ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет патентной заявки США номер 61/163222, поданной 25 марта 2009 года, содержание которой включено в настоящий документ в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится, главным образом, к области молекулярной биологии. Более конкретно, изобретение относится к антителам против FGFR3 и к их применению.
УРОВЕНЬ ТЕХНИКИ
Факторы роста фибробластов (FGF) и их рецепторы (FGFR) играют основную роль в процессе эмбрионального развития, гомеостаза тканей и метаболизма (1-3). У человека имеется 22 FGF (FGF1-14, FGF 16-23) и четыре рецептора FGF с тирозинкиназным доменом (FGFR1-4). FGFR состоят из внеклеточной области, связывающей лиганд, имеющий два или три иммуноглобулин-подобных домена (IgD1-3), трансмембранной области, однократно пронизывающей мембрану, и цитоплазматического разделенного тирозинкиназного домена. Каждый из FGFR1, 2 и 3 имеет две основных альтернативно сплайсированных изоформы, обозначаемых как IIIb и IIIc. Эти изоформы отличаются приблизительно 50 аминокислотами во второй половине IgD3 и имеют разную локализацию в тканях и отличную специфичность в отношении лиганда. Как правило, изоформа IIIb встречается в эпителиальных клетках, в то время как IIIc экспрессируется в мезенхимных клетках. При связывании FGF в тесном взаимодействии с гепаринсульфатными протеогликанами, FGFR димеризуются и фосфорилируются по конкретным остаткам тирозина. Это способствует привлечению основных адапторных белков, таких как субстрат FGFR 2α (FRS2α), что приводит к активации множества каскадов передачи сигнала, включая каскады митоген-активируемой протеинкиназы (MAPK) и PI3K-AKT (1, 3, 4). Следовательно, FGF и их рецепторы регулируют широкий набор клеточных процессов, включая пролиферацию, дифференцировку, миграцию и выживание, зависимым от условий образом.
FGFR с нарушенной активацией вовлечены в конкретные злокачественные опухоли человека (1, 5). В частности, хромосомная транслокация t(4;14) (p16.3;q32) происходит приблизительно у 15-20% пациентов с множественной миеломой, что приводит к сверхэкспрессии FGFR3, и коррелирует с более короткой общей выживаемостью (6-9). FGFR3 также вовлечен в придание хеморезистентности клеточным линиям миеломы в культуре (10), что согласуется с плохим клиническим ответом пациентов t(4;14)+ на общепринятую химиотерапию (8). Сверхэкспрессия мутационно активированного FGFR3 является достаточной для индукции онкогенной трансформации в кроветворных клетках и фибробластах (11-14, 15), трансгенных моделях на мышах (16) и на моделях трансплантации костного мозга у мышей (16, 17). Таким образом, FGFR3 был предложен в качестве возможной терапевтической мишени для лечения множественной миеломы. Действительно, несколько низкомолекулярных ингибиторов, нацеленных на FGFR, хотя и не селективных в отношении FGFR3 и имеющих перекрестную ингибиторную активность в отношении некоторых других киназ, продемонстрировали цитотоксичность против положительных по FGFR3 клеток миеломы в культуре и в моделях на мышах (18-22).
Сверхэкспрессия FGFR3 была описана в высокой доле случаев рака мочевого пузыря (23, 24). Более того, соматические активирующие мутации в FGFR3 были идентифицированы в 60-70% папиллярных и 16-20% мышечно-инвазивных карцином мочевого пузыря (24, 25). В экспериментах на клеточной культуре, РНК-интерференция (11, 26) или одноцепочечный Fv-фрагмент антитела против FGFR3 ингибировали пролиферацию клеток рака мочевого пузыря (27). В недавних исследованиях было показано, что конъюгат антитело против FGFR3-токсин снижает рост ксенотрансплантата клеточной линии рака мочевого пузыря через опосредуемую FGFR3 доставку токсинов в опухоли (28). Однако остается неясным, действительно ли передача сигнала FGFR3 является онкогенным пусковым фактором роста опухолей мочевого пузыря in vivo. Более того, не была определена терапевтическая мощность нацеливания на FGFR3 при раке мочевого пузыря на основе моделей in vivo. Публикации, касающиеся FGFR3 и антител против FGFR3 включают публикацию патента США No 2005/0147612; Rauchenberger et al., J Biol Chem 278 (40):38194-38205 (2003); WO2006/048877; Martinez-Torrecuadrada et al., (2008) Mol. Cancer Ther 7(4): 862-873; WO2007/144893; Trudel et al. (2006) 107(10): 4039-4046; Martinez-Torrecuadrada et al. (2005) Clin Cancer Res 11 (17): 6280-6290; Gomez-Roman et al. (2005) Clin Cancer Res 11: 459-465; Direnzo, R et al., (2007) Proceedings of AACR Annual Meeting, Abstract No. 2080; WO2010/002862.
Очевидно, что продолжает существовать необходимость в средствах, которые обладают клиническими признаками, оптимальными для разработки в качестве лекарственных средств. Изобретение, описанное в настоящем документе, удовлетворяет эту потребность и обеспечивает другие преимущества.
Все ссылки, цитированные в настоящем документе, включая патентные заявки и публикации, включены в качестве ссылок в полном объеме.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение частично основано на идентификации различных связывающих FGFR3 агентов (таких как антитела и их фрагменты). FGFR3 представляет собой важную и преимущественную терапевтическую мишень, и изобретение относится к композициям и способам, основанным на связывании этих агентов с FGFR3. Связывающие FGFR3 агенты по изобретению, как описано в настоящем документе, обеспечивают важные терапевтические и диагностические средства для использования для нацеливания на патологические состояния, связанные с экспрессией и/или активностью каскадов передачи сигнала FGFR3. Таким образом, изобретение относится к способам, композициям, наборам и изделиям, имеющим отношение к связыванию FGFR3.
Настоящее изобретение относится к антителам, которые связываются с FGFR3. В одном из аспектов изобретение относится к выделенному антителу, которое связывает FGFR3. В некоторых вариантах осуществления антитело связывает изоформу FGFR3 IIIb и/или изоформу FGFR3 IIIc. В некоторых вариантах осуществления антитело связывает мутантный FGFR3 (например, один или несколько из R248C, S249C, G372C, Y375C, K652E FGFR3 IIIb, и/или один или несколько из R248C, S249C, G370C, Y373C, K650E FGFR3 IIIc). В некоторых вариантах осуществления антитело связывает мономерный FGFR3 (например, мономерные изоформы FGFR3 IIIb и/или IIIc). В некоторых вариантах осуществления антитело обеспечивает образование мономерного FGFR3, например, путем стабилизации мономерной формы FGFR3 относительно димерной формы FGFR3.
В одном из аспектов изобретение относится к выделенному антителу против FGFR3, где полноразмерная форма IgG антитела связывает FGFR3 человека с Kd 1×10-7 или прочнее. Как известно в данной области, аффинность связывания лиганда с его рецептором можно определять любым из множества анализов и выражать в виде различных количественных величин. Таким образом, в одном из вариантов осуществления аффинность связывания выражают как величины Kd и они отражают собственно аффинность связывания (например, с минимизированными эффектами авидности). Как правило, и предпочтительно, аффинность связывания измеряют in vitro, как в бесклеточных условиях, так и в условиях на основе клеток. Для получения показателей аффинности связывания можно использовать любой из множества анализов, известных в данной области, включая анализы, описанные в настоящем документе, в том числе, например, Biacore, радиоиммунный анализ (RIA) и ELISA. В некоторых вариантах осуществления полноразмерная IgG-форма антитела связывает FGFR3 человека с Kd 1×10-8 или прочнее, с Kd 1×10-9 или прочнее, или с Kd 1×10-10 или прочнее.
Главным образом, антитела против FGFR3 по настоящему изобретению представляют собой антитела-антагонисты. Таким образом, в одном из аспектов антитела против FGFR3 ингибируют активность FGFR3 (например, активность FGFR3-IIIb и/или FGFR3-IIIc). В некоторых вариантах осуществления антитело против FGFR3 (главным образом, в двухвалентной форме) не обладает существенной функцией агониста FGFR3. В некоторых вариантах осуществления антитело-антагонист против FGFR3 (главным образом, в двухвалентной форме) обладает небольшой функцией агониста FGFR3 или не обладает ей. В одном из вариантов осуществления антитело по изобретению (главным образом, в двухвалентной форме) не проявляет уровень активности агониста FGFR3, который превышает фоновый уровень и который обладает статистической значимостью.
В одном из аспектов связывание антитела с FGFR3 может ингибировать димеризацию рецептора с другой молекулой рецептора, причем активация рецептора ингибируется (вследствие, по меньшей мере частично, отсутствия димеризации рецептора). Ингибирование может быть прямым или непрямым.
В одном из аспектов изобретение относится к антителам против FGFR3, которые не обладают существенной апоптотической активностью (например, не индуцируют апоптоз клетки, например, клетки карциномы переходных клеток или клетки множественной миеломы, такие как клетка множественной миеломы, содержащая транслокацию FGFR3, такую как транслокация t(4;14)). В некоторых вариантах осуществления антитело против FGFR3 обладает небольшой апоптотической функцией или не обладает ей. В некоторых вариантах осуществления антитела против FGFR3 не обладают апоптотической функцией, которая превышает фоновый уровень и которая является статистически значимой.
В одном из аспектов изобретение относится к антителам против FGFR3, которые не индуцируют существенное подавление FGFR3. В некоторых вариантах осуществления антитело против FGFR3 индуцирует небольшое подавление рецептора или не индуцирует его. В некоторых вариантах осуществления антитела против FGFR3 не индуцируют подавление рецептора, которое превышает фоновый уровень и которое является статистически значимым.
В одном из аспектов изобретение относится к антителам против FGFR3, которые обладают эффекторной функцией. В одном из вариантов осуществления эффекторная функция включает антителозависимую клеточную цитотоксичность (ADCC). В одном из вариантов осуществления антитело против FGFR3 (в некоторых вариантах осуществления «голое» антитело против FGFR3) способно вызывать гибель клетки, в некоторых вариантах осуществления, клеток множественной миеломы (например, клеток множественной миеломы, содержащих транслокацию, например, транслокацию t(4;14)). В некоторых вариантах осуществления антитело против FGFR3 способно вызывать гибель клетки, которая экспрессирует приблизительно 10000 молекул FGFR3 на клетку или более (например, приблизительно 11000, приблизительно 12000, приблизительно 13000, приблизительно 14000, приблизительно 15000, приблизительно 16000, приблизительно 17000, приблизительно 18000 или более FGFR3 молекул на клетку). В других вариантах осуществления клетка экспрессирует приблизительно 2000, приблизительно 3000, приблизительно 4000, приблизительно 5000, приблизительно 6000, приблизительно 7000, приблизительно 8000 или более молекул FGFR3 на клетку.
В одном из аспектов антитело против FGFR3 по изобретению ингибирует конститутивную активность FGFR3. В некоторых вариантах осуществления конститутивная активность FGFR3 представляет собой зависимую от лиганда конститутивную активность FGFR3. В некоторых вариантах осуществления конститутивная активность FGFR3 представляет собой независимую от лиганда конститутивную активность FGFR3.
В одном из аспектов антитело против FGFR3 ингибирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbR248C. Как используется в настоящем документе, термин "содержащий мутацию, соответствующую FGFR3-IIIbR248C," включает FGFR3-IIIbR248C и FGFR3-IIIcR248C, а также дополнительные формы FGFR3, содержащие мутацию R на C в положении, соответствующем R248 FGFR3-IIIb. Специалисту в данной области понятно, каким образом провести выравнивание последовательности FGFR3 для идентификации соответствующих остатков между соответствующими последовательностями FGFR3, например, выравнивание последовательности FGFR3-IIIc с последовательностью FGFR3-IIIb для идентификации положения в FGFR3, соответствующего положению R248 в FGFR3-IIIb. В некоторых вариантах осуществления антитело против FGFR3 ингибирует FGFR3-IIIbR248C и/или FGFR3-IIIcR248C.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3, который содержит мутацию, соответствующую FGFR3-IIIbK652E. Как используется в настоящей заявке термин "содержащий мутацию, соответствующую FGFR3-IIIbK652E," включает FGFR3-IIIbK652E и FGFR3-IIIcK650E, а также дополнительные формы FGFR3, содержащие мутацию K на E в положении, соответствующем K652 FGFR3-IIIb. Специалисту в данной области понятно, каким образом провести выравнивание последовательности FGFR3 для идентификации соответствующих остатков между соответствующими последовательностями FGFR3, например, выравнивание последовательности FGFR3-IIIc с последовательностью FGFR3-IIIb для идентификации положения в FGFR3, соответствующего положению K652 в FGFR3-IIIb. В некоторых вариантах осуществления антитело против FGFR3 ингибирует FGFR3-IIIbK652E и/или FGFR3-IIIcK650E.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbS249C. Как используется в настоящей заявке термин "содержащий мутацию, соответствующую FGFR3-IIIbS249C," включает FGFR3-IIIbS249C и FGFR3-IIIcS249C, а также дополнительные формы FGFR3, содержащие мутацию S на C в положении, соответствующем S249 FGFR3-IIIb. В некоторых вариантах осуществления антитело против FGFR3 ингибирует FGFR3-IIIbS249C и/или FGFR3-IIIcS249C.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbG372C. Как используется в настоящей заявке термин "содержащий мутацию, соответствующую FGFR3-IIIbG372C," включает FGFR3-IIIbG372C и FGFR3-IIIcG370C, а также дополнительные формы FGFR3, содержащие мутацию G на C в положении, соответствующем G372 FGFR3-IIIb. В некоторых вариантах осуществления антитело против FGFR3 ингибирует FGFR3-IIIbG372C и/или FGFR3-IIIcG370C.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbY375C. Как используется в настоящей заявке термин "содержащий мутацию, соответствующую FGFR3-IIIbY375C," включает FGFR3-IIIbY375C и FGFR3-IIIcY373C, а также дополнительные формы FGFR3, содержащие мутацию S на C в положении, соответствующем S249 FGFR3-IIIb. В некоторых вариантах осуществления антитело против FGFR3 ингибирует FGFR3-IIIbY375C и/или FGFR3-IIIcY373C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIbK652E и (b) один или несколько из FGFR3-IIIbR248C, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3IIIbG372C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIcK650E и (b) один или несколько из FGFR3-IIIcR248C, FGFR3-IIIcY373C, FGFR3-IIIcS249C и FGFR3IIIcG370C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIbR248C и (b) один или несколько из FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIcR248C и (b) один или несколько из FGFR3-IIIcK650E, FGFR3-IIIcY373C, FGFR3-IIIcS249C и FGFR3-IIIcG370C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIbG372C и (b) один или несколько из FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbR248C.
В одном из аспектов антитела против FGFR3 ингибируют (a) FGFR3-IIIcG370C и (b) один или несколько из FGFR3-IIIcK650E, FGFR3-IIIcY373C, FGFR3-IIIcS249C и FGFR3-IIIcR248C.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3-IIIbR248C, FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
В одном из аспектов антитела против FGFR3 ингибируют FGFR3-IIIcR248C, FGFR3-IIIcK650E, FGFR3-IIIcY373C, FGFR3-IIIcS249C и FGFR3-IIIcG370C.
В одном из аспектов изобретение относится к выделенному антителу против FGFR3, содержащему:
(a) по меньшей мере одну, две, три, четыре или пять последовательностей гипервариабельных областей (HVR), выбранных из:
(i) HVR-L1, содержащей последовательность A1-A11, где A1-A11 представляет собой RASQDVDTSLA (SEQ ID NO:87),
(ii) HVR-L2, содержащей последовательность B1-B7, где B1-B7 представляет собой SASFLYS (SEQ ID NO:88),
(iii) HVR-L3, содержащей последовательность C1-C9, где C1-C9 представляет собой QQSTGHPQT (SEQ ID NO:89),
(iv) HVR-H1, содержащей последовательность D1-D10, где D1-D10 представляет собой GFTFTSTGIS (SEQ ID NO:84),
(v) HVR-H2, содержащей последовательность E1-E18, где E1-E18 представляет собой GRIYPTSGSTNYADSVKG (SEQ ID NO:85), и
(vi) HVR-H3, содержащей последовательность F1-F20, где F1-F20 представляет собой ARTYGIYDLYVDYTEYVMDY (SEQ ID NO:86); и
(b) по меньшей мере один вариант HVR, где последовательность варианта HVR содержит модификацию по меньшей мере одного остатка (по меньшей мере двух остатков, по меньшей мере трех или более остатков) последовательности SEQ ID NO:1-18, 48-131 и 140-145. Желательно, чтобы модификация представляла собой замену, вставку или делецию.
В некоторых вариантах осуществления вариант HVR-L1 содержит 1-6 (1, 2, 3, 4, 5 или 6) замен в любой комбинации из следующих положений: A5 (V или D), A6 (V или I), A7 (D, E или S), A8 (T или I), A9 (A или S) и A10 (V или L). В некоторых вариантах осуществления вариант HVR-L2 содержит 1-2 (1 или 2) замены в любой комбинации из следующих положений: B1 (S или G), B4 (F или S или T) и B6 (A или Y). В некоторых вариантах осуществления вариант HVR-L3 содержит 1-6 (1, 2, 3, 4, 5 или 6) замен в любой комбинации из следующих положений: C3 (G или S или T), C4 (T или Y или A), C5 (G или S или T или A), C6 (A или H или D или T или N), C7 (Q или P или S) и C8 (S или Y или L или P или Q). В некоторых вариантах осуществления вариант HVR-H1 содержит 1-3 (1, 2 или 3) замены в любой комбинации из следующих положений: D3 (S или T), D5 (W или Y или S или T), D6 (S или G или T). В некоторых вариантах осуществления вариант HVR-H2 содержит 1-6 (1, 2, 3, 4, 5 или 6) замен в любой комбинации из следующих положений: E2 (R или S), E6 (Y или A или L или S или T), E7 (A или Q или D или G или Y или S или N или F), E8 (A или D или G), E9 (T или S), E10 (K или F или T или S), E11 (Y или H или N или I).
В одном из аспектов изобретение относится к выделенному антителу против FGFR3, содержащему:
(a) по меньшей мере одну, две, три, четыре или пять последовательностей гипервариабельных областей (HVR), выбранных из:
(i) HVR-L1, содержащей последовательность RASQX1X2X3X4X5X6A, где X1 представляет собой V или D, X2 представляет собой V или I, X3 представляет собой D, E или S, X4 представляет собой T или I, X5 представляет собой A или S, и X6 представляет собой V или L (SEQ ID NO:146),
(ii) HVR-L2, содержащей последовательность X1ASFLX2S, где X1 представляет собой S или G и X2 представляет собой A или Y (SEQ ID NO:147),
(iii) HVR-L3, содержащей последовательность QQX1X2X3X4X5X6T, где X1 представляет собой G, S или T, X2 представляет собой T, Y или A, X3 представляет собой G, S, T или A, X4 представляет собой A, H, D, T или N, X5 представляет собой Q, P или S, X6 представляет собой S, Y, L, P или Q (SEQ ID NO:148),
(iv) HVR-H1, содержащей последовательность GFX1FX2X3TGIS, где X1 представляет собой S или T, X2 представляет собой W, Y, S или T, X3 представляет собой S, G или T (SEQ ID NO:149),
(v) HVR-H2, содержащей последовательность GRIYPX1X2X3X4X5X6YADSVKG, где X1 представляет собой Y, A, L, S или T, X2 представляет собой A, Q, D, G, Y, S, N или F, X3 представляет собой A, D или G, X4 представляет собой T или S, X5 представляет собой K, F, T или S, X6 представляет собой Y, H, N или I (SEQ ID NO:150), и
(vi) HVR-H3, содержащей последовательность ARTYGIYDLYVDYTEYVMDY (SEQ ID NO:151).
В некоторых вариантах осуществления HVR-L1 содержит последовательность RASQX1VX2X3X4VA, где X1 представляет собой V или D, X2 представляет собой D, E или S, X3 представляет собой T или I, X4 представляет собой A или S (SEQ ID NO:152). В некоторых вариантах осуществления HVR-L3 содержит последовательность QQX1X2X3X4X5X6T, где X1 представляет собой S, G или T, X2 представляет собой Y, T или A, X3 представляет собой T или G, X4 представляет собой T, H или N, X5 представляет собой P или S, X6 представляет собой P, Q, Y или L (SEQ ID NO:153). В некоторых вариантах осуществления HVR-H2 содержит последовательность GRIYPX1X2GSTX3YADSVKG, где X1 представляет собой T или L, X2 представляет собой N, Y, S, G, A или Q; X3 представляет собой N или H (SEQ ID NO:154).
В другом аспекте изобретение относится к выделенному антителу против FGFR3, которое содержит одну, две, три, четыре, пять или шесть HVR, где каждая HVR содержит последовательность, выбранную из SEQ ID NO:1-18, 48-131 и 140-145, состоит или по существу состоит из них, и где SEQ ID NO:1, 7, 13, 48, 54, 60, 66, 72, 78, 84, 90, 96, 102, 108, 114, 120, 126 или 143 соответствует HVR-H1, SEQ ID NO:2, 8, 14, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127 или 144 соответствует HVR-H2, SEQ ID NO:3, 9, 15, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128 или 145 соответствует HVR-H3, SEQ ID NO:4, 10, 16, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129 или 140 соответствует HVR-L1, SEQ ID NO:5, 11, 17, 52, 58, 64, 70, 76, 82, 88, 94, 100, 106, 112, 118, 124, 130 или 141 соответствует HVR-L2, и SEQ ID NO:6, 12, 18, 53, 59, 65, 71, 77, 83, 89, 95, 101, 107, 113, 119, 125, 131 или 142 соответствует HVR-L3.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему HVR-H1, содержащую последовательность SEQ ID NO:1, 7, 13, 48, 54, 60, 66, 72, 78, 84, 90, 96, 102, 108, 114, 120, 126 или 143.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему HVR-H2, содержащую последовательность SEQ ID NO:2, 8, 14, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127 или 144.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему HVR-H3, содержащую последовательность SEQ ID NO:3, 9, 15, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128 или 145.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему область HVR-L1, содержащую последовательность SEQ ID NO:4, 10, 16, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129 или 140.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему область HVR-L2, содержащую последовательность SEQ ID NO:5, 11, 17, 52, 58, 64, 70, 76, 82, 88, 94, 100, 106, 112, 118, 124, 130 или 141.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему область HVR-L3, содержащую последовательность SEQ ID NO:6, 12, 18, 53, 59, 65, 71, 77, 83, 89, 95, 101, 107, 113, 119, 125, 131 или 142.
В одном из аспектов антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:1, 2, 3, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:4, 5, 6.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:7, 8, 9, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:10, 11, 12.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:13, 14, 15, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:16, 17, 18.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:48, 49, 50, и/или вариабельную область легкой цепи HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:51, 52, 53.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:54, 55, 56, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:57, 58, 59.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:60, 61, 62, 63, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:63, 64, 65.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:66, 67, 68, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:69, 70, 71.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:72, 73, 74, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:75, 76, 77.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:78, 79 80, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:81, 82, 83.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:84, 85, 86, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2 и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:87, 88, 89.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:90, 91, 92, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:93, 94, 95.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:96, 97, 98, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:99, 100, 101.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:102, 103, 104, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:105, 106, 107.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:108, 109, 110, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:111, 112, 113.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:114, 115, 116, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:117, 118, 119.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:120, 121, 122, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:123, 124, 125.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:126, 127, 128, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:129, 130, 131.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую HVR-H1, HVR-H2, HVR-H3, где каждая из них, по порядку, содержит SEQ ID NO:140, 141, 142, и/или вариабельную область легкой цепи, содержащую HVR-L1, HVR-L2, и HVR-L3, где каждая из них, по порядку, содержит SEQ ID NO:143, 144, 145.
Аминокислотные последовательности SEQ ID NO:1-18, 48-131 и 140-145 пронумерованы относительно отдельных HVR (т.е., H1, H2 или H3), как указано на фигуре 1, причем нумерация соответствует системе нумерации Кабат, как описано ниже.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:132 и вариабельную область легкой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область легкой цепи, содержащую SEQ ID NO:133, и вариабельную область тяжелой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:132, и вариабельную область легкой цепи, содержащую SEQ ID NO:133.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:134, и вариабельную область легкой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область легкой цепи, содержащую SEQ ID NO:135, и вариабельную область тяжелой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область легкой цепи, содержащую SEQ ID NO:139, и вариабельную область тяжелой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:134, и вариабельную область легкой цепи, содержащую SEQ ID NO:135.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:136, и вариабельную область легкой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область легкой цепи, содержащую SEQ ID NO:137, и вариабельную область тяжелой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:136, и вариабельную область легкой цепи, содержащую SEQ ID NO:137.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:138, и вариабельную область легкой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область легкой цепи, содержащую SEQ ID NO:139, и вариабельную область тяжелой цепи.
В другом аспекте антитело против FGFR3 содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:138, и вариабельную область легкой цепи, содержащую SEQ ID NO:139.
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему: по меньшей мере одну, две, три, четыре, пять и/или шесть последовательностей гипервариабельной области (HVR), выбранных из группы, состоящей из:
(a) HVR-L1, содержащей последовательность SASSSVSYMH (SEQ ID NO:155), SASSSVSYMH (SEQ ID NO:156) или LASQTIGTWLA (SEQ ID NO:157),
(b) HVR-L2, содержащей последовательность TWIYDTSILAS (SEQ ID NO:158), RWIYDTSKLAS (SEQ ID NO:159) или LLIYAATSLAD (SEQ ID NO:160),
(c) HVR-L3, содержащей последовательность QQWTSNPLT (SEQ ID NO:161), QQWSSYPPT (SEQ ID NO:162) или QQLYSPPWT (SEQ ID NO:163),
(d) HVR-H1, содержащей последовательность GYSFTDYNMY (SEQ ID NO:164), GYVFTHYNMY (SEQ ID NO:165) или GYAFTSYNMY (SEQ ID NO:166),
(e) HVR-H2, содержащей последовательность WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:167), WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:168) или WIGYIDPYIGGTSYNQKFKG (SEQ ID NO:169), и
(f) HVR-H3, содержащей последовательность ASPNYYDSSPFAY (SEQ ID NO:170), ARGQGPDFDV (SEQ ID NO:171) или ARWGDYDVGAMDY (SEQ ID NO:172).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему: по меньшей мере одну, две, три, четыре, пять и/или шесть последовательностей гипервариабельной области (HVR), выбранных из группы, состоящей из:
(a) HVR-L1, содержащей последовательность SASSSVSYMH (SEQ ID NO:155),
(b) HVR-L2, содержащей последовательность TWIYDTSILAS (SEQ ID NO:158),
(c) HVR-L3, содержащей последовательность QQWTSNPLT (SEQ ID NO:161),
(d) HVR-H1, содержащей последовательность GYSFTDYNMY (SEQ ID NO:164),
(e) HVR-H2, содержащей последовательность WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:167), и
(f) HVR-H3, содержащей последовательность ASPNYYDSSPFAY (SEQ ID NO:170).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему: по меньшей мере одну, две, три, четыре, пять и/или шесть последовательностей гипервариабельной области (HVR), выбранных из группы, состоящей из:
(a) HVR-L1, содержащей последовательность SASSSVSYMH (SEQ ID NO:156),
(b) HVR-L2, содержащей последовательность RWIYDTSKLAS (SEQ ID NO:159),
(c) HVR-L3, содержащей последовательность QQWSSYPPT (SEQ ID NO:162),
(d) HVR-H1, содержащей последовательность GYVFTHYNMY (SEQ ID NO:165),
(e) HVR-H2, содержащей последовательность WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:168), и
(f) HVR-H3, содержащей последовательность ARGQGPDFDV (SEQ ID NO:171).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему: по меньшей мере одну, две, три, четыре, пять и/или шесть последовательностей гипервариабельной области (HVR), выбранных из группы, состоящей из:
(a) HVR-L1, содержащей последовательность LASQTIGTWLA (SEQ ID NO:157),
(b) HVR-L2, содержащей последовательность LLIYAATSLAD (SEQ ID NO:160),
(c) HVR-L3, содержащей последовательность QQLYSPPWT (SEQ ID NO:163),
(d) HVR-H1, содержащей последовательность GYAFTSYNMY (SEQ ID NO:166),
(e) HVR-H2, содержащей последовательность WIGYIDPYIGGTSYNQKFKG (SEQ ID NO:169), и
(f) HVR-H3, содержащей последовательность ARWGDYDVGAMDY (SEQ ID NO:172).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему (a) легкую цепь, содержащую (i) HVR-L1, содержащую последовательность SASSSVSYMH (SEQ ID NO:155); (ii) HVR-L2, содержащую последовательность TWIYDTSILAS (SEQ ID NO:158); и (iii) HVR-L3, содержащую последовательность QQWTSNPLT (SEQ ID NO:161); и/или (b) тяжелую цепь, содержащую (i) HVR-H1, содержащую последовательность GYSFTDYNMY (SEQ ID NO:164); (ii) HVR-H2, содержащую последовательность WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:167); и (iii) HVR-H3, содержащую последовательность ASPNYYDSSPFAY (SEQ ID NO:170).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему (a) легкую цепь, содержащую (i) HVR-L1, содержащую последовательность SASSSVSYMH (SEQ ID NO:156); (ii) HVR-L2, содержащую последовательность RWIYDTSKLAS (SEQ ID NO:159); и (iii) HVR-L3, содержащую последовательность QQWSSYPPT (SEQ ID NO:162); и/или (b) тяжелую цепь, содержащую (i) HVR-H1, содержащую последовательность GYVFTHYNMY (SEQ ID NO:165); (ii) HVR-H2, содержащую последовательность WIGYIEPYNGGTSYNQKFKG (SEQ ID NO:168); и (iii) HVR-H3, содержащую последовательность ARGQGPDFDV (SEQ ID NO:171).
В одном из аспектов изобретение относится к антителу против FGFR3, содержащему (a) легкую цепь, содержащую (i) HVR-L1, содержащую последовательность LASQTIGTWLA (SEQ ID NO:157); (ii) HVR-L2, содержащую последовательность LLIYAATSLAD (SEQ ID NO:160); и (iii) HVR-L3, содержащую последовательность QQLYSPPWT (SEQ ID NO:163); и/или (b) тяжелую цепь, содержащую (i) HVR-H1, содержащую последовательность GYAFTSYNMY (SEQ ID NO:166); (ii) HVR-H2, содержащую последовательность WIGYIDPYIGGTSYNQKFKG (SEQ ID NO:169); и (iii) HVR-H3, содержащую последовательность ARWGDYDVGAMDY (SEQ ID NO:172). Некоторые варианты осуществления антител по изобретению содержат вариабельный домен легкой цепи гуманизированного антитела 4D5 (huMAb4D5-8) (HERCEPTIN®, Genentech, Inc., South San Francisco, CA, USA) (также указываемого в патенте США No. 6407213 и Lee et al., J. Mol. Biol. (2004), 340(5): 1073-1093), как представлено в SEQ ID NO:173 ниже:
1 Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys 107 (SEQ ID NO:173)
(остатки HVR подчеркнуты)
В одном из вариантов осуществления последовательность вариабельного домена легкой цепи huMAb4D5-8 модифицирована в одном или нескольких положениях 30, 66 и 91 (Asn, Arg и His, как указано полужирным шрифтом/курсивом выше, соответственно). В конкретном варианте осуществления, модифицированная последовательность huMAb4D5-8 содержит Ser в положении 30, Gly в положении 66, и/или Ser в положении 91. Таким образом, в одном из вариантов осуществления антитело по изобретению содержит вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:174 ниже:
1 Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys 107 (SEQ ID NO:174)
(остатки HVR подчеркнуты)
Остатки, замененные в huMAb4D5-8, указаны полужирным шрифтом/курсивом.
Антитела по изобретению могут содержать любую подходящую каркасную последовательность вариабельного домена, при условии, что активность связывания с FGFR3 по существу сохраняется. Например, в некоторых вариантах осуществления антитела по изобретению содержат консенсусную каркасную последовательность тяжелой цепи человека подгруппы III. В одном из вариантов осуществления этих антител, консенсусная каркасная последовательность содержит замену в положении 71, 73 и/или 78. В некоторых вариантах осуществления этих антител, положение 71 представляет собой A, 73 представляет собой T и/или 78 представляет собой A. В одном из вариантов осуществления эти антитела содержат каркасные последовательности вариабельного домена тяжелой цепи huMAb4D5-8 (HERCEPTIN®, Genentech, Inc., South San Francisco, CA, США) (также указываемого в патентах США No. 6407213 и 5821337, и Lee et al., J. Mol. Biol. (2004), 340(5): 1073-1093). В одном из вариантов осуществления эти антитела, кроме того, содержат консенсусную каркасную последовательность легкой цепи kI человека. В конкретном варианте осуществления эти антитела содержат последовательности HVR легкой цепи huMAb4D5-8, как описано в патентах США No. 6407213 и 5821337). В одном из вариантов осуществления эти антитела содержат последовательности вариабельных доменов легкой цепи huMAb4D5-8 (HERCEPTIN®, Genentech, Inc., South San Francisco, CA, США) (также казываемого в патентах США No. 6407213 и 5821337, и Lee et al., J. Mol. Biol. (2004), 340(5): 1073-1093).
В одном из вариантов осуществления антитело по изобретению содержит вариабельный домен тяжелой цепи, где каркасная последовательность содержит последовательность SEQ ID NO:19 и 203-205, 20 и 206-208, 21 и 209-211, 22 и 212-214, 23 и 215-217, 24 и 218-220, 25 и 221-223, 26 и 224-226, 27 и 227-229, 28 и 230-232, 29 и 233-235, 30 и 236-238, 31 и 239-241, 32 и 242-244, 33 и 245-247, 34 и 248-250, 35 и 251-253, 36 и 254-256, и/или 37 и 257-259, и последовательности H1, H2 и H3 HVR представляют собой SEQ ID NO:13, 14 и/или 15, соответственно. В другом варианте осуществления каркасная последовательность содержит последовательность SEQ ID NO:19 и 203-205, 20 и 206-208, 21 и 209-211, 22 и 212-214, 23 и 215-217, 24 и 218-220, 25 и 221-223, 26 и 224-226, 27 и 227-229, 28 и 230-232, 29 и 233-235, 30 и 236-238, 31 и 239-241, 32 и 242-244, 33 и 245-247, 34 и 248-250, 35 и 251-253, 36 и 254-256, и/или 37 и 257-259, и последовательности H1, H2 и H3 HVR представляют собой SEQ ID NO:48, 49 и/или 50, соответственно. В другом варианте осуществления, каркасная последовательность содержит последовательность SEQ ID NO:19 и 203-205, 20 и 206-208, 21 и 209-211, 22 и 212-214, 23 и 215-217, 24 и 218-220, 25 и 221-223, 26 и 224-226, 27 и 227-229, 28 и 230-232, 29 и 233-235, 30 и 236-238, 31 и 239-241, 32 и 242-244, 33 и 245-247, 34 и 248-250, 35 и 251-253, 36 и 254-256, и/или 37 и 257-259, и последовательности H1, H2 и H3 HVR представляют собой SEQ ID NO:84, 85 и/или 86, соответственно. В другом варианте осуществления, каркасная последовательность содержит последовательность SEQ ID NO:19 и 203-205, 20 и 206-208, 21 и 209-211, 22 и 212-214, 23 и 215-217, 24 и 218-220, 25 и 221-223, 26 и 224-226, 27 и 227-229, 28 и 230-232, 29 и 233-235, 30 и 236-238, 31 и 239-241, 32 и 242-244, 33 и 245-247, 34 и 248-250, 35 и 251-253, 36 и 254-256, и/или 37 и 257-259, и последовательности H1, H2 и H3 HVR представляют собой SEQ ID NO:108, 109 и/или 110, соответственно.
В конкретном варианте осуществления антитело по изобретению содержит вариабельный домен легкой цепи, где каркасная последовательность содержит последовательность SEQ ID NO:38 и 260-262, 39 и 263-265, 40 и 266-268 и/или 41 и 269-271, и последовательности L1, L2 и L3 HVR представляют собой SEQ ID NO:16, 17 и/или 18, соответственно. В другом варианте осуществления антитело по изобретению содержит вариабельный домен легкой цепи, где каркасная последовательность содержит последовательность SEQ ID NO:38 и 260-262, 39 и 263-265, 40 и 266-268, и/или 41 и 269-271, и последовательности L1, L2 и L3 HVR представляют собой SEQ ID NO:51, 52 и/или 53, соответственно. В дополнительном варианте осуществления антитело по изобретению содержит вариабельный домен легкой цепи, где каркасная последовательность содержит последовательность SEQ ID NO:38 и 260-262, 39 и 263-265, 40 и 266-268, и/или 41 и 269-271, и последовательности L1, L2 и L3 HVR представляют собой SEQ ID NO:87, 88 и/или 89, соответственно. В другом варианте осуществления антитело по изобретению содержит вариабельный домен легкой цепи, где каркасная последовательность содержит последовательность SEQ ID NO:38 и 260-262, 39 и 263-265, 40 и 266-268, и/или 41 и 269-271, и последовательности L1, L2 и L3 HVR представляют собой SEQ ID NO:111, 112 и/или 113, соответственно.
В другом аспекте антитело по изобретению содержит вариабельный домен тяжелой цепи, содержащий последовательность SEQ ID NO:132, и/или вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:133. В другом аспекте антитело по изобретению содержит вариабельный домен тяжелой цепи, содержащий последовательность SEQ ID NO:134, и/или вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:135. В другом аспекте антитело по изобретению содержит вариабельный домен тяжелой цепи, содержащий последовательность SEQ ID NO:136, и/или вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:137. В другом аспекте антитело по изобретению содержит вариабельный домен тяжелой цепи, содержащий последовательность SEQ ID NO:138, и/или вариабельный домен легкой цепи, содержащий последовательность SEQ ID NO:139.
В одном из аспектов изобретение относится к антителу против FGFR3, которое связывает полипептид, содержащий следующую аминокислотную последовательность: LAVPAANTVRFRCPA (SEQ ID NO:179) и/или SDVEFHCKVYSDAQP (SEQ ID NO:180), по существу состоящий или состоящий из нее.
В некоторых вариантах осуществления антитело связывает полипептид, содержащий аминокислоты номер 164-178 и/или 269-283 аминокислотной последовательности зрелого FGFR3 человека, по существу состоящий или состоящий из них.
В одном из вариантов осуществления антитело против FGFR3 по изобретению специфично связывает аминокислотную последовательность, имеющую по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 98% идентичность последовательности или сходство с последовательностью LAVPAANTVRFRCPA (SEQ ID NO:179) и/или SDVEFHCKVYSDAQP (SEQ ID NO:180).
В одном из аспектов антитело против FGFR3 по настоящему изобретению связывается по меньшей мере с одним, двумя, тремя, четырьмя или любым количеством вплоть до всех из остатков 154, 155, 158, 159, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 177, 202, 205, 207, 210, 212, 214, 216, 217, 241, 246, 247, 248, 278, 279, 280, 281, 282, 283, 314, 315, 316, 317 и/или 318 полипептида FGFR3 IIIb, или эквивалентных остатков полипептида FGFR3 IIIc. Специалисту в данной области понятно, как провести выравнивание последовательности FGFR3 для идентификации соответствующих остатков между соответствующими последовательностями FGFR3. Сочетания двух или более остатков могут включать любые из остатков 154, 155, 158, 159, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 177, 202, 205, 207, 210, 212, 214, 216, 217, 241, 246, 247, 248, 278, 279, 280, 281, 282, 283, 314, 315, 316, 317 и/или 318 полипептида FGFR3 IIIb, или эквивалентных остатков полипептида FGFR3 IIIc. В некоторых вариантах осуществления антитело против FGFR3 связывается по меньшей мере с одним, двумя, тремя, четырьмя или любым количеством вплоть до всех из остатков 158, 159, 169, 170, 171, 173, 175, 205, 207 и/или 315 полипептида FGFR3 IIIb, или эквивалентных остатков полипептида FGFR3 IIIc. В некоторых вариантах осуществления антитело против FGFR3 связывается по меньшей мере с одним, двумя, тремя, четырьмя или любым количеством вплоть до всех из остатков 158, 170, 171, 173, 175 и/или 315 полипептида FGFR3 IIIb, или эквивалентных остатков полипептида FGFR3 IIIc.
В одном из аспектов изобретение относится к антителу против FGFR3, которое конкурирует с любым из указанных выше антител за связывание с FGFR3. В одном из аспектов изобретение относится к антителу против FGFR3, которое связывается с тем же или аналогичным эпитопом на FGFR3, что и любое из упомянутых выше антител.
Как известно в данной области и как более подробно описано в настоящем документе ниже, обозначение аминокислотного положения/границ гипервариабельной области антитела может изменяться, в зависимости от контекста и различных определений, известных в данной области (как описано ниже). Некоторые положения в вариабельном домене могут рассматриваться как гибридные гипервариабельные положения, поскольку эти положения можно считать находящимися в пределах гипервариабельной области согласно одному набору критериев, и их можно считать находящимися вне гипервариабельной области согласно другому набору критериев. Одно или несколько из этих положений также могут быть обнаружены в удлиненных гипервариабельных областях (как дополнительно определено ниже).
В некоторых вариантах осуществления антитело представляет собой моноклональное антитело. В других вариантах осуществления антитело представляет собой поликлональное антитело. В некоторых вариантах осуществления антитело выбрано из группы, состоящей из химерного антитела, прошедшего стадию созревания аффинности антитела, гуманизированного антитела и антитела человека. В определенных вариантах осуществления антитело представляет собой фрагмент антитела. В некоторых вариантах осуществления антитело представляет собой Fab, Fab', Fab'-SH, F(ab')2 или scFv.
В некоторых вариантах осуществления антитело против FGFR3 представляет собой антитело с одним плечом (т.е., вариабельный домен тяжелой цепи и вариабельный домен легкой цепи образуют одно антиген-связывающее плечо), содержащее Fc-область, где Fc-область содержит первый и второй полипептиды Fc, где первый и второй полипептиды Fc присутствуют в комплексе и образуют Fc-область, которая увеличивает стабильность указанного фрагмента антитела по сравнению с молекулой Fab, содержащей указанное антиген-связывающее плечо. См., например, WO2006/015371.
В одном из вариантов осуществления антитело представляет собой химерное антитело, например, антитело, содержащее антиген-связывающие последовательности из не являющегося человеком донора, пересаженные в гетерологичную последовательность, не являющуюся последовательностью человека, последовательность человека или гуманизированную последовательность (например, каркасные последовательности и/или последовательности константного домена). В одном из вариантов осуществления не являющимся человеком донором является мышь. В другом варианте осуществления антиген-связывающая последовательность является искусственной, например, полученной мутагенезом (например, при скрининге способом фагового дисплея, и тому подобное). В конкретном варианте осуществления химерное антитело по изобретению имеет V-области мыши и C-область человека. В одном из вариантов осуществления V-область легкой цепи мыши слита с легкой цепью каппа человека. В другом варианте осуществления V-область тяжелой цепи мыши слита с C-областью IgG1 человека.
Гуманизированные антитела по изобретению включают антитела, которые имеют аминокислотные замены в каркасной области (FR), и варианты, прошедшие стадию созревания аффинности, с изменениями в пересаженных CDR. Замененные аминокислоты в CDR или FR не ограничиваются аминокислотами, представленными в донорном или реципиентном антителе. В других вариантах осуществления антитела по изобретению, кроме того, содержат изменения в аминокислотных остатках в Fc-области, которые обеспечивают улучшение эффекторной функции, в том числе усиление функции CDC и/или ADCC и уничтожение B-клеток. Другие антитела по изобретению включают антитела, имеющие определенные изменения, которые повышают стабильность. В других вариантах осуществления антитела по изобретению содержат изменения в аминокислотных остатках в Fc-области, которые приводят к снижению эффекторной функции, например, снижению функции CDC и/или ADCC и/или сниженному уничтожению B-клеток. В некоторых вариантах осуществления антитела по изобретению характеризуются снижением связывания (таким как отсутствие связывания) с фактором комплемента C1q человека и/или Fc-рецептором человека на природных киллерных (NK) клетках. В некоторых вариантах осуществления антитела по изобретению характеризуются снижением связывания (например, отсутствием связывания) с FcγRI, FcγRIIA и/или FcγRIIIA. В некоторых вариантах осуществления антитела по изобретению представляют собой антитела класса IgG (например, IgG1 или IgG4) и содержат по меньшей мере одну мутацию в положениях E233, L234, G236, D265, D270, N297, E318, K320, K322, A327, A330, P331 и/или P329 (нумерация согласно индексу EU). В некоторых вариантах осуществления антитела содержат мутации L234A/L235A или D265A/N297A.
Если антитело содержит Fc-область, то может быть изменен углевод, присоединенный к нему. Например, антитела со зрелой углеводной структурой, которая лишена фукозы, присоединенной к Fc-области антитела, описаны в патентной заявке США US 2003/0157108 (Presta, L.). См. также US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Антитела с биссекторным N-ацетилглюкозамином (GlcNAc) в углеводе, присоединенном к Fc-области антитела, указаны в WO 2003/011878, Jean-Mairet et al., и патенте США No. 6602684, Umana et al. Антитела по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области антитела, описаны в WO 1997/30087, Patel et al. См. также WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.), относящиеся к антителам с измененным углеводом, который присоединен к их Fc-области. Также см. US 2005/0123546 (Umana et al.), где описаны антиген-связывающие молекулы с модифицированным гликозилированием. В одном из аспектов изобретение относится к связывающим FGFR3 полипептидам, содержащим любую антиген-связывающую последовательность, представленную в настоящем документе, где связывающие FGFR3 полипептиды специфично связываются с FGFR3, например, с FGFR3 человека и/или яванского макака и/или мыши.
Антитела по изобретению связывают (например, специфично связывают) FGFR3 (например, FGFR3-IIIb и/или FGFR3-IIIc), и в некоторых вариантах осуществления могут модулировать (например, ингибировать) один или несколько аспектов передачи сигнала FGFR3 (таких как фосфорилирование FGFR3) и/или нарушение какого-либо биологически значимого каскада FGFR3 и/или лиганда FGFR3, и/или лечение и/или профилактику опухоли, клеточно-пролиферативного нарушения или злокачественной опухоли; и/или лечение или профилактику нарушения, связанного с экспрессией и/или активностью FGFR3 (таких как повышенная экспрессия и/или активность FGFR3). В некоторых вариантах осуществления антитело против FGFR3 специфично связывается с полипептидом, состоящим или по существу состоящим из FGFR3 (например, FGFR3 человека или мыши). В некоторых вариантах осуществления антитело специфично связывает FGFR3 с Kd 1×10-7 M или прочнее.
В некоторых вариантах осуществления антитело против FGFR3 по изобретению не является антителом против FGFR3, описанным в публикации патента США 2005/0147612 (например, антитело MSPRO2, MSPRO12, MSPRO59, MSPRO11, MSPRO21, MSPRO24, MSPRO26, MSPRO28, MSPRO29, MSPRO43, MSPRO55), антителом, описанным в Rauchenberger et al., J Biol. Chem. 278 (40): 38194-38205 (2003); антителом, описанным в публикации PCT WO2006/048877 (например, антитело PRO-001), антителом, описанным в Martinez-Torrecuadrada et al., Mol. Cancer Ther (2008) 7(4): 862-873 (например, scFvαFGFR3 3C), антителом, описанным в Direnzo, R et al. (2007) Proceedings of AACR Annual Meeting, Abstract No. 2080 (например, D11), или антителом, описанным в WO 2010/002862 (например, антитела 15D8, 27H2, 4E7, 2G4, 20B4).
В одном из аспектов изобретение относится к композициям, содержащим одно или несколько антител по изобретению и носитель. В одном из вариантов осуществления носитель является фармацевтически приемлемым.
В другом аспекте изобретение относится к нуклеиновым кислотам, кодирующим антитело против FGFR3 по изобретению.
В другом аспекте изобретение относится к векторам, содержащим нуклеиновую кислоту по изобретению.
В другом аспекте изобретение относится к композициям, содержащим одну или несколько нуклеиновых кислот по изобретению и носитель. В одном из вариантов осуществления носитель является фармацевтически приемлемым.
В одном из аспектов изобретение относится к клеткам-хозяевам, содержащим нуклеиновую кислоту или вектор по изобретению. Вектор может быть любого типа, например, рекомбинантный вектор, такой как вектор экспрессии. Можно использовать любую клетку-хозяин из существующего разнообразия. В одном из вариантов осуществления клетка-хозяин представляет собой прокариотическую клетку, например, E. coli. В другом варианте осуществления клетка-хозяин представляет собой эукариотическую клетку, например, клетку млекопитающего, такую как клетка яичника китайского хомяка (CHO).
В другом аспекте изобретение относится к способам получения антитела по изобретению. Например, изобретение относится к способам получения антитела против FGFR3 (которое, как определено в настоящем документе, включает полноразмерное антитело и его фрагменты), причем указанный способ предусматривает экспрессию в подходящей клетке-хозяине рекомбинантного вектора по изобретению, кодирующего антитело, и выделение антитела. В некоторых вариантах осуществления способ предусматривает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, так что экспрессируется нуклеиновая кислота. В некоторых вариантах осуществления способ дополнительно предусматривает выделение антитела из культуры клеток-хозяев. В некоторых вариантах осуществления антитело выделяют из культуральной среды клеток-хозяев. В некоторых вариантах осуществления способ дополнительно предусматривает объединение выделенного антитела с фармацевтически приемлемым носителем, эксципиентом или носителем с получением фармацевтического состава, содержащего гуманизированное антитело.
В одном из аспектов изобретение относится к изделию, содержащему контейнер; и композиции, содержащейся в контейнере, где композиция содержит одно или несколько антител против FGFR3 по изобретению. В одном из вариантов осуществления композиция содержит нуклеиновую кислоту по изобретению. В другом варианте осуществления композиция, содержащая антитело, дополнительно содержит носитель, который в некоторых вариантах осуществления является фармацевтически приемлемым. В одном из вариантов осуществления изделие по изобретению, кроме того, содержит инструкцию по применению композиции (например, антитела) индивидуумом (например, инструкцию по любому из способов, описанных в настоящем документе).
В другом аспекте изобретение относится к набору, содержащему первый контейнер, содержащий композицию, содержащую одно или несколько антител против FGFR3 по изобретению; и второй контейнер, содержащий буфер. В одном из вариантов осуществления буфер является фармацевтически приемлемым. В одном из вариантов осуществления композиция, содержащая антитело, кроме того, содержит носитель, который в некоторых вариантах осуществления является фармацевтически приемлемым. В другом варианте осуществления набор, кроме того, содержит инструкции по применению композиции (например, антитела) индивидуумом.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется в качестве лекарственного средства.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется для лечения или профилактики нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется для лечения или профилактики нарушения, такого как болезни скелета. В некоторых вариантах осуществления нарушение представляет собой ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется для снижения пролиферации клеток.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется для уничтожения клетки. В некоторых вариантах осуществления клетка представляет собой клетку множественной миеломы. В некоторых вариантах осуществления гибель клеток вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
В другом аспекте изобретение относится к антителу против FGFR3 по изобретению, которое используется для уменьшения количества клеток, таких как клетки множественной миеломы. В некоторых вариантах осуществления гибель клеток вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
В другом аспекте изобретение относится к применению антитела против FGFR3 по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезни скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В одном из аспектов изобретение относится к применению нуклеиновой кислоты по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезни скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В другом аспекте изобретение относится к применению вектора экспрессии по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезни скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В другом аспекте изобретение относится к применению клетки-хозяина по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезнь скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В другом аспекте изобретение относится к применению изделия по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезнь скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В одном из аспектов изобретение также относится к применению набора по изобретению для получения лекарственного средства для терапевтического и/или профилактического лечения нарушения, такого как патологическое состояние, связанное с активацией и/или экспрессией (в некоторых вариантах осуществления сверхэкспрессией) FGFR3. В некоторых вариантах осуществления нарушение представляет собой злокачественную опухоль, опухоль и/или клеточно-пролиферативное нарушение. В некоторых вариантах осуществления злокачественная опухоль, опухоль и/или клеточно-пролиферативное нарушение представляет собой множественную миелому или рак мочевого пузыря (например, карциному переходных клеток), рак молочной железы или рак печени. В некоторых вариантах осуществления нарушение представляет собой болезнь скелета, например, ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В другом аспекте изобретение относится к применению антитела против FGFR3 по изобретению для получения лекарственного средства для ингибирования пролиферации клеток. В другом аспекте изобретение относится к применению антитела против FGFR3 по изобретению для получения лекарственного средства для уничтожения клеток. В некоторых вариантах осуществления клетка представляет собой клетку множественной миеломы. В некоторых вариантах осуществления гибель клетки вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
В другом аспекте изобретение относится к применению антитела против FGFR3 по изобретению для получения лекарственного средства для уменьшения количества клеток, таких как клетки множественной миеломы. В некоторых вариантах осуществления гибель клетки вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
Изобретение относится к способам и композициям, подходящим для модулирования нарушений, связанных с экспрессией и/или передачей сигнала FGFR3, таких как повышенная экспрессия и/или передача сигнала или нежелательная экспрессия и/или передача сигнала.
Способы по изобретению можно использовать при любом подходящем патологическом состоянии. Характерные нарушения описаны в настоящем документе и включают злокачественную опухоль, выбранную из группы, состоящей из немелкоклеточного рака легкого, рака яичника, рака щитовидной железы, рака яичка, рака эндометрия, рака головы и шеи, злокачественной опухоли головного мозга (например, нейробластомы или менингиомы), рака кожи (например, меланомы, базально-клеточной карциномы или плоскоклеточной карциномы), рака мочевого пузыря (например, карциномы переходных клеток), карциномы молочной железы, рака желудка, рака ободочной и прямой кишки (CRC), печеночно-клеточной карциномы, рака шейки матки, рака легкого, рака поджелудочной железы, рака предстательной железы и гематологических злокачественных опухолей (например, T-клеточного острого лимфобластного лейкоза (T-ALL), B-клеточного острого лимфобластного лейкоза (B-ALL), острого миелогенного лейкоза (AML), B-клеточных злокачественных опухолей, лимфомы Ходжкина и множественных миелом). В некоторых вариантах осуществления нарушение представляет собой инвазивную карциному переходных клеток. В некоторых вариантах осуществления нарушение представляет собой множественную миелому. Другие характерные нарушения включают болезни скелета, такие как ахондроплазия, гипохондроплазия, карликовость, летальная дисплазия (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
В определенных вариантах осуществления злокачественная опухоль экспрессирует FGFR3, амплифицированный FGFR3, транслоцированный FGFR3 и/или мутантный FGFR3. В определенных вариантах осуществления злокачественная опухоль экспрессирует активированный FGFR3. В определенных вариантах осуществления злокачественная опухоль экспрессирует транслоцированный FGFR3 (например, с транслокацией t(4;14)). В определенных вариантах осуществления злокачественная опухоль экспрессирует конститутивный FGFR3. В некоторых вариантах осуществления конститутивный FGFR3 содержит мутацию в тирозинкиназном домене и/или юкстамембранном домене и/или лиганд-связывающем домене. В определенных вариантах осуществления злокачественная опухоль экспрессирует независимый от лиганда FGFR3. В некоторых вариантах осуществления злокачественная опухоль экспрессирует лиганд-зависимый FGFR3.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbS248C. В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbS248C и/или FGFR3-IIIcS248C.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbK652E. В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbK652E и/или FGFR3-IIIcK650E.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbS249C. В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbS249C и/или FGFR3-IIIcS249C.
В одном из аспектов злокачественная опухоль экспрессирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbG372C. В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbG372C и/или FGFR3-IIIcG370C.
В одном из аспектов злокачественная опухоль экспрессирует FGFR3, содержащий мутацию, соответствующую FGFR3-IIIbY375C. В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbY375C и/или FGFR3-IIIcY373C.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует (a) FGFR3-IIIbK652E и (b) один или несколько из FGFR3-IIIbR248C, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует (a) FGFR3-IIIbR248C и (b) один или несколько из FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует (a) FGFR3-IIIbG372C и (b) один или несколько из FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbR248C.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует FGFR3-IIIbR248C, FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
В определенных вариантах осуществления злокачественная опухоль экспрессирует повышенные уровни фосфо-FGFR3, фосфо-FRS2 и/или фосфо-MAPK по сравнению с контрольным образцом (например, с образцом нормальной ткани) или уровня.
В некоторых вариантах осуществления злокачественная опухоль экспрессирует (например, на поверхности клетки) приблизительно 10000 молекул FGFR3 на клетку или более (например, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000 или более рецепторов FGFR3). В некоторых вариантах осуществления злокачественная опухоль экспрессирует приблизительно 13000 молекул FGFR3. В других вариантах осуществления злокачественная опухоль экспрессирует приблизительно 5000, 6000, 7000, 8000 или более молекул FGFR3. В некоторых вариантах осуществления злокачественная опухоль экспрессирует менее чем приблизительно 4000, 3000, 2000, 1000 или меньше молекул FGFR3. В некоторых вариантах осуществления злокачественная опухоль экспрессирует менее чем приблизительно 1000 молекул FGFR3.
В одном из вариантов осуществления клетка, на которую нацелен способ по изобретению, представляет собой злокачественную клетку. Например, злокачественная клетка может представлять собой клетку, выбранную из группы, состоящей из клетки рака молочной железы, клетки рака ободочной и прямой кишки, клетки рака легкого (например, клетки немелкоклеточного рака легкого), клетки рака щитовидной железы, клетки множественной миеломы, клетки рака яичка, клетки папиллярной карциномы, клетки рака толстого кишечника, клетки рака поджелудочной железы, клетки рака яичника, клетки рака шейки матки, клетки злокачественной опухоли центральной нервной системы, клетки остеогенной саркомы, клетки карциномы почки, клетки печеночно-клеточной карциномы, клетки рака мочевого пузыря (например, клетки карциномы переходных клеток), клетки карциномы желудка, клетки плоскоклеточной карциномы головы и шеи, клетки меланомы, клетки лейкоза, клетки множественной миеломы (например, клетки множественной миеломы, содержащие транслокацию t(4:14) FGFR3) и клетки аденомы толстого кишечника. В одном из вариантов осуществления клетка, на которую нацелен способ по изобретению, представляет собой гиперпролиферативную и/или гиперпластическую клетку. В другом варианте осуществления клетка, на которую нацелен способ по изобретению, представляет собой диспластическую клетку. В другом варианте осуществления клетка, на которую нацелен способ по изобретению, представляет собой метастатическую клетку.
В одном из аспектов изобретение относится к способам ингибирования пролиферации клеток у индивидуума, причем способ предусматривает введение индивидууму эффективного количества антитела против FGFR3 для снижения пролиферации клеток.
В одном из аспектов изобретение относится к способам уничтожения клетки у индивидуума, причем способ предусматривает введение индивидууму эффективного количества антитела против FGFR3 для уничтожения клетки. В некоторых вариантах осуществления клетка представляет собой клетку множественной миеломы. В некоторых вариантах осуществления гибель клетки вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
В одном из аспектов изобретение относится к способам уменьшения количества клеток (таких как клетки множественной миеломы) у индивидуума, причем способ предусматривает введение индивидууму эффективного количества антитела против FGFR3 для уничтожения клетки. В некоторых вариантах осуществления гибель клетки вызвана ADCC. В некоторых вариантах осуществления антитело представляет собой «голое» антитело. В некоторых вариантах осуществления клетка сверхэкспрессирует FGFR3.
В одном из аспектов изобретение относится к способам лечения или профилактики болезней скелета. В некоторых вариантах осуществления болезнь представляет собой ахондроплазию, гипохондроплазию, карликовость, летальную дисплазию (TD; клинические формы TDI и TDII) или синдром краниосиностоза.
Способы по изобретению могут, кроме того, предусматривать дополнительные стадии лечения. Например, в одном из вариантов осуществления способ дополнительно предусматривает стадию, на которой клетку и/или ткань, на которые осуществляют нацеливание (например, злокачественную клетку), подвергают радиационной обработке или на которую воздействуют химиотерапевтическим средством.
В одном из аспектов изобретение относится к способам, предусматривающим введение эффективного количества антитела против FGFR3 в сочетании с эффективным количеством другого лекарственного средства (такого как средство против ангиогенеза, другое антитело, химиотерапевтическое средство, цитотоксическое средство, иммунодепрессивное средство, пролекарство, цитокин, цитотоксическая лучевая терапия, кортикостероид, противорвотное средство, противораковая вакцина, анальгетик или средство, ингибирующее рост). Например, антитела против FGFR3 используют в сочетаниях с противораковым средством или антиангиогенным средством для лечения различных неопластических или ненеопластических состояний. В конкретных примерах антитела против FGFR3 используют в сочетании с велкейдом, ревлимидом, тамоксифеном, лектрозолом, экземестаном, анастрозолом, иринотеканом, цетуксимабом, фулвестрантом, винорелбином, бевацизумабом, винкристином, цисплатином, гемцитабином, метотрексатом, винбластином, карбоплатином, паклитакселом, доцетакселом, пеметрекседом, 5-фторурацилом, доксорубицином, бортезомибом, леналидомидом, дексаметазоном, мелфалином, преднизоном, винкристином и/или талидомидом.
В зависимости от конкретной злокачественной опухоли, на которую направлено лечение, комбинированную терапию по изобретению можно сочетать с дополнительными лекарственными средствами, такими как химиотерапевтические средства, или дополнительными способами лечения, такими как лучевая терапия или хирургическая операция. В случае комбинированной терапии по изобретению можно использовать различные известные химиотерапевтические средства. Предпочтительно, используют химиотерапевтические средства, которые являются стандартными для лечения конкретных показаний. Дозировка или частота введения каждого используемого лекарственного средства в комбинированной терапии, предпочтительно, является такой же или меньше дозировки или частоты введения соответствующего средства в случае использования без другого средства(средств).
В другом аспекте изобретение относится к любому антителу против FGFR3, описанному в настоящем документе, причем антитело против FGFR3 содержит детектируемую метку.
В другом аспекте изобретение относится к комплексу любого из антител против FGFR3, описанных в настоящем документе, и FGFR3. В некоторых вариантах осуществления комплекс представляет собой комплекс in vivo или in vitro. В некоторых вариантах осуществления комплекс содержит злокачественную клетку. В некоторых вариантах осуществления антитело против FGFR3 мечено детектируемой меткой.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
ФИГУРЫ 1A, 1B и 1C: Последовательности HVR-петель тяжелой цепи и легкой цепи антител против FGFR3. На фигурах представлены последовательности HVR тяжелой цепи, H1, H2 и H3, и последовательности HVR легкой цепи, L1, L2 и L3. Нумерация последовательностей является следующей:
Клон 184.6 (HVR-H1 представляет собой SEQ ID NO:1; HVR-H2 представляет собой SEQ ID NO:2; HVR-H3 представляет собой SEQ ID NO:3; HVR-L1 представляет собой SEQ ID NO:4; HVR-L2 представляет собой SEQ ID NO:5; HVR-L3 представляет собой SEQ ID NO:6);
Клон 184.6.1 (HVR-H1 представляет собой SEQ ID NO:7; HVR-H2 представляет собой SEQ ID NO:8; HVR-H3 представляет собой SEQ ID NO:9; HVR-L1 представляет собой SEQ ID NO:10; HVR-L2 представляет собой SEQ ID NO:11; HVR-L3 представляет собой SEQ ID NO:12);
Клон 184.6.58 (HVR-H1 представляет собой SEQ ID NO:13; HVR-H2 представляет собой SEQ ID NO:14; HVR-H3 представляет собой SEQ ID NO:15; HVR-L1 представляет собой SEQ ID NO:16; HVR-L2 представляет собой SEQ ID NO:17; HVR-L3 представляет собой SEQ ID NO:18);
Клон 184.6.62 (HVR-H1 представляет собой SEQ ID NO:48; HVR-H2 представляет собой SEQ ID NO:49; HVR-H3 представляет собой SEQ ID NO:50; HVR-L1 представляет собой SEQ ID NO:51; HVR-L2 представляет собой SEQ ID NO:52; HVR-L3 представляет собой SEQ ID NO:53);
Клон 184.6.21 (HVR-H1 представляет собой SEQ ID NO:54; HVR-H2 представляет собой SEQ ID NO:55; HVR-H3 представляет собой SEQ ID NO:56; HVR-L1 представляет собой SEQ ID NO:57; HVR-L2 представляет собой SEQ ID NO:58; HVR-L3 представляет собой SEQ ID NO:59);
Клон 184.6.49 (HVR-H1 представляет собой SEQ ID NO:60; HVR-H2 представляет собой SEQ ID NO:61; HVR-H3 представляет собой SEQ ID NO:62; HVR-L1 представляет собой SEQ ID NO:63; HVR-L2 представляет собой SEQ ID NO:64; HVR-L3 представляет собой SEQ ID NO:65);
Клон 184.6.51 (HVR-H1 представляет собой SEQ ID NO:66; HVR-H2 представляет собой SEQ ID NO:67; HVR-H3 представляет собой SEQ ID NO:68; HVR-L1 представляет собой SEQ ID NO:69; HVR-L2 представляет собой SEQ ID NO:70; HVR-L3 представляет собой SEQ ID NO:71);
Клон 184.6.52 (HVR-H1 представляет собой SEQ ID NO:72; HVR-H2 представляет собой SEQ ID NO:73; HVR-H3 представляет собой SEQ ID NO:74; HVR-L1 представляет собой SEQ ID NO:75; HVR-L2 представляет собой SEQ ID NO:76; HVR-L3 представляет собой SEQ ID NO:77);
Клон 184.6.92 (HVR-H1 представляет собой SEQ ID NO:78; HVR-H2 представляет собой SEQ ID NO:79; HVR-H3 представляет собой SEQ ID NO:80; HVR-L1 представляет собой SEQ ID NO:81; HVR-L2 представляет собой SEQ ID NO:82; HVR-L3 представляет собой SEQ ID NO:83);
Клон 184.6.1.N54S (HVR-H1 представляет собой SEQ ID NO:84; HVR-H2 представляет собой SEQ ID NO:85; HVR-H3 представляет собой SEQ ID NO:86; HVR-L1 представляет собой SEQ ID NO:87; HVR-L2 представляет собой SEQ ID NO:88; HVR-L3 представляет собой SEQ ID NO:89);
Клон 184.6.1.N54G (HVR-H1 представляет собой SEQ ID NO:90; HVR-H2 представляет собой SEQ ID NO:91; HVR-H3 представляет собой SEQ ID NO:92; HVR-L1 представляет собой SEQ ID NO:93; HVR-L2 представляет собой SEQ ID NO:94; HVR-L3 представляет собой SEQ ID NO:95);
Клон 184.6.1.N54A (HVR-H1 представляет собой SEQ ID NO:96; HVR-H2 представляет собой SEQ ID NO:97; HVR-H3 представляет собой SEQ ID NO:98; HVR-L1 представляет собой SEQ ID NO:99; HVR-L2 представляет собой SEQ ID NO:100; HVR-L3 представляет собой SEQ ID NO:101);
Клон 184.6.1.N54Q (HVR-H1 представляет собой SEQ ID NO:102; HVR-H2 представляет собой SEQ ID NO:103; HVR-H3 представляет собой SEQ ID NO:104; HVR-L1 представляет собой SEQ ID NO:105; HVR-L2 представляет собой SEQ ID NO:106; HVR-L3 представляет собой SEQ ID NO:107);
Клон 184.6.58.N54S (HVR-H1 представляет собой SEQ ID NO:108; HVR-H2 представляет собой SEQ ID NO:109; HVR-H3 представляет собой SEQ ID NO:110; HVR-L1 представляет собой SEQ ID NO:111; HVR-L2 представляет собой SEQ ID NO:112; HVR-L3 представляет собой SEQ ID NO:113);
Клон 184.6.58.N54G (HVR-H1 представляет собой SEQ ID NO:114; HVR-H2 представляет собой SEQ ID NO:115; HVR-H3 представляет собой SEQ ID NO:116; HVR-L1 представляет собой SEQ ID NO:117; HVR-L2 представляет собой SEQ ID NO:118; HVR-L3 представляет собой SEQ ID NO:119);
Клон 184.6.58.N54A (HVR-H1 представляет собой SEQ ID NO:120; HVR-H2 представляет собой SEQ ID NO:121; HVR-H3 представляет собой SEQ ID NO:122; HVR-L1 представляет собой SEQ ID NO:123; HVR-L2 представляет собой SEQ ID NO:124; HVR-L3 представляет собой SEQ ID NO:125);
Клон 184.6.58.N54Q (HVR-H1 представляет собой SEQ ID NO:126; HVR-H2 представляет собой SEQ ID NO:127; HVR-H3 представляет собой SEQ ID NO:128; HVR-L1 представляет собой SEQ ID NO:129; HVR-L2 представляет собой SEQ ID NO:130; HVR-L3 представляет собой SEQ ID NO:131);
Клон 184.6.1.NS D30E (HVR-H1 представляет собой SEQ ID NO:143; HVR-H2 представляет собой SEQ ID NO:144; HVR-H3 представляет собой SEQ ID NO:145; HVR-L1 представляет собой SEQ ID NO:140; HVR-L2 представляет собой SEQ ID NO:141; HVR-L3 представляет собой SEQ ID NO:142).
Аминокислотные положения пронумерованы согласно системе нумерации Кабат, как описано ниже.
ФИГУРА 2A и 2B: представлены (A) аминокислотные последовательности вариабельных областей тяжелой цепи и вариабельных областей легкой цепи антител против FGFR3: 184.6.1.N54S, 184.6.58 и 184.6.62; и (B) гипервариабельные области антител против FGFR3: 1G6, 6G1 и 15B2.
ФИГУРЫ 3A, 3B и 4: представлены характерные акцепторные консенсусные каркасные последовательности человека для использования на практике настоящего изобретения со следующими идентификаторами последовательностей:
Консенсусные каркасные области вариабельной области тяжелой цепи (VH) (ФИГ. 3A, 3B)
консенсусная каркасная область VH человека подгруппы I минус CDR по Кабат (SEQ ID NO:19 и 203-205);
консенсусная каркасная область VH человека подгруппы I минус удлиненные гипервариабельные области (SEQ ID NO:20 и 206-208, 21 и 209-211, 22 и 212-214);
консенсусная каркасная область VH человека подгруппы II минус CDR по Кабат (SEQ ID NO:23 и 215-217);
консенсусная каркасная область VH человека подгруппы II минус удлиненные гипервариабельные области (SEQ ID NO:24 и 218-220, 25 и 221-223, 26 и 224-226);
консенсусная каркасная область VH человека подгруппы II минус удлиненные
консенсусная каркасная область VH человека подгруппы III минус CDR по Кабат (SEQ ID NO:27 и 227-229);
консенсусная каркасная область VH человека подгруппы III минус удлиненные гипервариабельные области (SEQ ID NO:28 и 230-232, 29 и 233-235, 30 и 236-238);
акцепторная каркасная область VH человека минус CDR по Кабат (SEQ ID NO:31 и 239-241);
акцепторная каркасная область VH человека минус удлиненные гипервариабельные области (SEQ ID NO:32 и 242-244, 33 и 2245-247);
каркасная область акцептора 2 VH человека минус CDR по Кабат (SEQ ID NO:34 и 248-250);
каркасная область акцептора 2 VH человека минус удлиненные гипервариабельные области (SEQ ID NO:35 и 251-253, 36 и 254-256, 37 и 257-259);
Консенсусные каркасные области вариабельной области легкой цепи (VL) (ФИГ. 4)
консенсусная каркасная область VL-каппа человека подгруппы I (SEQ ID NO:38 и 260-262);
консенсусная каркасная область VL-каппа человека подгруппы II (SEQ ID NO:39 и 263-265);
консенсусная каркасная область VL-каппа человека подгруппы III (SEQ ID NO:40 и 266-268);
консенсусная каркасная область VL-каппа человека подгруппы IV (SEQ ID NO:41 и 269-271);
ФИГУРА 5: представлены последовательности каркасной области легкой (SEQ ID NO:42-45) и тяжелой цепей (SEQ ID NO:46, 47, 175, 176) huMAb4D5-8. Номера надстрочными знаками/полужирным шрифтом указывают на положения аминокислот согласно Кабат.
ФИГУРА 6: представлены модифицированные/вариантные последовательности каркасной области легкой (SEQ ID NO:42, 43, 177, 45) и тяжелой цепей (SEQ ID NO:46, 47, 178 и 176) huMAb4D5-8. Номера надстрочными знаками/полужирным шрифтом указывают на положения аминокислот согласно Кабат.
ФИГУРА 7: нокдаун FGFR3 в клетках рака мочевого пузыря RT112 ингибирует пролиферацию и индуцирует остановку клеточного цикла в фазе G1 in vitro, и подавляет рост опухоли in vivo. Три различных shРНК для FGFR3 клонировали в индуцируемый Tet вектор экспрессии. Клетки RT112, стабильно экспрессирующие shРНК для FGFR3 или контрольную shРНК, получали с помощью селекции пуромицином. (A) Характерные блоты, на которых показана экспрессия FGFR3 в выбранных клонах, обработанных доксицилином или не обработанных им (Dox, 0, 0,1 и 1 мкг/мл, слева направо). (B) Включение [3H]-тимидина в стабильные клетки RT112. Стабильные клоны RT112 культивировали с 1 мкг/мл доксицилина или без него в течение 3 суток, а затем инкубировали в течение 16 часов с [3H]-тимидином (1 мкКи на лунку). Количество импульсов для включенного [3H]-тимидина нормализовали с количеством импульсов для клеток без индукции доксицилином. Величина ошибки соответствует SEM. (C) Гистограммы проточной цитометрии с флуоресценцией ДНК для стабильных клеток RT112. Клоны RT112, экспрессировавшие контрольную shРНК или shРНК4 для FGFR3, культивировали с 1 мкг/мл доксицилина или без него в течение 72 часов, и ядра окрашивали йодидом пропидия (PI). Для shРНК2 и 6 FGFR3 были получены аналогичные результаты (фигура 16). (D) рост клеток RT112, экспрессировавших контрольную shРНК (n=9 на группу введения) или shРНК4 для FGFR3 (n=11 на группу введения) у мышей. Мышам вводили 5% сахарозу без добавления или с добавлением 1 мг/мл доксицилина, и размер опухоли измеряли два раза в неделю. Величина ошибки соответствует SEM. Для shРНК2 и 6 FGFR3 были получены аналогичные результаты (фигура 16). Нижняя панель: экспрессия белка FGFR3 в лизатах опухолей, экстрагированных из тканей ксенотрансплантатов стабильных клеток, обработанных контрольной shРНК или shРНК4 для FGFR3.
ФИГУРА 8: R3Mab блокирует взаимодействие FGF/FGFR3. (A) Селективное связывание FGFR3 человека с помощью R3Mab. Химерные белки Fc и FGFR1-4 человека иммобилизовывали и инкубировали увеличивающимся количеством R3Mab. Обнаружение специфического связывания проводили с помощью антитела против Fab человека. (B-C) Блокирование связывания FGF1 с FGFR3-IIIb (B) или IIIc (C) человека с помощью R3Mab. Обнаружение специфического связывания проводили с использованием биотинилированного специфичного к FGF1 поликлонального антитела. (D-E) Блокирование связывания FGF9 с FGFR3-IIIb (D) или IIIc (E) человека с помощью R3Mab. Обнаружение специфического связывания проводили с использованием биотинилированного специфичного к FGF9 поликлонального антитела. Величина ошибки соответствует стандартной ошибке среднего значения (SEM) и иногда меньше символов.
ФИГУРА 9: R3Mab ингибирует пролиферацию клеток Ba/F3, индуцируемую FGFR3 дикого типа и мутантным FGFR3. (A) Ингибиторный эффект R3Mab на жизнеспособность клеток Ba/F3, экспрессирующих FGFR3-IIIb человека дикого типа. Клетки культивировали в среде без FGF1 (нет FGF1), или в присутствии 10 нг/мл FGF1 вместе с 10 мкг/мл гепарина отдельно (FGF1), или в сочетании с контрольным антителом (контроль) или R3Mab. Жизнеспособность клеток оценивали с помощью CellTiter-Glo (Promega) после инкубации в течение 72 ч с антителами. (B) Ингибирование фосфорилирования FGFR3 и MAPK с помощью R3Mab в стабильных клетках Ba/F3-FGFR3-IIIbWT. Клетки обрабатывали смесью 15 нг/мл FGF1 и 10 мкг/мл гепарина (+) или только гепарином (-) в течение 10 минут после преинкубации с контрольным Ab (контроль), снижающимися количествами R3Mab (1, 0,2, 0,04 мкг/мл соответственно) в PBS, или PBS отдельно (имитация) в течение 3 часов. Лизаты подвергали иммуноблоттингу для оценки фосфорилирования FGFR3 и p44/42 MAPK с антителами к pFGFRY653/654 и pMAPKThr202/Tyr204, соответственно. (C) Схематичное представление горячих точек мутации FGFR3 и их частоты при раке мочевого пузыря (нумерация представленных последовательностей основана на аминокислотной последовательности изоформы FGFR3 IIIb), в соответствие с опубликованными данными (32). TM, трансмембранный домен; TK1 и TK2, тирозинкиназный домен 1 и 2. (D-H) Ингибиторный эффект R3Mab на жизнеспособность клеток Ba/F3, экспрессирующих связанные со злокачественной опухолью мутанты FGFR3. G372C происходит из изоформы IIIc, а остальные мутанты происходят из изоформы IIIb. Нумерация последовательностей всех мутантов основана на аминокислотной последовательности изоформы FGFR3 IIIb (включая мутант G372C, который был бы пронумерован как G370C, исходя из аминокислотной последовательности изоформы FGFR3 IIIc). Жизнеспособность клеток оценивали после инкубирования в течение 72 часов с антителами, как описано в (A). Величина ошибки соответствует SEM.
ФИГУРА 10: Картирование эпитопа для R3Mab и кристаллическая структура комплекса между Fab-фрагментом R3Mab и IgD2-D3 FGFR3-IIIb человека. (A) Эпитоп, определенный связыванием 13 пептидов, охватывающих IgD2-D3 FGFR3 человека, с R3Mab. Каждый биотинилированный пептид иммобилизововывали на покрытые стрептавидином лунки микропланшета для титрования и инкубировали с R3Mab. Обнаружение специфично связанного R3Mab проводили с помощью антитела козы против IgG человека. (B) Выравнивание последовательностей пептидов FGFR3 человека 3 (LAVPAANTVRFRCPA (SEQ ID NO:179) и 11 (SDVEFHCKVYSDAQP (SEQ ID NO:180) с внеклеточными сегментами FGFR1 человека (пептид 3: HAVPAAKTVKFKCPS (SEQ ID NO:181); пептид 11: SNVEFMCKVYSDPQP (SEQ ID NO:182)). Остатки FGFR1, вовлеченные в первичное взаимодействие FGF2-FGFR1, связывание гепарина и ассоциацию рецептор-рецептор, показаны полужирным шрифтом, курсивом и подчеркиванием, соответственно. Функциональный анализ остатков FGFR1 основан на Plotnikov et al. (34). (C) Структура Fab R3Mab (показанная с помощью ленточно-спиральной структуры, легкая цепь: серый, тяжелая цепь: черный) в комплексе с IgD2-D3 FGFR3 человека (показанном в виде молекулярной поверхности, белый). Остатки рецептора, участвующие в связывание лиганда и димеризацию, окрашены серым цветом/заштрихованы и темно-серым цветом соответственно, на основе Plotnikov et al. (34). (D) Приближение кристаллической структуры показывает, что CDR-H3 и -H2 из Fab являются основными участками взаимодействия с IgD2 и IgD3 в FGFR3. (E) Наложение комплекса FGFR3-IIIc-FGF1 (код PDB 1RY7) и комплекса FGFR3-IIIb-Fab. FGFR3-IIIc и FGF1 окрашены серым цветом и темно-серым цветом, соответственно. FGFR3-IIIb показан белым цветом и Fab показан светло-серым цветом для легкой цепи, темно-серым цветом для тяжелой цепи. IgD2 использовали как основу для наложения. Было отмечено, что IgD2 обеих структур имели высокую степень совпадения, а IgD3 из FGFR3-IIIb принимали новую конформацию при их связывании с R3Mab. (F) Другое представление наложения комплекса FGFR3-IIIc-FGF1 (код PDB 1RY7) и комплекса FGFR3-IIIb-Fab. FGFR3-IIIc и FGF1 представлены в качестве молекулярных поверхностей, которые имеют серый цвет/ячеистую текстуру и темно-серый цвет/точечную текстуру, соответственно. FGFR3-IIIb показан белым цветом и Fab показан серым цветом для легкой цепи, черным цветом для тяжелой цепи. IgD2 использовали в качестве основы для наложения. Следует отметить высоко совпадающие IgD2 из обеих структур и новую конформацию, принимаемую IgD3 из FGFR3-IIIb, когда они связаны R3Mab.
ФИГУРА 11: R3Mab ингибирует пролиферацию, клональный рост и передачу сигнала FGFR3 в клетках рака мочевого пузыря, экспрессирующих FGFR3S249C дикого типа или мутантный FGFR3S249C. (A) Ингибирование включения [3H]-тимидина с помощью R3Mab в клеточной линии рака мочевого пузыря RT112. Величина ошибки соответствует SEM. (B) Блокирование активированной FGF1 передачи сигнала FGFR3 с помощью R3Mab (15 мкг/мл) в клеточной линии рака мочевого пузыря RT112 по сравнению со средой для обработки отдельно (имитация) или контрольным антителом (контроль). Клеточные лизаты подвергали иммунопреципитации с антителом против FGFR3 и оценивали их фосфорилирование FGFR3 с антителом против фосфотирозина (4G10). Лизаты подвергали иммуноблотннигу для обнаружения фосфорилирования AKT (pAKTS473) и p44/42 MAPK (pMAPKThr202/Tyr204). (C) Ингибирование клонального роста с помощью R3Mab (10 мкг/мл) в клеточной линии рака мочевого пузыря UMUC-14 (содержащей FGFR3S249C) по сравнению со средой для обработки отдельно (имитация) или контрольным антителом (контроль). (D) Количественное определение в исследовании (C), указывающее на число колоний, диаметр которых превышал 120 мкм из 12 повторяющихся лунок. Величина ошибки соответствует SEM. P<3,4×10-9 против имитации или контроля. (E) Ингибирование фосфорилирования FGFR3 в клетках UMUC-14 с помощью R3Mab (15 мкг/мл). Фосфорилирование FGFR3 анализировали как в (B). Следует отметить конститутивное фосфорилирование FGFR3 в этой клеточной линии.
ФИГУРА 12: R3Mab снижает уровень в стационарном состоянии связанного дисульфидной связью димера FGFR3S249C путем смещения равновесия димер-мономер в сторону мономерного состояния. (A) Эффект R3Mab на димер FGFR3S249C в клетках UMUC-14. Клетки инкубировали с R3Mab (15 мкг/мл) или с контрольным антителом (контроль) в течение 3 часов и лизаты цельных клеток анализировали с помощью иммуноблоттинга в невосстанавливающих и восстанавливающих условиях. (B) Эффект блокатора свободных сульфгидрильных групп DTNB на равновесие димер-мономер FGFR3S249C в клетках UMUC-14. Клетки UMUC-14 обрабатывали увеличивающейся концентрацией DTNB в течение 3 часов, и клеточные лизаты анализировали, как в (A). (C) Эффект R3Mab на очищенный рекомбинантный димер FGFR3S249C in vitro. Димер FGFR3S249C, состоящий из IgD2-D3, очищали через колонку для гель-фильтрации и инкубировали с PBS (имитация), контрольным антителом (контроль) или R3Mab при 37°C. В указанное время брали образцы для иммуноблоттинга в невосстанавливающих условиях. Обнаружение димера-мономера FGFR3 проводили с помощью антитела против гибридомы FGFR3 6G1 (A-C).
ФИГУРА 13: R3Mab ингибирует рост ксенотрансплантата клеток рака мочевого пузыря и рост аллотрансплантата Ba/F3-FGFR3S249C. (A) Эффект R3Mab на рост предварительно полученных ксенотрансплантатов рака мочевого пузыря RT112 по сравнению с контролем в виде носителя, n=10 на группу. (B) Ингибирование передачи сигнала FGFR3 в опухолевых тканях RT112 с помощью R3Mab. В отдельном эксперименте ксенотрансплантаты опухолей RT112, которые обрабатывали 15 мг/кг контрольного антитела (контроль) или R3Mab в течение 48 часов или 72 часов собирали (n=3 на группу), гомогенизировали и анализировали иммуноблоттингом в отношении активации FRS2α и MAPK. (C) Эффект R3Mab на рост предварительно полученных аллотрансплантатов Ba/F3-FGFR3S249C, n=10 на группу. (D) Эффект R3Mab на рост предварительно полученных ксентрансплантатов рака мочевого пузыря UMUC-14, n=10 на группу. (E) Эффект R3Mab на димер FGFR3S249C и передачу сигнала в опухолевых тканях UMUC-14. Ксенотрансплантаты опухолей UMUC-14, которые обрабатывали 30 мг/кг контрольного антитела (контроль) или R3Mab в течение 24 часов или 72 часов собирали (n=3 на группу), гомогенизировали и анализировали иммуноблоттингом в отношении димера-мономера FGFR3S249C, а также активации MAPK. Обнаружение димера-мономера FGFR3 проводили с использованием поликлонального антитела кролика против FGFR3 sc9007, чтобы избежать взаимного влияния с IgG мыши в лизатах опухоли. Величина ошибки соответствует SEM.
ФИГУРА 14: ADCC приводит к противоопухолевой эффективности R3Mab в моделях положительной по t(4;14) множественной миеломы. (A-B) Эффект R3Mab на рост предварительно полученных ксенотрансплантатов миеломы OPM2 (A) и KMS11 (B), n=10 на группу. (C-F) Цитолиз клеточных линий миеломы OPM2 (C) и KMS11 (D), или клеточных линий рака мочевого пузыря RT112 (E) и UMUC-14 (F), индуцированный R3Mab в клеточной культуре. Клетки миеломы или рака мочевого пузыря инкубировали со свежевыделенными PBMC человека в присутствии R3Mab или контрольного антитела. Цитотоксичность определяли путем измерения высвобождения LDH в супернатант. (G-H) Эффект R3Mab или его мутанта DANA на рост предварительно полученных ксенотрансплантатов миеломы OPM2 (G) и KMS11 (H), n=10 на группу. Величина ошибки соответствует SEM и иногда являются меньшими, чем символы.
ФИГУРА 15: Нокдаун FGFR3 с помощью siРНК ингибирует пролиферацию клеток клеточной линии рака мочевого пузыря. Genentech было разработано и синтезировано от шести до семи различных siРНК против FGFR3 и три неспецифических контрольных siРНК. Клеточные линии рака мочевого пузыря RT112 (A), SW780 (B), RT4 (C) и UMUC-14 (D) высевали в 96-луночный планшет (3000 клеток на лунку) и позволяли им прикрепляться в течение ночи и подвергали временной трансфекции 25 нМ siРНК в комплексе с РНКiMax (Invitrogen). Через 72 ч после трансфекции в культуру добавляли [3H]-тимидин (1 мкКи на лунку) (A, C и D) для инкубации в течение дополнительных 16 часов. Количественное определение включенного [3H]-тимидина проводили с помощью TopCount. Данные нормализовывали с данными клеток, трансфицированных РНКiMax отдельно (Mock). Величина ошибки соответствует SEM. Нижняя панель: характерные блоты, демонстрирующие экспрессию FGFR3 в клетках, трансфицированных siРНК. (B) Жизнеспособность клеток измеряли с помощью CellTiter-Glo (Promega) через 96 часов после трансфекции. Величина ошибки соответствует SEM.
ФИГУРА 16: Нокдаун FGFR3 в клеточной линии рака мочевого пузыря RT112 индуцирует остановку клеточного цикла в фазе G1 in vitro и подавляет рост опухоли in vivo. Было разработано три различных РНК против FGFR3 и клонировано в индуцируемый Tet экспрессирующий shРНК ретровирусный вектор. Стабильные клоны RT112, экспрессирующие shРНК FGFR3 или контрольную shРНК, получали селекцией пуромицином. (A) Гистограммы проточной цитометрии с флуоресценцией ДНК для окрашенных йодидом пропидия (PI) ядер, полученных для стабильных клеток RT112, экспрессировавших shРНК2 или shРНК6 для FGFR3, после обработки 1 мкг/мл доксицилина и без него в течение 72 часов. (B) Рост стабильных клеток RT112, экспрессирующих shРНК2-4 против FGFR3 (n=11 на группу введения) или shРНК6-16 FGFR3 (n=10 на группу введения) у мышей nu/nu. Мышам, обладавшим опухолями, вводили только 5% сахарозу (закрашенная окружность) или 5% сахарозу с 1 мг/мл доксицилина (закрашенный квадрат), и опухоли измеряли с помощью штангерциркуля два раза в неделю. Величина ошибки соответствует SEM.
ФИГУРА 17: Эффект гибридомных антител против FGFR3 16G, 6G1 и 15B2 на пролиферацию клеток Ba/F3, запускаемую FGFR3 дикого типа и мутантным FGFR3. Гибридомные антитела против FGFR3 получали иммунизацией мышей BALB/c химерой FGFR3-IIIb человека/Fc или FGFR3-IIIc человека/Fc. Селекцию слитых гибридомных клеток проводили с использованием селекции на гипоксантине-аминоптерине-тимидине в среде D из набора для селекции гибридом ClonaCell® (StemCell Technologies, Inc., Vancouver, BC, Canada). Затем гибридомные антитела скринировали на способность связываться с FGFR3-IIIb и FGFR3-IIIc с помощью ELISA и распознавать FGFR3 клеточной поверхности с помощью FACS. Затем отобранные гибридомы клонировали способом лимитирующих разведений. 16G, 6G1 и 15B2 представляют собой клоны, использованные для оценки эффекта в отношении пролиферации клеток Ba/F3, экспрессировавших FGFR3 дикого типа или мутантный FGFR3 аналогично тому, как описано на фигуре 9A. Величина ошибки соответствует SEM.
ФИГУРА 18: Сравнение эпитопов R3Mab, определенных пептидным картированием и анализом кристаллической структуры. (A) Эпитоп, выявленный по структуре Fab-фрагмента R3Mab в комплексе с внеклеточным сегментом IgD2-D3 из FGFR3 человека. Остатки FGFR3, контактирующие с тяжелой цепью и легкой цепью Fab, окрашены черным и серым цветом, соответственно. (B) Расположение пептидов 3 и 11 на FGFR3.
ФИГУРА 19: R3Mab ингибирует пролиферацию и передачу сигнала FGFR3 в клетках рака мочевого пузыря, содержащих FGFR3S249C дикого типа и мутантный FGFR3S249C. (A) Ингибирование жизнеспособности клеток с помощью R3Mab в клеточной линии рака мочевого пузыря RT4. Жизнеспособность клеток оценивали с помощью CellTiter-Glo (Promega) через 96 ч после инкубации с антителом. Величина ошибки соответствует SEM. (B) Блокирование активируемой FGF1 передачи сигнала FGFR3 с помощью R3Mab (15 мкг/мл) в клеточной линии рака мочевого пузыря RT4. (C) Ингибирование включения [3H]-тимидина с помощью R3Mab в клеточной линии рака мочевого пузыря RCC-97-7 (содержавшей FGFR3S249C). Величина ошибки соответствует SEM. (D) Ингибирование фосфорилирования FGFR3 в клетках TCC-97-7 с помощью R3Mab (15 мкг/мл). (E) Снижение димера FGFR3S249C в клетках TCC-97-7 после инкубации в течение 3 часов с R3Mab (15 мкг/мл) по сравнению с контрольным антителом (контроль).
ФИГУРА 20: Эффект ингибиторов эндоцитоза на интернализацию R3Mab и димера FGFR3S249C в клетках UMUC-14. (A) Эффект ингибиторов эндоцитоза на интернализацию R3Mab. Клетки UMUC-14, предварительно обработанные различными ингибиторами эндоцитоза или DMSO в течение 1 часа при 37°C, инкубировали с R3Mab (15 мкг/мл) в течение 3 часов при 37°C для осуществления интернализации. Промывку при низком значении pH использовали для удаления R3Mab с клеточной поверхности для визуализации интернализованного антитела. Клетки фиксировали и окрашивали с помощью меченного Alexa 488 IgG против антител человека. Изображение получали с использованием конфокальной микроскопии. (B) Эффект ингибиторов эндоцитоза на димер FGFR3S249C в клетках UMUC-14, обработанных R3Mab. Клетки UMUC-14, предварительно обработанные различными ингибиторами эндоцитоза или DMSO в течение 1 часа при 37°C, инкубировали с ложным антителом (дорожка 1), контрольным антителом (дорожка 2) или R3Mab (15 мкг/мл, дорожка 3) в течение 3 часов при 37°C. Клеточные лизаты анализировали на белок FGFR3 в невосстанавливающих или восстанавливающих условиях с помощью иммуноблоттинга. Следует отметить, что хлорпромазин (ингибитор опосредуемого клатрином эндоцитоза) и генистеин (общий ингибитор эндоцитоза) блокировали интернализацию R3Mab, но не оказывали эффекта на индуцируемое R3Mab снижение димера FGFR3S249C.
ФИГУРА 21: Чувствительность обнаружения для различных антител против FGFR3 в отношении мономерного и димерного FGFR3S249C в невосстанавливающих условиях. Клетки UMUC-14 подвергали лизису после обработки R3Mab (дорожка 1), контрольным IgG1 (дорожка 2) или PBS (дорожка 3) в течение 3 часов, и клеточные лизаты подвергали анализу иммуноблоттингом в восстанавливающих или невосстанавливающих условиях. Следует отметить, что с помощью 6G1 (гибридомное антитело мыши, полученное в Genentech) можно обнаружить как димер, так и мономер, FGFR3S249C, в то время как с помощью sc9007 (поликлональное антитело кролика, Santa Cruz Biotechnology) или sc13121 (гибридомное антитело мыши, Santa Cruz Biotechnology), предпочтительно, можно обнаружить димерный FGFR3S249C.
ФИГУРА 22: Эффект R3Mab на пролиферацию t(4;14)+ клеток множественной миеломы. (A) Ингибиторный эффект R3Mab на включение [3H]-тимидина клетками UTMC-2. Клетки UTMC-2 выращивали в среде, содержавшей R3Mab или контрольное антитело, в присутствии 25 нг/мл FGF9 и 5 мкг/мл гепарина или гепарина отдельно (без FGF9). Через 6 суток после инкубирования добавляли [3H]-тимидин для инкубирования в течение 16 ч. Данные нормализовывали с данными для клеток, выращенных в отсутствие FGF9 и антитела. (B-C) Эффект R3Mab на включение [3H]-тимидина клетками OPM2 (B) и KMS11 (C). Клетки, которые выращивали в среде с 1,5% FBS, обрабатывали R3Mab или контрольным антителом в течение 6 суток. Данные нормализовывали с данными, полученными для необработанных клеток. Величина ошибки соответствует SEM.
ФИГУРА 23: Уровни экспрессии на клеточной поверхности FGFR3 в клетках миеломы и рака мочевого пузыря. (A) Экспрессия на клеточной поверхности FGFR3 в клетках миеломы и клетках рака мочевого пузыря, оцененная FACS-анализом. Клетки окрашивали конъюгированным с фикоэритрином mAb мыши против FGFR3 человека (FAB766P, R&D Systems) или конъюгированным с фикоэритрином изотипическим контрольным IgG1 мыши (BD Pharmingen). (B) Анализ Скэтчарда плотности FGFR3 в клетках миеломы и клетках рака мочевого пузыря. R3Mab подвергали радиоактивному йодированию и инкубировали с клетками в суспензии с избытком немеченого антитела. После инкубации при комнатной температуре в течение 2 часов клетки осаждали центрифугированием и промывали два раза. Определяли специфично связанный 125I. Плотность рецепторов и аффинность связывания (Kd) соответствуют среднему значению для двух экспериментов по связыванию.
ФИГУРА 24: Эффект R3Mab или его мутанта DANA на рост ксенотрансплантата клеток карциномы мочевого пузыря. (A) Эффект R3Mab и его мутанта DANA (по 50 мг/кг каждого) на рост предварительно полученных опухолей RT112. (B) Эффект R3Mab и его мутанта DANA (по 50 мг/кг каждого) на рост предварительно полученных опухолей UMUC-14. Величина ошибки соответствует SEM.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение, раскрытое в настоящем документе, относится к антителам против FGFR3, которые могут использоваться, например, для лечения или профилактики заболеванй, связанных с экспрессией и/или активностью FGFR3, такой как повышенная экспрессия и/или активность или нежелательная экспрессия и/или активность. В некоторых вариантах осуществления антитела по изобретению могут быть использованы для лечения опухоли, рака и/или клеточно-пролиферативного нарушения.
В другом аспекте антитела против FGFR3 по изобретению могут быть использованы в качестве реагентов для обнаружения и/или выделения FGFR3, как например, обнаружение FGFR3 в различных тканях и типах клеток.
Кроме того, изобретение относится к способам получения и применения антител против FGFR3, и полинуклеотидов, кодирующих антитела против FGFR3.
Основные способы
Способы и процедуры, описанные или цитируемые в настоящем документе, в основном, хорошо известны и широко применяются с помощью общепринятых методик специалистами в данной области, например, таких как широко используемые методики, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al. eds., (2003)); серии METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE (R. I. Freshney, ed. (1987)).
Определения
"Выделенное" антитело представляет собой антитело, которое идентифицировано и отделено и/или извлечено из компонента его природного окружения. Загрязняющие компоненты его природного окружения представляют собой вещества, которые препятствуют диагностическому или терапевтическому применению антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело является очищенным (1) до более чем 95% по массе антитела, как определяют способом Лоури, и, наиболее предпочтительно, до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности при SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) в восстанавливающих или невосстанавливающих условиях с использованием кумасси синего или, предпочтительно, окрашивания серебром. Выделенное антитело включает антитело в рекомбинантных клетках in situ, поскольку в этом случае отсутствует по меньшей мере один компонент природного окружения антитела. Однако, как правило, выделенное антитело получают с помощью по меньшей мере одной стадии очистки.
"Выделенная" молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена по меньшей мере от одной загрязняющей молекулы нуклеиновой кислоты, с которой она обычно связана в природном источнике нуклеиновой кислоты. Форма или условия для выделенной молекулы нуклеиновой кислоты отличается от природной формы или условий. Таким образом, выделенная молекула нуклеиновой кислоты отличается от природной молекулы нуклеиновой кислоты клеток. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно экспрессируют нуклеиновую кислоту (например, нуклеиновую кислоту, кодирующую антитело), где, например, молекула нуклеиновой кислоты находится в области на хромосоме, отличающейся от области хромосомы, в которой она находится в клетках в природе.
Термины "нумерация остатков вариабельного домена как в Кабат" или "нумерация положения аминокислоты как в Кабат" и их варианты, относятся к системе нумерации, используемой для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи совокупности антител, представленных в Кабат et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991). С использованием этой системы нумерации истинная линейная аминокислотная последовательность может содержать меньшее количество аминокислот или она может содержать дополнительные аминокислоты, соответствующие укорочению или вставке в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать вставку из одной аминокислоты (остаток 52a согласно Кабат) после остатка 52 в H2 и встроенные остатки (например, остатки 82a, 82b и 82c, и тому подобное, согласно Кабат) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Кабат можно определять для данного антитела путем выравнивания областей гомологии последовательности антитела со "стандартной" пронумерованной по Кабат последовательностью.
Выражение "по существу аналогичный" или "по существу такой же", как используется в настоящем документе, обозначает достаточно высокую степень сходства между двумя числовыми величинами (как правило, одна из них связана с антителом по изобретению, а другая связана с эталонным/сравниваемым антителом), так чтобы специалист в данной области считал отличие между этими двумя величинами небольшими или не обладающими биологической и/или статистической значимостью в контексте биологической характеристики, измеренной с помощью указанных величин (например, величин Kd). Отличие между указанными двумя величинами, предпочтительно, составляет менее чем приблизительно 50%, предпочтительно, менее чем приблизительно 40%, предпочтительно, менее чем приблизительно 30%, предпочтительно, менее чем приблизительно 20%, предпочтительно, менее чем приблизительно 10% как функция величины для эталонного/сравниваемого антитела.
Термин "аффинность связывания" обычно относится к силе всех суммарных нековалентных взаимодействий между отдельным участком связывания молекулы (например, антитела) и его связывающим партнером (например, антигеном). Если нет иных указаний, как используется в настоящем документе, "аффинность связывания" относится к присущей аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y обычно можно представить константой диссоциации (Kd). Желательно, Kd составляет 1×10-7, 1×10-8, 5×10-8, 1×10-9, 3×10-9, 5×10-9 или даже 1×10-10 или прочнее. Аффинность можно измерять обычными способами, известными в данной области, включая способы, описанные в настоящем документе. Низкоаффинные антитела обычно связываются с антигеном медленно, и легко диссоциируют, тогда как высокоаффинные антитела обычно связываются с антигеном быстрее и дольше остаются в связанном состоянии. Специалистам известны различные способы измерения аффинности связывания, причем любой из них можно использовать для целей настоящего изобретения. Конкретные характерные варианты осуществления описаны далее.
В одном из вариантов осуществления "Kd" или "величину Kd" в соответствии с настоящим изобретением измеряют с помощью анализа связывания меченного радиоактивной меткой антигена (RIA), проводимого с Fab-версией представляющего интерес антитела и его антигеном, как описано в следующем анализе. Аффинность связывания Fab с антигеном в растворе измеряют путем уравновешивания Fab минимальной концентрацией (125I)-меченного антигена в присутствии серии титров немеченного антигена, с последующей фиксацией связанного антигена на планшете, покрытом антителом против Fab (Chen, et al., (1999) J. Mol. Biol 293:865-881). Для определения условий для анализа, на микропланшеты для титрования (Dynex) наносят в течение ночи 5 мкг/мл фиксирующего антитела против Fab (Cappel Labs) в 50 мМ растворе карбоната натрия (рH 9,6), а затем блокируют с использованием 2% (масс./об.) бычьего сывороточного альбумина в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В не адсорбирующем планшете (Nunc #269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями представляющего интерес Fab (например, в соответствии с оценкой антитела против VEGF, Fab-12, в Presta et al., (1997) Cancer Res. 57:4593-4599). Затем представляющий интерес Fab инкубируют в течение ночи; однако инкубацию можно продолжать в течение более длительного периода (например, 65 часов), чтобы быть уверенным в том, что равновесие достигнуто. После этого смеси переносят на фиксирующий планшет для инкубирования при комнатной температуре (например, в течение 1 часа). Затем раствор удаляют, и планшет промывают восемь раз 0,1% Tween-20 в PBS. После того, как планшеты высыхают, добавляют 150 мкл/лунка сцинтиллятора (MicroScint-20; Packard), и проводят подсчет в планшетах с использованием счетчика гамма-импульсов Topcount (Packard) в течение десяти минут. Концентрации каждого из Fab, которые обеспечивают связывание, меньшее или равное 20% от максимального связывания, выбирают для использования в конкурентных анализах связывания. В соответствии с другим вариантом Kd или величину Kd измеряют с использованием анализа способом поверхностного плазмонного резонанса. В таких анализах можно использовать BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами CM5 с иммобилизованным на них антигеном при ~10 единицах ответа (RU). Коротко, биосенсорные чипы из карбоксиметилированного декстрана (CM5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/минута для достижения приблизительно 10 единиц ответа (RU) связанного белка. После инъекции антигена путем инъекции вводят 1 M этаноламин для блокирования не прореагировавших групп. Для определения показателей кинетики, с помощью инъекции вводят двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS с 0,05% Tween 20 (PBST) при 25°C со скоростью потока приблизительно 25 мкл/мин. В некоторых вариантах осуществления используют следующие модификации способа анализа поверхностным плазмонным резонансом: антитело иммобилизуют на биосенсорных чипах CM5 для достижения приблизительно 400 RU, и для определения кинетики двукратные серийные разведения белка-мишени (например, FGFR3-IIIb или -IIIc) (начиная с 67 нМ) инъецируют в буфере PBST при 25°С со скоростью потока 30 мкл/минута. Константы скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания Ленгмюра "один к одному" (BIAcore Evaluation Software version 3.2) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как соотношение koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. Если константа ассоциации антитела превышает 106 M-1 с-1 в анализе способом поверхностного плазмонного резонанса, указанном выше, тогда константу ассоциации можно определять с использованием способа тушения флуоресценции, в котором измеряют повышение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) 20 нМ антитела (форма Fab) в PBS, pH 7,2, при 25°С, в присутствии возрастающих концентраций антигена при измерении в спектрофотометре, таком как спектрофотометр с останавливаемой струей (Aviv Instruments) или спектрофотометр SLM-Aminco серии 8000 (Thermo Spectronic) с перемешиваемой кюветой.
"Скорость связывания" или "скорость ассоциации" или "скорость соединения" или "kon" по изобретению также можно определять с помощью указанного способа поверхностного плазмонного резонанса, описанного выше, с использованием biacoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С с чипами CM5 с иммобилизованным антигеном при ~10 единицах ответа (RU). Коротко, биосенсорные чипы из карбоксиметилированного декстрана (CM5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/минута для достижения приблизительно 10 единиц ответа (RU) связанного белка. После инъекции антигена путем инъекции вводят 1 M этаноламин для блокирования не вступивших в реакцию групп. Для определения показателей кинетики, путем инъекции вводят двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS с 0,05% Tween 20 (PBST) при 25°C со скоростью потока приблизительно 25 мкл/мин. В некоторых вариантах осуществления используют следующие модификации способа анализа поверхностным плазмонным резонансом: антитело иммобилизуют на биосенсорных чипах CM5 для достижения приблизительно 400 RU, и для определения кинетики двукратные серийные разведения белка-мишени (например, FGFR3-IIIb или -IIIc) (начиная с 67 нМ) инъецируют в буфере PBST при 25°С со скоростью потока 30 мкл/минута. Константы скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания Ленгмюра "один к одному" (BIAcore Evaluation Software version 3.2) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как соотношение koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. Если константа ассоциации антитела превышает 106 M-1 с-1 в анализе способом поверхностного плазмонного резонанса, указанном выше, тогда константу ассоциации можно определять с использованием способа тушения флуоресценции, в котором измеряют повышение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) 20 нМ антитела против антигена (форма Fab) в PBS, pH 7,2, при 25°С, в присутствии возрастающих концентраций антигена при измерении в спектрофотометре, таком как спектрофотометр с останавливаемой струей (Aviv Instruments) или спектрофотометр SLM-Aminco серии 8000 (Thermo Spectronic) с перемешиваемой кюветой.
Термин "вектор", как используется в настоящем документе, относится к молекуле нуклеиновой кислоты, способной осуществлять транспорт другой нуклеиновой кислоты, с которой она связана. Одним типом вектора является "плазмида", которая относится к кольцевой двухцепочечной петле ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут встраиваться в геном клетки-хозяина при введении в клетку-хозяина, и, тем самым, они реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы называют в настоящем документе "рекомбинантными векторами экспрессии" или просто "векторами экспрессии". Как правило, векторы экспрессии, используемые в способах рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании термины "плазмида" и "вектор" могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора.
Термины "полинуклеотид" или "нуклеиновая кислота", используемые в настоящем документе взаимозаменяемо, относятся к полимерам нуклеотидов любой длины, и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой, или с помощью синтетической реакции. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. До и после сборки полимера может происходить модификация нуклеотидной структуры, если она происходит. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Кроме того, полинуклеотид может быть модифицирован после полимеризации, например, конъюгацией с осуществляющим мечение компонентом. Другие типы модификаций включают, например, "кэпы", замену одного или нескольких природных нуклеотидов аналогом, межнуклеотидные модификации, например, такие как модификации незаряженными связями (например, метилфосфонатами, фосфотриэфирами, фосфоамидатами, карбаматами и тому подобное) и заряженными связями (например, фосфоротиоатами, фосфородитиоатами, и тому подобное), модификации выступающими группами, например, такими как белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и тому подобное), модификации интеркаляторами (например, акридином, псораленом, и тому подобное), модификации хелаторами (например, металлами, радиоактивными металлами, бором, окислительными металлами и тому подобное), модификации алкилирующими соединениями, модификации модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и тому подобное), а также немодифицированные формы полинуклеотида(ов). Кроме того, любые гидроксильные группы, обычно присутствующие в сахарах, могут быть заменены, например, фосфонатными группами, фосфатными группами, они могут быть защищены стандартными защитными группами или активированы для образования дополнительных связей с дополнительными нуклеотидами, или они могут быть конъюгированы с твердыми или полутвердыми подложками. 5'- и 3'-концевой OH может быть фосфорилирован или замещен аминами или органическими кэпирующими группами из от 1 до 20 атомов углерода. Другие гидроксилы также могут быть преобразованы в стандартные защитные группы. Полинуклеотиды также могут содержать аналогичные формы сахаров рибозы или дезоксирибозы, которые, главным образом, известны в данной области, включая, например, 2'-O-метил-, 2'-O-аллил-, 2'-фтор- или 2'-азидорибозу, карбоциклические аналоги сахаров, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, сахара пиранозы, сахара фуранoзы, седогептулозы, ациклические аналоги и основные нуклеозидные аналоги, такие как метилрибозид. Одна или несколько фосфодиэфирных связей могут быть заменены альтернативными линкерными группами. Эти альтернативные линкерные группы включают, но ими не ограничиваются, варианты осуществления где фосфат заменен на P(O)S ("тиоат"), P(S)S ("дитиоат"), (O)NR2 ("амидат"), P(O)R, P(O)OR', CO или CH2 ("формацеталь"), где каждый R или R' независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно содержащий простую эфирную связь (-O-), арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не обязательно, чтобы все связи в полинуклеотиде были идентичными. Представленное выше описание применимо ко всем полинуклеотидам, указанным в настоящем документе, включая РНК и ДНК.
"Олигонуклеотид", как используется в настоящем документе, относится, главным образом, к коротким, как правило, одноцепочечным, как правило, синтетическим полинуклеотидам длиной, как правило, но не обязательно, менее 200 нуклеотидов. Термины "олигонуклеотид" и "полинуклеотид" не являются взаимоисключающими. Представленное выше описание полинуклеотидов в равной степени и полностью может использоваться для олигонуклеотидов.
"Процентную (%) идентичность аминокислотных последовательностей" в отношении пептидной или полипептидной последовательности, определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в последовательности специфической пептидной или полипептидной последовательности, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей, и не считая какие-либо консервативные замены частью идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей можно проводить различными способами, которые находятся в пределах знаний специалиста, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, требуемые для достижения максимального выравнивания на протяжении всей длины сравниваемых последовательностей. Однако для целей, представленных в настоящем документе, значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2, где полный исходный текст для программы ALIGN-2 приведен в Таблице 2, ниже. Компьютерная программа для сравнения последовательностей ALIGN-2 была составлена в Genentech, Inc. и исходный текст программы был представлен с пользовательской документацией в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован под U.S. Copyright Registration No. TXU510087. Программа ALIGN-2 является общедоступной через Genentech, Inc., South San Francisco, California или ее можно составить из исходного текста, представленного в WO 2007/001851. Программа ALIGN-2 должна быть составлена для использования на операционной системе UNIX, предпочтительно, цифровой UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не изменяются.
В случаях, когда ALIGN-2 используют для сравнений аминокислотных последовательностей, % идентичность аминокислотных последовательностей для этой аминокислотной последовательности A к, с или против данной аминокислотной последовательности B (что альтернативно может быть сформулировано как данная аминокислотная последовательность A, которая обладает определенной % идентичностью аминокислотной последовательности, содержит % идентичность аминокислотной последовательности, к, с или против данной аминокислотной последовательности B), или вычисляют следующим образом:
100 умножить на частное X/Y
где X представляет собой число аминокислотных остатков, оцененных как идентичные совпадения программой для выравнивания последовательностей ALIGN-2 при выравнивании этой программой A и B, и где Y представляет собой общее количество аминокислотных остатков в B. Понятно, что если длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичность аминокислотных последовательностей A к B не равна % идентичности аминокислотных последовательностей B к A.
В некоторых вариантах осуществления две или более аминокислотных последовательности, идентичные по меньшей мере на 50%, 60%, 70%, 80% или 90%. В некоторых вариантах осуществления две или более аминокислотных последовательности являются по меньшей мере на 95%, 97%, 98%, 99% или даже на 100% идентичными. Если конкретно не указано иное, все величины % идентичности аминокислотных последовательностей, используемые в настоящем документе, получают, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
Термин "FGFR3", как используется в настоящем документе, относится, если конкретно или контекстуально не указано иное, к любому нативному или вариантному (как нативному, так синтетическому) полипептиду FGFR3 (например, изоформе FGFR3-IIIb или изоформе FGFR3-IIIc). Термин "нативная последовательность" конкретно включает природные укороченные формы (например, последовательность внеклеточного домена или последовательность трансмембренной субъединицы), природные формы вариантов (например, альтернативно сплайсированные формы) и природные аллельные варианты. Термин "FGFR3 дикого типа" относится, главным образом, к полипептиду, содержащему аминокислотную последовательность природного белка FGFR3. Термин "последовательность FGFR3 дикого типа", главным образом, относится к аминокислотной последовательности, встречающейся в природном FGFR3.
Термин "лиганд FGFR3" (взаимозаменяемо называемый "FGF"), как используется в настоящем документе, относится, если конкретно не указано и не следует из контекста иного, к любому нативному или вариантному (как нативному, так и синтетическому) полипептиду лиганда FGFR3 (например, FGF1, FGF2, FGF4, FGF8, FGF9, FGF17, FGF18, FGF23). Термин "нативная последовательность" конкретно включает природные укороченные формы (например, последовательность внеклеточного домена или последовательность трансмембранной субъединицы), природные формы вариантов (например, альтернативно сплайсированные формы) и природные аллельные варианты. Термин "лиганд FGFR3 дикого типа" относится, главным образом, к полипептиду, содержащему аминокислотную последовательность природного белка лиганда FGFR3. Термин "последовательность лиганда FGFR3 дикого типа" относится, главным образом, к аминокислотной последовательности, встречающейся в природном лиганде FGFR3.
"Активация FGFR3" относится к активации или фосфорилированию рецептора FGFR3. Как правило, активация FGFR3 приводит к передаче сигнала (например, вызываемой внутриклеточным киназным доменом рецептора FGFR3, фосфорилирующего остатки тирозина в FGFR3 или субстратном полипептиде). Активация FGFR3 может опосредоваться связыванием лиганда FGFR с представляющим интерес рецептором FGFR3. Связывание лиганда FGFR3 (например, такого как FGF1 или FGF9) с FGFR3 может активировать киназный домен FGFR3 и, тем самым, приводить к фосфорилированию остатков тирозина в FGFR3 и/или фосфорилированию остатков тирозина в дополнительном субстратном полипептиде(ах).
Термин "конститутивный", как используется в настоящем документе, например, используемый в отношении активности рецепторной киназы, относится к постоянной активности рецептора в отношении передачи сигнала, которая не зависит от присутствия лиганда или других активирующих молекул. В зависимости от природы рецептора, вся активность может быть конститутивной или активность рецептора может, кроме того, увеличиваться путем связывания с другими молекулами (например, лигандами). Клеточные события, которые приводят к активации рецепторов, хорошо известны специалистам в данной области. Например, активация может включать олигомеризацию, например, димеризацию, тримеризацию и тому подобное, в рецепторные комплексы более высокого порядка. Комплексы могут содержать одну разновидность белка, т.е., они могут представлять собой гомомерный комплекс. Альтернативно, комплексы могут содержать по меньшей мере две различных разновидности белков, т.е., они могут представлять собой гетеромерный комплекс. Образование комплекса может быть вызвано, например, сверхэкспрессией нормальной и мутантных форм рецептора на поверхности клетки. Образование комплекса также может быть вызвано определенной мутацией или мутациями в рецепторе.
Термин "лиганд-независимый", как используется в настоящем документе, используемый, например, в отношении активности передачи сигнала рецептором, относится к активности передачи сигнала, которая не зависит от присутствия лиганда. Рецептор, обладающий лиганд-независимой киназной активностью, необязательно исключает связывание лиганда с этим рецептором для обеспечения дополнительной активации киназной активности.
Термин "лиганд-зависимый", как используется в настоящем документе, используемый, например, в отношении активности передачи сигнала рецептором, относится к активности передачи сигнала, которая зависит от присутствия лиганда.
Выражение "амплификация гена" относится к процессу, с помощью которого в конкретной клетке или клеточной линии образуется множество копий гена или фрагмента гена. Область дупликации (участок амплифицированной ДНК) часто называют "ампликоном". Обычно количество образующейся матричной РНК (мРНК), т.е., уровень экспрессии гена, также возрастает пропорционально количеству копий конкретного экспрессируемого гена.
"Тирозинкиназный ингибитор" представляет собой молекулу, которая ингибирует в некоторой степени тирозинкиназную активность тирозинкиназы, такой как рецептор FGFR3.
Образец злокачественной опухоли или биологический образец, который "обладает экспрессией, амплификацией или активацией FGFR3", представляет собой образец, который в диагностическом тесте экспрессирует (в том числе сверхэкспрессирует) FGFR3, имеет амплифицированный ген FGFR3 и/или демонстрирует активацию или фосфорилирование FGFR3 иным образом.
Образец злокачественной опухоли или биологический образец, который "проявляет активацию FGFR3" представляет собой образец, который в диагностическом тесте демонстрирует активацию или фосфорилирование FGFR3. Такую активацию можно определять напрямую (например, путем определения фосфорилирования FGFR3 с помощью ELISA) или косвенно.
Образец злокачественной опухоли или биологический образец, который "проявляет конститутивную активацию FGFR3", представляет собой образец, который в диагностическом тесте демонстрирует конститутивную активацию или фосфорилирование FGFR3. Такую активацию можно определять напрямую (например, путем определения фосфорилирования C-FGFR3 с помощью ELISA) или косвенно.
Образец злокачественной опухоли или биологический образец, который "проявляет амплификацию FGFR3", представляет собой образец, который в диагностическом тесте имеет амплифицированный ген FGFR3.
Образец злокачественной опухоли или биологический образец, который "проявляет транслокацию FGFR3", представляет собой образец, который в диагностическом тесте имеет транслоцированный ген FGFR3. Примером транслокации FGFR3 является транслокация t(4;14), которая встречается в некоторых опухолях множественной миеломы.
"Анализ фосфо-ELISA" в настоящем документе представляет собой анализ, в котором оценивают фосфорилирование одного или нескольких FGFR3, субстрата или нижерасположенных молекул в каскаде передачи сигнала, в твердофазном иммуноферментном анализе (ELISA) с использованием реагента, обычно антитела, для обнаружения фосфорилированных FGFR3, субстрата или нижерасположенной молекулы в каскаде передачи сигнала. В некоторых вариантах осуществления используют антитело, которое осуществляет обнаружение фосфорилированных FGFR3 или pMAPK. Анализ можно проводить на клеточных лизатах, предпочтительно, из свежих или замороженных биологических образцов.
Образец злокачественной опухоли или биологический образец, который "проявляет лиганд-независимую активацию FGFR3", представляет собой образец, который в диагностическом тесте демонстрирует лиганд-независимую активацию или фосфорилирование FGFR3. Такую активацию можно определять напрямую (например, путем определения фосфорилирования FGFR3 с помощью ELISA) или косвенно.
Образец злокачественной опухоли или биологический образец, который "проявляет лиганд-зависимую активацию FGFR3", представляет собой образец, который в диагностическом тесте демонстрирует лиганд-зависимую активацию или фосфорилирование FGFR3. Такую активацию можно определять напрямую (например, путем определения фосфорилирования FGFR3 с помощью ELISA) или косвенно.
Образец злокачественной опухоли или биологический образец, который "проявляет лиганд-независимую активацию FGFR3", представляет собой образец, который в диагностическом тесте демонстрирует лиганд-независимую активацию или фосфорилирование FGFR3. Такую активацию можно определять напрямую (например, путем определения фосфорилирования FGFR3 с помощью ELISA) или косвенно.
Злокачественная клетка со "сверхэкспрессией или амплификацией FGFR3" представляет собой клетку, которая имеет значительно более высокие уровни белка или гена FGFR3 по сравнению с незлокачественной клеткой того же типа ткани. Такая сверхэкспрессия может быть вызвана амплификацией гена или увеличенной транскрипцией или трансляцией. Сверхэкспрессию или амплификацию FGFR3 можно определять в диагностическом или прогностическом анализе путем оценки повышенных уровней белка FGFR3, присутствующего на поверхности клетки (например, в иммуногистохимическом анализе; IHC). Альтернативно или дополнительно, можно измерять уровни кодирующей FGFR3 нуклеиновой кислоты в клетке, например, с помощью флуоресцентной гибридизации in situ (FISH; см. WO98/45479, опубликованную в октябре 1998 года), саузерн-блоттинга или технологий полимеразной цепной реакции (ПЦР), таких как количественная ПЦР в реальном времени (qRT-PCR). Помимо указанных выше анализов квалифицированному специалисту доступны различные анализы in vivo. Например, можно подвергать клетки в организме пациента действию антитела, которое необязательно является меченым меткой, способной к обнаружению, например, радиоактивным изотопом, и можно оценивать связывание антитела с клетками у пациента, например, путем внешнего сканирования в отношении радиоактивности или с помощью анализа биоптата, полученного от пациента, которого ранее подвергали воздействию антитела.
Термин "мутация", как используется в настоящем документе, указывает на отличие аминокислотной последовательности или последовательности нуклеиновых кислот конкретного белка или нуклеиновой кислоты (гена, РНК) от белка или нуклеиновой кислоты дикого типа, соответственно. Мутантный белок или нуклеиновая кислота могут экспрессироваться или встречаться на одном аллеле (гетерозиготный) или обоих аллелях (гомозиготный) гена, и они могут быть соматическими или эмбриональными. В настоящем изобретении мутации, главным образом, являются соматическими. Мутации включают реорганизацию последовательности, такую как вставки, делеции и точковые мутации (включая однонуклеотидные/аминокислотные полиморфизмы).
"Ингибировать" означает уменьшать или снижать активность, функцию и/или количество по сравнению с эталоном.
Средство обладает "активностью или функцией агониста", когда средство имитирует по меньшей мере один из функциональных видов активности представляющего интерес полипептида (например, лиганда FGFR, такого как FGF1 или FGF9).
"Антитело-агонист", как используется в настоящем документе, представляет собой антитело, которое имитирует по меньшей мере один из функциональных видов активности представляющего интерес полипептида (например, лиганда FGFR, такого как FGF1 или FGF9).
"Экспрессия" белка относится к преобразованию информации, кодируемой геном, в матричную РНК (мРНК), а затем в белок.
В настоящем документе образец или клетка, которые "экспрессируют" представляющий интерес белок (такой как рецептор FGF или лиганд рецептора FGF), представляет собой образец или клетку, для которых определяют, что в них присутствуют мРНК, кодирующая белок, или белок, включая их фрагменты.
"Иммуноконъюгат" (используемый взаимозаменяемо с "конъюгатом антитело-лекарственное средство" или "ADC") означает антитело, конъюгированное с одним или несколькими цитотоксическими средствами, такими как химиотерапевтическое средство, лекарственное средство, ингибирующее рост средство, токсин (например, белковый токсин, ферментативно активный токсин происхождением из бактерий, грибов, растений или животных, или их фрагменты), или радиоактивный изотоп (т.е., радиоконъюгат).
Термин "Fc-область", как используется в настоящем документе, главным образом, относится к димерному комплексу, содержащему C-концевые полипептидные последовательности тяжелой цепи иммуноглобулина, где С-концевая полипептидная последовательность представляет собой последовательность, получаемую расщеплением папаином интактного антитела. Fc-область может содержать нативные или вариантные последовательности Fc. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут изменяться, Fc-область тяжелой цепи IgG человека обычно определяют как отрезок от аминокислотного остатка в положении Cys226, или от Pro230, до C-конца последовательности Fc. Последовательность Fc иммуноглобулина, как правило, содержит два константных домена, CH2-домен и CH3-домен, и необязательно содержит CH4-домен. C-концевой лизин (остаток 447 в системе нумерации EU) Fc-области может быть удален, например, в процессе очистки антитела, или при рекомбинантном конструировании нуклеиновой кислоты, кодирующей антитело. Соответственно, композиция, содержащая антитело, имеющее Fc-область по этому изобретению, может содержать антитело с K447, антитело, где все K447 удалены, или смесь антител с остатком K447 или без него.
Под "полипептидом Fc" в настоящем документе понимают один из полипептидов, которые образуют Fc-область. Полипептид Fc можно получать из любого подходящего иммуноглобулина, такого как подтипы IgG1, IgG2, IgG3 или IgG4, IgA, IgE, IgD или IgM. В некоторых вариантах осуществления полипептид Fc содержит часть последовательности шарнирной области дикого типа (как правило, на ее N-конце) или все эту последовательность. В некоторых вариантах осуществления полипептид Fc не содержит функциональную последовательность шарнирной области или последовательность шарнирной области дикого типа.
"Блокирующее" антитело или антитело-"антагонист" представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, которую оно связывает. Предпочтительные блокирующие антитела или антитела-антагонисты полностью ингибируют биологическую активность антигена.
"«Голое» антитело" представляет собой антитело, которое не конъюгировано с гетерологичной молекулой, такой как цитотоксическая группа или радиоактивная метка.
Антитело, обладающее "биологической характеристикой" указанного антитела, представляет собой антитело, которое обладает одной или несколькими из биологических характеристик этого антитела, которые отличают его от других антител, которые связываются с тем же антигеном.
Для скрининга антител, которые связывают эпитоп на антигене, связанном представляющим интерес антителом можно проводить общепринятый анализ перекрестного блокирования, такой как анализ, описанный в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow и David Lane (1988).
Для увеличения времени полужизни антител или полипептидов, содержащих аминокислотные последовательности по настоящему изобретению, можно присоединять к антителу (особенно фрагменту антитела) связывающий рецептор спасения эпитоп, как описано например, в патенте США 5739277. Например, молекула нуклеиновой кислоты, кодирующая связывающий рецептор спасения эпитоп, может быть связана в рамке считывания с нуклеиновой кислотой, кодирующей полипептидную последовательность по этому изобретению, так чтобы слитый белок, экспрессируемый со сконструированной молекулы нуклеиновой кислоты содержал связывающий рецептор спасения эпитоп и полипептидную последовательность по этому изобретению. Как используется в настоящем документе, термин "связывающий рецептор спасения эпитоп" относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за увеличение времени полужизни молекулы IgG в сыворотке in vivo (например, Ghetie et al., Ann. Rev. Immunol. 18:739-766 (2000), таблица 1). Антитела с заменами в Fc-области и увеличенным временем полужизни также описаны в WO00/42072, WO 02/060919; Shields et al., J. Biol. Chem. 276:6591-6604 (2001); Hinton, J. Biol. Chem. 279:6213-6216 (2004)). В другом варианте осуществления время полужизни в сыворотке также увеличивают, например, путем присоединения других полипептидных последовательностей. Например, антитела или другие полипептиды, подходящие в способах по изобретению, можно присоединять к сывороточному альбумину или части сывороточного альбумина, которая связывается с рецептором FcRn или связывающим сывороточный альбумин пептидом, так чтобы сывороточный альбумин связывался с антителом или полипептидом, например, такие полипептидные последовательности описаны в WO 01/45746. В одном из предпочтительных вариантов осуществления присоединяемый пептид сывороточного альбумина содержит аминокислотную последовательность DICLPRWGCLW (SEQ ID NO:183). В другом варианте осуществления время полужизни Fab возрастает при использовании этих способов. Также см. Dennis et al. J. Biol. Chem. 277:35035-35043 (2002) для связывающих сывороточный альбумин пептидных последовательностей.
Под "фрагментом" подразумевают часть полипептида или молекулы нуклеиновой кислоты, которая содержит, предпочтительно, по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или более от всей длины эталонной молекулы нуклеиновой кислоты или полипептида. Фрагмент может содержать 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100, 200, 300, 400, 500, 600 или более нуклеотидов или 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 120, 140, 160, 180, 190, 200 аминокислот или более.
Выражение "от небольшой функции агониста до ее отсутствия" в отношении антитела по изобретению, как используется в настоящем документе, означает, что антитело не вызывает биологически значимого уровня активности агониста, например, при введении индивидууму. Как было бы понятно в данной области, уровень активности можно определять количественно или качественно, при условии, что сравнение между антителом по изобретению и эталонным аналогом может быть проведено. Активность можно измерять или обнаруживать любым анализом или способом, известным в данной области, включая, например, анализы и способы, описанные в настоящем документе. Уровень активности для антитела по изобретению и его эталонного аналога можно определять в параллельных или отдельных анализах. В некоторых вариантах осуществления двухвалентное антитело по изобретению не обладает существенной функцией агониста.
Термины "апоптоз" и "апоптотическая активность" используют в широком значении и они относятся к упорядоченной или контролируемой форме гибели клеток у млекопитающих, которая, как правило, сопровождается одним или несколькими характерными изменениями клеток, включая конденсацию цитоплазмы, потерю микроворсинок плазматической мембраны, сегментацию ядра, деградацию хромосомной ДНК или утрату функции митохондрий. Эту активность можно определять и измерять с использованием способов, известных в данной области, например, анализом жизнеспособности клеток, FACS-анализом или электрофорезом ДНК, и, более конкретно, по связыванию аннексина V, фрагментации ДНК, сморщиванию клеток, расширению эндоплазматической сети, фрагментации клеток и/или образованию мембранных везикул (называемых апоптотическими тельцами).
Термины "антитело" и "иммуноглобулин" используют взаимозаменяемо в наиболее широком значении, и они включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, одновалентные антитела, поливалентные антитела, полиспецифические антитела (например, биспецифические антитела, при условии, что они обладают требуемой биологической активностью), а также могут включать определенные фрагменты антител (как более подробно описано в настоящем документе). Антитело может быть антителом человека, гуманизированным и/или полученным созреванием аффинности.
Под термином "вариабельный" понимают последовательности определенных сегментов вариабельных доменов, которые значительно отличаются у разных антител, и которые используются в связывании и определяют специфичность каждого конкретного антитела в отношении его конкретного антигена. Однако вариабельность последовательности встречается на протяжении вариабельных доменов антител неравномерно. Она сконцентрирована в трех сегментах, называемых определяющими комплементарность областями (CDR) или гипервариабельными областями в вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высоко консервативные участки вариабельных доменов называют каркасными областями (FR). Вариабельные домены каждой нативной тяжелой и легкой цепи содержат четыре FR-области, в основном принимающие конфигурацию β-слоя, связанные тремя CDR, которые формируют петли, соединяющие структуры β-слоя в некоторых случаях образующих их часть. CDR в каждой цепи удерживаются рядом FR-областями и, вместе с CDR из другой цепи, обеспечивают образование антиген-связывающего центра антител (см. Каbаt et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены не участвуют непосредственно в связывание антитела с антигеном, однако они проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности.
Расщепление антител папаином приводит к образованию двух идентичных антиген-связывающих фрагментов, называемых "Fab"-фрагментами, каждый с одним антиген-связывающим участком и остаточного "Fc"-фрагмента, название которого отражает его способность легко кристаллизоваться. Обработка антитела пепсином приводит к одному крупному F(ab')2-фрагменту, который имеет два антиген-связывающих участка, и, тем не менее, способен к перекрестному связыванию антигена.
"Fv" представляет собой минимальный фрагмент антитела, который содержит полный антиген-связывающий центр. В двухцепочечном типе Fv область состоит из димера из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочной нековалентной связью. В молекулах одноцепочечных Fv (scFv), один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи могут быть ковалентно связаны гибким пептидным линкером, так чтобы легкая и тяжелая цепи могли связываться в "димерный" структурный аналог молекул двухцепочечных Fv. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют, определяя антиген-связывающий участок на поверхности димера VH-VL. В совокупности, шесть CDR Fv обеспечивают антиген-связывающую специфичность антитела. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем целый участок связывания.
Fab-фрагмент содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конце CH1-домена тяжелой цепи, включая один или несколько цистеинов из шарнирной области антитела. Fab'-SH в настоящем документе представляет собой обозначение для Fab', в котором остаток(ки) цистеина константных доменов несет свободную тиольную группу. F(ab')2-фрагменты антитела исходно были получены в качестве пар Fab'-фрагментов, между которыми имеются остатки цистеина шарнирной области. Также известны другие химические соединения фрагментов антител.
"Легкие цепи" антител (иммуноглобулинов) из любых видов позвоночных могут быть отнесены к одному из двух довольно отличающихся типов, называемых каппа (k) и лямбда (λ), исходя из аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей, антитела (иммуноглобулины) могут быть отнесены к различным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть далее подразделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называют α, δ, ε, γ и µ, соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. "Фрагменты антитела" содержат только часть интактного антитела, где часть сохраняет по меньшей мере одну, несколько и большинство или все функции, обычно связанные с этой частью, если она присутствует в интактном антителе. Примеры фрагментов антител включают Fab-, Fab'-, F(ab')2- и Fv-фрагменты; диатела, линейные антитела; одноцепочечные молекулы антител и полиспецифические антитела, образованные фрагментами антител. В одном из вариантов осуществления фрагмент антитела содержит антиген-связывающую область интактного антитела и, таким образом, сохраняет способность связывать антиген. В другом варианте осуществления фрагмент антитела, например, фрагмент, который содержит Fc-область, сохраняет по меньшей мере одну из биологических функций, обычно связанных с Fc-областью, когда она присутствует в интактном антителе, такую как связывание FcRn, модулирование времени полужизни антитела, функция ADCC и связывание комплемента. В одном из вариантов осуществления, фрагмент антитела представляет собой одновалентное антитело, которое обладает временем полужизни in vivo, по существу аналогичным с интактным антителом. Например, такой фрагмент антитела может содержать антиген-связывающее плечо, связанное с последовательностью Fc, способной придавать фрагменту стабильность in vivo.
Термины "гипервариабельная область", "HVR" или "HV", как используются в настоящем документе, относятся к участкам вариабельного домена антитела, которые являются гипервариабельными по последовательности и/или образуют структурно определенные петли. Обычно антитела включают шесть гипервариабельных областей: три в VH (H1, H2, H3), и три в VL (L1, L2, L3). Ряд делинеаризированных гипервариабельных областей могут использоваться и входят в состав настоящей заявки. Определяющие комплементарность области (CDR) по Кабат основаны на вариабельности последовательностей и используются наиболее часто (Каbаt et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Chothia вместо этого рассматривает расположение структурных петель (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). Гипервариабельные области AbM представляют собой компромисс между CDR по Кабат и структурными петлями по Chothia, и они используются в программном обеспечении моделирования антител Oxford Molecular's AbM. "Контактные" гипервариабельные области основаны на анализе доступных комплексных кристаллических структур. Остатки из каждой из указанных гипервариабельных областей указаны ниже.
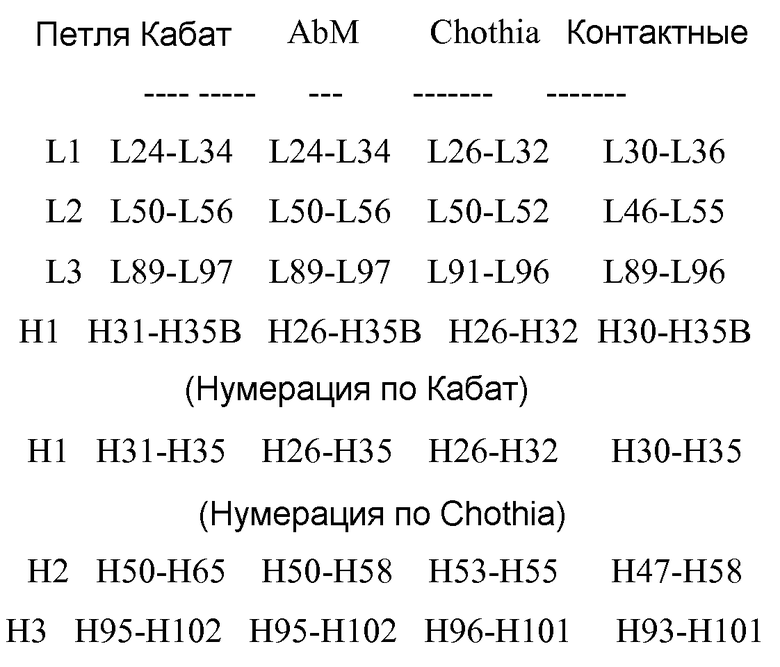
Гипервариабельные области могут включать следующие "удлиненные гипервариабельные области": 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (H2) и 93-102, 94-102 или 95-102 (H3) в VH. Остатки вариабельных доменов пронумерованы согласно Кабат et al., выше, для каждого из этих определений.
Остатки "каркасной области" или "FR" представляют собой остатки вариабельного домена, отличные от остатков HVR, как определено в настоящем документе.
"Гуманизированные" формы не являющихся человеческими антител (например, грызунов) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. В основном, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области видов, не относящихся к человеку, (донорное антитело), таких как мышь, крыса, кролик или не относящиеся к человеку приматы, которые обладают требуемой специфичностью, аффинностью и емкостью. В некоторых случаях, остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками, не являющиеся человеческими. Более того, гуманизированные антитела могут содержать остатки, которые отсутствуют в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения параметров антитела. Как правило, гуманизированное антитело содержит по существу все по меньшей мере из одного, и, как правило, из двух, вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют гипервариабельным петлям иммуноглобулина, не являющегося человеческим, и все или по существу все FR-области представляют собой FR-области из последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно содержит по меньшей мере участок константного домена (Fc) иммуноглобулина, как правило, константного домена иммуноглобулина человека. Для более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). Также см. следующие обзорные статьи и ссылки, цитированные в них: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
"Химерные" антитела (иммуноглобулины) имеют участок тяжелой и/или легкой цепи, идентичный или гомологичный соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) идентична или гомологична соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (патент США No 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). Гуманизированные антитела, как используется в настоящем документе, представляют собой подгруппу химерных антител.
"Одноцепочечные Fv" или "scFv" представляют собой фрагменты антител, которые содержат VH и VL-домены антитела, соединенные в единую полипептидную цепь. Как правило, полипептид sFv также необязательно содержит полипептидный линкер между VH и VL-доменами, который обеспечивает возможность формирования в sFv структуры, требуемой для связывания антигена. Для обзора sFv, см. Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994).
"Антиген" представляет собой заданный антиген, с которым антитело может селективно связываться. Антиген-мишень может представлять собой полипептид, углевод, нуклеиновую кислоту, липид, гаптен или другое природное или синтетическое соединение. Предпочтительно, антиген-мишень представляет собой полипептид.
Термин "диатела" относится к небольшим фрагментам антитела с двумя антиген-связывающими участками, причем эти фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL). С использованием линкера, который является слишком коротким для возможности спаривания двух доменов одной цепи, домены вынуждают образовывать пары с комплементарными доменами другой цепи с образованием двух антиген-связывающих центров. Диатела описаны более подробно, например, в EP 404097; WO93/1161; и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993).
"Антитело человека" представляет собой антитело, которое содержит аминокислотную последовательность, соответствующую последовательности антитела, продуцируемой у человека и/или полученной с использованием любых способов получения антител человека, как описано в настоящем документе. Это определение антитела человека, в частности, не включает гуманизированное антитело, содержащее антиген-связывающие остатки не человека.
Антитело, "прошедшее стадию созревания аффинности" представляет собой антитело с одним или несколькими изменениями в одной один или нескольких его CDR, которые приводят к увеличению аффинности антитела к антигену, по сравнению с исходным антителом, которое не имеет этого изменения(ий). Предпочтительные антитела, прошедшие стадию "созревания аффинности", имеют наномолярную или даже пикомолярную аффинность к антигену-мишени. Антитела, прошедшие стадию созревания аффинности, получают способами, известными в данной области. В Marks et al., Bio. Technology 10:779-783 (1992) описано созревание аффинности способом "перетасовки" VH- и VL-доменов. Случайный мутагенез CDR и/или каркасных остатков описан: Barbas et al., Proc Nat. Acad. Sci. USA 91:3809-3813 (1994); Schier et al., Gene 169:147-155 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995); и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
"Эффекторные функции" антитела относятся к таким видам биологической активности, которые являются свойственными Fc-области (Fc-области с нативной последовательностью или варианта Fc-участка по аминокислотной последовательности) антитела, и варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточно-опосредуемую цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности и активацию B-клеток.
"Антителозависимая клеточно-опосредуемая цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретируемые Ig, связанные с Fc-рецепторами (FcR), находящимися на определенных цитотоксических клетках (например, природных киллерных (NK) клетках, нейтрофилах и макрофагах), вызывают специфичное для этих цитотоксических эффекторных клеток связывание с несущей антиген клеткой-мишенью, а затем уничтожают клетки-мишени с помощью цитотоксинов. “Плечи” антител цитотоксических клеток являются абсолютно необходимыми для такого уничтожения. Основные клетки для осуществления ADCC, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках представлена в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991). Для оценки активности представляющей интерес молекулы в отношении ADCC можно проводить анализ ADCC in vitro, такой как анализ, описанный в патенте США 5500362 или 5821337 или в патенте США 6737056, выданном Presta. Подходящие для таких анализов эффекторные клетки включают периферические мононуклеарные клетки крови (PBMC), природные киллерные (NK) клетки. Альтернативно или дополнительно, активность представляющей интерес молекулы в отношении ADCC можно оценивать in vivo, например, в модели на животных, такой как описана в Clynes et al. PNAS (USA) 95:652-656 (1998).
"Эффекторные клетки человека" представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. Предпочтительно, клетки экспрессируют меньшей мере FcγRIII и выполняют эффекторную функцию ADCC. Примеры лейкоцитов человека, которые осуществляют ADCC, включают периферические мононуклеарные клетки крови (PBMC), природные киллерные (NK) клетки, моноциты, цитотоксические T-клетки и нейтрофилы; при этом предпочтительными являются PBMC и NK-клетки. Эффекторные клетки можно выделять из их природных источников, например, из крови.
Термины "Fc-рецептор" или "FcR" используют для описания рецептора, который связывается с Fc-областью антитела. Предпочтительно, FcR представляет собой нативную последовательность FcR человека. Более того, предпочтительный FcR представляет собой FcR, который связывает IgG-антитело (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно-сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые обладают аналогичными аминокислотными последовательностями, отличающимися, главным образом, своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA в своем цитоплазматическом домене содержит иммунорецепторный тирозин-связывающий активирующий мотив (ITAM). Ингибирующий рецептор FcγRIIB в своем цитоплазматическом домене содержит иммунорецепторный тирозин-связывающий ингибирующий мотив (ITIM), (см. обзор M. в Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). FcR описаны в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). К термину "FcR" в настоящем описании относятся другие FcR, том числе FcR, которые будут выявлены в будущем. Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регулирует гомеостаз иммуноглобулинов. В WO 00/42072 (Presta) описаны варианты антител с увеличенным или сниженным связыванием с FcR. Содержание каждой патентной публикации включено в настоящий документ в качестве ссылки в полном объеме. Также см. Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001).
Способы измерения связывания с FcRn известны (см., например, Ghetie 1997, Hinton 2004). Связывание с FcRn человека in vivo и время полужизни в сыворотке высокоаффинных связывающих полипептидов FcRn человека можно анализировать, например, у трансгенных мышей или в трансфицированных клеточных линиях человека, экспрессирующих FcRn человека, или у приматов, которым вводили варианты полипептидов Fc.
"Комплемент-зависимая цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического каскада комплемента начинается со связывания первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), связанными с распознаваемым им антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
Варианты полипептидов с измененными аминокислотными последовательностями Fc-областью и повышенной или сниженной способностью связывать C1q описаны в патенте США 6194551B1 и WO99/51642. Содержание этих патентных публикаций включено в настоящий документ в качестве ссылки в полном объеме. См. также, Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Термин "содержащий Fc-область полипептид" относится к полипептиду, такому как антитело или иммуноадгезин, который включает Fc-область. C-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области может быть удален, например, во время очистки полипептида или путем рекомбинантного конструирования нуклеиновой кислоты, кодирующей полипептид. Таким образом, композиция, содержащая полипептид, имеющий Fc-область, в соответствии с настоящим изобретением, может включать полипептиды с K447, полипептиды, из которых полностью удален K447, или смесь полипептидов, содержащих и не содержащих остаток K447.
"Акцепторная каркасная область человека" для целей настоящего описания представляет собой каркасную область, содержащую аминокислотную последовательность каркасной области VL или VH, происходящую из каркасной области иммуноглобулина человека или консенсусной последовательности каркасной области человека. Акцепторная каркасная область человека, "происходящая из" каркасной области иммуноглобулина человека или консенсусной последовательности каркасной области человека, может содержать такую же как и в них аминокислотную последовательность, или она может содержать предсуществующие изменения в аминокислотных последовательностях. Если предсуществующие изменения присутствуют, то предпочтительно, присутствует не более 5 и, предпочтительно, 4 или менее, 3 или менее предсуществующих изменений аминокислот. Если предсуществующие аминокислотные изменения присутствуют в VH, то предпочтительно, чтобы указанные изменения существовали только в трех, двух или одном из положений 71H, 73H и 78H; например, аминокислотные остатки в указанных положениях могут представлять собой 71A, 73T и/или 78A. В одном из вариантов осуществления акцепторная каркасная область VL человека идентична по последовательности каркасной области VL иммуноглобулина человека или консенсусной последовательности каркасной области человека.
"Консенсусная каркасная область человека" представляет собой каркасную область, которая представляет собой наиболее часто встречающиеся аминокислотные остатки при выборе каркасных последовательностей VL или VH иммуноглобулина человека. Обычно выбор последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельных доменов. Обычно подгруппа последовательностей представляет собой подгруппу согласно Кабат et al., выше. В одном из вариантов осуществления для VL подгруппа представляет собой подгруппу каппа I по Кабат et al., выше. В одном из вариантов осуществления для VH подгруппа представляет собой подгруппу III по Кабат et al., выше.
"Консенсусная каркасная область VH подгруппы III" включает консенсусную последовательность, образованную аминокислотными последовательностями вариабельных областей тяжелой цепи подгруппы III по Кабат et al., выше. В одном из вариантов осуществления аминокислотная последовательность консенсусной каркасной области VH подгруппы III включает по меньшей мере часть или все из каждой из следующих последовательностей: EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:184)-H1-WVRQAPGKGLEWV (SEQ ID NO:185)-H2-RFTISRDNSKNTLYLQMNSLRAEDTAVYYC (SEQ ID NO:186)-H3-WGQGTLVTVSS (SEQ ID NO:187).
"Консенсусная каркасная область VL подгруппы I" включает консенсусную последовательность, образованную аминокислотными последовательностями вариабельных областей легкой цепи каппа подгруппы I по Кабат et al., выше. В одном из вариантов осуществления аминокислотная последовательность консенсусной каркасной области VH подгруппы I включает по меньшей мере часть или все из каждой из следующих последовательностей: DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:188)-L1-WYQQKPGKAPKLLIY (SEQ ID NO:189)-L2-GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:190)-L3-FGQGTKVEIK (SEQ ID NO:191).
Как используется в настоящем документе, "мутант антитела" или "вариант антитела" относится к варианту аминокислотной последовательности антитела, где один или несколько из аминокислотных остатков зависимого от вида антитела модифицированы. Такие мутанты обязательно имеют менее чем 100% идентичность или сходство последовательности с зависимым от вида антителом. В одном из вариантов осуществления мутант антитела имеет аминокислотную последовательность, которая по меньшей мере на 75% идентична или аналогична по аминокислотной последовательности аминокислотной последовательности вариабельного домена либо тяжелой цепи, либо легкой цепи, зависимого от вида антитела, более предпочтительно, по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, более предпочтительно, по меньшей мере на 90%, и, наиболее предпочтительно, по меньшей мере на 95%. Идентичность или сходство в отношении этой последовательности определяют в настоящем документе как процент аминокислотных остатков в последовательности-кандидате, которые являются идентичными (т.е. тот же остаток) или аналогичными (т.е. аминокислотный остаток из той же группы на основе общих свойств боковой цепи, см. ниже) с остатками зависимого от вида антитела, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Ни одно из N-концевых, C-концевых или внутренних удлинений, делеций или вставок в последовательности антитела вне вариабельного домена не следует считать влияющими на идентичность или сходство последовательностей.
"Нарушение" или "заболевание" представляет собой любое состояние, при котором может быть эффективно лечение веществом/молекулой или способом по изобретению. Они включают хронические и острые нарушения или заболевания, включая патологические состояния, которые предрасполагают млекопитающего к рассматриваемому нарушению. Неограничивающие примеры нарушений, подлежащих лечению согласно настоящему описанию, включают злокачественные и доброкачественные опухоли; карциному, бластому или саркому.
"Лечение" относится как к терапевтическому лечению, так и к профилактическим или предупреждающим мерам. Нуждающиеся в лечении включают тех, кто уже имеет доброкачественную, предзлокачественную или неметастазирующую опухоль, а также тех, для предупреждения возникновение или рецидива злокачественной опухоли.
Термин "терапевтически эффективное количество" относится к количеству терапевтического средства для лечения или профилактики заболевания или нарушения у млекопитающего. В случае злокачественной опухоли, терапевтически эффективное количество лекарственного средства может снижать количество злокачественных клеток; уменьшать размер опухоли; ингибировать (т.е., замедлять до некоторой степени и, предпочтительно, останавливать) инфильтрацию злокачественных клеток в периферические органы; ингибировать (т.е., замедлять до некоторой степени и, предпочтительно, останавливать) метастазирование опухоли; ингибировать, до некоторой степени, рост опухоли; и/или смягчать до некоторой степени один или несколько симптомов, связанных с нарушением. В зависимости от того, может ли лекарственное средство препятствовать росту и/или уничтожать существующие злокачественные клетки, оно может быть цитостатическим и/или цитотоксическим. Для лечения злокачественной опухоли эффективность in vivo можно измерять, например, путем оценки длительности выживания, времени до прогрессирования заболевания (TTP), показателей ответа (RR), длительности ответа и/или качества жизни.
Термины "злокачественная опухоль", "рак" и "злокачественный" относятся к физиологическому состоянию у млекопитающих или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым ростом клеток. Это определение включает доброкачественные и злокачественные опухоли. Под "злокачественной опухолью на ранней стадии" или "опухолью на ранней стадии" понимают злокачественную опухоль, которая не является инвазивной или метастазирующей или классифицируется как злокачественная опухоль стадии 0, I или II. Примеры злокачественной опухоли включают, но ими не ограничиваются, карциному, лимфому, бластому (включая медуллобластому и ретинобластому), саркому (включая липосаркому и синовиальноклеточную саркому), нейроэндокринные опухоли (включая карциноидные опухоли, гастриному и злокачественную опухоль островковой ткани), мезотелиому, шванному (включая невриному слухового нерва), менингиому, аденокарциному, меланому и лейкоз или лимфоидные злокачественные опухоли. Более конкретные примеры таких злокачественных опухолей включают плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, включая мелкоклеточный рак легкого (SCLC), немелкоклеточный рак легкого (NSCLC), аденокарциному легкого и плокскоклеточную карциному легкого, злокачественную опухоль брюшины, печеночноклеточный рак, гастральный рак или рак желудка, включая желудочно-кишечный рак, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы (включая метастатический рак молочной железы), рак толстого кишечника, рак прямой кишки, рак ободочной и прямой кишки, карциному эндометрия или матки, карциному слюнной железы, рак почки или ренальный рак, рак предстательной железы, рак женских наружных половых органов, рак щитовидной железы, карциному печени, анальную карциному, карциному полового члена, рак яичка, рак пищевода, опухоли желчных путей, а также рак головы и шеи и множественную миелому.
Термин "предзлокачественный" относится к состоянию или росту, которые, как правило, предшествуют злокачественной опухоли или развиваются в злокачественную опухоль. "Предзлокачественный" рост имеет клетки, которые характеризуются нарушенной регуляцией клеточного цикла, пролиферацией или дифференцировкой, которые можно определять с помощью маркеров регуляции клеточного цикла, пролиферации клеток или дифференцировки.
Под "дисплазией" понимают любой аномальный рост или развитие ткани, органа или клеток. Предпочтительно, дисплазия представляет собой дисплазию высокой степени или предзлокачественную дисплазию.
Под "метастазом" понимают распространение злокачественной опухоли от ее первичной области в другие области организма. Злокачественные клетки могут отрываться от первичной опухоли, проникать в лимфатические и кровеносные сосуды, циркулировать через кровоток и расти в отделенном очаге (метастазировать) в нормальных тканях в другой области организма. Метастаз может быть местным или отдаленным. Метастазирование представляет собой последовательный процесс, зависящий от отделения опухолевых клеток от первичной опухоли, миграции через кровоток и остановки в отдаленной области. В новой области клетки обеспечивают подпитку кровью и могут расти с образованием угрожающей жизни массы.
Это поведение могут регулировать как стимулирующие, так и ингибиторные молекулярные каскады в опухолевой клетке, а также имеют значения взаимодействия между опухолевой клеткой и клетками хозяина в отдаленной области.
Под "неметастазирующей" понимают злокачественную опухоль, которая является доброкачественной или которая остается в первичной области и не проникает в лимфатическую или кровеносную систему или в ткани, отличные от первичной области. Как правило, неметастазирующая злокачественная опухоль представляет собой любую злокачественную опухоль, которая представляет собой злокачественную опухоль стадии 0, I или II и иногда злокачественную опухоль стадии III.
Под "первичной опухолью" или "первичной злокачественной опухолью" понимают исходную злокачественную опухоль, а не метастатический очаг, расположенный в другой ткани, органе или области организма индивидуума.
Под "доброкачественной опухолью" или "доброкачественным раком" понимают опухоль, которая остается локализованной в области ее происхождения и не обладает способностью к инфильтрации, вторжению в отдаленную область или метастазированию в нее.
Под "опухолевой массой" понимают количество злокачественных клеток, размер опухоли или величину злокачественной опухоли в организме. Опухолевую массу также называют опухолевой нагрузкой.
Под "числом опухолей" понимают количество опухолей.
Под "индивидуумом" понимают млекопитающего, включая, но ими не ограничиваясь, человека или не являющегося человеком млекопитающего, такого как бычьи, лошадиные, собачьи, овечьи или кошачьи. Предпочтительно, индивидуумом является человек.
Термин "средство против злокачественной опухоли" относится к терапии, подходящей для лечения злокачественной опухоли. Примеры лекарственных средств включают, но ими не ограничиваются, например, химиотерапевтические средства, ингибирующие рост средства, цитотоксические средства, средства, используемые в лучевой терапии, антиангиогенные средства, апоптотические средства, средства против тубулина и другие средства для лечения злокачественной опухоли, антитела против CD20, ингибиторы тромбоцитарного фактора роста (например, GleevecTM (иматиниба мезилат)), ингибитор COX-2 (например, целекоксиб), интерфероны, цитокины, антагонисты (например, нейтрализующие антитела), которые связываются с одной или несколькими из следующих мишеней: ErbB2, ErbB3, ErbB4, PDGFR-бета, BlyS, APRIL, BCMA или рецептор(ы) VEGF, TRAIL/Apo2 и другие биологически активные и органические химические агенты и тому подобное. Также в изобретение включаются их комбинации.
Термин "цитотоксическое средство", как используется в настоящем документе, относится к веществу, которое ингибирует функционирование клеток, или препятствует ему, и/или вызывает разрушение клеток. Подразумевают, что термин включает радиоактивные изотопы (например, I131, I125, Y90 и Re186), химиотерапевтические средства и токсины, такие как ферментативно активные токсины из бактерий, грибов, растений или животных, или их фрагменты.
"Химиотерапевтическое средство" представляет собой химическое соединение, подходящее для лечения злокачественной опухоли. Примеры химиотерапевтических средств включают химическое соединение, подходящее для лечения злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие средства, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бисульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогинины (особенно буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги на основе адозелезина, карзелезина и бизелезина); криптофицины (в частности криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CB1-TM1); элеутеробин; панкриастатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, иприт урацила; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как эндииновые антибиотики (например, калихеамицин, особенно калихеамицин гамма1I и калихеамицин омега1I (см., например, Agnew, Chem Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин A; бисфосфонаты, такие как клодронат; эсперамицин; а также хромофор на основе неокарциностатина и аналогичные хромопротеиновые хромофоры на основе эндииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, декторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин ADRIAMYCIN® (включая морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марселломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; антиадренальные средства, такие как аминоглутетимид, митотан, трилостан; средства для восполнения фолиевой кислоты, такие как фролиновая кислота; ацеглатон; альдофосфамид гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демекольцин; диазиквон; эльфорнитин; ацетат эллиптиния; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновая кислота; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2"-трихлортриэтиламин; трихотецены (особенно токсин T-2, верракурин A, роридин A и ангвидин); уретан; виндезин; декарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); циклофосфамид; тиотепу; таксоиды, например, паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), ABRAXANETM, не содержащий кремофоров, сконструированный с альбумином состав паклитаксела в виде частиц (American Pharmaceutical Partners, Schaumberg, Illinois), и доксетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин GEMZAR®; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин NAVELBINE®; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; иринотекан (Camptosar, CPT-11) (включая режим лечения иринотеканом с 5-FU и лейковорином); ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин, бортезомиб VELCADE; леналидомид REVLIMID; лейковорин (LV); оксалиплатин, включая режим лечения оксалиплатином (FOLFOX); ингибиторы PKC-альфа, Raf, H-Ras, EGFR (например, эрлотиниб (TarcevaTM)) и VEGF-A, которые снижают пролиферацию клеток, и фармацевтические приемлемые соли, кислоты или производные любого из указанных выше.
Также к этому определению относятся антигормональные средства, которые действуют, регулируя или ингибируя действие гормонов на опухоль, такие как антиэстрогены и селективные модулярторы рецепторов эстрогена (SERM), включая, например, тамоксифен (в том числе тамоксифен NOLVADEX®), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен FARESTON®; ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует продукцию эстрогена в надпочечниках, например, такие как 4(5)-имидазолы, аминоглутетимид; ацетат магестрола MEGASE®, экземестан AROMASIN®, форместан, фадрозол, ворозол RIVISOR®, лектрозол FEMARA® и анастрозол ARIMIDEX® и антиандрогены, такие как флутамид, нилутамид, бикалутамид, леупролид и госерелин; а также троксацитабин (нуклеозидный аналог цитозина 1,3-диоксолан); антисмысловые олигонуклеотиды, в частности, олигонуклеотиды, которые ингибируют экспрессию генов в каскадах передачи сигнала, вовлеченных в нарушенную пролиферацию клеток, например, такие как PKC-alpha, Raf и H-Ras; рибозимы, такие как ингибитор экспрессии VEGF (например, рибозим ANGIOZYME®) и ингибитор экспрессии HER2; вакцины, такие как вакцина для генной терапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; PROLEUKIN® rIL-2; ингибитор топоизомеразы 1 LURTOTECAN®; rmRH ABARELIX®; винорелбин и эсперамицины (см патент США 4675187) и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше.
Термин "пролекарство", как используется в настоящей заявке, относится к форме предшественника или производного фармацевтически активного вещества (например, низкомолекулярного соединения), которая является менее цитотоксической для клеток по сравнению с исходным лекарственном средством и может быть ферментативно или гидролитически активирована или преобразована в более активную исходную форму. См., например, Wilman, "Prodrugs in Cancer Chemotherapy" Biochemical Society Transactions, 14, pp. 375-382, 615th Meeting Belfast (1986) и Stella et al., "Prodrugs: A Chemical Approach to Targeted Drug Delivery," Directed Drug Delivery, Borchardt et al., (ed.), pp. 247-267, Humana Press (1985). Пролекарства по настоящему изобретению включают, но ими не ограничиваются, содержащие фосфат пролекарства, содержащие тиофосфат пролекарства, содержащие сульфат пролекарства, содержащие пептид пролекарства, пролекарства с модифицированными D-аминокислотами, гликозилированные пролекарства, содержащие β-лактам пролекарства, содержащие необязательно замещенный феноксиацетамид пролекарства, содержащие необязательно замещенный фенилацетамид пролекарства, пролекарства с 5-фторцитозином и другие пролекарства с 5-фторуридином, которые могут превращаться в более активное цитотоксическое свободное лекарственное средство. Примеры цитотоксических лекарственных средств, которые могут быть преобразованы в форму пролекарства для использования в настоящем изобретении, включают, но ими не ограничиваются, химиотерапевтические средства, описанные выше.
Под "лучевой терапией" понимают использование направленных гамма-лучей или бета-лучей для индукции достаточного повреждения клетки, чтобы ограничить ее способность нормально функционировать или разрушить клетку целиком. Понятно, что существует много известных в данной области способов определить дозировку и длительность лечения. Характерное лечение проходит в виде однократного облучения и характерные дозировки находятся в диапазоне от 10 до 200 единиц (Грей) в сутки.
Термин "биологический образец" (взаимозаменяемо называемый "образцом" или "образцом ткани или клетки") включает различные типы образцов, получаемые от индивидуума, и их можно использовать для диагностического анализа или мониторинга. Определение включает образцы крови и других жидкостей биологического происхождения, образцы солидных тканей, такие как биоптат или культуры тканей или клеток, полученные из него и их потомки. Также термин включает образцы, которые подвергали обработке любым образом после их извлечения, например, обработке реагентами, солюбилизации или обогащению определенным компонентами, такими как белки или полинуклеотиды, или погружению в полутвердый или твердый матрикс для целей получения срезов. Термин "биологический образец" включает клинический образец, а также включает клетки в культуре, клеточные супернатанты, клеточные лизаты, сыворотку, плазму, биологическую жидкость и образцы тканей. Источником биологического образца может быть любая солидная ткань, такая как ткань из свежего, замороженного и/или законсервированного органа, или образец ткани или биоптат или аспират; кровь или любые компоненты крови; жидкости организма, такие как спинномозговая жидкость, амниотическая жидкость перитонеальная жидкость или интерстициальная жидкость, клетки с любого момента беременности или развития индивидуума. В некоторых вариантах осуществления биологический образец получают из первичной или метастатической опухоли. Биологический образец может содержать соединения, которые не смешаны с тканью в природе, такие как консерванты, антикоагулянты, буферы, фиксажи, пищевые добавки, антибиотики или аналогичные с ними.
Для целей, представленных в настоящем документе, "срез" образца ткани означает отдельную часть или фрагмент образца ткани, например, тонкий срез ткани или клеток, вырезанный из образца ткани. Понятно, что в соответствии с настоящим изобретением можно получать и подвергать анализу множество срезов образцов тканей. В некоторых вариантах осуществления один срез образца ткани анализируют как на морфологическом, так и на молекулярном уровнях, или его анализируют в отношении как белка, так и нуклеиновой кислоты.
Термин "метка", как используется в настоящем документе, относится к обнаруживаемому соединению или композиции, которые конъюгированы или слиты непосредственно или косвенно с реагентом, таким как зонд нуклеиновой кислоты или антителом и способствуют обнаружению реагента, с которым они конъюгированы или слиты. Метка сама может быть обнаруживаемой (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, она может катализировать химическое изменение соединения или композиции субстрата, которое поддается обнаружения.
Композиции и способы по изобретению
Настоящее изобретение относится к композиции, в том числе к фармацевтическим композициям, содержащим антитело против FGFR3; и к полинуклеотидам, содержащим последовательности, кодирующие антитело против FGFR3. Как используют в настоящем документе, композиции содержат одно или несколько антител, которые связываются с FGFR3, и/или один или несколько полинуклеотидов, содержащих последовательности, кодирующие одно или несколько антител, которые связываются с FGFR3. Эти композиции, кроме того, могут содержать подходящие носители, такие как фармацевтически приемлемые эксципиенты, включая буферы, которые хорошо известны в данной области.
Также изобретение относится к вариантам осуществления выделенных антител и полинуклеотидов. Также изобретение относится к вариантам осуществления по существу чистых антител и полинуклеотидов.
Также изобретение относится к способу лечения нарушения, например, множественной миеломы или карциномы переходной стадии (например, инвазивной карциномы переходной стадии) с использованием антитела против FGFR3 (как описано в настоящем документе или как известно в данной области).
Композиции
Антитела против FGFR3 по изобретению, предпочтительно, являются моноклональными. Также объем изобретения включает фрагменты Fab, Fab', Fab'-SH и F(ab')2 антител против FGFR3, представленных в настоящем документе. Эти фрагменты антител могут быть получены обычными способами, такими как ферментативное расщепление, или их можно получать рекомбинантными способами. Такие фрагменты антител могут быть химерными или гуманизированными. Эти фрагменты подходят для диагностических и терапевтических целей, указанных ниже.
Моноклональные антитела получают из совокупности по существу гомогенных антител, т.е., отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных природных мутаций, которые могут быть представлены в небольших количествах. Таким образом, определение "моноклональные" указывает на свойство антитела, что оно не является смесью отдельных антител.
Моноклональные антитела против FGFR3 по изобретению можно получать способом гибридом, впервые описанного Kohler et al., Nature, 256:495 (1975), или их можно получать способами рекомбинантных ДНК (патент США 4816567).
В способе гибридом, мышь или другое подходящее животное-хозяина, такое как хомяк, иммунизируют для индукции лимфоцитов, которые продуцируют или способны продуцировать антитела, которые специфично связываются с белком, используемым для иммунизации. Антитела против FGFR3 можно индуцировать у животных многократными подкожными (sc) или внутрибрюшинными (ip) инъекциями FGFR3 и адъюванта. FGFR3 можно получать с использованием способов, хорошо известных в данной области, некоторые из которых дополнительно описаны в настоящем документе. Например, рекомбинантная продукция FGFR3 человека и мыши описана ниже. В одном из вариантов осуществления животных иммунизируют FGFR3, слитым с Fc-частью тяжелой цепи иммуноглобулина. В предпочтительном варианте осуществления животных иммунизируют слитым белком FGFR3-IgG1. Животных обычно иммунизируют против иммунногенных конъюгатов или производных FGFR3 с монофосфориллипидом A (MPL)/дикриномиколятом трегалозы (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT) и раствор инъецируют внутрикожно в несколько областей. Через две недели животным делают вспомогательную инъекцию. Через от 7 до 14 суток у животных проводят взятие крови и сыворотку анализируют в отношении титра антител против FGFR3. Животным делают вспомогательные инъекции до достижения титрами плато.
Альтернативно, лимфоциты можно иммунизировать in vitro. Затем лимфоциты подвергают слиянию с клетками миеломы с использованием подходящего реагента для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Гибридомные клетки, полученные таким способом, высевают и, выращивают в подходящей культуральной среде, которая предпочтительно, содержит одно или несколько веществ, которые ингибируют рост или выживание неслитых исходных клеток миеломы. Например, если исходные клетки миеломы лишены фермента гипоксантингуанинфосфорибозилтрансферазы (HGPRT или HPRT), культуральная среда для гибридом, как правило, включает гипоксантин, аминоптерин и тимидин ("среда HAT"), которые препятствуют росту дефицитных по HGPRT клеток.
Предпочтительные клеточные линии миеломы представляют собой клеточные линии, которые эффективно подвергаются слиянию, поддерживают стабильную экспрессию антитела на высоком уровне выбранными антителопродуцирующими клетками, и являются чувствительными к среде, такой как среда HAT. Среди них, предпочтительными клеточными линиями миеломы являются линии миеломы мыши, такие как линии, полученные из опухолей мыши MOPC-21 и MOPC-11, доступные, например, от Salk Institute Cell Distribution Center, San Diego, California USA и клетки SP-2 или Х63-Ag8-653, доступные от American Type Culture Collection, Rockville, Maryland USA. Также для продукции моноклональных антител человека описаны клеточные линии миеломы человека и гетеромиеломы мыши-человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., MONOCLONAL ANTIBODY PRODUCTION TECHNIQUES AND APPLICATIONS, pp. 51-63 (Marcel Dekker, Inc.: New York, 1987).
Культуральную среду, в которой культивируют гибридомные клетки, затем можно анализировать на присутствие моноклональных антител, направленных против FGFR3. Предпочтительно, специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, можно определять с помощью иммунопреципитации или анализа связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA).
Аффинность связывания моноклонального антитела можно определять, например, анализом Скэтчарда, Munson et al., Anal. Biochem., 107:220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с желаемыми специфичностью, аффинностью и/или активностью, клоны можно субклонировать способами лимитирующих разведений и выращивать стандартными способами (Goding, Monoclonal Antibodies: Principles и Practice, pp.59-103 (Academic Press, 1986)). Подходящие для этой цели культуральные среды включают, например, D-MEM и среду RPMI-1640. Кроме того, гибридомные клетки можно выращивать in vivo у животных в качестве асцитных опухолей.
Моноклональные антитела, секретируемые субклонами, подходящим образом выделяют из культуральной среды, асцитной жидкости или сыворотки общепринятыми способами очистки антител, например, такими как хроматография белка на A-сефарозе, хроматография с гидроксилапатитом, гель-электрофорез, диализ или аффинная хроматография.
Антитела против FGFR3 по изобретению могут быть получены с использованием комбинаторных библиотек для скрининга синтезированных клонов антител с требуемой активностью или видами активности. По существу, клоны синтетических антител повергают селекции с помощью библиотек фагового дисплея, которые содержат фаг, который экспонирует различные фрагменты вариабельной области (Fv) антитела, слитые с белком оболочки фага. Такие фаговые библиотеки сортируют с помощью аффинной хроматографии против требуемого антигена. Клоны, экспрессирующие Fv-фрагменты, способные связываться с требуемым антигеном, адсорбируются на антиген и, таким образом, их отделяют от несвязывающих клонов в библиотеке. Затем связывающие клоны элюируют от антигена, и их содержание можно далее повышать с помощью дополнительных циклов адсорбции/элюирования антигена. Путем разработки подходящего способа скрининга с помощью антигена для селекции представляющего интерес клона фага с последующим конструированием клона полноразмерного антитела против FGFR3 с использованием последовательностей Fv из представляющего интерес фагового клона и подходящих последовательностей константной области (Fc), описанных в Каbаt et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3, можно получать любые из антител против FGFR3 по изобретению.
Антиген-связывающий домен антитела образован из двух вариабельных (V) областей приблизительно из 110 аминокислот, по одной из легкой (VL) и тяжелой (VH) цепей, в обеих из которых присутствуют три гипервариабельные петли или определяющие комплементарность области (CDR). Вариабельные домены могут функционально экспонироваться на фаге, либо в качестве одноцепочечных Fv-фрагментов (scFv), в которых VH и VL ковалентно связаны через короткий подвижный пептид, либо в качестве Fab-фрагментов, в которых они обе являются слитыми с константным доменом и взаимодействуют нековалентно, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Как используется в настоящем документе, фаговые клоны, кодирующие scFv, и фаговые клоны, кодирующие Fab, совместно называют "фаговыми клонами Fv" или "клонами Fv".
Репертуары генов VH и VL можно по отдельности клонировать полимеразной цепной реакцией (ПЦР) и подвергать случайной рекомбинации в фаговых библиотеках, в которых затем проводят поиск антиген-связывающих клонов, как описано в Winter et al., Ann. Rev. Immunol, 12: 433-455 (1994). Библиотеки из иммунизированных источников обеспечивают высокоаффинные антитела к иммуногену без необходимости конструирования гибридом. Альтернативно, можно клонировать наивный репертуар для обеспечения одного источника антител человека к широкому диапазону несобственных, а также собственных антигенов без иммунизации, как описано Griffiths et al., EMBO J, 12: 725-734 (1993). В заключение, библиотеки с наивным репертуаром также можно получать синтетически путем клонирования не подвергнутых реаранжировке сегментов V-генов из стволовых клеток, и использования праймеров для ПЦР, содержащих случайную последовательность, для кодирования высоко вариабельных областей CDR3 и для проведения реаранжировки in vitro, как описано Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992).
Для экспонирования фрагментов антител используют нитевидные фаги, осуществляя слияние с минорным белком оболочки pIII. Фрагменты антител могут быть экспонированы в виде одноцепочечных Fv-фрагментов, в которых VH- и VL-домены связаны на одной полипептидной цепи подвижным полипептидным спейсером, например, как описано в Marks et al., J. Mol. Biol, 222: 581-597 (1991), или в качестве Fab-фрагментов, в которых одна цепь является слитой с pIII, а другая секретируется в периплазму бактериальной клетки-хозяина, где собирается структура Fab-белок оболочки, которая экспонируется на поверхности фага путем вытеснения некоторых из белков оболочки дикого типа, например, как описано в Hoogenboom et al., Nucl. Acids Res., 19: 4133-4137 (1991).
Как правило, нуклеиновые кислоты, кодирующие фрагменты гена антитела, получают из иммунных клеток, полученных от человека или животных. Если является желательной библиотека, смещенная в сторону клонов, направленных против FGFR3, то индивидуума иммунизируют FGFR3 для получения антительного ответа, и выделяют клетки селезенки и/или циркулирующие B-клетки, и другие лимфоциты периферической крови человека (PBL) для конструирования библиотеки. В предпочтительном варианте осуществления получают библиотеку фрагментов гена антитела против FGFR3 человека, смещенную в сторону клонов, направленных против FGFR3, путем обеспечения ответа антител против FGFR3 у трансгенной мыши, обладающей функциональным набором генов иммуноглобулинов человека (и лишенной функциональной эндогенной системы продукции антител), так что иммунизация с помощью FGFR3 приводит к возникновению B-клеток, продуцирующих антитела человека против FGFR3. Создание продуцирующих антитела человека трансгенных мышей описано ниже.
Дополнительное повышение содержания популяций клеток, реактивных против FGFR3, можно обеспечивать с использованием подходящего способа скрининга для выделения B-клеток, экспрессирующих специфичное к FGFR3 мембраносвязанное антитело, например, путем разделения клеток с использованием аффинной хроматографии с FGFR3 или адсорбции клеток на меченный флуорохромом FGFR3 с последующей активированной флуоресценцией сортировкой клеток (FACS).
Альтернативно применение клеток селезенки и/или B-клеток или других PBL от неиммунизированного донора обеспечивает лучшее представление возможного репертуара антител, а также позволяет конструирование библиотеки антител с использованием любого вида животных (человека или не относящегося к человеку), в котором FGFR3 не является антигенным. Для библиотек, включающих конструирование генов антител in vitro, собирают стволовые клетки индивида для получения нуклеиновых кислот, кодирующих не подвергнутые реаранжировке сегменты генов антител. Представляющие интерес иммунные клетки можно получать из различных видов животных, таких как человек, мышь, крыса, виды зайцеобразных, волчьих, собачьих, кошачьих, свиней, крупного рогатого скота, лошадей и птиц и тому подобное.
Нуклеиновую кислоту, кодирующую вариабельные сегменты генов (включая сегменты VH и VL) выделяют из представляющих интерес клеток и амплифицируют. В случае подвергнутых реаранжировке библиотек генов VH и VL, требуемую ДНК можно получать путем выделения геномной ДНК или мРНК из лимфоцитов с последующей полимеразной цепной реакцией (ПЦР) с праймерами, совпадающими с 5'- и 3'-концами подвергнутых реаранжировке генов VH и VL, как описано в Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86: 3833-3837 (1989), тем самым получая разнообразные репертуары V-генов для экспрессии. V-гены можно амплифицировать из кДНК и геномной ДНК с помощью обратных праймеров на 5'-конце экзона, кодирующего зрелый V-домен, и прямых праймеров, располагающихся в J-сегменте, как описано в Orlandi et al. (1989) и Ward et al., Nature, 341: 544-546 (1989). Однако для амплификации из кДНК, обратные праймеры также могут располагаться в лидерном экзоне, как описано в Jones et al., Biotechnol., 9: 88-89 (1991), а прямые праймеры могут располагаться в константной области, как описано в Sastry et al., Proc. Natl. Acad. Sci. (USA), 86: 5728-5732 (1989). Для максимального повышения комплементарности, в праймеры можно включать вырожденность, как описано в Orlandi et al. (1989) или Sastry et al. (1989). Предпочтительно, разнообразие библиотеки максимально увеличивают с помощью праймеров ПЦР, направленных на каждое семейство V-генов для амплификации всех доступных расположений VH и VL, присутствующих в образце нуклеиновой кислоты иммунных клеток, например, как описано в способе Marks et al., J. Mol. Biol., 222: 581-597 (1991) или как описано в способе Orum et al., Nucleic Acids Res., 21: 4491-4498 (1993). Для клонирования амплифицированной ДНК в векторы экспрессии, в праймер для ПЦР можно вносить редкие участки рестрикции в качестве метки на одном конце, как описано в Orlandi et al. (1989), или путем дальнейшей амплификации с помощью ПЦР с праймером с меткой, как описано в Clackson et al., Nature, 352: 624-628 (1991).
Репертуары подвергнутых синтетической реаранжировке V-генов можно получать in vitro из сегментов V-генов. Большинство из сегментов VH-генов человека были клонированы и секвенированы (как описано в Tomlinson et al., J. Mol. Biol, 227: 776-798 (1992)), и картированы (описано в Matsuda et al., Nature Genet., 3: 88-94 (1993); эти клонированные сегменты (включая все главные конформации петли H1 и H2) можно использовать для получения разнообразных репертуаров VH-генов с помощью праймеров для ПЦР, кодирующих петли H3 с разнообразной последовательностью и длиной, как описано в Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992). Также можно получать репертуары VH, где все разнообразие последовательности сфокусировано в длинной петле H3 с единой длиной, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 89: 4457-4461 (1992). Сегменты Vκ и Vλ человека были клонированы и секвенированы (описано в Williams and Winter, Eur. J. Immunol., 23: 1456-1461 (1993)) и их можно использовать для получения синтетических репертуаров легких цепей. Синтетические репертуары V-генов, на основе ряда сборок VH и VL, и длин L3 и H3, кодируют антитела со значительным структурным разнообразием. После амплификации ДНК, кодирующей V-ген, сегменты V-генов зародышевой линии можно подвергать реаранжировке in vitro в соответствии со способами Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992).
Репертуары фрагментов антител можно конструировать, комбинируя репертуары генов VH и VL различными путями. Каждый репертуар можно создавать в отдельных векторах, и векторы подвергать рекомбинации in vitro, например, как описано в Hogrefe et al., Gene, 128: 119-126 (1993), или in vivo путем комбинаторной инфекции, например, с помощью системы loxP, описанной в Waterhouse et al., Nucl. Acids Res., 21: 2265-2266 (1993). В подходе рекомбинации in vivo используется двухцепочечная структура Fab-фрагментов для преодоления ограничения размера библиотеки, определяемого эффективностью трансформации E. coli. Наивные репертуары VH и VL клонируют по отдельности, один в плазмиду, а другой в фаговый вектор. Затем две библиотеки комбинируют с помощью инфекции фагом содержащих фагмиду бактерий, так что каждая клетка содержит отличающееся сочетание, и размер библиотеки ограничивается только числом присутствующих клеток (приблизительно 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo, так что гены VH и VL рекомбинируют в один репликон и совместно упаковываются в фаговые вирионы. Эти огромные библиотеки дают большие количества разнообразных антител с высокой аффинностью (Kd -1 приблизительно 10-8 M).
Альтернативно, репертуары можно клонировать последовательно в один вектор, например, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 88: 7978-7982 (1991), или объединять с помощью ПЦР, а затем клонировать, например, как описано в Clackson et al., Nature, 352: 624-628 (1991). Сборку с помощью ПЦР также можно использовать для связывания ДНК VH и VL с ДНК, кодирующей подвижный пептидный спейсер, с образованием репертуаров одноцепочечных Fv (scFv). В другом способе используют "сборку с помощью ПЦР в клетках" для объединения генов VH и VL в лимфоцитах с помощью ПЦР, а затем клонирования репертуаров связанных генов, как описано в Embleton et al., Nucl. Acids Res., 20: 3831-3837 (1992).
Антитела, продуцируемые наивными библиотеками (либо природными, либо синтетическими) могут иметь умеренную аффинность (Kd -1 приблизительно от 106 до 107 M-1), однако созревание аффинности также можно имитировать in vitro с помощью конструирования и повторной селекции из вторичных библиотек, как описано в Winter et al. (1994), выше. Например, мутацию можно вносить случайным образом in vitro с использованием полимеразы с пониженной точностью (описано Leung et al., Technique, 1: 11-15 (1989)) в способе Hawkins et al., J. Mol. Biol., 226: 889-896 (1992) или в способе Gram et al., Proc. Natl. Acad. Sci USA, 89: 3576-3580 (1992). Кроме того, созревание аффинности можно проводить путем внесения мутации случайным образом в одну или несколько CDR, например, с использованием ПЦР с праймерами, имеющими случайную последовательность, охватывающую представляющую интерес CDR, в выбранных отдельных клонах Fv, и скринингом на клоны с наиболее высокой аффинностью. В WO 9607754 (опубликованной 14 марта 1996 года) описан способ индукции мутагенеза в определяющей комплементарность области легкой цепи иммуноглобулина для создания библиотеки генов легкой цепи. Другим эффективным подходом является рекомбинация VH- или VL-доменов, отобранных с помощью фагового дисплея, с репертуарами природных вариантов V-доменов, полученными из неиммунизированных доноров, и скрининг на наиболее высокую аффинность в нескольких раундах повторной перетасовки цепей, как описано в Marks et al., Biotechnol., 10: 779-783 (1992). Этот способ позволяет продукцию антител и фрагментов антител с аффинностью приблизительно 10-9 M.
Последовательности нуклеиновых кислот и аминокислотные последовательности FGFR3 известны в данной области. Последовательность нуклеиновой кислоты, кодирующую FGFR3, можно конструировать с использованием аминокислотной последовательности желаемой области FGFR3. Как хорошо известно в данной области, существует две основных изоформы FGFR3 по сплайсингу: FGFR3 IIIb и FGFR3 IIIc. Последовательности FGFR3 хорошо известны в данной области и могут включать последовательность с регистрационным номером UniProKB/Swiss-Prot P22607 (FGFR3 IIIc) или P22607_2 (FGFR3 IIIb). Мутации FGFR3 идентифицированы и хорошо известны в данной области и включают следующие мутации (относительно последовательностей, представленных в UniProKB/Swiss-Prot под регистрационным номером P22607 (FGFR3 IIIc) или P22607_2 (FGFR3 IIIb):

