Родственная заявка
По настоящей заявке испрашивается приоритет предварительной заявки на патент США № 61/351195, поданной 3 июня 2010 г., которая включена в настоящее описание изобретения в качестве ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам против STEAP-1 и их иммуноконъюгатам. Настоящее изобретение далее относится к способам применения антител против STEAP-1 и их иммуноконъюгатов.
Уровень техники
Рак предстательной железы является одной из наиболее часто диагностируемых злокачественных новообразований у мужчин и второй по распространенности причиной смерти людей от рака. По оценке Американского онкологического общества к 2000 г. будет диагностировано 180400 новых случаев рака предстательной железы и данное заболевание станет причиной смерти 31900 людей. На поздних стадиях развития рак предстательной железы метастазирует в кость. Несмотря на достижения, связанные с ранней диагностикой и лечением локальных опухолей, рак предстательной железы становится неизлечимым после образования метастазов. У индивидов с метастазирующим раком предстательной железы, проходящих курс лечения гормональными препаратами, в конце концов возникает резистентность к андрогену (андрогеннезависимое состояние), что вызывает прогрессирование болезни и смерть. В настоящее время широко используемым маркером опухоли для скрининга, диагностики и мониторинга рака предстательной железы является простатоспецифический антиген (PSA). Однако широкое применение PSA в качестве инструмента для скрининга является весьма спорным, так как PSA не позволяет точно отличить доброкачественное и злокачественное заболевание предстательной железы.
В зависимости от стадии рака предстательной железы и мочевого пузыря лечение включает один способ или комбинацию нижеследующих способов лечения: хирургическую операцию для удаления раковой ткани, лучевую терапию, химиотерапию, депривацию андрогена (например, гормональную терапию) в случае рака предстательной железы. Хотя хирургическая операция или лучевая терапия в значительной степени способствуют выживанию индивидов на ранних стадиях заболевания, возможности лечения очень ограничены на поздних стадиях заболевания, особенно в случае рецидива опухоли после устранения гормона. У большинства индивидов, прошедших курс гормональной терапии, продолжает развиваться андрогеннезависимое заболевание. В настоящее время отсутствует эффективное лечение для 20-40% индивидов с раком предстательной железы, у которых возникает рецидив заболевания после хирургической операции или лучевой терапии и у которых рак уже метастазировал в момент постановки диагноза. Химиотерапия вызывает токсические побочные эффекты, особенно у пожилых индивидов. Разработка новых форм лечения, особенно для заболевания, резистентного к отсутствию андрогена, является насущной потребностью при лечении рака предстательной железы.
В научной литературе (см. патент США № 6329503) был описан новый антиген на поверхности клетки, STEAP-1. STEAP-1 относится к трансмембранным антигенам змеевидного типа на поверхности клетки. Указанный антиген экспрессирован главным образом в раке предстательной железы, поэтому члены данного семейства получили название ”STEAP” (шесть трансмембранных эпителиальных антигенов предстательной железы). Белки STEAP человека характеризуются высокой степенью структурного консерватизма в данном семействе и не обладают значительной структурной гомологией с любыми известными белками человека. STEAP-1 является мембранным белком типа IIIa, экспрессированным главным образом в клетках предстательной железы нормальных тканей человека. В структурном отношении белок STEAP-1 состоит из 339 аминокислот, имеет шесть трансмембранных доменов и N- и С-концы внутри клетки, из чего следует, что укладка цепи данного белка происходит по ”змеевидному” принципу, образуя три внеклеточные и две внутриклеточные петли. Белок STEAP-1 высоко экспрессирован на разных стадиях рака предстательной железы. STEAP-1 сверхэкспрессирован в других раках человека, таких как рак легкого и рак ободочной кишки. Были созданы антитела мыши к фрагментам STEAP-1 человека, которые, как было показано, связываются с STEAP-1 на поверхности клетки (см. заявку на патент США № 20040253232А1).
Терапия на основе антител оказалась очень эффективной при лечении разных типов рака. Например, препараты герцептин® и ритуксан® (компании Genentech, S. San Francisco) были успешно использованы при лечении соответственно рака молочной железы и неходжкинской лимфомы. Герцептин® является гуманизированным моноклональным антителом, выделенным из рекомбинантной ДНК, которое избирательно связывается с внеклеточным доменом протоонкогена рецептора эпидермального фактора роста 2 человека (HER2). Сверхэкспрессия белка HER2 наблюдается в 25-30% случаев первичного рака молочной железы. Ритуксан® является генетически созданным химерным моноклональным антителом мыши/человека против антигена CD20, обнаруженного на поверхности нормальных и злокачественных В-лимфоцитов. Оба указанных антитела продуцированы в клетках СНО.
Использование конъюгатов антитела с лекарственным средством для местной доставки цитотоксических или цитостатических агентов, то есть лекарственных средств для уничтожения или ингибирования опухолевых клеток при лечении рака (Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drg. Del. Rev. 26:151-172; патент США № 4975278), позволяет осуществлять направленную доставку лекарственной части в опухоли и накапливать лекарственное средство внутри клетки, когда системная доставка неконъюгированных лекарственных средств может вызвать неприемлемые уровни токсичности для нормальных клеток наряду с опухолевыми клетками, подлежащими устранению (Baldwin et al., (1986) Lancet pp. (Mar. 15, 1986):603-05; Thorpe, (1985) “Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review,” in Monoclonal Antibodies ’84: Biological And Clinical Applications, A. Pinchera et al. (ed.s), pp. 475-506). При данном способе введения достигается максимальная эффективность при минимальной токсичности. Как отмечалось в научной литературе, в указанных способах лечения могут быть использованы как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother., 21:183-87). Лекарственные средства, используемые в указанных способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., Cancer Immunol. Immunother. 21:183-87 (1986)). Токсины, используемые в антитело-токсиновых конъюгатах, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Kerr et al. (1997) Bioconjugate Chem. 8(6):781-784; Mandler et al. (2000) Journal of the Nat. Cancer Inst. 92(19):1573-1581; Mandler et al. (2000) Bioorganic & Med. Chem. Letters 10:1025-1028; Mandler et al. (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (европейский патент 1391213; Liu et al., (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихеамицин (Lode et al. (1998) Cancer Res. 58:2928; Hinman et al. (1993) Cancer Res. 53:3336-3342). Токсины могут оказывать цитотоксическое и цитостатическое действие при помощи механизмов, включающих связывание тубулина, связывание ДНК или ингибирование топоизомеразы (Meyer, D.L. and Senter, P.D. “Recent Advances in Antibody Drug Conjugates for Cancer Therapy” in Annual Reports in Medicinal Chemistry, Vol. 38 (2003) Chapter 23, 229-237). Некоторые цитотоксические лекарственные средства становятся неактивными или менее активными при конъюгировании с крупными антителами или лигандами рецепторов белка.
Зевалин® (ибритумомаб тиуксетан, Biogen/Idec) является конъюгатом антитела с радиоактивным изотопом, который состоит из мышиного моноклонального антитела IgG1 с каппа-цепью, направленного против антигена CD20, обнаруженного на поверхности нормальных и злокачественных В-лимфоцитов, и радиоактивного изотопа 111In или 90Y, связанного хелатообразователем с тиомочевинным линкером (Wiseman et al. (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al. (2002) Blood 99(12):4336-42; Witzig et al. (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al. (2002) J. Clin. Oncol. 20(15):3262-69). Хотя зевалин активно воздействует на В-клеточную неходжкинскую лимфому (NHL), при его введении у большинства индивидов возникает тяжелая и продолжительная цитопения. Милотарг™ (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитела с лекарственным средством, состоящий из антитела человека против CD33-клеток, связанного с калихеамицином, был утвержден в 2000 г. для лечения острого миелоидного лейкоза путем инъекций (Drugs of the Future (2000) 25(7):686; патенты США №№ 4970198, 5079233, 5585089, 5606040, 5693762, 5739116, 5767285, 5773001). Кантузумаб мертанзин (Immunogen, Inc.), конъюгат антитела с лекарственным средством, состоящий из антитела C242 человека, связанного при помощи дисульфидного линкера SPP с майтанзиноидом, DM1, разработан для лечения злокачественных новообразований, экспрессирующих антиген CanAg, таких как рак ободочной кишки, поджелудочной железы, желудка и других. MLN-2704 (Millennium Pharm., BZL Biologics, Immunogen Inc.), конъюгат антитела с лекарственным средством, состоящий из моноклонального антитела против простатоспецифического мембранного антигена (PSMA), связанного с майтанзиноидом, DM1, находится в стадии разработки предположительно для лечения опухолей предстательной железы. Такой же майтанзиноид, DM1, был связан при помощи линкера SMCC, отличного от дисульфидного линкера, с моноклональным антителом мыши ТА.1 (Chari et al. (1992) Cancer Research 52:127-131). В научной литературе отмечалось, что указанный конъюгат является в 200 раз менее активным, чем соответствующий конъюгат с дисульфидным линкером. Линкер SMCC был признан ”нерасщепляемым”.
Из морского моллюска Dolabella auricularia было выделено несколько короткоцепных пептидных соединений, у которых была обнаружена биологическая активность (Pettit et al. (1993) Tetrahedron 49:9151; Nakamura et al. (1995) Tetrahedron Letters 36:5059-5062; Sone et al. (1995) Journal Org. Chem. 60:4474). Также были получены аналоги указанных соединений, при этом было установлено, что некоторые из них обладают биологической активностью (обзор таких соединений представлен в публикации Pettit et al. (1998) Anti-Cancer Drug Design 13:243-277). Например, ауристатин Е (патент США № 5635483) является синтетическим аналогом морского природного продукта доластатина 10, средства, ингибирующего полимеризацию тубулина благодаря связыванию с тем же сайтом тубулина, что и противораковое лекарственное средство винкристин (G.R. Pettit, (1997) Prog. Chem. Org. Nat. Prod. 70:1-79). Доластатин 10, ауристатин РЕ и ауристатин Е являются линейными пептидами, содержащими четыре аминокислоты, три из которых являются уникальными для соединений класса доластатина, и С-концевой амид.
Пептиды ауристатина, ауристатин Е (АЕ) и монометилауристатин (ММАЕ), синтетические аналоги доластатина, были конъюгированы с: (i) химерными моноклональными антителами cBR96 (специфичными к антигену Левиса Y на карциномах); (ii) сАС10, специфичным к CD30 на гематологических злокачественных новообразованиях (Klussman, et al. (2004), Bioconjugate Chemistry 15(4):765-773; Doronina et al. (2003) Nature Biotechnology 21(7):778-784; “Monomethylvaline Compounds Capable of Conjugation to Ligands”; Francisco et al. (2003) Blood 102(4):1458-1465; заявка на патент США 2004/0018194; (iii) антителами против CD20-клеток, такими как ритуксан® (ритуксимаб) (WO 04/032828), предназначенными для лечения CD20-экспрессирующих злокачественных новообразований и иммунных нарушений; (iv) антителами против EphB2 2Н9 и антителами против IL-8, предназначенными для лечения колоректального рака (Mao et al. (2004) Cancer Research 64(3):781-788; (v) антителом Е-селектином (Bhaskar et al. (2003) Cancer Res. 63:6387-6394) и (vi) другими антителами против CD30-клеток (WO 03/043583). Монометилауристатин (ММАЕ) был также конъюгирован с антителом 2Н9 против EphB2R, который является рецептором тирозинкиназы ТМ типа 1, характеризующимся близкой гомологией у мыши и человека, который сверхэкспрессирован в клетках колоректального рака (Mao et al. (2004) Cancer Res. 64:781-788).
Монометилауристатин MMAF, вариант ауристатина Е (ММАЕ), с фенилаланином у С-конца (патент США № 5767237, патент США № 6124431), как сообщалось в научной литературе, является менее активным, чем ММАЕ, но становится более эффективным при конъюгировании с моноклональными антителами (Senter et al., Proceedings of the American Association for Cancer Research, Volume 45, Abstract Number 623, presented March 28, 2004). Ауристатин F фенилендиамин (AFP), фенилаланиновый вариант ММАЕ, был связан с моноклональным антителом против CD70-клеток, 1F6, у С-конца 1F6 при помощи фенилендиаминового спейсера (Law et al., Proceedings of the American Association for Cancer Research, Volume 45, Abstract Number 625, presented Match 28, 2004).
В данной области существует потребность в дополнительных лекарственных средствах для лечения разных злокачественных новообразований, таких как раки и метастазы раков предстательной железы, легкого и ободочной кишки. Особенно полезными лекарственными средствами для указанной цели являются конъюгаты антитела против STEAP-1 с лекарственным средством, направленно воздействующие на клетки предстательной железы, легкого или ободочной кишки, которые обладают значительно более низкой токсичностью и приемлемой терапевтической эффективностью. Целью настоящего изобретения является устранение вышеуказанных и других ограничений и проблем, имевших место в прошлом.
Молекулярная визуализация является важным инструментом разработки и оценки новых фармацевтических средств. Иммуно-позитрон-эмиссионная томография (иммуно-ПЭТ) является способом быстрой визуализации для отслеживания и количественного определения моноклональных антител (mAb) in vivo, так как данный способ эффективно сочетает высокую чувствительность ПЭТ с высокой специфичностью моноклональных антител. Иммуно-ПЭТ является клиническим способом неинвазивной диагностики, обеспечивающим ”всестороннее иммуногистохимическое окрашивание in vivo” (van Dongen GA et al. “Immuno-PET: a navigator in monoclonal antibody development and applications” Oncologist 2007; 12:1379-89; Williams et al. (2001) Cancer Biother. Radiopharm. 16:25-35; Hollinger et al. (2005) Nat. Biotechnol. 23:1126-36)). Создание иммуно-ПЭТ на основе 89Zr сделало возможным измерение экспрессии мишени и биораспределения антител у мышей и человека; клиническое применение иммуно-ПЭТ с 89Zr-трастузумабом позволяет увидеть гетерогенную структуру метастазов в печени и кости (Dijkers et al., Nature Vol. 87 Number 5 (May 2010).
Системы визуализации ПЭТ создают изображения на основе распределения позитрон-излучающих изотопов в ткани индивида. Изотопы обычно вводят индивиду путем инъекции молекул зонда, содержащих позитрон-излучающий изотоп, такой как F-18, C-11, N-13 или О-15, ковалентно связанный с молекулой, которая легко метаболизируется или локализуется в организме (например, глюкоза), или химически связываются с сайтами рецепторов в организме. В некоторых случаях изотоп вводят индивиду в виде ионного раствора или путем ингаляции. Мелкие агенты, визуализируемые при помощи иммуно-ПЭТ, такие как Fab-фрагменты антитела (50 кДа) или диатела, спаренные димеры ковалентно связанной VH-VK области моноклонального антитела, 55 кДа (Shively et al. (2007) J. Nucl. Med. 48:170-2), могут быть особенно полезны, характеризуясь коротким временем полужизни в кровотоке, высокой проникаемостью в ткани и оптимальным соотношением радиоактивности между опухолью и фоном в течение двух-четырех часов после инъекции, что облегчает использование изотопов с коротким периодом полураспада, таких как широко используемый 18F (109,8 мин).
Любые ссылки, приведенные в настоящей заявке, не являются признанием того, что данная ссылка служит прототипом данной заявки. Все ссылки, приведенные в настоящей заявке, включая патенты, заявки на патенты и публикации, включены в настоящее описание изобретения в качестве ссылки в полном объеме.
Сущность изобретения
Настоящее изобретение относится к антителам против STEAP-1 и способам их применения.
В одном из аспектов настоящее изобретение относится к антителу, связывающемуся с STEAP-1, которое содержит вариабельный домен легкой цепи, содержащий аминокислотную последовательность, показанную на фигуре 2А (SEQ ID NO:6), или вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, показанную на фигуре 2В (SEQ ID NO:9). В одном из аспектов настоящее изобретение относится к антителу, связывающемуся с STEAP-1, которое содержит вариабельный домен легкой цепи, содержащий аминокислотную последовательность, показанную на фигуре 2А (SEQ ID NO:6), и вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, показанную на фигуре 2В (SEQ ID NO:9).
В одном из аспектов настоящее изобретение относится к антителу, связывающемуся с STEAP-1, которое содержит вариабельный домен тяжелой цепи, последовательность которого по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO:9 или 10. В одном из вариантов осуществления изобретения антитело содержит вариабельный домен тяжелой цепи SEQ ID NO:9 или 10. В одном из вариантов осуществления изобретения антитело содержит остовную область 1 вариабельного домена тяжелой цепи SEQ ID NO:25, остовную область 2 вариабельного домена тяжелой цепи SEQ ID NO:75, 76 или 77, или остовную область 3 вариабельного домена тяжелой цепи SEQ ID NO:78 или 79.
В одном из аспектов настоящее изобретение относится к антителу, содержащему вариабельный домен легкой цепи, последовательность которого по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO:6. В одном из вариантов осуществления изобретения антитело содержит вариабельный домен легкой цепи SEQ ID NO:6.
В одном из аспектов настоящее изобретение относится к антителу, связывающемуся с STEAP-1, которое содержит вариабельный домен тяжелой цепи, последовательность которого по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO:9 или 10. В одном из вариантов осуществления изобретения антитело содержит вариабельный домен тяжелой цепи SEQ ID NO:9 или 10. В одном из вариантов осуществления изобретения антитело содержит остовную область 1 вариабельного домена тяжелой цепи SEQ ID NO:25, остовную область 2 вариабельного домена тяжелой цепи SEQ ID NO:75, 76 или 77, или остовную область 3 вариабельного домена тяжелой цепи SEQ ID NO:78 или 79. В одном из вариантов осуществления изобретения антитело далее содержит вариабельный домен легкой цепи, последовательность которого по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO:6. В одном из вариантов осуществления изобретения антитело содержит вариабельный домен легкой цепи SEQ ID NO:6.
Определенные варианты осуществления изобретения относятся к полинуклеотиду, кодирующему любое из вышеуказанных антител. Один из вариантов осуществления изобретения относится к вектору, содержащему указанный полинуклеотид. Один из вариантов осуществления изобретения относится к клетке-хозяину, содержащей указанный вектор. В одном из вариантов осуществления изобретения клетка-хозяин является эукариотической клеткой. В одном из вариантов осуществления изобретения клетка-хозяин является клеткой яичника китайского хомячка (СНО). В одном из вариантов осуществления изобретение относится к способу получения антитела против STEAP-1, который включает культивирование клетки-хозяина в условиях, пригодных для экспрессии полинуклеотида, кодирующего антитело, и выделение указанного антитела.
В одном из аспектов настоящее изобретение относится к антителу, связывающемуся с STEAP-1, экспрессированным на поверхности клетки. В одном из вариантов осуществления изобретения антитело связывается с эпитопом в области STEAP-1 человека или мыши. В одном из вариантов осуществления изобретения клетка является клеткой млекопитающего. В одном из вариантов осуществления изобретения клетка является клеткой человека. В одном из вариантов осуществления изобретения клетка является раковой клеткой. В одном из вариантов осуществления изобретения клетка является клеткой предстательной железы, легкого или ободочной кишки. В одном из вариантов осуществления изобретения раковая клетка является раковой клеткой предстательной железы. В другом варианте осуществления изобретения клетка является клеткой из метастаза первичного рака предстательной железы, легкого или ободочной кишки.
В определенных вариантах осуществления изобретения любое из вышеуказанных антител является моноклональным антителом. В одном из вариантов осуществления изобретения антитело является фрагментом антитела, выбираемым из Fab, Fab'-SH, Fv, scFv или (Fab')2 фрагмента. В одном из вариантов осуществления изобретения антитело является гуманизированным антителом. В одном из вариантов осуществления изобретения антитело является антителом человека.
В одном из аспектов настоящее изобретение относится к способу обнаружения STEAP-1 в биологическом образце, который включает приведение в контакт биологического образца с любым из вышеуказанных антител в условиях, допускающих связывание антитела с STEAP-1 и обнаружение комплекса, образованного между антителом и STEAP-1. В одном из вариантов осуществления изобретения биологический образец включает клетки предстательной железы. В одном из вариантов осуществления изобретения биологический образец получают у млекопитающего с подозрением на наличие или с наличием нарушения клеток предстательной железы и/или с пролиферативным нарушением клеток или тканей, которое включает, не ограничиваясь ими, рак предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника и саркому Юинга, а также метастазы первичного рака предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника и саркомы Юинга. См., например, патент США № 6329503 и публикацию Rodeberg, D.A. et al., Clin. Cancer Res. 11(12):4545-4552 (2005)).
В одном из аспектов настоящее изобретение относится к способу диагностики пролиферативного нарушения клеток, связанного с повышенной экспрессией STEAP-1, который включает приведение в контакт исследуемой клетки с любым из вышеуказанных антител, определение уровня экспрессии STEAP-1 путем обнаружения связывания антитела с белком STEAP-1 и сравнение уровня экспрессии STEAP-1 в исследуемой клетке с уровнем экспрессии STEAP-1 в контрольной клетке, при этом более высокий уровень экспрессии STEAP-1 в исследуемой клетке по сравнению с контрольной клеткой указывает на наличие пролиферативного нарушения клетки, связанного с повышенной экспрессией STEAP-1. В одном из вариантов осуществления изобретения исследуемая клетка является клеткой индивида с подозрением на наличие пролиферативного нарушения клеток, такого как пролиферативное нарушение предстательной железы. В одном из вариантов осуществления изобретения пролиферативное нарушение клеток выбирают из нарушений клеток предстательной железы, которые включают, не ограничиваясь ими, рак предстательной железы. В одном из вариантов осуществления изобретения указанный способ включает определение уровня экспрессии STEAP-1 на поверхности исследуемой клетки (такой как, например, раковая клетка предстательной железы) и сравнение уровня экспрессии STEAP-1 на поверхности исследуемой клетки с уровнем экспрессии STEAP-1 на поверхности контрольной клетки (такой как, например, нормальная клетка предстательной железы в отличие от анормально пролиферирующей клетки предстательной железы).
В одном из аспектов настоящее изобретение относится к способу диагностики пролиферативного нарушения клеток, связанного с увеличением в клетках, таких как клетки предстательной железы, экспрессии STEAP-1, который включает приведение в контакт исследуемых клеток в биологическом образце с любым из вышеуказанных антител, определение уровня антитела, связанного с исследуемыми клетками в образце, путем обнаружения связывания антитела с STEAP-1 и сравнение уровня антитела, связанного с клетками, в контрольном образце, при этом уровень связанного антитела нормализован относительно числа клеток, экспрессирующих STEAP-1, в исследуемом и контрольном образцах, при этом более высокий уровень связанного антитела в исследуемом образце по сравнению с контрольным образцом указывает на наличие пролиферативного нарушения клетки, связанного с клетками, экспрессирующими STEAP-1.
В одном из аспектов настоящее изобретение относится к способу обнаружения растворимого STEAP-1 в крови или сыворотке, который включает приведение в контакт исследуемого образца крови или сыворотки, полученного у млекопитающего с подозрением на пролиферативное нарушение клеток предстательной железы, с антителом против STEAP-1 по настоящему изобретению и обнаружение увеличения растворимого STEAP-1 в исследуемом образце по сравнению с контрольным образцом крови или сыворотки, полученным у здорового млекопитающего. В одном из вариантов осуществления изобретения указанный способ обнаружения может быть использован в качестве способа диагностики пролиферативного нарушения клеток предстательной железы, связанного с увеличением растворимого STEAP-1 в крови или сыворотке млекопитающего.
В одном из аспектов настоящее изобретение относится к сконструированным антителам с цистеиновой заменой, в которых одна или несколько аминокислот материнского антитела заменены свободной аминокислотой цистеином, как описано в публикации WO2006/034488 (которая включена в настоящее описание изобретения в качестве ссылки в полном объеме). Сконструированное антитело с цистеиновой заменой содержит одну или несколько свободных аминокислот, являющихся цистеином, которые имеют значение реакционной способности тиоловой группы в пределах 0,6-1,0. Свободная аминокислота является остатком цистеина, который был введен в материнское антитело и не является частью дисульфидной мостиковой связи. Сконструированные антитела с цистеиновой заменой могут быть использованы для присоединения цитотоксических и/или визуализирующих соединений на сайте введенного цистеина, например, при помощи имида малеиновой кислоты или галогенацетила. Нуклеофильное взаимодействие функциональной тиоловой группы остатка Cys с группой имида малеиновой кислоты примерно в 1000 раз выше по сравнению с любой другой функциональной группой аминокислоты в белке, такой как аминогруппа остатков лизина или N-концевая аминогруппа. Специфическая функциональная тиоловая группа в иодацетиле и имиде малеиновой кислоты может взаимодействовать с аминогруппами, но для этого необходимо более высокое значение рН (>9,0) и более продолжительное время взаимодействия (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London).
Сконструированные антитела с цистеиновой заменой могут быть использованы при лечении рака и включают антитела, специфичные к поверхности клетки, трансмембранным рецепторам и опухолеспецифическим антигенам (ТАА). Такие антитела могут быть использованы в качестве “голых” антител (неконъюгированных с лекарственным средством или меткой) или в виде конъюгатов антитела с лекарственным средством (ADC). Сконструированные антитела с цистеиновой заменой по настоящему изобретению могут быть сайтспецифически и эффективно связаны с реагентом, взаимодействующим с тиоловой группой. Реагент, взаимодействующий с тиоловой группой, может быть многофункциональным линкером, иммобилизованной меткой, флуорофором или промежуточным соединением лекарственного средства с линкером. Сконструированное антитело с цистеиновой заменой может быть помечено детектируемой меткой, иммобилизовано на твердой подложке и/или конъюгировано с лекарственной частью. Реакционная способность тиоловой группы может быть сообщена любому антителу, в котором возможна замена аминокислот реакционноспособными остатками цистеина в легкой цепи в положениях: от L-10 до L-20, от L-38 до L-48, от L-105 до L-115, от L-139 до L-149, от L-163 до L-173; в тяжелой цепи в положениях: от Н-35 до Н45, от Н-83 до Н-93, от Н-114 до Н-127, от Н-170 до Н-184 и в Fc-области в положениях от Н-268 до Н-291, от Н-319 до Н-344, от Н-370 до Н-380 и от Н-395 до Н-405, где нумерация положений аминокислот начинается с положения 1 по системе нумерации Kabat (Kabat et al. (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD) и продолжается последовательно, как описано в публикации WO 2006/034488. Реакционная способность тиоловой группы может быть также сообщена определенным доменам антитела, таким как константный домен легкой цепи (CL) и константные домены тяжелой цепи, СН1, СН2 и СН3. Замены аминокислот цистеином с получением значений реакционной способности тиоловой группы, равных 0,6 и выше, могут быть произведены в константных доменах тяжелой цепи α, δ, ε, γ и μ интактных антител: соответственно, IgA, IgD, IgE, IgG и IgM, включая подклассы IgG: IgG1, IgG2, IgG3, IgG4, IgA и IgA2. Такие антитела и их применения описаны в публикации WO 2006/034488.
Сконструированное антитело с цистеиновой заменой по настоящему изобретению, предпочтительно, сохраняют антигенсвязывающую способность материнского антитела дикого типа. Таким образом, сконструированные антитела с цистеиновой заменой могут связываться, предпочтительно, специфически, с антигенами. Такие антигены включают, например, опухолеспецифические антигены (ТАА), белки рецепторов на поверхности клетки и другие молекулы на поверхности клетки, трансмембранные белки, сигнальные белки, факторы, регулирующие выживание клетки, факторы, регулирующие пролиферацию клетки, молекулы, связанные с развитием или дифференцировкой тканей (которые, как известно или предположительно функционально способствуют развитию или дифференцировке тканей), лимфокины, цитокины, молекулы, участвующие в регулировании клеточного цикла, молекулы, участвующие в образовании или развитии сосудов, и молекулы, связанные с развитием кровеносных сосудов (которые, как известно или предположительно функционально способствуют развитию кровеносных сосудов).
Антитело по настоящему изобретению может быть конъюгировано с другими агентами, взаимодействующими с тиоловыми группами, в которых реакционноспособная группа, например, представляет собой малеимид, иодацетамид, пиридилдисульфид или другие партнеры, взаимодействующие с тиоловыми группами (Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671). Указанный партнер может быть цитотоксическим агентом (например, токсином, таким как доксорубицин или коклюшевый токсин), флуорофором, таким как флуоресцентный краситель, подобный флуоресцеину или родамину, хелатообразователем для визуализирующего или радиотерапевтического металла, пептидильной или непептидильной меткой или детектирующей меткой, модификатором клиренса, таким как разные изомеры полиэтиленгликоля, пептидом, связывающимся с третьим компонентом, или другим углеводным или липофильным агентом.
В одном из аспектов настоящее изобретение относится к антителам по настоящему изобретению, конъюгированным с любой меткой, которая может быть ковалентно присоединена к антителу при помощи реакционноспособной части, активированной части или реакционноспособной тиоловой группы цистеина (Singh et al. (2002) Anal. Biochem. 304:147-15; Harlow E. and Lane, D. (1999) Using Antibodies: A Laboratory Manual, Cold Springs Harbor Laboratory Press, Cold Spring Harbor, NY; Lundblad R.L. (1991) Chemical Reagents for Protein Modification, 2nd ed. CRC Press, Boca Raton, FL). Присоединенная метка может: (i) подавать детектируемый сигнал; (ii) взаимодействовать со второй меткой с целью изменения детектируемого сигнала, подаваемого первой или второй меткой, например, осуществлять FRET (перенос энергии резонанса флуоресценции); (iii) стабилизировать взаимодействие или повышать аффинность связывания с антигеном или лигандом; (iv) влиять на подвижность, например, электрофоретическую подвижность или проницаемость клетки с помощью заряда, гидрофобности, формы или других физических параметров, или (v) быть иммобилизованной частью, используемой для модуляции аффинность лиганда, связывания антитела/антигена или ионного комплексообразования.
Меченые сконструированные антитела с цистеиновой заменой могут быть использованы при выполнении диагностических анализов, например, для обнаружения экспрессии представляющего интерес антигена в определенных клетках, тканях или сыворотке. Антитело, предназначенное для использования в диагностике, обычно метят детектируемой частью. Существует много меток, которые можно сгруппировать в следующие категории:
Радиоактивные изотопы (радионуклиды), такие как 3Н, 11С, 14С, 18F, 32Р, 35S, 64Cu, 68Ga, 86Y, 89Zr, 99Tc, 111In, 123I, 124I, 125I, 131I, 133Xe, 177Lu, 211At или 213Bi. Антитела, меченные радиоактивными изотопами, могут быть использованы в экспериментах по визуализации рецепторов-мишеней. Антитела могут быть помечены лигандами, которые связываются, образуют хелаты или в противном случае комплексы радиоактивных изотопов металлов и взаимодействуют с тиоловой группой цистеина, введенного в антитело, при помощи способов, описанных в публикации Current Protocols in Immunology. Volumes 1 and 2, Coligen et al., Ed. Wiley-Interscience, New York, NY, Pubs. (1991). Хелатообразующие лиганды, которые могут образовывать комплекс с ионом металла, включают DOTA, DOTP, DOTMA, DTPA и TETA (Macrocyclics, Dallas, TX). Радионуклиды могут оказывать направленное воздействие в результате образования комплекса с конъюгатами антитела с лекарственным средством по настоящему изобретению (Wu et al. (2005) Nature Biotechnology 23(9):1137-1146).
Линкеры, такие как DOTA-малеимид (4-малеимидобутирамидобензил-DOTA) могут быть получены в результате взаимодействия аминобензил-DOTA с 4-малеимидомасляной кислотой (Fluka), активированной изопропилхлорформиатом (Aldrich), способом, описанным в публикации Axworthy et al. (2000) Proc. Natl. Acad. Sci. USA 97(4):1802-1807). Реагенты, представляющие собой DOTA-малеимид, взаимодействуют со свободным цистеином в сконструированных антителах с цистеиновой заменой и образуют на антителе лиганд, образующий комплекс с металлом (Lewis et al. (1998) Bioconj. Chem. 9:72-86). Хелатообразующие линкеры, меченные реагентами, такие как DOTA-NHS (моно(N-гидроксисукцинимидоэфир 1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусной кислоты), могут быть приобретены коммерческим путем (Macrocyclics, Dallas, TX). Визуализация рецепторов-мишеней при помощи антител, меченных радиоактивными изотопами, позволяет определить активацию пути в результате обнаружения и количественного определения последовательного накопления антител в опухолевой ткани (Albert et al. (1998) Bioorg. Med. Chem. Lett. 8:1207-1210). Конъюгированные радиоактивные металлы могут оставаться внутри клетки после расщепления лизосом.
Хелатные комплексы металлов, которые могут быть использованы в качестве меток антител в экспериментах по визуализации, описаны в патентах США № 5342606, 5428155, 5316757, 5480990, 5462725, 5428139, 5385893, 5739294, 5750660, 5834456 и публикациях Hnatowich et al. (1983) J. Immunol. Methods 65:147-157; Meares et al. (1984) Anal. Biochem. 142:68-78; Mirzadeh et al. (1990) Bioconjugate Chem. 1:59-65; Meares et al. (1990) J. Cancer1990, Suppl. 10:21-26; Izard et al. (1992) Bioconjugate Chem. 3:346-350; Nikula et al. (1995) Nucl. Med. Biol. 22:387-90; Camera et al. (1993) Nucl. Med. Biol. 20:955-62; Kukis et al. (1998) J. Nucl. Med. 39:2105-2110; Verel et al. (2003) J. Nucl. Med. 44:1663-1670; Camera et al. (1994) J. Nucl. Med. 21:640-646; Ruegg et al. (1990) Cancer Res. 50:4221-4226; Verel et al. (2003) J. Nucl. Med. 44:1663-1670; Lee et al. (2001) Cancer Res. 61:4474-4482; Mitchell, et al. (2003) J. Nucl. Med. 44:1105-1112; Kobayashi et al. (1999) Bioconjugate Chem. 10:103-111; Miederer et al. (2004) J. Nucl. Med. 45:129-137; DeNardo et al. (1998) Clinical Cancer Research 4:2483-90; Blend et al. (2003) Cancer Biotherapy & Radiopharmaceuticals 18:355-363; Nikula et al. (1999) J. Nucl. Med. 40:166-76; Kobayashi et al. (1998) J. Nucl. Med. 39:829-36; Mardirossian et al. (1993) Nucl. Med. Biol. 20:65-74; Roselli et al. (1999) Cancer Biotherapy & Radiopharmaceuticals, 14:209-20.
(b) Флуоресцентные метки, такие как хелаты редкоземельных металлов (хелаты европия), флуоресцеин разных типов, включающие FITC, 5-карбоксифлуоресцеин, 6-карбоксифлуоресцеин, родамин разных типов, включающий TAMRA, данзил, лиссамин, цианины, фикоэритрины, техасский красный и их аналоги. Флуоресцентные метки могут быть конъюгированы с антителами способами, описанными, например, в приведенной выше публикации Current Protocols in Immunology. Флуоресцентные красители и флуоресцентные метки включают реагенты, которые могут быть приобретены коммерческим путем в компаниях Invitrogen/Molecular Probes (Eugene, OR) и Pierce Biotechnology, Inc. (Rockford, IL).
(с) Разные ферментные субстраты, используемые в качестве меток и описанные в научных публикациях (патент США № 4275149). Фермент обычно катализирует химическое изменение хромогенного субстрата, которое может быть измерено разными способами. Например, фермент может катализировать изменение цвета субстрата, которое можно измерить спектрофотометрическими способами. Альтернативно фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Способы количественного определения изменений флуоресценции описаны выше. Хемилюминесцентный субстрат испытывает электронное возбуждение в результате химического взаимодействия и может излучать свет, который может быть измерен (например, при помощи хемилюминометра), или передавать энергию акцептору флуоресценции. Примеры ферментативных меток включают люциферазы (например, люциферазу светляков и бактериальную люциферазу; патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уреазу, пероксидазу, такую как пероксидаза из хрена (HRP), щелочную фосфатазу (АР), β-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы (например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат-дегидрогеназу), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и тому подобные. Способы конъюгации ферментов с антителами описаны в публикации O’Sullivan et al. (1981) “Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay”, in Methods in Enzym. (ed. J. Langone & H. Van Vunakis), Academic Press, New York, 73:147-166.
Примеры комбинаций ферментных субстратов включают, например:
(i) пероксидазу из хрена (HRP) с гидропероксидазой в качестве субстрата, в котором гидропероксидаза окисляет предшественник красителя (например, ортофенилендиамин (OPD) или 3,3',5,5'-тетраметилбензидингидрохлорид (ТМВ));
(ii) щелочную фосфатазу (АР) с паранитрофенилфосфатом в качестве хромогенного субстрата и
(iii) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, паранитрофенил-β-D-галактозидазу) или 4-метилумбеллиферил-β-D-галактозидазу с флуорогенным субстратом.
В данной области существует много других комбинаций ферментных субстратов. Для ознакомления с общим обзором см. патенты США №№ 4275149 и 4318980.
Метка может быть косвенно конъюгирована с боковой цепью аминокислоты, боковой цепью активированной аминокислоты, сконструированным антителом с цистеиновой заменой и тому подобными. Например, антитело может быть конъюгировано с биотином и любая метка из вышеуказанных трех широких категорий может быть конъюгирована с авидином или стрептавидином или наоборот. Биотин избирательно связывается со стрептавидином и, таким образом, метка может быть конъюгирована с антителом таким непрямым способом. Альтернативно для косвенного конъюгирования метки с полипептидным вариантом такой полипептидный вариант конъюгируют с мелким гаптеном (например, дигоксином) и одну из вышеуказанных меток разных типов конъюгируют с полипептидным вариантом против гаптена (например, с антителом против дигоксина). Таким образом, может быть произведено косвенное конъюгирование метки с полипептидным вариантом (Hermanson, G. (1996) in Bioconjugate Techniques Academic Press, San Diego).
Антитело по настоящему изобретению может быть использовано при выполнении любого известного анализа, такого как ELISA, конкурентно-связывающих анализов, прямых и непрямых сэндвич-анализов и анализов способом иммунопреципитации (Zola, (1987) Monoclonal Antibodies: A Manual of Techniques, pp. 147-158, CRC Press, Inc.).
Детектирующая метка может быть использована для локализации, визуализации и количественного определения связывания или распознавания. Меченые антитела по настоящему изобретению могут обнаруживать рецепторы на поверхности клетки. Другим применением меченых антител является способ иммуноулавливания на гранулах, который включает конъюгирование гранулы с антителом, меченным флуоресцентным красителем, и обнаружение сигнала флуоресценции при связывании лиганда. В подобных способах обнаружения связывания использован эффект резонанса поверхностных плазмонов (SPR), позволяющий измерить и обнаружить взаимодействие антитела с антигеном.
Детектирующие метки, такие как флуоресцентные красители и хемилюминесцентные красители (Briggs et al. (1997) “Synthesis of Functionalised Fluorescent Dyes and Their Coupling to Amines and Amino Acids,” J. Chem. Soc., Perkin-Trans. 1:1051-1058), образуют детектируемый сигнал и обычно пригодны для мечения антител, предпочтительно, обладающих следующими свойствами: (i) меченое антитело должно формировать очень сильный сигнал при слабом фоне, чтобы можно было обнаружить небольшие количества антител при выполнении бесклеточных и клеточных анализов; и (ii) меченое антитело должно быть фотохимически устойчивым с тем, чтобы флуоресцентный сигнал можно было обнаружить, контролировать и зарегистрировать без существенного фотообесцвечивания. Метки, используемые в применениях, включающих связывание меченого антитела с мембранами или поверхностями клеток, особенно живых клеток, предпочтительно, (iii) должны обладать хорошей водорастворимостью для достижения эффективных концентраций конъюгата и чувствительности обнаружения и (iv) должны быть нетоксичными для живых клеток, то есть не должны нарушать нормальные процессы обмена в клетках или вызывать гибель недозрелых клеток.
Прямое количественное определение интенсивности флуоресценции клеток и нумерация событий, меченных флуоресцентным красителем, например, связывание на поверхности клетки конъюгатов пептида с красителем, может быть выполнено в системе (FMAT® 8100 HTS System, Applied Biosystems, Foster City, Calif.), которая позволяет автоматически выполнять нерадиоактивные анализы с живыми клетками или гранулами (Miraglia, “Homogeneous cell- and bead-based assays for high throughput screening using fluorometric microvolume assay technology”, (1999) J. of Biomolecular Screening 4:193-204). Меченые антитела также могут быть использованы в анализах связывания рецепторов на поверхности клетки, анализах иммуноулавливания, твердофазных иммуноферментных анализах с флуоресцентным усилением (FLISA), анализах расщепления каспазы (Zheng, “Caspase-3 controls both cytoplasmic and nuclear events associated with Fas-mediated apoptosis in vivo”, (1998) Proc. Natl. Acad. Sci. USA 95:618-23; патент США № 6372907), анализах апоптоза (Vermes, ”A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V” (1995) J. Immunol. Methods 184:39-51) и анализах цитотоксичности. Флуорометрический анализ микрообъема может быть использован для определения увеличения или уменьшения количества молекулы, направленно доставляемой к поверхности клетки (Swartzman, ”A homogeneous and multiplexed immunoassay for high-throughput screening using fluorometric microvolume assay technology”, (1999) Anal. Biochem. 271:143-51).
Меченые антитела по настоящему изобретению могут быть использованы в качестве визуализирующих биомаркеров и зондов в разных способах биомедицинской и молекулярной визуализации, таких как (i) MRI (магнитно-резонансная томография); (ii) MicroCT (компьютерная томография); (iii) SPECT (фотон-эмисионная компьютерная томография); (iv) позитрон-эмиссионная томография (РЕТ) или иммуно-позитрон-эмиссионная томография (иммуно-ПЭТ) (см. публикацию van Dongen GA, et al. “Immuno-PET: a navigator in monoclonal antibody development and applications” Oncologist 2007; 12:1379-89); (v) биолюминесценция; (vi) флуоресценция и (vii) ультразвук. Иммуносцинтиграфия является способом визуализации, в котором животному или человеку вводят антитела, меченные радиоактивными веществами, и регистрируют картину мест локализации антител в организме (патент США № 6528624). Визуализирующие биомаркеры могут быть объективно измерены и оценены в качестве индикатора нормальных биологических процессов, патогенных процессов или фармакологических реакций на терапевтическое воздействие. Биомаркеры могут быть нескольких типов: маркеры типа 0 являются естественными маркерами болезни и коррелируют с известными клиническими индексами, например, с оценкой способом MRI синовиального воспаления в случае ревматоидного артрита; маркеры типа I определяют эффект вмешательства в соответствии с механизмом действия, даже если указанный механизм не связан с клиническим результатом; маркеры типа II прогнозируют клиническую пользу на основании изменений или полученных сигналов для ”обоснования” конечной реакции, такой как измерение эрозии кости при помощи компьютерной томографии в случае ревматоидного артрита. Таким образом, визуализирующие биомаркеры могут предоставлять фармакодинамическую (PD) терапевтическую информацию о: (i) экспрессии белка-мишени, (ii) связывании лекарственного средства с белком-мишенью, то есть избирательности, и (iii) фармакокинетические данные о выведении и времени полужизни в кровотоке. Преимуществами визуализирующих биомаркеров in vivo по сравнению с лабораторными биомаркерами являются: неинвазивное воздействие, количественная оценка всего организма, повторное дозирование и оценка, то есть в течение нескольких временных периодов, и потенциально переносимые результаты доклинического (мелкие животные) на клиническое исследование (человека). В некоторых применениях визуализирующие биомаркеры позволяют отменить или свести к минимуму количество экспериментов на животных в доклинических исследованиях.
Хорошо известны способы мечения пептидов. См. публикации Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, (1997) Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Glazer et al. (1975) Chemical Modification of Proteins. Laboratory Techniques in Biochemistry and Molecular Biology (T. S. Work and E. Work, Eds.) American Elsevier Publishing Co., New York; Lundblad, R. L. and Noyes, C. M. (1984) Chemical Reagents for Protein Modification, Vols. I and II, CRC Press, New York; Pfleiderer, G. (1985) “Chemical Modification of Proteins”, Modern Methods in Protein Chemistry, H. Tschesche, Ed., Walter DeGryter, Berlin and New York; and Wong (1991) Chemistry of Protein Conjugation and Cross-linking, CRC Press, Boca Raton, Fla.); De Leon-Rodriguez et al. (2004) Chem.Eur. J. 10:1149-1155; Lewis et al. (2001) Bioconjugate Chem. 12:320-324; Li et al. (2002) Bioconjugate Chem. 13:110-115; Mier et al. (2005) Bioconjugate Chem. 16:240-237.
Пептиды и белки, меченные двумя частями, флуоресцентным репортером и гасителем, расположенными достаточно близко друг от друга, испытывают перенос энергии резонанса флуоресценции (FRET). Репортерные группы обычно являются флуоресцентными красителями, возбуждаемыми светом при определенной длине волны, которые передают энергию акцепторной или гасящей группе с соответствующим смещением Стоукса для эмиссии при максимальной яркости. Флуоресцентные красители включают молекулы с повышенной ароматичностью, такие как флуоресцеин и родамин, и их производные. Флуоресцентный репортер может быть частично или полностью погашен гасящей частью в интактном пептиде. При расщеплении пептида пептидазой или протеазой может быть измерено детектируемое увеличение флуоресценции (Knight, C. (1995) “Fluorimetric Assays of Proteolytic Enzymes”, Methods in Enzymology, Academic Press, 248:18-34).
Меченые антитела по настоящему изобретению могут быть также использованы в качестве очищающего агента. При выполнении указанного процесса меченое антитело иммобилизуют на твердой подложке, такой как смола сефадекс или фильтровальная бумага, способами, хорошо известными в данной области. Иммобилизованное антитело вводят в соприкосновение с образцом, содержащим очищаемый антиген, после чего подложку промывают соответствующим растворителем, удаляющим по существу все вещество в образце за исключением очищаемого антигена, связанного с иммобилизованным вариантом полипептида. И наконец, подложку промывают другим приемлемым растворителем, таким как глициновый буфер, рН 5,0, который позволяет выделить антиген из полипептидного варианта.
Реагенты, используемые для мечения, обычно содержат реакционноспособную функциональную группу, которая может взаимодействовать (i) непосредственно с тиоловой группой цистеина в сконструированном антителе с цистеиновой заменой с образованием меченого антитела, (ii) с линкером с образованием промежуточного соединения линкера с меткой или (iii) с линкерным антителом с образованием меченого антитела. Реакционноспособные функциональные группы реагентов, используемых для мечения, включают: малеимид, галогенацетил, сукцинимидиловый эфир иодацетамида (например, NHS, N-гидроксисукцинимид), изотиоцианат, сульфонилхлорид, 2,6-дихлортриазинил, пентафторфениловый эфир и фосфорамидит, хотя могут быть также использованы другие функциональные группы.
Характерной реакционноспособной функциональной группой является N-гидроксисукцинимидиловый эфир (NHS) заместителя карбоксильной группы детектируемой метки, например, биотин или флуоресцентный краситель. N-гидроксисукцинимидиловый эфир метки может быть заранее получен, выделен, очищен и/или исследован либо указанный эфир может быть получен in situ и подвергнут взаимодействию с нуклеофильной группой антитела. Карбоксильная форма метки обычно активируется в результате взаимодействия с определенной комбинацией карбодиимида, например, дициклогексилкарбодиимида, диизопропилкарбодиимида, или урония, например, TSTU (тетрафторборат O-(N-сукцинимидил)-N,N,N’,N’-тетраметилурония), HBTU (гексафторфосфат О-бензотриазол-1-ил)-N,N,N',N'-тетраметилурония) или HATU (гексафторфосфат О-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония), активатора, такого как 1-гидроксибензотриазол (HOBt), и N-гидроксисукцинимида с образованием NHS эфира метки. В некоторых случаях метка может быть связана с антителом путем активации метки in situ и осуществления взаимодействия с антителом с образованием конъюгата антитела с меткой в результате выполнения одной стадии. Другие активирующие и связывающие реагенты включают TBTU (гексафторфосфат 2-(1Н-бензотриазо-1-ил)-1,1,3,3-тетраметилурония), TFFH (2-фторгексафторфосфат N,N’,N”,N’’’-тетраметилурония), РуВОР (гексафторфосфат бензотриазол-1-ил-окси-трис-пирролидинофосфония), EEDQ (2-этокси-1-этоксикарбонил-1,2-дигидрохинолин), DCC (дициклогексилкарбодиимид), DIPCDI (диизопропилкарбодиимид), MSNT (1-(мезитилен-2-сульфонил)-3-нитро-1Н-1,2,4-триазол) и арилсульфонилгалогениды, например, триизолпропилбензолсульфонилхлорид.
Альбуминсвязывающие соединения пептида с Fab-фрагментом по настоящему изобретению
В одном из аспектов настоящее изобретение относится к антителу, слитому с альбуминсвязывающим белком. Связывание белка плазмы может быть эффективным способом улучшения фармакокинетических свойств нестойких молекул. Альбумин является широко распространенным белком плазмы. Сывороточные альбуминсвязывающие пептиды (АВР) могут изменять фармакодинамику слитых белков с активным доменом, включая изменение поглощения тканью, проникновение в ткань и диффузию. Указанные фармакодинамические параметры можно модулировать путем специального выбора соответствующей последовательности сывороточного альбуминсвязывающего пептида (заявка на патент США № 20040001827). Целый ряд альбуминсвязывающих пептидов был идентифицирован способом отображения на фаге (Dennis et al. (2002) “Albumin Binding As A General Strategy For Improving The Pharmacokinetics Of Proteins” J. Biol. Chem. 277:35035-35043; WO 01/45746). Соединения по настоящему изобретению включают последовательности АВР, представленные в публикациях: (i) Dennis et al. (2002) J. Biol. Chem. 277:35035-35043, таблицы III и IV, стр. 35038; (ii) заявка на патент США № 20040001827, SEQ ID NO:9-22 и (iii) WO 01/45746, стр. 12-13, которые включены в настоящее описание изобретения в качестве ссылки. Соединения альбуминсвязывающего пептида с Fab-фрагментом получают в результате гибридизации альбуминсвязывающего пептида, например, с С-концом тяжелой цепи Fab-фрагмента в стехиометрическом соотношении 1:1 (1 АВР / 1 Fab-фрагмент). Установлено, что связывание указанных соединений АВР-Fab с альбумином увеличивает время полужизни антитела у кроликов и мышей более чем в 25 раз. Поэтому рассмотренные выше реакционноспособные остатки Cys могут быть введены в указанные соединения АВР-Fab и использованы для сайтспецифического конъюгирования с цитотоксическими лекарственными средствами при выполнении исследований на животных in vivo.
Характерные последовательности альбуминсвязывающих пептидов включают, не ограничиваясь ими, аминокислотные последовательности SEQ ID NO:80-84.
Конъюгаты антитела с лекарственным средством
В другом аспекте настоящее изобретение относится к иммуноконъюгатам или конъюгатам антитела с лекарственным средством (ADC), содержащим антитело, конъюгированное с цитотоксическим агентом, таким как химиотерапевтическое средство, лекарственное средство, ингибитор роста, токсин (например, ферментативно активный токсин бактериального, грибного, растительного или животного происхождения или их фрагменты) или радиоактивный изотоп (то есть, радиоконъюгат). В другом аспекте настоящее изобретение относится к способам использования иммуноконъюгатов. Один из аспектов настоящего изобретения относится к иммуноконъюгату, содержащему любое из вышеуказанных антител против STEAP-1, ковалентно связанное с цитотоксическим агентом или детектируемым агентом.
Применение конъюгатов антитела с лекарственным средством для местной доставки цитотоксических или цитостатических агентов, то есть лекарственных средств, для уничтожения или ингибирования опухолевых клеток при лечении рака (Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drg. Del. Rev. 26:151-172; патент США № 4975278) делает возможной направленную доставку лекарственной части к опухолям и накопление ее внутри клетки, когда системное введение указанных неконъюгированных лекарственных средств может привести к неприемлемым уровням токсичности в нормальных клетках наряду с подлежащими уничтожению опухолевыми клетками (Baldwin et al., (1986) Lancet pp. (Mar. 15, 1986):603-05; Thorpe, (1985) “Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review,” in Monoclonal Antibodies ’84: Biological And Clinical Applications, A. Pinchera et al. (ed.s), pp. 475-506). Таким образом достигается максимальная эффективность при минимальной токсичности. В научной литературе отмечено, что для указанной цели могут быть использованы как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother., 21:183-87). Лекарственные средства, используемые при выполнении указанных способов, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al. (1986), см. выше). Токсины, используемые в конъюгатах антитела с токсином, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al. (2000) Jour. of the Nat. Cancer Inst. 92(19):1573-1581; Mandler et al. (2000) Bioorganic & Med. Chem. Letters 10:1025-1028; Mandler et al. (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (европейский патент № 1391213; Liu et al. (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихеамицин (Lode et al. (1998) Cancer Res. 58:2928; Hinman et al. (1993) Cancer Res. 53:3336-3342). Токсины могут оказывать цитотоксическое и цитостатическое действие в соответствии с механизмами, включающими связывание тубулина, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства становятся неактивными или менее активными при конъюгировании с большими антителами или лигандами рецепторов белка.
Зевалин® (ZEVALIN®, ибритумомаб тиуксетан Biogen/Idec) является конъюгатом антитела с радиоактивным изотопом, состоящим из моноклонального антитела IgG мыши с каппа-цепью, направленного против антигена CD20, обнаруженного на поверхности нормальных и злокачественных В-лимфоцитов, и радиоактивного изотопа 111In или 90Y, связанного хелатообразователем с тиомочевинным линкером (Wiseman et al. (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al. (2002) Blood 99(12):4336-42; Witzig et al. (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al. (2002) J. Clin. Oncol. 20(15):3262-69). Хотя зевалин активно воздействует на В-клеточную неходжкинскую лимфому (NHL), введение указанного средства вызывает у большинства индивидов тяжелую и длительную цитопению. Милотарг™ (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитела с лекарственным средством, состоящий из антитела huCD33, связанного с калихеамицином, был утвержден в 2000 г. для лечения острого миелоидного лейкоза путем инъекций (Drugs of the Future (2000) 25(7):686; патенты США №№ 4970198, 5079233, 5585089, 5606040, 5693762, 5739116, 5767285, 5773001). Кантузумаб мертанзин (Immunogen, Inc.) конъюгат антитела с лекарственным средством, состоящий из антитела huC242, связанного при помощи дисульфидного линкера SPP с мантанзиноидной лекарственной частью, DM1, проходит клинические испытания на стадии II с целью лечения злокачественных новообразований, экспрессирующих CanAg, таких как рак ободочной кишки, поджелудочной железы, желудка и других. MLN-2704 (Millennium Pharm., BZL Biologics, Immunogen Inc.), конъюгат антитела с лекарственным средством, состоящий из моноклонального антитела против простатоспецифического мембранного антигена (PSMA), связанного с майтанзиноидной лекарственной частью, DM1, находится на стадии разработки предположительно для лечения раков предстательной железы. Ауристатиновые пептиды, ауристатин Е (АЕ) и монометилауристатин (ММАЕ), синтетические аналоги доластатина, были конъюгированы с химерными моноклональными антителами cBR96 (специфичным к антигену Левиса Y на карциномах) и сАС10 (специфичным к CD30 на гематологических злокачественных новообразованиях) (Doronina et al. (2003) Nature Biotechnology 21(7):778-784) и в настоящее время находятся на стадии терапевтической разработки.
В настоящем описании изобретения рассмотрены химиотерапевтические средства, используемые для создания иммуноконъюгатов. Ферментативно активные токсины и их фрагменты, которые могут быть использованы для создания иммуноконъюгатов, включают цепь дифтерийного токсина А, несвязывающиеся активные фрагменты дифтерийного токсина, цепь экзотоксина А (из Pseudomonas aeruginosa), цепь рицина А, цепь абрина А, цепь модекцина А, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, заявку WO 93/21232, опубликованную 28 октября 1993 г. Существует целый ряд радионуклидов, используемых для создания радиоконъюгированных антител. Примеры таких радионуклидов включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела с цитотоксическим агентом получают, используя бифункциональные связывающиеся с белком агенты, такие как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как гидрохлорид диметиладипимидата), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(параазидобензоил)гександиамин), производные бисдиазония (такие как бис(парадиазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен в соответствии с описанием, приведенным в публикации Vitetta et al. (1987) Science, 238:1098. Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен-триаминопентауксусная кислота (МХ-DTPA) является типичным хелатообразователем, предназначенным для конъюгирования радионуклеотида с антителом (WO 94/11026).
В объем настоящего изобретения также входят конъюгаты антитела с одним или несколькими низкомолекулярными токсинами, такими как калихеамицин, майтанзиноиды, доластатины, ауристатины, трихотецен, СС1065 и производные указанных токсинов, обладающие активностью токсина.
Майтанзин и майтанзиноиды
В некоторых вариантах осуществления изобретения иммуноконъюгат содержит антитело (полноразмерное антитело или его фрагменты) по настоящему изобретению, конъюгированное с одной или несколькими молекулами майтанзиноида.
Майтанзиноиды являются митотическими ингибиторами, которые ингибируют полимеризацию тубулина. Майтанзин был впервые выделен из восточно-африканского кустарника Maytenus serrata (патент США № 3896111). Затем было обнаружено, что определенные микроорганизмы также продуцируют майтанзиноиды, такие как майтанзинол и сложные эфиры майтанзинола С-3 (патент США № 4151042). Синтетический майтанзинол, его производные и аналоги описаны, например, в патентах США № 4137230, 4248870, 4256746, 4260608, 4265814, 4294757, 4307016, 4308268, 4308269, 4309428, 4313946, 4315929, 4317821, 4322348, 4331598, 4361650, 4364866, 4424219, 4450254, 4362663 и 4371533.
Майтанзиноиды являются весьма привлекательными лекарственными частями для создания конъюгатов антитела с лекарственным средством благодаря (i) относительной доступности для получения путем ферментации или химической модификации, создания производных соединений продуктов ферментации, (ii) возможности создания производных соединений с функциональными группами, пригодными для конъюгирования с антителами при помощи линкеров, не являющихся дисульфидными линкерами, (iii) устойчивости в плазме и (iv) эффективному воздействию на разные линии опухолевых клеток.
Соединения майтанзина, пригодные для использования в качестве майтанзиноидных лекарственных частей, хорошо известны в данной области и могут быть выделены из природных источников известными способами или созданы способами генной инженерии (см. публикацию Yu et al. (2002) PNAS 99:7968-7973), либо майтанзинол и аналоги майтанзинола могут быть синтезированы известными способами.
Характерные майтанзиноидные лекарственные части включают модифицированное ароматическое кольцо, такое как С-19-дехлор (патент США № 4256746) (полученный путем восстановления ансамитоцина Р2 алюмогидридом лития); С-20-гидрокси (или С-20-деметил) +/-С-19-дехлор (патенты США №№ 4361650 и 4307016) (полученный путем деметилирования с использованием Streptomyces или Actinomyces или дехлорирования с использованием LAH) и С-20-деметокси, С-20-ацилокси (-OCOR), +/-дехлор (патент США № 4294757) (полученный путем ацилирования с использованием ацилхлоридов), и части, модифицированные в других положениях.
Типичные майтанзиноидные лекарственные части также включают такие модификации как: С-9-SH (патент США № 4424219) (полученный в результате взаимодействия майтанзинола с H2S или P2S5), С-14-алкоксиметил (деметокси/CH2OR) (патент США № 4331598), С-14-гидроксиметил или ацилоксиметил (СН2ОН или СН2ОАс) (патент США № 4450254) (полученный из Nocardia), C-15-гидрокси/ацилокси (патент США № 4364866) (полученный в результате конверсии майтанзинола видом Streptomyces), С-15-метокси (патенты США №№ 4313946 и 4315929) (выделенный из Trewia nudlflora), С-18-N-деметил (патенты США №№ 4362663 и 4322348) (полученный путем деметилирования майтанзинола видом Streptomyces) и 4,5-дезокси (патент США № 4371533) (полученный путем восстановления майтанзинола трихлоридом титана/LAH).
Характерные варианты майтанзиноидных лекарственных частей включают: DM1, DM3 и DM4, имеющие структуры:
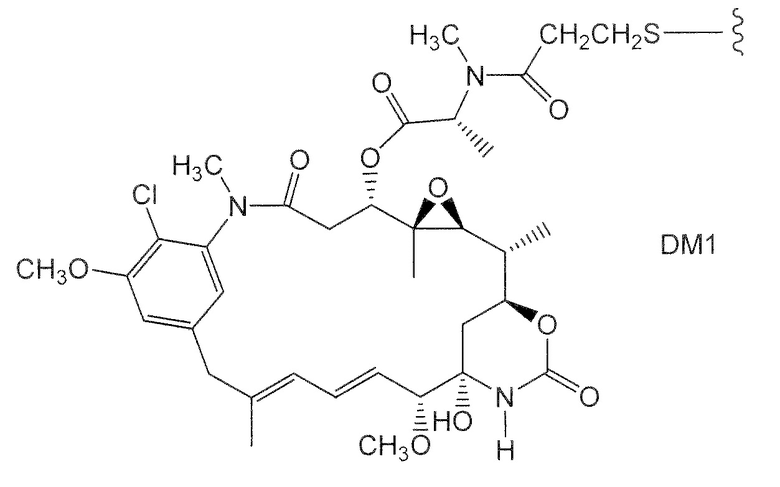
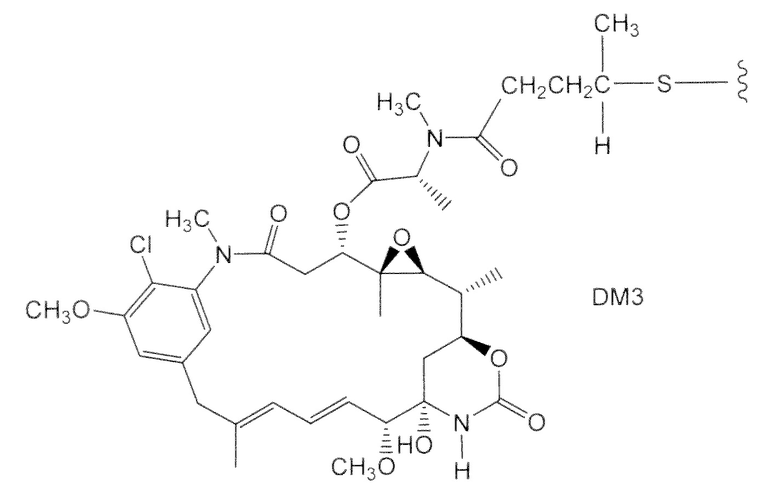

где волнистой линией показано ковалентное присоединение атома серы лекарственной части к линкеру (L) конъюгата антитела с лекарственным средством. В научной литературе описан герцептин® (HERCEPTIN®, трастузумаб), присоединенный при помощи SMCC к DM1 (публикация WO 2005/037992, которая включена в настоящее описание изобретения в качестве ссылки в полном объеме). Конъюгат антитела с лекарственным средством по настоящему изобретению может быть получен способами, рассмотренными в настоящем описании изобретения.
Другие типичные конъюгаты антитела с майтанзиноидом имеют следующие структуры и обозначены соответствующими аббревиатурами (где Ab означает антитело и р является числом от 1 до около 8):
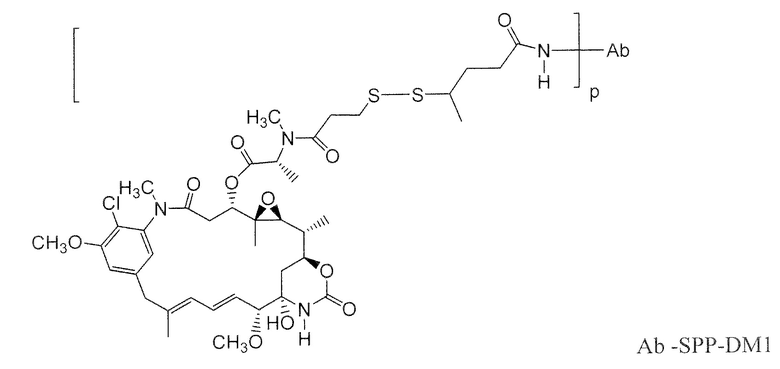
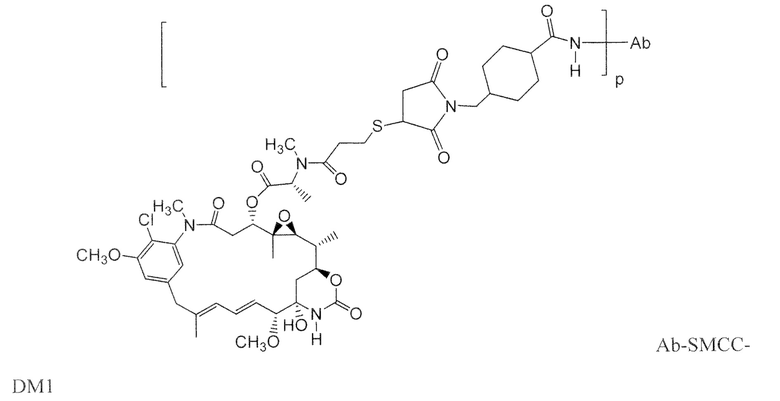
Типичные конъюгаты антитела с лекарственным средством, в которых DM1 присоединен при помощи линкера BMPEO к тиоловой группе антитела, имеют нижеследующую структуру и обозначены соответствующими аббревиатурами:
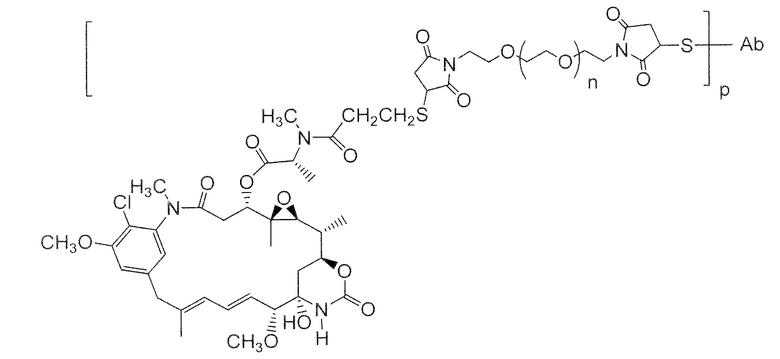
где Ab означает антитело; n равно 0, 1 или 2 и р равно 1, 2, 3 или 4.
Иммуноконъюгаты, содержащие майтанзиноиды, способы получения указанных иммуноконъюгатов и их терапевтическое применение описаны, например, в патентах США №№ 5208020, 5416064, 6441163 и в европейском патенте № 0425235 В1, которые включены в настоящее описание изобретения в качестве ссылки. В публикации Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описаны иммуноконъюгаты, содержащие майтанзиноид, обозначенный DM1, который связан с моноклональным антителом С242 против колоректального рака человека. Было установлено, что указанный конъюгат является высокотоксичным для культивируемых клеток рака ободочной кишки и обладает противоопухолевой активности при выполнении анализа роста опухоли in vivo. В публикации Chari et al., Cancer Research 52: 127-131 (1992) описаны иммуноконъюгаты, в которых майтанзиноид был конъюгирован при помощи дисульфидного линкера с антителом мыши А7, связывающимся с антигеном линий клеток рака ободочной кишки человека, или с другим мышиным моноклональным антителом ТА.1, связывающимся с онкогеном HER-2/neu. Цитотоксичность конъюгата антитела ТА.1 с майтанзиноидом исследовали in vitro с использованием линии клеток рака молочной железы человека SK-BR-3, экспрессирующей 3×105 антигенов HER-2 на поверхности каждой клетки. Конъюгат с лекарственным средством достигал такой же степени цитотоксичности, что и лекарственное средство на основе свободного майтанзиноида, которая могла быть повышена путем увеличения числа молекул майтанзиноида на молекулу антитела. Конъюгат антитела А7 с майтанзиноидом характеризовался низкой системной токсичностью у мышей.
Конъюгаты антитела против STEAP-1 с майтанзиноидом получают путем химического связывания антитела с молекулой майтанзиноида без существенного ослабления биологической активности антитела или молекулы майтанзиноида. См., например, патент США № 5208020 (который включен в настоящее описание изобретения в качестве ссылки в полном объеме). Цитотоксичность в отношении клеток-мишеней была увеличена без отрицательного влияния на функцию или растворимость антитела при использовании в среднем 3-4 молекул майтанзиноида на одну молекулу антитела, хотя, предположительно, даже одна молекула токсина/антитела повышает токсичность по сравнению с использованием “голого” антитела. Майтанзиноиды хорошо известны в данной области и могут быть синтезированы известными способами или получены из природных источников. Приемлемые майтанзиноиды описаны, например, в патенте США № 5208020, в других патентах и непатентных публикациях, на которые была сделана ссылка выше. Предпочтительными майтанзиноидами являются майтанзинол и аналоги майтанзинола, модифицированные в ароматическом кольце или в других положениях молекулы майтанзинола, такие как разные сложные эфиры майтанзинола.
В данной области известно много связующих групп, используемых для получения конъюгатов антитела с майтанзиноидом, которые включают, например, связующие группы, описанные в патентах США № 5208020, 6441163 или в европейском патенте № 0425235 В1, в публикации Chari et al., Cancer Research 52:127-131 (1992) и заявке на патент США 2005/0169933 А1, которые включены в настоящее описание изобретения в качестве ссылки в полном объеме. Конъюгаты антитела с майтанзиноидом, содержащие линкер SMCC, могут быть получены способом, описанным в заявке на патент США № 11/141344, поданной 31 мая 2005 г, “Antibody Drug Conjugates and Methods”. Связующие группы включают дисульфидные группы, тиоэфирные группы, кислотолабильные группы, фотолабильные группы, пептидазалабильные группы или эстеразалабильные группы, описанные в вышеуказанных патентах. В настоящем описании изобретения представлены дополнительные связующие группы.
Конъюгаты антитела с майтанзиноидом могут быть получены при использовании целого ряда агентов, связывающих бифункциональные белки, такие как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как гидрохлорид диметиладипимидата), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бисазидосоединения (такие как бис(параазидобензоил)-гександиамин), производные бисдиазония (такие как бис(парадиазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Особенно предпочтительными связующими агентами являются N-сукцинимидил-3-(2-пиридилдитио)-пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP), образующие дисульфидную связь.
Линкер может быть присоединен к молекуле майтанзиноида в разных положениях в зависимости от типа связи. Например, сложноэфирная связь может быть образована в результате осуществления взаимодействия с гидроксильной группой стандартными способами сочетания. Указанное взаимодействие может происходить в положении С-3, имеющего гидроксильную группу, положении С-14, модифицированного гидроксиметилом, положении С-15, модифицированного гидроксильной группой, и положении С-20, содержащего гидроксильную группу. В предпочтительном варианте осуществления изобретения указанная связь образована в положении С-3 майтанзинола или аналога майтанзинола.
В одном из вариантов осуществления изобретения любое антитело по настоящему изобретению (полноразмерное или его фрагмент) конъюгировано с одной или несколькими молекулами майтанзиноида. В одном из вариантов иммуноконъюгата цитотоксический агент D является майтанзиноидом DM1. В одном из вариантов иммуноконъюгата линкер выбран из группы, состоящей из SPDP, SMCC, IT, SPDP и SPP.
Ауристатины и долостатины
В некоторых вариантах осуществления изобретения иммуноконъюгат содержит антитело по настоящему изобретению, конъюгированное с доластатинами или пептидными аналогами и производными долостатина, ауристатинами (патенты США №№ 5635483, 5780588). Установлено, что доластатины и ауристатины нарушают динамику микротрубочек, препятствуют гидролизу GTP и делению ядер и клеток (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают противораковой (патент США № 5663149) и противогрибковой активностью (Pettit et al. (1998) Antimicrob. Agents Chemother. 42:2961-2965). Доластатиновая или ауристатиновая лекарственная часть может быть присоединена к антителу у N (амино) конца или С (карбоксильного) конца пептидной лекарственной части (WO 02/088172).
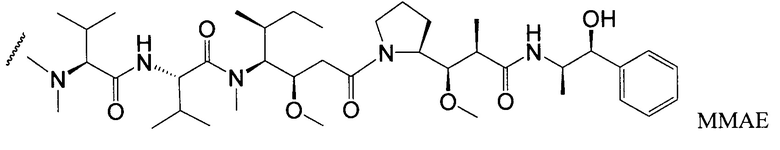
Типичные варианты ауристатина включают связанные с N-концом монометилауристатиновые лекарственные части DE и DF, описанные в публикации ”Senter et al., Proceedings of the American Association for Cancer Research, Volume 45, Abstract Number 623, presented Match 28, 2004, которая включена в настоящее описание изобретения в качестве ссылки в полном объеме.
Типичным вариантом ауристатина является ММАЕ (в котором волнистой линией показано положение ковалентного присоединения к линкеру (L) конъюгата антитела с лекарственным средством).
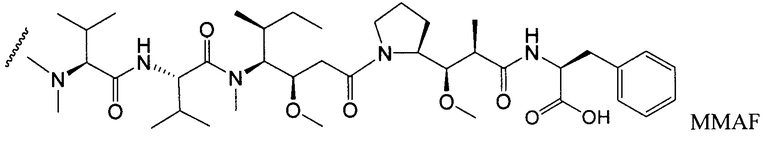
Другим типичным вариантом ауристатина является MMAF, в котором волнистой линией показано положение ковалентного присоединения к линкеру (L) конъюгата антитела с лекарственным средством (заявка на патент США 2005/0238649).
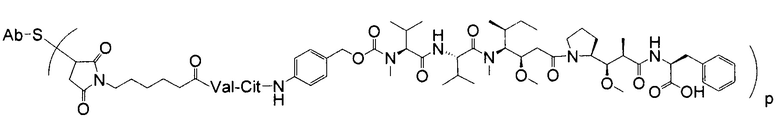
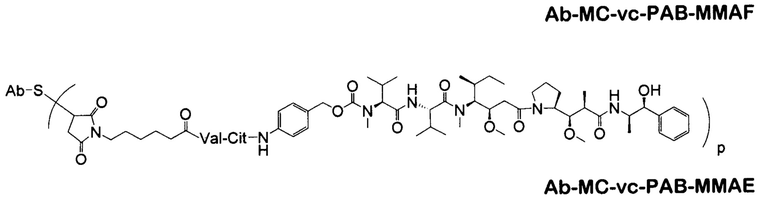
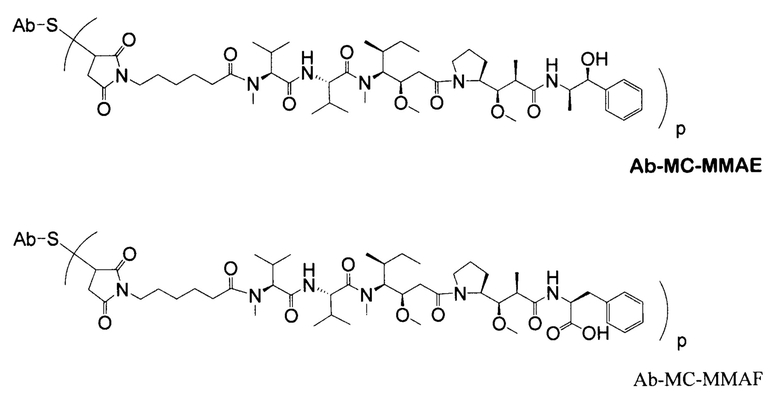
Дополнительные типичные варианты, включающие ММАЕ или MMAF и разные линкеры (описанные выше), имеют следующие структуры и обозначены соответствующими аббревиатурами (где Ab означает антитело и р является числом от 1 до около 8).
Лекарственные части на основе пептида могут быть получены путем образования пептидной связи между двумя или более аминокислотами и/или фрагментами пептидов. Такие пептидные связи могут быть получены, например, способом синтеза в жидкой фазе (см. публикацию Schroder and K. Lubke, “The Peptides”, volume 1, pp. 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Ауристатиновые/доластатиновые лекарственные части могут быть получены способами, описанными в патентах США №№ 5635483, 5780588 и публикациях Pettit et al. (1989) J. Am. Chem. Soc. 111:5463-5465, Pettit et al. (1998) Anti-Cancer Drug Design 13: 243-277, Pettit, G.R., et al. Synthesis, 1996, 719-725, Pettit et al. (1996) J. Chem. Soc. Perkin Trans. 15:859-863, и Doronina (2003) Nat. Biotechnol. 21(7):778-784.
Калихеамицин
В других вариантах осуществления изобретения иммуноконъюгат содержит антитело по настоящему изобретению, конъюгированное с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицинов способны разрывать двухцепочечную ДНК в субпикомолярных концентрациях. Получение конъюгатов с семейством калихеамицинов описано в патентах США №№ 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все указанные патенты принадлежат American Cyanamid Company). Структурные аналоги калихеамицина, которые могут быть использованы в настоящем изобретении, включают, не ограничиваясь ими, γ1I, α2I, α3I, N-ацетил-γ1I, PSAG и θI1 (Hinman et al., Cancer Research 53:3336-3342 (1993), Lode et al., Cancer Research 58:2925-2928 (1998) и вышеуказанные патенты США, принадлежащие компании American Cyanamid). Другим противоопухолевым средством, с которым может быть конъюгировано антитело, является QFA, представляющее собой антифолат. Как калихеамицин, так и QFA действуют внутри клетки и плохо проникают через плазматическую мембрану. Поэтому поглощение клеткой указанных средств благодаря опосредованной антителом интернализации значительно усиливает их цитотоксическое действие.
Другие цитотоксические агенты
Другие противоопухолевые средства, которые могут быть конъюгированы с антителами по настоящему изобретению, включают BCNU, стрептозоицин, винкристин и 5-фторурацил, семейство агентов, известных как комплекс LL-E33288, описанный в патентах США №№ 5053394, 5770710, а также эсперамицины (патент США № 5877296).
Ферментативно активные токсины и их фрагменты, которые могут быть использованы в настоящем изобретении, включают цепь дифтерийного токсина А, несвязывающиеся активные фрагменты дифтерийного токсина, цепь экзотоксина А (из Pseudomonas aeruginosa), цепь рицина А, цепь абрина А, цепь модекцина А, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, заявку WO 93/21232, опубликованную 28 октября 1993 г.
Настоящее изобретение далее относится к иммуноконъюгату, образованному антителом и соединением с нуклеолитической активностью (например, рибонуклеазой или ДНК-эндонуклеазой, такими как дезоксирибонуклеаза; ДНКаза).
Для избирательного разрушения опухоли антитело может включать радиоактивный атом. Существует целый ряд радиоактивных изотопов, предназначенных для создания радиоконъюгированных антител. Примеры таких радиоактивных изотопов включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Конъюгат, используемый для обнаружения, может включать радиоактивный атом для сцинтиграфического исследования, например, tc99m или I123, или спиновую метку для ядерно-магнитно-резонансной (ЯМР) томографии (также известной как магнитно-резонансная томография, MRI), такую как иод-123, иод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки могут быть введены в конъюгат известными способами. Например, можно синтезировать пептид способом биосинтеза или химического аминокислотного синтеза с использованием соответствующих предшественников аминокислот, включающих, например, фтор-19 вместо водорода. Такие метки как tc99m или I123, Re186, Re188 и In111 могут быть присоединены при помощи остатка цистеина в пептиде. Иттрий-90 может быть присоединен при помощи остатка лизина. Иод-123 может быть введен способом IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80:49-57. В публикации “Monoclonal Antibodies in Immunoscintigraphy” (Chatal, CRC Press, 1989) подробно описаны другие способы.
