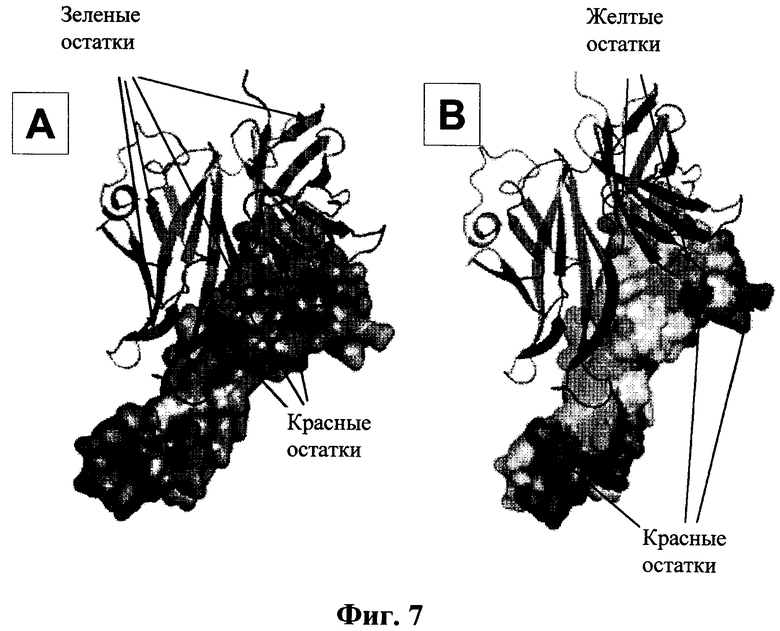
На Фиг.7 приведена структура fHbp:fH67 комплекса. На Фиг.7А fHbp показан в виде динамического изображения (остатки зеленого цвета сохраняются среди семейств, красные не сохраняются), а fH67 показан в виде плоскостной модели. На Фиг.7В остатки, которые варьируются между fH мыши и человека, показаны красным; наше исследование показало, что желтые остатки влияют на связывание fH.
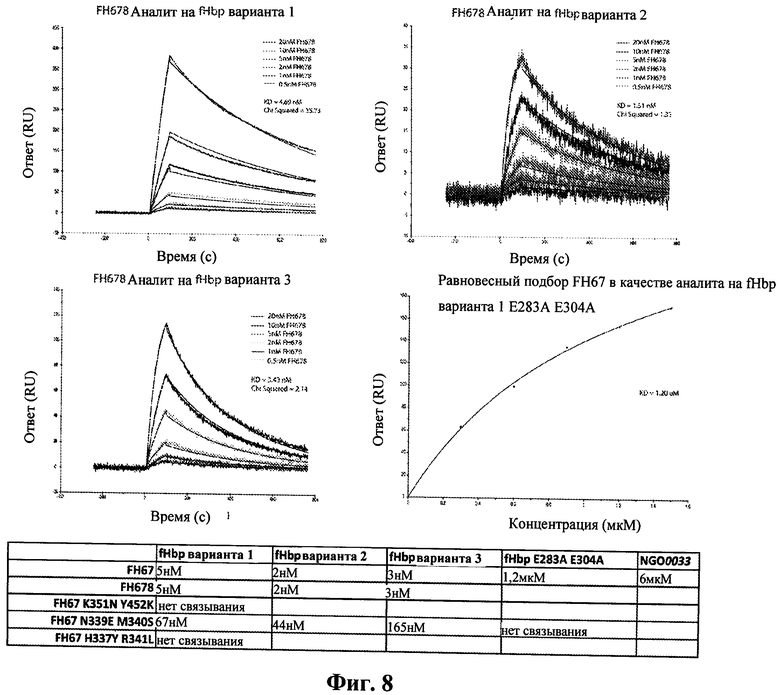
На Фиг.8 показано связывание fHbp варианта 1, 2 и 3 с fH678, как определено с помощью SPR. Взаимодействия с гонококковым гомологом NGO0033 также были изучены.
