Изобретение относится к области биотехнологии, биоинженерии, биохимии и медицинской микробиологии.
Основными областями применения ДНК-аптамеров являются медицинские и биологические исследования, создание высокоспецифических лигандов к любым органическим молекулам, включая малые, инструментов для клинической диагностики, в том числе создание высокочувствительных и высокоэффективных систем детекции мишеней белковой природы, а также терапия заболеваний различной этиологии.
Шига-токсины первого и второго типов (Stx1 и Stx2), распространенные, в частности, у бактерий Е. coli 0157:Н7, представляют собой двухкомпонентные бактериальные токсины, состоящие из рецепторного и эффекторного компонентов. Рецепторный компонент токсинов, представленный гомопентамером, связывается с рецепторами на поверхности клеток и обеспечивает создание поры, через которую происходит транслокация эффекторного компонента. Последний как в случае Stx1, так и в случае Stx2, связывает глоботриазилцерамид Gb3, что в силу различных механизмов приводит к инициации целого каскада событий сигнальной трансдукции, выражающихся в разнообразных физиологических последствиях - апоптозе, связывании лейкоцитов с клетками эндотелия, выбросе цитокинов, тромбообразовании, микроангиопатическом гемолизе [Psotka М.А., Obata F. с соавт. // Infect. Immun. - 2009. - V. 77. - №3. - P. 959-969].
Таким образом, шига-токсины относятся к подгруппе АВ5 двухкомпонентных токсинов. Альтернативные названия - «цитотоксины», «веротоксины» (на основе обнаруженной гиперчувствительности к данным токсинам, клеток линии Vero [Gentry М.K., Dalrymple J.M. // J. Clin. Microbiol. - 1980. - V. 12. - №3.- P. 361-366]). «Истинные» шига-токсины относятся к токсинам Shigella dysinteriae (Flexnerii) [Bergan J., Dyve Lingelem A.B. с соавт. // Toxicon. - 2012. - V. 60. - №6. - P. 1085-1107], в то время, как токсины энтерогеморрагической кишечной палочки обозначаются как «шига-подобные токсины», хотя гомология между соответствующими типами токсинов шигелл и Е. coli составляет более 90 %. Токсины 1 и 2 типов имеют между собой около 50-60 % гомологии [Flagler M.J., Mahajan S.S. с соавт. // Biochemistry. - 2010. - V. 49. - №8. - P. 1649-5167], при этом, как обнаружено в экспериментах на животных, токсичность Stx2 примерно в 400 раз превышает токсичность Stx1 по отношению к клеткам млекопитающих [Samuel J.E., Perera L.P. с соавт. // Infect Immun. - 1990. - V. 58. - №3. - P. 611-618].
Механизм действия обоих типов токсинов состоит в прерывании белкового синтеза путем модификации 28S РНК в 60S субъединице рибосомы [Sandvig K., Bergan J. с соавт. // Toxicon. - 2010. - V. 56. - Suppl. 7. - P. 1181-1185]. Эффекторная субъединица токсинов является N-гликозидазой, которая расщепляет специфическую молекулу аденина в составе РНК. В высокой концентрации при оверпродукции токсин также ингибирует рост бактерий.
Рецептором для токсина являются гликолипиды, в частности глоботриазилцерамид Gb3 [Samuel J.E., Perera L.P. с соавт. // Infect Immun. - 1990. - V. 58. - №3. - P. 611-618]. Концентрация Gb3 по невыясненным причинам особенно высока в клетках почечного эпителия. Поэтому один из основных синдромов тяжелой интоксикации шига-токсинами так называемый гемолитико-уремический синдром (ГУС), который приводит к тяжелой инвалидизации, а иногда и к смерти. В частности, при вспышке атипичной инфекции, вызванной несущим шига-токсин штаммом Е. coli О104:Н4 в Европе в 2011 году, из более 1500 заболевших 45 человек погибло несмотря на высокоэффективную медицину стран, охваченных инфекцией (Франция, Германия) [Beutin L., Martin A. J. с соавт.// Food Prot. -2012. - V. 75. - №2. - P. 408-418].
Инфекция токсигенной кишечной палочки и шигелл - зооантропонозы, основным переносчиком их являются свиньи и крупный рогатый скот [Pruimboom-Brees I.M., Morgan T.W. с соавт. // Proc. Natl. Acad. Sci. USA. - 2000. -V. 97. - №19. - P. 10325-10329]. Однако у этих млекопитающих отсутствуют рецепторы шига-токсина на поверхности клеток, поэтому инфекция у взрослых животных протекает практически бессимптомно, создавая природный резервуар токсигенных бактерий, в котором их элиминация и блокировка передачи к людям и контаминации продуктов питания серьезно затруднена.
Ранняя диагностика шигеллезной инфекции осложняется неспецифичностью симптомов (существует целый ряд кишечных инфекций как бактериальной, так ивирусной природы, имеющих сходные ранние симптомы - рвота, диарея), а также ввиду недавнего обнаружения природных атипичных штаммов кишечной палочки, несущих шига-токсин [Borges С.Α., Beraldo L.G. с соавт. // Foodborne Pathog. Dis. - 2012. - V. 9. - №12. - P. 1119-1125]. Полиморфизм самих генов шига-токсинов может вносить вклад в сложность ПЦР-диагностики, равно как и присутствие ингибиторов ПЦР, серьезно затрудняющих детекцию токсинов в образцах продуктов питания [Auvray F., Lecureuil С. с соавт. // Lett. Appl. Microbiol. - 2009. - V. 48. - №5 - P. 554-559]. Считается, что шига-токсины являются компонентом генома профага, а не основного генома Е. coli или шигелл. Мобилизация профага, которая может происходить в том числе in trans [Smith D.L., Rooks D.J. с соавт. // BMC Genomics. - 2012. - V. 13. - P. 311], может приводить к переносу генов токсина в другие штаммы кишечной палочки, поэтому специфическая детекция шига-подобных токсинов играет важнейшую роль в профилактике пищевых инфекций и предотвращении их распространения. Важно отметить, что если ареал встречаемости шигелл ограничен, в основном, тропической и субтропической зонами [Zaidi М.В., Estrada-Garcia Т. // Curr. Trop.Med. Rep. - 2014. - V. 1. - №2.- P. 81-87], ареал распространения токсигенных Ε. coli ограничен практически только распространением животноводства.