Конъюгаты антитела с цитотоксическим агентом могут быть получены при использовании разных агентов, связывающих бифункциональный белок, таких как N-сукцинимидил-3-(2-пиридилтио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как гидрохлорид диметиладипимидата), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бисазидосоединения (такие как бис(параазидобензоил)-гександиамин), производные бисдиазония (такие как бис(парадиазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен в соответствии с описанием, приведенным в публикации Vitella et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминопентауксусная кислота (МХ-DTPA) является типичным хелатообразователем, предназначенным для конъюгирования радионуклеотида с антителом. См. WO 94/11026. Линкер может быть ”расщепляемым линкером”, облегчающим высвобождение цитотоксического средства в клетку. Например, может быть использован кислотолабильный линкер, пептидаза-чувствительный линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Research 52:127-131 (1992); патент США № 5208020).
Соединения по настоящему изобретению включают, не ограничиваясь им, конъюгат антитела с лекарственным средством (ADC), полученный с использованием перекрестносшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, сульфо-SMPB и SVSB (сукцинимидил(4-винилсульфон)-бензоат), которые можно приобрести коммерческим путем (например, в компании Pierce Biotechnology, Inc., Rockford, IL., U.S.A.). См. страницы 467-498 Справочника и каталога заявок за 2003-2004 гг.
Получение конъюгатов антитела с лекарственным средством
В конъюгатах антитела с лекарственным средством (ADC) по настоящему изобретению антитело (Ab) конъюгировано с одной или несколькими лекарственными частями (D), например, примерно с 1-20 лекарственными частями на одно антитело, при помощи линкера (L). В одном из вариантов осуществления изобретения число лекарственных частей (D) на одно антитело составляет от около 1 до около 5, альтернативно от около 2 до около 6, альтернативно от около 2 до около 5, альтернативно от около 3 до около 4 лекарственных частей на одно антитело. Поскольку число лекарственных частей на одно антитело обычно является средним числом для всех конъюгатов в совокупности конъюгатов антитела с лекарственным средством, то число лекарственных частей на одно антитело может не быть целым числом. ADC формулы I можно получить несколькими способами с использованием реакций органической химии, условий и реагентов, известных специалистам в данной области, которые включают: (1) взаимодействие нуклеофильной группы антитела с двухвалентным линкером с образованием Ab-L, связанного ковалентной связью, и последующее взаимодействие с лекарственной частью D и (2) взаимодействие нуклеофильной группы лекарственной части с двухвалентным линкером с образованием D-L, связанного ковалентной связью, и последующее взаимодействие с нуклеофильной группой антитела. В настоящем описании изобретения представлены дополнительные способы получения ADC.
Ab-(L-D)р (формула I)
Линкер может состоять из одного или нескольких компонентов. Типичные компоненты линкера включают 6-малеимидокапроил (“MC”), малеимидопропаноил (“MP”), валин-цитруллин (“val-cit”), аланин-фенилаланин (“ala-phe”), парааминобензилоксикарбонил (“PAB”), N-сукцинимидил-4-(2-пиридилтио)пентаноат (“SPP”), N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (“SMCC”) и N-сукцинимидил(4-иодацетил)аминобензоат (“SIAB”). В одном из вариантов осуществления изобретения линкер представляет собой валин-цитруллин-парааминобензилоксикарбонил (“vc-PAB”). В данной области известны дополнительные компоненты линкеров, и некоторые из них рассмотрены в настоящем описании изобретения.
В некоторых вариантах осуществления изобретения линкер может содержать аминокислотные остатки. Типичные аминокислотные компоненты линкера включают дипептид, трипептид, тетрапептид или пентапептид. Типичные дипептиды включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Типичные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, являющиеся аминокислотным компонентом линкера, включают природные аминокислоты, а также минорные аминокислоты и не встречающиеся в природе аналоги аминокислот, такие как цитруллин. Могут быть созданы аминокислотные компоненты линкера с оптимизированной избирательностью ферментативного расщепления определенными ферментами, например, опухолеспецифической протеазой, катепсином В, С и D или плазминпротеазой.
Ниже показаны структуры типичных компонентов линкера (где волнистой линией показаны места ковалентного присоединения к другим компонентам ADC):
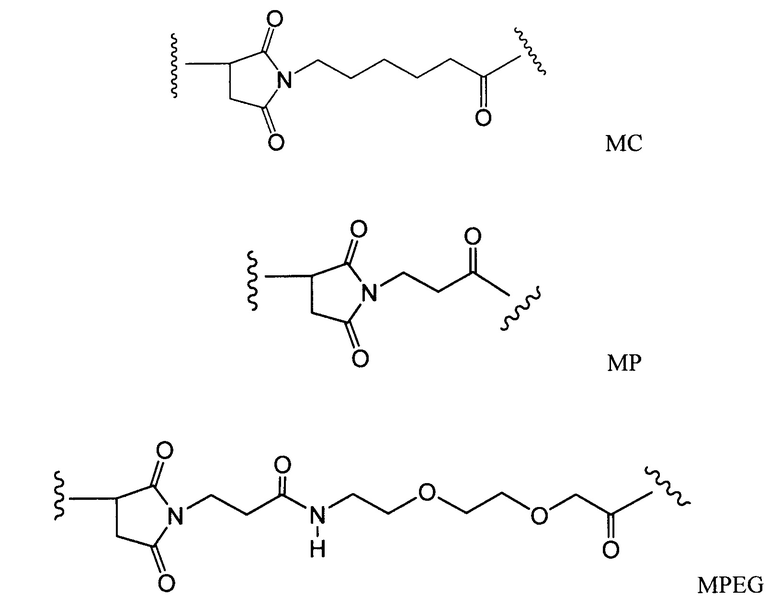
Дополнительные типичные компоненты линкера и обозначающие их аббревиатуры включают (где показаны антитело (Ab) и линкер и р является числом от 1 до около 8):
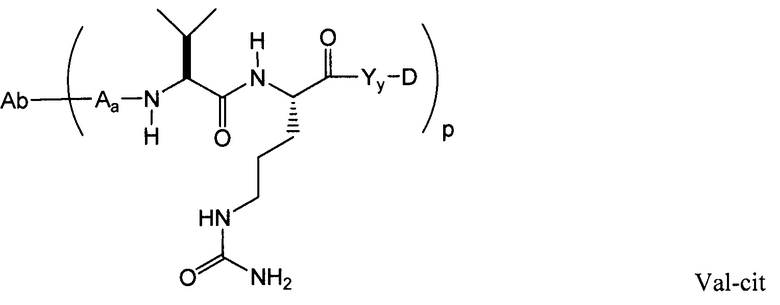
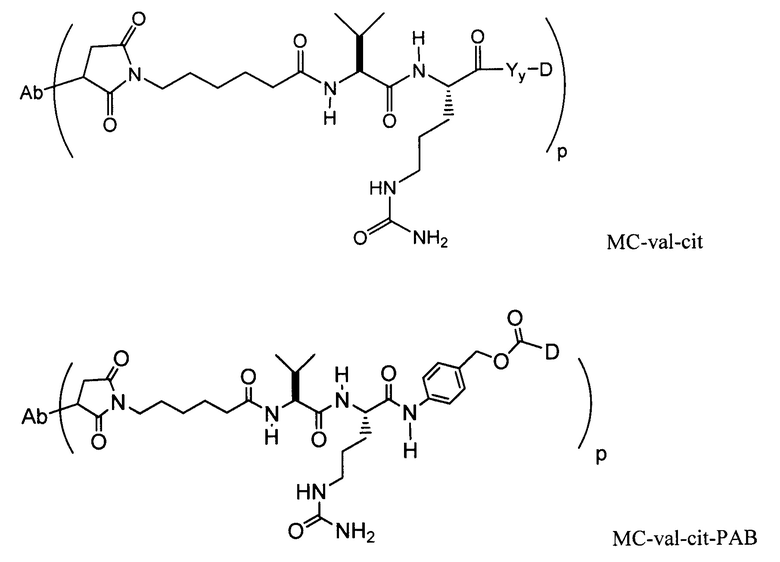
Нуклеофильные группы антител включают, не ограничиваясь ими: (i) N-концевые аминогруппы, (ii) аминогруппы боковой цепи, например, лизин, (iii) тиоловые группы боковой цепи, например, цистеин, и (iv) гидроксильные или аминогруппы сахара, когда антитело является гликозилированным. Аминогруппы, тиоловые и гидроксильные группы являются нуклеофильными группами, способными взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных частей и линкерных реагентов, включающих: (i) активные сложные эфиры, такие как HNS эфиры, HOBt эфиры, галогенформиаты и галогенангидриды кислот, (ii) алкилгалогениды и бензилгалогениды, такие как галогенацетамиды, (iii) альдегиды, кетоны, карбоксильные группы и группы имида малеиновой кислоты. Определенные антитела имеют восстанавливаемые межцепные дисульфидные связи, то есть цистеиновые мостики. Антителам может быть сообщена реакционная способность для конъюгирования с линкерными реагентами путем обработки восстановителем, таким как DTT (дитиотреитол). Таким образом, каждый цистеиновый мостик теоретически будет образовывать две реакционноспособные тиоловые нуклеофильные группы. Дополнительные нуклеофильные группы могут быть введены в антитела в результате осуществления взаимодействия лизинов с 2-иминотиоланом (реагент Траута), вследствие чего амин превращается в тиол. Реакционноспособные тиоловые группы могут быть введены в антитело (или его фрагмент) путем введения одного, двух, трех, четырех или более остатков цистеина (например, при получении мутантных антител, включающих один или несколько аминокислотных остатков цистеина).
Конъюгаты антитела с лекарственным средством по настоящему изобретению могут быть также получены в результате модификации антитела путем введения электрофильных частей, способных взаимодействовать с нуклеофильными заместителями в линкерном реагенте или лекарственном средстве. Сахара гликозилированных антител могут быть окислены, например, такими оксидантами как периодат, с образованием альдегидных или кетоновых групп, способных взаимодействовать с аминогруппой линкерных реагентов или лекарственных частей. Полученные иминогруппы шиффова основания могут образовывать устойчивую связь или могут быть восстановлены, например, борогидридом, с образованием устойчивых аминовых связей. В одном из вариантов осуществления изобретения в результате взаимодействия углеводной части гликозилированного антитела с галактозооксидазой или метапериодатом натрия в белке могут быть получены карбонильные (альдегидные или кетоновые) группы, способные взаимодействовать с соответствующими группами лекарственного средства (Hermanson, Bioconjugate Techniques). В другом варианте осуществления изобретения белки, содержащие N-концевые остатки серина или треонина, могут взаимодействовать с метапериодатом натрия с образованием альдегида вместо первой аминокислоты (Geoghegan & Stroh (1992) Bioconjugate Chem. 3:138-146; патент США № 5362852). Такой альдегид может быть подвергнут взаимодействию с лекарственной частью или нуклеофильной группой линкера.
Аналогичным образом нуклеофильные группы лекарственной части включают, не ограничиваясь ими, амин, тиол, гидроксил, гидразид, оксим, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид, способные взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных частей и линкерных реагентов, включающих: (i) активные эфиры, такие как HNS эфиры, HOBt эфиры, галогенформиаты и галогенангидриды кислот, (ii) алкилгалогениды и бензилгалогениды, такие как галогенацетамиды, (iii) альдегиды, кетоны, карбоксильные группы и группы имида малеиновой кислоты.
Способы конъюгирования линкерных и лекарственных частей с белками, направленно воздействующими на клетки, такими как антитела, иммуноглобулины или их фрагменты, описаны, например, в патентах США № 5208020 и 6441163, заявках на патент WO2005037992, WO2005081711 и WO2006/034488, которые включены в настоящее описание изобретения в качестве ссылки в полном объеме.
Альтернативно слитый белок, содержащий антитело и цитотоксический агент, может быть получен, например, способами рекомбинантных ДНК или синтеза пептидов. ДНК может включать соответствующие области, кодирующие две части конъюгата, расположенные рядом друг с другом или отделенные областью, кодирующей линкерный пептид, который не ухудшает требуемые свойства конъюгата.
В еще одном варианте осуществления изобретения антитело может быть конъюгировано с ”рецептором” (таким как стрептавидин) для предварительного направленного воздействия на опухоль, когда индивиду вводят конъюгат антитела с рецептором, затем удаляют несвязанный конъюгат из кровотока при помощи очищающего агента и затем вводят ”лиганд” (например, авидин), конъюгированный с цитотоксическим агентом (например, радионуклеотидом).
В одном из вариантов иммуноконъюгата цитотоксический агент D является ауристатином формулы DE или DF
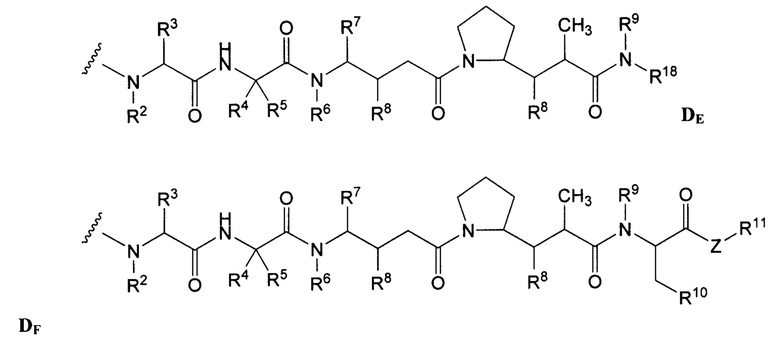
где R2 и R6 означают метил, R3 и R4 означают изопропил, R7 означает втор-бутил, каждый R8 независимо выбирают из СН3, О-СН3, ОН и Н; R9 означает Н; R10 означает арил; Z означает -О- или -NH-; R11 означает Н, С1-С8 алкил или -(СН2)2-О-(СН2)2-О-(СН2)2-О-СН3; и R18 означает -С(R8)2-C(R8)2-арил; и
(d) р является числом от около 1 до 8.
Нижеследующие варианты осуществления изобретения относятся к любому из вышеуказанных иммуноконъюгатов. В одном из вариантов осуществления изобретения иммуноконъюгат обладает киллерной активностью в отношении клеток in vitro или in vivo. В одном из вариантов осуществления изобретения линкер присоединен к антителу при помощи тиоловой группы антитела. В одном из вариантов осуществления изобретения линкер расщепляется протеазой. В одном из вариантов осуществления изобретения линкер включает дипептид val-cit. В одном из вариантов осуществления изобретения линкер включает парааминобензильное звено. В одном из вариантов осуществления изобретения парааминобензильное звено расположено между лекарственным средством и сайтом расщепления протеазой в линкере. В одном из вариантов осуществления изобретения парааминобензильное звено является парааминобензилоксикарбонилом (РАВ). В одном из вариантов осуществления изобретения линкер включает 6-малеимидокапроил. В одном из вариантов осуществления изобретения 6-малеимидокапроил расположен между антителом и сайтом расщепления протеазой в линкере. Вышеуказанные варианты осуществления изобретения могут быть осуществлены отдельно или в любой комбинации друг с другом.
В одном из вариантов осуществления изобретения лекарственное средство выбирают из ММАЕ и MMAF. В одном из вариантов осуществления изобретения иммуноконъюгат имеет формулу:

где Ab означает любое из вышеуказанных антител против STEAP-1, S означает атом серы и р является числом от около 2 до около 5. В одном из вариантов осуществления изобретения иммуноконъюгат имеет формулу:
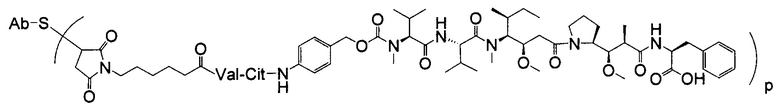
где Ab означает любое из вышеуказанных антител против STEAP-1, S означает атом серы и р является числом от около 1 до около 6, от около 2 до около 5, от около 2 до около 6, от около 2 до около 4, от около 2 до около 3, от около 3 до около 4, от около 3 до около 5, от около 3 до около 6 или от около 4 до около 6.
Способы визуализации меченого антитела
В другом варианте осуществления изобретения сконструированные антитела с цистеиновой заменой могут быть помечены с помощью тиоловой группы цистеина, содержащей радионуклиды, флуоресцентными красителями, субстратами, запускающими биолюминесценцию, субстратами, запускающими хемилюминесценцию, ферментами и другими детектируемыми метками в экспериментах по визуализации в диагностических, фармакодинамических и терапевтических применениях. Меченое сконструированное антитело с цистеиновой заменой, то есть ”биомаркер” или “зонд”, вводят при помощи инъекции, вливания или приема внутрь в живой организм, например, человеку, грызуну или другому мелкому животному, в перфузированный орган или образец ткани. Перемещение зонда обнаруживают через определенный период времени и представляют в виде изображения.
Реагенты, производящие мечение цирконием
В одном из вариантов осуществления изобретения создание иммуно-ПЭТ на основе 89Zr позволило измерять экспрессию мишени и биораспределение антитела у мышей и людей (Dijkers et al., Nature Vol. 87 Number 5 (May 2010)). Иммуно-ПЭТ в качестве инструмента для измерения экспрессии мишени и биораспределения антитела имеет важное значение в определении распределения мишеней ADC в местах первичного и метастазирующего поражений, особенно в случае рака предстательной железы и других злокачественных новообразований.
Радиоактивный изотоп или другие метки могут быть введены в конъюгат известными способами (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80:49-57; “Monoclonal Antibodies in Immunoscintigraphy” Chatal, CRC Press (1989)). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминопентауксусная кислота (МХ-DTPA) является типичным хелатообразователем, используемым для конъюгирования радионуклида с антителом (WO 94/11026).
Изделия
Еще один вариант осуществления изобретения относится к изделию или ”набору”, содержащему вещества, используемые для лечения вышеописанных нарушений. Изделие включает емкость и этикетку на емкости или вкладыш, прилагаемый к емкости. Приемлемые емкости включают, например, пузырьки, ампулы, шприцы, блистерную упаковку и т.д. Указанные емкости могут быть изготовлены из разных материалов, таких как стекло или пластик. В емкости со стерильным доступом (например, емкость может представлять собой мешок для внутривенного раствора или пузырек с пробкой, проницаемой для иглы для подкожных инъекций) находится конъюгат антитела с лекарственным средством (ADC), позволяющий эффективно лечить указанное состояние. По меньшей мере одним активным агентом в композиции является ADC. На этикетке или вкладыше должно быть указано, что данная композиция предназначена для лечения соответствующего состояния, такого как рак. Альтернативно или дополнительно изделие может включать вторую (или третью) емкость, содержащую фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), физиологический раствор с фосфатным буфером, раствор Рингера и раствор декстрозы. Изделие может далее включать другие материалы, требуемые с коммерческой точки зрения и с точки зрения потребителей, которые включают буферы, разбавители, фильтры, иглы и шприцы.
Фармацевтические композиции
В одном из аспектов настоящее изобретение относится к фармацевтической композиции, содержащей любой из вышеуказанных иммуноконъюгатов и фармацевтически приемлемый носитель. В одном из аспектов настоящее изобретение относится к способу лечения пролиферативного нарушения клеток предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или саркомы Юинга, который включает введение индивиду фармацевтической композиции. В одном из вариантов осуществления изобретения рак предстательной железы, легкого, ободочной кишки и яичника и саркома Юинга представляют собой метастазирование первичного рака предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или саркомы Юинга. В одном из вариантов осуществления изобретения пролиферативное нарушение клеток связано с повышенной экспрессией STEAP-1 на поверхности клетки.
В одном из аспектов настоящее изобретение относится к способу ингибирования пролиферации клетки, который включает воздействие на клетку любого из вышеуказанных иммуноконъюгатов в условиях, обеспечивающих связывание иммуноконъюгата с STEAP-1. В одном из вариантов осуществления изобретения клетка предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или саркомы Юинга является опухолевой клеткой. В одном из вариантов осуществления изобретения указанная опухолевая клетка является опухолевой клеткой предстательной железы, легкого, ободочной кишки, мочевого пузыря или яичника, или клеткой саркомы Юинга у млекопитающего с наличием или с подозрением на наличие пролиферативного нарушения предстательной железы, легкого, ободочной кишки, мочевого пузыря или саркомы Юинга, которое включает, не ограничиваясь им, метастазирование первичной раковой опухоли предстательной железы, легкого, ободочной кишки, мочевого пузыря или саркомы Юинга. В одном из вариантов осуществления изобретения клетка предстательной железы, легкого, ободочной кишки, мочевого пузыря или саркомы Юинга является ксенотрансплантатом. В одном из вариантов осуществления изобретения воздействие производят in vitro. В одном из вариантов осуществления изобретения воздействие производят in vivo.
В одном из аспектов настоящее изобретение относится к способу использования антитела против STEAP-1 по настоящему изобретению, который включает анализ растворимого в сыворотке STEAP-1, полученного у млекопитающего, страдающего пролиферативным нарушением клеток предстательной железы, легкого или ободочной кишки (или метастазированием первичной опухоли), измерение клинического прогрессирования или регрессии указанных заболеваний, или оценку опухолевой массы, или рецидива.
Краткое описание чертежей
В файле с данными о патентах или заявках находится по меньшей мере один чертеж, выполненный в цвете. Копии публикации данного патента или заявки на патент с цветными чертежами будут предоставлены по требованию и после оплаты необходимых пошлин.
На фигуре 1 изображена аминокислотная последовательность STEAP-1 человека (SEQ ID NO:1) в сравнении с аминокислотными последовательностями STEAP-1 мыши и обезьяны циномолгус (cyno) (соответственно, SEQ ID NO:2 и 3). Внеклеточные домены 1, 2 и 3 отмечены темными рамками.
Фигуры 2А-2В. На фигуре 2А изображена аминокислотная последовательность вариабельной области легкой цепи антитела мыши 120.545 против STEAP-1 в сравнении с последовательностью химерного антитела (120 chimera) и гуманизированного антитела (120 graft) и в сравнении с последовательностью подгруппы III человека. CDR-области указаны в рамках (CDR-L1, CDR-L2 и CDR-L3). Последовательности, охватывающие CDR-области, являются последовательностями остовных областей (FR-L1-FR-L4). Последовательности пронумерованы в соответствии с нумерацией Kabat. CDR-области по системе Kabat, Chotia и контактной системе указаны над CDR-областями в рамках. На фигуре 2В изображена аминокислотная последовательность вариабельной области тяжелой цепи антитела мыши против STEAP-1 (120.545) в сравнении с химерным антителом (120 chimera) и гуманизированным антителом (120 graft) и в сравнении с последовательностью человека каппа-цепи I. Гуманизированные варианты 24, 37, 48, 67 и 37/48, 67, 71 и 78 были получены в результате выполнения следующих замен аминокислот: A24V, V37I, V48M, F67I и L78F в тяжелой цепи антитела 120 graft. CDR-области указаны в рамках. Последовательности FR-H1, FR-H2, FR-H3 и FR-H4 охватывают CDR-области (CDR-H1, CDR-H2 и CDR-H3). Последовательности пронумерованы в соответствии с нумерацией Kabat. CDR-области по системе Kabat, Chotia и контактной системе указаны над CDR-областями в рамках.
На фигурах 3А и 3В показаны типичные последовательности консенсусных остовных областей вариабельного домена тяжелой цепи (VH) акцептора человека, используемые при осуществлении настоящего изобретения, со следующими идентификаторами последовательностей, где последовательности остовных областей указаны в следующем порядке FR-H1, FR-H2, FR-H3, FR-H4:
- консенсусная остовная область ”A” подгруппы I VH человека без CDR-областей по Kabat (SEQ ID NO:26, 27, 28, 29);
- консенсусные остовные области ”B”, “C” и ”D” подгруппы I VH человека без удлиненных гипервариабельных участков (SEQ ID NO:30, 31, 28, 29; SEQ ID NO:30, 31, 32, 29 и SEQ ID NO:30, 31, 33, 29);
- консенсусная остовная область ”A” подгруппы II VH человека без CDR-областей по Kabat (SEQ ID NO:34, 35, 36, 29);
- консенсусные остовные области ”B”, “C” и ”D” подгруппы II VH человека без удлиненных гипервариабельных участков (SEQ ID NO:37, 38, 36, 29; SEQ ID NO:37, 38, 39, 29 и SEQ ID NO:37, 38, 40, 29);
- консенсусная остовная область ”A” подгруппы III VH человека без CDR-областей по Kabat (SEQ ID NO:41, 42, 43, 29);
- консенсусные остовные области ”B”, “C” и ”D” подгруппы III VH человека без удлиненных гипервариабельных участков (SEQ ID NO:44, 45, 43, 29; SEQ ID NO:44, 45, 46, 29 и SEQ ID NO:44, 45, 46, 29);
- остовная область ”A” VH акцептора 1 человека без CDR-областей по Kabat (SEQ ID NO:48, 42, 49, 29);
- остовные области ”B” и “C” VH акцептора человека без удлиненных гипервариабельных участков (SEQ ID NO:44, 45, 49, 29 и SEQ ID NO:44, 45, 50, 29);
- остовная область ”A” VH акцептора 2 человека без CDR-областей по Kabat (SEQ ID NO:48, 42, 51, 29);
- остовные области ”В”, “C” и “D” VH акцептора 2 человека без удлиненных гипервариабельных участков (SEQ ID NO:44, 45, 51, 29; SEQ ID NO:44, 45, 52, 29 и SEQ ID NO:44, 45, 53, 29).
На фигурах 4А и 4В показаны типичные последовательности консенсусных остовных областей вариабельного домена легкой цепи (VL) акцептора человека, используемые при осуществлении настоящего изобретения, со следующими идентификаторами последовательностей:
- консенсусная остовная область подгруппы I-1 VL каппа-цепи человека (κv1-1): SEQ ID NO:54, 55, 56, 57;
- консенсусная остовная область подгруппы I VL каппа-цепи человека (κv1): SEQ ID NO:54, 58, 56, 57;
- консенсусная остовная область подгруппы II VL каппа-цепи человека (κv2):SEQ ID NO:69, 59, 60, 57;
- консенсусная остовная область подгруппы III VL каппа-цепи человека (κv3): SEQ ID NO:61, 62, 63, 57;
- консенсусная остовная область подгруппы IV VL каппа-цепи человека (κv4): SEQ ID NO:64, 65, 66, 57.
На фигуре 5 показан сравнительный анализ нативных последовательностей Fc-области IgG человека, humIgG1 (аллотип, отличный от А, SEQ ID NO:85, и аллотип А, где аминокислотная последовательность SREEM (SEQ ID NO:67) в SEQ ID NO:85 заменена последовательностью SRDEL (SEQ ID NO:68), humIgG2 (SEQ ID NO:86), humIgG3 (SEQ ID NO:87) и humIgG4 (SEQ ID NO:88), при этом отличия в последовательностях отмечены звездочками. Числа над последовательностями представляют систему нумерации EU. Также показана типичная константная область каппа-цепи.
На фигурах 6А-6D показан анализ FACS, нормализованный для уровня отображения каждого антитела или варианта на фаге. На фигуре 6А показаны изменения, выявленные FACS в клетках, экспрессирующих STEAP-1 (LB50), для четырех типичных антител. На фигуре 6В показаны изменения, выявленные FACS в клетках, не экспрессирующих STEAP-1, (S408) для нескольких антител, указанных на фигуре и в примере 1. На фигурах 6С и 6D показаны сравнительные анализы изменений, выявленных FACS, после нормализации уровней отображения на фаге.
На фигурах 7А-7F графически изображены анализы FACS, показывающие связывание антитела мыши, химерного антитела и гуманизированного варианта 24 антитела против STEAP-1 с белком STEAP-1 человека, экспрессированным по поверхности клетки. На фигурах 7А-7С показано, что антитело мыши 120, химерное антитело 120 и гуманизированное антитело 120v.24 против STEAP-1 связываются с белком STEAP-1 человека и обезьяны циномолгус и не связывается с белком STEAP-1 мыши. На фигурах 7D-7F изображены графики анализа FACS, на которых показано связывание мышиного антитела 120, химерного антитела 120 и гуманизированного антитела 120v.24 (клон 67) c белком STEAP-1, экспрессированным на поверхности клетки. Экзогенный белок STEAP-1 устойчиво экспрессирован в клетках 293 (клетки, обозначенные как LB50) и клетках РС3 (клетки, обозначенные как PS5.4) (фигуры 7D и 7Е) и эндогенно экспрессирован в клетках LNCaP BR (фигура 7F).
Фигуры 8А и 8В. На фигуре 8А изображен график, показывающий, что введение мышиного антитела против STEAP-1 120-МС-vc-PAB-MMAE в дозе 3 мг/кг оказывало эффективное воздействие в модели ксенотрансплантата опухоли предстательной железы (клетки LNCaP-Ner). См. пример 4. На фигуре 8В изображен график, показывающий, что введение однократной дозы гуманизированного антитела против STEAP-1 120v.24-MC-vc-PAB-MMAE (3 мг/кг), 120v.24-MC-MMAF (6 мг/кг), 120v.24-MC-MMAF (12 мг/кг) и химерного антитела против STEAP-1 120 chimera-MC-vc-PAB-MMAE (3 мг/кг) оказывало эффективное воздействие в модели ксенотрансплантата опухоли предстательной железы в клетках LNCaP. См. пример 4.
На фигуре 9 изображен график, показывающий, что введение химерного антитела против STEAP-1 120 chimera-МС-vc-PAB-MMAE (сокращенно обозначенное как антитело vcMMAE против STEAP) в дозе 3 мг/кг или химерного антитела против STEAP-1 120 chimera-МС-MMAF (сокращенно обозначенное как антитело mcMMAF против STEAP) в дозе 6 мг/кг оказывало эффективное воздействие в модели ксенотрансплантата рака предстательной железы у кастрированных мышей SCID-beige, которым были трансплантированы клетки LNCaP. См. пример 4.
На фигуре 10 изображен график, показывающий, что введение химерного антитела против STEAP-1 120 chimera-МС-vc-PAB-MMAE (сокращенно обозначенное как антитело vcMMAE против STEAP) в дозе 3 мг/кг оказывало эффективное воздействие в модели ксенотрансплантата рака предстательной железы у самцов мышей SCID-beige (андрогензависимые), которым были трансплантированы клетки LuCap 77. См. пример 4.
На фигуре 11 изображен график, показывающий, что введение гуманизированного антитела против STEAP-1 120v.24-MC-vc-PAB-MMAE в дозе 3 мг/кг и гуманизированного антитела против STEAP-1 120v.24-MC-MMAF в дозе 6 мг/кг и 12 мг/кг кастрированным мышам SCID-beige, которым была трансплантирована опухоль предстательной железы LuCap35V, оказывало эффективное воздействие по сравнению с контрольными антителами. См. пример 4.
На фигуре 12 изображена диаграмма, на которой показан белок STEAP-1 в клеточной мембране. Связывание антитела 120 против STEAP-1 зависит от конформации, при этом указанное антитело не узнает линейный эпитоп STEAP-1.
На фигурах 13А-13D показан белок STEAP-1, экспрессированный на поверхности клеток, который был обнаружен при помощи иммуногистохимии. На фигуре 13А показано иммуногистохимическое окрашивание клеток 293, экспрессирующих экзогенный белок STEAP-1 на поверхности клетки. На фигуре 13В показано иммуногистохимическое окрашивание клеток РС3, экспрессирующих экзогенный белок STEAP-1 на поверхности клетки. На фигуре 13С показано иммуногистохимическое окрашивание клеток LNCaP, экспрессирующих эндогенный белок STEAP-1 на поверхности клетки. На фигуре 13D показано иммуногистохимическое окрашивание клеток LuCAP 77, экспрессирующих эндогенный белок STEAP-1 на поверхности клетки.
На фигурах 14А-14Е изображены графики, показывающие относительную эффективность антитела против STEAP-1 120v.24-MC-MMAF и антитела против STEAP-1 120v.24-MC-vc-PAB-MMAE по уничтожению клеток, экспрессирующих STEAP-1, in vitro. Клетки PS5.4 (фигура 14А) являются клетками РС3, трансформированными вектором, кодирующим STEAP-1 с достижением экспрессии указанного белка на поверхности клетки. Клетки LB50 (фигура 14В) являются клетками 293, трансформированными вектором, кодирующим STEAP-1 с достижением экспрессии указанного белка на поверхности клетки. Клетки LNCaP (фигура 14С) экспрессируют STEAP-1 эндогенно. Обозначения ”PC3 vec” (фигура 14D) и “293 vec” (фигура 14Е) относятся, соответственно, к клеткам 293 и РС3, трансформированным контрольным вектором.
На фигуре 15 показаны конъюгаты антитела против STEAP-1 с введенным цистеином с лекарственным средством (ADC), в которых лекарственная часть присоединена к группе введенного цистеина в: легкой цепи (LC-ADC), тяжелой цепи (НС-ADC) и Fc-области (Fc-ADC).
На фигуре 16 показаны следующие стадии: (i) восстановление дисульфидных аддуктов цистеина и межцепных и внутрицепных дисульфидных связей в антителе против STEAP-1 с введенным цистеином (ThioMab) восстановителем ТСЕР (гидрохлорид трис(2-карбоксиэтил)фосфина); (ii) неполное окисление, то есть переокисление с повторным образованием межцепных и внутрицепных дисульфидных связей, при помощи dhAA (дегидроаскорбиновая кислота) и (iii) конъюгирование повторно окисленного антитела с промежуточным соединением лекарственного средства с линкером с образованием конъюгата антитела против STEAP-1 с введенным цистеином с лекарственным средством (ADC).
На фигурах 17А-С показаны сайты замен аминокислот, выполненных с целью создания антител против STEAP-1 с введенным цистеином (тио-mAb). На фигуре 17А показан вариант тио-LC V205C с соответствующей последовательной нумерацией и стандартизированной нумерацией по системе Kabat. На фигуре 17А показаны все последовательности в виде SEQ ID NO:70. На фигуре 17В показан вариант тио-НС А118С с соответствующей последовательной нумерацией и стандартизированной нумерацией по системе EU. На фигуре 17В показаны, соответственно, последовательности SEQ ID NO:71-74 и 102-108 в порядке появления. На фигуре 17С показан вариант тио-Fc S400C с соответствующей последовательной нумерацией и стандартизированной нумерацией по системе EU. На фигуре 17С показаны все последовательности в виде SEQ ID NO:109.
На фигурах 18А-F представлены анализы FACS, показывающие, что конъюгаты тиоантитела против STEAP-1 с лекарственным средством (TDC) сохраняют способность связываться с белком STEAP-1, экспрессированным на поверхности клетки. На фигурах 18А-18С изображены графики результатов анализов FACS, показывающие связывание конъюгатов тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (LCV205C) (сокращенно обозначенное как huSteap1 TDC (L205C) vcE и тиоантитела человека 120-vc-PAB-MMAE (HCA118C) (сокращенно обозначенное как huSteap1 TDC (HCA118C) vcE) с белком STEAP-1 человека, экспрессированным на поверхности клетки. Экзогенный белок STEAP-1 был устойчиво экспрессирован в клетках 293 (клетки, обозначенные как LB50) и клетках РС3 (клетки, обозначенные как PS5.4) (фигуры 18А и 18В) и эндогенно экспрессирован в клетках LNCaP BR (фигура 18С). На фигурах 18D, 18E и 18F показаны сравнительные анализы изменений, выявленных FACS, которые представлены, соответственно, на фигурах 7А, 7В и 7С.
На фигурах 19А-С показана относительная эффективность конъюгатов тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (LCV205C) (сокращенно обозначенное как huSteap1 TDC (L205C) vcE) и тиоантитела человека 120-vc-PAB-MMAE (HCA118C) (сокращенно обозначенное как huSteap1 TDC (HCA118C) vcE) с лекарственным средством (TDC) по уничтожению клеток, экспрессирующих STEAP-1, in vitro. Клетки LB50 (фигура 19А) являются клетками 293, трансформированными вектором, кодирующим STEAP-1 с достижением экспрессии указанного белка на поверхности клетки. Клетки PS5.4 (фигура 19В) являются клетками РС3, трансформированными вектором, кодирующим STEAP-1 с достижением экспрессии указанного белка на поверхности клетки. Клетки LNCaP (фигура 19С) экспрессируют STEAP-1 эндогенно.
На фигуре 20 изображен график, показывающий, что введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (HCA118C) (сокращенно обозначенное как hu Steap1 HC TDC vcE) в дозе 3 мг/кг оказывает эффективное воздействие по сравнению с контрольными антителами в модели ксенотрансплантата рака предстательной железы у самцов мышей SCID-beige (андрогензависимые мыши), которым были трансплантированы клетки LNCaP. См. пример 8.
На фигуре 21 изображен график, показывающий, что введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (HCA118C) (сокращенно обозначенное как hu Steap1 HC TDC vcE) в дозе 3 мг/кг или тиоантитела человека 120-МС-MMAF (HCA118C) (сокращенно обозначенное как hu Steap1 HC TDC mcF) в дозе 1, 3 или 6 мг/кг оказывало эффективное воздействие по сравнению с контрольными антителами в модели ксенотрансплантата рака предстательной железы у самцов мышей SCID-beige (андрогензависимые мыши), которым были трансплантированы клетки LNCaP. См. пример 8.
На фигуре 22 изображен график, показывающий, что введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (HCA118C) (сокращенно обозначенное как hu Steap1 HC TDC vcE) в дозе 3 мг/кг или тиоантитела человека 120-МС-MMAF (HCA125C) (сокращенно обозначенное как hu Steap1 HC TDC mcF) в дозе 3 или 6 мг/кг оказывало эффективное воздействие по сравнению с контрольными антителами в модели ксенотрансплантата рака предстательной железы у кастрированных мышей SCID-beige, которым были трансплантированы опухолевые клетки предстательной железы LuCaP 35V. См. пример 8.
На фигуре 23 показаны сайты замен аминокислот, выполненных с целью создания антитела против STEAP-1 с введенным цистеином (тио-mAb), обозначенного ”Simmons IV” или просто ”SGIV”. Аминокислотная последовательность легкой цепи SGIV (SEQ ID NO:90) показана в сравнении с легкой цепью мышиного антитела mu 120 (SEQ ID NO:85) и антитела 120.v24 (SEQ ID NO:91). Вариант тио-LC SGIV с соответствующей последовательной нумерацией и стандартизированной нумерацией по системе Kabat показан в сравнении с материнским антителом mu 120 и с вариантом тио-LC 120.v24 с соответствующей последовательной нумерацией и стандартизированной нумерацией по системе Kabat. CDR-области показаны в рамках (CDR-L1, CDR-L2 и CDR-L3). Последовательности, охватывающие CDR-области, являются последовательностями остовных областей (FR-L1 - FR-L4). Последовательности пронумерованы в соответствии с нумерацией Kabat. CDR-области по системе Kabat, Chotia и контактной системе указаны над CDR-областями в рамках. См. пример 9.
На фигуре 24 показаны сайты замен аминокислот в остовной области, выполненных с целью создания разных вариантов антител против STEAP-1 с введенным цистеином (тио-mAb) SGIV и 120v.24, показанных на фигуре 23А. Аминокислотная последовательность легкой цепи SGIV показана со стандартизированной нумерацией по системе Kabat в сравнении с вариантами LS.VLVH1 (SEQ ID NO:92), LS.VLVH2 (SEQ ID NO:93), LS.Q (SEQ ID NO:94) и LS.CH1 (SEQ ID NO:95). Аминокислотная последовательность легкой цепи варианта 120.v24 со стандартизированной нумерацией по системе Kabat показана в сравнении с вариантами ED.FW1 (SEQ ID NO:96), ED.FW2 (SEQ ID NO:97), ED.FW3 (SEQ ID NO:98), ED.all (SEQ ID NO:99), ED.Pro (SEQ ID NO:100) и ED.p1 (SEQ ID NO:101). CDR-области показаны в рамках. Последовательности пронумерованы в соответствии с нумерацией Kabat. См. пример 9. На фигуре 24 показаны последовательности вариантов ”Simmons IV” и ”120v.24” соответственно в виде SEQ ID NO:110-111.
На фигуре 25 изображены графики анализа Скэтчарда связывания антитела с белком STEAP-1, экспрессированным на поверхности клеток LNCaP.BR. Дублированные образцы исследовали, используя антитело 120.v24 (фигуры 25(А)-(D)) и вариант SGIV (фигуры 25(Е)-(Н)). См. пример 9.
На фигуре 26 изображены графики анализа Скэтчарда связывания антитела с белком STEAP-1, экспрессированным на поверхности клеток 293.LB50. Дублированные образцы исследовали, используя антитело 120.v24 (фигуры 26(А)-(D)) и вариант SGIV (фигуры 26(Е)-(Н)). См. пример 9.
На фигуре 27 представлена таблица, в которой сравниваются средние значения аффинности связывания, измеренные при выполнении анализа Скэтчарда, для антитела 1789 мыши, антитела 120 мыши, химерного антитела с Fc-областью, гуманизированного антитела 120.v24, тиоантитела 120.v24 и тиоантитела SGIV в клетках РС-3-PS5.4, 293-LB50 и LNCaP-BR, а также в клетках 293, временно экспрессирующих STEAP-1. См. пример 9.
На фигуре 28 представлены анализы FACS, показывающие изменения, выявленные с помощью FACS в клетках, устойчиво трансфицированных STEAP-1 (293 STEAP-1 LB48, 293 STEAP-1 LB50 и 293 STEAP-1 LB53), при использовании образцов антител SGIV и 120.v24. См. пример 9.
На фигуре 29 показан титр антител, имевший место в разных сборах из клеток, продуцирующих антитело SGIV или 120.v24.
На фигуре 30 показана визуализация способом ПЭТ с цирконием-89 у животных, которым вводили антитело против STEAP-1, меченное ZR89 с радиоактивностью 100 мкКи, и выполняли томографию через 4 дня, при этом поглощение Zr89-STEAP-1 было самым высоким в клетках Lucap35V и самым низким в клетках Lucap 96.1, что соответствовало результатам исследования in vitro.
Подробное описание вариантов осуществления изобретения
Настоящее изобретение относится к выделенным антителам, связывающимся с белком STEAP-1. Настоящее изобретение также относится к иммуноконъюгатам, содержащим антитела против STEAP-1. Антитела и иммуноконъюгаты по настоящему изобретению могут быть использованы, например, для диагностики или лечения нарушений, связанных с измененной экспрессией, например, повышенной экспрессией STEAP-1. В определенных вариантах осуществления изобретения антитела или иммуноконъюгаты по настоящему изобретению используют для диагностики или лечения пролиферативного нарушения клеток, такого как опухоль или рак. В определенных вариантах осуществления изобретения белок STEAP-1 экспрессирован в опухоли или раке ткани предстательной железы, легкого или ободочной кишки. В определенных вариантах осуществления изобретения антитела или иммуноконъюгаты по настоящему изобретению используют для обнаружения белка STEAP-1, например, STEAP-1, экспрессированного на поверхности клетки. В определенных вариантах осуществления изобретения антитела или иммуноконъюгаты по настоящему изобретению используют для обнаружения экспрессии STEAP-1 на поверхности нормальных и/или опухолевых или раковых клеток ткани предстательной железы, легкого или ободочной кишки.
Настоящее изобретение относится к полинуклеотидам, кодирующим антитела против STEAP-1. Настоящее изобретение также относится к векторам, содержащим полинуклеотиды, кодирующие антитела против STEAP-1, и к клеткам-хозяевам, содержащим такие векторы. Настоящее изобретение также относится к композициям, в том числе к фармацевтическим препаратам, содержащим один или несколько полинуклеотидов, антител против STEAP-1 или иммуноконъюгатов по настоящему изобретению.
Настоящее изобретение относится к способам лечения пролиферативных нарушений клеток, которые включают, не ограничиваясь ими, опухоль или рак, при помощи антитела против STEAP-1, конъюгата антитела с лекарственным средством или иммуноконъюгата. Такие способы включают, не ограничиваясь ими, лечение опухоли или рака в предстательной железе, легком или ободочной кишке млекопитающего. Настоящее изобретение относится к способам обнаружения экспрессии STEAP-1 на клетке ткани при помощи антитела против STEAP-1, конъюгата антитела с лекарственным средством или иммуноконъюгата. Такие способы включают, не ограничиваясь ими, обнаружение экспрессии STEAP-1 на нормальной клетке, опухолевой клетке или раковой клетке предстательной железы, легкого или ободочной кишки, указанных в качестве неограничивающих примеров.
Общие способы
Способы и процедуры, рассмотренные в настоящем описании изобретения, являются хорошо известными и широко применяемыми специалистами в данной области и включают, например, обычно используемые способы, описанные в публикациях Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Current Protocols in Molecular Biology (F. M. Ausubel, et al. eds., (2003)); the series Methods in Enzymology (Academic Press, Inc.): Pcr 2: A Practical Approach (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, A Laboratory Manual, and Animal Cell Culture (R. I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M. J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J. P. Mather and P. E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths, and D. G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D. M. Weir and C. C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J. E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C. A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); and Cancer: Principles and Practice of Oncology (V. T. DeVita et al., eds., J.B. Lippincott Company, 1993). 89Zr Immuno-PET: Comphrehensive Procedures for the Production of 89Zr-Labeled Monoclonal Antibodies (Verel I., et al. J Nucl Med; 44:1271-1281, 2003).
Определения терминов и аббревиатуры
Определения терминов
“Выделенное” антитело является антителом, которое было идентифицировано, отделено и/или выделено из компонента естественного окружения. Загрязняющие компоненты естественного окружения являются веществами, которые могут мешать исследованию, диагностике или терапевтическому применению антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах осуществления изобретения антитело очищают (1) до более чем 95 мас.% антитела при определении, например, способом Лоури и в некоторых вариантах осуществления изобретения до более чем 99 мас.%; (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности, например, при помощи секвенатора с центрифужной чашей, или (3) до гомогенного состояния способом SDS-PAGE в восстановительных или невосстановительных условиях, например, с окрашиванием кумассии голубым или серебром. Выделенное антитело представляет собой антитело in situ в рекомбинантных клетках, так как будет отсутствовать по меньшей мере один компонент естественного окружения антитела. Однако выделенное антитело обычно получают в результате выполнения по меньшей мере одной стадии очистки.
“Выделенная” молекула нуклеиновой кислоты является молекулой нуклеиновой кислоты, отделенной по меньшей мере от одной другой молекулы нуклеиновой кислоты, с которой данная молекула обычно связана, например, в естественном окружении. Выделенная молекула нуклеиновой кислоты далее включает молекулу нуклеиновой кислоты, находящуюся в клетках, которые обычно экспрессируют данную молекулу нуклеиновой кислоты, но при этом указанная молекула нуклеиновой кислоты находится вне хромосомы или в таком месте хромосомы, которое отличается от естественной локализации данной молекулы в хромосоме.
Термин ”очищенный” означает, что молекула присутствует в образце в концентрации, составляющей по меньшей мере 95 мас.% или по меньшей мере 98 мас.% образца, содержащего данную молекулу.
Термин ”по существу подобный” или “по существу такой же”, как используется в настоящем описании, означает достаточно высокую степень подобия двух числовых значений (например, одно из которых связано с антителом по настоящему изобретению и другое связано с эталонным/сравниваемым антителом), поэтому специалист в данной области может считать различие между двумя значениями незначительным или биологически и/или статистически незначимым для биологической характеристики, измеренной указанными значениями (например, значения Kd). Различие между двумя указанными значениями составляет, например, менее примерно 50%, менее примерно 40%, менее примерно 30%, менее примерно 20% и/или менее примерно 10% от эталонного/сравниваемого значения.
Фраза ”по существу меньший” или “по существу отличающийся”, как используется в настоящем описании, означает достаточно высокую степень отличия двух числовых значений (одно из которых обычно связано с исследуемой молекулой и другое связано с эталонной/сравниваемой молекулой), поэтому специалист в данной области может считать различие двух значений статистически значимым для биологической характеристики, измеренной указанными значениями (например, значения Kd). Различие между двумя указанными значениями составляет, например, более примерно 10%, более примерно 20%, более примерно 30%, более примерно 40% и/или более примерно 50% от значения эталонной/сравниваемой молекулы.
Термин ”вектор”, как используется в настоящем описании, означает молекулу нуклеиновой кислоты, способную переносить другую соединенную с ней нуклеиновую кислоту. Одним типом вектора является ”плазмида”, которая представляет собой кольцевую двухцепочечную ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Еще одним типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в геном вируса. Определенные векторы способны автономно реплицировать в клетке-хозяине, в которую указанные векторы были введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут быть встроены в геном клетки-хозяина в процессе введения в клетку-хозяина и будут реплицировать вместе с геномом хозяина. Кроме того, определенные векторы могут управлять экспрессией генов, с которыми они функционально связаны. Такие векторы определяются в настоящем описании изобретения как ”рекомбинантные векторы экспрессии” или просто ”векторы экспрессии”. Как правило, экспрессирующие векторы, используемые в способах рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании изобретения ”плазмида” и “вектор” могут иметь взаимозаменяемые значения, так как плазмида является наиболее распространенной формой вектора.
Термины ”полинуклеотид” или “нуклеиновая кислота”, имеющие взаимозаменяемые значения в настоящем описании изобретения, означают полимеры нуклеотидов любой длины и включают ДНК, и РНК. Указанные нуклеотиды могут быть дезоксирибонуклеотидами, рибонуклеотидами, модифицированными нуклеотидами или основаниями, и/или их аналогами, или любым субстратом, который может быть введен в полимер ДНК- или РНК-полимеразой или в результате выполнения реакции синтеза. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Структура нуклеотида может быть модифицирована в случае необходимости до или после сборки полимера. Последовательность нуклеотидов может прерываться компонентами, не являющимися нуклеотидами. Полинуклеотид может содержать модификации, произведенные после синтеза, такие как конъюгирование с меткой. Модификации других типов включают, например, ”кэпы”, замену одного или нескольких природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, нуклеотиды с неполяризованными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и с поляризованными связями (например, фосфортиоаты, фосфордитиоаты и т.д.), нуклеотиды, включающие боковые части, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальыне пептиды, ply-L-лизин и т.д.), нулеотиды с интеркаляторами (например, акридин, псорален и т.д.), нуклеотиды, содержащие хелатообразователи (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), нуклеотиды, содержащие алкилирующие агенты, нуклеотиды с модифицированными связями (например, аномерные нуклеиновые альфа-кислоты и т.д.), а также немодифицированные формы полинуклеотидов. Кроме того, любые гидроксильные группы, обычно присутствующие в сахарах, могут быть заменены, например, фосфонатными группами, фосфатными группами, защищены стандартными защитными группами или активированы для получения дополнительных связей с дополнительными нуклеотидами или могут быть конъюгированы с твердыми или полутвердыми подложками. 5'- или 3'-концевая группа ОН может быть фосфорилирована или замещена аминами или органическими кэппирующими группами, содержащими 1-20 атомов углерода. Другие гидроксилы также могут быть использованы для получения стандартных защитных групп. Полинуклеотиды могут также содержать аналогичные формы рибозы или дезоксирибозы, известные в данной области, которые включают, например, 2'-О-метил-, 2'-О-аллил-, 2'-фтор- или 2’-азидорибозу, карбоциклические аналоги сахаров, α-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозы, фуранозы, седогептулозы, ациклические аналоги и основные аналоги нуклеотидов, такие как метилрибозид. Одна или несколько фосфодиэфирных связей могут быть заменены альтернативными связующими группами. Указанные альтернативные связующие группы включают, не ограничиваясь ими, варианты, в которых фосфат заменен Р(О)S (”тиоат”), P(S)S (”дитиоат”), (О)NR2 (”амидат”), P(O)R, P(O)OR', CO или СН2 (”формацеталь”), где каждый элемент R или R' независимо означает Н, замещенный или незамещенный алкил (1-20 атомов С), необязательно содержащий простую эфирную (-О-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не все связи в полинуклеотиде должны быть идентичными. Настоящее изобретение относится ко всем полинуклеотидам, рассмотренным в настоящем описании изобретения, включая РНК и ДНК.
Термин ”олигонуклеотид” в использованном в настоящем описании значении означает короткие, одноцепочечные синтетические полинуклеотиды, которые обычно, но не обязательно, состоят из менее примерно 200 нуклеотидов в длину. Термины ”олигонуклеотид” и “полинуклеотид” не являются взаимоисключающими. Приведенное выше описание полинуклеотидов в равной степени и полностью относится к олигонуклеотидам.
Выражение “процентное значение (%) идентичности аминокислотной последовательности” в отношении эталонной полипептидной последовательности определяется как процентное значение аминокислотных остатков в последовательности-кандидате, идентичных аминокислотным остаткам в эталонной полипептидной последовательности после выполнения сравнительного анализа последовательностей и введения, при необходимости, разрывов для достижения максимального процентного значения идентичности последовательности, при этом при определении идентичности последовательности не учитываются любые консервативные замены. Сравнительный анализ с целью определения процентного значения идентичности аминокислотной последовательности может быть выполнен разными способами, известными в данной области, например, при помощи общедоступных компьютерных программ, таких как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры, требуемые для выполнения сравнительного анализа последовательностей, в том числе любые алгоритмы, необходимые для достижения максимального совмещения сравниваемых последовательностей по всей длине. Однако в соответствии с целями настоящего изобретения процентное значение идентичности аминокислотной последовательности определяют при помощи компьютерной программы сравнения последовательностей ALIGN-2. Компьютерная программа сравнения последовательностей ALIGN-2 создана в компании Genentech, Inc., и исходный код был направлен вместе с документацией пользователя в Ведомство по охране авторских прав США, Вашингтон, округ Колумбия, 20559, где указанная программа зарегистрирована под регистрационным номером Ведомства по охране авторских прав США TXU510087. Программа ALIGN-2 может быть приобретена в компании Genentech, Inc., South San Francisco, California, или может быть компилирована из исходного кода. Программу ALIGN-2 необходимо компилировать для использования в операционной системе UNIX, предпочтительно, в цифровой операционной системе UNIX V4.0D. В программе ALIGN-2 заданы все параметры сравнения последовательностей, которые не могут быть изменены.
При использовании программы ALIGN-2 для сравнения аминокислотных последовательностей процентное значение идентичности данной аминокислотной последовательности А относительно данной аминокислотной последовательности В (другими словами данной аминокислотной последовательности А, которая обладает определенным процентным значением идентичности относительно данной аминокислотной последовательности В) вычисляют следующим образом:
100 × результат деления Х/Y
где Х означает число аминокислотных остатков, являющихся идентичными парами по результатам сравнительного анализа последовательностей А и В, выполненного программой сравнения последовательностей ALIGN-2, и Y означает общее число аминокислотных остатков в В. Следует отметить, что, если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то процентное значение идентичности аминокислотной последовательности А по отношению к В не будет равно процентному значению идентичности аминокислотной последовательности В по отношению к А. За исключением особо оговоренных случаев все процентные значения идентичности аминокислотных последовательностей, использованные в настоящем описании изобретения, получены в соответствии с описанием, приведенным в предшествующем абзаце, при помощи компьютерной программы ALIGN-2.
Термин ”STEAP-1”, как используется в настоящем описании, означает любой нативный белок STEAP-1, выделенный у любого позвоночного, включая млекопитающих, таких как приматы (например, человек, обезьяна циномолгус (cyno)) и грызуны (например, мыши и крысы), за исключением особо оговоренных случаев. В определение данного термина входит наивный белок STEAP-1, а также любая форма STEAP-1, полученная в результате процессирования в клетке. В определение данного термина также входят природные варианты STEAP-1, например, сплайсированные варианты, аллельные варианты и изоформы. Аминокислотная последовательность белка STEAP-1 человека показана на фигуре 1 (SEQ ID NO:1). В одном из вариантов осуществления изобретения STEAP-1 экспрессирован на поверхности клетки, например, на поверхности нормальной клетки предстательной железы, легкого или ободочной кишки и характеризуется повышенной экспрессией в раковых клетках предстательной железы, легкого или ободочной кишки или в метастазах таких раковых клеток. На фигуре 1 также показана аминокислотная последовательность STEAP-1, выделенная у мыши и обезьяны циномолгус (соответственно SEQ ID NO:2 и 3).
“Антитела” (Ab) и “иммуноглобулины” (Ig) являются гликопротеинами с подобными характеристиками структуры. Если антитела обладают специфичностью связывания к определенному антигену, то иммуноглобулины включают как антитела, так и другие антителоподобные молекулы, у которых обычно отсутствует специфичность к антигену. Полипептиды последнего типа, например, продуцируются в небольших количествах лимфатической системой и на высоких уровнях в миеломах.
Термины ”антитело” и “иммуноглобулин” имеют взаимозаменяемые значения в самом широком смысле слова и включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, моновалентные антитела, мультивалентные антитела, мультиспецифические антитела (например, биспецифические антитела, если они обладают требуемой биологической активностью), а также могут включать определенные фрагменты антител (более подробно рассмотренные в настоящем описании изобретения). Антитело может быть химерным антителом, антителом человека, гуманизированным антителом и/или антителом с созревшей аффинностью.
Термин ”антитело против STEAP-1” или “антитело, связывающееся с STEAP-1” означает антитело, способное связываться с STEAP-1 с достаточной аффинностью, при этом указанное антитело может быть использовано в качестве диагностического и/или терапевтического агента для направленного воздействия на STEAP-1. Степень связывания антитела против STEAP-1 с неродственным белком, отличным от STEAP-1, составляет менее примерно 10% от связывания указанного антитела с STEAP-1, измеренного, например, при помощи радиоиммуноанализа (RIA). В определенных вариантах осуществления изобретения антитело, связывающееся с STEAP-1, имеет константу диссоциации (Kd) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ. В определенных вариантах осуществления изобретения антитело против STEAP-1 связывается с эпитопом STEAP-1, сохранившимся в STEAP-1 разных видов.
Термин ”вариабельная область” или “вариабельный домен” антитела означает аминоконцевые домены тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может быть обозначен как ”VH”. Вариабельный домен легкой цепи может быть обозначен как ”VL”. Указанные домены обычно являются самыми вариабельными частями антитела и содержат антигенсвязывающие центры.
Термин ”вариабельный” означает, что определенные части вариабельных доменов имеют разные последовательности и используются при связывании и определении специфичности определенного антитела к определенному антигену. Однако вариабельность распределена неравномерно в вариабельных доменах антител. Вариабельность сконцентрирована в трех сегментах, именуемых определяющими комплементарность областями (CDR) или гипервариабельными участками (HVR) в вариабельных доменах легкой и тяжелой цепей. Наиболее консервативные части вариабельных доменов именуются остовными областями (FR). Вариабельные домены нативных тяжелых и легких цепей включают четыре остовные области, имеющие в основном бета-складчатую конфигурацию, соединенные тремя CDR-областями, образующими петли, соединяющие и в некоторых случаях образующие часть бета-складчатой структуры. CDR-области в каждой цепи удерживаются в непосредственной близости друг от друга остовными областями и вместе с CDR-областями из другой цепи способствуют образованию антигенсвязывающего центра антител (см. публикацию Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены не участвуют непосредственно в связывании антитела с антигеном, но выполняют разные эффекторные функции, например, определяют антителозависимую клеточную токсичность.
“Легкие цепи” антител (иммуноглобулинов) любого позвоночного могут быть отнесены к одному из двух четко выраженных типов, именуемых каппа (κ) и лямбда (λ), на основании аминокислотных последовательностей константных доменов.
В зависимости от аминокислотных последовательностей константных доменов тяжелых цепей антитела (иммуноглобулины) могут быть отнесены к разным классам. Существуют пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из указанных классов могут быть далее разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют разным классам иммуноглобулинов, именуются соответственно α, δ, ε, γ и µ. Структуры субъединиц и трехмерные конфигурации разных классов иммуноглобулинов хорошо известны и описаны, например, в публикации Abbas et al., Cellular and Mol. Immunology, 4th ed. (2000). Антитело может быть частью более крупной слитой молекулы, образованной путем ковалентного или нековалентного связывания антитела с одним или несколькими другими белками или пептидами.
Термины ”непроцессированное антитело”, “интактное антитело” и “полное антитело” имеют взаимозаменяемые значения и означают антитело по существу в интактной форме, а не фрагменты антитела, описанные ниже. Указанные термины, в частности, служат для определения антитела, тяжелые цепи которого содержат Fc-область.
Термин ”фрагменты антитела” означает только часть интактного антитела, которая сохраняет по меньшей мере одну, большинство или все функции, связанные с указанной частью, находящейся в интактном антителе. В одном из вариантов осуществления изобретения фрагмент антитела включает антигенсвязывающий центр интактного антитела и, таким образом, сохраняет способность связываться с антигеном. В другом варианте осуществления изобретения фрагмент антитела, например, фрагмент, включающий Fc-область, сохраняет по меньшей мере одну из биологических функций, обычно связанных с Fc-областью, находящейся в интактном антителе, такую как связывание FcRn, модуляция времени полужизни антитела в кровотоке, функция ADCC и связывание комплемента. В одном из вариантов осуществления изобретения фрагмент антитела является моновалентным антителом, у которого время полужизни in vivo по существу соответствует времени полужизни интактного антитела. Например, такой фрагмент антитела может включать антигенсвязывающий домен, связанный с последовательностью Fc-области, способной сообщать устойчивость указанному фрагменту in vivo.
В результате расщепления антител папаином образуются два идентичных антигенсвязывающих фрагмента, именуемых ”Fab”-фрагментами, каждый из которых имеет один антигенсвязывающий центр, и остаточный ”Fc”-фрагмент, в названии которого отражена способность легко кристаллизоваться. В результате обработки пепсином образуется F(ab)2 фрагмент, который имеет два антигенсвязывающих центра и способен осуществлять перекрестное сшивание антигена.
“Fv” является минимальным фрагментом антитела, содержащим полный антигенсвязывающий центр. В одном из вариантов осуществления изобретения двухцепочечный Fv фрагмент состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, находящихся в тесной нековалентной ассоциации. В одноцепочечном Fv (scFv) фрагменте один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи могут быть ковалентно связаны гибким пептидным линкером, в результате чего легкая и тяжелая цепи образуют ”димерную” структуру, аналогичную структуре двухцепочечного Fv фрагмента. В указанной конфигурации три CDR-области каждого вариабельного домена взаимодействуют друг с другом, образуя антигенсвязывающий центр на поверхности VH-VL димера. Все шесть CDR-областей сообщают антителу антигенсвязывающую специфичность. Однако даже один вариабельный домен (или половина Fv-фрагмента, содержащая только три CDR-области, специфичные к антигену) способен распознавать и связывать антиген, хотя и с меньшей аффинностью по сравнению с полным антигенсвязывающим центром.
Fab фрагмент содержит вариабельные домены тяжелой и легкой цепей, а также константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab’ фрагменты отличаются от Fab фрагментов добавлением нескольких остатков к карбоксильному концу СН1 домена тяжелой цепи, включающих один или несколько остатков цистеина из шарнирной области антитела. Fab’-SH является обозначением Fab’ фрагмента, в котором один или несколько остатков цистеина константных доменов содержат свободную тиоловую группу. F(ab’)2 фрагменты антитела первоначально были получены в виде пар Fab' фрагментов, между которыми находились остатки цистеина из шарнирной области. Также известны другие химические сочетания фрагментов антитела.
“Одноцепочечный Fv” фрагмент или “scFv” фрагмент антитела содержит VH и VL домены антитела, находящиеся в одной полипептидной цепи. Полипептид scFv фрагмента обычно содержит полипептидный линкер между VH и VL доменами, позволяющий scFv фрагменту образовывать требуемую структуру для связывания антигена. Для ознакомления с scFv фрагментом см. публикацию Pluckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rodenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Термин ”диатела” означает небольшие фрагменты антитела с двумя антигенсвязывающими центрами, которые содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL). При использовании линкера, который является слишком коротким для спаривания двух доменов в одной цепи, домены принудительно спариваются с комплементарными доменами другой цепи и образуют два антигенсвязывающих центра. Диатела могут быть двухвалентными или биспецифическими. Диатела более полно описаны, например, в европейском патенте № 404097, WO93/1161, публикациях Hudson et al. (2003) Nat. Med. 9:129-134 и Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993). В публикации Hudson et al. (2003) Nat. Med. 9:129-134 также описаны триатела и тетратела.
Термин ”моноклональное антитело”, как используется в настоящем описании, означает антитело, полученное из популяции по существу гомогенных антител, то есть отдельные антитела, образующие популяцию, являются идентичными за исключением возможных мутаций, например, природных мутаций, которые могут присутствовать в незначительных количествах. Таким образом, определение “моноклональный” показывает, что указанное антитело не является смесью разных антител. В определенных вариантах осуществления изобретения такое моноклональное антитело обычно является антителом, включающим полипептидную последовательность, связывающуюся с мишенью, которая была получена способом, включающим отбор одной полипептидной последовательности, связывающейся с мишенью, из множества полипептидных последовательностей. Например, процесс отбора может включать выбор уникального клона из множества клонов, таких как пул клонов гибридом, клонов фагов или клонов рекомбинантных ДНК. Следует отметить, что выбранная последовательность, связывающаяся с мишенью, может быть далее изменена, например, для улучшения аффинности к мишени, гуманизации последовательности, связывающейся с мишенью, улучшения продуцирования в культуре клеток, уменьшения иммуногенности in vivo, создания мультиспецифического антитела и т.д., при этом антитело, включающее измененную последовательность, связывающуюся с мишенью, также является моноклональным антителом по настоящему изобретению. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных антигенных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено против одной антигенной детерминанты антигена. Помимо специфичности, преимуществом препаратов моноклональных антител является то, что они обычно не загрязнены другими иммуноглобулинами.
Определение ”моноклональный” указывает на характер антитела, полученного из по существу гомогенной популяции антител, и не ограничивает получение такого антитела каким-либо конкретным способом. Моноклональные антитела по настоящему изобретению могут быть получены разными способами, включающими, например, способ гибридом (см., например, публикации Kohler et al., Nature, 256:495 (1975); Harlow et al., Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981)), способы рекомбинантных ДНК (см., например, патент США № 4816567), способы отображения на фаге (см., например, публикации Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101 (34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2): 119-132 (2004) и способы продуцирования животными антител человека или человекоподобных антител, содержащих часть или все локусы иммуноглобулина человека или гены, кодирующие последовательности иммуноглобулина человека (см., например, публикации WO98/24893; WO96/34096; WO96/33735; WO91/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA 90: 2551 (1993); Jakobovits et al., Nature 362: 255-258 (1993); Bruggemann et al., Year in Immunol. 7:33 (1993); патенты США № 5545807; 5545806; 5569825; 5625126; 5633425; 5661016; Marks et al., Bio.Technology 10: 779-783 (1992); Lonberg et al., Nature 368: 856-859 (1994); Morrison, Nature 368: 812-813 (1994); Fishwild et al., Nature Biotechnol. 14: 845-851 (1996); Neuberger, Nature Biotechnol. 14: 826 (1996) and Lonberg and Huszar, Intern. Rev. Immunol. 13: 65-93 (1995).
В определение моноклональных антител входят ”химерные” антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, выделенных из определенного вида или принадлежащих к определенному классу или подклассу антител, в то время как остальная часть цепи идентична или гомологична соответствующим последовательностям в антителах, выделенных из другого вида или принадлежащих к другому классу или подклассу антител, а также фрагментам таких антител, при сохранении требуемой биологической активности (патент США № 4816567 и публикация Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
“Гуманизированные” формы антител, отличных от антител человека (например, мышиных), являются химерными антителами, содержащими минимальную последовательность, выделенную из иммуноглобулина, отличного от человеческого. В одном из вариантов осуществления изобретения гуманизированное антитело является иммуноглобулином человека (реципиентное антитело), в котором остатки из гипервариабельного участка реципиента заменены остатками из гипервариабельного участка вида, отличного от человека (донорское антитело), такого как мышь, крыса, кролик или примат кроме человека, обладающего требуемой специфичностью, аффинностью и/или антигенсвязывающей способностью. В некоторых случаях остатки остовной области (FR) иммуноглобулина человека заменяют соответствующими остатками, отличными от человеческих. Кроме того, гуманизированные антитела могут включать остатки, не обнаруженные в реципиентном антителе или донорском антителе. Для дальнейшего улучшения характеристик антитела могут быть произведены указанные модификации. Как правило, гуманизированное антитело может содержать по существу все за исключением по меньшей мере одного и обычно двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют петлям иммуноглобулина, отличного от человеческого, и все или по существу все остовные области являются остовными областями последовательности иммуноглобулина человека. Гуманизированное антитело может также необязательно содержать по меньшей мере часть константной области (Fc) иммуноглобулина, обычно иммуноглобулина человека. Для дальнейшего ознакомления с данным вопросом см. публикации Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332: 323-329 (1988) и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). См. также следующие обзорные статьи и приведенные в них ссылки: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
“Антитело человека” является антителом, аминокислотная последовательность которого соответствует аминокислотной последовательности, продуцированной человеком и/или полученной любым способом создания антител человека, рассмотренным в настоящем описании изобретения. Из определения антитела человека исключено гуманизированное антитело, содержащее антигенсвязывающие остатки, отличные от человеческих.
Термин ”гипервариабельный участок”, “HVR” или “HV”, использованный в настоящем описании изобретения, означает участки вариабельного домена антитела, имеющие гипервариабельную последовательность и/или образующие структурно определенные петли. Как правило, антитела содержат шесть гипервариабельных участков: три в VH (Н1, Н2, Н3) и три в VL (L1, L2, L3). В нативных антителах Н3 и L3 характеризуются наибольшим разнообразием шести гипервариабельных участков, при этом считается, что Н3 играет уникальную роль в сообщении антителам точной специфичности. Xu et al. (2000) Immunity 13:37-45; Johnson and Wu (2003) in Methods in Molecular Biology 248:1-25 (Lo, ed., Human Press, Totowa, NJ). Действительно, природные антитела, состоящие только из тяжелой цепи, являются функциональными и устойчивыми при отсутствии легкой цепи. Hamers-Casterman et al. (1993) Nature 363:446-448; Sheriff et al. (1996) Nature Struct. Biol. 3:733-736.
Существует несколько классификаций гипервариабельных участков, которые входят в объем настоящего изобретения. Наиболее часто используется классификация, включающая определяющие комплементарность области (CDR) Kabat, идентифицируемые на основании вариабельности последовательностей (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Классификация Chotia относится к локализации структурных петель (Chotia and Lesk, J. Mol. Biol. 196:901-917 (1987)). Гипервариабельные участки AbM представляют собой компромисс между CDR-областями Kabat и структурными петлями Chotia, и данная классификация использована в программном обеспечении моделирования антител AbM Oxford Molecular. “Контактная” классификация гипервариабельных участков основана на анализе существующих сложных кристаллических структур. Ниже приведены остатки всех гипервариабельных участков.
Гипервариабельные участки могут содержать ”удлиненные гипервариабельные участки”, в частности: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 или 89-96 (L3) в VL и 26-35 или 26-35А (Н1), 50-65 или 49-65 (Н2) и 93-102, 94-102 или 95-102 (Н3) в VH. Остатки вариабельных доменов во всех классификациях пронумерованы в соответствии с нумерацией, принятой в приведенной выше публикации Kabat et al. Гипервариабельные участки HVR-H1, HVR-H2 и HVR-H3 гуманизированных антител против STEAP-1 120v.24 по настоящему изобретению представляют собой Н26-Н35А, Н49-Н6 и Н95-Н102 по нумерации Kabat. Гипервариабельные участки HVR-L1, HVR-L2 и HVR-L3 гуманизированных антител против STEAP-1 120v.24 по настоящему изобретению представляют собой L24-34, L50-56 и L89-97 по нумерации Kabat. Как используется в настоящем описании, термины ”HVR” и “CDR” имеют взаимозаменяемые значения.
Остатки ”остовной области” или ”FR” представляют собой остатки вариабельных доменов, отличные от остатков гипервариабельных участков, рассмотренных в настоящем описании изобретения.
Термин ”нумерация остатков вариабельного домена по Kabat” или ”нумерация положений аминокислот по Kabat” и его варианты означают систему нумерации, используемую для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи компиляции антител, описанной в публикации Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). При использовании данной системы нумерации фактическая линейная аминокислотная последовательность может содержать меньше или больше аминокислот, соответствующих укорачиванию или инсерции в остовную область (FR) или гипервариабельный участок (HVR) вариабельного домена. Например, вариабельный домен тяжелой цепи может включать одну вставку аминокислоты (остаток 52а по нумерации Kabat) после остатка 52 Н2 и остатки (например, остатки 82а, 82b, 82с и т.д. по нумерации Kabat), вставленные после остатка 82 остовной области тяжелой цепи. Нумерация остатков по Kabat может быть определена для данного антитела в результате выполнения сравнительного анализа гомологичных областей последовательности антитела со ”стандартной” последовательностью, пронумерованной по системе Kabat.
Антитело с ”созревшей аффинностью” является антителом с одним или несколькими изменениями в одном или нескольких HVR, вызывающими повышение аффинности антитела к антигену по сравнению с материнским антителом, в котором отсутствуют такие изменения. В одном из вариантов осуществления изобретения антитело с созревшей аффинностью обладает наномолярной или даже пикомолярной аффинностью к антигену-мишени. Антитела с созревшей аффинностью получают способами, известными в данной области. В публикации Marks et al., Bio/Technology 10:779-783 (1992) описано созревание аффинности путем перестановки VH и VL доменов. Неспецифический мутагенез остатков HVR и/или остовной области описан в публикациях: Barbas et al., Proc. Nat. Acad. Sci., USA 91:3809-3813 (1994); Schier et al., Gene 169:147-155 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995); и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
“Блокирующее” антитело или антитело-”антагонист” является таким антителом, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается. Определенные блокирующие антитела или антитела-антагонисты существенно или полностью ингибируют биологическую активность антигена.
”Антитело-агонист”, как используется в настоящем описании, является антителом, которое имитирует по меньшей мере одну из функциональных активностей представляющего интерес полипептида.
“Эффекторные функции” антитела представляют собой биологические активности Fc-области (Fc-области нативной последовательности или вариантной Fc-области аминокислотной последовательности) антитела и изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антителозависимую клеточно-опосредованную цитотоксичность (ADCC); фагоцитоз; уменьшение экспрессии рецепторов на поверхности клетки (например, В-клеточного рецептора) и активацию В-клеток.
“Fc-рецептор” или “FcR” является рецептором, связывающимся с Fc-областью антитела. В некоторых вариантах осуществления изобретения FcR является нативным FcR человека. В некоторых вариантах осуществления изобретения FcR является рецептором, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно сплайсированные формы указанных рецепторов. Рецепторы FcγRII включают FcγRIIA (”активирующий рецептор”) и FcγRIIB (”ингибирующий рецептор”), которые имеют подобные аминокислотные последовательности, отличающиеся главным образом цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в цитоплазматическом домене активирующий фрагмент на основе тирозина иммунорецептора (ITAM). Ингибирующий рецептор FcγRIIB содержит в цитоплазматическом домене ингибирующий фрагмент на основе тирозина иммунорецептора (ITIM). (См. публикацию Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). Fc-рецепторы рассмотрены, например, в публикациях Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие Fc-рецепторы, в том числе рецепторы, которые могут быть обнаружены в будущем, также входят в определение термина “FcR”, приведенного в настоящем описании изобретения.
Термин ”Fc-рецептор” или “FcR” также включает неонатальный рецептор, FcRn, который отвечает за перенос IgG матери в плод (Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994)) и регуляцию гомеостаза иммуноглобулинов. Известны способы измерения связывания с FcRn (см., например, публикации Ghetie 1997, Hinton 2004). Связывание с FcRn человека in vivo и время полужизни в кровяном русле полипептидов, связывающихся с высокой степенью аффинности с FcRn человека, можно исследовать, например, в трансгенных мышах, линиях трансфицированных клеток человека, экспрессирующих FcRn человека, или в приматах, которым вводят полипептиды с вариантной Fc-областью.
В публикации WO00/42072 (Presta) описаны варианты антител с повышенным или пониженным связыванием с Fc-рецепторами. Указанная публикация патента включена в настоящее описание изобретения в качестве ссылки. См. также публикацию Shields et al., J. Biol. Chem. 9(2):6591-6604 (2001).
“Эффекторные клетки человека” являются лейкоцитами, которые экспрессируют один или несколько Fc-рецепторов и выполняют эффекторные функции. В определенных вариантах осуществления изобретения указанные клетки экспрессируют по меньшей мере FcγRIII и выполняют одну или несколько эффекторных функций ADCC. Примеры лейкоцитов человека, опосредующих ADCC, включают мононуклеарные клетки периферической крови (РВМС), естественные клетки-киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы. Эффекторные клетки могут быть выделены из естественного источника, например, из крови.
“Антителозависимая клеточно-опосредованная цитотоксичность” или “ADCC” означает форму цитотоксичности, в соответствии с которой секретированный Ig, связанный Fc-рецепторами (FcR), присутствующими на определенных цитотоксических клетках (например, естественных клетках-киллерах (NK), нейтрофилах и макрофагах), позволяет указанным цитотоксическим эффекторным клеткам специфически связываться с несущей антиген клеткой-мишенью и затем уничтожать указанную клетку-мишень цитотоксинами. Основные клетки, опосредующие ADCC, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гемопоэтических клетках суммирована в таблице 3 на странице 464 публикации Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Для оценки активности ADCC представляющей интерес молекулы может быть выполнен анализ ADCC in vitro, описанный в патенте США № 5500362 или 5821337 или в патенте США № 6737056, выданном Преста. Эффекторные клетки, пригодные для выполнения таких анализов, включают мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). Альтернативно или дополнительно активность ADCC представляющей интетрес молекулы можно определить in vivo, например, в животной модели, аналогичной рассмотренной в публикации Clynes et al., PNAS (USA) 95:652-656 (1998).
Термин ”комплементзависимая цитотоксичность” или “CDC” означает лизис клетки-мишени в присутствии комплемента. Классический путь активации комплемента инициируется в результате связывания первого компонента системы комплементов (C1q) с антителами (соответствующего подкласса), связанными с родственным антигеном. Для определения активации комплемента может быть выполнен анализ CDC, описанный, например, в публикации Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или пониженной C1q-связывающей способностью описаны, например, в патенте США № 6194551 В1 и WO99/51642. Указанные публикации патентов включены в настоящее описание изобретения в качестве ссылки. См. также, публикацию Idusogie et al., J. Immunol. 164:4178-4184 (2000).
Термин ”полипепитид, содержащий Fc-область” означает полипептид, такой как антитело или иммуноадгезин, содержащий Fc-область. С-концевой лизин (остаток 447 по системе нумерации EU) Fc-области может быть удален, например, в процессе очистки полипептида или создания способами рекомбинантных ДНК нуклеиновой кислоты, кодирующей данный полипептид. Поэтому композиция, содержащая полипептид, имеющий Fc-область по настоящему изобретению, может содержать полипептиды с остатком К447, без остатка К447 или смесь полипептидов с остатком К447 и без указанного остатка.
“Остовная область акцептора человека” в соответствии с целями настоящего изобретения означает остовную область, содержащую аминокислотную последовательность остовной области VL или VH, выделенную из остовной области иммуноглобулина человека или консенсусной остовной области человека. Остовная область акцептора человека, ”выделенная из” остовной области иммуноглобулина человека или консенсусной остовной области человека, может содержать такую же аминокислотную последовательность или может содержать ранее выполненные замены в аминокислотной последовательности. В некоторых вариантах осуществления изобретения число ранее выполненных замен аминокислот равно 10 или меньше, 9 или меньше, 8 или меньше, 7 или меньше, 6 или меньше, 5 или меньше, 4 или меньше, 3 или меньше либо 2 или меньше. Ранее выполненные замены аминокислот в VH, предпочтительно, находятся только в трех, двух или одном из положений 71Н, 73Н и 78Н; например, аминокислотными остатками в указанных положениях могут быть 71А, 73Т и/или 78А. В одном из вариантов осуществления изобретения остовная область VL акцептора человека идентична последовательности остовной области VL иммуноглобулина человека или консенсусной остовной области человека.