Нуклеиновые кислоты, кодирующие FGFR3, можно получать множеством способов, известных в данной области. Эти способы предусматривают, но ими не ограничиваются, химический синтез любым из способов, описанных в Engels et al., Agnew. Chem. Int. Ed. Engl, 28: 716-734 (1989), таких как способы сложных триэфиров, фосфитов, фосфорамидитов и H-фосфонатов. В одном из вариантов осуществления для конструирования ДНК, кодирующей FGFR3, используют кодоны, предпочтительные для экспрессируюущей клетки-хозяина. Альтернативно, ДНК, кодирующую FGFR3, можно выделять из библиотеки геномной ДНК или кДНК.
После конструирования молекулы ДНК, кодирующей FGFR3, молекула ДНК является функционально связанной с последовательностью контроля экспрессии в векторе экспрессии, таком как плазмида, где последовательность контроля распознается клеткой-хозяином, трансформированной вектором. Как правило, плазмидные векторы содержат последовательности для репликации и контрольные последовательности, которые происходят из видов, совместимых с клеткой-хозяином. Вектор обычно несет участок репликации, а также последовательности, которые кодируют белки, способные обеспечивать фенотипическую селекцию в трансформированных клетках. Подходящие векторы для экспрессии в прокариотических и эукариотических клетках-хозяевах известны в данной области и некоторые, кроме того, описаны в настоящем документе. Можно использовать эукариотические организмы, такие как дрожжи, или клетки, происходящие из многоклеточных организмов, таких как млекопитающие.
Необязательно, ДНК, кодирующая FGFR3, функционально связана с секреторной лидерной последовательностью, приводящей к секреции продукта экспрессии клеткой-хозяином в культуральную среду. Примеры секреторных лидерных последовательностей включают stII, экотин, lamB, GD вируса герпеса, lpp, щелочную фосфатазу, инвертазу и альфа-фактор. Также для использования в настоящем изобретении подходит лидерная последовательность белка A из 36 аминокислот (Abrahmsen et al., EMBO J., 4: 3901 (1985)).
Клетки-хозяева трансфицируют и, предпочтительно, трансформируют описанными выше векторами для экспрессии или клонирования по этому изобретению и культивируют в общепринятой питательной среде, модифицированной соответствующим образом для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих желаемые последовательности.
Трансфекция относится к захвату вектора экспрессии клеткой-хозяином независимо от того, экспрессируются ли в действительности кодирующие последовательности. Специалистам в данной области известны многочисленные способы трансфекции, например, преципитация с CaPO4 и электропорация. Успешную трансфекцию как правило определяют, если в клетке-хозяине возникает какой-либо признак действия этого вектора. Способы трансфекции хорошо известны в данной области, и некоторые из них, кроме того, описаны в настоящем документе.
Трансформация означает внесение ДНК в организм, так чтобы эта ДНК реплицировалась, либо в качестве внехромосомного элемента, либо после встраивания в хромосому. В зависимости от используемой клетки-хозяина, трансформацию проводят с использованием стандартных способов, подходящих для таких клеток. Способы трансформации хорошо известны в данной области и некоторые из них, кроме того, описаны в настоящем документе.
Прокариотические клетки-хозяева, используемые для продукции FGFR3, можно культивировать, как описано, главным образом, в Sambrook et al., выше.
Клетки-хозяева млекопитающих, используемые для продукции FGFR3, можно культивировать в различных средах, которые хорошо известны в данной области и некоторые из которых описаны в настоящем документе.
Клетки-хозяева, упоминаемые в настоящем описании, охватывают клетки в культуре in vitro, а также клетки, которые находятся в животном-хозяине.
Очистку FGFR3 можно проводить с использованием общепринятых способов, некоторые из которых описаны в настоящем документе.
Очищенный FGFR3 можно связывать с подходящей матрицей, такой как агарозные гранулы, акриламидные гранулы, стеклянные гранулы, целлюлоза, различные акриловые сополимеры, гидроксилметакрилатные гели, полиакриловые и полиметакриловые сополимеры, нейлон, нейтральные и ионные носители и тому подобное, для использования в разделении способами аффинной хроматографии клонов фагового дисплея. Присоединение белка FGFR3 к матрице можно проводить способами, описанными в Methods in Enzymology, vol. 44 (1976). Широко используемый способ для присоединения белковых лигендов к полисахаридным матрицам, например, агарозе, декстрану или целлюлозе, вовлекает активацию носителя цианогенгалогенидами и последующее присоединение первичных алифатических или ароматических аминов пептидных лигандов к активированной матрице.
Альтернативно, FGFR3 можно использовать для нанесения на лунки адсорбционных планшетов, экспрессировать на клетках-хозяевах, прикрепленных к адсорбционным планшетам или использовать в сортировке клеток, или конъюгировать с биотином для улавливания с помощью покрытых стрептавидином гранул, или использовать в любом другом известном в данной области способе пэннинга библиотек фагового дисплея.
Образцы для фагового дисплея приводят в контакт с иммобилизованным FGFR3 в условиях, подходящих для связывания по меньшей мере части фаговых частиц с адсорбентом. Обычно выбирают условия, включая pH, ионную силу, температуру и тому подобное, имитирующие физиологические условия. Фаги, связанные с твердой фазой, промывают, а затем элюируют кислотой, например, как описано в Barbas et al., Proc. Natl. Acad. Sci USA, 88: 7978-7982 (1991), или щелочью, например, как описано в Marks et al., J. Mol. Biol., 222: 581-597 (1991), или с помощью конкуренции с антигеном FGFR3, например, в способе, аналогичным со способом конкуренции с антигеном в Clackson et al., Nature, 352: 624-628 (1991). За один раунд селекции фаги можно обогащать в 20-1000 раз. Более того, обогащенные фаги можно выращивать в бактериальной культуре и подвергать последующим раундам селекции.
Эффективность селекции зависит от множества факторов, включая кинетику диссоциации при промывании, и возможность одновременной встречи множества фрагментов антител на одном фаге с антигеном. Антитела с быстрой кинетикой диссоциации (и слабой аффинностью связывания) можно задерживать с использованием короткого промывания, поливалентного фагового дисплея и высокой плотности нанесения антигена на твердую фазу. Высокая плотность не только стабилизирует фаг при поливалентных взаимодействиях, но также способствует повторному связыванию фага, который диссоциировал. Селекцию антител с низкой кинетикой диссоциации (и высокой аффинностью связывания) можно обеспечивать, используя длительные промывания и одновалентный фаговый дисплей, как описано в Bass et al., Proteins, 8: 309-314 (1990) и в WO 92/09690, и низкую плотность нанесения антигена, как описано в Marks et al., Biotechnol., 10: 779-783 (1992).
Селекцию фаговых антител с различной аффинностью к FGFR3 можно проводить даже при аффинности, которая отличается незначительно. Однако, случайная мутация выбранного антитела (например, как происходит в некоторых способах созревания аффинности, описанных выше) может привести к возникновению многих мутантов, большинство из которых связываются с антигеном, и немногие из которых обладают более высокой аффинностью. При ограничении FGFR3, редкий высокоаффинный фаг может конкурировать. Для сохранения всех мутантов с более высокой аффинностью, фаги можно инкубировать с избытком биотинилированного FGFR3, но концентрация биотинилированного FGFR3 должна иметь более низкую молярность, чем предполагаемая молярная аффинность, постоянная для FGFR3. Затем связывающиеся с высокой аффинностью фаги можно улавливать покрытыми стрептавидином парамагнитными гранулами. Такое "равновесное улавливание" позволяет селекцию антител согласно их аффинности связывания с чувствительностью, которая позволяет выделение мутантных клонов с аффинностью, большей только в два раза, из большого избытка фагов с более низкой аффинностью. Также можно изменять условия, используемые для промывания фагов, связанных с твердой фазой, для различения на основе кинетики диссоциации.
Клоны, направленные против FGFR3, можно отбирать на основе активности. В одном из вариантов осуществления изобретение относится к антителам против FGFR3, которые блокируют связывание между рецептором FGFR3 и его лигандом (таким как FGF1 и/или FGF9). Клоны Fv, соответствующие таким антителам против FGFR3, можно подвергать селекции с помощью (1) выделения клонов, направленных против FGFR3, из фаговой библиотеки, как описано выше, и необязательно, увеличения количества выделенной популяции фаговых клонов путем выращивания популяции в подходящем бактериальном хозяине; (2) селекции FGFR3 и второго белка, против которого является желательной блокирующая и неблокирующая активность, соответственно; (3) адсорбции фаговых клонов, направленных против FGFR3, на иммобилизованный FGFR3; (4) применения избытка второго белка для элюирования каких-либо нежелательных клонов, которые распознают связывающие FGFR3 детерминанты, которые перекрываются или являются общими со связывающими детерминантами второго белка; и (5) элюирования клонов, которые остаются адсорбированными после стадии (4). Необязательно, клоны с требуемыми блокирующими/неблокирующими свойствами можно далее обогащать путем повторения процессов селекции, описанных в настоящем документе, один или несколько раз.
ДНК, кодирующую образованные из гибридомы моноклональные антитела или клоны Fv фагового дисплея по изобретению, легко выделять и секвенировать с использованием общепринятых способов (например, с использованием олигонуклеотидных праймеров, сконструированных для специфичной амплификации представляющих интерес кодирующих участков тяжелой и легкой цепей из гибридомы или ДНК-матрицы фага). После выделения ДНК можно помещать в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьяны, клетки яичника китайского хомяка (CHO), или клетки миеломы, которые в ином случае не продуцируют белок иммуноглобулина, обеспечивая синтез требуемых моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитело, включают Skerra et al., Curr. Opinion in Immunol., 5: 256 (1993) и Pluckthun, Immunol. Revs, 130: 151 (1992).
ДНК, кодирующую клоны Fv по изобретению, можно комбинировать с известными последовательностями ДНК, кодирующими константные участки тяжелой цепи и/или легкой цепи (например, соответствующие последовательности ДНК можно получать в соответствии с Кабат et al., выше) с образованием клонов, кодирующих полноразмерные или неполные тяжелые и/или легкие цепи. Понятно, что для этой цели можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получать от человека или из любого вида животных. Клон Fv, образованный из ДНК вариабельного домена одного вида животных (такого как человек), а затем слитый с ДНК константной области другого вида животных с образованием кодирующей последовательности(ей) "гибридной" полноразмерной тяжелой цепи и/или легкой цепи, включен в определение "химерного" и "гибридного" антитела, как используется в настоящем документе. В предпочтительном варианте осуществления клон Fv, образованный из ДНК вариабельной области человека, является слитым с ДНК константной области человека с образованием кодирующей последовательности(ей) для полноразмерной или неполной тяжелой и/или легкой цепей.
ДНК, кодирующую антитело против FGFR3, продуцируемую гибридомой, также можно модифицировать, например, путем замены кодирующей последовательности константными доменами тяжелой и легкой цепей вместо гомологичных последовательностей мыши, происходящих из клона гибридомы (например, как в способе Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)). ДНК, кодирующую продуцируемое гибридомой или происходящей из Fv-клона антитело или фрагмент, далее можно модифицировать ковалентным связыванием с кодирующей последовательностью иммуноглобулина всей кодирующей последовательности не являющегося иммуноглобулином полипептида, или ее части. Таким образом, получают "химерные" или "гибридные" антитела, которые обладают специфичностью связывания происходящих из Fv-клона или клона гибридомы антител по изобретению.
Фрагменты антител
Настоящее изобретение относится к фрагментам антител. В некоторых обстоятельствах использование фрагментов антител имеет преимущества относительно использования интактных антител. Меньший размер фрагментов позволяет быстрое выведение и может обеспечивать улучшенный доступ к солидным опухолям.
Для получения фрагментов антител были разработаны различные способы. Обычно, эти фрагменты получали протеолитическим расщеплением интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако эти фрагменты в настоящее время можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Fab-, Fv- и scFv-фрагменты антител могут быть экспрессированы и секретированы в E. coli, таким образом, позволяя простой способ получения больших количеств этих фрагментов. Фрагменты антител можно выделять из фаговых библиотек антител, описанных выше. Альтернативно, фрагменты Fab'-SH можно выделять непосредственно из E. coli и химически связывать с получением фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-167 (1992)). В соответствии с другим подходом фрагменты F(ab')2 можно выделять непосредственно из рекомбинантной культуры клеток-хозяев. Fab- и F(ab')2-фрагмент с повышенным временем полужизни in vivo, содержащий остатки связывающего рецептор спасения эпитопа, описан в патенте США 5869046. Другие способы получения фрагментов антител будут очевидны квалифицированному специалисту. В других вариантах осуществления предпочтительное для выбора антитело представляет собой одноцепочечный Fv-фрагмент (scFv) (см., например, WO 93/16185; патент США 5571894; и патент США 5587458). Fv и sFv являются единственными типами с интактными антиген-связывающими участками, которые лишены константных участков; таким образом, они подходят для снижения неспецифического связывания в ходе применения in vivo. Можно конструировать слитые с sFv белки с получением эффекторного белка либо на N-, либо на С- конце sFv. См. Antibody Engineering, ed. Borrebaeck, выше. Фрагмент антитела также может представлять собой "линейное антитело", например, как описано в патенте США 5641870. Такие линейные фрагменты антител могут быть моноспецифическими или биспецифическими.
Гуманизированные антитела
Настоящее изобретение включает гуманизированные антитела. В данной области известны различные способы гуманизации антител, не являющихся человеческими. Например, гуманизированное антитело может обладать одним или несколькими встроенными в него аминокислотными остатками из источника, который не является человеческим. Эти не являющиеся человеческими аминокислотные остатки часто называют "импортными" остатками, которые, как правило, взяты из "импортного" вариабельного домена. Гуманизацию, главным образом, можно проводить в соответствии со способом Winter и коллег (Jones et al. (1986) Nature, 321:522-525; Riechmann et al. (1988) Nature, 332:323-327; Verhoeyen et al. (1988) Science, 239:1534-1536), с помощью замены CDR или последовательностями CDR крысы соответствующих последовательностей антитела человека. Таким образом, такие "гуманизированные" антитела представляют собой химерные антитела (патент США 4816567), где по существу менее интактной вариабельной области человека замещено соответствующей последовательностью вида, не относящегося к человеку. На практике, гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки гипервариабельной области и, возможно, некоторые остатки FR заменены остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека, как легких, так и тяжелых цепей, для использования при получении гуманизированных антител является очень важным для снижения антигенности. Согласно так называемому способу "наилучшего соответствия", последовательность вариабельного домена антитела грызуна анализируют относительно целой библиотеки известных последовательностей вариабельных доменов человека. Последовательность человека, которая является наиболее сходной с последовательностью грызуна, затем принимают в качестве каркасной области человека для гуманизированного антитела (Sims et al. (1993) J. Immunol, 151:2296; Chothia et al. (1987) J. Mol. Biol, 196:901). В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека из конкретной подгруппы легких или тяжелых цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al. (1992) Proc. Natl. Acad. Sci USA, 89:4285; Presta et al. (1993) J. Immunol, 151:2623).
Кроме того, обычно желательно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других положительных биологических свойств. Для достижения этой цели, в соответствии с предпочтительным способом гуманизированные антитела получают процессом анализа исходных последовательностей и различных предполагаемых гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широко доступными и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и выводят на экран возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Исследование этих выведенных на экран данных позволяет проводить анализ возможной роли остатков в функционировании предполагаемых последовательностей иммуноглобулинов, т.е., анализ остатков, которые влияют на способность иммуноглобулина-кандидата связывать свой антиген. Таким образом, можно отобрать остатки FR и комбинировать их из реципиентной и импортной последовательностей таким образом, чтобы были достигнуты требуемые свойства антител, такие как повышенная аффинность к антигену(ам)-мишени. Как правило, остатки гипервариабельной области оказывают непосредственное и наиболее значительное влияние на связывание антигена.
Антитела человека
Антитела человека против FGFR3 по изобретению можно конструировать путем комбинирования последовательности(ей) вариабельного домена клона Fv, выбранной из библиотек фагового дисплея, полученных из последовательностей человека, с известной последовательностью(ями) константного домена человека, как описано выше. Альтернативно, моноклональные антитела человека против FGFR3 по изобретению можно получать способом гибридом. Клеточные линии миеломы человека и гетеромиеломы мыши-человека для продукции моноклональных антител человека описаны, например, Kozbor J. Immunol, 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol, 147: 86 (1991).
В настоящее время возможным является получение трансгенных животных (например, мышей), которые при иммунизации способны продуцировать полный набор антител человека в отсутствии продукции эндогенных иммуноглобулинов. Например, было описано, что гомозиготная делеция гена участка соединения тяжелых цепей антител (JH) у химерных мышей и мутантных мышей зародышевой линии приводит к полному ингибированию продукции эндогенных антител. Перенос системы генов иммуноглобулинов зародышевой линии человека в таких мутантных мышей зародышевой линии приводит к продукции антител человека при антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255 (1993); Bruggermann et al., Year in Immuno., 7:33 (1993).
Также для получения антител человека из не являющихся человеческими антител, например, антител грызунов, можно использовать перетасовку генов, где антитело человека имеет аффинность и специфичность, аналогичные с исходным не являющимся человеческим антителом. Согласно этому способу, который также называют "импринтингом эпитопов", либо вариабельную область тяжелой цепи, либо вариабельную область легкой цепи, не являющегося человеческим фрагмента антитела, полученного способами фагового дисплея, как описано в настоящем документе, заменяют набором генов V-доменов человека, создавая популяцию химер scFv или Fab с не являющейся человеческой цепью/являющейся человеческой цепью. Селекция с антигеном приводит к выделению химерного scFv или Fab с не являющееся человеческой цепью/являющейся человеческой цепью, где человеческая цепь восстанавливает антиген-связывающий центр, разрушенный при удалении соответствующей не являющейся человеческой цепи в первичном клоне фагового дисплея, т.е. эпитоп определяет (осуществляет импринтинг) выбор цепи человека-партнера. Когда этот процесс повторяют для замены оставшейся не являющейся человеческой цепи, получают антитело человека (см. PCT WO 93/06213, опубликованную 1 апреля 1993 года). В отличие от традиционной гуманизации не являющихся человеческими антител путем пересадки CDR, этот способ обеспечивает полностью человеческие антитела, которые не имеют остатков FR или CDR происхождением не из человека.
Биспецифические антитела
Биспецифические антитела представляют собой моноклональные антитела, предпочтительно, антитела человека или гуманизированные антитела, которые обладают специфичностью связывания в отношении по меньшей мере двух различных эпитопов. В данном случае одна из специфичностей связывания направлена на FGFR3, а другая направлена на любой другой антиген. Характерные биспецифические антитела могут связываться с двумя различными эпитопами FGFR3. Биспецифические антитела также можно использовать для направления цитотоксических средств к клеткам, которые экспрессируют FGFR3. Эти антитела обладают FGFR3-связывающим плечом и плечом, которое связывает цитотоксическое средство (например, сапорин, антитело против интерферона-α, алкалоид барвинка, A-цепь рицина, метотрексат или гаптен радиоактивного изотопа). Биспецифические антитела могут быть получены как полноразмерные антитела или фрагменты антител (например, биспецифические антитела F(ab')2).
Способы получения биспецифических антител известны в данной области. Обычно, рекомбинантный способ получения полноразмерных биспецифических антител основан на совместной экспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулинов, где две цепи обладают различной специфичностью (Milstein and Cuello, Nature, 305:537 (1983)). Вследствие случайной сборки тяжелых и легких цепей иммуноглобулинов, такие гибридомы (квадромы) могут продуцировать смесь из 10 различных молекул антител, из которых только одна обладает правильной биспецифичной структурой. Очистка правильных молекул, которую обычно проводят с помощью стадий аффинной хроматографии, является довольно трудоемкой, и выход продукта является низким. Аналогичные способы описаны в WO 93/08829, опубликованной 13 мая 1993 г. и в Traunecker et al., EMBO J., 10:3655 (1991).
В соответствии с другим подходом проводят слияние вариабельных доменов антитела с требуемой специфичностью связывания (участками связывания антитело-антиген) с последовательностями константных доменов иммуноглобулинов. Слияние проводят, например, с константным доменом тяжелой цепи Ig, содержащим по меньшей мере часть шарнирной области, CH2 и CH3. Предпочтительно, наличие первой константной области тяжелой цепи (CH1), содержащей участок, необходимый для связывания легкой цепи, присутствующей по меньшей мере в одной из подвергаемых слиянию конструкций. ДНК, кодирующие предназначенные для слияния компоненты тяжелых цепей иммуноглобулинов и, если желательно, легкой цепи иммуноглобулина, встраивают в отдельные векторы экспрессии, и котрансфицируют в подходящий организм-хозяин. Это обеспечивает значительную гибкость при регулировании соотношений трех полипептидных фрагментов друг с другом в вариантах осуществления, где неравные соотношения трех полипептидных цепей в конструкции обеспечивает оптимальный выход. Однако возможным является встраивание двух или всех трех полипептидных цепей в один вектор экспрессии, если экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к высокому выходу продукта или если соотношения не имеют существенного влияния.
В предпочтительном варианте осуществления этого подхода, биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с одной специфичностью связывания на одном плече, и пары тяжелая цепь-легкая цепь гибридного иммуноглобулина (обладающей другой специфичностью связывания) на другом плече. Было обнаружено, что такая ассиметричная структура облегчает разделение требуемого биспецифического соединения и нежелательных сочетаний цепей иммуноглобулинов, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ разделения. Этот подход описан в WO 94/04690. Для более подробного описания получения биспецифических антител см., например, Suresh et al., Methods in Enzymology, 121:210 (1986).
В соответствии с другим подходом, для максимального увеличения процентного количества гетеродимеров, которые выделяют из рекомбинантных клеточных культур, можно сконструировать область контакта между парой молекул антител. Предпочтительная область контакта содержит по меньшей мере часть CH3-домена константного домена антитела. В этом способе одну или несколько небольших боковых цепей аминокислот из области контакта первой молекулы антитела заменяют на более крупные боковые цепи (например, тирозин или триптофан). В области контакта второй молекулы антитела создают компенсирующие "полости" идентичного или сходного с крупной боковой цепью(ями) размера с помощью замены крупных боковых цепей аминокислот на цепи меньших размеров (например, аланин или треонин). Это обеспечивает механизм для повышения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают поперечно-сшитые антитела или "гетероконъюгаты" антител. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое с биотином. Такие антитела, например, были предложены для нацеливания клеток иммунной системы против нежелательных клеток (патент США 4676980), и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/00373 и EP 03089). Гетероконъюгаты антител можно получать с использованием любого подходящего способа поперечного сшивания. Подходящие средства для поперечного сшивания хорошо известны в данной области, и описаны в патенте США 4676980, совместно с рядом способов поперечного сшивания.
Способы получения биспецифических антител из фрагментов антител также описаны в литературе. Например, биспецифические антитела могут быть получены с помощью химических связей. В Brennan et al., Science, 229:81 (1985) описан способ, где интактные антитела протеолитически расщепляют с получением F(ab')2-фрагментов. Эти фрагменты восстанавливают в присутствии вещества, образующего комплексы с дитиолами, арсенита натрия, для стабилизации соседних дитиолов и предотвращения образования межмолекулярного дисульфида. Затем полученные Fab'-фрагменты преобразуют в производные с тионитробензоатом (TNB). Одно из производных Fab'-TNB затем повторно преобразуют в Fab'-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB с получением биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве средств для селективной иммобилизации ферментов.
Последние достижения упростили прямое выделение из E. coli фрагментов Fab'-SH, которые можно химически связывать с получением биспецифических антител. В Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описано получение полностью гуманизированной молекулы F(ab')2 биспецифического антитела. Осуществляли секрецию из E. coli каждого Fab'-фрагмента по отдельности и их подвергали прямому химическому соединению in vitro с образованием биспецифического антитела. Биспецифическое антитело, полученное таким образом, обладало способностью связываться с клетками, сверхэкспрессирующими рецептор HER2, и нормальными T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против мишеней, представляющих собой рак молочной железы человека.
Также были описаны различные способы получения и выделения фрагментов биспецифических антител непосредственно из рекомбинантной клеточной культуры. Например, были получены биспецифические антитела с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды лейциновой молнии белков Fos и Jun связывали с Fab'-участками двух различных антител с помощью слияния генов. Гомодимеры антител восстанавливали в шарнирной области с получением мономеров, а затем повторно окисляли с получением гетеродимеров антител. Этот способ также можно использовать для получения гомодимеров антител. Способ "диател", описанный Hollinger et al., Proc. Natl. Acad. Sci USA, 90:6444-6448 (1993) обеспечивает альтернативный механизм получения биспецифических фрагментов антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) с помощью линкера, который является слишком коротким для возможности образования пары между двумя доменами на одной и той же цепи. Таким образом, домены VH и VL одного фрагмента вынуждены образовывать пару с комплементарными доменами VL и VH другого фрагмента, таким образом, образуя два антиген-связывающих центра. Также описан другой поход для получения фрагментов биспецифических антител с использованием димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol, 152: 5368 (1994).
Также предусмотрены антитела более чем c двумя валентностями. Например, можно получать триспецифические антитела. Tutt et al. J. Immunol. 147: 60 (1991).
Поливалентные антитела
Поливалентное антитело может интернализовываться (и/или катаболизироваться) быстрее, чем двухвалентное антитело, клеткой, экспрессирующей антиген, с которым антитела связываются. Антитела по настоящему изобретению могут быть поливалентными антителами (которые могут относиться к классу, отличному от класса IgM) с тремя или более антиген-связывающими участками (например, четырехвалентные антитела), которые могут быть легко получены с помощью рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Поливалентное антитело может содержать домен димеризации и три или более антиген-связывающих участков. Предпочтительный домен димеризации содержит (или состоит из них) Fc-участок или шарнирную область. В этом случае антитело будет содержать Fc-область и три или более антиген-связывающих участка со стороны N-конца Fc-участка. Предпочтительное поливалентное антитело, представленное в настоящем документе, содержит (или состоит из них) от трех до приблизительно восьми, однако, предпочтительно, четыре, антиген-связывающих участка. Поливалентное антитело содержит по меньшей мере одну полипептидную цепь (и предпочтительно, две полипептидные цепи), где полипептидная цепь(и) содержит два или более вариабельных доменов. Например, полипептидная цепь(и) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидную цепь Fc-участка, X1 и X2 представляют собой аминокислоту или полипептид, и n равно 0 или 1. Например, полипептидная цепь(и) может содержать: цепь VH-CH1-подвижный линкер-VH-CH1-Fc-участок; или цепь VH-CH1-VH-CH1-Fc-участок. Поливалентное антитело, представленное в настоящем документе, кроме того, предпочтительно, содержит два (и, предпочтительно, четыре) полипептида вариабельного домена легкой цепи. Поливалентное антитело, представленное в настоящем документе, например, может содержать от приблизительно двух до приблизительно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, рассматриваемые в настоящем документе, содержат вариабельный домен легкой цепи и, необязательно, дополнительно содержат CL-домен.
Варианты антител
В некоторых вариантах осуществления предусмотрена модификация(и) аминокислотной последовательности антител, описанных в настоящем документе. Например, может быть желательным повышение аффинности связывания и/или других биологических свойств антитела. Варианты антител по аминокислотной последовательности могут быть получены путем внесения соответствующих изменений в нуклеотидную последовательность, кодирующую антитело, или с помощью пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Для достижения конечного продукта можно проводить любую комбинацию из делеции, вставки и замены, при условии, что конечная конструкция обладает желаемыми характеристиками. Аминокислотные изменения можно вносить в рассматриваемую аминокислотную последовательность антитела во время получения этой последовательности.
Подходящий способ идентификации определенных остатков или участков антитела, которые являются предпочтительными участками для мутагенеза, называют "сканирующим аланином мутагенезом" как описано Cunningham and Wells (1989) Science, 244:1081-1085. В данном случае, выявляют остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют на нейтральную или отрицательно заряженную аминокислоту (наиболее предпочтительно, аланин или полиаланин) для нарушения взаимодействия аминокислот с антигеном. Те положения аминокислот, которые демонстрируют функциональную чувствительность к замещению, затем подтверждают с помощью внесения дополнительных или других вариантов в участки замещения, или вместо них. Таким образом, поскольку участок для внесения изменения в аминокислотную последовательность является предопределенным, нет необходимости в непосредственной предопределенности характера мутации. Например, для анализа действия мутации в данной области, в кодоне-мишени или участке-мишени проводят сканирующий ala или случайный мутагенез и экспрессированные иммуноглобулины подвергают скринингу в отношении требуемой активности.
Вставки в аминокислотную последовательность включают N- и/или C-концевые слитые последовательности, обладающие длиной в диапазоне от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или нескольких аминокислотных остатков внутрь последовательности. Примеры концевых вставок включают антитело с N-концевым метионильным остатком или антитело, слитое с цитотоксическим полипептидом. Другие варианты вставок в молекулу антитела включают слияние N- или C-конца антитела с ферментом (например, ADEPT) или полипептидом, который повышает время полужизни антитела в сыворотке.
Гликозилирование полипептидов, как правило, является либо N-связанным, либо O-связанным. N-связанное гликозилирование относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту, за исключением пролина, представляют собой последовательности, распознаваемые для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, обычно, к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования к антителу удобно проводить, изменяя аминокислотную последовательность, чтобы происходило внесение одной или нескольких из описанных выше трипептидных последовательностей (для участков N-связанного гликозилирования). Также изменение можно проводить с помощью добавления или замены одного или нескольких остатков серина или треонина в последовательности исходного антитела (для участков О-связанного гликозилирования).
Если антитело содержит Fc-область, то можно изменять углевод, связанный с ним. Например, антитела со зрелой углеводной структурой, которая лишена фукозы, присоединенной к Fc-области антитела, описаны в патентной заявке США No US 2003/0157108 (Presta, L.). Также см. US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Антитела с биссекторным N-ацетилглюкозамином (GlcNAc) в углеводе, присоединенном к Fc-области антитела, упоминаются в WO 2003/011878, Jean-Mairet et al. и патенте США 6602684, Umana et al. Антитела по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области антитела, описаны в WO 1997/30087, Patel et al. Также см. WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.), касающиеся антител с измененным углеводом, присоединенным к их Fc-области. Также см. US 2005/0123546 (Umana et al.) в отношении антиген-связывающих молекул с модифицированным гликозилированием.
Предпочтительный для настоящего изобретения вариант по гликозилированию содержит Fc-область, где углеводная структура, присоединенная к Fc-области, лишена фукозы. Такие варианты могут иметь улучшенную функцию ADCC. Необязательно, Fc-область, кроме того, содержит в ней одну или несколько аминокислотных замен, которые далее повышают ADCC, например, замены в положениях 298, 333 и/или 334 Fc-области (нумерация остатков Eu). Примеры публикаций, относящихся к "дефукозилированным" или "дефицитным по фукозе" антителам, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают клетки Lec13 CHO, дефицитные по фукозилированию белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США No US 2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., особенно в примере 11), и клеточные линии с нокаутом, такие как клетки CHO с нокаутом генов альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004)).
Другой тип варианта представляет собой вариант с аминокислотной заменой. Эти варианты имеют по меньшей мере один (по меньшей мере два, по меньшей мере три, по меньшей мере 4 или более) аминокислотный остаток в молекуле антитела, замененный отличающимся остатком. Представляющие наибольший интерес участки для мутагенеза с заменами включают гипервариабельные области, однако также предусмотрены изменения FR. Консервативные замены представлены в таблице 1 под заголовком "Предпочтительные замены". Если такие замены приводят к изменению биологической активности, тогда можно вносить более существенные изменения, обозначенные как "характерные замены" в таблице 1 или как дополнительно описано ниже в отношении классов аминокислот, и продукты подвергать скринингу.
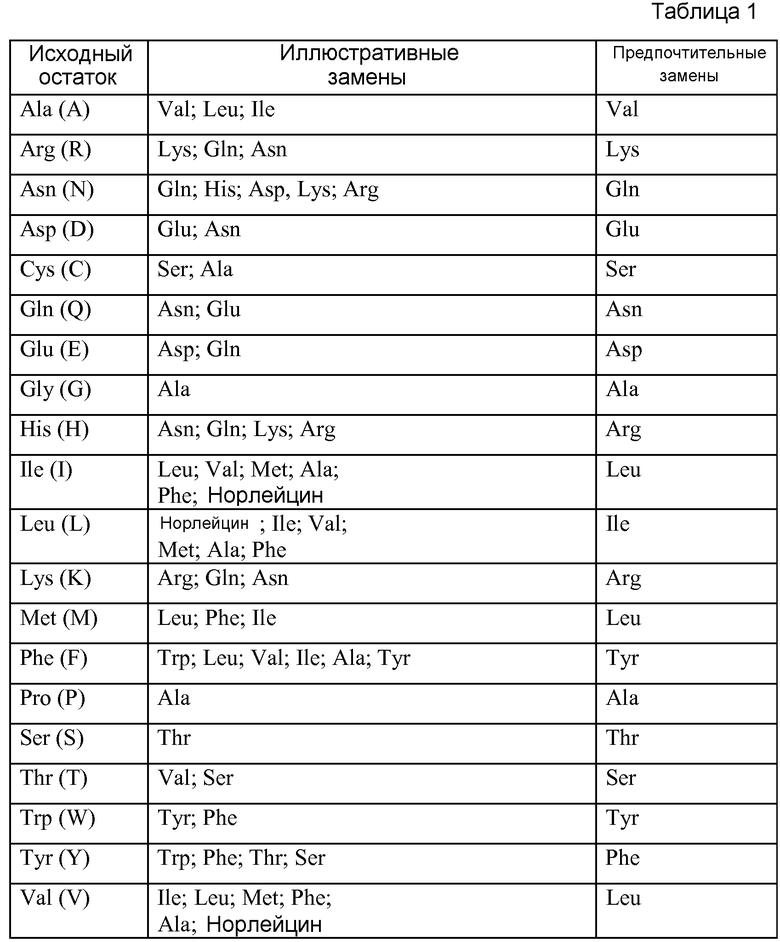
Существенные модификации биологических свойств антитела проводят, выбирая замены, которые значительно отличаются по их эффекту на поддержание (a) структуры полипептидного остова в области замены, например, такой как конформация в виде слоя или спирали, (b) заряда или гидрофобности молекулы в участке-мишени, или (c) объема боковой цепи. Природные остатки можно разделить на группы на основе общих свойств их боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr, asn, gln;
(3) кислые: asp, glu;
(4) основные: his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены включают замену члена одного из этих классов членом из другого класса.
Один из типов варианта с заменами включает замену одного или нескольких остатков гипервариабельного участка исходного антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный вариант(ы), выбранный для дальнейшей разработки, будет обладать улучшенными биологическими свойствами относительно исходного антитела, из которого они получены. Удобный путь получения таких вариантов с заменами вовлекает созревание аффинности с использованием фагового дисплея. Коротко, несколько областей гипервариабельной области (например, 6-7 областей) подвергают мутации с получением всех возможных аминокислотных замен в каждом участке. Полученные таким образом варианты антитела экспонируются на частицах нитевидных фагов в качестве молекул, слитых по меньшей мере с продуктом гена III фага M13, упакованных в каждую частицу. Затем экспонированные на фаге варианты подвергают скринингу в отношении их биологической активности (например, аффинности связывания), как описано в настоящем документе. В целях идентификации участков гипервариабельной области-кандидатов для модификации можно проводить сканирующий аланином мутагенез, идентифицируя остатки гипервариабельной области, вносящих значительный вклад в связывание антигена. Альтернативно или дополнительно, может быть целесообразным анализ кристаллической структуры комплекса антиген-антитело в целях идентификации точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки являются кандидатами для замены в соответствии со способами, известными в данной области, включая способы, приведенные в настоящем документе. После получения таких вариантов панель вариантов подвергают скринингу с использованием способов, известных в данной области, включая способы, описанные в настоящем документе, и варианты с наилучшими свойствами в одном или нескольких соответствующих анализах можно отбирать для дальнейшей разработки.
Молекулы нуклеиновых кислот, кодирующие варианты антитела по аминокислотной последовательности, получают различными способами, известными в данной области. Эти способы включают, но ими не ограничиваются, выделение из природного источника (в случае природных вариантов аминокислотной последовательности) или получение олигонуклеотид-опосредуемым (или сайт-направленным) мутагенезом, мутагенезом с помощью PCR, и кассетным мутагенезом ранее полученного варианта или не являющейся вариантом версии антитела.
Может быть желательным внесение одной или нескольких аминокислотных модификаций в Fc-область антител по изобретению, тем самым получая вариант по Fc-области. Вариант по Fc-области может включать последовательность Fc-области человека (например, Fc-области IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях, включая положение цистеина шарнирной области.
Согласно описанию и указаниям в данной области предусматривается, что в некоторых вариантах осуществления антитело по изобретению может содержать одно или несколько изменений по сравнению с альтернативным антителом дикого типа, например, в Fc-области. Эти антитела, тем не менее, сохраняют по существу те же характеристики, которые требуются для терапевтической применимости по сравнению с их аналогом дикого типа. Например, полагают, что в Fc-область можно вносить определенные изменения, которые могут привести к измененному (т.е., либо повышенному, либо сниженному) связыванию C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в WO99/51642. Также см. Duncan & Winter, Nature 322:738-40 (1988); патент США 5648260; патент США 5624821; и WO94/29351, касающиеся других примеров вариантов Fc-области. В WO00/42072 (Presta) и WO 2004/056312 (Lowman) описаны варианты антител с улучшенным или сниженным связыванием с FcR. Содержание этих патентных публикаций конкретно включено в настоящий документ в качестве ссылки. Также см., Shields et al. J. Biol. Chem. 9(2): 6591-6604 (2001). Антитела с увеличенным временем полужизни и повышенным связыванием с неонатальным Fc-рецептором (FcRn), которые отвечают за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US2005/0014934A1 (Hinton et al.). Эти антитела содержат Fc-область с одной или несколькими заменами в ней, которые повышают связывание Fc-области с FcRn. Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной способностью связывать C1q описаны в патенте США 6194551B1, WO99/51642. Содержание этих патентных публикаций конкретно включено в настоящий документ в качестве ссылки. Также см., Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Производные антител
Антитела по настоящему изобретению можно дополнительно модифицировать так, чтобы они содержали дополнительные небелковые группы, которые известны в данной области и легко доступны. Предпочтительно, группы, подходящие для получения производного антитела, представляют собой растворимые в воде полимеры. Неограничивающие примеры растворимых в воде полимеров включают, но ими не ограничиваются, полиэтиленгликоль (PEG), сополимеры этиленгликоль/пропиленгликоль, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилен/малеиновый ангидрид, полиаминокислоты (либо гомополимеры, либо случайные сополимеры), и декстран или поли(н-винилпирролидон)полиэтиленгликоль, пропропиленгликоль гомополимеры, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества при получении, благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу, и он может быть разветвленным или неразветвленным. Количество полимеров, связанных с антителом, может изменяться, и если связано более одного полимера, то они могут представлять собой одинаковые или разные молекулы. Как правило, количество и/или тип полимеров, используемых для получения производного, можно определять, исходя из соображений, включающих, но ими не ограничивающихся, конкретные свойства или функции антитела, подлежащего усовершенствованию, и того, будет ли производное антитела использовано в терапии в определенных условиях и тому подобное.
Скрининг антител с желаемыми свойствами
Антитела по настоящему изобретению могут быть охарактеризованы по их физическим/химическим свойствам и биологическим функциям с помощью различных анализов, известных в данной области (некоторые из которых описаны в настоящем документе). В некоторых вариантах осуществления антитела характеризуются любым одним или несколькими из снижения или блокирования связывания FGF (такого как FGF1 и/или FGF9), снижения или блокирования активации FGFR3, снижения или блокирования следующей после FGFR3 молекулярной передачи сигнала, нарушения или блокирования связывания FGFR3 с лигандом (например, FGF1, FGF9), уменьшения или блокирования димеризации FGFR3, стимуляции образования мономерного FGFR3, связывания с мономерным FGFR3, и/или лечения и/или профилактики опухоли, клеточно-пролиферативного нарушения или злокачественной опухоли; и/или лечения или профилактики нарушения, связанного с экспрессией и/или активностью FGFR3 (такой как увеличенная экспрессия и/или активность FGFR3). В некоторых вариантах осуществления антитела подвергают скринингу в отношении повышенной активации FGFR3, повышенной следующей после FGFR3 молекулярной передачи сигнала, апоптотической активности, подавления FGFR3 и эффекторной функции (например, активности ADCC).
Очищенные антитела, кроме того, могут быть охарактеризованы с помощью ряда анализов, включая, но ими не ограничиваясь, N-концевое секвенирование, анализ аминокислот, неденатурирующую эксклюзионную высокоэффективную жидкостную хроматографию (ВЭЖХ), масс-спектрометрию, ионообменную хроматографию и расщепление папаином.
В определенных вариантах осуществления изобретения антитела, полученные по настоящему изобретению, анализируют на их биологическую активность. В некоторых вариантах осуществления антитела по настоящему изобретению тестируют на их антиген-связывающую активность. Анализы связывания антигена, которые известны в данной области и которые можно использовать для настоящего изобретения, включают, но ими не ограничиваются, любые прямые или конкурентные анализы связывания с использованием способов, таких как вестерн-блоттинг, радиоиммунные анализы, ELISA (твердофазный иммуноферментный анализ), "сэндвич"-имммуноанализы, анализы иммунопреципитации, флуоресцентные иммуноанализы и иммуноанализы с белком A. Характерный анализ связывания антигена и другие анализы представлены ниже в разделе "Примеры".
Если желательно, то антитело против FGFR3, которое ингибирует рост клеток, антитело-кандидат можно тестировать в анализах in vitro и/или in vivo, которые определяют ингибирование роста клеток. Если желательно, то антитело против FGFR3, которое стимулирует или не стимулирует апоптоз, антитело-кандидат можно тестировать в анализах, которые определяют апоптоз. Способы исследования роста и/или пролиферации злокачественной клетки или определения апоптоза злокачественной клетки хорошо известны в данной области и некоторые из них описаны и проиллюстрированы в настоящем документе. Характерные способы определения роста и/или пролиферации и/или апоптоза клеток включают, например, анализ включения BrdU, анализ с MTT, анализ включения [3H]-тимидина (например, анализ TopCount (PerkinElmer)), анализы жизнеспособности клеток (например, CellTiter-Glo (Promega)), анализы фрагментации ДНК, анализы активации каспазы, анализы исключения триптанового синего, анализы морфологии хроматина и тому подобное.
В одном из вариантов осуществления настоящее изобретение относится к антителу, которое обладает эффекторными функциями. В определенных вариантах осуществления измеряют активность Fc антитела. Для подтверждения снижения/устранения видов активности CDC и/или ADCC можно проводить анализы цитотоксичности in vitro и/или in vivo. Например, анализы связывания Fc-рецепторов (FcR) можно проводить, чтобы убедиться, что антитело не связывает FcγR (таким образом, вероятно не имеет активности ADCC), однако сохраняет способность связывать FcRn. Главные опосредующие ADCC клетки, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках обобщенно представлена в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Пример анализа in vitro для оценки активности ADCC представляющей интерес молекулы описан в патентах США 5500362 или 5821337. Анализ для обнаружения активности ADCC также проиллюстирован в настоящем документе. Подходящие эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и природные киллерные (NK) клетки. Альтернативно или дополнительно, активность ADCC представляющей интерес молекулы можно оценивать in vivo, например, в модели на животных, такой как модель, описанная в Clynes et al. Proc. Nat'l Acad. Sci. USA 95:652-656 (1998). Также можно проводить анализы связывания C1q для подтверждения того, что антитело неспособно связывать C1q и, таким образом, лишено активности CDC. Для оценки активации комплемента можно проводить анализ CDC, например, как описано Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Также можно проводить определение связывания FcRn и выведения/времени полужизни in vivo с использованием способов, известных в данной области, например, способов, описанных в разделе "Примеры".
Если желательно, то антитело против FGFR3, которое связывает мономерный FGFR3, антитело-кандидат можно тестировать в анализах (таких как анализы in vitro), которые измеряют связывание с мономерным FGFR3 и стимуляцию образования мономерного FGFR3. Такие анализы известны в данной области и некоторые из анализов описаны и проиллюстрированы в настоящем документе.
Если желательно, то антитело против FGFR3, которое ингибирует димеризацию FGFR3, антитело-кандидат можно тестировать в анализах димеризации, например, как описано и проиллюстрировано в настоящем документе.
В некоторых вариантах осуществления определяют функцию антитела-кандидата в качестве агониста FGFR3. Способы оценки функции или активности в качестве агониста у антител против FGFR3 известны в данной области и некоторые из них также описаны и проиллюстрированы в настоящем документе.
В некоторых вариантах осуществления способность антитела против FGFR3 стимулировать подавление рецептора FGFR3 определяют, например, с использованием способов, описанных и проиллюстрированных в настоящем документе. В одном из вариантов осуществления антитело против FGFR3 инкубируют с подходящими тестируемыми клетками, например, клеточными линиями рака мочевого пузыря (например, RT112), и после подходящего периода времени клеточные лизаты собирают и исследуют в отношении общих уровней FGFR3. Для исследования уровней поверхностного рецептора FGFR3 после инкубации с антителами-кандидатами против FGFR3 также можно использовать FACS-анализ.
Векторы, клетки-хозяева и рекомбинантные способы
Для рекомбинантной продукции антитела по изобретению нуклеиновую кислоту, кодирующую антитело, выделяют и встраивают в реплицирующийся вектор для последующего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, можно легко выделять и секвенировать с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелую и легкую цепи антитела). Доступно множество векторов. Выбор вектора частично зависит от используемой клетки-хозяина. Как правило, предпочтительными клетками-хозяевами является клетки либо прокариотического, либо эукариотического (как правило, из млекопитающих) происхождения. Понятно, что для этой цели можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области могут быть получены от человека или любого вида животного.
a. Получение антител с использованием прокариотических клеток-хозяев:
i. Конструирование векторов
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела, могут быть получены с помощью стандартных рекомбинантных способов. Требуемые полинуклеотидные последовательности можно выделять из продуцирующих антитело клеток, таких как клетки гибридомы, и секвенировать. Альтернативно, полинуклеотиды можно синтезировать с использованием устройства для синтеза нуклеотидов или способами ПЦР. После получения, последовательности, кодирующие полипептиды, встраивают в рекомбинантный вектор, способный реплицировать и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. Для целей настоящего изобретения можно использовать множество векторов, которые доступны и известны в данной области. Выбор соответствующего вектора зависит, главным образом, от размера нуклеиновых кислот, подлежащих встраиванию в вектор, и конкретной клетки-хозяина, подлежащей трансформации с помощью вектора. Каждый вектор содержит различные компоненты, в зависимости от его функции (амплификация или экспрессия гетерологичного полинуклеотида, или обе из них) и его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты векторов обычно включают, но ими не ограничиваются: точку начала репликации, ген селективного маркера, промотор, участок связывания рибосом (RBS), сигнальную последовательность, вставку из гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
Как правило, применительно к этим хозяевам можно использовать плазмидные векторы, содержащие репликон и последовательности контроля из видов, совместимых с клеткой-хозяином. Вектор обычно содержит участок репликации, а также маркирующие последовательности, которые способны обеспечивать фенотипическую селекцию в трансформированных клетках. Например, E. coli обычно трансформируют с использованием pBR322, плазмиды, происходящей из вида E. coli. pBR322 содержит гены, кодирующие устойчивость к ампициллину (Amp) и тетрациклину (Tet) и, таким образом, обеспечивает простые способы идентификации трансформированных клеток. pBR322, ее производные, или другие плазмиды микроорганизмов или бактериофаг также могут содержать, или быть модифицированными, чтобы они содержали, промоторы, которые могут использоваться микроорганизмом для экспрессии эндогенных белков. Примеры производных pBR322, используемые для экспрессии конкретных антител, подробно описаны в Carter et al., патент США 5648237.
Кроме того, применительно к этим хозяевам, в качестве трансформируемых векторов можно использовать фаговые векторы, содержащие репликон и последовательности контроля, которые совместимы с микроорганизмом-хозяином. Например, при получении рекомбинантного вектора можно использовать бактериофаг, такой как λGEM.TM.-11, который можно использовать для трансформации восприимчивых к нему клеток-хозяев, таких как E. coli LE392.
Вектор экспрессии по изобретению может содержать две или более пар промотор-цистрон, кодирующих каждый из полипептидных компонентов. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5') цистрона, которая модулирует его экспрессию. Прокариотические промоторы, как правило, относятся к одному из двух классов: индуцибельные и конститутивные. Индуцибельный промотор представляет собой промотор, который инициирует повышение уровней транскрипции цистрона, находящегося под его контролем, в ответ на изменение условий культивирования, например, на наличие или отсутствие питательных веществ или изменение температуры.
Хорошо известно большое количество промоторов, распознаваемых различными потенциальными клетками-хозяевами. Выбранный промотор можно функционально связывать с цистронной ДНК, кодирующей легкую или тяжелую цепь, путем удаления промотора из ДНК, являющейся его источником, с помощью расщепления ферментом рестрикции, и встраивания выделенной последовательности промотора в вектор по изобретению. Для направления амплификации и/или экспрессии генов-мишеней можно использовать как нативную промоторную последовательность, так и множество гетерологичных промоторов. В некоторых вариантах осуществления используют гетерологичные промоторы, поскольку они позволяют более высокую транскрипцию и более высокий выход экспрессируемого гена-мишени по сравнению с нативным промотором полипептида-мишени.
Промоторы, подходящие для использования с прокариотическими хозяевами, включают промотор PhoA, промоторные системы β-галактамазы и лактозы, триптофановую (trp) промоторную систему и гибридные промоторы, такие как промотор tac или trc. Однако также подходят другие промоторы, которые являются функциональными в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, тем самым позволяя квалифицированному специалисту функционально лигировать их с цистронами, кодирующими требуемые легкие и тяжелые цепи (Siebenlist et al. (1980) Cell 20: 269) с использованием линкеров или адапторов для обеспечения каких-либо требуемых участков рестрикции.
В одном из вариантов осуществления изобретения каждый цистрон в рекомбинантном векторе содержит компонент, представляющий собой секреторную сигнальную последовательность, который направляет перемещение экспрессированных полипептидов через мембрану. Как правило, сигнальная последовательность может быть компонентом вектора, или она может быть частью ДНК полипептида-мишени, которую встраивают в вектор. Сигнальная последовательность, выбранная для целей этого изобретения, должна представлять собой последовательность, которая распознается клеткой-хозяином и подвергается процессингу (т.е. отщепляется сигнальной пептидазой) в ней. Для прокариотических клеток-хозяев, которые не распознают и не осуществляют процессинг сигнальных последовательностей, нативных для гетерологичных полипептидов, сигнальную последовательность заменяют прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Ipp или термостабильного энтеротоксина II (STII), LamB, PhoE, Pe1B, OmpA и MBP. В одном из вариантов осуществления изобретения, сигнальные последовательности, используемые в обоих цистронах экспрессирующей системы, представляют собой сигнальные последовательности STII или их варианты.
В другом аспекте продукция иммуноглобулинов по изобретению может происходить в цитоплазме клетки-хозяина, и, таким образом, она не требует присутствия секреторных сигнальных последовательностей в каждом цистроне. В этом отношении, легкие и тяжелые цепи иммуноглобулинов экспрессируются, сворачиваются и собираются с образованием функциональных иммуноглобулинов в цитоплазме. Определенные штаммы-хозяева (например, штаммы trxB- E. coli) обеспечивают условия в цитоплазме, которые способствуют образованию дисульфидных связей, тем самым обеспечивая правильное сворачивание и сборку экспрессированных субъединиц белков. Proba and Pluckthun Gene, 159:203 (1995).
Прокариотические клетки-хозяева, подходящие для экспрессии антител по изобретению, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры подходящих бактерий включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, виды Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления используют грамотрицательные клетки. В одном из вариантов осуществления, в качестве хозяев для изобретения используют клетки E. coli. Примеры штаммов E. coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D. C: American Society for Microbiology, 1987), pp. 1190-1219; депонированный образец ATCC 27325) и его производные, включая штамм 33D3, имеющий генотип W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTΔ (nmpc-fepE) degP41 kanR (патент США 5639635). Также подходят другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coliλ 1776 (ATCC 31537) и E. coli RV308 (ATCC 31608). Эти примеры являются иллюстративными, но не ограничивающими. Способы конструирования производных любой из указанных выше бактерий, имеющих определенные генотипы, известны в данной области и описаны, например, в Bass et al., Proteins, 8: 309-314 (1990). Как правило, необходимо выбирать соответствующие бактерии, учитывая способность к репликации репликона в клетках бактерий. Например, виды E. coli, Serratia или Salmonella можно подходящим образом использовать в качестве хозяев, когда для предоставления репликона используют хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Как правило, клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и может быть желательным добавление в клеточную культуру дополнительных ингибиторов протеаз.
ii. Продукция антител
Клетки-хозяева трансформируют описанными выше векторами экспрессии и культивируют в общепринятых питательных средах, модифицированных соответствующим образом для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности.
Трансформация означает внесение ДНК в прокариотического хозяина, чтобы ДНК была реплицируемой, либо в качестве внехромосомного элемента, либо в качестве элемента, встраивающегося в хромосому. В зависимости от используемой клетки-хозяина, трансформацию проводят с использованием стандартных способов, подходящих для таких клеток. Как правило, для бактериальных клеток, которые содержат плотные барьеры в виде клеточных стенок, применяют обработку кальцием с использованием хлорида кальция. В другом способе трансформации используют полиэтиленгликоль/DMSO. Другим используемым способом является электропорация.
Прокариотические клетки, используемые для получения полипептидов по изобретению, выращивают в средах, известных в данной области и подходящих для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают бульон Луриа (LB) вместе с необходимыми питательными веществами. В некоторых вариантах осуществления, среда также содержит агент для селекции, выбранный, исходя из конструкции вектора экспрессии, для селективного обеспечения роста прокариотических клеток, содержащих вектор экспрессии. Например, ампициллин добавляют в среду для выращивания клеток, экспрессирующих ген устойчивости к ампициллину.
Также, помимо источников углерода, азота и неорганического фосфата, можно добавлять любые необходимые добавки в соответствующих концентрациях, добавляемые отдельно или в качестве смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно, культуральная среда может содержать один или несколько восстановителей, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритрита и дитиотреитола.
Прокариотические клетки-хозяева культивируют при подходящих температурах. Для выращивания E. coli, например, предпочтительная температура находится в диапазоне от приблизительно 20°С до приблизительно 39°C, более предпочтительно, от приблизительно 25°C до приблизительно 37°C, более предпочтительно, она составляет приблизительно 30°С. pH среды может быть любым в диапазоне от приблизительно 5 до приблизительно 9, в зависимости, главным образом, от организма хозяина. Для E. coli, pH, предпочтительно, составляет от приблизительно 6,8 до приблизительно 7,4, и более предпочтительно, приблизительно 7,0.
Если в векторе экспрессии по изобретению используют индуцибельный промотор, то экспрессию белка индуцируют в условиях, подходящих для активации промотора. В одном из вариантов осуществления изобретения для контроля транскрипции полипептидов используют промоторы PhoA. Таким образом, для индукции трансформированные клетки культивируют в среде с ограниченным содержанием фосфата. В определенных вариантах осуществления среда с ограниченным содержанием фосфата представляет собой среду C.R.A.P (см., например, Simmons et al., J. Immunol. Methods (2002), 263:133-147). Можно использовать множество других индуцирующих веществ, в соответствии с используемой векторной конструкцией, как известно в данной области.
В одном из вариантов осуществления экспрессируемые полипептиды по настоящему изобретению секретируются в периплазму клетки-хозяина и из нее их выделяют. Выделение белка, как правило, включает разрушение микроорганизма, как правило, такими способами, как осмотический шок, обработка ультразвуком или лизис. После разрушения клеток, клеточный дебрис или целые клетки можно удалять центрифугированием или фильтрацией. Кроме того, белки можно очищать, например, хроматографией с аффинной смолой. Альтернативно, белки могут транспортироваться в культуральную среду и могут быть выделены из нее. Клетки можно удалять из культуры и культуральный супернатант фильтровать и концентрировать для последующей очистки продуцированных белков. Экспрессированные полипептиды можно далее выделять и идентифицировать с использованием широко известных способов, таких как электрофорез в полиакриламидном геле (PAGE) и вестерн-блот анализ.
В одном из аспектов изобретения продукцию антитела проводят в больших количествах путем процесса ферментации. Для продукции рекомбинантных белков доступны различные крупномасштабные способы ферментации с подпиткой. Крупномасштабная ферментация имеет емкость по меньшей мере 1000 литров, предпочтительно, приблизительно от 1000 до 100000 литров. В этих ферментерах используются перемешивающие лопасти для распределения кислорода и питательных веществ, особенно глюкозы (предпочтительного источника углерода/энергии). Мелкомасштабная ферментация относится, главным образом, к ферментации в ферментере, который имеет емкость не более чем приблизительно 100 литров, и она может находиться в диапазоне от приблизительно 1 литра до приблизительно 100 литров.
В процессе ферментации индукцию экспрессии белка, как правило, начинают после того, как клетки вырастают в подходящих условиях до требуемой плотности, например, OD550 приблизительно 180-220, на той стадии, когда клетки находятся в ранней стационарной фазе. Можно использовать различные индукторы в соответствии с используемой конструкцией вектора, как известно в данной области и как описано выше. Перед индукцией клетки можно выращивать в течение более коротких периодов времени. Клетки, как правило, индуцируют в течение приблизительно 12-50 часов, хотя можно использовать более длительную или более короткую индукцию.
Для повышения выхода продукции и качества полипептидов по изобретению, можно модифицировать различные условия ферментации. Например, для улучшения надлежащей сборки и сворачивания секретируемых полипептидов антитела, можно использовать дополнительные векторы, сверхэкспрессирующие белки-шапероны, такие как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил-цис,транс-изомераза с активностью шаперона) для котрансформации прокариотических клеток-хозяев. Было показано, что белки-шапероны способствуют правильному сворачиванию и растворимости гретерологичных белков, продуцируемых в бактериальных клетках-хозяевах. Chen et al. (1999) J Bio Chem 274: 19601-19605; Georgiou et al., патент США 6083715; Georgiou et al., патент США 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275:17100-17105; Ramm and Pluckthun (2000) J. Biol. Chem. 275:17106-17113; Arie et al. (2001) Mol. Microbiol. 39: 199-210.
Для минимизации протеолиза экспрессируемых гетерологичных белков (особенно белков, которые являются чувствительными к протеолизу), для настоящего изобретения можно использовать определенные штаммы-хозяева с дефицитом протеолитических ферментов. Например, штаммы клеток-хозяев можно модифицировать для обеспечения генетической мутации(ий) в генах, кодирующих известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их комбинации. Доступны некоторые штаммы E. coli с дефицитом протеаз и они описаны, например, в Joly et al. (1998), выше; Georgiou et al., патент США 5264365; Georgiou et al., патент США 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном из вариантов осуществления в качестве клеток-хозяев в экспрессирующей системе по изобретению используют штаммы E. coli с дефицитом протеолитических ферментов и трансформированные плазмидами, сверхэкспрессирующими один или несколько белков-шаперонов.
(iii) Очистка антитела
Можно использовать стандартные способы очистки белка, известные в данной области. Примерами способов очистки являются следующие способы: фракционирование на иммуноаффинных или ионообменных колонках, осаждение с этанолом, обращено-фазовая ВЭЖХ, хроматография на диоксиде кремния или на катионообменной смоле, такое как DEAE, хроматофокусирование, SDS-PAGE, осаждение с сульфатом аммония и гель-фильтрация с использованием, например, Sephadex G-75.
В одном из аспектов для иммуноаффинной очистки продуктов антитела по изобретению используют белок A, иммобилизованный на твердой фазе. Белок A представляет собой белок клеточной стенки массой 41 кДа из Staphylococcus aureas, который связывается с высокой аффинностью с Fc-областью антител. Lindmark et al., (1983) J. Immunol. Meth. 62: 1-13. Твердая фаза, на которую иммобилизуют белок A, может представлять собой колонку, содержащую поверхность из стекла или диоксида кремния, или она может представлять собой колонку с контролируемым размером пор или колонку с кремниевой кислотой. В некоторых вариантах применениях колонка покрыта реагентом, таким как глицерин, для возможного предотвращения неспецифического прикрепления примесей.
В качестве первой стадии очистки, препарат, полученный из клеточной культуры, как описано выше, можно наносить на твердую фазу с иммобилизованным белком A для обеспечения специфического связывания представляющего интерес антитела с белком A. Затем твердую фазу можно промывать для удаления примесей, неспецифично связавшихся с твердой фазой. В конце представляющее интерес антитело отделяют от твердой фазы путем элюирования.
b. Получение антител с использованием эукариотических клеток-хозяев
Компоненты вектора, как правило, включают, но ими не ограничиваются, один или несколько из следующих: сигнальная последовательность, точка начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
(i) Компонент - сигнальная последовательность
Вектор для использования в эукариотической клетке-хозяине также может содержать сигнальную последовательность или другой полипептид, обладающий участком для специфичного расщепления на N-конце зрелого белка или представляющего интерес полипептида. Выбранная гетерологичная сигнальная последовательность, предпочтительно, представляет собой последовательность, которая распознается и преобразуется (т.е., расщепляется сигнальной пептидазой) в клетке-хозяине. Для экспрессии в клетках млекопитающих, доступны сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, сигнальная последовательность gD вируса простого герпеса.
ДНК для такого предшествующего участка лигируют в рамке считывания с ДНК, кодирующей антитело.
(ii) Точка начала репликации
Как правило, для векторов экспрессии млекопитающих компонент, представляющий собой точку начала репликации, не требуется. Например, как правило, можно использовать точку начала SV40, только потому, что он содержит ранний промотор.
(iii) Компонент - ген селективного маркера
Векторы экспрессии и клонирующие векторы, как правило, содержат ген для селекции, также называемый селективным маркером. Типичные гены для селекции кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например, ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефекты ауксотрофов или (c) обеспечивают основные питательные вещества, которые не доступны из комплексных сред.
В одном из примеров схемы селекции используют лекарственное средство для остановки роста клетки-хозяина. Клетки, которые успешно трансформированы гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному средству и, таким образом, выживают после режима селекции. Примерами такой доминантной селекции являются применение лекарственных средств неомицина, микофеноловой кислоты и гигромицина.
Другим примером подходящих селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентификацию клеток, способных захватывать нуклеиновую кислоту антитела, такие как гены DHFR, тимидинкиназы, металлотионеина-I и -II, предпочтительно, гены металлотионеина приматов, аденозиндезаминазы, орнитиндекарбоксилазы и тому подобное.
Например, клетки, трансформированные геном для селекции DHFR, сначала идентифицируют культивированием всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. Подходящей клеткой-хозяином, когда используют DHFR дикого типа, является клеточная линия яичника китайского хомяка (CHO), дефицитная по активности DHFR (например, ATCC CRL-9096).
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, которые содержат эндогенный DHFR), трансформированные или котрансформированные последовательностями ДНК, кодирующими антитело, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), можно отбирать выращиванием клеток на среде, содержащей агент для селекции по селективному маркеру, такой как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См. патент США 4965199.
(iv) Промоторный компонент
Векторы экспрессии и клонирующие векторы, как правило, содержат промотор, который распознается организмом хозяина и функционально связан с последовательностью нуклеиновой кислоты, кодирующей представляющий интерес полипептид антитела. Промоторные последовательности эукариот хорошо известны. Практически все эукариотические гены обладают AT-богатой областью, расположенной приблизительно от 25 до 30 оснований выше участка инициации транскрипции. Другая последовательность, находящаяся от 70 до 80 оснований выше точки начала транскрипции множества генов, представляет собой участок CNCAAT, где N может представлять собой любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может представлять собой сигнал для добавления поли-A хвоста к 3'-концу кодирующей последовательности. Все из этих последовательностей можно встраивать в эукариотические векторы экспрессии.
Транскрипция с векторов полипептида антитела в клетках-хозяевах млекопитающих контролируется, например, промоторами, получаемыми из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и вирус обезьяны 40 (SV40), гетерологичными промоторами млекопитающих, например, промотором актина или промотором иммуноглобулина, промоторами теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 обычно получают в качестве рестрикционного фрагмента SV40, который также содержит точку начала репликации вируса SV40. Предранний промотор цитомегаловируса человека обычно получают в качестве рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в хозяевах, относящихся к млекопитающим, с использованием вируса папилломы крупного рогатого скота в качестве вектора, описана в патенте США 4419446. Модификация этой системы описана в патенте США 4601978. Альтернативно, в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
(v) Компонент - энхансерный элемент
Транскрипцию ДНК, кодирующей полипептид антитела по изобретению, у высших эукариот часто повышают благодаря встраиванию в вектор энхансерной последовательности. В настоящее время известно множество энхансерных последовательностей из генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако, как правило, используют энхансер из вируса эукариотической клетки. Примеры включают энхансер SV40 на поздней стороне точки начала репликации (п.о. 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне точки начала репликации и энхансеры аденовируса. См. также Yaniv, Nature 297: 17-18 (1982) в отношении элементов энхансера для активации эукариотических промоторов. Энхансер может быть присоединен в векторе к кодирующей полипептид антитела последовательности в положении 5' или 3', однако, предпочтительно, он расположен в участке, расположенном в 5'-направлении от промотора.
(vi) Компонент для терминации транскрипции
Векторы экспрессии, используемые в эукариотических клетках-хозяевах, также могут содержать последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно находятся на 5'- и, иногда, на 3'-нетранслируемых участках эукариотических или вирусных ДНК или кДНК. Эти участки содержат нуклеотидные фрагменты, транскрибируемые в качестве полиадинилированных фрагментов в нетранслируемом участке мРНК, кодирующей антитело. Одним подходящим компонентом для терминации транскрипции является участок полиаденилирования бычьего гормона роста. См. WO94/11026 и описанный в ней вектор экспрессии.
(vii) Селекция и трансформация клеток-хозяев
Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах, описанных в настоящем документе, включают клетки высших эукариот, описанные в настоящем документе, включая клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (культура ткани) стало общепринятым способом. Примерами подходящих клеточных линий млекопитающих являются линия почки обезьяны CV1, трансформированная SV40 (COS-7, ATCC CRL 1651); линия почки эмбриона человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки детенышей хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печения крысы буффало (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линия гепатомы человека (Hep G2).
Клетки-хозяева трансформируют описанными выше векторами экспрессии или клонирующими векторами для продукции антитела и культивируют в общепринятых питательных средах, модифицированных подходящим образом для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих желаемые последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продукции антитела по настоящему изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев подходящими являются коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда ((MEM), (Sigma)), RPMI-1640 (Sigma), и среда Игла, модифицированная способом Дульбекко, ((DMEM), Sigma). Кроме того, для клеток-хозяев в качестве культуральной среды можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal Biochem. 102:255 (1980), патентах США 4767704; 4657866; 4927762; 4560655 или 5122469; WO90/03430; WO 87/00195; или патенте США Re. 30985. Любые их этих сред можно дополнить при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCIN™), микроэлементами (определяемыми как неорганические соединения, обычно представленные в конечных концентрациях микромолярного диапазона) и глюкозой или эквивалентными источниками энергии. Также можно добавлять любые другие необходимые дополнительные вещества в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и тому подобное, представляют собой условия, которые ранее использовали для выбранных для экспрессии клеток-хозяев, и они будут понятны специалисту в данной области.
(ix) Очистка антитела
При использовании рекомбинантных способов антитело может продуцироваться внутриклеточно или непосредственно секретироваться в среду. Если антитело продуцируется внутриклеточно, на первой стадии, дебрис в виде частиц либо клеток-хозяев, либо лизированных фрагментов, удаляют, например, с помощью центрифугирования или ультрафильтрации. Когда антитело секретируется в среду, супернатанты из таких экспрессирующих систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например, элемента для ультрафильтрации Amicon или Millipore Pellicon. На любой из предшествующих стадий можно включать ингибитор протеаз, такой как PMSF, для ингибирования протеолиза, и для предотвращения роста ненужных контаминирующих организмов можно добавлять антибиотики.
Композицию антитела, полученную из клеток, можно очищать с использованием, например, хроматографии с гидроксилапатитом, гель-электрофореза, диализа и аффинной хроматографии, причем аффинная хроматография является предпочтительным способом очистки. Возможность использовать белок A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который присутствует в антителе. Белок A можно использовать для очистки антител, которые основаны на тяжелых цепях Y1, Y2 или Y4 человека (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендован для всех изотипов мыши и для и Y3 человека (Guss et al., EMBO J. 5: 15671575 (1986)). Матрицей, к которой присоединяют аффинный лиганд, наиболее часто является агароза, однако доступны другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол позволяют более быстрые скорости потока и более короткое время обработки, чем скорости потока и время обработки, которых можно достигать с помощью агарозы. Когда антитело содержит CH3-домен, для очистки подходит смола Bakerbond ABXTM (J. T. Baker, Phillipsburg, NJ). Также доступны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на гепарин-СЕФАРОЗЕTM, хроматография на анионо- или катионообменной смоле (такая как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония, в зависимости от подлежащего выделению антитела.
После предварительной стадии(ий) очистки, смесь, содержащую представляющее интерес антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия с низким значением pH с использованием буфера для элюирования при pH между приблизительно 2,5-4,5, предпочтительно, проводимой при низких концентрациях соли (например, от приблизительно 0-0,25 M соли).
Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам (также взаимозаменяемо называемым "конъюгатами антитело-лекарственное средство" или "ADC"), содержащим любое из антител против FGFR3, описанных в настоящем документе, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, лекарственным средством, ингибирующим рост средством, токсином (например, ферментативно активным токсином бактерий, грибов, растений или животных, в том числе его фрагменты и/или варианты), или с радиоактивным изотопом (т.е., радиоконъюгатом).
Применение конъюгатов антитело-лекарственное средство для локальной доставки цитотоксических средств или цитостатических средств, т.е. лекарственных средств, которые уничтожают или ингибируют клетки опухоли, при лечении злокачественной опухоли (Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drug Del. Rev. 26: 151-172; патент США 4975278) позволяет направленную доставку группы лекарственного средства в опухоли и внутриклеточное накопление в ней, при этом системное введение этих неконъюгированных лекарственных средств может приводить к неприемлемым уровням токсичности в отношении нормальных клеток, наряду с опухолевыми клетками, подлежащими устранению (Baldwin et al. (1986) Lancet (Mar. 15, 1986) pp. 603-05; Thorpe (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", Monoclonal Antibodies '84: Biological And Clinical Applications (A. Pinchera et al., eds) pp. 475-506). Таким образом, является желаемой максимальная эффективность при минимальной токсичности. В качестве подходящих в этих стратегиях описаны как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother. 21:183-87). Лекарственные средства, используемые в этих способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986), выше). Токсины, используемые в конъюгатах антитело-токсин, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al., (2000) J. Nat. Cancer Inst. 92(19):1573-1581; Mandler et al., (2000) Bioorganic & Med. Chem. Letters 10:1025-1028; Mandler et al., (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (EP 1391213; Liu et al., (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихеамицин (Lode et al., (1998) Cancer Res. 58:2928; Hinman et al., (1993) Cancer Res. 53:3336-3342). Токсины могут оказывать их цитотоксическое или цитостатическое действие через механизмы, включающие связывание тубулина, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства имеют тенденцию к инактивации или снижению активности при конъюгации с крупными антителами или лигандами белкового рецептора.
ZEVALIN® (ибритумомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело-радиоизотоп, состоящий из моноклонального антитела IgG1 каппа мыши, направленного против антигена CD20, находящегося на поверхности нормальных и злокачественных B-лимфоцитов, и радиоизотопа 111In или 90Y, связанного линкером-хелатором на основе тиомочевины (Wiseman et al., (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al., (2002) Blood 99(12):4336-42; Witzig et al., (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al., (2002) J. Clin. Oncol. 20(15):3262-69). Хотя ZEVALIN обладает активностью против B-клеточной неходжкинской лимфомы (NHL), его введение приводит к тяжелым и длительным цитопениям у большинства пациентов. MYLOTARGTM (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитело-лекарственное средство, состоящий из антитела huCD33, связанного с калихеамицином, был одобрен в 2000 для лечения острого миелоидного лейкоза путем инъекции (Drugs of the Future (2000) 25(7):686; патенты США 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Кантузумаб мертансин (Immunogen, Inc.), конъюгат антитело-лекарственное средство, состоящий из антитела huC242, связанного через дисульфидный линкер SPP с группой майтанзиноидного лекарственного средства, DM1, проходит испытание фазы II для лечения злокачественных опухолей, которые экспрессируют CanAg, таких как злокачественные опухоли толстого кишечника, поджелудочной железы, желудка и другие злокачественные опухоли. MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen Inc.), конъюгат антитело-лекарственное средство, состоящий из моноклонального антитела против простатспецифического мембранного антигена (PSMA), связанного с группой майтанзиноидного лекарственного средства, DM1, находится на стадии разработки для возможного лечения злокачественных опухолей предстательной железы. Пептиды ауристатина, ауристатин E (AE) и монометилауристатин (MMAE), синтетические аналоги доластатина, были конъюгированы с химерными моноклональными антителами cBR96 (специфичными к Lewis Y на карциноме) и cAC10 (специфичными к CD30 на гематологических злокачественных опухолях) (Doronina et al., (2003) Nature Biotechnol. 21(7):778-784) и находятся на стадии терапевтической разработки.
Химиотерапевтические средства, подходящие для получения таких иммуноконъюгатов, описаны в настоящем документе (например, выше). Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки-диантины, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. См., например, WO 93/21232, опубликованную 28 октября 1993 года. Для получения радиоконъюгированных антител доступны различные радионуклиды. Их примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического средства могут быть получены с помощью множества бифункциональных связывающих белки веществ, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как 2,6-диизоцианат толуола), и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина может быть получен, как описано в Vitetta et al., Science 238: 1098 (1987). Меченая углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой характерный пример хелатирующего агента для конъюгации радионуклеотида с антителом. См. WO94/11026.
Также в настоящем документе предусмотрены конъюгаты антитела и одного или нескольких низкомолекулярных токсинов, таких как калихеамицин, майтанзиноиды, доластатины, ауростатины, трихотецен и CC1065, и производные этих токсинов, которые обладают активностью токсинов.
i. Майтанзин и майтанзиноиды
В некоторых вариантах осуществления иммуноконъюгат содержит антитело (полноразмерное или фрагменты) по изобретению, конъюгированное с одной или несколькими молекулами майтанзиноида.
Майтанзиноиды представляют собой ингибиторы митоза, которые действуют, ингибируя полимеризацию тубулина. Майтанзин впервые был выделен из восточно-африканского кустарника Maytenus serrata (патент США 3896111). Впоследствии было открыто, что определенные микроорганизмы также продуцируют майтанзиноиды, такие как майтанзинол и C-3 сложные эфиры майтанзинола (патент США 4151042). Синтетический майтанзинол и его производные и аналоги описаны, например, в патентах США 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533.
Майтанзиноидные группы лекарственного средства являются привлекательными группами лекарственного средства в конъюгатах антитело-лекарственное средство, поскольку они: (i) относительно доступны для получения ферментацией или химической модификацией или преобразованием в производное продуктов ферментации, (ii) поддаются преобразованию в производные с помощью функциональных групп, подходящих для конъюгации через дисульфидные и недисульфидные линкеры с антителами, (iii) являются стабильными в плазме, и (iv) являются эффективными против различных опухолевых клеточных линий.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения, и их терапевтическое применение описаны, например, в патентах США 5208020, 5416064 и патенте Европы EP 0 425 235 B1, описания которых включены в настоящий документ в качестве ссылок в полном объеме. Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описали иммуноконъюгаты, содержащие майтанзиноид, обозначаемый как DM1, связанный с моноклональным антителом C242, направленным против рака ободочной и прямой кишки человека. Было выявлено, что конъюгат является высоко цитотоксичным в отношении культивируемых клеток рака толстого кишечника, и он продемонстрировал противоопухолевоую активность в анализе роста опухоли in vivo. В Chari et al., Cancer Research 52: 127-131 (1992) описаны иммуноконъюгаты, в которых майтанзиноид конъюгирован через дисульфидный линкер с антителом мыши A7, связывающимся с антигеном на клеточных линиях рака толстого кишечника человека или с другим моноклональным антителом мыши TA.1, которое связывает онкоген HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид тестировали in vitro на линии клеток рака молочной железы человека SK-BR-3, которая экспрессирует 3×105 поверхностных антигенов HER-2 на клетку. Конъюгат лекарственного средства достигал степени цитотоксичности, аналогичной со свободным майтанзиноидным лекарственным средством, которую можно повышать путем увеличения количества молекул майтанзиноида на молекулу антитела. Конъюгат A7-майтанзиноид продемонстрировал низкую системную цитотоксичность у мышей.
Конъюгаты антитело-майтанзиноид получают химическим связыванием антитела с молекулой майтанзиноида без существенного снижения биологической активности либо антитела, либо молекулы майтанзиноида. См., например, патент США 5208020 (описание которого включено в настоящий документ в качестве ссылки в полном объеме). В среднем 3-4 молекулы майтанзиноида, конъюгированных с одной молекулой антитела, продемонстрировали эффективность в отношении повышения цитотоксичности клеток-мишеней без отрицательного влияния на функцию или растворимость антитела, хотя можно ожидать, что даже одна молекула токсина/антитело повысит цитотоксичность относительно применения “голого" антитела. Майтанзиноиды хорошо известны в данной области и их можно синтезировать известными способами или выделять из природных источников. Подходящие майтанзиноиды описаны, например, в патенте США 5208020 и в других патентах и непатентных публикациях, указанных в настоящем документе выше. Предпочтительными майтанзиноидами являются майтанзинол и аналоги майтанзинола, модифицированные в ароматическом кольце или в других положениях молекулы майтанзинола, такие как различные сложные эфиры майтанзинола.
Для получения конъюгатов антитело-майтанзиноид существует множество линкерных групп, известных в данной области, включая, например, линкерные группы, описанные в патенте США 5208020 или в патенте EP 0425235 B1; Chari et al. Cancer Research 52:127-131 (1992); и в патентной заявке США 10/960602, поданной 8 октября 2004 года, описания которых включены в настоящий документ в качестве ссылок в полном объеме. Конъюгаты антитело-майтанзиноид, содержащие линкерный компонент SMCC, можно получать, как описано в патентной заявке США 10/960602, поданной 8 октября 2004 года. Линкерные группы включают дисульфидные группы, простые тиоэфирные группы, неустойчивые к действию кислот группы, фотолабильные группы, неустойчивые к пептидазам группы, или неустойчивые к эстеразе группы, как описано в указанных выше патентах, причем дисульфидные и тиоэфирные группы являются предпочтительными. Дополнительные линкеры описаны и проиллюстрированы в настоящем документе.
Конъюгаты антитела и майтанзиноида можно получать с использованием множества бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат), и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). В определенных вариантах осуществления связывающий агент представляет собой N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) или N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP), обеспечивающие дисульфидную связь.
Линкер можно связывать с молекулой майтанзиноида в различных положениях, в зависимости от типа связи. Например, сложноэфирную связь можно образовывать с помощью реакции с гидроксильной группой общепринятыми способами присоединения. Реакция может протекать в положении C-3, имеющем гидроксильную группу, положении C-14, модифицированном гидроксиметилом, положении C-15, модифицированном гидроксильной группой, и положении C-20, имеющем гидроксильную группу. В предпочтительном варианте осуществления связь образована в положении C-3 майтанзинола или аналога майтанзинола.
ii. Ауристатины и доластатины
В некоторых вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с доластатином или с пептидными аналогами и производными доластатина, ауристатинами (патенты США 5635483; 5780588). Было показано, что доластатины и ауристатины нарушают динамику микротрубочек, гидролиз GTP и деление ядер и клеток (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают активностью против злокачественной опухоли (патент США 5663149) и противогрибковой активностью (Pettit et al., (1998) Antimicrob. Agents Chemother. 42:2961-2965). Группу лекарственного средства в виде доластатина или ауристатина можно связывать с антителом через N(амино)-конец или C(карбоксильный)-конец пептидной группы лекарственного средства (WO 02/088172).
Характерные варианты осуществления ауристатина включают связывающиеся через N-конец группы лекарственного средства в виде монометилауристатина DE и DF, описанные в "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номером 10/983340, поданная 5 ноября 2004 года, описание которых включено в настоящий документ в качестве ссылки в полном объеме.
Как правило, группы лекарственного средства на основе пептидов можно получать за счет образования пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получать, например, способом жидкостного синтеза (см. E. Schroder and K. Lubke, "The Peptides", volume 1, pp 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Группы лекарственного средства в виде ауристатина/доластатина можно получать способами: патентов США 5635483 и 5780588; Pettit et al., (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al., (1998) Anti-Cancer Drug Design 13:243-277; Pettit, G.R., et al., Synthesis, 1996, 719-725 и Pettit et al., (1996) J. Chem. Soc. Perkin Trans. 1 5:859-863. Также см. Doronina (2003) Nat. Biotechnol. 21(7): 778-784; "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номером 10/983340, поданную 5 ноября 2004 года, включенной в настоящий документ в качестве ссылки в полном объеме (где описаны, например, линкеры и способы получения соединений монометилвалина, таких как MMAE и MMAF, конъюгированных с линкерами).
iii. Калихеамицин
В других вариантах осуществления иммуноконъюгат содержит антитело, конъюгированное с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицина способны образовывать двухцепочечные разрывы ДНК при субпикомолярных концентрациях. Для получения конъюгатов семейства калихеамицина, см. патенты США 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296 (все выданы American Cyanamid Company). Структурные аналоги калихеамицина, которые можно использовать, включают, но ими не ограничиваются, γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θI 1 (Hinman et al., Cancer Research 53: 3336-3342 (1993), Lode et al., Cancer Research 58:2925-2928 (1998) и упомянутые выше патенты США, выданные American Cyanamid). Другим противоопухолевым лекарственным средством, с которым можно конъюгировать антитело, является QFA, который представляет собой антифолат. Как калихеамицин, так и QFA, обладают внутриклеточным действием, и они не легко проходят через плазматическую мембрану. Таким образом, клеточный захват этих веществ с помощью опосредуемой антителом интернализации значительно повышает их цитотоксические эффекты.
iv. Другие цитотоксические средства
Другие противоопухолевые средства, которые можно конъюгировать с антителом, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство средств, совокупно известных как комплекс LL-E33288, описанный в патентах США 5053394 и 5770710, а также эсперамицины (патент США 5877296).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки-диантаны, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. См., например, WO 93/21232, опубликованную 28 октября, 1993.
Кроме того, настоящее изобретение относится к иммуноконъюгату, образованному между антителом и соединением с нуклеолитической активностью (например, рибонуклеазой или эндонуклеазой ДНК, такой как дезоксирибонуклеаза; ДНКаза).
Для селективного разрушения опухоли антитело может содержать высоко радиоактивный атом. Для продукции радиоактивных конъюгированных антител доступны различные радиоактивные изотопы. Их примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. В случае применения конъюгата для обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, tc99m или I123, или спиновую метку для получения изображения ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как также йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки можно встраивать в конъюгат известными способами. Например, пептид может быть биологически синтезированным или его можно синтезировать с помощью химического синтеза аминокислот с использованием подходящих предшественников аминокислот, включающих, например, фтор-19 вместо водорода. Метки, такие как tc99m или I123, Re186, Re188 и In111 можно присоединять через остаток цистеина в пептиде. Иттрий-90 можно присоединять через остаток лизина. Способ с IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80: 49-57) можно использовать для встраивания йода-123. В "Monoclonal Antibodies in Immunoscintigraphy" (Chatal, CRC Press 1989) подробно описаны другие способы.
Конъюгаты антитела и цитотоксического средства можно получать с использованием различных бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в Vitetta et al., Science 238: 1098 (1987). Характерными примерами хелатирующих агентов для конъюгации радионуклида с антителом является меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA). См. WO94/11026. Линкер может представлять собой "расщепляемый линкер", способствующий высвобождению цитотоксического лекарственного средства в клетке. Например, можно использовать неустойчивый к действию кислот линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al., Cancer Research 52:127-131 (1992); патент США 5208020).
Соединения напрямую предусматривают, но ими не ограничиваются, ADC, полученные с помощью поперечно-сшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A). См. стр. 467-498, 2003-2004 Applications Handbook and Catalog.
v. Получение конъюгатов антитело-лекарственное средство
В конъюгатах антитело-лекарственное средство (ADC) по изобретению, антитело (Ab) конъюгировано с одной или несколькими группами лекарственных средств (D), например, от приблизительно 1 до приблизительно 20 групп на антитело, через линкер (L). ADC формулы I можно получать несколькими путями с использованием реакций, условий и реагентов органической химии, известных специалистам в данной области, включая: (1) реакцию нуклеофильной группы антитела с двухвалентным линкерным реагентом с образованием Ab-L через ковалентную связь, с последующей реакцией с группой лекарственного средства D; и (2) реакцию нуклеофильной группы в группе лекарственного средства с двухвалентным линкерным реагентом, с образованием D-L, через ковалентную связь, с последующей реакцией с нуклеофильной группой антитела. Дополнительные способы получения ADC описаны в настоящем документе.
Линкер может состоять из одного или нескольких линкерных компонентов. Характерные линкерные компоненты включают 6-малеимидокапроил ("MC"), малеимидопропаноил ("MP"), валин-цитруллин ("val-cit" или "vc"), аланин-фенилаланин ("ala-phe"), п-аминобензилоксикарбонил ("PAB"), N-сукцинимидил-4-(2-пиридилтио)пентаноат ("SPP"), N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат ("SMCC"), N-сукцинимидил-(4-йодоацетил)аминобензоат ("SIAB"). Дополнительные линкерные компоненты известны в данной области и некоторые из них описаны в настоящем документе. Также см. "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номером 10/983340, поданную 5 ноября 2004 года, описание которой включено в настоящий документ в качестве ссылки в полном объеме.
В некоторых вариантах осуществления линкер может содержать аминокислотные остатки. Характерные примеры аминокислотных линкерных компонентов включают дипептид, трипептид, тетрапептид или пентапептид. Характерные примеры дипептидов включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Характерные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, которые содержат компонент в виде аминокислотного линкера, включают природные аминокислотные остатки, а также минорные аминокислоты и не природные аналоги аминокислот, такие как цитруллин. Аминокислотные линкерные компоненты можно конструировать и оптимизировать по их селективности к ферментативному расщеплению конкретными ферментами, например, связанной с опухолью протеазой, катепсином B, C и D или протеазой плазмина.
Нуклеофильные группы на антителах включают, но ими не ограничиваются: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, лизина, (iii) тиольные группы боковых цепей, например, цистеина, и (iv) гидроксилы или аминогруппы сахаров, где антитело является гликозилированным. Амино, тиольная и гидроксильная группы являются нуклеофильными и способны вступать в реакцию с образованием ковалентных связей с электрофильными группами на линкерных группах и линкерных реагентах, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) группы альдегидов, кетонов, карбоксильные и малеинимидные группы. Определенные антитела обладают восстановливаемыми дисульфидами между цепями, т.е. цистеиновыми мостиками. Реакционную способность антитела для конъюгации с линкерными реагентами можно обеспечивать с помощью обработки восстановителем, таким как DTT (дитиотреитол). Каждый цистеиновый мостик, таким образом, может образовывать, теоретически, два реакционноспособных тиольных нуклеофила. Дополнительные нуклеофильные группы можно вносить в антитела с помощью реакции лизина с 2-иминотиоланом (реагентом Траута), приводящей к превращению амина в тиол. Реакционноспособные тиольные группы можно вносить в антитело путем встраивания одного, двух, трех, четырех или более остатков цистеина (например, путем получения вариантов антител, содержащих один или несколько ненативных аминокислотных остатка цистеина).
Конъюгаты антитело-лекарственное средство также могут быть получены путем модификации антитела внесением электрофильных групп, которые могут реагировать с нуклеофильными заместителями на линкерном реагенте или лекарственном средстве. Сахара гликозилированных антител могут быть окисленными, например, окислителями на основе перйодатов, с образованием групп альдегидов или кетонов, которые могут реагировать с аминогруппой линкерных реагентов или групп лекарственного средства. Полученные иминогруппы шифова основания могут образовывать стабильную связь, или их можно восстанавливать, например, боргидридными реагентами, с образованием стабильных связей через амин. В одном из вариантов осуществления реакция углеводной части гликозилированного антитела либо с галактозооксидазой, либо с метаперйодатом натрия, может приводить к карбонильным группам (группам альдегидов и кетонов) в белке, которые могут взаимодействовать с соответствующими группами на лекарственном средстве (Hermanson, Bioconjugate Techniques). В другом варианте осуществления белка, содержащие N-концевые остатки серина или треонина могут взаимодействовать с метаперйодатом натрия, что приводит к образованию альдегида вместо первой аминокислоты (Geoghegan & Stroh, (1992) Bioconjugate Chem. 3:138-146; US 5362852). Такой альдегид можно подвергать реакции с группой лекарственного средства или нуклеофилом линкера.
Аналогично, нуклеофильные группы на группе лекарственного средства включают, но ими не ограничиваются: аминогруппу, тиольную, гидроксильную группы, группы гидразида, оксима, гидразина, тиосемикарбазона, гидразина карбоксилата и арилгидразида, способные вступать в реакции с образованием ковалентных связей с электрофильными группами на линкерных группах и линкерных реагентах, включающих: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеинимидные группы.
Альтернативно, слитый белок, содержащий антитело и цитотоксическое средство, может быть получен, например, рекомбинантными способами или пептидным синтезом. Отрезок ДНК может содержать соответствующие области, кодирующие две части конъюгата, либо соседние друг с другом, либо разделенные областью, кодирующей линкерный пептид, который на ухудшает желаемые свойства конъюгата.
В другом варианте осуществления антитело может быть конъюгировано с "рецептором" (таким как стрептавидин) для предварительного нацеливания на опухоль, где конъюгат антитело-рецептор вводят пациенту, а затем из кровотока удаляют не связавшийся конъюгат с использованием средства для удаления, с последующим введением "лиганда" (например, авидина), который конъюгирован с цитотоксическим средством (например, радионуклеотидом).
Способы с использованием антител против FGFR3
Настоящее изобретение относится к применению антитела против FGFR3 в качестве части определенного режима лечения, предназначенного для обеспечения благоприятного эффекта вследствие активности этого лекарственного средства. Настоящее изобретение особенно подходит для лечения злокачественных опухолей различных типов на различных стадиях.
Термин "злокачественная опухоль", "рак" включает совокупность пролиферативных нарушений, включая, но ими не ограничиваясь, предзлокачественный рост, доброкачественные опухоли и злокачественные опухоли. Доброкачественные опухоли локализируются в области их возникновения и не могут инфильтрировать, вторгаться или метастазировать в отдаленные области. Злокачественные опухоли вторгаются в другие ткани вокруг них и повреждают их. Также они могут приобретать способность отделяться от исходной области и распространяться в другие части организма (метастазировать), обычно через кровоток или через лимфатическую систему, где расположены лимфатические узлы. Первичные опухоли классифицируют по типу ткани, из которой они появляются; метастатические опухоли классифицируют по типу ткани, из которой злокачественные клетки происходят. С течением времени клетки злокачественной опухоли становятся более аномальными и становятся все менее похожими на нормальные клетки. Это изменение внешнего вида злокачественной опухоли называют степенью опухоли, и злокачественные клетки описывают как высоко дифференцированные (низкой степени), умеренно-дифференцированные, слабо-дифференцированные или недифференцированные (высокой степени). Высоко дифференцированные клетки имеют совершенно нормальный внешний вид и напоминают нормальные клетки, из которых они происходят. Недифференцированные клетки представляют собой клетки, которые стали настолько аномальными, что более невозможно определить источник этих клеток.
Системы определения стадий злокачественной опухоли описывают, насколько злокачественная опухоль распространилась анатомически, и предпринимают попытку поместить пациентов с аналогичным прогнозом и лечением в группу одной стадии. Для облегчения определения стадии злокачественной опухоли можно проводить несколько тестов, включая биопсию и определенные тесты по визуализации, такие как рентген грудной клетки, маммограмма, сканирование костей, КТ-сканирование и МРТ-сканирование. Также для оценки общего здоровья пациентов и обнаружения того, распространилась ли злокачественная опухоль на определенные органы, проводят анализы крови и клиническую оценку.
Для определения стадии злокачественной опухоли American Joint Committee on Cancer сначала относит злокачественную опухоль, в частности, солидные опухоли, к буквенной категории с использованием системы классификации TNM. Злокачественные опухоли обозначают буквами T (размер опухоли), N (пальпируемые узлы), и/или M (метастазы). T1, T2, T3 и T4 описывают растущий размер первичного очага; N0, N1, N2, N3 указывают на постепенно прогрессирующее вовлечение узлов; и M0 и М1 отражают отсутствие или присутствие отдаленных метастазов.
Во втором способе определения стадии, также известном как "Общее подразделение на стадии" или "Определение стадии с помощью римских чисел", злокачественные опухоли подразделяют на стадии от 0 до IV, включающие размер первичных очагов повреждения, а также присутствие распространения в узлы и отдаленных метастазов. В этой системе случаи подразделяют на четыре стадии, обозначаемые римскими цифрами с I по IV, или классифицируют как "рецидивирующие". Для некоторых злокачественных опухолей стадия 0 обозначают как "in situ" или "Tis", как например, карцинома протоков in situ или долевая карцинома in situ для рака молочной железы. Аденомы высокой степени также можно классифицировать как стадию 0. Как правило, злокачественные опухоли стадии I представляют собой небольшие локализованные злокачественные опухоли, которые обычно являются излечимыми, в то время как стадия IV обычно соответствует неоперабельной или метастазирующей злокачественной опухоли. Злокачественные опухоли стадии II и III обычно являются локально развернутыми и/или проявляют вовлечение местных лимфатических узлов. Как правило, более высокие номера стадии указывают на более развернутое заболевание, включая более высокий размер и/или распространение злокачественной опухоли в ближайшие лимфатические узлы и/или органы, соседние с первичной опухолью. Эти стадии определяют точно, однако определение отличается для каждого типа злокачественной опухоли и известно специалисту в данной области.
Во многих реестрах злокачественных опухолей, таких как NCI's Surveillance, Epidemiology, and End Results Program (SEER), используется суммарное определение стадии. Эту систему используют для всех типов злокачественной опухоли. Она подразделяет случаи злокачественной опухоли на пять основных категорий:
In situ представляет собой раннюю злокачественную опухоль, которая присутствует только в том слое клеток, в котором она возникла.
Локализованная представляет собой злокачественную опухоль, которая ограничена органом, в котором она возникла, без признаков распространения.
Регионарная представляет собой злокачественную опухоль, которая распространяется за пределы исходной (первичной) области в ближайшие лимфатические узлы или органы и ткани.
Отдаленная представляет собой злокачественную опухоль, которая распространилась из первичной области в отдаленные органы или в отдаленные лимфатические узлы.
Неизвестная - используют для описания случаев, для которых отсутствует достаточно информации для указания стадии.
Кроме того, для злокачественной опухоли является частым возвращение через месяцы или годы после удаления первичной опухоли. Злокачественную опухоль, которая вновь возникает после полного визуального устранения опухоли, называют рецидивирующим заболеванием. Заболевание, которое рецидивирует в области первичной опухоли, является локально рецидивирующим, а заболевание, которое рецидивирует с помощью метастазов, называют отдаленным рецидивом.
Опухоль может представлять собой солидную опухоль, или несолидную опухоль или опухоль мягких тканей. Примеры опухолей мягких тканей включают лейкоз (например, хронический миелогенный лейкоз, острый миелогенный лейкоз, взрослый острый лимфобластный лейкоз, острый миелогенный лейкоз, острый лимфобластный лейкоз зрелых B-клеток, хронический лимфоцитарный лейкоз, полилимфоцитарный лейкоз или волосатоклеточный лейкоз) или лимфому (например, неходжскинскую лимфому, Т-клеточную лимфому кожи или болезнь Ходжкина). Солидная опухоль включает любую злокачественную опухоль тканей организма, отличных от крови, костного мозга или лимфатической системы. Солидные опухоли могут быть далее подразделены на опухоли происхождением из эпителиальных клеток и опухоли происхождением из неэпителиальных клеток. Примеры солидных опухолей эпителиальных клеток включают опухоли желудочно-кишечного тракта, толстого кишечника, молочной железы, предстательной железы, легкого, почки, печени, поджелудочной железы, яичника, головы и шеи, полости рта, желудка, двенадцатиперстной кишки, тонкого кишечника, толстого кишечника, ануса, желчного пузыря, губы, носоглотки, кожи, матки, мужских половых органов, органов мочевыделения, мочевого пузыря и кожи. Солидные опухоли неэпителиального происхождения включают саркомы, опухоли головного мозга и опухоли костей. Другие примеры опухолей описаны в разделе "Определения".
В некоторых вариантах осуществления пациента подвергают диагностическому тесту, например, до и/или в процессе и/или после терапии. Как правило, если проводят диагностический тест, образец можно получать от пациента, нуждающегося в лечении. Когда индивидуум имеет злокачественную опухоль, образец может представлять собой образец опухоли или другой биологический образец, такой как биологическая жидкость, включая, но ими не ограничиваясь, кровь, мочу, слюну, асцитную жидкость, или ее производные, такое как сыворотка крови и плазма крови и тому подобное.
Биологический образец, описанный в настоящем документе, может представлять собой фиксированный образец, например, фиксированный в формалине, погруженный в парафин (FFPE) образец, или замороженный образец.
Различные способы определения экспрессии мРНК или белка включают, но ими не ограничиваются, определение профиля экспрессии генов, полимеразную цепную реакцию (ПЦР), включая количественную ПЦР в реальном времени (qRT-PCR), анализ на микрочипах, серийный анализ экспрессии генов (SAGE), MassARRAY, анализ экспрессии генов массивным параллельным секвенированием сигнатур (MPSS), протеомику, иммуногистохимию (IHC) и тому подобное. Предпочтительно, мРНК определяют количественно. Такой анализ мРНК, предпочтительно, проводят с использованием способа полимеразной цепной реакции (ПЦР) или анализом на микрочипах. Когда используют ПЦР, предпочтительной формой ПЦР является количественная ПЦР в реальном времени (qRT-PCR). В одном из вариантов осуществления экспрессию одного или нескольких из указанных выше генов считают положительной экспрессией, если она находиться на срединном уровне или выше, например, по сравнению с другими образцами того же типа опухоли. Срединный уровень экспрессии можно определять по существу одновременно с измерением экспрессии гена, или он может быть определен ранее.
Стадии характерного протокола определения профиля экспрессии генов с использованием фиксированных, погруженных в парафин тканей в качестве источника РНК, включая выделение, очистку мРНК, удлинение праймеров и амплификацию, приведены в различных опубликованных журнальных статьях (например: Godfrey et al. J. Molec. Diagnostics 2: 84-91 (2000); Specht et al., Am. J. Pathol. 158: 419-29 (2001)). Коротко, характерный процесс начинается с нарезания срезов погруженных в парафин образцов опухолевой ткани толщиной приблизительно 10 микрометров. Затем РНК экстрагируют и белок и ДНК удаляют. После анализа концентрации РНК, можно включать стадии репарации и/или амплификации РНК, если необходимо, и РНК подвергать обратной транскрипции с использованием специфичных к гену промоторов с последующей ПЦР. Наконец, данные анализируют для идентификации наилучшего варианта(ов) лечения, доступного для пациента, исходя из характерного паттерна экспрессии генов, идентифицированного в исследованном образце опухоли.
Обнаружение экспрессии гена или белка можно проводить непосредственно или косвенно.
Можно определять экспрессию или транслокацию или амплификацию FGFR3 в злокачественной опухоли (непосредственно или косвенно). Для этого доступны различные диагностические/прогностические анализы. В одном из вариантов осуществления сверхэкспрессию FGFR3 можно анализировать с помощью IHC. Погруженные в парафин срезы тканей из биоптата опухоли можно подвергать анализу IHC и согласовывать со следующими критериями интенсивности окрашивания белка FGFR3:
Показатель 0 - окрашивания не наблюдают или наблюдают окрашивание мембраны в менее чем 10% опухолевых клеток.
Показатель 1+ слабое/едва заметное окрашивание мембраны выявляется в более чем 10% опухолевых клеток. Клетки окрашиваются только в части их мембраны.
Показатель 2+ полное окрашивание мембраны от слабого до умеренного наблюдают в более чем 10% опухолевых клеток.
Показатель 3+ полное окрашивание мембраны от умеренного до сильного наблюдают в более чем 10% опухолевых клеток.
В некоторых вариантах осуществления опухоли с показателями 0 или 1+ в отношении оценки сверхэкспрессии FGFR3 могут быть охарактеризованы как не сверхэкспресирующие FGFR3, в то время как опухоли с показателями 2+ или 3+ могут быть охарактеризованы как сверхэкспрессирущие FGFR3.
В некоторых вариантах осуществления опухоли, сверхэкспрессирующие FGFR3, можно оценивать с помощью иммуногистохимических показателей, соответствующих числу копий молекул FGFR3, экспрессированных на клетку, и его можно определять биохимически:
0 = 0-90 копий/клетка,
1+ = по меньшей мере приблизительно 100 копий/клетка,
2+ = по меньшей мере приблизительно 1000 копий/клетка,
3+ = по меньшей мере приблизительно 10000 копий/клетка.
Альтернативно или дополнительно, можно проводить анализы FISH на фиксированной в формалине погруженной в парафин опухолевой ткани для определения присутствия или и/или степени (в случае присутствия) амплификации или транслокации FGFR3 в опухоли.
Активацию FGFR3 можно определять напрямую (например, тестированием с помощью фосфо-ELISA, или другими способами обнаружения фосфорилированного рецептора) или косвенно (например, с помощью обнаружения активированных последующих компонентов каскада передачи сигнала, обнаружения димеров рецепторов (например, гомодимеров, гетеродимеров), обнаружения профилей экспрессии генов и тому подобное.
Аналогично, обнаружение конститутивного FGFR3 и/или лиганд-независимого или лиганд-зависимого FGFR3 можно проводить непосредственно или косвенно (например, с помощью обнаружения мутаций рецептора с конститутивной активностью, с помощью обнаружения амплификации рецептора, коррелирующей с конститутивной активностью и тому подобное).
Способы обнаружения мутаций нуклеиновых кислот хорошо известны в данной области. Часто, хотя и не обязательно, нуклеиновую кислоту-мишень в образце амплифицируют, обеспечивая желаемое количество материала для определения того, имеется ли мутация. Способы амплификации хорошо известны в данной области. Например, амплифицированный продукт может охватывать или может не охватывать всю последовательность нуклеиновой кислоты, кодирующей представляющий интерес белок, при условии, что амплифицированный продукт содержит конкретное положение в аминокислотной последовательности/последовательности нуклеиновой кислоты, где предположительно имеется мутация.
В одном из примеров присутствие мутации можно определять путем контактирования нуклеиновой кислоты из образца с зондом нуклеиновой кислоты, который способен специфично гибридизоваться с нуклеиновой кислотой, кодирующей мутантную нуклеиновую кислоту, и обнаружения указанной гибридизации. В одном из вариантов осуществления зонд является меченым поддающейся обнаружения меткой, например, радиоизотопом (3H, 32P, 33P и тому подобное), флуоресцентным агентом (родамин, флуоресцеин и тому подобное) или хромогенным агентом. В некоторых вариантах осуществления зонд представляет собой антисмысловой олигомер, например, PNA, морфолинофосфорамидаты, LNA или 2'-алкоксиалкокси. Зонд может составлять от приблизительно 8 нуклеотидов до приблизительно 100 нуклеотидов, или от приблизительно 10 до приблизительно 75, или от приблизительно 15 до приблизительно 50, или от приблизительно 20 до приблизительно 30. В другом аспекте зонды нуклеиновой кислоты по изобретению предоставляют в наборе для идентификации мутаций FGFR3 в образце, причем указанный набор содержит олигонуклеотид, который специфично гибридизуется с участком мутации или рядом с участком мутации в нуклеиновой кислоте, кодирующей FGFR3. Кроме того, набор может содержать инструкции по лечению пациентов, имеющих опухоли, которые содержат мутации FGFR3, антагонистом FGFR3, исходя из результата теста гибридизации с использованием набора.
Обнаружение мутаций также можно проводить путем сравнения электрофоретической подвижности амплифицированной нуклеиновой кислоты с электрофоретической подвижностью соответствующей нуклеиновой кислоты, кодирующей FGFR3 дикого типа. Отличие в подвижности указывает на присутствие мутации в амплифицированной последовательности нуклеиновой кислоты. Электрофоретическую подвижность можно определять любым подходящим способом молекулярного разделения, например, на полиакриламидном геле.
Для обнаружения мутаций нуклеиновые кислоты также можно анализировать с использованием ферментативного обнаружения мутаций (EMD) (Del Tito et al., Clinical Chemistry 44:731-739, 1998). В EMD используется резольваза бактериофага T4 эндонуклеаза VII, которая сканирует двухцепочечную ДНК до тех пор, пока не выявляет и не расщепляет структурные искажения, вызываемые несовпадением спаривания оснований вследствие изменений нуклеиновой кислоты, таких как точковые мутации, инсерции и делеции. Обнаружение двух коротких фрагментов, образованных расщеплением резольвазой, например, с помощью гель-электрофореза, указывает на присутствие мутации. Преимуществом способа EMD является единый протокол для идентификации точковых мутаций, делеций и инсерций, анализируемых сразу после реакции амплификации, устранение необходимости очистки образца, укорочение времени гибридизации и увеличение соотношения сигнала к шуму. Можно анализировать смешанные образцы, содержащие вплоть до 20-кратного избытка нормальных нуклеиновых кислот и фрагментов размером вплоть до 4 т.п.н. Однако сканирование, EMD не идентифицирует конкретные изменения оснований, которые происходят в положительных по мутациям образцах, таким образом, часто требуя дополнительных процедур секвенирования для идентификации конкретной мутации, если необходимо. Фермент CEL I фермент можно использовать аналогично резольвазе T4, эндонуклеазе VII, как продемонстрировано в патенте США. 5869245.
Другим простым набором для обнаружения мутаций является тест-полоска для обратной гибридизации, аналогичная с Haemochromatosis StripAssayTM (Viennalabs http://www.bamburghmarrsh.com/pdf/4220.pdf) для обнаружения множества мутаций в генах HFE, TFR2 и FPN1, вызывающих гемохроматоз. Такой анализ основан на специфичной по последовательности гибридизации с последующей амплификацией с помощью ПЦР. Для анализов единичных мутаций можно использовать систему обнаружения на основе микропланшета, в то время как для анализа множества мутаций, можно использовать тест-полоски в качестве "макрочипов". Наборы могут включать готовые для использования реагенты для приготовления образца, амплификации и обнаружения мутаций. Мультиплексные протоколы амплификации обеспечивают удобство и позволяют тестирование образцов с очень ограниченными объемами. С использованием прямого формата StripAssay, можно проводить тестирование двадцати и более мутаций менее чем за пять часов без дорогостоящего оборудования. Из образца выделяют ДНК и нуклеиновую кислоту-мишень амплифицируют in vitro (например, с помощью ПЦР) и метят биотином, как правило, в одной ("мультиплексной") реакции амплификации. Затем продукты амплификации селективно гибридизуют с олигонуклеотидными зондами (специфичных к дикому типу и мутациям), иммобилизованными на твердой подложке, такой как тест-полоска, на которой иммобилизованы зонды в качестве параллельных линий или полос. Обнаружение связанных биотинилированных ампликонов проводят с использованием стрептавидина-щелочной фосфатазы и цветных субстратов. Такой анализ может обнаруживать все мутации по изобретению или любую их подгруппу. Что касается конкретной мутантной полосы зонда, возможен один из трех характеров подачи сигнала: (i) полоса только для зонда дикого типа, которая указывает на нормальную последовательность нуклеиновой кислоты, (ii) полоса для зонда дикого типа, так и для мутантного зонда, которая указывает на гетерозиготный генотип, и (iii) полоса только для мутантного зонда, которая указывает на гомозиготный мутантный генотип. Таким образом, в одном из аспектов изобретение относится к способу обнаружения мутаций по изобретению, включающему выделение и/или амплификацию последовательности нуклеиновой кислоты-мишени FGFR3 из образца, так чтобы продукт амплификации содержал лиганд, приведение в контакт продукта амплификации с зондом, который содержит обнаруживаемый партнер по связыванию для лиганда и способен специфично гибридизоваться с мутацией по изобретению, а затем обнаружения гибридизации указанного зонда с указанным продуктом амплификации. В одном из вариантов осуществления лиганд представляет собой биотин и партнер по связыванию содержит авидин или стрептавидин. В одном из вариантов осуществления партнер по связыванию содержит конъюгат стрептавидин-щелочная фосфатаза, который поддается обнаружению с помощью цветных субстратов. В одном из вариантов осуществления зонды иммобилизованы, например, на тест-полоске, где зонды, комплементарные различным мутациям, отделяют друг от друга. Альтернативно, амплифицированную нуклеиновую кислоту метят радиоизотопом, в случае которого зонд не должен содержать поддающуюся обнаружения метку.
Изменения гена дикого типа охватывают все формы мутаций, такие как вставки, инверсии, делеции и/или точковые мутации. В одном из вариантов осуществления мутации являются соматическими. Соматические мутации представляют собой мутации, которые встречаются только в определенных тканях, например, в опухолевой ткани, и не наследуются в зародышевой линии. Мутации зародышевой линии могут быть встречаться в любой из тканей организма.
Образец, содержащий нуклеиновую кислоту-мишень, можно получать способами, хорошо известными в данной области и которые подходят для конкретного типа и расположения опухоли. Для получения характерного фрагмента опухолевой ткани часто используют биопсию ткани. Альтернативно, опухолевые клетки можно получать косвенно в форме тканей/жидкостей, которые, как известно или предполагают, содержат представляющие интерес опухолевые клетки. Например, образцы очагов рака легкого можно получать резекцией, бронхоскопией, аспирационной диагностической пункцией, бронхиальным соскабливанием, или из мокроты, плевральной жидкости или крови. Обнаружение мутантных генов или продуктов генов можно проводить из опухоли или из других образцов организма, таких как моча, мокрота или сыворотка. Способы, рассмотренные выше для обнаружения мутантных генов-мишеней или продуктов генов в образцах опухоли, можно применять к другим образцам организма. Злокачественные клетки слущиваются с опухолей и появляются в таких образцах организма. С помощью скрининга таких образцов тканей можно достигать простой ранней диагностики для заболеваний, таких как злокачественная опухоль. Кроме того, мониторинг прогресса терапии можно легко проводить путем тестирования таких образцов организма в отношении мутантных генов-мишеней или продуктов генов.
Способы обогащения препарата ткани опухолевыми клетками известны в данной области. Например, ткань можно выделять из парафиновых или криостатных срезов. Злокачественные клетки также можно отделять от нормальных клеток проточной цитометрией или микродиссекцией с лазерным улавливанием. Эти, а также другие способы отделения опухолевых клеток от нормальных, хорошо известны в данной области. Если опухолевая ткань высоко контаминирована нормальными клетками, обнаружение мутаций может быть более трудной, хотя известны способы минимизации контаминации и/или ложноположительных/ложноотрицательных результатов, и некоторые из них описаны в настоящем документе ниже. Например, образец также можно оценивать на присутствие биомаркера (включая мутацию), известного тем, что он связан с интересующей опухолевой клеткой, но не с соответствующей нормальной клеткой, или наоборот.
Обнаружение точковых мутаций в нуклеиновых кислотах-мишенях можно проводить с помощью молекулярного клонирования нуклеиновых кислот-мишеней и секвенирования нуклеиновых кислот с использованием способов, хорошо известных в данной области. Альтернативно можно использовать способы амплификации, такие как полимеразная цепная реакция (ПЦР), для амплификации последовательностей нуклеиновых кислот-мишеней непосредственно из препарата геномной ДНК из опухолевой ткани. Затем можно определять последовательность нуклеиновой кислоты амплифицированных последовательностей и в них можно идентифицировать мутации. Способы амплификации хорошо известны в данной области, например, полимеразная цепная реакция, как описано в Saiki et al., Science 239:487, 1988; патенты США 4683203 и 4683195.
Следует отметить, что конструирование и выбор соответствующих праймеров хорошо известны в данной области.
Также для амплификации последовательностей нуклеиновых кислот-мишеней можно использовать лигазную цепную реакцию, которая известна в данной области. См., например, Wu et al., Genomics, Vol. 4, pp. 560-569 (1989). Кроме того, также можно использовать способ, известный как аллелеспецифическая ПЦР. См., например, Ruano and Kidd, Nucleic Acids Research, Vol. 17, p. 8392, 1989. Согласно этому способу используют праймеры, которые гибридизуются на их 3'-концах с конкретной мутацией нуклеиновой кислоты-мишени. Если конкретная мутация отсутствует, продукт амплификации не наблюдают. Также можно использовать амплификационную систему рефракторных мутаций (ARMS), как описано в публикации патентной заявки Европы 0332435, и в Newton et al., Nucleic Acids Research, Vol. 17, p.7, 1989. Обнаружение инсерций и делеций генов также можно проводить клонированием, секвенированием и амплификацией. Кроме того, для оценки изменения аллеля или инсереции в полиморфный фрагмент можно использовать зонды полиморфизма длин рестрикционных фрагментов (RFLP) для гена или окружающих маркерных генов. Анализ полиморфизма одноцепочечной конформации (SSCP) также можно использовать для обнаружения вариантов аллеля с изменением основания. См., например, Orita et al., Proc. Natl. Acad. Sci. USA Vol. 86, pp. 2766-2770, 1989, и Genomics, Vol. 5, pp. 874-879, 1989. Также можно использовать другие способы обнаружения инсерций и делеций, известные в данной области.
Обнаружение изменения генов дикого типа также можно проводить, исходя из изменения продукта экспрессии гена дикого типа. Такие продукты экспрессии включают как мРНК, так и белковый продукт. Обнаружение точковых мутаций можно проводить с помощью амплификации и секвенирования мРНК или с помощью молекулярного клонирования кДНК, полученной из мРНК. Последовательность клонированной кДНК можно определять с использованием способов секвенирования ДНК, которые хорошо известны в данной области. кДНК также можно секвенировать с помощью полимеразной цепной реакции (ПЦР).
Неправильно спаренные основания представляют собой гибридизованные дуплексы нуклеиновых кислот, которые не являются на 100% комплементарными. Отсутствие полной комплементарности может быть следствием делеций, инсерций, инверсий, замен или мутаций со сдвигом рамки считывания. Обнаружение неправильно спаренных оснований можно использовать для обнаружения точковых мутаций в нуклеиновой кислоте-мишени. Хотя эти способы могут быть менее чувствительными, чем секвенирование, они являются более простыми для проведения на большом количестве образцов ткани. Примером способа расщепления неправильно спаренных оснований является способ защиты с РНК-азой, который подробно описан в Winter et al., Proc. Natl. Acad. Sci. USA, Vol. 82, p. 7575, 1985, и Meyers et al., Science, Vol. 230, p. 1242, 1985. Например, способ по изобретению может включать использование меченного рибонуклеотидного зонда, который комплементарен нуклеиновой кислоте-мишени человека дикого типа. Рибонуклеотидный зонд и нуклеиновая кислота-мишень, происходящие из образца ткани, подвергают отжигу (гибридизуют) вместе, а затем расщепляют ферментом РНК-азой A, который способен выявлять некоторые неправильно спаренные основания в дуплексной структуре РНК. Если РНК-аза A выявляет неправильно спаренные основания, она расщепляет участок неправильного спаривания. Таким образом, когда подвергнутый отжигу препарат РНК разделяют на электрофоретической гелевой матрице, то, если неправильно спаренное выявлено и расщеплено РНК-азой A, будет виден продукт РНК, который является меньшим, чем полноразмерная дуплексная РНК для рибонуклеотидного зонда и мРНК или ДНК. Рибонуклеотидный зонд не должен быть полноразмерным относительно мРНК или гена нуклеиновой кислоты-мишени, а он может быть частью нуклеиновой кислоты-мишени, при условии, что он включает положение, предположительно являющееся мутантным. Если рибонуклеотидный зонд содержит только сегмент мРНК или гена нуклеиновой кислоты-мишени, может быть желательным использование нескольких таких зондов для скрининга всей последовательности нуклеиновой кислоты-мишени в отношении неправильно спаренных оснований, если желательно.
Аналогично, для обнаружения неправильно спаренных оснований можно использовать ДНК-зонды, например, с помощью ферментативного или химического расщепления. См., например, Cotton et al., Proc. Natl. Acad. Sci. USA, Vol. 85, 4397, 1988; и Shenk et al., Proc. Natl. Acad. Sci. USA, Vol. 72, p. 989, 1975. Альтернативно, обнаружение неправильно спаренных оснований можно проводить по сдвигам электрофоретической подвижности неправильно спаренных дуплексов относительно правильно спаренных дуплексов. См., например, Cariello, Human Genetics, Vol. 42, p. 726, 1988. С использованием либо рибонуклеотидных зондов, либо ДНК-зондов, мРНК или ДНК нуклеиновой кислоты-мишени, которые могут содержать мутацию, можно амплифицировать перед гибридизацией. Обнаружение изменений ДНК нуклеиновой кислоты-мишени также можно проводить с использованием саузерн-гибридизации, особенно если изменения представляют собой крупные перестройки, такие как делеции и инсерции.
Амплифицированные последовательности ДНК нуклеиновой кислоты-мишени также можно подвергать скринингу с использованием аллелеспецифических зондов. Эти зонды представляют собой олигомеры нуклеиновых кислот, каждый из которых содержит область гена нуклеиновой кислоты-мишени, содержащую известную мутацию. Например, один олигомер может иметь длину приблизительно 30 нуклеотидов, соответствующую части последовательности гена-мишени. С использованием набора таких аллелеспецифических зондов продукты амплификации нуклеиновой кислоты-мишени можно подвергать скринингу для идентификации присутствия ранее идентифицированной мутации в гене-мишени. Гибридизацию аллелеспецифических зондов с амплифицированными последовательностями нуклеиновых кислот можно проводить, например, на нейлоновом фильтре. Гибридизация с конкретным зондом в строгих условиях гибридизации указывает на присутствие этой мутации в опухолевой ткани, как и в аллелеспецифическом зонде.
Обнаружение изменения генов-мишеней дикого типа также можно проводить путем скрининга изменения соответствующего белка дикого типа. Например, для скрининга ткани можно использовать моноклональные антитела, иммунореактивные в отношении продукта гена-мишени, например, антитело, которое известно тем, что оно связывается с конкретным мутантным положением продукта гена (белка). Например, антитело, которое используют, может представлять собой антитело, которое связывается с делетированным экзоном или которое связывается с конформационным эпитопом, содержащим делетированную часть белка-мишени. Отсутствие собственного антигена указывает на мутацию. Антитела, специфичные к продуктам мутантных аллелей, также можно использовать для обнаружения продукта мутантного гена. Антитела можно идентифицировать из библиотек фагового дисплея. Такие иммунологические анализы можно проводить в любом удобном формате, известном в данной области. Они включают вестерн-блоты, иммуногистохимические анализы и анализы ELISA. Для обнаружения изменения генов-мишеней дикого типа можно использовать любые способы обнаружения измененного белка.
Для определения нуклеотидной последовательности нуклеиновой кислоты-мишени с использованием способов амплификации нуклеиновых кислот, таких как полимеразная цепная реакция, подходят пары праймеров. Пары одноцепочечных ДНК-праймеров можно подвергать отжигу с последовательностями в последовательности нуклеиновой кислоты-мишени или вокруг нее для запуска амплификации последовательности-мишени. Также можно использовать аллелеспецифические праймеры. Такие праймеры подвергаются отжигу только с мутантной последоательностью-мишенью, и, таким образом, амплифицируют продукт только в присутствии мутантной последовательности-мишени в качестве матрицы. Для способствования последующему клонированию амплифицированных последовательностей, праймеры могут иметь последовательности участков для ферментов рестрикции, присоединенные к их концам. Такие ферменты и участки хорошо известны в данной области. Сами праймеры можно синтезировать с использованием способов, которые хорошо известны в данной области. Как правило, праймеры можно получать с использованием устройств для синтеза олигонуклеотидов, которые являются коммерчески доступными. Конструирование конкретных праймеров находится в пределах квалификации специалиста в данной области.
Зонды нуклеиновых кислот подходят для ряда целей. Их можно использовать в саузерн-гибридизации с геномной ДНК и в способе защиты с РНК-азой для обнаружения точковых мутаций, уже описанных выше. Зонды можно использовать для обнаружения продуктов амплификации нуклеиновой кислоты-мишени. Также их можно использовать для обнаружения неправильно спаренных оснований с геном или мРНК дикого типа с использованием других способов. Обнаружение неправильно спаренных оснований можно проводить с использованием ферментов (например, S1-нуклеаза), химических реагентов (например, гидроксиламин или тетроксид осмия и пиперидин) или изменений электрофоретической подвижности неправильно спаренных гибридов по сравнению с полностью правильно спаренными гибридами. Эти способы известны в данной области. См. Novack et al., Proc. Natl. Acad. Sci. USA, Vol. 83, p. 586, 1986. Как правило, зонды комплементарны последовательностям вне киназного домена. Полный набор зондов нуклеиновой кислоты можно использовать для оставления набора для обнаружения мутаций в нуклеиновых кислотах-мишенях. Набор позволяет гибридизацию с большой областью представляющей интерес последовательности-мишени. Зонды могут перекрываться друг с другом или быть смежными.
Если рибонуклеотидный зонд используют для обнаружения неправильно спаренных оснований с мРНК, он, как правило, комплементарен мРНК гена-мишени. Таким образом, рибонуклеотидный зонд представляет собой антисмысловой зонд, который не кодирует соответствующий продукт гена, поскольку он комплементарен смысловой цепи. Рибонуклеотидный зонд, как правило, метят радиоактивным, колориметрическим или флуориметрическим материалом, что можно осуществлять любыми способами, известными в данной области. Если рибонуклеотидный зонд используют для обнаружения неправильно спаренных оснований в ДНК, он может иметь любую полярность: смысловую или антисмысловую. Аналогично, для обнаружения неправильно спаренных оснований можно использовать ДНК-зонды.
В некоторых случаях злокачественная опухоль сверхэкспрессирует FGFR3 или не сверхэкспрессирует его. Сверхэкспрессию рецептора можно определять в диагностическом или прогностическом анализе с помощью оценки увеличенных уровней рецепторного белка, присутствующего на поверхности клетки (например, с помощью иммуногистохимического анализа; IHC). Альтернативно или дополнительно, можно измерять уровни кодирующей рецептор нуклеиновой кислоты в клетке, например, с помощью флуоресцентной гибридизации in situ (FISH; см. WO98/45479, опубликованная в октябре 1998 года), саузерн-блоттинга или способов полимеразной цепной реакции (ПЦР), таких как количественная ПЦР в реальном времени (RT-PCR). Помимо указанных выше анализов, квалифицированному специалисту доступны различные анализы in vivo. Например, можно подвергать клетки в организме пациента действию антитела, которое необязательно является меченным поддающейся обнаружения меткой, например, радиоактивным изотопом, и можно оценивать связывание антитела с клетками у пациента, например, путем внешнего сканирования в отношении радиоактивности или путем анализа биоптата, взятого от пациента, ранее подвергнутого воздействию антитела.
Химиотерапевтические средства
Комбинированная терапия по изобретению, кроме того, может включать одно или несколько химиотерапевтическое средство(средств). Комбинированное введение включает совместное введение или одновременное введение с использованием отдельных составов или одного фармацевтического состава, и последовательное введение в любом порядке, где, предпочтительно, существует период времени, когда оба (или все) активных средства одновременно проявляют их виды биологической активности.
Химиотерапевтическое средство, если его вводят, обычно вводят в дозировках, известных для них, или, необязательно, их снижают вследствие комбинированного действия лекарственных средств или отрицательных побочных эффектов, свойственных введению антиметаболического химиотерапевтического средства. Получение и схемы дозирования для таких химиотерапевтических средств можно использовать согласно инструкциям изготовителя или как эмпирически определит специалист в данной области.
Различные химиотерапевтические средства, которые можно комбинировать, описаны в настоящем документе.
В некоторых вариантах осуществления химиотерапевтические средства, подлежащие комбинированию, выбирают из группы, состоящей из леналидомида (REVLIMID), ингибиторов протеосом (таких как бортезомиб (VELCADE) и PS342), таксоида бора (включая доцетаксел и паклитаксел), алкалоидов барвинка (таких как винорелбин или винбластин), соединения платины (такого как карбоплатин или цисплатин), ингибитора ароматазы (такого как лектрозол, анастразол или экземестан), антиэстрогена (например, фулвестрант или тамоксифен), этопозида, тиотепы, циклофосфамида, пеметрекседа, метотрексата, липосомального доксорубицина, пегилированного липосомального доксорубицина, капецитабина, гемцитабина, мелталина, доксорубицина, винкристина, ингибитора COX-2 (например, целекоксиб), или стероида (например, дексаметазон и преднизон). В некоторых вариантах осуществления (например, вариантах осуществления, вовлекающих лечение t(4;14)+ множественной миеломы), комбинируют дексаметазон и леналидомид, или дексаметазон, или бортезомиб, или винкристин, доксорубицин и дексаметазон, или талидомид и дексаметазон, или липосомальный доксорубицин, винкристин и дексаметазон, или леналидомид и дексаметазон, или бортезомиб и дексаметазон, или бортезомиб, доксорубицин, и дексаметазон. В некоторых вариантах осуществления (например, варианты осуществления, вовлекающие рак мочевого пузыря), комбинируют гемцитабин и цисплатин, или таксан (например, паклитаксел, доцетаксел), или пеметрексед, или метотрексат, винбластин, доксорубицин и цисплатин, или карбоплатин, или митомицин C в сочетании с 5-Фторурацил, или цисплатин, или цисплатин и 5-фторурацил.
Составы, дозировки и введение
Лекарственные средства, используемые для изобретения, можно составлять, дозировать и вводить так, чтобы это согласовывалось с "надлежащей медицинской практикой". Факторы, рассматриваемые в этом контексте, включают конкретное подвергаемое лечению нарушение конкретного подвергаемого лечению индивидуума, клиническое состояние конкретного пациента, причину нарушения и область доставки средства, способ введения, схему введения, взаимодействие лекарственное средство-лекарственное средство для комбинируемых средств, и другие факторы, известные медицинским практикующим специалистам.
Терапевтические составы получают с использованием стандартных способов известных в данной области, путем смешения активного ингредиента, имеющего желаемую степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences (20th edition), ed. A. Gennaro, 2000, Lippincott, Williams & Wilkins, Philadelphia, PA). Приемлемые носители включают физиологический раствор или буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные (менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; спирты сахаров, такие как манит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEEN®, PLURONICS® или полиэтиленгликоль (PEG).
Необязательно, но предпочтительно, состав содержит фармацевтически приемлемую соль, предпочтительно, хлорид натрия, и, предпочтительно, в приблизительно физиологических концентрациях. Необязательно, составы по изобретению могут содержать фармацевтически приемлемый консервант. В некоторых вариантах осуществления концентрация консерванта находится в диапазоне от 0,1 до 2,0%, как правило, об./об. Подходящие консерванты включают консерванты, известные в фармацевтической области. Предпочтительными консервантами являются бензиловый спирт, фенол, м-крезол, метилпарабен и пропилпарабен. Необязательно, составы по изобретению могут включать фармацевтически приемлемое поверхностно-активное вещество в концентрации от 0,005 до 0,02%.
Состав, представленный в настоящем документе, также может содержать более одного активного соединения, если необходимо для конкретного показания, подлежащего лечению, предпочтительно, соединения с дополняющими видами активностями, которые не оказывают неблагоприятного эффекта друг на друга. Такие молекулы соответственно присутствуют в сочетании в количествах, которые эффективны для поставленной цели.
Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, например, способами коацервации или межфазной полимеризации, например, микрокапсулы на основе гидроксиметилцеллюлозы или желатина и микрокапсулы на основе полиметилметакрилата, соответственно, в коллоидные системы для доставки лекарственного средства (например, липосомы, микросферы на основе альбумина, микроэмульсии, наночастицы и нанокапсулы), или в микроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences, выше.
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие антитело, которые представлены в виде имеющих форму изделий, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочная кислота-гликолевая кислота, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как этилен-винилацетат и молочная кислота-гликолевая кислота обеспечивают высвобождение молекулы в течение свыше 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. При инкапсулировании антитела остаются в организме в течение длительного времени, они могут подвергаться денатурации или образовывать агрегаты под воздействием влажности при 37°C, что приводит к потере биологической активности и возможным изменениями иммуногенности. В зависимости от вовлеченного механизма можно разрабатывать рациональные стратегии для стабилизации. Например, если открыто, что механизмом агрегации является межмолекулярное образование S-S связи через тио-дисульфидный обмен, стабилизации можно достигать модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, использованием соответствующих добавок и разработкой определенных полимерных композиций матриц.
Терапевтические средства по изобретению вводят пациенту-человеку хорошо известными способами, такими как внутривенное введение в качестве болюса или непрерывной инфузии в течение периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, внутрисуставным, интрасиновиальным, интратекальным, пероральным, местным или ингаляционным путями. Также для терапевтического использования можно использовать стратегию ex vivo. Стратегии ex vivo вовлекают трансфекцию или трансдукцию клеток, полученных от индивидуума, полинуклеотидом, кодирующим антагонист FGFR3. Затем трансфицированные или трансдуцированные клетки возвращают индивидууму. Клетки могут представлять собой любые из широкого диапазона типов, включая, но ими не ограничиваясь, гемопоэтические клетки (например, клетки костного мозга, макрофаги, моноциты, дендритные клетки, T-клетки или B-клетки), фибробласты, эпителиальные клетки, эндотелиальные клетки, кератиноциты или мышечные клетки.
Например, если антигонист FGFR3 представляет собой антитело, антитело вводят любым подходящим способом, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и интраназальное, и, если желательно для местного иммуносупрессивного лечения, введение в очаг повреждения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, антитело подходящим образом вводят путем импульсной инфузии, в частности, со снижающимися дозами антитела. Предпочтительно, дозирование проводят путем инъекций, наиболее предпочтительно, внутривенных или подкожных инъекций, в зависимости, частично, от того, является ли введение кратковременным или длительным.
В другом примере, соединение антигониста FGFR3 вводят локально, например, путем прямых инъекций, когда нарушение или область опухоли позволяют, и инъекции можно периодически повторять. Антагонист FGFR3 также можно доставлять системно индивидууму или напрямую в опухолевые клетки, например, в опухоль или в ложе опухоли после хирургического удаления опухоли, для профилактики или снижения локального рецидива или метастазирования.
Введение лекарственных средств в сочетании, как правило, проводят в течение определенного периода времени (обычно минут, часов, суток или недель, в зависимости от выбранной комбинации). Подразумевают, что комбинированная терапия включает введение этих лекарственных средств последовательно, т.е. где каждое лекарственное средство вводят в отличающееся время, а также введение этих лекарственных средств, или по меньшей мере двух из лекарственных средств, по существу одновременно.
Лекарственное средство можно вводить одним путем или различными путями. Например, антитело против FGFR3 в сочетании можно вводить путем внутривенной инъекции, в то время как химиотерапевтическое средство в сочетании можно вводить перорально. Альтернативно, например, оба лекарственных средства можно вводить перорально или оба лекарственных средства можно вводить путем внутривенной инъекции, в зависимости от конкретных лекарственных средств. Последовательность, в которой вводят лекарственные средства, также варьирует, в зависимости от конкретных средств.
В зависимости от типа и тяжести заболевания, приблизительно начальная предполагаемая дозировка для введения пациенту составляет от 1 мкг/кг до 100 мг/кг каждого лекарственного средства, например, с помощью одного или нескольких отдельных введений, или с помощью непрерывной инфузии. Типичная суточная дозировка может находиться в диапазоне от приблизительно 1 мкг/кг до приблизительно 100 мг/кг или более, в зависимости от факторов, упомянутых выше. Для повторяющихся введений в течение нескольких суток или более, в зависимости от состояния, лечение продолжают до тех пор, пока не вылечат злокачественную опухоль, при определении способами, описанными выше. Однако могут подходить другие режимы дозирования.
Настоящая заявка относится к введению антитела против FGFR3 с помощью генной терапии. См., например, WO96/07321, опубликованную 14 марта 1996 года, касающуюся применения генной терапии для получения внутриклеточных антител.
Изделия
В другом аспекте изобретения предусмотрено изделие, содержащее материалы, подходящие для лечения, профилактики и/или диагностики нарушений, описанных выше. Изделие содержит контейнер и ярлык на контейнере или вкладыш в упаковку, прилагаемый к контейнеру. Подходящие контейнеры включают, например, бутыли, флаконы, шприцы и тому подобное. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. В контейнере содержится композиция, которая сама по себе или при комбинировании с другой композицией(ями) эффективна для лечения, профилактики и/или диагностики состояния и может иметь отверстие для стерильного доступа (например, контейнер может представлять собой мешок для внутривенного раствора или флакон, имеющий пробку, проницаемую для иглы для подкожных инъекций). По меньшей мере одно активное вещество в композиции представляет собой антитело по изобретению. На ярлыке или вкладыше в упаковку указано, что композицию применяют для лечения выбранного состояния, такого как злокачественная опухоль. Более того, изделие может содержать (a) первый контейнер с композицией, содержащейся в нем, где композиция содержит антитело по изобретению; и (b) второй контейнер с композицией, содержащейся в нем, где композиция содержит дополнительное цитотоксическое средство. В этом варианте осуществления изобретения изделие может дополнительно содержать вкладыш в упаковку, на котором указано, что первую и вторую композиции антител можно применять для лечения конкретного состояния, например, злокачественной опухоли. Альтернативно, или дополнительно, изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфат-буферный солевой раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может включать другие материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Ниже представлены примеры способов и композиций по изобретению. Понятно, что можно применять на практике различные другие варианты осуществления, учитывая общее описание, представленное выше.
ПРИМЕРЫ
Материалы и способы
Клеточные линии и клеточная культура
Клеточная линия RT4 была получена от American Type Cell Culture Collection. Клеточные линии RT112, OPM2 и Ba/F3 были приобретены от German Collection of Microorganisms and Cell Cultures (DSMZ, (Германия)). Клеточная линия множественной миеломы KMS11 была любезно предоставлена Dr. Takemi Otsuki at Kawasaki Medical School (Япония). Клеточная линия рака мочевого пузыря TCC-97-7 была щедрым подарком Dr. Margaret Knowles at St James's University Hospital (Leeds, Великобритания). Клеточная линия UMUC-14 была получена от Dr. H. B. Grossman (в настоящее время в University of Texas M. D. Anderson Cancer Center, TX). Клетки поддерживали с помощью среды RPMI, дополненной 10% эмбриональной телячьей сывороткой (FBS) (Sigma), 100 Е/мл пенициллина, 0,1 мг/мл стрептомицина и L-глутамином в условиях 5% CO2 при 37°C.
Исследования с димеризацией FGFR3 S249C
Клетки UMUC-14 выращивали в не содержащей цистеин среде, обрабатывали R3Mab или DTNB в течение 3 ч, и клеточные лизаты подвергали иммуноблот-анализу в восстанавливающих и невосстанавливающих условиях. Для исследований димеризации in vitro, FGFR3-IIIbS249C (остатки 143-374) клонировали в вектор pAcGP67A и экспрессировали в клетках T.ni Pro. Рекомбинантный белок очищали через колонку Ni-NTA, а затем через колонку Superdex S200. Димерный FGFR3S249C элюировали в 25 мМ Tris (pH 7,5) и 300 мМ NaCl. R3Mab (1 мкМ) инкубировали с димером FGFR3S249C (0,1 мкМ) при 37°C в следующих условиях: 100 мМ KH2PO4 (pH 7,5), 25 мкМ DTT, 1 мМ ЭДТА и 0,75 мг/мл BSA. Аликвоты реакционной смеси отбирали в указанные моменты времени и реакцию останавливали добавлением буфера для образца без β-меркаптоэтанола. Димер-мономер анализировали с помощью иммуноблоттинга.
Исследования с ксенотрансплантатами
Все исследования были одобрены Genentech's Institutional Animal Care and Use Committee. Самок мышей nu/nu или мышей с тяжелым комбинированным иммунодефицитом (SCID) CB17 в возрасте 6-8 недель приобретали от Charles River Laboratory (Hollister, CA). Самки бестимусных мышей nude были получены от National Cancer Institute-Frederick Cancer Center. Мышей поддерживали в определенных условиях без патогенов. Стабильные клетки RT112 shРНК (7×106), RT112 (7×106), Ba/F3-FGFR3S249C (5×106), OPM2 (15×106), или KMS11 (20×106) имплантировали подкожно в бок мышей в объеме 0,2 мл в HBSS/матригеле (1:1 об./об., BD Biosciences). Клетки UMUC-14 (5×106) имплантировали без матригеля. Опухоли измеряли два раза в неделю с использованием штангенциркуля и объем опухоли вычисляли с использованием формулы: V=0,5 a×b2, где a и b представляют собой длину и ширину опухоли, соответственно. Когда средний объем опухоли достигал 150-200 мм3, мышей случайным образом разделяли на группы по 10 и проводили им два раза в неделю внутрибрюшинную (i.p) инъекцию R3Mab (0,3-50 мг/кг), или контрольного IgG1 человека, разбавленного в HBSS. Контрольным животным вводили носитель (HBSS) отдельно.
Статистика
Собранные данные выражают в качестве среднего значения +/- SEM. Для сравнения между двумя группами использовали непарные t-критерии Стьюдента (2-сторонние). Статистически значимой считали величину P<0,05 во всех экспериментах.
Получение стабильных клеток с shРНК для FGFR3
Три независимых shРНК для FGFR3 клонировали в вектор pHUSH, как описано (1). Последовательности для shРНК для FGFR3, использованных в исследованиях являются следующими: shРНК2:
5'GATCCCCGCATCAAGCTGCGGCATCATTCAAGAGATGATGCCGCAGCTTGATGCTTTTTTGGAAA (SEQ ID NO:192); shРНК4: 5'-GATCCCCTGCACAACCTCGACTACTATTCAAGAGATAGTAGTCGAGGTTGTGCATTTTTTGGAAA-3' (SEQ ID NO:193); shРНК6: 5'-GATCCCCAACCTCGACTACTACAAGATTCAAGAGATCTTGTAGTAGTCGAGGTTTTTTTTGGAAA-3' (SEQ ID NO:194). Все конструкции подтверждали секвенированием. Контрольная shРНК для EGFP описана в предыдущем исследовании автором изобретения (50). shРНК, содержащую ретровирус, получали котрансфекцией упаковочных клеток GP2-293 (Clontech Laboratories, Mountain View, CA) с конструкциями VSV-G (Clontech Laboratories) и pHUSH-FGFR3 shРНК, и супернатанты вирусов собирали через 72 ч после трансфекции, и отчищали от клеточного дебриса центрифугированием для эксперимента по трансдукции.
Клетки RT112 поддерживали в среде RPMI1640, содержавшей FBS, не содержащую тетрациклина (Clontech Laboratories), и трансдуцировали ретровирусным супернатантом в присутствии 4 мкг/мл полибрена. Через 72 часа после инфекции к среде добавляли 2 мкг/мл пуромицина (Clontech Laboratories) для селекции стабильных клонов, экспрессирующих shРНК. Стабильные клетки выделяли, обрабатывали 0,1 или 1 мкг/мл доксицилина (Clontech Laboratories) в течение 4 суток, и индуцибельный нокдаун экспрессии белка FGFR3 оценивали вестерн-блот анализом. Анализы клеточного цикла проводили, как описано (51).
Селекция фаговых антител, специфичных к FGFR3
Фаговые библиотеки антител человека с синтетическим разнообразием в выбранных определяющих комплементарность областях (H1, H2, H3, L3), имитирующим природное разнообразие репертуара IgG человека, использовали для пэннинга. Fab-фрагменты экспонировали бивалентно на поверхности частиц бактериофага M13 (52). В качестве антигенов использовали меченные His IgD2-D3 FGFR3-IIIb и IIIc. На 96-луночные иммунопланшеты MaxiSorp (Nunc) наносили в течение ночи при 4°C белок FGFR3-IIIb-His или белок FGFR3-IIIC-His (10 мкг/мл) и блокировали в течение 1 часа буфером PBST (PBS с 0,05% Tween 20), дополненным 1% BSA. Добавляли фаговые библиотеки антител и инкубировали их в течение ночи при комнатной температуре (RT). Планшеты промывали буфером PBST и связавшийся фаг элюировали с помощью 50 мМ HCl и 500 мМ NaCl в течение 30 минут и нейтрализовывали равным объемом 1 M основания Tris. Выделенные фаги амплифицировали в клетках E.coli XL-1 blue. В процессе последующих раундов селекции время инкубации фаговых антител снижали до 2 часов и строгость промывания планшетов постепенно увеличивали (53). Уникальные и специфические фаговые антитела, которые связываются как с изоформой IIIb, так и с изоформой IIIc FGFR3, идентифицировали фаговым ELISA и секвенированием ДНК. Из 400 подвергнутых скринингу клонов четыре были отобраны для изменения формата на полноразмерные IgG путем клонирования областей VL и VH отдельных клонов в векторы LPG3 и LPG4, соответственно, временно экспрессированные в клетках млекопитающих и очищенные с помощью колонок с белком A (54). Для созревания аффинности был отобран клон 184.6.
Для созревания аффинности фагмида, экспонирующая одновалентный Fab на поверхности бактериофага M13 (52), служила в качестве матрицы библиотеки для пересадки вариабельных доменов легкой цепи (VL) и тяжелой цепи (VH) фага Ab. В CDR-L3 были включены стоп-кодоны. Для созревания аффинности была выбрана стратегия мягкой рандомизации (53). Для рандомизации были отобраны две различных комбинации петель CDR, H1/H2/L3, H3/L3 или L1/L2/L3. Для селекции прошедших стадию созревания аффинности клонов, фаговые библиотеки сортировали против белка FGFR3 IIIb или IIIc-His, подвергали сортировке на планшете на первом раунде, а затем четырем раундам сортировке в растворе, как описано (52). После пяти раундов пэннинга использовали высокопроизводительный одноточечный конкурентный фаговый ELISA для быстрого скрининга высокоаффинных клонов, как описано (55). Клоны с низким соотношением поглощения при 450 нм в присутствии 10 нМ FGFR3-His к поглощению в отсутствие FGFR3-His были выбраны для дальнейшей охарактеризации.
Клоны 184.6.1, 184.6.21, 184.6.49, 184.6.51, 184.6.58, 184.6.62 и 184.6.92 значительно снижали жизнеспособность клеточных линий Ba/F3-FGFR3-IIIb, Ba/F3-FGFR3-IIIc и Ba/F3-FGFR3-S249C и клон 184.6.52 значительно снижал жизнеспособность клеточной линии Ba/F3-FGFR3-S249C. Увеличенная ингибиторная активность от приблизительно 50 раз (клон 184.6.52) до приблизительно 100 раз (клоны 184.6.1, 184.6.21, 184.6.49, 184.6.51, 184.6.58, 184.6.62 и 184.6.92) превышала ингибиторную активность исходного клона 184.6., в зависимости от анализированной клеточной линии. Кинетика связывания клонов 184.6.1, 184.6.58 и 184.6.62 с FGFR3-IIIb и FGFR3-IIIc была определена с использованием BIAcore следующим образом:

Клоны 184.6.1, 184.6.58 и 184.6.62 также продемонстрировали увеличенное ингибирование следующей после FGFR3 передачи сигнала в клетках Ba/F3-FGFR3, клетках RT112 и клетках OPM2.
Был отобран клон 184.6.1. Для улучшения технологичности в HVR H2 в остатке 54 вносили модификацию последовательности, N54S, получив клон 184.6.1N54S. Клоны 184.6.1 и 184.6.1N54S проявляли сравнимую кинетику связывания (измеренную в анализах Biacore) и сравнимую активность в анализе жизнеспособности клеток Ba/F3. Были получены дополнительные варианты HVR H2: N54S вносили в клон 184.6.58, и N54G, N54A, или N54Q были внесены в клон 184.6.1 и 184.6.58. Эти клоны продемонстрировали сравнимую активность в анализе жизнеспособности клеток Ba/F3 с исходными клонами 184.6.1 или 184.6.58.
Другую модификацию последовательности, D30E, вносили в HVR L1 клона 184.6.1N54S, создавая клон 184.6.1NSD30E. Клон 184.6.1NSD30E и клон 184.6.1N54S продемонстрировали сравнимую кинетику связывания и сравнимую активность в анализе жизнеспособности клеток BA/F3 с исходными клонами 184.6.1 или 184.6.58.
Как используют в настоящем документе "R3Mab" относится к клонам антитела против FGFR3 184.6.1N54S, 184.6.1, или 184.6. Клон 184.6.1N54S использовали на фигурах и в экспериментах, обозначая его как "R3Mab", за исключением экспериментов, приведших к результатам, представленным на следующих фигурах (для которых использованное антитело указано в скобках): фигуры 9B (клон 184.6.1), 10 (клон 184.6), 11A и B (клон 184.6), 13 (клон 184.6.1), 14A (клон 184.6.1), 14B, G и H (клон 184.6), 19 (клон 184.6.1), и 22B и C (клон 184.6.1).
Анализ BIAcore/поверхностным плазмонным резонансом (SRP) для определения аффинности связывания антител
Аффинность связывания R3Mab с FGFR3 измеряли с помощью Biacore/SRP с использованием устройства BIAcoreTM-3000, как описано (52), со следующими модификациями. R3Mab непосредственно наносили на биосенсорные чипы CM5 для достижения приблизительно 400 единиц ответа (RU). Для измерения кинетики двукратные серийные разведения белка FGFR3-IIIb или IIIc-His (начиная с 67 нМ) инъецировали в буфере PBST при 25°C со скоростью потока 30 мкл/минута. Константы ассоциации (Kon, в моль/с) и константы диссоциации (Koff, в с) вычисляли с использованием простой модели связывания Ленгмюра "один к одному" (BIAcore Evaluation Software version 3.2). Равновесную константу диссоциации (Kd, в моль) вычисляли как соотношение Koff/Kon.
Аффинность связывания антител гибридомы мыши с FGFR3 измеряли с помощью Biacore/SRP следующим образом. FGFR3-IIIb или IIIc человека связывали с тремя различными проточными ячейками (FC), FC2, FC3 и FC4, сенсорного чипа BIACORETM CM5 для достижения единиц ответа (RU) приблизительно 50 RU. Иммобилизации достигали случайным присоединением через аминогруппы с использованием протокола, предоставленного изготовителем. Сенсограммы регистрировали в отношении связывания происходящего из гибридомы IgG против FGFR3 или его Fab-фрагмента с этими поверхностями при 25°С путем инъекции серии растворов в диапазоне от 250 нМ до 0,48 нМ с 2-кратным шагом со скоростью потока 30 мкл/мин. Между каждой инъекцией, 10 мМ глицин-HCl, pH 1,7, служил в качестве буфера для регенерации сенсорного чипа. Сигнал от эталонной ячейки (FC1) вычитали из сенсограммы, наблюдаемой на FC2, FC3 и FC4. Кинетические константы вычисляли путем нелинейной регрессионной аппроксимации данных согласно модели связывания Ленгмюра 1:1 с использованием BIAcore Evaluation Software (версии 3.2), предоставляемого изготовителем.
Исследования связывания с помощью ELISA
кДНК, кодирующие внеклеточные домены (ECD) FGFR1-IIIb, IIIc, FGFR2-IIIb и IIIc, FGFR3-IIIb и IIIc и FGFR4 человека, клонировали в вектор на основе pRK для получения химерных белков FGFR человека Fc человека. Рекомбинантные белки продуцировали с помощью временной трансфекции клеток яичника китайского хомячка (CHO) и очищали аффинной хроматографией с белком A. Для тестирования связывания антител с FGFR человека на 96-луночные планшеты Maxisorp (Nunc) наносили в течение ночи при 4°C 50 мкл химерных белков ECD FGFR-Fc человека в концентрации 2 мкг/мл. После блокирования фосфатно-солевым буфером (PBS)/3% BSA добавляли антитело против FGFR3 и инкубировали при RT в течение 2 часов. Обнаружение специфично связанного антитела против FGFR3 проводили с использованием конъюгированного с HRP Fab против антител человека и колоригенный субстрат пероксидазы TMB (KPL, Gaithersburg, MD).
Для тестирования эффекта антител против FGFR3 на взаимодействие FGF/FGFR3, химерные белки FGFR3-Fc иммобилизовывали на планшете Maxisorp, покрытом антителом, специфичным к Fcγ-фрагменту иммуноглобулина человека (Jackson Immunoresearch, West Grove, PA). После промывания в планшет добавляли возрастающие количества антитела против FGFR3 и инкубировали в течение 30 минут. Затем добавляли FGF1 или FGF9 и гепарин для инкубации при RT в течение 2 часов. Планшеты промывали и инкубировали в течение 1 часа с биотинилированным специфичным к FGF1 поликлональным антителом (BAF232) или биотинилированным антителом против FGF9 (BAF273, R&D Systems), с последующим обнаружением с помощью стрептавидин-HRP и TMB.
Получение стабильных клеток Ba/F3-FGFR3
кДНК, кодирующую полноразмерный FGFR3 IIIb или IIIc человека, клонировали в вектор pQCXIP (Clontech Laboratories, Mountain View, CA) с получением pQCXIP-FGFR3-IIIb или IIIc. Конкретные мутации, т.е. R248C, S249C, G372C, Y375C и K652E, вносили в кДНК с помощью QuickChange (Stratagene, La Jolla, CA). Для получения стабильных клеток Ba/F3, экспрессирующих FGFR3 дикого типа или мутантный FGFR3, различные конструкции pQCXIP-FGFR3 котрансфицировали в упаковочные клетки GP2-293 с плазмидой VSV-G (Clontech Laboratories). После селекции с помощью 2 мкг/мл пуромицина в течение двух недель, клетки, экспрессировавшие FGFR3 дикого типа или мутантный FGFR3, окрашивали конъюгированным с фикоэритрином mAb против FGFR3 человека (FAB766P, R&D Systems), и отбирали с помощью активированной флуоресценцией сортировки клеток (FACS) для функциональных анализов. Для анализа пролиферации клеток в 96-луночном микропланшете для титрования использовали следующую плотность клеток: для клеток, экспрессирующих FGFR3-IIIb дикого типа и FGFR3-K652E: 5000 клеток/лунка; для остальных: 10000 клеток/лунка. Клетки высевали в среду RPMI1640, дополненную 10% эмбриональной телячьей сывороткой, 10 нг/мл FGF1 вместе с 10 мкг/мл гепарина (Sigma-Aldrich, St. Louis, MO). Добавляли R3Mab в указанной концентрации и антитела против FGFR3 гибридомы мыши добавляли в концентрации от 2000 до 0,49 нг/мл (четырехкратными серийными разведениями) в эксперименте с FGFR3-IIIb и в концентрации от 5000 до 1,2 нг/мл (четырехкратными серийными разведениями) в эксперименте с FGFR3-IIIc. После инкубации в течение 72 часов оценивали жизнеспособность клеток с помощью CellTiter-Glo (Promega, Madison, WI).
Анализ пролиферации клеток
Для анализов пролиферации клеток RT112, RT4 и TCC-97-7, 3000 клеток/лунка высевали в 96-луночный микропланшет для титрования и позволяли им прикрепляться в течение ночи. Затем среду заменяли средой с низким содержанием сыворотки (0,5% FBS) с контрольным антителом или R3Mab в указанных концентрациях. После инкубации в течение 4 суток в каждую лунку добавляли 1 мкКи [метил-3H]тимидина (PerkinElmer, Waltham, MA) и инкубировали в течение дополнительных 16 часов. Клетки переносили в UniFilters с использованием Packard Filtermate Harvester, и измеряли [3H]-тимидин, включенный в геномную ДНК растущих клеток, с использованием TopCount (PerkinElmer). В некоторых случаях жизнеспособность клеток оценивали с помощью CellTiter-Glo (Promega) после инкубации с антителами в течение 4 суток. Величины представлены в качестве средних значений +/- SE для четырех повторений.
Анализ клонального роста
Эффект R3Mab на клоногенность клеток оценивали согласно ранее описанному протоколу (50). Коротко, 400 клеток UMUC-14 высевали в 6-ячеечный планшет в среде DMEM, дополненной 10% эмбриональной телячьей сывороткой для обеспечения адгезии в течение ночи. Затем добавляли R3Mab или контрольное антитело, разбавленные в 0,1% среде BSA, до конечной концентрации 10 мкг/мл. В качестве другого контроля использовали равный объем 0,1% среды BSA отдельно (Mock). Клетки инкубировали в течение приблизительно 12 суток до тех пор, пока клетки в контрольных группах не образовывали достаточно крупные колонии. Колонии окрашивали 0,5% кристаллическим фиолетовым, и число и размер колоний количественно определяли с использованием GelCount (Oxford, Великобритания). Количество колоний крупнее 120 мкм в диаметре представляли в качестве среднего значения +/- SEM (n=12).
Анализы иммунопреципитации и иммуноблоттинга
Для исследования эффекта антител на передачу сигнала FGFR3 клетки подвергали голоданию в бессывороточной среде в течение ночи перед началом обработки. Клетки инкубировали либо с антителами, разбавленными в среде RPMI1640 с 0,1% BSA (масс./об.), либо со средой с 0,1% BSA отдельно (имитация). После 3 часов при 37°C к половине образцов добавляли FGF1 (конечная концентрация 15 нг/мл) и гепарин (конечная концентрация 5-10 мкг/мл). В качестве контролей к другой половине образцов добавляли ангалогичный объем гепарина отдельно. Инкубацию продолжали в течение 10 мин. Супернатанты извлекали аспирированием и клетки промывали ледяным PBS, затем лизировали в буфере RIPA (Upstate, Charlottesville, VA), дополненном 1 мМ ортованадатом натрия и полным коктейлем ингибиторов протеаз (Roche Applied Science, Indianapolis, IN). Лизаты очищали от нерастворимых материалов центрифугированием.
FGFR3 подвергали иммунопреципитации с использованием поликлонального антитела кролика (sc-123, Santa Cruz Biotechnology, Santa Cruz, CA) и анализировали с помощью натрий додецил-полиакриламидного гель-электрофореза (SDS-PAGE) и вестерн-блоттинга. Фосфорилированный FGFR3 оценивали с помощью моноклонального антитела против фосфотирозина (4G10, Upstate). Общий FGFR3 анализировали с помощью моноклонального антитела против FGFR3 (sc-13121, Santa Cruz Biotechnology). Фосфорилирование и активацию передачи сигнала FGFR3 исследовали с использованием следующих антител: антитело против FGFRY653/654, антитело против FRS2αY196, антитело против фосфо-p44/42 MAPKT202/Y204, антитело против общей p44/42 MAPK и антитело против AKTS473 были приобретены от Cell Signaling Technology (Danvers, MA); и антитело против общего FRS2α (sc-8318) было приобретено от Santa Cruz Biotechnology (Santa Cruz, CA). Блоты визуализировали с использованием хемилюминесцентного субстрата (ECL Plus, Amersham Pharmacia Biotech, Piscataway, NJ).
Картирование эпитопов антител
Для определения эпитопа R3Mab, 13 перекрывающихся пептидов, каждый длиной по 15 аминокислот, синтезировали, чтобы они охватывали внеклеточный домен FGFR3 человека от остатка 138 до остатка 310. Пептиды биотинилировали на C-конце, и иммобилизовывали на планшеты со стрептавидином (Pierce, Rockford, IL) в течение ночи. После блокирования с помощью PBS/3% BSA планшеты инкубировали с R3Mab и подвергали обнаружения с использованием конъюгированного с HRP IgG против антител человека (Jackson Immunoresearch) и колоригенного субстрата пероксидазы TMB (KPL, Gaithersburg, MD).
Антитела гибридомы мыши против FGFR3 человека 1G6, 6G1 и 15B2 тестировали в анализе ELISA для идентификации связываемых ими эпитопов. 1G6, 6G1 и 15B2 связываются с IgD2-IgD3 FGFR3 человека (как изоформы IIIb, так и изоформы IIIc), в то время как 5B8 связывает только IgD2-IgD3 FGFR3-IIIb человека. В конкурентном анализе 1G6, 6G1 и 15B2 конкурировали друг с другом за связывание FGFR3 человека, указывая на то, что 1G6, 6G1 и 15B2 имеют перекрывающиеся эпитопы. Ни одно из гибридомных антител не конкурировало с фаговым антителом 184.6. указывая на то, что гибридомные антитела имеют эпитоп(ы), отличающийся от эпитопа 184.6.
Получение и молекулярное клонирование антител мыши против FGFR3 1G6, 6G1 и 15B2
Мышей BALB/c иммунизировали 12 раз с помощью 2,0 мкг FGFR3-IIIb (химера rhFGFR3 (IIIB)/Fc, от R&D Systems, каталожный номер # 1264-FR, партия # CYH025011), или с помощью 2,0 мкг FGFR3-IIIc (химера rhFGFR3 (IIIc)/Fc, от R&D Systems, каталожный номер # 766-FR, партия # CWZ055041), ресуспендированных в адъюванте монофосфориллипид A/дикориномиколат трегалозы (Corixa, Hamilton, MT) в каждую из подушечек лап задних конечностей два раза в неделю. Через трое суток после последнего вспомогательного введения, подколенные лимфатические узлы подвергали слиянию с клеточной линией миеломы мыши P3X63Ag.U.1, с помощью электрослияния (Hybrimune, Cyto Pulse Sciences, Glen Burnie, Mаryland). Слитые гибридомные клетки отбирали из неслитых клеток подколенных узлов или клеток миеломы с использованием селекции с помощью гипоксантин-аминоптерин-тимидина (HAT) в среде D из набора для селекции гибридом ClonaCell® (StemCell Technologies, Inc., Vancouver, BC, Canada). Культуральные супернатанты первоначально подвергали скринингу в отношении их способности связываться с FGFR3-IIIb и FGFR3-IIIc в ELISA, а затем представляющие интерес гибридомы подвергали в отношении их способности к окрашиванию в FACS на трансфицированных FGFR3-IIIb клетках Ba/F и контрольных клетках Ba/F, а также в отношении блокирующей активности антитела. Затем отобранные гибридомы клонировали способом лимитирующих разведений.
Общую РНК экстрагировали из гибридомных клеток, продуцировавших моноклональное антитело мыши против FGFRIII человека 1G6 и 15B2, с использованием набора RNeasy Mini Kit (Qiagen, Germany). Вариабельные домены легкой цепи (VL) и вариабельные домены тяжелой цепи (VH) амплифицировали с использованием RT-PCR с помощью следующих вырожденных праймеров:
1G6:
Прямой для легкой цепи (LC): 5'-GTCAGATATCGTKCTSACMCARTCTCCWGC-3' (SEQ ID NO:195)
Прямой для тяжелой цепи (HC): 5'-GATCGACGTACGCTGAGATCCARYTGCARCARTCTGG-3' (SEQ ID NO:196)
6G1:
Прямой для легкой цепи (LC): 5'-GTCAGATATCGTGCTGACMCARTCTCC-3' (SEQ ID NO:197)
Прямой для тяжелой цепи (HC): 5'- GATCGACGTACGCTGAGATCCARYTGCARCARTCTGG-3' (SEQ ID NO:198)
15B2:
Прямой для легкой цепи (LC): 5'-GTACGATATCCAGATGACMCARTCTCC-3' (SEQ ID NO:199)
Прямой для тяжелой цепи (HC): 5'-GATCGACGTACGCTGAGATCCARYTGCARCARTCTGG-3' (SEQ ID NO:200)
Обратные праймеры для легкой цепи и тяжелой цепи для всех трех клонов являются следующими:
Обратный легкой цепи: 5'-TTTDAKYTCCAGCTTGGTACC-3' (SEQ ID NO:201)
Обратный тяжелой цепи: 5'-ACAGTGGGCCCTTGGTGGAGGCTGMRGAGACDGTGASHRDRGT-3' (SEQ ID NO:202).
Прямые праймеры были специфичными к N-концевой аминокислотной последовательности VL- и VH-области. Обратные праймеры LC и HC конструировали, чтобы они подвергались отжигу с областью в константном домене легкой цепи (CL) и константном домене 1 тяжелой цепи (CH1), соответственно, которые являются высоко консервативными среди видов.
Амплифицированный VL клонировали в вектор экспрессии клеток млекопитающих pRK (Shields et al., (2000) J. Biol. Chem. 276:659), содержащий константный домен каппа человека. Амплифицированый VH встраивали в вектор экспрессии клеток млекопитающих pRK, кодирующий полноразмерный константный домен IgG1 человека. Последовательность тяжелой и легкой цепей определяли с использованием общепринятых способов.
Кристаллизация, определение и уточнение структуры
ECD FGFR3-IIIb человека (остатки 143-374) клонировали в вектор pAcGP67A (BD Bioscience, San Jose, CA), продуцированный в клетках T.ni Pro, и очищали с использованием колонки Ni-NTA, а затем с помощью эксклюзионной хроматографии. Fab R3Mab экспрессировали в E.coli и очищали последовательно на аффинной колонке с белком G, на колонке с SP-сефарозой и на колонке с Superdex 75. Комплекс Fab-FGFR3 получали с помощью инкубации Fab с избытком ECD FGFR3, а затем комплекс дегликозилировали и очищали над колонкой с ограничением размера Superdex-200 в 20 мМ TrisCl, pH 7,5, и 200 мМ буфере NaCl. Содержащие комплекс фракции объединяли и концентрировали до 20 мг/мл и использовали в испытаниях по кристаллизации. Кристаллы, использованные в определении структуры, выращивали при 4°C при следующих условиях: 0,1 M какодилат натрия, pH 6,5, 40% MPD и 5% PEG8000 с использованием способа паровой диффузии. Данные обрабатывали с использованием HKL2000 и Scalepack (56). Структуру разрешали с помощью молекулярной замены с использованием программы Phaser (57) и координат 1RY3 (FGFR3) и 1N8Z (Fab-фрагмент). Модель завершали с использованием программы Coot (58) и структуру уточняли до R/Rfree 20,4%/24,3% с помощью программы Refmac (59). Координаты и показатели структуры депонировали в Protein Data Bank под регистрационным кодом 3GRW и они также описаны в USSN 61/163222, поданной 25 марта 2009 года, содержание которой включено в настоящий документ в качестве ссылки.
Анализ ADCC
PBMC человека выделяли центрифугированием в градиенте Ficoll гепаринизированной крови, и ADCC измеряли с использованием клеточных линий множественной миелома OPM2 или KMS11 или клеточных линий рака мочевого пузыря RT112 или UMUC-14 в качестве клеток-мишеней и PBMC в качестве эффекторных клеток в соотношении мишень:эффектор 1:100. Клетки-мишени (10000 клеток/лунка) обрабатывали R3Mab или контрольным IgG1 человека в течение 4 часов при 37°C. Цитотоксичность определяли путем измерения высвобождения LDH с использованием анализа целостности гомогенных мембран CytoTox-ONE согласно инструкциям изготовителя (Promega, Madison, WI). Результаты выражены в качестве процента специфичного цитолиза с использованием формулы: Цитотоксичность (%) = [(экспериментальный лизис - экспериментальный спонтанный лизис)/ (максимальный лизис мишени - спонтанный лизис мишени)]×100, где спонтанный лизис представляет собой неспецифичный цитолиз в отсутствие антитела, и максимальный лизис мишени индуцирован 1% Triton X-100.
Результаты
Индуцибельный нокдаун shРНК для FGFR3 снижает рост рака мочевого пузыря in vivo
В качестве вступления для оценки важности FGFR3 для роста опухоли мочевого пузыря in vivo, авторы изобретения исследовали эффект нокдауна FGFR3 in vitro. Несколько малых интерферирующих (si)РНК для FGFR3 эффективно подавляли FGFR3 в клеточных линиях рака мочевого пузыря, экспрессирующих либо FGFR3 WT (RT112, RT4, SW780), либо мутантный FGFR3 (UMUC-14, мутация S249C). Нокдаун FGFR3 во всех четырех клеточных линиях значительно подавлял пролиферацию в культуре (фигура 15). Далее, авторы настоящего изобретения получили стабильные клеточные линии RT112, экспрессирующие индуцибельную доксицилином shРНК для FGFR3. Индукция трех независимых shРНК для FGFR3 доксицилином снижала экспрессию FGFR3, в то время как индукция контрольной shРНК, нацеленной на EGFP, не имела эффекта (фигура 7A). В отсутствие экзогенного FGF обработка доксицилином снижала включение [3H]-тимидина клетками, экспрессирующими различные shРНК для FGFR3, но не контрольную shРНК (фигура 7B), подтверждая, что нокдаун FGFR3 ингибирует пролиферацию. Кроме того, анализ экспоненциально растущих клеток RT112 показал, что нокдаун FGFR3 при обработке доксицилином в течение 72 ч значительно и специфично снижал процент клеток в фазах S и G2 клеточного цикла при одновременном увеличении количества клеток в фазе G1 (фигура 7C). Аналогичный эффект наблюдали для двух других shРНК для FGFR3 (фигура 16A). Не было выявлено значимых количеств клеток с субдиплоидным содержанием ДНК, указывая на отсутствие изменения в уровнях апоптоза. Таким образом, ингибиторный эффект нокдауна FGFR3 на пролиферацию клеток RT112, главным образом, является следствием подавления прогрессирования клеточного цикла.
Далее авторы настоящего изобретения оценивали эффект нокдауна FGFR3 на рост предварительно установленных ксенотрансплантатов опухолей RT112 у мышей. Нокдаун FGFR3 значительно и специфично подавлял рост опухоли (фигура 7D, верхние панели, и фигура 16B). Анализ образцов опухоли на 45 сутки подтвердил эффективный нокдаун FGFR3 при индукции доксицилином shРНК для FGFR3 по сравнению с контрольной shРНК (фигура 7D, нижние панели). Эти результаты демонстрируют, что FGFR3 является крайне важным как in vitro, так и in vivo, для роста клеток рака мочевого пузыря RT112.
Получение блокирующего моноклонального антитела против FGFR3
Для дальнейшего исследования важности FGFR3 в росте опухоли и для исследования потенциала этого рецептора в качестве терапевтической мишени, авторы настоящего изобретения разработали антагонистическое моноклональное антитело против FGFR3 (дублированное R3Mab) с использованием подхода фагового дисплея. Авторы настоящего изобретения выбрали это конкретное антитело, исходя из его способности блокировать как связывание лиганда, так и димеризацию, с помощью FGFR3, и его уникальной способности ингибировать не только FGFR3 WT, но также наиболее распространенные связанные со злокачественной опухолью мутанты этого рецептора (см. ниже). R3Mab нацелено на внеклеточные домены IgD2 и IgD3 в FGFR3, что необходимо и достаточно для связывания FGF (4). R3Mab связывало как изоформу IIIb, так и изоформу IIIc FGFR3 человека, но не продемонстрировало поддающегося обнаружения связывания с FGFR1, FGFR2 или FGFR4 (фигура 8A). Анализ Biacore показал, что R3Mab имело аналогичную кажущуюся аффинность к FGFR3-IIIc мыши, яванского макака и человека (данные не представлены). Аффинность R3Mab к FGFR3 человека представлена в таблице 2.
Далее авторы настоящего изобретения тестировали способность R3Mab блокировать связывание FGFR3 с FGF1 и FGF9. R3Mab сильно ингибировало связывание FGF1 с FGFR3-IIIb и -IIIc, с полумаксимальными ингибиторными концентрациями (IC50) 0,3 нМ и 1,7 нМ, соответственно (фигура 8B,C). Аналогично, R3Mab эффективно блокировало связывание FGF9 с FGFR3-IIIb и -IIIc, с IC50 1,1 нМ и 1,3 нМ, соответственно (фигура 8D,E).
R3Mab ингибирует FGFR3 WT и его наиболее распространенные связанные со злокачественной опухолью мутантные варианты
Для исследования того, ингибирует ли R3Mab пролиферацию клеток, запускаемую FGFR3 WT или мутантным FGFR3, авторы настоящего изобретения воспользовались наблюдением, что эктопическая экспрессия FGFR3 в про-B-клетках Ba/F3 мыши обеспечивает интерлейкин (IL)-3-независимую, FGF1-зависимую пролиферацию и выживание (29). В отсутствие FGF1 и IL-3, клетки Ba/F3, стабильно экспрессировавшие FGFR3 WT, не были жизнеспособными, в то время как FGF1 значительно усиливал их пролиферацию (фигура 9A). R3Mab специфично блокировало стимулируемую FGF1 пролиферацию клеток Ba/F3-FGFR3 дозозависимым образом (фигура 9A). Далее авторы настоящего изобретения оценивали влияние R3Mab на передачу сигнала FGFR3 в этих клетках. FGF1 индуцировал фосфорилирование и активацию FGFR3 и сопутствующую активацию p44/42 MAPK, в то время как R3Mab эффективно подавляло активацию обеих молекул (фигура 9B).
При раке мочевого пузыря, соматические активирующие мутации в FGFR3 кластеризуются в линкерной области между IgD2 и IgD3, внеклеточном юкстамембранном домене или киназном домене (фигура 9C). Внеклеточные миссенс-замены наиболее часто приводят к неспаренному цистеину, что приводит к лиганд-независимой димеризации FGFR3. Эти мутации вызывают значительно отличающиеся уровни конститутивной активации FGFR3, возможно вследствие дифференциального влияния на ориентацию цитоплазматического киназного домена (30, 31). Наиболее частыми мутациями являются S249C, Y375C, R248C, G372C и K652E, которые совместно составляют 98% от всех мутаций FGFR3 при раке мочевого пузыря (32). Авторы настоящего изобретения пришли к выводу, что оптимальное лекарственное средство должно блокировать не только белок FGFR3 WT, который сверхэкспрессируется в определенных злокачественных опухолях, но также наиболее распространенные связанные с опухолью мутанты FGFR3. Для дальнейшей оценки R3Mab, авторы создали клеточные линии Ba/F3, стабильно экспрессирующие каждый из пяти наиболее распространенных мутантных вариантов FGFR3. Все мутанты экспрессировались на аналогичных уровнях на клеточной поверхности, и мутанты по цистеину димеризовывались спонтанно без лиганда (данные не представлены). Клеточные линии, экспрессировавашие различные мутанты по цистеину, проявляли различные ростовые ответы на FGF1, что согласуется с более ранними данными (30, 31). Как описано ранее (33), клетки, экспрессирующие FGFR3R248C, проявляют конститутивную лиганд-независимую пролиферацию и не отвечают на FGF1 (фигура 9D). Аналогично, наиболее частая мутация, FGFR3S249C, обеспечивала лиганд-независимую пролиферацию (фигура 9E). Примечательно, что R3Mab подавляло конститутивную пролиферацию, запускаемую каждым из мутантов (фигура 9 D,E). Для клеток, экспрессировавших мутации в юкстамембранном домене FGFR3G372C (фигура 9F) или FGFR3Y375C (фигура 9G) требовался FGF1 для пролиферации, и их рост полностью блокировался с помощью R3Mab. Клетки, экспрессировавшие FGFR3K652E, продемонстрировали слабую лиганд-независимую пролиферацию и значительный рост в ответ на FGF1 (33). R3Mab не влияло на слабую базальную активность FGFR3K652E (данные не представлены), но практически устраняло лиганд-индуцируемую пролиферацию, опосредуемую этим мутантом (фигура 9H). Таким образом, R3Mab обладает уникальной способностью ингибировать как WT, так и преобладающие связанные со злокачественной опухолью мутанты FGFR3. Более того, R3Mab не проявляло поддающейся обнаружения активности агониста.
В качестве отдельной попытки авторы настоящего изобретения получили и охарактеризовали несколько гибридомных антител мыши против FGFR3 человека. Ни одно из гибридомных антител не могло ингибировать все связанные со злокачественной опухолью мутанты FGFR3, которые протестировали авторы настоящего изобретения (фигура 17), также они не обладали общими перекрывающимися эпитопами с R3Mab.
Более того, все из гибридомных антител продемонстрировали активность агонистов, сильно стимулируя пролиферацию связанных со злокачественной опухолью мутантов по FGFR3 R248C и S249C, и демонстрируя некоторую стимуляцию пролиферации мутантов по Y375C и G370C. Гибридомные антитела продемонстрировали различные уровни антагонизма и агонизма, в зависимости от протестированного мутанта FGFR3, следующим образом:
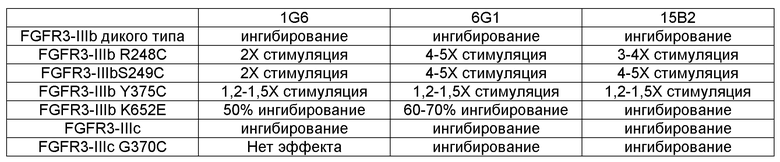
Таким образом, гибридомные антитела продемонстрировали непредсказуемый дифференциальный эффект на пролиферацию клеток Ba/F3, запускаемую различными мутантами FGFR3.
Охарактеризация гибридомных антител мыши против FGFR3 человека
Гибридомные антитела мыши против FGFR3 человека далее охарактеризовывали следующим образом:
(1) В анализе для тестирования способности гибридомных антител мыши против FGFR3 ингибировать связывание FGF1 с изоформами FGFR3-IIIb и IIIc человека, антитела 1G6, 6G1 и 15B2 были способны блокировать связывание FGF1 с изоформами FGFR3-IIIb и IIIc человека дозозависимым образом. При тестировании при диапазоне концентраций антител приблизительно от 2000 до 0,49 нг/мл, антитела 1G6, 6G1 и 15B2 блокировали связывание FGF1 с FGFR3-IIIb с величинами IC50 0,69, 0,87 и 0,72 нМ. При тестировании в диапазоне концентраций приблизительно от 5000 до 1,2 нг/мл, антитела 1G6, 6G1 и 15B2 блокировали связывание FGF1 с FGFR3-IIIc с величинами IC50 0,57, 3,4 и 0,7 нМ, соответственно.
(2) В анализе для тестирования способности гибридомных антител мыши против FGFR3 ингибировать связывание FGF9 с изоформами FGFR3-IIIb и IIIc человека, антитела 1G6, 6G1 и 15B2 эффективно блокировали связывание FGF1 с изоформами FGFR3-IIIb и IIIc человека дозозависимым образом. При тестировании при диапазоне концентраций антител приблизительно от 2000 до 0,49 нг/мл, антитела 1G6, 6G1 и 15B2 блокировали связывание FGF9 с FGFR3-IIIb с величинами IC50 0,13, 0,16 и 0,07 нМ, соответственно. При тестировании в диапазоне концентраций приблизительно от 5000 до 1,2 нг/мл, антитела 1G6, 6G1 и 15B2 блокировали связывание FGF9 с FGFR3-IIIc с величинами IC50 0,13, 0,11 и 0,07 нМ, соответственно.
(3) Аффинность связывания полноразмерных гибридомных антител мыши против FGFR3 1G6, 6G1 и 15B2 определяли с использованием анализа Biacore. Результаты этого анализа представлены в таблице 3.
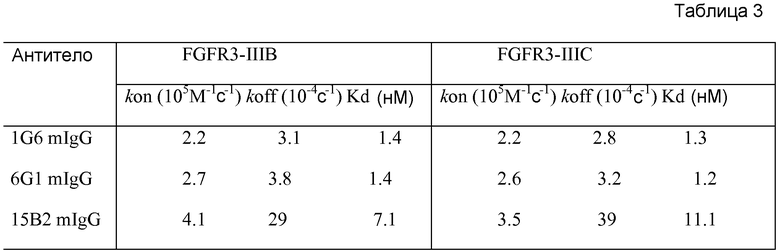
(4) В анализе для тестирования способности гибридомных антител мыши против FGFR3 ингибировать пролиферацию клеток Ba/F3, запускаемую FGFR3-IIIb или IIIc человека, антитела 1G6, 6G1 и 15B2 были способны блокировать пролиферацию клеток Ba/F3, запускаемую FGFR3-IIIb или IIIc человека, дозозависимым образом. При тестировании в диапазоне концентраций антитела приблизительно от 0,01 до 100 мкг/мл, антитела 1G6, 6G1 и 15B2 блокировали пролиферацию клеток Ba/F3, запускаемую FGFR3-IIIb, с величинами IC50 3-5 нМ, 3 нМ, и 6-8 нМ, соответственно, и блокировали пролиферацию клеток Ba/F3, запускаемую FGFR3-IIIc, с величинами IC50 10-35 нМ, 24 нМ и 60 нМ, соответственно.
(5) В анализе для тестирования способности гибридомных антител мыши против FGFR3 ингибировать индуцируемую FGF1 передачу сигнала в клетках Ba/F3, экспрессирующих FGFR3-IIIb человека, антитела 1G6, 6G1 и 15B2 были способны блокировать индуцируемую FGF1 передачу сигнала в клетках Ba/F3, экспрессировавших FGFR3-IIIb человека дозозависимым образом при тестировании в диапазоне концентраций антител приблизительно от 0,25 до 6,75 мкг/мл. В этом эксперименте использовали 25 нг/мл FGF1. В отсутствие FGF1 обработка антителом не имела эффекта на активацию FGFR3.
(6) В анализе для тестирования способности гибридомных антител мыши против FGFR3 ингибировать индуцируемую FGF1 передачу сигнала в клетках Ba/F3, экспрессировавших FGFR3-IIIc человека, антитела 1G6, 6G1 и 15B2 были способны блокировать индуцируемую FGF1 передачу сигнала в клетках Ba/F3, экспрессирующих FGFR3-IIIc человека дозозависимым образом при тестировании в диапазоне концентраций антител приблизительно от 0,25 до 6,75 мкг/мл. В этом эксперименте использовали 25 нг/мл FGF1. В отсутствие FGF1, обработка антителом не имела эффекта на активацию FGFR3.
Структурная основа для взаимодействия R3Mab с FGFR3
Для увеличения понимания способа взаимодействия R3Mab с FGFR3 авторы настоящего изобретения синтезировали панель из 13 перекрывающихся пептидов, охватывающих области IgD2 и D3 FGFR3-IIIb, и протестировали их связывание с R3Mab. Пептиды 3 (остатки 164-178) и 11 (остатки 269-283) продемонстрировали специфическое связывание с R3Mab, причем пептид 3 обладал более сильным взаимодействием (фигура 10A), указывая на то, что соответствующие области на FGFR3 являются критичными для распознавания R3Mab. Предшествующие кристаллографические исследования FGFR1 в комплексе с FGF2 идентифицировали критичные остатки рецептора, вовлеченные в непосредственное связывание с FGF и гепарином, а также в димеризацию рецептора (34). Выравнивание пептидов FGFR3 3 и 11 с функционально значимыми участками в FGFR1 показало, что эти пептиды охватывают соответствующие остатки FGFR1, необходимые для непосредственного связывания FGF2, димеризации рецептора, а также для взаимодействия с гепарином (фигура 10B). Эти данные указывают на то, что эпитоп R3Mab на FGFR3 перекрывается с остатками на рецепторе, вовлеченными в связывание лиганда и взаимодействие рецептор-рецептор.
Далее авторы изобретения кристаллизовали комплекс между Fab-фрагментом R3Mab и внеклеточной областью IgD2-D3 FGFR3-IIIb человека, и определили рентгеновскую структуру при разрешении 2,1 Å (фигура 10 C, D; таблица 4). В этом комплексе приблизительно 1400 Å2 и 1500 Å2 доступной для растворителя поверхности являются погруженными на FGFR3 и Fab, соответственно. Приблизительно 80% погруженной поверхности контакта вовлекает IgD2, а остальная часть включает линкер и IgD3 области. На Fab-стороне комплекса приблизительно 40% погруженной поверхности контакта вовлекает определяющую комплементарность область (CDR)-Н3, 20% CDR-H2, 20% CDR-L2, и небольшой вклад вносят остатки других CDR и каркасные остатки. Примечательно, что аминокислоты (а.к.) из CDR-H3 образуют две β-цепи, которые продолжают β-слой IgD2 (фигура 10D). Fab взаимодействует с а.к., которые составляют участок связывания FGF в FGFR3, а также остатки, которые образуют границу контакта димеризации рецептора, как ранее идентифицировано в различных димерных комплексах FGF:FGFR (например, код PDB - 1CVS, (34); и фигура 10C, области серого цвета/заштрихованные и темно-серого цвета). Поверхность взаимодействия, идентифицированная кристаллографией, полностью согласовывалась с данными на основе пептидов (фигура 18 A, B). Взятые вместе, эти результаты показывают, как R3Mab ингибирует связывание лиганда, и, кроме того, указывают на то, что связывание R3Mab с FGFR3 может предотвращать димеризацию рецептора. Аминокислоты FGFR3, которые контактируют с R3Mab, представлены в таблице 5. Кристаллографические координаты для этой структуры депонированы в Protein Data Bank под кодом доступа 3GRW.
bЧисла в скобках относятся к наиболее высокой оболочке разрешения.
сR = ∑|FO-FC|/∑F0. Rfree вычисляют как R, но за исключением 5% отражений, исключенных из всего уточнения.

