Область техники
Изобретение относится к области биотехнологии, а именно к технологии получения биологически активных веществ (БАВ) методами генной инженерии, точнее к методам получения рекомбинантных белков в культивируемых клетках млекопитающих.
Предшествующий уровень техники
Получение рекомбинантных белков в культивируемых клетках млекопитающих позволяет производить не менее половины лекарственных средств биологического происхождения, существенную часть вакцин, а также многие препараты для ветеринарии и реагенты для клинической диагностики. Среди линий клеток млекопитающих, используемых в биофармацевтическом производстве, наиболее распространена линия клеток яичника китайского хомячка (Cricetulus griseus) CHO (Chinese hamster ovary cells) и ее производные. Некоторые сублинии CHO отличаются инактивацией гена дигидрофолатредуктазы (DHFR) и позволяют, путем добавления ингибиторов DHFR, относительно легко проводить амплификацию введенных в геном клетки генетических конструкций, содержащих миниген DHFR. В частности, путем мутагенеза химическими агентами из базовой линии СНО-K1 была получена сублиния CHO-DG44, не содержащая активных аллелей DHFR (Urlaub G, Chasin LA. Isolation of Chinese hamster cell mutants deficient in dihydrofolate reductase activity. Proc Natl Acad Sci USA. 1980 Jul;77(7):4216-4220) и пригодная для получения стабильных линий-продуцентов рекомбинантных белков (Derouazi M et al, Genetic characterization of CHO production host DG44 and derivative recombinant cell lines. Biochem Biophys Res Commun. 2006 Feb 24;340(4):1069-1077).
Основным фактором, влияющим на продуктивность создаваемой промышленной линии-продуцента, является уровень мРНК целевого гена, определяемый, в свою очередь, силой промотора целевого гена, контекстом точки (или точек) интеграции целевого гена в хромосомы и числом копий целевого гена на клетку.
Стабильность линии-продуцента при последовательных пересевах определяется устойчивостью промотора целевого гена к инактивации метилированием остатков цитидина (Brooks AR e al Transcriptional silencing is associated with extensive methylation of the CMV promoter following adenoviral gene delivery to muscle. J Gene Med. 2004 Apr; 6(4):395-404), общей устойчивостью целевого гена и генетической кассеты к модификации ДНК (делециям, инсерциям и точечным мутациям) и интенсивности процессов хромосомной транслокации в областях интеграции копий целевого гена (Kim M et al, A mechanistic understanding of production instability in CHO cell lines expressing recombinant monoclonal antibodies. Biotechnol Bioeng. 2011 May 2. doi: 10.1002/bit.23189. Epub ahead of print). При этом устойчивость промотора к инактивации метилированием определяется его последовательностью (промоторы клеток устойчивее промоторов вирусов), устойчивость к модификации целевого гена и кассеты в целом определяется ее длиной (чем короче ген и чем ближе селекционный маркер к целевому гену, тем менее вероятна его модификация), а вероятность делеции целевого гена при хромосомных транслокациях определяется положениями точек интеграции и может быть проверена только экспериментально. Поскольку процессы метилирования ДНК, возникновения мутаций и транслокации участков хромосом идут в культивируемых клетках непрерывно и практически не поддаются внешнему контролю, "среднюю" стабильность линий-продуцентов можно увеличивать уменьшением числа генераций от момента трансфекции клеток генетической конструкцией до получения маточного клеточного банка.
Таким образом, при создании промышленной линии-продуцента рекомбинантного белка должны быть одновременно решены 2 различные технические задачи - максимизация уровня мРНК целевого гена и ограничение числа последовательных пересевов трансфицированной культуры до определения конечного клона-продуцента и его масштабирования.
Для максимизации уровня мРНК целевого гена в современной практике используют 3 общих подхода:
1) использование как можно более сильных промоторов и регуляторных участков ДНК, способствующих эухроматинизации области интеграции;
2) проведение селективной или направленной интеграции генетических конструкций в транскрипционно активные области генома;
3) амплификацию в геноме интегрированных генетических конструкций.
Применение первого и, в особенности, второго подхода позволяет минимизировать продолжительность пассирования культуры до момента получения конечного клона-продуцента. В то же время, использование третьего подхода позволяет увеличивать уровень мРНК целевого гена практически неограниченно, но требует увеличения времени культивирования.
Техническое решение, позволяющее использовать преимущества всех трех вышеуказанных подходов, должно представлять большой практический интерес. Такое техническое решение может быть основано на применении плазмидной ДНК, обладающей следующими основными свойствами:
1) сильный промотор целевого гена, минимально чувствительный к инактивации метилированием;
2) наличие регуляторных участков ДНК, препятствующих гетерохроматинизации целевой ДНК;
3) повышенная частота интеграции генетической конструкции в геном клеток;
4) наличие селекционного маркера, позволяющего вести первичный отбор стабильно трансфицированных клеток при различном уровне селекционного давления;
5) возможность осуществления амплификации целевых генов после получения первичной популяции стабильно трансфицированных клеток. Известно техническое решение, позволяющее вести экспрессию целевых генов в клетках СНО под контролем сильного промотора клетки-хозяина, при этом данный промотор находится в контексте собственных регуляторных последовательностей, расположенных в 5' и 3'-нетранскрибируемых областях его исходного гена - фактора элонгации трансляции 1 альфа (CHEF1) (Running Deer J, Allison DS. High-level expression of proteins in mammalian cells using transcription regulatory sequences from the Chinese hamster EF-1 alpha gene. Biotechnol Prog. 2004 May-Jun; 20(3):880-889). Использование данного промотора позволило увеличить уровень секреции нескольких модельных белков в 6-35 раз по сравнению с аналогичными конструкциями на основе известных промоторов CMV и EF1 человека, при этом полученные линии сохраняли продуктивность при культивировании в течение нескольких месяцев. Особенностью данного технического решения является расположение селекционного маркера - минигена DHFR мыши перед 3'-нетранскрибируемой областью (НТО) гена CHEF1, но после сигнала полиаденилирования и под контролем отдельного вирусного промотора SV40 и терминатора транскрипции SV40.
Следствием такого расположения функциональных участков является невысокая генетическая сцепленность целевого гена и гена селекционного маркера, существенно ограничивающая возможность амплификации целевого гена. Предположительно, при проведении амплификации построенных по данной схеме экспрессионных кассет под действием возрастающих концентраций ингибитора DHFR метотрексата (МТХ) будет возможна инактивация или делеция целевого гена и последующая амплификация в геноме клетки хозяина только области селекционного маркера, включающей его вирусный промотор, миниген DHFR и терминатор транскрипции. Опубликованные данные о стабильности генетических кассет на основе CHEF1 и DHFR при амплификации под действием МТХ отсутствуют.
Известны способы увеличения сцепленности амплифицируемого гена целевого белка и селекционного маркера - использование полученного в процессе трансляции гибридного белка, разделяющегося на целевой белок и белковый продукт селекционного маркера или бицистронной мРНК. Первый способ может быть реализован путем соединения полипептидных последовательностей целевого белка и продукта селекционного маркера пептидом 2А (Doronina VA et al. Site-specific release of nascent chains from ribosomes at a sense codon. Mol Cell Biol. 2008 Jul; 28(13):4227-4239). Такой способ имеет неустранимый недостаток - появление небольших количеств гибридного белка и целевого белка с неотделенным коротким пептидом р2В и не может быть использован для получения биофармацевтических белков. Второй способ может быть реализован путем соединения кодирующих областей целевого гена и гена DHFR внутренним сайтом связывания рибосом (internal ribosome entry site, IRES); он описан в работе (Lucas BK, Giere LM, Demarco RA, Shen A, Chisholm V, Crowley CW: High level production of recombinant proteins in CHO cells using a dicistronic DHFR intron expression vector. Nucleic Acids Res 1996, 24:1774-1779). Происхождение и эффективность работы IRES не имеют существенного значения для проведения амплификации, при конструировании данного функционального участка экспрессионного вектора может быть использован, например, IRES вируса энцефаломиокардита (Parks, G.D., G.M. Duke, and A.C. Palmenberg. Encephalomyocarditis 3C protease: efficient cell-free expression from clones which link viral 5' non-coding sequences to the P3 region. J. Virol. 1986, 60:376-384).
Практическая применимость экспрессионных векторов большой длины при получении эукариотических продуцентов ограничена низкой эффективностью их трансфекции в животные клетки (Kreiss P et al, Plasmid DNA size does not affect the physicochemical properties of lipoplexes but modulates gene transfer efficiency. Nucleic Acids Res. 1999 Oct 1;27(19):3792-3798) и низкой вероятностью успешной интеграции экспресионной кассеты в геном клетки. Поскольку небольшая часть нетрансфицированных клеток спонтанно приобретает устойчивость к действию селекционного агента и продолжает делиться в его присутствии, вероятность обнаружения клонов-продуцентов после первичного отбора становится незначительной при низком уровне интеграции экспрессионной кассеты.
Одним из способов увеличения вероятности интеграции является включение в экспрессионный вектор сигнальных участков ДНК вирусов, отвечающих за интеграцию провируса в геном клетки-хозяина. Известно техническое решение, в котором в последовательность экспрессионного вектора, содержащего промотор CMV и миниген DHFR, был вставлен фрагмент мультимеризованного концевого повтора вируса Эппштейн-Барра (US6180108 В1). Для полученного вектора, содержащего вставки, кодирующие фармацевтически значимые белки различного размера, было продемонстрировано увеличение вероятности получения стабильных трансфектантов, то есть увеличение числа колоний, растущих в селективной среде, в 3-10 раз. Фрагмент мультимеризованного концевого повтора вируса Эппштей-Барра (EBV-TR) не вызывал значимого изменения уровня продукции целевых белков. Для некоторых полученных стабильных трансфектантов была продемонстрирована возможность проведения амплификации целевого гена под действием возрастающих концентраций МТХ, однако влияние EBV-TR на ход амплификации и его общая эффективность не изучались. Фактически использованный в процитированной работе фрагмент EBV-TR содержал соединенные между собой половины единиц концевого повтора из состава конкатемера.
Подробное описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание вектора для получения высокопродуктивных и стабильных систем экспрессии рекомбинантных белков в клетках млекопитающих, в частности в клетках СНО, а также создание способа получения высокопродуктивных и стабильных при последовательном пассировании линий клеток млекопитающих, трансфицированных указанным вектором.