Термин ”консенсусная остовная область человека” означает остовную область, содержащую наиболее часто встречающиеся аминокислотные остатки при выборе последовательностей остовной области VL или VH иммуноглобулина человека. Последовательности VL или VH иммуноглобулина человека обычно выбирают из подгруппы последовательностей вариабельных доменов. Указанная подгруппа последовательностей обычно является подгруппой, описанной в публикации Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda MD (1991). В одном из вариантов осуществления изобретения подгруппа для VL является подгруппой каппа-цепи I, описанной в приведенной выше публикации Kabat et al. В одном из вариантов осуществления изобретения подгруппа для VH является подгруппой III, описанной в приведенной выше публикации Kabat et al.
”Консенсусная остовная область подгруппы III VH” включает консенсусную последовательность, полученную из аминокислотных последовательностей в подгруппе III вариабельного домена тяжелой цепи, описанной в приведенной выше публикации Kabat et al. В одном из вариантов осуществления изобретения аминокислотная последовательность консенсусной остовной области подгруппы III VH содержит по меньшей мере часть или всю любую из нижеследующих последовательностей: EVQLVESGGGLVQPGGSLRLSCAAS (FR-H1, SEQ ID NO:21)-HVR-H1-WVRQAPGKGLEWV (FR-H2, SEQ ID NO:22)-HVR-H2-RFTISRDTSKNTLYLQMNSLRAEDTAVYYCAR (FR-H3, SEQ ID NO:23)-HVR-H3-WGQGTLVTVSS (FR-H4, SEQ ID NO:24).
“Консенсусная остовная область подгруппы I VL” включает консенсусную последовательность, полученную из аминокислотных последовательностей в подгруппе I легкой каппа-цепи вариабельного домена, описанной в приведенной выше публикации Kabat et al. В одном из вариантов осуществления изобретения аминокислотная последовательность консенсусной остовной области подгруппы I VH включает по меньшей мере часть или всю любую из нижеследующих последовательностей: DIQMTQSPSSLSASVGDRVTITC (FR-L1, SEQ ID NO:17)-HVR-L1-WYQQKPGKAPKLLIY (FR-L2, SEQ ID NO:18)-HVR-L2-GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (FR-L3, SEQ ID NO:19)-HVR-L3-FGQGTKVEIKR (FR-L4, SEQ ID NO:20).
Термин ”сигнальная последовательность секреции” или “сигнальная последовательность” означает последовательность нуклеиновой кислоты, кодирующую короткий сигнальный пептид, который может быть использован для переноса представляющего интерес вновь синтезированного белка через клеточную мембрану, обычно внутреннюю мембрану или через внутреннюю и наружную мембраны прокариотических клеток. Представляющий интерес белок, такой как полипептид легкой или тяжелой цепи иммуноглобулина, секретируется в периплазму прокариотических клеток-хозяев или в культуральную среду. Сигнальный пептид, кодированный сигнальной последовательностью секреции, может быть эндогенным или экзогенным для клеток-хозяев, включая сигнальные пептиды, нативные для экспрессируемого полипептида. Сигнальные последовательности секреции, обычно локализованные у аминоконца экспрессируемого полипепитида, удаляются ферментами между биосинтезом и секрецией полипептида из цитоплазмы. Таким образом, сигнальный пептид обычно отсутствует в зрелом белковом продукте.
Термин “аминокислота свободного цистеина” означает аминокислотный остаток цистеина, введенный в материнское антитело, который содержит функциональную тиоловую группу (-SH), не спарен и не является частью внутримолекулярной или межмолекулярной дисульфидной мостиковой связи.
Термин ”значение реакционной способности тиоловой группы” является количественной характеристикой реакционной способности аминокислот свободного цистеина. Реакционная способность тиоловой группы определяется процентным значением аминокислоты свободного цистеина в сконструированном антителе с цистеиновой заменой, взаимодействующей с реагентом, активным в отношении тиола, которое может быть преобразовано в максимальное значение, равное 1. Например, аминокислота свободного цистеина в сконструированном антителе с цистеиновой заменой, которая взаимодействует со 100% выходом с реагентом, активным в отношении тиола, таким как реагент, состоящий из биотина и имида малеиновой кислоты, с образованием антитела, меченного биотином, имеет значение реакционной способности тиоловой группы, равное 1,0. Другая аминокислота цистеина, введенная в то же или другое материнское антитело, которая взаимодействует с 80% выходом с реагентом, активным в отношении тиола, имеет значение реакционной способности тиоловой группы, равное 0,8. Другая аминокислота цистеина, введенная в то же или другое материнское антитело, которая вообще не взаимодействует с реагентом, активным в отношении тиола, имеет значение реакционной способности тиоловой группы, равное 0. Значение реакционной способности тиоловой группы определенного цистеина можно определить при помощи анализа ELISA, масс-спектроскопии, жидкостной хроматографии, авторадиографии или других количественных аналитических способов. Реагенты, взаимодействующие с тиолом, которые позволяют удавливать сконструированные антитела с цистеиновой заменой и производить сравнение и количественное определение реакционной способности цистеина, включают биотин-РЕО-малеимид ((+)-биотинил-3-малеимидопропионамидил-3,6-диоксаоктаиндиамин, Oda et al. (2001) Nature Biotechnology 19:379-382, Pierce Biotechnology, Inc.), биотин-ВМСС, РЕО-иодацетилбиотин, иодацетил-LC-биотин и биотин-HPDP (Pierce Biotechnology, Inc.) и Nα-(3-малеимидилпропионил)биоцитин (MPB, Molecular Probes, Eugene, OR). Другие коммерческие реагенты для биотинилирования, бифункциональные и многофункциональные линкерные реагенты могут быть приобретены в компаниях Molecular Probes, Eugene, OR, и Sigma, St. Louis, MO.
“Материнское антитело” является антителом, содержащим аминокислотную последовательность, в которой один или несколько аминокислотных остатков заменены одним или несколькими остатками цистеина. Материнское антитело может содержать нативную последовательность или последовательность дикого типа. Материнское антитело может включать ранее выполненные модификации аминокислотной последовательности (такие как добавления, делеции и/или замены) по сравнению с другим нативным антителом, антителом дикого типа или модифицированными формами антитела. Материнское антитело может быть направлено против представляющего интерес антигена-мишени, например, биологически важного полипептида. В объем настоящего изобретения также входят антитела против антигенов, не являющихся полипептидами, (таких как опухолеспецифические гликолипидные антигены; см. патент США № 5091178).
Термин “аффинность связывания” обычно означает силу суммарных общих нековалентных взаимодействий между одним связывающим сайтом молекулы (например, антитела) и его партнером связывания (например, антигеном). За исключением особо оговоренных случаев в значении, использованном в настоящем изобретении, термин “аффинность связывания” означает собственную аффинность связывания, которая отражает взаимодействие с отношением 1:1 между членами связывающейся пары (например, антитела и антигена). Аффинность молекулы Х к партнеру Y обычно может быть представлена константой диссоциации (Kd). Аффинность можно измерить обычными способами, известными в данной области, в том числе способами, рассмотренными в настоящем описании изобретения. Низкоаффинные антитела медленно связываются с антигеном и легко диссоциируются, в то время как высокоаффинные антитела связываются с антигеном быстрее и остаются связанными в течение более длительного времени. В данной области известен целый ряд способов измерения аффинности связывания, которые могут быть использованы в соответствии с целями настоящего изобретения. Ниже представлены конкретные иллюстративные варианты осуществления изобретения.
В одном из вариантов осуществления изобретения “Kd” или “значение Kd” по настоящему изобретению измеряют при помощи анализа связывания антигена, меченного радиоактивным изотопом (RIA), выполняемого при использовании Fab-фрагмента представляющего интерес антитела и его антигена в соответствии с нижеследующим описанием. Аффинность связывания Fab-фрагментов с антигеном в растворе измеряют, уравновешивая Fab-фрагмент с (125I)-меченым антигеном в минимальной концентрации в присутствии серии титров немеченого антигена и улавливая связанный антиген на планшете, сенсибилизированном антителом против Fab-фрагмента (Chen et al. (1999) J. Mol. Biol. 293:865-881). С целью создания условий для выполнения данного анализа титрационные микропланшеты (Dynex) сенсибилизируют в течение ночи 5 мкг/мл улавливающего антитела против Fab-фрагмента (Cappel Labs) в 50 мМ карбоната натрия (рН 9,6) и затем блокируют 2% (мас./об.) бычьего сывороточного альбумина в PBS в течение двух-пяти часов при комнатной температуре (примерно 23°С). На неадсорбирующем планшете (Nunc № 269620) 100 пМ или 26 пМ [125I]-меченого антигена смешивают с серийно разведенным представляющим интерес Fab-фрагментом (например, в соответствии с оценкой антитела против VEGF, Fab-12, представленной в публикации Presta et al. (1997) Cancer Res. 57:4593-4599). Представляющий интерес Fab-фрагмент затем инкубируют в течение ночи; однако инкубация может быть произведена в течение более продолжительного времени (например, в течение примерно 65 часов) для гарантии достижения равновесия. Затем смесь переносят на улавливающий планшет для инкубации при комнатной температуре (например, в течение одного часа). Раствор удаляют и планшет восемь раз промывают 0,1% твина-20 в PBS. После высыхания планшетов добавляют 150 мкл/лунку сцинтиллятора (MicroScint-20™, Packard) и производят подсчет на планшетах счетчиком гамма-излучения Topcount (Packard) в течение десяти минут. В конкурентно-связывающих анализах используют концентрации Fab-фрагментов, которые обеспечивают менее 20% или соответствуют 20% максимального связывания.
В соответствии с другим вариантом осуществления изобретения Kd или значение Kd измеряют при помощи анализов резонанса поверхностных плазмонов в устройстве BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С на чипах с иммобилизованным антигеном СМ5 при ~10 единицах реактивности (RU). Биосенсорные чипы с карбоксиметилированным декстраном (СМ5, BIAcore®, Inc.) активируют гидрохлоридом N-этил-N’-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетата натрия, рН 4,8, до 5 мкг/мл (~0,2 мкМ) и вводят со скоростью потока 5 мкл/минуту до достижения примерно 10 единиц реактивности (RU) связанного белка. После введения антигена вводят 1 М этаноламина для блокирования непрореагировавших групп. Для измерений кинетики вводят Fab-фрагмент в виде двукратных серийных разведений (от 0,78 нМ до 500 нМ) в PBS с 0,05% твина-20 (PBST) при 25°С со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют при помощи простой модели связывания 1:1 Лангмюра (программа оценки BIAcore®, версия 3.2) путем одновременного совмещения сенсограмм ассоциации и диссоциации. Константу равновесной диссоциации (Kd) вычисляют в виде отношения koff/kon. См., например, публикацию Chen, Y. et al. (1999) J. Mol. Biol. 293:865-881). Скорость ассоциации, превышающую 106 М-1 s-1 при выполнении анализа резонанса поверхностных плазмонов, можно определить способом гашения флуоресценции, позволяющим измерить увеличение или уменьшение интенсивности флуоресценции (возбуждение = 295 нм; излучение = 340 нм, полоса пропускания 16 нм) при 25°С с использованием 20 нМ антитела против антигена (в виде Fab-фрагмента) в PBS, рН 7,2, в присутствии возрастающих концентраций антигена, выполняя измерение в спектрометре, таком как спектрометр, оснащенный прерывателем потока, (Aviv Instruments) или спектрофотометр SLM-Aminco серии 8000 (ThermoSpectronic) с перемешиваемой кюветой.
“Скорость ассоциации” или “kon” по настоящему изобретению можно также определить в соответствии с описанием, приведенным выше для использования системы BIAcore™-2000 или BIAcore™-3000 (BIAcore®, Inc., Piscataway, NJ).
“Нарушение” является любым состоянием или заболеванием, которое может быть улучшено в результате лечения веществом/молекулой или способом по настоящему изобретению. В определение данного термина входят хронические и острые нарушения или заболевания, в том числе патологические состояния, провоцирующие возникновение у млекопитающего данного нарушения. Неограничивающими примерами нарушений, подлежащих лечению способом по настоящему изобретению, являются раковые состояния, такие как рак или метастазы предстательной железы, легкого и ободочной кишки.
Термины ”пролиферативное нарушение клеток” и “пролиферативное нарушение” означают нарушения, обусловленные некоторой степенью аномальной пролиферации клеток. В одном из вариантов осуществления изобретения пролиферативным нарушением клеток является рак.
Термин ”опухоль”, как используется в настоящем описании, означает рост и пролиферацию всех неопластических клеток, как злокачественных, так и доброкачественных, и все предраковые и раковые клетки и ткани. Термины ”злокачественное новообразование”, “раковый”, “пролиферативное нарушение клеток”, “пролиферативное нарушение” и “опухоль” не являются взаимоисключающими, как используются в настоящем описании.
Термины ”злокачественное новообразование” и “раковый” означают или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется нерегулируемым ростом клеток. Примеры злокачественных новообразований включают, не ограничиваясь ими, карциному, лимфому, бластому, саркому и лейкоз и лимфоидные злокачественные новообразования. Более конкретные примеры таких злокачественных новообразований включают плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, в том числе мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточный рак легкого, рак брюшины, печеночно-клеточный рак, рак желудка, в том числе желудочно-кишечный рак, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевых путей, гепатому, рак молочной железы, рак ободочной кишки, рак прямой кишки, колоректальный рак, рак эндометрия или матки, рак слюнной железы, рак почки, рак предстательной железы, рак женских наружных половых органов, рак щитовидной железы, карциному печени, рак заднего прохода, рак полового члена, меланому, множественную меланому и В-клеточную лимфому, рак головного мозга, а также рак головы и шеи и связанные метастазы.
“STEAP-1-экспрессирующая клетка” является клеткой, экспрессирующей эндогенный или трансфицированный белок STEAP-1 на поверхности клетки. “STEAP-1-экспрессирующий рак” является рак, включающий клетки, на поверхности которых присутствует белок STEAP-1. “STEAP-1-экспрессирующий рак” продуцирует на поверхности клеток STEAP-1 в количестве, достаточном для того, чтобы антитело против STEAP-1 могло связываться с указанным белком и оказывать терапевтическое воздействие на рак. Рак, который ”сверхэкспрессирует” STEAP-1, является таким раком, в котором STEAP-1 присутствует на поверхности клетки в гораздо большем количестве по сравнению с нераковой клеткой ткани того же типа. Такая сверхэкспрессия может быть вызвана амплификацией гена или более высокой транскрипцией или трансляцией. Сверхэкспрессию STEAP-1 можно определить при выполнении диагностического или прогностического анализа, позволяющего выявить повышенные уровни белка STEAP-1 на поверхности клетки (например, при помощи иммуногистохимического анализа, анализа FACS). Альтернативно или дополнительно можно измерить уровни нуклеиновой кислоты, кодирующей STEAP-1, или мРНК в клетке, например, при помощи флуоресцентной гибридизации in situ (FISH, см. заявку WO98/45479, опубликованную в октябре 1998 г.), саузерн-блоттинга, нозерн-блоттинга или способами полимеразной цепной реакции (ПЦР), такими как количественная ПЦР в реальном времени (RT-ПЦР). Также можно исследовать сверхэкспрессию STEAP-1, измеряя ”слущивающийся” антиген в биологической жидкости, такой как сыворотка, например, при помощи анализов с использованием антитела (см. также, например, патент США № 4933294, выданный 12 июня 1990 г.; заявку WO91/05264, опубликованную 18 апреля 1991 г.; патент США № 5401638, выданный 28 марта 1995 г, и публикацию Sias et al., J. Immunol. Methods 132:73-80 (1990)). Помимо вышеуказанных анализов специалистам известны разные анализы in vivo. Например, можно воздействовать на клетки в организме индивида антителом, необязательно меченным детектируемой меткой, такой как радиоактивный изотоп, и определить связывание антитела с клетками индивида, например, путем наружного сканирования радиоактивности или анализа биопсийного образца, полученного у индивида до введения антитела. STEAP-1-экспрессирующим раком является рак предстательной железы, легкого и ободочной кишки.
Как используется в настоящем описании, термин ”лечение” (и варианты данного термина, такие как ”лечить”) означает клиническое вмешательство в попытке изменить естественное развитие заболевания у подвергаемого лечению индивида и может проводиться для профилактики или в процессе клинической патологии. Желаемые эффекты лечения включают, не ограничиваясь ими, предотвращение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любых прямых или непрямых патологических последствий заболевания, предотвращение возникновения метастазов, уменьшение интенсивности развития заболевания, улучшение или временное ослабление проявления болезни и ремиссию или лучший прогноз. В некоторых вариантах осуществления изобретения антитела по настоящему изобретению используют для задержки или замедления развития заболевания или нарушения.
Вышеуказанные показатели оценки успешного лечения или уменьшения проявления болезни можно легко измерить обычными способами, известными врачам. Эффективность лечения рака можно измерить, оценивая время до прогрессирования заболевания (ТТР) и/или определяя скорость реакции (RR). В случае рака предстательной железы эффективность лечения можно оценить обычными способами, измеряя уровни простатоспецифического антигена (PSA) в сыворотке; чем выше уровни PSA в крови, тем более распространенным является рак. Существуют коммерческие анализы для обнаружения PSA, например, наборы для анализа PSA Tandem-E и Tandem-R Hybitech, поликлональный анализ Yang ProsCheck (Yang Labs, Bellevue, WA), Abbott Imx (Abbott Labs, Abbott Park, IL) и другие. Метастазирование можно определить при помощи анализов определения стадии заболевания, сканирования костей и исследования уровня кальция и других ферментов с целью определения проникновения метастазов в кости. Также может быть выполнено сканирование с помощью компьютерной томографии для обнаружения проникновения метастазов в таз и лимфатические узлы. Рентгеновское исследование грудной клетки и измерение уровней ферментов в печени известными способами выполняют для обнаружения метастазов, соответственно, в легких и печени. Другие стандартные способы контроля за развитием болезни включают ректальную ультразвуковую эхографию (TRUS) и ректальную пункционную биопсию (TRNB).
“Индивид” является позвоночным. В определенных вариантах осуществления изобретения позвоночное является млекопитающим. Млекопитающие включают, но ими не ограничиваясь, сельскохозяйственных животных (таких как коровы), спортивных животных, домашних животных (таких как кошки, собаки и лошади), приматов, мышей и крыс. В определенных вариантах осуществления изобретения млекопитающее является человеком.
Термин ”эффективное количество” означает количество лекарственного средства, которое при вводимых дозах и в течение необходимых периодов времени позволяет достичь требуемого терапевтического или профилактического результата. “Терапевтически эффективное количество” вещества/молекулы по настоящему изобретению может изменяться в зависимости от таких факторов как состояние болезни, возраст, пол, масса тела индивида и способности данного вещества или молекулы вызывать желаемую реакцию у индивида. Терапевтически эффективным количеством является такое количество, в котором любые токсические или вредные эффекты данного вещества/молекулы перевешиваются терапевтически благоприятным воздействием. ”Профилактически эффективное количество” означает количество, позволяющее при вводимых дозах и в течение необходимых периодов времени достичь желаемого профилактического результата. Поскольку профилактическая доза используется для введения индивидам до возникновения заболевания или на ранней стадии заболевания, то профилактически эффективное количество должно быть меньше терапевтически эффективного количества. В случае рака терапевтически эффективное количество лекарственного средства может уменьшить число раковых клеток, сократить объем опухоли, ингибировать (то есть замедлить в некоторой степени или, предпочтительно, прекратить) инфильтрацию раковых клеток в периферические органы, ингибировать (то есть замедлить в некоторой степени или, предпочтительно, прекратить) метастазирование опухоли, подавить в некоторой степени рост опухоли и/или ослабить в некоторой степени один или несколько симптомов, связанных с раком. См. предыдущее определение термина ”лечение”. Лекарственное средство может быть цитостатическим и/или цитотоксическим в зависимости от степени, в которой оно может предотвращать рост и/или уничтожать существующие раковые клетки.
“Постоянное” введение означает введение одного или нескольких средств в продолжительном режиме, в отличие от экстренного режима введения, с целью сохранения первоначального терапевтического эффекта (активности) в течение продолжительного периода времени. “Периодическое” введение является такой формой лечения, которое проводится с перерывом и является циклическим по своему характеру.
Введение ”в комбинации с” одним или несколькими другими терапевтическими средствами включает одновременное и последовательное введение в любом порядке.
Термин “носители”, как используется в настоящем описании, включают фармацевтически приемлемые носители, наполнители или стабилизаторы, которые являются нетоксичными для клетки или млекопитающего в используемых дозах и концентрациях. Физиологически приемлемым носителем часто является водный забуференный раствор. Примеры физиологически приемлемых носителей включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту; низкомолекулярный (менее примерно 10 остатков) полипептид; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатообразователи, такие как EDTA; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, полиэтиленгликоль (PEG) и PLURONICS™.
Термин ”метка”, как используется в настоящем описании, означает детектируемое соединение или композицию, прямо или косвенно конъюгированную с антителом с образованием ”меченого” антитела. Метка может быть детектируемой сама по себе (например, радиоактивные изотопы или флуоресцентные метки) или, в случае ферментной метки, может катализировать химическое изменение детектируемого соединения или композиции субстрата.
Термин “иммуно-ПЭТ” означает позитрон-эмиссионную томографию (ПЭТ) с использованием антител, меченных радиоактивным изотопом, в том числе конъюгатов антитела с лекарственным средством, таких как ADC антитела против STEAP-1 с vc-ММАЕ или конъюгатов моноклонального тиоантитела с лекарственным средством (TDC). Термин ”89Zr-STEAP-1 иммуно-ПЭТ” означает визуализацию STEAP-1 при помощи ПЭТ с использованием антитела, меченного радиоактивным цирконием.
Термин ”меченый эпитоп”, как используется в настоящем описании, означает химерный полипептид, содержащий полипептид антитела против PSCA, слитый с “полипептидом-меткой”. Полипептид-метка содержит достаточное количество остатков для образования эпитопа, против которого может быть создано антитело, и в то же время является достаточно коротким, чтобы не мешать активности слитого полипептида Ig. Полипептид-метка также, предпочтительно, является достаточно уникальным, чтобы антитело не вступало в перекрестную реакцию с другими эпитопами. Приемлемые полипептиды-метки имеют по меньшей мере шесть аминокислотных остатков, обычно от около 8 до 50 аминокислотных остатков (предпочтительно, от около 10 до 20 аминокислотных остатков).
”Мелкая молекула” в значении, как используется в настоящем описании, имеет молекулярную массу меньше примерно 500 Дальтон.
Термин ”вкладыш в упаковку” означает инструкции, обычно вкладываемые в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, использовании, дозировке, введении, противопоказаниях и/или предупреждения, относящиеся к использованию таких терапевтических продуктов.
“Выделенная нуклеиновая кислота” является нуклеиновой кислотой, например, РНК, ДНК или смешанным полимером, которая по существу отделена от других последовательностей геномной ДНК, а также от белков и комплексов, таких как рибосомы и полимеразы, которые в естественных условиях сопровождают нативную последовательность. В определение данного термина входит последовательность нуклеиновой кислоты, которая была удалена из естественного окружения и включает изоляты рекомбинантной или клонированной ДНК и химически синтезированные аналоги или аналоги, биологически синтезированные гетерологичными системами. По существу чистая молекула включает выделенные формы молекулы.
Термин “вектор” означает челночные и векторы экспрессии. Плазмидная конструкция обычно также включает ориджин репликации (например, ориджин репликации ColE1) и селектируемый маркер (например, устойчивый к ампициллину или тетрациклину) для репликации и селекции плазмид в бактериях. Термин ”вектор экспрессии” означает вектор, содержащий необходимые регуляторные последовательности или регуляторные элементы для экспрессии антител, в том числе фрагмента антитела по настоящему изобретению, в бактериальных или эукариотических клетках. Приемлемые векторы рассмотрены ниже.
Клетка, продуцирующая антитело против STEAP-1 по настоящему изобретению, включает материнскую клетку гибридомы, например, гибридом, депонированные в АТСС, а также бактериальные и эукариотические клетки-хозяева, в которые введена нуклеиновая кислота, кодирующая указанные антитела. Приемлемые клетки-хозяева рассмотрены ниже.
Термин ”ингибитор роста”, как используется в настоящем описании, означает соединение или композицию, ингибирующую рост клетки, в частности, раковой клетки, экспрессирующей PSCA, in vitro или in vivo. Таким образом, ингибитор роста может быть веществом, которое значительно уменьшает процент PSCA-экспрессирующих клеток в S-фазе. Примеры ингибиторов роста включают агенты, блокирующие развитие клеточного цикла (помимо S-фазы) и вызывающие остановку развития в G1 и М-фазе. Классические блокаторы М-фазы включают препараты барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Агенты, вызывающие остановку развития в G1-фазе, также вызывают прекращение развития в S-фазе, например, ДНК-алкилирующие агенты, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ара-С. Дополнительная информация по данному вопросу представлена в публикации The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, entitled “Cell cycle regulation, oncogenes, and antineoplastic drugs” by Murakami et al. (WB Saunders: Philadelphia, 1995), especially p.13. Таксаны (паклитаксел и доцетаксел) являются противораковыми средствами, выделенными из тиса. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), выделенный из европейского тиса, является полусинтетическим аналогом паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки, предотвращая деполимеризацию, что вызывает ингибирование митоза в клетках.
Термин ”цитотоксический агент”, как используется в настоящем описании, означает вещество, ингибирующее или подавляющее функцию клетки и/или вызывающее гибель или разрушение клетки. В определение данного термина входят радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu), химиотерапевтические средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие средства, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины и ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, в том числе их фрагменты и/или варианты, токсины, ингибиторы роста, лекарственные части и разные противоопухолевые или противораковые средства, рассмотренные ниже. Другие цитотоксические агенты описаны ниже. Агенты, уничтожающие опухолевые клетки, вызывают разрушение опухолевых клеток.
“Токсин” является любым веществом, способным оказывать вредное воздействие на рост или пролиферацию клетки.
“Химиотерапевтическое средство” является химическим соединением, предназначенным для лечения рака. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включающие альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в частности, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (в том числе синтетический аналог топотекан (HYCAMTIN®), СРТ-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (в том числе его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (в том числе синтетические аналоги, KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урамустин, нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности, калихеамицин гамма 1I и калихеамицин омега I1 (см., например, публикацию Agnew, Chem. Intl. Ed. Engl., 33:183-186 (1994)); динемицин, в том числе динемицин А, эсперамицин, а также хромофор неокарциностатин и родственные хромофоры энедииновых антибиотиков хромопротеина), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицин, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин ADRIAMYCIN® (в том числе морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофенольная кислота, ногаламицин, оливомицины, пепломицин, порфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; ангалоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадренергические средства, такие как аминоглутетимид, митотан, трилостан; восполнитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамид-гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бизантрен; эдатраксат; дефофамин; демекольцин; диазиквон; элфорнитин; ацетат эллиптиния; эпофилон; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидаимин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; комплекс полисахарида PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2’,2”-трихлортриэтиламин; трихотецены (в частности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид (”Ara-C”); тиотепа; таксоиды, например, паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), препарат паклитаксела из наночастиц альбумина без кремофора ABRAXANE® (American Pharmaceutical Partners, Schaumberg, Illinois) и доцетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин (GEMZAR®); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (VELBAN®); платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин (ONCOVIN®); оксалиплатин; лейкововин; винорелбин (NAVELBINE®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин (XELODA®); фармацевтически приемлемые соли, кислоты или производные вышеуказанных средств, а также комбинации двух или более вышеуказанных средств, такие как СНОР, аббревиатура комбинированной терапии, включающей циклофосфамид, доксорубицин, винкристин и преднизолон, и FOLFOX, аббревиатура схемы лечения, включающей оксалиплатин (ELOXATIN™) в сочетании с 5-FU и лейкововином.
В определение данного термина также входят антигормональные средства, регулирующие, уменьшающие, блокирующие или ингибирующие действие гормонов, способных стимулировать рост рака, которые часто назначают в виде системного или общего лечения. Указанные средства могут быть самими гормонами. Примеры таких средств включают антиэстрогены и селективные модуляторы рецепторов эстрогенов (SERM), включающие, например, тамоксифен (в том числе тамоксифен NOLVADEX®), ралоксифен EVISTA®, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен FARESTON®; антипрогестеронные средства; средства, снижающие уровень рецепторов эстрогена (ERD); средства, подавляющие или блокирующие функцию яичников, например, агонисты релизинг-фатора лютеинизирующего гормона (LHRH), такие как ацетат лейпролида LUPRON® и ELIGARD®, ацетат госерелина, ацетат бусерелина и триптерелин; другие антиандрогенные средства, такие как флутамид, нилутамид и бикалутамид; и ингибиторы ароматазы, ингибирующие фермент ароматазу, который регулирует продуцирование эстрогенов в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, ацетат мегестрола MEGASE®, эксеместан AROMASIN®, форместан, фадрозол, ворозол RIVISOR®, летрозол FEMARA® и анастрозол ARIMIDEX®. Кроме того, в такое определение химиотерапевтических средств входят бисфосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат DIDROCAL®, NE-58095, золедроновая кислота/золедронат ZOMETA®, алендронат FOSAMAX®, памидронат AREDIA®, тилудронат SKELID® или ризедронат ACTONEL®, а также троксацитабин (аналог 1,3-диоксоланнуклеозидцитозина); антисмысловые олигонуклеотиды, ингибирующие экспрессию генов в сигнальных путях, определяющих пролиферацию аберрантных клеток, такие как, например, РКС-альфа, Raf, H-Ras, и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генотерапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 LURTOTECAN®; rmRH ABARELIX®; дитозилат лапатиниба (двойной низкомолекулярный ингибитор тирозинкиназы, включающий ErbB-2 и EGFR, известный также как GW572016); и фармацевтически приемлемые соли, кислоты или производные вышеуказанных средств.
Термин ”ингибитор роста”, как используется в настоящем описании, означает соединение или композицию, ингибирующую рост клетки (в частности, клетки, экспрессирующей STEAP-1), in vitro или in vivo. Таким образом, ингибитор роста может быть веществом, которое значительно уменьшает процент клеток (например, клеток, экспрессирующих STEAP-1) в S-фазе. Примеры ингибиторов роста включают агенты, блокирующие развитие клеточного цикла (помимо S-фазы) и вызывающие остановку развития в G1 и М-фазе. Классические блокаторы М-фазы включают препараты барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Агенты, вызывающие остановку развития в G1-фазе, также вызывают прекращение развития в S-фазе, например, ДНК-алкилирующие агенты, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ара-С. Дополнительная информация по данному вопросу представлена в публикации The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, entitled “Cell cycle regulation, oncogenes, and antineoplastic drugs” by Murakami et al. (WB Saunders: Philadelphia, 1995), especially p.13. Таксаны (паклитаксел и доцетаксел) являются противораковыми средствами, выделеннымии из тиса. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), выделенный из европейского тиса, является полусинтетическим аналогом паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки, предотвращая деполимеризацию, что вызывает ингибирование митоза в клетках.
Термин ”внутриклеточный метаболит” означает соединение, образовавшееся в результате процесса метаболизма или взаимодействия внутри клетки с участием конъюгата антитела с лекарственным средством (ADC). Процесс метаболизма или взаимодействие могут быть ферментативным процессом, таким как протеолитическое расщепление пептидного линкера ADC или гидролиз функциональной группы, такой как гидразон, сложный эфир или амид. Внутриклеточные метаболиты включают, не ограничиваясь ими, антитела и свободное лекарственное средство, подвергшиеся внутриклеточному расщеплению после проникновения, диффузии, поглощения или переноса в клетку.
Термины ”расщепленный внутри клетки” и “внутриклеточное расщепление” означают процесс метаболизма или взаимодействие внутри клетки с участием конъюгата антитела с лекарственным средством (ADC), в результате которого разрушается ковалентная связь, то есть линкер, между лекарственной частью (D) и антителом (Ab) и свободное лекарственное средство диссоциируют из антитела в клетку. Расщепленные части ADC являются внутриклеточными метаболитами.
Термин ”биологическая доступность” означает системную доступность (то есть на уровнях крови/плазмы) данного количества лекарственного средства, введенного индивиду. Биологическая доступность является абсолютным термином, который позволяет измерить время (скорость) и общее количество (степень) лекарственного средства, поступившего в кровоток из введенной лекарственной формы.
Термин ”цитотоксическая активность” означает уничтожающее клетку, цитостатическое или ингибирующее рост действие конъюгата антитела с лекарственным средством или внутриклеточного метаболита конъюгата антитела с лекарственным средством. Цитотоксическая активность может быть выражена в виде значения IC50, которое означает концентрацию (молярную или массовую) на единицу объема, при которой выживает половина клеток.
”Алкил” является С1-С18 углеводородом, содержащим нормальные, вторичные, третичные или циклические атомы углерода. Примеры алкилов включают метил (Ме, -СН3), этил (Et, -СН2СН3), 1-пропил (n-Pr, н-пропил, -СН2СН2СН3), 2-пропил (i-Pr, изопропил, -СН(СН3)2), 1-бутил (n-Bu, н-бутил, -СН2СН2СН2СН3), 2-метил-1-пропил (i-Bu, изобутил, -СН2СН(СН3)2), 2-бутил (s-Bu, втор-бутил, -СН(СН3)СН2СН3), 2-метил-2-пропил (t-Bu, трет-бутил, -С(СН3)3), 1-пентил (н-пентил, -СН2СН2СН2СН2СН3), 2-пентил (-СН(СН3)СН2СН2СН3), 3-пентил (-СН(СН2СН3)2), 2-метил-2-бутил (-С(СН3)2СН2СН3), 3-метил-2-бутил (-СН(СН3)СН(СН3)2), 3-метил-1-бутил (-СН2СН2СН(СН3)2), 2-метил-1-бутил (-СН2СН(СН3)СН2СН3), 1-гексил (-СН2СН2СН2СН2СН2СН3), 2-гексил (-СН(СН3)СН2СН2СН2СН3), 3-гексил (-СН(СН2СН3)(СН2СН2СН3)), 2-метил-2-пентил (-С(СН3)2СН2СН2СН3), 3-метил-2-пентил (-СН(СН3)СН(СН3)СН2СН3), 4-метил-2-пентил (-СН(СН3)СН2СН(СН3)2), 3-метил-3-пентил (-С(СН3)(СН2СН3)2), 2-метил-3-пентил (-СН(СН2СН3)СН(СН3)2), 2,3-диметил-2-бутил (-С(СН3)2СН(СН3)2), 3,3-диметил-2-бутил (-СН(СН3)С(СН3)3).
Термин ”C1-C8 алкил”, как используется в настоящем описании, означает насыщенный или ненасыщенный углеводород с прямой или разветвленной цепью, содержащий 1-8 атомов углерода. Типичные ”C1-C8 алкильные” группы включают, не ограничиваясь ими, -метил, -этил, -н-пропил, -н-бутил, -н-пентил-, -н-гексил, -н-гептил, -н-октил, -н-нонил и -н-децил; С1-С8 алкилы с разветвленной цепью включают, не ограничиваясь ими, -изопропил, -втор-бутил, -изобутил, -трет-бутил, -изопентил, 2-метилбутил, ненасыщенные С1-С8 алкилы включают, не ограничиваясь ими, -винил, -аллил, -1-бутенил, -2-бутенил, -изобутиленил, -1-пентенил, -2-пентенил, -3-метил-1-бутенил, -2-метил-2-бутенил, -2,3-диметил-2-бутенил, 1-гексил, 2-гексил, 3-гексил, -ацетиленил, -пропинил, -1-бутинил, -2-бутинил, -2-пентинил, -2-пентинил, -3-метил-1-бутинил, метил, этил, пропил, изопропил, н-бутил, изобутил, втор-бутил, трет-бутил, н-пентил, изопентил, неопентил, н-гексил, изогексил, 2-метилпентил, 3-метилпентил, 2,2-диметилбутил, 2,3-диметилбутил, 2,2-диметилпентил, 2,3-диметилпентил, 3,3-диметилпентил, 2,3,4-триметилпентил, 3-метилгексил, 2,2-диметилгексил, 2,4-диметилгексил, 2,5-диметилгексил, 3,5-диметилгексил, 2,4-диметилпентил, 2-метилгептил, 3-метилгептил, н-гептил, изогептил, н-октил и изооктил. С1-С8 алкильная группа может быть не замещена или замещена одной или несколькими группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -С(О)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2-NHC(O)R', -SO3R', -S(O)2R', -S(O)R’, -OH, -галоген, -N3, -NH2, NH(R'), -N(R')2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила и арила.
“Алкенил” является С2-С18 углеводородом, содержащим нормальные, вторичные, третичные или циклические атомы углерода при наличии по меньшей мере одного центра ненасыщенности, то есть углерод-углеродной, sp2 двойной связи. Примеры алкенилов включают, не ограничиваясь ими, этилен или винил (-СН=СН2), аллил (-СН2СН=СН2), циклопентенил (-С5Н7) и 5-гексенил (-СН2СН2СН2СН2СН=СН2).
“Алкинил” является С2-С18 углеводородом, содержащим нормальные, вторичные, третичные или циклические атомы углерода при наличии по меньшей мере одного центра ненасыщенности, то есть углерод-углеродной, sp тройной связи. Примеры алкинилов включают, не ограничиваясь ими, ацетилен (-С≡СН) и пропаргил (-СН2С≡СН).
“Алкилен” означает насыщенный углеводородный радикал с разветвленной или прямой цепью или циклической структурой, содержащий 1-18 атомов углерода и имеющий два центра одновалентных радикалов, полученных в результате удаления двух атомов водорода в положении одинаковых или двух разных атомов углерода исходного алкана. Типичные алкиленовые радикалы включают, не ограничиваясь ими, метилен (-СН2-), 1,2-этил (-СН2СН2-), 1,3-пропил (-СН2СН2СН2-), 1,4-бутил (-СН2СН2СН2СН2-) и тому подобные.
“С1-С10 алкилен” является насыщенной углеводородной группой с прямой цепью формулы -(СН2)1-10-. Примеры С1-С10 алкилена включают метилен, этилен, пропилен, бутилен, пентилен, гексилен, гептилен, оцитилен, нонилен и декален.
“Алкенилен” означает ненасыщенный углеводородный радикал с разветвленной или прямой цепью или циклической структурой, содержащий 2-18 атомов углерода и имеющий два центра одновалентных радикалов, полученных в результате удаления двух атомов водорода в положении одинаковых или двух разных атомов углерода исходного алкена. Типичные алкиленовые радикалы включают, не ограничиваясь ими, 1,2-этилен (-СН=СН-).
”Алкинилен” означает ненасыщенный углеводородный радикал с прямой или разветвленной цепью или циклической структурой, содержащий 2-18 атомов углерода и имеющий два центра одновалентных радикалов, полученных в результате удаления двух атомов водорода в положении одинаковых или двух разных атомов углерода исходного алкина. Типичные алкиленовые радикалы включают, не ограничиваясь ими, ацетилен (-С≡С-), пропаргил (-СН2С≡С-) и 4-пентинил (-СН2СН2СН2С≡С-).
“Арил” означает карбоциклическую ароматическую группу. Примеры арильных групп включают, не ограничиваясь ими, фенил, нафтил и антраценил. Карбоциклическая ароматическая группа или гетероциклическая ароматическая группа может быть не замещена или замещена одной или несколькими группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -С(О)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R’), -N(R’)2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила или арила.
“Арилен” является арильной группой, которая имеет две ковалентные связи и может иметь орто-, мета- или пара-конфигурацию, показанную следующими формулами:
где фенильная группа может быть не замещена или замещена одной-четырьмя группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2-NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила и арила.
“Арилалкил” означает ациклический алкильный радикал, в котором один из атомов водорода, связанный с атомом углерода, обычно с концевым или sp3 атомом углерода, заменен арильным радикалом. Типичные арилалкильные группы включают, не ограничиваясь ими, бензил, 2-фенилэтан-1-ил, 2-фенилэтен-1-ил, нафтилметил, 2-нафтилэтан-1-ил, 2-нафтилэтен-1-ил, нафтобензил, 2-нафтофенилэтан-1-ил и тому подобные. Арилалкильная группа включает 6-20 атомов углерода, например, алкильная часть, включая алканильные, алкенильные или алкинильные группы, арилалкильной группы, содержит 1-6 атомов углерода и арильная часть содержит 5-14 атомов углерода.
”Гетероарилалкил” означает ациклический алкильный радикал, в котором один из атомов водорода, связанный с атомом углерода, обычно с концевым или sp3 атомом углерода, заменен гетероарильным радикалом. Типичные гетероарилалкильные группы включают, не ограничиваясь ими, 2-бензимидазолилметил, 2-фурилэтил и тому подобные. Гетероарилалкильная группа включает 6-20 атомов углерода, например, алкильная часть, включая алканильные, алкенильные или алкинильные группы, гетероарилалкильной группы содержит 1-6 атомов углерода и гетероарильная часть содержит 5-14 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S. Гетероарильная часть гетероарилалкильной группы может быть моноциклической группой, содержащей 3-7 членов в кольце (2-6 атомов углерода), или бициклической группой, содержащей 7-10 членов в кольце (4-9 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S), например, бицикло [4,5],[5,5],[5,6] или [6,6] системой.
Термины “замещенный алкил”, “замещенный арил” и “замещенный арилалкил” означают соответственно алкил, арил и арилалкил, в которых один или несколько атомов водорода независимо замещены заместителем. Типичные заместители включают, не ограничиваясь ими, -X, -R, -O-, -OR, -SR, -S-, -NR2, -NR3, =NR, -CX3, -CN, -OCN, -SCN, -N=C=O, -NCS, -NO, -NO2, =N2, -N3, NC(=O)R, -C(=O)R, -C(=O)NR2, -SO3-, -SO3H, -S(=O)2R, -OS(=O)2OR, -S(=O)2NR, -S(=O)R, -OP(=O)(OR)2, -P(=O)(OR)2, -PO-3, -PO3H2, -C(=O)R, -C(=O)X, -C(=S)R, -CO2R, -СО2-, -C(=S)OR, -C(=O)SR, -C(=S)SR, -C(=O)NR2, -C(=S)NR2, -C(=NR)NR2, где каждый элемент Х независимо означает галоген: F, Cl, Br или I, и каждый элемент R независимо означает -Н, С2-С18 алкил, С6-С20 арил, С3-С14 гетероцикл, защитную группу или пролекарство. Вышеописанные алкиленовые, алкениленовые и алкиниленовые группы могут быть также замещены аналогичным образом.
Термины ”гетероарил” и “гетероцикл” означают кольцевую систему, в которой один или несколько атомов в кольце являются гетероатомами, например, атомами азота, кислорода и серы. Гетероциклический радикал включает 1-20 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S. Гетероцикл может быть моноциклом, имеющим 3-7 членов в кольце (2-6 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S), или бициклом, имеющим 7-10 членов в кольце (4-9 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S), например: бицикло [4,5], [5,5], [5,6] или [6,6] системой.
Гетероциклы описаны в публикациях Paquette, Leo A., “Principles of Modern Heterocyclic Chemistry” (W.A. Benjamin, New York, 1968), в частности, главы 1, 3, 4, 6, 7 и 9; “The Chemistry of Heterocyclic Compounds, A series of Monographs” (John Wiley & Sons, New York, 1950 to present), в частности, тома 13, 14, 16, 19 и 28; и J. Am. Chem. Soc. (1960) 82:5566.
Примеры гетероциклов включают, не ограничиваясь ими, пиридил, дигидропиридил, тетрагидропиридил (пиперидил), тиазолил, тетрагидротиофенил, тетрагидротиофенил с окисленной серой, пиримидинил, фуранил, тиенил, пирролил, пиразолил, имидазолил, тетразолил, бензофуранил, тианафталенил, индолил, индоленил, хинолинил, изохинолинил, бензимидазолил, пиперидинил, 4-пиперидонил, пирролидинил, 2-пирролидонил, пирролинил, тетрагидрофуранил, бис-тетрагидрофуранил, тетрагидропиранил, бис-тетрагидропиранил, тетрагидрохинолинил, тетрагидроизохинолинил, декагидрохинолинил, октагидроизохинолинил, азоцинил, триазинил, 6Н-1,2,5-тиадиазинил, 2Н,6Н-1,5,2-дитиазинил, тиенил, тиантренил, пиранил, изобензофуранил, хроменил, ксантенил, феноксатинил, 2Н-пирролил, изотиазолил, изоксазолил, пиразинил, пиридазинил, индолизинил, изоиндолил, 3Н-индолил, 1Н-индазолил, пуринил, 4Н-хинолизинил, фталазинил, нафтиридинил, хиноксалинил, хиназолинил, циннолинил, птеридинил, 4аН-карбазолил, карбазолил, β-карболинил, фенантридинил, акридинил, пиримидинил, фенантролинил, феназинил, фенотиазинил, фуразанил, феноксазинил, изохроманил, хроманил, имидазолидинил, имидазолинил, пиразолидинил, пиразолинил, пиперазинил, индолинил, изоиндолинил, хинуклидинил, морфолинил, оксазолидинил, бензотриазолил, бензизоксазолил, оксиндолил, бензоксазолинил и изатиноил.
В качестве примера, не ограничивающего объем изобретения, можно привести гетероциклы, связанные углеродной связью в положении 2, 3, 4, 5 или 6 пиридина, в положении 3, 4, 5 или 6 пиридазина, в положении 2, 4, 5 или 6 пиримидина, в положении 2, 3, 5 или 6 пиразина, в положении 2, 3, 4 или 5 фурана, тетрагидрофурана, тиофурана, тиофена, пиррола или тетрагидропиррола, в положении 2, 4 или 5 оксазола, имидазола или тиазола, в положении 3, 4 или 5 изоксазола, пиразола или изотиазола, в положении 2 или 3 азиридина, в положении 2, 3 или 4 азетидина, в положении 2, 3, 4, 5, 6, 7 или 8 хинолина или в положении 1, 3, 4, 5, 6, 7 или 8 изохинолина. Связанные углеродной связью гетероциклы в еще более типичных случаях включают 2-пиридил, 3-пиридил, 4-пиридил, 5-пиридил, 6-пиридил, 3-пиридазинил, 4-пиридазинил, 5-пиридазинил, 6-пиридазинил, 2-пиримидинил, 4-пиримидинил, 5-пиримидинил, 6-пиримидинил, 2-пиразинил, 3-пиразинил, 5-пиразинил, 6-пиразинил, 2-тиазолил, 4-тиазолил или 5-тиазолил.
В качестве примера, не ограничивающего объем изобретения, можно привести гетероциклы, связанные азотной связью в положении 1 азиридина, азетидина, пиррола, пирролидина, 2-пирролина, 3-пирролина, имидазола, имидазолидина, 2-имидазолина, 3-имидазолина, пиразола, пиразолина, 2-пиразолина, 3-пиразолина, пиперидина, пиперазина, индола, индолина, 1Н-индазола, в положении 2 изоиндола или изоиндолина, в положении 4 морфолина и в положении 9 карбазола или β-карболина. Связанные азотной связью гетероциклы в еще более типичных случаях включают 1-азиридил, 1-азетедил, 1-пирролил, 1-имидазолил, 1-пиразолил и 1-пиперидинил.
“C3-C8 гетероцикл” означает ароматическое или неароматическое С3-С8 углеродное кольцо, в котором один-четыре атома углерода в кольце независимо замещены гетероатомом из группы, состоящей из О, S и N. Типичные примеры С3-С8 гетероцикла включают, не ограничиваясь ими, бензофуранил, бензотиофен, индолил, бензопиразолил, кумаринил, изохинолинил, пирролил, тиофенил, фуранил, тиазолил, имидазолил, пиразолил, триазолил, хинолинил, пиримидинил, пиридинил, пиридонил, пиразинил, пиридазинил, изотиазолил, изоксазолил и тетразолил. С3-С8 гетероцикл может быть не замещен или замещен одной-семью группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила и арила.
“С3-С8 гетероцикло” означает вышеуказанную С3-С8 гетероциклическую группу, в которой один из атомов водорода гетероциклической группы заменен связью. C3-C8 гетероцикло может быть не замещен или замещен одной-шестью группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила и арила.
“Углеродное кольцо” означает насыщенное или ненасыщенное кольцо, содержащее 3-7 атомов углерода в виде моноцикла или 7-12 атомов углерода в виде бицикла. Моноциклические углеродные кольца имеют 3-6 атомов в кольце, обычно 5-6 атомов в кольце. Бициклические углеродные кольца имеют 7-12 атомов в кольце, например, расположенные в виде бицикло [4,5],[5,5],[5,6] или [6,6] системы, либо 9 или 10 атомов в кольце, расположенных в виде бицикло [5,6] или [6,6] системы. Примеры моноциклических углеродных колец включают циклопропил, циклобутил, циклопентил, 1-циклопент-1-енил, 1-циклопент-2-енил, 1-циклопент-3-енил, циклогексил, 1-циклогекс-1-енил, 1-циклогекс-2-енил, 1-циклогекс-3-енил, циклогептил и циклооктил.
“С3-С8 углеродное кольцо” является 3-, 4-, 5-, 6-, 7- или 8-членным насыщенным или ненасыщенным неароматическим карбоциклическим кольцом. Типичные С3-С8 углеродные кольца включают, не ограничиваясь ими, -циклопропил, -циклобутил, -циклопентил, -циклопентадиенил, -циклогексил, -циклогексенил, -1,3-циклогексадиенил, -1,4-циклогексадиенил, -циклогептил, -1,3-циклогептадиенил, -1,3,5-циклогептатриенил, -циклооктил и -циклооктадиенил. С3-С8 карбоциклическая группа может быть не замещена или замещена одной или несколькими группами, которые включают, не ограничиваясь ими, -С1-С8 алкил, -О-(С1-С8 алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN, где каждый элемент R' независимо выбирают из Н, -С1-С8 алкила и арила.
“С3-С8 карбоцикло” означает вышеуказанную С3-С8 карбоциклическую группу, в которой один из атомов водорода заменен связью.
“Линкер” означает химическую часть, включающую ковалентную связь или цепь атомов, которая ковалентно присоединяет антитело к лекарственной части. В разных вариантах осуществления изобретения линкеры включают двухвалентный радикал, такой как алкилдиил, арилдиил, гетероарилдиил, части, такие как -(CR2)nO(CR2)n-, повторяющиеся звенья алкилокси (например, полиэтиленокси, PEG, полиметиленокси) и алкиламино (например, полиэтиленамино, джеффамин™); сложный эфир и амиды двухосновных кислот, в том числе сукцинат, сукцинамид, дигликолят, малонат и капроамид.
Термин ”хиральный” относится к молекулам, обладающим свойством неналожения партнера, являющегося зеркальным отображением, в то время как термин ”ахиральный” относится к молекулам, которые могут налагаться на партнера, являющегося зеркальным отображением.
Термин ”стереоизомеры” означает соединения, которые имеют идентичное химическое строение, но отличаются расположением атомов или групп в пространстве.
“Диастереомер” означает стереоизомер с двумя или более центрами хиральности, молекулы которого не являются зеркальными отображениями друг друга. Диастереомеры обладают другими физическими свойствами, например, температурами плавления, температурами кипения, спектральными свойствами и реакционной способностью. Смеси диастереомеров можно разделить аналитическими способами с высоким разрешением, такими как электрофорез и хроматография.
“Энантиомеры” означают два стереоизомера соединения, которые являются неналагаемыми зеркальными отображениями друг друга.
Определения и условные обозначения стереохимических терминов, использованных в настоящем описании изобретения, как правило, соответствуют терминам, приведенным в публикациях S.P. Parker, Ed., McGraw-Hill Dictionary of Chemical Terms (1984) McGraw-Hill Book Company, New York; и Eliel, E. and Wilen, S., Stereochemistry of Organic Compounds (1994) John Wiley & Sons, Inc., New York. Многие органические соединения существуют в оптически активных формах, то есть такие соединения обладают способностью вращать плоскость плоскополяризованного света. При описании оптически активного соединения используются префиксы D и L, или R и S для обозначения абсолютной конфигурации молекулы вокруг ее хирального центра. Префиксы d и l или (+) и (-) служат для обозначения знака вращения плоскополяризованного света соединением, при этом (-) или l означает, что соединение является левовращающим. Соединение с префиксом (+) или d является правовращающим. Указанные стереоизомеры являются идентичными для данной химической структуры за исключением того, что они являются зеркальными отображениями друг друга. Конкретный стереоизомер может быть также определен как энантиомер, и смесь таких изомеров часто именуется энантиомерной смесью. Смесь энантиомеров с соотношением 50:50 является рацемической смесью или рацематом, которая может возникнуть при отсутствии стереоселекции или стереоспецифичности в химической реакции или процессе. Термины ”рацемическая смесь” и “рацемат” означают эквимолярную смесь энантиомеров двух видов, не обладающих оптической активностью.
“Удаляемая группа” означает функциональную группу, которая может быть замещена другой функциональной группой. Определенные удаляемые группы хорошо известны в данной области, и примеры таких групп включают, не ограничиваясь ими, галогенид (например, хлорид, бромид, иодид), метансульфонил (мезил), паратолуолсульфонил (тозил), трифторметилсульфонил (трифлат) и трифторметилсульфонат.
Аббревиатуры
Линкерные компоненты
МС = 6-малеимидокапроил
Val-Cit или “vc” = валин-цитруллин (типичный дипептид в расщепляемом протеазой линкере)
Цитруллин = 2-амино-5-уреидопентановая кислота
РАВ = парааминобензилоксикарбонил (пример ”жертвующего собой” линкерного компонента)
Me-Val-Cit = N-метил-валин-цитруллин (в котором пептидная связь линкера модифицирована для предотвращения его расщепления катепсином В)
МС(PEG)6-OH = малеимидокапроилполиэтиленгликоль (может быть присоединен к цистеинам антитела)
SPP = N-сукцинимидил-4-(2-пиридилтио)пентаноат
SPDP = N-сукцинимидил-3-(2-пиридилтио)пропионат
SMCC = сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат
IT = иминотиолан
Цитотоксические лекарственные средства
ММАЕ = монометилауристатин Е (молекулярная масса 718)
MMAF = вариант ауристатина Е (ММАЕ) с фенилаланином у С-конца лекарственного средства (молекулярная масса 731,5)
MMAF-DMAEA = MMAF с DMAEA (диметиламиноэтиламин) в амидной связи с С-концевым фенилаланином (молекулярная масса 801,5)
MMAF-TEG = MMAF с тетраэтиленгликолем, этерифицированным до фенилаланина
MMAF-NtBu = N-трет-бутил, присоединенный в виде амина к С-концу MMAF
DM1 = N(2’)-деацетил-N(2’)-(3-меркапто-1-оксопропил)майтанзин
DM3 = N(2’)-деацетил-N2-(4-меркапто-1-оксопентил)майтанзин
DM4 = N(2’)-деацетил-N2-(4-меркапто-4-метил-1-оксопентил)майтанзин
Ниже приведены другие аббревиатуры: АЕ означает ауристатин Е, Вос означает N-(трет-бутоксикарбонил), cit означает цитруллин, dap означает долапроин, DCC означает 1,3-дициклогексилкарбодиимид, DCM означает дихлорметан, DEA означает диэтиламин, DEAD означает диэтилазодикарбоксилат, DEPC означает диэтилфосфорилцианидат, DIAD означает диизопропилазодикарбоксилат, DIEA означает N,N-диизопропилэтиламин, dil означает долаизолейцин, DMA означает диметилацетамид, DMAP означает 4-диметиламинопиридин, DME означает диметиловый эфир этиленгликоля (или 1,2-диметоксиэтан), DMF означает N,N-диметилформамид, DMSO означает диметилсульфоксид, doe означает долафенин, dov означает N,N-диметилвалин, DTNB означает 5,5’-дитиобис(2-нитробензойную кислоту), DTPA означает диэтилентриаминпентауксусную кислоту, DTT означает дитиотреитол, EDCI означает гидрохлорид 1-(3-диметиламинопропил)-3-этилкарбодиимида, EEDQ означает 2-этокси-1-этоксикарбонил-1,2-дигидрохинолин, ES-MS означает электрораспылительную масс-спектрометрию, EtOAc означает этилацетат, Fmoc означает N-(9-флуоренилметоксикарбонил), gly означает глицин, HATU означает гексафторфосфат О-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония, HOBt означает 1-гидроксибензотриазол, HPLC означает жидкостную хроматографию высокого давления, ile означает изолейцин, lys означает лизин, MeCN(CH3CN) означает ацетонитрил, МеОН означает метанол, Mtr означает 4-анизилдифенилметил (или 4-метокситритил), nor означает (1S,2R)-(+)-норэфедрин, PBS означает физиологический раствор с фосфатным буфером (рН 7,4), PEG означает полиэтиленгликоль, Ph означает фенил, Pnp означает паранитрофенил, МС означает 6-малеимидокапроил, phe означает L-фенилаланин, PyBrop означает гексафторфосфат бром-трис-пирролидинофосфония, SEC означает вытеснительную хроматографию, Su означает сукцинимид, TFA означает трифторуксусную кислоту, TLC означает тонкослойную хроматографию, UV означает ультрафиолетовое излучение и val означает валин.
Композиции и способы их получения
Настоящее изобретение относится к антителам, связывающимся с белком STEAP-1. Настоящее изобретение также относится к иммуноконъюгатам, содержащим антитела против STEAP-1. Антитела и иммуноконъюгаты по настоящему изобретению пригодны, например, для диагностики или лечения нарушений, связанных с измененной экспрессией, например, повышенной экспрессией, белка STEAP-1. В определенных вариантах осуществления изобретения антитела или иммуноконъюгаты по настоящему изобретению пригодны для диагностики или лечения пролиферативного нарушения клеток, такого как рак.
Антитела против STEAP-1
В одном из аспектов настоящее изобретение относится к антителам, связывающемся с белком STEAP-1. Некоторые варианты осуществления изобретения относятся к антителам, связывающимся со зрелой формой STEAP-1 человека и обезьяны циномолгус (cyno). В одном таком варианте осуществления изобретения зрелая форма STEAP-1 человека имеет аминокислотную последовательность SEQ ID NO:1 (фигура 1). STEAP-1 обезьяны циномолгус имеет аминокислотную последовательность SEQ ID NO:3 (фигура 1). В некоторых вариантах осуществления изобретения антитело к STEAP-1 связывается со зрелой формой белка STEAP-1, экспрессированного на поверхности клетки. В некоторых вариантах осуществления изобретения антитело, связывающееся со зрелой формой STEAP-1, экспрессированного на поверхности клетки, ингибирует рост указанной клетки. В некоторых вариантах осуществления изобретения антитело против STEAP-1 связывается со зрелой формой STEAP-1, экспрессированного на поверхности клетки, и ингибирует пролиферацию клетки. В определенных вариантах осуществления изобретения антитело против STEAP-1 связывается со зрелой формой STEAP-1, экспрессированного на поверхности клетки, и вызывает гибель клетки. В некоторых вариантах осуществления изобретения антитело против STEAP-1 связывается со зрелой формой STEAP-1, экспрессированного на поверхности раковых клеток. В некоторых вариантах осуществления изобретения антитело против STEAP-1 связывается со зрелой формой STEAP-1, сверхэкспрессированного на поверхности раковых клеток по сравнению с нормальными клетками такой же ткани. В некоторых вариантах осуществления изобретения антитело против STEAP-1 конъюгировано с цитотоксином или детектируемой меткой и связывается с STEAP-1 на поверхности клетки. В некоторых вариантах осуществления изобретения конъюгат антитела с токсином ингибирует рост клетки. В некоторых вариантах осуществления изобретения конъюгат антитела с детектируемой меткой позволяет обнаружить in vitro или in vivo клетку, экспрессирующую на своей поверхности STEAP-1.
В одном из аспектов изобретения антитело против STEAP-1 является моноклональным антителом. В одном из аспектов изобретения антитело против STEAP-1 является фрагментом антитела, например, Fab, Fab'-SH, Fv, scFv или (Fab')2 фрагментом. В одном из аспектов изобретения антитело против STEAP-1 является химерным антителом, гуманизированным антителом или антителом человека. В одном из аспектов изобретения антитела против STEAP-1 по настоящему изобретению являются очищенными антителами.
Настоящее изобретение относится к типичным моноклональным антителам, выделенным из библиотеки фагов. В качестве антигена для скрининга библиотеки был использован полипептид, имеющий аминокислотную последовательность SEQ ID NO:28 или SEQ ID NO:30, соответствующую внеклеточным доменам (ECD) бета- и альфа-цепей STEAP-1. Антитела, полученные в результате скрининга библиотеки, являются антителами с созревшей аффинностью.
В одном из аспектов настоящее изобретение относится к моноклональным антителам, конкурирующим с антителом мыши 120.545, антителом 120 graft и гуманизированным антителом 120v.24 за связыванием с STEAP-1. Настоящее изобретение также относится к моноклональным антителам, которые связываются с тем же эпитопом, что и мышиное антитело 120.545, антитело 120 graft и гуманизированное антитело 120v.24.
В одном из аспектов настоящее изобретение относится к полинуклеотидам, кодирующим антитела против STEAP-1. Определенные варианты осуществления изобретения относятся к векторам, содержащим полинуклеотиды, кодирующие антитела против STEAP-1. Определенные варианты осуществления изобретения относятся к клеткам-хозяевам, включающим такие векторы. Другой аспект настоящего изобретения относится к композициям, содержащим антитела против STEAP-1 или полинуклеотиды, кодирующие антитела против STEAP-1. В определенных вариантах осуществления изобретения композиция по настоящему изобретению является фармацевтическим препаратом, предназначенным для лечения пролиферативного нарушения клеток, указанного в настоящем описании изобретения.
Ниже приведено подробное описание типичных антител против STEAP-1.
1. Специфические варианты антител против STEAP-1
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему вариабельный домен тяжелой цепи, содержащий последовательность SEQ ID NO:9 или 10, показанную на фигуре 2В. В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:6, показанную на фигуре 2А.
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему тяжелую цепь, содержащую последовательность SEQ ID NO:9, в которой была произведена одна или несколько следующих замен аминокислотных остатков в указанном положении по системе Kabat: A24V, V37I, V48M, F67I и L78F. В одном из вариантов осуществления изобретения тяжелая цепь содержит остовную область тяжелой цепи, выбираемую из последовательностей SEQ ID NO:25, 75, 76, 77, 78 и 79. В настоящем описании изобретения остовные области тяжелой цепи обозначены “FR-H1-H4” или “HC-FR1-FR4” и остовные области легкой цепи обозначены “FR-L1-L4” или “LC-FR1-FR4”. В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему легкую цепь, содержащую последовательность SEQ ID NO:6.
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему 1, 2, 3, 4, 5 или 6 последовательностей HVR антитела 120.v24, показанного на фигурах 2А и 2В.
Антитело против STEAP-1 может содержать любую приемлемую последовательность вариабельного домена остовной области при условии сохранения антителом способности связываться с STEAP-1. Например, в некоторых вариантах осуществления изобретения антитела против STEAP-1 по настоящему изобретению содержат консенсусную последовательность остовной области тяжелой цепи подгруппы III человека. В одном из вариантов указанных антител консенсусная последовательность остовной области тяжелой цепи включает замены в положении 24, 37, 48, 67 и/или 78. В одном из вариантов указанных антител в положении 24 находится А или V, в положении 37 находится I или V, в положении 48 находится М или V, в положении 67 находится I или F и/или в положении 78 находится F или L. В одном из вариантов осуществления изобретения указанные антитела включают последовательность остовной области вариабельного домена тяжелой цепи антитела huMab4D5-8, например, SEQ ID NO:21, 22, 23 и 24 (соответственно, FR-H1, FR-H2, FR-H3, FR-H4). Антитело huMab4D5-8 коммерчески известно как антитело против HER2 HERCEPTIN®, Genentech, Inc., South San Francisco, CA, USA; указанное антитело также описано в патентах США № 6407213 и 5821337 и в публикации Lee et al., J. Mol. Biol. (2004), 340(5):1073-93. В одном таком варианте осуществления изобретения указанные антитела далее включают консенсусную последовательность остовной области легкой κI-цепи. В одном таком варианте осуществления изобретения указанные антитела включают последовательность остовной области вариабельного домена легкой цепи антитела huMAb4D5-8, например, SEQ ID NO:17, 18, 19 и 20 (FR-L1, FR-L2, FR-L3, FR-L4).
В одном из вариантов осуществления изобретения антитело против STEAP-1 содержит вариабельный домен тяжелой цепи, содержащий последовательность остовной области и гипервариабельные участки, при этом последовательность остовной области содержит последовательности FR-H1-FR-H4 соответственно SEQ ID NO:21 или 25 (FR-H1), 22 (FR-H2), 23 (FR-H3) и 24 (FR-H4); HVR-H1 содержит аминокислотную последовательность SEQ ID NO:14, HVR-H2 содержит аминокислотную последовательность SEQ ID NO:15 и HVR-H3 содержит аминокислотную последовательность SEQ ID NO:16. В одном из вариантов осуществления изобретения антитело против STEAP-1 содержит вариабельный домен легкой цепи, содержащий последовательность остовной области и гипервариабельные участки, при этом последовательность остовной области содержит последовательности FR-L1-FR-L4 соответственно SEQ ID NO:17, 18, 19 и 20; HVR-L1 содержит аминокислотную последовательность, выбираемую из SEQ ID NO:11, 12 и 13. В одном из вариантов указанных антител вариабельный домен тяжелой цепи содержит SEQ ID NO:9 или 10 и вариабельный домен легкой цепи включает SEQ ID NO:6.
Некоторые варианты осуществления изобретения относятся к антителу против STEAP-1, содержащему вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO:9 или 10. В некоторых вариантах осуществления изобретения аминокислотная последовательность, характеризующаяся по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью, содержит замены, инсерции или делеции по сравнению с эталонной последовательностью, но антитело, включающее указанную аминокислотную последовательность, сохраняет способность связываться с STEAP-1. В некоторых вариантах осуществления изобретения в последовательности SEQ ID NO:9, 10, 14, 15, 16, 21, 22, 23, 24, 25, 75, 76, 77, 78 и/или 79 заменены, вставлены или удалены в общей сложности 1-10 аминокислот. В некоторых вариантах осуществления изобретения замены, инсерции или делеции находятся в областях за пределами гипервариабельных участков (HVR) (то есть в остовных областях). В некоторых вариантах осуществления изобретения антитело против STEAP-1 включает вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбираемую из SEQ ID NO:9 или 10.
Некоторые варианты осуществления изобретения относятся к антителу против STEAP-1, содержащему вариабельный домен тяжелой цепи, показанный на фигуре 2В (SEQ ID NO:9 или 10).
В некоторых вариантах осуществления изобретения последовательности HVR и FR тяжелой цепи содержат следующие последовательности:
HVR-H1 (GYSITSDYAWN, SEQ ID NO:14 )
HVR-H2 (GYISNSGSTSYNPSLKS, SEQ ID NO:15)
HVR-H3 (ERNYDYDDYYYAMDY, SEQ ID NO:16)
FR-H1 (EVQLVESGGGLVQPGGSLRLSCAAS, SEQ ID NO:21)
FR-H1 (EVQLVESGGGLVQPGGSLRLSCAVS, SEQ ID NO:25)
FR-H2 (WVRQAPGKGLEWV, SEQ ID NO:22)
FR-H2 (WIRQAPGKGLEWV, SEQ ID NO:75)
FR-H2 (WVRQAPGKGLEWM, SEQ ID NO:76)
FR-H2 (WIRQAPGKGLEWM, SEQ ID NO:77)
FR-H3 (RFTISRDNSKNTLYLQMNSLRAEDTAVYYCAR, SEQ ID NO:23)
FR-H3 (RITISRDNSKNTLYLQMNSLRAEDTAVYYCAR, SEQ ID NO:78)
FR-H3 (RFTISRDNSKNTFYLQMNSLRAEDTAVYYCAR, SEQ ID NO:79)
FR-H4 (WGQGTLVTVSS, SEQ ID NO:24)
Некоторые варианты осуществления изобретения относятся к антителу против STEAP-1, содержащему вариабельный домен легкой цепи, показанный на фигуре 2А (SEQ ID NO:6).
В некоторых вариантах осуществления изобретения последовательности HVR легкой цепи содержат следующие последовательности:
HVR-L1(KSSQSLLYRSNQKNYLA, SEQ ID NO:11)
HVR-L2 (WASTRES, SEQ ID NO:12)
HVR-L3 (QQYYNYPRT, SEQ ID NO:13).
В некоторых вариантах осуществления изобретения последовательности остовной области легкой цепи содержат следующие последовательности:
FR-L1 (DIQMTQSPSSLSASVGDRVTITC, SEQ ID NO:17);
FR-L2 (WYQQKPGKAPKLLIY, SEQ ID NO:18);
FR-L3 (GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC, SEQ ID NO:19)
FR-L4 (FGQGTKVEIKR, SEQ ID NO:20).
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему вариабельный домен легкой цепи, содержащий аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO:6. В некоторых вариантах осуществления изобретения аминокислотная последовательность, характеризующаяся по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью, содержит замены, добавления или делеции по сравнению с эталонной последовательностью, но антитело, содержащее указанную аминокислотную последовательность, сохраняет способность связываться с STEAP-1. В некоторых вариантах осуществления изобретения в последовательности, выбираемой из SEQ ID NO:6, 11, 12, 13, 17, 18, 19 и 20 заменены, вставлены или удалены в общей сложности 1-10 аминокислот. В некоторых вариантах осуществления изобретения замены, инсерции или делеции находятся в областях за пределами гипервариабельных участков (то есть в остовных областях). В некоторых вариантах осуществления изобретения антитело против STEAP-1 содержит вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO:6.
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему (а) вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности, выбираемой из SEQ ID NO:9 и 10; и (b) вариабельный домен легкой цепи, содержащий аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO:6. В некоторых вариантах осуществления изобретения аминокислотная последовательность, характеризующаяся по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью, содержит замены, добавления или делеции по сравнению с эталонной последовательностью, но антитело, включающее указанную аминокислотную последовательность, сохраняет способность связываться с STEAP-1. В некоторых вариантах осуществления изобретения в эталонной последовательности, которая включает, не ограничиваясь ими, последовательности, выбираемые из SEQ ID NO:9, 10, 14, 15, 16, 21, 22, 23, 24, 25, 75, 76, 77, 78, 79, заменены, вставлены или удалены в общей сложности 1-10 аминокислот. В некоторых вариантах осуществления изобретения замены, инсерции или делеции находятся в областях за пределами гипервариабельных участков (то есть в остовных областях). В некоторых вариантах осуществления изобретения антитело против STEAP-1 включает вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO:9 или 10, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбираемую из SEQ ID NO:6.
В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему (а) один, два или три гипервариабельных участка вариабельного домена тяжелой цепи (VH), выбираемых из HVR, показанных на фигуре 2В, и/или (b) один, два или три гипервариабельных участка вариабельного домена легкой цепи (VL), выбираемых из HVR, показанных на фигуре 2А. В одном из аспектов настоящее изобретение относится к антителу против STEAP-1, содержащему вариабельный домен тяжелой цепи, выбираемый из вариабельных доменов, показанных на фигуре 2В, и вариабельный домен легкой цепи, выбираемый из вариабельных доменов, показанных на фигуре 2А.
2. Фрагменты антител
Настоящее изобретение относится к фрагментам антител. Фрагменты антител могут быть получены традиционными способами, такими как расщепление ферментами, или способами рекомбинантных ДНК. В определенных условиях, предпочтительно, использовать фрагменты антител, а не полные антитела. Меньший размер фрагментов делает возможным быстрый клиренс и может обеспечить лучший доступ к солидным опухолям. Обзор определенных фрагментов антител приведен в публикации Hudson et al. (2003) Nat. Med. 9:129-134.
Были разработаны разные способы продуцирования фрагментов антител. Указанные фрагменты традиционно получают путем протеолитического расщепления интактных антител (см., например, публикации Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992) и Brennan et al., Science, 229:81 (1985)). Однако указанные фрагменты теперь могут быть продуцированы непосредственно рекомбинантными клетками-хозяевами. Все Fab, Fv и scFv фрагменты антител могут быть экспрессированы и секретированы из E. coli, благодаря чему можно легко продуцировать указанные фрагменты в больших количествах. Фрагменты антител могут быть выделены из вышеописанных библиотек отображения антител на фаге. Альтернативно Fab'-SH фрагменты могут быть выделены из E. coli и химически связаны с образованием F(ab')2 фрагментов (Carter et al., Bio/Technology 10:163-167 (1992)). В соответствии с другим способом F(ab')2 фрагменты могут быть выделены из культуры рекомбинантных клеток-хозяев. Fab и F(ab’)2 фрагменты с более продолжительным временем полужизни in vivo, включающие остатки эпитопа, связывающего восстанавливающий рецептор, описаны в патенте США № 5869046. Специалисту в данной области должны быть известны другие способы продуцирования фрагментов антител. В определенных вариантах осуществления изобретения антитело является одноцепочечным Fv фрагментом (scFv). См. WO 93/16185; патенты США №№ 5571894 и 5587458. Fv и scFv фрагменты являются единственными фрагментами с интактными объединяющими сайтами, у которых отсутствуют константные области; таким образом, указанные фрагменты антител могут быть использованы для менее активного неспецифического связывания при использовании in vivo. Гибридные белки scFv могут быть созданы для осуществления гибридизации эффекторного белка у аминоконца или карбоксильного конца scFv. См. приведенную выше публикацию Antibody Engineering, ed. Borrebaeck. Фрагмент антитела также может быть ”линейным антителом”, например, описанным в патенте США № 5641870. Такие линейные антитела могут быть моноспецифическими или биспецифическими антителами.
3. Гуманизированные антитела
Настоящее изобретение относится к гуманизированным антителам. В данной области известны разные способы гуманизации антител, отличных от человеческих. Например, гуманизированное антитело может иметь один или несколько аминокислотных остатков, введенных в него из источника, отличного от человека. Указанные аминокислотные остатки, отличные от человеческих, часто определяются как ”импортные” остатки, которые обычно получают из ”импортного” вариабельного домена. Гуманизация может быть выполнена способом, описанным в публикациях Винтера с соавторами (Jones et al. (1986) Nature, 321:522-525; Riechmann et al. (1988) Nature, 332:323-327; Verhoeyen et al. (1988) Sience, 239:1534-1536), путем замены соответствующих последовательностей антитела человека последовательностями гипервариабельных участков. Таким образом, такие ”гуманизированные” антитела являются химерными антителами (патент США № 4816567), в которых часть интактного вариабельного домена человека заменена соответствующей последовательностью вида, отличного от человека. На практике гуманизированные антитела обычно являются антителами человека, в которых некоторые остатки гипервариабельного участка и, возможно, некоторые остатки FR заменены остатками из аналогичных сайтов в антителах грызунов.
Выбор вариабельных доменов человека как легкой, так и тяжелой цепей, используемых для создания гуманизированных антител, имеет важное значение для уменьшения антигенности. В соответствии с так называемым способом ”наибольшего соответствия” последовательность вариабельного домена антитела грызуна сравнивают со всей библиотекой известных последовательностей вариабельных доменов человека. Последовательность человека, которая ближе всего соответствует последовательности грызуна, выбирают в качестве остовной области человека для гуманизированного антитела (Sims et al. (1993) J. Immunol., 151:2296; Chothia et al. (1993) J. Mol. Biol., 196:901). В другом способе используют конкретную остовную область, выделенную из консенсусной последовательности всех антител человека, относящихся к определенной подгруппе легких или тяжелых цепей. Одна и та же остовная область может быть использована для нескольких разных гуманизированных антител (Carter et al. (1992) Proc. Natl. Acad. Sci. USA, 89:4285; Presta et al. (1993) J. Immunol., 151:2623).
Обычно желательно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других благоприятных биологических свойств. Для достижения указанной цели в соответствии с одним способом получения гуманизированных антител выполняют анализ исходных последовательностей и разных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулина являются общедоступными и известны специалистам в данной области. Существуют компьютерные программы, иллюстрирующие и отображающие вероятные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Исследование указанных изображений позволяет анализировать вероятную роль остатков в функционировании последовательности иммуноглобулина-кандидата, то есть анализировать остатки, влияющие на способность иммуноглобулина-кандидата связываться с антигеном. Таким образом остатки остовной области (FR) могут быть выбраны и объединены с реципиентными и импортными последовательностями с возможностью достижения требуемого свойства антитела, такого как более высокая аффинность к антигену-мишени. Как правило, остатки гипервариабельного участка непосредственно и в большой степени влияют на связывание антигена.
4. Антитела человека
Антитела человека против STEAP-1 по настоящему изобретению могут быть созданы путем объединения последовательностей вариабельного домена клона Fv-фрагментов, выбранных из библиотек отображения на фаге, полученных у человека, с вышеописанными известными последовательностями константного домена человека. Альтернативно, моноклональные антитела человека против STEAP-1 по настоящему изобретению могут быть созданы способом гибридом. Линии клеток миеломы человека и гетеромиеломы мыши-человека, используемые для продуцирования моноклональных антител человека, описаны, например, в публикациях Kozbor, J. Immunol. 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147:86 (1991).
В настоящее время могут быть созданы трансгенные животные (например, мыши), способные в результате иммунизации продуцировать полный спектр антител человека при отсутствии продуцирования эндогенного иммуноглобулина. Например, в научной литературе было отмечено, что гомозиготная делеция гена (JH) соединительной области тяжелой цепи антитела у химерных и гаметических мутантных мышей вызывает полное ингибирование продуцирования эндогенных антител. Введение гаметического гена иммуноглобулина человека таким гаметическим мутантным мышам вызывает продуцирование антител человека при антигенной стимуляции. См., например, публикации Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255 (1993); Bruggermann et al., Year in Immunol., 7:33 (1993).
Для получения антител человека из антител, отличных от человеческих, например, из антител грызунов, в которых антитело человека обладает такой же аффинностью и специфичностью, что и исходное антитело, отличное от человеческого, может быть также использован способ перестановки генов. В соответствии с указанным способом, который также именуется ”импринтингом эпитопа”, вариабельную область тяжелой или легкой цепи фрагмента антитела, отличного от человеческого, полученного способами отображения на фаге, рассмотренными в настоящем описании изобретения, заменяют набором генов вариабельного домена человека, создавая таким образом популяцию химерных scFv или Fab фрагментов с цепью, отличной от человеческой/человеческой цепью. В результате отбора с использованием антигена выделяют химерные scFv или Fab фрагменты с цепью, отличной от человеческой/человеческой цепью, в которых цепь человека восстанавливает антигенсвязывающий центр, разрушенный в результате удаления соответствующей цепи, отличной от человеческой, в первичном клоне отображения на фаге, то есть указанный эпитоп определяет (запечатлевает) выбор цепи-партнера человека. При повторении указанного процесса в целью замены остальной цепи, отличной от человеческой, получают антитело человека (см. заявку РСТ WO 93/06213, опубликованную 1 апреля 1993 г.). В отличие от традиционного способа гуманизации антител, отличных от человеческих, путем трансплантации CDR указанный способ позволяет получить полностью антитела человека, в которых отсутствуют остатки FR или CDR с происхождением, отличным от человеческого.
5. Биспецифические антитела
Биспецифические антитела являются моноклональными антителами, обладающими специфичностью связывания в отношении по меньшей мере двух разных антигенов. В определенных вариантах осуществления изобретения биспецифические антитела являются антителами человека или гуманизированными антителами. В определенных вариантах осуществления изобретения одна из специфичностей связывания относится к STEAP-1 и другая специфичность связывания относится к любому другому антигену. В определенных вариантах осуществления изобретения биспецифические антитела могут связываться с двумя разными эпитопами STEAP-1. Биспецифические антитела могут быть также использованы для доставки цитотоксических агентов в клетки, экспрессирующие STEAP-1. Указанные антитела имеют STEAP-1-связывающий домен и домен, связывающийся с цитотоксическим агентом, таким как, например, сапорин, антитело против α-интерферона, алкалоид барвинка, цепь рицина А, метотрексат или радиоактивный изотоп гаптен. Биспецифические антитела могут быть получены в виде полноразмерных антител или фрагментов антител (например, в виде F(ab')2 фрагмента биспецифических антител).
