ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к моноклональным антителам, которые связываются с белком «anexelekto» (неконтролируемый), к средствам, содержащим указанные антитела в качестве активного ингредиента, а также к способам применения указанных антител.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Ген «anexelekto» (также называемый AXL, UFO, ARK, или TYRO7; в настоящей заявке обозначен как AXL), клонированный у пациентов с хроническим миелоидным лейкозом, представляет собой онкоген, способный трансформировать клетки NIH3T3 мышей при сверхэкспрессии (непатентные документы 1 и 2). Белок AXL представляет собой рецептор, связанный с тирозинкиназой, имеющий молекулярную массу 140 кДа (непатентный документ 3), и считается, что он отвечает за передачу сигнала по сигнальному пути за счет автофосфорилирования, которое происходит при связывании с лигандом Gas6 (продуктом гена 6 блокировки роста) (непатентный документ 4). Рецепторы, связанные с тирозинкиназой, такие как Sky, Mer и AXL, известны как рецепторы, связанные с тирозинкиназой, лигандом которых является Gas6 (непатентный документ 5).
Полагают, что AXL обладает молекулярными функциями, связанными с усилением роста клеток, подавлением апоптоза, миграцией клеток и клеточной адгезией. Экспериментально наблюдавшиеся явления в клетках, обработанных белком Gas6, подтвердили это предположение. Опубликованные экспериментальные данные включают подавление клеточной смерти и усиление роста гладкомышечных клеток сосудов крысы (непатентные документы 6 и 7); ускорение роста клеток NIH3T3 мыши и подавление клеточной смерти после выращивания клеток на бессывороточной среде (непатентные документы 8 и 9); усиление клеточного роста фибробластов сердца мыши (непатентный документ 10); усиление роста раковых клеток рака простаты человека (непатентный документ 11); усиление клеточного роста и инфильтрации, а также подавление клеточной смерти клеток карциномы желудка человека (непатентный документ 12); усиление способности к миграции гладкомышечных клеток сосудов человека и крысы (непатентный документ 13); способности к усилению миграции клеток нейронов мыши (непатентный документ 14) и агрегацию клеток, сверхэкспрессирующих AXL мыши (непатентный документ 15).
Аналогичным образом, молекулярные анализы внутриклеточных сигналов после обработки белком Gas6 показали, что путь PI3K-Akt и путь МАРК действуют как метаболические пути сигнальной трансдукции, опосредованной AXL, (непатентный документ 5). Анализ на дрожжах способом двойных гибридов с использованием внутриклеточной области AXL в качестве затравки подтвердил прямое молекулярное взаимодействие на указанные метаболические пути. В результате был идентифицирован путь GrbB2/PI3K/p55γ/SOCS-1/NcK2/RanBP2/C1-TEN (непатентный документ 16). Взаимодействия этих молекул позволяют предположить существование внутриклеточных путей передачи сигнала, следующих после AXL. Кроме того, наблюдаемые взаимодействия подтверждают предположение, что AXL действует в процессах усиления клеточного роста, подавления апоптоза, клеточной миграции и адгезии клеток. AXL также был идентифицирован как ген, сверхэкспрессирующий при появлении TNFα-индуцированной клеточной смерти фибробластов мыши подавлена с помощью IL-15. Подавление TNFα-индуцированной клеточной смерти прекращалось при подавлении экспрессии AXL, а фосфорилирование рецепторов IL-15 и AXL усиливалось при воздействии IL-15. Эти экспериментальные данные также показывают, что передача сигнала опосредована комплексом AXL и рецептора IL-15 (непатентный документ 17).
Сообщалось, что возникновение опухолей у “голых” мышей прекращалась при ингибировании Gas6-зависимого фосфорилирования AXL в глиомных линиях человека, сверхэкспрессирующих доминант-негативную форму AXL (непатентный документ 18). Однако не было никаких сообщений или предположений, и было неясно, существует ли антитело против AXL, которое ингибирует фосфорилирование.
AXL представляет собой рецептор, связанный с однократно пронизывающей мембрану тирозинкиназой, внеклеточная область которого состоит из двух иммуноглобулин-подобных доменов (называемых IgD1 и IgD2) и двух типа III фибронектиновых доменов (называемых FND1 и FND2) на N-концевой стороне (непатентный документ 3). Хотя известно, что FND экспрессируются в таких молекулах, как молекулы адгезии нервных клеток и нефрины, участвующие в межклеточной адгезии, более полная информация об активности FND-доменов в AXL отсутствует (непатентный документ 19).
AXL был идентифицирован как онкоген, который сохраняет характерную способность трансформировать клетки, его также рассматривали в качестве молекулы, связанной с канцерогенезом. Были представлены данные различных анализов экспрессии AXL на уровне белка и мРНК. Сообщалось о сверхэкспрессии белка AXL в опухолевых тканях и опухолевых клетках человека, в том числе рака легких (непатентный документ 20), рака молочной железы (непатентный документ 21), рака яичников (непатентный документ 22), рака щитовидной железы (непатентный документ 23), меланомы (непатентный документ 23), рака почки (непатентный документ 24), рака желудка (непатентный документ 12) и глиомы (непатентный документ 25). Кроме того, сверхэкспрессия белка AXL вызывается высокими уровнями мРНК AXL при раке пищевода (непатентный документ 26), раке толстой кишки (непатентный документ 27), остром миелобластном лейкозе (непатентный документ 28). Также имеются сообщения об ингибировании образования полостей клеток (люменов) при подавлении AXL с помощью иРНК в клетках HUVEC (непатентный документ 29), о пониженной способности раковых клеток рака молочной железы человека формировать опухоли у мышей в результате конститутивной супрессии AXL (непатентный документ 29), и о пониженной способности глиомных клеток человека формировать опухоли у мышей в результате конститутивной сверхэкспрессии у доминант-негативных мутантов (непатентный документ 25). Имеется твердое предположение об участии AXL на уровне его молекулярной активности в процессе роста опухолей.
Сообщалось, что поликлональные антитела к внеклеточному домену AXL оказывают ингибирующее действие на миграцию в клеточных линиях высокоинвазивного рака молочной железы (патентный документ 1). Однако доклинические исследования показали, что лекарственные средства, демонстрирующие ингибирующее действие на миграцию раковых клеток, не обязательно обладают противоопухолевую активность. Например, известно, что матричная металлопротеаза (далее сокращенно «MMP») способствует опухолевой инфильтрации и миграции. Таким образом, все внимание было сосредоточено на различных ингибиторах матричной металлопротеазы, которые ингибируют ферментативную активность MMP, и были проведены клинические исследования различных фармацевтических препаратов, таких как батимастат, маримастат и приномастат. Однако противоопухолевых эффектов в клинических испытаниях не наблюдалось (непатентный документ 30).
Соответственно, не существовало никаких сообщений или предположений, и остается неизвестно, обладает ли антитело против AXL какими-либо противоопухолевыми эффектами, в частности, in vivo, могут ли антитела подавлять ангиогенез, и могут ли они подавлять рак.
Патентный документ 1: WO 2004/008147
Непатентный документ 1: Liu, et al, Proc. Natl Acad. Sci. U.S.A. (1988) 85, 1952-6
Непатентный документ 2: Janssen, et al., Oncogene (1991) 6, 2113-20
Непатентный документ 3: O'Bryan, et al., Mol. Cell. Biol. (1991) 11, 5016-31
Непатентный документ 4: Varnum, et al., Nature (1995) 373, 623-626
Непатентный документ 5: Hafizi, et al., FEBS J. (2006) 273, 5231-5244
Непатентный документ 6: Nakano, et al., FEBS Lett. (1996) 387, 78-80
Непатентный документ 7: Nakano, et al., J. Biol. Chem. (1995) 270, 5702-5
Непатентный документ 8: Goruppi, et al., Mol. Cell. Biol. (1997) 17, 4442-53
Непатентный документ 9: Bellosta, et al., Oncogene (1997) 15, 2387-97
Непатентный документ 10: Stenhoff, et al., Biochem. Biophys. Res. Commun. (2004) 319, 871-8
Непатентный документ 11: Sainaghi, et al., J. Cell. Physiol. (2005) 204, 36-44
Непатентный документ 12: Sawabu, et al., Mol. Carcinog. (2007) 46, 155-164
Непатентный документ 13: Fridell, et al., J. Biol. Chem. (1998) 273, 7123-6
Непатентный документ 14: Allen, et al., Mol. Cell. Biol. (2002) 22, 599-613
Непатентный документ 15: McCloskey, et al., J. Biol. Chem. (1997) 272, 23285-91
Непатентный документ 16: Hafizi, et al., Biochem. Biophys. Res. Commun. (2002) 299, 793-800
Непатентный документ 17: Budagian et al., Embo J. (2005) 24, 4260-70
Непатентный документ 18: Vajkoczy P et al., Proc. Natl Acad. Sci. U.S.A. (2006) 103, 5799-804
Непатентный документ 19: Yamagata et al., Curr. Opin. Cell. Biol. (2003) 15, 621-632
Непатентный документ 20: Shieh, et al., Neoplasia (2005) 7, 1058-1064
Непатентный документ 21: Meric, et al., Clin. Cancer Res. (2002) 8, 361-367
Непатентный документ 22: Sun, et al., Oncology (2004) 66,450-457
Непатентный документ 23: Ito, et al., Thyroid (2002) 12, 971-975
Непатентный документ 24: Chung, et al., DNA Cell Biol. (2003) 22, 533-540
Непатентный документ 25: Vajkoczy, et al., Proc. Natl. Acad. Sci. U.S.A. (2006) 103, 5799-804
Непатентный документ 26: Nemoto, et al., Pathobiology (1997) 65, 195-203
Непатентный документ 27: Craven, et al., Int. J. Cancer (1995) 60, 791-797
Непатентный документ 28: Neubauer, et al., Blood (1994) 84, 1931-1941
Непатентный документ 29: Holland, et al., Cancer Res. (2005) 65, 9294-9303
Непатентный документ 30: Pavlaki et al., Cancer Metastasis Rev. (2003) 22, 177-203
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[Проблемы, которые решаются данным изобретением]
Целью настоящего изобретения является создание антител против AXL и их использование. Более конкретно, целью настоящего изобретения является создание способов ингибирования ангиогенеза с помощью антител против AXL, способов подавления клеточного роста, способов ингибирования функции AXL, способов ускорения функции AXL и способов снижения уровня экспрессии AXL. Другой целью настоящего изобретения является создание антител против AXL с новыми эффектами.
[Средства решения проблем]
В результате проведения тщательных исследований, авторам настоящего изобретения удалось получить антитела против AXL со специфическими функциями, и обнаружить, что эти антитела обладают подавляющим ангиогенез эффектом и противоопухолевым эффектом, и тем самым было сделано настоящее изобретение. Более конкретно, настоящее изобретение включает:
[1] моноклональное антитело, которое связывается с AXL;
[2] антитело по п. [1], которое обладает активностью, подавляющей клеточный рост;
[3] антитело по п. [1], которое подавляет рост раковых клеток;
[4] антитело по любому из пп. [1]-[3], которое связывается с FND1;
[5] антитело, полученное с использованием в качестве антигена пептида, содержащего полноразмерный FND1 или последовательность, содержащую по меньшей мере пять или более последовательных аминокислот указанного пептида;
[6] антитело по любому из пп. [1]-[5], которое обладает агонистической активностью в отношении AXL;
[7] антитело по любому из пп. [1]-[5], которое обладает антагонистической активностью в отношении AXL;
[8] антитело по п. [7], полученное путем отбора антитела, у которого не обнаруживался фосфорилированный тирозин в AXL при контакте с клеткой, экспрессирующей AXL и лиганда AXL;
[9] антитело по любому из пп. [1]-[8], которое обладает активностью, снижающей уровень экспрессии AXL;
[10] антитело по любому из пп. [1]-[9], которое обладает активностью, ингибирующей ангиогенез;
[11] антитело по любому из следующих пп. (a)-(j):
(a) антитело (Ax285), полученное из гибридомы с депозитарным номером FERM BP-10858;
(b) антитело (Ax292), полученное из гибридомы с депозитарным номером FERM BP-10859;
(c) антитело (Ax223), полученное из гибридомы с депозитарным номером FERM BP-10853;
(d) антитело (Ax96), полученное из гибридомы с депозитарным номером FERM BP-10852;
(e) антитело (Ax258), полученное из гибридомы с депозитарным номером FERM BP-10856;
(f) антитело (Ax284), полученное из гибридомы с депозитарным номером FERM BP-10857;
(g) антитело (Ax7), полученное из гибридомы с депозитарным номером FERM BP-10850;
(h) антитело (Ax51), полученное из гибридомы с депозитарным номером FERM BP-10851;
(i) антитело (Ax225), полученное из гибридомы с депозитарным номером FERM BP-10854; и
(j) антитело (Ax232), полученное из гибридомы с депозитарным номером FERM BP-10855;
[12] антитело, которое связывается с тем же эпитопом, что и эпитоп, связанный с любым антителом по п. [11];
[13] антитело, которое содержит последовательность CDR, идентичную последовательности CDR, находящейся в любом из антител по п. [11];
[14] антитело, в котором последовательности CDR 1, 2 и 3 тяжелой цепи представляет собой SEQ ID NO: 4, 5 и 6, соответственно;
[15] антитело, которое содержит CDR тяжелой цепи, включая аминокислотную последовательность CDR тяжелой цепи антитела по п. [14] с заменой, делецией, вставкой и/или добавлением одной или нескольких аминокислот, и которое функционально эквивалентно антителу по п. [14];
[16] антитело, в котором последовательности CDR 1, 2 и 3 легкой цепи представляют собой SEQ ID NO: 8, 9 и 10 соответственно;
[17] антитело, которое содержит CDR легкой цепи, включая аминокислотную последовательность CDR легкой цепи антитела по п. [16] с заменой, делецией, вставкой и/или добавлением одной или нескольких аминокислот, и которое функционально эквивалентно антителу по п. [16];
[18] антитело по любому из пп. [13]-[17], которое является химерным антителом;
[19] антитело по любому из пп. [13]-[17], которое является гуманизированным антителом;
[20] гибридома по любому из следующих пп. (a)-(j):
(a) гибридома с депозитарным номером FERM BP-10858 (Ax285);
(b) гибридома с депозитарным номером FERM BP-10859 (Ax292);
(c) гибридома с депозитарным номером FERM BP-10853 (Ax223);
(d) гибридома с депозитарным номером FERM BP-10852 (Ax96);
(e) гибридома с депозитарным номером FERM BP-10856 (Ax258);
(f) гибридома с депозитарным номером FERM BP-10857 (Ax284);
(g) гибридома с депозитарным номером FERM BP-10850 (Ax7);
(h) гибридома с депозитарным номером FERM BP-10851 (Ax51);
(i) гибридома с депозитарным номером FERM BP-10854 (Ax225); и
(j) гибридома с депозитарным номером FERM BP-10855 (Ax232);
[21] ингибитор ангиогенеза, содержащий антитело против AXL в качестве активного ингредиента;
[22] ингибитор ангиогенеза по п. [21], где антитело является антителом по любому из пп. [1]-[19];
[23] ингибитор клеточного роста, содержащий антитело против AXL в качестве активного ингредиента;
[24] ингибитор роста по п. [23], где клетки являются раковыми клетками;
[25] ингибитор роста по п. [23], где антитело является антителом по любому из пп. [1]-[19];
[26] ингибитор роста по п. [23], где антитело против AXL является антителом, связывающимся с FND1;
[27] ингибитор роста по п. [23], который содержит в качестве активного ингредиента антитело, которое связывается с IgD2 и обладает активностью ингибирования фосфорилирования;
[28] индуктор фосфорилирующей AXL активности, который содержит в качестве активного ингредиента антитело против AXL;
[29] индуктор по п. [28], где антитело против AXL является антителом, связывающимся с IgD;
[30] индуктор по п. [28], где антитело является антителом по п. [6];
[31] ингибитор активности фосфорилирующей AXL, который содержит антитело против AXL в качестве активного ингредиента;
[32] ингибитор по п. [31], где антитело против AXL является антителом, связывающимся с IgD2;
[33] ингибитор по п. [31], где антитело является антителом по п. [7] или п. [8];
[34] средство, снижающее уровень экспрессии AXL, которое содержит антитело против AXL в качестве активного ингредиента;
[35] средство, снижающее уровень экспрессии по п. [34], где антитело против AXL является антителом, связывающимся с FND1;
[36] средство, снижающее уровень экспрессии по п. [34], где антитело является антителом по п. [9];
[37] способ индукции фосфорилирования AXL с использованием антитела против AXL;
[38] способ снижения уровня экспрессии AXL с использованием антитела против AXL;
[39] способ ингибирования фосфорилирования AXL с использованием антитела против AXL;
[40] противораковое средство, содержащее антитело против AXL в качестве активного ингредиента;
[41] противораковое средство по п. [40], где антитело является антителом по любому из пп. [1]-[19];
[42] противораковое средство по п. [40], содержащее в качестве активного ингредиента антитело, которое связывается с IgD2 и обладает активностью ингибирования фосфорилирования;
[43] противораковое средство по п. [40], где рак является раком поджелудочной железы, раком желудка, раком легких, остеосаркомой, раком толстой кишки, раком предстательной железы, меланомой, раком эндометрия, раком яичников, лейомиомой матки, раком щитовидной железы, раком стволовых клеток, раком молочной железы, раком мочевого пузыря, раком почки, глиомой, нейробластомой или раком пищевода;
[44] противораковое средство по п. [42], где рак является глиомой, раком желудка, раком эндометрия, немелкоклеточным раком легкого, аденокарциномой поджелудочной железы или раком молочной железы;
[45] противораковое средство по п. [43], где рак является аденокарциномой поджелудочной железы или раком молочной железы;
[46] антитело по п. [1], которое обладает активностью ингибирования фосфорилирования;
[47] способ ингибирования ангиогенеза с использованием антитела против AXL;
[48] способ применения антитела против AXL при получении ингибитора ангиогенеза;
[49] подавления клеточного роста с использованием антитела против AXL;
[50] способ лечения и/или профилактики рака с использованием антитела против AXL;
[51] способ применения антитела против AXL при получении ингибитора клеточного роста;
[52] способ применения антитела против AXL при получении противоракового средства;
[53] способ применения антитела против AXL при получении индуктора фосфорилирования;
[54] способ применения антитела против AXL при получении ингибитора фосфорилирования;
[55] способ применения антитела против AXL при получении средства для снижения уровня экспрессии AXL; и
[56] способ получения антитела, специфичного в отношении AXL, включающий:
(a) иммунизацию животного, не человека, пептидом, содержащим полноразмерный FND1 или последовательность, содержащую по меньшей мере пять или более последовательных аминокислот указанного пептида; и
(b) сбор антител у животного, не человека, по п. (а) или сбор продуцирующих антитело клеток для сбора антитела, продуцируемого клетками, продуцирующими антитело.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1A представляет собой фотографию, на которой представлена активность моноклонального антитела против AXL (Ax292) по настоящему изобретению в отношении индукции фосфорилирования AXL в раковых клетках. Показано, что антитело индуцирует фосфорилирование киназного домена AXL.
Фигура 1B представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax258) по настоящему изобретению в отношении индукции фосфорилирования AXL в раковых клетках. Показано, что антитело индуцирует фосфорилирование киназного домена AXL.
Фигура 1C представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax285) по настоящему изобретению в отношении индукции фосфорилирования AXL в раковых клетках. Показано, что антитело индуцирует фосфорилирование киназного домена AXL.
Фигура 1D представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax223) по настоящему изобретению в отношении индукции фосфорилирования AXL в раковых клетках. Показано, что антитело индуцирует фосфорилирование киназного домена AXL.
Фигура 1E представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax96) по настоящему изобретению в отношении индукции фосфорилирования AXL в раковых клетках. Показано, что антитело индуцирует фосфорилирование киназного домена AXL.
Фигура 2A представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax51) по настоящему изобретению в отношении ингибирования лиганд-зависимого фосфорилирования AXL в клетке. Показано, что антитело ингибирует лиганд-зависимое фосфорилирование киназного домена AXL.
Фигура 2B представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax7) по настоящему изобретению в отношении ингибирования лиганд-зависимого фосфорилирования AXL в клетке. Показано, что антитело ингибирует лиганд-зависимое фосфорилирование киназного домена AXL.
Фигура 3A представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax292) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3B представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax284) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3C представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax285) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3D представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax223) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3E представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax7) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3F представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax225) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3G представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax96) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 3H представляет собой фотографию, на которой показана активность моноклонального антитела против AXL (Ax258) по настоящему изобретению в отношении индукции деактивации AXL в раковых клетках. Показано, что антитело индуцирует деактивацию белка AXL.
Фигура 4 представляет собой рисунок и фотографию, на которых показаны активности моноклональных антител против AXL по настоящему изобретению (Ax232, Ax292, Ax285 и Ax284) в отношении ингибирования ангиогенеза in vitro. Показано, что антитела обладают активностью, ингибирующей ангиогенез in vitro.
Фигура 5 представляет собой рисунок, на котором показаны противоопухолевые эффекты моноклональных антител против AXL по настоящему изобретению (Ax223, Ax285, Ax96, Ax292, Ax258, Ax7, Ax51, Ax284 и Ax225) у ксенотрансплантатных моделей на мышах аденокарциномы поджелудочной железы человека.
Фигура 6 представляет собой таблицу, в которой показаны противоопухолевые эффекты моноклональных антител против AXL по настоящему изобретению (Ax223, Ax285, Ax96, Ax292, Ax258, Ax7, Ax51, Ax284 и Ax225) (2) у ксенотрансплантатных моделей на мышах от аденокарциномы поджелудочной железы человека, и в которой сведены связывающие домены и активности ингибирования фосфорилирования каждого антитела.
Фигура 7 представляет собой рисунок, на котором показаны противоопухолевые эффекты моноклонального антитела против AXL (Ax225) по настоящему изобретению у ксенотрансплантатных моделей на мышах рака молочной железы человека.
ЛУЧШИЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новому антителу против AXL. Кроме того, настоящее изобретение относится к новому применению антитела против AXL.
Не существует никаких конкретных ограничений в отношении антитела против AXL по настоящему изобретению при условии, что оно связывается с AXL, и не существует также никаких особых ограничений в отношении источника его получения (например, человек, мышь, крыса, кролик или курица), тип (поликлональное антитело или моноклональное антитело), форму (например, измененное антитело, модифицированное антитело, фрагмент антитела, или мини-антитело [антитело с низким молекулярным весом]), и тому подобное. Хотя никаких особых ограничений на антитело против AXL по настоящему изобретению не существует, предпочтительно, антитело специфически связывается с AXL и, предпочтительно, является моноклональным антителом.
Антитело против AXL по настоящему изобретению также, предпочтительно, обладает подавляющей клеточный рост активностью.
Предпочтительным вариантом осуществления антител против AXL по настоящему изобретению являются антитела против AXL, связывающие FND1.
Как видно из примеров, описанных ниже, антитела, которые связываются с FND1, в частности, обладают более высокой in vivo противоопухолевой активностью по сравнению с активностью других антител.
Связывающая активность антител против AXL с FND1 может быть оценена способом, известным специалистам в данной области, например, одним из описанных ниже способов. Связывающую активность антитела против AXL с FND1 подтверждали электрофорезом FND1 и вестерн-блоттингом с антителом против AXL.
Антитело против AXL с агонистической в отношении AXL активностью является примером предпочтительного варианта осуществления антитела против AXL по настоящему изобретению. Антитело против AXL с агонистической в отношении AXL активностью имеет отношение к индукции фосфорилирования при участии AXL и, в частности, к индукции реакции фосфорилирования тирозина, когда антитело против AXL связывается с AXL. Хотя не существует никаких особых ограничений в отношении мишени в реакции фосфорилирования, которая вызвана антителом против AXL с агонистической активностью, пример включает автофосфорилирование AXL.
Обладает ли антитело против AXL агонистической активностью можно определить способом, известным специалистам в данной области, например, одним из описанных ниже способов. Исследуемое антитело против AXL приводили в контакт с клетками, экспрессирующими AXL (такими, как Calu-1, MDA-MB-231 или DU-145 клетки), и AXL затем извлекали из клеток. То, что тирозин в извлеченном AXL был фосфорилированным подтверждали, используя антитело против фосфотирозина. Более конкретнее, тот факт, что антитело против AXL обладает агонистической активностью, можно подтвердить способами, описанными в примерах.
Примеры антител против AXL с агонистической активностью включают антитела по пп. (a)-(g) ниже:
(а) антитело, полученное из гибридомы с депозитарным номером FERM BP-10858 (Ax285);
(b) антитело, полученное из гибридомы с депозитарным номером FERM BP-10852 (Ax96);
(c) антитело, полученное из гибридомы с депозитарным номером FERM BP-10856 (Ax258);
(d) антитело, полученное из гибридомы с депозитарным номером FERM BP-10859 (Ax292);
(е) антитело, полученное из гибридомы с депозитарным номером FERM BP-10853 (Ax223);
(f) антитело, распознающее тот же эпитоп, что и эпитоп, распознаваемый антителом по любому из пп. (a)-(e); и
(g) антитело с последовательностью CDR, идентичной последовательности CDR антитела по любому из пп. (a)-(e).
Антитело, распознающее тот же эпитоп, что и антитело, описанное выше, может быть получено, например, в соответствии со способом, описанным ниже.
Имеет ли исследуемое антитело и какое-либо еще антитело общий эпитоп, может быть подтверждено конкуренцией двух антител за этот эпитоп. Конкуренцию между антителами обнаруживают с помощью эпитоп-перекрестного конкурентного анализа и тому подобное. Конкурентный анализ ELISA, например, является предпочтительнее эпитоп-перекрестного конкурентного анализа. В частности, в эпитоп-перекрестном конкурентном анализе белок AXL, нанесенный на лунки обычного микропланшета, преинкубируют в присутствии или в отсутствии возможного конкурентного антитела, затем добавляют отмеченное выше антитело против AXL. Количество вышеупомянутого антитела против AXL, связанного с белком AXL в лунках, косвенным образом коррелирует со связывающей способностью возможного конкурентного антитела (исследуемого антитела), конкурирующего за связывание с тем же эпитопом. Таким образом, чем больше аффинность исследуемого антитела к тому же эпитопу, тем больше сокращается количество вышеупомянутого антитела против AXL, связавшегося с лунками, покрытыми белком AXL, и тем больше увеличивается количество исследуемого антитела, связанного с лунками, покрытыми белком AXL.
Количество антитела, связанного с лунками, легко может быть измерено посредством предварительного мечения антитела. Например, меченое биотином антитело можно измерить с помощью авидин-пероксидазного конъюгата и подходящего субстрата. Эпитоп-перекрестный конкурентный анализ, который использует ферментную метку, такую как пероксидаза, называется конкурентным анализом ELISA. Антитела могут быть помечены другими маркирующими веществами, которые можно детектировать и измерять. В частности, известны радиоактивные метки и флуоресцентные метки.
Если исследуемое антитело имеет константную область, происходящую от иных биологических видов, чем отмеченное выше антитело против AXL, то количество антитела, связанного с лунками, также может быть измерено с помощью меченого антитела, распознающего константную область этого антитела. В противном случае, если антитело происходит от того же биологического вида, но принадлежит другому классу, то количество антитела, связанного с лунками, может быть измерено с помощью антител, распознающих любой класс.
Если возможное конкурентное антитело может блокировать связывание антитела против AXL, по меньшей мере на 20%, предпочтительно, по меньшей мере на 20%-50%, еще более предпочтительно, по меньшей мере на 50% по сравнению со связывающей активностью, полученной в контрольном тесте, проведенном при отсутствии возможного конкурентного антитела, то возможное конкурентное антитело является антителом, которое в значительной степени связывается с тем же эпитопом, или которое конкурирует с вышеупомянутым антителом против AXL за связывание с тем же эпитопом.
Определение последовательности CDR для получения антитела с той же последовательностью CDR, что и у конкретных антител, может быть выполнено специалистом в данной области известными способами. Например, последовательность CDR может быть определена путем определения полноразмерной аминокислотной последовательности антитела или аминокислотной последовательности вариабельной области, и изучения ее гомологии, используя установленную аминокислотную последовательность в базе данных аминокислотных последовательностей антител, созданную Кабатом и сотрудниками (Kabat et al., «Sequence of Proteins of Immunological Interest», US Dept. of Health and Human Services, 1983). Порядковые номера в рамке считывания и порядковые номера в последовательностях CDR могут быть определены в соответствии с границами, указанными Кабатом (Kabat, A.E. et al., US Dept. of Health and Human Services, US Government Printing Offices, 1991).
Полноразмерная аминокислотная последовательность антитела или аминокислотная последовательность вариабельной области может быть определена специалистом в данной области известными способами.
Антитело с той же последовательностью CDR, что и у конкретного антитела, может иметь идентичную последовательность, по крайней мере, одного из шести CDR, которые присутствуют в антителе. Однако антитело, предпочтительно, имеет одинаковую последовательность всех трех CDR, присутствующих в тяжелой цепи, или имеет одинаковую последовательность всех трех CDR, присутствующих в легкой цепи, и даже, более предпочтительно, антитело имеет одинаковую последовательность всех шести CDR, присутствующих в антителе.
Антитела с той же последовательностью CDR, что и у конкретного антитела, включают химерные антитела и гуманизированные антитела. Химерные антитела и гуманизированные антитела будут описаны ниже.
Примером другого предпочтительного варианта осуществления антитела против AXL по настоящему изобретению является антитело против AXL с антагонистической активностью в отношении AXL. Антитело против AXL с антагонистической активностью в отношении AXL представляет собой антитело с активностью, ингибирующей реакцию фосфорилирования при участии AXL, индуцированную связыванием лиганда AXL (такого, как Gas6) с AXL, и, конкретно, реакцию фосфорилирования тирозина. Ингибирование реакции фосфорилирования может быть осуществлено путем ингибирования связывания лиганда AXL и AXL, или другим способом. Хотя не существует никаких особых ограничений в отношении мишеней реакции ингибирования фосфорилирования, индуцированной антителом против AXL с антагонистической активностью, примеры включают автофосфорилирование AXL.
Обладает ли антитело против AXL антагонистической активностью можно определить способом, известным специалистам в данной области, например, описанным ниже способом. Исследуемое антитело приводили в контакт с клетками, экспрессирующими AXL (такими, как Calu-1, MDA-MB-231 или DU-145 клетки), вместе с лигандом AXL, и AXL затем извлекали из клеток. Было подтверждено, что фосфорилированный тирозин в извлеченном AXL невозможно детектировать, используя антитело против фосфотирозина. Конкретнее, то, что антитело против AXL обладает антагонистической активностью, можно подтвердить с помощью способов, описанных в примерах.
Примеры антител против AXL с антагонистической активностью включают антитела по пп. (a)-(d) ниже:
(a) антитело, полученное из гибридомы с депозитарным номером FERM BP-10850 (Ax7);
(b) антитело, полученное из гибридомы с депозитарным номером FERM BP-10851 (Ax51);
(с) антитело, распознающее тот же эпитоп, что и эпитоп, распознаваемый антителом по любому из пп. (a)-(b); и
(d) антитело, обладающее последовательностью CDR, идентичной последовательности CDR антитела по любому из пп. (a)-(c).
Антитело, распознающее тот же эпитоп, и антитело, обладающее идентичной последовательностью CDR, могут быть получены с помощью ранее описанных способов.
Антитело с антагонистической активностью пригодно для ингибирования ангиогенеза, подавления клеточного роста, и тому подобное.
Примером другого предпочтительного варианта осуществления антитела по настоящему изобретению является антитело с активностью, снижающей уровень экспрессии AXL. В настоящем изобретении снижение уровня экспрессии AXL может означать уменьшение количества уже присутствующего AXL путем деградации AXL или тому подобное, либо может означать уменьшение количества вновь экспрессированного AXL путем подавления экспрессии AXL. Снижается ли уровень экспрессии AXL, можно определить способом, известным специалистам в данной области, и например, описанным ниже способом. Исследуемое антитело против AXL приводили в контакт с клетками, экспрессирующими AXL (такими, как Calu-1, MDA-MB-231 или DU-145 клетки), и количество AXL в клетках затем детектировали иммуноблоттингом или тому подобное. Затем сравнивали количество обнаруженного AXL, если исследуемое антитело находилось в контакте, и количество обнаруженного AXL, если исследуемое антитело в контакте не находилось. Более конкретно, это можно подтвердить способами, описанными в примерах.
Примеры антител против AXL с активностью, снижающей уровни экспрессии AXL, включают антитела по пп. (a)-(j) ниже:
a) антитело (Ax285), полученное из гибридомы с депозитарным номером FERM BP-10858;
(b) антитело (Ax96), полученное из гибридомы с депозитарным номером FERM BP-10852;
(с) антитело (Ax258), полученное из гибридомы с депозитарным номером FERM BP-10856;
(d) антитело (Ax7), полученное из гибридомы с депозитарным номером FERM BP-10850;
(e) антитело (Ax292), полученное из гибридомы с депозитарным номером FERM BP-10859;
(f) антитело (Ax223), полученное из гибридомы с депозитарным номером FERM BP-10853;
(g) антитело (Ax225), полученное из гибридомы с депозитарным номером FERM BP-10854;
(h) антитело (Ax284), полученное из гибридомы с депозитарным номером FERM BP-10857;
(i) антитело, распознающее тот же эпитоп, что и эпитоп, распознаваемый антителом по любому из пп. (a)-(h); и
(j) антитело, обладающее последовательностью CDR, идентичной последовательности CDR антитела по любому из пп. (a)-(i).
Антитело, распознающее тот же эпитоп, и антитело, обладающее идентичной последовательностью CDR, можно получить ранее описанным способом.
Антитело с активностью, снижающей уровень экспрессии AXL, может использоваться для ингибирования ангиогенеза, подавления клеточного роста, и тому подобное.
Примером другого предпочтительного варианта осуществления антитела по настоящему изобретению является антитело с эффектом ингибирования ангиогенеза. Хотя не существует никаких особых ограничений в отношении эффекта ингибирования ангиогенеза по настоящему изобретению, при условии, что новообразование кровеносных сосудов подавляется, примеры включают ингибирующий эффект на миграционную активность сосудистых эндотелиальных клеток, апоптоз-индуцирующее действие на сосудистые эндотелиальные клетки и ингибирующее действие на морфогенез сосудов сосудистых эндотелиальных клеток. Предпочтительным примером антитела с эффектом ингибирования ангиогенеза является антитело с эффектом ингибирования ангиогенеза опухолевых тканей. Не существует никаких особых ограничений на опухолевые ткани, и примеры включают ткань рака поджелудочной железы (такие, как ткань аденокарциномы поджелудочной железы), ткань рака желудка, ткань рака легких (ткань мелкоклеточного рака легкого, немелкоклеточного рака легкого, и т.д.), ткань остеосаркомы, ткань рака прямой кишки, ткань рака простаты, ткань меланомы, ткань рака эндометрия, ткань рака яичников, ткань лейомиомы матки, ткань рака щитовидной железы, ткань рака стволовых клеток, ткань рака молочной железы, ткань рака мочевого пузыря, ткань рака почек, ткань глиомы, ткань нейробластомы и ткань рака пищевода. Предпочтительными тканями являются ткань глиомы, ткань рака желудка, ткань рака эндометрия, ткань немелкоклеточного рака легкого, ткань аденокарциномы поджелудочной железы и ткань рака молочной железы, особенно ткань аденокарциномы поджелудочной железы и ткань рака молочной железы.
Обладает ли антитело эффектом ингибирования ангиогенеза можно подтвердить способом, известным специалисту в данной области, и например, это можно подтвердить, используя имеющийся в продаже ангиогенезный набор. Конкретнее, это можно подтвердить способами, описанными в примерах.
Конкретные примеры антител с эффектами ингибирования ангиогенеза включают ранее описанные антитела.
Примером другого предпочтительного варианта осуществления антитела по настоящему изобретению является антитело с подавляющей клеточный рост активностью.
Хотя не существует никаких особых ограничений в отношении клеток, чей рост подавляется антителом против AXL, предпочтительно, ими являются клетки, связанные с заболеванием, более предпочтительно, раковые клетки. Если клетки являются раковыми клетками, то не существует никаких особых ограничений на тип рака, и примеры включают рак поджелудочной железы (например, аденокарциному поджелудочной железы), рак желудка, рак легкого (мелкоклеточный рак легкого, немелкоклеточный рак легкого, и т.д.), остеосаркому, рак толстой кишки, рак предстательной железы, меланому, рак эндометрия, рак яичников, лейомиому матки, рак щитовидной железы, рак стволовых клеток, рак молочной железы, рак мочевого пузыря, рак почек, глиому, нейробластому и рак пищевода. Предпочтительным типом рака является глиома, рак желудка, рак эндометрия, немелкоклеточный рак легкого, аденокарцинома поджелудочной железы и рак молочной железы, особенно - аденокарцинома поджелудочной железы и рак молочной железы.
Не существует никаких особых ограничений в отношении механизма подавления клеточного роста антителом по настоящему изобретению, и рост клеток может быть подавлен с помощью любого механизма, такого как ингибирование ангиогенеза, ингибирование фосфорилирования, индукция фосфорилирования или снижение уровня экспрессии AXL.
Для оценки или измерения эффектов подавления клеточного роста, предпочтительно, используют следующие способы, основанные на антителе против AXL.
В качестве способа оценки или измерения подавляющей клеточный рост активности in vitro использовали способ, при котором поглощение жизнеспособными клетками [3H]-меченого тимидина, добавленного в их среду, измеряется как показатель способности репликации ДНК. Как более простой способ используют способ выделения красителя, в котором показатель исключения красителя, такого как трипановый синий, в окружающую среду из клеток измеряется под микроскопом, или используется способ МТТ. Последний оценивает способность живых клеток превращать МТТ (3-(4,5-диметилтиазол-2-ил)-2,5-дифенил тетразолия бромид), соль тетразолия, в синий формазановый продукт. Конкретно, исследуемое антитело добавляют к культуральному раствору тестовых клеток вместе с лигандом, а через заданный период времени добавляют раствор МТТ в культуральный раствор, и поддерживают в течение определенного периода, чтобы позволить МТТ включиться в клетки. МТТ, который представляет собой соединение желтого цвета, превращается в синее соединение с помощью сукцинатдегидрогеназы в митохондриях клеток. После растворения этого синего продукта и окрашивания, измеряют поглощение, которое используют в качестве показателя числа жизнеспособных клеток. Кроме МТТ, в продаже имеются такие реагенты, как MTS, XTT, WST-1 и WST-8 (Nacalai Tesque, и тому подобное), и они могут быть использованы надлежащим образом. При измерении этой активности может быть использовано контрольное антитело.
Для оценки или измерения подавляющей клеточный рост активности in vivo могут быть использованы модели опухолей на мышах. Например, после того, как раковые клетки, экспрессирующие AXL, трансплантируют тестовым животным, не человеку, внутрикожно или подкожно, исследуемое антитело вводят внутривенно или внутрибрюшинно, начиная со дня трансплантации или на следующий день, ежедневно или с интервалом в несколько дней. Подавляющая клеточный рост активность может быть оценена путем измерения размеров опухоли с течением времени. Аналогичным оценке in vitro образом, подавляющую клеточный рост активность можно определить введением контрольного антитела и наблюдая, становится ли размер опухоли в группе, которой вводят антитело против AXL, значительно меньше, чем размер опухоли в контрольной группе, которой вводят контрольное антитело. Если в качестве исследуемого животного, не человека, используют мышей, соответствующим образом можно использовать “голых” мышей (nu/nu), у которых утеряна функция Т-лимфоцитов из-за генетического дефекта в тимусе. Использование этих мышей позволяет устранить участие Т-лимфоцитов у тестовых животных во время оценки и измерения подавляющей клеточный рост активности введенного антитела.
Примеры антител против AXL с эффектами подавления клеточного роста включают ранее описанные антитела.
Моноклональное антитело против AXL по настоящему изобретению может быть получено известными способами. Моноклональные антитела, полученные у млекопитающих, особенно предпочтительны в качестве антитела против AXL по настоящему изобретению. Моноклональные антитела, полученные у млекопитающих, включают продуцируемые гибридомами, а также продуцируемые хозяином, трансформированным экспрессионным вектором, содержащим гены антитела, используя методы генной инженерии.
Гибридому, продуцирующую моноклональное антитело, можно получить с помощью известной технологии, например, описанной ниже. Сначала белок AXL используется в качестве сенсибилизирующего антигена для иммунизации в соответствии с обычными способами иммунизации. Иммунные клетки, полученные у иммунизированных животных, затем гибридизируют с известными родительскими клетками в соответствии с обычными способами гибридизации клеток с получением гибридом. Гибридому, продуцирующую антитело против AXL, можно выбрать из этих гибридом путем скрининга клеток, продуцирующих мишеневые антитела обычными способами скрининга.
Более конкретно, моноклональные антитела можно получить, например, способом, описанным ниже. Сначала белок AXL, используемый в качестве сенсибилизирующего антигена для получения антител, можно получить, экспрессируя ген AXL. Нуклеотидная последовательность гена AXL человека уже известна (GenBank, входящий № M76125). После введения последовательности гена, кодирующего AXL, в известный вектор экспрессии, с помощью которого трансформируют подходящие клетки-хозяева, интересующий белок AXL человека может быть очищен из клеток-хозяев или из культурального супернатанта с использованием известных способов. Очищенный природный белок AXL также можно использовать таким же образом. Очистку можно проводить с помощью нескольких хроматографий, таких как традиционная ионная хроматография и аффинная хроматография, выполненные один раз или несколько раз, либо в комбинации, либо отдельно. Слитый белок, в котором желательный частичный полипептид белка AXL гибридизировали с различными полипептидами, также можно использовать в качестве иммуногена. Fc фрагмент антитела или пептидный маркер и тому подобное могут быть использованы для получения слитого белка для использования в качестве иммуногена. Вектор, экспрессирующий слитый белок, можно получить гибридизацией двух или более видов желательных генов, кодирующих полипептидные фрагменты в рамке считывания, и вставкой гибридных генов в экспрессионный вектор, как описано выше. Способы получения слитых белков описаны во втором издании руководства «Молекулярное клонирование» (Sambrook, J. et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Laboratory Press, 1989). Очищенный таким образом белок AXL можно использовать в качестве сенсибилизирующего антигена для иммунизации млекопитающих.