Специфическая терапия интоксикации шига-подобными токсинами также практически отсутствует. Целый ряд антибактериальных препаратов может повысить уровень выброса токсина в кровь [Bielaszewska Μ., Idelevich Ε.А. с соавт. // Antimicrob. Agents Chemother - 2012. - V. 56. - №6 - P. 3277-3282]. Возможности моноклональных терапевтических антител, в последнее время разрабатываемых для специфической терапии бактериальных токсикоинфекций, ограничиваются внеклеточным пространством, поскольку антитела, как правило, не способны проникать внутрь клетки в интактном состоянии, а миниантитела, созданные на базе классических антител, как правило, не обладают достаточной стабильностью в плазме крови и быстро метаболизируются, не обеспечивая терапевтический эффект [Kubetzko S., Balic Ε. с соавт. // J.Biol. Chem. - 2006. - V. 281. - №46. - P. 35186-35201].
Одной из новых платформ для создания средств высокоэффективной диагностики и терапии являются аптамеры на основе нуклеиновых кислот. Наиболее рациональным вариантом нуклеиновых аптамеров по сумме характеристик (удобство синтеза, стабильность при хранении и в организме) являются ДНК-аптамеры и их производные. ДНК-аптамеры - одноцепочечные олигнуклеотиды, которые благодаря образованию сложных трехмерных структур и пространственному расположению зарядов в молекуле имеют высокую аффинность к молекулам-мишеням. Направленный отбор аптамеров против выбранных мишеней осуществляется по технологии SELEX ("Systematic Evolution of Ligands by Exponential Enrichment") [US Patent 5567588] с применением различных модификаций технологии, таких как микромагнитная селекция в микрожидкостных каналах (M-SELEX) [Lou X., Qian J., Xiao Y., Viel L., Gerdon A.E., Lagally E.Т., … &Soh Η.T. // Proceedings of the National Academy of Sciences. - 2009. - V. 106. - №. 9. - P. 2989-2994], использование капиллярного элекрофореза (ECEEM) [Drabovich Α., Berezovski Μ., Krylov S. Ν. // Journal of the American Chemical Society. - 2005. - V. 127. - №. 32. - P. 11224-11225] и др.
Известны различные варианты молекул, специфически ингибирующих токсическое действие шига-токсинов, в частности известна разработка малых ингибирующих молекул, нейтрализующих действие шига-токсина [Dasgupta S., Kitov P.I. с соавт. // Angew. Chem. Int. Ed. Engl. - 2014. - V. 53. - №6. - P. 1510-1515]. Однако указанная работа описывает лишь подходы к получению ингибирующих молекул, не описывая природу и эффективность действия подобных молекул в условиях in vivo.
Известны полноразмерные человеческие или гуманизированные моноклональные антитела [López E.L., Contrini Μ.Μ. с соавт. // Antimicrob. Agents Chemother. - 2010. - V. 54. - №1. - P. 239-243] или однодоменные антитела (VHH) [Tremblay J.M., Mukherjee J. с соавт. // Infect Immun. - 2013. - V. 81. - №12. - P. 4592-4603], разработанные с целью прямого ингибирования шига-токсина. При применении данных антител наблюдается смягчение или предотвращение ГУС в экспериментах с лабораторными животными. Однако попытки использовать однодоменные антитела для создания лекарств проводятся с 1998 года, и в отличие от классических антител ни одно из них не было утверждено в качестве терапевтического препарата [Lauwereys Μ. с соавт. // Тhе ЕМВО journal. - 1998. - V. 17. - №. 13. - Р. 3512-3520]. Однако, во-первых, ни одно из указанных антител не достигло III стадии клинических испытаний несмотря на многолетние (10 лет в случае VHH) исследования, а во-вторых, ни одно из описанных антител не способно нейтрализовать внутриклеточно локализованный токсин.
Известно также моноклональное терапевтическое антитело, блокирующее терминальные стадии активации комплемента (экулизумаб, [Delmas Y., Vendrely В. с соавт. // Nephrol. Dial. Transplant. - 2014. - V. 29. - №3. - P. 565-572]), характеризующееся непрямым действем, направленным на уменьшение эффекта интоксикации шига-токсинами при ГУС. Однако экулизумаб не способен предотвратить развитие интоксикации и гемолитико-уремического синдрома, поскольку это антитело не действует непосредственно на шига-токсин и не блокирует его взаимодействие с клеточными рецепторами или каталитическую активность. Оценка стоимости курса терапии моноклональными антителами, рассчитанная на основе протокола применения экулизумаба, в рамках которого пациенту вводится до нескольких граммов антител, составляет более 20 тысяч долларов США. Стоимость курса терапии аптамерами с учетом сравнимых с моноклональными антителами характеристик ингибирования действия токсина в пересчете на одну молекулу может составить несколько сотен долларов США, а с учетом инфраструктуры хранения и сроков годности общая разница по затратам будет еще значительнее.
Известны различные молекулярные системы для детекции шига-токсина на основе ПЦР [Belanger S.D., Boissinot Μ. с соавт. // J.Clin. Microbiol - 2002. - V. 40. - №4. - P. 1436-1440], иммуно-ПЦР [Не X., Qi W. с соавт. // Appl.Environ. Microbiol. - 2011. - V. 77. - №11. - P. 3558-3564], сенсорных технологий [Xing J.Z., Zhu L. с соавт. // J.Appl.Microbiol. - 2012. - V. 113. - №2. - P. 429-437], иммуноферментного анализа [Gavin P.J., Peterson L.R. ссоавт. // J.Clin. Microbiol - 2004. - V. 42. - №4 - P. 1652-1656]. Однако использование прямого метода ПЦР в образцах продуктов питания, которые зачастую являются источниками вспышки, образцах из окружающей среды, а также в клинических образцах бывает затруднительным ввиду присутствия ингибиторов ПЦР. Чувствительность иммуноферментного анализа составляет максимум 0,1 нг/мл, что недостаточно для ранней диагностики интоксикации [Gyles CL. // J.Anim.Sci. - 2007. - V. 85. - 13 Suppl.:E45-62]. Сенсорные технологии обеспечивают высокую чувствительность, однако для их использования необходимо нестандартное оборудование, не применяющееся в клинической практике и часто доступное только в виде экспериментальных образцов, работа с которыми требует высокой и специфической квалификации оператора. Иммуно-ПЦР может обеспечить требуемую чувствительность детекции и не требует от персонала дополнительной квалификации, кроме той, что необходима для постановки ПЦР и ИФА, однако требует подбора очень высокоэффективной пары моноклональных антител с весьма низким уровнем неспецифического взаимодействия с материалами, используемыми в тест-системе. Модификация антител с целью минимизации неспецифического связывания весьма трудоемка и дорогостояща, и ее эффективность невозможно предсказать заранее, поэтому использование классического иммуно-ПЦР до сих пор не получило того распространения, которое должен был бы заслуживать данный метод на основании его чувствительности, простоты постановки и устойчивости к ингибиторам ПЦР.