В данной области известны способы создания биспецифических антител. Продуцирование биспецифических антител способами рекомбинантных ДНК традиционно основано на соэкспрессии двух пар тяжелых цепей-легких цепей иммуноглобулина, в которых две тяжелые цепи обладают разными специфичностями (Milstein and Cuello, Nature, 305:537 (1983)). Вследствие произвольной реаранжировки тяжелых и легких цепей иммуноглобулина указанные гибридомы (квадромы) продуцируют потенциальную смесь 10 разных молекул антител, из которых только одна молекула имеет правильную биспецифическую структуру. Выделение правильной молекулы, обычно производимое при помощи аффинной хроматографии, является достаточно сложным процессом, при этом выход продукта является низким. Аналогичные способы описаны в заявке WO 93/08829, опубликованной 13 мая 1993 г. и в публикации Traunecker et al., EMBO J., 10:3655 (1991).
В соответствии с другим способом вариабельные домены антитела, обладающие требуемыми специфичностями связывания (сайты, объединяющие антитело с антигеном), сливают с последовательностями константного домена иммуноглобулина. Например, сливают с константным доменом тяжелой цепи иммуноглобулина, включающим по меньшей мере часть шарнирной области, СН2 и СН3 областей. В определенных вариантах осуществления изобретения первая константная область тяжелой цепи (СН1), содержащая сайт, необходимый для связывания с легкой цепью, присутствует по меньшей мере в одном из гибридов. ДНК, кодирующие гибриды тяжелой цепи иммуноглобулина и при желании легкой цепи иммуноглобулина, вводят в отдельные экспрессирующие векторы и котрансфицируют в приемлемый организм-хозяин. Таким образом достигается большая гибкость регулирования взаимных пропорций трех полипептидных фрагментов, когда неравные соотношения трех полипептидных цепей, используемых в данной конструкции, обеспечивают оптимальный выход. Однако кодирующие последовательности для двух или всех трех полипептидных цепей можно ввести в один вектор экспрессии, когда экспрессия по меньшей мере двух полипептидных цепей в одинаковых соотношениях обеспечивает высокий выход или когда соотношения компонентов не имеют существенного значения.
В соответствии с одним вариантом данного способа биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина, в которой первая специфичность связывания находится в одном домене и гибридная пара тяжелой цепи-легкой цепи иммуноглобулина (обеспечивающая вторую специфичность связывания) находится в другом домене. Было установлено, что указанная асимметричная структура облегчает отделение требуемого биспецифического соединения от нежелательных комбинаций цепей иммуноглобулина, так как наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы способствует отделению. Указанный способ описан в заявке WO 94/04690. Более подробно с способами создания биспецифических антител можно ознакомиться, например, в публикации Suresh et al., Methods in Enzymology, 121:210 (1986).
В соответствии с другим способом между двумя молекулами антител может быть создана область контакта, позволяющая максимально увеличить процентное количество гетеродимеров, выделяемых из культуры рекомбинантных клеток. Область контакта включает по меньшей мере часть СН3 домена константного домена антитела. В соответствии с указанным способом одну или несколько коротких боковых цепей аминокислот в области контакта первой молекулы антитела заменяют более длинными боковыми цепями (например, тирозином или триптофаном). В области контакта второй молекулы антитела создают компенсирующие ”полости”, соответствующие размеру длинных боковых цепей, заменяя длинные боковые цепи аминокислот меньшими боковыми цепями (например, аланином или треонином). Таким образом создается механизм увеличения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают перекрестносшитые или ”гетероконъюгированные” антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое с биотином. Такие антитела были предложены, например, для направленной доставки клеток иммунной системы к нежелательным клеткам (патент США № 4676980) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/00373 и европейский патент № 03089)). Гетероконъюгатные антитела могут быть получены любым известным способом перекрестного сшивания. Приемлемые перекрестносшивающие агенты хорошо известны в данной области и описаны в патенте США № 4676980, наряду с целым рядом способов перекрестного сшивания.
В научной литературе также описаны способы создания биспецифических антител из фрагментов антител. Например, биспецифические антитела могут быть получены способом химического связывания. В публикации Brennan et al., Science, 229:81 (1985) описан способ, в соответствии с которым интактные антитела протеолитически расщепляют с образованием F(ab’)2 фрагментов. Указанные фрагменты восстанавливают в присутствии агента, образующего комплекс с дитиолом, арсенита натрия для стабилизации соседних дитиолов и предотвращения образования межмолекулярных дисульфидных связей. Созданные Fab' фрагменты затем превращают в производные тионитробензоата (TNB). Одно из производных Fab'-TNB затем снова превращают в Fab'-тиол путем восстановления меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB с образованием биспецифического антитела. Полученные биспецифические антитела могут быть использованы в качестве агентов для селективной иммобилизации ферментов.
Прогресс, достигнутый за последнее время, позволил облегчить прямое выделение Fab'-SH фрагментов из E. coli, которые могут быть химически связаны с образованием биспецифических антител. В публикации Shalaby et al., J. Exp. Med., 175:217-225 (1992) описано получение F(ab')2 молекулы полностью гуманизированного биспецифического антитела. Каждый Fab' фрагмент был отдельно секретирован из E. coli и подвергнут направленному химическому связыванию in vitro с образованием биспецифического антитела. Полученное таким образом биспецифическое антитело обладало способностью связываться с клетками, сверхэкспрессирующими рецептор HER2, и нормальными Т-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против опухолей молочной железы человека.
В научной литературе также описаны разные способы получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела были получены с использованием лейциновых застежек-молний. Kostelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды лейциновой застежки-молнии из Fos и Jun белков были связаны с Fab' частями двух разных антител путем сливания генов. Гомодимеры антител восстанавливали в шарнирной области с образованием мономеров и затем повторно окисляли с образованием гетеродимеров антител. Указанный способ можно также использовать для получения гомодимеров антител. Способ ”диател”, описанный в публикации Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993), является альтернативным механизмом создания фрагментов биспецифических антител. Указанные фрагменты включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) при помощи линкера, который является слишком коротким для спаривания двух доменов в одной цепи. Поэтому VH и VL домены одного фрагмента принудительно спариваются с комплементарными VL и VH доменами другого фрагмента с образованием двух антигенсвязывающих центров. В научной литературе также описан другой способ получения фрагментов биспецифических антител путем использования димеров одноцепочечных Fv (sFv) фрагментов. См. публикацию Gruber et al., J. Immunol. 152:5368 (1994).
В объем настоящего изобретения входят антитела с более чем двумя валентностями. Например, могут быть получены триспецифические антитела. Tutt et al., J. Immunol., 147:60 (1991).
6. Мультивалентные антитела
Мультивалентное антитело может быть интернализировано (и/или катаболизировано) быстрее, чем бивалентное антитело, клеткой, экспрессирующей антиген, с которым связываются указанные антитела. Антитела по настоящему изобретению могут быть мультивалентными антителами (отличными от класса IgM) с тремя или более антигенсвязывающими центрами (например, четырехвалентные антитела), которые могут быть легко продуцированы в результате рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Мультивалентное антитело может содержать домен димеризации и три или более антигенсвязывающих центров. В определенных вариантах осуществления изобретения домен димеризации содержит (или состоит из) Fc-область или шарнирную область. Таким образом, антитело будет включать Fc-область и три или более антигенсвязывающих центров у аминоконца Fc-области. В определенных вариантах осуществления изобретения мультивалентное антитело включает (или состоит из) от трех до около восьми антигенсвязывающих центров. В одном таком варианте осуществления изобретения мультивалентное антитело включает (или состоит из) четыре антигенсвязывающих центра. Мультивалентное антитело содержит по меньшей мере одну полипептидную цепь (например, две полипептидные цепи), которые содержат два или более вариабельных доменов. Например, полипептидная цепь может включать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 означает первый вариабельный домен, VD2 означает второй вариабельный домен, Fc означает одну полипептидную цепь Fc-области, Х1 и Х2 означают аминокислоту или полипептид и n равно 0 или 1. Например, полипептидная цепь может включать: VH-CH1-гибкий линкер-VH-CH1-цепь Fc-области или VH-CH1-VH-CH1-цепь Fc-области. Мультивалентное антитело может далее содержать по меньшей мере два (например, четыре) полипептида вариабельного домена легкой цепи. Мультивалентное антитело может, например, содержать от около двух до около восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, входящие в объем настоящего изобретения, включают вариабельный домен легкой цепи и далее необязательно содержат домен CL.
7. Однодоменные антитела
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению является однодоменным антителом. Однодоменное антитело имеет одну полипептидную цепь, содержащую весь и часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах осуществления изобретения однодоменное антитело является однодоменным антителом человека (Domantis, Inc., Waltham, MA; см., например, патент США № 6248516 В1). В одном из вариантов осуществления изобретения однодоменное антитело состоит из всего или части вариабельного домена тяжелой цепи антитела.
8. Варианты антител
Некоторые варианты осуществления изобретения относятся к модификациям антител по настоящему изобретению. Например, может быть желательно улучшить аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела могут быть получены путем внесения соответствующих изменений в нуклеотидную последовательность, кодирующую антитело, или способом пептидного синтеза. Такие модификации включают, например, делеции, инсерции и/или замены остатков в аминокислотных последовательностях антитела. Может быть произведена любая комбинация делеций, инсерций и замен для достижения желаемой конечной конструкции при условии, что такая конечная конструкция обладает требуемыми характеристиками. Изменения могут быть внесены в аминокислотную последовательность антитела в процессе создания последовательности.
Приемлемый способ идентификации определенных остатков или областей антитела, которые являются предпочтительными местами для мутагенеза, получил название ”мутагенез со сканированием аланином” и описан в публикации Cunningham and Wells (1989) Science, 244:1081-1085. Остаток или группу остатков-мишеней идентифицируют (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) для достижения взаимодействия аминокислот с антигеном. В указанные положения аминокислот, функционально восприимчивые к заменам, затем вводят дополнительные или другие варианты на сайтах замен. Таким образом, хотя сайт для введения варианта аминокислотной последовательности определен заранее, характер самой мутации заранее не определен. Например, для анализа продуктивности мутации на данном сайте выполняют сканирование ala или неспецифический мутагенез в кодоне или области-мишени и экспрессированные иммуноглобулины исследуют в отношении требуемой активности.
Инсерции аминокислотной последовательности включают аминоконцевые и/или карбоксиконцевые слияния длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также инсерции одного или нескольких аминокислотных остатков внутри последовательности. Примеры концевых инсерций включают антитело с N-концевым остатком метионила. Другие инсерцинные варианты молекулы антитела включают слияние N- или С-конца антитела с ферментом (например, для ADEPT) или полипептидом, увеличивающим время полужизни антитела в кровяном русле.
В определенных вариантах осуществления изобретения антитело по настоящему изобретению изменяют для увеличения или уменьшения степени гликозилирования антитела. Гликозилирование полипептидов обычно происходит по N-связи или О-связи. Гликозилирование по N-связи означает присоединение углеводной части к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х означает любую аминокислоту за исключением пролина, являются распознаваемыми последовательностями для ферментативного присоединения углеводной части к боковой цепи аспарагина. Таким образом, наличие любой из указанных трипептидных последовательностей в полипептиде вызывает образование потенциального сайта гликозилирования. Гликозилирование по О-связи означает присоединение одного из сахаров, включающих N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, чаше всего к серину или треонину, хотя также может быть использован 5-гидроксипролин или 5-гидроксилизин.
Добавление или делеция сайтов гликозилирования в антителе обычно осуществляется в результате такого изменения аминокислотной последовательности, при котором образуется или удаляется одна или несколько вышеуказанных трипептидных последовательности (для сайтов гликозилирования по N-связи). Изменение может быть также произведено путем добавления, делеции или замены одного или нескольких остатков серина или треонина в последовательности исходного антитела (для сайтов гликозилирования по О-связи).
В антителе, содержащем Fc-область, может быть изменен присоединенный к нему углевод. Например, антитела с углеводом, имеющим зрелую структуру, в которой отсутствует фукоза, присоединенная к Fc-области антитела, описаны в заявке на патент США № 2003/0157108 (Presta, L.). См. также заявку на патент США № 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd.). Антитела, содержащие N-ацетилглюкозоамин (GlcNAc) в углеводе, присоединенном к Fc-области антитела, описаны в публикации WO 2003/011878, Jean-Mairet et al., и в патенте США № 6602684, Umana et al. Антитела, содержащие по меньшей мере один остаток галактозы в олигосахариде, присоединенном к Fc-области антитела, описаны в публикации WO 1997/30087, Patel et al. См. также публикации WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.), относящиеся к антителам с измененным углеводом, присоединенным к Fc-области. См. также заявку на патент США № 2005/123546 (Umana et al.), относящуюся к антигенсвязывающим молекулам с модифицированным гликозилированием.
В определенных вариантах осуществления изобретения гликозилированный вариант содержит Fc-область с присоединенным углеводом, в котором отсутствует фукоза. Такие варианты характеризуются улучшенной функцией ADCC. Fc-область далее необязательно содержит одну или несколько замен аминокислот, которые еще больше улучшают функцию ADCC, например, замены в положениях 298, 333 и/или 334 Fc-области (нумерация остатков по системе EU). Примеры публикаций, относящихся к ”дефукозилированным” антителам или антителам ”с отсутствием фукозы” включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры линий клеток, продуцирующих дефукозилированные антитела, включают клетки СНО Lec13, в которых отсутствует фукозилирование белка, (Ripka et al., Arch. Biochem. Biophys. 249:533-545 (1986); заявка на патент США № 2003/0157108 A1, Presta L.; и WO 2004/056312 A1, Adams et al., в частности, в примере 11) и линии клеток с отсутствием гена, такие как клетки СНО с отсутствием гена альфа-1,6-фукозил-трансферазы, FUT8 (Yamane-Ohnuki et al. Biotech. Bioeng. 87:614 (2004)).
В одном из вариантов осуществления изобретения антитело изменено для увеличения времени полужизни в сыворотке. Для увеличения времени полужизни антитела в сыворотке в антитело (в частности, фрагмент антитела) можно ввести эпитоп, связывающий восстанавливающий рецептор, описанный в патенте США № 5739277. Как используется в настоящем описании, термин ”эпитоп, связывающий восстанавливающий рецептор” означает эпитоп Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за увеличение времени полужизни молекулы IgG в кровяном русле in vivo (заявка на патент США № 2003/0190311, патенты США №№ 6821505, 6165745, 5624821, 5648260, 6165745, 5834597).
Другим типом варианта является вариант с заменой аминокислот. Указанные варианты содержат в молекуле антитела по меньшей мере один аминокислотный остаток, заменяемый другим остатком. Представляющие интерес сайты для заместительного мутагенеза включают гипервариабельные участки, но также возможны изменения остовной области. Консервативные замены показаны в таблице 1 под заголовком “предпочтительные замены”. Если такие замены приводят к желаемому изменению биологической активности, тогда могут быть произведены более существенные замены, озаглавленные ”типичные замены” в таблице 1 или описанные ниже со ссылкой на классы аминокислот.
Существенные модификации биологических свойств антитела выполняют, выбирая замены, которые значительно отличаются по своему воздействию на сохранение (а) структуры полипептидного остова в области замены, например, β-складчатой или спиральной конформации, (b) заряда или гидрофобности молекулы на сайте-мишени или (с) боковой цепи. Аминокислоты могут быть классифицированы по сходству свойств боковых цепей (в публикации A.L. Lehninger, in Biochemistry, second ed., pp. 73-75, Worth Publishers, New York (1975)):
(1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M);
(2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (C), Tyr (Y), Asn (N), Gln (Q);
(3) кислотные: Asp (D), Glu (E);
(4) основные: Lys (K), Arg (R), His (H).
Альтернативно природные остатки можно классифицировать на основании общих свойств боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены предполагают переход члена из одного из указанных классов в другой класс. Такие замещенные остатки могут быть также введены в сайты консервативных замен или оставшиеся (неконсервативные) сайты.
Один тип заместительного варианта включает замену одного или нескольких остатков в гипервариабельном участке материнского антитела (например, гуманизированного антитела или антитела человека). Полученный вариант, отобранный для дальнейшей разработки, будет обладать модифицированными (например, улучшенными) биологическими свойствами по сравнению с материнским антителом, из которого данный вариант был получен. Традиционный способ создания таких заместительных вариантов включает созревание аффинности с отображением на фаге. Несколько сайтов гипервариабельного участка (например, 6-7 сайтов) мутируют для выполнения всех возможных замен аминокислот на каждой сайте. Полученные таким образом антитела отображают на частицах нитчатого фага в виде гибридов по меньшей мере с частью белка оболочки фага (например, продукт гена III М13), упакованного в каждой частице. Отображенные на фаге варианты исследуют в отношении их биологической активности (например, аффинность связывания). Для идентификации используемых для модификации сайтов-кандидатов гипервариабельного участка, может быть выполнен мутагенез со сканированием (например, со сканированием аланином) с целью выявления остатков гипервариабельного участка, в значительной степени способствующих связыванию антигена. Альтернативно или дополнительно можно произвести анализ кристаллической структуры комплекса антиген-антитело для выявления точек контактирования антитела с антигеном. Такие контактные остатки и соседние остатки являются кандидатами для замены способами, известными в данной области, включая способы, рассмотренные в настоящем описании изобретения. После создания таких вариантов панель вариантов подвергают скринингу способами, известными в данной области, включая способы, рассмотренные в настоящем описании изобретения, и для дальнейшей разработки отбирают антитела с лучшими свойствами по результатам одного или нескольких выполненных анализов.
Молекулы нуклеиновой кислоты, кодирующей варианты аминокислотной последовательности антитела, получают разными способами, известными в данной области. Указанные способы включают, не ограничиваясь ими, выделение из природного источника (в случае природных вариантов аминокислотной последовательности) или получение при помощи сайтнаправленного мутагенеза с использованием олигонуклеотидов, мутагенеза с ПЦР и кластерного мутагенеза ранее полученного варианта или антитела, не являющегося вариантом.
Может быть желательно ввести одну или несколько модификаций аминокислот в Fc-область антител по настоящему изобретению с образованием варианта Fc-области. Вариант Fc-области может содержать последовательность Fc-области человека (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую модификацию аминокислот (например, замену) в одном или нескольких положениях, включая положение цистеина в шарнирной области.
В соответствии с настоящим описанием изобретения и данными, имеющимися в данной области, можно предположить, что в некоторых вариантах осуществления изобретения антитело по настоящему изобретению может содержать одно или несколько изменений по сравнению с антителом дикого типа, например, в Fc-области. Тем не менее указанные антитела в основном сохраняют характеристики, необходимые для терапевтического применения, по сравнению с антителом дикого типа. Например, считается, что в Fc-области могут быть произведены определенные замены, которые могут привести к изменению (то есть улучшению или ухудшению) связывания C1q и/или комплементзависимой цитотоксичности (CDC), как описано, например, в WO99/51642. См. также публикацию Duncan & Winter, Nature 322:738-40 (1988); патент США № 5648260, патент США № 5624821 и WO94/29351 для ознакомления с другими примерами вариантов Fc-области. В публикациях WO00/42072 (Presta) и WO 2004/056312 (Lowman) описаны варианты антител с повышенным или пониженным связыванием с FcR. Указанные публикации патентов включены в настоящее описание изобретения в качестве ссылки. См. также публикацию Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001). Антитела с увеличенным временем полужизни в кровяном русле или с улучшенным связыванием с неонатальным Fc-рецептором (FcRn), отвечающим за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117-587 (1976) and Kim et al., J. Immunol. 24:249 (1994), описаны в заявки на патент США US2005/0014934A1 (Hinton et al.). Указанные антитела содержат Fc-область с одной или несколькими заменами, улучшающими связывание Fc-области с FcRn. Полипептидные варианты с измененными аминокислотными последовательностями Fc-области и увеличенной или уменьшенной способностью связывания C1q описаны в патенте США № 6194551В1, WO99/51642. Указанные публикации патентов включены в настоящее описание изобретения в качестве ссылки. См., также публикацию Idusogie et al., J. Immunol. 164: 4178-4184 (2000).
В одном из аспектов настоящее изобретение относится к антителам, содержащим модификации в области контактов полипептидов Fc-области, которые облегчают и/или стимулируют гетеродимеризацию. Указанные модификации включают введение выступа в первый полипептид Fc-области и углубления во второй полипептид Fc-области, при этом выступ входит в углубление, стимулируя образование комплекса первого и второго полипептидов Fc-области. В данной области известны способы создания антител с указанными модификациями, например, описанные в патенте США № 5731168.
9. Производные антитела
Антитела по настоящему изобретению могут быть далее модифицированы с включением дополнительных небелковых частей, известных в данной области и легко доступных. Указанные части, пригодные для получения производного антитела, предпочтительно, являются водорастворимыми полимерами. Неограничивающие примеры водорастворимых полимеров включают, не ограничиваясь ими, полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля является более предпочтительным для производства благодаря устойчивости в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может изменяться, и в случае присоединения нескольких полимеров такие полимеры могут быть одинаковыми или разными молекулами. Как правило, число и/или тип полимеров, используемых для получения производного антитела, можно определить с учетом конкретных свойств или функций подлежащего улучшению антитела, применения производного антитела в терапии в определенных условиях и т.д.
Другой вариант осуществления изобретения относится к конъюгатам антитела с небелковой частью, которые могут быть избирательно нагреты под воздействием излучения. В одном из вариантов осуществления изобретения небелковая часть является углеродной нанотрубочкой (Kam et al., Proc. Natl. Acad. Sci. 102:11600-11605 (2005)). Излучение может иметь любую длину волны и включает, не ограничиваясь ими, длины волн, которые не причиняют вреда нормальным клеткам, но нагревают небелковую часть до температуры, при которой гибнут клетки, находящиеся рядом с небелковой частью антитела.
Некоторые способы создания антител
1. Некоторые способы на основе гибридом
Моноклональные антитела против STEAP-1 по настоящему изобретению могут быть получены способом гибридом, который впервые был описан в публикации Kohler et al., Nature, 256:495 (1975), или способами рекомбинантных ДНК (патент США № 4816567).
В соответствии со способом гибридом мышь или другое приемлемое животное-хозяина, такое как хомячок, иммунизируют для образования лимфоцитов, продуцирующих или способных продуцировать антитела, которые специфически связываются с белком, используемым для иммунизации. Антитела против STEAP-1 обычно образуются у животных в результате нескольких подкожных (sc) или внутрибрюшинных (ip) инъекций STEAP-1 и адъюванта. STEAP-1 можно получить способами, хорошо известными в данной области, некоторые из которых представлены далее в настоящем описании изобретения. Например, STEAP-1 может быть получен способами рекомбинантных ДНК. В одном из вариантов осуществления изобретения животных иммунизируют производным STEAP-1, содержащим внеклеточную часть STEAP-1, слитую с Fc-частью тяжелой цепи иммуноглобулина. В одном из вариантов осуществления изобретения животных иммунизируют слитым белком STEAP-1-IgG1. В одном из вариантов осуществления изобретения животных иммунизируют иммуногенными производными STEAP-1 в растворе с монофосфорильным липидом А (MPL)/дикриномиколятом трегалозы (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT), после чего полученный раствор инъецируют внутрикожно в несколько участков тела. Через две недели животных повторно иммунизируют. Через семь-четырнадцать дней у животных берут пробу крови и анализируют сыворотку в отношении титра антител против STEAP-1. Животных иммунизируют до получения положительного титра.
Альтернативно, лимфоциты могут быть иммунизированы in vitro. Затем лимфоциты сливают с миеломными клетками, используя приемлемый слитый агент, такой как полиэтиленгликоль, для образования клетки гибридомы (Goding, Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)).
Полученные таким образом клетки гибридом высевают и выращивают в приемлемой культуральной среде, например, в среде, содержащей одно или несколько веществ, ингибирующих рост или выживание неслитых, материнских миеломных клеток. Например, если в материнской миеломной клетке отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), культуральная среда для гибридом обычно включает гипоксантин, аминоптерин и тимидин (среда НАТ), которые предотвращают рост клеток с отсутствием HGPRT.
В определенных вариантах осуществления изобретения миеломные клетки эффективно гибридизируют и поддерживают устойчивое продуцирование антитела на высоком уровне отобранными антителопродуцирующими клетками и являются чувствительными к среде, такой как среда НАТ. Типичные миеломные клетки включают, не ограничиваясь ими, линии миеломных клеток мышей, выделенных, например, из мышиных опухолей МОРС-21 и МРС-11, которые могут быть приобретены в центре Salk Institute Cell Distribution Center, San Diego, California, USA, и из клеток SP-2 или X63-Ag8-653, которые можно приобрести в Американской коллекции типовых культур, Rockville, Maryland, USA. Миеломные клетки человека и линии гетеромиеломных клеток мыши-человека также могут быть использованы для продуцирования моноклональных антител человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, используемую для выращивания клеток гибридом, анализируют в отношении продуцирования моноклональных антител, связывающихся с STEAP-1. Специфичность связывания моноклональных антител, продуцированных клетками гибридом, предпочтительно, определяют способом иммунопреципитации или при помощи анализа связывания in vitro, такого как радиоиммуноанализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Аффинность связывания моноклонального антитела, например, можно определить при помощи анализа Скэтчарда, описанного в публикации Munson et al., Anal. Biochem., 107:220 (1980).
После идентификации клеток гибридом, продуцирующих антитела с требуемой специфичностью, аффинностью и/или активностью, клоны могут быть субклонированы способами ограничивающего разведения и выращены стандартными способами (Goding, Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)). Приемлемые культуральные среды, предназначенные для указанной цели, включают, например, среду DMEM или RPMI-1640. Кроме того, клетки гибридом могут быть выращены in vivo в виде асцитических опухолей у животного. Моноклональные антитела, секретированные субклонами, выделяют из культуральной среды, асцитической жидкости или сыворотки стандартными способами очистки иммуноглобулинов, такими как, например, хроматография на колонке с протеином А-сефарозой, хроматография на колонке с гидроксилапатитом, гель-электрофорез, диализ или аффинная хроматография.
2. Некоторые способы скрининга библиотек
Антитела против STEAP-1 по настоящему изобретению могут быть получены при использовании комбинаторных библиотек для скрининга антител с требуемой активностью или активностями. Например, в данной области известны разные способы создания библиотек отображения на фаге и скрининга таких библиотек в отношении антител, обладающих требуемыми характеристиками связывания. Такие способы описаны в публикации Hoogenboom et al. (2001) in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ) и в некоторых вариантах осуществления изобретения, представленных в публикации Lee et al., (2004) J. Mol. Biol. 340:1073-1093.
Как правило, синтезированные клоны антител отбирают путем скрининга библиотек, содержащих фаг, отображающий разные фрагменты вариабельной области антитела (Fv), гибридизированные с белком оболочки фага. Такие библиотеки фагов исследуют при помощи аффинной хроматографии против требуемого антигена. Клоны, экспрессирующие Fv-фрагменты, способные связываться с требуемым антигеном, адсорбируются на антигене и, таким образом, отделяются от несвязывающихся клонов в библиотеке. Связывающиеся клоны затем элюируют из антигена и далее обогащают, выполняя дополнительные циклы адсорбции на антигене/элюирования. Любые антитела против STEAP-1 по настоящему изобретению могут быть получены при помощи соответствующего способа скрининга антигена с целью отбора представляющего интерес фагового клона с последующим созданием клона полноразмерных антител против STEAP-1 с использованием последовательностей Fv-фрагментов из представляющего интерес фагового клона и приемлемых последовательностей константной области (Fc), описанных в публикации Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda, MD (1991), vols. 1-3.
В определенных вариантах осуществления изобретения антигенсвязывающий домен антитела образован двумя вариабельными (V) областями, состоящими примерно из 110 аминокислот, которые были выделены из легкой (VL) и тяжелой (VH) цепей, и содержащими три гипервариабельные петли (HVR) или определяющие комплементарность области (CDR). Вариабельные домены могут быть функционально отображены на фаге в виде одноцепочечных Fv (scFv) фрагментов, в которых VH и VL ковалентно связаны при помощи короткого, гибкого пептида, или в виде Fab-фрагментов, в которых VH и VL гибридизированы с константным доменом и взаимодействуют без ковалентной связи, как описано в публикации Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). В значении, как используется в настоящем описании, scFv-кодирующие фаговые клоны и Fab-кодирующие фаговые клоны совместно определяются как “фаговые клоны Fv-фрагментов” или “клоны Fv-фрагментов”.
Наборы генов VH и VL могут быть отдельно клонированы при помощи полимеразной цепной реакции (ПЦР) и произвольно объединены в библиотеках фагов, которые затем могут быть исследованы в отношении антигенсвязывающих клонов, как описано в публикации Winter et al., Ann. Rev. Immunol., 12:433-455 (1994). Библиотеки, созданные из иммунизированных источников, позволяют получить высокоаффинные антитела к иммуногену без необходимости создания гибридом. Альтернативно набор наивных антител может быть клонирован с целью создания одного источника антител человека к целому ряду несобственных, а также собственных антигенов без какой-либо иммунизации, как описано в публикации Griffiths et al., EMBO J., 12:725-734 (1993). И наконец, библиотеки наивных антител могут быть синтезированы путем клонирования нереаранжированных сегментов V-гена из стволовых клеток и использования праймеров для ПЦР, содержащих произвольную последовательность, для кодирования гипервариабельных участков CDR3 и реаранжирования in vitro, как описано в публикации Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992).
В определенных вариантах осуществления изобретения использован нитчатый фаг для отображения фрагментов антитела путем гибридизации с минорным белком оболочки pIII. Фрагменты антитела могут быть отображены в виде одноцепочечных Fv-фрагментов, в которых домены VH и VL соединены в одной полипептидной цепи при помощи гибкого полипептидного спейсера, например, в соответствии с описанием, приведенным в публикации Marks et al., J. Mol. Biol., 222:581-597 (1991), или в виде Fab-фрагментов, в которых одна цепь гибридизирована с белком pIII, а другая цепь секретирована в периплазму бактериальной клетки-хозяина, где происходит сборка структуры белка оболочки с Fab-фрагментом, отображаемой на поверхности фага путем вытеснения некоторых белков оболочки дикого типа, например, как описано в публикации Hoogenboom et al., Nucl. Acids Res., 19:4133-4137 (1991).
Как правило, нуклеиновые кислоты, кодирующие фрагменты генов антитела, получают из иммунокомпетентных клеток, выделенных у человека или животных. Если желательно смещение библиотеки в сторону клонов против STEAP-1, индивида иммунизируют белком STEAP-1 с целью определения реакции на антитело и выделяют клетки селезенки и/или циркулирующие в кровотоке В-клетки и другие лимфоциты периферической крови (PBL) для создания библиотеки. В предпочтительном варианте осуществления изобретения библиотеку фрагментов генов антитела человека, смещенную в сторону клонов антител против STEAP-1, получают путем возбуждения реакции на антитело против STEAP-1 у трансгенных мышей, несущих ряд функциональных генов иммуноглобулина человека (при отсутствии системы продуцирования функциональных эндогенных антител), в результате чего иммунизация белком STEAP-1 вызывает образование В-клеток, продуцирующих антитела человека против STEAP-1. Создание трансгенных мышей, продуцирующих антитела человека, описано ниже.
Популяции клеток, продуцирующих антитела против STEAP-1, могут быть дополнительно обогащены при помощи приемлемого способа скрининга, позволяющего выделить В-клетки, экспрессирующие STEAP-1-специфическое антитело, связанной с мембраной, например, путем разделения клеток способом аффинной хроматографии или адсорбции клеток на белке STEAP-1, меченном флуорохромом, с последующей сортировкой клеток с возбуждением флуоресценции (FACS).
Альтернативно использование клеток селезенки и/или В-клеток или других PBL, полученных у неиммунизированного донора, позволяет лучше представить спектр возможных антител и создать библиотеку антител с использованием животных любого вида (человека или животного, отличного от человека), в котором STEAP-1 не является антигеном. Для получения библиотек, включающий создание гена антитела in vitro, из стволовых клеток, полученных у индивида, выделяют нуклеиновые кислоты, кодирующие нереаранжированные сегменты гена антитела. Представляющие интерес иммунокомпетентные клетки могут быть получены у животных разных видов, таких как человек, мышь, крыса, зайцеобразные, волк, собака, кошка, свинья, бык, лошадь, птицы и т.д.
Нуклеиновую кислоту, кодирующую вариабельные сегменты гена антитела (включая сегменты VH и VL) выделяют из представляющих интерес клеток и амплифицируют. В случае библиотек реаранжированных генов VH и VL требуемую ДНК можно получить путем выделения геномной ДНК или мРНК из лимфоцитов с последующим выполнением полимеразной цепной реакции (ПЦР) c праймерами, соответствующими 5’- и 3’-концам реаранжированных генов VH и VL, как описано в публикации Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86:3833-3837 (1989), получая таким образом разнообразные наборы V-генов для экспрессии. V-гены могут быть амплифицированы из кДНК или геномной ДНК при помощи нижних праймеров у 5’-конца экзона, кодирующего зрелый V-домен, и верхних праймеров, находящихся в J-сегменте, как описано в публикациях Orlandi et al. (1989) и Ward et al., Nature, 341: 544-546 (1989). Однако для амплификации из кДНК нижние праймеры могут также находиться в лидерном экзоне, как описано в публикации Jones et al., Biotechnol., 9:88-89 (1991), и верхние праймеры могут находиться в константной области, как описано в публикации Sastry et al., Proc. Natl. Acad. Sci. (USA), 86:5728-5732 (1989). Для максимального увеличения комплементарности праймерам может быть сообщена вырожденность, как описано в публикациях Orlandi et al. (1989) или Sastry et al. (1989). В определенных вариантах осуществления изобретения разнообразие библиотек максимально увеличивается в результате использования праймеров для ПЦР, направленно воздействующих на каждое семейство V-генов с целью амплификации всех возможных расположений VH и VL, присутствующих в образце нуклеиновой кислоты иммунокомпетентных клеток, например, как описано в публикации Marks et al., J. Mol. Biol., 222:581-597 (1991) или в публикации Orum et al., Nucleic Acids Res., 21:4491-4498 (1993). Для клонирования амплифицированной ДНК в векторы экспрессии в праймеры для ПЦР могут быть введены редкие сайты рестрикции в виде метки у одного конца, как описано в публикации Orlandi et al. (1989), или может быть произведена дальнейшая амплификация при помощи ПЦР с использованием меченого праймера, как описано в публикации Clackson et al., Nature, 352:624-628 (1991).
Наборы синтетически реаранжированных V-генов могут быть получены in vitro из сегментов V-гена. Большинство сегментов VH-гена человека были клонированы и секвенированы (см. публикацию Tomlinson et al., J. Mol. Biol., 227:776-798 (1992)) и картированы (см. публикацию Matsuda et al., Nature Genet., 3:88-94 (1993); указанные клонированные сегменты (включая все основные конформации Н1 и Н2 петли) могут быть использованы для создания разнообразных наборов VH-генов с использованием праймеров для ПЦР, кодирующих Н3 петли различных последовательностей, как описано в публикации Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992). Наборы VH-генов могут быть также получены с использованием всего разнообразия последовательностей в длинной Н3 петле одинаковой длины, как описано в публикации Barbas et al., Proc. Natl. Acad. Sci. USA, 89:4457-4461 (1992). Vκ и Vλ сегменты человека были клонированы и секвенированы (см. публикацию Williams and Winter, Eur. J. Immunol., 23:1456-1461 (1993) и могут быть использованы для создания синтетических наборов легких цепей. Синтетические наборы V-генов на основе целого ряда складок VH и VL и длин L3 и Н3 кодируют антитела со значительным структурным разнообразием. После амплификации V-гена, кодирующего ДНК, сегменты гаметического V-гена могут быть реаранжированы in vitro способами, описанными в публикации Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992).
Наборы фрагментов антител могут быть созданы путем объединения наборов генов VH и VL несколькими способами. Каждый набор может быть создан в разных векторах, после чего векторы объединяют in vitro, например, как описано в публикации Hogrefe et al., Gene, 128:119-126 (1993), или in vivo путем комбинированного инфицирования, например, системой loxP, описанной в публикации Waterhouse et al., Nucl. Acids Res., 21:2265-2266 (1993). В процессе рекомбинации in vivo используют двухцепочечную природу Fab-фрагментов для преодоления ограничения, налагаемого на размер библиотеки эффективностью трансформации E. coli. Наивные наборы VH и VL клонируют отдельно, один в фагомид и другой в вектор на основе фага. Затем обе библиотеки объединяют, инфицируя фагом фагомидсодержащие бактерии, в результате чего каждая клетка содержит разную комбинацию и размер библиотеки ограничивается только числом присутствующих клеток (примерно 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo, благодаря чему гены VH и VL объединяются в один репликон и совместно упаковываются в вирионы фага. Указанные огромные библиотеки позволяют получить большое число разнообразных антител с хорошей аффинностью (Kd-1 около 10-8 М).
Альтернативно, наборы генов могут быть клонированы последовательно в один вектор, например, как описано в публикации Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или собраны вместе при помощи ПЦР и затем клонированы, например, как описано в публикации Clackson et al., Nature, 352:624-628 (1991). Сборка при помощи ПЦР также может быть использована для соединения ДНК VH и VL с ДНК, кодирующей гибкий пептидный спейсер, для создания наборов одноцепочечных Fv (scFv) фрагментов. В соответствии с другим способом используют ”сборку при помощи ПЦР в клетке” для объединения генов VH и VL в лимфоцитах при помощи ПЦР и затем клонируют наборы связанных генов в соответствии с описанием, приведенным в публикации Embleton et al., Nucl. Acids Res., 20:3831-3837 (1992).
Антитела, продуцированные наивными библиотеками (природными или синтетическими), могут обладать средней аффинностью (Kd-1 около 106-107 М-1), но созревание аффинности может быть также имитировано in vitro путем создания вторичных библиотек и выполнения повторного отбора, как описано в приведенной выше публикации Winter et al. (1994). Например, может быть введена произвольная мутация in vitro в результате использования подверженной ошибкам полимеразы (см. публикацию Leung et al., Technique, 1:11-15 (1989)) при выполнении способа, описанного в публикации Hawkins et al., J. Mol. Biol., 226:889-896 (1992), или способа, описанного в публикации Gram et al., Proc. Natl. Acad. Sci. USA, 89:3576-3580 (1992). Созревание аффинности может быть дополнительно выполнено путем произвольной мутации одной или нескольких CDR-областей, например, при помощи ПЦР с праймерами, содержащими произвольную последовательность, заполняющую представляющую интерес CDR, в выбранных отдельных клонах Fv-фрагментов, и скрининга клонов с большой аффинностью. В заявке WO 9607754 (опубликованной 14 марта 1996 г.) описан способ индукции мутагенеза в определяющей комплементарность области легкой цепи иммуноглобулина с целью создания библиотеки генов легкой цепи. Другим эффективным способом является рекомбинация доменов VH или VL, отобранных способом отображения на фаге, с набором природных вариантов V доменов, полученных у неиммунизированных доноров, и скрининг большей аффинности при выполнении нескольких циклов перестановки цепи, как описано в публикации Marks et al., Biotechnol., 10:779-783 (1992). Указанный способ позволяет продуцировать антитела и фрагменты антител с аффинностью около 10-9 М или меньше.
Скрининг библиотек может быть выполнен разными способами, известными в данной области. Например, STEAP-1, экспрессированный на клетках-хозяевах, иммобилизованных на адсорбционных планшетах, можно использовать для сенсибилизации лунок адсорбционных планшетов, STEAP-1 можно также использовать для сортировки клеток либо конъюгировать с биотином для улавливания гранулами, покрытыми стрептавидином, или использовать при выполнении любого другого способа скрининга библиотек отображения на фаге.
Образцы библиотеки фагов вводят в соприкосновение с иммобилизованным STEAP-1 в условиях, пригодных для связывания по меньшей мере части фаговых частиц с адсорбентом. Условия, включая рН, ионную силу, температуру и тому подобные, обычно выбирают с возможностью имитации физиологических условий. Фаги, связанные с твердой фазой, промывают и элюируют кислотой, например, в соответствии с описанием, приведенным в публикации Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или щелочью, например, в соответствии с описанием, приведенным в публикации Marks et al., J. Mol. Biol., 222:581-597 (1991), или способом конкурентного связывания с антигеном STEAP-1, например, аналогичным способу, описанному в публикации Clackson et al., Nature, 352: 624-628 (1991). Фаги могут быть обогащены в 20-1000 раз при выполнении одного цикла селекции. Кроме того, обогащенные фаги могут быть выращены в бактериальной культуре и подвергнуты дальнейшим циклам отбора.
Эффективность отбора зависит от многих факторов, включающих кинетику диссоциации во время промывки и способность нескольких фрагментов антитела на одном фаге одновременно связываться с антигеном. Антитела с быстрой кинетикой диссоциации (и слабой аффинностью связывания) могут быть сохранены в результате выполнения коротких промывок, отображения на мультивалентных фагах и благодаря высокой плотности оболочки антигена в твердой фазе. Высокая плотность не только стабилизирует фаг в процессе многократных взаимодействий, но также способствует повторному связыванию диссоциированного фага. Отбор антител с медленной кинетикой диссоциации (и хорошей аффинностью связывания) можно стимулировать с помощью продолжительных промывок, отображения на моновалентном фаге, как описано в публикации Bass et al., Proteins, 8:309-314 (1990) и в заявке WO 92/09690, и низкой плотности оболочки антигена, как описано в публикации Marks et al., Biotechnol., 10:779-783 (1992).
Можно выбирать антитела на фаге с разной аффинностью, даже с незначительно отличающейся аффинностью к STEAP-1. Однако неспецифическая мутация выбранного антитела (например, выполняемая в некоторых способах созревания аффинности), по-видимому, вызывает образование многих мутантов, большинство которых связывается с антигеном, но лишь небольшая часть связывается с более высокой степенью аффинности. Ограничивая количество белка STEAP-1, можно обнаружить редкий фаг с высокой степенью аффинности. Для сохранения всех мутантов с более высокой степенью аффинности фаги можно инкубировать с избытком биотинилированного STEAP-1, используемого в концентрации с более низкой молярностью по сравнению с аффинной константой молярности мишени для STEAP-1. Фаги, связывающиеся с высокой аффинностью, затем можно улавливать парамагнитными гранулами, покрытыми стрептавидином. Такое ”равновесное улавливание” позволяет отбирать антитела в соответствии с их аффинностью связывания, при этом могут быть выделены мутантные клоны по меньшей мере с двукратной аффинностью связывания из огромного количества фагов с более низкой аффинностью. Условия, используемые для промывки фагов, связанных с твердой фазой, также могут быть изменены для установления различий на основании кинетики диссоциации.
Клоны антител против STEAP-1 могут быть отобраны на основании активности. Определенные варианты осуществления изобретения относятся к антителам против STEAP-1, которые связываются с живыми клетками, экспрессирующими STEAP-1 в естественных условиях. Один вариант осуществления изобретения относится к антителам против STEAP-1, которые блокируют связывание лиганда STEAP-1 с белком STEAP-1, но не блокируют связывание лиганда STEAP-1 со вторым белком. Клоны Fv-фрагментов, соответствующие таким антителам против STEAP-1, могут быть отобраны в результате (1) выделения клонов антител против STEAP-1 из вышеописанной библиотеки фагов и необязательной амплификации выделенной популяции фаговых клонов путем выращивания популяции в приемлемом бактериальном хозяине; (2) отбора STEAP-1 и второго белка, на которые должна быть направлена соответственно блокирующая и неблокирующая активность; (3) адсорбции фаговых клонов антител против STEAP-1 на иммобилизованном STEAP-1; (4) использования избытка второго белка для элюирования нежелательных клонов, узнающих сайты связывания STEAP-1, которые перекрывают или совместно используются с сайтами связывания второго белка; и (5) элюирования клонов, которые остаются адсорбированными после выполнения стадии (4). Клоны, обладающие требуемыми блокирующими/неблокирующими свойствами, могут быть необязательно обогащены в результате однократного или многократного повторения процедур отбора антител, рассмотренных в настоящем описании изобретения.
ДНК, кодирующую моноклональные антитела, выделенные из гибридомы, или клоны Fv-фрагментов, отображенные на фаге, по настоящему изобретению можно легко выделить и секвенировать стандартными способами (например, используя олигонуклеотидные праймеры, созданные для специфической амплификации представляющих интерес кодирующих областей тяжелой и легкой цепей из матричной ДНК гибридомы или фага). Выделенную ДНК можно ввести в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьян, клетки яичника китайского хомячка (СНО) или миеломные клетки, которые в противном случае не продуцируют белок иммуноглобулина, для синтеза требуемых моноклональных антител в рекомбинантных клетках-хозяевах. Рекомбинантная экспрессия ДНК, кодирующей антитело, в бактериальных клетках описана в обзорных статьях Skerra et al., Curr. Opinion in Immunol., 5:256 (1993) и Pluckthun, Immunol. Revs., 130:151 (1992).
ДНК, кодирующая клоны Fv-фрагментов по настоящему изобретению, может быть объединена с известными последовательностями ДНК, кодирующими константные области тяжелой цепи и/или легкой цепи (например, соответствующие последовательности ДНК могут быть взяты из приведенной выше публикации Kabat et al.), для создания клонов, кодирующих полные или неполные тяжелые и/или легкие цепи. Следует отметить, что для указанной цели могут быть использованы константные области любого изотипа, в том числе константные области IgG, IgM, IgA, IgD и IgE, которые могут быть получены у человека или животных любых видов. Клон Fv-фрагментов, выделенный из ДНК вариабельного домена животного одного вида (такого как человек) и затем гибридизированный с ДНК константной области животного другого вида, с образованием одной или нескольких кодирующих последовательности для ”гибридной” полноразмерной тяжелой цепи и/или легкой цепи, входит в определение “химерного” или “гибридного” антитела, как используется в настоящем описании. В определенных вариантах осуществления изобретения клон Fv-фрагментов, выделенный из ДНК вариабельного домена человека, гибридизируют с ДНК константной области человека с образованием одной или нескольких кодирующих последовательностей для полных или неполных тяжелых и/или легких цепей.
ДНК, кодирующая антитело против STEAP-1, выделенное из гибридомы по настоящему изобретению, может быть также модифицирована, например, путем введения кодирующей последовательности константных доменов тяжелой и легкой цепи человека вместо гомологичных последовательностей мыши, выделенных из клона гибридом (как, например, в способе, описанном в публикации Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). ДНК, кодирующая антитело или фрагмент, выделенные из гибридомы или клона Fv-фрагментов, может быть далее модифицирована путем присоединения при помощи ковалентной связи к кодирующей последовательности иммуноглобулина или к кодирующей последовательности полипептида, не являющегося иммуноглобулином. Таким образом получают ”химерные” или “гибридные” антитела, обладающие специфичностью связывания клона Fv-фрагментов или антител по настоящему изобретению, выделенных из клона гибридом.
3. Векторы, клетки-хозяева и способы рекомбинантных ДНК
Для рекомбинантного продуцирования антитела по настоящему изобретению нуклеиновую кислоту, кодирующую указанное антитело, выделяют и вводят в реплицирующий вектор для дальнейшего клонирования (амплификации ДНК) или экспрессии. ДНК, кодирующую данное антитело, можно легко выделить и секвенировать стандартными способами (например, при помощи олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Существует много векторов. Выбор вектора частично зависит от используемой клетки-хозяина. Клетки-хозяева являются прокариотическими или эукариотическими (обычно клетки млекопитающих). Следует отметить, что для указанной цели могут быть использованы константные области любого изотипа, в том числе константные области IgG, IgM, IgA, IgD и IgE, которые могут быть получены у человека или животного любого вида.
Создание антител с использованием прокариотических клеток-хозяев
Конструкция вектора
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела по настоящему изобретению, могут быть получены стандартными способами рекомбинантных ДНК. Требуемые полинуклеотидные последовательности могут быть выделены и секвенированы из антителопродуцирующих клеток, таких как клетки гибридом. Альтернативно полинуклеотиды могут быть синтезированы в синтезаторе нуклеотидов или способами ПЦР. Полученные последовательности, кодирующие полипептиды, вводят в рекомбинантный вектор, способный реплицировать и экспрессировать гетерологичные полинуклеотиды в прокариотических клетках-хозяевах. Для достижения целей настоящего изобретения могут быть использованы многие существующие векторы, известные в данной области. Выбор соответствующего вектора зависит главным образом от размера нуклеиновых кислот, вводимых в вектор, и конкретной клетки-хозяина, трансформируемой указанным вектором. Каждый вектор содержит разные компоненты в зависимости от его функции (амплификация или экспрессия гетерологичного полинуклеотида либо обе функции) и совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора обычно включают, не ограничиваясь ими, ориджин репликации, ген селектируемого маркера, промотор, сайт связывания рибосомы (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
Плазмидные векторы, содержащие репликон и регуляторные последовательности, выделенные из вида, совместимого с клеткой-хозяином, используют вместе с указанными хозяевами. Вектор обычно содержит сайт репликации, а также маркерные последовательности, способные производить фенотипический отбор в трансформированных клетках. Например, E. coli обычно трансформируют плазмидой pBR322, выделенной из вида E. coli. Плазмида pBR322 содержит гены, кодирующие устойчивость к ампициллину (Amp) и тетрациклину (Tet), и, таким образом, позволяет легко идентифицировать трансформированные клетки. Плазмида pBR322 и ее производные, плазмиды других микроорганизмов или бактериофаг могут также содержать, или могут быть модифицированы с возможностью содержания, промоторы, которые могут быть использованы микроорганизмом для экспрессии эндогенных белков. Примеры производных плазмиды pBR322, используемых для экспрессии определенных антител, подробно рассмотрены в патенте США № 5648237, Carter et al.
Кроме того, векторы на основе фага, содержащие репликон и регуляторные последовательности, совместимые с микроорганизмом-хозяином, могут быть использованы в качестве трансформирующих векторов в указанных хозяевах. Например, такой бактериофаг как λGEM.TM.-11 может быть использован для создания рекомбинантного вектора, которым трансформируют восприимчивые клетки-хозяева, такие как E. coli LE392.
Вектор экспрессии по настоящему изобретению может содержать две или большее число пар промотора-цистрона, каждая из которых кодирует компоненты полипептида. Промотор является нетранслированной регуляторной последовательностью, локализованной вверху (5') от цистрона, модулирующего ее экспрессию. Прокариотические промоторы обычно делятся на два класса, индуцибельные и конститутивные. Индуцибельный промотор является промотором, который инициирует более высокую транскрипцию цистрона под своим управлением в ответ на изменение условий в культуре, например, наличие или отсутствие питательного вещества или изменение температуры.
Известно большое число промоторов, распознаваемых разными потенциальными клетками-хозяевами. Выбранный промотор может быть функционально связан с ДНК цистрона, кодирующей легкую или тяжелую цепь, в результате удаления промотора из исходной ДНК путем расщепления рестрикционным ферментом и введения выделенной последовательности промотора в вектор по настоящему изобретению. Для управления амплификацией и/или экспрессией генов-мишеней может быть использована как последовательность нативного промотора, так и многие гетерологичные промоторы. В некоторых вариантах осуществления изобретения использованы гетерологичные промоторы, так как такие промоторы обычно обеспечивают большую транскрипцию и более высокие выходы экспрессированного гена-мишени по сравнению с нативным промотором полипептида-мишени.
Промоторы, пригодные для использования с прокариотическими хозяевами, включают промотор PhoA, системы промоторов β-галактамазы и лактозы, систему промоторов триптофана (trp) и гибридные промоторы, такие как tac или trc промотор. Однако также могут быть использованы другие промоторы, функционирующие в бактериях (например, другие известные бактериальные или фаговые промоторы). Нуклеотидные последовательности таких промоторов опубликованы в научной литературе, благодаря чему специалист в данной области может функционально лигировать их с цистронами, кодирующими легкие и тяжелые цепи мишени (Siebenlist et al. (1980) Cell 20:269), при помощи линкеров или адапторов для получения любых необходимых сайтов рестрикции.
В одном из аспектов настоящего изобретения каждый цистрон в рекомбинантном векторе содержит секретирующий компонент сигнальной последовательности, который управляет транслокацией экспрессированных полипептидов через мембрану. Как правило, сигнальная последовательность может быть компонентом вектора или частью ДНК полипептида мишени, введенной в вектор. Сигнальная последовательность, выбранная в соответствии с целями настоящего изобретения, должна быть распознаваемой и процессируемой (то есть расщепляемой сигнальной пептидазой) клеткой-хозяином. В случае прокариотических клеток-хозяев, которые не узнают и не процессируют сигнальные последовательности, нативные для гетерологичных полипептидов, сигнальную последовательность заменяют прокариотической сигнальной последовательностью, выбираемой, например, из группы, состоящей из щелочной фосфатазы, пенициллиназы, Ipp или лидерных последовательностей термостойкого энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и МВР. В одном из вариантов осуществления изобретения сигнальные последовательности, используемые в обоих цистронах экспрессирующей системы, являются сигнальными последовательностями STII или их вариантами.
В одном из аспектов изобретения иммуноглобулины по настоящему изобретению могут быть продуцированы в цитоплазме клетки-хозяина и поэтому не требуют наличия сигнальных последовательностей секреции в каждом цистроне. В данном случае легкие и тяжелые цепи иммуноглобулина могут быть экспрессированы, уложены и собраны с образованием функциональных иммуноглобулинов в цитоплазме. Определенные штаммы-хозяева (например, штаммы trxB E. coli) создают условия в цитоплазме, благоприятные для образования дисульфидной связи, что позволяет произвести правильную укладку цепи и сборку экспрессированных субъединиц белка. Proba and Pluckthun, Gene, 159:203 (1995).
Антитела по настоящему изобретению могут быть также продуцированы при помощи экспрессирующей системы, в которой количественное соотношение экспрессированных компонентов полипептида может быть модулировано для максимального увеличения выхода секретированных и правильно собранных антител по настоящему изобретению. Такая модуляция по меньшей мере частично достигается в результате одновременной модуляции интенсивности трансляции для компонентов полипептида.
Один способ модуляции интенсивности трансляции описан в патенте США № 5840523, Simmons et al. В данном способе использованы варианты области инициации трансляции (TIR) в цистроне. Для данной TIR может быть создана серия вариантов аминокислотных последовательностей или последовательностей нуклеиновых кислот с разной интенсивностью трансляции, что позволяет регулировать указанный фактор для требуемого уровня экспрессии конкретной цепи. Варианты TIR могут быть созданы стандартными способами мутагенеза, вызывающими замены кодонов, которые могут изменить аминокислотную последовательность. В определенных вариантах осуществления изобретения замены в нуклеотидной последовательности являются молчащими. Изменения в TIR могут включать, например, изменения числа или размещения последовательностей Шайна-Далгарно наряду с изменениями сигнальной последовательности. Одним способом создания мутантных сигнальных последовательностей является создание ”банка кодонов” в начале кодирующей последовательности, который не изменяет аминокислотную последовательность сигнальной последовательности (то есть замены являются молчащими). Данный процесс может быть выполнен путем изменения положения третьего нуклеотида в каждом кодоне; дополнительно в банк могут быть добавлены некоторые аминокислоты, такие как лейцин, серин и аргинин, которые имеют множество первых и вторых положений. Указанный способ мутагенеза подробно описан в публикации Yansura et al. (1992) METHODS: A Companion to Methods in Enzymol. 4:151-158.
В одном из вариантов осуществления изобретения создают набор векторов с разной интенсивностью TIR для каждого цистрона. Такой ограниченный набор позволяет сравнивать уровни экспрессии каждой цепи, а также выход требуемых антител при наличии разных комбинаций интенсивности TIR. Интенсивность TIR можно определить путем количественного измерения уровня экспрессии гена-репортера, как подробно описано в патенте США № 5840523, Simmons et al. На основании сравнения интенсивности трансляции отбирают требуемые TIR для объединения в конструкциях векторов экспрессии по настоящему изобретению.
Прокариотические клетки-хозяева, пригодные для экспрессии антител по настоящему изобретению, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры полезных бактерий включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, вид Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления изобретения использованы грамотрицательные клетки. В одном из вариантов осуществления изобретения в качестве хозяев по настоящему изобретению использованы клетки E. coli. Примеры штаммов E. coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219; депозит АТСС № 27325) и их производные, в том числе штамм 33D3, имеющий генотип W3110 ∆fhuA (∆tonA) ptr3 lac Iq lacL8 ∆ompT∆(nmpc-fepE) degP41 kanR (патент США № 5639635). Также могут быть использованы другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coli λ 1776 (ATCC 31537) и E. coli RV308 (АТСС 31608). Приведенные примеры являются иллюстративными и не ограничивают объем изобретения. Способы создания производных любых вышеуказанных бактерий, имеющих определенные генотипы, известны в данной области и описаны, например, в публикации Bass et al., Proteins, 8:309-314 (1990). Соответствующие бактерии обычно выбирают с учетом реплицируемости репликона в клетках бактерий. Например, виды E. coli, Serratia или Salmonella могут быть использованы в качестве хозяина, когда репликон доставляется хорошо известными плазмидами, такими как pBR322, pBR325, pACYC177 или pKN410. Клетка-хозяин обычно должна секретировать минимальные количества протеолитических ферментов, при этом в культуру клеток могут быть введены дополнительные ингибиторы протеазы.
Продуцирование антител
Клетки-хозяева трансформируют вышеуказанными векторами экспрессии и культивируют в стандартных питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности.
Трансформация означает введение ДНК в прокариотическую клетку-хозяина, в котором ДНК реплицируется в качестве внехромосомного элемента или элемента, встроенного в хромосому. В зависимости от используемой клетки-хозяина трансформацию выполняют стандартными способами, приемлемыми для таких клеток. Бактериальные клетки, имеющие плохопроницаемые барьеры в виде оболочки клетки, обычно обрабатывают хлоридом кальция. В другом способе трансформации используют полиэтиленгликоль/ДМСО. В еще одном способе используют электропорацию.
Прокариотические клетки, используемые для продуцирования полипептидов по настоящему изобретению, выращивают в средах, известных в данной области и пригодных для культивирования выбранных клеток-хозяев. Примеры приемлемых сред включают бульон Луриа (LB), содержащий необходимые питательные добавки. В некоторых вариантах осуществления изобретения среда также содержит селектирующий агент, выбираемый с учетом конструкции вектора экспрессии, который обеспечивает избирательный рост прокариотических клеток, содержащих вектор экспрессии. Например, для роста клеток, экспрессирующих ген устойчивости к ампициллину, в среду добавляют ампициллин.
Любые необходимые добавки помимо углерода, азота и источников неорганического фосфата могут быть введены в соответствующих концентрациях отдельно или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Культуральная среда может необязательно содержать один или несколько восстановителей, выбираемых из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при приемлемых температурах. В определенных вариантах осуществления изобретения температуры выращивания E. coli находятся в пределах от около 20°С до около 39°С, от около 25°С до около 37°С или около 30°С. Значение рН среды может быть любым от около 5 до около 9 в зависимости главным образом от организма-хозяина. В определенных вариантах осуществления изобретения значение рН для E. coli находится в пределах от около 6,8 до около 7,4 или около 7,0.
При использовании индуцибельного промотора в векторе экспрессии по настоящему изобретению экспрессию белка индуцируют в условиях, пригодных для активации промотора. В одном из аспектов настоящее изобретение относится к промоторам PhoA, используемым для регулирования транскрипции полипептидов. Поэтому трансформированные клетки-хозяева культивируют в среде для индукции с ограниченным содержанием фосфата. В определенных вариантах осуществления изобретения средой с ограниченным содержанием фосфата является среда C.R.A.P. (см., например, публикацию Simmons et al., J. Immunol. Methods (2002), 263:133-147). Могут быть использованы разные другие индукторы в зависимости от применяемой конструкции вектора, как хорошо известно в данной области.
В одном из вариантов осуществления изобретения экспрессированные полипептиды по настоящему изобретению выделяют из периплазмы клеток-хозяев, в которую секретируются указанные полипептиды. Выделение белка обычно предполагает разрушение микроорганизма такими способами как осмотический шок, обработка ультразвуком или лизис. После разрушения клеток клеточный дебрис или цельные клетки могут быть удалены центрифугированием или фильтрацией. Белки затем могут быть очищены, например, при помощи аффинной хроматографии на смоле. Альтернативно белки могут быть перенесены в культуральную среду и выделены из нее. Клетки удаляют из культуры, а супернатанта культуры подвергают фильтрованию и концентрированию для дальнейшей очистки продуцированных белков. Экспрессированные полипептиды могут быть выделены и идентифицированы известными способами, такими как электрофорез в полиакриламидном геле (PAGE) и анализ способом вестерн-блоттинга.
В соответствии с одним аспектом изобретения антитело продуцируют в большом количестве при помощи процесса ферментации. Рекомбинантные белки могут быть получены в результате выполнения разных крупномасштабных периодических процессов ферментации. Производительность крупномасштабной ферментации составляет по меньшей мере 1000 литров и в определенных вариантах осуществления изобретения от около 1000 до 100000 литров. В указанных ферментерах использованы лопастные мешалки для распределения кислорода и питательных веществ, в частности, глюкозы (которая является предпочтительным источником углерода/энергии). Ферментация в небольшом масштабе обычно выполняется в ферментере, производительность которого не превышает примерно 100 литров и может составлять от около 1 литра до около 100 литров.
В процессе ферментации экспрессию белка обычно индуцируют после выращивания клеток в приемлемых условиях до требуемой плотности, например, до OD550, равной примерно 180-220, когда клетки находятся в ранней стационарной фазе. Могут быть использованы разные индукторы в зависимости от применяемой конструкции вектора, как известно в данной области и рассмотрено в настоящем описании изобретения. Клетки могут выращиваться до индукции в течение более коротких периодов времени. Клетки обычно индуцируют в течение примерно 12-50 часов, хотя могут быть использованы более продолжительные или более короткие периоды индукции.
Для улучшения выхода и качества полипептидов по настоящему изобретения могут быть модифицированы разные условия ферментации. Например, для улучшения правильной сборки и укладки цепи секретированных полипептидов антитела могут быть использованы дополнительные векторы, сверхэкспрессирующие белки-шапероны, такие как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпропил-цис,транс-изомераза с активностью шаперона), для котрансформации прокариотических клеток-хозяев. Установлено, что белки-шапероны облегчают правильную укладку цепи и улучшают растворимость гетерологичных белков, продуцированных в бактериальных клетках-хозяевах. Chen et al. (1999) J. Biol. Chem. 274:19601-19605; Georgiou et al., U.S. Patent No. 6,083,715; Georgiou et al., патент США № 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275:17100-17105; Ramm and Pluckthun (2000) J. Biol. Chem. 275:17106-17113; Arie et al. (2001) Mol. Microbiol. 39:199-210.
Чтобы свести к минимуму протеолиз экспрессированных гетерологичных белков (в частности, белков, подверженных протеолизу) в настоящем изобретении могут быть использованы определенные штаммы клеток-хозяев с отсутствием протеолитических ферментов. Например, штаммы клеток-хозяев могут быть модифицированы с достижением генетических мутаций в генах, кодирующих известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и комбинации указанных протеаз. Некоторые штаммы E. coli с отсутствием протеазы описаны, например, в приведенной выше публикации Joly et al. (1998); в патенте США № 5264365, Georgiou et al.; патенте США № 5508192, Georgiou et al.; и публикации Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном из вариантов осуществления изобретения в качестве клеток-хозяев в экспрессирующей системе по настоящему изобретению используются штаммы E. coli с отсутствием протеолитических ферментов, которые были трансформированы плазмидами, сверхэкспрессирующими один или несколько белков-шаперонов.
Очистка антитела
В одном из вариантов осуществления изобретения продуцированный белок антитела подвергают дальнейшей очистке для получения по существу гомогенных препаратов, предназначенных для последующих анализов и применений. Могут быть использованы стандартные способы очистки белков, известные в данной области. Нижеследующие способы являются примерами приемлемых способов очистки: фракционирование при помощи иммуноаффинной хроматографии или хроматографии на ионообменных колонках, преципитация этанолом, ВЭЖХ с обращенной фазой, хроматография на кремнеземе или катионообменной смоле, такой как DEAE, хроматографическое фокусирование, SDS-PAGE, преципитация сульфатом аммония или гель-фильтрация, например, через сефадекс G-75.
В соответствии с одним аспектом изобретения для очистки продуктов антитела по настоящему изобретению способом иммуноаффинной хроматографии используют протеин А, иммобилизованный на твердой фазе. Протеин А является белком клеточной оболочки с молекулярной массой 41 кДа, выделенным из Staphylococcus aureas, который связывается с высокой аффинностью с Fc-областью антител. Lindmark et al. (1983) J. Immunol. Meth. 62:1-13. Твердая фаза, на которой иммобилизован протеин А, может быть колонкой со стеклянной или кремнеземной поверхностью, стеклянной колонкой с контролируемым размером пор или колонкой с кремниевой кислотой. В некоторых применениях колонку покрывают реагентом, таким как глицерин, для возможного предотвращения неспецифического сцепления загрязняющих примесей.
В качестве первой стадии очистки препарат, выделенный из вышеописанной культуры клеток, наносят на твердую фазу с иммобилизованным протеином А для достижения специфического связывания представляющего интерес антитела с протеином А. Твердую фазу затем промывают для удаления загрязняющих примесей, неспецифически связанных с твердой фазой. И наконец, представляющее интерес антитело выделяют из твердой фазы элюированием.
Создание антител с использованием эукариотических клеток-хозяев
Вектор, предназначенный для использованием в эукариотической клетке-хозяине, обычно включает один или несколько нижеследующих неограничивающих компонентов: сигнальную последовательность, ориджин репликации, один или несколько генов-маркеров, энхансер, промотор и последовательность терминации транскрипции.
Сигнальная последовательность
Вектор, предназначенный для использования в эукариотической клетке-хозяине, может также содержать сигнальную последовательность или другой полипептид, имеющий специфический сайт расщепления у N-конца зрелого белка или представляющего интерес полипептида. Выбранная гетерологичная сигнальная последовательность может быть последовательностью, распознаваемой и процессируемой (например, расщепляемой сигнальной пептидазой) клеткой-хозяином. Для экспрессии в клетках млекопитающих могут быть использованы сигнальные последовательности млекопитающих, а также секреторные лидерные последовательности вирусов, например, сигнальная последовательность вируса простого герпеса gD. ДНК для такой области-предшественника лигируют в рамке считывания с ДНК, кодирующей антитело.
Ориджин репликации
Ориджин репликации обычно не нужен для векторов экспрессии млекопитающих. Например, ориджин SV40 может быть использован только потому, что он содержит ранний промотор.
Селектирующий ген
Векторы экспрессии и клонирующие векторы могут содержать селектирующий ген, также именуемый селектируемым маркером. Типичные селектирующие гены кодируют белки, которые (а) сообщают устойчивость к антибиотикам или другим токсинам, таким как ампициллин, неомицин, метотрексат или тетрациклин, (b) вызывают ауксотрофный дефицит комплемента или (с) предоставляют необходимые питательные вещества, отсутствующие в комплексной среде.
Одним примером схемы селекции является использование лекарственного средства для прекращения роста клетки-хозяина. Клетки, которые были успешно трансформированы гетерологичным геном, продуцируют белок, сообщающий лекарственную устойчивость и, таким образом, выживают в режиме селекции. Примерами таких лекарственных средств являются неомицин, микофенольная кислота и гигромицин.
Другим примером приемлемых селектируемых маркеров для клеток млекопитающих являются маркеры, позволяющие идентифицировать клетки, поглощающие нуклеиновую кислоту антитела, такие как DHFR, тимидинкиназа, металлотионеин-I и -II, предпочтительно, гены металлотионеина приматов, аденозиндеаминаза, орнитиндекарбоксилаза и т.д.
Например, в некоторых вариантах осуществления изобретения клетки, трансформированные селектирующим геном DHFR, сначала идентифицируют, культивируя все трансформанты в культуральной среде, содержащей метотрексат (Mtx), конкурентный антагонист DHFR. В некоторых вариантах осуществления изобретения клетка-хозяин при использовании DHFR дикого типа является линией клеток яичника китайского хомячка (СНО) с отсутствием активности DHFR (например, АТСС CRL-9096).
Альтернативно, клетки-хозяева (в частности, клетки-хозяева дикого типа, содержащие эндогенный белок DHFR), трансформированные или котрансформированные последовательностями ДНК, кодирующими антитело, белок DHFR дикого типа и другой селектируемый маркер, такой как аминогликозид-3'-фосфотрансфераза (АРН), могут быть выбраны при выращивании клеток в среде, содержащей селектирующий агент для селектируемого маркера, такой как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См. патент США № 4965199.
Промотор
Векторы экспрессии и клонирующие векторы обычно содержат промотор, распознаваемый организмом-хозяином и функционально связанный с нуклеиновой кислотой, кодирующий представляющий интерес полипептид (например, антитело). В данной области известны промоторные последовательности, используемые для эукариотических клеток. Например, фактически все эукариотические гены имеют область с большим содержанием АТ, находящуюся на расстоянии примерно 25-30 оснований вверху от сайта инициации транскрипции. Другая последовательность, обнаруженная на расстоянии 70-80 оснований вверху от начала транскрипции многих генов, является областью CNCAAT, где N может быть любым нуклеотидом. У 3'-конца большинства эукариотических генов находится последовательность ААТААА, которая может быть сигналом для добавления поли-А конца к 3'-концу кодирующей последовательности. В определенных вариантах осуществления изобретения любые или все указанные последовательности могут быть введены в эукариотические векторы экспрессии.
Транскрипцию векторов в клетках-хозяевах млекопитающих регулируют, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус оспы, аденовирус (такой как аденовирус 2), вирус папилломы коров, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и обезьяний вирус 40 (SV40), из гетерологичных промоторов млекопитающих, таких как, например, промотор актина или промотор иммуноглобулина, из промоторов термического шока, при условии, что такие промоторы совместимы с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 могут быть получены в виде рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Предранний промотор цитомегаловируса человека может быть получен в виде рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в клетках-хозяевах млекопитающих с использованием вируса папилломы коров в виде вектора описана в патенте США № 4419446. Модификация указанной системы описана в патенте США № 4601978. См. также публикацию Reyes et al., Nature 297:598-601 (1982), в которой описана экспрессия кДНК β-интерферона человека в клетках мыши, регулируемая промотором тимидинкиназы из вируса простого герпеса. Альтернативно в качестве промотора может быть использован длинный концевой повтор вируса саркомы Рауса.
Энхансерный элемент
Транскрипция ДНК, кодирующей антитело по настоящему изобретению в высших эукариотах, часто увеличивается в результате введения в вектор энхансерной последовательности. В настоящее время известны многие энхансерные последовательности из генов млекопитающих (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Однако обычно используют энхансер из вируса эукариотической клетки. В качестве примеров можно привести энхансер SV40 на поздней стороне ориджина репликации (100-270 п.о.), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне ориджина репликации и энхансеры аденовируса. См. также публикацию Yaniv, Nature 297:17-18 (1982), в которой описаны энхансерные элементы, используемые для активации эукариотических промоторов. Энхансер может быть введен в вектор в положении 5'- или 3'-конца по отношению к кодирующей полипептид последовательности антитела, но обычно находится в положении 5'-концевого сайта от промотора.
Компонент терминации транскрипции
Векторы экспрессии, используемые в эукариотических клетках-хозяевах, могут также содержать последовательности, необходимые для терминации транскрипции и стабилизации мРНК. Такие последовательности обычно находятся в 5'-концевой и иногда в 3’-концевой нетранслированных областях эукариотической или вирусной ДНК или кДНК. Указанные области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслированной части мРНК, кодирующей антитело. Одним приемлемым компонентом терминации транскрипции является область полиаденилирования гормона роста коров. См. WO94/11026 и вектор экспрессии, описанный в данной заявке.
Отбор и трансформация клеток-хозяев
Приемлемые клетки-хозяева, предназначенные для клонирования или экспрессии ДНК в векторах, включают клетки высших эукариотов по настоящему изобретению, в том числе клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (культура ткани) является обычной процедурой. В качестве примеров приемлемых линий клеток-хозяев млекопитающих можно привести линию клеток CV1 почки обезьяны, трансформированную SV40 (COS-7, ATCC CRT 1651); линию клеток эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59 (1997)); клетки почки детеныша хомячка (ВНК, ATCC CCL 10); клетки яичника китайского хомячка/DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC XRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени буйволовой крысы (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4 и линию клеток гепатомы человека (Hep G2).
Клетки-хозяева трансформируют вышеописанными векторами экспрессии или клонирующими векторами для продуцирования антител и культивирования в известной питательной среде, модифицированной соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности.
Культивирование клеток-хозяев
Клетки-хозяева, используемые для продуцирования антитела по настоящему изобретению, можно культивировать в разных средах. Для культивирования клеток-хозяев пригодны коммерчески доступные среды, такие как среда F10 Хэма (Sigma), минимальная поддерживающая среда ((МЕМ), (Sigma), RPMI-1640 (Sigma) и модифицированная по способу Дульбекко среда Игла ((DMEM), Sigma). Кроме того, в качестве культуральных сред для клеток-хозяев могут быть использованы любые среды, описанные в публикации Ham et al., Metrh. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), в патентах США № 4767704, 4657866, 4927762, 4560655 или 5122469; WO 90/03430; WO 87/00195 или патенте США № 30985. Любые указанные среды при необходимости могут дополнительно содержать гормоны и/или другие факторы роста (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия, фосфат кальция и магния), буферы (такие как HEPES), нуклеотиды (такие как аденозин и тимидин), антибиотики (такие как лекарственное средство GENTAMYCIN™), микроэлементы (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозу или эквивалентный источник энергии. Могут быть использованы любые другие добавки в соответствующих концентрациях, известных специалистам в данной области. Условия культивирования, такие как температура, рН и тому подобные, аналогичны ранее используемых для культивирования клетки-хозяина, выбранной для экспрессии, и хорошо известны специалистам в данной области.
Очистка антитела
При использовании способов рекомбинантных ДНК антитело может быть продуцировано внутри клетки или секретировано прямо в среду. Если антитело продуцировано внутри клетки, то на первой стадии удаляют дебрис клеток-хозяев или лизированных фрагментов способами центрифугирования или ультрафильтрации. Если антитело секретировано в среду, супернатанты из таких экспрессирующих систем сначала концентрируют, используя коммерчески доступный фильтр для концентрирования белков, например, ультрафильтрующее устройство Amicon или Millipore Pellicon. На любой из вышеуказанных стадий может быть использован ингибитор протеазы, такой как PMSF, для ингибирования протеолиза и антибиотики для предотвращения роста побочных загрязняющих примесей.
Антитело, полученное из клеток, может быть очищено, например, при помощи хроматографии на колонке с гидроксилапатитом, гель-электрофореза, диализа и аффинной хроматографии, причем стандартным способом очистки является аффинная хроматография. Пригодность протеина А в качестве аффинного лиганда зависит от вида и изотипа Fc домена любого иммуноглобулина, присутствующего в антителе. Протеин А может быть использован для очистки антител, созданных на основе тяжелых γ1, γ2 или γ4 цепей человека (Lindmark et al., J. Immunol. Methods 62:1-13 (1983)). Протеин G рекомендован для всех изотипов мышей и для γ3 изотипа человека (Guss et al., EMBO J. 5:15671575 (1986)). В качестве матрицы, к которой присоединен аффинный лиганд, может быть использована агароза, но существуют и другие матрицы. Матрицы, устойчивые к механическому воздействию, такие как стекло с контролируемым размером пор или полистиролдивинилбензол, обеспечивают более высокие скорости потока и более короткое время обработки по сравнению с агарозой. Для очистки антитела, содержащего СН3 домен, используют смолу ABX™ Bakerbond (J.T. Baker, Phillipsburg, N.J.). В зависимости от выделяемого антитела также могут быть использованы другие способы очистки белка, такие как фракционирование на ионообменной колонке, преципитация этанолом, ВЭЖХ с обращенной фазой, хроматография на кремнеземе, хроматография на гепарине SEPHAROSE™, хроматография на анионообменной или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматографическое фокусирование, SDS-PAGE и преципитация сульфатом аммония.
После выполнения любых стадий предварительной очистки смесь, включающая представляющее интерес антитело и загрязняющие примеси, может быть подвергнута дальнейшей очистке, например, при помощи гидрофобной хроматографии с низким значением рН с использованием элюирующего буфера при рН, равном примерно 2,5-4,5, предпочтительно, при низких концентрациях соли (например, около 0-0,25 М соли).
В данной области хорошо известны разные способы получения антител, предназначенных для исследований, тестирования и клинического применения, которые совместимы с вышеописанными способами и/или по мнению специалиста в данной области пригодны для получения конкретного антитела.
Иммуноконъюгаты
Настоящее изобретение также относится к иммуноконъюгатам (определяемым также как “конъюгаты антитела с лекарственным средством” или “ADC”), содержащим любые антитела против STEAP-1 по настоящему изобретению, конъюгированные с одним или несколькими цитотоксическими агентами, такими как химиотерапевтическое средство, лекарственное средство, ингибитор роста, токсин (например, ферментативно активный токсин бактериального, грибного, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (то есть радиоактивный конъюгат).
В определенных вариантах осуществления изобретения иммуноконъюгат содержит антитело против STEAP-1 и химиотерапевтическое средство или другой токсин. В настоящем описании изобретения рассмотрены (например, приведенные выше) химиотерапевтические средства, используемые для получения иммуноконъюгатов. Также могут быть использованы рассмотренные в настоящем описании изобретения ферментативно активные токсины и их фрагменты.
В определенных вариантах осуществления изобретения иммуноконъюгат содержит антитело против STEAP-1 и один или несколько низкомолекулярных токсинов, которые включают, не ограничиваясь ими, низкомолекулярные лекарственные средства, такие как калихеамицин, майтанзиноид, доластатин, ауристатин, трихотецен и СС1065, и производные указанных лекарственных средств, обладающие цитотоксической активностью. Такие иммуноконъюгаты более подробно рассмотрены ниже.
1. Типичные иммуноконъюгаты
Иммуноконъюгат (или ”конъюгат антитела с лекарственным средством” (“ADC”)) по настоящему изобретению может иметь нижеследующую формулу I, в которой антитело против STEAP-1 конъюгировано (то есть ковалентно связано) с одной или несколькими лекарственными частями (D) при помощи необязательного линкера (L).
Ab-(L-D)p (Формула I)
Таким образом, антитело против STEAP-1 может быть конъюгировано с лекарственным средством прямо или при помощи линкера. В формуле I р означает среднее число лекарственных частей на одно антитело, которое может составлять, например, от около 1 до около 20 лекарственных частей на одно антитело и в определенных вариантах осуществления изобретения от 1 до около 8 лекарственных частей на одно антитело.
Типичные линкеры
В настоящем описании изобретения рассмотрены типичные линкеры и лекарственные части. Линкер может состоять из одного или нескольких линкерных компонентов. Типичные линкерные компоненты включают 6-малеимидокапроил (”МС”), малеимидопропаноил (“МР”), валин-цитруллин (“val-cit” или “vc”), аланин-фенилаланин (“ala-phe”), парааминобензилоксикарбонил (“PAB”), N-сукцинимидил-4-(2-пиридилтио)пентаноат (“SPP”), N-сукцинимидил-4-(N-малеимидометил)-циклогексан-1-карбоксилат (“SMCC”), N-сукцинимидил(4-иодацетил)-аминобензоат (“SIAB”) и этиленокси-СН2СН2О- в качестве одного или нескольких повторяющихся звеньев (“ЕО” или “РЕО”). В данной области известны разные линкерные компоненты, некоторые из которых рассмотрены ниже.
Линкер может быть ”расщепляемым линкером”, облегчающим высвобождение лекарственного средства в клетке. Например, может быть использован кислотолабильный линкер (например, гидразон), протеазачувствительный (например, пептидазачувствительный) линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al., Cancer Research 52:127-131 (1992); патент США № 5208020).
В одном из вариантов осуществления изобретения линкер L конъюгата антитела с лекарственным средством (ADC) имеет формулу:
-Aa-Ww-Yy-
где:
-А- означает удлиняющее звено, ковалентно связанное с тиоловой группой цистеина в антителе (Ab);
а равно 0 или 1;
каждый элемент -W- независимо означает аминокислотное звено;
w независимо является целым числом от 0 до 12;
-Y- означает спейсерное звено, ковалентно связанное с лекарственной частью; и
у равно 0, 1 или 2.
Удлиняющее звено
Удлиняющее звено (-А-), при его наличии, способно связывать антитело с аминокислотным звеном (-W-). Антитело (Ab) содержит тиоловую группу свободного цистеина, которая может образовывать связь с электрофильной функциональной группой удлиняющего звена. Типичные удлиняющие звенья в конъюгатах формулы I представлены формулами II и III, где Ab-, -W-, -Y-, -D, w и у имеют указанные выше значения и R17 означает двухвалентный радикал, выбранный из (СН2)r, С3-С8 карбоциклила, О-(СН2)r, арилена, (СН2)r-арилена, -арилен-(СН2)r-, (СН2)r-(С3-С8 карбоциклила), (С3-С8 карбоциклил)-(СН2)r, С3-С8 гетероциклила, (СН2)r-(С3-С8 гетероциклила), -(С3-С8 гетероциклил)-(СН2)r-, -(СН2)rC(О)NRb(CH2)r-, -(CH2CH2O)r-, -(CH2CH2O)r-CH2-, -(CH2)rC(O)NRb(CH2CH2O)r-, -(CH2)rC(O)NRb(CH2CH2O)r-CH2-, -(CH2CH2O)rC(O)NRb(CH2CH2O)r-, -(CH2CH2O)rC(O)NRb(CH2CH2O)r-CH2- и -(CH2CH2O)rC(O)NRb(CH2)r-, где Rb означает Н, С1-С6 алкил, фенил или бензил, и r независимо означает целое число от 1 до 10.
Арилен включает двухвалентные ароматические углеводородные радикалы с 6-10 атомами углерода, полученные в результате удаления двух атомов водорода из ароматической кольцевой системы. Типичные ариленовые группы включают, не ограничиваясь ими, радикалы, полученные из бензола, замещенного бензола, нафталина, антрацена, бифенила и тому подобных.
Гетероциклильные группы включают кольцевую систему, в которой один или несколько атомов в кольце являются гетероатомами, например, атомами азота, кислорода и серы. Гетероциклильный радикал включает 1-20 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S. Гетероцикл может быть моноциклом, имеющим 3-7 членов в кольце (2-6 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S), или бициклом, имеющим 7-10 членов в кольце (4-9 атомов углерода и 1-3 гетероатома, выбираемых из N, O, P и S), например: бицикло [4,5], [5,5], [5,6] или [6,6] система. Гетероциклы описаны в публикациях Paquette, Leo A.; “Principles of Modern Heterocyclic Chemistry” (W.A. Benjamin, New York, 1968), particularly Chapters 1, 3, 4, 6, 7, and 9; “The Chemistry of Heterocyclic Compounds, A series of Monographs” (John Wiley & Sons, New York, 1950 to present), in particular Volumes 13, 14, 16, 19, and 28; and J. Am. Chem. Soc. (1960) 82:5566.
Примеры гетероциклов включают в качестве примера, не ограничивающего объем изобретения, пиридил, дигидропиридил, тетрагидропиридил (пиперидил), тиазолил, тетрагидротиофенил, тетрагидротиофенил с окисленной серой, пиримидинил, фуранил, тиенил, пирролил, пиразолил, имидазолил, тетразолил, бензофуранил, тианафталенил, индолил, индоленил, хинолинил, изохинолинил, бензимидазолил, пиперидинил, 4-пиперидонил, пирролидинил, 2-пирролидонил, пирролинил, тетрагидрофуранил, бис-тетрагидрофуранил, тетрагидропиранил, бис-тетрагидропиранил, тетрагидрохинолинил, тетрагидроизохинолинил, декагидрохинолинил, октагидроизохинолинил, азоцинил, триазинил, 6Н-1,2,5-тиадиазинил, 2Н,6Н-1,5,2-дитиазинил, тиенил, тиантренил, пиранил, изобензофуранил, хроменил, ксантенил, феноксатинил, 2Н-пирролил, изотиазолил, изоксазолил, пиразинил, пиридазинил, индолизинил, изоиндолил, 3Н-индолил, 1Н-индазолил, пуринил, 4Н-хинолизинил, фталазинил, нафтиридинил, хиноксалинил, хиназолинил, циннолинил, птеридинил, 4Ah-карбазолил, карбазолил, β-карболинил, фенантридинил, акридинил, пиримидинил, фенантролинил, феназинил, фенотиазинил, фуразанил, феноксазинил, изохроманил, хроманил, имидазолидинил, имидазолинил, пиразолидинил, пиразолинил, пиперазинил, индолинил, изоиндолинил, хинуклидинил, морфолинил, оксазолидинил, бензотриазолил, бензизоксазолил, оксиндолил, бензоксазолинил и изатиноил.
Карбоциклильные группы включают насыщенное или ненасыщенное кольцо, имеющее 3-7 атомов углерода в виде моноцикла или 7-12 атомов углерода в виде бицикла. Моноциклические углеродные кольца имеют 3-6 атомов в кольце, обычно 5 или 6 атомов в кольце. Бициклические углеродные кольца имеют 7-12 атомов в кольце, например, расположенных в виде бицикло [4,5], [5,5], [5,6] или [6,6] системы, либо 9 или 10 атомов в кольце, расположенных в виде бицикло [5,6] или [6,6] системы. Примеры моноциклических углеродных колец включают циклопропил, циклобутил, циклопентил, 1-циклопент-1-енил, 1-циклопент-2-енил, 1-циклопент-3-енил, циклогексил, 1-циклогекс-1-енил, 1-циклогекс-2-енил, 1-циклогекс-3-енил, циклогептил и циклооктил.
Из всех приведенных типичных вариантов ADC формулы I, в частности, имеющих формулы II-VI, должно быть понятно, что даже при отсутствии точного указания к антителу присоединены 1-4 лекарственные части (р = 1-4) в зависимости от числа введенных остатков цистеина.
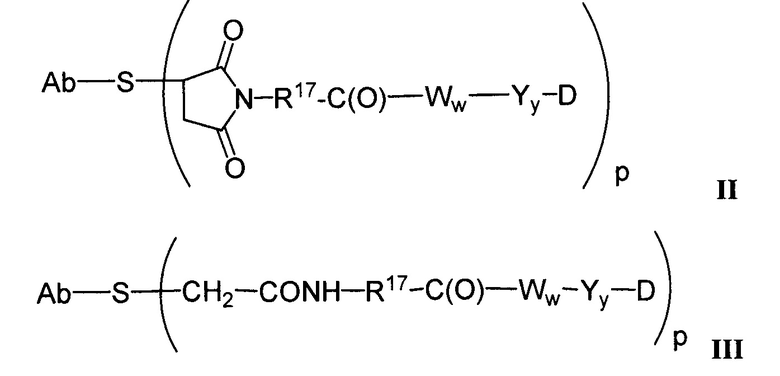
Иллюстративное удлиняющее звено формулы II получено из малеимидокапроила (МС), где R17 означает -(СН2)5-:

Иллюстративное удлиняющее звено формулы II получено из малеимидопропаноила (МР), где R17 означает -(СН2)2-:
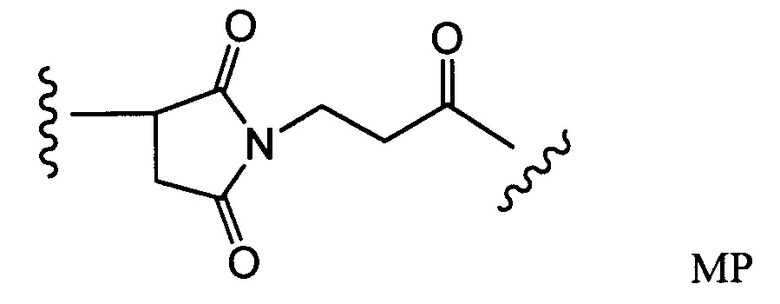
Другое иллюстративное удлиняющее звено формулы II, в котором R17 означает -(СН2СН2О)r-СН2- и r равно 2:
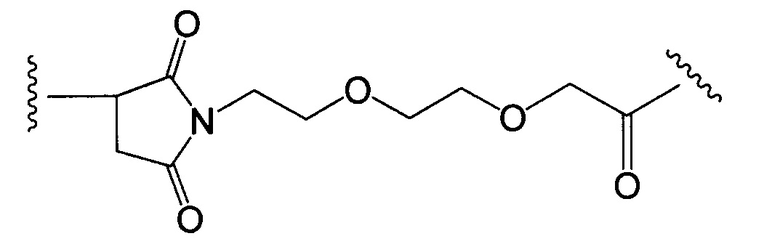
Еще одно иллюстративное удлиняющее звено формулы II, в котором R17 означает -(СН2)rC(О)NRb(CH2CH2O)r-CH2, где Rb означает Н и каждый r равен 2:
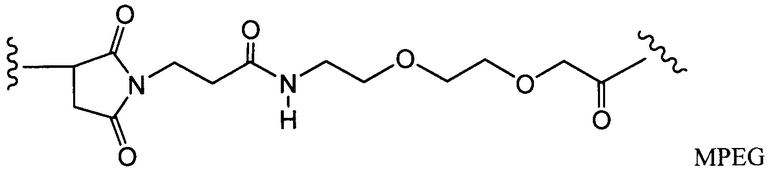
Иллюстративное удлиняющее звено формулы III, в котором R17 означает -(СН2)5-:
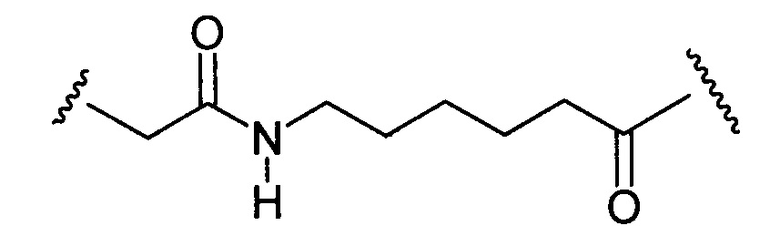
В другом варианте осуществления изобретения удлиняющее звено связано с антителом против STEAP-1 с введенным цистеином при помощи дисульфидной связи между атомом серы в сконструированном антителе с цистеиновой заменой и атомом серы в удлиняющем звене. Типичное удлиняющее звено по данному варианту осуществления изобретения представлено формулой IV, где R17, Ab-, -W-, -Y-, -D, w и у имеют указанные выше значения.
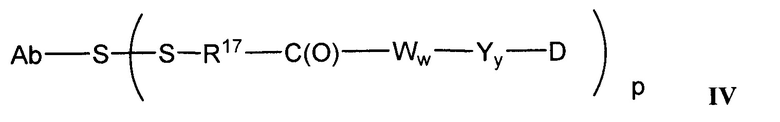
В другом варианте осуществления изобретения реакционноспособная группа удлиняющего звена содержит взаимодействующую с тиолом функциональную группу, которая может образовывать связь с тиоловой группой свободного цистеина в антителе. Примеры взаимодействующих с тиолом функциональных групп включают, не ограничиваясь ими, малеимид, α-галогенацетил, активированные сложные эфиры, такие как сложные эфиры сукцинимида, 4-нитрофениловые эфиры, пентафторфениловые эфиры, тетрафторфениловые эфиры, ангидриды, хлорангидриды кислот, сульфонилхлориды, изоцианаты и изотиоцианаты. Типичные удлиняющие звенья по данному варианту осуществления изобретения представлены формулами Va и Vb, где -R17-, Ab-, -W-, -Y-, -D, w и у имеют указанные выше значения.
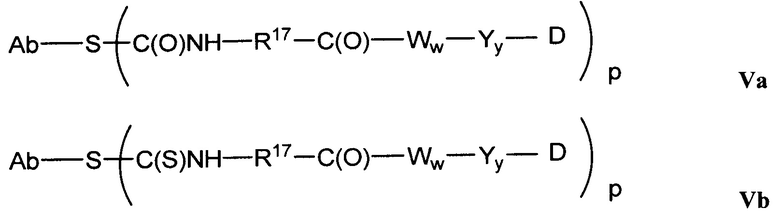
В другом варианте осуществления изобретения линкер может быть линкером дендритного типа, предназначенным для ковалентного присоединения к антителу более одной лекарственной части при помощи разветвленной многофункциональной линкерной части (Sun et al. (2002) Bioorganic & Medicinal Chemistry Letters 12:2213-2215; Sun et al. (2003) Bioorganic & Medicinal Chemistry 11:1761-1768; King (2002) Tetrahedron Letters 43:1987-1990). Дендритные линкеры могут увеличивать молярное отношение лекарственного средства к антителу, то есть нагрузку, определяющую действенность ADC. Таким образом, к сконструированному антителу с цистеиновой заменой, содержащему только одну реакционноспособную тиоловую группу цистеина, при помощи дендритного линкера можно присоединить несколько лекарственных частей.
Аминокислотное звено
Линкер может содержать аминокислотные остатки. Аминокислотное звено (-Ww-), при его наличии, связывает антитело (Ab) с лекарственной частью (D) в конъюгате сконструированного антитела с цистеиновой заменой с лекарственным средством по настоящему изобретению.
-Ww- означает дипептидное, трипептидное, тетрапептидное, пентапептидное, гексапептидное, гептапептидное, октапептидное, нонапептидное, декапептидное, ундекапептидное или додекапептидное звено. Аминокислотные остатки, образующие аминокислотное звено, включают природные аминокислоты, минорные аминокислоты и неприродные аналоги аминокислот, такие как цитруллин. Каждое -Ww- звено имеет формулу, приведенную ниже в квадратных скобках, и w является целым числом 0-12:
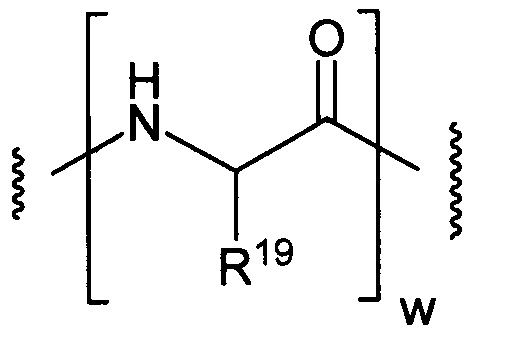
где R19 означает водород, метил, изопропил, изобутил, втор-бутил, бензил, п-гидроксибензил, -СН2ОН, -СН(ОН)СН3, -CH2CH2SCH3, -CH2CONH2, -CH2COOH, -CH2CH2CONH2, -CH2CH2COOH, -(CH2)3NHC(=NH)NH2, -(CH2)3NH2, -(CH2)3NHCOCH3, -(CH2)3NHCHO, -(CH2)4NHC(=NH)NH2, -(CH2)4NH2, -(CH2)4NHCOCH3, -(CH2)4NHCHO, -(CH2)3NHCONH2, -(CH2)4NHCONH2, -CH2CH2CH(OH)CH2NH2, 2-пиридилметил-, 3-пиридилметил-, 4-пиридилметил-, фенил, циклогексил,
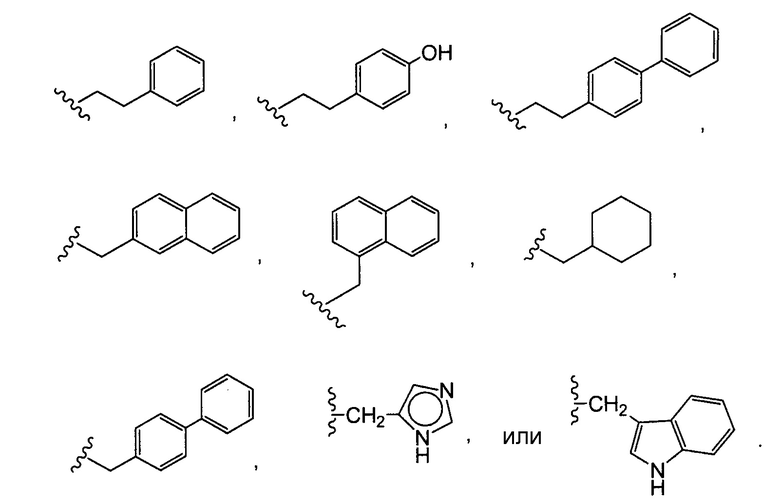
Когда R19 не является водородом, атом углерода, к которому присоединен R19, является хиральным атомом. Каждый атом углерода, к которому присоединен R19, независимо находится в (S) или (R) конфигурации или рацемической смеси. Таким образом, аминокислотные звенья могут быть энантиомерно чистыми, рацемическими или диастереомерными.
Типичные аминокислотные звенья -Ww- включают дипептид, трипептид, тетрапептид или пентапептид. Типичные дипептиды включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Типичные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, образующие аминокислотный линкер, включают природные аминокислоты, минорные аминокислоты и неприродные аналоги аминокислот, такие как цитруллин.
Аминокислотное звено может быть расщеплено одним или неколькими ферментами, включающими опухолеспецифическую протеазу, для высвобождения лекарственной части (-D), которая в одном из вариантов осуществления изобретения протонируется in vivo с образованием лекарственного средства (D). Компоненты аминокислотного линкера могут быть созданы с оптимизированной избирательностью к ферментативному расщеплению определенными ферментами, например, опухолеспецифической протеазой, катепсином В, С и D, или плазминпротеазой.
Спейсерное звено
Спейсерное звено (-Yy-), при его наличии, (у = 1 или 2) связывает аминокислотное звено (-Ww-) с лекарственной частью (D) при наличии аминокислотного звена (w = 1-12). Альтернативно спейсерное звено связывает удлиняющее звено с лекарственной частью при отсутствии аминокислотного звена. Спейсерное звено также связывает лекарственную часть с антителом при отсутствии как аминокислотного звена, так и спейсерного звена (w, y = 0). Спейсерные звенья относятся к двум общим типам: саморазрушающиеся и несаморазрушающиеся. В несаморазрушающемся спейсерном звене часть или все спейсерное звено остается связанным с лекарственной частью после отщепления, в частности, ферментами, аминокислотного звена от конъюгата антитела с лекарственным средством или от лекарственной части с линкером. Когда ADC, содержащий глицин-глициновое спейсерное звено или глициновое спейсерное звено, подвергается ферментативному расщеплению под воздействием протеазы, связанной с опухолевой клеткой, протеазы, связанной с раковой клеткой, или протеазы, связанной с лимфоцитом, глицин-глицин-лекарственная часть или глицин-лекарственная часть отщепляется от Ab-Aa-Ww-. В одном из вариантов осуществления изобретения в клетке-мишени происходит независимая реакция гидролиза, в результате которой расщепляется связь глицин-лекарственной части и высвобождается лекарственное средство.
В другом варианте осуществления изобретения -Yy- означает парааминобензилкарбамоильное (РАВ) звено, в котором фениленовая часть замещена Qm, где Q означает -С1-С8 алкил, -О-(С1-С8 алкил), -галоген, -нитро или -циано и m является целым числом 0-4.
Типичными вариантами несаморазрушающегося спейсерного звена (-Y-) являются: -Gly-Gly-, -Gly-, -Ala-Phe-, -Val-Cit-.
Один вариант осуществления изобретения относится к соединению лекарственной части с линкером или ADC с отсутствием спейсерного звена (у = 0), его фармацевтически приемлемой соли или сольвату.
Альтернативно ADC, содержащий саморазрушающееся спейсерное звено, может высвобождать -D. В одном из вариантов осуществления изобретения -Y- означает группу РАВ, связанную с -Ww- при помощи атома азота аминокислоты группы РАВ и присоединенную непосредственно к -D при помощи карбонатной, карбаматной или простой эфирной группы, при этом ADC имеет типичную структуру:
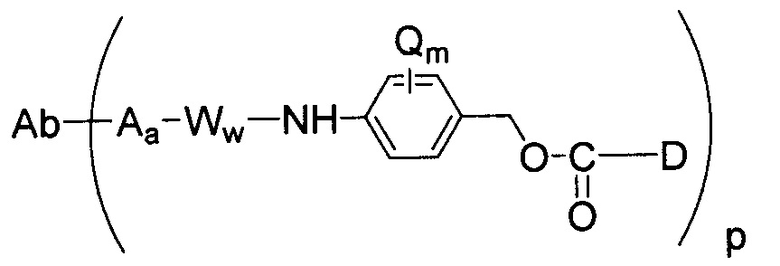
где Q означает -С1-С8 алкил, -О-(С1-С8 алкил), -галоген, -нитро или -циано, m является целым число от 0 до 4, и р равен 1-4.
Другие примеры саморазрушающихся спейсеров включают, не ограничиваясь ими, ароматические соединения, которые в электронном отношении подобны группе РАВ, такие как производные 2-аминоимидазол-5-метанола (Hay et al. (1999) Bioorg. Med. Chem. Lett. 9:2237), гетероциклические аналоги РАВ (заявка на патент США № 2005/0256030), бета-глюкуронид (WO 2007/011968) и орто- или парааминобензилацетали. Могут быть использованы спейсеры, подвергающиеся циклизации в результате гидролиза амидной связи, такие как замещенные и незамещенные амиды 4-аминомасляной кислоты (Rodrigues et al. (1995) Chemistry Biology 2:223), должным образом замещенные бицикло[2.2.1] и бицикло[2.2.2] кольцевые системы (Storm et al. (1972) J. Amer. Chem. Soc. 94:5815) и амиды 2-аминофенилпропионовой кислоты (Amsberry et al. (1990) J. Org. Chem. 55:5867). Примером саморазрушающегося спейсера, используемого в ADC, также является выделение аминосодержащих лекарственных средств, замещенных в положении глицина (Kingsbury et al. (1984) J. Med. Chem. 27:1447).
Типичные спейсерные звенья (Yy-) представлены формулами Х-XII:
Дендритные линкеры
В другом варианте осуществления изобретения линкер L может быть линкером дендритного типа, предназначенным для ковалентного присоединения к антителу более одной лекарственной части при помощи разветвленной многофункциональной линкерной части (Sun et al. (2002) Bioorganic & Medicinal Chemistry Lettrers 12:2213-2215; Sun et al. (2003) Bioorganic & Medicinal Chemistry 11:1761-1768). Дендритные линкеры могут увеличивать молярное отношение лекарственного средства к антителу, то есть нагрузку, определяющую действенность ADC. Таким образом, к сконструированному антителу с цистеиновой заменой, содержащему только одну реакционноспособную тиоловую группу цистеина, при помощи дендритного линкера можно присоединить несколько лекарственных частей. Типичными вариантами разветвленных дендритных линкеров являются дендримерные звенья 2,6-бис(гидроксиметил)-п-крезол и 2,4,6-трис(гидроксиметил)фенол (WO 2004/01993; Szalai et al. (2003) J. Amer. Chem. Soc. 125: 15688-15689; Shamis et al. (2004) J. Amer. Chem. Soc. 126:1726-1731; Amir et al. (2003) Angew. Chem. Int. Ed. 42:4494-4499).
В одном из вариантов осуществления изобретения спейсерное звено является разветвленным бис(гидроксиметил)стиролом (BHMS), который может быть использован для введения и высвобождения нескольких лекарственных средств и имеет структуру:
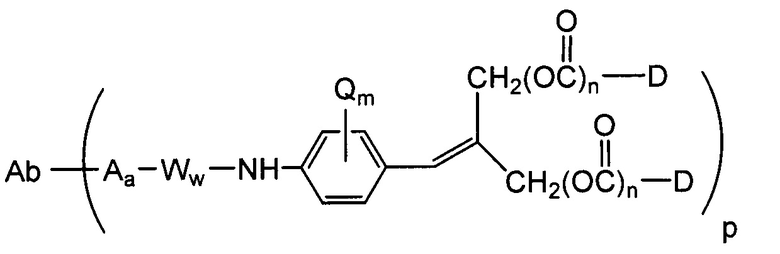
включающую дендримерное звено 2-(4-аминобензилиден)пропан-1,3-диол (WO 2004/043493; de Groot et al. (2003) Angew. Chem. Int. Ed. 42:4490-4494), где Q означает -С1-С8 алкил, -О-(С1-С8 алкил), -галоген, -нитро или -циано, m является целым числом от 0 до 4, n равен 0 или 1 и р является целым числом от 1 до 4.
Типичные варианты конъюгатов антитела с лекарственным средством формулы I включают соединения XIIIa (MC), XIIIb (val-cit), XIIIc (MC-val-cit) и XIIId (MC-val-cit-PAB):
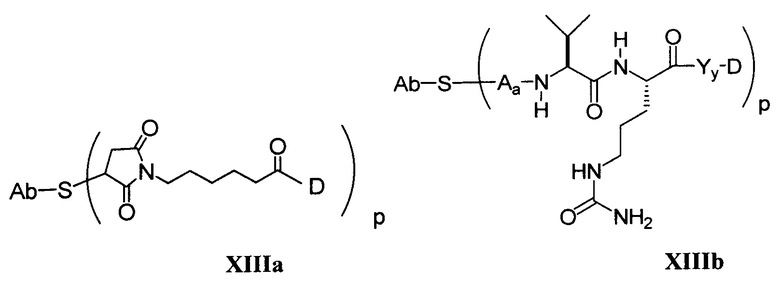

Другие типичные варианты конъюгатов антитела с лекарственным средством формулы Ia включают соединения XIVa-e:
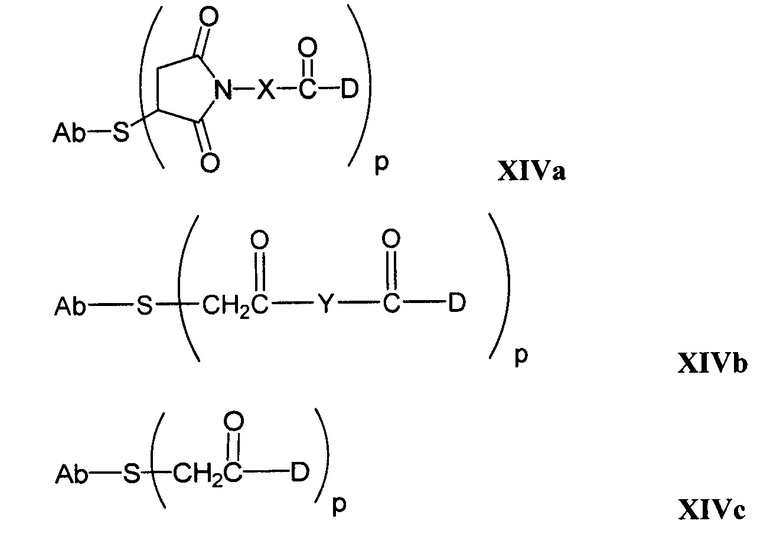
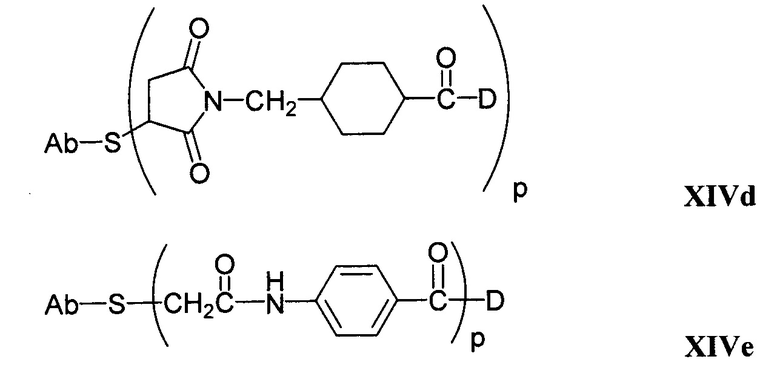
где Х означает
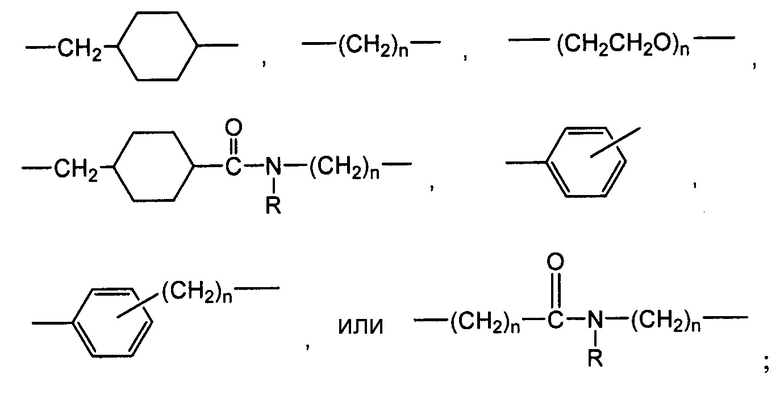
Y означает
R независимо означает Н или С1-С6 алкил и n является целым числом 1-12.
В другом варианте осуществления изобретения линкер имеет реакционноспособную функциональную группу, которая является нуклеофильной группой, взаимодействующей с электрофильной группой, присутствующей на антителе. Приемлемые электрофильные группы антитела включают, не ограничиваясь ими, карбонильные группы альдегида и кетона. Гетероатом нуклеофильной группы линкера может взаимодействовать с электрофильной группой антитела и образовывать ковалентную связь с компонентом антитела. Приемлемые нуклеофильные группы линкера включают, не ограничиваясь ими, гидразид, оксим, амино, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. Электрофильная группа антитела является удобным местом присоединения к линкеру.
Линкеры пептидного типа могут быть получены в результате образования пептидной связи между двумя или более аминокислотами и/или фрагментами пептидов. Такие пептидные связи могут быть получены, например, способом синтеза в жидкой фазе (E. Schroder and K. Lubke (1965) “The Peptides”, volume 1, pp.76-136, Academic Press), который хорошо известен в области химии пептидов. Промежуточные соединения линкера могут быть собраны в результате выполнения любой комбинации или последовательности реакций, включающих спейсер, удлиняющее звено и аминокислотные звенья. В спейсере, удлиняющем звене и аминокислотных звеньях могут быть использованы функциональные электрофильные, нуклеофильные группы или свободные радикалы. Реакционноспособные функциональные группы включают, не ограничиваясь ими, карбоксилы, гидроксилы, паранитрофенилкарбонат, изотиоцианат и удаляемые группы, такие как О-мезил, О-тозил, -Cl, -Br, -I, или малеимид.
В другом варианте осуществления изобретения линкер может быть замещен группами, модулирующими растворимость или реакционную способность. Например, заряженный заместитель, такой как сульфонат (-SO3-) или аммоний, может увеличивать растворимость реагента в воде и облегчать связывание линкерного реагента с антителом или лекарственной частью или связывание Ab-L (промежуточное соединение антитела с линкером) с D или D-L (промежуточное соединение лекарственного средства с линкером) с Ab в зависимости от схемы синтеза, используемой для получения ADC.
Типичные лекарственные части
Майтанзин и майтанзиноиды
В некоторых вариантах осуществления изобретения иммуноконъюгат включает антитело по настоящему изобретению, конъюгированное с одной или несколькими молекулами майтанзиноида. Майтанзиноиды являются митотическими ингибиторами, подавляющими полимеризацию тубулина. Майтанзин было впервые выделен из восточно-африканского кустарника Maytenus serrata (патент США № 3896111). Затем было обнаружено, что определенные микроорганизмы также продуцируют майтанзиноиды, такие как майтанзинол и сложные эфиры майтанзинола С-3 (патент США № 4151042). Синтетический майтанзинол, его производные и аналоги описаны, например, в патентах США №№ 4137230, 4248870, 4256746, 4260608, 4265814, 4294757, 4307016, 4308268, 4308269, 4309428, 4313946, 4315929, 4317821, 4322348, 4331598, 4361650, 4364866, 4424219, 4450254, 4362663 и 4371533.
Майтанзиноиды являются весьма привлекательными лекарственными частями для создания конъюгатов антитела с лекарственным средством, благодаря (i) относительной доступности для получения путем ферментации или химической модификации, создания производных соединений продуктов ферментации, (ii) возможности создания производных соединений с функциональными группами, пригодными для конъюгирования с антителами при помощи линкеров, не имеющих дисульфидных связей, (iii) устойчивости в плазме и (iv) эффективному воздействию на разные линии опухолевых клеток.
Соединения майтанзина, пригодные для использования в качестве майтанзиноидных лекарственных частей, хорошо известны в данной области и могут быть выделены из природных источников известными способами или созданы способами генной инженерии (см. публикацию Yu et al. (2002) PNAS 99:7968-7973), либо майтанзинол и аналоги майтанзинола могут быть синтезированы известными способами.
Типичные варианты майтанзиноидных лекарственных частей включают: DM1, DM3 и DM4, рассмотренные в настоящем описании изобретения.
Ауристатины и доластатины
В некоторых вариантах осуществления изобретения иммуноконъюгат содержит антитело по настоящему изобретению, конъюгированное с доластатином, пептиднымм аналогом и производным долостатина, например, ауристатином (патенты США №№ 5635483, 5780588). Установлено, что доластатины и ауристатины нарушают динамику микротрубочек, препятствуют гидролизу GTP и делению ядер и клеток (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают противораковой (патент США № 5663149) и противогрибковой активностью (Pettit et al. (1998) Antimicrob. Agents Chemother. 42:2961-2965). Доластатиновая или ауристатиновая лекарственная часть может быть присоединена к антителу у N (амино) конца или С (карбоксильного) конца пептидной лекарственной части (WO 02/088172).
Типичные варианты ауристатина включают связанные с N-концом монометилауристатиновые лекарственные части DE и DF, описанные в публикации Senter et al., Proceedings of the American Association for Cancer Research, Volume 45, Abstract Number 623, presented Match 28, 2004, которая включена в настоящее описание изобретения в качестве ссылки в полном объеме.
Пептидная лекарственная часть может быть выбрана из нижеследующих формул DE и DF:
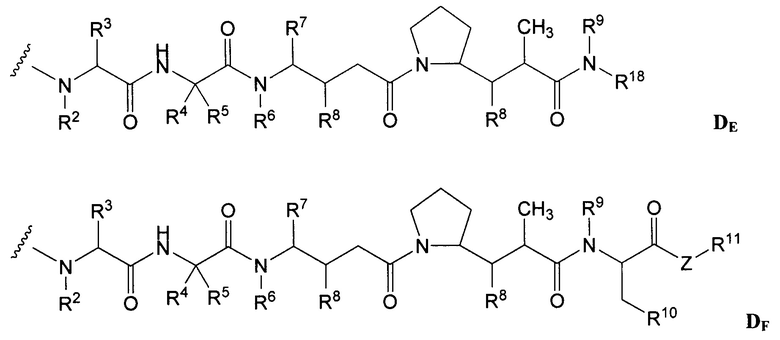
где волнистыми линиями в DE и DF показано место ковалентного присоединения к антителу или компоненту антитела с линкером независимо в каждом положении:
R2 выбирают из Н и С1-С8 алкила;
R3 выбирают из Н, С1-С8 алкила, С3-С8 карбоцикла, арила, С1-С8 алкиларила, С1-С8 алкил(С3-С8 карбоцикла), С3-С8 гетероцикла и С1-С8 алкил(С3-С8 гетероцикла);
R4 выбирают из Н, С1-С8 алкила, С3-С8 карбоцикла, арила, С1-С8 алкиларила, С1-С8 алкил(С3-С8 карбоцикла), С3-С8 гетероцикла и С1-С8 алкил(С3-С8 гетероцикла);
R5 выбирают из Н и метила;
или R4 и R5 вместе образуют карбоциклическое кольцо и имеют формулу -(CRaRb)n-, где Ra и Rb независимо выбирают из Н, С1-С8 алкила и С3-С8 карбоцикла и n выбирают из 2, 3, 4, 5 и 6;
R6 выбирают из Н и С1-С8 алкила;
R7 выбирают из Н, С1-С8 алкила, С3-С8 карбоцикла, арила, С1-С8 алкиларила, С1-С8 алкил(С3-С8 карбоцикла), С3-С8 гетероцикла и С1-С8 алкил(С3-С8 гетероцикла);
каждый R8 независиммо выбирают из Н, ОН, С1-С8 алкила, С3-С8 карбоцикла и О-(С1-С8 алкила);
R9 выбирают из Н и С1-С8 алкила;
R10 выбирают из арила или С3-С8 гетероцикла;
Z означает О, S, NH или NR12, где R12 является С1-С8 алкилом;
R11 выбирают из Н, С1-С20 алкила, арила, С3-С8 гетероцикла, -(R13O)m-R14 или -(R13O)m-CH(R15)2;
m является целым числом от 1 до 1000;
R13 означает С2-С8 алкил;
R14 означает Н или С1-С8 алкил;
каждый R15 независимо означает Н, СООН, -(CH2)n-N(R16)2, -(CH2)n-SO3H или -(CH2)n-SO3-C1-C8 алкил;
каждый R16 независимо означает Н, С1-С8 алкил или -(СН2)n-СООН;
R18 выбирают из -С(R8)2-C(R8)2-арила, -C(R8)2-C(R8)2-(C3-C8 гетероцикла) и -С(R8)2-C(R8)2-(C3-C8 карбоцикла); и
n является целым числом от 0 до 6.
В одном из вариантов осуществления изобретения R3, R4 и R7 независимо означают изопропил или втор-бутил и R5 означает -Н или метил. В типичном варианте осуществления изобретения R3 и R4 означают изопропил, R5 означает -Н и R7 означает втор-бутил.
В другом варианте осуществления изобретения R2 и R6 означают метил и R9 означает -Н.
В еще одном варианте осуществления изобретения каждый R8 означает -ОСН3.
В типичном варианте осуществления изобретения R3 и R4 означают изопропил, R2 и R6 означают метил, R5 означает -Н, R7 означает втор-бутил, каждый R8 означает -ОСН3 и R9 означает -Н.
В одном из вариантов осуществления изобретения Z означает -О- или -NH-.
В одном из вариантов осуществления изобретения R10 означает арил.
В типичном варианте осуществления изобретения R10 означает -фенил.
В типичном варианте осуществления изобретения, когда Z означает -О-, R11 означает -Н, метил или трет-бутил.
В одном из вариантов осуществления изобретения, когда Z означает -NH, R11 означает -СН(R15)2, где R15 означает -(СН2)n-N(R16)2 и R16 означает -С1-С8 алкил или -(СН2)n-СООН.
В другом варианте осуществления изобретения, когда Z означает -NH, R11 означает -СН(R15)2, где R15 означает -(СН2)n-SO3H.
Типичным вариантом ауристатина формулы DE является ММАЕ, где волнистой линией показано место ковалентного присоединения конъюгата антитела с лекарственным средством к линкеру (L):
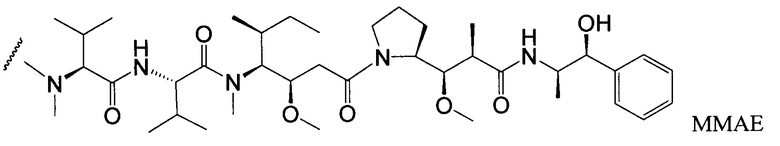
Типичным вариантом ауристатина формулы DF является MMAF, где волнистой линией показано место ковалентного присоединения конъюгата антитела с лекарственным средством к линкеру (L) (см. заявку на патент США № 2005/0238649 и публикацию Doronina et al. (2006) Bioconjugate Chem. 17:114-124):
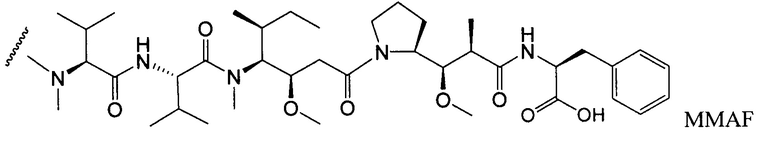
Другие лекарственные части включают следующие производные MMAF, где волнистой линией показано место ковалентного присоединения конъюгата антитела с лекарственным средством к линкеру (L):

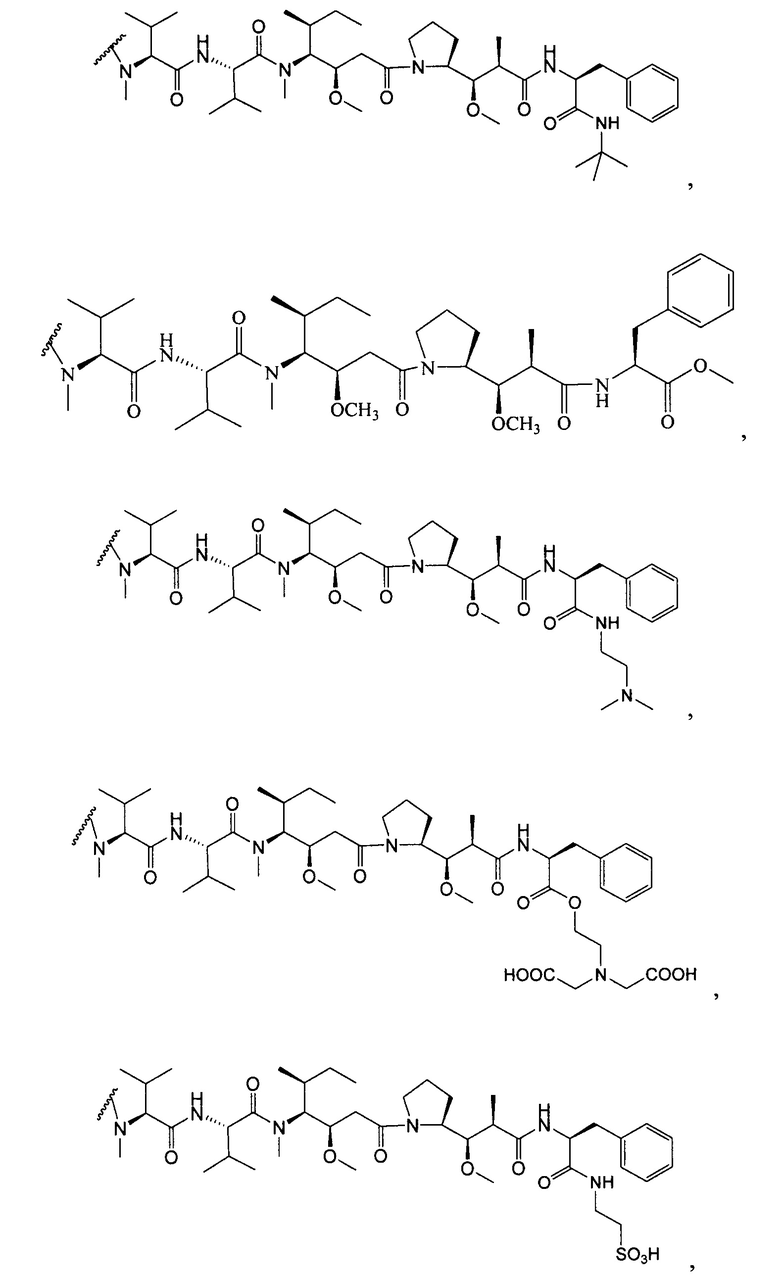
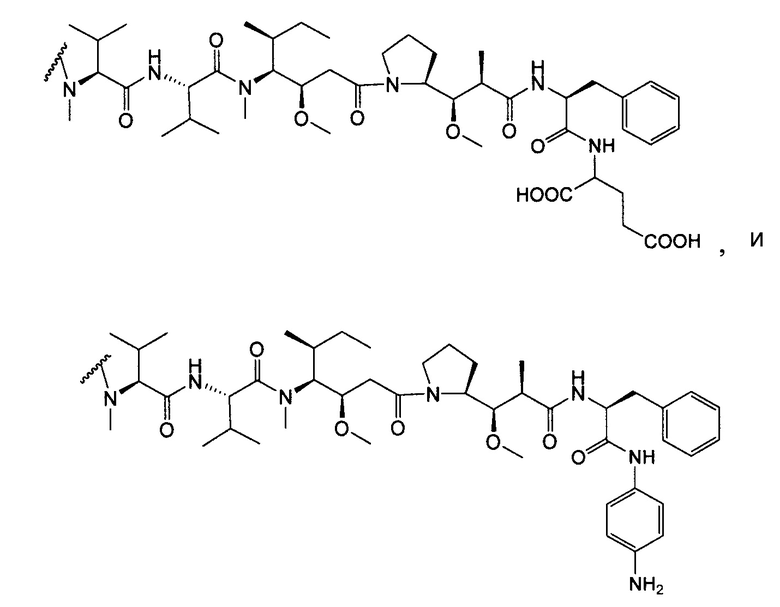
В одном из аспектов изобретения гидрофильные группы, которые включают, не ограничиваясь ими, сложные эфиры триэтиленгликоля (TEG), представленные выше, могут быть присоединены к лекарственной части в положении R11. Не ограничивая себя какой-либо конкретной теорией, можно отметить, что гидрофильные группы способствуют интернализации и препятствуют агломерации лекарственной части.
Типичные варианты ADC формулы I, включающие ауристатин/доластатин или их производные, описаны в заявке на патент США № 2005-0238649 А1 и в публикации Doronina et al. (2006) Bioconjugate Chem. 17: 114-124, которая включена в настоящее описание изобретения в качестве ссылки. Типичные варианты ADC формулы I, включающие ММАЕ или MMAF и разные линкерные компоненты, имеют следующие структуры и обозначены следующими аббревиатурами (где ”Ab” означает антитело, р является числом от 1 до около 8, ”Val-Cit” означает валин-цитруллиновый дипептид и “S” означает атом серы:
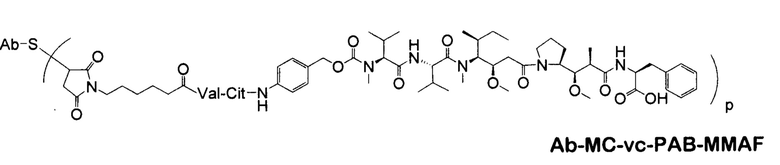
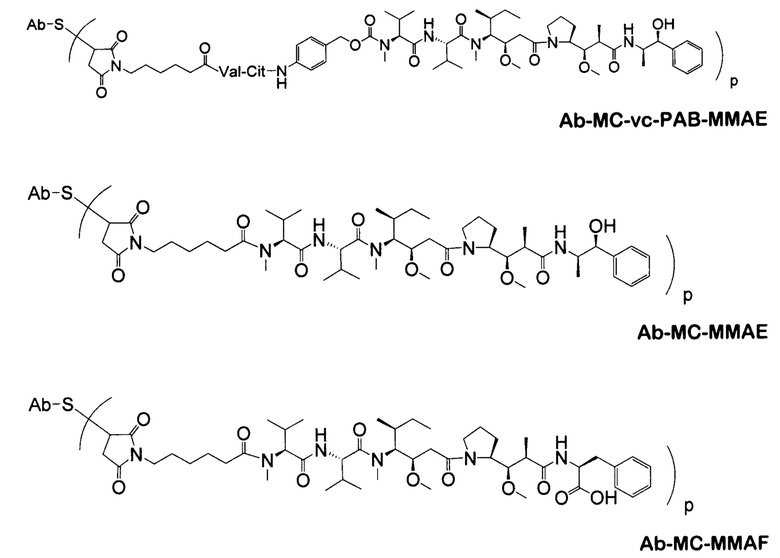
Типичными вариантами ADC формулы I, включающими MMAF и разные линкерные компоненты, далее являются Ab-MC-PAB-MMAF и Ab-PAB-MMAF. Интересно отметить, что иммуноконъюгаты, включающие MMAF, присоединенный к антителу линкером, который протеолитически не расщепляется, обладают активностью, сравнимой с иммуноконъюгатами, в которых MMAF присоединен к антителу протеолитически расщепляемым линкером. См. публикацию Doronina et al. (2006) Bioconjugate Chem. 17:114-124. В таких случаях считается, что на высвобождение лекарственного средства влияет разрушение антитела в клетке. Там же.
Лекарственные части на основе пептидов могут быть получены путем образования пептидной связи между двумя или более аминокислотами и/или фрагментами пептидов. Такие пептидные связи могут быть получены, например, способом синтеза в жидкой фазе (см. E. Schroder and K. Lubke, “The Peptides”, volume 1, pp.76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Лекарственные части ауристатина/доластатина могут быть получены способами, представленными в заявке на патент США 2005/0238649 А1; в патентах США №№ 5635483 и 5780588; в публикациях Pettit et al. (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al. (1998) AntiCancer Drug Design 13:243-277; Pettit, G.R., et al. Synthesis, 1996, 719-725; Pettit et al. (1996) J. Chem. Soc. Perkin Trans. 15:859-863; и Doronina (2003) Nat. Biotechnol. 21(7):778-784.
Лекарственные части ауристатина/доластатина формулы DF, такие как MMAF и его производные, могут быть получены способами, описанными в заявке на патент США № 2005-0238649 А1 и в публикации Doronina et al. (2006) Bioconjugate Chem. 17:114-124. Лекарственные части ауристатина/доластатина формулы DE, такие как ММАЕ и его производные, могут быть получены способами, описанными в публикации Doronina et al. (2003) Nat. Biotech. 21:778-784. Части лекарственного средства с линкером МС-MMAF, MC-MMAE, MC-vc-PAB-MMAF и MC-vc-PAB-MMAE могут быть успешно синтезированы стандартными способами, например, описанными в публикации Doronina et al. (2003) Nat. Biotech. 21:778-784 и в публикации заявки на патент США № 2005/0238649 A1, и затем конъюгированы с представляющим интерес антителом.
Лекарственная нагрузка
Лекарственная нагрузка обозначена буквой ”р” и представляет собой среднее число лекарственных частей на одно антитело в молекуле формулы I. Лекарственная нагрузка может быть равна 1-20 лекарственным частям (D) на одно антитело. ADC формулы I включают наборы антител, конъюгированных с лекарственными частями в количестве от 1 до 20. Среднее число лекарственных частей на одно антитело в препаратах ADC, полученных в результате выполнения реакций конъюгирования, можно определить стандартными способами, такими как масс-спектроскопия, анализ ELISA и ВЭЖХ. Также можно определить количественное распределение числа р в ADC. В некоторых случаях отделение, очистку и исследование гомогенного ADC, в котором р является определенным значением для ADC с другими лекарственными нагрузками, выполняют при помощи ВЭЖХ с обращенной фазой или электрофореза.
В некоторых конъюгатах антитела с лекарственным средством ”р” может быть ограничено числом мест присоединения на антителе. Например, если местом присоединения является тиоловая группа цистеина, как указано в вышеописанным типичных вариантах осуществления изобретения, антитело может иметь только одну или несколько тиоловых групп цистеина или может иметь только одну или несколько достаточно реакционноспособных тиоловых групп, с помощью которых может быть присоединен линкер. В определенных вариантах осуществления изобретения более высокая лекарственная нагрузка, например, р>5, может вызвать агрегацию, нерастворимость, токсичность или плохое проникновение в клетку определенных конъюгатов антитела с лекарственным средством. В определенных вариантах осуществления изобретения лекарственная нагрузка для ADC по настоящему изобретению находится в пределах от 1 до около 8, от около 2 до около 6, от около 3 до около 5, от около 3 до около 4, от около 3,1 до около 3,9, от около 3,2 до около 3,8, от около 3,2 до около 3,7, от около 3,2 до около 3,6, от около 3,3 до около 3,8, от около 3,3 до около 3,7. Действительно установлено, что для определенных ADC оптимальное отношение лекарственных частей к антителу может быть менее 8 и может составлять от около 2 до около 5. См. заявку на патент США 2005-0238649 А1 (которая включена в настоящее описание изобретения в качестве ссылки в полном объеме).
В определенных вариантах осуществления изобретения с антителом конъюгируют меньше лекарственных частей по сравнению с теоретическим максимумом. Антитело может содержать, например, остатки лизина, которые не взаимодействуют с промежуточным соединением лекарственного средства с линкером или линкерным реагентом, описанным ниже. Антитела обычно не содержат много свободных и реакционноспособных тиоловых групп цистеинов, которые могут быть связаны с лекарственной частью; действительно, большинство тиоловых групп цистеина находятся в антителах в виде дисульфидных мостиков. В определенных вариантах осуществления изобретения антитело может быть восстановлено восстановителем, таким как дитиотреитол (DTT) или трикарбонилэтилфосфин (ТСЕР) в условиях частичного или полного восстановления для создания реакционноспособных тиоловых групп цистеина. В определенных вариантах осуществления изобретения антитело денатурируют для выявления реакционноспособных нуклеофильных групп, таких как лизин или цистеин.
Нагрузку (соотношение лекарственного средства/антитела) ADC можно регулировать разными способами, например, (i) ограничивая молярный избыток промежуточного соединения лекарственного средства с линкером или линкерного реагента по отношению к антителу, (ii) ограничивая время или температуру реакции конъюгирования, (iii) частично ограничивая восстановительные условия для модификации тиоловой группы цистеина, (iv) создавая способами рекомбинантных ДНК такую аминокислотную последовательность антитела, в которой число и положения остатков цистеина модифицированы с возможностью регулирования числа и/или положения мест присоединения линкера-лекарственной части (с получением тиоMab или тиоFab, рассмотренных в настоящем описании изобретения и в публикации WO2006/034488 (которая включена в настоящее описание изобретения в качестве ссылки в полном объеме)).
Следует отметить, что при взаимодействии нескольких нуклеофильных групп с промежуточным соединением лекарственного средства с линкером или линкерным реагентом с последующим взаимодействием с лекарственной частью полученный продукт представляет собой смесь соединений ADC, содержащую одну или несколько лекарственных частей, присоединенных к антителу. Среднее число лекарственных частей на одно антитело можно вычислить в смеси при помощи двойного анализа антитела способом ELISA, специфичного к антителу и лекарственному средству. Отдельные молекулы ADC могут быть идентифицированы в смеси при помощи масс-спектроскопии или выделены при помощи ВЭЖХ, например, гидрофобной хроматографии (см., например, публикации Hamblett, K.J., et al. “Effect of drug loading on the pharmacology, pharmacokinetics, and toxicity of an anti-CD30 antibody-drug conjugate,” Abstract No. 624, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of the AACR, Volume 45, March 2004; Alley, S.C., et al. “Controlling the location of drug attachment in antibody-drug conjugates,” Abstract No. 627, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of the AACR, Volume 45, March 2004). В определенных вариантах осуществления изобретения гомогенный ADC с одной нагрузкой можно выделить из конъюгированной смеси электрофорезом или хроматографией.
Некоторые способы получения иммуноконъюгатов
ADC формулы I может быть получен несколькими способами при использовании реакций органической химии, условий и реагентов, известных специалистам в данной области, которые включают: (1) осуществление взаимодействия нуклеофильной группы антитела с двухвалентным линкерным реагентом с образованием Ab-L при помощи ковалентной связи с последующим осуществлением взаимодействия с лекарственной частью D; и (2) осуществление взаимодействия нуклеофильной группы лекарственной части с двухвалентным линкерным реагентом с образованием D-L при помощи ковалентной связи с последующим осуществлением взаимодействия с нуклеофильной группой антитела. Типичные способы получения ADC формулы I по последней схеме описаны в публикации заявки на патент США № 20050238649 А1, которая включена в настоящее описание изобретения в качестве ссылки.
Нуклеофильные группы антитела включают, не ограничиваясь ими: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, лизин, (iii) тиоловые группы боковых цепей, например, цистеин, и (iv) гидроксильные или аминогруппы сахара, когда антитело является гликозилированным. Аминогруппы, тиоловые и гидроксильные группы являются нуклеофильными группами и могут взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных частей и линкерных реагентов, включающих: (i) активные сложные эфиры, такие как NHS сложные эфиры, HOBt сложные эфиры, галогенформиаты и галогенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные группы и группы имида малеиновой кислоты. Некоторые антитела имеют восстанавливаемые дисульфидные связи между цепями, то есть цистеиновые мостики. Антителам может быть сообщена реакционная способность для конъюгирования с линкерными реагентами путем обработки таким восстановителем как DTT (дитиотреитол) или трикарбонилэтилфосфин (ТСЕР), в результате чего антитело становится полностью или частично восстановленным. Каждый цистеиновый мостик теоретически образуют две реакционноспособные тиоловые нуклеофильные группы. Дополнительные нуклеофильные группы могут быть введены в антитела в результате модификации остатков лизина, например, при осуществлении взаимодействия остатков лизина с 2-иминотиоланом (реагент Траута), что позволяет превратить амин в тиол. Реакционноспособные тиоловые группы могут быть введены в антитело путем введения одного, двух, трех, четырех или большего числа остатков цистеина (например, в результате получения вариантных антител, включающих один или несколько ненативных аминокислотных остатков цистеина).
Конъюгаты антитела с лекарственным средством по настоящему изобретению могут быть также получены в результате осуществления взаимодействия электрофильной группы антитела, такой как карбонильная группа альдегида или кетона, с нуклеофильной группой линкерного реагента или лекарственного средства. Приемлемые нуклеофильные группы линкерного реагента включают, не ограничиваясь ими, гидразид, оксим, амино, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. В одном из вариантов осуществления изобретения антитело модифицируют для введения электрофильных групп, способных взаимодействовать с нуклеофильными заместителями линкерного реагента или лекарственного средства. В другом варианте осуществления изобретения сахара гликозилированных антител могут быть окислены, например, периодатными оксидантами, с образованием альдегидных или кетонных групп, способных взаимодействовать с аминогруппой линкерных реагентов или лекарственных частей. Полученные иминогруппы шиффовых оснований могут образовывать устойчивую связь или могут быть восстановлены, например, борогидридными реагентами, с образованием устойчивых аминных связей. В одном из вариантов осуществления изобретения в результате осуществления взаимодействия углеводной части гликозилированного антитела с галактозооксидазой или метапериодатом натрия в антителе могут образоваться карбонильные группы (альдегида и кетона), которые могут взаимодействовать с соответствующими группами лекарственного средства (Hermanson, Bioconjugate Techniques). В другом варианте осуществления изобретения антитела, содержащие N-концевые остатки серина или треонина, могут взаимодействовать с метапериодатом натрия с образованием альдегида вместо первой аминокислоты (Geoghegan & Stroh (1992) Bioconjugate Chem. 3:138-146; патент США № 5362852). Такой альдегид может взаимодействовать с нуклеофильной группой лекарственной части или линкера.
Нуклеофильные группы лекарственной части включают, не ограничиваясь ими: амин, тиол, гидроксил, гидразид, оксим, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид, способные взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных частей или линкерных реагентов, включающих: (i) активные сложные эфиры, такие как NHS сложные эфиры, HOBt сложные эфиры, галогенформиаты и галогенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные группы и группы имида малеиновой кислоты.
Соединения по настоящему изобретению включают, не ограничиваясь ими, ADC, полученные при использовании следующих перекрестносшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, сульфо-SMPB и SVSB (сукцинимидил(4-винилсульфон)бензоат), которые являются коммерчески доступными реагентами (например, могут быть приобретены в компании Pierce Biotechnology, Inc., Rockford, IL., U.S.A.; see pages 467-498, 2003-2004 Applications Handbook and Catalog).
Иммуноконъюгаты, содержащие антитело и цитотоксический агент, могут быть также получены с использованием разных агентов, связывающих бифункциональный белок, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)-циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как гидрохлорид диметиладипимидата), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(параазидобензоил)гександиамин), производные бисдиазония (такие как бис(парадиазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен в соответствии с описанием, приведенным в публикации Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминопентауксусная кислота (МХ-DTPA) является типичным хелатообразователем, используемым для конъюгирования радионуклеотида с антителом. См. публикацию WO94/11026.
Альтернативно, слитый белок, содержащий антитело и цитотоксический агент, может быть получен, например, способами рекомбинантных ДНК или синтеза пептидов. Молекула рекомбинантной ДНК может включать области, кодирующие антитело и цитотоксические части конъюгата, расположенные рядом друг с другом или разделенные областью, кодирующей линкерный пептид, который не оказывает отрицательного влияния на требуемые свойства конъюгата.
В еще одном варианте осуществления изобретения антитело может быть конъюгировано с ”рецептором” (таким как стрептавидин), предназначенным для предварительного направленного воздействия на опухоль, когда индивиду вводят конъюгат антитела с рецептором, затем удаляют несвязанный конъюгат из кровотока при помощи очищающего агента и вводят ”лиганд” (например, авидин), конъюгированный с цитотоксическим агентом (например, радионуклеотидом).
Получение антител против STEAP-1 с введенным цистеином
Антитела против STEAP-1 с введенным цистеином, способные взаимодействовать с электрофильной функциональной группой, могут быть получены с помощью способов создания, отбора и получения по настоящему изобретению. Указанные способы также позволяют получить конъюгаты антитела, такие как конъюгат антитела с лекарственным средством (ADC), в которых молекулы лекарственного средства находятся в предназначенных для указанной цепи сайтах. Реакционноспособные остатки цистеина на поверхности антитела делают возможным специфическое конъюгирование лекарственной части при помощи реакционноспособной тиоловой группы, такой как малеимид или галогенацетил. Нуклеофильная реакционная способность функциональной тиоловой группы остатки Cys в отношении группы имида малеиновой кислоты примерно в 1000 раз больше по сравнению с любой другой аминокислотной функциональной группой в белке, такой как аминогруппа остатков лизина или N-концевая аминогруппа. Специфическая функциональная тиоловая группа в иодацетильных и малеимидных реагентах может взаимодействовать с аминогруппами, но для этого необходимо более высокое значение рН (>9,0) и более продолжительное время реакции (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academid Press, London). Количество свободного тиола в белке можно определить при помощи стандартного анализа Эллмана. Иммуноглобулин М является примером пентамера с дисульфидной связью, и иммуноглобулин G является примером белка с внутренними дисульфидными мостиками, связывающими структурные субъединицы вместе. В таких белках восстановление дисульфидных связей таким реагентом как дитиотреитол (DTT) или селенол (Singh et al. (2002) Anal. Biochem. 304:147-156) необходимо для создания реакционноспособного свободного тиола. Указанный способ может привести к утрате третичной структуры антитела и антигенсвязывающей специфичности.
Анализ PHESELECTOR (фаговый анализ способом ELISA для отбора реакционноспособных тиоловых групп) позволяет обнаружить реакционноспособные группы цистеина в антителах при помощи анализа ELISA в формате отображения на фаге, что способствует созданию сконструированных антител с цистеиновой заменой (WO 2006/034488). Сконструированное антитело с цистеиновой заменой наносят на поверхности лунок, инкубируют с фаговыми частицами, добавляют HRP-меченое вторичное антитело и детектируют оптическую плотность. Мутантные белки, отображенные на фаге, можно исследовать быстрым, надежным и высокопроизводительным способом. Можно создать библиотеки сконструированных антител с цистеиновой заменой, которые подвергают отбору в отношении связывания для идентификации соответствующих реакционноспособных сайтов для введения свободного цистеина (Cys) из произвольных библиотек отображения на фаге антител или других белков. Указанный способ включает взаимодействие мутантных белков цистеина, отображенных на фаге, с аффинным реагентом или репортерной группой, которая также взаимодействует с тиолом.
Анализ PHESELECTOR позволяет исследовать реакционноспособные тиоловые группы в антителах. В качестве типичного примера можно привести идентификацию варианта А121С указанным способом. Можно эффективно исследовать всю молекулу Fab-фрагмента для идентификации дополнительных вариантов тиоFab с реакционноспособными тиоловыми группами. Для идентификации тиоловых групп и количественного определения доступности растворителя к аминокислотным остаткам в полипептиде был использован параметр фракциональной доступности поверхности. Доступность поверхности может быть выражена в виде площади поверхности (Å2), которая может контактировать с молекулой растворителя, например, воды. Пространство, занимаемое водой, примерно равно сфере с радиусом 1,4 Å. Можно свободно приобрести или лицензировать программное обеспечение (Secretary to CCP4, Daresbury Laboratory, WarringEon, WA44AD, United Kingdon, Fax (+44) 1925603825, или по интернету: www.ccp4.ac.uk/dist/html/INDEX.html) в виде набора ССР4 программ по кристаллографии, которые включают алгоритмы для вычисления доступности поверхности каждой аминокислоты белка с координатами, установленными при помощи рентгеновской кристаллографии (“The CCP4 Suite: Programs for Protein Crystallograhy”) (1994) Acta. Cryst. D50:760-763). Двумя типичными модулями программного обеспечения, вычисляющими доступность поверхности, являются ”AREAIMOL” и “SURFACE”, созданные на основании алгоритмов, представленных в публикации B.Lee and F.M.Richards (1971) J. Mol. Biol. 55:379-400. Программа AREAIMOL определяет доступную для растворителя поверхность белка в виде локуса в центре сферы зонда (представляющей молекулу растворителя) по мере ее прохождения над поверхностью Van der Waals белка. Программа AREAIMOL вычисляет площадь доступной для растворителя поверхности, определяя точки поверхности на удлиненной сфере около каждого атома (на расстоянии от центра атома, равном сумме радиусов атома и зонда) и удаляя точки, лежащие в эквивалентных сферах, связанных с соседними атомами. Программа AREAIMOL находит доступную для растворителя площадь атомов в файле координат PDB и суммирует доступную площадь для остатка, цепи и всей молекулы. Доступные площади (или разности площадей) для отдельных атомов могут быть введены в выходной файл псевдо-PDB. Программа AREAIMOL принимает один радиус для каждого элемента и узнает только ограниченное число разных элементов.
Программы AREAIMOL и SURFACE сообщают значения абсолютной доступности, то есть число квадратных ангстремов (Å). Дробное значение доступности поверхности вычисляют со ссылкой на стандартное состояние аминокислоты в полипепитиде. Эталонное состояние представлено трипептидом Gly-X-Gly, где Х означает представляющую интерес аминокислоту, при этом эталонное состояние должно иметь ”удлиненную” конформацию, то есть подобную конформации в бета-цепях. Удлиненная конформация максимально увеличивает доступность аминокислоты Х. Вычисленную доступную площадь делят на доступную площадь в трипептиде Gly-X-Gly и сообщают результат, который является дробным значением доступности. Процентное значение доступности является дробным значением доступности, умноженным на 100. Другой типичный алгоритм для вычисления доступности поверхности создан на основе модуля SOLV программы xsae (Broger, C., F. Hoffman-LaRoche, Basel), которая вычисляет дробное значение доступности аминокислотного остатка для водной сферы на основании координат рентгеновского излучения полипептида. Дробное значение доступности поверхности для каждой аминокислоты в антителе может быть вычислена с использованием доступной информации о кристаллической структуре (Eigenbrot et al. (1993) J. Mol. Biol. 229:969-995).
ДНК, кодирующую сконструированные антитела с цистеиновой заменой, можно легко выделить и секвенировать стандартными способами (например, при помощи олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи зрелых антител). Источником такой ДНК являются клетки гибридом. Выделенную ДНК можно ввести в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьян, клетки яичника китайского хомячка (СНО) или другие клетки млекопитающих, такие как миеломные клетки (патент США № 5807715, заявка на патент США № 2005/0048572, заявка на патент США № 2004/0229310), которые в противном случае не продуцируют белок антитела, для синтеза моноклональных антител в рекомбинантных клетках-хозяевах.
Созданные и отобранные сконструированные антитела с цистеиновой заменой, например, тиоFab, содержащие неспаренные остатки цистеина (Cys) с сильной реакционной способностью, могут быть продуцированы в результате: (i) экспрессии в бактериальной системе, например, E. coli (Skerra et al. (1993) Curr. Opinion in Immunol. 5:256-262; Pluckthun (1992) Immunol. Revs. 130:151-188) или в культуре клеток млекопитающих (WO 01/00245), например, в клетках яичника китайского хомячка (СНО); и (ii) очистки стандартными способами очистки белков (Lowman et al. (1991) J. Biol. Chem. 266(17):10982-10988).
Тиоловые группы веденного цистеина (Cys) взаимодействуют с электрофильными линкерными реагентами и промежуточными соединениями лекарственного средства с линкером с образованием конъюгатов сконструированного антитела с цистеиновой заменой с лекарственным средством и других меченых сконструированных антител с цистеиновой заменой. Остатки Cys в сконструированных антителах с цистеиновой заменой и материнских антителах, которые спарены и образуют межцепные и внутрицепные дисульфидные связи, не имеют реакционноспособных тиоловых групп (если не будут обработаны восстановителем) и не взаимодействуют с электрофильными линкерными реагентами или промежуточными соединениями лекарственного средства с линкером. Вновь введенный остаток Cys может оставаться неспаренным и способным взаимодействовать, то есть конъюгировать с электрофильным линкерным реагентом или промежуточным соединением лекарственного средства с линкером, таким как лекарственное средство-малеимид. Типичные промежуточные соединения лекарственного средства с линкером включают: МС-ММАЕ, МС-MMAF, MC-vc-PAB-MMAE и MC-vc-PAB-MMAF. Положения введенных остатков Cys в структуре тяжелых и легких цепей пронумерованы в соответствии с системой последовательной нумерации. Система последовательной нумерации согласуется с системой нумерации Kabat (Kabat et al. (1991) Sequences of Proteins of Immunological Interest, 5th. Ed. Public Health Service, National Institutes of Health, Bethesda, MD), начинаясь у N-конца, и отличается от схемы нумерации Kabat (нижний ряд) инсерциями, обозначенными а, b, c. При использовании системы нумерации Kabat действительная линейная аминокислотная последовательность может содержать меньше или больше аминокислот, соответствующих делециям или инсерциям в FR или CDR вариабельного домена. Сайты варианта тяжелой цепи с введенным цистеином идентифицированы в соответствии с системами последовательной нумерации и нумерации Kabat.
В одном из вариантов осуществления изобретения антитело против STEAP-1 с введенным цистеином получают способом, включающим:
(а) замену одного или нескольких аминокислотных остатков в материнском антителе против STEAP-1 цистеином; и
(b) определение реакционной способности тиоловой группы в антителе против STEAP-1 с введенным цистеином при осуществлении взаимодействия антитела с введенным цистеином с реагентом, взаимодействующим с тиолом.
Сконструированное антитело с цистеиновой заменой может обладать большей реакционной способностью в отношении реагента, взаимодействующего с тиолом, чем материнское антитело.
Аминокислотные остатки свободного цистеина могут находиться в тяжелых или легких цепях либо в константных или вариабельных доменах. Фрагменты антител, например, Fab, также могут быть созданы с одной или несколькими аминокислотами цистеина, заменяющими аминокислоты фрагмента антитела, с образованием фрагментов сконструированного антитела с цистеиновой заменой.
Другой вариант осуществления изобретения относится к способу получения (создания) антитела против STEAP-1 с введенным цистеином, который включает:
(а) введение одной или нескольких аминокислот цистеина в материнское антитело против STEAP-1 с целью создания антитела против STEAP-1 с введенным цистеином; и
(b) определение реакционной способности тиола в сконструированном антителе с цистеиновой заменой в отношении реагента, взаимодействующего с тиолом;
при этом сконструированное антитело с цистеиновой заменой обладает большей реакционной способность в отношении реагента, взаимодействующего с тиолом, по сравнению с материнским антителом.
Стадия (а) способа получения сконструированного антитела с цистеиновой заменой может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновой заменой;
(ii) экспрессию сконструированного антитела с цистеиновой заменой; и
(iii) выделение и очистку сконструированного антитела с цистеиновой заменой.
Стадия (b) способа получения сконструированного антитела с цистеиновой заменой может включать экспрессию сконструированного антитела с цистеиновой заменой на вирусной частице, выбранной из фага или фагомидной частицы.
Стадия (b) способа получения сконструированного антитела с цистеиновой заменой может также включать:
(i) взаимодействие сконструированного антитела с цистеиновой заменой с аффинным реагентом, взаимодействующим с тиолом, с целью получения сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению; и
(ii) измерение связывания сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению, с улавливающей средой.
Другой вариант осуществления изобретения относится к способу скрининга сконструированных антитела с цистеиновой заменой при помощи высоко активных, неспаренных аминокислот цистеина с целью определения реакционной способности тиола, который включает:
(а) введение одной или нескольких аминокислот цистеина в материнское антитело с целью создания сконструированного антитела с цистеиновой заменой;
(b) взаимодействие сконструированного антитела с цистеиновой заменой с аффинным реагентом, взаимодействующим с тиолом, с целью создания сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению; и
(с) измерение связывания сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению, с улавливающей средой; и
(d) определение реакционной способности тиола в сконструированном антителе с цистеиновой заменой в отношении реагента, взаимодействующего с тиолом.
Стадия (а) способа скрининга сконструированного антитела с цистеиновой заменой может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновой заменой;
(ii) экспрессию сконструированного антитела с цистеиновой заменой; и
(iii) выделение и очистку сконструированного антитела с цистеиновой заменой.
Стадия (b) способа скрининга сконструированных антитела с цистеиновой заменой может включать экспрессию сконструированных антитела с цистеиновой заменой на вирусной частицы, выбираемой из фага или фагомидной частицы.
Стадия (b) способа скрининга сконструированных антител с цистеиновой заменой может также включать:
(i) взаимодействие сконструированного антитела с цистеиновой заменой с аффинным реагентом, взаимодействующим с тиолом, с целью получения сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению; и
(ii) измерение связывания сконструированного антитела с цистеиновой заменой, подвергнутого аффинному мечению, с улавливающей средой.
Меченые антитела против STEAP-1 с введенным цистеином
Антитела против STEAP-1 с введенным цистеином могут быть сайтспецифически и эффективно связаны с реагентом, взаимодействующим с тиолом. Реагент, взаимодействующий с тиолом, может быть многофункциональным линкерным реагентом, например, улавливающим реагентом с афинной меткой (например, биотиновым линкером), детектирующей меткой (например, флуорофором), твердофазным иммобилизующим реагентом (например, SEPHAROSE™, полистиролом или стеклом) или промежуточным соединением лекарственного средства с линкером. Одним примером реагента, взаимодействующего с тиолом, является N-этилмалеимид (NEM). В типичном варианте осуществления изобретения в результате взаимодействия тиоFab с биотиновым линкером образуется биотинилированный тиоFab, с помощью которого можно обнаружить присутствие остатка введенного цистеина и измерить реакционную способность указанного остатка. В результате взаимодействия тиоFab с многофункциональным линкерным реагентом получают тиоFab с функциональным линкером, который может быть далее подвергнут взаимодействию с лекарственной частью или другой меткой. В результате взаимодействия тиоFab с промежуточным соединением лекарственного средства с линкером получают конъюгат тиоFab с лекарственным средством.
Типичные способы, рассмотренные в настоящем описании изобретения, обычно относятся к идентификации и продуцированию антител и, в более широком смысле, других белков благодаря применению стадий создания и скрининга, рассмотренных в настоящем описании изобретения.
Такой способ применим к конъюгированию других реагентов, взаимодействующих с тиолом, в которых реакционноспособная группа является, например, имидом малеиновой кислоты, иодацетамидом, пиридилдисульфидом или другим партнером конъюгирования, взаимодействующим с тиолом (Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671). Реагент, взаимодействующий с тиолом, может быть лекарственной частью, флуорофором, таким как флуоресцентный краситель, подобный флуоресцеину или родамину, хелатообразователем для металла, применяемого в томографии или лучевой терапии, пептидильной или непептидильной меткой или детектирующей меткой, агентом, модифицирующим клиренс, таким как разные изомеры полиэтиленгликоля, пептидом, связывающимся с третьим компонентом, или другим углеводом или липофильным агентом.
Применение антител против STEAP-1 с введенным цистеином
Антитела против STEAP-1 с введенным цистеином и их конъюгаты могут быть использованы в качестве терапевтических и/или диагностических агентов. Настоящее изобретение далее относится к способам предотвращения, устранения, лечения или ослабления одного или нескольких симптомов, обусловленных нарушением, связанным с белком STEAP-1. Настоящее изобретение, в частности, относится к способам предотвращения, устранения, лечения или ослабления одного или нескольких симптомов, обусловленных пролиферативным нарушением клеток, таким как рак, например, рак предстательной железы, рак легкого, рак ободочной кишки, рак мочевого пузыря, рак яичника и саркома Юинга. Настоящее изобретение далее относится к способам диагностики нарушения, связанного с STEAP-1, или предрасположенности к возникновению такого нарушения, а также к способам идентификации антител и антигенсвязывающих фрагментов антител, которые, предпочтительно, связываются с клеточными полипептидами STEAP-1.
Другой вариант осуществления настоящего изобретения относится к использованию антитела против STEAP-1 с введенным цистеином для получения лекарственного средства, пригодного для лечения состояния, восприимчивого к нарушению, связанному с белком STEAP-1.
Получение конъюгатов антитела против STEAP-1 с введенным цистеином с лекарственным средством
ADC формулы I может быть получен несколькими способами с использованием реакций органической химии, условий реакций и реагентов, известных специалистам в данной области, которые включают: (1) взаимодействие цистеиновой группы сконструированного антитела с цистеиновой заменой с линкерным реагентом, с образованием промежуточного соединения антитела с линкером Ab-L при помощи ковалентной связи и последующее выполнение взаимодействия с активированной лекарственной частью D; и (2) взаимодействие нуклеофильной группы лекарственной части с линкерным реагентом с образованием промежуточного соединения D-L лекарственного средства с линкером при помощи ковалентной связи и последующее выполнение взаимодействия с цистеиновой группой сконструированного антитела с цистеиновой заменой. Способы конъюгации (1) и (2) могут быть использованы с разными сконструированными антителами с цистеиновой заменой, лекарственными частями и линкерами для получения конъюгатов антитела с лекарственным средством формулы I.
Тиоловые группы цистеина в антителе являются нуклеофильными и могут взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных реагентов и промежуточных соединений лекарственного средства с линкером, включающими (i) активные сложные эфиры, такие как NHS сложные эфиры, HOBt сложные эфиры, галогенформиаты и галогенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные группы и группы имида малеиновой кислоты; и (iv) дисульфиды, в том числе пиридилдисульфиды, в результате сульфидного обмена. Нуклеофильные группы лекарственной части включают, не ограничиваясь ими: амин, тиол, гидроксил, гидразид, оксим, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид, способные взаимодействовать с образованием ковалентных связей с электрофильными группами линкерных частей и линкерных реагентов.
Сконструированным антителам с цистеиновой заменой может быть сообщена реакционная способность для конъюгирования с линкерными реагентами в результате обработки восстановителем, таким как DTT (реагент Клеланда, дитиотреитол) или ТСЕР (гидрохлорид трис(2-карбоксиэтил)фосфина; Getz et al. (1999) Anal. Biochem. Vol. 273:73-80; Soltec Ventures, Beverly, MA), с последующим переокислением для повторного образования межцепных и внутрицепных дисульфидных связей (пример Х). Например, полноразмерные моноклональные сконструированные антитела с цистеиновой заменой (тиоMab), экспрессированные в клетках СНО, восстанавливают примерно 50-кратным избытком ТСЕР в течение 3 часов при 37°С для восстановления дисульфидных связей в аддуктах цистеина, которые могут образоваться между вновь введенными остатками цистеина и цистеином, присутствующим в культуральной среде. Восстановленное тиоMab разводят, вводят в колонку HiTrap S в 10 мМ ацетата натрия, рН 5, и элюируют PBS, содержащим 0,3 М хлорида натрия. Дисульфидные связи были восстановлены между остатками цистеина, присутствующими в материнском моноклональном антителе, разведенном в (200 нМ) водного раствора сульфата меди (CuSO4), при комнатной температуре в течение ночи. Альтернативно может быть использована дегидроаскорбиновая кислота (DHAA), которая является эффективным оксидантом для восстановления внутрицепных дисульфидных групп сконструированного антитела с цистеиновой заменой после восстановительного расщепления аддуктов цистеина. Могут быть использованы другие оксиданты, то есть окислители, и условия окисления, хорошо известные в данной области. Окисление окружающей атмосферой также является эффективным. Указанная стадия мягкого, частичного повторного окисления вызывает эффективное образование внутрицепных дисульфидов с высокой степенью достоверности и сохраняет тиоловые группы вновь введенных остатков цистеина. Примерно 10-кратный избыток промежуточного соединения лекарственного средства с линкером, например, МС-vc-PAB-MMAT, добавляли, смешивали и оставляли выстаиваться в течение примерно одного часа при комнатной температуре для конъюгирования и образования конъюгата антитела с лекарственным средством. Конъюгированную смесь фильтровали через гель, вводили в колонку HiTrap S и элюировали для удаления избытка промежуточного соединения лекарственного средства с линкером и других загрязняющих примесей.