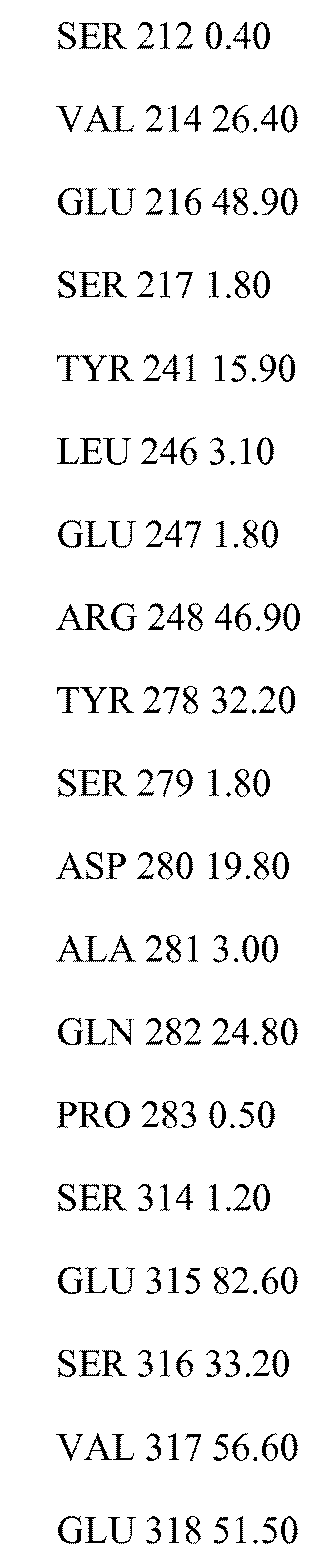
Авторы настоящего изобретения сравнили структуру R3Mab-FGFR3 с ранее опубликованной структурой FGFR3-IIIс в комплексе с FGF1 (4, 35) (фигура 10E, 10F). Наложение комплексов антитело-рецептор и лиганд-рецептор показало, что не существует значительных конформационных отличий в отдельных доменах рецептора, за исключением области, которая отличает FGFR3-IIIc от FGFR3-IIIb; однако ориентация IgD3 относительно IgD2 отличалась значительно (фигура 10E, белый и серый цвет; фигура 10F, белый и серый-зернистый). Поскольку относительные положения IgD2 и IgD3 являются важными для связывания лиганда, альтернативная конформация, принимаемая IgD3 при связывании R3Mab, может обеспечить дополнительный механизм для предотвращения взаимодействия лиганда с FGFR3.
R3Mab ингибирует эндогенный FGFR3 WT и мутантный FGFR3 в клетках рака мочевого пузыря
Для оценки того, может ли R3Mab подавлять функцию FGFR3 в клетках рака мочевого пузыря, авторы изобретения сначала исследовали клеточные линии RT112 и RT4, которые экспрессируют FGFR3 WT. R3Mab сильно ингибировало включение [3H]-тимидина клетками RT112 (фигура 11A) и проявляло значительное, хотя и более умеренное, подавление пролиферации клеток RT4 (фигура 19A). Для исследования эффекта R3Mab на активацию FGFR3, авторы изобретения исследовали фосфорилирование FGFR3 в клетках RT112. В соответствии с результатами в клетках Ba/F3-FGFR3 (фигура 9B), R3Mab значительно снижало индуцируемое FGF1 фосфорилирование FGFR3 (фигура 11B). Далее авторы изобретения исследовали фосфорилирование FRS2α, AKT и p44/42 MAPK, трех нижерасположенных медиаторов передачи сигнала FGFR3. FGF1 сильно активировал эти молекулы в клетках RT112, в то время как R3Mab значительно снижало эту активацию (фигура 11B). Аналогично, R3Mab подавляло индуцируемое FGF1 фосфорилирование FGFR3 и MAPK в клетках RT4 (фигура 19B).
Далее авторы изобретения исследовали, может ли R3Mab ингибировать активацию эндогенного мутанта FGFR3 в клетках рака мочевого пузыря человека. S249C является наиболее распространенной мутацией FGFR3 при раке мочевого пузыря (фигура 9C). Две доступных клеточных линии, UMUC-14 и TCC-97-7, несут мутантный аллель FGFR3S249C (ссылка 36 и данные не представлены). Хотя R3Mab не влияло на экспоненциальный рост клеток UMUC-14 в культуре (данные не представлены), оно значительно снижало клональный рост этих клеток (фигура 11C). Конкретно, R3Mab снижало число колоний, диаметр которых превышал 120 мкм, приблизительно на 77% по сравнению с контрольным антителом (фигура 11D). Более того, R3Mab ингибировало включение [3H]-тимидина клетками TCC-97-7 в культуре (фигура 19C).
Описано, что мутация S249C приводит к лиганд-независимой активации FGFR3 (26, 30). Действительно, FGFR3S249C был конститутивно фосфорилирован, независимо от обработки FGF1, в клетках UMUC-14 и клетках TCC-97-7, в то время как R3Mab снижало конститутивное фосфорилирование FGFR3S249C по сравнению с контрольным антителом в обеих клеточных линиях (фигуры 11E, 19D).
R3Mab ингибирует образование димеров FGFR3 S249C
Способность R3Mab ингибировать конститутивную передачу сигнала FGFR3S249C и пролиферацию в клетках рака мочевого пузыря была неожиданной, учитывая, что этот мутант может претерпевать лиганд-независимую димеризацию с образованием дисульфидных связей (26, 30). Для исследования того, как R3Mab ингибирует FGFR3S249C, авторы изобретения исследовали эффект R3Mab на олигомерное состояние этого мутанта в клетках UMUC-14. В восстанавливающих условиях FGFR3S249C мигрировал в качестве единой полосы размером ~97 кДа, что согласуется с мономерным размером (фигура 12A). В невосстанавливающих условиях в клетках, обработанных контрольным антителом, большая фракция FGFR3S249C выглядела в качестве полосы размером ~200 кДа, независимо от добавления FGF1, что указывает на конститутивное димерное состояние (фигура 12A). Обработка R3Mab существенно снижала количество димеров при одновременном увеличении количества мономеров (фигура 12A). Соответственно, R3Mab снижало уровень димеров FGFR3S249C в клетках TCC-97-7, независимо от обработки FGF1 (фигура 19E).
Каким образом R3Mab снижает уровни димера FGFR3S249C в клетках рака мочевого пузыря? Одним возможным объяснением является то, что оно может разрушать димер FGFR3S249C путем индуцируемой антителом интернализации FGFR3 и транспорта с помощью эндосом или лизосом. Авторы настоящего изобретения тестировали эту возможность с помощью фармакологического вмешательства в эндоцитоз. R3Mab, тем не менее, снижало количество димера в клетках UMUC-14, предварительно обработанных различными ингибиторами эндоцитоза, несмотря на существенную блокаду интернализации FGFR3S249C (фигура 20 A, B). Таким образом, разрушение димера с помощью R3Mab не зависит от эндоцитоза. Другое возможное объяснение состоит в том, что клеточный FGFR3S249C может существовать в динамическом равновесии мономер-димер; таким образом, связывание R3Mab с мономерным FGFR3S249C может предотвратить образование димера и, тем самым, сдвинуть равновесие в сторону мономерного состояния. Для исследования этой возможности авторы настоящего изобретения использовали не проникающий в клетку агент 5,5'-дитиобис-2-нитробензойную кислоту (DTNB), которая селективно реагирует с сульфгидрильными группами неспаренных остатков цистеина (37) и блокирует их. Обработка клеток UMUC-14 с помощью DTNB привела к накоплению мономеров FGFR3S249C за счет димеров (фигура 12B), указывая на то, что FGFR3S249C находится в динамическом равновесии между мономерами и димерами.
Для тестирования того, влияет ли R3Mab на это равновесие, авторы настоящего изобретения получили растворимый рекомбинантный белок, содержащий домены IgD2-D3 FGFR3S249C и выделили димеры эксклюзионной хроматографией. Авторы настоящего изобретения инкубировали димеры с буфером или антителами в присутствии очень низкой концентрации восстановителя (25 мкМ DTT), и анализировали олигомерное состояние рецептора с помощью SDS-PAGE в невосстанавливающих условиях. R3Mab значительно ускоряло появление полосы размером ~25 кДа, соответствующей мономерному FGFR3S249C, за счет димера размером ~50 кДа, по сравнению с имитацией или контрольным антителом (фигура 12C); действительно, через 2 ч снижение количества димеров было по существу более полным в присутствии R3Mab. Эти результаты указывают на то, что R3Mab сдвигает равновесие между мономерным и димерным состояниями FGFR3S249C в пользу мономера.
R3Mab не стимулирует подавление FGFR3
Авторы настоящего изобретения исследовали эффект R3Mab (клон 184.6.1) и гибридомных антител против FGFR3 на подавление FGFR3 с помощью анализа интернализации и деградации FGFR3 в клетках, обработанных антителом против FGFR3. Клеточные линии рака мочевого пузыря, экспрессирующие FGFR3 дикого типа (RT112) или мутантный FGFR3 (S249C в TCC97-7), обрабатывали R3Mab или гибридомными антителами 1G6 или 6G1 в течение от 4 до 24 часов, затем клеточные лизаты собирали для вестерн-блот-анализа общих уровней FGFR3. Обработка R3Mab не снижала уровни FGFR3, в то время как гибридомные mab 1G6 и 6G1 значительно снижали уровни FGFR3. Эти результаты показали, что R3Mab не стимулировало подавление FGFR3, в то время как mab 1G6 и 6G1 не стимулировали интернализацию и подавление рецептора FGFR3. В отдельном эксперименте, исследовали поверхностные уровни FGFR3 с использованием FACS-анализа. После обработки с помощью R3Mab (клон 184.6.1) клеток UMUC-14 (содержавших мутацию S249C FGFR3) в течение 24 часов, уровни FGFR3 на клеточной поверхности немного возрастали. Эти результаты демонстрируют, что обработка R3Mab не стимулировала подавление FGFR3.
R3Mab ингибирует рост и FGFR3 передачу сигнала во множестве моделей опухолей
Далее авторы настоящего изобретения исследовали эффект R3Mab на рост клеток рака мочевого пузыря in vivo. Авторы настоящего изобретения инъецировали мышам nu/nu клетки RT112 (которые экспрессируют FGFR3 WT), позволяли опухолям вырасти до среднего объема ~150 мм3, и животным два раза в неделю вводили носитель или R3Mab. По сравнению с контролем в виде носителя на 27 сутки, обработка с помощью R3Mab в дозе 5 или 50 мг/кг подавляла рост опухоли приблизительно на 41% или 73%, соответственно (фигура 13A). Анализ лизатов опухоли, собранных через 48 ч или 72 ч после обработки, показал, что R3Mab значительно снижало уровень фосфорилированного FRS2α (фигура 13B). Интересно, что общие уровни белка FRS2α также были ниже в опухолях, обработанных R3Mab, указывая на то, что ингибирование FGFR3 может, кроме того, привести к подавлению FRS2α. R3Mab также снижало количество фосфорилированной MAPK в опухолях без влияния на общие уровни MAPK (фигура 13B). Таким образом, R3Mab ингибирует рост ксенотрансплантатов опухолей RT112 совместно с блокированием передачи сигнала FGFR3 WT.
Далее авторы настоящего изобретения исследовали эффект R3Mab на рост ксенотрансплантатов, экспрессировавших мутантный FGFR3. Обработка R3Mab значительно ослабляла прогрессирование опухолей Ba/F3-FGFR3S249C (фигура 13C). Более того, R3Mab значительно ингибировало рост ксенотрансплантатов карциномы мочевого пузыря UMUC-14 (фигура 13D). Для оценки того, влияет ли R3Mab на активацию FGFR3S249C in vivo, авторы настоящего изобретения оценили уровень димера FGFR3S249C в лизатах опухолей, собранных через 24 ч или 72 ч после обработки. В невосстанавливающих условиях количество димера FGFR3S249C было по существу более низким, чем в опухолях, обработанных R3Mab, по сравнению с контрольной группой, в то время как общие уровни FGFR3S249C, исходя из количества, выявленного в восстанавливающих условиях, продемонстрировали небольшое изменение (фигура 13E). В лизатах опухолей не наблюдали заметного накопления мономера FGFR3S249C, в противоположность результатом в клеточной культуре (фигура 13E против 12A). Это могло быть следствием слабой чувствительности обнаружения для мономерного FGFR3 в невосстанавливающих условиях под действием поликлонального антитела кролика против FGFR3, использованного в этом исследовании (фигура 21). Важно, что R3Mab также значительно ингибировало фосфорилирование и активацию MAPK в опухолях UMUC-14 (фигура 13E), указывая на то, что R3Mab ингибирует активность FGFR3S249C in vivo. Авторы настоящего изобретения не наблюдали какого-либо значительного снижения массы тела или других крупных нарушений в каком-либо из исследований in vivo. Более того, в исследовании безопасности, проведенном у мышей, R3Mab, которое связывается с аналогичной аффинностью с FGFR3 как человека, так и мыши, не проявляло какой-либо заметной токсичности на какие-либо органы, включая мочевой пузырь (данные не представлены). Взятые вместе, эти данные указывают на то, что многократное воздействие R3Mab хорошо переносилось мышами.
Противоопухолевая активность R3Mab в моделях ксенотрансплантатов множественной миеломы вовлекает ADCC
Для оценки того, может ли R3Mab обладать терапевтическим потенциалом в отношении множественной миеломы, авторы настоящего изобретения сначала протестировали эффект R3Mab на пролиферацию и выживание трех клеточных линий t(4;14)+ в культуре. Клетки UTMC-2 обладают FGFR3 WT, в то время как OPM2 и KMS11 содержат замену K650E и Y373C, соответственно (7). В культуре R3Mab полностью устраняло индуцируемую FGF9 пролиферацию клеток UTMC-2 (фигура 22A). R3Mab умеренно ингибировало рост клеток OPM2, но не имело заметного эффекта на пролиферацию клеток KMS11 (фигура 22 B, C). Поскольку клетки UTMC-2 не образуют опухоли у мышей, авторы настоящего изобретения оценивали эффективность R3Mab против опухолей OPM2 и KMS11. R3Mab практически полностью устраняло рост ксенотрансплантатов опухолей обеих клеточных линий (фигура 14 A, B).
Выраженное отличие в активности R3Mab против опухолевых клеток OPM2 и KMS11 in vitro и in vivo указало на возможность того, что R3Mab может быть способно поддерживать Fc-опосредуемую иммунную эффекторную функцию против этих опухолей, сверхэкспрессирующих FGFR3. Обе клеточных линии экспрессируют высокие уровни CD55 и CD59 (данные не представлены), два ингибитора каскада комплемента; таким образом, не наблюдали комплементзависимой цитотоксичности (данные не представлены). Затем авторы настоящего изобретения сосредоточились на ADCC. ADCC происходит, когда антитело связывается с его антигеном на клетке-мишени, и, через его Fc-область, связывает Fcγ-рецепторы (FcγR), экспрессированные на иммунных эффекторных клетках (38). Для тестирования ADCC in vitro, авторы настоящего изобретения инкубировали клетки KMS11 или OPM2 со свежевыделенными мононуклеарными клеткими периферической крови (PBMC) человека в присутствии R3Mab или контрольного антитела. R3Mab опосредовало значительную цитолитическую активность PBMC в отношении обеих миеломных клеточных линии (фигура 14 C, D). Напротив, R3Mab не обеспечивало цитолиз клеток рака мочевого пузыря RT112 или UMUC-14 (фигура 14 E, F). При измерении в анализе Скэтчарда клетки множественной миеломы экспрессируют по существу больше FGFR3 на клеточной поверхности, чем клеточные линии карциномы мочевого пузыря (~ в 5-6 раз больше рецепторов на клетку; фигура 23 A, B).
Для установления вклада ADCC в активность R3Mab in vivo авторы вносили в Fc-домен антитела ранее охарактеризованную мутацию D265A/N297A (DANA). Эта двойная замена в Fc-домене антитела устраняет его связывание с FcγR (39), препятствуя привлечению иммунных эффекторных клеток. Мутация DANA не изменяла связывание R3Mab с FGFR3 или ингибирование активности FGFR3 in vitro, также она не изменяла фармакокинетику R3Mab у мышей (данные не представлены); однако она по существу устраняла активность in vivo против ксенотрансплантатов OPM2 или KMS11 (фигура 14 G, H). Напротив, мутация DANA не изменяла противоопухолевую активность R3Mab в отношении ксенотрансплантатов рака мочевого пузыря RT112 и UMUC-14 (фигура 24 A, B). Взятые вместе, эти результаты указывают на то, что Fc-зависимая ADCC играет важную роль в эффективности R3Mab против ксенотрансплантатов множественной миеломы OPM2 и KMS11.
Дополнительные исследования с ксенотрансплантатами
R3Mab (клон 184.6.1N54S) далее было охарактеризовано следующим образом:
(a) R3Mab тестировали в отношении эффективности in vivo с использованием модели ксенотрансплантата опухоли на основе клеточной линии рака печени (Huh7), по существу как описано выше. При тестировании при концентрации антитела 5 мг/кг и 30 мг/кг, R3Mab значительно ингибировало рост опухоли in vivo. Рост опухоли ингибировался приблизительно на 50% по сравнению с ростом опухоли у контрольных животных.
(b) R3Mab тестировали в отношении эффективности in vivo с использованием модели ксенотрансплантата опухоли, основанного на клеточной линии рака молочной железы (Cal-51), которая экспрессировала FGFR3, по существу как описано выше. Результаты этого исследования эффективности показали, что R3Mab было способно ингибировать опухоли in vivo при тестировании в диапазоне концентраций антител приблизительно от 1 мг/кг до 100 мг/кг. Рост опухоли ингибировался приблизительно на 30% по сравнению с ростом опухоли у контрольных животных.
Обсуждение
Ассоциация сверхэкспрессии FGFR3 с плохим прогнозом у пациентов с t(4;14)+ множественной миеломой и трансформирующей активностью активированного FGFR3 в нескольких экспериментальных моделях установила FGFR3 в качестве важного онкогенного пускового фактора, и, таким образом, потенциальной терапевтической мишени в этой гематологической злокачественной опухоли. Напротив, несмотря на сообщения о высокой частоте мутации и/или сверхэкспрессии FGFR3 в карциноме мочевого пузыря (24, 25, 40), критическая роль передачи сигнала FGFR3 в этой эпителиальной злокачественной опухоли не была установлена in vivo. Более того, терапевтическую мощность ингибирования FGFR3 при раке мочевого пузыря еще предстоит определить. В настоящем документе авторы настоящего изобретения демонстрируют, что генетическое или фармакологическое вмешательство в FGFR3 ингибирует рост нескольких ксенотрансплантатов рака мочевого пузыря человека у мышей. Эти результаты демонстрируют, что функция FGFR3 является критической для роста опухоли в этих условиях, подчеркивая потенциальную важность этого рецептора в качестве онкогенного пускового фактора и терапевтической мишени при раке мочевого пузыря. Блокада функции FGFR3 ингибировала рост ксенотрансплантатов, экспрессирующих либо FGFR3 WT, либо мутантный FGFR3, сходным образом, указывая на то, что обе формы рецептора могут вносить значительный вклад в прогрессирование опухоли мочевого пузыря. Хотя и реже чем при раке мочевого пузыря, мутации или сверхэкспрессия были идентифицированы в других злокачественных опухолях, включая карциному шейки матки (40), печеночно-клеточную карциному (41) и немелкоклеточный рак легкого (42, 43), указывая на потенциальный вклад FGFR3 в эти дополнительные типы эпителиальной злокачественной опухоли.
Кажущееся вовлечение FGFR3 в различные злокачественные опухоли идентифицирует этот рецептор в качестве интересного кандидата для направленной терапии. Хотя описаны низкомолекулырные соединения, которые могут ингибировать киназную активность FGFR3 (18-22, 44), тесная гомология киназных доменов в семействе FGFR препятствовала разработке селективных ингибиторов FGFR3. Отсутствие селективности описанных ингибиторов делает трудным отличение относительного вклада FGFR3 в биологию конкретных типов злокачественных опухолей; кроме того, возможно препятствование с точки зрения безопасности, ограничивая максимальные уровни доз и, таким образом, лимитируя оптимальное ингибирование FGFR3. Таким образом, для достижения селективного и специфичного нацеливания FGFR3, авторы настоящего изобретения обратились к стратегии на основе антител. Авторы настоящего изобретения заключили, что оптимальное терапевтическое антитело должно быть способно блокировать не только FGFR3 WT, но также преобладающие связанные со злокачественной опухоли мутанты FGFR3. Более того, учитывая, что димеризация FGFR3 является критичной для его активации, наилучшим было бы антитело, которое не только блокирует связывание лиганда, но также препятствует димеризации рецептора. Дополнительные желательные свойства могут включать способность поддерживать опосредуемую Fc эффекторную функцию и длительное время полужизни в сыворотке, обеспечиваемые природной каркасной областью полноразмерного антитела. Авторы настоящего изобретения сфокусировали их попытки скрининга и конструирования на идентификации молекулы антитела, которая сочетает все из этих признаков, что привело к получению R3Mab. Исследования связывания продемонстрировали способность R3Mab конкурировать с лигандами FGF за взаимодействие как с изоформой IIIb, так и с изоформой IIIc FGFR3. Дальнейшие эксперименты с трансфицированными клеточными линиями BaF/3 подтвердили примечательную способность R3Mab блокировать как WT, так и преобладающие связанные со злокачественной опухолью мутанты FGFR3. Кроме того, R3Mab проявляло значительную противоопухолевую активность в нескольких моделях с ксенотрансплантатом рака мочевого пузыря, экспрессирующим либо FGFR3 WT, либо FGFR3S249C, который является наиболее распространенным мутантом рецептора при этом заболевании. Фармакодинамические исследования указывают на то, что противоопухолевая активность R3Mab в этих моделях основана на ингибировании передачи сигнала FGFR3, о котором свидетельствует сниженное фосфорилирование его нижеследующих медиаторов FRS2α и MAPK. Эти данные, кроме того, подтверждают заключение, что FGFR3 требуется для прогрессирования опухоли мочевого пузыря, как демонстрируется исследованиями авторов настоящего изобретения с shРНК для FGFR3.
Мутации FGFR3 при раке мочевого пузыря представляют собой одно из наиболее частых изменений протеинкиназы в солидных злокачественных опухолях, напоминающее распространенную мутацию B-Raf при меланоме (45). Большинство из активирующих мутаций в FGFR3 приводят к неспаренному цистеину, что приводит к независимой от лиганда димеризации рецептора и к различным степеням конститутивной активации. В предыдущем исследовании с использованием одновалентного Fab-фрагмента против FGFR3 показана дифференциальная ингибиторная активность против определенных мутантов FGFR3 (46); однако молекулярная основа для этого переменного эффекта не была изучена. По сравнению с одновалентными фрагментами антител, двухвалентные антитела способны индуцировать кластеризацию антигенов, и, в случае рецепторных тирозинкиназ, могут вызывать олигомеризацию и активацию рецептора. Несмотря на полноразмерную двухвалентную конфигурацию R3Mab проявляло универсальное ингибирование FGFR3 WT и широкого спектра мутантов FGFR3, включая варианты, которые являются лиганд-зависимыми (FGFR3G372C, FGFR3Y375C), конститутивно активными (FGFR3R248C, FGFR3S249C) или и теми, и другими (FGFR3K652E). Эти результаты поднимают вопрос: каким образом R3Mab осуществляет антагонизм как FGFR3 WT, так и различных мутантов FGFR3, включая варианты с дисульфидными связями?
На основе выравнивания последовательности с FGFR1, пептидный эпитоп, распознаваемый R3Mab, перекрывается с остатками FGFR3, вовлеченными в связывание с лигандом и гепарином, а также в димеризацию рецептора. Это заключение было подтверждено кристаллографическими исследованиями комплекса между R3Mab и внеклеточными областями FGFR3. Рентгеновская структура показала, что антитело связывается с областями IgD2 и IgD3, которые являются критичными для взаимодействия лиганд-рецептор, а также контакта рецептор-рецептор. Таким образом, R3Mab может блокировать FGFR3 WT как с помощью конкуренции за связывание лиганда, так и с помощью предотвращения димеризации рецептора. R3Mab может использовать сходный механизм для ингибирования FGFR3K652E, который имеет низкую конститутивную активность, но требует лиганда для полной активации. Более того, связывание R3Mab изменяет относительную ориентацию IgD3 FGFR3 в отношении IgD2. Эти данные обеспечивают формальную возможность того, что антитело также может ингибировать активацию рецептора, вынуждая образовывать конформацию, которая не приводит к передаче сигнала - это представление требует дальнейшего исследования.
Для лучшего понимания того, как R3Mab блокирует варианты FGFR3, обладающие неспаренным цистеином, авторы настоящего изобретения анализировали наиболее распространенный мутант, FGFR3S249C, более подробно. Эксперименты с блокатором свободных сульфгидрилов DTNB указали на динамическое равновесие между мономерным и димерным состоянием FGFR3S249C. Аналогичное равновесие между окисленным и восстановленным состоянием, модулированным эндогенными окислительно-восстановительными регуляторами, было описано для рецепторов NMDA (46). Инкубация клеток рака мочевого пузыря, экспрессирующих FGFR3S249C, с R3Mab, привела к снижению количества димеров рецептора и одновременному увеличению уровня мономеров. Более того, очищенный фрагмент IgD2-D3 FGFR3S249C образовывал димеры в растворе; при инкубации с R3Mab димеры неизменно исчезали, в то время как мономерный FGFR3S249C накапливался. Взятые вместе со структурным анализом, эти результаты указывают на то, что R3Mab захватывает мономерный FGFR3 и препятствует его димеризации. С течением времени R3Mab сдвигает равновесие в сторону мономерного состояния, блокируя конститутивную активность рецептора. Этот механизм также может объяснить, каким образом R3Mab ингибирует другие мутанты FGFR3 по цистеину.
Другим важным открытием этого исследования была высокая противоопухолевая активность R3Mab против клеточных линий t(4;14)+ множественной миеломы OPM2 и KMS11 in vivo. Напротив, R3Mab имело от умеренного до минимального влияния на пролиферацию или выживание этих клеток в культуре. Клетки OPM2 и KMS11 экспрессируют относительно высокие уровни FGFR3 на клеточной поверхности (в 5-6 раз выше, чем клетки карциномы мочевого пузыря RT112 и UMUC-14). Эта более высокая плотность антигена может позволить R3Mab поддерживать эффективное привлечение несущих FcγR иммунных эффекторных клеток и активацию ADCC. Действительно, в присутствии PBMC человека R3Mab опосредовало цитолиз клеток OPM2 и KMS11, но не клеток рака мочевого пузыря RT112 или UMUC-14. Более того, мутантная версия R3Mab DANA, которая неспособна связывать FcγR, не имела эффекта на рост KMS11 или OPM2 in vivo, но, тем не менее, подавляла рост опухолей RT112 и UMUC-14 аналогично R3Mab. Взятые вместе эти данные указывают на то, что R3Mab обладает двойным механизмом противоопухолевой активности: (a) в клетках, экспрессирующих более низкие уровни FGFR3 WT или мутантного FGFR3 на поверхности, оно блокирует лиганд-зависимую или конститутивную передачу сигнала; (b) в клетках, экспрессирующих относительно высокие уровни FGFR3 на поверхности, оно индуцирует ADCC.
Результаты авторов настоящего изобретения также поднимают несколько новых вопросов. Во-первых, неизвестно, почему клеточные линии рака мочевого пузыря, протестированные в этом исследовании, проявляют переменную чувствительность к R3Mab. Такой дифференциальный ответ, который является распространенным для направленной терапии, может быть отражением отдаленного генетического характера отдельных опухолей. Действительно, Her2-положительные клетки рака молочной железы демонстрируют переменную чувствительность к антителу против Her2 (48), также как и различные злокачественные клетки в ответ на антитело против EGFR (49). В этом контексте срочно требуется разработка дополнительных моделей in vivo для рака мочевого пузыря с FGFR3 WT и мутантным FGFR3 для оценки чувствительности к молекулам FGFR3 у животных. Более того, установление прогнозирующих биомаркеров может помочь идентифицировать пациентов, которые оптимально могут получить пользу от нацеленной на FGFR3 терапии. Во-вторых, поскольку R3Mab не индуцировало регрессию опухоли в моделях, исследованных авторами изобретения, в будущих исследованиях должно быть изучено, может ли R3Mab действовать вместе с общепринятыми лекарственными средствами.
В заключение, данные авторов настоящего изобретения вовлекают как FGFR3 WT, так и мутантный FGFR3, в качестве важных факторов для роста рака мочевого пузыря, таким образом, расширяя онкогенное вовлечение in vivo этого рецептора в злокачественные опухоли от гематологических до эпителиальных. Более того, результаты авторов настоящего изобретения демонстрируют, что как на FGFR3 WT, так и на мутантный FGFR3, в опухолях можно эффективно осуществлять нацеливание с помощью полноразмерного антитела, которое сочетает в себе способность блокировать связывание лиганда, димеризацию рецептора и передачу сигнала, а также стимулировать лизис опухолевых клеток с помощью ADCC. Эти результаты обеспечивают убедительное обоснование для нацеленных на FGFR3 способов терапии на основе антител в различных злокачественных опухолях, связанных с этим рецептором.
Неполный список ссылок
1. Eswarakumar, V.P., Lax, I., and Schlessinger, J. 2005. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev 16:139-149.
2. L'Hote, C.G., and Knowles, M.A. 2005. Cell responses to FGFR3 signalling: growth, differentiation and apoptosis. Exp Cell Res 304:417-431.
3. Dailey, L., Ambrosetti, D., Mansukhani, A., and Basilico, C. 2005. Mechanisms underlying differential responses to FGF signaling. Cytokine Growth Factor Rev 16:233-247.
4. Mohammadi, M., Olsen, S.K., and Ibrahimi, O.A. 2005. Structural basis for fibroblast growth factor receptor activation. Cytokine Growth Factor Rev 16:107-137.
5. Grose, R., and Dickson, C. 2005. Fibroblast growth factor signaling in tumorigenesis. Cytokine Growth Factor Rev 16:179-186.
6. Chang, H., Stewart, A.K., Qi, X.Y., Li, Z.H., Yi, Q.L., and Trudel, S. 2005. Immunohistochemistry accurately predicts FGFR3 aberrant expression and t(4;14) in multiple myeloma. Blood 106:353-355.
7. Chesi, M., Nardini, E., Brents, L.A., Schrock, E., Ried, T., Kuehl, W.M., and Bergsagel, P.L. 1997. Frequent translocation t(4;14)(p16.3;q32.3) in multiple myeloma is associated with increased expression and activating mutations of fibroblast growth factor receptor 3. Nat Genet 16:260-264.
8. Fonseca, R., Blood, E., Rue, M., Harrington, D., Oken, M.M., Kyle, R.A., Dewald, G.W., Van Ness, B., Van Wier, S.A., Henderson, K.J., et al. 2003. Clinical and biologic implications of recurrent genomic aberrations in myeloma. Blood 101:4569-4575.
9. Moreau, P., Facon, T., Leleu, X., Morineau, N., Huyghe, P., Harousseau, J.L., Bataille, R., and Avet-Loiseau, H. 2002. Recurrent 14q32 translocations determine the prognosis of multiple myeloma, especially in patients receiving intensive chemotherapy. Blood 100:1579-1583.
10. Pollett, J.B., Trudel, S., Stern, D., Li, Z.H., and Stewart, A.K. 2002. Overexpression of the myeloma-associated oncogene fibroblast growth factor receptor 3 confers dexamethasone resistance. Blood 100:3819-3821.
11. Bernard-Pierrot, I., Brams, A., Dunois-Larde, C., Caillault, A., Diez de Medina, S.G., Cappellen, D., Graff, G., Thiery, J.P., Chopin, D., Ricol, D., et al. 2006. Oncogenic properties of the mutated forms of fibroblast growth factor receptor 3b. Carcinogenesis 27:740-747.
12. Agazie, Y.M., Movilla, N., Ischenko, I., and Hayman, M.J. 2003. The phosphotyrosine phosphatase SHP2 is a critical mediator of transformation induced by the oncogenic fibroblast growth factor receptor 3. Oncogene 22:6909-6918.
13. Ronchetti, D., Greco, A., Compasso, S., Colombo, G., Dell'Era, P., Otsuki, T., Lombardi, L., and Neri, A. 2001. Deregulated FGFR3 mutants in multiple myeloma cell lines with t(4;14): comparative analysis of Y373C, K650E and the novel G384D mutations. Oncogene 20:3553-3562.
14. Chesi, M., Brents, L.A., Ely, S.A., Bais, C., Robbiani, D.F., Mesri, E.A., Kuehl, W.M., and Bergsagel, P.L. 2001. Activated fibroblast growth factor receptor 3 is an oncogene that contributes to tumor progression in multiple myeloma. Blood 97:729-736.
15. Plowright, E.E., Li, Z., Bergsagel, P.L., Chesi, M., Barber, D.L., Branch, D.R., Hawley, R.G., and Stewart, A.K. 2000. Ectopic expression of fibroblast growth factor receptor 3 promotes myeloma cell proliferation and prevents apoptosis. Blood 95:992-998.
16. Chen, J., Williams, I.R., Lee, B.H., Duclos, N., Huntly, B.J., Donoghue, D.J., and Gilliland, D.G. 2005. Constitutively activated FGFR3 mutants signal through PLCgamma-dependent and -independent pathways for hematopoietic transformation. Blood 106:328-337.
17. Li, Z., Zhu, Y.X., Plowright, E.E., Bergsagel, P.L., Chesi, M., Patterson, B., Hawley, T.S., Hawley, R.G., and Stewart, A.K. 2001. The myeloma-associated oncogene fibroblast growth factor receptor 3 is transforming in hematopoietic cells. Blood 97:2413-2419.
18. Trudel, S., Ely, S., Farooqi, Y., Affer, M., Robbiani, D.F., Chesi, M., and Bergsagel, P.L. 2004. Inhibition of fibroblast growth factor receptor 3 induces differentiation and apoptosis in t(4;14) myeloma. Blood 103:3521-3528.
19. Trudel, S., Li, Z.H., Wei, E., Wiesmann, M., Chang, H., Chen, C., Reece, D., Heise, C., and Stewart, A.K. 2005. CHIR-258, a novel, multitargeted tyrosine kinase inhibitor for the potential treatment of t(4;14) multiple myeloma. Blood 105:2941-2948.
20. Chen, J., Lee, B.H., Williams, I.R., Kutok, J.L., Mitsiades, C.S., Duclos, N., Cohen, S., Adelsperger, J., Okabe, R., Coburn, A., et al. 2005. FGFR3 as a therapeutic target of the small molecule inhibitor PKC412 in hematopoietic malignancies. Oncogene 24:8259-8267.
21. Paterson, J.L., Li, Z., Wen, X.Y., Masih-Khan, E., Chang, H., Pollett, J.B., Trudel, S., and Stewart, A.K. 2004. Preclinical studies of fibroblast growth factor receptor 3 as a therapeutic target in multiple myeloma. Br J Haematol 124:595-603.
22. Grand, E.K., Chase, A.J., Heath, C., Rahemtulla, A., and Cross, N.C. 2004. Targeting FGFR3 in multiple myeloma: inhibition of t(4;14)-positive cells by SU5402 and PD173074. Leukemia 18:962-966.
23. Gomez-Roman, J.J., Saenz, P., Molina, M., Cuevas Gonzalez, J., Escuredo, K., Santa Cruz, S., Junquera, C., Simon, L., Martinez, A., Gutierrez Banos, J.L., et al. 2005. Fibroblast growth factor receptor 3 is overexpressed in urinary tract carcinomas and modulates the neoplastic cell growth. Clin Cancer Res 11:459-465.
24. Tomlinson, D.C., Baldo, O., Harnden, P., and Knowles, M.A. 2007. FGFR3 protein expression and its relationship to mutation status and prognostic variables in bladder cancer. J Pathol 213:91-98.
25. van Rhijn, B.W., Montironi, R., Zwarthoff, E.C., Jobsis, A.C., and van der Kwast, T.H. 2002. Frequent FGFR3 mutations in urothelial papilloma. J Pathol 198:245-251.
26. Tomlinson, D.C., Hurst, C.D., and Knowles, M.A. 2007. Knockdown by shRNA identifies S249C mutant FGFR3 as a potential therapeutic target in bladder cancer. Oncogene 26:5889-5899.
27. Martinez-Torrecuadrada, J., Cifuentes, G., Lopez-Serra, P., Saenz, P., Martinez, A., and Casal, J.I. 2005. Targeting the extracellular domain of fibroblast growth factor receptor 3 with human single-chain Fv antibodies inhibits bladder carcinoma cell line proliferation. Clin Cancer Res 11:6280-6290.
28. Martinez-Torrecuadrada, J.L., Cheung, L.H., Lopez-Serra, P., Barderas, R., Canamero, M., Ferreiro, S., Rosenblum, M.G., and Casal, J.I. 2008. Antitumor activity of fibroblast growth factor receptor 3-specific immunotoxins in a xenograft mouse model of bladder carcinoma is mediated by apoptosis. Mol Cancer Ther 7:862-873.
29. Ornitz, D.M., and Leder, P. 1992. Ligand specificity and heparin dependence of fibroblast growth factor receptors 1 and 3. J Biol Chem 267:16305-16311.
30. d'Avis, P.Y., Robertson, S.C., Meyer, A.N., Bardwell, W.M., Webster, M.K., and Donoghue, D.J. 1998. Constitutive activation of fibroblast growth factor receptor 3 by mutations responsible for the lethal skeletal dysplasia thanatophoric dysplasia type I. Cell Growth Differ 9:71-78.
31. Adar, R., Monsonego-Ornan, E., David, P., and Yayon, A. 2002. Differential activation of cysteine-substitution mutants of fibroblast growth factor receptor 3 is determined by cysteine localization. J Bone Miner Res 17:860-868.
32. Knowles, M.A. 2008. Novel therapeutic targets in bladder cancer: mutation and expression of FGF receptors. Future Oncol 4:71-83.
33. Naski, M.C., Wang, Q., Xu, J., and Ornitz, D.M. 1996. Graded activation of fibroblast growth factor receptor 3 by mutations causing achondroplasia and thanatophoric dysplasia. Nat Genet 13:233-237.
34. Plotnikov, A.N., Schlessinger, J., Hubbard, S.R., and Mohammadi, M. 1999. Structural basis for FGF receptor dimerization and activation. Cell 98:641-650.
35. Olsen, S.K., Ibrahimi, O.A., Raucci, A., Zhang, F., Eliseenkova, A.V., Yayon, A., Basilico, C., Linhardt, R.J., Schlessinger, J., and Mohammadi, M. 2004. Insights into the molecular basis for fibroblast growth factor receptor autoinhibition and ligand-binding promiscuity. Proc Natl Acad Sci U S A 101:935-940.
36. Jebar, A.H., Hurst, C.D., Tomlinson, D.C., Johnston, C., Taylor, C.F., and Knowles, M.A. 2005. FGFR3 and Ras gene mutations are mutually exclusive genetic events in urothelial cell carcinoma. Oncogene 24:5218-5225.
37. Ellman, G.L. 1959. Tissue sulfhydryl groups. Arch Biochem Biophys 82:70-77.
38. Adams, G.P., and Weiner, L.M. 2005. Monoclonal antibody therapy of cancer. Nat Biotechnol 23:1147-1157.
39. Gong, Q., Ou, Q., Ye, S., Lee, W.P., Cornelius, J., Diehl, L., Lin, W.Y., Hu, Z., Lu, Y., Chen, Y., et al. 2005. Importance of cellular microenvironment and circulatory dynamics in B cell immunotherapy. J Immunol 174:817-826.
40. Cappellen, D., De Oliveira, C., Ricol, D., de Medina, S., Bourdin, J., Sastre-Garau, X., Chopin, D., Thiery, J.P., and Radvanyi, F. 1999. Frequent activating mutations of FGFR3 in human bladder and cervix carcinomas. Nat Genet 23:18-20.
41. Qiu, W.H., Zhou, B.S., Chu, P.G., Chen, W.G., Chung, C., Shih, J., Hwu, P., Yeh, C., Lopez, R., and Yen, Y. 2005. Over-expression of fibroblast growth factor receptor 3 in human hepatocellular carcinoma. World J Gastroenterol 11:5266-5272.
42. Cortese, R., Hartmann, O., Berlin, K., and Eckhardt, F. 2008. Correlative gene expression and DNA methylation profiling in lung development nominate new biomarkers in lung cancer. Int J Biochem Cell Biol 40:1494-1508.
43. Woenckhaus, M., Klein-Hitpass, L., Grepmeier, U., Merk, J., Pfeifer, M., Wild, P., Bettstetter, M., Wuensch, P., Blaszyk, H., Hartmann, A., et al. 2006. Smoking and cancer-related gene expression in bronchial epithelium and non-small-cell lung cancers. J Pathol 210:192-204.
44. Xin, X., Abrams, T.J., Hollenbach, P.W., Rendahl, K.G., Tang, Y., Oei, Y.A., Embry, M.G., Swinarski, D.E., Garrett, E.N., Pryer, N.K., et al. 2006. CHIR-258 is efficacious in a newly developed fibroblast growth factor receptor 3-expressing orthotopic multiple myeloma model in mice. Clin Cancer Res 12:4908-4915.
45. Davies, H., Bignell, G.R., Cox, C., Stephens, P., Edkins, S., Clegg, S., Teague, J., Woffendin, H., Garnett, M.J., Bottomley, W., et al. 2002. Mutations of the BRAF gene in human cancer. Nature 417:949-954.
46. Trudel, S., Stewart, A.K., Rom, E., Wei, E., Li, Z.H., Kotzer, S., Chumakov, I., Singer, Y., Chang, H., Liang, S.B., et al. 2006. The inhibitory anti-FGFR3 antibody, PRO-001, is cytotoxic to t(4;14) multiple myeloma cells. Blood 107:4039-4046.
47. Gozlan, H., and Ben-Ari, Y. 1995. NMDA receptor redox sites: are they targets for selective neuronal protection? Trends Pharmacol Sci 16:368-374.
48. Hudziak, R.M., Lewis, G.D., Winget, M., Fendly, B.M., Shepard, H.M., and Ullrich, A. 1989. p185HER2 monoclonal antibody has antiproliferative effects in vitro and sensitizes human breast tumor cells to tumor necrosis factor. Mol Cell Biol 9:1165-1172.
49. Masui, H., Kawamoto, T., Sato, J.D., Wolf, B., Sato, G., and Mendelsohn, J. 1984. Growth inhibition of human tumor cells in athymic mice by anti-epidermal growth factor receptor monoclonal antibodies. Cancer Res 44:1002-1007.
50. Pai, R., Dunlap, D., Qing, J., Mohtashemi, I., Hotzel, K., and French, D.M. 2008. Inhibition of fibroblast growth factor 19 reduces tumor growth by modulating beta-catenin signaling. Cancer Res 68:5086-5095.
51. Pegram, M., Hsu, S., Lewis, G., Pietras, R., Beryt, M., Sliwkowski, M., Coombs, D., Baly, D., Kabbinavar, F., and Slamon, D. 1999. Inhibitory effects of combinations of HER-2/neu antibody and chemotherapeutic agents used for treatment of human breast cancers. Oncogene 18:2241-2251.
52. Lee, C.V., Liang, W.C., Dennis, M.S., Eigenbrot, C., Sidhu, S.S., and Fuh, G. 2004. High-affinity human antibodies from phage-displayed synthetic Fab libraries with a single framework scaffold. J Mol Biol 340:1073-1093.
53. Liang, W.C., Dennis, M.S., Stawicki, S., Chanthery, Y., Pan, Q., Chen, Y., Eigenbrot, C., Yin, J., Koch, A.W., Wu, X., et al. 2007. Function blocking antibodies to neuropilin-1 generated from a designed human synthetic antibody phage library. J Mol Biol 366:815-829.
54. Carter, P., Presta, L., Gorman, C.M., Ridgway, J.B., Henner, D., Wong, W.L., Rowland, A.M., Kotts, C., Carver, M.E., and Shepard, H.M. 1992. Humanization of an anti-p185HER2 antibody for human cancer therapy. Proc Natl Acad Sci U S A 89:4285-4289.
55. Sidhu, S.S., Li, B., Chen, Y., Fellouse, F.A., Eigenbrot, C., and Fuh, G. 2004. Phage-displayed antibody libraries of synthetic heavy chain complementarity determining regions. J Mol Biol 338:299-310.
56. Otwinowski, Z.a.M., W. 1997. Processing of X-ray diffraction data collected in oscillation mode. Methods in Enzymology 276:307-326.
57. McCoy, A.J., Grosse-Kunstleve, R.W., Storoni, L.C., and Read, R.J. 2005. Likelihood-enhanced fast translation functions. Acta Crystallogr D Biol Crystallogr 61:458-464.
58. Emsley, P., and Cowtan, K. 2004. Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr 60:2126-2132.
59. Murshudov, G.N., Vagin, A.A., and Dodson, E.J. 1997. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr D Biol Crystallogr 53:240-255
Хотя представленное выше изобретения более подробно описано с помощью иллюстраций и примеров для простоты понимания, описание и примеры не следует истолковывать как ограничивающие объем изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ AXL И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2577986C2 |
| АНТИТЕЛА ПРОТИВ BV8 И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2559542C2 |
| АНТИТЕЛА ПРОТИВ NOTCH2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2580029C2 |
| АНТИТЕЛА ПРОТИВ NRR Notch1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2476443C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ АКСИАЛЬНОГО СВЯЗЫВАНИЯ PD-1 И VEGF АНТАГОНИСТОВ | 2013 |
|
RU2689760C2 |
| АНТИТЕЛА ПРОТИВ EphB4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2450020C2 |
| АНТИ-ТАТ226 АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ | 2007 |
|
RU2448980C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ CD79b И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ПРИМЕНЕНИЯ | 2008 |
|
RU2557319C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2825390C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2013 |
|
RU2639506C2 |

















































































