Область техники
Изобретение относится к области биотехнологии, а именно к технологии получения биологически активных веществ (БАВ) методами генной инженерии, точнее к методам получения рекомбинантных белков в культивируемых клетках млекопитающих.
Предшествующий уровень техники
Некоторые секретируемые белки позвоночных животных содержат кластеры близкорасположенных гамма-карбоксилированных остатков глутаминовой кислоты, обычно обозначаемые как Gla-домен. Большинство известных белков, содержащих Gla-домен, являются зимогенами сериновых протеаз и участвуют в каскаде свертывания крови. Функции самого Gla-домена могут быть описаны как координирование ионов кальция и Са-зависимое связывание с фосфолипидной мембраной (Nelsestuen et al. (1976) Role of gamma-carboxyglutamic acid. Cation specificity ofprothrombin and factor X-phospholipid binding. J. Biol. Chem., 251, 6886-6893).
В настоящее время известно более 10 белков человека, содержащих Gla-домен, - факторы свертывания крови, такие как протромбин или фактор II, факторы VII, IX, X; антикоагулянты, такие как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранные Gla-белки (TMGPs). Для всех указанных белков гамма-карбоксилирование необходимо для проявления их функциональной активности (Furie et al. (1999) Vitamin K-dependent biosynthesis of gamma-carboxyglutamic acid. Blood, 93, 1798-1808).
Гамма-карбоксилирование остатков глутаминовой кислоты (Glu→Gla) происходит пост-трансляционно, локализовано в эндоплазматическом ретикулуме клеток и производится ферментом витамин-К зависимой гамма-глутамин карбоксилазой (GGCX, ЕС 4.1.1.90) (Berkner, K.L. (2000) The vitamin K-dependent carboxylase. J. Nutr., 130, 1877-1880). Источником присоединяемой карбоксильной группы в данной реакции является растворенный углекислый газ, а кофактором GGCX выступает восстановленная дигидрохиноновая форма витамина K, превращающаяся при прохождении реакции в окисленную форму 2,3-эпоксида хинона. Таким образом, реакция гамма-карбоксилирования требует постоянного поддержания существенной концентрации дигидрохиноновой формы витамина К в люмене эндоплазматического ретикулума.
Восстановление эпокси-формы витамина К до дигидрохинона в клетке производится ферментным комплексом VKOR или VKORC (vitamin К oxireductase complex), основным компонентом которого является интегральный белок "субъединица 1 комплекса 2,3-эпоксиредуктазы витамина K" (VKORC1, ЕС 1.1.4.1). Ген VKORC1 и основные свойства рекомбинантного белка VKORC1 человека описаны в патентах США 7482141 и 7524665. Известно, что некоторые мутации в этом гене вызывают устойчивость к ингибитору гамма-карбоксилирования белков варфарину, а также редкое наследственное заболевание "множественный дефицит факторов свертываемости крови тип 2" [Rost, Fregin, Ivaskevicius, Conzelmann, Hortnagel, Pelz, Lappegard, Seifried, Scharrer, Tuddenham, Muller, Strom and Oldenburg, Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2 // Nature, v.427, p.537-41 (2004)].
Поскольку белки системы гемостаза должны иметь полностью (или почти полностью) гамма-карбоксилированный Gla-домен для проявления функциональной активности, гетерологичная экспрессия их генов в культивируемых клетках млекопитающих может приводить к секреции неактивных форм белков. Этот нежелательный эффект наиболее выражен для систем экспрессии рекомбинантного фактора VII свертываемости крови и проявляется как невозможность получения существенных количеств функционально активного рекомбинантного фактора VII в клетках линии СНО, при этом клетки линий HepG2 или ВНК позволяют получать функционально активный белок при сходном уровне секреции. В ряде работ было установлено, что увеличение количества GGCX не приводит к улучшению гамма-карбоксилирования молекул фактора VII, в то время как оверэкспрессия гена VKORC1 человека в клетках линии СНО позволяет получить функционально активный продукт и существенно повысить скорость секреции клетками фактора VII (Bolt, G., Т.D. Steenstrup, et al. (2007). "All post-translational modifications except propeptide cleavage are required for optimal secretion of coagulation factor VII." Thromb Haemost 98(5): 988-997). Сходные данные были получены для рекомбинантного фактора свертываемости крови IX, секретируемого клетками линии ВНК [Wajih, Hutson, Owen and Wallin, Increased production of functional recombinant human clotting factor IX by baby hamster kidney cells engineered to overexpress VKORC1, the vitamin К 2,3-epoxide-reducing enzyme of the vitamin К cycle // J Biol Chem, v.280, p.31603-7 (2005)]. Для известных систем экспрессии фактора IX в клетках СНО, лимитированных уровнем процессинга пропептида целевого белка, уровень гамма-карбоксилирования, обеспечиваемый эндогенным ферментом VKORC1, по-видимому, достаточен для получения фактора IX с полной функциональной активностью [McGrath, Factor IX (protease zymogen) // Directory of therapeutic enzymes, p.209-238 (2005)]. В то же время уровень секреции фактора IX при оверэкспрессии гена VKORC1 человека увеличивается в 1,2-1,4 раза (патент США 7375084). Таким образом, оверэкспрессия гена VKORC1 в клетках-продуцентах позволяет увеличить как уровень секреции BK3-белков, так и долю их функционально активных форм.
Вследствие того, что исходно были охарактеризованы ген и кДНК VKORC1 человека, в большинстве работ для оверэкспрессии VKORC1 в культивируемых клетках были использованы плазмиды, содержащие кДНК VKORC1 человека. Известно, что изолированная каталитическая эффективность VKORC1 для различных видов млекопитающих отличается примерно в 4 раза [Wilson, Sauer, Carlson, Wallin, Ward and Hooser, Species comparison of vitamin K1 2,3-epoxide reductase activity in vitro: kinetics and warfarin inhibition // Toxicology, v.189, p.191-8 (2003)]. Поскольку удельная активность фермента VKORC1 может быть точно и непосредственно измерена только при разобщении витамин К оксиредуктазного комплекса и использовании дитиотрейтола в качестве донора электронов, собственная удельная активность VKORC1 не может охарактеризовать его фактическую каталитическую эффективность в живой клетке. Такая каталитическая эффективность, то есть способность поддерживать достаточную концентрацию восстановленной дигидрохиноновой формы витамина K в эндоплазматическом ретикулуме клеток, в которых происходит оверэкспрессия Gla-белка, может быть оценена только косвенным образом, а именно по уровню секреции гамма-карбоксилированной формы оверэкспрессируемого белка. Поскольку белки, переносящие электроны на мембранно-связанный фермент VKORC1, в настоящий момент неизвестны [Wilson, Sauer, Carlson, Wallin, Ward and Hooser, Species comparison of vitamin K1 2,3-epoxide reductase activity in vitro: kinetics and warfarin inhibition // Toxicology, v.189, p.191-8 (2003)], можно предположить, что их сродство к различным ортологам VKORC1 может существенно различаться. Тогда наилучшим вариантом VKORC1 для оверэкспрессии в клетках-продуцентах мог бы быть их собственный VKORC1, поскольку его достаточное сродство к белкам-донорам электронов клетки-хозяина не может вызывать сомнения. В случае клеток линии СНО такой аутологичный вариант VKORC1 не был описан, и его ферментативная активность не была охарактеризована ни прямыми, ни косвенными методами.
По-видимому, естественный уровень экспрессии гена VKORC1 в клетках СНО относительно низок, поскольку данный ген преимущественно экспрессируется в гепатоцитах, а линия клеток СНО получена из яичника. В частности, в работе [de Castilho Femandes, Fontes, Gonsales, Swiech, Picanco-Castro, Faca and Covas, Stable and high-level production of recombinant Factor IX in human hepatic cell line // Biotechnol Appi Biochem, v.58, p.243-9 (2011)] было установлено, что уровень экспрессии функционально активного фактора IX человека выше в линии клеток, полученных из печени (HepG2), чем в линиях клеток, полученных из других тканей; при этом во всех случаях использовался одинаковый экспрессионный вектор. В то же время, уровень экспрессии неустановленных белков, являющихся донорами электронов для VKORC1, в различных тканях неизвестен, также неизвестной остается и необходимость поддержания повышенного уровня экспрессии генов этих белков.
Краткое описание изобретения.
Целью настоящего изобретения является получение клеток китайского хомячка - продуцентов белков с Gla-доменом и предоставление способа получения белков с Gla-доменом с использованием полученных клеток.
Эта цель была достигнута путем того, что авторами настоящего изобретения впервые была клонирована и секвенирована область открытой рамки (ОРС) считывания кДНК VKORC1, полученной из клеток линии СНО DG-44. При оверэкспрессии ОРС аутологичного VKORC1 китайского хомячка, находящегося под контролем промотора CHEF1, в линии клеток СНО DG-44 было обнаружено резкое увеличение ферментативной активности VKORC1. Оверэкспрессия ОРС VKORC1 человека в таком же экспрессионном векторе приводила к значительно меньшему увеличению ферментативной активности VKORC1. Таким образом, ко-экспрессия аутологичного гена VKORC1 китайского хомячка под контролем сильного промотора может увеличить уровень секреции рекомбинантных Gla-белков, что представляет значительный интерес для биотехнологии и потенциально позволяет существенно уменьшить затраты при получении рекомбинантных факторов свертывания крови Vila, IX, X, протромбина, а также белка С, использующихся в качестве лекарственных средств.
Таким образом, объектом настоящего изобретения является субъединица 1 комплекса 2,3-эпоксиредуктазы витамина К (VKORC1) китайского хомячка, представленная в Перечне последовательностей под номером SEQ ID NO:2.
Также объектом настоящего изобретения является фрагмент ДНК, кодирующий VKORC1, описанную выше.
Также объектом настоящего изобретения является фрагмент ДНК, описаный выше, представленный в Перечне последовательностей под номером SEQ ID NO:1.
Также объектом настоящего изобретения является плазмида, обладающая способностью к экспрессии в клетке китайского хомячка, содержащая фрагмент ДНК, описанный выше, под контролем промотора, функционирующего в указанной клетке.
Также объектом настоящего изобретения является плазмида, описанная выше, в которой указанным промотором является сильный промотор.
Также объектом настоящего изобретения является плазмида, описанная выше, в которой указанной плазмидой является плазмида p1.2-Zeo-CHO-VKORC1, представленная в Перечне последовательностей под номером SEQ ID NO:5.
Также объектом настоящего изобретения является клетка китайского хомячка - продуцент белка с Gla-доменом, в которой повышена активность VKORC1 по п.1.