На фигуре 16 показан общий процесс получения сконструированного антитела с цистеиновой заменой, экспрессированного в культуре клеток для конъюгирования. При наличии в среде культуры клеток цистеина дисульфидные аддукты могут образовываться между вновь введенной аминокислотой цистеина и цистеином из среды. Указанные аддукты цистеина, показанные в виде кружков в типичном тиоMab (слева) на фигуре 12, необходимо восстановить для получения сконструированного антитела с цистеиновой заменой, обладающих реакционной способностью для конъюгирования. Аддукты цистеина, главным образом, с разными межцепными дисульфидными связями, расщепляются в восстановительных условиях с образованием восстановленной формы антитела при взаимодействии с восстановителем, таким как ТСЕР. Межцепные дисульфидные связи между спаренными остатками цистеина повторно образуются в условиях частичного окисления сульфатом меди, DHAA или под воздействием окружающего кислорода. Вновь введенные и неспаренные остатки цистеина продолжают оставаться доступными для взаимодействия с линкерными реагентами или промежуточными соединениями лекарственного средства с линкером с образованием конъюгатов антитела по настоящему изобретению. Моноклональные тиоантитела (тиоMab), экспрессированные в линиях клеток млекопитающих, вызывают образование конъюгированного аддукта цистеина (Cys) с введенным цистеином в результате образования -S-S- связи. Очищенные тиоMab подвергают восстановлению и повторному окислению, как описано в примере Х, для получения реакционноспособных тиоМаb. Указанные тиоMab используют для конъюгирования с лекарственными средствами, содержащими малеимид, флуорофорами и другими метками.
На фигуре 15 показаны варианты конъюгатов антитела против STEAP-1 с введенным цистеином с лекарственным средством, в которых лекарственная часть ауристатин присоединена к группе введенного цистеина в: легкой цепи (LC-ADC), тяжелой цепи (НС-ADC) и Fc-области (Fc-ADC).
Фармацевтические препараты
Введение конъюгатов антитела с лекарственным средством
Конъюгаты антитела с лекарственным средством (ADC) по настоящему изобретению могут быть введены индивиду любым способом, соответствующим состоянию, подлежащему лечению. ADC обычно вводят парентерально, то есть в виде подкожного, внутримышечного, внутривенного, чрезкожного, интратекального и эпидурального вливания.
В одном из вариантов осуществления изобретения для лечения рака, например, рака предстательной железы, легкого и/или ободочной кишки, конъюгат антитела с лекарственным средством вводят посредством внутривенного вливания. Доза, вводимая посредством вливания, составляет от около 1 мкг/м2 до около 10000 мкг/м2 в виде одной дозы, обычно при введении одной дозы в неделю, в общей сложности одной, двух, трех или четырех доз. Альтернативно доза составляет от около 1 мкг/м2 до около 1000 мкг/м2, от около 1 мкг/м2 до около 800 мкг/м2, от около 1 мкг/м2 до 600 мкг/м2, от около 1 мкг/м2 до около 400 мкг/м2, от около 10 мкг/м2 до около 500 мкг/м2, от около 10 мкг/м2 до около 300 мкг/м2, от около 10 мкг/м2 до около 200 мкг/м2 и от около 1 мкг/м2 до около 200 мкг/м2. Указанная доза может быть введена один раз в сутки, один раз в неделю, несколько раз в неделю, но реже одного раза в сутки, несколько раз в месяц, но реже одного раза в сутки, несколько раз в месяц, но реже одного раза в неделю, один раз в месяц или периодически для устранения или ослабления симптомов заболевания. Введение можно продолжать в соответствии с любым из указанных интервалов до достижения ремиссии опухоли или ослабления симптомов лимфомы или лейкоза. Введение можно продолжать после ремиссии или ослабления симптомов, если такая ремиссия или ослабление симптомов продлевается в результате такого постоянного введения.
Настоящее изобретение также относится к способу лечения рака предстательной железы, легкого и/или ободочной кишки и/или метастазов такого рака, который включает введение индивиду, страдающему раком предстательной железы, легкого или ободочной кишки, терапевтически эффективного количества гуманизированного антитела 120v.24 по любому из предшествующих вариантов осуществления изобретения, не конъюгированного с цитотоксической молекулой или детектируемой молекулой. Указанное антитело обычно вводят в дозе от около 1 мкг/м2 до около 1000 мг/м2.
Настоящее изобретение также относится к способу лечения рака предстательной железы, легкого и/или ободочной кишки и/или метастазов такого рака, который включает введение индивиду, страдающему раком предстательной железы, легкого или ободочной кишки, терапевтически эффективного количества гуманизированного антитела 120v.24 по любому из предшествующих вариантов осуществления изобретения, конъюгированного с цитотоксической молекулой или детектируемой молекулой. Указанное антитело обычно вводят в дозе от около 1 мкг/м2 до около 1000 мг/м2.
В одном из аспектов настоящее изобретение далее относится к фармацевтическим препаратам, содержащим по меньшей мере одно антитело против STEAP-1 по настоящему изобретению и/или по меньшей мере один иммуноконъюгат, и/или по меньшей мере один конъюгат антитела против STEAP-1 с лекарственным средством по настоящему изобретению. В некоторых вариантах осуществления изобретения фармацевтический препарат содержит 1) антитело против STEAP-1 и/или конъюгат антитела против STEAP-1 с лекарственным средством и/или его иммуноконъюгат и 2) фармацевтически приемлемый носитель. В некоторых вариантах осуществления изобретения фармацевтический препарат содержит 1) антитело против STEAP-1 и/или его иммуноконъюгат и необязательно 2) по меньшей мере одно дополнительное терапевтическое средство.
Фармацевтические препараты, содержащие антитело, иммуноконъюгат по настоящему изобретению или конъюгат антитела с лекарственным средством по настоящему изобретению получают с возможностью хранения путем смешивания антитела или конъюгата антитела с лекарственным средством, имеющего требуемую степень чистоты, с необязательными физиологически приемлемыми носителями, наполнителями или стабилизаторами (Remington's Pharmaceurtical Sciences, 16th edition, Osol, A. Ed. (1980)) в виде водных растворов, лиофилизованных или других высушенных препаратов. Приемлемые носители, наполнители или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают буферы, такие как фосфат, цитрат, гистидин и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония, хлорид бензетония); феноловый, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатообразователи, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы) и/или неионогенные поверхностно-активные вещества, такие как твин™, плюроникс™ или полиэтиленгликоль (PEG). Фармацевтические препараты, предназначенные для введения in vivo, обычно являются стерильными. Стерильность препаратов легко достигается фильтрованием через стерильные фильтрующие мембраны.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, способами коацервации или межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы в коллоидных системах доставки лекарственных средств (таких как, например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы), или в макроэмульсиях. Такие способы описаны в публикации Remington's Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980).
Могут быть получены препараты пролонгированного действия. Приемлемые примеры препаратов пролонгированного действия включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело или иммуноконъюгат по настоящему изобретению, которые представляют собой формованные изделия, например, пленки или микрокапсулы. Примеры матриц пролонгированного действия включают полиэфиры, гидрогели (например, поли-2-гидроксиэтилметакрилат или поливиниловый спирт), полиактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразрушаемый этиленвинилацетат, разрушаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида) и поли-D-(-)-3-гидроксимасляная кислота. Если полимеры, такие как сополимер этилена и винилацетата и сополимер молочной кислоты и гликолевой кислоты, высвобождают молекулы в течение более 100 дней, то определенные гидрогели высвобождают белки в течение более коротких периодов времени. Инкапсулированные антитела или иммуноконъюгаты, остающиеся в теле в течение длительного времени, могут денатурировать или агрегировать под воздействием влаги при 37°С, что вызывает утрату биологической активности и возможные изменения иммуногенности. Могут быть разработаны рациональные способы стабилизации в зависимости от действующего механизма. Например, если механизм агрегации заключается в образовании межмолекулярных S-S связей в результате тио-дисульфидного обмена, препарат может быть стабилизирован путем модификации сульфгидрильных остатков, лиофилизации из кислотных растворов, регулирования влажности, использования соответствующих добавок и создания специальных композиций полимерных матриц.
Лечение конъюгатом антитела с лекарственным средством
Предполагается, что конъюгаты антитела с лекарственным средством (ADC) по настоящему изобретению могут быть использованы для лечения разных заболеваний или нарушений, например, характеризующихся сверхэкспрессией опухолевого антигена. Типичные состояния или гиперпролиферативные нарушения включают доброкачественные или злокачественные опухоли, лейкоз и лимфоидные злокачественные новообразования. Другие заболевания включают нейронные, глиальные, астроцитальные, гипоталамусные, железистые, макрофаговые, эпителиальные, стромальные, бластоцелевые, воспалительные, ангиогенные и иммунологические, в том числе аутоиммунные, нарушения. Другие заболевания включают рак предстательной железы, легкого и ободочной кишки.
Соединения ADC, идентифицированные в животных моделях и при выполнении клеточных анализов, могут быть далее исследованы при введении высшим приматам, имеющим опухоли, и в клинических испытаниях с участием людей. Клинические испытания с участием людей могут быть направлены на исследование эффективности моноклонального антитела против STEAP-1 или иммуноконъюгата по настоящему изобретению у индивидов, страдающих пролиферативными нарушениями клеток предстательной железы, легкого или ободочной кишки, которые включают, не ограничиваясь ими, рак предстательной железы, легкого и ободочной кишки и метастазы таких злокачественных новообразований. Целью клинического испытания может быть оценка эффективности ADC в комбинациях с известными схемами лечения, такими как лучевая терапия и/или химиотерапия с использованием известных химиотерапевтических и/или цитотоксических агентов.
Рак может включать STEAP-1-экспрессирующие клетки, поэтому ADC по настоящему изобретению может связываться с раковыми клетками. Экспрессию STEAP-1 в раковых клетках можно обнаружить при помощи разных диагностических/прогностических анализов. В одном из вариантов осуществления изобретения сверхэкспрессия STEAP-1 может быть обнаружена при помощи иммуногистохимического анализа (IHC). Погруженные в парафин срезы ткани, полученные в результате биопсии опухоли, могут быть исследованы с помощью IHC анализа, при этом белок STEAP-1 определяют на основании интенсивности окрашивания и содержания опухолевых клеток.
Соответствующая доза ADC, используемая для профилактики или лечения заболевания, зависит от типа подлежащего лечению заболевания, тяжести и развития заболевания, введения молекулы в профилактических или терапевтических целях, предшествующего лечения, истории болезни индивида, реакции на антитело и решения лечащего врача. Указанную молекулу вводят индивиду однократно или в виде серии процедур. В зависимости от типа и тяжести заболевания начальная доза, вводимая индивиду, составляет от 1 мкг/кг до 15 мг/кг (например, 0,1-20 мг/кг), при этом указанную дозу вводят в виде одной или нескольких отдельных инъекций или в виде непрерывного вливания. Типичная суточная доза может составлять от около 1 мкг/кг до 100 мг/кг или больше в зависимости от вышеуказанных факторов. Типичная доза ADC, вводимая индивиду, составляет от около 0,1 до около 10 мг/кг массы тела индивида.
При повторном введении в течение нескольких дней или более продолжительного периода времени лечение продолжают в зависимости от состояния до требуемого подавления симптомов заболевания. Типичная схема введения предполагает введение начальной дозы, равной примерно 4 мг/кг, с последующим введением еженедельной поддерживающей дозы, равной примерно 2 мг/кг антитела против STEAP-1. Могут быть использованы другие схемы введения. Эффективность указанного лечения можно легко контролировать стандартными способами и анализами.
Комбинированная терапия
Конъюгат антитела с лекарственным средством (ADC) по настоящему изобретению может быть объединен в комбинированном фармацевтическом препарате или в схеме введения, применяемой в качестве комбинированной терапии, по меньшей мере с одним дополнительным соединением, обладающим противораковыми свойствами. По меньшей мере одно дополнительное соединение в комбинированном фармацевтическом препарате или схеме введения, предпочтительно, обладает активностью, комплементарной ADC в комбинации, благодаря чему такие соединения не оказывают вредного влияния друг на друга.
По меньшей мере одно дополнительное соединение может быть химиотерапевтическим средством, цитотоксическим агентом, цитокином, ингибитором роста, противогормональным средством и/или кардиозащитным средством. Такие молекулы могут присутствовать в комбинации в эффективных количествах для достижения указанной цели. Фармацевтическая композиция, содержащая ADC по настоящему изобретению, также может содержать терапевтически эффективное количество химиотерапевтического средства, такого как ингибитор образования тубулина, ингибитор топоизомеразы или ДНК-связывающий агент.
В соответствии с одним аспектом изобретения первым соединением является ADC против STEAP-1 по настоящему изобретению и по меньшей мере одним дополнительным соединением является терапевтическое антитело, отличное от антитела против STEAP-1 (“голого” антитела или ADC). В одном из вариантов осуществления изобретения по меньшей мере одним дополнительным соединением является антитело против PSCA. В одном из вариантов осуществления изобретения по меньшей мере одним дополнительным соединением является антитело против HER2, трастузумаб (например, герцептин®, Genentech, Inc., South San Francisco, CA). В одном из вариантов осуществления изобретения по меньшей мере одним дополнительным соединением является антитело против HER2, пертузумаб (омнитарг®, Genentech, Inc., South San Francisco, CA, см. патент США № 6949245). В одном из вариантов осуществления изобретения по меньшей мере одним дополнительным соединением является антитело против VEGF (например, авастин®, Genentech, Inc.). Во всех случаях по меньшей мере одно соединение является “голым” антителом или ADC. В одном из вариантов осуществления изобретения по меньшей мере одно дополнительное соединение является антителом (“голым” антителом или ADC), и дополнительное антитело является вторым, третьим, четвертым, пятым, шестым антителом и так далее, при этом комбинация, включающая второе, третье, четвертое, пятое, шестое или последующие антитела (“голые” или в виде ADC), позволяет эффективно лечить пролиферативное заболевание клеток в ткани, экспрессирующей STEAP-1.
Другие схемы лечения могут быть объединены с введением противоракового средства, соответствующего настоящему изобретению, которое может включать, не ограничиваясь ими, лучевую терапию, трансплантацию костного мозга и переливание периферической крови и/или введение цитотоксического агента, химиотерапевтического средства, или ингибитора роста. В одном таком варианте осуществления изобретения химиотерапевтическое средство является одним средством или комбинацией средств, таких как, например, циклофосфамид, гидроксидаунорубицин, адриамицин, доксорубицин, винкристин (онковин™), преднизолон, СНОР, CVP или СОР, иммунотерапевтические средства, такие как антитело против PSCA, антитело против HER2 (например, герцептин®, омнитарг™) или антитело против VEGF (например, авастин®). Комбинированная терапия может проводиться в соответствии с одновременной или последовательной схемой введения. При последовательном введении комбинацию можно вводить в виде двух или более введений. Объединенное введение предполагает совместное введение при использовании отдельных препаратов или одного фармацевтического препарата и последовательное введение в любом порядке, но, предпочтительно, в течение периода времени, когда оба (или все) активные агенты одновременно проявляют свою биологическую активность.
В одном из вариантов осуществления изобретения лечение ADC предполагает объединенное введение противоракового средства, указанного в настоящем описании изобретения, и одного или нескольких химиотерапевтических средство или ингибиторов роста, включая совместное введение коктейля из разных химиотерапевтических средств. Химиотерапевтические средства включают таксаны (такие как паклитаксел и доцетаксел) и/или антрациклиновые антибиотики. Схемы приготовления и дозирования таких химиотерапевтических средств могут быть определены в соответствии с инструкциями производителя или эмпирически квалифицированным лечащим врачом. Схемы приготовления и дозирования таких химиотерапевтических средств также описаны в публикации ”Chemotherapy Service”, (1992) Ed., M.C. Perry, Williams & Wilkins, Baltimore, Md.
Приемлемые дозы любых из вышеуказанных совместно вводимых средств соответствуют используемым в настоящее время и могут быть уменьшены благодаря объединенному действию (синергии) вновь созданного средства и других химиотерапевтических средств или способов лечения.
Комбинированная терапия может оказывать ”синергическое действие”, то есть действие, достигаемое при совместном использовании активных ингредиентов, оказывается больше суммарного действия, достигаемого при отдельном применении указанных соединений. Синергическое действие может быть достигнуто, когда активные ингредиенты: (1) вводят или доставляют одновременно в комбинированном унифицированном препарате; (2) доставляют поочередно или параллельно в отдельных препаратах или (3) вводят в соответствии с какой-либо другой схемой введения. При поочередной доставке синергическое действие достигается в том случае, когда соединения вводят или доставляют последовательно, например, в виде отдельных инъекций из разных шприцев. Как правило, при поочередной терапии эффективную дозу каждого активного ингредиента вводят последовательно, в то время как в комбинированной терапии эффективные дозы двух или более активных ингредиентов вводят одновременно.
Метаболиты конъюгатов антитела с лекарственным средством
В объем настоящего изобретения также входят продукты обмена веществ in vivo соединений ADC по настоящему изобретению, если такие продукты являются новыми и не были известны ранее. Такие продукты могут возникать, например, в результате окисления, восстановления, гидролиза, амидирования, этерификации, расщепления ферментами и подобных процессов, которым подвергается введенное соединение. Таким образом, настоящее изобретение относится к новым и неизвестным соединениям, образующимся в процессе контактирования соединения по настоящему изобретению с организмом млекопитающего в течение периода времени, достаточного для образования продукта обмена веществ.
Продукты обмена веществ обычно идентифицируют, получая меченный радиоактивным изотопом (например, 14С или 3Н) ADC, который вводят парентерально в детектируемой дозе (например, более примерно 0,5 мг/кг) животному, такому как крыса, мышь, морская свинка, обезьяна, или человеку, контролируют обмен веществ в течение достаточного периода времени (обычно от около 30 секунд до 30 часов) и выделяют продукты обмена веществ из мочи, крови или других биологических образцов. Указанные продукты можно легко выделить благодаря мечению (другие продукты выделяют при помощи антител, способных связываться с эпитопами, имеющимися в метаболите). Структуры метаболитов определяют стандартными способами, например, при помощи МС, ЖХ/МС или ЯМР. Как правило, метаболиты анализируют способами исследования метаболизма лекарственных средств, хорошо известными специалистам в данной области. Продукты обмена веществ, в случае их обнаружения in vivo, используют в диагностических анализах для определения терапевтических доз соединений ADC по настоящему изобретению.
Другие способы применения антител против STEAP-1 и иммуноконъюгатов
Диагностические способы и способы обнаружения
В одном из аспектов изобретения антитела против STEAP-1 и иммуноконъюгаты по настоящему изобретению используют для обнаружения STEAP-1 в биологическом образце. Термин ”обнаружение”, как используется в настоящем описании, означает количественное или качественное обнаружение. В определенных вариантах осуществления изобретения биологическим образцом является клетка или ткань. В определенных вариантах осуществления изобретения такие ткани включают нормальные и/или раковые ткани, экспрессирующие STEAP-1 на более высоких уровнях по сравнению с другими тканями, например, предстательной железы, легкого и ободочной кишки.
В одном из аспектов настоящее изобретение относится к способу обнаружения STEAP-1 в биологическом образце. В определенных вариантах осуществления изобретения указанный способ включает приведение в контакт биологического образца с антителом против STEAP-1 в условиях, допускающих связывание антитела против STEAP-1 с белком STEAP-1, и обнаружение комплекса, образованного между антителом против STEAP-1 и белком STEAP-1.
В одном из аспектов настоящее изобретение относится к способу диагностики нарушения, связанного с повышенной экспрессией STEAP-1. В определенных вариантах осуществления изобретения указанный способ включает приведение в контакт исследуемой клетки с антителом против STEAP-1, определение уровня экспрессии (в количественном или качественном отношении) STEAP-1 исследуемой клеткой путем обнаружения связывания антитела против STEAP-1 с белком STEAP-1 и сравнение уровня экспрессии STEAP-1 исследуемой клеткой с уровнем экспрессии STEAP-1 контрольной клеткой (например, нормальной клеткой из той же ткани, что и исследуемая клетка, или клеткой, экспрессирующей STEAP-1 на уровнях, сравнимых с аналогичными уровнями нормальной клетки), при этом более высокий уровень экспрессии STEAP-1 исследуемой клеткой по сравнению с контрольной клеткой указывает на наличие нарушения, связанного с повышенной экспрессией STEAP-1. В определенных вариантах осуществления изобретения исследуемую клетку получают у индивида с подозрением на нарушение, связанное с повышенной экспрессией STEAP-1. В определенных вариантах осуществления изобретения нарушение является пролиферативным нарушением клеток, таким как рак или опухоль.
Типичные пролиферативные нарушения клеток, которые могут быть диагностированы при помощи антитела по настоящему изобретению, включают рак предстательной железы, легкого и ободочной кишки или метастазы таких злокачественных новообразований.
В определенных вариантах осуществления изобретения способ диагностики или обнаружения, аналогичный вышеописанным способам, включает обнаружение связывания антитела против STEAP-1 с белком STEAP-1, экспрессированным на поверхности клетки или в мембранном препарате, полученном из клетки, экспрессирующей STEAP-1 на своей поверхности. В определенных вариантах осуществления изобретения указанный способ включает приведение в контакт клетки с антителом против STEAP-1 в условиях, допускающих связывание антитела против STEAP-1 с белком STEAP-1, и обнаружение комплекса, образованного между антителом против STEAP-1 и белком STEAP-1 на поверхности клетки. Типичным анализом для обнаружения связывания антитела против STEAP-1 с белком STEAP-1, экспрессированным на поверхности клетки, является анализ FACS.
Для обнаружения связывания антитела против STEAP-1 с белком STEAP-1 могут быть использованы некоторые другие способы. Такие способы включают, не ограничиваясь ими, антигенсвязывающие анализы, хорошо известные в данной области, такие как вестерн-блоттинг, радиоиммуноанализы, ELISA (твердофазный иммуноферментный анализ), иммуноферментные сэндвич-анализы, анализы способом иммунопреципитации, флуоресцентные иммуноанализы, иммуноанализы с использованием протеина А, иммуно-ПЭТ и иммуногистохимический анализ (IHC).
В определенных вариантах осуществления изобретения иммуно-ПЭТ с 89Zr-меченым антителом против STEAP-1, выполняемая in vitro или in vivo, может быть использована для количественного определения поглощения и биологического распределения антитела в первичных поражениях опухолью предстательной железы и для идентификации изменений профилей опухолевых антигенов при метастазировании первичных опухолей. Иммуно-ПЭТ требует использования позитрон-излучающего радиоактивного изотопа (такого как цирконий (89Zr); Verel I., J. Nucl. Med. 2003: 44:1271-1281; “89Zr Immuno-PET: Comprehensive Procedures for the Production of 89Zr-Labeled Monoclonal Antibodies”. При выполнении иммуно-ПЭТ комбинация ПЭТ с моноклональными антителами (mAb) является привлекательным вариантом более эффективного обнаружения опухоли и количественного определения с помощью моноклональных антител. Долгоживущий излучатель позитронов 89Zr обладает идеальными физическими характеристиками для иммуно-ПЭТ, такими как период полувыведения, равный 3,27 дня, который сравним со временем, необходимым для достижения интактными моноклональными антителами оптимального соотношения между опухолью и неопухолевыми тканями.
Для образования комплекса 89Zr с антителами, включающими моноклональные антитела (mAb), используют типичные бифункциональные реагенты на основе десферриоксамина В (Df). Десферриоксамин В (N'-{5-[ацетил(гидрокси)амино]пентил}-N-[5-({4-[(5-аминопентил)-(гидрокси)амино]-4-оксобутаноил}амино)пентил]-N-гидроксисукцинамид (регистрационный номер CAS 70-51-9), также известный как дефероксамин, десфероксамин В, DFO-B, DFOA, DFB или десферал) является бактериальный сидерофором, продуцируемым актинобактериями Streptomyces pilosus (фигура 20, вверху). Десферриоксамин В применяется в медицине в качестве хелатообразователя, используемого для удаления избытка железа из организма (Miller, Marvin J. “Synthesis and therapeutic potential of hydroxamic acid based siderophores and analogs” (1989) Chemical Reviews 89 (7):1563-1579). Мезилат DFO-B является коммерчески доступным продуктом. Первые эксперименты были выполнены с использованием N-(S-ацетил)тиоацетил-Df (SATA-Df) и моноклонального антитела, содержащего группы имида малеиновой кислоты, 4-[N-малеимидометил]-циклогексан-1-карбоксилат (mAb-SMCC), присоединенные к аминогруппе в боковой цепи лизина (Meijs W.E. et al. “Zirconuim-labeled monoclonal antibodies and their distribution in tumor-bearing nude mice” (1997) J. Nucl. Med. 38:112-8; Meijs W.E. et al. “A facile method for the labeling of proteins with zirconium isotopes” (1996) Nucl. Med. Biol. 23:439-48).
Другие типичные взаимодействующие с аминогруппой бифункциональные хелатообразователи на основе Df, модифицированного янтарным ангидридом (Suc), были использованы для превращения аминогруппы Df в карбоновую кислоту и последующей активации в виде 2,3,5,6-тетрафторфенилового эфира (TFP). TFP-N-Suc-Df (фигура 20, в центре) связывали с аминогруппами лизина в mAb и очищенный mAb-N-Suc-Df использовали для получения хелатного комплекса с 89Zr. Полученный комплекс 89Zr-mAb-N-Suc-Df был устойчив в физиологических условиях, и его биологическое распределение было сравнимо с распределением mAb-SMCC-SATA-Df у мышей (Verel I. et al. “89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies” (2003) J. Nucl. Med. 44:1271-81). Однако процесс получения TFP-N-Suc-Df требует защиты гидроксиамидных групп в виде комплекса Fe(III). Железо удаляют в результате обработки EDTA до образования хелатного комплекса с 89Zr, но указанный многостадийный способ является сложным, при этом возникает опасность неполного удаления железа из десферриоксамина и/или неполного удаления EDTA из буфера для конъюгирования, что может отрицательно влиять на образование хелатного комплекса с 89Zr. Поэтому недавно был разработан гетеробифункциональный реагент, взаимодействующий с аминогруппами, параизотиоцианатобензил-десферриоксаимин (Df-Bz-NCS), предназначенный для введения Df в белки при помощи тиомочевинной связи (Perk LR et al. “Facile radiolabeling of monoclonal antibodies and other proteins with zirconium-89 or gallium-68 for PET Imaging using p-isothiocyanatobenzyl-desferrioxamine” (2008) Nature Protocols, published online:DOI:10.1038/nprot.2008.22; Perk LR et al. “p-Isothiocyanatobenzyl-desferrioxamine: a new bifunctional chelate for facile radiolabeling of monoclonal antibodies with zirconium-89 for immuno-PET imaging” (2009) European Journal Of Nuclear Medicine And Molecular Imaging).
Конъюгаты антитела, полученные с использованием Df-Bz-NCS, характеризуются свойствами устойчивости и визуализации, сравнимыми с эталонными конъюгатами, полученными при использовании TFP-N-Suc-Df. После создания надежных способов связывания 89Zr с антителами при помощи аминогрупп лизина быстро растет число доклинических и клинических исследований с помощью иммуно-ПЭТ при использовании 89Zr-меченых антител (Verel I, et al. “Long-lived positron emitters zirconium-89 and iodine-124 for scouting of therapeutic radioimmunoconjugates with PET” (2003) Cancer Biother Radiopharm. 18:655-61; Nagengast WB et al. “In vivo VEGF imaging with radiolabeled bevacizumab in a human ovarian tumor xenograft” (2007) J Nucl Med. 48:1313-9; Perk LR, et al. “(89)Zr as a PET surrogate radioisotope for scouting biodistribution of the therapeutic radiometals (90)Y and (177)Lu in tumor-bearing nude mice after coupling to the internalizing antibody cetuximab” (2005) J Nucl Med. 46:1898-906; Perk LR et al. “Quantitative PET imaging of Met-expressing human cancer xenografts with (89)Zr-labelled monoclonal antibody DN30” (2008) European Journal Of Nuclear Medicine And Molecular Imaging 35:1857-67; Perk LR et al. “Preparation and evaluation of (89)Zr-Zevalin for monitoring of (90)Y-Zevalin biodistribution with positron emission tomography” (2006) European Journal Of Nuclear Medicine And Molecular Imaging 33:1337-45; Borjesson PK et al. “Performance of immuno-positron emission tomography with zirconium-89-labeled chimeric monoclonal antibody U36 in the detection of lymph node metastases in head and neck cancer patients” (2006) Clin Cancer Res. 12:2133-40; Aerts HJ et al. “Disparity between in vivo EGFR expression and 89Zr-labeled cetuximab uptake assessed with PET” (2009) J Nucl Med. 50:123-31; Dijkers EC et al. “Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging” (2009) J Nucl Med 50(6):974-981).
Варианты циркониевых комплексов также включают связывающиеся с цирконием (хелатообразующие) лиганды, такие как DTPA (регистрационный номер CAS 67-43-6), DOPA (1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусная кислота) (Liu, Shuang (2008) Advanced Drug Delivery Reviews 60(12): 1347-1370), циклопентадиенильные и аллильные группы (см. публикации Erker, G. (1991) Pure and Applied Chemistry 63(6): 797-806; Erker, G. (1990) Jour. of Organometallic Chem. 400(1-2):185-203), которые включены в настоящее описание изобретения в качестве ссылки.
Циркониевые комплексы (Z) и другие радионуклиды могут быть конъюгированы с антителами (Ab), включая моноклональные антитела (mAb), при помощи аминогруппы в боковой цепи лизина или тиоловой группы цистеина. Так как для конъюгирования в моноклональном антителе имеется примерно 40 боковых цепей лизина (Wang L. et al. “Structural characterization of the maytansinoid-monoclonal antibody immunoconjugate, huN901-DM1, by mass spectometry” (2005) Protein Sci. 14:2436-46) или 8 цистеинов (Hamblett K.J. et al. “Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate” (2004) Clin. Cancer. Res. 10:7063-70), оба способа обеспечивают гетерогенность соотношений конъюгатов mAb и места конъюгации. Модификация остатка лизина в месте связывания может уменьшить биологическую активность конъюгата (Cai W et al. “PET imaging of colorectal cancer in xenograft-bearing mice by use of an 18F-labeled T84.66 anti-carcinoembryonic antigen diabody” (2007) J Nucl Med. 48:304-10; Shively JE. “18F labeling for immuno-PET: where speed and contrast meet” (2007) J Nucl Med. 48:170-2; Tait JF et al. “Improved detection of cell death in vivo with annexin V radiolabeled by site-specific methods” (2006) J Nucl Med. 47:1546-53; Schellenberger EA et al. “Optical imaging of apoptosis as a biomarker of tumor response to chemotherapy” (2003) Neoplasia (New York, N.Y) 5:187-92), а модификация цистеинов в шарнирной области уменьшает время полужизни в плазме (Hamblett K.J. et al. “Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate” (2004) Clin. Cancer Res. 10:7063-70). Указанных ограничений можно избежать, используя генетически созданные моноклональные антитела, содержащие цистеин, избирательно расположенный в соответствии с целями сайтспецифического конъюгирования, наряду с выполнением биохимического анализа PHESELECTOR (патент США № 7521541; публикация Junutula J.R. et al. “Rapid identification of reactive cystein residues for site-specific labeling of antibody-Fabs” J. Immunol. Methods 2008; 332:41-52) для быстрой идентификации предпочтительных аминокислот в антителе для мутации в цистеин. Полученное антитело (THIOMAB) затем отбирают химическими способами и сайтспецифически конъюгируют с цитотоксическими средствами без какой-либо утраты аффинности связывания или вредного влияния на устойчивость антитела (Junutula J.R. et al. “Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index” (2008) Nat. Biotechnol. 26:925-32).
Что касается визуализации, то для достижения оптимального качества изображения необходима высокая аффинность к мишени и минимальное неспецифическое поглощение. Поэтому антитела с введенным цистеином, сайтспецифически меченные радиоактивными изотопами, (THIOMAB) могут быть радиоактивными индикаторами с неизменяемой аффинностью связывания и устойчивой структурой, которые могут свести к минимуму неспецифическое поглощение метаболитов за пределами ткани-мишени. Один аспект настоящего изобретения относится к способу сайтспецифического мечения THIOMAB радиоактивными изотопами с использованием новых взаимодействующих с тиолом бифункциональных реагентов на основе Df малеимидоциклогексилдесферриоксамина (Df-Chx-Mal), бромацетилдесферриоксамина (Df-Bac) и иодацетилдесферриоксамина (Df-Iac). В типичных вариантах осуществления изобретения указанные реагенты сайтспецифически конъюгированы с THIOMAB трастузумабом (тиотрастузумаб), образовали хелатный комплекс с 89Zr и оценены in vitro и in vivo.
В определенных вариантах осуществления изобретения антитела против STEAP-1 являются мечеными. Метки включают, не ограничиваясь ими, непосредственно обнаруживаемые метки или части (такие как флуоресцентные, хромофорные, электроноплотные, хемилюминесцентные и радиоактивные метки), а также такие части как ферменты или лиганды, обнаруживаемые опосредованно, например, в результате ферментативной реакции или молекулярного взаимодействия. Типичные метки включают, не ограничиваясь ими, радиоактивные изотопы 32Р, 14С, 125I, 3H и 131I, флуорофоры, такие как хелаты редкоземельных металлов, флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светляка и бактериальную люциферазу (патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу из хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат-дегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, связанные с ферментом, использующим пероксид водорода для окисления предшественника красителя, такого как HRP, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, устойчивые свободные радикалы и тому подобные.
В определенных вариантах осуществления изобретения антитела против STEAP-1 иммобилизованы на нерастворимой матрице. Иммобилизация позволяет отделить антитело против STEAP-1 от любого белка STEAP-1, остающегося в растворе. Указанной цели обычно достигают, делая антитело против STEAP-1 нерастворимым до выполнения анализа путем адсорбции на не растворимой в воде матрице или поверхности (патент США № 3720760, Bennich et al.) или путем ковалентного связывания (например, при помощи перекрестного сшивания с глутаровым альдегидом) или делая антитело против STEAP-1 нерастворимым после образования комплекса антитела против STEAP-1 с белком STEAP-1, например, способом иммунопреципитации.
Любой из вышеуказанных вариантов диагностики или обнаружения может быть выполнен с использованием иммуноконъюгата по настоящему изобретению вместо антитела против STEAP-1 или дополнительно к указанному антителу.
Терапевтические способы
Антитело или иммуноконъюгат по настоящему изобретению может быть использован в терапевтических способах in vitro, ex vivo и in vivo. Одним объектом настоящего изобретения является способ ингибирования роста или пролиферации клеток in vivo или in vitro, который включает воздействие на клетку антителом против STEAP-1 или его иммуноконъюгатом в условиях, допускающих связывание иммуноконъюгата с белком STEAP-1. Термин ”ингибирование роста или пролиферации клетки” означает уменьшение роста или пролиферации клетки по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 100% и индукцию гибели клетки. В определенных вариантах осуществления изобретения клетка является опухолевой клеткой. В определенных вариантах осуществления изобретения клетка является клеткой предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или клеткой саркомы Юинга. В определенных вариантах осуществления изобретения клетка является ксенотрансплантатом, например, представленным в настоящем описании изобретения.
В одном из аспектов изобретения антитело или иммуноконъюгат по настоящему изобретению используют для лечения или профилактики пролиферативного нарушения клетки предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или клетки саркомы Юинга. В определенных вариантах осуществления изобретения пролиферативное нарушение клетки связано с повышенной экспрессией и/или активностью STEAP-1. Например, в определенных вариантах осуществления изобретения пролиферативное нарушение клетки предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или клетки саркомы Юинга связано с повышенной экспрессией STEAP-1 на поверхности клетки предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или клетки саркомы Юинга. В определенных вариантах осуществления изобретения пролиферативное нарушение клетки предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или клетки саркомы Юинга является опухолью или злокачественным новообразованием или метастазами такого злокачественного новообразования.
В одном из аспектов настоящее изобретение относится к способу лечения пролиферативного нарушения клетки предстательной железы, легкого или ободочной кишки, который включает введение индивиду эффективного количества антитела против STEAP-1 или его иммуноконъюгата. В определенных вариантах осуществления изобретения способ лечения пролиферативного нарушения клетки предстательной железы, легкого или ободочной кишки включает введение индивиду эффективного количества фармацевтического препарата, включающего антитело против STEAP-1 или иммуноконъюгат антитела против STEAP-1 и необязательно по меньшей мере одно дополнительное терапевтическое средство из средств, представленных в настоящем описании изобретения.
В одном из аспектов изобретения по меньшей мере некоторые антитела или иммуноконъюгаты по настоящему изобретению могут связываться с белком STEAP-1 вида, отличного от человека. Поэтому антитела или иммуноконъюгаты по настоящему изобретению могут быть использованы для связывания белка STEAP-1, например, в культуре клеток, содержащей STEAP-1, в организме человека или других млекопитающих, содержащих STEAP-1, с которым перекрестно взаимодействует антитело или иммуноконъюгат по настоящему изобретению (например, шимпанзе, павиан, игрунка, циномолгус и макак-резус, собака, свинья, крыса или мышь). В одном из вариантов осуществления изобретения антитело против STEAP-1 или иммуноконъюгат могут быть использованы для направленного воздействия на белок STEAP-1 на клетках предстательной железы, легкого или ободочной кишки в результате осуществления контактирования антитела или иммуноконъюгата с белком STEAP-1 с образованиеми комплекса антитела или иммуноконъюгата с антигеном, благодаря чему конъюгированный цитотоксин иммуноконъюгата проникает внутрь клетки. В одном из вариантов осуществления изобретения белок STEAP-1, с которым связывается антитело против STEAP-1, является белком STEAP-1 человека. В одном из вариантов осуществления изобретения белок STEAP-1, с которым связывается антитело против STEAP-1, является белком STEAP-1 обезьяны циномолгуса. В одном из вариантов осуществления изобретения гуманизированное антитело против STEAP-1 связывается с белком STEAP-1 человека и/или обезьяны циномолгуса.
В одном из вариантов осуществления изобретения антитело против STEAP-1 или иммуноконъюгат может быть использован в способе связывания STEAP-1 в организме индивида, страдающего нарушением, связанным с повышенной экспрессией и/или активностью STEAP-1, который включает введение индивиду антитела или иммуноконъюгата с возможностью связывания STEAP-1 в организме индивида. В одном из вариантов осуществления изобретения связанное антитело или иммуноконъюгат интернализован в клетку предстательной железы, легкого, ободочной кишки, мочевого пузыря, яичника или в клетку саркомы Юинга, экспрессирующую STEAP-1. В одном из вариантов осуществления изобретения STEAP-1 является белком STEAP-1 человека и индивид является человеком. Альтернативно индивид может быть млекопитающим, экспрессирующим STEAP-1, с которым связывается антитело против STEAP-1. Далее индивид может быть млекопитающим, которому ввели белок STEAP-1 (например, в результате введения STEAP-1 или экспрессии трансгена, кодирующего STEAP-1).
Антитело против STEAP-1 или иммуноконъюгат можно вводить человеку в терапевтических целях. Кроме того, антитело против STEAP-1 или иммуноконъюгат можно вводить млекопитающему, отличному от человека, экспрессирующему STEAP-1, с которым перекрестно взаимодействует антитело (например, примат, собака, свинья, крыса или мышь), в ветеринарных целях или в качестве животной модели болезни человека. Такие животные модели могут быть использованы для оценки терапевтической эффективностм антител или иммуноконъюгатов по настоящему изобретению (например, для исследования доз и курсов введения).
Антитела или иммуноконъюгаты по настоящему изобретению могут быть использованы отдельно или в комбинации с другими композициями, применяемыми в терапии. Например, антитело или иммуноконъюгат по настоящему изобретению могут быть введены вместе по меньшей мере с одним дополнительным терапевтическим агентом и/или адъювантом. В определенных вариантах осуществления изобретения дополнительный терапевтический агент является цитотоксическим агентом, химиотерапевтическим средством или ингибитором роста. В одном таком варианте осуществления изобретения химиотерапевтическое средство представляет собой средство или комбинацию средств, таких как, например, циклофосфамид, гидроксидаунорубицин, адриамицин, доксорубицин, винкристин (онковин™), преднизолон, СНОР, CVP или СОР, иммунотерапевтические средства, такие как антитело против PSCA (см., например, патент США № 6824780), антитело против VEGF (например, авастин®, Genentech, Inc.), антитело против HER2 (например, герцептин®, омнитарг™, Genentech, Inc.) или антитело против HER2 в комбинации с таксолом® (см., например, публикацию BioWorld Today, November 17, 1999, page 1), предназначенные для использования в комбинированной терапии для лечения пролиферативных нарушений клеток, раков и/или метастазов раков предстательной железы, легкого и/или ободочной кишки.
Вышеуказанная комбинированная терапия включает объединенное введение (когда два или более терапевтических средств входят в состав одного или разных препаратов) и раздельное введение, когда антитело или иммуноконъюгат по настоящему изобретению вводят до, одновременно и/или после введения дополнительного терапевтического средства и/или адъюванта. Антитела или иммуноконъюгаты по настоящему изобретению могут быть также использованы в комбинации с лучевой терапией.
Антитело или иммуноконъюгат по настоящему изобретению (и любое дополнительное терапевтическое средство или адъювант) могут быть введены любым приемлемым способом, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и назальное введение и при желании местное введение и введение в место поражения. Парентеральные вливания включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, антитело или иммуноконъюгат можно вводить в виде импульсного вливания, в частности, в виде уменьшающихся доз антитела или иммуноконъюгата. Дозирование может быть произведено любым приемлемым способом, например, в виде инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или постоянным.
Антитела или иммуноконъюгаты по настоящему изобретению могут быть получены, дозированы и введены способом, совместимым с хорошей медицинской практикой. Факторы, учитываемые в данной связи, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние индивида, причину нарушения, место доставки лекарственного средства, способ введения, схему введения и другие факторы, известные лечащим врачам. Антитело или иммуноконъюгат не должны, но необязательно могут быть получены в виде одного препарата с одним или несколькими средствами, используемыми в настоящее время для профилактики или лечения данного нарушения. Эффективное количество других таких лекарственных средств зависит от количества антитела или иммуноконъюгата в препарате, типа нарушения или лечения и других факторов, рассмотренных выше. Антитело или иммуноконъюгат обычно используют в дозах, указанных в настоящем описании изобретения, или в количестве 1-99% от доз, указанных в настоящем описании изобретения, или в любых дозах и в соответствии с любым способом введения, которые эмпирически/клинически признаны правильными.
Соответствующая доза антитела или иммуноконъюгата (при отдельном использовании или в комбинации с одним или несколькими другими дополнительными терапевтическими средствами, такими как химиотерапевтические средства), используемая для профилактики или лечения заболевания, зависит от типа заболевания, подлежащего лечению, типа антитела или иммуноконъюгата, тяжести и развития заболевания, введения антитела или иммуноконъюгата в профилактических, или терапевтических целях, предшествующего лечения, истории болезни индивида, реакции на антитело или иммуноконъюгат и решения лечащего врача. Антитело или иммуноконъюгат вводят индивиду один или несколько раз. В зависимости от типа и тяжести заболевания начальная доза, вводимая индивиду, составляет от 1 мкг/кг до 100 мг/кг (например, 0,1-20 мг/кг) антитела или имммуноконъюгата, при этом указанную дозу вводят в виде одной или нескольких отдельных инъекций или в виде непрерывного вливания. Типичная суточная доза может составлять от около 1 мкг/кг до 100 мг/кг или больше в зависимости от вышеуказанных факторов. При повторном введении в течение нескольких дней или более продолжительного периода времени в зависимости от состояния лечение обычно продолжают до достижения требуемого подавления симптомов заболевания. Одна типичная доза антитела или иммуноконъюгата составляет от около 0,05 мг/кг до около 10 мг/кг. Таким образом, индивиду может быть введена одна или несколько доз антитела или иммуноконъюгата в количестве примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любая комбинация указанных доз). Такие дозы можно вводить периодически, например, один раз в неделю или один раз в три недели (при этом индивид получает от около двух до около двадцати доз или, например, около шести доз антитела или иммуноконъюгата). После начальной более высокой дозы можно ввести одну или несколько более низких доз. Типичная схема введения включает введение начальной дозы, равной примерно 4 мг/кг, и затем еженедельной поддерживающей дозы, равной примерно 2 мг/кг антитела. Однако могут быть использованы другие схемы введения. Эффективность указанного лечения можно легко контролировать стандартными способами и анализами.
Анализы
Физические/химические свойства и/или биологические активности антител против STEAP-1 и иммуноконъюгатов по настоящему изобретению могут быть исследованы в результаты выполнения разных анализов, известных в данной области.
Анализы активности
В одном из аспектов настоящее изобретение относится к анализам, позволяющим идентифицировать антитела против STEAP-1 или их иммуноконъюгаты, обладающие биологической активностью. Биологическая активность может включать, например, способность ингибировать рост или пролиферацию клеток (например, активность по ”уничтожению клеток”) или способность индуцировать гибель клетки, включая запрограммированную гибель клетки (апоптоз). Настоящее изобретение также относится к антителам или иммуноконъюгатам, обладающим такой биологической активаностью in vivo и/или in vitro.
В определенных вариантах осуществления изобретения антитело против STEAP-1 или его иммуноконъюгат исследуют в отношении способности ингибировать рост или пролиферацию клеток in vitro. Анализы ингибирования роста или пролиферации клеток хорошо известны в данной области. Некоторые анализы пролиферации клеток, определяемые как анализы ”уничтожения клеток”, позволяют измерить жизнеспособность клеток. Одним таким анализом является анализ жизнеспособности люминесцентных клеток CellTiter-Glo™, который является коммерчески доступным в компании Promega (Madison, WI). Указанный анализ позволяет определить число жизнеспособных клеток в культуре на основании количественного определения имеющегося АТР, который является показателем метаболически активных клеток. См. публикацию Crouch et al. (1993) J. Immunol. Meth. 160:81-88, патент США № 6602677. Указанный анализ может быть выполнен на 96-луночном или 384-луночном планшете, что позволяет производить автоматический высокопроизводительный скрининг (HTS). См. публикацию Cree et al. (1995) AntiCancer Drugs 6:398-404. Указанный анализ предполагает добавление одного реагента (реагент CellTiter-Glo®) непосредственно в культивируемые клетки. В результате этого происходит лизис клеток и образование люминесцентного сигнала в результате взаимодействия с люциферазой. Люминесцентный сигнал пропорционален количеству присутствующего АТР, которое прямо пропорционально числу жизнеспособных клеток в культуре. Данные могут быть зарегистрированы люминомером или визуализирующим устройством с камерой CCD. Выход люминесценции выражают в относительных световых единицах (RLU).
Другим анализом пролиферации клеток является анализ ”MTT”, колориметрический анализ, с помощью которого измеряют окисление бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия в формазан под воздействием митохондриальной редуктазы. Подобно анализу CellTiter-Glo™ указанный анализ позволяет определить число метаболически активных клеток в культуре клеток. См., например, публикации Mosmann (1983) J. Immunol. Meth. 65:55-63 и Zhang et al. (2005) Cancer Res. 65:3877-3882.
В одном из аспектов изобретения антитело против STEAP-1 исследуют в отношении способности индуцировать гибель клетки in vitro. Анализы индукции гибели клетки хорошо известны в данной области. В некоторых вариантах осуществления изобретения такие анализы позволяют измерить, например, утрату целостности мембраны, о чем свидетельствует поглощение иодида пропидия (PI), трипанового синего (см. публикацию Moore et al. (1995) Cytotechnology, 17:1-11) или 7AAD. В соответствии с типичным анализом поглощения PI клетки культивируют в модифицированной по способу Дульбекко среде Игла (D-MEM) и среде Хэма F-12 (50:50), содержащей 10% термоинактивированного FBS (Hyclone) и 2 мМ L-глутамина. Таким образом, указанный анализ выполняют при отсутствии комплемента и иммунокомпетентных эффекторных клеток. Клетки высевают с плотностью 3×106 на одну чашку в чашки размером 100×200 мм и оставляют для прикрепления в течение ночи. Среду удаляют и заменяют свежей средой без добавок или средой, содержащей антитело или иммуноконъюгат в разных концентрациях. Клетки инкубируют в течение 3 дней. После обработки монослои промывают PBS и отделяют, обрабатывая трипсином. Затем клетки центрифугируют со скоростью 1200 оборотов/минуту в течение 5 минут при 4°С, дебрис ресуспендируют в 3 мл холодного Са2+-связывающего буфера (10 мМ Hepes, рН 7,4, 140 мМ NaCl, 2,5 мМ CaCl2) и переносят аликвоту в 35 мм пробирки 12×75 мм с крышкой (1 мл в каждую пробирку, 3 пробирки на одну экспериментальную группу) для удаления скоплений клеток. Затем в пробирки добавляют PI (10 мкг/мл). Образцы анализируют в проточном цитометре FACSCAN™ при помощи программного обеспечения CellQuest FACSCONVERT™ (Becton Dickinson). Антитела или иммуноконъюгаты, индуцирующие гибель клеток на статистически значимых уровнях, идентифицируют по поглощению PI.
В одном из аспектов изобретения антитело против STEAP-1 или иммуноконъюгат исследуют в отношении способности индуцировать апоптоз (запрограммированную смерть клетки) in vitro. Типичным анализом антител или иммуноконъюгатов, индуцирующих апоптоз, является анализ связывания аннексина. При выполнении типичного анализа связывания аннексина клетки культивируют и высевают в чашки, описанные в предыдущем абзаце. Среду удаляют и заменяют свежей средой без добавок или средой, содержащей 0,001-10 мкг/мл антитела или иммуноконъюгата. После инкубации в течение трех дней монослои промывают PBS и отделяют, обрабатывая трипсином. Затем клетки центрифугируют, ресуспендируют в Са2+-связывающем буфере и переносят аликвоту в пробирки, описанные в предыдущем абзаце. Затем в пробирки добавляют меченый аннексин (например, аннексин V-FITC) (1 мкг/мл). Образцы анализируют в проточном цитометре FACSCAN™ при помощи программного обеспечения CellQuest FACSCONVERT™ (BD Biosciences). Идентифицируют антитела или иммуноконъюгаты, которые индуцируют связывание аннексина на статистически значимых уровнях по сравнению с контрольным антителом. Другим типичным анализом антител или иммуноконъюгатов, индуцирующих апоптоз, является колориметрический анализ ДНК гистона способом ELISA, позволяющий обнаружить межнуклеосомное разрушение геномной ДНК. Такой анализ может быть выполнен с использованием набора для обнаружения гибели клеток способом ELISA (Roche, Palo Alto, CA).
Клетки, используемые в любых вышеуказанных анализах in vitro, включают клетки или линии клеток, естественно экспрессирующих STEAP-1, или линии клеток, специально созданные для экспрессии STEAP-1. Такие клетки включают опухолевые клетки, сверхэкспрессирующие STEAP-1 по сравнению с нормальными клетками из такой же ткани. Такие клетки также включают линии клеток (в том числе линии опухолевых клеток), экспрессирующих STEAP-1, и линии клеток, которые не экспрессируют STEAP-1 в естественных условиях, но были трансфицированы нуклеиновой кислотой, кодирующей STEAP-1.
В одном из аспектов изобретения антитело против STEAP-1 или его иммуноконъюгат исследуют в отношении способности ингибировать рост или пролиферацию клеток in vivo. В определенных вариантах осуществления изобретения антитело против STEAP-1 или его иммуноконъюгат исследуют в отношении способности ингибировать рост опухоли in vivo. Для такого исследования могут быть использованы модельные системы in vivo, такие как модели ксенотрансплантатов. В типичной системе ксенотрансплантатов опухолевые клетки человека вводят животному, отличному от человека, с нарушенной иммунологической реактивностью, например, мыши SCID. Антитело и иммуноконъюгат по настоящему изобретению вводят животному. Измеряют способность антитела или иммуноконъюгата ингибировать или уменьшать рост опухоли. В определенных вариантах вышеуказанной системы ксенотрансплантатов опухолевые клетки человека являются опухолевыми клетками, полученными у больного человека. Такие клетки, пригодные для создания моделей ксенотрансплантатов, включают линии опухолевых клеток предстательной железы, легкого или ободочной кишки человека, которые включают, не ограничиваясь ими, клетки РС3, экспрессирующие экзогенный белок STEAP-1, и клетки, экспрессирующие STEAP-1 в естественных условиях, которые включают, не ограничиваясь ими, клетки LnCAP (Southern Research Institute, Birmingham, AL), клетки LuCAP77 и клетки LuCAP35V (University of Washington, Seattle, WA). В определенных вариантах осуществления изобретения опухолевые клетки человека вводят животному, отличному от человека, с нарушенной иммунологической реактивностью при помощи подкожной инъекции или трансплантации в приемлемый участок тела, такой как жировое тело молочной железы.
Анализы связывания и другие анализы
В одном из аспектов изобретения антитело против STEAP-1 исследуют в отношении антигенсвязывающей активности. Например, в определенных вариантах осуществления изобретения, антитело против STEAP-1 исследуют в отношении способности связываться с экзогенным или эндогенным белком STEAP-1, экспрессированным на поверхности клетки. Такое исследование может быть выполнено с помощью анализа FACS.
В одном из аспектов изобретения для идентификации моноклонального антитела, конкурирующего за связывание с белком STEAP-1 с антителом 120 graft или его гуманизированными вариантами, которые включают, не ограничиваясь им, антитело 120v.24, могут быть использованы конкурентные анализы. В определенных вариантах осуществления изобретения такое конкурирующее антитело связывается с тем же эпитопом (например, пептидом линейного эпитопа или конформационным эпитопом, образованным экспрессией STEAP-1 на поверхности клетки), с которым связывается антитело 120 graft или гуманизированное антитело 120 graft, включая вариант 120v.24 гуманизированного антитела против STEAP-1. Типичные конкурентные анализы включают, не ограничиваясь ими, стандартные анализы, описанные в публикации Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Типичные способы картирования эпитопа, с которым связывается антитело, подробно описаны в публикации Morris (1996) ”Epitope Mapping Protocols,” in Methods in Molecular Biology, vol. 66 (Humana Press, Totowa, NJ). Считается, что два антитела связываются с одним и тем же эпитопом, если каждое антитело блокирует связывание другого антитела на 50% или больше.
При выполнении типичного конкурентного анализа иммобилизованный белок STEAP-1 инкубируют в растворе, содержащем первое меченое антитело, связывающееся с STEAP-1, (например, антитело мыши 120.545, антитело 120 graft или гуманизированное антитело 120v.24) и второе немеченое антитело, исследуемое в отношении способности конкурировать с первым антителом за связывание с белком STEAP-1. Второе антитело может присутствовать в супернатанте гибридом. В качестве контрольного образца иммобилизованный белок STEAP-1 инкубируют в растворе, содержащем первое меченое антитело и не содержащем второе немеченое антитело. После инкубации в условиях, допускающих связывание первого антитела с белком STEAP-1, удаляют избыток несвязанного антитела и измеряют количество меток, связанных с иммобилизованным белком STEAP-1. Значительно меньшее количество меток, связанных с иммобилизованным STEAP-1, в исследуемом образце по сравнению с контрольным образцом указывает на то, что второе антитело конкурирует с первым антителом за связывание с STEAP-1. В определенных вариантах осуществления изобретения имммобилизованный STEAP-1 присутствует на поверхности клетки или в мембранном препарате, полученном из клетки, экспрессирующей STEAP-1 на своей поверхности.
В одном из аспектов изобретения очищенные антитела против STEAP-1 могут быть далее исследованы при выполнении серии анализов, которые включают, не ограничиваясь ими, секвенирование N-конца, аминокислотный анализ, неденатурирующую вытеснительную жидкостную хроматографию высокого давления (ВЭЖХ), масс-спектрометрию, ионообменную хроматографию и расщепление папаином.
Один вариант осуществления изобретения относится к измененному антителу, обладающему некоторыми, но не всеми эффекторными функциями, что делает такое антитело желаемым кандидатом для многих применений, в которых время полужизни антитела in vivo имеет важное значение, а определенные эффекторные функции (такие как комплемент и ADCC) являются ненужными или вредными. В определенных вариантах осуществления изобретения активность Fc-области антитела изменяют, чтобы удостовериться в сохранении только требуемых свойств. Анализы цитотоксичности in vitro и/или in vivo могут быть выполнены для подтверждения ослабления/устранения активностей CDC и/или ADCC. Например, анализы связывания с Fc-рецептором могут быть выполнены для гарантии того, что данное антитело не связывается с FcγR (вероятно, вследствие отсутствия активности ADCC), но сохраняет способность связываться с FcRn. Главные клетки, опосредующие ADCC, NK-клетки, экспрессируют только Fc(RIII, в то время как моноциты экспрессируют Fc(RI, Fc(RII и Fc(RIII. Экспрессия FcR гемопоэтическими клетками суммирована в таблице 3 на странице 464 публикации Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Пример анализа in vitro для оценки активности ADCC представляющей интерес молекулы приведен в патенте США № 5500362 или 5821337. Приемлемыми эффекторными клетками для таких анализов являются мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). Альтернативно или дополнительно активность ADCC представляющей интерес молекулы можно оценить in vivo, например, в животной модели, представленной в публикации Clynes et al. PNAS (USA) 95:652-656 (1998). Также могут быть выполнены анализы связывания C1q для подтверждения того, что данное антитело не способно связываться с C1q и, следовательно, не обладает активностью CDC. Для оценки активации комплемента может быть выполнен анализ CDC, например, описанный в публикации Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Связывание FcRn и клиренс/время полужизни in vivo также можно определить способами, известными в данной области.
Примеры
Ниже приведены примеры способов и композиций по настоящему изобретению. Предполагается, что на основании приведенного выше общего описания могут быть выполнены разные другие варианты осуществления изобретения.
Пример 1. Получение гуманизированных антител против STEAP-1
Молекулы нуклеиновых кислот, кодирующих варианты аминокислотной последовательности антитела, фрагмента антитела, VL домена или VH домена получают разными способами, известными в данной области. Указанные способы включают, не ограничиваясь ими, выделение из природного источника (в случае вариантов природной аминокислотной последовательности) или получение при помощи мутагенеза с использованием олигонуклеотидов (или сайтнаправленного мутагенеза), мутагенеза способом ПЦР и кластерного мутагенеза ранее полученного варианта антитела или антитела, не являющегося вариантом, фрагмента антитела, VL домена или VH домена. Например, могут быть созданы библиотеки для направленного воздействия на положения доступных аминокислот в VH домене и необязательно в одной или нескольких CDR-областях с целью замены аминокислот вариантными аминокислотами по способу Кункела. См., например, публикацию Kunkel et al., Methods Enzymol. (1987), 154:367-382 и примеры, приведенные в указанной публикации. В разделе ”Примеры” также описано создание рандомизированных последовательностей.
Последовательность олигонуклеотидов включает один или несколько созданных наборов кодонов для определенного положения в CDR-области (HVR) или остовной области (FR) полипептида по настоящему изобретению. Набор кодонов представляет собой набор разных последовательностей нуклеотидных триплетов, используемых для кодирования требуемых вариантных аминокислот. Наборы кодонов могут быть обозначены символами для определения соответствующих нуклеотидов или эквимолярных смесей нуклеотидов, как показано ниже согласно коду IUB.
Коды IUB
G Гуанин
А Аденин
Т Тимин
С Цитозин
R (A или G)
Y (C или Т)
М (А или С)
К (G или Т)
S (C или G)
W (А или Т)
Н (А, С или Т)
В (С, G или Т)
V (А, С или G)
D (А, G или Т)
N (А, С, G или Т)
Например, в наборе кодонов DVK D может быть нуклеотидами А, G или Т; V может быть А, G или С; и К может быть G или Т. Указанный набор кодонов может представлять 18 разных кодонов и может кодировать аминокислоты Ala, Trp, Tyr, Lys, Thr, Asn, Lys, Ser, Arg, Asp, Glu, Gly и Cys.
Наборы олигонуклеотидов или праймеров могут быть синтезированы стандартными способами. Набор олигонуклеотидов может быть синтезирован, например, при помощи твердофазного синтеза, и может содержать последовательности, представляющие все возможные комбинации нуклеотидных триплетов, образуемые данным набором кодонов, и кодирующие требуемую группу аминокислот. Синтез олигонуклеотидов с ”вырожденностью” выбранных нуклеотидов в определенных положениях хорошо известен в данной области. Такие наборы нуклеотидов, содержащие определенные наборы кодонов, могут быть синтезированы в коммерческих синтезаторах нуклеиновых кислот (например, компании Applied Biosystems, Foster City, CA) или могут быть приобретены коммерческим путем (например, в компании Life Technologies, Rockville, MD). Поэтому набор синтезированных олигонуклеотидов, содержащих определенный набор кодонов, обычно включает множество олигонуклеотидов с разными последовательностями, при этом различия определяются набором кодонов во всей последовательности. Олигонуклеотиды по настоящему изобретению имеют последовательности, способные гибридизировать с матрицей нуклеиновой кислоты вариабельного домена, а также могут включать сайты рестрикционных ферментов в целях клонирования.
В соответствии с одним способом по настоящему изобретению последовательности нуклеиновых кислот, кодирующие вариантные аминокислоты, могут быть созданы сайтнаправленным мутагенезом с использованием олигонуклеотидов. Указанный способ хорошо известен в данной области и описан в публикации Zoller et al., 1987, Nucleic Acids Res. 10:6487-6504. Последовательности нуклеиновых кислот, кодирующие вариантные аминокислоты, создают путем гибридизации набора олигонуклеотидов, кодирующих требуемые наборы кодонов, с матрицей ДНК, при этом указанная матрица является одноцепочечной формой плазмиды, содержащей последовательность матрицы нуклеиновой кислоты вариабельной области. После гибридизации используют ДНК-полимеразу для синтеза всей второй комплементарной цепи матрицы, которая включает олигонуклеотидный праймер и будет содержать наборы кодонов, предоставленные набором олигонуклеотидов.
Обычно используют олигонуклеотиды, состоящие по меньшей мере из 25 нуклеотидов. Оптимальный олигонуклеотид содержит 12-15 нуклеотидов, которые полностью комплементарны матрице с любой стороны нуклеотида, кодирующего мутации. Указанный способ гарантирует, что олигонуклеотид будет правильно гибридизировать с молекулой матрицы одноцепочечной ДНК. Олигонуклеотиды можно легко синтезировать способами, известными в данной области, которые описаны в публикации Crea et al., Proc. Nat’l. Acad. Sci. USA, 75:5765 (1978).
Матрица ДНК образована векторами, выделенными из векторов на основе бактериофага М13 (пригодны коммерчески доступные векторы M13mp18 и M13mp19), или векторами, содержащими ориджин репликации одноцепочечного фага, которые описаны в публикации Viera et al., Meth. Enzymol., 153:3 (1987). Таким образом, подлежащая мутации ДНК может быть введена в один из указанных векторов с целью создания одноцепочечной матрицы. Создание одноцепочечной матрицы описано в разделах 4.21-4.41 приведенной выше публикации Sambrook et al.
Для изменения нативной последовательности ДНК олигонуклеотид гибридизируют с одноцепочечной матрицей в приемлемых условиях гибридизации. Затем добавляют ДНК-полимеризующий фермент, обычно ДНК-полимеразу Т7 или фрагмент Кленова ДНК-полимеразы I, для синтеза комплементарной цепи матрицы с использованием олигонуклеотида в качестве праймера для синтеза. Гетеродуплексная молекула получена таким образом, что одна цепь ДНК кодирует мутированную форму гена 1, а другая цепь (исходная матрица) кодирует нативную, неизмененную последовательность гена 1. Указанную гетеродуплексную молекулу затем переносят в приемлемую клетку-хозяина, обычно в прокариотическую клетку, такую как E. coli JM101. Выращенные клетки культивируют на агарозных пластинках и исследуют с использованием олигонуклеотидного праймера, меченного радиоактивным изотопом 32-фосфата, для идентификации колоний бактерий, содержащих мутированную ДНК.
Вышеописанный способ может быть модифицирован с созданием гомодуплексной молекулы, в которой обе цепи плазмиды содержат мутации. Модификации выполняют следующим образом. Одноцепочечный олигонуклеотид гибридизируют с одноцепочечной матрицей, как было описано выше. Смесь трех дезоксирибонуклеотидов, а именно дезоксирибоаденозина (dATP), дезоксирибогуанозина (dGTP) и дезоксириботимидина (dTT), объединяют с модифицированным тиодезоксирибоцитозином, именуемым dCTP-(aS) (который может быть приобретен в компании Amersham). Указанную смесь вводят в коплекс матрицы с олигонуклеотидом. После добавления к указанной смеси ДНК-полимеразы образуется цепь ДНК, идентичная матрице, за исключением мутированных оснований. Кроме того, новая цепь ДНК будет содержать dCTP-(aS) вместо dCTP, что позволяет защитить ДНК от расщепления рестрикционной эндонуклеазой. После лигирования цепи матрицы двухцепочечного гетеродуплекса с соответствующим рестрикционным ферментом указанная цепь матрицы может быть расщеплена нуклеазой ExoIII или другой приемлемой нуклеазой после области, содержащей один или несколько сайтов, подвергаемых мутагенезу. Затем взаимодействие прекращают, получая при этом молекулу, которая лишь частично является одноцепочечной. Затем создают гомодуплекс полной двухцепочечной ДНК с использованием ДНК-полимеразы в присутствии трифосфатов всех четырех дезоксирибонуклеотидов, АТР и ДНК-лигазы. Молекула гомодуплекса затем может быть введена в приемлемую клетку-хозяина.
Как было указано выше, последовательность набора олигонуклеотидов имеет достаточную длину для гибридизации с нуклеиновой кислотой матрицы и может также, хотя и необязательно, содержать сайты рестрикции. Матрица ДНК может быть создана векторами, полученными из векторов на основе бактериофага М13 или векторов, содержащих ориджин репликации одноцепочечного фага, как описано в публикации Viera et al. (1987) Meth. Enzymol., 153:3. Таким образом, ДНК, которая должна быть мутирована, необходимо ввести в один из указанных векторов для создания одноцепочечной матрицы. Создание одноцепочечной матрицы описано в разделах 4.21-4.41 приведенной выше публикации Sambrook et al.
В соответствии с другим способом может быть создана библиотека, включающая наборы верхних и нижних олигонуклеотидов, при этом каждый набор содержит множество олигонуклеотидов с разными последовательностями, определяемыми наборами кодонов в последовательности олигонуклеотидов. Наборы верхних и нижних олигонуклеотидов наряду с последовательностью нуклеиновой кислоты матрицы вариабельного домена могут быть использованы при выполнении полимеразной цепной реакции для создания ”библиотеки” продуктов ПЦР. Продукты ПЦР могут быть определены как ”кассеты нуклеиновых кислот”, так как они могут быть гибридизированы с другими родственными или неродственными последовательностями нуклеиновых кислот, например, с белками оболочки вируса и доменами димеризации при помощи известных способов молекулярной биологии.
Наборы олигонуклеотидов могут быть использованы в полимеразной цепной реакции с использованием последовательности матрицы нуклеиновой кислоты вариабельного домена в качестве матрицы для создания кассеты нуклеиновых кислот. Последовательность матрицы нуклеиновой кислоты вариабельного домена может быть любой частью тяжелых цепей иммуноглобулина, содержащих последовательности-мишени нуклеиновых кислот (то есть последовательности нуклеиновых кислот, кодирующие аминокислоты, предназначенные для замены). Последовательность матрицы нуклеиновой кислоты вариабельной области является частью молекулы двухцепочечной ДНК, содержащей цепь первой нуклеиновой кислоты и комплементарную цепь второй нуклеиновой кислоты. Последовательность матрицы нуклеиновой кислоты вариабельного домена содержит по меньшей мере часть вариабельного домена и имеет по меньшей мере одну CDR-область. В некоторых случаях последовательность матрицы нуклеиновой кислоты вариабельного домена содержит несколько CDR-областей. Верхняя и нижняя части последовательности матрицы нуклеиновой кислоты вариабельного домена могут быть гибридизированы с членами набора верхних олигонуклеотидов и набора нижних олигонуклеотидов.
Первый олигонуклеотид набора верхних праймеров может гибридизровать с цепью первой нуклеиновой кислоты и второй олигонуклеотид набора нижних праймеров может гибридизировать с цепью второй нуклеиновой кислоты. Олигонуклеотидные праймеры могут включать один или несколько наборов кодонов и могут гибридизировать с частью последовательности матрицы нуклеиновой кислоты вариабельной области. С помощью указанных олигонуклеотидов можно ввести два или более наборов кодонов в продукт ПЦР (то есть кассета нуклеиновых кислот) после выполнения ПЦР. Олигонуклеотидный праймер, гибридизирующая с областями последовательности нуклеиновой кислоты, кодирующей вариабельный домен антитела, включает части, кодирующие остатки CDR, предназначенные для замены аминокислот.
Кроме того, могут быть синтезированы наборы верхних и нижних олигонуклеотидов, включающие сайты рестрикции в олигонуклеотидной последовательности. Указанные сайты рестрикции могут облегчить введение кассет нуклеиновых кислот (то есть продуктов реакции ПЦР) в вектор экспрессии, содержащий дополнительную последовательность антитела. В одном из вариантов осуществления изобретения сайт рестрикции предназначен для облегчения клонирования кассет нуклеиновых кислот без введения инородных последовательностей нуклеиновых кислот или удаления последовательностей нуклеиновых кислот исходной CDR-области или остовной области.
Кассеты нуклеиновых кислот могут быть клонированы в любой приемлемый вектор для экспрессии части или всей последовательности легкой или тяжелой цепи, содержащей замены аминокислот, выполненные при помощи ПЦР. В соответствии с способами, подробно описанными в настоящем изобретении, кассету нуклеиновых кислот клонируют в вектор, позволяющий создать часть или всю последовательность легкой или тяжелой цепи, гибридизированную со всем или частью белка оболочки вируса (то есть создание слитого белка) и отображенную на поверхности частицы или клетки. Хотя существуют векторы нескольких типов, которые могут быть использованы при осуществлении настоящего изобретения, фагомидные векторы являются более предпочтительными векторами, так как указанные векторы могут быть относительно легко созданы и легко амплифицированы. Фагомидные векторы обычно содержат разные компоненты, включающие промоторы, сигнальные последовательности, гены фенотипической селекции, сайты ориджина репликации и другие необходимые компоненты, известные специалистам в данной области.
Когда необходимо экспрессировать определенную комбинацию вариантных аминокислот, кассета нуклеиновых кислот содержит последовательность, способную кодировать весь или часть вариабельного домена тяжелой или легкой цепи и комбинации вариантных аминокислот. Для получения антител, содержащих указанные вариантные аминокислоты или комбинации вариантных аминокислот, так же, как в библиотеке, кассеты нуклеиновых кислот могут быть введены в вектор экспрессии, содержащий дополнительную последовательность антитела, например, все или части вариабельных или константных доменов вариабельных областей легкой и тяжелой цепей. Указанные дополнительные последовательности антитела могут быть также гибридизированы с другими последовательностями нуклеиновых кислот, которые кодируют белки оболочки вируса и поэтому позволяют продуцировать слитый белок.
В настоящем описании изобретения рассмотрен процесс гуманизации антитела мыши против STEAP-1 человека.
Материалы и способы
Номера остатков соответствуют нумерации Kabat (Kabat et al., Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Для обозначения аминокислот использованы однобуквенные аббревиатуры. Вырожденность ДНК представлена с использованием кода IUB (N=A/C/G/T, D=A/G/T, V=A/C/G, B=C/G/T, H=A/C/T, K=G/T, M=A/C, R=A/G, S=G/C, W=A/T, Y=C/T).
Клонирование вариабельных доменов антитела мыши 120 и создание химерного антитела 120. В результате экстрагирования полной РНК из клеток гибридом стандартными способами было получено антитело М2-120.545 (определяемое в настоящем описании изобретения как ”мышиное антитело 120” ил “mu120”). Вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH) амплифицировали при помощи RT-ПЦР с вырожденными праймерами к тяжелой и легкой цепям. Верхние праймеры были специфичны к N-концевой аминокислотной последовательности VL и VH областей. Соответственно, нижние праймеры к LC и HC были созданы для гибридизации с областью в константном домене легкой цепи (CL) и константном домене 1 тяжелой цепи (СН1), которые являются наиболее консервативными во всех видах. Амплифицированные VL и VH клонировали в векторы экспрессии млекопитающих. Полинуклеотидную последовательность вставок определяли стандартными способами секвенирования. Аминокислотные последовательности VL и VH антитела М2-120.545 (”mu 120”) показаны соответственно на фигурах 2А и 2В.
Создание мышиного химерного антитела 120. Химерное антитело против STEAP-1 было получено путем гибридизации вариабельных областей тяжелой цепи (VH) и вариабельных областей легкой цепи (VL) мышиного антитела 120 с константными доменами IgG человека. Полученное антитело определяется в настоящем описании изобретения как ”120 chimera”, “chimera 120” “химерный IgG 120” или “Fc chimera”.
Прямое введение трансплантатов гипервариабельных участков в консенсусную остовную область акцептора человека. Варианты, созданные в процессе гуманизации мышиного антитела 120, оценивали в виде белка в форме IgG или Fab-фрагмента, отображенного на фаге.
Фагомид, использованный в данном исследовании, является вектором для отображения моновалентного Fab-g3 и состоит из двух открытых рамок считывания, контролируемых промотором phoA. Первая открытая рамка считывания состоит из сигнальной последовательности stII, слитой с VL и CHI доменами легкой цепи акцептора, и вторая открытая рамка считывания состоит из сигнальной последовательности stII, слитой с VH и CH1 доменами тяжелой цепи акцептора, и минорного белка Р3 оболочки фага.
VL и VH домены из мышиного антитела 120 сравнивали с консенсусными последовательностями каппа-цепи I VL (huKI) и подгруппы III VH человека (huIII). Для создания трансплантатов CDR гипервариабельные участки из мышиного антитела 120 вводили в остовные области акцептора huKI и huIII.
Гипервариабельные участки из мышиного антитела 120 (mu120) были введены в консенсусную остовную область акцептора человека для создания прямого CDR-трансплантата (обозначенного в настоящем описании изобретения как ”120 graft” или ”graft 120”). В VL домене следующие области были трансплантированы в консенсусную основную область акцептора человека положения 24-34 (L1), 50-56 (L2) и 89-97 (L3). В VH домене трансплантат был введен в положения 26-35а (Н1), 49-65 (Н2) и 95-102 (H3). Последовательности вариабельных областей легкой и тяжелой цепей антитела 120 graft показаны на фигурах 2А-2В. CDR-области (также обозначенные в настоящем описании как HVR) показаны в рамках (фигуры 2А-2В). Указанные определения CDR включают положения, определяемые гипервариабельностью последовательностей (публикация Kabat), структурной локализацией (публикация Chotia) и их участием в контактировании антигена с антителом (MacCallum et al. J. Nol. Biol. 262:732-745 (1996)).
Прямые трансплантированные варианты, экспрессированные в виде Fab-фрагмента, отображенного на фаге, или в виде IgG, были созданы при помощи мутагенеза Кункела с использованием отдельного олигонуклеотида для каждого гипервариабельного участка. Правильные клоны определяли способом секвенирования ДНК.
Создание гуманизированных вариантов антитела 120 на фаге. Гуманизированные варианты антитела 120 были созданы в виде Fab-фрагмента, отображенного на фаге, при помощи мутагенеза Кункела. Фосфорилированный олигонуклеотид добавляли к 300 нг матрицы Кункела в 50 мМ трис-буфера, рН 7,5, 10 мМ MgCl2 в конечном объеме, равном 10 мкл. Смесь гибридизировали при 90°С в течение 2 минут, при 50°С в течение 5 минут и затем охлаждали на льду. Гибридизированную матрицу заполняли, добавляя 0,5 мкл 10 мМ АТР, 0,5 мкл 10 мМ dNTP (по 10 мМ каждого dATP, dCTP, dGTP и dTTP), 1 мкл 100 мМ DTT, 1 мкл 10-кратного буфера ТМ (0,5 М трис-буфера, рН 7,5, 0,1 М MgCl2), 80 ед. лигазы Т4 и 4 ед. полимеразы Т7 в общем объеме, равном 20 мкл, в течение 2 часов при комнатной температуре. Заполненный и лигированный продукт трансфицировали в клетки XL1-blue (Stratagene). Правильные клоны идентифицировали способом секвенирования ДНК.
Правильные фаговые клоны выращивали в 25 мл среды 2YT, содержащей 50 мкг/мл карбенациллина и фага-помощника М13/КО7 (MOI 10), в течение ночи при 37°С.
Оценка гуманизированных вариантов антитела 120. Гуманизированные варианты, экспрессированные в виде IgG, оценивали при помощи анализа FACS с использованием Steap1-положительных (293 Steap1 NT LB50) и Steap1-отрицательных (293 vector S408) линий клеток.
Гуманизированные варианты, экспрессированные в виде Fab-фрагмента, отображенного на фаге, также оценивали при помощи анализа FACS. У вариантов в виде Fab-фрагментов, отображенных на фаге, сначала оценивали уровень отображения Fab-фрагмента при помощи анализа ELISA, позволяющего обнаружить метку FLAG, гибридизированную с легкой цепью Fab-фрагмента. Титрационные микропланшеты MaxSorp сенсибилизировали антителом против gD 1766 в количестве 10 мкг/мл в PBS в течение ночи и затем блокировали казеиновым блокатором. Фаг из супернатантов культуры последовательно разводили в PBST, содержащем 0,5% BSA, на титрационном микропланшете с культурой ткани и переносили в сенсибилизированные лунки на 1 час для улавливания Fab-отображающего фага. Планшет промывали PBST, добавляли HRP-конъюгированное антитело против М13 (Amersham Pharmacia Biotech) (1:5000 в PBST, содержащем 0,5% BSA) и оставляли на 40 минут. Планшет промывали PBST и проявляли, добавляя тетраметилбензидиновый субстрат (Kirkegaard and Perry Laboratories, Gaithersburg, MD). Уровень отображения Fab-фрагмента на поверхности фага определяли на основании оптической плотности при длине волны 405 нм. Фаговые препараты нормализовали в отношении отображения разведением. При выполнении анализа FACS использовали фаг с низкой отображающей способностью (например, химеру).
При выполнении анализа связывания фага способом FACS клетки удаляли с планшета, используя 2 мМ EDTA, собирали в 15 мл пробирку с коническим основанием и центрифугировали. Клетки (5×105 клеток в одном образце) ресуспендировали в 100 мкл фага (нормализованного в отношении уровня отображения) в буфере для FACS (1% FBS, PBS c 2 мМ EDTA) и инкубировали в течение 1-2 часов на льду. Образцы дважды промывали буфером для FACS и центрифугировали. Добавляли контрольное антитело против М13 5G7 (Genentech, Inc. South San Francisco, CA) в количестве 2 мкг/мл и инкубировали на льду в течение по меньшей мере 45 минут. Образцы дважды промывали буфером для FACS и центрифугировали. Добавляли антитело против РЕ мыши (R-фикоэритрин козы против Fcy-фрагмента IgG мыши), разведенное в отношении 1:200 и инкубировали на льду в течение 30 минут. Образцы еще раз дважды промывали буфером для FACS, центрифугировали и анализировали способом FACS.
