Изобретение относится к области биотехнологии, биоинженерии, биохимии и диагностической медицинской микробиологии.
Основными областями применения ДНК-аптамеров являются медицинские и биологические исследования, клиническая диагностика заболеваний, в том числе создание высокочувствительных и высокоэффективных систем детекция мишеней белковой природы, а также терапия заболеваний различной природы.
Одной из актуальных проблем медицинской практики является быстрая и точная идентификация инфекционных возбудителей, обеспечивающая постановку правильного диагноза, что позволяет избежать развития различных осложнений и переход заболевания в хроническую форму.
Инфекция Legionella pneumophila по-прежнему ошибочно считается экзотической болезнью. Диагностирование инфекции затруднено вследствие неопределенного характера клинических симптомов, которые не позволяют отличить болезнь легионеров от прочих пневмоний, и ограниченного количества доступных и эффективных диагностических тестов на рынке. Невозможность своевременного определения этиологического агента пневмонии приводит к назначению неэффективной терапии, а также не позволяет проводить качественный эпидемиологический надзор за распространением и изменением изолятов L. pneumophila, циркулирующих в конкретных стационарах или регионах.
Основным методом, позволяющим осуществлять раннюю диагностику и мониторинг легионеллезной инфекции, является определение легионеллезного антигена в моче иммуно-хроматографическим или иммуноферментным методом. Эти тесты основаны, прежде всего, на выявлении ЛПС L. pneumophilla серогруппы I, т.к. именно этот возбудитель чаще всего вызывает легионеллезную инфекцию. Вместе с тем, последние исследования показывают, что доля легионеллезной инфекции, вызванной L. pneumophilla серогруппы I, варьирует от года к году и составляет от 50 до 91 %, что было подтверждено лабораторными исследованиями [St-Martin G. // ISRN Epidemiol. - 2013. - Vol. 2013. - Article ID 847283. - doi: 10.5402/2013/847283; Bruin J.P. // Eur J Clin Microbiol Infect Dis. - 2013. - Vol. 32, N 3. - P. 333-334. - doi:10.1007/s10096-012-1745-0.]. Более того, показатели смертности для всех групп пациентов, не входящих в группу заболевших от инфицирования L. pneumophilla серогруппы I, гораздо выше и составляют около 27 % [Benin A.L. // Clin Infect Dis. - 2002. - Vol. 35, N 9. - P. 1039-1046. - doi:10.1086/342903.]. L. pneumophila, L. micdadei и L. bozemanii были причиной 30-55% заболеваний легионеллезной инфекцией в Австралии и Новой Зеландии. Регистрировались также инфекции, вызванные L. jordanis, L. oakridgensis, L. anisa, L. gormanii и L. sainthelensi [Murdoch D.R. // Clin Infect Dis. - 2013. - Vol. 57, N 9. - P. 1275-1281. - doi:10.1093/cid/cit504; NNDSS, Annual Report Writing Group. Australia's notifiable disease status, 2011: annual report of the National Notifiable Diseases Surveil-lance System. Communicable diseases intelligence quarterly report. - 2013. - Vol. 37, N 4. - E313.].
Приведенные данные свидетельствуют о необходимости создания диагностических тестов, позволяющих выявлять инфекцию, вызванную любыми патогенными бактериями рода Legionella.
Известны достаточно эффективные диагностические тесты, позволяющие определять подавляющее большинство изолятов патогенной L. pneumophila. Тесты основаны на определении высококонсервативного родоспецифического пептидогликан-ассоциированного липопротеина (ПАЛ), известного также как урин-антиген, который определяется в моче больных легионеллезом, начиная с 3-его дня манифестации заболевания [Reller L. B. // Clinical Infectious Diseases. - 2003. - V. 36. - №. 1. - P. 64-69. - doi:10.1086/345529].
Однако при достаточно высокой специфичности существующих тест-систем, существует проблема недостаточной их чувствительности. Чувствительность некоторых достаточно экспрессных иммунологических тестов составляет около 70%. Существуют также тест-системы на основе ПЦР, которые при всей их эффективности обладают недостатками, присущими всем ПЦР-системам - невозможность определять антигены напрямую при отсутствии в образце ДНК возбудителя, ингибирование реакции амплификации и др. [Reller L. B. // Clinical Infectious Diseases. - 2003. - V. 36. - №. 1. - P. 64-69. - doi:10.1086/345529]. Поэтому существует необходимость разработки новых чувствительных тестов, позволяющих определять белок-антиген ПАЛ в образцах мочи.
Для высокоэффективной клинической диагностики легионеллезной инфекции могут быть успешно использованы специфические амплифицируемые лиганды - ДНК-аптамеры. Это одноцепочечные олигнуклеотиды, которые благодаря образованию сложных трехмерных структур и пространственному расположению зарядов в молекуле имеют высокую аффинность в отношении заданных молекул. Направленный отбор аптамеров против выбранных мишеней осуществляется по технологии SELEX ("Systematic Evolution of Ligands by Exponential Enrichment") [US Patent 5567588] с применением различных модификаций технологии, таких как микромагнитная селекция в микрожидкостных каналах (M-SELEX) [Lou, X., Qian, J., Xiao, Y., Viel, L., Gerdon, A. E., Lagally, E. T., … & Soh, H. T. // Proceedings of the National Academy of Sciences. - 2009. - V. 106. - №. 9. - P. 2989-2994.], использование капиллярного элекрофореза (ECEEM) [Drabovich A., Berezovski M., Krylov S. N. // Journal of the American Chemical Society. - 2005. - V. 127. - №. 32. - P. 11224-11225.] и др.
Ни в одной из существующих на сегодня баз данных селектированных и валидированных аптамеров не предложены последовательности, специфичные к пептидогликан-ассоциированному липопротеину легионелл.
Техническим результатом изобретения является селекция ДНК-аптамеров к пептидогликан-ассоциированному липопротеину патогенных микроорганизмов рода Legionella, обладающих высокой аффинностью и специфичностью в отношении ПАЛ.
Технический результат достигается тем, что предложена последовательность одноцепочечного ДНК-аптамера AptPAL_23 длиной 81 нуклеотид, с молекулярной массой 28 кДа, с первичной последовательностью CATACGTTCGACTGCTACGTGCATTCTTCTCCTGGGGGGGCTGCCGTCTCTACAGGGCATTATGTGAGATGTACAGACTAG, содержащая области для связывания с праймерами длиной 18 нуклеотидов в крайних положениях последовательности, образующая устойчивые специфичные связи с ПАЛ Legionella, имеющая константу равновесной диссоциации (KD) в отношении ПАЛ не менее 5×10-8 М.
Технический результат достигается применением оптимизированной системы селекции аптамеров, основанной на применении слитного рекомбинантного полипептида, позволяющего селектировать аптамеры с высокой специфичностью к мишени за малое количество раундов селекции [Патент RU №2558295]. Состав применяемого в системе селекции слитного рекомбинантного полипептида позволяет закрепить его на твердой фазе, а после связывания аптамеров с мишенью отделить мишень вследствие расщепления уникальной пептидной последовательности, распознаваемой и расщепляемой сайт-специфической протеазой. Такая структура слитного рекомбинантного полипептида для селекции аптамеров позволяет в мягких условиях элюции отделить мишень, несущую специфически связанные аптамеры, не элюируя при этом последовательности, связавшиеся с другими компонентами системы отбора и твердой фазой.
Также технический результат достигается путем клонирования селектированных последовательностей ДНК в плазмидный вектор, обеспечивающий селекцию, их секвенирования и определения структуры, проверки на специфическую активность в отношении легионеллезного ПАЛ.
Техническим результатом изобретения является Техническое решение предлагает применение слитного рекомбинантого белка для осуществления этапов позитивной селекции с применением этапа сайт-специфического протеолиза, что позволяет получить при отборе только последовательности, связанные с отщепленной мишенью (ПАЛ L. pneumophila).
Также техническое решение предлагает использовать иммобилизацию слитного рекомбинантного белка на поверхности магнитных микросфер, что позволяет повысить эффективность выделения комплекса аффинных аптамеров и белка-мишени за счет пространственного разделения белка-мишени, несущего специфические олигонуклеотиды, и остального комплекса системы селекции.
Важным преимуществом предлагаемой системы позитивной селекции является возможность получения пула высокоаффинных ДНК-аптамеров с минимальным содержанием низкоаффинных молекул, применив малое количество раундов селекции.
Изобретение иллюстрируют следующие графические материалы:
Фиг. 1. Последовательности ДНК, используемые для селекции молекул ДНК, для обогащения селектированного пула и для проведения амплификации селектированных последовательностей.
1) Последовательность олигонуклеотидной библиотеки, используемой в селекции, где N45 - вариабельная область, const1 и const2 - константные области для отжига специфических праймеров;
2) - 6) Последовательности праймеров, используемых в реакциях обогащения библиотеки, для клонирования и наращивания количества последовательностей.
7) - 8) Последовательности праймеров, используемых для получения последовательности ПАЛ L. pneumophila.
Фиг. 2. Пример результатов расщепления обратной цепи ДНК при обработке двуцепочечного фрагмента 5'-экзонуклеазой фага лямбда.
1. Маркер молекулярной массы (50 bp DNA Ladder, New England BioLabs, США);
2. Двуцепочечный фрагмент ДНК;
3. Отрицательный контроль (амплификация без внесения целевой ДНК);
4. Одноцепочечный фрагмент ДНК, полученный после обработки ПЦР-продукта 5'-экзонуклеазой фага лямбда.
Изображение получено с использованием лазерного сканера Typhoon FLA 9500.
Фиг. 3. Последовательность gst-apt-pal в составе плазмидной конструкции и экспрессируемый пептид GST/APT/PAL.
Фиг. 4. Результат расщепления пептида GST/APT/PAL в присутствии протеазы летального фактора B. anthracis.
1. Маркер молекулярной массы (PageRuler™ Prestained Protein Ladder #SM0671, Fermentas, США);
2. Химерный белок GST/APT/PAL без добавления летального фактора B. anthracis;
3. Белок протеаза летальный фактор B. anthracis;
4. Химерный белок GST/APT/PAL после обработки летальным фактором B. anthracis.
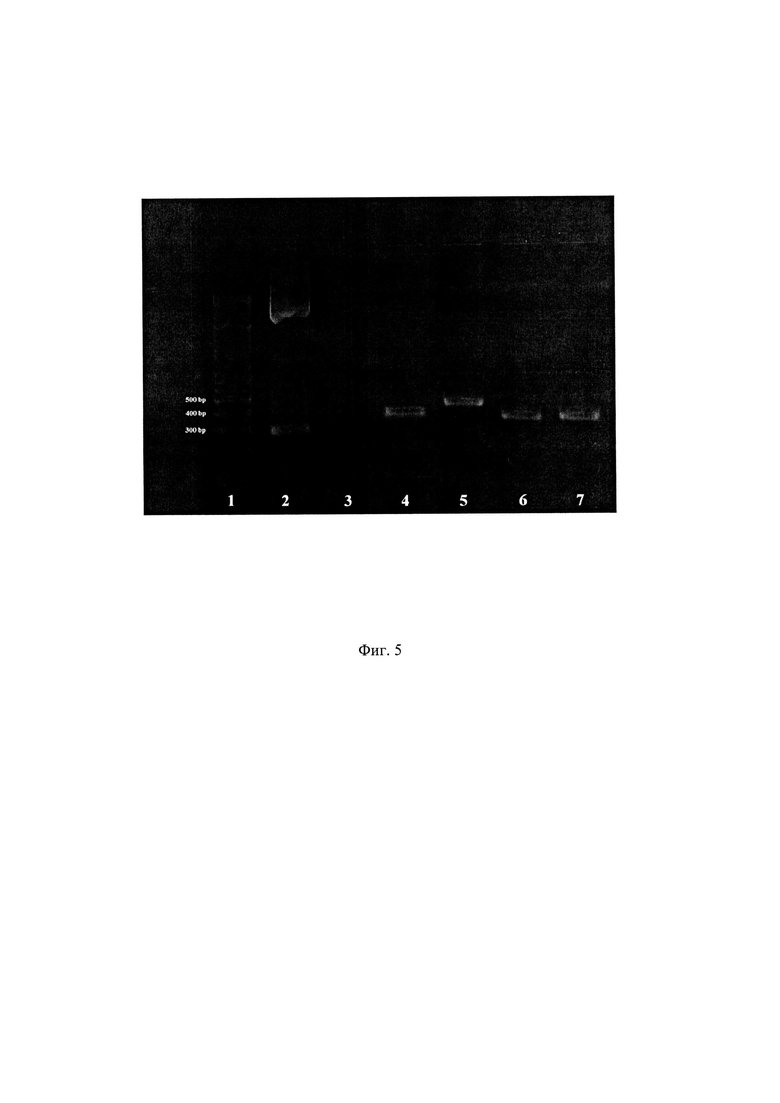
Фиг. 5. Типичные результаты ПЦР со стандартными праймерами M13/pUC отдельных выделенных колоний после клонирования селектированного пула ДНК в вектор pUC19 DNA.
1. Маркер молекулярной массы (O’RangeRuler 100+500 bp DNA Ladder, Thermo Scientific, США);
2. Положительный контроль (амплификация исходной плазмиды pUC19 DNA со стандартными праймерами M13/pUC);
3. Отрицательный контроль (амплификация без внесения целевой ДНК);
4, 6, 7. Результаты амплификации клонов, содержащих одну копию вставки, соответствующей 81 нуклеотиду.
5. Результат амплификации клона, содержащего две копии вставки.
Изображение получено с использованием лазерного сканера Typhoon FLA 9500.
Фиг. 6. Типичные результаты скрининга некоторых аптамеров по параметру эффективности связывания с белком ПАЛ L. pneumophila.
Фиг. 7А. Принципиальная схема проведения анализа для скрининга специфичности аптамеров.
Фиг. 7В. Типичные результаты скрининга специфичности аптамера AptPAL_23
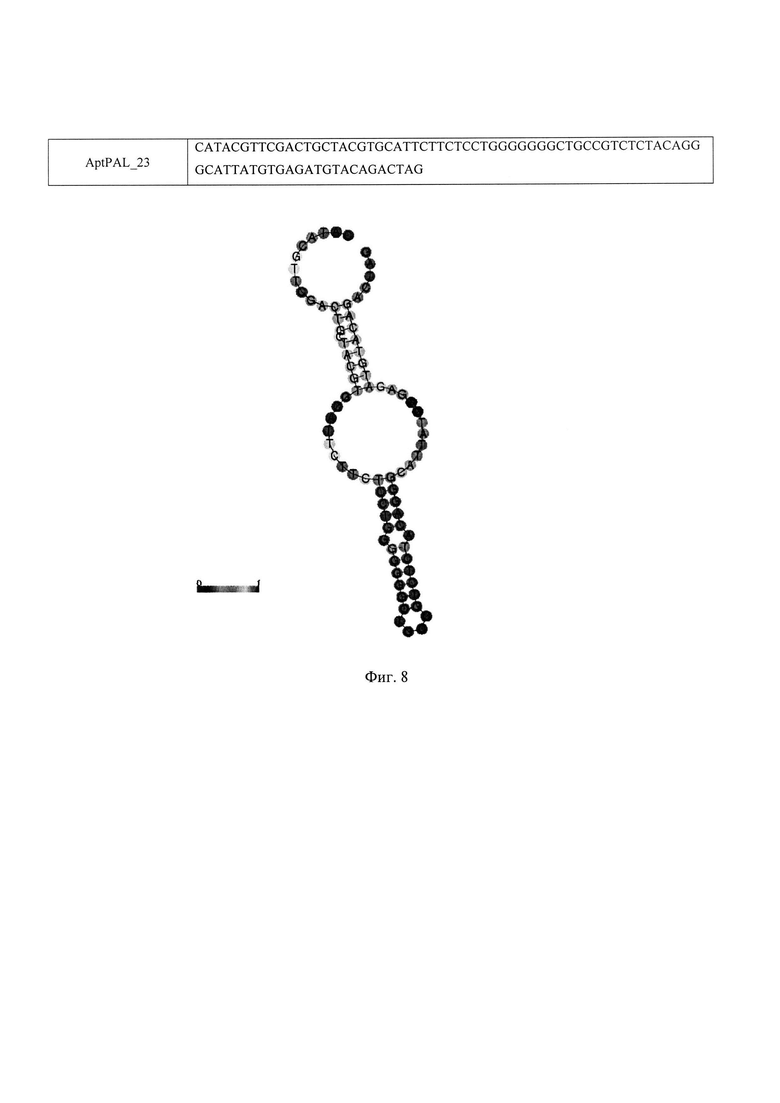
Фиг. 8. Последовательность и расчетная вторичная структура ДНК-аптамера AptPAL_23, специфичного к ПАЛ L. pneumophila.
Изобретение осуществляют следующим образом:
Синтезируют комбинаторную библиотеку олигонуклеотидов (ЗАО «Евроген», Россия; по заказу ФБУН ГНЦ ПМБ) вида 5'-const1-N45-const2, где N45 - вариабельная область, состоящая из 45 нуклеотидов, а const1 и const2 - константные фланкирующие области длиной по 18 нуклеотидов, которые служат для связывания со специфическими праймерами, позволяющими амплифицировать пул олигонуклеотидов после этапов селекции. Таким образом, олигонуклеотидная библиотека содержит 445 вариантов последовательности, содержащей 81 нуклеотид (Фиг. 1).
Селекцию проводят с использованием рекомбинантного слитного белка GST/APT/PAL, экспрессируемого с последовательности gst-apt-pal (Фиг. 3) в составе плазмидной конструкции pGST/APT/PAL, созданной на основе конструкта pGST/APT/X [Патент RU №2558295]. Для этого последовательность, кодирующую ПАЛ Legionella pneumophila, полученную прямой амплификацией с генома L. pneumophila, обрабатывают эндонуклеазами рестрикции BamHI и SalI Thermo, США) и клонируют в плазмиду pGST/APT/Х, обработанную эндонуклеазами рестрикции BamHI и XhoI (Thermo, США). Полученная плазмида pGST/APT/PAL кодирует экспрессионную конструкцию для синтеза химерного слитного белка, содержащего последовательность фрагмента глютатион-S-трансферазы, пептид RRKKVYPYPME, являющийся специфическим субстратом для протеолиза летальным фактором B. anthracis, in vivo биотинилируемый пептид, а также пептидогликан-ассоциированный липопротеин L. pneumophila.
Получают продуцент слитного рекомбинантного полипептида GST/APT/PAL, трансформируя компетентные клетки E. coli BL21(DE3) плазмидой pGST/APT/PAL. Проводят накопление полипептида в течение 5 часов после индукции экспрессии добавлением ИПТГ. По окончании продукции клеточную биомассу собирают центрифугированием, суспендируют в лизирующем буфере, суспензию бактерий обрабатывают лизоцимом и ДНКазой и центрифугируют для получения осветленного лизата, содержащего полипептид GST/APT/PAL в растворимой форме.
Очистку белка GST/APT/PAL из лизата проводят на колонке с иммобилизованным глутатионом, элюируют целевой белок раствором восстановленного глутатиона. Дополнительную очистку белка и перевод его в буфер для расщепления протеазой летальным фактором B. anthracis осуществляют при помощи эксклюзионной хроматографии на колонке с Superdex 200. Чистоту препарата белка GST/APT/PAL анализируют денатурирующим электрофорезом в полиакриламидном геле.
Проверяют эффективность отщепления белка-мишени от химерного протеина, обрабатывая белок GST/APT/PAL протеазой летальным фактором B. anthracis. Анализ продуктов расщепления проводят гель-электрофорезом в денатурирующих условиях в полиакриламидном геле (Фиг. 4).
Раунд селекции аптамеров осуществляют следующим образом: подготавливают магнитные микросферы, несущие глутатион, иммобилизируя на их поверхности слитный белок GST/APT/PAL за счет наличия глутатион-S-трансферазы в составе белка; инкубируют подготовленные магнитные микросферы с библилотекой одноцепочечных ДНК-олигонуклеотидов; промывают магнитные частицы от несвязавшихся олигонуклеотидов потоком отмывающего раствора в трубке, пропущенной сквозь магнит; отделяют аптамеры, специфически связанные с белком-мишенью ПАЛ L. pneumophila, от конструкта GST/APT/PAL и микросфер, удерживаемых магнитом, для чего инкубируют суспензию микросфер с рекомбинантным белком летальным фактором Bacillus anthracis; фиксируют магнитные микросферы при помощи магнита, собирают раствор, содержащий белок-мишень ПАЛ, несущий специфически связанные ДНК-олигонуклеотиды, и белок летального фактора Bacillus anthracis; отобранный пул олигонуклеотидов отделяют от белков методом фенол-хлороформной экстракции и концентрируют осаждением в этаноле, после чего проводят обогащение пула аптамеров амплификацией с использованием праймеров: прямого ForFAM, меченного FAM по 5'-концу цепи, и обратного RevP, несущего на 5'-конце остаток фосфорной кислоты (Фиг. 1) (отжиг при 54°С, элонгация 15 секунд). Полученный двуцепочечный ПЦР-продукт обрабатывают 5'-экзонуклеазой фага лямбда для получения одноцепочечного олигонуклеотида. Успешность расщепления обратной цепи контролируют электрофорезом в ПААГ визуально по наличию в электрофореграмме только одной, окрашенной FAM, полосы продукта (Фиг. 2). Проводят пять раундов селекции.
Получают индивидуальные последовательности аффинных к ПАЛ L. pneumophila аптамеров. Для этого пул олигонуклеотидов, прошедший все раунды селекции, амплифицируют с применением прямого ForP и обратного RevP праймеров (Фиг. 1) (отжиг при 55°С, элонгация 15 секунд), фосфорилированных по 5'-концу. Двуцепочечный ПЦР-продукт лигируют в вектор pUC19 DNA (Invitrogen, США), расщепленный эндонуклеазой рестрикции SmaI (Thermo, США) и обработанный щелочной фосфатазой (Invitrogen, США). Полученной плазмидой трансформируют электрокомпетентные клетки E. coli DH12S. Клетки высевают на плотную питательную среду, содержащую ампициллин, X-Gal и ИПТГ. По принципу бело-голубой селекции выбирают и отсевают колонии, содержащие в составе плазмиды вставку. Проверяют наличие искомой вставки в ПЦР со стандартными праймерами M13/pUC (Фиг. 5).
Колонии, содержащие вставку, соответствующую по размеру единичному аптамеру, подвергают амплификации с собственных праймеров ForFAM-Bio и RevP, тем самым увеличивая количество копий клонов аптамеров. Получают одноцепочечные фрагменты методом, описанным выше.
Проводят скрининг индивидуальных последовательностей аффинных к ПАЛ L. pneumophila аптамеров по параметру эффективности связывания с белком-мишенью. Для этого полученные биотинилированные по 5'-концу индивидуальные последовательности инкубируют в лунках 96-луночного иммунологического планшета, на поверхности лунок которого иммобилизирован рекомбинантный белок ПАЛ L. pneumophila. После инкубации отмывают несвязавшиеся последовательности высокосолевым раствором, а в лунки помещают раствор стрептавидина, конъюгированного с пероксидазой хрена. По завершении инкубации отмывают и визуализируют взаимодействие цветной реакцией. Оптическую плотность раствора в лунках считывают на планшетном спектрофотометре (Фиг. 6).
Последовательности, показавшие лучшие результаты в анализе эффективности связывания с белком-мишенью, анализируют по параметру специфичности взаимодействия с белком-мишенью, используя для этого панель гетерологичных белков. Для этого на поверхности лунок 96-луночного ПЦР-планшета иммобилизируют рекомбинантные белки: ПАЛ L. pneumophila, протеолитический домен BoNT/A (LC BoNT/A), рецепторсвязывающий домен тяжелой цепи BoNT/A(HC 50 BoNT/A), летальный фактор В. anthracis (LF), протективный антиген В. anthracis (PA), дифтерийный токсин (DT), стафилококковые энтеротоксины А и В (SEA и SEB), термолабильный токсин E. coli (LT) (ФБУН ГНЦ ПМБ, Оболенск). Индивидуальные последовательности подготавливают к анализу. Для этого амплифицируют их при помощи праймеров For(P-) и RevP, получают одноцепочечные последовательности как описано выше. Инкубируют одноцепочечные олигнуклеотиды в лунках планшета, отмывают высокосолевым буфером, после чего вносят в лунки планшета амплификационную смесь, содержащую интеркалирующий краситель, проводят ПЦР-РВ (Фиг. 7).
Для последовательности с наиболее выраженными свойствами чувствительности и специфичности определяют константу аффинности в отношении рекомбинантного ПАЛ L. pneumophila на приборе ProteOn™ XPR36 Protein Interaction Array System (Bio-Rad, США).
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Получение рекомбинантного слитного полипептида GST/APT/PAL.
Последовательность ДНК, кодирующую пептидогликан-ассоциированный липопротеин, получают ПЦР-амплификацией с геномной ДНК бактерий Legionella pneumophila (Государственная коллекция патогенных микроорганизмов ФБУН ГНЦ ПМБ, Оболенск) с помощью ПЦР с использованием праймеров PAL_FBam и PAL_RSal (Фиг.1). Температура отжига праймеров составляет 58°С, время элонгации 40 секунд. Анализируют полученный ПЦР-продукт при помощи электрофоретического разделения в 1% агарозном геле. Продукт апмлификации обрабатывают эндонуклеазами рестрикции BamHI и SalI, затем клонируют в плазмиду pGST/APT/Х [Патент RU №2558295], обработанную эндонуклеазами BamHI и XhoI. Полученная экспрессионная конструкция pGST/APT/PAL кодирует слитный белок, содержащий аминокислотную последовательность глютатион-S-трансферазы, пептид RRKKVYPYPME, являющийся субстратом летального фактора B. anthracis, in vivo биотинилируемый пептид и последовательность, кодирующую ПАЛ L. pneumophila.
Продуцент слитного рекомбинантного полипептида GST/APT/PAL получают электротрансформацией (прибор Eppendorf Electroporator 2510 (Eppendorf, Германия) режим 1,7 КB) компетентных клеток E. coli BL21(DE3) плазмидой pGST/APT/PAL. Трансформанты высевают на чашки с 2xYT-агаром, содержащим 100 мкг/мл ампициллина и 1 % глюкозы, выращивают в течение ночи при 37°C.
Единичные колонии-продуценты белка GST/APT/PAL выращивают в течение ночи при 37°С в жидкой среде 2хYT c добавлением 100 мкг/мл ампициллина, переносят 10 мл ночной культуры в 1000 мл среды 2xYT, содержащей 0,1% глюкозы и 100 мкг/мл ампициллина, и выращивают при 37°С также в течение ночи. Полученную ночную культуру объемом 1 л помещают в ферментер (Labfors 5, Infors, Швейцария), содержащий 9 л жидкой питательной среды 2хYT с добавлением 100 мкг/мл ампициллина и 0,1 % глюкозы. Наращивают культуру до оптической плотности 1 ОЕ, после чего культуру охлаждают до 30°С, добавляют ИПТГ до 0,2 мМ и проводят экспрессию белка в течение 5 часов при 30°С. По окончании ферментации клеточную биомассу собирают центрифугированием, суспендируют в буфере нанесения на сорбент состава 50 мМ Трис-HCl, 100 мМ NaCl, pH 7.4. Полученную суспензию бактерий обрабатывают лизоцимом, тритон Х-100 и ДНКазой, затем центрифугируют для получения осветленного лизата, содержащего полипептид GST/APT/PAL в растворимой форме.
Очистку рекомбинантного слитного белка GST/APT/PAL из осветленного лизата проводят на колонке с иммобилизованным глутатионом. Элюируют белок тем же буферным раствором для нанесения на сорбент, содержащим 25 мМ глутатиона в восстановленной форме. Дополнительную очистку белка и перевод его в буфер для расщепления протеазой летальным фактором B. anthracis (50 мМ Трис-HCl, 50 мМ NaCl, рН 7.4) осуществляют при помощи эксклюзионной хроматографии на колонке с Superdex 200. Чистоту препарата белка GST/APT/PAL анализируют электрофоретическим разделением белков в полиакриламидном геле в денатурирующих условиях с последующим окрашиванием (по Лэммли).
Эффективность расщепления белка GST/APT/PAL проверяют, добавляя к слитному белку GST/APT/PAL (~44 кДа) протеазу летальный фактор возбудителя сибирской язвы в соотношении 10/1, соответственно. Контролируют отщепление белка-мишени ПАЛ (~17 кДа), анализируя продукты расщепления гель-электрофорезом в денатурирующих условиях в полиакриламидном геле (Фиг. 4).
Пример 2. Проведение раунда селекции последовательностей олигонуклеотидов против ПАЛ Legionella pneumophila.
Слитный рекомбинантный полипептид GST/APT/PAL иммобилзируют на поверхности магнитных микросфер, покрытых глутатионом. Для этого 100 мкл суспензии частиц Pierce™ Glutathione Magnetic Beads (Thermo, США) в пробирке объемом 1,5 мл помещают на магнитный штатив DynaMag™-2 Magnet (Invitrogen, США), отбирают жидкость. Сняв пробирку со штатива, ресуспендируют микрочастицы в 300 мкл буфера состава 125 мМ Трис-HCl, 150 Мм NaCl, pH 8.0. Вновь помещают пробирку в магнитный штатив и отбирают жидкость. Повторяют трижды, отмывая тем самым микрочастицы от буфера хранения.
Получают раствор полипептида GST/APT/PAL объемом 300 мкл с конечной концентрацией 1 мг/мл в растворе 125 мМ Трис-HCl, 150 Мм NaCl, pH 8.0. Добавляют суспензию белка GST/APT/PAL к предварительно осушенным на магнитном стенде магнитным микрочастицам. Инкубируют суспензию со встряхиванием на вортексе при комнатной температуре в течение часа. По окончании инкубации помещают пробирку в магнитный штатив и отбирают супернатант. Отмывают частицы тем же раствором пятикратно в объеме не менее 1000 мкл. Ресуспендируют частицы в 300 мкл буфера. При необходимости хранят при +4°С, добавив к суспензии натрия азид до конечной концентрации 0,02 %.
Перед добавлением пула аптамеров к магнитным микрочастицам помещают пробирку в магнитный штатив и отбирают всю жидкость. Добавляют 20 мкг (4х1014 молекул) исходной комбинаторной библиотеки олигонуклеотидов. Инкубируют суспензию на протяжении часа при температуре 37°С при периодическом перемешивании на вортексе. Отмывают магнитные частицы от несвязавшихся молекул ДНК потоком буферного раствора объемом 300 мл состава 20 мМ Трис-HCl, 250 мМ NaCl, 5 мМ ЭДТА, pH 7.4, используя систему проточной магнитной сепарации MiniMACS™ Separator (Miltenyi Biotec, Германия). Прекращают магнитное воздействие на частицы, несущие на своей поверхности химерный полипептид GST/APT/PAL, связанный с аптамерами, смывая, собирают микросферы в чистую пробирку объемом 1,5 мл. Фиксируя магнитные микросферы на стенке пробирки при помощи магнитного штатива, отбирают супернатант. Добавляют 300 мкл раствора протеазы летального фактора B. anthracis [патент РФ №2355769] с содержанием 10 мкг протезы в буфере 50 мМ Трис-HCl, pH 7.4. Выдерживают в течение часа при температуре 30°С, периодически встряхивая. Фиксируют магнитные частицы на стенке пробирки при помощи магнитного штатива, отбирают супернатант, содержащий суспензию белка ПАЛ L. pneumophila, несущего специфически связанные аптамеры. Пул олигонуклеотидов отделяют от белка и очищают методом фенол-хлороформной экстракции нуклеиновых кислот и концентрируют преципитацией этанолом в присутствии копреципитанта Pellet Paint® Co-Precipitant (Novagen, Германия).
Обогащают селектированный пул одноцепочечных коротких фрагментов ДНК амплификацией с использованием праймеров: прямого ForFAM, несущего в положении 5' флуоресцентную метку карбоксифлуоресцеин, и обратного RevP, фосфорилированного по 5'-концу последовательности (температура отжига праймеров 54°С, время элонгации 15 секунд) (Фиг. 1). В результате проведенной ПЦР получают двуцепочечные фрагменты, одна из цепей которого мечена FAM.
Для получения обогащенного пула одноцепочечных фрагментов ПЦР-продукт обрабатывают 5'- экзонуклеазой фага лямбда, расщепляя обратную фосфорилированную цепь двуцепочечного ПЦР-фрагмента [Higuchi R. G., Ochman H. // Nucleic Acids Research. - 1989. - V. 17. - N. 14. - P. 5865.]. Эффективность расщепления цепи контролируют проведением электрофореза в 10%-ом полиакриламидном геле, содержащем мочевину, с последующим окрашиванием бромистым этидием, при этом наличие в пробе только полосы, флуоресцирующей зеленым, свидетельствует об успешном расщеплении обратной цепи фрагмента, а присутствие в пробе второй полосы с красным цветом флуоресценции свидетельствует о неуспешном или неполном расщеплении фосфорилированной цепи ДНК (Фиг. 2).
Полученные одноцепочечные ДНК очищают и концентрируют, используя фенол-хлороформную экстракцию и преципитацию этанолом. Пул обогащенных олигонуклеотидов используют для осуществления дальнейших раундов селекции.
Всего проводят пять раундов селекции.
Пример 3. Получение индивидуальных последовательностей олигонуклеотидов, подвергшихся негативной селекции.
Для получения индивидуальных последовательностей аптамеров из пула селектированных в пятом раунде селекции олигонуклеотидов при помощи ПЦР с использованием прямого и обратного фосфорилированных праймеров ForP и RevP (отжиг праймеров при 55°С, элонгация 15 секунд) (Фиг. 1) получают двуцепочечные, фосфорилированные по 5'-концам фрагменты ДНК. Полученные фрагменты клонируют в коммерческий вектор pUC19 DNA (Invitrogen, США). Для этого плазмиду pUC19 обрабатывают эндонуклеазой рестрикции SmaI в буфере Tango (Thermo, США), после чего обрабатывают щелочной фосфатазой (Invitrogen, США) для предотвращения лигирования внутри вектора. Фосфорилированные двуцепочечные фрагменты ДНК лигируют в подготовленную плазмиду.
Электропорацией (прибор Eppendorf Electroporator 2510 (Eppendorf, Германия) режим 1,7 КB) трансформируют клетки E. coli DH12S, используя подготовленную ранее плазмиду pUC19, содержащую вставки селектированных последовательностей. Трансформирманты высевают на плотную среду с 2xYT-агаром, содержащим 50 мкг/мл ампициллина, 80 мкг/мл X-Gal и 20 мМ ИПТГ, выращивают в течение ночи при 37°С.
По принципу бело-голубой селекции [Ullmann A, Jacob F, Monod J. // J Mol Biol. - 1967. - V. 24. - N. 2. - P. 339-343.] отбирают единичные колонии, потенциально содержащие вектор со вставкой, и анализируют наличие встроенного фрагмента ДНК искомой длины в плазмиде с помощью ПЦР со стандартными прямым и обратным праймерами M13/pUC (Fermentas, США) (Фиг. 5). Увеличивают количество копий индивидуальных последовательностей клонированных фрагментов ДНК амплификацией с использованием праймеров ForFAM-Bio и RevP (Фиг. 1). Получают одноцепочечный продукт для каждого из клонов обработкой 5'-экзонуклеазой фага лямбда с визуальным контролем после электрофоретического разделения (Фиг. 2).
Пример 4. Скрининг индивидуальных селектированных последовательностей по параметру эффективности связывания с ПАЛ L. pneumophila.
На поверхности лунок 96-луночного иммунологического планшета иммобилизируют рекомбинантный белок ПАЛ L. pneumophila, для этого вносят раствор белка ПАЛ в количестве 1 мкг/лунку в 100 мкл 10 мМ фосфатно-солевого буфера (ФСБ; 137 мМ NaCl, 2,7 мМ KCl, рН 7.4), инкубируют планшет в течение 3 часов при 37°С с перемешиванием при 500 об/мин на планшетном шейкере-инкубаторе. Отмывают лунки планшета трижды с использованием ФСБ с добавлением 0,05% твин-20 (ФСБ-Тв). Свободные центры связывания в лунках планшета блокируют обезжиренным (м.д.ж. не более 0,5%) безлактозным молоком в течение 40 минут. Промывают лунки планшета трижды при помощи ФСБ-Тв.
В лунки планшета в двукратных повторностях вносят индивидуальные одноцепочечные селектированные последовательности, биотинилированные по 5'-концу цепи, в количестве 0,2 мкг/лунку в 100 мкл раствора 20 мМ Трис-HCl, 50 мМ NaCl, 5 мМ ЭДТА (pH 7.4). В качестве отрицательного контроля используют лунку, в которую вносят 100 мкл чистого буфера, в качестве положительного контроля - биотинилированные мышиные моноклональные антитела против ПАЛ L. pneumophila (ФБУН ГНЦ ПМБ, Оболенск) в количестве 0,5 мкг антител на лунку в ФСБ. Инкубируют планшет в течение часа при температуре 37°С и перемешивании. Отмывают четырехкратно буферным раствором 20 мМ Трис-HCl, 250 мМ NaCl, 5 мМ ЭДТА (pH 7.4). Вносят во все лунки планшета раствор стрептавидина, конъюгированного с пероксидазой хрена (Sigma, США) в разведении, рекомендуемом производителем. Инкубируют в течение 40 минут в условиях, описанных выше. Отмывают шестикратно ФСБ-Тв. Визуализируют взаимодействие в лунках планшета цветной реакцией с использованием раствора тетраметилбензидина. Реакцию останавливают добавлением 2N раствора серной кислоты. Считывают результат на планшетном спектрофотометере при длине волны 450 нм (Фиг. 6).
Пример 5. Скрининг специфичности взаимодействия индивидуальных селектированных последовательностей с ПАЛ L. pneumophila.
Индивидуальные селектированные последовательности ДНК-аптамеров, показавшие наилучшие результаты в тесте на эффективность связывания с целевой мишенью белком ПАЛ L. pneumophila, исследуют на предмет специфичности взаимодействия с белком-мишенью.
Для этого на поликарбонатном 96-луночном ПЦР-планшете с высокосвязывающей поверхностью (Corning, США) иммобилизируют белки: ПАЛ L. pneumophila, протеолитический домен BoNT/A (LC BoNT/A), рецепторсвязывающий домен тяжелой цепи BoNT/A(HC 50 BoNT/A), летальный фактор B. anthracis (LF), протективный антиген B. anthracis (PA), дифтерийный токсин (DT), стафилококковые энтеротоксины А и В (SEA и SEB), термолабильный токсин E. coli (LT) (ФБУН ГНЦ ПМБ, Оболенск). В анализе задействуют 60 лунок планшета, ограниченных лунками B2-G2-B11-G11. Второй вертикальный ряд планшета (6 лунок B2-G2) не содержит белки и его лунки служат в качестве отрицательных контрольных проб. Начиная с третьего вертикального ряда вносят рекомбинатные белки из исследуемой панели в количестве 0,5 мкг/лунку в фосфатно-солевом буфере, pH 7.4 (ФСБ) в объеме 50 мкл. Инкубируют планшет на протяжении часа при 37°С при встряхивании на орбитальном шейкере. По окончании инкубации отмывают лунки планшета ФСБ-Тв трижды. Свободные валентности планшета блокируют буфером, состоящим из обезжиренного (м.д.ж. не более 0,5 %) безлактозного молока с добавлением 3 % БСА, в объеме 200 мкл/лунку в течение 40 минут при 37°С на орбитальном шейкере. Отмывку производят трижды ФСБ-Тв.
Индивидуальные селектированные последовательности, отобранные на предыдущем этапе (пример 4), амплифицируют с использованием праймеров For(P-) и RevP, получают одноцепочечные последовательности обработкой 5'-экзонуклеазой фага лямбда как было описано выше.
Амплифицированные одноцепочечные последовательности ДНК вносят в лунки горизонтальных рядов планшета в объеме 50 мкл и количестве 0,2 мкг/лунку. В первый и второй горизонтальные ряды в качестве контроля сравнения вносят исходную комбинаторную библиотеку в том же количестве. В последующие горизонтальные ряды вносят клоны аптамеров (каждый в 2 ряда по 10 лунок). Таким образом, каждая проба исследуется в двух повторностях (Фиг. 7А). Буфер для разведения ДНК содержит 20 мМ Трия-HCl, 50 мМ NaCl, 5 мМ ЭДТА (pH 7.4). Планшет заклеивают пленкой. Инкубируют планшет с ДНК в течение часа при 37°С. Отмывку от несвязавшихся молекул ДНК производят раствором, содержащим 20 мМ Трис-HCl, 250 мМ NaCl, 5 мМ ЭДТА (pH 7.4) пятикратно.
Контролируют реакцию связывания ДНК с белком в ПЦР в режиме реального времени. Для этого в лунки планшета вносят амплификационную смесь, содержащую флуоресцентный интеркалирующий краситель SYBR-Green I (ЗАО "Синтол", Россия) с добавлением праймеров ForP и RevP (Фиг. 1). Амплификацию проводят в режиме: температура отжига праймеров 55°С, время элонгации 10 секунд, детекцию сигнала проводят по каналу SYBR 1(FAM) (прибор IQ™5, Bio-Rad, США) (Фиг. 7В).
По результатам скрининга была выделена последовательность индивидуального клона аптамера AptPAL_23 (Фиг. 8), проявившая наибольшую специфическую активность в отношении ПАЛ L. pneumophila и вместе с тем не выявляющая неспецифические взаимодействия с гетерологичными белками, присутствующими в использованной скрининговой панели.
Пример 6. Характеризация аптамера AptPAL_23.
Для выбранной последовательности AptPAL_23 определяют константу равновесной диссоциации KD с использованием системы ProteOn™ XPR36 Protein Interaction Array System (Bio-Rad, США), которая составляет 4,3×10-8 М.
Определение первичной структуры аптамера AptPAL_23 проводят методом капиллярного секвенирования. Моделирование вторичной структуры осуществляют с использованием программы RNAFold (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi). Аптамер AptPAL_23 имеет нуклеотидную последовательность CATACGTTCGACTGCTACGTGCATTCTTCTCCTGGGGGGGCTGCCGTCTCTACAGGGCATTATGTGAGATGTACAGACTAG и предположительную вторичную структуру, представленные на Фиг. 8.
--->
Перечень последовательностей - коррекция
<110> ФБУН ГНЦ ПМБ; FSIS SRCAM&B
<120> Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila
<140> 2019136227 (071501)
<141> 12.11.2019
<160> SEQ ID NO: 1
<170> Patent-In 3.5
<210> 1
<211> 1234
<212> ДНК; DNA
<213> Искусственная последовательность; Artificial Sequence
<220>
<223> Фрагмент последовательности плазмиды pGST/APT/PAL, кодирующий слитный полипептид GST/APT/PAL
<400> 1
atg tcc cct ata cta ggt tat tgg aaa att aag ggc ctt gtg caa ccc 48
Met Ser Pro Ile Leu Gly Tyr Trp Lys Ile Lys Gly Leu Val Gln Pro
1 5 10 15
act cga ctt ctt ttg gaa tat ctt gaa gaa aaa tat gaa gag cat ttg 96
Thr Arg Leu Leu Leu Glu Tyr Leu Glu Glu Lys Tyr Glu Glu His Leu
20 25 30
tat gag cgc gat gaa ggt gat aaa tgg cga aac aaa aag ttt gaa ttg 144
Tyr Glu Arg Asp Glu Gly Asp Lys Trp Arg Asn Lys Lys Phe Glu Leu
35 40 45
ggt ttg gag ttt ccc aat ctt cct tat tat att gat ggt gat gtt aaa 192
Gly Leu Glu Phe Pro Asn Leu Pro Tyr Tyr Ile Asp Gly Asp Val Lys
50 55 60
tta aca cag tct atg gcc atc ata cgt tat ata gct gac aag cac aac 240
Leu Thr Gln Ser Met Ala Ile Ile Arg Tyr Ile Ala Asp Lys His Asn
65 70 75 80
atg ttg ggt ggt tgt cca aaa gag cgt gca gag att tca atg ctt gaa 288
Met Leu Gly Gly Cys Pro Lys Glu Arg Ala Glu Ile Ser Met Leu Glu
85 90 95
gga gcg gtt ttg gat att aga tac ggt gtt tcg aga att gca tat agt 336
Gly Ala Val Leu Asp Ile Arg Tyr Gly Val Ser Arg Ile Ala Tyr Ser
100 105 110
aaa gac ttt gaa act ctc aaa gtt gat ttt ctt agc aag cta cct gaa 384
Lys Asp Phe Glu Thr Leu Lys Val Asp Phe Leu Ser Lys Leu Pro Glu
115 120 125
atg ctg aaa atg ttc gaa gat cgt tta tgt cat aaa aca tat tta aat 432
Met Leu Lys Met Phe Glu Asp Arg Leu Cys His Lys Thr Tyr Leu Asn
130 135 140
ggt gat cat gta acc cat cct gac ttc atg ttg tat gac gct ctt gat 480
Gly Asp His Val Thr His Pro Asp Phe Met Leu Tyr Asp Ala Leu Asp
145 150 155 160
gtt gtt tta tac atg gac cca atg tgc ctg gat gcg ttc cca aaa tta 528
Val Val Leu Tyr Met Asp Pro Met Cys Leu Asp Ala Phe Pro Lys Leu
165 170 175
gtt tgt ttt aaa aaa cgt att gaa gct atc cca caa att gat aag tac 576
Val Cys Phe Lys Lys Arg Ile Glu Ala Ile Pro Gln Ile Asp Lys Tyr
180 185 190
ttg aaa tcc agc aag tat ata gca tgg cct ttg cag ggc tgg caa gcc 624
Leu Lys Ser Ser Lys Tyr Ile Ala Trp Pro Leu Gln Gly Trp Gln Ala
195 200 205
acg ttt ggt ggt ggc gac cat cct ccg aaa tct ggc gaa gat ctg ggt 672
Thr Phe Gly Gly Gly Asp His Pro Pro Lys Ser Gly Glu Asp Leu Gly
210 215 220
ggc ggt agc ggt cgc cgt aag aaa gtt tac ccg tat ccg atg gaa ggt 720
Gly Gly Ser Gly Arg Arg Lys Lys Val Tyr Pro Tyr Pro Met Glu Gly
225 230 235 240
ggc ggt agc ggt ggc ctg aac gac atc ttc gaa gct cag aag atc gaa 768
Gly Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu
245 250 255
tgg cac gag gat ccg gcc tgt tct aaa acc cca ggc agt gcg gat ggt 816
Trp His Glu Asp Pro Ala Cys Ser Lys Thr Pro Gly Ser Ala Asp Gly
260 265 270
ggg gct gcg gta ggt gat ggt gat gct act gcc caa ggt ttg ggg caa 864
Gly Ala Ala Val Gly Asp Gly Asp Ala Thr Ala Gln Gly Leu Gly Gln
275 280 285
atg act cac ttt gct gga caa gag cct ggt gag tcc tat aca acg cag 912
Met Thr His Phe Ala Gly Gln Glu Pro Gly Glu Ser Tyr Thr Thr Gln
290 295 300
gca ccg cat aat caa ttg tat ttg ttt gcg tat gat gac agc act ttg 960
Ala Pro His Asn Gln Leu Tyr Leu Phe Ala Tyr Asp Asp Ser Thr Leu
305 310 315 320
gca tca aaa tat tta cct tca gtt aac gcg cag gca gaa tat tta aaa 1008
Ala Ser Lys Tyr Leu Pro Ser Val Asn Ala Gln Ala Glu Tyr Leu Lys
325 330 335
acc cac cca ggt gct cga gtg atg ata gct gga cat act gat gaa cgt 1056
Thr His Pro Gly Ala Arg Val Met Ile Ala Gly His Thr Asp Glu Arg
340 345 350
ggt agt cgt gaa tat aac gtt gct ctt ggt gaa cgc cgt gct gat act 1104
Gly Ser Arg Glu Tyr Asn Val Ala Leu Gly Glu Arg Arg Ala Asp Thr
355 360 365
gta gct gaa att ctg cgt atg gca ggt gtt agc aga cga caa att cga 1152
Val Ala Glu Ile Leu Arg Met Ala Gly Val Ser Arg Arg Gln Ile Arg
370 375 380
gta gtt agc tac ggc aaa gag cgc ccg gct aat tat ggc cat gat gaa 1200
Val Val Ser Tyr Gly Lys Glu Arg Pro Ala Asn Tyr Gly His Asp Glu
385 390 395 400
gcg tcg cat gcg cag aat cgt cgc gtc gag ttt atc tac gag gcg acc 1248
Ala Ser His Ala Gln Asn Arg Arg Val Glu Phe Ile Tyr Glu Ala Thr
405 410 415
cgt taa 1254
Arg
<110> ФБУН ГНЦ ПМБ; FSIS SRCAM&B
<120> Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila
<140> 2019136227 (071501)
<141> 12.11.2019
<160> SEQ ID NO: 2
<170> Patent-In 3.5
<210> 2
<211> 417
<212> ПРТ; PRT
<213> Искусственная последовательность; Artificial Sequence
<220>
<223> Аминокислотная последовательность слитного полипептида GST/APT/PAL
<400> 2
Met Ser Pro Ile Leu Gly Tyr Trp Lys Ile Lys Gly Leu Val Gln Pro
1 5 10 15
Thr Arg Leu Leu Leu Glu Tyr Leu Glu Glu Lys Tyr Glu Glu His Leu
20 25 30
Tyr Glu Arg Asp Glu Gly Asp Lys Trp Arg Asn Lys Lys Phe Glu Leu
35 40 45
Gly Leu Glu Phe Pro Asn Leu Pro Tyr Tyr Ile Asp Gly Asp Val Lys
50 55 60
Leu Thr Gln Ser Met Ala Ile Ile Arg Tyr Ile Ala Asp Lys His Asn
65 70 75 80
Met Leu Gly Gly Cys Pro Lys Glu Arg Ala Glu Ile Ser Met Leu Glu
85 90 95
Gly Ala Val Leu Asp Ile Arg Tyr Gly Val Ser Arg Ile Ala Tyr Ser
100 105 110
Lys Asp Phe Glu Thr Leu Lys Val Asp Phe Leu Ser Lys Leu Pro Glu
115 120 125
Met Leu Lys Met Phe Glu Asp Arg Leu Cys His Lys Thr Tyr Leu Asn
130 135 140
Gly Asp His Val Thr His Pro Asp Phe Met Leu Tyr Asp Ala Leu Asp
145 150 155 160
Val Val Leu Tyr Met Asp Pro Met Cys Leu Asp Ala Phe Pro Lys Leu
165 170 175
Val Cys Phe Lys Lys Arg Ile Glu Ala Ile Pro Gln Ile Asp Lys Tyr
180 185 190
Leu Lys Ser Ser Lys Tyr Ile Ala Trp Pro Leu Gln Gly Trp Gln Ala
195 200 205
Thr Phe Gly Gly Gly Asp His Pro Pro Lys Ser Gly Glu Asp Leu Gly
210 215 220
Gly Gly Ser Gly Arg Arg Lys Lys Val Tyr Pro Tyr Pro Met Glu Gly
225 230 235 240
Gly Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu
245 250 255
Trp His Glu Asp Pro Ala Cys Ser Lys Thr Pro Gly Ser Ala Asp Gly
260 265 270
Gly Ala Ala Val Gly Asp Gly Asp Ala Thr Ala Gln Gly Leu Gly Gln
275 280 285
Met Thr His Phe Ala Gly Gln Glu Pro Gly Glu Ser Tyr Thr Thr Gln
290 295 300
Ala Pro His Asn Gln Leu Tyr Leu Phe Ala Tyr Asp Asp Ser Thr Leu
305 310 315 320
Ala Ser Lys Tyr Leu Pro Ser Val Asn Ala Gln Ala Glu Tyr Leu Lys
325 330 335
Thr His Pro Gly Ala Arg Val Met Ile Ala Gly His Thr Asp Glu Arg
340 345 350
Gly Ser Arg Glu Tyr Asn Val Ala Leu Gly Glu Arg Arg Ala Asp Thr
355 360 365
Val Ala Glu Ile Leu Arg Met Ala Gly Val Ser Arg Arg Gln Ile Arg
370 375 380
Val Val Ser Tyr Gly Lys Glu Arg Pro Ala Asn Tyr Gly His Asp Glu
385 390 395 400
Ala Ser His Ala Gln Asn Arg Arg Val Glu Phe Ile Tyr Glu Ala Thr
405 410 415
Arg
<110> ФБУН ГНЦ ПМБ; FSIS SRCAM&B
<120> Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila
<140> 2019136227 (071501)
<141> 12.11.2019
<160> SEQ ID NO: 3
<170> Patent-In 3.5
<210> 3
<211> 81
<212> ДНК; DNA
<213> Искусственная последовательность; Artificial Sequence
<220>
<223> Последовательность ДНК-аптамера AptPAL_23
<400> 3
catacgttcg actgctacgt gcattcttct cctggggggg ctgccgtctc tacagggcat 60
tatgtgagat gtacagacta g 81
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДУПРЕЖДЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2017 |
|
RU2766586C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| Рекомбинантный слитый белок | 2019 |
|
RU2732795C1 |
| МУКОАДГЕЗИВНЫЙ МИКРООРГАНИЗМ | 2017 |
|
RU2762940C2 |
| Генетическая конструкция, кодирующая предшественник белка YB-1 человека, штамм Escherichia coli - продуцент предшественника белка YB-1 человека, способ микробиологического синтеза этого предшественника | 2019 |
|
RU2728237C1 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| ПРОДУКТЫ И СПОСОБЫ ДЛЯ УХОДА ЗА ПОЛОСТЬЮ РТА, СОДЕРЖАЩИЕ HLP | 2016 |
|
RU2759134C2 |
| Способ микробиологического синтеза прохимозина быка с использованием рекомбинантного штамма Pichia pastoris, содержащего синтетический ген варианта препрохимозина с модифицированной сигнальной последовательностью секреции | 2020 |
|
RU2779307C2 |
| СПОСОБ ПОЛУЧЕНИЯ ХИМИЧЕСКИХ ПРОДУКТОВ ТОНКОГО СИНТЕЗА С ПОМОЩЬЮ CORYNEBACTERIUM, СЕКРЕТИРУЮЩЕЙ МОДИФИЦИРОВАННЫЕ α-1,6-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2763317C2 |






Изобретение относится к области биотехнологии и представляет собой последовательность ДНК-аптамера, связывающуюся с пептидогликан-ассоциированным липопротеином Legionella pneumophila, представляющую собой последовательность одноцепочечного ДНК-аптамера AptPAL_23 длиной 81 нуклеотид, с молекулярной массой 28 кДа, с первичной последовательностью CATACGTTCGACTGCTACGTGCATTCTTCTCCTGGGGGGGCTGCCGTCTCTACAGGGCATTATGTGAGATGTACAGACTAG, содержащую области для связывания с праймерами длиной 18 нуклеотидов в крайних положениях последовательности, формирующую трехмерную структуру, образующую устойчивые специфичные связи с ПАЛ Legionella, имеющую константу равновесной диссоциации (KD) в отношении ПАЛ не менее 5×10-8 М. Изобретение позволяет детектировать легионеллезный антиген ПАЛ в составе тест-систем в качестве специфической амплифицируемой матрицы. 8 ил., 6 пр.
Последовательность ДНК-аптамера, связывающаяся с пептидогликан-ассоциированным липопротеином Legionella pneumophila, представляющая собой последовательность одноцепочечного ДНК-аптамера AptPAL_23 длиной 81 нуклеотид, с молекулярной массой 28 кДа, с первичной последовательностью CATACGTTCGACTGCTACGTGCATTCTTCTCCTGGGGGGGCTGCCGTCTCTACAGGGCATTATGTGAGATGTACAGACTAG, содержащая области для связывания с праймерами длиной 18 нуклеотидов в крайних положениях последовательности, формирующая трехмерную структуру, образующая устойчивые специфичные связи с ПАЛ Legionella, имеющая константу равновесной диссоциации (KD) в отношении ПАЛ не менее 5×10-8 М.
| LOU, X | |||
| et al | |||
| Proceedings of the National Academy of Sciences | |||
| V | |||
| Светоэлектрический измеритель длин и площадей | 1919 |
|
SU106A1 |
| Ветряный двигатель | 1925 |
|
SU2989A1 |
| RELLER L | |||
| B | |||
| et al | |||
| Clinical Infectious Diseases | |||
| - V | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| Нефтяной конвертер | 1922 |
|
SU64A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGST/ART/X, КОДИРУЮЩАЯ СЛИТНЫЙ ХИМЕРНЫЙ ПОЛИПЕПТИД GST/ART/X ДЛЯ ВЫСОКОСПЕЦИФИЧНОЙ СЕЛЕКЦИИ АПТАМЕРОВ К БЕЛКУ-МИШЕНИ X В СОСТАВЕ ПОЛИПЕПТИДА | 2014 |
|
RU2558295C1 |

