Область техники.
Изобретение относится к области биотехнологии, а именно к технологии получения биологически активных веществ (БАВ) методами генной инженерии, точнее к методам получения фактора свертываемости крови VIII человека.
Предшествующий уровень техники.
Дефицит фактора свертывания крови VIII человека (фVIII) является причиной гемофилии А. Врожденная гемофилия А, вызываемая повреждениями в гене фVIII, наблюдается с частотой 1:5000-7000 мужчин. Ген фактора VIII расположен в X-хромосоме. Его общая длина составляет около 190 Кб, ген включает 26 экзонов размером от 69 п. о. до 3106 п. о. (Toole, Knopf et al., Nature, 1984, Nov, 312 (5992): 342-7). Экспрессия гена фактора VIII локализована в клетках печени, ген фактора VIII (фVIII) содержит регуляторные последовательности гепатоцитов. Продукт гена фVIII после удаления секреционного лидерного пептида длиной 19 аминокислот состоит из доменов А1-А2-В-А3-С1-С2, общая длина 2332 аминокислот.
Поскольку фVIII - секретируемый белок, он подвергается ряду посттрансляционных модификаций в эндоплазматическом ретикулуме и аппарате Гольджи. Наиболее существенные модификации фVIII - N-гликозилирование (около 25 сайтов, преимущественно в домене В), сульфатирование 6 остатков тирозина, О-гликозилирование и конверсия в двухцепочечную форму специфическим протеолизом на границе доменов В и A3. Зрелый фVIII, выделяемый из плазмы крови, практически полностью конвертирован в двухцепочечную форму, состоящую из тяжелой цепи (А1-А2-В) размером от 90 до 200 кДа и легкой цепи размером 80 кДа. Цепи фVIII соединены нековалентно, их взаимодействие опосредовано ионами меди, координированными в доменах А1 и A3. Фактор VIII в кровотоке образует комплекс со специфическим шапероном - фактором фон Виллебрандта (vWF) - и покидает этот комплекс при активации. Гетерогенность размеров тяжелой цепи обусловлена распадом фVIII по многим сайтам в составе В-домена, минимальная форма тяжелой цепи фVIII 90 кДа вообще не содержит остатков В-домена. Нормальная концентрация фVIII в кровотоке (1 международная единица (МБ) фVIII в 1 мл) составляет около 200 нг/мл, период полувыведения фVIII у человека около 20 ч.
Поскольку эффективная терапия гемофилии А основана на регулярных внутривенных инъекциях препаратов фVIII в дозе 250-2000 ME, мировое потребление фVIII является весьма значительным. В терапии гемофилии применяются: 1) препараты природного фактора VIII, 2) препараты рекомбинантного полноразмерного фVIII, 3) препараты рекомбинантного фактора VIII с делетированным В-доменом, вариант BDD SQ (делеция области 746-1639, отсчет по зрелому полипептиду).
Лекарственные препараты природного фVIII обычно получают из фракции криопреципитата донорской плазмы при помощи нескольких стадий хроматографической очистки. Часть препаратов природного фVIII очищают иммунноаффинной хроматографией, при этом происходит удаление vWF. Лекарственные препараты рекомбинантного фVIII также не содержат vWF и проходят многостадийную очистку, включающую иммунноаффинную хроматографию (или эквивалентную стадию - очистку на иммобилизованном пептиде), а также проходят процедуру вирус-инактивации.
Гетерологичная экспрессия гена фVIII крайне малоэффективна. Вследствие того что мРНК фVIII нестабильна и содержит транскрипционные сайленсеры в области открытой рамки считывания белка, уровень трансляции белка-предшественника фVIII снижен в десятки раз по сравнению с другими белками сходной длины. Около 90% предшественника фVIII не покидает эндоплазматический ретикулум вследствие связывания с шаперном BiP и последующей деградации. Зрелый секретированный фVIII также подвергается интенсивной деградации внеклеточными протеазами. Делеционный вариант фVIII BDD SQ в меньшей степени подвержен удерживанию в эндоплазматическом ретикулуме и протеолитической инактивации в культуральной среде, однако до 90% секретированного продукта подвергается обратному связыванию с мембраной клеток-продуцентов, в основном за счет специфического взаимодействия с фосфатидилсерином. Вследствие этих особенностей мРНК и белка фVIII известные промышленно пригодные системы получения рекомбинантного фVIII отличаются весьма низкой продуктивностью - от 0,5 до 10 МЕ/мл, т.е. не более 1 мг/л, при этом продуктивность систем экспрессии делеционного варианта фVIII в целом выше, чем систем экспрессии полноразмерного фVIII. Вариант делеции В-домена BDD SQ был использован при получении зарегистрированного лекарственного препарата фVIII "мороктоког альфа" (Lind, Larsson et al., Eur J Biochem., 1995, Aug 15, 232 (1): 19-27), другие варианты делеции В-домена в качестве лекарственных средств не использовались.
Исходный метод создания клеток-продуцентов делеционного варианта фVIII BDD SQ описан в патенте США US 4868112. Метод включает в себя конструирование плазмиды, содержащей область открытой рамки считывания (ОРС) фVIII с делецией области B-домена под контролем немедленного раннего промотора цитомегаловируса (CMV) и ген дигидрофолатредуктазы мыши (DHFR) под контролем промотора SV40, последующей трансфекции данной плазмиды в культивируемые клетки линий COS-1 (обезьяна) или СНО DUKX-B11 (китайский хомячок, инактивация одного аллеля гена DHFR), получению колоний стабильно трансфицированных клеток в среде, не содержащей гипоксантин и тимидин, и последующей селекции наиболее продуктивных клонов в среде, содержащей метотрексат (ингибитор DHFR). При использовании данного метода генерации линий-продуцентов максимальная опубликованная скорость секреции целевого белка в культуральную среду (удельная продуктивность) составляла 2 МЕ/млн клеток/день. Культивация клеток производилась в присутствии сыворотки крупного рогатого скота, содержащей vWF быка, плотно связывающийся с фVIII и защищающий его от захвата на мембрану и протеолитической деградации. Метод получения продуцента без использования сыворотки животных не был предложен.
Известные способы увеличения продуктивности линий-продуцентов фVIII BDD SQ имеют в своей основе внесение точечных аминокислотных замен или замену отдельных участков фVIII человека на гомологичные фрагменты фVIII млекопитающих. В частности, в патентах США US 6376463, US 7122634 описано внесение точечных мутаций в домены A и создание слитных белков фактора VIII человека и фактора VIII свиньи. Данные способы не имеют практической пригодности при создании лекарственных препаратов фVIII, хотя и могут быть основой решений для генотерапии гемофилии А, поскольку внесение мутаций в ОРС гена фVIII может привести к расширению спектра алло- и ауто-антител у больных гемофилией, в том числе за счет развития вторичной резистентности к фVIII свиньи (Parker, Craddock et al., J Thromb Haemost., 2004, Apr; 2 (4): 605-11.).
Другие известные способы улучшения клеток-продуцентов, не связанные с изменением структуры целевого белка, основаны на инсерции в геном культивируемых клеток генетических конструкций, кодирующих бицистронную РНК, то есть РНК с двумя транслируемыми областями - целевого гена и селекционного маркера. Популяцию стабильно трансфицированных такими генетическими конструкциями клеток подвергали продолжительной селекции в присутствии метотрексата (МТХ), что должно было приводить к амплификации в геноме трансфицированной генетической конструкции. Данный подход был описан в патенте США US 6114146 для клеток линий 293 и SK-HEP1 и плазмиды pCMVF/EDHPro, содержащей ОРС фVIII BDD SQ и ОРС DHFR, соединенных участком ДНК, содержащим сайт внутреннего связывания рибосом (IRES) вируса энцефаломиокардита. При культивировании колоний стабильно трансфицированных клеток в присутствии возрастающих концентраций МТХ и последующего получения клональных линий методом предельных разведений были созданы линии-продуценты, секретирующие фVIII BDD SQ со скоростью от 0,1 до 3 МЕ/млн клеток/день. Как и в описанных выше случаях, получение продуцентов по предлагаемой в патенте методике требует применения эмбриональной сыворотки крупного рогатого скота, что приводит к образованию плотного комплекса фактора VIII и vWF быка и существенного завышения измеряемой концентрации фVIII в культуральной среде (по сравнению с концентрацией фVIII в бессывороточной среде).
