Перекрестная ссылка на родственную заявку
По настоящей заявке испрашивается приоритет предварительной патентной заявки США No. 60/815257, поданной 21 июня 2006 г. Полное содержание указанной выше заявки включено в настоящее описание в качестве ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителу или его фрагменту, которое способно связывать гомолог 10 белка Frizzled (FZD10), такому как моноклональное антитело мыши, гибридное антитело и гуманизированное антитело. Настоящее изобретение также относится к способу лечения и/или профилактики заболевания, связанного с FZD10; способу диагностики или прогнозирования заболевания, связанного с FZD10; и способу визуализации FZD10 у субъекта in vivo.
Предшествующий уровень техники
Моноклональные антитела против молекул, специфичных для рака, как доказано, пригодны для лечения рака (Harris, M. (2004). Lancet Oncol, 5, 292-302). В дополнение к успешным примерам клинического применения гуманизированных или гибридных антител, таких как трастузумаб (Baselga, J. (2004). Oncology, 61, Supl 2 14-21), ритуксимаб (Maloney, D.G., et al. (1997). Blood, 90, 2188-95) и бевацизумаб (Ferrara, N., et al. (2004). Nat Rev Drug Discov, 3, 391-400) при раке молочной железы, злокачественной лимфоме и раке ободочной кишки, ряд моноклональных антител против других молекулярных мишеней находится в разработке и оценивается в отношении их противоопухолевой активности. Эти моноклональные антитела, как предполагается, дадут надежду больным с опухолями, не поддающимися эффективному лечению. Одним из других важных достоинств этих моноклональных антител является достижение избирательных терапевтических эффектов на раковые клетки без тяжелых токсических эффектов, обусловленных их специфическим взаимодействием с клетками, экспрессирующими молекулы-мишени (Crist, W.M., et al. (2001). J Clin Oncol, 19, 3091-102. Wunder, J.S., et al. (1998). J Bone Joint Surg Am, 80, 1020-33; Ferguson, W.S. and Goorin, A.M. (2001) Cancer Invest, 19, 292-315).
Помимо саркомы мягких тканей, остеосаркома, саркома Юинга и рабдомиосаркома чувствительны к химиотерапии и эти заболевания могут хорошо поддаваться лечению с помощью химиотерапии. С другой стороны, веретеноклеточные саркомы устойчивы к химиотерапии и облучению, и имеющие их больные характеризуются плохим прогнозом. Для синовиальной саркомы (SS) эффективным является хирургическое лечение больных на ранней стадии, но для больных на поздней стадии эффективное терапевтическое лекарство не доступно. Следовательно, разработка новых терапевтических способов воздействия, как ожидается, изменит прогноз у больных в лучшую сторону.
Широкий геномный анализ экспрессии генов в опухолях дает полезную информацию для идентификации новых молекулярных мишеней в целях разработки новых противораковых лекарственных средств и маркеров опухолей. В предшествующем исследовании авторы настоящего изобретения проанализировали профиль экспрессии генов ряда сарком мягких тканей с применением широкого геномного анализа с микроматрицей кДНК, состоящей из 23040 генов, и продемонстрировали, что гомолог 10 Frizzled (FZD10) (GenBank NO. доступа AB027464 (SEQ ID NO:1) и BAA84093 (SEQ ID NO:2)) специфично и часто положительно регулировался в SS (Nagayama, S., et al. (2002) Cancer Res, 62, 5859-66; и WO2004/020668). Продукт гена FZD10 представляет собой член семейства Frizzled и предполагаемый рецептор, передающий сигналы WNT (Koike, J., et al. (1999). Biochem Biophys Res Commun, 262, 39-43). Дополнительный анализ показал, что FZD10 специфично экспрессируется в SS, и не экспрессируется или экспрессируется на почти неопределимом уровне в других нормальных органах, за исключением плаценты, что позволяет предположить, что лекарственные агенты, направленные на эту молекулу, не должны вызывать или будут вызывать мало побочных эффектов (Nagayama, S., et al. (2002). Cancer Res, 62, 5859-66). Эксперименты с иРНК показали, что FZD10 в значительной степени вовлечен в рост опухоли SS (WO2006/013733). Более того, авторы настоящего изобретения разработали поликлональное антитело кролика против внеклеточного домена FZD10 (FZD10-ECD) и обнаружили, что это антитело обладает противоопухолевой активностью в мышиной модели ксенотрансплантата SS (Nagayama, S., et al. (2005). Oncogene, 24, 6201-12; и WO2005/004912). В целом, лечение антителами против FZD10 может, как ожидается, улучшить клинический исход SS.
Краткое изложение сущности изобретения
В настоящем описании ниже сообщается, что созданы моноклональные антитела мыши против FZD10 с помощью метода клеточной иммунизации для возможного клинического применения. Активность этих антител в отношении связывания с опухолью in vivo оценивалась с использованием систем визуализации флуоресценции in vivo с флуоресценцией в ближней инфракрасной части спектра в дополнение к традиционному методу с радиоизотопами. В настоящем описании авторы установили специфичность связывания моноклональных антител против FZD10 как in vitro, так и in vivo, а также интернализацию этих антител в клетках, экспрессирующих FZD10, и обнаружили, что у мышей, несущих ксенотрансплантат SYO-1, обработанных однократно через хвостовую вену Mab против FZD10, меченным 90Y в дозе 100 мкКи, наблюдался существенный противоопухолевый эффект.
Основываясь на представленных выше данных, авторы настоящего изобретения пришли к заключению, что мышиные моноклональные антитела против FZD10 обладают терапевтическим потенциалом при лечении и диагностике SS и других опухолей, сверхэкспрессирующих FZD10.
Следовательно, в первом аспекте в настоящем изобретении предлагается антитело или его фрагмент, которое включает V (вариабельную) область H (тяжелой) цепи, включающую область, определяющую комплементарность (CDR), имеющую аминокислотные последовательности, представленные в SEQ ID NO: 15, 17 и 19, или CDR, функционально эквивалентную ей, и V (вариабельную) область L (легкой) цепи, включающую CDR, имеющую аминокислотные последовательности, представленные в SEQ ID NO: 23, 25 и 27, CDR, функционально эквивалентную ей, которое способно связывать гомолог 10 белка Frizzled (FZD10) или его частичный пептид.
В одном варианте осуществления антитело или его фрагмент выбраны из группы, состоящей из антитела мыши, гибридного антитела, гуманизированного антитела, фрагмента антитела и одноцепочечного антитела.
В предпочтительном варианте осуществления антитело представляет собой антитело мыши. Предпочтительно, антитело мыши включает цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 57, и/или цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 59. Например, антитело мыши может продуцироваться клоном гибридомы 92-13 (FERM BP-10628).
В альтернативном предпочтительном варианте осуществления антитело представляет собой гибридное антитело. Предпочтительно, гибридное антитело включает область V цепи H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 13, например, гибридное антитело может включать цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 46. Предпочтительно, гибридное антитело включает область V цепи L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 21, например, гибридное антитело может включать цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 48.
Более предпочтительно, гибридное антитело включает область V цепи H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 13, и область V цепи L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 21. Например, гибридное антитело включает цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 46, и цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 48.
В одном варианте осуществления гибридное антитело дополнительно включает C (константную) область антитела человека.
В альтернативном предпочтительном варианте осуществления антитело представляет собой гуманизированное антитело. В одном варианте осуществления гуманизированное антитело дополнительно включает FR (каркасную) область антитела человека и/или C-область антитела человека.
Во втором аспекте в настоящем изобретении предлагается антитело или его фрагмент, которое включает V (вариабельную) область H (тяжелой) цепи, включающую область, определяющую комплементарность (CDR), имеющую аминокислотные последовательности, представленные в SEQ ID NO: 31, 33 и 35, или CDR, функционально эквивалентную ей, и V-область L (легкой) цепи, включающую CDR, имеющую аминокислотные последовательности, представленные в SEQ ID NO: 39, 41 и 43, или CDR, функционально эквивалентную ей, которое способно связывать гомолог 10 белка Frizzled (FZD10) или его частичный пептид.
В одном варианте осуществления антитело или его фрагмент выбраны из группы, состоящей из антитела мыши, гибридного антитела, гуманизированного антитела, фрагмента антитела и одноцепочечного антитела.
В предпочтительном варианте осуществления антитело представляет собой антитело мыши. Предпочтительно, антитело мыши включает цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 61, и/или цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 63. Например, антитело мыши может быть продуцировано клоном гибридомы 93-22 (FERM BP-10620).
В альтернативном предпочтительном варианте осуществления антитело представляет собой гибридное антитело. Предпочтительно, гибридное антитело включает область V цепи H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 29, например, гибридное антитело включает цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 50. Предпочтительно, гибридное антитело включает область V цепи L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 37, например, гибридное антитело включает цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 52.
Более предпочтительно, гибридное антитело включает область V цепи H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 29, и область V цепи L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 37. Например, гибридное антитело включает цепь H, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 50, и цепь L, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 52.
В одном варианте осуществления гибридное антитело дополнительно включает C (константную) область антитела человека.
В альтернативном предпочтительном варианте осуществления антитело представляет собой гуманизированное антитело. В одном варианте осуществления гуманизированное антитело дополнительно включает FR (каркасную) область антитела человека и/или C-область антитела человека.
В еще одном альтернативном варианте осуществления антитело или его фрагмент может быть помечено радиоизотопной меткой или флуоресцентной меткой. Такая радиоизотопная метка включает 90иттрий (90Y), 125йод (125I) и 111индий (111In).
В третьем аспекте в настоящем изобретении предлагается клон гибридомы 92-13 (FERM BP-10628), который продуцирует моноклональное антитело 92-13 мыши.
В четвертом аспекте в настоящем изобретении предлагается клон гибридомы 93-22 (FERM BP-10620), который продуцирует моноклональное антитело 93-22 мыши.
В пятом аспекте в настоящем изобретении предлагается способ лечения или профилактики заболевания, которое связано с гомологом 10 белка Frizzled (FZD10) у субъекта, включающий введение субъекту эффективного количества антитела или его фрагмента. В одном варианте осуществления заболевание, которое связано с FZD10, выбрано из синовиальной саркомы (SS), колоректального рака, рака желудка, хронического миелоидного лейкоза (CML) и острого миелоидного лейкоза (AML).
В шестом аспекте в настоящем изобретении предлагается способ диагностики или прогнозирования заболевания, которое связано с гомологом 10 белка Frizzled (FZD10), или предрасположенности к развитию заболевания у субъекта, включающий
(a) контактирование образца или препарата от субъекта с антителом или фрагментом, указанными выше;
(b) определение белка FZD10 в образце или препарате; и
(c) решение о том, страдает или нет субъект заболеванием или подвержен риску его развития, на основе относительного содержания белка FZD10 по сравнению с контролем.
В одном варианте осуществления заболевание, которое связано с FZD10, выбрано из синовиальной саркомы (SS), колоректального рака, рака желудка, хронического миелоидного лейкоза (CML) и острого миелоидного лейкоза (AML).
В седьмом аспекте в настоящем изобретении предлагается способ визуализации in vivo гомолога 10 белка Frizzled (FZD10) у субъекта, включающий введение субъекту эффективного количества антитела или фрагмента, указанных выше.
В восьмом аспекте в настоящем изобретении предлагается фармацевтическая композиция для лечения или профилактики заболевания, связанного с гомологом 10 Frizzled (FZD10), содержащая антитело или фрагмент, указанные выше, и фармацевтически приемлемый носитель или наполнитель.
В девятом аспекте в настоящем изобретении предлагается набор для диагностики или прогнозирования заболевания, связанного с гомологом 10 Frizzled (FZD10), содержащий антитело или фрагмент, указанные выше.
В десятом аспекте в настоящем изобретении предлагается фармацевтическая композиция для визуализации in vivo гомолога 10 белка Frizzled (FZD10), содержащая антитело или фрагмент, указанные выше.
В одиннадцатом аспекте в настоящем изобретении предлагается применение антитела или фрагмента, указанных выше, для получения набора для диагностики или прогнозирования заболевания, связанного с гомологом 10 Frizzled (FZD10).
В двенадцатом аспекте в настоящем изобретении предлагается применение антитела или фрагмента, указанных выше, для получения композиции для профилактики или лечения заболевания, связанного с гомологом 10 Frizzled (FZD10).
Термин «заболевание, которое связано с FZD10» (связанное с FZD10 заболевание), относится к заболеванию, которое связано со сверхэкспрессией белка FZD10. Такие заболевания включают, но не ограничиваются этим, синовиальную саркому (SS), колоректальный рак, рак желудка, хронический миелоидный лейкоз (CML) и острый миелоидный лейкоз (AML).
Термин «фрагмент» обозначает любой фрагмент антитела, который может быть получен из антитела против белка FZD10, и содержит определенные CDR. Такие фрагменты включают, но не ограничиваются этим, Fab фрагмент, F(ab')2 фрагмент и Fv фрагмент.
Термин «модифицированное антитело» обозначает любое антитело, которое может происходить из антитела против FZD10, и содержит определенные CDR. Такое модифицированное антитело включает, но не ограничивается этим, ПЭГ-модифицированное антитело. Фрагмент антитела или модифицированный фрагмент может легко узнаваться специалистом в данной области техники и продуцироваться с использованием любых методов, известных в данной области техники.
Термин «субъект» в настоящем описании относится к субъекту, который страдает заболеванием, связанным с FZD10, и также к субъекту, предположительно имеющему заболевание, связанное с FZD10. Субъектом по настоящему изобретению могут быть животные, включая млекопитающих и птиц. Например, млекопитающие включают человека, мышей, крыс, обезьян, кроликов и собак.
Краткое описание чертежей
На фиг. с 1a по 1f представлена характеристика специфичности связывания двух моноклональных антител против FZD10.
На фиг. 1a представлен анализ с помощью проточной цитометрии четырех антител, 39-2 и 39-10 (раскрытых в патенте WO2005/004912), 92-13 и 93-22, с использованием пяти линий SS (SYO-1, YaFuSS, HS-SY-2, Fuji и 1973/99) и одной линии клеток колоректального рака (LoVo). Сплошными линиями показана интенсивность флуоресценции, определяемая с помощью каждого из mAb; прерывистыми линиями изображена интенсивность флуоресценции клеток, проинкубированных с неиммунными IgG мыши в качестве отрицательного контроля.
На фиг. 1b представлена полуколичественная ОТ-ПЦР FZD10 в тех же линиях опухолевых клеток, что и используемые на фиг. 1a. Экспрессия гена β2-микроглобулина (β2-MG) служила в качестве внутреннего контроля.
На фиг. 1c представлен анализ с помощью проточной цитометрии антител 92-13 (верхние панели) и 93-22 (нижние панели) против экзогенного FZD10. Линию клеток рака ободочной кишки SNU-C5 трансфицировали пустым вектором pCAGGS (левые панели) или pCAGGS-FZD10-myc/His (правые панели) и анализировали через 48 часов после трансфекции. Сплошными линиями показана интенсивность флуоресценции, определяемая с помощью каждого из mAb; прерывистыми линиями изображена интенсивность флуоресценции клеток, проинкубированных с неиммунными IgG мыши в качестве отрицательного контроля.
На фиг. 1d представлено связывание меченных 125I 39-10, 39-2, 92-13 и 93-22 с нормальными клетками крови человека. Радиоактивно меченные Mab инкубировали каждое со свежевыделенными нормальными клетками крови человека от трех субъектов (A, B и C) в отсутствие (незакрашенный столбик) или в присутствии (закрашенный столбик) идентичных немеченых антител.
На фиг. 1e представлена связывающая активность меченных 125I Mab. Постоянное количество радиоактивно меченных Mab инкубировали с клеткой SYO-1 и возрастающим количеством немеченых антител. Наносили процент радиоактивности, связанной с клетками, против количества немеченого антитела. Закрашенный кружок; 92-13, незакрашенный кружок; 93-22.
На фиг. 1f представлен анализ с помощью проточной цитометрии аутоблока и перекрестного блока. Alexa-488-меченные 92-13 (верхние панели) и 93-22 (нижние панели) инкубировали с клеткой SYO-1 в (i) PBS, или в присутствии 100 мкг (ii) немеченых 92-13 и (iii) немеченых 93-22. На штрихованной гистограмме представлена интенсивность флуоресценции, определенная с помощью каждого из Alexa-488-меченных Mab; прерывистые линии изображают интенсивность флуоресценции клеток, инкубированных с PBS в качестве отрицательного контроля.
На фиг. 2 представлен иммуногистохимический анализ в SS и на нормальных замороженных срезах тканей человека без антитела (a, d, g, j и m), 92-13 (b, e, h, k и n) и 93-22 (c, f, i, l и o). (a-c), синовиальная саркома; (d-f), почка; (g-i), печень, (j-l), сердце; (m-o), мозг. Исходное увеличение: ×100.
На фиг. 3 представлено биологическое распределение 111In-меченных и 125I-меченных антител. 10 кВк (a), 111In-меченных 92-13, (b), 125I-меченных 92-13, (c), 111In-меченных 93-22 и (d), 125I-меченных 93-22 вводили внутривенно голым BALB/c мышам, несущим опухоль SYO-1. Органы и опухоль вырезали через 1 час (незакрашенный столбик), 24 часа (заштрихованный столбик) и 48 часов (закрашенный столбик) и измеряли радиоактивность. Представленные данные являются репрезентативными данными двух независимых экспериментов.
На фиг. 4a представлена визуализация in vivo флуоресценции мышей, несущих опухоль SYO-1, после введения Alexa 647-меченных 92-13 или 93-22. Флуоресцентно меченные Mab вводили в дозе 20 мкг на мышь внутрибрюшинно. Все флуоресцентные изображения получали при 60-секундном времени экспозиции (f/стоп = 2) перед введением, непосредственно после введения (0 часов), через 24, 48 и 96 часов. Стрелки указывают на расположение опухоли. S.C. опухоль локализована дорзально для 92-13 (верхние панели) и туловищно для 93-22 (нижние панели). Флуоресцентный сигнал от Alexa647 был псевдоокрашен в соответствии с цветным столбиком, представленным справа. В случае 93-22 (нижняя панель) головки стрелок указывают на место введения.
На фиг. 4b и 4c представлена иллюстративная визуализация извлеченных органов и опухолей от мышей, представленных на фиг. 4a, 4b; 92-13, и 4c; 93-22. i, опухоль SYO-1; ii, печень; iii, селезенка; iv, почка; v, поджелудочная железа; vi, ободочная кишка.
На фиг. 5a представлена визуализация in vivo флуоресценции мышей, несущих опухоль LoVo, после введения Alexa647-меченных 92-13 или 93-22. Флуоресцентно меченные Mab вводили как на фиг. 4. Все флуоресцентные изображения получали при 60-секундном времени экспозиции (f/стоп = 2) непосредственно после введения (0 часов), через 48, 72, 96 и 120 часов (h). Стрелка указывает на расположение опухоли. S.c. опухоль локализована в правом предплечье как для 92-13 (верхние панели), так и для 93-22 (нижние панели).
