По настоящей заявке испрашивается приоритет заявки США, имеющей серийный № 12/612912 и поданной 5 ноября 2009 г, являющейся частичным продолжением заявки США, имеющей серийный № 12/399241 и поданной 6 марта 2009 г, являющейся продолжением заявки США, имеющей серийный № 11/233258 и поданной 22 сентября 2005 г., в настоящее время заявка США 7521541, поданная 21 апреля 2009 г., а также по пункту 35 USC § 119(e) испрашивается приоритет Предварительной патентной заявки США, имеющей серийный № 60/612468 и поданной 23 сентября 2004 г., и Предварительной патентной заявки США, имеющей серийный № 60/696353 и поданной 30 июня 2005 г., полное содержание каждой из которых приведено в настоящем документе в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится, главным образом, к сконструированным антителам с реакционноспособными остатками цистеина, и более конкретно, к антителам с терапевтическими или диагностическими применениями. Сконструированные антитела с цистеиновыми заменами можно конъюгировать с химиотерапевтическими средствами, токсинами, аффинными лигандами, такими как биотин, и метками для детекции, такими как радиоактивные изотопы и флуорофоры. Изобретение также относится к способам использования антител и соединений - конъюгатов антитело-лекарственное средство для диагностики или лечения клеток млекопитающих или связанных с ними патологических состояний in vitro, in situ и in vivo.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Получение изображений на молекулярном уровне является важным инструментом для разработки и оценки новых лекарственных средств. Иммуно-позитронная эмиссионная томография (ИммуноПЭТ) представляет собой быстро развивающийся способ отслеживания и количественной оценки моноклональных антител (mAb) in vivo, поскольку в нем эффективно скомбинирована высокая чувствительность ПЭТ с высокой чувствительностью mAb. ИммуноПЭТ приближается к тому, чтобы стать клиническим способом выбора для неинвазивной диагностики, обеспечивающим «всеобъемлющее иммуногистохимическое окрашивание in vivo» (van Dongen GA, et al. «ImmunoPET: a navigator in monoclonal antibody development and applications» Oncologist 2007;12: 1379-89). Поскольку для ИммуноПЭТ необходимо присоединение позитронно-активного радиоизотопа к намеченной специфической молекуле, необходимо, чтобы биологическое время полужизни молекулы совпадало со временем полужизни радиоактивного изотопа (Verel I, et al. «The promise of immuno-PET in radioimmunotherapy» J Nucl Med 2005; 46 Suppl 1: 164S-71S). Хотя антитела (~150 кДа) обладают временем полужизни в сыворотке в диапазоне от суток до недель, получение изображений, как правило, обеспечивает максимальные соотношения сигнала к фону через 2-6 суток после введения метки на основе антител, что требует использования радиоактивных изотопов, таких как 89Zr и 124I со временем полужизни 3,3 суток и 4,2 суток, соответственно. К сожалению, время полужизни легко доступного 64Cu (12,7 ч) является слишком коротким для получения изображений с хорошим контрастом в эти временные рамки.
Разработка средств на основе Mab для получения изображений при позитронно-эмиссионной томографии (ПЭТ) (Иммуно-ПЭТ) продолжает являться многообещающим инструментом для локализации и количественного определения молекулярных мишеней и может улучшать клиническую диагностику патологических состояний (van Dongen et al (2007) Oncologist 12;1379-89; Williams et al (2001) Cancer Biother Radiopharm 16:25-35; Holliger et al (2005) Nat Biotechnol 23: 1126-36). ПЭТ представляет собой способ получения изображений на молекулярном уровне, все больше используемый для детекции заболеваний. Системы получения изображений ПЭТ создают изображения на основе распределения позитронно-активных изотопов в ткани пациента. Изотопы, как правило, вводят пациенту посредством инъекции молекул-зондов, содержащих позитронно-активный изотоп, такой как F-18, C-11, N-13 или O-15, ковалентно присоединенный к молекуле, поддающейся легкому метаболизму или локализации в организме (например, глюкоза), или химически связывающейся с участками рецепторов внутри организма. В некоторых случаях изотоп вводят пациенту в виде ионного раствора или посредством ингаляции. Небольшие средства для получения изображения иммуно-ПЭТ, такие как Fab-фрагменты антител (50 кДа) или диатела, спаренные димеры ковалентно связанной VH-VL области Mab, 55 кДа (Shively et al (2007) J Nucl Med 48: 170-2), могут являться особенно пригодными, поскольку они обладают коротким временем полужизни, высоким проникновением в ткани, и достигают оптимального соотношения опухоли к фону между двумя и четырьмя часами после инъекции, облегчая использование изотопов с коротким временем полужизни, таких как широкодоступный 18F (109,8 мин).
Иод 124 (124I) присоединяли к антителу 3F9 и использовали для дозиметрической оценки радиммунотерапии нейробластомы (Larson SM, et al «PET scanning of iodine-124-3F9 as an approach to tumor dosimetry during treatment planning for radioimmunotherapy in a child with neuroblastoma» J Nucl Med 1992;33:2020-3). Позже, с возникновением более сложных инструментов ПЭТ и улучшенных способов мечения радиоактивным иодом, 124I использовали в многочисленных исследованиях иммуноПЭТ (Verel I, et al «High-quality 124I-labelled monoclonal antibodies for use as PET scouting agents prior to 131I-radioimmunotherapy» European journal of nuclear medicine и molecular imaging 2004; 31:1645-52; Lee FT et al «Immuno-PET of human colon xenograft-bearing BALB/c nude mice using 124I-CDR-grafted humanized A33 monoclonal antibody» J Nucl Med 2001;42:764-9; Sundaresan G, et al. «124I-labeled engineered anti-CEA minibodies and diabodies allow high-contrast, antigen-specific small-animal PET imaging of xenografts in athymic mice» J Nucl Med 2003; 44:1962-9; Jain M and Batra SK. «Genetically engineered antibody fragments and PET imaging: a new era of radioimmunodiagnosis» J Nucl Med 2003; 44:1970-2; Gonzalez Trotter DE et al. «Quantitation of small-animal (124)I activity distributions using a clinical PET/CT scanner» J Nucl Med 2004; 45:1237-44; Robinson MK, et al. «Quantitative immuno-positron emission tomography imaging of HER2-positive tumor xenografts with an iodine-124 labeled anti-HER2 diabody» Cancer Res 2005; 65:1471-8; Jayson GC et al. «Molecular imaging and biological evaluation of HuMV833 anti-VEGF antibody: implications for trial design of antiangiogenic antibodies» J Natl Cancer Inst 2002; 94:1484-93; Divgi CR, et al. «Preoperative characterisation of clear-cell renal carcinoma using iodine-124-labelled antibody chimeric G250 (124I-cG250) and PET in patients with renal masses: a phase I trial» Lancet Oncol 2007; 8:304-10). Несмотря на относительно простые способы мечения радиоактивным иодом, доступные для присоединения 124I к mAb, важные ограничения замедляют повсеместное доклиническое использование этого радиоактивного изотопа. Следует отметить, что схема распада комплекса включает позитроны с высокой энергией (β+ макс. 1,5 и 2,1 МэВ), что отрицательно влияет на разрешение микроПЭТ для мелких животных. Кроме того, интернализованные меченные иодом белки подвергаются ферментативному деиодированию с быстрым выведением свободного иодида из клеток-мишеней, что приводит к тому, что изображения ПЭТ не отражают действительного поглощения mAb «Perera RM et al. «Internalization, intracellular trafficking, and biodistribution of monoclonal antibody 806: a novel anti-epidermal growth factor receptor antibody» Neoplasia (New York, N.Y 2007;9: 1099-110). Использование 89Zr преодолевает эти недостатки, поскольку позитроны, испускаемые при распаде 89Zr (β макс. 897 кэВ), обеспечивают разрешение микроПЭТ, сравнимое с 18F и 11C (приблизительно 1 мм). Метаболиты интернализованных 89Zr-mAb отслеживают также внутри клеток в лизосомах, обеспечивая лучшую корреляцию действительного поглощения mAb с получением изображений ПЭТ (van Dongen GA, et al. «Immuno-PET: a navigator in monoclonal antibody development and applications» Oncologist 2007;12: 1379-89).
Общепринятые средства присоединения, т.е., связывания посредством ковалентных связей, метки, такой как радиоактивный изотоп, флуоресцентный краситель или группа лекарственного средства, к антителу, как правило, приводит к гетерогенной смеси молекул, где группы метки присоединены к ряду участков антитела. Например, цитотоксические средства как правило, конъюгируют с антителами через многочисленные остатки лизина в антителе, получая гетерогенную смесь конъюгата антитело-лекарственное средство. В зависимости от условий реакции, гетерогенная смесь, как правило, содержит распределение антител с присоединенными от 0 до приблизительно 8 или более групп лекарственного средства. Кроме того, в пределах каждой подгруппы конъюгатов с определенным целочисленным соотношением групп лекарственного средства к антителу, присутствует потенциально гетерогенная смесь, где группа лекарственного средства присоединена к различным участкам антитела. Аналитические и препаративные способы являются неадекватными для разделения и характеризации видов молекул конъюгата антитело-лекарственное средство внутри гетерогенной смеси, полученной в результате реакции конъюгации. Антитела представляют собой большие, сложные и структурно разнообразные биомолекулы, часто со множеством реакционноспособных функциональных групп. Их реакционная способность по отношению к линкерным реагентам и промежуточным соединениям лекарственное средство-линкер зависит от таких факторов, как pH, концентрация, концентрация соли и сорастворители. Более того, способ многоступенчатой конъюгации может являться невоспроизводимым из-за сложностей в контролировании условий реакции и характеризации реагентов и промежуточных соединений.
Тиолы цистеина являются реакционноспособными при нейтральном pH, в отличие от большинства аминов, которые являются протонированными и менее нуклеофильными при pH около 7. Поскольку свободные тиоловые (RSH, сульфгидрильные) группы являются относительно реакционноспособными, белки с остатками цистеина часто существуют в их окисленной форме в виде связанных дисульфидными мостиками олигомеров или имеют дисульфидные группы, образующие внутренние мостики. Внеклеточные белки, как правило, не имеют свободных тиолов (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London, at page 55). Количество свободного тиола в белке можно оценивать общепринятым анализом Элмана. Иммуноглобулин M является примером пентамера, связанного дисульфидными связями, в то время как иммуноглобулин G является примером белка с внутренними дисульфидными мостиками, связывающими субъединицы вместе. В белках, подобных этим, необходимо восстановление дисульфидных связей с помощью такого реагента, как дитиотреитол (DTT) или селенол (Singh et al (2002) Anal. Biochem. 304: 147-156), для получения реакционноспособного свободного тиола. Этот способ может приводить к потере третичной структуры и антигенсвязывающей специфичности антитела.
Тиоловые группы остатков цистеина антител, как правило, являются более реакционноспособными, т.е. более нуклеофильными, по отношению к электрофильным реагентам для конъюгации, чем аминогруппы или гидроксильные группы антител. Остатки цистеина вводили в белки способами генной инженерии для формирования ковалентных присоединений к лигандам или для формирования новых внутримолекулярных дисульфидных связей (Better et al (1994) J. Biol. Chem. 269(13):9644-9650; Bernhard et al (1994) Bioconjugate Chem. 5: 126-132; Greenwood et al (1994) Therapeutic Immunology 1:247-255; Tu et al (1999) Proc. Natl. Acad. Sci USA 96:4862-4867; Kanno et al (2000) J. of Biotechnology, 76:207-214; Chmura et al (2001) Proc. Nat. Acad. Sci. USA 98(15):8480-8484; US 6248564). Однако, конструирование тиоловых групп цистеина посредством мутации различных аминокислотных остатков белка до цистеиновых аминокислот потенциально является проблематичным, в частности, в случае неспаренных (свободных Cys) остатков или остатков, относительно доступных для реакции или окисления. В концентрированных растворах белка, в периплазме E. coli, культуральных супернатантах, или частично или полностью очищенном белке, неспаренные остатки Cys на поверхности белка могут спариваться и окисляться с формированием межмолекулярных дисульфидных мостиков, и таким образом, димеров или мультимеров белка. Формирование дисульфидных димеров делает новые Cys нереакционноспособными для конъюгации с лекарственным средством, лигандом или другой меткой. Более того, если в белке с помощью окисления образуется внутримолекулярная дисульфидная связь между новым сконструированным Cys и существующим остатком Cys, обе группы Cys недоступны для участия в активном участке и взаимодействий. Более того, белок может становиться неактивным или неспецифическим из-за неправильного сворачивания или потери третичной структуры (Zhang et al (2002) Anal. Biochem. 311:1-9).
Сайт-специфическая конъюгация является предпочтительной по сравнению с модификацией случайных аминогрупп, поскольку она позволяет химическую модификацию в участке, отдаленном от участка связывания, обеспечивая полное сохранение биологической активности и позволяя контроль над возможным количеством добавленных простетических групп. Сконструированные антитела с цистеиновыми заменами разработаны в форме FAB-фрагментов антител (тиоFab) и экспрессированы в форме полноразмерных, моноклональных (тиоMab) антител IgG. См.: US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008; 332:41-52; Junutula JR et al. «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol. 26:925-32, содержание которых приведено в качестве ссылки. Антитела ТиоFab и ТиоMab конъюгировали посредством линкеров на тиолах нового введенного цистеина с помощью вступающих в реакцию с тиолом линкерных реагентов и реагентов лекарственное средство-линкер для получения конъюгатов с лекарственным средством сконструированного антитела с цистеиновыми заменами (Тио ADC) с противораковыми свойствами, включая анти-MUC16 (US 2008/0311134), анти-CD22 (US 2008/0050310), анти-ROBO4 (US 2008/0247951), анти-TENB2 (US 2009/0117100), анти-CD79B (US 2009/0028856; US 2009/0068202) Тио ADC.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, где одна или несколько аминокислот исходного антитела заменены на свободную аминокислоту цистеин. Сконструированное антитело с цистеиновыми заменами содержит одну или несколько свободных цистеиновых аминокислот, обладающих значением реакционной способности тиола в диапазоне от 0,6 до 1,0. Свободная аминокислота цистеин представляет собой остаток цистеина, введенный при конструировании в исходное антитело и не являющийся частью дисульфидного мостика.
Сконструированные антитела с цистеиновыми заменами могут являться пригодными для диагностики и лечения злокачественных опухолей и включают антитела, специфические для рецепторов поверхности клеток и трансмембранных рецепторов, и для опухолеассоциированных антигенов (TAA). Такие антитела можно использовать в форме голых антител (не конъюгированных с группой лекарственного средства или метки) или в форме конъюгатов антитело-цирконий (AZC).
Варианты осуществления способов получения и скрининга сконструированных антител с цистеиновыми заменами включают варианты, где исходное антитело представляет собой фрагмент антитела, такой как hu4D5Fabv8. Исходное антитело может представлять собой также слитый белок, содержащий альбуминсвязывающую пептидную последовательность (ABP). Исходное антитело может представлять собой также гуманизированное антитело, выбранное из huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (трастузумаб).
Сконструированные антитела с цистеиновыми заменами по изобретению можно сайт-специфическим и эффективным образом присоединять к вступающему в реакцию с тиолом реагенту. Вступающий в реакцию с тиолом реагент может представлять собой содержащий радиоактивный изотоп реагент, мультифункциональный линкерный реагент, связывающий метку реагент, содержащий флуорофор реагент или промежуточное соединение лекарственное средство-линкер.
Сконструированное антитело с цистеиновыми заменами можно метить поддающейся детекции меткой, иммобилизовывать на твердофазной подложке и/или конъюгировать с группой лекарственного средства.
Другой аспект изобретения относится к меченному цирконием сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающему формулой I:

где p представляет собой 1-4.
Другой аспект изобретения относится к меченному десферриоксамином сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающему формулой II:

где L-Df выбран из:
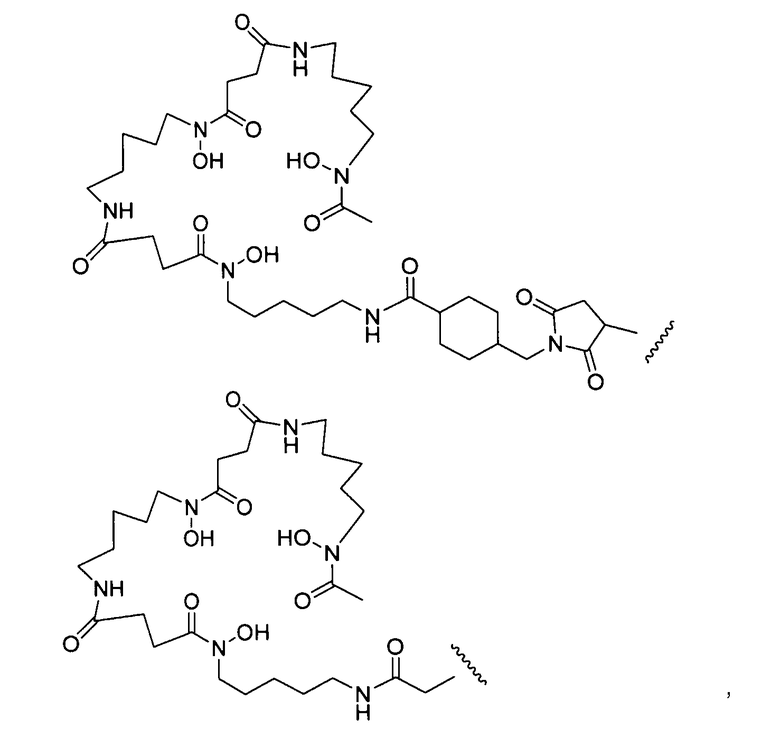
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
Другой аспект изобретения относится к реагенту для мечения десферриоксамином, выбранному из структур:
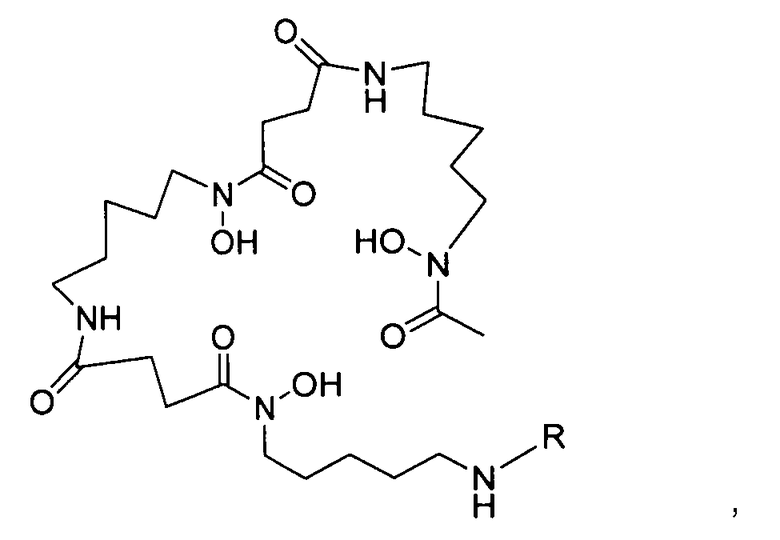
где R выбран из:
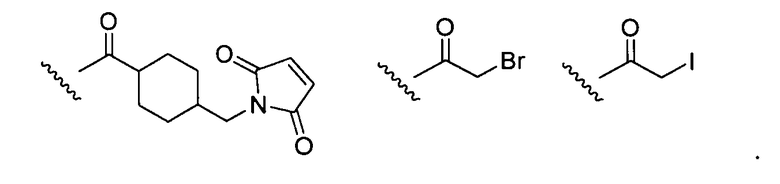 .
.
Другой аспект изобретения относится к способу получения меченного десферриоксамином сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающего формулой II:

где L-Df выбран из:
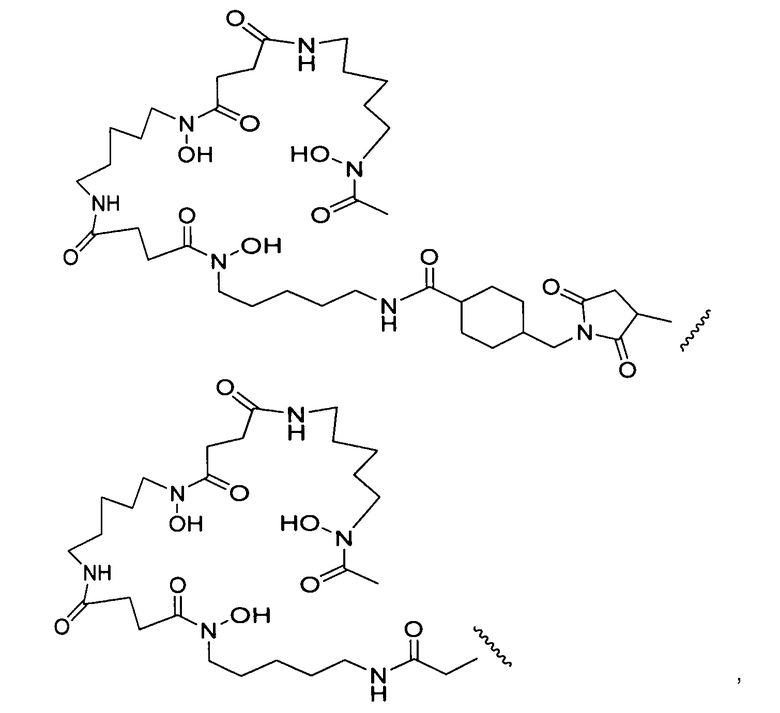
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
где способ включает реакцию композиции, выбранной из структур:
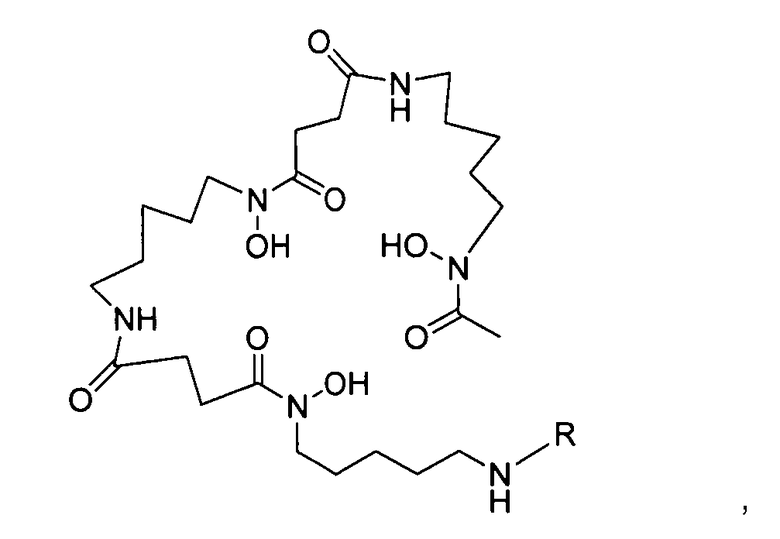
где R выбран из:
со сконструированным антителом с цистеиновыми заменами, имеющим одну или несколько свободных цистеиновых аминокислот, посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения меченного цирконием сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающего формулой I:

где p представляет собой 1-4;
где способ включает образование комплекса содержащего цирконий реагента с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, содержащим сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающим формулой II:

где L-Df выбран из:
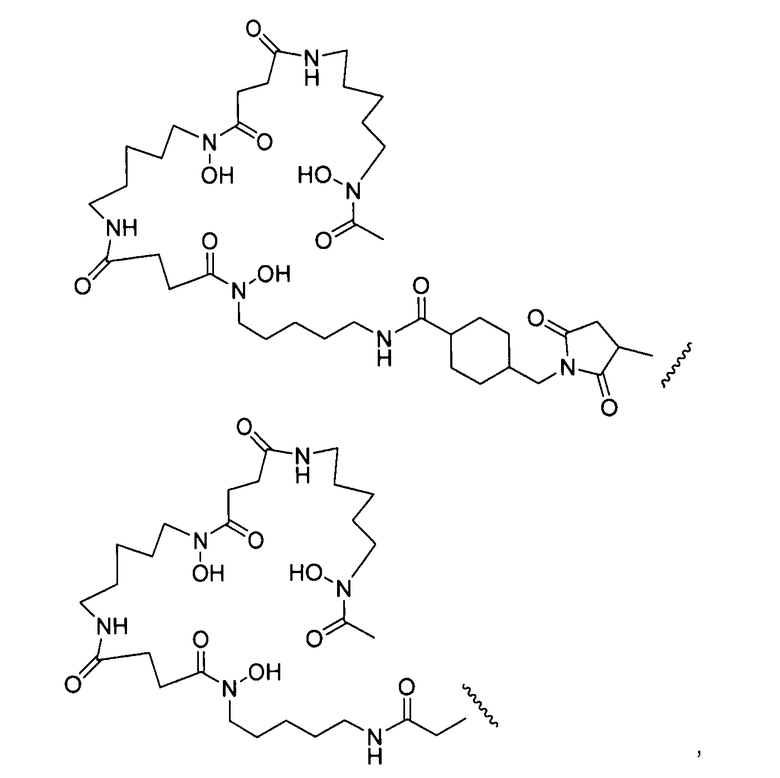
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения изображения, включающему:
введение меченного цирконием сконструированного антитела с цистеиновыми заменами животному; и
детекцию in vivo присутствия меченного цирконием сконструированного антитела с цистеиновыми заменами посредством получения изображений,
где меченное цирконием сконструированное антитело с цистеиновыми заменами содержит сконструированное антитело с цистеиновыми заменами (Ab), имеющее одну или несколько свободных цистеиновых аминокислот, конъюгированных с одним или несколькими комплексами циркония (Z) посредством линкера (L) и имеющее формулу I:

где p представляет собой 1-4.
Другой аспект изобретения относится к диагностическим применениям соединений и композиций, описанных в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1A показано трехмерное представление фрагмента антитела hu4D5Fabv7 в координатах кристалла, полученных с помощью рентгеновского излучения. Положения в структуре иллюстративных сконструированных остатков Cys тяжелой и легкой цепей пронумерованы (в соответствии с последовательной системой нумерации).
На фигуре 1B показана схема последовательной нумерации (верхний ряд), начиная с N-конца, по сравнению со схемой нумерации Kabat (нижний ряд) для 4D5v7fabH. Вставки с нумерацией по Kabat обозначены a, b, c.
На фигурах 2A и 2B показаны измерения связывания с детекцией оптической плотности при 450 нм для hu4D5Fabv8 и Cys мутантных (ТиоFab) фаговых вариантов hu4D5Fabv8: (A) небиотинилированного фаг-hu4D5Fabv8 и (B) биотинилированного фаг-hu4D5Fabv8 (B) посредством анализа PHESELECTOR для взаимодействий с BSA (незакрашенный столбец), HER2 (заштрихованный столбец) или стрептавидином (закрашенный столбец).
На фигурах 3A и 3B показаны измерения связывания с детекцией оптической плотности при 450 нм для hu4D5Fabv8 (слева) и Cys мутантных (ТиоFab) вариантов hu4D5Fabv8: (A) небиотинилированного фаг-hu4D5Fabv8 и (B) биотинилированного фаг-hu4D5Fabv8 посредством анализа PHESELECTOR для взаимодействий с: BSA (незакрашенный столбец), HER2 (заштрихованный столбец) и стрептавидином (незакрашенный столбец). Варианты легкой цепи расположены на левой стороне, а варианты тяжелой цепи расположены на правой стороне. Реакционная способность тиола = OD450нм для связывания стрептавидина ÷ OD450нм для связывания HER2 (антитела).
На фигуре 4A показаны значения относительной доступности поверхности остатков в hu4D5Fabv8 дикого типа. Участки легкой цепи расположены на левой стороне, а участки тяжелой цепи расположены на правой стороне.
На фигуре 4B показаны измерения связывания с детекцией оптической плотности при 450 нм для вариантов биотинилированного hu4D5Fabv8 (слева) и Cys мутанта (ТиоFab) hu4D5Fabv8 при взаимодействиях с HER2 (сутки 2), стрептавидином (SA) (сутки 2), HER2 (сутки 4) и SA (сутки 4). Cys варианты фаг-hu4D5Fabv8 выделяли и хранили при 4°C. На сутки 2 или сутки 4 проводили конъюгацию с биотином с последующими анализами PHESELECTOR для мониторирования их взаимодействия с Her2 и стрептавидином, как описано в примере 2, и тестировали стабильность реакционноспособных тиоловых групп на сконструированных вариантах ТиоFab.
На фигуре 5 показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированного с биотин-малеимидом-hu4D5Fabv8 (A121C) и небиотинилированного hu4D5Fabv8 дикого типа при связывании со стрептавидином и HER2. Каждый Fab тестировали при 2 нг и 20 нг.
На фигуре 6 показан анализ ELISA с детекцией оптической плотности при 450 нм для биотинилированного ABP-hu4D5Fabv8 дикого типа (wt), и цистеиновых мутантов ABP-hu4D5Fabv8 V110C и A121C при связывании с кроличьим альбумином, стрептавидином (SA) и HER2.
На фигуре 7 показан анализ ELISA с детекцией оптической плотности при 450 нм для цистеиновых мутантов (вариантов ТиоFab) биотинилированного ABP-hu4D5Fabv8: (слева направо) варианты с одним Cys ABP-V110C, ABP-A121C и варианты с двойным Cys ABP-V110C-A88C и ABP-V110C-A121C при связывании с кроличьим альбумином, HER2 и стрептавидином (SA), и использовании в качестве зондов Fab-HRP или SA-HRP.
На фигуре 8 показано связывание фага с биотинилированным ТиоFab и антитела против фага с HRP с HER2 (вверху) и стрептавидином (внизу).
На фигуре 13A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2, со связыванием меченного HRP вторичного антитела для детекции оптической плотности.
На фигуре 13B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотином-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании с иммобилизованным HER2. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг.
На фигуре 14A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2, со связыванием биотина с анти-IgG-HRP для детекции оптической плотности.
На фигуре 14B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотин-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании с иммобилизованным стрептавидином. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг.
На фигуре 15 показан общий способ подготовки сконструированного антитела с цистеиновыми заменами (ТиоMab), экспрессированного из культуры клеток, для конъюгации.
На фигуре 16 показан анализ электрофорезом в невосстанавливающем (вверху) и восстанавливающем (внизу) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab Fc (слева направо, дорожки 1-9): A339C; S337C; S324C; A287C; V284C; V282C; V279C; V273C, и 2H9 дикого типа после очистки на иммобилизованном белке A. Дорожка справа представляет собой маркер размера, указывающий на то, что интактные белки составляют приблизительно 150 кДа, фрагменты тяжелой цепи приблизительно 50 кДа, и фрагменты легкой цепи приблизительно 25 кДа.
На фигуре 17A показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab варианты (слева направо, дорожки 1-4): L-V15C; S179C; S375C; S400C, после очистки на иммобилизованном белке A.
На фигуре 17B показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле вариантов 2H9 и 3A5 ТиоMab, после очистки на иммобилизованном белке A.
На фигуре 18 показан анализ вестерн-блоттингом вариантов биотинилированного Тио-IgG. Варианты ТиоMab 2H9 и 3A5 анализировали электрофорезом в восстанавливающем денатурирующем полиакриламидном геле, белки переносили на нитроцеллюлозную мембрану. В качестве зондов для анализа присутствия антитела и конъюгированного биотина служили анти-IgG-HRP (вверху) и стрептавидин-HRP (внизу), соответственно. Дорожка 1: 3A5 H-A121C. Дорожка 2: 3A5 L-V110C. Дорожка 3: 2H9 H-A121C. Дорожка 4: 2H9 L-V110C. Дорожка 5: 2H9 дикого типа.
На фигуре 19 показан анализ ELISA связывания вариантов биотинилированного 2H9 со стрептавидином посредством использования в качестве зонда анти-IgG-HRP и измерения оптической плотности при 450 нм (верхняя столбчатая диаграмма). Нижняя схематическая диаграмма изображает дизайн эксперимента, используемый для анализа ELISA.
На фигуре 20 показаны бифункциональные реагенты для присоединения хелатирующего агента для 89Zr десферриоксамина B (Df, вверху) к белкам с использованием реакционноспособных по отношению к аминогруппам линкеров, TFP-N-SucDf и Df-Bz-NCS (в центре), и реакционноспособных по отношению к тиолу линкеров, Df-Chx-Mal, Df-Bac и Df-lac (внизу).
На фигуре 21 показано получение Df-Chx-Mal, Df-Bac, Df-Iac и конъюгация с тио-трастузумабом через остатки Cys, включенные в тяжелую цепь Fab. Условия реакции: i. DIEA, DMF/H2O (10:1), RT, 0,5-1 ч; ii. DIEA, DMF, 0°C, 4 ч; iii. pH 7,5, RT, 1 ч; iv. pH 9, RT, 5 ч; v. pH 9, RT, 2 ч.
На фигуре 22 показано образование хелатов оксалата циркония-89 с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, таких как варианты Df-линкер-трастузумаба, содержащие четыре линкера: N-Suc, Bz-SCN, Chx-малеимид (CHx-Mal), или ацетил (Ac).
На фигуре 23 показан масс-спектрометрический анализ восстановленных антител, показывающий отдельные сигналы от легкой и тяжелой цепей. A: тио-трастузумаб, B: Df-Ac-тио-трастузумаб (с использованием Df-Bac), и C: Df-Ac-тио-трастузумаб (с использованием Df-Iac), и D: Df-Chx-Mal-тио-трастузумаб.
На фигуре 24 показана стабильность 89Zr-Chx-Mal-тио-трастузумаба (незакрашенные круги) и 89Zr-Df-Ac-тио-трастузумаба (закрашенные круги) в сыворотке мыши при 37°C (n=3).
На фигуре 25 показаны репрезентативные изображения целого организма (проекция максимальной интенсивности), полученные через 96 часов после болюсной инъекции в хвостовую вену 100 мКи 89Zr-трастузумаба, полученного с использованием четырех различных линкеров (Bz-SCN, N-Suc, Chx-Mal и Ac).
На фигуре 26 показано поглощение in vivo в избранных тканях через 24, 96 и 144 часа после инъекции, как измерено посредством ПЭТ.
ПОДРОБНОЕ ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
В настоящее время приведены подробные ссылки на конкретные варианты осуществления изобретения, примеры которых проиллюстрированы в сопровождающих структурах и формулах. В то время как изобретение описано в отношении перечисленных вариантов осуществления, понятно, что они не предназначены для ограничения изобретения этими вариантами осуществления. С другой стороны, изобретение предназначено, чтобы включать все альтернативы, модификации и эквиваленты, которые могут быть включены в объем настоящего изобретения, как определено формулой изобретения.
Специалисту в данной области известно множество способов и материалов, подобных или эквивалентных описанным в настоящем документе, которые можно использовать в практике настоящего изобретения. Настоящее изобретение никаким образом не ограничено описанными способами и материалами.
Если не определено иначе, технические и научные термины, используемые в настоящем документе, обладают таким же значением, какое является общепринятым для специалиста в области, к которой относится это изобретение, и соответствует: Singleton et al (1994) Dictionary of Microbiology and Molecular Biology, 2nd Ed., J. Wiley & Sons, New York, NY; и Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immunobiology, 5th Ed., Garland Publishing, New York.
ОПРЕДЕЛЕНИЯ
Если не указано иначе, следующие термины и фразы, как применяют в настоящем документе, предназначены, чтобы обладать следующими значениями:
При использовании торговых наименований в настоящем документе, авторы настоящего изобретения подразумевают независимое включение состава продукта под торговым наименованием, аналога патентованного лекарственного средства и активного фармацевтического ингредиента(ингредиентов) продукта под торговым наименованием.
Термин «антитело» в настоящем документе используют в самом широком смысле, и он конкретно охватывает моноклональные антитела, поликлональные антитела, димеры, мультимеры, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антитела, пока они обладают желательной биологической активностью (Miller et al (2003) Jour. of Immunology 170:4854-4861). Антитела могут являться мышиными, человеческими, гуманизированными, химерными или полученными из других видов. Антитело представляет собой белок, образуемый иммунной системой, способный узнавать и связывать специфический антиген. (Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immuno Biology, 5th Ed., Garland Publishing, New York). Антиген-мишень, как правило, обладает несколькими участками связывания, называемыми также эпитопами, узнаваемыми CDR множества антител. Каждое антитело, которое специфически связывается с отличным эпитопом, обладает отличной структурой. Таким образом, один антиген может иметь более одного соответствующего антитела. Антитело включает полноразмерную молекулу иммуноглобулина или иммунологически активную часть полноразмерной молекулы иммуноглобулина, т.е., молекулы, содержащей антигенсвязывающий участок, содержащий антигенсвязывающий участок, иммуноспецифически связывающий антиген интересующей мишени или его часть, где такие мишени включают в качестве неограничивающих примеров, клетку или клетки злокачественных опухолей, продуцирующие аутоиммунные антитела, ассоциированные с аутоиммунным заболеванием. Иммуноглобулин, описанный в настоящем документе, может относиться к любому типу (например, IgG, IgE, IgM, IgD, и IgA), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекулы иммуноглобулина. Иммуноглобулины могут происходить из любых видов. В одном аспекте, однако, иммуноглобулин имеет человеческое, мышиное или кроличье происхождение.
«Фрагменты антител» содержат часть полноразмерного антитела, как правило, его антигенсвязывающую или вариабельную область. Примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv фрагменты; диатела; линейные антитела; миниантитела (Olafsen et al (2004) Protein Eng. Design & Sel. 17(4):315-323), фрагменты, полученные из экспрессирующей библиотеки Fab, анти-идиотипические (анти-Id) антитела, CDR (определяющую комплементарность область) и эпитоп-связывающие фрагменты любого из вышеуказанного, специфически связывающиеся с антигенами клеток злокачественных опухолей, вирусными антигенами или антигенами микроорганизмов, молекулы одноцепочечных антител; и мультиспецифические антитела, сформированные из фрагментов антител.
Термин «моноклональное антитело», как применяют в настоящем документе, относится к антителу, полученному из популяции по существу однородных антител, т.е., отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, являясь направленными против одного антигенного участка. Более того, в отличие от препаратов поликлональных антител, включающих различные антитела, нацеленные против различных детерминант (эпитопов), каждое моноклональное антитело нацелено против отдельной детерминанты антигена. В дополнение к их специфичности, моноклональные антитела являются преимущественными в том смысле, что их можно синтезировать без контаминации другими антителами. Определение «моноклональные» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует истолковывать как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела для использования в соответствии с настоящим изобретением можно получать способом гибридомы, впервые описанным Kohler et al (1975) Nature 256:495, или их можно получать способами рекомбинантной ДНК (см., например: US 4816567; US 5807715). Моноклональные антитела можно выделять также из фаговых библиотек антител с использованием способов, описанных, например, в Clackson et al (1991) Nature, 352:624-628; Marks et al (1991) J. Mol. Biol, 222:581-597.
Моноклональные антитела в настоящем документе конкретно включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как остаток цепи(цепей) является идентичным или гомологичным соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител, так же как фрагменты таких антител, пока они обладают желательной биологической активностью (US 4816567; и Morrison et al (1984) Proc. Natl. Acad. Sci. USA, 81:6851-6855). Интересующие химерные антитела в настоящем документе включают «приматизированные» антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученные из не относящегося к человеку примата (например, обезьяны Старого Света, обезьяны и т.д.), и человеческие последовательности константной области.
«Интактное антитело» в настоящем документе представляет собой антитело, содержащее домены VL и VH, а также константный домен (CL) легкой цепи и константные домены тяжелой цепи CH1, CH2 и CH3. Константные домены могут представлять собой константные домены с природной последовательностью (например, константные домены с человеческой природной последовательностью) или вариант их аминокислотной последовательности. Интактное антитело может обладать одной или несколькими «эффекторными функциями», которые относятся к тем видам биологической активности, которую можно приписать константной области Fc (природной последовательности Fc-области или варианту аминокислотной последовательности Fc-области) антитела. Примеры эффекторных функций антител включают связывание C1q; комплементзависимую цитотоксичность; связывание Fc рецептора; антителозависимую опосредуемую клетками цитотоксичность (ADCC); фагоцитоз; и понижающую регуляцию рецепторов поверхности клеток, таких как B-клеточный рецептор и BCR.
В зависимости от аминокислотных последовательностей константных доменов их тяжелых цепей, интактные антитела можно отнести к различным «классам». Существует пять главных классов интактных иммуноглобулинов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно далее разделить на «подклассы» (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. Формы Ig включают модификации шарнира или бесшарнирные формы (Roux et al (1998) J. Immunol. 161:4083-4090; Lund et al (2000) Eur. J. Biochem. 267:7246-7256; US 2005/0048572; US 2004/0229310).
«Рецептор ErbB» представляет собой рецепторную протеинтирозинкиназу, принадлежащую к семейству рецепторов ErbB, члены которого являются важными медиаторами роста, дифференцировки и выживаемости клеток. Семейство рецепторов ErbB включает четыре отдельных члена, включая рецептор эпидермального фактора роста (EGFR, ErbB1, HER1), HER2 (ErbB2 или p185neu), HER3 (ErbB3) и HER4 (ErbB4 или tyro2). Панель анти-ErbB2 антител охарактеризована с использованием линии клеток опухоли молочной железы человека SKBR3 (Hudziak et al (1989) Mol. Cell. Biol. 9(3):1165-1172. Максимальное ингибирование получено с антителом, называемым 4D5, ингибирующим пролиферацию клеток на 56%. Другие антитела в панели в этом анализе уменьшали пролиферацию клеток в меньшей степени. Кроме того, обнаружено, что антитело 4D5 сенсибилизирует линии клеток опухоли молочной железы со сверхэкспрессией ErbB2 к цитотоксическим эффектам TNF-α (US 5677171). Анти-ErbB2 антитела, обсуждаемые в Hudziak et al., дополнительно охарактеризованы в Fendly et al (1990) Cancer Research 50: 1550-1558; Kotts et al. (1990) In Vitro 26(3):59A; Sarup et al. (1991) Growth Regulation 1:72-82; Shepard et al. J. (1991) Clin. Immunol. 11(3):117-127; Kumar et al. (1991) Mol. Cell. Biol. 11(2):979-986; Lewis et al. (1993) Cancer Immunol. Immunother. 37:255-263; Pietras et al. (1994) Oncogene 9: 1829-1838; Vitetta et al. (1994) Cancer Research 54:5301-5309; Sliwkowski et al. (1994) J. Biol. Chem. 269(20): 14661-14665; Scott et al. (1991) J. Biol. Chem. 266: 14300-5; D'souza et al. Proc. Natl. Acad. Sci. (1994) 91:7202-7206; Lewis et al. (1996) Cancer Research 56:1457-1465; и Schaefer et al. (1997) Oncogene 15:1385-1394.
Рецептор ErbB, как правило, содержит внеклеточный домен, который может связывать лиганд ErbB; липофильный трансмембранный домен; консервативный внутриклеточный домен тирозинкиназы; и карбоксил-концевой сигнальный домен, несущий несколько остатков тирозина, которые могут быть фосфорилированы. Рецептор ErbB может представлять собой рецептор ErbB с «природной последовательностью» или «вариант его аминокислотной последовательности». Предпочтительно, рецептор ErbB представляет собой рецептор ErbB человека с природной последовательностью. Соответственно, «член семейства рецепторов ErbB» представляет собой EGFR (ErbB1), ErbB2, ErbB3, ErbB4 или любой другой рецептор ErbB, который известен в настоящее время или который будет идентифицирован в будущем.
Термины «ErbB1», «рецептор эпидермального фактора роста», «EGFR» и «HER1» используют в настоящем документе взаимозаменяемо, и они относятся к EGFR, как описано, например, в Carpenter et al (1987) Ann. Rev. Biochem., 56:881-914, включая их встречающиеся в природе мутантные формы (например, делеционный мутант EGFR, как в Humphrey et al (1990) Proc. Nat. Acad. Sci. (USA) 87:4207-4211). Термин erbB1 относится к гену, кодирующему белковый продукт EGFR. Антитела против HER1 описаны, например, в Murthy et al (1987) Arch. Biochem. Biophys., 252:549-560 и в WO 95/25167.
Термин «ERRP», «родственный EGF-рецептору белок», «родственный EGFR белок» и «родственный рецептору эпидермального фактора роста белок» используют в настоящем документе взаимозаменяемо, и они относятся к ERRP, как описано, например, в US 6399743 и в Публикации США № 2003/0096373.
Выражения «ErbB2» и «HER2» используют в настоящем документе взаимозаменяемо, и они относятся к белку HER2 человека, описанному, например, в Semba et al (1985) Proc. Nat. Acad. Sci. (USA) 82:6497-6501 и Yamamoto et al (1986) Nature, 319:230-234 (номер доступа в Genebank X03363). Термин «erbB2» относится к гену, кодирующему ErbB2 человека, и «neu» относится к гену, кодирующему p185neu крысы. Предпочтительным ErbB2 является природная последовательность ErbB2 человека.
«ErbB3» и «HER3» относятся к полипептиду рецептора, как описано, например, в Патентах США №№ 5183884 и 5480968, а также в Kraus et al (1989) Proc. Nat. Acad. Sci. (USA) 86:9193-9197. Антитела против ErbB3 известны в данной области и описаны, например, в Патентах США №№ 5183884, 5480968 и в WO 97/35885.
Термины «ErbB4» и «HER4» в настоящем документе относятся к полипептиду рецептора, как описано, например, в Патентной заявке EP № 599274; Plowman et al (1993) Proc. Natl. Acad. Sci. USA 90: 1746-1750; и Plowman et al (1993) Nature 366:473-475, включая их изоформы, например, как описано в WO 99/19488. Антитела против HER4 описаны, например, в WO 02/18444.
Антитела против рецепторов ErbB являются коммерчески доступными из ряда источников, включая, например, Santa Cruz Biotechnology, Inc., California, USA.
Термин «вариант аминокислотной последовательности» относится к полипептидам, обладающим аминокислотными последовательностями, отличающимися до некоторой степени от полипептида с природной последовательностью. Обычно варианты аминокислотной последовательности обладают по меньшей мере приблизительно 70% идентичностью последовательности по меньшей мере с одним связывающим рецептор доменом природного лиганда ErbB или по меньшей мере с одним связывающим лиганд доменом природного рецептора ErbB, и предпочтительно, они по меньшей мере приблизительно на 80%, более предпочтительно, по меньшей мере приблизительно на 90% гомологичны по последовательности таким связывающим рецептор или лиганд доменам. Варианты аминокислотной последовательности имеют замены, делеции и/или вставки в конкретных положениях внутри аминокислотной последовательности природной аминокислотной последовательности. Аминокислоты обозначают общепринятыми наименованиями, в однобуквенном и трехбуквенном кодах.
«Идентичность последовательности» определяют как процент остатков в варианте аминокислотной последовательности, являющихся идентичными после выравнивания последовательностей и введения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Способы и компьютерные программы для выравнивания хорошо известны в данной области. Одной такой компьютерной программой является «Align 2», авторство которой принадлежит Genentech, Inc. и подано с пользовательской документацией в Бюро регистрации авторских прав США, Washington, DC 20559, 10 декабря 1991 г.
«Антителозависимая опосредуемая клетками цитотоксичность» и «ADCC» относятся к опосредованной клетками реакции, при которой неспецифические цитотоксические клетки, экспрессирующие Fc-рецепторы (FcR) (например, клетки естественные киллеры (NK), нейтрофилы, и макрофаги) узнают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Первичные клетки, опосредующие ADCC, NK клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках обобщена в таблице 3 на странице 464 в Ravetch and Kinet, (1991) «Annu. Rev. Immunol.» 9:457-92. Для оценки активности ADCC интересующей молекулы можно проводить анализ ADCC in vitro, такой как описан в US 5500362 и US 5821337. Пригодные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и естественные клетки-киллеры (NK). Альтернативно или дополнительно, активность ADCC интересующей молекулы можно оценивать in vivo, например, в модели на животных, такой, как описана в Clynes et al (1998) PROC. NAT. ACAD. SCI. (USA) (USA) 95:652-656.
«Эффекторные клетки человека» представляют собой лейкоциты, которые экспрессируют один или несколько рецепторов константной области (FcR) и выполняют эффекторные функции. Предпочтительно, клетки экспрессируют по меньшей мере FcyRIII и выполняют эффекторную функцию(функции) ADCC. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), естественные клетки-киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы; где PBMC и NK клетки являются предпочтительными. Эффекторные клетки можно выделить из их природного источника, например, из крови или PBMC, как описано в настоящем документе.
Термины «Fc рецептор» или «FcR» используют для описания рецептора, который связывается с константной областью Fc антитела. Предпочтительный FcR представляет собой FcR человека с природной последовательностью. Более того, предпочтительным FcR является FcR, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcyRII, и FcγRIII, включая аллельные варианты и формы альтернативного сплайсинга этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), обладающие сходными аминокислотными последовательностями, которые отличаются, в первую очередь, в их цитоплазматических доменах. Активирующий рецептор FcγRIIA содержит иммунорецепторный активирующий мотив, основанный на тирозине (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный ингибирующий мотив, основанный на тирозине (ITIM), в его цитоплазматическом домене. (См. обзор M. в Daлron, «Annu. Rev. Immunol.» 15:203-234 (1997)). Обзор FcR приведен в Ravetch and Kinet, «Annu. Rev. Immunol»., 9:457-92 (1991); Capel et al (1994) Immunomethods 4:25-34; и de Haas et al (1995) J. Lab. Clin. Med. 126:330-41. Другие FcR, включая те, которые будут идентифицированы в будущем, включены в термин «FcR» в настоящем документе. Термин включает также неонатальный рецептор, FcRn, который является ответственным за перенос материнских IgG к плоду (Guyer et al (1976) J. Immunol, 117:587 и Kim et al (1994) J. Immunol. 24:249).
«Комплементзависимая цитотоксичность» или «CDC» относится к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента запускается связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе с родственными ей антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al J. Immunol. Methods, 202: 163 (1996).
«Нативные антитела» обычно представляют собой гетеротетрамерные гликопротеины приблизительно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как количество дисульфидных связей меняется среди тяжелых цепей различных изотипов иммуноглобулинов. В каждой тяжелой и легкой цепи также равномерно расположены внутрицепьевые дисульфидные мостики. Каждая тяжелая цепь на одном конце имеет вариабельный домен (VH) с последующими несколькими константными доменами. Каждая легкая цепь на одном конце имеет вариабельный домен (VL) и на другом конце - константный домен. Константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Считают, что конкретные аминокислотные остатки формируют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи.
Термин «вариабельный» относится к факту, что конкретные части вариабельных доменов сильно отличаются по последовательности среди антител и используются для связывания и специфичности каждого конкретного антитела для его конкретного антигена. Однако вариабельность не является равномерно распределенной на всем протяжении вариабельных доменов антител. Она сконцентрирована в трех участках, называемых гипервариабельными областями, в вариабельных доменах как на легкой цепи, так и на тяжелой цепи. Наиболее высоко консервативные части вариабельных доменов называют каркасными областями (FR). Каждый из вариабельных доменов природных тяжелых и легких цепей содержит четыре FR, преимущественно принимающие конфигурацию β-листа, соединенные тремя гипервариабельными областями, формирующими петли, соединяющие структуры β-листа, и в некоторых случаях формирующие их часть. Гипервариабельные области на каждой цепи удерживаются вместе в непосредственной близости посредством FR и, вместе с гипервариабельными областями из другой цепи, участвуют в формировании антигенсвязывающего участка антител (см. Kabat et al (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD). Константные домены не являются непосредственно вовлеченными в связывание антитела с антигеном, но обладают различными эффекторными функциями, такими как участие антитела в антителозависимой клеточной токсичности (ADCC).
Термин «гипервариабельная область» при использовании в настоящем документе относится к аминокислотным остаткам антитела, ответственным за связывание антигена. Гипервариабельная область обычно содержит аминокислотные остатки из «определяющей комплементарность области» или «CDR» (например, остатки 24-34 (LI), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al выше) и/или остатки из «гипервариабельной петли» (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (HI), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk (1987) J. Mol. Biol, 196:901-917). Остатки «каркасной области» или «FR» представляют собой остатки вариабельного домена, отличные от остатков гипервариабельной области, как определено в настоящем документе.
Расщеплением антител папаином получают два идентичных антигенсвязывающих фрагмента, называемых «Fab» фрагментами, каждый с одним антигенсвязывающим участком, и оставшийся «Fc» фрагмент, наименование которого отражает его способность легко кристаллизоваться. Обработкой пепсином получают F(ab')2 фрагмент, обладающий двумя антигенсвязывающими участками и еще способный к перекрестному связыванию антигена.
«Fv» представляет собой минимальный фрагмент антитела, содержащий полный узнающий антиген и антигенсвязывающий участок. Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи в тесной, нековалентной связи. В этой конфигурации три гипервариабельные области каждого вариабельного домена взаимодействуют для определения антигенсвязывающего участка на поверхности димера VH-VL. Шесть гипервариабельных областей совместно придают антигенсвязывающую специфичность антителу. Однако, даже отдельный вариабельный домен (или половина Fv, содержащая только три гипервариабельные области, специфичные для антигена) обладает способностью узнавать и связывать антиген, хотя и с более низкой аффинностью, чем целый связывающий участок.
Fab-фрагмент содержит также константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на карбокси-конце CH1 домена тяжелой цепи, включая один или несколько остатков цистеина из шарнирной области антитела. Fab'-SH в настоящем документе является обозначением Fab', в котором остаток(остатки) цистеина константных доменов несут по меньшей мере одну свободную тиоловую группу. F(ab')2 фрагменты антитела первоначально были получены в форме пар Fab'-фрагментов с шарнирными цистеинами между ними. Известны также другие химические соединения фрагментов антител.
«Легкие цепи» антител из любых видов позвоночных можно отнести к одному из двух явно различимых типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотных последовательностей их константных доменов.
«Одноцепочечные Fv» или «scFv» фрагменты антител содержат VH и VL домены антитела, где эти домены присутствуют на одной полипептидной цепи. Предпочтительно, полипептид Fv дополнительно содержит полипептидный линкер между VH и VL доменами, позволяющий scFv формировать желательную структуру для связывания антигена. Обзор scFv см. в Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994). scFv фрагменты анти-ErbB2 антитела описаны в WO 93/16185; Патентах США №№ 5571894; и 5587458.
Термин «диатела» относится к малым фрагментам антитела с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). С использованием линкера, слишком короткого, чтобы позволить спаривание двух доменов одной и той же цепи, домены вынуждают спариваться с комплементарными доменами другой цепи и формировать два антигенсвязывающих участка. Диатела более подробно описаны, например, в EP 404,097; WO 93/11161; и Hollinger et al (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448.
«Гуманизированные» формы не относящихся к человеку антител» (например, грызунов) представляют собой химерные антитела, содержащие минимальную последовательность, полученную из не относящегося к человеку иммуноглобулина. Гуманизация является способом переноса информации для связывания антигена от мыши к неиммуногенному акцепторному антителу человека, и приводит к получению множества терапевтически пригодных лекарственных средств. Способ гуманизации, как правило, начинается с переноса всех шести мышиных определяющих комплементарность областей (CDR) в каркас человеческого антитела (Jones et al, (1986) Nature 321:522-525). Эти CDR-привитые антитела, как правило, не сохраняют исходную аффинность для связывания антигена, и фактически, аффинность часто серьезно нарушена. Помимо CDR, избранные каркасные остатки не относящегося к человеку антитела также необходимо включать для поддержания правильной конформации CDR (Chothia et al (1989) Nature 342:877). Показано, что перенос ключевых мышиных остатков в человеческий акцептор для поддержания структурной конформации привитых CDR сохраняет связывание антигена и аффинность (Riechmann et al (1992) J. Mol. Biol. 224, 487-499; Foote and Winter, (1992) J. Mol. Biol. 224:487-499; Presta et al (1993) J. Immunol. 151, 2623-2632; Werther et al (1996) J. Immunol. Methods 157:4986-4995; и Presta et al (2001) Thromb. Haemost. 85:379-389). По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (антитело-реципиент), в котором остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области не относящихся к человеку видов (антитела-донора), таких как мышь, крыса, кролик или не относящийся к человеку примат, обладающими желаемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими не присутствующими у человека остатками. Более того, гуманизированные антитела могут содержать остатки, не обнаруженные в антителе-реципиенте или в антителе-доноре. Эти модификации выполняют для дальнейшего усовершенствования характеристик антитела. Как правило, гуманизированное антитело содержит в основном все из по меньшей мере одного, и как правило, двух вариабельных доменов, в которых все или в основном все гипервариабельные петли соответствуют петлям не относящегося к человеку иммуноглобулина, и все или в основном все FR являются FR с последовательностью человеческого иммуноглобулина. Гуманизированное антитело, необязательно, может содержать также по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Более подробно см. US 6407213; Jones et al (1986) Nature, 321:522-525; Riechmann et al (1988) Nature 332:323-329; и Presta, (1992) Curr. Op. Struct. Biol, 2:593-596.
«Свободная аминокислота цистеин» относится к остатку аминокислоты цистеина, который введен при конструировании в исходное антитело, имеет тиоловую функциональную группу (-SH), и не является спаренным в качестве внутримолекулярного или межмолекулярного дисульфидного мостика.
Термин «значение реакционной способности тиола» представляет собой количественную характеристику реакционной способности свободных цистеиновых аминокислот. Значение реакционной способности тиола представляет собой процент свободной аминокислоты цистеина в сконструированном антителе с цистеиновыми заменами, вступающий в реакцию с реакционноспособным по отношению к тиолу реагентом, и преобразованный до максимального значения 1. Например, свободная аминокислота цистеин в сконструированном антителе с цистеиновыми заменами, которая вступает в реакцию со 100% выходом с реакционноспособным по отношению к тиолу реагентом, таким как биотин-малеимидный реагент, с формированием меченного биотином антитела, имеет значение реакционной способности тиола 1,0. Другая аминокислота цистеин, введенная при конструировании в такое же или другое исходное антитело, которая вступает в реакцию с 80% выходом с реакционноспособным по отношению к тиолу реагентом, имеет значение реакционной способности тиола 0,8. Другая аминокислота цистеин, введенная при конструировании в такое же или другое исходное антитело, которая совсем не вступает в реакцию с реакционноспособным по отношению к тиолу реагентом, имеет значение реакционной способности тиола 0. Определение значения реакционной способности тиола конкретного цистеина можно проводить анализом ELISA, масс-спектроскопией, жидкостной хроматографией, радиоавтографией или другими количественными аналитическими тестами.
«Исходное антитело» представляет собой антитело, содержащее аминокислотную последовательность, в которой один или несколько аминокислотных остатков заменяют на один или несколько остатков цистеина. Исходное антитело может содержать природную последовательность или последовательность дикого типа. Исходное антитело может обладать предсуществующими модификациями аминокислотной последовательности (такими как добавления, делеции и/или замены) по сравнению с другими природными формами, формами дикого типа или модифицированными формами антитела. Исходное антитело может быть нацелено против интересующего антигена-мишени, например, биологически важного полипептида. Предусмотрены также антитела, нацеленные против неполипептидных антигенов (таких как опухолеассоциированные гликолипидные антигены; см. US 5091178).
Иллюстративные исходные антитела включают антитела, обладающие аффинностью и избирательностью для рецепторов поверхности клеток и трансмембранных рецепторов, и опухолеассоциированных антигенов (TAA).
Другие иллюстративные исходные антитела включают антитела, выбранные, и без ограничения, из антитела против рецептора эстрогена, антитела против рецептора прогестерона, анти-p53 антитела, анти-HER-2/neu антитела, анти-EGFR антитела, антитела против катепсина D, анти-Bcl-2 антитела, антитела против E-кадгерина, анти-CA125 антитела, анти-CA15-3 антитела, анти-CA19-9 антитела, анти-c-erbB-2 антитела, антитела против P-гликопротеина, анти-CEA антитела, антитела против белка ретинобластомы, антитела против онкобелка ras, антитела против Льюиса X, анти-Ki-67 антитела, анти-PCNA антитела, анти-CD3 антитела, анти-CD4 антитела, анти-CD5 антитела, анти-CD7 антитела, анти-CD8 антитела, анти-CD9/p24 антитела, анти-CD10 антитела, анти-CD11c антитела, анти-CD13 антитела, анти-CD14 антитела, анти-CD15 антитела, анти-CD19 антитела, анти-CD20 антитела, анти-CD22 антитела, анти-CD23 антитела, анти-CD30 антитела, анти-CD31 антитела, анти-CD33 антитела, анти-CD34 антитела, анти-CD35 антитела, анти-CD38 антитела, анти-CD41 антитела, анти-LCA/CD45 антитела, анти-CD45RO антитела, анти-CD45RA антитела, анти-CD39 антитела, анти-CD100 антитела, анти-CD95/Fas антитела, анти-CD99 антитела, анти-CD106 антитела, антитела против убиквитина, анти-CD71 антитела, анти-c-myc антитела, антитела против цитокератинов, антитела против виментинов, антитела против белков HPV, антитела против легких цепей каппа, антитела против легких цепей лямбда, антитела против меланосом, антитела против специфического антигена простаты, анти-S-100 антитела, антитела против антигена tau, антитела против фибрина, антитела против кератинов и антитела против Tn-антигена.
«Выделенное» антитело представляет собой антитело, которое идентифицировано и отделено и/или выделено из компонента его природного окружения. Загрязняющие компоненты его натурального окружения представляют собой материалы, которые могут мешать исследовательским, диагностическим или терапевтическим применениям антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело является очищенным (1) до более чем 95% по массе антитела, как определено способом Лоури, и наиболее предпочтительно, до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности при использовании секвенатора с вращающимся стаканом, или (3) до гомогенности в SDS-PAGE в невосстанавливающих или восстанавливающих условиях с использованием окраски Кумасси синим или, предпочтительно, серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, с тех пор, когда перестает присутствовать по меньшей мере один из компонентов природного окружения антитела. Обычно, однако, выделенное антитело получают посредством по меньшей мере одной стадии очистки.
Антитело, «которое связывает» интересующую молекулярную мишень или антиген, например, антиген ErbB2, представляет собой антитело, способное связывать этот антиген с достаточной аффинностью, так что антитело является пригодным для нацеливания на клетку, экспрессирующую антиген. Когда антитело представляет собой антитело, связывающее ErbB2, оно обычно предпочтительно связывает ErbB2 в отличие от других рецепторов ErbB, и может представлять собой антитело, которое не вступает в значительную перекрестную реакцию с другими белками, такими как EGFR, ErbB3 или ErbB4. В таких вариантах осуществления степень связывания антитела с этими не относящимися к ErbB2 белками (например, связывания поверхности клеток с эндогенным рецептором) составляет менее 10%, как определено анализом активированной флуоресценцией сортировки клеток (FACS) или радиоиммунопреципитацией (RIA). Иногда анти-ErbB2 антитело не вступает в значительную перекрестную реакцию с белком neu крысы, например, как описано в Schecter et al. (1984) Nature 312:513 и Drebin et al (1984) Nature 312:545-548.
Молекулярные мишени для антител, включенные в настоящее изобретение, включают CD белки и их лиганды, такие как, в качестве неограничивающих примеров: (i) CD3, CD4, CD8, CD19, CD20, CD22, CD34, CD40, CD79α (CD79a) и CD79β (CD79b); (ii) члены семейства рецепторов ErbB, такие как рецептор EGF, рецептор HER2, HER3 или HER4; (iii) молекулы клеточной адгезии, такие как LFA-1, Macl, p150,95, VLA-4, ICAM-1, VCAM и αν/β3 интегрин, включая их альфа- или бета-субъединицы (например, анти-CD11a, анти-CD18 или анти-CD11b антитела); (iv) факторы роста, такие как VEGF; IgE; антигены группы крови; рецептор flk2/flt3; рецептор ожирения (OB); рецептор mpl; CTLA-4; белок C, BR3, c-met, тканевой фактор, β7 и т.д.; и (v) опухолеассоциированный антигены (TAA) поверхности клеток и трансмембранные опухолеассоциированные антигены (TAA).
Если нет иных указаний, термин «моноклональное антитело 4D5» обозначает антитело, обладающее антигенсвязывающими остатками мышиного антитела 4D5 (ATCC CRL 10463), или полученное из него. Например, моноклональное антитело 4D5 может представлять собой мышиное моноклональные антитело 4D5 или его вариант, такой как гуманизированное 4D5. Иллюстративные гуманизированные антитела 4D5 включают huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (трастузумаб, ГЕРЦЕПТИН®), как в Патенте США № 5821337.
«Фаговый дисплей» представляет собой способ, посредством которого варианты полипептидов экспонируют в виде белков, слитых с белком оболочки, на поверхности частиц фага, например, нитевидного фага. Применимость фагового дисплея основана на факте, что большие библиотеки рандомизированных вариантов белка можно быстро и эффективно отсортировать по тем последовательностям, которые связывают антиген-мишень с высокой аффинностью. Экспонирование пептидных и белковых библиотек на фаге использовали для скрининга миллионов полипептидов для отбора полипептидов со специфическими свойствами связывания. Способы поливалентного фагового дисплея использовали для экспонирования малых случайных пептидов и малых белков, как правило, посредством слияния с pIII или pVIII нитевидного фага (Wells and Lowman, (1992) Curr. Opin. Struct. Biol, 3:355-362, и процитированные в этом документе ссылки). В моновалентном фаговом дисплее белковую или пептидную библиотеку сливают с белком оболочки фага или его частью, и экспрессируют на низких уровнях в присутствии белка дикого типа. Эффекты авидности снижены по сравнению с поливалентным фагом, так что отбор проходит на основе собственной аффинности лиганда, и используют фагмидные векторы, упрощающие манипуляции с ДНК. Lowman and Wells, Methods: A companion to Methods in Enzymology, 3:205-0216 (1991). Фаговый дисплей включает способы получения антителоподобных молекул (Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immunobiology, 5th Ed., Garland Publishing, New York, p.627-628; Lee et al ).
«Фагмида» представляет собой плазмидный вектор, обладающий бактериальной точкой начала репликации, например, ColE1, и копией межгенной области бактериофага. Фагмиду можно использовать для любого известного бактериофага, включая нитевидный бактериофаг и лямбдоидный бактериофаг. Плазмида также, как правило, содержит селективный маркер для устойчивости к антибиотику. Фрагменты ДНК, клонированные в эти векторы, можно размножать в форме плазмид. Когда клетки, несущие эти векторы, обеспечивают всеми генами, необходимыми для продукции фаговых частиц, тип репликации плазмиды меняется на репликацию по типу катящегося кольца для получения копий одной цепи плазмидной ДНК и упаковки фаговых частиц. Фагмиды могут формировать инфекционные или неинфекционные фаговые частицы. Этот термин включает фагмиды, содержащие ген белка оболочки фага или его фрагмент, связанный с геном гетерологичного полипептида в форме слитого гена, так что гетерологичный полипептид экспонирован на поверхности фаговых частиц.
«Линкер», «линкерное звено», или «связующее звено» обозначает химическую группу, содержащую ковалентную связь или цепь атомов, которая ковалентно присоединяет антитело к группе лекарственного средства. В различных вариантах осуществления линкер обозначают как L. Линкеры включают двухвалентный радикал, такой как алкилдииловые, ариленовые, гетероариленовые группы, такие как: -(CR2)nO(CR2)n-, повторяющиеся звенья алкилокси (например, полиэтиленокси, PEG, полиметиленокси) и алкиламино (например, полиэтиленамино, Джеффамин™); и сложные эфиры и амиды двухосновной кислоты, включая сукцинат, сукцинамид, дигликолят, малонат и капроамид.
Термин «метка» обозначает любую группу, которую можно ковалентно присоединять к антителу и которая функционирует для: (i) обеспечения поддающегося детекции сигнала; (ii) взаимодействия со второй меткой для модификации поддающегося детекции сигнала, обеспечиваемого первой или второй меткой, например, FRET (резонансный перенос энергии флуоресценции); (iii) стабилизации взаимодействий или увеличения аффинности связывания с антигеном или лигандом; (iv) влияния на подвижность, например, электрофоретическую подвижность, или клеточную проницаемость, посредством заряда, гидрофобности, формы или других физических параметров, или (v) предоставления связывающей группы, для модуляции аффинности лиганда, связывания антитело/антиген или образования комплексных ионов.
Стереохимические определения и обозначения, используемые в настоящем документе, в основном, следуют S. P. Parker, Ed., McGraw-Hill Dictionary of Chemical Terms (1984) McGraw-Hill Book Company, New York; и Eliel, E. and Wilen, S., Stereochemistry of Organic Compounds (1994) John Wiley & Sons, Inc., New York. Многие органические соединения существуют в оптически активных формах, т.е., они обладают способностью вращать плоскость плоскополяризованного света. При описании оптически активного соединения, префиксы D и L или R и S, используют для обозначения абсолютной конфигурации молекулы вокруг ее хирального центра(центров). Префиксы d и l или (+) и (-) используют для обозначения знака вращения плоскополяризованного света соединением, где (-) или l обозначает, что соединение является левовращающим. Соединение с префиксом (+) или d является правовращающим. Для данной химической структуры, эти стереоизомеры являются идентичными, за исключением того, что они являются зеркальными отражениями друг друга. Конкретный стереоизомер можно обозначать также, как энантиомер, и смесь таких изомеров часто называют энантиомерной смесью. Смесь энантиомеров 50:50 обозначают рацемической смесью или рацематом, которые могут возникать в отсутствие стереоизбирательности или стереоспецифичности химической реакции или процесса. Термины «рацемическая смесь» и «рацемат» относятся к эквимолярной смеси двух видов энантиомеров, лишенной оптической активности.
Фраза «фармацевтически приемлемая соль», как применяют в настоящем документе, относится к фармацевтически приемлемым органическим или неорганическим солям AZC. Иллюстративные соли включают соли сульфат, цитрат, ацетат, оксалат, хлорид, бромид, иодид, нитрат, бисульфат, фосфат, кислый фосфат, изоникотинат, лактат, салицилат, кислый цитрат, тартрат, олеат, таннат, пантотенат, битартрат, аскорбат, сукцинат, малеат, гентизинат, фумарат, глюконат, глюкуронат, сахарат, формат, бензоат, глутамат, метансульфонат, этансульфонат, бензолсульфонат, п-толуолсульфонат и памоат (т.е., 1,1'-метилен-бис-(2-гидрокси-3-нафтоат)), но не ограничены ими. Фармацевтически приемлемая соль может предполагать включение другой молекулы, такой как ион ацетата, ион сукцината или другой противоион. Противоион может представлять собой любую органическую или неорганическую группу, стабилизирующую заряд исходного соединения. Более того, фармацевтически приемлемая соль может иметь в структуре более одного заряженного атома. В случаях, когда несколько заряженных атомов составляют часть фармацевтически приемлемой соли, она может иметь несколько противоионов. Таким образом, фармацевтически приемлемая соль может иметь один или несколько заряженных атомов и/или один или несколько противоионов.
«Фармацевтически приемлемый сольват» обозначает ассоциацию одной или нескольких молекул растворителя и AZC. Растворители, формирующие фармацевтически приемлемые сольваты, включают в качестве неограничивающих примеров, воду, изопропанол, этанол, метанол, DMSO, этилацетат, уксусную кислоту и этаноламин.
Следующие аббревиатуры использованы в настоящем документе и имеют следующие определения: BME представляет собой бета-меркаптоэтанол, Boc представляет собой N-(трет-бутоксикарбонил), cit представляет собой цитруллин (2-амино-5-уреидопентановая кислота), dap представляет собой долапроин, DCC представляет собой 1,3-дициклогексилкарбодиимид, DCM представляет собой дихлорметан, DEA представляет собой диэтиламин, DEAD представляет собой диэтилазодикарбоксилат, DEPC представляет собой диэтилфосфорилцианидат, DIAD представляет собой диизопропилазодикарбоксилат, DIEA представляет собой N,N-диизопропилэтиламин, dil представляет собой долаизолейцин, DMA представляет собой диметилацетамид, DMAP представляет собой 4-диметиламинопиридин, DME представляет собой диметиловый эфир этиленгликоля (или 1,2-диметоксиэтан), DMF представляет собой N,N-диметилформамид, DMSO представляет собой диметилсульфоксид, doe представляет собой долафенин, dov представляет собой N,N-диметилвалин, DTNB представляет собой 5,5'-дитиобис(2-нитробензойную кислоту), DTPA представляет собой диэтилентриаминпентауксусную кислоту, DTT представляет собой дитиотреитол, EDCI представляет собой гидрохлорид 1-(3-диметиламинопропил)-3-этилкарбодиимида, EEDQ представляет собой 2-этокси-1-этоксикарбонил-1,2-дигидрохинолин, ES-MS представляет собой масс-спектрометрию с электрораспылением, EtOAc представляет собой этилацетат, Fmoc представляет собой N-(9-флуоренилметоксикарбонил), gly представляет собой глицин, HATU представляет собой гексафторфосфат O-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония, HOBt представляет собой 1-гидроксибензотриазол, HPLC представляет собой высокоэффективную жидкостную хроматографию, ile представляет собой изолейцин, lys представляет собой лизин, MeCN (CH3CN) представляет собой ацетонитрил, MeOH представляет собой метанол, Mtr представляет собой 4-анизилдифенилметил (или 4-метокситритил), nor представляет собой (1S,2R)-(+)-норэфедрин, PAB представляет собой п-аминобензилкарбамоил, PBS представляет собой фосфатно-солевой буфер (pH 7), PEG представляет собой полиэтиленгликоль, Ph представляет собой фенил, Pnp представляет собой п-нитрофенил, MC представляет собой 6-малеимидокапроил, phe представляет собой L-фенилаланин, PyBrop представляет собой гексафторфосфат бром-трис-пирролидинофосфония, SEC представляет собой эксклюзионную хроматографию, Su представляет собой сукцинимид, TFA представляет собой трифторуксусную кислоту, TLC представляет собой тонкослойную хроматографию, UV представляет собой ультрафиолет, и val представляет собой валин.
СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, в которых одна или несколько аминокислот антитела дикого типа или исходного антитела заменены на аминокислоту цистеин. Любую форму антитела можно модифицировать таким образом, т.е. подвергать мутации. Например, исходный Fab-фрагмент антитела можно модифицировать для получения сконструированного Fab с цистеиновыми заменами, обозначенного в настоящем документе как «ТиоFab». Подобным образом, исходное моноклональное антитело можно модифицировать для получения «ТиоMab». Следует отметить, что мутация в одном участке приводит к одному сконструированному остатку цистеина в ТиоFab, в то время как мутация в одном участке приводит к двум сконструированным остаткам цистеина в ТиоMab, из-за димерной природы антитела IgG. Мутанты с замененными («сконструированными») остатками цистеина (Cys) оценивали по реакционной способности вновь введенных сконструированных тиоловых групп цистеина. Значение реакционной способности тиола является относительным числовым термином в диапазоне от 0 до 1,0, и его можно измерять для любого сконструированного антитела с цистеиновыми заменами. Значения реакционной способности тиола сконструированных антител с цистеиновыми заменами по изобретению лежат в диапазонах 0,6-1,0; 0,7-1,0; или 0,8-1,0.
Способы дизайна, отбора и получения по изобретению предоставляют сконструированные антитела с цистеиновыми заменами, реакционноспособные по отношению к электрофильной функциональной группе. Эти способы, кроме того, предоставляют соединения конъюгата с антителом, такие как соединения конъюгата антитело-цирконий (AZC) с атомами циркония в обозначенных сконструированных избранных участках. Реакционноспособные остатки цистеина на поверхности антитела позволяют специфическую конъюгацию группы циркония через реакционноспособную по отношению к тиолу группу, такую как малеимид или галоацетил. Реакционная способность нуклеофильной тиоловой функциональной группы остатка Cys по отношению к малеимидной группе приблизительно в 1000 выше по сравнению с любой другой функциональной группой аминокислот в белке, такой как аминогруппа остатков лизина или N-концевая аминогруппа. Специфические для тиола функциональные группы в иодацетиловых и малеимидных реагентах могут вступать в реакцию с аминогруппами, однако, необходимы более высокий pH (>9,0) и более длительные периоды времени реакции (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London).
Сконструированные антитела с цистеиновыми заменами по изобретению предпочтительно сохраняют антигенсвязывающую способность эквивалентных им антител дикого типа, исходных антител. Таким образом, сконструированные антитела с цистеиновыми заменами способны связываться, предпочтительно, специфически, с антигенами. Такие антигены включают, например, опухолеассоциированные антигены (TAA), рецепторные белки поверхности клеток и другие молекулы поверхности клеток, трансмембранные белки, белки передачи сигнала, регуляторные факторы выживаемости клеток, регуляторные факторы пролиферации клеток, молекулы, ассоциированные с развитием или дифференцировкой тканей (например, как известно или как предполагают, вносящие в них функциональный вклад), лимфокины, цитокины, молекулы, вовлеченные в регуляцию клеточного цикла, молекулы, вовлеченные в васкулогенез, и молекулы, ассоциированные с ангиогенезом (например, как известно или как предполагают, вносящие в него функциональный вклад). Опухолеассоциированный антиген может представлять собой фактор кластера дифференцировки (т.е., CD белок). Антиген, с которым способно связываться сконструированное антитело с цистеиновыми заменами, может являться членом подгруппы одной из вышеупомянутых категорий, где другая подгруппа(подгруппы) указанной категории содержит другие молекулы/антигены, обладающие отличной характеристикой (по отношению к интересующему антигену).
Исходное антитело может представлять собой также гуманизированное антитело, выбранное из huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (Трастузумаб, ГЕРЦЕПТИН®), как описано в таблице 3 US 5821337, включенной в настоящее описание в качестве ссылки в полном объеме; антитела гуманизированное 520C9 (WO 93/21319) и гуманизированное 2C4, как описано в настоящем описании.
Сконструированные антитела с цистеиновыми заменами по изобретению можно сайт-специфически и эффективно присоединять к реакционноспособному по отношению к тиолу реагенту. Реакционноспособный по отношению к тиолу реагент может представлять собой мультифункциональный линкерный реагент, реагент с меткой связывания, т.е. аффинности, (например, биотин-линкерный реагент), метку для детекции (например, реагент с флуорофором), реагент для иммобилизации на твердой фазе (например, SEPHAROSE™, полистироле или стекле), или промежуточное соединение цирконий-линкер. Одним из примеров реакционноспособного по отношению к тиолу реагента является N-этилмалеимид (NEM). В иллюстративном варианте осуществления реакция ТиоFab с реагентом биотин-линкер предоставляет биотинилированное ТиоFab, посредством которого можно детектировать и измерять присутствие и реакционную способность сконструированного остатка цистеина. Реакция ТиоFab с мультифункциональным линкерным реагентом предоставляет ТиоFab с функционализированным линкером, который далее может вступать в реакцию с реагентом с группой циркония или с другой меткой. Реакция ТиоFab с промежуточным соединением цирконий-линкер предоставляет конъюгат ТиоFab с цирконием.
Иллюстративные способы, описанные в настоящем документе, можно широко использовать для идентификации и продукции антител, и более широко, для других белков, посредством использования стадий разработки и скрининга, описанных в настоящем документе.
Такой способ можно использовать для конъюгации других реакционноспособных по отношению к тиолу средств, в которых реакционноспособная группа представляет собой, например, малеимид, иодацетамид, пиридилдисульфид, или другой реакционноспособный по отношению к тиолу партнер для конъюгации (Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671). Партнер может представлять собой цитотоксическое средство (например, токсин, такой как доксорубицин или токсин коклюша), флуорофор, такой как флуоресцентный краситель, подобный флуоресцеину или родамину, хелатирующий агент для получения изображений или металлсодержащее радиотерапевтическое средство, пептидильную или непептидильную метку или метку для детекции, или модифицирующее выведение средство, такое как различные изомеры полиэтиленгликоля, пептид, связывающий третий компонент, или другой углевод или липофильное средство.
Участки, идентифицированные в иллюстративном фрагменте антитела, hu4D5Fabv8, в настоящей работе, находятся в первую очередь в константном домене антитела, который является достаточно консервативным во всех видах антител. Эти участки можно широко использовать для других антител, без дальнейшей необходимости структурной разработки или знаний о структурах конкретных антител и без влияния на свойства связывания антигена, присущие вариабельным доменам антитела.
Сконструированные антитела с цистеиновыми заменами, которые могут являться пригодными для лечения злокачественной опухоли, включают в качестве неограничивающих примеров, антитела против рецепторов поверхности клеток и опухолеассоциированных антигенов (TAA). Такие антитела можно использовать в форме голых антител (не конъюгированных с группой метки) или в форме конъюгатов антитело-цирконий (AZC) формулы I. Опухолеассоциированные антигены известны в данной области и могут быть получены для использования в получении антител с использованием способов и информации, хорошо известных в данной области. В попытках открыть эффективные клеточные мишени для диагностики и терапии злокачественных опухолей, исследователи стремились к идентификации трансмембранных или иным образом ассоциированных с опухолями полипептидов, специфически экспрессирующихся на поверхности одного или нескольких конкретных типа(типов) клеток злокачественных опухолей по сравнению с одной или несколькими нормальными незлокачественными клеткой(клетками). Часто, такие опухолеассоциированные полипептиды более сильно экспрессированы на поверхности клеток злокачественных опухолей по сравнению с поверхностью незлокачественных клеток. Идентификация таких опухолеассоциированных антигенных полипептидов на поверхности клеток привела к возможности специфического нацеливания на клетки злокачественных опухолей для их разрушения способами терапии на основе антител.
TAA включают в качестве неограничивающих примеров TAA (1)-(36), перечисленные ниже. Для удобства, информация относительно этих антигенов, все из которых известны в данной области, перечислена ниже и включает наименования, альтернативные наименования, номера доступа в Genbank и основополагающую ссылку(ссылки), следующие идентификационным обозначениям последовательностей нуклеиновых кислот и белков Национального центра биотехнологической информации (NCBI). Последовательности нуклеиновых кислот и белков, соответствующие TAA (1)-(36), доступны в публичных базах данных, таких как GenBank. Опухолеассоциированные антигены, являющиеся мишенями для антител, включают все варианты и изоформы аминокислотной последовательности, обладающие по меньшей мере приблизительно 70%, 80%, 85%, 90% или 95% идентичностью последовательности по отношению к последовательности, идентифицированной в процитированных ссылках, или обладающие по существу такими же биологическими свойствами или характеристиками, как TAA, обладающие последовательностью, обнаруженной в процитированных ссылках. Например, TAA, обладающие вариантом последовательности, как правило, способны специфически связываться с антителом, специфически связывающимся с TAA с соответствующими перечисленными последовательностями. Последовательности и описание в ссылке, конкретно процитированной в настоящем документе, в явной форме приведены в качестве ссылки.
ОПУХОЛЕАССОЦИИРОВАННЫЕ АНТИГЕНЫ (1)-(36):
(1) BMPR1B (рецептор костного морфогенного белка типа IB, номер доступа в Genbank NM_001203)
ten Dijke,P., et al Science 264 (5155): 101-104 (1994), Oncogene 14 (11): 1377-1382 (1997)); WO2004063362 (пункт формулы изобретения 2); WO2003042661 (пункт формулы изобретения 12); US2003134790-A1 (страницы 38-39); WO2002102235 (пункт формулы изобретения 13; страница 296); WO2003055443 (страницы 91-92); WO200299122 (Пример 2; страницы 528-530); WO2003029421 (пункт формулы изобретения 6); WO2003024392 (пункт формулы изобретения 2; Фиг.112); WO200298358 (пункт формулы изобретения 1; страница 183); WO200254940 (страницы 100-101); WO200259377(страницы 349-350); WO200230268 (пункт формулы изобретения 27; страница 376); WO200148204 (Пример; Фиг.4)
NP_001194 bone morphogenetic protein receptor, type IB /pid=NP_001194.1
Перекрестные ссылки: MIM:603248; NP_001194.1; AY065994
(2) E16 (LAT1, SLC7A5, номер доступа в Genbank NM_003486)
Biochem. Biophys. Res. Commun. 255 (2), 283-288 (1999), Nature 395 (6699):288-291 (1998), Gaugitsch, H.W., et al (1992) J. Biol. Chem. 267 (16): 11267-11273); WO2004048938 (Пример 2); WO2004032842 (Пример IV); WO2003042661 (пункт формулы изобретения 12); WO2003016475 (пункт формулы изобретения 1); WO200278524 (Пример 2); WO200299074 (пункт формулы изобретения 19; страницы 127-129); WO200286443 (пункт формулы изобретения 27; страницы 222, 393); WO2003003906 (пункт формулы изобретения 10; страницы 293); WO200264798 (пункт формулы изобретения 33; страницы 93-95); WO200014228 (пункт формулы изобретения 5; страницы 133-136); US2003224454 (Фиг.3); WO2003025138 (пункт формулы изобретения 12; страница 150);
NP_003477 solute carrier family 7 (cationic amino acid transporter, y+ system), member 5 /pid=NP_003477.3 - Homo sapiens
Перекрестные ссылки: MIM:600182; NP_003477.3; NM_015923; NM_003486_1
(3) STEAP1 (эпителиальный антиген простаты, содержащий шесть трансмембранных доменов, номер доступа в Genbank NM_012449)
Cancer Res. 61 (15), 5857-5860 (2001), Hubert, R.S., et al (1999) Proc. Natl. Acad. Sci. U.S.A. 96 (25): 14523-14528); WO2004065577 (пункт формулы изобретения 6); WO2004027049 (Фиг.1L); EP1394274 (Пример 11); WO2004016225 (пункт формулы изобретения 2); WO2003042661 (пункт формулы изобретения 12); US2003157089 (Пример 5); US2003185830 (Пример 5); US2003064397 (Фиг.2); WO200289747 (Пример 5; страницы 618-619); WO2003022995 (Пример 9; Фиг.13A, Пример 53; страница 173, Пример 2; Фиг.2A);
NP_036581 six transmembrane epithelial antigen of the prostate
Перекрестные ссылки: MIM:604415; NP_036581.1; NM_012449_1
(4) 0772P (CA125, MUC16, номер доступа в Genbank AF361486)
J. Biol. Chem. 276 (29):27371-27375 (2001)); WO2004045553 (пункт формулы изобретения 14); WO200292836 (пункт формулы изобретения 6; Фиг.12); WO200283866 (пункт формулы изобретения 15; страницы 116-121); US2003124140 (Пример 16); Перекрестные ссылки: GI:34501467; AAK74120.3; AF361486_1
(5) MPF (MPF, MSLN, SMR, фактор активации мегакариоцитов, мезотелин, номер доступа в Genbank NM_005823) Yamaguchi, N., et al Biol. Chem. 269 (2), 805-808 (1994), Proc. Natl. Acad. Sci. U.S.A. 96 (20): 11531-11536 (1999), Proc. Natl. Acad. Sci. U.S.A. 93 (1): 136-140 (1996), J. Biol. Chem. 270 (37):21984-21990 (1995)); WO2003101283 (пункт формулы изобретения 14); (WO2002102235 (пункт формулы изобретения 13; страницы 287-288); WO2002101075 (пункт формулы изобретения 4; страницы 308-309); WO200271928 (страницы 320-321); WO9410312 (страницы 52-57); Перекрестные ссылки: MIM:601051; NP_005814.2; NM_005823_1
(6) Napi3b (NAPI-3B, NPTIIb, SLC34A2, семейство переносчиков растворенных веществ 34 (фосфат натрия), член 2, натрий-зависимый переносчик фосфата 3b, типа II, номер доступа в Genbank NM 006424)
J. Biol. Chem. 277 (22): 19665-19672 (2002), Genomics 62 (2):281-284 (1999), Feild, J.A., et al (1999) Biochem. Biophys. Res. Commun. 258 (3):578-582); WO2004022778 (пункт формулы изобретения 2); EP1394274 (Пример 11); WO2002102235 (пункт формулы изобретения 13; страница 326); EP875569 (пункт формулы изобретения 1; страницы 17-19); WO200157188 (пункт формулы изобретения 20; страница 329); WO2004032842 (Пример IV); WO200175177 (пункт формулы изобретения 24; страницы 139-140);
Перекрестные ссылки: MIM:604217; NP_006415.1; NM_006424_1
(7) Sema 5b (FLJ10372, KIAA1445, Mm.42015, SEMA5B, SEMAG, Семафорин 5b Hlog, домен sema, семь повторов тромбоспондина (типа 1 и подобный типу 1), трансмембранный домен (TM) и короткий цитоплазматический домен, (семафорин) 5B, номер доступа в Genbank AB040878)
Nagase T., et al (2000) DNA Res. 7 (2): 143-150); WO2004000997 (пункт формулы изобретения 1); WO2003003984 (пункт формулы изобретения 1); WO200206339 (пункт формулы изобретения 1; страницы 50); WO200188133 (пункт формулы изобретения 1; страницы 41-43, 48-58); WO2003054152 (пункт формулы изобретения 20); WO2003101400 (пункт формулы изобретения 11);
Номера доступа: Q9P283; EMBL; AB040878; BAA95969.1. Genew; HGNC: 10737;
(8) PSCA hlg (2700050C12Rik, C530008O16Rik, кДНК RIKEN 2700050C12, кДНК RIKEN гена 2700050C12, номер доступа в Genbank AY358628); Ross et al (2002) Cancer Res. 62:2546-2553; US2003129192 (пункт формулы изобретения 2); US2004044180 (пункт формулы изобретения 12); US2004044179 (пункт формулы изобретения 11); US2003096961 (пункт формулы изобретения 11); US2003232056 (Пример 5); WO2003105758 (пункт формулы изобретения 12); US2003206918 (Пример 5); EP1347046 (пункт формулы изобретения 1); WO2003025148 (пункт формулы изобретения 20);
Перекрестные ссылки: GI:37182378; AAQ88991.1; AY358628_1
(9) ETBR (рецептор эндотелина типа B, номер доступа в Genbank AY275463);
Nakamuta M., et al Biochem. Biophys. Res. Commun. 177, 34-39, 1991; Ogawa Y., et al Biochem. Biophys. Res. Commun. 178, 248-255, 1991; Arai H., et al Jpn. Circ. J. 56, 1303-1307, 1992; Arai H., et al J. Biol. Chem. 268, 3463-3470, 1993; Sakamoto A., Yanagisawa M., et al Biochem. Biophys. Res. Commun. 178, 656-663, 1991; Elshourbagy N.A., et al J. Biol. Chem. 268, 3873-3879, 1993; Haendler B., et al J. Cardiovasc. Pharmacol. 20, s1-S4, 1992; Tsutsumi M., et al Gene 228, 43-49, 1999; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99, 16899-16903, 2002; Bourgeois C, et al J. Clin. Endocrinol. Metab. 82, 3116-3123, 1997; Okamoto Y., et al Biol. Chem. 272, 21589-21596, 1997; Verheij J.B., et al Am. J. Med. Genet. 108, 223-225, 2002; Hofstra R.M.W., et al Eur. J. Hum. Genet. 5, 180-185, 1997; Puffenberger E.G., et al Cell 79, 1257-1266, 1994; Attie T., et al, Hum. Mol. Genet. 4, 2407-2409, 1995; Auricchio A., et al Hum. Mol. Genet. 5:351-354, 1996; Amiel J., et al Hum. Mol. Genet. 5, 355-357, 1996; Hofstra R.M.W., et al Nat. Genet. 12, 445-447, 1996; Svensson P.J., et al Hum. Genet. 103, 145-148, 1998; Fuchs S., et al Mol. Med. 7, 115-124, 2001; Pingault V., et al (2002) Hum. Genet. 111, 198-206; WO2004045516 (пункт формулы изобретения 1); WO2004048938 (Пример 2); WO2004040000 (пункт формулы изобретения 151); WO2003087768 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO200261087 (Фиг.1); WO2003016494 (Фиг.6); WO2003025138 (пункт формулы изобретения 12; страницы 144); WO200198351 (пункт формулы изобретения 1; страницы 124-125); EP522868 (пункт формулы изобретения 8; Фиг.2); WO200177172 (пункт формулы изобретения 1; страницы 297-299); US2003109676; US6518404 (Фиг.3); US5773223 (пункт формулы изобретения 1a; кол. 31-34); WO2004001004;
(10) MSG783 (RNF124, гипотетический белок FLJ20315, номер доступа в Genbank NM_017763);
WO2003104275 (пункт формулы изобретения 1); WO2004046342 (Пример 2); WO2003042661 (пункт формулы изобретения 12); WO2003083074 (пункт формулы изобретения 14; страница 61); WO2003018621 (пункт формулы изобретения 1); WO2003024392 (пункт формулы изобретения 2; Фиг.93); WO200166689 (Пример 6);
Перекрестные ссылки: LocusID:54894; NP_060233.2; NM_017763_1
(11) STEAP2 (HGNC_8639, IPCA-1, PCANAP1, STAMP1, STEAP2, STMP, ассоциированный с раком предстательной железы ген 1, ассоциированный с раком предстательной железы белок 1, эпителиальный антиген простаты 2, содержащий шесть трансмембранных доменов, белок простаты, содержащий шесть трансмембранных доменов, номер доступа в Genbank AF455138)
Lab. Invest. 82 (11): 1573-1582 (2002)); WO2003087306; US2003064397 (пункт формулы изобретения 1; Фиг.1); WO200272596 (пункт формулы изобретения 13; страницы 54-55); WO200172962 (пункт формулы изобретения 1; Фиг.4B); WO2003104270 (пункт формулы изобретения 11); WO2003104270 (пункт формулы изобретения 16); US2004005598 (пункт формулы изобретения 22); WO2003042661 (пункт формулы изобретения 12); US2003060612 (пункт формулы изобретения 12; Фиг.10); WO200226822 (пункт формулы изобретения 23; Фиг.2); WO200216429 (пункт формулы изобретения 12; Фиг.10);
Перекрестные ссылки: GI:22655488; AAN04080.1; AF455138_1
(12) TrpM4 (BR22450, FLJ20041, TRPM4, TRPM4B, катионный канал транзиторного рецепторного потенциала, подсемейство M, член 4, номер доступа в Genbank NM_017636)
Xu, X.Z., et al Proc. Natl. Acad. Sci. U.S.A. 98 (19): 10692-10697 (2001), Cell 109 (3):397-407 (2002), J. Biol. Chem. 278 (33):30813-30820 (2003)); US2003143557 (пункт формулы изобретения 4); WO200040614 (пункт формулы изобретения 14; страницы 100-103); WO200210382 (пункт формулы изобретения 1; Фиг.9A); WO2003042661 (пункт формулы изобретения 12); WO200230268 (пункт формулы изобретения 27; страница 391); US2003219806 (пункт формулы изобретения 4); WO200162794 (пункт формулы изобретения 14; Фиг.1A-D);
Перекрестные ссылки: MIM:606936; NP_060106.2; NM_017636_1
(13) CRIPTO (CR, CR1, CRGF, CRIPTO, TDGF1, полученный из тератокарциномы фактор роста, номер доступа в Genbank NP 003203 или NM_003212)
Ciccodicola, A., et al EMBO J. 8 (7): 1987-1991 (1989), Am. J. Hum. Genet. 49 (3):555-565 (1991)); US2003224411 (пункт формулы изобретения 1); WO2003083041 (Пример 1); WO2003034984 (пункт формулы изобретения 12); WO200288170 (пункт формулы изобретения 2; страницы 52-53); WO2003024392 (пункт формулы изобретения 2; Фиг.58); WO200216413 (пункт формулы изобретения 1; страницы 94-95, 105); WO200222808 (пункт формулы изобретения 2; Фиг.1); US5854399 (Пример 2; кол. 17-18); US5792616 (Фиг.2);
Перекрестные ссылки: MIM:187395; NP_003203.1; NM_003212_1
(14) CD21 (CR2 (рецептор комплемента 2) или C3DR (рецептор C3d/вируса Эпштейна-Барр) или Hs.73792 номер доступа в Genbank M26004)
Fujisaku et al (1989) J. Biol. Chem. 264 (4):2118-2125); Weis J.J., et al J. Exp. Med. 167, 1047-1066, 1988; Moore M., et al Proc. Natl. Acad. Sci. U.S.A. 84, 9194-9198, 1987; Barel M., et al Mol. Immunol. 35, 1025-1031, 1998; Weis J.J., et al Proc. Natl. Acad. Sci. U.S.A. 83, 5639-5643, 1986; Sinha S.K., et al (1993) J. Immunol. 150, 5311-5320; WO2004045520 (Пример 4); US2004005538 (Пример 1); WO2003062401 (пункт формулы изобретения 9); WO2004045520 (Пример 4); WO9102536 (Фиг.9.1-9.9); WO2004020595 (пункт формулы изобретения 1);
Номера доступа: P20023; Q13866; Q14212; EMBL; M26004; AAA35786.1.
(15) CD79b (CD79B, CD79β, IGb (иммуноглобулин-ассоциированная молекула бета), B29, номер доступа в Genbank NM_000626 или 11038674)
Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (7):4126-4131, Blood (2002) 100 (9):3068-3076, Muller et al (1992) Eur. J. Immunol. 22 (6): 1621-1625); WO2004016225 (пункт формулы изобретения 2, Фиг.140); WO2003087768, US2004101874 (пункт формулы изобретения 1, страница 102); WO2003062401 (пункт формулы изобретения 9); WO200278524 (Пример 2); US2002150573 (пункт формулы изобретения 5, страница 15); US5644033; WO2003048202 (пункт формулы изобретения 1, страницы 306 и 309); WO 99/558658, US6534482 (пункт формулы изобретения 13, Фиг.17A/B); WO200055351 (пункт формулы изобретения 11, страницы 1145-1146);
Перекрестные ссылки: MIM:147245; NP_000617.1; NM_000626_1
(16) FcRH2 (IFGP4, IRTA4, SPAP1A (содержащий SH2 домен фосфатазный якорный белок 1a), SPAP1B, SPAP1C, номер доступа в Genbank NM_030764, AY358130)
Genome Res. 13 (10):2265-2270 (2003), Immunogenetics 54 (2):87-95 (2002), Blood 99 (8):2662-2669 (2002), Proc. Natl. Acad. Sci. U.S.A. 98 (17):9772-9777 (2001), Xu, M.J., et al (2001) Biochem. Biophys. Res. Commun. 280 (3):768-775; WO2004016225 (пункт формулы изобретения 2); WO2003077836; WO200138490 (пункт формулы изобретения 5; Фиг.18D-1-18D-2); WO2003097803 (пункт формулы изобретения 12); WO2003089624 (пункт формулы изобретения 25);
Перекрестные ссылки: MIM:606509; NP_110391.2; NM_030764_1
(17) HER2 (ErbB2, номер доступа в Genbank M11730)
Coussens L., et al Science (1985) 230(4730): 1132-1139); Yamamoto T., et al Nature 319, 230-234, 1986; Semba K., et al Proc. Natl. Acad. Sci. U.S.A. 82, 6497-6501, 1985; Swiercz J.M., et al J. Cell Biol. 165, 869-880, 2004; Kuhns J.J., et al J. Biol. Chem. 274, 36422-36427, 1999; Cho H.-S., et al Nature 421, 756-760, 2003; Ehsani A., et al (1993) Genomics 15, 426-429; WO2004048938 (Пример 2); WO2004027049 (Фиг.1I); WO2004009622; WO2003081210; WO2003089904 (пункт формулы изобретения 9); WO2003016475 (пункт формулы изобретения 1); US2003118592; WO2003008537 (пункт формулы изобретения 1); WO2003055439 (пункт формулы изобретения 29; Фиг.1A-B); WO2003025228 (пункт формулы изобретения 37; Фиг.5C); WO200222636 (Пример 13; страницы 95-107); WO200212341 (пункт формулы изобретения 68; Фиг.7); WO200213847 (страницы 71-74); WO200214503 (страницы 114-117); WO200153463 (пункт формулы изобретения 2; страницы 41-46); WO200141787 (страница 15); WO200044899 (пункт формулы изобретения 52; Фиг.7); WO200020579 (пункт формулы изобретения 3; Фиг.2); US5869445 (пункт формулы изобретения 3; Кол. 31-38); WO9630514 (пункт формулы изобретения 2; страницы 56-61); EP1439393 (пункт формулы изобретения 7); WO2004043361 (пункт формулы изобретения 7); WO2004022709; WO200100244 (Пример 3; Фиг.4);
Номера доступа: P04626; EMBL; M11767; AAA35808.1. EMBL; М11761; AAA35808.1.
(18) NCA (CEACAM6, номер доступа в Genbank M18728);
Barnett T., et al Genomics 3, 59-66, 1988; Tawaragi Y., et al Biochem. Biophys. Res. Commun. 150, 89-96, 1988; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99: 16899-16903, 2002; WO2004063709; EP1439393 (пункт формулы изобретения 7); WO2004044178 (Пример 4); WO2004031238; WO2003042661 (пункт формулы изобретения 12); WO200278524 (Пример 2); WO200286443 (пункт формулы изобретения 27; страница 427); WO200260317 (пункт формулы изобретения 2);
Номера доступа: P40199; Q14920; EMBL; M29541; AAA59915.1. EMBL; M18728;
(19) MDP (DPEP1, номер доступа в Genbank BC017023)
Proc. Natl. Acad. Sci. U.S.A. 99 (26): 16899-16903 (2002)); WO2003016475 (пункт формулы изобретения 1); WO200264798 (пункт формулы изобретения 33; страницы 85-87); JP05003790 (Фиг.6-8); W09946284 (Фиг.9);
Перекрестные ссылки: MIM:179780; AAH17023.1; BC017023_1
(20) IL20Rα (IL20Ra, ZCYTOR7, номер доступа в Genbank AF184971); Clark H.F., et al Genome Res. 13, 2265-2270, 2003; Mungall A.J., et al Nature 425, 805-811, 2003; Blumberg H., et al Cell 104, 9-19, 2001; Dumoutier L., et al J. Immunol. 167, 3545-3549, 2001; Parrish-Novak J., et al J. Biol. Chem. 277, 47517-47523, 2002; Pletnev S., et al (2003) Biochemistry 42: 12617-12624; Sheikh F., et al (2004) J. Immunol. 172, 2006-2010; EP1394274 (Пример 11); US2004005320 (Пример 5); WO2003029262 (страницы 74-75); WO2003002717 (пункт формулы изобретения 2; страница 63); WO200222153 (страницы 45-47); US2002042366 (страницы 20-21); WO200146261 (страницы 57-59); WO200146232 (страницы 63-65); W09837193 (пункт формулы изобретения 1; страницы 55-59);
Номера доступа: Q9UHF4; Q6UWA9; Q96SH8; EMBL; AF184971; AAF01320.1.
(21) Бревикан (BCAN, BEHAB, номер доступа в Genbank AF229053)
Gary S.C., et al Gene 256, 139-147, 2000; Clark H.F., et al Genome Res. 13, 2265-2270, 2003; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99, 16899-16903, 2002; US2003186372 (пункт формулы изобретения 11); US2003186373 (пункт формулы изобретения 11); US2003119131 (пункт формулы изобретения 1; Фиг.52); US2003119122 (пункт формулы изобретения 1; Фиг.52); US2003119126 (пункт формулы изобретения 1); US2003119121 (пункт формулы изобретения 1; Фиг.52); US2003119129 (пункт формулы изобретения 1); US2003119130 (пункт формулы изобретения 1); US2003119128 (пункт формулы изобретения 1; Фиг.52); US2003119125 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO200202634 (пункт формулы изобретения 1);
(22) EphB2R (DRT, ERK, Hek5, EPHT3, Tyro5, номер доступа в Genbank NM_004442)
Chan,J. и Watt, V.M., Oncogene 6 (6), 1057-1061 (1991) Oncogene 10 (5):897-905 (1995), Annu. Rev. Neurosci. 21:309-345 (1998), Int. Rev. Cytol. 196: 177-244 (2000)); WO2003042661 (пункт формулы изобретения 12); WO200053216 (пункт формулы изобретения 1; страница 41); WO2004065576 (пункт формулы изобретения 1); WO2004020583 (пункт формулы изобретения 9); WO2003004529 (страницы 128-132); WO200053216 (пункт формулы изобретения 1; страница 42);
Перекрестные ссылки: MIM:600997; NP_004433.2; NM_004442_1
(23) ASLG659 (B7h, номер доступа в Genbank AX092328)
US20040101899 (пункт формулы изобретения 2); WO2003104399 (пункт формулы изобретения 11); WO2004000221 (Фиг.3); US2003165504 (пункт формулы изобретения 1); US2003124140 (Пример 2); US2003065143 (Фиг.60); WO2002102235 (пункт формулы изобретения 13; страница 299); US2003091580 (Пример 2); WO200210187 (пункт формулы изобретения 6; Фиг.10); WO200194641 (пункт формулы изобретения 12; Фиг.7b); WO200202624 (пункт формулы изобретения 13; Фиг.1A-1B); US2002034749 (пункт формулы изобретения 54; страницы 45-46); WO200206317 (Пример 2; страницы 320-321, пункт формулы изобретения 34; страницы 321-322); WO200271928 (страницы 468-469); WO200202587 (Пример 1; Фиг.1); WO200140269 (Пример 3; страницы 190-192); WO200036107 (Пример 2; страницы 205-207); WO2004053079 (пункт формулы изобретения 12); WO2003004989 (пункт формулы изобретения 1); WO200271928 (страницы 233-234, 452-453); WO 0116318;
(24) PSCA (Предшественник антигена стволовых клеток простаты, номер доступа в Genbank AJ297436)
Reiter R.E., et al Proc. Natl. Acad. Sci. U.S.A. 95, 1735-1740, 1998; Gu Z., et al Oncogene 19, 1288-1296, 2000; Biochem. Biophys. Res. Commun. (2000) 275(3):783-788; WO2004022709; EP1394274 (Пример 11); US2004018553 (пункт формулы изобретения 17); WO2003008537 (пункт формулы изобретения 1); WO200281646 (пункт формулы изобретения 1; страница 164); WO2003003906 (пункт формулы изобретения 10; страница 288); WO200140309 (Пример 1; Фиг.17); US2001055751 (Пример 1; Фиг.1b); WO200032752 (пункт формулы изобретения 18; Фиг.1); WO9851805 (пункт формулы изобретения 17; страница 97); W09851824 (пункт формулы изобретения 10; страница 94); WO9840403 (пункт формулы изобретения 2; Фиг.1B);
Номер доступа: O43653; EMBL; AF043498; AAC39607.1.
(25) GEDA (номер доступа в Genbank AY260763);
AAP14954 lipoma HMGIC fusion-partner-like protein /pid=AAP14954.1 - Homo sapiens Species: Homo sapiens (human)
WO2003054152 (пункт формулы изобретения 20); WO2003000842 (пункт формулы изобретения 1); WO2003023013 (Пример 3, пункт формулы изобретения 20); US2003194704 (пункт формулы изобретения 45);
Перекрестные ссылки: GI:30102449; AAP14954.1; AY260763_1
(26) BAFF-R (рецептор фактора активации B-клеток, BLyS рецептор 3, BR3, номер доступа в Genbank AF116456); BAFF receptor /pid=NP_443177.1 - Homo sapiens
Thompson, J.S., et al Science 293 (5537), 2108-2111 (2001); WO2004058309; WO2004011611; WO2003045422 (Пример; страницы 32-33); WO2003014294 (пункт формулы изобретения 35; Фиг.6B); WO2003035846 (пункт формулы изобретения 70; страницы 615-616); WO200294852 (Кол. 136-137); WO200238766 (пункт формулы изобретения 3; страница 133); WO200224909 (Пример 3; Фиг.3);
Перекрестные ссылки: MIM:606269; NP_443177.1; NM_052945_1; AF132600
(27) CD22 (изоформа B-клеточного рецептора CD22-B, BL-CAM, Lyb-8, Lyb8, SIGLEC-2, FLJ22814, номер доступа в Genbank AK026467);
Wilson et al (1991) J. Exp. Med. 173: 137-146; WO2003072036 (пункт формулы изобретения 1; Фиг.1);
Перекрестные ссылки: MIM:107266; NP_001762.1; NM_001771_1
(28) CD79a (CD79A, CD79α, иммуноглобулин-ассоциированная молекула альфа, специфический для B-клеток белок, ковалентно взаимодействующий с Ig бета (CD79B) и формирующий комплекс на поверхности с молекулами IgM, передает сигнал, вовлеченный в дифференцировку B-клеток), pI: 4,84, MW: 25028 TM: 2 [P] Хромосома гена: 19q13.2, номер доступа в Genbank NP_001774.10)
WO2003088808, US20030228319; WO2003062401 (пункт формулы изобретения 9); US2002150573 (пункт формулы изобретения 4, страницы 13-14); W09958658 (пункт формулы изобретения 13, Фиг.16); WO9207574 (Фиг.1); US5644033; Ha et al (1992) J. Immunol. 148(5): 1526-1531; Mueller et al (1992) Eur. J. Biochem. 22: 1621-1625; Hashimoto et al (1994) Immunogenetics 40(4):287-295; Preud'homme et al (1992) Clin. Exp. Immunol. 90(1): 141-146; Yu et al (1992) J. Immunol. 148(2) 633-637; Sakaguchi et al (1988) EMBO J. 7(11):3457-3464;
(29) CXCR5 (Рецептор лимфомы Беркитта 1, связанный с G белком рецептор, активируемый хемокином CXCL13, функционирует в миграции лимфоцитов и гуморальной защите, играет роль в инфекции HIV-2 и, возможно, развитии СПИД, лимфомы, миеломы и лейкоза); 372 ак, pI: 8,54 MW: 41959 TM: 7 [P] Хромосома гена: 11q23.3, номер доступа в Genbank NP_001707.1)
WO2004040000; WO2004015426; US2003105292 (Пример 2); US6555339 (Пример 2); WO200261087 (Фиг.1); WO200157188 (пункт формулы изобретения 20, страница 269); WO200172830 (страницы 12-13); WO200022129 (Пример 1, страницы 152-153, Пример 2, страницы 254-256); W09928468 (пункт формулы изобретения 1, страница 38); US5440021 (Пример 2, кол. 49-52); W09428931 (страницы 56-58); W09217497 (пункт формулы изобретения 7, Фиг.5); Dobner et al (1992) Eur. J. Immunol. 22:2795-2799; Barella et al (1995) Biochem. J. 309:773-779;
(30) HLA-DOB (Бета-субъединица молекулы MHC класса II (Ia антиген), связывающая пептиды и представляющая их CD4+ T-лимфоцитам); 273 ак, pI: 6,56 MW: 30820 TM: 1 [P] Хромосома гена: 6p21.3, номер доступа в Genbank NP_002111.1)
Tonnelle et al (1985) EMBO J. 4(11):2839-2847; Jonsson et al (1989) Immunogenetics 29(6):411-413; Beck et al (1992) J. Mol. Biol. 228:433-441; Strausberg et al (2002) Proc. Natl. Acad. Sci USA 99: 16899-16903; Servenius et al (1987) J. Biol. Chem. 262:8759-8766; Beck et al (1996) J. Mol. Biol. 255: 1-13; Naruse et al (2002) Tissue Antigens 59:512-519; W09958658 (пункт формулы изобретения 13, Фиг.15); US6153408 (Кол. 35-38); US5976551 (кол. 168-170); US6011146 (кол. 145-146); Kasahara et al (1989) Immunogenetics 30(1):66-68; Larhammar et al (1985) J. Biol. Chem. 260(26): 14111-14119;
(31) P2X5 (управляемый лигандом пуринергического рецептора P2X ионный канал 5, ионный канал, управляемый внеклеточным ATP, может быть вовлечен в передачу возбуждения по синапсам и нейрогенез, недостаточность может вносить вклад в патофизиологию идиопатической нестабильности детрузора); 422 ак), pI: 7,63, MW: 47206 TM: 1 [P] Хромосома гена: 17p13.3, номер доступа в Genbank NP_002552.2)
Le et al (1997) FEBS Lett. 418(1-2): 195-199; WO2004047749; WO2003072035 (пункт формулы изобретения 10); Touchman et al (2000) Genome Res. 10: 165-173; WO200222660 (пункт формулы изобретения 20); WO2003093444 (пункт формулы изобретения 1); WO2003087768 (пункт формулы изобретения 1); WO2003029277 (страница 82);
(32) CD72 (антиген дифференцировки B-клеток CD72, Lyb-2) БЕЛКОВАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ Полная maeaity...tafrfpd (1..359; 359 ак), pI: 8,66, MW: 40225 TM: 1 [P] Хромосома гена: 9p13.3, номер доступа в Genbank NP_001773.1)
WO2004042346 (пункт формулы изобретения 65); WO2003026493 (страницы 51-52, 57-58); WO200075655 (страницы 105-106); Von Hoegen et al (1990) J. Immunol. 144(12):4870-4877; Strausberg et al (2002) Proc. Natl. Acad. Sci USA 99: 16899-16903;
(33) LY64 (Лимфоцитарный антиген 64 (RP105), мембранный белок типа I семейства богатых лейцином повторов (LRR), регулирует активацию и апоптоз B-клеток, потеря функции ассоциирована с увеличенной активностью заболевания у пациентов с системной красной волчанкой); 661 ак, pI: 6,20, MW: 74147 TM: 1 [P] Хромосома гена: 5q12, номер доступа в Genbank NP_005573.1)
US2002193567; WO9707198 (пункт формулы изобретения 11, стр. 39-42); Miura et al (1996) Genomics 38(3):299-304; Miura et al (1998) Blood 92:2815-2822; WO2003083047; W09744452 (пункт формулы изобретения 8, страницы 57-61); WO200012130 (страницы 24-26);
(34) FcRH1 (подобный Fc-рецептору белок 1, предположительный рецептор для Fc-домена иммуноглобулинов, содержащий Ig-подобный домен типа C2 и домен ITAM, может играть роль в дифференцировке B-лимфоцитов); 429 ак, pI: 5,28, MW: 46925 TM: 1 [P] Хромосома гена: 1q21-1q22, номер доступа в Genbank NP_443170.1)
WO2003077836; WO200138490 (пункт формулы изобретения 6, Фиг.18E-1-18-E-2); Davis et al (2001) Proc. Natl. Acad. Sci USA 98(17):9772-9777; WO2003089624 (пункт формулы изобретения 8); EP1347046 (пункт формулы изобретения 1); WO2003089624 (пункт формулы изобретения 7);
(35) IRTA2 (ассоциированный с транслокацией рецептор 2 суперсемейства иммуноглобулинов, предположительный иммунорецептор с возможной ролью в развитии B-клеток и лимфомагенезе; нарушение регуляции гена посредством транслокации происходит при некоторых злокачественных новообразованиях B-клеток); 977 ак, pI: 6,88 MW: 106468 TM: 1 [P] Хромосома гена: 1q21, номер доступа в Genbank Человек: AF343662, AF343663, AF343664, AF343665, AF369794, AF397453, AK090423, AK090475, AL834187, AY358085; Мышь: AK089756, AY158090, AY506558; NP_112571.1
WO2003024392 (пункт формулы изобретения 2, Фиг.97); Nakayama et al (2000) Biochem. Biophys. Res. Commun. 277(1): 124-127; WO2003077836; WO200138490 (пункт формулы изобретения 3, Фиг.18B-1-18B-2);
(36) TENB2 (TMEFF2, томорегулин, TPEF, HPP1, TR, предположительный трансмембранный протеогликан, родственный семейству факторов роста EGF/херегулина и фоллистатину); 374 ак, Номера доступа в NCBI: AAD55776, AAF91397, AAG49451, NCBI RefSeq: NP_057276; Ген в NCBI: 23671; OMIM: 605734; SwissProt Q9UIK5; номер доступа в Genbank AF179274; AY358907, CAF85723, CQ782436
WO2004074320 (SEQ ID NO 810); JP2004113151 (SEQ ID NO 2, 4, 8); WO2003042661 (SEQ ID NO 580); WO2003009814 (SEQ ID NO 411); EP1295944 (страницы 69-70); WO200230268 (страница 329); WO200190304 (SEQ ID NO 2706); US2004249130; US2004022727; WO2004063355; US2004197325; US2003232350; US2004005563; US2003124579; Horie et al (2000) Genomics 67: 146-152; Uchida et al (1999) Biochem. Biophys. Res. Commun. 266:593-602; Liang et al (2000) Cancer Res. 60:4907-12; Glynne-Jones et al (2001) Int J Cancer. Oct 15;94(2): 178-84;
(37) PMEL17 (гомолог silver; SILV; D12S53E; PMEL17; (SI); (SIL); ME20; gp100) BC001414; BT007202; M32295; M77348; NM_006928; McGlinchey,R.P. et al (2009) Proc. Natl. Acad. Sci. U.S.A. 106 (33), 13731-13736; Kummer,M.P. et al (2009) J. Biol. Chem. 284 (4), 2296-2306;
(38) TMEFF1 (трансмембранный белок с EGF-подобным доменом и двумя фоллистатин-подобными доменами 1; Томорегулин-1; H7365; C9orf2; C9ORF2; U19878; X83961) NM_080655; NM_003692; Harms, P.W. (2003) Genes Dev. 17 (21), 2624-2629; Gery, S. et al (2003) Oncogene 22 (18):2723-2727;
(39) GDNF-Ra1 (рецептор альфа 1 семейства GDNF; GFRA1; GDNFR; GDNFRA; RETL1; TRNR1; RET1L; GDNFR-alpha1; GFR-ALPHA-1; U95847; BC014962; NM_145793)
NM_005264; Kim, M.H. et al (2009) Mol. Cell. Biol. 29 (8), 2264-2277; Treanor, J.J. et al (1996) Nature 382 (6586): 80-83;
(40) Ly6E (комплекс лимфоцитарного антигена 6, локус E; Ly67,RIG-E,SCA-2,TSA-1)
NP_002337.1; NM_002346.2; de Nooij-van Dalen, A.G. et al (2003) Int. J. Cancer 103 (6), 768-774; Zammit, D.J. et al (2002) Mol. Cell. Biol. 22 (3):946-952;
(41) TMEM46 (гомолог shisa 2 (Xenopus laevis); SHISA2) NP_001007539.1;
NM_001007538,1; Furushima,K. et al (2007) Dev. Biol. 306 (2), 480-492; Clark,H.F. et al (2003) Genome Res. 13 (10):2265-2270;
(42) Ly6G6D (комплекс лимфоцитарного антигена 6, локус G6D; Ly6-D, MEGT1) NP_067079.2; NM_021246.2; Mallya, M. et al (2002) Genomics 80 (1): 113-123; Ribas, G. et al (1999) J. Immunol. 163 (1):278-287;
(43) LGR5 (содержащий богатые лейцином повторы связанный с белком G рецептор 5; GPR49, GPR67) NP_003658.1; NM_003667.2; Salanti,G. et al (2009) Am. J. Epidemiol. 170 (5):537-545; Yamamoto,Y. et al (2003) Hepatology 37 (3):528-533;
(44) RET (протоонкоген ret; MEN2A; HSCR1; MEN2B; MTC1; (PTC); CDHF12; Hs.168114; RET51; RET-ELE1) NP_066124.1; NM_020975.4; Tsukamoto, H. et al (2009) Cancer Sci. 100 (10): 1895-1901; Narita,N. et al (2009) Oncogene 28 (34):3058-3068;
(45) LY6K (комплекс лимфоцитарного антигена 6, локус K; LY6K; HSJ001348; FLJ35226) NP_059997.3; NM_017527.3; Ishikawa,N. et al (2007) Cancer Res. 67 (24): 11601-11611; de Nooij-van Dalen,A.G. et al (2003) Int. J. Cancer 103 (6):768-774;
(46) GPR19 (связанный с белком G рецептор 19; Mm.4787) NP_006134.1; NM_006143.2;
Montpetit, A. and Sinnett,D. (1999) Hum. Genet. 105 (1-2): 162-164; O'Dowd, B.F. et al (1996) FEBS Lett. 394 (3):325-329;
(47) GPR54 (рецептор KISS1; KISS1R; GPR54; HOT7T175; AXOR12) NP_115940.2; NM_032551.4; Navenot, J.M. et al (2009) Mol. Pharmacol. 75 (6): 1300-1306; Hata, K. et al (2009) Anticancer Res. 29 (2):617-623;
(48) ASPHD1 (содержащий домен аспартат-бета-гидроксилазы белок 1; LOC253982) NP_859069.2; NM_181718.3; Gerhard, D.S. et al (2004) Genome Res. 14 (10B):2121-2127;
(49) Тирозиназа (TYR; OCAIA; OCA1A; тирозиназа; SHEP3) NP_000363.1; NM_000372.4; Bishop, D.T. et al (2009) Nat. Genet. 41 (8):920-925; Nan, H. et al (2009) Int. J. Cancer 125 (4):909-917;
(50) TMEM118 (белок с «безымянным пальцем», трансмембранный белок 2; RNFT2; FLJ14627)
NP_001103373.1; NM_001109903.1; Clark, H.F. et al (2003) Genome Res. 13 (10):2265-2270; Scherer,S.E. et al (2006) Nature 440 (7082):346-351
(51) GPR172A (связанный с белком G рецептор 172A; GPCR41; FLJ11856; D15Ertd747e)
NP_078807.1; NM_024531.3; Ericsson, T.A. et al (2003) Proc. Natl. Acad. Sci. U.S.A. 100 (11):6759-6764; Takeda, S. et al (2002) FEBS Lett. 520 (1-3):97-101.
Исходное антитело может представлять собой также слитый белок, содержащий последовательность альбуминсвязывающего пептида (ABP) (Dennis et al. (2002) «Albumin Binding As A General Strategy For Improving The Pharmacokinetics Of Proteins» J Biol Chem. 277:35035-35043; WO 01/45746). Антитела по изобретению включают слитые белки с последовательностями ABP, объясняемые в: (i) Dennis et al (2002) J Biol Chem. 277:35035-35043 в таблицах III и IV, страница 35038; (ii) US 20040001827 в SEQ ID NO: 9-22; и (iii) WO 01/45746 на страницах 12-13, в SEQ ID NO: z1-z14, и содержание всех из них приведено в настоящем документе в качестве ссылки.
МУТАГЕНЕЗ
ДНК, кодирующую вариант аминокислотной последовательности исходного полипептида, получают множеством способов, известных в данной области. Эти способы включают, без ограничения, получение посредством сайт-специфического (или опосредованного олигонуклеотидом) мутагенеза, ПЦР-мутагенеза и кассетного мутагенеза ранее полученной ДНК, кодирующей полипептид. Варианты рекомбинантных антител можно также конструировать посредством манипуляций с рестрикционными фрагментами или посредством ПЦР с удлинением перекрывающихся фрагментов с использованием синтетических олигонуклеотидов. Праймеры для мутагенеза кодируют замену(замены) кодонов на кодоны для цистеина. Общепринятые способы мутагенеза можно использовать для получения ДНК, кодирующей такие мутантные сконструированные антитела с цистеиновыми заменами. Общее руководство можно найти в Sambrook et al Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989; и Ausubel et al Current Protocols in Molecular Biology, Greene Publishing and Wiley-Interscience, New York, N.Y., 1993.
Сайт-специфический мутагенез является одним из способов получения вариантов с заменами, т.е. мутантных белков. Этот способ хорошо известен в данной области (см., например, Carter (1985) et al Nucleic Acids Res. 13:4431-4443; Ho et al (1989) Gene (Amst.) 77:51-59; и Kunkel et al (1987) Proc. Natl. Acad. Sci. USA 82:488). Кратко, при проведении сайт-специфического мутагенеза ДНК, исходную ДНК изменяют, сначала проводя гибридизацию олигонуклеотида, кодирующего желательную мутацию, с одной цепью такой исходной ДНК. После гибридизации используют ДНК-полимеразу для синтеза полной второй цепи с использованием прогибридизовавшегося олигонуклеотида в качестве праймера, и с использованием одной цепи исходной ДНК в качестве матрицы. Таким образом олигонуклеотид, кодирующий желательную мутацию, включают в полученную двухцепочечную ДНК. Сайт-специфический мутагенез можно проводить в гене, экспрессирующем белок, подлежащий мутагенезу, в экспрессирующей плазмиде, и полученную плазмиду можно секвенировать для подтверждения вставки желательных мутаций замены на цистеин (Liu et al (1998) J. Biol. Chem. 273:20252-20260). Способы и форматы сайт-специфического мутагенеза включают коммерчески доступные способы и форматы, например, набор для множественного сайт-специфического мутагенеза QuikChange® (Stratagene, La Jolla, CA).
ПЦР-мутагенез также пригоден для получения вариантов аминокислотной последовательности исходного полипептида. См. Higuchi, (1990) in PCR Protocols, pp.177-183, Academic Press; Ito et al (1991) Gene 102:67-70; Bernhard et al (1994) Bioconjugate Chem. 5: 126-132; и Vallette et al (1989) Nuc. Acids Res. 17:723-733. Кратко, когда малые количества ДНК-матрицы используют в качестве исходного материала для ПЦР, праймеры, которые немного отличаются по последовательности от соответствующей области в ДНК-матрице, можно использовать для получения относительно больших количеств специфического фрагмента ДНК, отличающегося от последовательности матрицы только в положениях, в которых праймеры отличаются от матрицы.
Другой способ получения вариантов, кассетный мутагенез, основан на способе, описанном Wells et al (1985) Gene 34:315-323. Исходный материал представляет собой плазмиду (или другой вектор), содержащий ДНК исходного полипептида, подлежащего мутагенезу. Идентифицируют кодон(ы) в исходной ДНК, подлежащие мутагенезу. Должны присутствовать участки для уникальной рестрикционной эндонуклеазы с каждой стороны идентифицированного участка(участков) мутации. Если таких участков рестрикции не существует, их можно получить с использованием вышеописанного способа опосредованного олигонуклеотидами мутагенеза для введения их в подходящие положения ДНК исходного полипептида. Плазмидную ДНК разрезают в этих участках, чтобы линеаризовать ее. Двухцепочечный олигонуклеотид, кодирующий последовательность ДНК между участками рестрикции, но содержащий желательную мутацию(мутации), синтезируют с использованием общепринятых способов, когда две цепи олигонуклеотида синтезируют по отдельности и затем гибридизуют вместе с использованием общепринятых способов. Этот двухцепочечный олигонуклеотид обозначают кассетой. Эту кассету конструируют с наличием 5'- и 3'-концов, совместимых с концами линеаризованной плазмиды, так что ее можно напрямую лигировать в плазмиду. Эта плазмида теперь содержит последовательность ДНК с мутацией. Мутантную ДНК, содержащую закодированные цистеиновые замены, можно подтверждать секвенированием ДНК.
Отдельные мутации получают также направляемым олигонуклеотидами мутагенезом с использованием двухцепочечной плазмидной ДНК в качестве матрицы, посредством основанного на ПЦР мутагенеза (Sambrook and Russel, (2001) Molecular Cloning: A Laboratory Manual, 3rd edition; Zoller et al (1983) Methods Enzymol. 100:468-500; Zoller, M.J. and Smith, M. (1982) Nucl. Acids Res. 10:6487-6500).
По настоящему изобретению, hu4D5Fabv8, экспонированное на фаге M13 (Gerstner et al (2002) «Sequence Plasticity In The Antigen-Binding Site Of A Therapeutic Anti-HER2 Antibody», J Mol Biol. 321:851-62) использовали во всех экспериментах в качестве модельной системы. Цистеиновые мутации вводили в конструкции hu4D5Fabv8-фаг, hu4D5Fabv8 и ABP-hu4D5Fabv8. Выделения hu4D5-ТиоFab-фага проводили с использованием способа преципитации полиэтиленгликолем (PEG), как описано ранее (Lowman, Henry B. (1998) Methods in Molecular Biology (Totowa, New Jersey) 87 (Combinatorial Peptide Library Protocols) 249-264).
Олигонуклеотиды получают способом фосфорамидитного синтеза (US 4415732; US 4458066; Beaucage, S. and Iyer, R. (1992) «Advances in the synthesis of oligonucleotides by the phosphoramidite approach», Tetrahedron 48:2223-2311). Фосфорамидитный способ включает циклическое добавление нуклеотидных мономерных звеньев с реакционноспособной 3'-фосфорамидитной группой к олигонуклеотидной цепи, растущей на твердой подложке, состоящей из стекла с контролируемым размером пор или полистирола с высокой степенью сшивки, и, наиболее часто, в направлении от 3' к 5', где 3'-концевой нуклеозид присоединяют к твердой подложке в начале синтеза (US 5047524; US 5262530). Способ обычно осуществляют на практике с использованием автоматических, коммерчески доступных синтезаторов (Applied Biosystems, Foster City, CA). Олигонуклеотиды можно химически метить с помощью неизотопных групп для детекции, захвата, стабилизации или других целей (Andrus, A. «Chemical methods for 5' non-isotopic labelling of PCR probes and primers» (1995) in PCR 2: A Practical Approach, Oxford University Press, Oxford, pp. 39-54; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671; Keller, G. and Manak, M. in DNA Probes Second Edition (1993), Stockton Press, New York, pp. 121-23).
АНАЛИЗ PHESELECTOR
Анализ PHESELECTOR (фаговый ELISA для отбора реакционноспособных тиолов) позволяет детекцию реакционноспособных групп цистеина в антителах в формате фагового ELISA (US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008; 332:41-52). Способ покрытия интересующим белком (например, антителом) поверхностей лунок, последующей инкубации с фаговыми частицами и затем с меченным HRP вторичным антителом с детекцией оптической плотности подробно описан в примере 2. Можно проводить скрининг мутантных белков, экспонированных на фагах, быстрым, точным и высоко производительным способом. Библиотеки сконструированных антител с цистеиновыми заменами можно получать и подвергать отбору по связыванию с использованием такого же способа для идентификации подходящих реакционноспособных участков включения свободного Cys из случайных белково-фаговых библиотек антител или других белков. Этот способ включает реакцию мутантных по цистеину белков, экспонированных на фаге, с аффинным реагентом или репортерной группой, которая также является реакционноспособной по отношению к тиолу. Фигура 8 иллюстрирует анализ PHESELECTOR посредством схематического представления, изображающего связывание Fab или ТиоFab с HER2 (вверху) и биотинилированного ТиоFab со стрептавидином (внизу).
ЭКСПРЕССИЯ И ОЧИСТКА БЕЛКА
ДНК, кодирующую сконструированные антитела с цистеиновыми заменами, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Клетки гибридомы служат источником такой ДНК. После выделения ДНК можно помещать в экспрессирующие векторы, которыми затем трансфицируют клетки-хозяева, такие как клетки E. coli, клетки обезьяны COS, клетки яичников китайского хомяка (CHO) или другие клетки-хозяева млекопитающих, такие как клетки миеломы (US 5807715; US 2005/0048572; US 2004/0229310), которые в ином случае не продуцируют белка антитела, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Выходы сконструированных антител с цистеиновыми заменами hu4D5Fabv8 являлись сходными с выходами hu4D5Fabv8 дикого типа. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитело, включают Skerra et al (1993) Curr. Opinion in Immunol. 5:256-262 и Pluckthun (1992) Immunol. Revs. 130: 151-188.
После конструирования и отбора сконструированные антитела с цистеиновыми заменами, например, ТиоFab, с неспаренными остатками Cys с высокой реакционной способностью, можно получать посредством: (i) экспрессии в бактериальной системе, например, E. coli, или в системе культуры клеток млекопитающих (WO 01/00245), например, клеток яичников китайского хомяка (CHO); и (ii) очистки с использованием общепринятых способов очистки белка (Lowman et al (1991) J. Biol. Chem. 266(17): 10982-10988).
ТиоFab экспрессировали при индукции в 34B8, несупрессорном штамме E. coli (Baca et al (1997) Journal Biological Chemistry 272(16): 10678-84). См. Пример 3a. Собранный осадок клеток ресуспендировали в PBS (фосфатно-солевом буфере), общий лизис клеток проводили пропусканием через микрофлюидайзер, и ТиоFab очищали аффинной хроматографией на SEPHAROSE™ с белком G (Amersham). ТиоFab конъюгировали с биотин-PEO-малеимидом, как описано выше, и биотинилированые-ТиоFab дополнительно очищали гель-фильтрационной хроматографией на Superdex-200™ (Amersham), отсекающей свободный биотин-PEO-малеимид и олигомерную фракцию ТиоFab.
АНАЛИЗ МАСС-СПЕКТРОСКОПИИ
Масс-спектрометрический анализ жидкостной хроматографией с ионизацией электрораспылением (LC-ESI-MS) использовали для точного определения молекулярной массы конъюгированных с биотином Fab (Cole, R.B. Electro Spray Ionization Mass Spectrometry: Fundamentals, Instrumentation And Applications. (1997) Wiley, New York). Аминокислотную последовательность биотинилированного пептида hu4D5Fabv8 (A121C) определяли расщеплением трипсином с последующим анализом LC-ESI-тандемной MS (таблица 4, пример 3b).
Fab-фрагмент антитела hu4D5Fabv8 содержит приблизительно 445 аминокислотных остатков, включая 10 остатков Cys (пять на легкой и пять на тяжелой цепи). Определена структура с высоким разрешением вариабельного фрагмента гуманизированного 4D5 (Fv4D5), см.: Eigenbrot et al «X-Ray Structures Of The Antigen-Binding Domains From Three Variants Of Humanized Anti-P185her2 Antibody 4D5 And Comparison With Molecular Modeling» (1993) J Mol Biol. 229:969-995). Все остатки Cys присутствуют в форме дисульфидных связей, таким образом, эти остатки не имеют каких-либо реакционноспособных тиоловых групп для конъюгации с цирконий-малеимидом (если только не обработаны восстанавливающим агентом). Таким образом, вновь сконструированный остаток Cys может оставаться неспаренным, и способным вступать в реакцию, т.е., образовывать конъюгат с электрофильным линкерным реагентом или промежуточным соединением цирконий-линкер, таким как цирконий-малеимид. На фигуре 1A показано трехмерное представление фрагмента антитела hu4D5Fabv8 в координатах кристалла, полученных с помощью рентгеновского излучения. Положения в структуре сконструированных остатков Cys тяжелой и легкой цепей пронумерованы в соответствии с последовательной системой нумерации. Эта последовательная система нумерации коррелирует с системой нумерации Kabat (Kabat et al., (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD) для варианта 4d5v7fabH трастузумаба в соответствии с фигурой 1B, на которой показана схема последовательной нумерации (верхний ряд), начиная с N-конца, отличающаяся от схемы нумерации по Kabat (нижний ряд) вставками, обозначенными a,b,c. При использовании системы нумерации Kabat, действительная линейная аминокислотная последовательность может содержать меньше аминокислот или может содержать дополнительные аминокислоты, соответствующие укорочению FR или CDR вариабельного домена или вставке в них. Участки сконструированного варианта тяжелой цепи с цистеиновыми заменами идентифицированы по схеме последовательной нумерации и схеме нумерации Kabat в следующей таблице:

Для M13 фагмиды-Cys мутантных Fab (фигуры 3A и 3B) можно проводить быстрый скрининг по сравнению с Fab белками. Связывание фагмиды-ТиоFab с антигеном и со стрептавидином можно тестировать покрытием планшетов для ELISA HER2 и стрептавидином, соответственно, с последующим использованием в качестве зонда анти-Fab-HRP (Пероксидаза хрена), как описано в примере 2 и изображено на фигуре 8. Этот способ позволяет одновременное мониторирование эффекта на связывание антигена и на реакционную способность тиоловой группы сконструированного остатка Cys/конъюгированной молекулы биотина. Способ можно использовать также для скрининга реакционноспособных тиоловых групп любого белка, экспонированного на фаге M13. Конъюгированные или неконъюгированные фагмиды-ТиоFab очищают простой преципитацией PEG.
Антигенсвязывающий фрагмент гуманизированного 4D5 (hu4D5Fab) хорошо экспрессируется в E. Coli и был экспонирован на бактериофаге (Garrard et al (1993) Gene 128: 103-109). Fab-фрагмент антитела hu4D5Fabv8 был экспонирован на фаге M13 в качестве модельной системы в анализе на основе ELISA для тестирования реакционной способности тиола. Фигура 8 представляет собой графическое изображение анализа PHESELECTOR, показывающее связывание биотинилированного ТиоFab фага и антитела против фага с HRP с HER2 (вверху) и стрептавидином (внизу). Пять аминокислотных остатков (L-Ala43, H-Ala40, H-Ser119, H-Ala121 и H-Ser122) первоначально выбраны по информации о кристаллической структуре как удаленные от антигенсвязывающей поверхности (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Рентгеновская кристаллическая структура из базы данных белков обозначена 1FVC. Остатки Cys сконструированы в этих положениях посредством сайт-специфического мутагенеза. Препараты ТиоFab-фага выделяли и подвергали реакции с реагентом для биотинилирования.
Конъюгированные и неконъюгированные с биотином варианты тестировали по связыванию с HER2 и стрептавидином с использованием анализа PHESELECTOR на основе ELISA (фигура 8, пример 2) с HRP (пероксидаза хрена)-конъюгированным антителом против фага. Взаимодействие небиотинилированного фаг-hu4D5Fabv8 (фигура 2A) и биотинилированного фаг-hu4D5Fabv8 (Фигура 2B) с BSA (незакрашенный прямоугольник), HER2 (серый прямоугольник) или стрептавидином (закрашенный прямоугольник) мониторировали с помощью антитела анти-M13-пероксидаза хрена (HRP) посредством проявления стандартной реакции HRP и измерения оптической плотности при 450 нм. Оптическую плотность, полученную при превращении колориметрического субстрата, измеряли при 450 нм. Реакционная способность ТиоFab по отношению HER2 является показателем связывания антигена. Реакционная способность ТиоFab по отношению к стрептавидину является показателем степени биотинилирования. Реакционная способность ТиоFab по отношению к BSA является отрицательным контролем для неспецифического взаимодействия. Как видно на фигуре 2A, все варианты ТиоFab-фага обладали сходным связыванием с HER2 по сравнению со связыванием hu4D5Fabv8-фага дикого типа. Более того, конъюгация с биотином не влияла на связывание ТиоFab с HER2 (Фигура 2B).
Удивительно и неожиданно, для образцов ТиоFab-фагов показали изменяющиеся уровни активности связывания стрептавидина. Из всех протестированных фаг-ТиоFab, для сконструированного антитела с цистеиновыми заменами A121C показали максимальную реакционную способность тиола. Даже несмотря на то, что hu4D5Fabv8-фаг дикого типа инкубировали с такими же количествами биотин-малеимида, этот фаг обладал небольшим связыванием стрептавидина, что указывает на то, что предсуществующие остатки цистеина (вовлеченные в формирование дисульфидных связей) из hu4D5Fabv8 и белков оболочки фага M13 не мешают сайт-специфической конъюгации биотин-малеимида. Эти результаты показывают, что анализ фагового ELISA можно успешно использовать для скрининга реакционноспособных тиоловых групп на поверхности Fab.
Анализ PHESELECTOR позволяет скрининг реакционноспособных тиоловых групп в антителах. Идентификация варианта A121C этим способом является иллюстративной. Можно проводить эффективный поиск по целой молекуле Fab для идентификации большего числа вариантов ТиоFab с реакционноспособными тиоловыми группами. Параметр относительную доступность поверхности использовали для идентификации и количественной оценки доступности для растворителя аминокислотных остатков в полипептиде. Доступность поверхности можно выражать как площадь поверхности (Е2), которая может вступать в контакт с молекулой растворителя, например, воды. Пространство, занимаемое водой, составляет сферу радиусом приблизительно 1,4 Е. Программное обеспечение является свободно доступным или лицензируемым (Secretary to CCP4, Daresbury Laboratory, Warrington, WA44 AD, United Kingdom, Fax: (+44) 1925 603825, или доступным в интернет: www.ccp4.ac.uk/dist/html/INDEX.html) как пакет программ для кристаллографии CCP4, использующий алгоритмы для расчета доступности поверхности каждой аминокислоты белка с известными координатами, полученными рентгеновской кристаллографией («The CCP4 Suite: Programs for Protein Crystallography» (1994) Acta. Cryst. D50:760-763). Два иллюстративных модуля программного обеспечения, осуществляющие расчеты доступности поверхности «AREAIMOL» и «SURFACE», основаны на алгоритмах B.Lee and F. M. Richards (1971) J. Mol. Biol. 55:379-400.
AREAIMOL определяет поверхность белка, доступную для растворителя, как локус в центре сферы зонда (представляющей молекулу растворителя), по мере того, как она катится по Ван-дер-Ваальсовой поверхности белка. AREAIMOL рассчитывает площадь поверхности, доступной для растворителя, посредством получения точек поверхности на расширенной сфере вокруг каждого атома (на расстоянии от центра атома, равном сумме радиусов атома и зонда), и исключения точек, лежащих внутри одних и тех же сфер, ассоциированных с соседними атомами. AREAIMOL находит площадь атомов, доступных для растворителя, в файле координат PDB и суммирует доступную площадь по остатку, по цепи и по целой молекуле. Доступные площади (или разности площадей) для отдельных атомов можно записать в файл выходных данных псевдо-PDB. AREAIMOL присваивает один радиус каждому элементу и распознает только ограниченное количество различных элементов. Неизвестным типам атомов (т.е., не присутствующим во внутренней базе данных AREAIMOL) по умолчанию присваивается радиус 1,8 Å. Список распознаваемых атомов представляет собой:
AREAIMOL и SURFACE выдают абсолютные значения доступности, т.е. количество квадратных ангстрем (Å). Относительную доступность поверхности рассчитывают на основе стандартного состояния, соответствующего аминокислоте внутри полипептида. Стандартное состояние представляет собой трипептид Gly-X-Gly, где X представляет собой интересующую аминокислоту, и стандартное состояние должно представлять собой «растянутую» конформацию, т.е. подобную конформации в бета-цепях. Растянутая конформация максимизирует доступность X. Рассчитанную доступную площадь делят на доступную площадь в стандартном состоянии трипептида Gly-X-Gly и регистрируют частное, представляющее собой относительную доступность. Процент доступности представляет собой относительную доступность, умноженную на 100.
Другой иллюстративный алгоритм для расчета доступности поверхности основан на модуле SOLV программы xsae (Broger, C, F. Hoffman-LaRoche, Basel), рассчитывающем относительную доступность аминокислотного остатка для водяной сферы на основании рентгеновских координат полипептида.
Относительную доступность поверхности для каждой аминокислоты в hu4D5Fabv7 рассчитывали с использованием информации о кристаллической структуре (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Значения относительной доступности поверхности для аминокислот легкой цепи и тяжелой цепи hu4D5Fabv7 показаны в нисходящем порядке в таблице 1.
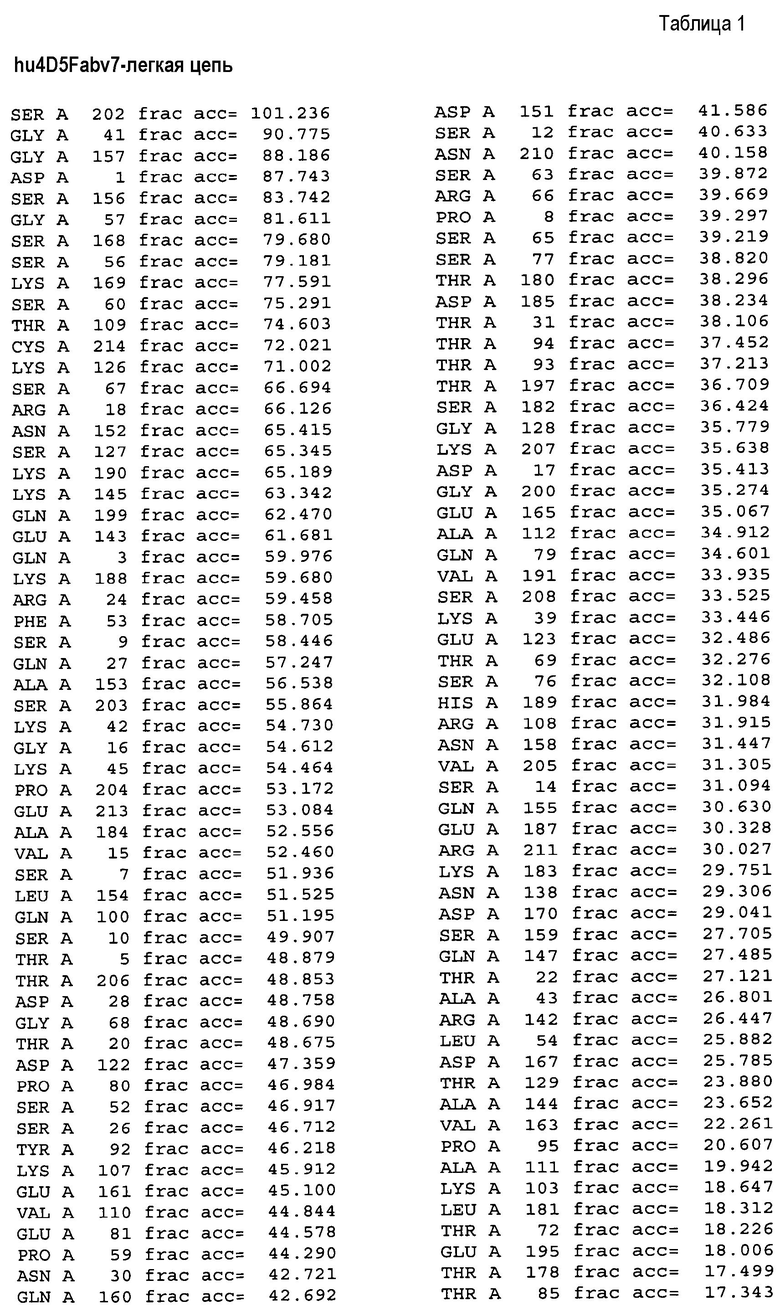
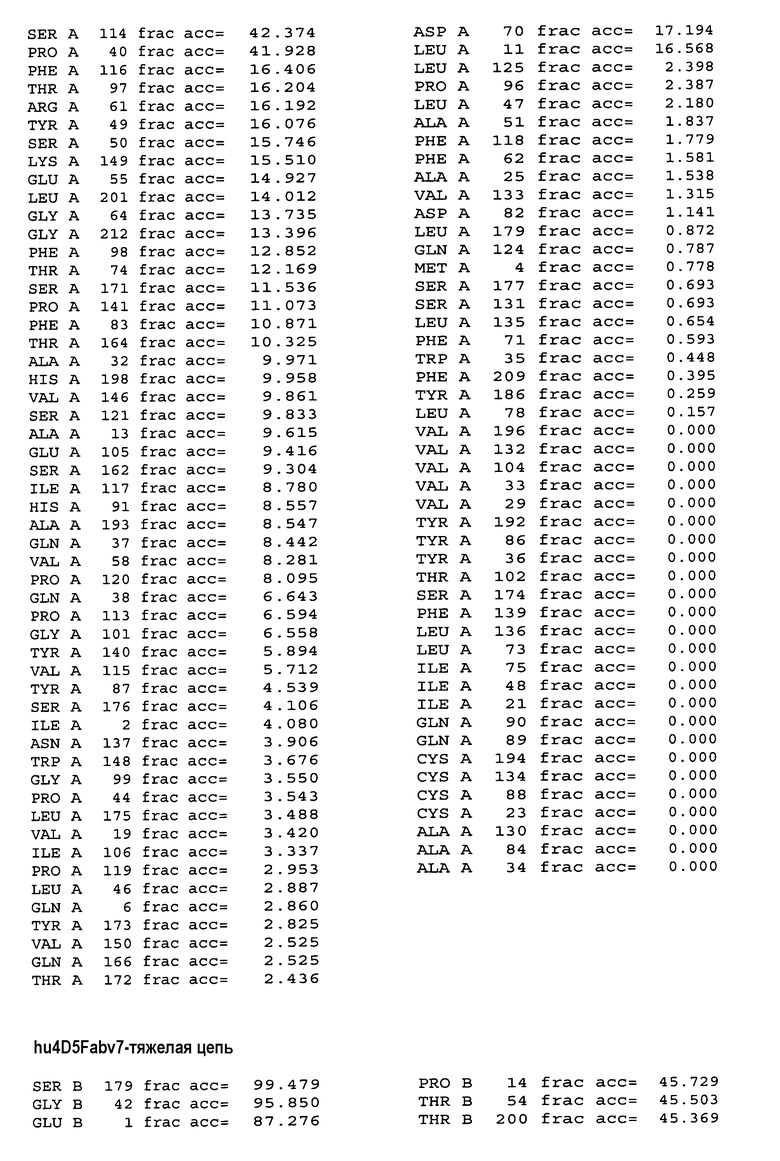
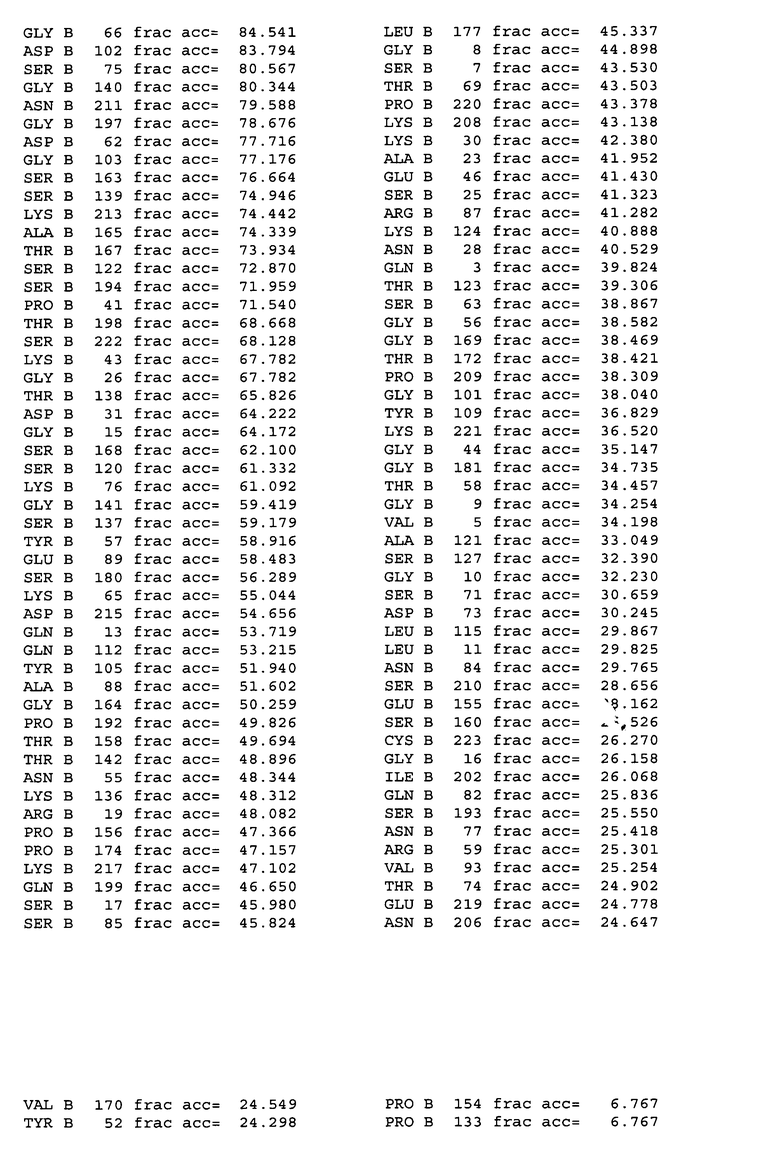
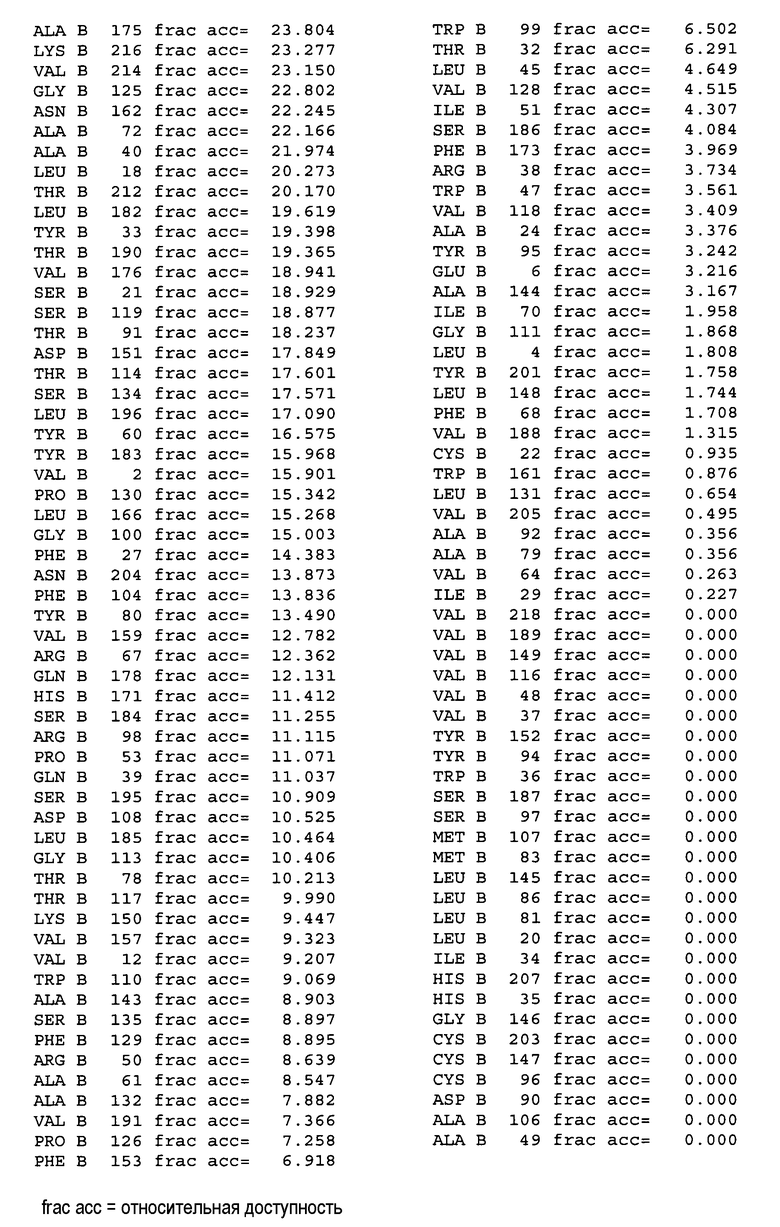
Следующие два критерия использовали для идентификации остатков hu4D5Fabv8, которые можно конструировать с заменой на остатки Cys:
1. Исключены аминокислотные остатки, которые полностью скрыты, т.е. менее чем с 10% относительной доступностью поверхности. В таблице 1 показано, что существует 134 (легкая цепь) и 151 (тяжелая цепь) остатков в hu4D5Fabv8, доступных более чем на 10% (относительная доступность поверхности). Выбрано десять наиболее доступных остатков Ser, Ala и Val из-за их близкого структурного свойства с Cys по сравнению с другими аминокислотами, так что вновь сконструированный Cys вносит только минимальные структурные ограничения в антитело. Можно также проводить скрининг других участков замены на цистеин, и они могут быть пригодны для конъюгации.
2. Остатки сортируют на основании их роли в функциональных и структурных взаимодействиях Fab. Затем отбирали остатки, не вовлеченные во взаимодействия с антигеном и удаленные от существующих дисульфидных связей. Вновь сконструированные остатки Cys должны быть удалены от участка связывания антигена, и не должны ни мешать связыванию антигена, ни ошибочно спариваться с остатками цистеина, вовлеченными в формирование дисульфидных связей.
Следующие остатки hu4D5Fabv8 удовлетворяли вышеуказанным критериям и были выбраны для замены на Cys: L-V15, L-A43, L-V110, L-A144, L-S168, H-A88, H-A121, H-S122, H-A175 и H-S179 (показаны на фигуре 1).
Реакционную способность тиола можно распространять на любое антитело, в котором можно выполнять замену аминокислот на реакционноспособные цистеиновые аминокислоты в пределах легкой цепи, выбранных из: L-10 - L-20; L-38 - L-48; L-105 - L-115; L-139 - L-149; L-163 - L-173; и в пределах тяжелой цепи, выбранных из: H-35 - H-45; H-83 - H-93; H-114 - H-127; и H-170 - H-184, и в пределах Fc-области, выбранных из H-268 - H-291; H-319 - H-344; H-370 - H-380; и H-395 - H-405.
Реакционную способность тиола можно распространять также на конкретные домены антитела, такие как константный домен легкой цепи (CL) и константные домены тяжелой цепи, CH1, CH2 и CH3. Замены на цистеин, приводящие к значению реакционной способности тиолов 0,6 и выше, можно выполнять в константных доменах тяжелых цепей α, δ, ε, γ и μ интактных антител: IgA, IgD, IgE, IgG и IgM, соответственно, включая подклассы IgG: IgG1, IgG2, IgG3, IgG4, IgA и IgA2.
Из данных кристаллической структуры очевидно, что выбранные 10 мутантов Cys находятся далеко от антигенсвязывающего участка, такого как поверхность контакта с HER2 в этом случае. Эти мутанты можно тестировать экспериментально по непрямым эффектам на функциональные взаимодействия. Реакционную способность тиола всех Cys вариантов Fab измеряли и рассчитывали, как описано в примерах 1 и 2, и представлено в таблице 2. Остатки L-V15C, L-V110C, H-A88C и H-A121C имели реакционноспособные и стабильные тиоловые группы (Фигуры 3A и 3B). Мутанты V15C, V110C, A144C, S168C представляют собой Cys варианты легкой цепи. Мутанты A88C, A121C, A175C, S179C представляют собой Cys варианты тяжелой цепи. Удивительно и неожиданно, что участки с высокой относительной доступностью поверхности не обладают наиболее высокой реакционной способностью тиола, как рассчитано посредством анализа PHESELECTOR (таблица 2). Иными словами, относительная доступность поверхности (таблицы 1, 2) не коррелирует с реакционной способностью тиола (таблица 2). Фактически, остатки Cys, сконструированные в участках с умеренной доступностью поверхности 20%-80% (фигура 4A, таблица 1), или частично экспонированные участки, подобные остаткам Ala или Val, обладают лучшей реакционной способностью тиола, т.е. >0,6, (фигура 3B, таблица 2), чем Cys, введенный вместо остатков Ser, таким образом, необходимость использования анализа PHESELECTOR для скрининга реакционноспособных участков тиола только по информации о кристаллической структуре является недостаточной для отбора этих участков (фигура 3B и 4A).
Данные по реакционной способности тиола показаны на фигурах 3A и 3B для аминокислотных остатков Cys мутантов 4D5 ТиоFab: (3A) небиотинилированный (контроль) и (3B) биотинилированные фаг-ТиоFab. Реакционноспособные тиоловые группы на поверхности антитела/Fab идентифицировали анализами PHESELECTOR по взаимодействию небиотинилированного фаг-hu4D5Fabv8 (3A) и биотинилированного фаг-hu4D5Fabv8 (3B) с BSA (незакрашенный прямоугольник), HER2 (серый прямоугольник) или стрептавидином (закрашенный прямоугольник). Анализ проводили, как описано в примере 2. Варианты легкой цепи расположены на левой стороне, и варианты тяжелой цепи расположены на правой стороне. Связывание небиотинилированных Cys мутантов 4D5 ТиоFab является низким, как ожидалось, однако, сильное связывание с HER2 сохраняется. Соотношение связывания со стрептавидином и с HER2 биотинилированных Cys мутантов 4D5 ТиоFab дает значение реакционной способности тиолов в таблице 2. Фоновая оптическая плотность при 450 нм или малые количества неспецифического связывания белка Cys мутантов биотинилированного 4D5 ТиоFab с BSA также видны на фигуре 3B. Значения относительной доступности поверхности избранных аминокислотных остатков, замененных на остаток Cys, показаны на фигуре 4A. Относительная доступность поверхности рассчитана по доступной структуре hu4D5Fabv7 и показана в таблице 1 (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Конформационные параметры структур hu4D5Fabv7 и hu4D5Fabv8 хорошо согласуются и позволяют определение любой корреляции между относительной доступностью поверхности hu4D5Fabv7 и реакционной способностью тиола цистеиновых мутантов hu4D5Fabv8. Измеренная реакционная способность тиола остатков Cys на фаге с ТиоFab, введенных в частично экспонированные остатки (Ala или Val), показывает лучшую реакционную способность тиола по сравнению с остатками Cys, введенными в остатки Ser (Таблица 2). В таблице 2 можно видеть, что для Cys мутантов ТиоFab корреляция между значениями реакционной способности тио- и относительной доступностью поверхности мала или отсутствует.
Аминокислоты в положениях L-15, L-43, L-110, L-144, L-168, H-40, H-88, H-119, H-121, H-122, H-175 и H-179 антитела можно, как правило, подвергать мутации (заменять) с помощью свободных цистеиновых аминокислот. Диапазоны в пределах приблизительно 5 аминокислотных остатков с каждой стороны от этих положений также можно заменять на свободные цистеиновые аминокислоты, т.е. L-10 - L-20; L-38 - L-48; L-105 - L-115; L-139 - L-149; L-163 - L-173; H-35 - H-45; H-83 - H-93; H-114 - H-127; и H-170 - H-184, так же как в диапазонах Fc-области, выбранных из H-268 - H-291; H-319 - H-344; H-370 - H-380; и H-395 - H-405, для получения сконструированных антител с цистеиновыми заменами по изобретению.
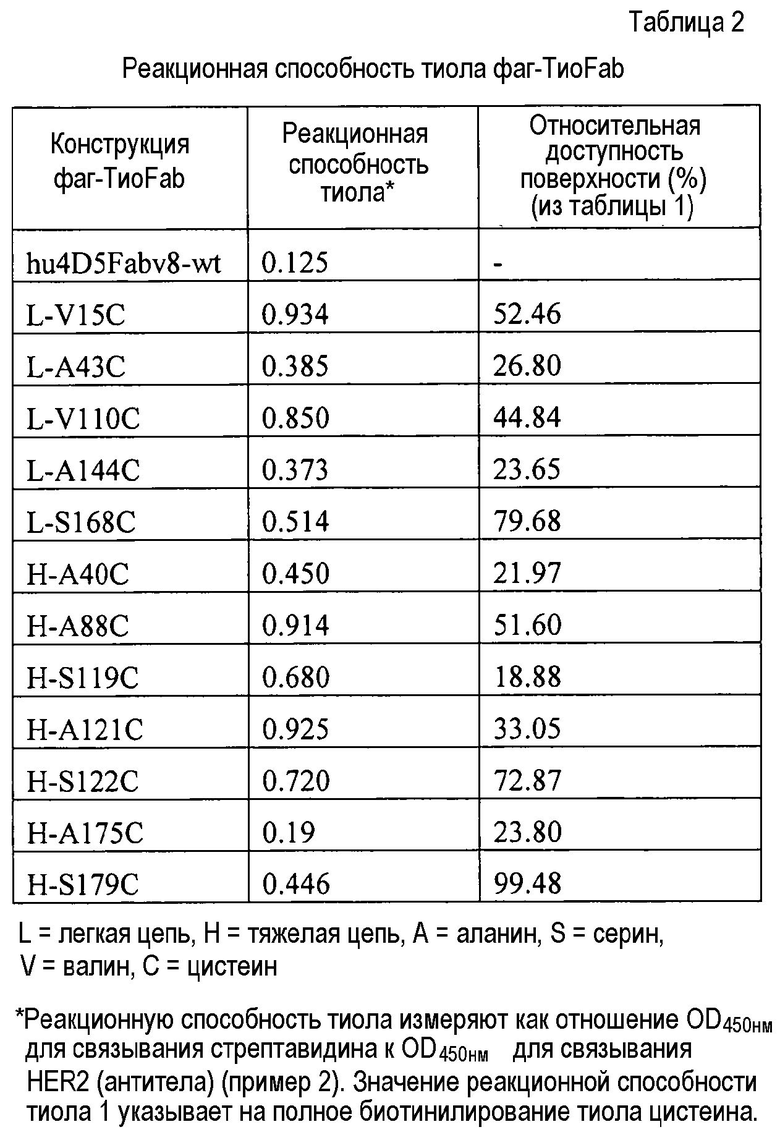
Два Cys варианта для легкой цепи (L-V15C и L-V110C) и два для тяжелой цепи (H-A88C и H-A121C) выбраны для дальнейшего анализа, поскольку для этих вариантов показана наивысшая реакционная способность тиола (таблица 2).
В отличие от очистки фага, получение Fab может требовать 2-3 суток, в зависимости от масштаба продукции. На протяжении этого времени, тиоловые группы могут терять реакционную способность из-за окисления. Для тестирования стабильности тиоловых групп на hu4D5Fabv8-фаге, измеряли реакционную способность тиола фаг-тиоFab (фигура 4B). После выделения ТиоFab-фага, на сутки 1, сутки 2 и сутки 4, все образцы конъюгировали с биотин-PEO-малеимидом и тестировали в анализе фагового ELISA (PHESELECTOR) для тестирования связывания HER2 и стрептавидина. L-V15C, L-V110C, H-A88C и H-A121C сохраняли значительные уровни реакционной способности тиола по сравнению с другими вариантами ТиоFab (фигура 4B).
СПОСОБЫ ПОЛУЧЕНИЯ СКОНСТРУИРОВАННЫХ АНТИТЕЛ С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, в которых одна или несколько аминокислот исходного антитела заменены на свободную аминокислоту цистеин. Сконструированное антитело с цистеиновыми заменами содержит одну или несколько свободных цистеиновых аминокислот, обладающих значением реакционной способности тиола в диапазоне 0,6-1,0. Свободная аминокислота цистеин представляет собой остаток цистеина, введенный при конструировании в исходное антитело и не являющийся частью дисульфидного мостика.
В одном аспекте сконструированное антитело с цистеиновыми заменами получено способом, включающим:
(a) замену одного или нескольких аминокислотных остатков исходного антитела на цистеин; и
(b) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами посредством реакции сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу реагентом.
Сконструированное антитело с цистеиновыми заменами может являться более реакционноспособным, чем исходное антитело, по отношению к реакционноспособному по отношению к тиолу реагенту.
Остатки свободной аминокислоты цистеина могут быть локализованы в тяжелой или легкой цепях, или в константном или вариабельном доменах. Фрагменты антитела, например, Fab, также можно конструировать с заменой аминокислот фрагментов антитела на одну или нескольких цистеиновых аминокислот для получения сконструированных фрагментов антитела с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения (изготовления) сконструированного антитела с цистеиновыми заменами, включающему:
(a) введение одной или нескольких цистеиновых аминокислот в исходное антитело для получения сконструированного антитела с цистеиновыми заменами; и
(b) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами с помощью реакционноспособного по отношению к тиолу реагента;
где сконструированное антитело с цистеиновыми заменами является более реакционноспособным, чем исходное антитело, по отношению к реакционноспособному по отношению к тиолу реагенту.
Стадия (a) способа получения сконструированного антитела с цистеиновыми заменами может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
Стадия (b) способа получения сконструированного антитела с цистеиновыми заменами может включать экспрессию сконструированного антитела с цистеиновыми заменами на вирусной частице, выбранной из частицы фага или фагмиды.
Стадия (b) способа получения сконструированного антитела с цистеиновыми заменами может включать также:
(i) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(ii) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой.
Другой аспект по изобретению относится к способу скрининга сконструированных антител с цистеиновыми заменами с высоко реакционноспособными, неспаренными цистеиновыми аминокислотами по реакционной способности тиола, включающему:
(a) введение одной или нескольких цистеиновых аминокислот в исходное антитело для получения сконструированного антитела с цистеиновыми заменами;
(b) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(c) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой; и
(d) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами с помощью реакционноспособного по отношению к тиолу реагента.
Стадия (a) способа скрининга сконструированных антител с цистеиновыми заменами может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
Стадия (b) способа скрининга сконструированного антитела с цистеиновыми заменами может включать экспрессию сконструированного антитела с цистеиновыми заменами на вирусной частице, выбранной из частицы фага или фагмиды.
Стадия (b) способа скрининга сконструированного антитела с цистеиновыми заменами может включать также:
(i) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(ii) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой.
МЕЧЕНЫЕ СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Сконструированные антитела с цистеиновыми заменами по изобретению можно конъюгировать с любой группой метки, которую можно ковалентно присоединять к антителу посредством тиоловой группы цистеина (Singh et al (2002) Anal. Biochem. 304: 147-15; Harlow E. и Lane, D. (1999) Using Antibodies: A Laboratory Manual, Cold Springs Harbor Laboratory Press, Cold Spring Harbor, NY; Lundblad R.L. (1991) Chemical Reagents for Protein Modification, 2nd ed. CRC Press, Boca Raton, FL). Присоединенная метка может функционировать для: (i) обеспечения поддающегося детекции сигнала; (ii) взаимодействия со второй меткой для модификации поддающегося детекции сигнала, обеспечиваемого первой или второй меткой, например, для получения FRET (резонансного переноса энергии флуоресценции); (iii) стабилизации взаимодействий или увеличения аффинности связывания с антигеном или лигандом; (iv) влияния на подвижность, например, электрофоретическую подвижность, или клеточную проницаемость, посредством заряда, гидрофобности, формы или других физических параметров, или (v) предоставления связывающей группы, для модуляции аффинности лиганда, связывания антитело/антиген, или образования комплексных ионов.
Меченые сконструированные антитела с цистеиновыми заменами могут являться полезными для диагностических анализов, например, для детекции экспрессии интересующего антигена в специфических клетках, тканях или сыворотке. Для диагностических применений антитело, как правило, метят поддающейся детекции группой.
Доступно множество меток, которые можно в общем сгруппировать по следующим категориям:
(a) Радиоизотопы (радиоактивные изотопы), такие как 3H, 11C, 14C, 18F, 32P, 35S, 64Cu, 68Ga, 86Y, 89Zr, 99Tc, 111In, 123I, 124I, 125I, 131I, 133Xe, 177LU, 211At или 213Bi. Меченные радиоизотопами антитела являются полезными в экспериментах получения изображений, направленных на рецепторы. Антитело можно метить с помощью реагентов с лигандами, которые связывают, образуют хелат или другой комплекс с радиоизотопом металла, где реагент является реакционноспособным по отношению к сконструированному тиолу антитела, с использованием способов, описанных в Current Protocols in Immunology, Volumes 1 и 2, Coligen et al, Ed. Wiley-Interscience, New York, NY, Pubs. (1991). Хелатирующие лиганды, которые могут образовывать комплексы с ионом металла, включают DOTA, DOPA, DOTP, DOTMA, DTPA и TETA (Macrocyclics, Dallas, TX). Радиоактивные изотопы можно нацеливать посредством образования комплексов с сконструированными антителами с цистеиновыми заменами в форме конъюгатов антитело-цирконий по изобретению (Wu et al (2005) Nature Biotechnology 23(9): 1137-1146).
Металл-хелатные комплексы, пригодные в качестве меток для антитела для экспериментов по получению изображений, описаны в: US 5342606; US 5428155; US 5316757; US 5480990; US 5462725; US 5428139; US 5385893; US 5739294; US 5750660; US 5834456; Hnatowich et al (1983) J. Immunol. Methods 65: 147-157; Meares et al (1984) Anal. Biochem. 142:68-78; Mirzadeh et al (1990) Bioconjugate Chem. 1:59-65; Meares et al (1990) J. Cancerl990, Suppl. 10:21-26; Izard et al (1992) Bioconjugate Chem. 3:346-350; Nikula et al (1995) Nucl. Med. Biol. 22:387-90; Camera et al (1993) Nucl. Med. Biol. 20:955-62; Kukis et al (1998) J. Nucl. Med. 39:2105-2110; Verel et al (2003) J. Nucl. Med. 44:1663-1670; Camera et al (1994) J. Nucl. Med. 21:640-646; Ruegg et al (1990) Cancer Res. 50:4221-4226; Verel et al (2003) J. Nucl. Med. 44:1663-1670; Lee et al (2001) Cancer Res. 61:4474-4482; Mitchell, et al (2003) J. Nucl. Med. 44:1105-1112; Kobayashi et al (1999) Bioconjugate Chem. 10:103-111; Miederer et al (2004) J. Nucl. Med. 45:129-137; DeNardo et al (1998) Clinical Cancer Research 4:2483-90; Blend et al (2003) Cancer Biotherapy & Radiopharmaceuticals 18:355-363; Nikula et al (1999) J. Nucl. Med. 40:166-76; Kobayashi et al (1998) J. Nucl. Med. 39:829-36; Mardirossian et al (1993) Nucl. Med. Biol. 20:65-74; Roselli et al (1999) Cancer Biotherapy & Radiopharmaceuticals, 14:209-20.
(b) Флуоресцентные метки, такие как хелаты редкоземельных элементов (хелаты европия), типы флуоресцеина, включая FITC, 5-карбоксифлуоресцеин, 6-карбоксифлуоресцеин; типы родамина, включая TAMRA; данзил; лиссамин; цианины; фикоэритрины; Техасский Красный; и их аналоги. Флуоресцентные метки можно конъюгировать с антителами с использованием способов, описанных, например, в Current Protocols in Immunology, выше. Реагенты с флуоресцентными красителями и флуоресцентной меткой включают реагенты, коммерчески доступные из Invitrogen/Molecular Probes (Eugene, OR) и Pierce Biotechnology, Inc. (Rockford, IL).
(c) Различные фермент-субстратные метки доступны или описаны (US 4275149). Фермент, как правило, катализирует химическое изменение хромогенного субстрата, которое можно измерять с использованием различных способов. Например, фермент может катализировать изменение окраски субстрата, которое можно измерять спектрофотометрически. Альтернативно, фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Способы количественного определения изменения флуоресценции описаны выше. Хемилюминесцентный субстрат становится электронно-возбужденным посредством химической реакции и может затем испускать свет, который можно измерять (например, с использованием хемилюминометра) или передавать энергию акцептору флуоресценции. Примеры ферментых меток включают люциферазы (например, люциферазу светлячков и бактериальную люциферазу; US 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уриназу, пероксидазу, такую как пероксидаза хрена (HRP), щелочную фосфатазу (AP), β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов (например, оксидазу глюкозы, оксидазу галактозы и глюкоза-6-фосфатдегидрогеназу), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.п. Способы конъюгации ферментов с антителами описаны в O'Sullivan et al (1981) «Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay», in Methods in Enzym. (ed J. Langone & H. Van Vunakis), Academic Press, New York, 73:147-166.
Примеры комбинаций фермент-субстрат включают, например:
(i) пероксидазу хрена (HRP) с перекисью водорода в качестве субстрата, где перекись водорода окисляет предшественник красителя (например, ортофенилендиамин (OPD) или 3,3',5,5'-тетраметилбензидингидрохлорид (TMB));
(ii) щелочную фосфатазу (AP) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и
(iii) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, п-нитрофенил-β-D-галактозидазой) или флуорогенным субстратом 4-метилумбеллиферил-β-D-галактозидазой.
Множество других комбинаций фермент-субстрат доступно специалистам в данной области. Общий обзор см. в US 4275149 и US 4318980.
Метку можно опосредованно конъюгировать с сконструированным антителом с цистеиновыми заменами. Например, антитело можно конъюгировать с биотином и любую метку из трех широких категорий, указанных выше, можно конъюгировать с авидином или стрептавидином, или наоборот. Биотин избирательно связывается со стрептавидином, и, таким образом, метку можно конъюгировать с антителом данным опосредованным образом. Альтернативно для достижения опосредованной конъюгации метки с вариантом полипептида, вариант полипептида конъюгируют с небольшим гаптеном (например, дигоксином) и одну из различных типов меток, указанных выше, конъюгируют с полипептидным вариантом против гаптена (например, антителом против дигоксина). Таким образом можно достигать опосредованной конъюгации метки с вариантом полипептида (Hermanson, G. (1996) in Bioconjugate Techniques Academic Press, San Diego).
Вариант полипептида по настоящему изобретению можно использовать в любом известном способе анализа, таком как ELISA, анализы конкурентного связывания, прямом и непрямом сэндвич-анализах, и анализах иммунопреципитации (Zola, (1987) Monoclonal Antibodies: A Manual of Techniques, pp.147-158, CRC Press, Inc.).
Метка для детекции может являться пригодной для локализации, визуализации и количественной оценки события связывания или узнавания. Меченые антитела по изобретению могут детектировать рецепторы поверхности клеток. Другим применением для меченных поддающейся детекции меткой антител является способ иммунного связывания с помощью бусин, включающий конъюгацию бусин с флуоресцентно меченным антителом и детекцию сигнала флуоресценции при связывании лиганда. В сходных способах детекции связывания используют эффект поверхностного плазмонного резонанса (SPR) для измерения и детекции взаимодействий антитело-антиген.
Такие метки для детекции, как флуоресцентные красители и хемилюминесцентные красители (Briggs et al (1997) «Synthesis of Functionalised Fluorescent Dyes and Their Coupling to Amines and Amino Acids», J. Chem. Soc, Perkin-Trans. 1: 1051-1058), обеспечивают поддающийся детекции сигнал и являются общепринятыми для мечения антитела, предпочтительно, со следующими свойствами: (i) меченое антитело должно производить очень сильный сигнал с низким фоном, так что небольшие количества антител можно с большой чувствительностью детектировать как в бесклеточных, так и в основанных на клетках анализах; и (ii) меченое антитело должно являться фотостабильным, так чтобы флуоресцентный сигнал можно было наблюдать, мониторировать и регистрировать без значительного фотообесцвечивания. Для применений, включающих связывание на поверхности клетки меченого антитела с мембранами или поверхностями клеток, особенно живых клеток, метки предпочтительно (iii) обладают хорошей растворимостью в воде для достижения эффективных концентраций конъюгата и чувствительности детекции и (iv) являются нетоксичными для живых клеток, так чтобы не нарушать нормальных процессов метаболизма клеток или не вызывать преждевременной гибели клеток.
Прямую количественную оценку интенсивности клеточной флуоресценции и подсчет событий флуоресцентного мечения, например, связывания на поверхности клеток конъюгатов пептид-краситель, можно проводить в системе (FMAT® 8100 HTS System, Applied Biosystems, Foster City, Calif.), автоматизирующей нерадиоактивные анализы смешивания и считывания с живыми клетками или бусинами (Miraglia, «Homogeneous cell- and bead-based assays for high throughput screening using fluorometric microvolume assay technology», (1999) J. of Biomolecular Screening 4: 193-204). Применения меченых антител включают также анализы связывания с рецептором на поверхности клеток, анализы иммунного связывания, твердофазные иммунофлуоресцентные анализы (FLISA), анализы расщепления каспазой (Zheng, «Caspase-3 controls both cytoplasmic and nuclear events associated with Fas-mediated apoptosis in vivo», (1998) Proc. Natl. Acad. Sci. USA 95:618-23; US 6372907), анализы апоптоза (Vermes, «A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V» (1995) J. Immunol. Methods 184:39-51) и анализы цитотоксичности. Способ флуориметрического анализа в микрообъемах можно использовать для идентификации повышающей или понижающей регуляции посредством молекулы, нацеленной на поверхность клеток (Swartzman, «A homogeneous and multiplexed immunoassay for high-throughput screening using fluorometric microvolume assay technology», (1999) Anal. Biochem. 271:143-51).
Меченые сконструированные антитела с цистеиновыми заменами по изобретению являются пригодными в качестве биомаркеров и зондов для получения изображений различными методами и способами получения биомедицинских и молекулярных изображений, такими как: (i) МРТ (магнитно-резонансная томография); (ii) МикроКТ (компьютерная томография); (iii) SPECT (однофотонная эмиссионная компьютерная томография); (iv) ПЭТ (позитронная эмиссионная томография) Chen et al (2004) Bioconjugate Chem. 15:41-49; (v) биолюминесценция; (vi) флуоресценция; и (vii) ультразвук. Иммуносцинтиография представляет собой способ получения изображений при котором антитела, меченные радиоактивными веществами, вводят пациенту - животному или человеку, и получают изображение в участках организма, где локализовано антитело (US 6528624). Биомаркеры для получения изображений можно объективно измерять и оценивать в качестве показателя нормальных биологических процессов, патогенных процессов или фармакологических ответов на терапевтическое вмешательство. Биомаркеры могут принадлежать к нескольким типам: маркеры типа 0 представляют собой маркеры естественного течения заболевания и по длительности проявления коррелируют с известными клиническими показателями, например, оценкой ЯМР синовиального воспаления при ревматоидном артрите; маркеры типа I фиксируют эффект вмешательства вне зависимости от механизма действия, даже несмотря на то, что механизм может не быть ассоциирован с клиническим исходом; маркеры типа II функционируют как суррогатные конечные точки, где по изменению биомаркера или по сигналу от биомаркера прогнозируют клиническое преимущество для «подтверждения» намеченного ответа, например, измерение эрозии костной ткани при ревматоидном артрите посредством КТ. Биомаркеры для получения изображений таким образом могут предоставлять фармакодинамическую (PD) терапевтическую информацию относительно: (i) экспрессии белка-мишени, (ii) связывания лекарственного средства с белком-мишенью, т.е., избирательности, и (iii) фармакокинетических данных о выведении и времени полужизни. Преимущества биомаркеров для получения изображений in vivo по сравнению с биомаркерами для лабораторных исследований включают: неинвазивную обработку, количественную оценку целого организма, повторяющиеся введение и оценку, т.е. множественные временные точки, и эффекты, потенциально переносимые от доклинических результатов (для мелких животных) к клиническим результатам (для человека). Для некоторых применений получение изображений биологических объектов заменяет эксперименты на животных или минимизирует количество экспериментов на животных в доклинических исследованиях.
Метки для получения изображений с радиоактивными изотопами включают такие радиоактивные изотопы, как 3H, 11C, 14C, 18F, 32P, 35S, 64Cu, 68Ga, 86Y, 89Zr, 99Tc, 111In, 123I, 124I, 125I, 131I, 133Xe, 177Lu, 211At или 213Bi. Можно получать комплекс иона радиоактивного изотопа металла с хелатирующим линкером, таким как DOTA. Линкерные реагенты, такие как DOTA-малеимид (4-малеимидобутирамидобензил-DOTA), можно получать реакцией аминобензил-DOTA с 4-малеимидомасляной кислотой (Fluka), активированной изопропилхлорформатом (Aldrich), с последующим использованием способа Axworthy et al (2000) Proc. Natl. Acad. Sci. USA 97(4): 1802-1807). DOTA-малеимидные реагенты вступают в реакцию со свободными цистеиновыми аминокислотами сконструированных антител с цистеиновыми заменами и обеспечивают наличие на антителе образующего комплексы с металлами лиганда (Lewis et al (1998) Bioconj. Chem. 9:72-86). Реагенты для мечения с хелатирующим линкером, такие как DOTA-NHS (сложный моно(N-гидроксисукцинимидный) эфир 1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусной кислоты), являются коммерчески доступными (Macrocyclics, Dallas, TX). Нацеленное на рецепторы получение изображений с помощью меченных радиоактивным изотопом антител может обеспечивать маркер активации пути посредством детекции и количественной оценки возрастающего накопления антител в ткани опухоли (Albert et al (1998) Bioorg. Med. Chem. Lett. 8: 1207-1210). Конъюгированные радиоактивные металлы могут оставаться внутри клеток после лизосомальной деградации.
Способы мечения пептидов хорошо известны. См. Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, (1997) Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Glazer et al (1975) Chemical Modification of Proteins. Laboratory Techniques in Biochemistry and Molecular Biology (T. S. Work и E. Work, Eds.) American Elsevier Publishing Co., New York; Lundblad, R. L. and Noyes, C. M. (1984) Chemical Reagents for Protein Modification, Vols. I and II, CRC Press, New York; Pfleiderer, G. (1985) «Chemical Modification of Proteins», Modern Methods in Protein Chemistry, H. Tschesche, Ed., Walter DeGryter, Berlin and New York; и Wong (1991) Chemistry of Protein Conjugation and Cross-linking, CRC Press, Boca Raton, Fla.); De Leon-Rodriguez et al (2004) Chem.Eur. J. 10: 1149-1155; Lewis et al (2001) Bioconjugate Chem. 12:320-324; Li et al (2002) Bioconjugate Chem. 13: 110-115; Mier et al (2005) Bioconjugate Chem. 16:240-237.
Пептиды и белки, меченные двумя группам, флуоресцентным репортером и тушителем в достаточной близости, подвергаются резонансному переносу энергии флуоресценции (FRET). Репортерные группы, как правило, представляют собой флуоресцентные красители, возбуждаемые светом определенной длины волны и переносящие энергию на группу акцептора, или тушителя, с соответствующим сдвигом Стокса для излучения с максимальной яркостью. Флуоресцентные красители включают молекулы с большими ароматическими группами, такие как флуоресцеин и родамин, и их производные. Флуоресцентный репортер может подвергаться частичному или полному тушению группой тушителя в интактном пептиде. При расщеплении пептида пептидазой или протеазой можно измерять поддающееся детекции увеличение флуоресценции (Knight, C. (1995) «Fluorimetric Assays of Proteolytic Enzymes», Methods in Enzymology, Academic Press, 248: 18-34).
Меченые антитела по изобретению можно использовать также в качестве средства для аффинной очистки. По этому способу, меченое антитело иммобилизуют на твердой фазе, такой как смола Sephadex или фильтровальная бумага, с использованием способов, хорошо известных в данной области. Иммобилизованное антитело приводят в контакт с образцом, содержащим антиген, подлежащий очистке, и затем подложку промывают подходящим растворителем, удаляющим в основном весь материал образца, за исключением антигена, подлежащего очистке, который является связанным с иммобилизованным вариантом полипептида. Наконец, подложку промывают другим подходящим растворителем, таким как глициновый буфер, pH 5,0, высвобождающим антиген с варианта полипептида.
Реагенты для мечения, как правило, несут реакционноспособную функциональную группу, которая может вступать в реакцию (i) неспосредственно с тиолом цистеина сконструированного антитела с цистеиновыми заменами с образованием меченого антитела, (ii) с линкерным реагентом с образованием промежуточного соединения линкер-метка, или (iii) с имеющим линкер антителом с образованием меченого антитела. Реакционноспособные функциональные группы реагентов для мечения включают: малеимид, галоацетил, сложный эфир иодацетамида и сукцинимидила (например, NHS, N-гидроксисукцинимид), изотиоцианат, сульфонилхлорид, 2,6-дихлортриазинил, сложный эфир пентарфторфенила и фосфорамидит, хотя можно использовать также другие функциональные группы.
Иллюстративной реакционноспособной функциональной группой является N-гидроксисукцинимидиловый сложный эфир (NHS) карбоксильной группы заместителя поддающейся детекции метки, например, биотина или флуоресцентного красителя. NHS сложный эфир метки может являться заранее полученным, выделенным, очищенным и/или охарактеризованным, или он может образовываться in situ и вступать в реакцию с нуклеофильной группой антитела. Как правило, карбоксильную форму метки активируют посредством реакции с некоторой комбинацией карбодиимидного реагента, например, дициклогексилкарбодиимида, диизопропилкарбодиимида или урониевого реагента, например, TSTU (тетрафторборат (O-(N-сукцинимидил)-N,N,N',N'-тетраметилурония), HBTU (гексафторфосфат O-бензотриазол-1-ил)-N,N,N',N'-тетраметилурония) или HATU (гексафторфосфат O-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония), активатора, такого как 1-гидроксибензотриазол (HOBt), и N-гидроксисукцинимида для получения NHS сложного эфира метки. В некоторых случаях, метку и антитело можно связывать активацией метки in situ и реакцией с антителом с образованием конъюгата метка-антитело в одну стадию. Другие реагенты для активации и связывания включают TBTU (гексафторфосфат 2-(1H-бензотриазо-1-ил)-1-1,3,3-тетраметилурония), TFFH (2-фтор-гексафторфосфат N,N',N'',N'''-тетраметилурония), PyBOP (гексафторфосфат бензотриазол-1-ил-окси-трис-пирролидинофосфония), EEDQ (2-этокси-1-этоксикарбонил-1,2-дигидрохинолин), DCC (дициклогексилкарбодиимид); DIPCDI (диизопропилкарбодиимид), MSNT (1-(мезитилен-2-сульфонил)-3-нитро-1H-1,2,4-триазол и арилсульфонилгалогениды, например, триизопропилбензолсульфонилхлорид.
КОНЪЮГАЦИЯ БИОТИН-МАЛЕИМИДА С ТИОFAB
Вышеописанные свойства ТиоFab определены в присутствии фага, поскольку слияние Fab с белком оболочки фага потенциально может изменять доступность или реакционную способность тиола Cys. Таким образом, конструкции ТиоFab клонировали в экспрессирующий вектор под промотором щелочной фосфатазы (Chang et al (1987) Gene 55: 189-196), и экспрессию ТиоFab индуцировали посредством выращивания клеток E. coli в бесфосфатной среде. ТиоFab очищали на колонке с SEPHAROSE™ с белком G и анализировали в восстанавливающих и невосстанавливающих гелях для SDS-PAGE. Эти анализы позволяют оценку того, сохранили ли ТиоFab свою реакционноспособную тиоловую группу или сделаны неактивными посредством формирования внутримолекулярных или межмолекулярных дисульфидных связей. ТиоFab L-V15C, L-V110C, H-A88C и H-A121C экспрессировали и очищали хроматографией на колонке с SEPHAROSE™ с белком-G (подробно см. в разделе методы). Очищенные белки анализировали в геле для SDS-PAGE в восстанавливающих (с DTT) и невосстанавливающих (без DTT) условиях. Другие восстанавливающие средства, такие как BME (бета-меркаптоэтанол), можно использовать в геле для расщепления межцепочечных дисульфидных групп. Из анализа SDS-PAGE в геле очевидно, что главная (~90%) фракция ТиоFab присутствует в мономерной форме, в то время как hu4D5Fabv8 дикого типа присутствует в основном в мономерной форме (47 кДа).
ТиоFab (A121C) и hu4D5Fabv8 дикого типа инкубировали со 100-кратным избытком биотин-малеимида в течение 3 часов при комнатной температуре, и биотинилированные Fab наносили на колонку для гель-фильтрации Superdex-200™. Эта стадия очистки являлась пригодной для отделения мономерных Fab от олигомерных Fab, а также от избыточного свободного биотин-малеимида (или свободного циркониевого реагента).
На фигуре 5 показано подтверждение свойств вариантов ТиоFab в отсутствие фагового окружения. Белки без слияния с фагом, hu4D5Fabv8 и hu4D5Fabv8-A121C (ТиоFab-A121C), экспрессировали и очищали с использованием бусин с агарозой с белком-G с последующей инкубацией со 100-кратным молярным избытком биотин-малеимида. Сравнивали связывание стрептавидина и HER2 с биотинилированным сконструированным ТиоFab с cys заменами и небиотинилированным Fab дикого типа. Степень конъюгации с биотином (взаимодействие со стрептавидином) и способность связывания с HER2 мониторировали посредством анализов ELISA. Каждый Fab тестировали при 2 нг и 20 нг.
Биотинилированный A121C ТиоFab сохранял связывание с HER2, сравнимое со связыванием hu4D5Fabv8 дикого типа (фигура 5). Fab дикого типа и A121C-ТиоFab очищали гель-хроматографией на колонке. Два образца тестировали по связыванию HER2 и стрептавидина посредством ELISA с использованием анти-Fab-HRP козы в качестве вторичного антитела. Как Fab дикого типа (незакрашенный прямоугольник), так и ТиоFab (прямоугольник с точками) обладают сходным связыванием с HER2, но только ТиоFab сохранял связывание со стрептавидином. Только фоновый уровень взаимодействия со стрептавидином наблюдали для небиотинилированного hu4D5Fabv8 дикого типа (фигура 5). Масс-спектральный (LC-ESI-MS) анализ биотинилированного-ТиоFab (A121C) приводил к главному пику 48294,5 дальтон в отличие от hu4D5Fabv8 дикого типа (47737 дальтон). Разница в 537,5 дальтон между двумя молекулами точно соответствует одному биотин-малеимиду, конъюгированному с ТиоFab. Результаты масс-спектрального секвенирования белка (LC-ESI-тандемного масс-спектрального анализа) дополнительно подтвердили, что конъюгированная молекула биотина присутствует на вновь сконструированном остатке Cys (таблица 4, пример 3).
САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ БИОТИН-МАЛЕИМИДА С АЛЬБУМИНСВЯЗЫВАЮЩИЙ ПЕПТИД (ABP)-ТИОFAB
Связывание с белком плазмы может являться эффективным способом улучшения фармакокинетических свойств короткоживущих молекул. Альбумин является наиболее распространенным белком в плазме. Связывающие сывороточный альбумин пептиды (ABP) могут изменять фармакодинамику слитых с ними активных доменов белков, включая изменение поглощения, проникновения и диффузии в тканях. Эти фармакодинамические параметры можно регулировать специфическим выбором подходящей последовательности связывающего сывороточный альбумин пептида (US 20040001827). Серию альбуминсвязывающих пептидов идентифицировали скринингом фагового дисплея (Dennis et al. (2002) «Albumin Binding As A General Strategy For Improving The Pharmacokinetics Of Proteins» J Biol Chem. 277:35035-35043; WO 01/45746). Соединения по изобретению включают ABP последовательности, объясняемые в: (i) Dennis et al (2002) J Biol Chem. 277:35035-35043 в таблицах III и IV, страница 35038; (ii) US 20040001827 в SEQ ID NO: 9-22; и (iii) WO 01/45746 на страницах 12-13, SEQ ID NO: z1-z14, и содержание всех из них приведено в настоящем документе в качестве ссылки.
Альбуминсвязывающие белки (ABP)-Fab конструировали слиянием альбуминсвязывающего пептида с C-концом тяжелой цепи Fab в стехиометрическом соотношении 1:1 (1 ABP/1 Fab). Показано, что связывание этих ABP-Fab с альбумином увеличивало время их полужизни более чем в 25 раз у кроликов и мышей. Вышеописанные реакционноспособные остатки Cys можно, таким образом, вводить в эти ABP-Fab и использовать для сайт-специфической конъюгации с циркониевыми реагентами с последующими исследованиями на животных in vivo.
Последовательности альбуминсвязывающих пептидов включают в качестве неограничивающих примеров аминокислотные последовательности, перечисленные в SEQ ID NO: 1-5:

Последовательности альбуминсвязывающих пептидов (ABP) связывают альбумин множества видов (мыши, крысы, кролика, быка, макака-резуса, бабуина и человека) с Kd (кролик) = 0,3 мкМ. Альбуминсвязывающий пептид не конкурирует с лигандами, как известно, связывающими альбумин и обладает временем полужизни (T½) у кролика 2,3 ч. Белки ABP-ТиоFab очищали на BSA-SEPHAROSE™ с последующей конъюгацией с биотин-малеимидом и очисткой хроматографией на колонке Superdex-S200, как описано в предыдущих разделах. Очищенные биотинилированные белки являлись гомогенными и лишенными каких-либо олигомерных форм (пример 4).
На фигуре 6 показаны свойства вариантов альбуминсвязывающий пептид (ABP)-ТиоFab. Анализы ELISA проводили для тестирования способности ABP-hu4D5Fabv8-wt, ABP-hu4D5Fabv8-V110C и ABP-hu4D5Fabv8-A121C связываться с кроличьим альбумином, стрептавидином и HER2. Биотинилированные ABP-ТиоFab способны связывать альбумин и HER2 с аффинностью, сходной с аффинностью ABP-hu4D5Fabv8 дикого типа, как подтверждено ELISA (фигура 6) и анализом кинетики BIAcore (таблица 3). Планшет для ELISA покрывали альбумином, HER2 и SA, как описано. Для связывания биотинилированного ABP-ТиоFab с альбумином, HER2 и SA в качестве зонда использовали анти-Fab HRP. Биотинилированные ABP-ТиоFab были способны связываться со стрептавидином по сравнению с небиотинилированным контрольным ABP-hu4D5Fabv8-wt, что указывает на то, что ABP-ТиоFab являлись конъюгированными с биотин-малеимидом, подобно ТиоFab, сайт-специфическим образом, поскольку одни и те же мутанты Cys использовали для обоих вариантов (фигура 6).
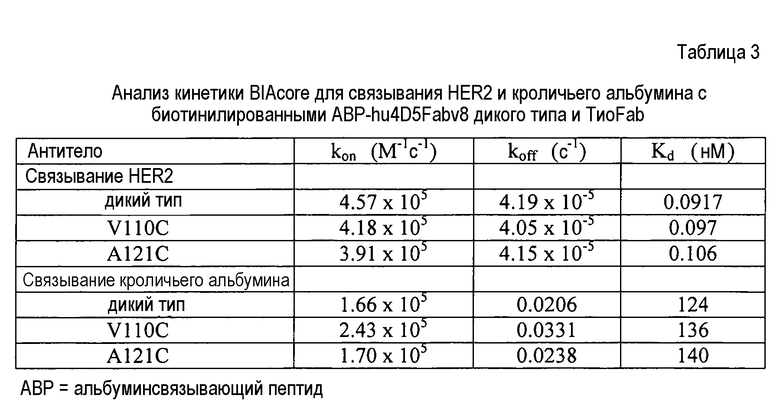
Альтернативно, альбуминсвязывающий пептид можно связывать с антителом посредством ковалентного присоединения через линкерную группу.
КОНСТРУИРОВАНИЕ ABP-ТИОFAB С ДВУМЯ СВОБОДНЫМИ ТИОЛОВЫМИ ГРУППАМИ НА FAB
Вышеуказанные результаты показывают, что все четыре варианта (L-V15C, L-V110C, H-A88C и H-A121C) тиоFab (сконструированных Fab антител с цистеиновыми заменами) обладают реакционноспособными тиоловыми группами, которые можно использовать для сайт-специфической конъюгации с реагентом для мечения, линкерным реагентом или промежуточным соединением цирконий-линкер. L-V15C можно экспрессировать и очищать, но с относительно низкими выходами. Однако, выходы экспрессии и очистки вариантов L-V110C, H-A88C и H-A121C являлись сходными с выходами hu4D5Fabv8. Таким образом, эти мутанты можно использовать для дальнейшего анализа и рекомбинации для получения более одной тиоловой группы на Fab. Для этой цели конструировали одну тиоловую группу на легкой и одну на тяжелой цепи для получения двух тиоловых групп на молекулу Fab (L-V110C/H-A88C и L-V110C/H-A121C). Эти два варианта с двойным Cys экспрессировали в экспрессирующей системе E. coli и очищали. Обнаружено, что гомогенность очищенных биотинилированных ABP-ТиоFab является сходной с гомогенностью вариантов с отдельным Cys.
Исследовали эффекты конструирования двух реакционноспособных остатков Cys на Fab (фигура 7). Присутствие второго биотина тестировали посредством исследования связывания биотинилированного ABP-ТиоFab с SA с использованием стрептавидин-HRP (фигура 7). Для анализа HER2/Fab планшет для ELISA покрывали HER2, и в качестве зонда использовали анти-Fab HRP. Для анализа SA/Fab планшет для ELISA покрывали SA, и в качестве зонда использовали анти-Fab HRP. Для анализа SA/SA планшет для ELISA покрывали SA, и в качестве зонда использовали SA-HRP. Фигура 7. Анализы ELISA взаимодействия cys вариантов биотинилированного ABP-hu4D5Fabv8 с HER2, стрептавидином (SA). Для HER2/Fab, SA/Fab и SA/SA показали, что их взаимодействия выявлены посредством анти-Fab-HRP, SA-HRP, соответственно. SA/Fab выявляет присутствие одного биотина на Fab, и более одного биотина на Fab выявляют анализом SA/SA. Связывание HER2 с мутантами с двойным cys является сходным со связыванием с вариантами с одним Cys (фигура 7). Однако, степень биотинилирования мутантов с двойным Cys была выше по сравнению с вариантами с одним Cys благодаря более одной свободной тиоловой группе на молекулу Fab (фигура 7).
КОНСТРУИРОВАНИЕ ТИО-IgG ВАРИАНТОВ ТРАСТУЗУМАБА
Цистеин вводили в конкретные остатки полноразмерного моноклонального антитела, трастузумаба (ГЕРЦЕПТИН®, Genentech Inc.). Мутанты трастузумаба с одним cys H-A88C, H-A121C и L-V110C, и мутанты трастузумаба с двойным cys V110C-A121C и V110C-A121C экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Последовательность мутантной тяжелой цепи A88C (450 ак) представляет собой SEQ ID NO:6. Последовательность мутантной тяжелой цепи A121C (450 ак) представляет собой SEQ ID NO:7. Последовательность мутантной легкой цепи V110C (214 ак) представляет собой SEQ ID NO:8.