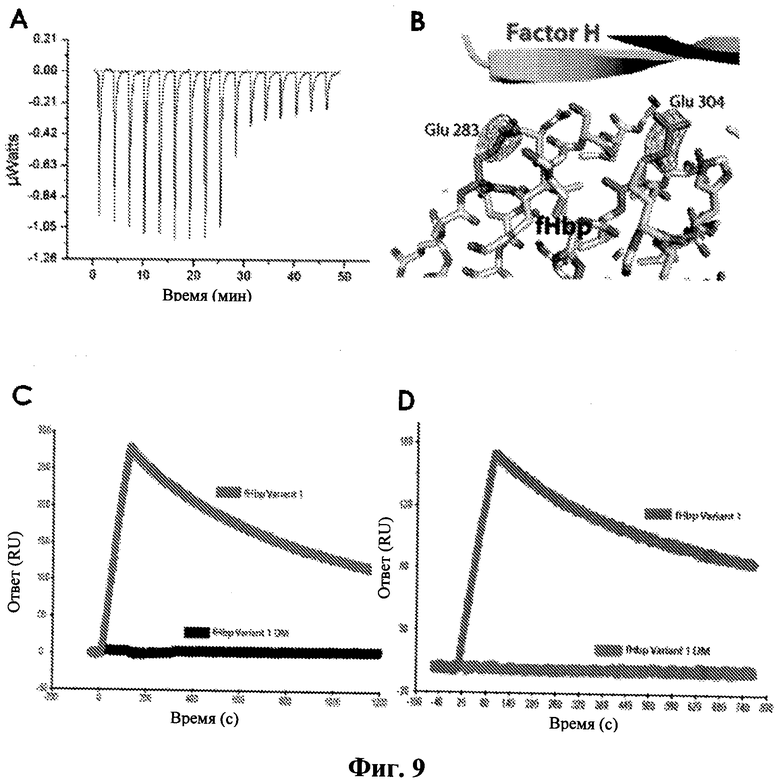
Фиг.9А, изотермическая калориметрия титрования подтверждает, что fHbp/fH взаимодействие характеризуется наномолярной KD в растворе. На Фиг.9В приведена структура двойного Glu - Ala мутанта fHbp, которая подтверждает, что в мутированных боковых цепях существуют только структурные изменения (красным показана разность электронной плотности в FO-FC, αС карта. На Фиг.9С показан ответ на поток 5нМ доменов 5, 6 и 7 фактора Н по поверхности дикого типа и мутанта fHbp (верхняя кривая приведена для fHbp варианта 1, нижняя кривая приведена для fHbp варианта 1 DM (двойной мутант)). На Фиг.9D показан ответ на поток 20 нМ интактного фактора Н по поверхности дикого типа и мутанта fHbp (верхняя кривая приведена для fHbp варианта 1, нижняя кривая приведена для fHbp варианта 1 DM).
Функция белка, связывающего фактор Н, последовательность которого приведена на Фиг.1, была уже ранее изучена путем подразделения белков на ряд областей, обозначенных "А", "В" и "С", покрывающих весь внеклеточный домен (Giuliani et al. (2005) Infect Immun 73, 1151-60). Вестерн-блот-анализ показывает, что все три области необходимы для высокой аффинности взаимодействий между фактором Н и белком, связывающим фактор Н (Фиг.1а), таким образом можно сделать вывод, что белок, связывающий фактор Н, имеет расширенный сайт распознавания для фактора Н по всей его поверхности.
Для того чтобы определить, какие 20 ССР доменов фактора Н опосредуют взаимодействие с белком, связывающим фактор Н, проводились следующие исследования: Фар-Вестерн-анализ, FACS-анализ (Фиг.1b) и поверхностный плазменный резонанс (Фиг.1с). Полученные результаты демонстрируют, что ключевая область фактора Н, распознаваемая белком, связывающим фактор Н, представляет собой шестой домен, ССР6, и что конструкции, содержащие ССР6, способны ингибировать зависимое от белка, связывающего фактор Н, взаимодействие между фактором Н и N. meningitidis (Фиг.1d). Квантификация взаимодействия демонстрирует, что константа диссоциации составляет около 5нМ (Фиг.1с, 1е и 1f) для любой конструкции, содержащей ССР6. Это взаимодействие не разрушается в высокосоляных условиях (1М NaCl, не показано) или в интервале рН от 4 до 8 (не показано), обеспечивающих дополнительную поддержку высокоаффинной природы связывающего элемента.
Как показано на Фиг.9A-D, поверхностный плазменный резонанс демонстрирует, что fHbp двойной мутант связывает домены 6 и 7 фактора Н с KD на два порядка выше. Мутант также связывает более длинные конструкции (содержащие домены 5, 6 и 7) и фактор Н полной длины Н (очищенный от сыворотки) намного слабее, чем fHbp дикого типа.