На фиг. 5b и 5c представлена иллюстративная визуализация извлеченных органов и опухолей мышей, представленных на фиг. 5a, 5b; 92-13, и 5c; 93-22. i, опухоль LoVo; ii, печень; iii, селезенка; iv, почка; v, поджелудочная железа; vi, ободочная кишка.
На фиг. 6 представлена интернализация 92-13 и 93-22, оцениваемая с помощью конфокальной микроскопии. Клетки обрабатывали PBS (a, d и g), 50 мкг/мл 92-13 (b, e и h) или 93-22 (c, f, i) в течение 3 часов при 37oC, 5% CO2. Антитела, связанные с клеточной поверхностью, десорбировали в кислой среде 0,1 М глициновым буфером (рН 2,5). Клетки фиксировали, пермиабилизировали и затем блокировали 3% BSA. Внутриклеточные антитела определяли с помощью козьих антимышиных IgG-Alexa488, и ядро окрашивали DAPI. (a - c), SYO-1; (d - f), YaFuSS; (g - i) LoVo.
На фиг. 7 представлено влияние 90Y-меченных 92-13 на рост опухоли. Когда опухоли прививались (0,4-2,7см3), мышам вводили однократно в хвостовую вену 100 мкКи 90Y-меченных 92-13.
На фиг. 8 представлена индуцируемая гибридными как 92-13, так и 93-22, ADCC конкретно в отношении к гиперэкспрессирующим FZD10 клеткам SYO-1. 1 мкг/мл гибридного антитела 93-22 (ch93-22) или гибридного антитела 92-13 (ch92-13) добавляли в различном соотношении эффектор:мишень. В качестве эффекторной клетки использовали PBMC от различных людей доноров; (a), (c) ADCC гибридных 92-13 против клетки SYO-1 с пятью здоровыми людьми донорами PBMC. (b), (d) ADCC гибридных 93-22 против клетки LoVo с двумя здоровыми людьми донорами PBMC. Количественная оценка цитотоксичности по активности LDH описана в (Nagayama, S., et al. Oncogene, 24, 6201-12).
Подробное описание и предпочтительные варианты осуществления настоящего изобретения
Гомолог 10 Frizzled (FZD10) представляет собой член семейства Frizzled, который является рецептором, проводящим сигнал от Wnt. Как описано в настоящем изобретении ниже, авторы успешно создали моноклональные антитела мыши и гибридные антитела против белка FZD10, которые могут быть пригодны для использования в медицине.
Моноклональные антитела мыши, специфичные для FZD10 (92-13 и 93-22 Mab), созданы путем иммунизации мышей клетками, трансфицированными FZD10. Как показано путем анализа с использованием проточной цитометрии (FACS), Mab, как 92-13, так и 93-22, обладают специфической связывающей активностью против FZD10 в линии клеток SS, клетках SYO-1 и клетках COS7, трансфицированными FZD10. Для подтверждения специфической связывающей активности этих антител in vivo авторы настоящего изобретения вводили флуоресцентно меченные Mab внутрибрюшинно или внутривенно мышам, несущим ксенотрансплантаты SS, и нашли с помощью использования флуоресцентной системы визуализации in vivo и радиоактивности, что эти Mab связываются с опухолями, экспрессирующими FZD10, а не с любыми другими нормальными тканями мышей. Последующий иммуногистохимический анализ с Mab подтвердил отсутствие или едва выявляемый уровень белка FZD10 в нормальных органах человека за исключением плаценты. Более того, интересно, что авторы настоящего изобретения нашли, используя конфокальную лазерную сканирующую микроскопию, что Mab интернализуются в клеточной линии SS, SYO-1, но не в негативную в отношении FZD10 линию клеток, LoVo. Неожиданно оказалось, что у мышей, несущих ксенотрансплантат SYO-1, с однократным введением в хвостовую вену 90Y-меченного Mab (92-13) против FZD10 в дозе 100 мкКи, наблюдался существенный противоопухолевый эффект. Суммируя вышеизложенное, авторы пришли к заключению, что эти специфические Mab против FZD10 могут быть использованы в качестве нового диагностического маркера или для лечения SS с минимальным риском побочных реакций или их отсутствием.
Часто очень трудно выработать антитела против белков с семью трансмембранными доменами из-за сложной структуры этих белков. В предшествующем исследовании авторы настоящего изобретения показали, что FZD10 образует гомоолигомер (Nagayama, S., et al. (2005). Oncogene, 24, 6201-12). После многих неудачных попыток создания моноклональных антител против FZD10, которые могут узнавать природную форму FZD10, путем использования полноразмерных или частично рекомбинантных белков FZD10 авторы настоящего изобретения наконец использовали иммунизацию путем инъекции живых клеток COS-7, гиперэкспрессирующих FZD10, в подушечки лап мышей Balb/c и успешно получили две гибридомы, продуцирующие антитела против FZD10, которые обладали способностью узнавать природную форму FZD10 в живых клетках, как показано FACS анализом. Так как эти антитела не определяют белок FZD10 при вестерн-блоттинге, авторы настоящего изобретения предположили, что эти Mab узнают третичную структуру FZD10.
Для исследования распределения in vivo Mab 92-13 и 93-22 авторы настоящего изобретения использовали два метода; один основывался на радиоизотопных методах с использованием 125I и 111In-меченных антител, а другой основывался на флуоресцентной визуализации с применением антител, меченных в ближней инфракрасной части спектра (Alexa647). Флуоресцирующий в ближней инфракрасной части спектра, главным образом индоцианиновый краситель, сейчас широко используется при визуализации in vivo для диагностической цели, так как свет этой длины волны проникает в живую ткань вполне эффективно (Chen, X., et al. (2004). Cancer Res, 64, 8009-14). Результаты, полученные с помощью двух подходов, очень согласовывались друг с другом и указывали на то, что 92-13 и 93-22 связываются с опухолевыми клетками SYO-1, а не с другими нормальными тканями. Для подтверждения того, могут ли эти антитела использоваться для клинического применения, авторы настоящего изобретения дополнительно проверили связывающую активность антител против нормальных клеток крови. Связывающая активность 125I-меченных 92-13 и 93-22 против нормальных клеток крови человека была не определима у всех трех индивидуальных доноров (фиг. 1d). Эти результаты согласовывались с полученными при анализе FACS с использованием мононуклеарной клетки периферической крови человека (данные не показаны), что предполагает клиническую применимость этих двух антител с небольшой возможностью побочного эффекта у больных SS, благодаря высокоспецифичному связывающему сродству к молекуле FZD10. Более того, эксперименты in vitro с использованием конфокальной микроскопии выявили, что специфическое связывание Mab 92-13 и 93-22 с FZD10 клеточной поверхности индуцирует интернализацию антител (фиг. 6). Как описано ранее (Stein, R., et al. (2001). Criti Rev Oncol Hematol, 39, 173-80; Stein, R., et al. (2005). Clin Cancer Res, 11, 2727-34), если меченые Mab интернализуются после связывания, 125I-меченное антитело метаболизируется в лизосомах и диффундирует из опухолевой клетки-мишени, тогда как 111In-меченное антитело остается в лизосомах. Как видно на фиг. 3, радиоактивность 111In-меченного антитела и 125I-меченного антитела в опухолях существенно различалась (фиг. 3, a и b, c и d). Эти данные предполагают, что Mab 92-13 (и 93-22) могут специфически интернализовываться в клетках SS через белок FZD10.
Когда антитела используются для лечения рака, следующие три механизма, как считается, индуцируют противоопухолевую активность; (i) в случае, если молекула-мишень вовлечена в стимуляцию роста, нейтрализация антителами должна блокировать передачу ростового сигнала и затем подавлять рост опухолевых клеток; (ii) второй возможностью является эффекторная активность в плане индукции антитело-зависимой клеточной цитотоксичности (ADCC) или зависимой от комплемента цитотоксичности (CDC). (iii) Третьим случаем являются радиоактивные изотопы или противоопухолевое лекарство, которые конъюгируются с антителами и эффективно доставляются к опухолевым клеткам-мишеням. Хотя авторы настоящего изобретения ранее показали, что молекула-мишень FZD10 вовлечена в рост опухоли SS, ни Mab 92-13, ни 93-22 не проявляли нейтрализующего эффекта in vitro при добавлении к клеточным культуральным средам (данные не показаны) или in vivo при введении мышам опухоленосителям (данные не показаны).
Конъюгированные с антителами радиоактивный изотоп или противораковое лекарство, такие как зевалин (антитело против CD20, конъюгированное с 90иттрием) и милотарг (антитело против CD33, конъюгированное с калихеамицином), как доказано, являются высокоэффективными в плане придания цитотоксичности антителам (Wiseman, G.A. and Witzig, T.E. (2005). Cancer Biother Radiopharm, 20, 185-8; van der Velden, V.H., et al. (2001). Blood, 97, 3197-204; Carter, P. (2001). Nat Rev Cancer, 1, 118-29). Милотарг проявляет свою противоопухолевую активность путем высвобождения противоопухолевого лекарства, калихеамицина, в опухолевой клетке после его интернализации (van der Velden, V.H., et al. (2001). Blood, 97, 3197-204). В примерах для терапевтических экспериментов создан конъюгат 90иттрий-DTPA-92-13 и исследована его противоопухолевая активность. В модели ксенотрансплантата мыши опухоли быстро уменьшались после обработки 90иттрий-DTPA-92-13 (фиг. 7). Замечено, что опухоли, включая опухоль более крупного объема (> 1 см3), не проявляли рефракции до 34 дней после введения, и не наблюдалось сильной токсичности. Так как антитела 92-13 и 93-22 против FZD10, очевидно, эффективно интернализовывались в антиген-позитивных клетках, как показано на фиг. 6, конъюгация противоракового лекарства с обоими Mab, 92-13 и 93-22, также, как ожидается, проявит сильный противораковый эффект на клетки SS. Относительно эффекторной активности, как гибридные 92-13, так и 93-22, индуцировали ADCC специфично в отношении FZD10-гиперэкспрессирующих клеток SYO-1 (фиг. 8, a и c), но не в отношении FZD10-негативных клеток LoVo (фиг. 8, b и d). Особенно гибридные 92-13 проявляли более высокую индукцию цитотоксичности по сравнению с гибридными 93-22, однако, их активность зависела от донора эффекторных клеток, возможно из-за полиморфизма рецептора Fc. В заключение авторы настоящего изобретения успешно получили моноклональные антитела, которые способны специфически связываться с FZD10 на FZD10-гиперэкспрессирующих опухолевых клетках in vitro и in vivo. Резюмируя, авторы настоящего изобретения уверены, что моноклональные антитела против FZD10 обладают большим потенциалом в плане разработки новой лекарственной терапии для лечения SS и других опухолей, которые гиперэкспрессируют FZD10.
1. Получение антитела
Антитела, которые могут быть использованы в настоящем изобретении, специфично направлены против белка FZD10, получаемого при заболевании, связанном с FZD10. Используемый в настоящем описании термин «антитело» означает молекулу антитела, как целиком, так и в виде ее фрагментов, таких как Fab фрагменты, F(ab')2 фрагменты и Fv фрагменты, которая может связываться с белком или его пептидными фрагментами в качестве антигенов. Антитело может представлять собой либо поликлональное антитело, либо моноклональное антитело. Оно может быть также гуманизированным или гибридным антителом, или одноцепочечным Fv (scFv) антителом. Антитела (поликлональные антитела и моноклональные антитела) для использования в настоящем изобретении могут быть получены, например, с помощью следующего способа.
(1) Моноклональное антитело
Исходно получают антиген для продукции антитела, пригодного для настоящего изобретения. Белок FZD10 или его частичный пептид может быть использован в качестве иммуногенного белка. Альтернативно, клетка, экспрессирующая белок FZD10 или его частичный пептид, может быть также использована в качестве иммуногена. Аминокислотная последовательность белка FZD10, используемого в качестве иммуногена в настоящем изобретении, и последовательность кДНК, кодирующая белок, есть в открытом доступе в GenBank под No. доступа BAA84093 (SEQ ID NO: 1) и AB027464 (SEQ ID NO: 2), соответственно. Белок FZD10 или его частичный пептид для использования в качестве иммуногена могут быть получены синтетически в соответствии с процедурой, известной в данной области техники, такой как метод твердофазного пептидного синтеза, с использованием доступной информации об аминокислотной последовательности. Пептидные фрагменты белка FZD10 включают, но не ограничиваются этим, пептид, содержащий остатки 1-225 аминокислотной последовательности, представленной в SEQ ID NO: 1, что соответствует N-концевому внеклеточному домену белка FZD10 (FZD10-ECD).
Белок или его частичный пептид, или экспрессирующая их клетка могут быть получены при использовании информации о последовательности кДНК, кодирующей белок FZD10 или его частичный пептид, в соответствии с известной процедурой рекомбинации генов. Получение белка или его пептидного фрагмента, а также экспрессирующей их клетки в соответствии с такой процедурой генной рекомбинации будет проиллюстрировано ниже.
Рекомбинантный вектор для продукции белка может быть получен путем сшивки указанной выше последовательности кДНК с подходящим вектором. Трансформант может быть получен путем введения рекомбинантного вектора для продукции белка в хозяина, так, чтобы белок-мишень FZD10 или его частичный пептид могли экспрессироваться.
В качестве вектора используют фаг или плазмиду, которые способны автономно реплицироваться в используемом хозяине. Примеры плазмидной ДНК включают pCAGGS, pET28, pGEX4T, pUC118, pUC119, pUC18, pUC19 и другие плазмидные ДНК, происходящие из Escherichia coli; pUB110, pTP5 и другие плазмидные ДНК, происходящие из Bacillus subtilis; и YEp13, YEp24, YCp50 и другие плазмидные ДНК, происходящие из дрожжей. Примеры ДНК фагов включают фаги лямбда, такие как λgt11 и λZAP. Кроме того, могут быть использованы векторы вирусов животных, такие как ретровирусный вектор и вектор коровьей оспы и могут быть также использованы векторы вирусов насекомых, такие как бакуловирусный вектор.
ДНК, кодирующую белок FZD10 или его частичный пептид (обозначаемую в настоящем описании далее как ДНК FZD10), вставляют в вектор, например, с помощью следующего метода. В этом методе очищенную ДНК расщепляют с помощью подходящего рестрикционного фермента и вставляют в сайт рестрикции ферментом или в сайт множественного клонирования подходящей ДНК вектора для лигирования в вектор.
В дополнение к промотору и ДНК FZD10 любые энхансеры и другие цис-элементы, сигналы сплайсинга, сигнал добавления поли-A, селективные маркеры, сайт рибосомального связывания (RBS) и другие элементы могут быть лигированы в рекомбинантный вектор для продукции белка для использования в клетках млекопитающих, если это желательно.
Для лигирования фрагмента ДНК с фрагментом вектора может быть использована известная лигаза ДНК. Фрагмент ДНК и фрагмент вектора отжигают и лигируют, получая тем самым рекомбинантный вектор для продукции белка.
Хозяин для использования при трансформации специально не ограничивается до тех пор, пока он позволяет экспрессироваться в нем белку FZD10 или его частичному пептиду. Примеры хозяина включают бактерии, например E. coli и Bacillus; дрожжи, например Saccharomyces cerevisiae; клетки животных, например клетки COS, клетки яичника китайского хомячка (CHO) и клетки насекомых.
Например, когда в качестве хозяина используют бактерию, рекомбинантный вектор для продукции белка должен предпочтительно обладать способностью к автономной репликации в бактерии-хозяине и включать промотор, сайт рибосомального связывания, ДНК FZD10 и последовательность терминации транскрипции. Рекомбинантный вектор может дополнительно включать ген для регуляции промотора. Пример Escherichia coli включает Escherichia coli BRL, и примером Bacillus является Bacillus subtilis. В настоящем описании может быть использован любой промотор, который может экспрессироваться в хозяине, таком как Escherichia coli.
Рекомбинантный вектор может быть введен в бактерию-хозяина с помощью любого из методов, известных в данной области техники. Такие методы включают, например, метод с использованием ионов кальция и электропорацию. Когда в качестве хозяина используют клетку дрожжей, клетку животных или клетку насекомых, трансформант может быть получен в соответствии с методом, известным в данной области техники, и затем белок FZD10 или его частичный пептид может быть получен в хозяине (трансформанте).
Белок FZD10 или его частичный пептид для использования в качестве иммуногена в настоящем изобретении может быть получен из культуры созданного выше трансформанта. «Культура» относится к любому культуральному супернатанту, культивируемым клеткам, культивируемым микроорганизмам и их гомогенатам. Трансформант культивируется в культуральной среде с помощью традиционного метода культивирования хозяина.
Культуральная среда для культивирования трансформанта, полученного с использованием Escherichia coli, дрожжей или других микроорганизмов в качестве хозяев, может быть либо природной средой, либо синтетической средой, до тех пор, пока она включает источник углерода, источник азота, неорганические соли и другие компоненты, используемые микроорганизмом, и позволяет трансформанту эффективно расти.
Трансформант обычно культивируют путем встряхивания культуры или аэрации культуры с перемешиванием в аэробных условиях при от 25°С до 37°С в течение от 3 до 6 часов. В процессе культивирования рН поддерживают на уровне приблизительно нейтрального путем доведения, например, неорганической или органической кислотой и щелочным раствором. В процессе культивирования к среде могут быть добавлены антибиотики, такие как ампициллин или тетрациклин, в соответствии с селективным маркером, включенным в рекомбинантный экспрессионный вектор, если это необходимо.
После культивирования, когда белок FZD10 или его частичный пептид продуцируется в микроорганизме или клетке, белок или его частичный пептид экстрагируют путем гомогенизации микроорганизма или клетки. Когда белок FZD10 или его частичный пептид секретируется из микроорганизма или клетки, культуральная среда используется, как она есть, или остатки микроорганизма или клетки удаляются из культуральной среды, например центрифугированием. После этого белок FZD10 или его частичный пептид может быть выделен из культуры и очищен с помощью традиционного биохимического метода для выделения и очистки белков, такого как осаждение сульфатом аммония, гель-хроматография, ионообменная хроматография и аффинная хроматография, либо по отдельности, либо в сочетании.
Получен или нет белок FZD10 или его частичный пептид, может быть подтверждено, например, электрофорезом в полиакриламидном геле с SDS.
Затем полученный белок FZD10 или его частичный пептид или трансформант растворяют в буфере для получения иммуногена. Когда это необходимо, туда может быть добавлен адъювант для эффективной иммунизации. Такие адъюванты включают, например, имеющийся в продаже полный адъювант Фрейнда и неполный адъювант Фрейнда. Любой из этих адъювантов может быть использован по отдельности или в сочетании.
Приготовленный таким образом иммуноген вводят млекопитающему, такому как кролик, крыса или мышь. Иммунизацию осуществляют главным образом с помощью внутривенного, подкожного или внутрибрюшинного введения. Интервал иммунизации не ограничивается специально, и млекопитающее иммунизируется от одного до 3 раз в интервалы в диапазоне от нескольких дней до недель. Клетки, продуцирующие антитела, собирают от 1 до 7 дней после последней иммунизации. Примеры клеток, продуцирующих антитела, включают клетки селезенки, клетки лимфатических узлов и клетки периферической крови.