Известны наиболее близкие к объекту изобретения аптамеры, специфичные к шига-токсину 2 типа, созданные на основе рибонуклеиновой кислоты (РНК) [Challa S., Tzipori S., Sheoran Α. // J. Nucl. Acids. - 2014:214929. doi: 10.1155/2014/214929]. Однако поскольку эти аптамеры получены на основе РНК, которая неустойчива к факторам внешней среды, их достаточно сложно химически трансформировать в устойчивую молекулу без потери аффинности [Sun Η., Zhu X. с соавт. // Mol. Ther. Nucl. Acids.- 2014. - 3:e182. doi: 10.1038/mtna.2014.32]. Кроме того, данные аптамеры не ингибируют токсический эффект шига-токсина по отношению к модельным эукариотическим клеткам. Получение данных аптамеров проводилось с использованием архаичной методики, не учитывающей значительное связывание неспецифических аптамеров с твердой фазой и не предлагающей методов негативной селекции таких молекул. Также не проводились количественные исследования констант связывания полученных аптамеров и исследование чувствительности детекции шига-токсина с помощью полученных аптамеров.
Задачей изобретения является получение аптамеров, обладающих аффинностью и специфичностью к рецепторной субъединице шига-подобного токсина Stx2, локализующегося в геноме энтерогеморрагических штаммов Е. coli (таких как 0157:Н7 или О104:Н4), и способностью к ингибированию токсического действия шига-подобного токсина Stx2.
Поставленная задача решается путем селекции аптамеров против химерного рекомбинантного полипептида, несущего в своем составе пептидную последовательность для иммобилизации данного полипептида на твердой фазе, отделенную от пептидной последовательности рецептор-связывающей субъединицы Stx2 уникальной пептидной последовательностью, распознаваемой и расщепляемой сайт-специфической протеазой.
Такая структура химерного рекомбинантного полипептида позволяет провести в мягких условиях элюцию мишени со специфически связанными с ней аптамерами, не элюируя одновременно молекулы аптамеров, связанные непосредственно с твердой фазой и с другими компонентами, помимо мишени находящимися на твердой фазе. Указанная процедура позволяет получить высокоспецифические аптамеры, используя небольшое число раундов селекции.
Также поставленная задача решается путем клонирования селектированных последовательностей ДНК в плазмидный вектор, определения их первичной структуры и проверки индивидуальных последовательностей ДНК на связывание с шига-токсином типа 2 при помощи ПЦР в режиме реального времени.
Также поставленная задача решается за счет тестирования защитных свойств ДНК-аптамеров на клетках Vero in vitro по отношению к шига-токсину Stx2n идентификации аптамеров с токсин-нейтрализующей активностью.
Техническим результатом изобретения является одноцепочечный ДНК-аптамер AptStx2B17 длиной 81 нуклеотид с молекулярной массой 27 кДа с первичной последовательностью
CATACGTTCGACTGCTACCTTAGCCTGGTCTTATGATGCCTGTTTGATCCTGATGGCT AATATGTGAGATGTACAGACTAG,
специфически связывающий рецепторную субъединицу Stx2 с константой связывания не менее 109 М-1 и ингибирующий токсичность Stx2 по отношению к эукариотическим клеткам Vero в концентрации ID50=3 мкМ.
Данное техническое решение использует специфическую элюцию белка-мишени с твердой фазы с использованием сайт-направленного протеолитического фермента для осуществления этапов позитивной селекции, что позволяет выделить при отборе исключительно те аптамеры, которые связаны с мишенью (Stx2B).
Также предлагаемое техническое решение предусматривает использование дополнительных раундов негативной селекции против твердой фазы, на которой иммобилизуется мишень, что дополнительно снижает процент отобранных последовательностей, не связывающихся с мишенью и способных амплифицироваться в процессе селекции.
Также техническое решение предполагает использование иммобилизации химерного белка на поверхности магнитных микросфер, что позволяет повысить эффективность выделения комплекса аффинных аптамеров и белка-мишени.
Также техническое решение предполагает использование иммуно-аптамерной ПЦР для анализа связывающей активности ДНК-аптамеров по отношению к шига-токсину Stx2.
Также техническое решение предусматривает тестирование токсин-нейтрализующей активности ДНК-аптамеров на клетках линии Vero.
Важным преимуществом заявленных последовательностей ДНК-аптамеров, специфически связывающих шига-токсин Stx2, является возможность использования их в качестве детектирующих молекул для определения низких концентраций токсина и в качестве кандидатных молекул при разработке медицинских препаратов для экстренной терапии шигеллезной интоксикации.
Изобретение осуществляют следующим образом:
Синтезируют комбинаторную библиотеку ДНК-олигонуклеотидов вида 5′-const1-N45-const2, где N45 - вариабельная область, состоящая из 45 нуклеотидов, a const1 и const2 - константные фланкирующие области длиной по 18 нуклеотидов, которые служат для связывания со специфическими праймерами, позволяющими амплифицировать пул олигонуклеотидов после этапов селекции (фиг. 1).
На основе рекомбинантной конструкции pGST/APT/X [Заявка на получение патента RU 2013147132] получают химерный белок GST/APT/STX2B. Для этого последовательность, кодирующую В-субъединицу шига-токсина 2, получают ПЦР-амплификацией с геномной ДНК бактерий Е. coli штамма 0157:Н7 (Государственная коллекция патогенных микроорганизмов ФБУН ГНЦ ПМБ, Оболенск) с помощью праймеров STX2ForBamHI и STX2RevXhoI, температура отжига праймеров составляет 56°C, время элонгации 20 с. Продукт апмлификации обрабатывают эндонуклеазами рестрикции BamHI и XhoI и клонируют в плазмидур GST/APT/X, обработанную теми же эндонуклеазами. Полученная экспрессионная плазмидар GST/APT/STX2B кодирует химерный слитный белок, содержащий последовательность фрагмента гена глютатион-S-трансферазы, пептид RRKKVYPYPME, являющийся субстратом летального фактора В. anthracis, in vivo биотинилируемый пептид и последовательность, кодирующую В-субъединицу шига-токсина 2.
Трансформируют плазмидой pGST/APT/STX2B клетки Ε. coli BL21(DE3), получая тем самым продуцент химерного пептида GST/APT/STX2B. Проводят продукцию полипептида в течение 3 ч после индукции жидкой культуры добавлением IPTG. После окончания продукции клеточную биомассу собирают центрифугированием, суспендируют в лизирующем буфере, суспензию бактерий обрабатывают лизоцимом и ДНКазой и центрифугируют для получения осветленного лизата, содержащего полипептид GST/APT/STX2B в растворимой форме.
Очистку белка GST/APT/STX2B из лизата проводят на колонке с иммобилизованным глутатионом, белок элюируют добавлением буфера, содержащего глутатион. Дополнительную очистку белка и перевод его в буфер для расщепления протеазой летальным фактором В. anthracis осуществляют при помощи эксклюзионной хроматографии на колонке с Superdex 200. Чистоту препарата белка GST/APT/STX2B анализируют денатурирующим электрофорезом в полиакриламидном геле.
Химерный белок GST/APT/STX2B иммобилизуют на магнитных частицах, покрытых глутатионом, за счет взаимодействия глутатион-8-трансферазы в составе химерного белка с глутатионом на поверхности магнитных частиц; проводят инкубацию подготовленных магнитных частиц, несущих химерную конструкцию, с пулом одноцепочечных олигонуклеотидов, подвергшихся предварительно негативной селекции; промывают магнитные частицы от несвязавшихся олигонуклеотидов потоком отмывающего раствора в трубке, пропущенной сквозь магнит; отделяют аптамеры, специфически связанные с белком-мишенью В-субъединицей Stx2, от иммобилизованного белка и микросфер, удерживаемых магнитом, в раствор обработкой комплекса протеазой летальным фактором Bacillus anthracis; отобранный пул олигонуклеотидов очищают методом фенол-хлороформной экстракции и концентрируют осаждением в этаноле, после чего проводят обогащение пула аптамеров амплификацией с использованием праймеров прямого ForFAM, меченого FAM по 5′-концу цепи, и обратного RevP, несущего на 5′-конце остаток фосфорной кислоты (фиг. 1). Двуцепочечный ПЦР-продукт обрабатывают 5′-экзонуклеазой фага лямбда для получения одноцепочечных ДНК-олигонуклеотидов. Деградацию цепи, фосфорилированной по 5′-концу, визуально контролируют электрофорезом в ПААГ по наличию в электрофореграмме единичной меченой FAM полосы одноцепочечной ДНК. Таким образом проводят пять раундов позитивной селекции.
Получают индивидуальные последовательности аффинных к В-субъединице Stx2 аптамеров. Для этого пул олигонуклеотидов, прошедший все раунды селекции, амплифицируют с применением прямого ForP и обратного RevP праймеров (фиг. 1), фосфорилированных по 5'-концу. Двуцепочечный ПЦР-продукт лигируют в вектор pBluescriptll SK(-) (Agilent Technologies, США), расщепленный эндонуклеазой рестрикции Smal (Thermo, США) и обработанный щелочной фосфатазой (Invitrogen, США). Полученной плазмидой трансформируют электрокомпетентные клетки Е. coli DH12S. Клетки высевают на плотную питательную среду, содержащую ампициллин, X-Gal и IPTG. По принципу бело-голубой селекции выбирают и отсевают колонии, содержащие вставку в составе плазмиды. Проверяют наличие искомой вставки в ПЦР со стандартными праймерами M13/pUC. Колонии, содержащие вставку, соответствующую по размеру единичному аптамеру, подвергают амплификации с собственных праймеров ForFAM и RevP и получают одноцепочечные фрагменты методом, описанным выше.
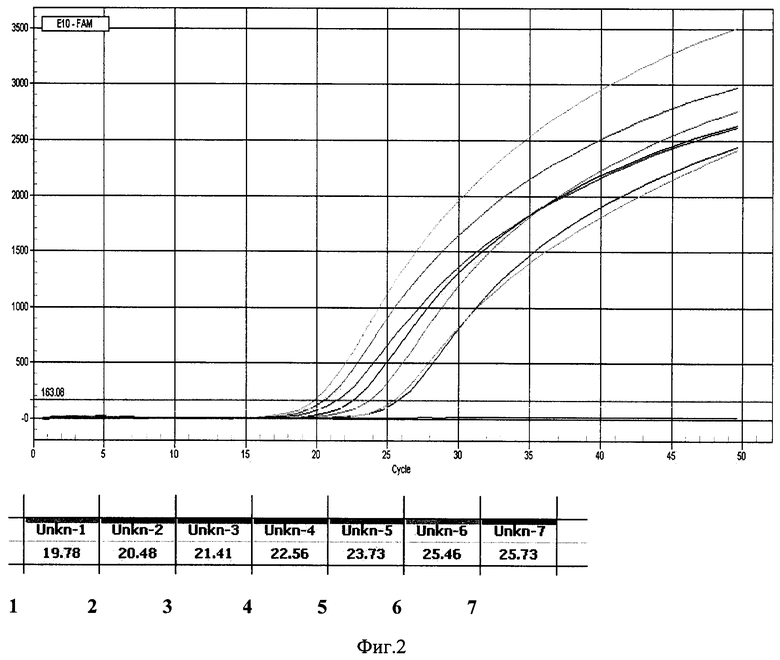
Проводят скрининг индивидуальных последовательностей аптамеров, аффинных к В-субъединице Stx2 в отношении чувствительности и специфичности связывания с белком-мишенью и контрольной панелью рекомбинантных белков токсинов по методу ПЦР в режиме реального времени. Для этого индивидуальные последовательности инкубируют в лунках ПЦР-планшета, на поверхности которых в различных концентрациях (от 10-8 до 10-12 г/мл) иммобилизованы следующие белки: В-субъединицы шига-токсинов энтерогеморрагической Е. coli 0157:Н7 (Stx1 и Stx2), протеолитический домен BoNT/A (LCBoNT/A), рецептор-связывающий домен тяжелой цепи BoNT/A (НС 50 BoNT/A), рецептор-связывающий домен тяжелой цепи BoNT/B (НС 50 BoNT/B), летальный фактор В. anthracis (LF), протективный антиген В. anthracis (РА), дифтерийный токсин (DT), стафилококковые энтеротоксины А и В (SEA и SEB), термолабильный токсин Е. coli (LT). Лунки планшета отмывают от свободных молекул ДНК, затем проводят ПЦР в режиме реального времени с применением интеркалирующего флуоресцентного красителя, на основании результатов которого отбирают наиболее эффективно связывающиеся с В-субъединицей токсина ДНК-аптамеры (фиг. 2).
Для последовательностей с наиболее выраженными свойствами чувствительности и специфичности определяют константу аффинности на приборе ProteOn™ XPR36 Protein Interaction Array System (Bio-Rad, США) по отношению к рекомбинантной В-субъединице шига-токсина Stx2.