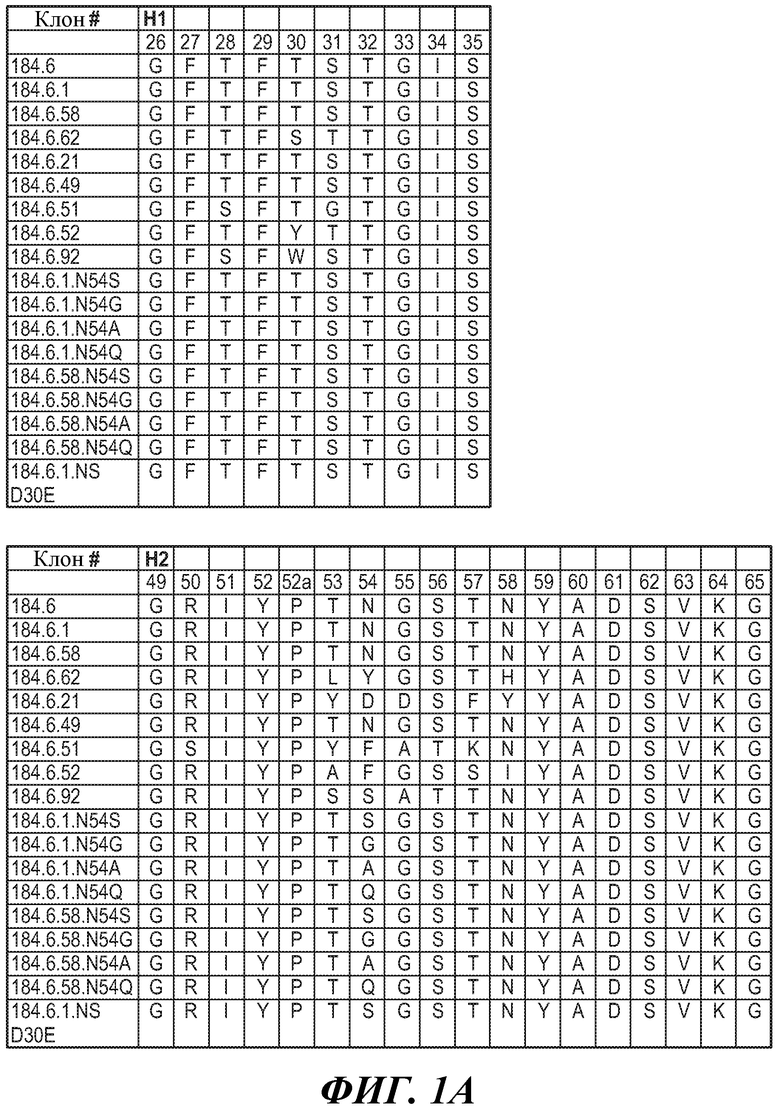
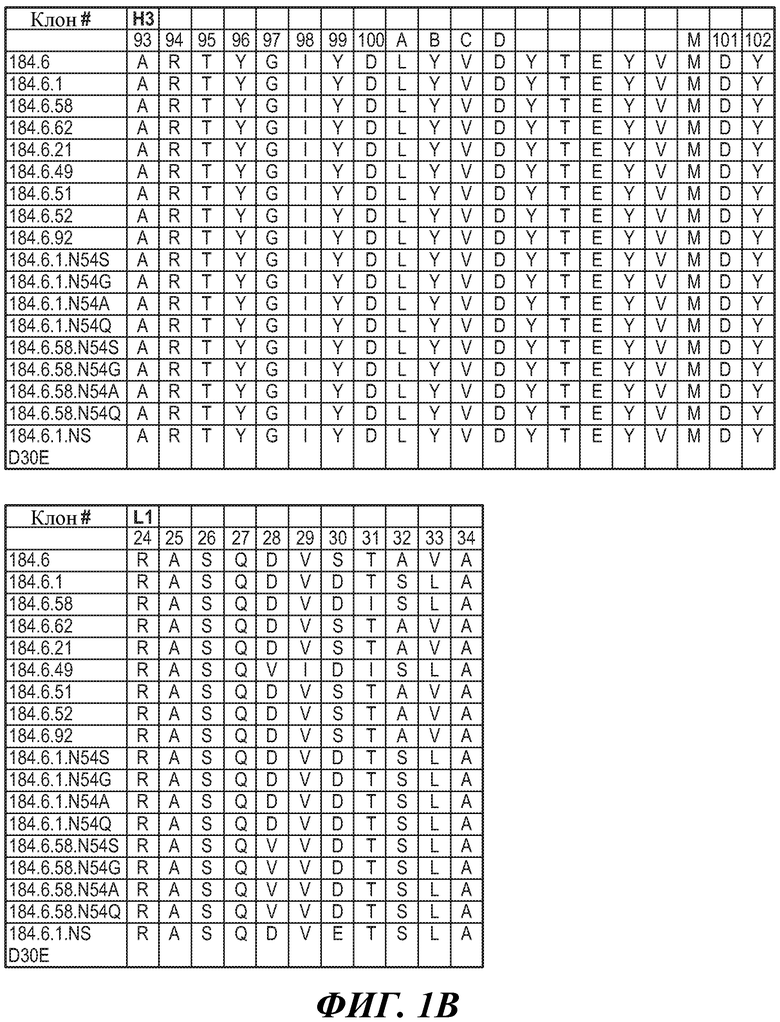
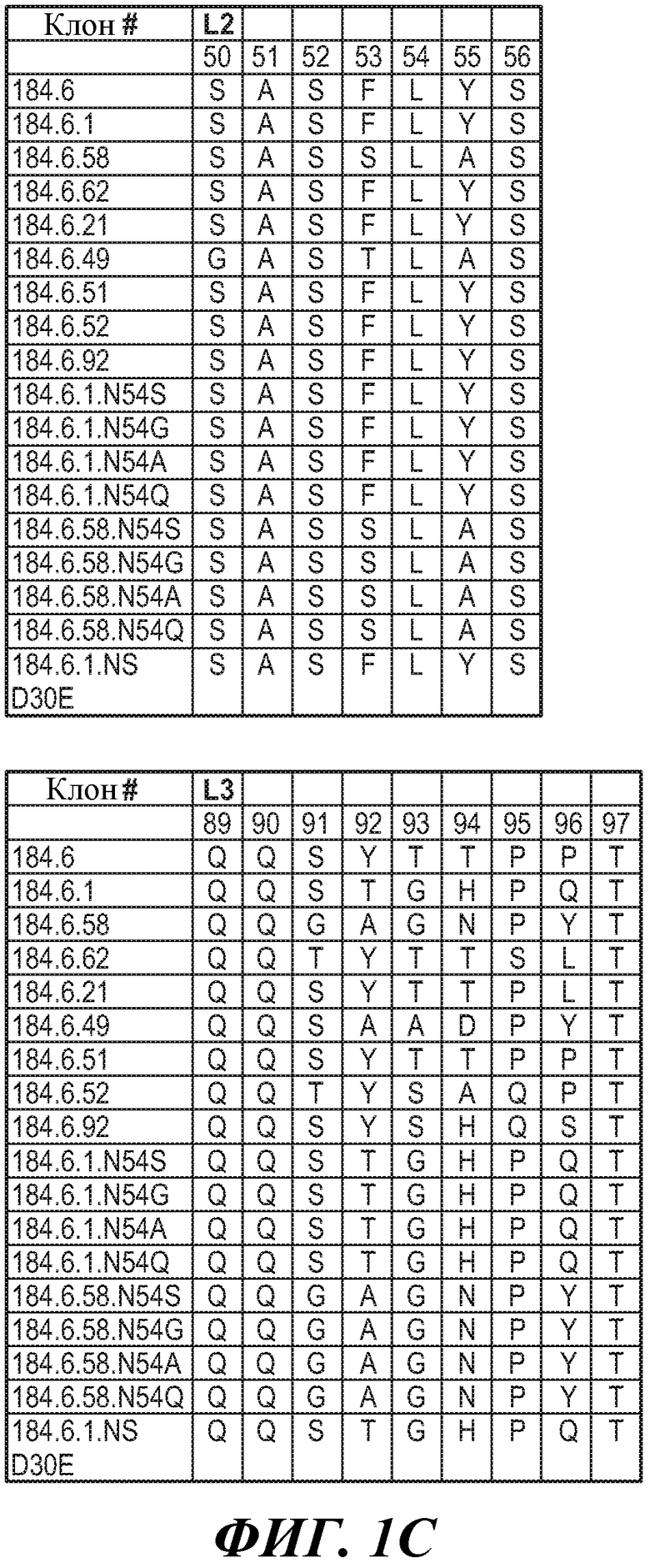