Получение линий-продуцентов фVIII BDD SQ при помощи бицистронного вектора с промотором CMV и селекционным маркером DHFR описано в патенте РФ RU 2429294, в данном случае используется безбелковая ростовая среда, но уровень продукции фVIII не превышает 0,5 МЕ/мл для суспензионной культуры с плотностью 1,2 млн клт/мл.
Наиболее близкий по технической реализации аналог настоящего изобретения - это описанная в патенте США US 6358703 система экспрессии гена фVIII, включающая трансфекцию бицистронного плазмидного вектора pCIS25DTR, кодирующего ген фVIII с делецией В-домена и ген DHFR, в клетки линии НКВ11. Полученные первичные олигоклональные линии продуцентов в дальнейшем подвергали селекции в присутствии возрастающих концентраций МТХ, клонированию методом предельных разведений и адаптации к бессывороточной среде, содержащей раствор белков плазмы человека (то есть vWF человека). Использование полученной данным способом клональной линии-продуцента для культивации в 15-литровом перфузионном биореакторе позволило достичь суммарной продуктивности 4 млн МЕ/день, что соответствует удельной продуктивности 10-30 МЕ/млн клт/день. Несмотря на заявленные очень высокие показатели продуктивности, данное промышленное решение имет ряд существенных недостатков, а именно основано на применении гибридной линии клеток, полученной слиянием культивируемых клеток человека линии 293S и раковых клеток лимфомы Беркитта, активно секретирующих иммуноглобулин и культивируемых в присутствии эмбриональной сыворотки крупного рогатого скота в составе культуральной среды. Культивационная среда для выращивания в биореакторе не содержала сыворотку, однако была снабжена раствором белков плазмы крови человека, которые могут быть контаминированы вирусами человека и прионами. Кроме того, раковые культивируемые клетки человека также могут быть источниками вирусных инфекций и не являются оптимальными предшественниками для получения промышленных линий-продуцентов фармацевтических белков (патентная заявка США US 2006/0099685 A1 п. 0058).
Краткое описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание высокопродуктивной технологии получения рекомбинантного фактора свертываемости крови VIII человека с делецией В-домена для биофармацевтического производства.
Технический результат достигался путем создания новой экспрессионной плазмидной ДНК p1.1-F8BDD, кодирующей открытую крамку считывания белка фактора свертываемости крови VIII человека с делецией В-домена, и создания на ее основе клональной клеточной линии-продуцента.
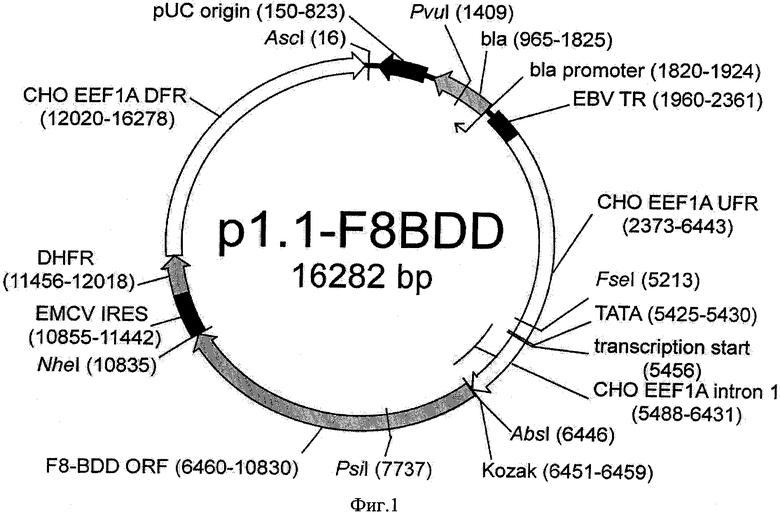
В основе данной технологии лежит разработанная плазмидная ДНК p1.1-F8BDD длиной 16282 п. о. (SEQ ID NO: 1, Фиг. 1), состоящая из:
- фрагмента ДНК длиной 4389 п. о., содержащего последовательность 4371 п. о. (SEQ ID NO: 1 6460-10830), кодирующую ОРС фактора свертываемости крови VIII человека с делецией В-домена и синтетическую последовательность Козак (сайт связывания рибосом, SEQ ID NO: 1 6451-6459), обеспечивающую инициацию трансляции мРНК,
- фрагмента 11893 п. о. плазмиды p1.1 (SEQ ID NO: 2, Фиг. 2), содержащего регуляторные элементы, обеспечивающие экспрессию целевого белка:
- 5' и 3' нетранслируемые области (НТО) гена фактора элонгации 1 альфа китайского хомячка, включающие соответственно функциональный промотор (5' НТО) и терминатор и сигнал полиаденилирования (3'НТО) этого гена, а также фланкирующие нетранскрибируемые области этого гена, обеспечивающие эухроматинизацию сайтов инсерции и транскрипционную активность генетической кассеты в геноме СНО;
- последовательность фактора устойчивости трансфецированных клеток к воздействию метотрексата - дигидрофолатредуктазы (DHFR) и регуляторные элементы для экспрессии DHFR - последовательность IRES вируса энцефаломиокардита. (EMCV), обеспечивающая бицистронную экспрессию в животных клетках и сигнал полиаденилирования;
- область начала репликации плазмиды pUC, открытую рамку считывания бета-лактамазы (bla) и прокариотический промотер гена bla, обеспечивающей устойчивость бактерий к антибиотику ампициллину, позволяющие проводить препаративную наработку плазмиды в E.coli.
Плазмида p1.1-F8-BDD содержит уникальные сайты узнавания эндонуклеазами рестрикции AscI (16), PvuI (1409), FseI (5213), AbsI (6446), PsiI (7737), NheI (10835).
Плазмида была введена в клетки СНО DG-44 методом липосомальной трансфекции (Straus S.E. et al, J. Virol., 1981, Jul; 39 (1): 290-4) при помощи культивирования в безбелковой среде, не содержащей гипоксантин и тимидин и дополнительно содержащей 50 нМ ингибитора DHFR метотрексата (МТХ), были получены олигоклональные линии стабильно трансфицированных клеток. Для отобранной олигоклональной линии с высокой продуктивностью была проведена амплификация кассеты в геноме путем последовательных культивирований в присутствии возрастающих концентраций метотрексата. Была получена поликлональная линия клеток, устойчивых к высокой концентрации метотрексата, и продуцирующая увеличенные количества целевого белка, и использована для клонирования методом предельного разведения. Полученные клоны клеток-продуцентов были проанализированы методами иммуноферментного анализа и коагулометрии и были отобраны клоны, дающие максимальный уровень экспрессии целевого белка. Среди них был выбран клон DG-P1.1-F8BDD-S3, при культивировании которого в суспензионной культуре в бессывороточной среде конечная активность В-домен делетированного фактора свертываемости крови VIII составляла 40±9 МЕ/мл при концентрации клеток 1,7 млн/мл. Полученная клеточная линия депонирована в Российской Коллекции Культур Клеток позвоночных Института Цитологии РАН, регистрационный номер №РККК(П) 756D.
Целью настоящего изобретения является предоставление плазмиды для экспрессии рекомбинантного фактора свертываемости крови VIII человека с делетированным В-доменом, в частности плазмиды p1.1-F8BDD. Также целью настоящего изобретения является предоставление моноклональной линии клеток млекопитающего, в частности клеток яичника китайского хомячка, - продуцентов рекомбинантного фактора свертываемости крови VIII человека с делетированным B доменом, содержащей в геноме множественные копии экспрессионной кассеты, соответствующей линеаризованной экспрессионной плазмиде, в частности плазмиде p1.1-F8BDD. Иллюстративным примером указанной моноклональной линии клеток являются клетки яичника китайского хомячка клона DG-P1.1-F8BDD-S3.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного фактора свертываемости крови VIII человека с делетированным В доменом с использованием указанных клеток.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание технологии высокопродуктивного получения рекомбинантного фактора свертываемости крови VIII с делецией В-домена для биофармацевтического производства с использованием культивируемых клеток яичника китайского хомячка, адаптированных к суспезионному культивированию в безбелковой среде, содержащих в геноме множественные копии генетической кассеты, представляющей собой линеаризованную экспрессионную плазмиду, содержащую фрагмент ДНК, кодирующий фактор свертываемости крови VIII с делецией В-домена под контролем промотора и регуляторных элементов, функционирующих в эукариотической клетке.
Термин «экспрессионная плазмида» («экспрессионная плазмидная ДНК») означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии внедренного в него гена, например, такие как промотор, терминатор, сигнал полиаденилирования. Конкретным примером генетических элементов, необходимых для экспрессии рекомбинантного фактора свертываемости крови VIII с делецией В-домена в составе экспрессионной кассеты согласно настоящему изобретению, является, но не ограничивается им, промотор гена фактора элонгации 1 альфа китайского хомячка.
Фрагментом ДНК, кодирующим рекомбинантный фактор свертываемости крови VIII с делецией В-домена согласно настоящему изобретению, является ген, кодирующий ОРС полипептида фактора VIII с делецией В-домена, который может быть получен, например, как указано в Примере 1. Также указанный фрагмент ДНК может быть получен, например, с использованием технологии клонирования фирмы Sloning BioTechnology, описанной в заявке PCT WO 2005071077.
Последовательность гена, кодирующего ОРС рекомбинантного фактора свертываемости крови VIII с делецией В-домена человека согласно настоящему изобретению, представлена в Перечне последовательностей под номером SEQ ID NO: 1 (нуклеотиды 6460-10830). Аминокислотная последовательность секретируемого фактора свертываемости крови VIII человека с делецией В-домена согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO: 3, и приведена на Фигуре 4, и представляет собой продукт трансляции нуклеотидов 6460-10830 последовательности ОРС SEQ ID NO: 1 без 19 первых аминокислот секреционного лидерного пептида. В-домен фактора VIII заменен на линкерный пептид длиной 14 аминокислот, частично отделяющийся в ходе пост-трансляционного процессинга фVIII BDD (SEQ ID NO: 3, аминокислоты 741-754), в котором первые пять аминокислот представляют собой аминокислотную последовательность N-концевого участка В-домена фVIII, а последующие девять аминокислот представляют собой аминокислотную последовательность С-концевого участка В-домена фVIII. Часть молекул зрелого секретированного фактора свертываемости крови VIII с делецией В-домена человека согласно настоящему изобретению может содержать один или несколько протеолитических разрывов полипептидной цепи, преимущественно после остатка Arg740, и представлять собой двухцепочечную форму фактора свертываемости крови VIII. Степень превращения одноцепочечной формы в двухцепочечную не влияет на биологическую активность фактора свертываемости крови VIII, для целей настоящего изобретения эти формы полностью эквивалентны.
Чтобы обеспечить эффективную трансляцию гена в клетках китайского хомячка, предпочтительно, чтобы ОРС предварялась последовательностью для кэп-зависимой инициации трансляции (последовательность Козак), например, синтетической.
Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO: 1 нуклеотиды 6460-10830), например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации. Фрагменты ДНК, которые кодируют по существу такой же функциональный белок, могут быть выявлены путем экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке и установления активности экспрессируемого продукта.
Фактор свертываемости крови VIII человека - это неэнзиматический кофактор системы свертывания крови, образующий плотный нековалентный комплекс с активрованным фактором IX и ускоряющий реакцию протеолитического расщепления фактора X до активированного фактора X. Природный фактор свертываемости крови VIII человека в кровотоке преимущественно циркулирует в двухцепочечной форме, то есть представляет собой гетеродимер тяжелой и легкой цепи, при этом тяжелая цепь может частично или полностью содержать область В-домена. Основные варианты тяжелой цепи природного фактора VIII имеют массу от 90 до 150 кДа, легкая цепь - около 80 кДа. Рекомбинантный фактор свертываемости крови VIII человека с делецией В-домена в одноцепочечной форме имеет массу около 170 кДа, в двухцепочечной - около 90 кДа у тяжелой цепи и около 80 кДа у легкой цепи. Рекомбинантный фактор свертываемости крови VIII с делетированным В-доменом полностью сохраняет прокоагулянтную активность, при этом его удельная прокоагулянтная активность, выраженная в международных единицах на один миллиграмм чистого белка, выше на 20-40% из-за меньшей молекулярной массы делетированного варианта.
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами рекомбинантного фактора свертываемости крови VIII человека, определяются по его способности уменьшать время свертывания (коагуляции) плазмы крови человека с удаленным фактором VIII. Так, например, активность рекомбинантного фактора свертываемости крови VIII человека можно детектировать по уменьшению времени образования сгустка в плазмы крови с удаленным фактором VIII после смешивания исследуемого раствора фактора VIII, плазмы крови с удаленным фактором VIII, контактного активатора свертывания эллаговой кислоты и раствора хлорида кальция. Время свертывания плазмы, смешанной с несколькими разведениями исследуемого раствора, и сопоставляют измеренные времена образования сгустка с временами образования сгустка, полученными для нескольких разведений стандарта активности фактора VIII - нормальной плазмы крови или раствора изолированного фактора VIII. Концентрацию фактора VIII человека определяют при помощи иммуноферментного анализа в сравнении со стандартом - плазмой крови или раствором очищенного фактора VIII с известным содержанием. Удельную активность фактора VIII определяют как отношение прокоагулянтной активности и концентрации фактора VIII и выражают в МЕ/мг. Считается, что вариант белка обладает свойствами рекомбинантного фактора VIII свертываемости крови человека при условии, что удельная активность указанного варианта составляет не ниже 1% от удельной активности природного фактора VIII человека, то есть не менее 50 МЕ/мг.
Экспрессионная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий фактор свертываемости крови VIII с делецией В-домена под контролем промотора и регуляторных элементов, функционирующих в эукариотической клетке. В качестве рекомбинантной плазмиды согласно настоящему изобретению могут использоваться различные плазмиды, обладающие способностью к экспрессии в клетке-реципиенте, такие как плазмиды pcDNA3.1, pCMV-Myc, pDEF38 и подобные им, но список плазмид не ограничивается ими.
Конкретным вариантом реализации настоящего изобретения является плазмида p1.1-F8BDD длиной 16282 п. о. (SEQ ID NO 1), которая содержит следующие функциональные элементы, перечисленные в порядке их расположения:
1. область начала репликации плазмиды pUC (150-283), открытую рамку считывания бета-лактамазы (bla) и прокариотический промотер гена bla (965-1924), позволяющие проводить препаративную наработку плазмиды в Е. coli;
2. участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), представляющий собой фрагмент конкатемера терминальных повторов вируса Эпштейна-Барр (1960-2361), обеспечивающей увеличение частоты интеграции генетической кассеты в геном клеток СНО и увеличение скорости амплификации кассеты в геноме;
3. фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «выше по течению» (2373-6443), включающий нетранскрибируемые области этого гена, промотор этого гена, первый интрон этого гена, обеспечивающий конститутивную экспрессию гена фактора VIII человека в геноме клеток СНО;
4. синтетическую последовательность Козак (сайт связывания рибосом) (6451-6459), обеспечивающую кэп-зависмую инициацию трансляции мРНК в животных клетках;
5. последовательность, кодирующую открытую рамку считывания фактора свертываемости крови VIII человека с делецией В-домена (6460-10830);
6. последовательность внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV (10855-11442), обеспечивающая кэп-независимую инициацию трансляции второго цистрона бицистронной РНК в животных клетках;
7. последовательность, кодирующую открытую рамку считывания дигидрофолатредуктазы (11456-12018) - фактора устойчивости трансфецированных клеток к воздействию метотрексата;
8. фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «ниже по течению» (12020-16278), включающий терминатор и сигнал полиаденилирования, а также нетранскрибируемые области этого гена, обеспечивающий конститутивную экспрессию гена фактора VIII человека в геноме клеток СНО.
Плазмида p1.1-F8-BDD содержит уникальные сайты узнавания эндонуклеазами рестрикции AscI (16), PvuI (1409), FseI (5213), AbsI (6446), PsiI (7737), NheI (10835).
