По настоящей заявке испрашивается приоритет на основании свода законов США 35 §119(е) по предварительной заявке на патент США US 60/736680, поданной 15 ноября 2005 г., полное содержание которой включено в настоящее описание в виде ссылки, несмотря на то, что оно полностью изложено ниже.
В соответствии со сводом законов США 35 §202(с) признается, что правительство США обладает бесспорными правами на настоящее изобретение, которое было выполнено совместно с Национальным институтом здоровья, грант PO1 HL-74124-01.
Область техники, к которой относится изобретение
Настоящее изобретение относится к области медицины и гематологии. Более конкретно, настоящее изобретение предусматривает новые коагулирующие агенты и способы их применения для модулирования каскада коагуляции у пациентов, нуждающихся в этом.
Уровень техники
В описании настоящего изобретения цитируются публикации и патентные материалы для описания состояния области, к которой относится настоящее изобретение. Каждая из цитируемых работ включена в настоящее изобретение в виде ссылки на ее сущность.
Коагулирующими ферментами являются ферменты типа трипсина, которые принадлежат семейству протеаз пептидазы S1, которые несут складчатость, подобную складчатости химотрипсина. Коагулирующие протеазы содержат каталитические домены, которые в высокой степени гомологичны друг другу и родовым сериновым протеазам пищеварения. Структурная гомология/идентичность настолько велика (>70%), что остатки в каталитических доменах коагулирующих ферментов пронумерованы в соответствии с соответствующими остатками в химотрипсиногене.
Ферменты коагуляции циркулируют в крови в виде неактивных предшественников, зимогенов, которым требуется протеолитическое расщепление для активирования. Зимогены обладают ~10000-кратной или меньшей протеолитической активностью при сопоставлении с сериновыми протеазами, вырабатываемыми в ходе активирования. Инициация коагуляции в месте повреждения сосуда приводит к сериям реакций, в которых зимоген конвертируется в протеазу через специфическое протеолитическое расщепление и формирует фермент для последующей реакции. В результате достигается активирование клеток крови и конверсия растворимого фибриногена в нерастворимый фибрин, после чего формируется сгусток. Избыток протеаз удаляется реакцией с циркулирующими в кровяном русле ингибиторами протеаз, выступающими в качестве «самоубийственных» субстратов, или с теми, которые распознают действующие ферменты. Таким образом, протеолитическое активирование зимогенов коагуляции является ключевым моментом в регуляции каскада коагуляции.
Хотя некоторых зимогены коагуляции расщепляются по двум или нескольким сайтам в соответствующих реакциях активирования, формирование протеаз требует расщепления по одному сайту. Расщепление по такому сайту и его структурные последствия оцениваются наиболее простым способом, используя систему гомологической нумерации, основанную на химотрипсиногене, причем интенсивная работа по структуре была проделана с трипсиногеном и трипсином. Конверсия зимогена в сериновую протеазу требует расщепления после Arg15 (обычно расщепление связи между Arg15 и ILe16), что обычно приводит к активированию пептида и проявлению нового N-конца в каталитическом домене, начинающемся с Ile16. В трипсине и факторе Ха новая N-концевая последовательность начинается с Ile16-Val17-Gly19-Gly19. Для ферментов коагуляции новая N-концевая последовательность является вариацией на ту же тему. N-концевая последовательность затем сворачивается в каталитический домен и встраивается в N-концевую связующую щель способом, зависящим от специфичности последовательности, который называется «молекулярной сексуальностью». См. фиг.2. Соответственно варианты с измененными N-концевыми последовательностями предположительно не претерпевают молекулярную сексуальность по сопоставимому пути. N-концевые инсерции приводят к формированию соляного мостика между группой α-NH2 остатков Ile16 и Asp194 внутри каталитического домена. Формирование солевого мостика связано с многочисленными изменениями в структуре каталитического домена, включая: реаранжировки так называемых доменов активирования (фиг.3), формирование оксианионной щели, необходимой для катализа и формирование субстратного сайта связывания. Эти изменения приводят к созреванию активной сериновой протеазы. Ключевое значение специфических по последовательностям взаимодействий нового N-конца через формирование молекулярной сексуальности и солевого мостика для созревания активной протеазы доказывается следующими фактами: бактериальные протеазы, не требующие расщепления для активации, имеют другую боковую цепь в каталитическом домене к солевому мостику с Asp194; трипсиноген может быть активирован в протеиназа-подобную конформацию без расщепления, но в чрезвычайно высоких концентрациях дипептида Ile-Val, который встраивают в щель, хотя и весьма неэффективно; дипептид Val-Ile и другие варианты гораздо менее эффективны; кроме того, есть два примера бактериальных белков, которые активируют зимогены коагуляции в отсутствии расщепления путем разрушения механизма активации через обеспечение их собственных N-концов, которые встраивают в N-концевую связывающую щель.
Структурные изменения, описанные выше, дают молекулярное объяснение для конверсии предшественника зимогена в действующую сериновую протеазу. Однако в отличие от трипсина, который в полной мере активен после расщепления по сайту Arg15, многие коагулирующие ферменты действуют очень слабо, находясь на соответствующих протеиновых субстратах. Даже если бы они располагали полностью функционально активными сайтами и могли расщеплять низкомолекулярные пептидильные субстраты, эффективное расщепление биологического субстрата часто требует кофакторного белка (фиг.2). В этих случаях кофакторные белки увеличивают скорость расщепления белкового субстрата в несколько тысяч раз. Хотя механизм, с помощью которого функция кофакторных белков по-прежнему может быть разрешена, они вряд ли действуют, придавая протеазе более ферментоподобный вид и, следовательно, делая более эффективной. Ключевой момент, за одним исключением, заключается в том, что кофакторы избирательно связывают протеазу и не связывают соответствующий зимоген. Например, фактор Ха с высоким сродством связывается с мембрано-связанным FVa, несмотря на то, что зимогенный фактор Х не связывает FVa.
В зависимости от состояния пациента может быть желательным создание белков измененного каскада коагуляции, которые проявляют повышенную или пониженную функцию коагуляции. Это объект настоящего изобретения, предусматривающий такие белки для применения в качестве лекарственных средств.
Краткое описание изобретения
В соответствие с настоящим изобретением композиции и способы предусмотрены для воздействия на регуляторные сайты в метаболическом пути транзиции FX зимогена → протеазы, тем самым вызывая выработку более «зимоген-подобных» видов FXa. Композиции и способы настоящего изобретения эффективны для модуляции гемостаза у пациентов, нуждающихся в этом.
В одном из вариантов осуществления настоящего изобретения предусмотрен вариант фактора Х/фактора Ха зимогена/протеазы, который модулирует гемостаз. Предпочтительно вариант зимогена/протеазы кодируется последовательностью SEQ ID NO:2, в которой нуклеотиды 1684-1695 могут соответствовать какой-либо аминокислоте, при условии, что нуклеотиды 1684-1686 не кодируют Val или Ala. Более предпочтительно вариант зимогена/протеазы содержит по меньшей мере одну модификацию в последовательности SEQ ID NO:1, выбранную из группы, состоящей из а) Не в положении 16 является Leu, Phe, Asp или Gly, б) Val в положении 17 является Leu, Ala или Gly, в) Asp в положении 194 является Asn или Glu. Нуклеиновые кислоты, кодирующие вариант зимогена/протеазы настоящего изобретения и способы их применения также описаны в настоящем изобретении. Такие нуклеотиды могут необязательно кодировать внутриклеточный сайт расщепления РАСЕ/фурина.
В другом варианте осуществления настоящего изобретения предусмотрена нуклеиновая кислота с последовательностью SEQ ID NO:2, в которой нуклеотиды в положениях 1684-1695 кодируют аминокислоты, выбранные из группы, включающей Leu-Val-Gly, Gly-Val-Gly, Ile-Ala-Gly, Phe-Val-Gly и Ile-Gly-Gly, причем указанная нуклеиновая кислота необязательно включает нуклеотиды в положениях 2233-2235, которые кодируют аминокислоту, выбранную из группы, состоящей из Asn или Glu.
Также предусмотрена фармацевтическая композиция, включающая вариант фактора Ха настоящего изобретения в биологически совместимом носителе. К другому предпочтительному объекту настоящего изобретения относятся способы лечения расстройств, связанных с гемостазом, у пациентов, нуждающихся в этом, заключающихся во введении в организм терапевтически эффективного количества фармацевтических композиций, содержащих вариант фактора Х/Ха зимогена/протеазы, описанный в настоящем изобретении. Такие способы могут быть эффективны при лечении расстройств, в которых требуется прокоагулянт и к которым без ограничения относятся: гемофилия А, гемофилия В, гемофилия А и гемофилия В, ассоциированные с ингибирующими антителами, недостаточность фактора коагуляции, недостаточность витамин К-эпоксидредуктазы С1, недостаточность гамма-карбоксилазы, кровотечение, связанное с травмой, повреждением, тромбозом, тромбоцитопенией, инсультом, коагулопатией, диссеминированной внутрисосудистой коагуляцией;
расстройства, связанные с избыточным антикоагуляционным лечением, синдром Бернара-Сулье, тромбоцитастения Гланцманна и недостаточность кровяного депо.
Некоторые варианты зимогена/протеазы могут быть применимы для лечения расстройств в тех случаях, когда желательна антикоагуляция. К таким расстройствам без ограничений относятся тромбоз, тромбоцитопения, инсульт и коагулопатия.
К другому объекту настоящего изобретения относятся клетки-хозяева, экспрессирующие вариант зимогена/протеазы настоящего изобретения, для выработки его больших количеств.
Краткое описание фигур
Фиг.1. Процессинг фактора X. Фактор Х (FX) синтезируется с сигнальной последовательностью и пропептидом, которые удаляются до его секреции. Фактор Х является зимогеном и не обладает ферментной активностью. FX конвертируется в фактор Ха после расщепления связи Arg15-Ile16, что приводит к высвобождению пептида активации.
Фиг.2. Конверсия зимогена в протеазу. Переход зимогена в протеазу для фактора Х и сборка фактора Ха в протромбиназу (FXa, FVa, фосфолипид и ионы кальция). Этот фермент конвертирует протромбин (II) в тромбин (IIa).
Фиг.3. Рентгеноструктура FXa. Каталитический домен FXa в стандартной ориентации. Отмечены структурные области наряду с важными остатками. По публикации Brandstetter и др., J. Biol. Chem. 271, 1996, cc.29988-29992.
Фиг.4. SDS-PAGE анализ вариантов FX/Xa. 4-12% SDS-PAGE гели прогоняют или в невосстановительных, или в восстановительных условиях, и затем окрашивают красителем кумасиновым синим.
Фиг.5. Аминокислотная последовательность (SEQ ID NO:1) и последовательность нуклеиновой кислоты (SEQ ID NO:2) фактора Ха. Сайты и положения аминокислот в желательных модификациях последовательности SEQ ID NO:1 выделены жирным шрифтом.
Фиг.6. Активность фактора Ха в плазме при гемофилии В. Дикого типа FXa или FXaI16L (2 нМ) вносили в плазму больного гемофилией В и через определенные интервалы времени образцы разводили (0,1 нМ) и исследовали по коагуляции с учетом АПТВ - активированного частичного (парциального) тромбопластинового времени.
Фиг.7. Коррекция АПТВ. Фактор Xa-I16L (200 мкг/кг; n=7 мышей) или фосфатно-солевой буфер (ФСБ) вводят инъекцией мышам с гемофилией В (C57BL/6) в хвостовую вену. После инъекции через 5 и 30 мин отбирают кровь и проводят исследование методом АПТВ. Красная пунктирная линия представляет величину АПТВ нормальных животных линии C57D1/6.
Фиг.8. Оценка гемостаза с последующим анализом методом зажима хвоста у мышей с гемофилией В (ГВ). Потеря крови измеряется по содержанию гемоглобина в физиологическом растворе (ФСБ) с помощью А525 после нанесения раны. Число мышей (Balb с): дикого типа (n=7), с ГВ-ФСБ (n=6), ГВ-FXaI16L (n=7).
Подробное описание изобретения
Протеолиз является важным фактором коагуляции крови и лежит в основе многих механизмов, регулирующих нормальный гемостаз. Прокофакторы и зимогены не могут в какой-либо значительной степени участвовать в их соответствующих макромолекулярных ферментных комплексах. Это свидетельствует о том, что протеолитическая активация должна привести к соответствующим структурным изменениям и к экспрессии сайтов, которые наделяют фермент, субстрат и кофактор связывающими способностями. Хотя активация прокофактора и зимогена интенсивно исследовалась, взаимосвязь между протеолизом и экспрессией сайтов связывания, которая обусловливает функцию, полностью не установлена. В настоящем изобретении предусмотрены модельные композиции и системы, которые разъясняют молекулярные механизмы, лежащие в основе экспрессии макромолекулярных связывающих взаимодействий, сопровождающих транзиции от состояния зимогена к ферменту.
Фактор Ха (FX)1 является витамин K-зависимым двухцепочечным гликопротеином, который играет главную роль в коагуляции крови (фиг.1). Зимоген сериновой протеазы является субстратом и для внешних (тканевый фактор/FVIIa), и для внутренних (FVIIIa/FIXa) ферментных комплексов, расщепляющих неустойчивую химическую связь Arg15-Ile16 в FX, высвобождая состоящий из 52 аминокислот активирующий пептид, который генерирует FXa. Фактор Ха является протеазой, ответственной за конверсию протромбина в тромбин (фиг.2), Хотя фактор Ха является полностью компетентной протеазой и осуществляет каталитический механизм расщепления протромбина, он весьма слабый катализатор для данной реакции. Его тесное связывающее взаимодействие с кофактором, фактором Va, на поверхности мембраны сильно увеличивает степень формирования тромбина без существенного воздействия на другие реакции, катализируемые фактором Ха. Изменения N-концевой последовательности (Ile-Val-Gly) после расщепления сайта Arg15, которые приводят к субоптимальной молекулярной сексуальности, предположительно вызывает формирование «зимоген-подобного» производного Ха, которое обладает пониженной, или даже нулевой, протеолитической активностью. Такие производные предположительно не чувствительны к подавлению ингибиторами плазматической протеазы, например, антитромбином III, и не участвуют в инициации коагуляции с последующим сосудистым поражением, поскольку предположительно они не связываются очень сильно с TFPI. Фактор Ха плотно связывает фактор Va в отличие от зимогенного фактора X. Таким образом, ожидается, что зимоген-подобные формы фактора Ха связываются с Va слабее, но полностью восстанавливаются при достаточно высоких концентрациях кофактора и эффективно катализируют формирование тромбина. Зимоген-подобные формы фактора Ха с подобными свойствами предположительно действуют в качестве долгоживущих в кровяном русле протеаз, которые с другой стороны разрушаются, но сохраняют способность катализировать формирование тромбина при связывании с фактором Va. Они могут быть терапевтическими прокоагулянтами, которые шунтируют недостаточности других факторов свертывания крови в каскаде без вредных воздействий, связанных с инфузией полностью функционального фактора дикого типа FXa.
I. Определения:
Различные термины, связанные с биологическими молекулами настоящего изобретения, используют в описании и формуле настоящего изобретения.
Понятие «вариант зимогена/протеазы» означает модифицированный зимоген FX или протеазу FXa, которые была изменены генетически таким образом, что их протеазная активность при конверсии в FXa снижена или «зимоген-подобна» в отсутствии специфических кофакторов (например, связывающее сродство для активного сайта ниже, чем у молекулы дикого типа). Важно отметить, что такое сродство/активность восстанавливается в присутствии характерных кофакторов, к которым относится без ограничений фактор Va. К предпочтительным сайтам аминокислотных изменений в исходной молекуле FX относятся замещение изолейцина в положении 16, замещение валина в положении 17 и замещение аспарагиновой кислоты в положении 194, при условии, что аминокислота в положении 16 не является валином или аланином.
Фраза «расстройства, связанные с гемостазом» подразумевает расстройства, связанные с кровотечением, например, гемофилию А и В, пациентов с гемофилией А и В с ингибирующими антителами, недостаточность факторов коагуляции, например, факторов VII, IX, X, XI, V, XII, II, фактора фон Виллебранда, комбинированная недостаточность факторов FV/FVIII, недостаточность витамин K-эпоксидредуктазы С1, недостаточность гамма-карбоксилазы; кровотечение, связанное с травмой, повреждением, тромбозом, тромбоцитопенией, инсультом, коагулопатией, диссеминированной внутрисосудистой коагуляцией; избыточная антикоагуляция, связанная с гепарином, низкомолекулярным гепарином, пентасахаридом, варфарином, низкомолекулярными антитромботическими факторами (т.е. ингибиторами FXa); расстройства, связанные с тромбоцитами, например, синдром Бернара-Сулье, тромбоцитастения Гланцманна и недостаточность кровяного депо. К расстройствам, связанным с гемостазом, также относятся кровотечения, связанные с тромботическими расстройствами, например, тромбозом глубоких вен, тромбозом, связанным с сердечнососудистым заболеванием или с онкологическим заболеванием, тромбозом, возникающим в результате установки постоянных катетеров или других инвазивных хирургических процедур, а также с тромбозом, ассоциированным с аутоиммунным заболеванием, например, красной волчанкой. Варианты зимогена/протеазы также могут обеспечить необходимый гемостаз пациентам с диссеминированной внутрисосудистой коагуляцией или коагулопатией потребления, которые возникают при различных болезненных состояниях.
Применительно к нуклеиновым кислотам настоящего изобретения иногда используется понятие «выделенная нуклеиновая кислота». Это понятие применительно к ДНК означает, что молекула ДНК отделена от последовательностей, с которыми она немедленно конъюгирует (в 5' и 3' направлениях) в нативном геноме организма, от которого она происходит. Например, «выделенная нуклеиновая кислота» может быть молекулой ДНК или кДНК, инсертированной в состав вектора, например, вирусного или плазмидного вектора, или может быть интегрирована в ДНК прокариота или эукариота.
Относительно молекул РНК настоящего изобретения понятие «выделенная нуклеиновая кислота» в первую очередь относится к молекуле РНК, кодируемой выделенной молекулой ДНК, описанной выше. В другом варианте это понятие может относиться к молекуле РНК, которая в существенной степени отделена от молекул РНК, с которыми она может быть связана в естественных условиях (т.е. в клетках или тканях), и в результате находится в «существенно чистой» форме (понятие «существенно чистая» форма характеризуется ниже).
В контексте настоящего изобретения применительно к белкам иногда употребляется понятие «выделенный белок» или «выделенный и очищенный белок». Это понятие преимущественно относится к белку, вырабатываемому в результате экспрессии выделенной молекулы нуклеиновой кислоты настоящего изобретения. В другом варианте это понятие относится к белку, который существенным образом отделен от других белков, с которыми он может быть естественным образом ассоциирован, при этом белок находится в «существенно чистой» форме.
Понятие «промоторная область» относится к областям регуляции транскрипции гена, которые могут располагаться с 5'- или 3'-конца кодирующей области, или внутри кодирующей области, или внутри интронов.
Понятие «вектор» относится к несущей молекуле ДНК небольшого размера, в состав которой может быть встроена последовательность ДНК для последующей интродукции в клетку-хозяина, в которой она может реплицироваться. Понятие «экспрессирующий вектор» означает специальный вектор, который содержит ген или последовательность нуклеиновой кислоты с необходимыми регуляторными областями, требуемыми для экспрессии в клетке-хозяине.
Понятие «оперативно связанный» означает, что регуляторные последовательности, необходимые для экспрессии кодирующей последовательности, расположены в молекуле ДНК в определенных положениях относительно кодирующей последовательности таким образом, чтобы влиять на экспрессию кодирующей последовательности. То же определение иногда используют применительно к сборке кодирующих последовательностей и элементов контроля транскрипции (например, промоторов, энхансеров, элементов терминации) в составе экспрессирующего вектора. Это определение иногда также применяют к сборке последовательностей нуклеиновых кислот из первой и второй молекулы нуклеиновой кислоты, в результате чего возникает гибридная молекула.
Понятие «существенно чистое» относится к препарату, включающему по меньшей мере 50-60 мас.% целевого соединения (например, нуклеиновой кислоты, олигонуклеотида, белка и т.д.). Более предпочтительно препарат включает по меньшей мере 75 мас.%, наиболее предпочтительно 90-99 мас.% целевого соединения. Чистоту определяют методами, применимыми для целевого соединения (т.е. хроматографическими методами, гель-электрофорезом в агарозе или полиакриламиде, ВЭЖХ и др.).
Фраза «в значительной степени состоящая из» применительно к определенной аминокислотной или нуклеотидной последовательности означает последовательность, имеющей свойства данной последовательности SEQ ID NO:. Например, при ссылке на аминокислотную последовательность эта фраза относится к последовательности самой по себе, а также к молекулярным модификациям, которые не влияют на основные и новые свойства последовательности.
Понятие «олигонуклеотид» в контексте настоящего изобретения относится к праймерам и зондам настоящего изобретения и представляет молекулу нуклеиновой кислоты, включающую два или более рибо- или дезоксирибонуклеотидов, предпочтительно более трех. Точный размер олигонуклеотида может зависеть от различных факторов и от конкретного назначения, для которого олигонуклеотид применяют.
Понятие «зонд» в контексте настоящего изобретения относится к олигонуклеотиду, полинуклеотиду или нуклеиновой кислоте, или РНК, или ДНК, которые либо естественным образом присутствуют после ферментного расщепления или вырабатываются синтетически, и способны к отжигу или специфической гибридизации с последовательностями нуклеиновых кислот, комплементарными зонду. Зонд может быть одноцепочечным или двухцепочечным. Реальная длина зонда может зависеть от многих факторов, включая температуру, источник зонда и способ применения. Например, для диагностики, в зависимости от сложности последовательности-мишени, олигонуклеотидный зонд обычно содержит 15-25 или более нуклеотидов, хотя также может быть меньше нуклеотидов.
В контексте настоящего изобретения зонды выбирают таким образом, что они «существенно» комплементарны разным цепям конкретной последовательности нуклеиновой кислоты-мишени. Это означает, что зонды должны быть существенно комплементарными, чтобы осуществлять «специфическую гибридизацию» или отжиг с их соответствующими цепями-мишенями при ряде заранее определенных условий. Следовательно, последовательность зонда не должна точно отражать комплементарную последовательность мишени. Например, некомплементарный нуклеотидный фрагмент может быть присоединен к 5'- или 3'-концу зонда, а оставшаяся последовательность зонда является комплементарной цепи-мишени. В другом варианте некомплементарные основания или более длинные последовательности могут быть распределены в зонде, при условии, что последовательность зонда в значительной степени комплементарна последовательности нуклеиновой кислоты-мишени, что позволяет специфически осуществлять отжиг.
Понятие «специфическая гибридизация» относится к ассоциации между двумя однонитевыми молекулами нуклеиновой кислоты с существенно комплементарными последовательностями для осуществления такой гибридизации при заранее определенных условиях, обычно применяемых в данной области (иногда употребляют понятие «существенная комплементарность»). В частности, это понятие относится к гибридизации олигонуклеотида с существенно комплементарной последовательностью, присутствующей в однонитевой молекуле ДНК или РНК настоящего изобретения, для реального исключения гибридизации олигонуклеотида с однонитевыми нуклеиновыми кислотами с некомплементарной последовательностью.
В контексте настоящего изобретения понятие «праймер» означает олигонуклеотид, или из РНК, или из ДНК, либо однонитевой, либо двунитевой, полученный из биологической системы, или выработанный в результате обработки ферментами рестрикции, или полученный синтетически, который при помещении в определенную среду может функционировать в качестве инициатора матричного синтеза нуклеиновой кислоты. При наличии соответствующей матрицы нуклеиновой кислоты, необходимых нуклеозидтрифосфатов - предшественников нуклеиновых кислот, фермента полимеразы, соответствующих кофакторов и условий, например, определенной температуры и рН, праймер может удлиняться со своего 3'-конца добавлением нуклеотидов за счет действия полимеразы или близкого действия для получения праймер-протяженного продукта.
Праймер может варьировать в длину в зависимости от определенных условий и потребностей. Например, для диагностики олигонуклеотидный праймер обычно содержит 15-25 или более нуклеотидов в длину. Праймер должен обладать достаточной комплементарностью по отношению к целевой матрице для индукции синтеза целевого наращиваемого продукта, т.е. быть способным к отжигу с целевой цепью матрицы способом, достаточным для обеспечения 3'-гидроксильной части молекулы праймера при соответствующем сопоставлении для применения в инициации синтеза полимеразой или близким по действию ферментом. Необязательно, чтобы последовательность праймера представляла точную копию целевой матрицы. Например, некомплементарная нуклеотидная последовательность может быть присоединена к 5'-концу другого комплементарного праймера. В другом варианте некомплементарные основания могут быть распределены в составе олигонуклеотидной последовательности праймера, при условии, что последовательность праймера обладает достаточной комплементарностью по отношению к последовательности целевой матричной цепи для функционального обеспечения комплекса матрицы-праймера для синтеза наращиваемого продукта.
В контексте настоящего изобретения понятие «процент идентичности» относится к сравнению последовательностей нуклеиновых кислот или аминокислотных последовательностей. Последовательности нуклеиновых кислот и аминокислот часто сравнивают, используя компьютерные программы, которые сопоставляют последовательности нуклеиновых кислот или аминокислот и выявляют разницу между двумя последовательностями. Для целей настоящего изобретения сравнение последовательностей нуклеиновых кислот проводят, используя программу GCG Wisconsin Package, версия 9.1, которую можно получить в Компьютерной генетической группе в Мэдисоне, штат Висконсин. Для удобства параметры, принимающие значение по умолчанию (пенальти создания гэпа = 12, пенальти протяженности гэпа = 4), устанавливаемые данной программой, предназначены для применения в настоящем изобретении для сравнения идентичности последовательностей. В другом варианте может применяться программа Blastn 2.0, предоставляемая Национальным центром информации в области биотехнологии (на сайте http://www.ncbi.nlm.nih.gov/blast/; Altschul и др., J Mol Biol 215, 1990, cc.403-410), использующая выверку гэпов с параметрами, принимающими значение по умолчанию, для определения уровня идентичности и близости между последовательностями нуклеиновой кислоты и последовательностями аминокислот.
II. Приготовление молекул нуклеиновой кислоты, кодирующей вариант зимогена-протеазы, и полипептидов
А. Молекулы нуклеиновой кислоты
Молекулы нуклеиновой кислоты, кодирующие зимоген/протеазы настоящего изобретения, могут быть получены с помощью методов рекомбинантной ДНК. Благодаря информации о нуклеотидной последовательности приготовление молекул отдельных нуклеиновых кислот настоящего изобретения возможно разными способами. Например, последовательности нуклеиновых кислот, кодирующие полипептид зимогена/протеазы, могут быть выделены из соответствующих биологических источников с помощью стандартных протоколов, хорошо известных в данной области.
Нуклеиновые кислоты настоящего изобретения могут поддерживаться в виде ДНК в каком-либо обычном клонирующем векторе. В предпочтительном варианте осуществления настоящего изобретения клоны поддерживают в плазмидном клонирующем/экспрессирующем векторе, например, pBluescript (фирма Stratagene, La Jolla, Калифорния), который размножается в соответствующих клетках-хозяевах Е. coli. В другом варианте нуклеиновые кислоты могут поддерживаться в векторе, пригодном для экспрессии в клетках млекопитающего. В тех случаях, когда пост-трансляционные модификации воздействуют на функцию зимогена/протеазы (например, фактор Ха), предпочтительно экспрессировать молекулу в клетках млекопитающего.
В одном из вариантов осуществления настоящего изобретения нуклеиновые кислоты, кодирующие варианты зимогена фактора X, могут быть дополнительно модифицированы за счет инсерции внутриклеточного сайта протеолитического расщепления. Для экспрессии «активированных» зимоген-подобных вариантов FXa в клетках млекопитающих внутриклеточный сайт протеолитического расщепления может быть встроен между положениями Arg15 и Arg16 в варианте зимогена FX. К таким сайтам расщепления относятся: Arg-Lys-Arg или Arg-Lys-Arg-Arg-Lys-Arg. Эти сайты расщепления эффективно распознаются протеазами (РАСЕ|фурин-подобными ферментами) в клетках и выводятся. Это приводит к процессированному варианту РХ(а), в котором тяжелая цепь молекулы теперь начинается с положения 16. Интродукция данного сайта расщепления в указанное положение может привести к внутриклеточной конверсии FX в FXa.
В другом варианте осуществления настоящего изобретения целый состоящий из 52 аминокислот активирующий пептид может быть удален и внутриклеточный сайт расщепления протеазой может быть встроен в место, которое приведет к формированию варианта FXa.
В конечном счете, эти типы модификаций позволяют секретироваться «активно» процессированной форме варианта РХ из клеток, экспрессирующих модифицированный вариант FX. Секреция фактора расщепления устраняет необходимость в протеолитическом расщеплении во время свертывания крови или при последующем выделении белка.
К вариантам молекул нуклеиновых кислот, кодирующих вариант зимогена/протеазы настоящего изобретения, относятся кДНК, геномная ДНК, РНК, их фрагменты, которые могут быть одно- или двунитевыми. Таким образом, настоящее изобретение предусматривает олигонуклеотиды (смысловые или антисмысловые цепи ДНК или РНК), чьи последовательности могут гибридизироваться по меньшей мере с одной последовательностью молекулы нуклеиновой кислоты настоящего изобретения. Такие олигонуклеотиды применимы в качестве зондов для выявления экспрессии зимогена/протеазы.
Б. Белки
Полипептид настоящего изобретения зимогена/протеазы полной длины или в виде варианта, может быть приготовлен разными способами, известными в данной области. Белок может быть очищен из соответствующих источников, например, из трансформированных бактерий или культивируемых животных клеток или тканей, которые экспрессируют зимоген/протеазу, методом иммуноаффинной очистки. Однако этот способ не является предпочтительным из-за низкого количества белка, присутствующего в клетках данного типа в данное время.
Доступность молекул нуклеиновой кислоты, кодирующих вариант полипептида зимогена/протеазы, позволяет вырабатывать зимоген/протеазу, используя in vitro методы экспрессии, известные в данной области. Например, кДНК или ген могут быть клонированы в соответствующем векторе транскрипции in vitro, например, pSP64 или pSP65 для транскрипции in vitro, с последующей трансляцией в бесклеточной системе в соответствующем бесклеточной системе трансляции, например, в лизатах проростков пшеницы или в лизатах ретикулоцитов кролика. Системы трансляции и транскрипции in vitro коммерчески доступны, например, в фирме Promega Biotech, Мэдисон, Висконсин, или в фирме BRL, Роквилл, Мэриленд.
В другом варианте согласно предпочтительному варианту осуществления настоящего изобретения могут быть получены повышенные количества зимогена/протеазы путем экспрессии в приемлемой прокариотической или эукариотической системе экспрессии. Например, часть или все молекулы ДНК, кодирующие вариант фактора Ха, могут быть встроены в плазмидный вектор, адаптированный для экспрессии в бактериальных клетках, например, Е. coli, или в клетках млекопитающих, например, клетках СНО или Hela. В другом варианте предпочтительного осуществления настоящего изобретения могут быть получены меченые гибридные белки, включая зимоген/протеазу. Такие зимоген/протеаза-меченые гибридные белки кодируются частью или целой молекулой ДНК, лигированной в корректный кодон рамки считывания в нуклеотидной последовательности, кодирующей часть или всю метку целевого полипептида, которая встроена в плазмидный вектор, адаптированный для экспрессии в бактериальных клетках, например, Е. coli, или в эукариотических клетках, например, дрожжевых или клетках млекопитающих, но ими перечень не ограничивается. Векторы, например, описанные выше, включают регуляторные элементы, необходимые для экспрессии ДНК в клетке-хозяине, расположенные таким образом, чтобы допустить экспрессию ДНК в клетке-хозяине. Такие регуляторные элементы, необходимые для экспрессии, включают, но ими не ограничиваются, промоторные последовательности, последовательности инициации транскрипции и энхансерные последовательности.
Варианты белков зимогена/протеазы, вырабатываемых генной экспрессией в рекомбинантной прокариотической или эукариотической системе, могут быть очищены способами, известными в данной области. В предпочтительном варианте осуществления настоящего изобретения может быть использована коммерчески доступная система экспрессии/секреции, с помощью которой рекомбинантный белок экспрессируется и впоследствии секретируется из клеток-хозяев, и которая способствует легкой очистке от окружающей среды, Если не используют векторы экспрессии/секреции, применяют другой подход, предусматривающий очистку рекомбинантного белка аффинным разделением, например, иммунологическим взаимодействием с антителами, которые специфически связываются с рекомбинантным белком или никелевой колонкой для выделения рекомбинантных белков, меченых 6-8 остатками гистидина с N-или С-конца. Другие метки могут включать эпитоп FLAG, GST или эпитоп гемагглютинина. Такие методы часто используются опытными специалистами.
Белки зимогена/протеазы, приготовленные указанными выше способами, могут анализироваться по стандартным методикам. Например, такие белки могут быть подвергнуты анализу аминокислотной последовательности по известным в данной области методикам.
Выше указывалось, что обычный способ получения полипептида настоящего изобретения заключается в экспрессии нуклеиновой кислоты, кодирующей его, при использовании этой нуклеиновой кислоты в системе экспрессии. Разные системы экспрессии, используемые в настоящем изобретении, хорошо известны специалистам в данной области.
Таким образом, настоящее изобретение также предусматривает способ получения полипептида (согласно описанному), способ, включающий экспрессию с нуклеиновой кислоты, кодирующей полипептид (в общем смысле с нуклеиновой кислоты). Этого можно достичь культивированием клеток-хозяев, содержащих такой вектор, в соответствующих условиях, которые вызывают или допускают выработку полипептида. Полипептиды также могут вырабатываться в системах in vitro, например, в лизатах ретикулоцитов.
III. Применение белков зимогена/протеазы и зимоген/протеаза-кодирующих нуклеиновых кислот
Варианты зимогенных/протеазных нуклеиновых кислот, кодирующие полипептиды с измененной протеазной активностью, могут быть использованы согласно настоящему изобретению, например, в качестве терапевтических и/или профилактических агентов (белка или нуклеиновой кислоты), которые модулируют каскад коагуляции крови. В настоящем изобретении установлено, что фактор Х/Ха зимогена/протеазы может повысить коагуляцию и обеспечить эффективный гемостаз.
А. Вариант полипептидов зимогена/протеазы
В предпочтительном варианте осуществления настоящего изобретения вариант полипептидов зимогена/протеазы может вводиться пациенту инфузией в биологически совместимом носителе, предпочтительно внутривенной инъекцией. Вариант зимогена/протеаз настоящего изобретения может необязательно быть заключен в липосомы или перемешан с другими фосфолипидами или мицеллами для повышения стабильности молекул. Зимоген/протеаза может вводиться отдельно или в комбинации с другими агентами, известными в качестве модуляторов гемостаза (например, фактором V, фактором Va и их производными). Соответствующая композиция для введения полипептидов зимогена/протеазы может быть определена специалистом-медиком на основании различных физиологических переменных, включая, но, не ограничиваясь ими, состояние пациента и гемодинамическое состояние. Различные композиции, соответствующие различным применениям и способам введения, хорошо известны в данной области и описаны в настоящем изобретении ниже.
Препарат, содержащий очищенный аналог фактора Х/Ха, включает физиологически приемлемый матрикс и предпочтительно перерабатывается в виде фармацевтического препарата. Препарат может быть составлен, используя известные в данной области методы, он может быть перемешан с буферными солями, например, NaCl, CaCl2, аминокислотами, например, глицином и/или лизином, при диапазоне рН 6-8. До применения очищенный препарат, содержащий фактор Х/Ха аналог, может храниться в форме готового раствора или в лиофильно высушенной форме. Предпочтительно препарат хранят в лиофилизированной форме и растворяют в визуально прозрачный раствор, используя соответствующий восстанавливающий раствор.
В другом варианте препарат по настоящему изобретению также может быть готовым к применению в виде жидкого препарата или в виде жидкости в состоянии глубокой заморозки.
Препарат по настоящему изобретению обладает стабильностью, т.е. он может сохраняться в растворенной форме на протяжении пролонгированного времени до применения.
Препарат по настоящему изобретению, который содержит аналог фактора Х в комбинации с фактором XIa или его производным, который способен активировать аналог фактора Х в фактор Ха или аналог фактора Ха, может быть доступен в форме комбинированного препарата, включающего контейнер, который содержит фактор XIa, иммобилизованный на матрице, возможно в форме миниатюрной колонки или шприца с добавлением протеазы, и контейнера, содержащего фармацевтический препарат с аналогом фактора X. Для активирования аналога фактора Х раствор, содержащий аналог фактора X, например, может быть выдавлен поверх иммобилизованной протеазы. Во время хранения препарата раствор, содержащий аналог фактора X, предпочтительно в пространственном отношении отделяется непроницаемой перегородкой, которая может быть легко удалена перед введением препарата. Растворы также могут храниться в отдельных контейнерах и приводятся в соприкосновение только непосредственно перед введением.
Аналог фактора Х может быть активирован в фактор Ха непосредственно перед применением, т.е. перед введением пациенту. Активирование может быть выполнено путем соприкосновения аналога фактора Х с иммобилизованной протеазой или путем перемешивания растворов, содержащих протеазу с одной стороны, и аналог фактора Х с другой стороны. Таким образом, можно раздельно поддерживать два компонента в растворе и смешивать их с помощью соответствующего устройства для инфузии, в котором компоненты приходят в соприкосновение друг с другом по мере прохождения через устройство, и таким образом происходит активирование в фактор Ха или в аналог фактора Ха. Таким образом, пациент получает смесь фактора Ха и, дополнительно, сериновую протеазу, ответственную за активацию. В этом контексте особенно важно уделить пристальное внимание дозировке, поскольку дополнительное введение сериновой протеазы также активирует эндогенный фактор X, который может сократить время коагуляции.
Препарат по настоящему изобретению может быть в виде фармацевтического препарата с активностью фактора Ха в форме однокомпонентного препарата или в комбинации с другими факторами в форме многокомпонентного препарата.
Перед переработкой очищенного белка в фармацевтический препарат, очищенный белок подвергают обычному контролю качества и формируют в лекарственный препарат. В частности, при получении рекомбинанта очищенный препарат проверяют на отсутствие нуклеиновых кислот клетки, а также тех нуклеиновых кислот, которые происходят от вектора экспрессии, предпочтительно используя способ, описанный в ЕР 0714987.
Другое свойство настоящего изобретения связано с получением доступного препарата, содержащего аналог фактора Ха с высокой стабильностью и структурной целостностью, который, в частности, является свободной формой неактивных промежуточных продуктов аналога фактора Х/Ха и продуктов аутопротеолитического разрушения, и который может быть получен путем активирования аналога фактора Х типа, описанного выше, и переработки его в соответствующий препарат.
Фармацевтический препарат может содержать дозировки 10-1000 мкг/кг, более предпочтительно примерно 10-250 мкг/кг и наиболее предпочтительно 10-75 мкг/кг, причем особенно предпочтительно, если вариант полипептида фактора Х содержится в количестве 40 мкг/кг. Пациенты могут подвергаться лечению непосредственно после поступления в клинику с кровотечением. В другом варианте пациенты могут получать болюсную инфузию один раз в 1-3 ч, или, в случае значительного улучшения, инфузию раз в сутки варианта фактора Ха, описанного в настоящем изобретении.
Б. Зимоген/протеаза-кодирующие нуклеиновые кислоты
Зимоген/протеаза-кодирующие нуклеиновые кислоты могут применяться для разных целей в соответствии с настоящим изобретением. В предпочтительном варианте осуществления настоящего изобретения предусмотрено средство доставки нуклеиновой кислоты (т.е, экспрессирующий вектор) для модулирования коагуляции крови, причем экспрессирующий вектор включает последовательность нуклеиновой кислоты, кодирующую вариант полипептида зимогена/протеазы, или его функциональный фрагмент, описанные выше. Введение зимоген/протеаза-кодирующих экспрессирующих векторов пациенту приводит к экспрессии полипептида зимоген/протеазы, который изменяет каскад коагуляции. В соответствии с настоящим изобретением последовательность нуклеиновой кислоты, кодирующей зимоген/протеазу, может кодировать полипептид зимогена/протеазы, описанный в настоящем изобретении, экспрессия которого повышает гемостаз. В предпочтительном варианте осуществления настоящего изобретения последовательность нуклеиновой кислоты зимогена/протеазы кодирует тромбин человека или вариант полипептида фактора Ха.
Экспрессирующие векторы, содержащие последовательности нуклеиновой кислоты варианта зимогена/протеазы, могут вводиться отдельно или в комбинации с другими молекулами, и применимы для модулирования гемостаза. Согласно настоящему изобретению Экспрессирующие векторы или комбинация терапевтических агентов могут вводиться пациенту отдельно или в фармацевтически приемлемых или в биологически совместимых композициях.
В предпочтительном варианте осуществления настоящего изобретения экспрессирующий вектор, включающий последовательности нуклеиновой кислоты, кодирующие варианты зимогена/протеазы, является вирусным вектором. Вирусные векторы, которые могут быть использованы в настоящем изобретении, включают, но ими не ограничиваются, аденовирусные векторы (с/без тканеспецифичными промоторами/энхансерами), адено-ассоциированные вирусные векторы (adeno-associated virus - AAV) множественных серотипов (например, AAV-2, AAV-5, AAV-7 и AAV-8) и гибридные векторы AAV векторы, лентивирусные векторы и псевдопитированные лентивирусные векторы [например, вируса Эболы, вирус везикулярного стоматита (vesicular stomatitis virus - VSV) и вирус кошачьего иммунодефицита (feline immunodeficiency virus -FIV)], вирус герпес-симплекс, векторы на основе вируса осповакцины и ретровирусные векторы.
В предпочтительном варианте осуществления настоящего изобретения предусмотрены способы введения вирусного вектора, включающие последовательности нуклеиновой кислоты, кодирующие вариант зимогена/протеазы или его функциональный фрагмент. Аденовирусные векторы, применяемые в способах настоящего изобретения, предпочтительно включают по меньшей мере существенные части ДНК аденовирусного вектора, В настоящем изобретении описано, что экспрессия полипептида варианта зимогена/протеазы после введения такого аденовирусного вектора служит для модулирования гемостаза. В контексте описанного в настоящем изобретении варианта фактора Ха такое введение повышает прокоагулирующую активность протеазы.
Рекомбинантные аденовирусные векторы нашли широкое применение для различных целей генной терапии. Их применение для таких целей связано преимущественно с высокой эффективностью in vivo генного переноса, достигаемого в разных органах.
Аденовирусные частицы могут применяться в качестве носителя для адекватной доставки гена. Такие вирионы обеспечивают ряд полезных свойств для такого применения, включая: структурные свойства, связанные с вирусной двуцепочечной ДНК, не заключенной в вирусную оболочку, и биологические свойства, например, тропизм в отношении дыхательной системы человека и желудочно-кишечного тракта. Кроме того, известно, что аденовирусы инфицируют широкий круг типов клеток in vivo и in vitro благодаря рецептор-опосредованному эндоцитозу. Подтверждая общую безопасность аденовирусных векторов, аденовирусная инфекция приводит к минимальному проявлению заболевания у людей, включая слабовыраженные симптомы, напоминающие грипп.
Из-за крупного размера (~36000 оснований) геномы аденовирусов хорошо подходят для применения в генной терапии в качестве переносчиков, потому что они могут осуществить инсерцию чужеродной ДНК после удаления аденовирусных генов, важных для репликации, а также незначительных областей. Такие замены компенсируют повреждения вирусного вектора, связанные с функциями репликации и способностью к инфицированию. Известно, что аденовирусы применяются в качестве векторов для генной терапии и для экспрессии гетерологичных генов.
Более подробное обсуждение применения аденовирусных векторов для генной терапии см. в работах Berkner, Biotechniques 6, 1988, сс.616-629, и Trapnell, Advanced Drug Delivery Reviews 12, 1993, сс.185-199.
Желательно интродуцировать вектор, который может обеспечить, например, большое количество копий целевого гена и, следовательно, повышенные количества продукта этого гена. Усиленные аденовирусные векторы и способы получения таких векторов были подробно описаны в ряде публикаций, патентов, патентных заявок, включая: Mitani, Kubo, Curr Gene Ther. 2 (2), 2002, сс.135-44, Olmsted-Davis и др., Hum Gene Ther. 13 (11), 2002, сс.1337-1347, Reynolds и др., Nat Biotechnol. 19 (9), 2001, сс.838-842, патенты US 5998205 (описаны опухолеспецифичные реплицирующие векторы, включающие много копий ДНК), 6228646 (описаны не содержащие хелпера, полностью дефектные аденовирусные векторы), 6093699 (предусмотрены векторы и способы генной терапии), 6100242 (в котором успешно применяли трансгенно-инсертированный дефектный по репликации аденовирусный вектор для генной терапии in vivo при болезни периферийных сосудов и болезни сердца) и международные патентные заявки WO 94/17810 и WO 94/23744.
Для некоторых применений экспрессирующая конструкция может дополнительно включать регуляторные элементы, необходимые для индукции экспрессии в клетках или тканях определенного типа. Такие регуляторные элементы известны специалистам в данной области и подробно обсуждаются в работах Sambrook и др. (1989) и Ausubel и др. (1992). Внедрение тканеспецифичных регуляторных элементов в экспрессирующие конструкции настоящего изобретения обеспечивает по меньшей мере частичный тканевый тропизм для экспрессии варианта зимогена/протеаз или их функциональных фрагментов. Например, Е1 делегированный тип 5 аденовирусного вектора, включающий последовательности аминокислот, кодирующие вариант зимогена/протеазы под контролем промотора цитомегаловируса может применяться для получения преимущества у способов настоящего изобретения.
Примеры способов получения аденовирусных векторов
Аденовирусные векторы для экспрессии рекомбинантного гена были получены в линии эмбриональных клеток почки человека 293 (Graham и др., J. Gen. Virol. 36, 1997, сс.59-72). Эта линия клеток допускает рост аденовируса 2 (Ad2) и мутантов аденовируса 5, дефектных по функции Е1, поскольку она включает левый конец генома аденовируса 5 и, следовательно, экспрессирует белки Е1. Гены Е1, интегрированные в клеточный геном клеток линии 293, экспрессируются на уровнях, которые облегчают применение этих клеток в качестве системы экспрессии, в которой вирусные векторы для амплификации этих генов были удалены. Клетки линии 293 интенсивно использовались для выделения и размножения Е1 мутантов, для хелпер-независимого клонирования, а также для экспрессии аденовирусных векторов. Таким образом, системы экспрессии, например, линия клеток 293, обеспечивают важные вирусные функции, связанные с переносом, и тем самым обеспечивают размножение вирусных векторов, в которых экзогенные последовательности нуклеиновой кислоты были замещены на гены Е1. См. Young и др. в кн.: «The Adenoviruses», 1984, под ред. Ginsberg, изд-во Plenum Press, Нью-Йорк и Лондон, сс.125-172.
Другие системы экспрессии, хорошо пригодные для размножения аденовирусных векторов, известны специалистам в данной области (например, клетки HeLa) и описаны во многих публикациях.
В настоящем изобретении также предусмотрен способ модулирования гемостаза, включающий получение клеток индивидуума с высвобождением нуклеиновой кислоты, кодирующей полипептид варианта зимогена/протеазы и позволяющей клеткам расти в условиях, при которых экспрессируется полипептид зимогена/протеазы.
Из последующей дискуссии можно видеть, что полипептиды зимогена/протеазы и экспрессирующие векторы с нуклеиновой кислотой зимогена/протеазы могут применяться для лечения расстройств, связанных с аберрантной коагуляцией крови.
В. Фармацевтические композиции
Экспрессирующие векторы настоящего изобретения могут быть инкорпорированы в фармацевтические композиции, которые могут быть доставлены субъекту таким образом, что позволяют вырабатывать биологически активный белок (например, полипептид варианта зимогена/протеазы или его функциональный фрагмент или производное). В одном из вариантов осуществления настоящего изобретения фармацевтические композиции, включающие достаточный генетический материал, позволяют реципиенту вырабатывать терапевтически эффективное количество полипептида варианта зимогена/протеазы, влияющего на гемостаз субъекта. Композиции могут быть введены отдельно или в комбинации по меньшей мере с одним из агентов, например, стабилизирующим соединением, которое может быть введено в каком-либо стерильном биосовместимом фармацевтическом носителе, включая, но, не ограничиваясь ими, физиологический раствор, буферный физиологический растров, глюкозу и воду. Композиции могут быть введены пациенту отдельно или в комбинации с другими агентами, влияющими на гемостаз.
В предпочтительных вариантах осуществления настоящего изобретения фармацевтические композиции также содержат фармацевтически приемлемый эксципиент. К таким эксципиентам относится фармацевтический агент, который не индуцирует иммунный ответ, опасный в случае индивидуального введения композиции, и который может вводиться без индукции токсичности. К фармацевтически приемлемым эксципиентам относятся, но ими не ограничиваются, жидкости, например, вода, физиологический раствор, глицерин, растворы сахаров и этанол. К фармацевтически приемлемым эксципиентам также могут быть отнесены, например, соли минеральных солей, например, гидрохлориды, гидробромиды, фосфаты, сульфаты и др., а также соли органических кислот, например, ацетаты, пропионаты, малонаты, бензоаты и др. Кроме того, дополнительные вещества, например, увлажняющие или эмульгирующие агенты, буферные вещества и др., могут входить в такие растворы. Доскональное изложение фармацевтически приемлемых эксципиентов приводится в кн.: «Remington's Pharmaceutical Sciences», 1990, 18-е изд., изд-во Mack Pub. Co., Easton, Пасадена.
Фармацевтические составы, приемлемые для парентерального введения, могут быть переработаны в водные растворы, предпочтительно в физиологически совместимых буферах, например, в растворе Хэнкса, растворе Рингера или в физиологически буферном солевом растворе. Инъекции водных суспензий могут содержать вещества, которые повышают вязкость суспензии, например, натриевую соль карбоксиметилцеллюлозы, сорбит или декстран. Кроме того, суспензии действующих соединения могут быть приготовлены в виде соответствующих инъекционных суспензий на масляной основе. К приемлемым липофильным растворителям или наполнителям относятся растительные масла, например, кунжутное масло, или сложные эфиры синтетических жирных кислот, например, этилолеат или триглицериды, или липосомы. Необязательно суспензия также может содержать приемлемые стабилизаторы или агенты, которые повышают растворимость соединений, что способствует приготовлению высококонцентрированных растворов.
Фармацевтическая композиция может быть предусмотрена в виде соли и может быть солью многих кислот, включая, но, не ограничиваясь ими, соляную, серную, уксусную, молочную, винную, яблочную, янтарную кислоты и др. Соли могут быть более растворимыми в водных или других протонных растворителях, чем в форме свободных оснований. В других случаях предпочтительным препаратом может быть лиофилизированный порошок, который может содержать какой-либо или все из следующих компонентов: 1-50 мМ гистидин, 0,1%-2% сахарозу и 2-7% маннит при диапазоне рН 4,5-5,5, которые перед применением объединяют с буфером.
После приготовления фармацевтические композиции могут быть помещены в соответствующий контейнер и снабжают этикеткой для применения. Для введения зимоген/протеаза-содержащих векторов такие этикетки должны включать количество, частоту и способ введения.
К фармацевтическим композициям, пригодным для применения в настоящем изобретении, относятся композиции, в которых действующие ингредиенты содержатся в эффективном количестве для достижения поставленной терапевтической цели. Специалист в данной области может определить терапевтически эффективную дозу, используя методы, описанные в настоящем изобретении. Терапевтические дозы могут зависеть наряду с другими факторами от возраста и общего состояния субъекта, тяжести фенотипа аберрантной коагуляции крови и концентрации контрольных последовательностей, регулирующих уровни экспрессии полипептида варианта зимогена/протеазы. Таким образом, у людей терапевтически эффективное количество может оказаться в относительно широком диапазоне, который может быть определен практикующим врачом на основании ответа конкретного пациента на лечение вектором, включающим зимоген/протеазу.
Г. Введение в организм
Полипептиды варианта фактора X, отдельно или в комбинации с другими агентами, могут быть непосредственно введены пациенту в соответствующем биологическом носителе согласно описанному выше. Векторы экспрессии настоящего изобретения, включающие последовательности нуклеиновой кислоты, кодирующие вариант зимогена/протеазы, или их функциональные фрагменты, могут быть введены пациенту различными способами (см. ниже) для достижения и поддержания профилактически и/или терапевтически эффективных уровней зимогенного/протеазного полипептида. Специалист в данной области может легко определить специфические протоколы для применения зимоген/протеаза-кодирующих экспрессирующих векторов настоящего изобретения для терапевтического применения конкретным пациентам. Протоколы для получения аденовирусных векторов и введения пациентам описаны в патентах US 5998205, 6228646, 6093699, 6100242 и в международных патентных заявках WO 94/17810 и WO 94/23744, сущность которых включена в настоящее изобретение в виде ссылок.
Вариант зимогена/протеазы, кодируемый аденовирусными векторами настоящего изобретения, может быть введен пациенту каким-либо известным способом. Прямое высвобождение фармацевтических композиций in vivo в основном выполняется инъекцией с помощью обычного шприца, хотя другие способы введения, например, конвекционно-повышенное высвобождение, рассматриваются в качестве допустимых (см., например, патент US 5720720). В связи с этим композиции могут вводиться подкожно, наноситься на кожу, вводиться внутрикожно, интратекально, вводиться внутриглазным способом, в слизистую оболочку, внутрибрюшинно, внутривенно, внутриартериально, перорально, внутрипеченочно или внутримышечно. К другим способам введения относится введение внутрь и введение в легкие, введение с помощью суппозиториев и чрескожных аппликаций. Клиническая специализация на лечении пациентов с расстройствами коагуляции крови может определить оптимальный способ введения аденовирусных векторов, включающих последовательности нуклеиновой кислоты, кодирующие зимоген/протеазу, основанный на ряде критерий, включая, но, не ограничиваясь ими: состояние пациента и цели лечения (например, повышенная или пониженная коагуляция крови).
Настоящее изобретение также включает векторы AAV, включающие последовательность нуклеиновой кислоты, кодирующей полипептид варианта зимогена/протеазы.
Также предусмотрены лентивирусные или псевдотипические лентивирусные векторы, включающие последовательность нуклеиновой кислоты, кодирующей полипептид варианта зимогена/протеазы.
В настоящем изобретении также предусмотрены «голые» плазмиды или экспрессирующие векторы, включающие последовательность нуклеиновой кислоты, кодирующую полипептид варианта зимогена/протеазы.
Примеры
Пример 1. Вариант фактора Ха зимогена/протеазы
Протеолитический процессинг предшественников белков плазмы для изменения активации является признаком коагуляции крови. Примером данного типа механизма активации является переход зимогена в протеазу в семействе химотрипсин-подобных сериновых протеиназ. Расщепление связи по высоко консервативному сайту (Arg15-Ile16, система нумерации хемотрипсина) раскрывает новый N-конец, который выступает в качестве внутримолекулярного лиганда для Asp 194 (фиг.2). Этот новый солевой мостик приводит к конформационному изменению, или связан с ним, и упорядочивает так называемый «домен активирования», поверхностные петли, состоящие из специфического кармана S1, оксианионной щели, автолизной петли и сайта связывания натрия (фиг.3). Достоверно установлено, что в системе трипсина формирование внутреннего Ile16-Asp194 солевого мостика аллостерически связано со специфическим сайтом S1; изменения сайтов влияют друг на друга.
Основные принципы зимогена по активации транзиции фермента для сериновых протеаз на структурном уровне хорошо подтверждены экспериментально, особенно для хемотрипсина и трипсина, и эти примеры выступают в качестве образца представителя семейства сериновой протеазы. Основные свойства могут быть суммированы следующим образом (см, фиг.2): 1) структура (~80-85%) зимогена относительно близка структуре протеазы, 2) перенос инициируется после высвобождения высоко консервативного N-конца (например, Ile16-Val-Gly-Gly19), 3) новая свободная α-аминогруппа (Ile16) становится погруженной в гидрофобную среду и ее α-аминный азот формирует внутренний солевой мостик с Asp194, 4) положение Asp194 существенно изменяется при активированном зимогеном повороте ~170°, 5) этот внутренний солевой мостик приводит к конформационному изменению, или связан с ним, в так называемом «домене активации», и поверхностно расположенные петли содержат остатки 16-19, 142-153, 184-194 и 216-223; и частично включающие специфический сайт S1 (номенклатура Schechter, Berger, Biochem. Biophys. Res. Comm., 436 19676 cc.694-702) и оксианионное отверстие. В различных исследованиях установлено, что зимоген и зрелый фермент находятся в равновесном состоянии с Keq=~108 в пользу зимогена. Bode и др. в изящном эксперименте показали, что трипсиноген может принимать активную трипсиноподобную структуру к сильному лигандному связыванию со специфическим карманом S1 или к соответствующим лигандам с высоким сродством к щели Ile16. Дополнительными примерами такой индукции без расщепления связи Arg/Lys15-Ile16 являются связывание стрептокиназы с плазминогеном, стафилокоагулазы с протромбином, и недавно описанное аутоантитело к протромбину (Madoiwa и др., Blood, 97, 2001, cc.3783-3789). Совместно эти исследования показывают, что сериновые протеазы, даже в форме зимогенов, могут принимать протеаза-подобные функции в зависимости от различных условий окружающего пространства, т.е. жесткое лигандное связывание с зимогеном.
Хорошо известно, что активирование FX приводит к крупным конформационным изменениям в домене сериновой протеазы, которые сопровождаются способностью протеазы связываться с намного большим сродством к S1-направленными зондами и мембраносвязанным фактором FVa (1-6). Общие молекулярные механизмы (механизм), управляющие транзицией сериновой протеазы, обычно соответствуют системе трипсина. Однако может не быть однородности. Одноцепочечный транспортный активатор плазминогена (ТАП) проявляет разную молекулярную стратегию для поддержания его зимоген-подобного статуса (8-11). Анализ пар зимоген/протеаза, участвующих в коагуляции крови, особенно FVII/FVIIa, показывает, что имеется несколько различий в такой транзиции по сравнению с системой трипсина (12). Хотя несколько структурных детерминант на FXa являются частью так называемого домена активирования, в настоящее время не установлено, связано ли формирование этих сайтов непосредственно с транзицией зимогена в протеазу. Ранее описанная модель зимогенного FX подтверждает, однако, что некоторые из этих элементов могут быть разупорядочены в зимоген (13). Сравнение модели зимогена с активным ферментом показывает, что остатки, укладывающие Са (Asp70-Glu80), Na+ (Ala183-Asp194, Gly219-Gly226) и аутолитическую (Thr144-Arg150) петли, претерпевают крупные изменения в их каркасном расположении при транзиции зимогена в протеазу. Поскольку уже достоверно установлено по меньшей мере для трипсиногена/трипсина, что S1 специфичный сайт и формирование Ile16-Asp194 аллостерически связаны, резонно предположить, что другие элементы домена активирования также связаны с транзицией зимогена в протеазу. В настоящем примере конструируют эксперименты для проверки гипотезы, что дестабилизация Ile16-Asp194 внутреннего солевого мостика путем изменений по положениям 16, 17 или 194 изменяет щель активного сайта, делая образующий вариант «зимоген-подобным». Мы также предполагаем, что эти изменения могут аллостерически модулировать FVa связывание.
Материалы и методы
Экспрессия фактора Ха
Хотя в литературе имеется несколько сообщений об экспрессии rFX, большинство основано на неубедительных версиях или не представляют адекватного описания (15-20). В первоначальных попытках экспрессии rFX в клетках НЕК 293 были получены уровни экспрессии в диапазоне 1-2 мг rFX/л кондиционированной среды, однако установлено, что только 10-40% полученного материала полностью γ-карбоксилировано (21). Оставшийся материал не обладает γ-карбоксилированием. На основании преимущества разного связывающего сродства витамин К-зависимых пропептидов карбоксилазы предполагают, что поскольку пропептид протромбина проявляет наименьшее сродство к карбоксилазе, измененный пропептид FX (наивысшая афинность) с таким же сродством, что и у протромбина, может повысить γ-карбоксилирование за счет допущения, что оборот субстрата повышен (22, 23). Используя этот новый вектор, отбирают стабильные трансфектанты клеток линии НЕК 293, размножают и очищают rFX иммунноафинной хроматографией. Применяют элюцию фосфатом с гидроксиапатита для разделения карбоксилированного материала от некарбоксилированного материала. Результаты, полученные в настоящее время от более чем 30 стабильных клеточных линий, показывают, что в среднем ~80-90% rFX полностью γ-карбоксилировано по сравнению с 10-40%, используя нативный пропептид FX. Эти результаты были опубликованы ранее, после чего данная стратегия была применена по меньшей мере еще в одной лаборатории (24, 25). Таким образом, используя данную новую систему экспрессии, получают миллиграммовые количества (15-25 мг полностью γ-карбоксилированного rFX из ~10 л L кондиционированных сред) белка для детального исследования структуры/функции.
Исследования ферментов
Концентрации ферментов могут быть определены титрованием активного сайта с помощью ρ-нитрофенила-ρ-гуанидинобензоата (IIa) или флуоресцеин-моно-ρ-гуанидингобензоата (FXa) (26, 27). Активность FXa хромогенного субстрата, в присутствии различных ингибиторов или без них, может быть измерена с начальных степеней гидролиза спектрозима FXa, S-2222 или S-2765, согласно ранее описанному (14). Кинетические параметры могут быть определены методом подбора наименьших квадратов данных начальной степени соответствующих уравнений.
Получение вариантов FXa
Мутанты FX получают, используя набор для сайт-направленного мутагенеза Quick-change (фирма Stratagene), и целую кДНК FX секвенируют для проверки идентичности продукта. Различные плазмиды временно трансфецируют в клетки НЕК 293, используя продукт липофектамин-2000. Через 48 ч после трансфекции среды собирают и определяют уровни антигена FX, используя FX-специфический анализ ELISA, а активность FXa оценивают хромогенным исследованием перед активированием с применением RVV-X или тканевого фактора FVIIa.
Результаты
Получение видов FXa, распределенных по метаболическому пути транзиции зимогена-протеазы выделены в табл.1. Результаты по временной трансфекции показывают, что получена серия вариантов FXa с переменными количествами активности, варьирующими от ~25% до <1%. Высказывают предположение, что эти различия в активности, вероятно, отражают варианты FX, которые в разной степени смещены в соответствии с транзицией зимогена в протеазу. При разных состояниях Ile16-Asp194 внутренний солевой мостик стабилизируется до различных степеней, в зависимости от аминокислот в положениях 16, 17 или 194. Для дальнейшего описания выбирают три из этих вариантов (rFXI16L, FXaI16G, FXaV17A).
Устанавливают стабильные клеточные линии в клетках НЕК293 и очищают зимоген из 10 л кондиционированной среды (14, 24). Варианты активируют RVV-X и затем очищают гель-фильтрационной хроматографией (14, 24). SDS-PAGE и rFXI16L, и rFXaI16L показаны на фиг.4.
Первоначально исследования были сосредоточены на оценке изменений пространства вокруг активного сайта каждого из вариантов, проводившейся с использованием специфических зондов, которые нацеливают данную область FXa. Кинетические исследования с применением пептидильных субстратов и активных сайт-направленных зондов, показывают, что FXaI16L и FXaV17A обладают повышенной способностью связывать эти зонды (от 15- до 25-кратного повышения Km или Ki), хотя степень катализа (kcat) снижается трехкратно по сравнению с FXa дикого типа (плазма-производный и рекомбинантный) (табл.2 и 3). Фактор XaI16G не ингибируется каким-либо из исследованных зондов и его хромогенная активность сильно ухудшилась (500-1000 раз), предотвращая подсчет кинетических параметров. Эти результаты согласуются с предположением, что дестабилизация формирования внутреннего солевого мостика (Ile16-Asp194) влияет на связывание специфического сайта S1. В противоположность этим результатам сборка FXaI16L и FXaV17A в протромбиназу почти полностью восстанавливает Km для пептидильных субстратов, хотя величина kcat уменьшается в 3 раза, показывая, что связывание FVa может сохранить связывание активного сайта (табл.2 и 3). Неожиданно даже величина Km для I16G была почти полностью восстановлена (трехкратное увеличение по сравнению с FXa дикого типа) при сборке в протромбиназу; однако установлено 60-кратное снижение величины kcat.
Для экспериментов, в которых применяют свободный фактор Ха, 2,0 нМ дикого типа или 6,0 нМ мутантного фактора Ха инкубируют с возрастающими концентрациями спектрозима FXa и для экспериментов, в которых Протромбиназа использует 5,0 нМ дикого типа или мутантного фактора Ха, инкубируют с 30 нМ фактора Va, 50 мкг PCPS и повышенными концентрациями субстрата (10-500 мкМ). Хромогенную активность оценивают мониторингом повышения поглощения при 405 нм на протяжении времени. Ошибки величин установленных констант соответствуют 95% доверительным пределам.
Согласно этим данным кинетические исследования с применением протромбина показали, что величины Km, полученные для каждого из этих вариантов, объединенных в протромбиназу, были в существенной степени эквивалентны ферменту дикого типа, хотя величины kcat были снижены до сходной степени, что и для хромогенных субстратов (табл.4). Взятые вместе полученные результаты показывают, что транзиция зимогена в протеазу для FX не только влияют на формирование сайта S1, но также принимают существенное участие в формировании FVa связывающего сайта. Поскольку прямое связывание таких РХа вариантов с FVa восстанавливает связывание по сайту S1, вероятно существует аллостерическая связь между этими сайтами. Совместно эти исследования иллюстрируют уникальный путь модификации транзиции зимогена в протеазу и выявляют возможный путь развития зимоген-подобных форм ферментов, которые «активируются» последующим связыванием жесткого лиганда, например, кофакторных белков,
Результаты, полученные с хромогенным субстратом и активные сайт-направленные ингибиторы, показывают, что зимоген-подобные варианты FXa связываются с зондами активных сайтов с пониженной афинностью. Однако сборка таких вариантов в протромбиназу существенно улучшает сродство с зондами активных сайтов, подтверждая, что FVa связывание может спасти связывание по активному сайту. На следующем этапе исследуют, является ли реверсия также истинной, и что занятие зимоген-подобного активного сайта влияет на связывание с FVa. Для оценки этой гипотезы измеряют константы связывания между FVa и FXaI16L и FXaV17A. Для достижения этого инкубируют FVa с флуоресцирующим производным неактивного wtFXa в присутствии синтетических фосфолипидных пузырьков и ионов Са2+. Формирование комплекса приводит к повышению сигнала флуоресценции относительно флуоресценции только одного FXa. Затем вносят повышенные концентрации нефлуоресцирующего FGXa, который, при связывании с FVa, смещает флуоресценцию FXa, что выражается в снижении сигнала флуоресценции. Для контроля вносят S195A FXa в качестве конкурента. Этот мутант неактивен, потому что в каталитической триаде недостает Ser, но он обладает высоким сродством в отношении FVa (табл.5). Напротив, при внесении либо FXaI16L, либо FXaV17A, сродство этих зимоген-подобных вариантов к FXa существенно снижено по сравнению с FXaS195A (табл.5). Затем исследуют, может ли ковалентное заполнение активного сайта FXaI16L восстановить связывание с FVa. Для этого модифицируют активный сайт дикого типа Fxa и FXaI16L с необратимым ингибитором (EGR-хлорметилкетоном) и затем повторяют эксперимент. Полученные данные показывают, что данный активный сайт блокирует FXaI16L связанных FVa-мембран с той же аффинностью, что и блокирует активный сайт дикого типа у FXa. Это означает, что заполнение зимоген-подобного FXa-активного сайта непосредственно воздействует на связывание с FVa.
Основываясь на том наблюдении, что зимоген-подобные FXa производные обладают плохой реакционной способностью по отношению к направленным на активный сайт зондам и ингибиторам в отсутствии FVa, но проявляют практически нормальную активность при сборке вариантов в протромбиназу, на следующем этапе исследуют активность FXaI16L в среде плазмы. Плазму больных гемофилией А или В берут под наблюдение вместе с FXa дикого типа для коррекции времени коагуляции (аРТТ) этих образцов плазмы; 0,1 нМ wtFXa показывает время коагуляции ~32 сек. Добавление той же концентрации FXaI16L приводит к времени коагуляции, равному ~42 сек, которое составляет ~50-70% активности относительно wtFXa, подтверждая, что зимоген-подобный вариант проявляет почти нормальную коагулирующую активность в плазме. Затем подвергают мониторингу период полураспада дикого типа FXa и FXaI16L в плазме больных гемофилией В. Белки вносят в плазму больных гемофилией В (ГВ) и в разное время отбирают аликвоты и исследуют методом на основе АПТВ (активированного частичного парциального тромбопластинового времени). Результаты с плазмой больных гемофилией В (ГВ) показывают, что относительная остаточная активность дикого типа FXa ингибируется очень быстро (<2 мин) (фиг.6). Напротив, активность FXaI16L сохраняется на гораздо большее время при периоде полураспада >2 ч. Сходные результаты были получены на плазме больных гемофилией А. Эти результаты означают, что можно модулировать свойства фермента таким образом, что он начинает обладать длительным периодам полураспада в плазме и может корректировать время коагуляции плазмы больных гемофилией.
На следующем этапе оценивают способность зимоген-подобного FXaI16L модулировать гемостаз на модельных грызунах с гемофилией (Schlachterman и др., J. Tromb. Haemost., 3, 2005, сс.2730-2737). Величина активированного частичного парциального тромбопластинового времени (АПТВ) мышей с гемофилией В (C57BL/6) примерно составляет 50-55 сек. Фактор XaI16L (200 мкг/кг, n=7) или ФСБ (n=4) вводят инъекцией в хвостовую вену мышам с гемофилией В. В определенное время (5 и 30 сек) отбирают кровь и во всех образцах определяют аРТТ. На фиг.7 показано, что инфузия FXaI16L приводит к полной коррекции аРТТ до уровней, наблюдаемых у здоровых животных. Этот эффект длится по меньшей мере на протяжении 30 мин, показывая, что молекула имеет относительно длинный период полураспада in vivo. Инфузия ФСБ обладает только предельным эффектом. Эти данные согласуются с данными экспериментов на плазме in vitro и показывают, что FXaI16L, а возможно другие зимоген-подобные FXa варианты, в самом деле, могут эффективно модулировать гемостаз in vivo.
Для дальнейшего исследования эффективности FXaI16L in vivo исследуют, может ли данная молекула корректировать время кровотечения у мышей с гемофилией В после травмы хвоста (Schlachterman и др., J. Tromb. Haemost., 3, 2005, сс.2730-2737). Потерю крови измеряют на протяжении 10 мин после надсечения дистальной стороны хвоста. В данном типе исследования потеря крови минимальна у здоровых мышей дикого типа линии Bald-c (n=7) и чрезвычайно устойчива у мышей с гемофилией В (мыши линии Bald с), которым вводили инъекцией ФСБ (n=6) после повреждения хвоста (фиг.8). Напротив, инъекция 450 мкг/кг FXaI16L существенно снижает объем кровопотери из раны на хвосте животного (n=7). Суммарный анализ полученных данных доказывает, что FXaI16L способен улучшить гемостаз у пациентов с гемофилией А и гемофилией В.
СПИСОК ЛИТЕРАТУРЫ
1. Furie В., Furie В.С. Spectral changes in bovine factor X associated with activation by the venom coagulant protein Vipera russelli. J. Biol. Chem. 251, 1976, сс.6807-6814.
2. Robison D., Furie В., Furie В.С., Bing D.H. Active site of bovine factor X. Characterization using substituted benzamidines as competitive inhibitors and affinity-labeling reagents. J. Biol. Chem. 255, 1980, сс.2014-2021.
3. Keyt В., Furie В.С. Furie В. Structural transitions in bovine factor X associated with metal binding and zymogen activation. Studies using conformational-specific antibodies. J. Biol. Chem. 257, 1982. сс.8687-8695.
4. Persson E., Valcarce C., Stenflo J. The γ-carboxyglutamic acid and epidermal growth factor-like domains of factor X. Effect of isolated domains on prothrombin activation and endothelial cell binding of factor X. J Biol Chem 266, 1991, с.2458.
5. Persson E., Hogg P.J., Stenflo J. Effects of Ca2+ binding on the protease module of factor Xa and its interaction with factor Va: evidence for two Gla-independent Ca2+ binding sites in factor Xa. J Biol Chem 268, 1993, сс.22531-22539.
6. Dahlbäck В., Stenflo J. Binding of bovine coagulation factor Xa to platelets. Biochemistry 17, 1978, сс.4938-4945.
7. Miletich J.P., Jackson С.M., Majerus P.W. Properties of the factor Xa binding site on human platelets. J. Biol. Chem. 253, 1978, сс.6908-6916.
8. Madison E., Kobe A., Gething M., Sambrook J.F., Goldsmith E. Converting tissue plasminogen activator to a zymogen: A regulatory triad of Asp-His-Ser. Science 262, 1993, сс.419-421.
9. Tachias К., Madison E. Converting tissue-type plasminogen activator into a zymogen. J. Biol. Chem. 271, 1996, сс.28749-28752.
10. Tachias К., Madison E. Converting tissue type plasminogen activator into a zymogen. Important role of Lys156 J. Biol. Chem. 272, 1997, сс.28-31.
11. Renatus M., Engh R. A., Stubbs M.Т., Huber R., Fischer S., Kohnert U., Bode W. Lysine 156 promotes the anomalous proenzyme activity of tPA: X-ray crystal structure of single-chain human tPA. EMBO J 16, 1997, сс.4797-4805.
12. Eigenbrot С., Kirchhofer D., Dennis M.S., Santell L., Lazarus R.A., Stamos J., Ultsch M.H. The factor VII zymogen structure reveals reregistration of beta strands during activation. Structure 9, 2001, сс.627-636.
13. Venkateswarlu D., Perera L., Darden Т., Pedersen L.G. Structure and dynamics of zymogen human blood coagulation factor X. Biophys. J. 82, 2002, сс.1190-1206.
14. Camire R. M. Prothrombinase assembly and S1 site occupation restore the catalytic activity of FXa impaired by mutation at the sodium-binding site. J. Biol. Chem. 277, 2002, сс.37863-37870.
15. Rezaie A.R., Neuenschwander P.F., Morrissey J.H., Esmon С.Т. Analysis of the functions of the first epidermal growth factor-like domain of factor X. J Biol Chem 268, 1993, сс.8176-8180.
16. Rezaie A.R., Esmon С.Т. Asp-70 to Lys mutant of factor X lacks high affinity Ca2+ binding site yet retains function. J. Biol. Chem. 269, 1994, сс.21495-21499.
17. Rezaie A.R., Esmon С.Т. Contribution of residue 192 in factor Xa to enzyme specificity and function. J. Biol. Chem. 270, 1995, сс.16176-16181.
18. Rezaie A.R. Role of residue 99 at the S2 subsite of factor Xa and activated protein С in enzyme specificity. J. Biol. Chem. 271, 1996, сс.23807-23814.
19. Rezaie A.R. Identification of basic residues in the heparin-binding exposit of factor Xa critical for heparin and factor Va binding. J. Biol. Chem. 275, 2000, сс.3320-3327.
20. Rezaie A.R., He X. Sodium binding site of factor Xa: Role of sodium in the prothrmombinase complex. Biochemistry 39, 2000, сс.1817-1825.
21. Larson P.J., Camire R.M., Wong D., Fasano N.C., Monroe D.M., Tracy P.В., High K.A. Structure/function analyses of recombinant variants of human factor Xa: Factor Xa incorporation into prothrombinase on the activated platelet surface is not mimicked by synthetic phospholipids vesicles. Biochemistry 37, 1998, сс.5029-5038.
22. Stanley Т.В., Jin D.Y., Lin P., Stafford D.W. The propeptides of the vitamin K-dependent proteins possess different affinities for the vitamin K-dependent carboxylase. J. Biol. Chem. 274, 1999, сс.16940-16944.
23. Stanley Т.В., Humphries J., High К.A., Stafford D.W. Amino acids responsible for the reduced affinities of vitamin K-dependent propeptides for the carboxylase. Biochemistry 38, 1999, сс.15681-15687.
24. Camire R.M., Larson Р.J., Stafford D.W., High K.A. Enhanced γ-carboxylation of recombinant factor X using a chimeric construct containing the prothrombin propeptide. Biochemistry 39, 2000, сс.14322-14329.
25. Zhong D., Bajaj M.S., Schmidt A.E., Bajaj S.P. The N-terminal EGF-like domain in factor IX and factor X represents an important recognition motif for binding to tissue factor. J. Biol. Chem. 277, 2001, сс.3622-3631.
26. Chase Т., Shaw E. Comparison of the esterase activities of trypsin, plasmin, and thrombin on guanidinobenzoate esters. Titration of the enzymes. Biochemistry. 8, 1969, cc.2212-2224.
27. Bock P.E., Craig P.A., Olson S.Т., Singh P. Isolation of human blood coagulation α-factor Xa by soybean-trypsin inhibitor-Sepharose chromatography and its active-site titration with fluorescein mono-r-guanidinobenzoate. Arch. Bioch. Biophys. 273, 1989, cc.375-388.
Хотя конкретные варианты из числа предпочтительных вариантов осуществления настоящего изобретения были описаны выше, это не означает, что ими ограничивается настоящее изобретение. Также могут быть произведены различные модификации, не уклоняющиеся от области охвата и духа настоящего изобретения, изложенных ниже в формуле настоящего изобретения.
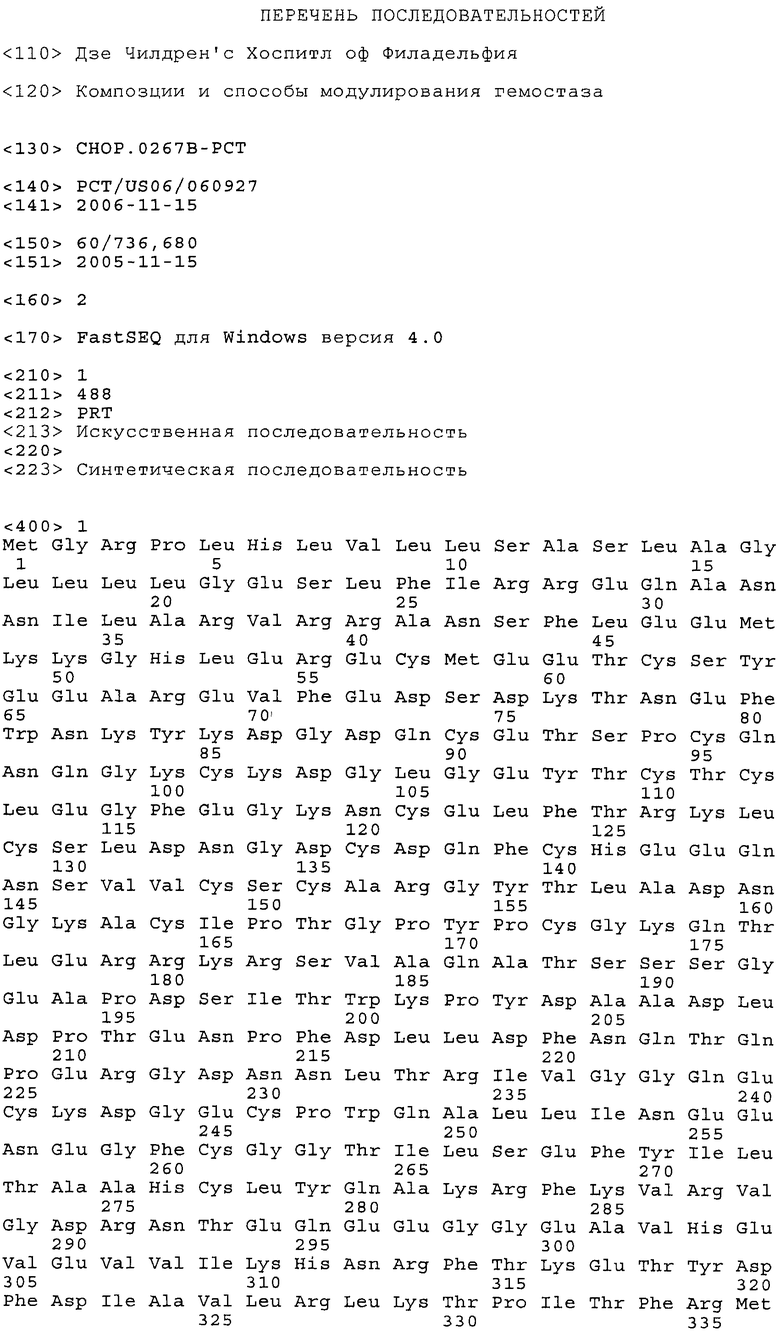

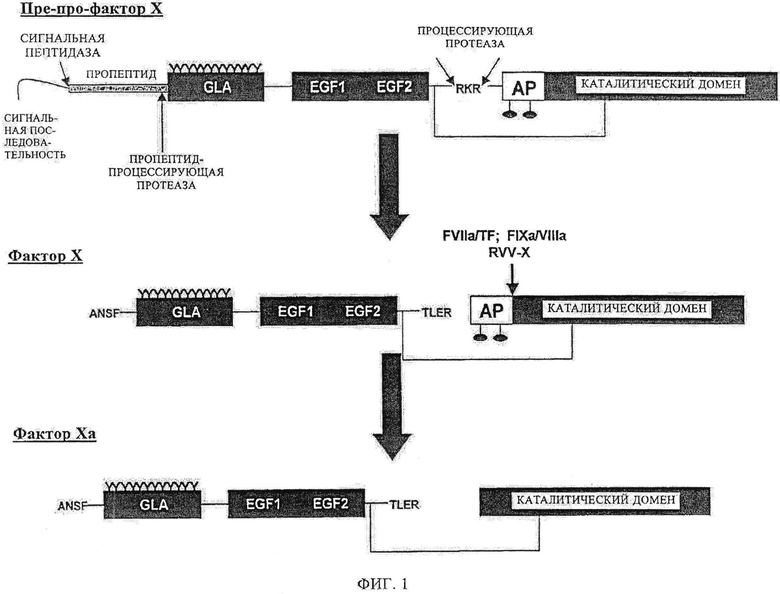
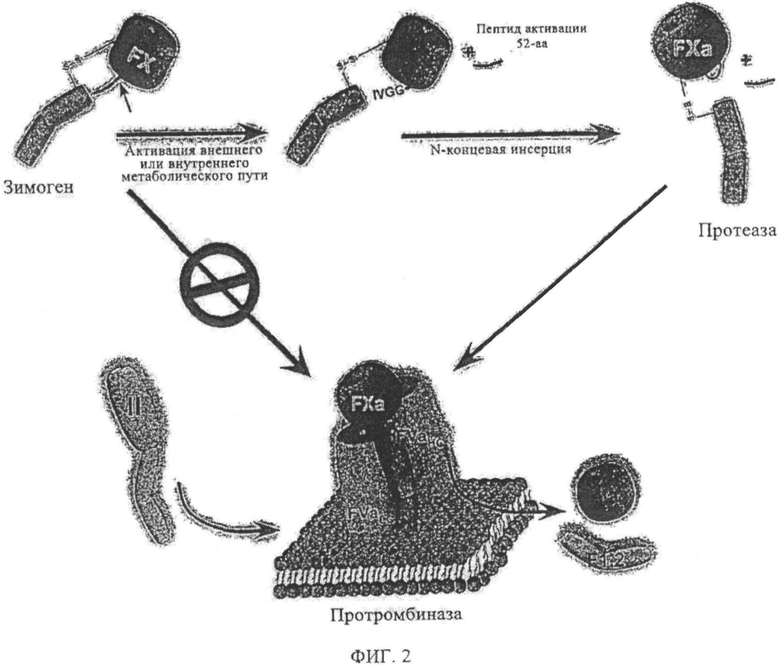
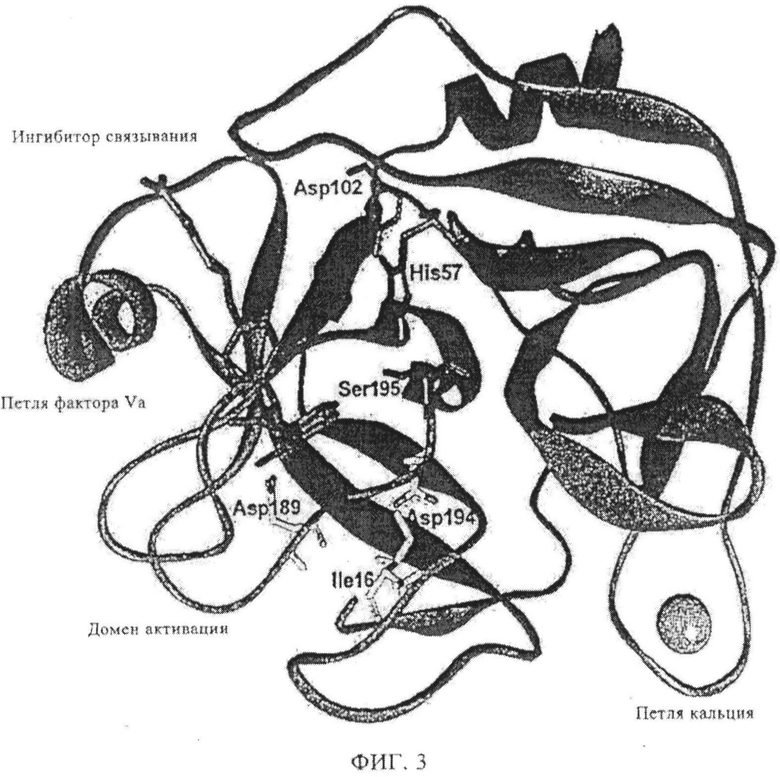
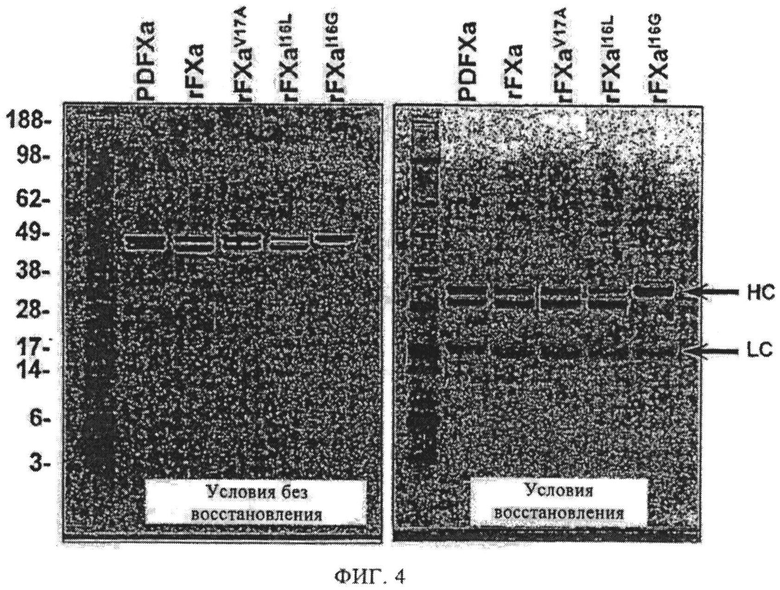



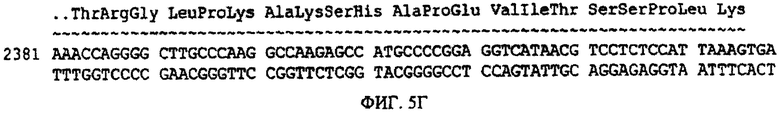
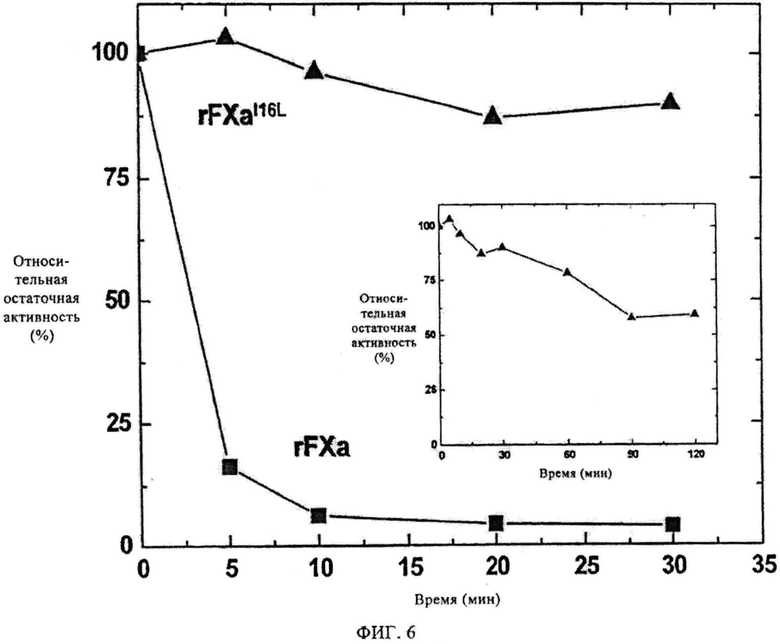
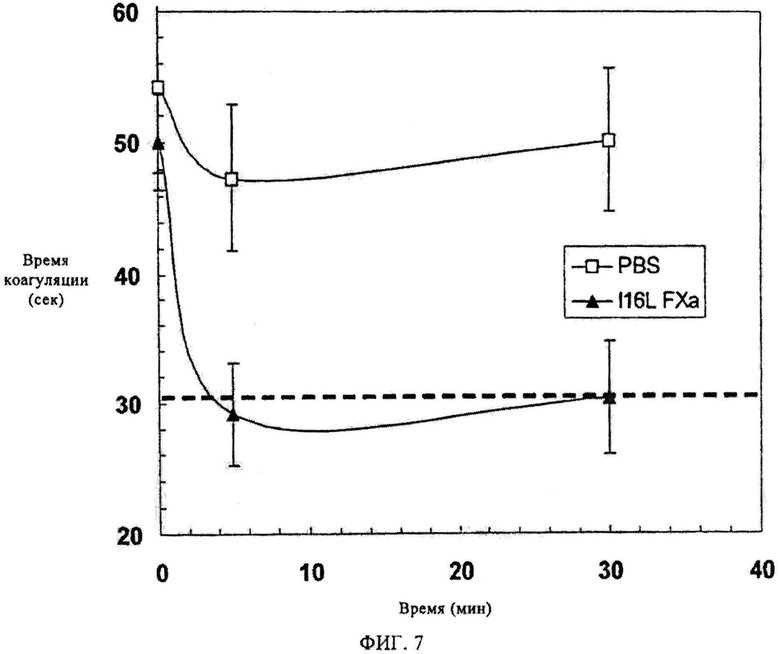
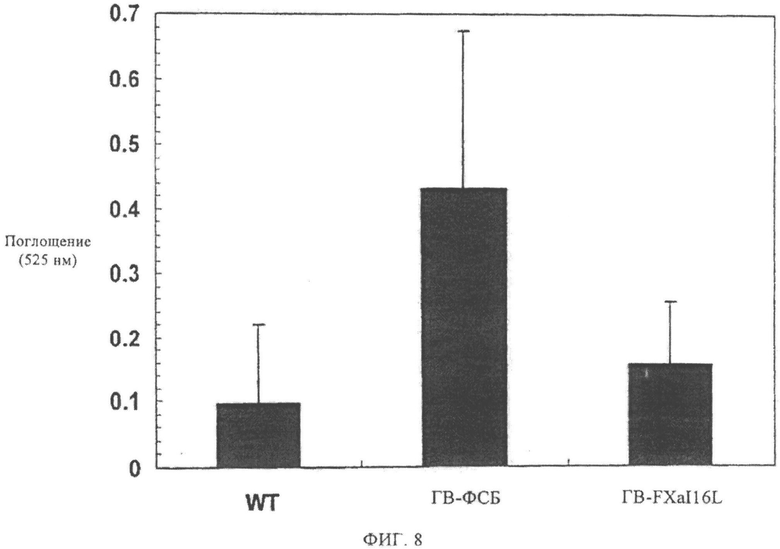
Изобретение относится к области биохимии, в частности к увеличивающим гемостаз вариантам фактора Ха, кодирующим их нуклеиновым кислотам, получению и применению фактора Xa для лечения расстройств, связанных с гемостазом. Заявленный фактор Xa включает аминокислотную замену в последовательности фактора X дикого типа в положении 16, 17 или 194 в системе нумерации химотрипсина. Изобретение позволяет эффективно увеличивать гемостаз. 7 н. и 14 з.п. ф-лы, 11 ил., 5 табл., 1 пр.
1. Фактор Ха, который увеличивает гемостаз, включающий замену в аминокислотной последовательности фактора X дикого типа, выбранную из группы, состоящей из:
а) Ile в положении 16 в системе нумерации химотрипсина, где Ile замещен аминокислотой, которая выбрана из группы, состоящей из Leu, Phe, Asp и Gly,
б) Val в положении 17 в системе нумерации химотрипсина, где Val замещен аминокислотой, которая выбрана из группы, состоящей из Leu, Ala и Gly, и
в) Asp в положении 194 в системе нумерации химотрипсина, где Asp замещен Asn или Glu,
причем связывание субстрата активным сайтом фактора Ха слабее по сравнению с фактором Ха дикого типа и сильнее, когда указанный фактор Ха связывается с помощью фактора Va в протромбиназном комплексе.
2. Фактор Ха по п.1, где Ile в положении 16 замещен Leu или Gly, или Val в положении 17 замещен Ala.
3. Фактор Ха по п.1, в котором последовательности в положениях 16-18 выбраны из группы, состоящей из Leu-Val-Gly, Gly-Val-Gly, Ile-Ala-Gly, Phe-Val-Gly и Ile-Gly-Gly.
4. Фактор Ха по п.1, где Ile в положении 16 замещен Leu.
5. Фактор Ха по п.1, где фактор Ха имеет более длинный период полужизни в плазме, чем фактор Ха дикого типа.
6. Фактор Ха по п.1, где указанный фактор Ха включает аминокислоты 41-179 и аминокислоты 235-488 SEQ ID NO: 1, где Ile в положении 16 (положение 235 SEQ ID NO: 1) замещен Leu.
7. Фактор Ха по п.1, где указанный фактор Ха состоит из аминокислот 41-179 и аминокислот 235-488 SEQ ID NO: 1, где Ile в положении 235 SEQ ID NO: 1 (положение 16 в системе нумерации химотрипсина) замещен Leu.
8. Фармацевтическая композиция для лечения связанных с гемостазом расстройств, включающая фактор Ха по любому из пп.1-7 в биологически совместимом носителе.
9. Применение фактора Ха по любому из пп.1-7 для лечения расстройств, связанных гемостазом.
10. Применение фактора Ха по п.9, где указанное расстройство выбрано из группы, состоящей из гемофилии А, гемофилии В, недостатка фактора коагуляции, кровотечения, обусловленного повреждением, травмы, тромбоза, тромбоцитопении, инсульта, коагулопатии, рассеянной внутриваскулярной коагуляции (DIC).
11. Выделенная молекула нуклеиновой кислоты для получения фактора Ха по любому из пп.1-7, где указанная молекула нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую человеческий полипептид фактора X (FX), имеющий замены, выбранные из группы, состоящей из
а) Ile в положении 16 в системе нумерации химотрипсина замещен аминокислотой, которая выбрана из группы, состоящей из Leu, Phe, Asp и Gly,
б) Val в положении положение 17 в системе нумерации химотрипсина замещен аминокислотой, которая выбрана из группы, состоящей из Leu, Ala и Gly, или
в) Asp в положении 194 в системе нумерации химотрипсина, замещен Asn или Glu, и
где указанная нуклеиновая кислота кодирует внутриклеточный сайт расщепления, где указанный внутриклеточный сайт расщепления находится в положении между 15 и 16 в системе нумерации химотрипсина или заменен белком активации, а расщепление внутриклеточного сайта расщепления в экспрессируемом факторе X приводит к получению фактора Ха.
12. Молекула нуклеиновой кислоты по п.11, где указанный FX полипептид включает аминокислоты 41-179 и 235-488 SEQ ID NO: 1.
13. Молекула нуклеиновой кислоты по п.11, где Ile в положении 16 замещен Leu или Gly, или Val в положении 17 замещен Ala.
14. Молекула нуклеиновой кислоты по п.11, где Ile в положении 16 замещен Leu.
15. Молекула нуклеиновой кислоты по п.11, где указанный FX полипептид включает последовательность пропептида.
16. Молекула нуклеиновой кислоты по п.11, где указанный внутриклеточный протеолитический сайт расщепления является сайтом расщепления РАСЕ/фурин.
17. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по любому из пп.11-16, функционально связанную с регуляторной последовательностью.
18. Вектор экспрессии по п.17, выбранный из группы, состоящей из аденовирусного вектора, связанного с аденовирусом вектора, ретровирусного вектора, плазмиды и лентивирусного вектора.
19. Клетка-хозяин для получения фактора Ха, включающая молекулу нуклеиновой кислоты по любому из пп.11-16, функционально связанную с регуляторной последовательностью.
20. Клетка-хозяин по п.19, где указанные клетки-хозяева являются клетками СНО.
21. Способ получения активированного фактора X (FXa), включающий инкубацию клетки-хозяина по п.19 и очистку продуцируемого и секретируемого ею FXa.
| US 20030181381 A1, 25.09.2003 | |||
| RODNEY M.C., Prothrombinase Assembly and S1 Site Occupation Restore the Catalytic Activity of FXa Impaired by Mutation at the Sodium-binding Site, J | |||
| Biol | |||
| Chem., 2002, vol.277(40), pp.37863-37870 | |||
| MANITHODY C | |||
| et al., Role of Basic Residues of the Autolysis Loop in the Catalytic Function of Factor Xa, Biochemistry, 2002, vol.41 no.21, pp.6780-6788 | |||
| RU 99120393 A, 27.06.2001. |