В одном из вариантов осуществления сконструированные антитела с цистеиновыми заменами тио-трастузумаб содержат одну или несколько из следующих последовательностей вариабельной области тяжелой цепи со свободной аминокислотой цистеином (SEQ ID NO: 9-16).

В другом варианте осуществления сконструированные антитела с цистеиновыми заменами тио-трастузумаб содержат одну или несколько из следующих последовательностей вариабельных областей легкой цепи со свободной аминокислотой цистеином (SEQ ID NO: 17-27).

Полученные полноразмерные варианты IgG тио-трастузумаба анализировали по реакционной способности тиола и активности связывания HER2. На фигуре 13A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2 и меченным HRP вторичным антителом для детекции оптической плотности. На фигуре 13B показаны измерения связывания с иммобилизованным HER2 с детекцией оптической плотности при 450 нм (слева направо): небиотинилированный трастузумаб дикого типа (Wt), конъюгированные с биотином-малеимидом варианты тио-трастузумаба V110C (один cys), A121C (один cys) и V110C-A121C (двойной cys). Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг. Измерения показывают, что биотинилированные анти-HER2 ТиоMab сохраняют активность связывания HER2.
На фигуре 14A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2 со связыванием биотина с анти-IgG-HRP для детекции оптической плотности. На фигуре 14B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотином-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании со стрептавидином. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио IgG и исходный трастузумаб тестировали при 1, 10 и 100 нг. Измерения показывают, что ТиоMab против HER2 обладают высокой реакционной способностью тиола.
Цистеин вводили в конкретные остатки полноразмерного анти-EphB2R антитела 2H9. Мутант с одним cys H-A121C 2H9 экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Мутантная последовательность тяжелой цепи A121C 2H9 (450 ак) представляет собой SEQ ID NO:28.

Сконструированные тио-2H9 антитела с цистеиновыми заменами содержат следующие последовательности константной области тяжелой цепи Fc со свободной аминокислотой цистеином (SEQ ID NO: 29-38).

На фигуре 16 показан анализ электрофорезом в невосстанавливающем (вверху) и восстанавливающем (внизу) денатурирующем SDS-PAGE (электрофорезом в полиакриламидном геле) вариантов 2H9 ТиоMab Fc (слева направо, дорожки 1-9): A339C; S337C; S324C; A287C; V284C; V282C; V279C; и V273C, вместе с 2H9 дикого типа, после очистки на иммобилизованном белке A. Дорожка справа представляет собой маркер размера, указывающий на то, что интактные белки составляют приблизительно 150 кДа, фрагменты тяжелой цепи приблизительно 50 кДа и фрагменты легкой цепи приблизительно 25 кДа. На фигуре 17A показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (справа) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab (слева направо, дорожки 1-4): L-V15C; S179C; S375C; S400C, после очистки на иммобилизованном белке A. На фигуре 17B показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле дополнительных вариантов ТиоMab 2H9 и 3A5 после очистки на иммобилизованном белке A. Варианты ТиоMab 2H9 (в области Fab, так же как в Fc-области) экспрессировали и очищали, как описано. Как показано на фигурах 16, 17A и 17B, все белки являются гомогенными в SDS-PAGE после процедуры восстановления и окисления из примера 11 для получения реакционноспособных ТиоMab для конъюгации (пример 12).
Цистеин вводили в конкретные остатки полноразмерного анти-MUC16 антитела 3A5. Мутант 3A5 с одним cys H-A121C экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Мутантная последовательность тяжелой цепи A121C 3A5 (446 ак) содержит SEQ ID NO:39.

Сконструированные антитела с цистеиновыми заменами тио-3A5 анти-MUC16 содержат следующие последовательности вариабельной области тяжелой цепи со свободной аминокислотой цистеином (SEQ ID NO: 40-44).
Сконструированные тио-3A5 анти-MUC16 антитела с цистеиновыми заменами содержат следующие последовательности вариабельной области легкой цепи со свободной аминокислотой цистеином (SEQ ID NO: 45-49).
РЕАКЦИОННАЯ СПОСОБНОСТЬ ТИОЛА ТИОMAB
Реакционную способность тиола полноразмерных сконструированных антител с цистеиновыми заменами IgG (ТиоMab) измеряли по биотинилированию и связыванию стрептавидина. Анализ вестерн-блоттингом разрабатывали для скрининга ТиоMab, специфически конъюгированного с биотин-малеимидом. В этом анализе антитела анализировали восстанавливающим SDS-PAGE, и присутствие биотина специфически тестировали инкубацией со стрептавидином-HRP. Как видно на фигуре 18, наблюдают взаимодействие стрептавидина-HRP с тяжелой цепью или легкой цепью в зависимости от того, какой сконструированный с заменами cys вариант используют, и не наблюдают взаимодействия для дикого типа, что указывает на специфическую конъюгацию вариантов ТиоMab с биотином на сконструированном остатке Cys. На фигуре 18 показан анализ электрофорезом в денатурирующем геле восстановленных биотинилированных вариантов Тио-IgG после связывания с иммобилизованным анти-IgG-HRP (верхний гель) и стрептавидином-HRP (нижний гель). Дорожка 1: 3A5 H-A121C. Дорожка 2: 3A5 L-V110C. Дорожка 3: 2H9 H-A121C. Дорожка 4: 2H9 L-V110C. Дорожка 5: исходное анти-EphB2R 2H9, дикого типа. Каждый мутант (дорожки 1-4) связывался с анти-IgG с детекцией HRP (вверху), что указывает на сохранение избирательности и аффинности. Связывание с иммобилизованным стрептавидином с детекцией HRP (внизу) подтвердило локализацию биотина на тяжелой и легкой цепях. Локализацией цистеиновой мутации на сконструированных антителах с цистеиновыми заменами в дорожках 1 и 3 является тяжелая цепь. Локализацией цистеиновой мутации на сконструированных антителах с цистеиновыми заменами в дорожках 2 и 4 является легкая цепь. Участок цистеиновой мутации подвергается конъюгации с биотин-малеимидным реагентом.
Анализ ТиоMab сконструированных антител с цистеиновыми заменами на фигуре 18 и варианта 2H9 V15C посредством LC/MS дает количественные показатели реакционной способности тиола (Таблица 5).
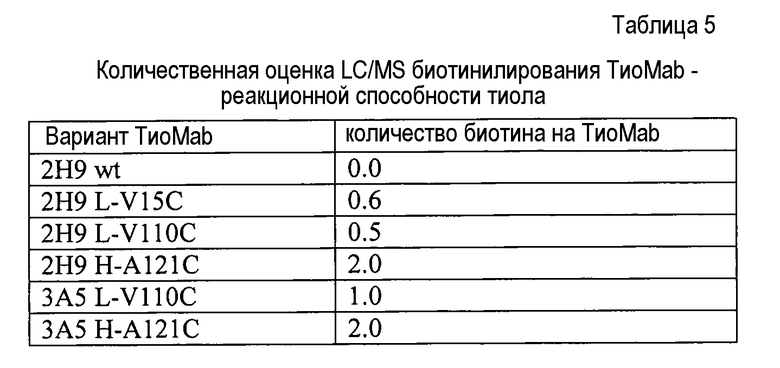
Конструирование цистеиновых замен проводили в константном домене, т.е. Fc-области, IgG антител. Множество аминокислотных участков переводили в цистеиновые участки, и экспрессированные мутанты, т.е. сконструированные антитела с цистеиновыми заменами, оценивали по реакционной способности тиола.
Варианты биотинилированного Fc ТиоMab 2H9 оценивали по реакционной способности тиола посредством количественной оценки с помощью HRP посредством связывания с иммобилизованным стрептавидином в анализе ELISA (фигура 19). Анализ ELISA разрабатывали для быстрого скрининга остатков Cys с реакционноспособными тиоловыми группами. Как показано на схематической диаграмме на фигуре 19, взаимодействие стрептавидин-биотин мониторировали с использованием в качестве зонда анти-IgG-HRP с последующим измерением оптической плотности при 450 нм. Эти результаты подтвердили, что 2H9-ТиоFc варианты V282C, A287C, A339C, S375C и S400C обладали реакционной способностью тиола, от средней до наиболее высокой. Степень конъюгации с биотином вариантов 2H9 ТиоMab Fc количественно оценивали анализом LS/MS, как указано в таблице 6. Анализ LS/MS подтвердил, что варианты A282C, S375C и S400C обладали 100% конъюгацией с биотином, и V284C и A339C обладали 50% конъюгацией, что указывает на присутствие реакционноспособной тиоловой группы цистеина. Другие варианты ТиоFc, и исходное 2H9 дикого типа либо обладали очень малым биотинилированием, либо не были биотинилированы.
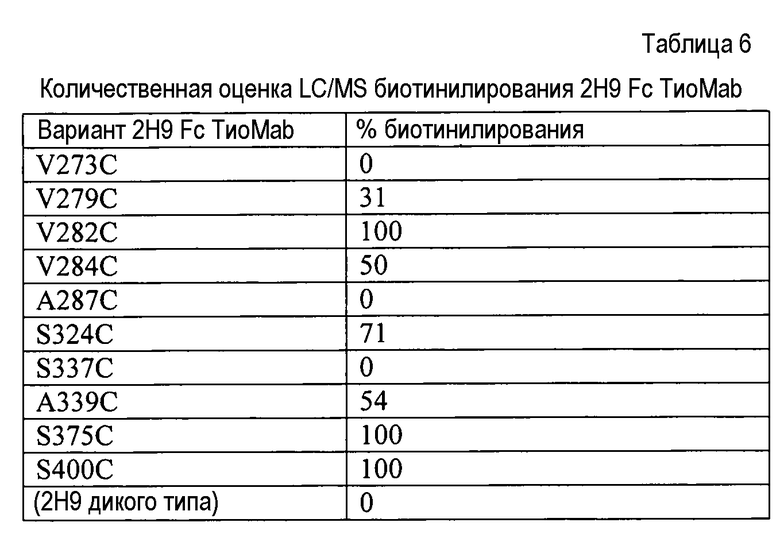
РЕАКЦИОННАЯ СПОСОБНОСТЬ ТИОЛА ВАРИАНТОВ ЛЕГКОЙ ЦЕПИ ТИО-4D5 FAB
Скринингом множества сконструированных вариантов легкой цепи Fab анти-ErbB2 антитела 4D5 с цистеиновыми заменами получили ряд вариантов с значением реакционной способности тиола 0,6 и выше (таблица 7), как измерено анализом PHESELECTOR на фигуре 8. Значения реакционной способности тиола из таблицы 7 нормализованы по варианту тяжелой цепи 4D5 ТиоFab (HC-A121C), который принимали за 100%, принимая полное биотинилирование варианта HC-A121C, и представлены как значения в процентах.
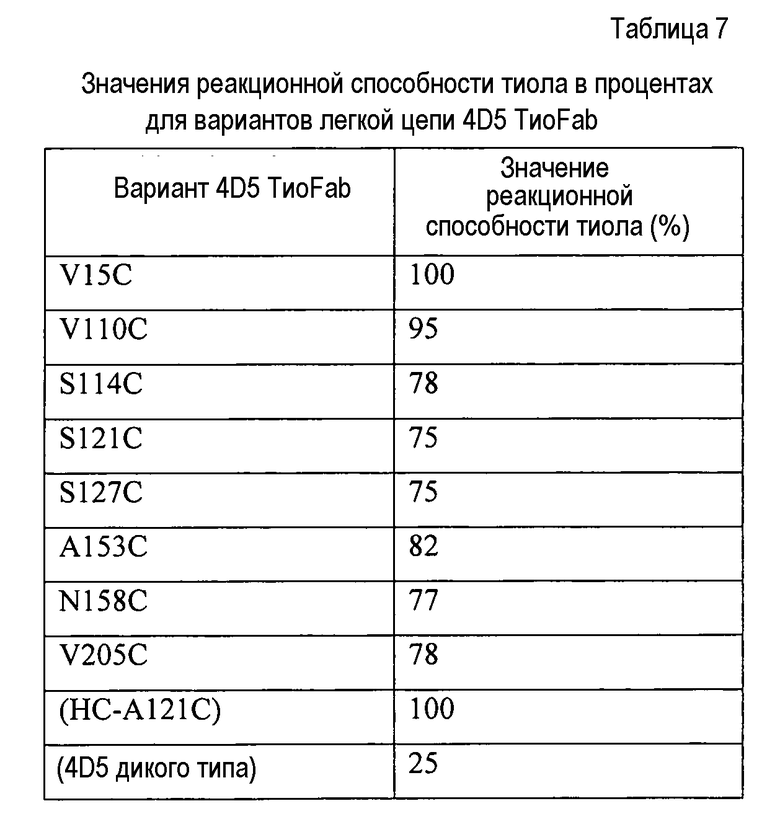
РЕАГЕНТЫ ДЛЯ МЕЧЕНИЯ ЦИРКОНИЕМ
Иллюстративные бифункциональные реагенты на основе десферриоксамина B (Df) использовали для образования комплексов 89Zr с антителами, включая моноклональные антитела (mAb). Десферриоксамин B (Ν'-{5-[ацетил(гидрокси)амино]пентил}-N-[5-({4-[(5-аминопентил)(гидрокси)амино]-4-оксобутаноил}амино)пентил]-N-гидроксисукцинамид (CAS Reg. No. 70-51-9); и известный также как дефероксамин, десфероксамин B, DFO-B, DFOA, DFB или десферал), представляет собой бактериальный сидерофор, продуцируемый actinobacter Streptomyces pilosus (фигура 20, вверху). Десферриоксамин B имеет применение в медицине в качестве хелатирующего агента, используемого для удаления избытка железа из организма (Miller, Marvin J. «Syntheses and therapeutic potential of hydroxamic acid based siderophores and analogs» (1989) Chemical Reviews 89 (7): 1563-1579). Соль мезилат DFO-B является коммерчески доступной. Первоначальные эксперименты проводили с N-(S-ацетил)тиоацетил-Df (SATA-Df) и mAb с добавленными малеимидными группами, 4-[N-малеимидометил]циклогексан-1-карбоксилатом (mAb-SMCC), присоединенными к ε-аминогруппе в боковой цепи лизина (Meijs WE et al. «Zirconium-labeled monoclonal antibodies and their distribution in tumor-bearing nude mice» (1997) J. Nucl. Med. 38: 112-8; Meijs WE et al. «A facile method for the labeling of proteins with zirconium isotopes» (1996) Nucl Med Biol. 23:439-48). Однако, полученные тиоэфирные конъюгаты (mAb-SMCC-SATA-Df) были нестабильными в сыворотке человека при 37°C (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med 44: 1271-81). Другие иллюстративные реакционноспособные по отношению к аминогруппам бифункциональные хелаторы на основе Df, модифицированного янтарным ангидридом (Suc), использовали для превращения аминогруппы Df в карбоновую кислоту и последующей активации в форме сложного эфира 2,3,5,6-тетрафторфенила (TFP). TFP-N-Suc-Df (фигура 20, в центре) присоединяли к ε-аминогруппам лизина mAb, и получали хелаты очищенного mAb-N-Suc-Df с 89Zr. Полученный 89Zr-mAb-N-Suc-Df являлся стабильным в физиологических условиях, и его биораспределение сравнивали с mAb-SMCC-SATA-Df у мышей (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med. 44: 1271-81). Однако, получение TFP-N-Suc-Df требует защиты гидроксамидных групп в форме комплекса Fe(III). Железо удаляют обработкой ЭДТА до образования хелатов с 89Zr, но многоступенчатый способ является трудоемким и подвержен опасности неполного удаления железа из десферриоксамина и/или неполного удаления ЭДТА из буфера для конъюгации, что может отрицательно влиять на выход образования 89Zr-хелатов. Таким образом, гетеробифункциональный реакционноспособный по отношению к аминогруппам реагент, п-изотиоцианатбензил-десферриоксамин (Df-Bz-NCS), недавно разработан для включения Df в белки посредством связи с тиомочевиной, фигура 20, в центре (Perk LR et al. «Facile radiolabeling of monoclonal antibodies and other proteins with zirconium-89 or gallium-68 for PET Imaging using p-isothiocyanatobenzyl-desferrioxamine» (2008) Nature Protocols, published online:DOI: 10.1038/nprot.2008.22; Perk LR et al. «p-Isothiocyanatobenzyl-desferrioxamine: a new bifunctional chelate for facile radiolabeling of monoclonal antibodies with zirconium-89 for immuno-PET imaging» (2009) European Journal Of Nuclear Medicine And Molecular Imaging). Для конъюгатов антител, полученных с использованием Df-Bz-NCS, показали стабильность и свойства получения изображений, сравнимые с контрольными конъюгатами, полученными с использованием TFP-N-Suc-Df. Поскольку разработаны надежные способы присоединения 89Zr к антителам через ε-аминогруппы лизина, число опубликованных доклинических и клинических исследований иммуноПЭТ с меченными 89Zr антителами быстро увеличивается (Verel I, et al. «Long-lived positron emitters zirconium-89 and iodine-124 for scouting of therapeutic radioimmunoconjugates with PET» (2003) Cancer Biother Radiopharm. 18:655-61; Nagengast WB et al. «In vivo VEGF imaging with radiolabeled bevacizumab in a human ovarian tumor xenograft» (2007) J Nucl Med. 48:1313-9; Perk LR, et al. «(89)Zr as a PET surrogate radioisotope for scouting biodistribution of the therapeutic radiometals (90)Y and (177)Lu in tumor-bearing nude mice after coupling to the internalizing antibody cetuximab» (2005) J Nucl Med. 46: 1898-906; Perk LR et al. «Quantitative PET imaging of Met-expressing human cancer xenografts with (89)Zr-labelled monoclonal antibody DN30» (2008) European Journal Of Nuclear Medicine And Molecular Imaging 35:1857-67; Perk LR et al. «Preparation and evaluation of (89)Zr-Zevalin for monitoring of (90)Y-Zevalin biodistribution with positron emission tomography» (2006) European Journal Of Nuclear Medicine And Molecular Imaging 33:1337-45; Borjesson PK et al. «Performance of immuno-positron emission tomography with zirconium-89-labeled chimeric monoclonal antibody U36 in the detection of lymph node metastases in head and neck cancer patients» (2006) Clin Cancer Res. 12:2133-40; Aerts HJ et al. «Disparity between in vivo EGFR expression and 89Zr-labeled cetuximab uptake assessed with PET» (2009) J Nucl Med. 50:123-31; Dijkers EC et al. «Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging» (2009) J Nucl Med 50(6):974-981).
Варианты осуществления комплексов циркония включают также связывающие цирконий (образующие с ним хелаты) лиганды, такие как DTPA (CAS Reg. No. 67-43-6), DOPA (1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусная кислота) (Liu, Shuang (2008) Advanced Drug Delivery Reviews 60(12):1347-1370), циклопентадиенильные и аллильные группы (Erker, G. (1991) Pure and Applied Chemistry 63(6):797-806; Erker, G. (1990) Jour, of Organometallic Chem. 400(1-2):185-203), содержание каждого из которых приведено в настоящем документе в качестве ссылки.
Комплексы циркония (Z) и других радиоактивных изотопов можно конъюгировать с антителами (Ab), включая моноклональные антитела (mAb), через ε-аминогруппу боковой цепи лизина или через тиоловую группу цистеина. Поскольку приблизительно 40 боковых цепей лизина (Wang L et al «Structural characterization of the maytansinoid-monoclonal antibody immunoconjugate, huN901-DM1, by mass spectrometry» (2005) Protein Sci. 14:2436-46) или 8 остатков цистеина (Hamblett KJ et al. «Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate» (2004) Clin Cancer Res. 10:7063-70) доступны для конъюгации в mAb, оба способа обеспечивают гетерогенность по отношению к соотношениям конъюгатов mAb и участкам конъюгации. Модификация остатка лизина внутри участка связывания может уменьшать биологическую активность конъюгата (Cai W et al. «PET imaging of colorectal cancer in xenograft-bearing mice by use of an 18F-labeled T84.66 anti-carcinoembryonic antigen diabody» (2007) J Nucl Med. 48:304-10; Shively JE. «18F labeling for immuno-PET: where speed and contrast meet» (2007) J Nucl Med. 48: 170-2; Tait JF et al «Improved detection of cell death in vivo with annexin V radiolabeled by site-specific methods» (2006) J Nucl Med. 47: 1546-53; Schellenberger EA et al «Optical imaging of apoptosis as a biomarker of tumor response to chemotherapy» (2003) Neoplasia (New York, N.Y) 5: 187-92), в то время как модификация остатков цистеина в шарнирной области обеспечивает уменьшенное время полужизни в плазме (Hamblett KJ et al. «Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate» (2004) Clin Cancer Res. 10:7063-70). Этих ограничений можно избежать с использованием mAb, сконструированных с содержанием цистеина, расположенного избирательно, с целью сайт-специфической конъюгации, с помощью биохимического анализа PHESELECTOR (US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008;332:41-52) для быстрой идентификации предпочтительных аминокислот в антителе для мутации с заменой на цистеин. Полученное антитело (ТИОMAB) затем химически избирательно и сайт-специфически конъюгируют с цитотоксическими лекарственными средствами без какой-либо потери аффинности связывания или неблагоприятных эффектов на стабильность каркаса антитела (Junutula JR et al. «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol. 26:925-32).
С точки зрения получения изображений, высокая аффинность к мишени и минимальное неспецифическое поглощение необходимы для оптимального качества изображения. Соответственно, сайт-специфически радиоактивно меченные сконструированные антитела с цистеиновыми заменами (ТИОMAB) могут обеспечивать метки с неизмененной аффинностью связывания и стабильностью каркаса, которые могут минимизировать неспецифическое поглощение метаболитов вне ткани-мишени. Один аспект настоящего изобретения относится к способу сайт-специфического радиоактивного мечения ТИОMAB с использованием новых реакционноспособных по отношению к тиолу бифункциональных реагентов на основе Df малеимидоциклогексил-десферриоксамина (Df-Chx-Mal), бромацетил-десферриоксамина (Df-Bac) и иодацетил-десферриоксамина (Df-Iac) (фигура 20). Иллюстративные варианты осуществления включают варианты осуществления, в которых эти реагенты сайт-специфически конъюгируют с ТИОMAB трастузумабом (тио-трастузумаб), получают хелаты с 89Zr и оценивают in vitro и in vivo.
Одним из метастабильных изомеров циркония является 89Zr со временем полужизни 78,4 часов с типами распада бета (испускание электронов), испускания позитронов (бета плюс) и гамма-излучения.
Радиоактивный изотоп или другие метки можно включать в конъюгат известными способами (Fraker et al (1978) Biochem. Biophys. Res. Commun. 80: 49-57; «Monoclonal Antibodies in Immunoscintigraphy» Chatal, CRC Press 1989). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой иллюстративный хелатирующий агент для конъюгации радиоактивного изотопа с антителом (WO 94/11026).
ЛИНКЕРЫ
«Линкер» (L) представляет собой бифункциональную или мультифункциональную группу, которую можно использовать для соединения одной или нескольких групп комплекса с цирконием (Z) и молекулы антитела (Ab) с образованием конъюгатов антитело-цирконий (AZC) формулы I. Конъюгаты антитело-цирконий (AZC) можно удобно получать с использованием линкера, обладающего реакционноспособной функциональной группой для связывания с цирконием и с антителом. Тиол цистеина сконструированного антитела с цистеиновыми заменами (Ab) может образовывать связь с функциональной группой линкерного реагента, группой циркониевой метки или промежуточным соединением цирконий-линкер.
В одном аспекте линкер имеет реакционноспособный участок с электрофильной группой, являющейся реакционноспособной по отношению к нуклеофильному цистеину, присутствующему в антителе. Тиол цистеина антитела является реакционноспособным по отношению к электрофильной группе линкера и образует ковалентную связь с линкером. Пригодные электрофильные группы включают в качестве неограничивающих примеров малеимидные и галоацетамидные группы.
Сконструированные антитела с цистеиновыми заменами могут вступать в реакцию с линкерными реагентами или промежуточными соединениями цирконий-линкер, с электрофильными функциональными группами, такими как малеимид или α-галокарбонил, согласно способу конъюгации на странице 766 в Klussman, et al (2004), Bioconjugate Chemistry 15(4):765-773, и согласно способам из примеров 17-19.
В другом варианте осуществления группы Z являются одинаковыми.
В другом варианте осуществления группы Z являются различными.
Иллюстративные варианты осуществления соединений конъюгатов антитело-цирконий (AZC) формулы I включают:
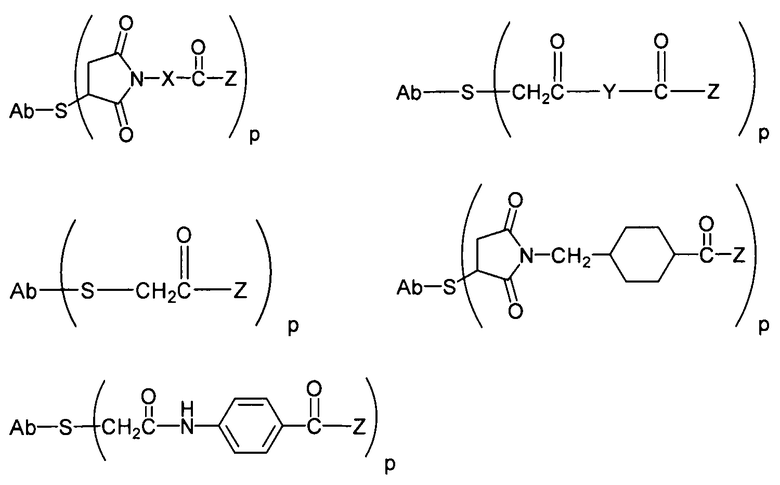
где X представляет собой:
где Y представляет собой:
R независимо представляет собой H или C1-C6 алкил; и n представляет собой 1-12.
В другом варианте осуществления линкер обладает реакционноспособной функциональной группой, обладающей нуклеофильной группой, реакционноспособной по отношению к электрофильной группе, присутствующей в антителе. Пригодные электрофильные группы антитела включают в качестве неограничивающих примеров, карбонильные группы альдегидов и кетонов. Гетероатом нуклеофильной группы линкера может вступать в реакцию с электрофильной группой антитела и образовывать ковалентную связь с молекулой антитела. Пригодные нуклеофильные группы линкера включают в качестве неограничивающих примеров, гидразид, оксим, амино, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. Электрофильная группа антитела обеспечивает удобный участок для присоединения к линкеру.
В другом варианте осуществления линкер может быть замещен группами, модулирующими растворимость или реакционную способность. Например, заряженный заместитель, такой как сульфонат (-SO3 -) или аммоний, может увеличивать растворимость реагента в воде и облегчать реакцию присоединения линкерного реагента к антителу или группе циркония, или облегчать реакцию присоединения Ab-L (промежуточного соединения антитело-линкер) к Z или Z-L (промежуточного соединения цирконий-линкер) к Ab, в зависимости от способа синтеза, используемого для получения AZC.
Соединения по изобретению конкретно включают, но без ограничения, AZC, полученные с помощью линкерных реагентов: BMPEO, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), и включая бис-малеимидные реагенты: DTME, BMB, BMDB, BMH, BMOE, BM(PEO)2 и BM(PEO)43, коммерчески доступные от Pierce Biotechnology, Inc., Customer Service Department, P.O. Box 117, Rockford, IL. 61105 U.S.A. Бис-малеимидные реагенты позволяют присоединение тиоловой группы сконструированного антитела с цистеиновыми заменами к содержащей тиол циркониевой группе, метке или линкерному промежуточному соединению, последовательным или одновременным образом. Другие функциональные группы помимо малеимида, реакционноспособные по отношению к тиоловой группе сконструированного антитела с цистеиновыми заменами, циркониевой группе, метке или линкерному промежуточному соединению, включают иодацетамид, бромацетамид, винилпиридин, дисульфид, пиридилдисульфидизоцианат и изотиоцианат.
Пригодные линкерные реагенты можно получать также из других коммерческих источников, таких как Molecular Biosciences Inc. (Boulder, CO), или синтезировать способами, описанными в Toki et al (2002) J. Org. Chem. 67: 1866-1872; Walker, M.A. (1995) J. Org. Chem. 60:5352-5355; Frisch et al (1996) Bioconjugate Chem. 7: 180-186; US 6214345; WO 02/088172; US 2003130189; US2003096743; WO 03/026577; WO 03/043583; и WO 04/032828.
Иллюстративные линкерные реагенты включают:
где n представляет собой целое число в диапазоне 1-10, и T представляет собой -H или -SO3Na;
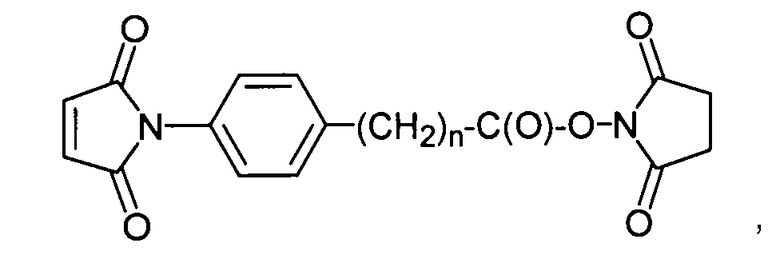
где n представляет собой целое число в диапазоне 0-3;
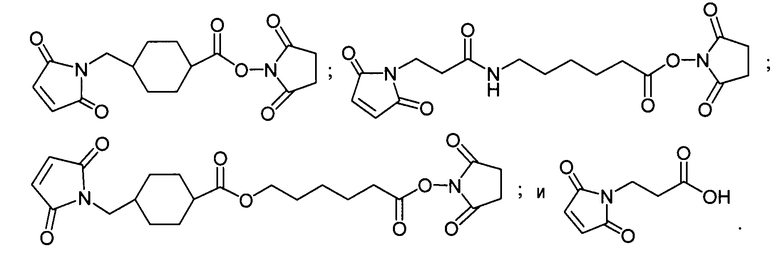
В другом варианте осуществления линкер L может представлять собой линкер древовидного типа для ковалентного присоединения более одной группы циркония через разветвленную, мультифункциональную линкерную группу к антителу (Sun et al (2002) Bioorganic & Medicinal Chemistry Letters 12:2213-2215; Sun et al (2003) Bioorganic & Medicinal Chemistry 11:1761-1768). Линкеры древовидного типа могут увеличивать молярное соотношение циркония к антителу, т.е. нагрузку AZC. Таким образом, когда сконструированное антитело с цистеиновыми заменами несет только одну реакционноспособную тиоловую группу цистеина, множество циркониевых групп можно присоединять через линкер древовидного типа.
Следующие иллюстративные варианты осуществления линкерных реагентов древовидного типа позволяют конъюгацию вплоть до девяти содержащих циркониевую группу нуклеофильных реагентов посредством реакции с содержащими хлорэтил и азотистый иприт функциональными группами:
Другие варианты осуществления разветвленных, древовидных линкеров включают линкеры с самоуничтожающимися дендримерными звеньями 2,6-бис(гидроксиметил)-п-крезола и 2,4,6-трис(гидроксиметил)-фенола (WO 2004/01993; Szalai et al (2003) J. Amer. Chem. Soc. 125: 15688-15689; Shamis et al (2004) J. Amer. Chem. Soc. 126: 1726-1731; Amir et al (2003) Angew. Chem. Int. Ed. 42:4494-4499).
МЕЧЕННЫЕ ДЕСФЕРРИОКСАМИНОМ СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
В одном аспекте изобретение относится к меченному десферриоксамином сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающему формулой II:

где L-Df выбран из:
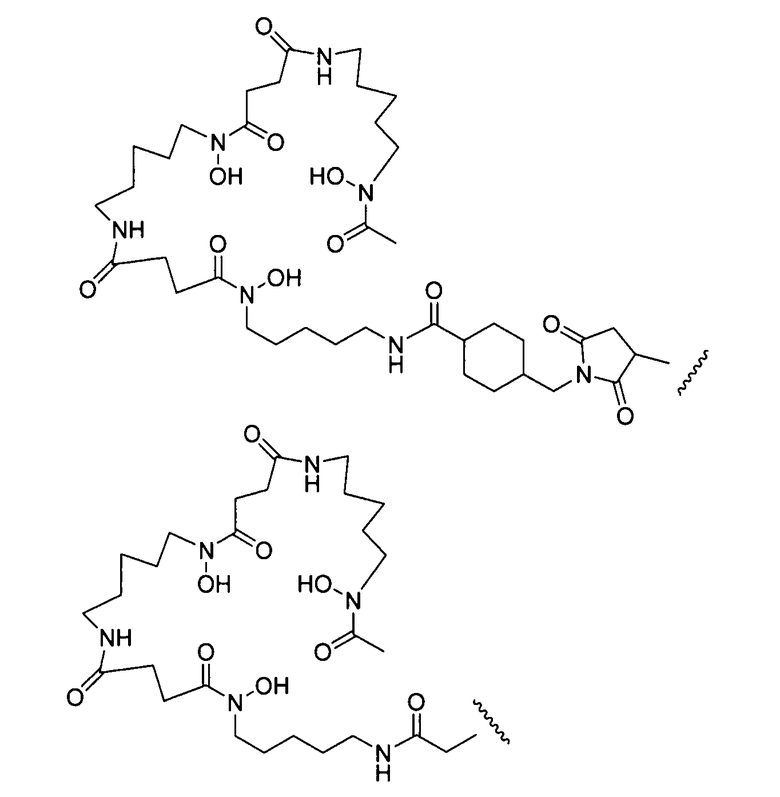
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
ПОЛУЧЕНИЕ КОНЪЮГАТОВ АНТИТЕЛО-ЦИРКОНИЙ
Конъюгаты антитело-цирконий (AZC) формулы I можно получать несколькими способами с использованием реакций, условий и реагентов органической химии, известных специалистам в данной области, включая: (1) реакцию цистеиновой группы сконструированного антитела с цистеиновыми заменами с линкерным реагентом для образования промежуточного соединения антитело-линкер Ab-L, посредством ковалентной связи, с последующей реакцией с активированной группой циркониевой метки Z; и (2) реакцию нуклеофильной группы циркониевой группы с линкерным реагентом для образования промежуточного соединения циркониевая метка-линкер Z-L, посредством ковалентной связи, с последующей реакцией с цистеиновой группой сконструированного антитела с цистеиновыми заменами. Способы конъюгации (1) и (2) можно использовать со множеством сконструированных антител с цистеиновыми заменами, групп циркониевой метки и линкеров для получения конъюгатов антитело-цирконий формулы I.
Тиоловые группы цистеина антитела являются нуклеофильными и способными вступать в реакцию для формирования ковалентных связей с электрофильными группами линкерных реагентов и промежуточных соединений цирконий-линкер, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил и бензилгалиды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы; и (iv) дисульфиды, включая пиридилдисульфиды, посредством сульфидного обмена. Нуклеофильные группы на группе циркониевой метки включают в качестве неограничивающих примеров: группы амина, тиола, гидроксила, гидразида, оксима, гидразина, тиосемикарбазона, гидразинкарбоксилата и арилгидразида, способные вступать в реакцию для формирования ковалентных связей с электрофильными группами линкерных групп и линкерных реагентов.
В конкретных условиях сконструированные антитела с цистеиновыми заменами можно сделать реакционноспособными для конъюгации с линкерными реагентами посредством обработки восстанавливающим агентом, таким как DTT (реактив Клеланда, дитиотреитол) или TCEP (трис(2-карбоксиэтил)фосфингидрохлорид; Getz et al (1999) Anal. Biochem. Vol 273:73-80; Soltec Ventures, Beverly, MA). Полноразмерные сконструированные моноклональные антитела с цистеиновыми заменами (ТиоMab), экспрессированные в клетках CHO, восстанавливали с помощью приблизительно 50-кратного избытка TCEP в течение 3 часов при 37°C для восстановления дисульфидных связей, которые могут формироваться между вновь введенными остатками цистеина и цистеином, присутствующим в культуральной среде. Восстановленное ТиоMab разводили и наносили на колонку HiTrap S в 10 мМ ацетате натрия, pH 5, и элюировали с помощью PBS, содержащего 0,3 М хлорид натрия. Дисульфидные связи вновь образовывались между остатками цистеина, присутствующими в исходном Mab, с помощью разведенного (200 нМ) водного сульфата меди (CuSO4) при комнатной температуре, в течение ночи. Можно использовать другие окислители, т.е. окисляющие агенты и окисляющие условия, известные в данной области. Окисление атмосферным воздухом также является эффективным. Эта стадия мягкого, частичного повторного окисления формирует внутрицепочечные дисульфиды эффективно с высокой точностью. Приблизительно 10-кратный избыток промежуточного соединения цирконий-линкер добавляют, перемешивают и оставляют приблизительно на один час при комнатной температуре для осуществления конъюгации и образования конъюгата ТиоMab антитело-цирконий. Смесь после конъюгации подвергают гель-фильтрации, наносят на колонку HiTrap S и проводят элюцию для удаления избытка промежуточного соединения цирконий-линкер и других примесей.
На фигуре 15 показан общий способ подготовки сконструированного антитела с цистеиновыми заменами, экспрессированного из культуры клеток, для конъюгации. Цистеиновые аддукты, предположительно с различными межцепьевыми дисульфидными связями, подвергают восстановительному отщеплению для получения восстановленной формы антитела. Межцепочечные дисульфидные связи между спаренными остатками цистеина вновь формируют в условиях частичного окисления, таких как воздействие атмосферного кислорода. Вновь введенные сконструированные и неспаренные остатки цистеина остаются доступными для реакции с линкерными реагентами или промежуточными соединениями цирконий-линкер для образования конъюгатов антител по изобретению. С ТиоMab, экспрессированных в линиях клеток млекопитающих, получали внешний аддукт Cys, конъюгированный с конструированным Cys посредством формирования -S-S- связи. Таким образом, очищенные ТиоMab необходимо обрабатывать способами восстановления и окисления, как описано в примере 11, для получения реакционноспособных ТиоMab. Эти ТиоMab используют для конъюгации с содержащими малеимид радиоактивными метками, цитотоксическими лекарственными средствами, флуорофорами и другими метками.
ПОЛУЧЕНИЕ И АНАЛИЗ КОНЪЮГАТОВ 89 Zr-Df-ТРАСТУЗУМАБА
Защищенный активный сложный эфир TFP-N-SucDf-Fe получали согласно описанному ранее способу (Verel I et al «89Zr Immuno-PET: Comprehensive Procedures For The Production Of 89Zr-Labeled Monoclonal Antibodies» (2003) J Nucl Med 44: 1271-81) и конъюгировали с трастузумабом с использованием 5-кратного молярного избытка TFP-N-SucDf-Fe для получения N-SucDf-трастузумаба в среднем с 1,6 молекулами десферриоксамина (Таблица 8). Df-Bz-SCN-трастузумаб получали объединением с 8-кратным молярным избытком Df-Bz-SCN при pH 8,5 (Perk LR, et al. «Facile radiolabeling of monoclonal antibodies and other proteins with zirconium-89 or gallium-68 for PET Imaging using p-isothiocyanatobenzyl-desferrioxamine» (2008) Nature Protocols; published online:DOI: 10.1038/nprot.2008.22). Реакция приводит к Df-Bz-SCN-трастузумабу с присоединенными в среднем 2,4 молекулами десферриоксамина (Таблица 1).
Новый реакционноспособный по отношению к тиолу бифункциональный линкер на основе малеимида Df-Chx-Mal получали из эквимолярных количеств мезилата десферриоксамина и SMCC (фигура 21, пример 13). Реакция завершалась через 30 мин при комнатной температуре, и продукт выделяли преципитацией при добавлении воды с 45% выходом и более чем 95% чистотой. Реакцией 8,5-кратного молярного избытка Df-Chx-Mal со свежеприготовленым тио-трастузумабом (фигура 21, пример 17) получали конъюгат Df-Chx-Mal-тио-трастузумаб точно с 2 молекулами десферриоксамина за 1 час (таблица 1, фигура 21). Бромацетилдесферриоксамин (BDf-Bac) получали реакцией с эквимолярными количествами мезилата десферриоксамина и бромацетилбромида при 0°C (пример 14). Продукт получали с 14% выходом после очистки HPLC. Алкилированием свежеполученного тио-трастузумаба (фигура 21, пример 16) с помощью 12-кратного молярного избытка Df-Bac получали конъюгат (Df-Ac-тио-трастузумаб) с 1,8 молекулами Df на антитело через 5 часов (таблица 8, фигура 21, пример 18). Низкая реакционная способность бромида побудила авторов настоящего изобретения исследовать более реакционноспособное производное иодацетила (Df-Iac). Df-Iac получали с 53% выходом реакцией мезилата десферриоксамина с небольшим избытком иодацетата N-гидроксисукцинимидила (фигура 21, пример 15). Продукт получали более чем с 95% чистотой посредством преципитации из реакционной смеси. Последующей реакцией с 11-кратным избытком Df-Iac получали Df-Ac-тио-трастузумаб с 1,8 молекулами Df через 2 часа (таблица 1, фигура 21, пример 19). На основании опыта авторов настоящего изобретения, Df-Chx-Mal является предпочтительным реагентом из трех исследованных соединений. Следует отметить, что реакция Df-Chx-Mal завершалась при среднем pH в пределах 1 часа, в отличие от более высокого pH и более длительного времени реакции, необходимых для алкилирования тиоловых групп с помощью Df-Bac и Df-Iac. Кроме того, более низкая реакционная способность галогенацетамидов может вызывать неполную загрузку обеих доступных остатков цистеина тио-трастузумаба.
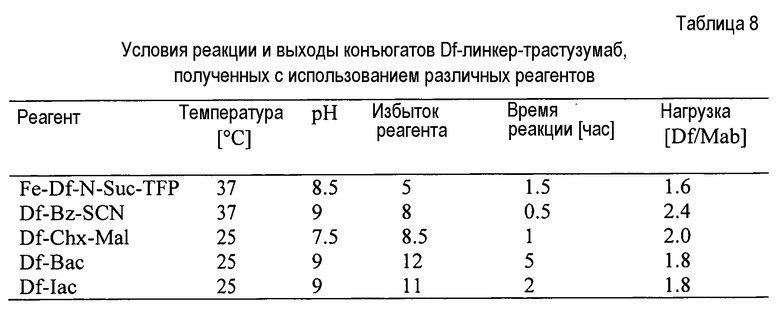
89Zr образовывал хелаты в форме оксалата 89-циркония со всеми четырьмя вариантами Df-трастузумаба с использованием ранее описанного экспериментального способа (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med. 44: 1271-81). Белки с радиоактивной меткой очищали на обессоливающей колонке, и конечный раствор концентрировали до необходимого объема мембранной фильтрацией. Выход, чистота и конечная сецифическая активность конъюгатов 89Zr обобщены в таблице 9. Как правило, выход хелатов превышал 80%, за исключением Df-N-Suc линкера, полученного с низким выходом, предположительно, из-за более низкого количества Df на молекулу антитела и/или неполного удаления Fe(III), используемого для защиты хелатирующего агента в ходе активации и конъюгации. После очистки вариантов Df-трастузумаба с использованием обессоливающей колонки, чистота продукта составляла более 90% с малым количеством (1-6%) высокомолекулярных агрегатов, детектированных в каждом образце. Df-Chx-Mal-тио-трастузумаб обеспечивал комплекс 89Zr с чистотой 99% (Таблица 9), в отличие от конъюгата Df-Ac, загрязненного приблизительно 8% низкомолекулярных загрязнений и 2% высокомолекулярных агрегатов. Загрязнения выдерживали удаление с использованием колонки NAP-10, но удаление являлось возможным с использованием повторяющейся смены буфера на фильтре Amicon.
Выходы, специфическая активность и чистота меченного радиоактивной меткой 89Zr-Df-линкер-трастузумаба
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ КОНЪЮГАТОВ 89 Zr-Df-ТРАСТУЗУМАБА
Биологическую активность вновь полученных сайт-специфических конъюгатов Df-линкер-тио-трастузумаб определяли с использованием анализа Скэтчарда, анализа связывания с линией клеток рака молочной железы BT474. Полученные значения KD сравнивали с немодифицированным трастузумабом (0,91±0,20 нМ). KD для конгъюгата тио-трастузумаба, содержащего Chx-Mal линкер, составляла 0,93±0,15 нМ, и значения для конъюгатов, содержащих Ac линкер, составляли 1,22±0,22 нМ для конъюгата, полученного с использованием Df-Bac, и 0,87±0,15 нМ для конъюгата, полученного с использованием Df-Iac. Результаты анализов биологической активности указывают на то, что модификация тио-трастузумаба не влияет на аффинность связывания антитела с HER2.
СТАБИЛЬНОСТЬ СЫВОРОТКИ IN VITRO
Ранее опубликованные конъюгаты Df-антитела с линкерами N-Suc и Bz-SCN, содержащие связи амида или тиомочевины, являлись стабильными in vitro в течение периода 6 суток в сыворотке при 37°C (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» J Nucl Med 2003;44: 1271-81; Perk LR et al, (2009) European Journal Of Nuclear Medicine And Molecular Imaging 35(10): 1857-1867). Стабильность конъюгатов 89Zr-тио-трастузумаба с Chx-Mal и Ac линкерами определяли в сыворотке мыши при 37°C. Значительной потери связанного с антителом 89Zr не наблюдали в течение периода 5 суток. Оба тио-конъюгата являлись стабильными со средней потерей связанного с антителом 89Zr 1,8% в сутки для 89Zr-Df-Chx-Mal-тио-трастузумаба и 1,4% в сутки для 89Zr-Df-Ac-тио-трастузумаба (Фигура 24). Медленное формирование высокомолекулярных молекул, предположительно, агрегатов, наблюдали для обоих линкеров Df-Chx-Mal и Df-Ac.
ПОЛУЧЕНИЕ ИЗОБРАЖЕНИЙ МИКРОПЭТ IN VIVO
Двадцати животным (5 животных на группу), несущим подкожные ксенотрансплантаты BT474M1 (размер ~200 мм3), инъецировали внутривенно 89Zr-трастузумаб. Количество антитела, инъецированного на животное, составляло 1,4±0,29 мг/кг. Изображения для проекции максимальной интенсивности для репрезентативных животных (через 96 ч p.i.) показаны на фиг.3. Поглощение 89Zr-трастузумаба избранными тканями обобщено на фиг.4. В изображениях через 1 час (не показаны) преобладало сильное поглощение в кровяных депо, за исключением Df-Bac, где быстрое выделение липофильных загрязнений из печени и желчных путей приводило к увеличенному поглощению в кишечнике. Загрязнения полностью выводились в течение первых 24 часов, и увеличенное поглощение в тонком и толстом кишечнике не детектировали через 24 часа или в более позднее время после инъекции метки. Хотя поглощение тканями конъюгата Df-Ac в результате являлось немного меньшим (~8%), на соотношения опухоли к крови (таблица 10) не влияла потеря инъецированной радиоактивности. В изображениях через 96 часов преобладало сильное поглощение в опухолях с небольшими различиями, обнаруженными среди четырех различных вариантов 89Zr-трастузумаба (фигура 25). Поглощение в опухоли было идентичным для каждой метки, достигая максимальных значений через 24 часа после инъекции и максимальных соотношений для опухоли и крови через 144 часа из-за выведения из крови (таблица 10). Для конъюгата на основе тиола 89Zr-Df-Chx-Mal-тио-трастузумаба показали увеличенное поглощение в костной ткани (P<0,05) по сравнению с конъюгатами на основе амина (Df-Bz-SCN и Df-N-Suc) через 96 и 144 часов p.i. Поглощение в костной ткани Df-Ac-тио-трастузумаба не было значимо повышенным (P=0,20) по сравнению с линкерами Df-Bz-SCN и Df-N-Suc, но может становиться значимым при коррекции на 8% потери радиоактивности в течение первых 24 часов. Поглощение в почках для каждой метки являлось низким (фигура 26), как ожидали для меток на основе антитела, но для 89Zr-Df-Chx-Mal-тио-трастузумаба было немного выше по сравнению с другими линкерами через 24, 96 и 144 часа (P<0,05).

Для ксенотрансплантов BT474 (3+ уровень экспрессии HER2) показали более низкое абсолютное поглощение метки (15 %ID/г), чем ранее измерено Dijkers et al для SKOV3 (3+ уровень экспрессии HER2) 33,4±7,7%ID/г (Dijkers EC, et al. «Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging» (2009) J Nucl Med 50(6):974-981). Однако, соотношение опухоли к крови 5,7-7,1 (таблица 10) является сравнимым со значением, полученным для SKOV3 (соотношение опухоли к крови 7,6). Различия в поглощении в опухоли можно отнести к модели опухоли и общей дозы трастузумаба. Использовали материал с более высокой специфической активностью, таким образом, инъецировали значительно меньше антитела (35 мкг, 1,4 мг/кг) по сравнению с исследованием Dijkers et al с SKOV3 (100 мкг, 4 мг/кг). Различия в специфической активности также можно отнести к более низкому поглощению в костях свободного 89Zr в эксперименте в настоящем документе 2-3 %ID/г по сравнению с моделью SKOV3 (5-10 %ID/г). К сожалению, Dijkers et al не приводят объяснений относительно повышенного поглощения в костной ткани. Известно, что цирконий связывается с белками плазмы (Mealey J, Jr. «Turn-over of carrier-free zirconium-89 in man» (1957) Nature 179:673-4 и позже депонируется в костных минералах (Fletcher CR. «The radiological hazards of zirconium-95 and niobium-95» (1969) Health Phys. 16:209-20; Shiraishi Y and Ichikawa R. «Absorption and retention of 144 Ce and 95 Zr-95 Nb in newborn, juvenile and adult rats» (1972) Health Phys. 22:373-8). Поскольку инъецированный материал не содержит свободного 89Zr, поглощение в костной ткани может происходить из-за разрушения 89Zr антитела или из-за 89Zr, неспецифически ассоциированного с антителом, который может затем образовывать транс-хелаты с белками плазмы в отличие от 89Zr, связанного с Df.
Три специфичных для тиола реагента приведены в настоящем документе в качестве примера для хемоселективной конъюгации десферриоксамина (Df) с моноклональными антителами через тиоловую группу цистеина сконструированного антитела с цистеиновыми заменами. Тиол-специфические Df-реагенты получали ацилированием аминогруппы десферриоксамина B с выходами 14% (Df-Bac), 53% (Df-Iac) и 45% (Df-Chx-Mal), и конъюгировали с тио-трастузумабом, получая сайт-специфическую модификацию обоих сконструированных остатков цистеина за 1-5 ч. Активность связывания сайт-специфических конъюгатов тио-трастузумаба с HER2 были идентичны активности немодифицированного трастузумаба. Для Df-модифицированного тио-трастузумаба (Df-Ac-тио-трастузумаб и Df-Chx-Mal-тио-трастузумаб) получали хелаты с 89Zr (фигура 22) с выходами, превышающими 80%, в течение 1 часа, что сравнимо с лизиновыми конъюгатами, полученными с использованием ранее описанных линкеров Df-Bz-SCN и Df-N-Suc. Как для 89Zr-Df-Ac-тио-трастузумаба, так и для 89Zr-Df-Chx-Mal-тио-трастузумаба показали сравнимую стабильность в сыворотке мыши. Для обоих соединений показали также возможность получения изображений ПЭТ на модели рака молочной железы BT474M1, что сравнимо с лизиновыми конъюгатами, достигающими поглощения в опухоли 10-15% ID/г с соотношением опухоли к крови в диапазоне 6,1-7,1. В целом, новые реагенты являются легко доступными, обладая хорошей реакционной способностью по отношению к тиоловым группам белка, и обладают очень хорошими хелатирующими свойствами по отношению к 89Zr. 89Zr-меченные антитела являлись стабильными в сыворотке и обладали отличными свойствами получения изображений ПЭТ. Df-Chx-Mal является пригодным реагентом для конъюгации Df с антителами через боковую цепь цистеина и обладает несколькими преимуществами по сравнению с Df-Bac и Df-Iac. Во-первых, умеренный pH 7,5 необходим для полной конъюгации Df-Chx-Mal в течение 1 часа по сравнению с pH 9 и 2 или 5 часами, необходимыми для Df-Bac и Df-Iac. Кроме того, сайт-специфически меченные 89Zr конъюгаты сконструированного ТИОMAB можно использовать сходным образом с 18F меченными конъюгатами ТИОFAB (Gill HS, et al. «A modular platform for the rapid site-specific radiolabeling of proteins with 18F exemplified by quantitative positron emission tomography of human epidermal growth factor receptor 2» (2009) Jour, of Med. Chem. 52:5816-25) в качестве полезных инструментов для применений получения изображений ПЭТ в биомедицинских исследованиях.
ВВЕДЕНИЕ КОНЪЮГАТОВ АНТИТЕЛО-ЦИРКОНИЙ
Конъюгаты антитело-цирконий (AZC) по изобретению можно вводить любым способом, подходящим для состояния, подлежащего лечению. AZC, как правило, вводят парентерально, т.е. инфузией, подкожно, внутримышечно, внутривенно, внутрикожно, интратекально и эпидурально.
ФАРМАЦЕВТИЧЕСКИЕ СОСТАВЫ
Фармацевтические составы диагностических конъюгатов антитело-цирконий (AZC) по изобретению, как правило, получают для парентерального введения, т.е. болюсной, внутривенной, внутриопухолевой инъекции вместе с фармацевтически приемлемым парентеральным носителем и в пригодной для инъекции единичной лекарственной форме. Конъюгат антитело-цирконий (AZC), обладающий желательной степенью чистоты, необязательно смешивают с фармацевтически приемлемыми разбавителями, носителями, наполнителями или стабилизаторами (Remington's Pharmaceutical Sciences (1980) 16th edition, Osol, A. Ed.), в форме лиофилизированного состава или водного раствора.
Приемлемые разбавители, носители, наполнители и стабилизаторы в используемых дозах и концентрациях не токсичны для реципиентов и включают буферы, такие как фосфатный, цитратный и относящиеся к другим органическим кислотам; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметэтония; хлорид бензалкония, хлорид бензэтония; феноловый, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); полипептиды с низкой молекулярной массой (менее чем приблизительно 10 остатков); белки, сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металла (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как ТВИН™, ПЛЮРОНИК™ или полиэтиленгликоль (PEG). Например, лиофилизированные составы анти-ErbB2 антитела описаны в WO 97/04801, содержание которой приведено в настоящем документе в качестве ссылки в полном объеме.
Активные фармацевтические ингредиенты можно заключать также в микрокапсулы, полученные, например, способами коацервации или межповерхностной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметацилатные) микрокапсулы, соответственно, в коллоидные системы для доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Пригодные примеры препаратов с замедленным высвобождением включают содержащие AZC полупроницаемые матрицы из твердых гидрофобных полимеров, где матрицы находятся в виде изделий с заданной формой, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат), или поливиниловый спирт)), полилактиды (US 3773919), сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, неразлагаемый ацетат этиленвинила, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (пригодные для инъекции микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту.
Подлежащие использованию для введения in vivo составы должны быть стерильными, что легко осуществить фильтрацией через стерильные мембраны для фильтрации.
Составы включают составы, пригодные для вышеуказанных способов введения. Составы можно для удобства предоставлять в единичной дозированной форме, и их можно получать любыми способами, хорошо известными в данной области фармацевтики. Способы и составы в основном можно найти в Remington's Pharmaceutical Sciences (Mack Publishing Co., Easton, PA). Такие способы включают стадию объединения активного ингредиента с носителем, представляющим собой один или несколько вспомогательных ингредиентов. Как правило, составы получают посредством однородного и тесного объединения активного ингредиента с жидкими носителями или тонкоизмельченными твердыми носителями, или и с теми, и с другими, и затем, при необходимости, формования продукта.
Водные суспензии по изобретению содержат активные вещества в смеси с наполнителями, пригодными для изготовления водных суспензий. Такие наполнители включают суспендирующее вещество, такое как карбоксиметилцеллюлоза натрия, кроскармеллоза, повидон, метилцеллюлоза, гидроксипропилметилцеллюлоза, альгинат натрия, поливинилпирролидон, трагакантовая камедь и гуммиарабик, и диспергирующие или увлажняющие средства, такие как природный фосфатид (например, лецитин), продукт конденсации алкиленоксида с жирной кислотой (например, полиоксиэтиленстеарат), продукт конденсации этиленоксида с алифатическим спиртом с длинными цепями (например, гептадекаэтиленоксицетанол), продукт конденсации этиленоксида с неполным сложным эфиром, полученным из жирной кислоты и ангидрид гексита (например, моноолеат полиоксиэтиленсорбитана). Водная суспензия может содержать также один или несколько консервантов, таких как этил- или н-пропил-п-гидроксибензоат, одно или несколько окрашивающих средств, одно или несколько придающих вкус средств и один или несколько подсластителей, таких как сахароза или сахарин.
Фармацевтические композиции AZC могут присутствовать в форме стерильного пригодного для инъекции препарата, такого как стерильная пригодная для инъекции водная или масляная суспензия. Эту суспензию можно составлять, как известно в данной области, с использованием тех пригодных диспергирующих или увлажняющих средств и суспендирующих средств, которые упомянуты выше. Стерильный пригодный для инъекции препарат может представлять собой также стерильные пригодные для инъекции раствор или суспензию в нетоксичном пригодном для парентерального введения разбавителе или растворителе, например, раствор в 1,3-бутандиоле, или может представлять собой лиофилизированный порошок. Среди пригодных носителей и растворителей, которые можно использовать, присутствуют вода, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, стерильные жирные масла являются общепринятыми в качестве растворителя или суспендирующего вещества. Для этой цели можно применять любое легкое жирное масло, включая синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, можно подобным образом использовать для получения пригодных для инъекции препаратов.
Количество активного ингредиента, которое можно объединять с материалом носителя для получения единичной лекарственной формы, меняется в зависимости от обрабатываемого хозяина и конкретного способа введения. Например, водный раствор, предназначенный для внутривенной инфузии, может содержать приблизительно от 3 до 500 мкг активного ингредиента на миллилитр раствора, чтобы могла происходить инфузия подходящего объема со скоростью приблизительно 30 мл/ч.
Составы, пригодные для парентерального введения, содержат водные и неводные стерильные растворы для инъекции, которые могут содержать анти-окислители, буферы, бактериостатические средства и растворенные вещества, делающие состав изотоническим с кровью предполагаемого реципиента; и водные и неводные стерильные суспензии, которые могут включать суспендирующие средства и загустители.
Хотя пероральное введение белковых лекарственных средств нежелательно из-за гидролиза или денатурации в пищеварительном тракте, составы AZC, пригодные для перорального введения, можно получать в форме единичных лекарственных форм, таких как капсулы, саше или таблетки, каждая из которых содержит предопределенное количество AZC.
Составы можно упаковывать в контейнеры для единичных доз или множественных доз, например, герметично закрытые ампулы и флаконы, и можно хранить в высушенном замораживанием (лиофилизированном) состоянии, требующем только добавления стерильного жидкого носителя, например, воды, для инъекции, непосредственно перед использованием. Приготовленные для немедленного использования растворы и суспензии для инъекций получают из стерильных порошков, гранул и таблеток описанного ранее вида. Предпочтительными единичными дозированными составами являются составы, содержащие ежесуточную дозу активного ингредиента или единичную часть ежесуточной дозы, как перечислено в настоящем документе выше, или ее подходящую часть.
Изобретение, кроме того, относится к ветеринарным композициям, содержащим по меньшей мере один активный ингредиент, как определено выше, вместе с ветеринарным носителем, соответственно. Ветеринарные носители представляют собой материалы, пригодные для цели введения композиции, и могут представлять собой твердые, жидкие или газообразные материалы, которые в ином случае являются инертными или приемлемыми в области ветеринарии и являются совместимыми с активным ингредиентом. Эти ветеринарные композиции можно вводить парентерально, перорально или любым другим желательным способом.
СПОСОБЫ ПОЛУЧЕНИЯ ИЗОБРАЖЕНИЙ МЕЧЕНОГО АНТИТЕЛА
В другом варианте осуществления изобретения сконструированные антитела с цистеиновыми заменами могут являться меченными через тиол цистеина радиоактивными изотопами, флуоресцентными красителями, превращаемыми под действием биолюминесценции субстратными группами, превращаемыми под действием хемилюминесценции субстратными группами, ферментами и другими детектируемыми метками для экспериментов по получению изображений для диагностических, фармакодинамических и терапевтических применений. Как правило, меченое сконструированное антитело с цистеиновыми заменами, т.е. «биомаркер» или «зонд», вводят посредством инъекции, перфузии или перорального приема внутрь в живой организм, например, человеку, грызуну или другому мелкому животному, в перфузируемый орган или образец ткани. Распределение зонда детектируют с течением времени и представляют в виде изображения.
ПРИМЕРЫ
Препараты растворителей и химикатов закупали в Aldrich (Milwaukee, WI), если не указано иначе. Следующие системы для обращеннофазовой HPLC использовали для анализа и очистки продуктов. Система A: Phenomenex BioSep-SEC-S 3000 (300×4,60 мм, 5 мкм) 50 мМ PBS 0,5 мл/мин, оборудованная детектором УФ-поглощения и радиоактивности (PMT); Система B Altima C-18 (100×22,0 мм, 5 мкм) 0,05% TFA+10-50% ацетонитрил, 0-30 мин, 24 мл/мин, оборудованная УФ-детектором. Масс-спектрометрический анализ низкомолекулярных продуктов проводили в системе PE Sciex API 150EX LCMS, оборудованной колонкой Onyx Monolithic C18. LCMS анализ белков проводили на тройном квадрупольном масс-спектрометре TSQ Quantum с расширенным диапазоном массовых чисел (Thermo Electron, Thermo Fisher Scientific Inc., USA). Образцы белка для анализа LCMS восстанавливали обработкой 20 мМ дитиотреитолом (DTT) при 37°C в течение 1 часа для разделения тяжелых и легких цепей. Образцы подвергали хроматографии на микроколонке PRLP-S 1000 A (50×2,1 мм, Polymer Laboratories, Varian Inc., USA), нагретой до 75°C. Использовали линейный градиент от 30-40% B (растворитель A, 0,05% TFA в воде; растворитель B, 0,04% TFA в ацетонитриле), и элюат напрямую ионизировали с использованием источника электрораспыления. Данные собирали в системе данных Xcalibur и деконволюцию проводили с использованием программного обеспечения ProMass (Novatia, Monmouth Junction, NJ). Спектры ЯМР регистрировали на спектрометре Bruker Avance II 400 при 298K, и регистрировали химические сдвиги относительно TMS. Концентрации белка измеряли при 280 нм с использованием Eppendorf BioPhotometer (Westbury, NY). 89Zr получали из Memorial Sloan-Kettering Cancer Center (New York, NY) в форме оксалата 89Zr(IV) в 1M растворе щавелевой кислоты с удельной активностью 470-1195 Ки/ммоль (Holland JP, et al (2009) «Standardized methods for the production of high specific-activity zirconium-89» Nucl Med Biol. 36:729-39). Гетеробифункциональный линкер сукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (SMCC) закупали в Pierce (Rockford, IL) и иодацетат N-гидроксисукцинимидила получали из Indofine Chemical Company (Hillsborough, NJ). Колонки NAP-10 получали от GE Healthcare, USA и фильтры для ультрацентрифугирования Amicon Ultra-4 (10000 MWCO) от Millipore (Billerica, MA). Df-Bz-SCN закупали в Macrocyclics (Dallas, TX).
Пример 1 Получение биотинилированного ТиоFab-фага
Проводили реакцию ТиоFab-фага (5×1012 фаговых частиц) с 150-кратным избытком биотин-PEO-малеимида ((+)-биотинил-3-малеимидопропионамидил-3,6-диоксаоктандиамин, Oda et al (2001) Nature Biotechnology 19:379-382, Pierce Biotechnology, Inc.) в течение 3 часов при комнатной температуре. Избыток биотин-PEO-малеимида удаляли из биотин-конъюгированного фага повторяющимися преципитациями PEG (3-4 раза). Можно использовать другие коммерчески доступные реагенты для биотинилирования с электрофильными группами, реакционноспособными по отношению к тиоловым группам цистеина, включая биотин-BMCC, PEO-иодацетил-биотин, иодацетил-LC-биотин и биотин-HPDP (Pierce Biotechnology, Inc.), и N α-(3-малеимидилпропионил)биоцитин (MPB, Molecular Probes, Eugene, OR). Другие коммерческие источники биотинилированных, бифункциональных и мультифункциональных линкерных реагентов включают Molecular Probes, Eugene, OR, и Sigma, St. Louis, MO.
Пример 2 Анализ PHESELECTOR
Бычьим сывороточным альбумином (BSA), внеклеточным доменом erbB2 (HER2) и стрептавидином (100 мкл при 2 мкг/мл) по отдельности покрывали 96-луночные планшеты Maxisorp. После блокирования с помощью 0,5% Твин-20 (в PBS), биотинилированный и небиотинилированный hu4D5Fabv8-ТиоFab-фаг (2×1010 фаговых частиц) инкубировали в течение 1 часа при комнатной температуре с последующей инкубацией с меченным пероксидазой хрена (HRP) вторичным антителом (антитело против белка оболочки фага M13, белка pVIII). Фигура 8 иллюстрирует анализ PHESELECTOR посредством схематического представления, изображающего связывание Fab или ТиоFab с HER2 (вверху) и биотинилированного ТиоFab со стрептавидином (внизу).
Проводили стандартную реакцию с HRP и измеряли оптическую плотность при 450 нм. Реакционную способность тиола измеряли, рассчитывая соотношение OD450 для стрептавидина/OD450 для HER2. Значение реакционной способности тиола 1 указывает на полное биотинилирование тиола цистеина. В случае измерений связывания белка Fab, использовали hu4D5Fabv8 (2-20 нг) с последующей инкубацией с меченными HRP поликлональными анти-Fab антителами козы.
Пример 3a Экспрессия и очистка ТиоFab
ТиоFab экспрессировали при индукции в 34B8, несупрессорном штамме E. coli (Baca et al (1997) Journal Biological Chemistry 272(16): 10678-84). Собранный осадок клеток ресуспендировали в PBS (фосфатно-солевом буфере), общий лизис клеток проводили пропусканием через микрофлюидайзер, и ТиоFab очищали аффинной хроматографией на SEPHAROSE™ с белком G (Amersham).
ТиоFab L-V15C, L-V110C, H-A88C и H-A121C экспрессировали и очищали хроматографией на колонке с SEPHAROSE™ с белком G. Олигомерный-Fab присутствовал в фракциях 26-30, и большая часть мономерной формы находилась в фракциях 31-34. Фракции, состоящие из мономерной формы, объединяли и анализировали посредством SDS-PAGE вместе с hu4D5Fabv8 дикого типа, и анализировали в геле для SDS-PAGE в восстанавливающих (с DTT или BME) и невосстанавливающих (без DTT или BME) условиях. Фракции после гель-фильтрации A121C-ТиоFab анализировали невосстанавливающим SDS-PAGE.
ТиоFab конъюгировали с биотин-PEO-малеимидом, как описано выше, и биотинилированные-ТиоFab дополнительно очищали гель-фильтрационной хроматографией на Superdex-200™ (Amersham), отсекающей свободный биотин-PEO-малеимид и олигомерную фракцию ТиоFab. Каждый из hu4D5Fabv8 дикого типа и hu4D5Fabv8 A121C-ТиоFab (в количестве 0,5 мг) по отдельности инкубировали со 100-кратным молярным избытком биотин-PEO-малеимида в течение 3 часов при комнатной температуре и наносили на колонку для гель-фильтрации Superdex-200 для отделения свободного биотина, так же как олигомерных Fab, от мономерной формы.
Пример 3b Анализ ТиоFab
Фрагменты после ферментативного расщепления биотинилированного hu4D5Fabv8 (A121C) ТиоFab и hu4D5Fabv8 дикого типа анализировали масс-спектрометрическим анализом жидкостной хроматографией с ионизацией электрораспылением (LS-ESI-MS). Различие между исходной массой 48294,5 биотинилированного hu4D5Fabv8 (A121C) и исходной массой 47737,0 hu4D5Fabv8 дикого типа составляло 557,5 единиц массы. Этот фрагмент указывает на присутствие одной биотин-PEO-малеимидной группы (C23H36N5O7S2). В таблице 4 показано распределение значений фрагментации, подтверждающее последовательность.
LC-ESI-Масс-спектрометрический анализ биотинилированного hu4D5Fabv8 ТиоFab A121C после расщепления трипсином
До и после гель-фильтрации на Superdex-200 проводили анализы SDS-PAGE в гелях, с восстановлением и без восстановления посредством DTT или BME, биотинилированного ABP-hu4D5Fabv8-A121C, биотинилированного ABP-hu4D5Fabv8-V110C, биотинилированного ABP-hu4D5Fabv8-(V110C-A88C) с двойным Cys и биотинилированного ABP-hu4D5Fabv8-(V110C-A121C) с двойным Cys.
Масс-спектрометрический анализ (MS/MS) hu4D5Fabv8-(V110C)-BMPEO-DM1 (после очистки гель-фильтрацией на Superdex-200): Fab+1 51607,5, Fab 50515,5. Эти данные показывают 91,2% конъюгацию. Анализ MS/MS hu4D5Fabv8-(V110C)-BMPEO-DM1 (восстановленного): LC 23447,2, LC+1 24537,3, HC (Fab) 27072,5. Эти данные показывают, что вся конъюгация DM1 произошла на легкой цепи Fab.
Пример 11 Восстановление/окисление ТиоMab для конъюгации
Полноразмерные сконструированные моноклональные антитела с цистеиновыми заменами (ТиоMab), экспрессированные в клетках CHO, восстанавливали с помощью приблизительно 50-кратного избытка TCEP (трис(2-карбоксиэтил)фосфингидрохлорида; Getz et al (1999) Anal. Biochem. Vol 273:73-80; Soltec Ventures, Beverly, MA) в течение 3 часов при 37°C. Восстановленный ТиоMab (Фигура 15) разводили и наносили на колонку HiTrap S в 10 мМ ацетате натрия, pH 5, и элюировали с помощью PBS, содержащего 0,3 M хлорид натрия. Элюированный восстановленный ТиоMab обрабатывали 200 нМ водным сульфатом меди (CuSO4) при комнатной температуре, в течение ночи. Окисление окружающим воздухом также являлось эффективным.
Пример 13 N-[4-(N-малеимидометил)циклогексан-1-карбоксил]десферриоксамин (Df-Chx-Mal)
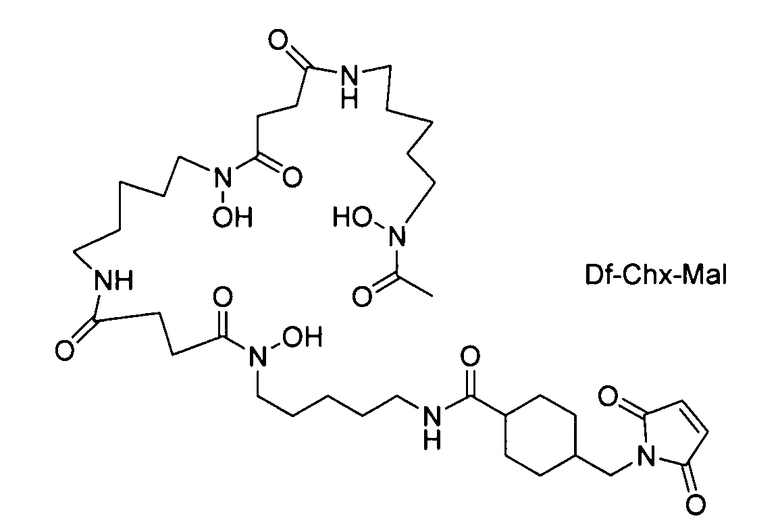
4-[N-малеимидометил]циклогексан-1-карбоксилат (SMCC, 40 мг, 0,12 ммоль), мезилат десферриоксамина (78 мг, 0,12 ммоль) и N,N-диизопропилэтиламин (22 мкл, 0,13 ммоль) растворяли в смеси DMF (2,0 мл) и 0,2 мл воды (фигура 21). Полученный мутный раствор перемешивали при комнатной температуре в течение 30 мин. Добавляли воду (8 мл), и преципитированный продукт выделяли посредством фильтрации, промывали водой и высушивали под пониженным давлением с получением 42 мг (45%) N-[4-(N-малеимидометил)циклогексан-1-карбоксил]десферриоксамина (Df-Chx-Mal) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 0,87-0,90 (м, 2H), 1,20-1,26 (м, 8Н), 1,35-1,41 (м, 6Н), 1,45-1,55 (м, 8H), 1,60-1,70 (м, 4H), 1,97 (с, 3H, ацетил), 2,26-2,29 (м, 4H), 2,56-2,60 (м, 4H), 2,95-3,05 (м, 6H), 3,24 (д, J=7,0 Гц, 2H), 3,44-3,48 (м, 6H), 7,00 (с, 2H, малеимид), 7,62 (т, J=5,4 Гц, 1H, амид), 7,75 (м, 2H, амид), 9,59 (с, 2H, гидроксил), 9,64 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C37H62N7O11 780,44; обнаружено 780.6.
Пример 14 N-бромацетилдесферриоксамин (Df-Bac)
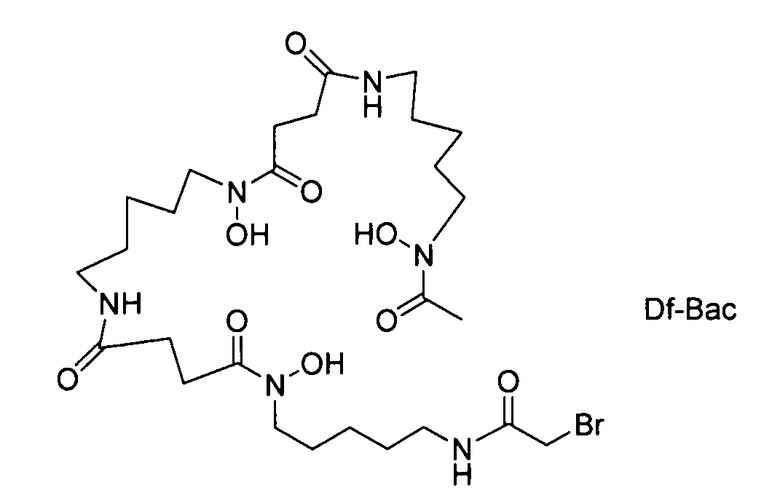
Раствор бромацетилбромида (27 мкл, 0,30 ммоль) в DMF (1 мл) добавляли по каплям за 5 минут в охлажденную (0°C) смесь мезилата десферриоксамина (200 мг, 0,30 ммоль) и N,N-диизопропилэтиламина (106 мкл, 0,60 ммоль) в DMF (5 мл), после чего реакционную смесь перемешивали при 0°C в течение 4 часов (фигура 21). Добавляли воду (10 мл), и продукт выделяли с использованием HPLC (Система B, время удержания 7,5 мин) с получением 29 мг (14%) N-бромацетилдесферриоксамина (Df-Bac) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 1,18-1,26 (м, 6H), 1,35-1,42 (м, 6H), 1,45-1,55 (м, 6H), 1,97 (с, 3Н, ацетил), 2,24-2,30 (м, 4H), 2,54-2,59 (м, 4Н), 2,96-3,07 (м, 6Н), 3,44-3,47 (м, 6Н), 3,82 (c, 2Н, бромацетил), 7,74 (м, 2H, амид), 8,21 (т, 1H, амид), 9,59 (с, 2H, гидроксил), 9,63 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C27H50BrN6O9 681,27, 683,27; обнаружено 681,1, 683,0.
Пример 15 N-иодацетилдесферриоксамин (Df-Iac)
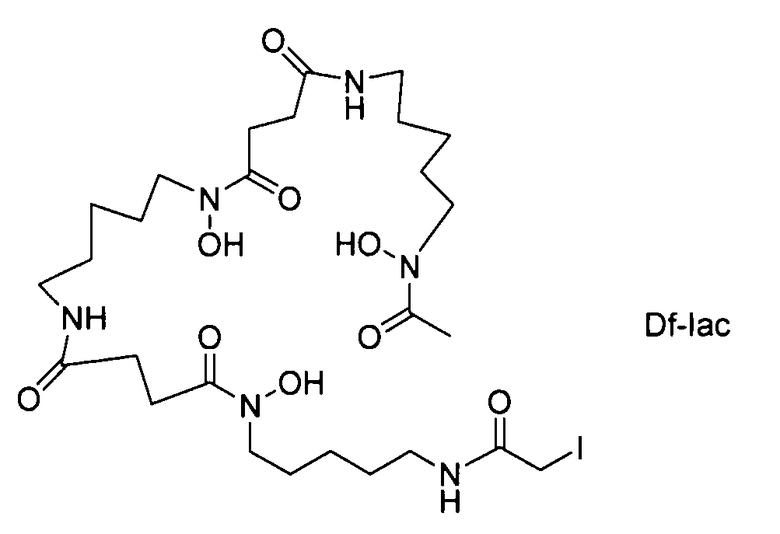
Мезилат десферриоксамина (200 мг, 0,30 ммоль) и N,N-диизопропилэтиламин (53 мкл, 0,30 ммоль) смешивали в DMF (4 мл) и воде (0,4 мл). Добавляли иодацетат N-гидроксисукцинимидила (93 мг, 0,33 ммоль) и полученную смесь перемешивали при комнатной температуре в течение 1 часа (фигура 21). Добавляли воду (8 мл) и преципитированный продукт отделяли, промывали водой и высушивали под пониженным давлением с получением 115 мг (53%) N-иодацетилдесферриоксамина (Df-Iac) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 1,20-1,25 (м, 6H), 1,35-1,42 (м, 6H), 1,47-1,54 (м, 6H), 1,97 (с, 3H, ацетил), 2,25-2,29 (м, 4H), 2,56-2,59 (м, 4Н), 2,98-3,03 (м, 6Н), 3,45 (м, 6Н), 3,61 (с, 2Н, иодацетил), 7,75 (м, 2H, амид), 8,17 (т, 1H, амид), 9,57 (с, 2H, гидроксил), 9,61 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C27H50IN6O9 729,26; обнаружено 729,1.
Пример 16 Тио-трастузумаб
Конструирование, экспрессия и очистка ТИОMAB с заменой Cys вместо Ala114 (нумерация Kabat) в тяжелой цепи описаны ранее (Junutula JR, et al «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol 26:925-32). Выделенный тио-трастузумаб подготавливали для конъюгации способом восстановления и повторного окисления для удаления дисульфидных аддуктов, связанных с Cys114. Сначала белок восстанавливали в течение 24 часов обработкой 40-кратным молярным избытком DTT и 2 мМ ЭДТА в 88 мМ Трис-буфере pH 7,5. Для удаления DTT перед повторным окислением, pH раствора тио-трастузумаба доводили до 5 добавлением 10 мМ буфера сукцината натрия. Затем раствор наносили на ионообменную колонку (HiTrap SP FF, GE Healthcare), которую стерилизовали и уравновешивали с помощью 10 мМ буфера сукцината натрия pH 5. Колонку промывали 10 мМ буфером сукцинатом натрия (10 мл), и затем тио-трастузумаб элюировали 3 мл буфера 50 мМ Трис, 150 мМ NaCl с pH 7,5. Повторное окисление тио-трастузумаба осуществляли обработкой 25-кратным молярным избытком дегидроаскорбиновой кислоты (100 мМ в N,N-диметилацетамиде (DMA)) в буфере 75 мМ Трис, 150 мМ NaCl pH 7,5 при 25°C в течение 3,5 часов. После повторного окисления тио-трастузумаб конъюгировали с десферриоксамином без дополнительной очистки. MS ESI (m/z): обнаружено для легкой цепи 23440,0, для тяжелой цепи 50627,3.
Пример 17 Df-Chx-Mal-тио-трастузумаб
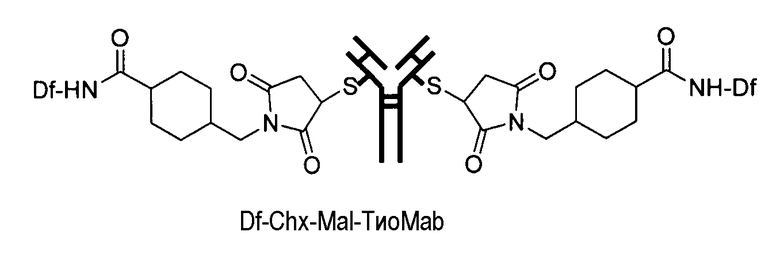
2 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Chx-Mal (1,5 мг, 2 мкмоль) в 1:1 смеси (1 мл) DMF и DMA посредством нагревания при 44°C в течение 30 мин, затем исходный раствор разделяли на аликвоты и хранили при -80°C (фигура 21). Затем аликвоту исходного раствора (220 мкл, 0,440 мкмоль) добавляли к раствору тио-трастузумаба (7,5 мг, 52 нмоль) в буфере 50 мМ Трис, 150 мМ NaCl pH 7,5 (1,5 мл) и инкубировали при комнатной температуре в течение 1 часа. Затем раствор подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,25 M ацетат натрия с получением 1 мл раствора конъюгата Df-Chx-Mal-тио-трастузумаба в концентрации 6 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,2, для тяжелой цепи 51407,3 (фигура 23, D).
Пример 18 Df-Ac-тио-трастузумаб с использованием Df-Bac
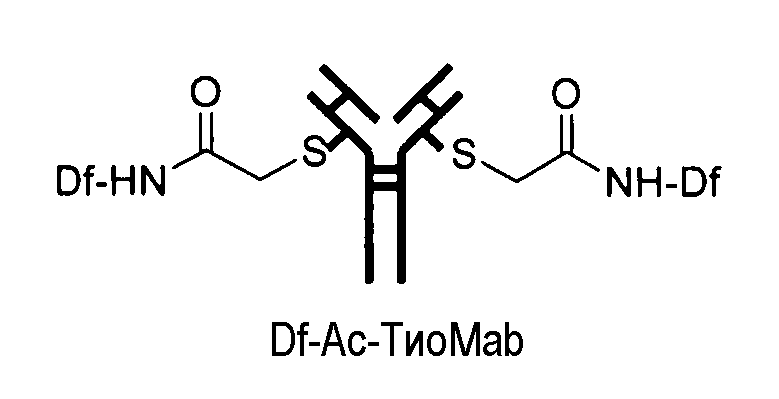
12 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Bac (8 мг, 12 мкмоль) в 1 мл DMA (фигура 21). Затем исходный раствор разделяли на аликвоты и хранили при -80°C. Повторно окисленный тио-трастузумаб подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,05 M борат натрия pH 9. Аликвоту исходного раствора Df-Bac (35 мкл, 0,410 мкмоль) добавляли к раствору тио-трастузумаба (4,9 мг, 34 нмоль) в буфере 0,05 M борате натрия pH 9 (1 мл) и инкубировали при комнатной температуре в течение 5 часов. Реакционную смесь наносили на колонку NAP-10, и Df-Ac-тио-трастузумаб элюировали с помощью 1,5 мл буфера 0,25 M ацетата натрия с получением продукта в концентрации 3,2 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,1, для тяжелой цепи 51228,1 (фигура 23, B).
Пример 19 Df-Ac-тио-трастузумаб с использованием Df-Iac
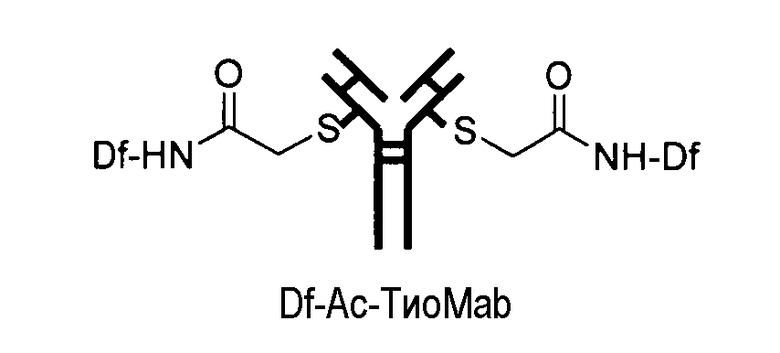
11 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Iac (8 мг, 11 мкмоль) в DMSO (1 мл), затем исходный раствор разделяли на аликвоты и хранили при -80°C (фигура 21). pH раствора тио-трастузумаба (3,2 мл) доводили до 9 добавлением 0,5 мл 0,1 M карбоната натрия. Затем аликвоту исходного раствора (110 мкл, 1,20 мкмоль) добавляли к раствору тио-трастузумаба (16 мг, 110 нмоль) в буфере 50 мМ Трис, 150 мМ NaCl, 0,0125 M карбонат натрия с pH 9 (4 мл) и инкубировали при комнатной температуре в течение 2 часов. Затем раствор подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,25 M ацетат натрия буфер с получением 1 мл раствора конъюгата Df-Ac-тио-трастузумаба в концентрации 8 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,1, для тяжелой цепи 51228,3 (фигура 23, C).
Пример 20 Общий способ получения хелатов 89Zr
Раствор оксалата 89Zr(IV) (2-4 мКи, 100 мкл) в 1 M щавелевой кислоте смешивали с 2 M раствором Na2CО3 (45 мкл) и инкубировали при комнатной температуре в течение 3 мин, после чего добавляли буфер 0,5 M HEPES (0,15 мл) (фигура 22). Конъюгат Df-тио-трастузумаба (1 мг, 7 нмоль) разводили 0,25 M ацетатом натрия/0,5% гентизиновой кислотой до конечного объема 0,356 мл и добавляли к раствору 89Zr. Наконец, добавляли вторую часть буфера HEPES (0,350 мл), получая общий объем 1 мл. Смесь инкубировали при комнатной температуре в течение 1 часа. Для удаления свободного 89Zr белок с радиоактивной меткой очищали с использованием обессоливающей колонки NAP-10. Колонку NAP-10 уравновешивали с помощью 20 мл 0,25 M ацетата натрия/0,5% гентизиновой кислоты. Реакционную смесь наносили на колонку NAP-10, и 89Zr-Df-тио-трастузумаб элюировали с помощью 1,5 мл буфера 0,25 M ацетата натрия/0,5% гентизиновой кислоты (1,5 мл). При необходимости 89Zr-Df-тио-трастузумаб концентрировали с использованием фильтра Amicon Ultra-4 до желательного объема. Продукт анализировали посредством SEC HPLC (Система A).
Пример 21 Стабильность в сыворотке in vitro
Раствор конъюгата 89Zr-Df-тио-трастузумаба 0,5-1,5 мКи (1 мг) в буфере 0,25 M ацетате натрия/0,5% гентизиновой кислоте (0,1 мл) добавляли к свежей сыворотке мыши (0,9 мл) и инкубировали при 37°C в течение 0-96 часов. Образцы (20 мкл) раствора сыворотки анализировали с использованием SEC HPLC (Система A), результаты показаны на фигуре 24.
Пример 22 Модели на животных
Бежевых голых мышей XID в возрасте 6-8 недель получали из Harlan Sprague Dawley (Livermore, CA). За трое суток до инокуляции клеток, мышам имплантировали (s.c, левый бок) 0,36 мг частиц 17β-эстрадиола с замедленным высвобождением в течение 60-суток (Innovative Research of America) для поддержания уровня эстрогена в сыворотке. Мышам инокулировали в жировую прослойку молочной железы 5×106 клеток BT474M1 в 50% матригеле без содержания фенола красного. BT474M1 представляет собой субклон линии клеток опухоли молочной железы человека BT474, полученный из California Pacific Medical Center. При содержании животных и манипуляциях с ними следовали протоколам, одобренным Genentech's Institutioned Animal Care and Use Committee, аккредитованным Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC).
Пример 23 Получение изображений микроПЭТ
Мышей подвергали анестезии приблизительно 3% севофлураном до получения эффекта и инъецировали мышам i.v. через хвостовую вену приблизительно 0,1 мКи меченного радиоактивным 89Zr трастузумаба в изотоническом растворе (100-130 мкл) и возвращали их в клетку для восстановления. Получение изображений ПЭТ (фигура 25) проводили на сканере Inveon PET/CT через 1, 24, 96 и 144 часа после инъекции метки следующим образом. Животных после подвергания анестезии севофлураном располагали в положении лежа, головой вперед на площадке сканера и проводили статическое сканирование 15 или 30 мин. Температуру тела измеряли ректальным зондом и поддерживали с помощью теплого воздуха. Реконструкцию повторяющихся изображений целого организма получали с использованием алгоритма максимальной апостериорной вероятности (MAP, гиперпараметр бета (β) 0,05) и корректировали на затухание сигнала с использованием плотности тканей, полученной при КТ. Проекции получали с помощью программного обеспечения ASIPro (Siemens Preclinical Solutions) и использовали для получения количественных уровней активности в каждом интересующем органе с использованием анализа интересующих областей.
Статистический анализ: Графики на фигуре 26 получали с помощью программного обеспечения R версии 2.4.1 (R Foundation for Statistical Computing, Vienna, Austria). Статистическую значимость определяли с использованием двустороннего критерия Стьюдента или ANOVA, и значения P менее 0,05 считали значимыми; данные представлены как среднее ± s.d., если не указано иначе.
Объем настоящего изобретения не является ограниченным конкретными вариантами осуществления, описанными в примерах, которые предназначены в качестве иллюстраций некоторых аспектов изобретения, и любые варианты осуществления, являющиеся функционально эквивалентными, входят в объем этого изобретения. Действительно, различные модификации по изобретению в дополнение к показанным и описанным в настоящем документе очевидны специалистам в данной области и предназначены для включения в объем прилагаемой формулы изобретения.
Содержание всех патентов, патентных заявок и ссылок, процитированных на протяжении описания, конкретно приведено в качестве ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕННЫЕ С ПОМОЩЬЮ ГЕННОЙ ИНЖЕНЕРИИ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ И ИХ КОНЪЮГАТЫ | 2011 |
|
RU2626537C2 |
| ПОЛУЧЕННЫЕ С ПОМОЩЬЮ ГЕННОЙ ИНЖЕНЕРИИ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ И ИХ КОНЪЮГАТЫ | 2011 |
|
RU2755066C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, АНАЛОГИ АНТИТЕЛ, КОМПОЗИЦИИ И СПОСОБЫ | 2010 |
|
RU2580038C2 |
| АНТИТЕЛА ПРОТИВ TENB2, СКОНСТРУИРОВАННЫЕ С ЦИСТЕИНОМ, И КОНЪЮГАТЫ АНТИТЕЛО - ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2008 |
|
RU2505544C2 |
| КОНЪЮГАТЫ ПРОИЗВОДНОГО АНТРАЦИКЛИНА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ПРОТИВООПУХОЛЕВЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2523419C2 |
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ, ВИЗУАЛИЗИРУЕМЫЕ ПРИ ПОМОЩИ ИММУНО-ПОЗИТРОН-ЭМИССИОННОЙ ТОМОГРАФИИ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2613886C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2553566C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2511410C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2791984C2 |
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2436796C9 |

