Структура плазмиды p1.1-F8-BDD приведена на Фигуре 1.
Плазмида p1.1-F8-BDD сконструирована на базе экспрессионной плазмиды p1.1, разработанной авторами настоящего изобретения ранее и подробно описанной в патентной заявке РФ 2011146243, целиком включенной в настоящее описание посредством ссылки. Плазмида p1.1 содержит функциональные промотор и терминатор гена фактора элонгации 1 альфа китайского хомячка, обеспечивающие конститутивную экспрессию целевого гена в клетках СНО, фланкированные 5' и 5' НТО этого гена, обеспечивающими эухроматинизацию сайтов интеграции экспрессионной кассеты в геном СНО; участок для клонирования открытых рамок считывания целевых белков; внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV IRES), обеспечивающий реинициацию трансляции на бицистронной мРНК в клетках млекопитающих и полное генетическое сцепление уровней продукции целевого белка и DHFR; ОРС DHFR мыши, экспрессирующуюся в составе бицистронной мРНК вместе с целевым геном (IRES DHFR) и обеспечивающую устойчивость стабильно трансфицированных клеток к отсутствию в среде тимидина и дозозависимую устойчивость к воздействию метотрексата (МТХ), что позволяет вести направленную селекцию высокопродуктивных клонов с множественными копиями экспрессионной кассеты в геноме СНО; участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), обеспечивающий повышенный уровень интеграции кассет в геном клеток СНО и ускорение процесса амплификации амплификации целевого гена в геноме под действием МТХ. Использование плазмиды p1.1 позволяет осуществлять высокочастотную интеграцию и ускоренную амплификацию экспрессионной кассеты в клетках млекопитающих, а также получать линии-продуценты с высоким уровнем продукции целевого белка и высокой стабильностью.
При помощи созданной плазмиды согласно настоящему изобретению, в частности плазмиды p1.1-F8-BDD, можно трансформировать эукариотическую клетку, предпочтительно иммортализованную клетку китайского хомячка. Выбор конкретной линии клеток не является критическим, поскольку методология и приемы трансформации хорошо известны специалисту в данной области техники. После проведения одного или нескольких раундов селекции и клонирования популяции трансформированных клеток могут быть получены клональные линии-продуценты рекомбинантного фактора свертываемости крови VIII человека. Методология и приемы селекции и клонирования клеток известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования и селекции трансформантов уровень экспрессии фактора свертываемости крови VIII человека может варьироваться, факт транзиентной экспрессии целевого белка будет иметь место при условии успешной трансформации клетки- реципиента, факт стабильной экспрессии целевого белка будет иметь место при успешном проведении раунда селекции популяции клеток-трансформантов.
«Трансформация клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Методы трансформации включают любые стандартные методы, известные специалисту в данной области техники, например электропорацию, использование трансфекционных агентов, например метод, описанный в техническом документе компании Invitrogen, Inc. "Lipofectamine® 2000 Reagent" Pub. No. MAN0000995 Rev. Date 20 July 2012.
Согласно настоящему изобретению «клеточная линия - продуцент фактора свертывания крови VIII человека с делетированным В-доменом» означает клональную линию клеток млекопитающих, обладающую способностью к продукции и секреции фактора свертывания крови VIII человека с делетированным В-доменом, когда она согласно настоящему изобретению выращивается в указанной питательной среде. Используемый здесь термин «клеточная линия - продуцент фактора свертывания крови VIII человека с делетированным В-доменом» также означает линию клеток, которые способна секретировать фактор свертывания крови VIII человека с делетированным В-доменом в количестве не менее чем 5 МЕ/мл при концентрации клеток 1-2 млн/мл, более предпочтительно не менее чем 20 МЕ/мл при концентрации клеток 1-2 млн/мл при культивировании в суспензионной культуре в бессывороточной среде. Указанный фактор свертывания крови VIII человека с делетированным В-доменом секретируется указанными клетками в культивационную среду.
Среди клеток млекопитающих предпочтительно использование клеток китайского хомячка (Cricetulus griseus), предпочтительно клеток яичника (СНО). В качестве примера предпочтительных клеток яичника китайского хомячка может быть приведена линия СНО DG44 (Invitrogen cGMP banked, США; Mol Cell Biol 5, 1750-1759 Kaufman RJ et al. 1985). Круг сублиний СНО не ограничен каким-либо образом, например, могут быть использованы сублинии СНО CHOZN DHFR, СНО DUKX B11 и подобные им.
Конкретным примером линии для получения продуцента фактора свертывания крови VIII человека с делетированным В-доменом согласно настоящему изобретению является линия СНО DG44, но спектр линий клеток не ограничиваются ей.
Линия СНО DG44 характеризуется следующими культурально-морфологическими, физиолого-биохимическими признаками и генетическими признаками:
- Данные по видовой принадлежности: китайский хомячок Cricetulus griseus, яичник.
- Контроль видовой идентичности: кариологический, изоферментный (ЛДГ и Г6ФДГ).
- Маркерные признаки и методы их оценки: иммунологические, цитогенетические, биохимические, физиологические:
- Морфология: эпителиоподобная.
- Кариология: 2n=22, пределы изменчивости по числу хромосом 10-28, модальное число хромосом 22, псевдодиплоид, имеются микрохромосомы, количество полиплоидов 9,0%.
- Биохимия: Дигидрофолатредуктазная активность по резистентности к метотрексату.
- Условия выращивания: среда - бессывороточная, определенного химического состава, CD Opti СНО (Invitrogen, США), температура - +37°C, концентрация CO2 - 8%, скорость перемешивания - 120-130 об/мин.
- Культуральные свойства: способ культивирования - суспензионный, посевная доза - 300000 на мл., кратность рассева - 1:2-1:6, процедура пересева - центрифугирование суспензии на скорости 1200 об/мин, 5 мин, ресуспендирование в новой ростовой среде. Оптимальная плотность 1,0-1,2×106 клеток/мл.
Трансформация клеток линии СНО DG44 плазмидой p1.1-F8BDD с последующей селекцией и клонированием приводит к получению клеточных линий-продуцентов рекомбинантного фактора VIII человека с делетированным В-доменом.
Линия-продуцент DG-P1.1-F8BDD-S3 обеспечивает синтез и секрецию рекомбинантного фактора VIII человека с делетированным В-доменом в количестве 40±9 ME конечной активности В-домен делетированного фактора свертываемости крови VIII на 1 мл среды при концентрации клеток 1,7 млн/мл при культивировании в суспензионной культуре в бессывороточной среде. Линия-продуцент DG-P1.1-F8BDD-S3 депонирована в Российской Коллекции Культур Клеток позвоночных Института Цитологии РАН, регистрационный номер №РККК(П) 756D.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного фактора VIII человека с делетированным В-доменом в клетках млекопитающих, включающего культивирование в питательной среде описанных выше клеток млекопитающих - продуцентов рекомбинантного фактора VIII человека с делетированным В-доменом, и выделение полученного целевого рекомбинантного белка из культуральной жидкости.
Выращивание клеток, выделение и очистка целевого белка из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам культивирования, в которых рекомбинантный белок продуцируется с использованием клеток млекопитающих.
Питательная среда, используемая для культивирования, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста клеток. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза или сахароза и другие органические кислоты. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Для получения фармацевтически значимого рекомбинантного фактора VIII человека с делетированным В-доменом предпочтительно использование бессывороточной питательной среды для культивирования.
Выращивание может осуществляться в аэробных условиях, предпочтительно с повышенным содержанием CO2 (8%), таких как перемешивание культуральной жидкости в колбах, при температуре в пределах от 20 до 40°C, предпочтительно в пределах от 30 до 38°C. Обычно, выращивание в течение от 12 часов до 4 дней приводит к накоплению целевого рекомбинантного белка в культуральной среде или в цитоплазме клетки.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования, а затем целевой белок может быть выделен и очищен методами хроматографии и/или концентрирования.
Особенности созданной экспрессионной плазмиды, линии клеток и результаты практического применения изобретения приведены на следующих фигурах.
Краткое описание Фигур.
