ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
По данной заявке испрашивается приоритет предварительной заявки США No. 61/101917, поданной 1 октября 2008 года, содержание которой включено в настоящий документ в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится, главным образом, к области молекулярной биологии. Более конкретно, изобретение относится к антителам против Notch, в том числе к антителам против отрицательной регуляторной области (NRR) Notch2, и к их применению. Также представлены антитела против NRR Notch1 и способы их применения.
УРОВЕНЬ ТЕХНИКИ
Семейство рецепторов Notch представляет собой класс эволюционно консервативных трансмембранных рецепторов, которые передают сигналы, влияющие на развитие организмов, настолько разнообразных, как морские ежи и человек. Рецепторы Notch и их лиганды, Delta и Serrate (у млекопитающих известных как Jagged), представляют собой трансмембранные белки с крупными внеклеточными доменами, которые содержат подобные эпидермальному фактору роста (EGF) повторы. Количество паралогов Notch отличается между видами. Например, существует четыре рецептора Notch у млекопитающих (Notch1-Notch4), два у Caenorhabditis elegans (LIN-12 и GLP-1) и один у Drosophila melanogaster (Notch). Также рецепторы Notch в процессе транспорта к поверхности клетки подвергаются протеолитическому процессингу фурин-подобной протеазой в участке S1 на N-концевой стороне трансмембранного домена, с образованием внеклеточной субъединицы Notch (ECN) и трансмембранной субъединицы Notch (NTM). Эти две субъединицы остаются нековалентно связанными и формируют зрелый гетеродимерный рецептор клеточной поверхности. Рецепторы Notch и каскад передачи сигнала Notch рассмотрены, например, в Aster et al., Annu. Rev. Pathol. Mech. Dis. 3:587-613, 2008, и Bolos et al., Endocrine Reviews 28:339-363, 2007.
Субъединицы Notch2 ECN содержат 36 N-концевых EGF-подобных повторов, за которыми следуют три тандемно повторяющихся модуля Lin 12/Notch Repeat (LNR), которые предшествуют участку S1. Каждый модуль LNR содержит три дисульфидных связи и группу консервативных кислотных и полярных остатков, которые, как предполагают, координируют ион кальция. В области EGF-повтора находятся связывающие участки для активирующих лигандов.
Notch2 NTM содержит внеклеточную область (которая содержит участок расщепления S2), трансмембранный сегмент (который содержит участок расщепления S3) и крупную внутриклеточную часть, которая включает домен RAM23, шесть анкириновых повторов, домен трансактивации и C-концевую последовательность PEST. Стабильная ассоциация субъединиц ECN и NTM зависит от домена гетеродимеризации (HD), содержащего C-конец ECN (называемый HD-N) и внеклеточный N-конец NTM (называемый HD-C). Перед индуцируемой лигандом активацией Notch поддерживается в покоящейся конформации с помощью отрицательной регуляторной области (NRR), которая содержит три LNR и HD-домен. Кристаллическая структура NRR Notch2 описана в Gordon et al, (2007) Nature Structural & Molecular Biology 14:295-300, 2007.
Связывание лиганда Notch с субъединицей ECN инициирует два последовательных протеолитических расщепления, которые происходят путем регулируемого внутримембранного протеолиза. Первое расщепление металлопротеиназой (ADAM17) в участке S2 делает трансмембранную субъединицу Notch чувствительной ко второму расщеплению в участке S3 вблизи внутреннего слоя плазматической мембраны. Расщепление в участке S3, которое катализируется мультибелковым комплексом, содержащим пресенилин и никастрин и стимулирующим активность γ-секретазы, высвобождает внутриклеточную часть трансмембранной субъединицы Notch, позволяя ей перемещаться в ядро и активировать транскрипцию генов-мишеней. (Для обзора протеолитического расщепления Notch, см., например, Sisodia et al., Nat. Rev. Neurosci. 3:281-290, 2002.)
У человека идентифицировано пять лигандов Notch классов Jagged и Delta-подобных белков (Jagged1 (также называемый Serrate1), Jagged2 (также называемый Serrate2), Delta-подобный 1 (также называемый DLL1), Delta-подобный 3 (также называемый DLL3) и Delta-подобный 4 (также называемый DLL4)). Каждый из этих лигандов представляет собой однократно пронизывающий мембрану трансмембранный белок с консервативным N-концевым мотивом Delta, Serrate, LAG-2 (DSL), необходимым для связывания Notch. Серии EGF-подобных модулей, находящихся со стороны C-конца мотива DSL, предшествуют трансмембранному сегменту. В отличие от рецепторов Notch, лиганды имеют на C-конце короткие цитоплазматические хвостовые части из 70-215 аминокислот. Кроме того, описаны другие типы лигандов (например, DNER, NB3 и F3/контактин). (Для обзора лигандов Notch и опосредуемой лигандами активации Notch, см., например, D'Souza et al., Oncogene 27:5148-5167, 2008.)
Каскад Notch функционирует в ходе различных связанных с развитием и физиологических процессов, включая процессы, влияющие на нейрогенез у мух и позвоночных. Как правило, передача сигнала Notch вовлечена в латеральное ингибирование, определение направления дифференцировки и установление границ между группами клеток (см., например, Bray, Molecular Cell Biology 7:678-679, 2006). Было показано, что множество заболеваний человека, включая злокачественные опухоли и нейродегенеративные нарушения, являются следствием мутаций в генах, кодирующих рецепторы Notch или их лиганды (см., например, Nam et al., Curr. Opin. Chem. Biol. 6:501-509, 2002). Связь между неограниченной передачей сигнала Notch и злокачественной опухолью впервые была признана, когда была идентифицирована рекуррентная t(7;9)(q34;q34,3) хромососмная транслокация, которая приводит к укороченному конститутивно активному варианту Notch1 человека в подгруппе острых лимфобластных лейкозов (T-ALL) человека (см., например, Aster et al., Annu. Rev. Pathol. Mech. Dis. 3:587-613, 2008). В моделях на мышах было показано, что передача сигнала Notch1 является необходимой для развития T-клеток и что опосредуемые Notch1 сигналы стимулируют развитие T-клеток за счет развития B-клеток (см., например, Wilson et al., J. Exp. Med. 194:1003-1012, 2001).
Notch2 также вовлечен в некоторые злокачественные опухоли. В частности, Notch2 сверхэкспрессируется при B-клеточном хроническом лимфоцитарном лейкозе (B-CLL), что в свою очередь приводит к сверхэкспрессии CD23, характерного признака клеток B-CLL. (См. Hubmann et al., Blood 99:3742-3747, 2002.) Как Notch1, так и Notch2, высоко экспрессируются в клетках множественной миеломы (злокачественных плазматических B-клетках), и стимуляция лигандом резко увеличивает рост опухолевых клеток. (См. Jundt et al., Blood 103:3511-3515, 2004.) Notch2 и последующие эффекторы сверхэкспрессируются при меланоме (см. Hoek et al., Cancer Res. 64:5270-5282, 2004; Seykora et al., Am J Dermatopathol 25:6-11, 2003), и локус Notch2 рекуррентно амплифицируется в клеточных линиях меланомы (Jonsson et al., Oncogene, 26:4738- 4748, 2007). Кроме того, во множестве исследований выявлена связь нарушенной передачи сигнала Notch2 с раком молочной железы и другими солидными опухолями (рассмотрено Leong and Karsay, Blood 107:2223-2233, 2006). Notch2 также требуется для развития B-клеточной маргинальной зоны. (См. Pillai et al., Annu. Rev. Immunol. 23:161-196, 2005.)
С учетом вовлечения передачи сигнала Notch в широкое множество заболеваний человека, очевидно, что продолжает существовать потребность в средствах, которые регулируют передачу сигнала Notch и которые имеют клинические качества, подходящие для их разработки в качестве лекарственных средств. Изобретение, описанное в настоящем документе, удовлетворяет эту потребность и обеспечивает другие преимущества.
Все ссылки, цитированные в настоящем документе, включая патентные заявки и публикации, включены в качестве ссылок в полном объеме.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к антителам против Notch и способам их применения.
В одном аспекте представлено моноклональное антитело, которое связывается с NRR Notch2. В одном варианте осуществления антитело ингибирует активность Notch2. В другом варианте осуществления антитело по существу не связывается с представителями семейства Notch, отличными от Notch2. В другом варианте осуществления антитело связывается с NRR Notch2 мыши и NRR Notch2 человека. В другом варианте осуществления антитело связывается с NRR Notch2 с Kd ≤ 10 нМ.
В следующем варианте осуществления представлено моноклональное антитело, которое связывается с NRR Notch2, где антитело содержит:
(a) HVR-H1, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:3;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:4;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:5;
(d) HVR-L1, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 10;
(e) HVR-L2, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 14; и
(f) HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 19.
В одном из таких вариантов осуществления антитело содержит HVR-H1, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 1-2; HVR-L1, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 6-9; HVR-L2, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 15-18. В одном варианте осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO: 1, HVR-L1 содержит аминокислотную последовательность SEQ ID NO: 6, HVR-L2 содержит аминокислотную последовательность SEQ ID NO: 11, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO: 15. В другом варианте осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO: 2, HVR-L1 содержит аминокислотную последовательность SEQ ID NO: 7, HVR-L2 содержит аминокислотную последовательность SEQ ID NO: 11 и HVR-L3 содержит аминокислотную последовательность SEQ ID NO: 16. В другом варианте осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO: 2, HVR-L1 содержит аминокислотную последовательность SEQ ID NO: 8, HVR-L2 содержит аминокислотную последовательность SEQ ID NO: 12 и HVR-L3 содержит аминокислотную последовательность SEQ ID NO: 17. В другом варианте осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO: 2, HVR-L1 содержит аминокислотную последовательность SEQ ID NO: 9, HVR-L2 содержит аминокислотную последовательность SEQ ID NO: 13 и HVR-L3 содержит аминокислотную последовательность SEQ ID NO: 18. В любом из указанных выше вариантов осуществления антитело, кроме того, содержит по меньшей мере одну каркасную область, выбранную из каркасной области акцептора 2 VH человека и консенсусной каркасной области подгруппы I VL каппа человека.
В другом аспекте представлено моноклональные антитело, которое связывается с NRR Notch2, где антитело содержит вариабельный домен тяжелой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 20-21, и вариабельный домен легкой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 22-25. В одном варианте осуществления антитело содержит вариабельный домен тяжелой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 20 и вариабельный домен легкой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 22. В одном из таких вариантов осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 20, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 22. В другом варианте осуществления антитело содержит вариабельный домен тяжелой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 21, и вариабельный домен легкой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 23-25. В одном из таких вариантов осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 23-25.
В другом аспекте представлено выделенное антитело, которое связывается с тем же эпитопом, что и антитело, выбранное из антитела D, антитела D-1, антитела D-2 или антитела D-3. В другом аспекте представлено выделенное антитело, которое связывается с доменом LNR-A и с доменом HD-C Notch2.
В другом аспекте антитело против NRR Notch2 представляет собой фрагмент антитела, выбранный из Fab-, Fab'-SH-, Fv-, scFv- или (Fab')2-фрагмента. В другом аспекте антитело против NRR Notch2 представляет собой гуманизированное, химерное антитело или антитело человека.
Любой из указанных выше вариантов осуществления может существовать отдельно или в комбинации.
В другом аспекте представлен способ ингибирования активности Notch2, включающий воздействие на клетку, которая экспрессирует Notch2, антитела согласно любому из указанных выше вариантов осуществления. В другом аспекте представлен способ лечения нарушения, ассоциированного с повышенной экспрессией или активностью Notch2, включающий введение индивидууму, нуждающемуся в этом, эффективного количества антитела согласно любому из указанных выше вариантов осуществления. В другом аспекте представлен способ лечения B-клеточной злокачественной опухоли, включающий введение индивидууму, нуждающемуся в этом, эффективного количества антитела согласно любому из указанных выше вариантов осуществления. В другом аспекте представлен способ лечения меланомы, включающий введение индивидууму, нуждающемуся в этом, эффективного количества антитела согласно любому из указанных выше вариантов осуществления.
В другом аспекте представлен способ лечения нарушения, ассоциированный с повышенной экспрессией или активностью Notch1, включающий совместное введение индивидууму, нуждающемуся в этом, эффективного количества антитела против NRR Notch1 и лекарственного средства, выбранного из дексаметазона и тамоксифена, где лекарственное средство снижает изменение дифференцировки клеток кишечника, вызываемое антителом. В одном из таких вариантов осуществления нарушение представляет собой T-клеточную злокачественную опухоль.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фигуре 1 представлены последовательности гипервариабельных областей (HVR) H1, H2 и H3 тяжелых цепей моноклональных антител против NRR Notch2, обозначаемых как антитело D, антитело D-1, антитело D-2 и антитело D-3, как описано в примере B(1). Аминокислотные положения пронумерованы согласно системе нумерации Kabat, как описано ниже.
На фигуре 2 представлены последовательности HVR L1, L2 и L3 легких цепей моноклональных антител против NRR Notch2, обозначаемых как антитело D, антитело D-1, антитело D-2 и антитело D-3, как описано в примере B(1). Аминокислотные положения пронумерованы согласно системе нумерации Kabat, как описано ниже.
На фигуре 3 представлено выравнивание последовательностей вариабельных областей тяжелых цепей антитела D, антитела D-1, антитела D-2 и антитела D-3. HVR заключены в рамки, как описано в примере B(1).
На фигуре 4 представлено выравнивание последовательностей вариабельных областей легких цепей антитела D, антитела D-1, антитела D-2 и антитела D-3. HVR заключены в рамки.
На фигурах 5A и 5B представлены иллюстративные консенсусные каркасные последовательности вариабельных областей тяжелых цепей (VH) человека для применения для осуществления на практике настоящего изобретения. Идентификаторы последовательностей являются следующими:
- консенсусная каркасная область "A" подгруппы I VH человека минус CDR по Kabat (SEQ ID NO: 32, 33, 34, 35).
- консенсусные каркасные области "B," "C" и "D" подгруппы I VH человека минус удлиненные гипервариабельные области (SEQ ID NO: 36, 37, 34, 35; SEQ ID NO: 36, 37, 38, 35; и SEQ ID NO: 36, 37, 39, 35).
- консенсусная каркасная область "A" подгруппы II VH человека минус CDR по Kabat (SEQ ID NO: 40, 41, 42, 35).
- консенсусные каркасные области "B," "C" и "D" подгруппы II VH человека минус удлиненные гипервариабельные области (SEQ ID NO: 43, 44, 42, 35; SEQ ID NO: 43, 44, 45, 35; и SEQ ID NO: 43, 44, 46, и 35).
- консенсусная каркасная область "A" подгруппы III VH человека минус CDR по Kabat (SEQ ID NO: 47, 48, 49, 35).
- консенсусные каркасные области "B," "C" и "D" подгруппы III VH человека минус удлиненные гипервариабельные области (SEQ ID NO: 50, 51, 49, 35; SEQ ID NO: 50, 51, 52, 35; и SEQ ID NO: 50, 51, 53, 35).
- каркасная область акцептора "A" VH человека минус CDR по Kabat (SEQ ID NO: 54, 48, 55, 35).
- каркасные области акцептора "B" и "C" VH человека минус удлиненные гипервариабельные области (SEQ ID NO: 50, 51, 55, 35; и SEQ ID NO: 50, 51, 56, 35).
- каркасная область акцептора 2 "A" VH человека минус CDR по Kabat (SEQ ID NO: 54, 48, 57, 35).
- каркасные области акцептора 2 "B", "C" и "D" VH человека минус удлиненные гипервариабельные области (SEQ ID NO: 50, 51, 57, 35; SEQ ID NO: 50, 51, 58, 35; и SEQ ID NO: 50, 51, 59, 35).
На фигуре 6 представлены иллюстративные акцепторные консенсусные каркасные последовательности вариабельной области легкой цепи (VL) человека для применения для осуществления на практике настоящего изобретения. Идентификаторы последовательностей являются следующими:
- консенсусная каркасная область подгруппы I VL каппа человека (κv1): SEQ ID NO: 60, 61, 62, 63
- консенсусная каркасная область подгруппы II VL каппа человека (κv2): SEQ ID NO: 64, 65, 66, 63
- консенсусная каркасная область подгруппы III VL каппа человека (κv3): SEQ ID NO: 67, 68, 69, 63
- консенсусная каркасная область подгруппы IV VL каппа человека (κv4): SEQ ID NO: 70, 71, 72, 63
На фигуре 7 показаны каркасные последовательности легкой и тяжелой цепей huMAb4D5-8. Числа в виде надстрочных знаков/выделенные полужирным шрифтом указывают на аминокислотные положения по Kabat.
На фигуре 8 показаны каркасные последовательности легкой и тяжелой цепей huMAb4D5-8 с указанными модификациями. Числа в виде надстрочных знаков/выделенные полужирным шрифтом указывают на аминокислотные положения по Kabat.
На фигурах 9A и 9B показано, что антитела против NRR1 и против NRR2 специфично связываются с их рецепторами, как описано в примере B(1). (A) Анализ ELISA для измерения связывания антитела против NRR1 (левая панель) и антитела против NRR2 (правая панель) с очищенными фрагментами белка NRR из каждого из четырех рецепторов Notch человека (h) и мыши (m). Связывание, показанное как A560 на оси y, представлено на графике против титров антитела против NRR1 или антитела против NRR2. (B) Анализ FACS для определения связывания антитела против NRR1 (панели 1-6) или антитела против NRR2 (панели 7-12) с нетрансфицированными клетками K1-CHO (панели 1, 4, 7 и 10), клетками K1-CHO, стабильно трансфицированными N-myc-Notch1 (панели 2, 5, 8 и 11) или клетками K1-CHO, стабильно трансфицированными N-myc-Notch2 (панели 3, 6, 9 и 12). Трансгены N-myc-Notch экспрессировали под контролем индуцибельного промотора tet; нижний ряд, контрольная экспрессия в отсутствии индукции доксицилином (-Dox); верхний ряд, индуцированная экспрессия после добавления доксицилина (+Dox); следует отметить, что линия K1-CHO эндогенно экспрессирует Notch2, который выявляется антителом против NRR2 в присутствии и отсутствии доксицилина (например, панели 7 и 10 для сравнения).
На фигурах 10A-C показано, что антитело против NRR1 и антитело против NRR2 специфично ингибирует передачу сигнала от его рецепторов-мишеней, включая рецепторы, несущие активирующие мутации, как описано в примерах B(2) и B(3). (A) Анализ совместной культуры для определения ингибирования антителом против NRR1 передачи сигнала Notch1. Клетки NIH-3T3, стабильно трансфицированные Jag1, использовали для индукции передачи сигнала Notch в клетках NIH-3T3, стабильно трансфицированных Notch1 (за исключением "-Jag1", где вместо клеток, экспрессирующих Jag1, использовали нетрансфицированные клетки NIH-3T3). Передачу сигнала Notch определяли с использованием репортерного гена Notch (CSL-зависимый промотор, запускающий экспрессию люциферазы светляка), и она выражена относительно экспрессии контрольного гена (конститутивный промотор, запускающий экспрессию люциферазы Renilla), нормализованной к условиям с DAPT (определенным как величина 1). +Jag1, стандартный анализ с совместным культивированием; DMSO, носитель DAPT отдельно; DAPT, 5 мкм в DMSO; α-gD, изотипическое контрольное антитело в концентрации 2000 нг/мл; α-NRR1, антитело против NRR1 в указанных концентрациях; последние четыре анализа включали 80 нг/мл либо α-NRR1, либо α-gD, вместе с очищенными белковыми фрагментами NRR либо Notch1, либо Notch2, как указано (+NRR1 или +NRR2). (B) Анализ совместной культуры для определения ингибирования антителом против NRR2 передачи сигнала Notch2. Клетки NIH-3T3, стабильно трансфицированные Jag1, использовали для индукции передачи сигнала Notch в клетках U87MG, которые экспрессируют высокие уровни Notch2. Анализ проводили, как описано в (A). (C) Анализ совместной культуры для определения ингибирования антителом против NRR1 передачи сигнала Notch1 с рецепторов Notch1 дикого типа или с мутантных рецепторов Notch1. Анализ проводили как в (A) за исключением того, что экспрессирующие рецептор клетки получали путем временной трансфекции плазмид, экспрессирующих указанные рецепторы Notch1. WT, Notch1 дикого типа; ΔPEST, Notch1, лишенный домена PEST; L1594P, Notch1, несущий указанную конститутивно активирующую точечную мутацию; E25, 625 нг/мл изотипического контрольного антитела; α-NRR1, 625 нг/мл антитела против NRR1; DMSO, носитель GSI отдельно; DAPT, 5 мкм в DMSO; CmpE, 1 мкМ соединение E в DMSO.
На фигурах 11A-D показано, что антитело против NRR1 и антитело против NRR2 функционируют в качестве рецептор-специфичных ингибиторов in vivo, как описано в примере B(4). Мышам Balb/c инъецировали четыре раза каждые четверо суток 5 мг/кг изотипического конрольного α-gD, α-NRR1 или α-NRR2, и на 13 сутки, через одни сутки после четвертой дозы, собирали клетки из тимуса или селезенки. (A) Измерение массы тимуса. Масса тимуса (в мг) выражена относительно общей массы тела (в г). Величины соответствуют среднему значению плюс стандартное отклонение для трех мышей на группу. (B) Количество клеток в тимусе. (C) FACS на CD4 и CD8 для идентификации двойных положительных по CD4+/CD8+ T-клеток. Числа соответствуют проценту клеток тимуса в двойных отрицательных, одинарных положительных и двойных положительных популяциях. Относительно контрольного антитела против gD (77,5%), антитело против NRR1 резко снижало процент клеток в популяции CD4+/CD8+ (5,89%) в то время как антитело против NRR2 (80%) не имело значительного эффекта. (D) FACS на CD21 и CD23 для идентификации B-клеток маргинальной зоны. Количества соответствуют средним процентам ± стандартное отклонение (для трех животных) клеток в области ворот MZB, которая заключена в рамку; также указаны значения p; представлены типичные точечные графики для одного из трех животных в каждой группе. Относительно контрольного антитела против gD (6,61%), антитело против NRR2 практически устраняло клетки MZB (0,97%), в то время как антитело против NRR1 (6%) не имело значительного эффекта.
На фигурах 12A-D структура размером 2,2 Ǻ совместного кристалла Fab против NRR1/NRR1 указывает на то, что антитело против NRR1 одновременно контактирует с доменами LNR-A, LNR-B и HD-C, как описано в примере B(5). (A) В таблице обобщенно представлено связывание α-NRR1 или α-NRR2 с фрагментами химерного белка Notch1-NRR или Notch2-NRR. Указанные фрагменты белка NRR (темно-синий, последовательности Notch1; голубой, последовательности Notch2) экспрессировались в качестве секретируемых белков, слитых с щелочной фосфатазой, для обеспечения быстрого определения связывания антитела в анализе на планшетах. После использования активности щелочной фосфатазы для нормализации по экспрессии и секреции NRR, культуральную среду, содержащую указанные химерные белки NRR, добавляли в 96-луночный планшет, на который было нанесено α-NRR1, α-NRR2 или изотипическое контрольное антитело (использованное для оценки фонового связывания, не показано). Связывание антитела оценивали путем измерения активности щелочной фосфатазы, которая оставалась связанной с планшетом. Y, сильное связывание; N, связывание не выявлено; W, выявлено слабое связывание. (B) Структура NRR1 человека. NRR1 представлен в качестве C-альфа изображения. Три иона кальция в мотивах LNR показаны в качестве сфер. Положение участка расщепления S2 указано стрелкой. (C) Наложение NRR1 на NRR2 (цепь A из pdb, код 2OO4), исходя из структурно консервативных атомов. NRR1 (закрашенный) и NRR2 (белый) показаны в качестве C-альфа изображений. (D) Вид по типу "открытая книга" для поверхности контакта между NRR1 (слева) и α-NRR1 Fab (справа, граница между тяжелой и легкой цепями показана черной пунктирной линией). Указана степень, с которой доступная для растворителя площадь поверхности погружается при образовании комплекса. Указаны остатки, которые погружены по меньшей мере на 50%, и идентичные остатки NRR в Notch1 и Notch2 обозначены черным шрифтом (см. также фигуру 18).
На фигурах 13A-C показано, что антитело против NRR1 вызывает гиперветвление эндотелиальных клеток, как описано в примере B(6). (A) Анализ ветвления эндотелиальных клеток in vitro. HUVEC наносили на гранулы Cytodex и совместно культивировали с фибробластами кожи. Культуры либо обрабатывали имитирующей обработкой (контроль), либо их обрабатывали 1 мкМ DBZ, 5 мкг/мл α-NRR1 или 5 мкг/мл α-Dll4. Планка масштаба = 100 мкм. (B) Измерение длины ветвлений в культурах из (A). (C) Анализ сетчатки новорожденных мышей в отношении ветвления эндотелиальных клеток и ангиогенеза. Новорожденным мышам инъецировали на P1 и P3 указанные антитела, и на P5 сетчатки приготавливали для визуализирующей перфузии изолектина или маркера пролиферации Ki67. Панели I и II, планки масштаба = 1 мм. Панели III-VI представляют собой увеличения частей панелей I и II, планка масштаба = 0,2 мм.
На фигурах 14A-E показано, что селективное антитело, блокирующее передачу сигнала Notch1, нарушает ангиогенез опухоли и ингибирует рост опухоли, как описано в примере B(7). На графиках для трех моделей с ксенотрансплантатами показан объем опухоли (среднее значение +/- SEM) против времени после введения указанных антител: антитела против агглютинина амброзии (отрицательный контроль), α-VEGF или α-NRR1. Значения P показаны для сравнения антитела против агглютинина амброзии против α-NRR1. (A) Модель Calu6. (B) Модель HM7. (C) Модель HM7 с титрованием дозы α-NRR1. (D) Окрашивание эндотелиальных клеток в репрезентативных срезах опухолей из модели Calu6, показанной на (A). Антитела, использованные в моделях с ксенотрансплантатами, показаны сверху. DAPI и α-CD31 использовали для окрашивания ДНК и эндотелиальных клеток, соответственно. Нижний ряд панелей демонстрирует объединенные изображения. Планка масштаба = 50 мкм. (E) Количественное определение окрашивания на CD31, представленного в (D). С использованием изображения J для количественного определения окрашивания на CD31 и с помощью DAPI в изображениях, сходных с изображениями, показанными в (D), относительное окрашивание на CD31 (окрашивание на CD31, нормализованное к окрашиванию DAPI) наносили на график для каждого из трех введений антител относительно контрольного антитела против агглютинина амброзии, которое было принято в качестве значения 1; данные соответствуют среднему значению +/- стандартное отклонение на протяжении 8 изображенных полей.
На фигурах 15A-C показано, что селективное антитело, блокирующее Notch1, является достаточным для изменения участи клеток в тонком кишечнике, как описано в примере B(8). (A) Общее изменение массы тела (среднее значение +/- стандартное отклонение) против времени. Мышам проводили введение, как описано для фигуры 11, контрольного изотипического антитела против gD, LTβR-Fc, антитела против NRR1 или антитела против NRR2. Стрелками указаны сутки дозирования. (B) Иммуногистохимические анализы тонкого кишечника. Мышам вводили указанные концентрации контрольного изотипического антитела против gD, DBZ или антитела против NRR1 на 0, 2 и 6 сутки и на 7 сутки тонкие кишечники подготавливали для иммуногистохимического анализа. В каждом ряду, как указано, представлено окрашивание гематоксилином и эозином (H & E), альциановым синим на муцин и для маркирования секреторных бокаловидных клеток, окрашивание на лизоцим для маркирования клеток Панета, окрашивание на Ki-67 для определения пролиферации и на Hes1 в качестве нижерасположенной мишени Notch. См. фигуры 19 и 20 для анализов толстого кишечника на 7 сутки и тонкого кишечника на 2 сутки, соответственно. Планка масштаба = 40 мкм. (C) Сравнение специфического ингибирования Notch1 против специфического ингибирования Notch2 в отношении дифференцировки клеток тонкого кишечника. Тонкие кишечники из исследования, описанного в (A), приготавливали для окрашивания альциановым синим и окрашивания на Ki-67 через одни сутки после конечного дозирования α-gD, α-NRR1 или α-NRR2, как указано. Планка масштаба = 50 мкм.
На фигуре 16 показано, что антитело против NRR1 является сильным ингибитором передачи сигнала Notch1, индуцируемой множеством лигандов, как описано в примере B(2). Анализы передачи сигнала Notch1 в совместной культуре проводили, как описано на фигуре 10A, за исключением того, что лиганд-экспрессирующие клетки экспрессировали Jag1, Jag2 или Dll1, как указано.
На фигуре 17 показаны химерные белковые конструкции NRR Notch1/NRR Notch2 с низким выходом, как описано в примере B(5). Таблица дополняет фигуру 12A, и в ней приведены химерные белки NRR, которые приводили к низкой активности щелочной фосфатазы или к ее отсутствию, указывая на то, что химеры не правильно сворачивались или были иным образом нестабильными. Для фрагментов, которые приводили к слабой, но поддающейся детекции активности щелочной фосфатазы, обобщенно представлено связывание α-NRR1 или α-NRR2. ---, нет экспрессии, и связывание не поддается тестированию; N, отсутствует экспрессия или связывание; W, слабая экспрессия или связывание.
На фигуре 18 показана консервативность остатков NRR1, с которыми контактировало антитело против NRR1, как описано в примере B(1) и B(5). Представлено выравнивание аминокислотных последовательностей NRR-домена Notch1 человека (SEQ ID NO: 26), NRR-домена Notch1 мыши (SEQ ID NO: 27), NRR-домена Notch2 человека (SEQ ID NO: 28) и NRR-домена Notch2 мыши (SEQ ID NO: 29), и границы субдомнов представлены справа. NRR Notch1 человека приведен в качестве эталонной последовательности, и остатки в других последовательностях NRR, которые идентичны остаткам в Notch1 человека, указаны полужирным шрифтом. Остатки, которые по меньшей мере на 25% погружены в структуру Fab против NRR1/Notch1-NRR (фигура 12), подчеркнуты, причем сплошные линии против пунктирных линий отражают повышение степени, с которой остатки погружены. Из 21 аминокислоты, которые по меньшей мере на 25% погружены в последовательности Notch1 человека, все 21 являются идентичными в последовательности Notch1 мыши, но только шесть из них идентичны в последовательностях Notch2 человека и мыши (плюс седьмое консервативное отличие, T вместо S, в S1712 Notch1 человека); среди этих идентичных и "погруженных" остатков в последовательностях Notch2, ни один из них не относится к классу погруженных >75%. Это сравнение последовательностей согласуется с (a) сильным связыванием антитела против NRR1 (приблизительно равные аффинности) с Notch1 как человека, так и мыши и (b) отсутствием связывания антитела против NRR1 с Notch2 человека и мыши. Более того, все погруженные остатки находятся в пределах LNR-A, LNR-B и HD-C, что согласуется с экспериментом по обмену доменов, представленным на фигурах 11A и 17.
На фигуре 19 показано, что селективное антитело, блокирующее Notch1, является достаточным для изменения участи клеток в толстом кишечнике, как описано в примере B(8). Гистопатологический анализ образцов толстого кишечника, взятых на 7 сутки эксперимента, описан на фигуре 15B.
На фигуре 20 показано, что изменения участи клеток кишечника развиваются на 2 сутки после блокирования Notch1, как описано в примере B(8). Гистопатологический анализ образцов тонкого кишечника, взятых на 2 сутки эксперимента, описан на фигуре 15B, на которой показаны образцы, взятые на 7 сутки.
На фигуре 21 показано, что антитело против NRR1 не блокирует индуцируемую Notch2 передачу сигнала in vitro, как описано в примере B(2). Анализ совместной культуры в клетках U87MG, как описано на фигуре 10B. Хотя антитело против NRR1 активно ингибирует передачу сигнала Notch1 in vitro (фигура 10A) и in vivo (фигуры 11A-C), оно не влияет на передачу сигнала Notch2, даже при использовании в концентрации 1000 нг/мл, что более чем в 100 раз превышает IC50. Этот результат согласуется со специфическим связыванием антитела против NRR1 с Notch1 (фигуры 9 и 12), а также с отсутствием ингибирования Notch2 антителом против NRR1 in vivo (фигура 11D).
На фигуре 22 показаны возможные синергические эффекты антитела против NRR1 и антитела против NRR2 на дифференцировку клеток кишечника, как описано в примере B8.
На фигуре 23 показано, что дексаметазон приводит по меньшей мере к частичному восстанавливаю фенотипа кишечника от фенотипа, вызванного антителом против NRR1, как описано в примере B(9).
На фигурах 24A и B показано, что селективное блокирование либо Notch1, либо Notch2 минимизирует или предотвращает метаплазию бокаловидных клеток, ассоциированную с тотальным ингибированием Notch, в то время как блокирование как Notch1, так и Notch2, вызывает тяжелую метаплазию бокаловидных клеток. (A) Как описано в примере B8, мышам вводили дозы 5 мг/кг антитела против NRR1, антитела против NRR2, или оба, или отрицательное контрольное антитело против gD на сутки, отмеченные стрелками; показано общее изменение массы тела (среднее значение +/- стандартное отклонение) против времени. (B) Иммуногистохимические анализы тонкого кишечника от мышей, которым проводили введение согласно (A), с использованием альцианового синего, окрашивающего муцин, для мечения секреторных бокаловидных клеток.
На фигурах 25A и 25B показано, что антитело против NRR2 (обозначаемое как "антитело против N2") ингибирует рост клеточных линий меланомы человека SK23 и LOX-IMVI in vitro, как описано в примере B(10).
На фигуре 26 показан эффект антитела против NRR2 (обозначаемого как "антитело против Notch2") на пять клеточных линий диффузных крупноклеточных B-клеточных лимфом (DLBCL) (приведенных справа). Как описано в примере B(11), рост одной из клеточных линий, "DB" строго ингибировался обработкой антителом против NRR2.
На фигуре 27 показан эффект антитела против NRR2 (называемого "антителом против Notch2") на рост клеточной линии DB DLBCL с течением времени, как описано в примере B(11).
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Изобретение относится к выделенным антителам, которые связываются с Notch, и к способам их применения, например, для диагностики или лечения заболеваний, ассоциированных с экспрессией или активностью Notch.
I. ОБЩИЕ СПОСОБЫ
Способы и процедуры, описанные или цитированные в настоящем документе, обычно хорошо понятны и широко применяются с использованием общепринятых технологий специалистами в данной области, например, таких как широко используемые технологии, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Current Protocols in Molecular Biology (F. M. Ausubel, et al. eds., (2003)); серия Methods in Enzymology (Academic Press, Inc.): PCR 2: A Practical Approach (M. J. MacPherson, B. D. Hames и G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, A Laboratory Manual, and Animal Cell Culture (R. I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M. J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J. P. Mather and P. E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths, and D. G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D. M. Weir and C. C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al, eds., 1994); Current Protocols in Immunology (J. E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C. A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (V. T. DeVita et al., eds., J. B. Lippincott Company, 1993).
II. ОПРЕДЕЛЕНИЯ
Для интерпретации описания применяются следующие определения, и, когда это приемлемо, термины, используемые в единственном числе, также включают множественное число и наоборот. В случае, когда любое указанное ниже определение противоречит документу, включенному в настоящий документ в качестве ссылки, следует руководствоваться определением, указанным ниже.
Термин "антитело" используют в настоящем документе в наиболее широком значении и, конкретно, он охватывает моноклональные антитела, поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), образованные из по меньшей мере двух целых антител, и фрагменты антител, при условии, что они проявляют требуемую биологическую активность.
"Выделенное" антитело представляет собой антитело, которое идентифицировано и отделено и/или извлечено из компонента его естественных окружающих условий. Загрязняющие компоненты его естественных окружающих условий представляют собой вещества, которые препятствуют исследовательскому, диагностическому или терапевтическому применению антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело является очищенным (1) до более чем 95% по массе антитела, как определяют способом Лоури, и наиболее предпочтительно до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности при SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием кумасси синего или, предпочтительно, окрашивания серебром. Выделенное антитело включает антитело в рекомбинантных клетках in situ, поскольку в этом случае отсутствует по меньшей мере один компонент условий естественного окружения антитела. Однако, как правило, выделенное антитело получают посредством по меньшей мере одной стадии очистки.
"Нативные антитела" обычно представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как количество дисульфидных связей варьирует среди тяжелых цепей различных изотипов иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет межцепочечные дисульфидные мостики, находящиеся на правильных расстояниях. Каждая тяжелая цепь имеет на ее одном конце вариабельный домен (VH), за которым следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на ее другом конце; причем константный домен легкой цепи находится на одном уровне с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи находится на одном уровне с вариабельным доменом тяжелой цепи. Полагают, что поверхность контакта между вариабельными доменами легкой цепи и тяжелой цепи формируют конкретные аминокислотные остатки.
Термин "антитело против NRR Notch1" или "антитело, которое связывается с NRR Notch1" относится к антителу, которое способно связывать NRR Notch1 с достаточной аффинностью, чтобы антитело было пригодным в качестве диагностического и/или лекарственного средства при нацеливании на Notch1. Предпочтительно, степень связывания антитела против NRR Notch1 с неродственным не являющимся Notch белком составляет менее чем приблизительно 10% от связывания антитела с NRR Notch1 при определении, например, радиоиммунным анализом (RIA). В определенных вариантах осуществления антитело, которое связывается с NRR Notch1, имеет константу диссоциации (Kd)≤1 мкм, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ. В определенных вариантах осуществления антитело против NRR Notch1 связывается с эпитопом Notch, который является консервативным среди Notch из различных видов, например, грызунов (мышей, крыс) и приматов.
Термин "антитело против NRR Notch2" или "антитело, которое связывается с NRR Notch2" относится к антителу, которое способно связывать NRR Notch2 с достаточной аффинностью, чтобы антитело было пригодным в качестве диагностического и/или лекарственного средства при нацеливании на Notch2. Предпочтительно, степень связывания антитела против NRR Notch2 с неродственным не являющимся Notch белком составляет менее чем приблизительно 10% от связывания антитела с NRR Notch2 при определении, например, радиоиммунным анализом (RIA). В определенных вариантах осуществления антитело, которое связывается с NRR Notch2, имеет константу диссоциации (Kd) ≤1 мкм, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ. В определенных вариантах осуществления антитело против NRR Notch2 связывается с эпитопом Notch, который является консервативным среди Notch из различных видов, например, грызунов (мышей, крыс) и приматов.
"Вариабельная область" или "вариабельный домен" антитела относится к N-концевым доменам тяжелой и легкой цепи антитела. Вариабельный домен тяжелой цепи может быть обозначен как "VH". Вариабельный домен легкой цепи может быть обозначен как "VL". Эти домены, как правило, являются наиболее вариабельными частями антитела и содержат антигенсвязывающие центры.
Термин "вариабельный" относится к тому факту, что последовательности определенных сегментов вариабельных доменов значительно отличаются среди антител. Однако вариабельность не является равномерно распределенной на протяжении вариабельных доменов антител. Она сконцентрирована в трех сегментах, называемых гипервариабельными областями (HVR) в вариабельных доменах как легких цепей, так и тяжелых цепей. Наиболее высоко консервативные участки вариабельных доменов называются каркасными областями (FR). Все вариабельные домены природных тяжелых и легких цепей содержат четыре области FR, главным образом, приминающих конфигурацию бета-слоев, соединенных тремя HVR, которые формируют петли, соединяющие структуры бета-слоев, и в некоторых случаях формирующие их часть. HVR в каждой цепи расположены вместе в непосредственной близости от FR и, совместно с HVR другой цепи, участвуют в формировании антигенсвязывающего центра антител (см. Kabat et al., Sequences of Protein of Immunological Interest, 5th Ed. National Institutes of Health, Bethesda, MD. (1991)). Константные домены не вовлечены непосредственно в связывание антитела с антигеном, но они проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности.
"Легкие цепи" антител (иммуноглобулинов) любого вида позвоночных могут быть отнесены к одному из двух типов, называемых каппа (κ) и лямбда (λ), отчетливо различающихся на основе аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей, антитела (иммуноглобулины) могут быть отнесены к различным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно подразделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны и описаны, главным образом, например, в Abbas et al. Cellular and Mol. Immunology, 4th ed. (W.B. Saunders, Co., 2000). Антитело может быть частью более крупной слитой молекулы, образованной путем ковалентной или нековалентной ассоциации антитела с одним или несколькими другими белками или пептидами.
Термины "полноразмерное антитело", "интактное антитело" и "целое антитело" используют в настоящем документе взаимозаменяемо для обозначения антитела в его по существу целой форме, а не фрагментов антитела, определенных ниже. Эти термины, в частности, относятся к антителу с тяжелыми цепями, которые содержат Fc-область.
"Простое антитело" для целей настоящего описания представляет собой антитело, которое не конъюгировано с цитотоксической группой или радиоактивной меткой.
"Фрагменты антитела" содержат часть целого антитела, предпочтительно его антигенсвязывающую область. Примеры фрагментов антител включают Fab-, Fab'-, F(ab')2- и Fv-фрагменты, антитела-димеры, линейнные антитела, одноцепочечные молекулы антител, и полиспецифические антитела, образованные фрагментами антител.
Расщепление антител папаином приводит к образованию двух идентичных антигенсвязывающих фрагментов, называемых "Fab"-фрагментами, каждый из которых содержит один антигенсвязывающий центр, и остаточного "Fc"-фрагмента, название которого отражает его способность легко кристаллизоваться. Обработка пепсином приводит к одному крупному F(ab')2-фрагменту, который имеет два антигенсвязывающих центра и все еще способен перекрестно связывать антиген.
"Fv" представляет собой минимальный фрагмент антитела, который содержит полный антигенсвязывающий центр. В одном варианте осуществления двухцепочечный тип Fv состоит из димера, состоящего из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочной нековалентной связью. В одноцепочечных типах Fv (scFv), один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи могут быть ковалентно связаны подвижным пептидным линкером, так чтобы легкая и тяжелая цепи могли ассоциировать в "димерные" структурные аналоги, аналогичные двухцепочечным типам Fv. Именно в этой конфигурации три HVR каждого вариабельного домена взаимодействуют, определяя антигенсвязывающий центр на поверхности димера VL-VH. В совокупности шесть HVR придают антигенсвязывающую специфичность антителу. Однако даже один вариабельный домен (или полвина Fv, содержащая только три HVR, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем целый участок связывания.
Fab-фрагмент состоит из вариабельных доменов тяжелой и легкой цепей, а также он содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов наличием дополнительных нескольких остатков на C-конце домена CH1, включая один или несколько цистеинов из шарнирной области антитела. Fab'-SH представляет собой обозначение для Fab', в котором остаток(ки) цистеина константных доменов обладает свободной тиольной группой. F(ab')2-фрагменты антитела исходно были получены в качестве пар Fab'-фрагментов, которые обладают шарнирными цистеинами между ними. Также известно другое химическое связывание фрагментов антител.
"Одноцепочечные Fv" или "scFv", представляют собой фрагменты антител, которые содержат VH- и VL-домены антитела, где эти домены присутствуют в единой полипептидной цепи. Как правило, полипептид sFv также необязательно содержит полипептидный линкер между VH- и VL-доменами, который обеспечивает формирование в sFv структуры, требуемой для связывания антигена. Для обзора sFv см. Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York, 1994), pp. 269-315.
Термин "антитела-димеры" относится к фрагментам антитела с двумя антигенсвязывающими участками, причем эти фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL). С использованием линкера, который является слишком коротким для возможности спаривания двух доменов одной цепи, домены вынуждают образовывать пары с комплементарными доменами другой цепи с образованием двух антигенсвязывающих центров. Антитела-димеры могут быть двухвалентными или биспецифическими. Антитела-димеры описаны более подробно, например, в EP 404097; WO93/1161; Hudson et al., Nat. Med. 9:129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Антитела-тримеры и антитела-тетрамеры также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
В настоящем документе, термин "моноклональное антитело" относится к антителу из совокупности по существу однородных антител, т.е. отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных мутаций, например, природных мутаций, которые могут быть представлены в небольших количествах. Таким образом, определение "моноклональный" указывает на тот признак антитела, что оно не является смесью различных антител. В определенных вариантах осуществления такое моноклональное антитело, как правило, включает антитело, содержащее полипептидную последовательность, которая связывает мишень, где связывающая мишень полипептидная последовательность получена посредством процесса, который включает селекцию единичной связывающей мишень полипептидной последовательности из множества полипептидных последовательностей. Например, процесс селекции может представлять собой селекцию единичного клона из множества клонов, таких как пул гибридомных клонов, фаговых клонов или клонов рекомбинантных ДНК. Следует понимать, что отобранную связывающую мишень последовательность можно дополнительно изменять, например, для повышения аффинности к мишени, для гуманизации связывающей мишень последовательности, для повышения ее продукции в клеточной культуре, для снижения ее иммуногенности in vivo, для создания полиспецифичного антитела, и т. д., и что антитело, содержащее измененную связывающую мишень последовательность, также является моноклональным антителом по настоящему изобретению. В противоположность препаратам поликлональных антител, которые как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной детерминанты на антигене. В дополнение к их специфичности, препараты моноклонального антитела является преимущественными в том, что они, как правило, не содержат примесей других иммуноглобулинов.
Определение "моноклональный" указывает на тот признак антитела, что его получают из по существу гомогенной совокупности антител, и не подразумевает того, что антитело должно быть получено каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, можно получать множеством способов, включая, например, способ гибридом, (например, Kohler and Milstein, Nature, 256:495-97 (1975); Hongo et al, Hibridoma, 14 (3): 253-260 (1995); Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in Monoclonal Antibodies and T-Cell Hybridomas 563-681, (Elsevier, N.Y., 1981)), способы рекомбинантных ДНК (см., например, патент США No. 4816567), технологии фагового дисплея (см., например, Clackson et al., Nature, 352:624-628 (1991); Marks et al., J. Mol. Biol., 222:581-597 (1991); Sidhu et al., J. Mol. Biol. 338(2):299-310 (2004); Lee et al., J. Mol. Biol. 340(5):1073-1093 (2004); Fellouse, Proc. Nat. Acad. Sci. USA 101(34):12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2):119-132 (2004), и технологии продуцирования антитела человека или антител, подобных антителам человека, у животных, которые имеют части локусов или генов иммуноглобулинов человека, кодирующие последовательности иммуноглобулинов, или все эти локусы или гены (см., например, WO 1998/24893; WO 1996/34096; WO 1996/33735; WO 1991/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immuno., 7:33 (1993); патенты США No. 5545807; 5545806; 5569825; 5625126; 5633425; и 5661016; Marks et al., Bio/Technology, 10: 779-783 (1992); Lonberg et al., Nature, 368:856-859 (1994); Morrison, Nature, 368:812-813 (1994); Fishwild et al., Nature Biotechnology, 14:845-851 (1996); Neuberger, Nature Biotechnology, 14:826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol., 13:65-93 (1995).
Моноклональные антитела в настоящем документе конкретно включают "химерные" антитела, в которых участок тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) идентична или гомологична соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (см, например, патент США No 4816567; и Morrison et al., Proc. Natl. Acad. Sci USA, 81:6851-6855 (1984)). Химерные антитела включают PRIMATIZED®-антитела, где антигенсвязывающая область антитела происходит из антитела, продуцируемого, например, путем иммунизации макак представляющим интерес антигеном.
"Гуманизированные" формы не являющихся человеческими антител (например, грызунов) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. В одном осуществлении, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области видов, не относящихся к человеку, (донорное антитело), таких как мышь, крыса, кролик или не относящиеся к человеку приматы, которые обладают требуемой специфичностью, аффинностью и емкостью. В некоторых случаях, остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками, не являющиеся человеческими. Более того, гуманизированные антитела могут содержать остатки, которые отсутствуют в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения параметров антитела. Как правило, гуманизированное антитело содержит по существу все по меньшей мере из одного, и, как правило, из двух, вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют гипервариабельным петлям иммуноглобулина, не являющегося человеческим, и все или по существу все FR-области представляют собой FR-области из последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно содержит по меньшей мере участок константной области (Fc) иммуноглобулина, как правило, константного домена иммуноглобулина человека. Для более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). Также см. следующие обзорные статьи и ссылки, цитированные в них: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994) и патенты США No. 6982321 и 7087409.
"Антитело человека" представляет собой антитело, которое содержит аминокислотную последовательность, соответствующую последовательности антитела, продуцируемой у человека и/или полученной с использованием любых способов получения антител человека, как описано в настоящем документе. Это определение антитела человека, в частности, не включает гуманизированное антитело, содержащее антигенсвязывающие остатки не человека. Антитела человека можно получать с использованием различных способов, известных в данной области, включая библиотеки фагового дисплея. Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol, 222:581 (1991). Также для получения моноклональных антител человека доступны способы, описанные в Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol, 147(1): 86-95 (1991). Также см. van Dijk and van de Winkel, Curr. Opin. Pharmacol, 5: 368-74 (2001). Антитела человека можно получить путем введения антигена трансгенному животному, модифицированному для продукции таких антител в ответ на нагрузку антигеном, но имеющему поврежденные эндогенные локусы, например, иммунизированным ксеномышам (см., например, патенты США No. 6075181 и 6150584 в отношении технологии XENOMOUSETM). Также см, например, Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006) в отношении антител человека, полученных технологией B-клеточных гибридом человека.
Термины "гипервариабельная область", "HVR" или "HV" в настоящем документе относятся к участкам вариабельного домена антитела, которые являются гипервариабельными по последовательности и/или образуют структурно определенные петли. Обычно антитела включают шесть гипервариабельных областей: три в VH (H1, H2, H3), и три в VL (L1, L2, L3). В нативных антителах H3 и L3 проявляют наибольшее разнообразие среди шести HVR, и, в частности, полагают, что H3 играет уникальную роль в обеспечении высокой специфичности антител. См., например, Xu et al., Immunity 13:37-45 (2000); Johnson and Wu, Methods in Molecular Biology 248:1-25 (Lo, ed., Human Press, Totowa, NJ, 2003). Действительно, природные антитела верблюжьих, состоящие только из тяжелых цепей, являются функциональными и стабильными в отсутствии легких цепей. См., например, Hamers-Casterman et al., Nature 363:446-448 (1993); Sheriff et al., Nature Struct. Biol. 3:733-736 (1996).
Используют несколько способов обозначения гипервариабельных областей, и они включены в рассматриваемое описание. Определяющие комплементарность области (CDR) по Kabat основаны на вариабельности последовательностей и используются наиболее часто (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). Chothia вместо этого рассматривает расположение структурных петель (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). HVR AbM представляют собой компромисс между HVR по Kabat и структурными петлями по Chothia, и они используются в программном обеспечении моделирования антител Oxford Molecular's AbM. "Контактные" HVR основаны на анализе доступных комплексных кристаллических структур. Остатки из каждой из указанных HVR указаны ниже.
HVR могут включать следующие "удлиненные HVR": 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 (L3) в VL и 26-35B (H1), 50-65, 47-65 или 49-65 (H2) и 93-102, 94-102 или 95-102 (H3) в VH. Остатки вариабельных доменов пронумерованы согласно Kabat et al., выше, для каждого из этих определений.
Остатки "каркасной области" или "FR" представляют собой остатки вариабельного домена, отличные от остатков HVR, как определено в настоящем документе.
Термины "нумерация остатка вариабельного домена по Kabat" или "нумерация аминокислотного положения по Kabat", и их варианты, относятся к системе нумерации, которую используют для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи в совокупности антител, представленной в Kabat et al.,выше. С использованием этой системы нумерации, истинная линейная аминокислотная последовательность может содержать меньшее количество или дополнительное количество аминокислот, что соответствует укорочению FR или HVR вариабельного домена или встраиванию в них. Например, вариабельный домен тяжелой цепи может включать одну вставку представляющей интерес аминокислоты (остаток 52a по Kabat) после остатка 52 в H2 и встроенные остатки (например, остатки 82a, 82b, и 82c, и т.д. по Kabat) после остатка 82 FR тяжелой цепи. Нумерацию по Kabat для остатков можно определить для конкретного антитела путем выравнивания по участкам гомологии последовательности антитела со "стандартной" последовательностью, пронумерованной по Kabat.
Систему нумерации Kabat обычно используют при указании на остаток в вариабельном домене (приблизительно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). "Систему нумерации EU" или "индекс EU" обычно используют при указании на остаток в константной области тяжелой цепи иммуноглобулина (например, индекс EU, указанный в Kabat et al., выше). "Индекс EU по Kabat" относится к нумерации остатков антитела IgG1 человека EU. Если в настоящем документе нет иных указаний, указания на номера остатков в вариабельных доменах антител означают нумерацию остатков в соответствии с системой нумерации Kabat. Если в настоящем документе нет иных указаний, указания на номера остатков в константном домене антител означают нумерацию остатков в соответствии с системой нумерации EU. (например, см. публикацию патентной заявки No. US 2008/0181888 A1, фигуры для нумерации EU).
Антитело, полученное "созреванием аффинности", представляет собой антитело с одним или несколькими изменениями в одной один или нескольких его HVR, которые приводят к повышению аффинности антитела к антигену, по сравнению с исходным антителом, которое не имеет этого изменения(ий). Предпочтительные антитела, полученные "созреванием аффинности", имеют наномолярную или даже пикомолярную аффинность к антигену-мишени. Антитела, полученные созреванием аффинности, получают способами, известными в данной области. В Marks et al., Bio./Technology 10:779-783 (1992) описано созревание аффинности способом "перетасовки" VH- и VL-доменов. Случайный мутагенез HVR и/или каркасных остатков описан: Barbas et al., Proc Nat. Acad. Sci. USA 91:3809-3813 (1994); Schier et al., Gene 169:147-155 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995); и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
"Блокирующее антитело" или "антитело-антагонист" представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, который оно связывает. Предпочтительные блокирующие антитела или антитела-антагонисты частично или полностью ингибируют биологическую активность антигена.
"Антитело-агонист" в настоящем документе представляет собой антитело, которое частично или полностью имитирует по меньшей мере один из видов функциональной активности представляющего интерес полипептида.
"Ингибирующие рост" представляют собой антитела, которые препятствуют пролиферации или снижают пролиферацию клетки, экспрессирующей антиген, с которым антитело связывается. Например, антитело может препятствовать пролиферации злокачественных клеток in vitro и/или in vivo, снижать ее.
Антитела, которые "индуцируют апоптоз", представляют собой антитела, которые индуцируют запрограммированную гибель клеток при определении стандартными анализами апоптоза, такими как связывание аннексина V, фрагментация ДНК, сморщивание клеток, расширение эндоплазматической сети, фрагментация клетки и/или образование мембранных везикул (называемых апоптотическими тельцами).
"Эффекторные функции" антитела относятся к таким видам биологической активности, которые являются свойственными Fc-области (Fc-области с нативной последовательностью или варианта Fc-области по аминокислотной последовательности) антитела, и варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антителозависимую клеточно-опосредуемую цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора; BCR) и активацию B-клеток.
Термин "Fc-область" используют в настоящем документе для определения C-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативной последовательностью и варианты Fc-областей. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи IgG человека обычно определяют как отрезок от аминокислотного остатка в положении Cys226, или от Pro230, до его C-конца. C-концевой лизин (остаток 447 в системе нумерации EU) Fc-области может быть удален, например, в процессе получения или очистки антитела, или при рекомбинантном конструировании нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, композиция целых антител может включать совокупности антител, во всех из которых удален остаток K447, совокупности антител, в которых не удален остаток K447, и совокупности антител, содержащие смесь антител, содержащих и не содержащих остаток K447.
"Функциональная Fc-область" обладает "эффекторной функцией" Fc-области с нативной последовательностью. Примеры "эффекторных функций" включают связывание C1q; CDC; связывание Fc-рецептора; ADCC; фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, рецептора В-клеток; BCR) и т.д. Такие эффекторные функции обычно требуют, чтобы Fc-область была объединена со связывающим доменом (например, вариабельным доменом антитела) и их можно оценить, используя различные анализы, как описано, например, в разделе "Определения", настоящего документа.
"Fc-область с нативной последовательностью" включает аминокислотную последовательность, идентичную нативной аминокислотной последовательности Fc-области. Fc-области человека с нативной последовательностью включают Fc-область IgG1 человека с нативной последовательностью (аллотипы не-A и A); Fc-область IgG2 человека с нативной последовательностью; Fc-область IgG3 человека с нативной последовательностью; и Fc-область IgG4 человека с нативной последовательностью, а также их природные варианты.
"Вариант Fc-области" включает аминокислотную последовательность, которая отличается от Fc-области с нативной последовательностью тем, что содержит по меньшей мере одну модификацию аминокислоты, предпочтительно одну или несколько аминокислотную замену(замен). Предпочтительно, вариант Fc-области включает по меньшей мере одну аминокислотную замену по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например от приблизительно одной до приблизительно десяти аминокислотных замен, и предпочтительно от приблизительно одной до приблизительно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. Вариант Fc-области в настоящем документе предпочтительно обладает по меньшей мере приблизительно 80% гомологией с Fc-областью с нативной последовательностью и/или Fc-областью исходного полипептида, и наиболее предпочтительно он обладает гомологией с ними по меньшей мере приблизительно 90%, более предпочтительно он обладает гомологией с ними по меньшей мере приблизительно 95%.
Термины "Fc-рецептор" или "FcR" обозначают рецептор, который связывается с Fc-областью антитела. В некоторых вариантах осуществления FcR представляет собой FcR человека с нативной последовательностью. В некоторых вариантах осуществления FcR представляет собой FcR, который связывает антитело IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно-сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые обладают сходными аминокислотными последовательностями, отличающимися, главным образом, своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA в своем цитоплазматическом домене содержит иммунорецепторный тирозин-связывающий активирующий мотив (ITAM). Ингибирующий рецептор FcγRIIB в своем цитоплазматическом домене содержит иммунорецепторный тирозин-связывающий ингибирующий мотив (ITIM), (см. обзор M. в Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). FcR описаны в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). К термину "FcR" в настоящем описании относятся другие FcR, в том числе FcR, которые будут выявлены в будущем.
Термин "Fc рецептор" или "FcR" также включает неонатальный рецептор, FcRn, который ответствен за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регуляцию гомеостаза иммуноглобулинов. Способы определения связывания с FcRn известны (см., например, Ghetie and Ward., Immunol. Today 18(12):592-598 (1997); Ghetie et al, Nature Biotechnology, 15(7):637-640 (1997); Hinton et al, J. Biol. Chem. 279(8):6213-6216 (2004); WO 2004/92219 (Hinton et al).
Связывание с FcRn человека in vivo и период полужизни в сыворотке для полипептидов с высокой аффинностью связывания FcRn человека можно анализировать, например, у трансгенных мышей или в трансфицированных клеточных линиях человека, экспрессирующих FcRn человека, или у приматов, которым вводят полипептиды с вариантом Fc-области. В WO 2000/42072 (Presta) описаны варианты антител с повышенным или сниженным связыванием с FcR. См. также, например, Shields et al. J. Biol. Chem. 9(2):6591-6604 (2001).
"Эффекторные клетки человека" представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. В некоторых вариантах осуществления клетки экспрессируют по меньшей мере FcγRIII и выполняют эффекторную функцию ADCC. Примеры лейкоцитов человека, которые осуществляют ADCC, включают периферические мононуклеарные клетки крови (PBMC), естественные киллерные (NK) клетки, моноциты, цитотоксические T-клетки и нейтрофилы; при этом предпочтительными являются PBMC и NK-клетки. Эффекторные клетки можно выделять из их природных источников, например, из крови.
"Антителозависимая клеточно-опосредуемая цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретируемые Ig, связанные с Fc-рецепторами (FcR), находящимися на определенных цитотоксических клетках (например, естественных киллерных (NK) клетках, нейтрофилах и макрофагах), вызывают специфичное для этих цитотоксических эффекторных клеток связывание с несущей антиген клеткой-мишенью, а затем уничтожают клетки-мишени посредством цитотоксинов. Основные клетки для осуществления ADCC, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках представлена в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991). Для оценки активности представляющей интерес молекулы в отношении ADCC можно проводить анализ ADCC in vitro, такой как анализ, описанный в патенте США No. 5500362 или 5821337 или в патенте США No.6737056 (Presta). Пригодные для таких анализов эффекторные клетки включают PBMC и NK-клетки. Альтернативно, или дополнительно, активность представляющей интерес молекулы в отношении ADCC можно оценивать in vivo, например, в модели на животных, такой как описана в Clynes et al. PNAS (USA) 95:652-656 (1998).
"Комплементзависимая цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического каскада комплемента начинается со связывания первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), связанными с распознаваемым им антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Варианты полипептидов с измененными аминокислотными последовательностями Fc-областью и повышенной или сниженной способностью связывать C1q описаны в патенте США No. 6194551B1 и WO99/51642. См. также, Idusogie et al. J. Immunol. 164:4178-4184 (2000).
Термин "содержащее Fc-область антитело" относится к антителу, которое включает Fc-область. C-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области может быть удален, например, во время очистки антитела или путем рекомбинантного конструирования антитела. Таким образом, композиция, включающая содержащее Fc-область антитело в соответствии с настоящим изобретением, может включать антитело, содержащее K447, антитело, из которого полностью удален K447, или смесь антител, содержащих и не содержащих остаток K447.
"Аффинность связывания", как правило, относится к силе полной суммы нековалентных взаимодействий между единичным участком связывания молекулы (например, антитела) и ее партнера по связыванию (например, антигена). Если нет иных указаний, "аффинность связывания" в настоящем документе, относится к присущей аффинности, которая отражает взаимодействие 1:1 между членами связывающей пары (например, антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y, как правило, может быть выражена константой диссоциации (Kd). Аффинность можно измерять общепринятыми способами, известными в данной области, включая способы, описанные в настоящем документе. Низкоаффинные антитела, как правило, связывают антиген медленно и имеют тенденцию к легкой диссоцации, в то время как высокоаффинные антитела, как правило, связывают антиген быстрее и имеют тенденцию к тому, чтобы дольше оставаться связанными. В данной области известно множество способов измерения аффинности связывания, любые из которых можно использовать для целей настоящего изобретения. Конкретные иллюстративные и типичные варианты осуществления для измерения аффинности связывания описаны ниже.
В одном варианте осуществления "Kd" или "величину Kd" в соответствии с настоящим изобретением измеряют с помощью анализа связывания меченного радиоактивной меткой антигена (RIA), проводимого с Fab-версией представляющего интерес антитела и его антигеном, как описано в следующем анализе, в котором измеряют аффинность связывания Fab с антигеном в растворе путем уравновешивания Fab минимальной концентрацией (125I)-меченного антигена в присутствии серии титров немеченого антигена, с последующей фиксацией связанного антигена на планшете, покрытом антителом против (см. Chen, et al., J. Mol. Biol 293:865-881 (1999)). Чтобы установить условия для анализа, на многолуночные планшеты MICROTITER® (Dynex) наносят в течение ночи 5 мкг/мл фиксирующего антитела против Fab (Cappel Labs) в 50 мМ растворе карбоната натрия (pH 9,6), а затем блокируют с использованием 2% (масс./об.) бычьего сывороточного альбумина в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В неадсорбирующем планшете (Nunc #269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями представляющего интерес Fab (например, в соответствии с антитела против VEGF, Fab-12, в Presta et al., Cancer Res. 57:4593-4599) (1997). Затем представляющий интерес Fab инкубируют в течение ночи; однако, инкубацию можно продолжать в течение более длительного периода (например, 65 часов), чтобы убедиться в том, что равновесие достигнуто. После этого смеси переносят на фиксирующий планшет для инкубации при комнатной температуре (например, в течение 1 часа). Затем раствор удаляют, и планшет промывают восемь раз 0,1% TWEEN-20TM в PBS. После того, как планшеты высыхают, добавляют 150 мкл/лунка сцинтиллятора (MICROSCINT-20TM; Packard), и проводят подсчет в планшетах с использованием счетчика гамма-импульсов TOPCOUNTTM (Packard) в течение десяти минут. Концентрации каждого из Fab, которые обеспечивают связывание, меньшее или равное 20% от максимального связывания, выбирают для использования в конкурентных анализах связывания.
В соответствии с другим вариантом осуществления Kd или величину Kd измеряют с использованием анализа способом поверхностного плазмонного резонанса с использованием BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами CM5 с иммобилизованным на них антигеном при ~10 единицах ответа (RU). В кратком изложении, биосенсорные чипы из карбоксиметилированного декстрана (CM5, BIACORE Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/минута для достижения приблизительно 10 единиц ответа (RU) связанного белка. После инъекции антигена инъецируют 1 M этаноламин для блокирования не вступивших в реакцию групп. Для определения показателей кинетики, инъецируют двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS с 0,05% TWEEN-20TM (PBST) при 25°C со скоростью потока приблизительно 25 мкл/мин. Константу ассоциации (kon) и константу диссоциации (koff) вычисляют с использованием простой модели связывания Ленгмюра "один к одному" (BIACORE® Evaluation Software версии 3.2) посредством одновременного приведения в соответствие сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как соотношение koff/kon. См., например, Chen et al., J. Mol. Biol. 293:865-881 (1999). Если константа ассоциации антитела превышает 106 M-1 с-1 в анализе способом поверхностного плазмонного резонанса, указанным выше, тогда константу ассоциации можно определять с использованием способа тушения флуоресценции, в котором измеряют повышение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 25°С 20 нМ антитела (форма Fab) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена при измерении в спектрофотометре, таком как спектрофотометр с останавливаемой струей (Aviv Instruments) или спектрофотометр SLM-AMINCOTM серии 8000 (ThermoSpectronic) с перемешиваемой кюветой.
"Скорость связывания" или "скорость ассоциации" или "скорость соединения" или "kon согласно настоящему изобретению также можно определять, как описано выше с использованием систем BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Piscataway, NJ).
Выражение "по существу сходный" или "по существу такой же" в настоящем документе обозначает достаточно высокую степень сходства между двумя числовыми значениями (как правило, одно связано с антителом по изобретению, а другое связано с эталонным/сравнительным антителом), так что специалист в данной области может считать различие между двумя значениями небольшим или не имеющим биологической и/или статистической значимости в контексте биологического признака, измеряемого указанными значениями (например, значениями Kd). Различие между указанными двумя значениями предпочтительно составляет менее чем приблизительно 50%, предпочтительно менее чем приблизительно 40%, предпочтительно менее чем приблизительно 30%, предпочтительно менее чем приблизительно 20%, и/или менее чем приблизительно 10% в качестве функции от значения для эталонного/сравнительного антитела.
Выражение "значимо сниженный" или "значимо отличающийся" в настоящем документе обозначает значимую высокую степень отличия между двумя числовыми значениями (как правило, одно связано с молекулой, а другое связано с эталонной/сравнительной молекулой), так что специалист в данной области может считать различие между двумя значениями статистически значимым в контексте биологического признака, измеряемого указанными значениями (например, значениями Kd). Различие между указанными двумя значениями предпочтительно составляет более чем приблизительно 10%, более чем приблизительно 20%, более чем приблизительно 30%, более чем приблизительно 40%, и/или более чем приблизительно 50% в качестве функции от значения для эталонной/сравнительной молекулы.
"Акцепторная каркасная область человека" или "человеческая каркасная область" для целей настоящего описания представляет собой каркасную область, содержащую аминокислотную последовательность каркасной области VL или VH, образованной каркасной областью иммуноглобулина человека или консенсусной последовательностью каркасной области человека. Акцепторная каркасная область человека, "образованная" каркасной областью иммуноглобулина человека или консенсусной последовательностью каркасной области человека, может содержать такую же как и в них аминокислотную последовательность, или она может содержать предсуществующие изменения в аминокислотных последовательностях. В некоторых вариантах осуществления количество предсуществующих аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. Когда предсуществующие аминокислотные изменения присутствуют в VH, предпочтительно, чтобы указанные изменения существовали только в трех, двух или одном из положений 71H, 73H и 78H; например, аминокислотные остатки в указанных положениях могут представлять собой 71A, 73T и/или 78A. В одном варианте осуществления акцепторная каркасная область VL человека идентична по последовательности каркасной области VL иммуноглобулина человека или консенсусной последовательности каркасной области человека.
"Консенсусная каркасная область человека" представляет собой каркасную область, которая представляет собой наиболее часто встречающиеся аминокислотные остатки при выборе каркасных последовательностей VL или VH иммуноглобулина человека. Обычно выбор последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельных доменов. Обычно подгруппа последовательностей представляет собой подгруппу согласно Kabat et al, выше. В одном варианте осуществления для VL подгруппа представляет собой подгруппу каппа I по Kabat et al, выше. В одном варианте осуществления для VH подгруппа представляет собой подгруппу III по Kabat et al, выше.
"Консенсусная каркасная область VH подгруппы III" включает консенсусную последовательность, образованную аминокислотными последовательностями вариабельных областей тяжелой цепи подгруппы III по Kabat et al, выше. В одном варианте осуществления аминокислотная последовательность консенсусной каркасной области VH подгруппы III включает по меньшей мере часть или все из каждой из следующих последовательностей: (SEQ ID NO: 50)-H1-(SEQ ID NO: 51)-H2-(SEQ ID NO: 57 или 59)-H3-(SEQ ID NO: 35). В некоторых вариантах осуществления последний остаток (S11) SEQ ID NO: 35 заменен аланином.
"Консенсусная каркасная область VL подгруппы I" включает консенсусную последовательность образованную аминокислотными последовательностями вариабельных областей легкой цепи каппа подгруппы I по Kabat et al., выше. В одном варианте осуществления аминокислотная последовательность консенсусной каркасной области VH подгруппы I включает по меньшей мере часть или все из каждой из следующих последовательностей: (SEQ ID NO: 60)-L1-(SEQ ID NO: 61)-L2-(SEQ ID NO: 62)-L3-(SEQ ID NO: 63).
"Очищенный" означает, что молекула присутствует в образце в концентрации по меньшей мере 95% по массе, или по меньшей мере 98% по массе образца, в котором она содержится.
"Выделенная" молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая отделена по меньшей мере от одной другой молекулы нуклеиновой кислоты, с которой она обычно ассоциирована, например, в ее природном окружении. Выделенная молекула нуклеиновой кислоты, кроме того, включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно экспрессируют молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты находится вне хромосомы или в области хромосомы, которая отличается от ее природного расположения на хромосоме.
Термин "вектор" в настоящем документе относится к молекуле нуклеиновой кислоты, способной осуществлять транспорт другой нуклеиновой кислоты, с которой она связана. Одним типом вектора является "плазмида", которая относится к кольцевой двухцепочечной петле ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут встраиваться в геном клетки-хозяина при введении в клетку-хозяина, и, тем самым, они реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы называют в настоящем документе "рекомбинантными экспрессирующими векторами" или просто "экспрессирующими векторами". Как правило, экспрессирующие векторы, применяемые в способах рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании термины "плазмида" и "вектор" могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора.
Термин "полинуклеотид" или "нуклеиновая кислота", используемые в настоящем документе взаимозаменяемо, относится к полимерам нуклеотидов любой длины, и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой, или посредством синтетической реакции. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. До и после сборки полимера может происходить модификация нуклеотидной структуры, если она происходит. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Кроме того, полинуклеотид может быть модифицирован после полимеризации, например, конъюгацией с осуществляющим мечение компонентом. Другие типы модификаций включают, например, "кэпы", замену одного или нескольких природных нуклеотидов аналогом, межнуклеотидные модификации, например, такие как модификации незаряженными связями (например, метилфосфонатами, фосфотриэфирами, фосфоамидатами, карбаматами и т.д.) и заряженными связями (например, фосфоротиоатами, фосфородитиоатами, и т.д.), модификации выступающими группами, например, такими как белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), модификации интеркаляторами (например, акридином, псораленом, и т.д.), модификации хелаторами (например, металлами, радиоактивными металлами, бором, окислительными металлами и т.д.), модификации алкилирующими соединениями, модификации модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т. д.), а также немодифицированные формы полинуклеотида(ов). Кроме того, любые гидроксильные группы, обычно присутствующие в сахарах, могут быть заменены, например, фосфонатными группами, фосфатными группами, они могут быть защищены стандартными защитными группами или активированы для образования дополнительных связей с дополнительными нуклеотидами, или они могут быть конъюгированы с твердыми подложками. 5'- и 3'-концевой OH может быть фосфорилирован или замещен аминами или органическими кэпирующими группами из от 1 до 20 атомов углерода. Другие гидроксилы также могут быть преобразованы в стандартные защитные группы. Полинуклеотиды также могут содержать аналогичные формы сахаров рибозы или дезоксирибозы, которые, главным образом, известны в данной области, включая, например, 2'-O-метил-, 2'-O-аллил-, 2'-фтор- или 2'-азидорибозу, карбоциклические аналоги сахаров, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, сахара пиранозы, сахара фуранозы, седогептулозы, ациклические аналоги и абазические нуклеозидные аналоги, такие как метилрибозид. Одна или несколько фосфодиэфирных связей могут быть заменены альтернативными линкерными группами. Эти альтернативные линкерные группы включают, но не ограничиваются ими, варианты осуществления где фосфат заменен на P(O)S ("тиоат"), P(S)S ("дитиоат"), "(O)NR2 ("амидат"), P(O)R, P(O)OR', CO или CH2 ("формацеталь"), где каждый R или R' независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно содержащий простую эфирную связь (-O-), арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не обязательно, чтобы все связи в полинуклеотиде были идентичными. Представленное выше описание применимо ко всем полинуклеотидам, указанным в настоящем документе, включая РНК и ДНК.
"Олигонуклеотид" в настоящем документе относится, главным образом, к коротким, как правило, одноцепочечным, как правило, синтетическим полинуклеотидам длиной, как правило, но не обязательно, менее 200 нуклеотидов. Термины "олигонуклеотид" и "полинуклеотид" не являются взаимоисключающими. Представленное выше описание полинуклеотидов в равной степени и полностью применимо к олигонуклеотидам.
Термин "Notch" в настоящем документе относится к любому нативному Notch (Notch1-4) из любого являющегося его источником позвоночного, включая млекопитающих, таких как приматы (например, люди) и грызунов (например, мышей и крыс), если нет иных указаний. Термин охватывает "полноразмерный" непроцессированный Notch, а также любую форму Notch, которая является следствием процессинга в клетке. Термин также охватывает природные варианты Notch, например, варианты по сплайсингу или аллельные варианты.
Термин "Notch1" в настоящем документе относится к любому нативному Notch1 из любого являющегося источником позвоночного, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если нет иных указаний. Термин охватывает "полноразмерный" непроцессированный Notch1, а также любую форму Notch1, которая является следствием процессинга в клетке. Термин также охватывает природные варианты Notch1, например, варианты по сплайсингу или аллельные варианты.
Термин "Notch2" в настоящем документе относится к любому нативному Notch2 из любого являющегося источником позвоночного, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если нет иных указаний. Термин охватывает "полноразмерный" непроцессированный Notch2, а также любую форму Notch2, которая является следствием процессинга в клетке. Термин также охватывает природные варианты Notch2, например, варианты по сплайсингу или аллельные варианты.
Термин "активность Notch2" относится к передаче сигнала Notch2. Средство (например, антитело), которое "ингибирует активность Notch2" в значительной степени снижает передачу сигнала Notch2 относительно уровня передачи сигнала Notch2, наблюдаемого у соответствующего контроля по существу в идентичных условиях. В определенных вариантах осуществления активность Notch2 можно оценивать с помощью пригодного репортерного анализа, как описано в настоящем документе в примере B(2). В определенных вариантах осуществления активность Notch2 можно оценивать путем измерения образования клеток маргинальной B-зоны, как описано в настоящем документе в примере B(4). В определенных вариантах осуществления снижение передачи сигнала Notch2 по меньшей мере в 2, 3, 4, 5 или 10 раз ниже уровня, наблюдаемого в контроле.
Термин "NRR Notch2" относится к области Notch2, состоящей из трех модулей LNR (LNR-A, LNR-B и LNR-C) и HD-домена (HD-N и HD-C). Иллюстративные последовательности NRR Notch2 человека и мыши представлены на фигуре 18 (SEQ ID NO: 28 и 29, соответственно). NRR Notch2 может состоять из нековалентно связанных фрагментов, например, которые являются результатом процессинга Notch2 в S1, а также из единой непрерывной полипептидной последовательности. В качестве примера и ссылаясь на фигуру 18, NRR Notch2 человека может состоять из аминокислот 1422-1677 Notch2 человека (SEQ ID NO: 75), или альтернативно аминокислот 1422-1608 SEQ ID NO: 75, нековалентно связанных с аминокислотами 1609-1677 SEQ ID NO: 75.
"Процентную (%) идентичность аминокислотных последовательностей" в отношении последовательности эталонного полипептида определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в последовательности эталонного полипептида, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей, и не считая какие-либо консервативные замены частью идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей можно проводить различными способами, которые находятся в пределах специальных знаний, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, требуемые для достижения максимального выравнивания на протяжении всей длины сравниваемых последовательностей. Однако для целей, представленных в настоящем документе, значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была составлена в Genentech, Inc. и исходный текст был представлен с пользовательской документацией в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован под U.S. Copyright Registration No. TXU510087. Программа ALIGN-2 является общедоступной через Genentech, Inc., South San Francisco, California или ее можно составить из исходного текста, представленного в таблице 1, ниже. Программа ALIGN-2 должна быть составлена для применения на операционной системе UNIX, предпочтительно цифровой UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не изменяются.
В случаях, когда ALIGN-2 используют для сравнений аминокислотных последовательностей, % идентичность аминокислотных последовательностей для данной аминокислотной последовательности A к, с или против данной аминокислотной последовательности B (что альтернативно может быть сформулировано как данная аминокислотная последовательность A, которая обладает определенной % идентичностью аминокислотной последовательности, содержит % идентичность аминокислотной последовательности, к, с или против данной аминокислотной последовательности B), или вычисляют следующим образом:
100 умножить на частное X/Y
где X представляет собой число аминокислотных остатков, оцененных как идентичные совпадения программой для выравнивания последовательностей ALIGN-2 при выравнивании этой программой A и B, и где Y представляет собой общее количество аминокислотных остатков в B. Понятно, что когда длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичность аминокислотных последовательностей A к B не равна % идентичности аминокислотных последовательностей B к A. Если нет иных конкретных указаний, все величины % идентичности аминокислотных последовательностей, использованные в настоящем документе, получены, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
"Нарушение" представляет собой любое состояние, при котором может быть полезным лечение композицией или способом по изобретению. Оно включает хронические и острые нарушения или заболевания, включая патологические состояния, которые предрасполагают млекопитающего к рассматриваемому нарушению. Неограничивающие примеры нарушений, подлежащих лечению согласно настоящему описанию, включают такие состояния, как злокачественная опухоль.
"B-клеточные злокачественные опухоли", также называемые "B-клеточными новообразованиями", включают, но не ограничиваются ими, болезнь Ходжкина (например, болезнь Ходжкина с преобладанием лимфоцитов (LPHD)); неходжскинскую лимфому (NHL); лимфомы фоллекулярных центральных клеток (FCC); острый лимфоцитарный лейкоз (ALL, также называемый B-ALL); лейкоз предшественников B-лимфобластов; хронический лимфоцитарный лейкоз (CLL, также называемый B-CLL); волосатоклеточный лейкоз; лимфомы B-клеток маргинальной зоны (включая узелковый, MALT и селезеночный типы); множественную миелому (плазмацитому, миелому плазматических клеток); лимфомы мелких клеток с нерасщепленным ядром (например, лимфома Беркитта); крупноклеточные лимфомы (включая диффузную крупноклеточную (B-клеточную) лимфому, диффузную лимфому смешанных клеток и иммунобластную лимфому); лимфома из клеток мантийной зоны; мелколимфоцитарную лимфому; связанную со СПИД лимфому и макроглобулинемию Вальденстрема. Вялотекущая лимфома представляет собой медленно растущее неизлечимое заболевание, при котором в среднем пациент выживает между шестью и 10 годами после многочисленных периодов ремиссии и обострения; агрессивные лимфомы представляют собой быстрорастующие формы лимфомы, охватывающие высокозлокачественные и некоторые категории среднезлокачественных NHL.
Термин "меланома" относится к злокачественной опухоли меланоцитов, которые встречаются, главным образом, в коже, однако которые также встречаются в глазу (увеальная меланома) или в кишечнике.
В настоящем документе "лечение" (и его варианты, такие как "лечить" или "проведение лечения") относится к клиническому вмешательству в попытках изменения естественного течения у индивидуума или в клетке, подвергаемых лечению, и его можно проводить либо для профилактики, либо в ходе клинической патологии. Желаемые эффекты лечения включают предотвращение возникновения или рецидива заболевания, смягчение симптомов, уменьшение любого прямого или непрямого патологического последствия заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, смягчение или временное облегчение болезненного состояния и ремиссию или улучшенный прогноз. В некоторых вариантах осуществления антитела по изобретению используют для замедления развития заболевания или нарушения или для замедления прогрессирования заболевания или нарушения.
"Индивидуум", "субъект" или "пациент" представляет собой позвоночное. В определенных вариантах осуществления позвоночным является млекопитающее. Млекопитающие включают, но не ограничиваются ими, сельскохозяйственных животных (таких как коровы), спортивных животных, домашних животных (таких как кошки, собаки и лошади), приматов, мышей и крыс. В определенных вариантах осуществления млекопитающим является человек.
Термин "фармацевтический состав" относится к препарату, который имеет такую форму, чтобы обеспечить эффективность биологической активности активного ингредиента, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для индивидуума, которому состав будут вводить. Такие составы могут быть стерильными.
"Стерильный" состав является асептическим или не содержит живых микроорганизмов и их спор.
"Эффективное количество" антитела относится к количеству, эффективному, в необходимых дозировках и в течение необходимых периодов времени, для осуществления желаемой терапевтической или профилактической цели.
"Терапевтически эффективное количество" вещества/молекулы по изобретению может варьировать, в зависимости от таких факторов, как болезненное состояние, возраст, пол и масса индивидуума и способность вещества/молекулы вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество охватывает количество, в котором над любыми токсическими или вредными эффектами вещества/молекулы преобладают терапевтически полезные эффекты. "Профилактически эффективное количество" относится к количеству, эффективному, в необходимых дозировках и в течение необходимых периодов времени, для достижения требуемого профилактического результата. Как правило, но не обязательно, поскольку профилактическую дозу используют у субъектов до заболевания или на его ранних стадиях, профилактически эффективное количество будет меньшим, чем терапевтически эффективное количество.
Термин "цитотоксическое средство" в настоящем документе относится к веществу, которое ингибирует функционирование клеток, или препятствует ему, и/или вызывает разрушение клеток. Подразумевают, что термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические средства, например метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин C, хлорамбуцил, даунорубицин или другие интеркалирующие вещества, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины из бактерий, грибов, растений или животных, включая их фрагменты и/или варианты, и различные противоопухолевые средства или средства против злокачественной опухоли, как описано ниже. Другие цитотоксические средства описаны ниже. Уничтожающее опухолевые клетки средство вызывает разрушение опухолевых клеток.
"Токсин" представляет собой любое вещество, способное оказывать вредный эффект на рост или пролиферацию клетки.
"Химиотерапевтическое средство" представляет собой химическое соединение, полезное при лечении злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие средства, такие как тиотепа и циклофосфамид (CYTOXAN®); алкилсульфонаты, такие как бисульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, в том числе альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогинины (особенно буллатацин и буллатацинон); дельта-9-тетрагидроканнабиол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (в том числе синтетический аналог топотекан (HYCAMTIN®), CPT-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; CC-1065 (в том числе его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (в том числе синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкрастатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлорэтамин, мехлорэтамина оксида гидрохлорид, мелфалан, новэмбихин, фенестерин, преднемустин, трофосфамид, иприт урацила; нитрозомочневина, такая как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как эндииновые антибиотики (например, калихеамицин, особенно, калихеамицин гамма1I и калихеамицин омегаI1 (см., например, Agnew, Chem Intl. Ed. Engl., 33: 183-186 (1994)); CDP323, пероральный ингибитор альфа-4 интегрина, динемицин, в том числе динемицин A; эсперамицин; а также хромофор неокарциностатин и сходные хромофоры хромопротеиновых эндииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (в том числе ADRIAMYCIN®, морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин, липосомную инъекцию доксорубицин-HCl (DOXIL®), липосомальный доксорубицин TLC D-99 (MYOCET®), пегилированный липосомальный доксорубицин (CAELYX®), и деоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марселломицин, митомицины, такие как митомицин C, микофеноловавую кислотау, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (GEMZAR®), тегафур (UFTORAL®), капецитабин (XELODA®), эпотилон и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как даноптерин, метотрексат, птероптерин, триметрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидеоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан,тестолактон, антиадреналовые средства, такие как аминоглутетимид, митотан, трилостан; освежающий раствор на основе фолиевой кислоты, такой как фролиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демекольцин; диазиквон; эльфорнитин; эллиптиния ацетат; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2"-трихлортриэтиламин; трихотецены (особенно токсин T-2, верракурин A, роридин A и ангвидин); уретан; виндезин (ELDISINE®, FILLDISINE®), декарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); тиотепу; таксоид, например, паклитаксел (TAXOL®), сконструированный на основе альбумина препарат из наночастиц паклитаксела (ABRAXANETM) и доцетаксел (TAXOTERE®); хлорамбуцил; 6-тиогуанин; меркаптопурин; метотрексат; средства на основе платины, такие как цисплатин, оксаплатин (например, ELOXATIN®) и карбоплатин; средства на основе барвинка, которые предотвращают полимеризацию тубулина при формировании микротрубочек, в том числе винбластин (VELBAN®), винкристин (ONCOVIN®), виндезин (ELDISINE®, FILDESIN®) и винорелбин (NAVELBINE®); этопзид (VP-16); ифосфамид; митоксантрон; лейковорин; новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота, в том числе бексаротен (TARGRETIN®); бифосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат (DIDROCAL®), NE-58095, золедроновая кислота/золедронат (ZOMETA®), алендронат (FOSAMAX®), памидронат (AREDIA®), тилудронат (SKELID®) или ризедронат (ACTONEL®); троксацитабин (1,3-диоксолановый нуклеозидный аналог цитозина); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, которые ингибируют экспрессию генов сигнальных путей, вовлеченных в абберантную клеточную пролиферацию, таких как, например, PKC-alpha, Raf, H-Ras и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN®, и вакцина VAXID®; ингибитор топоизомеразы 1 (например, LAUROTECAN®); rmRH (например, ABARELIX®); BAY439006 (сорафениб; Bayer); SU-11248 (Pfizer); перифозин, ингибитор COX-2 (например, целекоксиб или эторикоксиб), ингибитор протеасом (например, PS341); бортезомиб (VELCADE®); CCI-779; типифарниб (R11577); орафениб, ABT510; ингибитор Bcl-2, такой как облимерсен натрия (GENASENSE®); пиксантрон; ингибиторы EGFR (см. определение ниже); ингибиторы тирозинкиназы (см. определение ниже); ингибиторы серин-треонинкиназы, такие как рапамицин (сиролимус, RAPAMUNE®); ингибиторы фарнезилтрансферазы, такие как лонафарниб (SCH 6636, SARASARTM); и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше; а также сочетания двух или более из указанных выше, такие как CHOP, сокращение для комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращение для режима лечения оксалиплатином (ELOXATINTM) в сочетании с 5-FU и лейковорином.
Химиотерапевтические средства, как определено в настоящем документе, включают "антигормональные средства" или "эндокринные лекарственные средства", которые действуют посредством регуляции, уменьшения, блокирования или ингибирования воздействия гормонов, которые могут стимулировать рост опухоли. Они сами по себе могут представлять собой гормоны, включая, но не ограничиваясь ими, антиэстрогены со смешанным профилем агонист/антагонист, в том числе, тамоксифен (NOLVADEX®), 4-гидрокситамоксифен, торемифен (FARESTON®), идоксифен, дролоксифен, ралоксифен (EVISTA®), триоксифен, кеоксифен и селективные модуляторы рецепторов эстрогена (SERM), такие как SERM3; полные антиэстрогены без свойств агонистов, такие как (FASLODEX®) и EM800 (такие средства могут блокировать димеризацию рецепторов эстрогенов (ER), ингибировать связывание ДНК, повышать обновление ER, и/или снижать уровень ER); ингибиторы ароматазы, в том числе стероидные ингибиторы ароматазы, такие как форместан и экземестан (AROMASIN®) и нестероидные ингибиторы ароматазы, такие как анастразол (ARIMIDEX®), лектрозол (FEMARA®) и аминоглутетимид, и другие ингибиторы ароматазы, в том числе ворозол (RIVISOR®), ацетат магестрола (MEGASE®), фадрозол, и 4(5)-имидазолы; агонисты лютеинизирующего рилизинг-гормона, в том числе леупролид (LUPRON® и ELIGARD®), гозерелин, бузерелин и триптерелин; половые стероидные гормоны, включающие прогестины, такие как ацетат магестрола и ацетат медроксипрогестерона, эстрогены, такие как диэтилстилбестрол и премарин, и андрогены/ретиноиды, такие как флуоксиместерон, полностью трансретиноевая кислота и фенретинид; онапристон; анти-прогестероны; ингибиторы рецепторов эстрогена (ERD); антиандрогены, такие как флутамид, нилутамид и бикалутамид; и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше; а также комбинации двух или более из указанных выше.
"Ингибирующее рост средство", в настоящем документе, относится к соединению или композиции, которые ингибируют рост клетки (такой как клетка, экспрессирующая Notch), либо in vitro, либо in vivo. Таким образом, ингибирующее рост средство может представлять собой средство, которое значительно снижает процентное количество клеток (таких как клетка, экспрессирующая Notch) в S-фазе. Примеры ингибирующих рост средств включают средства, которые блокируют прогрессирование клеточного цикла (в месте, отличном от S-фазы), такие как средства, которые индуцируют остановку в G1-фазе и остановку M-фазе. Классические блокаторы M-фазы включают алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Те средства, которые вызывают остановку в G1-фазе, также вовлекают остановку в S-фазе, например, алкилирующие ДНК средства, такие как тамоксифен, преднизон, декарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, entitled "Cell cycle regulation, oncogenes, and antineoplastic drugs", Murakami et al. (WB Saunders: Philadelphia, 1995), especially p. 13. Таксаны (паклитаксел и доцетаксел) представляют собой лекарственные средства против злокачественной опухоли, полученные из тисового дерева. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из тиса европейского, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел обеспечивают сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки, препятствуя деполимеризации, что приводит к ингибированию митоза в клетках.
III. КОМПОЗИЦИИ И СПОСОБЫ
Изобретение основано, частично, на идентификации связывающих Notch средств, например, антител и их фрагментов. В конкретных вариантах осуществления изобретение относится к средствам, связывающим отрицательную регуляторную область Notch2 (NRR), таким как выделенные антитела, которые связывают NRR Notch2 и его фрагменты. Такие антитела пригодны, например, для диагностики или лечения нарушений, ассоциированных с экспрессией и/или активностью Notch2 (например, повышенной экспрессией или активностью). В конкретных вариантах осуществления антитела против NRR Notch2 пригодны для диагностики или лечения злокачественной опухоли, например, B-клеточных злокачественных опухолей и меланомы. Таким образом, изобретение относится к способам, композициям, наборам и изделиям, связанным со связыванием NRR Notch2.
A. Антитела против NRR Notch2
1. Иллюстративные антитела, полученные с помощью фаговой библиотеки
В настоящем документе представлены иллюстративные моноклональные антитела, которые были получены с помощью фаговой библиотеки, как описано в примере B. Было выделено антитело, обозначенное "антителом D", которое связывается с NRR Notch2. Это антитело подвергали созреванию аффинности с получением антител, обозначенных как антитело D-1, антитело D-2 и антитело D-3. Последовательности гипервариабельных областей тяжелой цепи и легкой цепи (HVR) антитела D, антитела D-1, антитела D-2 и антитела D-3, представлены на фигурах 1 и 2. Последовательности вариабельных доменов тяжелой и легкой цепей антитела D, антитела D-1, антитела D-2 и антитела D-3 представлены на фигурах 3 и 4. Следующие варианты осуществления антител против NRR Notch2 представлены ниже.
В одном аспекте представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из:
(a) HVR-H1, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 3;
(b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 4;
(c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 5;
(d) HVR-L1, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 10;
(e) HVR-L2, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 14; и
(f) HVR-L3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 19.
В следующем аспекте антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5 и по меньшей мере одну, две, три, четыре или пять HVR, выбранных из (a), (b), (d), (e) и (f), выше. В следующем аспекте, антитело содержит (a), (b), (c), (d), (e) и (f), выше. В отношении (a), (d), (e) и (f), предусматривается любой один или несколько из следующих вариантов осуществления: HVR-H1 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-2; HVR-L1 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 6-9; HVR-L2 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и HVR-L3 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 15-18.
В другом аспекте представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит HVR-H1, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 3, HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4, и HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5. В одном варианте осуществления HVR-H1 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-2.
В другом аспекте представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит HVR-L1, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 10, HVR-L2, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 14, и HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 19. Следующие варианты осуществления предусматриваются в любой комбинации: HVR-L1 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 6-9; HVR-L2 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и HVR-L3 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 15-18. В одном варианте осуществления антитело, которое связывается с NRR Notch2, содержит HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 6; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15. В другом варианте осуществления антитело, которое связывается с NRR Notch2, содержит HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 16. В другом варианте осуществления антитело, которое связывается с NRR Notch2, содержит HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 8; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 12; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 17. В другом варианте осуществления антитело, которое связывается с NRR Notch2, содержит HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 9; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 18.
В одном варианте осуществления представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 1;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 6;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
В другом варианте осуществления представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2;
(b) HVR-H2 содержащую аминокислотную последовательность SEQ ID NO: 4;
(c) HVR-H3 содержащую аминокислотную последовательность SEQ ID NO: 5;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 16.
В другом варианте осуществления представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 8;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 12; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 17.
В другом варианте осуществления представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит:
(a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4;
(c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5;
(d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 9;
(e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и
(f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 18.
В определенных вариантах осуществления любое из указанных выше антител, кроме того, содержит по меньшей мере одну каркасную область, выбранную из консенсусной каркасной области VH подгруппы III и консенсусной каркасной области VL подгруппы I.
В определенных вариантах осуществления антитело против NRR Notch2 является подвергнутым созреванию аффинности. Например, любую из представленных ниже замен в указанных положениях HVR (нумерация Kabat) можно проводить в любой комбинации:
- в HVR-H1 (SEQ ID NO: 1): S28T; T30S;
- в HVR-L1 (SEQ ID NO: 6): S28N; I29N или V; S30R или K; S31R; Y32F
- в HVR-L2 (SEQ ID NO: 11): G50R; S53I или T; A55E
- в HVR-L3 (SEQ ID NO: 15): S93I или R; L96W или H
Конкретные антитела, описанные в настоящем документе, т.е., антитело D, а также полученные созреванием аффинности формы антитела D (D-1, D-2 и D-3), могут подвергаться дальнейшему созреванию аффинности. Таким образом, представлены подвергнутые созреванию аффинности формы любых из антител, описанных в настоящем документе.
В определенных вариантах осуществления антитело против NRR Notch2, имеющее любую из указанных выше последовательностей HVR, может, кроме того, содержать любую пригодную последовательность каркасной области вариабельного домена, при условии по существу сохранения активности связывания с NRR Notch2. В определенных вариантах осуществления антитело против NRR Notch2 содержит консенсусную последовательность каркасной области вариабельного домена тяжелой цепи (VH) человека, как в любой из консенсусных последовательностей каркасных областей VH, показаных на фигурах 5A и 5B. В одном варианте осуществления консенсусная последовательность каркасной области VH содержит консенсусную последовательность каркасной области тяжелой цепи подгруппы III человека, например, как показано на фигурах 5A и 5B. В другом варианте осуществления консенсусная последовательность каркасной области VH содержит каркасную последовательность акцептора 2, например, как показано на фигурах 5A и 5B. В конкретном варианте осуществления консенсусная последовательность каркасной области VH содержит FR1-FR4 акцептора 2B или акцептора 2D, где FR4 содержит SEQ ID NO: 35 (фигуры 5A и 5B), причем последний остаток SEQ ID NO: 35 (S11) необязательно замещен аланином. В следующем конкретном варианте осуществления консенсусная последовательность каркасной области VH содержит последовательности SEQ ID NO: 50; 51; 57 или 59; и 35, где S11 SEQ ID NO: 35 необязательно замещен аланином.
В определенных вариантах осуществления антитело против NRR Notch2, имеющее любую из указанных выше последовательностей HVR, может, кроме того, содержать последовательность каркасной области вариабельного домена легкой цепи (VL) человека, как показано на фигурах 6A и 6B. В одном варианте осуществления консенсусная последовательность каркасной области VL содержит консенсусную последовательность каркасной области VL каппа подгруппы I человека (κv1), например, как показано на фигурах 6A и 6B. В другом варианте осуществления консенсусная последовательность каркасной области VL содержит FR1-FR4 huMAb4D5-8, как показано на фигурах 7 или 8. В конкретном варианте осуществления консенсусная последовательность каркасной области VL содержит последовательности SEQ ID NO: 60, 61, 62 и 63.
В другом аспекте антитело против NRR Notch2 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 20-21. В определенных вариантах осуществления последовательность VH, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность, содержит замены (например, консервативные замены), вставки или делеции относительно эталонной последовательности, однако антитело против NRR Notch2, содержащее эту последовательность, сохраняет способность связываться с NRR Notch2. В определенных вариантах осуществления всего 1 из 10 заменен, вставлена и/или удалена аминокислот в аминокислотной последовательности, выбранной из SEQ ID NO: 20-21. В определенных вариантах осуществления замены, вставки или делеции встречаются в областях вне HVR (т.е., в FR). В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 3, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 4, и (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 5. В одном из таких вариантов осуществления HVR-H1 содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-2.
В другом аспекте представлено антитело, которое специфично связывается с NRR Notch2, где антитело содержит вариабельный домен легкой цепи (VL), имеющий по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 22-25. В определенных вариантах осуществления последовательность VL, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность, содержит замены (например, консервативные замены), вставки или делеции относительно эталонной последовательности, однако антитело против NRR Notch2, содержащее эту последовательность, сохраняет способность связываться с NRR Notch2. В определенных вариантах осуществления всего 1 из 10 аминокислот заменен, вставлена и/или удалена в аминокислотной последовательности, выбранной из SEQ ID NO: 22-25. В определенных вариантах осуществления замены, вставки или делеции встречаются в областях вне HVR (т.е., в FR). В конкретном варианте осуществления VL содержит одну, две или три HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 10; (b) HVR-L2, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 14; и (c) HVR-L3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO: 19. В одном из таких вариантов осуществления VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 6-9; (b) HVR-L2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и (c) HVR-L3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 15-18. В одном из таких вариантов осуществления VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 6; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 11; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15. В другом из таких вариантов осуществления VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 11; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 16. В другом из таких вариантов осуществления VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 8; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 12; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 17. В другом из таких вариантов осуществления VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 9; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 13; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 18.
В определенных вариантах осуществления варианты последовательностей VH и VL, представленных выше, замены, вставки или делеции могут встречаться в HVR. В таких вариантах осуществления замены, вставки или делеции могут встречаться в пределах одной или нескольких HVR, при условии, что такие изменения по существу не снижают способность антитела связывать антиген. Например, в HVR можно вносить консервативные изменения, которые по существу не снижают аффинность связывания. В некоторых случаях, изменения в HVR могут в действительности повысить аффинность антитела. Такие изменения можно проводить в "горячих точках" HVR (т.е., в остатках, кодируемых кодонами, которые претерпевают мутацию с высокой частотой в ходе процессов соматического созревания) в целях повышения аффинности антитела. (см., например, Chowdhury, Способы Mol. Biol. 207:179-196, 2008.) В определенных вариантах осуществления вариантов последовательностей VH и VL, представленных выше, каждая HVR либо является консервативной (неизмененной), либо содержит не более одной аминокислотной замены, вставки или делеции.
В другом аспекте представлено антитело, которое специфично связывает NRR Notch2, где антитело содержит VH, как в любом из вариантов осуществления представленных выше, и VL, как в любом из вариантов осуществления представленных выше. В одном варианте осуществления антитело содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 20, и VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 22.
В одном из таких вариантов осуществления VH содержит одну, две или три HVR, выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 1, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 4, и (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 5, и VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 6; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 11; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15. В конкретном варианте осуществления антитело содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 20, и VL, содержащую аминокислотную последовательность SEQ ID NO: 22.
В другом варианте осуществления антитело против NRR Notch2, которое специфично связывает NRR Notch2, содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 21, и VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 23-25.
В одном из таких вариантов осуществления VH содержит одну, две или три HVR, выбранных из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 2, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 4, и (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 5, и VL содержит одну, две или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 7-9; (b) HVR-L2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и (c) HVR-L3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 16-18. В конкретных вариантах осуществления антитело содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 21, и VL, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 23-25.
В определенных вариантах осуществления представлена подвергнутая созреванию аффинности форма любого из указанных выше антител. В следующих вариантах осуществления представлен рекомбинантный белок, который специфично связывает NRR Notch2, где рекомбинантный белок содержит антигенсвязывающий центр(ы) любого из указанных выше антител. В одном из таких вариантов осуществления рекомбинантный белок содержит любую одну или несколько из HVR, представленных выше.
В определенных вариантах осуществления представлен полинуклеотид, кодирующий любое из указанных выше антител. В одном варианте осуществления представлен вектор, содержащий полинуклеотид. В одном варианте осуществления представлена клетка-хозяин, содержащая вектор. В одном варианте осуществления клетка-хозяин является эукариотической клеткой. В одном варианте осуществления клетка-хозяин представляет собой клетку CHO. В одном варианте осуществления представлен способ получения антитела против NRR Notch2, где способ включает культивирование клетки-хозяина в условиях, пригодных для экспрессии полинуклеотида, кодирующего антитело, и выделение антитела.
2. Дополнительные иллюстративные антитела
В одном варианте осуществления изобретение относится к выделенному антителу против NRR Notch2, которое ингибирует активность Notch2. Например, антитела по изобретению могут модулировать один или несколько аспектов передачи сигнала Notch2, включая нарушение любого биологически значимого каскада передачи сигнала Notch2.
В следующем варианте осуществления изобретение относится к выделенному антителу против NRR Notch2, которое связывает NRR Notch2 с Kd ≤100 нМ. В определенных вариантах осуществления антитело против NRR Notch2 связывает NRR Notch2 с Kd ≤10 нМ, ≤1 нМ или ≤0,1 нМ. Как описано в разделе "Примеры" настоящего документа, иллюстративное фаговое антитело D-3 связывается с Kd 5 нМ. Как общепринято в данной области, аффинность связывания лиганда с его рецептором можно определять с использованием любого из множества анализов, и ее можно выражать с помощью различных количественных величин. Таким образом, в одном варианте осуществления аффинность связывания выражают как величины Kd, и она отражает собственно аффинность связывания (например, с минизмизированными эффектами авидности). Как правило и предпочтительно, аффинность связывания измеряют in vitro, как в бесклеточных, так и в связанных с клетками условиях. Для получения показателей аффинности связывания можно использовать любое количество анализов, известных в данной области, включая анализы, описанные в настоящем документе, включая, например, Biacore, радиоиммунный анализ (RIA) и ELISA.
В другом варианте осуществления представлено выделенное антитело, которое связывается с NRR Notch2, где антитело по существу не связывается с представителем семейства Notch, отличным от Notch2 (т.е., Notch1, 3 и 4 у млекопитающих). Такое антитело можно идентифицировать с использованием анализов, представленных в примере B(I). В одном варианте осуществления антитело связывается с NRR Notch2 человека и NRR Notch2 из по меньшей мере одного другого не являющегося человеком вида, например, мыши.
В другом варианте осуществления представлено выделенное антитело, которое связывается с тем же эпитопом, что и антитело, представленное в настоящем документе. В одном варианте осуществления представлено выделенное антитело против NRR Notch2, которое связывается с тем же эпитопом, что и антитело, выбранное из антитела D, антитела D-1, антитела D-2 и антитела D-3. В другом варианте осуществления изобретение относится к антителу против NRR Notch2, которое конкурирует с антителом, выбранным из антитела D, антитела D-1, антитела D-2 и антитела D-3. В другом варианте осуществления представлено выделенное антитело, которое связывается по меньшей мере с одним доменом, выбранным из домена LNR-A и домена HD-C Notch2. В одном из таких вариантов осуществления антитело связывается как с доменом LNR-A и так и с доменом HD-C. В другом из таких вариантов осуществления антитело, кроме того, связывается с доменами LNR-B и/или HD-N. Эти домены указаны на фигуре 18.
Указанные выше варианты осуществления антител против NRR Notch2 могут существовать отдельно или в любой комбинации. Более того, любое антитело против NRR Notch2, описанное в настоящем документе, может обладать любым одним или несколькими из следующих признаков: в определенных вариантах осуществления антитело против NRR Notch2 представляет собой моноклональное антитело. В определенных вариантах осуществления антитело против NRR Notch2 представляет собой фрагмент антитела, выбранный из Fab-, Fab'-SH-, Fv-, scFv- или (Fab')2-фрагмента. В другом варианте осуществления антитело против NRR Notch2 представляет собой гуманизированное антитело, антитело человека или химерное антитело.
3. Фрагменты антител
Настоящее изобретение охватывает фрагменты антител. Фрагменты антител можно получать традиционными способами, такими как ферментативное расщепление, или рекомбинантными способами. В определенных обстоятельствах, преимущественным является применение фрагментов антител, а не целых антител. Меньший размер фрагментов позволяет быстрое выведение и может привести к повышенному доступу к солидным опухолям. Для обзора определенных фрагментов антител, см. Hudson et al. (2003) Nat. Med. 9:129-134.
Для получения фрагментов антител были разработаны различные способы. Традиционно, эти фрагменты получали протеолитическим расщеплением полноразмерных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако эти фрагменты в настоящее время можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Fab-, Fv- и scFv-фрагменты антител могут быть экспрессированы и секретированы в E. coli, таким образом, позволяя простой способ получения больших количеств этих фрагментов. Фрагменты антител можно выделять из фаговых библиотек антител, описанных выше. Альтернативно фрагменты Fab'-SH можно выделять непосредственно из E. coli и химически связывать с получением фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-167 (1992)). В соответствии с другим подходом фрагменты F(ab')2 можно выделять непосредственно из рекомбинантной культуры клеток-хозяев. Fab- и F(ab')2-фрагмент с повышенным временем полужизни in vivo, содержащий остатки связывающего рецептор спасения эпитопа, описан в патенте США No. 5869046. Другие способы получения фрагментов антител будут очевидны квалифицированному специалисту. В других вариантах осуществления предпочтительное для выбора антитело представляет собой одноцепочечный Fv-фрагмент (scFv). См. WO 93/16185; патент США No. 5571894; и патент США No. 5587458. Fv и sFv являются единственными типами с целыми антигенсвязывающими участками, которые лишены константных участков; таким образом, они пригодны для снижения неспецифического связывания в ходе применения in vivo. Можно конструировать слитые с sFv белки с получением эффекторного белка либо на N-, либо на С- конце sFv. См. Antibody Engineering, ed. Borrebaeck, выше. Фрагмент антитела также может представлять собой "линейное антитело", например, как описано в патенте США 5641870. Такие линейные фрагменты антител могут быть моноспецифическими или биспецифическими.
4. Гуманизированные антитела
Изобретение охватывает гуманизированные антитела. Способы гуманизации антител, не являющихся человеческими, описаны в данной области. Как правило, гуманизированное антитело обладает одним или несколькими встроенными в него аминокислотными остатками из источника, который не является человеческим. Такие не являющиеся человеческими аминокислотные остатки часто называют "импортными" остатками, которые, как правило, взяты из "импортной" вариабельной области. Гуманизацию, главным образом, можно проводить в соответствии со способом Winter и коллег [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)], посредством замены CDR или последовательностями CDR крысы соответствующих последовательностей антитела человека. Таким образом, такие "гуманизированные" антитела представляют собой химерные антитела (патент США No. 4816567), где по существу менее целой вариабельной области человека замещено соответствующей последовательностью вида, не относящегося к человеку. На практике, гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и возможно некоторые остатки FR заменены остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека, как легких, так и тяжелых цепей, для использования при получении гуманизированных антител является очень важным для снижения антигенности. Согласно так называемому способу "наилучшего соответствия", последовательность вариабельного домена антитела грызуна анализируют относительно целой библиотеки известных последовательностей вариабельных доменов человека. Последовательность человека, которая является наиболее сходной с последовательностью грызуна, затем принимают в качестве каркасной области человека для гуманизированного антитела. См., например, Sims et al. (1993) J. Immunol, 151:2296; Chothia et al. (1987) J. Mol. Biol, 196:901. В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека из конкретной подгруппы легких или тяжелых цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител. См., например, Carter et al. (1992) Proc. Natl. Acad. Sci USA, 89:4285; Presta et al. (1993) J. Immunol, 151:2623.
Кроме того, обычно желательно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других положительных биологических свойств. Для достижения этой цели, в соответствии с предпочтительным способом гуманизированные антитела получают с помощью процесса анализа исходных последовательностей и различных предполагаемых гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широко доступными и известны специалистам в данной области. Доступными являются компьютерные программы, которые иллюстрируют и выводят на экран возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Исследование этих выведенных на экран данных позволяет проводить анализ возможной роли остатков в функционировании предполагаемых последовательностей иммуноглобулинов, т.е., анализ остатков, которые влияют на способность иммуноглобулина-кандидата связывать свой антиген. Таким образом, можно отобрать остатки FR и комбинировать их из реципиентной и импортной последовательностей таким образом, чтобы были достигнуты требуемые свойства антител, такие как повышенная аффинность к антигену(ам)-мишени. Как правило, остатки гипервариабельной области оказывают непосредственное и наиболее значительное влияние на связывание антигена.
5. Антитела человека
Антитела человека по изобретению можно конструировать путем комбинирования последовательности(ей) вариабельного домена клона Fv, выбранную из библиотек фагового дисплея, полученных из последовательностей человека, с известной последовательностью(ями) константного домена человека, как описано выше. Альтернативно моноклональные антитела человека по изобретению можно получать способом гибридом способ. Клеточные линии миеломы человека и гетеромиеломы мыши-человека для продукции моноклональных антител человека описаны, например, Kozbor J. Immunol, 133: 3001 (1984); Brodeur et al, Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al, J. Immunol, 147: 86 (1991).
В настоящее время возможным является получение трансгенных животных (например, мышей), которые при иммунизации способны продуцировать полный набор антител человека в отсутствии продукции эндогенных иммуноглобулинов. Например, было описано, что гомозиготная делеция гена участка соединения тяжелых цепей антител (JH) у химерных мышей и мутантных мышей зародышевой линии приводит к полному ингибированию продукции эндогенных антител. Перенос системы генов иммуноглобулинов зародышевой линии человека в таких мутантных мышей зародышевой линии приводит к продукции антител человека при антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggermann et al., Year in Immuno., 7:33 (1993).
Также для получения антител человека из не являющихся человеческими антител, например антител грызунов, можно использовать перетасовку цепей, где антитело человека имеет аффинность и специфичность, сходные с исходным не являющимся человеческим антителом. Согласно этому способу, который также называют "импринтингом эпитопов", либо вариабельную область тяжелой цепи, либо вариабельную область легкой цепи, не являющегося человеческим фрагмента антитела, полученного способами фагового дисплея, как описано в настоящем документе, заменяют набором генов V-доменов человека, создавая популяцию химер scFv или Fab с не являющейся человеческой цепью/являющейся человеческой цепью. Селекция с антигеном приводит к выделению химерного scFv или Fab с не являющееся человеческой цепью/являющейся человеческой цепью, где человеческая цепь восстанавливает антигенсвязывающий центр, разрушенный при удалении соответствующей не являющейся человеческой цепи в первичном клоне фагового дисплея, т.е. эпитоп определяет (осуществляет импринтинг) выбор цепи человека-партнера. Когда этот процесс повторяют для замены оставшейся не являющейся человеческой цепи, получают антитело человека (см. PCT WO 93/06213, опубликованную 1 апреля 1993 года). В отличие от традиционной гуманизации не являющихся человеческими антител путем пересадки CDR, этот способ обеспечивает полностью человеческие антитела, которые не имеют остатков FR или CDR происхождением не из человека.
6. Биспецифические антитела
Биспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностям связывания по отношению к по меньшей мере двум различным антигенам. В определенных вариантах осуществления биспецифические антитела представляют собой антитела человека или гуманизированные антитела. В определенных вариантах осуществления одна из специфичностей связывания направлена на Notch2, а другая направлена на другой антиген. В определенных вариантах осуществления биспецифические антитела могут связываться с двумя различными эпитопами Notch2. Биспецифические антитела также можно использовать для локализации цитотоксических средств у клеток, которые экспрессируют Notch2. Эти антитела обладают Notch2-связывающим плечом и плечом, которое связывает цитотоксическое средство, например, такое как сапорин, антитело против интерферона-α, алкалоид барвинка, A-цепь рицина, метотрексат или гаптен радиоактивного изотопа. Биспецифические антитела можно получать в качестве полноразмерных антител или фрагментов антител (например, биспецифические антитела F (ab')2).
Способы получения биспецифических антител известны в данной области. Традиционно, рекомбинантный способ получения полноразмерных биспецифических антител основан на совместной экспрессии двух пар тяжелая цепь - легкая цепь иммуноглобулинов, где две цепи обладают различной специфичностью (Millstein and Cuello, Nature, 305:537-539 (1983)). Вследствие случайной сборки тяжелых и легких цепей иммуноглобулинов, такие гибридомы (квадромы) потенциально продуцируют смесь из 10 различных молекул антител, из которых только одна обладает правильной биспецифичной структурой. Очистка правильных молекул, которую обычно проводят посредством стадий аффинной хроматографии, является довольно трудоемкой, и выход продукта является низким. Аналогичные способы описаны в WO 93/08829 опублекованном 13 мая 1993 года, и в Traunecker et al., EMBO J., 10:3655-3659 (1991).
В соответствии с другим подходом проводят слияние вариабельных доменов антитела с требуемой специфичностью связывания (участками связывания антитело-антиген) с последовательностями константных доменов иммуноглобулинов. Слияние проводят, например, с константным доменом тяжелой цепи Ig, содержащим по меньшей мере часть шарнирной области, CH2 и CH3. В определенных вариантах осуществления по меньшей мере один из компонентов, предназначенных для слияния, обладает первым константным доменом тяжелой цепи (CH1), содержащим участок, необходимый для связывания легкой цепи. ДНК, кодирующие предназначенные для слияния компоненты тяжелых цепей иммуноглобулинов и, если желательно, легкой цепи иммуноглобулина, встраивают в отдельные экспрессирующие векторы, и совместно трансфицируют в пригодный организм-хозяин. Это обеспечивает значительную гибкость при регулировании соотношений трех полипептидных фрагментов друг с другом в вариантах осуществления, где неравные соотношения трех полипептидных цепей в конструкции обеспечивает оптимальный выход требуемого биспецифического антитела. Однако возможным является встраивание двух или всех трех полипептидных цепей в один экспрессирующий вектор, если экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к высокому выходу продукта или если соотношения не имеют существенного влияния на требуемое сочетание цепей.
В одном варианте осуществления этого подхода, биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с одной специфичностью связывания на одном плече, и пары тяжелая цепь - легкая цепь гибридного иммуноглобулина (обладающей другой специфичностью связывания) на другом плече. Было обнаружено, что такая ассиметричная структура облегчает разделение требуемого биспецифического соединения и нежелательных сочетаний цепей иммуноглобулинов, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифичной молекулы обеспечивает легкий способ разделения. Этот подход описан в WO 94/04690. Для более подробного описания получения биспецифических антител см., например, Suresh et al., Methods in Enzymology, 121:210 (1986).
В соответствии с другим подходом, для максимального увеличения процентного количества гетеродимеров, которые выделяют из рекомбинантных клеточных культур, можно сконструировать область контакта между парой молекул антител. Область контакта содержит по меньшей мере часть CH3-домена константного домена антитела. В этом способе одну или несколько небольших боковых цепей аминокислот из области контакта первой молекулы антитела заменяют на более крупные боковые цепи (например, тирозин или триптофан). В области контакта второй молекулы антитела создают компенсирующие "полости" идентичного или сходного с крупной боковой цепью(ями) размера посредством замены крупных боковых цепей аминокислот на цепи меньших размеров (например, аланин или треонин). Это обеспечивает механизм для повышения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают поперечно-сшитые антитела или "гетероконъюгаты" антител. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое с биотином. Такие антитела, например, были предложены для нацеливания клеток иммунной системы против нежелательных клеток (патент США No. 4676980), и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373, и EP 03089). Гетероконъюгаты антител можно получать с использованием любого пригодного способа поперечного сшивания. Пригодные средства для поперечного сшивания хорошо известны в данной области, и описаны в патенте США No. 4676980, совместно с рядом способов поперечного сшивания.
Способы получения биспецифических антител из фрагментов антител также описаны в литературе. Например, биспецифические антитела можно получать с использованием образования химических связей. В Brennan et al., Science, 229: 81 (1985) описан способ, где полноразмерные антитела протеолитически расщепляют с получением F(ab')2-фрагментов. Эти фрагменты восстанавливают в присутствии вещества, образующего комплексы с дитиолами, арсенита натрия, для стабилизации соседних дитиолов и предотвращения образования межмолекулярного дисульфида. Затем полученные Fab'-фрагменты превращают в производные с тионитробензоатом (TNB). Одно из производных Fab'-TNB затем повторно превращают в Fab'-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB с получением биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве средств для селективной иммобилизации ферментов.
Последние достижения упростили прямое выделение из E. coli фрагментов Fab'-SH, которые можно химически связывать с получением биспецифических антител. В Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описано получение полностью гуманизированной молекулы F(ab')2 биспецифического антитела. Осуществляли секрецию из E. coli каждого Fab'-фрагмента по отдельности и их подвергали прямому химическому соединению in vitro с образованием биспецифического антитела. Биспецифическое антитело, полученное таким образом, обладало способностью связываться с клетками, сверхэкспрессирующими рецептор HER2, и нормальными T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против мишеней, представляющих собой рак молочной железы человека.
Также были описаны различные способы получения и выделения фрагментов биспецифических антител непосредственно из рекомбинантной клеточной культуры. Например, были получены биспецифические антитела с использованием лейциновых молний. Kositelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды лейциновой молнии белков Fos и Jun связывали с Fab'-участками двух различных антител посредством слияния генов. Гомодимеры антител восстанавливали в шарнирной области с получением мономеров, а затем повторно окисляли с получением гетеродимеров антител. Этот способ также можно использовать для получения гомодимеров антител. Способ "димеров антител", описанный Hollinger et al., Proc. Natl. Acad. Sci USA, 90:6444-6448 (1993) обеспечивает альтернативный механизм получения биспецифических фрагментов антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким для возможности образования пары между двумя доменами на одной и той же цепи. Таким образом, домены VH и VL одного фрагмента вынуждены образовывать пару с комплементарными доменами VL и VH другого фрагмента, таким образом, формируя два антигенсвязывающих центра. Также описана другая стратегия для получения фрагментов биспецифических антител с использованием димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol, 152: 5368 (1994).
Также представлены антитела более чем c двумя валентностями. Например, можно получать триспецифические антитела. Tutt et al. J. Immunol. 147: 60 (1991).
7. Поливалентные антитела
Поливалентное антитело может интернализовываться (и/или катаболизироваться) быстрее, чем двухвалентное антитело, клеткой, экспрессирующей антиген, с которым антитела связываются. Антитела по настоящему изобретению могут представлять собой поливалентные антитела (которые могут относиться к классу, отличному от класса IgM) с тремя или более антигенсвязывающими участками (например, четырехвалентные антитела), которые можно легко получать посредством рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Поливалентное антитело может содержать домен димеризации и три или более антигенсвязывающих участков. В определенных вариантах осуществления домен димеризации содержит (или состоит из них) Fc-участок или шарнирную область. В этом случае антитело будет содержать Fc-область и три или более антигенсвязывающих участка со стороны N-конца Fc-участка. В определенных вариантах осуществления поливалентное антитело, представленное в настоящем документе, содержит (или состоит из) от трех до приблизительно восьми антигенсвязывающих участков. В одном из вариантов осуществления поливалентное антитело содержит четыре антигенсвязывающих участка. Поливалентное антитело содержит по меньшей мере одну полипептидную цепь (и предпочтительно две полипептидные цепи), где полипептидная цепь(и) содержит два или более вариабельных доменов. Например, полипептидная цепь(и) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидуную цепь Fc-участка, X1 и X2 представляют собой аминокислоту или полипептид, и n равно 0 или 1. Например, полипептидная цепь(и) может содержать: цепь VH-CH1-подвижный линкер-VH-CH1-Fc-участок; или цепь VH-CH1-VH-CH1-Fc-участок. Поливалентное антитело, представленное в настоящем документе, например, может дополнительно содержать от приблизительно двух (например, четырех) полипептидов вариабельного домена легкой цепи. Поливалентное антитело, представленное в настоящем документе, например, может содержать от приблизительно двух до приблизительно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, рассматриваемые в настоящем документе, содержат вариабельный домен легкой цепи и, необязательно, дополнительно содержат CL-домен.
8. Однодоменные антитела
В некоторых вариантах осуществления антитело по изобретению представляет собой однодоменное антитело. Однодоменное антитело представляет собой единичную полипептидную цепь, содержащую весь вариабельный домен тяжелой цепи антитела или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В определенных вариантах осуществления одндодоменное антитело представляет собой однодоменное антитело человека (Domantis, Inc., Waltham, MA; см., например, патент США No. 6248516 B1). В одном варианте осуществления однодоменное антитело состоит из всего вариабельного домена тяжелой цепи антитела или его части.
9. Варианты антител
В некоторых вариантах осуществления представлена модификация(и) аминокислотной последовательности антител, описанных в настоящем документе. Например, может быть желательным повышение аффинности связывания и/или других биологических свойств антитела. Варианты антител по аминокислотной последовательности можно получать путем внесения соответствующих изменений в нуклеотидную последовательность, кодирующую антитело, или с помощью пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Для достижения конечного продукта можно проводить любую комбинацию из делеции, вставки и замены, при условии, что конечная конструкция обладает желаемыми характеристиками. Аминокислотные изменения можно вносить в рассматриваемую аминокислотную последовательность антитела во время получения этой последовательности.
Пригодный способ идентификации определенных остатков или участков антитела, которые являются предпочтительными участками для мутагенеза, называют "сканирующим аланином мутагенезом", как описано Cunningham and Wells (1989) Science, 244:1081-1085. В данном случае, выявляют остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют на нейтральную или отрицательно заряженную аминокислоту (наиболее предпочтительно аланин или полиаланин) для нарушения взаимодействия аминокислот с антигеном. Те положения аминокислот, которые демонстрируют функциональную чувствительность к замещению, затем подтверждают посредством внесения дополнительных или других вариантов в участки замещения, или вместо них. Таким образом, поскольку участок для внесения изменения в аминокислотную последовательность является предопределенным, нет необходимости в непосредственной предопределенности характера мутации. Например, для анализа действия мутации в данной области, в кодоне-мишени или участке-мишени проводят сканирующий ala или случайный мутагенез и экспрессированные варианты антитела против сфинголипида подвергают скринингу в отношении наличия требуемой активности.
Вставки в аминокислотную последовательность включают N- и/или C-концевые слитые последовательности, обладающие длиной в диапазоне от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или нескольких аминокислотных остатков внутрь последовательности. Примеры концевых вставок включают антитело против сфинголипида с N-концевым метионильным остатком или антитело, слитое с эпитопной меткой. Другие варианты вставок в молекулу антитела включают слияние N- или C-конца антитела с ферментом (например, ADEPT) или полипептидом, который повышает время полужизни антитела в сыворотке.
В определенных вариантах осуществления антитело по настоящему изобретению изменяют для повышения или снижения степени гликозилирования антитела. Гликозилирование полипептидов, как правило, является либо N-связанным, либо O-связанным. N-связанное гликозилирование относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту, за исключением пролина, представляют собой последовательности, распознаваемые для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, обычно, к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования к антителу удобно проводить, изменяя аминокислотную последовательность, чтобы происходило внесение или удаление одной или нескольких из описанных выше трипептидных последовательностей (для участков N-связанного гликозилирования). Также изменение можно проводить посредством добавления, делеции или замены одного или нескольких остатков серина или треонина в последовательности исходного антитела (для участков О-связанного гликозилирования).
Когда антитело содержит Fc-область, можно изменять углевод, связанный с ним. Нативные антитела, продуцируемые клетками млекопитающих, как правило, содержат разветвленный биантенный олигосахарид, который, как правило, связан N-связью с Asn297 CH2-домена Fc-области. См., например, Wright et al. (1997) TIBTECH 15:26-32. Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в "стебле" биантенной олигосахаридной структуры. В некоторых вариантах осуществления модификации олигосахарида в антителе по изобретению можно проводить для получения вариантов антител с определенными улучшенными свойствами.
Например, представлены варианты антител, имеющие углеводную структуру, которая лишена фукозы, присоединенной (прямо или непрямо) к Fc-области. Такие варианты могут иметь улучшенную функцию ADCC. См., например, публикации патентов США No. US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, относящихся к "дефукозилированным" или "дефицитным по фукозе" вариантам антител включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO2002/031140; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al Biotech. Bioeng. 87: 614 (2004). Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают клетки Lee 13 CHO, дефицитные по фукозилированию белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США No US 2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., особенно в примере 11), и клеточные линии с нокаутом, такие как клетки CHO с нокаутом генов альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al, Biotechnol. Bioeng., 94(4):680-688 (2006); и WO2003/085107).
Кроме того, представлены варианты антител с олигосахаридами из двух частей, например, в которых биантенный олигосахарид, связанный с Fc-областью антитела, разделен пополам посредством GlcNAc. Такие варианты антител могут иметь сниженное фукозилирование и/или улучшенную функцию ADCC. Примеры таких вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet et al); патенте США No. 6602684 (Umana et al); и US 2005/0123546 (Umana et al). Также представлены варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Такие варианты антител могут иметь улучшенную функцию CDC. Такие варианты антител описаны, например, в WO 1997/30087 (Patel et al); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
В определенных вариантах осуществления вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые дополнительно улучшают ADCC, например, замены в положениях 298, 333, и/или 334 Fc-области (нумерация остатков Eu). Такие замены могут встречаться в комбинации с любым из вариантов, описанных выше.
В определенных вариантах осуществления изобретение относится к варианту антитела, который обладает некоторыми, но не всеми эффекторными функциями, которые делают его желаемым кандидатом для многих применений, в которых важно время полужизни антитела in vivo, но, тем не менее определенные эффекторные функции (такие как комплементзависимая цитотоксичность и ADCC) являются необязательными или вредными. В определенных вариантах осуществления активность Fc антитела определяют для того, чтобы убедиться, что сохранены только желаемые свойства. Анализы цитотоксичности in vitro и/или in vivo можно проводить для подтверждения снижения/устранения видов активности CDC и/или ADCC. Например, анализы связывания Fc-рецепторов (FcR) можно проводить, чтобы убедиться, что антитело не связывает FcγR (таким образом, вероятно не имеет активности ADCC), однако сохраняет способность связывать FcRn. Главные опосредующие ADCC клетки, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках обобщенно представлена в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Неограничивающие примеры анализов in vitro для оценки активности ADCC представляющей интерес молекулы описаны в патенте США No. 5500362 (см., например Hellstrom, L, et al. Proc. Nat'l Acad. Sci. USA 83:7059-7063 (1986)) и Hellstrom, I., et al., Proc. Natl Acad. Sci. USA 82:1499-1502 (1985); 5821337 (см. Bruggemann, M. et al., J. Exp. Med. 166:1351-1361 (1987)). Альтернативно можно использовать нерадиоактивные способы анализа (см., например, нерадиоактивный анализ цитотоксичности ACTITM для проточной цитометрии (CellTechnology, Inc. Mountain View, CA); и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Madison, WI). Пригодные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и естественные киллерные (NK) клетки. Альтернативно или дополнительно, активность ADCC представляющей интерес молекулы можно оценивать in vivo, например, в модели на животных, такой как модель, описанная в Clynes et al. Proc. Nat'l Acad. Sci. USA 95:652-656 (1998). Также можно проводить анализы связывания C1q для подтверждения того, что антитело неспособно связывать C1q и, таким образом, лишено активности CDC. Для анализа активации комплемента можно проводить анализ CDC (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996); Cragg, M.S. et al., Blood 101:1045-1052 (2003); и Cragg, M.S. и M.J. Glennie, Blood 103:2738-2743 (2004)). Также можно проводить определение связывания FcRn и выведения/времени полужизни in vivo с использованием способов, известных в данной области (см., например, Petkova, S.B. et al., Int'l. Immunol. 18(12):1759-1769 (2006)).
Представлены другие варианты антител, имеющие одну или несколько аминокислотных замен. Представляющие интерес участки для мутагенеза с заменами включают гипервариабельные области, однако также предусмотрены изменения FR. Консервативные замены представлены в таблице 1 под заголовком "Предпочтительные замены". Более существенные изменения, обозначенные как "иллюстративные замены" предоставлены в таблице 1, или как дополнительно описано ниже в отношении классов аминокислот. Аминокислотные замены можно вносить в представляющее интерес антитело и продукты можно подвергать скринингу, например, в отношении желательной активности, такой как улучшенное связывание антигена, сниженная иммуногенность, повышенная ADCC или CDC, и т.д.
Модификации биологических свойств антитела можно проводить, выбирая замены, которые влияют на (a) структуру полипептидного остова в области замены, например, такую как конформация в виде слоя или спирали, (b) заряд или гидрофобность молекулы в заданном участке, или (c) объем боковой цепи. Аминокислоты можно подразделить на группы на основе сходства свойств их боковых цепей: (A. L. Lehninger, in Biochemistry, second ed., pp. 73-75, Worth Publishers, New York (1975)):
(1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M)
(2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (C), Tyr (Y), Asn (N), Gln (Q)
(3) кислотные: Asp (D), Glu (E)
(4) основные: Lys (K), Arg (R), His(H)
Альтернативно природные остатки можно подразделить на группы, исходя из общих свойств боковых цепей:
(1) гидрофобные: Норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены вовлекают замену члена одного из этих классов членом из другого класса. Такие замещенные остатки также можно вносить в участки консервативных замен или, более предпочтительно, в остальные (неконсервативные) участки.
Один из типов варианта с заменами включает замену одного или нескольких остатков гипервариабельного участка исходного антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный вариант(ы), выбранный для дальнейшей разработки, будет обладать модифицированными (улучшенными) биологическими свойствами относительно исходного антитела, из которого они получены. Иллюстративным вариантом с заменами является антитело, полученное созреванием аффинности, которое можно удобным образом получать способами созревания аффинности с использованием фагового дисплея. В кратком изложении несколько областей гипервариабельной области (например, 6-7 областей) подвергают мутации с получением всех возможных аминокислотных замен в каждом участке. Полученные таким образом варианты антитела экспонируются на частицах нитевидных фагов в качестве молекул, слитых по меньшей мере с частью белка оболочки фага (например, продукта гена III фага M13), упакованных в каждую частицу. Затем экспонированные на фаге варианты подвергают скринингу в отношении их биологической активности (например, аффинности связывания). В целях идентификации участков гипервариабельной области-кандидатов для модификации можно проводить сканирующий мутагенез (например, сканирование аланином), идентифицируя остатки гипервариабельной области, вносящие значительный вклад в связывание антигена. Альтернативно или дополнительно, может быть целесообразным анализ кристаллической структуры комплекса антиген-антитело в целях идентификации точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки являются кандидатами для замены в соответствии со способами, известными в данной области, включая способы, приведенные в настоящем документе. После получения таких вариантов панель вариантов подвергают скринингу с использованием способов, известных в данной области, включая способы, описанные в настоящем документе, и варианты с наилучшими свойствами в одном или нескольких соответствующих анализах можно отбирать для дальнейшей разработки.
Молекулы нуклеиновых кислот, кодирующие варианты антитела по аминокислотной последовательности, получают различными способами, известными в данной области. Эти способы включают, но не ограничиваются ими, выделение из природного источника (в случае природных вариантов аминокислотной последовательности) или получение олигонуклеотид-опосредуемым (или сайт-направленным) мутагенезом, мутагенезом посредством PCR, и кассетным мутагенезом ранее полученного варианта или не являющейся вариантом версии антитела.
Может быть желательным внесение одной или нескольких аминокислотных модификаций в Fc-область антител по изобретению, тем самым получая вариант по Fc-области. Вариант по Fc-области может включать последовательность Fc-области человека (например, Fc-области IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях, включая положение цистеина шарнирной области.
Согласно описанию и указаниям в данной области предусматривается, что в некоторых вариантах осуществления антитело по изобретению может содержать одно или несколько изменений по сравнению с альтернативным антителом дикого типа, например в Fc-области. Эти антитела, тем не менее, сохраняют по существу те же характеристики, которые требуются для терапевтической применимости по сравнению с их аналогом дикого типа. Например, полагают, что в Fc-область можно вносить определенные изменения, которые могут привести к измененному (т.е., либо повышенному, либо сниженному) связыванию C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в WO99/51642. Также см. Duncan & Winter, Nature 322:738-40 (1988); патент США No. 5648260; патент США No. 5624821; и WO94/29351, касающиеся других примеров вариантов Fc-области. В WO00/42072 (Presta) и WO 2004/056312 (Lowman) описаны варианты антител с улучшенным или сниженным связыванием с FcR. Содержание этих патентных публикаций конкретно включено в настоящий документ в качестве ссылки. Также см., Shields et al. J. Biol. Chem. 9(2): 6591-6604 (2001). Антитела с увеличенным временем полужизни и повышенным связыванием с неонатальным Fc-рецептором (FcRn), которые отвечают за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al, J. Immunol. 24:249 (1994)), описаны в US2005/0014934A1 (Hinton et al.). Эти антитела содержат Fc-область с одной или несколькими заменами в ней, которые повышают связывание Fc-области с FcRn. Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной способностью связывать C1q описаны в патенте США No. 6194551B1, WO99/51642. Содержание этих патентных публикаций конкретно включено в настоящий документ в качестве ссылки. Также см., Idusogie et al. J. Immunol. 164:4178-4184 (2000).
В другом аспекте изобретение относится к антителам, содержащим модификации на границе контакта Fc-полипептидов, содержащих Fc-область, где модификации способствуют гетеродимеризации и/или запускают ее. Эти модификации включают внесение выступа в первый Fc-полипептид и полости во второй Fc-полипептид, где выступ может располагаться в полости так, чтобы обеспечить образование комплекса первого и второго Fc-полипептидов. Способы получения антител с этими модификациями известны в данной области, например, как описано в патенте США No. 5731168.
В другом аспекте, может быть желательным создание антител со встроенным способами инженерии цистеином, например, "тио-MAb", в которых один или несколько остатков антитела замещены остатками цистеина. В конкретных вариантах осуществления замещенные остатки находятся в доступных участках антитела. Путем замены этих остатков цистеином, в доступные участки антитела помещают реакционноспособные тиольные группы и их можно использовать для конъюгации антитела с другими группами, такими как группы лекарственных средств или группы линкер-лекарственное средство, как описано в настоящем документе далее. В определенных вариантах осуществления цистеином может быть заменен один или несколько из следующих остатков: V205 (нумерация по Kabat) легкой цепи; A118 (нумерация EU) тяжелой цепи; и S400 (нумерация EU) Fc-области тяжелой цепи.
10. Производные антител
Антитела по настоящему изобретению можно дополнительно модифицировать, чтобы они содержали дополнительные небелковые группы, которые известны в данной области и легкодоступны. Предпочтительно, группы, пригодные для получения производного антитела, представляют собой растворимые в воде полимеры. Неограничивающие примеры растворимых в воде полимеров включают, но не ограничиваются ими, полиэтиленгликоль (PEG), сополимеры этиленгликоль/пропиленгликоль, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилен/малеиновый ангидрид, полиаминокислоты (либо гомополимеры, либо случайные сополимеры), и декстран или поли(н-винил пирролидон)полиэтиленгликоль, пропропиленгликоль гомополимеры, сополимеры окись полипропилена/окись этилена, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества при изготовлении вследствие его стабильности в воде. Полимер может иметь любую молекулярную массу, и он может быть разветвленным или неразветвленным. Количество полимеров, связанных с антителом, может варьировать, и если связано более одного полимера, они могут представлять собой одинаковые или разные молекулы. Как правило, количество и/или тип полимеров, используемых для получения производного, можно определять, исходя из соображений, включающих, но не ограничивающихся ими, конкретные свойства или функции антитела, подлежащего усовершенствованию, применения производного антитела в терапии в определенных условиях и т.д.
В другом варианте осуществления представлены конъюгаты антитела и небелковой группы, которые можно селективно нагревать путем воздействия облучения. В одном варианте осуществления небелковая группа представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Облучение может быть любой длины волны, и оно включает, но не ограничивается ими, длины волн, которые не повреждают обычные клетки, но которые нагревают небелковую группу до температуры, при которой происходит уничтожение клеток вблизи конъюгата антитело-небелковая групп.
B. Определенные способы получения антител
1. Определенные способы на основе гибридом
Моноклональные антитела по изобретению можно получать с использованием способа гибридом, впервые описанного Kohler et al, Nature, 256:495 (1975), и, кроме того, описанного, например, в Hongo et al, Hybridoma, 14 (3): 253-260 (1995), Harlow et al, Antobodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al, in: Monoclonal Antibodies and T-Cell Hibridomas 563-681 (Elsevier, N.Y., 1981), и Ni, Xiandai Mianyixue, 26(4):265-268 (2006) в отношении гибридом человека-человека. Дополнительные способы включают способы, описанные, например, в патенте США No. 7189826 в отношении продукции моноклональных природных антител IgM человека из гибридомных клеточных линий. Технология гибридом человека (технология триом) описана в Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3):185-91 (2005).
Для различных других способов гибридом, см., например, US 2006/258841; US 2006/183887 (полностью человеческие антитела), US 2006/059575; US 2005/287149; US 2005/100546; US 2005/026229; и патенты США No. 7078492 и 7153507. Иллюстративный протокол для получения моноклональных антител с использованием способа гибридом описан далее. В одном варианте осуществления мышь или другое подходящее животное-хозяина, такое как хомячок, иммунизируют для индукции лимфоцитов, которые продуцируют или способны продуцировать антитела, которые специфично связываются с белком, используемым для иммунизации. Антитела индуцируют у животных путем множества подкожных (sc) или внутрибрюшинных (ip) инъекций полипептида, содержащего Notch2 или его фрагмент (например, NRR Notch2), и адъювант, такой как монофосфориллипид A (MPL)/дикриномиколят трегалозы (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT). Полипептид, содержащий Notch2 или его фрагмент, можно получать с использованием способов, хорошо известных в данной области, таких как рекомбинантные способы, некоторые из которых дополнительно описаны в настоящем документе. Сыворотку иммунизированных животных анализируют в отношении антител против Notch2, и необязательно проводят вспомогательные иммунизации. Лимфоциты животных, продуцирующих антитела против Notch2, выделяют. Альтернативно лимфоциты можно иммунизировать in vitro.
Затем лимфоциты подвергают слиянию с клетками миеломы с использованием пригодного средства для слияния, такого как полиэтиленгликоль, с образованием гибридомных клеток. См., например, Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986). Клетки миеломы можно использовать для эффективного слияния, поддержания стабильной продукции антитела на высоком уровне с помощью выбранных антителопродуцирующих клеток, и они являются чувствительными к среде, такой как среда HAT. Иллюстративные клетки миеломы включают, но не ограничиваются ими, линии миеломы мыши, такие как линии, происходящие из опухолей мыши MOPC-21 и MPC-11, доступные от Salk Institute Cell Distribution Center, San Diego, California USA, и клетки SP-2 или X63-Ag8-653, доступные от American Type Culture Collection, Rockville, Maryland USA. Также для продукции моноклональных антител человека описаны миеломные клеточные линии и линии гетеромиеломы мыши-человека (Kozbor, J. Immunol, 133:3001 (1984); и Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Полученные таким образом гибридомные клетки высевают и выращивают в пригодной культуральной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживание неслитых исходных клеток миеломы. Например, если исходные клетки миеломы лишены фермента гипоксантин-гуанин-фосфорибозил-трансферазы (HGPRT или HPRT), культуральная среда для гибридом, как правило, включает гипоксантин, аминоптерин и тимидин (среда HAT), которые препятствуют росту клеток, дефицитных по HGPRT. Предпочтительно, используют бессывороточные способы культивирования гибридомных клеток для уменьшения применения полученной от животного сыворотки, такой как эмбриональная телячья сыворотка, как описано, например, в Even et al., Trends in Biotechnology, 24(3), 105-108 (2006).
Олигопептиды в качестве инструментов повышения продуктивности гибридомных клеточных культур описаны в Franek, Trends in Monoclonal Antibody Research, 111-122 (2005). Конкретно, стандартную культуральную среду обогащают определенными аминокислотами (аланином, серином, аспарагином, пролином), или фракциями белкового гидролизата, и апоптоз можно значительно подавлять с помощью синтетических олигопептидов, сконструированных из от трех до шести аминокислотных остатков. Пептиды присутствуют в миллимолярных или более высоких концентрациях.
Культуральную среду, в которой выращивают гибридомные клетки, оценивают на продукцию моноклональных антител, которые связывают Notch2. Специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют посредством иммунопреципитации или анализа связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Аффинность связывания моноклонального антитела можно определять, например, посредством анализа Скэтчарда, описанного в Munson et al., Anal. Biochem., 107:220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с требуемой специфичностью, аффинностью и/или активностью, клоны можно субклонировать способами лимитирующих разведений и выращивать стандартными способами. См., например, Goding, выше. Пригодные для этой цели культуральные среды включают, например, среду D-MEM или RPMI-1640. Кроме того, гибридомные клетки можно выращивать in vivo у животных в качестве асцитных опухолей. Моноклональные антитела, секретируемые субклонами, пригодным образом выделяют из культуральной среды, асцитной жидкости или сыворотке с помощью общепринятых способов очистки иммуноглобулинов, например, таких как хроматография с белком A-Sepharose, хроматография с гидроксилапатитом, гель-электрофорез, диализ или аффинная хроматография. Один из способов выделения белков из гибридомных клеток описан в US 2005/176122 и патенте США No. 6919436. Способ включает применение минимальных солей, таких как лиотропные соли, в процессе связывания, а также предпочтительно с использованием небольших количеств органических растворителей в процессе элюирования.
2. Некоторые способы скрининга библиотек
Антитела по изобретению можно получать с использованием комбинаторных библиотек для скрининга антител с требуемой активностью или видами активности. Например, в данной области известны различные способы создания библиотек фагового дисплея и скрининга таких библиотек в отношении антител, обладающих требуемыми характеристиками связывания. Такие способы описаны, главным образом, в Hoogenboom et al., in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001). Например, один способ получения представляющих интерес антител осуществляют с использованием фаговой библиотеки антител, как описано в Lee et al. (2004) J. Mol. Biol. 340:1073-1093.
По существу, клоны синтетических антител повергают селекции посредством библиотек фагового дисплея, которые содержат фаг, который экспонирует различные фрагменты вариабельной области (Fv) антитела, слитые с белком оболочки фага. Такие фаговые библиотеки сортируют с помощью аффинной хроматографии против требуемого антигена. Клоны, экспрессирующие Fv-фрагменты, способные связываться с требуемым антигеном, адсорбируются на антиген и, таким образом, их отделяют от несвязывающих клонов в библиотеке. Затем связывающие клоны элюируют от антигена, и их содержание можно далее повышать посредством дополнительных циклов адсорбции/элюирования антигена. Путем разработки пригодного способа скрининга с помощью антигена для селекции представляющего интерес клона фага с последующим конструированием клона полноразмерного антитела с использованием последовательностей Fv из представляющего интерес фагового клона и пригодных последовательностей константной области (Fc), описанных в Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3, можно получать любые из антител по изобретению.
В определенных вариантах осуществления антигенсвязывающий домен антитела образован из двух вариабельных (V) областей приблизительно из 110 аминокислот, по одной из легкой (VL) и тяжелой (VH) цепей, в обеих из которых присутствуют три гипервариабельные петли (HVR) или определяющие комплементарность области (CDR). Вариабельные домены могут функционально экспонироваться на фаге, либо в качестве одноцепочечных Fv-фрагментов (scFv), в которых VH и VL ковалентно связаны через короткий подвижный пептид, либо в качестве Fab-фрагментов, в которых они обе являются слитыми с константным доменом и взаимодействуют нековалентно, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). В настоящем документе фаговые клоны, кодирующие scFv, и фаговые клоны, кодирующие Fab, совместно называют "фаговыми клонами Fv" или "клонами Fv".
Репертуары генов VH и VL можно по отдельности клонировать полимеразной цепной реакцией (ПЦР) и подвергать случайной рекомбинации в фаговых библиотеках, в которых затем проводят поиск антигенсвязывающих клонов, как описано в Winter et al., Ann. Rev. Immunol, 12: 433-455 (1994). Библиотеки из иммунизированных источников обеспечивают высокоаффинные антитела к иммуногену без необходимости конструирования гибридом. Альтернативно, можно клонировать наивный репертуар для обеспечения одного источника антител человека к широкому диапазону несобственных, а также собственных антигенов без иммунизации, как описано Griffiths et al., EMBO J, 12: 725-734 (1993). В заключение, библиотеки с наивным репертуаром также можно получать синтетически путем клонирования не подвергнутых реаранжировке сегментов V-генов из стволовых клеток, и использования праймеров для ПЦР, содержащих случайную последовательность, для кодирования высоко вариабельных областей CDR3 и для проведения реаранжировки in vitro, как описано Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992).
В определенных вариантах осуществления используют нитевидные фаги для экспонирования фрагментов антител путем слияния с минорным белком оболочки pIII. Фрагменты антител могут быть экспонированы в виде одноцепочечных Fv-фрагментов, в которых VH- и VL-домены связаны на одной полипептидной цепи подвижным полипептидным спейсером, например, как описано в Marks et al., J. Mol. Biol, 222: 581-597 (1991), или в качестве Fab-фрагментов, в которых одна цепь является слитой с pIII, а другая секретируется в периплазму бактериальной клетки-хозяина, где собирается структура Fab-белок оболочки, которая экспонируется на поверхности фага путем вытеснения некоторых из белков оболочки дикого типа, например, как описано в Hoogenboom et al., Nucl. Acids Res., 19: 4133-4137 (1991).
Как правило, нуклеиновые кислоты, кодирующие фрагменты гена антитела, получают из иммунных клеток, полученных от человека или животных. Если является желательной библиотека, смещенная в сторону клонов, направленных против Notch2 (клонов против NRR Notch2), индивидуума иммунизируют посредством Notch2 (или NRR Notch2) для обеспечения антительного ответа, и выделяют клетки селезенки и/или циркулирующие B-клетки, и другие лимфоциты периферической крови человека (PBL) для конструирования библиотеки. В предпочтительном варианте осуществления получают библиотеку фрагментов гена антитела человека, смещенную в сторону клонов, направленных против Notch2, путем обеспечения ответа антител против Notch2 у трансгенной мыши, обладающей функциональным набором генов иммуноглобулинов человека (и лишенной функциональной эндогенной системы продукции антител), так что иммунизация посредством Notch2 приводит к возникновению B-клеток, продуцирующих антитела человека против Notch2. Создание продуцирующих антитела человека трансгенных мышей описано ниже.
Дополнительное повышение содержания популяций клеток, реактивных против Notch2, можно обеспечивать с использованием пригодного способа скрининга для выделения B-клеток, экспрессирующих специфичное к Notch2 мембраносвязанное антитело, например, путем разделения клеток с использованием аффинной хроматографии с Notch2 или адсорбции клеток на меченный флуорохромом Notch2 с последующей активированной флуоресценцией сортировкой клеток (FACS).
Альтернативно применение клеток селезенки и/или B-клеток или других PBL от неиммунизированного донора обеспечивает лучшее представление возможного репертуара антител, и также позволяет конструирование библиотеки антител с использованием любого вида животных (человека или не относящегося к человеку), в котором Notch2 не является антигенным. Для библиотек, включающих конструирование генов антител in vitro, собирают стволовые клетки субъекта для получения нуклеиновых кислот, кодирующих не подвергнутые реаранжировке сегменты генов антител. Представляющие интерес иммунные клетки можно получать из различных видов животных, таких как человек, мышь, крыса, видов зайцеобразных, волчьих, собачьих, кошачьих, свиней, крупного рогатого скота, лошадей и птиц и т.д.
Нуклеиновую кислоту, кодирующую вариабельные сегменты генов (включая сегменты VH и VL) выделяют из представляющих интерес клеток и амплифицируют. В случае подвергнутых реаранжировке библиотек генов VH и VL, требуемую ДНК можно получать путем выделения геномной ДНК или мРНК из лимфоцитов с последующей полимеразной цепной реакцией (ПЦР) с праймерами, совпадающими с 5'- и 3'-концами подвергнутых реаранжировке генов VH и VL, как описано в Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86: 3833-3837 (1989), тем самым получая разнообразные репертуары V-генов для экспрессии. V-гены можно амплифицировать из кДНК и геномной ДНК с помощью обратных праймеров на 5'-конце экзона, кодирующего зрелый V-домен, и прямых праймеров, располагающихся в J-сегменте, как описано в Orlandi et al. (1989) и Ward et al., Nature, 341:544-546 (1989). Однако для амплификации из кДНК, обратные праймеры также могут располагаться в лидерном экзоне, как описано в Jones et al., Biotechnol., 9:88-89 (1991), а прямые праймеры могут располагаться в константной области, как описано в Sastry et al., Proc. Natl. Acad. Sci. (USA), 86: 5728-5732 (1989). Для максимального повышения комплементарности, в праймеры можно включать вырожденность, как описано в Orlandi et al. (1989) или Sastry et al. (1989). В определенных вариантах осуществления разнообразие библиотеки максимально увеличивают с использованием праймеров ПЦР, направленных на каждое семейство V-генов для амплификации всех доступных расположений VH и VL, присутствующих в образце нуклеиновой кислоты иммунных клеток, например, как описано в способе Marks et al., J. Mol. Biol., 222:581-597 (1991) или как описано в способе Orum et al., Nucleic Acids Res., 21:4491-4498 (1993). Для клонирования амплифицированной ДНК в экспрессирующие векторы, в праймер для ПЦР можно вносить редкие участки рестрикции в качестве метки на одном конце, как описано в Orlandi et al. (1989), или путем дальнейшей амплификации посредством ПЦР с праймером с меткой, как описано в Clackson et al., Nature, 352: 624-628 (1991).
Репертуары подвергнутых синтетической реаранжировке V-генов можно получать in vitro из сегментов V-генов. Большинство из сегментов VH-генов человека были клонированы и отсеквенированы (как описано в Tomlinson et al., J. Mol. Biol, 227: 776-798 (1992)), и картированы (описано в Matsuda et al., Nature Genet., 3: 88-94 (1993); эти клонированные сегменты (включая все главные конформации петли H1 и H2) можно использовать для получения разнообразных репертуаров VH-генов с помощью праймеров для ПЦР, кодирующих петли H3 с разнообразной последовательностью и длиной, как описано в Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992). Также можно получать репертуары VH, где все разнообразие последовательности сфокусировано в длинной петле H3 с единой длиной, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 89:4457-4461 (1992). Сегменты Vκ и Vλ человека были клонированы и отсеквенированы (описано в Williams and Winter, Eur. J. Immunol., 23:1456-1461 (1993)) и их можно использовать для получения синтетических репертуаров легких цепей. Синтетические репертуары V-генов, на основе ряда сборок VH и VL, и длин L3 и H3, кодируют антитела со значительным структурным разнообразием. После амплификации ДНК, кодирующей V-ген, сегменты V-генов зародышевой линии можно подвергать реаранжировке in vitro в соответствии со способами Hoogenboom and Winter, J. Mol. Biol, 227:381-388 (1992).
Репертуары фрагментов антител можно конструировать, комбинируя репертуары генов VH и VL различными путями. Каждый репертуар можно создавать в отдельных векторах, и векторы подвергать рекомбинации in vitro, например, как описано в Hogrefe et al., Gene, 128: 119-126 (1993), или in vivo путем комбинаторной инфекции, например, с помощью системы loxP, описанной в Waterhouse et al., Nucl. Acids Res., 21: 2265-2266 (1993). В подходе рекомбинации in vivo используется двухцепочечная структура Fab-фрагментов для преодоления ограничения размера библиотеки, определяемого эффективностью трансформации E. coli. Наивные репертуары VH и VL клонируют по отдельности, один в плазмиду, а другой в фаговый вектор. Затем две библиотеки комбинируют посредством инфекции фагом содержащих фагмиду бактерий, так что каждая клетка содержит отличающееся сочетание, и размер библиотеки ограничивается только числом присутствующих клеток (приблизительно 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo, так что гены VH и VL рекомбинируют в один репликон и совместно упаковываются в фаговые вирионы. Эти огромные библиотеки обеспечивают большие количества разнообразных антител с высокой аффинностью (Kd -1 приблизительно 10-8 M).
Альтернативно репертуары можно клонировать последовательно в один вектор, например, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или объединять посредством ПЦР, а затем клонировать, например как описано в Clackson et al., Nature, 352:624-628 (1991). Сборку посредством ПЦР также можно использовать для связывания ДНК VH и VL с ДНК, кодирующей подвижный пептидный спейсер, с образованием репертуаров одноцепочечных Fv (scFv). В другом способе используют "сборку посредством ПЦР в клетках" для объединения генов VH и VL в лимфоцитах посредством ПЦР, а затем клонирования репертуаров связанных генов, как описано в Embleton et al., Nucl. Acids Res., 20:3831-3837 (1992).
Антитела, продуцируемые наивными библиотеками (либо природными, либо синтетическими) могут иметь умеренную аффинность (Kd -1 приблизительно от 106 до 107 M-1), однако созревание аффинности также можно имитировать in vitro посредством конструирования и повторной селекции из вторичных библиотек, как описано в Winter et al. (1994), выше. Например, мутацию можно вносить случайным образом in vitro с использованием полимеразы с пониженной точностью (описано Leung et al., Technique, 1:11-15 (1989)) в способе Hawkins et al., J. Mol. Biol., 226:889-896 (1992) или в способе Gram et al., Proc. Natl. Acad. Sci USA, 89: 3576-3580 (1992). Кроме того, созревание аффинности можно проводить путем внесения мутации случайным образом в одну или несколько CDR, например, с использованием ПЦР с праймерами, имеющими случайную последовательность, охватывающую представляющую интерес CDR, в выбранных отдельных клонах Fv, и скринингом на клоны с наиболее высокой аффинностью. В WO 9607754 (опубликованной 14 марта 1996 года) описан способ индукции мутагенеза в определяющей комплементарность области легкой цепи иммуноглобулина для создания библиотеки генов легкой цепи. Другим эффективным подходом является рекомбинация VH- или VL-доменов, отобранных посредством фагового дисплея, с репертуарами природных вариантов V-доменов, полученными из неиммунизированных доноров, и скрининг на наиболее высокую аффинность в нескольких раундах повторной перетасовки цепей, как описано в Marks et al., Biotechnol., 10:779-783 (1992). Этот способ позволяет продукцию антител и фрагментов антител с аффинностью приблизительно 10-9 M или менее.
Скрининг библиотек можно проводить различными способами, известными в данной области. Например, Notch2 (или NRR Notch2) можно использовать для нанесения на лунки адсорбционных планшетов, экспрессировать на клетках-хозяевах, фиксированных на адсорбционных планшетах или использовать в сортировке клеток, или конъюгировать с биотином для улавливания покрытыми стрептавидином гранулами, или использовать в любом способе пэннинга библиотек фагового дисплея.
Образцы для фагового дисплея подвергают контактированию с иммобилизованным Notch2 в условиях, пригодных для связывания по меньшей мере части фаговых частиц с адсорбентом. Обычно выбирают условия, включая pH, ионную силу, температуру и т.п., имитирующие физиологические условия. Фаги, связанные с твердой фазой, промывают, а затем элюируют кислотой, например, как описано в Barbas et al., Proc. Natl. Acad. Sci USA, 88:7978-7982 (1991), или щелочью, например, как описано в Marks et al., J. Mol. Biol., 222:581-597 (1991), или посредством конкуренции с антигеном Notch2, например в способе, сходным со способом конкуренции с антигеном в Clackson et al., Nature, 352:624-628 (1991). За один раунд селекции фаги можно обогащать в 20-1000 раз. Более того, обогащенные фаги можно выращивать в бактериальной культуре и подвергать последующим раундам селекции.
Эффективность селекции зависит от множества факторов, включая кинетику диссоциации при промывании, и возможность одновременной встречи множества фрагментов антител с антигеном. Антитела с быстрой кинетикой диссоциации (и слабой аффинностью связывания) можно задерживать с использованием короткого промывания, поливалентного фагового дисплея и высокой плотности нанесения антигена на твердую фазу. Высокая плотность не только стабилизирует фаг при поливалентных взаимодействиях, но также способствует повторному связыванию фага, который диссоциировал. Селекцию антител с низкой кинетикой диссоциации (и высокой аффинностью связывания) можно обеспечивать, используя длительные промывания и одновалентный фаговый дисплей, как описано в Bass et al., Proteins, 8:309-314 (1990) и в WO 92/09690, и низкую плотность нанесения антигена, как описано в Marks et al., Biotechnol., 10:779-783 (1992).
Селекцию между фаговыми антителами с различной аффинностью к Notch2 можно проводить даже при аффинности, которая отличается немного. Однако случайная мутация выбранного антитела (например, как происходит в некоторых способах созревания аффинности) может привести к возникновению многих мутантов, большинство из которых связываются с антигеном, и немногие из которых обладают более высокой аффинностью. При ограничении Notch2, редкий высокоаффинный фаг может быть конкурентно устранен. Для сохранения всех мутантов с более высокой аффинностью, фаги можно инкубировать с избытком биотинилированного Notch2, но с биотинилированным Notch2 в концентрации с более низкой молярностью, чем предполагаемая молярная аффинность, постоянная для Notch2. Затем связывающие с высокой аффинностью фаги можно улавливать покрытыми стрептавидином парамагнитными гранулами. Такое "равновесное улавливание" позволяет селекцию антител согласно их аффинности связывания, с чувствительностью, которая позволяет выделение мутантных клонов с аффинностью, большей только в два раза, из большого избытка фагов с более низкой аффинностью. Также можно изменять условия, используемые для промывания фагов, связанных с твердой фазой, для различения на основе кинетики диссоциации.
Клоны, направленные против Notch2, можно отбирать на основе активности. В определенных вариантах осуществления изобретение относится к антителам против Notch2, которые связываются с живыми клетками, которые естественным образом экспрессируют Notch2. В одном варианте осуществления изобретение относится к антителам против Notch2, которые блокируют связывание между лигандом Notch2 и Notch2, но не блокируют связывание между лигандом Notch2 и вторым белком. Клоны Fv, соответствующие таким антителам против Notch2, можно подвергать селекции посредством (1) выделения клонов, направленных против Notch2, из фаговой библиотеки, как описано выше, и необязательно размножения выделенной популяции фаговых клонов путем выращивания популяции в пригодном бактериальном хозяине; (2) селекции Notch2 и второго белка, против которого является желательной блокирующая и неблокирующая активность, соответственно; (3) адсорбции фаговых клонов, направленных против Notch2, на иммобилизованный Notch2; (4) применения избытка второго белка для элюирования каких-либо нежелательных клонов, которые распознают связывающие Notch2 детерминанты, которые перекрываются или являются общими со связывающими детерминантами второго белка; и (5) элюирования клонов, которые остаются адсорбированными после стадии (4). Необязательно, клоны с требуемыми блокирующими/неблокирующими свойствами можно дополнительно обогащать путем повторения процессов селекции, описанных в настоящем документе, один или несколько раз.
ДНК, кодирующую образованные из гибридомы моноклональные антитела или клоны Fv фагового дисплея по изобретению, легко выделять и секвенировать с использованием общепринятых способов (например, с использованием олигонуклеотидных праймеров, сконструированных для специфичной амплификации представляющих интерес кодирующих участков тяжелой и легкой цепей из гибридомы или ДНК-матрицы фага). После выделения ДНК можно помещать в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьяны, клетки яичника китайского хомячка (CHO), или клетки миеломы, которые в ином случае не продуцируют белок иммуноглобулина, обеспечивая синтез требуемых моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитело, включают Skerra et al., Curr. Opinion in Immunol., 5:256 (1993) и Pluckthun, Immunol. Revs, 130:151 (1992).
ДНК, кодирующую клоны Fv по изобретению, можно комбинировать с известными последовательностями ДНК, кодирующими константные участки тяжелой цепи и/или легкой цепи (например, соответствующие последовательности ДНК можно получать в соответствии с Kabat et al., выше) с образованием клонов, кодирующих полноразмерные или неполные тяжелые и/или легкие цепи. Будет понятно, что для этой цели можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получать от человека или из любого вида животных. Клон Fv, образованный из ДНК вариабельного домена одного вида животных (такого как человек), а затем слитый с ДНК константной области другого вида животных с образованием кодирующей последовательности(ей) "гибридной" полноразмерной тяжелой цепи и/или легкой цепи, включен в определение "химирного" и "гибридного" антитела как используют в настоящем документе. В определенных вариантах осуществления клон Fv, образованный из ДНК вариабельной области человека, является слитым с ДНК константной области человека с образованием кодирующей последовательности(ей) для полноразмерной или неполной тяжелой и/или легкой цепей.
ДНК, кодирующую антитело против Notch2, происходящую из гибридомы, также можно модифицировать, например, путем замены кодирующей последовательности константными доменами тяжелой и легкой цепей вместо гомологичных последовательностей мыши, происходящих из клона гибридомы (например, как в способе Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). ДНК, кодирующую происходящее из гибридомы или Fv-клона антитело или фрагмент, можно дополнительно модифицировать ковалентным связыванием с кодирующей последовательностью иммуноглобулина всей кодирующей последовательности не являющегося иммуноглобулином полипептида, или ее части. Таким образом, получают "химерные" или "гибридные" антитела, которые обладают специфичностью связывания происходящих из Fv-клона или клона гибридомы антител по изобретению.
3. Векторы, клетки-хозяева и рекомбинантные способы
Также антитела можно получать с использованием рекомбинантных способов. Для рекомбинантной продукции антитела против Notch2 нуклеиновую кислоту, кодирующую антитело, выделяют и встраивают в реплицируемый вектор для последующего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, можно легко выделять и секвенировать с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелую и легкую цепи антитела). Доступно множество векторов. Компоненты векторов, как правило, включают, но не ограничиваются ими, один или несколько из следующих компонентов: сигнальная последовательность, ориджин (начало) репликации, один или несколько генов селективных маркеров, энхансерный элемент, промотор и последовательность терминации транскрипции.
a) Компонент - сигнальная последовательность
Антитело по изобретению можно получать рекомбинантными способами не только непосредственно, но также в качестве полипептида, слитого с гетерологичным полипептидом, который предпочтительно представляет собой сигнальную последовательность или другой полипептид, обладающий участком для специфичного расщепления на N-конце зрелого белка или полипептида. Выбранная гетерологичная сигнальная последовательность предпочтительно представляет собой последовательность, которая распознается и преобразуется (т.е., расщепляется сигнальной пептидазой) в клетке-хозяине. В случае прокариотических клеток-хозяев, которые не распознают и не преобразуют нативную сигнальную последовательность антитела, сигнальную последовательность можно заменять прокариотической сигнальной последовательностью, выбранной, например, из группы из лидерных последовательнотсей щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II. В случае секреции в дрожжах, нативную сигнальную последовательность можно заменять, например, лидерной последовательностью инвертазы дрожжей, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора Saccharomyces и Kluyveromyces) или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы C. albicans или сигнальной последовательностью, описанной в WO 90/13646. Для экспрессии в клетках млекопитающих, доступны сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, сигнальная последовательность gD вируса простого герпеса.
b) Ориджин репликации
Векторы как для экспрессии, так и для клонирования, содержат последовательность нуклеиновой кислоты, которая обеспечивает репликацию вектора в одной или нескольких выбранных клетках-хозяевах. Как правило, в векторах для клонирования эта последовательность представляет собой последовательность, которая обеспечивает репликацию вируса независимо от хромосомной ДНК хозяина, и включает ориджины репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для множества бактерий, дрожжей и вирусов. Ориджин репликации из плазмиды pBR322 является пригодным для большинства грамотрицательных бактерий, плазмидный ориджин 2µ пригоден для дрожжей, и различные вирусные ориджины (SV40, вируса полиомы, аденовируса, VSV или BPV) пригодны для клонирования векторов в клетках млекопитающих. Как правило, для экспрессирующих векторов млекопитающих компонент, представляющий собой ориджин репликации, не требуется (ориджин SV40, главным образом, можно использовать только потому, что он содержит ранний промотор).
с) Компонент - селективный ген
Экспрессирующие и клонирующие векторы могут содержать ген для селекции, также называемый селективным маркером. Типичные гены для селекции кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например, ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефекты ауксотрофов или (c) обеспечивают основные питательные вещества, которые не доступны из комплексных сред, например, ген, кодирующий D-аланинрацемазу для Bacilli.
В одном примере схемы селекции используют лекарственное средство для остановки роста клетки-хозяина. Клетки, которые успешно трансформированы гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному средству и, таким образом, выживают после режима селекции. Примерами такого доминантного вида селекции является применение лекарственных средств неомицина, микофеноловой кислоты и гигромицина.
Другим примером пригодных селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентификацию клеток, способных захватывать нуклеиновую кислоту, кодирующую антитело, такие как гены DHFR, глутаминсинтетазу (GS), тимидинкинизы, металлотеонеина-I и -II, предпочтительно гены металлотеонеина приматов, аденозиндезаминазы, орнитиндекарбоксилазы и т.д.
Например, клетки, трансформированные геном для селекции DHFR, сначала идентифицируют культивированием всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. В этих условиях ген DHFR амплифицируется вместе с любой другой совместно трансформированной нуклеиновой кислотой. При использовании DHFR дикого типа, пригодные клетки-хозяева представляют собой клеточную линию яичника китайского хомячка (CHO), дефицитную по активности DHFR (например, ATCC CRL-9096).
Альтернативно, клетки, трансформированные геном CS, выявляют культивированием трансформантов в культуральной среде, содержащей L-метионинсульфоксимин (Msx), ингибитор GS. В этих условиях ген GS амплифицируется вместе с любой другой совместно трансформированной нуклеиновой кислотой. Систему для селекции/амплификации GS можно использовать в комбинации с системой селекции/амплификации DHFR, описанной выше.
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, которые содержат эндогенный DHFR), трансформированные или совместно трансформированные с последовательностями ДНК, кодирующими антитело по изобретению, ген DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), можно отбирать выращиванием клеток на среде, содержащей вещество для селекции по селективному маркеру, такое как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См. патент США No. 4965199.
Пригодным геном для селекции для применения в дрожжах является ген trp1, находящийся в дрожжевой плазмиде YRp7 (Stinchcomb et al., Nature, 282:39 (1979)). Ген trp1 представляет собой селективный маркер для мутантного штамма дрожжей, у которого отсутствует способность расти в отсутствии триптофана, например, ATCC No. 44076 или PEP4-1. Jones, Genetics, 85:12 (1977). Затем наличие повреждения trp1 в геноме дрожжевой клетки-хозяина обеспечивает эффективные условия для определения трансформации по росту в отсутствии триптофана. Аналогично, дефицитные по Leu2 штаммы дрожжей (ATCC 20622 или 38626), комплементируют известными плазмидами, обладающими геном Leu2.
Кроме того, векторы, полученные из 1,6-мкм кольцевой плазмиды pKD1, можно использовать для трансформации дрожжей Kluyveromyces. Альтернативно описаны экспрессирующие системы для крупномасштабной продукции рекомбинантного химозина телят для K. lactis. Van den Berg, Bio/Technology, 8:135 (1990). Также описаны стабильные мультикопийные экспрессирующие векторы для секреции зрелого рекомбинантного сывороточного альбумина человека посредством промышленных штаммов Kluyveromyces. Fleer et al., Bio/Technology, 9:968-975 (1991).
d) Компонент - промотор
Экспрессирующие и клонирующие векторы, как правило, содержат промотор, который распознается организмом-хозяином и функционально связан с нуклеиновой кислотой, кодирующей антитело. Промоторы, пригодные для применения в прокариотических хозяевах, включают промотор phoA, промоторные системы β-лактамазы и лактозы, промотор щелочной фосфатазы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако пригодными являются другие бактериальные промоторы. Промоторы для применения в бактериальных системах также содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей антитело.
Известны промоторные последовательности для эукариот. Практически все эукариотические гены обладают AT-богатой областью, расположенной приблизительно от 25 до 30 оснований выше участка инициации транскрипции. Другая последовательность, находящаяся от 70 до 80 оснований выше точки начала транскрипции множества генов, представляет собой участок CNCAAT, где N может представлять собой любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может представлять собой сигнал для добавления поли-A хвоста к 3'-концу кодирующей последовательности. Все из этих последовательностей можно встраивать в эукариотические экспрессирующие векторы.
Примеры пригодных промоторных последовательностей для применения в дрожжевых хозяевах включают промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, таких как енолаза, глицеральдегид-3-фосфат дегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкоза-6-фосфат изомераза, 3-фосфоглицеромутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкоизомераза и глюкокиназа.
Другие дрожжевые промоторы, которые представляют собой индуцибельные промоторы, обладающие дополнительным преимуществом контролируемой условиями выращивания транскрипции, представляют собой промоторные участки алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, ферментов деградации, связанных с метаболизмом азота, металлотеонеина, глицеральдегид-3-фосфат дегидрогеназы и ферментов, ответственных за употребление мальтозы и галактозы. Кроме того, пригодные векторы и промоторы для применения для экспрессии в дрожжах описаны в EP 73657. Также предпочтительно с дрожжевыми промоторами используют дрожжевые энхансеры.
Транскрипция антитела с векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами, получаемыми из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и наиболее предпочтительно, вирус обезьяны 40 (SV40), гетерологичными промоторами млекопитающих, например, промотором актина или промотором иммуноглобулина, промоторами теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 обычно получают в качестве рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Предранний промотор цитомегаловируса человека обычно получают в качестве рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в хозяевах, относящихся к млекопитающим, с использованием вируса папилломы крупного рогатого скота в качестве вектора описана в патенте США No. 4419446. Модификация этой системы описана в патенте США No. 4601978. См. также Reyes et al., Nature 297:598-601 (1982) в отношении экспрессии кДНК интерферона человека в клетках мышей под контролем промотора тимидинкиназы из вируса простого герпеса. Альтернативно в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
e) Компонент - энхансерный элемент
Транскрипцию ДНК, кодирующей антитело по настоящему изобретению, у высших эукариот часто усиливают посредством встраивания в вектор энхансерной последовательности. Известно много энхансерных последовательностей из генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако, как правило, используют энхансер из вируса эукариотической клетки. Их примеры включают энхансер SV40 на поздней стороне ориджина репликации (п.о. 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне ориджина репликации и энхансеры аденовируса. См. также Yaniv, Nature 297:17-18 (1982) в отношении энхансеров для активации эукариотических промоторов. Энхансер может быть присоединен в векторе к кодирующей антитело последовательности в положении 5' или 3', однако предпочтительно он расположен в участке, расположенном в 5'-направлении от промотора.
f) Компонент для терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (в клетках дрожжей, грибов, насекомых, растений, животных, человека, или содержащих ядро клетках из других многоклеточных организмов) также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно находятся на 5'- и, иногда, на 3'-нетранслируемых участках эукариотических или вирусных ДНК или кДНК. Эти участки содержат нуклеотидные фрагменты, транскрибируемые в качестве полиадинилированных фрагментов в нетранслируемом участке мРНК, кодирующей антитело. Одним пригодным компонентом для терминации транскрипции является участок полиаденилирования бычьего гормона роста. См. WO94/11026 и описанный в ней экспрессирующий вектор.
g) Выбор и трансформация клеток-хозяев.
Пригодные клетки-хозяева для клонирования или экспрессии ДНК в векторах, представленных в настоящем документе, представляют собой клетки прокариот, дрожжей или высших эукариот, описанных выше. Пригодные для этой цели прокариоты включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанные в DD 266710, опубликованной 12 апреля 1989 года), Pseudomonas, такие как P. aeruginosa и Streptomyces. Одним предпочтительным хозяином E. coli для клонирования является E. coli 294 (ATCC 31446), хотя также пригодными являются другие штаммы, такие как E. coli B, E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Эти примеры являются иллюстративными, но не ограничивающими.
Полноразмерное антитело, слитые белки антитела и фрагменты антитела можно получать в бактериях, в частности, когда гликозилирование и Fc-эффекторная функция не требуются, например, когда терапевтическое антитело конъюгировано с цитотоксическим средством (например, токсином) и иммуноконъюгат самостоятельно демонстрирует эффективность в отношении разрушения клеток опухоли. Полноразмерные антитела обладают более высоким временем полужизни в кровотоке. Продукция в E. coli является более быстрой и более экономичной. Для экспрессии фрагментов антитела и полипептидов в бактериях, см., например, U.S. 5648237 (Carter et. al.), U.S. 5789199 (Joly et al.), и U.S. 5840523 (Simmons et al.), в которых описаны участок инициации трансляции (TIR) и сигнальные последовательности для оптимизации экспрессии и секреции. См. также Charlton, Methods in Molecular Biology. Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), pp. 245-254, в которой описана экспрессия фрагментов антител в E. coli. После экспрессии антитело выделяют из клеточной массы E. coli в растворимой фракции и их можно очищать, например, с помощью колонки с белком A или G, в зависимости от изотипа. Конечную очистку можно проводить, аналогично процессу очистки антитела, экспрессируемого, например, в клетках CHO.
В дополнение к прокариотам, пригодными хозяевами для клонирования или экспрессии кодирующих связывающего IgG антитела векторов являются эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи. Saccharomyces cerevisiae, или обычные пекарские дрожжи, наиболее распространены среди низших эукариотических микроорганизмов-хозяев. Однако широко доступным и пригодным здесь является ряд других родов, видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K. lactis, K.fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans, и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитевидные грибы, такие как, например, хозяева Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger. Для обзора применения дрожжей и нитчатых грибов для продукции терапевтических белков см., например, Gerngross, Nat. Biotech. 22:1409-1414 (2004).
Можно выбирать определенные штаммы грибов и дрожжей, в которых каскады гликозилирования являются "гуманизированными", что приводит к продукции антитела с частично или полностью человеческим паттерном гликозилирования. См., например, Li et al., Nat. Biotech. 24:210-215 (2006) (где описана гуманизация каскада гликозилирования в Pichia pastoris); и Gerngross et al., выше.
Пригодные клетки-хозяева для экспрессии гликозилированного антитела также получают из многоклеточных организмов (безпозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Выявлен ряд штаммов и вариантов бакуловирусов и соответствующих пермиссивных клеток-хозяев среди хозяев, таких как Spodoptera frugiperda (гусеница), Aedes aegypti (москит), Aedes albopictus (москит), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Множество штаммов вирусов для трансфекции являются широко доступными, например, вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV и такие вирусы можно использовать здесь в качестве вирусов в соответствии с настоящим изобретением, в частности, для трансфекции клеток Spodoptera frugiperda.
Также в качестве хозяев можно использовать растительные клеточные культуры хлопка, кукурузы, картофеля, сои, петунии, томата, ряски (Lemnaceae), люцерны (M. truncatula) и табака. См., например, патенты США No. 5959177, 6040498, 6420548, 7125978 и 6417429 (где описана технология PLANTIBODIESTM для продукции антител трансгенными растениями).
В качестве хозяев можно использовать клетки позвоночных, и размножение клеток позвоночных в культуре (культуре тканей) стало общепринятой методикой. Примерами пригодных хозяев, представляющих собой клеточные линии млекопитающих, являются линия клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); эмбриональная линия почки человека (клетки 293 или 293, субклонированные для выращивания в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки молодого хомячка (BHK, ATCC CCL 10); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки цервикльной карциномы человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы буффало (BRL 3A, ATCC CR11442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линия гепатомы человека (Hep G2). Другие пригодные клеточные линии млекопитающих включают клетки яичника китайского хомячка (CHO), включающие клетки DHFR- CHO (Urlaub et al, Proc. Natl. Acad. Sci. USA 11:4216 (1980)); и клеточные линии миеломы, такие как NSO и Sp2/0. Для обзора определенных клеточных линий млекопитающих, пригодных для продукции антител, см., например, Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), pp. 255-268.
Для продукции антитела клетки-хозяева трансформируют описанными выше экспрессирующими и клонирующими векторами и культивируют их в общепринятой среде, модифицированной соответствующим образом, для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности.
h) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продукции антитела по настоящему изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев пригодными являются коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда ((MEM), (Sigma)), RPMI-1640 (Sigma), и среда Игла, модифицированная способом Дульбекко, ((DMEM), Sigma). Кроме того, для клеток-хозяев в качестве культуральной среды можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal Biochem. 102:255 (1980), патентах США No. 4767704; 4657866; 4927762; 4560655 или 5122469; WO9003430; WO 87/00195; или патенте США Re. 30985. Любые их этих сред можно дополнить при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCINTM), микроэлементами (определяемыми как неорганические соединения, обычно представленные в конечных концентрациях микромолярного диапазона) и глюкозой или эквивалентными источниками энергии. Также можно добавлять любые другие необходимые дополнительные вещества в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, которые ранее использовали для выбранных для экспрессии клеток-хозяев, и они будут понятны специалисту в данной области.
i) Очистка антитела
При использовании рекомбинантных способов антитело может продуцироваться внутриклеточно, в периплазматическом пространстве или оно может непосредственно секретироваться в среду. При внутриклеточной продукции антитела в качестве первого этапа удаляют частицы дебриса, либо клетки-хозяева, либо лизированные фрагменты, например, посредством центрифугирования или ультрафильтрации. В Carter et al., Bio/Technology 10: 163-167 (1992) описан способ выделения антител, которые секретируются в периплазматическое пространство E. coli. В кратком изложении, клеточную массу размораживают в присутствии ацетата натрия (pH 3,5), EDTA и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 мин. Клеточный дебрис можно удалять центрифугированием. Когда антитело секретируется в среду, супернатанты из таких экспрессирующих систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например, элемента для ультрафильтрации Amicon или Millipore Pellicon. На любой из предшествующих стадий можно включать ингибитор протеаз, такой как PMSF, для ингибирования протеолиза, и для предотвращения роста ненужных контаминирующих организмов можно добавлять антибиотики.
Композицию антитела, полученную из клеток, можно очищать с использованием, например, хроматографии с гидроксилапатитом, хроматографии гидрофобного взаимодействия, гель-электрофореза, диализа и аффинной хроматографии, причем аффинная хроматография является предпочтительным способом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который находится в антителе. Белок A можно использовать для очистки антител, в основе которых лежат тяжелые цепи γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендован для всех изотипов мыши и для цепи γ3 человека (Guss et al., EMBO J. 5:15671575 (1986)). Матрица, к которой присоединен аффинный лиганд, наиболее часто представляет собой агарозу, однако доступными являются и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и более короткое время обработки, чем те, которых достигают с помощью агарозы. В случае, когда антитело содержит домен CH3, для очистки пригодна смола ABXTM (J. T. Baker, Phillipsburg, NJ). Также в зависимости от антитела, подлежащего выделению, пригодны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на SEPHAROSE™ с гепарином, хроматография на анионной или катионной обменной смоле (такая как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
После предварительной стадии(ий) очистки смесь, содержащую представляющее интерес антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия с низким значением pH с использованием буфера для элюирования при pH между приблизительно 2,5-4,5, предпочтительно проводимой при низких концентрациях соли (например, от приблизительно 0-0,25 M соли).
В общем, в данной области хорошо известны различные способы получения антител для применения в разработке, тестировании и клинике, которые согласуются с описанными выше способами и/или которые специалист в данной области сочтет пригодными для конкретного представляющего интерес антитела.
C. Иммуноконъюгаты
Это изобретение также относится к иммуноконъюгатам (также взаимозаменяемо называемым "конъюгатами антитело-лекарственное средство" или "ADC"), содержащим антитело, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, лекарственное средство, ингибирующее рост средство, токсин (например, белковый токсин, ферментативно активный токсин бактерий, грибов, растений или животных, или их фрагменты), или с радиоактивным изотопом (т.е., радиоконъюгатом).
Иммуноконъюгаты применяют для локальной доставки цитотоксических средств, т.е., лекарственных средств, которые уничтожают или ингибируют рост или пролиферацию клеток, при лечении злокачественной опухоли (Lambert, J. (2005) Curr. Opinion in Pharmacology 5:543-549; Wu et al (2005) Nature Biotechnology 23(9): 1137-1146; Payne, G. (2003) i 3:207-212; Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drug Deliv. Rev. 26: 151-172; патент США No. 4975278). Иммуноконъюгаты позволяют направленную доставку группы лекарственного средства к опухоли и ее внутриклеточное накопление, где системное введение неконъюгированных лекарственных средств может приводить к неприемлемым уровням токсичности в отношении нормальных клеток, наряду с опухолевыми клетками, подлежащими устранению (Baldwin et al., Lancet (Mar. 15, 1986) pp. 603-05; Thorpe (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", Monoclonal Antibodies '84: Biological And Clinical Applications (A. Pinchera et al., eds) pp. 475-506. В качестве пригодных в этих стратегиях описаны как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother. 21:183-87). Лекарственные средства, используемые в этих способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986), выше). Токсины, используемые в конъюгатах антитело-токсин, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al (2000) J. Nat. Cancer Inst. 92(19):1573-1581; Mandler et al (2000) Bioorganic & Med. Chem. Letters 10:1025-1028; Mandler et al (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (EP 1391213; Liu et al., (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихеамицин (Lode et al (1998) Cancer Res. 58:2928; Hinman et al (1993) Cancer Res. 53:3336-3342). Токсины могут оказывать их цитотоксическое действие через механизмы, включающие связывание тубулина, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства имеют тенденцию к инактивации или снижению активности при конъюгации с крупными антителами или лигандами белкового рецептора.
ZEVALIN® (ибритутомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело-радиоизотоп, состоящий из моноклонального антитела IgG1 каппа мыши, направленного против антигена CD20, находящегося на поверхности нормальных и злокачественных B-лимфоцитов, и радиоизотопа 111In или 90Y, связанного линкером-хелатором на основе тиомочевины (Wiseman et al (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al (2002) Blood 99(12):4336-42; Witzig et al (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al (2002) J. Clin. Oncol. 20(15):3262-69). Хотя ZEVALIN обладает активностью против B-клеточной неходжкинской лимфомы (NHL), его введение приводит к тяжелым и длительным цитопениям у большинства пациентов. MYLOTARGTM (гемтузумаб озагомицин, Wyeth Pharmaceuticals), конъюгат антитело-лекарственное средство, состоящий из антитела huCD33, связанного с калихеамицином, был одобрен в 2000 для лечения острого миелоидного лейкоза путем инъекции (Drugs of the Future (2000) 25(7):686; патенты США No. 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Кантузумаб мертансин (Immunogen, Inc.), конъюгат антитело-лекарственное средство, состоящий из антитела huC242, связанного через дисульфидный линкер SPP с группой майтанзиноидного лекарственного средства, DM1, проходит испытание фазы II для лечения злокачественных опухолей, которые экспрессируют CanAg, таких как злокачественные опухоли толстого кишечника, поджелудочной железы, желудка и другие злокачественные опухоли. MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen Inc.), конъюгат антитело-лекарственное средство, состоящий из моноклонального антитела против простатспецифического мембранного антигена (PSMA), связанного с группой майтанзиноидного лекарственного средства, DM1, находится на стадии разработки для возможного лечения злокачественных опухолей предстательной железы. Пептиды ауристатина, ауристатин E (AE) и монометилауристатин (MMAE), синтетические аналоги доластатина, были конъюгированы с химирными моноклональными антителами cBR96 (специфичными к Lewis Y на карциноме) и cAC10 (специфичными к CD30 на гематологических злокачественных опухолях) (Doronina et al (2003) Nature Biotechnol. 21(7):778-784) и находятся на стадии терапевтической разработки.
В определенных вариантах осуществления иммуноконъюгат содержит антитело и химиотерапевтическое средство или другой токсин. Химиотерапевтические средства, пригодные для получения таких иммуноконъюгатов, описаны в настоящем документе (например, выше). Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки-диантаны, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. См. например, WO 93/21232, опубликованную 28 октября 1993 года. Для получения радиоконъюгированных антител доступны различные радионуклиды. Их примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического средства можно получать с использованием множества бифункциональных связывающих белки веществ, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как 2,6-диизоцианат толуола), и вторичные активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина можно получать, как описано в Vitetta et al. Science 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой иллюстративный хелатирующий агент для конъюгации радионуклеотида с антителом. См. WO94/11026.
Также в настоящем документе представлены конъюгаты антитела и одного или нескольких низкомолекулярных токсинов, таких как калихеамицин, майтанзиноиды, доластатины, ауростатины, трихотецен и CC1065, и производные этих токсинов, которые обладают активностью токсинов.
1. Майтанзин и майтанзиноиды
В некоторых вариантах осуществления иммуноконъюгат содержит антитело (полноразмерное или фрагменты), конъюгированное с одной или несколькими молекулами майтанзиноида.
Майтанзиноиды представляют собой ингибиторы митоза, которые действуют, ингибируя полимеризацию тубулина. Майтанзин впервые был выделен из восточно-африканского кустарника Maytenus serrata (патент США No. 3896111). Впоследствии, было открыто, что определенные микроорганизмы также продуцируют майтанзиноиды, такие как майтанзинол и C-3 сложные эфиры майтанзинола (патент США No. 4151042). Синтетический майтанзинол и его производные и аналоги описаны, например, в патентах США No. 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533.
Майтанзиноидные группы лекарственного средства являются привлекательными группами лекарственного средства в конъюгатах антитело-лекарственное средство, поскольку они: (i) относительно доступны для получения ферментацией или химической модификацией или преобразованием в производное продуктов ферментации, (ii) поддаются преобразованию в производные с помощью функциональных групп, пригодных для конъюгации через дисульфидные и недисульфидные линкеры с антителами, (iii) являются стабильными в плазме, и (iv) являются эффективными против различных опухолевых клеточных линий.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения, и их терапевтическое применение описаны, например, в патентах США No. 5208020, 5416064 и патенте Европы EP 0 425 235 B1, описания которых включены в настоящий документ в качестве ссылок в полном объеме. Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описали иммуноконъюгаты, содержащие майтанзиноид, обозначаемый как DM1, связанный с моноклональным антителом C242, направленным против рака ободочной и прямой кишки человека. Было выявлено, что конъюгат является высоко цитотоксичным в отношении культивируемых клеток рака толстого кишечника, и он продемонстрировал противоопухолевоую активность в анализе роста опухоли in vivo. В Chari et al., Cancer Research 52: 127-131 (1992) описаны иммуноконъюгаты, в которых майтанзиноид конъюгирован через дисульфидный линкер с антителом мыши A7, связывающимся с антигеном на клеточных линиях рака толстого кишечника человека или с другим моноклональным антителом мыши TA.1, которое связывает онкоген HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид тестировали in vitro на клеточной линии рака молочной железы человека SK-BR-3, которая экспрессирует 3×105 поверхностных антигенов HER-2 на клетку. Конъюгат лекарственного средства достигал степени цитотоксичности, сходной со свободным майтанзиноидным лекарственным средством, которую можно повышать путем увеличения количества молекул майтанзиноида на молекулу антитела. Конъюгат A7-майтанзиноид продемонстрировал низкую системную цитотоксичность у мышей.
Конъюгаты антитело-майтанзиноид получают химическим связыванием антитела с молекулой майтанзиноида без существенного снижения биологической активности либо антитела, либо молекулы майтанзиноида. См., например, патент США No. 5208020 (описание которого включено в настоящий документ в качестве ссылки в полном объеме). В среднем 3-4 молекулы майтанзиноида, конъюгированных с одной молекулой антитела, продемонстрировали эффективность в отношении повышения цитотоксичности клеток-мишеней без отрицательного влияния на функцию или растворимость антитела, хотя можно ожидать, что даже одна молекула токсина/антитело повысит цитотоксичность относительно применения простого антитела. Майтанзиноиды хорошо известны в данной области и их можно синтезировать известными способами или выделять из природных источников. Пригодные майтанзиноиды описаны, например, в патенте США No. 5208020 и в других патентах и непатентных публикациях, указанных в настоящем документе выше. Предпочтительными майтанзиноидами являются майтанзинол и аналоги майтанзинола, модифицированные в ароматическом кольце или в других положениях молекулы майтанзинола, такие как различные сложные эфиры майтанзинола.
Для получения конъюгатов антитело-майтанзиноид существует множество линкерных групп, известных в данной области, включая, например, линкерные группы, описанные в патенте США No. 5208020 или в патенте EP 0 425 235 B1; Chari et al. Cancer Research 52:127-131 (1992); и US 2005/016993 A1, описания которых включены в настоящий документ в качестве ссылок в полном объеме. Конъюгаты антитело-майтанзиноид, содержащие линкерный компонент SMCC, можно получать, как описано в US 2005/016993 A1. Линкерные группы включают дисульфидные группы, простые тиоэфирные группы, неустойчивые к действию кислот группы, фотолабильные группы, неустойчивые к пептидазам группы, или неустойчивые к эстеразе группы, как описано в указанных выше патентах, причем дисульфидные и тиоэфирные группы являются предпочтительными. Дополнительные линкеры описаны и проиллюстрированы в настоящем документе.
Конъюгаты антитела и майтанзиноида можно получать с использованием множества бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат), и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). В определенных вариантах осуществления связывающий агент представляет собой N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) или N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP), обеспечивающие дисульфидную связь.
Линкер можно связывать с молекулой майтанзиноида в различных положениях, в зависимости от типа связи. Например, сложноэфирную связь можно образовывать посредством реакции с гидроксильной группой с использованием общепринятых способов присоединения. Реакция может протекать в положении C-3, имеющем гидроксильную группу, положении C-14, модифицированном гидроксиметилом, положении C-15, модифицированном гидроксильной группой, и положении C-20, имеющем гидроксильную группу. В предпочтительном варианте осуществления связь образована в положении C-3 майтанзинола или аналога майтанзинола.
2. Ауристатины и доластатины
В некоторых вариантах осуществления иммуноконъюгат содержит антитело, конъюгированное с доластатином или с пептидным аналогом или производным доластатина, например, ауристатином (патенты США No. 5635483; 5780588). Было показано, что доластатины и ауристатины нарушают динамику микротрубочек, гидролиз GTP и деление ядер и клеток (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают активностью против злокачественной опухоли (US 5663149) и противогрибковой активностью (Pettit et al. (1998) Antimicrob. Agents Chemother. 42:2961-2965). Группу лекарственного средства в виде доластатина или ауристатина можно связывать с антителом через N(амино)-конец или C(карбоксильный)-конец пептидной группы лекарственного средства (WO 02/088172).
Иллюстративные варианты осуществления ауристатина включают связывающиеся через N-конец группы лекарственного средства в виде монометилауристатина DE и DF, описанные в "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номером No. 10/983340, поданная 5 ноября 2004 года, описание которых включено в настоящий документ в качестве ссылки в полном объеме.
Как правило, группы лекарственного средства на основе пептидов можно получать образованием пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получать, например, способом жидкостного синтеза (см. E. Schroder and K. Lubke, "The Peptides", volume 1, pp 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Группы лекарственного средства в виде ауристатина/доластатина можно получать согласно способам: US 5635483; US 5780588; Pettit et al. (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al. (1998) Anti-Cancer Drug Design 13:243-277; Pettit, G.R., et al. Synthesis, 1996, 719-725 и Pettit et al. (1996) J. Chem. Soc. Perkin Trans. 1 5:859-863. Также см. Doronina (2003) Nat. Biotechnol. 21(7): 778-784; "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номеров No. 10/983340, поданная 5 ноября 2004 года, включенная в настоящий документ в качестве ссылки в полном объеме (где описаны, например, линкеры и способы получения соединений монометилвалина, таких как MMAE и MMAF, конъюгированных с линкерами)
3. Калихеамицин
В других вариантах осуществления иммуноконъюгат содержит антитело, конъюгированное с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицина способны образовывать двухцепочечные разрывы ДНК при субпикомолярных концентрациях. Для получения конъюгатов семейства калихеамицина, см. патенты США No. 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все выданы American Cyanamid Company). Структурные аналоги калихеамицина, которые можно использовать, включают, но не ограничиваются ими, γ1I, α2I, α3I, N-ацетил-γ1I, PSAG и θI1 (Hinman et al. Cancer Research 53: 3336-3342 (1993), Lode et al. Cancer Research 58: 2925-2928 (1998) и упомянутые выше патенты США, выданные American Cyanamid). Другим противоопухолевым лекарственным средством, с которым можно конъюгировать антитело, является QFA, который представляет собой антифолат. Как калихеамицин, так и QFA, обладают внутриклеточным действием, и они не легко проходят через плазматическую мембрану. Таким образом, клеточный захват этих веществ посредством опосредуемой антителом интернализации значительно повышает их цитотоксические эффекты.
c. Другие цитотоксические средства
Другие противоопухолевые средства, которые можно конъюгировать с антителом, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство средств, совокупно известных как комплекс LL-E33288, описанный в патентах США No. 5053394, 5770710, а также эсперамицины (патент США No. 5877296).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A-цепь дифтерийного токсина, несвязывющие активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки-диантаны, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. См., например, WO 93/21232, опубликованную 28 октября, 1993.
Кроме того, настоящее изобретение относится к иммуноконъюгату, образованному между антителом и соединением с нуклеолитической активностью (например, рибонуклеазй или эндонуклеазой ДНК, такой как дезоксирибонуклеаза; ДНКаза).
Для селективного разрушения опухоли антитело может содержать высоко радиоактивный атом. Для продукции радиоактивных конъюгированных антител доступны различные радиоактивные изотопы. Их примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. В случае применения конъюгата для дететкции, он может содержать радиоактивный атом для сцинтиграфических исследований, например tc99m или I123, или спиновую метку для получения изображения ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как также йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки можно встраивать в конъюгат известными способами. Например, пептид может быть биологически синтезированным или его можно синтезировать посредством химического синтеза аминокислот с использованием пригодных предшественников аминокислот, включающих, например, фтор-19 вместо водорода. Метки, такие как tc99m или I123, Re186, Re188 и In111 можно присоединять через остаток цистеина в пептиде. Иттрий-90 можно присоединять через остаток лизина. Способ с IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80: 49-57) можно использовать для встраивания йода-123. В "Monoclonal Antibodies in Immunoscintigraphy" (Chatal, CRC Press 1989) подробно описаны другие способы.
Конъюгаты антитела и цитотоксического средства можно получать с использованием различных бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бисдиазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в Vitetta et al., Science 238: 1098 (1987). Иллюстративным хелатирующим агентом для конъюгации радионуклида с антителом является меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA). См. WO94/11026. Линкер может представлять собой "расщепляемый линкер", способствующий высвобождению цитотоксического лекарственного средства в клетке. Например, можно использовать неустойчивый к действию кислот линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al, Cancer Research 52:127-131 (1992); патент США No. 5208020).
Соединения прямо предусматривают, но не ограничиваются ими, ADC, полученные с помощью поперечно-сшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A). См. стр. 467-498, 2003-2004 Applications Handbook and Catalog.
5. Получение конъюгатов антитело-лекарственное средство
В конъюгатах антитело-лекарственное средство (ADC) антитело (Ab) конъюгировано с одной или несколькими группами лекарственных средств (D), например от приблизительно 1 до приблизительно 20 групп на антитело, через линкер (L). ADC формулы I можно получать несколькими путями с использованием реакций, условий и реагентов органической химии, известных специалистам в данной области, включая: (1) реакцию нуклеофильной группы антитела с двухвалентным линкерным реагентом с образованием Ab-L через ковалентную связь, с последующей реакцией с группой лекарственного средства D; и (2) реакцию нуклеофильной группы в группе лекарственного средства с двухвалентным линкерным реагентом, с образованием D-L, через ковалентную связь, с последующей реакцией с нуклеофильной группой антитела. Дополнительные способы получения ADC описаны в настоящем документе.
Линкер может состоять из одного или нескольких линкерных компонентов. Иллюстративные линкерные компоненты включают 6-малеимидокапроил ("MC"), малеимидопропаноил ("MP"), валин-цитруллин ("val-cit" или "vc"), аланин-фенилаланин ("ala-phe" или "af"), п-аминобензилоксикарбонил ("PAB"), N-сукцинимидил-4-(2-пиридилтио)пентаноат ("SPP"), N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат ("SMCC"), N-сукцинимидил-(4-йодоацетил)аминобензоат ("SIAB"). Дополнительные линкерные компоненты известны в данной области и некоторые из них описаны в настоящем документе. Также см. "Monomethylvaline Compounds Capable of Conjugation to Ligands", US с серийным номеров No. 10/983340, поданную 5 ноября 2004 года, описание которой включено в настоящий документ в качестве ссылки в полном объеме.
В некоторых вариантах осуществления линкер может содержать аминокислотные остатки. Иллюстративные аминокислотные линкерные компоненты включают дипептид, трипептид, тетрапептид или пентапептид. Иллюстративные дипептиды включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Иллюстративные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, которые содержат компонент в виде аминокислотного линкера, включают природные аминокислотные остатки, а также минорные аминокислоты и неприродные аналоги аминокислот, такие как цитруллин. Аминокислотные линкерные компоненты можно конструировать и оптимизировать по их селективности к ферментативному расщеплению конкретными ферментами, например, ассоциированной с опухолью протеазой, катепсином B, C и D или протеазой плазмина.
Нуклеофильные группы на антителах включают, но не ограничиваются ими: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, лизина, (iii) тиольные группы боковых цепей, например, цистеина, и (iv) гидроксилы или аминогруппы сахаров, где антитело является гликозилированным. Амино, тиольная и гидроксильная группы являются нуклеофильными и способны вступать в реакцию с образованием ковалентных связей с электрофильными группами на линкерных группах и линкерных реагентах, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) группы альдегидов, кетонов, карбоксильные и малеинимидные группы. Определенные антитела обладают поддающимися восстановлению дисульфидами между цепями, т.е. цистеиновыми мостиками. Реакционную способность антитела для конъюгации с линкерными реагентами можно обеспечивать посредством обработки восстановителем, таким как DTT (дитиотреитол). Каждый цистеиновый мостик, таким образом, может образовывать, теоретически, два реакционноспособных тиольных нуклеофила. Дополнительные нуклеофильные группы можно вносить в антитела посредством модификации остатков лизина, например, реакцией остатков лизина с 2-иминотиоланом (реагентом Трота), приводящей к превращению амина в тиол. Реакционно способные тиольные группы можно вносить в антитело путем встраивания одного, двух, трех, четырех или более остатков цистеина (например, путем получения мутантных антител, содержащих один или несколько ненативных аминокислотных остатка цистеина).
Конъюгаты антитело-лекарственное средство также можно получать путем модификации антитела внесением электрофильных групп, которые могут реагировать с нуклеофильными заместителями на линкерном реагенте или лекарственном средстве. Сахара гликозилированных антител могут быть окисленными, например окислителями на основе периодатов, с образованием групп альдегидов или кетонов, которые могут реагировать с аминогруппой линкерных реагентов или групп лекарственного средства. Полученные иминогруппы шифового основания могут образовывать стабильную связь, или их можно восстанавливать, например, боргидридными реагентами, с образованием стабильных связей через амин. В одном варианте осуществления реакция углеводной части гликозилированного антитела либо с галактозооксидазой, либо с метаперйодатом натрия, может приводить к карбонильным группам (группам альдегидов и кетонов) в антителе, которые могут вступать в реакции с соответствующими группами на лекарственном средстве (Hermanson, Bioconjugate Techniques). В другом варианте осуществления антитела, содержащие N-концевые остатки серина или треонина могут реагировать с метаперйодатом натрия, что приводит к образованию альдегида вместо первой аминокислоты (Geoghegan & Stroh, (1992) Bioconjugate Chem. 3:138-146; US 5362852). Такой альдегид можно подвергать реакции с группой лекарственного средства или нуклеофилом линкера.
Аналогично, нуклеофильные группы на группе лекарственного средства включают, но не ограничиваются ими: аминогруппу, тиольную, гидроксильную группы, группы гидразида, оксима, гидразина, тиосемикарбазона, гидразина карбоксилата и арилгидразида, способные вступать в реакции с образованием связей с электрофильными группами на линкерных группах и линкерных реагентах, включающих: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеинимидные группы.
Альтернативно слитый белок, содержащий антитело и цитотоксическое средство, можно получать, например, рекомбинантными способами или пептидным синтезом. Отрезок ДНК может содержать соответствующие области, кодирующие две части конъюгата, либо соседние друг с другом, либо разделенные областью, кодирующей линкерный пептид, который на ухудшает желаемые свойства конъюгата.
В другом варианте осуществления антитело может быть конъюгировано с "рецептором" (таким как стрептавидин) для применения в предварительном нацеливании на опухоль, где конъюгат антитело-рецептор вводят пациенту, а затем из кровотока удаляют не связавшийся конъюгат с использованием средства для удаления, с последующим введением "лиганда" (например, авидина), который конъюгирован с цитотоксическим средством (например радионуклидом).
D. Способы
1. Диагностические способы и способы детекции
В одном аспекте антитела по изобретению пригодны для детекции наличия Notch2 в биологическом образце. Термин "детекция" в настоящем документе включает количественную или качественную детекцию. В определенных вариантах осуществления биологический образец включает клетку или ткань, такую как ткань злокачественной опухоли.
В одном аспекте изобретение относится к способу детекции наличия Notch2 в биологическом образце. В определенных вариантах осуществления способ включает контактирование биологического образца с антителом против Notch2 (например, с антителом против NRR Notch2) в условиях, позволяющих связывание антитела против Notch2 с Notch2, и детекцию образования комплекса между антителом против Notch2 и Notch2.
В одном аспекте изобретение относится к способу диагностики нарушения, ассоциированного с повышенной экспрессией Notch2. В определенных вариантах осуществления способ включает контактирование тестируемой клетки с антителом против Notch2; определение уровня экспрессии (либо количественно, либо качественно) Notch2 тестируемой клеткой путем детекции связывания антитела против Notch2 с Notch2; и сравнение уровня экспрессии Notch2 тестируемой клеткой с уровнем экспрессии Notch2 контрольной клеткой (например, нормальной клеткой, происходящей из той же ткани, что и тестируемая клетка, или клеткой, которая экспрессирует Notch2 на уровнях, сравнимых c уровнями для нормальной клетки или со средним уровнем экспрессии для множества контрольных клеток), где более высокий уровень экспрессии Notch2 тестируемой клеткой по сравнению с контрольной клеткой указывает на наличие нарушения, ассоциированного с повышенной экспрессией Notch2. В определенных вариантах осуществления тестируемую клетку получают от индивида, предположительно имеющего нарушение, ассоциированное с повышенной экспрессией Notch2. В определенных вариантах осуществления нарушение представляет собой клеточно-пролиферативное нарушение, такое как злокачественная опухоль или опухоль.
Иллюстративные нарушения, которые можно диагностировать с использованием антитела по изобретению, включают злокачественную опухоль, например, B-клеточные злокачественные опухоли, меланому, T-клеточные злокачественные опухоли (например, T-ALL), рак молочной железы, рак мозга, рак шейки матки, рак толстого кишечника и рак поджелудочной железы.
Для детекции связывания антител против Notch2 с Notch2 можно использовать некоторые другие способы. Такие способы включают, но не ограничиваются ими, анализы связывания антигена, которые хорошо известны в данной области, такие как вестерн-блоты, радиоиммунные анализы, ELISA (твердофазный иммуноферментный анализ), иммунные "сэндвич"-анализы, анализы иммунопреципитации, флуоресцентные иммунологические анализы, иммунологические анализы с белком A и иммуногистохимию (IHC).
В определенных вариантах осуществления антитела являются мечеными. Метки включают, но не ограничиваются ими, метки или группы, которые подвергают детекции прямо (такие как флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также такие группы, как ферменты или лиганды, которые подвергают детекции непрямо, например, посредством ферментативной реакции или молекулярного взаимодействия. Иллюстративные метки включают, но не ограничиваются ими, радиоизотопы 32P, 14C, 125I, 3H и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светляка и бактериальную люциферазу (патент США No. 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, связанные с ферментом, который использует пероксид водорода для окисления предшественника красителя, такого как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
В определенных вариантах осуществления антитела против Notch2 иммобилизуют на нерастворимой матрице. Иммобилизация включает отделение антитела против Notch2 от любого Notch2, который остается свободным в растворе. Ее осуществляют, либо обеспечивая нерастворимость антитела против Notch2 перед анализом, например, путем адсорбции на нерастворимой в воде матрице или поверхности (Bennich et al., U.S. 3720760) или путем ковалентного присоединения (например, с использованием поперечного сшивания глутаральдегидом), либо обеспечивая нерастворимость антитела против Notch2 после образования комплекса между антителом против Notch2 и Notch2, например, путем иммунопреципитации.
Понятно, что любой из указанных выше вариантов осуществления диагностики или детекции можно проводить с использованием иммуноконъюгата по изобретению вместо антитела против Notch2 или дополнительно к нему.
2. Способы лечения
Антитело по изобретению можно использовать, например, в способах лечения in vitro, ex vivo и in vivo. В одном аспекте изобретение относится к способам ингибирования активности Notch2, т.е. передачи сигнала Notch2, либо in vivo, либо in vitro, включающий воздействие на клетку антитела против NRR Notch2 по изобретению в условиях, позволяющих связывание иммуноконъюгата с Notch2. В определенных вариантах осуществления клетка представляет собой злокачественную клетку, например, злокачественную B-клетку или клетку меланомы. В одном варианте осуществления антитело против NRR Notch2 по изобретению можно использовать для ингибирования активности Notch2, включающий воздействие на Notch2 антитела против NRR Notch2 по изобретению, так чтобы активность Notch2 ингибировалась.
Антитело против NRR Notch2 по изобретению можно использовать, например, для лечения нарушений, ассоциированных с экспрессией и/или активностью Notch2, например, нарушений, ассоциированных с повышенной экспрессией или активностью Notch2, или нарушений, при которых экспрессия или активность Notch2 приводят к патогенному состоянию. "Нарушение, ассоциированное с повышенной экспрессей или активностью Notch2" относится к нарушению, при котором экспрессия или активность Notch2 значительно превышает норму.
В одном аспекте антитело по изобретению применяют для лечения или профилактики злокачественной опухоли, например, B-клеточных злокачественных опухолей, меланомы, T-клеточных злокачественных опухолей (например, T-ALL), рака молочной железы, злокачественной опухоли головного мозга, рака шейки матки, рака толстого кишечника, и рака поджелудочной железы. В определенных вариантах осуществления антитело против NRR Notch2 применяют для лечения злокачественной опухоли, такой как рак молочной железы, B-клеточная злокачественная опухоль или меланома.
В следующем аспекте антитело по изобретению применяют для лечения или профилактики злокачественной опухоли, конкретно, солидной опухоли, содержащей злокачественные стволовые клетки. Злокачественные стволовые клетки способны пролиферировать, давая начало дополнительным злокачественным стволовым клеткам, а также другим популяциям опухолевых клеток. См. Al-Hajj et al., Proc. Nat'l Acad. Sci. USA 100: 3983-3988 (2003). Полагают, что злокачественные стволовые клетки приводят к рецидивам злокачественных опухолей и устойчивости злокачественнйх опухолей к лекарственным средствам. Рецепторы Notch идентифицированы в качестве маркеров злокачественных стволовых клеток и в качестве мишени для устранения злокачественных стволовых клеток, ответственных за образование и рецидив солидных опухолей. См. WO 2008/091641. Такие солидные опухоли включают, но не ограничиваются ими, рак молочной железы, рак толстого кишечника, рак поджелудочной железы, рак предстательной железы, рак легкого, рак головы и шеи, рак прямой кишки и рак ободочной и прямой кишки.
В одном аспекте изобретение относится к способам лечения злокачественной опухоли, включающим введение индивидууму, нуждающемуся в этом, эффективного количества антитела по изобретению. В определенных вариантах осуществления способ лечения злокачественной опухоли включает введение индивидууму, нуждающемуся в этом, эффективного количества фармацевтического состава, содержащего антитело по изобретению и, необязательно, по меньшей мере одно дополнительное лекарственное средство, такое как средства, представленные ниже.
Антитела по изобретению можно использовать либо отдельно, либо в комбинации с другими композициями, в лечении. Например, антитело по изобретению можно вводить совместно по меньшей мере с одним дополнительным лекарственным средством и/или адъювантом. В определенных вариантах осуществления антитело против Notch1 (например, антитело против NRR Notch1) вводят с дополнительным лекарственным средством, которое ингибирует или снижает измененную дифференцировку клеток кишечника, которая в ином случае индуцировалась бы таким антителом против Notch1. В определенных вариантах осуществления дополнительным лекарственным средством является дексаметазон или тамоксифен. Такая комбинированная терапия может быть полезна, например, при лечении ангиогенных нарушений, включающих ассоциированный с опухолью ангиогенез, и злокачественной опухоли. В определенных вариантах осуществления антитело против Notch2 (например, антитело против NRR Notch2) вводят с дополнительным лекарственным средством, например, химиотерапевтическим средством. Такая комбинированная терапия может быть пригодна, например, для лечения злокачественной опухоли, такой как B-клеточные злокачественные опухоли и меланома.
Такие комбинированные способы лечения, описанные выше, включают комбинированное введение (где два или более лекарственных средств включены в один и тот же или в отдельные составы), и раздельное введение, в случае которого введение антитела по изобретению может происходить до, одновременно и/или после введения дополнительного лекарственного средства и/или адъюванта. Антитела по изобретению также можно использовать в комбинации с лучевой терапией.
В одном аспекте по меньшей мере некоторые из антител по изобретению могут связывать Notch2 отличных от человека видов. Таким образом, антитела по изобретению можно использовать для связывания Notch2, например, в культуре клеток млекопитающего, экспрессирующих эндогенный или рекомбинантный Notch2, у человека, или у других млекопитающих, имеющих Notch2, с которым антитело по изобретению перекрестно реагирует (например шимпанзе, бабуин, мартышка, яванский макак и макак-резус, свинья, крыса или мышь).
В одном варианте осуществления антитело по изобретению применяют в способе связывания Notch2 у индивидуума, страдающего нарушением, ассоциированным с повышенной экспрессией и/или активностью Notch2, включающий введение индивидууму антитела, так чтобы Notch2 у индивидуума был связан. В одном варианте осуществления Notch2 представляет собой Notch2 человека, и индивидуумом является человек. Альтернативно индивидуум может представлять собой любое млекопитающее, у которого экспрессируется Notch2, с которым связывается антитело по изобретению. Кроме того, индивидуумом может быть млекопитающее, которому вводят Notch2 (например, путем введения Notch2 или экспрессии трансгена, кодирующего Notch2).
Антитело по изобретению можно вводить человеку для терапевтических целей. Более того, антитело по изобретению можно вводить не являющемуся человеком млекопитающему, экспрессирующему Notch2, с которым антитело перекрестно реагирует (например, примату, свинье, крысе или мыши) для ветеринарных целей или в качестве модели на животных для заболевания человека. Что касается последнего, такие модели на животных могут быть пригодны для оценки терапевтической эффективности антител по изобретению (например, тестирования дозировок и длительности введения).
Антитело по изобретению (и любое дополнительное лекарственное средство или адъювант) можно вводить любым пригодным способом, включая парентеральный, подкожный, внутрибрюшинный, внутрилегочный и интраназальный, и, если желательно для местного лечения, введение в область повреждения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, антитело можно вводить импульсной инфузией, в частности, со снижающимися дозами антитела. Дозирование можно проводить любым пригодным способом, например путем инъекций, таких как внутривенные или подкожные инъекции, в зависимости отчасти от того, является ли введение кратковременным или длительным.
При получении и введении антитела можно учитывать расположение мишени для связывания антитела по изобретению. Когда мишень для связывания представляет собой внутриклеточную молекулу, некоторые варианты осуществления изобретения относится к антителу или его антигенсвязывающему фрагменту, подлежащему введению в клетку, где расположена мишень для связывания. В одном варианте осуществления антитело по изобретению можно экспрессировать внутриклеточно в качестве внутреннего антитела. Термин "внутреннее антитело" в настоящем документе относится к антителу или его антигенсвязывающей части, которые экспрессируются внутриклеточно и которые способны селективно связываться с молекулой мишенью, как описано, например, в Marasco, Gene Therapy 4: 11-15 (1997); Kontermann, Methods 34: 163-170 (2004); патентах США No. 6004940 и 6329173; публикации патентной заявки США No. 2003/0104402 и публикации PCT No. WO2003/077945. Также см, например, WO96/07321, опубликованную 14 марта 1996 года, в отношении применения генной терапии для получения внутриклеточных антител.
Внутриклеточной экспрессии внутреннего антитела можно достигать путем введения нуклеиновой кислоты, кодирующей желаемое антитело или его антигенсвязывающий фрагмент (лишенные лидерной последовательности дикого типа и секреторных сигналов, в норме ассоциированных с геном, кодирующим эти антитело или антигенсвязывающий фрагмент) в клетку-мишень. В клетку мишень можно доставлять одну или несколько нуклеиновых кислот, кодирующих все антитело по изобретению или его часть, так чтобы экспрессировались одно или несколько внутренних антител, которые способны связываться с внутриклеточным полипептидом-мишенью и модулировать активность полипептида-мишени. Можно использовать любой стандартный способ введения нуклеиновых кислот в клетку, включая, но не ограничиваясь ими, микроинъекцию, баллистическую инъекцию, электропорацию, осаждение с фосфатом кальция, липосомы и трансфекцию векторами на основе ретровируса, аденовируса, аденоассоциированного вируса и вируса осповакцины, содержащего представляющую интерес нуклеиновую кислоту.
В определенных вариантах осуществления нуклеиновую кислоту (необязательно содержащуюся в векторе) можно вводить в клетки пациента способами in vivo и ex vivo. В одном примере доставки in vivo нуклеиновую кислоту инъецируют прямо пациенту, например, в область, где требуется терапевтическое вмешательство. В следующем примере доставки in vivo нуклеиновую кислоту вводят в клетку с использованием трансфекции вирусных векторов (таких как аденовируса, вируса простого герпеса I или аденоассоциированного вируса) и систем на основе липидов (пригодными липидами для опосредуемого липидами переноса гена являются, например, DOTMA, DOPE и DC-Choi). Для обзора определенных протоколов генного маркирования и генной терапии см. Anderson et al, Science 256:808-813 (1992) и WO 93/25673 и ссылки, цитированные в них. В примере лечения ex vivo, клетки пациента извлекают, в эти выделенные клетки вводят нуклеиновую кислоту, и модифицированные клетки вводят пациенту либо прямо, либо, например, в инкапсулированном виде в пористых мембранах, которые имплантируют пациенту (см., например, патенты США No. 4892538 и 5283187). Обычно для доставки ex vivo нуклеиновой кислоты используют ретровирусный вектор.
В другом варианте осуществления представлены интернализующиеся антитела. Антитела могут обладать определенными характеристиками, которые усиливают доставку антител в клетку, или они могут быть модифицированы, чтобы они обладали такими характеристиками. Способы для достижения этого известны в данной области. Например, известно, что катионизация антитела способствует его захвату в клетки (см., например, патент США No. 6703019). Также для доставки антитела в клетки можно использовать липофекцию или липосомы. Когда используют фрагменты антител, преимущественным может быть наименьший ингибиторный фрагмент, который специфично связывается с белком-мишенью. Например, на основе последовательностей вариабельных областей антитела можно разработать пептидные молекулы, которые сохраняют способность связываться с белковой последовательностью-мишенью. Такие пептиды можно синтезировать химически и/или получать способами рекомбинантных ДНК. См., например, Marasco et al, Proc. Natl. Acad. Sci. USA, 90: 7889-7893 (1993).
Проникновение антител в клетки-мишени можно усиливать другими способами, известными в данной области. Например, определенные последовательности, такие как последовательности, происходящие из Tat ВИЧ или гомеодоменного белка Antennapedia, способны обеспечивать эффективный захват гетерологичных белков через клеточные мембраны. См., например, Chen et al., Proc. Natl. Acad. Sci. USA (1999), 96:4325-4329.
Когда мишень для связывания антитела расположена в головном мозге, определенные варианты осуществления изобретения относятся к антителу, проходящему через гематоэнцефалический барьер. Для транспорта молекул через гематоэнцефалический барьер существует несколько известных в данной области подходов, включая, но не ограничиваясь ими, физические способы, способы на основе липидов, способы на основе стволовых клеток и способы на основе рецепторов и каналов.
Физические способы транспорта антитела через гематоэнцефалический барьер включают, но не ограничиваются ими, полный обход гематоэнцефалического барьера или создание отверстий в гематоэнцефалическом барьере. Способы обхода включают, но не ограничиваются ими, прямую инъекцию в головной мозг (см., например, Papanastassiou et al., Gene Therapy 9:398-406 (2002)), интерстициальную инфузию/усиленную конвекцией доставку (см., например, Bobo et al., Proc. Natl. Acad. Sci. USA 91:2076-2080 (1994)), и имплантацию устройства для доставки в головной мозг (см., например, Gill et al., Nature Med. 9:589-595 (2003); и Gliadel WafersTM Guildford Pharmaceutical). Способы создания отверстий в барьере включают, но не ограничиваются ими, ультразвук (см., например, публикация патента США No. 2002/0038086), осмотическое давление (например, путем введения гипертонического маннита (Neuwelt, E. A., Implication of the Blood-Brain Barrier and its Manipulation, Vols 1 & 2, Plenum Press, N. Y. (1989)), повышение проницаемости, например, посредством брадикинина или усиливающего проницаемость средства A-7 (см., например, патенты США No. 5112596, 5268164, 5506206 и 5686416), и трансфекцию нейронов, которые проходят через гематоэнцефалический барьер, векторами, содержащими гены, кодирующие антитело (см., например, публикацию патента США No. 2003/0083299).
Способы транспорта антитела через гематоэнцефалический барьер на основе липидов включают, но не ограничиваются ими, инкапсулирование антитела в липосомы, которые связаны со связывающими фрагментами антител, которые связываются с рецепторами на эндотелии сосудов гематоэнцефалического барьера (см., например, публикацию патентной заявки США No. 20020025313), и нанесение антитела на частицы липопротеинов низкой плотности (см., например, публикацию патентной заявки США No. 20040204354) или аполипопротеин E (см., например, публикацию патентной заявки США No. 20040131692).
Способы транспорта антитела через гематоэнцефалический барьер на основе стволовых клеток охватывают генную инженерию нейрональных клеток-предшественников (NPC) для экспрессии представляющего интерес антитела, а затем имплантацию стволовых клеток в головной мозг индивидуума, подвергаемого лечению. См. Behrstock et al. (2005) Gene Ther. последняя он-лайн публикация 15 декабря 2005 года (где описано, что NPC, генетически модифицированные для экспрессии нейротрофического фактора GDNF, снижали симптомы болезни Паркинсона при имплантации в головной мозг в моделях на грызунах и приматах).
Способы транспорта антитела через гематоэнцефалический барьер на основе рецепторов и каналов включают, но не ограничиваются ими, применение глюкокортикоидных блокаторов для повышения проницаемости гематоэнцефалического барьера (см., например, публикации патентных заявок США No. 2002/0065259, 2003/0162695 и 2005/0124533); активацию калиевых каналов (см., например, публикацию патентной заявки США No. 2005/0089473), ингибирование ABC-переносчиков лекарственных средств (см., например, публикацию патентной заявки США No. 2003/0073713); нанесение на антитела трансферрина и модулирование активности одного или нескольких рецепторов трансферрина (см., например, публикацию патентной заявки США No. 2003/0129186) и катионизацию антител (см., например, патент США No. 5004697).
Антитела по изобретению можно изготавливать, дозировать и вводить способами, согласующимися с "Надлежащей медицинской практикой". Факторы, учитываемые в этом контексте, включают конкретное нарушение, подвергаемое лечению, конкретное млекопитающее, подвергаемое лечению, клиническое состояние отдельного пациента, причину нарушения, область доставки средства, способ введения, график введения и другие факторы, известные врачам. Антитело или иммуноконъюгат не должны быть изготовлены, однако их необязательно изготавливают, с одним или несколькими средствами, используемыми в настоящее время для профилактики и лечения рассматриваемого нарушения. Эффективное количество таких других средств зависит от количества антител по изобретению, присутствующего в составе, типа нарушения или лечения, и других факторов, рассмотренных выше. Их обычно используют в тех же дозировках и с теми же способами введения, которые описаны в настоящем документе выше или в количестве приблизительно от 1 до 99% от дозировок, описанных в настоящем документе, или в любой дозировке и с любым способом, которые эмпирически/клинически определены, как пригодные.
Для профилактики или лечения заболевания, дозировка и способ введения антитела (при использовании отдельно в сочетании с одним или несколькими дополнительными лекарственными средствами) могут зависеть от типа заболевания, подлежащего лечению, как определено выше, тяжести и течения заболевания, от профилактических или терапевтических целей введения молекулы, предшествующего лечения, клинического анамнеза пациента и ответа на антитело и от решения лечащего врача. Антитело подходящим образом вводят пациенту за один раз или на протяжении серии введений. В зависимости от типа и тяжести заболевания, исходная предполагаемая дозировка для введения пациенту составляет от приблизительно 1 мкг/кг до 15 мг/кг (например, приблизительно 0,1 мг/кг - 10 мг/кг) антитела, например, либо посредством одного или нескольких отдельных введений, либо посредством непрерывной инфузии. Одна из типичных суточных дозировок может находиться в диапазоне приблизительно от 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. В случае многократных введений в течение нескольких суток или более, в зависимости от состояния, введение повторяют до возникновения требуемого подавления симптомов заболевания. Одна иллюстративная дозировка антитела или иммуноконъюгат может находиться в диапазоне от приблизительно 0,05 мг/кг до приблизительно 10 мг/кг. Таким образом, пациенту можно вводить одну или несколько доз, составляющих приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любое их сочетание) антител. Такие дозы можно вводить периодически, например каждую неделю или каждые три недели (например, так чтобы пациент получал от приблизительно двух до приблизительно двадцати, или, например, приблизительно шесть доз антитела). Можно вводить первоначальную более высокую нагрузочную дозу, а затем одну или несколько более низких доз. Иллюстративная схема дозирования может включать введение исходной нагрузочной дозы, составляющей приблизительно 4 мг/кг, с последующей поддерживающей дозой антитела приблизительно 2 мг/кг каждую неделю. Однако могут быть пригодными другие схемы дозирования. Мониторинг хода этой терапии легко проводить посредством общепринятых способов и анализов.
Понятно, что любой из описанных выше способов лечения можно проводить с использованием иммуноконъюгата по изобретению вместо добавления антитела против Notch2.
3. Способы анализа
Антитела против Notch2 по изобретению могут быть охарактеризованы по их физическим/химическим свойствам и/или видам биологической активности различными анализами, известными в данной области.
a) Анализы активности
В одном аспекте представлены анализы для идентификации антител против Notch2 (например, антител против NRR Notch2), имеющих биологическую активность. Биологическая активность может включать, например, ингибирование или снижение активности Notch2, например, передачи сигнала Notch2. Также представлены антитела, имеющие такую биологическую активность in vivo и/или in vitro.
В определенных вариантах осуществления антитело против NRR Notch2 по изобретению тестируют в отношении его способности ингибировать образование B-клеток маргинальной зоны. Иллюстративный способ анализа предоставлен в разделе "Примеры". В некоторых других вариантах осуществления антитело по изобретению тестируют в отношении его способности ингибировать экспрессию репортерного гена, который отвечает за передачу сигнала Notch2. Иллюстративный способ анализа представлен в разделе "Примеры".
b) Анализы связывания и другие способы анализа
В одном аспекте антитело по изобретению тестируют в отношении его антигенсвязывающей активности, например, известными способами, такими как ELISA, вестерн-блоттинг и т.д. В другом аспекте можно использовать конкурентные анализы для идентификации антитела (например, моноклонального антитела), которое конкурирует с антителом по изобретению. В одном варианте осуществления антитело конкурирует с антителом D, антителом D-1, антителом D-2 или антителом D-3 за связывание с Notch2. В одном из таких вариантов осуществления конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), с которым связывается антитело D, антитело D-1, антитело D-2 или антитело D-3. В другом варианте осуществления конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), с которым связывается антитело против NRR1, как описано в примере B(5), т.е., с эпитопом, содержащим домены LNR-A, LNR-B и HD-C из NRR Notch1. Иллюстративные конкурентные анализы включают, но не ограничиваются ими, общепринятые анализы, такие как представлены в Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Подробное описание иллюстративных способов картирования эпитопа, с которым связывается антитело, представлены в Morris (1996) "Epitope Mapping Protocols", in Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ). Два антитела называют "связывающимися с одним эпитопом", если каждое из них блокирует связывание другого на 50% или более.
В иллюстративном конкурентном анализе иммобилизованный NRR Notch2 инкубируют в растворе, содержащем первое меченое антитело, которое связывается с NRR Notch2 (например, антитело D, антитело D-1, антитело D-2 или антитело D-3), и второе немеченое антитело, которое тестируют в отношении его способности конкурировать с первым антителом за связывание с NRR Notch2. Второе антитело может присутствовать в супернатанте гибридом. В качестве контроля иммобилизованный NRR Notch2 инкубируют в растворе, содержащем первое меченое антитело, но не второе немеченое антитело. После инкубации в условиях, позволяющих связывание первого антитела с NRR Notch2, избыток несвязанного антитела удаляют и измеряют количество метки, связанной с иммобилизованным NRR Notch2. Если количество метки, ассоциированной с иммобилизованным NRR Notch2, является по существу сниженным в тестируемом образце относительно контрольного образца, тогда это указывает на то, что второе антитело конкурирует с первым антителом за связывание с NRR Notch2.
В одном аспекте антитела по изобретению можно далее охарактеризовать с помощью серии анализов, включающих, но не ограничивающихся ими, N-концевое секвенирование, анализ аминокислот, неденатурирующую эксклюзионную высокоэффективную жидкостную хроматографию (ВЭЖХ), масс-спектрометрию, ионообменную хроматографию и расщепление папаином.
Понятно, что любой из описанных выше способов можно проводить с использованием иммуноконъюгата по изобретению вместо антитела против NRR Notch2 или в дополнение к нему.
E. Изделия
В другом аспекте изобретения представлено изделие, содержащее материалы, пригодные для лечения, профилактики и/или диагностики нарушений, описанных выше. Изделие содержит контейнер и ярлык на контейнере или вкладыш в упаковку, прилагаемый к контейнеру. Пригодные контейнеры включают, например, бутылки, флаконы, шприцы и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластмасса. В контейнере находится композиция, которая самостоятельно или в комбинации с другой композицией эффективна для лечения, профилактики и/или диагностики состояния и может иметь отверстие для стерильного доступа (например, контейнер может представлять собой мешок с внутривенным раствором или флакон, имеющий пробку, проницаемую для иглы для подкожной инъекции). По меньшей мере одно активное средство в композиции представляет собой антитело или иммуноконъюгат по изобретению. На ярлыке или вкладыше в упаковку указано, что композицию используют для лечения выбранного состояния. Более того, изделие может содержать (a) первый контейнер с содержащейся в нем композицией, где композиция содержит антитело или иммуноконъюгат по изобретению; и (b) второй контейнер с композицией, содержащейся в нем, где композиция содержит дополнительное цитотоксическое или иное лекарственное средство. В этом варианте осуществления изобретения изделие может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно использовать для лечения конкретного состояния. Альтернативно или дополнительно, изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекции (BWFI), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Кроме того, он может включать другие материалы, желательные с коммерческой точки зрения и с точки зрения потребителя, включая другие буферы, разбавители, фильтры, иглы и шприцы.
IV. ПРИМЕРЫ
Ниже предоставлены примеры способов и композиций по изобретению. Понятно, что можно осуществлять на практике различные другие варианты осуществления с учетом общего описания, представленного выше.
A. Материалы и способы
1. Получение антител против NRR Notch2
Сортировка библиотеки и скрининг для идентификации антител против NRR Notch2
Для пэннинга использовали фаговые библиотеки антител человека с разнообразными синтетическими выбранными определяющими комплементарность областями (H1, H2, H3, L3), имитирующими природное разнообразие набора IgG человека. Fab-фрагменты бивалентно экспонировали на поверхности частиц бактериофага M13. (См. Lee et al., J. Mol. Biol. 340:1073-1093 (2004).) Фрагменты NRR Notch2 экспрессировали в качестве секретируемых белков, слитых с эпитопными метками (FLAG или 6xHis) с использованием бакуловирусной экспрессирующей векторной системы или клеток 293T, очищенных до чистоты >90% с использованием аффинной хроматографии и тестированных в отношении отсутствия агрегации с использованием рассеяния света. Последовательности антигенов NRR Notch2 были следующими:
FLAG-NRR-Notch2-человека-6xHis:
KDDDDKGSGDVCPQMPCLNGGTCAVASNMPDGFICRCPPGFSGARCQSSCGQVK CRKGEQCVHTASGPRCFCPSPRDCESGCASSPCQHGGSCHPQRQPPYYSCQCAPPF SGSRCELYTAPPSTPPATCLSQYCADKARDGVCDEACNSHACQWDGGDCSLTME NPWANCSSPLPCWDYINNQCDELCNTVECLFDNFECQGNSKTCKY
DKYCADHFKDNHCNQGCNSEECGWDGLDCAADQPENLAEGTLVIVVLMPPEQLL QDARSFLRALGTLLHTNLRIKRDSQGELMVYPYYGEKSAAMKKQRM TRRSLPGEQEQEVAGSKVFLEIDNRQCVQDSDHCFKNTD AAAALLASHAIQGTLSY PLVSVVSESLTPERTEFGLVPRGSGHHHHHH (SEQ ID NO:73)
NRR-Notch2-мыши-FLAG:
ADVCPQKPCLNGGTCAVASNMPDGFICRCPPGFSGARCQSSCGQVKCRRGEQCIH TDSGPRCFCLNPKDCESGCASNPCQHGGTCYPQRQPPHYSCRCPPSFGGSHCELYT APTSTPPATCQSQYCADKARDGICDEACNSHACQWDGGDCSLTMEDPWANCTST LRCWEYINNQCDEQCNTAECLFDNFECQRNSKTCKYDKYCADHFKDNHCDQGCN SEECGWDGLDCASDQPENLAEGTLIIVVLLPPEQLLQDSRSFLRALGTLLHTNLRIK QDSQGALMVYPYFGEKSAAMKKQKMTRRSLPEEQEQEQEVIGSKIFLEIDNRQCV QDSDQCFKNTDAAAALLASHAIQGTLSYPLVSVFSELESPRNARRAGSGDYKDDD
DKENLYFQ (SEQ ID NO:74)
На 96-луночные иммунопланшеты Maxisorp Nunc immunoplates наносили в течение ночи при 4°C антиген-мишень (10 мкг/мл) и блокировали в течение 1 часа при комнатной температуре буфером для блокирования фага PBST (фосфатно-солевой буфер (PBS) и 1% (масс./об.) бычьим сывороточным альбумином (BSA) и 0,05% (об./об.) tween-20). В планшеты с антигеном по отдельности добавляли VH (см., например, Lee et al., J. Immunol. Meth. 284:119-132, 2004) и VH/VL (см. Liang et al., J. Mol. Biol. 366: 815-829, 2007) фаговых библиотек антител и инкубировали в течение ночи при комнатной температуре. На следующие сутки покрытые антигеном планшеты промывали десять раз посредством PBT (PBS с 0,05% Tween-20) и связавшийся фаг элюировали посредством 50 мМ HCl и 500 мМ NaCl в течение 30 минут и нейтрализовывали равным объемом 1 M основания Tris (pH7,5). Выделенный фаг амплифицировали в клетках E. coli XL-1 Blue. В ходе последующих раундов селекции инкубацию фага с антителом в покрытых антигеном планшетах снижали до 2-3 часов, и строгость промывания планшета постепенно повышали.
После 4 раундов пэннинга, наблюдали значительное обогащение. При сортировке VH и VH/VL было отобрано 96 клонов для определения того, связываются ли они специфически с NRR Notch2 человека и мыши. Вариабельные области этих клонов секвенировали способом ПЦР для идентификации уникальных по последовательности клонов.
Аффинность фаговых антител ранжировали с использованием точечного конкурентного ELISA. Величины IC50 фаговых антител далее определяли с использованием конкурентного ELISA связывания фага. Уникальные фаговые антитела, которые связывали NRR Notch2 как человека, так и мыши, выбирали и преобразовывали в полноразмерные IgG для оценки в клеточном анализе in vitro.
Представляющие интерес клоны преобразовывали в IgG путем клонирования областей VL и VH отдельных клонов в векторы LPG3 и LPG4, соответственно, временно экспрессируя их в клетках млекопитающих CHO, и очищая на колонке с белком A.
Конструирование библиотек для повышения аффинности антитела D
Фагмида pW0703 (полученная на основе фагмиды pV0350-2b (Lee et al, J. Mol. Biol 340, 1073-1093 (2004)), экспонирующей одновалентный Fab на поверхности бактериофага M13) служила в качестве матрицы библиотеки для пересадки вариабельных доменов легкой (VL) и тяжелой (VH) цепей антитела D из библиотеки VH/VL для созревания аффинности. Затем стоп-кодоны включали в CDR-L3 матрицы библиотеки. Для созревания аффинности использовали стратегию слабой рандомизации, которая обеспечивала скорость мутаций приблизительно 50% в выбранных положениях при использовании стратегии "испорченных" олигонуклеотидов со смесями 70-10-10-10 оснований в пользу нуклеотидов дикого типа (Gallop et al., Journal of Medicinal Chemistry 37: 1233-1251 (1994)). В частности, нацеливались на остатки в положениях 28-32 CDR-L1, 50 и 53-55 CDR-L2, 91-94 и 96 CDR-L3, 28-35 CDR-H1, 50-58 CDR-H2, 95-100 CDR-H3. Также конструировали три различных библиотеки с комбинациями слабо рандомизированных петель CDR, L1/L2/L3, L3/H1/H2 и L3/H3.
Стратегия фаговой сортировки для обеспечения повышения аффинности
Для селекции по повышению аффинности, фаговые библиотеки подвергали сортировке на планшетах на первом раунде, с последующими пятью раундами сортировки в растворе. Библиотеки сортировали против NRR Notch2 мыши и человека по отдельности. На первом раунде сортировки на планшетах три библиотеки сортировали против покрытого планшета (планшет NUNC Maxisorp) по отдельности с фагом, добавленным в количестве приблизительно 3 O.D./мл в 1% BSA и 0,05% Tween 20, в течение 2 часов при комнатной температуре. На следующем раунде сортировки в растворе, 1 O.D./мл фага, размножившегося на первом раунде сортировки в планшете, инкубировали с 50 нМ биотинилированного белка-мишени (концентрация основана на величине IC50 для исходного клона фага) в 100 мкл буфера, содержащего 1% Superblock (Pierce Biotechnology) и 0,05% Tween20 в течение 30 минут при комнатной температуре. Смесь далее разбавляли 10X 1% Superblock и 100 мкл/лунка добавляли в покрытые нейтравидином лунки (5 мкг/мл) в течение 15 минут при комнатной температуре при осторожном встряхивании для улавливания связавшегося с биотинилированной мишенью фага. Затем лунки промывали PBS-0,05% Tween20 десять раз. Для определения фонового связывания в контрольных лунках, содержащих фаг с мишенями, которые не были биотинилированными, проводили улавливание на покрытые нейтравидином планшеты. Связавшийся фаг элюировали 0,1 Н HCl в течение 20 минут, нейтрализовывали 1/10 объема 1 M Tris pH11, титровали и размножали для следующего раунда. Далее проводили четыре дополнительных раунда сортировки в растворе вместе с двумя способами повышения строгости селекции. Первым из этих способов является зависимая от уровня селекция путем снижения концентрации биотинилированного белка-мишени от 10 нМ до 0,5 нМ, и второй из которых представляет собой независимую от уровня селекцию путем добавления избыточных количеств небиотинилированного белка-мишени (избыток в 100-500) для устранения путем конкуренции более слабых связующих веществ при комнатной температуре. Также добавление фага снижали (0,1-0,5 O.D/мл) для снижения фонового связывания фага.
Высокопроизводительный ELISA со скринингом аффинности (конкуренция в одной точке)
Колонии отбирали после шестого раунда сортировки и выращивали в течение ночи при 37°С в 500 мкл/лунка среды 2YT с 50 мкг/мл карбенициллина и 1E10/мл KO7 в 96-луночном планшете (Falcon). Из того же планшета в качестве контроля отбирали колонию XL-1, инфицированных исходным фагом. На 96-луночные планшеты Maxisorp Nunc наносили 100 мкл/лунка белка NRR Notch2 человека и мыши (1 мкг/мл) по отдельности в PBS при 4°С в течение ночи или комнатной температуре в течение 2 часов. Планшеты блокировали 65 мкл 1% BSA в течение 30 мин и 40 мкл 1% Tween 20 в течение других 30 минут.
Супернатант фага разбавляли 1:5 в буфере для ELISA (твердофазный иммуноферментный анализ) (PBS с 0,5% BSA, 0,05% Tween20) с 10 нМ белком-мишенью в или без него в общем объеме 100 мкл и инкубировали в течение по меньшей мере 1 часа при комнатной температуре в F-планшете (NUNC). 75 мкл смеси с белком-мишенью или без него переносили параллельно в планшеты, покрытые белком-мишенью. Планшеты осторожно встряхивали в течение 15 мин для обеспечения улавливания не связавшегося фага с планшетом, покрытым белком-мишенью. Планшет промывали по меньшей мере пять раз посредством PBS-0,05% Tween 20. Связывание количественно определяли путем добавления конъюгированного с пероксидазой хрена (HRP) антитела против M13 в буфере для ELISA (1:5000) и инкубировали в течение 30 минут при комнатной температуре. Планшеты промывали посредством PBS-0,05% Tween 20 по меньшей мере пять раз. Затем в лунки добавляли 100 мкл/лунка субстрата пероксидазы, 3,3',5,5'-тетраметилбензидина (TMB), и раствора пероксидазы B (H2O2) (Kirkegaard-Perry Laboratories (Gaithersburg, MD)) в соотношении 1:1 и инкубировали в течение 5 минут при комнатной температуре. Реакцию останавливали добавлением в каждую лунку 100 мкл 1 M фосфорной кислоты (H3PO4) и реакционную смесь инкубировали в течение 5 минут при комнатной температуре. OD (оптическую плотность) для желтого цвета в каждой лунке определяли с использованием стандартного устройства для считывания планшетов для ELISA при 450 нм. Снижение OD (%) вычисляли с помощью следующего уравнения.
Снижение OD450нм(%)=[(OD450нм лунки с конкурентом)/(OD450нм лунки без конкурента)]*100
По сравнению со снижением OD450 нм (%) для лунки исходного фага (100%), клоны, которые имели снижение OD450 нм (%) ниже чем 50% для мишени как человека, так и мыши, отбирали для анализа последовательности. Уникальные клоны подвергали селекции для получения фага для определения аффинности связывания (IC50 для фага) против NRR Notch2 как человека, так и мыши, по сравнению с исходными клонами. Клоны с наибольшим повышением аффинности преобразовывали в IgG1 человека и экспрессировали в клетках млекопитающих.
Характеризация антител против NRR Notch2 (Biacore)
Аффинность связывания антител против NRR Notch2 измеряли путем поверхностного плазмонного резонанса (SRP) с использованием устройства BIAcoreTM-3000. IgG человека против NRR Notch2 улавливали с помощью антитела мыши против IgG человека, нанесенного на сенсорный чип CM5, до достижения приблизительно 200 единиц ответа (RU). Для определения кинетики двукратные серийные разведения NRR Notch2 человека и мыши (от 3,9 нМ до 500 нМ) инъецировали в буфер PBT (PBS с 0,05% Tween 20) при 25°C со скоростью потока 30 мкл/мин. Константы ассоциации (kon) и константы диссоциации (koff) вычисляли с использованием простой модели связывание Ленгмюра "один к одному" (BIAcore Evaluation Software версии 3.2). Равновесную константу диссоциации (KD) вычисляли в качестве соотношения koff/kon.
2. Получение антител против NRR Notch1
Получение и характеризация определенных антител против NRR Notch1 описаны ранее. См. публикацию патентной заявки США No. US 2009/0081238 A1.
3. ELISA
Антитело против Anti-Flag использовали для улавливания меченного Flag белка NRR из всех четырех рецепторов для Notch в планшете высокого связывания с черными лунками (Greiner). Антитело против NRR и контрольные антитела E25 связывали с белками NRR и отмывали PBS. Конъюгированное с щелочной фосфатазой вторичное антитело против (H+L) человека (Jackson ImmunoResearch) использовали для детекции связанного антитела против NRR и E25. Несвязанное вторичное антитело отмывали и проводили детекцию AP с субстратом PNPP (Pierce).
4. FACS
Получали экспрессирующие плазмиды, в которых внеклеточный домен и трансмембранные домены либо Notch1, либо Notch2, были мечены на N-конце посредством myc, и экспрессировали их под контролем отвечающего на тетрациклин элемента (TRE) выше минимального предраннего промотора CMV. Эти плазмиды стабильно трансфицировали в клетки CHO-K1 (Clonetech), сконструированные для экспрессии трансактиваторного белка в присутствии доксицилина, который связывает и активирует TRE в трансфицированной плазмиде. Клетки обрабатывали 1 мг/мл доксицилина или носителем в течение 48 ч, а затем окрашивали антителом против NRR, антителом против myc (9E10, Millipore) или контрольным антителом E25 и вторичными антителами козы против антител человека либо с Alexa 488, либо с Alexa 647 (Invitrogen). После окрашивания, клетки анализировали на устройстве FACScalibur (BD Biosciences) и накапливали 15000 событий и ворота диапазона были настроены для оценки положительно и отрицательно окрашенных популяций.
5. Репортерный анализ Notch
Клетки NIH-3T3, либо стабильно трансфицированные Notch1, либо временно трансфицированные плазмидами, содержащими другие рецепторы Notch, котрансфицировали отвечающей на Notch TP-1 (12X CSL), люциферазный репортер светляка и конститутивно активным люциферазным репортером Renilla (pRL-CMV, Promega) для контроля эффективности трансфекции. Клеткам позволяли восстановиться после трансфекции от 6 часов до периода в течение ночи. Для стимуляции рецепторных клеток используют обработку антителами и клетками NIH-3T3, стабильно трансфицированными лигандом. Через 20 часов люциферазу светляка и Renilla определяют с помощью системы для анализа люциферазы Dual Glo Luciferase Assay system (Promega). Для каждого условия анализировали восемь повторений путем деления сигнала от люциферазы светляка на сигнал от Renilla для контроля эффективности трансфекции. Среднее значение и стандартное отклонение вычисляли и величины нормализовывали к вычисленным значениям для совместной культуры, стимулированной клетками NIH-3T3 без трансфекции лиганда.
6. Картирование эпитопов
Конструировали синтетические последовательности NRR Notch1 и Notch2 с уникальными участками рестрикции в точках соединения между пятью доменами NRR - LNR-A; LNR-B; LNR-C; HD-N; и HD-C - и изготавливали их в Blue Heron Technology (Bothell, WA). Обмен различными доменами между этими двумя синтетическими последовательностями с использованием сконструированных участков рестрикции приводил к химерным белкам NRR. Химерные клоны собирали в модифицированный вектор pUC19 и переносили в вектор, содержащий SEAP, pCSC.AP, для экспрессии. Меченные AP белки NRR получали путем трансфекции клеток 293T. После культивирования в течение 72 ч, кондиционированные среды собирали, очищали центрифугированием и тестировали в отношении активности AP с использованием системы Phospha-Light System (Applied Biosystems, Bedford, MA). Активность AP в каждом образце нормализовывали и ELISA проводили для тестирования способности α-NRR1 или α-NRR2 связывать химерные белки NRR. ELISA проводили следующим образом: либо α-NRR1, либо α-NRR2, улавливали с помощью связанного с планшетом антитела a-Fc в 96-луночном формате и инкубировали с кондиционированной средой в течение ночи. Планшет промывали для удаления не связавшегося белка, и уловленный белок тестировали в отношении активности AP с использованием системы Phospha-Light System. Полученную активность AP использовали для количественного определения способности каждого антитела против α-NRR связывать конкретные химерные белки и установления важных эпитопов для связывания.
7. Анализ HUVEC на гранулах из фибринового геля
Клетки HUVEC выращивали на гранулах из фибринового геля, как описано ранее (Nakatsu et al., 2003). На гранулы Cytodex 3 (Amersham Pharmacia Biotech) наносили 350-400 HUVEC на гранулу в 2 мл среды EGM-2 (Clonetics). Приблизительно 200 гранул, покрытых HUVEC, погружали в фибриновый сгусток в одной лунке 12-луночного планшета для культивирования тканей. 6×104 фибробластных клеток Detroit 551 высевали сверху сгустка и среды, содержавшей либо 5 мкг/мл антитела против NRR1, либо 5 мкг/мл антитела против DLL4, либо 1 мкМ DBZ, либо контроль в виде носителя. Среду меняли каждые двое суток. После от семи до девяти суток, фибриновые гели фиксировали 4% параформальдегидом и эндотелиальные клетки окрашивали антителом против CD31 (R&D Systems AF806) с использованием конъюгированного с FITC вторичного антитела для детекции.
8. Анализ неонатальной сетчатки in vivo
Новорожденным CD1 инъецировали внутрибрюшинно на P1 и P3 либо 10 мг/кг полученного из амброзии контроля, либо 20 мг/кг Notch-1. Глаза собирали на P5 и фиксировали с помощью 4% PFA. Сетчатки вырезали, блокировали (5% BSA, 0,5% Triton X-100) течение одного часа, а затем проводили обработку в течение 10 мин 1 M цитратом натрия. Затем сетчатки инкубировали в течение ночи при 4°С с биотинилированным изолектином B4 (Sigma) и антителом Ki67 (NeoMarkers). Затем сетчатки промывали и окрашивали конъюгатом стрептавидин-Alexa 488 и антителом козы против антител кролика с Cy3 в 1%BSA, 0,5% Triton X-100. Сетчатки помещали на предметное стекло и визуализировали с использованием эпифлуоресцентного микроскопа.
9. Исследования опухоли in vivo
Опухолевые клеточные линии человека Calu-6 и HM7 выращивали в среде Хэма F12 и DMEM с низком содержанием 1:1 глюкозы, дополненной 10% об./об. FBS, 1% об./об. пенициллином/стрептомицином, 2 мМ L-Gln и 1 мкг/мл FungizoneTM (InvitrogenTM, CA). Клетки инкубировали при 37°С в атмосфере 95% воздух/5% CO2. Для экспериментов с ксенотрансплантатами мышей, опухолевые клетки суспендировали в концентрации 1×108 или 1×109 клеток/мл и подкожно инъецировали (100 мкл/мышь) в дорзальную боковую область мышей Balb-c nude (Harlan Sprague Dawley, IN). Когда опухоли достигали объема 400 мм3, случайным образом выбирали группу (n=10) в качестве контролей 0 суток. Оставшихся мышей разделяли на группы по 10 мышей, и фаговое антитело против Notch1 вводили внутрибрюшинно в дозе 10 мг/кг два раза в неделю. Трансплантированные опухоли измеряли два раза в неделю вдоль наибольшей оси и перпендикулярной оси, как описано. Для каждых суток, на которые измеряли опухоли, для каждой мыши вычисляли объем опухоли и средние объемы опухолей для каждого контрольного антитела (антитела против агглютинина амброзии), группу антител против VEGF сравнивали со средним объемом опухоли у мышей, которым вводили антитело против Notch1, с помощью критерия Даннетта, осуществленного в системе для статистического анализа JMPTM (версии 5.1 для Windows; SAS Institute, Cary, NC), на уровне P<0,05. Мышей умерщвляли, когда объем опухоли достигал 2000 мм3. С помощью криостата делали срезы опухолей размером 7 микрометров, их фиксировали ацетоном и окрашивали посредством DAPI (Invitrogen), антитела хомячка против CD31 (Serotech, Inc.) и вторичного антитела против антител хомячка с Cy3 (Jackson Immunolabs). Предметные стекла закрепляли флуоресцентной закрепляющей средой (Dako). Изображения получали с помощью микроскопа Zeiss Axioskop2 и анализировали посредством ImageJ в отношении площади окрашивания DAPI и окрашивания Cy3. Окрашивание Cy3 делили на площадь окрашивания DAPI для нормализации к количеству присутствующих клеток и величины нормализовали к величинам для опухоли, обработанной антителом против агглютинина амброзии. Все исследования на животных проводили согласно протоколам, одобренным Institutional Animal Care and Use Committee.
10. Анализ B-клеток маргинальной зоны
Мышам Balb-c (Jackson) в возрасте 12 недель внутрибрюшинно инъецировали два раза в неделю 5 мг/кг антитела против gD, антитела против NRR1 или антитела против NRR2. Спленоциты мышей выделяли и окрашивали для FACS посредством B220-PerCP, CD23-PE и CD21-FITC (BD Biosciences), как описано ранее (Saito, et al, Immunity, Vol. 18, 675-685, May 2003).
11. Гистология и иммуногистохимия ткани кишечника мыши
В шести группах по шесть мышей C57BL/6 в возрасте 10 недель (Jackson) вводили либо MCT раз в двое суток, либо DBZ в количестве 30 мкмоль/кг раз в двое суток, либо антитело против NRR1 в количестве 10, 2, или 0,4 мг/кг или контроля в виде антитела против gD (HSV-I) в количестве 10 мг/кг на 0, 2 и 6 сутки, или, в ином случае, как указано. Фиксированные в формалине и погруженные в парафин ткани тонкого кишечника мыши нарезали на срезы толщиной 3 микрометра. Гистохимическую идентификацию типов клеток кишечника проводили с помощью альцианового синего, как рекомендовано изготовителем (PolyScientific). Для окрашивания антителом против Ki67, срезы предварительно обрабатывали раствором для извлечения мишени (S1700, DAKO) и инкубировали с антителом кролика против Ki67 (1:200, клон SP6, Neomarkers). Вторичное антитело козы против антител кролика в концентрации 7,5 мкг/мл (Vector labs) подвергали детекции с помощью набора Vectastain ABC Elite Kit (Vector labs). Окрашенные на Ki67 срезы контрастно окрашивали гематоксилином Мейера. Для окрашивания антителом против лизоцима срезы обрабатывали на платформе Discovery XT (Ventana Medical Systems, Inc.) с использованием условий выделения эпитопа CC1m, детекции антител кролика с помощью OmniMap и гематоксилина Ventana II/с контрастным окрашиванием синим красителем. Для окрашивания с помощью HES-1, после антитела против HES-1 крысы (клон NM1, MBL, International) следовала обработка TSA-HRP.
B. Результаты
1. Получение синтетических антител, которые специфично нацелены на отрицательные по Notch1 и Notch2 регуляторные области
Для обеспечения независимого антагонизма передачи сигнала Notch1 и Notch2 у человека и мышей, авторы изобретения использовали фаговый дисплей для нацеливания на NRR и отбора синтетических антител человека с нижеследующими качествами. Во-первых, каждое антитело отбирали, чтобы оно эффективно ингибировало свою мишень - Notch1 или Notch2 - но не другие рецепторы Notch. Во-вторых, антитела отбирали по связыванию с последовательностями-мишенями человека для обеспечения терапевтического нацеливания. В-третьих, для установления биологических последствий системного ингибирования Notch1 или Notch2 in vivo и для исследования функций этих рецепторов в моделях на мышах, антитела совместно отбирали по связыванию с ортологичными последовательностями мыши. В-четвертых, для предотвращения иммунного ответа человека против антител определяющие комплементарность области клонировали в каркас иммуноглобулинов человека.
В соответствии с этим подходом, авторы выделили антитела против NRR Notch1 и против NRR Notch2 с каждым из этих желаемых свойств. Эти антитела имеют несколько преимуществ относительно "общих ингибиторов Notch", которые нацелены на множество рецепторов Notch, таких как ингибиторы γ-секретазы. Поскольку эти антитела отличают конкретные рецепторы Notch, они могут устранять различные функции этих отдельных рецепторов. Более того, общие ингибиторы Notch демонстрируют потенциально нежелательные побочные эффекты in vivo вследствие двойного ингибирования как Notch1, так и Notch2, и, таким образом, антитела, которые являются селективными в отношении либо Notch1, либо Notch2, могут быть более привлекательными кандидатами для терапевтических целей. Антитела против NRR Notch1 и против NRR Notch2, выделенные согласно указанному выше подходу, дополнительно описаны и охарактеризованы ниже.
Выделение и характеризация антител против Notch NRR1 рассмотрены в публикации патентной заявки США No. US 2009/0081238 A1. Одно из антител, описанных в этой заявке, "антитело A-2", было охарактеризовано в отношении его структуры и некоторых видов биологической активности. Некоторые из исследований, рассмотренные в этой заявке, обобщены в настоящем документе. Для удобства "антитело A-2" из публикации патентной заявки США No. US 2009/0081238 A1 называют в настоящем документе "антителом против NRR1".
Выделение и характеризация антител против NRR Notch2 описаны в настоящем документе. Было выделено антитело против NRR Notch2, называемое "антителом D". Это антитело подвергали созреванию аффинности, как описано выше, с получением антитела D-1, антитела D-2 и антитела D-3. На фигуре 1 показаны последовательности HVR-H1, HVR-H2 и HVR-H3 антитела D, антитела D-1, антитела D-2 и антитела D-3. На фигуре 2 показаны последовательности HVR-L1, HVR-L2 и HVR-L3 антитела D, антитела D-1, антитела D-2 и антитела D-3. На фигуре 3 показаны последовательности вариабельной области тяжелой цепи антитела D, антитела D-1, антитела D-2 и антитела D-3. На фигуре 4 показаны последовательности вариабельной области легкой цепи антитела D, антитела D-1, антитела D-2 и антитела D-3. Для удобства антитело D-3 называют в настоящем документе как "антитело против NRR2".
Измерение аффинности связывания с использованием поверхностного плазмонного резонанса (SPR) показало, что антитело против NRR1 связывалось со сходной и высокой аффинностью (Kd=3 нМ) с очищенными белками NRR1 с последовательностью как мыши, так человека. Авторы изобретения наблюдали сходную аффинность (Kd=5 нМ) в отношении антигенов NRR2 в тестах антител против NRR2. Связывание обоих антител оказалось высоко специфичным в отношении их рецепторов; не было выявлено связывания с последовательностями человека или мыши любого из других трех не являющихся мишенями рецепторов как для антитела против NRR1, так и для антитела против NRR2, с использованием различных способов, включающих SPR, твердофазный иммуноферментный анализ (ELISA) и активированную флуоресценцию сортировку клеток (FACS) (фигура 9 и данные не представлены). Например, результаты ELISA показали, что антитело против NRR1 связывалось в равной степени хорошо с очищенными белками NRR Notch1 человека и мыши, причем полумаксимальное связывание наблюдали при концентрации антитела приблизительно 0,1 мкг/мл. Антитело против NRR1 не связывалось на поддающемся детекции уровне с очищенным белками NRR Notch2, NRR Notch3 или NRR Notch4 человека или мыши, даже при наиболее высокой протестированной концентрации антитела (фигура 9A, левая панель), несмотря на тот факт, что последовательности NRR Notch1 и NRR Notch2 человека являются на 45% идентичными (фигура 18). Авторы настоящего изобретения выявили сходные результаты для антитела против NRR2, которое специфично связывалось с белками NRR Notch2, но не с белками NRR из других рецепторов Notch (фигура 9A, правая панель).
Для определения того, связывается ли антитело против NRR1 и антитело против NRR2 специфично с полноразмерными рецепторами, экспрессированными на поверхности клеток, авторы настоящего изобретения использовали активируемую флуоресценцией сортировку клеток (FACS). Авторы настоящего изобретения оценивали связывание либо с Notch1, либо с Notch2, каждый из которых содержит эпитопную метку myc на N-конце для того, чтобы отличать трансгенные рецепторы Notch от эндогенных рецепторов Notch (в частности Notch2, который экспрессировался клетками K1-CHO). Экспрессию рецептора индуцировали в клетках K1-CHO, которые либо оставались нетрансфицированными (фигура 9B, панели 1, 4, 7 и 10), либо были трансфицированы myc-Notch1 (фигура 9B, панели 2, 5, 8 и 11), либо были трансфицированы myc-Notch2 (фигура 9B, панели 3, 6, 9 и 12). Связывание антитела против NRR1 с myc-положительными клетками наблюдали в клетках myc-N1 (фигура 9B, панель 2), но не в исходных клетках K1-CHO (фигура 9B, панель 1) и только после индукции (фигура 9B, сравнить панели 2 и 5), указывая на то, что антитело против NRR1 специфично связывалось с Notch1. Напротив, хотя экспрессия myc-Notch2 очевидно индуцировалась в клетках myc-N2 (фигура 9B, сравни панели 3 и 6), антитело против NRR1 не связывалось с myc-Notch2 (фигура 9B, сравнить панели 2 и 3). Авторы изобретения провели сходный анализ с использованием антитела против NRR2 (фигура 9B, панели 7-12), хотя этот анализ был осложнен экспрессией myc-Notch2, происходящей в отсутствии индукции (фигура 9B, панели 6 и 12) и эндогенной экспрессией Notch2 хомячка в линии K1-CHO (фигура 9B, сравнить панели 7 и 10 с панелями 1 и 4). Тем не менее, антитело против NRR2 в значительной степени связывалось только с клетками, экспрессирующими myc-Notch2 после индукции, что согласуется со специфичным связыванием с Notch2 (фигура 9B, сравнить панель 9 с панелями 8 и 12). Взятые вместе, результаты авторов настоящего изобретения по связыванию при использовании очищенных белков NRR, а также экспрессии на клеточной поверхности полноразмерных рецепторов, демонстрируют, что антитело против NRR1 специфично связывается с NRR1 Notch1, но не с NRR Notch2, и аналогично, что антитело против NRR2 специфично связывается с NRR Notch2, но не с NRR Notch1. Эта специфичность далее подтверждена рядом структурных и функциональных исследований, описанных ниже.
2. Функционирование антитела против NRR1 и антитела против NRR2 в качестве сильных и специфичных ингибиторов передачи сигнала Notch1 и Notch2 in vitro
Для оценки того, влияют ли антитела против NRR на передачу сигнала Notch, авторы настоящего изобретения сначала использовали анализ совместной культуры, в котором использовали клеточные линии NIH-3T3, сконструированные для экспрессии Jag1, в качестве лиганда Notch, или либо Notch1 (фигура 10A), либо Notch2 (фигура 10B) в качестве рецептора Notch. Анализы точно отражали передачу сигнала Notch, поскольку сильный сигнал репортера Notch (люцифераза светляка) зависел от присутствия как клеток, экспрессирующих лиганд, так и клеток, экспрессирующих рецептор (фигуры 10A и B, сравнить -Jag1 с +Jag1), и уровень репортера снижался до фонового уровня, когда был включен GSI (ингибитор γ-секретазы) (фигуры 10A и B, сравнить DMSO с DAPT).
Добавление возрастающих количеств антитела против NRR1 ингибировало передачу сигнала в клетках Notch1, причем полное ингибирование наблюдалось при концентрации антитела между 80 и 400 нг/мл (фигура 10A). Контрольное антитело человека не ингибировало передачу сигнала (фигура 10A, сравнить титрование α-gD с α-NRR1), так же как и многочисленные другие антитела, направленные против антигенов, отличных от Notch1 (данные не представлены). Для дальнейшего тестирования того, отражает ли эта ингибиторная активность связывание NRR1, авторы настоящего изобретения решили установить, восстанавливает ли добавление очищенного антигена NRR1 передачу сигнала в присутствии ингибиторной концентрации (80 нг/мл) антитела против NRR1, путем конкуренции за связывание антитела с Notch1, экспрессируемым на передающих сигнал клетках. Добавление антигена NRR1, но не NRR2 (который оказался активным и надлежащим образом свернутым в других анализах; см. фигура 10B) восстановило передачу сигнала до контрольных уровней (фигура 10A, сравнить 80 нг/мл α-NRR1 + NRR1 с 80 нг/мл α-NRR1 отдельно и 80 нг/мл α-NRR1 + NRR2). В качестве контроля, ни антиген NRR1, ни антиген NRR2, не влияли на передачу сигнала в совместных культурах, которые содержали контрольное антитело, подтверждая, что фрагменты NRR сами по себе не оказывают прямого эффекта на передачу сигнала Notch (фигура 10A, α-gD+NRR1 и α-gD+NRR2). С использованием клеток, экспрессирующих Notch2, для анализа активности против NRR2, авторы настоящего изобретения получили сходные результаты для ингибирования антителом против NRR2 передачи сигнала Notch2 (фигура 10B). Оба антитела ингибировали передачу сигнала, индуцируемую всеми из лигандов Notch, которые авторы настоящего изобретения протестировали, а именно Jag1, Jag2, Dll1 и Dll4 (фигура 16 и данные не представлены). Взятые вместе, эти результаты демонстрируют, что антитело против NRRl и антитело против NRR2 являются сильными паралог-специфическими ингибиторами передачи сигнала Notch1 и Notch2, соответственно.
3. Антитело против NRR1 ингибирует передачу сигнала через оба основных класса мутантных рецепторов, встречающихся при T-ALL
Одним из механизмов, посредством которых, как было показано, передача сигнала Notch прямо влияет на онкогенез, является механизм через мутационную активацию передачи сигнала Notch1 при T-ALL. В частности, активирующие Notch1 мутации при T-ALL относятся к двум широким категориям: (1) мутации, которые укорачивают домен PEST, таким образом, стабилизируя ICD и повышая передачу сигнала лиганд-зависимым образом, и (2) мутации, которые дестабилизируют NRR, таким образом, обеспечивая расщепление ADAM и стимуляцию передачи сигнала независимым от лиганда образом. Для тестирования того, может ли антитело против NRR1 прекратить передачу сигнала через такие мутантные рецепторы, авторы настоящего изобретения модифицировали анализ совместной культуры для экспрессии Notch1-WT (Notch1 дикого типа), Notch1-ΔPEST (Notch1, лишенный домена PEST), или Notch1-L1594P (Notch1, имеющий мутацию L1594P в домене гетеродимеризации (HD) NRR). Как было выявлено для передачи сигнала Notch1 дикого типа (фигура 10C, верхняя панель), антитело против NRR1 полностью ингибировало зависимую от лиганда и независимую от лиганда передачу сигнала через Notch1-ΔPEST и Notch1-L1594P (фигура 10C, средняя и нижняя панели). Таким образом, антитело против NRR1 осуществляет антагонизм передачи сигнала через оба класса мутантных рецепторов, включая мутацию L1594P, которая находится в том же домене, на который нацелено антитело, и дестабилизирует его.
4. Антитело против NRR1 и антитело против NRR2 функционируют в качестве сильных и специфичных ингибиторов передачи сигнала Notch1 и Notch2 in vivo
Для определения того, функционирует ли антитело против NRR1 и антитело против NRR2 как специфичные к рецептору ингибиторы in vivo, авторы настоящего изобретения исследовали, каким образом антитела влияли на определение участи клеток, роль в котором генетическими способами была установлена для Notch1 или Notch2. Конкретно, зависимая от условий инактивация Notch1 или Notch2 ранее показала, что Notch1 является ключевым для определения участи T-клеток относительно B-клеток в ходе развития лимфоидной системы, в то время как Notch2 требуется для образования B-клеток маргинальной зоны (MZB) селезенки. В соответствии с функцией Notch1 в развитии T-клеток, авторы настоящего изобретения открыли, что введение мышам антитела против NRR1, но не антитела против NRR2, значительно снижало массу тимуса (фигуры 11A и B). Аналогично, антитело против NRR1, но не антитело против NRR2, значительно снижало общее число клеток в тимусе (фигура 11B) и практически полностью ингибировало образование двойных положительных CD4+/CD8+ T-клеток (фигура 11C).
Напротив, эффекты антител были обратными, когда исследовали клетки MZB. Введение антитела против NRR2 практически устраняло клетки MZB (0,97+/-0,45 по сравнению с контрольным значением, составляющим 6,61+/-0,25), снижая популяцию в еще более высокой степени, чем очищенный рекомбинантный слитый белок рецептор лимфотоксина-β (LTβR)-Fc слитый белок (3,48+/-0,06), который служил в качестве положительного контроля (фигура 11D). Этот ингибиторный эффект был специфичным к антителу против NRR2, поскольку антитело против NRR1 не снижало в значительной степени популяцию MZB (6,00+/-0,44) относительно контрольного антитела против gD (6,61+/-0,25; фигура 11D). Взятые вместе, эти исследования in vivo указывают на то, что оба антитела являются сильными ингибиторами их соответствующих мишеней, которые могут индуцировать биологически значимые изменения в определении участи клеток; более того, поскольку антитело против NRR1 влияет на образование T-клеток, но не MZB-клеток, в то время как антитело против NRR2 влияет на образование MZB-клеток, но не T-клеток, каждое антитело функционирует специфично в отношении мишени, для которой оно предназначено, in vivo.
5. Совместная кристаллическая структура показала, что антитело против NRR1 связывается с доменами LNR и HD-C и, вероятно, стабилизирует конформацию "off" NRR
Для установления специфичности антитела против NRR1 и антитела против NRR2 и повышения механистического понимания антагонистической активности антитела, авторы настоящего изобретения протестировали связывание обоих антител с рядом химерных белков NRR. Конкретно, авторы настоящего изобретения получили химерные NRR путем обмена каждого из субдоменов NRR (LNR-A, LNR-B, LNR-C, HD-N и HD-C) между Notch1 и Notch2, экспрессировали эти химеры в качестве секретируемых белков, слитых с щелочной фосфатазой для простоты детекции и анализировали связывание нормализованных количеств каждого химерного NRR с антителом против NRR1 или антителом против NRR2. Из 26 химерных NRR, которые экспрессировали авторы изобретения 17 эффективно выявлялись в качестве секретируемых белков (фигура 12A), 7 плохо поддавались детекции и 2 не поддавались детекции выше фонового уровня (фигура 17), указывая на то, что большинство, но не все, из химерных NRR экспрессировались, сворачивались надлежащим образом и секретировались. Обмен единичного домена LNR-A из NRR2 в каркас NRR1 нарушал связывание антитела против NRR1 (фигура 12A, химера BC.Hd и все другие с LNR-A из NRR2), указывая на то, что контакт антитела против NRR1 с субдоменом LNR-A в NRR1 является необходимым для связывания. Напротив, обмен доменами LNR-C и/или HD-N из NRR2 в NRR1 не повлиял на связывание антитела против NRR1 (фигура 12A, химеры AB.Hd, ABC.Hc и AB.Hc), демонстрируя, что ни один из этих доменов не является необходимым для специфичности связывания. Наконец, AB.Hc представлял собой химеру с наименьшим количеством доменов NRR1, которая, тем не менее, поддерживала полное связывание антитела против NRR1 (хотя с химерой AB наблюдали слабое связывание), указывая на то, что большинство, и, возможно, все, из участков контакта, требуемых для специфичности связывания антитела против NRR1, содержатся в доменах LNR-A, LNR-B и HD-C.
Характеризация антитела против NRR2 показала паттерн связывания, сходный с паттерном, определенным для антитела против NRR1, хоти и с некоторыми отчетливыми отличиями. Обмен доменом LNR-A из NRR1 в каркас NRR2 полностью нарушал связывание антитела против NRR2, в то время как обмен LNR-B или LNR-C не имел эффекта; таким образом, как было в случае антитела против NRR1, связывание антитела против NRR2 требует субдомена LNR-A из Notch2. Аналогично также был необходим домен HD-C из Notch2. Две различных химеры (C.Hn и BC) только с тремя судбоменами из NRR2 поддерживали полное связывание в этом анализе; обе из этих химер имели общие субдомены LNR-A и HD-C из NRR2 и, в соответствии с этими данными, химера, которая содержала только субдомены LNR-A и HD-C из NRR2 поддерживала слабое, но поддающееся детекции связывание с антителом против NRR2. Таким образом, при определении субдоменов, которые определяют специфичность связывания антитела против NRR2, эксперименты по обмену доменами: (a) показали, что LNR-A и HD-C являются наиболее важными (необходимыми и частично достаточными для специфичности связывания) и (b) указали на то, что участки контакта антитело-NRR2 в LNR-B и HD-N также могут играть роль.
Для лучшего понимания молекулярной основы антагонистической активности антитела против NRR1, авторы настоящего изобретения определи 2,2 Ǻ2 кристаллическую структуру Fab-фрагмента антитела, связанного с NRR1 человека. Эта структура показала, что NRR1 формирует компактную структуру, очень сходную со структурой NRR2 человека, где три связывающих Ca2+ модуля LNR обернуты вокруг центрального домена HD (фигуры 12B и C). Структура показала, что кажущимся эффектом связывания Fab является стабилизация взаимодействий LNR-HD, так чтобы S2 не был доступен для расщепления, таким образом, поддерживая каскад передачи сигнала Notch1 в состоянии покоя. Эта интерпретация подтверждается данными, что Fab не прямо закрывает участок S2, а вместо этого, связывается на поверхности контакта между доменами HD, LNR-A и LNR-B (фигура 12D), что согласуется с экспериментами по обмену доменами (фигура 12A). В этой поверхности контакта погружено ~2000 Ǻ доступной для растворителя площади поверхности, равно распределенной между fab и NRR1. Тяжелая цепь Fab контактирует с LNR-B, конечной спиралью в домене HD-C и периферией LNR-A. CDR H3 располагается в поверхности контакта между LNR-B и HD. В частности, R99 из H3 образует водородную связь с карбонилом Phe1501 основной цепи LNR-A, в то время как алифатическая часть боковой цепи взаимодействует с L1710 из домена HD. Легкая цепь составляет ~40% погруженной площади поверхности, обеспечиваемой Fab, и контактирует с соединительной петлей перед конечной спиралью в домене HD, а также LNR-A.
Эта структура также раскрывает основу Fab-специфичности для Notch1 относительно Notch2. Несмотря на ~45% идентичность последовательностей между доменами NRR Notch1 и Notch2, большинство из остатков, вовлеченных в поверхность контакта, отличаются между двумя белками. Например, из 21 остатка NRR, которые погружают по меньшей мере 25% доступной для растворителя площади поверхности при связывании с Fab, только шесть идентичны в Notch1 и Notch2. Эти шесть остатков вносят вклад менее чем в четверть площади поверхности в эпитопе Fab и распределены по всему эпитопу (фигуры 12D и 18).
6. Селективное ингибирование передачи сигнала Notch1 нарушает регуляцию ангиогенеза
Чтобы начать исследование специфичных к рецептору Notch антител в целях определения функциональной значимости отдельных рецепторов in vivo, авторы изобретения сначала решили установить, нарушает ли селективное блокирование передачи сигнала Notch1 антителом против NRR1 ангиогенез у млекопитающих. Многочисленные сообщения указывают на то, что каскад Notch функционирует ниже сосудисто-эндотелиального фактора роста (VEGF) и играет ключевую роль в ангиогенезе путем регуляции определения участи эндотелиальных клеток между концевыми клетками и клетками трубки. Эксперименты с использованием различных генетических и биохимических инструментов, включая специфичные к Dll4 блокирующие антитела, установили Dll4 в качестве ключевого лиганда Notch, вовлеченного в ангиогенную передачу сигнала Notch, хотя в ангиогенез в качестве рецепторов Dll4 вовлечены два рецептора Notch, Notch1 и Notch4.
Сначала авторы изобретения протестировали, влияет ли селективное ингибирование Notch1 на ветвление эндотелиальных клеток in vitro. Эндотелиальные клетки пупочной вены человека (HUVEC), совместно культивированные с фибробластами кожи человека, образуют ветвления со структурами, подобными просвету. В соответствие с предшествующей работой, авторы настоящего изобретения выявили, что общее ингибирование Notch с использованием DBZ, ингибитора γ-секретазы, а также селективного антитела, блокирующего Dll4, повышало как количество, так и длину ветвлений эндотелиальных клеток. Важно, что селективное блокирование передачи сигнала Notch1 с использованием антитела против NRR1 привело к сходному фенотипу (фигуры 13A и 13B).
Для определения того, влияет ли антитело против NRRl на ангиогенез in vivo, авторы настоящего изобретения использовали модель, основанную на развитии сосудов сетчатки у новорожденных мышей. Системная доставка антитела против NRR1 в значительной степени нарушала развитие сосудов сетчатки у новорожденных мышей, которым проводили введение (фигура 13C). Относительно введения контрольного антитела против агглютинина амброзии, введение против NRR1 приводило к плотной, компактной и на вид запутанной сосудистой сети (фигура 13C, сравнить панели I и III с панелями II и IV, соответственно). Это повышенное накопление эндотелиальных клеток коррелировало с повышенной пролиферацией, оцениваемой по мечению маркером пролиферации Ki67 (фигура 13C, сравнить панели V и VI). Все эти данные, включая повышенную плотность сосудов, накопление эндотелиальных клеток и повышенную пролиферацию, близко имитируют данные, выявленные после ингибирования Dll4 или γ-секретазы с использованием этой модели на новорожденных (данные не представлены) и согласуется с результатами других исследований, включая анализ сосудов опухоли (фигуры 13D и 13E) и анализ кармана роговицы у мышей (данные не представлены). Результаты, полученные авторами настоящего изобретения, демонстрируют не только то, что антитело против NRR1 обладает высокой биологической активностью, но то, что ингибирование Notch1 отдельно в противоположность Notch4 или комбинации Notch1 и Notch4, является достаточным для выраженного нарушения ангиогенеза у млекопитающих.
7. Селективное блокирование антителом Notch1 ингибирует рост опухоли в доклинических моделях
Для определения того, является ли достаточным селективное блокирование Notch1 с использованием антитела против NRR1 для ингибировании роста опухоли, авторы настоящего изобретения использовали доклинические модели опухоли и сравнивали рост установившихся опухолей после введения антитела против NRR1, антитела против VEGF или антитела против агглютинина амброзии. В обеих моделях на Calu6 и HM7, группы, в которых вводили антитело против NRR1, продемонстрировали значимое снижение размера опухоли относительно контрольных групп, даже в первые исследованные моменты времени, от трех до четырех суток после начальной дозы (фигуры 13A-C). В модели Calu6 группа антитела против NRR1 продемонстрировала снижение размера опухоли от приблизительно 250 мм3 до менее чем 100 мм3, аналогично снижению, наблюдаемому в группе антитела против VEGF; напротив, опухоли в контрольной группе антитела против агглютинина амброзии росли до размера более 400 мм (фигура 14A). Авторы настоящего изобретения наблюдали сходные результаты при использовании агрессивно растущих HM7; введение антитела против NRR1 приводило к тому, что размер опухоли оставался неизменным (фигура 14B) или практически неизменным (фигура 14C) в течение 12-13 суток, что значительно отличается от роста в течение шести суток от 250 мм3 до более чем 900 мм3, наблюдаемого в контрольной группе (фигуры 14B и 5C). Авторы настоящего изобретение наблюдали сходное ингибирование роста опухоли с использованием ряда концентраций доз антитела против NRR1, между 20 мг/кг и 2,5 мг/кг, хотя группы 10 и 20 мг/кг продемонстрировали тенденцию в отношении более сильного ингибирования относительно групп 2,5 и 5 мг/кг (фигура 14C).
Авторы настоящего изобретения исследовали модели опухолей Calu6 или HM7, поскольку их ответ на антиангиогенные средства документально подтвержден. Для прямого тестирования того, нарушает ли антитело против NRR1 ангиогенез в модели Calu6, авторы настоящего изобретения исследовали сосуды в репрезентативных срезах опухолей от мышей в трех группах введения. Путем номализации окрашивания на CD31, маркер эндотелиальных клеток, к окрашиванию ДНК, с использованием DAPI, авторы настоящего изобретения открыли, что антитело против NRR1 значимо повышало окрашивание на CD31 относительно других групп (фигура 14D). Это повышение было статистически значимым (p < 0,01), и постоянно выявлялось на множестве изображений (фигура 14E). В противоположность этому и как ожидалось, антитело против VEGF вызывало уменьшение окрашивания на CD31 (фигуры 14D и 14E). Таким образом, результаты селективного блокирования Notch1 отражают результаты, описанные для селективного блокирования Dll4, которое аналогично приводит к повышению окрашивания опухоли на CD31, а также к обеспечению плохого функционирования сосудов опухоли. Взятые вместе, эти результаты указывают на то, что противоопухолевые эффекты, оказываемые антителом против NRR1 в моделях Calu6 и HM7, главным образом, отражают нарушение ангиогенеза опухоли.
8. Ингибирование антителом как Notch1, так и Notch2, имело выраженные эффекты на участь клеток кишечника
Ингибиторы гамма-секретазы (GSI), которые представляют собой общие ингибиторы Notch, которые ингибируют множество рецепторов Notch, вызывают снижение массы тела и метаплазию бокаловидных клеток кишечника, отражая роль, которую Notch играет в определении участи клетки путем поддержания пролиферации клеток-предшественников крипт кишечника и препятствования дифференцировке в секреторные клетки. (См. van Es et al., Nature 435:959-963 (2005).) Более того, генетические исследования с использованием мышей с зависимым от условий нокаутом Notch указывают на то, что для обеспечения метаплазии бокаловидных клеток кишечника требуется нарушение как Notch1, так и Notch2. (См. Riccio et al., EMBO Rep. 9:377-383 (2008).) Авторы настоящего изобретения использовали антитело против NRR1 и антитело против NRR2 для определения эффектов селективного ингибирования Notch1 или Notch2, или обоих из них, на снижение массы тела и дифференцировку клеток кишечника.
Мыши, которым вводили антитело против NRR1, имели небольшое снижение общей массы тела в ходе дозирования антител. В первом эксперименте, прямо сравнивающем антитело против NRR1 и антитело против NRR2, авторы настоящего изобретения выявили, что антитело против NRR1 приводило к 5% снижению общей массы тела, в противоположность повышению массы тела, наблюдаемому в случае антитела против NRR2, а также контрольных групп антитела против gD и антитела против LTβR-Fc (фигура 15A). Во втором эксперименте селективное ингибирование Notch1 или Notch2 с использованием антитела против NRR1 или антитела против NRR2, соответственно, приводило небольшому эффекту на массу или отсутствию этого эффекта. Результаты этого эксперимента представлены на фигуре 24A, где мышам вводили 5 мг/кг антитела против NRR1 (квадраты), антитела против NRR2 (треугольники) или контрольного антитела ("α-gD", ромбы) на сутки, указанные стрелками. Однако у мышей, которым вводили антитело против NRR1 и антитело против NRR2 ("X" на фигуре 24A), масса тела снижалась практически на 20% от исходной уже на 7 сутки.
Авторы настоящего изобретения исследовали то, отражает ли небольшое снижение массы тела вследствие введения антитела против NRR1 отдельно или более значительное снижение массы тела вследствие введения как антитела против NRR1, так и антитела против NRR2, изменения в определении участи клеток кишечника. После дозирования мышам антитела против NRR1 или антитела против gD или DBZ в качестве контролей, срезы толстого (фигура 19) и тонкого кишечника анализировали на 2 сутки (фигура 20) и 7 сутки (фигура 15B) с использованием различных гистохимических красок. DBZ имел те же эффекты, которые описаны ранее для GSI (фигура 15B, столбец DBZ): ингибирование экспрессии Hes1 коррелировало со снижением пролиферации клеток-предшественников (как показано с помощью окрашивания на Ki-67) и с выраженной экспансией популяции бокаловидных клеток (как показано с помощью окрашивания на муцин альциановым синим). Окрашивание на лизоцим также показало, что DBZ повышал количество и/или активность клеток Панета, популяцию вторичных секреторных клеток (фигура 15B), и это предположение было далее подтверждено с использованием окрашивания азуром A-эозином B на гранулы клеток Панета (данные не представлены). Антитело против NRR1 в концентрации 10 мг/кг индуцировало изменения в кишечнике, неотличимые от изменений, индуцируемых DBZ (фигура 15 A, сравнить DBZ, 30 мкмоль/кг, с α-NRR1, 10 мг/кг). Эти изменения кишечника зависели от концентрации антитела против NRR1, так что наблюдали значительные изменения участи клеток при применении 10 мг/кг и 2,0 мг/кг но небольшие, при их наличии, заметные изменения наблюдали при применении 0,4 мг/кг. Толстый кишечник отвечал на введение DBZ и антитела против NRR1 сходным образом: введение обоих из них снижало передачу сигнала Notch (фигура 19, окрашивание Hes1), блокировало пролиферацию клеток-предшественников (фигура 19, Ki-67) и повышало количество бокаловидных клеток (фигура 19, альциановый синий и H&E). Изменение участи клеток в кишечнике только слабо поддавалось детекции в ранний момент времени на 2 сутки этого исследования (фигура 20; следует отметить небольшие, но поддающиеся детекции изменения, вызываемые как DBZ, так и антителом против NRR1, наблюдаемые при окрашивании на Ki-67 и альциановым синим).
Для определения того, может ли ингибирование передачи сигнала Notch2 сходным образом влиять на определение участи клетки, авторы настоящего изобретения прямо сравнили эффекты антитела против NRR1 и антитела против NRR2 на толстый (данные не представлены) и тонкий кишечник (фигура 15C, где показано окрашивание альциановым синим и окрашивание на Ki-67, и фигура 24B, где показано окрашивание альциановым синим). В то время как антитело против NRR1 вызывало метаплазию бокаловидных клеток, которая совпадала со снижением экспрессии Ki-67 в предшественниках крипт, антитело против NRR2 не вызывало каких-либо поддающихся детекции эффектов (фигуры 15C и 24B, сравнить антитело против NRR2 с контролем против gD). Поскольку антитело против NRR1 и антитело против NRR2 функционируют в качестве сильных и специфических ингибиторов передачи сигнала Notch1 и Notch2, соответственно, как in vivo (фигура 11), так и in vitro (фигуры 9, 10 и 21), эти результаты убедительно подтверждают, что селективное ингибирование Notch1, но не Notch2, является достаточным для изменения определения участи клеток кишечника.
Также исследовали эффект ингибирования как Notch1, так и Notch2, на определение участи клеток кишечника. На фигуре 22 представлены синергичные эффекты антитела против NRR1 и антитела против NRR2 на измененную дифференцировку клеток кишечника. Самкам мышей Balb/c вводили 5 мг/кг антитела против NRR1, 5 мг/кг антитела против NRR2 или 5 мг/кг антитела против NRR1 вместе с 5 мг/кг антитела против NRR2 на 0, 4, 7 и 11 сутки. Кишечники извлекали и окрашивали H&E для установления морфологии крипт. Для мышей, которым вводили антитело против NRR1 или антитело против NRR2 отдельно, кишечники извлекали на 12 сутки; для мышей, которым вводили оба антитела вместе, кишечники извлекали на 6 сутки, поскольку у мышей в этой группе быстро снижалась масса тела. Анализ кишечников показал, что комбинация антител приводила к более тяжелому фенотипу по сравнению с фенотипом, наблюдаемым после введения антитела против NRR1 отдельно, указывая на то, что Notch1 и Notch2 функционирую совместно и отчасти чрезмерно в кишечнике млекопитающего. Дальнейшее экспериментирование подтвердило это наблюдение. Как показано на фигуре 24B, кишечники от мышей, которым вводили антитело против NRR1 и антитело против NRR2, продемонстрировали тяжелую метаплазию бокаловидных клеток. Хотя введение антитела против NRR1 отдельно было достаточным для индукции некоторой метаплазии бокаловидных клеток, эффект антитела против NRR1 отдельно был мягким относительно эффекта как антитела против NRR1, так и антитела против NRR2, в комбинации.
Взятые вместе, результаты указывают на то, что Notch1 и Notch2 функционируют чрезмерно при дифференцировке клеток кишечника, хотя существенное ингибирование Notch1, но не Notch2, является достаточным для выявления неполного фенотипа. Авторы настоящего изобретения предположили, что их способность выявить этот неполный фенотип, не описанный ранее в генетических исследованиях, отражает то, что антитело против NRR1 обеспечивает более единообразное и сильное ингибирование передачи сигнала Notch1 в клетках крипт, которое может быть достигнуто после зависящего от условий нокаута, который может быть неполным. Важно, что путем существенного снижения или устранения метаплазии бокаловидных клеток, которые являются признаками общего ингибирования Notch, специфические ингибиторы в виде антител против Notch1 и Notch2, описанные в настоящем документе, представляют собой очевидное достижение по сравнению с существующими общими ингибиторами Notch, такими как GSI.
9. Восстановление фенотипа кишечника, вызванного антителом против NRR1
На фигуре 23 показано, что дексаметазон по меньшей мере частично восстанавливает фенотип кишечника, вызываемый антителом против NRR1. Антитела вводили внутрибрюшинно (IP) самкам мышей NCR.nude (три мыши на группу) следующим образом:
Группа A) комбинация носителей, MCT, один раз в сутки, контрольное антитело против gD в количестве 4 мг/кг, раз в четверо суток (не показано на фигуре);
Группа B) дексаметазон, 90 мг/кг, раз в сутки (не показано на фигуре);
Группа C) антитело против NRR1, 4 мг/кг, раз в четверо суток;
Группа D) дексаметазон, 90 мг/кг, раз в сутки и антитело против NRR1, 4 мг/кг, раз в четверо суток. Носитель представлял собой 0,5% (масс./об.) гидроксипропилметилцеллюлозу (Methocel E4M) и 0,1% (масс./об.) Tween 80 в воде (MCT), которые, как показано ранее, не вызывают неблагоприятных клинических эффектов.
Кишечники извлекали на 9 сутки и окрашивали антителом против Ki67 (маркер пролиферации) и альциановым синим (на муцин). Этот эксперимент показал, что мыши, которым вводили комбинацию дексаметазона вместе с антителом против NRR1, проявляли более мягкий фенотип кишечника относительно мышей, которым вводили антитело против NRR1 отдельно, таким образом, указывая на то, что дексаметазон защищает кишечник от индуцируемых антителом против NRR1 изменений дифференцировки.
10. Антитело против NRR2 подавляет рост клеточных линий меланомы
Исследовали эффект антитела против NRR1 и антитела против NRR2 на рост клеточных линий меланомы. Клеточные линии меланомы человека SK23 и LOX-IMVI высевали в среду, содержащую 2% FBS при низкой плотности (4000 клеток/лунка для SK23, 2500 клеток/лунка для LOX-IMVI) в 96-луночных планшетах, на которые было нанесено покрытие без (-jag-1) или с лигандом Jagged-1 (+jag-1) (R&D Systems, Minneapolis, MN). В культуры добавляли антитело против NRR1 (20 мг/мл), антитело против NRR2 (20 мг/мл) или ингибитор гамма-секретазы (GSI) DAPT (5 мкм) и после этого повторно добавляли раз в двое суток. Равный объем DMSO служил в качестве контроля в виде носителя, в то время как равная концентрация фагового антитела против gD HuB6 служила в качестве изотипического контрольного антитела. Через шесть суток после посева проводили анализы титра клеток Glo (Promega, Madison, WI). Результаты представлены на фигурах 25A и 25B (антитело против NRR1 и антитело против NRR2 обозначены как "антитело против N1" и "антитело против N2"). На фигурах 25A и 25B, жизнеспособность клеток (ось y) выражена относительно значений для контрольных лунок с носителем DMSO, -jag-1. Обе меланомных клеточных линии продемонстрировали снижение жизнеспособности в ответ на введение GSI (особенно в присутствии лиганда Jag1) и введение антитела против NRR2, но не на введение антитела против NRR1.
11. Антитело против NRR2 подавляет рост диффузной крупноклеточной B-клеточной лимфомы in vitro
Диффузная крупноклеточная B-клеточная лимфома (DLBCL) является наиболее распространенным типом неходжскинской лимфомы. Недавнее исследование злокачественных клеток от пациентов с DLBCL показало, что приблизительно 8% пациентов имеют мутацию в домене PEST Notch2, которая, как предполагают, продлевает передачу сигала Notch2 после активации лиганда. (См. Lee et al., Cancer Sci. 100:920-926, 2009.) Однако то, как клетки DLBCL будут отвечать на ингибирование Notch2, не было известно. Таким образом, авторы исследовали эффект антитела против NRR2 на пять клеточных линий DLBCL. Линии DLBCL, приведенные справа на фигуре 26, выращивали с повторами в лунках 384-луночных планшетов в течение трех суток. Клеточные культуры включали указанные концентрации антитела против NRR2 (обозначаемого как "антитело против Notch2" на фигурах 26 и 27), которые соответствовали трехкратным серийным разведениям, начиная с 10 мкг/мл. Рост оценивали с использованием анализов титра клеток Glo (Promega) и наносили на график как процент жизнеспособных клеток относительно введения изотипического контрольного антитела. Данные на фигуре 26 соответствуют средним величинам, полученным для отдельных лунок для каждой клеточной линии при указанных концентрациях антител. Как показано на фигуре 26, рост одной из клеточных линий, "DB", в значительной степени ингибировался при введении антитела против NRR2. На фигуре 27 показан рост клеточной линии DB с течением времени. Линию DB выращивали в ячейках 12-ячеечной чашки, и культуры обрабатывали DMSO, ингибитором гамма-секретазы DAPT, изотипическим контрольным антителом против gD или антителом против NRR2 в указанных концентрациях. Рост оценивали путем подсчета жизнеспособных клеток (Vi-CELL, Beckman Coulter, Fullerton, CA) на 2, 3 и 5 сутки после инокуляции и обработки культур. Величины соответствуют среднему значению для измерений в трех независимых культурах, плюс/минут стандартное отклонение. На фигуре 27 показано, что антитело против NRR2 подавляет рост клеточной линии DB DLBCL in vitro.
Хотя представленное выше изобретение описано с некоторыми деталями в качестве иллюстрации и примера для ясности понимания, описание и примеры не следует истолковывать как ограничивающие объем изобретения. Описание всей патентной и научной литературы, цитированной в настоящем документе, включено в настоящий документ в качестве ссылки в полном объеме.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ NRR Notch1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2476443C2 |
| АНТИТЕЛА ПРОТИВ NOTCH1 | 2011 |
|
RU2622083C2 |
| АНТИТЕЛА К JAGGED И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2666990C2 |
| АНТИТЕЛА ПРОТИВ DLL4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2415869C2 |
| КОМПОЗИЦИИ ГУМАНИЗИРОВАННЫХ СЛИТЫХ БЕЛКОВ NOTCH И СПОСОБЫ ЛЕЧЕНИЯ | 2008 |
|
RU2532830C2 |
| АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ Notch3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С Notch3 ЗАБОЛЕВАНИЙ | 2007 |
|
RU2493167C2 |
| АНТИТЕЛО К NOTCH4 ЧЕЛОВЕКА | 2016 |
|
RU2720280C2 |
| ОСНОВАННЫЕ НА Notch3 ЧЕЛОВЕКА ГИБРИДНЫЕ БЕЛКИ В КАЧЕСТВЕ ЛОВУШЕК-ИНГИБИТОРОВ ПЕРЕДАЧИ СИГНАЛА Notch3 | 2009 |
|
RU2567662C2 |
| АГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ NOTCH3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ NOTCH3-АССОЦИИРОВАННЫХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2461569C2 |
| ЛЕЧЕНИЕ ВИРУСА ГРИППА А H7N9 | 2015 |
|
RU2682049C2 |






































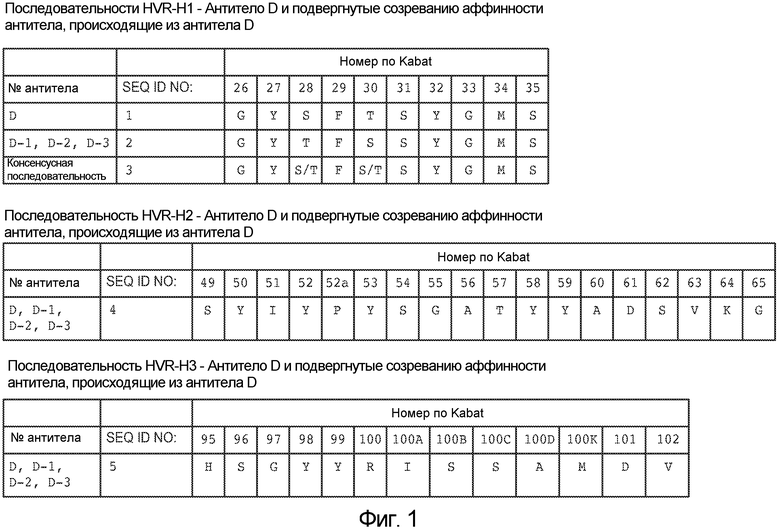
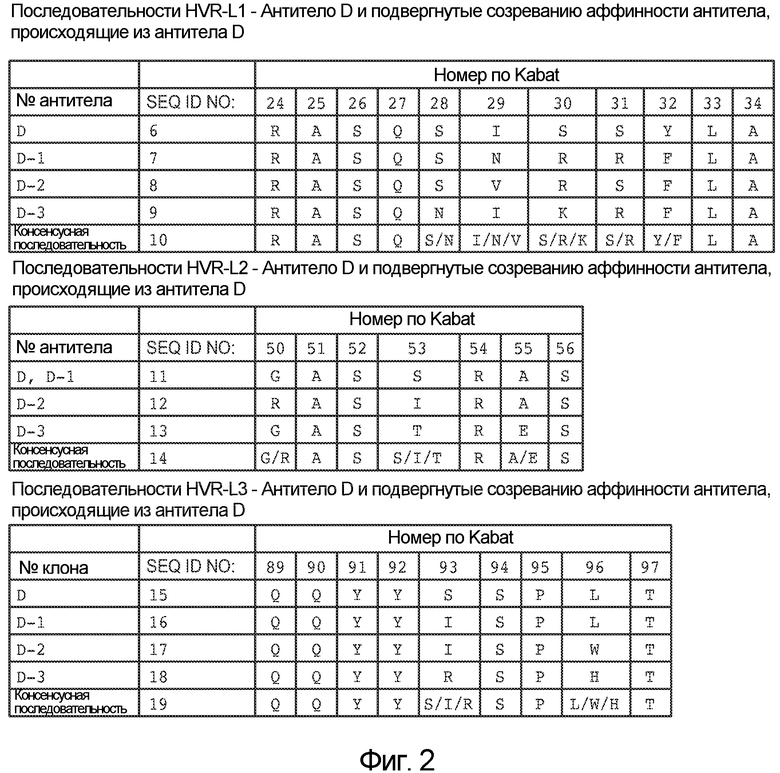


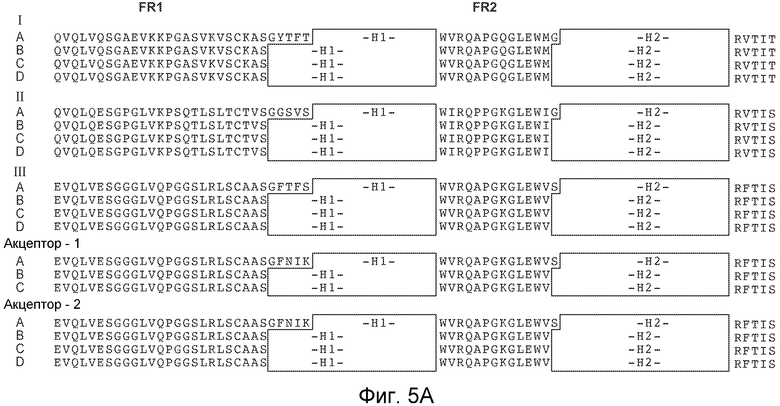
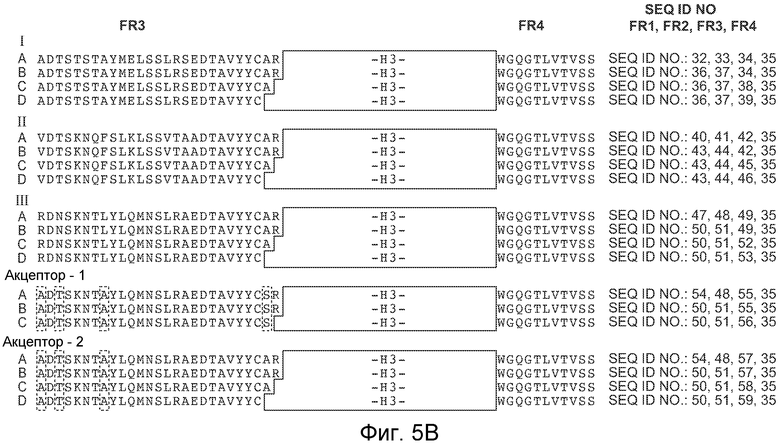
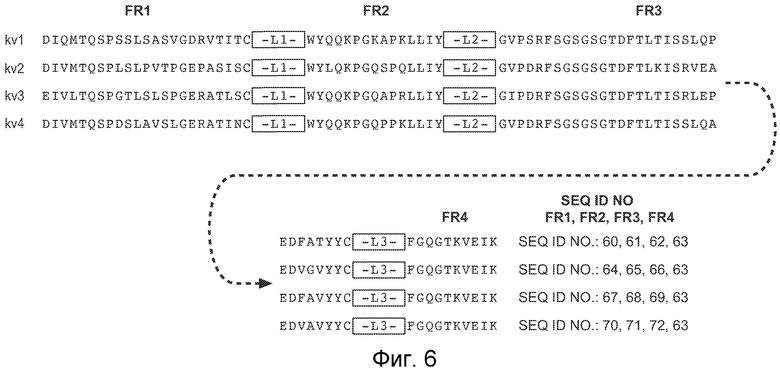


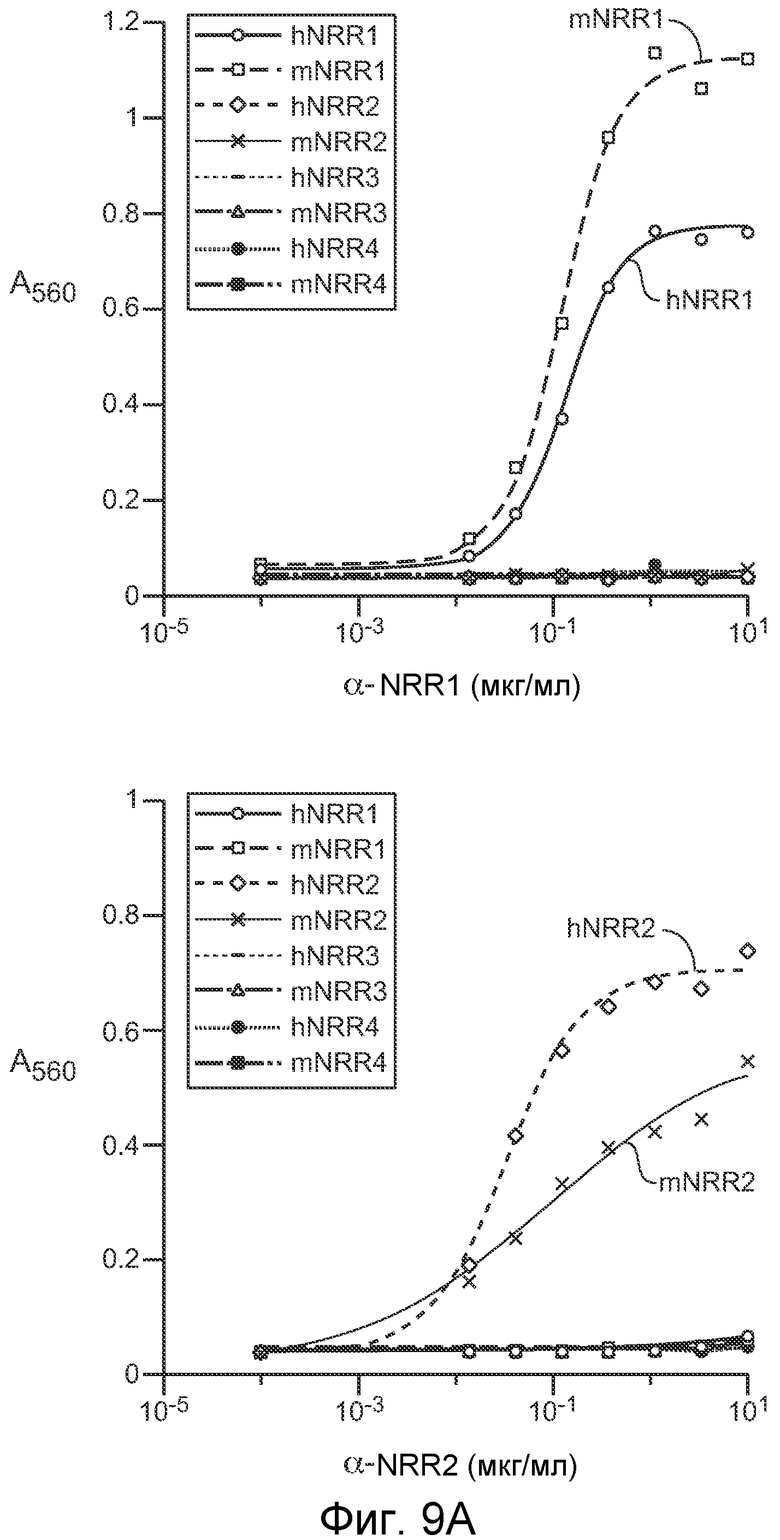
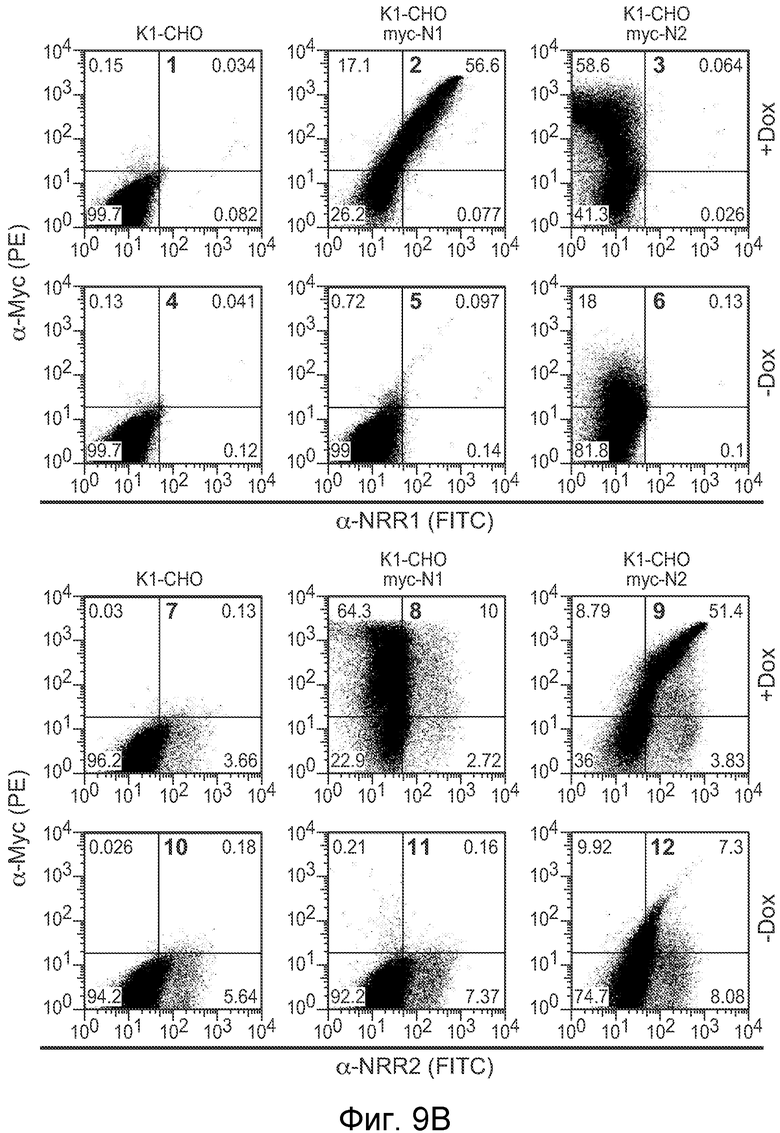
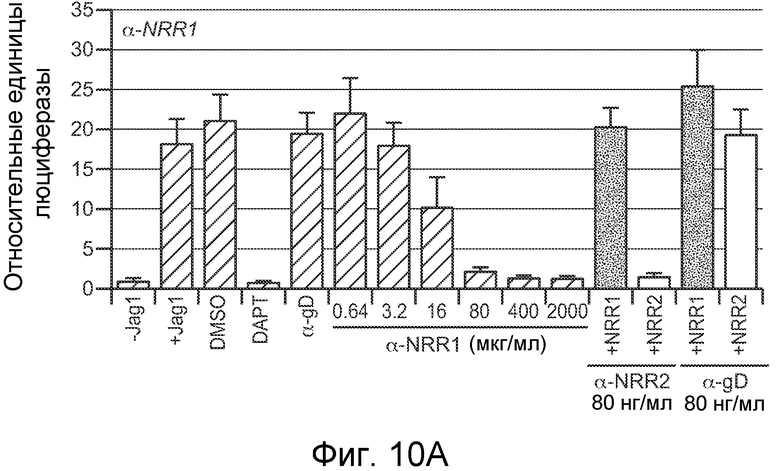
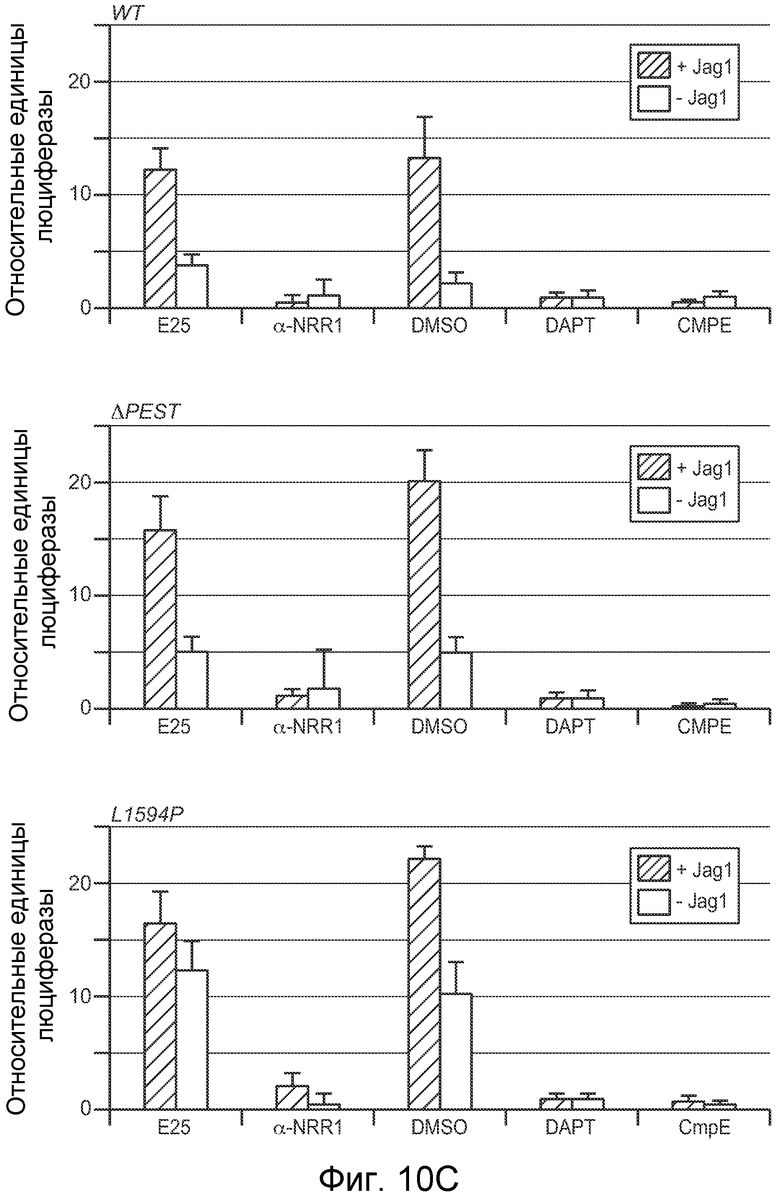
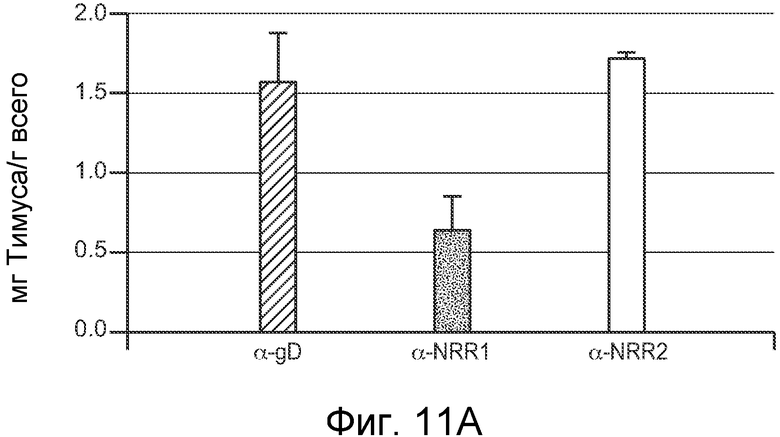
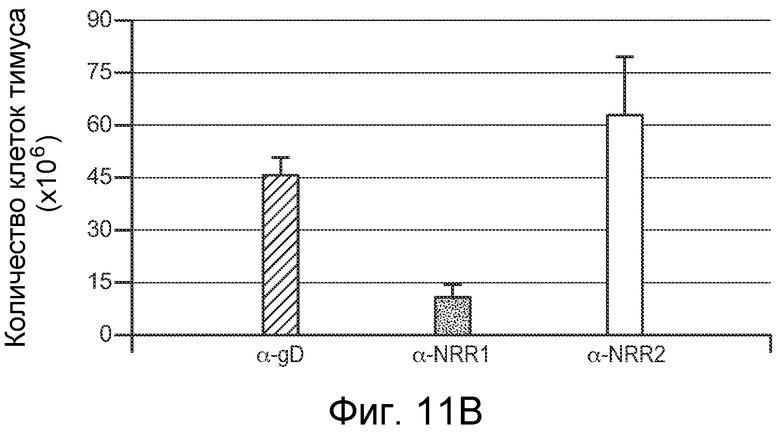
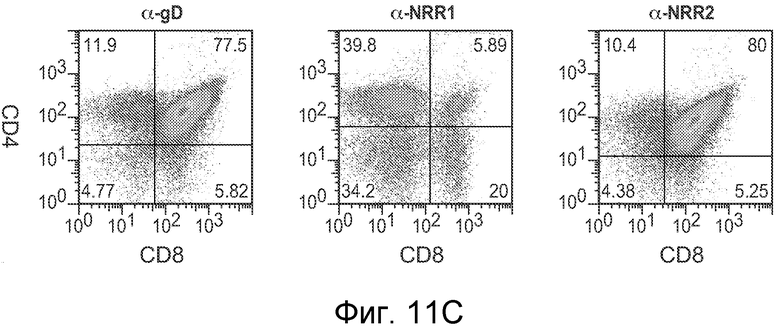
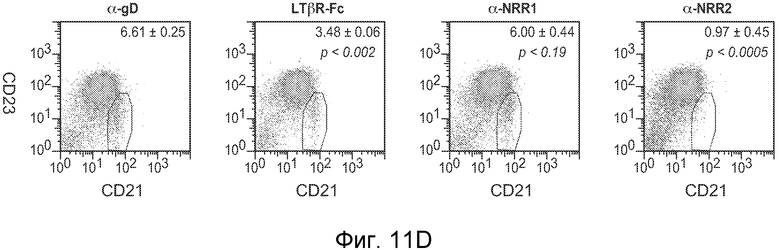
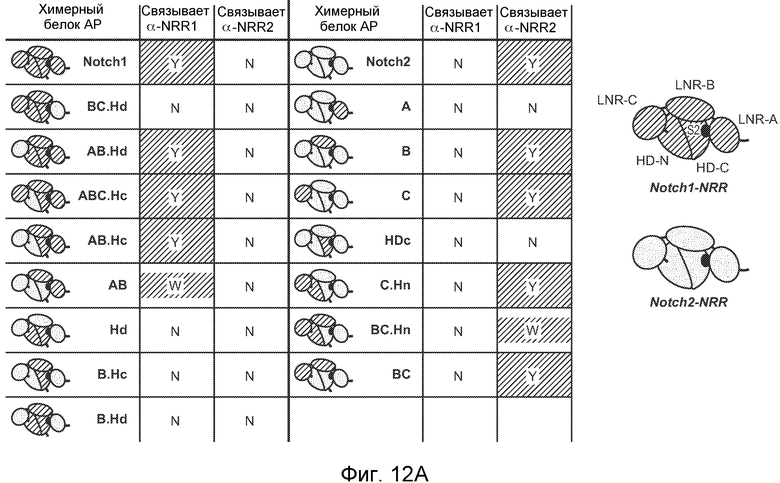
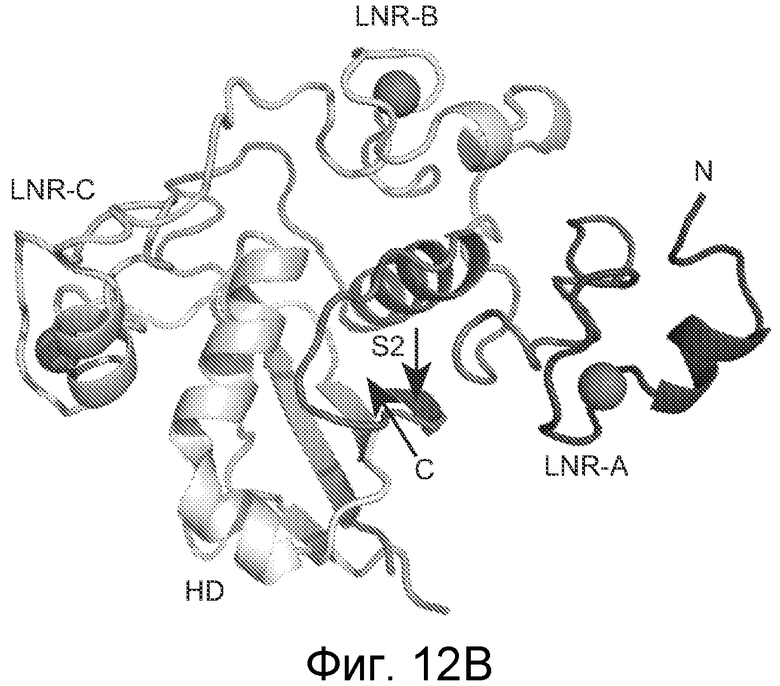

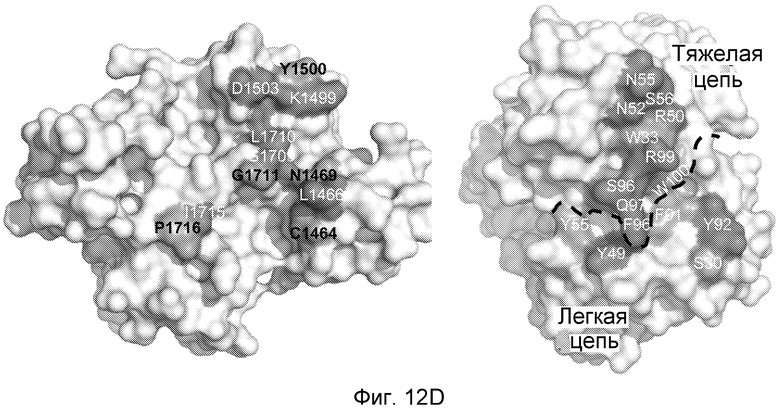
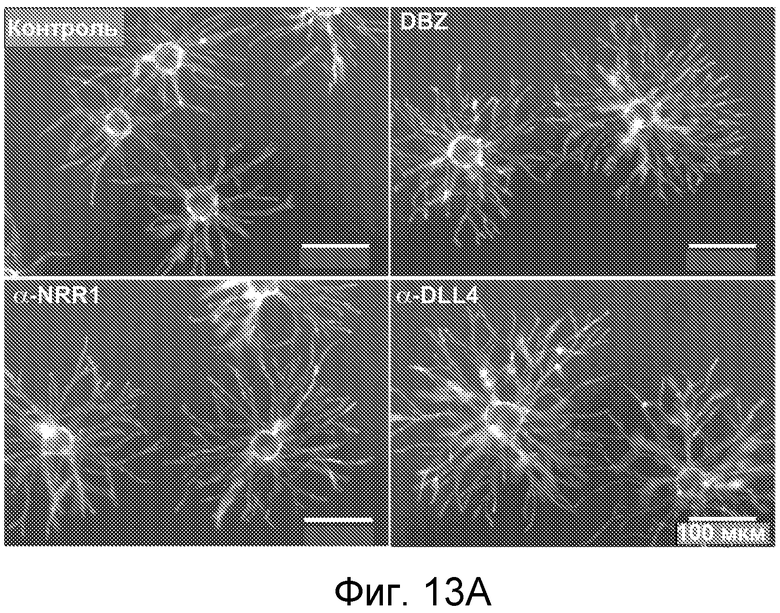
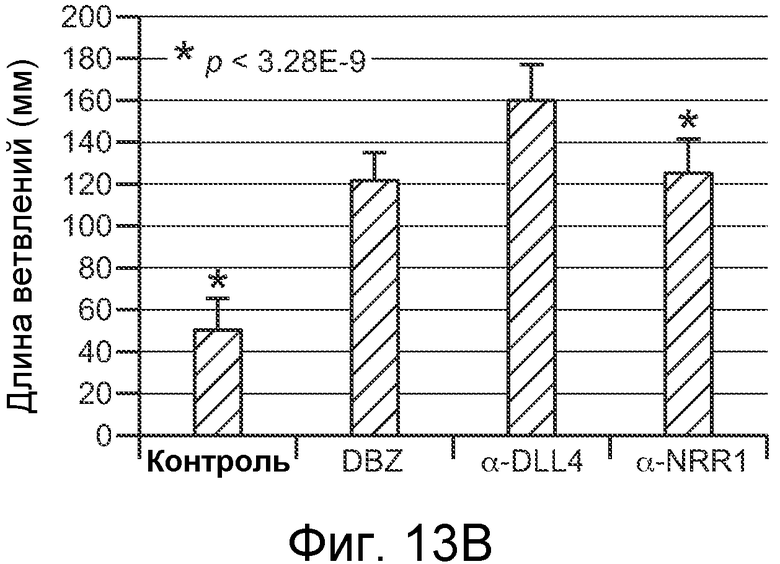
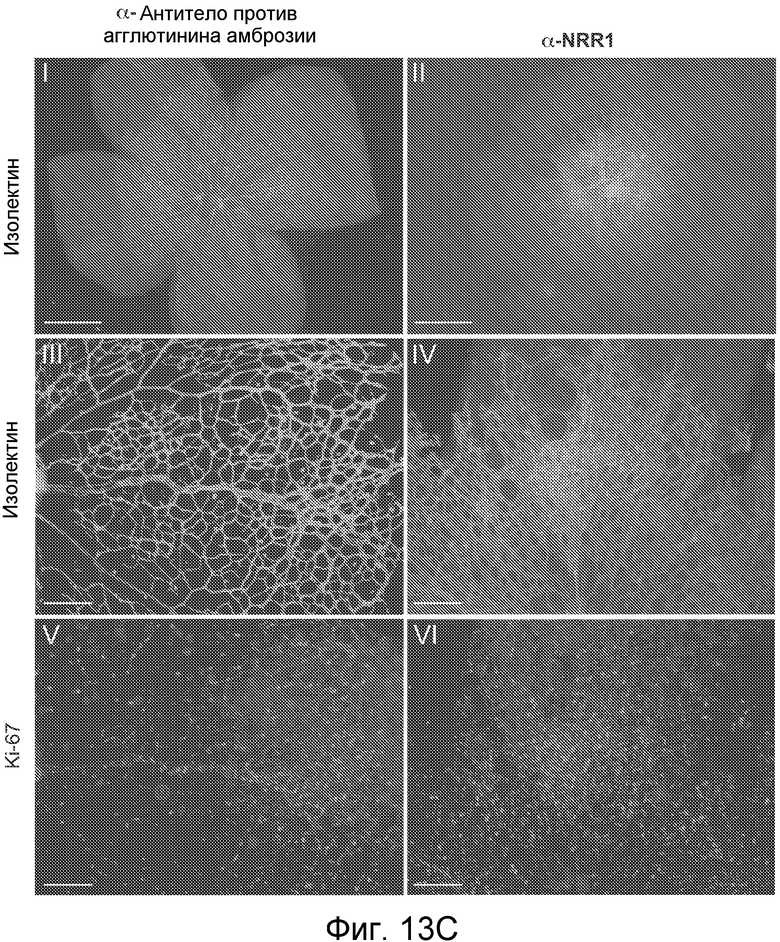
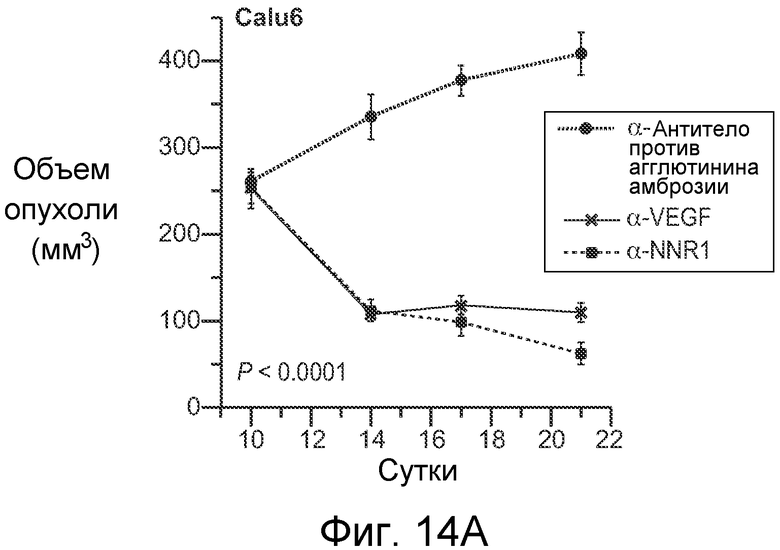
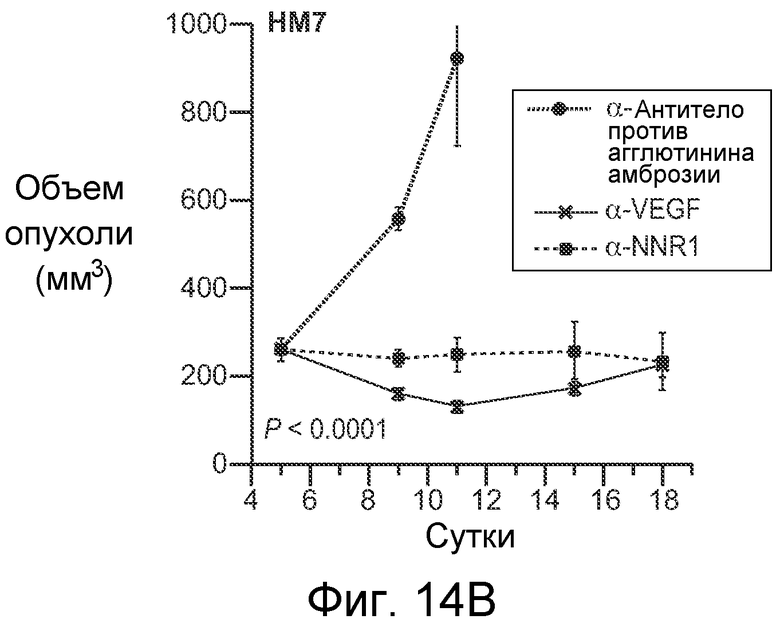
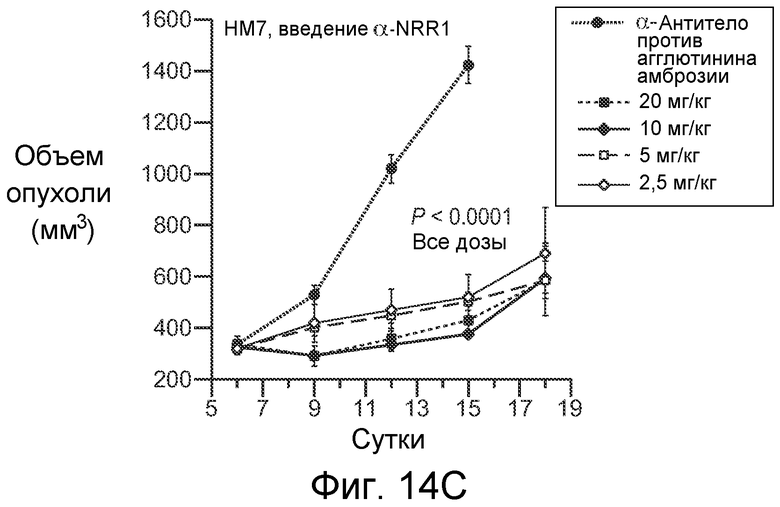
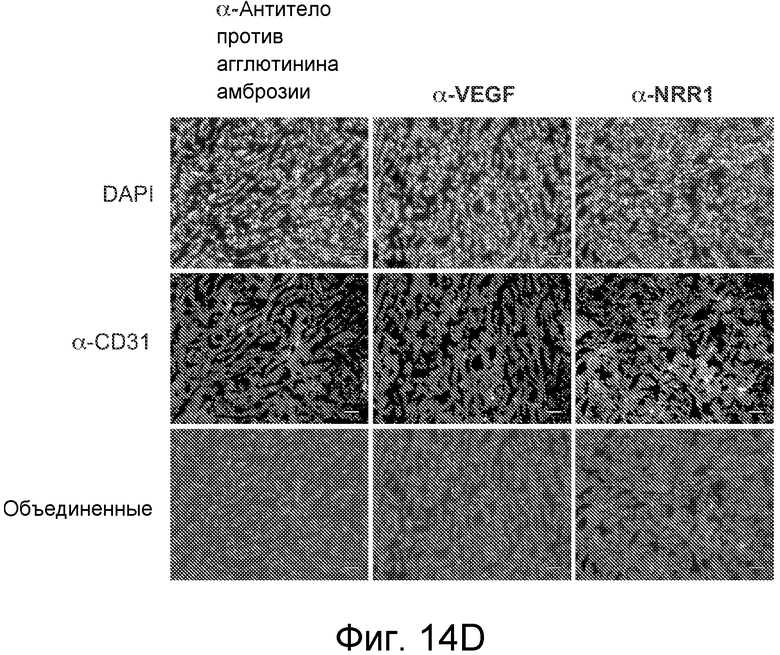
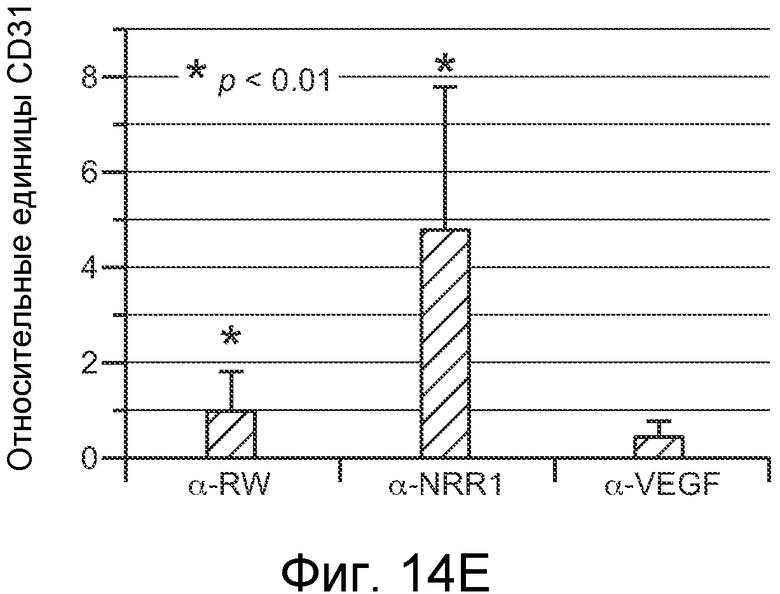
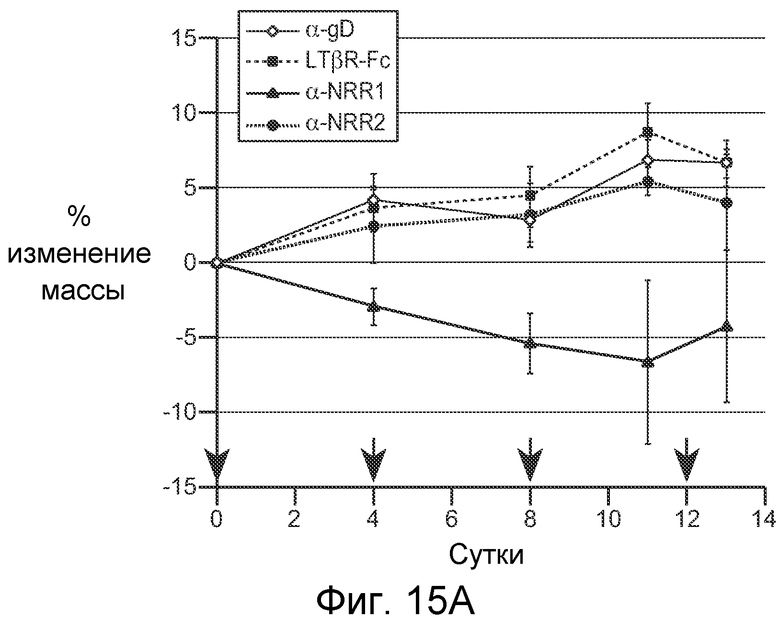
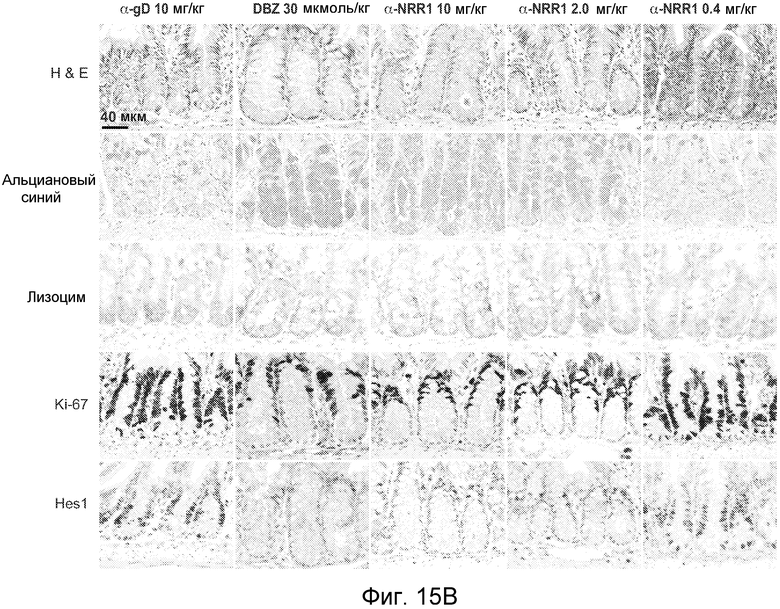
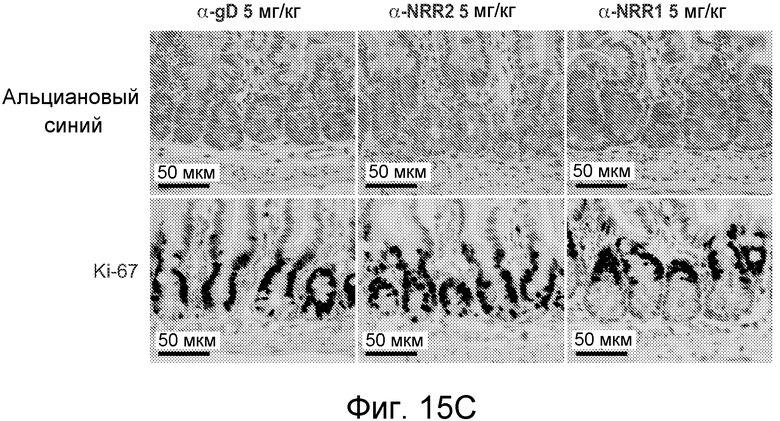
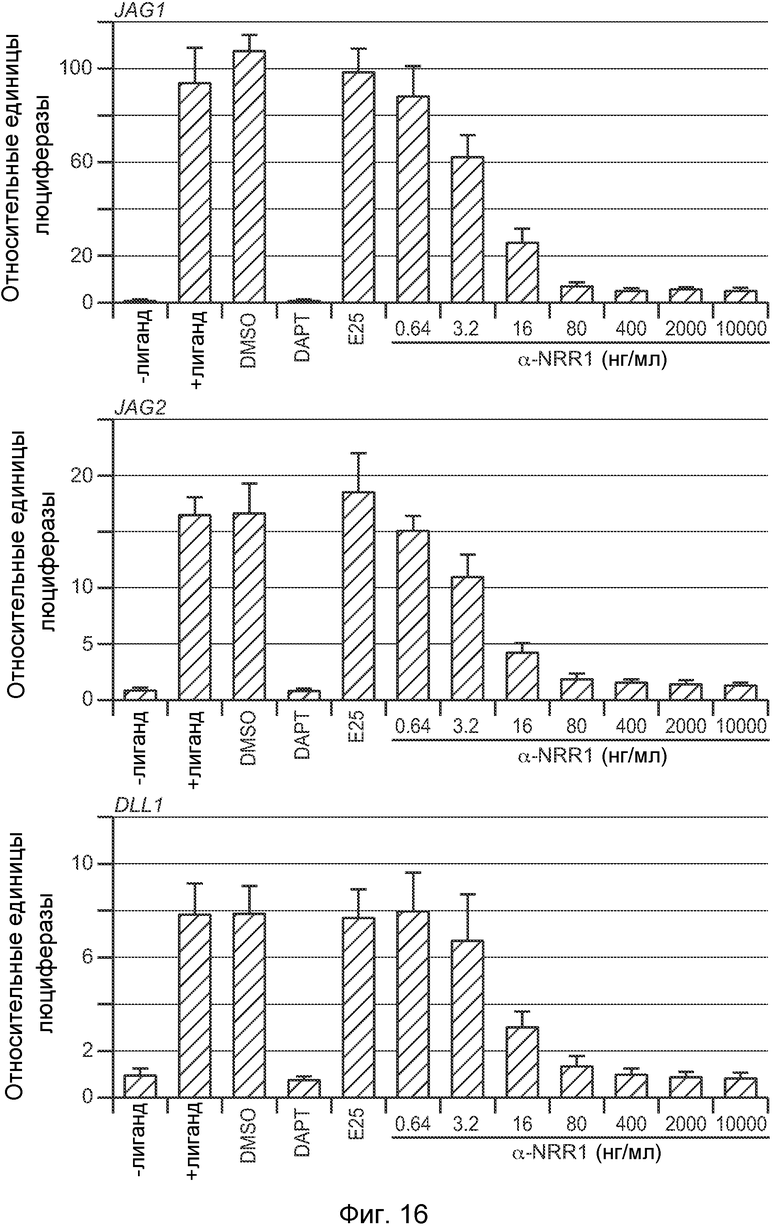
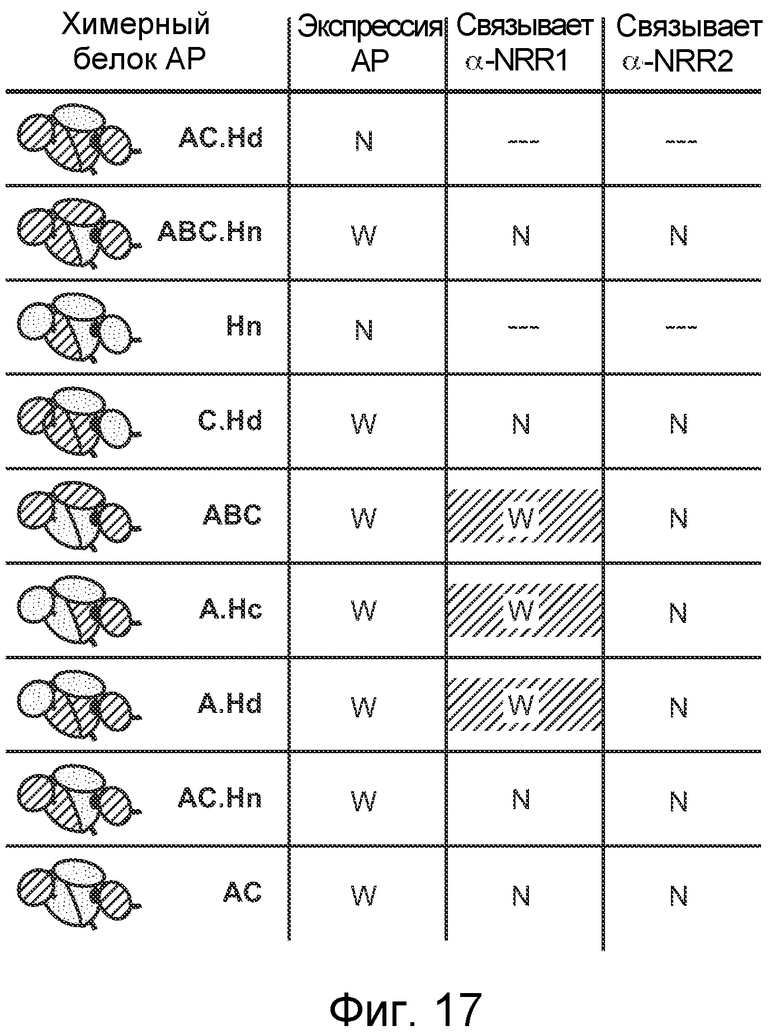
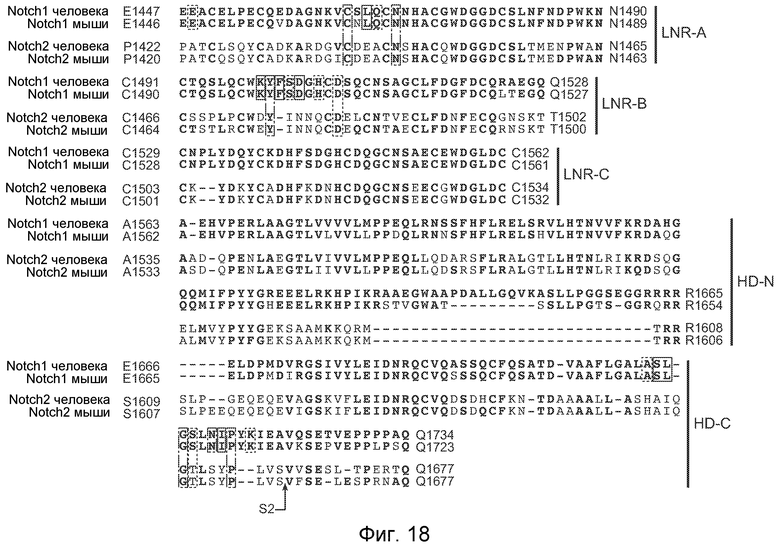
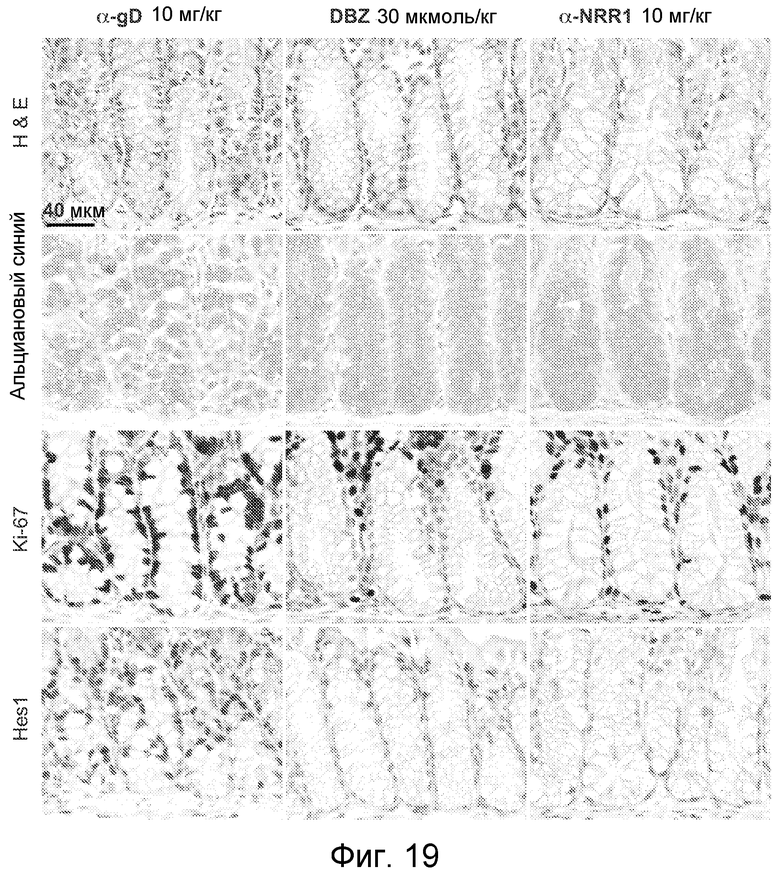
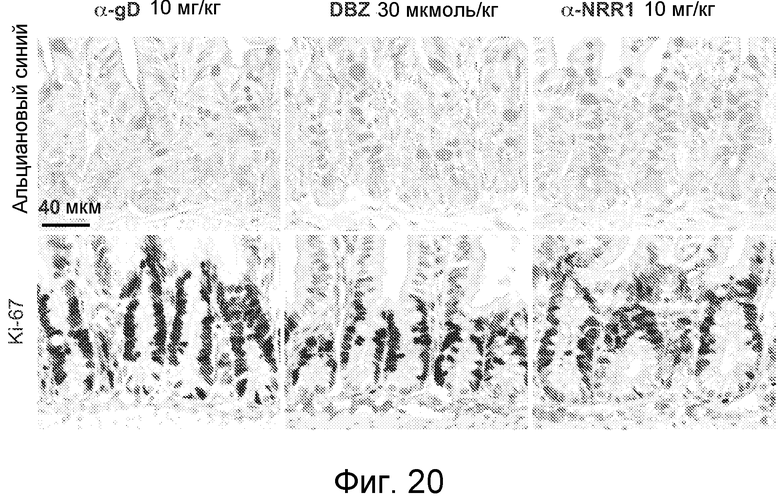
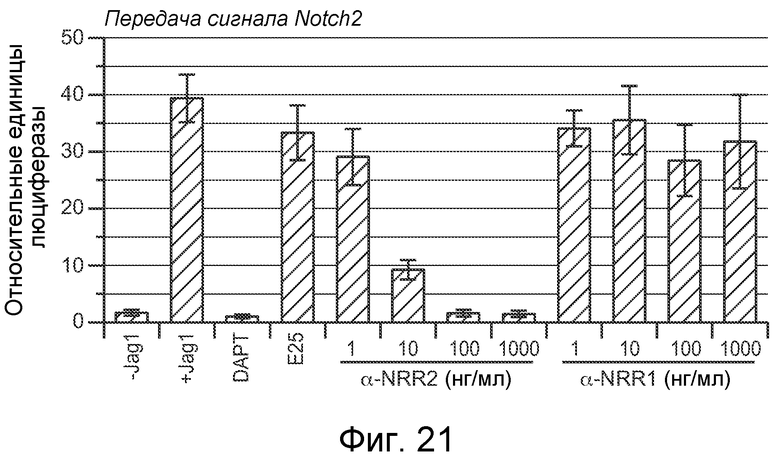
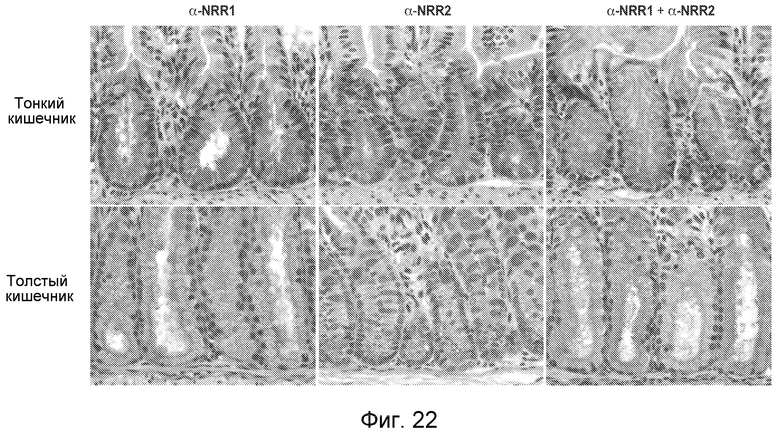
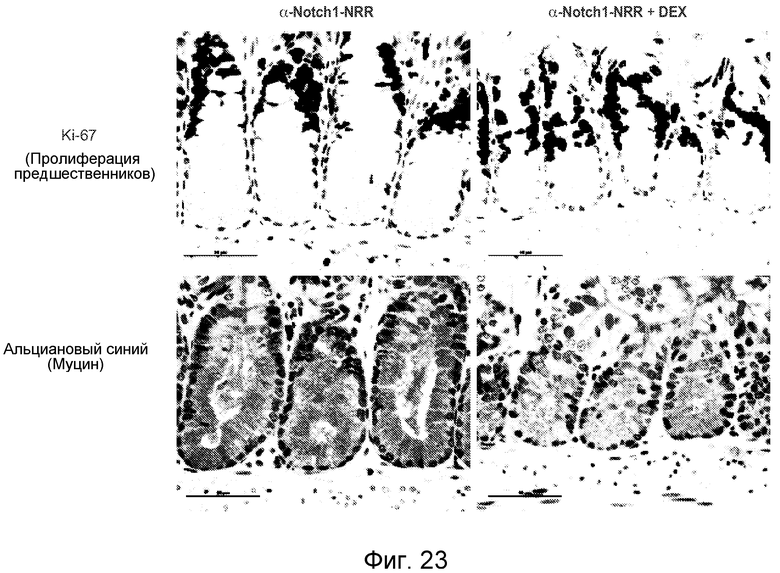
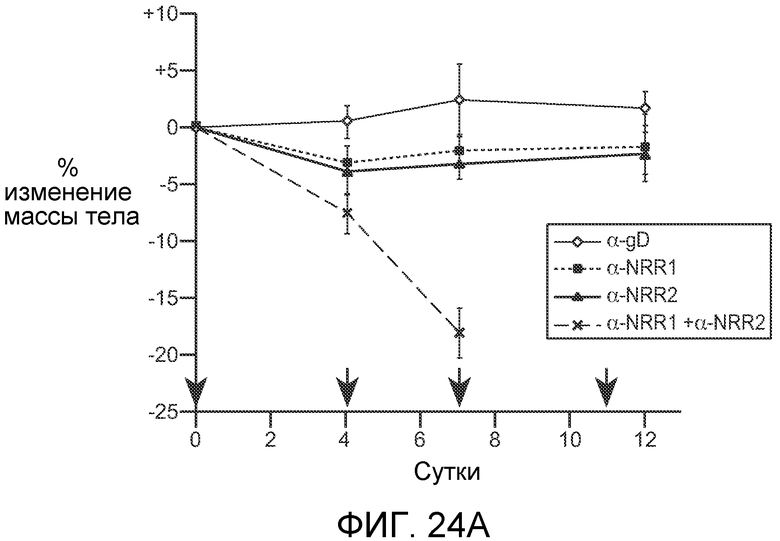
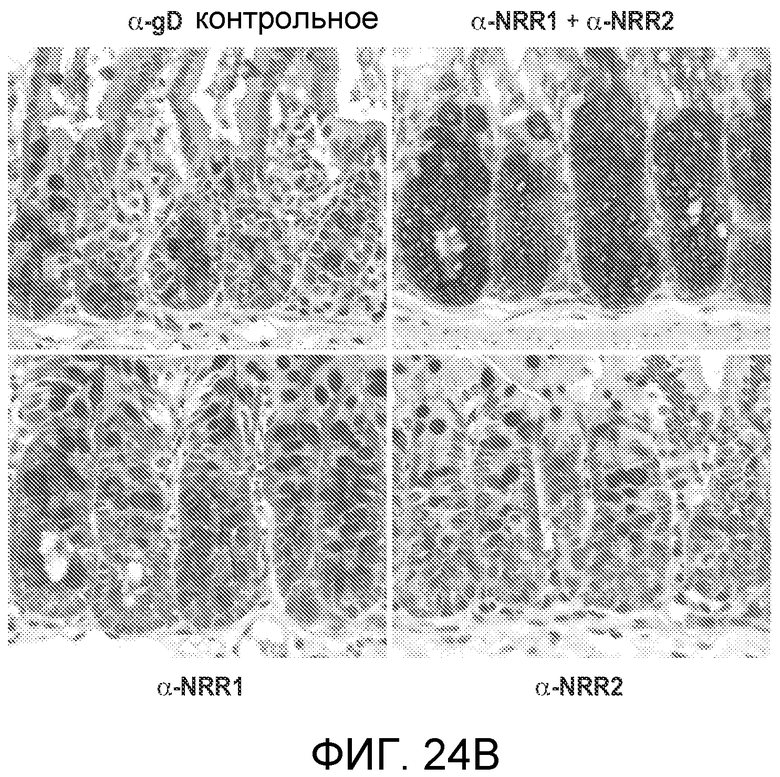
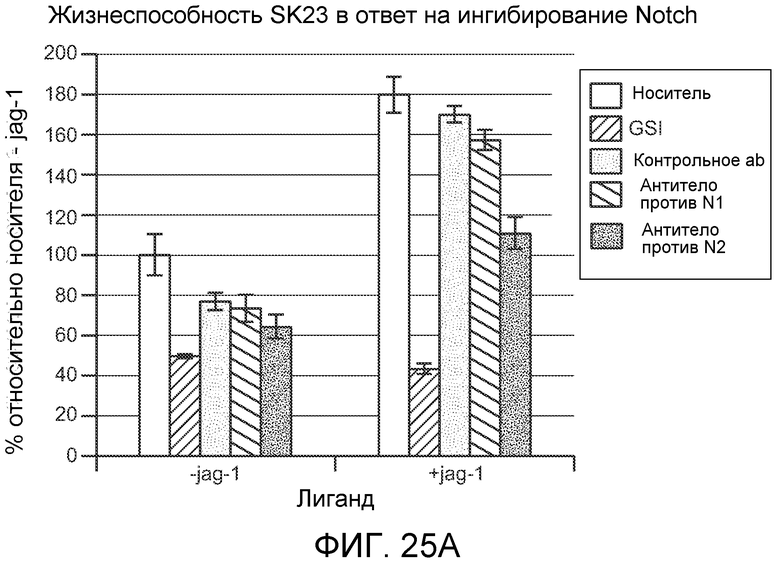
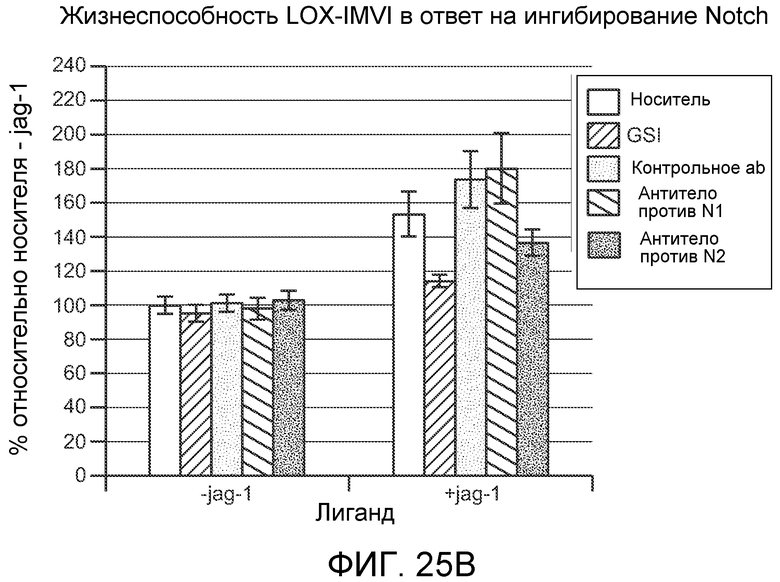
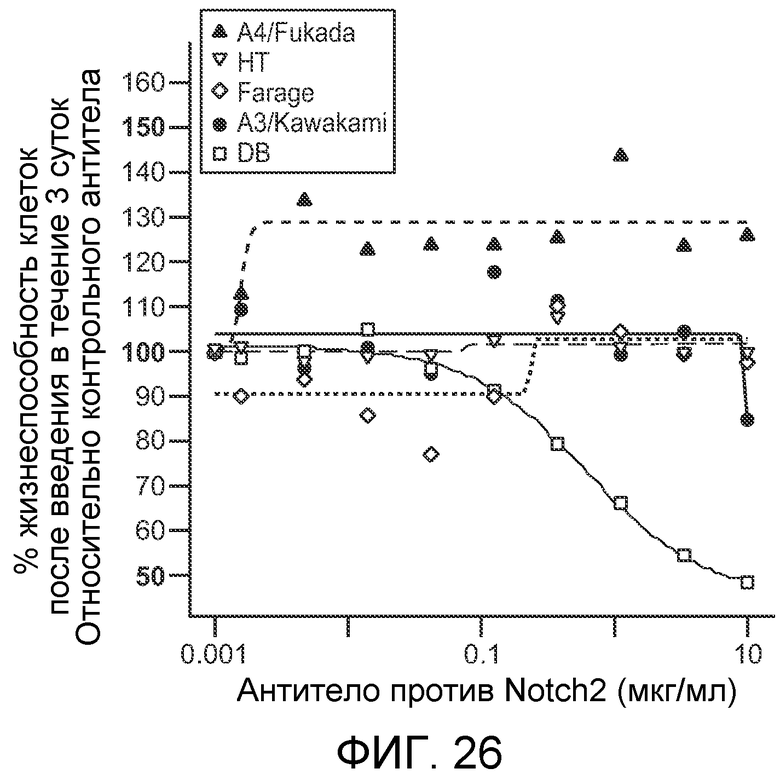
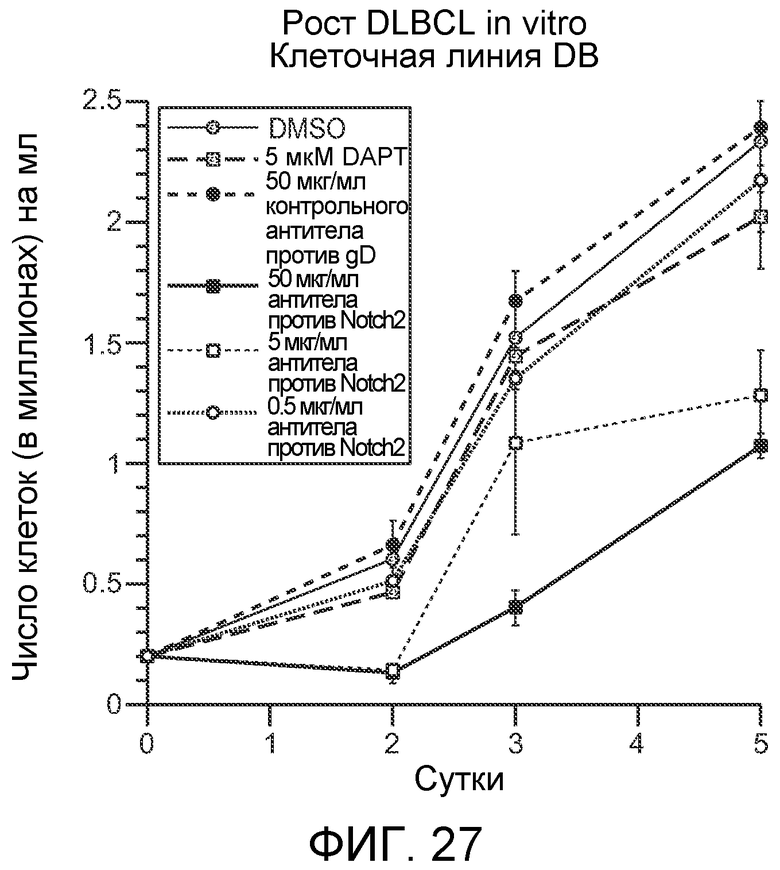
Изобретение относится к области биохимии, в частности к антителам, которые связывают Notch2. Заявлены моноклональные антитела, которые специфически связываются с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибируют передачу сигнала Notch2. Также изобретение относится к способам применения этих антител. Изобретение позволяет использовать антитела, связывающие Notch2, для лечения нарушения, ассоциированного с повышенной экспрессией или передачей сигнала Notch2, такого как злокачественная опухоль. 9 н. и 28 з.п. ф-лы, 47 ил., 1 табл.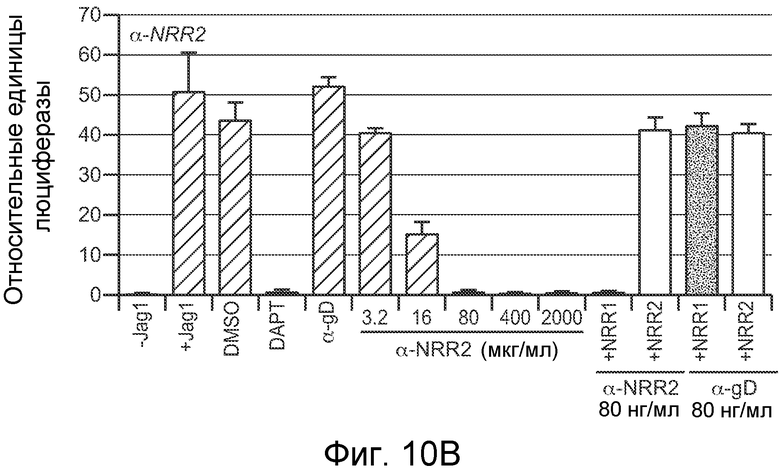
1. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, и где антитело содержит:
(a) гипервариабельную область тяжелой цепи HVR-H1, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 1-2;
(b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4; (с) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5; (d) гипервариабельную область легкой цепи HVR-L1, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 6-9; (е) HVR-L2, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 11-13; и (f) HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 15-18.
2. Антитело по п.1, где антитело снижает число В-клеток маргинальной зоны селезенки in vivo.
3. Антитело по п.2, где антитело снижает число В-клеток маргинальной зоны селезенки in vivo на ≥60%.
4. Антитело по п.1, где антитело снижает жизнеспособность клеток меланомы in vitro.
5. Антитело по п.1, где антитело подавляет рост клеток диффузной крупноклеточной В-клеточной лимфомы in vitro.
6. Антитело по п.1, где антитело связывается с полипептидом, содержащим аминокислоты 1-44 и 188-256 последовательности SEQ ID NO: 28.
7. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, и где антитело содержит: HVR-H1 содержащую аминокислотную последовательность SEQ ID NO: 1; HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4; HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 6; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
8. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, и где антитело содержит: HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2; HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4; HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 16.
9. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, и где антитело содержит: HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2; HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4; HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 8; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 12; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 17.
10. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации(HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, и где антитело содержит: HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 2; HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 4; HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 9; HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 13; и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 18.
11. Антитело по п.1, содержащее каркасную область акцептора 2 вариабельных областей тяжелых цепей (VH) человека.
12. Антитело по п.1, содержащее консенсусную каркасную область вариабельных областей легких цепей (VL) каппа подгруппы I человека.
13. Моноклональное антитело, которое специфически связывается с доменом Lin12/Notch Repeat (LNR)-А и доменом гетеродимеризации (HD)-С отрицательной регуляторной области (NRR) Notch2 и ингибирует передачу сигнала Notch2, где антитело связывается как с NRR Notch2 мыши, так и с NRR Notch2 человека с Kd≤10 нМ, где антитело содержит вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 20-21, и вариабельный домен легкой цепи, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 22-25.
14. Антитело по п.13, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 20, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 22.
15. Антитело по п.13, где антитело содержит вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 23-25.
16. Антитело по п.15, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 23.
17. Антитело по п.15, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 24.
18. Антитело по п.15, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 25.
19. Антитело по п.1, где антителом является IgG1 антитело.
20. Антитело по п. 1, где антитело представляет собой фрагмент антитела, выбранный из фрагментов Fab, Fab′-SH, Fv, scFv или (Fab′)2.
21. Антитело по п.1, где антитело представляет собой антитело человека, гуманизированное антитело или химерное антитело.
22. Антитело по п.1, где аффинность определяется с помощью анализа связывания меченного радиоактивной меткой антигена или анализа на основе иммунносорбента, связанного с ферментом.
23. Антитело по п.1, где аффинность определяется с помощью способа поверхностного плазмонного резонанса.
24. Антитело по п.1, где антитело связывается как с NRR Notch2 мыши, так и NRR Notch2 человека с Kd, равной 5 нМ.
25. Способ ингибирования передачи сигнала Notch2, включающий воздействие на клетку, которая экспрессирует Notch2, антителом по п.1.
26. Способ лечения нарушения, ассоциированного с повышенной экспрессией или передачей сигнала Notch2, включающий введение индивидууму, нуждающемуся в этом, эффективного количества антитела по п.1.
27. Способ по п.26, где нарушением, ассоциированным с повышенной экспрессией или передачей сигнала Notch2, является В-клеточная злокачественная опухоль.
28. Способ по п.26, где нарушением, ассоциированным с повышенной экспрессией или передачей сигнала Notch2, является меланома.
29. Способ по п.26, где нарушением является рак.
30. Способ по п.26, где антитело содержит вариабельный домен тяжелой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 21, и вариабельный домен легкой цепи, имеющий по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 23-25.
31. Способ по п.30, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 21, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 25.
32. Способ по п.26, где антитело вводят с по крайней мере одним терапевтическим средством.
33. Способ по п.32, где терапевтическим средством является химиотерапевтическое средство.
34. Способ по п.26, где антитело и терапевтическое средство вводят субъекту отдельно.
35. Применение антитела по любому из пп. 1-24 для получения лекарственного средства для лечения нарушения, ассоциированного с повышенной экспрессией или передачей сигнала Notch2.
36. Применение по п.35, где нарушением является В-клеточная злокачественная опухоль.
37. Применение по п.35, где нарушением является меланома.
38. Применение по п.35, где нарушением является рак.
| WO 2008057144 A2, 15.05.2008 | |||
| WO 2008091641 A2, 31.07.2008 | |||
| GORDON W.R | |||
| et al., Structural basis for autoinhibition of Notch, Nature Structural & Molecular Biology, 2007, vol.14 no.4, pp.295-300 | |||
| LI KANG et al., Modulation of Notch signaling by antibodies specific for the extracellular negative regulatory region of NOTCH3, JOURNAL OF BIOLOGICAL |