Также объектом настоящего изобретения является клетка, описанная выше, в которой экспрессия VKORC1 повышена за счет повышения экспрессии VKORC1 в указанной клетке.
Также объектом настоящего изобретения является клетка, описанная выше, в которой экспрессия VKORC1 повышена за счет трансформации указанной клетки плазмидой по п.4.
Также объектом настоящего изобретения является клетка, описанная выше, в которой экспрессия VKORC1 повышена за счет использования сильного промотора, функционирующего в указанной клетке.
Также объектом настоящего изобретения является клетка, описанная выше, при этом указанной клеткой является клетка СНО DG44.
Также объектом настоящего изобретения является клетка, описанная выше, в которой белок с Gla-доменом выбран из группы, состоящей из факторов свертывания крови, таких как (протромбин или фактор II, факторы VII, IX, X; антикоагулянтов, таких как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранных Gla-белков (TMGPs).
Также объектом настоящего изобретения является способ получения белка с Gla-доменом, включающий стадии культивирования описанной выше в питательной среде и выделения полученного белка с Gla-доменом из культуральной жидкости.
Также объектом настоящего изобретения является способ, описанный выше, в котором культивируют клетки СНО DG44/p1.2-Zeo-CHO-VKORC1.
Также объектом настоящего изобретения является способ, описанный выше, в котором белок с Gla-доменом выбран из группы, состоящей из факторов свертывания крови, таких как (протромбин или фактор II, факторы VII, IX, X; антикоагулянтов, таких как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранных Gla-белков (TMGPs).
Более подробно настоящее изобретение описано ниже.
Подробное описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание системы для оверэкспрессии белков с Gla-доменом в клетках китайского хомячка.
Технический результат достигался путем клонирования области открытой рамки считывания кДНК гена VKORC1 китайского хомячка, конструирования на ее основе новой экспрессионной плазмидной ДНК, в частности p1.2-Zeo-CHO-VKORC1, пригодной для трансфекции клеток-продуцентов белков с Gla-доменом и последующей селекции вспомогательного трансгена, позволяющей получать линии-продуценты белков с Gla-доменом с высоким уровнем продукции полностью гамма-карбоксилированного целевого белка и достаточной стабильностью основного и вспомогательного трансгенов в геноме.
В основе технического решения лежит разработанная авторами плазмидная ДНК p1.2-Zeo-CHO-VKORC1 длиной 12318 п.о., кодирующая ген VKORC1 китайского хомячка под контролем промотора гена CHEF1 (Chinese Hamster translation Elongation Factor 1 alpha), функционирующего в клетке китайского хомячка и фланкированного нетранскрибируемыми областями гена CHEF1, а также генетический элемент, обеспечивающий возможность селекции клонов с инсерциями экспрессионной кассеты в геноме клеток продуцентов. Функциональные промотор и терминатор гена CHEF1 обеспечивают конститутивную экспрессию целевого гена; последовательность кДНК гена VKORC1 китайского хомячка обеспечивает экспрессию полипептидной цепи фермента, последовательность, включающая промотор SV40 и ген фермента устойчивости к действию антибиотика зеоцина (Zeo) обеспечивает возможность селекции стабильно трансфицированных клеток СНО в присутствии зеоцина.
Термин «субъединица 1 комплекса 2,3-эпоксиредуктазы витамина K (VKORC1)» означает фермент, катализирующий следующую реакцию: 2-метил-3-фитил-1,4-нафтохинон+окисленный дитиотреитол⇔2,3-эпокси-2,3-дигидро-2-метил-3-фитил-1,4-нафтохинон+1,4-дитиотреитол. Указанную активность классифицируют как ЕС 1.1.4.1. Предыдущее название - витамин K-зависимый фактор дефицита свертывания крови 2 (vitamin K dependent clotting factors deficiency 2, VKCFD2). Наличие активности ацил-СоА-дегидрогеназы может быть установлено способом, описанным, например, Goodstadt L. и Ponting C.P. (Vitamin К epoxide reductase: homology, active site and catalytic mechanism. Trends Biochem Sci. 2004 Jun; 29(6):289-92) или способом, описанным в Примерах 5 или 7. В настоящий момент известны VKORC1 человека, мыши, крысы. Так, аминокислотная последовательность рекомбинантного фермента VKORC1 человека представлена в Перечне последовательностей под номером SEQ ID NO:4. Последовательность ОРС гена, кодирующего рекомбинантный фермент VKORC1 человека, представлена в Перечне последовательностей под номером SEQ ID NO:3. Примером VKORC1 китайского хомячка согласно настоящему изобретению является фермент, последовательность которого приведена в Перечне последовательностей под номером SEQ ID NO:2 или его функциональный вариант. Последовательность белка депонирована заявителем в GenBank (03-APR-2012) под номером AFG26681.1.
Термин "вариант белка", используемый в настоящем изобретении, означает белок с изменениями в последовательности, будь то делеции, вставки, добавления или замены аминокислот, в котором сохраняется активность указанного белка. Число изменений в варианте белка зависит от положения или типа аминокислотного остатка в третичной структуре белка. Оно может быть от 1 до 15, предпочтительно от 1 до 10, более предпочтительно от 1 до 5. Данные изменения в вариантах могут иметь место в областях, не критичных для функции белка. Данные изменения возможны потому, что некоторые аминокислоты имеют высокую гомологию друг другу, поэтому такие изменения не влияют на третичную структуру или активность. Следовательно, вариант белка может быть представлен белками с гомологией не менее 80%, предпочтительно, не менее 90%, и, наиболее предпочтительно, не менее 95%, по отношению к полной аминокислотной последовательности, приведенной в Перечне последовательностей, при условии, что активность белка такого сохраняется.
Гомология между двумя аминокислотными последовательностями может быть определена с использованием известных методов, например, компьютерной программы BLAST 2.0, которая считает три параметра: число аминокислот, идентичность и сходство.
Замена, делеция, вставка или добавление одной или нескольких аминокислотных остатков являются консервативной (-ыми) мутацией (-ями), если сохраняется активность. Типичным примером консервативной мутации является консервативная замена. Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Не на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, He или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp, и замену Val на Met, Ile или Leu.
Фрагментом ДНК, кодирующим VKORC1 китайского хомячка согласно настоящему изобретению, является любой (с точки зрения вырожденности генетического кода) фрагмент ДНК, кодирующий фермент из китайского хомячка, обладающий активностью VKORC1.
Примером последовательности ОРС гена, кодирующего фермент VKORC1 клеток китайского хомячка DG44 согласно настоящему изобретению, является последовательность, представленная в Перечне последовательностей под номером SEQ ID NO:1. Указанная последовательность депонирована 03.04.2012 заявителем в базу данных GenBank под номером JQ400047.1. Аминокислотная последовательность фермента VKORC1 клеток китайского хомячка DG44 согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO:2.
Фрагменты ДНК, которые кодируют, по существу, те же регуляторные элементы могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO:1), например, посредством метода сайт-направленного мутагенеза, так, что один или несколько нуклеотидов в определенном сайте будут делегированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Показатели функциональной активности, при которой считается, что полученный вектор позволяет получить высокопродуктивный клон клеток с увеличенным содержанием ферментов, осуществляющих гамма-карбоксилирование, определяются по ферментативной активности витамин-K эпоксиредуктазного комплекса в лизате клеточной массы (при этом удельная ферментативная активность определяется как отношение скорости восстановления 2,3-эпоксида витамина K1 лизатом клеток в присутствии избытка донора электронов, например дитиотреитола, и концентрации общего белка в исследуемом лизате). Измерение удельной ферментативной активности описано в Примерах 5 и 7.
Измерение степени превращения 2,3-эпоксида витамина K в восстановленную форму витамина К может быть проведено путем последовательного отбора аликвот из лизата исследуемых клеток, смешанных с донором электронов дитиотреитолом и субстратом реакции -2,3-эпоксидом витамина K1, экстракции всех форм витамина K1 смесью гексана и изопропанола, последующим отделением органической фазы, ее выпариванием в вакууме, растворением нелетучего осадка в минимальном объеме метанола, хроматографическом разделении окисленной и восстановленной форм витамина K1 на колонке для обращенно-фазовой хроматографии, расчета объемов пиков форм витамина K1 и вычисления доли восстановленной формы витамина K1.
Активность фермента в клетке может быть повышена за счет увеличения количества соответствующей мРНК, повышения количества соответствующего фермента, или увеличения удельной активности фермента. Количество соответствующей мРНК может быть увеличено за счет увеличения числа копий соответствующего гена или за счет усиления транскрипции соответствующего гена, достигаемого путем использования более сильного промотора, снятия репрессии, или за счет увеличения стабильности мРНК. Количество соответствующего фермента может быть повышено за счет усиления трансляции соответствующей мРНК, достигаемого модификацией нуклеотидной последовательности сайта связывания рибосом соответствующей мРНК или за счет увеличения его стабильности. Удельная активность фермента может быть увеличена за счет введения соответствующих мутаций в аминокислотную последовательность фермента.
Методы усиления экспрессии гена включают увеличение числа копий гена. Введение гена в вектор, способный функционировать в клетке согласно настоящему изобретению, увеличивает число копий гена.
Термин «плазмида, обладающая способностью к экспрессии в клетке китайского хомячка» означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии в клетке-хозяине внедренного в него гена, например, такие как промотор, терминатор, и элементов для отбора клонов с множественными копиями экспрессионной кассеты в геноме. Конкретным примером генетических элементов, согласно настоящему изобретению, является, но не ограничивается им, промотор гена CHEF1, и ген фермента, обеспечивающего устойчивость клеток к действию зеоцина.
