ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ НА ИЗОБРЕТЕНИЕ
В данной заявке на изобретение заявлено преимущество предварительной заявки на патент США №61/881834, поданной 24 сентября 2013 г., включенной в данное описание путем ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Перечень последовательностей, поданный одновременно с данной заявкой на изобретение в соответствии с 37 CFR §1.821 в машиночитаемой форме (CRF) через EFS-Web в виде файла с названием PC72049_SEQLIST_ST25.txt, включен в данное описание путем ссылки. Электронная копия перечня последовательностей создана 10 сентября 2014 г., где размер файла составляет 31 килобайт.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Эффективные способы терапии необходимы для того, чтобы контролировать чрезмерные кровотечения в ряде клинических состояний, при которых кровотечение нельзя адекватно контролировать путем медицинского или хирургического вмешательства. Данная неудовлетворенная потребность особенно критична для пациентов, страдающих от гемофилии, в особенности тех, у которых заместительная терапия в отношении фактора свертывания крови оказывается менее эффективной вследствие продукции ингибиторных антител.
Активированный фактор свертывания крови X (FXa) занимает центральную позицию в каскаде свертывания крови на схождении внутренних и внешних путей свертывания крови. Связанный с мембраной FXa в присутствии своего кофактора фактора Va (FVa) превращает протромбин в тромбин, который активирует тромбоциты и превращает фибриноген в фибрин с образованием тромба. В принципе, заместительная терапия путем прямого введения FXa может корректировать кровотечение. Тем не менее, терапевтический потенциал FXa ограничен вследствие его очень короткого периода полувыведения из плазмы крови и возможности вызывать чрезмерное свертывание крови вследствие активации других факторов свертывания крови.
В более ранней работе идентифицировали вариант FXa (вариант I16L), в котором лейцин заменял изолейцин по N-концу тяжелой цепи FXa дикого типа (позиция 16 в схеме нумерации химотрипсина). Эта замена приводила к варианту FXa, обладающему зимогеноподобными свойствами. Toso R. et al., The conformational switch from the factor X zymogen to protease state mediates exosite expression and prothrombinase assembly. J. Biol. Chem. 283, 18627-18635 (2008); Ivanciu L. et al., A zymogen-like factor Xa variant corrects the coagulation defect in hemophilia. Nat. Biotechnol. 29:1028-33 (2011).
Будучи не включенным в протромбиназный комплекс со своим кофактором, а именно фактором Va (FVa), вариант FXa I16L не обладал значимой каталитической активностью и был лучше защищен от инактивации ингибиторами сывороточной протеазы по сравнению FXa дикого типа. В результате этот вариант обладал более длительным периодом полувыведения из сыворотки крови по сравнению с FXa дикого типа. Тем не менее, связывание с FVa в протромбиназе приводило к переходу варианта от зимогеноподобного состояния к активной конформации, таким образом, восстанавливая способность этого варианта катализировать превращение протромбина в тромбин и, таким образом, его прокоагулянтную активность. В мышиных моделях гемофилии А и гемофилии В введение варианта FXa I16L до повреждения уменьшало кровопотерю после отсечения хвоста в зависимости от дозы. Результаты этих экспериментов свидетельствуют о том, что вариант FXa I16L может быть полезен для лечения неконтролируемого кровотечения у людей, страдающих от гемофилии.
Тем не менее, вариант FXa I16L, используемый в более ранних исследованиях, получали в небольших количествах из стабильно трансфицируемых клеток НЕК 293 с последующей активацией белка FX с использованием протеазы яда гадюки Рассела RWX. Toso R., Zhu H. & Camire R.M. The conformational switch from the factor X zymogen to protease state mediates exosite expression and prothrombinase assembly. J. Biol. Chem. 283, 18627-18635 (2008). Несмотря на пользу данного подхода для мелкомасштабных исследований, он не подходит для продукции больших количеств очищенного варианта белка FXa, необходимых для клинических исследований и конечной поставки пациентам. Соответственно, в области техники существует потребность в препаратах варианта белка FXa I16L, приготавливаемых в таком количестве и с такой чистотой, что они могут быть тестированы в клинических исследованиях, и после одобрения предложены субъектам, нуждающимся в гемостазе.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Описание настоящего изобретения относится к неудовлетворенной потребности в области техники, описанной выше, путем того, что предложены композиции вариантов белка FXa, получаемые с достаточной чистотой и в достаточном количестве, таким образом, что они могут доставляться субъектам нуждающимся в гемостазе. В различных воплощениях эти композиции содержат разные изоформы и посттрансляционные модификации вариантов белка FXa.
В одном из воплощений композиции содержат бета форму варианта белка FXa, в которой последовательности легкой и тяжелой цепей белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 386, аминокислот с 1 по 140 и со 146 по 384, аминокислот с 1 по 140 и со 146 по 386, аминокислот с 1 по 141 и со 146 по 384, аминокислот с 1 по 141 и со 146 по 386, аминокислот с 1 по 142 и со 146 по 384, аминокислот с 1 по 142 и со 146 по 386, аминокислот с 1 по 143 и со 146 по 384, и аминокислот с 1 по 143 и со 146 по 386, все из которых имеют аминокислотную последовательность в соответствии с SEQ ID NO: 1.
В еще одном воплощении композиции содержат альфа форму варианта белка FXa, в которой последовательности легкой и тяжелой цепей белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 398, аминокислот с 1 по 140 и со 146 по 398, аминокислот с 1 по 140 и со 146 по 399, аминокислот с 1 по 141 и со 146 по 398, аминокислот с 1 по 141 и со 146 по 399, аминокислот с 1 по 142 и со 146 по 398, и аминокислот с 1 по 142 и со 146 по 399, все из которых имеют аминокислотную последовательность в соответствии с SEQ ID NO: 1.
В любом одном или более чем одном из воплощений бета и альфа изоформы вариантов белков FXa, описанных выше, легкие цепи белков могут быть модифицированы таким образом, что включают β-гидрокси Asp63, О-связанную гексозу и 9, 10 или 11 остатков Gla. В некоторых воплощениях присутствует 9 остатков Gla. В других воплощениях присутствует 10 остатков Gla. И в других воплощениях присутствует 11 остатков Gla.
В любом одном или более чем одном воплощении бета изоформы вариантов белка FXa, описанных выше, тяжелые цепи могут быть модифицированы таким образом, что включают один или два О-связанных гликана ядра-1. В некоторых воплощениях только первый, только второй или оба О-связанных гликана ядра-1 могут быть не сиалилированы, моносиалилированы или дисиалилированы. В соответствии с некоторыми воплощениями отсутствуют какие-либо группы сиаловой кислоты. В других воплощениях присутствует одна группа сиаловой кислоты. В других воплощениях присутствует две группы сиаловой кислоты. В дополнительных воплощениях присутствует три группы сиаловой кислоты. И в других дополнительных воплощениях в общей сложности присутствует четыре группы сиаловой кислоты.
В соответствии с некоторыми воплощениями композиции варианта белка FXa включают по меньшей мере один вид варианта белка FXa, представленный в таблице 2 описания изобретения. В других воплощениях композиции в соответствии с описанием изобретения содержат по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 55 или по меньшей мере 60 видов варианта белка FXa, перечисленных в таблице 2.
В соответствии с другими воплощениями композиции варианта белка FXa включают по меньшей мере один вариант белка FXa, в котором легкая цепь представляет собой виды, перечисленные в таблице 3 описания изобретения. И в других воплощениях композиции варианта белка FXa включают по меньшей мере один вариант белка FXa, в котором тяжелая цепь представляет собой виды, перечисленные в таблице 4 описания изобретения.
В некоторых воплощениях любой один или более чем один вариант белка FXa в соответствии с описанием изобретения может быть представлен в композиции со средней частотой, которая варьирует относительно других видов, которые могут быть представлены. Например, конкретный вид может быть представлен с основной, минорной или следовой частотой по сравнению с другими представленными видами. Альтернативно, конкретный вид может быть представлен с высокой частотой, средней частотой, низкой частотой или очень низкой частотой по сравнению с другими представленными видами.
В описании настоящего изобретения также предложены нуклеиновые кислоты, векторы, клетки-хозяева и способы получения и очистки вариантов белка FXa.
Например представлены последовательности нуклеиновых кислот, кодирующие вариант белка FX, включающий замену нативной последовательности активационного пептида (АР) на сайт процессинга РАСЕ (фермент, расщепляющий белок в месте спаренных основных аминокислот), таким образом, давая возможность для внутриклеточной активации фактора свертывания крови. В некоторых воплощениях последовательность АР удаляют полностью и заменяют на аминокислотную последовательность Arg-Lys-Arg. В соответствии с некоторыми воплощениями в качестве последовательности кДНК (комплементарная ДНК), кодирующей вариант белка FX, предложена последовательность нуклеиновой кислоты SEQ ID NO: 4, и в качестве кодируемой ею аминокислотной последовательности предложена аминокислотная последовательность SEQ ID NO: 1. Родственные способы предложены для получения варианта белка FXa путем выращивания клеток-хозяев, коэкспрессирующих РАСЕ (или сходную протеазу) и вариант белка FX, и затем очистки варианта белка FXa, продуцируемого, процессируемого и секретируемого клетками-хозяевами. Также раскрыты варианты белка FXa, продуцируемые в соответствии с этими способами. В некоторых воплощениях этих способов клетки-хозяева представляют собой клетки СНО (яичника китайского хомячка).
Очистка вариантов белка FXa может быть осуществлена путем хроматографии, включающей пропускание раствора, содержащего белки, через колонку для хроматографии в смешанном режиме (ММС) (например, с использованием сред Capto ММС), с последующим промыванием и элюированием, затем пропусканием белков через первую колонку для ионообменной хроматографии (например, с использованием сред Q-Sepharose Fast Flow), с последующим промыванием и элюированием, и затем пропусканием белков через вторую отличающуюся от первой колонку для ионообменной хроматографии (например, с использованием сред Fractogel SO3-). с последующим промыванием и элюированием. Также возможны другие способы очистки вариантов белка FXa.
Способы очистки могут возможно включать стадию вирусной инактивации, а также стадии ультрафильтрования и диафильтрования. После очистки вариантов белка FXa, и в некоторых случаях их концентрирования, они могут быть разведены в фармацевтически приемлемом разбавителе, возможно содержащем другие ингредиенты, такие как буферы или эксципиенты.
В описании также предложены способы лечения субъектов, нуждающихся в гемостазе, путем введения гемостатически эффективного количества композиции, содержащей один или более чем один вариант белка FXa в соответствии с описанием изобретения. В других воплощениях субъектам вводят гемостатически эффективное количество композиции, содержащей один или более чем один вариант белка FXa в соответствии с описанием изобретения, до возникновения кровотечения для профилактического предупреждения неконтролируемого кровотечения у восприимчивого субъекта. В некоторых воплощениях субъекта лечат от гемофилии А. В других воплощениях субъекта лечат от гемофилии В. И в других воплощениях субъекта лечат от травмы или других типов неконтролируемого кровотечения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1А представлена аминокислотная последовательность (SEQ ID NO: 1) зрелого варианта белка FX, в котором изолейциновый остаток дикого типа в положении 146 заменен на лейцин, показанный жирным шрифтом. Положение 146 соответствует положению 16 в системе нумерации химотрипсина. Потенциальные остатки γ-карбоксиглутаминовой кислоты (Gla) подчеркнуты и выделены курсивом. Предсказанные внутрицепочечные и межцепочечные дисульфидные связи проиллюстрированы при помощи линий, проведенных между связанными цистеинами. Предсказанный сайт β-гидроксилирования заключен в рамку из сплошных линий. Сайт распознавания и расщепления РАСЕ, сконструированный для замены активационного пептида, заключен в рамку из пунктирных линий. Лизин, составляющий С-концевую аминокислоту бета формы тяжелой цепи (Lys386) заключен в овал из сплошной линии. На Фиг. 1В представлена предсказанная аминокислотная последовательность (SEQ ID NO: 2) легкой цепи зрелого варианта белка FX. На Фиг. 1С представлена предсказанная аминокислотная последовательность (SEQ ID NO: 3) тяжелой цепи варианта белка FXa. На Фиг. 1D представлена нуклеотидная последовательность (SEQ ID NO: 4) кДНК, кодирующей вариант белка FX, включающий сигнальную последовательность и пропептид.
На Фиг. 2 представлен масс-спектр пептидов, полученных в результате восстановления, алкилирования и протеолитического расщепления при помощи Lys-C очищенного препарата интактного варианта белка FXa. Обозначение пиков, основанное на масс-спектре после анализа пептидов при помощи RP-HPLC (высокоэффективная жидкостная хроматография на обращенной фазе)/ESI-QТОF MS (времяпролетная квадрупольная масс-спектрометрия с ионизацией электрораспылением), использовали для подтверждения аминокислотной последовательности белка и для идентификации некоторых посттрансляционных модификаций, описанных в таблице 1 и в настоящем описании. Маркировка пиков является следующей. "L" с последующим номером относится к конкретному пептиду, происходящему из легкой цепи. "Н" с последующим номером относится к конкретному пептиду, происходящему из тяжелой цепи. "Н201": С-концевая альфа-форма H цепи, оканчивающаяся Lys399, содержащая два дисиалилированых О-гликана ядра-1. "Н202": С-концевая альфа-форма H цепи, оканчивающаяся Lys399, содержащая один дисиалилированый О-гликан ядра-1. "Н203": С-концевая альфа-форма H цепи, оканичивающаяся Leu398, содержащая один дисиалилированый О-гликан ядра-1. “L41Gla”: пептид L цепи #4, содержащий 1 остаток γ-карбоксиглутаминовой кислоты. "L10glc": пептид L цепи #10, содержащий потенциально О-связанную глюкозу (гексозу). "L1Gla": пептид L цепи #1 с 2 остатками γ-карбоксиглутаминовой кислоты. "L36-8Gla": пептид L цепи #3 с 6, 7 или 8 остатками γ-карбоксиглутаминовой кислоты. "L8b-OH": пептид L цепи #8, содержащий β-гидрокси Asp. "d": деамидирование. "R": пик реагента, т.е. продукты буфера, соответствующие реагентному контролю. "*": сверхалкилирование.
На Фиг. 3 представлена хроматограмма HPLC очищенного препарата интактного варианта белка FXa и продемонстрировано присутствие в препарате различных изоформ варианта белка FXa.
На Фиг. 4 представлена хроматограмма HPLC очищенного препарата интактного варианта белка FXa и продемонстрировано присутствие в препарате различных изоформ варианта белка FXa. Препарат также анализировали при помощи масс-спектрометрии.
На Фиг. 5 представлен масс-спектр интактного варианта белка FXa из очищенного препарата. Масс-спектр подтверждает присутствие двух основных изоформ альфа и бета, а также обширную гетерогенность по массам, присущую вариабельному расщеплению по сайту расщепления РАСЕ, и присутствие различных типов посттрансляционной модификации. Основные пики идентифицированы на фигуре по их обнаруженной массе в дальтонах. Обозначение пиков, основанное на масс-спектре после анализа RP-HPLC/ESI-QTOF MS, использовали для подтверждения аминокислотной последовательности белковых изоформ и для идентификации некоторых посттрансляционных модификаций, описанных в таблице 2 и описании изобретения. Пики, обозначенные "*", указывают на присутствие 11 остатков Gla в домене Gla легкой цепи.
На Фиг. 6 представлена хроматограмма HPLC очищенного препарата варианта белка FXa после восстановления и алкилирования для разделения легкой и тяжелой цепей путем устранения межцепочечных и внутрицепочечных дисульфидных связей. Хроматограмма демонстрирует присутствие различных изоформ варианта белка FXa. Препарат также анализировали при помощи масс-спектрометрии.
На Фиг. 7А представлен масс-спектр легкой цепи варианта белка FXa после восстановления и алкилирования очищенного препарата белка. Масс-спектр демонстрирует обширную гетерогеннность по массам, присущую вариабельному расщеплению по сайту расщепления РАСЕ, и присутствие различных типов посттрансляционных модификаций. Основные пики идентифицированы на фигуре по их обнаруженной массе в дальтонах. Обозначение пиков, основанное на масс-спектре после анализа RP-HPLC/ESI-QTOF MS, использовали для подтверждения аминокислотной последовательности белковых изоформ и для идентификации некоторых посттрансляционных модификаций, описанных в таблице 3 и описании изобретения.