Технический результат достигался путем конструирования новой экспрессионной плазмидной ДНК p1.1, содержащей регуляторные элементы для гетерологичной экспрессии рекомбинантных белков, высокочастотной интеграции и ускоренной амплификации экспрессионной кассеты в клетках млекопитающих, а также выявления методов трансфекции, первичной селекции продуцентов и амплификации целевого гена, позволяющих получать линии-продуценты с высоким уровнем продукции целевого белка и достаточной стабильностью.
В основе технического решения лежит разработанный авторами плазмидный ДНК вектор, содержащий функциональные промотор и терминатор гена фактора элонгации 1 альфа китайского хомячка, обеспечивающие конститутивную экспрессию целевого гена в клетках СНО, фланкированные 5' и 3' НТО этого гена, обеспечивающими эухроматинизацию сайтов интеграции экспрессионной кассеты в геном СНО; участок для клонирования открытых рамок считывания целевых белков; внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV IRES), обеспечивающий реинициацию трансляции на бицистронной мРНК в клетках млекопитающих и полное генетическое сцепление уровней продукции целевого белка и DHFR; ОРС DHFR мыши, экспрессирующуюся в составе бицистронной мРНК вместе с целевым геном (IRES DHFR) и обеспечивающую устойчивость стабильно трансфицированных клеток к отсутствию в среде тимидина и дозозависимую устойчивость к воздействию метотрексата, что позволяет вести направленную селекцию высокопродуктивных клонов с множественными копиями экспрессионной кассеты в геноме СНО; участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), обеспечивающий повышенный уровень интеграции кассет в геном клеток СНО и ускорение процесса амплификации амплификации целевого гена в геноме под действием МТХ.
Термин «экспрессионный вектор» означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии внедренного в нее целевого гена, например, промотор и терминатор, и элементы для амплификации экспрессионной кассеты и отбора клонов с множественными копиями экспрессионной кассеты в геноме.
Конкретным примером генетических элементов для экспрессии согласно настоящему изобретению являются промотор и терминатор фактора элонгации 1 альфа китайского хомячка, но не ограничиваются ими.
Примером последовательности промотора гена фактора элонгации 1 альфа китайского хомячка, фланкированного 5' НТО этого гена, является последовательность, соответствующая нуклеотидам с 2377 по 6443 в последовательности SEQ ID NO:1.
Примером последовательности терминатора гена фактора элонгации 1 альфа китайского хомячка, фланкированного 3' НТО этого гена, является последовательность, соответствующая нуклеотидам с 7652 по 11909 в последовательности SEQ ID NO:1.
Нетранслируемая область (НТО, UTR - untranslated region) согласно настоящему изобретению представляет собой участок ДНК, не кодирующий открытых рамок считывания (ОРС), который при этом может содержать функциональные регуляторные элементы. Размер указанной нетранслируемой области варьируется в широких пределах и составляет 150 до 10000 нуклеотидов, предпочтительно 4000-5000 нуклеотидов, необходимых для обеспечения инициации и терминации транскрипции, процессинга транскрибируемой РНК, а также для эухроматинизации сайтов интеграции экспрессионной кассеты в геном СНО.
Последовательность участка для клонирования открытых рамок считывания целевых белков (полилинкер) может представлять представляют собой, например, последовательность, соответствующие нуклеотидам с 6444 по 6486 в последовательности SEQ ID NO:1. Указанный полилинкер содержит уникальные сайты узнавания эндонуклеазами рестрикции: AscI (20), PvuI (1413), FseI (5213), AbsI (6446), NheI (6467), HpaI (6484), AgeI (8861), SbfI (11764).
Последовательность внутреннего сайта связывания рибосом вируса энцефаломиокардита (EMCV IRES), представляет собой, например, последовательность, соответствующую нуклеотидам с 6487 по 7074 в последовательности SEQ ID NO:1.
Последовательность OPC DHFR мыши представляет собой, например, последовательность, соответствующую нуклеотидам с 7087 по 7650 в последовательности SEQ ID NO:1.
Фрагментом ДНК, кодирующим элемент для амплификации экспрессионной кассеты согласно настоящему изобретению, может являться, например, фрагмента конкатемера терминального повтора вируса Эпштейн-Барр человека (EBVTR). Указанный фрагмент ДНК может быть получен методом ПЦР (см. Пример 1, Фиг.1). Также указанный фрагмент ДНК может быть получен с использованием технологии клонирования фирмы Sloning BioTechnology, описанной в заявке РСТ WO 2005071077.
Последовательность участка фрагмента конкатемера терминального повтора вируса Эпштейн-Барр человека (EBVTR) представляет собой, например, последовательность, соответствующую нуклеотидам с 1964 по 2365 в последовательности SEQ ID NO:1.
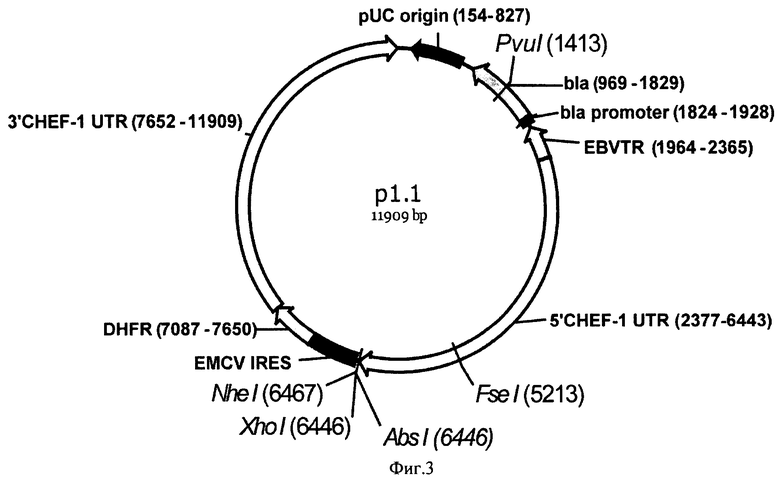
Конкретным воплощением вектора согласно настоящему изобретению является плазмида p1.1 длиной 11909 п.о., представленная в Перечне последовательностей под номером SEQ ID NO:1. Структура плазмиды pl.l приведена на Фиг.3.
Указанная плазмида размером 11909 п.о. состоит из:
1) 1-1964 участка, включающего
а. 154-827 (674 по) область начала репликации плазмиды pUC,
b. 969-1829 (861 по) открытую рамку считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину;
2) 1964-2365 (402 по) участка фрагмента конкатемера терминального повтора вируса Эпштейн-Барр человека (EBVTR);
3) 2377-6443 (4067 по) участка, содержащего функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5' НТО этого гена (5'CHEF-1 UTR);
4) 6444-6486 (43 по) участка для клонирования вставок целевых ОРС (полилинкер);
5) 6487-7074 (588 по) участка, содержащего внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV IRES);
6) 7087-7650 (564 по) участка, содержащего открытую рамку считывания дегидрофолатредуктазы мыши для селективного отбора и амплификации в эукариотических клетках;
7) 7652-11909 участка, содержащего сигнал полиаденилирования и функциональный терминатор и (7920-7925) гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3' НТО этого гена (3'CHEF-1 UTR).
Вектор был получен с использованием стандартных методов генной инженерии, коммерчески доступных плазмид и химически синтезированных олигонуклеотидов (Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. 2nd ed. New York, NY: Cold Spring Harbor Laboratory Press; 1989).
Элементы вектора перечислены в порядке их расположения. Взаимный порядок расположения функциональных элементов является существенным для эффективной работы вектора.
В качестве рекомбинантной плазмиды согласно настоящему изобретению могут использоваться различные плазмиды, обладающие способностью к интеграции в геном клетки-реципиента после трансфекции, содержащие конститутивный эукариотический промотор, экспрессионную бицистронную кассету и области, обеспечивающие эухроматинизацию района интеграции экспрессионный кассеты.
Предпочтительно, чтобы экспрессионный плазмидный вектор согласно настоящему изобретению дополнительно содержал консенсусную последовательность Козак. Последовательность Козак располагается вокруг старт-кодона целевого гена и способствует инициации трансляции мРНК целевого гена.
Последовательность экспрессионной плазмиды p1.1-eGFP несущей вставку гена репортера - зеленого флуоресцентного белка eGFP согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO:2.
Фрагменты ДНК, которые кодируют по существу те же регуляторные элементы могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO:1), например, посредством метода сайт-направленного мутагенеза, так, что один или несколько нуклеотидов в определенном сайте будут делегированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Показатели функциональной активности, при которой считается, что полученный вектор позволяет получить высокопродуктивный клон клеток с множественными копиями экспрессионной кассеты в геноме, определяются по концентрации секретируемого белка в культуральной среде или по концентрации несекретируемого белка в лизате клеточной массы или по специфическим функциональным свойствам целевого белка, в случае eGFP - по отношению интенсивности флуоресценции лизата клеточной массы к содержанию общего белка в клеточной массе. Измерение интенсивности флуоресценции и тотального клеточного белка описано в Примере 6, измерение концентрации секретируемого или несекретируемого целевого белка может быть проведено при помощи твердофазного ИФА, измерение функциональной активности целевого белка может быть проведено, например, коагулометрически для факторов свертываемости крови, спектрофотометрически для ферментов, при помощи твердофазного ИФА с использованием антигена на подложке - для иммуноглобулинов и т.д. Считается, что вероятность интеграции генетической кассеты в геном увеличена, если на 21 день после трансфекции доля лунок культуральных планшетов, в которых интенсивность флуоресценции eGFP достоверно выше фонового значения, более чем вдвое выше доли таких лунок для клеток, трансфицированных контрольной генетической конструкцией в одинаковых условиях. Считается, что частота амплификации целевого гена выше, чем для контрольной генетической конструкции, если средняя удельная интенсивность флуоресценции eGFP для пяти лунок культуральных планшетов, проанализированных через 15 дней после введения повышенной концентрации метотрексата и дающих максимальные интенсивности флуоресценции среди всех проанализированных лунок, более чем вдвое выше аналогичного показателя для культуры, трансфицированной контрольной генетической конструкцией и подвергнутой такой же обработке метотрексатом.