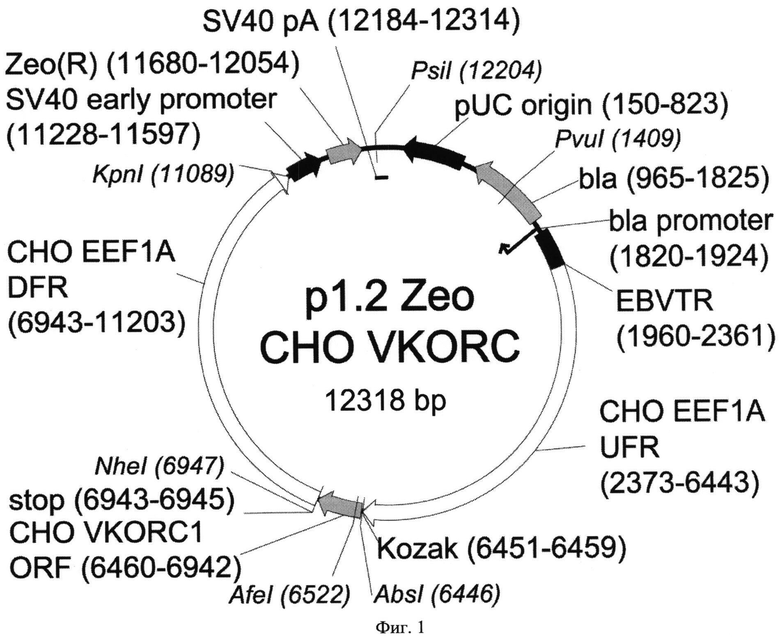
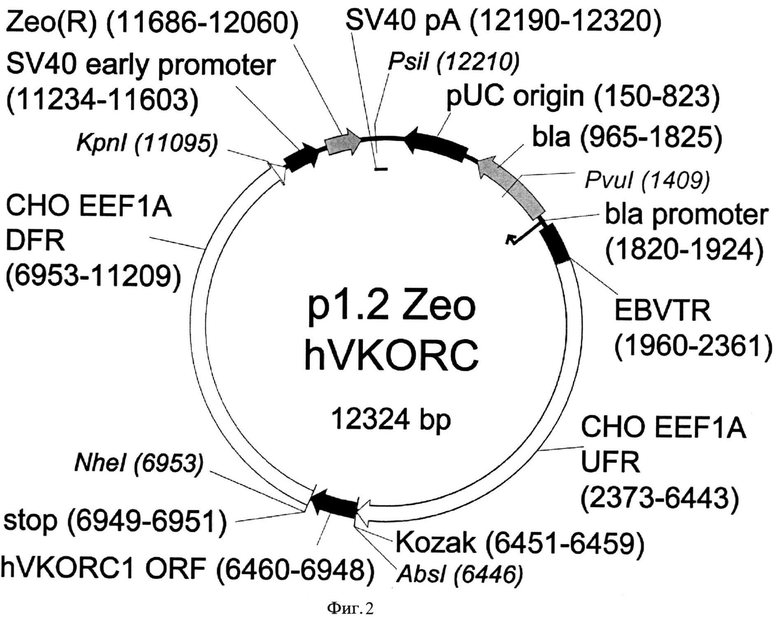
Конкретным примером такой плазмиды является плазмида p1.2-Zeo-CHO-VKORC1. Карта экспрессионной плазмиды p1.2-Zeo-CHO-VKORC1 согласно настоящему изобретению, приведена на Фиг. 1, последовательность плазмиды представлена в Перечне последовательностей под номером SEQ ID NO:5. Карта экспрессионной плазмиды p1.2-Zeo-hVKORC1, кодирующей VKORC1 человека, приведена на Фиг. 2, последовательность плазмиды представлена в Перечне последовательностей под номером SEQ ID NO:6.
Практические функциональные свойства плазмиды, содержащей ген VKORC1 китайского хомячка, p1.2-Zeo-CHO-VKORC1 (SEQ ID NO:5) можно оценить по сравнению с контрольной плазмидой p1.2-Zeo-hVKORC1 (SEQ ID NO:6), содержащей OPC VKORC1 человека.
Плазмида p1.2-Zeo-CHO-VKORC1 размером 12318 п.о. состоит из:
1) фрагмента размером 501 п.о. (6446-6946), включающего открытую рамку считывания VKORC1 китайского хомячка, последовательность Козак, обеспечивающую инициацию трансляции и стоп-кодон;
2) фрагмента векторной плазмиды p1.2-Zeo размером 11817 п.о. (6947-6445), содержащего элементы для экспрессии целевого белка в клетках млекопитающих (функциональный вирусный промотор, сигнал полиаденилирования и функциональный терминатор) и селекционный маркер для отбора в эукариотических клетках, а также элементы, необходимые для продукции плазмиды в бактериальной клетке - область начала репликации и селекционный маркер.
Указанная плазмида содержит уникальные сайты узнавания эндонуклеазами рестрикции: PvuI (1409), AbsI (6446), AfeI (6522), NheI (6947), KpnI (11089), PsiI (12204).
Плазмида была получена с использованием стандартных методов генной инженерии, коммерчески доступных плазмид и химически синтезированных олигонуклеотидов [Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. 2nd ed. New York, NY: Cold Spring Harbor Laboratory Press; 1989].
Усиление экспрессии гена может также быть достигнуто путем введения множества копий гена в геном. Для введения множества копий гена в геном выполняется гомологичная рекомбинация с использованием в качестве целевых последовательностей, присутствующих в хромосоме во множестве копий. Последовательности с множеством копий в геномной ДНК, включают, но не ограничиваются ими, повторяющиеся ДНК, или инвертированные повторы на концах транспонируемых элементов. Также возможно включить ген в состав транспозона и обеспечить его перенос для введения множества копий гена в хромосомную ДНК.
Усиление экспрессии гена также может быть достигнуто путем подстановки фрагмента ДНК согласно настоящему изобретению под контроль сильного промотора. Использование сильного промотора можно сочетать с увеличением копий гена.
С другой стороны, действие промотора может быть усилено, например, введением в промотор мутации, ведущей к увеличению уровня транскрипции локализованного за промотором гена. Кроме того, возможно ввести нуклеотидную замену в область промотора гена на хромосоме, результатом чего является усиление функции промотора.
Усиление трансляции может быть достигнуто путем введения во фрагмент ДНК согласно настоящему изобретению более эффективной последовательности Козак. Последовательность Козак располагается вокруг старт-кодона целевого гена и способствует инициации трансляции мРНК целевого гена. Использование сильного промотора можно сочетать с использованием более эффективной последовательности Козак.
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989). Согласно настоящему изобретению термин «клетка китайского хомячка - продуцент белка с Gla-доменом» означает клональную линию клеток китайского хомячка, обладающую способностью к продукции и секреции белка с Gla-доменом, когда она согласно настоящему изобретению выращивается в питательной среде. Используемый здесь термин «клетка китайского хомячка - продуцент белка с Gla-доменом» также означает линию клеток, которые способна секретировать белок с Gla-доменом в количестве не менее чем 0,1 мкг/мл, более предпочтительно, не менее чем 1 мкг/мл при культивировании в суспензионной культуре в бессывороточной среде. Указанный белок с Gla-доменом секретируется указанными клетками в культивационную среду.
В качестве примера предпочтительных клеток яичника китайского хомячка может быть приведена линия СНО DG44 (Invitrogen cGMP banked, США; Mol Cell Biol 5, 1750-1759 Kaufman RJ et al. 1985). Круг сублиний СНО не ограничен каким-либо образом, например, могут быть использованы сублинии СНО CHOZN DHFR, CHO DUKX B11 и подобные им.
Конкретным примером линии для получения продуцента фактора свертывания крови VIII человека с делегированным В-доменом согласно настоящему изобретению является линия СНО DG44, но спектр линий клеток не ограничиваются ей.
Белком с Gla-доменом является белок, выбранный из группы, состоящей из факторов свертывания крови, таких как (протромбин или фактор II, факторы VII, IX, X; антикоагулянтов, таких как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранных Gla-белков (TMGPs).
Также для реализации настоящего изобретения еще одной технической задачей явилось создание способа получения высокопродуктивных и стабильных при последовательном пассировании линий клеток млекопитающих-продуцентов белков с Gla-доменом с увеличенным содержанием ферментов, обеспечивающих гамма-карбоксилирование, с использованием эукариотической клетки (или эукариотической клетки-продуцента Gla-домен содержащих белков), трансфицированной экспрессионной плазмидой.
Также целью настоящего изобретения является предоставление способа получения белка с Gla-доменом в клетках китайского хомячка, включающего культивирование в питательной среде описанных выше клеток - продуцентов белка с Gla-доменом, и выделение полученного целевого рекомбинантного белка из культуральной жидкости.
Выращивание клеток, выделение и очистка целевого белка из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам культивирования, в которых рекомбинантный белок продуцируется с использованием клеток млекопитающих.
Питательная среда, используемая для культивирования, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста клеток. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза или сахароза и другие органические кислоты. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Выращивание может осуществляться в аэробных условиях, предпочтительно с повышенным содержанием СО2 (8%), таких как перемешивание культуральной жидкости в колбах, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. Обычно, выращивание в течение от 12 часов до 4 дней приводит к накоплению целевого рекомбинантного белка в культуральной среде или в цитоплазме клетки.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования, а затем целевой белок может быть выделен и очищен методами хроматографии и/или концентрирования.
Особенности настоящего изобретения и результаты его практического применения приведены на следующих Фигурах и Примерах.
Краткое описание Фигур.
На Фигуре 1 показана карта экспрессионной плазмиды p1.2-Zeo-CHO-VKORC1 (длина 12318 пар оснований). Используются следующие обозначения: pUC origin - область начала репликации плазмиды pUC; bla - открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; bla promoter - прокариотический промотор гена bla; EBVTR -участок терминального повтора вируса Эпштейна-Барр человека; СНО EEF1A UFR функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, 5′ нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; transcription start - точка начала транскрипции, Kozak - область, кодирующая последовательность Козак для кэп-зависимой инициации трансляции, СНО VKORC ORF - ОРС гена VKORC1 китайского хомячка, stop - стоп-кодон, СНО EEF1A DFR функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, 3′ нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; SV40 early promoter -промотор ранних генов вируса SV40, Zeo(R)- ген устойчивости к селекционному агенту - антибиотику зеоцину, SV40 pA - сигнал полиаденилирования и терминатор вируса SV40. Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
На Фигуре 2 показана карта экспрессионной плазмиды p1.2-Zeo-hVKORC1 (длина 12324 пар оснований). Обозначения аналогично Фигуре 1. hVKORC1 ORF - ОРС гена VKORC1 человека.