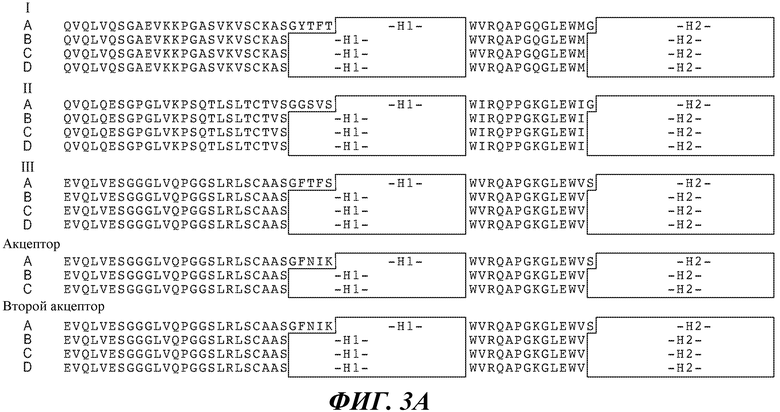
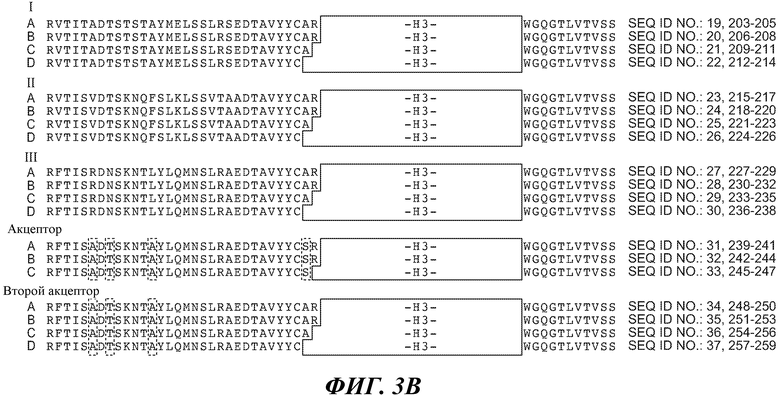
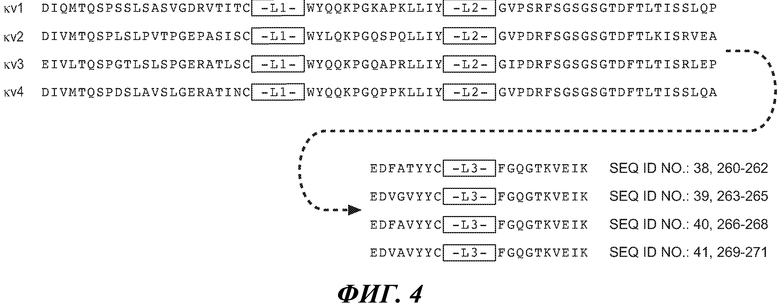