Были получены кристаллические структуры комплекса между CCPs 67 фактора Н (выше упоминается как fH67) и внеклеточной частью белка, связывающего фактор Н, содержащего области А, В и С, и модели для белка, связывающего фактор Н, и fH67 были построены с разрешением 2.35 Å в двух кристаллических формах с, в целом, семью независимыми копиями комплекса (Фиг.2а, b и с). Как можно увидеть на Фиг.2а, внеклеточная часть белка, связывающего фактор Н, укладывается с образованием β-цилиндров с N-концевым цилиндром, состоящим из "А" и части "В" областей, тогда как С-концевой цилиндр состоит из остатка "В" и "С" областей. Поиск в структурных базах данных по М-концевым цилиндрам не обнаружил никаких близких структурных гомологов, и различные топологии β-цилиндров (Фиг.2с) позволяют предположить, что они не возникают в результате дупликации гена.
Комплекс fH67:fHBP (фактор Н: белок, связывающий фактор Н) удерживается вместе благодаря повсеместным взаимодействиям между β-цилиндрами белка, связывающего фактор Н, и ССР6 фактора Н (Фиг.2b), это согласуется с изучениями связывания. В частности, спиральная вставка во вторую β-тяж ССР6 (необычный признак этого ССР домена) занимает в комплексе центральное положение. Анализ с помощью PISA сервера позволил получить все семь независимых комплексов с коэффициентом статистической значимости 1.0 (то есть весьма вероятна биологическая значимость), причем средняя площадь поверхности белка, связывающего фактор Н, находящегося в комплексе по изобретению (2860±177Å2), больше, чем площадь поверхности в случае большинства комплексов антитело: антиген. Более того, ΔG (-6 ккал/моль), как предсказано на основе структуры для образования комплекса, хорошо согласуется с аффинностью, полученной в результате изучения связывания (-11 ккал/моль, вычислено исходя из KD, приведенной на Фиг.1е), что обеспечивает дополнительную поддержку физиологической релевантности кристаллизованного комплекса. Поверхность взаимодействия показывает хорошую комплементарность формы с многочисленными электростатическими взаимодействиями (Фиг.2с), включающими многочисленные водородные связи и солевые мостики. Это согласуется с результатами изучения fHbp:fH взаимодействия, которые не показали каких-либо различий в связывании (Фиг.1е, Фиг.4 и Фиг.5) между 402His/Tyr изоформами фактора Н, связанными с возрастной макулярной дегенерацией. His402 только периферически участвует во взаимодействие с белком, связывающим фактор Н, и контактирует с гибкой петлей между тяжами 1 и 2 N-концевого β-цилиндра белка, связывающего фактор Н (Фиг.4).
Для следующего исследования воссоздавали поверхность взаимодействия дважды мутированных/замещенных форм обоих белков, в которых заряженные боковые цепи замещались маленьким гидрофобным остатком Аланином (А). В fH67 полученные мутации представляли собой R341A и Н337А, и в белке, связывающем фактор Н, полученные мутации представляли собой Е283А и Е304А. Применяя SPR для изучения взаимодействий между мутированными белками и их партнерами дикого типа, обнаружили, что аффинность обоих мутантных форм уменьшилась более чем на два порядка, так что практически не наблюдалось никакого взаимодействия при концентрациях аналита (~50 нМ) около десятикратной величины константы диссоциации дикого типа (Фиг.3а). При концентрации аналита в тысячу раз выше (микромолярный интервал) мутантные формы обоих белков не взаимодействовали с их партнерами дикого типа: fHbpE283A, E304A сохранил качественное сходство по скорости ассоциации, но скорость диссоциации возросла по сравнению с диким типом, в то время как fH67R341A, H337A был более схож с диким типом в отношении взаимодействия по скорости его диссоциации, но имел чрезвычайно уменьшенную скорость ассоциации (Фиг.1е).
fHbp, принадлежащие к различным семействам, имеют подобные характеристики связывания
Штаммы N. meningitidis экспрессируют один fHbp, принадлежащий к одному из трех возможных семейств, вариант 1, вариант 2 и вариант 3. Наши исследования были сфокусированы на fHbp семейства варианта 1, так как это наиболее преобладающее семейство среди изолятов, вызывающих заболевание. На Фиг.7 приведена вариация последовательности среди трех семейств, обозначенных на структуре комплекса, и показано, что, хотя уровень сохранения последовательности является высоким (>60%), существует значительная вариация около fH-связывающего сайта. Поэтому мы определили, является ли способ распознавания fH неизменным для различных семейств fHbp и являются ли одни и те же остатки fHbp и fH ключевыми для связывания. Это очень важная информация для получения вариантов fHbp с уменьшенным связыванием в целях дальнейшего развития вакцин.
В конце исследования мы клонировали, экспрессировали и очистили fHbp семейства варианта 2 и варианта 3 и охарактеризовали их взаимодействия с fH (Fig.8). Наши SPR исследования показали, что способы распознавания являются подобными; аффинность имеет значения одного порядка, а мутация одних и тех же остатков в fH оказывает критически важное влияние на связывание fHbps всех семейств (не показано). Известно, что гомолог fHbp из Neisseria gonorrhoeae (NG00033) имеет близкое родство к варианту 3 (91% идентичности аминокислот), и очищенный белок не взаимодействует с fH до какой-либо значительной степени (Фиг.8).
fHbpE283A, E304A мутант сохраняет его атомную структуру
Возможное объяснение неспособности fHbpE283A, E304A мутанта (двойной мутант, DM) связываться с fH67 при константе диссоциации (KD) в наномолярном диапазоне (нМ) заключается в том, что аминокислотные изменения разрушают общую структуру белка. В связи с этим была определена кристаллическая структура fHbpE283A, E304A мутанта в комплексе с fH67. Результаты подтвердили, что только структурные изменения в fHbp связаны с мутированными боковыми цепями (Фиг.9В). Дополнительно мы определили значимость этих мутаций для связывания с fH5,6,7 и Ж полной длины (Фиг.9С и D). Концентрации fH в 10 нМ диапазоне, как можно увидеть, взаимодействуют с белком дикого типа, что позволяет предположить, что аффинность fH полной длины находится в том же диапазоне, что и аффинность более маленьких фрагментов (например, fH5,6,7). Однако аффинность связывания fHbpE283A, E304A белка значительно снижена по сравнению с белком дикого типа.
Обозначение на структуре известных из опубликованных документов эпитопов белка, связывающего фактор Н, которые вызывают бактериальные антитела (Giuliani et al. (2005) Infect Immun 73, 1151-60; Cantini et al. (2006) J Biol Chem 281, 7220-7), демонстрирует, что ни один из сайтов, охарактеризованных на сегодняшний день, не лежит непосредственно в сайте распознавания фактора Н. Однако эпитопы, распознаваемые антителами, которые влияют на связывание фактора, расположены вблизи сайта распознавания и таким образом, вероятно, ингибируют связывание фактора Н по причине стерических препятствий
fHbp одинарные и двойные мутанты сохраняют свою иммуногенность.
Для того чтобы исследовать влияние Е283А и Е304А аминокислотных изменений, белки из семейства fHbp варианта 1 с этими замещениями, либо отдельно (то есть fHbpE283A и fHbpE304A), либо в комбинации (fHbpE283A,E304A), экспрессировали и очистили. fHbp дикого типа и эти белки применяли для иммунизации групп мышей (5 группа) и определили присутствие бактериальных антител в иммунной сыворотке. Результаты (Таблица 1) показаны как обратная величина от наибольшего разбавления сыворотки, которое приводит к гибели 50% или более N. meningitidis серогруппы В дикого типа (штамм Н44/76). Сыворотка до иммунизации не приводила к гибели.
Результаты показывают, что мутантные белки имеют, по меньшей мере, равную белкам дикого типа способность обеспечивать титры SBA против N. meningitidis серогруппы В.
Способы:
Вестерн-блот-анализ: Образцы белка разделили с помощью SDS-PAGE, затем переносили на PVDF мембраны при 15 В в течение 60 минут. Мембраны блокировали в 5% молока в PBS при 4°С всю ночь, один раз промыли в PBSTM (PBS, 0.05% Tween20, 0.5% снятого молока), затем инкубировали с первичными антителами или источником fH в течение 1-2 часов при комнатной температуре. После трех промывок в PBSTM мембраны инкубировали с козлиными политональными антителами (pAbs) к fH человека (при конечном разбавлении 1:1000) или мышиными поликлональными антителами (pAbs) к fHbp (при конечном разбавлении 1:10,000) в PBS. Связывание обнаружили с помощью антител к IgG кролика, конъюгированных с пероксидазой (Dakocytomation), которые применяли при разбавлении 1:500.