На Фигуре 1 показана карта экспрессионной плазмиды p1.1-F8BDD. Используются следующие обозначения: pUC origin - область начала репликации плазмиды pUC; bla - открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; bla promoter - прокариотический промотер гена bla; EBV TR - участок терминального повтора вируса Эпштейна-Барр человека; CHOEEF1AUFR функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, 5' нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; transcription start - точка начала транскрипции, TATA - ТАТА-бокс, СНО EEF1A intron 1 - первый интрон гена фактора элонгации 1 альфа китайского хомячка, EMCV IRES - внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV); DHFR - открытая рамка считывания дигидрофолатредуктазы мыши для селективного отбора и амплификации в эукариотических клетках; CHOEEF1ADFR - функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, 3' нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; Kozak - область, кодирующая последовательность Козак для кэп-зависимой инициации трансляции, F8-BDD ORF - ОРС фактора VIII с делетированным В-доменом. Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
На Фигуре 2 показана карта экспрессионного вектора p1.1. Используются обозначения аналогично фигуре 1.
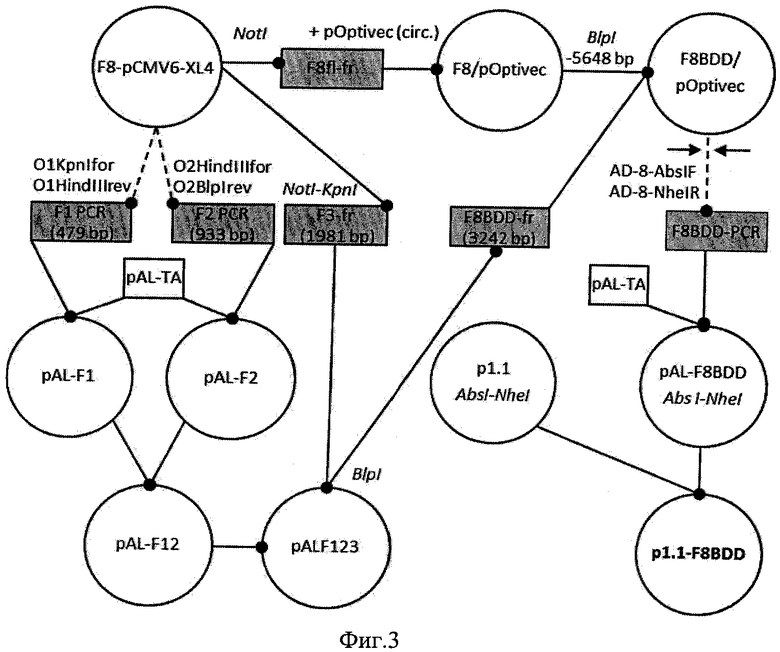
На Фигуре 3 показана схема получения плазмид p1.1-F8BDD. Обозначения: пунктирной линией обозначены стадии ПЦР, сплошной линией обозначены стадии рестрикции-лигирования, кольцевые плазмиды обозначены кружками, линейные фрагменты ДНК - прямоугольниками; названия использовавшихся для клонирования эндонуклеаз рестрикции, указаны курсивом, названия олигонуклеотидных праймеров использовавшихся при ПЦР указаны прямым шрифтом.
На Фигуре 4 показана аминокислотная последовательность полипептида, секретируемого клетками DG-P1.1-F8BDD-S3 (продукт экспрессии плазмиды с p1.1-F8BDD без секреционного лидерного пептида).
На Фигуре 5 приведены результаты иммуноблотинга секретированного и внутриклеточного фVIII BDD для линии 3-В8. Используются следующие обозначения: «heavy chain» и «light chain» - гибридизация с моноклональными антителами к тяжелой и легкой цепи фVIII соответственно; "CMV» и «CHEF1» - образцы, соответствующие клеткам линии DG-BDDFVIII-18 (промотор CMV, описана в патенте РФ RU 2429294) и 3-В8 (промотор CHEF1) соответственно. Приведенные для каждого из антител дорожки соответствуют одной мембране, разделение в 7.5% ДСН-ПААГ в восстанавливающих условиях, молекулярные массы указаны в кДа.
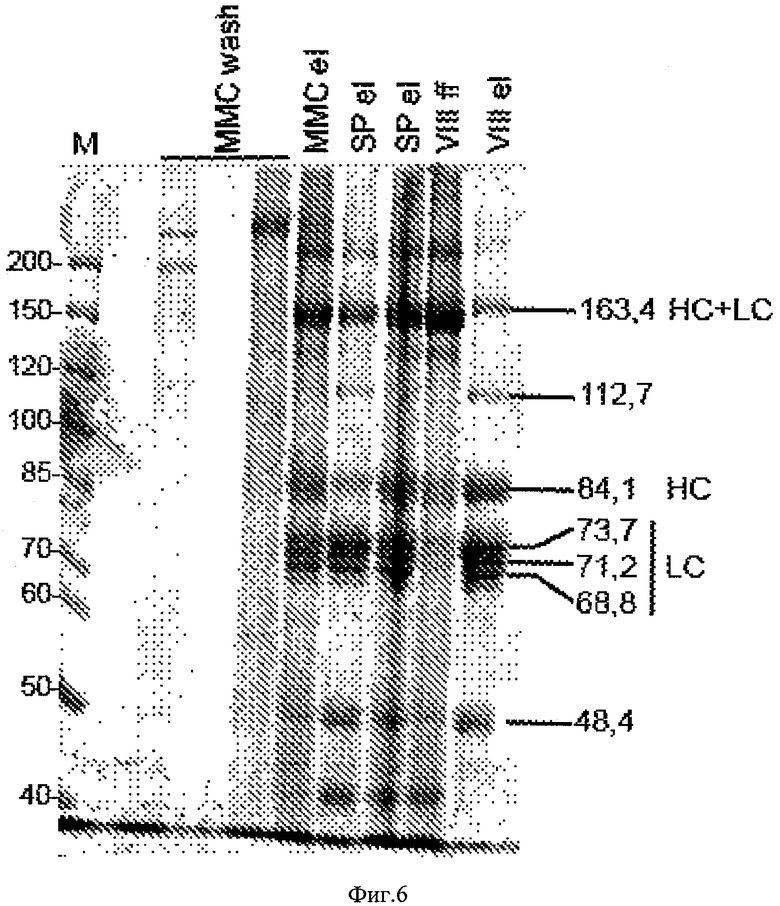
На Фигуре 6 приведена электрофореграмма белковых фракций промывок колонок и элюатов при очистке фVIII. Восстанавливающие условия, окраска Кумасси синим. Исследуемые фракции сконцентрированы и обессолены переосаждением белков трихлоруксусной кислотой. Используются следующие обозначения: полосы легкой цепи - LC, полоса тяжелой цепи - НС, полоса одноцепочечного предшественника - HC+LC. ММС wash - фракции промывки колонки Capto ММС, SP el - фракции элюции колонки Sepharose SP, VIII ff - фракция проскока для колонки VIII Select, VIII el - фракция элюции с колонки VIII Select, М - маркер молекулярных масс. Молекулярные массы указаны в кДа.
Практическая применимость настоящего изобретения иллюстрируется следующими примерами.
Пример 1. Получение генетической конструкции p1.1-F8BDD, кодирующей белок человеческого фактора свертываемости крови VIII с делецией В домена, для трансфекции линии клеток CHO-DG-44.
Получение экспрессионной конструкции проводили согласно общей схеме, представленной на Фиг. 3. Для получения ОРС фактора VIII с делегированным В-доменом методом ПЦР со специфических праймеров O1KpnIfor и O1HindIIIrev, 02HmdIIIfor и 02B1pIrev (SEQ ID NO:4-7, соответственно) были получены два ПЦР-продукта F1 и F2, фланкирующие область делеции. В качестве матрицы использовали коммерчески доступный клон pCMV6-XL4/NM_000132. Олигонуклеотиды были синтезированы ЗАО ″Евроген″, РФ. ПЦР проводили при помощи смеси Tersus polymerase mix (ЗАО ″Евроген″, РФ) на приборе РТС-100 Thermal Cycler (MJ Reseach, США).