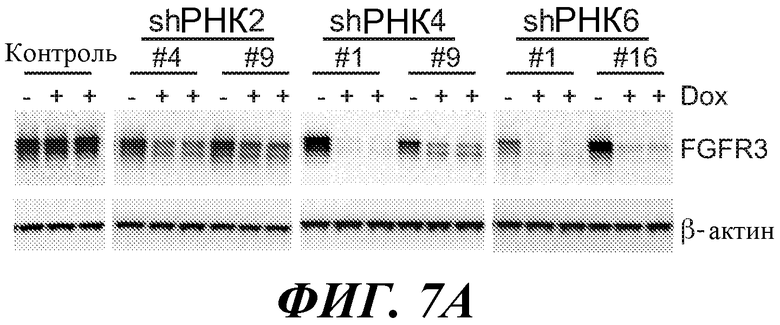

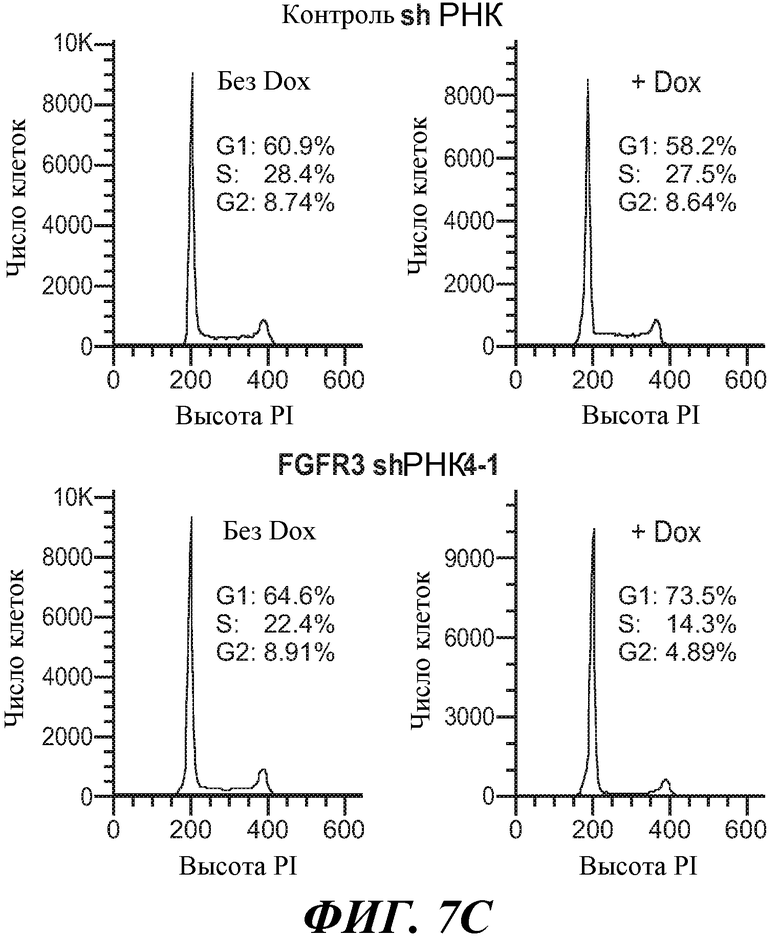
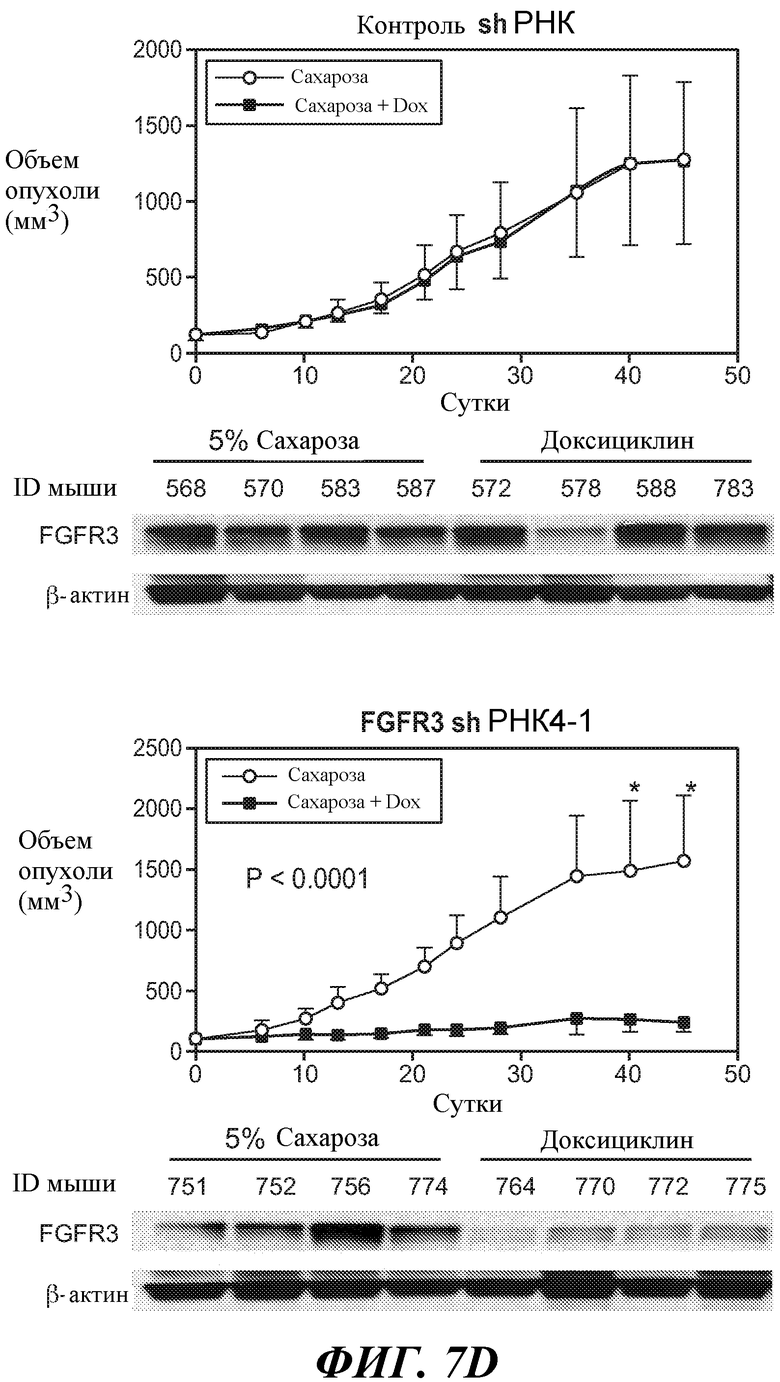
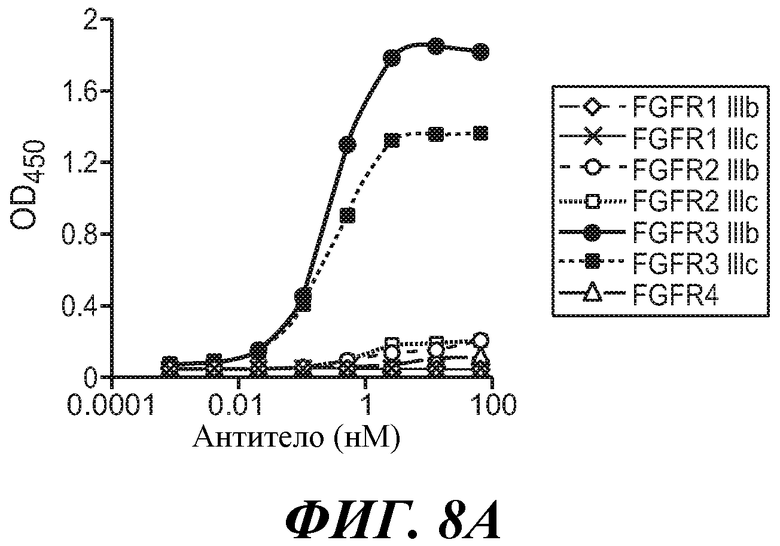

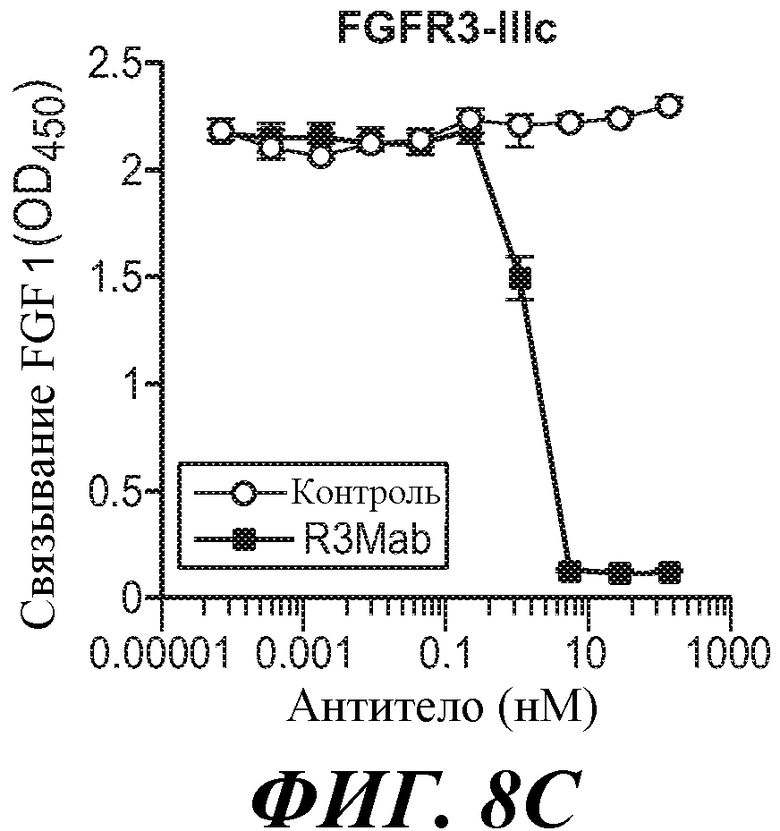
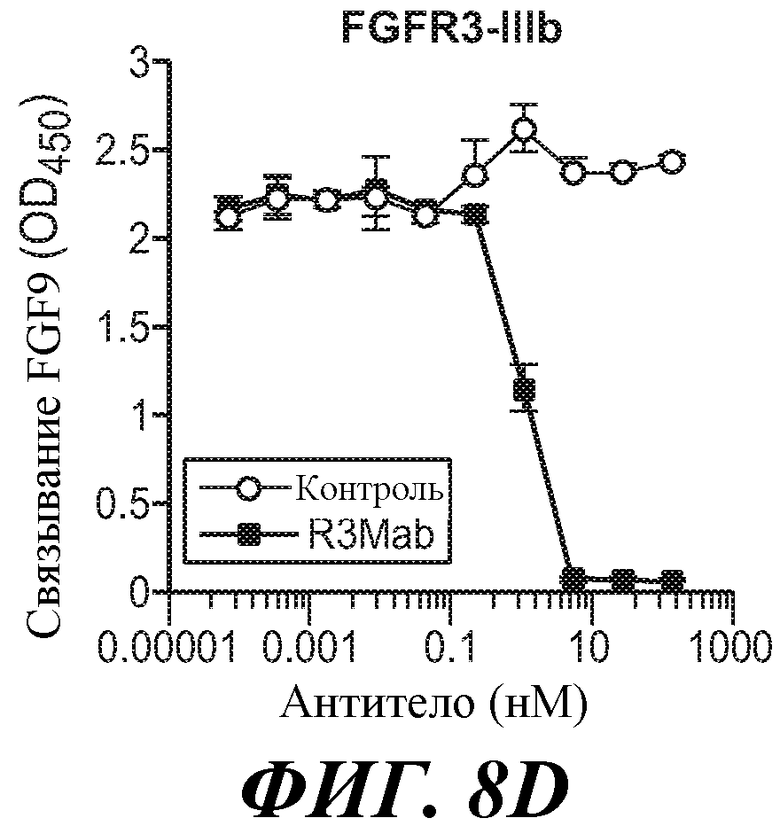
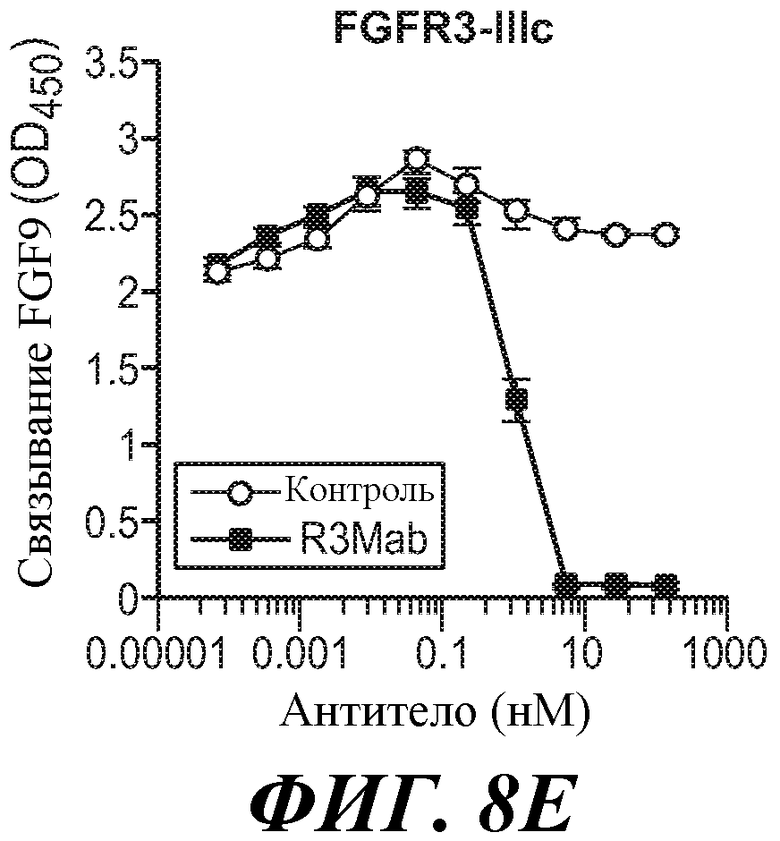
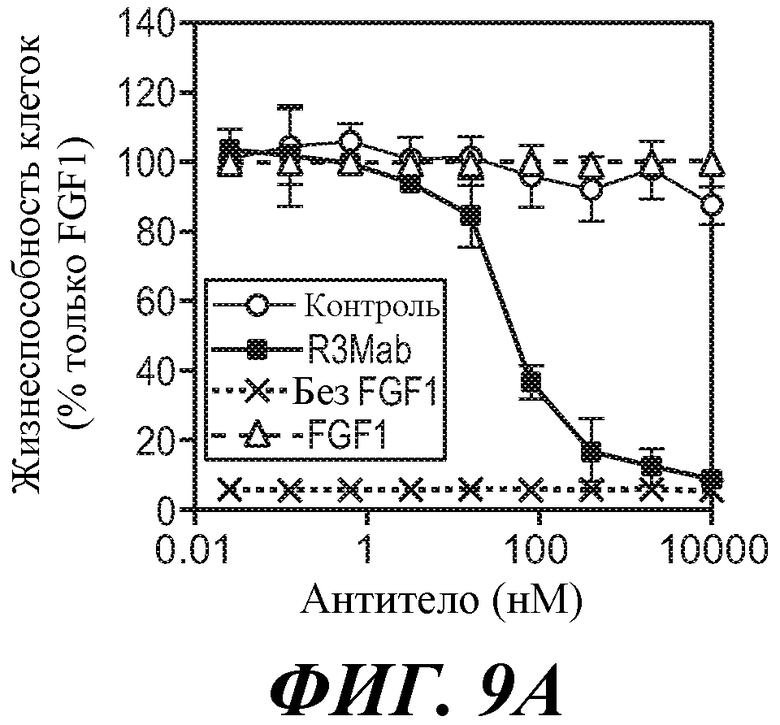
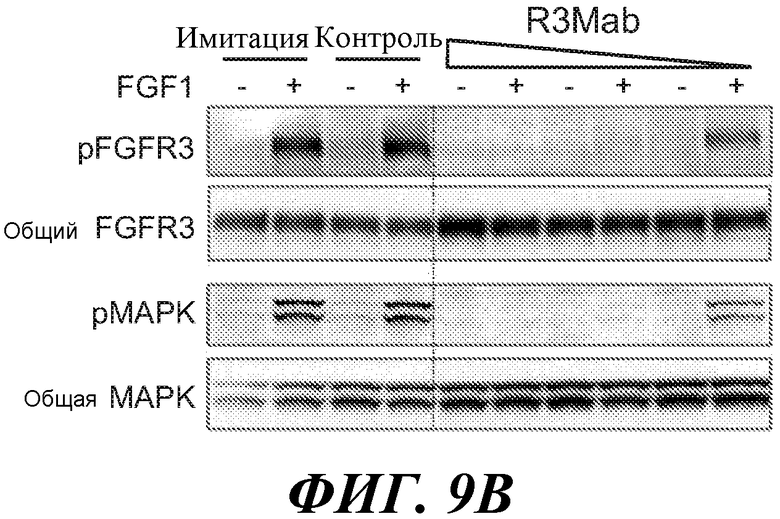
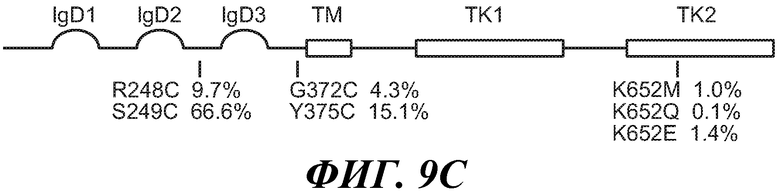
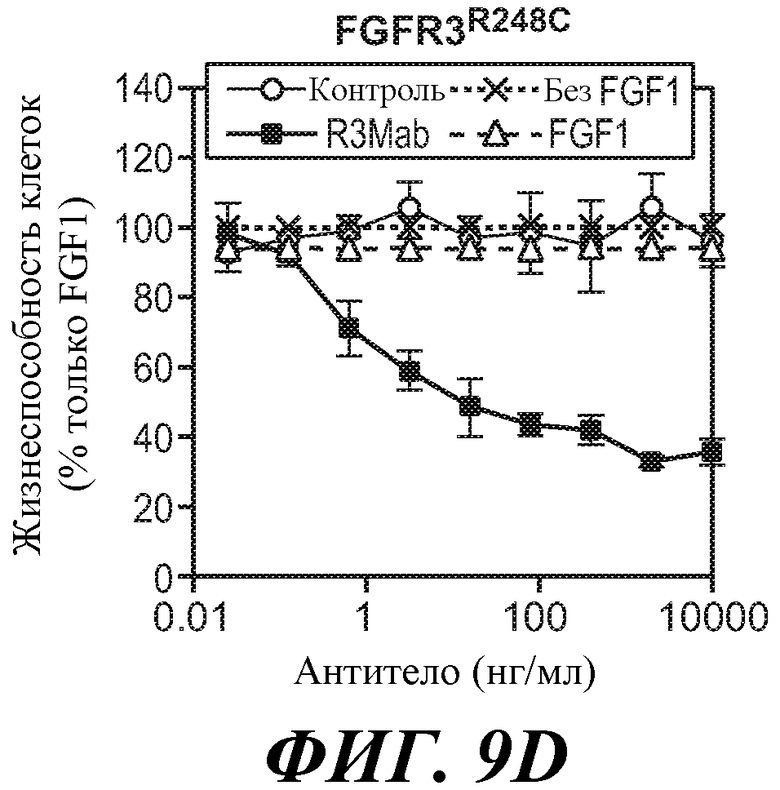

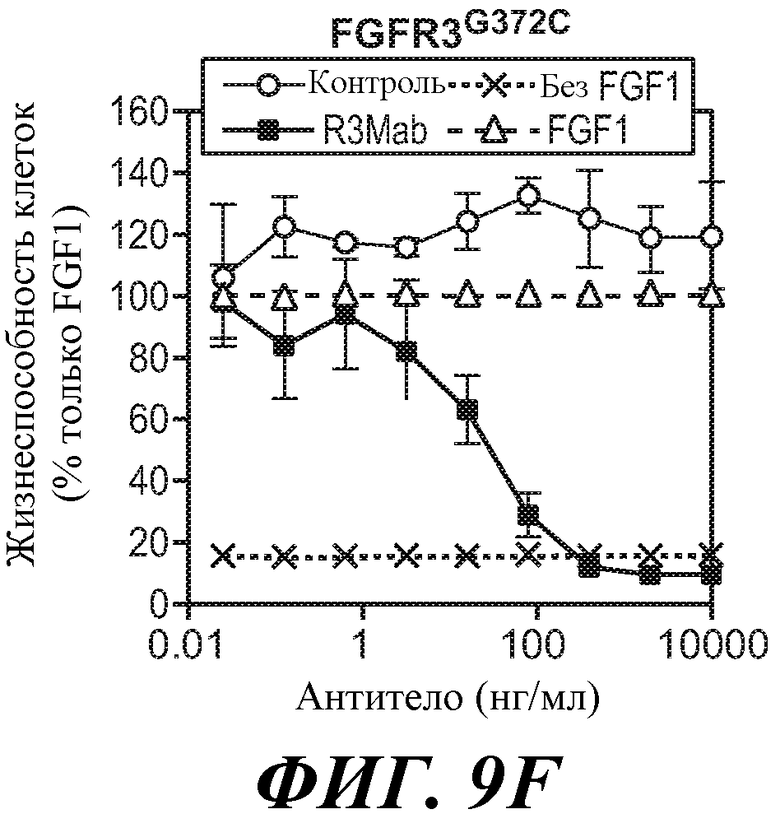
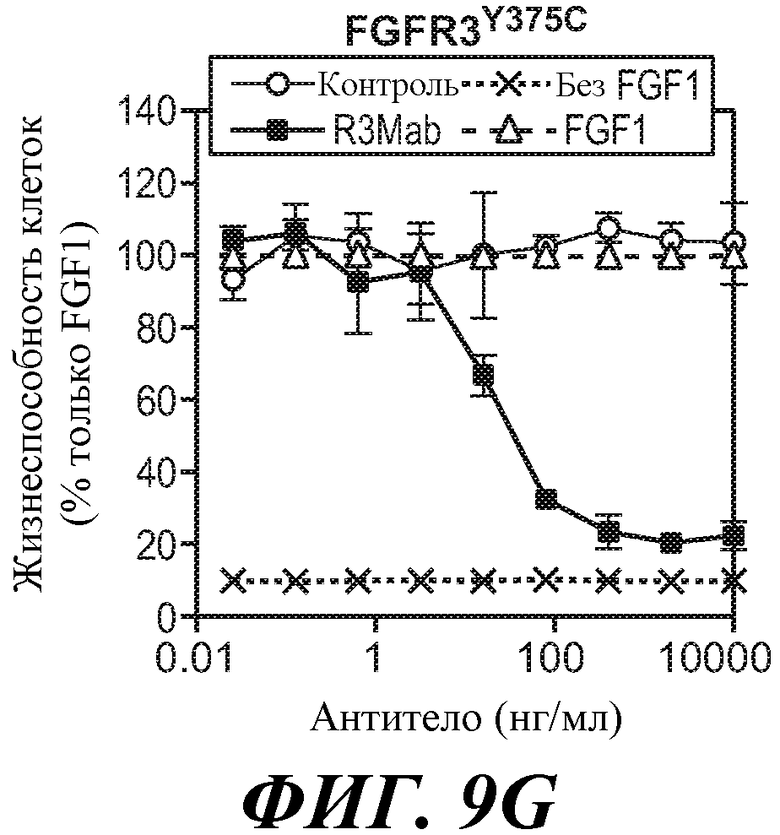
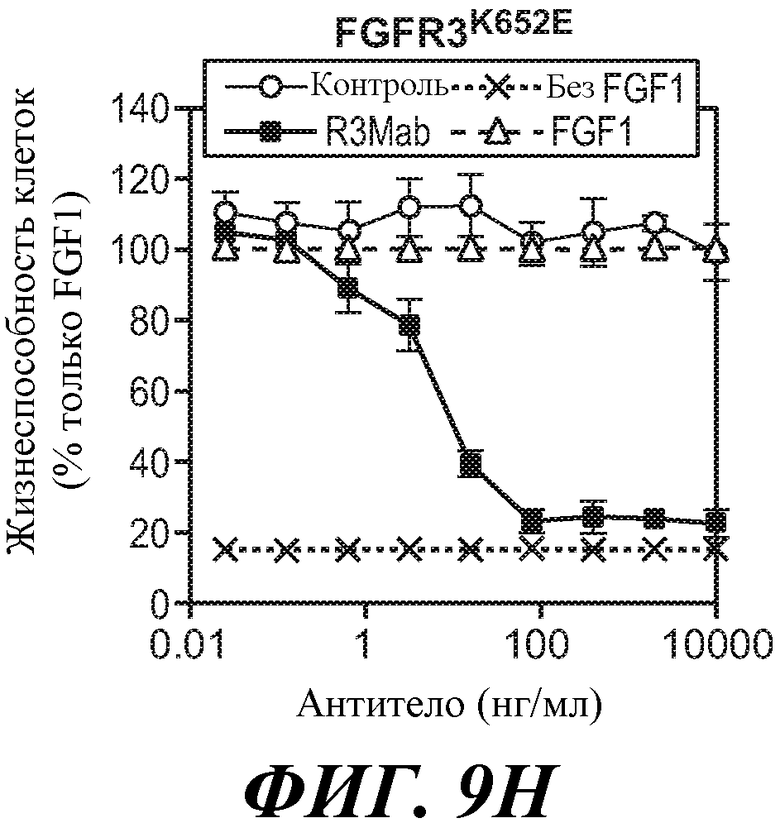
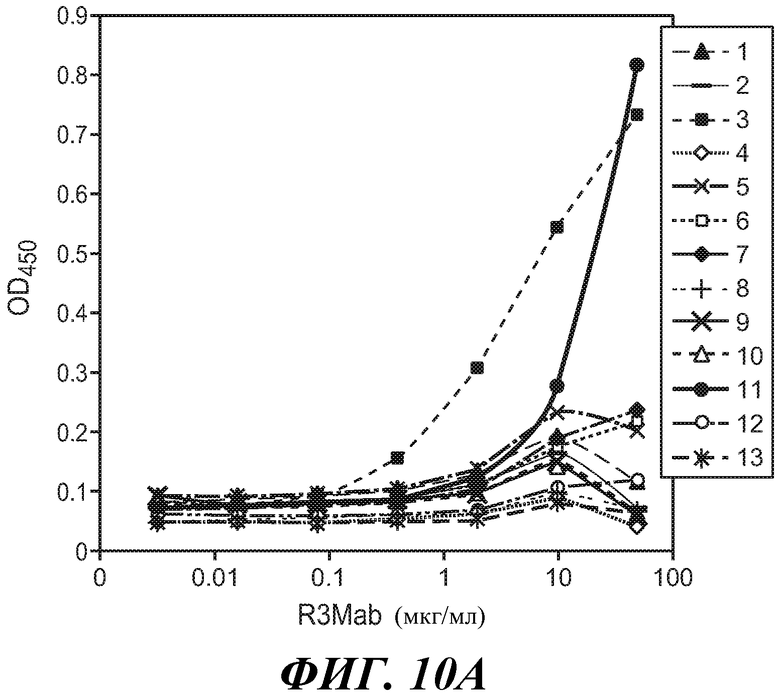
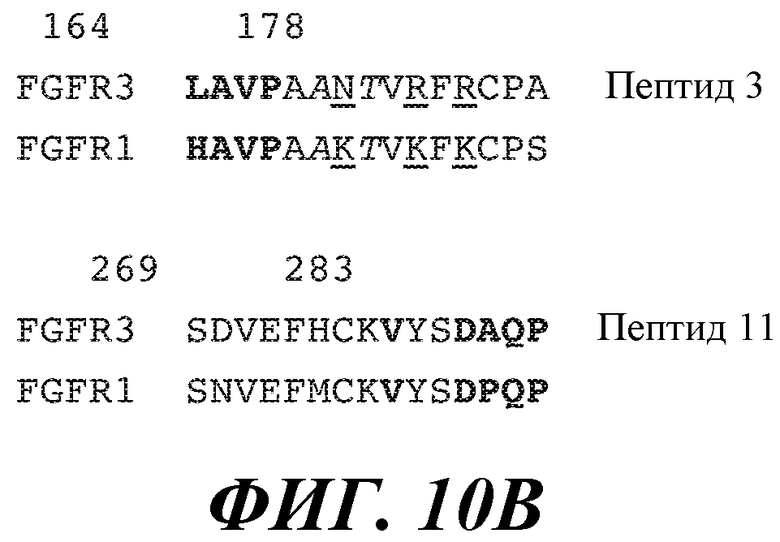
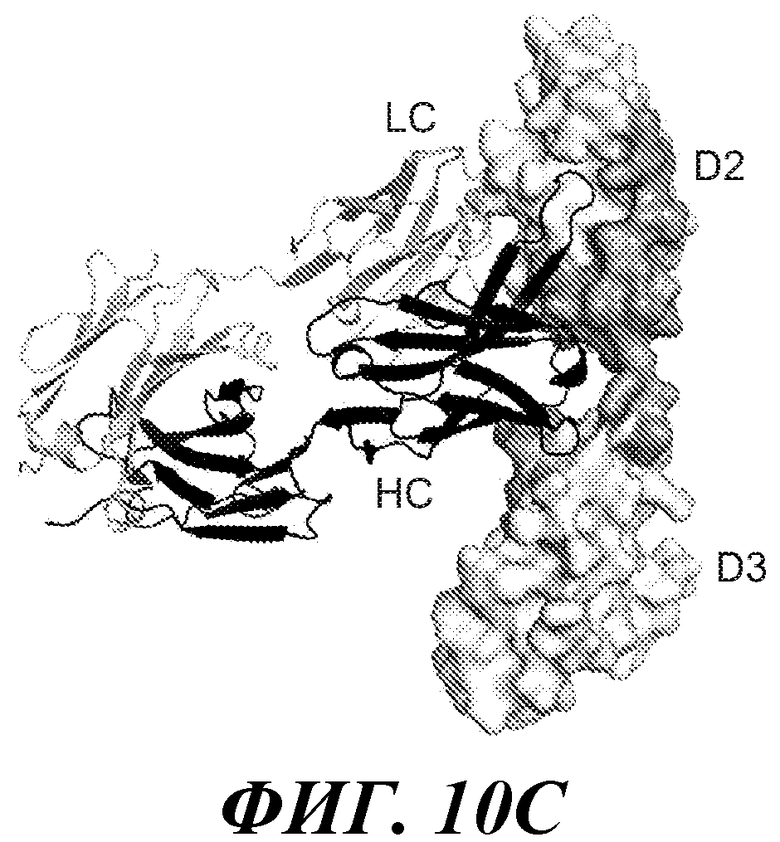

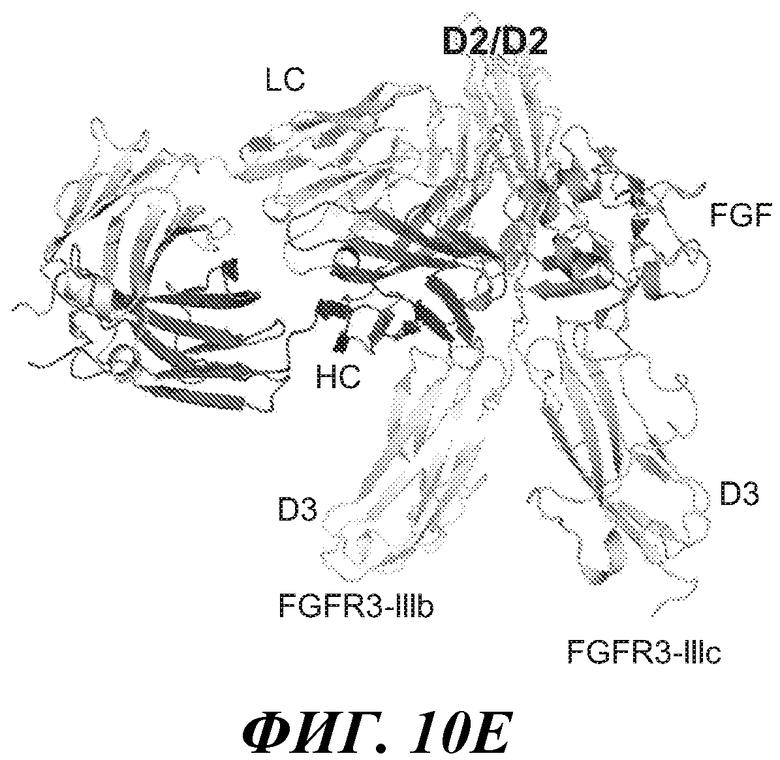

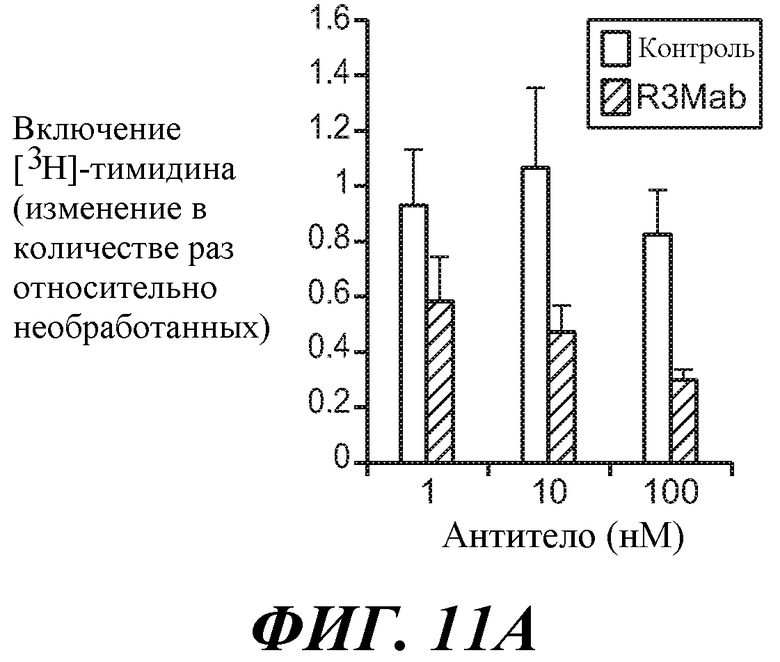

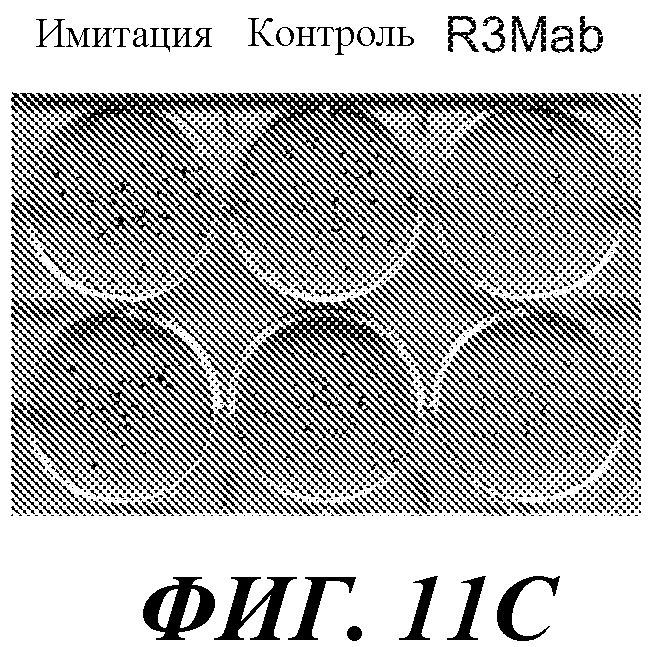
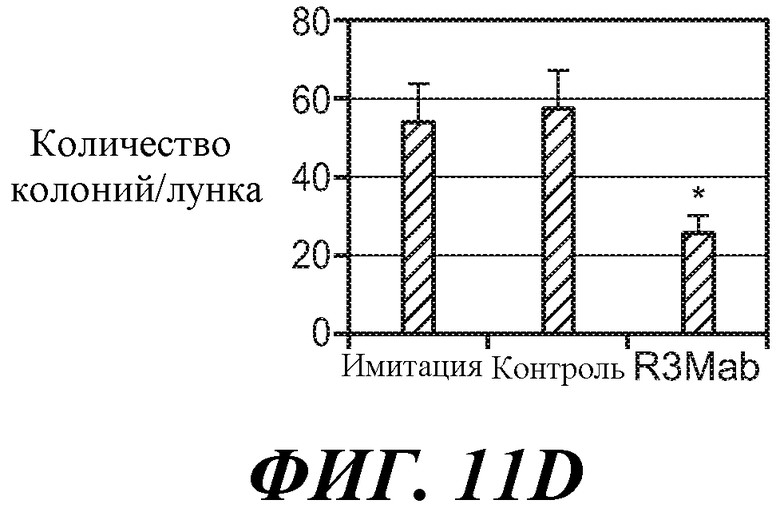
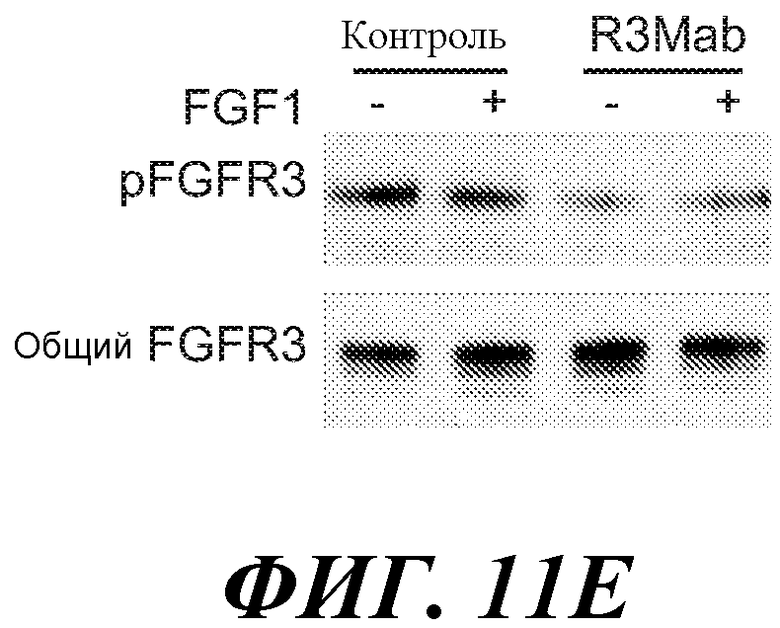
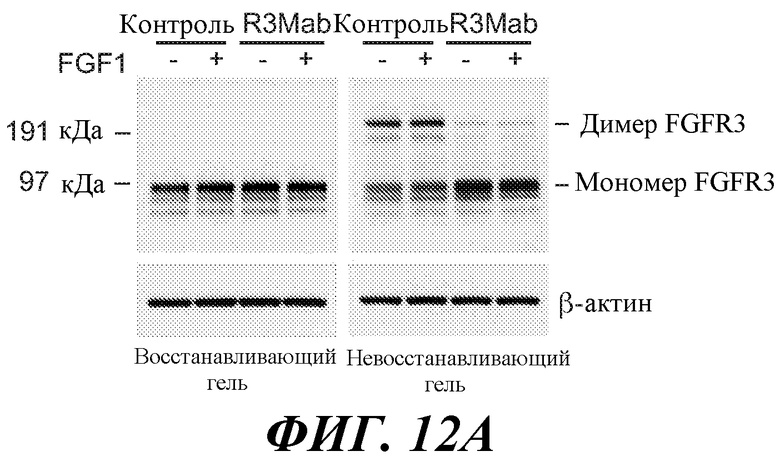

