Изобретение относится к области биотехнологии, биоинженерии, биохимии, молекулярной медицины и лабораторных медицинских исследований. Представляет собой способ получения пулов олигонуклеотидов, используемых для твердофазной селекции аптамеров к целевому белку-мишени, истощенных по содержанию структур, связывающихся с нецелевыми растворимыми белковыми компонентами, находящимися в составе комплекса, используемого для отбора специфических аптамеров, осуществляемый с помощью сепарации смесей нуклеиновая кислота (НК)-белок гель-фильтрационной хроматографией.
Основными областями применения способа направленного истощения олигонуклеотидных библиотек являются медицинские и биологические исследования, в том числе создание новых медицинских терапевтических препаратов на основе аптамеров, а также высокочувствительных и высокоэффективных систем детекции мишеней с использованием аптамеров на основе нуклеиновых кислот.
Аптамеры представляют собой одноцепочечные олигонуклеотиды (ДНК или РНК), обладающие трехмерной структурой, которую составляют петли, шпильки, псевдоузлы, триплексы, квадруплексы и др. [Stoltenburg R., Reinemann С, Strehlitz В. // Biomolecular engineering. - 2007. - V. 24. - №. 4. - P. 381-403.], способные, благодаря своей структуре и пространственному расположению зарядов в молекуле, образовывать высокоспецифичные и высокоаффинные связи с заданными молекулярными мишенями различной природы (белки, липиды, нуклеиновые кислоты и др.). Аптамеры часто рассматривают в качестве аналогов моноклональных антител в диагностике и терапии заболеваний [Rimmle М. // Chembiochem. - 2003. - V. 4. - №. 10. - Р. 963-971.]. Терапевтическое применение высокоаффинных олигонуклеотидов имеет ряд существенных преимуществ перед использованием в этих целях антител. Короткие последовательности нуклеиновых кислот (20-100 нуклеотидов) не обладают иммуногенностью, могут быть получены сравнительно быстро и недорого, могут быть отобраны к огромному спектру мишеней, включая неиммуногенные и токсичные компоненты, легко могут быть подвержены химическим модификациям для достижения большей стабильности без потери аффинных свойств. Кроме того, в составе, например, биосенсоров, аптамеры могут быть использованы многократно за счет способности нуклеиновых кислот денатурировать и ренатурировать без потери свойств молекулы [Лахин А.В., Тарантул В.З., Генинг Л.В. // Acta Naturae - 2013. - Т. 5. - №. 4 (19) - С. 37-48.].
Для осуществления направленного отбора аптамеров против выбранных мишеней используют технологию SELEX ("Systematic Evolution of Ligands by Exponential Enrichment") [US Patent 5567588] с применением различных модификаций технологии, таких как микромагнитная селекция в микрожидкостных каналах (М-SELEX) [Lou, X., Qian, J., Xiao, Y., Viel, L., Gerdon, A.E., Lagally, E. Т., … & Soh, H.T. // Proceedings of the National Academy of Sciences. - 2009. - V. 106. - №. 9. - P. 2989-2994], использование капиллярного элекрофореза (ECEEM) [Drabovich A., Berezovski М., Krylov S.N. // Journal of the American Chemical Society. - 2005. - V. 127. - №. 32. - P. 11224-11225] и др. В основе метода SELEX лежит серия повторяющихся раундов связывания пула олигонуклеотидной библиотеки с мишенью, удаления несвязавшихся олигонуклеотидов и амплификации связавшихся кандидатных аптамеров при помощи полимеразной цепной реакции (ПЦР) с применением праймеров к их констатным фланкирующим областям.
Негативная селекция аптамерных библиотек в отношении структур различной природы является одним из факультативных элементов стратегии SELEX (этап преселекции) и используется для элиминации из пула структур, специфичных к молекулам и компонентам системы отбора [Klug S. J., Famulok М. // Molecular biology reports. - 1994. - V. 20. - №. 2. - P. 97-107; Gopinath S. С.B. // Analytical and bioanalytical chemistry. - 2007. - V. 387. - №. 1. - P. 171-182; Stoltenburg R., Reinemann C., Strehlitz B. // Biomol. Engineering. - 2007. - V. 24. - N. 4. - P. 381-403].
Широкий набор мишеней для селекции аптамеров (очищенные рекомбинантные белки, экспрессируемые в эу- и прокариотических клетках белки, нативные молекулярные мишени на поверхности или в цитозоле клеток и тканей [Kanoatov М., Javaherian S., Krylov S.N. // Analytica chimica ac a. - 2010. - V. 681. - №. 1. - P. 92-97; Ye M. et al. // International journal of molecular sciences. - 2012. - V. 13. - №. 3. - P. 3341-3353.], опухолевые клетки in vivo [Mi J. et al. // Nature chemical biology. - 2010. - V. 6. - №. 1. - P. 22-24.] и др. [Лахин А.В., Тарантул В.3., Генинг Л.В. // Acta Naturae - 2013. - Т. 5. - №. 4 (19) - С. 37-48.]) обусловливает существующее многообразие протоколов отбора и отсутствие универсальных методов селекции и тестирования полученных последовательностей. Варьируют также и способы негативной селекции в рамках различных протоколов отбора аптамеров.
Известно использование негативной селекции при отборе аптамеров к клеткам или тканям, несущим на поверхности своей мембраны специфические молекулярные маркеры (например, в технологии Cell-SELEX). При таком подходе после 1-2 раундов позитивной селекции аптамеров к поверхности клеток-мишеней проводится негативный отбор в отношении контрольных клеток/тканей, не содержащих на своей поверхности таргетных молекул. Таким образом, удается удалить из селектированного пула подавляющую часть кандидатных аптамеров, аффинных к структурам и молекулам, присущим как клеткам-мишеням, так и контрольным клеткам [Ohuchi S.Р., Ohtsu Т., Nakamura Y. // Biochimie. - 2006. - V. 88. - №. 7. - P. 897-904; Sefah K. et al. // Nature protocols. - 2010. - V. 5. - №. 6. - P. 1169-1185; US Patent 1998/015619].
Известно истощение олигонуклеотидной библиотеки для отбора аптамеров против хроматографического сорбента, несущего мишень, иммобилизованную на хроматографической колонке [Klug S.J., Famulok М. // Molecular biology reports. - 1994. - V. 20. - №. 2. - P. 97-107], что позволяет избавиться от артефактов при проведении дальнейшей селекции с использованием того же сорбента. Тем не менее, истощение олигонуклеотидного пула исключительно против поверхности твердофазного носителя может являться недостаточным для исключения ко-селекции структур НК, аффинных, в частности, к нецелевым белковым компонентам системы, а, следовательно, и для обеспечения высокоспецифического отбора аптамеров к иммобилизованной мишени.
Известно проведение негативной селекции олигонуклеотидного пула против вспомогательных аминокислотных последовательностей, используемых для селекции в составе рекомбинантных белков. Иммобилизация на парамагнитных частицах или хроматографических сорбентах за счет вспомогательной аминокислотной последовательности (тэга), химеризованной с последовательностью белка, и способной эффективно связываться с молекулярной мишенью на поверхности сорбента или частиц, является наиболее доступной и часто употребимой стратегией отбора аптамеров к рекомбинантным водорастворимым белкам. В качестве тегов часто используются последовательность шести гистидинов (для иммобилизации мишени на металл-хелатном сорбенте) [Murphy М.В., Fuller S.T., Richardson P.M., Doyle S.A. // Nucleic Acids Res. - 2003. - V. 31. - №18. - P.e110], глутатион-S-трансферазы (для иммобилизации мишени на сорбентах, несущих глутатион) [Oktem Н.А., Bayramoglu G., Ozalp V.C., Arica M.Y. // Biotechnol. Prog. - 2007. - V. 23. - №1. - P. 146-154] и др. Однако применимо к селекции аптамеров было установлено, что металл-хелатные сорбенты, как правило, обладают высоким уровнем неспецифической адсорбции НК, варьирующим в зависимости от типа смолы и задействованного бивалентного катиона [Kowalska Е., Bartnicki F., Pels K., Strzalka W. // Anal. Bioanal. Chem. - 2014. - V. 406. - №22. - P. 5495-5499], что снижает эффективность селекции и увеличивает количество раундов отбора. Аналогично, результативное проведение процедур селекции аптамеров к мишеням, содержащим глутатион-S-трансферазу, включает этап удаления структур НК, способных связаться с твердофазным носителем с иммобилизованным белком глутатион-S-трансферазы [Weiss S., Proske D., Neumann M., Groschup M.H., Kretzschmar H.A., Famulok M., Winnacker E.L. // J. Virol. - 1997. - V.71. - №11. - Р. 8790-8797]. При отсутствии стадии негативной селекции требуется верификация кандидатных аптамеров конечного селектируемого пула на отсутствие связывание тегов. Получение химер тег-белок мишень неизбежно требует проведения ряда генно-инженерных манипуляций, продукции и очистки рекомбинантного белка, тогда как не все белки-мишени легко получить в достаточном количестве и в растворимом состоянии в виде химеры в гетерологичной системе экспрессии. Помимо этого, рекомбинантный белок, используемый для селекции, должен быть валидирован по функциональной активности, но даже этот этап не может исключить наличия в белке структурных изменений, которые могут повлиять на процесс селекции аптамеров и отразиться на эффективности их взаимодействия с нативным аналогом белка.
Известна альтернатива данному методу, заключающаяся в иммобилизации мишени, используемой для отбора, на сорбенте или парамагнитных частицах, несущих белки авидинового ряда (авидин, стрептавидин, нейтравидин и др.) Авидин и его аналоги обладают высоким (Kd=10-15M) сродством к биотину [Livnah О., Bayer Е., Wilchek М., Sussman J. // Proc. Natl. Acad. Sci. USA. - 1993. - V.90. - №11. - P. 5076-5080]. Таким образом, связывание биотинилированной мишени с белком авидинового ряда происходит крайне быстро. Успешный протокол отбора аптамеров в системах, использующих в качестве маркера для иммобилизации мишени биотин, был недавно представлен в работе [Mayer G., 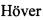 Т. // Meth. Mol. Biol. - 2009. - V. 535. - P. 19-32.]. Некоторые из данных систем задействуют этап негативной селекции, проводимый против носителя с иммобилизованным белком авидинового ряда (как правило, стрептавидином, в силу наличия коммерчески доступных парамагнитных частиц с этим белком, например, Dynabeads компании Life Technologies, США) [Faryammanesh R., Lange Т., Magbanua E., Haas S., Meyer C, Wicklein D., Schumacher U., Hahn U. // PLoS One. - 2014. - V.9. - №4. - Р.: е93173]. Однако, емкость частиц (даже в наиболее современных вариантах, например, Dynabeads MyOne компании Life Technologies, США) не превышает 1300 рМ биотина на мг частиц, что является достаточным для иммобилизации белка-мишени, но может оказаться недостаточным для удаления из раствора неспецифически связывающихся со стрептавидином структур НК. При проведении преселекции и селекции для блокировки свободных валентностей поверхности частиц и снижения неспецифической адсорбции исследователи вынуждены прибегать к использованию в буферных растворах дополнительных блокирующих агентов, в частности, бычьего сывороточного альбумина [Faryammanesh R., Lange Т., Magbanua Е., Haas S., Meyer С., Wicklein D., Schumacher U., Hahn U. // PLoS One. - 2014. - V. 9. - №4. - P.: e93173].
Т. // Meth. Mol. Biol. - 2009. - V. 535. - P. 19-32.]. Некоторые из данных систем задействуют этап негативной селекции, проводимый против носителя с иммобилизованным белком авидинового ряда (как правило, стрептавидином, в силу наличия коммерчески доступных парамагнитных частиц с этим белком, например, Dynabeads компании Life Technologies, США) [Faryammanesh R., Lange Т., Magbanua E., Haas S., Meyer C, Wicklein D., Schumacher U., Hahn U. // PLoS One. - 2014. - V.9. - №4. - Р.: е93173]. Однако, емкость частиц (даже в наиболее современных вариантах, например, Dynabeads MyOne компании Life Technologies, США) не превышает 1300 рМ биотина на мг частиц, что является достаточным для иммобилизации белка-мишени, но может оказаться недостаточным для удаления из раствора неспецифически связывающихся со стрептавидином структур НК. При проведении преселекции и селекции для блокировки свободных валентностей поверхности частиц и снижения неспецифической адсорбции исследователи вынуждены прибегать к использованию в буферных растворах дополнительных блокирующих агентов, в частности, бычьего сывороточного альбумина [Faryammanesh R., Lange Т., Magbanua Е., Haas S., Meyer С., Wicklein D., Schumacher U., Hahn U. // PLoS One. - 2014. - V. 9. - №4. - P.: e93173].
He являясь близкими аналогами данного изобретения, описанные системы отбора демонстрируют значимость проведения этапа негативной селекции библиотеки олигонуклеотидов для удаления структур НК, неспецифически связывающихся с компонентами системы отбора, для успешности процесса селекции. Описанных близких аналогов данного изобретения в базах данных объектов интеллектуальной собственности и научной литературе не обнаружено.
Задачей настоящего изобретения является создание универсального способа направленного истощения олигонуклеотидных библиотек для снижения уровня ко-селекции структур нуклеиновых кислот (НК), аффинных к вспомогательным элементам структуры рекомбинантных белков и иным белкам, используемым для твердофазной селекции аптамеров на основе нуклеиновых кислот.
Техническим результатом данного изобретения является способ направленного истощения исходного олигонуклеотидного пула, при котором библиотека олигонуклеотидов связывается с избытком молекулярной мишени и последующее отделение пула олигонуклеотидов, не связывающегося с мишенью, проводится при помощи гель-фильтрационной хроматографии.
Технический результат изобретения достигается за счет подготовки исходного пула олигонуклеотидной библиотеки, в течение которой получают разведение исходной библиотеки одноцепочечных коротких последовательностей НК до 1015 молекул/мл, что учитывает достаточную ее представленность для дальнейшей селекции и для равномерного и распределенного присутствия молекул в растворе.
Также технический результат изобретения достигается за счет прогревания предварительно разведенного пула олигонуклеотидов при 90°C для разрушения существующих двухцепочечных структур и плавления комплементарно связавшихся между собой, в том числе за счет высокой исходной концентрации, молекул ДНК и приведения всех олигонуклеотидов в состояние одиночной молекулы для максимальной представленности и вариабельности исходного пула молекул библиотеки.
Технический результат изобретения также достигается за счет медленного остывания подготовленного разведенного пула олигонуклеотидной библиотеки до 37°C для формирования в растворе индивидуальной пространственной структуры молекул олигонуклеотидов.
Технический результат изобретения достигается за счет связывания подготовленного исходного пула олигонуклеотидной библиотеки с мишенью, в отношении которой необходимо провести истощение пула олигонуклеотидов, проводимого в растворе.
Также технический результат изобретения достигается за счет присутствия избытка молекулы-мишени в растворе при инкубации ее с подготовленным пулом олигонуклеотидной библиотеки, при котором каждая молекула имеет потенциальную возможность взаимодействовать с мишенью и проявить аффинные свойства при их наличии.
Также технический результат изобретения достигается за счет оптимального времени инкубации пула олигонуклеотидной библиотеки с молекулой-мишенью, которое составляет от 60 до 120 минут, в течение которых происходит связывание с мишенью не только высокоаффинных структур НК, но и структур, обладающих более низким сродством к мишени, за счет чего удается элиминировать из пула олигонуклеотидов максимальное количество структур НК, аффинных к мишени, и представить к этапу позитивной селекции аптамеров на твердой фазе максимально истощенную по признаку связывания с предыдущей мишенью библиотеку.
Технический результат изобретения достигается также использованием высокоэффективной гель-фильтрационной хроматографии для эффективного разделения свободных от мишени последовательностей пула олигонуклеотидной библиотеки.
Наконец, технический результат изобретения достигается концентрированием истощенного по признаку связывания с заданной мишенью пула библиотеки олигонуклеотидов с использованием метода преципитации этанолом с дальнейшим растворением преципитата олигонуклеотидов в необходимом для дальнейшей работы объеме раствора.
Преимуществом изобретения является возможность истощения исходной олигонуклеотидной библиотеки против растворимых белков, находящихся в нативной конформации или конформации близкой к нативной, что позволяет олигонуклеотидам контактировать со всеми эпитопами молекулы-мишени, что невозможно при ориентированной иммобилизации мишени или химической модификации.
Важным достоинством предлагаемого изобретения является универсальность его применения для любых типов растворимых молекулярных мишеней.
Также преимуществом изобретения является применение гель-фильтрационной хроматографии на сорбенте, подходящем для эффективного разделения связавшихся и свободных последовательностей. Выбор типа сорбента зависит от молекулярной массы молекул, присутствующих в разделяемом составе.
Также преимуществом метода является простота, быстрота и доступность его осуществления. Проведение негативной селекции по данному методу не требует затрат на дорогостоящие носители для белка-мишени или парамагнитные частицы.
Также немаловажно, что совпадение молекулярной массы мишени и олигонуклеотидов библиотеки не препятствует удалению из смеси олигонуклеотидов, имеющих высокие и средние показатели аффинности к заданной мишени, поскольку такой комплекс эффективно отделится в процессе гель-фильтрации от несвязанных между собой компонентов смеси. В случае выхода свободных аптамеров и свободных молекул мишени в составе единого хроматографического пика, их разделение возможно провести методом фенол-хлороформной экстракции с дальнейшей преципитацией нуклеиновых кислот этанолом.
Также предлагаемое изобретение позволяет провести не только эффективное, но и быстрое направленное целевое истощение исходного олигонуклеотидного пула против растворимой мишени, занимающее один рабочий день.
Изобретение осуществляют следующим образом:
Синтезируют (ЗАО «Евроген», Россия; по заказу ФБУН ГНЦ ПМБ) комбинаторную библиотеку олигонуклеотидов, состоящую из вариабельной части размером в 45 нуклеотидов и фланкирующих константных участков длиной по 18 нуклеотидов: 5'-const1-N45-const2, где N45 - вариабельная область, a const1 и const2 -константные фланкирующие области, которые служат для связывания со специфическими праймерами, позволяющими амплифицировать пул олигонуклеотидов после проведения этапов селекции. Таким образом, исходная олигонуклеотидная библиотека содержит 445 вариантов последовательности, содержащей 81 нуклеотид (Фиг. 1).
Делают разведение исходной библиотеки олигонуклеотидов с содержанием 1015 молекул олигонуклеотидов в растворе, содержащем 20 мМ трис-HCl, 100 мМ хлорида натрия. Прогревают полученный раствор при 90°C в течение 10 минут, затем медленно охлаждают до 37°C. Приводят раствор к объему 500 мкл с использованием центрифужных концентраторов (Amicon Ultra-0,5, Millipore, США или аналогичных) и инкубируют в объеме 1 мл с не менее чем 5-кратным молярным избытком мишени при температуре 37°C и медленном перемешивании. На колонку, упакованную сорбентом для гель-фильтрационной хроматографии, способным по молекулярной массе и особенностям структуры отделить свободные олигонуклеотиды от комплекса мишени и связанных с ней олигонуклеотидов, уравновешенную буферным раствором, содержащим 20 мМ трис-HCl, 100 мМ хлорида натрия, наносят полученную смесь ДНК и белка. После отделения свободных молекул НК в отдельный пул, суспензию доочищают фенол-хлороформом и концентрируют, используя метод преципитации этанолом. Преципитат олигонуклеотидов доводят рабочим раствором до 500 мкл и повторяют раунд. Всего проводят три аналогичных раунда истощения пула олигонуклеотидной библиотеки по вышеописанной методике.
Полученный после истощения в отношении заданной мишени пул амплифицируют для обогащения при помощи ПЦР с использованием специфических к константным областям праймеров: прямого, меченного по 5'-концу карбоксифлуоресцеином (FAM метка), и обратного, несущего на 5'-конце остаток фосфорной кислоты. Фосфорилированную цепь расщепляют обработкой двуцепочечных фрагментов 5'-экзонуклеазой фага лямбда. Наличие FAM метки позволяет визуально контролировать полноту расщепления обратной цепи при проведении электрофореза в 10% полиакриламидном геле, содержащем мочевину (Фиг. 2). Истощенный и амплифицированный пул одноцепочечных последовательностей применяют для проведения твердофазной селекции аптамеров.
Проводят предварительный скрининг проявления аффинности полученного истощенного пула олигонуклеотидов в отношении мишени для истощения в формате ПЦР. Для этого на ПЦР-планшете иммобилизуют мишень, после чего инкубируют с исходным (контроль сравнения) и истощенным пулами библиотеки олигонуклеотидных последовательностей. Результат визуализируют в ПЦР в режиме реального времени с применением интеркалирующего красителя SYBR-Green I (ЗАО «Синтол», Россия). Количество анализируемых молекул нуклеиновых кислот в лунках с контролем и исследуемой пробой уравнивают.
Изобретение иллюстрируют следующие графические материалы:
Фиг. 1. Последовательности ДНК, используемые в процессе селекции и амплификации селектированных последовательностей.
1) Последовательность олигонуклеотидной библиотеки, используемой в селекции, где N45 - вариабельная область, const1 и const2 - константные области для отжига специфических праймеров;
2) Последовательности специфических праймеров, используемых в реакциях обогащения библиотеки и в реакциях амплификации при проведении ПЦР-РВ.
Фиг. 2. Расщепление обратной цепи ДНК при обработке двуцепочечного фрагмента 5'-экзонуклеазой фага лямбда.
1. Маркер молекулярной массы (50 bp DNA Ladder, New England BioLabs, США)
2. Двуцепочечный фрагмент ДНК
3. Отрицательный контроль (амплификация без внесения целевой ДНК)
4. Одноцепочечный фрагмент ДНК, полученный после обработки ПЦР-продукта 5'-экзонуклеазой фага лямбда.
Изображение получено с использованием лазерного сканера Typhoon FLA 9500.
Фиг. 3. Схема проведения раунда негативной селекции для получения истощенного пула олигонуклеотидной библиотеки с использованием разработанного способа.
Фиг. 4. Результаты скрининга в ПЦР-РВ эффективности истощения пулов олигонуклеотидной библиотеки в отношении глутатион-S-трансферазы и стрептавидина.
По оси абсцисс обозначены:
1) Результаты* ПЦР-РВ для пула исходной библиотеки ДНК-олигонуклеотидов;
2) Результаты* ПЦР-РВ для пула библиотеки ДНК-олигонуклеотидов после одного раунда истощения;
3) Результаты* ПЦР-РВ для пула библиотеки ДНК-олигонуклеотидов после двух раундов истощения;
4) Результаты* ПЦР-РВ для пула библиотеки ДНК-олигонуклеотидов после трех раундов истощения.
*представлен средний результат для трех повторностей.
Фиг. 5. Принципиальная схема проведения раунда позитивной селекции аптамеров к LF и LC BoNT/A с использованием магнитных микрочастиц.
А) Вариант селекции с использованием пула исходной олигонуклеотидной библиотеки.
В) Вариант селекции с использованием пула истощенной в отношении глутатион-S-трансферазы или стрептавидина олигонуклеотидной библиотеки.
Фиг. 6. Принципиальная схема проведения раунда позитивной селекции аптамеров к IV домену РА и HC50 BoNT/A с использованием хроматографического сорбента.
A) Вариант селекции с использованием пула исходной олигонуклеотидной библиотеки.
B) Вариант селекции с использованием пула истощенной в отношении глутатион-S-трансферазы или стрептавидина олигонуклеотидной библиотеки.
Фиг. 7. Результаты скрининга в ПЦР-РВ эффективности селекции пула специфичных олигонуклеотидов из исходной и истощенной библиотек в отношении белков LF, PA IV, LC BoNT/A и НС50 BoNT/А.
По оси абсцисс обозначены:
1. Результаты* ПЦР-РВ для пула исходной библиотеки ДНК-олигонуклеотидов до преселекции и селекции к рекомбинантным белкам;
2. Результаты* ПЦР-РВ для пула истощенной библиотеки ДНК-олигонуклеотидов: истощенной в отношении глутатион-S-трансферазы для белков LF и PA IV, истощенной в отношении стрептавидина для белков LC BoNT/A и НС50 BoNT/A;
3. Результаты* ПЦР-РВ для пула специфичных ДНК-олигонуклеотидов, селектированных из исходной библиотеки олигонуклеотидов после пяти раундов отбора против рекомбинантных белков;
4. Результаты* ПЦР-РВ для пула специфичных ДНК-олигонуклеотидов, селектированных из истощенной (в отношении глутатион-S-трансферазы для белков LF и PA IV, в отношении стрептавидина для белков LC BoNT/A и НС50 BoNT/A) библиотеки олигонуклеотидов после пяти раундов отбора против рекомбинантных белков.
*представлен средний результат для трех повторностей.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Проведение раунда направленного истощения пула олигонуклеотидной библиотеки на примере комбинаторной библиотеки последовательностей ДНК и мишеней: глутатион-S-трансферазы и стрептавидина.
К 45 мкг (1015 молекул) раствора исходного пула олигонуклеотидной комбинаторной библиотеки (Фиг. 1) добавляют буферный раствор, содержащий 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4), до общего объема 1000 мкл, тщательно перемешивают. Полученный раствор ДНК помещают в твердотельный термостат и прогревают при температуре 90°C в течение 10 минут. Устанавливают температуру твердотельного термостата в значении 37°C, и дожидаются охлаждения пробы до установленной температуры. Доводят объем полученного раствора до 500 мкл с использованием центрифужных концентраторов (Amicon Ultra-0,5, Millipore, США).
Белок глутатион-S-трансфераза, имеющий массу 26 кДа, в количестве 0,5 мг или стрептавидин с молекулярной массой 53 кДа в количестве 1 мг в буферном растворе 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (рН 7,4) и объеме не более 500 мкл добавляют к подготовленному раствору олигонуклеотидов. При необходимости доводят общий объем смеси до 1 мл. Таким образом, в смеси представлен молярный избыток белка по отношению к ДНК. Каждую смесь инкубируют в течение 1 часа при температуре 37°C с перемешиванием на орбитальном шейкере.
По окончании инкубации каждую из смесей разделяют при помощи гель-фильтрационной хроматографии на колонке, содержащей сорбент низкого давления Superdex 75™ (GE Healthcare, Великобритания) под контролем спектрофотометра при длине волны 260 нм, при скорости потока 0,5 мл/мин на фракции белка, несвязавшихся молекул ДНК и молекул ДНК, ассоциировавших с белком.
Пул отделенных несвязавшихся олигонуклеотидов очищают с использованием метода фенол-хлороформной экстракции нуклеиновых кислот, затем концентрируют преципитацией ДНК этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant (Novagen, Германия). Преципитат ДНК растворяют в 500 мкл буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4). Повторяют описанную выше процедуру дважды. Схема раунда истощения приведена на Фиг. 3.
Пример 2. Скрининг эффективности связывания истощенного пула олигонуклеотидов с мишенью.
Полученные истощенные пулы одноцепочечных коротких фрагментов ДНК амплифицируют для обогащения их молекулами ДНК, имеющими низкий показатель аффинности в отношении глутатион-S-трансферазы или стрептавидина, соответственно. Амплификацию каждого пула проводят с использованием праймеров: прямого ForFAM, несущего в положении 5' флуоресцентную метку карбоксифлуоресцеин, и обратного RevP, фосфорилированного по 5'-концу последовательности (температура отжига праймеров 54°C, время элонгации 15 секунд) (Фиг. 1). В результате получают двуцепочечные фрагменты, одна из цепей которых мечена FAM.
Для получения обогащенных пулов одноцепочечных фрагментов ПЦР-продукт обрабатывают 5'-экзонуклеазой фага лямбда, которая расщепляет обратную фосфорилированную цепь фрагмента [Higuchi R. G., Ochman Н. // Nucleic Acids Research. - 1989. - V. 17. - N. 14. - P. 5865]. Эффективность расщепления цепи контролируют проведением электрофореза в 10%-ом полиакриламидном геле, содержащем мочевину, с последующим окрашиванием бромистым этидием, при этом наличие в пробе только полосы, флуоресцирующей зеленым, свидетельствует об успешном расщеплении обратной цепи фрагмента, а присутствие в пробе второй полосы с красным цветом флуоресценции свидетельствует о неуспешном или неполном расщеплении фосфорилированной цепи ДНК (Фиг. 2). Полученные одноцепочечные ДНК очищают и концентрируют, используя фенол-хлороформную экстракцию и преципитацию этанолом.
На поверхности лунок поликарбонатного 96-луночного ПЦР-планшета (Corning Costar, США) иммобилизируют белок глутатион-S-трансферазу или стрептавидин в количестве 0,5 мкг/лун в 50 мкл буферного раствора (20 мМ Tris-HCl, 50 мМ NaCl, pH 7,4). Для этого инкубируют планшет с белком в течение 1 часа при 37°C. Проводят отмывку лунок планшета буфером состава 20 мМ Tris-HCl, 50 мМ NaCl, pH 7,4 с содержанием Tween-20 0,05% трижды. Свободные валентности пластика блокируют инертным раствором белков с составом 3% бычьего сывороточного альбумина, 1% α-казеина в буфере 20 мМ Tris-HCl, 50 мМ NaCl, pH 7,4 в течение часа при температуре 37°C, после чего проводят трехкратную отмывку планшета. В лунки планшета вносят пробы одноцепочечной ДНК в количестве 0,2 мкг/лун в 50 мкл буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4 - истощенный в отношении глутатион-S-трансферазы и истощенный по связыванию со стрепавидином пулы олигонуклеотидов наносят на планшет, покрытый глутатион-S-трансферазой и стрептавидином, соответственно. В качестве контроля сравнения используют разведения исходного пула олигонуклеотидов, в качестве отрицательного контроля амплификации используют лунки без внесения ДНК. Все пробы тестируют в трех повторностях. Инкубируют пробы ДНК в течение 1 часа при температуре 37°C и перемешивании на орбитальном шейкере. Промывку лунок планшета проводят пятикратно буфером, содержащим 20 мМ Tris-HCl, 250 мМ NaCl и 5 мМ EDTA, pH 7,4.
Контролируют реакцию связывания одноцепочечной ДНК с белком в ПЦР в режиме реального времени. Для этого в лунки планшета после их отмывки вносят амплификационную смесь, содержащую флуоресцентный интеркалирующий краситель SYBR-Green I (ЗАО "Синтол", Россия) с добавлением праймеров ForP и RevP (Фиг. 1). Проводят амплификацию в режиме: температура отжига праймеров 54°C, время элонгации 10 секунд, детекцию сигнала проводят по каналу SYBR 1 (FAM) (прибор IQ™5, Bio-Rad, США). Результаты скрининга представлены на Фиг. 4.
Пример 3. Проведение раунда позитивной селекции в отношении рекомбинантного белка летального фактора В. anthracis (LF), содержащего глутатион-S-трансферазу, с использованием исходного и истощенного пулов олигонуклеотидной библиотеки на твердой фазе, представленной магнитными микрочастицами.
Рекомбинантный белок летального фактора В. anthracis (LF), содержащий в своем составе в качестве дополнительного компонента глутатион-S-трансферазу, иммобилизируют на поверхности магнитных микрочастиц, несущих глутатион (Pierce™ Glutathione Magnetic Beads, Thermo, США), в соответствии с инструкцией производителя.
Помещают по 100 мкл суспензии магнитных частиц с иммобилизованным на их поверхности рекомбинантным белком LF, в две пластиковые пробирки объемом 1,5 мл, устанавливают пробирки на магнитный стенд DynaMag™-2 Magnet (Invitrogen, США), отбирают жидкость. К сухому осадку магнитных частиц добавляют пул ДНК олигонуклеотидов в объеме 1 мл: 1) пул, истощенный по признаку связывания с глутатион-S-трансферазой; 2) исходный пул олигонуклеотидной библиотеки, концентрацию которого уравнивают в соответствии с концентрацией истощенного пула одноцепочечной ДНК. Буфер для инкубации ДНК с магнитными частицами имеет состав 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4. Инкубацию проводят в течение 1 часа при температуре 37°C при постоянном перемешивании на орбитальном шейкере, а также каждые 10 минут тщательно перемешивают смесь вортексированием.
По окончании инкубации суспензию магнитных частиц переносят в хроматографический шланг с внутренним диаметром 0,5 мм, который пропущен через неодимовую трубку длиной 10 см с радиально направленным магнитным полем. Шланг подключают к перистальтическому насосу и промывают стерильным буфером, содержащим 20 мМ Tris-HCl pH 7,5, 100 мМ NaCl и 5 мМ EDTA, pH 7,4, в течение 10 минут при скорости потока 2 мл/мин. Останавливают насос, смещают неодимовую магнитную трубку к концу шланга, после чего трубку снимают, а суспензию магнитных частиц собирают в стерильную пробирку.
Собранные после промывки частицы собирают на стенке пробирки, помещая ее на магнитный штатив, отбирают промывочный раствор. Снимают пробирку с магнита и добавляют к частицам, несущим глутатион, 200 мкл раствора (20 мМ Tris-HCl pH 7,5, 50 мМ NaCl и 5 мМ EDTA, pH 7,4), содержащего 0,2 мМ восстановленного глутатиона. Инкубируют при постоянном перемешивании в течение 10 минут. Фиксируют магнитные частицы магнитным полем на штативе, отбирают жидкость в отдельную стерильную пробирку.
ДНК выделяют из раствора с использованием метода фенол-хлороформной экстракции нуклеиновых кислот, затем концентрируют преципитацией этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant. Преципитат ДНК растворяют в 1 мл буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4).
Амплификацию селектированных последовательностей и получение одноцепочечных молекул ДНК проводят по методике, описанной в Примере 2.
Всего проводят 5 раундов селекции. Принципиальная схема раунда селекции представлена на Фиг 5.
Пример 4. Проведение раунда позитивной селекции в отношении рекомбинантного белка IV домена протективного антигена В. anthracis (PA IV), содержащего глутатион-S-трансферазу, с использованием исходного и истощенного пулов олигонуклеотидной библиотеки на твердой фазе, представленной хроматографическим сорбентом глутатион сефарозой.
Рекомбинантным белком PA IV, содержащим глутатион-S-трансферазу, заполняют все активные центры предварительно уравновешенного рабочим буфером сорбента Glutathione Sepharose 4 FastFlow, помещенного в две колонки объемом 1 мл (GSTrap™ FF, GE Healthcare, Великобритания) в соответствии с характеристиками сорбента, заявленными производителем (10 мг рекомбинантного белка глутатион-S-трансферазы на 1 мл сорбента). Рабочий буфер представлен составом 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4).
Два варианта пула ДНК-олигонуклеотидов (истощенный пул и исходный пул олигонуклеотидной библиотеки) в равных концентрациях доводят до 10 мл рабочим буфером, набирают в шприцы с люэровским соединением. Подсоединяют шприцы к коннекторам колонок и прогоняют раствор ДНК через сорбент за время не более 3 секунд. Промывают сорбент потоком раствора 20 мМ Tris-HCl, 250 мМ NaCl и 5 мМ EDTA (pH 7,4) в объеме не менее 400 мл. Элюируют рекомбинантный белок PA IV со связавшимися молекулами ДНК буфером 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4), содержащим 0,2 мМ восстановленного глутатиона.
ДНК выделяют из раствора с использованием метода фенол-хлороформной экстракции нуклеиновых кислот, затем концентрируют преципитацией этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant. Преципитат ДНК растворяют в 1 мл буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4).
Амплификацию селектированных последовательностей и получение одноцепочечных молекул ДНК проводят по методике, описанной в Примере 2.
Всего проводят 5 раундов селекции. Принципиальная схема раунда селекции представлена на Фиг. 6.
Пример 5. Проведение раунда позитивной селекции в отношении рекомбинантного белка протеолитического домена (легкой цепи) ботулотоксина типа А (LC BoNT/A), содержащего in vivo биотинилируемый пептид, с использованием исходного и истощенного пулов олигонуклеотидной библиотеки на твердой фазе, представленной магнитными микрочастицами.
Рекомбинантный белок LC BoNT/A, содержащий в своем составе пептид, биотинилированный in vivo ферментом биотин-лигазой Е. coli, иммобилизируют на поверхности магнитных микрочастиц, несущих стрептавидин (Pierce™ Streptavidin Magnetic Beads, Thermo, США), в соответствии с инструкцией производителя.
Помещают по 100 мкл суспензии магнитных частиц с иммобилизованным на их поверхности рекомбинантным белком LC BoNT/A в две пластиковые пробирки объемом 1,5 мл, устанавливают пробирку на магнитный стенд DynaMag™-2 Magnet (Invitrogen, США), отбирают жидкость. К сухому осадку магнитных частиц добавляют пул ДНК олигонуклеотидов в объеме 1 мл: 1) пул, истощенный по признаку связывания со стрептавидином; 2) исходный пул олигонуклеотидной библиотеки, концентрацию которого уравнивают в соответствии с концентрацией истощенного пула одноцепочечной ДНК. Буфер для инкубации ДНК с магнитными частицами имеет состав 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4. Инкубацию проводят в течение 1 часа при температуре 37°C при постоянном перемешивании на орбитальном шейкере, а также каждые 10 минут тщательно перемешивают смесь вортексированием.
Отмывку магнитных частиц проводят по методике, описанной в Примере 3, с использованием перистальтического насоса и трубки, пропущенной сквозь неодимовый магнит.
Собранные после промывки частицы собирают на стенке пробирки, помещая ее на магнитный штатив, отбирают промывочный раствор. Снимают пробирку с магнита и добавляют к частицам 200 мкл раствора 8 М гуанидин-HCl, pH 1,5. Инкубируют при постоянном перемешивании в течение 5 минут. Фиксируют магнитные частицы магнитным полем на штативе, отбирают жидкость в отдельную стерильную пробирку, немедленно доводят pH раствора до значений 7,4-7,6. Диализуют отобранную пробу против буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4.
ДНК выделяют из раствора с использованием метода фенол-хлороформной экстракции нуклеиновых кислот, затем концентрируют преципитацией этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant. Преципитат ДНК растворяют в 1 мл буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,4).
Амплификацию селектированных последовательностей и получение одноцепочечных молекул ДНК проводят по методике, описанной в Примере 2.
Всего проводят 5 раундов селекции. Принципиальная схема раунда селекции представлена на Фиг 5.
Пример 6. Проведение раунда позитивной селекции в отношении химически модифицированного (биотинилированного) рекомбинантного белка рецепторсвязывающего домена тяжелой цепи ботулинического нейротоксина типа А (НС50 BoNT/A) с использованием исходного и истощенного пулов олигонуклеотидной библиотеки на твердой фазе, представленной хроматографическим сорбентом стрептавидин сефарозой.
Рекомбинантным биотинилированным белком НС50 BoNT/A заполняют все активные центры предварительно уравновешенного рабочим буфером сорбента Streptavidin Sepharose High Performance (GE Healthcare, Великобритания), помещенного в две колонки объемом 1 мл (HiTrap Streptavidin HP, GE Healthcare, Великобритания) в соответствии с характеристиками сорбента, заявленными производителем (>300 нМ биотина на 1 мл сорбента). Рабочий буфер представлен составом 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4.
Истощенный в отношении стрептавидина и исходный пулы олигонуклеотидной библиотеки в равных концентрациях доводят до 10 мл рабочим буфером. Наносят растворы ДНК на колонки с использованием шприцов с люэровским соединением за время не более 3 секунд. Промывают сорбент потоком раствора 20 мМ Tris-HCl, 250 мМ NaCl и 5 мМ EDTA (pH 7,4) в объеме не менее 400 мл. Элюируют рекомбинантный белок НС50 BoNT/A со связавшимися молекулами ДНК буфером 8 М гуанидин-НСl, pH 1,5. По окончании элюции немедленно доводят pH раствора до значений 7,4 - 7,6 и диализуют отобранную пробу против буфера 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA, pH 7,4.
Выделение ДНК и амплификацию селектированных последовательностей и получение одноцепочечных молекул ДНК проводят по методике, описанной выше.
Всего проводят 5 раундов селекции. Принципиальная схема раунда селекции представлена на Фиг. 6.
Пример 7. Сравнительный скрининг эффективности селекции аптамеров с применением истощенного и исходного пулов ДНК-олигонуклеотидов в отношении рекомбинантных белков LF, PA IV, LC BoNT/A и НС50 BoNT/А.
Скрининг эффективности проводят по схеме, аналогично описанной в Примере 2. Для скрининга используют пулы ДНК-олигонуклеотидов, подвергнутых позитивной селекции в отношении рекомбинантных мишеней. При этом сравнивают эффективность связывания для пулов, которые предварительно были истощены по признаку связывания с глутатион-S-трансферазой, входящей в состав рекомбинантных белков, или стрептавидином, находящимся на поверхности твердой фазы, на которой проводится селекция аптамеров, и для пулов олигонуклеотидов, не подвергавшихся преселекции.
На поверхности лунок поликарбонатного 96-луночного ПЦР-планшета иммобилизируют рекомбинантные белки - LF или PA IV, вместо глутатион-S-трансферазы содержащие полигистидиновую последовательность, или LC BoNT/A, или НС50 BoNT/A, - в количестве 500 мкг/лунку в 50 мкл буферного раствора (20 мМ Tris-HCl, 50 мМ NaCl, pH 7,4). Свободные активные валентности планшета блокируют раствором инертных белков. В лунки планшета вносят пробы одноцепочечных структур ДНК в количестве 0,1 мкг/лун в 50 мкл буфера. В качестве контроля сравнения используют разведения исходного пула олигонуклеотидов, в качестве отрицательного контроля амплификации используют лунки без внесения ДНК. Все пробы тестируют в трех повторностях. Инкубируют в течение 1 часа при температуре 37°C и перемешивании.
Контролируют реакцию связывания одноцепочечной ДНК с белком в ПЦР в режиме реального времени как описано в Примере 2. За положительный результат связывания ДНК с белком-мишенью принимают показания прибора для пороговых циклов, которые ниже пороговых циклов контроля сравнения не менее чем на пять циклов (Фиг. 7).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С ПРОТЕОЛИТИЧЕСКОЙ СУБЪЕДИНИЦЕЙ НЕЙРОТОКСИНА ТИПА A CLOSTRIDIUM BOTULINUM | 2014 |
|
RU2571210C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯ ШИГА-ТОКСИН ТИПА 2 | 2014 |
|
RU2566552C1 |
| Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila | 2019 |
|
RU2737829C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGST/ART/X, КОДИРУЮЩАЯ СЛИТНЫЙ ХИМЕРНЫЙ ПОЛИПЕПТИД GST/ART/X ДЛЯ ВЫСОКОСПЕЦИФИЧНОЙ СЕЛЕКЦИИ АПТАМЕРОВ К БЕЛКУ-МИШЕНИ X В СОСТАВЕ ПОЛИПЕПТИДА | 2014 |
|
RU2558295C1 |
| Способ термической диссоциации для проведения селекции ДНК-аптамеров | 2019 |
|
RU2723373C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, ПРИМЕНИМАЯ В КАЧЕСТВЕ ЗОНДА ДЛЯ ОБЕСПЕЧЕНИЯ МАКСИМАЛЬНОГО СООТНОШЕНИЯ "СИГНАЛ/ФОН" В ТЕСТ-СИСТЕМАХ НА ОСНОВЕ ИММУНО-ПЦР | 2014 |
|
RU2566550C1 |
| СПОСОБ СПЕЦИФИЧЕСКОГО ОТБОРА ВЫСОКОАФФИННЫХ МОЛЕКУЛ ДНК (ДНК-АПТАМЕРОВ) К РЕКОМБИНАНТНОМУ БЕЛКУ-МИШЕНИ | 2012 |
|
RU2513700C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯСЯ С БАКТЕРИЯМИ Escherichia coli O157:H7 | 2014 |
|
RU2566551C1 |
| АПТАМЕРЫ ДЛЯ ОЧИСТКИ И КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ ГЕЛЬЗОЛИНА (GELSOLIN) И ЕГО АНАЛОГОВ | 2015 |
|
RU2644191C1 |
| СПОСОБЫ РАЗДЕЛЕНИЯ СПЕРМАТОЗОИДОВ И ПРИМЕНЯЕМЫЕ ДЛЯ ЭТОГО КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АПТАМЕРЫ ИЛИ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВЫХ КИСЛОТ | 2007 |
|
RU2475539C2 |



Изобретение относится к области биотехнологии и касается способа направленного истощения олигонуклеотидных библиотек в отношении водорастворимых белковых мишеней глутатион-S-трансферазы и стрептавидина. Представленный способ включает подготовку комбинаторной олигонуклеотидной библиотеки в количестве 1015 молекул к отбору, разведение олигонуклеотидов библиотеки до концентрации 1015 молекул на 1 мл, денатурацию олигонуклеотидов при 90°С в течение 10 минут и отжиг олигонуклеотидов при 37°С. Связывание глутатион-S-трансферазы или стрептавидина в количестве 1 мг с подготовленным пулом исходной олигонуклеотидной библиотеки в буферном растворе, содержащем 20 мМ Tris-HCl рН 7,4, 50 мМ NaCl и 5 мМ EDTA, и разделение полученной смеси нуклеиновая кислота/белок-мишень гель-фильтрационной хроматографией на фракции. Изобретение позволяет элиминировать из пула олигонуклеотидов максимально возможное количество аффинных к заданной мишени молекул, а повторение раундов такой селекции гарантирует отсутствие в финальном пуле молекул олигонуклеотидов, имеющих значимую для дальнейшего применения аффинность к мишени. Изобретение может быть применено для получения выскоаффинных аптамеров, применимых в области биомедицины. 7 ил., 7 пр.
Способ направленного истощения олигонуклеотидных библиотек в отношении водорастворимых белковых мишеней глутатион-S-трансферазы и стрептавидина, включающий подготовку комбинаторной олигонуклеотидной библиотеки в количестве 1015 молекул к отбору, разведение олигонуклеотидов библиотеки до концентрации 1015 молекул на 1 мл, денатурацию олигонуклеотидов при 90°С в течение 10 минут и отжиг олигонуклеотидов при 37°С, связывание глутатион-S-трансферазы или стрептавидина в количестве 1 мг с подготовленным пулом исходной олигонуклеотидной библиотеки в буферном растворе, содержащем 20 мМ Tris-HCl рН 7,4, 50 мМ NaCl и 5 мМ EDTA, и разделение полученной смеси нуклеиновая кислота/белок-мишень гель-фильтрационной хроматографией на фракции, содержащие олигонуклеотиды, связавшиеся или не связавшиеся с белком-мишенью, для выделения пула олигонуклеотидов, не способных связываться с белковой мишенью в заданных условиях.
| STUART K ARCHER et al., Selective and flexible depletion of problematic sequences from RNA-seq libraries at the cDNA stage, BMC Genomics, 2014, 15:401, pp.1-9 | |||
| STUART K | |||
| ARCHER et al, Probe-Directed Degradation (PDD) for Flexible Removal of Unwanted cDNA Sequences from RNA-Seq Libraries, Curr Protoc Hum Genet., 2015 Apr 1 Vol.85, No.11.15, pp.1-36 | |||
| СПОСОБ СКРИНИНГА БИБЛИОТЕКИ ФАГОВОГО ДИСПЛЕЯ | 2005 |
|
RU2402777C2 |