На Фиг. 7В представлен масс-спектр тяжелой цепи варианта белка FXa после восстановления и алкилирования очищенного препарата белка. Масс-спектр демонстрирует обширную гетерогеннность по массам, присущую присутствию различных типов посттрансляционных модификаций. Основные пики идентифицированы на фигуре по их обнаруженной массе в дальтонах. Обозначение пиков, основанное на масс-спектре после анализа RP-HPLC/ESI-QTOF MS, использовали для подтверждения аминокислотной последовательности белковых изоформ и для идентификации некоторых посттрансляционных модификаций, описанных в таблице 4 и в описании изобретения. Пики, обозначенные "*", указывают на сверхалкилирование тяжелой цепи.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Здесь описаны композиции, содержащие вариант белка FXa, полученный с достаточной чистотой и в достаточном количестве, тестируемые в клинических исследованиях, и предложенные для субъектов, нуждающихся в гемостазе. Также описаны способы получения и очистки варианта белка FXa.
В некоторых воплощениях фраза "вариант белка FX", "вариант белка FXa" и похожие термины относятся (если из контекста не ясно иное) к человеческому фактору X (FX) или активированному фактору X (FXa), соответственно, в котором изолейцин (IIе или I) непосредственно за последовательностью активационного пептида в тяжелой цепи заменен на лейцин (Leu или L). На основе выравнивания последовательностей тяжелой цепи FXa и каталитического домена химотрипсина эту замену также называют мутацией путем замены I16L с использованием схемы нумерации химотрипсина. Эта позиция соответствует аминокислоте 146 в SEQ ID NO: 1 и аминокислоте 1 в SEQ ID NO: 3. Как обсуждалось выше, FXa, включающий эту мутацию, обладает зимогенными свойствами, которые дают ему возможность циркулировать в крови в течение более длительных периодов времени по сравнению с FXa дикого типа, а также способен расщеплять протромбин с высокой скоростью при включении в протромбиназный комплекс.
В соответствии с другими воплощениями вариант белка FX и вариант белка FXa относятся к FX или FXa, соответственно, в котором аминокислота, соответствующая положению 146 в SEQ ID NO: 1 (положение 16 в схеме нумерации химотрипсина) заменена на Phe (F), Asp (D) или Gly (G). В соответствии с другими воплощениями вариант белка FX и вариант белка FXa относятся к человеческому FX или FXa, соответственно, в котором валин (Val или V), соответствующий положению 147 в SEQ ID NO: 1 (положение 17 в схеме нумерации химотрипсина), заменен на Leu (L), Ala (А) или Gly (G). В других воплощениях вариант белка FX и вариант белка FXa относятся к человеческому FX или FXa, соответственно, в котором аминокислота, соответствующая положению 329 в SEQ ID NO: 1 (положение 194 в схеме нумерации химотрипсина), заменена на Asn (Ν) или Glu (Ε).
Фактор X дикого типа обычно циркулирует в крови в виде неактивных двух цепей зимогена, удерживаемых вместе при помощи дисульфидной связи. Двухцепочечный зимоген образуется из зрелого FX (т.е. лишенного сигнального пептида и пропептида) после протеолитического удаления пептида Arg-Lys-Arg, представленного в зрелом белке аминокислотами 140-142. Тем не менее, для активации требуется удаление активационного пептида (АР) фактором IXa или фактором VIIa до образования сгустка, или при помощи других протеаз, таких как RWX. АР соответствует аминокислотам 143-194 в зрелом одноцепочечном FX. В двухцепочечном зимогене он представлен по N-концу тяжелой цепи. М. Hertzberg, Biochemistry of factor X, Blood Rev. 8(1):56-62 (1994).
В примере воплощения последовательность варианта белка фактора X модифицирована путем удаления активационного пептида и его замены на аминокислотную последовательность Arg-Lys-Arg (RKR в однобуквенном коде) для создания сайта распознавания и расщепления для фермента, расщепляющего спаренные основные аминокислоты (РАСЕ; также называемого фурин). Вместо того, чтобы активировать FXa на отдельной стадии, замена АР дает возможность для внутриклеточной активации варианта белка FX, когда РАСЕ коэкспрессируется в клетках-хозяевах. Это позволяет избежать потребности в исходной очистке варианта FX, продуцируемого клетками, отдельной активации белка с использованием протеазы (такой как RWX) и затем очистки варианта FXa. Путем того, что избегают этих дополнительных стадий, вариант FXa может быть получен в более чистой форме и в больших количествах.
Аминокислотная последовательность зрелого варианта фактора X изображена на Фиг. 1А (SEQ ID NO: 1), в которой лейциновая замена I16L располагается по аминокислотному положению 146 (показана жирным шрифтом), последовательность RKR, заменяющая активационный пептид, располагается в положениях 143-145 и сайт процессинга РАСЕ располагается в положениях 140-145 (показан рамкой с пунктирной линией). На Фиг. 1В и Фиг. 1С изображена предсказанная аминокислотная последовательность легкой цепи (SEQ ID NO: 2) и тяжелой цепи (SEQ ID NO: 3) варианта FXa, соответственно, после расщепления по сайту РАСЕ.
Анализ множества препаратов варианта белка FXa, продуцируемых в соответствии со способами в соответствии с описанием изобретения, продемонстрировал неожиданную степень гетерогенности в отношении белковой последовательности и посттрансляционной модификации по сравнению с предсказанной структурой. Тем не менее, такие композиции, содержащие структурно гетерогенные популяции вариантов белка FXa способны действовать в качестве сильных прокоагулянтов в анализе in vitro. Как дополнительно описано ниже, гетерогенность присуща вариациям в аминокислотной последовательности легкой цепи и тяжелой цепи и посттрансляционным модификациям, представленным в одной, другой или обеих цепях.
Не желая быть связанными какой-либо конкретной теорией действия, полагают, что один источник структурной гетерогенности присущ вариабельному расщеплению по сайту распознавания и расщепления протеазой РАСЕ/фурин, сконструированному для замены активационного пептида. В не ограничивающем объем изобретения воплощении аминокислотная последовательность сайта РАСЕ представляет собой RKRRKR (в однобуквенном коде) (SEQ ID NO: 5), сконструированную в варианте белка FX путем замены нативной аминокислотной последовательности АР на остатки RKR, которые вместе с последними тремя остатками легкой цепи, образуют сайт протеолитического распознавания RKRRKR (SEQ ID NO: 5). Также могут быть использованы другие сайты распознавания РАСЕ (например, только RKR), а также сайты распознавания и расщепления протеазами, отличающимися от РАСЕ/фурина.
В некоторых воплощениях сайт РАСЕ протеолитически отщеплен для полного его удаления из легкой цепи, не оставляя остатки позади (ANS…TLER139/RKRRKR/) (SEQ ID NO: 6 и SEQ ID NO: 5, соответственно, легкая цепь и пептидный фрагмент). В предшествующем и следующих воплощениях, описанных в этом абзаце, косая черта ("I") указывает на возможные сайты расщепления между соседними аминокислотами, и надстрочный номер указывает на позицию С-концевой аминокислоты легкой цепи после расщепления (на основе Фиг. 1А; SEQ ID NO: 1). В других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя свой первый остаток по С-концу легкой цепи (ANS…TLERR140/KRRKR/) (SEQ ID NO: 7 и SEQ ID NO: 8, соответственно, легкая цепь и пептидный фрагмент). В других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя свои первые два остатка по С-концу легкой цепи (ANS…TLERRK141/RRKR/) (SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, легкая цепь и пептидный фрагмент). В других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя свои первые три остатка по С-концу легкой цепи (ANS…TLERRKR142/RKR/) (SEQ ID NO: 11). В других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя свои первые четыре остатка по С-концу легкой цепи (ANS…TLERRKRR143/KR/) (SEQ ID NO: 12). В других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя свои первые пять остатков по С-концу легкой цепи (ANS…TLERRKRRK144/R/) (SEQ ID NO: 13). И в других воплощениях сайт РАСЕ протеолитически расщеплен, оставляя всю свою последовательность по С-концу легкой цепи (ANS…TLERRKRRKR145/) (SEQ ID NO: 2).
В других воплощениях структурная гетерогенность в легкой цепи может также включать присутствие варьирующего количества остатков Gla в домене Gla, присущего вариациям степени γ-карбоксилирования некоторых остатков глутаминовой кислоты. В некоторых воплощениях количество остатков Gla составляет 9, 10 или 11. Потенциальные сайты γ-карбоксилирования глутаминовой кислоты в домене Gla идентифицированы на Фиг. 1А. В соответствии с другими воплощениями дополнительные потенциальные источники структурной гетерогенности легкой цепи включают присутствие или отсутствие β-гидроксилированного остатка аспарагиновой кислоты по Asp63 (Фиг. 1А; SEQ ID NO: 1) и присутствие или отсутствие О-связанной гексозы. В некоторых воплощениях О-связанная гексоза представляет собой глюкозу, но в других воплощениях она может представлять собой другую альдогексозу, или циклический полуацеталь, кетогексозу или другую О-связанную гексозу.
Тяжелая цепь также может демонстрировать структурную гетерогенность. В соответствии с некоторыми воплощениями вариация может возникать по длине тяжелой цепи. Например некоторые тяжелые цепи представляют собой более длинную альфа изоформу, оканчивающуюся по К399 (SEQ ID NO: 3) (надстрочный индекс указывает на положение С-концевой аминокислоты тяжелой цепи на основе Фиг. 1А; SEQ ID NO: 1), а в других случаях концевой лизин отсутствует, таким образом, что белок оканчивается по L398 (SEQ ID NO: 14). В других воплощениях тяжелая цепь представляет собой более короткую бета форму, оканчивающуюся по К386 или К384 (SEQ ID NO: 46 и SEQ ID NO: 45, соответственно).
Еще один источник гетерогенности в тяжелых цепях представляет собой степень О-связанного гликозилирования. Таким образом, в некоторых воплощениях тяжелые цепи не гликозилированы. В других воплощениях тяжелые цепи включают один или два О-гликана ядра-1. О-гликан ядра-1 представляет собой О-гликан, где N-ацетилгалактозамин (GalNAc) присоединен по серину или треонину, и галактоза (Gal) присоединена по GalNAc. В некоторых воплощениях изобретения представлен сахар GalNAc, но отсутствует сахар Gal. В соответствии с другими воплощениями каждый из двух О-связанных гликанов ядра-1 по отдельности может быть не сиалилирован, моносиалилирован или дисиалилирован, таким образом, что тяжелые цепи могут нести в общей сложности 0-4 группы сиаловой кислоты. Когда группы сиаловой кислоты присутствуют, тогда они могут быть присоединены по GalNAc или по Gal, или по обоим сахарам.
В соответствии с другими воплощениями возможны различные варианты образования пар легких цепей и тяжелых цепей, таким образом, давая возможность для получения большого комбинаторного набора двухцепочечных зимогенов с различными длинами цепи и посттрансляционными модификациями.
В других воплощениях различные виды вариантов белка FXa могут варьировать по своей относительной частоте. В одном из воплощений частота различных видов может быть определена путем сравнения высот пиков в масс-спектрах интактных вариантов белка FXa. Альтернативно, частота легкой и тяжелой цепей могут быть по отдельности проанализированы при помощи масс-спектрометрии после восстановления интактного белка и его алкилирования для устранения межцепочечных и внутрицепочечных дисульфидных связей. Как описано в приведенных ниже примерах, частота различных видов вариантов FXa может быть оценена как основная, минорная или следовая путем сравнения с самым высоким пиком в масс-спектрах.
Тем не менее, сравнение высоты пиков масс-спектров представляет собой не единственный способ измерения частоты. Другие способы измерения абсолютной или относительной частоты структурных видов находятся в пределах знаний специалистов в данной области техники. В не ограничивающем объем изобретения примере другого способа классификации относительной частоты виды рассматривают как имеющие высокую частоту в том случае, если высота их пиков в масс-спектре составляет по меньшей мере приблизительно 50% от наиболее частых видов (т.е. самый высокий пик в масс-спектре). Вид имеет среднюю частоту в том случае, если его пик составляет приблизительно 10-50% от наиболее частых видов. Виды обладают низкой частотой в том случае, если их пик составляет приблизительно 2-10% от наиболее частых видов, и обладают очень низкой частотой в том случае, если их пик составляет меньше чем приблизительно 2% от наиболее частых видов. Также возможны другие соглашения для сравнения относительной частоты видов.
Специфические не ограничивающие объем изобретения воплощения структурной гетерогенности в легких цепях, тяжелых цепях варианта белка FXa, и их комбинаций описаны ниже.
Воплощения изобретения включают варианты белка FXa, в которых С-концы легкой цепи оканчиваются по различным аминокислотам вследствие вариабельного расщепления по сайту РАСЕ. Родственные воплощения включают присутствие варьирующего числа остатков Gla и посттрансляционные модификации, такие как β-гидроксилирование остатка аспарагиновой кислоты и О-связанную гексозу. Некоторые из этих воплощений описаны ниже.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 139 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми из других вариантов белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 140 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 141 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 142 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 143 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
Воплощения в соответствии с описанием изобретения включают варианты белка FXa, в котором С-конец тяжелой цепи оканчивается по различным аминокислотам. Родственные воплощения включают варьирующие количества О-связанных гликанов, включающие варьирующие степени сиалилирования. Некоторые из этих воплощений описаны ниже.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. Композиции в соответствии с описанием изобретения могут дополнительно содержать любой из других вариантов белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. Композиции в соответствии с описанием изобретения могут дополнительно содержать любой из других вариантов белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
Воплощения в соответствии с описанием изобретения включают варианты белка FXa, в которых С-конец легкой цепи оканчивается по различным аминокислотам вследствие вариабельного расщепления по сайту РАСЕ и в которых С-конец тяжелой цепи также оканчивается по различным аминокислотам. Родственные воплощения включают варьирующие количества остатков Gla и посттрансляционные модификации, такие как β-гидроксилирование остатка аспарагиновой кислоты и О-связанную гексозу в легких цепях, и варьирующие количества О-связанных гликанов, включающие варьирующие степени сиалилирования в тяжелых цепях. Некоторые из этих воплощений описаны ниже.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 139 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 139 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 139 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 139 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра 1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 140 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 140 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 140 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 140 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 141 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 141 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 141 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 141 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 142 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 142 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 142 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 142 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 143 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 384 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 143 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 386 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 143 аминокислотной последовательности SEQ ID NO: 1 и тяжелая цепь состоит из аминокислот с 146 по 398 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В некоторых воплощениях композиции в соответствии с описанием изобретения содержат вариант белка FXa, в котором легкая цепь состоит из аминокислот с 1 по 143 аминокислотной последовательности SEQ ID NO: 1 и1 тяжелая цепь состоит из аминокислот с 146 по 399 аминокислотной последовательности SEQ ID NO: 1. В родственных воплощениях легкая цепь дополнительно содержит 9 остатков Gla, 10 остатков Gla или 11 остатков Gla. В каждом из этих воплощений легкая цепь может дополнительно содержать β-гидрокси Asp63, О-связанную гексозу или обе эти посттрансляционные модификации. В других родственных воплощениях тяжелая цепь дополнительно содержит один гликан ядра-1, который может быть не сиалилирован, моносиалилирован или дисиалилирован, или два гликана ядра-1, каждый из которых может быть независимо не сиалилирован, моносиалилирован или дисиалилирован. В других родственных воплощениях каждая возможная пермутация посттрансляционных модификаций легкой цепи комбинирована с каждой возможной пермутацией посттрансляционных модификаций тяжелой цепи. Композиции в соответствии с описанием изобретения могут содержать любые варианты белка FXa, описанные в этом абзаце, сами по себе или в комбинации с другими описанными, а также с любыми другими вариантами белка FXa в соответствии с описанием изобретения.
В описании настоящего изобретения дополнительно предложены выделенные нуклеиновые кислоты, содержащие последовательности нуклеиновых кислот, которые кодируют вариант белка FX. В соответствии с примером не ограничивающего объем изобретения воплощения последовательность комплементарной ДНК (кДНК), кодирующая вариант белка FX, здесь раскрыта как SEQ ID NO: 4. Как будет понятно специалистам в данной области техники, множество других последовательностей нуклеиновых кислот, кодирующих вариант белка FX, возможно в свете вырожденности генетического кода.
В некоторых воплощениях может быть предложена нуклеиновая кислота, кодирующая вариант белка FX, имеющая последовательность, кодирующую сигнальный пептид и/или пропептид (т.е. лидерные последовательности), располагающийся по N-концу белка FX, который, среди других потенциальных функций, направляет вновь синтезированный белок в секреторный компартмент. Посттрансляционный процессинг затем удаляет лидерную последовательность до секреции зрелого белка из клетки. В некоторых воплощениях эти последовательности происходят из нативного человеческого FX. Могут быть использованы ненативные лидерные последовательности, такие как последовательности из человеческого протромбина. Также могут быть использованы лидерные последовательности из других белков (включая белки не человеческого происхождения). В воплощении последовательности нуклеиновой кислоты в соответствии с SEQ ID NO: 4 N-концевая лидерная последовательность происходит из человеческого протромбина.
В соответствии с другими воплощениями нуклеиновая кислота, кодирующая вариант белка FX, может быть модифицирована таким образом, чтобы дать возможность для внутриклеточной активации фактора свертывания крови с получением вариантов белка FXa.
В соответствии с указанным выше, FX обычно циркулирует в виде двухцепочечного зимогена, в котором активационный пептид из 52 аминокислот расположен по N-концу тяжелой цепи FX. Для активации с образованием FXa, способного функционировать в качестве прокоагулянта в протромбиназном комплексе, требуется протеолитическое удаление АР протеазами из внутренней или внешней системы свертывания крови, такой как активированный FIX или активированный FVII. В предшествующих исследованиях использовали аналогичные способы активации. В частности, вариант белка FX экспрессировали в клетках и затем активировали на отдельной стадии путем обработки ядом гадюки Расселла (RWX), после чего вариант белка FXa очищали для последующего анализа.
Хотя активированный вариант белка FX в принципе может быть продуцирован в промышленном масштабе с использованием похожих способов, это может быть весьма неэффективным и дорогостоящим. Например может потребоваться тщательная очистка для устранения какого-либо остаточного RWX. Для того, чтобы избежать этих недостатков, нативный АР может быть удален и заменен на сайты распознавания и расщепления протеаз, способных экспрессироваться и функционировать внутриклеточно. Таким образом, возможно, чтобы вариант белка FX активировался внутриклеточно без необходимости в дополнительных стадиях процессинга. Активируемые варианты белка FX, секретируемые клетками-хозяевами в питательную среду, затем могут быть очищены.
Не ограничивающий объем изобретения пример последовательности нуклеиновой кислоты, кодирующей вариант FX, в котором последовательность АР заменена на сайт распознавания и расщепления РАСЕ, приведен в SEQ ID NO: 4. Белковая последовательность зрелого варианта белка FX, кодируемого этим примером последовательности кДНК, приведена в SEQ ID NO: 1. В этих воплощениях аминокислоты, составляющие АР (аминокислоты с 143 по 194 последовательности белка FX дикого типа), заменены на последовательность Arg-Lys-Arg (RKR в однобуквенном коде). Эта последовательность, комбинированная с последними тремя остатками последовательности легкой цепи (также RKR; аминокислоты 140-142 в SEQ ID NO: 1), формирует сайт распознавания и расщепления (т.е. RKRRKR) (SEQ ID NO: 5) для фермента, расщепляющего спаренные основные аминокислоты (РАСЕ). Таким образом, растворимый фермент РАСЕ, коэкспрессируемый в тех же самых клетках-хозяевах, экспрессирующих модифицированный вариант белка FX, может внутриклеточно активировать фактор свертывания крови.
В некоторых воплощениях, когда клетки-хозяева исходно экпрессируют фермент с сайтом распознавания, сходным с таковым у РАСЕ, может отсутствовать необходимость в коэкспрессии экзогенной протеазы. Наоборот, нативная протеаза может быть достаточна для активации варианта белка FX, экспрессируемого в клетках, таким образом, продуцируя и секретируя активированный вариант белка FX.
В других воплощениях сайты распознавания и расщепления для протеаз, отличающихся от РАСЕ, но также способных функционировать внутриклеточно, могут быть использованы для замены аминокислотной последовательности АР. Такие ферменты могут быть коэкспрессироваться в одних и тех же клетках-хозяевах вместе с вариантом белка FX для активации модифицированного фактора свертывания крови. Если нативные ферменты, способные расщеплять сконструированный сайт, продуцируются на достаточных уровнях в клетках-хозяевах, тогда коэкспрессия экзогенных протеаз может не потребоваться.
Последовательности нуклеиновых кислот, кодирующие варианты белка FX в соответствии с описанием настоящего изобретения могут быть включены в векторы с использованием способов, хорошо известных специалистам в данной области техники.
В некоторых воплощениях векторы включают плазмиды, как правило бактериальные плазмиды, эукариотические эписомы, дрожжевые искусственные хромосомы и вирусные геномы. Не ограничивающие объем изобретения примеры вирусов включают ретровирусы, аденовирусы, адено-ассоциированные вирусы (AAV) и вирусы растений, такие как вирус мозаики цветной капусты и вирус табачной мозаики. Возможны другие типы векторов. В некоторых воплощениях векторы способны к автономной репликации в подходящих хозяевах. В других воплощениях векторы поддерживаются в хозяевах экстрахромосомно или могут интегрироваться в геном хозяина, давая возможность для репликации вектора с геномом хозяина. Векторы, содержащие ген и контрольные последовательности, достаточные для поддержания транскрипции и трансляции гена, названы экспрессирующимися векторами. Векторы в соответствии с описанием настоящего изобретения могут быть выбраны или сконструированы в соответствии со знаниями специалиста в данной области техники таким образом, чтобы функционировать в любом типе клеток, способном поддерживать экспрессию варианта белка FX, включающем бактериальные клетки, другие прокариотические клетки, дрожжевые клетки, другие клетки грибов, клетки растений, клетки животных, клетки насекомых, клетки млекопитающих, клетки СНО и человеческие клетки или другие.
Векторы могут возможно содержать одну или более чем одну контрольную последовательность. Некоторые контрольные последовательности дают возможность для репликации, такие как ориджины репликации. Другие контрольные последовательности контролируют или модулируют транскрипцию, такие как промоторы, энхансеры и сайты терминации транскрипции. Не ограничивающие объем изобретения примеры промотора или энхансеров представляют собой промоторы и энхансеры, происходящие из ретровирусных LTR (длинных концевых повторов), цитомегалловируса (CMV), вируса обезьян 40 (SV40), аденовируса (например основанного на аденовирусе позднего промотора (AdMLP)) или вируса полиомы. Дополнительные примеры включают тканеспецифические промоторы и энхансеры, конститутивно действующие промоторы и энхансеры и индуцируемые промоторы и энхансеры. Также возможны другие промоторы и энхансеры. Другие контрольные последовательности контролируют или модулируют просттранскрипционный процесссинг РНК, такие как сигналы сплайсинга и полиаденилирования, или сигналы, которые увеличивают или уменьшают стабильность мРНК. Другие контрольные последовательности контролируют или модулируют трансляцию белка, такие как последовательности инициации трансляции (например консенсусная последовательность Козака), посттрансляционный процессинг или стабильность белка.
Векторы могут также включать селективные маркерные гены, дающие возможность для отбора клеток-хозяев, в которые встроены векторы. Не ограничивающие объем изобретения примеры включают селективные маркерные гены, которые придают резистентный к лекарственному средству фенотип, такие как ген дигидрофолатредуктазы (DHFR) (для применения в DHFR- клетках-хозяевах, дающий возможность для отбора с использованием метотрексата), ген neo (дающий возможность для отбора при помощи G418 или похожих лекарственных средств), ген hph (дающий возможность для отбора при помощи гигромицина В) и ген глутаматсинтетазы (дающий возможность для отбора при помощи метионинсульфоксимина).
Векторы, содержащие последовательности нуклеиновых кислот, кодирующих варианты белка FX в соответствии с описанием изобретения, могут быть введены в один или более чем один тип клеток-хозяев, способный поддерживать экспрессию варианта белка FX. Способы введения векторов в подходящие клетки-хозяева хорошо известны специалистам в данной области техники. Не ограничивающие объем изобретения примеры включают временную и стабильную трансфекцию, трансформацию, трансдукцию и инфицирование целевых клеток-хозяев вирусами. Другие примеры включают опосредованную декстраном трансфекцию, осаждение фосфатом кальция, опосредованную полибреном трансфекцию, слияние протопластов, электропорацию, инкапсулирование полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра. Способы трансформации клеток растений, бактериальных и дрожжевых клеток также хорошо известны в области техники.
FX или варианты белка FXa могут быть экспрессированы с другими белками для оптимизации экспрессии или посттрансляционной обработки FX или вариантов белка FXa. В не ограничивающем объем изобретения примере вектор, включающий ген, кодирующий γ-глутамилкарбоксилазу, может быть трансфицирован одновременно с вектором, кодирующим вариант белка FX, в клетки-хозяева для увеличения карбоксилирования глутаминовых кислот в домене Gla легкой цепи. Благодаря этому степень карбоксилирования и образования остатков Gla в домене Gla может быть увеличена, таким образом улучшая выход активного фактора свертывания крови, продуцируемого клетками. В еще одном описанном выше воплощении вариант белка FX, модифицированный таким образом, что нативная аминокислотная последовательность АР заменена на сайт расщепления РАСЕ, может быть экспрессирован вместе с растворимым ферментом РАСЕ в тех же самых клетках-хозяевах. Коэкспрессируемый РАСЕ может затем внутриклеточно расщеплять вариант белка FX с образованием активированного варианта белка FX, который затем секретируется из клетки. Как указано выше, этот подход позволяет избежать необходимости активировать вариант белка FX на отдельной стадии, что, в противном случае, может быть весьма неэффективным и дорогостоящим.
Клетки, способные экспрессировать варианты белка FX и другие белки, полезные для оптимизации экспрессии варианта белка FX в активной форме, включают клетки млекопитающих. Линии клеток млекопитающих, подходящие в качестве хозяев для экспрессии белка, известные в области техники, включают линии клеток, происходящие из людей, крыс, мышей и других млекопитающих. Не ограничивающие объем изобретения примеры включают некоторые иммортализованные клеточные линии, доступные из Американской коллекции типовых культур (АТСС) или других источников, включающие клетки яичников китайского хомячка (СНО), клетки NS0, клетки SP2, клетки НЕК-293Т, клетки NIH-3T3, клетки HeLa, клетки почки новорожденного хомяка (ВНК), клетки почки африканской зеленой мартышки (например, клетки COS, CV-1 или Vero), клетки человеческой гепатоклеточной карциномы (например, HepG2), клетки А549, клетки А431, клетки HeLa, клетки L, клетки ВНК21, клетки HL-60, клетки U937, клетки НаК, клетки Юркат и другие. Применение других клеток млекопитающих в качестве клеток-хозяев для экспрессии белка возможно в соответствии со знаниями специалистов в данной области техники.
В других воплощениях могут быть использованы клеточные линии из насекомых, растений, бактерий или грибов. Не ограничивающие объем изобретения примеры клеток насекомых включают клетки Sf9 или Sf21, которые часто используют в сочетании с системой экспрессии на основе бакуловирусного вектора. Не ограничивающие объем изобретения примеры клеток растений включают клетки видов табака, арабидопсиса, ряски, кукурузы, пшеницы и картофеля. Не ограничивающие объем изобретения примеры бактерий включают Escherichia coli, Bacillus subtilis, Salmonella typhimurium и штаммы Streptomyces. He ограничивающие объем изобретения примеры грибов включают Schizosaccharomyces pombe, Saccharomyces cerevisiae, Pichia pastoris, дрожжевые штаммы Kluyveromyces и дрожжевые штаммы Candida. Применение клеток других насекомых, растений, бактерий и грибов в качестве клеток-хозяев для белковой экспрессии возможно в соответствии со знаниями специалиста в данной области техники.
Способы, реагенты и условия для выращивания клеток-хозяев для продукции активированного варианта белка FX находятся в пределах знаний специалистов в данной области техники, и не предполагаются в качестве ограничивающих объем изобретения. В не ограничивающем объем изобретения примере варианты белка FXa могут быть продуцированы в 500 литрах культур бессывороточной среды, дополненной витамином К. В еще одном не ограничивающем объем изобретения примере варианты белка FXa в соответствии с описанием изобретения могут быть продуцированы в промышленном масштабе, например в ферментационных чанах объемом 2500 литров, или даже более крупных объемов, в условиях, разработанных для оптимизации экспрессии и выделения.
После выращивания клеток-хозяев в условиях выращивания, поддерживающих экспрессию и внутриклеточную активацию вариантов белка FX среда для выращивания клеток может быть обработана для выделения и очистки вариантов белка FXa, секретируемых во время клеточного роста. В других воплощениях, в частности, когда используют клетки-хозяева, отличающиеся от млекопитающих, вариант белка FXa, оставшийся в клетках, может быть высвобожден, например путем механического, ферментативного разрушения клеток, или разрушения детергентами.
Культуральная среда с клетками может быть центрифугирована и/или профильтрована для удаления клеток и дебриса, например путем глубинного фильтрования самого по себе или с последующим мембранным фильтрованием, например через мембранный фильтр со средним размером пор 0,45 мкм, 0,2 мкм или другим размером пор. После удаления клеточных остатков путем центрифугирования и/или фильтрования среда может быть обработана для дополнительной очистки вариантов белка FXa. В зависимости от последующих стадий очистки, может быть желательно разбавление и/или замена буфера в осветленном супернатанте или фильтрате.
Способы очистки белка хорошо известны из уровня техники. Примеры не ограничивающих объем изобретения способов очистки белка включают осаждение солью, гель-фильтрационную хроматографию, ионообменную хроматографию и аффинную хроматографию. Например, антитела, специфическим образом распознающие вариант белка FXa, могут быть иммобилизированы на колонках для очистки для обратимого захвата белка из окружающей среды или буфера. После промывания аффинной колонки для удаления примесей связанный вариант белка FXa может быть затем элюирован.
Частично очищенный вариант белка FXa может быть подвергнут стадиям обработки в соответствии с надлежащей производственной практикой или другими регулирующими требованиями, включающими, например удаление или инактивацию вирусов. Вирусы могут быть удалены путем ультрафильтрования или инактивированы путем обработки образца спиртом и/или детергентом в соответствии со знаниями специалиста в данной области техники. Например могут быть использованы вирусные фильтры марки Virosart®, доступные от Sartorius Stedim Biotech, а также фильтры других производителей. Возможны другие способы удаления или инактивации вирусов.
В некоторых воплощениях хроматография в смешанном режиме (ММС) может быть использована для очистки варианта белка FXa самого по себе или в комбинации с другими стадиями очистки. ММС, также названная мультимодальной хроматографией, относится к применению хроматографических сред, которые обеспечивают более чем один тип взаимодействия между лигандом и компонентами образца. Примеры типов взаимодействий включают электростатическое (анионо- или катионообменник), гидрофобное, взаимодействие пи-пи, образование водородных связей и тиофильное взаимодействие. Эти взаимодействия могут работать кооперативно или работать независимо. Специфические не ограничивающие объем изобретения примеры сред ММС включают Capto adhere, Capto adhere ImpRes, Capto ММС, Capto ММС ImpRes, Capto Core 700, все из которых дополнительно описаны в Multimodal Chromatography Handbook, опубликованной GE Healthcare Life Sciences (2013). Capto ММС, например, обеспечивает тиофильное, гидрофобное связывание, образование водородных связей, а также электростатические свойства катионообменника. Также возможно применение сред ММС из других источников.
В некоторых воплощениях ионообменная хроматография может быть использована для очистки варианта белка FXa самого по себе или в комбинации с другими стадиями очистки. Ионообменная хроматография относится к применению хроматографических сред, обеспечивающих разделение молекул на основе различий в их общем поверхностном заряде, который может варьировать в зависимости от рН, соли и других условий. При величине рН, превосходящей изоэлектрическую точку белка, последний будет связываться с положительно заряженной средой (анионообменник), тогда как при рН ниже изоэлектрической точки белка, последний будет связываться с отрицательно заряженной средой (катионообменник). Ионообменные среды и буферные условия (например, концентрация соли, рН и т.п.) могут быть выбраны таким образом, чтобы со средами предпочтительно связывался очищаемый белок (например, вариант белка FXa), давая возможность для отмывания от примесей, которые в тех же самых условиях связываются менее прочно, после чего интересующий белок может быть элюирован с хроматографической колонки. Дополнительная информация об ионообменной хроматографии описана в Ion Exchange Chromatography & Chromatofocusing Principles and Methods Handbook, опубликованной GE Healthcare (2010).
Ионообменники могут быть классифицированы на сильные или слабые, что указывает на степень, с которой они остаются заряженными при изменениях рН. Сильные обменники остаются заряженными в широком диапазоне рН, тогда как слабые обменники не остаются. Примеры функциональных групп анионообменника включают четвертичный аммоний (Q), который является сильным, и диэтиламиноэтил (DEAE) и диэтиламинопропил (ANX), которые рассматривают как слабые. Примеры функциональных групп катионообменника включают сульфопропил (SP) и метилсульфонат (S), оба из которых рассматривают как сильные, и карбоксиметил (СМ), который является слабым.
В одном из воплощений вариант белка FXa может быть очищен с использованием только стадии анионообменной хроматографии или с последующей стадией катионообменной хроматографии. В еще одном воплощении вариант белка FXa может быть очищен с использованием только стадии катионообменной хроматографии или с последующей стадией анионообменной хроматографии.
Специфические не ограничивающие объем изобретения примеры ионообменных сред включают Capto DEAE, Capto Q ImpRes, Capto SP ImpRes, Capto S, Capto Q, SOURCE 15Q, SOURCE 15S, SOURCE 30Q, SOURCE 30S, MacroCap SP, MacroCap Q, Mini Q PC, Mini S PC, Mini Q PE, Mini S PE, HR Mono Q, HR Mono S, PC Mono Q, PC Mono S, Mono Q GL, Mono S GL, DEAE Sephacel, CM Sephadex C-25, CM Sephadex C-50, DEAE Sephadex A-25, DEAE Sephadex A-50, QAE Sephadex A-25, QAE Sephadex A-50, SP Sephadex C-25, SP Sephadex C-50, ANX Sepharose 4 Fast Flow (High Sub), ANX Sepharose 4 Fast Flow (Low Sub), CM Sepharose Fast Flow, CM Sepharose High Performance, DEAE Sepharose CL-6B, DEAE Sepharose Fast Flow, Q Sepharose Big Beads, Q Sepharose Fast Flow, Q Sepharose High Performance, Q Sepharose XL, SP Sepharose Big Beads, SP Sepharose Big Beads Food Grade, SP Sepharose High Performance, SP Sepharose Fast Flow и SP Sepharose XL, имеющиеся в продаже от GE Healthcare. Другие ионообменные среды могут быть получены из Merck Millipore, включающие, например среды серий Fractogel® или серий Eshmuno®. Примеры в сериях Fractogel® включают смолу ТМАЕ, смолу ТМАЕ Hicap, смолу ТМАЕ Medcap, смолу DEAE, смолу DMAE, смолу SO3-, смолу SE Hicap и смолу СОО-. Примеры в сериях Eshmuno® включают смолы СРХ, S, Q и НСХ. Также могут быть использованы ионообменные среды из других источников.
После достаточной очистки варианта белка FXa очищенный белок может быть концентрирован с использованием ультрафильтрования и диафильтрования при приготовлении для упаковки или дополнительной обработки.
В некоторых воплощениях очищенные и концентрированные варианты белка FXa могут быть разведены в водных растворах, содержащих буферы и другие ингредиенты, подходящие для введения субъектам, и затем упакованы для длительного хранения до доставки. Могут быть использованы дополнительные стадии обработки. Например композиции, содержащие варианты белка FXa, эксципиенты, буферы и другие фармацевтически приемлемые ингредиенты, могут быть лиофилизированы для длительного хранения.
Варианты белка FXa в соответствии с описанием изобретения, продуцируемые в соответствии со способами, описанными в нижеприведенных примерах, или при помощи других способов, могут быть проанализированы для определения присутствия и степени структурной гетерогенности с использованием способов, знакомых специалистам в данной области техники.
Например, в некоторых воплощениях интактный белок или приготовленные из него протеолитические фрагменты могут быть проанализированы при помощи непосредственного белкового секвенирования, например с использованием деградации по Эдману. Белок или пептиды также могут быть проанализированы при помощи жидкостной хроматографии (LC), включающей обращенно-фазовую LC (RP-LC), а также масс-спектрометрии, включающей квадрупольную времяпролетную масс-спектрометрию с ионизацией электрораспылением (ESI-QTOF MS) для обнаружения вариации по аминокислотной последовательности и присутствия конкретных посттрансляционных модификаций. Дополнительно, присутствие заряженных групп, например γ-карбоксилированных остатков глутаминовой кислоты (Gla), может быть обнаружено с использованием анионообменной жидкостной хроматографии (AEX-HPLC).
Когда предполагают присутствие углеводных группировок, тогда их существование может быть подтверждено путем их удаления специфическими гликозидазами. После этого обработанный белок, который должен быть лишен углевода, удаляемого гликозидазой, может быть повторно проанализирован, например при помощи масс-спектрометрии, для подтверждения идентичности предполагаемого и присутствующего углевода.
Другие аналитические методики, с которыми знакомы специалисты в данной области техники, могут быть использованы для выявления степени и идентичности структурной гетерогенности в вариантах белка FXa.
Композиции, содержащие варианты белка FXa в соответствии с описанием изобретения, могут быть протестированы в отношении прокоагулянтной активности. В не ограничивающем объем изобретения примере коагулирующую активность измеряют при помощи одностадийного анализа коагулирующей активности крови с использованием в качестве субстрата плазмы крови людей с дефицитом фактора VIII. Один из таких анализов известен как анализ активированного частичного тромбопластинового времени (АРТТ), но другие анализы могут быть использованы в соответствии со знанием специалиста в данной области техники. Анализ АРТТ более подробно описан в Kamal, A.F., et al., How to interpret and pursue an abnormal prothrombin time, activated partial thromboplastin time, and bleeding time in adults, Mayo Clin Proc. 82(7):864-873 (2007).
В некоторых воплощениях композиции в соответствии с описанием изобретения включают "терапевтически эффективное количество" или "профилактически эффективное количество" вариантов белка FXa. "Терапевтически эффективное количество" относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого терапевтического результата, например гемостаза неконтролируемого кровотечения. Терапевтически эффективное количество варианта FXa может варьировать в зависимости от факторов, таких как болезненное состояние, возраст, пол и масса индивида, и способность варианта FXa вызывать желаемый ответ у индивида. Терапевтически эффективное количество также представляет собой количество, при котором любые токсические или вредные действия варианта FXa перевешиваются терапевтически благоприятными действиями. "Профилактически эффективное количество" относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого профилактического результата, например предупреждения неконтролируемого кровотечения у восприимчивого субъекта. Например, доза может быть введена до планируемого хирургического вмешательства. Примеры восприимчивых субъектов включают субъектов, страдающих от гемофилии А или В, включая субъектов, у которых продуцируются ингибирующие антитела против продуктов, заменяющих фактор.
Композиции в соответствии с описанием изобретения могут быть введены субъекту, включающему пациента-человека, нуждающемуся в0 лечении или предупреждении любого состояния, отличающегося недостаточным свертыванием крови или избыточным кровотечением. Не ограничивающие объем изобретения примеры таких состояний включают гемофилию А, гемофилию В, гемофилию А или В, ассоциирующуюся с ингибирующими антителами, дефицит фактора свертывания крови, дефицит эпоксидредуктазы витамина К С1, дефицит гамма-карбоксилазы, кровотечение, ассоциированное с травмой, повреждения, тромбоз, тромбоцитопению, инсульт, коагулопатию, диссеминированное внутрисосудистое свертывание (DIC), расстройства, связанные с избыточной антикоагуляционной терапией, синдром Бернара-Сулье, тромбастению Гланцмана и дефицит пула тромбоцитов. Также возможно лечение или предупреждение других расстройств, отличающихся недостаточным свертыванием крови или избыточным кровотечением.
Схемы введения доз могут быть скорректированы для обеспечения оптимальной желаемой реакции (например, терапевтической или профилактической реакции). Например может быть введен единичный болюс, несколько дробных доз могут быть введены с течением времени, или доза может быть пропорционально уменьшена или увеличена в соответствии с необходимостью терапевтической ситуации. В соответствии с некоторыми воплощениями композиции для парентерального введения готовят в стандартной лекарственной форме для легкого введения и однородности дозы. Стандартная лекарственная форма относится к физически дискретным единицам, служащим в качестве однократных доз для субъектов, которых лечат, где каждая единица содержит предопределенное количество активного соединения, рассчитанное таким образом, чтобы обеспечивать желаемое терапевтическое действие в ассоциации с подходящим фармацевтическим носителем.
В некоторых воплощениях терапевтически или профилактически эффективное количество вариантов белка FXa составляет от приблизительно 0,0001 до 50 мг/кг, от приблизительно 0,001 до 50 мг/кг, от приблизительно 0,001 до 5 мг/кг, от приблизительно 0,001 до 0,5 мг/кг, от приблизительно 0,001 до 0,05 мг/кг, от приблизительно 0,01 до 5 мг/кг или от приблизительно 0,01 до 0,5 мг/кг. В соответствии с родственными воплощениями терапевтически или профилактически эффективная концентрация в сыворотке крови вариантов белка FXa составляет от приблизительно 0,0003 до 300 нМ, от приблизительно 0,003 до 300 нМ, от приблизительно 0,03 до 300 нМ, от приблизительно 0,003 до 30 нМ, от приблизительно 0,03 до 30 нМ или от приблизительно 0,3 до 3 нМ. Концентрация в сыворотке крови варианта FXa может быть измерена при помощи любого способа, известного из уровня техники.
Стандартные дозы композиций в соответствии с описанием изобретения могут быть приготовлены для того, чтобы обеспечить удобное введение субъектам терапевтически или профилактически эффективных количеств вариантов белка FXa или для достижения желаемой концентрации в сыворотке крови таких белков у таких субъектов.
Для любого конкретного субъекта специфические схемы введения доз или диапазоны могут быть скорректированы по времени в соответствии с потребностями субъекта, которого лечат, и профессиональным суждением индивида, ответственного за введение, или контролирующего введение композиций. Соответственно, приведенные здесь схемы введения доз и диапазоны приведены только для примера, и не предполагается, что они ограничивают объем или практическое применение композиций в соответствии с описанием изобретения.
Дополнительно предложены наборы, включающие, например одну или более чем одну стандартную дозу композиции в соответствии с описанием изобретения. Такие стандартные дозы могут быть представлены в жидкой форме или в виде лиофилизата. Если композиция упакована в лиофлизированной форме, тогда наборы могут дополнительно включать контейнер, заполненный разбавителем, таким как стерильная вода или физиологический раствор, для ресуспендирования высушенного осадка. Наборы также могут включать изделия, используемые для введения, включающие, например, шприцы для подкожных инъекций, или трубки игольные или иглы-бабочки и т.п. Наборы могут дополнительно включать указания по применению, спиртовые салфетки для стерилизации кожи или другие компоненты.
Композиции, содержащие вариант белка FXa, могут быть введены однократно или множество раз до остановки кровотечения или достижения достаточного свертывания крови в соответствии со знанием специалиста в данной области техники. Когда используют множество введений, тогда они могут быть ежечасными, ежесуточными, еженедельными или производиться с другими интервалами. Введения могут осуществляться по схеме, такой как каждые 10 минут, каждые 15 минут, каждые 20 минут, каждые 30 минут, каждый час, каждые два часа, каждые три часа, каждые четыре часа, три раза в сутки, два раза в сутки, один раз в сутки, один раз каждые двое суток, один раз каждые трое суток и один раз в неделю. Вариант FXa также может быть введен непрерывно, например при помощи микронасоса. Вариант FXa может быть введен парентеральным путем (например, внутривенно, подкожно, внутрибрюшинно или внутримышечно). Вариант FXa может быть введен однократно, дважды или более чем дважды, или в течение по меньшей мере периода времени, требующегося для достижения эффективного свертывания крови.
В еще одном воплощении композиции, содержащие вариант белка FXa, могут быть введены вместе с еще одним прокоагулянтом, включающим отличающийся вариант FXa (например, вариант, имеющий замену по аминокислоте номер 146, отличающуюся от лейцина), Фактор IX, Фактор XIa, Фактор ХIIа, Фактор VIII, Фактор VIIa, FEIBA (шунтирующая активность при наличии ингибиторов к фактору VIII) или концентрат протромбинового комплекса (РСС).
Композиции, содержащие варианты белка FXa, могут дополнительно содержать фармацевтически приемлемый носитель или наполнитель, который может представлять собой растворители, диспергирующие среды, антибактериальные и противогрибковые агенты, изотонические агенты и агенты, замедляющие абсорбцию, и подобные физиологически совместимые агенты. Некоторые примеры фармацевтически приемлемых носителей, приведенные исключительно для иллюстрации, представляют собой воду, физиологический раствор, физиологический раствор, забуференный фосфатом, декстрозу, глицерин, этанол и т.п., а также их комбинации. В композицию также могут быть включены изотонические агенты, например соли (например, хлорид натрия), сахара (например, сахароза) или полиспирты (например, маннит или сорбит). Дополнительные примеры фармацевтически приемлемых веществ представляют собой увлажнители, эмульгаторы, консерванты или буферы, которые усиливают стабильность или другие свойства данных композиций.
Композиции для применения в соответствии с описанием изобретения могут находиться в любой подходящей форме для введения субъекту, например в виде жидкости для инъекции или для инфузии. Форма зависит от предполагаемого способа введения и терапевтического применения.
Терапевтические композиции обычно являются стерильными и стабильными в условиях приготовления и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарства. Стерильные инъецируемые растворы могут быть приготовлены путем включения вариантов белка FXa в соответствии с описанием изобретения в требуемое количество подходящего растворителя с одним ингредиентом или комбинацией ингредиентов, описанных выше, с последующей стерилизацией путем фильтрования. В случае стерильных порошков для приготовления стерильных инъецируемых растворов способы приготовления включают вакуумную сушку или сушку замораживанием (лиофлизацию), которые дают возможность получать порошок активного ингредиента с любым дополнительным желательным ингредиентом из его раствора, предварительно подвергнутого стерильной фильтрации.
Следующие примеры приведены только для иллюстративных задач, и их не следует рассматривать как ограничивающие каким-либо образом объем формулы изобретения или других раскрытых здесь изобретений.
ПРИМЕРЫ
ПРИМЕР 1: Экспрессия и очистка рекомбинантного варианта белка FXa
С использованием стандартных для молекулярной биологии способов и реагентов получали кДНК, кодирующую вариант белка FX, включающий мутацию I16L, в котором нативная аминокислотная последовательность активационного пептида фактора X заменена на аминокислотную последовательность RKR (однобуквенный код), которая вместе с последовательностью RKR на С-конце легкой цепи образовывала сайт протеолитического расщепления РАСЕ, имеющий аминокислотную последовательность RKRRKR (SEQ ID NO: 5). Последовательность кДНК и кодируемого ею зрелого белка описаны на Фиг. 1D (SEQ ID NO: 4) и Фиг. 1А (SEQ ID NO: 1), соответственно. Таким образом, предшественник FX варианта FXa может быть активирован путем внутриклеточного протеолитического расщепления ферментами РАСЕ, экспрессируемыми вместе с фактором свертывания крови. Дополнительно, нативная сигнальная последовательность и пропептид человеческого FX заменены на сигнальную последовательность и пропептид человеческого протромбина.
кДНК, кодирующая вариант FX, субклонировали в вектор экспрессии под контролем конститутивного промотора. Вектор также экспрессировал ген устойчивости к неомицину, обеспечивающий возможность для антибиотического отбора стабильных трансфектантов. Был создан второй вектор для экспрессии растворимой формы фермента РАСЕ.
Клетки-хозяева СНО К1 адаптировали для выращивания в бессывороточной суспензии и котрансфицировали путем липофекции линеаризованными векторами для экспрессии варианта FXa и растворимого РАСЕ. Трансфицированные клетки отбирали путем роста в средах, дополненных антибиотиком G418 и витамином К1. Резистентные клоны отбирали в отношении экспрессии и активности рекомбинантного варианта белка FXa. Клоны, демонстрирующие относительно высокую экспрессию и активность рекомбинантного варианта белка FXa, адаптировали для роста в бессывороточной суспензионной культуре. Один клон отбирали и использовали для создания предварительного банка клеток и затем основного банка клеток с использованием химически определенной среды, не содержащей компоненты животного или человеческого происхождения.
Для продукции лекарственного вещества для дополнительного исследования клетки, экспрессирующие вариант FXa, выращивали в масштабе 500 литров (л) или 2500 л с использованием химически определенных сред. Для запуска флаконы с клетками из клеточного банка размораживали, выращивали и постепенно увеличивали по массе путем выращивания в сосудах увеличивающегося объема с использованием химически определенной среды, не содержащей животные компоненты. После завершения увеличения массы культуры поддерживали в небольших биореакторах с перемешиванием содержимого резервуара. Для продукции выращивание продолжали в биореакторах объема 500 л или 2500 л до сбора материала. Для сбора клеточную культуральную среду центрифугировали для удаления клеток и дебриса. Среду дополнительно осветляли путем глубинного фильтрования и фильтрования через фильтр 0,2 мкм.
Профильтрованную среду затем разбавляли буфером и наносили на колонку для хроматографии в смешанном режиме (ММС) Capto (GE Healthcare Life Sciences). После промывания продукт элюировали с колонки. Частично очищенный продукт затем разводили в растворе детергента для инактивации вирусов. После инактивации вирусов продукт наносили на колонку для хроматографии с использованием Q-Sepharose Fast Flow (GE Healthcare Life Sciences), промывали и элюировали с колонки. Элюированный продукт затем разбавляли буфером и наносили на хроматографическую колонку Fractogel SO3" (Merck Millipore), промывали и элюировали с колонки. Элюированный продукт затем фильтровали с использованием фильтров Virosart® CPV (Sartorius Stedim Biotech) для удаления любого остаточного вируса, который может присутствовать. После фильтрования вируса очищенный продукт концентрировали с использованием ультрафильтрования и диафильтрования. Очищенное лекарственное вещество затем тестировали для того, чтобы охарактеризовать вариант белка FXa, продуцируемый клетками СНО.
ПРИМЕР 2: Концевая аминокислотная последовательность
N-концевое секвенирование варианта FXa путем деградации по Эдману использовали для подтверждения первых десяти остатков N-конца легкой цепи и N-конца тяжелой цепи. Анализ осуществляли путем автоматизированной деградации по Эдману после разделения варианта FXa при помощи SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) в невосстанавливающих и восстанавливающих условиях и подвергали электроблоттингу на поливинилидендифторидной (PVDF) мембране.
N-концевая последовательность легкой цепи представляла собой ANSFL(X)(X)MKK (однобуквенный код) (SEQ ID NO: 15). Никакая сигнальная последовательность не появляется по остаткам 6 и 7 (представленным как X), что согласуется с присутствием отстатков гамма-карбоксиглутаминовой кислоты (Gla) в этих позициях. N-концевая последовательность тяжелой цепи представляла собой LVGGQE(Z)KDG (SEQ ID NO: 16). Никакая сигнальная последовательность не обнаружена по остатку 7 (представленному как Z), что согласуется с присутствием цистеина в этой позиции. Эти результаты подтверждали ожидаемые N-концевые последовательности легкой цепи и тяжелой цепи по сравнению с теоретической последовательностью.
ПРИМЕР 3: Картирование пептида для подтверждения первичной структуры и идентичности посттрансляционных модификаций
Для подтверждения первичной аминокислотной последовательности и для обнаружения посттрансляционных модификаций вариант белка FXa, продуцируемый как описано в примере 1, анализировали путем пептидного картирования. В частности, белок восстанавливали, алкилировали и расщепляли лизилэндопептидазой Lys-C (протеаза 1 Achromobacter lyticus), после чего фрагменты анализировали путем обращенно-фазовой высокоэффективной жидкостной хроматографии/квадрупольной времяпролетной масс-спектрометрии с ионизацией электрораспылением (RP-HPLC/ESI-QTOF MS).
Пример получающегося в результате профиля пептидной карты представлен на Фиг. 2, в котором пептиды отмечены в соответствии с их предполагаемыми относительными позициями относительно теоретической последовательности легкой цепи, представленный на Фиг. 1В, и тяжелой цепи, представленной на Фиг. 1С. Пики, отмеченные буквой "L", указывают на пептид легкой цепи и пики, отмеченные буквой "Н", указывают на пептид тяжелой цепи. Минорные пики, представляющие продукты буфера, отмечены "R". Несколько пиков следового уровня, представляющих деамидированные (d) или сверхалкилированные пептиды, отмечены "d" и "*", соответственно.
Пептиды, обнаруженные путем пептидного картирования, их теоретические и обнаруженные массы и аминокислотные последовательности представлены в таблице 1 ниже. Нумерация аминокислот основана на Фиг. 1А (SEQ ID NO: 1).
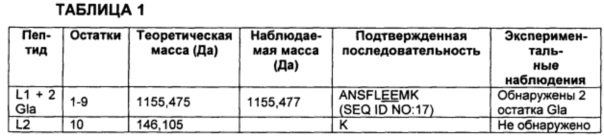
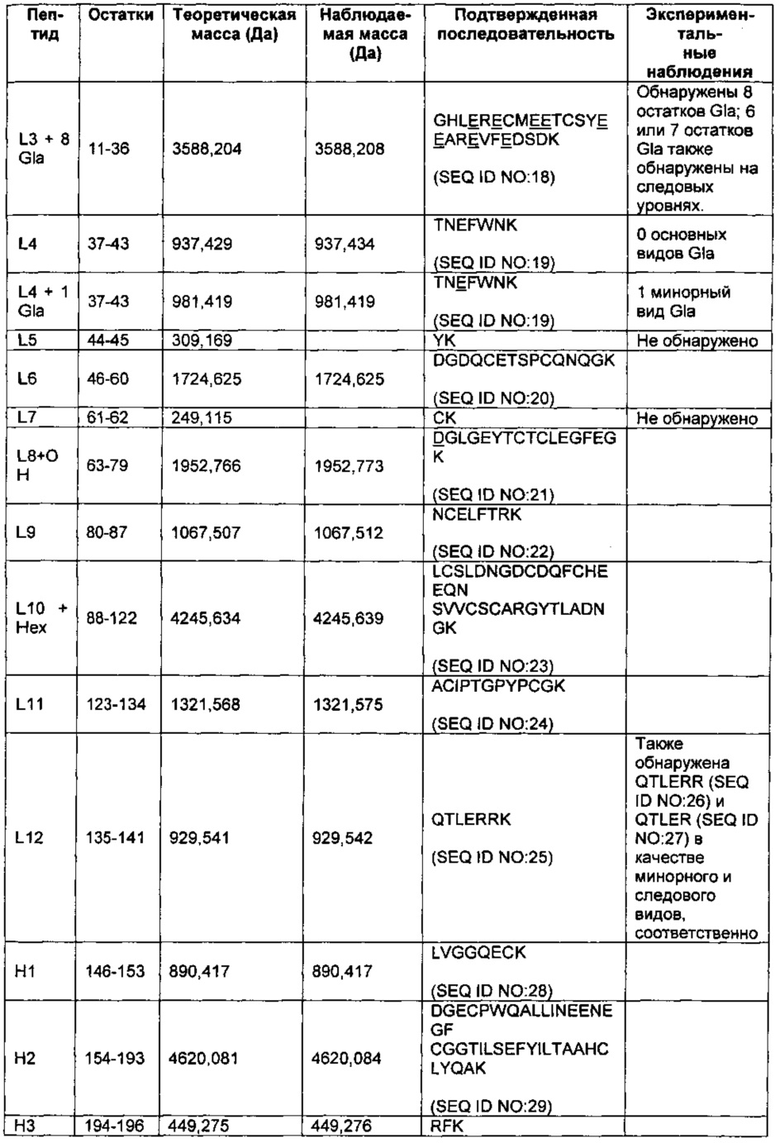
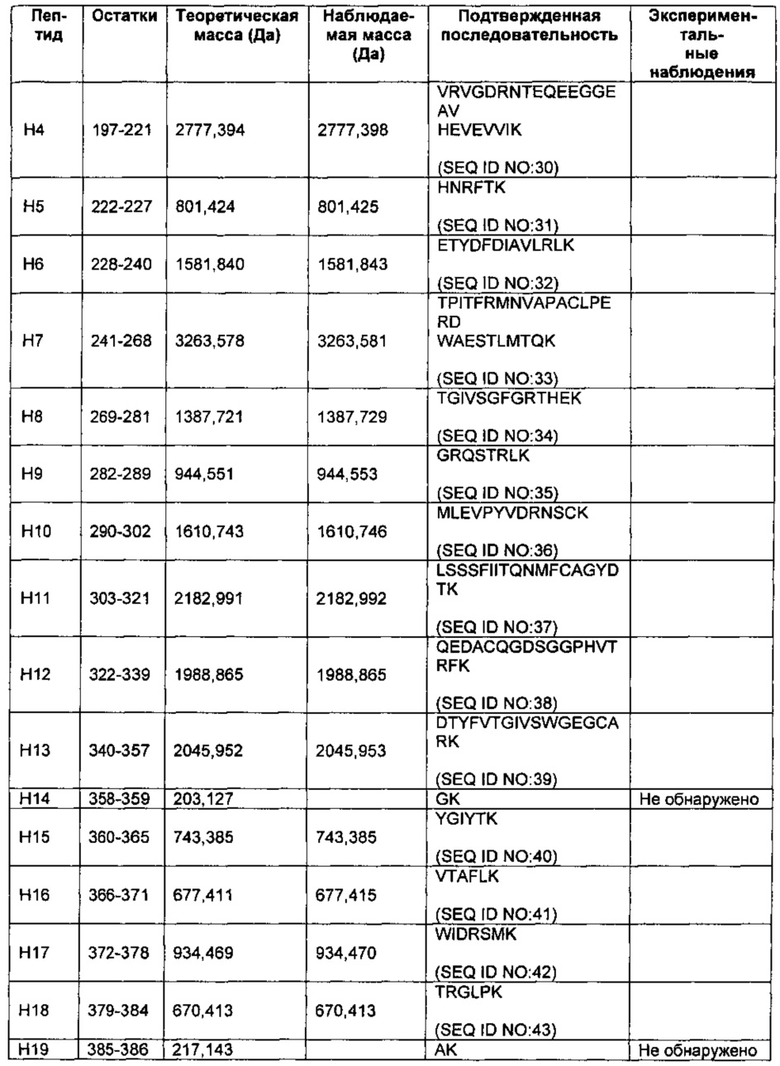

Теоретические массы рассчитывали при помощи PAWS (Genomic Solutions, Ann Arbor, MI). Наблюдаемые массы рассчитывали по наиболее частому несущему множественный заряд иону в масс-спектре. Все наблюдаемые массы согласовывались с теоретическими массами в пределах 10 млн-1. Покрытие последовательности среди пептидов для легкой цепи (LC) и тяжелой цепи (НС) составляло приблизительно 94 и 98%, соответственно. Три пептида из цепи L (L2, L5, L7) и два пептида из тяжелой цепи (Н14, Н19) не были обнаружены. Рассчитанные различия между теоретической и наблюдаемой массой пептида дали возможность сделать заключение о присутствии некоторых посттрансляционных модификаций. Эти остатки, как обнаружено, представляющие собой объект для модификации, подчеркнуты в столбце 5 таблицы 1.
N-концевой домен Gla в легкой цепи в общем содержал от 9 до 11 остатков гамма-карбоксилированной глутаминовой кислоты (Gla). Основные виды легкой цепи содержали 10 остатков Gla. Хотя пептид L1 был всегда карбоксилирован, гетерогенность обнаруживали в пептидах L3 и L4. В частности, пептид L3 обнаружен с 6, 7 или 8 остатками Gla, а пептид L4 обнаружен без остатков или с одним остатком Gla. См. таблицу 1.
Гетерогенность в отношении гамма-карбоксилирования подтверждена с использованием анионообменной высокоэффективной жидкостной хроматографии (AEX-HPLC) для измерения гетерогенности по заряду. Разделение при помощи AEX-HPLC варианта белка FXa позволило разделить изоформы, содержащие 9, 10 и 11 остатков гамма-карбоксиглутаминовой кислоты (Gla). Относительная частота количества остатков Gla в легкой цепи варианта FXa согласовывалась среди семи протестированных препаратов очищенного варианта белка FXa. В частности, на основе относительного распределения пиков частота появления 9 остатков Gla находилась в диапазоне от 10% до 17% (среднее значение равно 12,1%, SD (стандартное отклонение) равно 2,5%), частота появления 10 остатков Gla находилась в диапазоне от 41% до 46% (среднее значение равно 43,0%, SD равно 1,8%) и частота появления 11 остатков Gla находилась в диапазоне от 36% до 46% (среднее значение равно 42,1%, SD равно 3,3%).
Показано, что очищенные изоформы 10 и 11 Gla в равной степени активны в основанном на АРТТ анализе коагулирующей активности крови, тогда как изоформа 9 Gla обладала уменьшенной активностью (приблизительно 20% активности изоформ 10 и 11 Gla при нормализации по массе).
Также обнаружили другие посттрансляционные модификации легкой цепи. В частности, обнаружили, что масса пептида L8 согласуется с присутствием остатка бета-гидроксилированной аспарагиновой кислоты в положении 63 (Asp63) (SEQ ID NO: 1). Дополнительно, пептид L10 обнаружен с разностью масс, указывающей на добавление О-связанной гексозы.
Тяжелая цепь также подвергается О-связанному гликозилированию. На пептидной карте обнаружено, что основная форма С-конца альфа изоформы тяжелой цепи оканчивается по аминокислоте L398, и обнаружено, что минорная форма С-конца оканчивается по аминокислоте К399. Обнаружено, что оба этих пептида содержат один или два дисиалилированных О-гликана ядра-1, тогда как преобладающие виды обоих пептидов содержат один дисиалилированный О-гликан ядра-1. В профиле пептидной карты пик, помеченный как "Н203", соответствует пептиду Н20, оканчивающемуся по L398, а пик, помеченный как "Н202", соответствует пептиду Н20, оканчивающемуся по К399, каждый из которых содержит один О-связанный гликан. Пик, помеченный как "Н201", соответствует пептиду Н20, оканчивающемуся по К399 и содержащему два О-связанных гликана. Пептид Н20, оканчивающийся по L398, имеющий два О-связанных гликана, обнаруживали на следовых уровнях, и он не помечен на Фиг. 2. Местоположение О-гликанов в Н20 с Thr394 по Ser395 было определено путем бета-элиминирования с последующей фрагментацией путем MS (масс-спектрометрия).
ПРИМЕР 4: Структурная гетерогенность, определенная при помощи высокоэффективной жидкостной хроматографии
Обращеннофазовую высокоэффективную жидкостную хроматографию (RP-HPLC) использовали для измерения структурной гетерогенности в одном из препаратов интактного очищенного варианта белка FXa. Очищенный белок инжектировали на обращенно-фазовую колонку и элюировали градиентом подкисленного ацетонитрила. В соответствии с обнаруженным в результате анализа путем пептидного картирования вариант FXa содержал структурную гетерогенность по С-концу тяжелой цепи, приводя в результате к двум основным структурным изоформам, альфа (оканчивающейся по Lys399) и бета (оканчивающейся по Lys386). Обнаружена дополнительная минорная изоформа, названная гамма, вследствие обрезания тяжелой цепи после Lys281 или Arg283. Хроматограмма HPLC, полученная в этих экспериментах, представлена на Фиг. 3.
Относительная частота основных структурных изоформ согласовывалась среди семи протестированных препаратов очищенного варианта белка FXa. В частности, на основе относительного распределения пиков частота появления альфа изоформы находилась в диапазоне от 54% до 74% (среднее значение равно 66,9%, SD равно 7,0%), частота появления изоформы бета находилась в диапазоне от 24% до 42% (среднее значение равно 31,4%, SD равно 6,5%), и частота появления изоформы гамма находилась в диапазоне от 1% до 3% (среднее значение равно 2,0%, SD равно 0,8%).
Альфа и бета изоформы очищали с использованием гель-фильтрационной HPLC и тестировали в отношении активности в стандартном основанном на АРТТ анализе коагулирующей активности крови. Изоформа бета демонстрировала слегка меньшую активность (79%) по сравнению с альфа изоформой (нормализованной как 100%). Активность минорных видов гамма не тестировали.
ПРИМЕР 5: Структурная гетерогенность интактного белка, определенная при помощи масс-спектроскопии
Молекулярную массу (Мr) интактного варианта FXa из одного препарата очищенного белка анализировали при помощи RP-HPLC с последующей масс-спектрометрией с использованием гибридного квадрупольного времяпролетного масс-анализатора высокого разрешения (ESI-QTOF MS). Значения Мг определяли по масс-спектрам для нулевого заряда после обращенной свертки данных для множественного заряда при помощи программного обеспечения Waters MaxEnt-1. Все относительные ошибки между теоретическими и наблюдаемыми основными и минорными значениями масс находились в пределах ±60 млн-1, согласуясь с функциональными характеристиками масс-спектрометра Waters Q-ToF. Хроматограмма HPLC представлена на Фиг. 4. Масс-спектр интактного варианта белка FXa представлен на Фиг. 5, и обозначение пиков, основанное на масс-спектре, перечислено в таблице 2 ниже. Нумерация аминокислот основана на Фиг. 1А (SEQ ID NO: 1). 1ANS…LER139 соответствует SEQ ID NO: 6; 1ANS…ERR140 соответствует SEQ ID NO: 7; 1ANS…RRK141 соответствует SEQ ID NO: 9; 1ANS…RKR142 соответствует SEQ ID NO: 11; 1ANS…KRR143 соответствует SEQ ID NO: 12; 146LVG…LPK384 соответствует SEQ ID NO: 45; 146LVG…KAK386 соответствует SEQ ID NO: 46; 146LVG…SPL398 соответствует SEQ ID NO: 14; и 146LVG…PLK399 соответствует SEQ ID NO: 3.
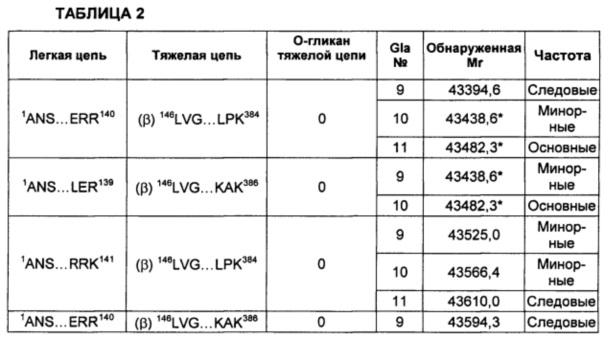
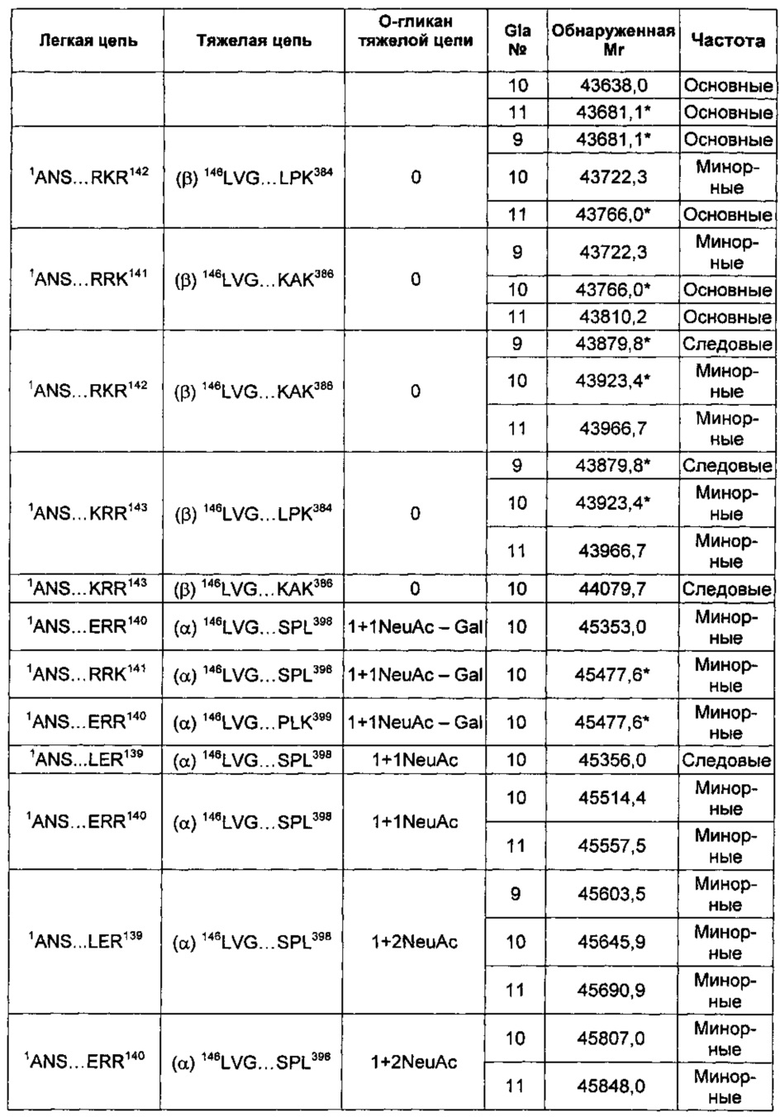
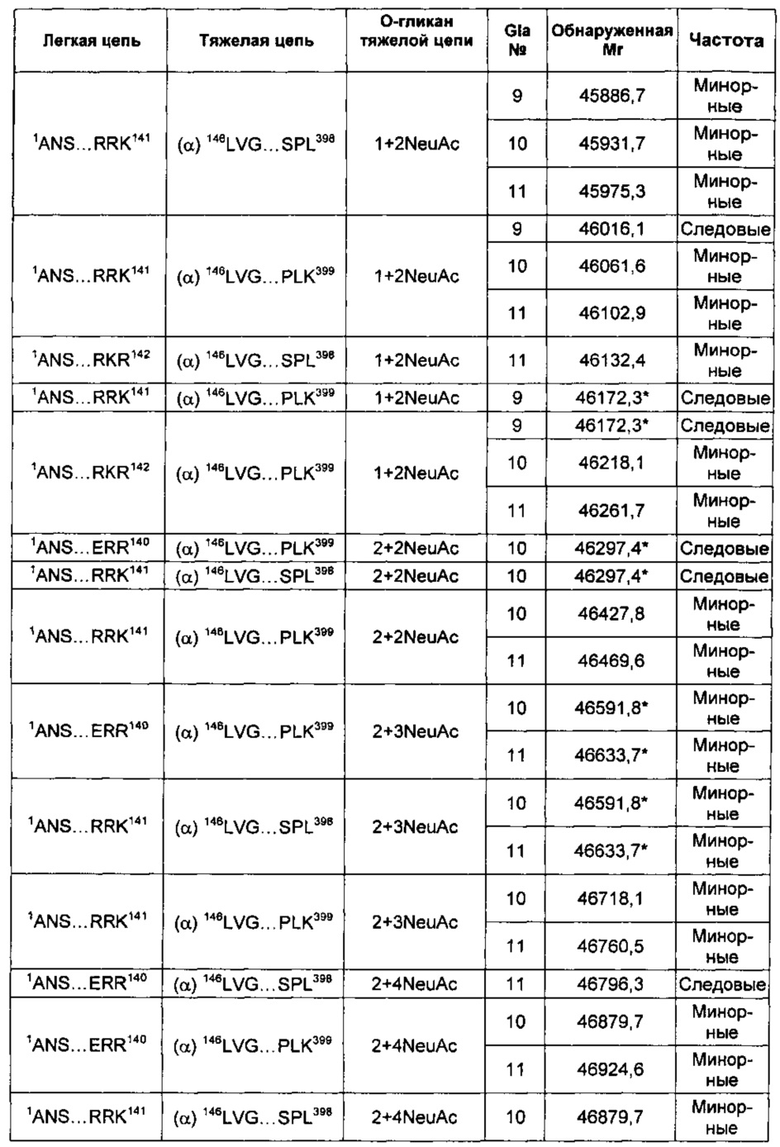
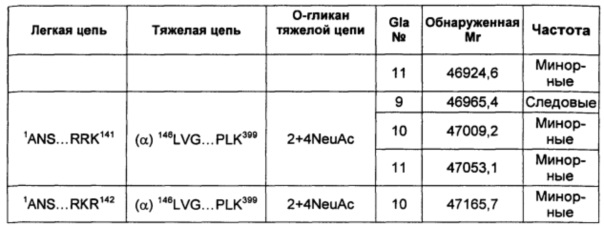
HPLC вновь продемонстрировала структурную гетерогенность варианта белка FXa (Фиг. 4). Пики, меченные А, соответствуют видам гамма. Пик, меченный В, соответствует альфа форме белка, включающей два О-гликана. Пик, меченный С, соответствует альфа форме белка, включающей один О-гликан. Пик, меченный D, соответствует бета форме белка, в котором тяжелая цепь оканчивается по К386. Пик, меченный Е, соответствует бета форме белка, в котором тяжелая цепь оканчивается по К384.
На масс-спектре обнаружены три области (Фиг. 5). Первая на низкомолекулярном конце масс-спектра (слева) соответствует интактному варианту белка FXa, содержащему бета изоформу тяжелой цепи. Эта область также содержит так называемые основные виды, которые появляются с большей частотой по сравнению с другими, на основе относительной интенсивности пиков. Основные виды включают виды, содержащие бета форму тяжелой цепи, оканчивающуюся по К386, в которой С-концевой остаток легкой цепи представлял собой R139, R140 или К141, по-видимому вследствие вариабельного расщепления по сайту расщепления РАСЕ. Виды, в которых легкая цепь оканчивается по R142, также, по-видимому, возникающие вследствие вариабельного расщепления, также представлены в виде минорных видов. Среди эти основных и минорных видов, легкая цепь включала β-гидрокси Asp63 и О-связанную гексозу, а также 10 или 11 остатков Gla.
В масс-спектре также появились две области с большей массой, обе из которых соответствуют минорным видам интактного варианта белка FXa, содержащим альфа изоформу тяжелой цепи (оканчивающуюся по L398 или К399). Одна из групп таких видов дополнительно включает единичную О-гликановую, а вторая группа дополнительно включает две О-гликановые посттрансляционные модификации (центр и правая часть на Фиг. 5, соответственно).
Обозначения, специально приведенные на Фиг. 5, соответствуют видам интактного варианта белка FXa, в которых легкая цепь содержит 10 остатков Gla, β-гидрокси Asp63 и О-связанную глюкозу. Соседние пики в спектре, помеченные "*", представляют виды, содержащие 11 остатков Gla, которые увеличивают массу на 44,0 Да.
В таблице 2 представлены основные и минорные виды, идентифицированные на описанном выше масс-спектре, а также многочисленные другие минорные и следовые виды, идентифицированные по данным масс-спектра. В таблице 2 виды, имеющее то же самое значение обнаруженной Мr, которое нельзя отличить от похожей изобарической изоформы в таблице, обозначены "*". Обозначение кажущейся частоты различных видов как основное, минорное и следовое основано на относительных интенсивностях пиков по данным масс-спектра. В столбце, озаглавленном "О-гликан", приведено состояние гликозилирования тяжелой цепи различных видов вариантов белка FXa, идентифицированных в таблице. В соответствии с обсуждаемым здесь легкая цепь содержит О-связанную гексозу. В таблице "0" обозначает отсутствие О-связанных гликанов в тяжелой цепи; "1+1NeuAc" указывает на присутствие одного моносиалилированного О-гликана в ядре-1; "1+1NeuAc - Gal" указывает на присутствие одного моносиалилированного О-гликана в ядре-1, в котором концевая галактоза (Gal) отсутствует; "1+2NeuAc" указывает на присутствие одного дисиалилированного О-гликана в ядре-1; "2+2NeuAc" указывает на присутствие двух О-гликанов в ядре-1, один из которых дисиалилирован или оба из которых моносиалилированы; "2+3NeuAc" указывает на присутствие двух О-гликанов в ядре-1, один из которых моносиалилирован, а другой из которых дисиалилирован; и "2+4NeuAc" указывает на присутствие двух О-гликанов в ядре-1, каждый из которых дисиалилирован.
ПРИМЕР 6: Структурная гетерогенность восстановленного и алкилированного белка, определенная при помощи масс-спектроскопии
Для подтверждения описанного выше анализа очищенный интактный вариант белка FXa восстанавливали и алкилировали для устранения межцепочечных и внутрицепочечных дисульфидных связей. Разделенные легкие и тяжелые цепи затем анализировали при помощи RP-HPLC/ESI-QTOF MS для определения их аминокислотной структуры и посттрансляционной модификации.
Хроматограмма HPLC представлена на Фиг. 6, в которой легкие и тяжелые цепи помечены. Альфа и бета изоформы тяжелой цепи элюируются в виде множественных хроматографических пиков. Следовые уровни видов гамма (тяжелая цепь бета, усеченная после аминокислоты К281 или R283) обнаружены при элюировании непосредственно перед тяжелой цепью на хроматограмме приблизительно на 48-50 минуте.
Масс-спектр легкой цепи для нулевого заряда представлен на Фиг. 7А, и виды, идентифицированные из спектра, включая их относительные частоты как основные, минорные или следовые, перечислены в таблице 3 ниже. Нумерация аминокислот основана на Фиг. 1А (SEQ ID NO: 1). 1ANS…LER139 соответствует SEQ ID NO: 6; 1ANS…ERR140 соответствует SEQ ID NO: 7; 1ANS…RRK141 соответствует SEQ ID NO: 9; 1ANS…RKR142 соответствует SEQ ID NO: 11; и 1ANS…KRR143 соответствует SEQ ID NO: 12.
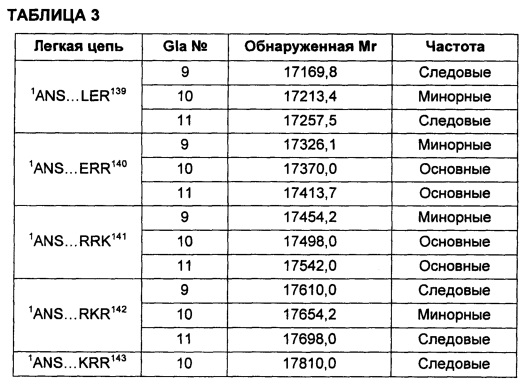
Как видно для интактного варианта белка FXa, обнаружено, что все виды легкой цепи включают β-гидроксилирование Asp63 и О-связанную гексозу. Карбоксилирование глутаминовой кислоты в домене Gla и расщепление РАСЕ, тем не менее, варьировали, приводя к образованию видов легкой цепи с 9, 10 или 11 остатками Gla и С-концевой гетерогенностью, возникающей в результате неаккуратного расщепления по сайту РАСЕ.
Масс-спектр тяжелой цепи для нулевого заряда представлен на Фиг. 7 В, и различные виды, идентифицированные из спектра, включая их относительные частоты как основные, минорные или следовые, перечислены в таблице 4 ниже. Нумерация аминокислот основана на Фиг. 1А (SEQ ID NO: 1). 146LVG…LPK384 соответствует SEQ ID NO: 45; 146LVG…KAK386 соответствует SEQ ID NO: 46; 146LVG…SPL398 соответствует SEQ ID NO: 14; и 146LVG…PLK399 соответствует SEQ ID NO: 3.