ПЦР продукты очищали используя набор реактивов ″Wizard SV Gel and PCR Clean-Up System″ (″Promega″, США) по протоколу производителя, и клонировали в вектор pAL-ТА (Евроген) ДНК-лигазой фага Т4 (Fermentas, Литва) по методике производителя фермента. Полученной лигазной смесью трансформировали клетки Е. coli штамма DH5α, с генотипом F-φ80lacZΔM15 Δ(lacZYA-argF) U169 recAl endAl hsdR17 (rk-, mk+) phoA supE44 λ- thi-1 gyrA96 relAl. Для этого к 200 мкл замороженной суспензии клеток Е. coli добавляли 5 мкл лигазной смеси, инкубировали на льду 30 минут для сорбции плазмидной ДНК, нагревали до 42°C на 60 секунд и инкубировали на льду 5 минут. После чего добавляли 800 мкл питательного бульона SOC (20 г/л триптона, 5 г/л дрожжевого экстракта, 0,5 г/л NaCl, 250 мМ KCl, 10 мМ MgCl2) и инкубировали при 37°C 60 минут. После инкубации переносили суспензию на чашку Петри с твердой агаризованной средой 2xYT-агар (16 г/л триптона, 10 г/л дрожжевого экстракта, 10 г/л NaCl, 18 г/л агара), содержащей ампициллин в концентрации 100 мкг/мл, и помещали в термостат на 37°C на 18 часов. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 37°C и выделяли плазмидную ДНК набором реактивов "Wizard Plus SV Minipreps" («Promega», США) по протоколу производителя. Полностью секвенировали области вставки полученных плазмид с праймеров Т7 и SP6 с использованием набора BigDye Terminator v. 3.1 cycle sequencing kit (Applied Biosystems, США) и капиллярного секвенатора ABI PRISM 3730 genetic analyzer (Applied Biosystems, США), анализ данных вели при помощи программы Chromas 1.45 (Technelysium Pty Ltd, Australia).
Фрагмент F3 fVIII получали без использования ПЦР, при препаративной рестрикции плазмиды pCMV6-XL4/NM_000132, эндонуклеазами NotI и KpnI. Продукты рестрикции смешивали 6:1 с буфером для нанесения «6х Loading dye» («Fermentas», Литва) и проводили разделение в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, визуализированную при помощи УФ, вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл, после чего проводили выделение ДНК, ноабором реактивов "Wizard SV Gel and PCR Clean-Up System" («Promega», США) согласно по протоколу производителя.
Сборку фрагментов F1-F3 проводили в векторе pAL-TA с образованием плазмиды pALTA/F123. Отбор клонов Е. coli проводился методом ПЦР с клонов с использованием праймера Odelf и стандартных праймеров к последовательности вектора T7prom (SEQ ID NO: 9) и SP6 (SEQ ID NO: 10). ПЦР с колоний Е. coli проводили с использованием набора реактивов «ScreenMix-HS» (ЗАО Евроген, Россия), по методике производителя, помещая образцы бактериальных клонов в пробирки с 10 мкл готовой смеси для ПЦР. Продукты ПЦР анализировали электрофорезом в 1% агарозном геле, позитивные клоны наращивали в 5 мл среды LB с добавлением ампициллина в концентрации 50 мкг/мл 18 часов при 37°C и выделяли плазмидную ДНК набором реактивов "Wizard Plus SV Minipreps" («Promega», США) по протоколу производителя. Полностью секвенировали области вставки полученных плазмид с праймеров T7prom (SEQ ID NO: 9) и SP6 (SEQ ID NO: 10) и специфического праймера Odelf (SEQ ID NO: 8), как описано выше.
Полученный исскуственный фрагмент гена В-домен делетированного фактора VIII лигировали с рестрицированной по сайту BlpI и дефосфорилированной плазмидой pOptivec/F8.
Для создания плазмиды pOptivec/F8 использовался вектор pOptivec, полученный путем рециркуляризации линейного вектора pOptivec-TOPO (Invitrogen, США) при инкубации в деионизованной воде 30 минут с последующим лигированием ДНК-лигазой фага Т4 (Fermentas, Литва) по методике производителя фермента. Плазмида pOptivec была рестрицирована эндонуклеазой NotI и лигирована с фрагментом NotI-NotI плазмиды pCMV6-XL4/NM_000132 (Origene, США) с образованием плазмиды pOptivec/F8. Фрагмент BlpI-BlpI pOptivec/FS был заменен на фрагмент BlpI-BlpI pALTA/F123 с образованием плазмиды pOptivec/F8BDD. ПЦР-анализ колоний вели с использованием двух пар специфических праймеров 8sq4f (SEQ ID NO: 11) и 8sq5r (SEQ ID NO: 12) или CMV for (SEQ ID NO: 13) и 8sq15r (SEQ ID NO: 14). При помощи праймеров AD-8-AbsF (SEQ ID NO: 15), и AD-ONheI-R (SEQ ID NO: 16), и pOptivec/F8BDD в качестве матрицы получили ПЦР-продукт, содержащий открытую рамку считывания фактора VIII свертываемости крови человека с делетированным В-доменом. Полученный продукт ПЦР клонировали в вектор PAL-TA и анализировали секвенированием области вставки, используя праймеры к последовательностям вектора T7prom (SEQ ID NO: 9) и SP6 (SEQ ID NO: 10), а также специфические праймеры к последовательности гена фактора VIII делетированным В-доменом - 8sq1f, 8sq2f, 8sq3f (SEQ ID NO: 17-19 соответственно), 8sq4f (SEQ ID NO: 11), 8sq5f, 8sq12f, 8sq13f, 8sq14f, 8sq15f (SEQ ID NO: 20-24 соответственно), 8sq5r (SEQ ID NO: 12), 8sq15r (SEQ ID NO: 14), 8sq2r, 8sq3r, 8sq4r, 8sq12r, 8sq13r, 8sq14r (SEQ ID NO: 25-30 соответственно). После обнаружения клонов с корректной нуклеотидной последовательностью области вставки плазмиду PAL-F8BDD рестрицировали AbsI и NheI и переносили фрагмент, содержащий ОРС F8BDD с участком консенсусной последовательности Козак в рестрицированный эндонуклеазами AbsI и NheI вектор p1.1 (SEQ ID NO: 2) с образованием экспрессионной плазмиды p1.1-F8BDD (SEQ ID NO: 1). Открытую рамку считывания делеционного варианта BDD-FVHI, а также области основных функциональных элементов экспрессионной конструкции секвенировали, как описано выше. Для секвенирования области ОРС и функциональных элементов экспрессионной плазмиды использовали специфические праймеры к последовательности гена фактора VIII делетированным В-доменом - 8sq1f, 8sq2f, 8sq3f (SEQ ID NO: 17-19 соответственно), 8sq4f (SEQ ID NO: 11), 8sq5f, 8sq12f, 8sq13f, 8sq14f, 8sq15f (SEQ ID NO: 20-24 соответственно), 8sq5r (SEQ ID NO: 12), 8sq15r (SEQ ID NO: 14), 8sq2r, 8sq3r, 8sq4r, 8sq12r, 8sq13r, 8sq14r (SEQ ID NO: 25-30 соответственно), а также праймеры к последовательностям вектора SQ-3CH1-R, SQ-5CH6-F, IRESArev (SEQ ID NO: 31-33 соответственно). Результаты сравнивали с теоретически предсказанными последовательностями гена фактора FVIII свертываемости крови человека с делетированным В-доменом и соответствующих регуляторных элементов, нуклеотидных замен не было обнаружено.
Полученная экспрессионная конструкиця фактора свертываемости VIII человека с делетированым В-доменом в векторе p1.1-F8BDD имеет размер 16282 п. о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции AscI (16), PvuI (1409), FseI (5213), AbsI (6446), PsiI (7737), NheI (10835). Данная конструкция предназначена для трансфекции эукариотических клеток геном фактора свертываемости VIII человека с делетированым В-доменом с последующим отбором и амплификацией по сцепленному признаку DHFR+. Сообщает клеткам-реципиентам следующие фенотипические признаки: при трансформации прокариот - устойчивость к ампицилину, карбенициллину; при трансфекции эукариот ограниченная устойчивость к метотрексату, увеличивающаяся при селективном отборе, экспрессия рекомбинантного фактора FVIII свертываемости крови человека с делетированным В-доменом.
Созданную генетическую конструкцию использовали для получения линии-продуцента.
Пример 2. Получение первичных поликлональных линий клеток СНО DG-44, стабильно трансфицированных плазмидой p1.1-F8BDD.
Экспрессионная плазмида p1.1-F8BDD, кодирующая рекомбинантный фактор свертываемости крови VIII человека с делецией В-домена по варианту SQ, была сконструирована по методике, приведенной в Примере 1, выращена в препаративных количествах в штамме E.coli ТОР 10 (Invitrogen, Ltd., США), очищена при помощи набора EndoFree Plasmid MaxiKit, (Qiagen, США), линеаризована эндонуклеазой рестрикции Pvul (Fermentas, Литва) с разрушением гена бета-лактамазы и стерилизована фильтрованием.
Клетки линии CHO-DG-44 (Invitrogen, Ltd., США) культивировали в безбелковой среде CD DG-44 (Gibco, США) с 8 тМ L-глутамина и 0,018% сурфактанта Pluronic F-68 (BASF Inc., США). Данная культуральная среда содержит достаточные количества гипоксантина и тимидина для поддержания роста клеток, дефектных по гену dhfr. Клетки выращивали в стерильных одноразовых колбах Эрленмеера (30 мл суспензии в колбах вместимостью 125 мл) на орбитальной качалке при 130 об/мин в атмосфере, содержащей 8% углекислого газа при температуре 37°C. Подсчет плотности клеточной суспензии и доли живых клеток проводится в счетной камере Горяева, при окрашивании трипановым синим. Пассирование культуры проводили по достижении плотности 1,2 млн клт/мл, посевная концентрация 0,3 млн клт/мл. За 48 ч и 24 ч до трансфекции клетки дополнительно пассировали до посевной концентрации 0,3 млн клт/мл.
Трансфекцию линеаризованной плазмиды p1.1-F8BDD проводили при помощи липосомного реагента Unfectin-56 (Unifect Group, Россия) в бессывороточной среде CD DG-44 (Invitrogen), используя 1,5×107 клеток в 30 мл среды в колбе Эрленмеера, 18 мкг плазмидной ДНК и 15 мкл трансфекционного агента. Смешивание ДНК и трансфекционного реагента проводили по методике производителя агента.
Культуру клеток выращивали 48 ч без смены среды, после чего отбирали аликвоту культуральной жидкости для определения концентрации фVIII (0,048 МЕ/мл по ИФА) и высевали клетки в лунки 96-луночных культуральных планшетов по 50 тыс. клеток на лунку в среду СНО A (Invitrogen) с 8 мМ аланил-глутамина («Invitrogen») и 50 нМ метотрексата, по 200 мкл среды в лунку и растили 14 дней при 37°C, 5% CO2. После прохождения селекции стабильно трансфицированных клеток, делящихся в отсутствие гипоксантина и тимидина и в присутствие ингибитора DHFR метотрексата, в лунках с крупными делящимися колониями клеток отбирали культуральную среду для определения концентрации секретированного фVIII методом иммуноферментного анализа. В 96 лунках, содержащих фVIII в максимальной концентрации (от 0,1 до 1 МЕ/мл), проводили замену среды и повторное тестирование через 4 дня. Клетки из 48 лунок с наибольшей концентрацией фVIII пересаживали в лунки 24-луночных планшетов, выращивали до покрытия монослоем не менее 50% поверхности в 80% лунок и повторно тестировали методом ИФА. Клетки из 18 лунок с наибольшей концентрацией фVIII переносили в лунки 6-луночных неадгезионных планшетов и вносили в лунки по 5 мл среды Opti СНО (Invitrogen) с 8 мМ аланил-глутамина («Invitrogen») и 50 нМ метотрексата. Поскольку среда Opti СНО содержит сурфактант Pluronic F-68, клетки теряют способность распластываться на дне лунок и образовывают неприкрепленный слой. Культуры растили до достижения концентрации клеток около 1 млн клт/мл, измеряли концентрацию секретированного фVIII и три наиболее продуктивные олигоклональные линии пересаживали в колбы Эрленмейера и в дальнейшем растили при постоянном перемешивании. Для дальнейшей работы была выбрана олигоклональная линия 3-В8 (Таблица 1), секретировавшая преимущественно корректно процессированные варианты цепей фVIII (Фигура 5).
Пример 3. Амплификация целевого гена под действием метотрексата и получение клональных линий-продуцентов фVIII.
Амплификацию целевого гена в геноме клеток отобранной линии 3-В8 проводили в суспензионной культуре, добавляя в культуральную среду метотрексат в возрастающих концентрациях до момента падения жизнеспособности культуры. При обнаружении падения жизнеспособности культуру культивировали при постоянной концентрации метотрексата до восстановления жизнеспособности >85%, после чего концентрацию метотрексата удваивали. Процесс амплификации считали законченным, когда жизнеспособность клеток переставала восстанавливаться после 20 дней культивирования. Ход амплификации и показатели продуктивности культуры приведены в Таблице 2.
Полученная поликлональная популяция клеток 3-В8 1K 4К была использована для получения клональных линий-продуцентов методом предельного разведения. Клетки клонировали в условиях адгезионного культивирования в среде CD СНО-А с 8 мМ аланил-глутамина («Invitrogen»), по 200 мкл среды в каждую лунку планшета при 37°C, 5% CO2 в течение 21 дня. Метотрексат не включали в состав среды при клонировании и не использовали при дальнейшем культивировании. Рост одиночных колоний наблюдали и документировали на 14-й день культивирования. В 10 планшетах было получено 361 одиночная колония. Выросшие колонии переносили в 24-луночные планшеты и при помощи ИФА осуществляли скрининг кондиционированной среды из лунок, содержащих активно растущие колонии. Клональные линии с наилучшими уровнями секреции продолжали растить в адгезионной культуре и затем реадаптировали к условиям суспензионного культивирования в течение трех последовательных пассажей в лунках 6-луночных планшетов, используя среду Pro СНО 4 (Lonza, США) с 8 мМ L-глутамина. Скрининг кондиционированной среды из лунок 6-луночных планшетов проводили при помощи ИФА и коагулометрического измерения активности фVIII. Три выбранные линии с высокой продуктивностью и временем деления менее 40 ч выращивали в условиях суспензионного культивирования в перемешиваемых колбах Эрленмейера. Основные параметры трех отобранных линий приведены в таблице 3.
В качестве основной линии-продуцента был выбран клон 11А4Н, обладавший максимальной удельной продуктивностью и временем деления клеток менее 30 ч. Линия-продуцент была обозначена как DG-P1.1-F8 BDD-S3, сохранена в форме исследовательского клеточного банка в криоампулах, депонирована в Специализированную коллекцию культур клеток позвоночных Российской коллекции клеточных культур 05.02.2013 г. под номером РККК (П) 756Д. Линия DG-P1.1-F8 BDD-S3 была протестирована на постоянство уровня секреции при продолжительном культивировании. Было установлено, что конечная активность фVIII в кондиционированной среде, определяемая в моменты пассирования культуры, существенно не меняется в течение 60 дней последовательного культивирования (данные не приводятся). Таким образом, полученная клеточная линия потенциально пригодна для промышленного применения, т.е. сохраняет постоянную продуктивность на период времени, достаточный для подготовки посевного материала и проведения 10 циклов периодического культивирования в биореакторе большого объема.
Пример 4. Выделение целевого белка из супернатанта культуры линии DG-P1.1-F8BDD.
Препаративное культивирование линии DG-P1.1-F8BDD вели в перемешиваемых колбах Эрленмейера с использованием бессывороточной среды Pro СНО 4 с добавлением 8 мМ L-глютамина. Проводили посев культуры на плотности 0,3 млн клт/мл и заменяли 7/8 объема культуральной суспензии каждые 96 ч на свежую культуральную среду.
Получаемые порции клеточной суспензии немедленно осветляли центрифугированием и наносили на колонку с сорбентом Capto ММС (GE Healthcare, США). Колонку промывали высокосолевым раствором и элюировали фVIII раствором, содержащим 50% этиленгликоля и 0,8 М аргинина гидрохлорида. Элюат разбавляли водой в 12 раз и наносили на колонку SP Sepharose FF (GE Healthcare), колонку промывали раствором, содержащим 0,15 М NaCl, и элюировали фVIII раствором, содержащим 0,4 М NaCl. К раствору полуочищенного целевого белка добавляли маннитол до конечной концентрации 0,2 М и полученный стабилизированный раствор замораживали и хранили для последующей очистки.
Объединенные порции замороженного раствора полуочищенного фVIII подвергали окончательной очистке при помощи аффинного сорбента VIII Select (GE Healthcare) по инструкции производителя сорбента. Элюированный с аффинной колонки раствор целевого белка разбавляли водой в 12 раз, наносили на колонку с сорбентом Capto Q (GE Healthcare), удаляли протеолитические фрагменты фVIII промывкой раствором, содержащим 0,3 М NaCl, после чего к выходу колонки присоединяли колонку с гель-фильтрационным сорбентом Superdex 200 (GE Healthcare), предварительно уравновешенную изотоническим раствором, содержащим все вспомогательные вещества для получения лекарственной формы фVIII (хлорид натрия, сахарозу, хлорид кальция, гистидин гидрохлорид, полисорбат-80). Проводили элюцию фVIII с колонки Capto Q при помощи раствора, содержащего 0,4 М NaCl, и продолжали подавать элюирующий раствор на тандемно соединенные колонки до выхода фракции мономера фVIII с гель-фильтрационной колонки. Полученный раствор очищенного мономерного фVIII разделяли на аликвоты и замораживали. В образцах элюата со всех стадий очистки определяли активность фVIII, содержание общего белка по методу Брэдфорд, содержание белков штамма-продуцента и ДНК штамма-продуцента. Основные параметры очистки фVIII приведены в Таблице 4, результаты гель-электрофореза фракций элюата и промывки колонок приведены на Фигуре 6.
Полученный прототип формулированного раствора активной фармацевтической субстанции фVIII обладал удельной активностью около 11 тыс. МЕ/мг общего белка при измерении одностадийным методом, что соответствует фармакопейным требованиям для лекарственных препаратов делеционного варианта фVIII.
Описанные в настоящих примерах методики применимы для создания линий клеток-продуцентов рекомбинантного фактора свертываемости крови VIII с делецией В-домена на основе любых эукариотических клеточных линий, имеющих генетический нокаут гена dhfr, как прикрепленных, так и суспензионных, а также трансфецированных как описанным методом, так и с использованием других липосомальных агентов, а также методами кальций-фосфатной трансфекции, нуклеофекции или микроинъекции.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.






























Изобретение относится к области биотехнологии и может быть использовано для рекомбинантного получения фактора свертываемости крови VIII (FVIII) человека с делецией В-домена. Конструируют плазмиду для экспрессии рекомбинантного FVIII человека с делетированным В-доменом, в клетках СНО, нокаутных по гену дигидрофолатредуктазы (DHFR), включающую область начала репликации плазмиды pUC, открытую рамку считывания бета-лактамазы (bla) и прокариотический промотер гена bla, участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), функциональные элементы гена фактора элонгации 1 альфа китайского хомячка, последовательность Козак; открытую рамку считывания гена FVIII человека с делетированным В-доменом; внутренний сайт связывания рибосом (IRES) вируса EMCV; открытую рамку считывания DHFR, экспрессирующуюся в составе бицистронной мРНК вместе с геном, кодирующим FVIII человека с делетированным В-доменом (IRES DHFR). Полученной плазмидой трансформируют клетку СНО с получением клетки-продуцента указанного фактора. Изобретение позволяет существенно повысить эффективность продукции в клетках СНО и сократить затраты на получение фармацевтически значимого рекомбинантного FVIII человека с делецией В-домена. 3 н. и 7 з.п. ф-лы, 6 ил., 4 табл., 4 пр.
1. Плазмида для экспрессии рекомбинантного фактора свертываемости крови VIII человека с делетированным В-доменом в клетках яичника китайского хомячка (СНО), нокаутных по гену дигидрофолатредуктазы, в следующей последовательности по существу состоящая из области начала репликации плазмиды pUC, открытой рамки считывания бета-лактамазы (bla) и прокариотического промотора гена bla; участка терминального повтора вируса Эпштейн-Барр человека (EBVTR), функционального промотора и первого интрона гена фактора элонгации 1 альфа китайского хомячка, фланкированного 5′ нетранслируемой областью этого гена; консенсусной последовательности Козак; открытой рамки считывания гена, кодирующего фактор свертываемости крови VIII человека с делетированным В-доменом; внутреннего сайта связывания рибосом (IRES) вируса энцефаломиокардита (EMCV); открытой рамки считывания дигидрофолатредуктазы (DHFR), экспрессирующуюся в составе бицистронной мРНК вместе с геном, кодирующим фактор свертываемости крови VIII человека с делетированным В-доменом (IRES DHFR); и функционального терминатора гена фактора элонгации 1 альфа китайского хомячка, фланкированного 3′ нетранслируемой областью этого гена.
2. Плазмида по п. 1, отличающаяся тем, что ген фактор свертываемости крови VIII человека кодирует полипептид фактора свертываемости крови VIII человека с делетированным В доменом, последовательность аминокислот которого приведена в SEQ ID NO: 3.
3. Плазмида по п. 1, отличающаяся тем, что участок терминального повтора вируса Эпштейн-Барр человека (EBVTR) представляет собой фрагмент конкатемера терминального повтора EBVTR.
4. Плазмида по п. 1, отличающаяся тем, что используют открытую рамку считывания дигидрофолатредуктазы (DHFR) мыши.
5. Плазмида по п. 1, отличающаяся тем, что указанной плазмидой является вектор p1.1-F8BDD, представленный в Перечне последовательностей под номером SEQ ID NO: 1.
6. Клетка яичника китайского хомячка (СНО) - продуцент рекомбинантного фактора свертываемости крови VIII человека с делетированным В-доменом, трансформированная плазмидой по пп. 1-5.
7. Клетка по п. 6, отличающаяся тем, указанной клеткой является клетка яичника китайского хомячка моноклональной линии DG-P1.1-F8BDD-S3, депонированной в Российской Коллекции Культур Клеток позвоночных Института Цитологии РАН под регистрационным номером №РККК (П) 756Д.
8. Способ получения рекомбинантного фактора свертываемости крови VIII с делетированным В-доменом, включающий следующие стадии:
- культивирование в питательной среде клеток по п. 6; и
- выделение полученного рекомбинантного белка из культуральной жидкости.
9. Способ по п. 8, отличающийся тем, что культивируют клетки яичника китайского хомячка моноклональной линии DG-P1.1-F8BDD-S3, депонированной в Российской Коллекции Культур Клеток позвоночных Института Цитологии РАН, регистрационный номер №РККК (П) 756Д.
10. Способ по п. 8, отличающийся тем, что питательная среда для культивирования является бессывороточной.
| ЭКСПРЕССИОННАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pOptivec/F8BDD, КОДИРУЮЩАЯ РЕКОМБИНАНТНЫЙ ФАКТОР СВЕРТЫВАЕМОСТИ КРОВИ VIII ЧЕЛОВЕКА С ДЕЛЕЦИЕЙ В-ДОМЕНА, И КЛЕТОЧНАЯ ЛИНИЯ DG-OV-F8BDD-18, ПРОДУЦИРУЮЩАЯ РЕКОМБИНАНТНЫЙ ФАКТОР СВЕРТЫВАЕМОСТИ КРОВИ VIII ЧЕЛОВЕКА С ДЕЛЕЦИЕЙ В-ДОМЕНА | 2009 |
|
RU2429294C2 |
| ORLOVA N.A | |||
| et al., Stable Expression of Recombinant Factor VIII in CHO Cells Using Methotrexate-Driven Transgene Amplification, Acta Naturae, 2012, v.4, n.1, p.93-100 | |||
| ОРЛОВА Н.А | |||
| и др., Высокопродуктивная линия-продуцент фактора свертывания крови VIII человека на основе вектора с нетранслируемыми областями гена фактора | |||