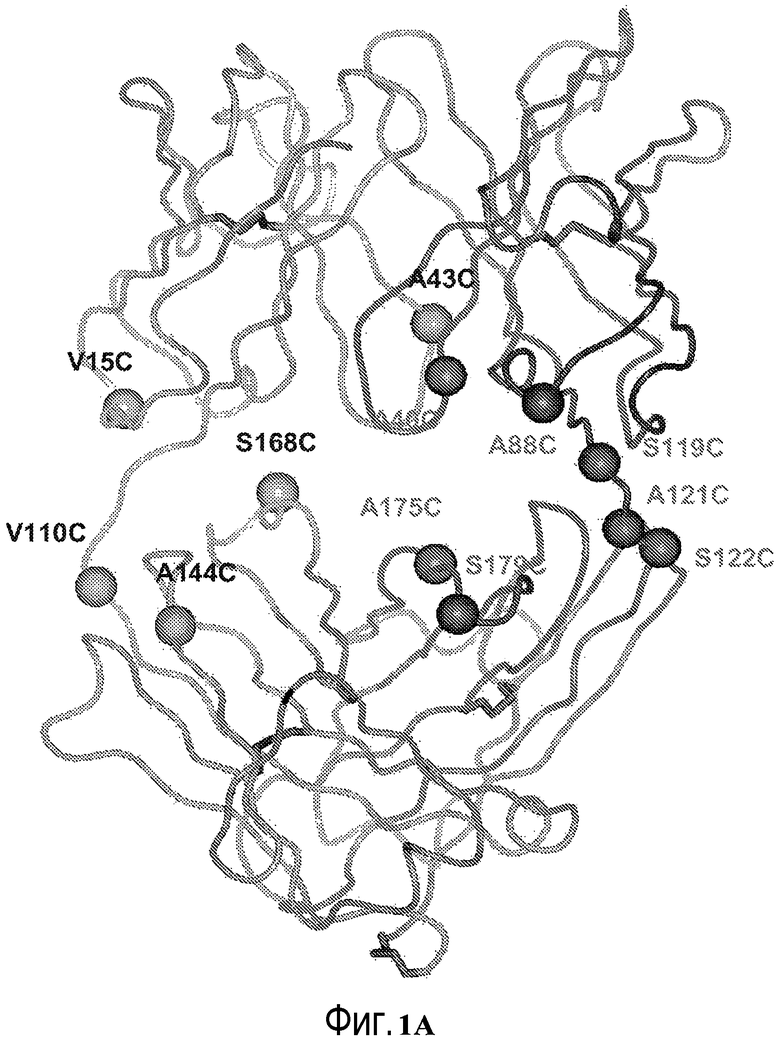
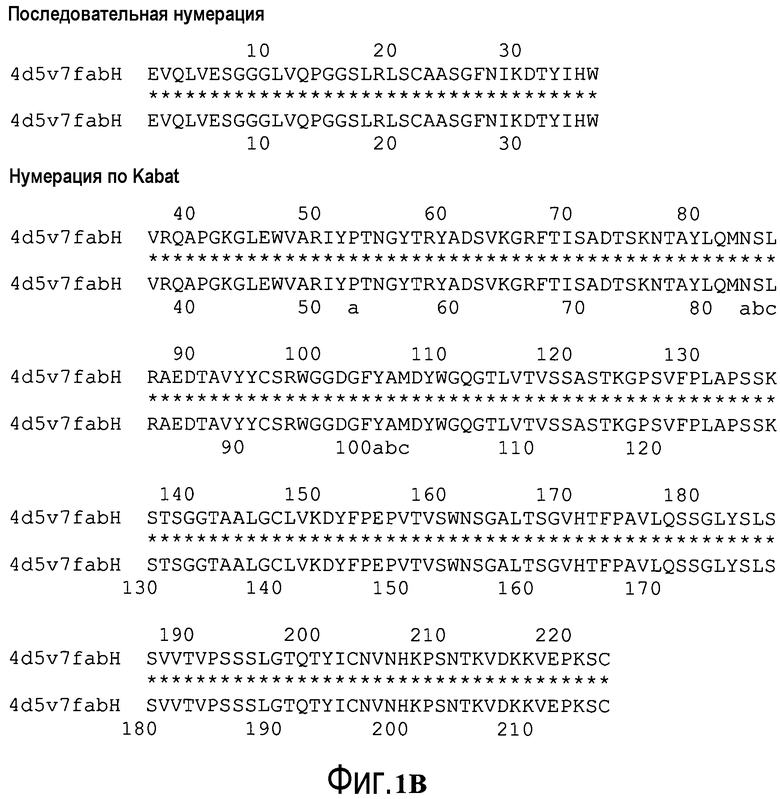
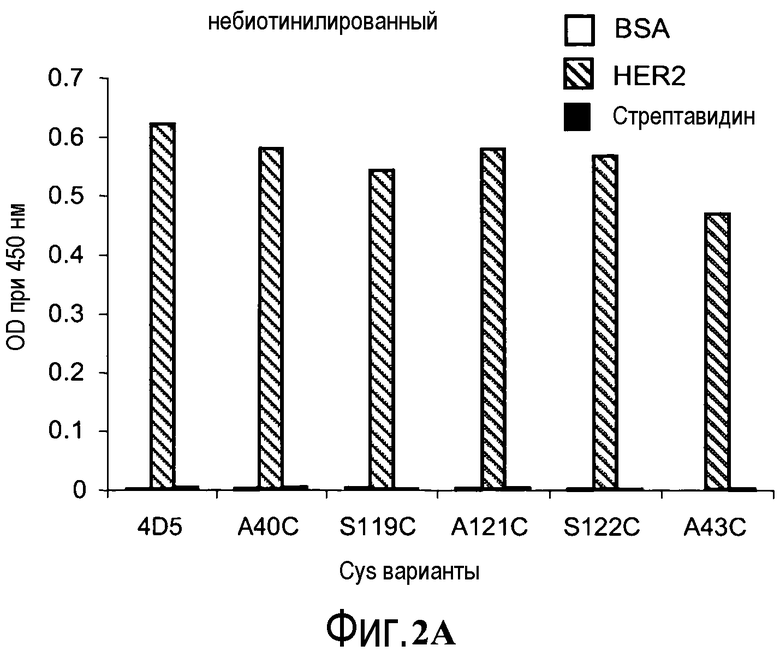
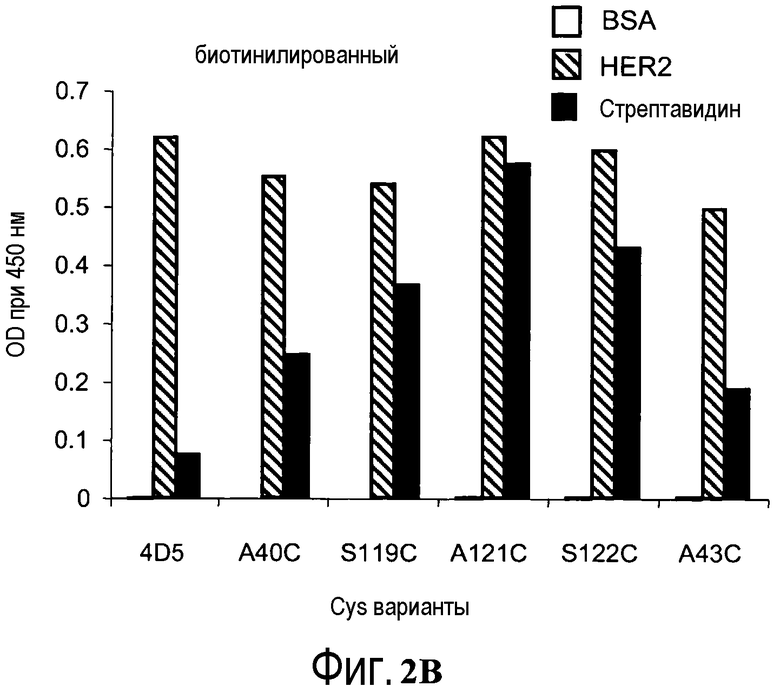
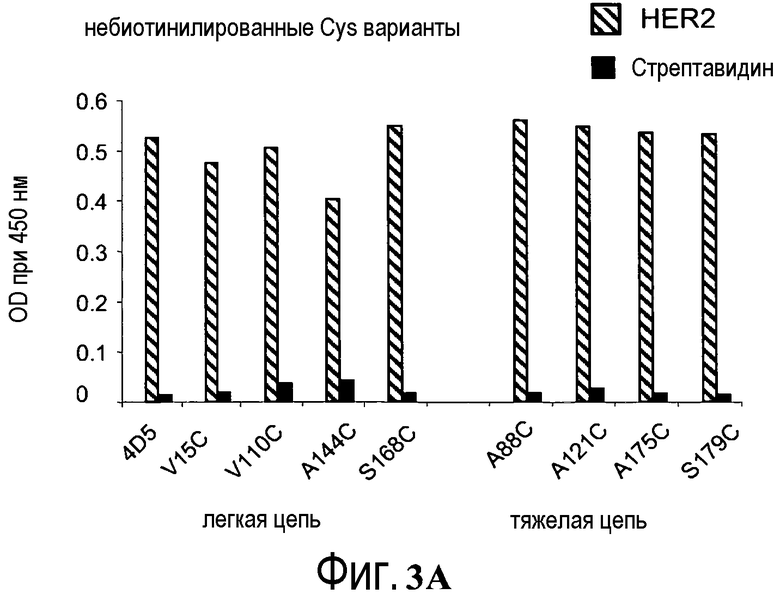
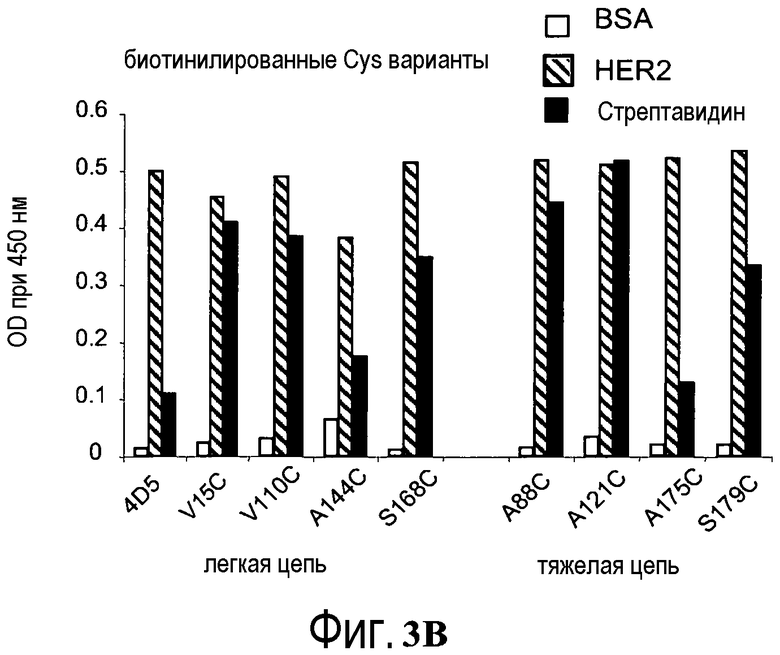
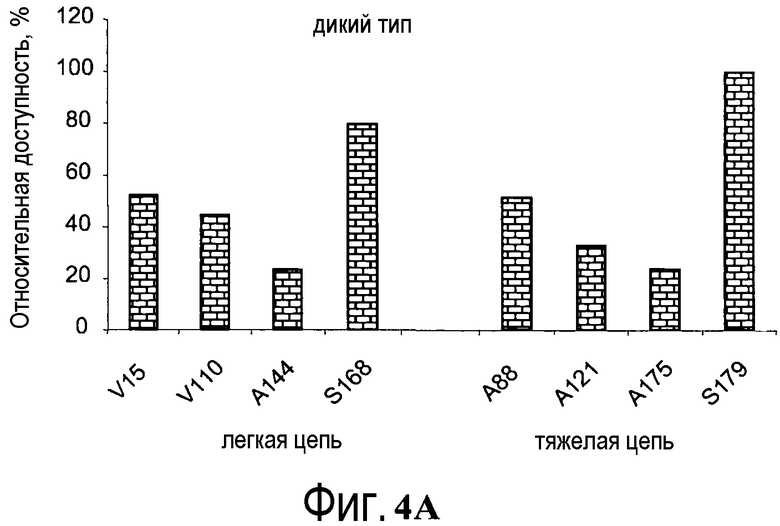
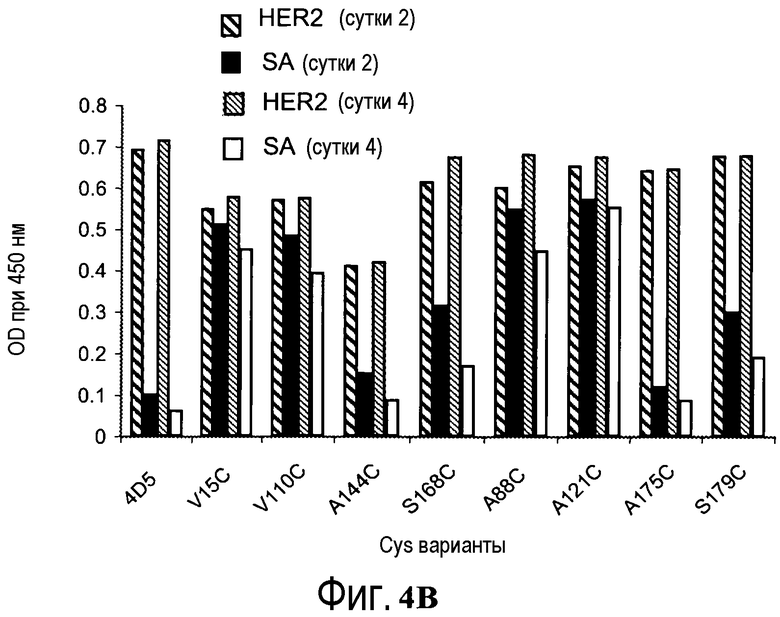
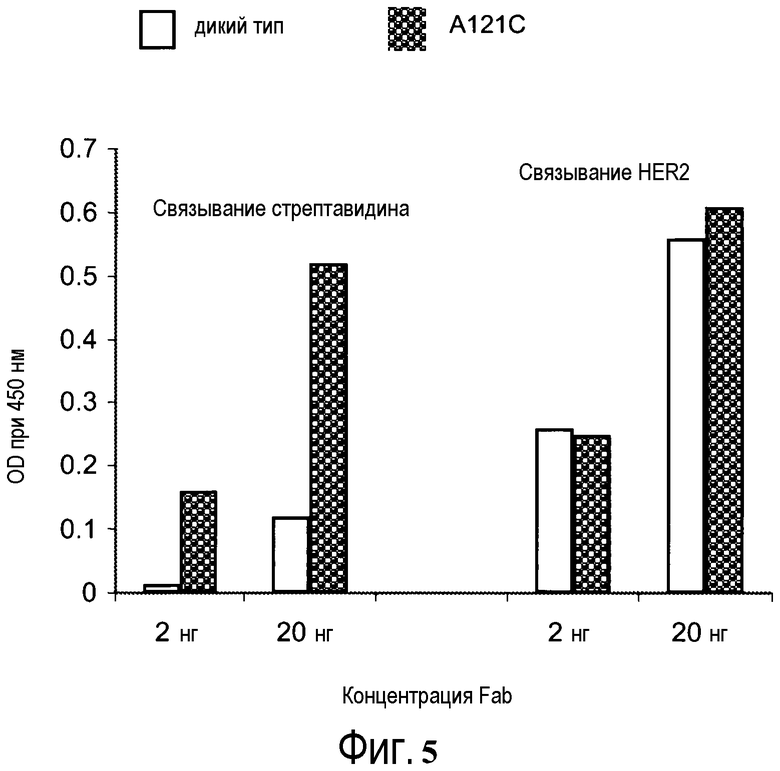
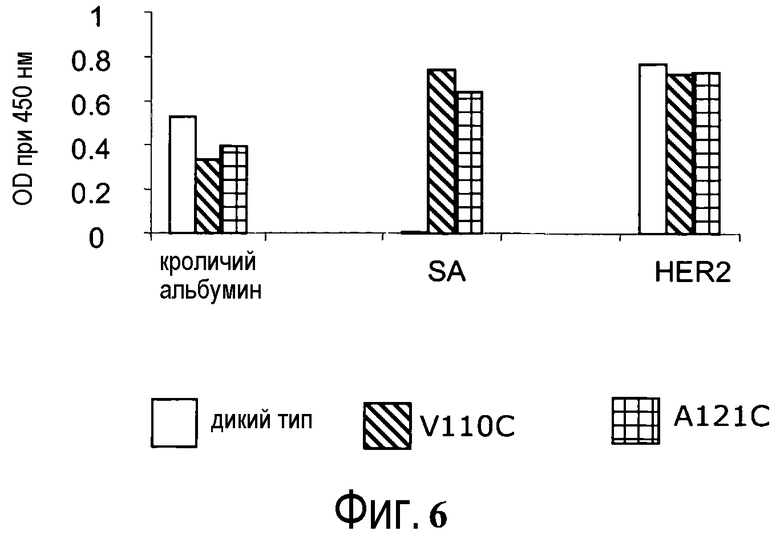
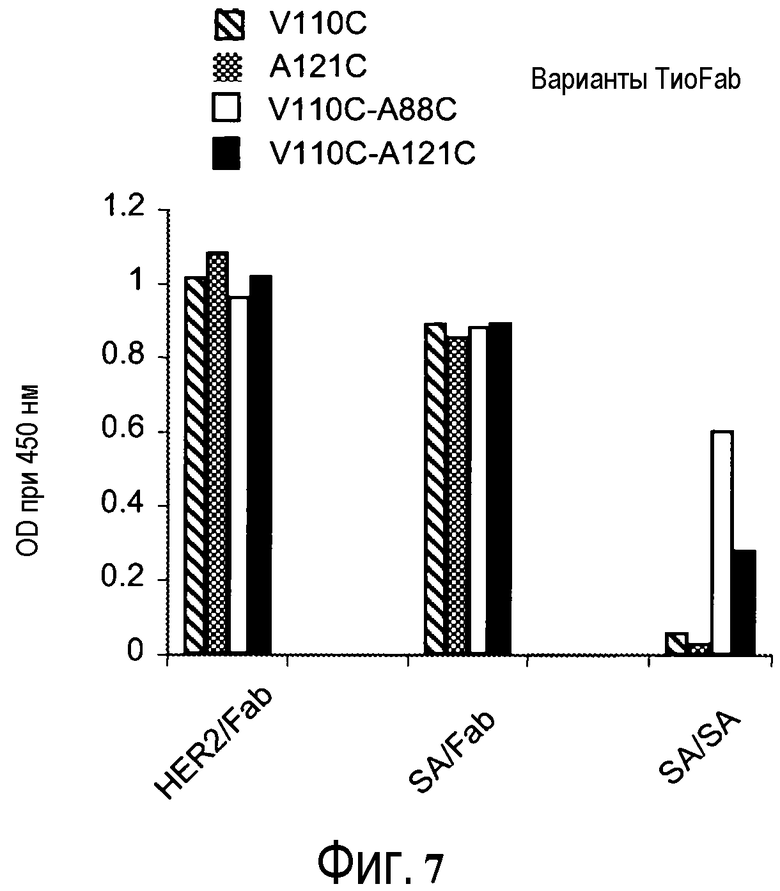
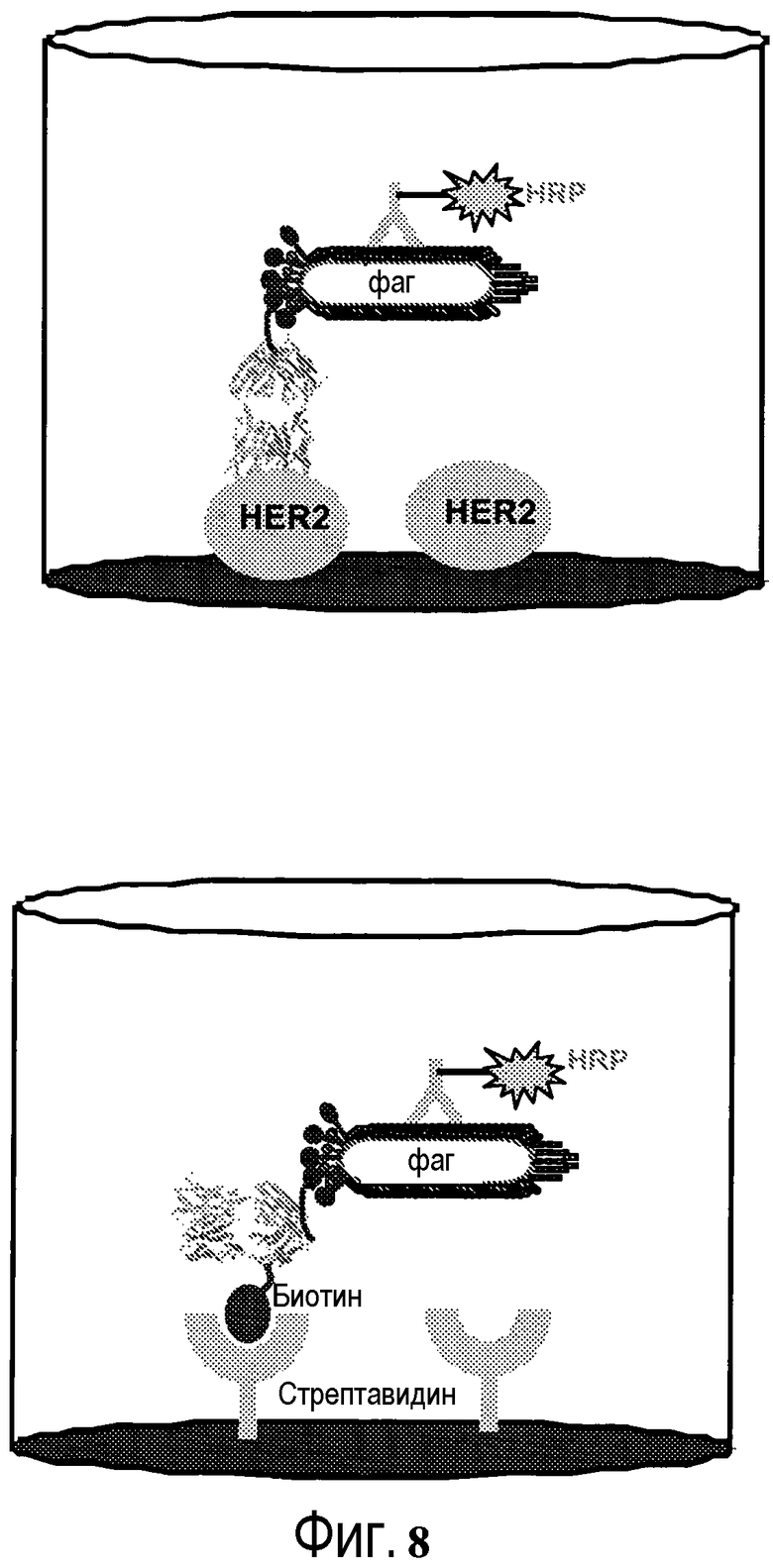
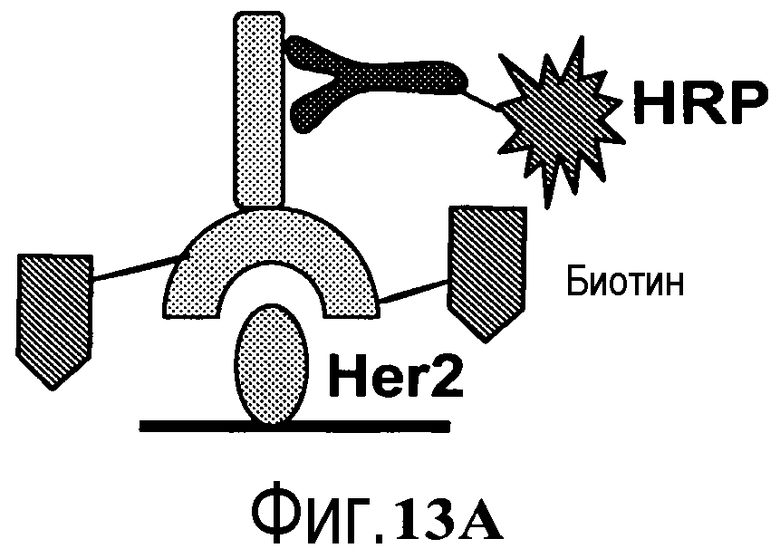
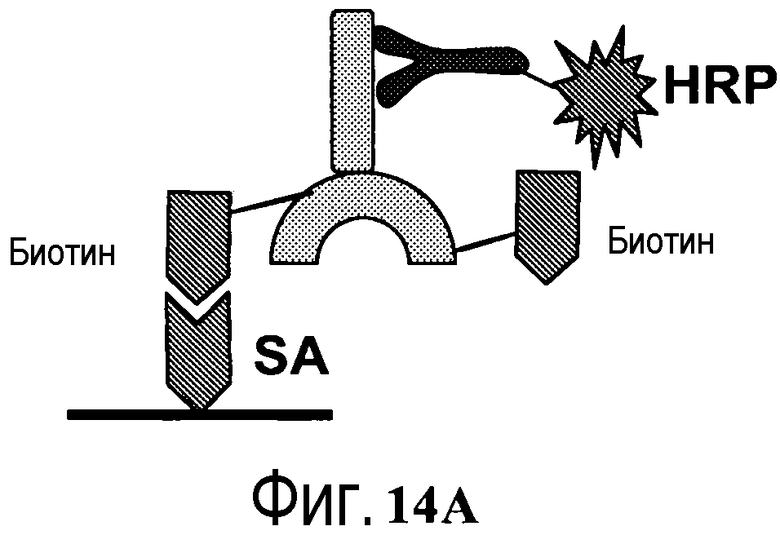
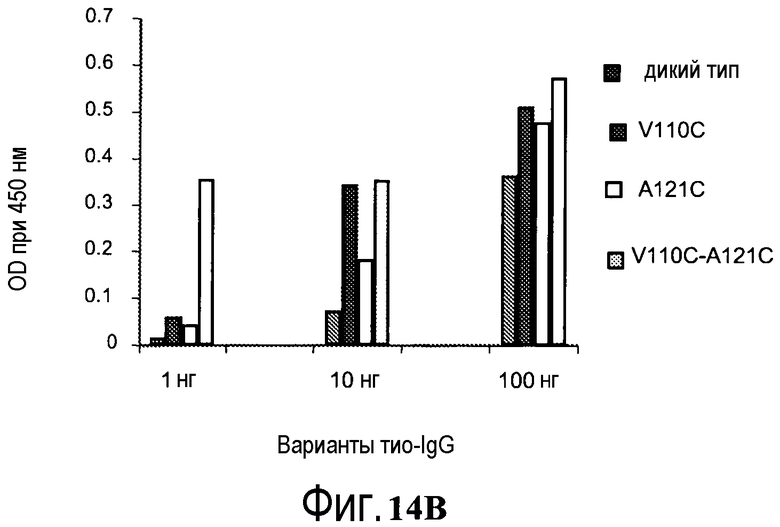
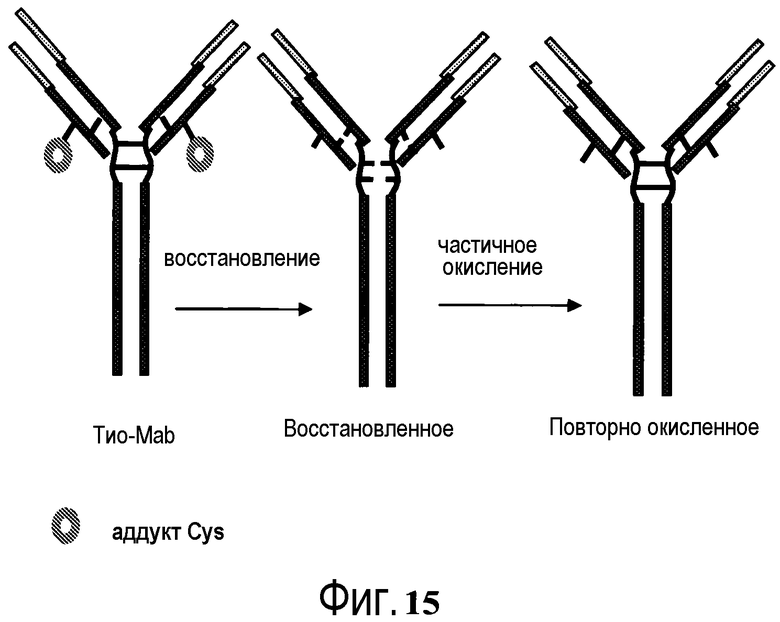
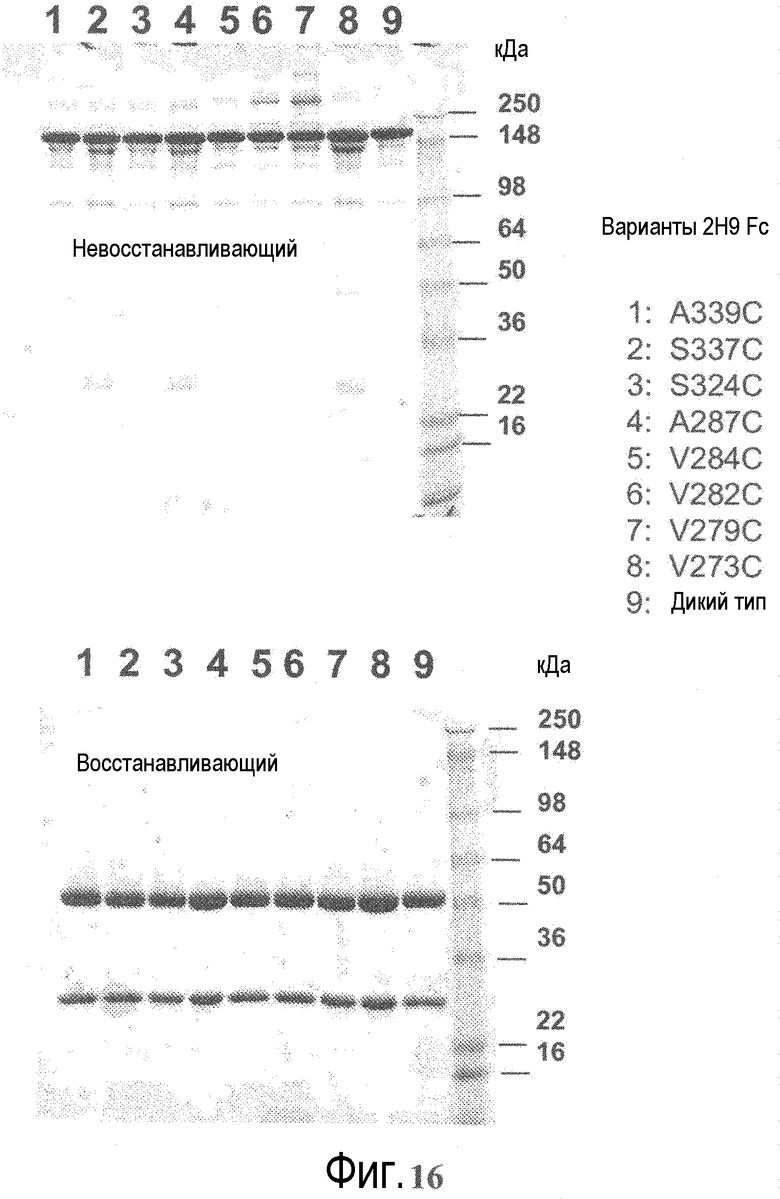
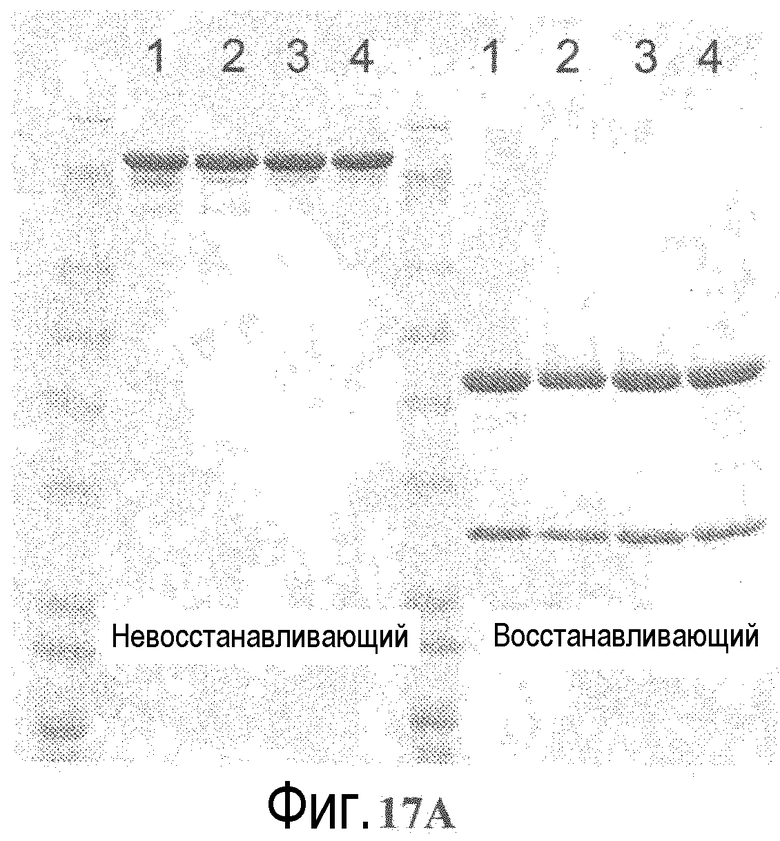
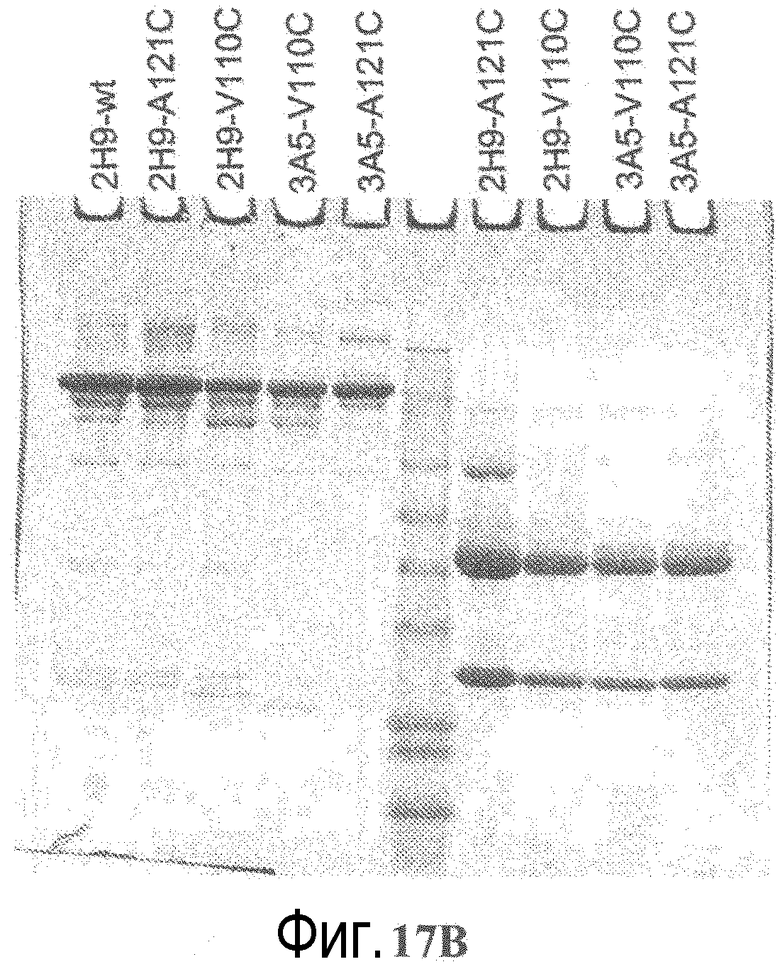
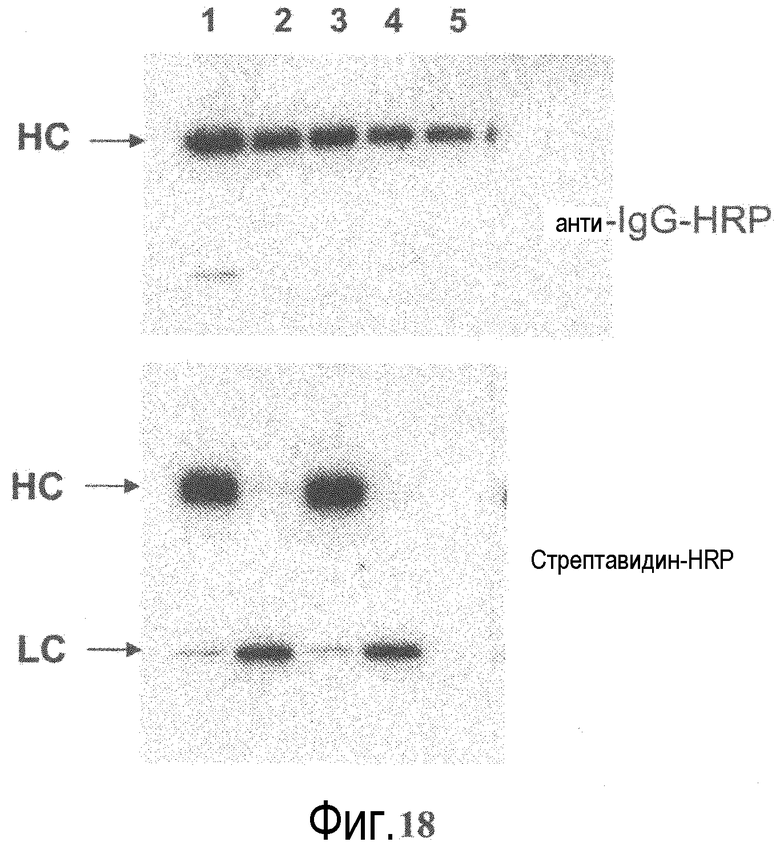
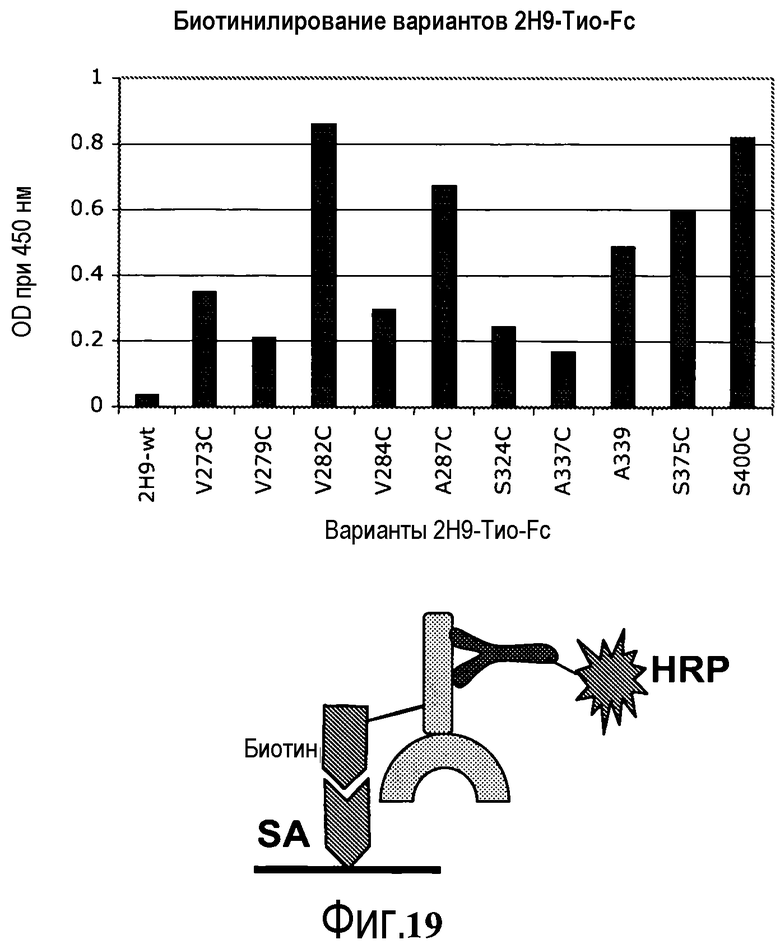
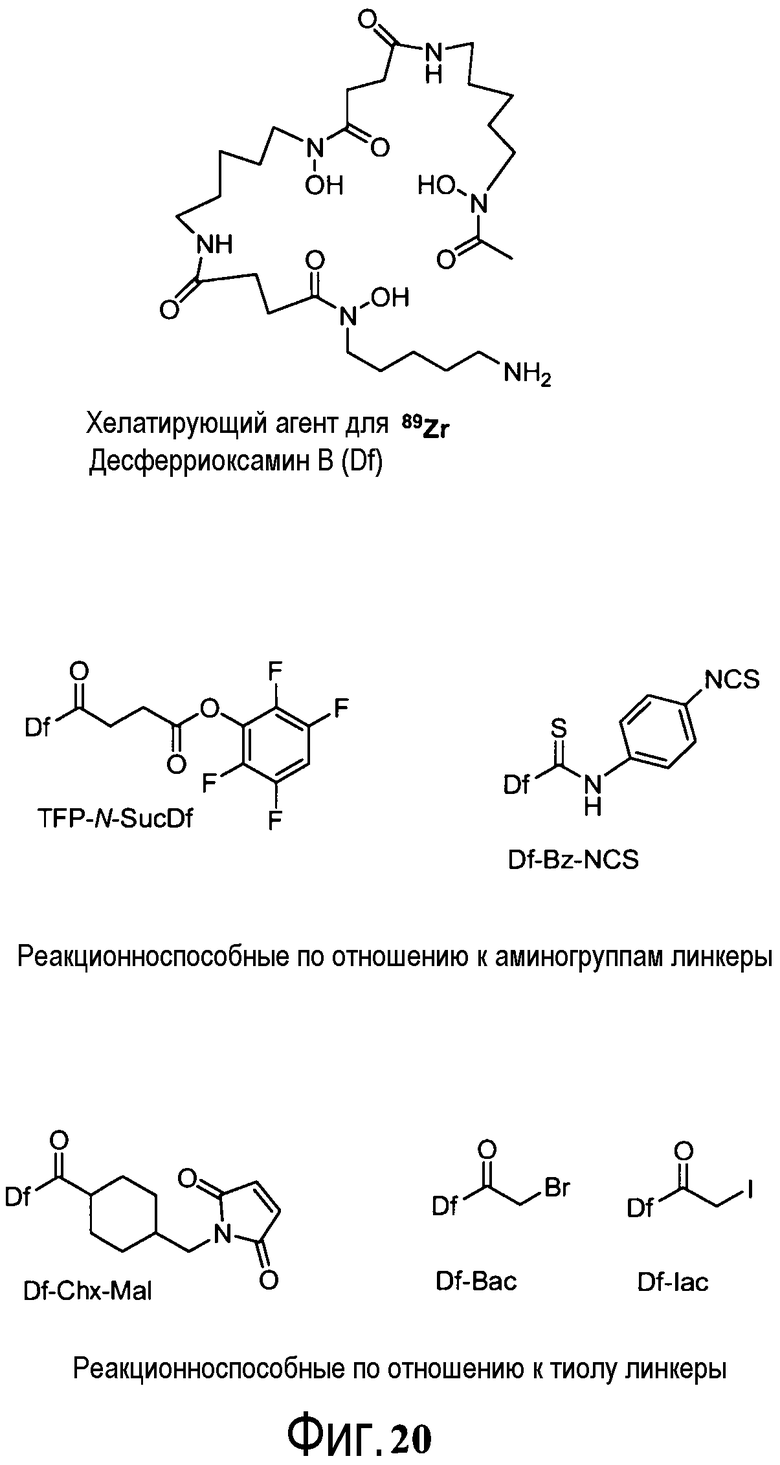
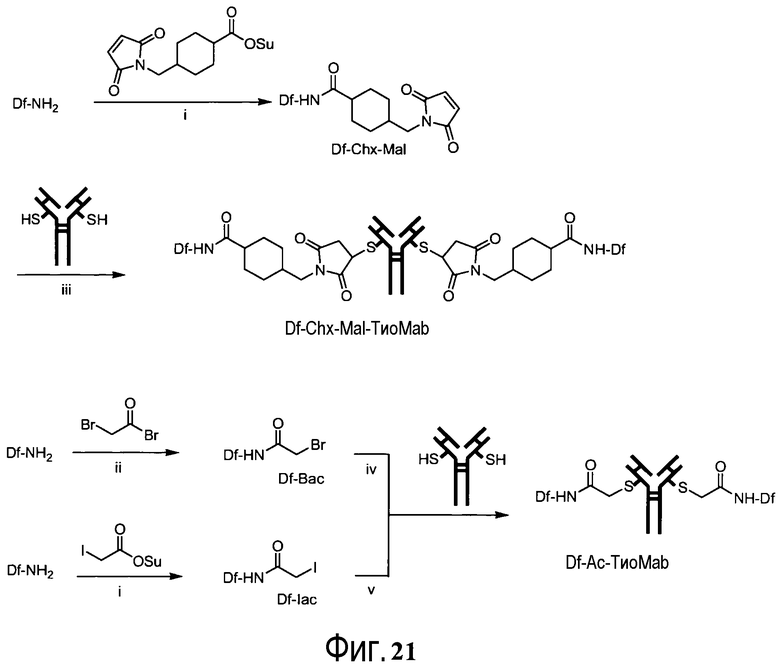