Проточная цитометрия: Всего 108 N. meningitidis инкубировали с 10 мкл очищенного fH (2 мкМ) или CCPs (Конечная концентрация, 3 мМ) в течение 30 минут при 37°С. После трех промывок с применением PBS-0.1% Tween20 (PBST) бактерии инкубировали в течение 30 минут с mAb MRC OX241 или с pAbs к фактору Н человека в PBS. После трех промывок в PBST клетки инкубировали с вторичными FITC-конъюгированными антителами в течение 30 минут в темноте при 4°С. Фактор Н, связанный с менингококками, обнаружили с помощью проточной цитометрии, применяя проточный цитофлуориметр FACS Calibur (Becton Dickinson). Для анализа ингибирования фактор Н полной длины инкубировали с бактериями вместе с возрастающими концентрациями CCPs 67, как указано.
Экспрессия белков и очищение комплекса: белок, связывающий фактор Н, и усеченные версии белка экспрессировали в Е.coli BL21(DE3) клетках-хозяевах в виде гибридных белков, помеченных полигистидином на их С-концах (Masignani et al (2003) J Exp Med Vol 197, No 6, 789-97). Культуры штаммов выращивали до середины логарифмического роста и экспрессию индуцировали изопропил-D-тиогалактозидом (IPTG, конечная концентрация 1 мМ). Белки очистили с помощью аффинной хроматографии на His-Trap колонке согласно инструкциям производителя (GE Healthcare) и элюировали с колонки с помощью 250 мМ имидазола. Конструкции фактора Н экспрессировались в Е.coli, как описано ранее (с рефолдингом, так чтобы было возможно правильное образование четырех дисульфидных связей внутри двух ССР фрагментов), и очищались либо с помощью аффинной очистки на гепариновой колонке, либо с помощью эксклюзионной хроматографии. После смешивания fHbp с fH67 (при большом мольном избытке) комплекс очищали с помощью эксклюзионной хроматографии (Superdex S-200, GE Healthcare Inc., буфер Tris pH 7.5, 150 мМ NaCl), где наблюдалось сильное взаимодействие между не находящимся в комплексе fH67 и матриксом колонки, что гарантировало хорошее отделение пика, содержащего комплекс, от пика, содержащего избыток fH67. Присутствие обоих белков в предполагаемом пике, соответствующем комплексу, подтверждалось с помощью SDS-PAGE и масс-спектрометрии (данные не приводятся).
Сайт-направленный мутагенез: Двойные мутанты получали последовательно, применяя набор QuikChange для сайт-направленного мутагенеза (Stratagene). R341A, Н337А двойные мутанты из fH67 получали с применением следующих праймеров:
Н337А прямой,
5'AAACATGGAGGTCTATATGCTGAGAATATGCGTAGACCATACTTTCC3' (SEQ ID NO:2);
Н337А обратный,
5'TTTGTACCTCCAGATACGACTCTTATACGCATCTGGTATGAAAGG3'(SEQ ID NO:3).
R341A прямой,
5'GGTCTATATCATGAGAATATGGCTAGACCATACTTTCCAGTAGC 3'(SEQ ID NO:4);
R341A обратный,
5'CCAGATATAGTACTCTTATACCGATCTGGTATGAAAGGTCATCG 3'(SEQ ID NO:5).
Введение мутации подтверждали секвенированием. Рекомбинантные фрагменты мутанта fH67 экспрессировались и подвергались рефолдингу под действием телец включения, как описано ранее (Prosser et al (2007) Acta Crystallogr Sect F Struct Biol Cryst Common Jun 1:63(PT6):480-3).
Для получения Е283А, E304A белка, связывающего фактор Н, применялись следующие праймеры:
Е283А прямой 5' ACAACCAAGCCGCGAAAGGCAGTTAC 3';
Е283А обратный, 5' GTAACTGCCTTTCGCGGCTTGGTTGT 3';
E304A прямой TGCCGGCAGCGCGGCAGTGAAAACCG 5';
E304A обратный 5' CGGTTTTCACTGCCGCGCTGCCGGCA 3'
и белок экспрессировали и очищали, как описано выше.
Поверхностный плазменный резонанс: Либо белок, связывающий фактор Н, либо конструкции фактора Н соединяют с поверхностью сенсора (СМ5 чипы), применяя стандартные протоколы аминного связывания. Соответствующий лиганд применялся в жидкой фазе при скорости течения потока 40 мкл/мин. Сильное взаимодействие не нарушалось ни при действии высокой/низкой соли (0-3 М NaCl), ни при экстремальном рН (интервал 4-8 проверен) и поэтому между последовательными инъекциями применялся увеличенный промежуток времени (60 минут). Полученные данные для более крупного белка, связывающего фактор Н, в жидкой фазе не могут быть удовлетворительными в случае подбора с применением простой модели 1:1 (предположительно из-за сложностей с движением более крупного белка в матрицу/из матрицы чипа или стерических затруднений в сайте связывания фактора Н, введенных связыванием более мелких белков с поверхностью чипа), грубый анализ почти равновесных значений позволил предположить, что величина KD лежит в интервале от десятых до сотых нМ (в зависимости от применяемой конструкции фактора Н). Если взаимодействие является обратимым (при fH на чипе), данные могут бать удовлетворительными при подборе с применением модели Ленгмюра 1:1 с χ2 около 3 (Фиг.1f), что указывает на то, что точная KD конструкций, содержащих ССР6, обладает значением порядка 5 нМ (смотрите Фиг.1f). Двойные мутанты белка, связывающего фактор Н, и fH67 также протестировали с fHbp партнером, связанным с поверхностью чипа (fHbpWT около 800 RU, связанный по каналу 1, fHbpE283A,Е304А около 1500 RU, связанный по каналу 2). Направляли поток fH67 (дикий тип и мутант) при концентрации 50 мкМ (около десятикратной нативной KD), в результате чего наблюдалось незначительно взаимодействие или не наблюдалось взаимодействие между мутантами и их партнерами дикого типа по сравнению со значением взаимодействия, наблюдаемого между парой дикого типа. Для того чтобы вычислить аффинность fH67R341A,H337A к fHbpWT, применяли ряд инъекций (в двойном выполнении), при постепенном увеличении концентрации от 250нМ до 20 мкМ, и подбор осуществляли с помощью BiaEvaluation программного обеспечения с получением значений, показанных на Фиг.1f.
Сбор данных с помощью кристаллизации и Х-лучей: кристаллы выращивали, применяя метод диффузии через газообразное состояние в варианте сидящей капли из 0.2 мкл комплекса + 0.2 мкл капель маточной жидкости (комплекс вычислили при концентрации 4.5 мг/мл, применяя предполагаемый коэффициент экстинкции 2.2, оптимальные характеристики маточного раствора были выбраны с помощью JCSG-plus условие 15, 20% полиэтиленгликоля 6000 MW, 0.1М Bicine pH 9.0). Собрали кристаллы и с помощью SDS-PAGE подтвердили, что присутствует и fH67, и белок, связывающий фактор Н (данные не приводятся). Кристаллы (обычно 50×10×5 мкм) подверглись сверхбыстрой заморозке в жидком азоте с последующей криозащитой с 15% этиленгликоля. Данные собраны при европейском центре синхротронного излучения ESRF и центре синхротронного излучения «Diamond» методом вращения, применяя диапазон колебаний 0.5 или 1 градус, при этом кристаллы хранятся при 120K. Данные для нативной кристаллической формы Р1 были собраны с помощью канала синхротронного излучения ID14eh4 (ESRF) при длине волны 0.9523 Å, наборы данных для нативной кристаллической формы С2 и Pt были собраны с помощью канала синхротронного излучения 103 (Diamond) при длине волны 0.9814 Å, Hg набор данных собрали с помощью канала синхротронного излучения ВМ14 (ESRF) с длиной волны 1.003 Å, и S-SAD собрали с помощью канала синхротронного излучения ID29 (ESRF) с длиной волны 1.8 Å. Данные подвергли интегрированию и масштабированию, применяя xia2 с 3d опцией для повышения эффективности программы XDS4, применяемой для интеграции, и программы Scala5, применяемой для масштабирования.
Определение структуры и оптимизация: структуру комплекса fh67/fHbp определили в С2 кристаллической форме с помощью комбинации способа молекулярного замещения структурой fh67 (заимствованной из более ранней структуры fH678 в комплексе с октасульфатом сахарозы, SOS, PDBID:2UWN) и MIRAS способа, применяя Pt, Hg производные и аномальное рассеяние, получаемое от 31 из 33 атомов серы, присутствующих в С2 асимметрической единице. Три копии fH67 были обнаружены, применяя MolRep7 (CCP4 пакет программ), и два ССР домена подверглись моделированию жесткими телами, применяя Buster-TNT без моделирования пропущенных атомов. Эти фазы затем были вовлечены в SHARP фазирование и применялись для определения местоположения сайтов тяжелых атомов в других кристаллах (6 Pt, 1 Hg и 31 S). Модель с тяжелыми атомами была оптимизирована с помощью SHARP (с В-фактором, связанным с коэффициентом Вильсона для каждого кристалла), применяя данные для 3.2 Å, и полученные фазы объединяли с фазами, полученными при молекулярном замещении в рамках SHARP с получением фаз с добротностью 0.54 (0.35 для наивысшего разрешения 3.3-3.2 Å). Показатели фазирования производных были следующими: (аномальный/изоформный) 0.7/2.4 для Pt, 0.2/0.3 для Hg и 0.3/0.0 для аномального сигнала серы. Карта, полученная с применением этих фаз, была подвергнута подавлению помех (solvent flattened) в рамках SHARP и усреднению в DM, применяя три отдельных оператора для белка, связывающего фактор Н, ССР6 области фактора Н и ССР7 области фактор Н. ЯМР-модель (Cantini et al (2006) J Biol Chem Mar 17:281(11):7220-7) для C-концевого цилиндра белка, связывающего фактор Н, не может применяться при такой электронной плотности (как видно, полосы искажаются), поэтому модель построили вручную, применяя программу Coot. Реконструирование и оптимизация с использованием данных в интервале 40 - 2.35 Å, обработанных с помощью Coot и Buster-TNT с некристаллографической симметрией, применялись для фиксации центра каждого домена, тогда как в отношении области петель допускалась вариация, пока R/Rfree составляли 25.5/27.1, с отсутствием остатков в запрещенных областях карты Рамачандрана (94.6% в наиболее предпочтительных, как определено с помощью MolProbity). Модель содержит все остатки fH67 и все остатки, за исключением пяти, на N-конце белка, связывающего фактор Н, и шесть гистидинов аффинного хвоста, которые неупорядочены во всех копиях молекулы. Молекулярное замещение отдельных доменов (ССР6 и ССР7фактора Н и белок, связывающий фактор Н), применяя программу MolRep, позволило определить Р1 кристаллическую форму (четыре независимые копии комплекса при различной упаковке). Второстепенное реконструирование областей петлей и оптимизация (90 - 2.35 Å), применяя Coot/Buster-TNT, привели к значениям R/Rfree, равным 23.2/26.3, с отсутствием остатков в запрещенных областях карты Рамачандрана (95.3% в наиболее предпочтительных). Структурные изображения были получены с помощью программы PyMol.
Протокол иммунизации
BALB/C мышей в возрасте семи недель иммунизировали путем подкожного введения очищенного fHbp или подкожного введения мутанта fHbp с адъювантом Фрейнда в день 0 и день 10. Каждое животное получило 25 мкг белка в PBS с равным объемом адъюванта с общим объемом 200 мкл. На 14 день животных умертвили, собрали сыворотку, объединили, затем аликвотировали и хранили при -80°С.
Бактерицидная активность сыворотки (SBA)
Штамм Н44/76 Neisseria meningitdis серогруппы В выращивали всю ночь на твердой среде, собрали в PBS и определили число колониеобразующих единиц. Серийные разбавления сыворотки добавляли к бактериям (104 колониеобразующих единиц в SBA буфере) и комплементу крольченка-сосунка (Pelfreeze, при конечном разбавлении 1 к 8) в течение одного часа и число выживших бактерий определили с помощью культивирования на твердой среде.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ФАКТОР Н-СВЯЗЫВАЮЩЕГО БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714248C2 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2665841C2 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ | 2018 |
|
RU2780425C2 |
| ВАРИАНТЫ ФАКТОР H-СВЯЗЫВАЮЩЕГО БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2811942C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2009 |
|
RU2475496C2 |
| ПЕПТИД СО СВОЙСТВАМИ АНТИГЕНА NEISSERIA MENINGITIDIS, КОДИРУЮЩИЙ ЕГО ПОЛИНУКЛЕОТИД, ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ ЗАБОЛЕВАНИЙ ИЛИ СОСТОЯНИЙ, ВЫЗВАННЫХ NEISSERIA MENINGITIDIS, (ВАРИАНТЫ), АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С УКАЗАННЫМ ПЕПТИДОМ | 2001 |
|
RU2252224C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРОТИВ NEISSERIA MENINGITIDIS (ВАРИАНТЫ) | 2014 |
|
RU2662968C2 |
| Композиции с Neisseria meningitidis и способы их применения | 2020 |
|
RU2839906C1 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2739504C2 |
| МУЛЬТИВАЛЕНТНАЯ ВАКЦИНА ИЗ НАТИВНЫХ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ МЕНИНГОКОККОВ, СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2009 |
|
RU2477145C2 |





Группа изобретений относится к области биотехнологии и включает иммуногенную и фармацевтическую композиции для их применения для лечения или профилактики инфекции или заболевания, вызываемых Neisseria meningitidis и/или Neisseria gonorrhoeae, а также набор для применения для индукции иммунного ответа. Композиция содержит по меньшей мере один модифицированный белок, связывающий фактор Н, и фармацевтически приемлемый носитель. Композиция способна вызвать иммунный ответ на Neisseria meningitides и Neisseria gonorrhoeae при введении человеку или не относящемуся к человеческому роду животному. Модифицированный белок, связывающий фактор Н, имеет последовательность Seq ID No: 1 за исключением измененной аминокислоты в по меньшей мере одном положении, что приводит к отсутствию связывания фактора Н или по меньшей мере пятикратному уменьшению связывания фактора Н по сравнению с белком, связывающим фактор Н, имеющим последовательность Seq ID No: 1. Измененный аминокислотный остаток находится в положении номер 106, как определено в последовательности Seq ID No:1. 4 н. и 7 з.п. ф-лы, 9 ил., 1 табл.
1. Иммуногенная композиция для лечения или профилактики инфекции или заболевания, вызываемых Neisseria meningitidis и/или Neisseria gonorrhoeae, содержащая по меньшей мере один модифицированный белок, связывающий фактор Н, и фармацевтически приемлемый носитель, где композиция способна вызвать иммунный ответ на Neisseria meningitides и Neisseria gonorrhoeae при введении человеку или не относящемуся к человеческому роду животному, где модифицированный белок, связывающий фактор Н, имеет последовательность Seq ID No:1 за исключением измененной аминокислоты в по меньшей мере одном положении, что приводит к отсутствию связывания фактора Н или по меньшей мере пятикратному уменьшению связывания фактора Н по сравнению с белком, связывающим фактор Н, имеющим последовательность Seq ID No: 1, где измененный аминокислотный остаток находится в положении номер 106, как определено в последовательности Seq ID No: 1.
2. Композиция по п. 1, где белок, связывающий фактор Н, имеет последовательность, по меньшей мере на 90% идентичную последовательности Seq ID No: 1.
3. Композиция по п. 1, где белок, связывающий фактор Н, происходит из Neisseria meningitides или Neisseria gonorrhoeae.
4. Композиция по п. 1, где модифицированный белок, связывающий фактор Н, включает две или более аминокислоты, которые были изменены по сравнению с Seq ID No: 1.
5. Композиция по п. 1, где одна или более аминокислот, которые меняются, представляют собой аминокислотные остатки, которые менее выставлены на поверхности, когда белок, связывающий фактор Н, находится в комплексе с фактором Н по сравнению с выделенным белком, связывающим фактор Н дикого типа.
6. Композиция по любому из пп. 1-3 или 5, где последовательность определяется как Seq ID No:1 с одной аминокислотной заменой в положении 106.
7. Композиция по п. 1, где композиция способна вызывать защитный иммунный ответ против Neisseria meningitidis и/или Neisseria gonorrhoeae.
8. Композиция по п. 1 для применения в качестве профилактической или терапевтической вакцины, направленной против N. meningitidis или Neisseria gonorrhoeae.
9. Фармацевтическая композиция для лечения или профилактики инфекции или заболевания, вызываемых Neisseria meningitidis и/или Neisseria gonorrhoeae, содержащая композицию по любому из предшествующих пунктов и при необходимости эксципиент.
10. Применение композиции по любому из пп. 1-9 для получения лекарственного средства для иммунизации человека или не принадлежащего к человеческому роду млекопитающего против инфекции, вызываемой N. meningitidisis или Neisseria gonorrhoeae.
11. Набор для применения для индукции иммунного ответа в организме для лечения или профилактики инфекции или заболевания, вызываемых Neisseria meningitidis и/или Neisseria gonorrhoeae, содержащий композицию по любому из пп. 1-9 и инструкции по введению.
| MASIGNANI V ET AL., Vaccination against Neisseria meningitides using three variants of the lipoprotein GNA1870, The Journal of experimental medicine, Rockefeller university press, vol.197, no.6, 17.03.2003, pages 789-799, abstract, figure 6 | |||
| BEERNINK PETER T ET AL., Prevalence of factor H-binding protein variants and NadA among meningococcal group |