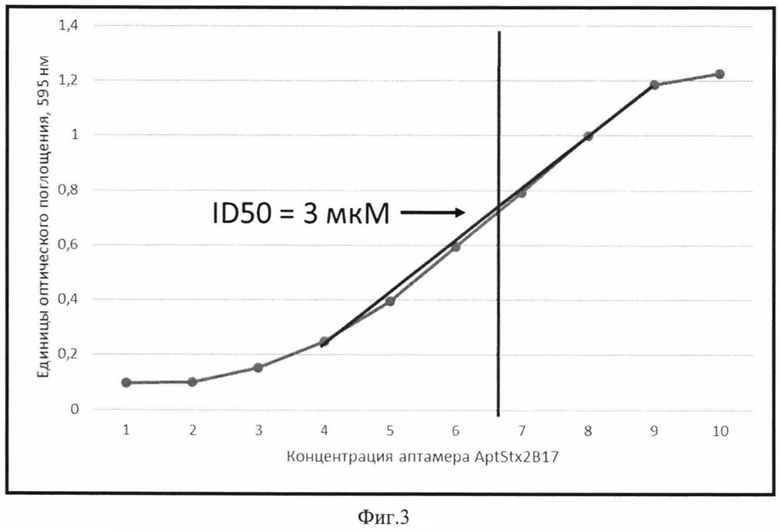
Выбранные ДНК-аптамеры тестируют на способность к нейтрализации токсического эффекта шига-токсина Stx2 на клетках перевиваемой линии Vero. Для аптамеров, способных к нейтрализации токсического эффекта Stx2, определяют ID50, которая не должна превышать 15 мкМ (фиг. 3).
Проводят определение первичных последовательностей аптамеров, связывающихся с В-субъедницей шига-токсина Stx2 и ингибирующих ее токсическую активность (фиг. 4).
Изобретение иллюстрируют следующие графические материалы:
Фиг. 1. Последовательности ДНК, используемые для отбора молекул ДНК, для обогащения селектированного пула, проведения амплификации и анализа полученных аптамерных последовательностей.
A. Последовательность олигонуклеотидной библиотеки, используемой в процессе селекции, где N45 - вариабельная область, const1 и const2 - константные области для отжига специфических праймеров;
Б. Последовательности праймеров, использованных для клонирования В-субъединицы шига-токсина Stx2;
B. Последовательности праймеров, используемых в реакциях обогащения библиотеки, для клонирования и наращивания количества последовательностей ДНК-аптамеров.
Фиг. 2. ДетекцияВ-субъединицы шига-токсина Stx2 аптамером AptStx2B17 по методу иммуно-аптамерной ПЦР.
Результаты ПЦР в режиме реального времени. Рекомбинантная В-субъединица Stx2, иммобилизована на планшете в концентрациях 10-8 г/мл (1), 10-9 г/мл (2), 10-10 г/мл (3), 10-11 г/мл (4), (5), 10-12 г/мл (6); 7 - отрицательный контроль (лунка, не содержащая иммобилизованного белка В-субъединицы Stx2).
Фиг. 3. Результат анализа ингибирования токсического эффекта шига-токсина Stx2 аптамером AptStx2B17.
По оси X представлена концентрация аптамера AptStx2B17, выраженная в мкМ: 1-0,005; 2-0,01; 3-0,05; 4-0,1; 5-0,5; 6-1; 7-5; 8-10; 9-50; 10-100. По оси Y представлено значение оптического поглощения при 600 нм как результат анализа жизнеспособности культуры в тесте с кристаллическим фиолетовым.
Фиг. 4. Первичная последовательность и результат моделирования вторичной структуры аптамера AptStx2B17, специфичного к В-субъединице шига-токсина Stx2.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Получение рекомбинантного полипептида GSТ/АРТ/8ТХ2В.
Последовательность ДНК, кодирующую В-субъединицу шига-токсина 2, получают ПЦР-амплификацией с геномной ДНК бактерий Е. coli штамма 0157:Н7 (Государственная коллекция патогенных микроорганизмов ФБУН ГНЦ ПМБ, Оболенск) с помощью ПЦР с использованием праймеров STX2ForBamHI и STX2RevXhoI (фиг. 1). Температура отжига праймеров составляет 56°C, время элонгации 20 с. Продукт апмлификации обрабатывают эндонуклеазами рестрикции BamHI и XhoI и клонируют в плазмиду pGST/APT/X [Заявка на получение патента RU 2013147132], обработанную теми же эндонуклеазами. Полученная экспрессионная плазмида pGST/APT/STX2B кодирует химерный слитный белок, содержащий аминокислотную последовательность глютатион-S-трансферазы, пептид RRKKVYPYPME, являющийся субстратом летального фактора В. anthracis, in vivo биотинилируемый пептид и последовательность, кодирующую В-субъединицу шига-токсина 2.
Продуцент слитного рекомбинантного полипептида GST/APT/STX2B получают электротрансформацией (прибор ВТХ 600, режим 129 Ом, 2,5 KB) электрокомпетентных клеток Е. coli BL21(DE3) плазмидой pGST/APT/STX2B. Трансформанты высевают на чашки с 2хYТ-агаром, содержащим 50 мкг/мл ампициллина и 2% глюкозы и выращивают в течение ночи при 37°С.
Для препаративной продукции белка GST/APT/STX2B единичные колонии-продуценты выращивают в течение ночи при 37°C в среде 2xYT с добавлением 2% глюкозы, переносят 10 мл ночной культуры в 1000 мл среды 2xΥΤ, содержащей 0,1% глюкозы и 50 мкг/мл ампициллина, и выращивают при 37°C до оптической плотности 1 ОЕ. Выросшую культуру охлаждают до 30°C, добавляют ИПТГ до 0,2 мМ и проводят экспрессию белка в течение 3 ч при 30°C. Уровень продукции белка определяют электрофорезом в полиакриламидном геле. Для проведения электрофореза в денатурирующих условиях в полиакриламидном геле отбирают аликвоты жидкой культуры продуцента, центрифугируют при 5000 об/мин 5 мин, осажденные клетки растворяют в 100 мкл лизирующего буфера с красителем бромфеноловым синим, обрабатывают 20 с ультразвуком, нагревают 3 мин при 100°C и наносят на гель. После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Shimadzu CS-930. По денситометрическим измерениям количество слитного химерного белка GST/APT/STX2B составляет 40% от общего клеточного белка. Трансформируют плазмидой pGST/APT/STX2B клетки Е. coli BL21(DE3), получая тем самым продуцент химерного пептида GST/APT/STX2B. Проводят продукцию полипептида в течение 3 ч после индукции жидкой культуры добавлением IPTG. После окончания продукции клеточную биомассу собирают центрифугированием, суспендируют в лизирующем буфере, суспензию бактерий обрабатывают лизоцимом и ДНКазой и центрифугируют для получения осветленного лизата, содержащего полипептид GST/APT/STX2B в растворимой форме.
Очистку белка GST/APT/STX2B из лизата проводят на колонке с иммобилизованным глутатионом, белок элюируют добавлением буфера, содержащего 25 мМ глутатиона. Дополнительную очистку белка и перевод его в буфер для расщепления протеазой летальным фактором В. anthracis осуществляют при помощи эксклюзионной хроматографии на колонке с Superdex 200. Чистоту препарата белка GST/APT/STX2B анализируют денатурирующим электрофорезом в полиакриламидном геле.
Пример 2. Проведение селекции олигонуклеотидов с В-субъединицей рекомбинантного шига-токсина Stx2, находящейся в составе рекомбинантного полипептида.
Очищенный из лизата культуры Е. coli BL21(DE3) растворимый химерный пептид GST/APT/STX2B иммобилизуют на поверхности магнитных микрочастиц, покрытых глутатионом. Для этого 100 мкл суспензии частиц Pierce™ Glutathione Magnetic Beads (Thermo, США) в пробирке объемом 1,5 мл помещают на магнитный штатив DynaMag™-2 Magnet (Invitrogen, США) и отбирают жидкость. Сняв пробирку со штатива, ресуспендируют микрочастицы в 300 мкл буфера состава 125 мМ Трис, 150 мМ NaCl, pH 8,0. Вновь помещают пробирку в магнитный штатив и отбирают жидкость. Повторяют трижды, отмывая тем самым микрочастицы от буфера хранения.
Белок химерного конструкта GST/APT/STX2B приводят к конечной концентрации 1 мг/мл и объему 300 мкл буфером 125 мМ Трис pH 8,0, 150 мМ NaCl. Добавляют суспензию белка к изолированным с помощью магнитного стенда магнитным микрочастицам. Инкубируют суспензию при комнатной температуре в течение 1 ч, встряхивая пробирку на вортексе каждые 15 мин. По окончании инкубации помещают пробирку в магнитный штатив и отбирают супернатант. Пятикратно отмывают частицы 500 мкл того же буферного раствора. Ресуспендируют частицы в 300 мкл буфера. При необходимости хранят при +4°C, добавив к суспензии натрия азид до конечной концентрации 0,02%.
Перед добавлением олигонуклеотидной библиотеки к магнитным микрочастицам помещают пробирку в магнитный штатив и отбирают супернатант.Добавляют суспензию олигонуклеотидовв объеме 300 мкл (буферный раствор состава 20 мМ Трис-НСl, 50 мМ NaCl, 5 мМ EDTA, pH 7,4) с содержанием ДНК в растворе 50 мкг. Инкубируют суспензию на протяжении 1 ч при комнатной температуре при периодическом перемешивании на вортексе. Отмывают магнитные частицы от несвязавшихся молекул ДНК потоком буферного раствора объемом 300 мл состава 20 мМ Трис-НСl, 250 мМ NaCl, 5 мМ EDTA, pH 7,4, используя систему проточной магнитной сепарации MiniMACS™ Separator (Miltenyi Biotec, Германия). Удалив магнитное воздействие на частицы, несущие на своей поверхности химерный полипептид GST/APT/STX2B, связанный с аптамерами, собирают микросферы в чистую пробирку объемом 1,5 мл. Фиксируя магнитные микросферы, отбирают супернатант. Добавляют 300 мкл раствора протеазы летального фактора В. anthracis [патент РФ №2355769] с содержанием 5 мкг протеазы в буфере 30 мМ Трис-НСl, pH 7,4. Выдерживают в течение 1 ч при температуре 30°С.Фиксируют частицы при помощи магнитного штатива, отбирают супернатант, содержащий отщепленный белок Stx2B, несущий специфически связанные аптамеры. Пул олигонуклеотидов очищают методом фенол-хлороформной экстракции и концентрируют преципитацией этанолом.
Наращивают селектированный пул аптамеров с помощью ПЦР-амплификации с использованием прямого праймера ForFAM и обратного праймера RevP (фиг. 1В). Получают одноцепочечный продукт обработкой 5′-экзонуклеазой фага лямбда, которая расщепляет обратную фосфорилированную цепь фрагмента. Контроль удаления комплементарной цепи ДНК осуществляют при помощи электрофореза в ПААГ. Полученный обогащенный на связывание с Stx2B пул ДНК-аптамеров используют для осуществления дальнейших раундов селекции. Всего проводят пять раундов позитивной селекции.
Пример 3. Получение индивидуальных аптамерных последовательностей.
Фрагменты для клонирования в коммерческий вектор pBluescriptII SK(-) (Agilent Technologies, США) нарабатывают на основе пула олигонуклеотидов, отобранных на связывание с белком Stx2B при помощи ПЦР с использованием прямого и обратного фосфорилированных праймеров ForP и RevP (отжиг праймеров при 54°C, элонгация 15 с) (фиг. 1). Плазмиду pBluescriptll SK(-) обрабатывают эндонуклеазой рестрикции Smal в буфере Tango (Thermo, США), после чего обрабатывают щелочной фосфатазой (Invitrogen, США). Фосфорилированные двуцепочечные фрагменты ДНК лигируют в подготовленную плазмиду.
Электрокомпетентные клетки Е. coli DH12S трансформируют подготовленной ранее плазмидой pBluescriptll SK(-), содержащей вставки селектированных последовательностей, применив электротрансформацию (прибор EppendorfElectroporator 2510 (Eppendorf, Германия), режим 1,7 кB). Трансформированные клетки высевают на плотную среду с 2хYТ-агаром, содержащим 50 мкг/мл ампициллина, 80 мкг/мл X-Gal и 20 мМ IPTG, и выращивают в течение ночи при 37°C.
Единичные колонии, потенциально содержащие вектор со вставкой (по принципу бело-голубой селекции [Ullmann A., Jacob F., Monod J. // J MolBiol. - 1967. - V. 24. - N. 2. -P. 339-343]) верифицируют на наличие единичного встроенного фрагмента ДНК искомой длины в плазмиде с помощью ПЦР со стандартными прямым и обратным праймерами M13/pUC (Fermentas, США). Наращивают количество копий индивидуальных последовательностей клонированных фрагментов ДНК амплификацией с использованием праймеров ForFAM и RevP (фиг. 1В). Получают одноцепочечный продукт для каждого из клонов обработкой 5′-экзонуклеазой фага лямбда с визуальным контролем в электрофорезе. В результате проведенных процедур получают клонированные одноцепочечные индивидуальные последовательности из 500 бактериальных клонов.
Пример 4. Скрининг чувствительности и специфичности индивидуальных селектированных последовательностей в отношении В-субъединицы шига-токсина 2 при помощи ПЦР в режиме реального времени.
На поликарбонатном 96-луночном ПЦР-планшете с высокосвязывающей поверхностью (Corning, США) иммобилизуют в различных концентрациях (от 10-8 до 10-12 г/мл) следующие рекомбинантные белки: шига-токсины Stx1 и Stx2 энтерогеморрагической Е. coli 0157:Н7, протеолитический домен BoNT/A (LC BoNT/A), рецептор-связывающий домен тяжелой цепи BoNT/A (НС 50 BoNT/A), рецептор-связывающий домен тяжелой цепи BoNT/B (НС 50 BoNT/B), летальный фактор В. anthracis (LF), протективный антиген В. anthracis (РА), дифтерийный токсин (DT), стафилококковые энтеротоксины А и В (SEA и SEB), термолабильный токсин Е. coli (LT). Первый вертикальный ряд планшета (8 лунок) не содержит белки, и его лунки служат в качестве отрицательных контрольных проб. Начиная со второго вертикального ряда вносят рекомбинатные белки из исследуемой панели в количестве 0,5 мкг/лунку в фосфатно-солевом буфере, pH 7,4 (ФСБ) в объеме 50 мкл. Инкубируют планшет на протяжении 1 ч при 37°C при встряхивании на орбитальном шейкере. По окончании инкубации трижды отмывают лунки планшета ФСБ, содержащим 0,05% Tween-20. Свободные валентности планшета блокируют буфером, состоящим из обезжиренного (содержание жира 0,5%) молока с добавлением 3% БСА, в объеме 200 мкл/лунку в течение 40 мин при 37°C на орбитальном шейкере. Отмывку производят трижды ФСБ с 0,05% Tween-20.
В лунки горизонтальных рядов планшета вносят одноцепочечную ДНК индивидуальных аптамеров в объеме 50 мкл и количестве 0,2 мкг/лунку. В первый и второй горизонтальные ряды в качестве контроля сравнения вносят исходную комбинаторную библиотеку. В последующие горизонтальные ряды вносят ДНК аптамеров (в 2 ряда по 12 лунок). Таким образом, каждая проба исследуется в двух повторностях, Буфер для разведения ДНК содержит 20 мМ Трис-НСl, 50 мМ NaCl, 5 мМ EDTA (pH 7,4). Инкубируют планшет с ДНК в течение 1 ч при 37°C. Отмывку от несвязавщихся молекул ДНК производят раствором, содержащим 20 мМ Трис-НСl, 300 мМ NaCl, 5 мМ EDTA (pH 7,4) пятикратно.
Контролируют реакцию связывания ДНК с белками в ПЦР в режиме реального времени. Для этого в лунки планшета вносят амплификационную смесь, содержащую флуоресцентный интеркалирующий краситель SYBR-GreenI (ЗАО "Синтол", Россия) с добавлением праймеров ForP и RevP (фиг. 1). Амплификацию проводят в режиме: температура отжига праймеров 54°C, время элонгации 10 с, детекцию сигнала проводят по каналу SYBR 1 (FAM) (прибор IQ™5, Bio-Rad, США) (фиг. 2).
По результатам проведенного скрининга выделяют аптамер (AptStx2B 17) (фиг. 4), проявивший наибольшую специфическую активность в отношении Stx2B и вместе с тем не обнаруживший неспецифического взаимодействия с другими рекомбинантными белками токсинов, присутствующими в использованной скрининговой панели.
Для аптамера AptStx2B17 определяют константу связывания с белком Stx2B с использованием системы ProteOn™ XPR36 Protein Interaction Array System (Bio-Rad, США), которая составляет 1,7·10-9 Μ.
Пример 6. Нейтрализация токсичности шига-токсина типа 2 по отношению к клеткам Vero с помощью аптамера AptStx2B17.
Для получения частично очищенного шига-токсина типа 2 клетки токсигенной Е. coli 0157:Н7 из Государственной коллекции микроорганизмов ФБУН ГНЦ ПМБ, продуцирующие только Stx2, выращивают в колбах в объеме 1 л в минимальной среде с добавлением 1% казаминовых кислот (М9СА, Difco), 2% глюкозы, 0,004% триптофана, и 0,004% никотиновой кислоты, в течение 72 ч ночи при температуре 37°C. По окончании роста клетки концентрируют центрифугированием, разрушают ультразвуком, удаляют дебрис центрифугированием при 5000×g, а среду осветляют высокоскоростным центрифугированием при 20000×g. Осветленный лизат концентрируют с использованием ультрафильтрационной мембраны с отсечкой молекулярной массы 30 кДа, к концентрированному лизату, содержащей холотоксин Stx2, добавляют ингибитор протеаз «Complete» (Roche) согласно инструкции производителя, стерилизуют, пропуская через фильтр с диаметром пор 0,22 мкм, и используют в тестах на цитотоксичность.
Клетки линии Vero, обладающие повышенной чувствительностью к действию Stx2, выращивают в среде DMEM, содержащей 10% фетальной сыворотки телят, при 37°C в 5% СO2 в культуральных флаконах 75 или 125 см2 в течение 24-48 ч. Выросшие клетки трипсинизируют, проводят подсчет и переносят в 96-луночные планшеты в количестве 103 клеток на лунку. После инкубации в течение 16 ч в лунки добавляют различные разведения культуральной жидкости, содержащей Stx2, для определения LD50.
Для определения LD50, серийные разведения токсина в DMEM добавляют в лунки в виде 50-мкл образцов, микропланшеты инкубируют дополнительно в течение 16 ч. Инкубируют также отрицательные контроли и образцы, в качестве положительного контроля цитотоксичности используют цисплатин, согласно ранее описанной методике [Rao M., Kumar M.M. с соавт. // J.Biochem. - 1999. - V. 125. - №2. - Р. 383-390].
По окончании инкубации с токсинами клетки, перешедшие в среду, токсин и среду удаляют, оставшиеся прикрепленными клетки фиксируют 2% раствором формальдегида в PBS, pH 7,2, фиксирующий агент удаляют, и клетки окрашивают 0,13% кристаллическим фиолетовым в PBS, содержащем 5% этанола и 2% формальдегида. Краситель отмывают с микропланшета 100 мкл 50% этанола, смешивают с 900 мкл PBS и измеряют адсорбцию при 595 нмс использованием микропланшетного мультиридера Themo Varioskan. Разведение токсина, вызывавшее отделение 50% клеток, определяют как LD50.
Для проверки нейтрализующего эффекта на культуре клеток аптамеры синтезируют с заблокированными спейсерными фосфороамидитами 3′- и 5′- концами для предотвращения деградации под действием экзонуклеаз. При проведении анализа нейтрализации токсинамодифицированные аптамеры в серийных разведениях (100-0,005 мкМ) смешивают с токсином, взятым в концентрации, соответствующей LD50. Образцы смеси наносят на микропланшеты с прикрепленными клетками Vеrо и определяют процент выживших клеток, как описано выше. Результаты анализа для аптамера AptStx2B17 соответствуют данным, приведенным на фиг. 3. Аптамер AptStx2B17 нейтрализует активность токсина Stx2 по отношению к клеткам перевиваемой линии Vero в концентрации ID50=3 мкМ.
Пример 7. Определение первичной и моделирование вторичной структуры ДНК-аптамера AptStx2B 17 к В-субъединице шига-токсина 2.
Определение первичной структуры аптамера AptStx2B17 проводят методом капиллярного секвенирования. Моделирование вторичной структуры осуществляют с использованием программы RNAFold (http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi). Аптамер AptStx2B17 имеет нуклеотидную последовательность и предположительную вторичную структуру, представленные на рисунке (фиг. 4).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С ПРОТЕОЛИТИЧЕСКОЙ СУБЪЕДИНИЦЕЙ НЕЙРОТОКСИНА ТИПА A CLOSTRIDIUM BOTULINUM | 2014 |
|
RU2571210C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGST/ART/X, КОДИРУЮЩАЯ СЛИТНЫЙ ХИМЕРНЫЙ ПОЛИПЕПТИД GST/ART/X ДЛЯ ВЫСОКОСПЕЦИФИЧНОЙ СЕЛЕКЦИИ АПТАМЕРОВ К БЕЛКУ-МИШЕНИ X В СОСТАВЕ ПОЛИПЕПТИДА | 2014 |
|
RU2558295C1 |
| Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila | 2019 |
|
RU2737829C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С БАКТЕРИЯМИ Escherichia coli O157:H7 | 2014 |
|
RU2566551C1 |
| СПОСОБ СПЕЦИФИЧЕСКОГО ОТБОРА ВЫСОКОАФФИННЫХ МОЛЕКУЛ ДНК (ДНК-АПТАМЕРОВ) К РЕКОМБИНАНТНОМУ БЕЛКУ-МИШЕНИ | 2012 |
|
RU2513700C1 |
| НЕТОКСИЧНЫЙ РЕКОМБИНАНТНЫЙ ШИГА ТОКСИН 2-ГО ТИПА (Stx2) | 2013 |
|
RU2573924C2 |
| Способ направленного истощения олигонуклеотидных библиотек для снижения неспецифической адсорбции при твердофазной селекции аптамеров на основе нуклеиновых кислот | 2015 |
|
RU2618872C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БОТУЛИНИЧЕСКОГО НЕЙРОТОКСИНА ТИПА А НА ОСНОВЕ ИММУНОДЕТЕКЦИИ, СОПРЯЖЕННОЙ С ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИЕЙ | 2013 |
|
RU2549463C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НАЛИЧИЯ БАКТЕРИЙ Escherichia coli O157:H7 В БИОЛОГИЧЕСКИХ И ПИЩЕВЫХ ОБРАЗЦАХ НА ОСНОВЕ ИММУНОДЕТЕКЦИИ, СОПРЯЖЕННОЙ С ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИЕЙ | 2014 |
|
RU2569196C1 |
| МИКРООРГАНИЗМ-НОСИТЕЛЬ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, КОДИРУЮЩИХ АНТИГЕНЫ И БЕЛКОВЫЕ ТОКСИНЫ | 2007 |
|
RU2447145C2 |

Изобретение относится к биотехнологии, а именно к последовательности ДНК-аптамеров. Указанный ДНК-аптамер связывается с шига-токсином типа 2 и представляет собой одноцепочечную ДНК AptStx2B17 длиной 81 нуклеотид и молекулярной массой 27 кДа. ДНК-аптамер специфически связывает В-субъединицу шига-подобного токсина 2 с константой связывания 1,7·109 М-1 и первичной последовательностью CATACGTTCGACTGCTACCTTAGCCTGGTCTTATGATGCCTGTTTGATCCTGATGGCTAATATGTGAGATGTACAGACTAG. Последовательность ДНК-аптамера отобрана в результате раундов позитивной селекции в отношении рекомбинантного белка В-субъединицы шига-токсина типа 2 на основе системы магносепарации с отделением белка-мишени, аффинно связанного со специфичными аптамерами, из состава химерного полипептида посредством расщепления химерного конструкта по сайту, содержащему субстрат протеазы летального фактора. ДНК-аптамер способен специфически детектировать В-субъединицу шига-подобного токсина 2 по методу ПЦР в режиме реального времени в концентрации до 10-11 г и нейтрализовать его активность по отношению к клеткам перевиваемой линии Vero в концентрации ID50=3 мкМ. Изобретение позволяет с высокой эффективностью детектировать В-субъединицу шига-токсина. 4 ил., 7 пр.
Последовательность ДНК-аптамеров, связывающая шига-токсин типа 2, представляет собой одноцепочечную ДНК AptStx2B17 длиной 81 нуклеотид и молекулярной массой 27 кДа, специфически связывает В-субъединицу шига-подобного токсина 2 с константой связывания 1,7·109 М-1, и первичной последовательностью CATACGTTCGACTGCTACCTTAGCCTGGTCTTATGATGCCTGTTTGATCCTGATGGCTAATATGTGAGATGTACAGACTAG, отобранная в результате раундов позитивной селекции в отношении рекомбинантного белка В-субъединицы шига-токсина типа 2 на основе системы магносепарации с отделением белка-мишени, аффинно связанного со специфичными аптамерами, из состава химерного полипептида посредством расщепления химерного конструкта по сайту, содержащему субстрат протеазы летального фактора, способна специфически детектировать В-субъединицу шига-подобного токсина 2 по методу ПЦР в режиме реального времени в концентрации до 10-11 г и нейтрализовать его активность по отношению к клеткам перевиваемой линии Vero в концентрации ID50=3 мкМ.
| CHALLA S | |||
| ET AL., Selective Evolution of Ligands by Exponential Enrichment to Identify RNA Aptamers against Shiga Toxins, J Nucleic Acids, 2014, 2014:214929 | |||
| SUN H | |||
| ET AL., Oligonucleotide aptamers: new tools for targeted cancer therapy, Mol Ther Nucleic Acids, 2014, 3:e182 | |||
| US 20120071639 A1, 22.03.2012 | |||
| АПТАМЕРНЫЙ ОЛИГОНУКЛЕОТИД - ПРЯМОЙ ИНГИБИТОР ТРОМБИНА | 2008 |
|
RU2401306C2 |