Частичный пептид AXL также можно использовать в качестве сенсибилизирующего антигена. Примеры частичных пептидов AXL включают пептид, химически синтезированный по аминокислотной последовательности AXL человека, пептид, полученный путем введения части гена AXL человека в экспрессионный вектор, и его экспрессии, и пептид, полученный протеолитической деградацией белка AXL человека. Не существует никаких особых ограничений в отношении области, используемой в качестве частичного пептида, и например, можно использовать внеклеточную область AXL.
Кроме того, пептид с полной последовательностью FND1 или содержащий по меньшей мере пять его последовательных аминокислот, можно предпочтительно использовать в качестве частичного пептида. Последовательности, содержащие по меньшей мере пять последовательных аминокислот относятся к тем, что содержат предпочтительно шесть или более, более предпочтительно, восемь или более последовательных аминокислот. Кроме того, последовательности, которые содержат по меньшей мере пять, или более последовательных аминокислот, относятся к аминокислотным последовательностям, обладающих антигеннностью.
Не существует никаких особых ограничений в отношении млекопитающих, иммунизированных сенсибилизирующим антигеном. Для получения моноклональных антител путем слияния клеток, предпочтительно, выбрать иммунизируемое животное после рассмотрения его совместимости с родительскими клетками, используемыми для слияния клеток. В целом, в качестве иммунизируемых животных предпочтительны грызуны. Конкретнее, в качестве иммунизируемых животных могут использоваться мыши, крысы, хомяки или кролики. Также могут использоваться в качестве иммунизируемых животных обезьяны и тому подобное.
Животных, описанных выше, можно иммунизировать сенсибилизирующим антигеном, используя известные способы. Например, в конкретном способе млекопитающее иммунизируют инъекцией сенсибилизирующего антигена внутрибрюшинно или подкожно. В частности, сенсибилизирующий антиген вводят млекопитающему несколько раз с интервалом от 4 до 21 дней. Сенсибилизирующий антиген используют для иммунизации после разбавления до подходящей степени разбавления в фосфатно-буферном растворе (PBS), физиологическом растворе, и тому подобное. Сенсибилизирующий антиген может быть введен с адъювантом. Например, сенсибилизирующий антиген может быть смешан с полным адъювантом Фрейнда и эмульгирован для использования в качестве сенсибилизирующего антигена. При иммунизации сенсибилизирующим антигеном можно использовать подходящий носитель. В частности, если в качестве сенсибилизирующего антигена для иммунизации используют частичный пептид с низким молекулярным весом, то желательно связать сенсибилизирующий антиген с белком-носителем, таким как альбумин, гемоцианин лимфы улитки и тому подобное.
После того как млекопитающие иммунизированы подобным образом, и было подтверждено, что уровень желательных антител в сыворотке крови увеличился, иммунные клетки млекопитающего собирали и использовали для слияния клеток. В частности, в качестве иммунных клеток предпочтительно можно использовать клетки селезенки.
Миеломные клетки млекопитающих используют в качестве клеток слияния с иммунными клетками. Миеломные клетки, предпочтительно, имеют соответствующий селекционный маркер для скрининга. Селекционный маркер имеет отношение к характерному свойству, которое позволяет клеткам выживать (или погибать) при определенных условиях культивирования. Известные селекционные маркеры включают дефицит гипоксантин-гуанин фосфорибозилтрансферазы (далее сокращенно «дефицит HGPRT») и дефицит тимидинкиназы (далее сокращенно «дефицит TK»). Клетки, дефицитные по HGPRT или TK являются гипоксантин-аминоптерин-тимидин чувствительными (далее сокращенно «HAT-чувствительность»). HAT-чувствительные клетки не способны синтезировать ДНК и погибают в HAT-селективной среде. Однако если их гибридизуют с нормальными клетками, то они могут продолжать синтезировать ДНК, используя «реутилизационный» путь нормальных клеток, и поэтому они начинают расти на HAT-селективной среде.
И HGPRT-дефицитные клетки, и TK-дефицитные клетки можно отбирать на среде, содержащей 6-тиогуанин, 8-азагуанин (далее сокращенно «8AG») или 5'-бромдезоксиуридин. Хотя нормальные клетки погибают в результате встраивания этих пиримидиновых аналогов в их ДНК, клетки, дефицитные по этим ферментам, не в состоянии встраивать эти пиримидиновые аналоги и, тем самым, способны выживать в селективной среде. Селекционный маркер, называемый G418-устойчивым, также придает устойчивость к антибиотикам типа 2-дезоксистрептамина (гентамициновые аналоги), поскольку является геном устойчивости к неомицину. Известны различные миеломные клетки, подходящие для слияния клеток, и примеры миеломных клеток, которые могут быть использованы, включают P3 (P3x63Ag8.653) (J. Immunol. (1979) 123, 1548-1550), P3x63Ag8U.1 (Current Topics in Microbiology and Immunology (1978) 81, 1-7), NS-1 (Kohler, G and Milstein, C., Eur. J. Immunol. (1976) 6, 511-519), MPC-11 (Margulies, D.H. et al., Cell (1976) 8, 405-415), SP2/0 (Shulman, M. et al., Nature (1978) 276,269-270), FO (de St. Groth, S.F. et al., J. Immunol. Methods (1980) 35, 1-21), S194 (Trowbridge, I.S., J. Exp. Med. (1978) 148, 313-323), и R210 клетки (Galfre, G et al., Nature (1979) 277, 131-133).
Слияние вышеупомянутых иммунных клеток и миеломных клеток может быть осуществлено известными способами, такими как способ Келера и Мильштейна (Kohler, G and Milstein, C., Methods Enzymol. (1981) 73, 3-46).
Более конкретно, вышеупомянутое слияние клеток может быть осуществлено в обычной питательной среде культивирования в присутствии активатора слияния клеток. Примеры активаторов слияния клеток, которые могут быть использованы, включают полиэтиленгликоль (PEG) и вирус Сендай (HVJ). Для дальнейшего повышения эффективности слияния при желании также может быть добавлено вспомогательное средство, такое как диметилсульфоксид.
Отношение, в котором используются иммунные клетки и миеломные клетки, может быть установлено произвольно. Например, предпочтительным является 1-10 кратное превышение иммунных клеток над миеломными клетками. Примеры питательных сред, которые могут быть использованы для слияния клеток, описанные выше, включают культуральные среды MEM и RPMI1640, предпочтительно используемые для роста вышеупомянутых линий миеломных клеток, а также для этого вида клеточной культуры использовали обычную питательную среду. Сывороточная добавка, такая как фетальная сыворотка теленка (FCS), также может быть добавлена в культуральную среду.
Слияние клеток осуществляется для формирования целевых гибридных клеток (гибридом) путем тщательного перемешивания заранее определенных количеств иммунных клеток и миеломных клеток в питательной среде, а затем перемешивания в растворе PEG, подогретом приблизительно до 37°C. Например, в процессе слияния клеток можно добавить PEG со средней молекулярной массой около 1000-6000, как правило, в концентрации 30%-60% (вес/об). После этого, средства слияния клеток и другие средства, не отвечающие за рост гибридом, удаляют последовательными повторными добавлениями подходящей среды культивирования, как указывалось выше, центрифугированием и удалением супернатанта.
Полученные таким образом гибридомы могут быть селектированы с помощью селективной культуральной среды, соответствующей селекционному маркеру, в зависимости от миеломы, используемой для слияния клеток. Например, HGPRT или TK-дефицитные клетки могут быть селектированы культивированием в культуральной среде HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Если для слияния клеток используют HAT-чувствительные миеломные клетки, то те клетки, которые успешно сливаются с нормальными клетками, могут быть селективно выращены на культуральной среде HAT. Культивирование на среде HAT продолжают достаточно долго, для того чтобы другие клетки, отличные от целевых гибридом (неслитые клетки), погибли. В частности, целевые гибридомы обычно селектируют культивированием в течение промежутка времени от нескольких дней до нескольких недель. Далее, скрининг и моноклонирование гибридомы, продуцирующей целевое антитело, можно выполнить обычным способом серийных разведений. Кроме того, распознающее AXL антитело может быть получено способом, описанным в международной публикации WO 03/104453.
Скрининг и моноклонирование целевого антитела предпочтительно проводят известным способом скрининга, основанном на реакции антиген-антитело. Например, антиген связывают с носителем, таким как полистирольные гранулы или имеющиеся в продаже 96-луночные планшеты для микротитрования, и проводят реакцию с культуральным супернатантом гибридомы. Носитель затем промывают, и проводят реакцию с меченым ферментом вторичным антителом и т.д. Если целевое антитело, которое вступает в реакцию с сенсибилизирующим антигеном, присутствует в культуральном супернатанте, вторичные антитела связывают с носителем посредством этого антитела. В итоге, присутствует ли целевое антитело в культуральном супернатанте, можно определить детекцией вторичных антител, связанных с носителем. Гибридому, продуцирующую желательные антитела, способные связывать антиген, можно клонировать таким способом, как серийные разведения. На данном этапе антиген, использованный для иммунизации, или в значительной степени эквивалентный белку AXL, может быть использован преимущественно в качестве антигена.
В дополнение к способу получения гибридом путем иммунизации отличных от человека животных антигеном, целевое антитело можно получить путем сенсибилизации антигеном лимфоцитов человека. В частности, сначала лимфоциты человека сенсибилизируют in vitro белком AXL. Иммуносенсибилизированные лимфоциты затем гибридизуют с подходящим партнером слияния. Например, миеломные клетки человеческого происхождения, способные делиться постоянно, могут быть использованы в качестве партнера слияния (см. заявку на патент Японии Kokoku Publication № (JP-B) H1-59878 (рассмотренная, утвержденная заявка на патент Японии, опубликованная для опротестования)). Антитело против AXL, полученное способом, является антителом человека со связывающей белок AXL активностью.
Антитело человека против AXL может быть получено путем введения белка AXL в качестве антигена трансгенным животным с полным репертуаром генов антител человека. Клетки иммунизированных животных, продуцирующих антитела, могут быть иммортализованы с помощью таких процедур, как гибридизация с подходящим партнером слияния или заражение вирусом Эпштейна-Барр. Антитело против AXL может быть получено путем выделения антитела человека, направленного против белка AXL, из иммортализованных клеток, полученных таким образом (см. международные публикации WO 94/25585, WO 93/12227, WO 92/03918 и WO 94/02602). Клетки, продуцирующие антитела с интересующей реакционной специфичностью, могут также быть клонированы путем клонирования иммортализованных клеток. Когда трансгенных животных используют в качестве иммунизируемых животных, иммунная система животного распознает AXL человека, как чужеродный. Таким образом, можно легко получить антитело человека, направленное против AXL человека. Гибридому, продуцирующую моноклональное антитело, полученную таким образом, можно повторно культивировать на обычной питательной среде. Гибридому также можно хранить в течение длительного периода времени в жидком азоте.
Гибридому можно культивировать в соответствии с обычными способами, чтобы получить целевые моноклональные антитела из культурального супернатанта. В противном случае, моноклональные антитела можно получать путем введения гибридомы млекопитающим, совместимым с ней, чтобы обеспечить рост гибридомы, используя полученную в результате асцитную жидкость в качестве источника моноклонального антитела. Вышеупомянутый способ подходит для получения особо чистых антител.
В настоящем изобретении также может быть использовано антитело, кодируемое генами антител, клонированными из продуцирующих антитела клеток. Клонированные гены антител можно экспрессировать в виде антител путем их вставки в соответствующий вектор и введения вектора хозяину. Способы выделения генов антител, вставки их в вектор и трасформации ими клеток-хозяев уже созданы (см., например, Vandamme, A.M. et al., Eur. J. Biochem. (1990) 192, 767-775).
Например, кДНК, кодирующую вариабельную область (V-область) антитела против AXL, можно получить из гибридомных клеток, продуцирующих антитело против AXL. Для достижения этого обычно сначала экстрагируют тотальную РНК из гибридом. Примеры способов экстракции мРНК из клеток включают ультрацентрифугирование в гуанидине (Chirgwin, J.M. et al., Biochemistry (1979) 18, 5294-5299) и способ AGPC (Chomczynski, P. et al., Anal. Biochem. (1987) 162, 156-159).
Экстрагированную мРНК можно очистить, используя набор для очистки мРНК (GE Healthcare Bio-sciences) и тому подобное. В качестве варианта, можно приобрести такие наборы, как набор QuickPrep для очистки мРНК (GE Healthcare Bio-sciences) для экстракции всех мРНК непосредственно из клеток. Эти наборы можно использовать для получения всех мРНК из гибридом. кДНК, кодирующую V-область антитела, можно синтезировать из полученных мРНК с помощью обратной транскриптазы. кДНК можно получить, например, с помощью набора для синтеза первой цепи кДНК с помощью обратной транскриптазы AMV (Seikagaku Corp.). Можно использовать набор для амплификации 5'-Ampli FINDER RACE Kit (Clontech) и 5'-RACE способ с использованием ПЦР (Frohman, M.A. et al., Proc. Natl Acad. Sci. U.S.A. (1988) 85, 8998-9002, Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932) для синтеза и амплификации кДНК. Подходящие сайты рестрикции, описанные ниже, также можно вставить на обоих концах кДНК в ходе синтеза кДНК.
Целевые фрагменты кДНК очищают от полученного продукта ПЦР и связывают с векторной ДНК. Таким образом, получают рекомбинантный вектор, и после его вставки в Escherichia coli или тому подобное и селекции колоний можно получить желательный рекомбинантный вектор из E. coli, которые образуют колонии. Имеет ли рекомбинантный вектор нуклеотидные последовательности целевой кДНК или нет, можно подтвердить известным способом, например, секвенированием дидезоксинуклеотидного обрыва цепи.
Чтобы получить ген, кодирующий вариабельную область, также можно использовать ПЦР с праймером, амплифицирующим вариабельную область гена. Сначала синтезируют кДНК, используя экстрагированную мРНК в качестве матрицы для создания библиотеки кДНК. Удобно использовать имеющийся в продаже набор для синтеза библиотек кДНК. Поскольку количество мРНК, полученное лишь из небольшого числа клеток, очень мало, его непосредственная очистка приводит к малому выходу конечного продукта. Таким образом, мРНК обычно очищают после добавления РНК носителя, который заведомо не содержит никаких генов антител. В противном случае, когда существует возможность экстрагировать определенное количество РНК, может быть эффективно экстрагирована РНК из клеток, продуцирующих только антитела. Например, добавление РНК носителя может не быть необходимым для экстракции РНК от 10 или более, 30 или более, или лучше 50 или более клеток, продуцирующих антитело.
Ген антитела затем амплифицируют посредством ПЦР, используя библиотеку кДНК, созданную таким образом, в качестве матрицы. Праймеры для амплификации генов антител посредством ПЦР известны. Например, можно разработать праймеры для амплификации генов антител человека на основании литературных источников (например, J. Mol. Biol. (1991) 222, 581-597). Эти праймеры имеют нуклеотидные последовательности, различные для каждого подкласса иммуноглобулинов. Таким образом, когда в качестве матрицы используют библиотеку кДНК неизвестного подкласса, ПЦР проводят с учетом всех возможностей.
Например, если необходимо получить ген, кодирующий IgG человека, можно использовать праймеры, амплифицирующие ген, кодирующий γ1-γ5 в качестве тяжелых цепей, а κ и λ цепи - в качестве легких цепей. Для амплификации вариабельной области гена IgG в качестве праймера на 3' стороне обычно используют праймер, гибридизующийся с последовательностью, соответствующей шарнирной области. С другой стороны, праймер, соответствующий каждому подклассу, можно использовать в качестве праймера на 5' стороне.
Продукты ПЦР, амплифицированные праймерами, амплифицирующими гены каждого подкласса тяжелых цепей и легких цепей, производят в независимых библиотеках. Использование библиотеки, синтезированной таким образом, позволяет реконструировать иммуноглобулины, состоящие из комбинации тяжелых цепей и легких цепей. Затем можно скринировать целевое антитело, используя в качестве индикатора связывающую с AXL активность реконструированных иммуноглобулинов.
После получения кДНК, кодирующей V-область целевого антитела против AXL, кДНК расщепляют рестриктазой, распознающей сайт рестрикции, вставленный по обоим концам кДНК. Предпочтительная рестриктаза распознает и расщепляет нуклеотидную последовательность, чего не может случиться с нуклеотидной последовательностью, составляющей ген антитела. Рестриктаза, создающая «липкий» конец, является предпочтительной при вставке одной копии расщепленного фрагмента в вектор в нужном направлении. Вектор, экспрессирующий антитело, можно получить путем вставки кДНК, кодирующей V-область антитела против AXL, расщепленной как описано выше, в подходящий вектор экспрессии. В настоящее время, можно производить химерные антитела путем слияния в рамке считывания генов, кодирующих константную область (C-область) антитела, и генов, кодирующих V-области, описанные выше. В документе «химерным антителом» называют антитело, содержащее константные и вариабельные области, полученные из различных организмов. Таким образом, ксеногенные химерные антитела, такие как человеческо-мышиные антитела и человеческо-человеческие аллогенные химерные антитела, включены в химерные антитела по настоящему изобретению. Вектор экспрессии химерного антитела можно также создать путем включения генов V-области в вектор экспрессии, который первоначально имел константные области.
В частности, последовательность узнавания рестриктазы, расщепляющей ген V-области, можно разместить на 5' стороне вектора экспрессии, оставляя в сохранности ДНК, кодирующую желательную константную область (C-область) антитела. Вектор экспрессии химерного антитела создают расщеплением двух из них с помощью той же комбинации рестриктаз, а затем сливая их в рамке считывания.
Гены антител можно вставить в вектор экспрессии для экспрессии под контролем области регуляции экспрессии, чтобы продуцировать антитело против AXL по настоящему изобретению. Область регуляции экспрессии для экспрессии антитела может включать, например, энхансер и промотор. Рекомбинантные клетки, экспрессирующие ДНК, кодирующую антитело против AXL, можно затем получить путем трансформации подходящих клеток-хозяев этим вектором экспрессии.
При экспрессии генов антител каждую из ДНК, кодирующих тяжелую цепь (H-цепь) и легкую цепь (L-цепи) антитела, можно вставить в отдельные векторы экспрессии. Векторы, включающие либо H-цепь, либо L-цепь, могут экспрессировать молекулу антитела с H-цепью и L-цепью после того, как векторы одновременно трансформируют (котрансфицируют) одну и ту же клетку-хозяина. И наоборот, ДНК, кодирующие H-цепь и L-цепь, можно включить в единый вектор экспрессии для трансформации клеток-хозяев (см. международную публикацию WO 94/11523).
Для получения антител известны многие комбинации хозяев и векторов экспрессии, когда сначала выделяют гены антител, а затем вставляют их подходящему хозяину. Все эти системы экспрессии применимы к настоящему изобретению. Животные клетки, клетки растений или грибковые клетки можно использовать, когда в качестве хозяев используют эукариотические клетки. Конкретные примеры животных клеток, которые можно использовать по настоящему изобретению, включают клетки млекопитающих (например, клетки CHO, COS, миеломные, BHK [почек детеныша хомяка], Hela и Vero), клетки амфибий (например, ооциты Xenopus) и клетки насекомых (например, клетки sf9, sf21 и Tn5).
Среди известных примеров растительных клеток, используемых в системах экспрессии генов антител, находятся клетки из рода Nicotiana, например, Nicotiana tabacum. Для трансформации клеток растений можно использовать клетки каллюсных культур.
Примеры грибковых клеток, которые можно использовать включают дрожжи (род Saccharomyces, например, Saccharomyces cerevisiae, и метанол-утилизирующие дрожжи рода Pichia, например, Pichia pastoris) и мицелиальные грибы (рода Aspergillus, например, Aspergillus niger).
Также известны системы экспрессии генов антител, использующие прокариотические клетки. Например, по настоящему изобретению можно использовать клетки таких бактерий, как E. coli или Bacillus subtilis.
При использовании клеток млекопитающих, можно создать вектор экспрессии, в котором между собой функционально связаны обычно используемый подходящий промотор, гены экспрессируемого антитела, и полиА сигнальная последовательность на 3' стороне в 5'-3' направлении. Примером промотора/энхансера является предранний промотор/энхансер цитомегаловируса человека.
Другие примеры промоторов/энхансеров, которые можно использовать для экспрессии антитела по настоящему изобретению, включают вирусные промоторы/энхансеры или промоторы/энхансеры клеток млекопитающих, такие как человеческий фактор элонгации 1α (HEF1α). Конкретные примеры вирусов, чьи промоторы/энхансеры находят применение, включают ретровирусы, полиомавирусы, аденовирусы и вирус обезьян 40 (SV40).
При использовании SV40 промотора/энхансера, можно использовать способ Mulligan et al. (Nature (1979) 277, 108). Также можно использовать промотор/энхансер HEF1α, чтобы удобно экспрессировать целевой ген способом Mizushima et al. (Nucleic Acids Res. (1990) 18, 5322).
При использовании E. coli, гены антитела можно экспрессировать, функционально связывая обычно используемый подходящий промотор, сигнальную последовательность секреции антитела, а также гены экспрессируемого антитела. Примеры промоторов включают lacZ промотор и araB промотор. При использовании lacZ промотора можно использовать способ Ward et al. (Nature (1989) 341, 544-546; FASEB J. (1992) 6, 2422-2427). В противном случае araB промотор можно использовать для экспрессии целевых генов согласно способу Better et al. (Science (1988) 240, 1041-1043).
Примером сигнальной последовательности секреции антитела, которую можно использовать для продукции в периплазму E. coli, является сигнальная последовательность pelB (Lei, S. P. et al., J. Bacteriol. (1987) 169, 4379). Антитело, продуцируемое в периплазму, отделяют, а затем так проводят структурный рефолдинг с использованием белкового денатуранта, например, гуанидинхлорида или мочевины, чтобы антитела имели желательную связывающую активность.
Примеры пригодных репликаторов, которые можно вставить в вектор экспрессии, включают те, которые происходят из SV40, полиомавирусов, аденовирусов и вируса папилломы крупного рогатого скота (BPV). Можно также вставить в вектор экспрессии селекционный маркер, чтобы амплифицировать количество генных копий в системе клетки-хозяина. Конкретные примеры селекционных маркеров, которые можно использовать, включают ген аминогликозидтрансферазы (APH), ген тимидинкиназы (TK), ген E. coli ксантингуанин-фосфорибозилтрансферазы (Ecogpt) и ген дигидрофолатредуктазы (dhfr).
Целевое антитело получают путем введения этих векторов экспрессии в клетки-хозяева и культивирования трансформированных клеток-хозяев in vitro или in vivo. Культивирование клеток-хозяев осуществляют в соответствии с известными способами. Примеры питательных сред, которые можно использовать, включают DMEM, MEM, RPMI1640 и IMDM, и их можно использовать в сочетании с сывороточной добавкой, такой как FCS.
Антитело, экспрессируемое и продуцируемое описанным выше образом, можно очистить, применяя известные способы, которые обычно используют для очистки белков, либо отдельно, либо в подходящем сочетании. Например, антитела можно разделить и очистить путем подходящего подбора и сочетания, например, аффинной колонки, например, колонки с белком A, хроматографической колонки, фильтра, ультрафильтрации, высаливания или диализа (Antibodies - A Laboratory Manual, Ed Harlow and David Lane, Cold Spring Harbor Laboratory, 1988).
В дополнение к клеткам-хозяевам, описанным выше, для получения рекомбинантных антител также можно использовать трансгенных животных. Целевое антитело можно получить из животного, в организм которого был введен ген, кодирующий целевое антитело. Например, гены антител можно сконструировать в виде химерных генов, включив их в ген, кодирующий белок, по своей природе производимый в молоке в рамке считывания. Например, в качестве этого белка, секретируемого в молоко, можно использовать β казеин козы. Фрагмент ДНК, содержащий химерные гены, в который были вставлены гены антител, вводят с помощью инъекции в козий эмбрион, и инъецированный эмбрион имплантируют козе-самке. Желательные антитела можно получить в форме слитого белка, слитого с молочным белком, из молока, производимого трансгенными козами (или их потомками), рожденными от коз, которым эмбрион был имплантирован. Трансгенной козе при необходимости можно назначить гормоны, чтобы увеличить количество молока, содержащего желательное антитело, продуцируемое ею (Ebert, K.M. et al., Bio/Technology (1994) 12, 699-702). C-область, возникшую в антителе животных, можно использовать для C-области рекомбинантного антитела по настоящему изобретению. Примеры подходящих C-областей H-цепи антител мыши включают Cγ1, Cγ2a, Cγ2b, Cγ3, Cµ, Cδ, Cα1, Сα2 и Cε, а примеры C-областей L-цепи включают Cκ и Cλ. Примеры подходящих антител животных, отличных от антител мыши, включают антитела крыс, кроликов, коз, овец, верблюдов и обезьян. Последовательности этих антител известны. C-область также можно модифицировать, чтобы улучшить стабильность антитела или его продукцию. В настоящем изобретении, при назначении антитела человеку можно сделать искусственно модифицированное рекомбинантное антитело, например, для того, чтобы снизить его ксеногенную антигеннность в организме человека. Примеры рекомбинантных антител включают химерные антитела и гуманизированные антитела.
Эти модифицированные антитела можно получить, используя известные способы. Химерными антителами называются антитела, в которых вариабельные области и константные области различного происхождения связаны между собой. Например, антитело с вариабельными областями тяжелой цепи и легкой цепи антитела мыши и константными областями тяжелой цепи и легкой цепи антитела человека является человеческо-мышиным ксеногенным химерным антителом. Рекомбинантный вектор, экспрессирующий химерное антитело можно изготовить путем связывания ДНК, кодирующей вариабельные области антител мыши с ДНК, кодирующей константную область антитела человека, а затем вставив это в вектор экспрессии. Рекомбинантные клетки, трансформированные вектором, культивируют, и встроенные ДНК экспрессируют, получая химерные антитела, продуцируемые в культуре. C-области антитела человека используют в качестве C-областей химерных антител и гуманизированных антител. Так, например, Cγ1, Сγ2, Cγ3, Cγ4, Gµ, Cδ, Cα1, Сα2 и Cε можно использовать для C-области в H-цепи. Cκ и Cλ можно использовать для C-области в L-цепи. Известны аминокислотные последовательности этих C-областей и нуклеотидные последовательности, кодирующие их. Можно также модифицировать C-область антитела человека, повышая стабильность антитела как таковую или продукцию антител.
Как правило, химерные антитела состоят из V-областей, происходящих от антител животного, не человека, и C-областей, происходящих от антител человека. В отличие от этого, гуманизированные антитела состоят из определяющих комплементарность областей (CDR), происходящих от антител животных, отличных от человека, каркасных областей (FR), происходящих от антител человека, и C-областей, происходящих от антител человека. Поскольку гуманизированные антитела имеют пониженную антигенность в организме человека, они подходят в качестве активного ингредиента терапевтического средства по настоящему изобретению.
Вариабельные области антител, как правило, состоят из трех CDR, фланкированных четырьмя FR. CDR по сути представляет собой область, определяющую специфичность связывания антитела. Аминокислотные последовательности CDR очень разнообразны. Напротив, аминокислотные последовательности, составляющие FR, часто демонстрируют высокую гомологию, даже среди антител с различными специфичностями связывания. Соответственно, как правило, считается, что специфичность связывания определенного антитела можно привить другому антителу путем пересадки CDR.
Гуманизированное антитело также называют «реконструированным» антителом человека. А именно, известны гуманизированные антитела, в которых CDR антител животного, не человека, такого как мышь, пересажены в антитела человека. Также известны общепринятые методы генной рекомбинации для получения гуманизированных антител.
Конкретный пример известного способа пересадки CDR антител мыши в FR человека представляет собой ПЦР с удлинением перекрывающихся фрагментов. В ПЦР с удлинением перекрывающихся фрагментов нуклеотидную последовательность, кодирующую CDR антитела мыши, которая будет пересажена, добавляют к праймерам для синтеза FR антитела человека. Праймеры изготавливают для каждой из четырех FR. В большинстве случаев, это считается выгодным с точки зрения поддержания функции CDR, чтобы выбрать FR человека с высокой гомологией к FR мыши, когда пересаживают CDR мыши в FR человека. Значит, как правило, предпочтительнее использовать FR человека с аминокислотной последовательностью, высоко гомологичной аминокислотной последовательности FR, прилегающей к пересаживаемому CDR мыши.
Связываемые нуклеотидные последовательности проектируют так, чтобы они соединялись в общей рамке считывания. FR человека синтезируют индивидуально с помощью специфичных наборов праймеров. В результате получают продукты, в которых ДНК, кодирующую CDR мыши, добавляют к каждой FR. Нуклеотидные последовательности, кодирующие CDR мыши продуктов, проектируют так, чтобы они перекрывали друг друга. Затем проводят реакцию синтеза комплементарной нити путем взаимной гибридизации перекрывающихся частей CDR продуктов, синтезированных с использованием гена антитела человека в качестве матрицы. В результате этой реакции FR человека связывают через последовательность CDR мыши.
Наконец, полноразмерный ген V-области, в котором связаны три CDR и четыре FR, амплифицируют с помощью праймеров, гибридизующихся с его 5' и 3' концами, и которые имеют подходящие добавленные последовательности узнавания рестриктазами. Затем можно изготовить вектор для экспрессии гуманизированного антитела посредством вставки ДНК, полученной описанным выше образом, и ДНК, кодирующей C-область антитела человека, в вектор экспрессии так, чтобы они были сплавлены в рамке считывания. Затем продуцируют гуманизированное антитело при культивировании клеток в культуре путем введения рекомбинантного вектора в клетку-хозяина, чтобы создать рекомбинантные клетки, за чем следует культивирование рекомбинантных клеток и экспрессия ДНК, кодирующей гуманизированное антитело (см. Европейскую патентную публикацию № EP 239400 и международную публикацию № WO 96/02576).
Предпочтительные FR антитела человека можно отобрать так, что CDR образуют подходящий антиген-связывающий участок, когда они соединены через CDR, с помощью качественного или количественного измерения и оценки его связывающей активности в отношении антигена гуманизированного антитела, полученного описанным выше образом. Аминокислотные остатки FR также в случае необходимости можно заместить так, чтобы CDR реконструированного антитела человека образовывали подходящий антиген-связывающий участок. Например, в FR можно ввести мутации аминокислотной последовательности, применяя ПЦР, используемый для пересадки CDR мыши к FR человека. В частности, мутацию частичной последовательности нуклеотидов можно ввести в праймер для гибридизации с FR. Мутированную нуклеотидную последовательность вставляют в FR, синтезированную с участием этого праймера. Можно отобрать мутантную последовательность FR с желательными свойствами, измеряя и оценивая связывающую активность мутантного антитела с аминокислотными замещениями в отношении антигена, с использованием описанного выше способа (Sato, K. et al., Cancer Res. (1993) 53, 851-856).
Известны также способы получения антител человека. Например, лимфоциты человека сенсибилизируют желательным антигеном или клетками, экспрессирующими желательный антиген in vitro. Далее, желательное антитело человека со связывающей активностью в отношении антигена можно получить путем слияния сенсибилизированных лимфоцитов с миеломными клетками человека (см. JP-B H1-59878). Например, в качестве миеломных клеток человека можно использовать клетки U266, исполняющие роль партнера по слиянию.
Желательное антитело человека можно также получить путем иммунизации желательным антигеном трансгенных животных с полным репертуаром генов антител человека (см. международные публикации WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096 и WO 96/33735). Кроме того, также известны технологии, с помощью которых антитела человека можно получить пэннингом, используя библиотеку антител человека. Например, V-область антитела человека можно экспрессировать на поверхности фага в виде одноцепочечного антитела (scFv), применяя способ фагового дисплея, таким образом, делая возможным селекцию фага, который связывается с антигеном. Анализируя гены отобранного фага, можно определить последовательность ДНК, кодирующую V-область антитела человека, которое связывается с антигеном. После определения последовательности ДНК scFv, которая связывается с антигеном, последовательность V-области гибридизуют в рамке считывания с последовательностью C-области желательного антитела человека, а затем вставляют в подходящий вектор экспрессии для получения вектора экспрессии. Антитело человека можно получить путем введения вектора экспрессии в предпочтительные экспрессирующие клетки, как описано выше, и экспрессируя ген, кодирующий антитело человека. Эти способы уже известны (международные публикации WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438 и WO 95/15388).
Антитела по настоящему изобретению включают в себя не только бивалентные антитела, представленные IgG, но также моновалентные антитела или поливалентные антитела, представленные IgM, если только они связываются с белком AXL. Поливалентные антитела по настоящему изобретению включают те, что имеют одинаковые антиген-связывающие участки, и те, у которых все или некоторые из антиген-связывающих участков различны. Антитело по настоящему изобретению не ограничивается целой молекулой антитела, но также может являться мини-антителом или модифицированным антителом такового, если только оно связывается с белком AXL.
Мини-антитела включают фрагменты антител, у которых удалена часть целого антитела (такого как целый IgG). Допускаются частичные делеции в молекулах антител, если только сохраняется способность связывать антиген AXL. Фрагмент антител по настоящему изобретению предпочтительно состоит из чего-либо одного из двух или обеих: вариабельной области (VH) тяжелой цепи и вариабельной области (VL) легкой цепи. Аминокислотные последовательности VH или VL могут содержать замены, делеции, добавления и/или инсерции. Кроме того, может быть удалена часть одного или обеих VH и VL, если только способность связывать антиген AXL сохраняется. Вариабельные области можно также химеризовать или гуманизировать. Конкретные примеры фрагментов антител включают, например, Fab, Fab', F(ab')2 и Fv. Конкретные примеры мини-антител включают Fab, Fab', F(ab')2, Fv, scFV (одноцепочечные Fv), диатело, sc(Fv)2 (одноцепочечные (Fv)2) и тому подобное. Полимеры этих антител (например, димеры, тримеры, тетрамеры или полимеры), также включены в мини-антитела по настоящему изобретению.
Фрагменты антител можно получить путем производства фрагмента антител, обрабатывая антитела ферментом. Известные примеры ферментов, используемых для получения фрагментов антител, включают папаин, пепсин, плазмин и тому подобное. Или же, гены, кодирующие эти фрагменты антител, можно создать, вставить в вектор экспрессии, а затем экспрессировать в подходящих клетках-хозяевах (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2698-2976; Better, M. and Horwitz, A.H., Methods in Enzymology (1989) 178, 476-496; Plueckthun, A. and Skerra, A., Methods in Enzymology (1989) 178, 476-496; Lamoyi, E., Methods in Enzymology (1989) 121, 652-663; Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-669; and Bird, R.E. et al., TIBTECH (1991) 9, 132-137).
Пищеварительные ферменты расщепляют фрагмент антитела по особым положениям, образуя фрагмент антитела со специфической структурой, как указано ниже. Произвольную часть антитела можно удалить, применяя методы генной инженерии к фрагменту антитела, ферментативно полученному таким образом.
Расщепление папаином: F(ab)2 или Fab
Расщепление пепсином: F(ab')2 или Fab'
Расщепление плазмином: Facb
«Диателом» называют бивалентные фрагменты антител, созданные слиянием генов (см. Holliger, P. et al., Proc. Natl Acad. Sci. U.S.A. (1993) 90, 6444-6448; EP 404,097; WO 93/11161, и тому подобное). Диатела представляют собой димеры, состоящие из двух полипептидных цепей. Как правило, VL и VH внутри одной и той же цепи полипептидных цепей, которые образуют димер, оба связаны линкерами. Линкеры в диателе, как правило, слишком коротки, чтобы дать возможность VL и VH связаться друг с другом. В конкретном плане, число аминокислотных остатков, которые образуют линкер, составляет, например, около пяти остатков. Таким образом, VL и VH, закодированные в одной и той же полипептидной цепи, не могут образовывать одноцепочечный фрагмент вариабельной области, но вместо этого образует димер с другим одноцепочечным фрагментом вариабельной области. В результате, диатело имеет два антиген-связывающих участка.
scFv получают связыванием V-области H-цепи и V-области L-цепи антитела. В scFv, V-область H-цепи и V-область L-цепи связаны через линкер, предпочтительно - пептидный линкер (Huston, J.S. et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 85, 5879-5883). V-область H-цепи и V-область L-цепи в scFv могут происходить из любого антитела, описанного в настоящем документе в качестве антитела. Не существует никаких особых ограничений на пептидные линкеры, связывающие V-области. Например, произвольный одноцепочечный пептид, содержащий приблизительно от 3 до 25 остатков, можно использовать в качестве линкера. V-области можно связать, например, описанным выше способом ПЦР. Чтобы связать V-области способом ПЦР, ДНК, кодирующую полную или желательную частичную аминокислотную последовательность последовательности ДНК, кодирующей H-цепь или V-область H-цепи вышеупомянутого антитела, и последовательность ДНК, кодирующую L-цепь или V-область L-цепи вышеупомянутого антитела, используют в качестве матрицы.
Как ДНК, кодирующую V-области H-цепи, так и ее же, кодирующую L-цепь, амплифицировали способом ПЦР, используя пары праймеров с последовательностями, соответствующими последовательностям на обоих концах ДНК, которую амплифицируют. Далее, изготавливали ДНК, кодирующую часть пептидного линкера. ДНК, кодирующую пептидный линкер, можно синтезировать способом ПЦР. Нуклеотидные последовательности, которые могут соединять продукты амплификации каждой отдельно синтезированной V-области, добавляют к 5' стороне праймеров, использованных в этой настройке. Далее, осуществляют реакцию ПЦР с помощью «ДНК V-области H-цепи», «ДНК пептидного линкера», и «ДНК V-области L-цепи», совместно с праймерами для сборочного ПЦР. Праймеры для сборочного ПЦР состоят из сочетания праймера, который гибридизуют с 5' стороной «ДНК V-области H-цепи», и праймера, который гибридизуют с 3' стороной «ДНК V-области L-цепи». Поэтому, праймеры для сборочного ПЦР состоят из набора праймеров, способных амплифицировать ДНК, кодирующую полную последовательность синтезируемой scFv. С другой стороны, нуклеотидные последовательности, которые можно присоединить к каждой ДНК V-области, добавляют к «ДНК пептидного линкера». В результате эти ДНК связаны друг с другом, а полноразмерную scFv, в конечном итоге, производят в виде продукта амплификации праймеров, используемых для сборочного ПЦР. После того, как получена ДНК, кодирующая scFv, обычными способами можно получить вектор экспрессии, содержащий ДНК, и рекомбинантные клетки, трансформированные вектором экспрессии. Также можно получить scFv, экспрессируя ДНК, кодирующую scFv, в культурах полученных рекомбинантных клеток.
sc(Fv)2 представляет собой мини-антитело, в котором два VH и два VL могут быть соединены с помощью линкера, либо так, чтобы образовать единую цепь (Hudson, et al., J. Immunol. Methods (1999) 231, 177-189). Например, sc(Fv)2 можно изготовить, соединяя два scFv линкером.
sc(Fv)2 предпочтительно является антителом, в котором два VH и два VL расположены в таком порядке: VH, VL, VH, VL (VH-линкер-VL-линкер-VH-линкер-VL) с использованием N-концевой стороны одноцепочечного полипептида в качестве исходной точки.
Любой произвольный пептидный линкер, который можно вставить посредством генной инженерии, линкер - синтетическое соединение (например, раскрытые в Protein Engineering, (1996) 9(3), 299-305) или тому подобное, можно использовать в качестве линкера, чтобы соединить вариабельные области антител. В настоящем изобретении пептидные линкеры предпочтительнее. Не существует никаких особых ограничений на длину пептидных линкеров, и длину может адекватно выбрать специалист в данной области в зависимости от цели использования. Обычно количество аминокислотных остатков в составе пептидного линкера колеблется от 1 до 100 аминокислот, предпочтительно, от 3 до 50 аминокислот, более предпочтительно, от 5 до 30 аминокислот, и особенно предпочтительно, от 12 до 18 аминокислот (например, 15 аминокислот).
Последовательность аминокислот, составляющих пептидный линкер, может быть произвольной последовательности, если только это не препятствует связывающей функции scFv.
Альтернативно, V-области можно соединить, используя синтетический химический линкер (химическое перекрестно-сшивающее средство). Перекрестно-сшивающие средства обычно используют для перекрестной сшивки пептидных соединений, и это можно использовать в настоящем изобретении. Примеры применимых перекрестно-сшивающих средств, включают:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидилпропионат) (DSP),
дитиобис(сукцинимидилпропионат) (DTSSP),
этиленгликольбис(сукцинимидилсукцинат) (EGS),
этиленгликольбис(сульфосукцинимидилсукцинат) (sulfo-EGS),
дисукцинимидилтартрат (DST),
дисульфосукцинимидилтартрат (sulfo-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES) и
бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (sulfo-BSOCOES).
Обычно необходимы три линкера, когда между собой связаны четыре вариабельные области антител. Используемые несколько линкеров могут быть одинаковыми или разными. Предпочтительным мини-антителом по настоящему изобретению является диатело или sc(Fv)2. Чтобы получить эти мини-антитела, антитело обрабатывали ферментом, таким как папаин или пепсин, чтобы получить фрагменты антител. В качестве варианта, создавали ДНК, кодирующую эти фрагменты антител, вставляли в вектор экспрессии, а затем экспрессировали в подходящих клетках-хозяевах (см., например, Co,M.S. et al., J. Immunol. (1994) 152, 2698-2976; Better,M. and Horwitz, A.H., Methods Enzymol. (1989) 178, 476-496; Plueckthun, A. and Skerra, A., Methods Enzymol. (1989) 178, 497-515; Lamoyi, E., Methods Enzymol. (1986) 121, 652-663; Rousseaux, J. et al., Methods Enzymol. (1986) 121, 663-669; и Bird, R.E. and Walker, B.W., Trends Biotechnol. (1991) 9, 132-137).
Антитело по настоящему изобретению также можно использовать в виде модифицированного антитела, связанного с различными молекулами, такими как полиэтиленгликоль (PEG) или цитотоксические вещества. Такие модифицированные антитела можно получить путем химической модификации антитела по настоящему изобретению. Способы модификации антитела уже широко известны в данной области.
Антитело по настоящему изобретению может также являться биспецифическим антителом. «Биспецифическим антителом» называется антитело, которое имеет вариабельные области, распознающие различные эпитопы внутри одной и той же молекулы антитела. Эпитопы могут находиться в разных молекулах или присутствовать в одной и той же молекуле. В настоящем изобретении, биспецифическое антитело может иметь антиген-связывающие сайты, распознающие различные эпитопы в молекуле AXL. Или же, посредством одного сайта узнавания биспецифическое антитело распознает AXL, а посредством другого сайта узнавания - цитотоксическое вещество. Эти антитела также включены в антитела по настоящему изобретению.
Биспецифическое антитело, распознающее антиген, отличный от AXL, в настоящем изобретении может быть комбинированным. Например, биспецифическое антитело, распознающее антиген, отличный от AXL, который специфически экспрессируется на поверхности целевых раковых клеток таким же образом, что и AXL, может быть комбинированным.
Способы получения биспецифических антител известны. Например, биспецифическое антитело можно получить путем соединения двух типов антител, распознающих различные антигены. Каждое из соединенных антител может представлять собой половину молекулы, с H и L цепями, или четверть молекулы, содержащей только H цепь. В качестве варианта, можно изготовить химерные клетки, продуцирующие биспецифические антитела, путем слияния гибридом, продуцирующих различные моноклональные антитела. Биспецифические антитела можно также изготовить методами генной инженерии.
Связывающая активность антитела
Известные средства можно использовать для определения антиген-связывающей активности антитела (Antibodies A Laboratory Manual, Ed Harlow and David Lane, Cold Spring Harbor Laboratory, 1988). Примеры способов, которые можно использовать, включают ELISA (ферментный иммуносорбентный анализ), EIA (иммуноферментный анализ), RIA (радиоиммунный анализ), люминесцентный иммуноферментный анализ, и тому подобное. Примеры средств измерения связывающей активности антитела с антигеном, экспрессируемым в клетках, включают способ, описанный на стр. 359-420 в «Antibodies A Laboratory Manual», упомянутом выше.
Способы с использованием проточного цитометра особенно предпочтительно применять для измерения связывания антигена, экспрессируемого на поверхности клеток, суспендированных в буфере и тому подобное, с антителом к этому антигену. Примеры используемых проточных цитометров включают FACSCanto™ II, FACSAria™, FACSArray™, FACSVantage™ SE и FACSCalibur™ (все от BD Biosciences), а также EPICS ALTRA HyPerSort, Cytomics FC 500, EPICS XL-MCL ADC, EPICS XL ADC и Cell Lab Quanta/Cell Lab Quanta SC (все от Beckman Coulter).
Примером предпочтительного способа измерения связывающей активности исследуемого антитела против AXL с антигеном является способ окрашивания вторичным антителом, меченным FITC, которое распознает исследуемое антитело, вступившее в реакцию с клетками, экспрессирующими AXL, а затем измерения с помощью FACSCalibur (BD Biosciences) и анализа интенсивности флуоресценции с помощью пакета программ CellQuest (BD Biosciences).
Гибридомы
Настоящее изобретение также предусматривает гибридомы, с депозитарным номером FERM BP-10858 (AX285), FERM BP-10859 (AX292), FERM BP-10853 (AX223), FERM BP-10852 (AX96), FERM BP-10856 (AX258), FERM BP-10857 (AX284), FERM BP-10850 (Ax7), FERM BP-10851 (Ax51), FERM BP-10854 (Ax225) и FERM BP-10855 (Ax232). Эти гибридомы продуцируют антитела против AXL с агонистической активностью, антитела против AXL с антагонистической активностью, антитела против AXL с активностью, снижающей уровень экспрессии AXL, антитела против AXL с ингибирующей ангиогенез активностью и/или антитела против AXL с ингибирующей клеточный рост активностью.
Ингибиторы ангиогенеза
Настоящее изобретение также предусматривает ингибиторы ангиогенеза, содержащие антитело против AXL. Не существует никаких особых ограничений на механизм, посредством которого ингибируется ангиогенез. Примеры механизма включают ингибирующее действие на миграционную активность сосудистых эндотелиальных клеток, апоптоз-индуцирующее действие на сосудистые эндотелиальные клетки и ингибирующее действие на морфогенез сосудов сосудистых эндотелиальных клеток. Ингибиторы ангиогенеза по настоящему изобретению предпочтительно ингибируют ангиогенез в раковых тканях. Не существует никаких особых ограничений на опухолевые ткани. Примеры этих раковых тканей включают ткани рака поджелудочной железы (ткани аденокарциномы поджелудочной железы и тому подобное), ткани рака желудка, ткани рака легких (ткани мелкоклеточного рака легкого, немелкоклеточного рака легкого и тому подобное), ткани остеосаркомы, ткани рака прямой кишки, ткани рака простаты, ткани меланомы, ткани рака эндометрия, ткани рака яичников, ткани лейомиомы матки, ткани рака щитовидной железы, ткани рака стволовых клеток, ткани рака молочной железы, ткани рака мочевого пузыря, ткани рака почек, ткани глиомы, ткани нейробластомы и ткани рака пищевода. Более предпочтительными тканями являются ткань глиомы, ткань рака желудка, ткань рака эндометрия, ткань немелкоклеточного рака легкого, ткань аденокарциномы поджелудочной железы и ткань рака молочной железы, особенно ткань аденокарциномы поджелудочной железы и ткань рака молочной железы.
Не существует никаких особых ограничений на антитела, используемые в качестве ингибиторов ангиогенеза по настоящему изобретению, если только они имеют ингибирующее ангиогенез действие. Например, можно использовать антитела, описанные выше (антитела с агонистической активностью, антитела с антагонистической активностью, антитела с активностью, снижающей уровень экспрессии AXL, и тому подобное).
Ингибиторы ангиогенеза, содержащие антитело против AXL по настоящему изобретению, можно представить как способы ингибирования ангиогенеза с использованием антитела против AXL. Ингибиторы ангиогенеза, содержащие антитело против AXL по настоящему изобретению, можно представить как применение антитела против AXL для получения ингибитора ангиогенеза.
Ингибиторы клеточного роста
Настоящее изобретение также предусматривает ингибиторы клеточного роста, содержащие антитела против AXL. Не существует никаких особых ограничений на механизм, по которому подавляется рост клеток. Примеры механизмов включают основанные на ингибирующем ангиогенез действии, основанные на цитотоксической активности антител и основанные на цитотоксическом веществе, связанным с антителом, но основанные на ингибирующем ангиогенез действии - предпочтительнее.
Не существует никаких особых ограничений на клетки, чей рост подавляется антителом против AXL. Клетками предпочтительно являются те, что связаны с болезнью, более предпочтительно, раковые клетки. Таким образом, примеры предпочтительных вариантов осуществления ингибиторов клеточного роста по настоящему изобретению включают противораковое средство, содержащее антитело против AXL. Когда клетки являются раковыми клетками, не существует никаких особых ограничений на тип рака, и данные типы включают рак поджелудочной железы (аденокарциному поджелудочной железы и тому подобное), рак желудка, рак легкого (мелкоклеточный рак легкого, немелкоклеточный рак легкого, и тому подобное), остеосаркому, рак толстой кишки, рак предстательной железы, меланому, рак эндометрия, рак яичников, лейомиому матки, рак щитовидной железы, рак стволовых клеток, рак молочной железы, рак мочевого пузыря, рак почек, глиому, нейробластому и рак пищевода. Более предпочтительными типами рака являются глиома, рак желудка, рак эндометрия, немелкоклеточный рак легкого, аденокарцинома поджелудочной железы и рак молочной железы, особенно - аденокарцинома поджелудочной железы и рак молочной железы.
Не существует никаких особых ограничений на антитела, используемые в качестве ингибиторов клеточного роста по настоящему изобретению, если только они имеют подавляющую клеточный рост активность. Например, можно использовать антитела, описанные выше (антитела с агонистической активностью, антитела с антагонистической активностью, антитела активностью, снижающей уровень экспрессии AXL, и тому подобное).
Ингибиторы клеточного роста, содержащие антитело против AXL по настоящему изобретению можно представить как способы подавления роста клеток с использованием антитела против AXL. Когда клетки, чей рост подавляется, являются раковыми клетками, противораковые средства, содержащие антитело против AXL по настоящему изобретению, можно представить как способы лечения и/или профилактики рака с использованием антитела против AXL. Ингибиторы клеточного роста, содержащие антитело против AXL по настоящему изобретению, можно представить как применение антитела против AXL для получения ингибитора клеточного роста. Когда клетки, чей рост подавляется, являются раковыми клетками, их можно представить как применение антитела против AXL для получения противоракового средства.
Индукторы фосфорилирования
Настоящее изобретение также предусматривает индукторы фосфорилирования, содержащие антитело против AXL. Индукторы фосфорилирования по настоящему изобретению обычно индуцируют фосфорилирование в клетках, экспрессирующих AXL. Хотя нет никаких особых ограничений на цели индукции фосфорилирования, данными целями, как правило, являются полипептиды, имеющие тирозин, и предпочтительно являющиеся AXL.
Не существует никаких особых ограничений на антитела, используемые в качестве индукторов фосфорилирования по настоящему изобретению. Например, можно использовать антитела с агонистической активностью, описанные выше.
Индукторы фосфорилирования, содержащие антитело против AXL по настоящему изобретению, можно представить как способы индукции фосфорилирования с использованием антитела против AXL. Индукторы фосфорилирования, содержащие антитело против AXL по настоящему изобретению, можно представить как применение антитела против AXL для получения индуктора фосфорилирования.
Ингибиторы фосфорилирования
Настоящее изобретение также предусматривает ингибиторы фосфорилирования, содержащие антитело против AXL. Ингибиторы фосфорилирования по настоящему изобретению обычно ингибируют фосфорилирование, индуцированное связыванием лиганда AXL (таким как Gas6) с AXL. Хотя нет никаких особых ограничений на цели ингибирования фосфорилирования, данными целями, как правило, являются полипептиды, имеющие тирозин, и предпочтительно являющиеся AXL.
Не существует никаких особых ограничений на антитела, используемые в качестве ингибиторов фосфорилирования по настоящему изобретению. Например, можно использовать антитела с антагонистической активностью, описанные выше.
Ингибиторы фосфорилирования, содержащие антитело против AXL по настоящему изобретению, можно представить как способы ингибирования фосфорилирования с использованием антитела против AXL. Ингибиторы фосфорилирования, содержащие антитело против AXL по настоящему изобретению, можно представить как применение антитела против AXL для получения ингибитора фосфорилирования.
Средства для снижения уровня экспрессии AXL
Настоящее изобретение также предусматривает средства, снижающие уровень экспрессии AXL, содержащие антитело против AXL. Средство, снижающее уровень экспрессии AXL, снижает уровень экспрессии AXL в клетках, экспрессирующих AXL. Не существует никаких особых ограничений на клетки, которые экспрессируют AXL. Примеры таких клеток включают раковые клетки (Calu-1, MDA-MB-231, DU-145, и тому подобное).
Снижение уровня экспрессии AXL может являться снижением уже присутствующего количества AXL, в результате деградации AXL, или тому подобное, либо может являться снижением количества вновь экспрессированного AXL в результате подавления экспрессии AXL.
Средства, снижающие уровень экспрессии AXL, содержащие антитело против AXL по настоящему изобретению, можно представить как способы снижения уровня экспрессии AXL с использованием антитела против AXL. Кроме того, средства, снижающие уровень экспрессии AXL, содержащие антитело против AXL по настоящему изобретению, можно представить как применение антитела против AXL для получения средства, снижающего уровень экспрессии AXL.
Фармацевтические композиции
Данные ингибиторы ангиогенеза, ингибиторы клеточного роста, индукторы фосфорилирования, ингибиторы фосфорилирования, или средства, снижающие уровень экспрессии AXL, по настоящему изобретению, можно вводить либо способами перорального введения, либо способами парентерального введения. Способы парентерального введения особенно предпочтительны. Конкретные примеры таких способов введения включают введение в форме инъекции, трансназальное введение, транспульмонарное введение а также чрескожное введение. Фармацевтические композиции по настоящему изобретению можно вводить системно или локально посредством введения в форме инъекции, например, внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции или тому подобное. Подходящие способы введения также можно выбрать в зависимости от возраста и симптомов больного. Дозировку можно выбрать, например, в диапазоне от 0,0001 мг до 1000 мг на килограмм массы тела за одно введение. В противном случае, дозировку можно выбрать, например, в пределах от 0,001 до 100000 мг/организм в расчете на одного пациента. Однако дозировка фармацевтических композиций по настоящему изобретению не ограничивается только приведенными.
Данные ингибиторы ангиогенеза, ингибиторы клеточного роста, индукторы фосфорилирования, ингибиторы фосфорилирования, или средства для снижения уровня экспрессии AXL по настоящему изобретению можно составить в соответствии с обычными способами (например, Remington's Pharmaceutical Science, самое последнее издание, Mark Publishing Company, Easton, USA), и может включать фармацевтически приемлемые основы или добавки. Примеры основ и добавок включают, без ограничений, поверхностно-активные вещества, наполнители, красители, отдушки, консерванты, стабилизаторы, буферы, суспендирующие средства, изотонические средства, связующие вещества, разрыхлители, смягчающие компоненты, загустители и ароматизаторы. При необходимости можно использовать другие, часто используемые основы. Конкретные примеры таких основ включают двуокись кремния низкой вязкости, лактозу, кристаллическую целлюлозу, маннит, крахмал, кальциевую соль кармеллозы, натриевую соль кармеллозы, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, поливинилацетальдиэтиламиноацетат, поливинилпирролидон, желатин, среднецепочечные триглицериды жирных кислот, полиоксиэтиленовое гидрированное касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганические соли и тому подобное.
Все ссылки ограничительной части патентной формулы, процитированные в данном документе, включены как неотделимая часть во всей полноте.
Примеры
Хотя настоящее изобретение будет более подробно пояснено следующими примерами, настоящее изобретение не ограничивается этими примерами.
[Пример 1]
1-1 Получение антигена
Клетки яичника китайского хомячка (CHO (dhfr-) клетки) трансфицировали вектором экспрессии слитого белка (hAXL-ECD-mIgG2aFc), в котором были гибридизированы внеклеточная область AXL человека и Fc-область IgG2a мыши, и линии клеток СНО, продуцирующие белок hAXL-ECD-mIgG2aFc клонировали с селекцией G418. Культуральный супернатант продуцирующих белок hAXL-ECD-mIgG2aFc линий клеток СНО, собранный с использованием бессывороточной среды (CHO-S-SFM II; Gibco) наносили на колонку с белком G (HiTrap Protein G HP, GE Healthcare), уравновешенную связывающим буфером (20 мМ фосфатный буфер, рН 7,0). После промывки несвязанных белков связывающим буфером, фракции hAXL-ECD-mIgG2aFc белка собирали в буфере элюции (100 мМ глицин-HCl, рН 2,7) в пробирки, содержащие нейтрализующий буфер (1 М Трис-HCl, рН 9,0). Затем, буфер очищенного белка заменяли на фосфат-буферный физиологический раствор (рН 7,35-7,65; Takara Bio) и очищенный белок концентрировали, применяя набор для ультрафильтрации фракций с молекулярной массой 10 кДа (Centricon®, Millipore). Концентрацию очищенного белка рассчитывали по оптической плотности при 280 нм с помощью молярного коэффициента поглощения, вычисленного по расчетной формуле Pace et al. (Prof. Sci. (1995) 4, 2411-2423).
1-2 Получение гибридомы, продуцирующей антитело против AXL
Четыре BALB/с мыши (самцы, 6 недель на начало иммунизации, Charles River Laboratories Japan) и две MRL/lpr мыши (самцы, 6 недель на начало иммунизации, Charles River Laboratories Japan) иммунизировали, как описано ниже, антигеном, полученным в предыдущем разделе (hAXL-ECD-mIgG2aFc белком). Антиген, эмульгированный в полном адъюванте Фрейнда (H37 Ra, Difco Laboratories) вводили подкожно в дозе 40 мкг/голову в качестве первичной иммунизации. Через две недели антиген, эмульгированный в неполном адъюванте Фрейнда (Difco Laboratories), вводили подкожно в дозе 40 мкг/голову. Животных последовательно иммунизировали еще три раза с интервалами в одну неделю. Повышение в сыворотке титра антител в ответ на антиген подтверждали с помощью ELISA, как указано в следующем разделе, за чем следовала окончательная иммунизация внутривенным введением антигена, разбавленного в фосфат-буферном физиологическом растворе (фосфат-буферный солевой раствор без ионов кальция и магния, PBS(-); Nissui Pharmaceutical) в дозе 10 мкг/голову. Через три дня после окончательной иммунизации, клетки селезенки мыши и миеломные клетки мыши P3X63Ag8U.1 (под названием P3U1, АТСС CRL-1597) гибридизировали в соответствии с обычными способами, используя PEG 1500 (Roche Diagnostics). Слитые клетки культивировали в среде RPMI1640 (Invitrogen), содержащей 10% FBS (Invitrogen) (далее обозначаемый как 10% FBS/RPMI1640). На следующий день после слияния слитые клетки суспендировали в полужидкой среде (StemCells) с последующим селективным культивированием и образованием колоний гибридом. Колонии гибридом отбирали из среды на девятый или десятый день после слияния и рассевали в 96-луночный планшет с селективной средой HAT (10% FBS/RPMI1640, 2% по объему HAT 50× концентрат [Dainippon Pharmaceutical] и 5% по объему BM-Condimed H1 [Roche Diagnostics]) по одной колонии на каждую лунку. После культивирования в течение трех-четырех дней супернатант собирали из каждой лунки, и гибридомы со связывающей внеклеточную область AXL человека активностью отбирали путем измерения их связывающей активности с вышеупомянутым антигеном и контрольным белком, слитым с Fc-областью IgG2a мыши, с помощью ELISA, как указано в следующем разделе.
Связывающие активности супернатантов отобранных гибридом приведены в таблице 1.
№ клона
AXL-mFc
FGFR2-mFc
AbΔ
AXL-His
IgG
Гибридомы, отобранные авторами настоящего изобретения, были депонированы в Международном патентном депозитарии организмов Национального института современной технической науки и технологии. Следующий раздел предусматривает описание содержания, точно определяющее депонирование.
(a) Название и адрес учреждения депозитария
Название: International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology
Адрес: Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan 305-8566
(b) Дата приемки (дата депонирования): 5 июля 2007
(c) Номер доступа
AXL № 7 #070402 (Ax7) (номер доступа FERM BP-10850)
AXL № 51 #070406 (Ax51) (номер доступа FERM BP-10851)
AXL № 232 #070406 (Ax232) (номер доступа FERM BP-10855)
AXL № 96 #070402 (Ax96) (номер доступа FERM BP-10852)
AXL № 223 #070402 (Ax223) (номер доступа FERM BP-10853)
AXL № 225 #070402 (Ax225) (номер доступа FERM BP-10854)
AXL № 258 #070402 (Ax258) (номер доступа FERM BP-10856)
AXL № 284 #070402 (Ax284) (номер доступа FERM BP-10857)
AXL № 285 #070402 (Ax285) (номер доступа FERM BP-10858)
AXL № 292 #070411 (Ax292) (номер доступа FERM BP-10859)
1-3 Связывающая активность с AXL человека
Антиген (hAXL-ECD-mIgG2aFc) разбавленный до 1 мкг/мл буфером для сенсибилизации поверхностей (100 мМ бикарбонат натрия [рН 9,6], 0,02% азид натрия) или контрольный белок, гибридизированный с Fc-областью IgG2a мыши, наносили в 96-луночный планшет (Nunc-Immuno™ 96 Micro Well™ MaxiSorp™ plate; Nalge Nunc International) по 80 мкл/лунку, а затем инкубировали, по меньшей мере в течение ночи при 4°C. После того, как трижды промывали фосфатно-буферным раствором, содержащим 0,05% по объему Твин® 20 (tPBS [-]), планшет блокировали по меньшей мере в течение ночи при 4°C буфером разведения (BlockingOne, разведение 1/5; Nacalai Tesque). После удаления буфера, антисыворотку мыши или супернатант гибридомной культуры, разбавленные буфером разведения, добавляли в планшет по 80 мкл/лунку, а затем инкубировали в течение 1 часа при комнатной температуре. После трехкратной промывки планшеты буфером tPBS(-), меченое HRP антитело мыши против IgG (Stressgen), разбавленное 1/5000 буфером разведения, добавляли по 80 мкл/лунку, а затем инкубировали в течение 1 часа при комнатной температуре. После пятикратной промывки планшеты буфером tPBS(-), хромогенный субстрат (Peroxidase Substrate, Kirkegaad & Perry Laboratories) добавляли по 80 мкл/лунку, а затем инкубировали в течение 20 минут при комнатной температуре. После добавления стоп-раствора пероксидазы (Peroxidase Stop Solution, Kirkegaad & Perry Laboratories) по 80 мкл/лунку, измеряли оптическую плотность при 405 нм с помощью микропланшетного ридера Microplate Reader Model 3550 (Bio-Rad Laboratories).
1-4 Очистка антитела из супернатанта культуры гибридом
Полученные в результате гибридомы, описанные выше, выращивали на избирательной среде HAT, применяя FBS с низким содержанием IgG (Invitrogen) в качестве FBS. Частицы с белком G (Protein G beads, Pharmacia), в котором растворитель замещали промывочным буфером (20 мМ натрий-ацетатный буфер, рН 5,0), добавляли к 20-50 мл культурального супернатанта по 50 мкл на 10 мл культурального супернатанта, а затем перемешивали переворачиванием в течение ночи при 4°C. После того как частицы белка G извлекали и промывали промывочным буфером, антитело элюировали буфером элюции (50 мМ натрий-ацетатный буфер, рН 3,3), вслед за чем немедленно нейтрализовали буфером нейтрализации (трис-HCl буфер, рН 7,8). Буфер замещали на фосфат-буферный физиологический раствор (рН 7,35-7,65; Nissui Pharmaceutical), и очищенное антитело концентрировали, применяя набор для ультрафильтрации фракций с молекулярной массой 10 кДа (Amicon®, Millipore), а затем стерилизовали с помощью 0,22 мкм фильтров для стерилизации (Millipore GV, Millipore).
[Пример 2] Анализ антителоиндуцированного фосфорилирования
Тестировали способность моноклональных антител против AXL, полученных в примере 1, индуцировать фосфорилирование AXL в раковых клетках. Клетки (линии Calu-1 клеток немелкоклеточного рака легких человека, линии MDA-MB-231 раковых клеток молочной железы человека, а также линии DU-145 раковых клеток рака предстательной железы человека) рассеивали в 6-луночные планшеты при плотности 4×105 клеток/лунку, и через 24 часа среду замещали средой, из которой была удалена сыворотка (бессывороточная среда), а затем клетки культивировали в течение ночи. Затем, полученное вышеуказанным образом моноклональное антитело против AXL добавляли до 2 мкг/мл, а рекомбинантные GAS6 (R&D) добавляли до 200 нг/мл в качестве положительного контроля, с последующей инкубацией в течение 30 минут при 37°C. Далее клетки промывали PBS(-) и лизировали на льду в течение 30 минут буфером лизиса клеток (137 мМ NaCl, 20 мМ Трис-HCl [рН 8,0], 10% глицерин, 2 мМ ЭДТА, 1 мМ натрий ванадат, 1% по объему NP-40, 1 мМ фенилметилсульфонил фторид [PMSF], 10 мкг/мл апротинин, 10 мкг/мл леупептин, 10 мкг/мл пепстатин). Смесь раствора клеток гомогенизировали ультразвуковым гомогенизатором (Tomy Seiko), а затем центрифугировали (20000×g) в течение 10 минут при 4°C. Супернатант смеси раствора клеток перемешивали в течение 30 минут с 0,05 объемами белок G-агарозы (Roche Diagnostics). После центрифугирования (2300×g) в течение 1 минуты при температуре 4°C 1,2 мкг моноклонального антитела против AXL (R&D) добавляли в супернатант, который встряхивали в течение 1 часа при 4°C. Затем добавляли 10 мкл белок G-агарозы, и раствор встряхивали еще один час при 4°C. После центрифугирования (2300×g) в течение 1 минуты при температуре 4°C иммунопреципитат промывали и суспендировали в буфере для образца NuPAGE-LDS (Invitrogen), а затем прогревали в течение 10 минут при 70°C. Иммунопреципитат подвергали электрофорезу течение одного часа при 150 вольт с использованием 7% NuPAGE (Invitrogen).
После иммунопреципитации и электрофореза на 7% NuPAGE, белок электрофоретически переносили на 0,45 мкм поливинилидендифторидный фильтр (Immobilon-FL, Millipore) в течение 1 часа при 30 мА с буфером переноса NuPAGE (Invitrogen) и буфером, содержащим 20% по объему метанола. Фильтр промывали TBS (50 мМ Трис-HCl [рН 7,6], 150 мМ NaCl), а затем блокировали инкубацией в течение ночи в блокирующем буфере Odyssey (Li-COR). Фильтр промывали 4 раза по 5 минут каждый буфере TEST (TBS, содержащий 0,05% по объему Твин® 20), а затем инкубировали в течение 2 часов при комнатной температуре с биотинилированным антителом 4G10 против фосфотирозина (разбавление буфером TEST 1:1000; Upstate) и с антителом против AXL (разбавление буфером TBST 1:15000; Santa Cruz). После того как фильтр промывали 4 раза по 5 минут каждый буфером TBST, фильтр инкубировали в течение 1 часа с Alexa 680-меченым стрептавидином (Invitrogen), разбавленным буфером TBST 1:10000, и IRDye 800-меченым вторичным антителом против козы (Rockland), разбавленным буфером TBST 1:10000. После того как фильтр промывали три раза по пять минут каждый буфером TBST, его снова промывали один раз в течение пяти минут буфером TBS, а затем сканировали инфракрасной системой формирования изображений Odyssey (Li-COR).
Полученную иммуноблоттингом иммунопреципитированного внутриклеточного AXL с антителом против AXL полосу и полученную иммуноблоттингом его же с антителом против фосфотирозина полосу накладывали друг на друга, и усиление полосы для фосфорилированного по тирозинам AXL наблюдали после добавления моноклональных антител против AXL Ax285, Ax292, Ax223, Ax96 и Ax258, а также после добавления рекомбинантных GAS6, использованных в качестве положительного контроля (фигуры 1 A, B, C, D и E). Таким образом, наблюдали усиление фосфорилирования тирозинов AXL, как результат добавления моноклональных антител против AXL, полученных авторами настоящего изобретения. Таким образом, эти моноклональные антитела против AXL могут индуцировать фосфорилирование киназного домена AXL.
[Пример 3] Анализ ингибирования лиганд-зависимого фосфорилирования антителом
Тестировали способность моноклонального антитела против AXL ингибировать лиганд-зависимое фосфорилирование в раковых клетках. Клетки (линии Calu-1 клеток немелкоклеточного рака легких человека, линии MDA-MB-231 раковых клеток рака молочной железы человека, а также линии DU-145 раковых клеток рака предстательной железы человека) рассеивали в 6-луночные планшеты при плотности 4×105 клеток/лунку, и через 24 часа среду замещали средой, из которой была удалена сыворотка (бессывороточная среда), а затем клетки культивировали в течение ночи. Затем, полученное в примере 1 моноклональное антитело против AXL добавляли до 2 мкг/мл, а затем рекомбинантные GAS6 (R&D) добавляли одновременно до 200 нг/мл и инкубировали в течение 30 минут при 37°C. Далее клетки промывали PBS(-) и белок экстрагировали из клеток ранее описанным буфером лизиса клеток. Продукты лизиса клеток, иммунопреципитированные имеющимся в продаже антителом против AXL (Santa Cruz™), разделяли на 7% NuPAGE (Invitrogen), подвергали иммуноблоттингу в форме вестерн-блоттинга и анализу фосфорилирования тирозинов, как описано выше. Иммунопреципитированный внутриклеточный AXL переносили на блот с антителом против фосфотирозина посредством обработки GAS6, которые являются его лигандом. Однако блот антитела против фосфотирозина ослаблялся моноклональными антителами против AXL Ax7 и Ax51 (фигуры 2 A и B). Таким образом, было подтверждено, что лиганд-зависимое фосфорилирование тирозинов AXL ингибируется под действием моноклональных антител против AXL, полученных авторами настоящего изобретения. Эти моноклональные антитела против AXL могут ингибировать лиганд-зависимое фосфорилирование киназного домена AXL.
[Пример 4] Анализ индукции деактивации белка AXL антителом
Тестировали способность моноклональных антител против AXL индуцировать деактивацию белка AXL в раковых клетках. Клетки (линии Calu-1 клеток немелкоклеточного рака легких человека, линии MDA-MB-231 раковых клеток рака молочной железы человека, а также линии DU-145 раковых клеток рака предстательной железы человека) рассеивали в 6-луночные планшеты при плотности 4×105 клеток/лунку, и через 24 часа среду замещали средой, из которой была удалена сыворотка (бессывороточная среда), а затем клетки культивировали в течение ночи. Затем, полученное вышеуказанным образом моноклональное антитело против AXL добавляли до 2 мкг/мл, а рекомбинантные GAS6 (R&D) добавляли до 200 нг/мл в качестве положительного контроля, с последующей инкубацией в течение 24 часов при 37°C. Далее клетки промывали PBS(-) и белок экстрагировали из клеток ранее описанным буфером лизиса клеток. Продукты лизиса клеток, иммунопреципитированные имеющимся в продаже антителом против AXL (Santa Cruz™), разделяли на 7% NuPAGE (Invitrogen), подвергали иммуноблоттингу в форме вестерн-блоттинга и анализу фосфорилирования тирозинов, как описано выше.
25 мкг каждого раствора белка суспендировали в буфере для образца NuPAGE-LDS (Invitrogen), прогревали в течение 10 минут при 70°C, и подвергали электрофорезу в течение одного часа при 150 вольт на 7% NuPAGE (Invitrogen). Гели, разделенные электрофорезом, электрофоретически переносили на 0,45 мкм поливинилидендифторидный фильтр (Immobilon-FL, Millipore) в течение 1 часа при 30 мА в буфере переноса NuPAGE (Invitrogen) и буфере, содержащем 20% по объему метанола. Фильтр промывали TBS (50 мМ Трис-HCl [рН 7,6], 150 мМ NaCl), а затем блокировали инкубацией в течение ночи в блокирующем буфере Odyssey (Li-COR). Фильтр промывали 4 раза по 5 минут каждый буфере TEST, а затем инкубировали в течение 2 часов при комнатной температуре с антителом против AXL (разбавление 1:15000 буфером TBST; Santa Cruz) и антителом против актина (разбавление 1:5000 буфером TBST). После того как фильтр промывали 4 раза по 5 минут каждый буфером TBST, его инкубировали в течение 1 часа с Alexa 680-меченым вторичным антителом против кролика (Invitrogen), разбавленным буфером TEST 1:10000, и IRDye 800-меченым вторичным антителом против козы (Rockland), разбавленным буфером TEST 1:10000. После того как фильтр промывали три раза по пять минут каждый буфером TEST, его снова промывали один раз в течение пяти минут буфером TBS, а затем сканировали инфракрасной системой формирования изображений Odyssey (Li-COR).
Наблюдали ослабление блотов с AXL после воздействия моноклональных антител против AXL Ax285, Ax292, Ax223, Ax96, Ax258, Ax284, Ax7, и Ax225 (фигуры 3 A, B, C, D, E, F, G и H). Следовательно, эти моноклональные антитела против AXL могут индуцировать деактивацию белка AXL.
[Пример 5] Ингибирующая ангиогенез in vitro активность антитела против AXL
Активность антитела против AXL ингибировать образование полостей клеток (люменов) у эндотелиальных клеток пупочной вены человека (HUVEC) измеряли с помощью ангиогенезного набора, полученного от Kurabo Industries. Порядок проведения эксперимента находился в соответствии с протоколом, приложенным к набору, и приводится ниже. HUVEC и фибробласты культивировали совместно, и 24-луночный планшет (входящий в набор), содержащий клетки на ростовой стадии раннего образования люмена помещали в инкубатор на 3 часа при температуре 37°C в атмосфере 5% CO2 и увлажненного воздуха. Крышки трех контейнеров, содержащих 25 мл среды специального назначения (входит в набор), ослабляли и помещали в термостат на 30 минут при 37°С в атмосфере 5% CO2 и увлажненного воздуха. Планшет затем удаляли из инкубатора, и покрывающий лунки лист снимают. Крышку планшета заменяли новой (входит в набор). Убеждались, что клетки нормальны, наблюдая их под микроскопом. Культуральную среду (> 12 мл/планшет), нагретую до 37°C, распределяли в пробирки Falcon, и VEGF-A (2 мкг/мл) добавляли к среде до конечной концентрации 10 нг/мл при 200-кратном разведении. Антитело против AXL, полученное, как описано выше, добавляли в пробирки до конечной концентрации 10 мкг/мл. PBS(-) использовали вместо антитела для отрицательного контроля. Среду в лунках 24-луночного планшета аккуратно отсасывали, и затем аккуратно добавляли 500 мкл содержащей лекарственное средство среды. Состояние клеток изучали микроскопированием, а затем их возвращали в инкубатор. Среду заменяли, используя ту же процедуру, в дни 4, 7 и 9, если считать день, в который добавляли антитело, как день 1.
Слой клеток фиксировали и окрашивали с помощью набора для окрашивания люмена (Kurabo) на 11-й день после добавления антитела. Процедуру проводили в соответствии с протоколом, поставляемым вместе с набором, и приводимым ниже. После наблюдения клеток под микроскопом, среду отсасывали и лунку промывали добавлением 1 мл промывочного буфера (PBS(-) рН 7,4; Sigma) в каждую лунку, а затем промывочный буфер удаляли отсасыванием. 1 мл ледяного фиксирующего раствора (70% этанол), добавляли в каждую лунку и выдерживали в течение 30 минут при комнатной температуре. Фиксирующий раствор затем удаляли, 1 мл блокирующего раствора добавляли в каждую лунку, хорошо промывали, и блокирующий раствор отсасывали. 0,5 мл первичного антитела, поставляемого вместе с набором, разводили в соответствии с протоколом, и добавляли в каждую лунку, с последующей инкубацией в течение 1 часа при 37°C. Первичное антитело отсасывали, и каждую лунку промывали 3 раза 1 мл блокирующего раствора (PBS(-), содержащий 1% БСА, рН 7,4; Sigma). 0,5 мл вторичного антитела, поставляемого вместе с набором и разбавленного в соответствии с протоколом, добавляли в каждую лунку, с последующей инкубацией в течение 1 часа при 37°C. Вторичное антитело отсасывали, и каждую лунку промывали 3 раза 1 мл дистиллированной воды. 0,5 мл раствора субстрата, поставляемого вместе с набором, добавляли в каждую лунку, с последующей инкубацией в течение 10-30 минут при 37°C, пока люмен не станет темно-пурпурным. Затем раствор субстрата отсасывали, и каждую лунку промывали 3 раза 1 мл дистиллированной воды и высушивали на воздухе. Микроскопические изображения каждой фиксированной клетки фиксировали по пяти позициям CCD-камерой (Nikon Digital Camera, dxm1200), а площади сосудов подсчитывали с использованием программного обеспечения для количественного описания ангиогенеза (версия 1.0, Kurabo).
Коэффициент снижения площади образовавшихся в люмене сосудов в лунках, к которым добавляли антитело против AXL, относительно площади образовавшихся в люмене сосудов в лунках, к которым добавляли PBS(-) в качестве отрицательного контроля, использовали как показатель ингибирующей активности антител, а Ax232, Ax292, Ax285 и Ax284 показали ингибирующие активности (фигура 4).
[Пример 6] Связывающая активность антитела против AXL с AXL мыши
После того, как внеклеточный домен AXL мыши (далее обозначаемый mAXL-ECD; R&D) растворяли в буфере для сенсибилизации поверхностей (100 мм натрий-бикарбонатный буфер, рН 9,6) до 2 мкг/мл, по 100 мкл разливали в 96-луночный планшет (Nunc-Immuno™ 96 MicroWell™ MaxiSorp™ планшеты; Nalge Nunc International). После того, как планшет помещали в холодильник на ночь, раствор антител в планшете удаляли, разливали по 200 мкл/лунку буфера для разбавления (BlockingOne; Nacalai Tesque), а затем блокировали в течение двух часов при комнатной температуре. После удаления буфера для разбавления, полученное ранее антитело против AXL разбавляли до 3 мкг/мл в буфере для разбавления, разливали по 100 мкл/лунку, и оставляли на 1,5 часа при комнатной температуре. После удаления раствора антител лунки промывали три раза с помощью tPBS(-). Меченую смесь антител, содержащую меченое щелочной фосфатазой антитело козы против IgG1 мыши, меченое щелочной фосфатазой антитело козы против IgG2a мыши и меченое щелочной фосфатазой антитело козы против IgG2b мыши (SouthernBiotech), получали с конечным разведением каждого антитела 1/2250:1/4000:1/4000, разливали по 100 мкл/лунку, и оставляли на 1 час при комнатной температуре. После удаления раствора антител лунки промывали три раза tPBS(-). Разливали по 100 мкл/лунку раствора хромогенного субстрата щелочной фосфатазы (BluePhos Microwell Phosphatase Substrate System, Kirkegaad & Perry Laboratories), с последующим развитием окрашивания при комнатной температуре. Оптическую плотность при 650 нм измеряли с помощью микропланшетного ридера (Emax, Molecular Devices).
Связывание с AXL мыши подтверждено для Ax96, Ax119, Ax223, Ax225 и Ax284.
[Пример 7] Ингибирующая активность in vitro роста раковой клетки антитела против AXL
Оценку проводили, используя HCT-116 (CCL-247), Calu-1 (HTB-54), DU-145 (HTB-81) и T-47D (HTB-133), приобретенные у ATCC, а также AsPC-1, MDA-MB-231 и PANC-1, приобретенные у Dainippon Sumitomo Pharma. Клетки поддерживали в соответствии с условиями, рекомендованными поставщиком каждой клетки. Изготавливали серии разведений антитела против AXL, как описано выше, с 10% FBS/RPMI1640, и разливали по 20 мкл в 96-луночный планшет (с плоским дном). Каждую из суспензий HCT-116, Calu-1, DU145, T-47D, AsPC-1, MDA-MB-231 и PANC-1 клеток изготавливали при 2000, 3000, 2000, 5000, 3000, 5000 и 3000 клеток на лунку, соответственно, и 180 мкл суспензии клеток добавляли в каждую лунку, а затем культивировали в термостате при 37°С в атмосфере 5% CO2. Четыре дня спустя, в каждую лунку добавляли по 10 мкл WST-8 (Cell Counting Kit-8, Dojindo Laboratories), и измеряли поглощение при 450 нм с помощью микропланшетного ридера (Model 3550-UV, Bio-Rad), в соответствии с протоколом, поставляемым вместе с набором. Клеточную ингибирующую активность антитела против AXL (в %) рассчитывали, присвоив значение 0% ингибирования измеряемой величине, когда не включено никакое исследуемое вещество, и присвоив значение 100% ингибирования измеряемой величине, когда не включены ни исследуемое вещество, ни клетки.
Ax51 продемонстрировали CGI активность (ингибирующую клеточный рост активность) 30% или более в отношении HCT116 клеткам.
[Пример 8] Измерение противоопухолевых эффектов антитела против AXL у модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека
1. Подготовка модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека
Линию клеток аденокарциномы поджелудочной железы человека PANC-1, приобретенную у Dainippon Pharmaceutical (в настоящее время - Dainippon Sumitomo Pharma), доводили до плотности 5×107 клеток/мл с помощью сбалансированного солевого раствора Хенкса (HBSS). 200 мкл суспензии клеток (1×107 клеток/мышь) подкожно трансплантировали в паховую область CAnN.Cg-Foxnl<nu>/CrlCrlj nu/nu (BALB-nu/nu) мышей, приобретенных у Charles River Laboratories, Япония. Мышь подвергали эксперименту, когда опухоль достигала объема около 210 мм3.
2. Подготовка и введение антитела
Антитела, приведенные в таблице 1, доводили до концентрации 2 мг/мл с помощью PBS и вводили два раза в неделю в течение 2 недель в дозе 20 мг/кг в брюшную полость мышей с трансплантированной аденокарциномой поджелудочной железы человека. В качестве отрицательного контроля таким же образом вводили PBS. Gemzar (Eli Lilly Japan) доводили до концентрации 12 мг/мл в физиологическом растворе, в качестве положительного контроля и вводили внутрибрюшинно по 120 мг/кг дважды в неделю в течение 2 недель.
3. Оценка противоопухолевых эффектов
Противоопухолевые эффекты у модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека рассчитывали как подавляющие рост опухоли эффекты путем сравнения роста опухолей в группе, обработанной антителом, с ростом опухолей в отрицательной контрольной группе через четыре дня после окончательного введения (фигура 5).
Подавляющий рост опухолей эффект (%)=(1-число роста опухолей в группе, обработанной антителом/число роста опухолей в контрольной группе)×100
4. Статистическая обработка
Объем опухоли представляли в виде среднего значения ± стандартное отклонение. Статистический анализ состоял в сопоставлении контрольной группы и опытной группы методом наименьшей достоверной разности (LSD), применяя пакет программ программного обеспечения для обработки статистических данных (SAS) доклинических исследований, версию 5.0. Достоверность 95% (*: р<0,05), определяли, как представляющее статическую значимость.
5. Результаты
Все антитела тормозили рост опухоли и продемонстрировали противоопухолевые эффекты (фигура 5).
[Пример 9] Измерение противоопухолевых эффектов антитела против AXL у модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека (2)
1. Подготовка модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека
Линию клеток аденокарциномы поджелудочной железы человека PANC-1, приобретенную у Dainippon Pharmaceutical (в настоящее время - Dainippon Sumitomo Pharma), доводили до плотности 5×107 клеток/мл с помощью HBSS. 200 мкл суспензии клеток (1×107 клеток/мышь) подкожно трансплантировали в паховую область CAnN.Cg-Foxnl<nu>/CrlCrlj nu/nu (BALB-nu/nu) мышей, приобретенных у Charles River Laboratories, Япония. Мышь подвергали эксперименту, когда опухоль достигала объема около 270 мм3.
2. Подготовка и введение антитела
Антитело против AXL доводили до концентрации 2 мг/мл с помощью PBS и вводили в брюшную полость мышей с трансплантированной аденокарциномой поджелудочной железы человека два раза в неделю в течение 2 недель в дозе 20 мг/кг. В качестве отрицательного контроля таким же образом вводили PBS. Gemzar (Eli Lilly Japan) доводили до концентрации 12 мг/мл в физиологическом растворе для использования в качестве положительного контроля и вводили внутрибрюшинно дважды в неделю в течение 2 недель по 120 мг/кг.
3. Оценка противоопухолевых эффектов
Противоопухолевые эффекты у модели на мышах с трансплантированной аденокарциномой поджелудочной железы человека рассчитывали как подавляющие рост опухоли эффекты путем сравнения роста опухолей в группе, обработанной антителом, с ростом опухолей в отрицательной контрольной группе через четыре дня после окончательного введения.
Подавляющий рост опухолей эффект (%)=(1-число роста опухолей в группе, обработанной антителом/число роста опухолей в контрольной группе)×100
4. Результаты
Результаты подавления роста опухоли показаны на фигуре 6. Подавляющий эффект роста опухолей (в %) меньший, чем 30%, обозначен как «-», около 30% или более - обозначен как «+», а около 60% или более - обозначен как «++». Результаты анализа ингибирования антителом лиганд-зависимого фосфорилирования из примера 3 также показаны на фигуре 6.
Антитела, которые связываются с FND-1, продемонстрировали 60% или более активности ингибирования роста опухолей (TGI), даже если введение начинали в то время, когда средние объемы опухолей составляли около 270 мм3. Этот результат, что антитела против AXL, которые связываются с FND-1, имеют такие значительные противоопухолевые эффекты in vivo, впервые обнаружен в этом исследовании, и был совершенно неожиданным.
Кроме того, существование антител против AXL, связывающихся с IgD2, что демонстрирует эффект ингибирования фосфорилирования и противоопухолевый эффект in vivo, как указано в примерах 3, 8 и 9, также обнаружен впервые в этом исследовании, и также был совершенно неожиданным.
[Пример 10] Связывающая активность с AXL-FND1 человека и с AXL-IgD2 человека
1. Связывающая активность с AXL-FND1 человека и с AXL-IgD2 человека
Тестировали связывающие активности моноклонального антитела против AXL с доменом 1 AXL-фибронектина типа 3 (AXL-FND1) и доменом 2 AXL из семейства иммуноглобулинов (AXL-IgD2).
2. Подготовка векторов экспрессии рекомбинантного AXL-FND1 человека и рекомбинантного AXL-IgD2 человека
Рекомбинантный AXL-FND1 человека получали амплификацией с помощью ПЦР эквивалента области с 225-й по 331-ю аминокислоту из полноразмерной кДНК AXL человека (O'Bryan, et al., Mol. Cell. Biol. (1991) 11, 5016-5031) (GenBank № NM 021913), клонированием амплифицированных продуктов в pET-41a(+) (Novagen) для экспрессии слитых белков с GST-таговым маркером, и созданием pET-AXL-FND1. Другие домены получали путем амплификацией с помощью ПЦР эквивалента области со 137-й по 224-ю аминокислоту, и клонированием амплифицированных продуктов в pET-41a(+) для экспрессии слитых белков с GST-таговым маркером. Каждый из полученных векторов (5 мкл) трансформировали в DHScc (Toyobo Co., Ltd., Cat. № DNA-903) способом теплового шока, а затем культивировали в среде SOC. Селекцию колоний проводили после культивирования в течение ночи при 37°C на чашках Петри с LB, содержащей канамицин.
3. Очистка рекомбинантного AXL-FND1 человека и рекомбинантного AXL-IgD2 человека
Каждую из полученных колоний пре-культивировали в течение ночи при 37°C в 20 мл LB среды, содержащей канамицин, а затем переводили в 500 мл среды. Каждую колонию культивировали до значения A600 около 0,5±0,05 и добавляли IPTG до концентрации 0,5 мМ. После культивирования в течение одного часа при температуре 37°C, бактериальные клетки собирали и суспендировали в буфере A (50 мМ Трис-HCl (рН 8,0), 1 мМ ЭДТА, 0,5 мМ PMSF и 1 мМ DTT). Замораживание и оттаивание повторяли дважды с использованием жидкого азота. Затем добавляли NP-40 до 0,5%, и клетки гомогенизировали с помощью ультразвукового гомогенизатора (30 сек × 5) и центрифугировали в течение 30 минут при 204000×g, а затем собирали супернатант.
Рекомбинантный AXL-FND1 человека очищали, описанным ниже образом, используя полученный в результате супернатант. Солюбилизированный супернатант E. coli смешивали с быстропроточной глутатион-сефарозой 4 (Glutathione Sepharose™ 4 Fast Flow, GE Healthcare) и перемешивали в течение 1 часа при 4°С с помощью ротатора. После центрифугирования в течение 5 минут при 500×g, супернатант отбрасывали, и глутатион-сефарозу 4B промывали, добавляя буфер А. Эту процедуру промывки повторяли три раза. После перенесения рекомбинантного AXL-FND1 человека с промытой быстропроточной глутатион-сефарозы 4 на мини-колонку, его отделили и элюировали из быстропроточной глутатион-сефарозы 4 с помощью 50 мМ Трис-HCl (рН 7,5) и 25 мМ глутатиона. Каждый из других доменов AXL экспрессировали, разделяли и элюировали таким же образом.
4. Оценка связывающей активности антитела против AXL с AXL-FND1 с помощью вестерн-блоттинга
Рекомбинантные AXL-FND1, отделенные и элюированные с быстропроточной глутатион-сефарозы 4, равно как и AXL-IgD1, AXL-IgD2, AXL-FND2, AXL-IgD1+IgD2, AXL-IgD2+FND1 и AXL-FND1+FND2 количественно анализировали с помощью белкового анализа BIO-RAD DC. По 1 мкг каждого смешивали с буфером для образца NuPAGE® Sample Buffer (Invitrogen), и подвергали электрофорезу в NuPAGE® 10% Bis-TrisGel. Электрофорезный гель переносили на PVDF мембрану Immobilon™-FL (Millipore). PVDF мембрану, содержащую перенесенный белок, блокировали буфером Odyssey® Blocking Buffer (LI-COR) и погружали в раствор первичных антител, в котором антитело против AXL разбавляли до 5 мкг/мл и инкубировали в течение ночи при 4°C. PVDF мембрану, содержащую перенесенный белок, и погруженную в раствор первичных антител, промывали 4 раза по 5 минут в 0,1% TBS-T (TBS (Tris-Buffered Saline (Takara)) с 0,1% твин-20). PVDF мембрану, погруженную в антитело против AXL, погружали в раствор вторичных меченых Alexa Fluor® 680 антител козы против IgG (H+L) мыши (Invitrogen), разбавленных до 80 нг/мл, и инкубировали в течение 1 часа при комнатной температуре. После промывки PVDF мембраны, погруженной в раствор вторичных антител, три раза по пять минут каждый в 0,1% TBS-T, мембраны промывали в течение пяти минут в TBS-T с 0,01% SDS, а затем промывали в течение пяти минут в TBS. Связывание промытой PVDF мембраны оценивали сканированием с помощью система формирования изображения в дальней инфракрасной области спектра Odyssey®.
5. Результаты
Результаты оценки представлены на фигуре 6.
Было показано, что антитело против AXL, продуцируемое гибридомой, с депозитарным номером FERM BP-10854 (Ax225), распознает FND1 из AXL (фигура 6). Было признано, что антитело против AXL, продуцируемое гибридомой, с депозитарным номером FERM BP-10857 (Ax284), распознает FND1 и IgD2 из AXL (фигура 6). Было показано, что антитело против AXL, продуцируемое гибридомой, с депозитарным номером FERM BP-10850 (Ax7), и антитело против AXL, продуцируемое гибридомой, с депозитарным номером FERM BP-10851 (Ax51), распознают IgD2 из AXL (фигура 6).
[Пример 11] Измерение противоопухолевых эффектов антитела против AXL у модели на мышах с трансплантированным раком молочной железы человека
1. Подготовка модели на мышах с трансплантированным раком молочной железы человека
Линию клеток рака молочной железы человека MDA-MB-435S, полученную от ATCC, доводили до плотности 5×107 клеток/мл с помощью HBSS. 200 мкл суспензии клеток (1×107 клеток/мышь) подкожно трансплантировали в паховую область CAnN.Cg-Foxnl<nu>/CrlCrlj nu/nu (BALB-nu/nu) мышей, приобретенных у Charles River Laboratories, Япония. Мышь использовали в эксперименте, когда опухоль достигала объема около 200 мм3.
2. Подготовка и введение антитела
Антитело против AXL доводили до концентрации 2 мг/мл с помощью PBS и вводили в брюшную полость мышей с трансплантированным раком молочной железы человека два раза в неделю в течение 2 недель в дозе 20 мг/кг. В качестве отрицательного контроля таким же образом вводили PBS.
3. Оценка противоопухолевых эффектов
Противоопухолевые эффекты у модели на мышах с трансплантированным раком молочной железы человека рассчитывали как подавляющие рост опухоли эффекты путем сравнения роста опухолей в группе, обработанной антителом, с ростом опухолей в отрицательной контрольной группе через четыре дня после окончательного введения.
Подавляющий рост опухолей эффект (%)=(1-число роста опухолей в группе, обработанной антителом/число роста опухолей в контрольной группе)×100
4. Статистическая обработка
Объем опухоли представляли в виде среднего значения ± стандартное отклонение. Статистический анализ состоял в сопоставлении контрольной группы и опытной группы методом LSD, применяя пакет программ программного обеспечения для обработки статистических данных (SAS) доклинических исследований, версию 5.0. Достоверность 95% (*: р<0,05), определяли, как представляющее статическую значимость.
5. Результаты
Использованные антитела против AXL подавляли рост опухолей и продемонстрировали противоопухолевые эффекты (фигура 7). Следовательно, ожидается, что антитела против AXL, связывающиеся с FND1, имеют противоопухолевые эффекты против различных опухолей.
[Пример 12] Анализ последовательности кДНК антитела
1. Получение векторов экспрессии химерного антитела
Из клеток гибридомы, с депозитарным номером FERM BP-10854 (Ax225), экстрагировали общую РНК, применяя набор RNeasy Mini Kit (Qiagen), и кДНК синтезировали, применяя набор для амплификации кДНК SMART RACE cDNA Amplification Kit (BD Biosciences). Ген вариабельной области антитела выделяли, проведя ПЦР с ДНК-полимеразой PrimeSTAR HS DNA Polymerase (Takara), используя следующие праймеры (H-цепь, MHCgl; L-цепь, MLCk), которые были настроены для соответствующих константных областей антител, а также 10-кратную смесь универсальных праймеров, поставляемых с набором SMART RACE cDNA Amplification Kit (BD Biosciences).
MHCg1: 5'-GGGCCAGTGGATAGACAGATG-3' (SEQ ID NO:1)
MLCk: 5'-GCTCACTGGATGGTGGGAAGATG-3' (SEQ ID NO:2)
Нуклеотидную последовательность каждого выделенного фрагмента ДНК определяли с использованием набора BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) с помощью автоматических секвенаторов ДНК ABI PRISM 3730×L DNA Sequencer или ABI PRISM 3700 DNA Sequencer (Applied Biosystems) в соответствии со способом, описанным в инструкции.
2. Результаты
Вариабельная область тяжелой цепи аминокислотной последовательности полученного в результате антитела мыши AXL225 показана в SEQ ID NO:3, CDR1 этой области показан в SEQ ID NO:4, CDR2 показан в SEQ ID NO:5, а CDR3 показан в SEQ ID NO:6. Вариабельная область легкой цепи аминокислотной последовательности полученного в результате антитела мыши AXL225 показана в SEQ ID NO:7, CDR1 этой области показан в SEQ ID NO:8, CDR2 показан в SEQ ID NO:9 и CDR3 показан в SEQ ID NO:10.
Промышленная применимость
Авторы настоящего изобретения впервые обнаружили, что антитела против AXL оказывают подавляющий ангиогенез эффект и эффект подавления рака. Антитело против AXL по настоящему изобретению является пригодным в качестве ингибитора ангиогенеза и в качестве ингибитора клеточного роста. Используя антитело по настоящему изобретению, также можно индуцировать или ингибировать фосфорилирование AXL. Кроме того, используя антитело по настоящему изобретению, можно снизить уровень экспрессии AXL.
| название | год | авторы | номер документа |
|---|---|---|---|
| AXL-АНТИТЕЛА | 2008 |
|
RU2506276C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО И ПРОДУЦИРУЮЩАЯ ЕГО ГИБРИДОМА | 2004 |
|
RU2312109C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ AXL | 2010 |
|
RU2571224C2 |
| НАПРАВЛЕННЫЕ НА ОПУХОЛЬ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ FZD10 И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2412203C2 |
| Антитело, связывающееся с карбоангидразой, и его применение | 2017 |
|
RU2727682C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2567657C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2603742C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2598258C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2597971C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2012 |
|
RU2610428C2 |





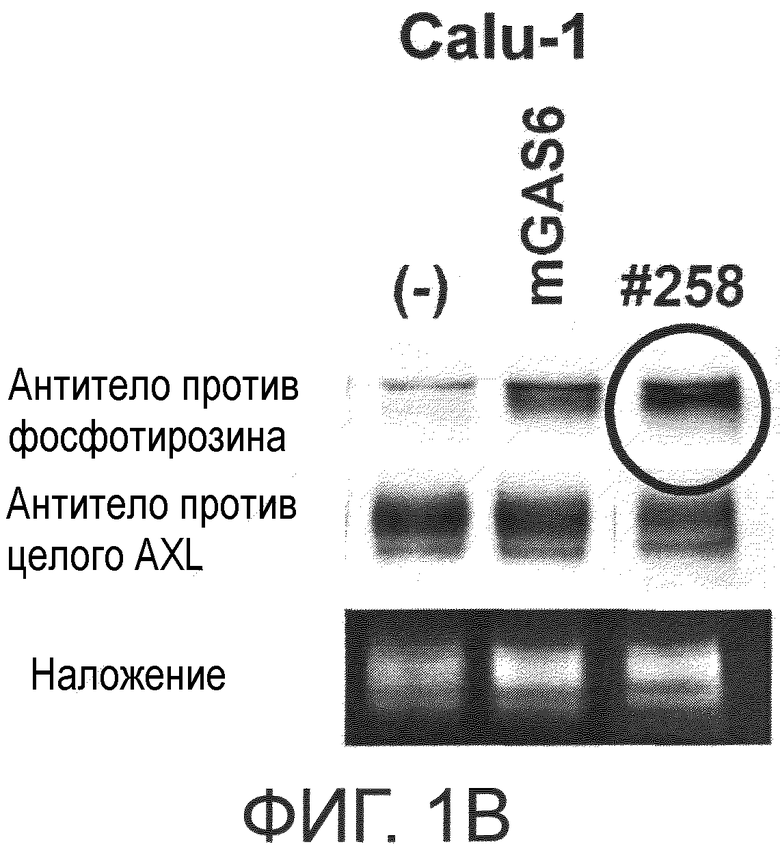
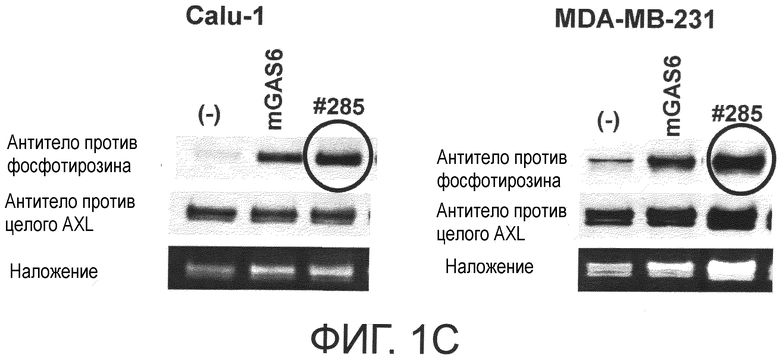
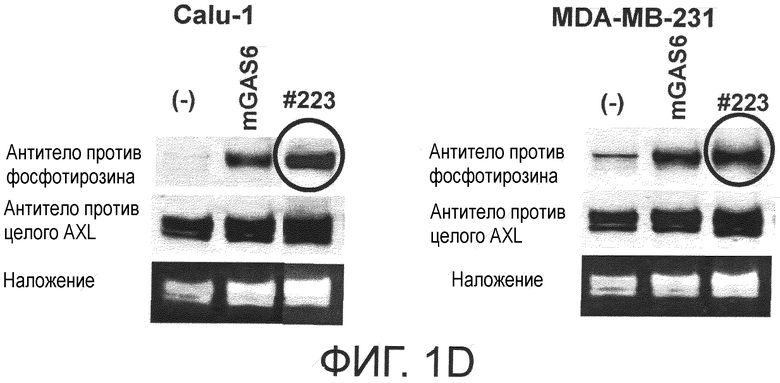
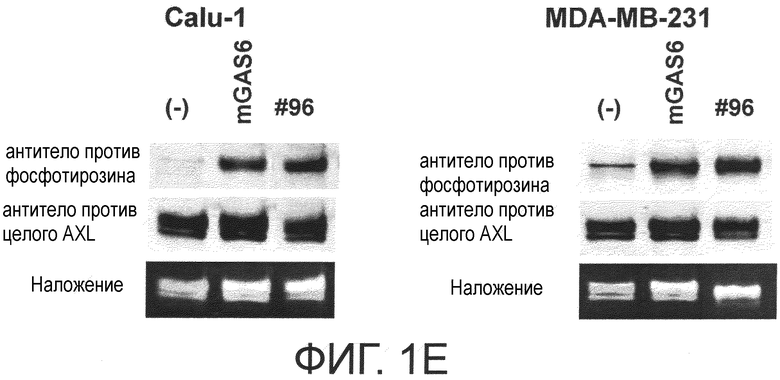
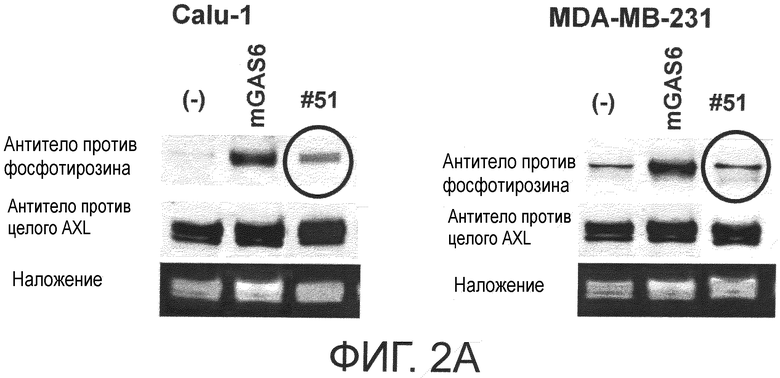
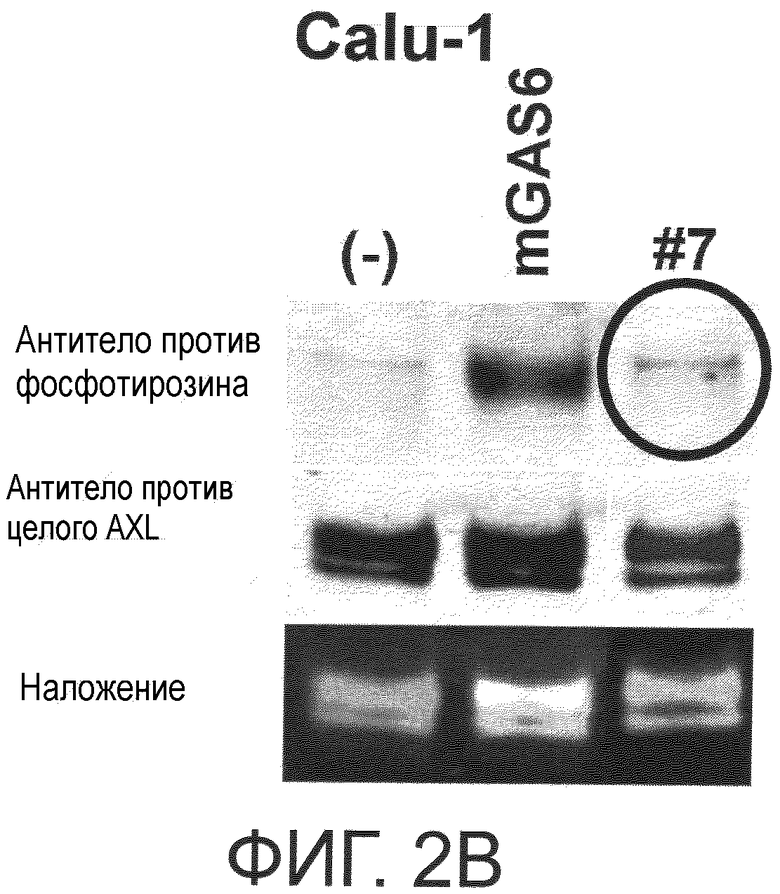
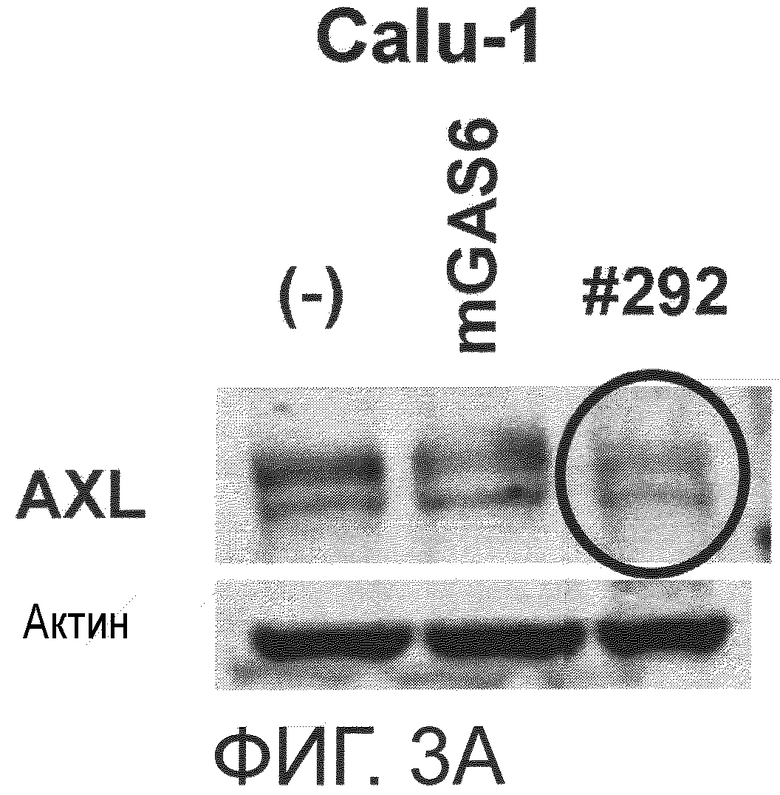
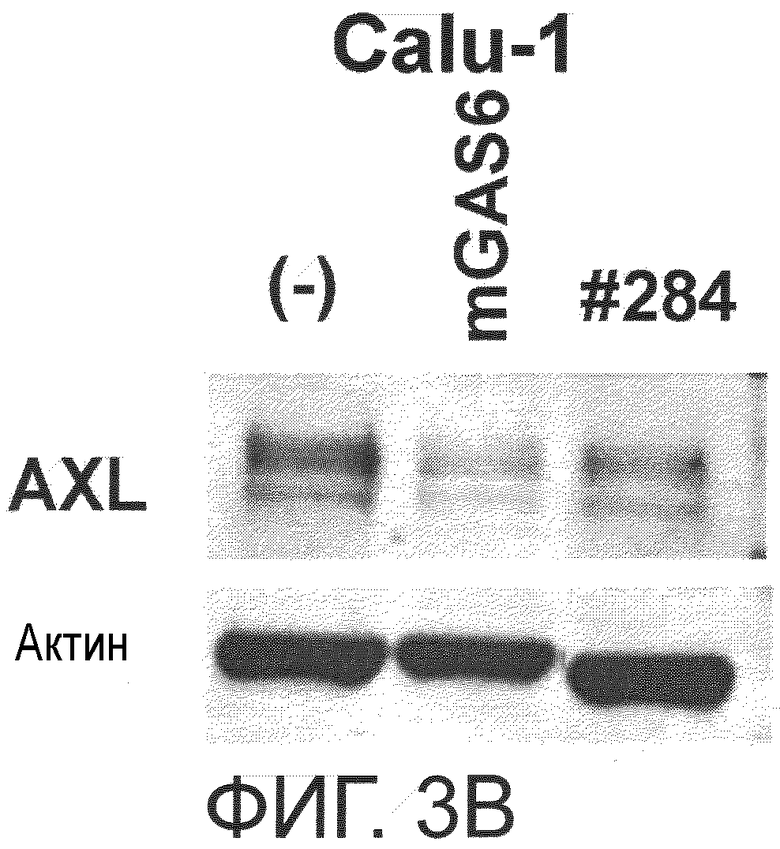
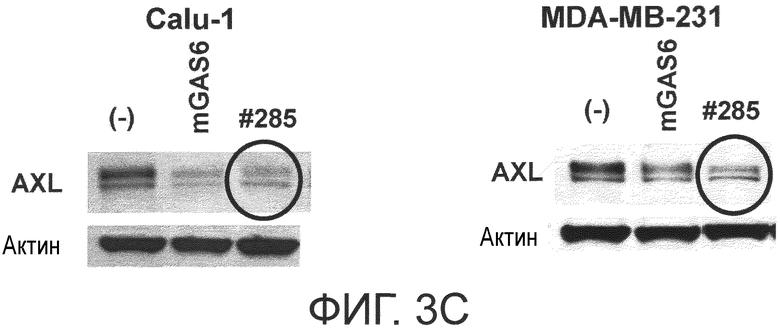
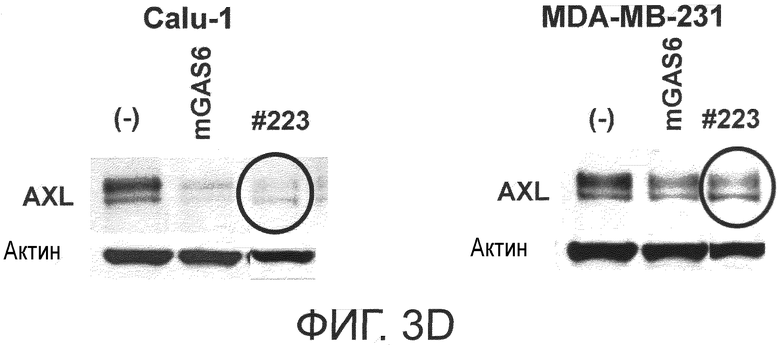
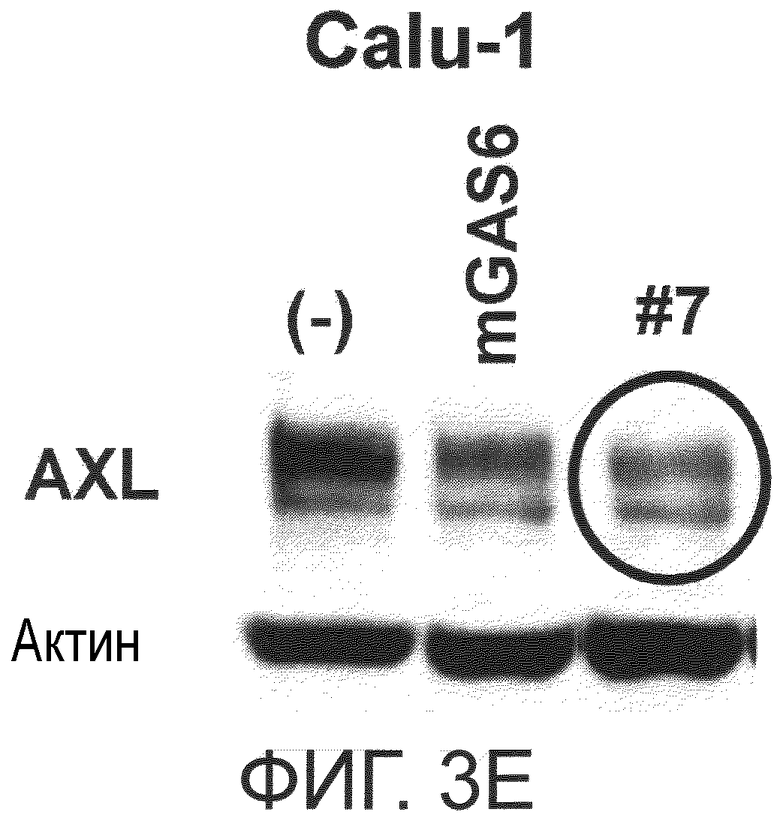
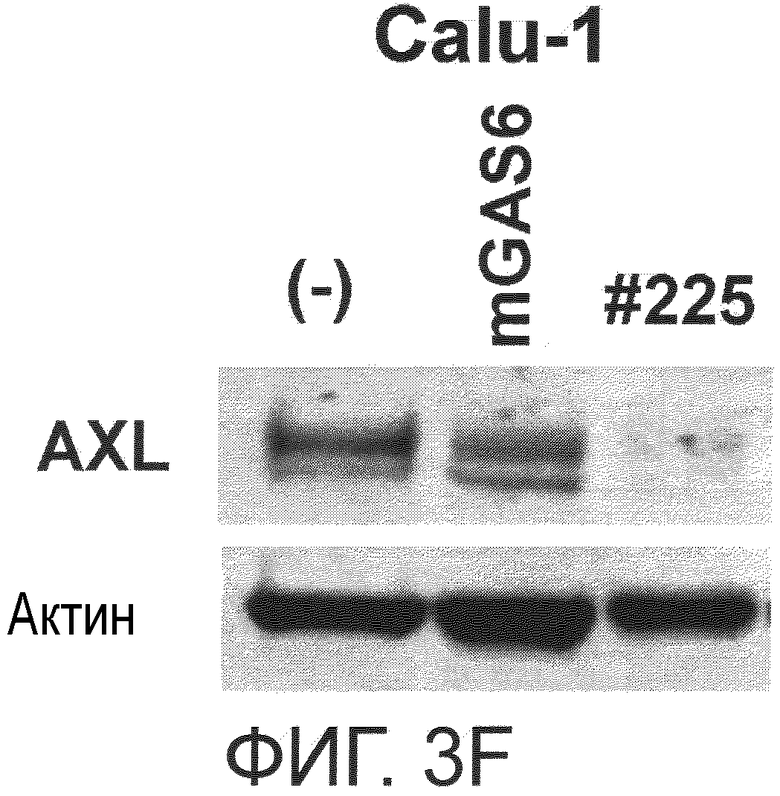
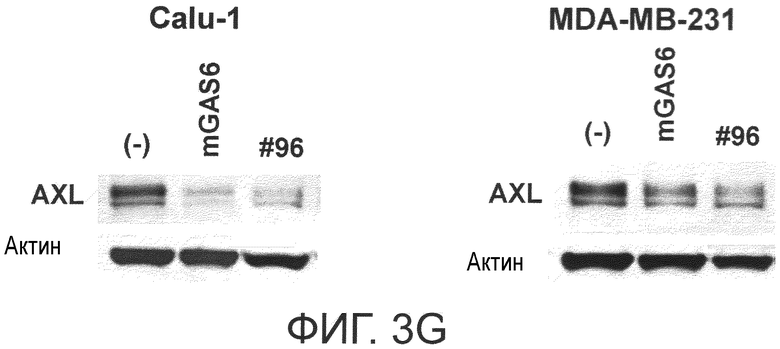
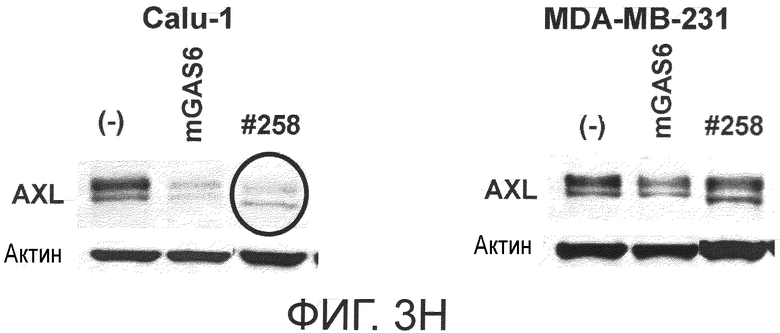

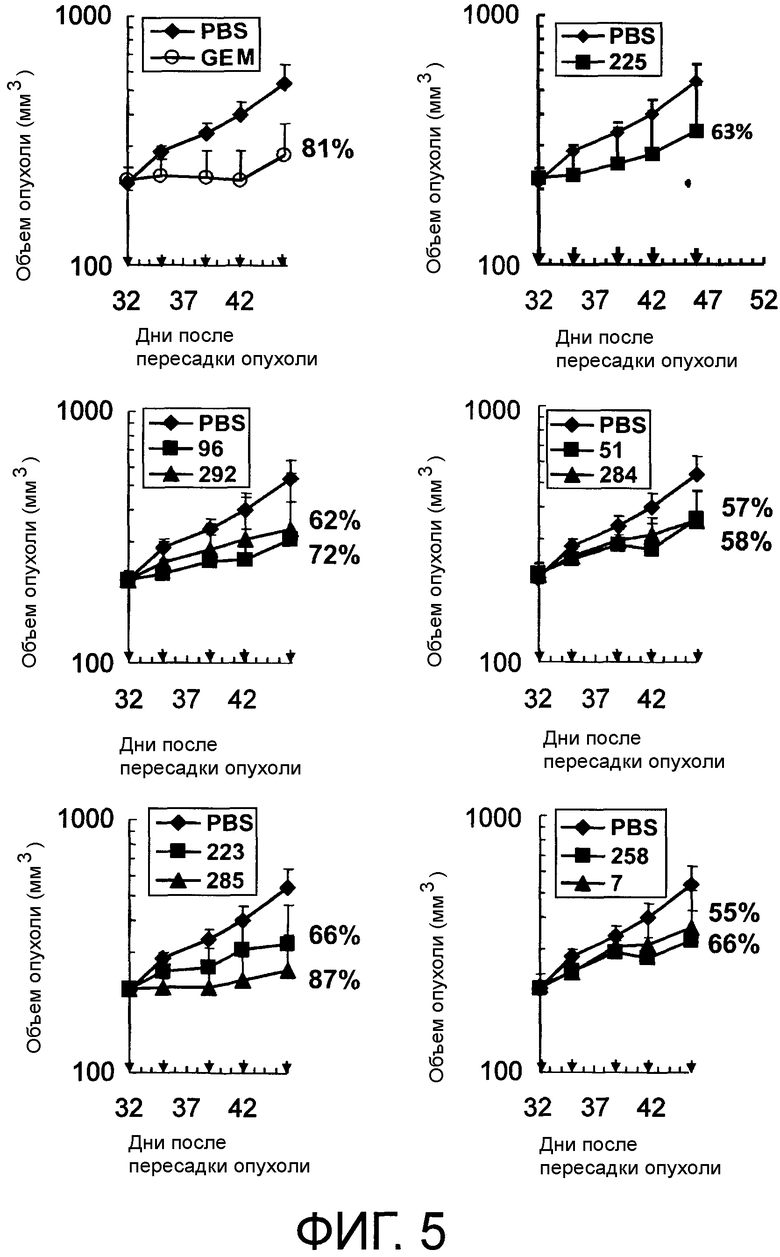
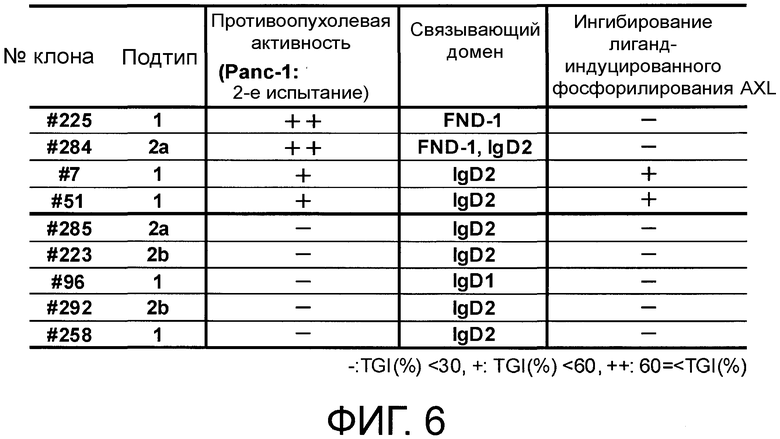
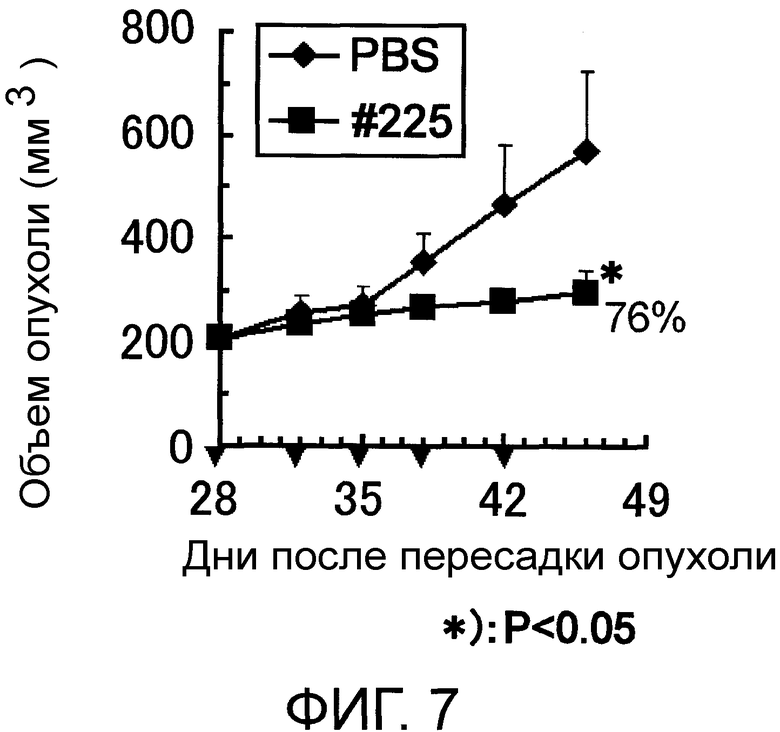
Данное изобретение относится к области биотехнологии. Предложены антитела, которые связываются с фибронектиновым доменом III типа 1 (FND1) белка AXL и вызывают деактивацию AXL, а также применение таких антител для подавления роста злокачественной опухоли, опосредованной AXL, продуцирующие их гибридомы и композиция для подавления роста злокачественной опухоли, опосредованной AXL, которая в качестве активного ингредиента содержит антитело по настоящему изобретению. 7 н. и 5 з.п. ф-лы, 7 ил., 2 табл., 12 пр.
1. Применение моноклонального антитела, которое связывается с фибронектиновым доменом III типа 1 (FND1) AXL и обладает активностью, которая индуцирует деактивацию белка AXL для подавления роста злокачественной опухоли, опосредованной AXL.
2. Антитело, которое связывается с FND1 AXL, по любому из следующих подпунктов (а)-(b):
(a) антитело (Ах284), полученное из гибридомы с депозитарным номером FERM BP-10857; и
(b) антитело (Ах225), полученное из гибридомы с депозитарным номером FERM BP-10854.
3. Применение антитела IgG против AXL, которое связывается с FND1 и блокирует связывание любого антитела по п. 2 с белком AXL на 50% по сравнению со связывающей активностью, полученной в контрольном тесте, проведенном в отсутствии заявленного антитела, для подавления роста злокачественной опухоли, опосредованной AXL.
4. Антитело, которое специфически связывается с FND1 AXL, в котором последовательности CDR1, 2 и 3 тяжелой цепи представляют собой SEQ ID NO: 4, 5 и 6, и последовательности CDR1, 2 и 3 легкой цепи представляют собой SEQ ID NO: 8, 9 и 10 соответственно.
5. Антитело по п.4, которое является химерным антителом.
6. Антитело по п.4, которое является гуманизированным антителом.
7. Гибридома, депонированная под депозитарным номером FERM ВР-10857 (Ах284), продуцирующая антитело против AXL, связывающееся с FND1.
8. Гибридома, депонированная под депозитарным номером FERM ВР-10854 (Ах225), продуцирующая антитело против AXL, связывающееся с FND1.
9. Композиция для подавления роста злокачественной опухоли, опосредованной AXL, которая в качестве активного ингредиента содержит антитело, описанное в любом из пп.1-6, где количество активного ингредиента составляет от 0,0001 мг до 1000 мг/кг.
10. Композиция для подавления роста злокачественной опухоли по п.9, где злокачественной опухолью является рак поджелудочной железы, рак желудка, рак легких, остеосаркома, рак толстой кишки, рак предстательной железы, меланома, рак эндометрия, рак яичников, лейомиома матки, рак щитовидной железы, рак стволовых клеток, рак молочной железы, рак мочевого пузыря, рак почки, глиома, нейробластома или рак пищевода.
11. Композиция для подавления роста злокачественной опухоли по п.9, где злокачественной опухолью является глиома, рак желудка, рак эндометрия, немелкоклеточный рак легкого, аденокарцинома поджелудочной железы или рак молочной железы.
12. Композиция для подавления роста злокачественной опухоли по п.9, где злокачественной опухолью является аденокарцинома поджелудочной железы или рак молочной железы.
| Кипятильник для воды | 1921 |
|
SU5A1 |
| LAY J.-D | |||
| et al., "Sulfasalazine Suppresses Drug Resistance and Invasiveness of Lung Adenocarcinoma Cells Expressing AXL", Cancer Res (15 апреля 2007); 67(8):3878-3887 | |||
| O'BRYAN J.P | |||
| et al., "The transforming receptor tyrosine kinase, Axl, is post-translationally regulated by proteolytic cleavage", J Biol Chem | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |