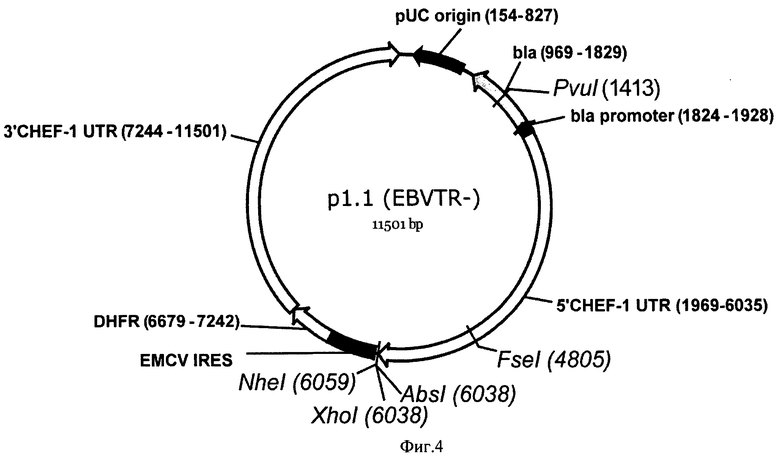
Практические функциональные свойства вектора p1.1 (SEQ ID NO:1) можно оценить по оценке экспрессии репортерного белка, ОРС которого с соответствующей последовательностью Козак клонирована в p1.1 (примером такой конструкции является плазмида p1.1-eGFP SEQ ID NO:2) по сравнению с контрольным вектором p1.1(EBVTR-) (SEQ ID NO:3), в который клонирована идентичная вставка (с образованием конструкции p1.1-eGFP (EBVTR-)(SEQ ID NO:4)).
Также целью настоящего изобретения является предоставление бицистронной мРНК, образующейся в ходе экспрессии описанной выше экспрессионной кассеты, содержащей открытую рамку считывания целевого рекомбинантного белка, внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV) и открытую рамку считывания дегидрофолатредуктазы (DHFR). Экспрессия целевого гена и гена селекционного маркера в составе указанной бицистронной мРНК обеспечивает их повышенную генетическую сцепленность. При использовании клеток мыши для получения рекомбинантных белков предпочтительно использование открытой рамки считывания дегидрофолатредуктазы (DHFR) мыши.
Также целью настоящего изобретения является предоставление способа получения стабильных линий клеток млекопитающих с множественными копиями экспрессионной кассеты в геноме, продуцирующих целевой белок с высокой эффективностью.
Указанный способ включает трансфекцию клеток млекопитающего, не содержащих активные аллели дегидрофолатредуктазы, экспрессионным вектором согласно настоящему изобретению, содержащим вставку с ОРС целевого гена, кодирующей целевой рекомбинантный белок; отбор жизнеспособных клеток на среде, не содержащей гипоксантин и тимидин, с последующим отбором клеток млекопитающих - продуцентов целевого белка на среде, содержащей метотрексат. Предпочтительная концентрация метотрексата в среде составляет 50 нМ, предпочтительно 100 нМ, предпочтительно 200 нМ.
В качестве клеток млекопитающего, не содержащих активные аллели дегидрофолатредуктазы, могут быть использованы любые традиционно применяемые линии клеток с инактивируемой дигидрофолатредуктазой. Предпочтительно использование клеток эпителия яичника китайского хомячка (СНО), нокаутных по гену дигидрофолатредуктазы (dhfr-/-), линия DG-44 (Invitrogen, США).
Целевым рекомбинантных белком согласно настоящему изобретению может быть любой белок, представляющий интерес в данной области техники. Примером таких белков является, например, зеленый флуоресцентный белок eGFP, но не ограничиваются ими.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором культивируют клетки СНО DG44, трансфицированные экспрессионным вектором согласно настоящему изобретению, содержащим вставку с ОРС целевого гена, кодирующей целевой рекомбинантный белок.
Также целью настоящего изобретения является предоставление способа получения рекомбинантных белков в клетках млекопитающих, включающего культивирование в питательной среде клеток млекопитающих - продуцентов целевого белка, полученных описанным выше способом, и выделение полученного целевого белка из культуральной жидкости.
Выращивание клеток, выделение и очистка целевого белка из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам культивирования, в которых рекомбинантный белок продуцируется с использованием клеток млекопитающих.
Питательная среда, используемая для культивирования, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста клеток. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза или сахароза и другие органические кислоты. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Выращивание может осуществляться в аэробных условиях, предпочтительно с повышенным содержанием CO2 (8%), таких как перемешивание культуральной жидкости в колбах, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. Обычно, выращивание в течение от 12 часов до 4 дней приводит к накоплению целевого рекомбинантного белка в культуральной среде.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования, а затем целевой белок может быть выделен и очищена методами хроматографии и/или концентрирования.
Особенности вектора и результаты его практического применения приведены на следующих чертежах.
Краткое описание чертежей:
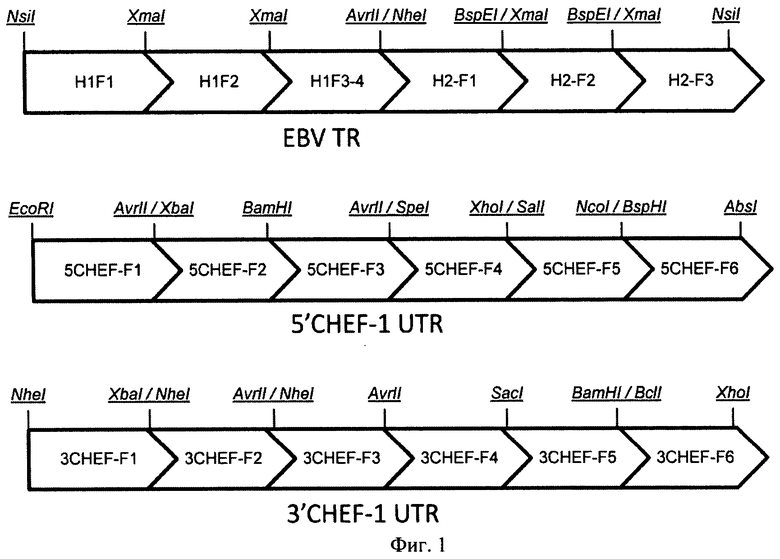
На Фигуре 1 показана схема сборки участка терминального повтора вируса EBV и областей промотора и терминатора гена фактора элонгации 1 альфа китайского хомячка с фланкирующими их областями. Обозначения: EBVTR - участок терминального повтора вируса Эпштейн-Барр человека (EBV); 5'CHEF-1 UTR функциональный промотор гена фактора элонгации 1 альфа китайского хомячка фланкированный 5' НТО этого гена; 3'CHEF-1 UTR функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка фланкированный 3' НТО этого гена.
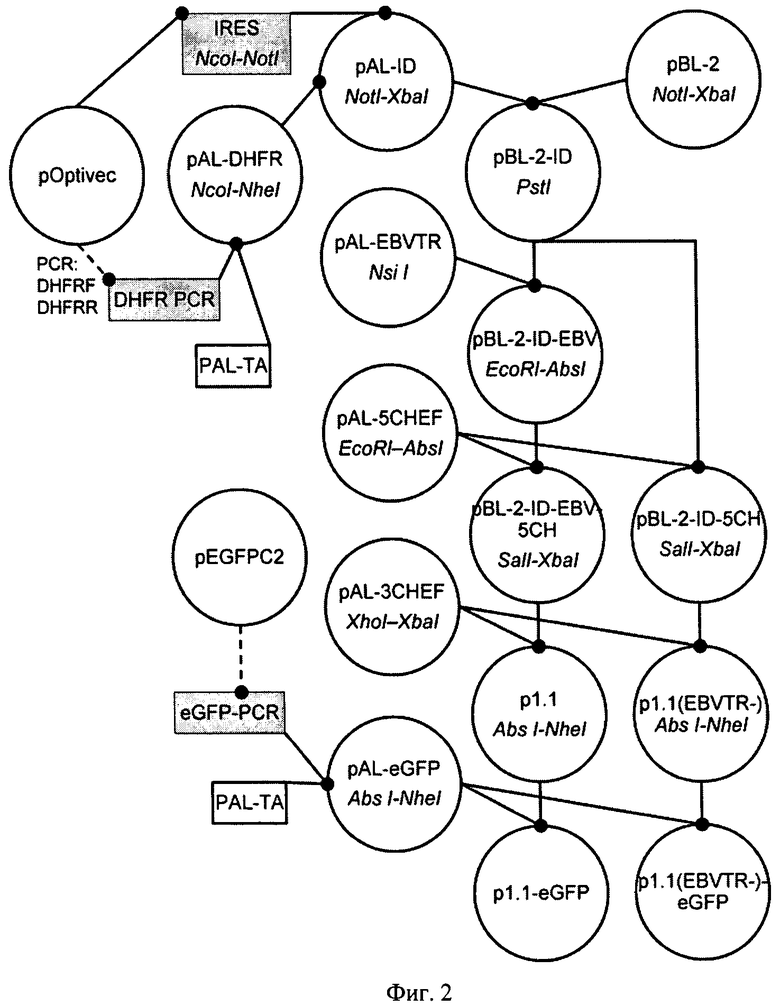
На Фигуре 2 показана схема получения плазмид p1.1, p1.1 (EBVTR-), p1.1-eGFP, p1.1(EBVTR-)-eGFP. Обозначения: пунктирной линией обозначены стадии ПЦР, сплошной линией обозначены стадии рестрикции-лигирования, эндонуклеазы рестрикции, использовавшиеся для клонирования, указаны под названиями плазмид.
На Фигуре 3 показана карта экспрессионной плазмиды p1.1 (длина 11909 пар оснований). Используются следующие обозначения: pUC origin - область начала репликации плазмиды pUC; bla - открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; bla promoter - прокариотический промотер гена bla; EBVTR - участок терминального повтора вируса Эпштейн-Барр человека (EBVTR); 5'CHEF-1 UTR функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5' НТО этого гена; EMCV IRES - внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV); DHFR - открытая рамка считывания дигидрофолатредуктазы мыши для селективного отбора и амплификации в эукариотических клетках; 3'CHEF-1 UTR функциональный терминатор и сигнал полиаденилирования (7920-7925) гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3' НТО этого гена; Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
На Фигуре 4 показана карта экспрессионной плазмиды p1.1 (EBVTR-) (длина 11501 пар оснований). Обозначения аналогично фигуре 3.
На Фигуре 5 показана карта экспрессионной плазмиды p1.1-eGFP (длина 12623 пар оснований). Обозначения аналогично фигуре 3, eGFP - открытая рамка считывания зеленого флуоресцентного белка с последовательностью Козак и стоп-код оном.
На Фигуре 6 показана карта экспрессионной плазмиды p1.1(EBVTR-)-eGFP (длина 12215 пар оснований). Обозначения аналогично фигуре 5.
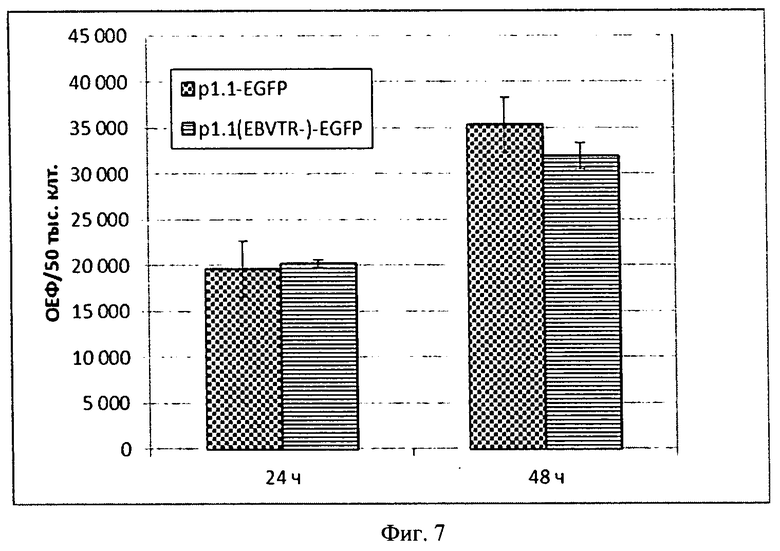
На Фигуре 7 показана диаграмма удельных интенсивностей флуоресценции белка eGFP при транзиентной трансфекции клеток СНО плазмидами p1.1-EGFP и p1.1(EBVTR-)-EGFP. По оси ординат отложена нормализованная интенсивность флуоресценции в условных единицах, линеаризация по данным калибровочного графика для раствора белка eGFP.
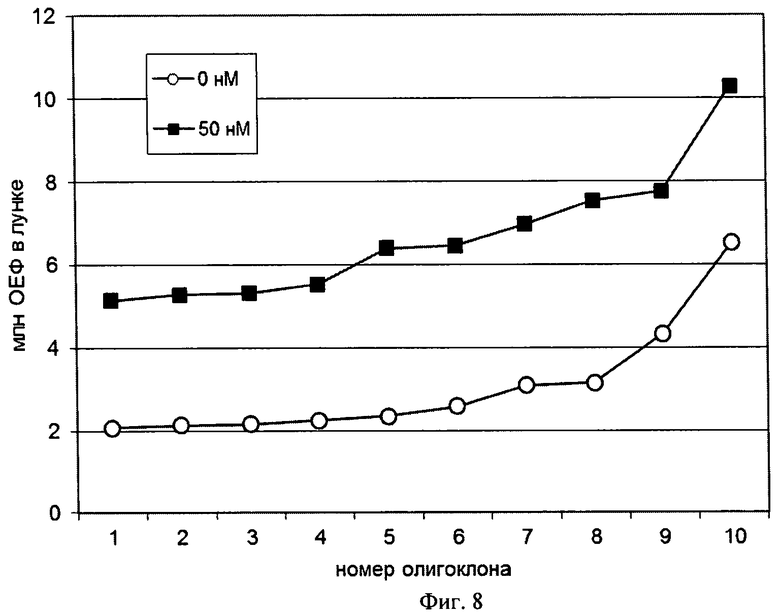
На Фигуре 8 показан график абсолютных интенсивностей флуоресценции EGFP для 10 наиболее продуктивных лунок, полученных при первичной селекции клонов в присутствии и отсутствии 50 нМ метотрексата.
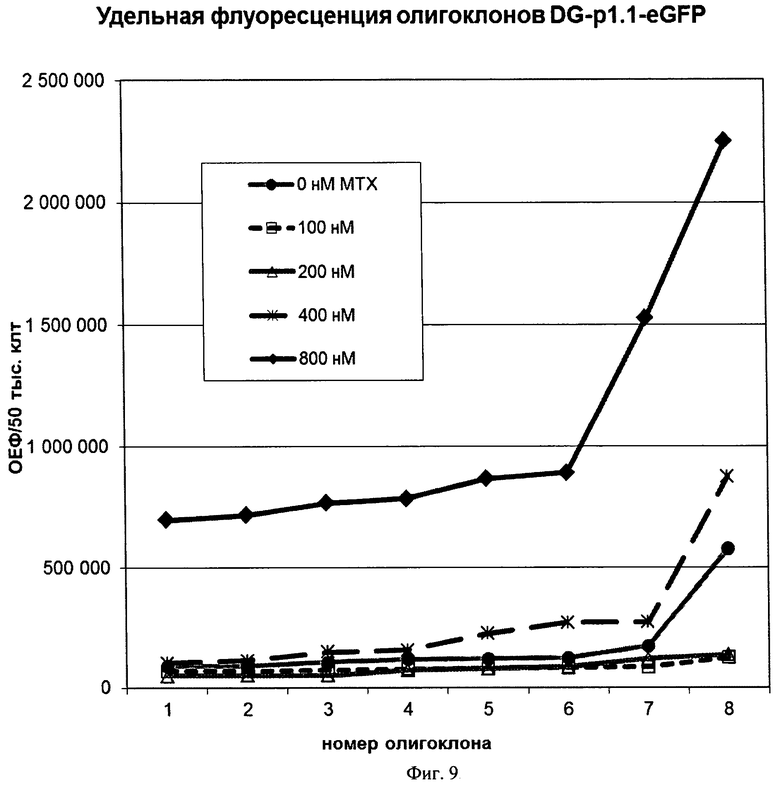
На Фигуре 9 показан график удельных интенсивностей флуоресценции EGFP для 8 наиболее продуктивных олигоклональных линий, полученных при амплификации целевого гена под действием различных концентраций метотрексата.
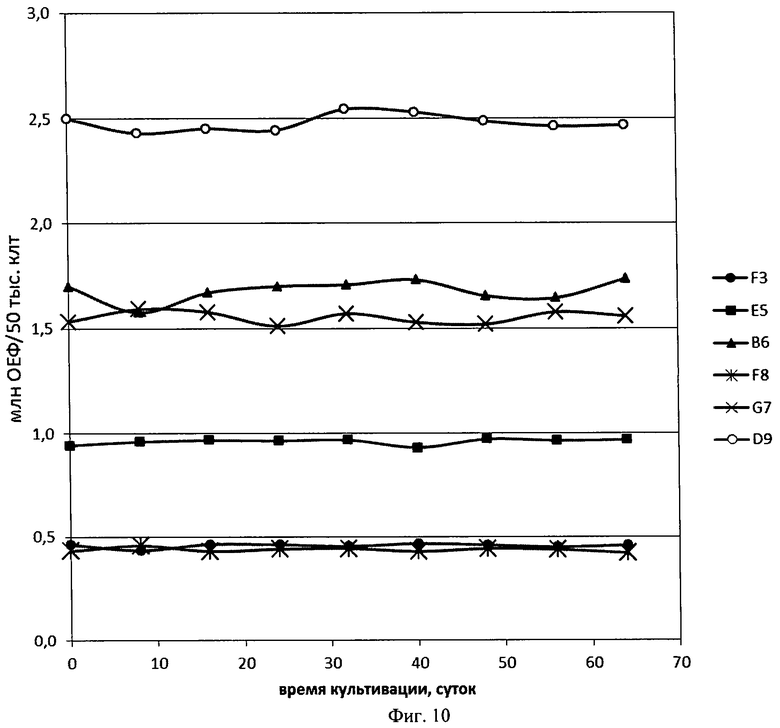
На Фигуре 10 показана динамика удельных интенсивностей флуоресценции EGFP для 6 олигоклональных линий с амплификацией целевого гена при их продолжительном культивировании.
Существо настоящего изобретения и его промышленная применимость в биотехнологии иллюстрируются следующими примерами.
ПРИМЕРЫ
Пример 1. Получение плазмиды PAL-EBV
ПЦР-продукт, содержащий терминальные повторы EBV (SEQ ID NO:5) собирали из синтетических олигонуклеотидов EBV-H1F1-F (SEQ ID NO:6); EBV-H1F1-R (SEQ ID NO:7); EBV-H1F2-F (SEQ ID NO:8); EBV-H1F2-R (SEQ ID NO:9); EBV-H1F3-F (SEQ ID NO:10); EBV-H1F3-R (SEQ ID NO:11); EBV-H1F4-F (SEQ ID NO:12); EBV-H1F4-R (SEQ ID NO:13); EBV-H2F1-F (SEQ ID NO:14); EBV-H2F1-R (SEQ ID NO:15); EBV-H2F2-F (SEQ ID NO:16); EBV-H2F2-R (SEQ ID NO:17); EBV-H2F3-F (SEQ ID NO:18); EBV-H2F3-R (SEQ ID NO:19); для аналитических ПЦР в процессе сборки использовали специфические праймеры EBV-H1F2-SHORTF (SEQ ID NO:20); EBV-H2F2-SHORTR (SEQ ID NO:21); EBV-H2F1-SHORTF (SEQ ID: NO:22); для анализа полученной конструкции PAL-EBV (SEQ ID NO:23) использовали стандартные праймеры к последовательностями вектора PAL-TA (ЗАО «Евроген», Россия) M13dir (SEQ ID NO:24), M13rev (SEQ ID NO:25); SP6 (SEQ ID NO:26), T7prom (SEQ ID NO:27).
Праймеры EBV-H1F1-F (SEQ ID NO:6) и EBV-H1F1-R (SEQ ID NO:7) фосфорилировали, отжигали и клонировали в вектор PAL-TA (ЗАО «Евроген», Россия).
Реакцию фосфорилирования олигонуклеотидов проводили в буфере трис-HCl, рН 7,5, содержащем 10 мМ MgCl2, 50 мМ дитиотреитола, 1 мМ АТФ и 100 пМ олигонуклеотида, 1 ед. полинуклеотидкиназы фага Т4 (Сибэнзим, Россия) в течение 30 минут при 37°С. После окончания реакции фермент инактивировали при 65°С 10 мин.
Для получения дуплекса вносили в пробирку по 100 пм каждого олигонуклеотида, нагревали до 95°С и медленно охлаждали до комнатной температуры и лигировли в вектор PAL-TA (ЗАО «Евроген», Россия) с использованием ДНК-лигазы фага Т4 и стандартного буфера (Fermentas, Литва). Полученной лигазной смесью трансформировали клетки Е.coli штамма DH5alpha, с генотипом F- φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rk-, mk+) phoA supE44 λ- thi-1 gyrA96 relA1. Для этого к 200 мкл замороженной суспензии клеток Е.coli добавляли 5 мкл лигазной смеси, инкубировали на льду 30 мин, нагревали до 42°С на 45 секунд и инкубировали на льду 5 минут, затем добавляли 800 мкл питательного бульона SOB, инкубировали при 37°С 60 минут, затем переносили суспензию на чашку Петри с твердой агаризованной средой, содержащей ампициллин в концентрации 100 мкг/1 мл агара и помещали в термостат на 37°С 18 часов. Колонии Е.coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов, с использованием праймеров EBV-H1F1-F (SEQ ID NO:6) и EBV-H1F1-R (SEQ ID NO:7), отобранные клоны наращивали в 5 мл среды 2xYT-Amp и выделяли плазмидную ДНК Получали плазмиду PAL-H1F1 при помощи набора Gene JET Plasmid Miniprep Kit (Fermentas, Литва) по протоколу производителя. После чего выделяли фрагмент PAL-H1F1/XmaI из 1,5% агарозного геля набором "Wizard SV Gel and PCR Clean-Up System" («Promega», США) по протоколу производителя.
Праймеры EBV-H1F3-F (SEQ ID NO:10), EBV-H1F3-R (SEQ ID NO:11) и EBV-H1F4-F (SEQ ID NO:12) и EBV-H1F4-R (SEQ ID NO:13) отжигали попарно (нагревали до 95°С и медленно охлаждали до комнатной температуры), фосфорилировали как описано выше и лигировали два дуплекса между собой ДНК-лигазой фага Т4 (Fermentas, Литва), после чего клонировали в вектор PAL-TA как описано выше. Получали плазмиду PAL-H1F3-4, обрабатывали рестриктазой XmaI и лигировали с фрагментом PAL-H1F1/XmaI, отбирали клоны и выделяли плазмиды как описано выше. Полученную плазмиду PAL-H1F1-3-4 обрабатывали эндонуклеазой XmaI и лигировали с стоженными, фосфорилированными частично комплиментарными праймерами EBV-H1F2-F (SEQ ID NO:8) и EBV-H1F2-R (SEQ ID NO:9), при отжиге образующими дуплекс с выступающими «липкими» 5'-концами комплиментарными липким концам, образующимся при рестрикции акцептора нуклеазой XmaI. Трансформацию проводили как описано выше.
Клоны PAL-H1 анализировали методом ПЦР с клонов при помощи праймеров EBV-H1F2-SHORTF (SEQ ID NO:20), EBV-H1F1-R (SEQ ID NO:7), M13dir (SEQ ID NO:24), M13rev (SEQ ID NO:25).
Праймеры EBV-H2F1-F (SEQ ID 14) и EBV-H2F1-R (SEQ ID NO:15) фосфорилировали, отжигали и клонировали в вектор PAL-TA (Евроген) как описано выше, полученную плазмиду PAL-H2F1 обрабатывали рестриктазой BspEI и подставляли двуцепочечный адаптер полученный как описано выше из праймеров EBV-H2F2-F (SEQ ID NO:16) и EBV-H2F2-R (SEQ ID NO:17), выступающие 5' концы которых коплиментарны липким концам, образующимся при обработке рестриктазой BspEI с исчезновением одного сайта. Полученную плазмиду PAL-H1F1-2 обрабатывали по сохранившемуся уникальному сайту рестриктазой BspEI и подставляли двуцепочечный адаптер, полученный как описано выше из праймеров EBV-H2F3-F (SEQ ID NO:18) и EBV-H2F3-R (SEQ ID NO:19).
Клоны PAL-H2 анализировали методом ПЦР с клонов при помощи праймеров EBV-H2F2-SHORTR (SEQ ID NO:21), EBV-H2F2-SHORTF (SEQ ID NO:22), M13dir (SEQ ID NO:24), M13rev (SEQ ID NO:25). Две половины целевой последовательности EBV-H1 и EBV-H2 собирали по сайтам рестрикции AvrII/NheI. Схема сборки фрагментов EBVTR представлена на Фиг.1
Полученная конструкция PAL-EBV представлена в списке последовательностей под номером SEQ ID NO:23. Ее секвенировали с использованием праймеров SP6 (SEQ ID NO:26), T7prom (SEQ ID NO:27).
Пример 2. Получение плазмиды PBL-2-ID и PBL-2-ID-EBV
ОРС, кодирующую дегидрофолатредуктазу (аминокислотная последовательнось SEQ ID NO:28), получали при помощи ПЦР с использованием праймеров AD-DHFR-F (SEQ ID NO:29), AD-DHFR-R (SEQ ID NO:30) и плазмиды pOptivec-circ (SEQ ID NO:31) в качестве матрицы. ПЦР проводили набором реактивов "Encycio PCR kit" (ЗАО Евроген, Россия) по инструкции производителя, ПЦР-продукт очищали из 1% агарозного геля набором "Wizard SV Gel and PCR Clean-Up System" («Promega», США) по протоколу производителя и лигировли в векторную плазмиду PAL-TA (ЗАО «Евроген», Россия) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора (Fermentas, Литва). Полученными лигазными смесями трансформировали клетки Е.coli штамма DH5 alpha и вели отбор клонов и выделение плазмид как описано в Примере 1. Полученная конструкция PAL-DHFR представлена в списке последовательностей под номером 32. Ее секвенировали с использованием праймеров SP6 (SEQ ID 26), T7prom (SEQ ID 27).
Фрагмент, кодирующий внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV), переносили рестрикцией из pOptivec-circ (SEQ ID NO:31). Для этого pOptivec-circ (SEQ ID NO:31), обрабатывали рестриктазами BamHI-BglII и лигировали в PAL-DHFR/BglII с образованием плазмиды PAL-ID (SEQ ID NO:34). Для ее секвенирования использовали праймеры IRESrev (SEQ ID NO:35) и IRESArev (SEQ ID NO:36) и AD-DHFR-F (SEQ ID NO:29).
В качестве основы для сборки целевого вектора использовали вектор PBL-2 (SEQ ID NO:37). Фрагмент, содержащий IRES-DHFR, вырезали из плазмиды PAL-ID (SEQ ID NO:34) рестриктазами NotI и XbaI, лигировали в PBL-2 (SEQ ID 37)/ NotI и XbaI с образованием плазмиды PBL-2-ID (SEQ ID NO:38). Анализ клонов вели с праймеров AD-DHFR-F (SEQ ID NO:29) и SQ-BB-R (SEQ ID NO:40). Для ее секвенирования использвали праймеры SQ-BlaN-F (SEQ ID NO:39) и SQ-BB-R (SEQ ID NO:40).
Фрагмент, содержащий EBVTR, вырезали из плазмиды PAL-EBV (SEQ ID NO:23) рестрикцией NsiI, и лигировали в PBL-2-ID (SEQ ID 38)/ PstI с образованием плазмиды PBL-2-ID-EBV (SEQ ID NO:41).
Пример 3. Получение векторов p1.1(EBVTR-) и p1.1, содержащих 5' и 3' НТО EF-1 китайского хомячка
Фрагмент, содержащий промотор фактора элонгации 1 альфа китайского хомячка и 5'НТО этого гена, соответствующий области 8532-12603 AY188393 (SEQ ID 42), получали методом ПЦР с использованием геномной ДНК выделенной из клеток СНО DG44 и праймеров AD-5CHEF1-R (SEQ ID NO:43); AD-5CHEF2-F (SEQ ID NO:44); AD-5CHEF2-R (SEQ ID NO:45); AD-5CHEF3-F (SEQ ID NO:46); AD-5CHEF3-R (SEQ ID NO:47); AD-5CHEF4-F (SEQ ID NO:48); AD-5CHEF4Xho-R (SEQ ID NO:49); AD-5CHEF1-F (SEQ ID NO:50); AD-5CHEF5-F (SEQ ID NO:51); AD-5CHEF5-R (SEQ ID NO:52); AD-5CHEF61-F (SEQ ID NO:53); AD-5CHEF61-R (SEQ ID NO:54); AD-5CHEF62-F (SEQ ID NO:55); AD-5CHEF62-R (SEQ ID NO:56).
Методом ПЦР с геномной ДНК с использованием праймеров AD-5CHEF1-F (SEQ ID NO:50) и AD-5CHEF1-R (SEQ ID NO:43) получали фрагмент 5CHEF-F1 (SEQ ID:57); с использованием праймеров AD-5CHEF2-F (SEQ ID NO:44) и AD-5CHEF2-R (SEQ ID NO:45) получали фрагмент 5CHEF-F2 (SEQ ID NO:58); с использованием праймеров AD-5CHEF3-F (SEQ ID NO:46) и AD-5CHEF3-R (SEQ ID NO:47) получали фрагмент 5CHEF-F3 (SEQ ID NO:59); с использованием праймеров AD-5CHEF4-F (SEQ ID NO:48) и AD-5CHEF4Xho-R (SEQ ID NO:49) получали фрагмент 5CHEF-F4 (SEQ ID NO:60); с использованием праймеров AD-5CHEF5-F (SEQ ID NO:51) и AD-5CHEF5-R (SEQ ID NO:52) получали фрагмент 5CHEF-F5 (SEQ ID NO:61). Фрагменты клонировали в вектор PAL-TA и секвенировали с использованием праймеров SP6 (SEQ ID NO:26), T7prom (SEQ ID NO:27). Фрагмент 5CHEF-F6 (SEQ ID NO:62) получали отжигом праймеров AD-5CHEF61-F (SEQ ID NO:53); AD-5CHEF61-R (SEQ ID NO:54); AD-5CHEF62-F (SEQ ID NO:55); AD-5CHEF62-R (SEQ ID NO:56) и также клонировали в вектор PAL-TA и секвенировали с использованием праймеров SP6 (SEQ ID NO:26) и SQ-5CH6-F(SEQ ID NO:64).
Схема сборки фрагментов представлена на Фиг.1
Акцептор PAL-5CHEF-F1/ NotI-AvrII, содержащий фрагмент 5CHEF-F1 (SEQ ID NO:57), лигировали с XbaI-PspOMI фрагментом PAL-5CHEF-F2, содержащим фрагмент 5CHEF-F2 (SEQ ID NO:58) и получали PAL-5CHEF-F12. Акцептор PAL-5CHEF-F12/ NotI-BamHI, содержащий фрагменты 5CHEF-F1-2, лигировали с BamHI - NotI фрагментом PAL-5CHEF-F3, содержащим фрагмент 5CHEF-F3 (SEQ ID NO:59) и получали PAL-5CHEF-F123. Акцептор PAL-5CHEF-F123/ NotI-AvrII, содержащий фрагменты 5CHEF-F1-2,-3 лигировали с SpeINotI фрагментом PAL-5CHEF-F4, содержащим фрагмент 5CHEF-F4 (SEQ ID NO:59) и получали PAL-5CHEF-F1234. Акцептор PAL-5CHEF-F5/ NcoI, содержащий фрагмент 5CHEF-F5 (SEQ ID NO:61), лигировали с BspHI-PciI фрагментом PAL-5CHEF-F6, содержащим фрагмент 5CHEF-F6 (SEQ ID NO:62) и получали PAL-5CHEF-F56. Акцептор PAL-5CHEF-F1234/ XhoI, содержащий фрагменты 5CHEF-F1-2-3-4, лигировали с SalI-SalI фрагментом, PAL-5CHEF-F56, содержащим фрагменты 5CHEF-F5-6, определяли ориентацию с использованием F5REV и F4FOR и получали PAL-5CHEF.
Полученная конструкция PAL-5CHEF с полной сборкой соответствующей области 8532-12603 AY188393 представлена в списке последовательностей под номером SEQ ID NO:63.
Общее количество нуклеотидных замен в сборке 5'CHEF НТО по сравнению с участком 8532-12603 последовательности AY188393, депонированной в публичной базе данных GeneBank, составило 6 нуклеотидов, в том числе 5 точечных делеций или инсерций. Вероятная причина выявленных расхождений - артефакты секвенирования ДНК в опубликованной последовательности.
Область 12603-12626 последовательности AY188393, соответствующая участку длиной 20 нуклеотидов (ACCGCTAATTCAA АТССААС), непосредственно предшествующих стартовому (ATG) кодону кодируемого природным геном белка фактора элонгации трансляции 1 альфа в базовом векторе p1.1 удалена. При подстановке в вектор вставок ОРС целевых белков эта последовательность должна быть частично восстановлена - стартовому кодону целевого белка в готовой экспрессионной плазмиде должна предшествовать последовательность из 14 нуклеотидов (tcgaggccgccacc), содержащая консенсусную последовательность Козак и фрагмент рестриктного сайта AbsI (cctcgagg), по которому производится клонирование вставок ОРС целевых белков в вектор p1.1 или p1.1 (EBVTR-). Порядок действий по клонированию вставки с ОРС модельного белка в вектор p1.1 приведен в Примере 2.
PAL-5CHEF (SEQ ID NO:63) расщепляли рестриктазой EcoRI и AbsI и полученный фрагмент лигировали в плазмиду PBL-2-ID-EBV (SEQ ID NO:41)/EcoRI-AbsI с образованием плазмиды PBL-2-ID-5CH (SEQ ID NO:65), а также в плазмиду PBL-2-ID (SEQ ID NO:38) /EcoRI-AbsI с образованием плазмиды PBL-2-ID-EBV-5CH (SEQ ID 66)
Фрагмент, содержащий терминатор и сигнал полиаденилирования фактора элонгации 1 альфа китайского хомячка и 3'НТО этого гена, соответствующий области 3'CHEF 14545-18794 AY188393 (SEQ ID NO:67), получали методом ПЦР с использованием геномной ДНК, выделенной из клеток СНО DG44 и адапторных праймеров AD-3CHEF1-NheIF(SEQ ID NO:68); AD-3CHEF1-R (SEQ ID NO:69); AD-3CHEF2-F (SEQ ID NO:70); AD-3CHEF2-R (SEQ ID NO:71); AD-3CHEF3-F (SEQ ID NO:72); AD-3CHEF3-R (SEQ ID NO:73); AD-3CHEF4-F (SEQ ID NO:74); AD-3CHEF4-R (SEQ ID NO:75); AD-3CHEF5-F (SEQ ID NO:76); AD-3CHEF5-R (SEQ ID NO:77); AD-5CHEF6-F (SEQ ID NO:78); AD-3CHEF6-XhoR (SEQ ID NO:79), несущих сайты узнавания рестриктаз для последующей сборки фрагментов.
С использованием праймеров AD-3CHEF1-NheIF (SEQ ID NO:68); AD-3CHEF1-R (SEQ ID NO:69) получали фрагмент 3CHEF-F1 (SEQ ID NO:80), фланкированный сайтами NheI и XbaI; с использованием праймеров AD-3CHEF2-F (SEQ ID NO:70); AD-3CHEF2-R (SEQ ID NO:71) получали фрагмент 3CHEF-F2 (SEQ ID NO:81), фланкированный сайтами NheI и AvrII; с использованием праймеров AD-3CHEF3-F (SEQ ID NO:72); AD-3CHEF3-R (SEQ ID NO:73) получали фрагмент 3CHEF-F3 (SEQ ID NO:82), фланкированный сайтами NheI и AvrII; с использованием праймеров AD-3CHEF4-F (SEQ ID NO:74); AD-3CHEF4-R (SEQ ID NO:75) получали фрагмент 3CHEF-F4 (SEQ ID NO:83), фланкированный сайтами AvrII и SacI; с использованием праймеров AD-3CHEF5-F (SEQ ID NO:76); AD-3CHEF5-R (SEQ ID NO:77) получали фрагмент 3CHEF-F5 (SEQ ID NO:84), фланкированный сайтами SacI и BamHI; с использованием праймеров AD-5CHEF6-F (SEQ ID NO:78); AD-3CHEF6-XhoR (SEQ ID NO:79) получали фрагмент 3CHEF-F6 (SEQ ID NO:85), фланкированный сайтами BclI. и XhoI. Фрагменты клонировали в PAL-TA (Евроген) и секвенировали с использованием праймеров SP6 (SEQ ID NO:26), T7prom (SEQ ID NO:27) как описано выше.
Схема сборки фрагментов представлена на Фиг.1
Акцептор PAL-3CHEF-F3, содержащий фрагмент 3CHEF-F3 (SEQ ID NO:82), расщепляли рестриктазой NheI, лигировали фрагмент NheI-AvrII из PAL-3CHEF-F2, содержащий фрагмент 3CHEF-F2 (SEQ ID NO:81), отбирали методом ПЦР клоны с правильной ориентацией вставки. Полученные конструкции рестрицировали NheI, и лигировали с фрагментом NheI-XbaI плазмиды PAL-3CHEF-F1, содержащей 3CHEF-F1 (SEQ ID NO:80), отбирали методом ПЦР клоны с правильной ориентацией вставки. Акцептор PAL-3CHEF-F5, содержащий фрагмент 3CHEF-F5 (SEQ ID NO:84), расщепляли рестриктазами PciI-Sac и лигировали фрагмент NcoI-SacI плазмиды PAL-3CHEF-F4, содержащий 3CHEF-F4 (SEQ ID NO:83). Затем расщепляли акцептор, содержащий собранные фрагменты 3CHEF-F1-3, эндонуклеазами AvrII - PspOMI и лигировали очищенные фрагменты 3CHEF-F4-5, полученные из PAL-3CHEF-F45 рестрикцией AvrII - BamHI, и 3CHEF-F6 (SEQ ID NO:85), полученный из PAL-3CHEF-F6/ BclI - NotI.
Полученная конструкция PAL-3CHEF с полной сборкой соответствующей области 14545-18794 AY188393 представлена в списке последовательностей под номером 86.
Общее количество нуклеотидных замен в общей сборке по сравнению с 14545-18794 AY188393 депонированной в публичной базе данных GeneBank составило 5, из них инсерций-делеций 2.
PAL-3CHEF (SEQ ID NO:86) расщепляли рестриктазами NheI и XhoI и полученный фрагмент лигировали в плазмиду PBL-2-ID-5CH (SEQ ID NO:65)/XbaI-SalI с образованием плазмиды p1.1(EBVTR-) (SEQ ID NO:3), а также в плазмиду PBL-2-ID-EBV-5CH (SEQ ID NO:66) /XbaI-SalI с образованием плазмиды p1.1 (SEQ ID 1).
Пример 4. Получение экспрессионных конструкций p1.1-EGFP и p1.1(EBVTR-)-EGFP, кодирующих зеленый флуоресцентный белок
С использованием праймеров AD-EG-AbsF(SEQ ID NO:87) и AD-EG-NheR (SEQ ID NO:88) и pEGFP-N2 (Clontech, США) в качестве матрицы получили ПЦР-продукт eGFP PCR (SEQ ID NO:89), содержащий ОРС зеленого флуоресцентного белка (аминокислотная последовательность SEQ ID NO:90). Полученный продукт ПЦР клонировали в вектор PAL-TA и анализировали секвенированием области вставки, используя праймеры SP6, после обнаружения клонов с корректной нуклеотидной последовательностью области вставки плазмиду PAL- eGFP расщепляли рестриктазами AbsI и NheI и переносили фрагмент, содержащий ОРС eGFP с участком консенсусной последовательности Козак в векторы p1.1 (SEQ ID NO:1) и p1.1(EBVTR-) (SEQ ID NO:3) с образованием p1.1- eGFP (SEQ ID 2) и eGFP-p1.1(EBVTR-) (SEQ ID 4), соответственно.
Пример 5. Получение сверхскрученной и линеаризованной плазмидной ДНК для трансфекции клеток СНО
Полученными в Примере 4 экспрессионными генетическими конструкциями p1.1-EGFP и p1.1(EBVTR-)-EGFP, а также контрольной плазмидой pEGFP-N2 (Clontech, США) трансформировали клетки Е.coli штамма ТОРР10 (Invitrogen, США), одиночными колониями трансформантов инокулировали пробирки, содержащие по 5 мл среды LB с 50 мкг/мл ампициллина, выращивали ночные культуры и инокулировали ими колбы, содержащие по 0,5 литра жидкой среды LB с 50 мкг/мл ампициллина. Выращивали культуры в течение 6 ч. Выделение сверхскрученной плазмидной ДНК проводили методом щелочного лизиса бактерий с использованием набора EndoFree Plasmid MaxiKit (Qaigen, США) по методике производителя. Получали от 0,3 до 2 мг сверхскрученной плазмидной ДНК, содержащей менее 5% релаксированной кольцевой и линейной форм и содержащей менее 50 ЭЕ/мл ЛАЛ-эндотоксина. Для аликвот растворов сверхскрученных плазмид, содержащих по 100 мкг ДНК, проводили получение линеаризованных форм путем расщепления эндонуклеазой рестрикции PvuI. Полученные линеаризованные формы содержали в обоих случаях разрыв кольца в области гена устойчивости к ампициллину bla. Дополнительную очистку линейных ДНК проводили переосаждением 70% этанолом. Сверхскрученные и линеаризованные формы экспрессионных плазмид p1.1-EGFP и p1.1(EBVTR-)-EGFP переводили в фосфатно-солевой буферный раствор и стерилизовали растворы плазмид при помощи одноразовых фильтров с порами 0,22 мкм (Millipore, США).
Пример 6. Транзиентная трансфекция
Клетки эпителия яичника китайского хомячка (СНО), нокаутные по гену дигидрофолатредуктазы (dhfr-/-), линия DG-44 (Invitrogen, США), посеянные в количестве 0,5 миллиона клеток на лунку 24-луночного планшета, трансфицировали очищенными суперскрученными плазмидами p1.1-EGFP и p1.1(EBVTR-)-EGFP и pCMV-EGFP (2 мкг/лунка для каждой плазмиды) при помощи трансфекционного реагента FugeneHD (Promega, США) по протоколу производителя реагента. Каждой из исследуемых плазмид трансфицировали по 3 лунки.
Через 48 часов после введения ДНК рассчитывали долю живых и долю трансфицированных клеток в культурах путем подсчета в камере Горяева с окрашиванием трипановым синим в проходящем свете и при помощи пары светофильтров для детекции флуоресцеинизотионата (таблица 1).
Через 24 и 48 часов после введения ДНК определяли уровень продукции целевого белка - eGFP. Для этого отбирали небольшие аликвоты исследуемых суспензионных культур, содержащие по 50 тыс. живых клеток (по данным подсчета в камере Горяева с окрашиванием трипановым синим), отделяли культуральную среду центрифугированием, осадок клеток лизировали в 100 мкл раствора 1% Тритона-Х100, 100 мМ Tris-HCl рН=8,0, 100 мМ NaCl, 10 мМ ЭДТА-Na, 1xProtein Inhibitor Cocktail №8820 (Sigma, США), понижали вязкость полученных лизатов обработкой ультразвуковым диспергатором, переносили полученные лизаты в гнезда светоизолирующего 96-луночного планшета и измеряли интенсивность флуоресценции в образцах, используя пару светофильтров 485 нм/520 нм. Результаты измерений представлены на Фигуре 7. Поскольку доля транзиентно трансфицированных клеток и общий уровень продукции eGFP не имел достоверных различий для случаев трансфекции клеток СНО DG-44 плазмидами сходной длины, содержащими или не содержащими участок EBVTR, было установлено, что EBVTR не оказывает непосредственного влияния на уровень экспрессии гена целевого белка и эффективность транзиентной трансфекции. Эффективность транзиентной трансфекции для плазмид на основе вектора p1.1 значительно ниже, чем для контрольной плазмиды pEGFP-N2 с минимальным вирусным промотором, что объясняется большей длиной вектора p1.1.
Пример 7. Получение пулов стабильно трансфицированных клеток
Клетки СНО DG-44, суспензионно культивирующиеся при 37°С и атмосфере 8% CO2 в безсывороточной среде DG-44 (Invitrogen, США), содержащей гипоксантин и тимидин (НТ+), были трансфицированы линеаризованными плазмидами p1.1-EGFP и p1.1(EBVTR-)-EGFP. Был использован трансфекционный реагент FugeneHD (Promega, США) и соотношение 15 млн клеток/60 мкг ДНК/180 мкл FugeneHD/30 мл культивационного объема. Смесь ДНК и трансфекционного реагента готовили по инструкции производителя трансфекционного реагента и добавляли в суспензионную культуру клеток в указанном выше соотношении, одинаковом для обеих плазмид. Через 48 часов культивации трансфицированных клеток в перемешиваемых колбах культуральную среду заменяли на не содержащую гипоксантин и тимидин селективную среду (НТ-) Opti СНО (Invitrogen, США) и меняли ее в дальнейшем каждые 3 дня центрифугированием/ресуспендированием осадка клеток. При этом оценивали количество клеток подсчетом в камере Горяева и их жизнеспособность по окрашиванию трипановым синим. При культивации в селективной безсывороточной среде Opti СНО происходит постепенная смерть клеток, не содержащих в геноме интегрированного функционального гена dhfr из-за блокирования их нуклеотидного метаболизма.
В общей динамике клеточной культуры смерть нетрансфицированных и транзиентно трансфицированных клеток выражается в прохождении культурой минимума общего количества клеток и минимума выживаемости (то есть доли жизнеспособных клеток) культуры. Культура, трансфицированная плазмидой p1.1-EGFP, содержащей участок EBVTR, проходила минимум клеточной плотности на 11-й день культивации в селективной среде, при этом минимальная плотность составила 147,5 тыс. клеток/мл при доле жизнеспособных клеток 27,7%. В случае плазмиды p1.1(EBVTR-)-EGFP, не содержащей участка EBVTR, минимум был достигнут на 13-ый день культивации в селективной среде, при этом минимальная плотность составила 42,5 тыс. клеток/мл при доле жизнеспособных клеток 3,6%. Таким образом, суммарное количество жизнеспособных клеток в колбах на точках минимума составляло 1225 тыс. и 45,9 тыс., соответственно, то есть эффективность интеграции кассет в геном при наличии в составе плазмиды участка EBVTR была увеличена в 27 раз. Следует отметить, что при предположительной скорости деления стабильно трансфицированных клеток 1 раз за 24 ч количество различных стабильно трансфицированных клеток, содержащихся в культуре на момент внесения селективной среды (день 1), должно составлять 600 шт на колбу для плазмиды, содержащей EBVTR и 6 шт на колбу для плазмиды без EBVTR. Прямое измерение числа различных клеток, в которых прошла интеграция экспрессионной кассеты в геном, невозможно.
Пулы стабильно трансфицированных клеток принято считать сформированными и пригодными для последующего клонирования или амплификации при достижении культурой плотности 1,2 млн клеток/мл и доле жизнеспособных клеток более 85%. При трансфекции клеток плазмидой p1.1-EGFP данные показатели были достигнуты через 22 дня после внесения селективной среды, а в случае плазмиды p1.1(EBVTR-)-EGFP - через 27 дней. Средний уровень экспрессии целевого гена eGFP для обоих полученных пулов не имел существенных различий - 61981 ОЕФ/50 тыс. клеток и 63804 ОЕФ/50 тыс. клеток, соответственно.
Таким образом, наличие в составе экспрессионной плазмиды участка EBVTR позволяет не только увеличить размер исходной популяции стабильно трансфицированных клеток, но и существенно сократить продолжительность культивирования трансфицированных клеток до момента получения плотного жизнеспособного пула. Участок EBVTR не влияет на средний уровень экспрессии целевого гена в стабильно трансфицированных клетках.
Пример 8. Получение олигоклональных линий-продуцентов без амплификации целевого гена
Для количественной оценки динамики интеграции была определена относительная эффективность колониеобразования для стабильно трансфицированных клеток. Для этого клетки СНО DG-44 трансфицировали линеаризованными плазмидами p1.1-EGFP и p1.1(EBVTR-)-EGFP, как описано в Примере 7. Через 48 ч после трансфекции заменяли культуральную среду на селективную безсывороточную среду без гипоксантина и тимидина (НТ-) СНО-А (Invitrogen, США) с добавлением рекомбинантного инсулина и трансферрина человека. Данная ростовая среда предназначена для получения адгезионных культур клеток СНО. Для каждой трансфицированной плазмиды засевали десять 96-луночных культуральных планшетов по 0,5 млн. клеток на планшет. Через 2 недели селективной культивации без смены среды при 37°С и 5% CO2 в лунках подсчитывали количество образовавшихся колоний. В качестве контроля использовали клетки DG-44, трансфицированные реагентом FugeneHD без добавления ДНК, для них не было зафиксировано лунок с колониями. Для исследуемых плазмид было определено число лунок с колониями, общее число колоний и число колоний, экспрессирующих eGFP по данным флуоресцентной микроскопии. Для нескольких отобранных визуально лунок с колониями с максимальной интенсивностью флуоресценции проводили лизис клеток и измерение удельной интенсивности флуоресценции. Результаты измерений приведены в Таблице 2.
При прямом получении олигоклональных линий-продуцентов eGFP трансфекция плазмидой, содержащей участок EBVTR, позволяет увеличить количество позитивных колоний в 40 раз, при этом уровень экспрессии целевого гена для самой продуктивной олигоклональной линии выше в 4,6 раза.
Пример 9. Получение олигоклональных линий-продуцентов при увеличенном селекционном давлении
Получение первичных олигоклональных линий-продуцентов в стандартных условиях селекции обычно приводит к появлению значительной доли колоний, не экспрессирующих целевой ген (Таблица 2), а среди позитивных колоний большая их часть демонстрирует относительно невысокий уровень экспрессии. Это явление вызывается существенной вероятностью интеграции генетической кассеты в транскрипционно неактивные участки генома или интеграции поврежденной генетической кассеты. Экспрессионные конструкции, в которых уровень экспрессии селекционного маркера жестко связан с уровнем экспрессии целевого гена, например, путем экспрессии единственной бицистронной РНК, позволяют проводить первичный отбор линий-продуцентов при повышенном селекционном давлении. При этом происходит уменьшение общего числа образующихся колоний клеток-продуцентов и одновременно с этим возможно сильное увеличение доли высокопродуктивных клонов.
Для проверки возможности обогащения набора олигоклональных линий-продуцентов высокопродуктивными линиями проводили трансфекцию клеток СНО DG-44 плазмидой p1.1-EGFP по условиям Примера 7 и получение олигоклональных линий-продуцентов по условиям Примера 8, по одному культуральному планшету для селекционной среды из Примера 8, дополнительно содержащей 50 нМ ингибитора DHFR метотрексата и среды из Примера 8, не содержащей метотрексат.
Параметры колониеобразования приведены в Таблице 3, уровни экспрессии целевого гена для 8 наиболее продуктивных линий приведены на Фиг.8.
Среднее число клеток на момент анализа, то есть скорость роста колоний, для 10 наиболее продуктивных линий, полученных в присутствии 50 нМ метотрексата, и 10 аналогичных линий, полученных в отсутствие метотрексата, не имело существенных различий, что указывает на отсутствие влияния медленного процесса амплификации целевого гена под действием метотрексата на продуктивность полученных линий. Генерация олигоклональных линий-продуцентов в присутствии низкой концентрации ингибитора селекционного маркера для плазмиды на основе экспрессионного вектора p1.1 позволяет существенно увеличить продуктивность наиболее продуктивных линий при незначительном падении общего числа колоний. Это указывает на высокое генетическое сцепление уровней экспрессии целевого гена и гена селекционного маркера и позволяет управлять продуктивностью получаемых олигоклональных линий-продуцентов.
Пример 10. Получение высокопродуктивных олигоклональных линий амплификацией целевого гена под действием метотрексата
Для получения высокопродуктивных линий-продуцентов eGFP проводили амплификацию целевого гена в геноме клеток под действием метотрексата. Для этого у культивируемых пулов клеток, стабильно трансфицированных плазмидами р1.1-EGFP и p1.1(EBVTR-)-EGFP как описано в примере 7, проводили замену суспензионной среды на культуральную среду для адгезионного культивирования СНО-А с добавлением рекомбинантного инсулина и трансферрина человека. Полученную суспензию клеток разделяли на аликвоты и добавляли к ним метотрексат до конечных концентраций 0, 50, 100, 200, 400, 800 нМ. Суспензию засевали в культуральные адгезионные 96-луночные планшеты, по 0,5 млн клеток /планшет, по 3 планшета на каждую концентрацию метотрексата для обоих пулов. Через 2 недели селективной культивации без смены среды при 37°С и 5% CO2 проводили инспекцию планшетов и смену среды, в дальнейшем среду заменяли каждые 4 дня. По достижению конфлюентности в 10% лунок для каждого планшета проводили прижизненную оценку интенсивности флюоресценции в лунках планшета, клетки из 16 наиболее интенсивно флуоресцирующих лунок переносили в 48-луночный планшет. Время культивации клеток в планшетах до пересева составило 15 дней для концентрации метотрексата 100 и 800 нм, 16 дней для концентрации метотрексата 200 и 400 нм. Продолжали культивирование отобранных олигоклональных культур, отбрасывая олигоклоны, не обладавшие достаточной жизнеспособностью при пассировании и не пригодные для отделения от подложки смывом. Сохраняли по 24 олигоклона для каждого уровня метотрексата, по 1 лунке 24 луночного планшета. Переносили ¼ конфлюентной культуры каждого олигоклона в новый 24-луночный планшет, остатки культуры лизировали и проводили измерение интенсивности флюоресценции и содержания общего белка (т.е. количества клеток). Удельная интенсивность флюоресценции для 8х6 олигоклонов, полученных при амплификации пула клеток, стабильно трансфицированных плазмидой p1.1-EGFP, приведена на Фиг.9.
При амплификации целевого гена под действием метотрексата для трансфектантов контрольной плазмиды p1.1(EBVTR-)-EGFP и использовании концентраций метотрексата 400 и 800 нМ не было получено ни одной жизнеспособной колонии, удельные интенсивности флуоресценции для остальных полученных олигоклональных линий не имели существенных отличий от удельных интенсивностей флуоресценции линий, полученных при культивировании трансфектантов в отсутствие метотрексата (данные не приводятся).
Таким образом, наличие участка EBVTR в плазмиде p1.1 существенно увеличивает скорость амплификации целевого гена и позволяет получать высокопродуктивные линии в короткие сроки (15 дней) под действием высоких концентраций метотрексата. Возможный механизм действия участка EBVTR при амплификации целевого гена - образование шпильки на одноцепочечной ДНК при транспозиции области целевого гена, ускорение амплификации целевых генов по механизму транспозиции описано в работе (Tanaka H et al. Short inverted repeats initiate gene amplification through the formation of a large DNA palindrome in mammalian cells. Proc Natl Acad Sci USA. 2002 Jun 25;99(13):8772-8777). Наиболее вероятно образование шпильки между участками GGGGGCCGCGGG (нуклеотиды 161-172) и CCCGCAGCCCCC (84-95) или CCCGCGGACCCC (100-111), образующими инвертированные повторы с одной некомплементарной парой нуклеотидов.
Взаимное расположение промотора целевого гена, участка внутренней инициации трансляции, ОРС селекционного маркера, а также сигнальных областей целевого гена позволяет увеличивать продуктивность нескольких наилучших олигоклональных линий приблизительно в 5 раз при выполнении однократной процедуры амплификации целевого гена.
Пример 11. Получение клональных высокопродуктивных линий-продуцентов и оценка их стабильности при культивировании
Для 10 полученных по условиям Примера 10 олигоклональных линий различной продуктивности была проведена генерация моноклональных линий-продуцентов eGFP. Использовали только олигоклональные линии, полученные от клеток, трансфицированных плазмидой p1.1(EBVTR-)-EGFP. Клонирование проводили методом предельных разведении, из расчета 1 клеток на лунку. Через 3 недели после внесения клеток в культуральные планшеты визуально и микроскопически определяли лунки с одиночными колониями и переносили эти колонии в 48-луночный планшет по достижении ими конфлюентности. Продолжали культивирование отобранных клонов до их переноса в 12-луночные планшеты. Для проверки стабильности уровня продукции линий при продолжительном культивировании отобрали 6 клонов с различным уровнем продукции целевого белка (различной удельной интенсивностью флуоресценции) и вели их культивацию в 12-луночном планшете, пассируя в разведении 1:5 каждые 3-4 дня. Динамика удельной интенсивности флуоресценции при наблюдениях в течение 64 дней приведена на Фиг.10, значимого изменения уровня продуктивности не наблюдалось ни в одной из линий. Таким образом, линии-продуценты, полученные при трансфекции клеток СНО DG-44 плазмидой на основе вектора p1.1 и последующей одностадийной амплификации целевого гена, сохраняют стабильный уровень экспрессии целевого гена в течение 60 дней. Такой срок сохранения неизменного уровня продукции целевого белка достаточен для промышленного использования линий-продуцентов на основе вектора p1.1.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.


Изобретение относится к биотехнологии и биофармацевтике. Описан экспрессионный плазмидный вектор для гетерологичной экспрессии рекомбинантных белков, высокочастотной интеграции и ускоренной амплификации экспрессионной кассеты в клетках млекопитающих. Вектор содержит функциональные промотор и терминатор гена фактора элонгации 1 альфа китайского хомячка, фланкированные 5' и 3' НТО этого гена; участок для клонирования открытых рамок считывания целевых белков; внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV); открытую рамку считывания дегидрофолатредуктазы (DHFR) мыши, экспрессирующуюся в составе бицистронной мРНК вместе с целевым геном (IRES DHFR); и участок терминального повтора вируса Эпштейн-Барр человека (EBVTR). Также описан способ получения стабильных линий продуцентов рекомбинантных белков с высокой копийностью экспрессионной кассеты в геноме с использованием указанного экспрессионного плазмидного вектора. 4 н. и 8 з.п. ф-лы, 10 ил., 3 табл., 11 пр.
1. Экспрессионный плазмидный вектор для гетерологичной экспрессии рекомбинантных белков, высокочастотной интеграции и ускоренной амплификации экспрессионной кассеты в клетках млекопитающих в следующей последовательности, по существу, содержащий область начала репликации плазмиды pUC; участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5' нетранслируемой областью этого гена; участок для клонирования открытой рамки считывания целевого белка (полилинкер); внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV); открытую рамку считывания дегидрофолатредуктазы (DHFR), экспрессирующуюся в составе бицистронной мРНК вместе с целевым геном (IRES DHFR); и функциональный терминатор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3' нетранслируемой областью этого гена.
2. Экспрессионный плазмидный вектор по п.1, отличающийся тем, что участок терминального повтора вируса Эпштейн-Барр человека (EBVTR) представляет собой фрагмент конкатемера терминального повтора EBVTR.
3. Экспрессионный плазмидный вектор по п.1, отличающийся тем, что используют открытую рамку считывания дегидрофолатредуктазы (DHFR) мыши.
4. Экспрессионный плазмидный вектор по п.1, отличающийся тем, что указанный вектор дополнительно содержит консенсусную последовательность Козак.
5. Экспрессионный плазмидный вектор по п.1, отличающийся тем, что указанным вектором является вектор p1.1, представленный в Перечне последовательностей под номером SEQ ID NO:1.
6. Бицистронная мРНК, образующаяся в процессе экспрессии вектора по п.1, содержащая открытую рамку считывания целевого рекомбинантного белка, внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV) и открытую рамку считывания дегидрофолатредуктазы (DHFR).
7. Бицистронная мРНК по п.6, отличающаяся тем, что используют открытую рамку считывания дегидрофолатредуктазы (DHFR) мыши.
8. Способ получения стабильной линии клеток млекопитающих с множественными копиями экспрессионной кассеты в геноме продуцентов рекомбинантного белка, включающий следующие стадии:
а) трансфекцию клеток млекопитающего, не содержащих активные аллели дегидрофолатредуктазы, экспрессионным вектором по п.1, содержащим вставку с открытой рамкой считывания гена, кодирующего целевой рекомбинантный белок;
б) отбор жизнеспособных клеток на среде, не содержащей гипоксантин и тимидин; и
в) отбор полученных на стадии б) клеток млекопитающих - продуцентов целевого белка на среде, содержащей метотрексат.
9. Способ по п.8, отличающийся тем, что концентрация метотрексата в среде составляет от 50 нМ до 200 нМ.
10. Способ по п.8, отличающийся тем, что используют клетки эпителия яичника китайского хомячка (СНО) DG-44, нокаутные по гену дигидрофолатредуктазы.
11. Способ получения рекомбинантного белка, включающий следующие стадии:
- культивирование в питательной среде клеток млекопитающих - продуцентов целевого белка, полученных способом по п.8; и
- выделение полученного целевого белка из культуральной жидкости.
12. Способ по п.11, отличающийся тем, что культивируют клетки СНО DG44, трансфицированные экспрессионным вектором по п.1, содержащим вставку с открытой рамкой считывания гена, кодирующего целевой рекомбинантный белок.
| RUNNING DEER J | |||
| et al., "High-level expression of proteins in mammalian cells using transcription regulatory sequences from the Chinese hamster EF-1 alpha gene", Biotechnol Prog | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| DORONINA V.A | |||
| et al., "Site-specific release of nascent chains from ribosomes at a sense codon", Mol Cell Biol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |