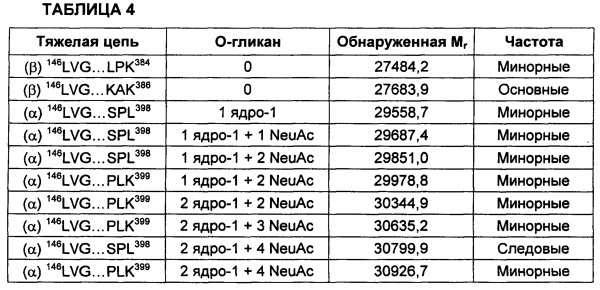
В масс-спектре тяжелая цепь обнаружена в виде трех преобладающих областей, соответствующих изоформам бета и альфа. Наблюдаемая Мг наиболее частой изоформы тяжелой цепи (27683,9 Да) хорошо сравнима с теоретической массой бета изоформы тяжелой цепи (27684,3 Да; 146LVG…KAK386) с алкилированием всех девяти цистеиновых остатков. Дополнительная гетерогеннность по массам возникает вследствие вариации по С-концевому остатку и О-гликозилирования альфа изоформ. В частности, некоторые пики соответствовали альфа изоформам, оканчивающимся по L398 или К399, тогда как другие пики соответствовали тяжелым цепям, несущим один или два О-гликана ядра-1, которые произвольно были не сиалилированы, моносиалилированы или дисиалилированы. На Фиг. 7В присутствие "*" указывает на сверхалкилирование тяжелой цепи, согласующееся с пиками, обнаруженными путем пептидного картирования варианта белка FXa (см. Пример 3).
Структурная гетерогенность, обнаруженная в легкой и тяжелой цепях после восстановления и алкилирования, согласуется с результатами, обнаруженными с использованием интактного варианта белка FXa.
Теоретические массы рассчитывали при помощи программного обеспечения PAWS (2000.06.08, Genomic Solutions, Ann Arbor, Ml). Обнаруженные массы определяли по масс-спектрам для нулевого заряда после обращенной свертки данных для множественного заряда при помощи программного обеспечения Waters MaxEnt-1. Все относительные ошибки между теоретическими и наблюдаемыми основными и минорными значениями масс составляли меньше 60 млн-1, согласуясь с функциональными характеристиками масс-спектрометра Waters Q-ToF для анализа интактного гликопротеина. Разделение видов на основные, минорные и следовые основано на соответствующих интенсивностях пиков, что коррелирует с частотой изоформы.
ПРИМЕР 7: Анализ in vitro коагулирующей активности крови
Коагулирующую активность для семи различных препаратов варианта белка FXa определяли с использованием одностадийного анализа коагулирующей активности крови с использованием в качестве субстрата человеческой плазмы, дефицитной по фактору VIII (анализ АРТТ). Специфическую активность определяли путем деления коагулирующей активности (мг/мл) на концентрацию белка (мг/мл). Средняя коагулирующая активность среди препаратов составляла 1,52 мг/мл при стандартном отклонении 0,09. Средняя специфическая активность составляла 101,3% при стандартном отклонении 5,9.
Описание настоящего изобретения не ограничивается объемом конкретных описанных здесь воплощений. Действительно, различные модификации описания изобретения дополнительно к описанным здесь понятны специалисту в данной области техники из предшествующего описания и сопутствующих графических материалов. Предполагается, что такие модификации оказываются в пределах объема формулы изобретения.
Здесь цитируются различные ссылки, включающие заявки на патенты, патенты и технические публикации. Описание каждого такого источника во всей его полноте включено в данное описание путем ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНЪЮГИРОВАННЫЕ МОЛЕКУЛЫ ФАКТОРА VIII | 2009 |
|
RU2573587C2 |
| ПОЛИПЕПТИДЫ НА ОСНОВЕ МОДИФИЦИРОВАННОГО ФАКТОРА VII И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571931C2 |
| Биофармацевтические композиции и связанные с ними способы | 2020 |
|
RU2824761C2 |
| ВАРИАНТЫ ДОМЕНА GLA ФАКТОРА VII ИЛИ VIIa | 2004 |
|
RU2373282C2 |
| МОДИФИЦИРОВАННЫЕ ВИТАМИН К-ЗАВИСИМЫЕ ПОЛИПЕПТИДЫ | 2005 |
|
RU2396347C2 |
| АНТИТЕЛА ПРОТИВ ИНГИБИТОРА МЕТАБОЛИЧЕСКОГО ПУТИ ТКАНЕВОГО ФАКТОРА | 2009 |
|
RU2562114C2 |
| МОДИФИЦИРОВАННЫЙ КОАГУЛИРУЮЩИЙ ФАКТОР VIIa С ПРОДЛЕННЫМ ВРЕМЕНЕМ ПОЛУЖИЗНИ | 2007 |
|
RU2466141C2 |
| НОВЫЕ АНТИТЕЛА К ТКАНЕВОМУ ФАКТОРУ В КАЧЕСТВЕ АНТИКОАГУЛЯНТОВ | 2003 |
|
RU2345789C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВИТАМИН К-ЗАВИСИМЫХ БЕЛКОВ | 2001 |
|
RU2321633C2 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |





















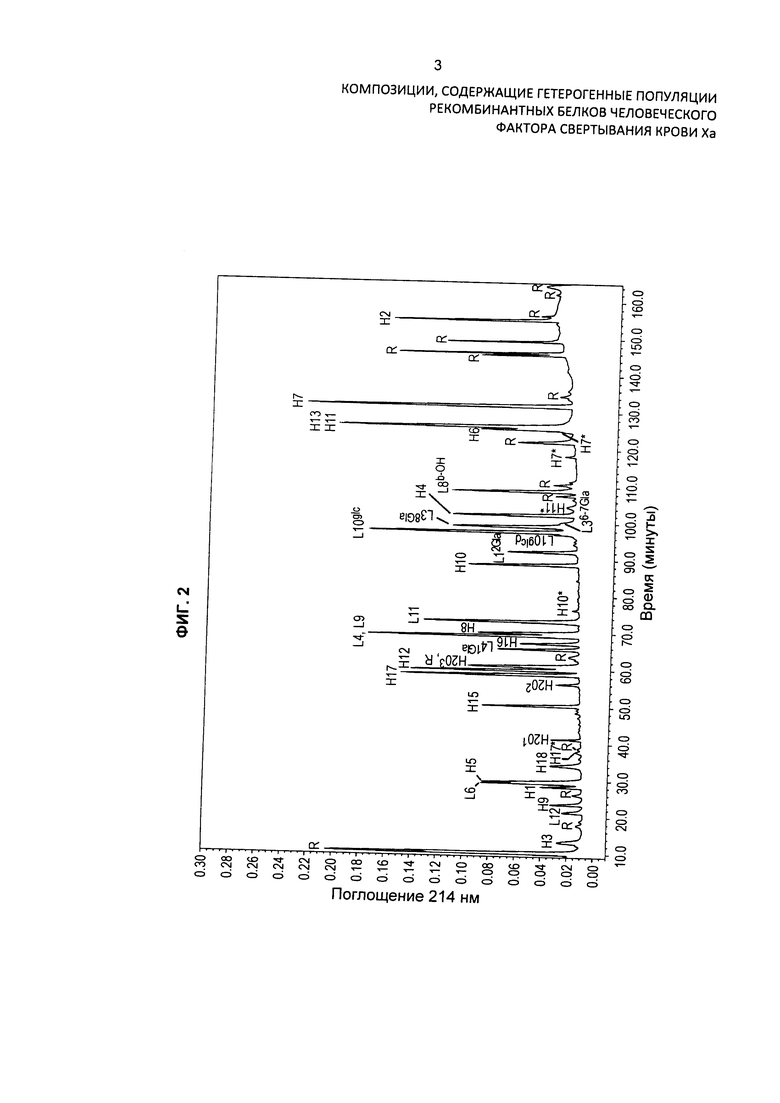
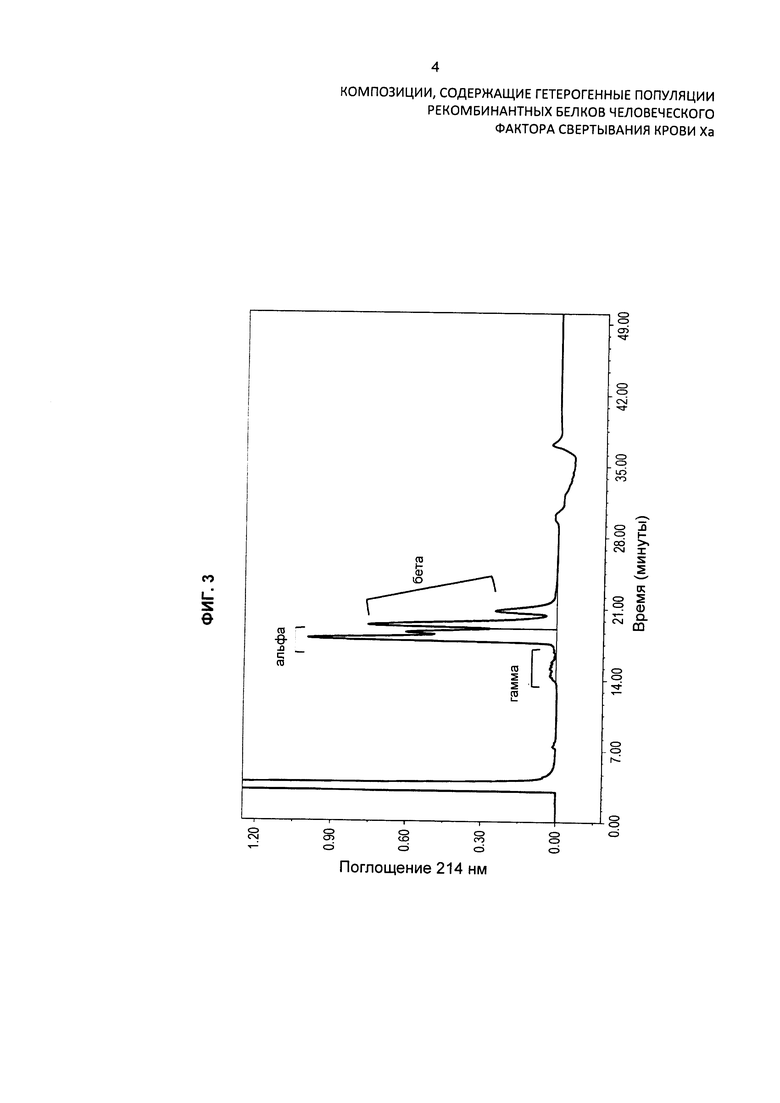
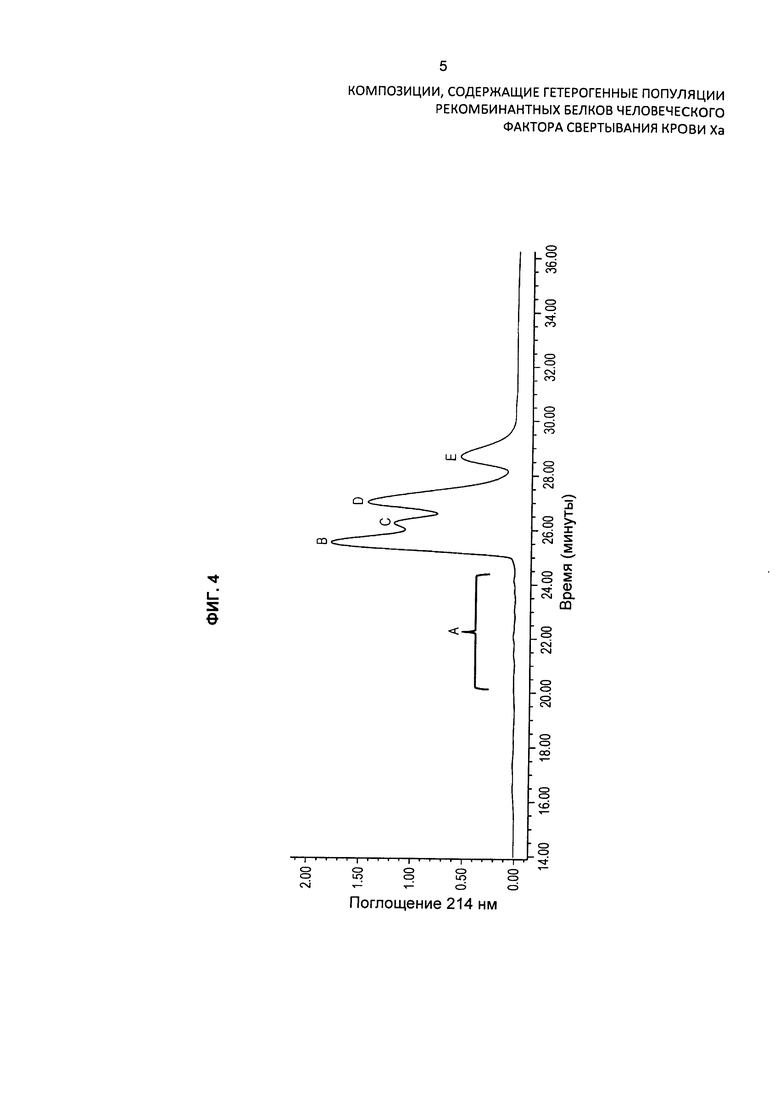
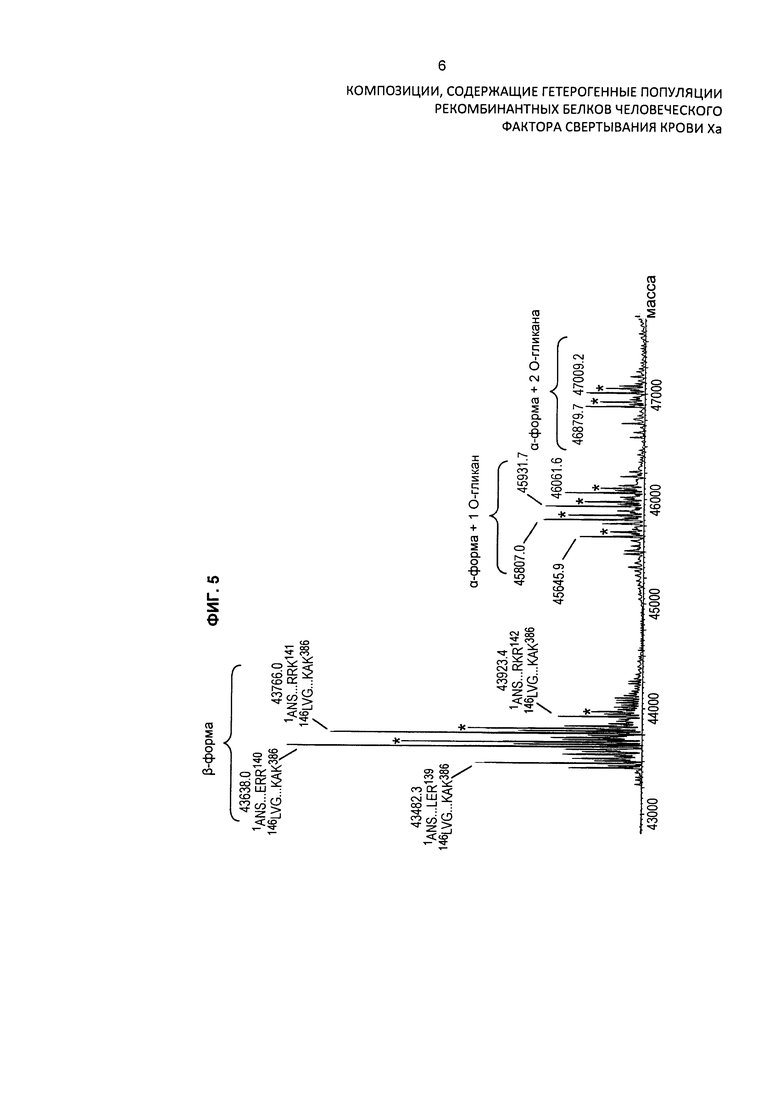
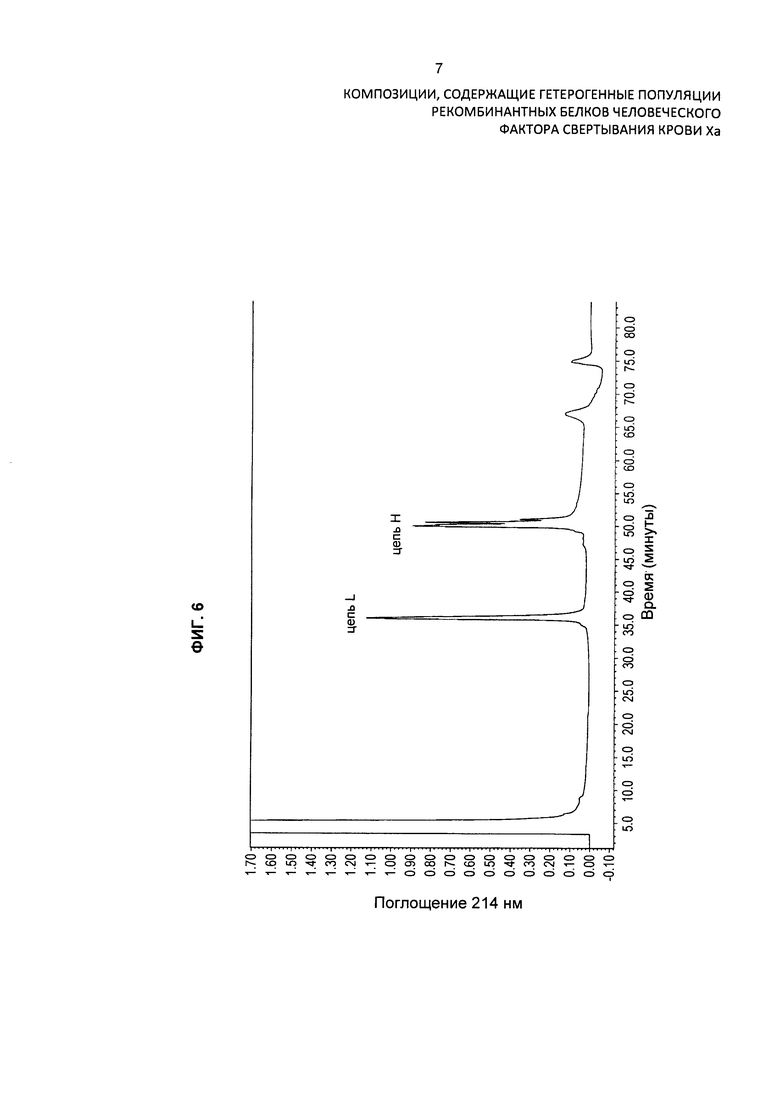
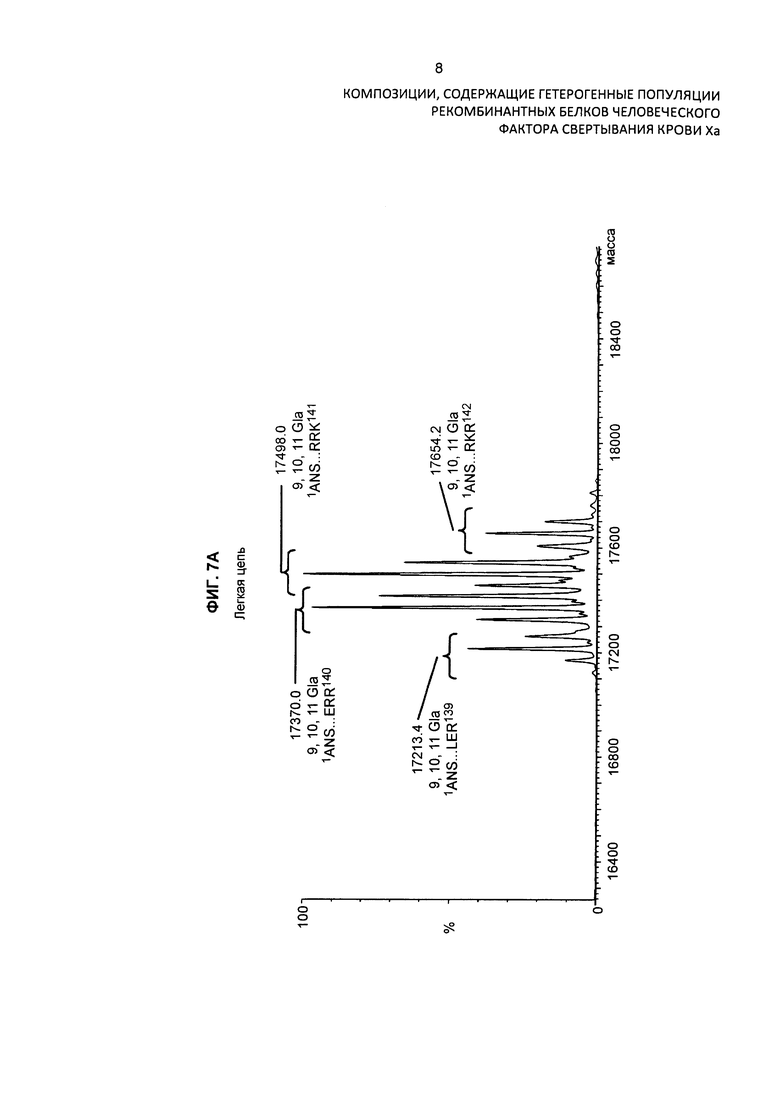
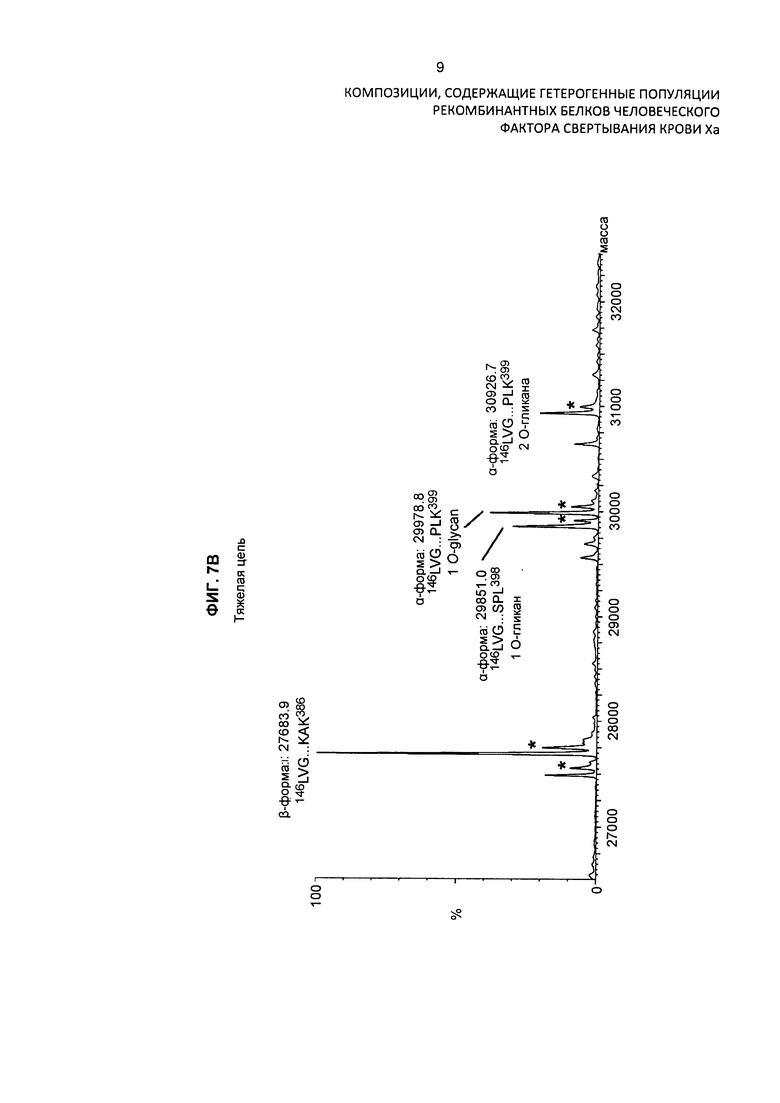
Изобретение относится к области биотехнологии, конкретно к композициям, содержащим рекомбинантные варианты человеческого фактора свертывания крови Ха, что может быть использовано в медицине. Рекомбинантным путем получают широкое разнообразие изоформ и посттрансляционных модификаций FXa. Изобретение позволяет осуществлять эффективную терапию гемостаза у нуждающихся в этом субъектов. 4 н. и 33 з.п. ф-лы, 7 ил., 4 табл., 7 пр.
1. Композиция для эффективной терапии субъекта, нуждающегося в гемостазе, содержащая в эффективном количестве два или более рекомбинантных вариантов белка FXa, выбранных из группы, состоящей из:
(а) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(б) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(в) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(г) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(д) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 143 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(е) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(ж) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(з) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1, и
(и) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 143 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
где легкие цепи указанных вариантов белка FXa возможно содержат модификацию, выбранную из группы, состоящей из β-гидроксилирования Asp63, включения О-связанной гексозы и включения 9, 10 или 11 остатков Gla.
2. Композиция для эффективной терапии субъекта, нуждающегося в гемостазе, содержащая в эффективном количестве два или более рекомбинантных варианта белка FXa, выбранных из группы, состоящей из:
(а) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(б) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(в) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(г) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(д) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1,
(е) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1, и
(ж) рекомбинантного варианта белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1,
где легкие цепи указанных вариантов белка FXa возможно содержат модификацию, выбранную из группы, состоящей из β-гидроксилирования Asp63, включения О-связанной гексозы и включения 9, 10 или 11 остатков Gla, и
где тяжелые цепи указанных вариантов белка FXa возможно содержат модификацию, представляющую собой включение одного или двух О-связанных гликанов ядра-1.
3. Композиция по п. 1 или 2, где легкие цепи указанных вариантов белка FXa включают β-гидрокси Asp63 и О-связанную гексозу.
4. Композиция по п. 1 или 2, где легкие цепи указанных вариантов белка FXa включают 9, 10 или 11 остатков Gla.
5. Композиция по п. 3, где легкие цепи указанных вариантов белка FXa дополнительно включают 9, 10 или 11 остатков Gla.
6. Композиция по п. 5, где легкие цепи указанных вариантов белка FXa включают 9 остатков Gla.
7. Композиция по п. 5, где легкие цепи указанных вариантов белка FXa включают 10 остатков Gla.
8. Композиция по п. 5, где легкие цепи указанных вариантов белка включают 11 остатков Gla.
9. Композиция по п. 2, где тяжелые цепи указанных вариантов белка FXa включают один или два О-связанных гликана ядра-1.
10. Композиция по п. 9, где один О-связанный гликан ядра-1 моносиалилирован или дисиалилирован.
11. Композиция по п. 10, где один О-связанный гликан ядра-1 моносиалилирован.
12. Композиция по п. 10, где один О-связанный гликан ядра-1 дисиалилирован.
13. Композиция по п. 9, где каждый из двух О-связанных гликанов ядра-1, если присутствуют, моносиалилирован.
14. Композиция по п. 9, где каждый из двух О-связанных гликанов ядра-1, если присутствуют, дисиалилирован.
15. Композиция по п. 9, где один из двух О-связанных гликанов ядра-1, если присутствуют, моносиалилирован, а другой дисиалилирован.
16. Композиция по п. 1, содержащая 3, 4, 5, 6, 7, 8 или все 9 рекомбинантных вариантов белка FXa, выбранных из указанной группы.
17. Композиция по п. 2, содержащая 3, 4, 5, 6 или все 7 рекомбинантных вариантов белка FXa, выбранных из указанной группы.
18. Композиция по п. 1, где относительная частота вариантов белка FXa (а), (б), (в) и (г), каждый из которых содержит 10 остатков Gla, составляет (б) больше (в), (в) больше (а), (а) больше (г), как определено с использованием масс-спектрометрии.
19. Композиция по п. 2, где относительная частота вариантов белка FXa (а), (б), (в) и (е), каждый из которых содержит 10 остатков Gla и один О-связанный гликан ядра-1, составляет (б) больше (в), (в) больше (е), (е) больше (а), как определено с использованием масс-спектрометрии.
20. Композиция рекомбинантных вариантов белка FXa для эффективной терапии субъекта, нуждающегося в гемостазе, содержащая в эффективном количестве:
(а) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(б) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(в) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(г) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(д) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 143 и со 146 по 386 аминокислотной последовательности SEQ ID NO: 1,
(е) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(ж) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(з) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(и) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 143 и со 146 по 384 аминокислотной последовательности SEQ ID NO: 1,
(к) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 139 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(л) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(м) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(н) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 398 аминокислотной последовательности SEQ ID NO: 1,
(о) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 140 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1,
(п) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 141 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1, и
(р) вариант белка FXa, где последовательности легкой цепи и тяжелой цепи белка соответственно состоят из аминокислот с 1 по 142 и со 146 по 399 аминокислотной последовательности SEQ ID NO: 1,
где легкие цепи указанных вариантов белка FXa возможно содержат модификацию, выбранную из группы, состоящей из β-гидроксилирования Asp63, включения О-связанной гексозы и включения 9, 10 или 11 остатков Gla, и
где тяжелые цепи указанных вариантов белка FXa возможно содержат модификацию, представляющую собой включение одного или двух О-связанных гликанов ядра-1.
21. Композиция по п. 20, где легкие цепи указанных вариантов белка FXa включают β-гидрокси Asp63 и О-связанную гексозу.
22. Композиция по п. 20, где легкие цепи указанных вариантов белка FXa включают 9, 10 или 11 остатков Gla.
23. Композиция по п. 21, где легкие цепи указанных вариантов белка FXa дополнительно включают 9, 10 или 11 остатков Gla.
24. Композиция по п. 23, где легкие цепи указанных вариантов белка FXa включают 9 остатков Gla.
25. Композиция по п. 23, где легкие цепи указанных вариантов белка FXa включают 10 остатков Gla.
26. Композиция по п. 23, где легкие цепи указанных вариантов белка включают 11 остатков Gla.
27. Композиция по п. 20, где тяжелые цепи указанных вариантов белка FXa, имеющих тяжелую цепь альфа формы, включают один или два О-связанных гликана ядра-1.
28. Композиция по п. 27, где один О-связанный гликан ядра-1 моносиалилирован или дисиалилирован.
29. Композиция по п. 28, где один О-связанный гликан ядра-1 моносиалилирован.
30. Композиция по п. 28, где один О-связанный гликан ядра-1 дисиалилирован.
31. Композиция по п. 27, где каждый из двух О-связанных гликанов ядра-1, если присутствуют, моносиалилирован.
32. Композиция по п. 27, где каждый из двух О-связанных гликанов ядра-1, если присутствуют, дисиалилирован.
33. Композиция по п. 27, где один из двух О-связанных гликанов ядра-1, если присутствуют, моносиалилирован, а другой дисиалилирован.
34. Способ лечения субъекта, нуждающегося в гемостазе, включающий стадию введения указанному субъекту количества композиции по любому из пп. 1-33, эффективного для того, чтобы вызывать гемостаз у указанного субъекта.
35. Способ по п. 34, где указанный субъект страдает от гемофилии А.
36. Способ по п. 34, где указанный субъект страдает от гемофилии В.
37. Способ по п. 34, где указанный субъект страдает от травмы.
| LACRAMIOARA I | |||
| A zymogen-like factor Xa variant corrects the coagulation defect in hemophilia, Nature Biotechnology, 2011, v.29, n.11, p.1028-1035 | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| YAN-MEI S | |||
| Vitamin K epoxide reductase significantly improves carboxylation in a cell line overexpressing factor X, Blood, 2005, v.106, numb | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| MIN-NING H., Characterization of the Human Blood Coagulation Factor X Promoter, The J | |||
| of Biol | |||
| Chem., 1992, v.267, n.22, p.15440-15446 | |||
| МОДИФИЦИРОВАННЫЕ ВИТАМИН К-ЗАВИСИМЫЕ ПОЛИПЕПТИДЫ | 2005 |
|
RU2396347C2 |