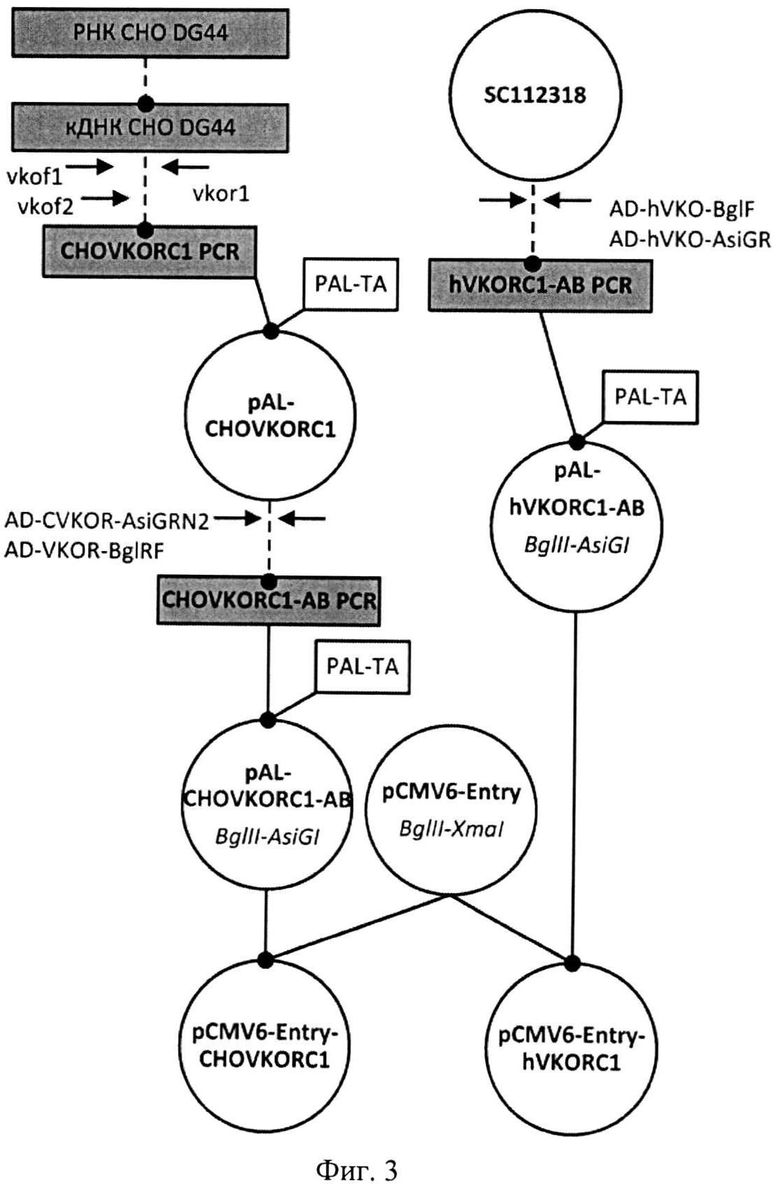
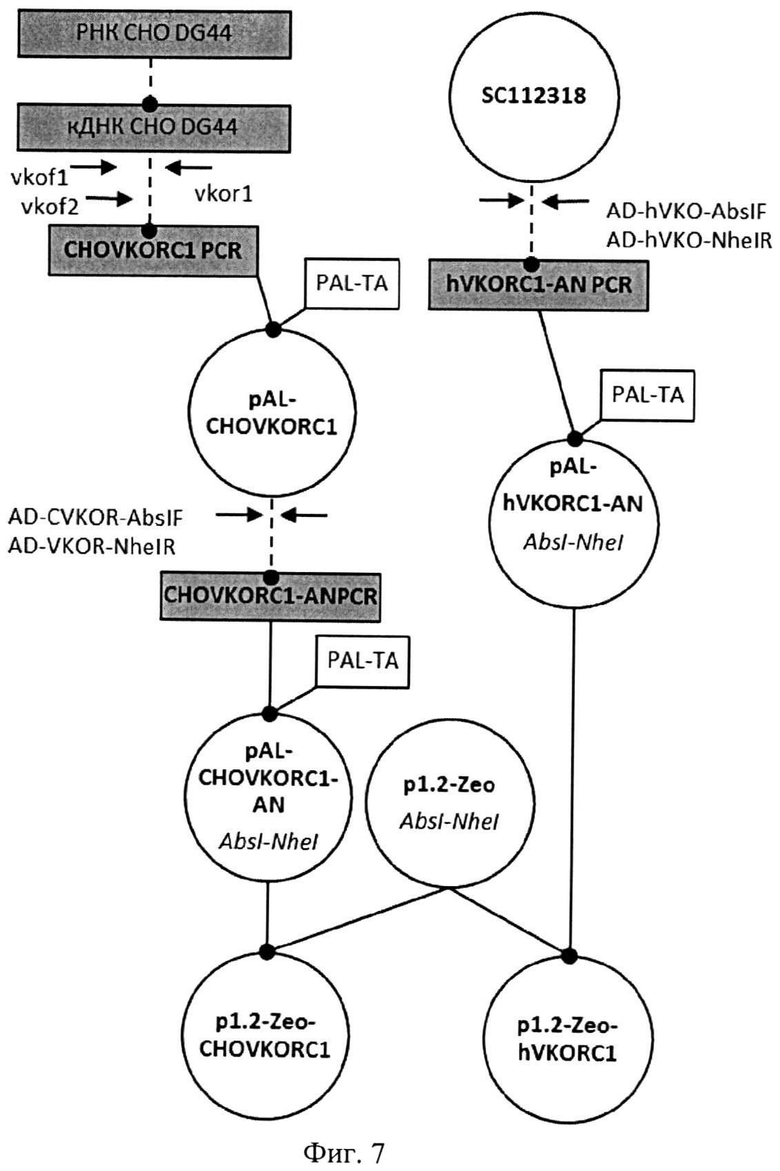
На Фигуре 3 показана схема получения экспрессионных плазмид pCMV6-Entry-CHO-VKORC1 и pCMV6-Entry-hVKORC1. Используются следующие обозначения: Линейные фрагменты ДНК изображены в виде прямоугольников, плазмиды - кругов. Пунктирной линией обозначены стадии ПЦР, сплошной линией обозначены стадии рестрикции-лигирования, названия эндонуклеаз рестрикции, использовавшихся для клонирования, указаны под названием плазмид курсивом, названия олигонуклеотидных праймеров показаны прямым шрифтом.
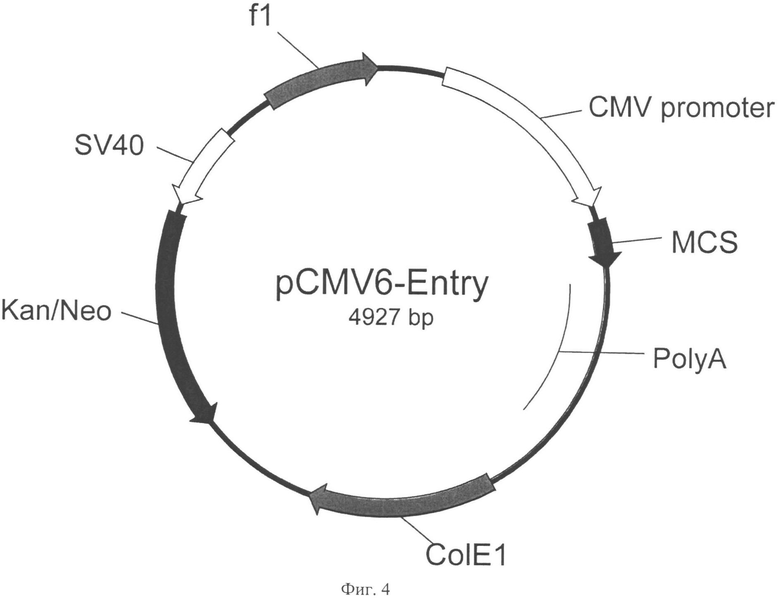
На Фигуре 4 показана карта экспрессионного вектора pCMV6-Entry (длина 4927 пар оснований). Используются следующие обозначения: CMV-promoter - цитомегаловирусный промотор, polyA - сигнал полиаденилирования гормона роста быка, ColE1 - бактериальная область начала репликации, f1 - область начала репликации бактериофага f1, Kan/Neo - последовательность, кодирующая аминогликозид-3'-фосфотрансферазу с бактериальным промотором, обеспечивающая устойчивость бактерий к канамицину, SV40 - ранний промотор полиомавируса SV40, обеспечивающий экспрессию аминогликозид-3'-фосфотрансферазы в эукариотических клетках, что позволяет вести их селекцию антибиотиком G418. Направления транскрипции генов указаны стрелками.
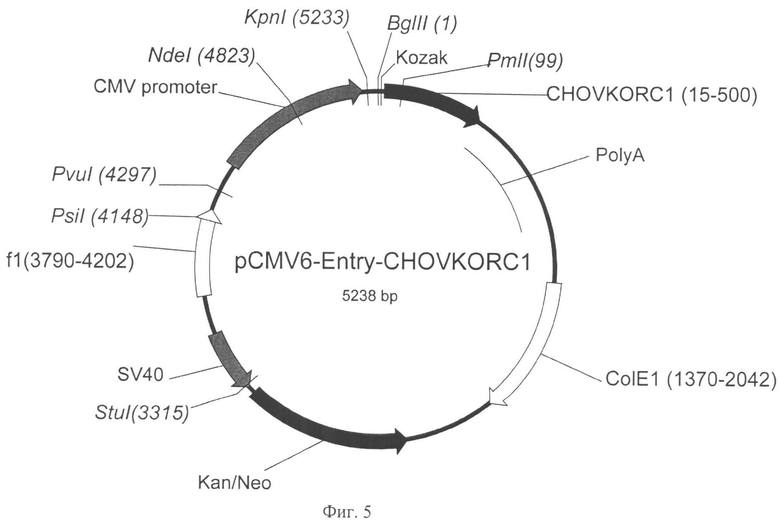
На Фигуре 5 показана карта экспрессионной плазмиды pCMV6-Entry-CHO-VKORC1 (длина 5238 пар оснований). Обозначения аналогично фигуре 4, а также: CHOVKORC1 - ОРС гена VKORC1 китайского хомячка, Kozak - последовательность Козак, обеспечивающая инициацию трансляции в клетках эукариот. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
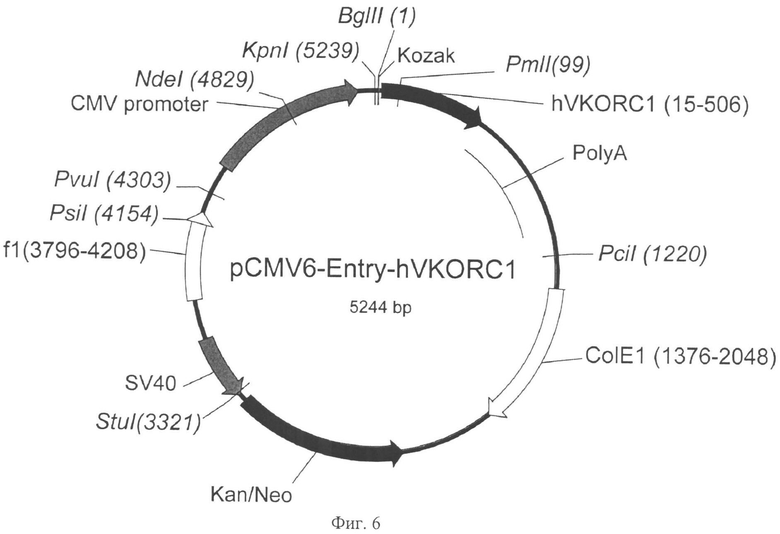
На Фигуре 6 показана карта экспрессионной плазмиды pCMV6-Entry-h VKORC1 (длина 5244 пар оснований). Обозначения аналогично Фигуре 5. hVKORC1 - ОРС гена VKORC1 человека.
На Фигуре 7 показана схема получения экспрессионных плазмид p1.2-Zeo-CHO-VKORC1 и р1.2-Zeo-hVKORC1. Обозначения аналогично Фигуре 3
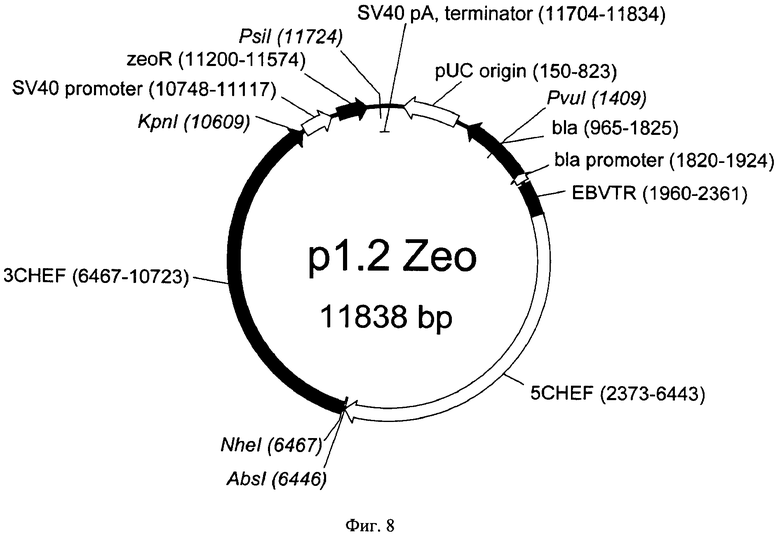
На Фигуре 8 показана карта экспрессионного вектора p1.2-Zeo (длина 11838 пар оснований). Используются следующие обозначения: pUC origin - область начала репликации плазмиды pUC; bla - открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; bla promoter - прокариотический промотер гена bla; EBVTR - участок терминального повтора вируса Эпштейн-Барр человека (EBVTR); 5CHEF функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5' НТО этого гена; 3CHEF функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3' НТО этого гена; SV40 promoter - область промотора вируса SV40; SV40 рА, terminator - терминатор и сигнал полиаденилирования вируса SV40; zeoR - ген устойчивости к зеоцину. Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
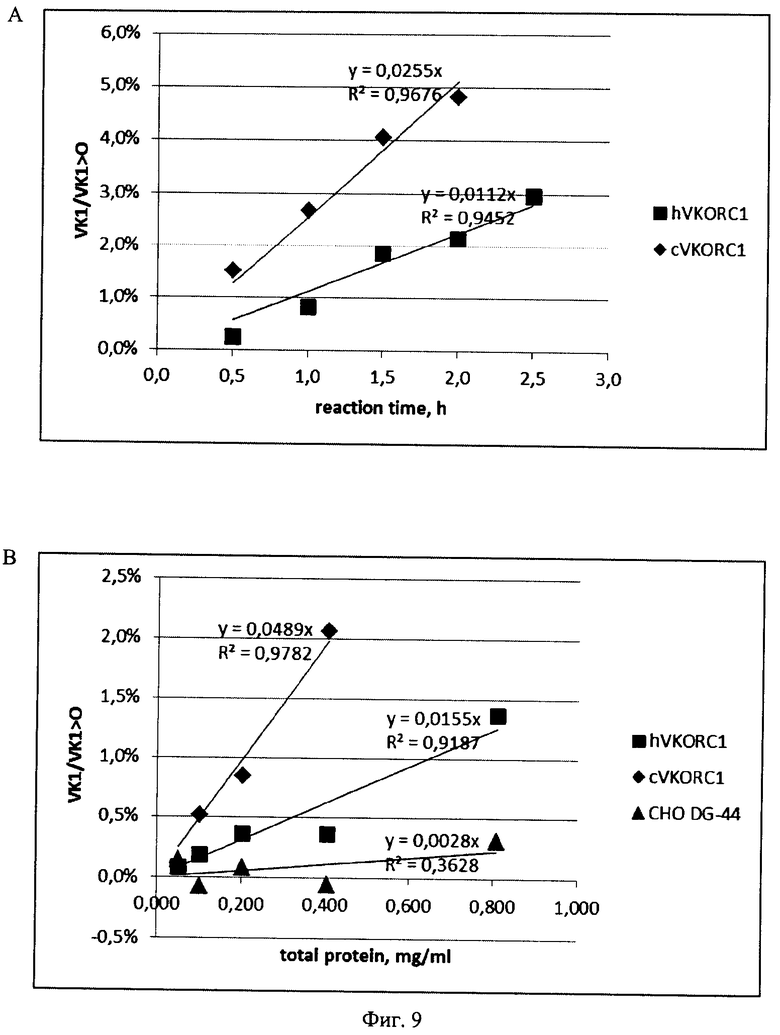
На Фигуре 9 показаны зависимости уровней восстановления эпоксида витамина K1 (VK1>0) от концентрации лизатов (А) и времени реакции(В). Концентрация общего белка для зависимости уровня превращения VK1>0 от времени реакции - 0,81 мг/мл, время реакции для зависимости уровня превращения VK1>0 от концентрации общего белка в лизатах - 3 ч. Все уровни восстановления скорректированы на величину фонового уровня восстановления VK1>0 в отсутствие лизатов. Обозначения: с VKORC1 - лизат клеток, стабильно трансфицированных плазидой p1.2-Zeo-CHO-VKORCl; hVKORC1 - лизат клеток, стабильно трансфицированных плазидой p1.2-Zeo-h VKORC1; СНО DG-44 - лизат нетрансфицированных клеток линии СНО DG-44.
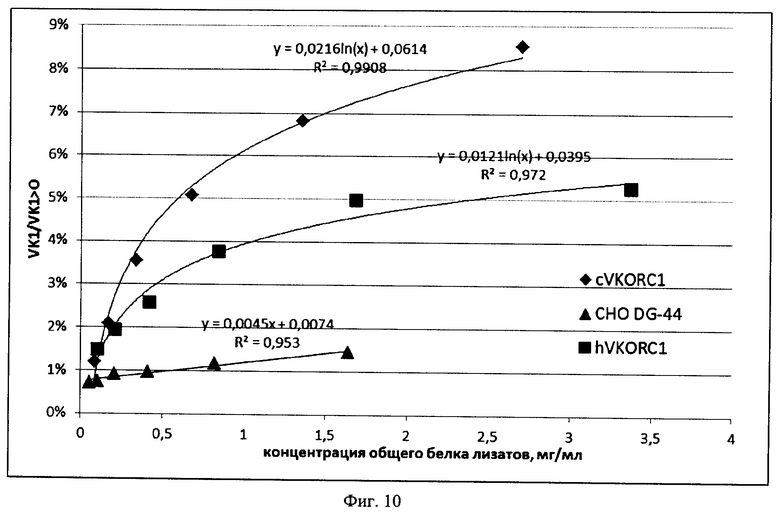
На Фигуре 10 показана оценка максимальной скорости реакции восстановления эпоксида витамина K1 (VK1>0), катализируемой VKORC1 китайского хомячка и человека. Время реакции - 3 ч. Обозначения аналогично Фиг.9.
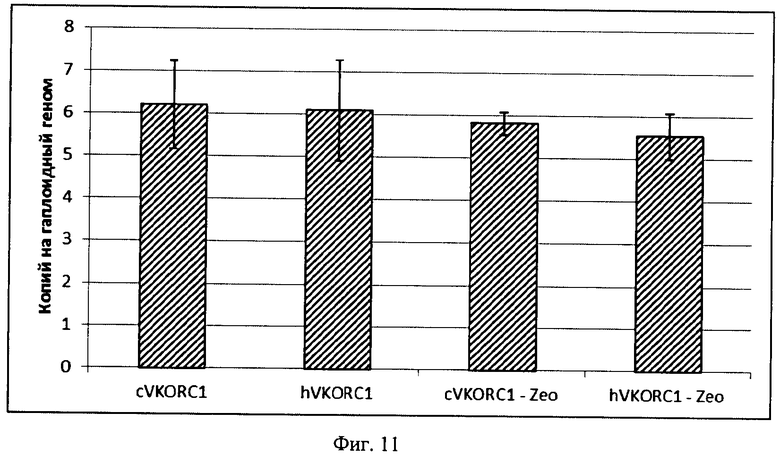
На Фигуре 11 показаны результаты определения числа копий экспрессионных кассет, содержащих ОРС VKORC1 китайского хомячка и человека. Копийность генов нормализована на копийность опорного гена китайского хомячка PPIB с известной копийностью 1. Планкой погрешности обозначены стандартные отклонения, не менее трех параллельных определений для каждого образца. Относительная погрешность определения числа копий опорного гена не более 30% для всех образцов. Обозначения: Zeo - область ОРС маркера устойчивости к действию зеоцина, остальные обозначения аналогично Фиг.9.
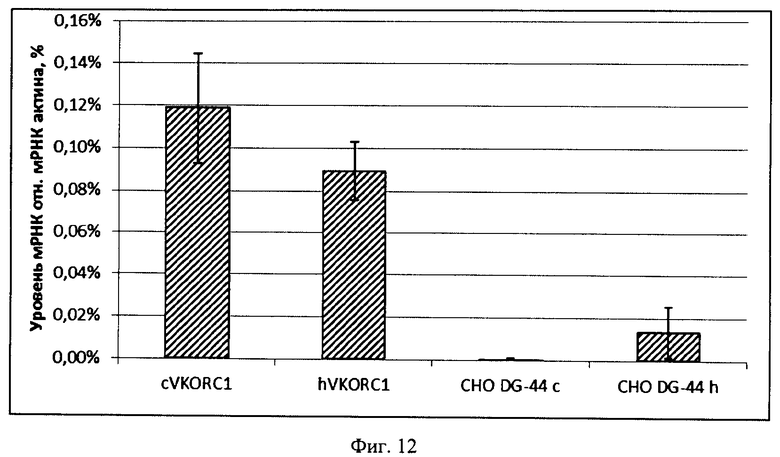
На Фигуре 12 показаны относительные уровни мРНК оверэкспрессированных ОРС VKORC1 китайского хомячка и человека, нормализованные на уровень мРНК бета-актина. Планкой погрешности обозначены стандартные отклонения, не менее трех параллельных определений для каждого образца. Относительная погрешность определения уровня мРНК бета-актина не более 30% для всех образцов. Обозначения; "СНО DG-44 с" - амплификация при помощи праймеров к искусственному гену VKORC1 китайского хомячка; "СНО DG-44 h" - амплификация при помощи праймеров к ОРС VKORC1 человека, комплементарных ОРС эндогенной мРНК VKORC1 китайского хомячка и ОРС VKORC1 человека, остальные обозначения аналогично Фиг.9.
Существо и промышленная применимость изобретения в биотехнологии иллюстрируются следующими примерами.
Пример 1. Получение плазмидной ДНК pAL-CHOVKORC1.
Из замороженной биомассы, содержащей 2 млн клеток CHO-DG44, реагентом TRI Reagent (MRC) была выделена РНК, растворена в воде с 0.5 мкг/мкл проверена на 1,2% ТАЕ агарозном геле. Синтез кДНК проводили набором MINT по протоколу производителя, используя 2 мкл РНК матрицы. Реакционная смесь 10 мкл была разбавлена в 2 раза водой и использована для амплификации кДНК набором Encycio PCR Kit по протоколу производителя. Состав реакционной смеси (10×PCR-буфер - 5 мкл; Encycio полимераза- 1 мкл, dNTP (по 10 мкМ каждого) - 1 мкл, M1-праймер (SEQ ID NO:7) - 10 мкM 2 мкл, первая цепь cDNA, разбавленная в 2 раза - 2 мкл, вода до 50 мкл). Режим амплификации: 95°С - 1 мин и 17 циклов: 95°С - 20 с/66°С - 15 с/72°С - 3 мин. кДНК очищали на колонке, элюировали 50 мкл 5 мМ tris-HCl pH 8.0, концентрация составила 25 нг/мкл. Амплифицированную кДНК перед использованием прогревали 65°С 1 мин, перемешивали, аликвоту разбавляли водой в 20 раз, использовали 1 мкл разбавленной кДНК на 25 мкл ПЦР-смеси. Вели ПЦР, используя праймеры vkof1, vkof2, vkor1 (SEQ ID NO:8-10).
ПЦР проводили при помощи смеси Tersus polymerase mix или Encycio PCR Kit no (ЗАО «Евроген») инструкции производителя на приборе РТС-100 Thermal Cycler («MJ Reseach», США). В параллельных экспериментах использовали реакционную смесь, содержавшую дополнительно 5% DMSO и не содержавшую DMSO. Режим амплификации: 1) 95°С - 3 мин и 39 циклов: 95°С - 20 с/градиент 52-65°С - 20 с/72°С -1 мин, финальная элонгация 72°С - 5 мин; 2) 1) 95°С - 3 мин, 30 циклов: 95°С - 20 с/ инкремент от 65°С с шагом 0,5°С/цикл - 20 с/72°С - 1 мин, 10 циклов: 95°С - 20 с/ 54°С - 20 с/72°С -1 мин, финальная элонгация 72°С - 5 мин.
Продукты ПЦР выделяли из 1% агарозного геля, используя набор реактивов «Wizard SV Gel and PCR Clean-Up System» («Promega», США) по протоколу производителя, после чего лигировали в Т-вектор PAL-TA («Евроген», Россия) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора («Fermentas», Литва). Лигирование вели в объеме 10 мкл при молярном соотношении вектора и вставки 1:10, в течение 2-20 часов при комнатной температуре. Полученными лигазными смесями трансформировали клетки Е. coli штамма DH5α, с генотипом F-φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 end A1 hsdR17 (rk-, mk+) phoA supE44 λ- thi-1 gyrA96 relA1. Для этого к 200 мкл замороженной суспензии клеток Е. coli добавляли 5 мкл лигазной смеси, инкубировали на льду 20 минут для сорбции плазмидной ДНК, нагревали до 42С в течение 45 секунд и инкубировали на льду в течение 5 минут. После чего добавляли 800 мкл питательного бульона SOB и инкубировали при 37°С в течение 60 минут. Затем переносили суспензию на чашку Петри с твердой агаризованной средой, содержащей ампициллин в концентрации 100 мкг/1 мл агара, и помещали в термостат на 18 часов при 37С. Колонии Е. coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов с праймеров SP6 и T7prom (SEQ ID NO:11-12). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Amp и проводили выделение плазмидной ДНК набором реактивов «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя, первичную нуклеотидную последовательность области вставки полученной плазмиды pAL-CHOVKORC 1 определяли методом ПЦР-секвенирования с использованием праймеров SP6 и T7prom (SEQ ID NO:11-12).
Последовательность ОРС гена, кодирующего фермент VKORC 1 клеток китайского хомячка DG44 согласно настоящему изобретению, представлена в Перечне последовательностей под номером SEQ ID NO:1. Последовательность депонирована 03.04.2012 заявителем в GenBank JQ400047.1 Аминокислотная последовательность фермента VKORC 1 клеток китайского хомячка DG44 согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO:2. Последовательность ОРС белка депонирована заявителем в GenBank (03-APR-2012) AFG26681.1.
Полученную плазмиду pAL-CHOVKORC 1, содержащую ОРС гена VKORC 1 клеток китайского хомячка DG44, использовали для получения экспрессионных конструкций.
Пример 2. Получение экспрессионной плазмидной ДНК pCMV6-Entry-CHO VKORC1.
При помощи адапторных праймеров AD-VKOR-BglRF и AD-CVKOR-AsiGRN2 (SEQ ID NO:13-14) и pAL-CHOVKORC1 в качестве матрицы получили ПЦР-продукт, содержащий ОРС VKORC1 китайского хомячка с добавленными рестриктными сайтами для субклонирования. Полученный продукт ПЦР клонировали в вектор PAL-TA как описано в примере 1 с образованием pAL-CHOVKORC1-АВ, анализировали методом ПЦР с клонов и секвенированием области вставки, используя праймеры SP6 и T7prom (SEQ ID NO:11-12).
Плазмиду pAL-CHOVKORC1-AB с подтвержденной первичной нуклеотидной последовательностью рестрицировали BglII. и AsiGI (изошизомер AgeI) переносили фрагмент, содержащий ОРС в вектор pCMV6-Entry (Кат №PS 100001 Origene, США, Фиг.4), рестрицированный BglII-XmaI, с образованием pCMV6-Entry-CHOVKORC1 (Фиг.5, SEQ ID NO:15). Для этого продукты рестрикции разделяли в 1% агарозном геле и выделяли набором реактивов Wizard SV Gel&PCR Clean-Up System no методике производителя. Реакцию лигирования очищенных фрагментов и трансформацию лигатами клеток Е. со/г штамма DHSa вели как описано в примере 1. Трансформантов высевали на агаризованную среду с канамицином (30 мкг/1 мл агара). Е. coli анализировали методом ПЦР с клонов с праймеров AD-VKOR-BglRF и AD-CVKOR-AsiGRN2(SEQ ID NO:13-14). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan (30 мкг/мл), проводили выделение плазмидной ДНК набором «Wizard Plus SV Minipreps». Нуклеотидную последовательность верифицировали при помощи ПЦР-секвенирования с праймеров AD-VKOR-BglRF и AD-CVKOR-AsiGRN2 (SEQ ID NO:13-14).
Плазмидную ДНК pCMV6-Entry-CHOVKORCl (SEQ ID NO:15) для трансфекции нарабатывали в клетках Е. соli (штамм Stbl4, #11635018 «Invitrogen»). Клетки культивировали в 0.5 л среды ТВ-Kan (30 мкг/мл) 18 ч, и выделяли плазмиды набором EndoFree Plasmid MaxiKit («Qiagen», США) по протоколу производителя.
Полученную экспрессионную плазмиду pCMV6-Entry-CHOVKORC1, содержащую ОРС гена VKORC 1 клеток китайского хомячка DG44, использовали для получения стабильно трансфицированных поликлональных линий клеток СНО.
Пример 3. Получение экспрессионной плазмидной ДНК pCMV6-Entry-hVKORC1.
При помощи праймеров AD-hVKO-BglF и AD-hVKO-AsiGR (SEQ ID NO:16-17) и Origene clone SC 112318 (Origene, США), соответствующему кДНК VKORC 1 человека (GenBank ID NM_024006.4), в качестве матрицы получили ПЦР-продукт, содержащий ОРС VKORC 1 человека человека с добавленными рестриктными сайтами для субклонирования. Полученный продукт ПЦР клонировали в вектор PAL-TA как описано в примере 1, с образованием pAL-hVKORC1-AB, анализировали методом ПЦР с клонов и секвенированием области вставки, используя праймеры SP6 и T7prom (SEQ ID NO:11-12).
Плазмиду pAL-h VKORC 1-АВ с подтвержденной первичной нуклеотидной последовательностью рестрицировали BglII и AsiGI (Agel) и переносили фрагмент, содержащий ОРС, в вектор pCMV6-Entry (Кат №PS 100001 Origene, США Фиг.4), рестрицированный BglII-XmaI, с образованием pCMV6-Entry-hVKORCl (Фиг.6, SEQ ID NO:18). Для этого продукты рестрикции разделяли в 1% агарозном геле и выделяли с помощью набора реактивов Wizard SV Gel&PCR Clean-Up System по методике производителя. Лигирование очищенных фрагментов и трансформацию лигатами клеток Е. coli штамма DH5α вели как описано в примере 1. Трансформантов высевали на агаризованную среду с канамицином (30 мкг/1 мл агара). Е. coli анализировали методом ПЦР с клонов с праймеров AD-hVKO-BglFn AD-hVKO-AsiGR (SEQ ID NO:16-17). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, проводили выделение плазмидной ДНК набором «Wizard Plus SV Minipreps». Нуклеотидную последовательность верифицировали при помощи ПЦР-секвенирования с праймеров AD-hVKO-BglFn AD-hVKO-AsiGR (SEQ ID NO:16-17).
Плазмидную ДНК pCMV6-Entry-hVKORC1 (SEQ ID N0:18) для трансфекции нарабатывали в клетках E. coli (штамм Stbl4, #11635018 «Invitrogen»). Клетки культивировали в 0.5 л среды ТВ-Кап (30 мкг/мл) в течение 18 ч, и выделяли плазмиды набором EndoFree Plasmid MaxiKit («Qiagen», США) по протоколу производителя.
Полученную экспрессионную плазмиду pCMV6-Entry-hVKORC1, содержащую ОРС гена VKORC 1 человека, использовали для получения стабильно трансфицированных поликлональных линий клеток СНО.
Пример 4. Получение стабильно трансфицированных экспрессионными плазмидами pCMV6-Entry-CHOVKORC1 и pCMV6-Entry-hVKORC1 поликлональных линий клеток СНО.
Клетки эпителия яичника китайского хомячка (СНО), линия DG-44 (Invitrogen, США), были трансфицированы очищенными суперскрученными плазмидами pCMV6-Entry-CHOVKORCl (SEQ ID N0:15), pCMV6-Entry-hVKORC1 (SEQ ID NO:18) и pEGFP-N2 (Clontech Кат. №6081-1, последовательность в GenBank U57608); плазмиду pEGFP-N2, кодирующую зеленый флюоресцентный белок, использовали в качестве отрицательного контроля. Был использован трансфекционный реагент FugeneHD (Promega, США) и соотношение 15 млн клеток/60 мкг ДНК/180 мкл FugeneHD/30 мл культивационного объема. Смесь ДНК и трансфекционного реагента готовили по инструкции производителя трансфекционного реагента и добавляли в суспензионную культуру клеток в указанном выше соотношении, одинаковом для обеих плазмид. Через 48 часов культивации трансфицированных клеток в перемешиваемых колбах культуральную среду заменили на селективную среду, содержавшую 750 мкг/мл антибиотика G418 (Sigma-Aldrich, США) и меняли ее в дальнейшем каждые 3 дня центрифугированием/ресуспендированием осадка клеток. При достижении культурами плотности 1,2 млн кл./мл и доле жизнеспособных клеток более 70% считали культуры установившимися, снижали концентрацию G418 до 250 мкг/мл и проводили отбор образцов клеточной массы для анализа.
Пример 5. Определение удельной активности вариантов VKORC1 китайского хомяка и человека, находящихся под контролем промотора CMV.
Измерение активности проводили по модифицированной методике [Rost S, Fregin A, Ivaskevicius V, Conzelmann E, Hortnagel К, Pelz HJ, Lappegard К, Seifried E, Scharrer I, Tuddenham EG, Muller CR, Strom TM, Oldenburg J. Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2. Nature. 2004 Feb 5; 427(6974):537-4L].
2,3-эпоксид витамина K1 синтезировали из хинона витамина K1 (Sigma Aldrich, США) окислением перекисью водорода и очищали согласно [Max Tishler, Louis F. Fieser, and Norman L. Wendler. Hydro, Oxido and Other Derivatives of Vitamin K1 and Related Compounds. Journal of the American Chemical Society 1940 62 (10), 2866-2871].
Для измерения активности готовили раствор для лизиса клеток состава 25 мМ имидазола рН=7.6, 0.5% CHAPS, 50% глицерина, 5 мМ бензамидина и раствор для проведения ферментативной реакции состава: 25 мМ имидазола рН=7.6, 0.5% CHAPS, 5 мМ CaCl2, 5 мМ DTT, 8 мкМ 2,3-эпоксида витамина K1. Проводили лизис образцов, содержащих по 5 млн клеток, в объеме 500 мкл раствора для лизиса на 1 пробирку, ресуспендировали клетки до исчезновения видимых глазом частиц, инкубировали 30 мин на льду и разрушали клетки при помощи гомогенизатора Даунса. Лизат осветляли центрифугированием 2 мин на 4 тыс. об/мин, отбирали по 10 мкл супернатанта для измерения содержания общего белка по Брэдфорд, в качестве стандарта концентрации белка использовали раствор бычьего сывороточного альбумина. Готовили серию двукратных разведений полученного осветленного лизата в растворе для лизиса и отрицательный контроль - пробирку с 30 мкл раствора для лизиса. В пробирки с разведениями лизата вносили по 500 мкл полного раствора для проведения ферментативной реакции, перемешивали, инкубировали 1 ч в термоблоке при температуре 30°С. После этого вели экстракцию, внося по 660 мкл раствора для экстракции состава изопропанол:гексан (3:2). Из верхней фазы отбирали по 450 мкл в новые пробирки, экстракт полностью выпаривали в вакуумной центрифуге при комнатной температуре, вносили в пробирки по 30 мкл метанола, перемешивали на вортексе и разделяли формы витамина K1 при помощи обращенно-фазовой хроматографии. Использовали колонку Symmetry С 18 75×4.6 mm (Waters, США), метанол в качестве подвижной фазы, скорость потока 1 мл/мин, длина волны детекции - 254 нм, температура колонки - 30°С, объем инъекции - 20 мкл. Для калибровки хроматографической системы и в качестве стандарта количества вещества использовали растворы 2,3-эпоксида и хинона витамина K1 в метаноле. Результаты представлены в таблице 1.
Выводы: при использовании плазмид с промотором CMV ферментативная активность VKORC1 в стабильно трансфицированных клетках практически не изменяется как для VKORC1 человека, так и для VKORC1 китайского хомячка.
Пример 6. Получение экспрессионных плазмидных ДНК p1.2-Zeo-CHO-VKORC1 и p1.2-Zeo-hVKORC1.
При помощи адапторных праймеров AD-CVKO-AbsIF и AD-CVKO-NheIR (SEQ ID NO:19-20) и pAL-СНО VKORC1 в качестве матрицы получили ПЦР-продукт, содержащий ОРС VKORC1 китайского хомячка с добавленными рестриктными сайтами для субклонирования. Полученный продукт ПЦР клонировали в вектор PAL-TA как описано в примере 1 с образованием pAL-CHOVKORC1-AN, анализировали методом ПЦР с клонов и секвенированием области вставки, используя праймеры SP6 и T7prom (SEQ ID N0:11-12).
Полученную плазмиду pAL-CHOVKORC1-AN с подтвержденной первичной нуклеотидной последовательностью рестрицировали AbsI и NheI переносили фрагмент, содержащий ОРС в вектор p1.2Zeo (OOO "ЛМБТ", заявка на патент RU2013122048, Фиг.8), рестрицированный AbsI и NheI, с образованием p1.2Zeo-CHOVKORC1 (Фиг.1, SEQ ID NO:5). Для этого продукты рестрикции разделяли в 1% агарозном геле и выделяли набором «Wizard SV Gel&PCR Clean-Up System». Реакцию лигирования очищенных фрагментов и трансформацию лигатами клеток Е. coli штамма DH5α вели как описано в примере 1. Бактерии после трансформации высевали на агаризованную среду с карбеницилином (50 мкг/1 мл агара). Е. coli анализировали методом ПЦР с клонов с праймеров AD-CVKO-AbsIF и AD-CVKO-NheIR (SEQ ID NO:19-20). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Carb (50 мкг/мл), проводили выделение плазмидной ДНК набором «Wizard Plus SV Minipreps». Нуклеотидную последовательность верифицировали при помощи ПЦР-секвенирования с праймеров AD-CVKO-AbsIF и AD-CVKO-NheIR (SEQ ID NO:19-20).
При помощи праймеров AD-hVKO-AbsIF и AD-hVKO-NheIR (SEQ ID NO:21-22) и плазмиды Origene clone SC 112318 (Origene, США), кодирующей кДНК VKORC1 человека (GenBank ID NM_024006.4) в качестве матрицы получили ПЦР-продукт, содержащий ОРС VKORC1 человека с рестриктными сайтами для субклонирования. Полученный продукт ПЦР клонировали в вектор PAL-TA как описано в примере 1, с образованием pAL-hVKORC1-AN, анализировали методом ПЦР с клонов и секвенированием области вставки, используя праймеры SP6 и T7prom (SEQ ID NO:11-12).
Плазмиду pAL-hVKORC1-AN с подтвержденной первичной нуклеотидной последовательностью рестрицировали AbsI и NheI, переносили фрагмент, содержащий ОРС в вектор pl.2Zeo (ЛМБТ, заявка на патент RU2013122048, Фиг.8), рестрицированный AbsI и NheI, с образованием p1.2Zeo-hVKORC1 (Фиг.2, SEQ ID NO:6). Для этого продукты рестрикции разделяли в 1% агарозном геле, выделяли набором «Wizard SV Gel&PCR Clean-Up System». Реакцию лигирования очищенных фрагментов и трансформацию лигатами клеток Е. coli штамма DH5a. вели, как описано в примере 1. Бактерии после трансформации высевали на агаризованную среду с канамицином (30 мкг/1 мл агара). Е. coli анализировали методом ПЦР с клонов с праймеров AD-hVKO-AbsIF и AD-hVKO-NheIR (SEQ Ю N0:21-22). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, проводили выделение плазмидной ДНК набором «Wizard Plus SV Minipreps». Нуклеотидную последовательность верифицировали при помощи ПЦР-секвенирования с праймеров AD-hVKO-AbsIF и AD-hVKO-NheIR (SEQ ID NO:21-22).
Плазмидные ДНК p1.2Zeo-CHOVKORC1 и p1.2Zeo-hVKORC1 для трансфекции нарабатывали в клетках E. coli (штамм Stbl4, #11635018 «Invitrogen»). Клетки культивировали в 0.5 л среды TB-Carb (50 мкг/ мл) в течение 18 ч, и выделяли плазмиды с использованием набора EndoFree Plasmid MaxiKit («Qiagen», США) по протоколу производителя.
Полученные экспрессионные плазмиды p1.2Zeo-CHOVKORC1 (SEQ ID NO:5) и p1.2Zeo-hVKORCl (SEQ ID NO:6), содержащие соответственно ОРС гена VKORC1 китайского хомячка DG44 и ОРС гена VKORC1 человека, использовали для получения стабильно трансфицированных поликлональных популяций клеток СНО DG-44.
Пример 7. Получение стабильно трансфицированных экспрессионными плазмидами р1.2-Zeo-CHO-VKORC1 и p1.2-Zeo-hVKORC1 поликлональных популяций клеток СНО DG-44, измерение активности VKORC1.
Трансфекцию клеток линии СНО DG-44 очищенными плазмидами p1.2-Zeo-CHO-VKORC1 и p1.2-Zeo-hVKORC1, кодирующими VKORC1 китайского хомячка и человека, соответственно, проводили, как указано в Примере 4. Получали стабильно трансфицированные культуры при помощи культивации в присутствии 400 мкг/мл антибиотика зеоцин до восстановления жизнеспособности клеток выше 85%. Измерение активности VKORC1 в лизате клеток проводили, как указано в Примере 5, с изменениями - выравнивали концентрацию общего белка в лизатах перед добавлением субстрата реакции и использовали несколько двукратных разведений лизата исследуемых клеток в буферном растворе для лизиса. Реакцию вели 3 ч, отбирали пробы с интервалом 30 мин. Результаты измерения активности VKORC1 при времени реакции 3 ч приведены в Таблице 2, зависимости скоростей восстановления эпоксида витамина K1 (VK1>0) от концентрации лизатов и времени реакции приведены на Фиг.9. Максимальную скорость VKORC1 китайского хомячка и человека оценивали, проводя реакцию восстановления VK1>0 с максимальными концентрациями лизатов клеток, результаты приведены на Фиг.10.
Выводы: Уровень ферментативной активности VKORC1 может быть повышен в 24 раза при оверэкспрессии аутологичного VKORC1 китайского хомячка и только в 8 раз при оверэкспрессии VKORC1 человека. Оба ортолога VKORC1 практически не подвержены термоинактивации или протеолитическому распаду в течение 3-х часов. Скорость восстановления VK1>0 линейно зависит от концентрации обоих ферментов в выбранном диапазоне концентраций общего белка в лизатах. Максимальная скорость реакции, развиваемая в присутствии избытка VKORC1 китайского хомячка приблизительно вдвое выше максимальной скорости реакции, катализируемой избытком VKORC1 человека.
Пример 8. Измерение числа копий экспрессионных плазмид p1.2-Zeo-CHO-VKORC1 и р1.2-Zeo-hVKORC1, введенных в геном клеток и уровней мРНК VKORC1 китайского хомячка и человека в трансфицированных клетках.
Количественный анализ копийности экспрессионной кассеты в геноме и уровня мРНК проводили методом ПНР в реальном времени (ПЦР-РВ) с использованием амплификатора iCycler iQ (Bio-Rad, США). Для ПЦР использовали готовую 5х реакционную смесь с горячим стартом, содержащую интеркалирующий краситель SYBR Green I, qPCRmix-HS SYBR («Евроген», Россия). Каждую реакцию повторяли 3 раза в объеме 25 мкл, в 3-5 повторах.
Геномную ДНК выделяли из 0,5-5 млн клеток набором «Wizard SV Genomic DNA Purification System» (Promega, США) по инструкции производителя. Суммарную РНК выделяли из 0,5-5 млн клеток набором «RNeasy Mini Kit» (Qiagen, США) по инструкции производителя. Для приготовления образцов кДНК использовали по 1 мкг суммарной РНК и набор реактивов «Mint» (Евроген, Россия).
Для определения копийности экспрессионной кассеты в геноме в качестве матрицы использовали геномную ДНК и олигонуклеотидные праймеры, не гомологичные геномным последовательностям китайского хомячка. Для VKORC1 СНО использовали праймеры к С-концевой области ОРС VKORC1 СНО и нетранслируемой области экспрессионной плазмиды р1.2-Zeo-CHO-VKORC1 - RT-cVKOspC-F и RT-cVKOspC-R (SEQ ID NO:23-24). Для VKORC1 человека использовали уникальные праймеры к гену человека - RT-hVKOR-f RT-hVKOR-r (SEQ ID NO:25-26).
Для определения уровня экспрессии мРНК генов VKORC1 в качестве матрицы использовали кДНК и уникальные олигонуклеотидные праймеры, не гомологичные последовательностям китайского хомячка.
Для VKORC1 СНО использовали праймеры RT-cVKOspN-F и RT-cVKOspN-R (SEQ ID NO:27-28), комплементарные участкам двух соседних экзонов искусственного гена, кодируемого плазмидой p1.2-Zeo-CHO-VKORC1. Таким образом, короткий продукт ПЦР мог образовываться только с кДНК-матрицы p1.2-Zeo-CHO-VKORC1, но не с геномных копий p1.2-Zeo-CHO-VKORC1 или природного гена VKORC1 или природной кДНК VKORC1.
Для VKORC1 человека использовали пару праймеров RT-hVKOR-f RT-hVKOR-r (SEQ ID NO:25-26), специфичных к последовательности гена человека, кодируемой плазмидой p1.2-Zeo-hVKORC1.
Для анализа геномной ДНК клеток обеих линий использовали пару праймеров к гену устойчивости к зеоцину - RT-Zeo-F и RT-Zeo-R (SEQ ID NO:29-30).
Для амплификации опорных генов, в случае ПЦР с геномной ДНК, использовали праймеры к области гена пептидил-пролил изомеразы В (циклофилина В), предположительно уникальной для генома СНО, RT-PPIB-F и RT-PPIB-R (SEQ ID NO:31-32), а в случае ПЦР с кДНК использовали праймеры к бета-актину СНО - RT-bACT-F и RT-bACT-R (SEQ ID NO:33-34).
Копийность экспрессионной кассеты определяли из калибровочной кривой, построенной для серийных разведений экспрессионных плазмид p1.2-Zeo-CHO-VKORC1 для VKORC1 китайского хомячка и p1.2-Zeo-hVKORC1 для VKORC1 человека. Результаты ПЦР сравнивали с результатами для контрольного ампликона, представленного в геноме клеток СНО только один раз по поисковой выдаче алгоритма BLAST из базы данных NCBI Nucleotide Collection. При анализе уровней экспрессии мРНК было установлено, что эффективность ПЦР составляет не менее 99% для всех использованных пар праймеров, вычисление относительных концентраций мРНК проводили по формуле C=2^(-CT), где СТ - пороговый цикл амплификации.
Выводы: Для обеих плазмид p1.2-Zeo-CHO-VKORC1 и p1.2-Zeo-hVKORC1 число копий экспрессионных кассет в геноме клеток составляет около 6 и не различается при амплификации областей ОРС VKORC1 и ОРС гена устойчивости к зеоцину (фиг.11). Уровень мРНК VKORC1 человека и китайского хомячка различается не более, чем в 1,33 раза (фиг.12). Таким образом, повышенная ферментативная активность оверэкспрессированного VKORC1 китайского хомячка определяется собственными свойствами фермента, а не большей эффективностью трансфекции или лучшим уровнем транскрипции искусственного гена.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Список последовательностей приведен в графической части.



























Настоящее изобретение относится к области биотехнологии и может быть использовано для получения рекомбинантных белков с Gla-доменом в клетках китайского хомячка. Сконструирована плазмида для экспрессии в клетке китайского хомячка, в следующей последовательности, по существу, содержащая следующие элементы: область начала репликации плазмиды pUC; открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; прокариотический промотор гена bla; участок терминального повтора вируса Эпштейна-Барр человека; функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, 5' нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; область, кодирующая последовательность Козак для кэп-зависимой инициации трансляции; ОРС гена субъединицы 1 комплекса 2,3-эпоксиредуктазы витамина К (VKORC1) китайского хомячка со стоп-кодоном; функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, 3' нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген; промотор ранних генов вируса SV40; ген устойчивости к селекционному агенту; и сигнал полиаденилирования и терминатор вируса SV40. Полученной плазмидой трансформируют клетки китайского хомячка - продуцента белка с Gla-доменом, которые используют в способе рекомбинантного получения белков с Gla-доменом. Изобретение позволяет повысить продуктивность указанных клеток за счет повышения активности нативного VKORC1. 3 н. и 9 з.п. ф-лы, 12 ил., 2 табл., 8 пр.
1. Плазмида для экспрессии в клетке китайского хомячка, в следующей последовательности, по существу, содержащая следующие элементы:
- область начала репликации плазмиды pUC;
- открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину;
- прокариотический промотор гена bla;
- участок терминального повтора вируса Эпштейна-Барр человека;
- функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, 5′ нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген;
- область, кодирующая последовательность Козак для кэп-зависимой инициации трансляции;
- ОРС гена субъединицы 1 комплекса 2,3-эпоксиредуктазы витамина К (VKORC1) китайского хомячка со стоп-кодоном;
- функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, 3′ нетранслируемая область этого гена и нетранскрибируемая область, фланкирующая этот ген;
- промотор ранних генов вируса SV40;
- ген устойчивости к селекционному агенту; и
- сигнал полиаденилирования и терминатор вируса SV40.
2. Плазмида по п.1, отличающаяся тем, что указанная субъединица VKORC1 китайского хомячка содержит последовательность аминокислот, представленную в Перечне последовательностей под номером SEQ ID NO:2.
3. Плазмида по п.1, отличающаяся тем, что указанной ОРС гена VKORC1 китайского хомячка является фрагмент ДНК, представленный в Перечне последовательностей под номером SEQ ID NO:1
4. Плазмида по п.1, отличающаяся тем, что указанным геном устойчивости к селекционному агенту является ген устойчивости к зеоцину.
5. Плазмида по п.1, отличающаяся тем, что указанной плазмидой является плазмида p 1.2-Zeo-CHO-VKORC1, представленная в Перечне последовательностей под номером SEQ ID NO:5.
6. Клетка китайского хомячка - продуцент белка с Gla-доменом, трансформированная плазмидой по п.1.
7. Клетка по п.7, отличающаяся тем, что указанной клеткой является клетка СНО DG44.
8. Клетка по п.7, отличающаяся тем, что указанный белок с Gla-доменом выбран из группы, состоящей из факторов свертывания крови, таких как (протромбин или фактор II, факторы VII, IX, X; антикоагулянтов, таких как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранных Gla-белков (TMGPs).
9. Клетка по п.7, отличающаяся тем, что указанной плазмидой является плазмида p 1.2-Zeo-CHO-VKORC1, представленная в Перечне последовательностей под номером SEQ ID NO:5.
10. Способ получения белка с Gla-доменом, включающий стадии культивирования клетки по п.7 в питательной среде и выделения полученного белка с Gla-доменом из культуральной жидкости.
11. Способ по п.10, отличающийся тем, что культивируют клетки СНО DG44/p 1.2-Zeo-CHO-VKORC1.
12. Способ по п.10, отличающийся тем, что указанный белок с Gla-доменом выбран из группы, состоящей из факторов свертывания крови, таких как (протромбин или фактор II, факторы VII, IX, X; антикоагулянтов, таких как белок С, белок S, белок Z; остеокальцин или Gla-белок кости, матриксный Gla-белок кости (MGP); белок регулятор клеточного цикла Gas6 (growth arrest specific gene 6 protein) и трансмембранных Gla-белков (TMGPs).
| XU X | |||
| et al., The genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line, Nature Biotechnology, v.29, n.8, p | |||
| РЕГЕНЕРАТИВНЫЙ РАДИОПРИЕМНИК | 1923 |
|
SU735A1 |
| УСТРОЙСТВО ДЛЯ ИСПЫТАНИЯ АППАРАТОВ, ИМЕЮЩИХ КЛАВИАТУРУ ИЛИ КНОПКИ | 0 |
|
SU400047A1 |
| US 20060194284 A1, 31.08.2006 | |||
| WO 2006067116 A1, 29.06.2006 | |||
| WO 2006063292 A1, 15.06.2006 | |||
| RU 2004136574 A, 27.05.2006. | |||