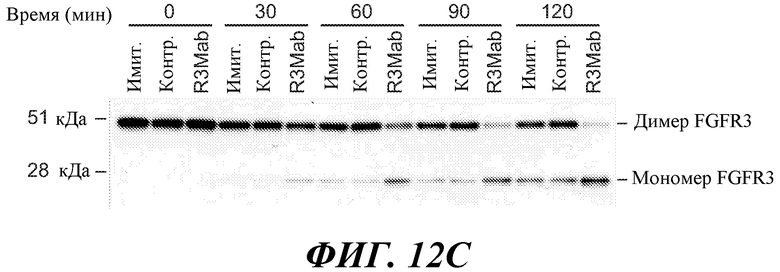
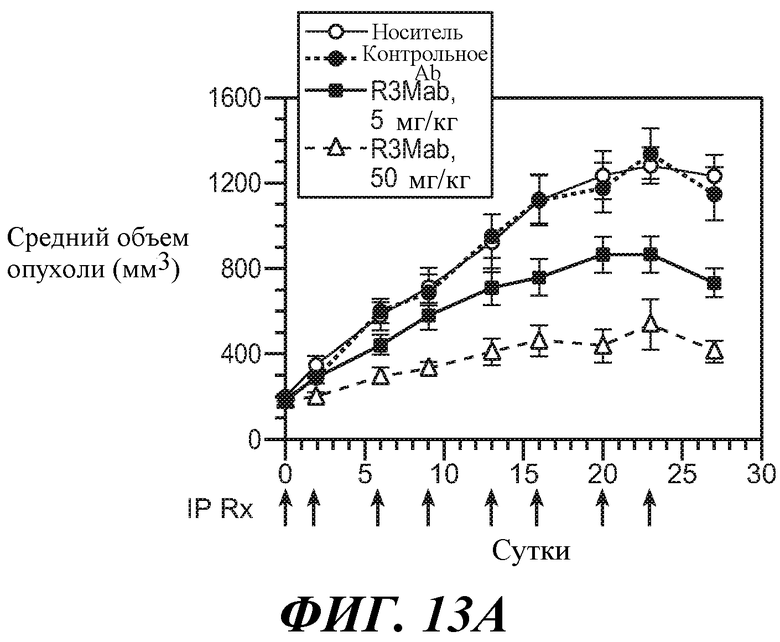
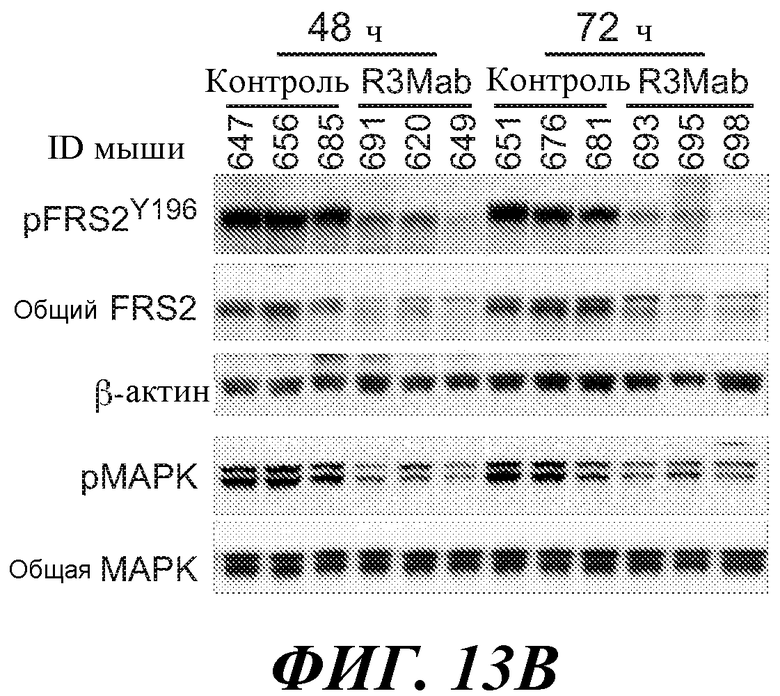

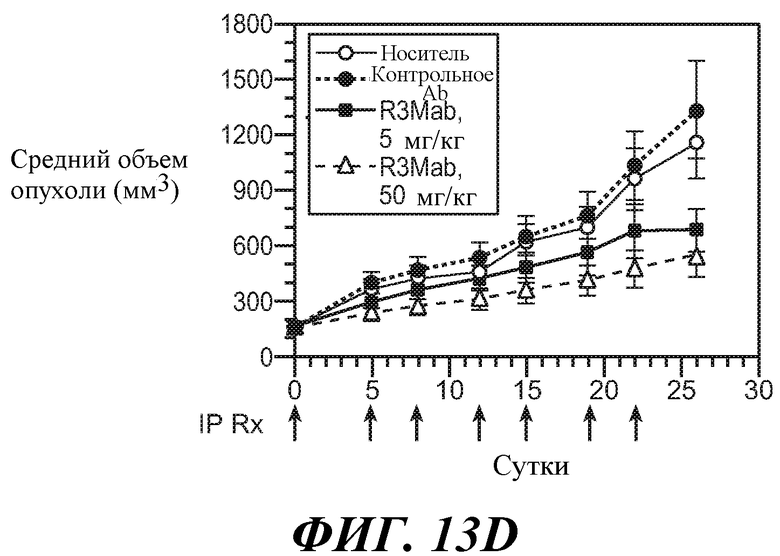
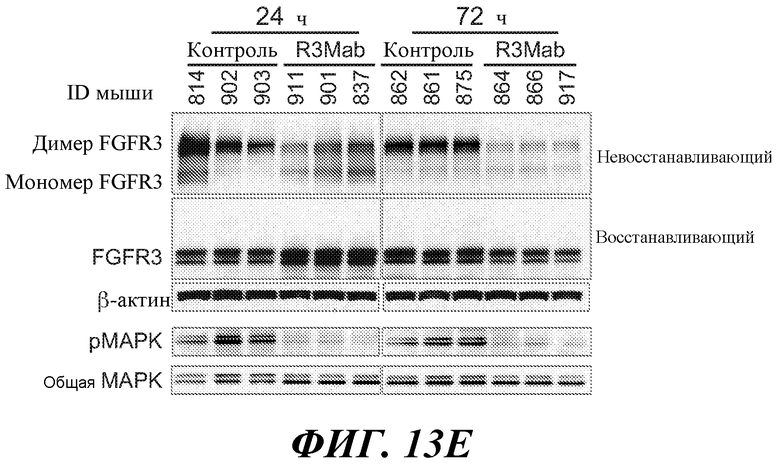
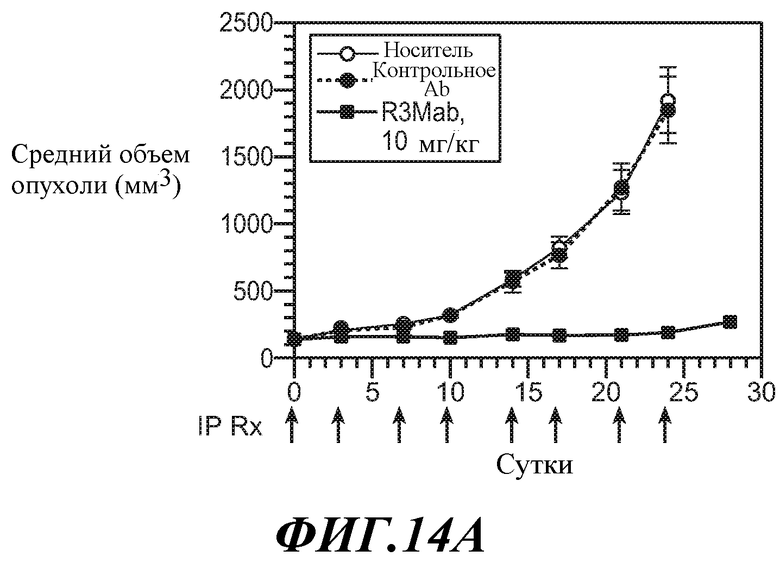
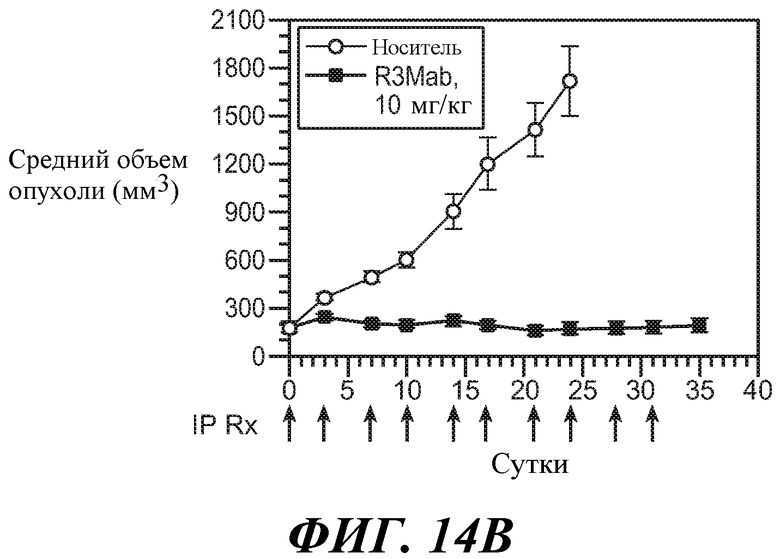
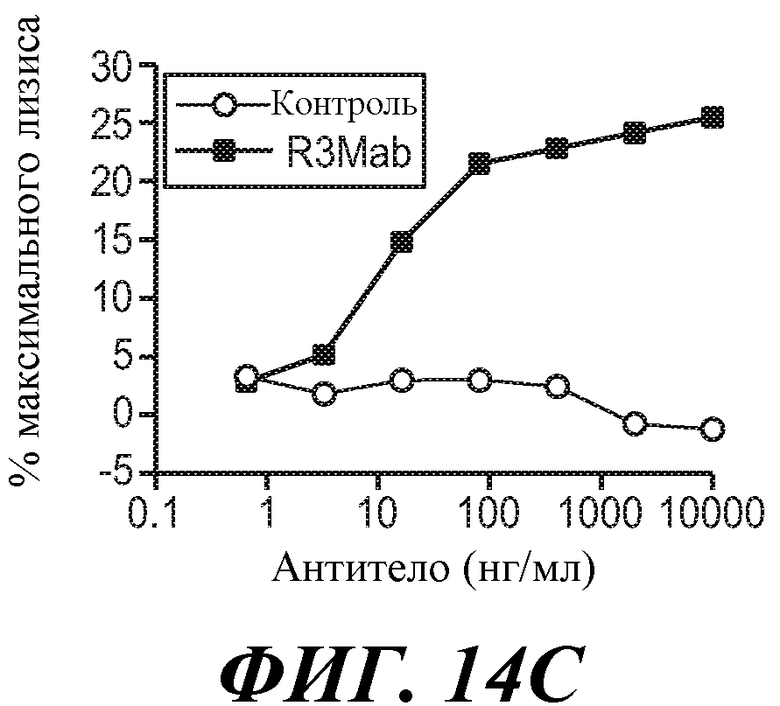
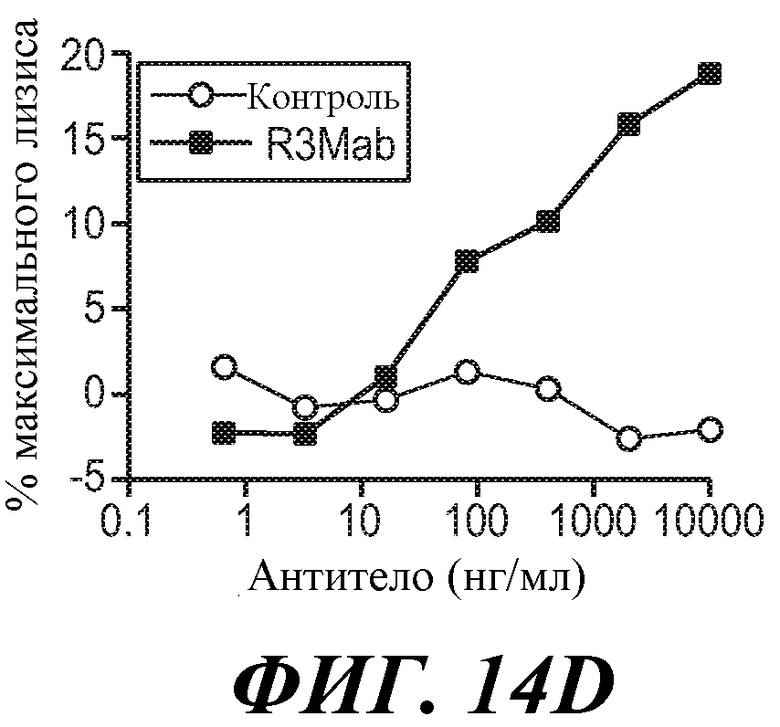
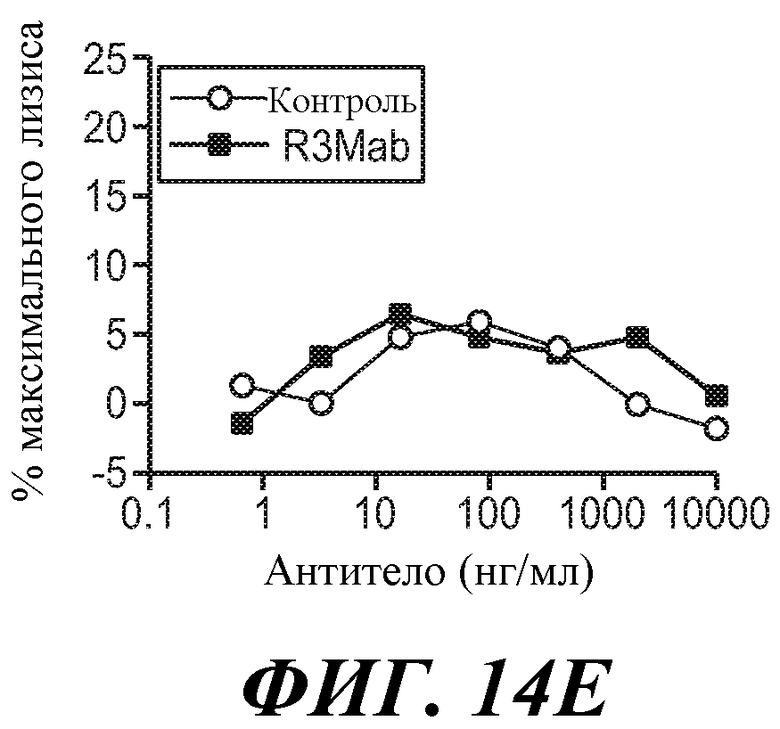
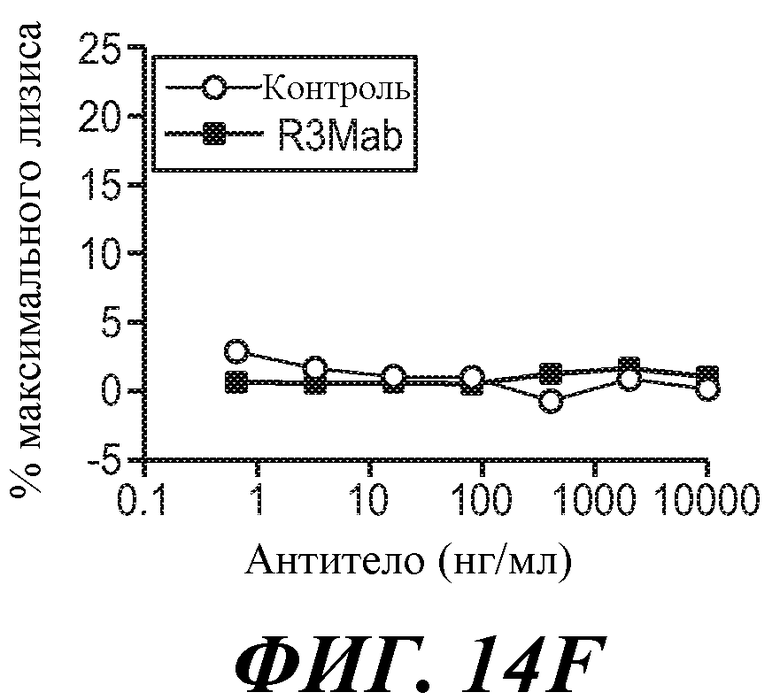

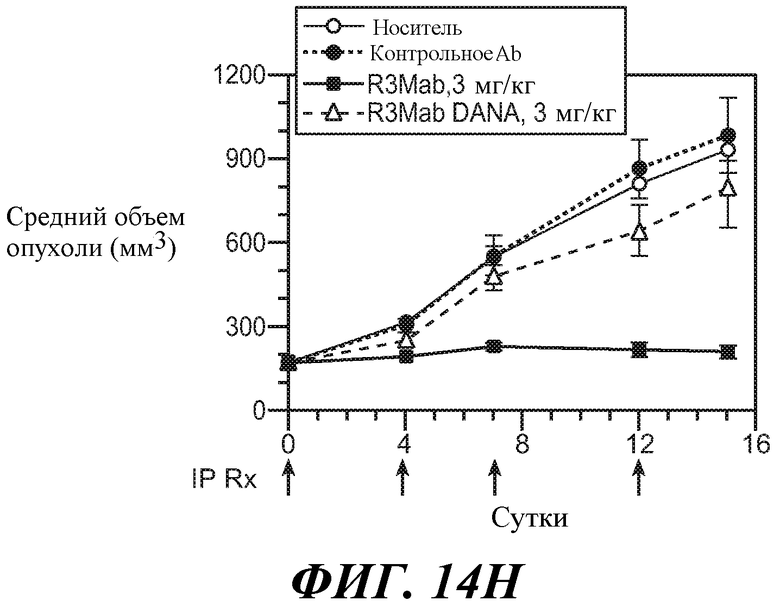
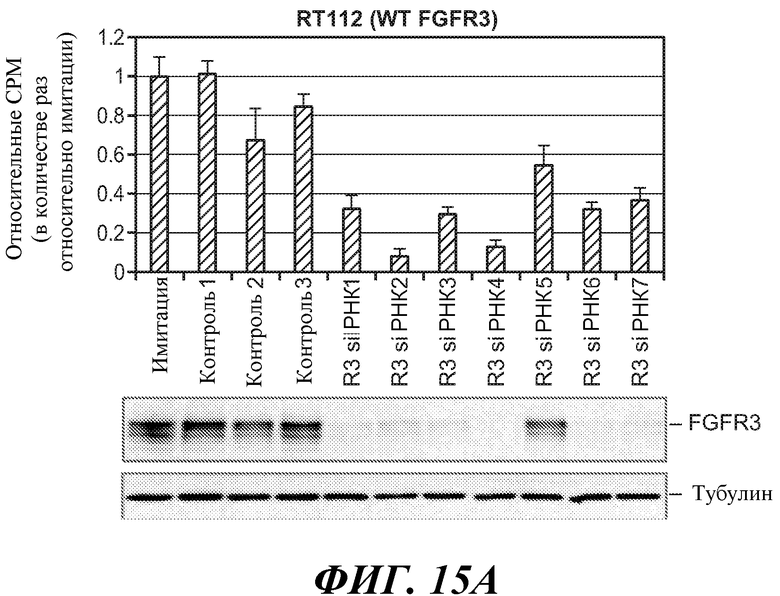
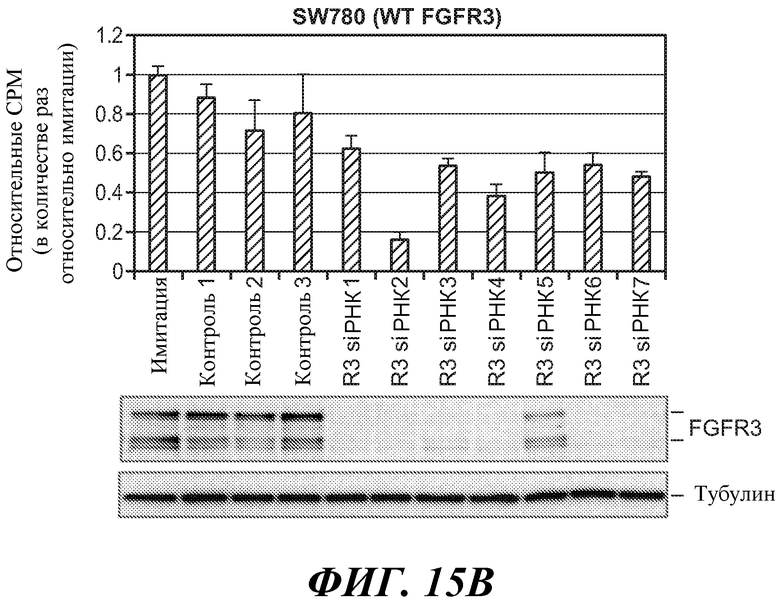
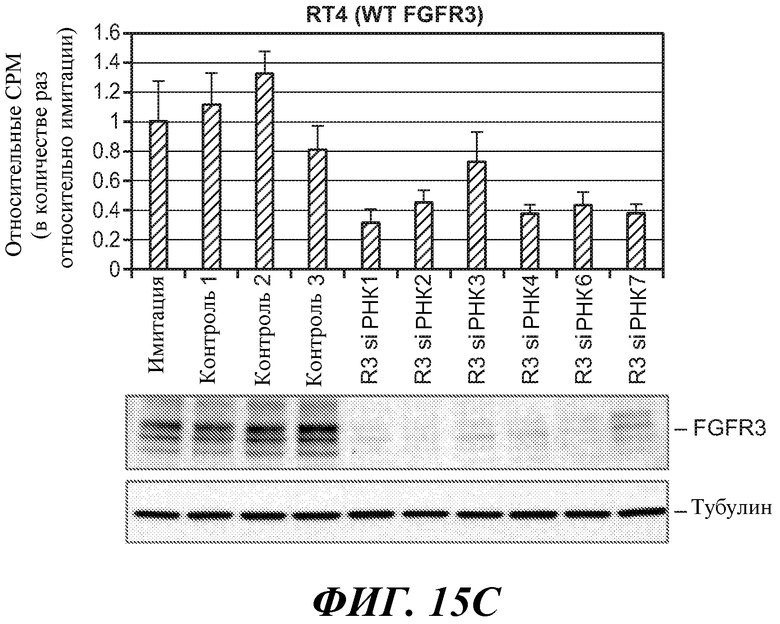
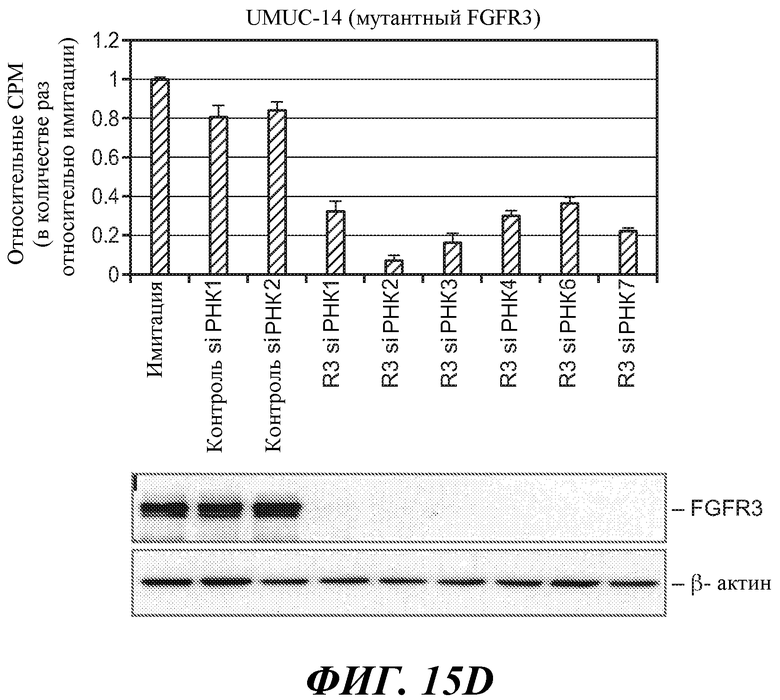
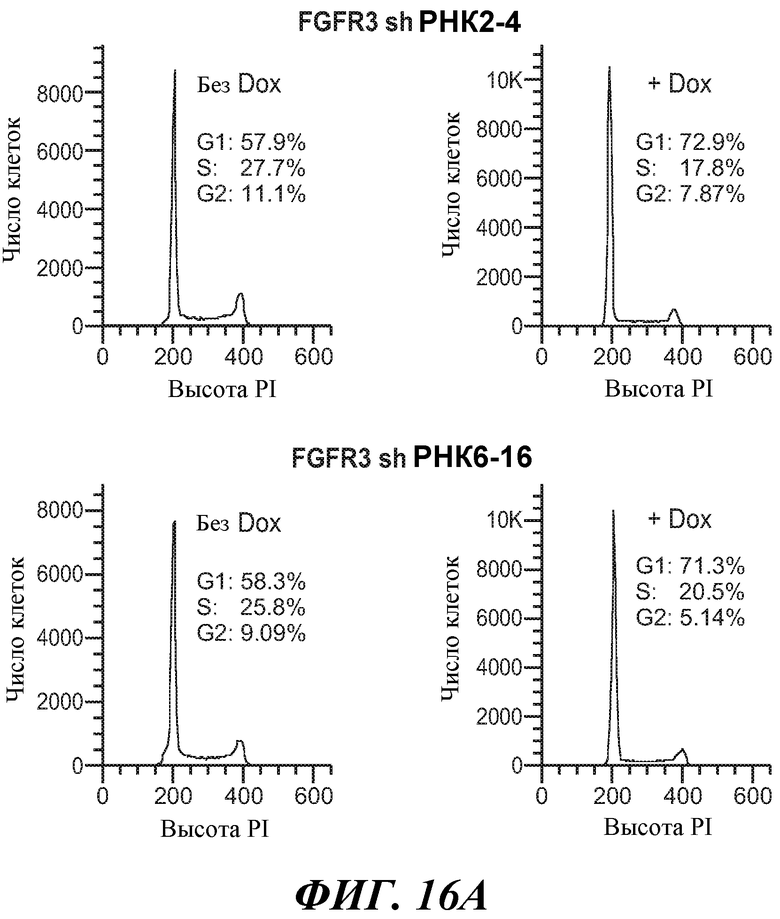
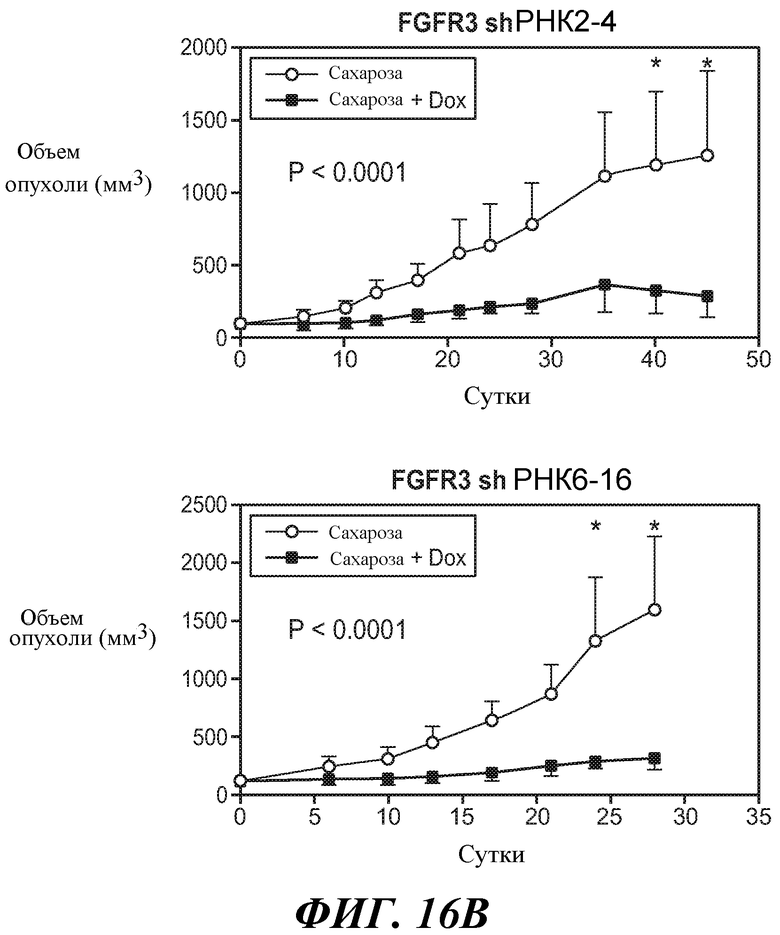
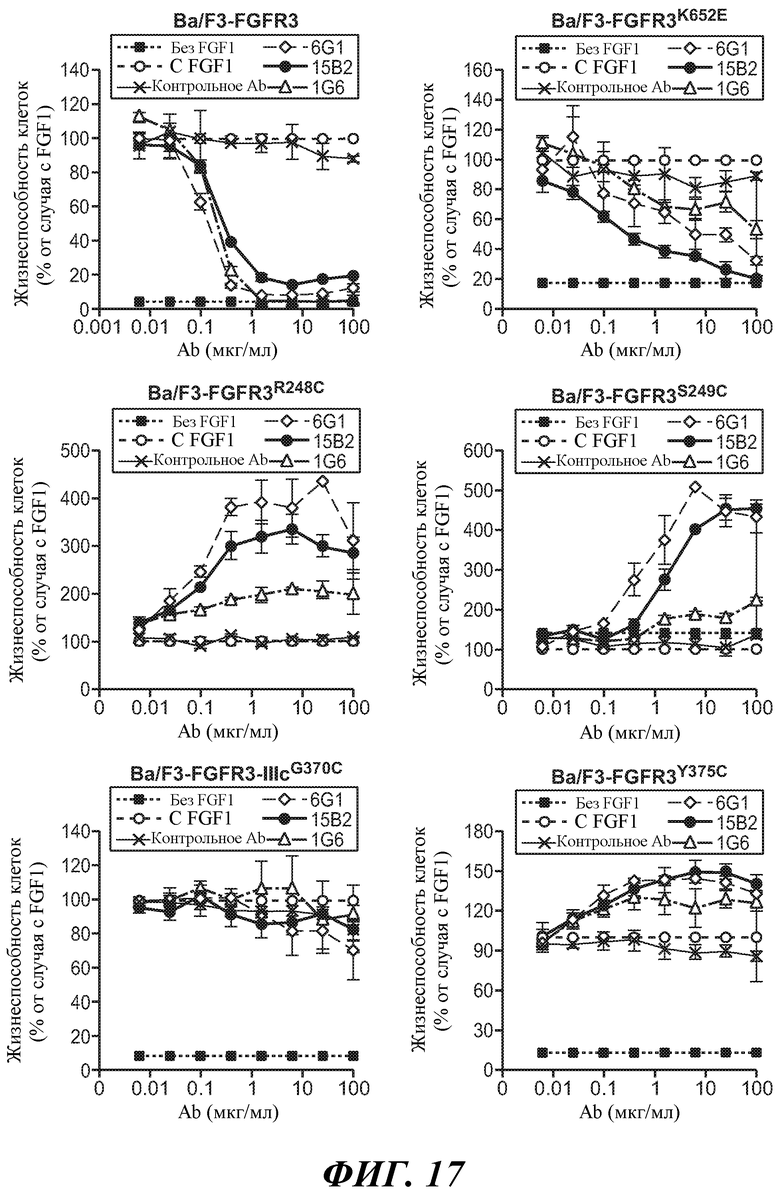
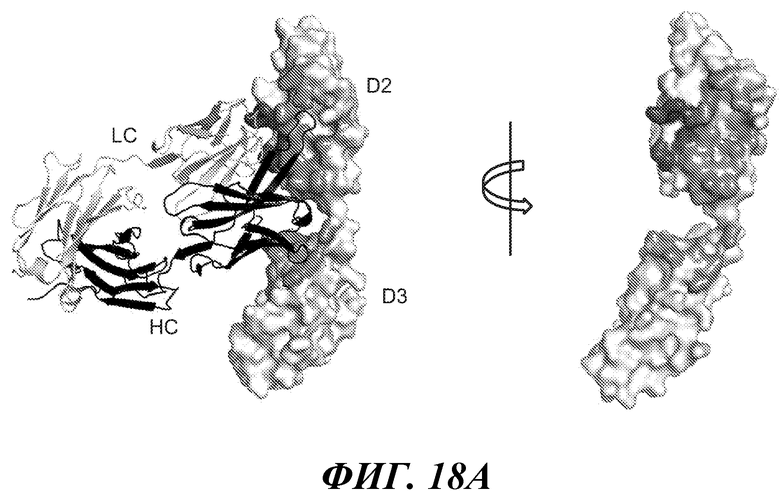
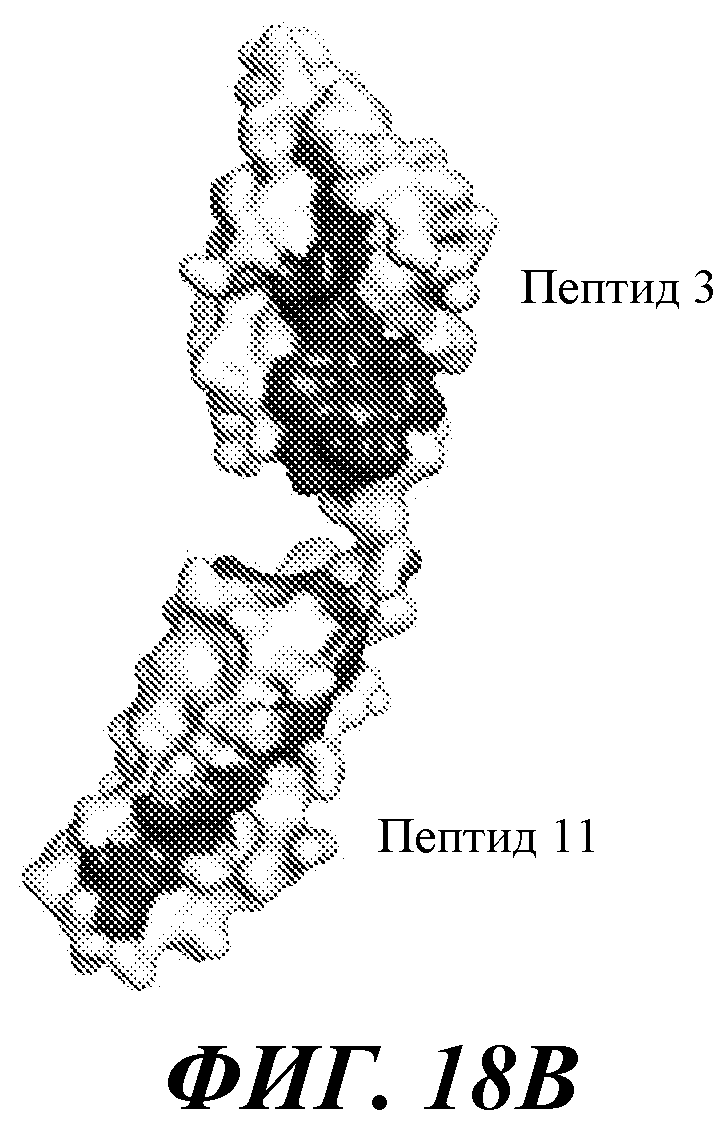
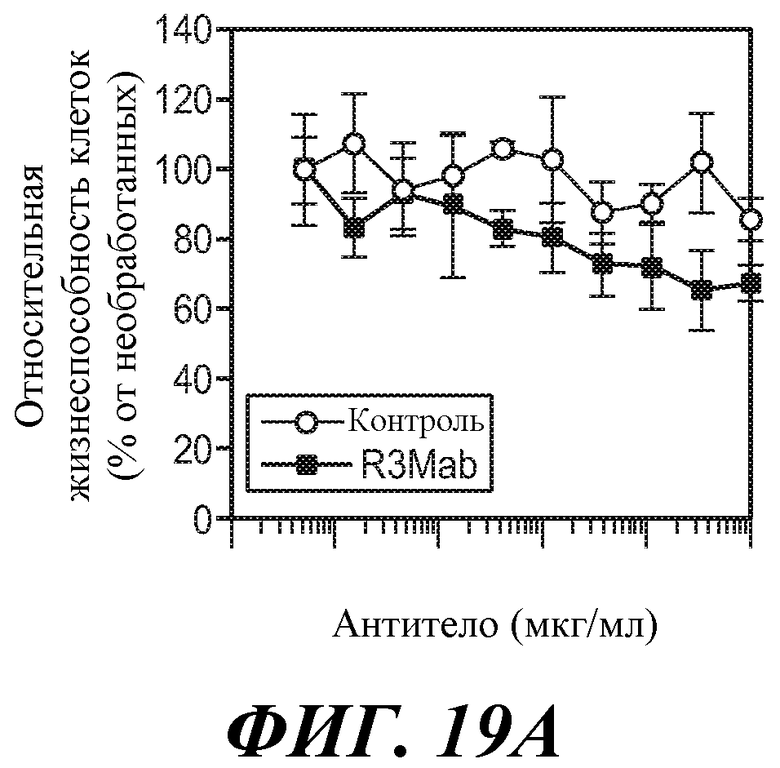
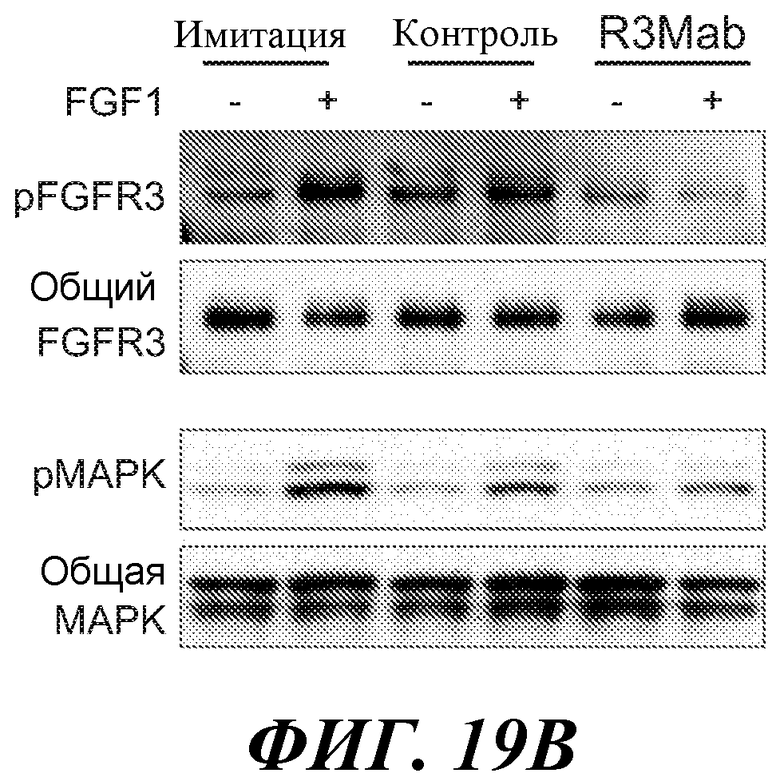
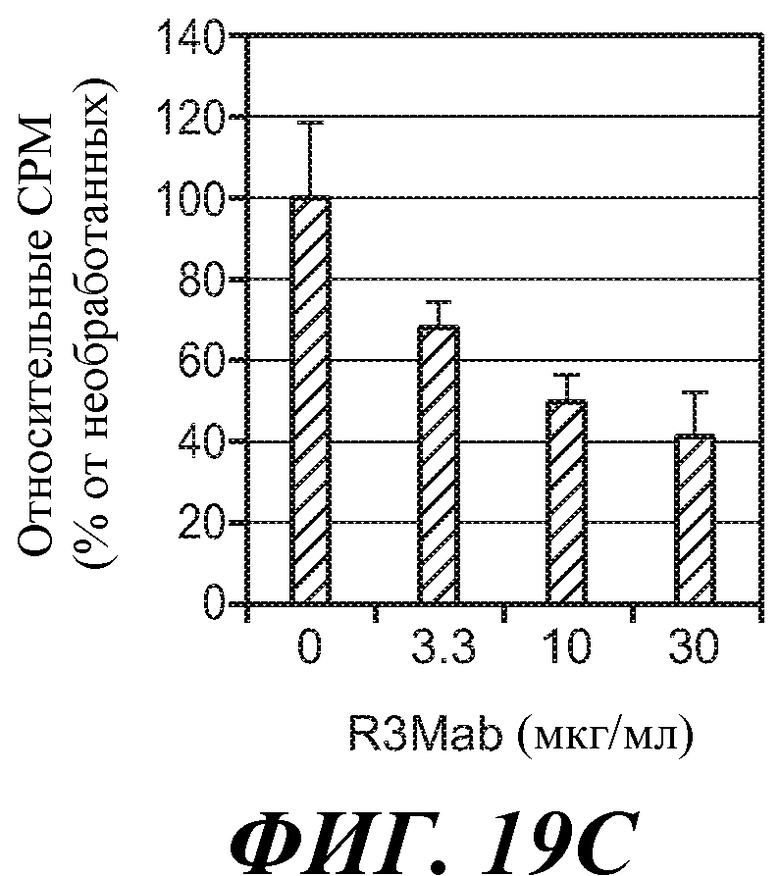
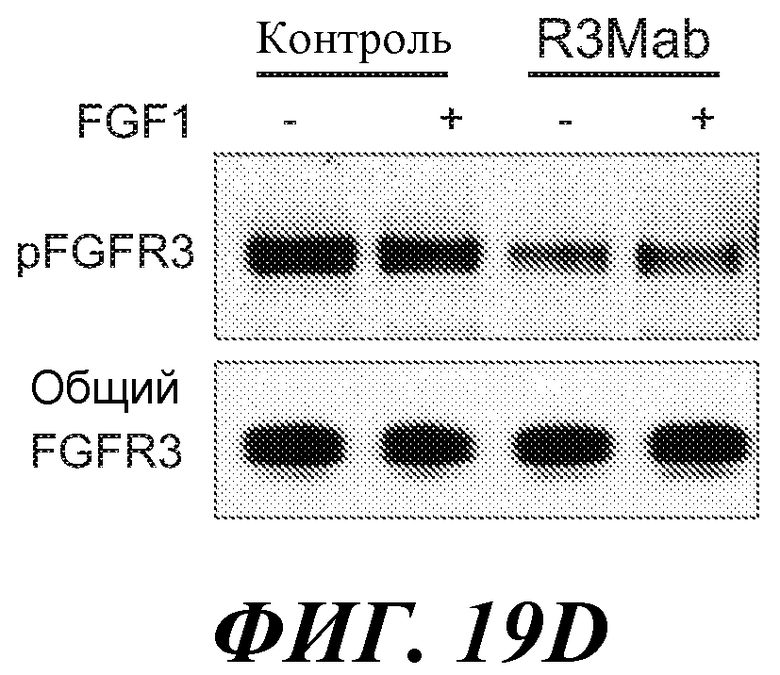
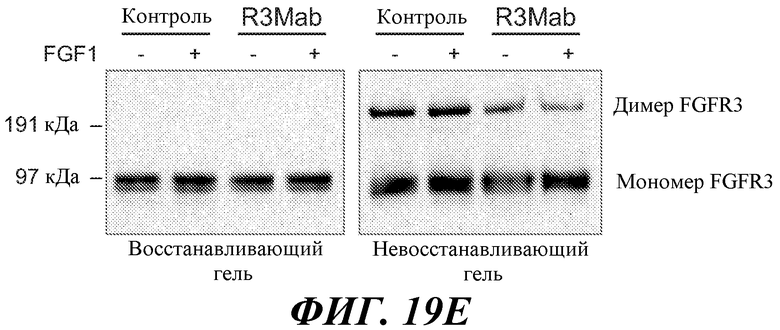
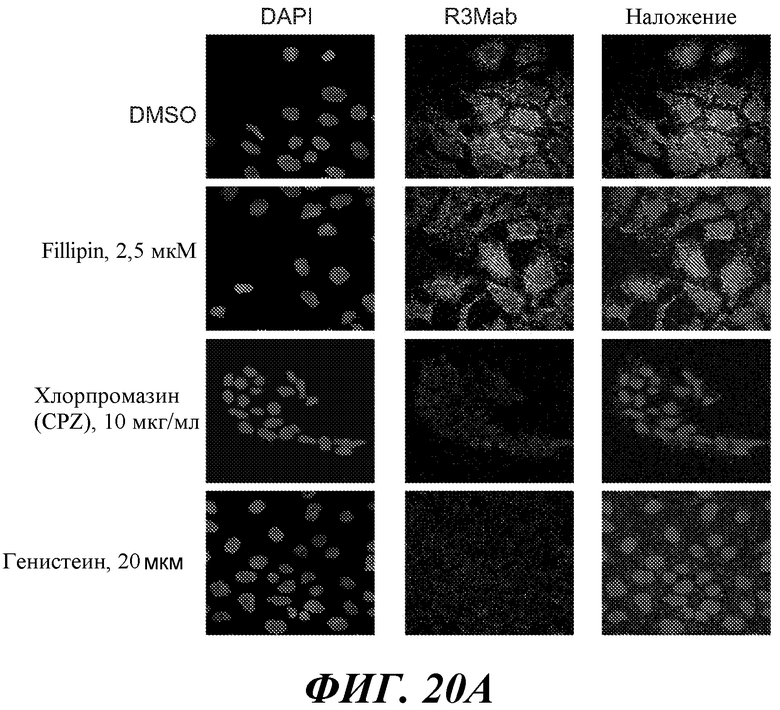
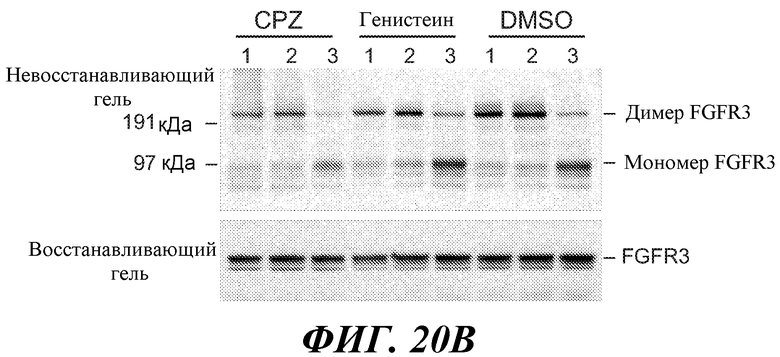
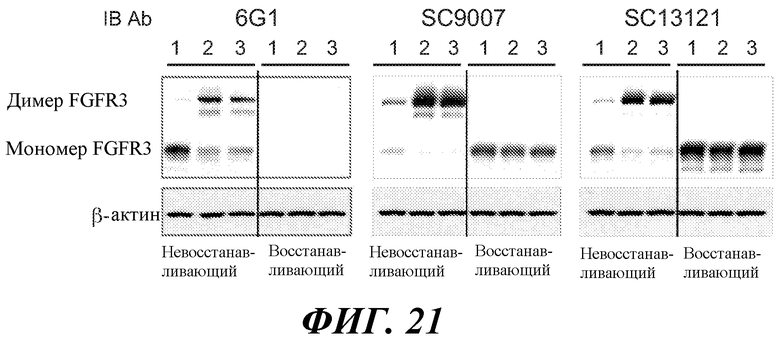


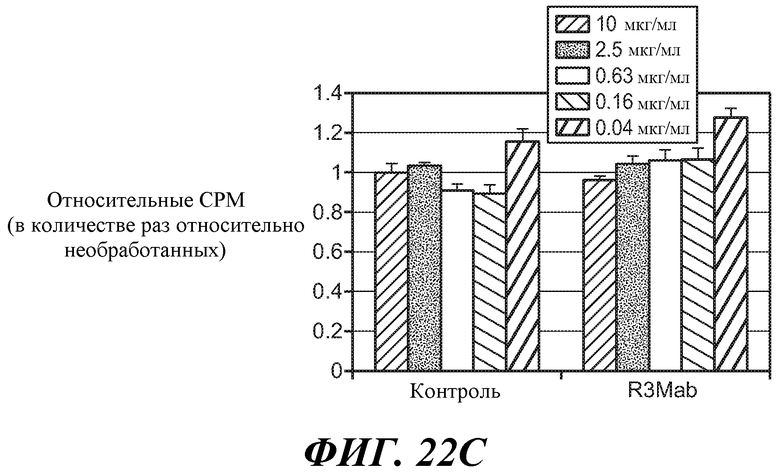
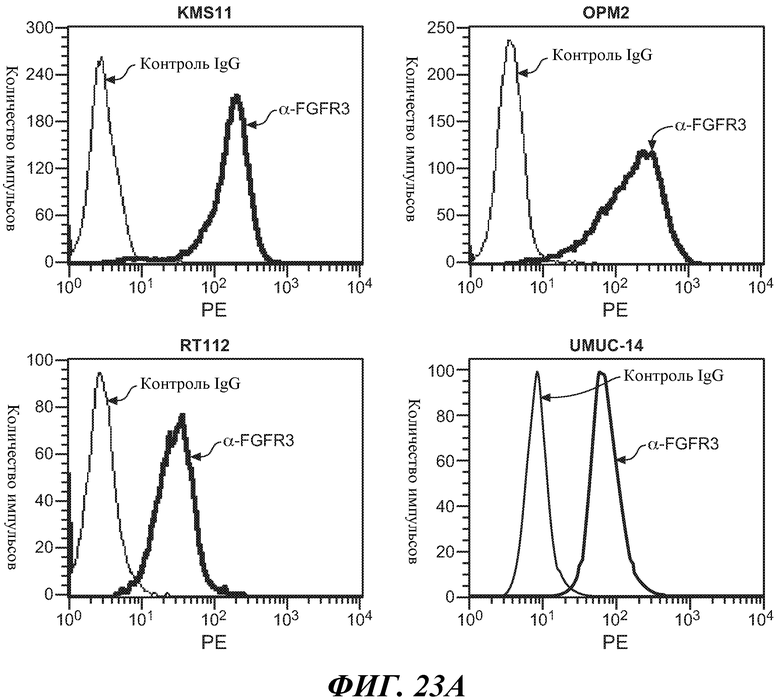
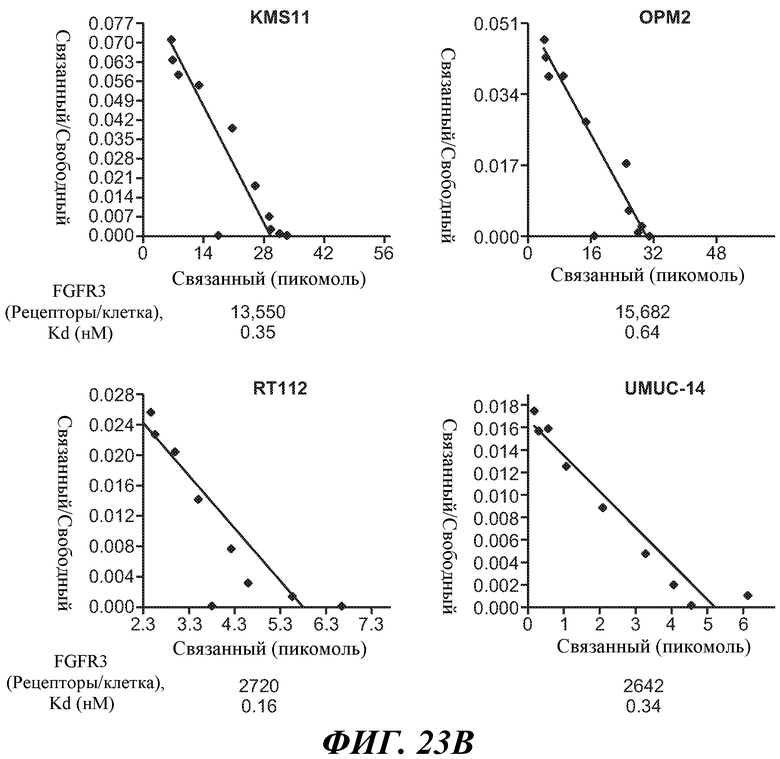
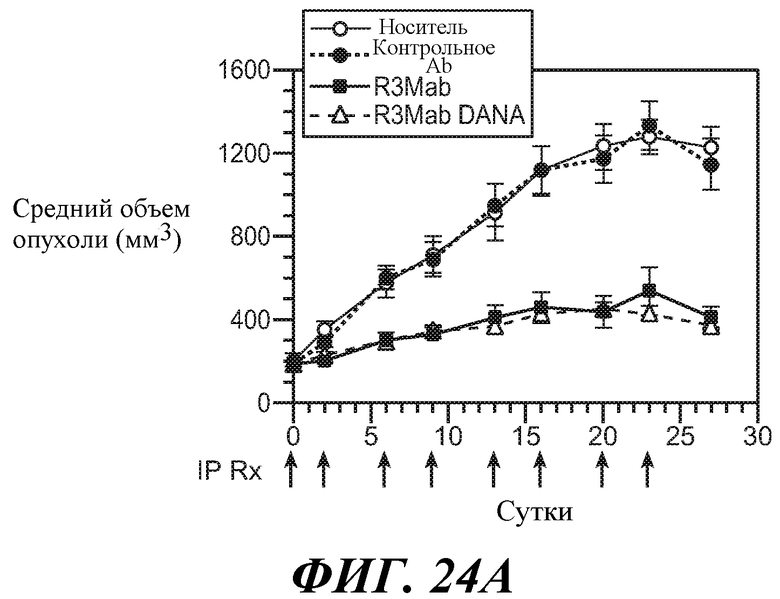
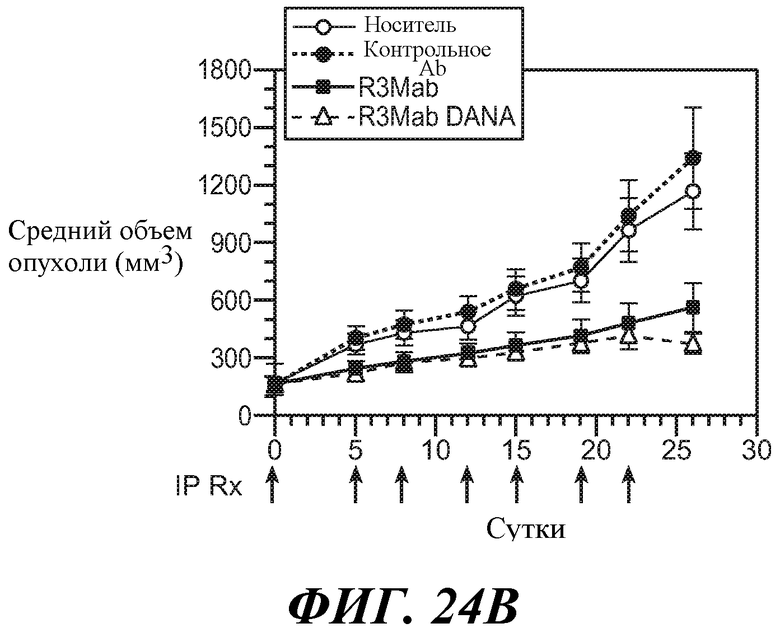
Изобретение относится к области иммунологии. Предложены выделенное антитело-антагонист против FGFR3 и его функциональный фрагмент, а также полинуклеотид, вектор экспрессии, клетка-хозяин и способ получения антитела против FGFR3 или его функционального фрагмента. Кроме того, рассмотрены фармацевтические композиции и применение антитела или его функционального фрагмента по изобретению для лечения или профилактики рака и болезни скелета, а также для ингибирования клеточной пролиферации и/или уменьшения количества раковых клеток. Данное изобретение может найти дальнейшее применение в терапии заболеваний, связанных с активацией и/или сверхэкспрессией FGFR3. 10 н. и 10 з.п. ф-лы, 24 ил., 5 табл.
1. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, содержащее:
(i) HVR-L1, содержащую последовательность RASQDVDTSLA (SEQ ID NO:87) ;
(ii) HVR-L2, содержащую последовательность SASFLYS (SEQ ID NO:88) ;
(iii) HVR-L3, содержащую последовательность QQSTGHPQT (SEQ ID NO:89);
(iv) HVR-H1, содержащую последовательность GFTFTSTGIS (SEQ ID NO:84) ;
(v) HVR-H2, содержащую последовательность GRIYPTSGSTNYADSVKG (SEQ ID NO:85); и
(vi) HVR-H3, содержащую последовательность ARTYGIYDLYVDYTEYVMDY (SEQ ID NO:86).
2. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1, содержащее вариабельную область тяжелой цепи, содержащую SEQ ID NO:132, и вариабельную область легкой цепи, содержащую SEQ ID NO:133.
3. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1 или 2, имеющее одно или несколько следующих свойств:
(a) способно связываться с FGFR3 человека с Kd, равным 1×10-8 или сильнее;
(b) способно связываться с полипептидом, содержащим аминокислотную последовательность, которая по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98% идентична или сходна аминокислотной последовательности LAVPAANTVRFRCPA (SEQ ID NO:179) и/или SDVEFHCKVYSDAQP (SEQ ID NO:180);
(c) способно ингибировать димеризацию рецептора FGFR3 с другой единицей рецептора, ингибируя, таким образом, активацию рецептора; и
(d) способно ингибировать (a) FGFR3-IIIbR248C и (b) один или несколько из FGFR3-IIIbK652E, FGFR3-IIIbY375C, FGFR3-IIIbS249C и FGFR3-IIIbG372C.
4. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1 или 2, которое имеет одно или несколько следующих свойств:
(a) обладает антитело-зависимой клеточной цитотоксичностью;
(b) не индуцирует значительное ингибирование FGFR3;
(c) бивалентная форма антитела не обладает существенной агонистической функцией;
(d) способно связываться по меньшей мере с одной, двумя, тремя, четырьмя или любым числом вплоть до всех остатков 154, 155, 158, 159, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 177, 202, 205, 207, 210, 212, 214, 216, 217, 241, 246, 247, 248, 278, 279, 280, 281, 282, 283, 314, 315, 316, 317 и/или 318 FGFR3 IIIb;
(e) способно ингибировать конститутивную лиганд-зависимую активность FGFR3; и
(f) способно ингибировать конститутивную лиганд-зависимую активность FGFR3.
5. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1 или 2, содержащее изменения в аминокислотных остатках Fc-области, что приводит к снижению эффекторной функции.
6. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1, содержащее каркасную последовательность, и по меньшей мере часть каркасной последовательности представляет собой консенсусную каркасную последовательность человека.
7. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1, содержащее консенсусную каркасную последовательность легкой цепи человека подгруппы κ и/или консенсусную каркасную последовательность тяжелой цепи человека подгруппы III.
8. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.5, где антитело относится к IgG1 классу и содержит мутации D265/N297A (нумерация согласно индексу EU).
9. Выделенное антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1 или 2, где антитело выбрано из группы, состоящей из моноклонального антитела, химерного антитела, гуманизированного антитела, прошедшего стадию созревания аффинности антитела, антитела человека и биспецифического антитела.
10. Полинуклеотид, кодирующий антитело, или его функциональный фрагмент, по пп.1 или 2.
11. Вектор экспрессии, содержащий полинуклеотид по п.10.
12. Клетка-хозяин in vitro, содержащая вектор по п.11, экспрессирующая антитело-антагонист против FGFR3, или его функциональный фрагмент, по п.1 или 2.
13. Способ получения антитела против FGFR3, или его функционального фрагмента, причем указанный способ предусматривает культивирование клетки-хозяина по п.12 таким образом, что экспрессируется антитело против FGFR3, или его функциональный фрагмент, и, необязательно, выделение его из культуры.
14. Фармацевтическая композиция, содержащая эффективное количество антитела, или его фрагмента, по п.1 или 2, и фармацевтически приемлемый носитель, для лечения или профилактики рака, связанного с активацией и/или сверхэкспрессией FGFR3.
15. Фармацевтическая композиция, содержащая эффективное количество антитела, или его фрагмента, по п.1 или 2, и фармацевтически приемлемый носитель, для лечения или профилактики болезни скелета, связанного с активацией и/или сверхэкспрессией FGFR3.
16. Применение антитела-антагониста против FGFR3, или его функционального фрагмента, по п.1 или 2 для лечения рака, связанного с активацией и/или сверхэкспрессией FGFR3.
17. Применение антитела-антагониста против FGFR3, или его функционального фрагмента, по п.1 или 2 для ингибирования клеточной пролиферации и/или уменьшения количества раковых клеток, связанных с активацией и/или сверхэкспрессией FGFR3.
18. Применение по п.16, где рак выбран из группы, состоящей из солидной опухоли, множественной миеломы, карциномы мочевого пузыря, немелкоклеточного рака легкого, рака яичника, рака щитовидной железы, рака головы и шеи, рака печени, карциномы молочной железы, рака желудка, карциномы переходных клеток и рака ободочной и прямой кишки.
19. Применение антитела-антагониста против FGFR3, или его функционального фрагмента, по п.1 или 2 для лечения или профилактики болезни скелета, связанной с активацией и/или сверхэкспрессией FGFR3.
20. Применение по п.19, где болезнь скелета выбрана из группы, состоящей из ахондроплазии, гипохондроплазии, карликовости, летальной дисплазии или синдрома краниосиностоза.
| RAUCHENBERGER R | |||
| et al., "Human combinatorial Fab library yielding specific and functional antibodies against the human fibroblast growth factor receptor 3", Journal of Biological Chemistry, 2003; 278(40): 38194-38205WO 2002/102973 А2, 27.12.2002WO 2007/144893 A2, 21.12.2007TRUDEL S | |||
| et al., "The inhibitory anti-FGFR3 antibody, PRO-001, is |