Для анализа IgG способом FACS клетки получали так же, как при выполнении анализа фага способом FACS. Каждый IgG добавляли в количестве 5 мкг/мл на льду и оставляли на 1 час. Образцы дважды промывали буфером для FACS, центрифугировали, добавляли конъюгат антитела против РЕ человека (R-фикоэритрин козы против Fcy-фрагмента IgG человека, Jackson Immunoresearch) и оставляли на 30 минут. Образцы еще раз промывали буфером для FACS, центрифугировали и анализировали способом FACS.
Получение IgG и определение аффинности. IgG очищали аффинной хроматографией с протеином G. Аффинность определяли при помощи анализа Скэтчарда в клетках 293 STEAP-1 NT LB50.
Результаты и их рассмотрение
Создание последовательностей вариабельного домена мышиного антитела 120 и трансплантата CDR. Остовная область акцептора человека, используемая для гуманизации антитела М2-120.545, получена на основе VL домена консенсусной последовательности каппа-цепи I человека и VH домена консенсусной последовательности подгруппы III человека. VL и VH домены мышиного антитела М2-120.545 сравнивали с доменами каппа-цепи I и подгруппы III; каждую область, определяющую комплементарность, (CDR) идентифицировали и трансплантировали в остовную область акцептора человека с целью создания трансплантата CDR, который мог быть отображен в виде Fab-фрагмента на фаге и экспрессирован в виде IgG. Последовательности 24 вариабельных областей гуманизированного варианта антитела против STEAP-1 показаны на фигурах 2А и 2В. Fab-фрагмент антитела 120 graft, отображенный на фаге, и IgG антитела 120 graft исследовали в отношении связывания с клетками (293 STEAP-1 NT LB50), экспрессирующими экзогенный белок STEAP-1, при помощи анализа способом FACS. Хотя IgG антитела 120 graft специфически связывался с STEAP-1-экспрессирующими клетками, сигнал FACS, полученный для IgG антитела 120 graft, был слабее сигнала, полученного для IgG химерного антитела 120, что указывает на утрату аффинности связывания. Фаг, отображающий Fab-фрагмент антитела 120 graft, также генерировал сигнал FACS, который имел место только в STEAP-1-экспрессирующих клетках. Указанное изменение было меньше изменения, наблюдаемого для IgG химерного антитела 120. Результат анализа Скэтчарда IgG антитела 120 graft также свидетельствовал о значительной (примерно 5-кратной) утрате аффинности связывания (KD = 36 нМ для антитела 120v.78; KD = 260 нМ для антитела 120 graft).
Гуманизация антитела М2-120.545. Было идентифицировано примерно 30 верньерных положений, влияющих на конформацию CDR и упаковку VL:VH доменов, и в процессе гуманизации антител необходимо учитывать различия в указанных положениях между остовными областями донора и человека (Foote, J. and Winter, G., J. Mol. Biol. 224(2): 487-499 (1992)). Сравнение мышиного антитела М2-120.545 с VL доменом консенсусной последовательностип каппа-цепи I человека и VH доменом консенсусной последовательности подгруппы III человека позволило выявить различия последовательностей в 6 основных верньерных положениях VH домена: 24, 37, 48, 67, 73, 78 (см. фигуру 2В). Для оценки влияния указанных положений мышиные остатки вводили в VH домен консенсусной последовательности подгруппы III Fab-фрагмента на фаге. Данный процесс предполагает выполнение следующих мутаций в Fab-фрагменте антитела 120 graft, отображенном на фаге: A24V (120.v24), V37I (120.v37), V48M (120.v48), F67I (120.v67) и L78F (120.v78). N73T не исследовали. Каждый вариант фага нормализовали путем разведения до эквивалентного уровня отображения Fab-фрагмента, определенного путем титрования метки эпитопа, слитой с легкой цепью, отображенной на фаге, и затем оценивали в отношении связывания с STEAP-1 при помощи анализа FACS в STEAP-1-экспрессирующих клетках (293 STEAP-1 NT LB50) и в клетках, не экспрессирующих STEAP-1 (293 vector S408). Термин ”2°” означает вторичное антитело в анализе FACS. Термин ”α-120” означает мышиное антитело 120 против STEAP-1. Термин ”α-10Н1” означает контрольное антитело. Термины ”24 Phage”, “37 Phage” и тому подобные означают гуманизированные варианты антитела против STEAP-1 по настоящему изобретению, отображенные на фаге. Термин “Ch 120 Phage” означает химерное антитело 120, отображенное на фаге, и термин ”120.graft Phage” означает антитело 120 graft, отображенное на фаге. (Фигура 6). Значение нормализации фаговых клонов на основании уровня отображения Fab-фрагмента проиллюстрировано анализом FACS антитела 120 graft при разных титрах фага: 7×1012 фага/мл на фигуре 6 и 2×1011 фага/мл на фигуре 6. После разведения до более низкой концентрации фага фаг с антителом 120 graft не вызывает изменения, детектируемого способом FACS. Таким образом, нормализация разных фаговых клонов в отношении уровня отображения была важной стадией оценки различий в аффинности к белку Steap1.
После нормализации в отношении уровней отображения Fab-фрагмента вариант антитела 120 graft, содержащий дополнительную мутацию A24V (120.v24), вызвал изменение показателей FACS, превосходящее другие варианты (фигура 6). При экспрессии в виде IgG антитело 120.v24 вызвало такое же изменение показателей FACS, что и химерное антитело 120 при всех исследованных концентрациях. В последующем анализе Скэтчарда антитела 120.v24 было получено значение Kd 2,2 нМ для связывания с клетками 293 STEAP-1 NT LB50, что является двукратным улучшением по сравнению с химерным антителом 120 и исходным мышиным антителом М2-120.545 (таблица 2).
Аффинность связывания антитела против STEAP-1 к белку STEAP-1 на поверхности клетки (Kd (нМ))
187256 сайтов в одной клетке
103204 сайта в одной клетке
301.100 сайтов в одной клетке
252892 сайта в одной клетке
264172 сайта в одной клетке
37207 сайтов в одной клетке
22021 сайт в одной клетке
Связывающую активность “голых” антител против STEAP-1, антитела 120 мыши и химерного антитела 120 также исследовали при помощи анализа FACS. Связывание сравнивали для экзогенного белка STEAP-1 в устойчивых клетках 293 STEAP-1 NT LB50 и устойчивых клетках РС3 STEAP-1 PS5.4 и для эндогенного белка STEAP-1 в клетках LNCaP. Результаты также показаны на фиг.7D-7F. Клетки NT LB50, экспрессирующие экзогенный белок STEAP-1 человека на поверхности клетки, были получены в результате устойчивой трансформации клеток 293 (АТСС CRL-1573) при помощи ДНК STEAP-1 человека. Клетки PS5.4, экспрессирующие экзогенный белок STEAP-1 человека на поверхности клетки, были получены в результате устойчивой трансформации РС3 (АТСС CLL-1435) при помощи ДНК STEAP-1 человека. Клетки LNCaP (АТСС CRL-1740) экспрессируют эндогенный STEAP-1.
Пример 2. Исследование антител против STEAP-1
Антитела против STEAP-1 (“голые” антитела и конъюгаты антитела с лекарственным средством, рассмотренные в настоящем описании изобретения) исследовали стандартными способами.
Анализы способом ELISA. Антитела против STEAP-1 исследовали при помощи анализа ELISA, выполняя все инкубации при комнатной температуре. Аналитические планшеты (Nunc Immunoplate) сенсибилизировали в течение 2 часов очищенным белком STEAP-1 в 50 мМ натрий-карбонатного буфера, рН 9,6, затем блокировали 0,5% бычьего сывороточного альбумина в физиологическом растворе с фосфатным буфером (PBS) в течение 30 минут и четыре раза промывали PBS, содержащим 0,05% твина-20 (PBST). Добавляли супернатанты исследуемого антитела и инкубировали, встряхивая, в течение двух часов, затем четыре раза промывали PBST. Планшеты проявляли, добавляя 100 мкл/лунку раствора, содержащего 10 мг дигидрохлорида о-фенилендиамина (Sigma, № Р8287) и 10 мкл 30% раствора пероксида водорода в 25 мл фосфат-цитратного буфера, рН 5,0, и инкубировали в течение 15 минут. Взаимодействие прекращали, добавляя 100 мкл/лунку 2,5 М раствора серной кислоты. Данные получены в результате считывания планшетов в автоматическом аппарате для чтения планшетов ELISA при оптической плотности, равной 490 нм.
Исследование связывания антитела против STEAP-1 при помощи анализа Скэтчарда
Аффинность связывания моноклонального антитела можно определить, например, при помощи анализа Скэтчарда, описанного в публикации Munson et al., Anal. Biochem., 107:220 (1980), выполняемого стандартными способами, хорошо известными в данной области. См. также публикацию Scatchard, G., Ann. N.Y. Acad. Sci. 51:660 (1947).
Пример 3. Получение конъюгатов антитела против STEAP-1 с лекарственным средством
Получение ADC антитела против STEAP-1 с ауристатином. ADC антитела против STEAP-1 были получены в результате конъюгирования антител против STEAP-1, включающих антитело мыши 120.545, химерное антитело 120, антитело 120 graft и гуманизированные варианты остовной области антитела 120, со следующими частями лекарственного средства-линкера: spp-DM1, smcc-DM1, MC-vc-PAB-MMAE, MC-vc-PAB-MMAF, MC-MMAE, MC-MMAF, vc-MMAE и vc-MMAF, которые наряду со способами их присоединения рассмотрены в настоящем описании изобретения, а также в заявке WO 2004/010957, опубликованной 5 февраля 2004 г., заявке WO 2006/034488, опубликованной 9 сентября 2005 г. и в публикации Doronina, S.O. et al., Nature Biotechnol. 21:778-784 (2003) (все публикации включены в настоящее описание изобретения в качестве ссылки в полном объеме). До конъюгирования антитела частично восстанавливали ТСЕР стандартными способами в соответствии с способикой, описанной в заявке WO 2004/010957. Частично восстановленные антитела конъюгировали с вышеуказанными частями лекарственного средства-линкера стандартными способами в соответствии с способикой, описанной в публикации Doronina et al. (2003) Nat. Biotechnol. 21:778-784 и в заявке на патент США 2005/0238649 А1. Частично восстановленные антитела объединяли с частями лекарственного средства-линкера с достижением конъюгирования указанных частей с остатками цистеина. Реакции конъюгирования гасили и конъюгаты антитела с лекарственным средством очищали. Лекарственную нагрузку (среднее число лекарственных частей в одном антителе) для каждого ADC определяли при помощи ВЭЖХ. В настоящем описании изобретения компонент линкера-лекарственного средства в ADC ”-MC-vc-PAB-MMAE” или “-MC-vc-PAB-MMAF” иногда обозначен аббревиатурой ”-vcMMAE” или “-vcMMAF” и компонент “-MC-MMAF” иногда обозначен аббревиатурой “MCMMAF” или “mcMMAF”.
Получение ADC антитела против STEAP-1 с майтанзиноидом. ADC антитела против STEAP-1 были получены в результате конъюгирования антител против STEAP-1, включающих мышиное антитело 120, химерное антитело 120 chimera, антитело 120 graft и гуманизированные варианты остовной области антитела 120, с частью линкера-лекарственного средства -smcc-DM1. Такое конъюгирование можно выполнить способом, описанным в заявке WO 2005/037992 для конъюгирования антитела против HER2 герцептина®.
Пример 4. Анализ уменьшения объема опухоли in vivo
Способность конъюгированных с токсином или неконъюгированных моноклональных антител против STEAP-1 эффективно уменьшать объем опухоли in vivo и in vitro исследовали в соответствии со следующим протоколом.
Линии клеток млекопитающих и ксенотрансплантаты опухоли человека: клетки 293 представляют собой линию иммортализованных клеток почки эмбриона человека (АТСС, № CRL1573), клетки РС-3 представляют собой линию клеток аденокарциномы предстательной железы (АТСС, № CRL1435) и клетки LNCaP представляют собой линию клеток карциномы предстательной железы (АТСС, № CRL1740). Все клетки выращивали в модифицированной по способу Дульбекко среде Игла с высоким содержанием глюкозы (50/50), среде Хэма F12, содержащей 10% фетальной телячьей сыворотки (FBS), 2 мМ глутамина, 1% пенициллина-стрептомицина и культивировали при 37°С в 5% СО2. Устойчивые линии клеток 293 и РС-3 трансфицировали (Fugene, Roche) вектором на основе цитомегаловируса, кодирующим полноразмерный белок STEAP-1, (соответственно, LB50 и PS5.4) или пустым вектором и производили отбор в 400 мкг/мл G418 (Geneticin, Life Technologies). Модели эксплантатов предстательной железы человека, LuCAP 77 и LuCAP 35V, были предоставлены Университетом Сиэтла.
Экспрессия экзогенного и эндогенного белка STEAP-1 на поверхности клетки была продемонстрирована при помощи иммуногистохимического анализа (IHC) и анализа FACS следующим образом. Антитела против STEAP-1 овцы и мыши (Agensys, Inc., Santa Monica, CA) были созданы против внутриклеточного аминоконцевого пептида STEAP-1 (см. публикацию Hubert, R.S., Vivanco, I. et al., PNAS 25:14523-14528 (1999)). Моноклональные антитела против внеклеточных доменов STEAP-1 (Agensys, Inc.) были созданы в результате иммунизации мышей клетками 293Т, временно трансфицированными белком STEAP-1. При выполнении IHC анализа для обнаружения использовали первичное антитело овцы против STEAP-1. При выполнении анализа FACS клетки выращивали до 90% слияния и удаляли с планшетов, используя 2 мМ EDTA в PBS. Клетки промывали, ресуспендировали в буфере для FACS (PBS с 1% BSA) и инкубировали в течение 60 минут с антителами против STEAP-1 при комнатной температуре и затем в течение 60 минут с соответствующим вторичным антителом, конъюгированным с фикоэритрином. Анализ выполняли в сканирующем устройстве для FACS (BD Biosciences). Для анализа иммунофлуоресценции клетки выращивали на предметных стеклах с лунками в течение ночи и затем инкубировали с первичным антителом при 37°С в течение 60 минут. Клетки фиксировали в параформальдегиде, блокировали в 1% BSA и инкубировали с соответствующим вторичным антителом, конъюгированным с флуоресцеином.
Модели ксенотрансплантатов рака предстательной железы in vivo использовали для исследования эффективности ADC антитела против STEAP-1. Указанные модели включали линию клеток человека LNCaP (АТСС CRL-1740 или Southern Research Institute, Birmingham, AL). Модели эксплантатов предстательной железы включали LuCaP 77 и LuCaP35V (University of Washington, Seattle, WA). Модели эксплантатов предстательной железы сохраняли путем последовательной трансплантации кастрированным (андрогеннезависимая модель, LuCAP 35V) или некастрированным (андрогензависимая модель, LuCAP 77) самцам мышей SCID-beige компании Charles River Lab. До имплантации некастрированным мышам вводили гранулу тестостерона и кастрировали мышей по меньшей мере за две недели до имплантации опухолей для максимального снижения уровня тестостерона. Когда у мышей-доноров объем опухоли достигал 800-1000 мм3, опухолевую ткань удаляли в асептических условиях и рассекали на мелкие кусочки имплантируемого размера (примерно 20 мм3) для введения экспериментальным животным. Опухоль помещали в карман на месте имплантации и кожу закрывали зажимами для ран. Для модели линии клеток LNCaP выращенные in vitro клетки LNCaP инъецировали подкожно в количестве 8-10 миллионов клеток на одну мышь в 50% матригеле самцам мышей SCID-beige, которым предварительно вводили гранулу тестостерона. Когда средний размер опухоли достигал 100-200 мм3, животных произвольно распределяли в десять групп по десять мышей в каждой и производили одно внутривенное введение ADC исследуемого антитела или контрольного антитела (“голое” или контрольное). В некоторых экспериментах вводили несколько доз исследуемого или контрольного антитела (см. фигуры 8А, 9 и 10). В некоторых экспериментах вводили однократную дозу исследуемого и контрольного антитела, как показано на фигурах 8В и 11. Когда моделью эксплантата предстательной железы была модель LuCap 77, гранулу тестостерона имплантировали мышам примерно за 3-7 дней до трансплантации экзогенной опухоли. Опухоли измеряли два раза в неделю в течение 4 недель, затем один или два раза в неделю на протяжении остального времени исследования или один раз в неделю в течение всего исследования. Значительно меньший объем опухоли у экспериментальных животных в течение указанного времени считался показателем эффективности антитела. В некоторых случаях объем опухоли значительно уменьшался от первоначального объема и оставался небольшим на протяжении всего исследования. Результаты представлены в виде графиков на фигурах 8-11.
Конъюгаты антитела против STEAP-1 с лекарственным средством ауристатином уменьшают объем опухоли предстательной железы in vivo
Введение мышиного антитела против STEAP-1 120-МС-vc-PAB-MMAE в количестве 3 мг/кг оказывало эффективное воздействие в модели ксенотрансплантата опухоли предстательной железы (клетки LNCaP-Ner). В качестве контрольных образцов использовали PBS и антитело против gp120-MC-vc-PAB-MMAE (3 мг/кг). Дозы вводили в 0-й, 7-й и 14-й день. См. фигуру 8А.
Введение гуманизированного антитела против STEAP-1 120v.24-MC-vc-PAB-MMAE (3 мг/кг), 120v.24-MC-MMAF (6 мг/кг), 120v.24-MC-MMAF (12 мг/кг) и химерного антитела против STEAP-1 120 chimera-MC-vc-PAB-MMAE (3 мг/кг) мышам SCID-beige, которым была трансплантирована опухоль LNCap-Ner (указанным мышам вводили гранулу тестостерона, как указано в настоящем описании изобретения), было признано эффективным. В качестве контрольных образцов использовали наполнитель, антитело против амброзии-MC-vc-PAB-MMAE (3 мг/кг) и антитело против амброзии-MC-MMAF (12 мг/кг). Дозы вводили в дни, указанные на фигуре 8. Результаты представлены в виде графика на фигуре 8В.
Введение химерного антитела против STEAP-1 120 chimera-MC-vc-PAB-MMAE (3 мг/кг) и химерного антитела против STEAP-1 120 chimera-MC-MMAF (6 мг/кг) было признано эффективным в модели ксенотрансплантата рака предстательной железы у мышей SCID-beige, которым были трансплантированы клетки LNCaP. Мышам вводили три дозы примерно в 15-й, 25-й и 30-й день в количестве 3 мг/кг (антитело против STEAP-vcMMAE) или 6 мг/кг (антитело против STEAP-mcMMAF). В качестве контрольных образцов использовали антитело против амброзии-MC-vc-PAB-MMAE (3 мг/кг) и антитело против амброзии-MC-MMAF (6 мг/кг). См. фигуру 9.
Введение гуманизированного антитела против STEAP-1 120 chimera-МС-vc-PAB-MMAE (3 мг/кг) было признано эффективным в модели ксенотрансплантата рака предстательной железы у самцом мышей SCID-beige (андрогензависимые), которым были трансплантированы клетки LuCap 77. В качестве контрольных образцов использовали наполнитель и антитело против амброзии-Mc-vc-PAB-MMAE. Вводили три дозы в количестве 3 мг/кг исследуемых и контрольных антител. См. фигуру 10.
Введение гуманизированного антитела против STEAP-1 120v.24-MC-vc-PAB-MMAE в дозе 3 мг/кг, антитела против STEAP-1 120v.24-MC-MMAF в дозе 6 мг/кг и 12 мг/кг кастрированным мышам SCID-beige, которым были трансплантированы опухолевые клетки предстательной железы LuCap35V, было признано эффективным по сравнению с контрольными антителами. Лекарственная нагрузка составляла 3,1 на одно антитело. В качестве контрольных антител использовали антитело против амброзии-MC-MMAF, вводимое в дозе 12 мг/кг, и антитело против gp120-MC-vc-PAB-MMAE, вводимое в дозе 6 мг/кг. См. фигуру 11.
Конъюгаты антитела против STEAP-1 с лекарственным средством ауристатином уменьшают объем опухоли предстательной железы in vitro
Анализы лизиса клеток in vitro выполняли для оценки эффективности конъюгатов антитела против STEAP-1 с лекарственным средством в отношении ингибирования роста и/или лизиса клеток, экспрессирующих STEAP-1. Клетки, экспрессирующие белок STEAP-1, культивировали в количестве примерно 2000 клеток/лунку на 96-луночном планшете и через 24 часа производили дублированную обработку конъюгатом антитела с лекарственным средством. Планшеты инкубировали в течение 5-7 дней при 37°С и проявляли при помощи набора для анализа жизнеспособности люминесцентных клеток CellTiter-Glo® (Promega, Madison, WI, USA). Исследуемые клетки включали PS5.4 (клетки РС3, экспрессирующие экзогенный белок STEAP-1), LB50 (клетки 293, экспрессирующие экзогенный белок STEAP-1), клетки РС3, трансфицированные только вектором, клетки 293, трансфицированные только вектором, и клетки LNCaP, экспрессирующие эндогенный белок STEAP-1. Исследуемые конъюгаты антитела с лекарственным средством включали контрольное антитело-MC-MMAF, контрольное антитело-vc-MMAE, антитело против STEAP-1 120-chimera-vc-MMAE, антитело против STEAP-1 120-chimera-МС-MMAF (две разные партии материалов) и антитело против STEAP-1 chimera-vc-MMAF. Результаты показаны на фигуре 14А-Е.
Пример 5. Получение антител против STEAP-1 с введенным цистеином для конъюгирования способом восстановления и повторного окисления
Полноразмерные моноклональные антитела против STEAP-1 с введенным цистеином (ThioMab), экспрессированные в клетках СНО, растворяли в 500 мМ бората натрия и 500 мМ хлорида натрия при рН около 8,0 и восстанавливали примерно 50-100-кратным избытком 1 мМ ТСЕР (гидрохлорид трис(2-карбоксиэтил)фосфина (Getz et al. (1999) Anal. Biochem. Vol. 273:73-80; Soltec Ventures, Beverly, MA) в течение примерно 1-2 часов при 37°С. Восстановленное антитело ThioMab разводили и вводили в колонку HiTrap S в 10 мМ ацетата антрия, рН 5, и элюировали PBS, содержащим 0,3 М хлорида натрия. Элюированное восстановленное антитело ThioMab обрабатывали 2 мМ дегидроаскорбиновой кислоты (dhAA) при рН 7 в течение 3 часов или 2 мМ водного раствора сульфата меди (CuSO4) в течение ночи при комнатной температуре. Окисление в условиях окружающей атмосферы также может быть эффективным. Буфер заменяли, производя элюирование через сефадекс G24, и элюировали PBS с 1 мМ DTPA. Содержание тиола в антителе проверяли, определяя концентрацию восстановленного антитела на основании оптической плотности раствора при 280 нм и концентрацию тиола определяли в результате взаимодействия с DTNB (Aldrich, Milwaukee, WI) и на основании оптической плотности при 412 нм.
Пример 6. Получение конъюгатов антитела против STEAP-1 с введенным цистеином с лекарственным средством путем конъюгирования антител против STEAP-1 с введенным цистеином с промежуточными соединениями лекарственного средства с линкером
После процедур восстановления и повторного окисления, описанных в примере 5, антитело против STEAP-1 с введенным цистеином растворяли в PBS (физиологический раствор с фосфатным буфером) и охлаждали на льду. Примерно 1,5 молярного эквивалента промежуточного соединения лекарственного средства ауристатина с линкером, такого как МС-ММАЕ (малеимидокапроилмонометилауристатин Е), МС-MMAF, MC-val-cit-PAB-MMAE или MC-val-cit-PAB-MMAF, в зависимости от количества введенных остатков цистеина в одном антителе и при использовании имида малеиновой кислоты в качестве взаимодействующей с тиолом функциональной группы, растворяли в ДМСО, разводили в ацетонитриле и воде и добавляли к охлажденному, восстановленному, повторно окисленному антителу в PBS. Примерно через один час добавляли избыток имида малеиновой кислоты, чтобы погасить реакцию и кэппировать любые непрореагировавшие тиоловые группы антитела. Реакционную смесь концентрировали ультрафильтрованием в центрифуге, конъюгат антитела против STEAP-1 с введенным цистеином с лекарственным средством очищали и обессоливали, элюируя через смолу G25 в PBS, фильтровали через 0,2 мкм фильтры в стерильных условиях и замораживали для хранения.
В соответствии с вышеописанным способом были получены следующие конъюгаты антитела против STEAP-1 с лекарственным средством (нумерация вариантов стандартизирована (нумерация Kabat для легкой цепи и нумерация EU для тяжелой цепи), как показано в настоящем описании изобретения и на фигуре 17):
конъюгат тиоантитела человека 120-МС-MMAF, полученный в результате конъюгирования тиоантитела человека 120 в положении V205C легкой цепи с МС-MMAF;