Для получения гибридомы клетку, продуцирующую антитела, и миеломную клетку сливают. В качестве миеломной клетки для слияния с клеткой, продуцирующей антитела, может быть использована обычно доступная созданная клеточная линия. Предпочтительно, чтобы используемая клеточная линия обладала бы селективностью к лекарствам и такими свойствами, чтобы она не могла выживать в HAT-селективной среде (содержащей гипоксантин, аминоптерин и тимидин) в неслитой форме и могла выживать только при слиянии с клеткой, продуцирующей антитела. Возможные миеломные клетки включают, например, линии мышиных миеломных клеток, такие как P3X63-Ag.8.U1 (P3U1) и NS-I.
Затем миеломную клетку и клетку, продуцирующую антитела, сливали. Для слияния эти клетки смешивали, предпочтительно в отношении клеток, продуцирующих антитела, к миеломным клеткам 5:1 в культуральной среде для животных клеток, которая не содержит сыворотки, такой как среда DMEM и RPMI-1640, и сливали в присутствии агента, стимулирующего слияние клеток, такого как полиэтиленгликоль (ПЭГ). Слияние клеток может быть также осуществлено с использованием имеющегося в продаже устройства для слияния клеток с применением электропорации.
Затем гибридому отбирают от клеток после указанной выше обработки для слияния. Например, суспензию клеток соответствующим образом разводят, например, средой RPMI-1640, содержащей эмбриональную телячью сыворотку, и затем высаживают на планшет для микротитрования. К каждой лунке добавляют селективную среду, и клетки культивируют с соответствующей заменой селективной среды. В результате в виде гибридомы могут быть получены клетки, которые растут приблизительно 30 дней после начала культивирования в селективной среде.
Затем проводят скрининг культурального супернатанта растущей гибридомы на присутствие антитела, которое взаимодействует с белком FZD10 или его частичным пептидом. Скрининг гибридомы может быть осуществлен в соответствии с общепринятой процедурой, например, с использованием твердофазного иммуноферментного анализа (ELISA), иммуноферментного анализа (ИФА) или радиоиммуноанализа (РИА). Слитые клетки клонируют путем серийного разведения для создания гибридомы, которая продуцирует интересующее моноклональное антитело.
Моноклональное антитело может быть собрано из созданной гибридомы, например, с помощью общепринятого метода культивирования клеток или путем продукции асцитов. Если это необходимо, антитело может быть очищено в описанном выше методе сбора антител в соответствии с известным методом, таким как осаждение сульфатом аммония, ионообменная хроматография, гель-фильтрация, аффинная хроматография или их сочетание.
Тип глобулина моноклональных антител, пригодных в настоящем изобретении, специально не ограничивается, до тех пор, пока они способны специфически связываться с белком FZD10, и они могут представлять собой любой из IgG, IgM, IgA, IgE и IgD. Среди них предпочтительными являются IgG и IgM.
В настоящем изобретении мышиные моноклональные антитела 93-22 и 92-13 успешно созданы и предпочтительно используются. Клон гибридомы 93-22, продуцирующий моноклональное антитело мыши 93-22, был сдан на международное хранение Shuichi Nakatsuru в IPOD International Patent Organism Depository of the National Institute of Advanced Industrial Science and Technology (AIST Tsukuba Central 6, 1-1, Higashi 1-chome, Tsukuba-shi, Ibaraki-Ken, 305-8566 Japan) 14 июня 2006 г. под регистрационным номером FERM BP-10620. Также клон гибридомы 92-13, продуцирующий моноклональное антитело мыши 92-13, был сдан на международное хранение Shuichi Nakatsuru в IPOD International Patent Organism Depository of the National Institute of AIST 28 июня 2006 г. под регистрационным номером FERM BP-10628. Моноклональное антитело, продуцируемое гибридомой, может предпочтительно использоваться в настоящем изобретении.
В настоящем изобретении также может использоваться моноклональное антитело рекомбинантного типа, которое может быть получено путем клонирования гена антитела из гибридомы, интеграции гена антитела в подходящий вектор, введения вектора в хозяина, продукции антитела хозяином в соответствии с общепринятыми способами рекомбинации генов (см., например, Vandamme, A. M. et al., Eur. J. Biochem. (1990) 192, 767-75).
Более конкретно, мРНК, кодирующую вариабельную (V) область моноклонального антитела мыши против FZD10, выделяют из гибридомы, продуцирующей антитела (например, такой, как описанные выше). Выделение мРНК осуществляют путем получения суммарной РНК с помощью известного метода, такого как метод ультрацентрифугирования в гуанидине (Chirgwin, J. M. et al., Biochemistry (1979) 18, 5294-9) и метод AGPC (Chomczynski, P. et al., Anal. Biochem. (1987) 162, 156-9), и затем получают желаемую мРНК из суммарной РНК с использованием набора для очистки мРНК (Pharmacia) или тому подобного. Альтернативно, мРНК может быть также получена прямо при использовании набора QuickPrep mRNA Purification (Pharmacia).
Затем из мРНК с помощью обратной транскриптазы синтезируют кДНК для V-области антитела. Синтез кДНК может быть осуществлен с использованием имеющегося в продаже набора, например Gene RacerTM (Invitrogen). кДНК может быть также синтезирована или амплифицирована с помощью метода 5'-RACE (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-32) с применением набора 5'-Ampli FINDER RACE (Clontech) в сочетании с методом ПЦР.
Аминокислотные последовательности H-цепи и L-цепи моноклонального антитела 92-13 мыши представлены в SEQ ID NO: 57 и 59, соответственно (кодируемые нуклеотидной последовательностью, как представлено в SEQ ID NO: 58 и 60, соответственно). Аминокислотные последовательности H-цепи и L-цепи моноклонального антитела 93-22 мыши представлены в SEQ ID NO: 61 и 63, соответственно (кодируемые нуклеотидной последовательностью, как представлено в SEQ ID NO: 62 и 64, соответственно). Основываясь на информации о последовательностях, с помощью общепринятого метода могут быть созданы праймеры, используемые для амплификации H-цепи или L-цепи интересующего моноклонального антитела.
Интересующий фрагмент ДНК выделяют и очищают из полученного с помощью ПЦР продукта и затем лигируют с ДНК вектора для получения рекомбинантного вектора. Рекомбинантный вектор вводят в хозяина, такого как E. coli, и отбирают колонию, содержащую желаемый рекомбинантный вектор. Нуклеотидную последовательность интересующей ДНК в рекомбинантном векторе подтверждают с использованием, например, автоматического секвенатора.
После получения ДНК, кодирующей V-область антитела против FZD10, ДНК интегрируют в экспрессионный вектор, содержащий ДНК, кодирующую константную (C) область антитела.
Для получения антитела против FZD10, используемого в настоящем изобретении, ген антитела интегрируют в экспрессионный вектор так, чтобы ген антитела мог экспрессироваться под контролем элементов, контролирующих экспрессию (например, энхансера, промотора). Клетка-хозяин трансформируется экспрессионным вектором для экспрессии антитела.
При экспрессии гена антитела ДНК, кодирующая тяжелую (H) цепь, и ДНК, кодирующая легкую (L) цепь антитела, могут быть интегрированы в отдельные экспрессионные векторы, и затем клетка-хозяин совместно трансформируется полученными рекомбинантными экспрессионными векторами. Альтернативно, как ДНК, кодирующая H-цепь, так и ДНК, кодирующая L-цепь антитела, могут быть интегрированы совместно в один экспрессионный вектор, и затем клетка-хозяин трансформируется полученным рекомбинантным экспрессионным вектором (например, в патенте WO 94/11523).
Ген антитела может быть экспрессирован с помощью известных методов. Для экспрессии в клетке млекопитающих могут быть оперативно связаны традиционно применяемый промотор, ген антитела для ожидаемой экспрессии и сигнал поли(A) (локализованный ниже 3'-конца гена антитела). Например, в качестве пригодной системы промотор/энхансер может быть использована система немедленного раннего промотора/энхансера цитомегаловируса чнловека.
Другие системы промотор/энхансер, например, происходящие от вирусов (например, ретровируса, вируса полиомы, аденовируса и вируса 40 обезьян (SV40) и происходящие от клеток млекопитающих (например, фактора 1 альфа элонгации человека (HEF1 альфа)), также могут использоваться для экспрессии антитела в настоящем изобретении.
Когда применяют систему промотор/энхансер SV40, экспрессия гена может легко осуществляться с помощью метода Mulligan et al. (Nature (1979) 277, 108-14). Когда применяют систему промотор/энхансер HEF1 альфа, экспрессия гена может легко осуществляться с помощью метода Mizushima et al. (Nucleic Acids Res. (1990) 18, 5322).
Для экспрессии в E. coli могут быть функционально связаны традиционно применяемый промотор, сигнальная последовательность для секреции интересующего антитела и ген антитела. В качестве промотора может быть использован lacZ промотор или araB промотор. Когда используют lacZ промотор, экспрессия гена может осуществляться с помощью метода Ward et al. (Nature (1098) 341, 544-6; FASBE J. (1992) 6, 2422-7), в то время как, когда используют araB промотор, экспрессия гена может осуществляться с помощью метода Better et al. (Science (1988) 240, 1041-3).
В отношении сигнальной последовательности для секреции антитела, когда интересующее антитело предназначено для секреции в периплазматическое пространство E. coli, может быть использована сигнальная последовательность pelB (Lei, S.P. et al., J. Bacteriol. (1987) 169, 4379-83). Антитело, секретируемое в периплазматическое пространство, выделяют и затем повторно укладывают так, чтобы антитело приняло соответствующую конфигурацию.
Может быть использован ориджин репликации, происходящий от вирусов (например, SV40, вируса полиомы, аденовируса, вируса папилломы коров (BPV)) или тому подобного. Для того чтобы увеличить количество копий гена в системе клетки-хозяина, экспрессионный вектор может дополнительно содержать ген селективного маркера, такой как ген аминогликозидфосфотрансферазы (APH), ген тимидинкиназы (TK), ген ксантин-гуанинфосфорибозилтрансферазы (Ecogpt) и ген дигидрофолатредуктазы (dhfr).
Для получения антитела, используемого в настоящем изобретении, может быть использована любая экспрессионная система, включая эукариотные и прокариотные клеточные системы. Эукариотная клетка включает признанные клеточные линии животных (например, млекопитающих, насекомых, плесневых грибов и грибов, дрожжей). Прокариотная клетка включает бактериальные клетки, такие как клетки E. coli. Предпочтительно, чтобы антитело, используемое в настоящем изобретении, экспрессировали в клетке млекопитающего, такой как клетка CHO, COS, миеломы, BHK, Vero и HeLa.
Затем трансформированную клетку-хозяина культивируют in vitro или in vivo для получения интересующего антитела. Культивирование клетки-хозяина может быть осуществлено с помощью любого известного метода. Культуральная среда, которая может быть использована в настоящем описании, может представлять собой среду DMEM, MEM, RPMI 1640 или IMDM. Культуральная среда может содержать добавку сыворотки, такой как фетальная телячья сыворотка (FCS).
При получении рекомбинантного антитела, кроме указанных выше клеток-хозяев, в качестве хозяина может быть также использовано трансгенное животное. Например, ген антитела вставляют в предварительно определенный сайт гена, кодирующего белок, которому свойственно продуцироваться в молоко животного (например, бета-казеин) для получения гибридного гена. Фрагмент ДНК, содержащий гибридный ген с введенным геном антитела, вводят эмбриону животного, не являющегося человеком, и эмбрион затем вносят в самку животного. Самка животного, несущая в себе эмбрион, несет трансгенное животное, не являющееся человеком. Антитело, представляющее интерес, секретируется в молоко трансгенного животного, не являющегося человеком, или его потомства. В целях увеличения количества молока, содержащего антитело, трансгенному животному может быть введен подходящий гормон (Ebert, K.M. et al., Bio/Technology (1994) 12, 699-702).
Антитело, экспрессируемое и получаемое как описано выше, может быть выделено из клеток или организма животного-хозяина и очищено. Выделение и очистка антитела, используемого в настоящем изобретении, могут быть осуществлены на аффинной колонке. Могут быть также использованы другие способы, традиционно применяемые для выделения и очистки антитела; таким образом, метод особенно не ограничивается. Например, для выделения и очистки антитела, представляющего интерес, могут быть использованы различные хроматографические методы, фильтрация, ультрафильтрация, высаливание и диализ, по одиночке или в сочетании (Antibodies A Laboratory Manual. Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988).
(2) Гибридное антитело и гуманизированное антитело
В настоящем изобретении может быть использовано искусственно модифицированное рекомбинантное антитело, включая гибридное антитело и гуманизированное антитело. Эти модифицированные антитела могут быть получены любым известным методом. Например, могут быть использованы способы, разработанные для получения «гибридных антител» (Morrison et al., 1984, Proc. Natl. Acad. Sci., 81: 6851-5; Neuberger et al., 1984, Nature, 312: 604-8; Takeda et al., 1985, Nature, 314: 452-4). Гибридное антитело представляет собой молекулу, в которой различные части происходят от различных видов животных, такую как молекула, обладающая вариабельной областью, происходящей от mAb мыши, и константной областью иммуноглобулина человека, например, «гуманизированные антитела».
Гибридное антитело по настоящему изобретению может быть получено с помощью лигирования ДНК, кодирующей V-область антитела, с ДНК, кодирующей C-область антитела человека, интеграции продукта лигирования в экспрессионный вектор и введения полученного рекомбинантного экспрессионного вектора в хозяина для получения гибридного антитела.
Гуманизированное антитело обозначается также как «антитело человека с восстановленной формой», в котором области, определяющие комплементарность (CDR), антитела млекопитающего, не являющегося человеком (например, мыши), соединены с таковыми антитела человека. Обычный способ генетической рекомбинации для получения такого гуманизированного антитела также известен (например, в патентах EP 125023; WO 96/02576).
Конкретно, создается последовательность ДНК, в которой CDR антитела мыши лигируются через каркасные области (FR), и синтезируется с помощью метода ПЦР с использованием нескольких олигонуклеотидов в качестве праймеров, которые создаются как обладающие областями, перекрывающими концевые области CDR и FR. Полученную ДНК лигируют с ДНК, кодирующей C-область антитела человека, и продукт лигирования интегрируют в экспрессионный вектор. Полученный рекомбинантный экспрессионный вектор вводят в хозяина, получая тем самым гуманизированное антитело (например, патент WO 96/02576).
FR, лигированные через CDR, отбирают так, чтобы CDR могли образовывать функциональный антигенсвязывающий сайт. Если это необходимо, аминокислота(ы) в FR V-области антитела могут быть замещены так, что CDR антитела человека с восстановленной формой могло образовывать соответствующий антигенсвязывающий сайт (Sato, K. et al., Cancer Res. (1993) 53, 851-6).
Гибридное антитело состоит из V-областей, происходящих от антитела млекопитающего, не являющегося человеком, и C-областей, происходящих от антитела человека. Гуманизированное антитело состоит из CDR, происходящих от антитела млекопитающего, не являющегося человеком, и FR и C-областей, происходящих от антитела человека. Гуманизированное антитело может быть пригодно для клинического применения, потому что антигенность этого антитела против организма человека снижена.
Конкретным примером гибридного антитела или гуманизированного антитела, используемого в настоящем изобретении, является антитело, в котором CDR происходят от моноклонального антитела мыши 92-13, или антитело, в котором CDR происходят от моноклонального антитела мыши 93-22. Метод получения таких гибридных антител и гуманизированных антител описан ниже.
Для клонирования ДНК, включающей нуклеотидную последовательность, кодирующую V-область мышиного моноклонального антитела против FZD10, из гибридом может быть выделена мРНК, и с использованием обратной транскриптазы может быть синтезирована каждая кДНК в V-областях L- и H-цепей, как описано выше. При синтезе кДНК может быть использован олиго-dT праймер или другой подходящий праймер, который гибридизуется с C-областью L- или H-цепи. Например, может использоваться, но не ограничиваться этим, CH1 (IgG2a) праймер, имеющий нуклеотидную последовательность, как представлено в SEQ ID NO: 3, для V-области Н-цепи, и CL1 (каппа) праймер, имеющий нуклеотидную последовательность, как представлено в SEQ ID NO: 4, для V-области L-цепи.
Амплификация кДНК как L-, так и Н-цепей, может быть осуществлена путем ПЦР (полимеразной цепной реакции) с применением имеющегося в продаже набора (например, набора GeneRacerTM от Invitrogen) или используя известный метод, включая метод 5'-RACE (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA, 85, 8998-9002, 1988; Belyavsky, A. et al., Nucleic Acids Res., 17, 2919-32, 1989).
Конкретные праймеры для амплификации ДНК V-областей мышиного моноклонального антитела 92-13 включают праймеры, имеющие нуклеотидные последовательности, представленные в SEQ ID NO: 5 и 6 для V-области Н-цепи, и праймеры, имеющие нуклеотидные последовательности, представленные в SEQ ID NO: 7 и 8 для V-области L-цепи. При использовании этих праймеров могут быть амплифицированы ДНК, кодирующая V-область Н-цепи, имеющая аминокислотную последовательность, как представлено в SEQ ID NO: 13, и ДНК, кодирующая V-область L-цепи, имеющая аминокислотную последовательность, как представлено в SEQ ID NO: 21. Конкретные праймеры для амплификации ДНК V-областей мышиного моноклонального антитела 93-22 включают праймеры, имеющие нуклеотидные последовательности, представленные в SEQ ID NO: 53 и 54 для V-области Н-цепи, и праймеры, имеющие нуклеотидные последовательности, представленные в SEQ ID NO: 55 и 56 для V-области L-цепи. При использовании этих праймеров могут быть амплифицированы ДНК, кодирующая V-область Н-цепи, имеющая аминокислотную последовательность, как представлено в SEQ ID NO: 29, и ДНК, кодирующая V-область L-цепи, имеющая аминокислотную последовательность, как представлено в SEQ ID NO: 37.
Затем амплифицированные продукты подвергают электрофорезу в агарозном геле в соответствии с общепринятыми методами, и интересующие фрагменты ДНК вырезают, выделяют и лигируют с ДНК вектора.
Полученную ДНК и ДНК вектора можно лигировать с использованием известного набора для лигирования для создания рекомбинантного вектора. ДНК вектора может быть получена с помощью известного метода: J. Sambrook, et al., "Molecular Cloning", Cold Spring Harbor Laboratory Press, 1989. ДНК вектора гидролизуют ферментом(тами) рестрикции, и нуклеотидная последовательность желаемой ДНК может быть определена известным методом или при использовании автоматического секвенатора.
После клонирования фрагментов ДНК, кодирующих V-области L- и Н-цепей моноклонального антитела мыши (в настоящем описании далее L- или Н-цепь антитела иногда может обозначаться как «L- или Н-цепь мыши» для антител мыши и «L- или Н-цепь человека» для антител человека), ДНК, кодирующие V-области мыши, и ДНК, кодирующие константные области антитела человека, лигируют и экспрессируют с получением гибридных антител.
Стандартный метод получения гибридных антител включает лигирование лидирующей последовательности мыши и последовательности V-области, присутствующей в клонированной кДНК, с последовательностью, кодирующей C-область антитела человека, уже присутствующей в экспрессионном векторе клетки млекопитающего. Альтернативно, лидирующую последовательность мыши и последовательность V-области, присутствующую в клонированной кДНК, лигируют с последовательностью, кодирующей C-область антитела человека, с последующим лигированием в экспрессионный вектор клетки млекопитающего.
Полипептид, включающий C-область антитела человека, может представлять собой любую H- или L-цепь C-областей антител человека, включая, например, C гамма 1, C гамма 2, C гамма 3 или C гамма 4 для Н-цепей человека или C лямбда или C каппа для L-цепей.
Для получения гибридного антитела сначала конструируют два экспрессионных вектора; это экспрессионный вектор, содержащий ДНК, кодирующие V-область L-цепи мыши и C-область L-цепи человека, под контролем элемента, контролирующего экспрессию, такого как система промотор/энхансер, и экспрессионный вектор, содержащий ДНК, кодирующие V-область Н-цепи мыши и C-область Н-цепи человека, под контролем элемента, контролирующего экспрессию, такого как система промотор/энхансер. Затем клетки-хозяева, такие как клетки млекопитающих (например, COS клетка), совместно трансформируются этими экспрессионными векторами, и трансформированные клетки культивируются in vitro или in vivo для получения гибридного антитела: см., например, патент WO91/16928.
Альтернативно, лидирующая последовательность мыши, присутствующая в клонированной кДНК, и ДНК, кодирующие V-область L-цепи мыши и C-область L-цепи человека, а также лидирующая последовательность мыши и ДНК, кодирующие V-область Н-цепи мыши и C-область Н-цепи человека, вводят в один экспрессионный вектор (см., например, патент WO94/11523), и указанный вектор используют для трансформации клетки-хозяина; затем трансформированного хозяина культивируют in vivo или in vitro для получения желаемого гибридного антитела.
Вектор для экспрессии Н-цепи гибридного антитела может быть получен путем введения кДНК, включающей нуклеотидную последовательность, кодирующую V-область Н-цепи мыши (в настоящем описании далее обозначается также как «кДНК V-области Н-цепи»), в подходящий экспрессионный вектор, содержащий геномную ДНК, включающую нуклеотидную последовательность, кодирующую C-область Н-цепи антитела человека (в настоящем описании далее обозначается также как «геномная ДНК C-области Н-цепи»), или кДНК, кодирующую указанную область (в настоящем описании далее обозначается также как «кДНК C-области Н-цепи»). C-область Н-цепи включает, например, области C гамма 1, C гамма 2, C гамма 3 или C гамма 4.
Экспрессионные векторы, имеющие геномную ДНК, кодирующую C-область Н-цепи, в частности те, которые кодируют область C гамма 1, включают, например, HEF-PMh-g гамма 1 (патент WO92/19759) и DHER- INCREMENT E-RVh-PM1-f (патент WO92/19759). Альтернативно, библиотека константных областей человека может быть получена, используя кДНК из PBMC человека (мононуклеарных клеток периферической крови), как описано ранее (Liu, A.Y. et al., Proc. Natl. Acad. Sci. USA, Vol.84, 3439-43, 1987; Reff, M.E. et al., Blood, Vol.83, No.2, 435-45, 1994).
Когда кДНК, кодирующую V-область Н-цепи мыши, вставляют в эти экспрессионные векторы, в указанную кДНК может быть введена подходящая нуклеотидная последовательность с помощью метода ПЦР. Например, ПЦР может быть проведена с использованием ПЦР праймера, который создается так, что указанная кДНК имеет на своем 5'-конце узнаваемую последовательность для подходящего фермента рестрикции и консенсусную последовательность Козака непосредственно перед ее кодоном инициации так, чтобы улучшить эффективность транскрипции, а также ПЦР праймер, который создается так, что указанная кДНК имеет на своем 3'-конце узнаваемую последовательность для подходящего фермента рестрикции и донор сайта сплайсинга для надлежащего сплайсинга продуктов первичной транскрипции геномной ДНК с получением мРНК, для введения этих подходящих нуклеотидных последовательностей в экспрессионный вектор.
Созданную таким образом кДНК, кодирующую V-область Н-цепи мыши, обрабатывают подходящим(ми) ферментом(тами) рестрикции, затем ее вставляют в указанный экспрессионный вектор для создания гибридного экспрессионного вектора Н-цепи, содержащего геномную ДНК, кодирующую C-область (область C гамма 1) Н-цепи.
Созданную таким образом кДНК, кодирующую V-область Н-цепи мыши, обрабатывают подходящим(ми) ферментом(тами) рестрикции, лигируют с кДНК, кодирующей C-область C гамма 1 Н-цепи, и вставляют в экспрессионный вектор, такой как pQCXIH (Clontech) для создания экспрессионного вектора, содержащего кДНК, кодирующую гибридную Н-цепь.
Вектор для экспрессии L-цепи гибридного антитела может быть получен с помощью лигирования кДНК, кодирующей V-область L-цепи мыши, и геномной ДНК или кДНК, кодирующих C-область L-цепи антитела человека, и введения в подходящий экспрессионный вектор. C-область L-цепи включает, например, каппа цепь и лямбда цепь.
При создании экспрессионного вектора, содержащего кДНК, кодирующую V-область L-цепи мыши, в указанный экспрессионный вектор методом ПЦР могут быть введены подходящие нуклеотидные последовательности, такие как узнаваемая последовательность или консенсусная последовательность Козака.
Полная нуклеотидная последовательность кДНК, кодирующей C-область цепи L лямбда человека, может быть синтезирована с помощью ДНК-синтезатора и сконструирована с помощью метода ПЦР. Известно, что C-область цепи L лямбда человека содержит, по меньшей мере, 4 разных изотипа, и каждый изотип может быть использован для конструирования экспрессионного вектора.
Сконструированная кДНК, кодирующая C-область цепи L лямбда человека, и указанная выше сконструированная кДНК, кодирующая V-область L-цепи мыши, могут быть лигированы между подходящими сайтами ферментов рестрикции и вставлены в экспрессионный вектор, такой как pQCXIH (Clontech), для конструирования экспрессионного вектора, содержащего кДНК, кодирующую цепь L лямбда гибридного антитела.
ДНК, кодирующая C-область цепи L каппа человека, предназначенная для лигирования с ДНК, кодирующей V-область L-цепи мыши, может быть сконструирована, например, из HEF-PM1k-gk, содержащей геномную ДНК (см. WO92/19759). Альтернативно, с помощью кДНК из PBMC (мононуклеарных клеток периферической крови) человека может быть получена библиотека константной области человека, как описано ранее (Liu, A.Y. et al., Proc. Natl. Acad. Sci. USA, Vol.84, 3439-43, 1987; Reff, M.E. et al., Blood, Vol.83, No.2, 435-45, 1994).
Узнаваемые подходящими ферментами рестрикции последовательности могут быть введены методом ПЦР в 5'- и 3'-концы ДНК, кодирующей C-область цепи L каппа, и ДНК, кодирующей V-область L-цепи мыши, сконструированной выше, и ДНК, кодирующая C-область цепи L каппа, может быть лигирована с каждой из них и вставлена в экспрессионный вектор, такой как pQCXIH (Clontech), для создания экспрессионного вектора, содержащего кДНК, кодирующую цепь L каппа гибридного антитела.
Для получения гуманизированного антитела, в котором CDR моноклонального антитела мыши соединена с антителом человека, желательно наличие высокой гомологии между FR моноклонального антитела мыши и FR антитела человека. Соответственно производится сопоставление между V-областями H и L-цепей моноклонального антитела мыши против FZD10 и V-областями всех известных антител, структура которых была выяснена, с помощью Protein Data Bank. Кроме того, их одновременно сравнивают с подгруппами антител человека (HSG: Human subgroup), классифицируемыми по Kabat et al. на основании длины FR антитела, аминокислотной гомологии и тому подобного: Kabat, E.A. et al., US Dep, Health and Human Services, US Government Printing Offices, 1991.
Первая стадия создания ДНК, кодирующей V-область гуманизированного антитела, заключается в выборе V-области антитела человека в качестве основы для конструирования. Например, для получения гуманизированного антитела может быть использована FR V-области антитела человека с гомологией более 80% с FR V-области антитела мыши.
В гуманизированном антителе C-область и каркасные (FR) области V-области указанного антитела происходят от человека, а области, определяющие комплементарность (CDR) V-области, происходят от мыши. Полипептид, включающий V-область гуманизированного антитела, может быть получен способом, называемым прививкой CDR, с помощью метода ПЦР при условии наличия фрагмента ДНК антитела человека в качестве матрицы. Термин «прививка CDR» относится к методу, при котором получают фрагмент ДНК, кодирующей CDR от мыши, и заменяют им CDR антитела человека в качестве матрицы.
Если фрагмент ДНК антитела человека, подлежащий использованию в качестве матрицы, недоступен, может быть синтезирована на синтезаторе ДНК нуклеотидная последовательность, имеющаяся в базе данных, и с помощью метода ПЦР может быть получена ДНК V-области гуманизированного антитела. Кроме того, когда в базе данных имеется лишь аминокислотная последовательность, полная нуклеотидная последовательность может быть выведена из аминокислотной последовательности на основе знаний об использовании кодонов в антителах, как сообщалось Kabat, E.A. et al. в US Dep. Health and Human Services, US Government Printing Offices, 1991. Нуклеотидную последовательность синтезируют на синтезаторе ДНК, и ДНК V-области гуманизированного антитела может быть получена с помощью метода ПЦР и введена в подходящего хозяина с последующей ее экспрессией для получения желаемого полипептида.
Общие процедуры прививки CDR с помощью метода ПЦР описаны ниже для случая, когда имеется фрагмент ДНК антитела человека в качестве матрицы.
Прежде всего синтезируют фрагменты ДНК, соответствующие интересующим CDR. CDR с 1 по 3 синтезируют на основе нуклеотидных последовательностей ранее клонированных V-областей H и L-цепей мыши. Например, когда гуманизированное антитело получают на основе моноклонального антитела мыши 92-13, последовательности CDR V-области Н-цепи могут представлять собой аминокислотные последовательности, представленные в SEQ ID NO: 15 (VH CDR1), 17 (VH CDR2) и 19 (VH CDR3); а последовательности CDR V-области L-цепи могут представлять собой аминокислотные последовательности, представленные в SEQ ID NO: 23 (VL CDR1), 25 (VL CDR2) и 27 (VL CDR3). Когда гуманизированное антитело получают на основе моноклонального антитела мыши 93-22, последовательности CDR V-области Н-цепи могут представлять собой аминокислотные последовательности, представленные в SEQ ID NO: 31 (VH CDR1), 33 (VH CDR2) и 35 (VH CDR3); а последовательности CDR V-области L-цепи могут представлять собой аминокислотные последовательности, представленные в SEQ ID NO: 39 (VL CDR1), 41 (VL CDR2) и 43 (VL CDR3).
ДНК V-области Н-цепи гуманизированного антитела может быть лигирована с ДНК любой C-области Н-цепи антитела человека, например, областью C гамма 1 Н-цепи человека. Как упоминалось выше, ДНК V-области Н-цепи может быть обработана подходящим ферментом рестрикции и лигирована с ДНК, кодирующей C-область Н-цепи человека, экспрессия которой управляется контролирующим элементом, таким как энхансерная/промоторная система для получения экспрессионного вектора, содержащего ДНК гуманизированной V-области Н-цепи и C-области Н-цепи человека.
ДНК V-области L-цепи гуманизированного антитела может быть лигирована с ДНК любой C-области L-цепи антитела человека, например, областью C лямбда L-цепи человека. ДНК V-области L-цепи может быть обработана подходящим ферментом рестрикции и лигирована с ДНК, кодирующей C-область L-цепи лямбда человека, экспрессия которой управляется контролирующим элементом, таким как энхансерная/промоторная система для получения экспрессионного вектора, содержащего ДНК гуманизированной V-области L-цепи и C-области L-цепи лямбда человека.
ДНК, кодирующая V-область Н-цепи гуманизированного антитела и C-области Н-цепи, и ДНК, кодирующая гуманизированную V-область L-цепи и C-область L-цепи человека, могут быть также введены в единый экспрессионный вектор, такой как раскрытый в патенте WO94/11523, причем указанный вектор может быть использован для трансформации клетки-хозяина, и трансформированный хозяин может культивироваться in vivo или in vitro с получением желаемого гуманизированного антитела.
Для получения гибридного или гуманизированного антитела следует получить два упомянутых выше экспрессионных вектора. Так, в случае гибридного антитела конструируют экспрессионный вектор, включающий ДНК, кодирующую V-область Н-цепи мыши и C-область Н-цепи человека, управляемую контролирующим элементом экспрессии, таким как энхансер/промотор, и экспрессионный вектор, включающий ДНК, кодирующую V-область L-цепи мыши и C-область L-цепи человека, управляемую контролирующим элементом экспрессии. В случае гуманизированного антитела конструируют экспрессионный вектор, включающий ДНК, кодирующую гуманизированную V-область Н-цепи и C-область Н-цепи человека, управляемую контролирующим элементом экспрессии, и экспрессионный вектор, включающий ДНК, кодирующую гуманизированную V-область L-цепи и C-область L-цепи человека, управляемую контролирующим элементом экспрессии.
После этого клетку-хозяина, такую как клетка млекопитающего (например, клетку COS), совместно трансформируют этими экспрессионными векторами, и полученную трансформированную клетку можно культивировать in vitro или in vivo с получением гибридного или гуманизированного антитела (см., например, патент WO91/16928).
Альтернативно, ДНК, кодирующая V и C-области Н-цепи, и ДНК, кодирующая V и C-области L-цепи, могут быть лигированы в единый вектор и трансформированы в подходящую клетку-хозяина с получением антитела. Так, при экспрессии гибридного антитела ДНК, кодирующая лидирующую последовательность мыши, находящуюся в клонированной кДНК, V-область Н-цепи мыши и C-область Н-цепи человека, а также ДНК, кодирующая лидирующую последовательность мыши, V-область L-цепи мыши и C-область L-цепи человека, могут быть введены в единый экспрессионный вектор, такой как, например, раскрытый в патенте WO94/11523. В случае экспрессии гуманизированного антитела ДНК, кодирующая гуманизированную V-область Н-цепи и C-область Н-цепи человека, и ДНК, кодирующая гуманизированную V-область L-цепи и C-область L-цепи человека, могут быть введены в единый экспрессионный вектор, такой как, например, раскрытый в патенте WO94/11523. Такой вектор используют для трансформации клетки-хозяина, и трансформированную клетку культивируют in vivo или in vitro с получением интересующего гибридного или гуманизированного антитела.
Для получения гибридного или гуманизированного антитела против белка FZD10 по настоящему изобретению может быть использована любая экспрессионная система. Например, эукариотные клетки включают клетки животных, такие как стабильные линии клеток млекопитающих, клетки грибов и клетки дрожжей; прокариотные клетки включают бактериальные клетки, такие как Escherichia coli. Предпочтительно, гибридное или гуманизированное антитело по настоящему изобретению экспрессируют в клетке млекопитающего, такой как клетка COS или CHO.
Могут быть использованы любые традиционные промоторы, пригодные для экспрессии в клетках млекопитающих. Например, предпочтительно используют немедленный ранний промотор цитомегаловируса человека (HCMV). Кроме того, промоторы для экспрессии генов в клетках млекопитающих могут включать вирусные промоторы, такие как промоторы ретровируса, вируса полиомы, аденовируса и обезьяньего вируса (SV) 40, и промоторы из клеток млекопитающих, такие как промотор фактора-1 альфа элонгации полипептидной цепи человека (HEF-1 alpha). Например, промотор SV40 может быть легко использован по способу Mulligan et al. (Nature, 277, 108-14, 1979); способ Mizushima, S. et al. (Nucleic Acids Research, 18, 5322, 1990) может быть легко использован с промотором HEF-1 alpha.
Ориджин репликации включает ориджины репликации из SV40, вируса полиомы, аденовируса и вируса папилломы крупного рогатого скота (BPV). Кроме того, экспрессионный вектор может включать ген фосфотрансферазы (APH(3') II или I (neo), тимидинкиназы (TK), ксантин-гуанинфосфорибозилтрансферазы E. coli (Ecogpt) или дигидрофолатредуктазы (DHFR) в качестве маркера селекции для повышения количества копий гена в системе клетки-хозяина.
Гибридное или гуманизированное антитело, представляющее интерес, которое получают таким образом путем культивирования трансформанта, трансформированного ДНК, кодирующей гибридное или гуманизированное антитело, может быть выделено из клетки и затем очищено.
Выделение и очистка гибридного или гуманизированного антитела, представляющего интерес, могут быть осуществлены с помощью колонки протеин A-агарозы, но могут быть также произведены с помощью любых способов, используемых для выделения и очистки белка, и, таким образом, не являются ограничивающими. Например, для выделения и очистки гибридного или гуманизированного антитела могут быть необязательно выбраны или использованы в сочетании хроматография, ультрафильтрация, высаливание или диализ.
После выделения гибридного антитела или гуманизированного антитела концентрация полученного очищенного антитела может быть определена с помощью ELISA.
Определение антигенсвязывающей активности или других типов активности гибридного антитела или гуманизированного антитела, включая связывающую активность в отношении нормальной клетки, может быть произведено с помощью любых известных способов (Antibodies A Laboratory Manual, Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988).
Что касается способа определения антигенсвязывающей активности антитела, могут быть использованы такие методы, как ELISA (твердофазный иммуноферментный анализ), ИФА (иммуноферментный анализ), РИА (радиоиммунный анализ) или флуоресцентный анализ.
(3) Фрагмент антитела и модифицированное антитело
Антитело, используемое в настоящем изобретении, может быть фрагментом антитела или модифицированным антителом, до тех пор пока оно связывает белок FZD10 и подавляет его активность. Например, фрагмент антитела включает Fab, F(ab')2, Fv или одноцепочечный Fv (scFv), состоящий из фрагмента Fv Н-цепи или фрагмента Fv L-цепи, соединенных друг с другом посредством подходящего линкера. Конкретно, такие фрагменты антитела могут быть получены путем расщепления антитела ферментом (например, папаином, пепсином) до фрагментов антитела или путем конструирования гена, кодирующего фрагмент антитела, и вставки гена в экспрессионный вектор и введения полученного рекомбинантного экспрессионного вектора в подходящую клетку-хозяина, экспрессирующую таким образом фрагмент антитела (см., например, Co, M. S., et al., J. Immunol. (1994), 152, 2968-76; Better, M. & Horwitz, A.H., Methods in Enzymology (1989), 178, 476-96, Academic Press, Inc.; Pluckthun, A. & Skerra, A., Methods in Enzymology (1989) 178, 497-515, Academic Press, Inc.; Lamoyi, E., Methods in Enzymology (1989) 121, 652-63; Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-9; и Bird, R.E. et al., Trends Biotechnol. (1991) 9, 132-7). Альтернативно, могут быть сконструированы экспрессионные библиотеки Fab (Huse et al., 1989, Science, 246: 1275-81) для обеспечения быстрой и простой идентификации моноклональных фрагментов Fab с желаемой специфичностью.
scFv может быть получен лигированием V-области Н-цепи с V-областью L-цепи через линкер, предпочтительно пептидный линкер (Huston, J. S. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 5879-83). V-область Н-цепи и V-область L-цепи в scFv могут происходить от любого из описанных в настоящем описании антител. Пептидный линкер, связывающий V-области, может быть любым одноцепочечным пептидом, например, состоящим из 12-19 аминокислотных остатков.
В качестве модифицированного антитела также могут быть использованы, например, антитело против FZD10 или его фрагмент, конъюгированные с любой молекулой (например, полиэтиленгликолем). Такие модифицированные антитела также охватываются термином «антитело» настоящего изобретения. Модифицированные антитела могут быть получены путем химических модификаций антител. Способы химической модификации, пригодные для этой цели, уже были установлены в данной области техники.
2. Терапевтическое применение
Ниже описаны способы и фармацевтические композиции для лечения и/или профилактики связанного с FZD10 заболевания с использованием антитела настоящего изобретения. Результатом лечения является, по меньшей мере, благотворное действие на подвергающегося лечению субъекта, которое в случае опухолей включает, но не ограничивается этим, ремиссию опухолей, облегчение симптомов опухолей и контроль над метастатическим распространением опухолей.
Конкретно, способ лечения и/или профилактики связанного с FZD10 заболевания у субъекта по настоящему изобретению включает введение нуждающемуся в этом субъекту антитела или фрагмента, описанного выше.
Термин «субъект» в настоящем описании относится к субъекту, который страдает связанным с FZD10 заболеванием, а также к субъекту, предположительно имеющему связанное с FZD10 заболевание. Субъектом в настоящем изобретении может быть животное, включая млекопитающих и птиц. Например, млекопитающие могут включать человека, мышей, крыс, обезьян, кроликов и собак.
Термин «связанное с FZD10 заболевание» в настоящем описании относится к заболеванию, связанному с избыточной экспрессией белка FZD10. Конкретно, связанные с FZD10 заболевания включают, но не ограничиваются этим, синовиальную саркому (SS), колоректальный рак, рак желудка, хронический миелоидный лейкоз (CML) и острый миелоидный лейкоз (AML).
Описанное в настоящем описании антитело или его фрагмент может специфически связываться с белком FZD10, так что при введении антитела или его фрагмента субъекту оно связывается с белком FZD10 у субъекта, и активность белка FZD10 может быть подавлена. Альтернативно, когда антитело или его фрагмент могут быть конъюгированы с терапевтической частью и введены субъекту, оно доставляется в область, которая экспрессирует белок FZD10 (т.е. страдающую область) субъекта, и терапевтическая часть может быть избирательно доставлена в страдающую область и воздействовать на нее. Такая терапевтическая часть может представлять собой любое известное лекарственное средство или средство, которое будет разработано в будущем, оказывающее лечебное действие на связанное с FZD10 заболевание и включающее, не ограничиваясь этим, радиоизотопную метку и химиотерапевтический агент. Радиоизотопная метка, которая может быть использована в качестве терапевтического агента, может быть выбрана в зависимости от множества факторов, включая энергию β-излучения и эффективность его эмиссии, наличие или отсутствие γ-излучения, его энергии и эффективности эмиссии, физического периода полужизни и процедуры промечивания. Обычно может быть использована радиоизотопная метка на основе иттрия (такого как 90Y) и йода (такого как 125I и 131I). Химиотерапевтический агент может представлять собой любой известный агент или агент, который будет создан в будущем, для лечения связанного с FZD10 заболевания и включающий, не ограничиваясь этим, метотрексат, таксол, меркаптопурин, тиогуанин, цисплатин, карбоплатин, митомицин, блеомицин, доксорубицин, идарубицин, даунорубицин, дактиномицин, винбластин, винкристин, винорелбин, паклитаксель и доцетаксель. Описанное в настоящем описании антитело или его фрагмент может избирательно связываться с белком FZD10 и не связываться с нормальной клеткой, в результате чего побочное действие, которое вызывается антителом или его фрагментом или радиоизотопным или химиотерапевтическим агентом, может быть эффективно нивелировано, и, следовательно, терапевтическая эффективность может быть высокой.
Антитело или его фрагмент, описанное в настоящем описании, может быть введено субъекту в эффективной дозе для лечения или профилактики связанного с FZD10 заболеванием. Эффективная доза означает такое количество антитела или его фрагмента, которое достаточно для обеспечения благотворного действия у подвергаемого лечению субъекта. Составы и способы введения, которые могут быть использованы в случаях, когда фармацевтическая композиция содержит антитело по настоящему изобретению, описаны ниже.
Фармацевтические композиции для применения по настоящему изобретению могут быть составлены традиционным способом с использованием одного или более фармацевтически приемлемых носителей или наполнителей.
Антитела или их фрагменты могут быть составлены для парентерального введения (т.е. внутривенного или внутримышечного) путем инъекции посредством, например, болюсной инъекции или продолжительной инфузии. Составы для инъекции могут быть представлены в виде единиц лекарственной формы, например ампул или контейнеров с множеством доз, с добавленным консервантом. Композиции могут принимать такие формы, как суспензии, растворы или эмульсии в масляных или водных носителях, и могут содержать обеспечивающие образование формы агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, антитело может находиться в форме лиофилизованного порошка для соединения с подходящим носителем, например стерильной, свободной от пирогенов водой, перед применением.
Токсичность и терапевтическая эффективность антитела или фрагмента или терапевтической части, присоединенной к ним, могут быть определены с помощью стандартных фармацевтических процедур на клеточных культурах или экспериментальных животных, например, для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Соотношение доз, вызывающих токсичное и терапевтическое действие, представляет собой терапевтический индекс и может быть выражено в виде отношения LD/ED.
Предпочтительны антитела или терапевтические части, проявляющие высокие терапевтические индексы. В случаях использования антител или частей, проявляющих токсичное побочное действие, следует обеспечить систему доставки, которая направляет такие антитела или части к участку поврежденной ткани для сведения к минимуму потенциального повреждения неинфицированных клеток и, тем самым, для снижения побочного действия.
Данные, полученные при анализе клеточных культур и в исследованиях на животных, можно использовать для формирования диапазонов доз для применения у человека. Доза таких антител предпочтительно находится в диапазоне концентраций в плазме циркулирующей крови, включающем ED50 с низкой или отсутствующей токсичностью. Доза может варьировать в пределах этого диапазона в зависимости от используемой формы дозы, используемого пути введения и типа и количества конъюгированной терапевтической части. Для любого антитела, используемого в способе изобретения, эффективная доза может быть установлена изначально на основании тестов в клеточных культурах. Доза может быть определена на животных моделях для обеспечения диапазона концентраций в плазме циркулирующей крови, включающего IC50 (т.е. концентрацию тестируемого антитела, которая обеспечивает полумаксимальное ингибирование симптомов), определенную в клеточной культуре. Эта информация может быть использована для более точного определения доз, пригодных для человека. Уровни в плазме могут быть измерены, например, с помощью высокоэффективной жидкостной хроматографии.
В зависимости от состояния и возраста субъекта и/или пути введения специалист в данной области техники может подобрать подходящую дозу фармацевтической композиции по настоящему изобретению. Например, фармацевтическую композицию по настоящему изобретению вводят в таком количестве, что антитело по настоящему изобретению вводится субъекту в количестве от приблизительно 3 до приблизительно 15 мкг на кг массы тела субъекта и предпочтительно от приблизительно 10 до приблизительно 15 мкг на кг массы тела субъекта. Интервал между введениями и время введения могут быть выбраны с учетом состояния и возраста субъекта, пути введения и реакции на фармацевтическую композицию. Например, фармацевтическую композицию можно вводить субъекту от одного до 5 раз, предпочтительно 1 раз в день на протяжении от 5 до 10 дней.
Фармацевтическую композицию можно вводить системно или местно. Предпочтительно ее введение способом направленной доставки для доставки активного компонента в область повреждения.
В отдельных вариантах осуществления способы и композиции по настоящему изобретению используют для лечения или профилактики связанного с FZD10 заболевания совместно с одним химиотерапевтическим агентом или сочетанием химиотерапевтических агентов, включающими, но не ограниченными этим, метотрексат, таксол, меркаптопурин, тиогуанин, цисплатин, карбоплатин, митомицин, блеомицин, доксорубицин, идарубицин, даунорубицин, дактиномицин, винбластин, винкристин, винорелбин, паклитаксель и доцетаксель.
Что касается рентгенотерапии, то может быть использован любой протокол рентгенотерапии в зависимости от типа подвергаемого лечению связанного с FZD10 заболевания. Например, но не ограничиваясь этим, может быть использовано рентгеновское облучение. Для экспозиции тканей могут быть также использованы испускающие гамма-излучение радиоизотопы, такие как радиоактивные изотопы радия, кобальта и других элементов.
В другом варианте осуществления химиотерапию или рентгенотерапию применяют на протяжении, по меньшей мере, часа, пяти часов, 12 часов, суток, недели, месяца и более, предпочтительно нескольких месяцев (например, до трех месяцев) после использования способов и композиций, содержащих антитело настоящего изобретения. Химиотерапия или рентгенотерапия, используемая до, одновременно или после лечения с применением способов и композиций по настоящему изобретению, может использоваться с помощью любого известного в данной области техники способа.
3. Диагностическое и прогностическое использование
Антитела против белка FZD10 или его фрагментов можно также использовать в качестве диагностических и прогностических агентов, как описано в настоящем описании. Такие диагностические способы можно использовать для выявления наличия или отсутствия связанного с FZD10 заболевания и риска развития такого заболевания. Способ диагностики и/или предсказания настоящего изобретения связанного с FZD10 заболевания включает иммунологическое выявление или определение в образце белка FZD10, появляющегося в результате заболевания, с помощью антитела или его фрагмента по настоящему изобретению. Конкретно, способ диагностики и/или предсказания связанного с FZD10 заболевания или предрасположенности субъекта к такому заболеванию по настоящему изобретению включает:
(a) введение в контакт образца от субъекта с антителом против белка FZD10 или его фрагмента;
(b) выявление белка FZD10 в образце; и
(c) принятие решения о том, страдает ли субъект от заболевания или имеет риск развития заболевания, на основании относительного избытка белка FZD10 по сравнению с контролем.
Способ диагностики и/или предсказания по настоящему изобретению может быть реализован на основе любых процедур, пока они представляют собой анализ с использованием антитела, т.е. иммунологический анализ. Таким образом, белок FZD10 может быть определен с помощью антитела или его фрагмента по настоящему изобретению в качестве антитела, используемого для анализа. Например, белок FZD10 может быть выявлен с помощью иммуногистохимического окрашивания, иммуноанализа, такого как иммуноферментный анализ (ELISA и ИФА), иммунофлуоресцентного анализа, радиоиммунологического анализа (РИА) или вестерн-блоттинга.
Образец, подлежащий тестированию способом диагностики и/или предсказания связанного с FZD10 заболевания по настоящему изобретению, не имеет особых ограничений до тех пор, пока он представляет собой биологический образец, который может содержать белок FZD10, появляющийся в результате связанного с FZD10 заболевания. Примеры образца включают экстракт клетки или органа и срезы ткани, а также кровь, сыворотку, плазму, супернатант культивируемых лимфоцитов, мочу, спинномозговую жидкость, слюну, пот и асциты. Избыток белка FZD10, выявленный в таких образцах, как опухолевая ткань, биопсия опухоли и ткань с метастазом, с помощью антитела или его фрагмента по настоящему изобретению, может быть особенно пригодным в качестве показателя связанного с FZD10 заболевания.
Например, антитела или их фрагменты, описанные в настоящем описании, могут быть использованы для количественного или качественного выявления белка FZD10. Антитела (или их фрагменты) по настоящему изобретению, кроме того, могут быть использованы для гистологии, например, иммуннофлуоресцентной или иммуноэлектронной микроскопии, для выявления белка FZD10 in situ. Выявление in situ может быть осуществлено отбором гистологического образца у субъекта, такого как заключенные в парафин срезы тканей (таких как образцы, полученные при хирургических операциях), и нанесения на них меченого антитела по настоящему изобретению. Антитело (или его фрагмент) предпочтительно наносят покрытием образца меченым антителом (или его фрагментом). Специалистам в данной области техники должно быть вполне понятно, что при использовании настоящего изобретения для достижения такого выявления in situ любой из множества гистологических методов (такой, как процедура окрашивания) может быть модифицирован.
Иммунологические анализы белка FZD10 обычно включают инкубацию подвергаемого исследованию образца от субъекта, такого как биологическая жидкость, экстракт ткани, свежесобранные клетки или лизаты клеток, инкубированных в клеточной культуре, в присутствии антитела по настоящему изобретению, меченого выявляемой меткой, и выявление связанного антитела с помощью любого из ряда способов, хорошо известных специалистам в данной области техники.
Образец может быть введен в контакт и иммобилизован на твердофазной подложке или носителе, такой как нитроцеллюлоза, или другой твердой подложке, которая способна иммобилизовывать клетки, клеточные частицы или растворимые белки. Подложку затем можно промыть подходящими буферами и затем обработать меченым выявляемой меткой антителом против FZD10. Твердофазную подложку можно затем промыть буфером второй раз для удаления несвязанного антитела. Затем можно определить с помощью традиционных средств количество связанной метки на твердой подложке.
Термин «твердофазная подложка или носитель» означает любую подложку, способную связывать антиген или антитело. Специалистам в данной области техники должны быть известны многие подходящие носители для связывания антител или антигенов, или специалисты должны быть способны выявить такие носители путем традиционного экспериментирования.
Связывающую активность данной партии антитела против FZD10 можно определить с помощью хорошо известных способов. Специалисты в данной области техники должны быть способны определить рабочие и оптимальные условия анализа для каждого определения с помощью традиционного экспериментирования.
Взаимодействие между антителом (или его фрагментом) по настоящему изобретению и белком FZD10, легко получаемым в образце из области, затронутой связанным с FZD10 заболеванием, можно выявить прямо путем промечивания антитела по настоящему изобретению или непрямо путем промечивания вторичного антитела. Последняя процедура непрямого выявления, такая как сэндвичный анализ или конкурентный анализ ELISA, предпочтительно используется в способе по настоящему изобретению благодаря улучшенной чувствительности.
Примерами меток для использования в настоящем описании являются следующие. В иммуноферментном анализе можно использовать пероксидазы (POD), щелочные фосфатазы, β-галактозидазу, уреазу, каталазу, глюкозооксидазу, лактатдегидрогеназу, амилазы и комплексы авидин-биотин. В иммунофлуоресцентном анализе можно использовать флуоресцеина изотиоцианат (FITC), тетраметилродамина изотиоцианат (TRITC), замещенный родамина изотиоцианат, дихлортриазина изотиоцианат и Alexa488. В радиоиммунологическом анализе можно использовать тритий, йод (такой как 125I и 131I) и индий (такой как 111In). В иммунолюминесцентном анализе можно использовать тест НАДH-ФМНH2-люцифераза, систему люминол-перекись водорода-POD, сложные эфиры акридина и диоксетановые соединения.
Метку можно присоединять к антителу с помощью традиционной процедуры. Например, метку можно присоединить к антителу с помощью глутаральдегидного способа, малеимидного способа, пиридилдисульфидного способа или перйодатного способа в иммуноферментном анализе и способа хлорамин-T или способа Болтона-Хантера в радиоиммунологическом анализе.
Анализ можно проводить в соответствии с известной процедурой (Ausubel, F.M. et al. Eds., Short Protocols in Molecular Biology, Chapter 11 "Immunology" John Wiley & Sons, Inc. 1995).
Например, когда антитело по настоящему изобретению прямо метят описанной выше меткой, образец вводят в контакт с меченым антителом с образованием в результате комплекса между белком FZD10 и антителом. Затем отделяют несвязанное меченое антитело, и уровень белка FZD10 в образце можно определить на основании количества связанного меченого антитела или количества несвязанного меченого антитела.
При использовании вторичного меченого антитела антителу по настоящему изобретению дают провзаимодействовать с образцом в первой реакции, и полученному комплексу дают провзаимодействовать с меченым вторичным антителом во второй реакции. Первая реакция и вторая реакция могут быть произведены в обратном порядке, одновременно или через некоторый промежуток времени между ними. Первая и вторая реакции дают комплекс [белок FZD10]-[антитело по изобретению]-[меченое вторичное антитело] или комплекс [антитело по изобретению]-[белок FZD10]-[меченое вторичное антитело]. Несвязанное меченое вторичное антитело затем отделяют, и уровень белка FZD10 в образце можно определить на основании количества связанного меченого вторичного антитела или количества несвязанного меченого вторичного антитела.
Согласно другому варианту осуществления антитело по настоящему изобретению метят радиоизотопом или флуоресцентной меткой, и меченое антитело вводят парэнтерально субъекту. Таким образом с помощью неинвазивного способа может быть быстро определена локализация первичной опухоли и связанной с ней метастазной опухоли связанного с FZD10 заболевания. Такой способ диагностики известен как визуализация опухоли in vivo, и специалист в данной области техники может легко понять его процедуры. Меченое антитело можно вводить субъекту системно или местно, предпочтительно парентеральным путем, таким как внутривенная инъекция, внутримышечная инъекция, внутрибрюшинная инъекция или подкожная инъекция.
Антитела по настоящему изобретению специфически взаимодействуют с белком FZD10, как упоминалось выше, и могут благодаря этому использоваться в наборах для диагностики и/или прогнозирования связанного с FZD10 заболевания.
Набор для диагностики и/или прогнозирования по настоящему изобретению включает антитело или его фрагмент, описанные выше. Путем определения белка FZD10 в образце от субъекта с подозрением на наличие связанного с FZD10 заболевания с использованием набора для диагностики и/или прогнозирования по настоящему изобретению можно быстро и легко определить, страдает ли субъект связанным с FZD10 заболеванием. Наборы для диагностики и/или прогнозирования заболеваний с использованием подобных иммунологических реакций получили широкую известность, и специалист в данной области техники может легко выбрать подходящие компоненты, отличные от антитела. Наборы для диагностики и/или прогнозирования по настоящему изобретению можно использовать любыми способами, пока они представляют собой способы иммунологического анализа.
ПРИМЕРЫ
Настоящее изобретение далее будет иллюстрировано следующими неограничивающими примерами.
Линии клеток и образцы тканей, использованные в следующих примерах, получали, как описано ниже. Конкретно, линии клеток, берущие начало от синовиальных сарком (HS-SY-2, YaFuSS, 1973/99, Fuji и SYO-1), раков ободочной кишки (LoVo, SNU-C4 и SNU-C5), клетки HEK293 и COS7 выращивали в виде монослоя в подходящих средах с добавлением 10% фетальной телячьей сыворотки и 1% раствора антибиотика/противогрибкового агента и поддерживали при 37°С в атмосфере воздуха, содержащего 5% CO2. Образцы первичной синовиальной саркомы (SS) получали после информированного согласия, мгновенно замораживали в жидком азоте сразу после резекции и хранили при -80°С.
Пример 1
Получение моноклональных антител против FZD10
(1) Получение моноклональных антител с помощью иммобилизации клеток
Моноклональные антитела мыши против FZD10 (Mab) получали путем иммунизации четырехнедельных самок мыши линии Balb/c в подушечки лап 2×107 клеток COS-7, трансфицированных 2×107 pCAGGS/neo-FZD10-myc/His (Medical and Biological Laboratories, Nagoya, Japan). Конструкция pCAGGS/neo-FZD10-myc/His была описана ранее (Nagayama, S., et al. (2005). Oncogene, 24, 6201-12), и она экспрессирует полную последовательность, кодирующую кДНК FZD10 и Myc и метки эпитопа His на C-конце. Мышей иммунизировали полным адъювантом Фрейнда (Mitsubishi Kagaku Iatron, Inc., Tokyo, Japan) за один день до иммунизации клетками. От иммунизированных мышей получали клетки селезенки, которые сливали с клетками миеломной линии. Гибридомы субклонировали и тестировали с помощью клеточного ELISA на способность секретировать иммуноглобулин, который связывается с внеклеточным доменом FZD10 (аминокислотные остатки 1-225 FZD10). Для клеточного ELISA клетки COS-7, экспрессирующие FZD10-myc/His (полная последовательность кДНК, кодирующей FZD10 и Myc и His эпитопные метки на его C-конце), высевали в 96-луночные планшеты. Затем в планшету добавляли по 50 мкл культуральных супернатантов, полученных из гибридом, и инкубировали в течение 30 минут при комнатной температуре. После промывки клеток добавляли козьи IgG-POD против мыши (Medical and Biological Laboratories, Nagoya, Japan) в разведении 1:10000, инкубировали в течение 30 минут при комнатной температуре. Связанные антитела выявляли по OD450-620 нм. Положительные клоны дополнительно анализировали на специфическую связывающую активность. Эти клоны включают: клоны 39-2 и 39-10 (раскрытые в патенте WO2005/004912, обозначенные как 5F2), а также 92-13 и 93-22. Все Mab имели изотип IgG2a, определенный с помощью набора IsoStrip для изотипирования моноклональных антител мыши (Roche). Mab подвергали аффинной очистке на протеин-G-сефарозе для дополнительной характеристики.
Гибридомный клон 93-22, продуцирующий моноклональное антитело мыши 93-22, был сдан на международное хранение Shuichi Nakatsuru в IPOD International Patent Organism Depository of the National Institute of Advanced Industrial Science and Technology (AIST Tsukuba Central 6, 1-1, Higashi 1-chome, Tsukuba-shi, Ibaraki-Ken, 305-8566 Japan) 14 июня 2006 г. с регистрационным номером FERM BP-10620. Также гибридомный клон 92-13, продуцирующий моноклональное антитело мыши 92-13, был сдан на международное хранение Shuichi Nakatsuru в IPOD International Patent Organism Depository of the National Institute of AIST, 28 июня 2006 г. с регистрационным номером FERM BP-10628.
(2) Промечивание антител радионуклидами
Меченные 125I Mab получали способом с хлорамином T (Arano, Y., et al. (1999). Cancer Res, 59, 128-34). 740 кБк/2 мкл Na125I добавляли к 10 мкг Mab в 100 мкл 0,3 М натрий-фосфатного буфера. Затем добавляли один мкг хлорамина T в 3 мкл 0,3 М натрий-фосфатного буфера, инкубировали в течение 5 мин при комнатной температуре. Меченое антитело очищали с помощью колонки 6 Biospin (Bio-Rad).
Для промечивания Mab 111In 1 мг Mab в 100 мкл 50 мМ боратного буфера (pH 8,5) конъюгировали с изотиоцианатбензилдиэтилентриаминпентауксусной кислотой (SCN-BZ-DTPA; Macrocyclics) в диметилформамиде в молярном отношении 1:3. После инкубации при 37°С в течение 20 часов конъюгаты Mab очищали с помощью колонки 6 Biospin. 40 мкл 111In инкубировали в 60 мкл 0,25 М уксуснокислого буфера (pH 5,5) и включали в 10 мкг/мкл конъюгатов Mab-DTPA в течение одного часа при комнатной температуре. Меченое антитело очищали с помощью колонки 6 Biospin.
Для получения 92-13, конъюгированного с 90Y, 92-13 конъюгировали с DTPA по остаткам лизина. DTPA-92-13 метили иттрием до удельной активности 100 мкКи/мг, и иммунореактивность 90Y-DTPA-92-13 составляла приблизительно 70%.
(3) Синтез Mab, меченных Alexa647
Промечивание Mab Alexa-Fluoro647 производили согласно инструкции производителя с использованием набора для промечивания Alexa647 моноклональных антител (Molecular Probes, Eugene, Oregon). Химически активный краситель Alexa647 содержит сукцинимидильную сложноэфирную часть, которая взаимодействует с первичными аминами белков, и полученные конъюгаты Mab-краситель очищали с помощью колонки с исключением по размеру.
Пример 2
Связывающая активность моноклональных антител против FZD10
Авторы настоящего изобретения использовали два способа оценки связывающего сродства моноклональных антител мыши; проточный цитометрический анализ с флуоресцентными красителями и измерение радиоактивности с использованием 125I.
(1) Проточный цитометрический (FACS) анализ
Для исследования сродства связывания с клеткой четырех антител, 39-2 и 39-10 (раскрытых в патенте WO2005/004912), 92-13 и 93-22 заявители проводили эксперименты с проточной цитометрией (FACS). Для проточного цитометрического анализа с непрямой флуоресценцией суспензии 5×106 клеток инкубировали с 10 мкг/мл Mab или IgG неиммунизированной мыши (Beckman Coulter) в течение 30 мин при 4°С. После промывки PBS добавляли 2 мкг флуоресцентного IgG козы против мыши (Alexa Fluor 488, Molecular Probes, Eugene, Oregon), и клеточную суспензию инкубировали в течение 30 мин при 4°С для анализа с помощью FACScan (Becton Dickinson, Franklin Lakes, NJ). Для прямого иммунофлуоресцентного анализа клетки инкубировали с 2 мкг Alexa488-Mab в присутствии или в отсутствие избытка (100 мкг) немеченых Mab в течение 30 мин при 4°С и подвергали анализу с помощью FACScan.
Для подтверждения экспрессии FZD10 в линиях клеток авторы проводили ОТ-ПЦР. Для экспериментов ОТ-ПЦР из клеточных линий экстрагировали суммарные РНК с помощью реагента TRIzol (Invitrogen, Carlsbad, CA, USA), и 3-мкл аликвоту каждой суммарной РНК подвергали обратной транскрипции. Амплификацию с помощью ПЦР проводили с использованием кДНК в качестве матриц со следующими праймерами: 5'-TATCGGGCTCTTCTCTGTGC-3' (SEQ ID NO: 9) и 5'-GACTGGGCAGGGATCTCATA-3' (SEQ ID NO: 10) для FZD10 и 5'-TTAGCTGTGCTCGCGCTACT-3' (SEQ ID NO: 11) и 5'-TCACATGGTTCACACGGCAG-3' (SEQ ID NO: 12) для β2-микроглобулина (β2MG), внутреннего контроля.
Как показано на фиг. 1a, все четыре Mab, 39-2, 39-10, 92-13 и 93-22 связываются с четырьмя экспрессирующими FZD10 SS клеточными линиями, SYO-1, YafuSS, HS-SY-2, и Fuji, зависимым от дозы FZD10 образом, но не связываются с двумя клеточными линиями, 1973/99 и LoVo, в которых транскрипт FZD10 не был выявлен. В таблице 1 ниже показана корреляция между относительной средней интенсивностью флуоресценцией (MFI) этих Mab и уровнем экспрессии FZD10, показанным на фиг. 1b. Кроме того, заявители, в частности, показали, что Mab 92-13 и 93-22 также связываются с SNU-C5, трансфицированными конструктом FZD10-myc/His, тогда как связывание не выявлялось при использовании клеток SNU-C5, трансфицированных пустым вектором (фиг. 1c), что предполагает специфическое связывание этих Mab 92-13 и 93-22 против белка FZD10.
(2) Связывающая активность в отношении нормальных клеток крови
Для подтверждения того, что эти антитела могут использоваться для клинической цели, авторы настоящего изобретения дополнительно проверяли связывающую активность антител в отношении нормальных клеток крови. Для оценки неспецифической связывающей активности Mab против нормальных клеток крови меченные 125I Mab инкубировали с 100 мкл свежей крови здорового донора. После инкубации в течение одного часа при комнатной температуре измеряли радиоактивность клеточного осадка, как описано выше.
Связывающая активность меченных 125I Mab 92-13 и 93-22 в отношении нормальных клеток крови не выявлялась у всех трех индивидуальных доноров, тогда как активность Mab 39-2 и 39-10 выявлялась у всех трех индивидуальных доноров (фиг. 1d). Эти результаты соответствуют результатам FACS анализа с использованием мононуклеарных клеток периферической крови человека (данные не показаны), что позволяет предполагать клиническую пригодность только антител 92-13 и 93-22 с малой вероятностью неблагоприятного действия на больных SS благодаря очень специфичному связывающему сродству к молекуле FZD10. Поэтому заявители сконцентрировали свое внимание лишь на антителах 92-13 и 93-22 для последующего анализа.
(3) Дополнительные анализы
Кроме того, тестирование связывания проводили с помощью меченных 125I Mab (см. пример 1 (2)) для оценки связывающего сродства в отношении молекул FZD10 на клеточной поверхности. Для радиоактивного анализа полученные в примере 1 (2) 0,5 кБк (0,001 мкг антитела) меченных 125I Mab добавляли к 100 мкл клеточной суспензии с различными количествами идентичных немеченых Mab. После инкубации в течение одного часа при комнатной температуре клеточную суспензию центрифугировали при 800×g. Супернатант удаляли, и измеряли радиоактивность клеточного осадка.
Результаты показали более высокое связывающее сродство антитела 92-13 по сравнению с антителом 93-22; при одинаковых условиях с клетками связывалось приблизительно 33% 92-13 и приблизительно 9% антитела 93-22 (фиг. 1e). Количество связанного антитела снижалось при добавлении немеченых антител зависимым от дозы образом.
Авторы затем провели конкурентный связывающий анализ Mab 92-13 и 93-22 с использованием проточной цитометрии. Связывание клетками обоих меченных Alexa488 антител полностью блокировалось большим количеством немеченых антител (фиг. 1f, ii и iii) при добавлении к каждому из них, что позволяет предполагать, что Mab 92-13 и 93-22, вероятно, узнают очень сходные или один и тот же эпитоп FZD10. Эти результаты позволяют предполагать, что данные Mab способны специфически узнавать FZD10, экспрессируемый на поверхности SS клеток.
Пример 3
Иммуногистохимия
Для оценки специфичности связывания 92-13 и 93-22 с тканями человека авторы провели иммуногистохимический анализ с использованием замороженных срезов тканей. Тканевые срезы замороженных нормальных органов взрослого человека (BioChain, Hayward, California) фиксировали 4% параформальдегидом при 4°С в течение 15 мин и инкубировали с 5 мкг/мл Mab в течение одного часа при комнатной температуре. Затем добавляли мышиный ENVISION Polymer Reagent (DAKO) и визуализировали субстратом пероксидазы (3,3'-диаминобензидина тетрагидрохлоридом).
Результаты показаны на фиг. 2. На фиг. 2 показан иммуногистохимический анализ в SS и нормальных замороженных срезов ткани и здорового человека без антитела (a, d, g, j и m), с 92-13 (b, e, h, k и n) и с 93-22 (c, f, i, l и o). (a-c), синовиальная саркома; (d-f), почки; (g-i), печень, (j-l), сердце; (m-o), мозг. Как и ожидалось, авторы наблюдали сильную иммунную реакцию в отношении FZD10 в образце SS (фиг. 2, a, b и c) и плаценте (данные не показаны), но не обнаруживали ее в нормальной почке, сердце, мозге и печени (фиг. 2, d-o), что соответствует результатам экспериментов нозерн-блоттинга и ОТ-ПЦР (Nagayama, S., et al. (2005). Oncogene, 24, 6201-6212).
Пример 4
Биораспределение Mab против FZD10 в модели ксенотрансплантата у мышей Balb/c
Распределение 92-13 и 93-22 в модели in vivo анализировали у мышей BALB/c с помощью двух независимых способов, радионуклидной визуализации и флуоресцентной визуализации.
(1) Радионуклидная визуализация in vivo
Эксперименты in vivo проводили в виварии согласно утвержденным правилам. Мышам BALB/cA Jcl-nu (самки в возрасте 7 недель) инъецировали подкожно (п/к) клетки опухоли SYO-1 (5×106 клеток) в 0,1 мл PBS по бокам. Для исследования биораспределения мышам с полностью развившимися опухолями вводили 10 кБк (0,5-1 мкг) меченных 125I Mab и 10 кБк (0,5-1 мкг) меченных 111In Mab через хвостовую вену. Через 1, 24, 48 часов животных умерщвляли, и измеряли массу и радиоактивность тканей. Распределение для всех образцов выражали в виде % от введенной дозы/г ткани. Для оптической визуализации биораспределения кроме мышей с опухолью SYO-1 использовали мышей, несущих опухоль LoVo. Опухолевые клетки LoVo (1×107 клеток) инъецировали п/к мышам BALB/cA Jcl-nu, как описано выше. После полного развития опухолей мышей подвергали визуализирующему исследованию.
Результаты на фиг. 3a показывают, что радиоактивность 111In-92-13, связанная с кровью, снижалась с 35% от инъецированной дозы на грамм (%ID/g) через один час после инъекции до 12% через 48 часов. Радиоактивность 111In-92-13, связанная с печенью, почкой, кишечником, селезенкой, поджелудочной железой, легким, сердцем, желудком и мышцей, оставалась без заметных изменений или снижалась в процессе наблюдения (фиг. 3a). Радиоактивность 111In-92-13, связанная с опухолью, накапливалась в ходе эксперимента с 2%ID/g через один час после инъекции до 11%ID/g через 48 часов. С другой стороны, на фиг. 3b показано, что радиоактивность меченного 125I 92-13, связанного с опухолью, не возрастала существенно, хотя радиоактивность, связанная с кровью, снижалась с 25% через один час до 7% через 48 часов, а радиоактивность, связанная с другими нормальными органами, оставалась постоянной. Возможно, меченные 125I антитела деградировали внутри клеток после интернализации. Меченное 111In 93-22 тоже аккумулировалось в опухоли SYO-1 через 48 часов после инъекции (фиг. 3c), а меченное 125I 93-22 показало плохую аккумуляцию (фиг. 3d), что позволяет предполагать интернализацию, как и 92-13.
(2) Флуоресцентная визуализация in vivo
Флуоресцентную визуализацию in vivo проводили с помощью визуализирующей системы серии 100 IVISTM (Xenogen, Alameda, CA). Для получения флуоресценции Alexa647-Mab in vivo использовали оптимизированный фильтр cy5.5. Мышам-носителям опухоли SYO-1 инъецировали 20 мкг меченных Alexa647 Mab внутрибрюшинно и подвергали флуоресцентной визуализации через разные промежутки времени. Мышей кормили пищей, которая не содержала alfalfa, в течение четырех дней перед инъекцией Mab для снижения фоновой флуоресценции. При получении изображений мышей анестезировали 2% изофлураном (Abbott Laboratories) и помещали в систему IVIS. Мышей умерщвляли через четыре дня после инъекции Mab, извлекали опухоль и основные органы и получали флуоресцентное изображение.
Как показано на фиг. 4a, значительная флуоресценция была выявлена в месте локализации опухоли через 24 часа после инъекции. Связанная с опухолью флуоресценция наблюдалась для обоих Mab, 92-13 и 93-22; сигналы достигали максимального уровня через приблизительно 48 часов после инъекции, и их можно было обнаружить через 96 часов после инъекции. Авторы настоящего изобретения умерщвляли этих мышей через 120 часов после инъекции и измеряли интенсивность флуоресценции в их опухоли и также важных нормальных органах (печени, селезенке, почке, поджелудочной железе, ободочной кишке) (фиг. 4b и 4c). Очень сильный флуоресцентный сигнал наблюдался в извлеченной опухоли, тогда как в нормальных органах флуоресцентный сигнал не обнаруживался. Для подтверждения специфичности связывания авторы настоящего изобретения создавали ксенотрансплантаты с помощью негативной по антигену клеточной линии LoVo у голых мышей и инъецировали меченные Alexa647 Mab с проведением флуоресцентного визуализирующего анализа. У мышей-носителей LoVo флуоресценция не была выявлена ни в месте локализации опухоли (фиг. 5a), ни в извлеченных опухоли или других органах (фиг. 5b и 5c). Эти результаты показывают, что данные Mab также способны специфически связываться с экспрессирующими FZD10 опухолевыми клетками in vivo.
Пример 5
Интернализация Mab против FZD10 в антиген-позитивные клетки
Для исследования молекулярного поведения этих Mab после связывания с клеточными поверхностями отслеживали их локализацию с помощью системы визуализации in vitro.
Клетки помещали в 8-луночные закрытые предметные стекла (Nalge Nunc International, Naperville, IL) при плотности 5×104 клеток на лунку. Клетки инкубировали с Mab в течение трех часов при 37°С в воздушной камере, содержащей 5% CO2. Mab, связанные с клеточной поверхностью, удаляли с помощью кислого удаляющего буфера (0,1 М глицин, 500 мМ NaCl, pH 2,5) при 4°С в течение 10 мин и нейтрализации 500 мМ трис (pH 7,5). Клетки затем фиксировали 3,7% формальдегидом в течение 15 мин при комнатной температуре и пермеабилизировали экспозицией с 0,2% тритоном X-100 в течение 10 мин с последующим блокированием 3% бычьим сывороточным альбумином в течение одного часа при комнатной температуре. Для выявления Mab, интернализированных в клетку, образцы инкубировали с меченным Alexa488 IgG козы против мыши (разведение 1:700) в течение одного часа при комнатной температуре. Стекла заключали c DAPI (Vectashield, Vector Laboratories, Burlingame, CA) и анализировали с помощью конфокальной оптики Leica TCS SP1.
Как показано на фиг. 6, по данным конфокальной микроскопической визуализации, производимой с использованием меченного Alexa488 IgG козы против мыши, оба Mab 92-13 и 93-22 эффективно включались в цитозоль клеток SYO-1 и клеток YaFuSS через 3 часа после инкубации Mab с клетками (фиг. 6, a-f). С другой стороны, флуоресцентные сигналы этих Mab едва выявлялись в клетках LoVo, не экспрессирующих FZD10 (фиг. 6, g-i), что показывает, что специфическое связывание Mab с FZD10 клеточной поверхности индуцировало интернализацию антител.
Пример 6
Специфическая цитотоксичность Mab
92-13 и 93-22 не влияли на рост опухолевых клеток при добавлении непосредственно к культивируемым клеткам (данные не показаны). Для терапевтических исследований опухоли SYO-1 выращивали у мышей BALB/cA Jcl-nu способом, сходным с описанным в примере 4. Диаметр опухолей измеряли с помощью кронциркуля, и объем опухолей определяли с использованием следующей формулы: 0,5 × (больший диаметр) × (меньший диаметр)2, как описано ранее (Nagayama, S., et al. (2005). Oncogene, 24, 6201-12). Когда объем опухоли достигал более 0,4-2,8 см3, мышей Balb/c-nude, несущих подкожную опухоль SYO-1, случайным образом распределяли в группы обработки, и им делали внутривенные инъекции 100 мкКи меченных 90Y Mab или контрольные Mab в хвостовую вену. Мышей взвешивали и фиксировали диаметры опухоли.
На фиг. 7 показано, что объемы опухоли значительно снижались сразу после лечения, почти до следовых значений в течение одной недели у всех мышей. При введении мышам 50 мкКи 90Y-DTPA-92-13 опухоли с объемом >1 см3 восстанавливались через две недели после лечения, хотя размер опухолей сразу после лечения значительно снижался. У мышей наблюдалось временное снижение массы (10-15%), но они выздоравливали в течение одной недели, и не наблюдалось явных признаков токсичности (данные не показаны).
Пример 7
Создание гибридных антител
Гибридные антитела ch92-13 и ch93-22, соответствующие антителам 92-13 и 93-22 мыши, были созданы путем переноса последовательности вариабельной области каждого антитела мыши на константную область IgG1 человека под контролем промотора CMV. Из гибридомных клонов 92-13 и 93-22 экстрагировали суммарные РНК. Из суммарной РНК синтезировали кДНК с помощью набора GeneRacerTM (Invitrogen). Последовательности вариабельных областей моноклональных антител амплифицировали с использованием прямого праймера (GeneRacerTM5'Primer) и обратного праймера; CH1 (IgG2a); 5'-AATTTTCTTGTCCACCTTGGTG-3' (SEQ ID NO: 3) для тяжелой цепи и CL1 (каппа); 5'-CTAACACTCATTCCTGTTGAAGCTCT-3' (SEQ ID NO: 4) для легкой цепи. Продукты ПЦР секвенировали, и были определены последовательности, кодирующие вариабельную область m92-13 и m93-22.
В результате были определены следующие аминокислотные последовательности вариабельных областей H-цепи и вариабельных областей L-цепи Ig:
92-13, вариабельная область H-цепи:
MKCSWVIFFLMAVVTGVNSEVQLQQSGAELVKPGASVKLSCTASGFNINDTYMHWVKQRPEQGLEWIGRIDPANGNTKYDPKFQGKATITADTSSNTAYLQLSSLTSEDTAVYYCARGARGSRFAYWGQGTLVTVSA (SEQ ID NO: 13), кодируемая нуклеотидной последовательностью SEQ ID NO: 14, и
92-13, вариабельная область L-цепи:
MSVPTQVLGLLLLWLTDARCDIQMTQSPASLSVSVGETVTITCRASENIYSNLAWYQQKQGKSPQLLVYVATNLADGVPSRFSGSGSGTQYSLKINSLQSEDFGSYYCQHFWGTPYTFGGGTKL (SEQ ID NO: 21), кодируемая нуклеотидной последовательностью SEQ ID NO: 22; и
93-22, вариабельная область H-цепи:
MGWSRIFLFLLSITAGVHCQVQLQQSGPELVKPGASVKISCKASGYAFSSSWMNWVKQRPGQGLEWIGRIYPGDGDTNYNGKFKGKATLTADKSSSTAYMQLSSLTSVDSAVYFCARGGNYGWFAYWGQGTLVTVSAGS (SEQ ID NO: 29), кодируемая нуклеотидной последовательностью SEQ ID NO: 30, и
93-22, вариабельная область L-цепи:
METDTLLLWVLLLWVPGSTGDIVLTQSPASLAVSLGQRATISCRASKSVSTSGYSYMHWYQQKPGQPPKLLIYLASNLESGVPARFSGSGSGTDFTLNIHPVEEEDAATYYCQHSRELYTFGGGTKLGS (SEQ ID NO: 37), кодируемая нуклеотидной последовательностью SEQ ID NO: 38. Подчеркиванием отмечены сигнальные последовательности.
Были определены следующие последовательности CDR (области, определяющей комплементарность) антител:
92-13, INDTYMH (SEQ ID NO: 15) в качестве VH CDR1, RIDPANGNTKYD (SEQ ID NO: 17) в качестве VH CDR2 и GSRFAY (SEQ ID NO: 19) в качестве VH CDR3, RASENIYSNLA (SEQ ID NO: 23) в качестве VL CDR1, VATNLAD (SEQ ID NO: 25) в качестве VL CDR2 и QHFWGTPY (SEQ ID NO: 27) в качестве VL CDR3; и
93-22, SSWMN (SEQ ID NO: 31) в качестве VH CDR1, RIYPGDGDTNYN (SEQ ID NO: 33) в качестве VH CDR2 и GGNYGWFAY (SEQ ID NO: 35) в качестве VH CDR3, RASKSVSTSGYSYMH (SEQ ID NO: 39) в качестве VL CDR1, LASNLES (SEQ ID NO: 41) в качестве VL CDR2 и QHSRELY (SEQ ID NO: 43) в качестве VL CDR3.
Кроме того, были определены следующие аминокислотные последовательности Н-цепей и L-цепей моноклональных антител мыши 92-13, 93-22 и 39-10:
92-13, Н-цепь: SEQ ID NO: 58 (кодируемая нуклеотидной последовательностью SEQ ID NO: 57);
92-13, L-цепь: SEQ ID NO: 60 (кодируемая нуклеотидной последовательностью SEQ ID NO: 59);
93-22, Н-цепь: SEQ ID NO: 62 (кодируемая нуклеотидной последовательностью SEQ ID NO: 61);
93-22, L-цепь: SEQ ID NO: 64 (кодируемая нуклеотидной последовательностью SEQ ID NO: 63);
39-10, Н-цепь: SEQ ID NO: 66 (кодируемая нуклеотидной последовательностью SEQ ID NO: 65);
39-10, L-цепь: SEQ ID NO: 68 (кодируемая нуклеотидной последовательностью SEQ ID NO: 67).
В соответствии с определенной последовательностью были сконструированы специфические праймеры для вариабельной области m92-13: 5'-AATAGCGGCCGCACCATGAAATGCAGCTGGGTTATCTT-3' (SEQ ID NO: 5) и 5'-AATAGCTAGCTGCAGAGACAGTGACCAGAGTCC-3' (SEQ ID NO: 6) для тяжелой цепи и 5'-AATAGCGGCCGCACCATGAGTGTGCCCACTCAGG-3' (SEQ ID NO: 7) и 5'-TTCCAGCTTGGTCCCCCC-3' (SEQ ID NO: 8) для легкой цепи. Кроме того, были сконструированы специфические праймеры для вариабельной области m93-22, 5'-AATAGCGGCCGCACCATGGGATGGAGCCGGATCTTT-3' (SEQ ID NO: 53) и 5'-AATAGGATCCTGCAGAGACAGTGACCAGAGTCCCTT-3' (SEQ ID NO: 54) для тяжелой цепи и 5'-AATAGCGGCCGCACCATGGAGACAGACACACTCCT-3' (SEQ ID NO: 55) и 5'-AATAGGATCCCAGCTTGGTCCCCCCTCCGAACGT-3' (SEQ ID NO: 56) для легкой цепи. Для конструирования экспрессионного вектора для гибридных антител было получено два кассетных вектора. Фрагмент ДНК, кодирующий IgG1 (CH1-CH3) человека, был вставлен в pQCXIH (Clontech) (pQCXCHIH), а фрагмент ДНК, кодирующий Igκ (CL1) человека, был вставлен в pQCXIP (pQCXCLIP). Для получения фрагментов ДНК, кодирующих IgG1 человека или Igκ человека, была получена библиотека константной области человека с использованием кДНК из PBMC (мононуклеарных клеток периферической крови) человека с помощью описанного ранее способа (Liu, A.Y. et al., Proc. Natl. Acad. Sci. USA, Vol.84, 3439-43, 1987; Reff, M.E. et al., Blood, Vol.83, No.2, 435-45, 1994). ДНК, кодирующие вариабельную область тяжелой цепи и легкой цепи m92-13 и m93-22, амплифицировали с помощью ПЦР, секвенировали и субклонировали в pQCXCHIH и pQCXCLIP, соответственно, с использованием сайтов NotI и BamHI. Эти векторы совместно трансфицировали в клетки CHO. Трансфицированные клетки культивировали в среде F-12, содержащей 500 мкг/мл гигромицина и 10 мкг/мл пуромицина. Когда клетки вырастали до состояния, близкого к конфлуентности, среду заменяли бессывороточной средой (CHO-S-SFM II; GIBCO), и гибридное антитело очищали из супернатанта с помощью аффинной колонки протеина A (GE Amersham) и секвенировали. Последовательность тяжелой цепи гибридного антитела ch92-13 включает SEQ ID NO: 46, кодируемую нуклеотидной последовательностью SEQ ID NO: 45; а последовательность легкой цепи гибридного антитела ch92-13 включает SEQ ID NO: 48, кодируемую нуклеотидной последовательностью SEQ ID NO: 47. Последовательность тяжелой цепи гибридного антитела ch93-22 включает SEQ ID NO: 49, кодируемую нуклеотидной последовательностью SEQ ID NO: 50; а последовательность легкой цепи гибридного антитела ch93-22 включает SEQ ID NO: 52, кодируемую нуклеотидной последовательностью SEQ ID NO: 51.
Пример 8
Связывающая активность гибридных антител
Антителозависимую клеточную цитотоксическую (ADCC) активность, индуцируемую гибридными 92-13 и 93-22, определяли по активности LDH, как описано ранее (Nagayama, S., et al. (2005). Oncogene, 24, 6201-6212). Свежие эффекторные клетки выделяли из гепаринизированной периферической крови здорового донора с помощью Ficoll-Plaque (Amersham Bioscience). Эффекторные клетки (E) и клетки-мишени (T) (каждые в количестве 5×103/лунку) совместно инкубировали в течение 6 час при 37°С в четырех параллелях при различных соотношениях E:T вместе с гибридным 92-13, гибридным 93-22 или IgG неиммунизированного человека в 0,1 мл не содержащей фенол красный среде RPMI 1640 с добавкой 5% FBS в 96-луночном планшете. LDH, выделенную в культуральный супернатант, определяли по поглощению при 490 нм. Процент специфической цитотоксичности рассчитывали в соответствии с инструкциями производителя.
Что касается эффекторной активности, оба гибридных 92-13 и 93-22 индуцировали ADCC специфично в отношении сверхэкспрессирующих FZD10 клеток SYO-1 (фиг. 8, a и c), но не в отношении FZD10-негативных клеток LoVo (фиг. 8, b и d). Конкретно, гибридное 92-13 вызывало большую индукцию цитотоксичности по сравнению с гибридным 93-22; однако их активность зависела от донора эффекторных клеток, возможно, из-за полиморфизма рецептора Fc.
Каждый из заявок и патентов, упомянутых в настоящем документе, и каждый документ, цитированный или на который имеется ссылка в упомянутых выше заявках и патентах, включен в настоящее описание в качестве ссылки. Более того, все цитированные в настоящем тексте документы и все документы, цитированные или на которые имеется ссылка в документах, цитированных в настоящем тексте, включены тем самым в настоящее описание в качестве ссылки.
Различные модификации и вариации описанных способов и системы изобретения в пределах объема и духа изобретения должны быть очевидны для специалистов в данной области техники. Хотя изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что изобретение, как оно заявлено, не следует неоправданно ограничивать такими конкретными вариантами осуществления и что в пределах объема изобретения в нем могут быть произведены разнообразные модификации и добавления. Действительно, различные модификации описанных способов для осуществления изобретения, которые очевидны для специалистов в области молекулярной биологии или смежных областей, рассматриваются как входящие в объем формулы изобретения. Более того, различные сочетания характерных черт следующих зависимых пунктов формулы изобретения могут быть произведены с характерными чертами независимых пунктов формулы изобретения без выхода за границы объема настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНОЕ ТЕРАПЕВТИЧЕСКОЕ АНТИТЕЛО НН1 ПРОТИВ CD-37 | 2012 |
|
RU2658438C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ FZD10, ЕГО ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2765431C2 |
| НОВЫЕ АНТИ-IL 13 АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2004 |
|
RU2487887C2 |
| АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА | 2002 |
|
RU2314317C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| НОВЫЕ ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ VAP-1 | 2008 |
|
RU2459832C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ TNFα | 2012 |
|
RU2549700C1 |
| АГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ NOTCH3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ NOTCH3-АССОЦИИРОВАННЫХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2461569C2 |

































































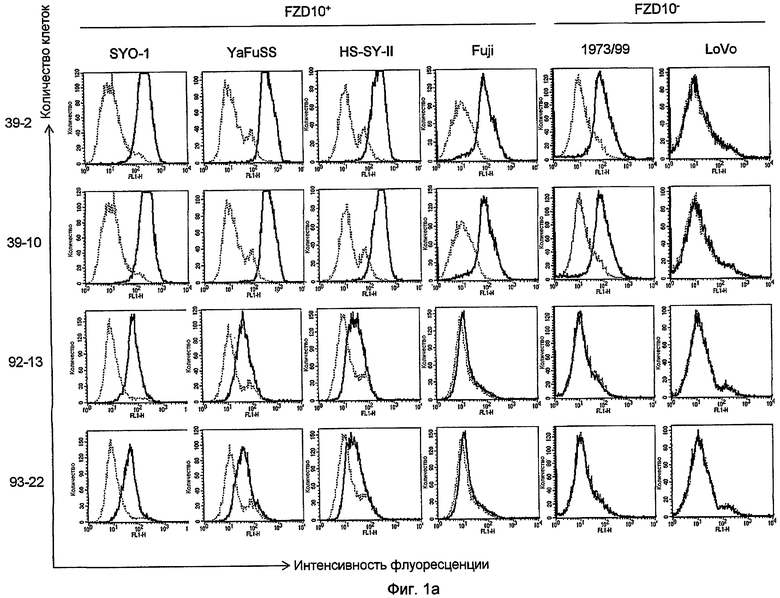
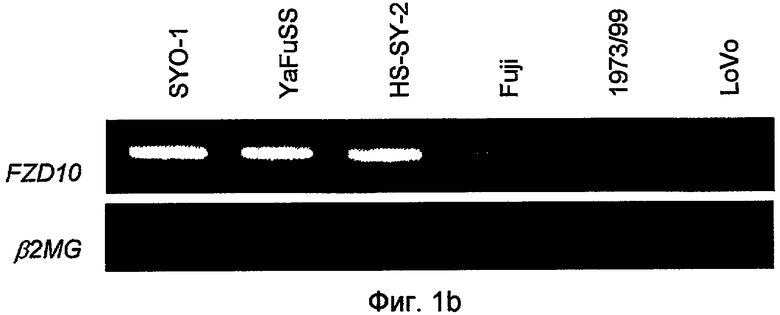
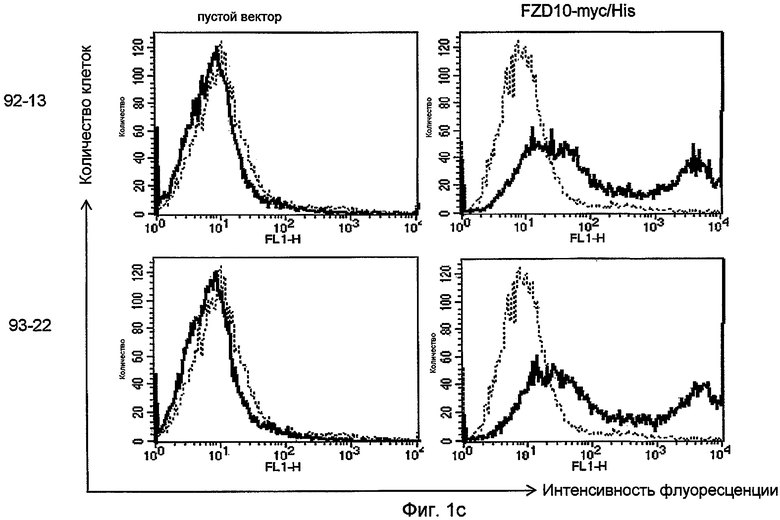
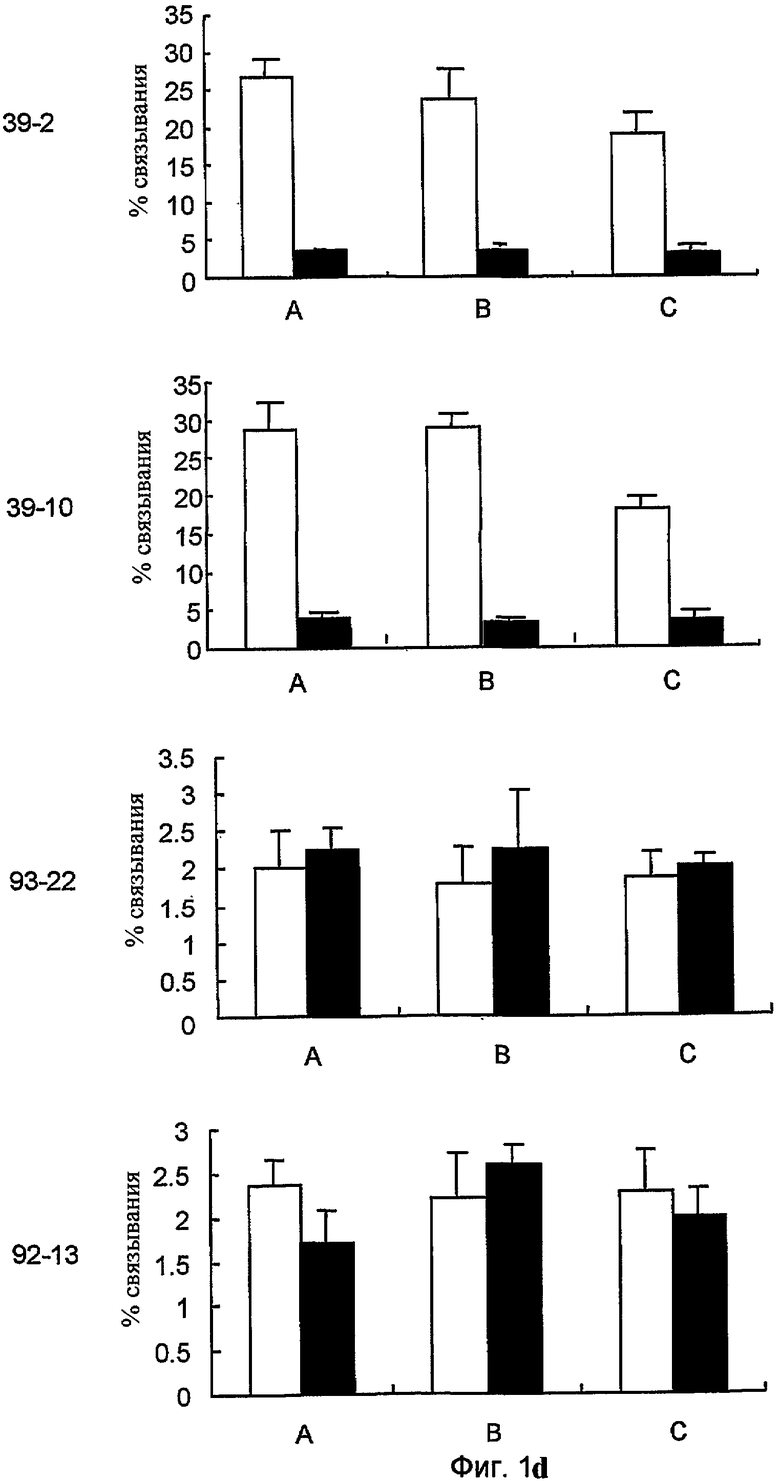
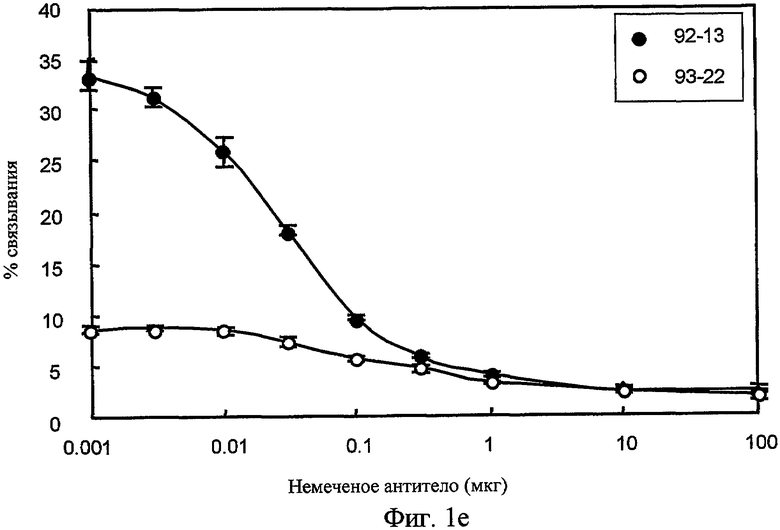

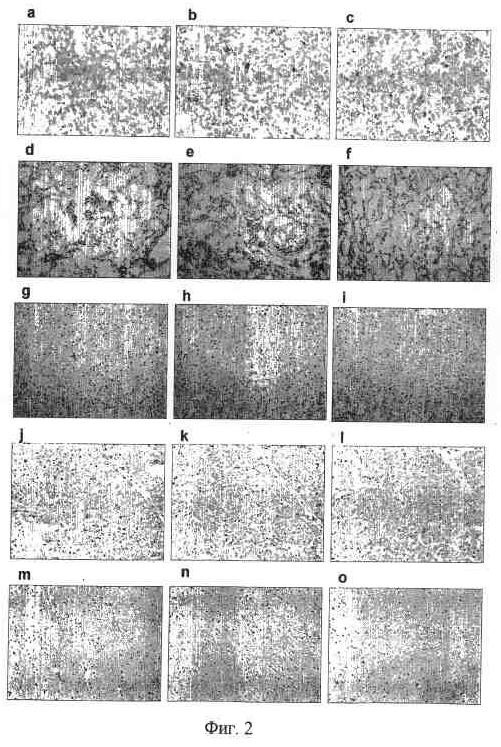
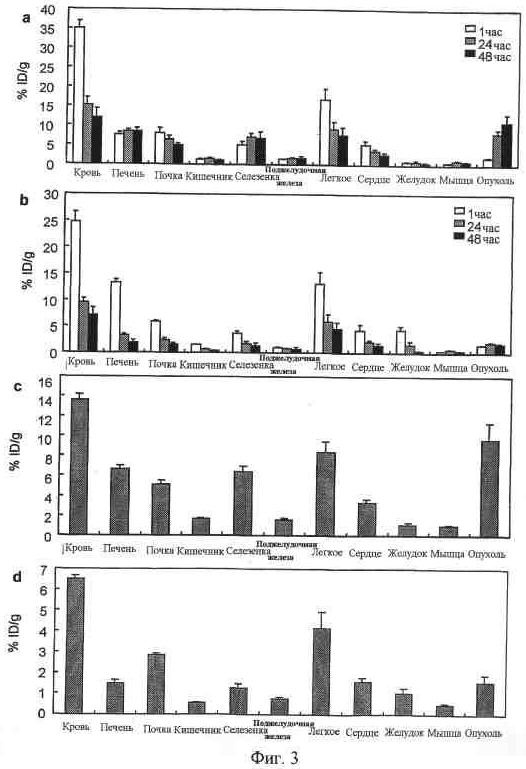
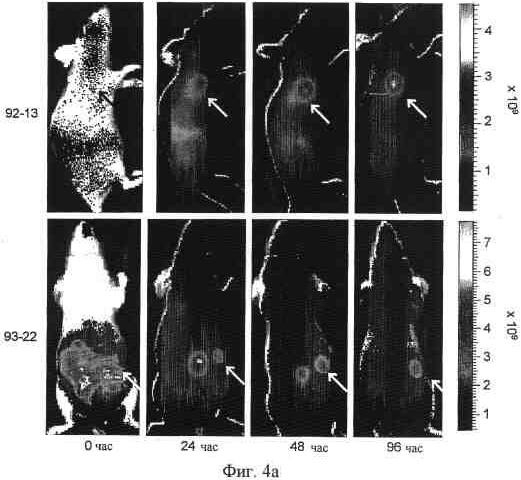
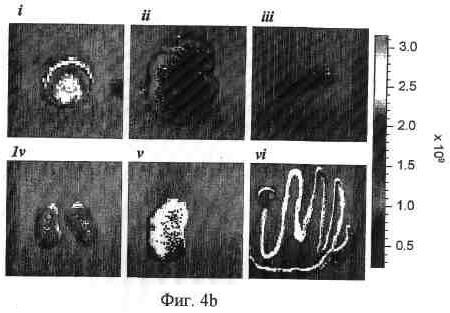
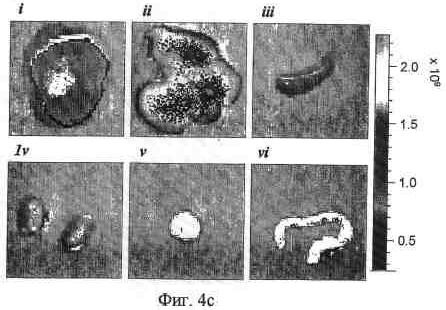
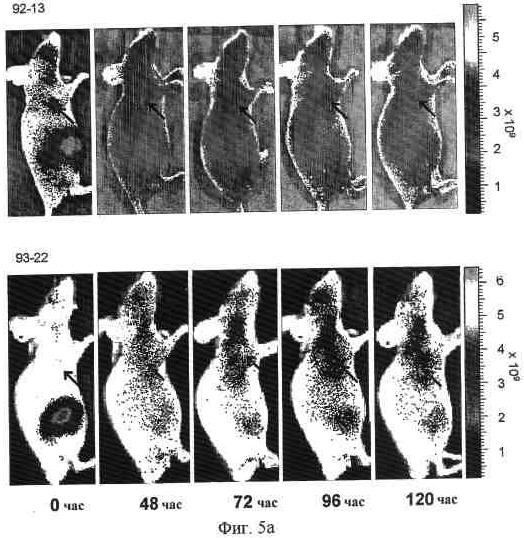
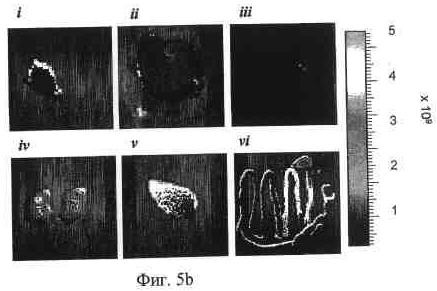
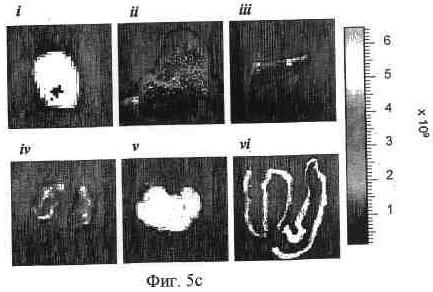

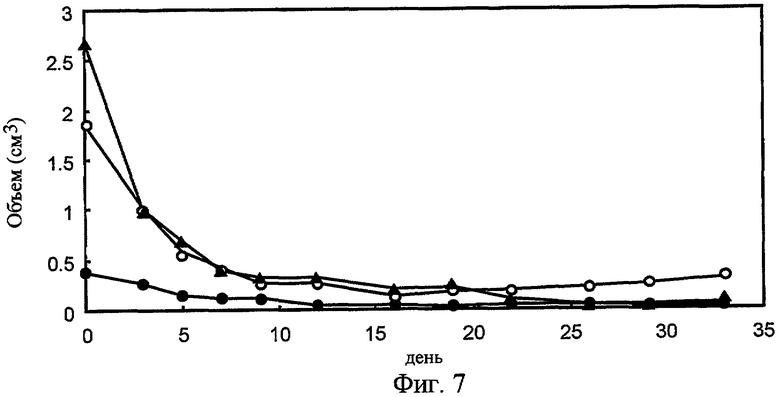
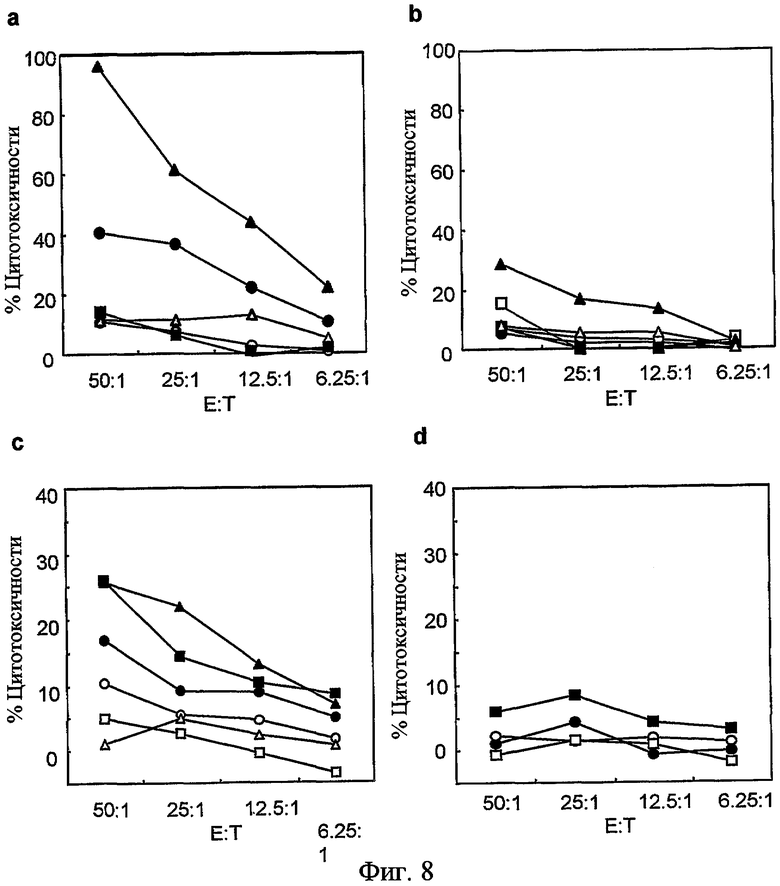
Изобретение относится к биотехнологии и представляет собой антитело или его фрагмент, которое способно связываться с гомологом 10 белка Frizzled (FZD10), такое как моноклональное антитело мыши, гибридное антитело, химерное и гуманизированное антитело. Также представлены клоны гибридомы, продуцирующие антитело, а также способ лечения или профилактики связанного с FZD10 заболевания; способ диагностики или прогнозирования связанного с FZD10 заболевания; способ визуализации in vivo FZD10 у субъекта; фармацевтическая композиция и набор, содержащие антитело. Лечение такими антителами позволяет улучшить клинический исход при заболеваниях, связанных с гомологом 10 белка Frizzled (FZD10). 11 н. и 33 з.п. ф-лы, 1 табл., 8 ил.
1. Антитело или его фрагмент, которое включает V (вариабельную) область Н (тяжелой) цепи, включающую область, определяющую комплементарность (CDR), имеющую аминокислотные последовательности, представленные в SEQ ID NO: 15, 17 и 19, и V-область L (легкой) цепи, включающую CDR, имеющую аминокислотные последовательности, представленные в SEQ ID NO: 23, 25 и 27, и которое способно связывать гомолог 10 белка Frizzled (FZD10) или его частичный пептид.
2. Антитело или его фрагмент по п.1, где антитело выбрано из группы, состоящей из антитела мыши, гибридного антитела, гуманизированного антитела, фрагмента антитела и одноцепочечного антитела.
3. Антитело или его фрагмент по п.1, где антитело представляет собой антитело мыши.
4. Антитело или его фрагмент по п.3, где антитело мыши включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 57, и/или L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 59.
5. Антитело или его фрагмент по п.3, где антитело мыши продуцируется клоном гибридомы 92-13 (FERM ВР-10628).
6. Антитело или его фрагмент по п.1, где антитело представляет собой гибридное антитело.
7. Антитело или его фрагмент по п.6, где гибридное антитело включает V-область Н-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 13.
8. Антитело или его фрагмент по п.6 или 7, где гибридное антитело включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 46.
9. Антитело или его фрагмент по п.6, где гибридное антитело включает V-область L-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 21.
10. Антитело или его фрагмент по п.6 или 9, где гибридное антитело включает L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 48.
11. Антитело или его фрагмент по п.6, где гибридное антитело включает V-область Н-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 13, и V-область L-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 21.
12. Антитело или его фрагмент по п.6 или 11, где гибридное антитело включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 46, и L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 48.
13. Антитело или его фрагмент по п.6, где гибридное антитело дополнительно включает С (константную) область антитела человека.
14. Антитело или его фрагмент по п.1, где антитело представляет собой гуманизированное антитело.
15. Антитело или его фрагмент по п.14, где гуманизированное антитело дополнительно включает FR (каркасную) область антитела человека и/или С-область антитела человека.
16. Антитело или его фрагмент, которое включает V (вариабельную) область Н (тяжелой) цепи, включающую область, определяющую комплементарность (CDR), имеющую аминокислотные последовательности, представленные в SEQ ID NO: 31, 33 и 35, и V-область L (легкой) цепи, включающую CDR, имеющую аминокислотные последовательности, представленные в SEQ ID NO: 39, 41 и 43, и которое способно связывать гомолог 10 белка Frizzled (FZD10) или его частичный пептид.
17. Антитело или его фрагмент по п.16, где антитело выбрано из группы, состоящей из антитела мыши, гибридного антитела, гуманизированного антитела, фрагмента антитела и одноцепочечного антитела.
18. Антитело или его фрагмент по п.16, где антитело представляет собой антитело мыши.
19. Антитело или его фрагмент по п.18, где антитело мыши включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 61, и/или L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 63.
20. Антитело или его фрагмент по п.18, где антитело мыши продуцируется клоном гибридомы 93-22 (FERM ВР-10620).
21. Антитело или его фрагмент по п.16, где антитело представляет собой гибридное антитело.
22. Антитело или его фрагмент по п.21, где гибридное антитело включает V-область Н-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 29.
23. Антитело или его фрагмент по п.21 или 22, где гибридное антитело включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 50.
24. Антитело или его фрагмент по п.21, где гибридное антитело включает V-область L-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 37.
25. Антитело или его фрагмент по п.21 или 24, где гибридное антитело включает L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 52.
26. Антитело или его фрагмент по п.21, где гибридное антитело включает V-область Н-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 29, и V-область L-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 37.
27. Антитело или его фрагмент по п.21 или 26, где гибридное антитело включает Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 50, и L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 52.
28. Антитело или его фрагмент по п.21, где гибридное антитело дополнительно включает С (константную) область антитела человека.
29. Антитело или его фрагмент по п.16, где антитело представляет собой гуманизированное антитело.
30. Антитело или его фрагмент по п.29, где гуманизированное антитело дополнительно включает FR (каркасную) область антитела человека и/или С-область антитела человека.
31. Химерное или гуманизированное антитело или его фрагмент, в котором определяющие комплементарность области (CDR) происходят от мышиного моноклонального антитела 92-13 (FERM ВР-10628).
32. Антитело или его фрагмент по п.31, представляющее собой химерное антитело, которое содержит:
(i) V-область Н-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 13, и/или V-область L-цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 21; или
(ii) Н-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 46, и/или L-цепь, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 48.
33. Антитело или его фрагмент по п.1 или 31, которое содержит радиоизотопную метку или флуоресцентную метку.
34. Антитело или его фрагмент по п.33, где радиоизотопная метка выбрана из 90иттрия (90Y), 125йода (125I) и 111индия (111In).
35. Клон гибридомы 92-13 (FERM BP-10628), который продуцирует моноклональное антитело 92-13 мыши.
36. Клон гибридомы 93-22 (FERM BP-10620), который продуцирует моноклональное антитело 93-22 мыши.
37. Способ лечения или профилактики заболевания, которое связано с гомологом 10 белка Frizzled (FZD10) у субъекта, включающий введение субъекту эффективного количества антитела или фрагмента по любому из пп.1-34.
38. Способ по п.37, где заболевание, которое связано с FZD10, выбрано из синовиальной саркомы (SS), колоректального рака, рака желудка, хронического миелоидного лейкоза (CML) и острого миелоидного лейкоза (AML).
39. Способ диагностики или прогнозирования заболевания, которое связано с гомологом 10 белка Frizzled (FZD10), или предрасположенности к развитию заболевания у субъекта, включающий
(a) контактирование образца или препарата от субъекта с антителом или фрагментом по любому из пп.1-34;
(b) выявление белка FZD10 в образце или препарате; и
(c) принятие решения о том, страдает ли субъект заболеванием или подвержен ли риску развития заболевания, на основе относительного содержания белка FZD10 по сравнению с контролем.
40. Способ по п.39, где заболевание, которое связано с FZD10, выбрано из синовиальной саркомы (SS), колоректального рака, рака желудка, хронического миелоидного лейкоза (CML) и острого миелоидного лейкоза (AML).
41. Способ визуализации in vivo гомолога 10 белка Frizzled (FZD10) у субъекта, включающий введение субъекту эффективного количества антитела или фрагмента по п.33 или 34.
42. Фармацевтическая композиция для лечения или профилактики заболевания, связанного с гомологом 10 Frizzled (FZD10), содержащая антитело или фрагмент по любому из пп.1-34 и фармацевтически приемлемый носитель или наполнитель.
43. Набор для диагностики или прогнозирования заболевания, связанного с гомологом 10 Frizzled (FZD10), содержащий антитело или фрагмент по любому из пп.1-34 и меченое вторичное антитело.
44. Фармацевтическая композиция для визуализации in vivo гомолога 10 Frizzled белка, (FZD10), содержащая антитело или фрагмент по п.33 или 34 и фармацевтически приемлемый носитель или наполнитель.
| NAGAYAMA S | |||
| et al | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Oncogene | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| KOIKE J | |||
| et al | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Biochem Biophys Res Commun | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| WO 2005004912, 20.01.2005 | |||
| WO 2004020668, 11.03.2004 | |||
| WO 2002092635, 21.11.2002. | |||