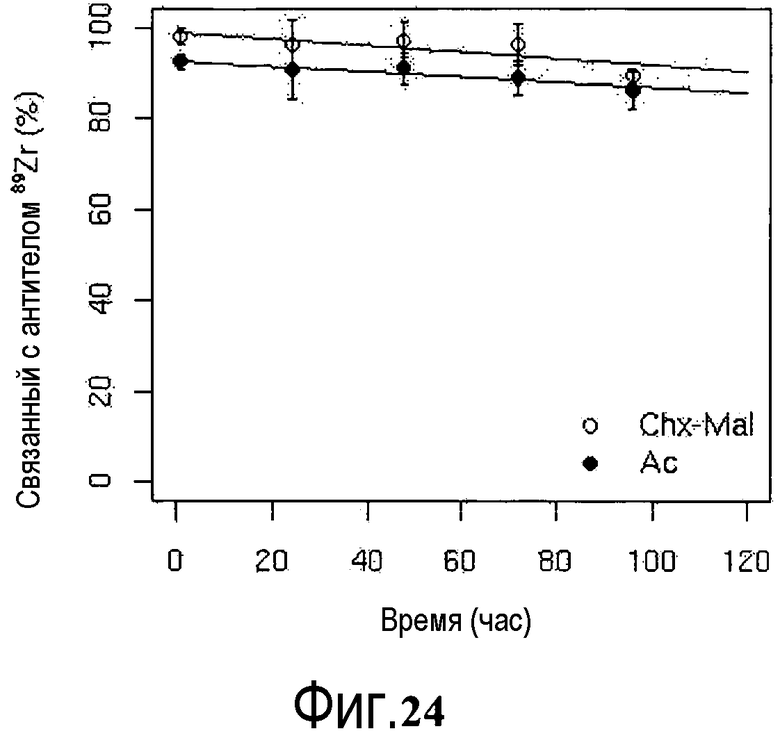
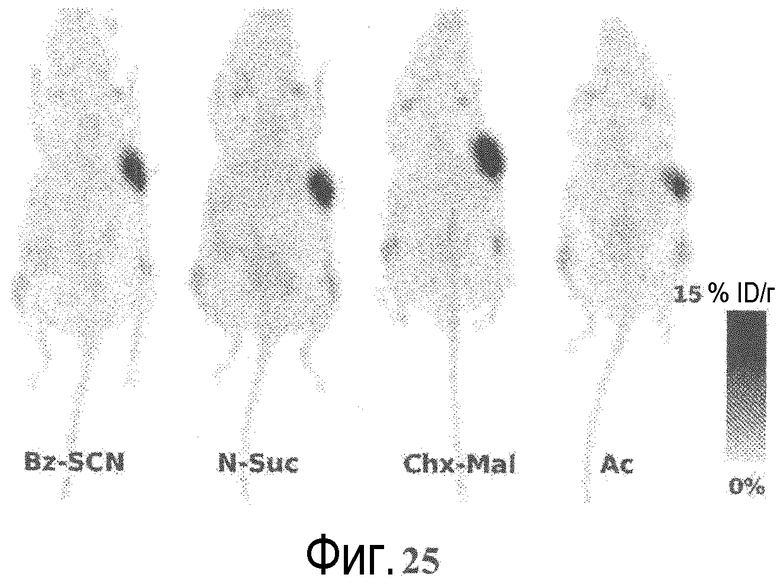
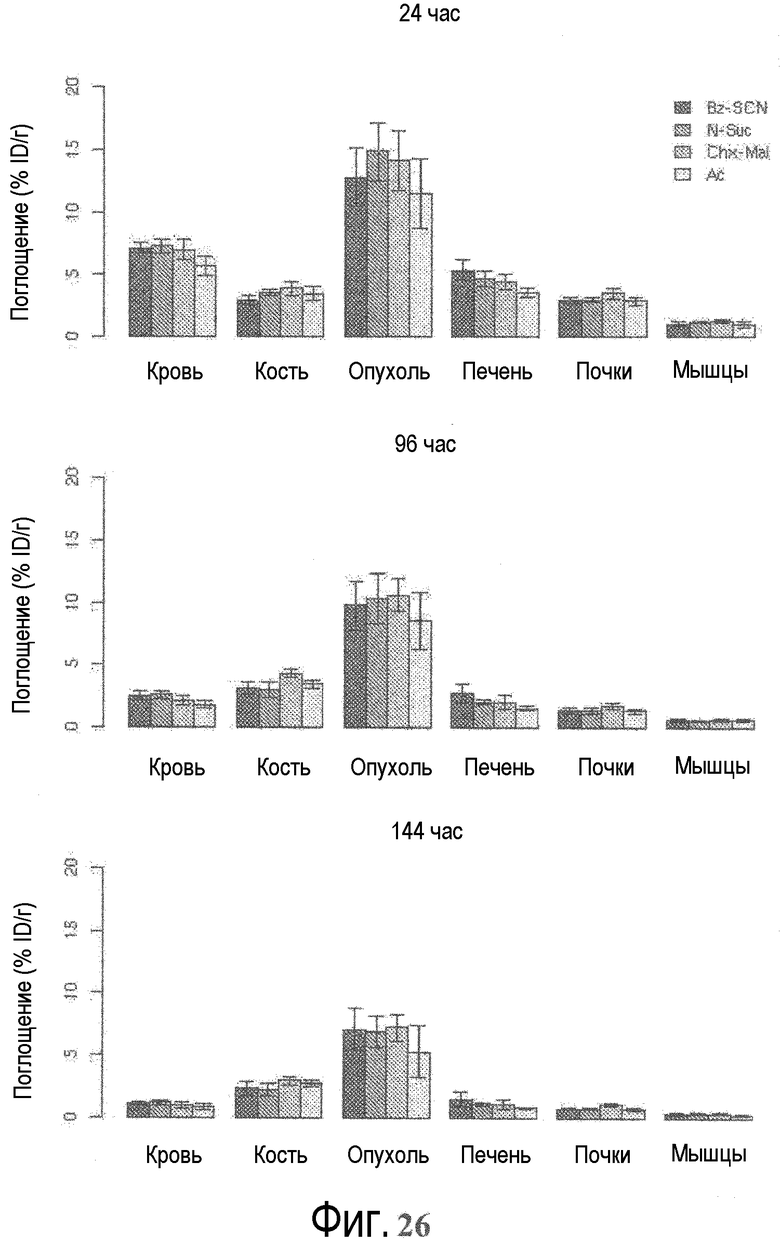
Изобретение относится к области биотехнологии, конкретно к получению меченых антител, и может быть использовано для диагностики. Антитела конструируют с заменой одной или нескольких аминокислот исходного антитела на не являющиеся перекрестно связанными, высокореакционноспособные цистеиновые аминокислоты. Сконструированные антитела с цистеиновыми заменами (Ab) конъюгируют с одной или несколькими метками с комплексом циркония (Z) через линкер (L) для получения конъюгатов меченных цирконием сконструированных антител с цистеиновыми заменами, обладающих формулой I: Ab-(L-Z)p, где p представляет собой 1-4. Изобретение позволяет повысить эффективность диагностики за счет использования меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами для получения изображений. 5 н. и 10 з.п. ф-лы, 29 ил., 12 табл., 23 пр.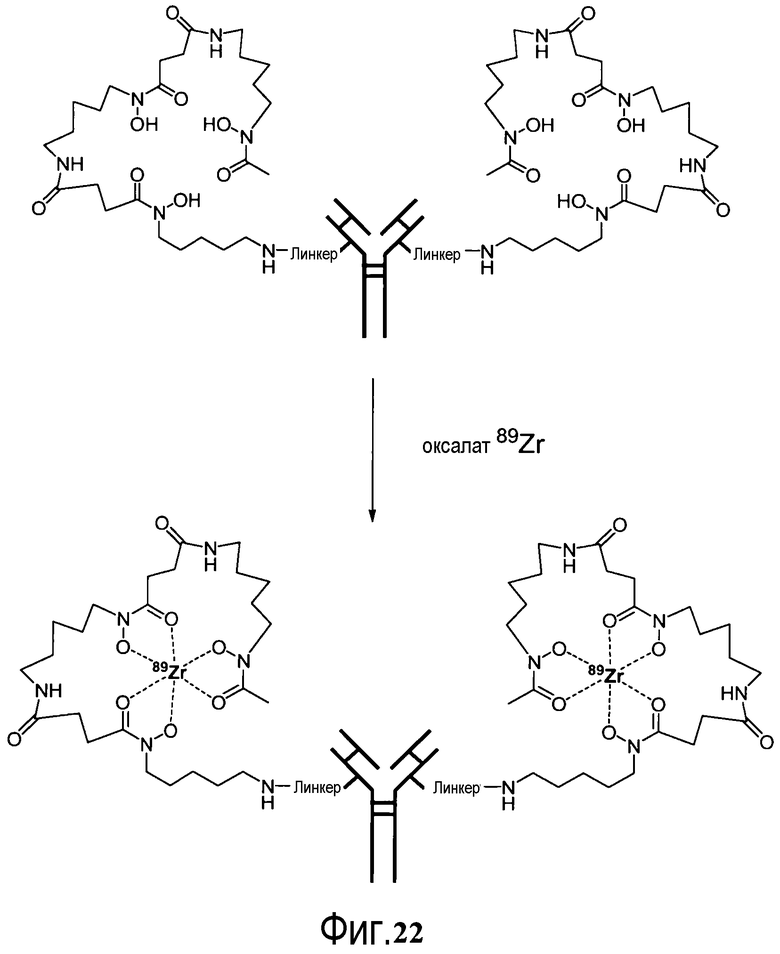
1. Меченное цирконием сконструированное антитело с цистеиновыми заменами, содержащее сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающее формулой I:

где p представляет собой 1-4;
где (L-Z) выбран из
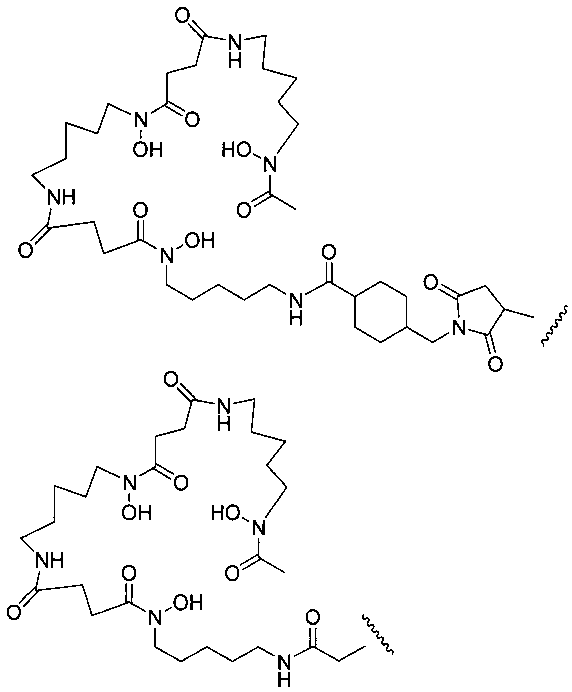 ,
,
где волнистая линия обозначает присоединение к антителу; и где цирконий образует комплекс с десферриоксаминовой группой; и
где сконструированное антитело содержит последовательность в тяжелой цепи, выбранную из SEQ ID NO: 11, 12, 13 и 15:

где цистеин в SEQ ID NO: 11, 12, 13 и 15 представляет собой свободную аминокислоту цистеин, где сконструированное антитело с цистеиновыми заменами в легкой цепи содержит последовательность, выбранную из SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46:

где цистеин в SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46 представляет собой свободную аминокислоту цистеин.
2. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где p равно 2.
3. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами получают способом, включающим:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
4. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами выбрано из моноклонального антитела, биспецифического антитела, химерного антитела, человеческого антитела, гуманизированного антитела и Fab-фрагмента.
5. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами представляет собой А121С тио-трастузумаб.
6. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами получают способом, включающим замену одного или нескольких аминокислотных остатков исходного антитела на одну или несколько свободных цистеиновых аминокислот, где исходное антитело избирательно связывается с антигеном и сконструированное антитело с цистеиновыми заменами избирательно связывается с тем же антигеном, что и исходное антитело.
7. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами или исходное антитело связывается с одним или несколькими из рецепторов (1)-(51):
(1) BMPR1B (рецептор костного морфогенного белка типа IB);
(2) Е16 (LAT1, SLC7A5);
(3) STEAP1 (эпителиальный антиген простаты, содержащий шесть трансмембранных доменов);
(4) 0772Р (СА125, MUC16);
(5) MPF (MPF, MSLN, SMR, фактор активации мегакариоцитов, мезотелин);
(6) Napi3b (NAPI-3B, NPTIIb, SLC34A2, семейство переносчиков растворенных веществ 34 (фосфат натрия), член 2, натрий-зависимый переносчик фосфата 3b, типа II);
(7) Sema 5b (FLJ10372, KIAA1445, Mm. 42015, SEMA5B, SEMAG, Семафорин 5b Hlog, домен sema, семь повторов тромбоспондина (типа 1 и подобный типу 1), трансмембранный домен (ТМ) и короткий цитоплазматический домен, (семафорин) 5В);
(8) PSCA hlg (2700050C12Rik, C530008O16Rik, кДНК RIKEN 2700050С12, кДНК RIKEN гена 2700050С12);
(9) ETBR (рецептор эндотелина типа В);
(10) MSG783 (RNF124, гипотетический белок FLJ20315);
(11) STEAP2 (HGNC_8639, IPCA-1, PCANAP1, STAMP1, STEAP2, STMP, ассоциированный с раком предстательной железы ген 1, ассоциированный с раком предстательной железы белок 1, эпителиальный антиген простаты 2, содержащий шесть трансмембранных доменов, белок простаты, содержащий шесть трансмембранных доменов);
(12) TrpM4 (BR22450, FLJ20041, TRPM4, TRPM4B, катионный канал транзиторного рецепторного потенциала, подсемейство М, член 4);
(13) CRIPTO (CR, CR1, CRGF, CRIPTO, TDGF1, полученный из тератокарциномы фактор роста);
(14) CD21 (CR2 (рецептор комплемента 2) или C3DR (рецептор C3d/вируса Эпштейна-Барр) или Hs. 73792);
(15) CD79b (CD79B, CD79β, IGb (иммуноглобулин-ассоциированная молекула бета), В29);
(16) FcRH2 (IFGP4, IRTA4, SPAP1A (содержащий SH2 домен фосфатазный якорный белок 1a), SPAP1B, SPAP1C);
(17) HER2;
(18) NCA;
(19) MDP;
(20) IL20Rα;
(21) Бревикан;
(22) EphB2R;
(23) ASLG659;
(24) PSCA;
(25) GEDA;
(26) BAFF-R (рецептор фактора активации В-клеток, BLyS рецептор 3, BR3;
(27) CD22 (изоформа B-клеточного рецептора CD22-B);
(28) CD79a (CD79A, CD79α, иммуноглобулин-ассоциированная молекула альфа, специфический для В-клеток белок, ковалентно взаимодействующий с Ig бета (CD79B) и формирующий комплекс на поверхности с молекулами IgM, передает сигнал, вовлеченный в дифференцировку В-клеток);
(29) CXCR5 (рецептор лимфомы Беркитта 1, связанный с G белком рецептор, активируемый хемокином CXCL13, функционирует в миграции лимфоцитов и гуморальной защите, играет роль в инфекции HIV-2 и, возможно, развитии СПИД, лимфомы, миеломы и лейкоза);
(30) HLA-DOB (бета-субъединица молекулы МНС класса II (1а антиген), связывающая пептиды и представляющая их CD4+ Т-лимфоцитам);
(31) Р2Х5 (управляемый лигандом пуринергического рецептора Р2Х ионный канал 5, ионный канал, управляемый внеклеточным АТР, может быть вовлечен в передачу возбуждения по синапсам и нейрогенез, недостаточность может вносить вклад в патофизиологию идиопатической нестабильности детрузора);
(32) CD72 (антиген дифференцировки В-клеток CD72, Lyb-2);
(33) LY64 (лимфоцитарный антиген 64 (RP105), мембранный белок типа I семейства богатых лейцином повторов (LRR), регулирует активацию и апоптоз В-клеток, потеря функции ассоциирована с увеличенной активностью заболевания у пациентов с системной красной волчанкой);
(34) FcRH1 (подобный Fc-рецептору белок 1, предположительный рецептор для Fc-домена иммуноглобулинов, содержащий Ig-подобный домен типа С2 и домен ITAM, может играть роль в дифференцировке В-лимфоцитов);
(35) IRTA2 (ассоциированный с транслокацией рецептор 2 суперсемейства иммуноглобулинов, предположительный иммунорецептор с возможной ролью в развитии В-клеток и лимфомагенезе; нарушение регуляции гена посредством транслокации происходит при некоторых злокачественных новообразованиях В-клеток);
(36) TENB2 (предположительный трансмембранный протеогликан, родственный семейству факторов роста EGF/херегулина и фоллистатину);
(37) PMEL17 (гомолог silver; SILV; D12S53E; PMEL17; (SI); (SIL); МЕ20; gp100);
(38) TMEFF1 (трансмембранный белок с EGF-подобным доменом и двумя фоллистатин-подобными доменами 1; Томорегулин-1; Н7365; C9orf2; C90RF2; U19878; Х83961;
(39) GDNF-Ra1 (рецептор альфа 1 семейства GDNF; GFRA1; GDNFR; GDNFRA; RETL1; TRNR1; RET1L; GDNFR-alpha1; GFR-ALPHA-1; U95847; ВС014962);
(40) Ly6E (комплекс лимфоцитарного антигена 6, локус Е; Ly67, RIG-E, SCA-2, TSA-1);
(41) ТМЕМ46 (гомолог shisa 2 (Xenopus laevis); SHISA2);
(42) Ly6G6D (комплекс лимфоцитарного антигена 6, локус G6D; Ly6-D, MEGT1);
(43) LGR5 (содержащий богатые лейцином повторы, связанный с белком G рецептор 5; GPR49, GPR67);
(44) RET (протоонкоген ret; MEN2A; HSCR1; MEN2B; МТС1; (РТС); CDHF12; Hs. 168114; RET51; RET-ELE1);
(45) LY6K (комплекс лимфоцитарного антигена 6, локус K; LY6K; HSJ001348; FLJ35226);
(46) GPR19 (связанный с белком G рецептор 19; Mm. 4787);
(47) GPR54 (рецептор KISS1; KISS1R; GPR54; НОТ7Т175; AXOR12);
(48) ASPHD1 (содержащий домен аспартат-бета-гидроксилазы белок 1; LOC253982);
(49) Тирозиназа (TYR; OCAIA; OCA1A; тирозиназа; SHEP3);
(50) TMEM118 (белок с «безымянным пальцем», трансмембранный белок 2; RNFT2; FLJ14627); и
(51) GPR172A (связанный с белком G рецептор 172А; GPCR41; FLJ11856; D15Ertd747e).
8. Меченное десферриоксамином сконструированное антитело с цистеиновыми заменами, содержащее сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающее формулой II:

где L-Df выбран из:
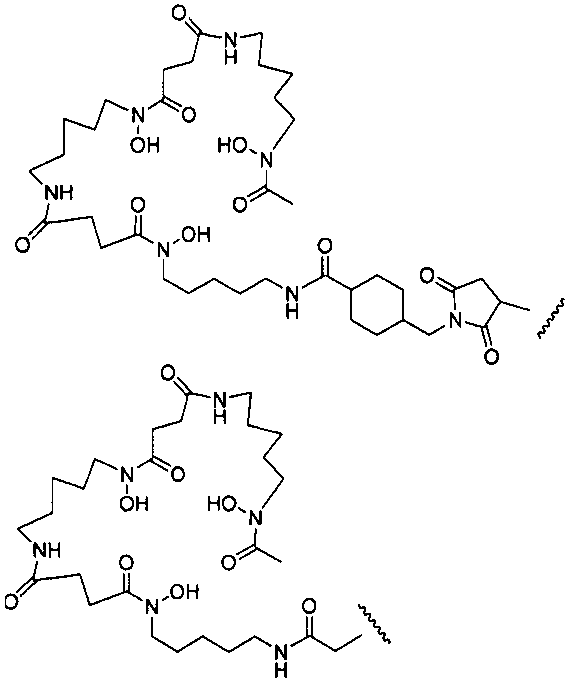
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
9. Способ получения меченного десферриоксамином сконструированного антитела с цистеиновыми заменами по п. 8,
где способ включает реакцию композиции, выбранной из структур:
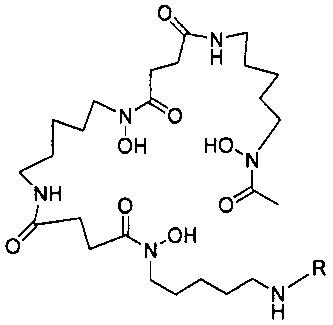
где R выбран из:
со сконструированным антителом с цистеиновыми заменами, имеющим одну или несколько свободных цистеиновых аминокислот, посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
10. Способ получения меченного цирконием сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающего формулой I:

где p представляет собой 1-4;
где способ включает образование комплекса циркониевого реагента с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, содержащим сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающим формулой II:

где L-Df выбран из:
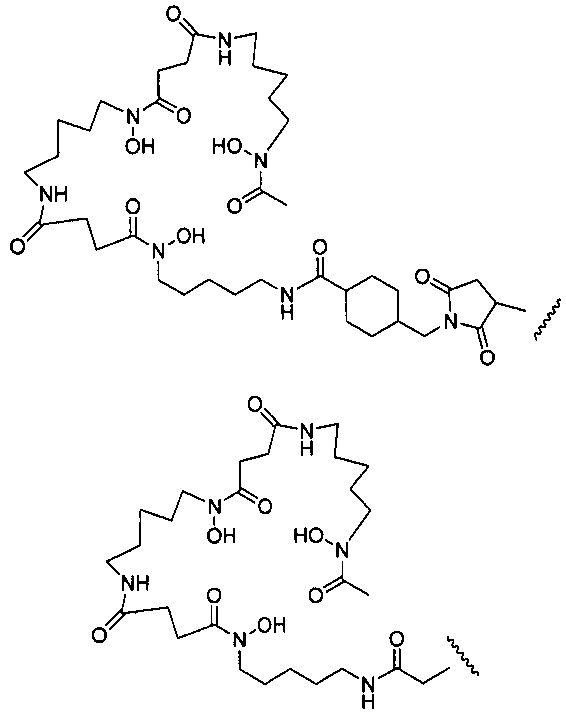
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
11. Способ по п. 10, где циркониевый реагент представляет собой оксалат 89циркония.
12. Способ получения изображений, включающий:
введение меченного цирконием сконструированного антитела с цистеиновыми заменами по п. 1 животному; и
детекцию in vivo присутствия меченного цирконием сконструированного антитела с цистеиновыми заменами посредством получения изображений.
13. Способ по п. 12, где Z содержит цирконий в комплексе с десферриоксамином В.
14. Способ по п. 12, где меченное цирконием сконструированное антитело с цистеиновыми заменами связывается с антигеном.
15. Способ по п. 12, где животное представляет собой модель ксенотрансплантата опухоли на мышах.
| WO2006034488 A2, 30.03.2006 | |||
| US2008311134 A1, 18.12.2008 | |||
| DIJKERS E.C.F | |||
| et al, Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging, The Journal of Nuclear Medicine, 2009, v | |||
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Шеститрубный элемент пароперегревателя в жаровых трубках | 1915 |
|
SU974A1 |
| GOVINDAN S.V | |||
| et al, Deferoxamine as a chelator for 67Ga in the preparation of antibody conjugates, 2005, | |||