конъюгат тиоантитела человека 120-МС-MMAF, полученный в результате конъюгирования тиоантитела человека 120 в положении А118С тяжелой цепи с МС-MMAF;
конъюгат тиоантитела человека 120-МС-val-cit-PAB-MMAE, полученный в результате конъюгирования тиоантитела человека 120 в положении V205C легкой цепи с MC-val-cit-PAB-MMAE; и
конъюгат тиоантитела человека 120-MC-val-cit-PAB-MMAE, полученный в результате конъюгирования тиоантитела человека в положении А118С тяжелой цепи с MC-val-cit-PAB-MMAE.
Пример 7. Исследование антител против STEAP-1 с введенным цистеином
Конъюгаты антитела против STEAP-1 с введенным цистеином с лекарственным средством (TDC), полученные в соответствии в приведенным выше описанием, анализировали для подтверждения сохранения указанными конъюгатами активности материнского антитела in vitro. TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (LCV205C) (обозначенный аббревиатурой huSteap1 TDC (L205C) vcE) и тиоантитела человека 120-vc-PAB-MMAE (HCA118C) (обозначенный аббревиатурой huSteap1 TDC (HCA118C) vcE) оценивали при помощи анализа FACS в отношении связывания с белком STEAP-1 на STEAP-1-экспрессирующих клетках (293 STEAP-1 NT LB50) и неэкспрессирующих (293 вектор S408) клетках. Термин ”2nd only” означает вторичное антитело при выполнении анализа FACS. Контрольный TDC (vcE) и стандартный контрольный ADC (vcE) являются контрольными конъюгатами соответственно тиоантитела и нетиоантитела с vc-PAB-MMAE. ADC huSteap1 (стандартный) является конъюгатом с лекарственным средством vc-PAB-MMAE, полученным из материнского антитела человека против STEAP-1. Как показано в настоящем описании изобретения, TDC вызывали изменения при выполнении анализа FACS, аналогичные изменениям исходного ADC huSteap1.
Также были выполнены анализы лизиса клеток in vitro для оценки эффективности конъюгатов антитела против STEAP-1 с введенным цистеином с лекарственным средством в отношении ингибирования роста и/или лизиса клеток, экспрессирующих STEAP-1. Клетки, экспрессирующие STEAP-1, культивировали в количестве примерно 2000 клеток/лунку на 96-луночном планшете и через 24 часа производили дублированную обработку конъюгатом антитела с лекарственным средством. Планшеты инкубировали в течение 5-7 дней при 37°С и проявляли при помощи набора для анализа жизнеспособности люминесцентных клеток CellTiter-Glo® (Promega, Madison, WI, USA). Исследуемые клетки включали PS5.4 (клетки РС3, экспрессирующие экзогенный белок STEAP-1), LB50 (клетки 293, экспрессирующие экзогенный белок STEAP-1) и клетки LNCaP, экспрессирующие эндогенный белок STEAP-1. Исследуемые конъюгаты антитела с лекарственным средством включали конъюгат контрольного антитела-vc-MMAE (стандартный контрольный ADC (vcE)), конъюгат контрольного тиоантитела-vc-MMAE (контрольный TDC (vcE)), TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (LCV205C) (обозначенный аббревиатурой TDC huSteap1 (L205C) vcE) и тиоантитела человека 120-vc-PAB-MMAE (HCA118C) (обозначенный аббревиатурой TDC huSteap1 (HCA118C) vcE) и ADC huStreap1 (стандартный), конъюгат с лекарственным средством vc-PAB-MMAE, полученный из материнского антитела человека против STEAP-1. Как показано на фигурах 19А-С, TDC антитела против STEAP-1 сохраняет активность исходного ADC in vitro.
Пример 8. Анализы уменьшения объема опухоли in vivo конъюгатами антитела против STEAP-1 с введенным цистеином с лекарственным средством
Для исследования эффективности ADC антитела против STEAP-1 с введенным цистеином были использованы модели ксенотрансплантатов рака предстательной железы in vivo. Модели и способы исследования аналогичны описанным в примере 4.
Введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (НСА118С) (обозначенный аббревиатурой huSteap1 HC TDC vcE) (3 мг/кг) мышам SCID-beige, которым была трансплантирована опухоль LNCap-Ner (вводили гранулу тестостерона в соответствии с настоящим описанием изобретения), было признано эффективным. В качестве контрольных образцов использовали наполнитель (PBS), конъюгат контрольного антитела-vc-MMAE (стандартный контрольный ADC vcE) и конъюгат контрольного тиоантитела-vc-MMAE (контрольный TDC HC vcE). Эффект TDC антитела против STEAP-1 также сравнивали с конъюгатом антитела человека против STEAP-1 120-MC-vc-PAB-MMAE (стандартный ADC hu Sreap1 vcE), используемым в качестве положительного контрольного образца. Вводили однократную дозу в 0-й день. Все антитела вводили в дозе 3 мг/кг. Результаты представлены в виде графика на фигуре 20.
На фигуре 21 показано, что введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (НСА118С) (обозначенный аббревиатурой huSteap1 HC TDC vcE) в дозе 3 мг/кг и TDC тиоантитела человека против STEAP-1 120-МС-MMAF (НСА118С) (обозначенный аббревиатурой huSteap1 HC TDC mcF) в дозе 1, 3 или 6 мг/кг было признано эффективным в модели ксенотрансплантата рака предстательной железы у мышей SCID-beige, которым были трансплантированы клетки LNCaP. Мышам вводили однократные дозы в 0-й день в количестве 0,3, 1 или 3 мг/кг (huStea1 HC TDC vcE) или 1, 3 или 6 мг/кг (huSteap1 HC TDC mcF). В качестве контрольных образцов использовали наполнитель (PBS), конъюгат контрольного антитела-vc-MMAE (стандартный контрольный ADC vcE) и конъюгат контрольного тиоантитела-vc-MMAE (контрольный TDC HC vcE).
На фигуре 22 показано, что введение TDC тиоантитела человека против STEAP-1 120-vc-PAB-MMAE (НСА118С) (обозначенный аббревиатурой huSteap1 HC TDC vcE) в дозе 3 мг/кг и TDC тиоантитела человека против STEAP-1 120-MC-MMAF (НСА118С) (обозначенный аббревиатурой huSteap1 HC TDC mcF) в дозе 3 или 6 мг/кг было признано эффективным в модели ксенотрансплантата рака предстательной железы у самцов мышей SCID-beige (андрогензависимые), которым были трансплантированы клетки LuCap 35V. Мышам вводили однократные дозы в 0-й день в количестве 0,3, 1 или 3 мг/кг (huSteap1 HC TDC vcE) или 1, 3 или 6 мг/кг (huSteap1 HC TDC mcF). В качестве контрольных образцов использовали наполнитель (PBS), конъюгат контрольного антитела-vc-MMAE (стандартный контрольный ADC vcE) и конъюгат контрольного тиоантитела-vc-MMAE (контрольный TDC HC vcE).
Пример 9. Получение антитела против STEAP-1 SGIV из варианта 24 антитела 120 и исследование указанного антитела
Был получен еще один вариант LC антитела против STEAP-1, в котором легкая цепь и остовные области были далее модифицированы для достижения более высоких уровней экспрессии антитела.
Материалы и способы
Номера остатков соответствуют нумерации Kabat (Kabat et al., Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Использованы однобуквенные аббревиатуры для обозначения аминокислот. Вырожденность ДНК представлена с использованием кода IUB (N=A/C/G/T, D=A/G/T, V=A/C/G, B=C/G/T, H=A/C/T, K=G/T, M=A/C, R=A/G, S=G/C, W=A/T, Y=C/T).
Получение варианта с измененной легкой цепью. Был создан и исследован вариант антитела 120.v24, обозначенный ”Simmons IV” или просто ”SGIV”. Аминокислотная последовательность легкой цепи SGIV представлена в SEQ ID NO:90. Указанная последовательность в сравнении с соответствующими областями мышиного антитела 120 (SEQ ID NO:89) и антитела 120.v24 (SEQ ID NO:91) показана на фигуре 23.
Оценка варианта SGIV по сравнению с вариантом 120.v24. Антитела SGIV и 120.v24, экспрессированные в виде IgG, оценивали при помощи анализа FACS с использованием устойчиво трансформированных Steap1-положительных линий клеток 293 Streap1 NT LB48, 293 Steap1 NT LB50 и 293 Steap1 NT LB53, а также клеток LNCaP, экспрессирующих эндогенный белок STEAP-1 (фигура 28). Клетки были получены в соответствии с описанием, приведенным в примере 1. Каждый IgG добавляли в количестве 5 мкг/кг на льду и оставляли на 1 час. Образцы дважды промывали буфером для FACS, центрифугировали, добавляли конъюгат антитела против РЕ человека (R-фикоэритрин козы против Fcy-фрагмента IgG человека, Jackson Immunoresearch), разведенный в отношении 1:200, и оставляли на 30 минут. Образцы еще раз дважды промывали буфером для FACS, центрифугировали и анализировали способом FACS.
Определение аффинности связывания антител SGIV и 120.v24 к белку STEAP-1 при помощи анализа Скэтчарда. Аффинность связывания антител 120.v24 и Simmons IV (”SGIV”) к белку STEAP-1 определяли при помощи анализа Скэтчарда стандартными способами. IgG очищали аффинной хроматографикей с протеином G. Аффинность определяли при помощи дублированного анализа Скэтчарда в клетках РС-3-PS5.4, 293-LB50 и LNCaP-BR. Графики результатов анализа Скэтчарда для антител 120.v24 и SGIV в клетках LNCaP BR и 293.LB50 показаны соответственно на фигурах 25 и 26. На фигуре 27 представлена таблица, в которой сравниваются средние значения аффинности связывания для мышиного антитела 1789, мышиного антитела 120, химерного антитела с Fc-областью, гуманизированного антитела 120.v24, тиоантитела 120.v24 и тиоантитела SGIV в клетах РС-3-PS5.4, 293-LB50 и LNCaP-BR, а также в клетках 293, временно экспрессирующих STEAP-1.
Сайтнаправленный мутагенез антител SGIV и 120.v24. Варианты антител SGIV и 120.v24 были получены стандартными способами мутагенеза, описаннымии выше. Варианты первого класса были получены при помощи сайтнаправленного мутагенеза, в результате которого определенные остатки антитела Simmons IV (”SGIV”) были заменены соответствующими остатками антитела 120.v24 для дальнейшего улучшения аффинности связывания. Как показано на фигуре 24, были получены следующие конкретные варианты:
(1) LS.VLVH1, в котором остатки 42 (”Q”) и 43 (”P”) были модифицированы, соответственно, в ”K” и ”A” (SEQ ID NO:92);
(2) LS.VLVH2, в котором остаток 3 (”V”) был модифицирован в ”Q”, остатки 42 (”Q”) и 43 (”Р”) были модифицированы, соответственно, в ”К” и ”А” и остаток 85 (”V”) был модифицирован в ”Т” (SEQ ID NO:93);
(3) LS.Q, в котором остаток 3 (”V”) был модифицирован в (”Q”) (SEQ ID NO:94);
(4) LS.CH1, в котором остаток 15 (”L”) был модифицирован в ”V” и остаток 83 (”V”) был модифицирован в ”F” (SEQ ID NO:95).
Варианты второго класса были созданы при помощи сайтнаправленного мутагенеза, в результате которого определенные остатки антитела 120.v24 былим заменены соответствующими остатками антитела Simmons IV (SGIV) с целью повышения уровней экспрессии антитела. Как показано на фигуре 24, были получены следующие конкретные варианты:
(1) ED.FW1, в котором остаток 3 (”Q”) был модифицирован в ”V”, остаток 9 (”S”) был модифицирован в ”D”, остаток 12 (”S”) был модифицирован в ”А”, остаток 13 (”А”) был модифицирован в ”V”, остаток 15 (”V”) был модифицирован в ”L”, остаток 17 (”D”) был модифицирован в ”E”, остаток 19 (”V”) был модифицирован в ”А” и остаток 22 (”Т”) был модифицирован в ”N” (SEQ ID NO:96);
(2) ED.FW2, в котором остатки 42 (”К”) и 43 (”А”) антитела 120.v24 были модифицированы, соответственно, в ”Q” и ”Р” (SEQ ID NO:97);
(3) ED.FW3, в котором остаток 60 (”S”) был модифицирован в ”D”, остаток 80 (”Р”) был модифицирован в ”А”, остаток 83 (”F”) был модифицирован в ”V” и остаток 85 (”Т”) был модифицирован в ”V” (SEQ ID NO:98);
(4) ED.all, в котором остаток 3 (”Q”) был модифицирован в ”V”, остаток 9 (”S”) был модифицирован в ”D”, остаток 12 (”S”) был модифицирован в ”А”, остаток 13 (”А”) был модифицирован в ”V”, остаток 15 (”V”) был модифицирован в ”L”, остаток 17 (”D”) был модифицирован в ”Е”, остаток 19 (”V”) был модифицирован в ”А”, остаток 22 (”Т”) был модифицированы в ”N”, остатки 42 (”К”) и 43 (”А”) антитела 120.v24 были модифицирован в ”Q” и ”Р”, остаток 60 (”S”) был модифицирован в ”D”, остаток 80 (”Р”) был модифицирован в ”А”, остаток 83 (”F”) был модифицирован в ”V” и остаток 85 (”Т”) был модифицирован в ”V” (SEQ ID NO:99);
(5) ED.Pro, в котором остаток 43 (”А”) был модифицирован в ”Р” и остаток 80 (”Р”) был модифицирован в ”А” (SEQ ID NO:100);
(6) ED.pl, в котором остаток 9 (”S”) был модифицирован в ”D”, остаток 42 (”К”) был модифицирован в ”Q” и остаток 60 (”S”) был модифицирован в ”D” (SEQ ID NO:101).
Результаты и их рассмотрение
Получение антитела SGIV. Последовательность вариабельной области антитела против STEAP-1 варианта 24 (120.v24) показана на фигурах 23 и 24 (SEQ ID NO:91). В результате выполнения сайтнаправленного мутагенеза вышеописанными стандартными способами был получен еще один вариант, получивший название ”Simmons IV” или просто ”SGIV”. На фигурах 23 и 24 показана последовательность легкой цепи SGIV в сравнении с мышиным антителом 120 и вариантом антитела 120.v24. Титры разных сборов антитела SGIV показаны на фигуре 29.
Сравнение связывания антител SGIV и 120.v24 с белком STEAP-1 при помощи анализа FACS. Способность обоих антител, 120.v24 и SGIV связываться с белком STEAP-1, экспрессированным на поверхности клетки, измеряли при помощи анализа FACS. Связывание антитела с линиями клеток, экспрессирующих экзогенный белок STEAP-1 (293 STEAP-1 LB48, 293 STEAP-1 LB50 и 293 STEAP-1 LB53) или эндогенный белок STEAP-1 (LNCaP.Br), измеряли в дублированных экспериментах; полученные результаты суммированы на фигуре 28. Как показано на фигуре 28, оба антитела связывались с белком STEAP-1 во всех четырех линиях клеток.
Аффинность связывания антитела SGIV к белку STEAP-1 и сравнение с антителом 120.v24. Аффинность связывания антител SGIV и 120.v24 к белку STEAP-1 исследовали при помощи анализа Скэтчарда. Графики результатов анализа Скэтчарда для антител 120.v24 и SGIV в клетках LNCap BR и 293.LB50 показаны, соответственно, на фигурах 25 и 26. На фигуре 27 представлена таблица, в которой сравниваются средние значения аффинности связывания для мышиного антитела 1789, мышиного антитела 120, антитела с химерным Fc-фрагментом, гуманизированного антитела 120.v24, тиоантитела 120.v24 и тиоантитела SGIV в клетках РС-3-PS5.4, 293-LB50 и LNCaP-BR, а также в клетках 293, временно экспрессирующих STEAP-1. Полученные результаты показывают, что аффинность связывания антитела 120.v24 в клетках 293-LB50 и LNCaP.BR в 1,5 раза превышает аналогичный показатель варианта SGIV.
Пример 10. Иммуно-ПЭТ с использованием конъюгата 89Zr-меченого антитела против STEAP-1-vc-MMAE
Иммуно-ПЭТ с использованием конъюгата меченного цирконием-89 антитела против STEAP-1-vc-MMAE выполняли для измерения уровней экспрессии STEAP-1-vc-MMAE в нескольких первичных опухолях предстательной железы у мышей для оценки интернализации антитела и его поглощения опухолью в мышиных моделях по сравнению с другими известными исследованиями и измерениями экспрессии рецепторов in vitro. Для исследования эффективности ADC антитела против STEAP-1 были использованы модели ксенотрансплантатов рака предстательной железы in vivo. Указанные модели включали линию клеток человека LNCaP (ATCC CRL-1740 или Southern Research Institute, Birmingham, AL). Модели эксплантатов предстательной железы включали LuCaP 77 и LuCaP35V (University of Washington, Seattle, WA). Каждую модель эксплантата предстательной железы сохраняли путем последовательной трансплантации кастрированным (андрогеннезависимая модель, LuCAP 35V) или некастрированным (андрогензависимая модель, LuCAP 77) самцам мышей. Животным вводили конъюгат 89Zr-антитела против STEAP-1-vc-MMAE с радиоактивностью 100 мкКи и производили визуализацию через четыре (4) дня. Первичные опухоли Lucap 96.1, Lucap 70, Lucap 77 и Lucap 35V исследовали аналогичным образом in vitro (см. пример 4, фигуры 8-11). Как показано на фигуре 30, поглощение 89Zr-STEAP-1-vc-MMAE было самым высоким в Lucap35V и самым низким в Lucap 96.1 [Lucap 96.1>Lucap 70>Lucap 77>Lucap 35V]. Исследования эффективности, выполняемые в течение многих недель, проводили параллельно с 3-дневными исследованиями поглощения опухолями [модель Lucap35V характеризовалась высоким поглощением STEAP-1-vc-MMAE, соответствующим высокой эффективности; модель Lucap 70 характеризовалась средним поглощением и средней эффективностью. Таким образом, 89Zr иммуно-ПЭТ является неинвазивным способом контроля за биологическим распределением антител в тканях и может быть использован для исследования разных видов млекопитающих от мышей до человека (как показано для поражений с положительной динамикой лечения герцептином 2 у индивидов с метастазирующим раком молочной железы в публикации Dijkers E.C. et al., Nature, Volume 87 Number 5 (May 2010). 89Zr иммуно-ПЭТ может быть использована для определения точки насыщения опухоли и облегчения выбора схемы лечения для будущих лекарственных препаратов.
Хотя настоящее изобретение подробно описано с использованием иллюстраций и примеров для лучшего понимания, указанные иллюстрации и примеры не ограничивают объем изобретения. Все патенты и научные публикации, приведенные в настоящем описании изобретения, включены в описание изобретения в качестве ссылки в полном объеме.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2639543C9 |
| АГЕНТЫ ПРОТИВ КЛЕТКИ-МИШЕНИ, НАЦЕЛЕННЫЕ НА CD138, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2537265C2 |
| СПОСОБЫ УЛУЧШЕНИЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА CD138-ЭКСПРЕССИРУЮЩИЕ ОПУХОЛЕВЫЕ КЛЕТКИ И АГЕНТЫ ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 2008 |
|
RU2486203C2 |
| АНТИ-ТАТ226 АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ | 2007 |
|
RU2448980C2 |
| АНТИТЕЛА ПРОТИВ ПОЛИУБИКВИТИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2603093C2 |
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2436796C9 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2511410C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2553566C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ CD79b И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ПРИМЕНЕНИЯ | 2008 |
|
RU2557319C2 |
| ПРОТИВООПУХОЛЕВЫЕ КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ КОНЪЮГАТЫ НА ОСНОВЕ АНТИТЕЛА К CEACAM5, ТРИФЛУРИДИН И ТИПИРАЦИЛ | 2021 |
|
RU2835019C1 |









































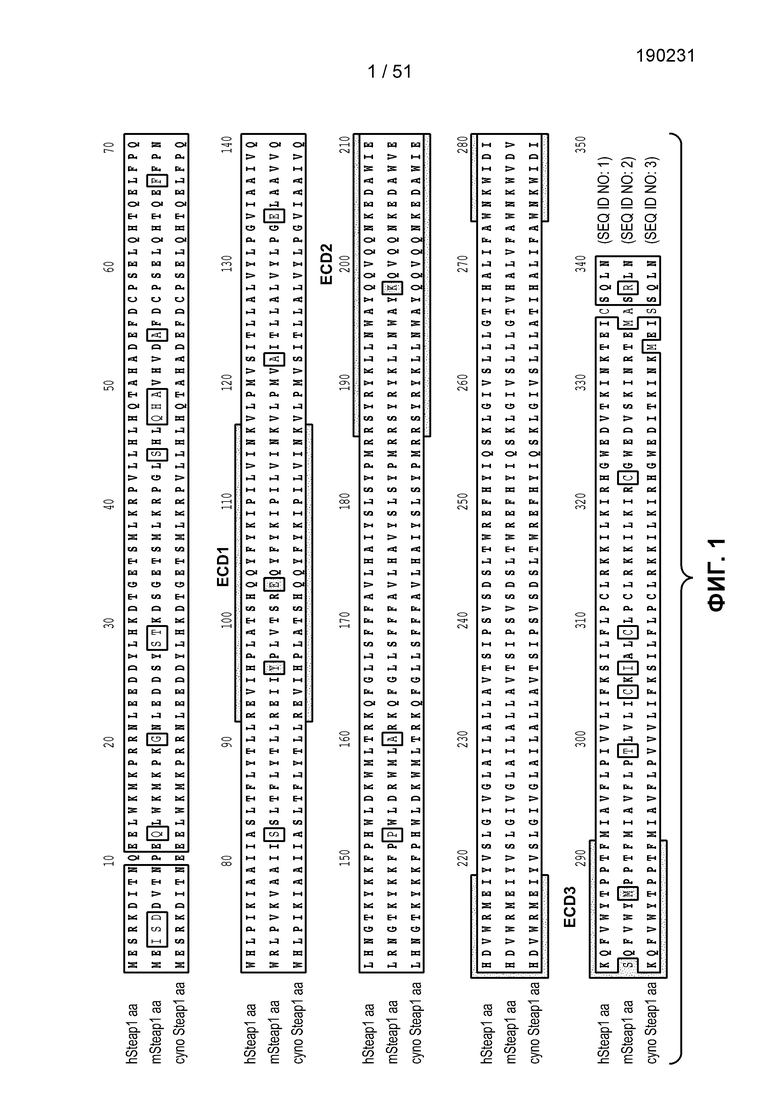
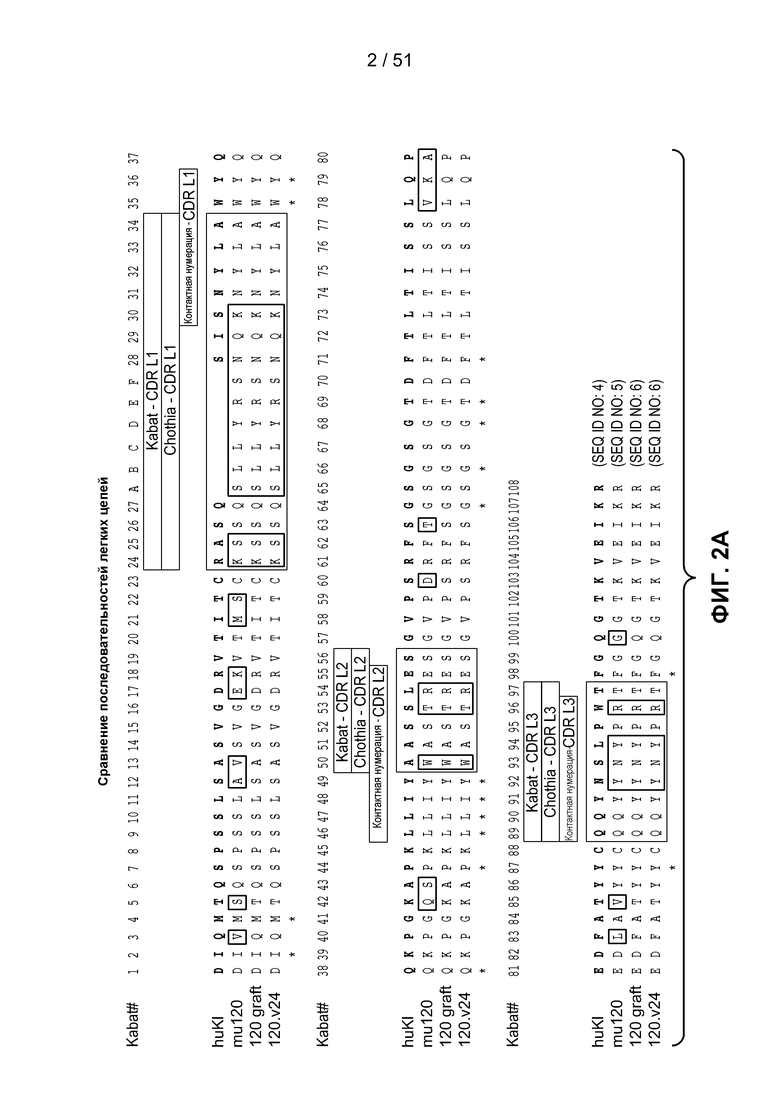
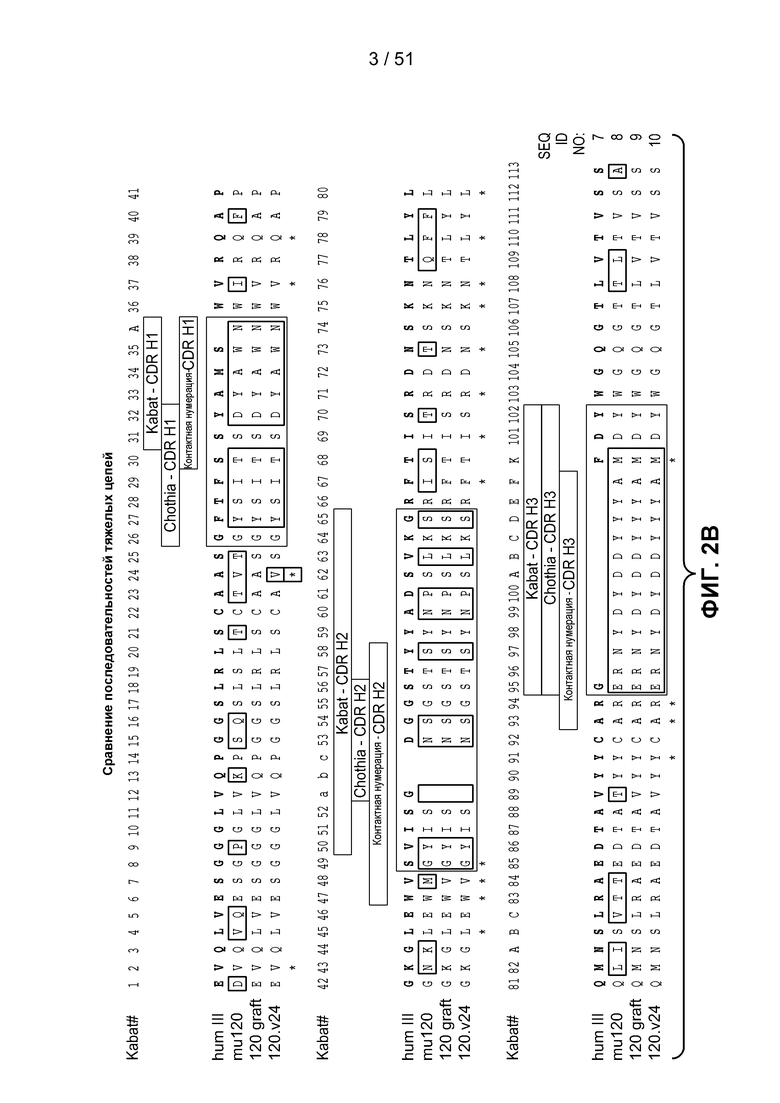
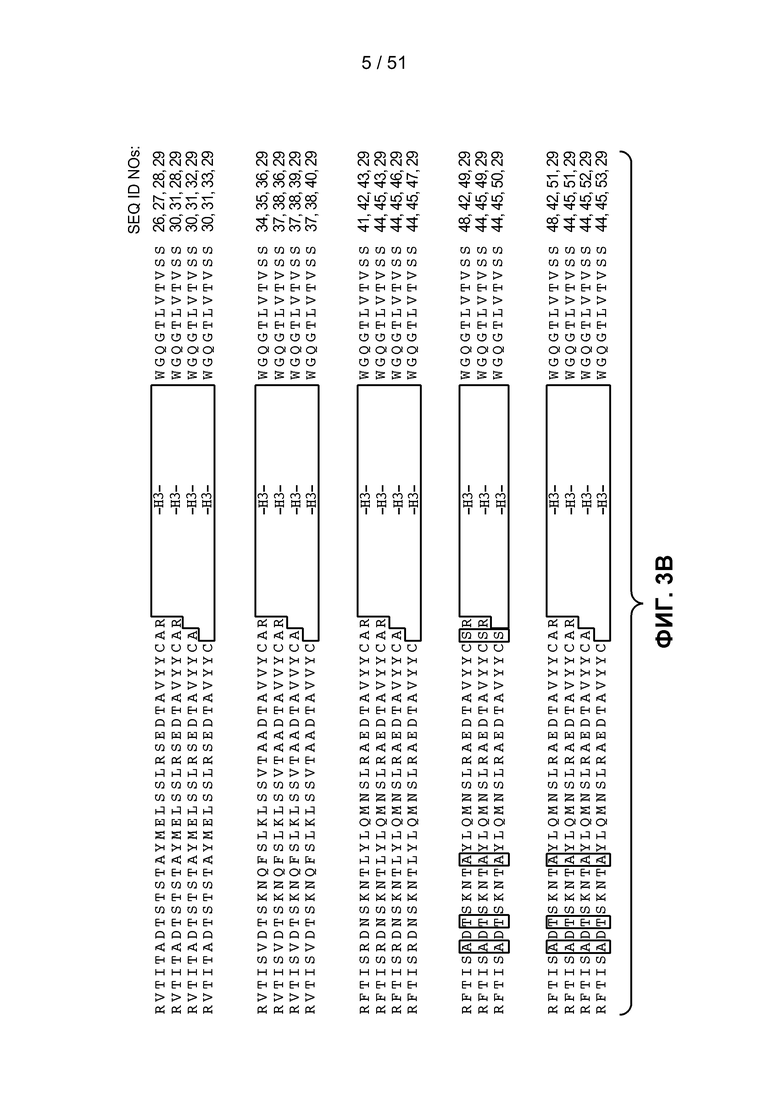
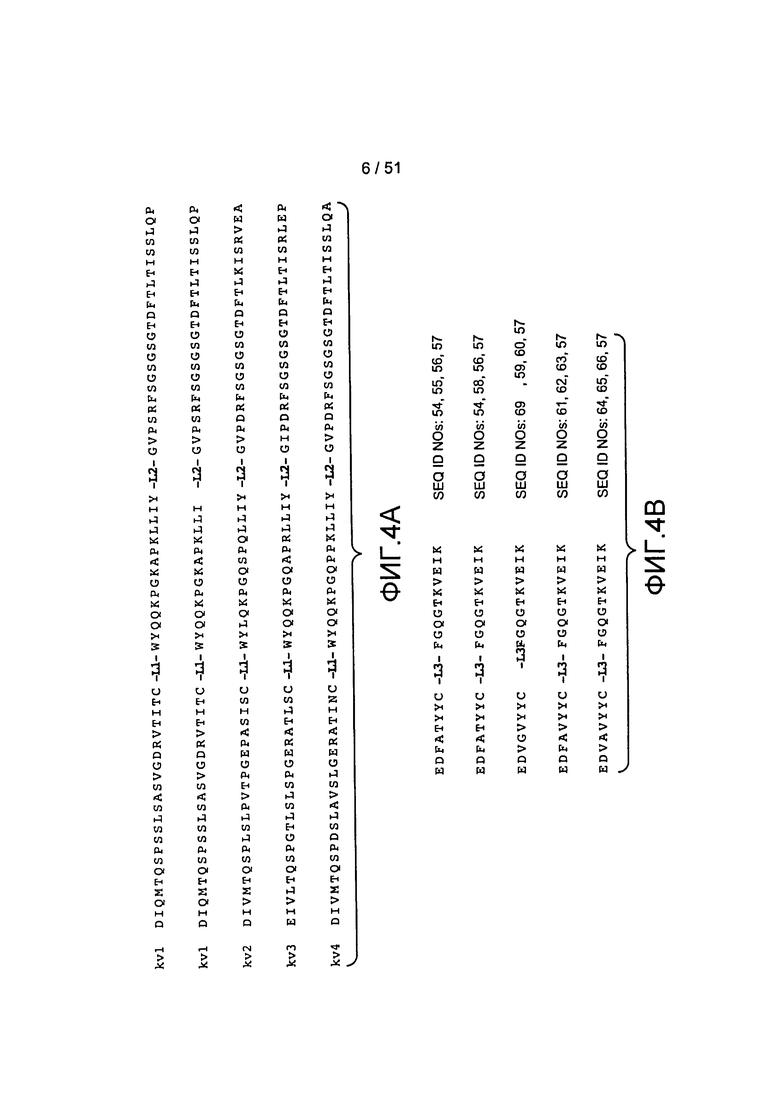

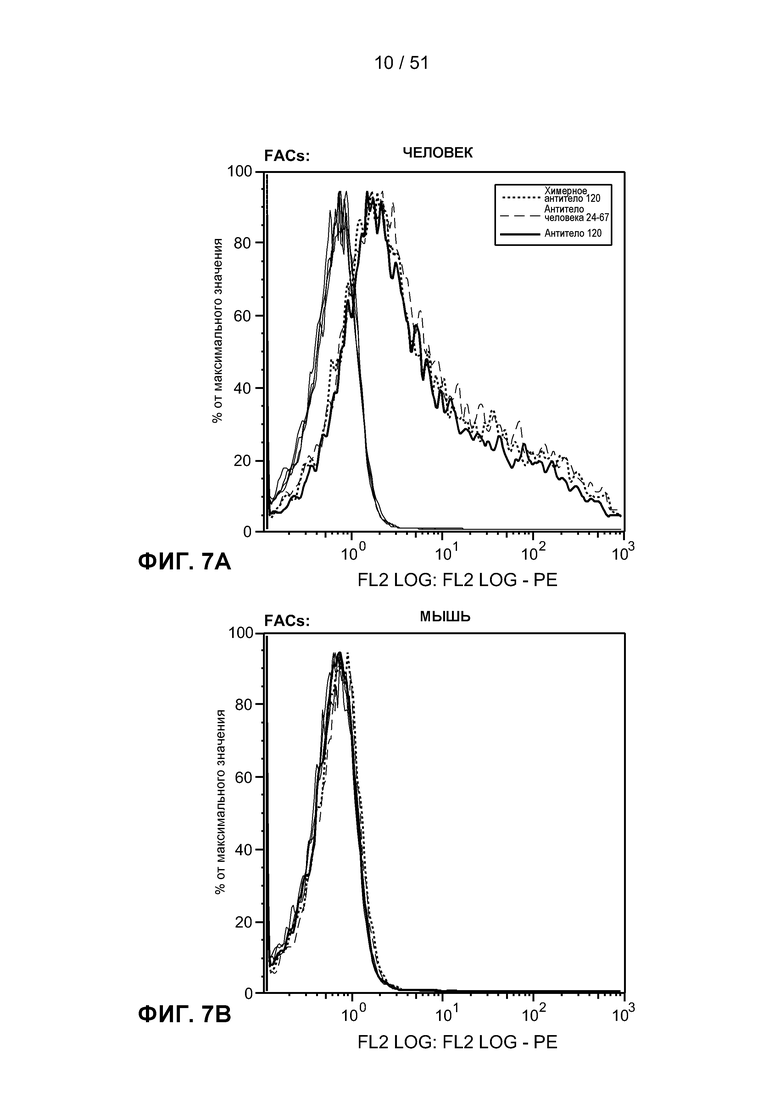
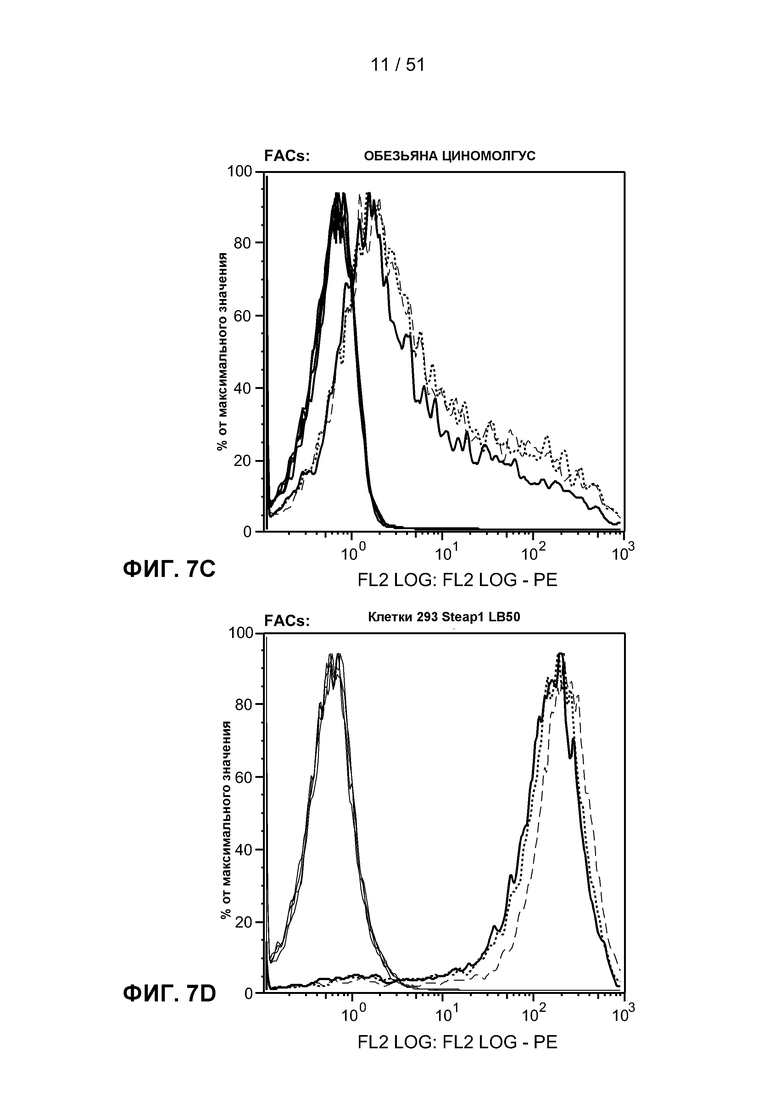
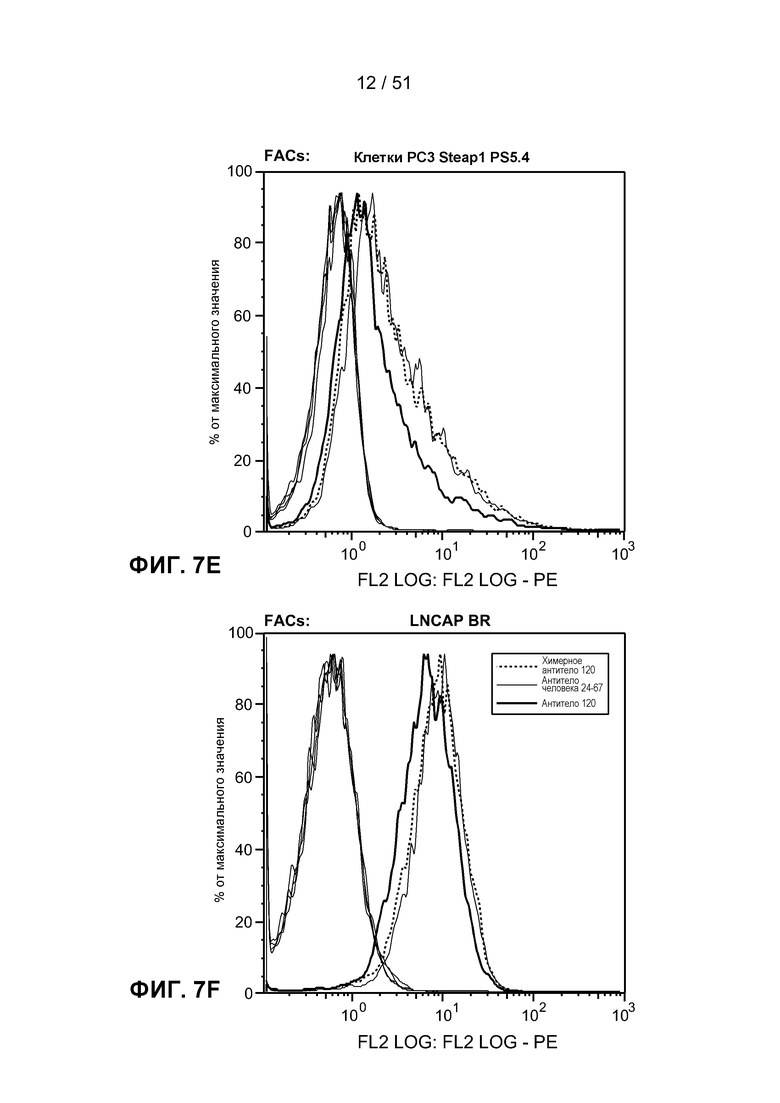
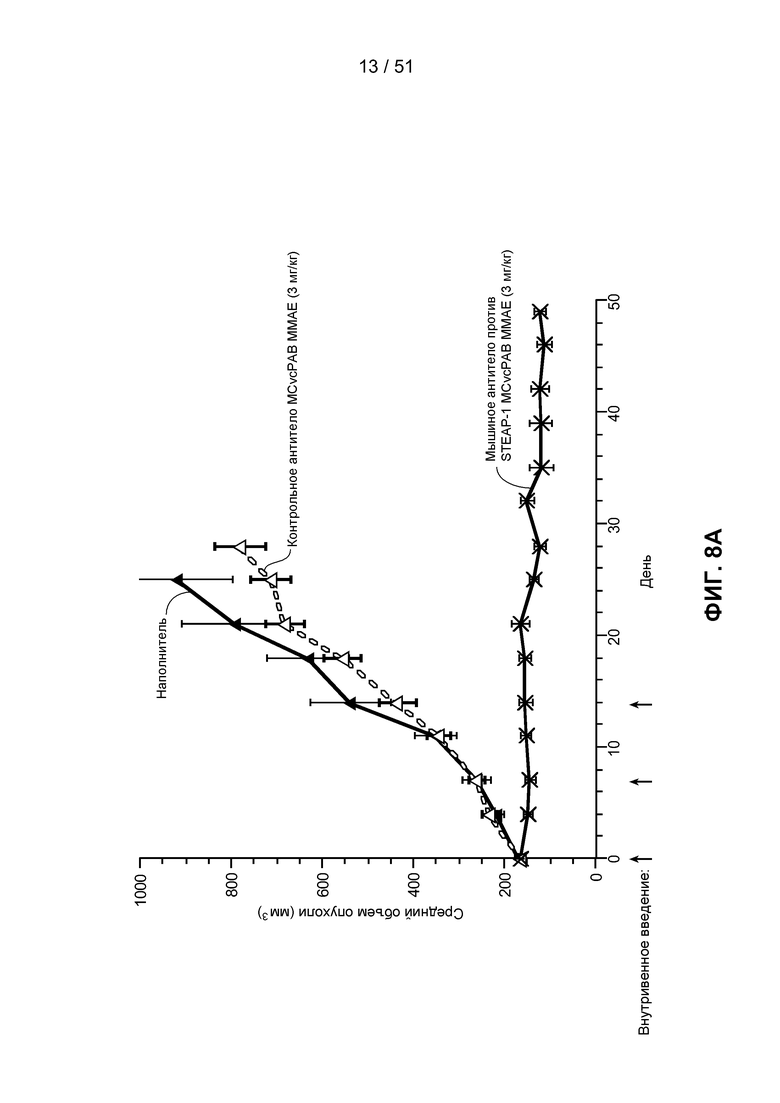
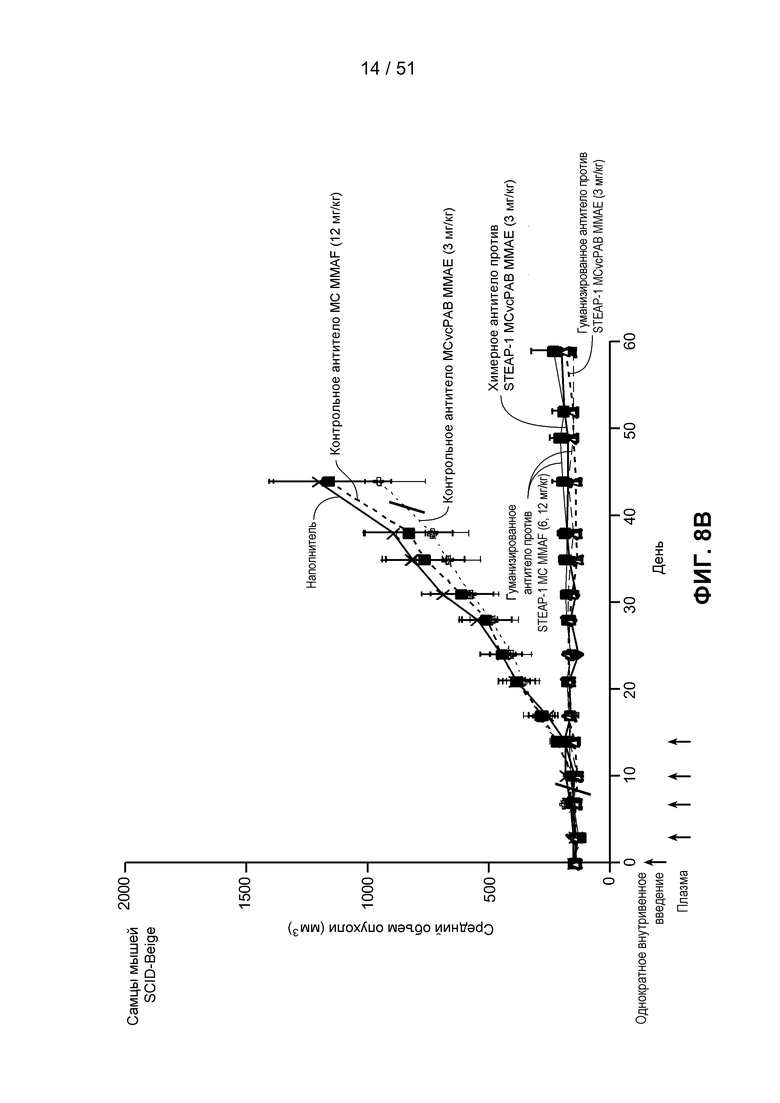
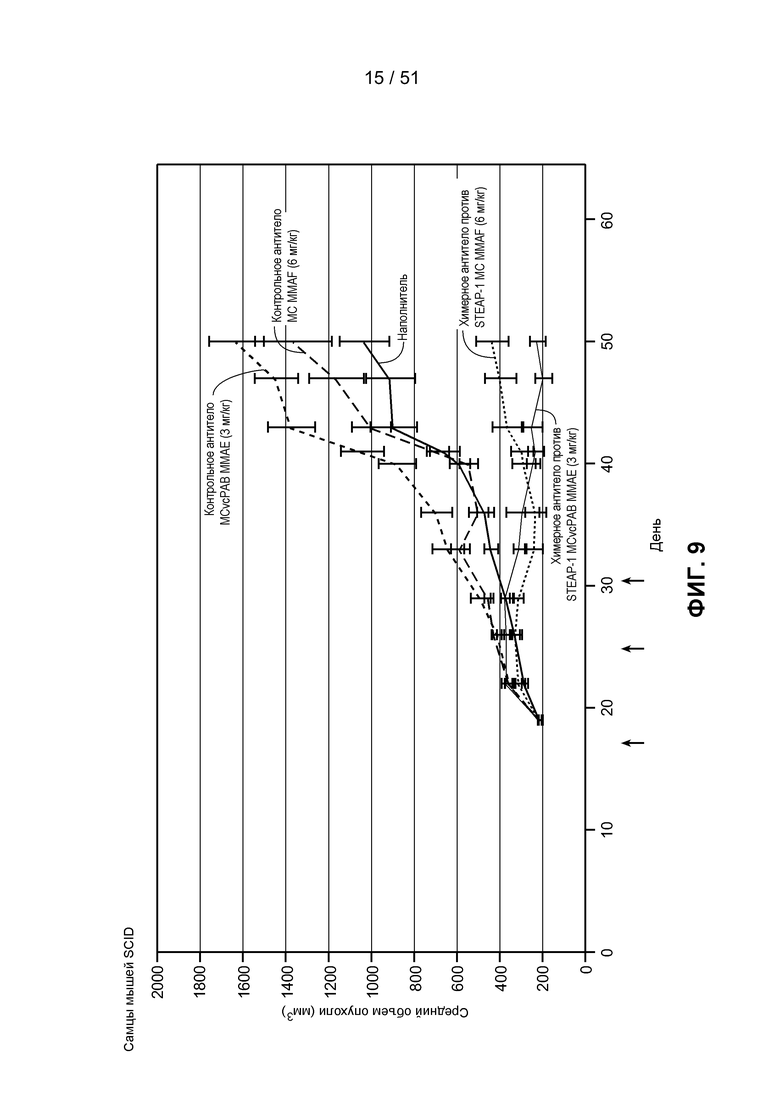
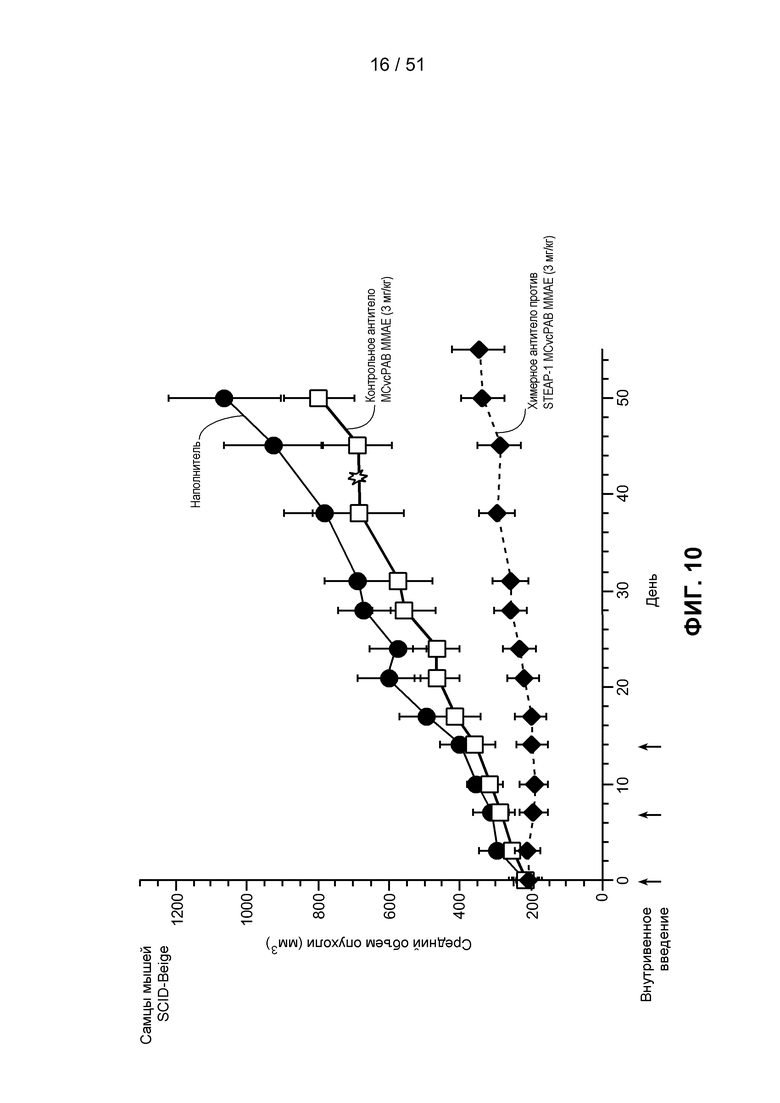
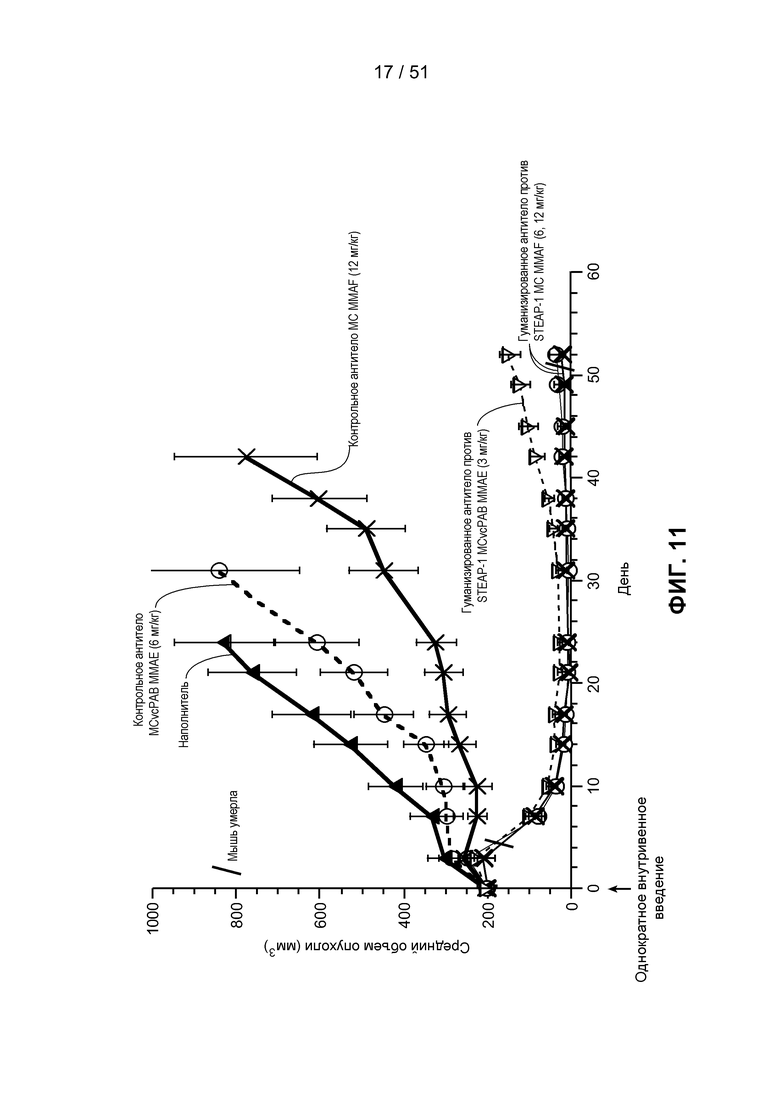
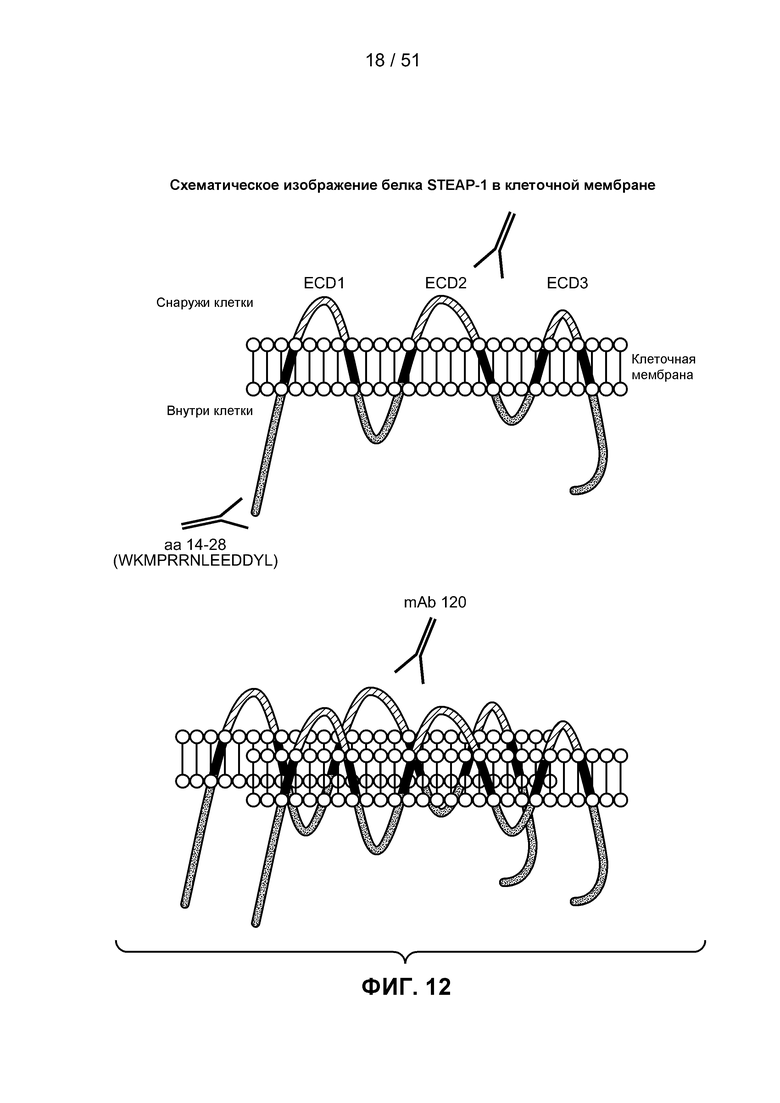

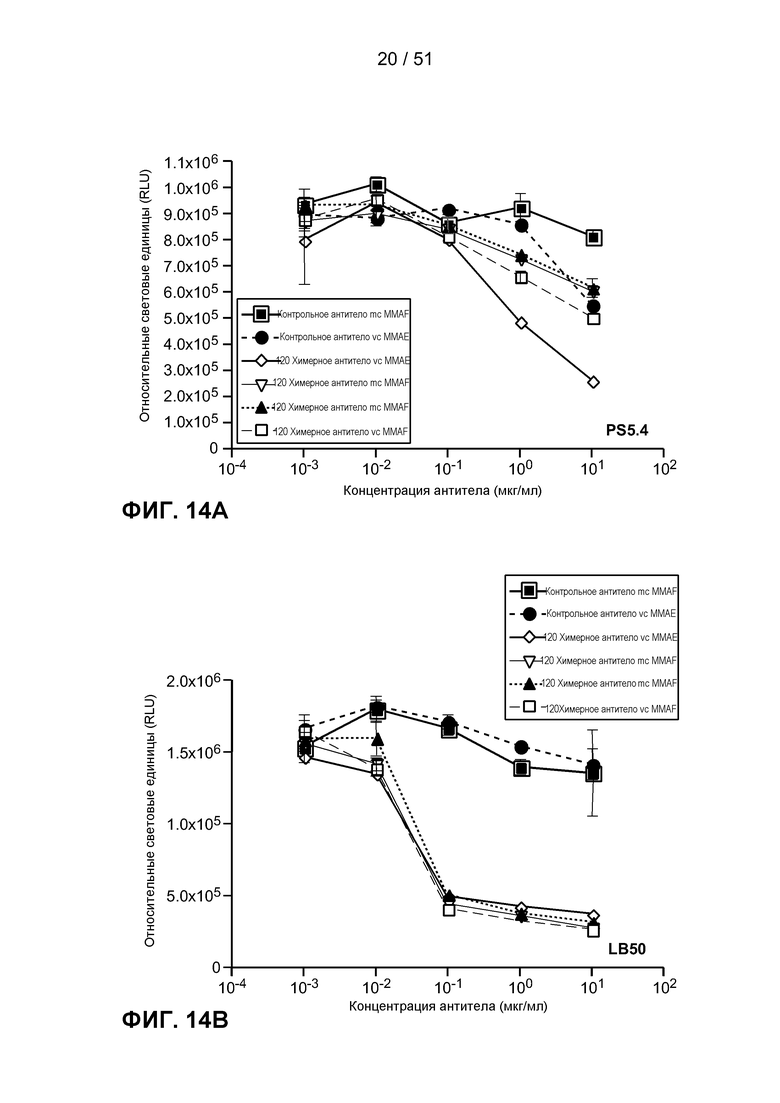
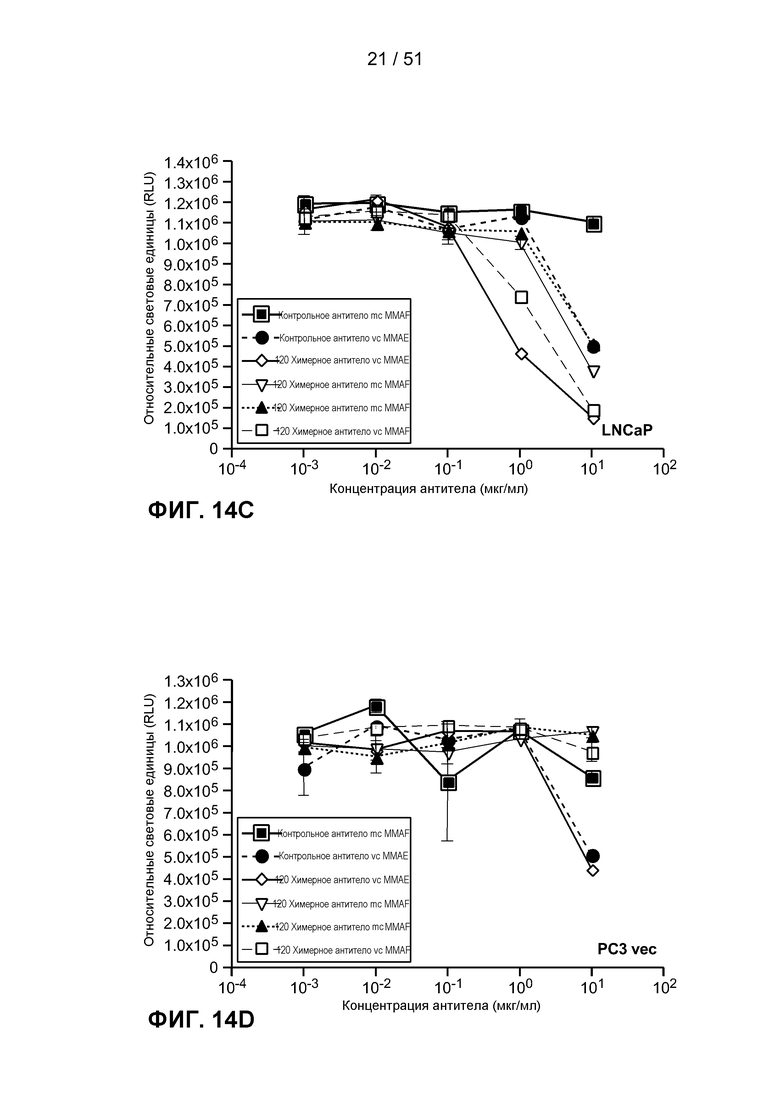
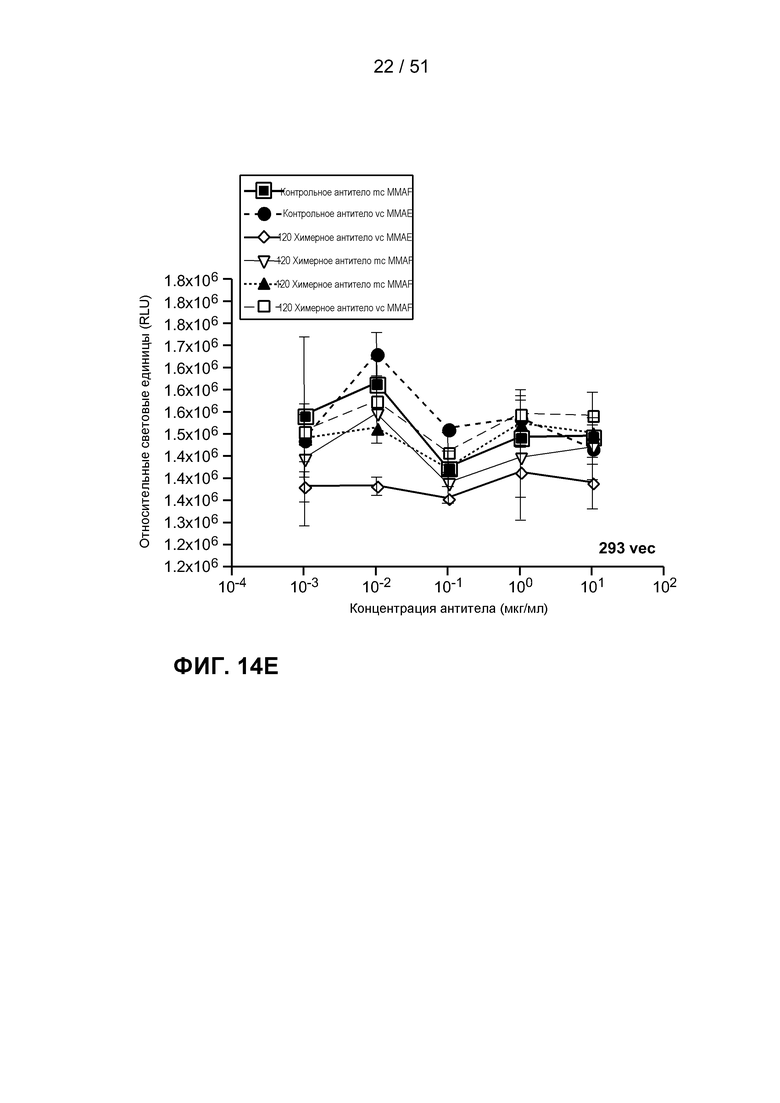
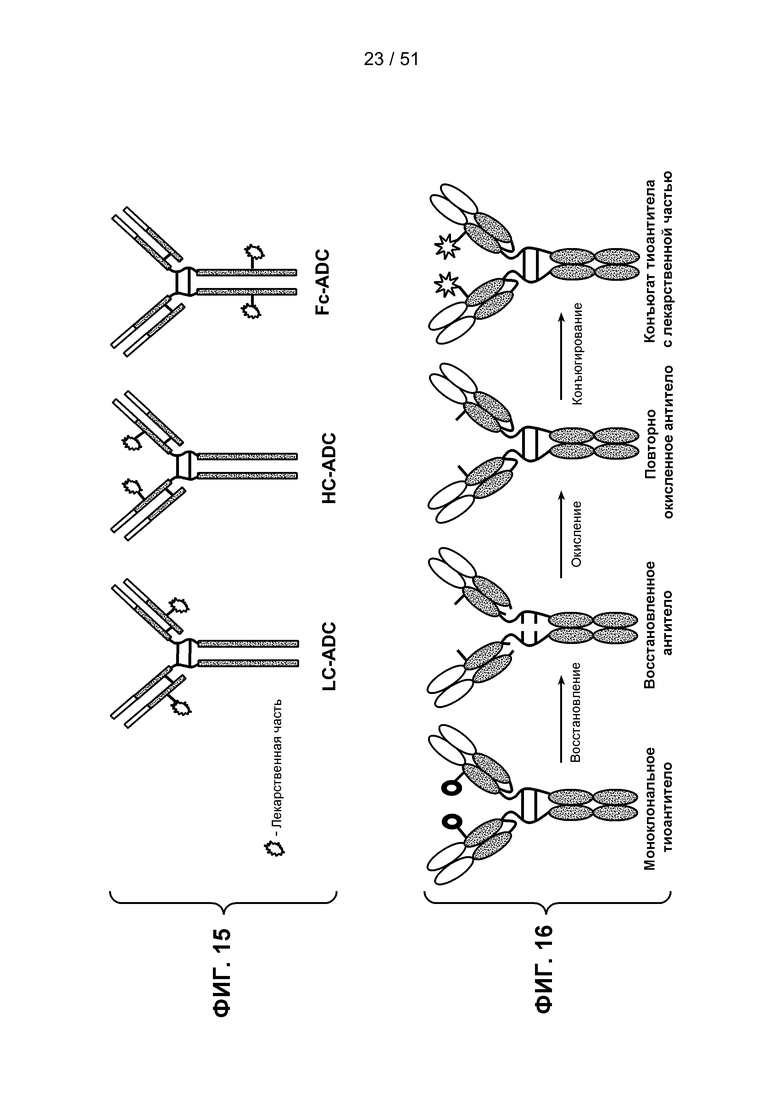

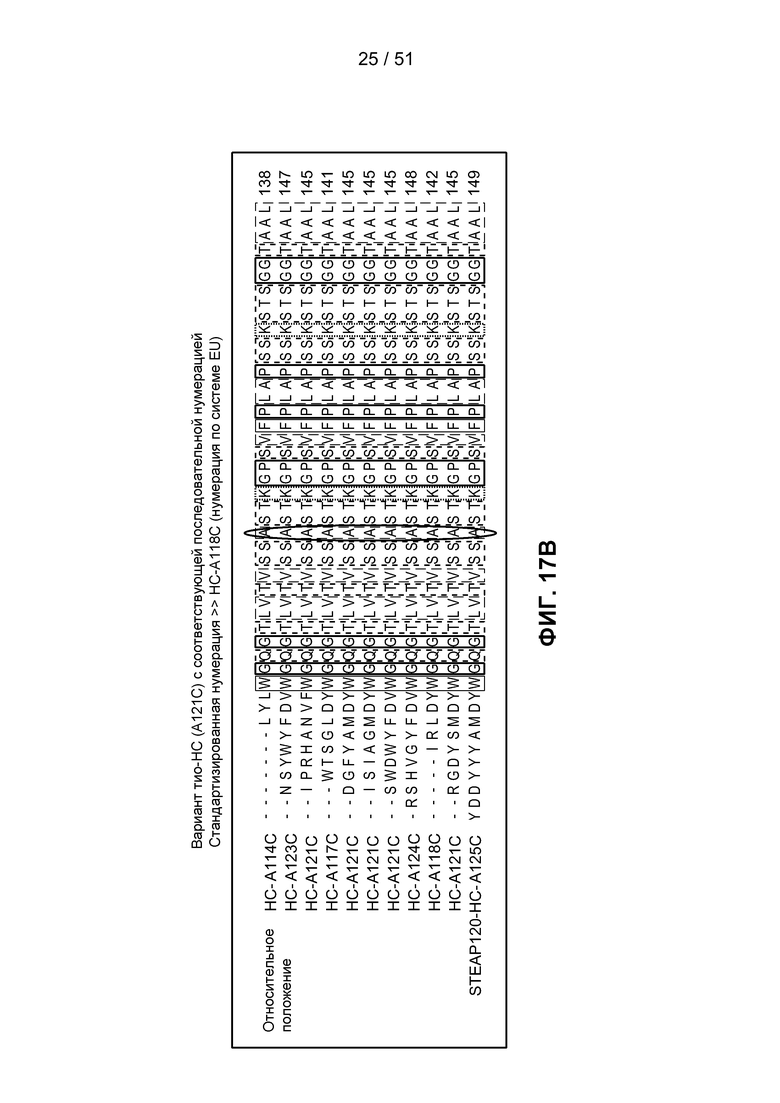

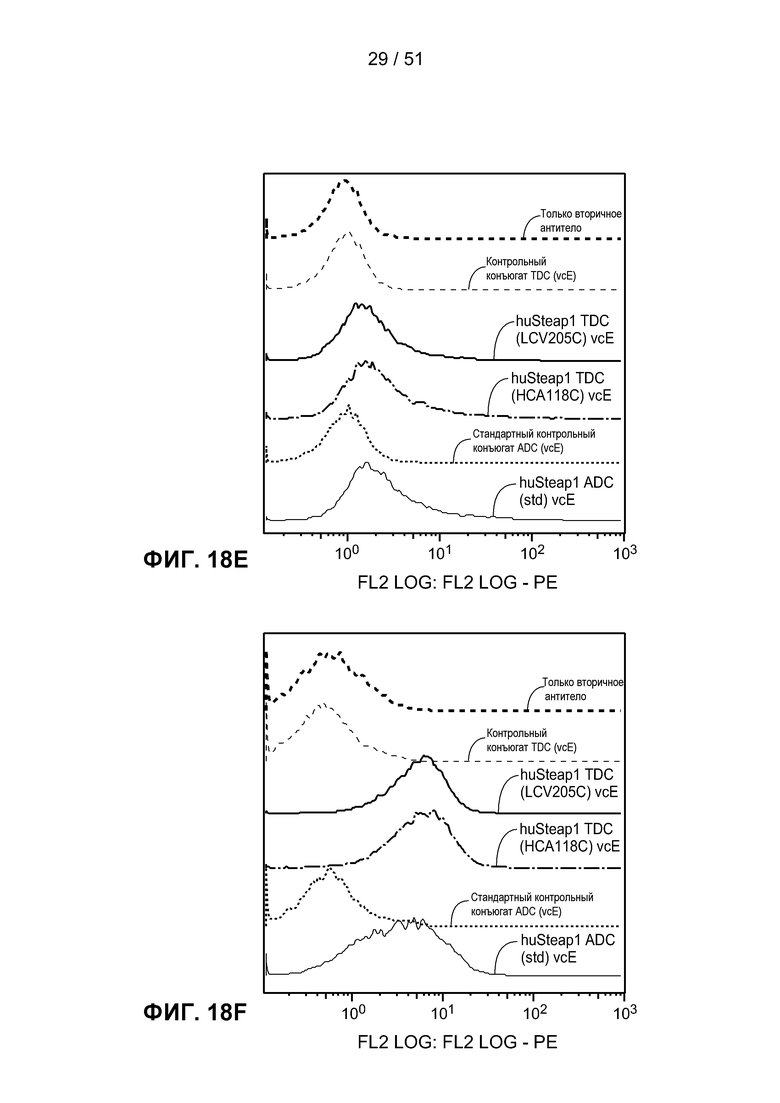
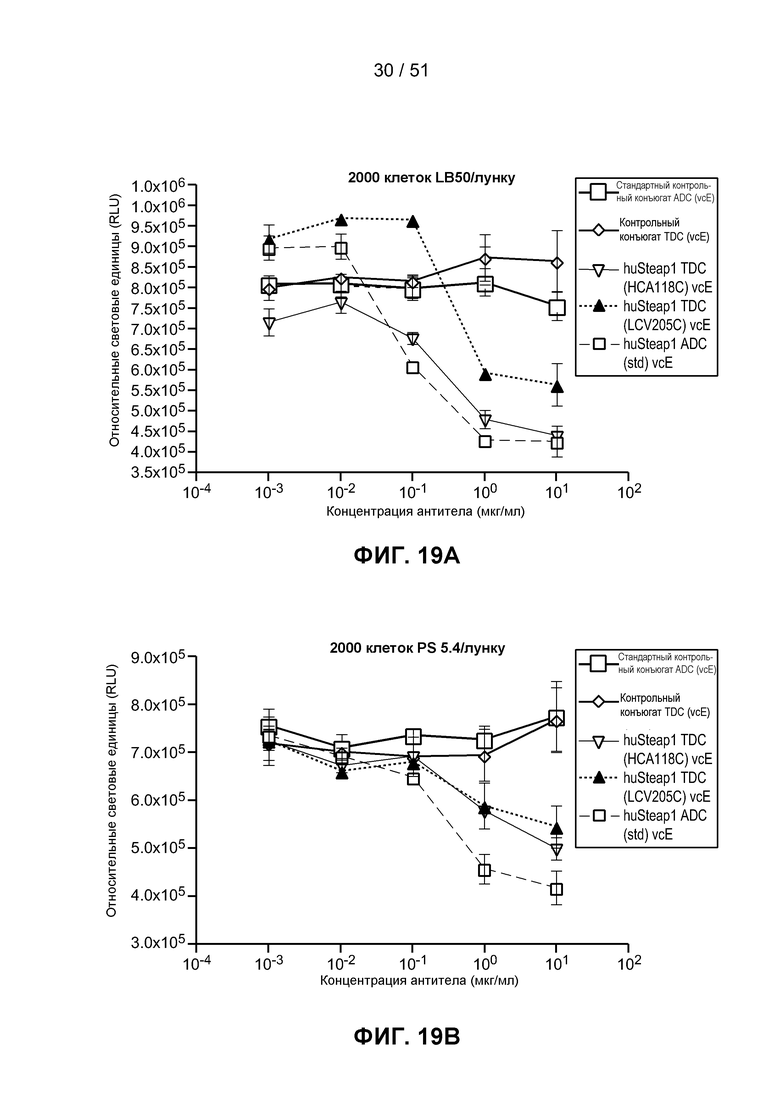
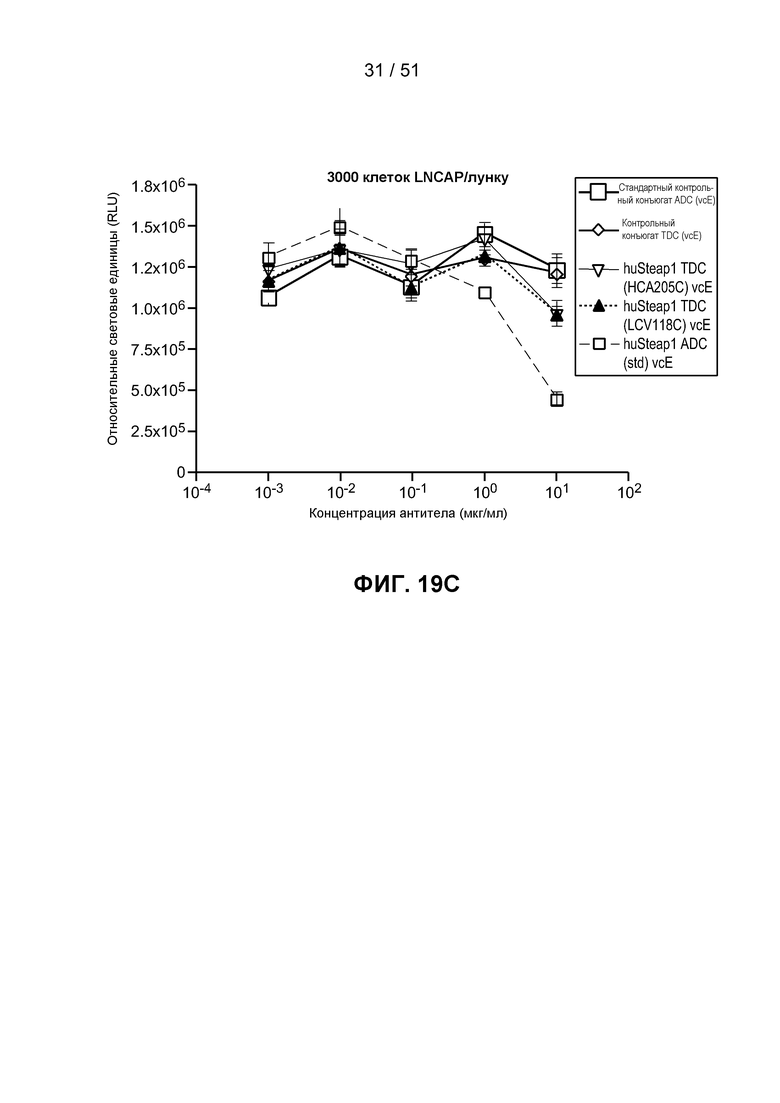

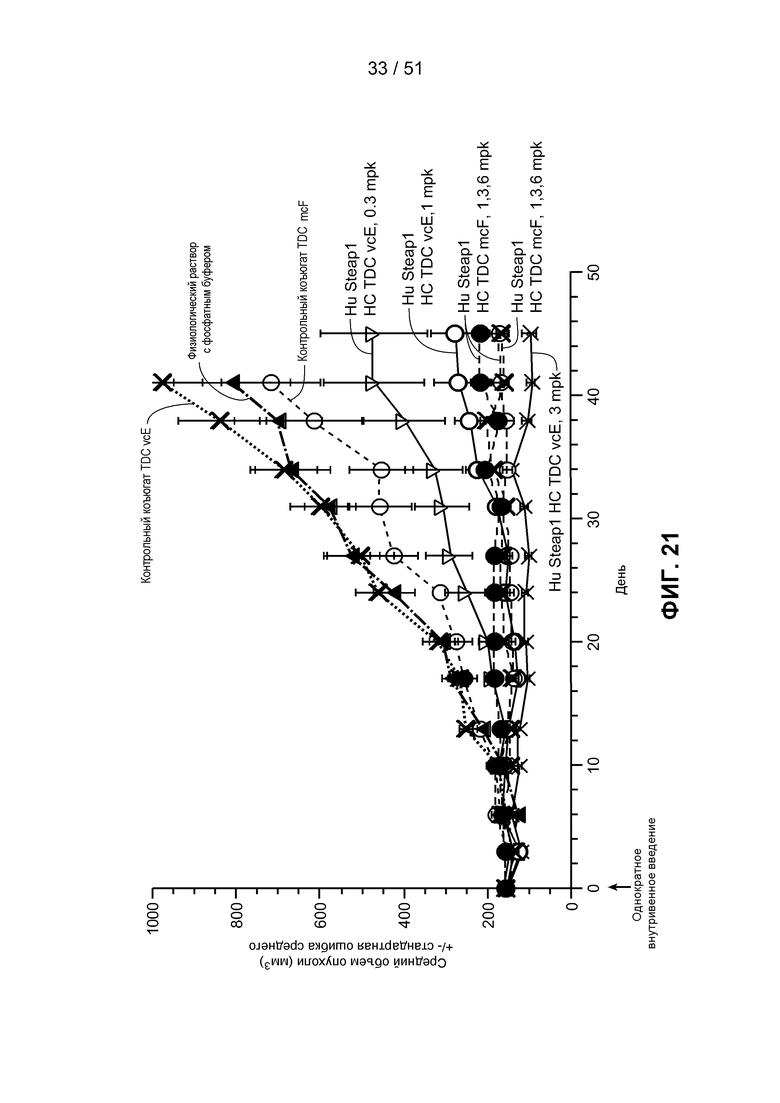
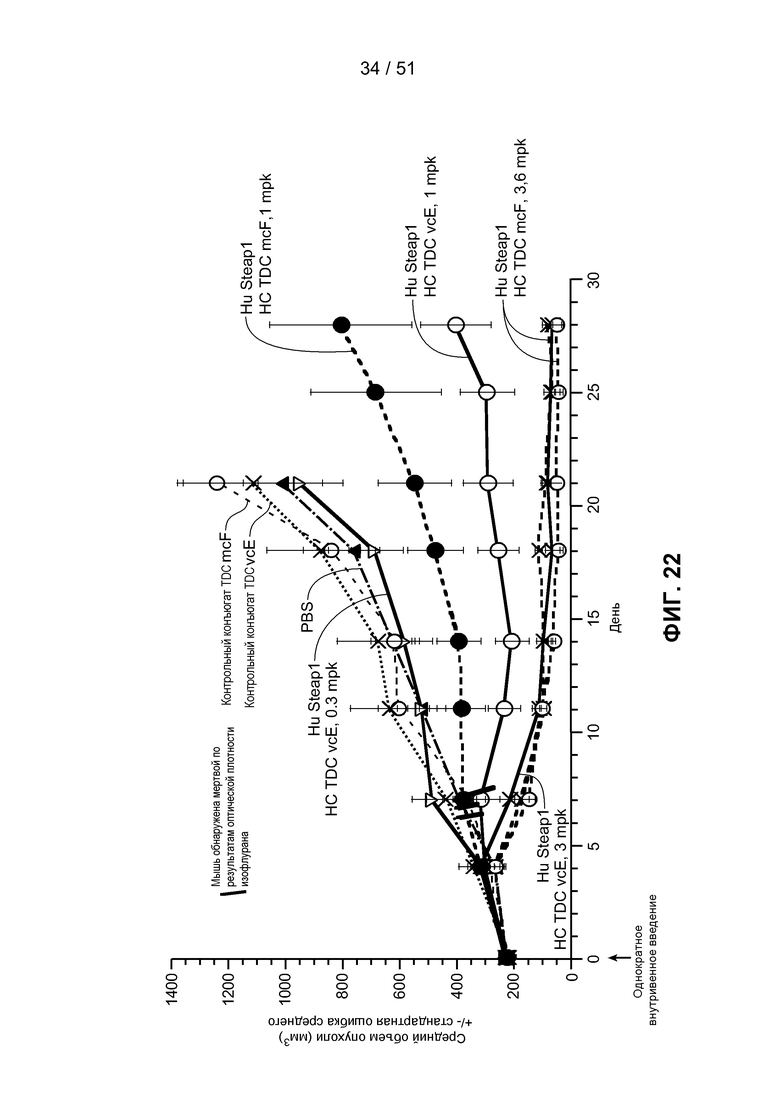
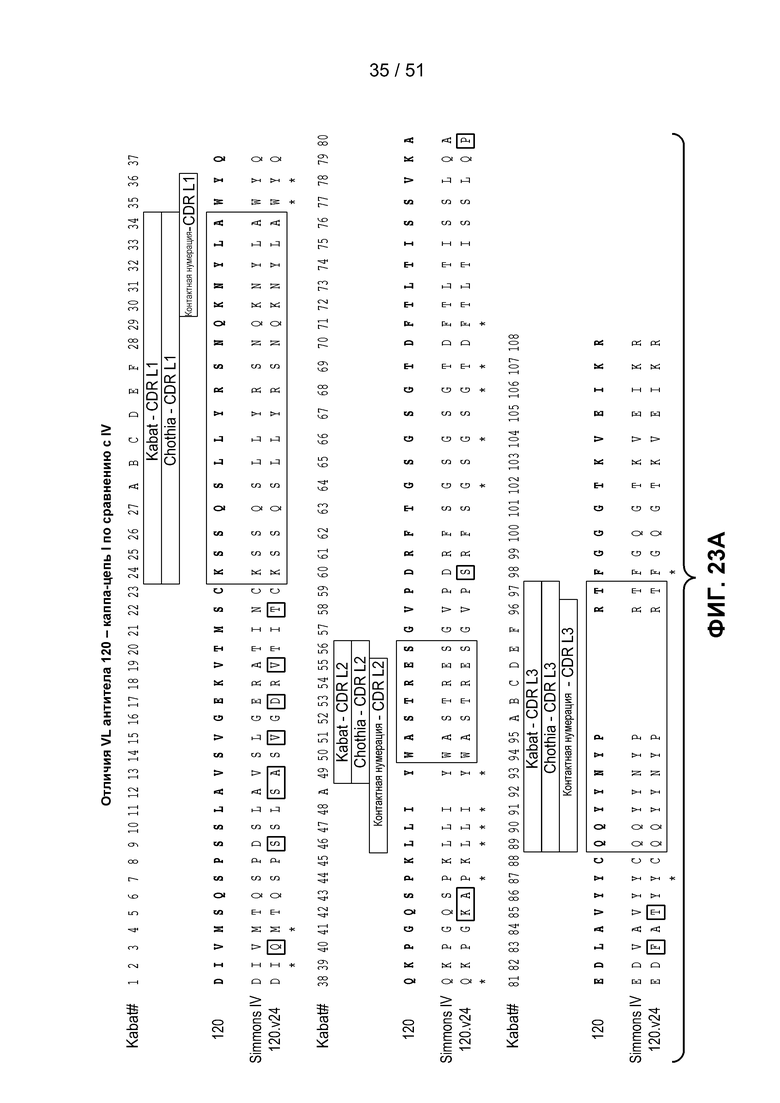
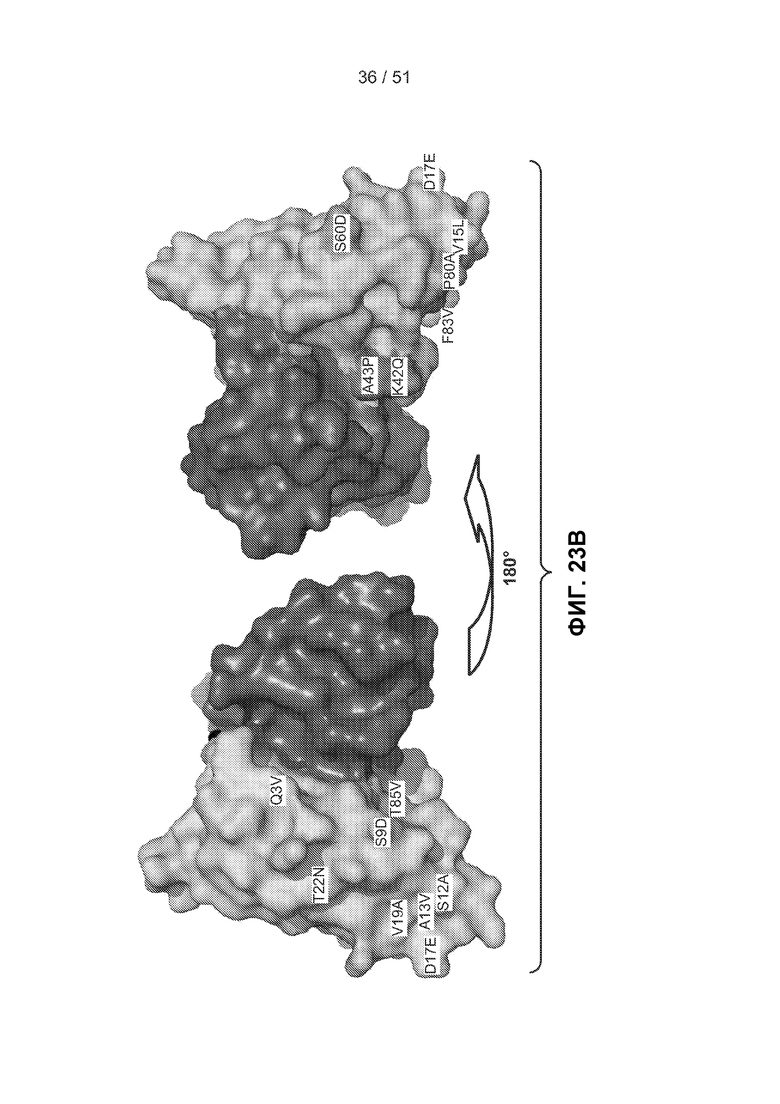
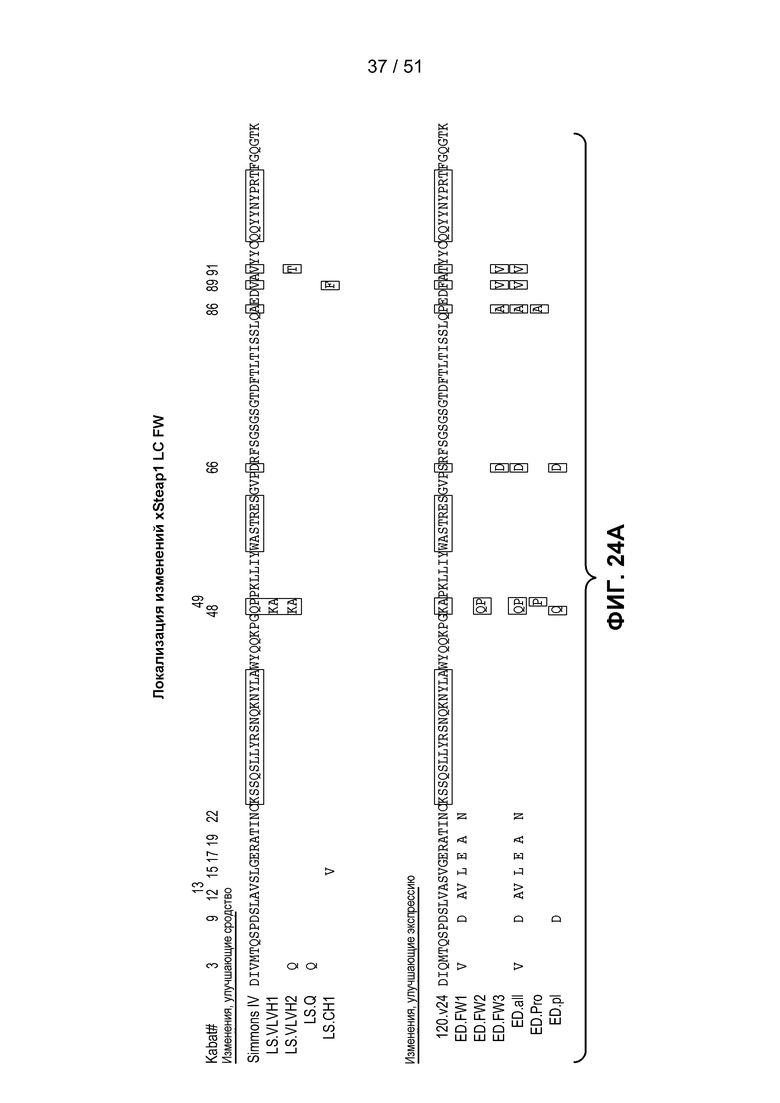
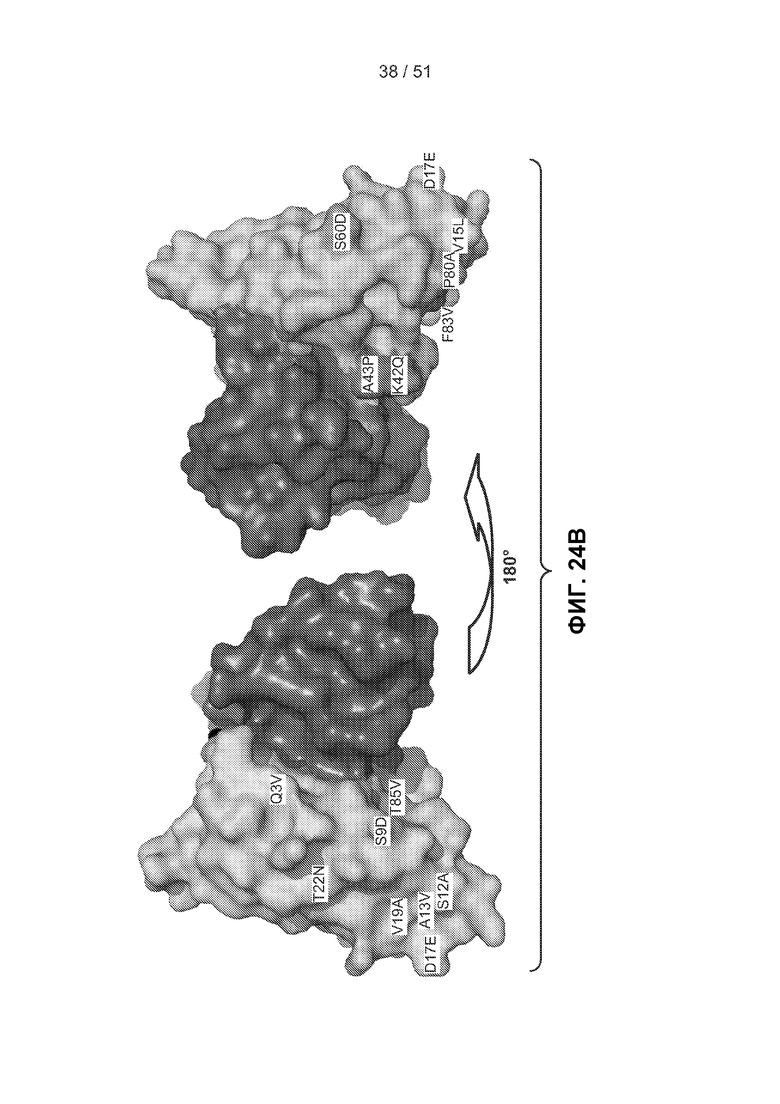
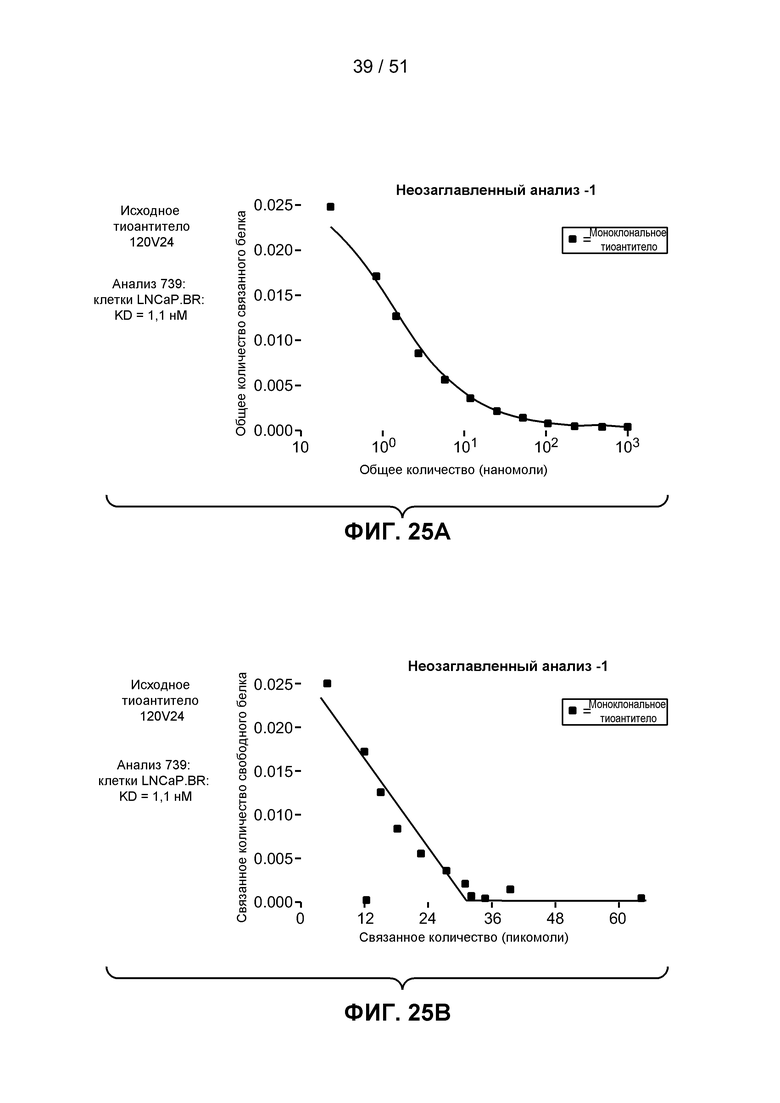
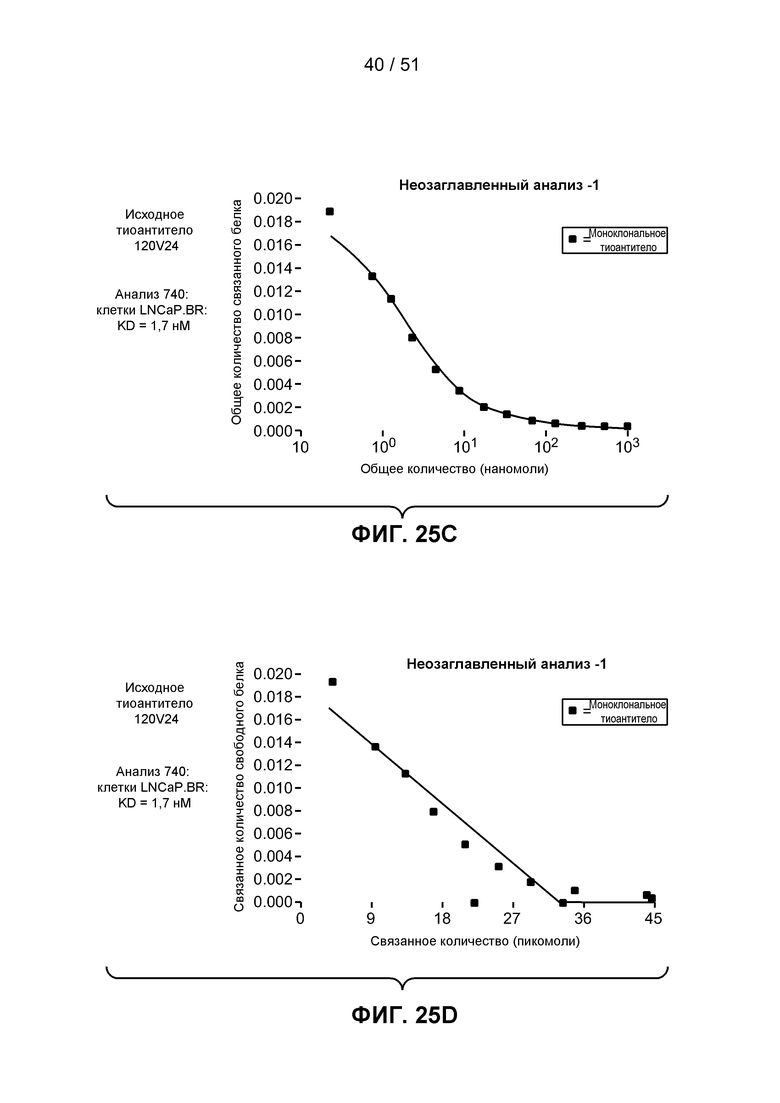
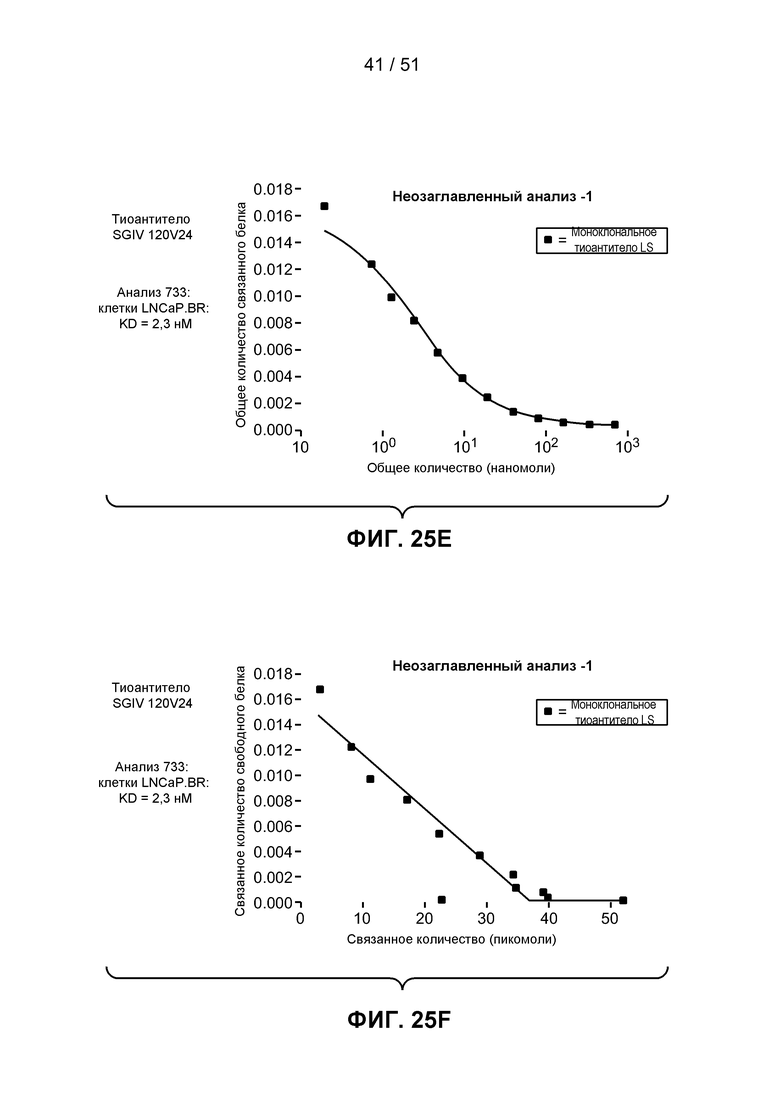
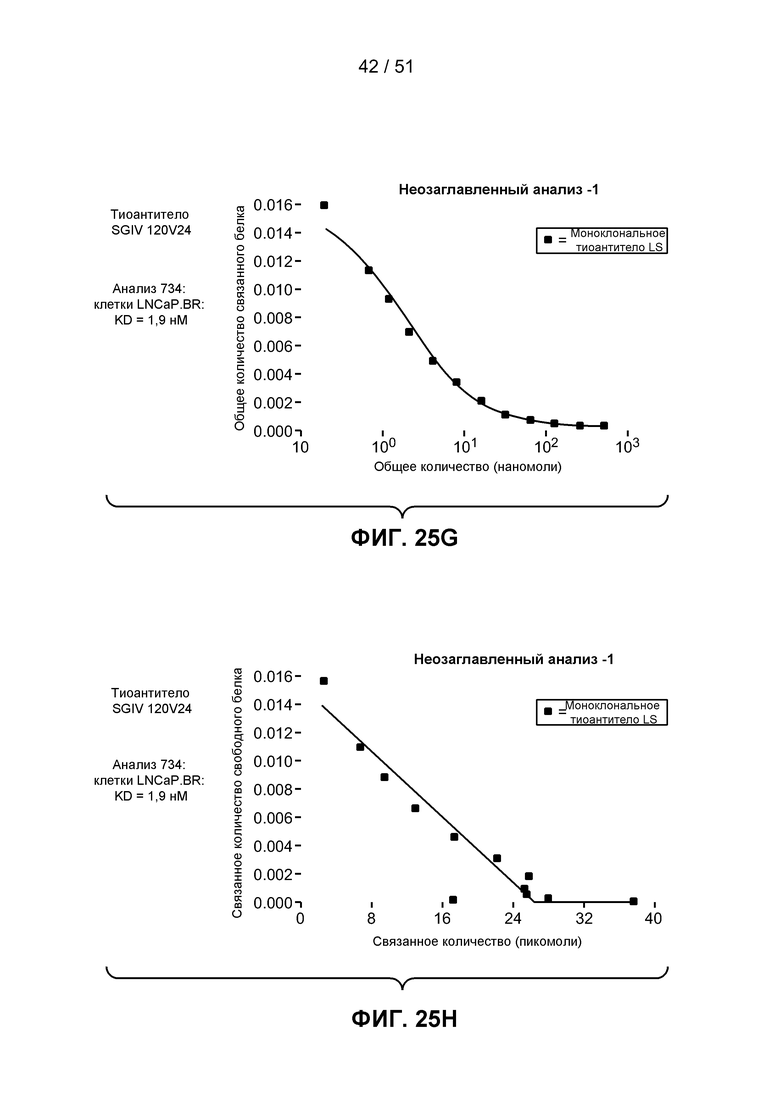
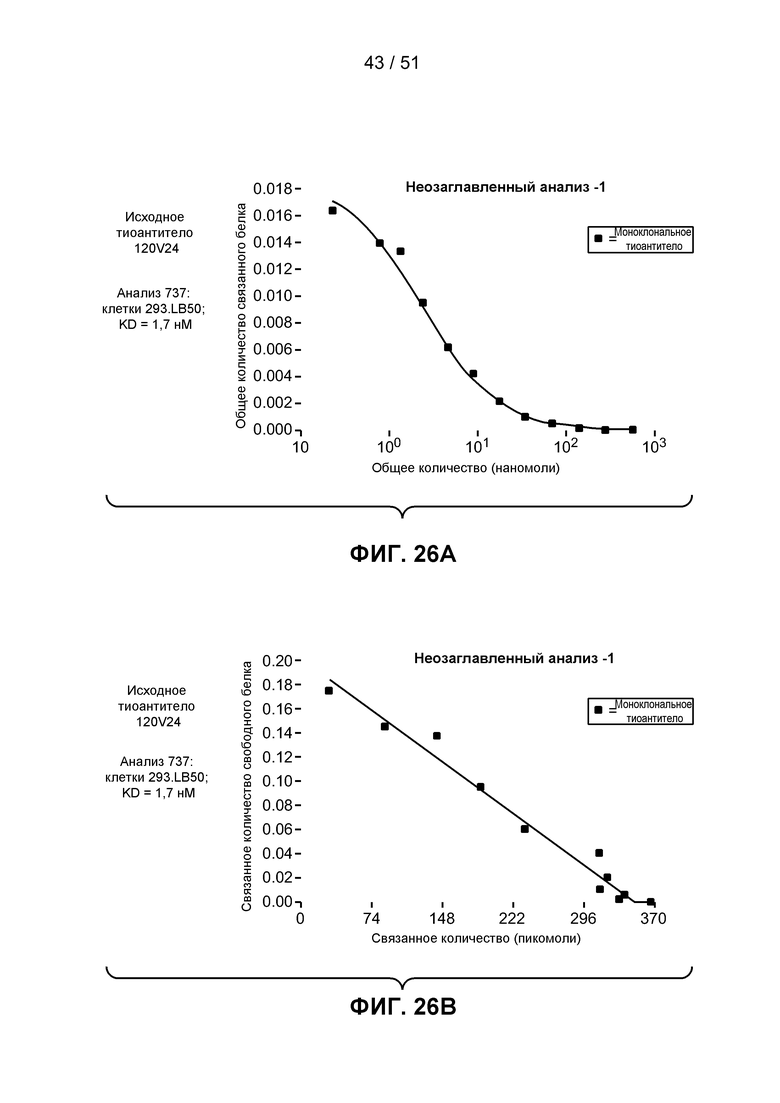
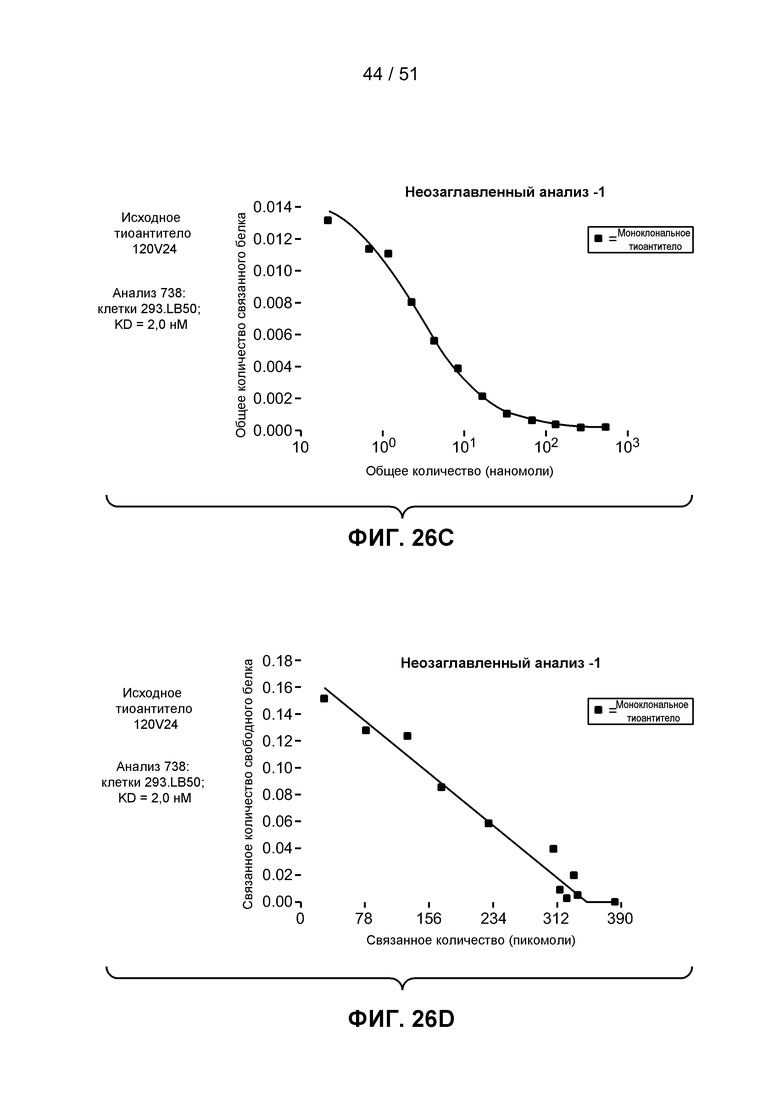
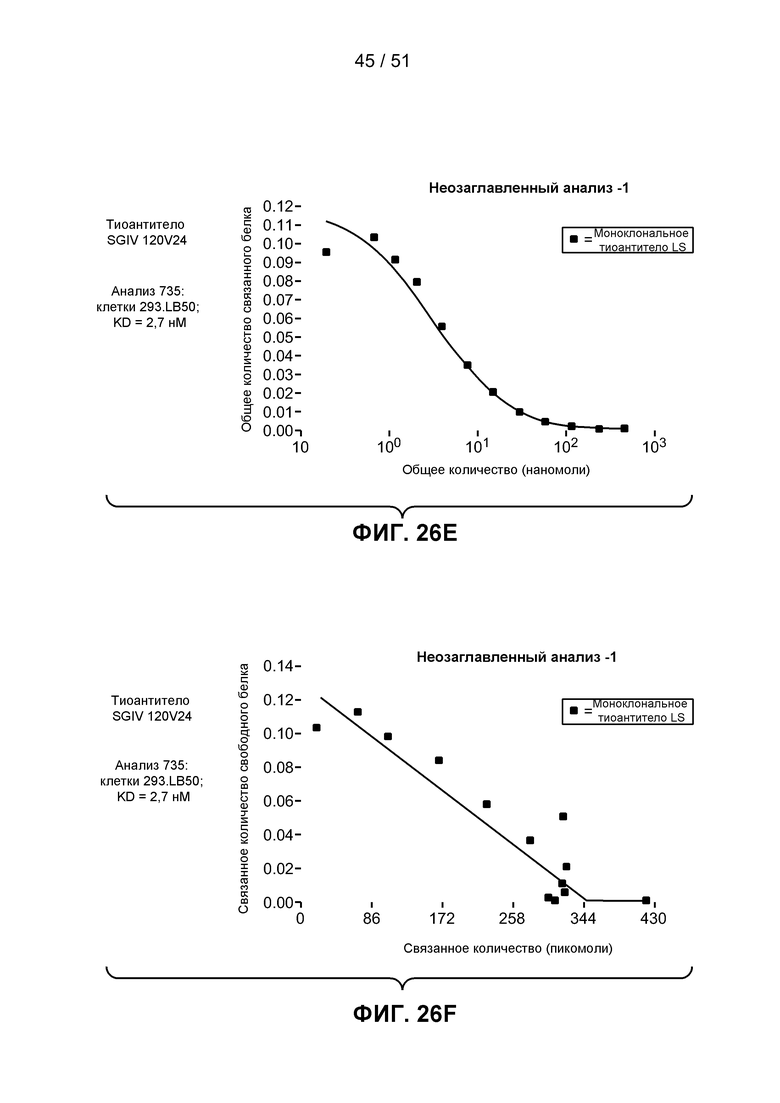
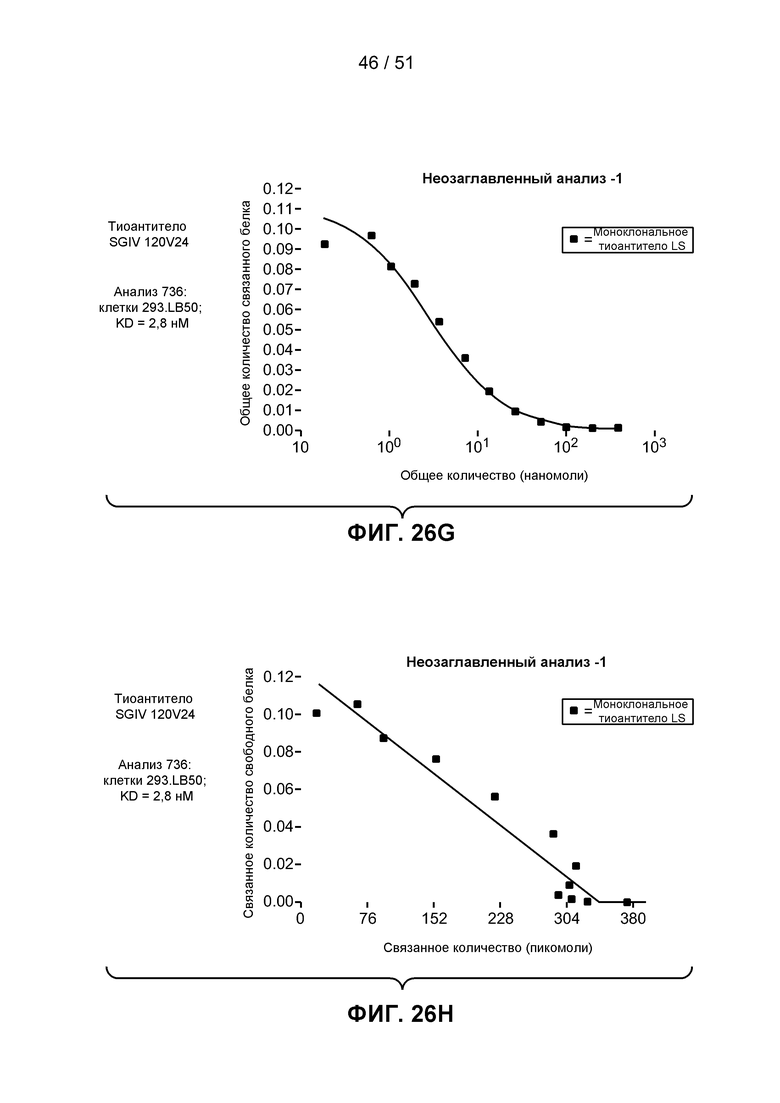
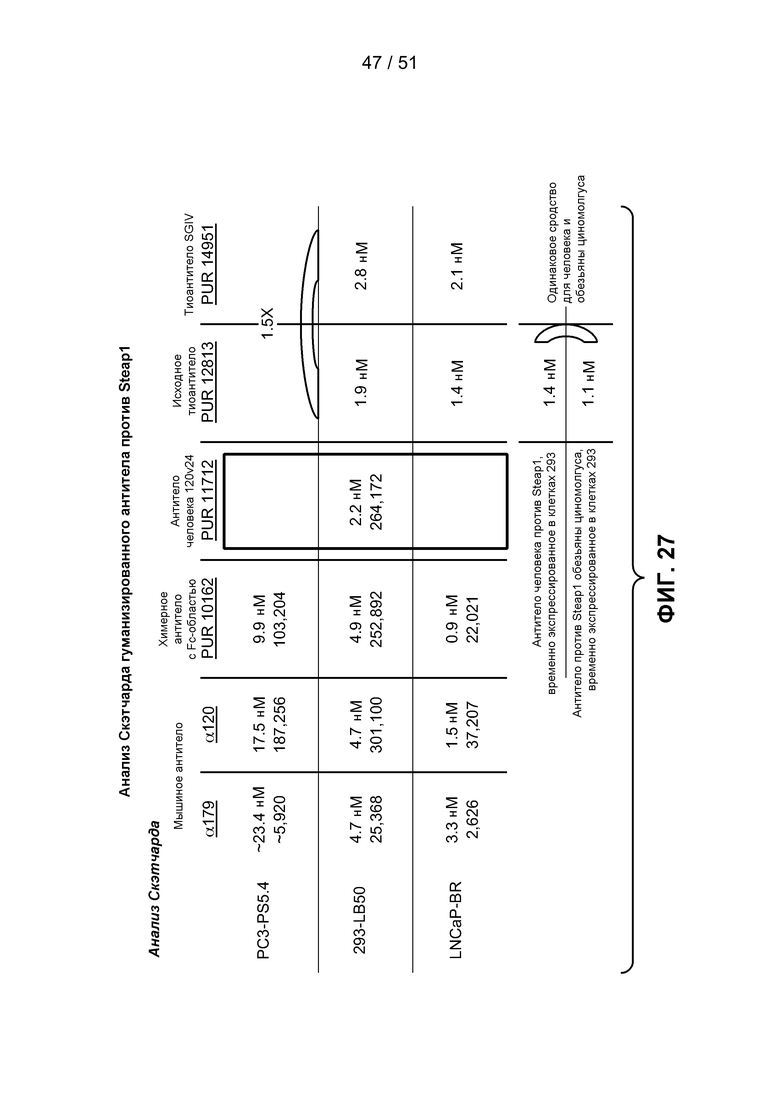
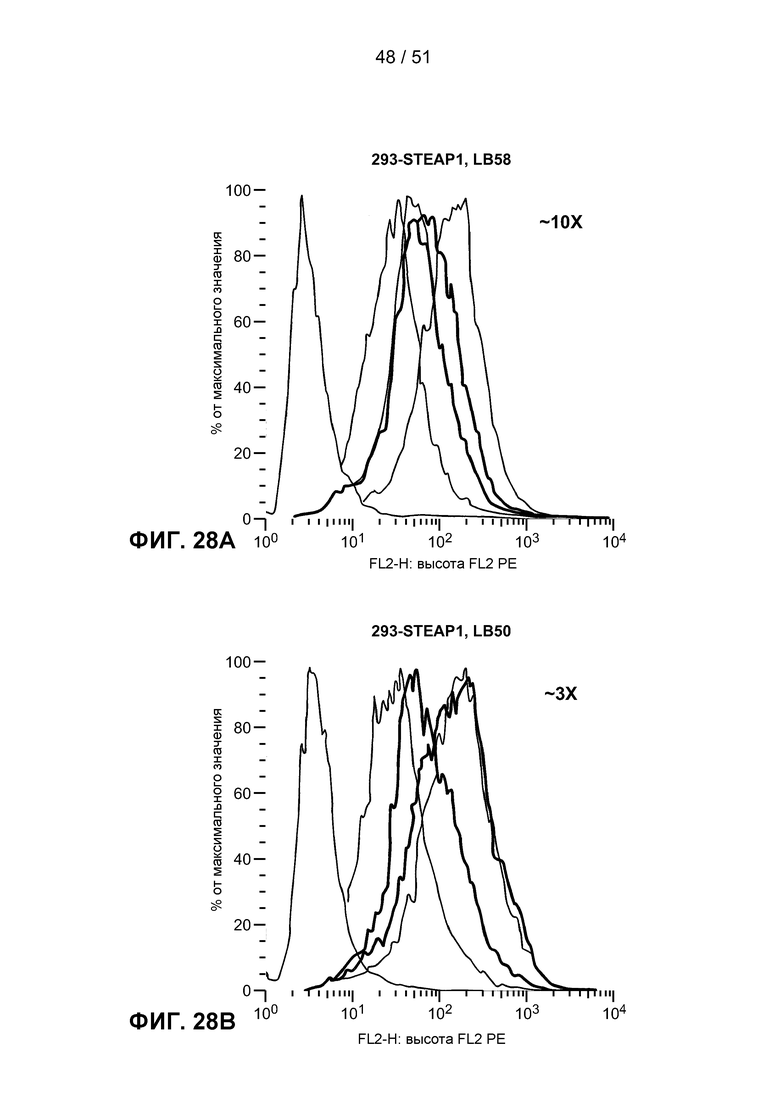
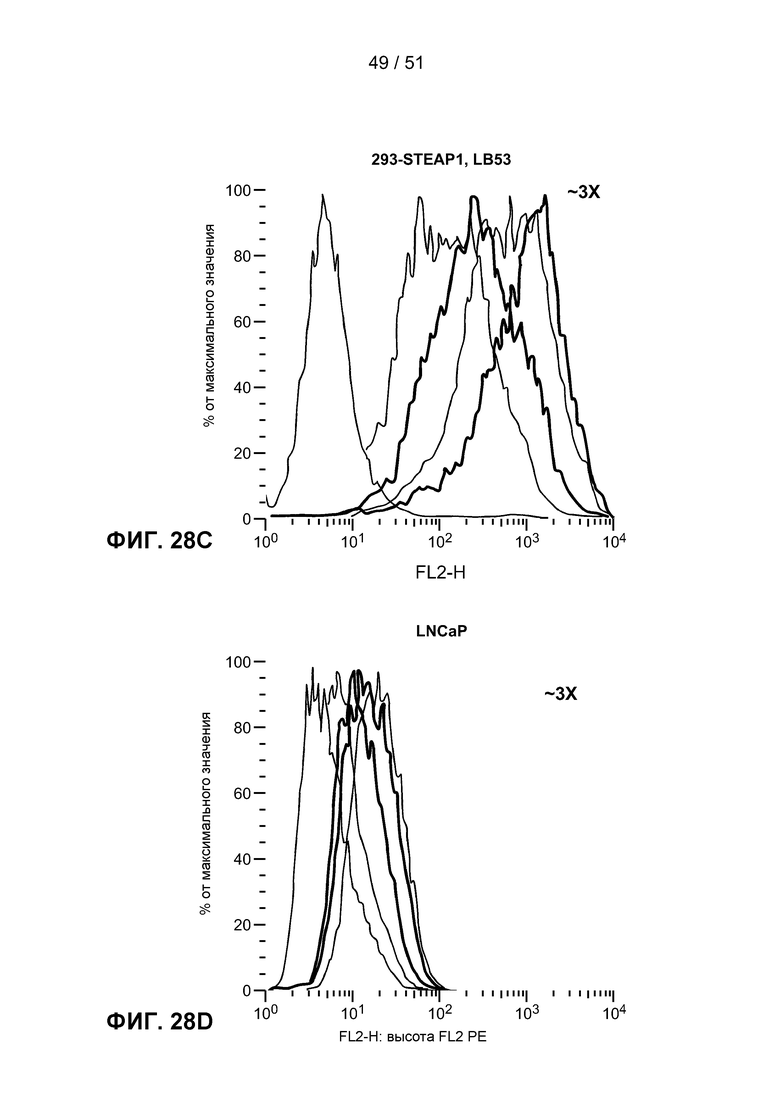
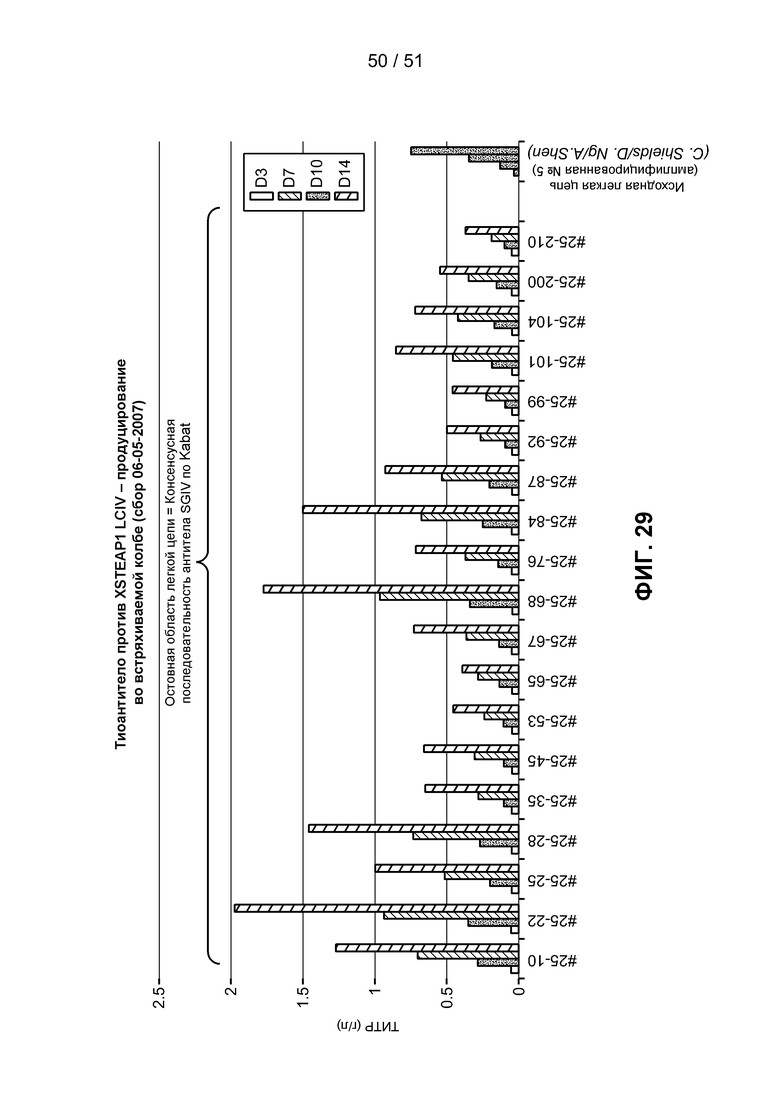
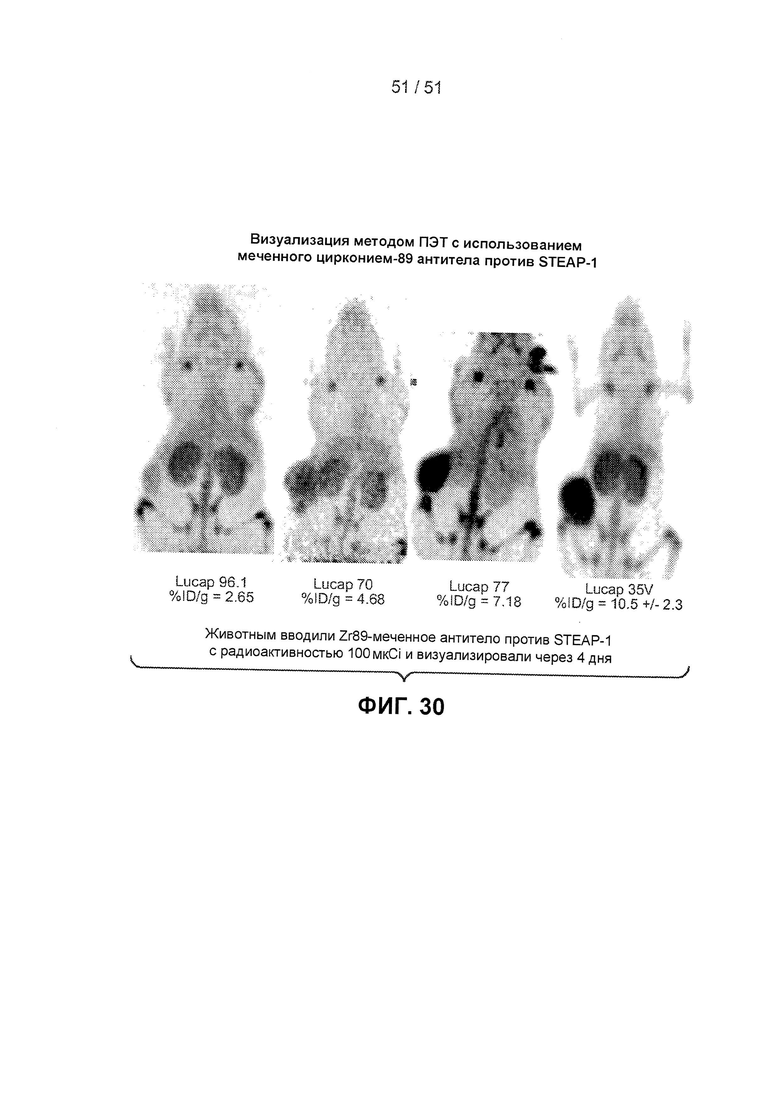
Изобретение относится к медицине и может быть использовано для определения присутствия белка STEAP-1 в биологическом образце, полученном от индивида, имеющего подозрение на метастатический рак предстательной железы. Для этого биологический образец подвергают воздействию меченного 89цирконием антитела против STEAP-1 и измеряют связывание меченного 89цирконием антитела против STEAP-1 с белком в биологическом образце. При этом связывание меченного 89цирконием антитела против STEAP-1 с белком является показателем присутствия белка STEAP-1 в образце. Меченное 89цирконием антитело против STEAP-1 содержит вариабельную область тяжелой цепи, содержащую гипервариабельную область CDR-H1, содержащую GYSITSDYAWN, CDR-H2, содержащую GYISNSGSTSYNPSLKS, и CDR-H3, содержащую ERNYDYDDYYYAMDY; и вариабельную область легкой цепи, содержащую CDR-L1, содержащую KSSQSLLYRSNQKNYLA, CDR-L2, содержащую WASTRES, и CDR-L3, содержащую QQYYNYPRT, и меченное 89цирконием антитело против STEAP-1 представляет собой 89цирконий-десферриоксамид (Df) антитело против STEAP-1. Изобретение позволяет определять присутствия белка STEAP-1 в биологическом образце, полученном от индивида, с метастатическим раком предстательной железы. 5 з.п. ф-лы, 2 табл., 30 ил., 10 пр.
1. Способ определения присутствия белка STEAP-1 в биологическом образце, полученном от индивида, имеющего подозрение на метастатический рак предстательной железы, с подозрением на содержание указанного белка STEAP-1, в котором указанный биологический образец подвергают воздействию меченного 89цирконием антитела против STEAP-1 и измеряют связывание указанного меченного 89цирконием антитела против STEAP-1 с указанным белком в указанном биологическом образце, при этом связывание указанного меченного 89цирконием антитела против STEAP-1 с указанным белком является показателем присутствия указанного белка STEAP-1 в указанном биологическом образце, где способ осуществляется у пациента с подозрением на метастатический рак простаты с метастазами в кости и где
(A) меченное 89цирконием антитело против STEAP-1 содержит
(a) вариабельную область тяжелой цепи, содержащую
(i) гипервариабельную область (CDR)-H1, содержащую GYSITSDYAWN,
(ii) CDR-H2, содержащую GYISNSGSTSYNPSLKS, и
(iii) CDR-H3, содержащую ERNYDYDDYYYAMDY, и
(b) вариабельную область легкой цепи, содержащую
(i) CDR-L1, содержащую KSSQSLLYRSNQKNYLA,
(ii) CDR-L2, содержащую WASTRES, и
(iii) CDR-L3, содержащую QQYYNYPRT, и
(B) меченное 89цирконием антитело против STEAP-1 представляет собой 89цирконий-десферриоксамид (Df) антитело против STEAP-1.
2. Способ по п. 1, который является способом визуализации при помощи иммуно-ПЭТ.
3. Способ по п. 1, где меченное 89цирконием антитело против STEAP-1 содержит 89цирконий, конъюгированный через аминогруппу в боковой цепи лизина антитела против STEAP-1.
4. Способ по п. 3, в котором индивид является человеком.
5. Способ по п. 1, в котором меченное 89цирконием антитело против STEAP-1 является 89Zr-меченым антителом против STEAP-1 с радиоактивностью 100 мкКи.
6. Способ по пп. 1-5, в котором меченное 89цирконием антитело против STEAP-1 содержит антитело, связывающееся с белком STEAP-1, при этом антитело, связывающееся с белком STEAP-1, содержит вариабельный домен тяжелой цепи, последовательность которого по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 9 или 10, и вариабельный домен легкой цепи, последовательность которого по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 6.
| US 2010111856 A1, 06.05.2010 | |||
| VAGHJIANI RJ et al., Six-transmembrane epithelial antigen of the prostate (STEAP1 and STEAP2)-differentially expressed by murine and human mesenchymal stem cells,Tissue Eng Part A | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| VEREL I et al., 89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies.J Nucl Med | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |

