Родственные заявки
Приоритет принадлежит заявке США №60/923,512, Edwin Madison, Christopher Thanos, Sandra Waugh Ruggles и Shaun Coughlin, озаглавленной "MODIFIED FACTOR VII POLYPEPTIDES AND USES THEREOF", зарегистрированной 13 апреля 2007 года.
Эта заявка близка соответствующей заявке США №12/082,662 Edwin Madison, Christopher Thanos, Sandra Waugh Ruggles и Shaun Coughlin, озаглавленной "MODIFIED FACTOR VII POLYPEPTIDES AND USES THEREOF", которая обладает приоритетом по отношению к заявке США №60/923,512.
Где это допустимо, предмет каждой из вышеупомянутых заявок включен в настоящий документ в полном объеме.
Область техники, к которой относится изобретение
Настоящим изобретением предусмотрены модифицированные белки для терапевтических целей. В частности, предусмотрены модифицированные полипептиды фактора VII, которые включают Фактор VIIa и другие формы фактора VII, а также их применение.
Предпосылки создания изобретения
Гемостаз представляет собой сложный физиологический процесс, который ведет к прекращению кровотечения. Тромбоциты, белки плазмы, кровеносные сосуды и эндотелиальные клетки являются тремя компонентами этого процесса, и каждый из них играет важную роль в событиях, которые сразу следуют за повреждением тканей и которые, в нормальных обстоятельствах, приводят к быстрому образованию сгустка. Центральным событием этого каскада свертывания является ряд протеолитических актов, в которых некоторые белки плазмы (или факторы свертывания крови) последовательно активируются в "каскаде" другими ранее активированными факторами свертывания, что приводит к быстрому образованию тромбина. Образованный в больших количествах в данном каскаде тромбин расщепляет фибриноген на пептиды фибрина, которые необходимы для формирования тромбов.
Факторы свертывания циркулируют в виде неактивных одноцепочечных зимогенов и активируются при расщеплении по одному или нескольким положениям для образования двухцепочечной активированной формы белка. Фактор VII (FVII), витамин К-зависимый белок плазмы, в исходном состоянии циркулирует в крови в виде зимогена. Зимоген FVII активируется протеолитическим расщеплением по одному положению, Arg152-Ile153, что приводит к образованию двухцепочечной протеазы, в которой цепи связаны одной дисульфидной связью (FVIIa). FVIIa связывает свой кофактор, тканевый фактор (TF) с образованием комплекса, в котором FVIIa может эффективно активировать фактор Х (FX) в FXa, тем самым запуская серию событий, которые приводят к образованию фибрина и гемостазу.
Хотя в большинстве случаев достигается нормальный гемостаз, дефекты в этом процессе могут привести к расстройствам, связанным с кровоточивостью, при которых увеличено время, необходимое для формирования тромбов. Такие расстройства могут быть врожденными или приобретенными. Например, гемофилии А и В представляют собой наследственные заболевания, характеризующиеся недостатком фактора VIII (FVIII) и фактора IX (FIX), соответственно. Заместительная терапия является традиционным способом лечения гемофилии А и В и включает внутривенное введение FVIII или FIX, либо полученных из человеческой плазмы, либо рекомбинантных белков. Во многих случаях, однако, у пациентов вырабатываются антитела (также известные как ингибиторы) против введенных белков, которые уменьшают или сводят на нет эффективность лечения. Для лечения гемофилии А или В у пациентов, у которых вырабатываются ингибиторы FVIII или FIX было одобрено применение рекомбинантных FVIIa (Novoseven ®), они также используются для остановки или предотвращения кровотечений, связанных с травмой и/или хирургическим вмешательством. Рекомбинантный FVIIa также был одобрен для лечения пациентов с врожденным дефицитом FVII, и все шире применяется для недокументированного использования, например для лечения кровотечений, связанных с другими врожденными или приобретенными расстройствами, связанными с кровотечениями, травмами, и в хирургии не страдающих гемофилией пациентов.
Использование рекомбинантного FVIIa для содействия формированию тромбов подчеркивает его растущую важность в качестве терапевтического агента. FVIIa терапия приводит к его значительной нехватке для медицинских целей. Например, по данным клинических испытаний, для остановки острых кровотечений у пациентов с гемофилией необходимо в среднем по 3 дозы FVIIa в течение 6 часовой или больший период. Для уменьшения требуемых количеств необходим более эффективный вариант FVIIa. Таким образом, одной из целей настоящего изобретения является предоставление модифицированных FVII-полипептидов, которые обладают улучшенными лечебными свойствами.
Краткое изложение сущности изобретения
В настоящем изобретении предусмотрены модифицированные полипептиды фактора VII (FVII). В частности, в настоящем изобретении предусмотрены модифицированные FVII-полипептиды, которые проявляют прокоагуляционную активность. В FVII-полипептидах модифицирована первичная последовательность по сравнению с немодифицированным FVII-полипептидом, и данная модификация может включать аминокислотные вставки, делеции и замены. Предусмотренные в настоящем изобретении модифицированные FVII-полипептиды включают FVII-полипептиды, которые проявляют повышенную устойчивость к влиянию ингибитора запускаемого тканевым фактором (TFPI) пути, повышенной устойчивостью к ингибиторному влиянию антитромбина-III (AT-III), сниженной способностью к связыванию Zn2+, обладают улучшенными фармакокинетическими свойствами, такими как увеличенное время полураспада, увеличенной каталитической активностью в присутствии и/или в отсутствие TF, и/или увеличенной способностью к связыванию с активированными тромбоцитами. Модифицированные полипептиды FVII могут содержать любую комбинацию предусмотренных настоящим изобретением модификаций, причем один или более видов активности или свойств полипептида изменены по сравнению с немодифицированным FVII-полипептидом. Как правило, модифицированные полипептиды FVII сохраняет прокоагуляционную активность. Кроме того, в настоящем документе предусмотрены нуклеиновой кислоты, векторы и клетки, которые кодируют / экспрессируют модифицированные FVII-полипептиды. Фармацевтические композиции, продукты производства, наборы и методы лечения также приводятся в настоящем документе. FVII-полипептиды включают аллельные и видовые варианты и полипептиды, и другие варианты модифицикации которых влияют на другие виды активности и/или свойства. В настоящее изобретение также включены активные фрагменты FVII-полипептидов, которые включают модификации, приведенные в данном документе. Примерами FVII-полипептидов являются полипептиды, которые включают последовательность аминокислот, изложенную в SEQ ID №3, а также ее варианты 60%, 65%, 70%, 75%, 80%, 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательности.
Полипептиды фактора VII (FVII) модифицированы по первичной последовательности по сравнению с немодифицированным полипептидом FVII и могут включать аминокислотные вставки, делеции и замены. Модифицированные FVII-полипептиды, предусмотренные в настоящем документе, включают FVII-полипептиды, которые демонстрируют повышенную устойчивость к ингибиторному воздействию TFPI, повышенную устойчивость к ингибирующему влиянию AT-III, сниженную способность к связыванию Zn2+, обладают улучшенными фармакокинетическими свойствами, такими как увеличенное время полураспада, увеличенной каталитической активностью в присутствии и/или в отсутствие TF, и/или увеличенной способностью к связыванию с активированными тромбоцитами. Модифицированные полипептиды FVII могут содержать любую комбинацию предусмотренных настоящим изобретением модификаций, причем один или более видов активности или свойств полипептида изменены по сравнению с немодифицированным FVII-полипептидом. FVII-полипептиды могут включать другие модификации, которые изменяют другие свойства и/или виды активности. Как правило, модифицированные FVII-полипептиды сохраняют прокоагуляционную активность. Кроме того, в настоящем документе предусмотрены нуклеиновые кислоты, векторы и клетки которые кодируют/экспрессируют модифицированные FVII-полипептиды. Фармацевтические композиции, продукты производства, наборы и методы лечения также приводятся в настоящем документе.
В частности, в настоящем документе предусмотрены модифицированные полипептиды фактора VII (FVII), их аллельные и видовые варианты, или их активные фрагменты, или их другие варианты. В настоящем документе предусмотрены FVII-полипептиды, включающие их видовые и аллельные варианты или их активные фрагменты, которые содержат модификацию в положениях, соответствующих положениям D196, К197 или К 199 в FVII-полипептиде, обладающие последовательностью аминокислот, приведенной в SEQ ID №3, или в соответствующих остатках в FVII-полипептиде, включая его видовые и аллельные варианты, активные фрагменты, а также другие FVII-полипептиды, модифицированы для других видов активности или свойств. FVII-полипептиды содержат по крайней мере одну модификацию в положениях, соответствующих положениям D196, К197 или К199 в FVII-полипептиде, содержащем последовательность аминокислот, приведенную в SEQ ID №3 или в соответствующих остатках в FVII-полипептиде. Модификация представляет собой вставку и/или замену на гидрофобную или кислую аминокислоту, выбранную из Val (V), Leu (L), Не (I), Phe (F), Trp (W), Met (M), Tyr (Y), Cys (C), Asp (D) и Glu (E). Если предусмотрен активный фрагмент, то он включает такие модификации. Например, если в документе предусмотрены полипептиды фактора VII (FVII), их аллельные и видовые варианты или активные фрагменты, или другие их варианты, то они также включают замены, выбранные из D196F, D196W, D196L, D196I, D196Y, К197Е, K197D, K197L, К197М, K197I, K197V. K197F, K197W, K199D, К199Е и K197Y.
Кроме того, предусмотренные модифицированные FVII-полипептиды могут содержать дополнительные модификации, включая замены аминокислоты, вставки или делеции, по другой позиции в FVII-полипептиде. В некоторых примерах, дополнительная модификация представляет собой замену аминокислоты по положению, соответствующему выбранному из D196, К197, К199, G237, Т239, R290 и К341, где первая модификация и вторая модификация происходят с различными аминокислотами. Эти модификации включают, но не ограничиваются D196K, D196R, D196A, D196Y, D196F, D196W, D196L, D196I, K197Y, К197А, К197Е, K197D, K197L, К197М, K197I, K197V, K197F, K197W, К199А, K199D, К199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, К341Е, K341R, K341N, К341М, K341D и K341Q. Другие примеры модификаций являются примерами вставок аминокислот, включая, но не ограничиваясь, любой из следующих вставок: G237Т238вставкаА, G237Т238вставка8, G237Т238вставкаV, G237Т238вставкаАS, G237Т238вставка8А, D196К197вставкаК, D196К197вставкаК, D196К197вставкаY, D196K197вставкаW, D196К197вставкаА, В196К197вставкаМ, К1971198вставкаЕ, К1971198вставкаУ, К1971198вставкаА и К197П98вставкаS. Другие примеры модифицированных полипептидов FVII включают следующие модификации: D196R/K197E/K199E, D196K/K197E/K199E, D196R/K197E/K199E/R290E, D196R/K197M/K199E, D196R/K197M/ K199E/R290E, D196K/K197L, D196F / K197L, D196L / K197L, D196M/K197L, D196W/K197L, D196F/K197E, D196W/K197E, K196V/K197E, K197E/K341Q, K197L/K341Q, K197E/G237V/K341Q, К197Е/К199Е, K197E/G237V, К199Е/ K341Q или K197E/K199E/K341Q.
Также предусмотрены модифицированные FVII-полипептиды, включающие их видовые и аллельные варианты или их активные фрагменты, или другие их варианты, которые также могут содержать модифицированные аминокислоты, соответствующие какой-либо одной или нескольким из D196R, D196Y, D196F, D196W, D196L, D196I, K197Y, К197Е, K197D, K197L, К197М, К197М, К1971, K197V, K197F, K197W, K199D, К199Е, G237W, G237I, G237V, R290M, R290V, К341М, K341D, G237Т238вставкаА, G237Т238вставкаS, G237Т238вставкаУ, С237Т238вставкаА8, G237Т238вставка8А, D196K197вставкаK, D196K197вставкаR, D196K197вставкаY, D196K197вставкаW, D196K197вставкаA, D196K197вставкаM, К1971198вставкаЕ, К1971198вставкаУ, К1971198вставкаА или К1971198вставка8 в FVII-полипептиде с последовательностью аминокислот, приведенной в SEQ ID №3 или в соответствующих остатках в полипептиде FVII. Кроме того, эти модифицированные полипептиды FVII могут содержать дополнительные модификации, в том числе замены аминокислот, вставки или делеции по другим положениям в полипептиде FVII. В некоторых примерах, дополнительная модификация может представлять собой замену аминокислоты в положениях, соответствующих положениям D196, К197, К199, G237, Т239, R290 и К341, где положение, по которому произошла дополнительная модификация, отличается от положения, по которому произошла первая модификация. Например, модифицированный полипептид FVII может дополнительно содержать замену аминокислоты, выбранную из D196K, D196R, D196A, D196Y, D196F, D196M, D196W, D196L, D196I, K197Y, К197А, К197Е, K197D, K197L, К197М, K197I, K197V, K197F, K197W, К199А, K199D, К199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, К341Е, K341R, K341N, К341М, K341D и K341Q. Примеры таких модифицированных полипептидов FVII включают модификации, выбранные из D196R/R290E, D196R/R290D, D196R/K197E/K199E, D196K/K197E/K199E, D196R/K197E/K199E/R290E, D196R/K197M/K199E, D196R/K197M/K199E/R290E, D196K/K197L, D196F/K197L, D196L/K197L, D196M/K197L, D196W/K197L, D196F/K197E, D196W/K197E, K197L/K341Q, G237V/K341Q, K197E/G237V/K341Q, К197Е/К199Е, K197E/G237V, K199E/K341Q, K197E/K199E/K341Q и K196V/K197E.
В некоторых случаях, модифицированный полипептид фактора VII (FVII), в том числе его видовые и аллельные варианты, или его активные фрагменты, или другие его варианты могут включать две или более модификаций в полипептиде FVII, где, по крайней мере, две модификации аминокислот соответствуют D196K, D196R, D196A, D196Y, D196F, D196M, D196W, D196L, D196I, K197Y, К197А, К197Е, K197D, K197L, К197М, K197I, K197V, K197F, K197W, К199А, K199D, К199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, К341Е, K341R, K341N, К341М, K341D, K341Q, G237Т238вставкаА, G237Т238вставкаS, G237Т238вставкаV, G237Т238вставкаАS, G237Т238вставкаSА, G196К197вставкаК, D196К197вставкаR, D196К197вставкаY, D196K197вставкаW, D196К197вставкаА, D196К197вставкаМ, К197I198вставкаЕ, К1971198вставкаY, К1971198вставкаА или К1971198вставкаS в полипептиде FVII, обладающем последовательностью, или включающем последовательность аминокислот, приведенную в SEQ ID №3 или в соответствующих остатках в полипептиде FVII. Полипептид может включать более двух модификаций, например 2, 3, 4, 5, 6 или 7 модификаций.
Примерами этого являются FVII-полипептиды, которые обладают модификациями, выбранными из D196R/R290E, D196K/R290E, D196R/R290D, D196R/K197E/K199E, D196K/K197E/K199E, D196R/K197E/K199E/R290E, D196R/K197M/K199E, D196R/K197M/K199E/R290E, D196K/K197L, D196F/K197L, D196L/K197L, D196M/K197L, D196W/K197L, D196F/K197E, D196W/K197E, D196V/K197E, K197E/K341Q, K197L/K341Q, G237V/K341Q, K197E/G237V/K341Q, К197Е/К199Е, K197E/G237V, K199E/K341Q, K197E/K199E/K341Q и K197E/G237V/M298Q.
Любой из вышеупомянутых модифицированных FVII-полипептидов может проявлять повышенную устойчивость к ингибитору запускаемого тканевым фактором пути (TFPI) по сравнению с немодифицированным FVII-полипептидом. В некоторых примерах, модифицированный полипептид FVII по крайней мере приблизительно или точно на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500% или более устойчив к TFPI. Кроме того, эти модифицированные полипептиды FVII также могут содержать гетерологичный Gla домен или его достаточную для связывания фосфолипидов часть.
Кроме того, в настоящем документе предусмотрены модифицированные FVII-полипептиды, содержащие гетерологичный Gla домен, или его достаточную часть, например 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или большую часть гетерологичного Gla домена, для связывания фосфолипидов. Все и каждый в отдельности из вышеупомянутых модифицированных FVII-полипептидов также могут включать замены Gla, в том числе замены Gla, как проиллюстрировано и описано в данном документе.
Гетерологичный Gla домен может быть выбран из Gla доменов фактора IX (FIX), фактора Х (FX), протромбина, протеина С, протеина S, остеокальцина, Gla протеина матрикса, белка Gas6 (Growth-arrest -specific) или белка Z. В некоторых примерах настоящего документа гетерологичный Gla домен в модифицированном полипептиде FVII обладает последовательностью аминокислот, приведенной в любой из Последовательностей SEQ ID №№: 110-118, 120 и 121, или его частью, достаточной для связывания фосфолипидов. Модификации в полипептиде FVII могут быть осуществлены удалением всего или непрерывной части нативного Gla домена FVII, которая может включать аминокислоты 1-45 в полипептиде FVII, обладающем последовательностью или включающем последовательность аминокислот, приведенную в SEQ ID №3, или в соответствующих остатках в полипептиде FVII, и заменой его на гетерологичный Gla домен или его часть, достаточную, чтобы повлиять на связывание фосфолипидов, например увеличить его. Такие модификации приводят к тому, что модифицированный полипептид FVII проявляет повышенную способность к связыванию фосфолипидов по сравнению с немодифицированным FVII-полипептидом. Например, модифицированный FVII-полипептид может проявлять, по крайней мере, приблизительно или точно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500% или более повышенную способность связывать фосфолипиды.
У модифицированных FVII-полипептидов может быть удален весь нативный FVII Gla домен или его непрерывная часть, и он может быть заменен на гетерологичный домен Gla, или его часть, достаточную для осуществления связывания фосфолипидов. Примерами таких полипептидов являются те, в которых нативный FVII Gla домен содержит аминокислоты 1-45 в полипептиде FVII, обладающем последовательностью или включающем последовательность аминокислот, приведенную в SEQ ID №3, или в соответствующих остатках в полипептиде FVII. Gla модификации включают, но не ограничиваются, Gla заменой FIX, Gla заменой FX, Gla заменой белка С, Gla заменой белка S и Gla заменой тромбина.
В силу замены Gla модифицированные полипептиды FVII могут проявлять повышенную способность связывать фосфолипиды. Такое увеличение может быть, по крайней мере, приблизительно или точно 0,5%, 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500% или более.
Модифицированные полипептиды FVII, содержащие гетерологичный Gla домен, могут содержать дополнительные модификации в позициях, которые приводят к или проявляют повышенную устойчивость к ингибитору пути, запускаемому тканевым фактором (TFPI) по сравнению с немодифицированным FVII-полипептидом. Примерами таких модификаций являются модификации(ия) одной или нескольких аминокислот по позициям, выбранным из D 196, К 197, К199, G237, Т239, R290 и К341 в полипептиде FVII, обладающем последовательностью или имеющем последовательность аминокислот, приведенную в SEQ ID №3 или в соответствующих остатках в полипептиде FVII. В частности, такие модификации могут включать одну или несколько модификаций аминокислот(ы), выбранных из D196K-, D196R, D196A, D196Y, D196F, D196M, D196W, D196L, D196I, K197Y, К197А, К197Е, K197D, K197L, К197М, K197I, K197V, K197F, K197W, К199А, K199D, К199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, К341Е, K341R, K341N, К341М, K341D, K341Q, G237Т238вставкаА, G237Т238вставкаS, G237Т238вставкаV, G237Т238вставкаАS, G237Т238вставкаSА, В196K197вставкаК, D196К197вставкаR, D196К197вставкаY, D196K197вставкаW, D196К197вставкаА, D196К197вставкаМ, К1971198вставкаЕ, К1971198вставкаУ, К1971198вставкаА и К197Н98вставкаS. Например, модифицированные FVII-полипептиды, содержащие гетерологичный Gla домен, также могут содержать аминокислотные замены и/или вставки, такие как D196R/R290E, D196K/R290E, D196R/R290D, D196R/K197E/K199E, D196K/K197E/K199E, D196R7K197E/K199E/R290E, D196R/K197M/K199E и D196R/K197M/K199E/R290E, но не ограничиваясь этим. В некоторых случаях такие дополнительные модификации приводят к повышенной устойчивости модифицированного полипептида к TFPI по сравнению с полипептидом FVII, не содержащим модификаций, т.е. с немодифицированным FVII-полипептидом.
Любые из описанных выше модифицированных FVII-полипептидов, в том числе те, которые демонстрируют повышенную устойчивость к TFPI или повышенную способность связывать фосфолипиды, или любая их комбинация, может дополнительно содержать другие модификации, в том числе любые описанные в уровне техники. Такая дальнейшая модификация аминокислот может повысить устойчивость к антитромбину-III (AT-III), повысить способность к связыванию и/или сродство к фосфолипидам, увеличить сродство к тканевому фактору (TF), повысить присущую полипептиду активность, изменить конформацию полипептида для изменения зимогенности, включая изменение конформации в более зимогенподобную форму или менее зимогенподобную форму, повысить устойчивость к протеазам, снизить гликозилирование, увеличить гликозилирование, снизить иммуногенность, повысить стабильность, и/или способствовать к связыванию химических групп. Например, предусмотренные в настоящем документе модифицированные полипептиды FVII могут содержать модификации(ию) в позициях Q176, М298 и Е296 в полипептиде FVII, обладающем последовательностью или имеющем последовательность аминокислот, приведенную в SEQ ID №3 или в соответствующих остатках в FVII-полипептиде. В некоторых примерах, модифицированные FVII-полипептиды могут дополнительно содержать модификации аминокислот, выбранные из Q176A, M298Q, E296V и/или Е296А. В других примерах, модифицированные FVII-полипептиды могут дополнительно содержать, в дополнение к замене Gla и/или другим указанным модификациям, одну или более из следующих дополнительных модификаций аминокислот (ы): S278C/V302C, L279C/N301C, V280C/V301C, S281C/V299C, вставка тирозина в положение 4, F4S, F4T, P10Q, Р10Е, P10D, P10N, Q21N, R28F, R28E, I30C, BOD, I30E, K32D, K32Q, К32Е, K32G, К32Н, К32Т, К32С, К32А, K32S, D33C, D33F, D33E, D33K, А34С, А34Е, A34D, A34I, A34L, А34М, A34V, A34F, A34W, A34Y, R36D, R36E, Т37С, T37D, Т37Е, К38С, К38Е, К38Т, K38D, K38L, K38G, К38А, K38S, K38N, К38Н, L39E, L39Q, L39H, W41N, W41C, W41E, W41 D, I42R, I42N, I42S, I42A, I42Q, I42N, I42S, I42A, I42Q, I42K, S43Q, S43N, Y44K, Y44C, Y44D, Y44E, S45C, S45D, S45E, D46C, A51N, S53N, G58N, G59S, G59T, К62Е, K62R, K62D, K62N, K62Q, К62Т, L65Q, L65S, L65N, F71D, F71Y, F71E, F71Q, F71N, P74S, Р74А, А75Е, A75D, Е77А, E82Q, E82N, E82S, Е82Т Т83К, N95S, N95T, G97S, G97T, Y101N, D104N, T106N, K109N, E116D, G117N, G124N, S126N, T128N, L141C, L141D, L141E, E142D, E142C, K143C, K143D, K143E, R144E, R144C, R144D, N145Y, N145G, N145F, N145M, N145S, N1451, N145L, N145T, N145V, N145P, N145K, N145H, N145Q, N145E, N145R, N145W, N145D, N145C, K157V, K157L, K157I, K157M, K157F, K157W, K157P, K157G, K157S, K157T, K157C, K157Y, K157N, K157E, K157R, K157H, K157D, K157Q, V158L, V158I, V158M, V158F, V158W. V158P, V158G, V158S, V158T, V158C, V158Y, V158N, V158E, V158R, V158K, V158H, V158D, V158Q, A175S, A175T, G179N, I186S, I186T, V188N, R202S, R202T, I205S, I205T, D212N, E220N, I230N, P231N, P236N, G237N. Q250C, V253N, E265N, T267N, E270N, A274M, A274L, A274K, A274R, A274D, A274V, A274I, A274F, A274W, A274P, A274G, A274T, A274C, A274Y, A274N, A274E, A274H, A274S, A274Q, F275H, R277N, F278S, F278, F278N, F278Q, F278G, L280N, L288K, L288C, L288D, D289C, D289K, L288E, R290C, R290G, R290A, R290S, R290T, R290K, R290D, R290E, G291E, G291D, G291C, G291N, G291K, A292C, A292K, A292D, A292E, T293K, E296V, E296L, E296I, E296M, E296F, E296W, E296P, E296G, E296S, E296T, E296C, E296Y, E296N, E296K, E296R, E296H, E296D, E296Q, M298Q, M298V, M298L, M298I, M298F, M298W, M298P, M298G, M298S, M298T, M298C, M298Y, M298N, M298K, M298R, M298H, M298E, M298D, P303S, P303ST, R304Y, R304F, R304L, R304M, R304G, R304T, R304A, R304S, R304N, L305V, L305Y, L305I, L305F, L305A, L305M, L305W, L305P, L305G, L305S, L305T, L305C, L305N, L305E, L305K, L305R, L305H, L305D, L305Q, M306D, M306N, D309S, D309T, Q312N, Q313K, Q313D, Q313E, S314A, S314V, S3141, S314M, S314F, S314W, S314P, S314G, S314L, S314T, S314C, S314Y, S314N, S314E, S314K, S314R, S314H, S314D, S314Q, R315K, R315G, R315A, R315S, R315T, R315Q, R315C, R315D, R315E, K316D, К316С, К316Е, V317C, V317K, V317D, V317E, G318N, N322Y, N322G, N322F, N322M, N322S, N3221, N322L, N322T, N322V, N322P, N322K, N322H, N322Q, N322E, N322R, N322W, N322C, G331N, Y332S, Y332A, Y332N, Y332Q, Y332G, D334G, D334E, D334A, D334V, D334I, D334M, D334F, D334W, D334P, D334L, D334T, D334C, D334Y, D334N, D334K, D334R. D334H, D334S, D334Q, S336G, S336E, S336A, S336V, S336I, S336M, S336F, S336W, S336P, S336L, S336T, S336C, S336Y, S336N, S336K, S336R, S336H, S336D, S336Q, K337L, K337V, K337I, К337М, K337F, K337W, К337Р, K337G, K337S, К337Т, К337С, K337Y, K337N, К337Е, K337R, К337Н, K337D, K337Q, К341Е, K341Q, K341G, К341Т, К341А, K341S, G342N, H348N, R353N, Y357N, I361N, F374P, F374A, F374V, F374I, F374L, F374M, F374W, F374G, F374S, F374T, F374C, F374Y, F374N, F374E, F374K, F374R, F374H, F374D, F374Q, V376N, R379N, L390C, L390K, L390D, L390E, M391D, М391С, М391К, M391N, М391Е, R392C, R392D, R392E, S393D, S393C, S393K, S393E, Е394К, Р395К, Е394С, P395D, Р395С, Р395Е, R396K, R396C, R396D, R396E, P397D, Р397К, Р397С, Р397Е, G398K, G398C, G398D, G398E, V399C, V399D, V399K, V399E, L400K, L401K, L401C, L401D, L401E, R402D, R402C, R402K, R402E, А403К, А403С, A403D, А403Е, Р404Е, P404D, Р404С, Р404К, F405K, Р406С, K32N/A34S, K32N/A34T, F31N/D33S, F31N/D33T, I30N/K32S, I30N/K32T, A34N/R36S, A34N/R36T, K38N/F40S, K38N/F40T, T37N/L39S, T37N/L39T, R36N/K38S, R36N/K38T, L39N/W41S, L39N/W41T, F40N/I42S, F40N/I42T, I42N/Y44S, I42N/Y44T, Y44N/D46S, Y44N/D46T, D46N/D48S, D46N/D48T, G47N/Q49S, G47N/Q49T, K143N/N145S, K143N/N145T, E142N/R144S, E142N/R144T, L141N/K143S, L141N/K143T, I140N/E142S/, I140N/E142T, R144N/A146S, R144N/A146T, A146N/K148S, A146N/K148T, S147N/P149S/, S147N/P149T, R290N/A292S, R290N/A292T, D289N/G291 S, D289N/G291T, L288N/R290S, L288N/R290T, L287N/D289S, L287N/D289, A292N/A294S, A292N/A294T, T293N/L295S, T293N/L295T, R315N/V317S, R315N/V317T, S314N /K316S, S314N /К316Т, Q313N/ R315S, Q313N/R315T, K316N/G318S, K316N/G318T, V317N/D319S, V317N/D319T, K341N/ D343S, K341N / D343T, S339N/K341S, S339N/K341T, D343N/G345S, D343N/G345T, R392N/E394S, R392N/E394T, L390N / R392S, L390N / R392T, K389N/M391S, K389N/M391T, S393N/P395S, S393N/P395T, E394N/R396S, E394N/R396T, P395N/P397S, P395N/P397T, R396N/G398S, R396N/G398T, P397N/V399S, P397N/V399T, G398N/L400S, G398N/L400T, V399N/L401S, V399N/L401T, L400N/R402S, L400N/R402T, L401N/A403S, L401N/A403T, R402N/P404S, R402N/P404T, A403N/F405S, A403N/F405T, P404N/P406S и P404N/P406T, V158D/G237V/E296V/M298Q, K197E/G237V/M298Q, K197E/G237V/M298Q/K341Q, K197E/K199E/G237V/M298Q/K341Q, G237V/M298Q, G237V/M298Q/K341Q, M298Q/Gla замена FIX, K197E/M298Q и M298Q/K341D.
Любой из предусмотренных в настоящем документе модифицированных FVII-полипептидов может содержать одну или несколько дополнительных модификаций аминокислот(ы), которые повышают устойчивость к антитромбину-III (AT-III), повышают способность к связыванию и/или сродство к фосфолипидам, увеличивают сродство к тканевому фактору (TF), повышают присущую полипептиду активность, изменяют конформацию полипептида, что приводит к изменению отношения активностей зимогена и активного фермента, повышают устойчивость к протеазам, снижают гликозилирование, увеличивают гликозилирование, снижают иммуногенность, повышают стабильность и/или способствуют связыванию химических групп. Например, изменение отношения активностей зимогена и активного фермента может привести к более зимогенподобной форме или менее зимогенподобной форме. Такие модифицированные FVII-полипептиды, например, могут включать замещение аминокислот по позициям 300-322, 305-322, 300-312, или 305-312 соответствующими аминокислотами из трипсина, тромбина или FX, или замещение аминокислот по позициям 310-329, 311-322 и 233-329 соответствующими аминокислотами из трипсина.
Модифицированные полипептиды, предусмотренные в настоящем документе, включают полипептиды, в которых немодифицированный полипептид FVII включает или содержит только последовательность аминокислот, приведенную в SEQ ID №3. Примеры модифицированных FVII-полипептидов представляют собой полипептиды, которые обладают последовательностями или включают последовательности аминокислот, приведенные в SEQ ID №№18-43, 125-150 и 206-250 или их аллельные или видовые варианты, или другие их варианты. Аллельные или видовые варианты, или другие варианты могут обладать 40%, 50%, 60%, 65%, 70%. 75%, 80%, 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательности по отношению к полипептиду, приведенному в SEQ ID №3, за исключением модификаций аминокислот(ы). Источники полипептидов FVII включают людей и другие виды. Полипептиды могут представлять собой зрелые полипептиды или предшественники полипептидов. В некоторых вариантах, изменяется только первичная последовательность полипептида FVII. Кроме того, FVII-полипептиды могут включать химические модификации или посттрансляционные модификации, включая, но не ограничиваясь, гликозилированием, карбоксилированием, гидроксилированием, сульфированием, фосфорилированием, присоединением альбумина или связыванием с другим фрагментом, например полиэтиленгликольным (PEG) фрагментом.
Модифицированные FVII-полипептиды могут быть представлены одноцепочечными полипептидами или в виде смеси одноцепочечных и двуцепочечных или мультицепочечных форм, или они могут быть представлены как двуцепочечные, трехцепочечные или другие мультицепочечные формы. Модифицированные FVII-полипептиды могут быть представлены неактивными или активированными полипептидами. Активация может быть произведена, например, самоактивационным протеолитическими расщеплением, расщеплением фактором IX (FIXa), расщеплением фактором Х (FXa), расщеплением фактором XII (FXIIa) или расщеплением тромбином.
Модифицированные FVII-полипептиды обычно сохраняют один или более видов активности или свойств немодифицированного FVII-полипептида. Модификации могут включать модификации по 1, 2, 3,4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50 или 60 аминокислотным позициям при условии, что полипептид сохраняет, по крайней мере, один вид FVII активности немодифицированного FVII-полипептида. Сохранение активности может быть не менее, приблизительно или точно 1%, 2%, 3%, 4%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500% или более активности немодифицированного FVII-полипептида. Виды активности включают, например, связывание тканевого фактора (TF), активацию фактора Х (FX), активацию фактора IX (FIX), связывание фосфолипидов и коагуляционную активность. Активности могут быть увеличены или уменьшены. Среди предусмотренных в настоящем документе модифицированных полипептидов представлены полипептиды, у которых увеличена коагуляционная активность. Активность может быть оценена в in vitro или in vivo.
Также в настоящем документе предусмотрены молекулы нуклеиновой кислоты, которые включают последовательность нуклеотидов, которая кодирует любые из модифицированных полипептидов FVII. Также в настоящем документе предусмотрены векторы, такие как прокариотические векторы или эукариотические векторы, в том числе векторы млекопитающих и вирусные векторы. Примеры вирусных векторов включают аденовирусный, аденосвязанный вирусный, ретровирусный векторы, вектор на основе вируса герпеса, лентивирусный, поксвирусный и цитомегаловирусный векторы. Также в настоящем документе предусмотрены клетки, содержащие указанные молекулы нуклеиновой кислоты или векторы. Клетки могут быть эукариотическими, например, клетки млекопитающих или дрожжей, или прокариотическими. Клетки млекопитающих включают, например, клетки почек детенышей хомяков (ВНК-21) или клетки культуры 293 и клетки СНО. Клетки могут быть выращены при определенных условиях, при которых экспрессируется модифицированный полипептид FVII. Он может быть выделен. В настоящем документе предусмотрен модифицированный полипептид FVII, продуцируемый такими клетками.
Также в настоящем документе предусмотрены композиции, содержащие модифицированные полипептиды FVII, предусмотренные в настоящем документе. В частности, предусмотрены фармацевтические композиции, которые содержат такие полипептиды. Композиции могут содержать терапевтически эффективные концентрации или количества модифицированного полипептида FVII, или молекул нуклеиновой кислоты, или вектора, либо предусмотренных в настоящем документе клеток с фармацевтически приемлемым носителем. Фармацевтические композиции могут быть предназначены для введения одной дозой или многократными дозами. Фармацевтическая композиция может быть в любой форме, такой как жидкая, гелеобразная или твердая, и находиться в капсулах, контейнерах и других подходящих формах. Она может быть составлена для разбавления перед введением или в любом удобном виде. Количество может зависеть от конкретного подвергающегося лечению расстройства и/или от индивидуума, подвергающегося лечению, и, при необходимости, может быть определено эмпирически. Фармацевтическая композиция может быть составлена для любого способа введения, в том числе, например, такого как локальное, системное, или местное введение, или составлена для орального, назального, легочного, буккального, трансдермального, подкожного, интрадуоденального, энтерального, парентерального, внутривенного или внутримышечного введения. Фармацевтические композиции могут быть составлены для управляемого высвобождения.
Также предусмотрены методы лечения и применения композиций для лечения. Фармацевтические композиции вводятся или составляются для введения субъекту, который имеет заболевание или состояние, которое подвергается лечению, введением FVII, включая лечение введением активного FVII (FVIIa). Лечение фармацевтической композицией улучшает или облегчает симптомы, связанные с заболеванием или состоянием. Введение может следовать за наблюдением, либо происходить в сочетании с наблюдением субъекта на предмет изменений в симптомах, связанных с FVII-опосредованными заболеваниями или состояниями. Заболевания или состояния, подвергающиеся лечению, включают, но не ограничиваются, нарушениями свертывания крови, гематологическими расстройствами, геморрагическими расстройствами, гемофилиями, дефицитом фактора VII и кровоточивостью. Примерами заболеваний являются гемофилия А или гемофилия В, или гемофилия С. Гемофилия может быть врожденной или приобретенной, например возникшей в связи с кровотечениями, являющимися осложнениями операции или травмы. Кровотечение может проявляться в виде острых гемартрозов, хронической гемофильной артропатии, гематом, гематурии, кровотечений в центральной нервной системе, желудочно-кишечных кровотечений, или кровоизлияния в мозг, и может быть результатом, например, удаления зубов или операций, такие как, например, ангиопластика, легочная хирургия, абдоминальная хирургия, спинальная хирургия, хирургия мозга, сосудистая хирургия, стоматологическая хирургия или трансплантация органов, такая как трансплантация костного мозга, сердца, легких, поджелудочной железы и печени. Субъект может иметь аутоантитела к фактору VIII или фактору IX. Лечение может сопровождаться последовательным или периодическим введением одного или более дополнительных факторов коагуляции, таких как, например, очищенные из плазмы или рекомбинантные факторы свертывания, прокоагулянты, такие как витамин К, производные витамина К, ингибиторы протеина С, плазма, тромбоциты, эритроциты и кортикостероиды. Фармацевтическая композиция может быть использована с композициями, которые содержат указанные другие факторы свертывания крови.
В настоящем документе предусмотрены изделия, которые включают упаковочные материалы и фармацевтическую композицию, содержащуюся в упаковочном материале, и, необязательно, инструкции по применению. Например, модифицированный FVII-полипептид в фармацевтической композиции для лечения FVII-опосредованных заболеваний или расстройств, и упаковочный материал, который может включать этикетку, на которой указано, что модифицированный FVII-полипептид используется для лечения FVII-опосредованных заболеваний или расстройств. Также предусмотрены наборы, содержащие фармацевтические композиции, и устройство для введения композиции и, необязательно, инструкции для введения.
Краткое описание фигур
На фигуре 1 изображен каскад коагуляции. На фигуре показаны внутренний и внешний пути коагуляции, приводящие к независимому получению FXa и схождению путей к общему пути выработки тромбина и фибрина для образования сгустка. Они являются взаимосвязанными путями. На фигуре изображен порядок участия молекул в активационном каскаде, в котором зимоген преобразуются в активированную протеазу путем расщепления одной или нескольких пептидных связей. Затем активированная протеаза служит в качестве активирующей протеазы для следующей молекулы зимогена в каскаде, в конечном итоге приводя к образованию сгустков.
На фигуре 2 изображена клеточная модель коагуляции (см., например, Hoffman и др. (2001) Thromb Haemost 85:958-965). На фигуре изображены акты коагуляции, разделенные на три фазы, где инициация коагуляции производится активацией FX в FXa под действием TF / FVIIa комплекса на TF-несущей клетке, что приводит к генерации небольшого количества тромбина после активации под действием FXa / FVa. Усиление происходит, когда тромбин связывается и активирует тромбоциты, а также инициирует активацию достаточного количества соответствующих факторов коагуляции с образованием FVIIIa / FIXa и FVa / FXa комплексов. Распространение коагуляции происходит на поверхности большого количества активированных тромбоцитов в месте повреждения, что приводит к резкому увеличению генерации тромбина, количество которого достаточно для генерации достаточного количества фибрина из фибриногена для создания сгустка в месте повреждения.
На фигуре 3 показаны механизмы, с помощью которых FVIIa может инициировать образование тромбина. Фигура иллюстрирует TF-зависимый путь FVIIa генерации тромбина, который происходит на поверхности TF-несущих клеток и включает образование комплекса FVIIa с TF до активации FX в FXa. На фигуре также изображен TF-независимый путь FVIIa генерации тромбина, в ходе которого FVIIa связывается с фосфолипидами на активированных тромбоцитах и активирует FX в FXa, который, в свою очередь, образует комплекс с FV, который расщепляет протромбин с образованием тромбина.
На фигуре 4 изображен четвертичный ингибиторный комплекс, который образуется, когда TFPI/FXa комплекс связывается TF/FVIIa комплексом. TFPI содержит три домена Кунитца. Домен Кунитца 2 (К-2) взаимодействует с FXa и ингибирует его, домен Кунитца 1 (К-1) взаимодействует и ингибирует FVIIa.
На фигуре 5 изображены аминокислотные последовательности первого домена Кунитца BPTI5L15 (аминокислоты с 1 по 55 позиции из SEQ ID №106) и TFPI-2 (аминокислоты 14-65 позиции из SEQ ID №105), и TFPI-1 (аминокислоты 26-76 позиции из SEQ ID №102), и TFPI-2 (аминокислоты 14-65 позиции из SEQ ID №105) и показаны консервативные аминокислоты.
На фигуре 6 изображена гомологичная модель, используемая для определения контактирующих остатков в области взаимодействия между FVIIa и TFPI. Структура домена Кунитца 1 (К-1) из TFPI-2 была получена из кристаллической структуры комплекса трипсин / TFPI и смоделирована на BPTI5L15 на TF/FVIIa/BPTI5L15 кристаллической структуре. Было проведено компьютерное моделирование мутагенеза для подгонки соответствующих аминокислот TFPI-1 К-1 в модели. Были определены контактирующие остатки FVIIa, которые, вероятно, участвуют во взаимодействии с TFPI в области белок-белкового контакта, и они были приняты в качестве кандидатов для мутагенеза в разработке TFPI-устойчивого TFPI полипептида.
На фигуре 7 изображено смоделированное взаимодействие между FVIIa и TFPI. В частности, на фигуре изображены контактирующие остатки FVII, которые находятся в области взаимодействия FVIIa и TFPI, и соответствующие контактные остатки TFPI, которые образуют дополнительные электростатические контакты.
Подробное описание
Оглавление
А. Определения
В. Обзор гемостаза
1. Адгезия и агрегация тромбоцитов
2. Каскад коагуляции
а. Инициация
b. Усиление
с. Распространение
3. Регуляция коагуляции
С.фактор VII (FVII)
1. Структура и организация FVII
2. Посттрансляционные модификации
3. Процессинг FVII
4. Активация FVII
5. Функции FVII
а. Зависимая от тканевого фактора активность FVIIa
b. Независимая от тканевого фактора активность FVIIa
6. FVII как биотехнологический лекарственный препарат
D. Модифицированные FVII-полипептиды
1. Устойчивость к ингибиторам
a. TFPI, модификации, приводящие к повышенной устойчивости к TFPI
b. Антитромбин III (AT-III), модификации, приводящие к повышенной устойчивости к AT-III
2. Связывание с активированными тромбоцитами, модификация путем введения гетерологичного Gla домена
3. Комбинации и дополнительные модификации
а. Модификации, которые увеличивают внутреннюю активность
b. Модификации, повышающие устойчивость к протеазам
с. Модификации, увеличивающие способность связывать фосфолипиды
d. Модификации, изменяющие гликозилирование
е. Модификации, содействующие связыванию химических групп
f. Примеры мутантных комбинаций FVII
Е. Способы и методы модификации FVII
1. Рациональные
2. Эмпирические (то есть скрининг)
а. Случайный мутагенез
b. Специфичный мутагенез
с. Скрининг
3. Выбор вариантов FVII
F. Производство полипептидов FVII
1. Векторы и клетки
2. Системы экспрессии
а. Прокариотическая экспрессия
b. Дрожжи
с. Насекомые и клетки насекомых
d. Клетки млекопитающих
е. Растения
2. Очистка
3. Гибридные белки
4. Модификации полипептидов
5. Нуклеотидные последовательности
G. Оценка активности модифицированных FVII-полипептидов
1. Анализы in vitro
а. Посттрансляционные модификации
b. Протеолитическая активность
с. Коагуляционная активность
d. Связывание и/или ингибирование другими белками
е. Связывание фосфолипидов
2. Модели на животных
3. Клинические анализы
Н. Фармацевтические композиции и способы введения
1. Фармацевтические композиции
а. Дозировка
b. Лекарственные формы
2. Введение модифицированных полипептидов FVII
3. Введение нуклеиновых кислот, кодирующих модифицированные FVII-полипептиды (генная терапия)
I. Терапевтическое применение
1. Врожденные нарушения свертывания крови
а. Гемофилия
b. Дефицит FVII
с. Другое
2. Приобретенные нарушения свертывания крови
а. Приобретенная в результате химиотерапии тромбоцитопения
b. Другие коагулопатии
с. Кровотечения при трансплантации
d. Вызванные антикоагулянтной терапией кровотечения
е. Приобретенная гемофилия
3. Кровотечения при травме и хирургии
J. Комбинированная терапия
К. Изделия и наборы
L. Примеры
А. Определения
Если не определено иное, все технические и научные термины, используемые в настоящем документе, имеют тот же смысл, как они обычно понимаются специалистом в области техники, к которой относится изобретение. Все патенты, патентные заявки, опубликованные заявки и публикации, Genbank последовательности, базы данных, веб-сайты и других печатные материалы, упомянутые в процессе раскрытия изобретения, если не указано иное, приведены в ссылках в полном объеме. В том случае, если для терминов, используемых в настоящем документе, есть множество определений, то превалируют значения, указанные в этом разделе. Когда делается ссылка на URL или другой такой идентификатор или адрес, то понимается, что такие идентификаторы могут измениться, и конкретная информация в Интернете может появляться и исчезать, но может быть найдена эквивалентная информация в Интернете. Ссылка на такую информацию свидетельствует о наличии и широком распространении такой информации.
В настоящем документе пути коагуляции или каскады коагуляции относятся к сериям актов активации, которые приводят к образованию нерастворимого сгустка фибрина. При коагуляционном каскаде или пути неактивные белки сериновых протеаз (также называемые зимогенами) преобразуются в активные протеазы путем расщепления одной или нескольких пептидных связей, а затем служат в качестве активирующих протеаз для следующей молекулы зимогена в каскаде. В конечной протеолитической стадии каскада фибриноген протеолитически расщепляется тромбином в фибрин, который затем сшивается в месте повреждения и формирует сгусток.
Используемый в настоящем документе термин "гемостаз" относится к остановке кровотечения или кровотока в органе или части тела. Термин гемостаз может охватить весь процесс свертывания крови для предотвращения потери крови после повреждения кровеносного сосуда и последующее разрушение сгустка крови, следующее за восстановлением ткани.
Используемые в настоящем документе термины "свертывание" или "коагуляция" относятся к образованию нерастворимого сгустка фибрина или процессу, в котором факторы свертывания крови взаимодействуют в каскаде коагуляции, что в конечном итоге приводит к образованию нерастворимого сгустка фибрина.
Используемый в настоящем документе термин "протеаза" относится к ферменту, который катализирует гидролиз пептидных ковалентных связей. Эти обозначения относятся к зимогенным формам и их активированным одно-, двух- и мультицепочечным формам. Для ясности, упоминание протеаз относится ко всем этим формам. Протеазы включают, например, сериновые протеазы, цистеиновые протеазы, аспарагиновые протеазы, треониновые и металлопротеазы в зависимости от каталитической активности их активного центра и механизма расщепления пептидных связей в целевом субстрате.
Используемые в настоящем документе термины «сериновая протеаза» или сериновая эндопептидаза относятся к классу пептидаз, которые характеризуются наличием остатков серина в активном центре фермента. Сериновые протеазы участвуют в широком диапазоне функций в организме, включая свертывание крови и воспаление, а также функционируют как пищеварительные ферменты у прокариотов и эукариотов. Механизм расщепления сериновыми протеазами основан на нуклеофильной атаке целевой пептидной связи серином. Цистеин, треонин или молекулы воды, связанные с аспартатом или металлами, также могут играть эту роль. Боковые цепи серина, гистидина и аспартата формируют каталитическую триаду общую для большинства сериновых протеаз.
Активный центр сериновых протеаз имеет форму расщелины, где связывается полипептидный субстрат.
Используемый в настоящем документе термин «фактор VII» (FVII, F7 также упоминается как фактор 7, фактор свертывания VII, сывороточный фактор VII, сывороточный ускоритель преобразования протромбина, SPCA, проконвертин и эптаког альфа) относится к сериновым протеазам, которые являются частью каскада коагуляции. FVII включает Gla домен, два EGF домена (EGF-1 и EGF-2), серин-протеазный домен (или пептидазный домен S1), который является высококонсервативным у всех членов семейства S1 пептидаз сериновых протеаз, таких как, например, у химотрипсина. Последовательность одного из предшественника FVII, имеющего сигнальный пептид и пропептид, приведена в SEQ ID №1. Пример зрелого полипептида FVII приведен в SEQ ID №3. FVII представлен в виде одноцепочечного зимогена, зимогеноподобного двуцепочечного полипептида и полностью активированной двуцепочечной формы. Полная активация, которая происходит через конформационный переход от зимогеноподобной формы, происходит при связывании полипептида со своим кофактором, тканевым фактором. Кроме того, могут быть введены мутации, которые приводят к изменению конформации в отсутствие тканевого фактора. Таким образом, FVII включает его одно- и двухцепочечные формы, включая зимогеноподобные и полностью активированные двухцепочечные формы.
Упоминаемые полипептиды FVII также включает полипептиды-предшественники и зрелые полипептиды FVII в одноцепочечной или двуцепочечной формах, в укороченных формах, которые обладают активностью, и включает аллельные и видовые варианты, варианты, кодирующиеся сплайсированными вариантами, и другие варианты, включая полипептиды, обладающие, по крайней мере, 40%, 45%, 50%, 55%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или большей идентичностью по отношению к последовательности полипептида-предшественника, приведенной в SEQ ID №1 или к его зрелым формам. Также включены модифицированные FVII-полипептиды, такие как приведены в SEQ ID №№18-43, 125-150 или 206-150, и их варианты. Также включены полипептиды, которые сохраняют, по меньшей мере, активность FVII, такую как связывание TF, связывание фактора X, связывание фосфолипидов, и/или коагуляционную активность FVII. При сохранении активности она может быть изменена, например, уменьшена или увеличена по сравнению с диким типом FVII настолько, пока уровень сохраненной активности достаточен для обнаруживаемого эффекта. FVII-полипептиды включают, но не ограничиваются, тканеспецифичными изоформами и их аллельными вариантами, синтетическими молекулами, полученными трансляцией нуклеиновых кислот, белками, полученными химическим синтезом, таким как сшивание коротких полипептидов, рекомбинантными методами, выделением белков из человеческих и нечеловеческих тканей и клеток, химерными полипептидами FVII и его модифицированными формами. FVII-полипептиды также включают фрагменты или части FVII, которые имеют достаточную длину или включают соответствующие районы для сохранения, по крайней мере, одного вида активности (при активации в случае необходимости) зрелого полипептида полной длины. FVII-полипептиды также включают полипептиды, которые содержат химические или посттрасляционные модификации, и те, которые не содержат химических или посттрасляционных модификаций. Такие модификации включают, но не ограничиваются, пегилированием, присоединением альбумина, гликозилированием, фарнизилированием, карбоксилированием, гидроксилированием, фосфорилированием, и другими известными в уровне техники модификациями полипептидов.
Примерами полипептидов FVII являются полипептиды млекопитающих, включая человека. Примеры аминокислотных последовательностей FVII человеческого происхождения приведены в SEQ ID №№1, 2 и 3. Примеры вариантов таких человеческих полипептидов FVII включают любые из предшественников полипептидов, приведенных в SEQ ID №№44-100. FVII-полипептиды включают также любые полипептиды нечеловеческого происхождения, включая, но не ограничиваясь, мышиными или крысиными, собачьими, кошачьими, заячьими, птичьими, бычьими, бараньими, свиными, лошадиными, рыбьими, лягушачьими и жабьими полипептидами и полипептидами фактора VII других приматов. Примеры FVII-полипептидов нечеловеческого происхождения включают, например, полипептиды коровы (Bos Taurus, последовательность SEQ ID №4), мыши (Mus musculus, последовательность SEQ ID №5), карликового шимпанзе (Pan paniscus, последовательность SEQ ID №6), шимпанзе (Pan troglodites, последовательность SEQ ID №7), кролика (Oryctolagus cuniculus, последовательность SEQ ID №8), крысы (Rattus.Norvegicus, последовательность SEQ ID №9), макаки резуса (Масаса mulatta, последовательность SEQ ID №10), свиньи (Sus Scrofa, последовательность SEQ ID №11), собаки (Canis familiaris, последовательность SEQ ID №12), полосатого данио (Brachydanio rerio, последовательность SEQ ID №13), японского иглобрюха (Fugu rubripes, SEQ ID №: 14), курицы (Gallus Gallus, последовательность SEQ ID №15), орангутанга (Pongo pygmaeus, последовательность SEQ ID №16) и гориллы (Gorilla Gorilla, последовательность SEQ ID №17).
Специалисту в данной области техники известно, что указанные позиции в зрелом полипептиде фактора VII (последовательность SEQ ID №3) отличаются на 60 аминокислотных остатков по сравнению с изоформой предшественника полипептида FVII, приведенной в SEQ ID №1, которая представляет собой изоформу полипептида фактора VII, содержащую сигнальный пептид и пропептидные последовательности. Таким образом, первый аминокислотный остаток в SEQ ID №3 "соответствует" шестьдесят первому (61) аминокислотному остатку в SEQ ID №1. Специалисту в данной области техники также известно, что указанные позиции зрелого полипептида фактора VII (последовательность SEQ ID №3) отличаются на 38 аминокислотных остатков по сравнению с предшественником FVII-полипептида, приведенным в SEQ ID №2, который является изоформой b полипептида фактора VII, содержащего сигнальный пептид и пропептидные последовательности. Таким образом, первый аминокислотный остаток в SEQ ID №3 "соответствует" тридцать девятому (39) аминокислотному остатку от SEQ ID №2.
В настоящем документе соответствующие остатки относится к остаткам, которые находятся в соответствующих выровненных локусах. Родственные или вариантные полипептиды выравниваются любым методом, известным специалисту в данной области. Такие методы обычно обеспечивают максимальное соответствие и включают такие методы, как ручное выравнивание или выравнивание с использованием многочисленных доступных программ выравнивания (например, BLASTP) и других программ, известных специалисту в данной области техники. С помощью выравнивания последовательностей полипептидов специалист в данной области может определить соответствующие остатки, используя консервативные и идентичные аминокислотные остатки в качестве ориентиров. Например, с помощью выравнивания последовательности полипептида фактора VII специалист в данной области может определить соответствующие остатки, используя консервативные и идентичные аминокислотные остатки в качестве ориентиров. Например, аланин в 1 позиции (А1) в SEQ ID №3 (зрелый фактор VII) соответствует аланину в позиции 61 (А61) в SEQ ID №1 и аланину в позиции 39 (A39) в SEQ ID №2. В других случаях могут быть идентифицированы соответствующие районы. Например, Gla домен соответствуют аминокислотным позициям с А1 до F45 в SEQ ID №3, аминокислотным позициям с А61 до S105 в SEQ ID №1 и аминокислотным позициям с А39 до S83 в SEQ ID №2. Специалист в данной области также может использовать консервативные аминокислотные остатки в качестве ориентиров, чтобы найти соответствующие аминокислотные остатки между человеческими и нечеловеческими последовательностями. Например, аминокислотные остатки S43 и Е163 в SEQ ID №3 (человеческий) соответствуют S83 и Е203 в SEQ ID №4 (бычий). Соответствующие позиции также могут быть основаны на структурном выравнивании, например, с помощью компьютерной имитации выравнивания структуры белка. В других случаях соответствующие районы могут быть идентифицированы.
Используемые в настоящем документе термины "прорегион", "пропептид" или "про-последовательность" относятся к району или сегменту, который расщепляется для получения зрелого белка. Эти термины могут обозначать сегменты, которые подавляют протеолитическую активность, маскируя каталитический механизм и предотвращая таким образом формирование каталитических интермедиатов (т.е. стерически перекрывая сайт связывания субстрата). Прорегион представляет собой последовательность аминокислот, расположенную на амино-конце зрелого биологически активного полипептида, и он может составлять всего лишь несколько аминокислот, или может иметь мультидоменную структуру.
Используемый в настоящем документе термин "зрелый фактор VII" относится к полипептидам FVII, у которых отсутствует сигнальная последовательность и пропептидная последовательность. Как правило, сигнальная последовательность направляет белок для секреции через эндоплазматический ретикулум и (ЭР)-Гольджи путь и отщепляется после проникновения в ЭР в процессе трансляции. Пропептидная последовательность обычно функционирует при посттрансляционных модификациях белков и отщепляется до выделения белка из клетки. Таким образом, зрелые полипептиды FVII обычно представляют собой секретированные белки. В одном примере зрелый человеческий полипептид FVII представлен SEQ ID №3. Аминокислотная последовательность, приведенная в SEQ ID №3, отличается от полипептида-предшественника, приведенного в SEQ ID №№1 и 2; в SEQ ID №3 отсутствует сигнальная последовательность, которая соответствует аминокислотным остаткам 1-20 в SEQ ID №№1 и 2, а также в ней отсутствует пропептидная последовательность, которая соответствует аминокислотным остаткам 21-60 в последовательности SEQ ID №1 и аминокислотным остаткам 21-38 в последовательности SEQ ID №2. Упоминаемый зрелый полипептид FVII охватывает одноцепочечную зимогенную форму и двухцепочечную форму.
Используемые в настоящем документе термины "дикого типа" или "нативный" совместно с FVII относятся к полипептиду FVII, кодируемому нативным или естественным FVII геном, включая аллельные варианты, которые присутствуют в организме, в том числе у человека и других животных, в природе. Упоминание фактора VII дикого типа без упоминания вида предназначено для охвата любых видов фактора VII дикого типа. Полипептиды FVII дикого типа включают закодированные предшественники полипептидов, их фрагменты, их процессированные формы, такие, как зрелые форме без сигнального пептида, а также их пре- или посттрансляционно процессированные или модифицированные формы. Также нативными полипептидами FVII являются те, которые модифицированы посттрансляционно, включая, но, не ограничиваясь, модифицированными гликозилированием, карбоксилированием и гидроксилированием. Нативные полипептиды FVII также включают одноцепочечные и двухцепочечные формы. Например, у людей экспрессируется нативный FVII. Аминокислотные последовательности FVII человека дикого типа приведены в SEQ ID №№1, 2, 3, а аллельные варианты и их зрелые формы приведены в SEQ ID №№44-100. Другие животные производят нативные FVII, включая, но, не ограничиваясь, коровой (Bos Taurus, последовательность SEQ ID №4), мышью (Mus musculus, последовательность SEQ ID №5), карликовым шимпанзе (Pan paniscus, последовательность SEQ ID №6), шимпанзе (Pan troglodites, последовательность SEQ ID №7), кроликом (Oryctolagus cuniculus, последовательность SEQ ID №8), крысой (Rattus №rvegicus, последовательность SEQ ID №9), макакой резусом (Масаса mulatta, последовательность SEQ ID №10), свиньей (Sus Scrofa, последовательность SEQ ID №11), собакой (Canis familiaris, последовательность SEQ ID №12), полосатым данио (Brachydanio rerio, последовательность SEQ ID №13), японским иглобрюхом (Fugu rubripes, SEQ ID №: 14), курицей (Gallus Gallus, последовательность SEQ ID №15), орангутангом (Pongo pygmaeus, последовательность SEQ ID №16) и гориллой (Gorilla Gorilla, последовательность SEQ ID №17).
Используемый в настоящем документе термин «видовые варианты» относится к вариантам полипептидов среди различных видов, включая различные виды млекопитающих, таких как мышь и человек.
Используемый в настоящем документе термин «аллельные варианты» относятся к вариантам белков у членов одного и того же вида.
Используемый в настоящем документе термин «сплайсинговые варианты» относится к вариантам, полученным дифференциальным процессингом первичных транскриптов геномной ДНК, что приводит к более чем одному типу мРНК.
Используемый в настоящем документе термин «зимоген» относится к протеазе, которая активируется протеолитическим расщеплением, в том числе расщеплением, приводящим к созреванию, таким как активационное расщепление, и/или сложной ассоциацией с другим белком (белками) и/или кофактором (кофакторами). Зимоген является неактивным предшественником протеолитического фермента. Такие предшественники, как правило, хотя и не обязательно, крупнее, чем активная форма. Со ссылкой на сериновые протеазы зимогены превращаются в активные ферменты специфическим расщеплением, в том числе каталитическим и автокаталитическим расщеплением, или связыванием активирующего кофактора, что генерирует активный фермент. Например, обычно, зимогены находятся в одноцепочечной форме. Зимогены, как правило, являются неактивными и могут быть преобразованы в зрелые активные полипептиды каталитическим или автокаталитическим расщеплением по одному или нескольким протеолитическим сайтам с образованием мультицепочечного, такого как двухцепочечного, полипептида. Зимоген, таким образом, представляет собой неактивный с ферментативной точки зрения белок, который преобразуется в протеолитический фермент под действием активаторов. Расщепление может быть осуществлено путем самоактивации. Множество коагуляционных белков представляют собой зимогены, они неактивны, но расщепляются и активируются после инициации системы коагуляции после повреждения сосуда. FVII-полипептиды существуют в плазме крови в виде зимогенов до расщепления протеазами, такими как, например, активированный фактор IX (FIXa), активированный фактор Х (FXa), активированный фактор XII (FXIIa), тромбин, или до самоактивации с образованием зимогеноподобных двухцепочечных форм, которые затем требуют дальнейших конформационных изменений для полной активности,
Используемый в настоящем документе термин "зимогеноподобные" белки или полипептиды относится к белкам, которые были активированы протеолитическим расщеплением, но все же проявляют свойства, ассоциируемые с зимогенами, такие как, например, низкая или нулевая активность, или конформация которых напоминает конформацию зимогенной формы белка. Например, в отсутствие связи с тканевым фактором двухцепочечная активированная форма FVII является зимогенподобным белком, она сохраняет конформацию, аналогичную конформации нерасщепленного зимогена FVII и поэтому проявляет очень низкую активность. После связывания с тканевым фактором двухцепочечная активированная форма FVII претерпевает конформационные изменения и приобретает свою полную активность в качестве фактора свертывания.
Используемый в настоящем документе термин «активационная последовательность» относится к последовательности аминокислот в зимогене, которая представляет собой сайт, необходимый для активационного расщепления или расщепления для созревания с формированием активной протеазы. Расщепление активационной последовательности может катализироваться активационными партнерами или быть автокаталитическим.
Используемый в настоящем документе термин «активизационное расщепление» относится к одному из видов созревания расщеплением, которое вызывает изменение конформации, необходимое для развития полной энзиматической активности. Это классический путь активации, например, для сериновых протеаз, в которых в результате расщепления появляется новый N-конец, который взаимодействует с косервативными регионами протеазы, такими как Asp 194 в химотрипсине, что индуцирует конформационные изменения, необходимые для проявления активности. Активация может привести к образованию мультицепочечных форм протеаз. В некоторых случаях одноцепочечные формы протеазы могут проявлять протеолитическую активность.
Используемые в настоящем документе термины «активированный фактор VII" или "FVIIa" относятся к любым двухцепочечным формам полипептида FVII. Двухцепочечные формы обычно являются результатом протеолитического расщепления, но могут быть получены синтетическим путем. Активированный фактор VII, таким образом, включает зимогеноподобную двухцепочечную форму с низкой коагуляционной активностью, полностью активированную форму (примерно с 1000 раз большей активностью), которая образуется при связывании с тканевым фактором, и мутантные формы, которые существуют в полностью активированной двухцепочечной форме или подвергаются конформационным изменениям в полностью активированную форму. Например, одноцепочечная форма полипептида FVII (см., например, последовательность SEQ ID №3) протеолитически расщепляется между аминокислотными остатками, R152 и 1153 зрелого полипептида FVII. Продукты расщепления, FVII тяжелая цепь и FVII легкая цепь, удерживающиеся вместе дисульфидной связью (между аминокислотными остатками 135С и 162С в FVII, SEQ ID №3), образуют двухцепочечный активированный FVII фермент. Протеолитическое расщепление может осуществляться, например, активированным фактором IX (FIXa), активированным фактором Х (FXa), активированным фактором XII (FXIIa), тромбином или путем самоактивации.
Используемый в настоящем документе термин "свойство" полипептида FVII относится к физическим или структурным свойствам, таким как трехмерная структура, р1, время полужизни, конформация и другим подобным физическим характеристикам.
Используемый в настоящем документе термин "активность" полипептида FVII относится к любой активности, проявляемой полипептидом фактора VII. Активность может быть определена in vitro и/или in vivo и включает, но не ограничивается, коагуляционной или коагулянтной активностями, прокоагуляционной активностью, протеолитической или каталитической активностями, такими как осуществление активации фактора Х (FX) или фактора IX (FIX);
иммуногенностью (способностью связываться или конкурировать с полипептидами за связывание с анти-FVII антителами); способностью связывать тканевой фактор, фактор Х или фактор IX; и/или способностью к связыванию с фосфолипидами. Активность может быть оценена in vitro или in vivo с использованием признанных методов анализа, например, путем измерения коагуляции in vitro или in vivo. Результаты таких анализов показывают, что полипептид проявляет активность, которая может быть соотнесена с активностью полипептида in vivo, где in vivo активность может быть соотнесена как биологическая активность. Способы анализа для определения функций или активности модифицированных форм FVII известны специалистам в области техники. Примеры анализов для оценки активности полипептида FVII включают анализ на тромбопластиновое время (РТ) или активированное частичное тромбопластиновое время (аРТТ) для оценки коагуляционной активности, или хромогенный анализ с использованием синтетических субстратов, таких как описано в примерах 4, 5 и 11, для оценки каталитической или протеолитической активности.
Используемые в настоящем документе термины "проявление, по крайней мере, одного вида активности" или "сохранение, по крайней мере, одного вида активности" относятся к активности, проявляемой модифицированным полипептидом FVII по сравнению с немодифицированным полипептидом FVII одной и той же формы в одних и тех же условиях. Например, модифицированный полипептид FVII в двухцепочечной форме сравнивается с немодифицированным полипептидом FVII в двухцепочечной форме в тех же экспериментальных условиях, когда единственным различием между двумя полипептидами является изучаемая модификация. В качестве другого примера, модифицированный полипептид FVII в одноцепочечной форме сравнивается с немодифицированным полипептидом FVII в одноцепочечной форме в тех же экспериментальных условиях, когда единственным различием между двумя полипептидами является изучаемая модификация. Как правило, модифицированный полипептид FVII сохраняет или проявляет, по крайней мере, один вид активности немодифицированного полипептида FVII того же вида, сохраняет активность в достаточной степени для того, чтобы при введении in vivo модифицированный полипептида FVII являлся терапевтически эффективным при прокоагулянтной терапии. Как правило, модифицированный полипептид FVII сохраняет терапевтическую эффективность в качестве прокоагулянта, и сохраненная активность составляет, приблизительно или точно 0,5%, 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более от активности немодифицированного полипептида FVII в той же форме, которая показывает терапевтическую эффективность в качестве прокоагулянта. Активность, которая необходима для поддержания терапевтической эффективности как прокоагулянта, может быть эмпирически определена в случае необходимости. Как правило, сохранение активности от 0,5% до 20%, от 0,5% до 10%, от 0,5% до 5% достаточно для сохранения терапевтической эффективности в качестве прокоагулянта in vivo.
Подразумевается, что проявляемая или сохраненная активность модифицированного полипептидна FVII может представлять собой любую активность, включая, но, не ограничиваясь, коагуляционной или коагулянтной активностями, прокоагулянтной активностью;
протеолитической или каталитической активностями, такими как активация фактора Х (FX) или фактора IX (FIX); иммуногенностью (способностью связываться или конкурировать с полипептидами за связывание с анти-FVII антителами); способностью связывать тканевой фактор, фактор Х или фактор IX; и/или способностью к связыванию с фосфолипидами. В некоторых случаях, модифицированный полипептид FVII может обладать активностью, которая увеличена по сравнению с немодифицированным FVII-полипептидом. В некоторых случаях, модифицированный полипептид FVII может сохранять активность, сниженную по сравнению с немодифицированным FVII-полипептидом. Активность модифицированного полипептида FVII может составлять любой процент от активности немодифицированного полипептида, где оба полипептида представлены в одной форме, включая, но не ограничиваясь, 1% от активности, 2%, 3%, 4%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500%, или более высокой активностью по сравнению с полипептидом, который не содержит модификаций. Например, модифицированный полипептид FVII может проявлять увеличенную или уменьшенную активность по сравнению с немодифицированным полипептидом FVII в той же форме. Например, он может сохранять, по меньшей мере, приблизительно или точно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или, по крайней мере, 99% от активности немодифицированного FVII-полипептида. В других вариантах, измененная активность увеличена, по меньшей мере, в 2 раза, 3 раза, в 4 раза, в 5 раз, 6 раз, 7 раз, 8 раз, в 9 раз, в 10 раз, 20 раз, 30, в 40 раз, 50 раз, в 60 раз, 70 раз, 80 раз, 90 раз, в 100 раз, 200 раз, 300 раз, 400 раз, 500 раз, 600 раз, 700 раз, 800 раз, 900 раз, в 1000 раз или в более раз по сравнению с немодифицированным FVII. Определенный сохраненный уровень зависит от предполагаемого использования полипептида и может быть эмпирически определен. Активность может быть измерена, например, с помощью in vitro и in vivo анализов, таких как описано в настоящем документе или в примерах ниже.
Используемые в настоящем документе термины "коагуляционная активность" или "коагулянтная активность" и "прокоагулянтная активность" относятся к способности полипептида осуществлять коагуляцию. Анализы для оценки коагулянтной активности известны специалистам в области техники и включают в себя тромбопластиновое время (РТ) или активированное частичное тромбопластиновое время (аРТТ).
Используемые в настоящем документе термины "каталитическая активность" или "протеолитическая активность" в отношении FVII относятся к способности белка FVII катализировать протеолитическое расщепление субстрата, а также используются как синонимы. Анализы для оценки такой активности известны в данной области техники. Например, протеолитическая активность FVII может быть измерена с помощью хромогенных субстратов, таких как Spectrozyme FVIIa (СН3SO2-D-СНА-But-Arg-pNA), где расщепление субстрата наблюдается по абсорбции и скорость гидролиза субстрата определяется по линейной регрессии.
Используемый в настоящем документе термин "собственная активность" в отношении FVII относится к каталитической, протеолитической и/или коагулянтной активности белка FVII в отсутствие тканевого фактора.
Используемый в настоящем документе термин «домен» (как правило, последовательность из трех или более, как правило, 5, или 7, или более аминокислот) относится к части молекулы, такой как белок или кодирующая его нуклеиновая кислота, которая структурно и/или функционально отлична от других частей молекулы и является идентифицируемой. Например, домены включают те участки полипептидной цепи, которые могут образовывать независимые складчатые структуры белка, содержащие один или более структурных мотивов и/или которые распознаются в силу их функциональной активности, такой как протеолитическая активность. Белок может обладать одним или более чем одним, отдельным доменом. Например, домен может быть определен, отделен или различен по гомологии последовательности в нем по сравнению с близкими членами семейства, например по гомологии мотивов, которые определяют протеазный домен или Gla домен. В другом примере, домен можно отличить по своей функции, например по протеолитической активности, или способности взаимодействовать с биомолекулой, такой как связывание ДНК, связывание лигандов и димеризация. Домен может проявлять биологическую функцию или активность, домен независимо или в соединении с другой молекулой может осуществлять активность, такую как, например, протеолитическую активность или связывание лигандов. Домен может представлять собой линейную последовательность или нелинейную аминокислотную последовательность. Многие полипептиды содержат множество доменов. Такие домены известны и могут быть определены специалистами в данной области техники. Для примера в настоящем документе приведены определения, однако подразумевается, что в уровне техники необходимо распознавать определенные домены по названию. Если необходимо, для определения доменов может быть использовано соответствующее программное обеспечение.
Используемый в настоящем документе термин «протеазный домен» относится к каталитически активной части протеазы. Упоминаемый протеазный домен протеазы включает одно-, двух- и мультицепочечные формы любого из этих белков. Протеазный домен белка обладает всеми необходимыми свойствами этого белка, требуемыми для его протеолитической активности, такими как, например, наличие каталитического центра. В отношении FVII протеазный домен объединен гомологией и структурными чертами с химотрипсин/трипсин семейством протеазных доменов, включая каталитическую триаду. Например, в зрелом полипептиде FVII, приведенном в последовательности SEQ ID №3, протеазный домен соответствует аминокислотным позициям с 153 по 392.
Используемый в настоящем документе термин «гамма-карбоксиглутаматный (Gla) домен» относится к части белка, например витамин К-зависимого белка, которая содержит посттрансляционно модифицированные остатки глутамата, как правило, большинство, но не все остатки глутамата, подвергнутые витамин К-зависимому карбоксилированию с образованием Gla. Gla домена отвечает за высокое сродство к связыванию ионов кальция и связывание отрицательно заряженных фосфолипидов. Как правило, домен Gla начинается на N-конце зрелой формы витамин К-зависимых белков и заканчивается консервативным ароматическим остатком. В зрелом FVII-полипептиде Gla домен соответствует аминокислотным позициям с 1 до 45 примера полипептида, приведенного в SEQ ID №3. Gla домены хорошо известны, и их локусы могут быть определены в конкретных полипептидах. Gla домены различных витамин К-зависимых белков обладают гомологией в последовательности, структуре и функциях, включая кластеризацию N-концевых гидрофобных остатков в гидрофобные участки, которые способствуют взаимодействию с отрицательно заряженными фосфолипидами на поверхности мембран клеток. Примеры других Gla-содержащих полипептидов, включают, но не ограничиваются, FIX, FX, протромбином, протеином С, протеином S, остеокальцином, протеином Gla, Gas6 белком (Growth-arrest-specific) и белком Z. Gla домены этих и других примеров белков приведены в любой из последовательностей SEQ ID №№110-121.
Используемые в настоящем документе термины "нативный" или "эндогенный" со ссылкой на домен Gla относятся к естественным Gla доменам и соотнесены со всеми или с частью полипептидов, имеющихся в домене Gla. Для целей настоящего изобретения нативный домен Gla упоминается со ссылкой на FVII-полипептид. Например, нативный домен Gla из FVII, приведенный в SEQ ID №119, соответствует аминокислотам 1-45 последовательности аминокислот, приведенной в SEQ ID №3.
Используемый в настоящем документе термин «гетерологичный домен Gla» относится к домену Gla полипептида, одного или разных видов, который не является Gla доменом FVII. Примеры гетерологичного Gla домена представляют собой Gla домены Gla-содержащих полипептидов, включая, но не ограничиваясь, FIX, FX, протромбином, протеином С, протеином S, остеокальцином, протеином Gla, Gas6 белком и белком Z. Gla домены этих и других примеров белков приведены в любой из последовательностей SEQ ID №№110-118, 120 и 121.
Используемый в настоящем документе термин «непрерывная часть домена Gla» относится, к, по крайней мере, двум или более соседним аминокислотам, как правило, 2, 3, 4, 5, 6, 8, 10, 15, 20, 30, 40 или более аминокислотам, вплоть до всех аминокислот, которые составляют домена Gla.
Используемый в настоящем документе термин "достаточная часть домена Gla, чтобы привести к связыванию фосфолипидов" относится к, по крайней мере, одной аминокислоте, как правило, 2, 3, 4, 5, 6, 8, 10, 15 или более аминокислотам домена, но менее чем ко всем аминокислотам, входящим в состав домена, при условии, что полипептид, содержащий такую часть, проявляет способность связывать фосфолипиды.
Используемые в настоящем документе термины "заменить" по отношению к домену Gla или "замена Gla домена" относится к процессу замены эндогенного домена Gla белка с помощью рекомбинантных, синтетических или других методов на домен Gla другого белка. В контексте "замены Gla домена", "Gla домен" представляет собой любые выбранные аминокислоты из домена Gla и прилегающих районов, которые достаточны для сохранения активности по связыванию фосфолипидов. Как правило, замена Gla домена заключается в замене 40-50 аминокислот эндогенного белка 40-50 аминокислотами другого белка, но может включать меньше или больше аминокислот.
Используемый в настоящем документе термин «домен эпидермального фактора роста» (ЭФР) (ЭФР-1 или ЭФР-2) относится к той части белка, которая обладает гомологией последовательности со специфической частью из 30 до 40 аминокислот в последовательности эпидермального фактора роста (ЭФР). Данный домен ЭФР включает шесть остатков цистеина, для которых было показано (в ЭФР) участие в образовании дисульфидных связей. Основная структура домена ЭФР состоит из двух бета-листов, включающих по две цепочки аминокислот, следуемой за ними петли, ведущей к короткому двуцепочечному листу на С-конце. FVII включает два ЭФР домена: ЭФР-1 и ЭФР-2. Эти домены соответствуют аминокислотным позициям 46-82 и 87-128, соответственно, в зрелом полипептиде FVII, приведенном в SEQ ID №3.
Используемые в настоящем документе термины "немодифицированный полипептид" или "немодифицированный FVII" и их грамматические вариации относятся к начальному полипептиду, который выбран в настоящем документе для модифицирования. Начальный полипептид может представлять собой полипептид естественного происхождения, полипептид дикого типа. Кроме того, начальный полипептид может быть изменен или мутирован, поэтому он может отличаться от нативной изоформы дикого типа, но тем не менее он будет упомянут в настоящем документе в качестве начального немодифицированного полипептида по отношению к последующим модифицированным полипептидам, полученным по настоящему изобретению. Таким образом, существующие белки, известные в уровне техники, которые были модифицированы для желаемого увеличения или уменьшения той или иной активности или свойства по сравнению с немодифицированным белком, могут быть выбраны и использованы в качестве начального немодифицированного полипептида. Например, белок, который был модифицирован из его нативной формы одной или более заменами аминокислот и обладает увеличением или уменьшением желаемого свойства, например заменой аминокислотных остатков или одного остатка для изменения гликозилирования, может упоминаться в этом документе как немодифицированный и подвергаться дальнейшей модификации для изменения того же или другого свойства. Примеры модифицированных FVII-полипептидов, известных в уровне техники, включают любые полипептиды FVII, описанные, например, в патентах США №№5580560, 6017882, 6693075, 6762286 и 6806063, опубликованных заявках США №№20030100506 и 20040220106 и международных заявках №№WO 1988010295, WO 200183725, WO 2003093465, WO 200338162, WO 2004083361, WO 2004108763, WO 2004029090, WO 2004029091, WO 2004111242 и WO 2005123916.
Используемые в настоящем документе термины "модифицированные полипептиды фактора VII " и "модифицированный фактор VII" относятся к FVII-полипептидам, которые обладают одним или несколькими аминокислотными различиями по сравнению с немодифицированным полипептидом фактора VII. Одно или более аминокислотных различий могут представлять собой мутации аминокислот, например одну или более аминокислотных замен (замещений), вставок и делеций, или представлять собой вставки или делеций целых доменов, и любые их комбинации. Как правило, модифицированный полипептид FVII обладает одной или несколькими модификациями в первичной последовательности по сравнению с немодифицированным FVII-полипептидом. Например, модифицированный полипептид FVII, предусмотренный в настоящем документе, может обладать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50 или более аминокислотными различиями по сравнению с немодифицированным FVII-полипептидом. Любые модификации предполагаются до тех пор, пока полученный полипептид проявляет, по крайней мере, один вид FVII активности, связанный с нативным полипептидом FVII, такой как, например, каталитическая активность, протеолитическая активность, способность связывать TF или способность связывать активированные тромбоциты.
Используемый в настоящем документе термин "ингибиторы коагуляции" относится к белкам или молекулам, которые ингибируют или предотвращают коагуляцию или образование сгустка. Ингибирование или предотвращение коагуляции может наблюдаться in vivo или in vitro и может анализироваться с помощью любого метода, известного в уровне техники, включая, но не ограничиваясь, анализом на тромбопластиновое время (РТ) или активированное частичное тромбопластиновое время (аРТТ).
Используемый в настоящем документе термин «ингибитор пути, запускаемого тканевым фактором» (TFPI, так же известный, как TFPI-1) является ингибитором Кунитц-типа, который вовлечен в формирование четвертичного TF/FVIIa/TFPI/FXa ингибиторного комплекса, в котором активность FVIIa ингибирована. TFPI экспрессируется в виде предшественников в двух разных формах, претерпевающих затем альтернативный сплайсинг, TFPIa (последовательность SEQ ID №101) и TFPIp (последовательность SEQ ID №103) предшественников, которые расщепляются во время секреции с образованием 276 аминокислотного (последовательность SEQ ID №102) и 223 аминокислотного фрагментов (последовательность SEQ ID №104) зрелых белков, соответственно. TFPI содержит 3 домена Кунитца, из которых Кунитц-1 домен отвечает за связывание и ингибирование FVIIa.
Используемый в настоящем документе термин «TFPI-2» (также известный как плацентарный белок 5 (РР5) и связанный с матриксом ингибитор сериновой протеазы (MSPI)) относится к гомологу TFPI. Зрелый белок TFPI-2 (последовательность SEQ ID №105) содержит 213 аминокислоты и три домена Кунитца, которые демонстрируют 43%, 35% и 53% идентичности первичной последовательностей с 1, 2 и 3 доменами Кунитца TFPI-1, соответственно. TFPI-2 играет важную роль в регуляции расщепления и перестройки внеклеточного матрикса и поэтому не считается важным фактором в пути коагуляции.
Используемый в настоящем документе термин «антитромбин III» (АТ-III) относится к ингибитору сериновой протеазы (серпину). AT-III синтезируется в виде белка-предшественника, содержащего 464 аминокислотных остатков (последовательность SEQ ID №122), который расщепляется ходе секреции с образованием содержащего 432 аминокислоты зрелого антитромбина (последовательность SEQ ID №123).
Используемый в настоящем документе термин «кофакторы» относится к белкам или молекулам, которые связываются с другими специфическими белками или молекулами с образованием активных комплексов. В некоторых примерах, связывание с кофактором необходимо для оптимальной протеолитической активности. Например, тканевый фактор (ТФ) является кофактором FVIIa. Связывание FVIIa с ТФ индуцирует конформационные изменения, что приводит к повышению протеолитической активности FVIIa по отношению к его субстратам, FX и FIX.
Используемый в настоящем документе термин «тканевый фактор» (ТФ) относится к содержащему 263 аминокислоты трансмембранному гликопротеину (последовательность SEQ ID №124), который функционирует в качестве кофактора для FVIIa. Он конститутивно экспрессируется в гладкомышечных клетках и фибробластах, а также помогает инициировать коагуляцию путем связывания FVII и FVIIa, когда эти клетки вступают в контакт с кровью после повреждения тканей.
Используемый в настоящем документе термин «активированные тромбоциты» относится к тромбоцитам, которые были активированы связыванием молекул, таких как коллаген, тромбоксан А2, АДФ и тромбин, что приводит к различным изменениям в их морфологии, фенотипе и функциях, что в конечном итоге содействует коагуляции. Например, у активированного тромбоцита была изменена форма на более аморфную с выступающими пальцеобразными выростами. Активированные тромбоциты также претерпевают "переворот" в клеточной мембране, так что фосфатидилсерин и другие отрицательно заряженные фосфолипиды, которые обычно присутствуют на внутренней стороне клеточной мембраны, перемещаются на внешнюю, плазма-ориентированную поверхность. Мембраны активированных тромбоцитов предоставляют поверхность, на которой осуществляются многие реакции каскада коагуляции. Активированные тромбоциты также выделяют пузырьки, содержащие такие прокоагулянтные факторы, как vWF (фактор Виллибранда), FV, тромбин, АДФ и тромбоксан А2, и они сливаются друг с другом с образованием тромбоцитной пробки, которая стабилизируются фибрином с образованием сгустка.
Используемый в настоящем документе термин «увеличенное связывание и/или аффинность к активированным тромбоцитам», и любые его грамматические модификации, означает повышение способности полипептида или белка, например полипептида FVII, связываться с поверхностью активированных тромбоцитов по сравнению со сравниваемым полипептидом или белком. Например, способность модифицированного полипептида FVII связываться с активированными тромбоцитами может быть больше, чем способность немодифицированного полипептида FVII связываться с активированными тромбоцитами. Связывание и/или аффинность полипептидов к активированным тромбоцитам может быть увеличена, по крайней мере, на, приблизительно, 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению со связыванием и/или аффинностью немодифицированного полипептида. Анализы для определения связывания и/или аффинности полипептида к активированным тромбоцитам известны в уровне техники. Связывание полипептида FVII с активированными тромбоцитами опосредовано взаимодействием аминокислот домена Gla полипептида FVII и отрицательно заряженных фосфолипидов, таких как фосфатидилсерин на активированных тромбоцитах. Таким образом, в методах анализа связывания полипептидов, таких как FVII-полипептиды, с активированными тромбоцитами используют мембраны и пузырьки, которые содержат фосфолипиды, такие как фосфатидилсерин. Например, способность полипептидов связываться с активированными тромбоцитами отражается на способности полипептидов связываться с фосфолипидными пузырьками, что может быть измерено по рассеянию света.
Используемый в настоящем документе термин «увеличение способности связывать и/или увеличение аффинности к фосфолипидам», и любые его грамматические вариации, означает повышение способности полипептида или белка связывать фосфолипиды по сравнению со сравниваемым полипептидом или белком. Фосфолипиды могут включать любые фосфолипиды, но особенно фосфатидилсерин. Связывание и/или аффинность полипептидов к фосфолипидам может быть увеличена, по крайней мере, приблизительно, на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению со связыванием и/или аффинностью немодифицированного полипептида. Анализы для определения аффинности и/или способности полипептида связывать фосфолипиды известны в уровне техники. Например, связывание FVII-полипептида с фосфолипидными везикулами может быть определено по относительному рассеянию света под углом 90° к падающему свету. Интенсивность рассеивания света одними фосфолипидными пузырьками и фосфолипидными пузырьками с FVII измеряется для определения константы диссоциации. Также для измерения аффинности полипептидов FVII к фосфолипидам мембран может быть использован плазменный резонанс на поверхности, такой как полученный на биосенсоре BIAcore.
Используемый в настоящем документе термин «повышенная устойчивость к ингибиторам» или "повышенная устойчивостью к TFPI" или "повышенная устойчивость к AT-III" относится к любому понижению чувствительности полипептида к действию ингибитора, такого как TFPI или AT-III, по сравнению со сравниваемым полипептидом, таким как немодифицированный FVII-полипептид. Например, TFPI в комплексе с FXa связывается с TF/FVIIa комплексом. Таким образом, он подавляет активность FVIIa. Таким образом, модифицированный полипептид FVII, который снижает связывание или препятствует связыванию TFPI с TF/FVIIa комплексом и соответственно уменьшает или препятствует TFPI-опосредованному ингибированию активности FVIIa, проявляет повышенную устойчивость к TFPI. Повышенная устойчивость к ингибиторам, таким как TFPI, может анализироваться на основе оценки связывания модифицированных полипептидов FVII с ингибитором. Повышенная устойчивость к ингибиторам, таким как TFPI, также может быть анализирована по измерению собственной активности или когулянтной активности полипептида FVII в присутствии TFPI. Анализы для определения связывания полипептида с ингибитором, таким как TFPI или AT-III, известны в данной области техники. Для нековалентных ингибиторов, таких как, например, TFPI, может быть измерена kj. Для ковалентных ингибиторов, таких как, например, AT-III, может быть измерена константа скорости ингибирования второго порядка. Кроме того, поверхностный плазменный резонанс, такой как выполненный на биосенсоре BIAcore, также может быть использован для измерения связывания полипептидов FVII с TFPI, AT-III и другими ингибиторами. Однако для ковалентных ингибиторов, таких как AT-III, с помощью BIAcore может быть измерена только скорость присоединения ингибитора. Анализы для определения ингибиторного эффекта, например TFPI на коагулянтную активность FVII или его собственную активность также известны в уровне техники. Например, может быть измерена способность модифицированного полипептида FVII расщеплять его субстрат FX в присутствии или в отсутствие TFPI, и может быть определена степень, с которой TFPI ингибирует реакцию. Результаты можно сравнить со способностью немодифицированного полипептида FVII расщеплять его субстрат FX в присутствии или отсутствие TFPI. Модифицированные полипептиды проявляют повышенную устойчивостью к ингибиторам, например повышенную на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%, 200%, 300%, 400%, 500% или более устойчивость к воздействию ингибиторов по сравнению с немодифицированным полипептидом.
Используемый в настоящем документе термин "биологическая активность" относится к активности соединения in vivo или физиологическим ответам, которые проявляются при введении in vivo соединения, композиции или другой смеси. Биологическая активность, таким образом, включает в себя терапевтический эффект и фармацевтическую активность таких соединений, композиций и смесей. Биологическая активность может наблюдаться в системах in vitro, предназначенных для тестирования или использования такой активности. Таким образом, для целей настоящего изобретения биологическая активность полипептидов FVII включает коагулянтную активность.
Используемый здесь термин "оценки", и его грамматические модификации, призван включать количественные и качественные определения для получения абсолютного значения активности полипептида, а также получения показателя, отношения, доли, визуального или другого ориентировочного значения уровня активности. Оценка может быть прямой или косвенной. Например, обнаружение расщепления субстрата полипептидом может быть достигнуто прямым измерением продукта или может быть определено косвенным путем определением активности расщепленного субстрата.
Используемый в настоящем документе термин "химотрипсиновая нумерация" относится к нумерации аминокислот в зрелом полипептиде химотрипсина, приведенном в SEQ ID №107. Выравнивание протеазного домена другой протеазы, такой как, например, протеазного домена фактора VII, может быть осуществлено по химотрипсину. В таком случае аминокислотам фактора VII, которые соответствуют аминокислотам химотрипсина, дается нумерация аминокислот химотрипсина. Соответствующие позиции могут быть определены с помощью такого выравнивания специалистом в данной области выравниванием вручную или с помощью многочисленных доступных программ выравнивания (например, BLASTP). Соответствующие позиции также могут быть определены с помощью структурного выравнивания, например с помощью компьютерной имитации выравнивания структуры белка. Перечисление аминокислот полипептида, соответствующих аминокислотам в раскрытой последовательности, относится к аминокислотам, определенным по выравниванию полипептида с раскрытой последовательностью по максимальной идентичности или гомологии (где выровнены соответствующие консервативные аминокислоты) с помощью стандартного алгоритма выравнивания, такого как алгоритм GAP. Соответствующие химотрипсиновые номера аминокислотных позиций с 153 по 406 полипептида FVII, приведенного в SEQ ID №3, приведены в таблице 1. Аминокислотные позиции в последовательности, приведенной в SEQ ID №3, указаны нормальным шрифтом, аминокислотные остатки на этих позициях выделены жирным шрифтом и соответствующие номера в химотрипсине выделены курсивом. Например, после выравнивания зрелого фактора VII (последовательность SEQ ID №3) со зрелыми химотрипсином (последовательность SEQ ID №107) изолейцину (I) в 153 аминокислотной позиции в факторе VII дается химотрипсиновый номер 116. Последующие аминокислоты пронумерованы соответственно. Например, глутаминовой кислоте (Е) в 210 аминокислотной позиции зрелого фактора VII (последовательность SEQ ID №3) соответствует аминокислотная позиция Е70 по химотрипсиновой нумерации. Если остаток присутствует в протеазе, но отсутствует в химотрипсине, аминокислотному остатку дается буквенное обозначение. Например, остатки в химотрипсине, которые являются частью петли с 60 аминокислотой по химотрипсиновой нумерации, но вставлены в последовательность фактора VII по сравнению с химотрипсином, обозначаются, например, К60а, 160b, К60с или N60d. Эти остатки соответствуют К197, I198, К199 и N200, соответственно, по нумерации последовательности зрелого фактора VII (последовательность SEQ ID №3).

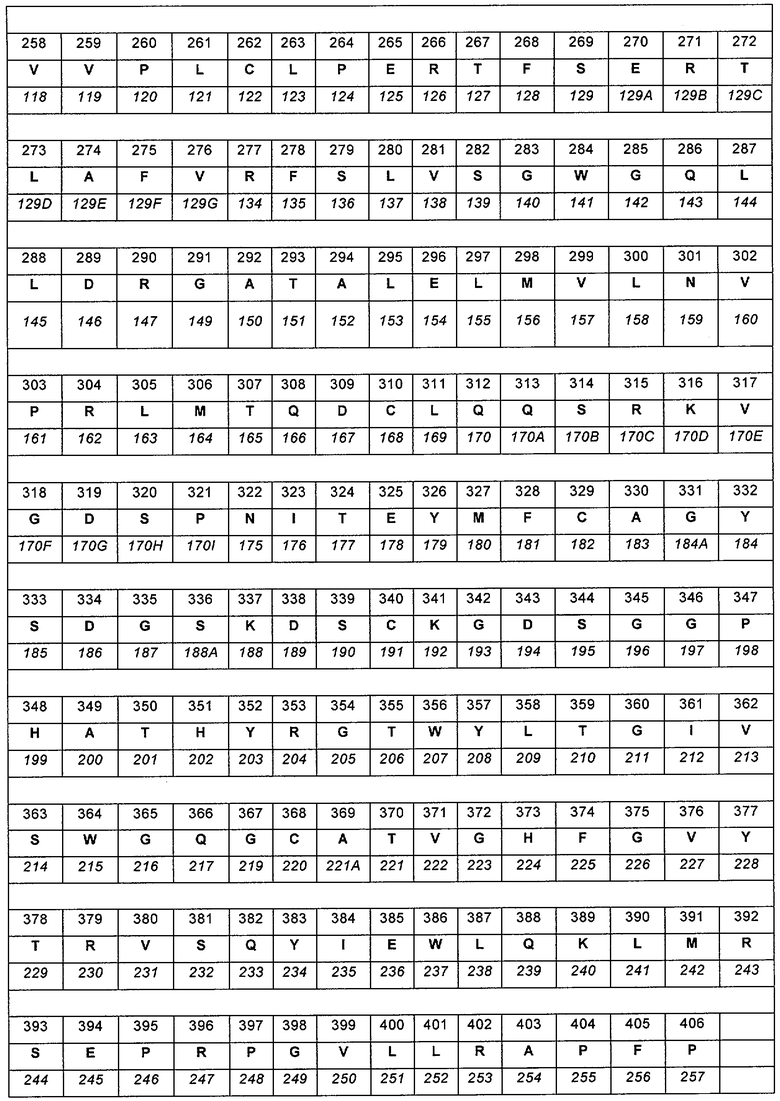
Используемый в настоящем документе термин «нуклеиновые кислоты» относится к ДНК, РНК и их аналогам, в том числе пептидным нуклеиновым кислотам (ПНК) и их смесям. Нуклеиновые кислоты могут быть одно- или двуцепочечными. В отношении зондов или праймеров, которые являются необязательно мечеными, например с обнаруживаемой меткой, такой как флуоресцентная или радиоактивная метка, рассматриваются как одноцепочечные молекулы. Такие молекулы, как правило, обладают такой длиной, что их мишень является статистически уникальной или представленной небольшим числом копий (обычно менее 5, как правило, менее 3) для зондирования или праймирования библиотеки. Как правило, зонд или праймер содержит непрерывную последовательность, по меньшей мере, из 14, 16 или 30 нуклеотидов, комплементарных или идентичных интересующему гену. Зонды и праймеры могут включать 10, 20, 30, 50, 100 или более остатков нуклеиновых кислот.
Используемый в настоящем документе термин «пептид» относится к полипептиду, который включает от 2 до 40 аминокислот.
Используемые в настоящем документе аминокислоты, которые включены в различные последовательности аминокислот, содержащихся в настоящем документе, определяются в соответствии с их известными, трехбуквенными или однобуквенными аббревиатурами (табл.2). Нуклеотиды, включенные в различные фрагменты нуклеиновых кислот, обозначены стандартными однобуквенными обозначениями, обычно используемыми в уровне техники.
Используемый в настоящем документе термин "аминокислоты" относится к органическим соединениям, содержащим аминогруппу и карбоксильную группу. Полипептид содержит две или более аминокислоты. Для целей настоящего изобретения, к аминокислотам относятся двадцать природных аминокислот, неприродные аминокислоты и аналоги аминокислот (например, аминокислоты, с боковой цепью у α-углеродного атома).
Аббревиатуры для аминокислотных остатков, соответствующие стандартной номенклатуре полипептидов, описанной в J. Biol. Chem., 243: 3552-3559 (1969), и принятой 37 C.F.R., §§ 1.821-1.822, приведены в Таблице 2:
Следует отметить, что все последовательности остатков аминокислот, представленные в настоящем документе в виде формул, ориентированы слева направо в обычном направлении от амино-конца к карбоксильному концу. Кроме того, фраза "аминокислотный остаток" представляет собой широкое определение, включающее аминокислоты, перечисленные в таблице соответствия, (таблица 2) модифицированные и необычные аминокислоты, такие как упомянутые в 37 C.F.R. §§ 1.821-1.822 и приведенные в настоящем документе в качестве ссылки. Кроме того, следует отметить, что тире в начале или в конце последовательности аминокислотных остатков обозначает пептидную связь с дополнительной последовательностью из одного или более аминокислотных остатков, с амино-концевой группой, такой как NН2 или с карбоксильной концевой группой, такой как СООН.
Используемый в настоящем документе термин "гидрофобная аминокислота" относится к любой из аминокислот, определенной как гидрофобная с помощью шкалы гидрофобности Айзенберга. Примерами природных гидрофобных аминокислот являются изолейцин, фенилаланин, валин, лейцин, триптофан, метионин, аланин, глицин, цистеин и тирозин (Eisenberg и др., (1982) Faraday Symp.Chem. Soc. 17:109-120). В настоящем документе также включены неприродные гидрофобные аминокислоты.
Используемый в настоящем документе термин "кислая аминокислота" относится к природным аминокислотам - остаткам аспарагиновой кислоты и глутаминовой кислоты. В настоящем документе также включены неприродные кислые аминокислоты.
Используемый в настоящем документе термин "природные аминокислоты" относится к 20 L-аминокислотам, которые присутствуют в полипептидах.
Используемый в настоящем документе термин "неприродные аминокислоты" относится к органическим соединениям, содержащим аминогруппу и карбоксильную группу, которые не являются природными аминокислотами, перечисленными в Таблице 2. К неприродным аминокислотам таким образом относятся, например, аминокислоты или аналоги аминокислот, отличающиеся от 20 природных аминокислот и включающие, но не ограничивающиеся, D-изостереомерами аминокислот. Примеры неприродных аминокислот известны специалисту в данной области и могут быть включены в модифицированный полипептид фактора VII.
Используемый в настоящем документе термин «ДНК-конструкция» относится к одно- или двухцепочечным, линейным или кольцевым молекулам ДНК, которые содержат сегменты ДНК, комбинированные и совмещенные в порядке, не встречающемся в природе. ДНК-конструкции существуют как результат манипуляций человека и включают клоны и другие копии созданных молекул.
Используемый в настоящем документе термин «сегмент ДНК» относится к части более крупной молекулы ДНК, имеющей определенные свойства. Например, сегмент ДНК, кодирующий определенный полипептид, является частью большей молекулы ДНК, такой как плазмида или фрагмент плазмиды, который при считывании в направлении от 5' к 3' концу кодирует последовательность аминокислот определенного полипептида.
Используемый здесь термин «полинуклеотид» означает одно- или двухцепочечный полимер дезоксирибонуклеотидных или рибонуклеотидных оснований, который считывается от 5' к 3' концу. Полинуклеотиды включают РНК и ДНК, могут быть выделены из природных источников, синтезированы in vitro или получены комбинацией природных и синтетических молекул. Длина молекул полинуклеотидов задается в настоящем документе в нуклеотидах (сокращенно "нт") или парах оснований (сокращенно "нп"). Термин нуклеотиды используется для одно- и двухцепочечных молекул, где позволяет контекст. Когда этот термин применяется к двухцепочечным молекулам, он используется для обозначения общей длины и эквивалентен парам оснований. Специалисту в данной области очевидно, что две цепи двухцепочечного полинуклеотида могут незначительно отличаться по длине, и что их концы могут быть несовмещенными, и поэтому не все нуклеотиды в двухцепочечной молекуле полинуклеотида могут быть сопряжены. Такие непарные концы, как правило, не превышают 20 нуклеотидов в длину.
Используемый в настоящем документе термин "первичная последовательность" относится к последовательности аминокислотных остатков полипептида.
Используемый в настоящем документе термин "сходство" между двумя белками или нуклеиновыми кислотами указывает на родство между последовательностями аминокислот в белках или нуклеотидными последовательностями нуклеиновых кислот. Сходство может быть основано на степени идентичности и/или гомологии последовательности остатков и остатков, содержащихся в белке. Методы оценки степени сходства белков или нуклеиновых кислот известны специалистам в данной области. Например, в одном методе оценки сходства последовательностей две последовательности аминокислот или нуклеотидов выравниваются таким образом, чтобы достичь максимального уровня идентичности между последовательностями. "Идентичность" относится к степени, в которой аминокислотные или нуклеотидные последовательности являются инвариантными. При выравнивании аминокислотных последовательностей и, в некоторой степени, нуклеотидных последовательностей также могут приниматься во внимание консервативные различия и/или частые замены аминокислот (или нуклеотидов). Консервативными различиями являются те, при которых сохраняются физико-химические свойства участвующих остатков. Выравнивание может быть глобальным (выравнивание сравниваемой последовательности по всей длине последовательности, а также по всем остаткам) или локальным (выравнивание части последовательности, которая включает только самый сходный район или районы).
Используемые здесь термины "гомология" и "идентичность" используются как синонимы, но гомология для белков может подразумевать консервативные замены аминокислот. В общем, чтобы определить соответствующие позиции, последовательности аминокислот выравниваются таким образом, чтобы достичь наибольшей степени совпадения (см., например. Computational Molecular Biology, Lesk, A.M., изд. Oxford University Press, New York, 1988; Biocomputing: Informatics and Ge.Nome Projects, Smith, DW, изд. Academic Press, New York, 1993; Computer Analysis of Sequence Data, часть I, Griffin, A.M., и Griffin, H. G., изд. Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., изд. М Stockton Press, New York, 1991; Carillo и др. (1988) SIAMJ Applied Math 48: 1073).
Используемый в настоящем документе термин идентичность последовательности относится к множеству идентичных аминокислот (или нуклеотидов) при сравнении исследуемого и указанного полипептидов или полинуклеотидов. Гомология полипептидов имеет отношение к ранее определенным идентичным или гомологичным аминокислотным остаткам. Гомология подразумевает консервативные замены аминокислот, а также идентичность остатков. Идентичность последовательности может быть определена с помощью программ со стандартными алгоритмами выравнивания со стандартными настройками, установленными каждым поставщиком программного обеспечения. Гомология молекул нуклеиновой кислоты имеет отношение к ранее определенному множеству идентичных или гомологичных нуклеотидов. Гомология подразумевает замены, которые не меняют закодированные аминокислоты (т.е. "молчащие замены"), а также идентичные остатки. Значительно гомологичные молекулы нуклеиновой кислоты, как правило, гибридизуются в умеренных условиях или в жестких условиях по всей длине нуклеиновой кислоты или, по меньшей мере, приблизительно на протяжении 70%, 80% или 90% от полной длины молекулы интересующей нуклеиновой кислоты. Также предусмотрены молекулы нуклеиновой кислоты, которые содержат вырожденные кодоны на месте кодонов в гибридизуемой молекуле нуклеиновой кислоты. (Для определения гомологии белков, консервативные аминокислоты, а также идентичные аминокислоты могут быть выровнены, в этом случае доля идентичности и процент гомологии отличаются). Нуклеотидные последовательности любых двух молекул нуклеиновой кислоты (или аминокислотные последовательности любых двух полипептидов), которые являются, по крайней мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% "идентичными", могут быть определены с помощью известных компьютерных алгоритмов, таких как программа "FASTA", с использованием, параметров, например, как указано Pearson и др. Proc. Nat. Acad. Sci. USA 85: 2444 (1988) (другие программы включают пакет программ GCG (Devereux J., и др. Nucleic Acids Research 12(1): 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F., и др., J. Molec. Biol. 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, изд.. Academic Press, San Diego (1994), и Carillo и др. SIAM J Applied Math 48: 1073 (1988)). Например, для определения идентичности может быть использована функция BLAST Национального центра биотехнологической информации. Другие коммерчески или публично доступные программы включают DNAStar программу "MegAlign" (Madison, WI) и программу "Gap" (Madison, WI) Genetics Computer Group (UWG) Университета штата Висконсин США. Доля гомологии или идентичности молекул белка и/или нуклеиновой кислоты может быть определена, например, путем сравнения информации о последовательностях с помощью компьютерной программы GAP (например, Needleman и др. J. Mol. Biol. 48: 443 (1970), как проверено Smith и Waterman (Adv. Appl. Math. 2: 482 (1981)). Программа GAP определяет сходство как количество выровненных символов (т.е. нуклеотидов или аминокислот), которые являются аналогичными, деленное на общее число символов в более короткой из двух последовательностей. Параметры для программы GAP могут включать: (1) унарную матрицу сравнения (содержащую значение 1 для идентичных и 0 для неидентичных аминокислот), взвешенную матрицу сравнения, см. Gribskov и др. Nucl. Acids Res. 14: 6745 (1986), как описано Schwartz и Dayhoff, изд.. Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, c. 353-358 (1979); (2) "штраф" в размере 3,0 для каждого гэпа и дополнительно 0,10 для каждого символа в каждом гэпе и (3) отсутствие штрафа для гэпов в конце последовательностей.
Используемый в настоящем документе термин "идентичность" относится к сравнению тестируемого и указанного полипептида или полинуклеотида. Например, фраза "по крайней мере, на 90% идентичен по сравнению с" относится к идентичности от 90 до 100% по отношению к указанному полипептиду. Например, идентичность 90% или более свидетельствует о том, что при сравнении тестируемого и указанного полипептидов длиной 100 аминокислот не более чем 10% (т.е. 10 из 100) аминокислот в тестируемом полипептиде отличается от указанного полипептида. Подобные сравнения можно провести между тестируемым и указанным полинуклеотидами. Такие различия могут быть представлены в виде точечных мутаций, хаотически распределенных по всей длине аминокислотной последовательности, или же они могут быть сгруппированы в одном или более районах различной длины вплоть до максимально допустимой, например 10/100 различий аминокислот (около 90% идентичности). Различия определяются как замещения, вставки и делеции в нуклеиновых кислотах или аминокислотах. При уровне гомологии или идентичности, превышающем, приблизительно, 85-90%, результат не должен зависеть от программы и гэп-параметров; такой высокий уровень идентичности можно легко выявить, зачастую не полагаясь на программное обеспечение.
Подразумевается, что используемый в настоящем документе термин "существенно идентичный" или "сходный" варьируется в зависимости от контекста, как известно или как может быть оценено специалистами в данной области.
Используемый в настоящем документе термин «выровненные последовательности» относится к использованию гомологии (сходства и/или идентичности) для выравнивания соответствующих позиций в последовательности нуклеотидов или аминокислот. Как правило, выравниваются две или более последовательности, которые обладают 50% или более идентичности. Множество выровненных последовательностей относится к 2 или более последовательностям, которые выровнены по соответствующим позициям и могут включать выровненные последовательности, полученные из РНК, таких как EST (Expressed sequence tags) и кДНК, выровненных с геномной последовательностью ДНК.
Используемый в настоящем документе термин "специфично гибридизуется" относится к отжигу на основе комплементарного спаривания оснований молекулы нуклеиновой кислоты (например, олигонуклеотида) с целевой молекулой нуклеиновой кислоты. Специалистам в данной области известны in vitro и in vivo параметры, которые влияют на специфическую гибридизацию, такие как длина и состав конкретной молекулы. Особенно значимые для in vitro гибридизации параметры также включают температуру отжига и отмывания, состав буфера и концентрацию солей. Примеры условий отмывания для удаления неспецифически связанных молекул нуклеиновой кислоты при высокой точности включают 0,1 (SSPE (раствор 180 мМ хлорида натрия, 10 мМ дигидрофосфата натрия и 1 мМ ЭДТА), 0,1% SDS (додецилсульфат натрия), 65°С, а при средней точности включают 0,2 (SSPE, 0,1% SDS, 50°C. Эквивалентные условия точности известны в уровне техники. Квалифицированный специалист может легко подобрать эти параметры для достижения специфической гибридизации молекулы нуклеиновой кислоты с целевой молекулой нуклеиновой кислоты, подходящей для конкретного применения.
Используемый в настоящем документе изолированный или очищенный полипептид или белок, или его биологически активная часть является практически свободным от клеточного материала или других загрязняющих белков из клеток ткани, из которой он был выделен, или практически свободным от химических веществ-предшественников и других химических веществ, если он был химически синтезирован. Препараты могут быть определены, как практически чистые, если они свободны от легко обнаруживаемых примесей, определяемых стандартными методами анализа, такими как тонкослойная хроматография (ТСХ), гель-электрофорез и высокоэффективная жидкостная хроматография (ВЭЖХ), которые используются специалистами для оценки чистоты; или препараты могут быть определены, как достаточно чистые, если дальнейшая очистка существенно не изменяет физические и химические свойства, такие как протеолитическая и биологическая активности вещества. Методы очистки соединений для получения существенно химически чистых соединений известны специалистам в данной области. Существенно химически чистые соединения, однако, могут быть смесью стереоизомеров. В таких случаях дальнейшая очистка может привести к увеличению удельной активности соединения.
Термин «практически свободный от клеточного материала» относится к препартам белков, в которых белок отделен от компонентов клетки, из которой его выделяют или получают рекомбинантно. В одном варианте термин свободный от клеточного материала относится к получению белков протеаз, содержащих менее чем, приблизительно, 30% (на сухой вес) непротеазных белков (также упоминаемых в этом документе как загрязняющие белки), как правило, менее 20% непротеазных белков, или менее 10% непротеазных белков, или менее чем, приблизительно, 5% непротеазных белков. Если протеазный белок или его активная часть получены рекомбинантно, они также являются практически свободными от культуральной среды, то есть культуральная среда представляет собой менее чем, или приблизительно, или точно 20%, 10% или 5% от объема препарата протеазного белка.
Используемый здесь термин «практически свободный от химических предшественников и других химических веществ» относится к препаратам протеазных белков, при котором белок отделен от химических предшественников или других химических веществ, которые участвуют в синтезе белка. Этот термин относится к препаратам протеазных белков, имеющих менее 30% (на сухой вес), 20%, 10%, 5%, или меньшее количество химических предшественников или непротеазных химических веществ или компонентов.
Используемый в настоящем документе термин «получение рекомбинантными методами с использованием метода рекомбинантной ДНК» относится к использованию известных методов молекулярной биологии для экспрессии белков, кодируемых клонированной ДНК.
Используемый в настоящем документе термин «вектор» (или плазмида) относится к дискретным элементам, которые используются, чтобы ввести гетерологичную нуклеиновую кислоту в клетки для ее экспрессии или репликации. Векторы обычно остаются эписомными, но могут быть разработаны для интеграции гена или его части в хромосому генома. Также предусмотрены векторы, которые являются искусственными хромосомами, такими как бактериальные искусственные хромосомы, искусственные хромосомы дрожжей и искусственные хромосомы млекопитающих. Выбор и использование таких носителей хорошо известны специалистам в данной области.
Используемый в настоящем документе термин «экспрессия» относится к процессу, посредством которого нуклеиновые кислоты транскрибируется в мРНК и транслируется в пептиды, полипептиды или белки. Если нуклеиновая кислота получена из геномной ДНК, экспрессия, при выборе соответствующей эукариотической клетки-хозяина или организма, может включать процессинг, такой как сплайсинг РНК.
Используемый в настоящем документе термин «векторы экспрессии» относится к векторам, способным экспрессировать ДНК, которая связана с регуляторными последовательностями, такими как промотерные районы, которые способны вызывать экспрессию таких фрагментов ДНК. Такие дополнительные сегменты могут включать промоторные и терминаторные последовательности и необязательно могут включать одну или более точку начала репликации, один или более селективный маркер, энхансер, сигнал полиаденилирования и тому подобное. Векторы экспрессии, как правило, получают из плазмидной или вирусной ДНК, или вектор может содержать элементы того и другого. Таким образом, вектор экспрессии представляет собой рекомбинантную ДНК или РНК конструкцию, например, плазмиду, фаг, рекомбинантный вирус или другой вектор, которая после введения в соответствующую клетку-хозяина приводит к экспрессии клонированной ДНК. Соответствующие векторы экспрессии хорошо известны специалистам в данной области, и включают векторы, которые реплицируются в эукариотических клетках и/или клетках прокариот и векторы, которые остаются эписомальными или интегрируются в геном клетки-хозяина.
Используемый в настоящем документе термин вектор относится также к "вирусным векторам". Вирусные векторы представляют собой рекомбинантные вирусы, которые оперативно связаны с экзогенными генами для передачи (как носители или челноки) экзогенных генов в клетки.
Используемый в настоящем документе термин «аденовирус» относится к любому из группы ДНК-содержащих вирусов, которые вызывают конъюнктивит и инфекции верхних дыхательных путей у человека.
Используемый в настоящем документе термин «голая ДНК» относится к свободной от гистонов ДНК, которая могут быть использована для вакцин и генной терапии. Голая ДНК представляет собой генетический материал, который передается от клетки к клетке во время процесса переноса гена, называемого трансформацией или трансфекцией. В процессе трансформации или трансфекции очищенная или голая ДНК, принятая клеткой-реципиентом, дает клетке-реципиенту новую характеристику или фенотип.
Используемый в настоящем документе термин «действенно или оперативно связанные», если речь идет о сегментах ДНК, означает, что сегменты расположены таким образом, что действуют согласно их назначению, например транскрипция инициируется на промоторе и проходит через кодирующий сегмент до терминатора.
Используемый в настоящем документе термин «агент», относится к агенту, модулирующему активность белка или экспрессию гена или нуклеиновой кислоты, уменьшая, или увеличивая, или иным образом изменяя активность белков, или, каким-либо образом, повышая, или уменьшая, или иным образом изменяя экспрессию нуклеиновых кислот в клетке.
Используемые в настоящем документе термины "химерный белок" или "гибридный белок" относятся к полипептиду, оперативно связанному с другим полипептидом. Химерные или гибридные белки, предусмотренные в настоящем документе, включают один или более FVII-полипептидов, или его части, и один или несколько других полипептидов, для каждого с одним или более сигналами управления транскрипции / трансляции, сигнальными последовательностями, метками для локализации, метками для очистки, частью домена иммуноглобулина G и/или агентами нацеливания. Химерные полипептиды FVII также включают полипептиды, имеющие внутренние домены или районы, обмененные с другим полипептидом. Эти химерные или гибридные белки включают белки, полученные рекомбинантными методами, такие как гибридные белки, полученные химическими методами, такими как химическое соединение, посредством, например, соединения сульфогидрильных групп, а также белки, полученные любым другим методом, посредством которого, по крайней мере, один полипептид (т.е. FVII), или его часть связана прямо или через линкер(ы) с другим полипептидом.
Используемый в настоящем документе термин «оперативно-связанные», когда речь идет о синтезе белка, относится к протеазным полипептидам и непротеазным полипептидам, которые гибридизованы в рамке друг с другом. Непротеазный полипептид может быть гибридизован с N-концом или С-концом протеазного полипептида.
Используемый в настоящем документе термин «агент позиционирования» относится к любому фрагменту, такому как белок или его эффективная часть, которая обеспечивает специфическое связывание с молекулами поверхности клетки, такими как рецепторы, расположенные на поверхности клетки, которые в некоторых случаях могут образовать конъюгат или его часть. Агенты позиционирования также могут способствовать или облегчать, например, аффинное отделение или очистку конъюгата; присоединение конъюгата к поверхности; или обнаружение конъюгатов или комплексов, содержащих этот конъюгат.
Используемые в настоящем документе термины «производное» или «аналог молекулы» относятся к части, полученной из молекулы, или к модифицированной молекуле.
Используемый в настоящем документе термин "заболевание или расстройство" относится к патологическим состояниям организма, вызванными причинами или состояниями, включающими инфекции, приобретенные состояния, генетические состояния, но не ограничивающимися ими, и характеризующимися идентифицируемыми симптомами. В настоящем документе рассматриваются заболевания и расстройства, затрагивающие коагуляцию, в том числе опосредованные белками, ответственными за коагуляцию, и те, в которых белки, ответственные за коагуляцию, играют определенную роль в этиологии и патологии. Заболевания и расстройства также включают те, которые вызваны отсутствием таких белков, например гемофилию, и особый интерес в настоящем документе представляют собой заболевания и расстройства, при которых коагуляция не происходит из-за дефицита или дефекта ответственного за коагуляцию белка.
Используемый в настоящем документе термин "прокоагулянт" означает любое вещество, которое способствует свертыванию крови.
Используемый в настоящем документе термин "антикоагулянт" означает любое вещество, которое препятствует свертыванию крови
Используемые в настоящем документе термин "гемофилия" относится к заболеванию, связанному с кровоточивостью, обусловленному дефицитом факторов свертывания крови. Гемофилия может быть результатом, например, отсутствия, уменьшения экспрессии или уменьшения активности фактора свертывания крови. Наиболее распространенным типом гемофилии является гемофилия А, обусловленная дефицитом фактора VIII. Вторым наиболее распространенным типом гемофилии является гемофилия В, которая обусловлена дефицитом фактора IX. Гемофилия С, также называемая FXI недостаточностью, является более легкой и менее распространенной формой гемофилии.
Используемый в настоящем документе термин "врожденная гемофилия" относится к тем типам гемофилии, которые передаются по наследству. Врожденная гемофилия является результатом мутации, делеции, вставки или иной модификации гена фактора свертывания крови, при которой образование фактора свертывания крови отсутствует, уменьшено или не приводит к функциональному продукту. Например, наследственные мутации в генах факторов свертывания крови, таких как фактор VIII и фактор IX, приводят к врожденным гемофилиям, гемофилиям А и В, соответственно.
Используемый в настоящем документе термин "приобретенная гемофилия" относится к типу гемофилии, которая развивается в зрелом возрасте в результате образования аутоантител, инактивирующих FVIII.
Используемый в настоящем документе термин "расстройство, связанное с кровоточивостью" означает состояние, при котором снижена способность контролировать кровотечение. Кровоточивость может быть наследственной или приобретенной, а также может быть результатом, например, дефектов или недостатков в пути свертывания, дефектов или недостатков в активности тромбоцитов, или дефектов сосудов.
Используемый в настоящем документе термин "приобретенное связанное с кровоточивостью расстройство" относится связанному с кровоточивостью расстройству, которое является результатом дефекта свертывания, вызванного такими состояниями, как заболевания печени, дефицит витамина К или результатом терапии кумадином (варфарином), или другой антикоагулянтной терапии.
Используемый в настоящем документе термин "лечение" субъекта, страдающего заболеванием или состоянием, означает, что полипептид, композиция или другой продукт, предусмотренный в настоящем изобретении, вводится этому субъекту.
Используемые в настоящем документе термины «терапевтический агент», «лечебный режим», «радиопротектант» или «химиотерапия» относятся к обычным препаратам и медикаментозной терапии, в том числе вакцинам, которые известны специалистами в данной области. Радиотерапевтические агенты хорошо известны в уровне техники.
Используемый в настоящем документе термин «лечение» относится к любой последовательности действий, в результате которой симптомы состояния, расстройства или заболевания улучшаются или иным образом благотворно изменяются. Поэтому лечение включает профилактику, терапию и/или лечение. Лечение также включает любое фармацевтическое использование композиций по настоящему изобретению. Лечение также охватывает любое фармацевтическое использование модифицированного FVII и описанных здесь композиций.
Используемый в настоящем документе термин «улучшение симптомов» конкретного заболевания или расстройства посредством лечения, например путем введения фармацевтической композиции или другого лекарственного препарата, относится к любому снижению, постоянному или временному, кратковременному или длительному, симптомам, которое можно объяснить или связать с введением композиции или лекарственного препарата.
Используемые в настоящем документе термины «предотвращение» или «профилактика» относятся к методам, посредством которых снижается риск развития заболевания или состояния. Профилактика включает уменьшение риска развития заболевания или состояния и/или предотвращение ухудшения симптомов или прогрессирования заболевания или снижение риска ухудшения симптомов или прогрессирования заболевания.
Используемый здесь термин «эффективное количество» соединения или композиции для лечения конкретного заболевания относится к количеству, которое достаточно для улучшения, или какого-либо уменьшения симптомов, связанных с этим заболеванием. Такое количество может быть введено в виде одной дозы или может вводиться в соответствии с режимом, при котором оно является эффективным. Указанное количество может вылечить заболевание, но, как правило, вводится в целях улучшения симптомов заболевания. Как правило, повторное введение является необходимым для достижения желаемого улучшения симптомов.
Используемые в настоящем документе термины "терапевтически эффективное количество" или "эффективная терапевтическая доза" относятся к агенту, соединению, материалу или композиции, содержащей соединение, которого, по меньшей мере, достаточно для получения терапевтического эффекта. Эффективное количество представляет собой количество терапевтических агентов, необходимое для предотвращения, лечения, облегчения, прекращения или частичного прекращения симптомов заболевания или расстройства.
Используемые в настоящем документе термины "больной", подвергающийся лечению, или "субъект", подвергающийся лечению, относятся к людям и/или животным, включая млекопитающих. Млекопитающие включают приматов, таких как люди, шимпанзе, гориллы и мартышки; одомашненных животных, например собак, лошадей, кошек, свиней, коз, коров, и грызунов, таких как мыши, крысы, хомяки и песчанки.
Используемый в настоящем документе термин «комбинация» относится к какой-либо ассоциации двух или более отдельных лекарственных объектов. Ассоциации могут быть пространственными или относиться к применению двух или более объектов для достижения общей цели.
Используемый в настоящем документе термин «композиция» относится к любой смеси двух или нескольких продуктов или соединений (например, агентов, модуляторов, регуляторов и т.д.). Это может быть раствор, суспензия, жидкость, порошок, паста, водная или неводная рецептура или любая их комбинация.
Используемый в настоящем документе термин "продукт" относится к продукту, который производится и продается. Используемый в настоящем документе термин должен охватывать модифицированные протеазные полипептиды и нуклеиновые кислоты, содержащиеся в упаковке.
Используемый в настоящем документе термин «жидкость» относится к любой композиции, которая может течь. Термин жидкость, таким образом, охватывает композиции в полутвердом виде, в виде паст, растворов, водных смесей, гелей, лосьонов, кремов и другие подобные композиции.
Используемый в настоящем документе термин "набор" относится к упакованной комбинации, необязательно включающей реагенты и другие продукты и/или компоненты для практического осуществления методов с использованием элементов комбинации. Например, в настоящем документе предусмотрены наборы, содержащие модифицированные полипептидные протеазы или молекулы нуклеиновой кислоты, предусмотренные в настоящем документе, и другой объект для целей, включающих, но не ограничивающихся введением, диагностикой и оценкой биологической активности или свойства. Наборы необязательно включают инструкции по использованию.
Используемый в настоящем документе термин «антитела» включает фрагменты антител, такие как фрагменты Fab, которые состоят из легкой цепи и вариабельной области тяжелой цепи.
Используемый в настоящем документе термин «рецептор» относится к молекуле, которая имеет сродство к конкретному лиганду. Рецепторы могут быть естественными или синтетическими молекулами. Рецепторы также могут упоминаться в уровне техники как анти-лиганды.
Используемый в настоящем документе термин «животные» включает любых животных, включая, но не ограничиваясь, приматами, включая человека, гориллами и мартышками; грызунами, такими как мыши и крысы, птицами, такими как куры; жвачными животными, такими как козы, коровы, олени, овцы; свиньями и другими животными. Термин животные, не включающие человека, исключает людей как животных. Предусмотренные в настоящем документе протеазы могут происходить из любого источника, животного, растения, прокариотического организма и гриба.
Используемый в настоящем документе термин «генная терапия» предполагает передачу гетерологичных нуклеиновых кислот, таких как ДНК, в определенные клетки, клетки-мишени млекопитающих, в частности, человека, страдающего расстройствами или состояниями, для которых необходима такая терапия. Нуклеиновые кислоты, такие как ДНК, вводятся в выбранные клетки-мишени, непосредственно, либо с помощью вектора или другого носителя, таким образом, чтобы гетерологичная нуклеиновая кислота, такая как ДНК, экспрессировалась и таким образом производился закодированный терапевтический продукт. С другой стороны, гетерологичная нуклеиновая кислота, такая как ДНК, может каким-либо образом способствовать экспрессии ДНК, которая кодирует терапевтический продукт, или она может кодировать продукт, такой как пептид или РНК, который каким-либо образом способствует, прямо или косвенно, экспрессии терапевтического продукта. Генная терапия также может быть использована для доставки нуклеиновых кислот, кодирующих продукты генов, которые заменяют дефектный ген или дополняют продукт гена, произведенный млекопитающим или клеткой, в которые введена нуклеиновая кислота. Введенная нуклеиновая кислота может кодировать терапевтическое соединение, такое как протеаза или модифицированная протеаза, которые обычно не производятся в организме млекопитающего или которые не производятся в терапевтически эффективных количествах или в течение терапевтически эффективного времени. Гетерологичные нуклеиновые кислоты, такие как ДНК, кодирующие терапевтические продукты, могут быть модифицированы до введения в клетки больного в целях повышения или иного изменения продукта или его экспрессии. Генная терапия также может включать поставку ингибитора, или репрессора, или другие модуляторы экспрессии генов.
Используемый в настоящем документе термин «гетерологичная нуклеиновая кислота» относится к нуклеиновой кислоте, которая обычно не производится in vivo клеткой, в которой она экспрессируется; или к нуклеиновой кислоте, которая производится клеткой, но в другом локусе, или иным образом экспрессируется; или которая способствует или кодирует медиаторы, которые изменяют экспрессиию эндогенных нуклеиновых кислот, таких как ДНК, воздействием на транскрипцию, трансляцию, или другой регулируемый биохимический процесс. Гетерологичная нуклеиновая кислота, как правило, не является эндогенной для клетки, в которую она введена, а получена из другой клетки или получена синтетически. Гетерологичная нуклеиновая кислота может быть эндогенной, но представлять собой нуклеиновую кислоту, которая экспрессируется в другом локусе, или нуклеиновую кислоту с измененной экспрессией. Как правило, хотя и не обязательно, такие нуклеиновые кислоты кодируют РНК и белки, которые обычно не производятся в клетке, или не производятся в том же виде в клетке, в которой она экспрессируется. Гетерологичная нуклеиновая кислота, такая как ДНК, также может быть отнесена к чужеродной нуклеиновой кислоте, такой как ДНК. Таким образом, гетерологичная нуклеиновая кислота или чужеродная нуклеиновая кислота включает молекулы нуклеиновой кислоты, не присутствующие в такой ориентации или позиции, как у аналогичной молекулы нуклеиновой кислоты, такой как ДНК, находящейся в геноме. Указанный термин также может относиться к молекуле нуклеиновой кислоты из другого организма или другого вида (то есть экзогенной).
Любая нуклеиновая кислота, такая как ДНК, которую специалист в данной области рассматривает или считает гетерологичной или чужеродной в клетке, в которой она экспрессируется, относится к гетерологичным нуклеиновым кислотам; гетерологичные нуклеиновые кислоты включают экзогенно добавленные нуклеиновые кислоты, которые также экспрессируются эндогенно. Примеры гетерологичных нуклеиновых кислот включают, но не ограничиваются, нуклеиновыми кислотами, которые кодируют обнаруживаемые белки-маркеры, такие как белки, которые обусловливают лекарственную устойчивость; нуклеиновыми кислотами, которые кодируют эффективные терапевтические вещества, такие как противораковые агенты, ферменты и гормоны; и нуклеиновыми кислотами, такими как ДНК, которые кодируют другие типы белков, такие как антитела. Антитела, закодированные гетерологичной нуклеиновой кислотой, могут секретироваться или экспрессироваться на поверхности клетки, в которую гетерологичная нуклеиновая кислота была введена.
Используемый в настоящем документе термин «терапевтически эффективный продукт генной терапии» относится к продукту, который кодируется гетерологичной нуклеиновой кислотой, как правило, ДНК, и который после введения нуклеиновой кислоты реципиенту экспрессируется и улучшает или полностью устраняют симптомы проявления наследственных или приобретенных заболеваний или что лечит заболевание. В настоящем изобретении также включены биологически активные молекулы нуклеиновой кислоты, такие как RNAi и антисмысловые РНК.
Используемый в настоящем документе оборот: полипептид "состоит по существу" из приведенной последовательности аминокислот, означает, что только приведенная часть или ее фрагмент присутствуют в полипептиде полной длины. Полипептид необязательно может и, обычно, будет включать дополнительные аминокислоты из другого источника, или может быть вставлен в другой полипептид.
Используемые в настоящем документе термины, употребляемые в единственном числе, подразумевают также их использование во множественном числе, если контекст четко не указывает иное. Так, например, ссылка на соединение, включающее "внеклеточный домен" включает соединения с одним или многими внеклеточными доменами.
Используемые в настоящем документе термины «диапазон» и «количество» могут быть переданы фразой "приблизительно" относительно индивидуального значения или диапазона. Термин приблизительно также относится к точному значению. Следовательно, "приблизительно 5 оснований" означает "приблизительно 5 оснований", а также "5 оснований".
Используемые в настоящем документе термины "необязательный" или "необязательно" означают, что описанные впоследствии события или обстоятельства, могут или произойти, или не произойти, и что описание включает случаи, когда указанное событие или обстоятельство имеет место, и случаи, когда этого не происходит. Например, фраза «необязательно замещенная группа» означает, что группа является незамещенной или замещенной.
Используемые в настоящем документе сокращения для любых защитных групп, аминокислот и других соединений, если не указано иное, соответствуют их обычно используемым, признанным сокращениям, или сокращениям, принятым IUPAC-IUB (Комиссией по биохимической номенклатуре (см., (1972) Biochem. 11: 1726)).
В. Гемостаз
В настоящем документе предусмотрены модифицированные полипептиды фактора VII (FVII). У указанных полипептидов FVII увеличена коагулянтная активность. Соответственно, эти полипептиды имеют множество вариантов использования и применения, например, в качестве лекарственного средства для модулирования гемостаза, и других родственных биологических процессов. Чтобы оценить модификации, предусмотренные в настоящем документе, и использование таких модифицированных молекул FVII, необходимо понимание системы гемостаза и каскада коагуляции. Последующее обсуждение обеспечивает такой фон, являющийся вступлением к обсуждению фактора VII и его модификаций.
Гемостаз является физиологическим механизмом, который останавливает кровотечение, которое является результатом повреждения сосудов. Нормальный гемостаз зависит от клеточных компонентов и растворимых белков плазмы и включает серию сигнальных событий, которые в конечном итоге приводят к образованию сгустка. Коагуляция быстро инициируется после травмы кровеносного сосуда и повреждения клеток эндотелия. На первичном этапе коагуляции активируются тромбоциты, что приводит к формированию гемостатической пробки в месте повреждения. На втором этапе гемостаза в процесс вовлекаются плазменные факторы коагуляции, которые действуют в протеолитическом каскаде, приводящем к образованию нитей фибрина, которые укрепляют тромбоцитную пробку.
После повреждения сосуда приток крови непосредственно к поврежденной области ограничивается сужением сосудов, что позволяет тромбоцитам присоединиться к экспонированным фибриллам коллагена субэндотелиальной соединительной ткани. Адгезия обусловлена фактором Виллебранда (vWF), который связывается с эндотелием в течение трех секунд с момента травмы, тем самым способствуя адгезии и агрегации тромбоцитов. Активация агрегированных тромбоцитов приводит к секреции целого ряда факторов, включая АДФ, АТФ, тромбоксан и серотонин. Также высвобождаются молекулы адгезии, фибриноген, фактор Виллебранда, тромбоспондин и фибронектин. Такая секреция дополнительно способствует адгезии и агрегации тромбоцитов, увеличению активации тромбоцитов и сужению кровеносных сосудов, а также экспозиции анионных фосфолипидов на поверхности тромбоцитов, которая служит платформой для сборки ферментных комплексов свертывание крови. Тромбоциты изменяют форму, что приводит к формированию псевдоподий, что еще больше упрощает агрегацию с другими тромбоцитами, результатом чего является неплотная тромбоцитарная пробка.
Одновременно инициирруется каскад пептидаз свертывания (каскад коагуляции). Каскад коагуляции включает целый ряд активационных актов, включающих протеолитическое расщепление. С помощью такого каскада неактивные белки сериновых протеаз (также называемые зимогенами) преобразуются в активные протеазы путем расщепления одной или более пептидных связей, который затем служат в качестве активирующих протеаз для следующих зимогенных молекул в каскаде, что в конечном итоге приводит к образованию сгустков путем сшивания фибрина. Например, каскад генерирует активированные молекулы, такие как тромбин (расщеплением протромбина), что еще больше активизирует тромбоциты, а также генерирует фибрин расщеплением фибриногена. Фибрин затем формирует сшитый полимер вокруг тромбоцитарной пробки для стабилизации сгустка. После репарации повреждений фибрин разрушается фибринолитической системой, основными компонентами которой являются плазминоген и активатор плазминогена тканевого типа (tPA). Оба эти белка включены в полимеризованный фибрин, где они взаимодействуют с образованием плазмина, который, в свою очередь, воздействует на фибрин с растворением образовавшегося сгустка. Во время образования сгустка ингибиторы фактора свертывания также циркулируют в крови для предотвращения формирования сгустков за пределами места повреждения.
Взаимодействия в системе от травмы до образования сгустка и последующего фибринолиза, описаны ниже.
1. Адгезия и агрегация тромбоцитов
Свертывание крови активно тормозится в нормальных условиях. Эндотелий кровеносных сосудов поддерживает расширение сосудов, препятствует адгезии и активации тромбоцитов, подавляет коагуляцию, усиливает расщепление фибрина и обладает противовоспалительными свойствами. Эндотелиальные клетки выделяют молекулы, такие как окись азота (№) и простациклин, которые ингибируют агрегацию тромбоцитов и расширяют кровеносные сосуды. Высвобождение этих молекул активирует растворимую гуанилатциклазу (SGC) и цГМФ-зависимую протеинкиназу I (cGKI) и увеличивает уровень циклического гуанозинмонофосфата (цГМФ), который вызывают релаксацию гладкой мускулатуры сосудистой стенки. Кроме того, эндотелиальные клетки экспрессируют АДФазы, расположенные на поверхности клетки, такие как CD39, которые контролируют активацию и агрегацию тромбоцитов путем преобразования АДФ, освобожденного из тромбоцитов, в адениннуклеотидные ингибиторы тромбоцитов. Эндотелий также играет важную роль в регуляции ферментов фибринолитического каскада. Эндотелиальные клетки непосредственно содействуют генерации плазмина через экспрессию рецепторов плазминогена (аннексина II) и урокиназы, а также выделению активаторов плазминогена тканевого и урокиназного типов, все они способствуют растворению сгустка. На последнем этапе протромботического регулирования эндотелиальные клетки играют активную роль в ингибировании каскада свертывания посредством образования гепаран сульфата, что увеличивает кинетику ингибирования тромбина и других факторов свертывания крови антитромбином III.
При острой сосудистой травме, однако, сосудосуживающие механизмы преобладают, и эндотелий становится протромботическим, прокоагулянтным и провоспалительным. Это достигается за счет уменьшения количества расширяющих эндотелий агентов: аденозина, №и простациклина, и за счет прямого действием АДФ, серотонина и тромбоксана на гладкомышечные клетки сосудов, чтобы вызывает их сокращение (Becker, Heindl и др., 2000). Главным толчком к изменению функции эндотелия, которое приводит к образованию гемостатических тромбов, является потеря эндотелиального клеточного барьера между кровью и компонентами внеклеточного матрикса (ЕСМ) (Ruggery (2002) Nat Med 8:1227-1234). Циркулирующие тромбоциты определяют и различают области поражения эндотелия и присоединяются к экспонированному субэндотелию. Их взаимодействия с различными тромбогенными субстратами и локально образованными или освобожденными агонистами приводят к активации тромбоцитов. Этот процесс описывается как протекающий в два этапа: 1) адгезия: первоначальное закрепление на поверхности, и 2) агрегация: тромбоцит-тромбоцит когезия (Savage и др., (2001) Curr Opin Hematol 8:270-276).
Адгезии тромбоцитов начинается, когда циркулирующие тромбоциты связываются с экспонированным коллагеном путем взаимодействия с коллаген связывающими белками на поверхности клеток, и со взаимодействия с vWF, также присутствующим на эндотелии. Белок vWF обладает мультимерной структурой различного размера, секретируется в двух направлениях эндотелием: базолатерально и в кровь. vWF также связывает фактор VIII, что играет важную роль в стабилизации фактора VIII и его сохранении при циркуляции.
Адгезия тромбоцитов и последующая активация достигаются при связывании vWF через домен А1 с GPIB (часть тромбоцитарного рецепторного гликопротеиного комплекса GPIB-IX-V). Взаимодействие между vWF и GPIB регулируется усилием сдвига, так что увеличение напряжения сдвига приводит к соответствующему увеличению сродства vWF к GPIB. Интегрин α1β2, известный также как присутствующий на лейкоцитах белок VLA-2, является основным коллагеновым рецептором тромбоцитов, и взаимодействие через этот рецептор генерирует внутриклеточные сигналы, которые способствуют активации тромбоцитов. Связывание с α1β2 способствует привлечению рецептора коллагена низкого сродства, GP VI. Он принадлежит к суперсемейству иммуноглобулинов и является рецептором, который генерирует наиболее мощный внутриклеточный сигнал для активации тромбоцитов. Активации тромбоцитов приводит к освобождению аденозиндифосфата (АДФ), который превращается в тромбоксан А2.
Активация тромбоцитов также приводит к экспрессии на поверхности тромбоцитов рецептора гликопротеина IIb-IIIa (GP IIb-IIIa), GP IIb-IIIa рецепторы позволяют тромбоцитам присоединяться друг к другу (то есть агрегировать) посредством молекул фибриногена, связывающих тромбоциты через эти рецепторы. Это приводит к образованию тромбоцитарной пробки в месте повреждения, что предотвращает дальнейшую потерю крови, пока поврежденная ткань сосуда высвобождает факторы, которые запускают каскад свертывания и образование стабилизирующей фибриновой сети вокруг тромбоцитарной пробки.
2. Каскад коагуляции
Путь коагуляции является протеолитическим путем, в котором каждый фермент присутствует в плазме в виде зимогена, или неактивной формы. Расщепление зимогенов регулирует высвобождение активных форм из молекул-предшественников. Кофакторы активированных протеаз, такие как гликопротеины FVIII и FV, также активизируются в каскаде реакций и играют определенную роль в образовании сгустка. Путь функционирует как ряд положительных и отрицательных обратных связей, которые контролируют процесс активации, где конечная цель состоит в образовании тромбина, который затем может преобразовать растворимый фибриноген в фибрин и сформировать сгусток. Факторам коагуляции обычно присваивают номера, записанные римскими цифрами, со строчной буквой "а" для указания активированной формы. В таблице 3 ниже приведен примерный перечень факторов, в том числе их обычные названия и их роли в каскаде коагуляции. Как правило, эти белки участвуют в свертывании крови через один или несколько путей, представленных внутренним, внешним или общим путями свертывания (см. Фиг.1). Как отмечено ниже, эти пути связаны между собой, и свертываемость крови, как полагают, осуществляется через клеточную модель активации, где фактор VII (FVII) является главным инициатором свертывания.
Факторы свертывания
Из таблицы (с изменениями) M.W.King (2006) из med.unibs.it/~Marchesi/blood.html
Генерация тромбина исторически была разделена на три пути:
внутренний (предположительно, все компоненты пути характерны для плазмы) и внешний (предположительно, один или более компонентов пути являются внешними для плазмы) пути, которые обеспечивают альтернативные пути для генерации активированного фактора Х (FXa), и окончательный общий путь, результатом которого является образование тромбина (фиг.1). Эти белки вместе участвуют во взаимосвязанном и взаимозависимом процессе коагуляции. Была разработана клеточная модель коагуляции, описывающая эти пути (фиг.2) (Hoffman и др., (2001) Thromb Haemost 85:958-965). В этой модели "внешний" и "внутренний" пути происходят на поверхностях различных клеток, несущих тканевый фактор (ТФ) клеток и тромбоцитов, соответственно. Процесс коагуляции разделяется на отдельные фазы, инициации, усиления и распространения, в ходе которых внешний и внутренний пути функционируют на различных этапах для получения большого количества тромбина, необходимого для преобразования достаточного количества фибриногена в фибрин для формирования сгустка.
А. Инициация
FVII считается фактором свертывания, ответственным за инициирование каскада коагуляции, которое зависит от его взаимодействия с ТФ. ТФ представляет собой трансмембранный гликопротеин, экспрессируемый различными клетками, такими как гладкомышечные клетки, фибробласты, моноциты, лимфоциты, гранулоциты, тромбоциты и эндотелиальные клетки. Миелоидные клетки и эндотелиальные клетки экспрессируют ТФ только при раздражении, например, провоспалительными цитокинами. Гладкомышечные клетки и фибробласты, однако, экспрессируют ТФ конститутивно. Соответственно, как только эти клетки вступают в контакт с кровью после повреждения тканей, каскад коагуляции быстро инициируется связыванием ТФ с фактором VII или FVIIa в плазме.
Как будет показано ниже, большая часть FVII в крови находится в форме зимогена с небольшим, примерно 1%, содержанием FVIIa. В отсутствие связывания с ТФ, однако, даже FVIIa имеет зимогенподобные характеристики и не проявляет значительную активность, пока не образует комплекс с ТФ. Таким образом, плазменный FVII требует активации протеолитическим расщеплением, а также дополнительных конформационных изменений при взаимодействии с ТФ для полноценной активности. Было показано, что ряд протеаз, в том числе факторы IХа, Ха, ХIIа и тромбин, способны расщепления FVII in vitro, и этот процесс ускоряется в присутствии ТФ. FVIIa сам по себе также может активировать FVII в присутствии ТФ, этот процесс называют самоактивацией. Небольшие количества FVIIa в крови обусловлены, вероятно, его активацией FXa и/или FIXa (Wildgoose и др., (1992) Blood 80:25-28 и Butenas и др. (1996) Biochemistry 35:1904-1910). Таким образом, TF / FVIIa комплексы могут образоваться прямым связыванием FVIIa с ТФ или связыванием FVII с TF с последующей активацией FVII в FVIIa протеазой плазмы, такой как FXa, FIXa, FXIIa, или самоактивацией FVIIa. TF / FVIIa комплекс остается связанным с ТФ-несущей клеткой, где он активирует небольшие количества FX в FXa, что известно как "внешний путь" коагуляции.
TF / FVIIa комплекс также расщепляет небольшие количества FIX с образованием FIXa. FXa также связывается со своим кофактором FVa с образованием комплекса на ТФ-несущей клетке, которая затем может преобразовывать протромбин в тромбин. Образуется небольшое количество тромбина, которого, однако, недостаточно для поддержания образования фибрина, необходимого для полного свертывания. Кроме того, циркулирующие в крови активные FXa и FIXa ингибируются антитромбином III (AT-III) и другими серпинами, которые будут рассмотрены более подробно ниже. Это, как правило, предотвращает формирование сгустков в циркулирующей крови. При наличии травмы, однако, повреждение сосуда приводит к агрегации тромбоцитов и активации в этом месте образования тромбина, позволяя усилить сигнал коагуляции.
В. Усиление
Усиление происходит, когда тромбин связывается с тромбоцитами и активирует их. Активированные тромбоциты высвобождают FV из их альфа-гранул, который активируется тромбином с образованием FVa. Тромбин также высвобождается и активирует FVIII из FVIII / vWF комплекса на тромбоцитарной мембране, а также расщепляет FXI с образованием FXIa. Эти реакции приводят к активированным тромбоцитам, которые несут FVa, FVIIIa и FIXa на поверхности, что закладывает основу для образования большого количества тромбина на этапе распостранения.
С. Распространение
Распространение свертывания происходит на поверхности большого числа тромбоцитов в месте повреждения. Как описано выше, активированные тромбоциты несут FXIa, FVIIIa и FVa на своей поверхности. Именно здесь осуществляется внешний путь. FXIa активизирует FIX с образованием FIXa, который затем может связаться с FVIIIa. Этот процесс, в дополнение к небольшому количеству FIXa, образованному расщеплением FIX ТФ / FVIIa комплексом на ТФ-несущей клетке, приводит к образование большого количества FXIa / FVIIIa комплекса, который, в свою очередь, может активировать значительные количества FX с образованием FXa. Молекулы FXa связывают молекулы FVa с образованием протромбиназного комплекса, который активирует протромбин в тромбин. Образование тромбина действует как положительная обратная связь для активации дополнительных тромбоцитов и вновь инициирует процессы, описанные для фазы усиления.
За очень короткое время образуется достаточное количество активированных тромбоцитов с соответствующими комплексами для создания всплеска тромбина, которого достаточно для образования достаточного количества фибрина из фибриногена для формирования гемостатического сгустка фибрина. Фибриноген представляет собой растворимый в плазме крови димер, который при расщеплении тромбином высвобождает фибринопептид А и фибринопептид В. Фибринопептид В затем расщепляется тромбином, и мономеры фибрина, образованные при этом втором протеолитическом расщеплении, спонтанно образуют нерастворимый гель. Полимеризованный фибрин удерживается нековалентными и электростатическими силами и стабилизируется трансамидирующим ферментным фактором XIIIa (FXIIIa), образующимся расщеплением FXIII тромбином. Тромбин также активизирует TAFI, который ингибирует фибринолиз за счет уменьшения образования плазмина на поверхности сгустка. Кроме того, тромбин сам включен в структуру сгустка для дополнительной стабилизации. Этот нерастворимый агрегат фибрина (сгусток) вместе с агрегированными тромбоцитами (тромб) блокирует поврежденный кровеносный сосуд и предотвращает дальнейшее кровотечение.
3. Регуляция коагуляции
При коагуляции каскад регулируется конститутивными и стимулируемыми процессами, ингибирующими дальнейшее образование сгустков. Есть несколько причин для таких механизмов регулирования. Во-первых, регулирование необходимо для ограничения ишемии тканей при образовании сгустка фибрина. Во-вторых, регулирование предотвращает распространенный тромбоз за счет локализации образования сгустков лишь в месте повреждения тканей.
Регуляция осуществляется катионами нескольких ингибиторных молекул. Например, антитромбин III (AT-III) и ингибитор пути тканевого фактора (TFPI) конститутивно ингибируют факторы каскада коагуляции. AT-III ингибирует тромбин, FIXa и FXa, a TFPI ингибирует FXa и FVIIa / ТФ комплекс. Дополнительный фактор, протеин С, который стимулируется активацией тромбоцитов, регулирует коагуляцию протеолитическим расщеплением и инактивацией FVa и FVIIIa. Протеин S повышает активность протеина С. Кроме того, еще одним фактором, который способствует ингибированию коагуляции, является интегральный мембранный белок тромбомодулин, который образуется эндотелиальными клетками и служит рецептором тромбина. Связывание тромбина тромбомодулином ингибирует прокоагулянтную активность тромбина, а также способствует активации протеина С.
Фибринолиз, распад сгустка фибрина, также является механизмом для регулирования коагуляции. Сшитые мультимеры фибрина в сгустке разрушаются до растворимых полипептидов плазмином, сериновой протеазой. Плазмин может образоваться из неактивного предшественника плазминогена и оказываться рядом со сгустком фибрина двумя способами:
за счет взаимодействия с тканевым активатором плазминогена (tPA) на поверхности сгустка фибрина, а также за счет взаимодействия с урокиназным активатором плазминогена (uРА) на поверхности клеток. Первый механизм представляется основным механизмом, ответственным за разрушение сгустков крови в сосудах. Второй механизм способствует растворению сгустка, а также может играть важную роль в ремоделировании тканей, миграции клеток и воспалении.
Растворение сгустка также регулируется двумя путями. Во-первых, эффективная активация плазмина и фибринолиз происходят только в комплексах, образованных на поверхности сгустка или на клеточной мембране, в то время как свободные белки в крови являются неэффективными катализаторами и быстро инактивируются. Во-вторых, активаторы плазминогена и плазмина инактивируются такими молекулами, как ингибитор активатора плазминогена 1 типа (PAI-I) и PAI-2, которые действуют на активаторы плазминогена, а также а2-антиплазмином и а2-макроглобулином, которые инактивируют плазмин. В нормальных условиях отрегулированное во времени равновесие между коагуляцией и фибринолизом приводит к формированию и эффективному разрушению сгустков после повреждения сосудов при одновременном предотвращении нежелательного тромбообразования или кровотечений.
Краткий обзор некоторых факторов свертывания, кофакторов и регуляторных белков, а также их активности приведены в таблице 4 ниже.
Таблица адаптирована из M.W.King (2006) med.unibs.it/~Marchesi/blood.html
С.фактор VII (FVII)
Фактор VII является витамин К-зависимой сериновой гликопротеиновой протеазой, которая синтезируется в организме животных, включая млекопитающих, в виде одноцепочечного зимогена в печени и выделяется в кровь. Как описано выше, FVII является коагуляционной протеазой, ответственной за инициирование каскада протеолитических событий, который приводит к образованию тромбина и отложению фибрина. Он является частью внешнего пути, хотя последующие эффекты его активности также существенно влияют на внутренний путь. Эта общая роль в формировании сгустка привлекает значительный интерес к FVII как мишени для антикоагулянтной и гемостатической терапии в клинике. Например, был разработан активированный рекомбинантный FVII (rFVIIa) в качестве кровоостанавливающего агента для субъектов с гемофилией и субъектов с другими состояниями, связанными с кровотечениями. В настоящем документе предусмотрены модифицированные FVII-полипептиды, которые обладают увеличенной коагуляционной активностью при активации, которые могут применяться как улучшенные лекарственные средства для лечения заболеваний и состояний, поддающихся терапии фактором VII.
1. Структура и организация FVII
Ген человека FVII (F7) находится в хромосоме 13 в 13q34 бэнде и обладает длиной 12,8 Кб и 9 экзонами. Ген FVII обладает значительным сходством организации с генами, кодирующими другие витамин-К-зависимые белки, такие как протромбин, факторов IX, фактор Х и белок С.мРНК FVII претерпевает альтернативный сплайсинг с получением двух транскриптов: вариант 1 (Genbank Accession №. NM_000131, последовательность, приведенная в SEQ ID №: 108) и вариант 2 (Genbank Accession №. NM_019616 последовательность, приведенная в SEQ ID №: 109). Транскрипт варианта 2, который является более распространенной формой в печени, не включает экзон 1b и кодирует более короткий полипептид-предшественник из 444 аминокислот (изоформа b предшественника FVII; последовательность SEQ ID №: 2) по сравнению с 466 аминокислотным полипептидом-предшественником, закодированным транскиптом варианта 1 (изоформа предшественника FVII; последовательность SEQ ID №: 1). Аминокислоты, которые не представлены в изоформе b предшественника FVII, соответствуют аминокислотным позициям с 22 до 43 изоформы предшественника FVII. Эти аминокислоты являются частью пропептидной последовательности, что приводит к укороченной изоформе b пропептида FVII. Полипептиды-предшественники состоят из следующих сегментов и доменов: гидрофобный сигнальный пептид (аминокислоты 1-20 из последовательностей SEQ ID №№: 1 и 2), пропептид (аминокислоты 21-60 из SEQ ID №: 1, аминокислоты 21-38 в SEQ ID №: 2), домен G1A (аминокислоты 39-83 в SEQ ID №1, аминокислоты 61-105 в SEQ ID №: 2), домен эпидермального фактора роста типа В (ЭФР-подобный домен 1, аминокислоты 84-120 в SEQ ID №: 1, аминокислоты 106-142 в SEQ ID №: 2), домен эпидермального фактора роста типа А (ЭФР-подобный домен 2, аминокислоты 125-166 в SEQ ID №: 1; и аминокислоты 147-188 в SEQ ID №: 2), сериновый протеазный домен (аминокислоты 191-430 в SEQ ID №: 1, аминокислоты 213-452 в SEQ ID №: 2).
406-аминокислотная зрелая форма полипептида FVII (SEQ ID №: 3) не имеет сигнального пептида и пропептидных последовательностей и идентична по длине и последовательности независимо от изоформы предшественника, из которого она была получена. В зрелой форме полипептида FVII соответствующие аминокислотные позиции для вышеуказанных доменов таковы: Gla домен (аминокислоты 1-45 из SEQ ID №: 3), ЭФР-подобный домен 1 (аминокислоты 46-82 из SEQ ID №: 3), ЭФР-подобный домен 2 (аминокислоты 87-128 из SEQ ID №: 3), сериновый протеазный домен (аминокислоты 153-392 из SEQ ID №: 3).
Домен Gla из FVII представляет собой связывающий мембраны фрагмент, который в присутствии ионов кальция взаимодействует с фосфолипидами мембран, которые включают фосфатидилсерин. Домен Gla также играет определенную роль в связывании с кофактором FVIIa, тканевым фактором (ТФ). В комплексе с ТФ домен Gla из FVIIa загружается семью ионами Са2+, выбрасывает три гидрофобные боковые цепи в направлении клеточной мембраны для взаимодействия с фосфолипидами на поверхности клеток, а также образует значительный контакт с С-концевым доменом ТФ. Домен Gla является консервативным у всех витамин-К-зависимых белков, таких как протромбин, факторы коагуляции VII, IX и X, белки С, S и Z. Этим белкам требуется витамин К для посттрасляционного синтеза γ-карбоксиглутаминовой кислоты, аминокислоты, сосредоточенной в N-концевом домене Gla этих белков. Все остатки глутаминовой кислоты в домене являются потенциальными сайтами карбоксилирования, и многие из них модифицируются карбоксилированием.
В дополнение к домену Gla, зрелый белок FVII также содержит два ЭФР-подобных домена. Первый ЭФР-подобный домен (ЭФР-подобный домен 1 или EGF1) является кальций-связывающим ЭФР доменом, в котором шесть консервативных коровых цистеинов образуют три дисульфидных моста. Домен EGF1 из FVII связывает только один ион Са2+, но со значительно более высоким сродством, чем домен G1A (Banner и др., (1996) Nature 380:41-46). Этот связанный ион Са2+способствует сильному взаимодействию между EGF1 доменом FVII и ТФ (Osterbund и др., (2000) Eur J Biochem 267:6204-621 1) Второй ЭФР-подобный домен (ЭФР-подобный домен 2 или EGF2) не является кальций-связывающим доменом, но формирует 3 дисульфидных мостика. Как и другие домены FVII, EGF2 домен взаимодействует с ТФ. Он также связан дисульфидными связями с протеазным доменом, с которым он обладает большой поверхностью контакта.
Наконец, сериновый протеазный домен из FVII является доменом, ответственным за протеолитическую активность FVIIa. Последовательность аминокислот в каталитическом домене FVII демонстрирует высокую идентичность последовательности и сходство третичной структуры с другими сериновыми протеазами, такими как трипсин и химотрипсин (Jin и др., (2001) J Mol. Biol, 307: 1503-1517). Например, эти сериновые протеазы обладают общей каталитической триадой Н57, D102, S195 по химотрипсиновой нумерации. В отличие от других сериновых протеаз, однако, расщепление FVIIa не является достаточным, чтобы завершить преобразование зимогена в полностью активный фермент.Вместо этого, как указано ниже, каталитическая функция FVIIa аллостерически активируется путем связывания с рецептором ТФ на клеточной поверхности, которое вызывает конформационные изменения в протеазном домене FVIIa, переводя его из зимогеноподобного неактивного состояния в каталитически активный фермент. Район спираль-петля между сайтом связывания кофактора и активным центром (т.е. аминокислотные позиции 305-321, соответствующие остаткам 163-1701 по химотрипсиновой нумерации) FVIIa имеет важное значение для аллостеричесокой и зимогенной активности FVIIa (Persson и др., (2004) Biochem J., 379: 497-503). Этот район состоит из короткой α-спирали (аминокислотные позиции с 307 до 312) и следующей за ней петли. N-концевая части спирали образует часть поверхности взаимодействия между протеазным доменом и ТФ, а также содержит ряд остатков, которые являются важными для протеолитической функции и оптимального связывания с ТФ. Сравнение кристаллической структуры FVIIa и комплекса FVIIa с ТФ указывает, что α-спираль претерпевает значительные конформационные изменения при связывании FVIIa с ТФ. α-Спираль FVIIa без ТФ выглядит искаженной, укороченной и ориентирована по-другому. Это влияет на прилегающую петлевую структуру, отодвигая ее от активного центра. Напротив, α-спираль FVIIa в комплексе с ТФ стабилизируется, и соседние петли расположены ближе к активному центру. Такая стабилизация осуществляется посредством механизмов, в которые вовлечена, по крайней мере, аминокислота метионин в аминокислотной позиции 306 (аминокислотный остаток Met164 по химотрипсиновой нумерации) FVII (Pike и др., (1999) PNAS 8925-8930).
2. Посттрансляционные модификации
Полипептид предшественника FVII (либо изоформа гена фактора VII) является мишенью в клетке для секреторного пути гидрофобного сигнального пептида, который проникает в эндоплазматический ретикулум (ЭР) для начала перемещения через мембрану. После перемещения белка через мембрану эндоплазматического ретикулума, 20 аминокислотный сигнальный пептид отщепляется сигнальной пептидазой в просвете ЭР, после чего полипептид подвергается дальнейшим посттрансляционным модификациям, в том числе N- и O-гликозилированию, витамин-К-зависимому карбоксилированию N-терминальной глутаминовой кислоты в γ-карбоксиглутаминовую кислоту и гидроксилированию аспарагиновой кислоты с образованием р-гидроксиаспарагиновой кислоты.
Пропептид несет сайт связывания витамин-К-зависимой карбоксилазы, которая распознает амфипатическую α-спираль из 10 остатков в пропептиде FVII. После связывания карбоксилаза γ-карбоксилирует 10 остатков глутаминовой кислоты в домене Gla полипептида FVII, образуя γ-карбоксиглутаминовые остатки на позициях Е66, Е67, Е74, Е76, Е79, Е80, Е85, Е86, Е89 и Е95 по аминокислотной последовательности FVII предшественника, приведенной в SEQ ID №: 2. Эти позиции соответствуют позициям Е6, Е7, Е14, Е19, Е20, Е25, Е26, Е29 и Е35 зрелого полипептида FVII, приведенного в SEQ ID №: 3. Для обеспечения оптимальной активности молекуле FVII требуется кальций, который связывает полипептид и облегчает конформационные изменения, необходимые для связывания FVIIa с ТФ и липидами. γ-карбоксилированный домен Gla связывает семь ионов Са2+с различным сродством, что индуцирует конформационные изменения, которые позволяют домену Gla взаимодействовать с С-концевым доменом ТФ, а также фосфатидилсеринами и другими отрицательно заряженными фосфолипидами на мембране тромбоцитов.
N-гликозилирование осуществляется переносом Glс3Маn9(GIсМАс) на два остатка аспарагина в полипептиде FVII на позиции, которые соответствуют аминокислотным остаткам 145 и 262 зрелого белка (последовательность SEQ ID №3). O-гликозилирование происходит по аминокислотным остаткам 52 и 60 зрелого полипептида, и гидроксилирование с образованием β-гидроксиаспарагиновой кислоты происходит по остатку аспарагиновой кислотой в положении 63. Эти O-гликозилированные остатки серина и β-гидроксилированный остаток аспарагиновой кислоты расположены в ЭФР-1 домене FVII. Эти модифицикации осуществляются в ЭР и аппарате Гольджи перед окончательным процессингом полипептида в его зрелую форму.
3. Процессинг FVII
Модифицированный пpo-FVII-полипептид транспортируется через аппарат Гольджи в транс-Гольджи компартмент, где пропептид расщепляется пропептидазой непосредственно перед секрецией белка из клетки. РАСЕ / фурин (где РАСЕ является аббревиатурой для фермента, расщепляющего белок в месте спаренных основных аминокислот) является эндопептидазой, локализованной в мембране аппарата Гольджи, которая расщепляет многие белки по карбоксильному концу последовательности Arg-[любой остаток]-(Lys или Arg)-Arg. Это пропептидаза расщепляет витамин-К-зависимые гликопротеины, такие как про-фактор IX и про-vWF полипептиды (Himmelspach и др., (2000) Thromb Research 97; 51-67), отделяя пропептид от зрелых белков. Включение соответствующего сайта распознавания РАСЕ / Фурин в предшественника рекомбинантного фактора VII способствует правильному процессингу и секреции рекомбинантного полипептида (Margaritas и др., (2004) dm Invest 113 (7): 1025-1031). PACE / Фурин или другой пропептидазный фермент сходного типа, скорее всего, ответственен за протеолитический процессинг про-FVII в FVII. Он может распознавать и связывать непрерывный -Arg-Arg-Arg-Arg- мотив, соответствующий аминокислотным позициям 35-38 последовательности, приведенной в SEQ ID №1, и 57-60 последовательности, приведенной в SEQ ID №2, расщепляя пропептид и высвобождая зрелый белок для секреции.
4. Активация FVII
Подавляющее большинство FVII находится в крови в виде неактивированных одноцепочечных зимогенов, хотя небольшое количество присутствует в виде двухцепочечной активированной формы. Активация FVII происходит при протеолитическом расщеплении связи Arg152-Ile153 (позиции по отношению к зрелому полипептиду FVII, приведенному в SEQ ID №3), которое приводит к двухцепочечному полипептиду, содержащему 152 аминокислотную легкую цепь (около 20 кДа), связанную дисульфидным мостиком с 254 аминокислотной тяжелой цепью (около 30 кДа). Легкая цепь FVIIa содержит домен Gla и ЭФР-подобный домен, в то время как тяжелая цепь содержит каталитическую или серин-протеазную часть молекулы. Преобразование одноцепочечного FVII в двухцепочечный FVIIa осуществляется расщеплением FIXa, FXa, FXIIa, тромбином, или происходит автокаталитически при участии эндогенного FVIIa (Butenas и др., (1996) Biochem 35:1904-1910; Nakagaki и др., (1991) Biochem 30:10819-10824). Следовые количества FVIIa, которые присутствуют в циркулирующей крови, скорее всего, возникают из-за действия FXa и FIXa.
Как говорилось выше, расщепление зимогенный формы FVII с образованием FVIIa не достаточно для полноценной активности. Для полноценной активности FVIIa необходимо соединиться с ТФ (Higashi и др., (1996) J. Biol. Chem. 271:26569-26574). Из-за этого FVIIa без ТФ приписаны зимогенподобные черты, отображающие зимогенный фолдинг и форму, а также проявление низкой активности. Эти зимогенподобные характеристики FVIIa в отсутствие его связи с ТФ делают его относительно устойчивьм к антитромбину III (AT-III) и другим серпинам, которые обычно действуют на активные формы сериновых протеаз, а не на зимогенные формы. Кроме того, TFPI, основной ингибитор активности ТФ / FVIIa также не связывает эффективно "неактивную" некомплексную форму FVIIa.
После комплексообразования с ТФ FVIIa претерпевает конформационные модификации, которые обусловливают полноценную активность молекулы. Все домены FVII участвуют во взаимодействии с ТФ, но конформационные изменения локализованы в протеазном домене FVIIa. Например, конформационные изменения, которые происходят при аллостерическом взаимодействии FVIIa и ТФ, включают создание обширного сайта связывания макромолекулярного субстрата. Этот сайт связывания значительно повышает FVII-опосредованную протеолитическую активацию фактора X.
Активность FVIIa дополнительно увеличивается (т.е. в тысячу раз) при взаимодействии FVIIa с экспрессированным на поверхности клеток ТФ. Это происходит потому, что фосфолипиды мембраны, среди которых присутствуют отрицательно заряженные фосфолипиды, такие как фосфатидилсерин, являются сайтам взаимодействия для других витамин-К-зависимых факторов свертывания, таких как FIX и FX, которые связываются с ними через домены G1A. Таким образом, локальная концентрация указанных витамин-К-зависимых белков высока на поверхности клеток, что способствует их взаимодействию с ТФ / FVIIa комплексом.
5. Функции FVII
Хотя FVIIa проявляет повышенную активность после аллостерической активации ТФ, есть свидетельства того, что существуют механизмы, посредством которых FVIIa один может инициировать коагуляцию. Следовательно, FVII может функционировать ТФ-зависимо и ТФ-независимо. Этот последний путь может играть гораздо меньшую роль в нормальном гемостазе, хотя он может быть существенным при рассмотрении нарушений свертываемости крови и их лечении.
А. Зависимая от тканевого фактора активность FVIIa
Циркулирующий в крови FVII связывается с ТФ на поверхности клетки и активируется FIXa, FXa, тромбином или автокаталитически эндогенным FVIIa, как описано выше. Кроме того, очень небольшое количество циркулирующего FVIIa может напрямую связать ТФ. Затем ТФ / FVIIa комплекс связывает небольшое количество плазменненного FX, и каталитический домен FVIIa расщепляет FX с образованием FXa. Тромбин, таким образом, образуется по внешнему пути на поверхности ТФ-несущей клетки, когда FXa комплексы с FVa активируют протромбин в тромбин (Фиг.3). FIX также активируется ТФ / FVIIa комплексом, что обеспечивает связь с внутренним путем, который действует на поверхности активированных тромбоцитов. Системы положительной обратной связи в каскаде свертывания, описанном выше, обеспечивают механизмы, с помощью которых производится большое количество тромбина, который расщепляет фибриноген в фибрин для образования сгустка.
В. Независимая от тканевого фактора активность FVIIa
В дополнение к ТФ-зависимому механизму активации FX в FXa, FVIIa также может активировать FX в отсутствие ТФ. Активированные тромбоциты перемещают фосфатидилсерины и другие отрицательно заряженные фосфолипиды на внешнюю, плазма-ориентированную поверхность клетки (Hemker и др., (1983) Blood Cells 9:303-317). Это обеспечивает альтернативные "рецепторы", через которые может связаться FVIIa, хотя и с относительно низким сродством, которое в 1000 раз меньше, чем сродство связывания FVIIa с ТФ (Моnrое и др., (1997) Br. J. Haematol. 99:542-7). Это взаимодействие опосредовано остатками в домене GlA (Harvey и др., (2003) 278:8363-8369). Затем FVIIa может преобразовывать FX в FXa и FIX в FIXa на поверхности активированных тромбоцитов (Hoffman и др., (1998) Blood Coagul Fibrinolysis 9: S61-S65). FXa остается связанным с поверхностью тромбоцитов, где он может связаться с FVa и образовать достаточное количество тромбина из протромбина, в то время как образованный FIXa объединяется с FVIIIa и катализирует активизацию дополнительного FX в FXa (Фиг.3). Гемостаз в отсутствие ТФ может быть достигнут положительными обратными связями и механизмами распространения, описанными выше. Примечательно, однако, что хотя FVIIIa может участвовать в процессе коагуляции на активированных тромбоцитах, их присутствие не обязательно для образования тромбина в ТФ-независимом механизме (Фиг.3). Таким образом, в отсутствие FVIII, например у больных гемофилией, FVIIa может инициировать и/или усиливать образование тромбина, благодаря этому второму механизму и вызывать формирование сгустка.
6. FVII как биофармацевтический лекарственный препарат
FVII инициирует свертывание крови. Рекомбинантный FVIIa (NovoSeven®; rFVIIa) предназначен для лечения кровотечений или профилактики кровотечений при хирургических или инвазивных процедурах у пациентов с гемофилией А или В с ингибиторами фактора VIII или фактора IX, а также у пациентов с врожденным дефицитом фактора VII. NovoSeven® является генно-инженерным препаратом фактора FVII, который производится с помощью системы экспрессии млекопитающих в клетках почек детенышей хомяков (ВНК). Агент почти идентичен фактору FVII плазмы по структуре и функции (Ratko и др., (2004), Р & Т, 29: 712-720).
Было показано, что введениt рекомбинантного FVIIa (rFVIIa) содействует свертыванию крови у больных гемофилией, и лечение FVIIa было признано безопасными и хорошо переносимым человеком. Как правило, rFVIIa применяют для лечения больных, у которых выработались ингибиторы (то есть аллоантитела) фактора VIII или фактора IX. Применение rFVIIa в качестве коагулянта было распространено на лечение других нарушений, связанных с кровоточивостью, например тромбостении Гланцманна, других состояний, связанных с обширными кровотечениями, например, возникших в результате травм или операций, включая, но не ограничиваясь, пересадкой печени, предстательной железы и геморрагической травмой; неонатальной коагулопатией, тяжелыми формами заболеваний печени; трансплантацией костного мозга, тромбоцитопенией и расстройствами функционирования тромбоцитов; немедленным обращением оральной антикоагуляции, врожденным дефицитом факторов V, VII, Х и XI; и болезнью фон Виллебранда с ингибиторами фактора Виллебранда.
Для достижения терапевтического эффекта необходимы высокие дозы rFVII. Дозы и режим, необходимые при введении rFVII, варьируются в зависимости от клинических указаний. Например, типичная доза rFVII для геморрагических эпизодов у пациентов с гемофилией А или гемофилией В с аллоантителами составляет 90 мкг / кг при введении внутривенной (ВВ) инъекции. Так как период полураспада rFVII составляет 2 часа, необходимо повторное введение. Дополнительная доза может вводиться через каждые два часа, пока не будет достигнут гемостаз. Величина дозы может быть изменена в зависимости от тяжести состояния. Например, эффективная доза может составлять 35-120 мкг / кг. Кроме того, доза и режим введения могут изменяться в зависимости от других показаний. Например, больным гемофилией А или гемофилией В, перенесшим операцию, начальная доза 90 мкг/кг может вводиться непосредственно перед операцией, с повторным введением каждые два часа во время и после операции. В зависимости от тяжести операции и эпизода кровотечения внутривенное введение болюса может продолжаться каждые два-шесть часов, пока не будет достигнут терапевтический эффект. Пациентам с врожденным дефицитом FVII rFVII, как правило, вводится для профилактики кровотечений при хирургии и других инвазивных процедурах в количестве 15-30 мкг / кг каждые 4-6 часов, пока не будет достигнут гемостаз.
Механизм инициирования гемостаза rFVII объясняет необходимость высоких доз. У больных гемофилией этап инициации коагуляции, при котором ТФ / FVIIa комплекс активирует FX в FXa, приводящий к образованию тромбина на ТФ-несущей клетке, протекает нормально. Впоследствии, однако, процесс свертывания нарушается, так как у больных гемофилией отсутствует FVIII (гемофилия А) или FIX (гемофилия В), и поэтому они не в состоянии сформировать FVIIIa / FIXa комплекс на поверхности активированных тромбоцитов, что обычно служит для активации большого количеств FX в FXa для фаз усиления и распространения, описанных ранее. В связи с наличием ингибиторов, таких как TFPI и AT-III, FXa, образованный на ТФ-несущей клетке после расщепления ТФ / FVIIa, не в состоянии легко диффундировать между клеточными поверхностями. В результате этого не происходит образования большого количества тромбина на поверхности активированных тромбоцитов, и сгусток не образуется.
Существуют доказательства того, что кровоостанавливающее действие высоких доз rFVII достигается с помощью ТФ-зависимого и/или ТФ-независимого образования FXa с помощью rFVIIa на активированных тромбоцитах (Фиг.3). ТФ-зависимое образование тромбина может быть очень быстро минимизировано насыщением молекул ТФ эндогенными FVIIa и rFVIIa. В некоторых случаях, высокие дозы rFVIIa могут связывать активированные тромбоциты и превращать FX в FXa. Связанный на поверхности FXa активизирует FV, что приводит к образованию достаточного количества тромбина для гемостаза. Так как rFVII связывается с поверхностью тромбоцитов с низким сродством, для образования тромбина могут потребоваться высокие дозы rFVII. Активация FXa на активированных тромбоцитах гарантирует, что rFVIIa-опосредованной гемостаз будет локализован в месте повреждения.
Приемы, приводящие к уменьшению дозы rFVII, могут повысить его полезность и эффективность в качестве лекарственного средства. В настоящем изобретении предусмотрены модифицированные FVII-полипептиды. Предусмотрены модифицированные FVII-полипептиды, которые демонстрируют повышенную устойчивость к TFPI, повышенную устойчивость к AT-III, улучшенные фармакокинетические свойства, такие как увеличение времени полужизни, увеличение каталитической активности в присутствии и/или в отсутствие ТФ, и/или увеличение связывания с активированными тромбоцитами. Эти FVII-полипептиды могут проявлять увеличенную коагулянтную активность. FVII-полипептиды, предусмотренные в настоящем документе, могут быть использованы в лечении для инициирования гемостаза по ТФ-зависимому и/или ТФ-независимому механизмам, вследствие чего образуется FXa и вырабатывается тромбин.
D. Модифицированные FVII-полипептиды
В настоящем изобретении предусмотрены модифицированные FVII-полипептиды. Модифицированные FVII-полипептиды отличаются по одному или более видам активности или свойствам по сравнению с немодифицированным FVII-полипептидом. Активности или свойства, которые могут быть изменены в результате модификаций, включают, но не ограничиваются, коагуляцией или коагулянтной активностью, прокоагулянтной активностью; протеолитической или каталитической активностью, такими как активация фактора Х (FX) или активация фактора IX (FIX); иммуногенность (способность к связыванию или конкурирование с полипептидами за связывание с анти-FVII антителами); способность связывать тканевой фактор, фактор Х или фактор IX, способность к связыванию с клеточной поверхностью активированных тромбоцитов; способность связывать фосфолипиды, время полужизни; трехмерная структура; р1 и/или конформация. Как правило, модифицированные полипептиды FVII проявляют прокоагулянтную активность. В настоящем изобретении предусмотрены модифицированные FVII-полипептиды, которые проявляют увеличенную коагулянтную активность при активации их одноцепочечной зимогенной формы. Такие модифицированные полипептиды FVII могут быть использованы в лечении расстройств, связанных с кровоточивостью, или состояний, таких как гемофилии или травмы, где FVII-полипептиды могут содействовать свертыванию крови. К числу таких модифицированных полипептидов FVII относятся те, которые обладают повышенной устойчивость к ингибиторам, таким как ингибитор пути, запускаемого тканевым фактором (TFPI), и антитромбин III (AT-III), а также полипептиды с увеличенной каталитической активностью в присутствии и/или в отсутствие ТФ; полипептиды, которые обладают улучшенными фармакокинетическими свойствами, такими как увеличенное время полужизни; и полипептиды, у которых увеличена способность связываться, и/или аффинность к поверхности тромбоцитов. В частности, такие модифицированные полипептиды FVII могут быть использованы при заболеваниях или состояниях, обеспечивая коагулянтную активность, без потребности в FVIIIa и FIXa. В одном примере, модифицированные FVII-полипептиды, предусмотренные в настоящем документе, могут быть использованы для лечения больных с гемофилией, обладающих аутоантителами к FVIIIa и FIXa. Таким образом, модифицированные полипептиды FVII, предусмотренные в настоящем документе, обладают преимуществами, включая уменьшение количества вводимого FVII, которое необходимо для поддержания достаточной для гемостаза концентрации активного FVII в сыворотке крови. Это может привести, например, к более низким дозам и/или частоте доз, необходимых для достижения сопоставимого биологического эффекта, быстрому наступлению терапевтического эффекта, быстрому улучшению при острых эпизодах кровотечения, повышению комфортности и перенесения лечения, а также ослаблению вторичных, побочных эффектов.
Модифицированные полипептиды FVII могут быть получены для любой формы полипептидов FVII, в том числе для видовых и аллельных вариантов, вариантов сплайсинга, вариантов, известных в уровне техники, или гибридных или химерных молекул FVII. Например, модификации, предусмотренные в настоящем документе, могут быть осуществлены в предшественниках полипептидов FVII, приведенных в SEQ ID №№1 или 2, зрелых полипептидах FVII, приведенных в SEQ ID №3, или в любых видовых, аллельных или модифицированных вариантах и их активных фрагментах, которые обладают 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, или 99% идентичностью последовательности по отношению к любому из FVII-полипептидов, приведенных в SEQ ID №№1-3. Аллельные варианты FVII включают, но не ограничиваются, любыми из этих полипептидов-предшественников с последовательностями аминокислот, приведенными в SEQ ID №№18-74. Примеры видовых вариантов для модификаций в настоящем документе включают, но не ограничиваются, человеческими и нечеловеческими полипептидами, включая FVII-полипептид коровы, мыши, карликового шимпанзе, шимпанзе, кролика, крысы, макаки резуса, свиньи, собаки, рыбы-зебры, иглобрюха, курицы, гориллы и орангутанга, последовательности которых приведены в SEQ ID №№4-17, соответственно. Модификации в полипептидах FVII могут быть осуществлены для FVII-полипептидов, которые также содержат другие модификации, такие как описано в уровне техники, в том числе модификации первичной последовательности и модификации не в первичной последовательности полипептида.
Модификация полипептидов FVII также включает модификацию полипептидов, которые являются гибридами различных полипептидов FVII, а также синтетическими FVII-полипептидами, полученными рекомбинантно, или синтезированными или сконструированными с помощью других методов, известных в уровне техники, на основе последовательностей известных полипептидов. Например, на основе выравнивания FVII с другими членами семейства факторов свертывания, такими как фактор IX (FIX) или фактор Х (FX), среди членов семейства легко идентифицировать гомологичные домены. Могут быть сконструированы химерные варианты полипептидов FVII, когда одна или более аминокислот или целых доменов в FVII аминокислотной последовательности заменяются с использованием последовательности аминокислот соответствующего члена семейства. Кроме того, химерные полипептиды FVII включают полипептиды, в которых одна или более аминокислот или целых доменов в аминокислотной последовательности человеческого FVII заменяются с использованием аминокислотной последовательности разных видов (см., например, Williamson и др. (2005) J Thromb Haemost 3:1250-6). Такие химерные белки могут быть использованы в качестве начальных, немодифицированных FVII-полипептидов в настоящей заявке.
Предусмотренные в настоящей заявке модификации начального немодифицированного указанного полипептида включают аминокислотные замены или замещения, добавления или делеции аминокислот или любые их комбинации. Например, модифицированные FVII-полипептиды включают полипептиды с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50 или более модифицированными позициями. Кроме того, в настоящей заявке предусмотрены модифицированные FVII-полипептиды с двумя или более модификациями по сравнению с начальным указанным полипептидом FVII. Модифицированные FVII-полипептиды включают полипептиды с 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50 или более модифицированными позициями. Любые предусмотренные в настоящей заявке модификации могут сочетаться с любыми другими известными специалистам в данной области модификациями, до тех пока полученный модифицированный полипептид FVII проявляет увеличенную коагуляционную активность, когда находится в двухцепочечной или активированной форме. Как правило, модифицированные полипептиды FVII проявляют повышенную коагуляционную активность. Активности или свойства, которые могут быть изменены в результате модификаций, включают, но не ограничиваются, коагуляцией или коагулянтной активностью, прокоагулянтной активностью; протеолитической или каталитической активностью, такой как активация фактора Х (FX) или активация фактора IX (FIX); иммуногенностью (способность к связыванию или конкуренции с полипептидами за связывание с анти-FVII антителами); способностью связывать тканевый фактор, ингибиторный фактор тканевого фактора (TFPI), антитромбин III, фактор Х или фактор IX; способностью связываться с поверхностью активированных тромбоцитов, способностью к связыванию с фосфолипидами; временем полураспада в сыворотке; трехмерной структурой; р1 и/или конформацией. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, включают те, которые обладают повышенной устойчивостью к ингибитору пути, опосредованного тканевым фактором (TFPI), повышенной устойчивостью к антитромбину III (AT-III), увеличенной каталитической активностью в присутствии и/или в отсутствие ТФ, улучшенными фармакокинетическими свойствами, такими как увеличение времени полураспада в сыворотке, увеличенной внутренней активностью и/или повышенным сродством и/или способностью связываться с активированными тромбоцитами.
В некоторых примерах, модификация может повлиять на два или более свойства или активности полипептида FVII. Например, модификация может привести к увеличенной устойчивости к TFPI и увеличенной каталитической активности модифицированного полипептида FVII по сравнению с немодифицированным FVII-полипептидом. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть исследованы по каждому свойству и активности для выявления множества эффектов модификации. Такие анализы известны в уровне техники и описаны ниже. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, также включают FVII-полипептиды, которые дополнительно модифицированы клеточными механизмами и включают, например, гликозилированные, γ-карбоксилатные и β-гидроксилированные полипептиды.
Модифиации, предусмотренные в настоящем документе для полипептида FVII, осуществлены для увеличения устойчивости к TFPI, увеличения устойчивости к AT-III, улучшения фармакокинетических свойств, такие как увеличенное времени полураспада в сыворотке, повышения каталитической активности в присутствии и/или в отсутствие ТФ, и/или увеличения сродства и/или связывания с активированными тромбоцитами. Например, полипептид FVII может включать модификацию (модификации), увеличивающие, одновременно, или отдельно, устойчивость к TFPI и связывание с тромбоцитами. В других случаях какие-либо модификации, предусмотренные в настоящем документе, могут сочетаться с любыми другим известными специалисту в данной области модификациями, до тех пор, пока полученный модифицированный полипептид FVII проявляет повышенную коагуляционную активность, когда находится в двухцепочечной форме. Как правило, такая повышенная коагуляционная активность связана с повышенной устойчивостью к TFPI, улучшенными фармакокинетическими свойствами, такими как увеличение периода полураспада в сыворотке, повышенной устойчивостью к AT-III, изменениями гликозилирования, увеличением каталитической активности и/или увеличением связываемоеTM и/или аффинности к фосфолипидам. В некоторых случаях, модификации, которые введены в полипептид FVII для изменения конкретного вида активности или свойства, вместе с желаемым изменением, или вместо желаемого изменения, могут повлиять на другую активность или другое свойство. Таким образом, предусмотренные в настоящем документе модификации могут повлиять на свойство или активность, на которые они были призваны повлиять, и на одно или более другое свойство или активность. Например, модификации, введенные в полипептид FVII для увеличения устойчивости к TFPI, также могут улучшить фармакокинетические свойства, такие как период полураспада в сыворотке. В некоторых примерах, одна модификация, такая как одна аминокислотная замена, изменяет 2, 3, 4 или более свойств или активностей полипептида FVII. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть исследованы по каждому свойству и активности для выявления множества эффектов модификации. Такие анализы известны в уровне техники и описаны ниже. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, также включают FVII-полипептиды, которые дополнительно модифицированы клеточными механизмами и включают, например, гликозилированные, γ-карбоксилированные и β-гидроксилированные полипептиды.
Модификации, предусмотренные в настоящем документе, могут быть осуществлены стандартными методами рекомбинантных ДНК, известные специалистам в данной области. Может быть использован любой метод, известный в области техники, который приводит к мутации одной или более аминокислот в целевом белке. Методы включают стандартный сайт-направленный мутагенез (используя, например, набор QuikChange, Stratagene) кодирующей молекулы нуклеиновой кислоты, или методы твердофазного синтеза полипептидов. Кроме того, предусмотренные в настоящем документе модифицированные химерные белки (например, с заменой домена Gla) могут быть получены с помощью обычной рекомбинантной ДНК технологии. Например, химерные полипептиды могут быть получены с помощью ферментов рестрикции и методик клонирования для рутинного субклонирования компонентов желаемого химерного полипептида.
Другие модификации, которые присутствуют или не присутствуют в первичной последовательности полипептида, также могут быть включены в модифицированные полипептиды FVII, либо их конъюгаты, включая, но не ограничиваясь, добавлением углеводного фрагмента, добавлением полиэтиленгликольного (ПЭГ) фрагмента, добавлением домена Fc и т.д. Например, такие дополнительные модификации могут быть осуществлены для повышения стабильности или времени полураспада белка.
Полученные модифицированные полипептиды FVII включают одноцепочечные зимогенные или активные полипептиды, или одноцепочечные или двухцепочечные зимогеноподобные полипептиды.
Например, любой модифицированный полипептид, предусмотренный в настоящем документе, который является одноцепочечным зимогенным полипептидом, может быть автоактивирован или активирован другими факторами свертывания для получения модифицированного FVII в двухцепочечной форме (т.е. FVIIa). Активность модифицированного полипептида FVII, как правило, проявляется в его двухцепочечной форме.
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут проявлять повышенную устойчивость к TFPI, повышенную устойчивость к AT-III, повышенную каталитическую активность в присутствии и/или в отсутствие ТФ, улучшенные фармакокинетические свойства, такие как повышенное время полураспада в сыворотке, и/или повышение связываемоеTM и/или аффинности к фосфолипидам. Как правило, такие свойства и/или активности модифицированных полипептидов FVII, предусмотренных в настоящем документе, достигаются при сохранении других активностей или свойств FVII, включая, но не ограничиваясь, связыванием ТФ и/или связыванием и активацией FX. Следовательно, модифицированные FVII-полипептиды, предусмотренные в настоящем документе, сохраняют способность связывать ТФ и/или связывать и активировать FX по сравнению с диким типом или начальной формой полипептида FVII. Как правило, такая активность существенно не меняется (изменяется менее 1%, 5%, 10%, 20% или 30%) по сравнению с диким типом или начальным белком. В других примерах активность модифицированного полипептида FVII увеличивается или уменьшается по сравнению с диким типом или начальным FVII-полипептидом. Активность может быть оценена in vitro, ex vivo или in vivo и может быть сравнена с активностью немодифицированного полипептида FVII, такого как, например, зрелый нативный полипептид FVII дикого типа (последовательность SEQ ID №3), предшественник FVII-полипептида дикого типа (последовательность SEQ ID №1 или 2), или любого другого полипептида FVII, известного специалистам в данной области, который используется в качестве исходного материала.
Следовательно, в силу модификаций, предусмотренных в настоящем документе, модифицированные полипептиды FVII могут проявлять увеличенную коагуляционную активность при ТФ-зависимой и/или ТФ-независимой активации. Как правило, повышенная коагуляционная активность модифицированных полипептидов FVII, предусмотренных в настоящем документе, может наблюдаться при соответствующих анализах in vitro, или in vivo, при введении субъекту, такому как человек или не человек. Повышение активности модифицированных полипептидов FVII может, по меньшей мере, составлять 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 1 10%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190%, 200%, 300%, 400%, 500% или более по сравнению с активностью начального или немодифицированного FVIIa полипептида.
1. Повышенная устойчивость к ингибиторам
Как и большинство биологических систем, каскад коагуляции регулируется положительными и отрицательными механизмами. Эти механизмы часто действуют путем ингибирования или повышения активности одного или нескольких факторов, участвующих в каскаде, прямо или косвенно. Таким образом, интерес представляют терапевтические факторы коагуляции, которые устойчивы к ингибирующим механизмам, негативно регулирующим их активность. В частности, интерес представляют модифицированные FVII-полипептиды, которые являются устойчивыми к ингибирующим молекулам, которые в нормальных условиях ингибируют активность FVII. Основным регулятором ТФ / FVIIa комплекса является ингибитор пути, опосредованного тканевым фактором (TFPI). Кроме того, антитромбин III (AT-III) подавляет активность FVIIa, хотя и в меньшей степени.
а. TFPI
Ингибитор пути, опосредованного тканевым фактором, представляет собой одноцепочечный полипептид, закодированный 9 экзонами, расположенными на хромосоме 2q31-q32.1. TFPI (также упоминаемый как TFPI-I) является ингибитором Кунитц-типа, который в первую очередь синтезируется эндотелиальными клетками и содержит отрицательно заряженный аминоконцевой регион, три тандемных ингибиторных доменов Кунитц-типа и высокоосновный карбокси-терминальный хвост (Wun и др., J. Biol. Chem. 263:6001 (1988)). С помощью альтернативного сплайсинга мРНК создаются две различные формы TFPI: TFPIa и Р. TFPIa является секретируемым белком с молекулярной массой около 46 кДа. Полипептид-предшественник содержит 304 аминокислоты (последовательность SEQ ID №101), и процессируется отщеплением 28 аминокислотного сигнального пептида, что приводит к секреции 276 аминокислотного зрелого гликопротеина (последовательность SEQ ID №102). Зрелый белок содержит 18 остатков цистеина, образующих 9 дисульфидных мостов при правильном фолдинге. Первичная последовательность белка также содержит три Asn-X-Ser/Thr N-связанных сайтов гликозилирования, аспарагиновые остатки которых находятся в позициях 145, 195 и 256. Кунитц-1 и Кунитц-2 домены ответственны за связывание и ингибирование ТФ / FVIIa комплекса и FXa, соответственно (см., например, Фиг.4). Было показано, что Кунитц-3 домен, который не обладает ингибиторной активностью к протеиназам, а также С-конец TFPIa принимают участие в его локализации на поверхности клетки (Piro и др., (2004) Circulation 110:3567-3572).
TFPIβ является формой TFPI, полученной альтернативным сплайсингом, в которой Кунитц-3 домен и С-концевая область TFPIa замещены неродственной С-концевой областью, которая направляет прикрепление гликозилфосфатидилинозотольного (GPI) якоря. Полипептид-предшественник (последовательность SEQ ID №103) процессируется в зрелый белок, содержащий 223 аминокислоты (последовательность SEQ ID №104). Масса белка TFPIβ (28 кДа) значительно меньше, чем TFPIα (36 кДа). Оба передвигаются как белки с одинаковыми кажущимися молекулярными массами (46 кДа) при электрофорезе в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), что указывает на различия в посттрансляционных модификациях. TFPIα и β связаны с поверхностью клетки через GPI. TFPIR содержит GPI-якорь на своем С-конце, в то время как TFPIα связан с еще не идентифицированным GPI-связанным белком (белками). Хотя TFPIβ составляет лишь 20% от всех поверхностных TFPI, на него приходится большая часть анти-ТФ/FVIIα активности, что свидетельствует о потенциальной альтернативной роли расположенного на клеточной поверхности TFPIα, такой как связывание и расщепление FXa (Piro и др., (2005) J Thromb Haemost 3:2677-2683).
TFPI ингибирует каскад коагуляции, по меньшей мере, двумя способами: путем связывания с активным центром FXa и инактивации ТФ / FVIIa комплекса (см., например, Фиг.4). Первый Кунитц-домен связывает FVIIa, в то время как второй домен связывает FXa (Girard и др., (1989) Nature 338:518-520). Свободный TFPI связывает FVIIa очень медленно в сравнении с его связыванием FXa. После связывания FXa, однако, TFPI / FXa комплекс демонстрирует высокую аффинность к ТФ / FVIIa комплексу. Их взаимодействие приводит к образованию гетеротетрамерного комплекса на поверхности клеток эндотелия, в котором FVIIa является каталитически неактивным (фиг.4). Это четвертичная структура тем самым препятствует ТФ / FVIIa-инициации каскада коагуляции.
У людей существует, по крайней мере, один другой гомолог TFPI, TFPI-2, также известный как плацентарный белок 5 (РР5) и связанный с матриксом ингибитор сериновой протеазы (MSPI). TFPI-2 представляет собой 32 кДа связанный с матриксом сериновый ингибитор протеиназ Кунитц-типа, который обладает ингибирующей активностью по отношению к широкому спектру протеиназ, в том числе включая трипсин, плазмин, химотрипсин, катепсин G, плазменный калликреин и фактор VII-комплекса тканевого фактора. 213 аминокислот зрелого TFPI-2 белка (SEQ ID №: 105) содержат три домена типа Кунитц, которые демонстрируют 43%, 35% и 53% идентичности первичной последовательности с доменами типа TFPI Кунитц 1, 2 и 3, соответственно. Первый домен Кунитц-типа человеческого TFPI-2 содержит все структурные элементы для ингибирования сериновых протеаз, хотя кинетические и молекулярные исследования показывают, что TFPI-2 является более эффективным ингибитором плазмина, чем некоторые другие сериновые протеиназы, в том числе TF / FVIIa (Чанд и др., (2004) J Biol Chem 279:17500-17507). Считается, что в регуляции внеклеточного матрикса расщепление и перестройка играют важную роль. В этой связи снижение синтеза TFPI-2 связывалось с многочисленными патофизиологическими процессами, такими как воспаление, ангиогенез, атеросклероз, дегенерация сетчатки и опухолевый рост / метастазирование (Чанд др. (2005) Thromb Haemost 94:1122-30). Таким образом, гомология TFPI, TFPI-2 не считается важным фактором в пути свертывания. Выравнивание последовательностей Кунитц-1 домена TFPI-1 и TFPI-2 показаны на Фиг.6b.
Иммуноистощение TFPI на модели увеличенной коагуляции у кролика (Warn-Cramer и др. (1993) Arteriosci Thromb 13:1551-1557, Ragni др. (2000) Circulation 102:113-117) показало, что нарушение формирования TF / FVIIa-TFPI/FXa четвертичного комплекса может ингибировать негативное регулирование TF / FVIIa начала коагуляции. Кроме того, нарушение взаимодействия с FVIIa TFPI может уменьшить расщепление FVIIa. Снижение расщепления может проявляться как повышение времени полужизни (half-life). Кроме того, связанный FVIIa с TF на клеточной поверхности может быть интернализован и разрушен, без расщепления TF на поверхности клеток. Эта форма интернализации и деградации значительно увеличивалась в присутствии TFPI / FXA, который образует четвертичный комплекс с TF / FVIIa клеточной поверхности. Интернализации и деградации, которая следует за связыванием, опосредуется белком, родственным рецептору липопротеинов низкой плотности, (LRP)- зависимого пути через окаймленные ямки (Iakhiaev др., (1999) J. Biol. Chem 274:36995-37003). TFPI также может помочь расщеплению FVIIa через другие механизмы, такие как печеночный или почечный механизмы клиренса.
Модификации, приводящие к повышенной устойчивости к TFPI
Предусмотрены измененные FVII-полипептиды, проявляющие повышенную устойчивость к TFPI. Такая устойчивость к TFPI может быть достигнута, например, в результате мутации одного или нескольких остатков в FVII, которые участвуют во взаимодействии и связывании с TFPI, уменьшая или предотвращая такое связывание, тем самым делая модифицированные FVII-полипептиды устойчивыми к естественным ингибирующим воздействиям TFPI в отношении инициации коагуляции. В опытах in vitro либо in vivo, таких как после введения прокоагулянтных терапевтических средств, модифицированные устойчивые к TFPI полипептиды FVII могут проявлять более высокую коагуляционную активность по сравнению с неизмененными полипептидами FVII. Среди мутантов FVIIa, которые показывают увеличенную к TFPI устойчивость, имеются мутанты, которые имеют увеличенное время полужизни (half-life). Например, мутант, описанный в примере 9, проявляет увеличенное время полужизни.
Предусмотрены измененные FVII-полипептиды, имеющие одну или несколько мутаций в FVII /TFPI контактных остатках или остатках в непосредственной близости от области взаимодействия. Такие контактные остатки включают остатки аспарагиновой кислоты в позиции 196 (D1 96, Asp196), остатки лизина в позиции 197 (KL 97, Lys197), остатки лизина в позиции 199 (KL 99, Lys199), остатки треонина в позиции 239 (Т239, Thr239), остатки аргинина в позиции 290 (R290, Arg2), а также остатки лизина в позиции 341 (К341, Lys341), с нумерацией, соответствующей аминокислоте зрелого полипептида FVII в SEQ ID №: 3. Когда определена нумерация химотрипсина, эти остатки соответствуют D60, К60а, К60с, Т99, R147 и К. 192 (Фиг.7). Остатком, который находится в непосредственной близости к области взаимодействия FVII / TFPI и который может быть модифицирован, чтобы вызвать стерические затруднения, является глицин в позиции 237 (G237, Gly237) в зрелом FVII, что соответствует G97 (Gly97) по нумерации для химотрипсина.
Выявленные остатки могут быть изменены перемещением, удалением или заменой аминокислоты. Кроме того, может быть использована вставка аминокислоты для модифицирования конформации целевых аминокислотных остатков или структуры белка в непосредственной близости от целевого аминокислотного остатка. Например, выявленные остатки могут быть перемещены или заменены другой аминокислотой с тем, чтобы нарушить взаимодействия между FVII и TFPI и препятствовать эффективному связыванию двух белков, генерируя тем самым устойчивые к TFPI модифицированные FVII-полипептиды. Например, возможна замена аминокислотой, которая пространственно препятствует взаимодействию FVII и TFPI (т.е. замещение GIy237 (GIy97) другой большей аминокислотой). В других вариантах, устойчивые к TFPI модифицированные полипептиды FVII могут быть получены путем замены контактных остатков альтернативными аминокислотами, так что происходит замена или нейтрализации заряда в определенных позициях. Такие мутации могут существенно влиять на электростатические вклады замещенных FVII аминокислот и нарушать нормальное взаимодействие контактных остатков с соответствующими контактными остатками TFPI. Например, отрицательно заряженные остатки аспарагиновой кислоты в FVII, которые могут обычно взаимодействовать с положительно заряженными остатками аргинина в TFPI, могут быть заменены на положительно заряженный лизин, чтобы не только ликвидировать первоначальное электростатическое притяжение, но также создать силы отталкивания между остатками и препятствовать эффективному связыванию белков. Эта мутация, следовательно, препятствует эффективному связыванию белков. Таким образом, отрицательно заряженные аминокислоты (например, кислые аминокислоты) такие, как аспарагиновая кислота (Asp, D) или глутаминовая кислота (Glu, E), могут быть заменены на положительно заряженные аминокислоты (т.е. основные аминокислоты) такие, как лизин (Lys, К), аргинин (Arg, R), или гистидин (His, H). Наоборот, основные аминокислоты такие, как лизин (Lys, К), аргинин (Arg, R) или гистидин (His, H), могут быть заменены на кислые аминокислоты такие, как аспарагиновая кислота (Asp, D) или глутаминовая кислота (Glu, E). Кроме того, любой контактный остаток может быть заменен на нейтральную аминокислоту такую, как аланин (Аlа, А), тирозин (Туr, Y), метионин (Met, М) или лейцин (Leu, L), изолейцин (Не, I), валин(VАl, V), фенилаланин (Phe, F), пролин (Pro, P) или глицин (Gly, G), серин (Ser, S), треонин (Thr, Т), аспарагин (Asn, N), глютамин (Gin, Q), триптофан (Trp, W) и цистеин (Cys, С). Примерами нейтральных аминокислот являются гидрофобные аминокислоты, к которым относятся изолейцин (Не, I), фенилаланин (Phe, F), валин (VAI, V), лейцин (Leu, L), триптофан (Trp, W), метионин (Met, М), аланин (Аlа, А), глицин (Gly, G), цистеин (Cys, С) и тирозин (Туr, Y).
В некоторых вариантах, глицин в позиции 237 зрелого FVII заменяют таким образом, что новая аминокислота приводит к стерическим затруднениям и тормозит взаимодействие между FVII и TFPI. Глицин является самой маленькой и простой аминокислотой, содержащей только один атом водорода в боковой цепи. Благодаря своим малым размерам, глицин может поместиться на небольшом пространстве и может принять необходимые конформации, что другие аминокислоты не могут. Замена глицина в позиции 237 на аминокислоту с большей боковой цепью может привести к стерическим затруднениям с другими аминокислотами в белке FVII, таким образом изменяя конформацию для района связывания TFPI и/или может причинить стерические затруднения с аминокислотами в TFPI белке. Обе ситуации могут привести к замедлению взаимодействия и связывания FVII и TFPI и тем самым увеличить устойчивость модифицированного полипептида FVII к ингибиторным эффектам TFPI. Любая из 19 природных аминокислот содержит большие боковые цепи, чем глицин, и может быть использована для модифицирования FVII-полипептида. Примерами являются триптофан (Trp, W), изолейцин (Не, I), валин (VAl, V) и треонин (Thr, Т). В других случаях, аминокислотные вставки, включенные в полипептид FVII вблизи выявленных остатков, мешают взаимодействию между TFPI и FVII. Один или более аминокислотных остатков могут быть включен в одну или несколько позиций в полипептид FVII. Например, тирозин может быть вставлен между D 196 и К197 (соответствует D60 и К60а, соответственно, в нумерации химотрипсина). Эта модификация представляет собой G196К197вставкаY (т.е. D196 и К 197 находятся в позициях, между которыми был вставлен тирозин (вставка Y)). В другом примере два аминокислотных остатка вставляют в позиции, которые считаются важными для взаимодействия с TFPI. Например, аланин и серии можно вставить между G237 и Т238 (соответствует Т98 и G97, соответственно, в нумерации химотрипсина), получая модификацию 0237Т238вставкаА8. Такие инсеционные мутации могут привести к замедленному взаимодействию и связыванию FVII и TFPI, тем самым увеличивая устойчивость модифицированного полипептида FVII к ингибиторным эффектам TFPI.
В таблице 5 дается не ограничивающий ряд примеров возможных аминокислотных замен в выявленных остатках, соответствующих позициям аминокислот зрелого полипептида FVII, отображенного в SEQ ID №: 3. Как уже отмечалось, такие полипептиды FVII предназначены для повышения устойчивости к TFPI путем ингибирования связывания FVII / TFPI и, следовательно, увеличения коагуляционной активности. В таких мутациях первой аминокислоте (однобуквенная аббревиатура) соответствует аминокислота, которая заменена, число соответствует позиции в зрелом FVII-полипептиде в последовательности SEQ ID №: 3, а вторая аминокислота (однобуквенная аббревиатура) соответствует выбранной аминокислоте, которая заменяет первую аминокислоту в этой позиции. Позиции аминокислот для мутации также приведены для нумерационной схемы химотрипсина. В таблице 5 ниже указаны номера последовательностей (SEQ ID №) модифицированных полипептидов FVII, a также любые полипептидные идентификационные номера.
В дальнейшем воплощении, могут быть получены сочетания мутантов. К числу таких комбинационных мутантов относятся полипептиды, имеющие две или более мутации в остатках D 196, К197, К199, G237, Т239, R290 или К341, основанные на SEQ ID №: 3 зрелого FVII (соответствует D60, К60а, К60с, G97, Т99, RL 47 и К192, соответственно, нумерация для химотрипсина). Например, модифицированный полипептид FVII может обладать аминокислотными заменами на 2, 3, 4, 5, 6 или 7 идентифицированных позициях. Как отмечалось выше, заменяют остатки так, что благоприятное (удобное) взаимодействие между FVIIa и TFPI исключается, и наблюдаются неблагоприятные взаимодействия или стерические затруднения в еще одной позиции. Таким образом, модифицированный полипептид может содержать 1, 2, 3, 4, 5, 6 или 7 мутаций, которые обеспечивают одно или более стерических затруднений, изменение заряда или его нейтрализацию или другие неблагоприятные взаимодействия, или их сочетание. Например, две или несколько мутаций могут быть сделаны, при которых отрицательно заряженные аминокислоты такие, как аспарагиновая кислота (Asp, D) или глутаминовая кислота (Glu, Е), могут быть заменены на положительно заряженные аминокислоты такие, как лизин (Lys, К), аргинин (Arg, R) или гистидин (His, H), или наоборот, или нейтральные замены могут быть сделаны заменой любой аминокислотой такой, как, например, аланин (Аlа, А), тирозин (Туr, Y), метионин (Met, M), или лейцин (Leu, L), изолейцин (Не, I), валин (VAl, V), фенилаланин (Phe, F), пролин (Pro, Р) или глицин (Gly, G), серии (Ser, S), треонин (Thr, Т), аспарагин (Asn, N), глютамин (Gin, Q), триптофан (Тrр, W) и цистеин (Cys, С) или любой их комбинацией. Примерами нейтральных аминокислот, которые являются гидрофобными, являются изолейцин (Не, I), фенилаланин (Phe, F), валин (VAl, V), лейцин (Leu, L), триптофан (Тrр, W), метионин (Met, M), аланин (Аlа, А), глицин (Gly, G), цистеин (Cys, С) и тирозин (Туr, Y). Две или более мутации могут быть сделаны, где, по крайней мере, осуществлена одна перемена заряда или его нейтрализация и другое замещение, которое заменяет глицин на аминокислоты, которые содержат большие боковые цепи, такие как триптофан (Тrр, W), изолейцин (Ilе, I), валин (VAl, V) и треонин (Thr, Т). В других примерах инсерционная мутация может быть включена в модифицированный полипептид FVII с другими инсерционными мутациями, либо с одной или более аминокислотной заменой, которые вносят неблагоприятные взаимодействия или стерические затруднения между FVII и TFPI. Любые изменения комбинаций могут быть включены в модифицированные полипептиды FVII, изложенные в таблице 5 выше. Например, FVII-полипептиды, содержащиеся в настоящем документе, могут содержать D196K и одно или несколько дополнительных изменений в позиции К197, К199, G237, Т239, R290 и/или К341, и/или инсерции (вставки) после D 196, К197 и/или G237. Специалист в данной области может легко генерировать всевозможные комбинации, используя модификации, изложенные в таблице 5. Например, FVII варианты включают в качестве одной из мутаций аминокислотную замену в позиции 197 зрелого FVII по SEQ ID №: 3. Любой аминокислотный остаток может быть введен вместо лизина, который занимает 197 позицию в немодифицированном FVII-полипептида. В некоторых вариантах, лизин заменен тирозином (Y), аланином (А), глутаминовой кислотой (Е), аспарагиновой кислотой (D), лейцином (L), метионином (M), изолейцином (I), валином (V), фенилаланином (F), или триптофаном (W). Например, пример модифицированного полипептида FVII содержит замещение K197L или К.197Е. Другой пример модифицированного в позиции 197 полипептида FVII (например, замена, удаление или добавление) также может включать в себя другую модификацию по другой позиции. Как правило, такие другие модификации, которые предполагают модулировать взаимодействия FVII с TFPI, включают удаление благоприятных взаимодействий, создание неблагоприятных взаимодействий, создание стерических препятствий или их сочетание. К числу таких дополнительных, описанных выше модификаций относят D196, К199, G237, Т239, R290 или К341. Возможны также другие дополнительные модификации, известные в данной области, такие как описано ниже. Таким образом, в дополнение к изменению в позиции 197, полипептид FVII может содержать 1, 2, 3, 4, 5, 6 или более аминокислотных модификаций. Например, примерами вариантов FVII, имеющих, по крайней мере, модификацию в позиции 197, относятся те, которые содержат 2 замены, такие как D196F и К197Е; те, которые содержат 3 замены, например, D196R, К197М и К199Е; те, которые содержат 4 замены, такие как D196R, К197Е, К199Е и R290E; те, которые содержат 5 замен, такие, как K197L, K199D, G237T, R290D и K341Q; те, которые содержат 6 замен, такие как D196F, K197L, К199Е, G237V, Т239А и R290D; и те, которые содержат 7 замен, такие как D196Y, K197L, К199А, G237T, Т239А, R290D и K341R. Примеры FVII комбинаций мутантов, которые включают в качестве одной из мутаций аминокислотную замену в позиции 197 в SEQ ID №: 3 зрелого FVII, предсталены в таблице 6.
Примеры FVII-полипептидов, которые содержат две или более аминокислотных замен D196, К197, К199, G237, Т239, R290 или К341 в SEQ ID №: 3 зрелого полипептида FVII, указаны в таблице 6. В таблице приведены также аминокислотные позиции мутаций для химотрипсиновой нумерационной схемы. Такие модификации направлены на увеличение устойчивости к TFPI путем ингибирования связывания FVII с TFPI и, следовательно, увеличения коагуляционной активности.
FVII-полипептиды, содержащие одно или более аминокислотных замещений в упомянутых выше остатках, также могут содержать дополнительные мутации, такие как описано в Международной патентной заявке №WO2004/083361, Neuenschwander и др., (1995) Biochemistry 34:8701-87, Чанг и др., (1999) Biochemistry 38:10940-10948 и Iakhiaev и др., (2001) Thromb. Haemost. 85:458-463. Таким образом, в одном варианте FVII-полипептиды по настоящей заявке могут содержать одну или более аминокислотных замещений на один или более аминокислотных остатков Q176, D196, К197, К199, G237, Т239, R290. Е296, К341 и Q366, что соответствует позиции аминокислоты зрелого полипептида FVII в SEQ ID №: 3. Например, одна или несколько дополнительных мутаций может включать, но не ограничиваться этим, любую одну или несколько замен из Q176A, D196N, G237L, Е296А, K341N, K341Q, K341Е, Q366A, Q366E и Q366G.
В. Антитромбин III (АТ-III)
Антитромбин III (также известен как антитромбин или AT-III) является важным серпин антикоагулянтом (сериновым ингибитором протеазы). AT-III синтезируется в виде предшественника белка, содержащего 464 аминокислотных остатка (SEQ ID №: 122). В ходе секреции 32 остатка сигнального пептида отщепляются, образуя 432 аминокислотный зрелый человеческий антитромбин (SEQ ID №: 123). 58 кДа гликопротеин AT-III циркулирует в крови и функционирует как сериновый ингибитор протеазы (серпин), ингибируя большое число сериновых протеаз свертывающей системы. Основными мишенями AT-III являются тромбин и фактор Ха, хотя AT-III также ингибирует активности FIXa, FXIa, FXIIa и, в меньшей степени, FVIIa. Действия AT-III значительно увеличиваются гликозаминогликанами, такими, как естественно присутствующими гепаран сульфатом или полученными из различных тканей гепаринами, которые широко используются в качестве антикоагулянтов в клинической практике. AT-III связывается с высокой специфичностью с уникальной пентасахаридной последовательностью в гепарине, что вызывает конформационное изменение реактивного центра петли. В такой конформации реактивный центр петли AT-III может более эффективно взаимодействовать с реактивным сайтом сериновой протеазы, и происходит эффект торможения.
AT-III обычно не ингибирует свободный FVIIa плазмы, даже в присутствии гепарина, вероятно, благодаря зимогенподобной конформации FVIIa, которая препятствует эффективному взаимодействию с AT-III. Тормозящее действие AT-III увеличивается, однако, как только FVIIa образует комплекс с ТФ. Связывание AT-III с ТФ / FVIIa комплексом может освободить FVIIa от ТФ и поддерживать его в неактивном комплексе с AT-III. Повышенное сродство AT-III к TF-связанному FVIIa по сравнению с FVIIa только предположительно отражает становление активного сайта FVIIa, когда он образует комплекс с ТФ, вследствие чего он становится подходящим для связывания с AT-III (RAO и др. (1993) Blood 81:2600-2607). Таким образом, влияние AT-III на FVIIa пропорционально собственной активности молекулы FVIIa. Хотя FVIIa сохраняет свою зимогенподобную конформацию, AT-III имеет незначительный эффект. Если, однако, FVIIa измененяет конформацию к более активной форме, например, путем связывания ТФ или путем конкретных модификаций in vitro, AT-III значительно увеличивает ингибирование. FVIIa модифицированные полипептиды с увеличенной внутренней активностью часто демонстрируют одновременное увеличение восприимчивости к AT-III ингибированию. Например, изменение одной или нескольких аминокислот в активационном кармане FVIIa, например, путем аминокислотных замен К337А, L305V, M298Q, V158D и E296V (по отношению к зрелым последовательностям FVII в SEQ ID №: 3), приводит к повышенной чувствительности полипептида FVIIa к AT-III, тем самым ингибируя активность FVIIa до 90% (Persson и др., (2001) 98:13583-13588 PNAS). В другом случае, индукция более выраженной зимогенподобной конформации путем модификации аминокислот, включенных в α-спираль FVIIa, приводит к повышению активности модифицированного белка FVIIa, а также повышает его восприимчивость к AT-III (Perssen и др., (2004) Biochem J 379:497-503).
Модификации, приводящие к повышенной устойчивости к AT-III
Модификации могут быть сделаны в полипептиде FVII, которые повышают его устойчивость к ингибированию AT-III. Как правило, такие измененные полипептиды FVII сохраняют, по крайней мере, один вид активности полипептида FVII. Как правило, такие модификации включают одну или более аминокислотных замен в любом положении полипептида FVII, которые непосредственно участвуют во взаимодействии FVIIa (или TF / FVIIa комплекса) с AT-III, или в других остатках, которые влияют на позицию или конформацию остатков, которые непосредственно участвуют во взаимодействиях между FVIIa (или TF / FVIIa комплекса) и AT-III. Модифицированные FVII-полипептиды, которые имеют повышенную устойчивость к AT-III, могут проявлять снижение степени ингибирования при определенных условиях или второй порядок константы скорости ингибирования AT-III, по крайней мере, на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению со степенью ингибирования или вторым порядком константы скорости ингибирования немодифицированного или дикого типа полипептида FVII в условиях in vivo, ex vivo, или in vitro. Модифицированные полипептиды FVII поэтому устойчивы к естественному ингибирующему действию AT-III в отношении инициирования свертывания. По результатам анализа in vitro или ex vivo, или in vivo после введения субъекту прокоагулянтных терапевтических средств модифицированные АТ-III-устойчивые полипептиды FVII показывают повышение коагуляционной активности по сравнению с неизмененными полипептидами FVII.
Согласно приведенному ниже описанию, специалист в данной области может эмпирическим или рациональным способом разработать модифицированные FVII-полипептиды, которые должны показывать повышенную устойчивостью к AT-III. Такие модифицированные полипептиды FVII могут быть исследованы, чтобы определить, что модифицированные полипептиды FVII показывают повышенную устойчивостью к AT-III. Например, константа ингибирования второго порядка AT-III может быть измерена для таких модифицированных полипептидов FVIIa. Как правило, модифицированный полипептид FVII, который имет увеличенную стабильность к AT-III, будет показывать уменьшенное связывание при определенных условиях (например, после введения фиксированного количества белка пациенту) и/или уменьшение ингибирования AT-III. Как правило, такие анализы проводятся на двухцепочных формах FVII, таких как активированная форма FVII (FVIIa). Кроме того, анализы по определению эффектов AT-III, как правило, производятся в присутствии гепарина и тканевого фактора, хотя такие анализы также могут быть выполнены в отсутствие одного или обоих кофакторов.
Модифицированные FVII-полипептиды, которые имеют повышенную устойчивость к AT-III, могут проявлять снижение степени ингибирования при определенных условиях или снижение константы скорости ингибирования второго порядка AT-III, по крайней мере, на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению с величинами ингибирования или константой скорости ингибирования второго порядка немодифицированного полипептида FVII или полипептида FVII дикого типа в условиях in vivo или in vitro. Увеличение устойчивости модифицированных полипептидов FVII к AT-III может быть установлено как увеличение коагуляционной активности в присутствии AT-III. Коагуляционная активность АТ-III-модифицированного полипептида FVII может быть увеличена, по крайней мере, на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению с коагуляционной активностью немодифицированного полипептида FVII или полипептида FVII дикого типа в условиях in vivo, ex vivo или in vitro.
2. Связывание с активированными тромбоцитами
Также могут быть сделаны модификации в полипептиде FVII, которые повышают его коагуляционную активность при ТФ-зависимых и/или ТФ-независимых инициациях коагуляции. Механизм ТФ-независимого начала коагуляции (свертывания) предполагает включение прямой активации FX и FIX FVIIa (то есть не в комплексе с ТФ), которая осуществляется на поверхности активированных тромбоцитов. FVIIa связывает активированные тромбоциты взаимодействием между остатками в Gla домене полипептида FVIIa и фосфатидилсеринами и другими отрицательно заряженными фосфолипидами, экспрессированными на поверхности тромбоцитов. Это взаимодействие является относительно слабым и, вероятно, не играет существенной роли в первоначальном выбросе тромбина, который обычно наблюдается при коагуляции, что является весьма вероятным результатом TF-зависимой активности. Как уже говорилось выше, TF-независимое инициирование коагуляции и относительно слабые взаимодействия FVIIa с поверхностью тромбоцитов может требовать введения высоких доз rFVIIa для достижения эффективности в клинике. Например, только около 1% от 10нМ FVII в плазме присутствует в виде TF / FVIIa, а терапевтическая доза rFVIIa составляет 90 мкг / кг, или около 50 нМ в плазме крови, что намного выше этого уровня. Таким образом, высокие дозы rFVIIa компенсируют низкое сродство связывания rFVIIa с активированными тромбоцитами, что приводит к достаточной коагуляционной активности терапевтических средств во время лечения. Таким образом, представляют терапевтический интерес модифицированные FVII-полипептиды, которые могут быть введены в более низких дозах из-за увеличения связывания и/или аффинности к активированным тромбоцитам, достаточных для осуществления начала коагуляции. Модифицированные FVII-полипептиды, предназначенью для этой цели, с увеличенным связыанием и/или аффинностью к активированным тромбоцитам получают посредством модификации домена GIA.
Взаимодействие звеньев в γ-карбоксилированном домене Gla витамин К-зависимых белков плазмы, таких как FVII, FIX, FX, протромбин, протеин С и протеин S, и отрицательно заряженных фосфолипидов на поверхности мембран имеет важное значение для гемостаза. Gla домены витамин К-зависимых белков плазмы обычно содержат около 45 аминокислот, из которых от 9 до 12 остатков глутаминовой кислоты посттрансляционно изменены витамин К-зависимым карбоксилированием с образованием γ-карбоксиглутамата (Gla). Аминокислоты, которые образуют домен Gla, размещены сразу после тех, которые формируют сигнальный пептид и пропептид белка, поэтому они расположены на N-конце после процессинга и отщепления прекурсорных полипептидов в зрелом белке. Например, аминокислоты, которые формируют Gla домен в FVII, находятся в позициях 39-83 предшественника полипептида с SEQ ID №: 1, в позициях 61-105 предшественника полипептида с SEQ ID №: 2 и в позициях с 1 по 45 зрелых полипептидов с SEQ ID №: 3. Из них 10 остатков глутаминовой кислоты на позициях Е6, Е7, Е14, Е19, Е20, Е25, Е26, Е29 и Е35 зрелого полипептида FVII с SEQ ID №: 3 модифицируются карбоксилированием с получением γ-карбоксиглутаматных (Gla) остатков.
Из-за того, что глутаминовая кислота является лишь слабым Са2+ хелатором, и γ-карбоксиглутамат (Gla) является гораздо более сильным хелатором, эта модификационный шаг витамин К карбоксилазы, значительно увеличивает Са2+ связывающую аффинность Gla домена белка. Кальций также связывает протеазный домен FVIIa, что облегчает конформационные изменения, необходимые для оптимальной активности. Через связывание с сайтам в Gla и протеазными доменами FVIIa кальций повышает связывание с ТФ и фосфолипидами, а также повышает каталитическую эффективность активации FX. γ-Карбоксилатный домен Gla связывает семь ионов Са2+ с вариабильной аффинностью, что ведет к конформационному изменению и что позволяет домену Gla взаимодействовать с С-концевым доменом ТФ. Са2+ связывание также способствует взаимодействию FVIIa с отрицательно заряженными фосфолипидами на мембранах тромбоцитов, основная часть которых являются фосфатидилсеринами. Фосфатидилсерин (PS) является ключевым компонентом отрицательно заряженных мембран, который поддерживает свертывание крови. В большинстве клеток, PS присутствует на внутренней стороне клеточной мембраны и поэтому не подвергается действию плазмы. Специфичные клеточные процессы, такие как активация тромбоцитов, приводят к транслокации PS на внешнюю, предназначенную для плазмы поверхность, презентируя сайт связывания для домена Gla витамин К-зависимых белков (Hemker и др. (1983) Blood Cells 9: 303-317).
В дополнение к участию во взаимодействии FVIIa с ТФ и активированными тромбоцитами, домен Gla FVIIa также причастен к связыванию и активации FX. Это может быть осуществлено через косвенные механизмы, когда домен Gla FVIIa помогает выравнивать Lys165 и Lys166 ТФ с Gla доменом РХ,что является важным шагом в активизации FX до FXa. Кроме того, домен Gla FVIIa непосредственно взаимодействует с доменом Gla FX для корректного выравнивания фермента и субстрата перед активацией (Neuenschwander др., (1994) J Biol Chem 269:8007-8013, Huang и др. (1996) J Biol Chem 271:21752-2175 7). Было показано, что аргинин в позиции 36 FVIIa важен для эффективной стыковки FX Gla-домена в отсутствии фосфолипида, что демонстрирует, что Gla-домен FVIIa участвует в белок-белковых взаимодействий с FX (Ruf др., (1999) Biochemistry 38:1957-1966).
Хотя Gla домены различных витамин К-зависимых белков плазмы показывают значительную гомологию, они связывают отрицательно заряженные фосфолипиды мембран с очень разным сродством, с константами диссоциации (Kd'S), различающимися более чем в 1000 раз. Человеческий FVII имеет однин из самых низких показателей аффинности к фосфолипидам мембран среди этих белков (McDonald и др. (1997) Biochemistry 36:5120-5127). Какие определенные взаимодействия ответственны за связывание домена Gla с PS не полностью понятно, хотя и вполне вероятно, что отдельные аминокислотные остатки в этом домене вносят свой вклад в наблюдающуюся различную аффинность. Более рание исследования показали, что существует корреляция между аминокислотами в позициях 10, 32 и 33 (в зрелом FVII белке) и общей мембранной аффинностью (McDonald др. (1991) Biochemistry 36:5120-5127). Последующий анализ мутаций выявил, что специфические остатки вносят вклад в аффинность связывания. Например, замена гистидина в позиции 10 зрелой формы белка С на пролин приводит к 3 кратному снижению мембранной аффинности (Shen др. (1997) Biochemistry 36:16025-16031). С другой стороны, мутации конкретного остатка в GlA домене витамин К-зависимых белков плазмы может дать белки с более высокой мембранной аффинностью и расширенной функцией. Эффекты мутации определенных позиций Gla домена, однако, не обязательно являются неизменными для различных белков. Например, мутации бычьего протеина С Q32E и N33D опосредует значительное увеличения мембранного связывания, в то время как соответствующие мутации в человеческом белке С показали минимальные изменения в связывании. Аналогично мутация человеческого FVII (K32E) приводит к 13-кратному увеличению аффинности связывания по сравнению с белком FVII дикого типа (Harvey и др. (2003) J Biol Chem 278:8363-8369).
А. Модификация введением гетерологичного Gla домена
Из-за относительно низкой аффинности связывания к активированным тромбоцитам домен Gla из FVII модифицируют с целью повышения взаимодействия между модифицированные FVII и фосфолипидами мембран, тем самым увеличивая коагуляционную активность. Изменение может быть осуществлено путем замены некоторых аминокислот, которые участвуют в этом взаимодействии (см., например, Shah и др.. PNAS 95:4429-4234, Harvey и др. (2003) J Biol Chem 278:8363-8369). Кроме того, изменение может быть осуществлено путем замены всего домена Gla доменом Gla другого витамин К-зависимого белка, т.е. обменной домена Gla. Такого рода модификации приводят к химерным белкам, подобные тем, которые получали, когда домен Gla белка С был заменен доменом Gla из FVII (Geng и др. (1997) Thromb Haemost 77:926-933).
Как правило, такие модификации включают интродукцию, например, путем присоединения или замещения гетерологичного домена Gla или его части для осуществления связывания фосфолипидов в зоне полипептида FVII с получением химерного модифицированного FVII-полипептида. Как правило, такой химерный FVII-полипептид сохраняет, по крайней мере, одну активность FVII. Связывание и/или аффинность Gla-модифицированных FVII-полипептидов с активированными тромбоцитами может быть увеличена по крайней мере на около 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению со связыванием и/или аффинностью немодифицированного FVII-полипептида или FVII-полипептида дикого типа in vivo или in vitro. Связывание и/или аффинность к активированным тромбоцитам модифицированных FVII-полипептидов также может проявляться как повышение активности коагуляции. Коагуляционная активность Gla модифицированных FVII-полипептидов может быть увеличена, по крайней мере, на или приблизительно на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению с коагуляционной активностью немодифицированного FVII-полипептида или FVII-полипептида дикого типа в условиях in vivo или in vitro.
GlA домен или достаточная его часть, содержащаяся в каком-либо полипептиде, может быть использована в качестве источника гетерологичного домена Gla для внедрения или замены участка полипептида FVII. Как правило, такой гетерологичный домен Gla демонстрирует аффинность к фосфолипидам, например фосфолипидам, присутствующим на поверхности активированных тромбоцитов. Как правило, выбор гетерологичного GlA домена основан на проявлении большего сродства к фосфолипидам по сравнению с аффинностью Gla домена FVII. Полный домена Gla, или достаточная его часть, используемые в качестве гетерологичного домена для модификация FVII-полипептида, могут быть рациональнным или эмпирическим образом определены. Примеры других Gla-содержащий полипептидов включают, но не ограничиваются, FIX, FX, протромбин, протеин С, протеин S, остеокальцин, матриксный GlA протеин, прекращающий рост специфичный белок 6 (Gas6), и белок Z. GlA домены этих протеинов описаны в SEQ ID №№: 110-118, 120 и 121. Например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или более смежных (непрерывных) аминокислот или весь домен Gla из гетерологичного домена Gla могут быть введены в FVII-полипептид. Кроме того, введение Gla домена в FVII-полипептид также может включать дополнительные аминокислоты, не являющиеся частью домена Gla из гетерологичного полипептида, в той мере пока дополнительные аминокислоты не оказывают существенного ослабления способности фосфолипидного связывания GlA домена.
В некоторых примерах введение представляет собой такое добавление Gla домена к FVII-полипептиду, что гетерологичный Gla домен вставляется в эндогенный Gla домен или в другой район или домен FVII-полипептида, так что модифицированный FVII-полипептид сохраняет, по крайней мере, одну активность FVII. В таких примерах, нативный Gla домен FVII-полипептида сохраняется в полипептиде, хотя в некоторых случаях последовательность аминокислот, входящих в состав нативного Gla домена, прерывается. В других примерах гетерологичный Gla домен или достаточная его часть вводится либо с N- или с С-конца нативного Gla домена так, что нативный Gla домен не прерывается. В других примерах, гетерологичный GlA домен, или достаточная его часть, вставляются в другой домен FVII-полипептида.
Кроме того, в настоящем документе раскрыт модифицирований Gla домен FVII-полипептида, когда все или смежные части эндогенного Gla домена FVII удаляются и заменяются гетерологичным Gla доменом или его частью для осуществления фосфолипидного связывания, при том, что модифицированный FVII-полипептид сохраняет, по крайней мере, одну активность FVII. Такая модификация также относится к GlA доменной замене. Например, весь или часть домена Gla FVII, как описано в SEQ ID №: 119, соответствующая 1-45 аминокислотная последовательность, описанная в SEQ ID №: 3, может быть удалялена из FVII-полипептида и заменена гетерологичным Gla доменом, или его частью. Гетерологичная часть включает в себя любой домен Gla, известный специалисту, который связывается с фосфолипидом, включая, но не ограничиваясь этим Gla доменом с последовательностью аминокислот из SEQ ID №№: 110-118, 120 и 121.В некоторых случаях, необходимая часть Gla домена представляет собой SEQ ID №№: 110-118, 120 и 121 для осуществления фосфолипидного связывания, которая может заменить эндогенный Gla домен FVII или его часть. Например, Gla домен FVII может быть заменен Gla доменом белка С с SEQ ID №: 113. В результате модифицированный FVII-полипептид содержит Gla домен из протеина С из собственного EGF-I, EGF-2 и домена серии протеазы.
В некоторых случаях, гетерологичный Gla домен содержит мутации, что дополнительно увеличивает связывание с активированными тромбоцитами за счет увеличения фосфолипидного связвания. Например, если белок С Gla домена вводится для получения модифицированного полипептида FVII, белок С Gla домена может содержать аминокислотные мутации, что увеличивает фосфолипидное связывание.
В других случаях гетерологичный Gla домен может содержать дальнейшие мутации, которые наделяют GIA домен FVII-подобными функциями. Например, как отмечалось выше, R36 из Gla домена FVII, в SEQ ID №: 119 может быть вовлечен во взаимодействий с FX. Следовательно, гетерологичный Gla домен может содержать дополнительные модификации, такие, которые необходимы для поддержания аргинина в позиции 36 зрелого FVII-полипептида, как указано в SEQ ID №: 3, или любые другие модификации, необходимые для поддержания FX-активационых свойств модифицированного FVII-полипептида (Ruf и др., (1999) Biochem 38:1957-1966). Так, в некоторых случаях, соответствующая мутация R36 может быть осуществлена в гетерологичном Gla домене. Соответствующая позиция может быть определена специалистом, например, путем выравнивания аминокислотных последовательностей.
Как описано здесь ниже, специалист в данной области может эмпирически или рационально сконструировать модифицированный FVII-полипептид, содержащий гетерологичный Gla домен, так что полученный FVII-полипептид сохранит, по крайней мере, одну активность FVII. Как правило, такие модифицированные полипептиды FVII сохраняют способность связывать и активировать FX. В некоторых случаях, такие модифицированные полипептиды FVII также сохраняют способность связывать и активировать FIX. Полученные Gla-модифицированные FVII-полипептиды включают те, которые сохраняют способность к связыванию с ТФ. Модифицированные FVII-полипептиды, с замененным Gla-доменом, также включают полипептиды с увеличенной внутренней активностью в отсутствие ТФ. Таким образом, полученные Gla модифицированные FVII-полипептиды включают полипептиды, которые не связываются с ТФ, или которые связывают ТФ плохо по сравнению с диким типом FVIIa. Как правило, такие модифицированные полипептиды FVII проявляют повышенную способность связывать фосфолипиды. В частности, такие модифицированные FVII-полипептиды проявляют повышенную способность связывать фосфатидилсерин. Повышенную способность связывать фосфолипиды можно оценить в анализах на изолированных фосфолипидах или на поверхности активированных тромбоцитов. Такие анализы, как правило, проводят на двухцепочечной форме FVII, такой как активированная форма FVII (FVIIa).
В настоящем документе предусмотрены модифицированные FVII-полипептиды, содержащие гетерологичный домен GlA. Примеры таких модифицированных полипептидов FVII включают полипептиды, в которых эндогенный домен Gla заменен полностью или частью домена Gla любого из белков: FIX (последовательность SEQ ID №110), FX (последовательность SEQ ID №111), тромбин (последовательность SEQ ID №112), белок С (последовательность SEQ ID №113) или белок S (последовательность SEQ ID №114). Такие модифицированные полипептиды FVII могут проявлять повышенную способность связывать активированные тромбоциты, что приводит к повышению коагулянтной активности. В таблице 7 приведены примеры модификаций, которые могут быть осуществлены в полипептиде FVII для замены эндогенного домена Gla доменами FIX, FX, белка С, белка S или тромбина. В таблице указывается название модификации (например, Gla замена FX) и подробная информация о модификации, в том числе аминокислотные позиции, по которым экзогенный домен Gla вставлен в FVII-полипептид, и последовательность экзогенного домена Gla. Также приведены номера последовательностей (последовательность SEQ ID №), под которыми приведены примеры аминокислотных последовательностей модифицированных полипептидов FVII, а также любые полипептидные идентификационные номера. Например, модификация "Gla замена FIX" (А1У44делеция вставка
YNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQY) предполагает исключение эндогенного Gla домена FVII делецией аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 45 аминокислотных остатков, которые соответствуют аминокислотным остаткам с Y1 до Y45 FIX домена Gla, приведенного в последовательности SEQ ID №110. Аналогичным образом, модификация «Gla замена FX» предполагает исключение аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 44 аминокислотных остатков, которые соответствуют остаткам с А1 до Y44 из FX Gla домена, приведенного в последовательности SEQ ID №111. Модификация «Gla замена Тромбин» предполагает исключение аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 44 аминокислотных остатков, которые соответствуют аминокислотным остаткам с Y1 до Y44 из Gla домена тромбина, приведенного в последовательности SEQ ID №112. Модификация «Gla замена белок С» предполагает исключение аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 44 аминокислотных остатков, которые соответствуют аминокислотным остаткам с А1 до Н44 из Gla домена белка С, приведенного в последовательности SEQ ID №113. Модификация «Gla замена белок S» предполагает исключение аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 44 аминокислотных остатков, которые соответствуют аминокислотным остаткам с Y1 до Y44 из Gla домена белка S, приведенного в последовательности SEQ ID №114.
Prot - белок
Swap - замена из
Delins - удаление-вставка
Модифицированные FVII-полипептиды, содержащие гетерологичный домен Gla, могут проявлять повышенную коагулянтную активность при более низких дозах по сравнению с FVII дикого типа, таким как NovoSeven®, за счет увеличения связывания и/или аффинности к активированным тромбоцитам. Коагулянтная активность Gla модифицированных FVII-полипептидов может быть увеличена, по крайней мере, или приблизительно на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200%, 300%, 400%, 500% или более по сравнению с коагулянтной активностью немодифицированного FVII-полипептида или полипептида дикого типа in vivo, ex vivo или in vitro.
3. Комбинации и дополнительные модификации
В дополнение к модифицированным FVII-полипептидам, обладающим повышенной устойчивостью к TFPI, повышенной устойчивостью к AT-III, повышенной ТФ-зависимой и/или ТФ-независимой каталитической активностью, улучшенными фармакокинетическими свойствами, такими как увеличение периода полураспада, и/или увеличенной способностью связывать и/или аффинностью к фосфолипидам мембраны, модифицированные FVII-полипептиды, предусмотренные в настоящем документе, также включают те, которые демонстрируют более одного из вышеупомянутых свойств. Например, полипептиды FVII могут быть модифицированы таким образом, что полученные полипептиды проявляют повышенную способность связывать и/или аффинность к фосфолипидам, а также проявляют повышенную устойчивость к TFPI. Соответственно, полипептиды FVII могут быть модифицированы введением гетерологичного домена Gla из других витамин-К-зависимых белков плазмы и замещением одной или нескольких аминокислот в позициях 196, 197, 199, 237, 239, 290 или 341 зрелого полипептида FVII, приведенного в последовательности SEQ ID №3.
Кроме того, любые предусмотренные в настоящем документе модифицированные полипептиды FVII также могут содержать одну или несколько других модификаций, описанных в уровне техники. Как правило, такими дополнительными модификации являются те, которые сами приводят к повышению коагулянтной активности модифицированного полипептида и/или повышению стабильности полипептида. Таким образом, полученные модифицированные полипептиды FVII проявляют увеличенную коагулянтную активность. Дополнительные модификации могут включать, например, любые аминокислотные замены, удаления или вставки, известные в уровне техники, как правило, увеличивающие коагулянтную активность и/или стабильность FVII-полипептида, Любые модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут содержать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более дополнительных аминокислотных модификаций, пока полученные модифицированные полипептиды FVII сохраняют активность FVII дикого типа или немодифицированного полипептида.
В одном примере, в последовательность FVII-полипептида могут быть внесены дополнительные модификации, которые изменяют его взаимодействия с другими факторами, молекулами и белками. Например, могут быть заменены аминокислотные остатки, которые участвуют во взаимодействии с тканевым фактором (ТФ), что увеличит сродство модифицированного полипептида FVII к ТФ. Другие модификации включают, но не ограничиваются, модификациями аминокислот, которые участвуют во взаимодействиях с фактором X, фактором IX, TFPI и AT-III.
Также могут быть осуществлены дополнительные модификации модифицированных полипептидов FVII, предусмотренных в настоящем документе, которые изменяют конформацию или фолдинг полипептида. К ним относятся, например, замены одной или нескольких аминокислот цистеином, что приводит к образованию новой дисульфидной связи, или модификации, которые стабилизируют конформацию а-спирали, тем самым придавая повышенную активность модифицированному FVII-полипептиду.
В полипептид FVII также могут быть внесены дополнительные модификации, влияющие на посттрансляционные модификации. Например, полипептид может быть модифицирован для включения дополнительных сайтов гликозилирования, так что полученный модифицированный полипептид FVII обладает повышенным гликозилированием по сравнению с немодифицированным FVII-полипептидом. Также могут быть осуществлены модификации, вводящие аминокислотные остатки, которые могут быть впоследствии связаны с химическими фрагментами для увеличения стабильности модифицированного полипептида FVII. Полипептид FVII также может быть изменен модифицированием потенциального сайта расщепления (сайтов) эндогенными протеазами, что приводит к уменьшению деградации варианта FVIIa и повышению стабильности модифицированного полипептида FVII.
Кроме того, замены, удаления или вставки аминокислот могут быть осуществлены в эндогенном или гетерологичном домене Gla так, что модифицированные полипептиды FVII проявляют повышенную способность связывать и/или аффинность к фосфолипидам мембран. В других примерах предусмотренные в настоящем документе модифицированные полипептиды FVII могут обладать делениями в эндогенном домене Gla, или заменами на позициях, которые в норме гамма-карбоксилированы (US20070037746).
В следующих разделах описаны неограничивающие примеры модификаций, описанных в уровне техники для повышения стабильности и/или коагулянтной активности полипептида FVII. Как отмечалось выше, такие модификации также могут быть дополнительно включены в любые модифицированные полипептиды FVII, предусмотренные в настоящем документе. Аминокислотные позиции, приведенные ниже, соответствуют зрелому полипептиду FVII, приведенному в последовательности SEQ ID № 3. Соответствующие мутации могут быть осуществлены и в других полипептидах FVII, таких как аллельные, видовые варианты или варианты сплайсинга зрелого полипептида FVII, приведенного в последовательности SEQ ID №3.
а. Модификации, которые увеличивают внутреннюю активность
В одном примере, могут быть осуществлены дополнительные модификации модифицированного полипептида фактора VII, предусмотренного в настоящем документе, что приводит к увеличению каталитической активности к фактору Х и/или фактору IX. Например, могут быть внесены модификации в аминокислоты, которые участвуют во взаимодействии с его кофактором, ТФ, такие, что полученные модифицированные полипептиды FVII обладают повышенной аффинностью к ТФ, тем самым показывая возросшую активность по отношению к FX и/или FIX. Модификации также могут быть осуществлены для активации кармана полипептида FVII, что повышает внутреннюю активность модифицированного полипептида FVII по отношению к FX и/или FIX по сравнению с активностью немодифицированного полипептида. Другая стратегия модификаций, которая приводит к увеличению активности, предполагает модификацию полипептида FVII, которая изменяет фолдинг и конформацию белка в более активную форму, или она предполагает смещение равновесия между высокоактивной и неактивной (или менее активной) конформациями в пользу высокоактивной конформации (конформаций). Получение более активного полипептида также может быть достигнуто путем модификации аминокислот, находящихся в р-тяжах полипептида FVII. Например, могут быть осуществлены аминокислотные замены, вводящие новые цистеиновые пары, которые могут образовывать новые дисульфидные связи, которые могут функционировать для "фиксирования" модифицированного полипептида FVII в более активной форме.
Примеры дополнительных модификаций, которые могут быть включены в модифицированные FVII-полипептиды, предусмотренные в настоящем документе, для увеличения внутренней активности модифицированного FVII-полипептида, включают, но не ограничиваются, описанными Persson и др., (2004) Biochem J. 379:497-503, Maun и др., (2005) Prot Sci 14:1171-1180, Persson и др., (2001) PNAS 98:13583-13588, Persson и др., (2002) Eur J Biochem 269:5950 - 5955, Soejima др., (2001) J Biol Chem 276:17229-17235, Soejima др., (2002) J Biol Chem 277:49027-49035, WO200183725, WO2002022776, WO2002038162, WO2003027147, WO200338162, WO2004029090, WO2004029091, WO2004108763 и WO2004111242. Неограничивающие примеры модификаций аминокислот, описанные в уровне техники, которые могут привести к усилению внутренней активности модифицированного полипептида FVII, включают одну или более из S278C/V302C, L279C/N301C, V280C/V301C, S281C/V299C, S314E, L39E, L39Q, L39H, I42R, S43Q, S53N, К62Е, K62R, K62D, K62N, K62Q, К62Т, L65Q, L65S, F71D, F71Y, F71E. F71Q, F71N, P74S, Р74А, А75Е, A75D, Е77А, E82Q, E82N, Т83К, E116D, K157V, K157L, K157I, К157М, K157F, K157W, К157Р, K157G, K157S, К157Т, К157С, K157Y, K157N, К157Е, K157R, К157Н, K157D, K157Q, V158L, V158I, V158M, V158F, V158W, V158P, V158G, V158S, V158T, V158C, V158Y, V158N, V158E, V158R, V158K, V158H, V158D, V158Q, А274М, A274L, А274К, A274R, A274D, A274V, A274I, A274F, A274W, А274Р, A274G, А274Т, А274С, A274Y, A274N, А274Е, А274Н, A274S, A274Q, F275H, E296V, E296L, E296I, Е296М, E296F, E296W, Е296Р, E296G, E296S, Е296Т, Е296С, E296Y, E296N, Е296К, E296R, Е296Н, E296D, E296Q, M298Q, M298V, M298L, M298I, M298F, M298W, М298Р, M298G, M298S, М298Т, М298С, M298Y, M298N, М298К, M298R, М298Н, М298Е, M298D, R304Y, R304F, R304L, R304M, L305V, L305Y, L305I, L305F, L305A, L305M, L305W, L305P, L305G, L305S. L305T, L305C, L305N, L305E, L305K, L305R, L305H, L305D, L305Q, M306D, M306N, D309S, D309T, S314A, S314V, S3141, S314M, S314F, S314W, S314P, S314G, S314L, S314T. S314C. S314Y, S314N, S314E, S314K, S314R, S314H, S314D, S314Q, D334G, D334E, D334A, D334V, D334I, D334M, D334F, D334W, D334P, D334L, D334T, D334C, D334Y, D334N, D334K, D334R, D334H, D334S, D334Q, S336G, S336E, S336A, S336V, S336I, S336M, S336F, S336W, S336P, S336L, S336T, S336C, S336Y, S336N, S336K, S336R, S336H, S336D, S336Q, K337L, K337V, K337I, К337М, K337F, K337W, К337Р, K337G, K337S, К337Т, К337С, K337Y, K337N, К337Е, K337R, К337Н, K337D, K337Q, F374P, F374A, F374V, F374I, F374L, F374M, F374W, F374G, F374S, F374T, F374C, F374Y, F374N, F374E, F374K, F374R, F374H, F374D, F374Q, и замен позиций 300-322, 305-322, 300-312, или 305-312 соответствующими аминокислотами трипсина, тромбина или FX, а также замещения позиций 310-329, 311-322 или 233-329 соответствующими аминокислотами трипсина.
b. Модификации, повышающие устойчивость к протеазам
Модифицированные FVII-полипептиды, предусмотренные в настоящем документе, также могут содержать дополнительные модификации, которые приводят к увеличению устойчивости полипептида к протеазам. Например, могут быть осуществлены аминокислотные замены, удаляющие один или более потенциальных сайтов протеолитического расщепления. Таким образом, могут быть получены более устойчивые к протеазам модифицированные полипептиды FVII с повышенной тем самым стабильностью и периодом полураспада.
Примеры дополнительных модификаций, которые могут быть включены в модифицированные полипептиды FVII, предусмотренные в настоящем документе, для увеличения устойчивости к протеазам, включают, но не ограничиваются, описанными в патенте США №US5580560 или международных заявках №№WO1988010295 и WO2002038162. Неограничивающие примеры модификаций, описанных в данной области техники, которые могут привести к увеличению устойчивости модифицированного полипептида FVII к ингибиторам и/или протеазам, включают одну или более из K32Q, К32Е, K32G, К32Н, К32Т, К32А, K32S, К38Т, K38D, K38L, K38G, К38А, K38S, K38N, К38Н, I42N, I42S, I42A, I42Q, Y44N, Y44S, Y44A, Y44Q, F278S, F278A, F278N, F278Q, F278G, R290G, R290A, R290S, R290T, R290K, R304G, R304T, R304A, R304S, R304N, R315G, R315A, R315S, R315T, R315Q, Y332S, Y332A, Y332N, Y332Q, Y332G, К341Е, K341Q, K341G, К341Т, К341А и K341S.
с. Модификации, увеличивающие аффинность к фосфолипидам
Коагулянтная активность FVII может быть повышена за счет увеличения связывания и/или аффинности полипептида к фосфолипидам, таким как экспрессированным на поверхности активированных тромбоцитов. Например, могут быть осуществлены дополнительные замены аминокислот в конкретных позициях в эндогенном домене Gla полипептида FVII, результатом которых являются модифицированные полипептидны FVII с повышенной способностью связывать фосфатидилсерин и другие отрицательно заряженные фосфолипиды. Таким образом, такие модификации также могут быть включены в модифицированные полипептиды FVII, предусмотренные в настоящем документе, при условии, что такие модифицированные полипептиды FVII обладают эндогенными доменами Gla, то есть еще не были подвергнуты модификациям, приводящим к замещению эндогенного домена Gla доменом другого витамин-К-зависимого белка.
Примеры дополнительных модификаций для увеличения связывания и/или аффинности к фосфолипидам, которые можно ввести в модифицированный полипептид FVII, предусмотренный в настоящем документе, который содержит эндогенный FVII Gla домен, включают, но не ограничиваются, описанными в Harvey и др., (2003) J Biol Chem 278:8363-8369, US20030100506, US20040220106, US20060240526, US6017882, US6693075, US6762286, WO200393465 и WO2004111242. Примеры таких модификаций включают одну или более из следующих модификаций: вставка тирозина в положении 4 или одна или более модификаций P10Q, Р10Е, P10D, PLON, R28F, R28E, К32Е, K32D, D33F, D33E, D33K А34Е, A34D, A34I, A34L, А34М. A34V, A34F, A34W, A34Y, R36D, R36E, К38Е и K38D.
d. Модификации, изменяющие гликозилирование
Изменение уровня гликозилирования белков описано в уровне техники как средство снижения иммуногенности, повышения стабильности, сокращения частоты введения и/или уменьшения неблагоприятных побочных эффектов, таких как воспаление. Обычно это достигается увеличением уровня гликозилирования. Сайт гликозилирования (сайты) представляет собой сайт прикрепления углеводного фрагмента к полипептиду, такой, что при введении полипептида в эукариотическую клетку, способную к гликозилированию, он гликозилируется.
В FVII присутствуют четыре естественных сайта гликозилирования, два сайта N-гликозилирования на N145 и N322 и два сайта O-гликозилирования на S52 и S60, соответствующих аминокислотным позициям в зрелом полипептиде FVII, приведенном в последовательности SEQ ID №3. В одном варианте, могут быть осуществлены дополнительные модификации в модифицированном полипептиде FVII, предусмотренном здесь, приводящие к нарушению гликозилирования по указанным выше сайтам. Это может привести к модифицированным полипептидам FVII с повышенной коагулянтной активностью (см., например, WO2005123916). Неограничивающие примеры модификаций, описанных в данной области техники, которые могут привести к снижению гликозилирования и увеличению активности модифицированного полипептида FVII по сравнению с немодифицированным полипептидом FVII, включают, но не ограничиваются, N145Y, N145G, N145F, N145M, N145S, N1451, N145L, N145T, N145V, N145P, N145K, N145H, N145Q, N145E, N145R, N145W, N145D, N145C, N322Y, N322G, N322F, N322M, N322S, N3221, N322L, N322T, N322V, N322P, N322K, N322H, N322Q, N322E, N322R, N322W и N322C.
В другом варианте, в аминокислотную последовательность модифицированных полипептидов FVII, предусмотренных в настоящем документе, могут быть внесены дополнительные модификации, которые вводят дополнительные сайты гликозилирования, повышая тем самым уровень гликозилирования модифицированного полипептида FVII по сравнению с немодифицированным FVII-полипептидом. Сайт гликозилирования может представлять собой N- или О-сайт гликозилирования. Примеры модификаций, которые могут быть осуществлены в полипептидах FVII для введения одного или нескольких новых сайтов гликозилирования, включают, но не ограничиваются примерами, описанными в US6806063 и WO2003 93465. Неограничивающие примеры модификаций, описанных в данной области техники, которые могут привести к увеличению гликозилирования модифицированного полипептида FVII по сравнению с немодифицированным полипептидом FVII, включают, но не ограничиваются, F4S, F4T, P10N, Q21N, W41N, S43N, A51N, G58N, L65N, G59S, G59T, E82S, Е82Т, N95S, N95T, G97S, G97T, Y101N, D104N, T106N, K109N, G117N, G124N, S126N, T128N, A175S, A175T, G179N, I186S, I186T, V188N, R202S, R202T, I205S, I205T, D212N, E220N, I230N, P231N, P236N, G237N, V253N, E265N, T267N, E270N, R277N, L280N, G291N, P303S, P303ST, L305N, Q312N, G318N, G331N, D334N, K337N, G342N, H348N, R353N, Y357N, I361N, V376N, R379N, M391N, K32N/A34S, K32N/A34T, F31N/D33S, F31N/D33T, I30N/K32S, I30N/K32T, A34N/R36S, A34N/R36T, K38N/F40S, K38N/F40T, T37N/L39S, T37N/L39T, R36N/K38S, R36N/K38T, L39N/W41S, L39N/W41T, F40N/I42S, F40N/I42T, I42N/Y44S, I42N/Y44T, Y44N/D46S, Y44N/D46T, D46N/D48S, D46N/D48T, G47N/Q49S, G47N/Q49T, K143N/N145S, K143N/N145T, E142N/R144S, E142N/R144T, L141N/K143S, L141N/K143T, I140N/E142S, I140N/E142T, R144N/A146S, R144N/A146T, A146N/K148S, A146N/K148T, S147N/P149S, S147N/P149T, R290N/A292S, R290N/A292T, D289N/G291S, D289N/G291T, L288N/R290S, L288N/R290T, L287N/D289S, L287N/D289, A292N/A294S, A292N/A294T, T293N/L295S, T293N/L295T, R315N/V317S, R315N/V317T, S314N/K316S, S314N/K316T, Q313N/R315S, Q313N/R315T, КЗ 16N/G318S, КЗ 16N/G318Т, V317N/D319S, V317N/D319Т, K341N/D343S, K341N/D343T, S339N/K341S, S339N/K341T, D343N/G345S, D343N/G345T, R392N/E394S, R392N/E394T, L390N/R392S, L390N/R392T, K389N/M391S, K389N/M391T, S393N/P395S, S393N/P395T, E394N/R396S, E394N/R396T, P395N/P397S, P395N/P397T, R396N/G398S, R396N/G398T, P397N/V399S, P397N/V399T, G398N/L400S, G398N/L400T, V399N/L401S, V399N/L401T, L400N/R402S, L400N/R402T, L401N/A403S, L401N/A403T, R402N/P404S, R402N/P404T, A403N/F405S, A403N/F405T, P404N/P406S и P404N/P406T.
е. Модификации для облегчения связывания химических групп
Дополнительные модификации модифицированных полипептидов FVII, предусмотренных в настоящем документе, также могут быть осуществлены с целью облегчения последующего связывания химических групп.Одна или более аминокислотных замен или вставок могут быть сделаны так, что химические группы могут связываться с модифицированным полипептидом FVII через замененные аминокислоты.
Например, в модифицированный полипептид FVII может быть вставлен цистеин, к которому может быть присоединен полиэтиленгликольный (PEG) фрагмент для большей стабильности и периода полураспада в сыворотке. Другие остатки, к которым осуществляется присоединение, включают остатки лизина, аспарагиновой кислоты и глутаминовой кислоты. В некоторых вариантах аминокислотные остатки заменяются для уменьшения числа потенциальных позиций для связывания. Например, может быть уменьшено количество лизинов. Примеры модификаций, которые могут быть введены в последовательность аминокислот полипептида FVII, которые могут облегчить последующее связывание с химическими группами, включают, но не ограничиваются, описанными в US20030096338, US20060019336, US6806063, WO200158935 и WO2002077218. Неограничивающие примеры модификаций полипептида FVII, которые могут облегчить последующее связывание с химическими группами, включают, но не ограничиваются, Q250C, R396C, Р406С, I42K, Y44K, L288K, D289K, R290K, G291K, А292К, Т293К, Q313K, S314K. R315K, V317K, L390K, М391К, R392K, S393K, Е394К, Р395К, R396K, Р397К, G398K, V399K, L400K, L401K, R402K, А403К, Р404К, F405K, I30C, К32С, D33C, А34С, Т37С, К38С, W41C, Y44C, S45C, D46C, L141C, Е142С, К143С, R144C, L288C, D289C, R290C, G291C, А292С, S314C, R315C, К316С, V317C, L390C, М391С, R392C, S393C, Е394С, Р395С, R396C, Р397С, G398C, V399C, L401C, R402C, А403С, Р404С, I30D, K32D, A34D, T37D, K38D, W41D, Y44D, S45D, D46C, L141D, E142D, K143D, R144D, L288D, R290D, G291D, A292D, Q313D, S314D, R315D, K316D, V317D, L390D, M391D, R392D, S393D, P395D, R396D, P397D, G398D, V399D, L401D, R402D, A403D, P404D, 130Е, К32Е, А34Е, Т37Е, К38Е, W41E, Y44E, S45E, D46C, L141E, Е142Е, К143Е, R144E, L288E, R290E, G291E, А292Е, Q313E, S314E, R315E, К316Е, V317E, L390E, М391Е, R392E, S393E, Р395Е, R396E, Р397Е, G398E, V399E, L401E, R402E, А403Е, Р404Е, Kl 8R, K32R, K38R, K62R, K85R, K109R, K137R, K143R, K148R, K157R, K161R, K197R, K199R, K316R, K337R, K341R, K389R. Кл 8Q, K32Q, K38Q, K62Q, K85Q, K109Q, K137Q, K143Q, K148Q, K157Q, K161Q, K197Q, K199Q, K316Q, K337Q, K341Q, K389Q, Kl 8N, K32N, K38N, K62N, K85N, K109N, K137N, K143N, K148N, K157N, K161N, K197N, K199N, K316N, K337N, K341N, K389N, К18Н, К32Н, К38Н, К62Н, К85Н, К109Н, К137Н, К143Н, К148Н, К157Н, К161Н, К197Н, К199Н, К316Н, К337Н, К341Н и К389Н.
f. Примеры комбинаций модификаций
В настоящем документе предусмотрены модифицированные FVII-полипептиды, которые обладают двумя или более модификациями, призванными воздействовать на одно или несколько свойств или активностей немодифицированного FVII-полипептида. В некоторых примерах две или более модификации изменяют два или более свойства или активности полипептида FVII. В FVII-полипептиды могут быть внесены модификации, изменяющие одно или более свойств, выбранных из устойчивости к TFPI, устойчивости к AT-III, внутренней активности, амидолитической активности, каталитической активности (в присутствии или отсутствии ТФ), способности связывания и/или аффинности к фосфолипидам, гликозилирования, устойчивости к протеазам, измененного времени полураспада и взаимодействия и/или активации других факторов или молекул, таких как FX и FIX. Как правило, комбинируются две или более модификации, так что полученный модифицированный полипептид FVII обладает повышенной коагулянтной активностью, повышенной продолжительностью коагулянтной активности и/или расширенным терапевтическим индексом по сравнению с немодифицированным FVII-полипептидом. Модифицированные полипептиды FVIIa могут характеризоваться более быстрым наступлением коагулянтной активности in vitro, ex vivo, или in vivo. Модификации могут включать замены, вставки или удаления аминокислот. Повышенная коагулянтная активность, увеличенная продолжительность коагулянтной активности, ускоренное наступление коагулянтной активности и/или расширение терапевтического индекса модифицированного полипептида FVII, содержащего две или более модификации, могут быть увеличены, по крайней мере, или приблизительно на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 1 10%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190%, 200%, 300%, 400%, 500% или более по сравнению с активностью начального или немодифицированного FVIIa полипептида.
В настоящем документе предусмотрены модифицированные FVII-полипептиды, которые содержат две или более модификации, которые вводятся в немодифицированный полипептид FVII для изменения двух или более видов активности или свойств. Модифицированные полипептиды FVII могут содержать 2, 3, 4, 5, 6 или более модификаций. Кроме того, каждая модификация может затрагивать один или более аминокислотных остатков. Например, модифицированный полипептид FVII может содержать две модификаци, каждая из которых представляет собой одну аминокислотную замену. В другом примере, модифицированный полипептид FVII может содержать две модификации, одна из которых представляет собой единичную аминокислотную замену, а другая предполагает делецию более чем одного аминокислотного остатка, и последующее включение более одного аминокислотного остатка. Например, модифицированный полипептид FVII, предусмотренный в настоящем документе, может содержать аминокислотную замену M298Q (остаток, соответствующий зрелому полипептиду FVII, приведенному в последовательности SEQ ID №3) для увеличения внутренней активности и модификацию «Gla замена FIX», которая предполагает исключение эндогенного Gla домена FVII, исключение аминокислотных остатков с А1 до Y44 (остатки соответствующего зрелого полипептида FVII, приведенного в последовательности SEQ ID №3) и вставку 45 аминокислотных остатков, которые соответствуют аминокислотным остаткам с Y1 по Y45 домена Gla FIX, приведенного в последовательности SEQ ID №110.
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут иметь две или более модификации, выбранные из тех, которые приведены в таблицах с 5 по 8. В других примерах, модифицированный полипептид FVII содержит две или более модификации, где одна или несколько модификаций выбраны из числа указанных в таблицах с 5 по 8, и одна или более модификации представляют собой дополнительные модификации, не указанные в таблицах с 5 по 8, такие как, например, модификации, известные в данной области. В некоторых примерах, одна или несколько дополнительных модификаций могут быть выбраны из модификаций, которые предусмотрены в разделе D.3.a-e выше. Например, модифицированный полипептид FVII может содержать модификации по одному или нескольким аминокислотным остаткам, выбранным из D196, К197, К199, G237, Т239, R290 или К341 по нумерации зрелого FVII, приведенной в последовательности SEQ ID №3 (соответствующим D60, КбОа, КбОс, G97, Т99, R147 и К 192 остаткам соответственно, по химотрипсиновой нумерации), которые могут увеличить устойчивость к TFPI, и модификации по одному или более аминокислотным остаткам, которые затрагивают внутреннюю активность, такую как, например, VI 58 и М298, (V21 и Ml 56 соответственно, по химотрипсиновой нумерации). Например, модифицированный полипептид FVII может содержать две аминокислотные замены, что повышает устойчивость к TFPI, такие как К197Е и G237V, и одну аминокислотную замену, которая увеличивает внутреннюю активность, такую как M298Q, что приводит к полипептиду FVII с повышенной коагулянтной активностью.
Неограничивающие примеры комбинаций модификаций приведены в таблице 9. Эти примеры комбинаций модификаций включают две или более модификации, которые призваны изменить два или более видов активности или свойств полипептида FVII, включая, но не ограничиваясь, устойчивостью к TFPI, устойчивостью к AT-III, внутренней активностью, амидолитической активностью, каталитической активностью (в присутствии и/или отсутствие ТФ), связывание и/или аффинность к фосфолипидам, гликозилирование, устойчивость к протеазам, время полураспада и взаимодействие и/или активация других факторов или молекул, таких как FX и FIX. Модифицированные FVII-полипептиды, содержащие такие комбинации модификаций, могут обладать увеличенной коагулянтной активностью, увеличенной продолжительностью коагулянтной активности, ускоренным наступлением коагулянтной активности и/или расширенным терапевтическим индексом. Модификации, которые указаны в таблице 8 ниже, приведены с использованием той же терминологии и системы нумерации, как описано для таблиц с 5 до 7 выше. Например, "модификация Gla замена FIX" включает исключение эндогенного домена Gla FVII делецией аминокислотных остатков с А1 до Y44 (остатки соответствуют зрелому полипептиду FVII, приведенному в последовательности SEQ ID №3) и вставку 45 аминокислотных остатков, соответствующих аминокислотным остаткам с Y1 до Y45 Gla домена FIX, приведенного в последовательности SEQ ID №110, как описано выше. Аминокислотные позиции соответствуют аминокислотным позициям зрелого полипептида FVII, приведенного в последовательности SEQ ID №3, а также упоминаются соответствующие обозначения по химотрипсиновой нумерации. В таблице 9 ниже определены номера последовательностей (SEQ ID №), под которыми приведены примеры аминокислотных последовательностей модифицированных полипептидов FVII, а также любые полипептидные идентификационные номера.
Swap - замена из
е. Принципы и методы модификации FVII
В настоящей статье предусмотрены модифицированные FVII-полипептиды. FVII-полипептиды модифицируются таким образом, что они могут отличаться по одному или более видам активности или свойствами по сравнению с немодифицированным FVII-полипептидом. Активности и свойства, которые могут быть изменены в результате модификаций, включают, но не ограничиваются, коагуляцией или коагулянтной активностью, прокоагулянтной активностью; протеолитической или каталитической активностями, такими как способность активировать фактор Х (FX) или фактор IX (FIX); антигенностью, например, оцениваемой по способности связываться или конкурировать с полипептидами за связывание с анти-FVII антителами; способностью связывать тканевый фактор, фактор Х или фактор IX, способностью к связыванию с фосфолипидами, временем полураспада; трехмерной структурой; р1 и/или конформацией. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, включают полипептиды с увеличенной коагулянтной активностью. Эти полипептиды включают те, у которых увеличение коагулянтной активности обусловлено или связано с повышенной устойчивостью модифицированных FVII к TFPI, повышенной устойчивостью к AT-III, улучшенными фармакокинетическими свойствами, такими как увеличение времени полураспада, увеличением каталитической активности и/или увеличением связывания с активированными тромбоцитами. Примеры методов выявления таких модифицированных полипептидов FVII, и такие модифицированные полипептиды приведены в настоящем документе. Например, модифицированные полипептиды FVII, которые проявляют повышенную устойчивостью к TFPI, повышенную устойчивость к AT-III и/или увеличение связывания с активированными тромбоцитами, могут быть получены рационально или эмпирически: (а) рационально отмечая сайты, которые, как предполагается, вовлечены в проявление свойств, или (б) эмпирически тестируя модифицированные FVII-полипептиды в функциональных анализах на стойкость к воздействию TFPI, устойчивость к AT-III, улучшенные фармакокинетические свойства, такие как увеличение периода полураспада, и/или увеличения связывания с фосфолипидами или активированными тромбоцитами. Во многих случаях сочетание рациональных и эмпирических подходов к конструированию может быть использовано для создания модифицированных FVII-полипептидов.
Как только домен, регион или аминокислота определены для модификации с помощью рационального или эмпирического подхода, может быть использован любой метод, известный в уровне техники, вызывающий мутации одной или более аминокислот в целевом белке. Методы включают стандартный сайт-направленный мутагенез (используя, например, комплект оборудования, такой как комплект QuikChange, доступный от Stratagene) кодирующих молекул нуклеиновой кислоты, или твердофазные методы синтеза полипептидов. Кроме того, предусмотренные в настоящем документе модифицированные химерные белки (например, с заменой домена Gla) могут быть получены обычной техникой рекомбинантной ДНК. Например, химерные полипептиды могут быть созданы с помощью ферментов рестрикции и методов клонирования для рутинного субклонирования компонентов желаемого химерного полипептида. Определенные и полученные модифицированные FVII-полипептиды могут быть оценены на предмет их активности, такой как каталитическая активность или коагулянтная активность, с использованием одного или более различных методов анализа, известных в уровне техники или описанных здесь ниже.
1. Рациональная модификация
В некоторых примерах, модифицированные полипептиды FVII, предусмотренные в настоящем документе, получаются с помощью рациональных модификаций для увеличения коагулянтной активности. В таком методе, домены, районы или аминокислоты, ответственные за активность полипептида FVII, известны или могут быть рационально определены. Например, различные базы данных в общественном доступе предоставляют последовательность и информацию о домене, относящегося к FVII дикого типа (см., например, сервер ExPASy Proteomics ca.expasy.org). Другая общедоступная информация может быть использована для определения аминокислот, районов и/или доменов в полипептидах FVII, участвующих в конкретном взаимодействии или активности. Такие источники включают, например, научные журналы, базы данных последовательностей и функций, или патентные базы данных. В некоторых случаях прямые данные могут быть использованы для определения влияния одной или нескольких аминокислот на данное взаимодействие или активность. В других случаях данные экстраполируются косвенно, например, из фактов или информации о родственных белках, которые проявляют аналогичные активности и/или взаимодействия. В таких примерах аминокислоты, которые ответственны за активность и взаимодействия в родственном белке, могут быть использованы для определения соответствующих аминокислот в полипептиде FVII по выравниванию родственного полипептида с полипептидом FVII, основанном на последовательности и/или структурном сходстве. Таким образом, могут быть определены аминокислоты в полипептиде FVII, которые могут быть вовлечены в данный вид активности или взаимодействие. Кроме того, моделирование гомологии может быть использовано для прогнозирования остатков, которые играют важную роль в формировании и/или стабилизации белок-белковых взаимодействий, и эта информация может быть использована для разработки вариантов с улучшенными свойствами.
После определения доменов, районов и/или аминокислот, ответственных за активность полипептида FVII, они могут быть модифицированы таким образом, чтобы модификации, согласно прогнозам, привели к увеличению коагулянтной активности полипептида. Это может быть осуществлено несколькими способами в зависимости от активности или взаимодействия, которое требуется изменить. В некоторых примерах могут быть осуществлены одна или более замен, вставок и удалений аминокислот, которые увеличат аффинность модифицированного FVII к другим белкам или молекулам.
В одном примере, модификации могут быть осуществлены в любом домене, районе, с любой аминокислотой (аминокислотами) таким образом, чтобы изменить свойства или взаимодействия модифицированных FVII с другими белками, например, увеличить или уменьшить взаимодействие в зависимости от его вида, выбранного для модифицирования. Для осуществления таких изменений домены, районы и/или аминокислоты, которые участвуют или предположительно участвуют в таком взаимодействии, должны быть известны или рационально определены. Например, модифицированный FVII может быть рационально сконструирован со сниженной способностью взаимодействовать с ингибиторным белком, таким как TFPI или AT-III, для увеличенной коагулянтной активности модифицированного FVII-полипептида. Методы рационального конструирования полипептидов FVII со сниженной способностью взаимодействовать с TFPI приведены в Примере 1. Аналогичные подходы могут быть применены для увеличения или уменьшения взаимодействия FVII с любым другим полипептидом, с которым FVII взаимодействует или связывается.
Например, FVII был модифицирован для повышения устойчивости к TFPI. TFPI в комплексе с FXa связывается с ТФ / FVIIa комплексом с образованием четвертичного комплекса, в котором каталитическая активность FVIIa подавлена. Первый домен Кунитца TFPI-I (TFPI-1 K1) является доменом, который отвечает за взаимодействие и ингибирование FVIIa. Определение аминокислотных остатков FVII, которые участвуют в этом взаимодействии, может содействовать конструированию полипептидов FVII, которые устойчивы к TFPI. Для определения того, какие аминокислотные остатки FVII участвуют в этом взаимодействии, могут быть использованы различные подходы, включая, но не ограничиваясь, мутагенезом, сканированием и рациональными подходами, такими как компьютерное моделирование. В качестве основы для определения контактирующих остатков может быть использована кристаллическая структура FVIIa в комплексе с TFPI. Кроме того, как описано в примере 1, если кристаллическая структура недоступна, может быть использовано компьютерное моделирование, основанное на выравнивании и экстраполяции известных структур и последовательностей белков и их взаимодействии друг с другом.
Результаты компьютерного моделирования могут быть использованы для выявления остатков, которые непосредственно вовлечены в контакт с желаемым белком, таким как, например, TFPI, и/или остатков, которые косвенно участвуют в процессе взаимодействия, например, благодаря близости к контактирующим остаткам. Например, с использованием результатов компьютерного моделирования, как описано в Примере 1, для выявления контактирующих остатков в TFPI-I было осуществлено выравнивание первого домена Кунитца TFPI-I и TFPI-2 (Фиг.5b). Замены аминокислот могут быть определены по изменению взаимодействий полипептида FVII с целевым белком. Заменяющие аминокислоты могут представлять собой все другие 19 естественных аминокислот, а также "неестественные" аминокислоты. Альтернативно, такие замены аминокислот могут быть определены in silico мутагенезом на основе рациональных предположений о взаимодействии. Пример 1 иллюстрирует in silico подходом к определению замен аминокислот, при котором анализ компьютерной модели указывает на остатки, участвующие в электростатических взаимодействиях факторов VII и TFPI-I K-1. Например, было выявлено, что отрицательно заряженный остаток D60 из FVII (по химотрипсиновой нумерации) участвует в электростатическом взаимодействии с положительно заряженным R20 из TFPI-I. Поэтому этот остаток является кандидатом для мутагенеза, приводящего к нарушению электростатического взаимодействия с R20 из TFPI, тем самым приводя к модифицированному полипептиду FVII с повышенной устойчивостью к TFPI. Это может быть осуществлено с помощью замен на остатки с другим зарядом или нейтрально заряженные остатки. Например, может быть осуществлена замена отрицательно заряженной аспарагиновой кислоты на положительно заряженный лизин. Такая замена с изменением заряда приводит к отталкиванию зарядов в этом сайте, а также к нарушению связывания и взаимодействия TFPI с FVIIa. В другом примере отрицательно заряженный D60 из FVII может быть заменен на основный остаток аргинина для нарушения электростатического взаимодействия с R20 из TFPI и введения неблагоприятного взаимодействия. Такие рационально просчитанные модификации могут быть осуществлены с одним или несколькими остатками FVII, для которых было установлено прямое или косвенное участие в контакте и взаимодействии с остатками TFPI.
В другом примере, рациональные модификации полипептида FVII могут быть осуществлены на основе известных функций доменов и районов полипептида. Например, домен Gla из FVII, как известно, ответственен за связывание FVIIa с фосфолипидами, такими как экспонированными на активированных тромбоцитах, хотя и очень слабое. Другие витамин-К-зависимые белки, такие как, например, FIX, FX, протромбин, белок С и белок S, также обладают Gla доменами, которые связывают белок с отрицательно заряженными фосфолипидами, во многих случаях с гораздо большей аффинностью, чем Gla домен FVII. Предполагается, что введение гетерологичного домена Gla в полипептид FVII увеличит сродство FVII к фосфолипидам. Например, FX (и FXa) проявляют относительно высокое сродство к фосфолипидам по сравнению с FVII (и FVIIa). Таким образом, в одном примере домен Gla из FX или достаточная для осуществления связывания фосфолипидов часть домена Gla из FX могут быть введены в полипептид FVII для создания химерного модифицированного FVII-полипептида.
При необходимости для разработки FVII-полипептидов могут быть использованы сочетания рациональных и эмпирических методов. Например, связывание содержащих Gla-домен полипептидов может быть оценено и проверено для определения, какие полипептиды демонстрируют высокое связывание и/или аффинность к фосфолипидам, таким как, например, расположенным на поверхности тромбоцитов. Альтернативно или дополнительно серия FVII-полипептидов, содержащих различные введенные гетерологичные Gla домены или достаточные их части, может быть проверена на наличие химерных полипептидов, проявляющих наибольшую аффинность и/или связывание с фосфолипидами.
2. Эмпирический подход (т.е. скрининг)
Модифицированные FVII-полипептиды также могут быть получены эмпирически, а затем тестированы или подвергнуты скринингу для выявления полипептидов, которые обладают конкретным видом активности или свойством. Например, модифицированные FVII-полипептиды могут быть получены мутацией одного или более аминокислотных остатков полипептида FVII с использованием любого метода, широко известного в данной области (см. также заявку США №2004/0146938). Такие модифицированные полипептиды FVII могут быть тестированы с помощью функциональных методов анализа на коагуляцию для определения результатов повышения коагулянтной активности. Полипептиды с увеличенной коагулянтной активностью могут быть далее тестированы для определения повышенной устойчивость к ингибиторам, таким как TFPI или АТ-III и/или повышенной связываемости с фосфолипидами, или улучшенние фармакокинетических свойств, таких как увеличение периода полураспада, с использованием любого из методов анализа, описанных в настоящем документе или известных специалистам в данной области. Примеры методов мутирования FVII включают методы, которые приводят к случайному мутагенезу всей последовательности, и методы, которые приводят к целенаправленному мутагенезу выбранного района или домена FVII. В одном примере, количество мутаций, введенных в полипептид составляет 1, 2, 3, 4, 5, 6,7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40 или более.
А. Случайный мутагенез
Может быть использован любой из ряда общих подходов для направленной модификации белка на основе мутагенеза. Для модификации полипептидов, таких как FVII, с целью достижения требуемого свойства, может быть использован любой из методов, отдельно или в комбинации. Такие методы включают случайный мутагенез, где аминокислоты в исходной белковой последовательности заменяются всеми (или группой) из 20 природных аминокислот (а также неприродными аминокислотами), одновременно заменяя одну или несколько аминокислот на различных аминокислотных позициях одной молекулы. Другой метод, ограниченный случайный мутагенез, вводит все или некоторые из 20 природных аминокислот (а также неприродные аминокислоты) или соответствующие изменения остатков ДНК. Изменение основывается на последовательности ДНК, а не на последовательности белка, осуществляется стохастическим или полустохастическим образом, соответственно, в пределах ограниченных или предопределенных районов белка, для которых заранее известно, что они принимают участие в изменяемой биологической активности. Примеры методов модификации протеаз описаны в заявке США №10/677,977, приведенной здесь в качестве ссылки в полном объеме. Кроме того, для модификации или изменения последовательности протеазного полипептида, такого как протеазный полипептид FVII, может быть использован любой метод, известный в уровне техники.
К методам случайного мутагенеза относятся, например, использование Е. coli XL I red, УФ-облучение, химическая модификация, например, с помощью дезаминирования, алкилирования, или использование аналогов оснований в качестве мутагенов, использование ПЦР методов, такого как ДНК шаффлинг, кассетный мутагенез, сайт-направленный случайный мутагенез, или ПЦР пониженной точности (см., например, заявку США №: 2006-0115874). Такие примеры включают, но не ограничиваются, химической модификацией гидроксиламином (Ruan, H., и др., (1997) Gene 188:35-39), использованием дНТФ аналогов (Zaccolo, М., и др. (1996) J. Mol. Biol. 255:589-603), или использованием коммерчески доступных наборов для случайного мутагенеза, таких как, например, основанный на ПЦР GeneMorph набор для случайного мутагенеза (Stratagene) или набор для диверсификации случайного мутагенеза (Clontech). Набор для диверсификации случайного мутагенеза позволяет выбрать желаемую частоту мутаций для данной последовательности ДНК (от 2 до 8 мутаций на 1000 пар оснований), варьируя количество марганца (Мn2+) или dGTP в реакционной смеси. Повышение уровня марганца сначала увеличивает частоту мутаций, а дальнейшее увеличение частоты мутаций обусловлено повышением концентрации dGTP. Еще более высокие уровни мутаций могут быть достигнуты путем проведения дополнительных циклов ПЦР.
b. Направленный мутагенез
Направленные мутации могут быть достигнуты введением одной или нескольких мутаций в заданный район последовательности гена, например в район протеазного домена, который обусловливает каталитическую активность, в район домена Gla, который связывает фосфолипиды, районы полипептида FVII, с которыми связываются ингибиторы или молекулы, такие как TFPI, AT-III и Zn+, как предусмотрено в настоящем документе, или в районы белка, которые взаимодействуют с факторами или рецепторами, участвующими в расщеплении FVIIa, такими как предусмотрено в настоящем документе. В одном примере одна или более аминокислот полипептида мутируют с использованием любого стандартного набора для единичного или множественного сайт-направленного мутагенеза, такого как, например, QuikChange (Stratagene). В другом примере одна или более аминокислот протеазы мутируют с помощью мутагенеза насыщения (Zheng и др. (2004) Nucl. Acids. Res. 32:115). В одном из примерных вариантов используется техника мутагенеза насыщения, при которой остаток (остатки) района или домена мутируют в каждую из 20 возможных природных аминокислот (а также неприродных аминокислот) (см., например, метод Kunkle, Current Protocols in Molecular Biology, John Wiley and Sons, Inc, Media Pa.). В такой технике может быть синтезирован мутагенный вырожденный олигонуклеотидный праймер, в котором содержатся рандомизированные нуклеотиды в нужном кодоне (кодонах), кодирующем выбранную аминокислоту (аминокислоты). Примеры схем рандомизации включают NNS- или NNK-рандомизацию, где N представляет собой любой нуклеотид, S представляет собой гуанин или цитозин и К представляет собой гуанин или тимин. Вырожденный мутагенный праймер отжигается на одноцепочечной матрице ДНК, и добавленная ДНК-полимераза синтезирует комплементарную матрице цепь. После лигирования двухцепочечная матрица ДНК переносится в Е. coli для амплификации.
В дополнительных примерах ориентированный мутагенез может быть ограничен аминокислотами, которые определены как горячие точки на начальных этапах скрининга. Например, после селекции модифицированных полипептидов FVII из комбинаторных библиотек случайного мутагенеза может наблюдаться непропорциональное количество мутаций в конкретных позициях или районах. Они обозначаются как "горячие точки", и их можно наблюдать после более чем одного цикла мутагенеза и скрининга. Затем могут быть проведены функциональные анализы модифицированных полипептидов FVII для установления, коррелируют ли мутации в горячих точках с одним или более желаемьм свойством или активностью. Если связь между горячими точками и желаемой активностью или свойством установлена, то ориентированный мутагенез может быть использована непосредственно для дальнейшего мутагенеза этих горячих точек. Эта стратегия позволяет достичь более разнообразного и глубокого мутагенеза в отдельных указанных позициях, в отличие от более поверхностного мутагенеза, который происходит при случайном мутагенезе полипептидной последовательности. Например, мутагенез насыщения может быть использован для мутирования "горячих точек" с помощью олигонуклеотидов, содержащих NNt / g или NNt / с на этих позициях.
с. Скрининг
Модифицированные полипептиды могут быть протестированы в скрининговых исследованиях индивидуально или могут быть протестированы в наборе, например в библиотеках. В одном примере варианты FVII-полипептидов случайно полученны мутагенезом и клонированы индивидуально. Затем осуществляется индивидуальная оценка активности по каждому отдельному мутанту белка после его экспрессии и измерения соответствующей активности. В некоторых примерах отдельные клоны могут анализироваться в определенных последовательностях, в которых они физически отделены друг от друга так, что идентичность каждого отдельного полипептида известна на основе его местонахождения в последовательности. Например, если каждый из них является единственным продуктом независимой реакции мутагенеза, конкретные мутации могут быть легко определены без необходимости секвенирования. Альтернативно, может быть осуществлено секвенирование полученных модифицированных полипептидов для выявления тех мутаций, которые ответственны за их активность.
В другом примере модифицированные полипептиды FVII могут подвергнуться скринингу в виде совокупностей или в библиотеке. Например, библиотека полипептидов FVII может быть проявлена с помощью генетического пакета для скрининга, включая, но не ограничиваясь, каким-либо репликативным вектором, например фагом, вирусом или бактерией, которые могут проявлять полипептидный фрагмент. Множество полипептидов проявляются с помощью генетического пакета посредством связывания и/или взаимодействия полипептидов с целевым полипептидом. Примеры генетических пакетов включают, но не ограничиваются, бактериофагами (см., например, Clackson др., (1991) Making Antibody Fragments Using Phage Display Libraries, Nature, 352:624-628; Glaser и др., (1992) Antibody Engineering by Condon-Based Mutagenesis in a Filamentous Phage Vector System, J. ImmoNol, 149:3903 3913; Hoogenboom др. (1991) Multi-Subunit Proteins on the Surfase of Filamentous Phage: Methodologies for Displaing Antibody (Fate)Heavy and 30 Light Chains, Nucleic Acid Res., 19:4133-41370), бакуловирусами (см., например, Boublik и др., (1995) Eukaryotic Virus Display: Engineering the Major Surface Glycoproteins of the Autographa California Nuclear Polyhedrosis Virus (ACNPV) for the Presentation of Foreign Proteins on the Virus Surface, Bio/Technology, 13:1079-1084), бактериями и другими векторами, подходящими для выявления белка, например выявления протеазы фагом. Примеры интересующих бактериофагов включают, но не ограничиваются, фагом Т4, фагом М13 фагом HI. Генетические пакеты могут быть необязательно амплифицированы, например, в бактериальном хозяине.
Метод фагового дисплея известен специалистам в данной области и описан, например, Ladner др., в патенте США №5223409; Rodi и др., (2002) Curr. Opin. Chem. Biol. 6:92-96; Smith (1985) Science 228:1315-1317; WO 92/18619; WO 91/17271; WO 92/20791; WO 92/15679; WO 93/01288; WO 92/01047; WO 92 / 09690; WO 90/02809; de Haard и др., (1999) J. Biol. Chem 274:18218-30; Hoogenboom и др., (1998) Immu-Notech.Nology 4:1-20; Hoogenboom и др., (2000) ImmoNol Today 2:371-8; Fuchs и др., (1991) Bio / Technology 9:1370-1372; Hey и др., (1992) Hum Antibod Hybridomas 3:81-85; Huse и др., (1989) Science 246:1275-1281; Griffiths и др., (1993) EMBO J 12:725-734; Hawkins и др., (1992) JMol Biol 226:889-896; Clackson и др., (1991) Nature 352:624-628; Gram и др., (1992) PNAS 89:3576-3580; Garrard и др., (1991) Bio/Technoogy 9:1373-1377; Rebar и др., (1996) Methods Enzymol. 267: 129-49; Hoogenboom и др., (1991) Nuc Acid Res 19:4133-4137; и Barbas и др., (1991) PNAS 88:7978-7982. Подходящие для фагового дисплея нуклеиновые кислоты, например, фаговые векторы, известны в уровне техники (см., например, Andris-Widhopf и др., (2000) J Immunol Methods, 28: 159-81, Armstrong и др., (1996) Academic Press, Kay и др., Ed. cc.35-53; Corey и др., (1993) Gene 128(1):129-34; Cwiria и др., (1990) Proc Nati Acad Sd USA 87(16):6378-82; Fowlkes и др., (1992) Biotechniques 13(3):422-8; Hoogenboom и др., (1991) Nuc Acid Res 19(15):4133-7; McCafferty и др., (1990) Nature 348(6301):552-4; McConnell и др. (1994) Gene 151 (1-2):115-8; Scott and Smith (1990) Science 249(4967):386-90).
Библиотеки вариантов полипептидов FVII для скрининга также могут быть экспрессированы на поверхности клеток, например прокариотических или эукариотических клеток. Примеры клеток для экспрессии на клеточной поверхности включают, но не ограничиваются, бактериями, дрожжами, клетками насекомых, клетками птиц, клетками растений и клетками млекопитающих (Chen и Georgiou (2002) Biotechnol Bioeng 79: 496-503). В одном примере бактериальные клетки для экспрессии представляют собой Escherichia Coli.
Варианты полипептидов могут быть экспрессированы как химерные белки с белком, который экспрессируется на поверхность клетки, таким как мембранный белок или белок, связанный с клеточной поверхностью. Например, вариант протеазы может быть экспрессирован в E.coli в виде химерного белка с белком наружной мембраны кишечной палочки (например, ОтрА), генно-инженерной гибридной молекулы основного липопротеина Е. coli (LPP) и белка внешней мембраны ОтрА или белка, связанного с клеточной поверхностью (например, субъединицы пилей и жгутиков). Как правило, выявление гетерологичных пептидов и белков при использовании бактериальных белков внешней мембраны достигается путем генетической вставки в разрешенные сайты белка-носителя. Экспрессия гетерологичного пептида или белка зависит от структурных особенностей вставленного домена белка, так как пептид или белок находится в менее естественной конформации при вставке в разрешенный сайт по сравнению с присоединением к N- или С-концу белка. Для улучшения экспрессии гибридного белка могут быть осуществлены модификации гибридного белка, такие как включение гибкой линкерной или спейсерной пептидной последовательности или модификации бактериального белка (например, в результате мутации, вставки или делеции в аминокислотной последовательности). Ферменты, такие как (β-лактамаза и Сех экзоглюконаза из Cellulomonas fimi, были успешно экспрессированы в виде Lpp-OmpA гибридного белка на поверхности кишечной палочки (Francisco J.A. и Georgiou G. Ann N Y Acad Sci 745:372-382 (1994) и Georgiou G. и др. Protein Eng. 9:239-247 (1996)). Другие пептиды, содержащие 15-514 аминокислот, были проявлены во второй, третьей и четвертой петлях на внешней поверхности OmpA (Samuelson и др., J. Biotechnol. 96: 129-154 (2002)). Таким образом, белки внешней мембраны могут нести и проявлять гетерологичные генные продукты на внешней поверхности бактерий.
Для скрининга библиотеки вариантов полипептида также возможно использование других форматов проявления. Примеры других форматов проявления включают слияние нуклеиновых кислот с белками, рибозим дисплей (см., например, Hanes и Pluckthun (1997) Proc. Natl. Acad. Sci. U.S.A. 13:4937-4942), дисплей с гранулами (Lam, К. S. и др.. Nature (1991) 354, 82-84;, К. S. и др. (1991) Nature, 354, 82-84; Houghten, R. А. и др. (1991) Nature, 354, 84-86; Furka, A. и др. (1991) Int. J. Peptide Protein Res. 37,487-493; Lam, К. S., и др., (1997) Chem. Rev., 97,41 1-448; U.S. Published Patent Application 2004-0235054) и белковые матрицы (см., например, Cahill (200L) J. 1 Immunol. Meth. 250:81-91, WO 01/40803, WO 99/51773, и US2002-0192673-A1).
В других конкретных случаях может быть выгодно присоединить варианты полипептидов, или фаговые библиотеки, или клетки, экспрессирующие варианты полипептидов, к твердой поддержке. Например, в некоторых примерах, клетки, экспрессирующие варианты полипептидов FVII, могут быть естественно адсорбированы на гранулах так, что совокупность гранул содержит одну клетку на гранулу (Freeman и др., Biotechnol. Bioeng. (2004) 86:196-200). После иммобилизации на стеклянной подложке микроколонии могут выращиваться, подвергаться скринингу с помощью хромогенного или флуорогенного субстрата. В другом примере варианты полипептидов FVII или фаговые библиотеки или клетки, экспрессирующие варианты протеазы, могут быть высеяны на титровальные планшеты и иммобилизированы.
Для идентификации модифицированных полипептидов FVII, которые демонстрируют повышенную коагулянтную активность, модифицированные FVII-полипептиды подвергаются скринингу по отдельности или в библиотеке и тестируются с помощью функциональных методов анализа для определения полипептидов, проявляющих повышенную устойчивость к ингибиторам, таким как TFPI и AT-III, повышенную способность связывать фосфолипиды активированных тромбоцитов, которые проявляют увеличенное время полураспада и/или увеличенную каталитическую активность. Такие анализы описаны в настоящем документе или известны специалистам в данной области. Например, модифицированные полипептиды FVII тестировались на протеолитическую активность. FVII-полипептиды, отдельно или в присутствии ТФ, инкубировали с хромогенным субстратом в различных концентрациях, таким как пептидный субстрат Spectrozyme FVIIa (CH3SO2-D-CHA-But-Arg-pNA.AcOH). Расщепление субстрата определяется по абсорбции, и скорость гидролиза субстрата определяется методом линейной регрессии с помощью доступного программного обеспечения. В другом примере устойчивость модифицированных полипептидов FVII к TFPI или AT-III оценивается после инкубации ингибитора с полипептидом FVII, который, в некоторых случаях, предварительно инкубировали с ТФ. Затем активность FVII измеряли с помощью одного или нескольких анализов на активность или коагуляцию, известных в уровне техники, и ингибирование TFPI или AT-III оценивали путем сравнения активности FVII-полипептидов, которые инкубировали с ингибитором, и с активностью FVII-полипептидов, которые не инкубировали с ингибитором.
Конкретные мутации полипептида могут быть определены с использованием обычных методов рекомбинантной ДНК, таких как секвенирование. В одном варианте комбинация удачных мутаций может быть осуществлена для дальнейшего улучшения свойств и/или коагулянтной активности модифицированного FVII-полипептида.
3. Выбор вариантов FVII
Варианты FVII-полипептидов, разработанные и идентифицированные по какому-либо одному или нескольким подходам, описанных выше, могут быть выбраны для определения кандидатов FVII-полипептидов, которые демонстрируют заданные свойства или активности. Выбор варианта полипептида FVII основан на: 1) во-первых, тестировании варианта полипептида на конкретные виды активности или свойства, которые были модифицированы (т.е. устойчивость к TFPI, устойчивость к AT-III, связывание Zn2+, каталитическая активность, время полураспада, связывание и/или аффинность к фосфолипидам, и 2) во-вторых, тестировании вариантов полипептида на сохранение активности FVII, необходимой для гемостаза и коагуляции. Активности FVII, необходимые для коагуляции, включают, например, ферментативную, протеолитическую или каталитическую активности, такие как осуществление активации фактора Х (FX) или активации фактора IX (FIX). Другие активности FVII, которые могут быть оценены, включают, но не ограничиваются, иммуногенностью, способностью связывать тканевый фактор, фактор Х или фактор IX, и способностью к связыванию с фосфолипидами. Таким образом, модифицированные полипептиды FVII, такие как любые содержащиеся в настоящем документе или идентифицированные по методам, представленным здесь, должны сохранить определенный уровень, повышенный или пониженный, активности FVII, необходимой для коагулянтной активности. Стандартные анализы для осуществления каждой из двух упомянутых выше оценок, известные в уровне техники или описанные здесь ниже, могут быть выполнены in vitro или in vivo. Например, протеолитическая и каталитическая активности FVII могут быть оценены с помощью различных методов, включая измерение расщепления синтетического субстрата или измерение активации фактора Х (см., например, примеры 4, 5 и 11 ниже), а устойчивость к TFPI или AT-III может быть оценена с помощью анализов на ингибирование TFPI или AT-III, соответственно (см., например, примеры 7,12 и 16). Кроме того, также могут быть использованы in vivo анализы на прокоагулянтную активность, такие как описано, например, в примерах 8 и 14.
Общим эффектом любого варианта полипептида FVII является проявление прокоагулянтной активности. Например, вариант полипептида может обладать повышенной устойчивостью к TFPI, устойчивостью к AT-III, периодом полураспада и/или способностью связывать и/или аффинностью к фосфолипидам, а также проявлять повышенную каталитическую активность. Такой полипептид FVII будет выбран в качестве кандидата варианта FVII с повышенной коагулянтной активностью.
В некоторых случаях, однако, полипептид FVII, модифицированный для улучшения коагулянтной активности за счет одной или нескольких модификаций, таких как повышенная устойчивость к TFPI, повышенная устойчивость к AT-III, увеличение периода полураспада или увеличение способности связывать и/или аффинности к фосфолипидам, может также проявлять в результате конкретной модификации сниженную каталитическую активность, или другую сниженную активность, необходимую для свертывания. Эти последствия могут возникнуть в результате, например, конформационных изменений в модифицированных полипептидах FVII, которые мешают связыванию с другой молекулой, конформационных изменений, которые приводят к изменению в активном сайте, или аминокислотных замен, которые непосредственно связаны с одним или более аминокислотными остатками, которые ответственны за взаимодействие с другой молекулой.
Таким образом, в другом примере, вариант полипептида, который обладает повышенной устойчивостью к TFPI, устойчивостью к AT-III, увеличенным периодом полураспада и/или аффинностью к фосфолипидам, может проявлять сопутствующее снижение каталитической активности. Уровень снижения активности FVII, такой как каталитическая активность, который был бы приемлем для улучшения коагулянтной активности, зависит от сопутствующего улучшения свойств, которые были модифицированы для улучшения (например, повышенной устойчивости к TFPI) и могут быть эмпирически определены. Таким образом, результаты такой оценки, приведенной выше, могут быть сбалансированы для идентификации вариантов полипептидов, которые демонстрируют улучшенные свойства, и в то же время сохраняют, по крайней мере, достаточную активность FVII, т.е. каталитическую активность, вызывающую коагуляцию. Обычно, чем значительнее улучшения свойства, подвергнутого модификации (т.е. чем больше увеличение устойчивости к TFPI), тем значительнее уменьшение протеолитической или каталитической активности.
Например, полипептид FVII, который проявляет 100-кратное увеличение устойчивости к TFPI или AT-III, может проявлять снижение протеолитической или каталитической активности, которая уменьшилась, точно или приблизительно, в 1,5 раза, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90 и более раз по сравнению с активностью немодифицированного полипептида FVII, и при этом является реальным кандидатом на повышение коагулянтной активности. И наоборот, если устойчивость к TFPI или AT-III увеличивается в 10 раз, уровень снижения каталитической активности для поддержания общей прокоагулянтной активности, как правило, будет составлять, точно или приблизительно, 1, 5, 2, 3, 4, 5, 6, 7, 8, 9 и более раз по сравнению с каталитической активностью немодифицированного полипептида. Специалист в данной области может оценить сопутствующие изменения устойчивости к TFPI, устойчивости к АТ-III, связывании с активированными тромбоцитами, периода полураспада и протеолитической и каталитической активности, или любой другой активности или свойства для определения полезности модифицированного полипептида FVII в качестве прокоагулянтного лекарственного средства, например, для лечения эпизодов кровотечения при больных гемофилией.
F. Получение FVII-полипептидов
FVII-полипептиды, в том числе модифицированные FVII-полипептиды, или домены FVII или домены других витамин-К-полипептидов могут быть получены методами очистки белков и рекомбинантной экспрессии белка, хорошо известными в уровне техники. Может быть использован любой метод, известный специалистам в данной области для идентификации нуклеиновых кислот, которые кодируют желаемые гены. Любой метод, известный в уровне техники, может быть использован для получения полной (например, охватывающей весь кодирующий район) к ДНК или клонов геномной ДНК, кодирующих полипептиды FVII или другие витамин-К-полипептиды, из клеточного или тканевого источников, таких как, например, печень. Модифицированные полипептиды FVII могут быть созданы, как описывалось выше, например, путем сайт-направленного мутагенеза.
FVII может быть клонирован или выделен с использованием любых имеющихся методов, известных в данной области для клонирования и выделения молекул нуклеиновой кислоты. Такие методы включают ПЦР-амплификацию нуклеиновых кислот и скрининг библиотек, включая скрининг гибридизацией нуклеиновых кислот, скрининг на основе антител и скрининг по активности.
Методы амплификации нуклеиновых кислот могут быть использованы для выделения молекул нуклеиновой кислоты, кодирующих полипептиды FVII, включая, например, полимеразную цепную реакцию (ПЦР). Материал, содержащий нуклеиновые кислоты, может быть использован в качестве исходного материала, из которого могут быть выделены кодирующие FVII молекулы нуклеиновой кислоты. Например, препараты ДНК и РНК, клеточные экстракты, экстракты тканей (например, печени), образцы жидкостей (например, крови, сыворотки, слюны), образцы, взятые у здоровых и/или больных субъектов, могут быть использованы в методах амплификации. Библиотеки нуклеиновых кислот также могут быть использованы в качестве источника исходного материала. Для амплификации кодирующих FVII молекул могут быть созданы праймеры. Например, праймеры могут быть разработаны на основе экспрессированных последовательностей, с которыми образуется FVII. Праймеры могут быть разработаны с помощью обратной трансляции аминокислотной последовательности FVII. Молекулы нуклеиновых кислот, образованные в результате амплификации, могут быть секвенированы для подтверждения кодирования полипептида FVII.
К кодирующей FVII молекуле нуклеиновой кислоты может быть присоединена дополнительная нуклеотидная последовательность, включая линкерную последовательность, содержащую сайты узнавания эндонуклеазы рестрикции для клонирования синтетических генов в векторе, например, векторе экспрессии белка или векторе, предназначенном для амплификации последовательности ДНК, кодирующей белок. Кроме того, с кодирующей FVII молекулой нуклеиновой кислоты могут быть оперативно связаны дополнительные нуклеотидные последовательности, такие как функциональные элементы ДНК. Примеры таких последовательностей включают, но не ограничиваются, промотерными последовательностями, предназначенными для осуществления внутриклеточной экспрессии белка, последовательностями секреции, предназначенными для осуществления секреции белков. Дополнительные нуклеотидные последовательности, например последовательности, определяющие связывающие сайты белка, также могут быть связаны с кодирующими FVII молекулами нуклеиновой кислоты. Такие районы включают, но не ограничиваются, последовательностями, предназначенными для содействия поглощению FVII конкретными клетками-мишенями или улучшения фармакокинетики синтетических генов иным образом.
Идентифицированные и выделенные молекулы нуклеиновой кислоты затем могут быть вставлены в соответствующий вектор клонирования. Может быть использовано большое количество систем вектор-хозяин, известных в данной области. Подходящие векторы включают, но не ограничиваются, плазмидами или модифицированными вирусами, но векторная система должна быть совместима с используемыми клетками-хозяевами. Такие векторы включают, но не ограничиваются, бактериофагами, такими как производные бактериофага лямбда, или плазмидами, такими как производные pBR322 или pUC или вектор Bluescript (Stratagene, La Jolla, CA). Включение в вектор клонирования может, например, быть достигнуто путем лигирования фрагментов ДНК в векторе клонирования, который имеет комплементарные липкие концы. Инсерция может осуществляться с использованием ТОРО клонирующего вектора (Invitrogen, Carlsbad, CA). Если комплементарные сайты рестрикции, используемые для фрагментации ДНК, не присутствуют в векторе клонирования, концы молекулы ДНК могут быть ферментативно модифицированы. Кроме того, любые желаемые сайты могут быть получены путем лигирования нуклеотидных последовательностей (линкеров) к концам ДНК, эти лигированные линкеры могут содержать специфические химически синтезированные олигонуклеотиды, кодирующие распознаваемые рестриктазой последовательности. В альтернативном методе расщепленный вектор и ген белка FVII могут быть модифицированы гомополимерным хвостом. Рекомбинантные молекулы могут быть введены в клетки хозяина с помощью, например, трансформации, трансфекции, инфекции, электропорации и сонопорации, так что генерируется большое количество копий последовательностей гена.
В конкретных вариантах, трансформация клеток хозяина молекулами рекомбинантной ДНК, которые включают изолированный ген белка FVII, кДНК или синтезированные последовательности ДНК, вызывает образование несколько копий этого гена. Таким образом, ген может быть получен в больших количествах в растущих трансформантах, рекомбинантные молекулы ДНК могут быть изолированы из трансформантов и, при необходимости, встроенный ген может быть извлечен из изолированной рекомбинантной ДНК.
1. Векторы и клетки
Для рекомбинантной экспрессии одного или нескольких FVII белков нуклеиновые кислоты, содержащие часть нуклеотидной последовательности, кодирующей белок FVII, или содержащие всю последовательность, могут быть вставлены в соответствующий вектор экспрессии, то есть вектор, который содержит все необходимые элементы для транскрипции и трансляции вставленной кодирующей белок последовательности. Пример такого вектора представляет собой любой вектор экспрессии в млекопитающих, такой как, например, pCMV. Необходимые транскрипционные и трансляционные сигналы также могут содержаться в нативном промотере FVII гена, и/или его фланкирующих районах.
Также предусмотрены векторы, содержащие нуклеиновые кислоты, кодирующие FVII или модифицированный FVII. Также предусмотрены клетки, содержащие векторы. Клетки включают эукариотические клетки и прокариотические клетки, а векторы представляют собой любые пригодные для использования в настоящем документе векторы.
Предусмотреные прокариотические и эукариотические клетки включают клетки эндотелия, содержащие векторы. Такие клетки включают бактериальные клетки, дрожжевые клетки, клетки грибов, археи, клетки растений, клетки насекомых и клетки животных. Клетки используются для получения полипептидов FVII или модифицированных полипептидов FVII посредством выращивания выше описанных клеток в условиях, в которых закодированный белок FVII экспрессируется в клетке, и выделения экспрессированного FVII белка. Для целей настоящего изобретения FVII может выделяться в среду.
В одном варианте предусмотрены векторы, содержащие последовательность нуклеотидов, которая кодирует полипептид, который обладает активностью FVII и содержит часть полипептида FVII, или весь полипептид, или несколько их копий. Векторы могут быть выбраны для экспрессии полипептида FVII или модифицированного полипептида FVII в клетке, или для экспрессии белка FVII в виде секретируемого белка. Когда FVII экспрессируется, нуклеиновая кислота связывается с нуклеиновой кислотой, кодирующей сигнал секреции, такой как сигнальная последовательность α-фактора спаривания Saccharomyces cerevisiae или ее частью, или с нативной сигнальной последовательностью.
Множество систем вектор-хозяин могут быть использованы для экспрессии последовательности, кодирующей белок. Они включают, но не ограничиваются, клеточными системами млекопитающих, зараженными вирусом (например, вирусом коровьей оспы, аденовирусом и другими вирусами); клеточными системами насекомых, зараженными вирусом (например, бакуловирусом);
микроорганизмами, такими как дрожжи, содержащими дрожжевые векторы; или бактериями, трансформированными бактериофагами, ДНК, плазмидной ДНК, или космидной ДНК. Экспрессируемые элементы векторов варьируют по преимуществам и особенностям. В зависимости от используемой системы хозяин-вектор может быть использован любой элемент из числа подходящих транскрипционных и трансляционных элементов.
Могут быть использованы любые методы, известные специалистам в данной области, для вставки фрагментов ДНК в вектор для создания векторов экспрессии, содержащих рекомбинантный ген, содержащий необходимые транскрипционные/трансляционные сигналы контроля и кодирующую белок последовательность. Эти методы могут включать in vitro рекомбинацию ДНК и синтетические методы, и in vivo рекомбинацию (генетическую рекомбинацию). Экспрессия последовательности нуклеиновой кислоты, кодирующей полипептид FVII или модифицированный полипептид FVII, или их домены, производные, фрагменты или гомологи, может регулироваться второй последовательностью нуклеиновой кислоты, так что гены или их фрагменты экспрессируются в хозяине, трансформированном рекомбинантной молекулой ДНК (молекулами). Например, экспрессия белка может контролироваться любым промотером/энхансером, известным в уровне техники. В конкретном варианте, промотер не является нативным для гена белка FVII. Промотеры, которые могут быть использованы, включают, но не ограничиваются, ранним промотером SV40 (Bemoist и Chambon, Nature, 290:304-310 (1981)), промотером, содержащимся в 3' протяженном терминальном повторе вируса саркомы Рауса (Yamamoto и др.. Cell 22:787-197 (1980)), промотером тимидинкиназы герпеса (Wagner и др., Proc. Natl. Acad. Sci. USA 75:1441-1445 (1981)), регуляторными последовательностями генов металлотионеина (Brinster и др.. Nature, 296:39-42 (1982)); промотором прокариотических векторов экспрессии, такими как промотер р-лактамазы (Jay и др., (1981) Proc. Natl. Acad. Sci. USA 75:5543) или tac промотера (DeBoer и др., Proc. Natl. Acad. Sci. USA 50:21-25 (1983)); см. также "Useful Proteins for Recombinant Bacteria" в Scientific American 242:79-94 (1980)); промоторами растительных векторов экспрессии, содержащими промотер нопалин-синтетазы (Herrar-Estrella и др., Nature, 303:209-213 (1984)) или промотор 35S РНК вируса мозаики цветной капусты (Garder др. Nucleic Acids Res. 9:2871 (1981)), и промотер фотосинтетического фермента рибулозобисфосфаткарбоксилазы (Herrera-Estrella и др., Nature 370:115-120 (1984)); промотерными элементами дрожжей и других грибов, такими как Gal4 промотер, промотер алкогольдегидрогеназы, промотер фосфоглиролкиназы, промотер щелочной фосфатазы, и следующие районы контроля транскрипции животных, которые демонстрируют тканеспецифичность и были использованы в трансгенных животных: район контроля гена эластазы I, который активен в ацинарных клетках поджелудочной железы (Swift и др., Cell 35:639-646 (1984); Omitz и др. Cold Spring Harbor Symp.Quant. Biol. 50:399-409 (1986); MacDonald, Hepatology 7:425-515 (1987)); район контроля гена инсулина, который активен в бета-клетках поджелудочной железы (Hanahan др., Nature 375:1 15-122 (1985)), район контроля гена иммуноглобулина, который активен в лимфоидных клетках (Grosschedl и др., Cell 35:647-658 (1984); Adams и др., Nature 375:533-538 (1985), Alexander и др., Mol. Cell Biol. 7:1436-1444 (1987)), контролирующим районом вируса опухоли молочных желез мышей, который активен в клетках яичек, молочной железы, лимфоидной ткани и тучных клетках (Leder и др. Cell 45:485-495 (1986)), контролирующим районом гена альбумина, который активен в печени (Pinckert и др., Genes and Devel. 7:268-276 (1987)), контролирующим районом гена альфа-фетопротеина, который активен в печени (Krumlauf и др., Mol Cell Biol 5:1639-1648 (1985); Hammer и др. Science 235:53-58 1987)), контролирующим районом гена альфа-1 антитрипсина, который активен в печени (Kelsey и др., Genes and Devel 7:161-171 (1987)), контролирующим районом гена бета глобина, который активен в миелоидных клетках (Mogram и др., Nature 375:338-340 (1985); Kollias др., Cell 45:89-94 (1986)), контролирующим районом гена основного белка миелина, который активен в олигодендроцитах головного мозга (Readhead и др., Cell 48:703-112 (1987)), контролирующим районом гена легкой цепи-2 миозина, который активен в скелетных мышцах (Sani, Nature 314:283-286 (1985)), и контролирующим районом гена гонадотропного рилизинг гормона, который активен в гонадотропном районе гипоталамуса (Mason и др. Science 234:1372-1378 (1986)).
В конкретном варианте используется вектор, содержащий промотор, оперативно связанный с нуклеиновой кислотой, кодирующей полипептид FVII или модифицированный полипептид FVII, или его домен, фрагмент, производное или гомолог, одну или более точек начала репликации, и, необязательно, один или более специфический маркер (например, ген устойчивости к антибиотикам). Векторы и системы для экспрессии полипептидов FVII включают хорошо известные векторы Pichia (доступен, например, от Invitrogen, San Diego, CA), в частности, предназначенные для секреции кодируемых белков. Примеры плазмидных векторов для экспрессии в клетках млекопитающих включают, например, pCMV. Примеры плазмидных векторов для трансформации клеток Е. coli включают, например, вектор экспрессии pQE (доступен Qiagen, Valencia, CA, см. также литературу, опубликованную Qiagen с описанием системы). pQE вектор обладает промотером фага Т5 (распознаваемого РНК-полимеразой Е. coli) и двойным lac операторным модулем репрессии, обеспечивающим жестко регулируемый, высокий уровень экспрессии рекомбинантных белков в E.coli, синтетический сайт связывания рибосомы (RBS II) для эффективной трансляции, последовательность, кодирующую 6xHis метку, t0 и Т1 терминаторы транскрипции, CoIEl точку начала репликации, а также ген бета-лактамазы для устойчивости к ампициллину. PQE векторы позволяют включить 6xHis метку на N- или С-конце рекомбинантного белка. Такие плазмиды включают pQE 32, pQE 30, и pQE 31, в которых предусмотрено множество сайтов клонирования для всех трех рамок считывания и предусмотрена возможность экспрессии N-терминально 6xHis-меченого белка. Другие примеры плазмидных векторов для трансформации клеток Е. coli, включают, например, рЕТ векторы экспрессии (см. патент США 4952496; доступный из NOVAGEN, Madison, WI; см. также литературу, опубликованную Novagen с описанием системы). Такие плазмиды включают рЕТ 11a, в которой содержится промотер Т7lас, Т7 терминатор, индуцируемый lac оператор Е. coli, и ген lac репрессора; рЕТ -12а-с, который содержит Т7 промотер, Т7 терминатор и Е. coli сигнал секреции ompT; и рЕТ-15b и pET-19b (Novagen, Madison, WI), которые содержат His-TagTM лидерную последовательность для использования в очистке с His колонкой и сайт расщепления тромбином, что позволяет осуществить расщепление после очистки на колонке, Т7-lас промотерный район и Т7 терминатор.
2. Системы экспрессии
FVII-полипептиды (модифицированные и немодифицированные) могут быть получены любыми методами, известными в данной области техники, для получения белка, в том числе in vitro и in vivo методами, например, введением молекул нуклеиновых кислот, кодирующих FVII в клетки-хозяева, в животного-хозяина и экспрессией молекул нуклеиновой кислоты, кодирующих FVII in vitro. FVII и модифицированные полипептиды FVII могут быть экспрессированы в любом организме, подходящем для создания требуемого количества и формы полипептида FVII, необходимые для введения и лечения. Экспрессирующие хозяева включают прокариотические и эукариотические организмы, такие как Е.соli, дрожжи, растения, клетки насекомых, клетки млекопитающих, включая человеческие клеточные линии и трансгенных животных. Экспрессирующие хозяева могут отличаться по уровням образования белка, а также типам посттрансляционных модификаций, которые присутствуют в экспрессированном белке. Выбор экспрессирующего хозяина может быть сделан на основе этих и других факторов, таких как нормативные ограничения и соображения безопасности, издержки производства, а также необходимость и методы очистки.
Экспрессия в эукариотических хозяевах может включать экспрессию в дрожжах, таких как Saccharomyces cerevisiae и Pichia Pastoria, клетках насекомых, таких как клетки дрозофилы и чешуекрылых, растениях и растительных клетках, таких как табак, кукуруза, рис, водоросли и ряска. Эукариотические клетки для экспрессии также включают линии клеток млекопитающих, такие как клетки яичника китайского хомяка (СНО), клетки почек детенышей хомяков (ВНК). Эукариотические экспрессирующие хозяева также включают трансгенных животных, например, включают получение белка в сыворотке, крови, молоке и яйцах. Трансгенные животные для получения полипептидов FVII дикого типа известны в уровне техники (опубликованные заявки США №№20020166130 и 20040133930) и могут быть адаптированы для производства модифицированного полипептида FVII, предусмотренного в настоящем документе.
Многие векторы экспрессии доступны и известны специалистам в данной области для экспрессии FVII. Выбор вектора экспрессии зависит от выбора хозяина системы экспрессии. Такой выбор вполне может быть осуществлен специалистом в данной области. В общем, вектор экспрессии может включать промотеры транскрипции и, необязательно, энхансеры, сигналы трансляции, и сигналы терминации транскрипции и трансляции. Векторы экспрессии, которые используются для стабильной трансформации, как правило, обладают маркером для отбора, который позволяет выбрать и эксплуатировать трансформированные клетки. В некоторых случаях точка начала репликации может быть использована для амплификации копий вектора в клетках.
FVII или модифицированные полипептиды FVII также могут быть использованы или экспрессированы как гибридные белки. Например, гибридизация может быть осуществлена для добавления дополнительных функций полипептиду. Примеры гибридных белков включают, но не ограничиваются, гибридами с сигнальными последовательностями, такими как метки для локализации, например, His6 метка или myc метка, или метки для очистки, например, GST, и последовательностями для направления секреции белка и/или встраивания в мембрану.
В одном варианте полипептид FVII или модифицированный полипептид FVII может быть экспрессирован в активной форме, в которой активация достигается самоактивацией полипептида после секреции. В другом варианте протеаза экспрессируется в неактивной, зимогенной форме.
Методы получения FVII-полипептидов могут включать коэкспрессию одного или нескольких дополнительных гетерологичных полипептидов, которые могут содействовать генерации полипептида FVII. Например, такие полипептиды могут участвовать в посттрансляционном процессинге полипептида FVII. Примеры полипептидов включают, но не ограничиваются, пептидазами, которые помогают расщеплять предшественников последовательностей FVII, такие как пропептидные последовательности, и ферментами, которые участвуют в модификации полипептидов FVII, например, путем гликозилирования, гидроксилирования, карбоксилирования, или фосфорилирования. Примеры пептидаз, которые могут коэкспрессироваться с FVII, включают РАСЕ / Фурин (или PACE-SOL), которая помогает расщеплять пропептидную последовательность FVII. Примеры белков, которые помогают карбоксилировать полипептид FVII, включают варфарин-чувствительный фермент витамин-К-2,3-эпоксидредуктазу (VKOR), которая образует восстановленный витамин К для использования витамин-К-зависимой у-карбоксилазой в качестве кофактора (Wajih и др., J. Biol. Chem. 280 (36) 31603-31607). Субъединица этого фермента, VKORC1 может быть коэкспрессирована с модифицированным полипептидом FVII для увеличения γ-карбоксилирования. Один или несколько дополнительных полипептидов могут быть экспрессированы с одного вектора экспрессии, с которого экспрессируется и полипептид FVII, или с другого вектора.
а. Прокариотическая экспрессия
Прокариоты, особенно Е. coli, обеспечивают систему для получения больших количеств FVII (см., например, Platis и др., (2003) Protein Exp.Purif. 31 (2): 222-30; и Khalizzadeh и др. (2004) J. Ind. Microbiol. Biotechnol. 31 (2): 63-69). Трансформация Е. coli, является простым и быстрым способом, хорошо известным специалистам в данной области. Векторы экспрессии для кишечной палочки могут содержать индуцибильные промотеры, которые являются полезными для индуцирования высоких уровней экспрессии белка и для экспрессии белков, которые демонстрируют некоторую токсичность для клеток хозяина. Примеры индуцибильных промотеров включают lac промотер, trp промотер, гибридный tac промотер, Т7 и SP6 РНК промотеры и регулируемый температурой λPL промотер.
FVII может быть экспрессирован в цитоплазматической среде Е. coli. Цитоплазма является восстановительной средой, и для некоторых молекул это может привести к образованию нерастворимых телец включения. Восстанавливающие агенты, такие как дитиотреитол и β-меркаптоэтанол, и денатуранты (например, такие как гуанидин-НСl и мочевина) могут быть использованы для ресолюбилизации белков. Альтернативный подход представляет собой экспрессию FVII в периплазматическом пространстве бактерий, в котором поддерживается окислительная среда и присутствуют шаперонин-подобные белки и дисульфидизомеразы, что приводит к получению растворимого белка. Как правило, лидерная последовательность, которая направляет белок в периплазматическое пространство, гибридизуется с белком, который будет экспрессирован. Лидерная последовательность затем удаляется сигнальной пептидазой внутри периплазматического пространства. Примеры лидерных последовательностей, обусловливающих периплазматическую ориентацию, включают реlВ лидер из гена пектат-лиазы и лидер, полученный из гена щелочной фосфатазы. В некоторых случаях, периплазматическая экспрессия приводит к утечке экспрессированного белка в культуральную среду. Секреция белка обеспечивает быструю и простую очистку от культурального супернатанта. Белки, которые не являются секретируемыми, могут быть получены из периплазмы с помощью осмотического лизиса. Как и при цитоплазматической экспрессии, в некоторых случаях белки могут стать нерастворимыми, и для облегчения растворения и рефолдинга могут быть использованы денатурат и восстановители. Температура индукции и роста также может повлиять на уровень экспрессии и растворимость. Как правило, используются температуры между 25°С и 37°С. Мутации также могут быть использованы для увеличения растворимости экспрессируемых белков. Как правило, бактерии производят агликозилированные белки. Таким образом, если белку для функционирования требуется гликозилирование, гликозилирование может быть осуществлено in vitro после очистки из клеток хозяина.
b. Дрожжи
Дрожжи, такие как Saccharomyces cerevisiae, Schizosaccharomyces pombe, Yarrowia lipolytica, Kluyveromyces Lactis и Pichia pastoris, являются полезными хозяевами для экспрессии FVII (см., например, Skoko и др., (2003) Biotechnol. Appl. Biochem. 38 (Pt3):257-65). Дрожжи можно трансформировать эписомальным репликативным вектором или стабильной хромосомной интеграцией путем гомологичной рекомбинации. Как правило, для регулирования экспрессии генов используются индуцибильные промотеры. Примеры таких промотеров включают GAL1, GAL7 и GAL5, и промотеры металлотионеина, такие как CUP1. Векторы экспрессии часто включают маркеры для отбора, такие как LEU2, TRPL, HIS3 и URA3 для селекции и сохранения трансформированных ДНК. Белки, экспрессируемые в дрожжах, зачастую являются растворимыми, и при совместной экспрессии с шаперонинами, такими как Bip, и протеиндисульфидизомеразами, может повыситься уровень экспрессии и растворимость. Кроме того, экспрессируемые в дрожжах белки могут быть направлены на секрецию с помощью гибридизации с сигнальными пептидами секреции, такими как сигнал секреции альфа-фактора типа спаривания дрожжей Saccharomyces cerevisiae и гибридизации с белками клеточной поверхности дрожжей, такой как гибридизация с Aga2p рецептором спаривания или глюкоамилазой из Arxula adeninivorans. Сайт расщепления протеазами (например, Кех-2 протеазой) может быть встроен для удаления гибридной последовательности от полипептида, когда он выходит на путь секреции. Дрожжи также способны к гликозилированию Asn-X-Ser/Thr мотива.
с. Насекомые и клетки насекомых
Насекомые и клетки насекомых, в частности, с применением бакуловирусной системы экспрессии являются полезными для экспрессии полипептидов, таких как FVII или его модифицированых форм (см., например, Muneta др., (2003) J. Vet. Med. Sci. 65 (2):219-23). Клетки насекомых и личинки насекомых, включая экспрессию в гемолимфе, экспрессируют высокие уровни белка и способны осуществлять большую часть посттрансляционных модификаций, используемых высшими эукариотами. Бакуловирусы имеют ограничительный диапазон хозяев, который повышает безопасность и уменьшает нормативные ограничения эукариотической экспрессии. Как правило, в векторах экспрессии используется промотор, такой как полигедриновый промотер бакуловируса для высокого уровня экспрессии. Обычно используемые бакуловирусные системы включают такие бакуловирусы, как вирус ядерного полиэдроза Autographa califomica (AcNPV) и вирус ядерного полиэдроза Bombyx mori (BmNPV) и клеточные линии насекомых, такие как Sf9, полученные из Spodoptera frugiperda, Pseudaletia unipuncta (A7S) и Danaus plexippus (DpNl). Для высокого уровня экспрессии нуклеотидная последовательность молекулы, которая будет экспрессирована, гибридизуется непосредственно за кодоном инициации транскрипции полигедрина этого вируса. Сигналы секреции млекопитающих точно процессируются в клетках насекомых и могут быть использованы для секреции экспрессируемого белка в культуральную среду. Кроме того, клеточные линии Pseudaletia unipuncta (A7S) и Danaus plexippus (DpNl) производят белки с паттернами гликозилирования, похожими на паттерны клеточных систем млекопитающих.
Альтернативные системы экспрессии в клетках насекомых относятся к использованию стабильно трансформированных клеток. Клеточные линии, такие как Schnieder 2 (S2), Кс клетки (Drosophila mela-Nogaster) и С7 клетки (Aedes albopictus), могут быть использованы для экспрессии. Промотер металлотионеина дрозофилы может быть использован для индуцирования высокого уровня экспрессии при индукции тяжелыми металлами в присутствии кадмия и меди. Векторы экспрессии, как правило, поддерживаются за счет использования маркеров для отбора, таких как неомицин и гигромицин.
d. Клетки млекопитающих
Системы экспрессии млекопитающих могут быть использованы для экспрессии FVII-полипептидов. Конструкции для экспрессии могут быть перенесены в клетки млекопитающих с помощью вирусной инфекции, такой как аденовирус, или прямой передачей ДНК, такой как с использованием липосом, фосфата кальция, DEAE-декстрана и физических средств, таких как электропорации и микроинъекции.
Векторы экспрессии для клеток млекопитающих, как правило, включают мРНК кэп сайт, ТАТА бокс, последовательность инициации трансляции (консенсусная последовательность Козака) и полиаденилированные элементы. Такие векторы часто включают транскрипционные промотер-энхансеры для высокого уровня экспрессии, например, промотер-энхансер SV40, промотер цитомегаловируса человека (ЦМВ), и протяженный терминальный повтор вируса саркомы Рауса (RSV). Эти промотер-энхансеры действуют во многих типах клеток. Ткане- и клеточного типа промотерные и энхансерные районы также могут быть использованы для экспрессии. Примеры промотер/энхансерных районов включают, но не ограничиваются промотер/энхансерными районами генов, таких как ген эластазы I, инсулина, иммуноглобулина, промотер/энхансерным районом вирус опухоли молочных желез мышей, гена альбумина, альфа-фетопротеина, альфа-1-антитрипсина, бета-глобина, основного белка миелина, легкой цепи-2 миозина, а также гонадотропного рилизинг гормона.
Маркеры для отбора могут быть использованы для выбора и для поддержания клеток с конструкцией для экспрессии. Примеры генов-маркеров для отбора включают, но не ограничиваются, гигромицин В фосфотрансферазой, аденозиндезаминазой, ксантин-гуанин фосфорибозил трансферазой, аминогликозид фосфотрансферазной, дигидрофолат редуктазой и тимидинкиназой. Гибридизация с сигнальными молекулами, обусловливающими локализацию на поверхности клетки, такими как TCR-ζ и FcεRI-γ, может направить экспрессию белка в активном состоянии на поверхность клеток.
Множество клеточных линий, доступных для экспрессии в млекопитающих, включают клеточные линии мыши, крысы, человека, обезьяны, курицы и хомяка. Примеры клеточных линий включают, но не ограничиваются, ВНК (т.е. ВНК-21 клетками), 293-F, СНО, Balb/3Т3, HeLa, MT2, мышь NSO (несекретирующая) и другими миеломными клеточными линиями, гибридомами и гибридомными клеточными линиями, лимфоцитами, фибробластами, SP2/0, COS, NIH3T3, НЕК293, 293S, 293Т, 2 В8, и НКВ клетками. Клеточные линии также можно адаптировать к бессывороточной среде, что облегчает очистку выделяемых белков от клеточной культуральной среды. Одним из таких примеров является бессывороточная EBNA-1 линия клеток (Pham и др., (2003) Biotechnol. Bioeng. 84:332-42). Экспрессия рекомбинантных полипептидов FVII, имеющих одинаковую структуру и посттрансляционные модификации, как полученных из плазмы FVII, известна в уровне техники (см., например, Jurlander и др., (2003) Semin Throm Hemost). Известны методы оптимизации экспрессии витамин-К-зависимых белков. Например, добавление витамина К в культуральную среду или коэкспрессия витамин-К-зависимой γ-карбоксилазы (Wajih и др., J. Biol. Chem. 280 (36) 31603-31607) может способствовать посттрансляционным модификациям витамин-К-зависимых белков, таких как полипептиды FVII.
е. Растения
Трансгенные растительные клетки и растения могут быть использованы для экспрессии FVII. Конструкции для экспрессии, как правило, переносятся в растения с использованием прямого переноса ДНК, такого как микропроекционная бомбардировка и PEG-опосредованная передача в протопласт, а также с помощью опосредованной агробактериями трансформации. Векторы экспрессии могут включать промотерные и энхансерные последовательности, элементы терминации транскрипции, а также элементы управления трансляцией. Векторы экспрессии и методы трансформации обычно делятся между двудольными хозяевами, такими как арабидопсис и табак, и однодольными хозяевами, такими как кукуруза и рис. Примеры растительных промотеров для экспрессии включают промотер вируса мозаики цветной капусты, промотер нопалин-синтазы, промотер рибозобисфосфаткарбоксилазы и промотеры убиквитина и UBQ3. Маркеры для отбора, такие как гигромицин, фосфоманнозизомераза и неомицинфосфотрансфераза, часто используются для упрощения отбора и поддержания трансформированных клеток. Трансформированные растительные клетки могут поддерживаться в культуре клеток, агрегатах (каллусной ткани) или регенерировать в целые растения. Так как растения имеют отличные от клеток млекопитающих паттерны гликозилирования, то это может повлиять на выбор этих хозяев для получения FVII. Трансгенные клетки растения также могут включать и водоросли, предназначенные для получения белка (см., например, Mayfield и др., (2003) 100:438-442 PNAS). Так как растения имеют отличные от клеток млекопитающих паттерны гликозилирования, то это может повлиять на выбор этих хозяев для получения FVII.
2. Очистка
Методы очистки полипептидов FVII из клеток хозяина зависят от выбранных клеток хозяина и системы экспрессии. Для секретируемых молекул белки, как правило, очищаются от культуральной среды после удаления клеток. При внутриклеточной экспрессии клетки могут быть лизированы и белки очищены от экстракта. Когда трансгенные организмы, такие как трансгенные растения и животные, используются для экспрессии, в качестве исходного материала для получения экстракта лизированных клеток могут быть использованы ткани или органы. Кроме того, получение в трансгенных животных может включать получение полипептидов в молоке или яйцах, которые могут быть собраны, и, при необходимости, далее белки могут быть экстрагированы и затем очищены с помощью стандартных методов, известных в уровне техники.
FVII могут быть очищены с использованием стандартных методов очистки белков, известных в данной области, включая, но не ограничиваясь, SDS-PAGE, фракционированием по размеру и хроматографией исключения размера (эксклюзионной), осаждением сульфатом аммония, хелатной хроматографией и ионообменной хроматографией. Например, FVII-полипептиды могут быть очищены анионобменной хроматографией. Примеры методов очистки FVII-полипептидов включают использование ионообменной колонки, которая способна связать любой полипептид, который обладает функциональным доменом Gla, и последующую элюцию в присутствии кальция (см., например, Пример 3). Методы аффинной очистки также могут быть использованы для повышения эффективности и чистоты препаратов. Например, антитела, рецепторы и другие молекулы, которые связывают FVII, могут быть использованы в аффинной очистке. В качестве другого примера, очистка также может быть улучшена с использованием аффинной колонки с растворимым ТФ (рТФ) (Maun и др., (2005) Prot Sci 14:1171-1180). Конструкции для экспрессии также могут быть сконструированы с добавлением аффинной метки, такой как MYC эпитоп, гибридизация с GST (глутатион трансфераза) или His6 и аффинно очищаться с помощью антител к myc, глутатионовой смолы и Ni-смолы, соответственно. Чистота может быть оценена любым методом, известным в уровне техники, в том числе гель-электрофорезом с окрашиванием и спектрофотометрическими методами.
Протеазы FVII могут быть экспрессированы и очищены в неактивной форме (зимогенной форме), либо, альтернативно, экспрессированные протеазы могут быть очищены в активной форме, например, полученной автокатализом. Например, FVII-полипептиды, которые были активированы с помощью протеолитического расщепления Arg152-Ile153, могут быть получены in vitro (т.е. FVIIa; двухцепочечной форме). FVII-полипептиды могут быть получены с помощью любого метода получения, описанного в настоящем документе, включая, но не ограничиваясь, получением в клетках млекопитающих с последующей очисткой. Расщепление полипептид ов FVII с получением активной формы протеазы, FVIIa, можно осуществить несколькими способами. Например, самоактивация во время инкубации с фосфолипидными везикулами в присутствии кальция может быть достигнута за 45 минут (Nelsestuen и др., (2001) J Biol Chem 276:39825-31). FVII-полипептиды также могут быть полностью активированы инкубацией с фактором Ха, фактор ХIIа или ТФ в присутствии кальция, в присутствии или без фосфолипидов (см., например, Пример 3, Broze и др., (1980) J Biol Chem 255:1242-1247, Higashi и др., (1996) J Biol Chem 271:26569-26574, Harvey и др., J Biol Chem 278:8363-8369).
3. Гибридные белки
Также предусмотрены гибридные белки, содержащие модифицированные полипептиды FVII и один или несколько других полипептидов. Предусмотрены фармацевтические композиции, содержащие такие гибридные белки, изготовленные для введения подходящим путем. Гибридные белки образуются путем объединения в любом порядке имодифицированного полипептида FVII и агента, такого как антитело или его фрагмент, фактор роста, рецептор, лиганд, и другие подобные агенты для облегчения очистки полипептида FVII, изменения фармакодинамических свойств полипептида FVII, например, направляя полипептид в целевые клетки или ткани, и/или для увеличения экспрессии или секреции полипептида FVII. Обычно любой гибридный белок FVII сохраняет, по меньшей мере, около 30%, 40%, 50%, 60%, 70%, 80%, 90%, или 95% коагулянтной активности по сравнению с негибридным полипептидом FVII, в том числе 96%, 97%, 98%, 99% или больше коагулянтной активности по сравнению с негибридным полипептидом.
Связь полипептида FVII с другим полипептидом может осуществляться непосредственно или опосредованно через линкер. В одном из примеров связь может представлять собой химическую связь, такую как через гетеробифункциональные агенты, или тиольные связи, или другие подобные связи. Гибридизация также может быть осуществлена с помощью рекомбинантных методов. Гибридизация полипептида FVII с другим полипептидом может осуществляться по N- или С-концу полипептида FVII. Неограничивающие примеры полипептидов, которые могут быть использованы в гибридных белках с полипептидами FVII, предусмотренными в настоящем документе, включают, например, GST (глутатион S-трансфераза) полипептид, Fc домен иммуноглобулина G, или гетерологичную сигнальную последовательность. Гибридные белки могут содержать дополнительные компоненты, такие как связывающий мальтозу белок Е. coli (MBP), которые помогают поглощению белка клетками (см. Международную заявку РСТ №WO 01/32711).
Гибридные белки могут быть получены стандартными рекомбинантными методами. Например, фрагменты ДНК, кодирующие различные полипептидные последовательности, могут быть лигированы вместе с учетом рамки считывания в соответствии с обычными методами, например, с использованием тупых или липких концов для лигирования, расщепления ферментами рестрикции для образования соответствующих концов, заполнения липких концов по мере необходимости, обработки щелочной фосфатазой, чтобы избежать нежелательного присоединения и ферментативного лигирования. В другом варианте, гибридные гены могут быть синтезированы обычными методами, включая использование автоматизированных синтезаторов ДНК. Альтернативно, могут осуществляться ПЦР-амплификации фрагментов гена с помощью якорных праймеров, которые приводят к комплементарным оверхэнгам между двумя последовательными генетическими фрагментами, которые могут быть впоследствии подвергнуты отжигу и реамплифицированы для создания химерных последовательностей генов (см., например, Ausubel и др., (eds.) Current Protocols in Molecular Biology, John Wiley & Sons, 1992). Кроме того, коммерчески доступны многие векторы экспрессии, которые уже кодируют гибридизуемый фрагмент (например, полипептид GST). FVII-кодирующая нуклеиновая кислота может быть клонирована в такой вектор, что гибридизуемый фрагмент связан в одной рамке считывания с протеазным белком.
4. Модификация полипептидов
Модифицированные полипептиды FVII могут быть получены в виде голых полипептидных цепей или в виде комплекса. Для некоторых целей может быть желательно получить FVII модифицированные полипептиды в "голой" форме без посттрансляционных или иных химических модификаций. Голые полипептидные цепи могут быть получены в соответствующих хозяевах, которые посттрансляционно не модифицируют FVII. Такие полипептиды также могут быть получены в in vitro системах и с использованием химического синтеза полипептидов. Для других целей могут быть желаемы специфические модификации, включая пегилирование, присоединение альбумина, гликозилирование, карбоксилирование, гидроксилирование, фосфорилирование, или другие известные модификации. Модификации могут быть осуществлены in vitro или, например, при получении модифицированных FVII в подходящем хозяине, которые производят такие модификации.
5. Нуклеотидные последовательности
В настоящем документе предусмотрены молекулы нуклеиновой кислоты, кодирующие FVII или модифицированные полипептиды FVII. Молекулы нуклеиновой кислоты включают аллельные варианты или варианты сплайсинга любых молекул, кодирующих FVII-полипептиды. Примеры молекул нуклеиновых кислот, предусмотренные здесь, включают любые молекулы, которые кодируют модифицированные полипептиды FVII, предусмотренные в настоящем документе, такие как любые, кодирующие полипептиды, приведенные в любой из SEQ ID №: 18-43, 125-150 или 206-250. В одном варианте, молекулы нуклеиновой кислоты, предусмотренные в настоящем документе, обладают, по меньшей мере, 50, 60, 65, 70,75, 80, 85, 90, 91, 92, 93, 94, 95, или 99% идентичностью последовательности, или гибридизуются в условиях средней или высокой жесткости, по крайней мере, на 70% от полной длины любой нуклеиновой кислоты, кодирующей полипептид FVII, предусмотренный в настоящем документе. В другом варианте, молекулы нуклеиновой кислоты могут включать последовательности с вырожденным кодоном, кодирующие любой из FVII-полипептидов, предусмотренных в настоящем документе.
G. Оценка активности модифицированных полипептидов FVII
Активности и свойства полипептидов FVII можно оценить in vitro и/или in vivo. Анализы для такой оценки известны специалистам в данной области, и известно, что активность при исследованиях, результаты терапии и in vivo активность коррелируют.В одном примере, варианты FVII могут быть оценены в сравнении с немодифицированным FVII и/или FVII дикого типа. В другом примере, активность модифицированных полипептидов FVII может быть оценена после воздействия TFPI или AT-III in vitro или in vivo по сравнению с активностью модифицированных полипептидов FVII, которые не обрабатывали TFPI или AT-III. In vitro анализы включают любые лабораторные анализы, известные специалистам в данной области, например, анализы с использованием клеток, включая анализы на коагуляцию, анализы на связывание, анализы белков, и молекулярнобиологические анализы. In vivo анализы включают анализы FVII на животных моделях, а также введение людям. В некоторых случаях активность FVII in vivo может быть определена на основе исследования крови, сыворотки, или других жидкостей организма на определенные параметры. Варианты FVII также могут быть исследованы in vivo для оценки активности или свойств, таких как терапевтический эффект.
Как правило, описанные здесь анализы относятся к двухцепочечной активированной форме FVII, т.е. FVIIa. Такие анализы также могут быть выполнены с одноцепочечной формой, например, для обеспечения негативного контроля, поскольку такая форма, как правило, не проявляет протеолитической или каталитической активности, необходимой для коагулянтной активности FVII. Кроме того, такие анализы также могут быть выполнены в присутствии кофакторов, таких как ТФ, которые в некоторых случаях увеличивают активность FVII.
1. In vitro анализы
Примеры in vitro анализов включают анализы для оценки модификаций и активности полипептида. Модификация может быть оценена с помощью in vitro анализов, которые оценивают у-карбоксилирование и другие посттрансляционные модифицикации анализов белка и конформационных анализов, известных в уровне техники. Анализы активности включают, но не ограничиваются, измерением взаимодействия FVII с другими факторами свертывания, такими как ТФ, фактор Х и фактор IX, протеолитическими анализами для определения протеолитической активности FVII-полипептидов, анализами для определения способности связывать и/или аффинности полипептидов FVII к фосфатидилсерину и другим фосфолипидам, и клеточные анализы для определения влияния полипептидов FVII на коагуляцию.
Концентрации модифицированных полипептидов FVII можно оценить по методам, хорошо известным в данной области, включая, но не ограничиваясь, фермент-связанным иммуносорбантным анализом (ELISA), SDS-PAGE;
Брэдфорд, Лоури, БСА методом; УФ абсорбцией, а также другими методами количественного измерения меченого белка, включая, но не ограничиваясь, иммунологическими, радиоактивными и флуоресцентными методами и связанными с ними методами.
Оценка продуктов расщепления, полученных в результате реакций протеолиза, в том числе расщепления полипептидов FVII, или продуктов, полученных в результате протеазной активности FVII, может быть выполнена с использованием методов, включающих, но не ограничивающихся, расщеплением хромогенного субстрата, ВЭЖХ, SDS-PAGE анализом, ELISA, Вестерн-блотом, иммуногистохимией, иммунопреципитацией, NН2-концевой секвенированием и мечением белка.
Структурные свойства модифицированных полипептидов FVII также могут быть оценены. Например, может быть выполнена рентгеноструктурная кристаллография, ядерный магнитный резонанс (ЯМР), криоэлектронная микроскопия (крио-ЭМ) модифицированных полипептидов FVII для оценки трехмерной структуры полипептидов FVII и/или других свойств полипептидов FVII, таких как связывание Са+ или кофактора.
Кроме того, наличие и степень деградации FVII могут быть измерены стандартными методами, такими как электрофорез с натрий додецилсульфатом в полиакриламидном геле (SDS-PAGE) и Вестерн-блоттинг подвергнутых электрофорезу FVII содержащих образцов. FVII-полипептиды, которые были обработаны протеазами, также могут быть подвергнуты N-концевому секвенированию для определения местонахождения или изменений в сайтах расщепления модифицированных полипептидов FVII.
а. Посттрансляционные модификации
FVII-полипептиды также могут быть оценены на предмет наличия посттрансляционных модифицикаций. Такие анализы известны в уровне техники и включают анализы для измерения гликозилирования, гидроксилирования и карбоксилирования. Примеры анализов на гликозилирование включают анализ на углеводы, например, SDS-PAGE анализ FVII-полипептидов, подвергнутых гидразинолизу или обработке эндогликозидазой. Гидразинолиз отделяет N- и O-связанные гликаны от гликопротеина при инкубации с безводным гидразином, а обработка эндогликозидазой PNGaзой E приводит к отделению большинства N-гликанов от гликопротеинов. Гидразинолиз или обработка эндогликозидазами полипептидов FVII приводит к образованию восстанавливающего конца, который может быть мечен флуорофорной или хромофорной меткой. Меченые FVII-полипептиды могут быть проанализированы с помощью флуорофор-ассоциированного электрофореза углеводов (FACE). Флуоресцентные метки для гликанов также могут быть использованы для анализа моносахаридов, профилирования или выявления характерных особенностей структуры сложных паттернов гликозилирования методом высокоэффективной жидкостной хроматографии. Примеры методов ВЭЖХ включают гидрофильную хроматографию, электронного взаимодействия, ионообменную, гидрофобную, а также хроматографию исключения размера. Примеры реагентов на гликаны включают, но не ограничиваются, 3-(ацетиламино)-6-аминоакридином (АА-АС) и 2-аминобензойной кислотой (2-АА). Углеводные фрагменты также могут быть обнаружены с использованием специфических антител, которые распознают гликозилированный FVII-полипептид. Примеры анализов для измерения β-гидроксилирования включают обратнофазную ВЭЖХ полипептидов FVII, которые были подвергнуты щелочному гидролизу (Przysiecki и др., (1987) PNAS 84:7856-7860). Карбоксилирование и γ-карбоксилирование полипептидов FVII может быть оценено с помощью масс-спектрометрии с ионизацией матрица-активированной лазерной десорбцией с измерением времени пролета (MALDI-TOF), как описано в уровне техники (см., например, Harvey и др., J Biol Chem 278:8363-8369, Maum и др., Prot Sci 14:1171-1180). Взаимодействие полипептида FVII, содержащего пропептид (Pro-FVII), с карбоксилазой, ответственной за посттрансляционное γ-карбоксилирование, также может быть оценено. Константа диссоциации (Kd) карбоксилазы после инкубации с флуоресцин-меченным про-FVII-полипептидом может быть измерена путем определения количества связанной карбоксилазы с помощью анизотропии (Lin и др., (2004) J Biol Chem 279:6560-6566).
В. Протеолитическая активность
Модифицированные полипептиды FVII могут быть проверены на протеолитическую активность. Протеолитическая активность FVII может быть измерена с помощью хромогенных субстратов, таких как Chromozym t-PA (MeSO2-D-Phe-Gly-Arg-pNA), S-2288 (H-D-Ile-Pro-Arg-pNA), S-2266 (H-D-Val-Leu-Arg-pNA), S-2765 (Z-D-Arg-Gly-Arg-pNA), Spectrozyme FXA и Spectrozyme FVIIa (CH3SO2-D-CHA-But-Arg-pNA). FVII-полипептиды, отдельно или в присутствии ТФ, инкубировали с различными концентрациями хромогенного субстрата. Расщепление субстрата может контролироваться с помощью измерения абсорбции, и скорость гидролиза субстрата определяется методом линейной регрессии с помощью доступного программного обеспечения.
Активация субстратов факторов свертывания, таких как FX, полипептидами FVII также может быть оценена. FVII-полипептиды, с предварительной инкубацией с ТФ или без инкубации, могут инкубироваться с очищенным FX (коммерчески доступным). Количество активного FXa, полученного в результате инкубации с полипептидом FVII, определяется как активность FXa по отношению к хромогенному субстрату, такому как S-2222 или Spectrafluor FXa (CH3SO2-D-СНА -Gly-Arg-AMC. AcOH), которая контролируется через оптическую плотность изменения абсорбции (Harvey и др., J Biol Chem 278:8363-8369, см. также Пример 5 ниже). Источник фосфолипидов также может быть включен при инкубации FVII и FX (Nelsestuen и др., (2001) J Biol Chem 276:39825-31).
с. Коагулянтная активность
FVII-полипептиды могут быть тестированы на коагулянтную активность с использованием анализов, хорошо известных в уровне техники. Например, некоторые анализы включают, но не ограничиваются, двухстадийным анализом на свертывание (Leibman и др., (1985) PNAS 82:3879-3883); анализом на протромбиновое время (РТ, которое может характеризовать ТФ-зависимую активность FVIIa во внешнем пути); анализами, которые являются модификациями теста РТ; активированным частичным тромбопластиновым временем (АЧТВ, которое характеризует ТФ-независимую активность FVIIa), активированным временем свертывания крови (ACT); рекальцифицированным активированным временем свертывания крови; время свертывания Ли-Вайта; или тромбоэластографией (ТЭГ) (Pusateri и др., (2005) Critical Care 9: S15-S24). Например, коагулянтная активность модифицированного полипептида FVII может быть определена анализом на основе РТ, при котором FVII разводят FVII-дефицитной плазмой и смешивают с реагентом для измерения протромбинового времени (рекомбинантный ТФ с фосфолипидами и кальцием), таким как доступный от loNovin TM, Dade Behring. Тромбообразование обнаруживают оптически и определяют время образования сгустка, сравнивая со временем для FVII-дефицитной плазмы.
d. Связывание с другими белками и/или ингибирование другими белками
Анализы на ингибирование могут быть использованы для измерения устойчивости модифицированных полипептидов FVII к FVII ингибиторам, таким как, например, TFPI и AT-III, или молекулам, таким как Zn2+. Оценка ингибирования другими ингибиторами также может быть оценена и включает, но не ограничивается, другими ингибиторами сериновой протеазы и FVII-специфическими антителами. Ингибирование может быть оценено инкубацией, например, TFPI, AT-III или Zn2+ с полипептидами FVII, которые были предварительно инкубированы с ТФ. Активность FVII может быть измерена с помощью определения одной или нескольких активностей или коагулянтных анализов, описанных выше, и ингибирование TFPI, AT-III и Zn2+ может быть оценено сравнением активности FVII-полипептидов, инкубированных с ингибитором, с активностью полипептидов FVII, которые не были инкубированы с ингибитором.
FVII-полипептиды могут тестироваться на связывание с другими факторами коагуляции и ингибиторами. Например, прямое и непрямое взаимодействия FVII с кофакторами, такими как ТФ, субстратами, такими как FX и FIX, а также ингибиторами, такими как TFPI, антитромбин III и гепарин, могут быть оценены с помощью любого анализа на связывание, известного в уровне техники, включая, но не ограничиваясь, иммунопреципитацией, очисткой на колонке, невосстанавливающим SDS-PAGE, BIAcore® анализами, поверхностным плазменным резонансом (SPR), флуоресцентным резонансным переносом энергии (FRET), флуоресцентной поляризацией (FP), калориметрией изотермического титрования (ITC), круговым дихроизмом (CD), структурным анализом комплементации фрагментов белка (РСА), ядерным магнитным резонансом (ЯМР), рассеянием света, седиментационным равновесием, малых зон гель-фильтрационной хроматографией, удерживанием в геле. Far-western блоттингом, поляризацией флуоресценции, гидроксил-радикальным футпринтингом белка, фаговым дисплеем, а также различными двухгибридными системами. В одном примере, связывание Zn2+ оценивали с помощью анализа равновесия (Petersen и др., (2000) Protein Science 9:859-866)
е. Аффинность к фосфолипидам
Способность модифицированного полипептида FVII связывать и/или аффинность к фосфатидилсерину (PS) и другим фосфолипидам можно определить с помощью анализов, хорошо известных в уровне техники. Фосфолипиды высокой чистоты (например, известные концентрации бычьего PS и яичного фосфатидилхолина (PC)), которые являются коммерчески доступными, например, от Sigma, в органическом растворителе могут быть использованы для получения малых однослойных фосфолипидных везикул. Связывание FVII-полипептидов с этими PS / PC может быть определено по относительному рассеянию света под углом 90° к падающему свету. Интенсивность рассеяния света PC / PS и PC / PS / FVII измеряется для определения константы диссоциации (Harvey и др., J Biol Chem 278:8363-8369). Поверхность плазменного резонанса, например, выполненного на биосенсоре BIAcore, также может быть использовано для измерения аффинности полипептидов FVII к фосфолипидам мембран (Sun и др., Blood 101:2277-2284).
2. Не относящиеся к человеку животные модели
Не относящиеся к человеку животные модели могут быть использованы для оценки активности, эффективности и безопасности модифицированных полипептидов FVII. Например, животные, не включающие человека, могут быть использованы для моделирования заболеваний или состояний. Животные, не включающие человека, могут инъецироваться веществом, вызывающим заболевание и/или фенотип, до введения вариантов FVII, таких как любые варианты FVII, приведенные в любой из SEQ ID №8: 18-43,125-150 или 206-250, для мониторинга воздействия на прогрессирование заболевания. Генетические модели также являются полезными. Могут быть созданы животные, такие как мыши, имитирующие заболевания или состояния по причине гиперэкспрессии, недостаточной экспрессии или нокаута одного или нескольких генов, такие как, например, фактор VIII нокаут-мыши, у которых проявляется гемофилия A (Bi и др., (1995) Nat Gen 10:119-121). Такие животные могут быть получены методами трансгенного получения животных, хорошо известными в уровне техники, или с использованием естественных или индуцированных мутантных штаммов. Примеры полезных не относящихся к человеку животных моделей заболеваний, связанных с FVII, включают, но не ограничиваются, моделями нарушения свертываемости крови, в частности, гемофилией или болезнями тромбообразования. Не относящиеся к человеку животные модели травм также могут быть использованы для оценки активности, такой как коагулянтная активность, полипептидов FVII. Эти не относящиеся к человеку животные модели могут быть использованы для определения активности вариантов FVII по сравнению с FVII-полипептидом дикого типа.
Животные модели также могут быть использованы для определения стабильности, периода полураспада и расщепления модифицированного полипептида FVII. Такие анализы полезны для сравнения модифицированных полипептидов FVII и для расчета доз и режима доз для дополнительных не связанных с человеком животных моделей и испытаний на человеке. Например, модифицированные полипептиды FVII, такие как любые варианты FVII, предусмотренные в настоящем документе, включая, например, любые последовательности, приведенные в любой из SEQ ID №8: 18-43,125-150 или 206-250, могут быть введены в хвостовую вену мыши. Затем образцы крови берутся через определенные временные интервалы после инъекции (например, минуты, часы и дни после инъекции), а затем определяется уровень модифицированных полипептидов FVII в образцах, включающих, но не ограничивающихся, сывороткой или плазмой, через определенные промежутки времени, например с помощью ELISA или радиоиммуноанализа. Образцы крови, взятые в различные моменты времени после введения FVII-полипептидов, также могут быть исследованы на коагулянтную активность с использованием различных методов, таких как это описано в Примерах 9 и 14. Эти типы фармакокинетических исследований могут предоставить информацию о периоде полураспада, расщеплении и стабильности полипептидов FVII, которые могут помочь в определении подходящих доз для введения вещества как прокоагулянта.
Модифицированные FVII-полипептиды, такие как любые приведенные в любой из SEQ ID №8: 18-43,125-150 или 206-250, могут быть проверены на терапевтическую эффективность для лечения гемофилии с использованием животных моделей. В одном из неограничивающих примеров могут быть использованы животные модели, например, мыши. Мышиные модели гемофилии доступны в уровне техники и могут быть использованы для проверки модифицированных FVII-полипептидов. Например, мышиная модель гемофилии А, полученная инъекцией анти-FVIII антител, может быть использована для оценки коагулянтной активности полипептидов FVII (см. примеры 8 и 14, Tranholm и др.. Blood (2003) 102:3615 -3620). Мышиная модель гемофилии В также может быть использована для тестирования полипептидов FVII (Margaritis и др., (2004) J Clin Invest 113:1025-1031). Также существуют немышиные модели нарушений, связанных с кровоточивостью. Активность FVII-полипептидов может быть оценена на крысах с вызванной варфарином или мелагатраном кровоточивостью (Diness и др., (1992) Thromb Res 67:233-241, Elg и др., (2001) Thromb Res 101:145-157), и на кроликах с гепарин-индуцированной кровоточивостью (Chan и др., (2003) J Thromb Haemost 1:7б0-7б5). Собаки с врожденной гемофилией А, гемофилией В и болезнью Виллебранда, у которых проявляется тяжелая кровоточивость, также могут быть использованы для исследований полипептидов FVII на животных (Brinkhous и др., (1989) 86:1382-1386 PNAS). Активность полипептидов FVII также может быть оценена на модели кровоточивости у кролика, в которой тромбоцитопения индуцируется комбинацией гамма-излучения и использования антител против тромбоцитов (Tranholm и др., (2003) Thromb Res 109:217-223).
В дополнение к животным с генерализованными расстройствами, связанными с кровоточивостью, модели ранений и травм также могут быть использованы для оценки активности полипептидов FVII, а также их безопасности и эффективности в качестве коагулянтных лекарственных средств. Неограничивающие примеры таких моделей включают модель с коронарным стенозом на кролике (Fatorutto и др., (2004) Can J Anaesth 51-672-679), модель с повреждением печени V класса на свинье (Lynn и др., (2002) J Trauma 52: 703-707), модель с повреждением печени V класса на свинье (MartiJ№witz и др., (2001) J Trauma 50:721-729), а также модель с аортотомией на свинье (Sondeem и др. (2004) Shock 22:163-168).
3. Клинические исследования
Доступно множество анализов для оценки активности FVII для клинического использования. Такие анализы могут включать оценку коагуляции, стабильности белков, периода полураспада in vivo и фенотипический анализ. Фенотипические анализы и анализы для оценки терапевтического эффекта FVII включают оценку уровня FVII в крови (например, измерение сывороточного уровня FVII до введения и через определенные промежутки времени после введений, в том числе, после первого введения, сразу после последнего введения, и через определенные промежутки времени между введениями, с поправками на индекс массы тела (ИМТ)), оценку свертывания крови in vitro с использованием методов, описанных выше, после лечения FVII (например, анализа РТ), и фенотипический ответ на лечение FVII, включая улучшение симптомов с течением времени по сравнению с субъектами, которым вводили немодифицированный FVII и/или FVII дикого типа, или плацебо. Пациенты, получавшие лечение полипептидами FVII, могут быть исследованы на потерю крови, потребность в переливании, и уровень гемоглобина. Пациенты могут исследоваться регулярно в течение определенного периода времени после обычного или неоднократного введения, или после введения в ответ на острые события, такие как кровотечение, травма или хирургическое вмешательство.
Н. Композиция и введение
В данном документе приводятся композиции для использования в лечении расстройств, связанных с кровоточивостью. Такие композиции содержат терапевтически эффективное количество полипептида фактора VII, как описано выше. Полипептиды FVII в эффективной концентрации или их фармацевтически приемлемые производные смешиваются с подходящим фармацевтическим носителем или наполнителем для системного, местного или наружного введения. Соединения включены в эффективном для лечения выбранного расстройстве количестве. Концентрация активного соединения в композиции будет зависеть от поглощения, инактивации, уровня выделения активных соединений, дозировки, а также введенного количества и других факторов, известных специалистам в данной области.
Фармацевтические носители или наполнители, подходящие для введения соединений, предусмотренных в настоящем документе, включают любые носители, известные специалистам в данной области как подходящие для конкретного режима введения. Фармацевтические композиции, которые включают терапевтически эффективные количества полипептидов FVII, описанных здесь, также могут представлять собой лиофилизированный порошок, восстановленный, например, стерильной водой непосредственно перед применением.
1. Композиции
Фармацевтические композиции, содержащие модифицированный FVII, могут быть получены любым обычным способом путем смешивания выбранного полипептида с одним или более физиологически приемлемым носителем или наполнителем. Выбор носителя или вспомогательного вещества определяется знаниями специалиста и может зависеть от ряда параметров. К ним относятся, например, способ введения (например, системное, оральное, назальное, легочное, местное, поверхностное, или любой другой способ введения) и заболевание. Фармацевтические композиции могут быть выполнены в виде простого дозового (прямое) введения или с возможностью разведения или других изменений. Концентрации соединений в композициях должны быть эффективными как для способа введения, так и для предполагаемого лечения. Как правило, композиции разработаны для простого дозового введения. Чтобы получить состав, массовую долю соединения или их смесь растворяют, суспендируют, диспергируют, или иным образом смешивают в выбранных условиях в эффективной концентрации, чтобы болезненное состояние облегчалось или улучшалось.
Модифицированные FVII-полипептиды по настоящему изобретению могут быть использованы для введения субъекту в виде двуцепочного FVIIa белка. Модифицированные полипептиды FVII могут быть активированы в композиции любым методом, известным в уровне техники. Например, FVII может быть самоактивирован при очистке ионообменной хроматографией (Jurlander др. (2001) Semin Thromb Hemost 27:373-384). Модифицированные FVII-полипептиды также могут быть активированы инкубацией с FXA, иммобилизованном на гранулах (Kemball-Cook и др. (1998) J Biol Chem 273:8516-8521), или любыми другими методами, известными в данной области (см. Пример 3 ниже). Включение кальция в эти процессы обеспечивает полную активацию и правильный фолдинг модифицированного FVIIa белка. Модифицированные FVII-полипептиды также могут быть использованы для введения в виде одноцепочного белка. Одноцепочные FVII-полипептиды могут быть очищены таким образом, чтобы не допустить расщепления (см., например, US6677440). Модифицированные FVII-полипептиды могут быть использованы в виде смеси одноцепочного и двухцепочного белков в фармацевтической композиции, в любом соотношении с соответствующим выбором средств для исключения или контроля самоактивации.
Соединение может быть суспендировано в микронизированной или другой подходящей форме или может быть модифицировано для получения более растворимых активных продуктов. Форма получаемой смеси зависит от ряда факторов, в том числе от предполагаемого способа введения и растворимости соединений в выбранном носителе или среде. Получающиеся смеси представляют собой растворы, суспензии, эмульсии и другие подобные смеси и могут представлять собой как неводные, так и водные смеси, кремы, гели, мази, эмульсии, растворы, эликсиры, лосьоны, суспензии, настойки, пасты, пены, аэрозоли, средства для орошения, спреи, свечи, бинты, или любые другие подходящие средства, удобные для системного, поверхностного или местного введения. Для местного внутреннего введения, такого как внутримышечное, парентеральное, или внутрисуставного введения, полипептиды могут быть использованы как суспензии в водной среде, такие как изотонический буферный раствор, или в сочетании с биологически совместимым носителем или биоадгезивом, предназначенным для внутреннего введения. Эффективная концентрация нацелена на смягчение целевого состояния и может быть эмпирически определена. Для получения состава массовую долю соединений растворяют, суспендируют, диспергируют или иным образом смешивают в эффективном количестве с выбранной средой, чтобы болезненное состояние облегчалось или улучшалось.
Как правило, фармацевтически приемлемые композиции готовятся с учетом утвержденных требований Регулирующих органов или с требованиями Фармакопеи для животных и людей. Фармацевтические композиции могут включать носители, такие как растворитель, адъвант, наполнитель или среду, с которой изоформ вводится. Такие фармацевтические носители могут быть стерильными жидкостями, такими как вода или масла, в том числе природные смеси жидких углеводородов животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральные масла и кунжутное масло. Вода является типичным носителем, когда фармацевтическую композицию вводят внутривенно. Солевые растворы и водные растворы глюкозы и глицерина также могут быть использованы в качестве жидких носителей, особенно для инъекционных растворов. Композиции могут содержать наряду с активным ингредиентом: разбавители, такие как лактоза, сахароза, двухкальциевый фосфат, или карбоксиметилцеллюлозу; смазки, такие как стеарат магния, кальция стеарат и тальк, связующее, такое как крахмал, природные смолы, такие как смола акации, глюкоза, патоки, поливинилпирролидон, целлюлозы и их производные, повидон, кросповидоны и другие известные связующие. Подходящие фармацевтические эксипиенты включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, глицерин моностеарат, тальк, натрия хлорид, сухое обезжиренного молоко, глицерин, пропиленгликоль, воду и этанол. Состав при желании также может содержать незначительные количества смачивателя или эмульгатора, или рН буферизующих агентов, например, ацетат, цитрат натрия, циклодекстриновые производные, сорбитан монолаурат, триэтаноламин ацетатнатрия, триэтаноламин олеат, и другие подобные агенты. Эти композиции могут иметь форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков и форм с замедленным высвобождением. Капсулы и картриджи, например желатин для использования в ингаляторе или инсуффлятор, могут содержать порошковую смесь терапевтического соединения и подходящего базового порошка, такого как лактоза или крахмал. Состав может быть сформулирован в виде свечи, с традиционными связующими и носителями, такими как триглицериды. Пероральные композиции могут включать в себя стандартные носители такие, как фармацевтически пригодные маннит, лактоза, крахмал, стеарат магния, натрия сахарин, целлюлоза, карбонат магния и другие подобные агенты. Композиции для перорального введения также могут содержать ингибиторы протеазы, такие как Bowman-Birk ингибитор, конъюгированный Bowman-Birk ингибитор, апротинин и камостат 1. Примеры подходящих фармацевтических носителей описаны в "Ремингтоне фармацевтических наук" E.W.Martin. Такие композиции могут содержать терапевтически эффективное количество соединения, как правило, в чистом виде, а также необходимое количество носителя, с тем чтобы обеспечить надлежащую форму для введения субъекту или пациенту.
Композиция должна соответствовать определенному способу введения. Например, модифицированный FVII может быть выполнен в виде инъекций (например, болюсной инъекции или непрерывной инфузии) для парентерального введения. Инъекционные композиции могут быть суспензиями, растворами или эмульсиями на основе липидных или водных сред. Стерильные инъекционные композиции также могут быть растворами или суспензиями в нетоксичных парентерально-приемлемых разбавителях или растворителях, например, в виде раствора в 1-4, бутандиоле. Стерильные, твердые масла являются условно используемыми в качестве растворителя или суспендирующей среды. Для этой цели любое легкое твердое масло может быть использовано, в том числе, но не ограничиваясь, синтетические моно-или диглицериды, жирные кислоты (в том числе олеиновая кислота), натуральные растительные масла, такие как кунжутное масло, кокосовое масло, арахисовое масло, хлопковое масло и другие масла, синтетические масляные среды, такие как этилолеат. Буфферы, консерванты, антиоксиданты и подходящие ингредиенты могут быть включены, если это требуется, или, напротив, могут содержаться в композиции.
Полипептиды могут быть единственными активными фармацевтическими ингредиентами в составе или могут сочетаться с другими активными ингредиентами. Полипептиды могут иметь целевую доставку, например, при конъюгации с векторным агентом, таким как антитела. Липосомальные суспензии, в том числе нацеленные на определенные ткани липосомы, также могут быть пригодны как фармацевтически приемлемые носители. Они могут быть приготовлены в соответствии с методами, известными специалистам в данной области. Например, липосомальные композиции могут быть приготовлены так, как описано в Патенте США №4522811. Липосомальные композиции также могут представлять собой системы с медленным высвобождением, в том числе фармацевтические матрицы такие, как коллагеновые гели и липосомы, модифицированные фибронектином (см., например, Weiner и др. (1985) J Pharm Sci. 74 (9): 922-5). Композиции также могут содержать один или более адъювантов, облегчающих высвобождение, таких как, но не ограничиваясь, инертные носители или коллоидные дисперсионные системы. Репрезентативные и неограничивающие примеры таких инертных носителей могут быть выбраны из воды, изопропилового спирта, газообразных фторуглеродов, этилового спирта, поливинилпирролидона, пропиленгликоля, формирующего гель материала, стеарилового спирта, стеариновой кислоты, спермацета, сорбита моноолеата, метилцеллюлозы, а также подходящих комбинаций двух или более указанных носителей. Активное соединение включают в фармацевтически приемлемый носитель в количестве, достаточном для проявления терапевтического эффекта в отсутствии нежелательных побочных эффектов при лечении субъекта. Терапевтически эффективные концентрации могут быть определены эмпирически путем тестирования соединений в известных in vitro и in vivo системах, таких, которые приведены в описании.
а. Дозы
Точное количество или дозы терапевтического средства зависят от конкретного FVII-полипептида, пути введения, а также других факторов, таких, как тяжесть болезни, вес и общее состояние пациента. Местное введение терапевтических агентов, как правило, требуют меньшей дозы, чем любой способ системного введения, хотя концентрация терапевтического агента для местного введения может в некоторых случаях быть выше, чем для безопасного системного введения. При необходимости, конкретные дозы и продолжительность лечения и протокол могут быть эмпирически определены или экстраполированы. Например, примерные дозы рекомбинантного и нативного FVII-полипептида могут быть использованы в качестве отправной точки для определения соответствующих доз. Например, рекомбинантные FVII (rFVIIa) полипептиды, которые были активированы в rFVIIa, Novo SEVEN ®, были введены пациентам с гемофилией А или гемофилией В, в состоянии кровотечения, в дозе 90 мкг/кг в виде болюсных вливаний в течение более 2 до 5 минут, с достижением эффективного циркулирующего уровня не менее 2 мкг/мл. Дозы вводили каждые 2 часа, пока не был достигнут гемостаз. Модифицированные FVII-полипептиды могут быть эффективными при сокращении количества доз и/или частоты по сравнению с рекомбинантным FVII. Например, модифицированные FVII-полипептиды могут быть введены в дозе 80 мкг/кг, 70 мкг/кг, 60 мкг/кг, 50 мкг/кг, 40 мкг/кг, 30 мкг/кг, 20 мкг/кг, 15 мкг/кг или менее. В некоторых вариантах, дозировки могут быть выше, например, 100 мкг/кг, 110 мкг/кг, 120 мкг/кг или выше. Продолжительность лечения и интервал между инъекциями будут различаться в зависимости от тяжести кровотечения и реакции пациента на лечение, и могут быть скорректированы соответствующим образом. Такие факторы, как уровень активности и время полужизни модифицированных FVII по сравнению с немодифицированным FVII, могут быть приняты во внимание при определении дозы. Отдельные дозировки и схемы лечения могут быть определены эмпирически.
В другом случае, рекомбинантный FVII (rFVIIa) полипептид, который был активирован в rFVIIa, No voSEVEN ®, был введен пациентам с врожденным дефицитом FVII, в состоянии кровотечения, в дозировке 15-30 мкг/кг болюсным вливанием в течение более 2 до 5 минут. Дозы повторялись каждые 4-6 часа до достижения гемостаза. Модифицированные полипептиды FVII могут быть эффективными при сокращенном количестве доз и/или частоте по сравнению с рекомбинантными FVII. Например, модифицированные полипептиды FVII, предусмотренных в настоящем изобретении, могут быть введены в дозе 20 мкг/кг, 15 мкг/кг, 10 мкг/кг, 5 мкг/кг, 3 мкг/кг или менее. В некоторых случаях, дозировки могут быть выше, такие, как 35 мкг/кг, 40 мкг/кг, 45 мкг/кг, или выше. Продолжительность лечения и интервал между инъекциями будут меняться в зависимости от тяжести кровотечения и реакции пациента на лечение, и могут быть скорректированы соответствующим образом. Такие факторы, как уровень активности и время полужизни модифицированных FVII по сравнению с неизмененным FVII, могут быть использованы при определении дозы. Например, модифицированный FVII-полипептид, который показывает большее время полужизни, чем немодифицированый FVII-полипептид, может быть введен при более низких дозах и/или менее часто, чем немодифицированный FVII-полипептид. Аналогичным образом, дозы, необходимые для терапевтического эффекта модифицированного FVII-полипептида, который показывает увеличенную коагуляционную активность по сравнению с немодифицированным FVII полипептидом, могут быть сокращены по частоте и по величине. Отдельные дозы и схемы могут быть определены специалистм эмпирически.
b. Дозированные лекарственные формы
Фармацевтические терапевтически активные соединения и их производные, как правило, получают и вводят в единичных лекарственных формах или мультидозированных формах. Композиции для введения людям и животным могут быть представлены в лекарственных формах, которые включают, но не ограничиваются таблетками, капсулами, пилюлями, порошками, гранулами, стерильными парентеральными растворами или суспензиями, оральными растворами или суспензиями, жировыми и водными эмульсиями, содержащими в подходящих количествах соединения или фармацевтически приемлемые их производные. Каждая единичная доза содержит заранее определенное количество терапевтически активных соединений, достаточное для получения желаемого терапевтического эффекта, в сочетании с необходимыми фармацевтическими носителями, средами или разбавителями. Примерами единичных дозированных форм являются ампулы и шприцы, индивидуально упакованные таблетки или капсулы. В некоторых примерах, единичные дозированные формы предоставляют собой лиофилизированный порошок, восстановленный до введения. Например, полипептид FVII может быть изготовлен в виде лиофилизированного порошка, воссозданного подходящим раствором для создания раствора в виде единой дозы для инъекций. В некоторых вариантах, лиофилизированный порошок может содержать полипептид FVII и дополнительные компоненты, такие как соли, такие, что восстановление стерильной дистиллированной водой полипептида FVII дает буферный или физиологический раствор полипептида. Единичные дозированные формы могут быть введены в долях или многократных долях. Множественные дозированные формы представляют собой множество идентичных дозированных лекарственных форм, упакованных в одном контейнере, для введения в сегрегационной единичной дозе. Примерами различных многодозовых форм являются флаконы, бутылки, наполненные таблетками или капсулами, или бутылки, наполненные галлонами и пинтами жидкости. Таким образом, многодозовые формы представляют собой множество единичных доз, которые не разделены в упаковке.
2. Введение модифицированных FIVI полипептидов
FVII-полипептиды по настоящему изобретению (т.е. активные соединения) могут быть введены in vitro, ex vivo или in vivo в жидкости тела или другой образец ткани. Например, при введении ех vivo жидкости тела или ткани субъекта контактируют с FVII-полипептидом, который нанесен на трубку или фильтр, такие как, например, труба или фильтр аппарата искусственного кровообращения. При введении in vivo, активные вещества могут вводиться любым известным способом, например, перорально, нозально, пульмонально, парентерально, внутривенно, внутрикожно, подкожно, интраартикулярно, интрацистенально, интраокулярно, интравентикулярно, интратекально, внутримышечно, внутрибрюшинно, интратрахеально или наружно, а также любой комбинацией любых двух или более способов, в жидкой, полужидкой или твердой форме таким образом, который подходит для каждого способа введения. Модифицированные FVII-полипептиды можно вводить один или более чем один раз, например, два, три, четыре раза, или любое количество раз, которые необходимы для достижения терапевтического эффекта. Несколько введений может быть произведено с помощью любого способа или комбинаций способов, а может осуществляться каждый час, каждые 2 часа, каждые три часа, каждые четыре часа и более.
Наиболее подходящий способ введения будет варьироваться в зависимости от болезненного состояния, на которое нужно воздействовать, например, локализации места кровотечения. Как правило, FVII-полипептиды вводятся внутривенной болюсной инъекцией со временем введения (инфузии) примерно 2-5 минут. В других случаях желаемый уровень FVII в крови может быть обеспечен путем непрерывной инфузии активного агента, длительность которой может быть установлена по плазме крови. Следует отметить, что лечащий врач должен знать, как и когда следует прекратить, прервать или изменить терапию для снижения дозировки из-за токсичности или дисфункций костного мозга, печени или почек. Наоборот, лечащий врач должен также знать, как и когда следует повысить уровень лечащей дозы, если клинический ответ не является адекватным (исключая токсичные побочные эффекты). В других случаях, место кровотечения может означать, что введение FVII нужно осуществлять через альтернативный путь введения. Например, местное введение, включая введение в мозг (например, интравентикулярно), может быть выполнено, когда пациент испытывает кровотечение в этом регионе. Аналогично, для лечения кровотечений в суставах, может быть использовано местное введение терапевтического агента в сустав (т.е. интраартикулярно, внутривенно или подкожно). В других случаях, поверхностное введение терапевтических агентов на кожу, например, в виде крема, геля или мази, или введение в легкие при вдыхании или интратрахеально может быть более целесообразно, если кровотечение локализовано в этих областях.
В случае, когда модифицированные FVII-полипептиды являются депо препаратами, длительно действующие композиции могут быть введены имплантацией (например, внутримышечно или подкожно) или путем внутримышечных инъекций. Так, например, терапевтические соединения могут быть совмещены с соответствующим полимерным или гидрофобным материалом (например, в виде эмульсии в приемлемом масле) или ионообменной смолой, или быть использованы в виде малорастворимых производных, например, малорастворимых солей.
Композиции, при желании, могут быть выполнены в виде блока, набора, или дозирующего устройства, которые могут содержать одну или более отдельных лекарственных форм, содержащих активный ингредиент. Блок, например, может содержать металлическую или пластмассовую пленку, такую, как в блистерной упаковке. Блок или дозирующее устройство могут содержать инструкции по введению. Композиции, содержащие активные вещества, могут быть упакованы в упаковочный материал и содержать этикетку, указывающую на расстройство, для которого предназначено лекарство.
3. Введения нуклеиновых кислот, кодирующих модифицированные FVII-полипептиды (генная терапия).
Также раскрываются композиции нуклеиновых кислот, кодирующих модифицированные FVII-полипептиды, и экспрессионные векторы, кодируещие их, пригодные для генной терапии. Вместо того, чтобы доставлять белки, нуклеиновые кислоты могут быть введены in vivo системно или другим образом, или ех vivo, например, путем забора клеток, в том числе лимфоцитов, введения нуклеиновых кислот в них, и обратное их введение хозяину или реципиенту.
Модифицированные FVII-полипептиды могут быть доставлены в клетки и ткани экспрессией молекул нуклеиновой кислоты.
Модифицированные FVII-полипептиды могут быть доставлены как молекулы нуклеиновой кислоты, кодирующие модифицированные FVII-полипептиды, в том числе технологиями ех vivo и методами экспрессии in vivo. Нуклеиновые кислоты могут быть доставлены в клетки и ткани любым методом, известным из уровня техники. Выделенные последовательности нуклеиновых кислот могут быть включены в векторы для дальнейших манипуляций. Используемые в настоящем документе вектор (или плазмиды) относятся к дискретным элементам, которые используются для гетерологичного введения ДНК в клетки для экспрессии или репликации. Выбор и использование таких средств доставки известен специалистам из уровня техники.
Методы введения модифицированных FVII-полипептидов, кодируемые молекулами нуклеиновых кислот, включают введение рекомбинантных векторов. Векторы могут быть введены эписомально, например, путем добавления источника репликации или могут быть интегрирированы в хромосомы клетки. Модифицированные FVII-полипептиды также могут быть использованы в ex vivo генной терапии с использованием не-вирусных векторов. Например, могут быть сконструированы клетки для экспрессии модифицированного FVII-полипептида, например, путем включения нуклеиновых кислот, кодирующих модифицированные FVII-полипептиды, в участки генома, оперативно связанные с регуляторными последовательностями, или в такие места, где он будет оперативно связан с регуляторными последовательностями геномного участка. Такие клетки затем можно вводить локально или системно эксипиенту, например, пациенту, нуждающемуся в лечении.
Вирусные векторы относятся, например, к аденовирусам, адено-ассоциированным вирусам (AAV), поксовирусам, вирусам герпеса, ретровирусам и др., подходящим для генной терапии. Векторы могут оставаться эписамальными или могут быть интегрированы в хромосомы. Модифицированные FVII-полипептиды могут быть экспрессированы вирусом, который был введен субъекту, нуждающемуся в лечении. Вирусные векторы, подходящие для генной терапии, относятся к аденовирусам, адено-ассоциированным вирусам (AAV), ретровирусам, лентивирусам, вакцинным вирусам и другим вирусам, указанным выше. Например, аденовирусные приемы экспрессии хорошо известны из уровня техники и такие способы получения введения также хорошо известны. Аденовирусные серотипы доступны, например, из American Type Culture Collection (ATCC, Rockville, MD). Аденовирус может быть использован ex vivo, например, когда выделенные клетки пациентов, нуждающихся в лечении, обрабатываются аденовирусным вектором, экспрессирующим модифицированные FVII-полипептиды. После соответствующего периода культивирования трансдуцированные клетки вводятся пациенту местно и/или системным образом. Альтернативно, модифицированные FVII-полипептиды, экспрессируемые аденовирусным вектором, могут быть выделены и совмещены с фармацевтически приемлемым носителем для ведения терапевтически эффективного количества для профилактики, лечения или смягчения симптомов болезни или состояния пациента. Как правило, аденовирус вводят в дозе от 1 до 1014 единиц на килограмм веса тем, как правило, от 106 или 108 единиц до 1012 единиц на килограмм веса субъекта. В некоторых случаях целесообразно обеспечить исходной нуклеиновой кислотой агентами, которые нацеливают клетки, такие как антитела, специфичные к поверхностному мембранному белку или к клетке-мишени, или лиганд к рецептору клетки-мишени. FVII также может быть нацелен для высвобождения в специфических типах клеток. Например, аденовирусный вектор, кодирующий FVII-полипептиды, может быть использован для стабильной экспрессии в неделящихся клетках, таких как клетки печени (Margaritis др. (2004) J din Invest 113:1025-1031). В другом варианте, вирусные или невирусные векторы, кодирующие FVII-полипептиды, могут быть трансдуцированы в изолированные клетки для последующей доставки. Другими типами клеток для экспрессии и доставки FVII могут быть, но не ограничиваясь этим, фибробласты и эндотелиальные клетки.
Молекулы нуклеиновой кислоты могут быть введены в искусственные хромосомы и другие не-вирусные векторы. Искусственные хромосомы, такие, как ACES (см., Lindenbaum и др. (2004) Acids Res. 32 (21); е172) могут быть сконструированы для кодирования и экспрессиии изоформы. Короче говоря, искусственные хромосомы млекопитающих (MACs) являются средством внедрения больших объемов генетической информации в клетку в автономной репликации, без интеграции. Уникальная среди MACs, сателитная ДНК млекопитающих, основанная на искусственной хромосомной экспресии (АСЕ), может быть репродуктивно воспроизведена de Novo в клеточных линиях различного вида и легко очищена от хромосом клеток хозяина. Очищенные MACs млекопитающих могут быть реиндуцированы в различные реципиентные клеточные линии, где они будут стабильно поддерживаться в течение длительного времени в отсутствии селективного давления, используя АСЕ Систему. Использование этого подхода позволило достичь мишени из одного или двух генов в LMTK (-) и СНО клетках. Другим методом введения нуклеиновой кислоты, кодирующей модифицированные FVII-полипептиды, является двушаговый способ генной замены в дрожжах, начиная с полного аденовирусного генома (Ad2; Ketner др.. (1994) PNAS 91: 6186-6190), клонированного в дрожжевой. Искусственной хромосоме (YAC) и плазмиде, содержащей аденовирусную последовательность, нацеленной на определенную область в YAC клоне, экспрессионную кассету интересующего гена и положительные и отрицательные селективные маркеры. YACs представляют особый интерес, поскольку они позволяют включать большие гены. Этот подход может быть использован для конструирования аденовирусных векторов, которые переносят нуклеиновые кислоты, кодирующие любой из описанных модифицированных FVII-полипептидов, в гены клеток млекопитающих или целых животных.
Нуклеиновые кислоты могут быть инкапсулированы в носителе, таком как липосомы, либо их вводят в клетки, такие как бактериальные клетки, особенно в ослабленные бактерии, или в вирусные векторы. Например, когда используются липосомы, можно использовать протеины, которые прикрепляются к мембранному белку клеточной поверхности, связанному с эндоцитозом, для нацеливания и/или облегчения поглощения, например, капсидные белки или их фрагменты, которые имеют сродство к определенному типу клеток, антитела к белкам, которые подвергаются интернализации, и белки, которые определяют внутриклеточную локализацию и повышают время полужизни внутри клетки.
Для ex vivo и in vivo методов, молекулы нуклеиновой кислоты, кодирующие модифицированные FVII-полипептиды, вводятся в клетки из подходящего донора или субъекта, подлежащие лечению. Клетки, в которые вводятся нуклеиновые кислоты в целях терапии, включают, например, любые необходимые, доступные типы клеток для соответствующей болезни или состояния, в том числе, но не ограничиваясь этим, эпителиальные клетки, эндотелиальные клетки, кератиноциты, фибробласты, клетки мышц, гепатоциты, клетки крови, такие как Т-лимфоциты, В-лимфоциты, моноциты, макрофаги, нейтрофилы, эозинофилы, мегакариоциты, гранулоциты, различные стволовые клетки и их предшественники и, в частности, гемопоэтические стволовые клетки или их предшественники или, например, такие, как стволовые клетки, полученные из костного мозга, пуповинной крови, периферической крови, фетальной печени, а также других источников.
Для лечения ех vivo, клетки донора, совместимые с пациентом, выделяются, нуклеиновые кислоты вводят в эти выделенные клетки, и модифицированные клетки вводятся пациенту. Лечение включает в себя прямое введение, такое как, например, инкапсулирование в пористые мембраны, которые имплантируются в организм пациента (см., например, патенты США № 4892538 и 5283187, каждый из которых включен здесь в качестве ссылки в полном объеме). Подходящие методы для ввода нуклеиновых кислот в клетки млекопитающих в условиях in vitro включают использование липосом и катионых липидов (например, DOTMA, DOPE и DC-Chol) электропорацией, микроинъекцией, слиянием клеток, DEAE-декстрановой трансфекцией, и методом осаждения фосфатом кальция. Способы доставки ДНК могут быть использованы для экспрессии модифицированных FVII-полипептидов в условиях in vivo. Такие методы включают липосомальную доставку нуклеиновых кислот и депротеинизированной ДНК, включая местные и системные доставки, с использованием электропорации, ультразвука и кальций-фосфатную доставку. Другие методы включают микроинъекции, слияние клеток, хромосом-опосредованный перенос генов, микроклеточный-опосредованный перенос генов и сферопластовое слияние.
In vivo экспрессия FVII модифицированных полипептидов может быть связана с экспрессией других дополнительных молекул. Например, экспрессия FVII модифицированных полипептидов может быть связана с экспрессией цитотоксических продуктов, например в сконструированном вирусе или цитотоксическом вирусе. Такие вирусы могут быть направлены на конкретный тип клеток, которые являются мишенью для терапевтического эффекта. Экспрессия модифицированных FVII-полипептидов может быть использована для повышения цитотоксичности вируса.
Экспрессия in vivo модифицированных FVII-полипептидов может включать в себя оперативное связывание молекул нуклеиновой кислоты, кодирующих модифицированные FVII-полипептиды, со специфичными регуляторными последовательностями, такими как клеточно-специфичные или ткане-специфичные промотеры. Модифицированные FVII-полипептиды также могут быть экспрессированы векторами, которые специально заражали, и/или реплецированы в целевых типовых клетках и/или тканях. Индуцибильные промотеры можно использовать для селективного регулирования экспрессии модифицированных FVII-полипептидов. Примером регулируемой системы экспрессии является доксициклин-индуцируемая система экспрессии гена, которая была использована для регулирования экспрессии рекомбинантного FVII (Srour et al.. (2003) Thromb Haemost. 90 (З): 398-405).
Молекулы нуклеиновых кислот, как депротеинизированная ДНК или векторы, искусственные хромосомы, липосомы и другие частицы, могут вводиться субъекту системно, наружно, местно и другими способами введения. Когда осуществляется введение системно и in vivo, молекула нуклеиновой кислоты или другие частицы, содержащие молекулы нуклеиновой кислоты, можгут быть нацелены на клетку.
Введение также может быть, например, путем введения вектора или клеток, которые обычно ориентированы на определенные клетки или ткани. Например, опухолевые и пролифирирующие клетки могут быть нацеленными клетками для экспрессии in vivo модифицированных FVII-полипептидов. Клетки, используемые для экспрессии in vivo модифицированных FVII-полипептидов, также включают аутологичные для пациента клетки. Такие клетки могут быть удалены у пациента, внесены нуклеиновые кислоты для экспресии модифицированных FVII-полипептидов, а затем введены пациенту например, путем инъекции или приживлением трансплантанта.
I. Терапевтическое использование
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы для лечения любых состояний, при которых применяется рекомбинантный FVII. Как правило, такое лечение включает увеличение коагуляции, такое как увеличение гемостатического ответа. Модифицированные полипептиды FVII обладают терапевтической активностью самостоятельно или в сочетании с другими агентами. Модифицированные полипептиды, предусмотренные в настоящем документе, созданы таким образом, чтобы сохранять терапевтическую активность и проявлять модифицированные свойства, в частности, повышенную устойчивость к TFPI, повышенную устойчивость к AT-III, повышенную каталитическую активность, увеличенное время полураспада и/или увеличенную способность связывать и/или аффинность к активированным тромбоцитам. Такие модифицированные свойства, например, могут повысить терапевтическую эффективность полипептидов посредством увеличения коагулянтной активности модифицированных полипептидов FVII. Этот раздел содержит примеры использования и методы введения. Эти описанные виды терапии приведены в качестве примеров и не ограничивают применение модифицированных полипептидов FVII.
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы для различных видов терапевтического применения, а также в диагностических методах, в которых применяется FVII. Такие методы включают, но не ограничиваются, методами лечения физиологических и медицинских состояний и описаны ниже. Модифицированные FVII-полипептиды, предусмотренные в настоящем документе, могут проявлять увеличение активности in vivo и терапевтического эффекта по сравнению с FVII дикого типа, включая более низкие дозы для достижения того же эффекта и другие улучшения в ведении и лечении, такие как меньшие и/или менее частые введения, уменьшение побочных эффектов и увеличенный терапевтический эффект. Модифицированные полипептиды FVII могут быть введены как зимогены FVII (т.е. в одноцепочечной форме), но, как правило, модифицированные полипептиды FVII, предусмотренные в настоящем документе, вводят в активированной двухцепочечной форме, например, после самоактивации или активации другими факторами коагуляции, например, во время очистки.
В частности, модифицированные полипептиды FVII предназначены для использования в терапевтических методах, в которых FVII использовали для лечения. Такие методы включают, но не ограничиваются, методами лечения заболеваний и расстройств, включающих, но не ограничивающихся, нарушениями свертывания крови, гематологическими расстройствами, геморрагическими расстройствами, гемофилиями, такими как гемофилия А, гемофилия В и дефицит фактора VII, и приобретенными заболеваниями крови, такими как приобретенная недостаточность фактора VII, вызванная болезнью печени. Модифицированные полипептиды FVII также могут быть использованы при лечении заболеваний, связанных с кровотечениями, и расстройств, включая, но не ограничиваясь, тромбоцитопенией (например, возникшей из-за химиотерапии), болезнью Виллебранда, наследственными тромбоцитарными заболеваниями (например, заболеваниями пула, такими как синдромы Чедиака-Хигаси и Германски-Пудлака, дисфункцией тромбоксана А2, тромбастенией Гланцманна и синдромом Бернарда-Сулье), гемолитическим уремическим синдромом, наследственной геморрагической телеангиэктазией, известной также как синдром Ренду-Ослера-Вебера, аллергической пурпурой (болезнью Шенлейна-Геноха) и диссеминированной интраваскулярной коагуляцией.
В некоторых вариантах, кровотечения, подвергающиеся лечению полипептидами FVII, происходят в таких органах, как мозг, внутренняя область уха, глаза, печень, легкие, опухолевые ткани, желудочно-кишечный тракт. В других вариантах, кровотечение является диффузным, таким как геморрагический гастрит и обильное маточное кровотечение. Пациенты с нарушениями свертываемости крови, такими как, например, гемофилиями А и В, часто подвергаются риску кровотечений во время операции или травмы. Такие кровотечения могут проявляться в виде острых гемартрозов (кровотечений в суставах), хронической гемофильной артропатии, гематом (например, мышечных, ретроперитонеальных, сублингвальных и ретрофарингеальных), гематурии (кровотечения из почечного тракта), кровотечений в центральной нервной системе, желудочно-кишечных кровотечений (например, кровотечений верхнего пищеварительного тракта) и кровотечений в мозг, которые также можно лечить с помощью модифицированных полипептидов FVII. Кроме того, любые кровотечения, связанные с хирургией (например, гепатэктомией) или удалением зубов, можно лечить с помощью модифицированных полипептидов FVII. В одном варианте, модифицированные полипептиды FVII могут быть использованы для лечения эпизодов кровотечений, возникших в результате травмы, или операции, или уменьшенного количества или активности тромбоцитов у субъекта. Примеры методов для пациентов, подвергающихся хирургическому вмешательству, включают лечение для предотвращения кровотечений и лечения до, во время и после таких операций, включая, но не ограничиваясь, сердечной хирургией, ангиопластикой, легочной хирургией, абдоминальной хирургией, хирургией позвоночника, сосудистой хирургией, стоматологической хирургией и трансплантацией органов, включая трансплантацию костного мозга, сердца, легкого, поджелудочной железы или печени.
Лечение заболеваний и состояний модифицированными полипептидами FVII может быть произведено их введением любым подходящим путем с использованием подходящих композиций, как описывалось выше, включая, но не ограничиваясь, инъекцией, легочным, оральным и трансдермальным введением. Лечение обычно осуществляется внутривенным введением болюса.
В случае необходимости, конкретная дозировка, продолжительность и протокол лечения могут быть эмпирически определены или экстраполированы. Примеры доз рекомбинантного и нативного полипептидов FVII могут быть использованы в качестве отправной точки для определения соответствующих дозировок. Например, рекомбинантные FVII (rFVIIa) полипептиды, которые были активированы в rFVIIa, Novoseven ®, были введены пациентам с гемофилией А или гемофилией В с эпизодом кровотечения в дозе 90 мкг/кг болюсной инфузией от 2 до 5 минут, с получением эффективного циркулирующего уровня не менее 2 мкг / мл, со средним периодом полураспада 2,7 часа. Дозы повторяют каждые 2 часа, пока не будет достигнут гемостаз. Модифицированные FVII-полипептиды, которые обладают повышенной коагулянтной активностью вследствие повышенной устойчивости к TFPI, повышенной устойчивости к AT-III, увеличенной каталитической активности, увеличенного периода полураспада и/или увеличенной способности связывать и/или аффинности к активированным тромбоцитам, могут быть эффективными при малых дозах и/или малой частоте введения по сравнению с рекомбинантным FVII. Дозировка для FVII дикого типа и немодифицированных полипептидов FVII может быть использована в качестве ориентира для определения доз для модифицированных полипептидов FVII. Такие факторы, как уровень активности и время полураспада модифицированных FVII по сравнению с немодифицированными FVII, могут быть использованы для определения таких доз. Определенные дозы и режимы могут быть эмпирически определены.
Дозы и режимы могут быть определены исходя из известных доз и режимов, и, в случае необходимости, могут быть экстраполированы на основе изменений свойств модифицированных полипептидов и/или могут быть определены эмпирически на основе целого ряда факторов. Такие факторы включают вес тела индивидуума, общее состояние здоровья, возраст, активность конкретного применяемого соединения, пол, диету, время введения, уровень экскреции, комбинацию лекарств, тяжесть и течение болезни, а также предрасположение пациента к болезни и решение лечащего врача. Активный ингредиент, полипептид, как правило, используется в сочетании с фармацевтически эффективным носителем. Количество активного ингредиента, который может быть комбинирован с носителями для производства единичных дозированных форм или мультидозированных форм, может изменяться в зависимости от реципиента и определенного способа введения.
Влияние полипептидов FVII на время свертывания крови можно контролировать с помощью любого из анализов на свертывание, известных в данной области, включая, но не ограничиваясь, протромбиновым временем цельной крови (РТ), активированным частичным тромбопластиновым временем (АЧТВ), активированным временем свертывания крови (АКТ), активированным рекальцифицированным временем свертывания крови, или временем свертывания Ли-Вайта.
После улучшения состояния пациента, если необходимо, может быть введена поддерживающая доза соединения или композиции; и дозировка, лекарственная форма, частота введения, или их сочетания могут быть изменены. В некоторых случаях субъекту может потребоваться прерывистое долгосрочное лечение при повторении симптомов заболевания или на основе запланированных доз. В других случаях, могут быть необходимы дополнительные введения в ответ на острые события, такие как кровотечение, травма или хирургическое вмешательство.
Ниже приведены некоторые примеры состояний, при которых FVII (вводится в виде FVIIa) использовали в качестве агента для лечения отдельно или в комбинации с другими агентами.
1. Врожденные нарушения свертывания крови
а. Гемофилия.
Врожденная гемофилия является рецессивным расстройством крови, при котором наблюдается снижение уровня факторов свертывания в плазме, что приводит к нарушению каскада коагуляции и увеличению времени свертывания крови. Гемофилия А, которая составляет примерно 85% всех случаев гемофилии, является результатом мутации (мутаций) в гене фактора VIII в Х-хромосоме, что приводит к дефициту или дисфункции белка FVIII. Гемофилия В вызвана дефицитом или дисфункцией фактора свертывания FIX, как правило, возникшими в результате точечных мутаций или делеций в гене FIX в Х хромосоме. Заболеваемость гемофилией А в мире составляет примерно 1 случай на 5000 мужчин, и 1 случай на 25000 мужчин для гемофилии В. Гемофилии А и В далее классифицируются как легкие, умеренные и тяжелые. Уровень 5%-25% нормального функционирующего фактора VIII или IX в плазме классифицируется как легкая гемофилия, 1%-5% является умеренной, и менее 1% является тяжелой. Гемофилия С, часто называемая недостаточностью FXI, является относительно легким и редким заболеванием, которое затрагивает 1 из 100000 человек, является аутосомно-рецессивным.
Гемофилии А и В различно проявляются клинически. В отличие от травм и операций, малые порезы и ссадины не приводят к чрезмерным кровотечениям. У пациента также будут многочисленные кровотечения в суставах и мышцах и легко проявляются кровоподтеки. Гемартроз или кровотечения в суставах являются одним из основных осложнений у больных гемофилией, и может происходить спонтанно или в ответ на травму. Блоковидные суставы, такие как коленный, локтевой и голеностопный суставы, затрагиваются чаще всего. Бедра и плечи страдают гораздо реже, так как вокруг шаровидных суставов больше мускулатуры, защищающей их от повреждений. Кровотечение может вызвать сильную острую боль, ограничивать движения и привести к осложнениям, включая синовиальную гипертрофию. Кроме того, повторяющиеся кровоизлияния в суставы могут вызвать хронический синовит, который может вызвать повреждения суставов, уничтожая синовиальную оболочку, хрящи и кости. Опасные для жизни кровотечения, такие как внутричерепные кровоизлияния и кровотечения в центральной нервной системе, также затрагивают субъектов с гемофилией. Внутричерепные кровоизлияния происходят приблизительно у 10% пациентов с тяжелой гемофилией, приводя к уровню смертности 30%. Гемофилия С является более мягкой. Спонтанные кровотечения редки, и кровоизлияния в суставы, мягкие ткани и мышцы также являются редкими. Кровотечения, как правило, лечат переливанием свежезамороженной плазмы (FFP), FXI заместительной терапией, или, для местного лечения, лечения внешних ран или удаления зубов, фибриновым клеем.
Наиболее частым видом лечения гемофилии А или В является заместительная терапия, при которой пациенту вводят FVIII или FIX. Композиции коммерчески доступны как полученные из плазмы или рекомбинантные продукты, рекомбинантные белки в настоящее время являются предпочтительным видом лечением для ранее не получавших лечения пациентов. Хотя эти виды терапии могут быть очень успешными, если пациент вырабатывает ингибиторы к введенным фактору VIII или фактору IX, возникают осложнения. Ингибиторы представляют собой антитела IgG, основном подкласса IgG4, которые взаимодействуют с FVIII или FIX и препятствуют прокоагулянтной функции. Ингибиторы затрагивают приблизительно 1 из 5 пациентов с тяжелой гемофилией А. Большинство субъектов вырабатывают эти ингибиторы вскоре после введения первой инъекции фактора VIII, которая часто происходит в раннем детстве, хотя субъекты вырабатывают их позже. Ингибиторы также затрагивают примерно 1 из 15 человек с легкой или умеренной гемофилией А. Эти ингибиторы обычно вырабатываются во взрослом возрасте и не только уничтожают вводимые экзогенные FVIII, но и эндогенные FVIII. В результате, легкая и умеренная гемофилии становятся тяжелыми. Клинически, пациенты с гемофилией А с ингибиторами делятся на респондентов с высоким и низким ответом в зависимости от силы анамнестического ответа, который они проявляют при повторном экспонировании с FVIII. Ингибиторы затрагивают примерно 1 из 100 пациентов с гемофилией В. В большинстве случаев, ингибиторы вырабатываются после первого введения терапевтического фактора IX и могут сопровождаться аллергическими реакциями.
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы для лечения пациентов с гемофилией, особенно пациентов с гемофилией, у которых вырабатываются ингибиторы. Рекомбинантный продукт FVIIa (Novoseven, Novo Nordisk) был одобрен и лицензирован для лечения эпизодов кровотечения при гемофилии А или В у пациентов с ингибиторами FVIII или FIX и для профилактики кровотечений при хирургических вмешательствах или инвазивных процедурах у пациентов с гемофилией А или В с ингибиторами FVIII или FIX. Лечение с использованием rFVIIa увеличивает образование тромбина в обход потребности в FVIIIa и/или FIXa. Коагуляция начинается в месте повреждения в результате взаимодействия rFVIIa с ТФ, что приводит к первоначальной активации FX, тромбина и активации тромбоцитов. Полная коагуляция с помощью rFVIIa может быть осуществлена по ТФ-зависимому и ТФ-независимому механизмам, где некоторое количество образованного тромбина может возникнуть в результате прямой активации FX на активированных тромбоцитах с помощью rFVIIa, который сам связывает активированные тромбоциты через взаимодействия низкой аффинности с фосфолипидами мембран.
Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы в терапии для лечения гемофилии, в том числе для лечения эпизодов кровотечения и предотвращения кровотечения при хирургических вмешательствах или инвазивных процедурах. Модифицированные FVII-полипептиды обладают повышенной устойчивостью к ингибитору ТФ/FVIIa комплекса, TFPI, повышенной устойчивостью к AT-III, увеличенной каталитической активностью, увеличенным временем полураспада и/или увеличенной способностью связывать и/или аффинностью к активированным тромбоцитам. Поэтому FVII-полипептиды отображают повышенную коагулянтную активность в ТФ-зависимом пути (например, через повышение устойчивости к TFPI), и/или ТФ-независимом пути (например, путем увеличения способности связывать и/или аффинности к активированным тромбоцитам). Таким образом, модифицированные полипептиды FVII могут быть использованы для обеспечения более активной терапии для лечения гемофилии. Примеры улучшений терапии с использованием модифицированных FVII-полипептидов включают, но не ограничиваются, более низкими дозами, меньшими и/или менее частыми введениями, снижением побочных эффектов, а также увеличенным терапевтическим эффектом.
Модифицированные полипептиды FVII, как правило, вводятся как активированные FVII (FVIIa) полипептиды. Модифицированные полипептиды FVII могут быть проверены на терапевтическую эффективность, например, с помощью животных моделей. Например, мыши с вызванной антителами гемофилией, или любая другая известная модель для гемофилии, могут подвергаться лечению модифицированными полипептидами FVII. Прогрессирование фенотипа и симптомов заболевания контролируется с целью оценки эффекта модифицированных полипептидов FVII. Модифицированные полипептиды FVII также могут быть введены субъекту, например, в ходе клинических испытаний для оценки эффективности in vivo по сравнению с плацебо-контролем и/или контролем с немодифицированными FVII.
b. FVII дефицит
Дефицит фактора VII является аутосомным рецессивным расстройством, связанным с кровоточивостью, которое затрагивает приблизительно 1 из 500000 человек. Дефицит FVII может быть клинически легким, умеренным или тяжелым, с легкой и умеренной недостаточностью, характеризующейся повышенной кровоточивостью после операций и травм. Пациенты с тяжелой недостаточностью FVII (менее 1% FVII активности) проявляют аналогичные гемофилии симптомы. Например, FVII-дефицитные субъекты склонны к кровотечениям в суставах, спонтанным носовым кровотечениям, желудочно-кишечным кровотечениям, кровотечениям из мочевыводящих путей. Также описаны внутримозговые кровоизлияния и кровотечения в мышцах, тогда как женщины могут столкнуться с тяжелой меноррагией (тяжелые менструальные кровотечения). Лечение может быть осуществлено путем проведения заместительной терапии. Рекомбинантный продукт FVIIa (Novoseven®, Novo Nordisk) был одобрен и лицензирован для лечения эпизодов кровотечений у пациентов с врожденным дефицитом FVII и для профилактики кровотечений при хирургических вмешательствах или инвазивных процедурах у пациентов с врожденной недостаточностью FVII. Таким образом, модифицированные полипептиды FVII по настоящему документу могут быть использованы аналогичным образом. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы при лечении эпизодов кровотечений и предотвращения кровотечений при хирургических вмешательствах или инвазивных процедурах у FVII-дефицитных пациентов. Например, новорожденным пациентам с тяжелым дефицитом FVII с внутричерепными кровоизлияниями внутривенно болюсом могут быть введены модифицированные FVII-полипептиды для достижения коагуляции и поддержания гемостаза. Обычно, модифицированные полипептиды FVII вводят как активированные FVII (FVIIa) полипептиды.
с. Другое
Другие заболевания, связанные с кровотечениями, можно лечить с помощью FVII-полипептидов, предусмотренных в настоящем документе для содействия коагуляции. Врожденные дефициты факторов V и Х также приводят к повышенному времени свертывания крови, и потенциально могут подвергаться лечению введением терапевтических доз FVII. Например, пациенту с дефицитом фактора Х может быть введен rFVIIa для контроля кровотечений, связанных со спленектомией (Boggio и др., (2001) Br J Haematol 112:1074-1075). Спонтанные и связанные с хирургией эпизоды кровотечения, связанные с болезнью Виллебранда (vWD), также можно лечить с помощью модифицированных полипептидов FVII, содержащихся в настоящем документе. VWD является заболеванием, связанным с кровотечением, обусловленным дефектом или дефицитом белка свертывания крови, фактора Виллебранда (vWF), и, по оценкам, затрагивает от 1% до 2% населения. У субъектов с vWD легко проявляются гематомы, происходят периодические кровотечения из носа, кровотечения после удаления зубов, удаления миндалин или других операций, и у женщин-пациентов могут быть увеличенные менструальные кровотечения. Модифицированные полипептиды FVII могут быть использованы для улучшения спонтанных и связанных с хирургией кровотечений у vWD пациентов (von Depka и др., (2006) Blood Coagul Fibrin 17:311-316). Другие связанные с тромбоцитами нарушения свертываемости крови, такие как, например, тромбастения Гланцманна и синдром Германски-Пудлака также связаны со сниженной эндогенной активностью свертывания. Избыточные спонтанные или связанные с операциями кровотечения у пациентов со связанными с тромбоцитами нарушениями также могут регулироваться терапевтическими дозами модифицированных полипептидов FVII. Например, пациента с тромбастенией Гланцманна, подвергающегося хирургической операции, можно лечить модифицированными полипептидами FVII до, во время и/или после операции для предотвращения серьезной потери крови (Van Buuren и др., (2002) Dig Dis Sci 47:2134-2136). Как правило, модифицированные полипептиды FVII вводят как активированные FVII (FVIIa) полипептиды.
2. Приобретенные расстройства, связанные с кровоточивостью
а. Приобретенная в результате химиотерапии тромбоцитопения
Расстройства, связанные с кровоточивостью, также могут быть приобретенными, а не врожденными. Так, например, химиотерапия, например, против лейкемии и других раковых заболеваний, может привести к тромбоцитопении. Она возникает, вероятно, из-за потери образования тромбоцитов в костном мозге больных, получающих химиотерапию, и обычно происходит после 6-10 дней после лечения. Лечение приобретенной тромбоцитопении обычно осуществляется тромбоцитами, эритроцитами или переливанием плазмы, что служит для предотвращения любых ненормальных спонтанных кровотечений, которые могут быть результатом дефицита тромбоцитов. Кровотечения у пациентов с вызванной химиотерапией тромбоцитопенией или любыми другими приобретенными или врожденными тромбоцитопениями, также могут регулироваться введением терапевтических количеств модифицированных полипептидов FVII, предусмотренных в настоящем документе. Например, пациентам с тромбоцитопенией с неконтролируемым кровотечением, например, в желудочно-кишечном тракте, может быть введен внутривенно болюс, содержащий терапевтическое количество полипептидов FVII для остановки кровотечения (Gerotziafas и др., (2002) Am J Hematol 69:219-222). Как правило, модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
и. Другие коагулопатии
Другие приобретенные коагулопатии можно лечить с помощью модифицированных полипептидов FVII, предусмотренных в настоящем документе. Коагулопатии могут быть следствием условий, включающих, но не ограничивающихся, фульминантной печеночной недостаточностью (FHF, такой как вызванной гепатотоксическими наркотиками, токсинами, метаболическими заболеваниями, инфекционными заболеваниями и ишемией), другими заболеваниями печени, включающими цирроз и заболевания, связанные с болезнью Вильсона, дефицит витамина К (например, вызванный лечением антибиотиками или диетой), гемолитический уремический синдром, тромбоцитопению (ТТК) и диссеминированную внутрисосудистую коагулопатию (DIC). Обычное лечение, как правило, представляет собой переливание плазмы, введение эритроцитов (RBC), или тромбоцитов, но может быть неудачным. В одном варианте, модифицированные полипептиды FVII могут быть введены пациенту с FHF, переносящему инвазивные процедуры для предотвращения кровотечения. Обычное лечение свежезамороженной плазмой (FFP) часто является неудачным и может потребовать большого количества плазмы, создавая объемную перегрузку и анасарку (генерализованную инфильтрацию отечной жидкости в подкожную соединительную ткань). Лечение терапевтическими количествами модифицированных полипептидов внутривенным введением болюса FVII во время, до и/или после инвазивной хирургии, такой как, например, биопсии печени или трансплантации печени, может предотвратить кровотечение и достигнуть гемостаза у больных FHF. Пациент может контролироваться по РТ крови для определения эффективности лечения (Shami и др., (2003) Liver Transpl 9:138-143). В другом варианте, FVII могут быть введены пациенту с тяжелым кровотечением, связанным с коагулопатией, таким как, например, тяжелым интраабдоминальным кровотечением после кесарева сечения, связанным с дисфункцией печени и DIC, которое не реагирует на обычное переливание или инфузию (Moscardo и др., (2001) Br J Haematol 113:174-176). Кроме того, модифицированные полипептиды FVII могут быть использованы для лечения коагулопатии у новорожденных и детей. В конкретном варианте, новорожденные и дети не реагируют на обычное лечение, такое как RBC и инфузия тромбоцитов. Например, новорожденным с тяжелой легочной геморрагией, связанной с увеличением РТ, которые не реагируют на RBC и трансфузию тромбоцитов, могут быть введены модифицированные FVII-полипептиды для уменьшения РТ и установления гемостаза (Olomu и др., (2002) J Permatol 22:672-674). Модифицированные полипептиды FVII, предусмотренные в настоящем документе, проявляют повышенную коагулянтную активность по сравнению с немодифицированными полипептидами FVII, и поэтому могут вводиться, например, в более низких дозах, реже и с меньшим количеством побочных эффектов. Модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
с. Кровотечения, приобретенные в результате трансплантации
Тяжелое кровотечение после трансплантации костного мозга (ВМТ) и трансплантации стволовых клеток (SCT) является относительно частым и опасным для жизни осложнением, связанным с этими процедурами, обусловленным снижением количества тромбоцитов. Например, диффузное альвеолярное кровоизлияние (DAH) является легочным осложнением ВМТ с оцениваемой встречаемостью 1-21% среди пациентов с пересадкой, смертность составляет 60-100%. Обычное лечение таких эпизодов кровотечения включает лечение кортикостероидами и переливанием плазмы, введением тромбоцитов и/или RBC, хотя эти виды лечения в значительной степени являются неудачными с общим уровнем смертности примерно 50% (Hicks и др., (2002) Bone Marrow Transpl 30:975-978). Для лечения DAH и установления гемостаза может быть осуществлено введение FVII внутривенным болюсом, с одновременным введением кортикостероидов и/или введением тромбоцитов или без них, (Hicks и др., (2002) Bone Marrow Transpl 30:975-978). Модифицированные полипептиды FVII, предусмотренные в настоящем изобретении, проявляют повышенную коагулянтную активность по сравнению с немодифицированным полипептидом FVII, и поэтому могут вводиться, например, в более низких дозах, реже и с меньшей продолжительностью лечения, с меньшим количеством побочных реакций при той же биологической активности и эффективности. Модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
d. Кровотечения, вызванные антикоагулянтной терапией
У пациентов, подвергающихся антикоагулянтной терапии для лечения состояний, таких как тромбоэмболия, могут проявляться эпизоды кровотечения при экстренном приеме антикоагулянтов, таких как варфарин, гепарин и фондапаринукс, или развиться геморрагические расстройства в результате такого длительного лечения. Лечение эпизодов кровотечений обычно включает введение прокоагулянтов, таких как витамин К, плазма, экзогенные FIX и протамины для нейтрализации гепарина. Для нейтрализации влияния антикоагулянтов, увеличения РТ, аРРТ и/или других маркеров коагуляции и установления гемостаза также может быть осуществлено введение экзогенного FVII, (Deveras и др., (2002) Ann Inten Med 137:884-888). Модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы для контроля эпизодов кровотечения у пациентов с приобретенными нарушениями свертываемости крови вследствие антикоагулянтной терапии. Модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
е. Приобретенная гемофилия
Ингибиторы фактора VIII могут спонтанно выработаться у во всем остальном здоровых людей, что приводит к состоянию, известному как "приобретенная гемофилия". Приобретенная гемофилия является редким заболеванием, ежегодная заболеваемость составляет 0,2-1,0 на миллион населения. Аутоантитела в основном представляют собой IgG4 антитела, которые, при связывании с FVIII, ингибируют активность FVIII, нарушая расщепление тромбина, взаимодействие с фактором Виллебранда и/или связывание фосфолипидов. Это приводит к опасной для жизни геморрагии у, приблизительно, 87% пациентов. Частые места кровотечения представляют собой кожу, слизистые оболочки, мышцы и забрюшинное пространство, в отличие от пациентов с наследственной гемофилией, у которых кровотечения происходят преимущественно в суставах и мышцах. Приобретенная гемофилия может подвергаться лечению концентратом активированного протромбинового комплекса или рекомбинантным активированным фактором VII (Novoseven®, Novo Nordisk) для борьбы с эпизодами кровотечения. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, проявляют повышенную коагулянтную активность по сравнению с немодифицированными полипептидами FVII, и поэтому могут вводиться, например, в более низких дозах, реже и при меньшей продолжительности лечения, с меньшим количеством побочных реакций при той же биологической активности и эффективности. Модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
3. Кровотечения при травме и хирургическом вмешательстве
FVII-полипептиды могут быть использованы в качестве терапии для лечения кровотечений, связанных с травматической и периоперационной кровопотерей у субъектов с нормальными системами коагуляции. Например, FVII-полипептиды могут быть введены пациенту для содействия свертыванию крови и снижения потери крови, связанной с хирургическим вмешательством и, кроме того, могут уменьшить потребность в переливании крови. В одном варианте, FVII-полипептиды могут быть введены субъектам, подвергающимся позадилонной (залобковой) простатэктомии. Позадилонная простатэктомия часто связана со значительной потерей крови и необходимостью последующего переливания. Субъектам, подвергающимся такой или подобной операции, может быть введен болюс, содержащий терапевтическое количество FVII, на раннем этапе операции для сокращения периоперационной потери крови путем усиления коагуляции в области операции. Сокращение потерь крови приводит к устранению необходимости переливания крови у этих больных (Friedrich и др., (2003) Lancet 361:201-205). FVII-полипептиды могут быть введены пациентам с нормальной коагуляцией, подвергающимся другим видам хирургического вмешательства, для достижения быстрого гемостаза и предотвращения потери крови. Неограничивающие примеры хирургических процедур, в которых в качестве терапии для сокращения периоперационной потери крови может быть использован FVII, как правило, вводимый в активированной форме (т. FVIIa), включают, но не ограничиваются, хирургией сердечных клапанов (Al Douri и др., (2000) Blood Coag Fibrinol 1 1: S121-S127), заменой клапанов аорты (Kastrup и др., (2002) Ann Thorac Surg 74:910-912), резекцией рекуррентной гемангиоперицитомы (Gerlach и др., (2002) J Newosurg 96:946-948), раковой хирургией (Sajdak и др., (2002) Eur J Gynaecol Oncol 23:325-326) и хирургией двенадцатиперстной кишки (Viot и др., (2000) Am J Med 108:421-423). Лечение FVII может способствовать гемостазу в области, подвергающейся операции, и сократить или предотвратить потерю крови, тем самым ослабляя или отменяя потребность в переливании крови. Модифицированные полипептиды FVII, предусмотренные в настоящем документе, разработаны для проявления повышенной коагулянтной активности по сравнению с немодифицированными полипептидами FVII, и поэтому могут вводиться, например, в более низких дозах, реже и с меньшим количеством побочных реакций. Модифицированные полипептиды FVII вводят как активированные FVII (FVIIa) полипептиды.
Полипептиды фактора VII также могут быть использованы для содействия коагуляции и предотвращения потери крови у субъектов с травмой. Травма определяется как повреждение живой ткани внешним агентом и является четвертой по значимости причиной смертности в Соединенных Штатах. Травмы классифицируются на тупые травмы (приводящие к внутреннему сжатию, повреждению органов и внутреннему кровоизлиянию) или проникающие травмы (приводящие к проникновению агента в тело и уничтожению тканей, сосудов и органов, приводящей к внешнему кровотечению). Травма может быть вызвана несколькими событиями, включая, но не ограничиваясь, дорожно-транспортными происшествиями (что приводит к тупой и/или проникающей травме), огнестрельными ранениями (приводящими к проникающим травмам), ножевыми ранениями (приводящими к проникающим травмам), производственными происшествиями (приводящими к проникающим и/или тупым травмам) и падениями со значительной высоты (приводящими к проникающим и/или тупым травмам). Неконтролируемое кровотечение в результате травмы ответственно за большинство связанных с травмами смертельных случаев. Диффузная коагулопатия является довольно частым осложнением у пациентов с травмой, встречающимся у 25-36% субъектов. Коагулопатия может развиваться вскоре после травмы в результате целого ряда факторов, таких как разведение и потребление факторов свертывания крови и тромбоцитов, фибринолиз, ацидоз и гипотермия. Обычные способы лечения включают заместительную терапию с переливанием свежезамороженных тромбоцитов плазмы (FFP), RBC и/или криопреципитата, коррекцию ацидоза и лечение гипотермии. Эти действия часто являются недостаточными для остановки кровотечения и предотвращения смертельного исхода. Лечение введением терапевтических количеств FVII может способствовать коагуляции и уменьшению кровопотери у пациентов с травмой. Например, пациентам с огнестрельными травмами при тяжелой потере крови, в дополнение к хирургическому вмешательству, для контроля коагулопатического кровотечения будут вводиться FVII (Kenet и др., (1999) Lancet 354:1879). Коагулянтная FVII терапия может эффективно понизить потерю крови и кровотечения у пациентов с тупой и проникающей травмой (Rizoli и др., (2006) Crit Care 10: R178). Модифицированные полипептиды FVII, предусмотренные в настоящем документе, созданы для проявления повышенной коагулянтной активности по сравнению с немодифицированными полипептидами FVII, и поэтому могут вводиться, например, в более низких дозах, реже и с меньшим количеством побочных реакций. Модифицированные полипептиды FVII вводятся как активированные FVII (FVIIa) полипептиды.
J. Комбинированная терапия
Любой из модифицированных описанных здесь полипептидов FVII может быть введен в сочетании с, до, с перерывами, или после других терапевтических агентов или процедур, включая, но не ограничиваясь, другими биопрепаратами, низкомолекулярными соединениями и хирургией. Для любого заболевания или состояния, включая те, которые были приведены в качестве примеров выше, для которых показан или был использован FVII (включая FVIIa и rFVIIa), и в отношении которых пригодны другие агенты и виды лечения, FVII может быть использован в сочетании с ними. Таким образом, модифицированные полипептиды FVII, предусмотренные в настоящем документе, могут быть использованы аналогичным образом. В зависимости от заболевания или состояния, подвергающегося лечению, примеры комбинаций включают, но не ограничиваются, комбинациями с другими очищенными из плазмы или рекомбинантными факторами коагуляции, прокоагулянтами, такими как витамин К, производными витамина К и ингибиторами протеина С, плазмой, тромбоцитами, эритроцитами и кортикостероидами.
К. Изделия и наборы
Фармацевтические препараты модифицированных полипептидов FVII или нуклеиновых кислот, кодирующих модифицированные FVII-полипептиды, или их производных или биологически активных частей могут быть упакованы в изделиях, содержащих упаковочный материал, фармацевтическую композицию, которая является эффективной для лечения заболеваний или расстройств гемостаза, и этикетку, которая указывает, что модифицированные полипептиды FVII или молекулы нуклеиновой кислоты предназначены для лечения заболевания или расстройства гемостаза.
Изделия, предусмотренные в настоящем документе, содержат упаковочный материал. Упаковочные материалы, используемые в упаковке фармацевтической продукции, хорошо известны специалистам в данной области. См., например, патенты США №№5323907, 5052558 и 5033352, каждый из которых включен здесь в полном объеме. Примеры фармацевтических упаковочных материалов включают, но не ограничиваются, блистерной упаковкой, бутылями, пробирками, ингаляторами, помпами, пакетами, флаконами, контейнерами, шприцами, бутылками и любым упаковочным материалом, подходящим для выбранной композиции и выбранного способа введения и лечения. Широкий спектр композиций соединений и композиций, предусмотренных в настоящем документе, рассматривается как множество возможных вариантов лечения для любого заболевания или расстройства гемостаза.
Модифицированные полипептиды FVII и молекулы нуклеиновых кислот также могут быть предусмотрены как наборы. Наборы могут включать фармацевтическую композицию, описанную выше, и изделие для введения. Например, модифицированные FVII могут поставляться вместе устройством для введения, таким как шприц, ингалятор, чашка для дозирования, пипетка или аппликатор. Набор необязательно включает инструкцию по применению, включающую дозировки препаратов, режимы дозирования и инструкции по способам введения. Наборы также могут включать фармацевтическую композицию, описанную выше, и изделие для установления диагноза. Например, такие наборы могут включать изделие для измерения концентрации, количества или активности FVII или FVII регулируемую систему субъекта.
Следующие примеры включены в иллюстративных целях и не ограничивают сферу применения изобретения.
L. Примеры
Пример 1
In silico получение вариантов фактора VII с повышенной устойчивостью к TFPI
А. Моделирование взаимодействия фактора VII и TFPI
Компьютерное моделирование взаимодействия фактора VII и его естественного ингибитора, первого домена Кунитца (К1) ингибитора пути, опосредованного тканевым фактором (TFP I)-I (TFPI-1 К-1), было осуществлено с целью определить контактирующие аминокислотные остатки на поверхности сайта взаимодействия. Для создания модели гомологии была использована публично доступная информация из банка данных белков (rcsb.org/pdb/). Пока еще не определены ни кристаллическая структура TFPI-1 К-1, ни структура четвертичного комплекса ТФ/FVIIa/TFPI-l/FXa. Вместо этого, компьютерное моделирование проводилось с использованием 2,1 А кристаллической структуры тройного комплекса между ТФ, FVII и 5L15 вариантом бычьего панкреатического ингибитора трипсина (BPTI5L15; PDB код IFAK_1) в качестве начальной модели процесса, после чего использовалась информация о кристаллической структуре комплекса трипсинЛТР1-2 (Фиг.6). BPTI5L15 представляет собой сериновую протеазу Кунитц-типа и проявляет гомологию с TFPI-1 и TFPI-2. Первый домен Кунитца (К1) BPTI5L15 (SEQ ID NO: 106) характеризуется 45% идентичностью первичной последовательности в первом домене Кунитца (К1) (фиг.5). Координаты для кристаллической структуры TFPI-2 К1 были взяты из файла 1TFX программы базы данных (pdb) из банка данных белков, который описывает кристаллическую структуру трипсинЛТР1-2 комплекса. Координаты TFPI-2 К1 были выровнены с BPTI5L15 трехмерными координатами кристаллической структуры ТФ/FVIIA/BPTI5L15 (pdb файл IFAK1) с использованием программы выравнивания, рассматривающей С-α скелет как твердое тело, в приложениях из пакета PyMol (pymol.sourceforge.net/). Анализ соответствия TFPI-2 К1 в модели путем измерения совпадений привел к среднеквадратичному отклонению (RMSD) менее 1 Å. Это указывает на точное выравнивание между двумя структурами и надежность модели FVII и TFPI-2 К-1 комплекса.
Боковые цепи TFPI-2 К-1, которые по данным модели находятся в тесном контакте с поверхностью FVII, были выявлены и подвергнуты in silico мутагенезу для соответствия боковым цепям в TFPI-1 K1. Результаты этого анализа показали наличие электростатической комплементарности между фактором VII и TFPI-1 K1 по нескольким остаткам в непосредственной близости от активного центра (т.е. остаткам 2-ой сферы). Остатки в FVII, участвующие во взаимодействии с TFPI-1 K1 по данным приведенного выше анализа на гомологию, включают D60, К60а, К60с, Т99, R147 и К 192 по химотрипсиновой нумерации, которые соответствуют D196, К197, К199, Т239, R290 и К341, соответственно, по нумерации зрелого FVII (Фиг.7). Рассмотрение модели взаимодействия показывает, что остаток глицина в позиции 97 по химотрипсиновой нумерации (G97), который соответствует G237 по нумерации зрелого FVII, находится в непосредственной близости от контактирующих остатков. Модификации по этому положению могут привести к стерическим препятствиям, которые нарушат взаимодействие FVII с TFPI.
В. In silico мутагенез фактора VII, направленный на обеспечение повышенной устойчивости к TFPI-1 K1
Аминокислотные остатки, расположенные рядом или непосредственно на взаимодействующих поверхностях FVII/TFPI-1 K1, были определены, как описано выше. Создаваемые варианты FVII с измененными аминокислотами обладали а) заменами комплементарных электростатических контактов между FVII и TFPI-1 на отталкивающие контакты между зарядами и/или b) нарушенными положительными электростатическими контактами вследствие замен заряженных остатков в FVII на нейтральные остатки и/или с) стерическими затруднениями, обусловленными заменами остатков, содержащих небольшие боковые цепи, остатками, содержащими большие боковые цепи. Примеры вариантов приведены в таблице 9.
Примеры вариантов фактора VII
Пример 2
Клонирование и экспрессия FVII
А. Клонирование FVII
Последовательность нуклеотидов, кодирующая 466-аминокислотную изоформу предшественника полипептида FVII человека (Р08709; приведенную в SEQ ID NO: 1), была клонирована в векторе экспрессии млекопитающих pCMV Script (Stratagene; SEQ ID NO: 151), который содержит промотер цитомегаловируса (CMV). Олигонуклеотиды СВО-125 (SEQ ID NO: 152) и СВО-126 (SEQ ID NO: 153) были использованы в качестве прямого и обратного праймеров, соответственно, для амплификации последовательности FVII методом ПЦР с использованием кДНК человеческого FVII (Invitrogen) в качестве матрицы. СВО-125 праймер содержит сайт рестрикции BamHI (выделен жирным шрифтом), последовательность Козака (подчеркнут два раза), за которой следуют 18 нуклеотидов, гомологичных 5' концу последовательности кДНК FVII (подчеркнута), включая стартовый кодон ATG. СВО-126 праймер содержит сайт рестрикции EcoRI (выделен жирным шрифтом), стоп-кодон (подчеркнут два раза) и 21 нуклеотид, гомологичные 3' концу последовательности кДНК FVII (подчеркнута).
СВО-125 прямой
праймер 5 'gcatcateacgtgacggatccgccaccatggtctcccaggccctc3'
СВО -126 обратный праймер
5' gatcgtaceatacgtgaattcctagggaaatggggctcgcapgag 3'
Были использованы стандартная реакция ПЦР и условия термоциклирования в сочетании с набором KoD HiFi PCR (EMD Biosciences), как рекомендовано изготовителем. Продукт ПЦР был расщеплен ферментами рестрикции ВатН I и EcoR I и лигирован по сайтам рестрикции ВатН I и EcoR I с pCMV Script вектором с помощью стандартных молекулярных методов. Вектор затем был трансфецирован в Escherichia Coli. Выбранные колонии были выращены, и бактериальные клетки собрали для очистки плазмид с использованием рутинных методов молекулярной биологии.
В. Получение вариантов FVII
Варианты FVII были получены с использованием набора QuikChange II XL Site-Directed Mutagenesis (Stratagene) в соответствии с инструкциями изготовителя, со специально разработанными олигонуклеотидами, которые служили праймерами, которые вносили конкретные мутации во вновь синтезированную ДНК. Метод QuikChange предполагает линейную амплификацию матрицы ДНК с помощью PfuUltra высокоточной ДНК-полимеразы. Комплементарные праймеры, которые включают желаемую мутацию, были удлинены во время циклирования с использованием очищенного двуспирального суперспирализованного вектора pCMV Scpipt, который содержал клонированную кДНК FVII в качестве матрицы. Удлинение праймеров привело к включению интересующей мутации во вновь синтезированные цепи и созданию мутированной плазмиды с одноцепочечными разрывами. После амплификации, нуклеиновые кислоты обрабатывали Dpn I, который расщепил dam-метилированные родительские цепи Е. coli pCMV Script вектора. Это привело к "отбору" вновь синтезированных мутированных плазмид, которые не были метилированы. Векторная ДНК, содержащая требуемую мутацию (мутации), была трансфецирована в XLlO-Gold ультракомпетентные клетки Е. coli, где бактериальная лигаза репарировала одноцепочечные разрывы, что привело к нормальной репликации.
Были созданы варианты FVII с единичными аминокислотными заменами по позициям D196, К197, К199, G237, Т239, R290 и К341, по нумерации зрелого FVII (соответствующим D60, К60а, К60с, G97, Т99, R147 и К192 по химотрипсиновой нумерации). FVII варианты с несколькими мутациями по позициям D196, К197 и К199, или мутациями по позициям D196 и К197, также были получены. Нуклеотидные последовательности одного из олигонуклеотидов каждой пары комплементарных праймеров, используемых для получения вариантов FVII, приведены в таблице №10. Соответствующие последовательности дикого типа также показаны для сравнения. Последовательность из 3 пар оснований, которая кодирует замененную аминокислоту, показана жирным шрифтом. Например, для создания варианта FVII, содержащего замену D196K (D60K по химотрипсиновой нумерации; СВ554, SEQ ID NO: 18), СВО-157 D60K олигонуклеотид и олигонуклеотид, комплементарный СВО-157 D60K, были использованы с набором QuickChange II XL Site-Directed Mutagenesis для замены 3 пар оснований "gac" последовательности дикого типа на последовательность 3 пар оснований "aag". Мутированный вектор, кодирующий вариант полипептида FVII, содержащий замену D196K, затем был трансфецирован в XLlO-Gold ультракомпетентные клетки Е. coli.
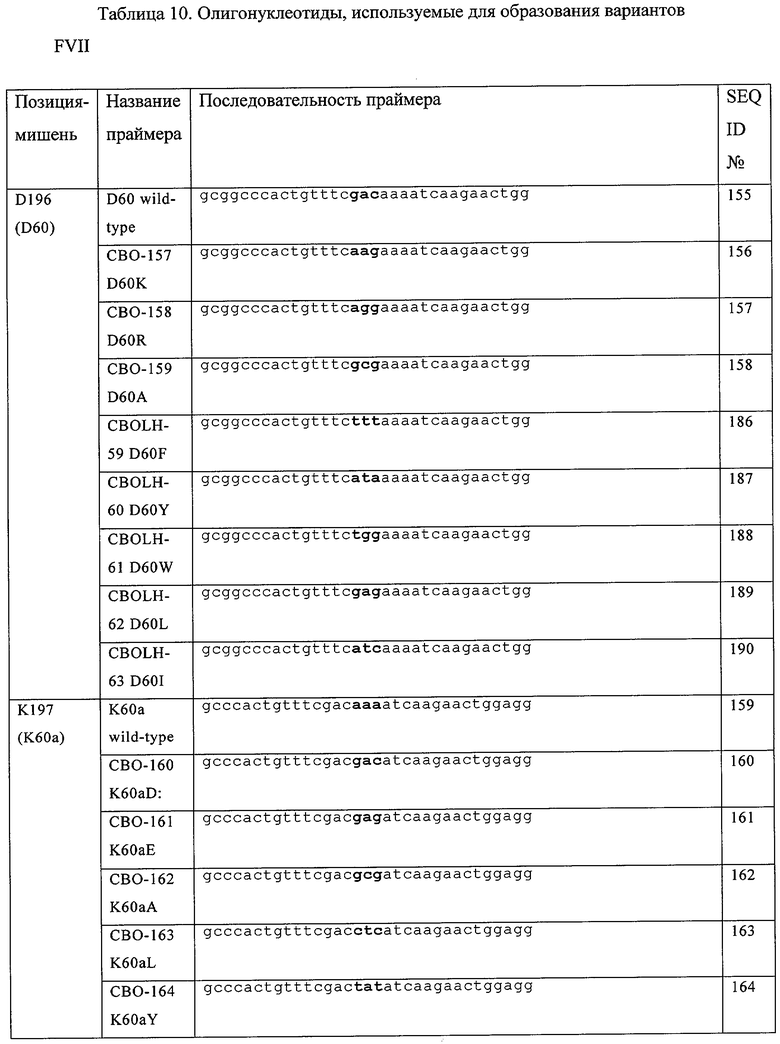
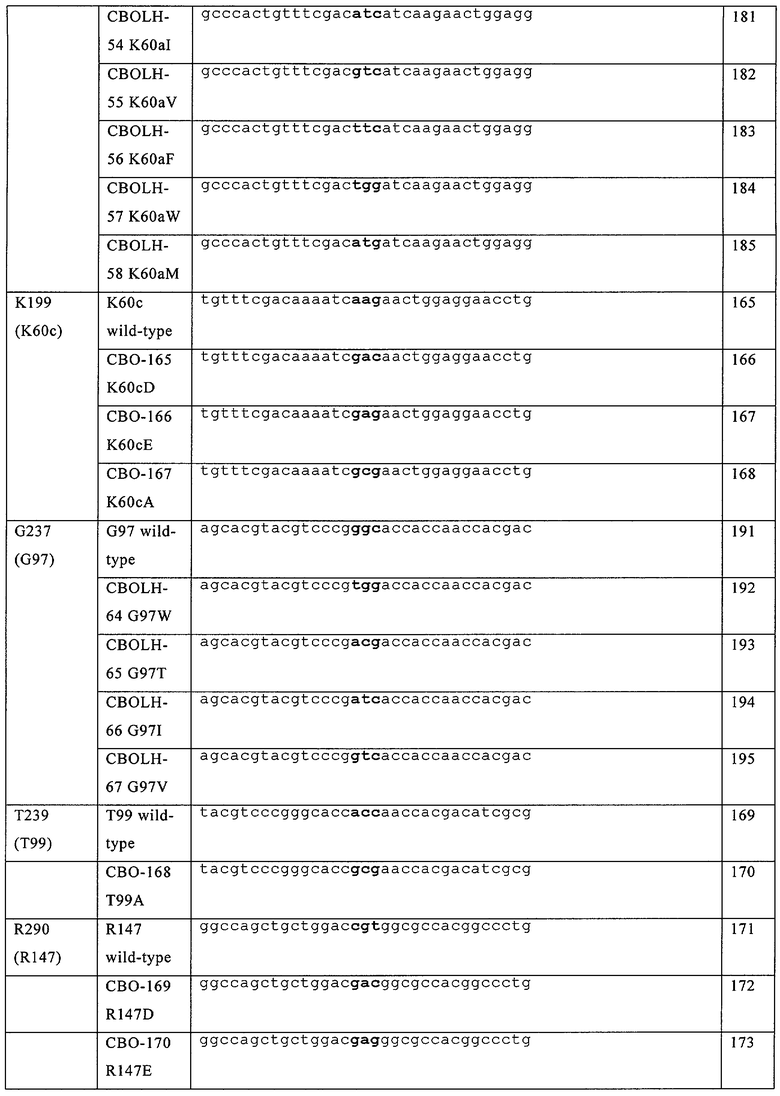
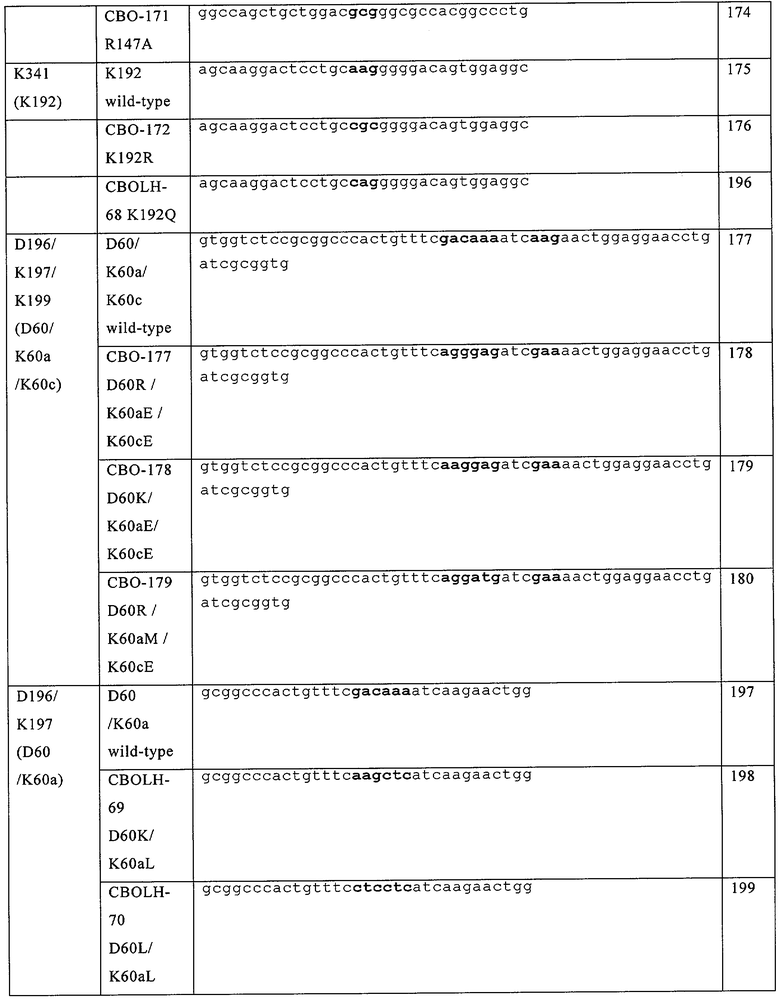
Wild type -дикий тип
С. Экспрессия полипептидов FVII
Для первоначального анализа экспрессии с помощью ELISA и вестерн-блоттинга, FVII-полипептиды экспрессировали в клетках ВНК-21. Для биохимических анализов, таких как описано ниже, FVII-полипептиды экспрессировали в клетках FreestyleTM 293-F (Invitrogen).
Полипептид фактора VII дикого типа (СВ553-02, SEQ ID NO: 3) и варианты полипептида FVII первоначально были экспрессированы в линии клеток почек детенышей хомяков ВНК-21 (АТСС CRL 1632). ВНК-21 клетки культивировали в минимальной среде Игла (ЕМЕМ, Invitrogen) с 10% эмбриональной телячьей сывороткой (FCS) в 100 мм культуральных чашках при 37°С и 5% СO2. После роста до, приблизительно, 90% покрытия, клетки трансфецировали 24 мкг FVII плазмидной ДНК с помощью набора Липофектамин 2000 (Invitrogen) в соответствии с инструкциями изготовителя. Культуральную среду заменили через 6 часов после трансфекции на бессывороточную ЕМЕМ, содержащую 1 мкг/мл витамина K1 (Sigma), и клетки инкубировали в течение еще 72 часов. Экспрессию FVII в среде с культурой клеток оценивали по ELISA или вестерн-блоттингу.
Для последующих биохимических анализов, полипептид фактора VII дикого типа (СВ553-02, SEQ ID NO: 3) и варианты полипептида FVII экспрессировали в клетках FreestyleTM 293-F (Invitrogen). Клетки культивировали в среде FreestyleTM 293 (Invitrogen) при 37°С и 8% СО2 в колбах Эрленмейера с вентилируемыми колпачками. Клетки трансфецировали по предложенному производителем протоколу. После роста до 1×106 клеток/мл, клетки центрифугировали и сменили среду. Затем клетки трансфецировали 240 мкг плазмидной ДНК FVII на каждые 240 мл клеток с использованием 293fectin (Invitrogen). Кроме того, к каждым 240 мл клеток было добавлено 50 мкл 1 мг/мл раствора витамина K1 (Sigma) в этаноле. Клетки растили в течение 5 дней, а затем собрали культуральный супернатант. Экспрессию FVII в среде культуры клеток оценивали с помощью ELISA.
1. ELISA
Иммуноанализ был использован для оценки количества человеческого FVII и FVIIa в образце. Поликлональные антитела к человеческому FVII использовали для захвата и обнаружения протеаз в растворе. Иммуноанализ может быть использован для определения концентрации белка в кондицированной среде или очищенном сырье, или для определения концентрации FVII в другом образце, например образце плазмы человека или мыши. Обычная концентрация для FVII в крови человека составляет около 50 нМ, а для энзиматически активной формы FVIIa составляет, приблизительно, 1 нМ.
Для определения количества человеческих FVII или FVIIa белков в образцах был выполнен ELISA сэндвич-типа. Девяностошестиячеечные планшеты Maxisorp immune (Nunc) с плоским дном были покрыты 100 мкл 5 нг/мкл раствора авидина на ячейку (NeutrAvidin, Pierce Biotech.). Планшеты закрыли и инкубировали при встряхивании в течение 1 часа при комнатной температуре (RT) с последующей четырехкратной промывкой PBS с 0,01% Твин-20 (PBST). Планшеты блокировали в течение не менее 1 часа при комнатной температуре при встряхивании инкубацией с 1% раствором бычьего сывороточного альбумина (БСА) (вес/объем) в PBS, добавляя в каждую ячейку по 200 мкл раствора. Блокированные планшеты затем хранили при 4°С до использования (до 2 недель).
Перед использованием планшеты промывали четыре раза PBST для удаления БСА, и в каждую ячейку добавляли 100 мкл 1 нг/мкл раствора биотинилированных анти-фактор VII антител (R & D Systems), и планшеты инкубировали при комнатной температуре в течение 45 минут при встряхивании для комплексообразования с авидиновым покрытием. Избыток свободных антител удалили промыванием планшета PBST (четыре раза).
Серию разведений стандарта FVII в два раза (American Diagnostica;
разбавлен PBST), начиная с 50 нг/мкл до 0,8 нг/мкл, добавили в планшеты по 100 мкл на ячейку. В качестве буферного контроля была включена ячейка, содержащая PBST без FVII. Для анализа очищенных образцов (до и после активации, см. Пример 3) FVII или FVIIa, пробу сначала разводили 1: 25 PBST, а затем делали серию 2-кратных разведений, так чтобы тестировать 25-кратно, 50-кратно, 100-кратно и 200-кратно разбавленные растворы. Разбавленные образцы добавляли в ячейки, по две ячейки на образец, по 100 мкл на одну ячейку. Для анализа образцов плазмы, содержащих FVII или FVIIa, образцы плазмы разводили 1:100 и 1:400 PBST и добавляли в ячейки, по две ячейки на образец, по 100 мкл на одну ячейку. Образцы плазмы без FVII или FVIIa также были включены для определения фонового уровня. Затем планшеты инкубировали в течение 30 минут при комнатной температуре при встряхивании, позволяя FVII или FVIIa в образце образовать комплекс с анти-FVII антителами.
После инкубации с образцом, планшеты 4 раза промывали PBST. Вторичные антитела лошади против FVII человека (American Diagnostica) разбавили 1:5000 в PBST и добавили в каждую ячейку в объеме 100 мкл. Планшеты инкубировали в течение 30 минут при комнатной температуре при встряхивании для связывания антител с FVII или FVII комплексами на планшете. Для удаления избытка вторичных антител планшеты промыли PBST (4 раза). Для выявления связанных вторичных антител в каждую ячейку добавили 100 мкл HRP-конъюгированных антител коза анти-лошадь при разведении 1:5000 в PBST. После инкубации в течение 30 минут при комнатной температуре при встряхивании, планшеты промыли четыре раза PBST и добавили раствор, содержащий 1:1 смеси ТМВ субстрата и раствора пероксида водорода (Pierce Biotech.), 100 мкл на ячейку. Планшеты встряхивали около 1 минуты при комнатной температуре до добавления 2М H2SО4,100 мкл на ячейку, для остановки реакции. Оптическая плотность при 450 нм была измерена с помощью планшет-ридера Molecular Device M5, и фоновое значение для планшета (измеренное для PBST) было вычтено из измеренного для каждой ячейки значения. Калибровочный график был построен отложением значений концентрации стандарта FVII против абсорбции. Диапазон калибровочной кривой около 0,2-50 нг/мл обычно определяли в соответствии с указанными выше условиями ELISA. Концентрации каждого образца определяли с помощью калибровочной кривой при умножении на коэффициент разбавления с определением среднего и стандартного отклонения.
2. Вестерн-блоттинг
Экспрессию FVII в культуральной среде оценивали с помощью вестерн-блоттинга. Аликвоты, содержащие неразбавленные образцы или два последовательных разведения в 2 раза в PBS культуральной среды трансфецированных FVII ВПК-21 клеток, отметили как конц. 1 (неразбавленный), конц. 2 (2-кратное разведение) и конц. 3 (4-кратное разведение). Образцы загрузили на SDS PAGE гель рядом с 10, 25 и 50 нг очищенного rFVII плазмы в качестве контроля (American Diagnostica, CB553-01). FVII белок, образованный ВПК-21 клетками, был обнаружен с помощью вестерн-блоттинга с первичными поликлональными лошадиными анти-FVII антителами (American Diagnostica;
используемыми в предложенной производителем концентрации) и HPR-конъюгированными вторичными антителами против лошадиного IgG (разведение 1:2000 1 мг/мл раствора от Zymed Laboratories). Сравнение уровня экспрессии проводилось с очищенным rFVII плазмы в качестве контроля. Результаты показали, что в аликвотах культуральной среды концентрация составляла от, приблизительно, 20 нг до более чем 50 нг FVII.
Пример 3
Очистка и активация полипептидов FVII
FVII-полипептиды очищали с помощью Q Sepharose Fast Flow (XKL 6) колонки, на которой адсорбировались FVII-полипептиды с функциональными доменами Gla, а затем элюировались с помощью кальция. 240 мл культурального супернатанта трансфецированных FreestyleTM 293 -F клеток разбавили в 2 раза раствором, содержащим 20 мМ Трис с рН 8,0 и 0,01% Твин-20, а затем к разбавленному образцу добавили 1,5 мл 500 мМ ЭДТА с рН 8,0. Образцы отфильтровали перед загрузкой на Q Sepharose Fast Flow (XKL 6) колонку, которая была предварительно уравновешена сначала буфером В (20 мМ Трис с рН 8,0, 1 М NaCl, 0,01% Твин-20), затем буфером А (20 мМ Трис с рН 8,0,0,15 М NaCl, 0,01% Твин-20) при 8 мл/мин. После загрузки колонку промывали буфером, пока оптическая плотность потока при 280 нм не достигла базового уровня. Буфер заменили буфером С (20 мМ Трис с рН 8,0,0,15 М NaCl, 0,01% Твин-20, 5 мМ CaCl2), и для полной замены буфера в колонке провели промывание насосом. После завершения промывки, буфер С пропускали через колонку со скоростью 8 мл/мин до элюирования FVII-полипептидов, которые собирали в 5 мл фракции в течение 60 минут. После элюции колонку промыли буфером В, в то же время собирая фракции, пока розовый пигмент (из культуральной среды) не смылся с колонки. Затем колонку промыли буфером А, чтобы подготовить ее для очистки FVII из следующего образца.
Элюированные фракции затем очистили с помощью Mono Q 5/5 колонки (содержащей 1 мл смолы), которую предварительно уравновесили сначала буфером В, а затем буфером А при 2 мл/мин 5 мл фракции с 5 по 8, собранные с буфером С, были объединены и разбавлены в 2 раза буфером А, а затем к ним добавили 1,6 мл 500 мМ ЭДТА, рН 8,0. На этой стадии для анализа, например, с помощью ELISA, по желанию могут быть отобраны аликвоты (100 мкл). Объединенные образцы загрузили на колонку Mono Q при 2 мл/мин, а затем промыли буфером А. Для элюирования связанных полипептидов FVII, через колонку пропускали градиент буфера В от 0% до 30% в течение 20 минут при 1 мл/мин, при этом собирали 0,5 мл фракции. Затем колонку промыли буфером В, а затем буфером А для подготовки к очистке FVII из следующего образца.
Очищенные FVII-полипептиды активировали в FVIIa с помощью биотинилированного фактора Ха из набора Restriction Protease Factor Xa Cleavage and Removal (Roche). Обычно, 7 фракций с Mono Q объединяли в 15 мл коническую пробирку и к раствору добавляли 388 мкл 500 мМ CaCl2, 38,9 мкл 10% БСА в дистиллированной воде и 3,2 мкг биотинилированного фактора Ха. После инкубации в течение 14-16 часов при температуре 37°С к образцу добавили 250 мкл иммобилизованного авидина (Pierce), и образец перемешивали при температуре 4°С в течение 30 минут. Полученный раствор отфильтровали через Econo-Pak колонку (Bio-Rad), и фильтрат смешали с 250 мкл иммобилизованного авидина и перемешивали в течение 30 минут. Раствор вновь отфильтровали, и фильтрат сконцентрировали до, приблизительно, 300-500 мкл с помощью Amicon Ultra-4 10 кДа центрифужного фильтра (Millipore). Концентрацию FVIIa определяли с помощью ELISA (как описано в Примере 2.С.1), и уровень активации фактора VII контролировали с помощью вестерн-блоттинга. Вестерн-блоттинг проводили, как описано в примере 2.С.2, но с использованием антител кролика против человеческого фактора VII (Haematologic Technologies, Inc) в разведении 1: 2000 в качестве первичных антител в течение 1 часа, а затем HRP-коза-анти-кролик IgG (H+L) (Invitrogen) в разведении 1:5000 в течение 30 минут.
Пример 4
Определение константы Михаэлиса-Ментен для амидолитической активности FVIIa к низкомолекулярным субстратам
Амидолитическую активность вариантов FVII оценивали путем измерения константы Михаэлиса-Ментен полипептидов FVIIa на пептидильном субстрате Spectrozyme FVIIa (CH3SO2-D-CHA-But-Arg-pNA.AcOH). Липидированный человеческий очищенный тканевый фактор (Innovin, Dade Behring, VWR Cat#68100-390) был включен в анализ для обеспечения оптимальной активности FVIIa. ТФ-FVIIa комплекс расщепляет Spectrozyme FVIIa как весьма специфичный хромогенный субстрат, высвобождая паранитроанилинный хромофор (pNA), уровень которого может контролироваться путем измерения поглощения при 405 нм. Ферментативная активность определяется путем измерения абсорбции свободного pNA при 405 нм как функции времени.
Реакции проводили при трех разных концентрациях фермента. Для реакции варианты FVIIa сначала разбавляли до 40 нМ в 1 × буфере для прямого анализа (100 мМ Трис с рН 8,4, 100 мМ NaCl, 5 мМ CaCl2, 0,01% БСА) в 1,7 мл пробирках (низкоадгезивные микроцентрифужные пробирки из ISC Bioexpress). Затем FVIIa разбавляли в присутствии ТФ (Innovin, Dade Behring) до 2 нМ в 12-ячеечной полипропиленовой емкости (Axygen) следующим образом: смешивали 720 мкл 5× прямого буфера (500 мМ Трис с рН 8,4, 500 мМ NaCl, 25 мМ СаСl2 0,05% БСА), 180 мкл 40 нМ FVIIa и 2700 мкл 2×ТФ (6 нМ раствор, восстановленный в 10 мл воды). Разбавленную протеазу инкубировали в течение 5 минут при комнатной температуре. 2 нМ исходного раствора FVIIa далее разбавляли в 2 раза для получения серии разведений из 1 нМ и 0,5 нМ растворов протеазы, соответственно, также в присутствии ТФ. Последовательные реакции разведения: сначала 1800 мкл 2 нМ раствора FVIIa/ТФ разбавили в 360 мкл 5× прямого буфера, 900 мкл 2 × ТФ и 540 мкл воды. Этот разбавленный раствор разбавли 1:1 1800 мкл 1х ТФ прямым буфером.
Был использован планшет с разведениями субстрата Spectrozyme FVIIa (American Diagnostica). Исходный расвор Spectrozyme FVIIa получили восстановлением 50 мкмолей в дистиллированной воде до 10 мМ, и хранили при температуре 4°С. Восемьдесят мкл 7,5 мМ Spectrozyme FVIIa раствора, полученного из 10 мМ Spectrozyme FVIIa, 60 мкл (10 мМ Spectrozyme FVIIa)+20 мкл воды, добавили в ячейки в две соседние колонки 96-ячеечных полипропиленовых планшетов для анализа (Costar). Содержимое двух ячеек последовательно разбавляли в 2 раза в каждой из 8 ячеек в соответствующей колонке для получения серии 10× концентраций субстрата от 10 мМ до 78 мкМ субстрата в ячейках в первой колонке и от 7,5 мМ до 58,6 мкМ субстрата в ячейках во второй колонке.
Пять мкл каждого разведения субстрата Spectrozyme FVIIa добавили в ячейки свободной половины 96-ячеечного планшета для анализа (Costar). Сорок пять мкл каждого из трех разведений FVIIa/ТФ добавили в три группы колонок с сериями разведений субстрата. На этом этапе были приняты меры, чтобы избежать введения пузырьков в ячейки для анализа. Если пузыри были введены, их убирали прокалыванием чистой иглой до начала каждого анализа. Содержимое планшетов перемешивали покачиванием. До начала анализа, длина оптического пути ячеек для анализа была измерена с помощью планшетного спектрофотометра Spectramax Gemini M5 (Molecular Devices) путем определения конечной точки и с помощью функции Pathcheck программного обеспечения SoftMax Pro (Molecular Devices). Увеличение абсорбции при 405 нм измеряли каждые 30 секунд в течение одного часа при 37°С.
Программное обеспечение SoftMax Pro было использовано для преобразования уровня абсорбции (миллиединиц/сек) в концентрацию свободного pNA (мкМ/сек) с помощью оптического пути и коэффициента экстинкции уходящей группы pNA при 405 нм, 9600 М-1см-1. Уравнение преобразования: уровень × (1/60×1000)×(1/9600 × длина оптического пути) × 100000. Результаты для каждой концентрации протеазы были отражены на графике с помощью программного обеспечения Graph Pad Prism с концентрацией субстрата на оси × и определяемой величиной мкМ/сек по оси у. С использованием программного обеспечения Graph Pad Prism 4 были определены Ymax и Км путем сопоставления данных с уравнением Михаэлиса-Ментен:
У=((kкатКM/1000000)Х[Е])/(1+(X+КM),
где: Х представляет собой концентрацию субстрата (мкМ),
У представляет собой активность фермента (мкМ / сек)
kкатКM является константой (М-1сек-1)
КM является константой Михаэлиса (мкМ)
Е представляет собой концентрацию фермента (мкМ)
Были заданы начальные значения Е=1, КM=Х при 0,5×Yмакс и kкатКM=1000.
Удельная активность (миллиединиц(оптической плотности)/ мин / нМ) каждого из вариантов FVIIa, FVIIa дикого типа (СВ553-02) была определена с помощью описанного выше анализа с 420 мкМ Spectrozyme FVIIa (табл.11). Активность вариантов FVIIa, как правило, была выше, чем активность FVIIa дикого типа (оценена в 2,7 миллиединиц / мин / нМ). Активность вариантов FVII сопоставили с активностью вариантов, описанных в литературе (СВ591-СВ594), и большинство вариантов FVII обладали сопоставимой или большей активностью, чем активность вариантов, которые ранее были описаны в литературе.
Удельная активность вариантов FVIIa
Пример 5. Определение каталитической активности FVIIa для его субстрата, фактора Х
Каталитическая активность вариантов FVIIa для их субстрата, фактора Х (FX), была оценена косвенно с помощью флуорогенных анализов на определение активности FXa, образованного в результате активации FVIIa, с помощью синтетического субстрата Spectrafluor FXa. Липидированная форма очищенного тканевого фактора (ТФ) была включена в анализ для обеспечения оптимальной активности FVIIa. Ферментативная активность FXa по отношению к Spectrafluor FXa (CH3S02-D-CHA-Gly-Arg-AMC.AcOH) определяли путем измерения увеличения абсорбции свободного флуорофора АМС, (7-амино-4-метилкумарина), как функции времени.
Варианты FVIIa сначала разбавили до 0,5 мкМ 1 × буфером для прямого анализа (100 мМ Трис с рН 8,4,100 мМ NaCl, 5 мМ CaCls и 0,01% БСА), затем дополнительно разбавили до 0,1 нМ буфером для прямого анализа. Липидированный ТФ с полной длиной цепи (Innovin; Dade Behring) восстановили в 20 мл воды с получением 3 нМ раствора и разбавили до 0,2 нМ 1 × буфером для прямого анализа. Четыреста мкл 0,1 нМ FVIIa смешали с 400 мкл 0,2 нМ ТФ и инкубировали при комнатной температуре в течение 5 минут. Раствор дополнительно разбавили 2 раза в 2 раза 1 × буфером для прямого анализа, содержащим 0,2 нМ ТФ для получения в общей сложности трех разведении FVIIa с 0,05 нМ, 0,025 нМ, или 0,0125 нМ FVIIa, каждое из которых содержит 0,2 нМ ТФ (FVIIa / ТФ растворы).
Субстрат, фактор Х (FX, American Diagnostica; 80 мкг) восстановили в 135,6 мкл дистиллированной воды с получением 10 мкМ раствора и хранили в аликвотах при температуре -80°С. Аликвоты не замораживались и размораживались более одного раза. Исходный раствор FX разбавили до 800 нМ в буфере для прямого анализа, затем последовательно разбавляли в 2 раза с получением растворов FX в диапазоне от 800 нМ до 50 нМ.
Spectrofluor Xa (American Diagnostica; 10 мкмолей) восстановили в дистиллированной воде до 5 мМ и хранили при 4°С. 5 мкл Spectrofluor Xa (American Diagnostica) добавили в каждую ячейку черного 96-ячеечного планшета для анализа с уменьшенной вдвое плошадью ячеек (Costar). Затем в каждую ячейку добавили 25 мкл раствора FX. В последний ряд ячеек планшета, в качестве негативного контроля, не было добавлено FX. В двух повторностях, три концентрации раствора ТФ/ FVIIa по 20 мкл добавили в ячейки соответствующих колонок планшета так, чтобы каждое разведение ТФ / FVIIa анализировали против разведения FX, с одним набором колонок, не содержащих ТФ / FVIIa (т.е. только FX). Планшеты перемешивали встряхиванием. Флуоресценцию измеряли во времени, причем спектрофлуориметр осуществлял измерения каждые 30 секунд в течение 1 часа при температуре 37°С (возбуждение: 380 нм, излучение: 450 нм, светотеневая граница: 435 нм), время отмечалось в единицах квадрата времени. После анализа для перехода от единиц флуоресценции к единицам молярности субстрата, высвобожденного в анализе, на том же планшет-ридере была построена стандартная кривая АМС флуоресценции. 1 мМ АМС в ДМСО (Invitrogen) разбавили до 0,02 мМ 1 × буфером для прямого анализа. Были получены шесть последовательных разведений АМС 1 × буфером для прямого анализа в два раза в диапазоне от 20 нМ до 0,625 нМ. Флуоресценцию АМС измеряли в тех же условиях анализа, как описано выше, и построили график флуоресценции против концентраци АМС. Был рассчитан наклон этой линии, который использовали как коэффициент пересчета из единиц флуоресценции в мкМ в последующих расчетах.
Константы кинетики активации FX с помощью FVIIa были рассчитаны с помощью линейной регрессии зависимости обратной концентрации субстрата от обратной скорости расщепления субстрата (в секундах2), где Vmax, Fviia рассчитывали как величину, обратную отрезку, отсекаемому но оси Y, Kmpviia рассчитывали как наклон прямой, отсекающей отрезок на оси Y, и Vmax/К.т, pviia как величину, обратную наклону. Значение Ккат получили по уравнению:
Ккат/Km, FVIIa=Vmax/Кm, FVIIa × 1/(0,5 × k2 × [FVIIa в мкМ] × (единицы флуоресценуии / коэффициент пересчета в мкМ)),
где: k2=([S] × Ккат, FXa) / (KM, FXa+[S]), Где Ккат, FXa и KM, FXa являются константами для FXa расщепления Spectrofluor Xa, определяемыми экспериментально для FXa cтандартов, Ккат, FXa=117 сек-1 и KM, FXa=164 мкМ.
С использованием указанных выше условий анализа значение кинетической константы k2 было определено как 88,1 сек-1.
КM и Ккат для каждого варианта FVIIa были определены для оценки каталитической активности, Ккат/КM (М-1сек-1), каждого варианта для субстрата FX (табл.12). Для FVIIa протеазы дикого типа (СВ553-02) также было определено, что она проявляет активность 1,8×107 М-1сек-1. Активация фактора Х фактором Vila, как определено Krishnaswamy и др. (J. Biol. Chem. (1998) 273:8 4378-86), составляет 2,9×107 М-1сек-1. Множество вариантов проявляли активность, сопоставимую с активностью немодифицированной протеазы. В некоторых случаях, варианты FVIIa проявляли увеличенную каталитическую активность по сравнению с немодифицированной протеазой. Например, СВ558, СВ561 и СВ563 проявляли каталитическую активность 5,2×107, 4,3×107, 5,9×107 M-1 сек-1.
Каталитическая активность вариантов FVIIa
Ориентировочное значение
Пример 6. Определение концентрации каталитически активной протеазы в растворе
Концентрацию каталитически активного FVIIa в растворе определяли титрованием комплекса фактора Vila и растворимого тканевого фактора (рТФ) необратимым пептидным ингибитором FVIIa, Phe-Phe-Arg-хлорметилкетоном (FFR-CMK). Ингибитор связывается с FVIIa, но не FVII. Продолжительная инкубация при высокой концентрации FVIIa (50 нМ) обеспечивает полное титрование протеазы. Для определения концентрации каталитически активного FVIIa в исходном растворе была измерена остаточная активность FVIIa / ТФ комплекса после инкубации с FFR-CMK.
А. 96-ячеечный формат планшета для анализа
В 96-ячеечный прозрачный планшет с вдвое уменьшенной площадью ячеек (Nunc) предварительно добавили 150 мкл/ячейку 1 × планшетного буфера (100 мМ Трис с рН 8,4,100 мМ NaCl, 0,01% БСА, 0,01% Твин-20) в каждую ячейку и инкубировали планшет при 37°С минимум в течение 1 часа. Буфер был полностью удален промоканием фильтровальной бумагой и центрифугированием планшета вверх дном для устранения любых остатков буфера, планшет высушивали на воздухе в течение 1 часа и хранили закрытым при комнатной температуре (RT).
Для приготовления FVIIa /рТФ/ FFR-CMK реакционной смеси, раствор FVIIa (American Diagnostica; разбавили до 5 мкМ 50% глицерином (объем / объем) и хранили в охлажденных аликвотах при температуре -20°С) или варианта FVIIa сначала разбавляли до 500 нМ, 1 × буфером для прямого анализа (100 мМ Трис с рН 8,4,100 мМ NaCl, 5 мМ СаСl2 0,01% БСА). Затем FVIIa / рТФ смесь приготовили смешиванием 90 мкл дистиллированной воды, 36 мкл 5 × буфера для прямого анализа, 18 мкл 500 нМ FVIIa и 18 мкл 5 мкМ рТФ (рекомбинантный человеческий фактор коагуляции III / растворимый тканевый фактор, R & D Systems; исходный раствор 19,6 мкМ в 50% глицерине, разбавляли до 5 мкМ в 1 × буфере для прямого анализа и хранили до двух недель при температуре 4°С). Затем компоненты соединяли с образованием комплекса в течение 5 минут при комнатной температуре.
Раствор 10 мМ FFR-CMK (Bachem) в ДМСО (хранится при температуре -20°С), разбавили водой до 3,5 мкМ. В одном ряду полипропиленового непрозрачного планшета (Costar # 3363) были приготовлены последовательные разведения в два раза в воде FFR-CMK в 11 ячейках 96-ячеечного непрозрачного планшета, при этом последняя ячейка ряда содержала только воду в качестве контроля. Получили 10 × последовательных растворов FFR-CMK ингибитора. В каждую ячейку ряда предварительно обработанного 96-ячеечного прозрачного планшета для анализа с уменьшенной вдвое площадью ячеек добавили 10,8 мкл FVIIa / рТФ смеси, а затем 1,2 мкл 10 × FFR-CMK растворов ингибитора. Растворы тщательно перемешали, и планшет центрифугировали при <3000 об/мин в течение 5 минут, чтобы удалить подтеки в ячейках. Планшет закрыли и инкубировали в течение 8 часов при 37°С.
Для определения остаточной активности FVIIa / ТФ комплекса сначала приготовили смесь субстрата Spectrozyme FVIIa (American Diagnostica, # 217L; восстановленный раствор 50 мкмоль в 5 мл дистиллированной воды до 10 мМ, хранится при температуре 4°С) и 5 × прямого буфера (500 мМ Трис с рН 8,4, 500 мМ NaCl, 25 мМ СаСl2 и 0,05% БСА), смешивая 360 мкл 5 × прямого буфера с 180 мкл 10 мМ раствора Spectrozyme FVIIa и 1080 мкл воды. В каждую ячейку планшета для анализа добавили 108 мкл приготовленного раствора субстрата. Содержимое ячеек перемешали, и планшеты инкубировали при 37°С. Увеличение абсорбции при 405 нм измеряли каждые 30 секунд в течение одного часа при 37°С на Spectramax Gemini M5 планшет-ридере от Molecular Devices.
Уровни абсорбции измерили с использованием SoftMax Pro Software (Molecular Devices), и относительную активность протеаз, инкубированных с ингибитором, определяли делением измеренного уровня на уровень для неингибированной протеазы. Относительные активности были отложены на графике против концентрации FFR-CMK, и точки, соответствующие >90% или <10% активности неингибированной протеазы, были отброшены. Затем через остальные точки была проведена линия для определения отсекаемого на оси Х отрезка, который представляет собой концентрацию активной протеазы в растворе. Были измерены значения в нескольких анализах, и было определено среднее и стандартное отклонение.
В. 384-ячеечный формат анализа
Для повышения точность и производимости титрования формат предыдущего анализа был изменен на 384-ячеечный. Инкубация проводилась в течение семи часов при высокой концентрации протеазы (250 нМ) с серией концентраций FFR-CMK, охватывающей диапазон от 400 нМ до 53 нМ. Остаточную активность измеряли добавлением субстрата для FVIIa (мезил-dFPR-АСС) и измерением изменения флуоресцентного сигнала во времени.
384-ячеечный черный планшет для анализа (Nunc) предварительно обработали, как описано в Примере 6. Затем приготовили FVIIa / рТФ раствор, смешивая 32,5 мкл 1 мкМ раствора FVIIa+19,5 мкл 5 мкМ рТФ+6,5 мкл 5 × AST буфера (100 мМ Na Hepes, рН 7,5, 750 мМ NaCl, 25 мМ СаСl2, 0,5% БСА, 0,5 PEG 8000) и 6,5 мкл dH2O. Инкубацию проводили при комнатной температуре в течение 5 минут. Половина 384-ячеечного планшета вмещает 5 анализов протеаз в трех повторностях и один контрольный образец для анализа. Во время FVIIa / рТФ инкубации 20 мМ FFR-CMK разбавили до 8 мкМ в 1 мМ НСl, затем приготовили 9 последовательных разведении в 1,25 раза в 96-ячеечном планшете. Остальные две ячейки в левом ряду заполнили 1 мМ НСl. Затем этот ряд разбавили в 10 раз 1 × AST буфером и перемешали для получения серии FFR-CMK концентраций от 800 нМ до 107 нМ. К каждому ряду 384-ячеечного планшета добавили 4 мкл FFR-CMK из серии растворов. Затем, 4 мкл FVIIa / рТФ раствора смешали с серией растворов ингибитора в одиннадцати столбцах планшета. Последний столбец наполнили 4 мкл 1 × AST буфера в качестве контроля. Планшет центрифугировали, закрыли и инкубировали при комнатной температуре в течение семи часов при встряхивании. Анализ начали с 72 мкл 110 мкМ мезил-dFPR-ACC, растворенного в 1 × буфере AST. Увеличение флуоресценции контролировали при возбуждении 380 нм и излучении 460 нм в течение 20 минут с измерениями каждые 45 секунд при температуре 37°С. Данные были проанализированы аналогично Примеру 6, со следующими изменениями. Во-первых, остаточная активность была ограничена значениями от 20% до 80% от активности протеазы без ингибитора. Во-вторых, отрезок, отсекаемый на оси Y линейной эмпирической кривой, должен попадать между 0,9 и 1,1. В-третьих, отрезок, отсекаемый на оси X, был ограничен значениями от 125 нм и 375 нм. Длины отрезков, отсекаемых на оси Х в трех повторностях для каждой протеазы, были усреднены, и были определены среднее и стандартное отклонение.
Пример 7. Определение IС50 для TFPI ингибирования FVIIa / ТФ
Взаимодействие между TFPI и FVIIa / ТФ комплексом оценивали по измерению уровня ингибирования различными концентрациями TFPI каталитической активности FVIIa / ТФ по отношению к субстрату, Spectrazyme Vila. Концентрация TFPI, которая требуется для 50% ингибирования (IС50), были рассчитана для каждого варианта FVII и стандарта FVIIa.
96-ячеечный прозрачный планшет для анализа с уменьшенной вдвое площадью ячеек (Nunc) был предварительно обработан добавлением 150 мкл/ячейку 1 × планшетного буфера (100 мМ Трис с рН 8,4,100 мМ NaCl, 0,01% БСА, 0,01% Твин-20) в каждую ячейку и инкубацией планшета при 37°С минимум в течение 1 часа. Буфер полностью удалили вытряхиванием и промоканием планшета, а также центрифугированием планшета вверх дном для удаления оставшегося буфера. Планшет высушивали на воздухе в течение 1 часа и хранили при комнатной температуре (RT).
В 1,7 мл пробирках для микроцентрифугирования (низкоадгезивные микроцентрифужные пробирки из Bioexpress ISC) была получена смесь FVIIa / ТФ с общим объемом 450 мкл путем смешивания 9 мкл 250 нМ FVIIa (American Diagnostica, CB553-02 или соответствующего варианта для тестирования) с 337,5 мкл 2 × ТФ (Innovin; Dade Behring; лиофилизированный продукт ресуспендировали в 10 мл дистиллированной воды для получения 2 × ТФ, что приблизительно эквивалентно 7 нМ липидированного ТФ), с 90 мкл 5 × буфера для анализа (500 мМ Трис с рН 8,4,500 мМ NaCl, 25 мМ CaCl2, 0,05% БСА) и 13,5 мкл воды, полученный раствор содержал 5 нМ FVIIa и 5,2 нМ ТФ. Смесь инкубировали при комнатной температуре в течение 5 минут, что позволило компонентам образовать комплекс. В каждую ячейку 2 колонок в предварительно обработанном 96-ячеечном прозрачном планшете добавили 25 мкл соответствующей FVIIa / рТФ смеси, и планшет закрыли для предотвращения испарения.
Человеческий рекомбинантный TFPI (R & D Systems) сначала растворили в 33 мкл 50% глицерина (объем / объем) с получением 10 мкМ раствора для хранения при -20°С. Раствор TFPI затем разбавили до 1,5 мкМ в 1 × прямом буфере (100 мМ Трис с рН 8,4,100 мМ NaCl, 5 мМ CaCl2, 0,01% БСА) в полипропиленовом планшете следующим образом: для каждого анализа протеазы было приготовлено 87,5 мкл 1,5 мкМ раствора TFPI смешиванием 13,1 мкл 10 мкМ TFPI с 17,5 мкл 5 × буфера для анализа и 56,9 мкл дистиллированной воды. Серия разведении раствора TFPI в 3 раза была приготовлена в 1 × буфере для анализа смешиванием 27,5 мкл TFPI с 55 мкл 1 × буфера для анализа, так что были получены растворы, содержащие 750 нМ, 250 нМ, 83,3 нМ, 27,8 нМ, 9,26 нМ, 3,1 нМ и 1,03 нМ TFPI. Последняя ячейка в серии содержит только 1 × прямой буфер в качестве контроля.
Двадцать пять мкл каждого разведения TFPI было добавлено в 2 ячейки (т.е. в двух повторностях) 2 колонок 96-ячеечного прозрачного планшета для анализа с уменьшенной вдвое площадью ячеек, содержащие FVIIa / ТФ смесь, так что протеазная смесь анализировалась в двух повторностях с каждым разведением TFPI. Раствор 1 × буфера для анализа без TFPI также добавили в 2 ячейки, содержащие FVIIa / ТФ смесь, в качестве негативного контроля. Планшет встряхивали непродолжительное время и затем центрифугировали при 3000 об/мин в течение 5 минут до инкубации при 37°С в течение 1,5 часов.
Исходный раствор Spectrazyme Vila (American Diagnostica) получили восстановлением 50 мкмолей в 5 мл дистиллированной воды до 10 мМ, и хранили при -40°С до использования. Непосредственно перед использованием раствор разбавили до 600 мкМ дистиллированной водой. После инкубации планшета для анализа, описанного выше, в каждую ячейку планшета для анализа добавили 10 мкл разбавленного Spectrazyme Vila. Содержимое перемешали, и планшет инкубировали при 37°С. Увеличение абсорбции при 405 нм измеряли каждые 30 секунд в течение одного часа при 37°С, уровни абсорбции рассчитывали с использованием SoftMax Pro Software (Molecular Devices).
Для определения степени ингибирования TFPI, уровни абсорбции протеазных проб, содержащих TFPI, сначала были разделены на уровень абсорбции проб, не содержащих TFPI (контрольная проба), для получения относительной активности, а также был определен log10 каждой концентрации TFPI. С использованием GraphPad Prism Software, log10[TFPI] был отложен против относительной активности для каждой протеазы, и была получена кривая доза-реакция с аппроксимированием, при котором учтено, что наибольшая и наименьшая активности принимаются равными 1 и 0 соответственно. Программное обеспечение было использовано для определения TFPI ингибирования как значения логарифма IC50 (pIC50), и абсолютного значения IС50 (ингибирование TFPI в нМ) для каждой протеазы, было определено их среднее и стандартное отклонение.
Уровни ингибирования TFPI каждого из вариантов FVIIa в комплексе с рТФ были определены и выражены в IС50 или рIС50 (табл.13). Степень, в которой TFPI препятствует активности немодифицированного FVIIa (CB553-02) в комплексе с рТФ, также была определена, и IС50 оказался равным 88 нМ. Девять из созданных вариантов FVIIa демонстрировали пониженную способность взаимодействовать с TFPI, что отразилось в увеличении значений IC50.
Ингибирование вариантов FVIIa TFPI
Пример 8. In vivo оценка прокоагулянтной активности FVIIa дикого типа
Мышиная модель гемофилии была создана с целью оценки прокоагулянтной активности полипептидов FVIIa. Гемофилию А вызывали у CD-1 мышей введением анти-FVIII антител, и последующего хирургического удаления кончиков хвостов для начала кровотечения. Затем мышей обрабатывали FVIIa полипептидом и измеряли время, необходимое для остановки кровотечения, и количество крови, потерянной за это время, для определения прокоагулянтной активности полипептидов FVIIa.
Самцов CD-1 мышей анестезировали внутрибрюшинным введением тиобарбитала натрия в дозе 100 мг/кг и кетамина в дозе 100 мг/кг. Лидокаин вводился подкожно в вентральную область шеи для снижения чувствительности. В трахею и сонные артерии были введены катетеры через небольшой разрез кожи в области шеи для облегчения дыхания и введения анти-фактор VIII антител, рекомбинантного человеческого фактора Vila (rhFVIIa) и/или модифицированных полипептидов FVII.
Мышам с катетерами вводили 3,76 мг овечьих антител к FVIII человека (Affinity Biologicals, Лот IG129R4, 612 мышей BU/мл) в 40 мкл. Эта доза была определена с помощью проведения предварительного эксперимента с антителами с использованием 0,376, 0,94,1,88 и 3,76 мг антител против человеческого FVIII, и оценки кровопотери и времени кровотечения. Через 20 минут, хвосты мышей помещали в 15 мл пробирки, содержащие фосфатный солевой буфер при 39°С (PBS), на 10 минут. На 30 минуте хвосты быстро удалили из раствора PBS, и последние 5 мм хвостов иссекали для начала кровотечения. Время, за которое началось кровотечение, было отмечено. Хвосты затем вновь поместили в пробирку, содержащую PBS при 39°С и позволили кровоточить в течение 5 минут (пред-кровотечение), чтобы убедиться, что на мышей подействовали анти-FVIII антитела. После предварительного кровотечения мышам вводили FVIIa полипептиды или носитель, в котором FVIIa белки были получены и введены. FVIIa полипептиды были разбавлены PBS или буфером, состоящим из 52 мМ хлорида натрия, 10,2 мм дегидратированного хлорида кальция, 9,84 мМ глицилглицина, 0,01% полисорбата 80 и 165 мМ маннита. FVIIa препараты вводили в дозах 1, 3 или 10 мг/кг, в объеме, эквивалентном 3 мл / кг, через катетер сонной артерии, и хвосты поместили в пробирки, содержащие свежий PBS при +39°С. Кровотечение контролировали в течение 20 минут, и время, при котором кровотечение остановилось, было отмечено. Общее время кровотечения было рассчитано как сумма длительности кровотечения в ходе предварительного кровотечения, и длительности кровотечения после введения полипептидов FVIIa, или PBS, или буфера.
Для определения количества крови, потерянной во время эпизода кровотечения, содержимое 15 мл пробирки анализировали на содержание гемоглобина. Тритон Х-100 разбавляли 1 к 4 в стерильной воде, и 100 мкл раствора добавляли к 1 мл образцов для гемолиза. Абсорбцию образцов измеряли при длине волны 546 нм. Для определения количества потерянной крови, абсорбцию отмечали на стандартной кривой, полученной измерением абсорбции при 546 нм известных объемов мышиной крови, разбавленной PBS и подвергнутой гемолизу, как описано выше, с Тритон Х-100.
Эксперимент был проведен сравнением rhFVIIa (CB533), полученным, как описано выше, с коммерчески доступным рекомбинантным человеческим FVIIa (NovoSeven®, Novo Nordisk), потери крови оценивали после введения 3 мг/кг каждого белка. Кровопотери в группе с носителем (буфер, N=15) составляли 671,9±57,89 мкл в течение 20-минутного периода. Кровопотеря была уменьшена при введении rhFVIIa, произведенного Catalist Biosciences, до 264,1±56,59 мкл и при введении rhFVIIa, произведенного NovoSeven®, до 273,7±53,93 мкл (N=14). Этот эксперимент продемонстрировал эквивалентность между двумя белками.
Пример 9. Фармакокинетический анализ FVIIa полипептидов
Фармакокинетические свойства полипептидов FVIIa оценивали путем измерения количества человеческого фактора Vila в плазме мышей. Для определения количества FVIIa в плазме были использованы два анализа. Для определения количества FVIIa белка в плазме крови мышей был использован ELISA, и для количественного определения коагулянтной активности полипептидов FVIIa в плазме был использован FVIIa-зависимый анализ свертывания (FVII: С).
А. Введение полипептидов FVIIa мышам
Модифицированные FVIIa полипептиды и немодифицированный рекомбинантный человеческий FVIIa (rhFVIIa) белок (NovoSeven®, Novo Nordisk) были оценены в фармакокинетических исследованиях. Для каждого исследования, 18 самцам CD-1 мышей вводили внутривенно болюсные дозы (0,1-3,0 мг/кг, в зависимости от анализа) rFVIIa. На 5,15, 30, 60, 120 и 240 минуте после инъекции, трех мышей, получивших одну инъекцию, эвтаназировали с помощью асфиксии СО2, и приблизительно 1,0 мл крови отбирали в 1,0 мл шприц через чрескожную сердечную пункцию. Каждый шприц был предварительно загружен достаточным количеством цитрата натрия для достижения конечной концентрации 3,2% в 1 мл крови. Образцы крови затем центрифугировали при 9000 об/мин в течение 8 мин при 4°С. Плазму перенесли в отдельные отмеченные 1,5 мл пробирки (эппендорфы), быстро заморозили в жидком азоте и хранили при -80°С. Кроме того, одна мышь на эксперимент была инъецирована носителем (симуляция), и плазма этой мыши была использована для определения фоновой активности FVIIa.
В. ELISA
Коммерчески доступный набор, IMUBIND® фактор VII ELISA (American Diagnostica), был использован для выявления FVII белков в сыворотке крови с помощью ELISA. В этом наборе используется планшет, предварительно покрытый анти-FVII/FVIIa антителами для связывания белка, и биотинилированные анти-FVII антитела для обнаружения с помощью стрептавидин-меченой пероксидазы хрена. Набор был использован в соответствии с указаниями производителей со следующими исключениями: во-первых, стандартная кривая была сужена для обеспечения линейного спектра во всем диапазоне концентраций, и охватывала концентрации от 0,88 нг/мл до 10 нг/мл; во-вторых, для стандартной кривой был использован очищенный вариант FVIIa, а не стандартные FVII, поставляемые в наборе из-за различий в аффинности антител. Опыты показали, что комплекс FVIIa с антитромбином III (ATIII), потенциальным ингибитором FVIIa в плазме, выявлен в 75% от уровня свободной протеазы, что показывает, что тест может определять весь FVIIa в образце плазмы, как в активной, так и в неактивной формах.
Образцы плазмы разморозили при комнатной температуре и разбавили в 10 раз в буфере для образцов (PBS+0,1% Тритон Х-100+1,0% БСА), затем разбавляли последовательно в 5,5 раза с получением четырех разведении. Четыре разбавленных образца были разбавлены в 2 раза на планшете для ELISA для окончательного разведения образцов в 20,110, 605 и 3327,5 раза. Эти четыре разведения плазмы мышей охватывали широкий круг концентраций протеазы от 33000 нг/мл до 20 нг/мл. Каждый образец измеряли на двух отдельных планшетах для анализа, и измерения в диапазоне стандартной кривой использовали для расчета концентрации вариантов FVIIa в образцах плазмы.
С. Анализ свертывания
Коммерчески доступный набор (STACLOT FVIIa-RTF, Diagnostica Stago, Parsippany, NJ) был использован для анализа свертывания. Для определения коагулянтной активности активных полипептидов FVIIa образы плазмы были исследованы с помощью F Vila-зависимого анализа на свертывание (FVII: С). Анализ проводили с использованием реагентов и инструкций, содержащихся в коммерческом наборе, и время свертывания крови измеряли с помощью электромеханического прибора обнаружения сгустка (STArt4, Diagnostica Stago, Parsippany, NJ). Набор был использован в соответствии с инструкциями производителей со следующими исключениями: во-первых, для стандартной кривой был использован очищенный вариант FVIIa, а не стандартные rhFVIIa, содержащийся в наборе, во-вторых, для рутинных фармакокинетических исследований были использованы коммерческие реагенты, которые дали сопоставимые результаты с реагентами из набора: растворимый тканевый фактор (Calbiochem, La Jolla, CA), смесь синтетических фосфолипидов (Avanti Polar Lipids, Alabaster, AL), TBSA буфер (Трис-NaCl, pH 7,5 с 1% БСА; DiaPharma, West Chester, Ohio) и 25 мкМ раствор хлорида кальция (Diagnostuca Stago, Parsippany, NJ).
Анализ свертывания проводили следующим образом. Замороженные образцы плазмы размораживали при комнатной температуре в течение примерно 45 минут и затем разбавляли 1:5000 буфером. Пятьдесят мкл разбавленной плазмы в сочетании с 50 мкл фактор VII-дефицитной плазмой человека и 50 мкл релипидированного тканевого фактора предварительно инкубировали в течение 180 секунд. После инкубации для инициации свертывания добавили 50 мкл раствора хлорида кальция (25 мкМ). Время свертывания крови определяли с помощью электромеханического обнаружения сгустков. Каждый образец плазмы анализировали в двух повторностях. Систему калибровали путем построения стандартной кривой с помощью времени свертывания серии разведении буфера, содержащих известные количества конкретного исследуемого варианта FVIIa. Концентрации FVIIa в образцах плазмы мыши были рассчитаны на основе линейной части стандартной кривой logFVIIa против времени свертывания. Способность образцов плазмы вызывать свертывание в фактор VII-дефицитной плазме описывалась как содержание FVIIa нг/мл в плазме мыши, с учетом фонового эндогенного FVIIa дикого типа в плазме контрольных мышей.
Период полураспада каждого белка FVII определяли в обычном порядке, аппроксимируя логарифм активности к прямой линии, и измеряя время, требующееся для уменьшения активности белка FVIIa вдвое. Для белков FVII с мультиэкспоненциальной кинетикой распада, период полураспада определяли по конечной части графика логарифма концентрации в плазме против времени. Дополнительные фармакокинетические параметры были рассчитаны с использованием коммерчески доступных программ (WinNonLin V5.1, Pharsight Corporation, Mountain View, CA).
D. фармакокинетические свойства СВ728, CB735 и СВ945
С использованием описанного выше протокола была проведена оценка фармакокинетических свойств FVIIa дикого типа и двух вариантов FVIIa: СВ728, CB735 и СВ945. Результаты приведены в таблице 14. CB735 и СВ945 проявляли улучшенные фармакокинетические параметры по сравнению с FVIIa дикого типа.
Фармакокинетические параметры вариантов FVIIa в мыши
Swap - замена
E. фармакокинетические свойства CB558
Период полураспада модифицированного полипептида FVIIa, который проявлял повышенную устойчивость к TFPI (CB-558; содержит мутацию K197E), был измерен и сравнен с периодом полураспада немодифицированного рекомбинантного человеческого FVIIa (rhFVIIa) белка (NovoSeven®, Novo Nordisk) с помощью фармакокинетического исследования, аналогичного описанному выше, с некоторыми модификациями. Пятнадцати самцам CD-1 мышей вводили 0,5 мг/кг внутримышечного болюса CB-558, и 15 мышам вводили 0,5 мг/кг внутривенного болюса rhFVIIa. На 5, 15, 30, 60 и 120 минуте после инъекции трех мышей из каждого протокола инъекций эвтаназировали асфиксией CO2, и приблизительно 1,0 мл крови отбирали в 1,0 мл шприц через чрескожную сердечную пункцию. Каждый шприц предварительно загружали достаточным количеством цитрата натрия для достижения конечной концентрации 3,2% в 1 мл крови. Образцы крови затем центрифугировали при 9000 об/мин в течение 8 мин при 4°C. Плазму переносили в отдельную меченую 1,5 мл пробирку (эппендорф), быстро замораживали в жидком азоте и хранили при -80°C.
Для определения коагулянтной активности и периода полураспада активных полипептидов FVIIa, образцы плазмы оценивали в Machaon Diagnostica, Inc (Oakland, CA) с использованием FVIIa-зависимого анализа на свертывание (FVII:C) и автоматизированного прибора для оценки гемостаза (AMAX 190plus, Trinity Biotech). Образцы замороженной плазмы размораживали при 37°C на водяной бане в течение 5 минут и затем разбавили 1:40 вероналовым буфером. Шесть мкл разбавленной плазмы добавили к 54 мкл вероналового буфера в кювету и смешали с металлическими шариками. В кюветы было добавлено равное количество (60 мкл) фактор VII-дефицитной плазмы человека (Precision Biologic), которые инкубировали при 37°C в течение 60 секунд при постоянном перемешивании.
Чтобы инициировать начало проведения реакции, в кюветы было добавлено 120 мкл тромбопластина (источник - мозг кролика, тромбопластин-DS; Pacific Hemostasis) при постоянном перемешивании. Реакцию свертывания контролировали фотооптически при 405 нм для обнаружения образования фибрина. Систему калибровали, анализируя время свертывания последовательных разведении указанной плазмы с опубликованными уровнями FVII (Precision Biologic), а затем анализируя время свертывания нормальной плазмы в качестве контроля (Precision Biologic) и абнормальной плазмы в качестве контроля (Precision Biologic).
Способность образцов плазмы индуцировать свертывание в фактор VII-дефицитной плазме была оценена в процентах наблюдаемого времени свертывания крови к времени для нормальной человеческой плазмы. Коагулянтная активность плазмы обработанных FVIIa (NovoSeven ®) и СВ-558 мышей снижалась с течением времени. Период полураспада каждого белка FVII был определен путем обычного аппроксимирования логарифма активности прямой линией, а также измерением времени, требующегося для уменьшения активности белка FVIIa вдвое. Определенный период полураспада СВ-558 оказался примерно в два раза больше, чем период полураспада FVIIa (NovoSeven®): 156 минут по сравнению с 67 минутами.
Пример 10. Образование дополнительных вариантов FVII
Был получен ряд дополнительных мутантов FVII для изменения одного или нескольких свойств и/или активностей FVII. В дополнение к модификациям для увеличения устойчивости к TFPI, были осуществлены модифицикации для увеличения устойчивости к AT-III, повышения каталитической активности, увеличения периода полураспада, и/или увеличения аффинности и/или связываемоеTM с фосфолипидами, такими как присутствующие на активированных тромбоцитах. Модификации включают замены аминокислот, вставки и делеции. Также были получены FVII варианты, содержащие две или более модификации. В таблице 23 приведены варианты FVII, которые были созданы и экспрессированы в Freestyle TM 293-F клетках. Аминокислотные замены были включены в FVII-полипептиды с использованием методов, описанных в примере 2.В выше, с соответствующим олигонуклеотидом.
В таблице 15 ниже изложены дополнительные варианты FVII, которые были получены. Были созданы FVII варианты с заменой Gla домена, в которых аминокислотные остатки с А1 до Y44 (по нумерации зрелого FVII) наК-конце белка FVII дикого типа были заменены аминокислотными остатками соответствующего домена Gla из FIX, FX, протеина С, протеина S или тромбина. Вариант FVII "Gla замена из FIX" (т.е. полипептид FVII, в котором эндогенный домен Gla заменен доменом Gla из FIX) содержит аминокислотные остатки с А2 до Y45 из SEQ ID NO: 110 на N-конце; вариант FVII "Gla замена из FX" содержит аминокислотные остатки с А1 до Y44 из SEQ ID NO: 111 HaN-конце; вариант FVII "Gla замена из протеина С" содержит аминокислотные остатки с А1 до Н44 из SEQ ID NO: 113 на N-конце; вариант FVII "Gla замена из протеина S" содержит аминокислотные остатки с А1 до Y44 из SEQ ID NO: 114 на N-конце, и вариант FVII "Gla замена из тромбина" содержит аминокислотные остатки с А1 до Y44 из SEQ ID NO: 115 на N-конце.
Примеры вариантов фактора VII
Ins - вставка
Prot - белок
Swap - замена
Thrombin - тромбин
Пример 11. Анализ каталитической активности вариантов FVIIa по отношению к их субстрату, фактору Х
Каталитическую активность вариантов FVIIa по отношению к их субстрату, фактору Х (FX), оценивали косвенно двумя типами хромогенных анализов путем измерения активности FXa, образованного активацией FVIIa, с помощью синтетического субстрата Spectrafluor FXa. Два анализа были выполнены в присутствии или в отсутствие липидированного тканевого фактора для оценки как ТФ-зависимой, так и ТФ-независимой активности. Варианты FVII экспрессировали, очищали и активировали в FVIIa, как описано выше в примерах 2 и 3. Хотя большинство FVII вариантов экспрессировали только в FreestyleTM 293 -F клетках, некоторые из вариантов также были экспрессированы в ВНК-21 клетках.
Непрямой анализ с липидированным тканевым фактором
Каталитическую активность вариантов FVIIa в присутствии тканевого фактора оценивали с помощью анализа, описанного в примере 5 выше, с незначительными изменениями. Одним из таких изменений является использование протеазы субстрата фактора X, которую обрабатывали ERG-CMK и FFR-CMK для снижения фоновой активности (Molecular Innvations). Проводили два типа анализа данных с использованием двух отдельных проб: линейный анализ спектра и гиперболический анализ. В линейном анализе использовали диапазон концентраций фактора Х между 0 и 150 нМ для обеспечения точных измерений кинетических констант в линейном диапазоне кривой доз. Напротив, в гиперболическом анализе использовали диапазон концентраций фактора Х от 0 до 1,44 мкМ для обеспечения точного измерения кинетических констант по кривой доз с насыщением (гиперболической).
Непрямой анализ с липидированным тканевым фактором с линейным анализом данных проводился, как описано в примере 5 выше, со следующими изменениями. Растворы варианта FVIIa / ТФ были получены как растворы 0,1 нМ FVIIa/0,4 ТФ, и их инкубировали в течение 30 минут, после чего разбавляли в два раза 0,4 нМ ТФ до раствора, содержащего 1,5625 пМ FVIIa / 0,4 нМ ТФ. Двадцать пять мкл FVIIa / ТФ раствора смешали с 25 мкл раствора субстрата, содержащего 1,0 ммоль Spectrofluor FXa (American Diagnostica) и 300 нМ, 200 мМ, 133,3 нМ, 88,9 нМ, 59,3, 39,5 нМ, 36,3 нМ или 0 нМ фактора Х (Molecular Innovations). Таким образом, окончательные концентрации для анализа составляли 0,8 пМ FVIIa, 0,2 нМ ТФ, 0,5 ммоль Spectrofluor FXa и 150 нМ, 100 мМ, 66,7 нМ, 44,4 нМ, 29,6 нМ, 19,8 нМ, 13,2 нМ или 0 нМ фактора Х (Molecular Innovations) в 50 мкл на ячейку. Стандартная кривая АМС, которая служила коэффициентом пересчета единиц флуоресценции в мкМ в последующих расчетах, была расширена за счет включения в диапазон доз, которые охватывают от 0 мкМ до 100 мкМАМС.
Непрямой анализ с липидированным тканевым фактором с гиперболическим анализом диапазона данных проводился, как описано в примере 5 выше, со следующими изменениями. Растворы вариантов FVIIa / ТФ были получены как 0,1 нМ FVIIa/0,4 нМ ТФ растворы, и их инкубировали в течение 30 минут, после чего разбавляли два раза 0,4 нМ ТФ до 1,5625 пМ (или 0,78 пМ для протеаз, для которых ожидается высокая активность) FVIIa / 0,4 нМ ТФ. Двадцать пять мкл FVIIa / ТФ раствора смешали с 25 мкл раствора субстрата, содержащего 1,0 ммоль Spectrofluor FXa (American Diagnostica) и 1440 нМ, 720 мМ, 360 нМ, 180 нМ, 90 нМ, 45 нМ, 22,5 нМ или 0 нМ фактора Х (Molecular Innovations). Таким образом, окончательные концентрации для анализа составляли 0,8 (или 0,39) пМ FVIIa, 0,2 нМ ТФ, 0,5 ммоль Spectrofluor FXa и 7 нМ, 720 мМ, 360 нМ, 180 нМ, 90 нМ, 45 нМ, 22,5 нМ, 11,25 нМ или 0 нМ фактора Х (Molecular Innovations) в 50 мкл на ячейку. Ккат и Km параметры вычисляли по гиперболическому уравнению Михаэлиса-Ментен вида (Vmax / (L+(Км / х))). Стандартная кривая АМС, которая служила коэффициентом пересчета единиц флуоресценции в мкМ в последующих расчетах, была расширена за счет включения в диапазон доз, которые охватывают от 0 мкМ до 100 мкМ АМС.
Для определения кинетических констант скоростей активации FX FVIIa или вариантами FVIIa сырые данные, собранные с применением SoftMax Pro (Molecular Devices), экспортировали как XML файлы. Дальнейшие линейные и нелинейные анализы данных проводили с помощью XLfit4 пакета программного обеспечения для автоматизированного аппроксимирования кривых и статистического анализа в таблицах Microsoft Excel (IDBS Software).
По данным, собранным с помощью линейного анализа, Ккат/КM (М-1сек-1) кинетические константы вычисляли непосредственно из наклона прямой линейной регрессии зависимости концентрации FX от скорости расщепления флуорогенного субстрата (в мкМ/сек2), где Ккат/КM=наклон / [FVIIa]×0,5×k2. Поправочный коэффициент k2=45 был определена по методу, описанному в примере 5, и кинетическим константам для FXa расщепления Spectrofluor FXa, Ккат, FXa=56 сек-1 и КM, FXa=126 нМ, определенным экспериментально с помощью активированного FX (FXa), который ранее был оттитрован по активному сайту АТ-III/гепарином. Исключение точек, которые приводили к значениям R2 менее 0,98, обеспечивало линейность наборов данных, используемых в рутинном аппроксимировании.
Анализ данных, собранных с помощью анализа гиперболического диапазона, был осуществлен из нелинейного регрессионного анализа зависимости концентрации FX от скорости расщепления флуорогенного субстрата (в мкМ/сек2). Индивидуальные параметры Ккат и КM рассчитывали как параметры аппроксимирования с помощью гиперболического уравнения Михаэлиса-Ментен вида (Vmax/(L+(Км/х))), где Ккат=Vmax / [FVIIa]×0,5×k2. Кинетическая константа, Ккат/КM был рассчитана из индивидуальных параметров аппроксимирования Ккат и КM.
Независимый от тканевого фактора непрямой анализ
Каталитическая активность вариантов FVIIa в присутствии тканевого фактора была оценена с помощью непрямого анализа, аналогичного описанному выше, за исключением того, что тканевый фактор не был включен в анализ. Таким образом, тест для оценки ТФ-независимой активности осуществляли, главным образом, как описано выше, со следующими изменениями. Растворы вариантов FVIIa разбавляли до 50 нМ. Двадцать пять мкл каждого раствора FVIIa смешивали с 25 мкл раствора субстрата, содержащего 1,0 ммоль Spectrofiuor FXa (American Diagnostica) и 1050 нМ, 700 мМ, 466,7 нМ, 311,1 нМ, 207,4 нМ, 138,3 нМ, 92,2 нМ или 0 нМ фактора Х (Molecular Innovations). Таким образом, окончательные концентрации для анализа составляли 25 нМ FVIIa, 0,5 ммоль Spectrofiuor FXa и 525 мМ, 350 нМ, 233,3 нМ, 155,6 нМ, 103,7 нМ, 69,1 нМ, 46,1 нМ или 0 нМ фактора Х (Molecular Innovations) в 50 мкл на ячейку. Анализ данных проводили, как описано для линейного анализа диапазона выше, без каких-либо изменений.
В таблицах 16-18 представлены результаты анализов, которые проводили для измерения каталитической активности вариантов FVIIa. В таблицах 15 и 16 приведены значения каталитической активности, измеренной в ТФ-зависимом непрямом анализе с использованием полипептидов FVIIa, экспрессированных в 293-F и клетках и ВНК-21 клетках, соответственно, а в таблице 17 приведена каталитическая активность, измеренная в ТФ-независимом анализе с использованием полипептидов FVIIa, экспрессированных в 293-F клетках и/или ВНК-21 клетках. Результаты представлены в виде кинетической константы каталитической активности, Ккат/КM (М-1сек-1), а также выражены в процентах активности FVIIa дикого типа, где активность является каталитической активностью, Ккат/КM (М-1сек-1) каждого варианта FVIIa по отношению к субстрату, FX. Для значений, представленных в таблицах, также указано использование анализа линейного или гиперболического диапазона данных. Не все варианты FVIIa анализировали в каждом тесте. Некоторые из вариантов FVII, которые анализировали в Примере 5 выше, проявляли немного уменьшенную или увеличенную каталитическую активность к FX в данном наборе анализов по сравнению с анализами, описанными в Примере 5. Разница в активностях белков в Примере 5, вероятно, связана с различной фоновой активностью остаточного FXa в субстрате FX, полученном от American Diagnostica. Несколько вариантов FVIIa проявляли увеличенную каталитическую активность по сравнению с FVIIa дикого типа. Например, CB760, в котором содержится мутация Q366V, обладает каталитической активностью от 1,5 до 5 раз выше, чем FVIIa дикого типа.
Linear - линейный
Giperbolic - гиперболический
Ins - вставка
Prot - белок
Swap - замена Thrombin - тромбин
Каталитическая активность вариантов FVIIa: ТФ-зависимый непрямой анализ с полипептидами FVIIa из ВНК-21 клеток
Linear - линейный
Ins - вставка
Swap - замена
WT - дикий тип
Пример 12. Определение ингибирования FVIIa / ТФ или FVIIa AT-III/гепарином
Возможность взаимодействия комплекса AT-III/ гепарин и FVIIa в присутствии или в отсутствие растворимого тканевого фактора (рТФ), т.е. ТФ-зависимого или ТФ-независимого взаимодействия, была оценена по измерению уровня ингибирования каталитической активности FVIIa /рТФ по отношению к субстрату, мезил-FRP-ACC, различными концентрациями AT-III. Значения К0,5 определяли для каждого тестируемого варианта FVIIa, они соответствуют концентрациям AT-III, которые требуются для 50% ингибирования (IC50) варианта FVIIa в 30 минутном анализе при комнатной температуре (25°С).
Были осуществлены два отдельных анализа, один с ТФ и один без ТФ. 2 мкМ раствор комплекса АТ-III/гепарин (окончательно 5 мМ гепарина) приготовили смешиванием 26,4 мкл 151,7 мкМ AT-III (очищенный AT-III плазмы человека; Molecular Innovations) с 50 мкл 0,2 мМ низкомолекулярного гепарина (Calbiochem), 400 мкл 5 × буфера для анализа (100 мМ HEPES, 750 MMNaCl, 25 мМ СаСl2, 0,05% БСА, 0,5% ПЭГ 8000, рН 7,4) и 1,523 мл воды, предназначенной для лабораторного использования. Этот раствор использовали как раствор с самой высокой концентрацией в ТФ-зависимом анализе. Раствор, содержащий 4 мкМ комплекса АТ-III/гепарин (окончательно 5 мМ гепарина) приготовили для использования в ТФ-независимом анализе смешиванием 52,8 мкл 151,7 мкМ АТ-III (Molecular Innovations) с 50 мкл 0,2 мМ низкомолекулярного гепарина (Calbiochem), 400 мкл 5 × буфера для анализа и 1,497 мл воды, предназначенной для использования в лаборатории. Раствор АТ-III/гепарина инкубировали в течение 5-10 минут при комнатной температуре и затем разбавляли в два раза в 96-ячеечном полипропиленовом планшете с глубокими ячейками до окончательного объема 1 мл, в которм содержалось 5 мкМ гепарина, что привело к разведениям до 2000, 1000, 500, 250, 125, 62,5, 31,25 и 0 нМ, или 4000, 2000, 1000, 500, 250, 125, 62,5 и 0 нМ. Варианты FVIIa и FVIIa дикого типа разбавляли до 250 нМ 1 × буфером для анализа (20 мМ HEPES, 150 мМ NaCl, 5 мМ СаСl2, 0,01% БСА, 0,1% ПЭГ 8000, рН 7,4). Для ТФ-зависимого анализа, 5 нМ FVIIa/50 нМ рТФ комплексы получали смешиванием 20 мкл FVIIa с 10 мкл 5 мкМ рТФ (R & D Systems человеческий фактор свертывания III: # 2339-PA), 200 мкл 5 × буфера для анализа и 770 мкл воды для использования в лаборатории, и раствор инкубировали в течение 10-15 минут при комнатной температуре. Для ТФ-независимого анализа 100 мкл FVIIa смешивали с 200 мкл 5 × буфера для анализа и 700 мкл воды для использования в лаборатории для приготовления 25 нМ раствора FVIIa. Для начала анализа, 25 мкл FVIIa / ТФ или только раствора FVIIa отдельно смешивали с 25 мкл каждого разведения АТ-III/гепарина в ячейках 96-ячеечного черного планшета для анализа с уменьшенной вдвое площадью ячеек (Nunc). Окончательные условия для ТФ-зависимого анализа включают 2,5 нМ FVIIa/25 нМ рТФ и AT-III / гепарин в концентрациях от 1000 нМ до 0 нМ. Для ТФ-независимого анализа концентрация FVIIa составляла 12,5 нМ, и концентрация АТ-III/гепарина варьировала от 2000 нМ до 0 нМ. Планшеты инкубировали в течение 30 минут при встряхивании при комнатной температуре (25°С).
Исходный раствор субстрата FVIIa (мезил-FPR-ACC) получили растворением субстрата в ДМСО до 20 мМ, 0,5 мМ рабочий раствор затем готовили на 1 × буфере для анализа. После инкубации описанного выше планшета для анализа, в каждую ячейку планшета для анализа добавили 50 мкл субстрата FVIIa. Содержимое ячеек перемешали, и остаточную активность FVIIa определяли после начального расщепления субстрата в течение 15 минут по флуоресценции при 30°С.
Чтобы определить степень ингибирования FVIIa или вариантов FVIIa АТ-III/гепарином, сырые данные, собранные с применением SoftMax Pro (Molecular Devices) экспортировали в виде XML файлов. Дальнейший нелинейный анализ данных проводили с помощью XLfit4 пакета программного обеспечения для автоматизированного аппроксимирования и статистического анализа в таблицах Microsoft Excel (IDBS Software). Электронные таблицы использовали для расчета последовательных разведении AT-III, соотношений AT-III к FVIIa и значений Vi / Vo для каждой повторности анализа FVIIa для каждой экспериментальной концентрации AT-III. Нелинейный регрессионный анализ зависимости остаточной активности FVIIa (обозначается как Vi / Vo) от концентрации AT-III осуществили с помощью XLfit4 и гиперболического уравнения ингибирования вида (С+(Amp*(L (X / (К0,5+Х))))); где С=смещение (установлено в 0 для экстраполяции данных, которые не достигают 100%-ного ингибирования в ходе анализа). Amp=амплитуда аппроксимирующей кривой и К0,5 соответствует концентрации AT-III, необходимой для половины максимального ингибирования в соответствии с условиями анализа. Для нескольких вариантов FVIIa, AT-III ингибирует активность менее чем на 20-25% от общей активности протеазы при самой высокой исследованной концентрации AT-III, представляющей верхний предел обнаружения для анализа. Варианты с менее чем 20-25% максимального ингибирования были оценены как обладающие нижним пределом значения К0,5 (5 мкМ для ТФ-зависимого и 10 мкМ для ТФ-независимого анализа), и в большинстве случаев, как ожидается, обладали устойчивостью к AT-III больше указанной величины.
В таблицах 19 и 20 представлены результаты анализов, которые проводились на вариантах FVIIa, экспрессированных в Freestyle ТМ 293-F клетках и/или ВНК-21 клетках, в присутствии и отсутствии ТФ, соответственно. Результаты представлены как параметры аппроксимирования К0,5 и как оценки степени устойчивости к AT-III для каждого варианта по сравнению с FVIIa дикого типа, выраженные как отношения их параметров аппроксимирования К0,5 (К0,5 варианта / К0,5 дикого типа). Несколько вариантов FVIIa проявляли повышенную устойчивость к AT-III по сравнению с FVIIa дикого типа.
Ингибирование вариантов FVIIa АТ-III/гепарином в присутствии ТФ
Ins - вставка
Пример 13 Определение устойчивости вариантов FVIIa к TFPI
Устойчивость различных вариантов FVIIa к TFPI оценивали с помощью анализов, описанных в примере 7 выше. В таблице 21 приведены результаты анализа. Результаты показаны как устойчивость каждого варианта FVIIa к TFPI по сравнению с FVIIa дикого типа.
Пример 14. In vivo оценка прокоагулянтной активности FVIIa полипептидов
Мышиная модель гемофилии была создана с целью оценки прокоагулянтной активности полипептидов FVIIa. Гемофилию вызывали у CD-1 мышей введением анти-FVIII антител, и затем хирургическим удалением кончиков хвостов инициировали кровотечения (аналогично описанному выше в Примере 8). Также были использованы мыши с недостаточностью FVIII (FVIII-/- мыши), но они не обрабатывались анти-FVIII антителами. Мышей затем обрабатывали FVIIa полипептидами, и количество крови, потерянной в течение 20 минут, измеряли для определения прокоагулянтной активности полипептидов FVIIa.
А. Анализ коагулянтной активности FVIIa на CD-I мышах с индуцированной гемофилией
Первоначальные эксперименты проводили с целью определения необходимых доз, времени и продолжительности эффекта антител против человеческого фактора FVIII при введении внутрибрюшинно для индуцирования гемофилии у CD-I мышей. Для первой партии анти-FVIII антител (партия 1; Affinity Biologicals, Лот IG129R4) дозы основывались на дозах, используемых в экспериментах при введении через катетер, описанных выше в Примере 8. Доза, вызывающая состояние гемофилии, составляла (неконтролируемое кровотечение за период анализа в 20 минут) 7,54 мг/мышь (80 мкл 94,25 мг/мл исходного раствора). Данная партия нейтрализовала свертывание у 612 мышей BU/мл. Для второй партии антител против человеческого FVIII (партия 2; Affinity Biologicals, Лот IG1577R2, нейтрализующая активность у 474 мышей BU/мл) применяемая доза составляла 11,98 мг/мышь (120 мкл 99,8 мг/мл исходного раствора), она вводилась за 6 часов до иссечения хвоста.
Для индуцирования гемофилии самцам CD-I мышей (25-35 г) внутрибрюшинно вводили партию 1 или партию 2 антител против FVIII до эксперимента. Самцов CD-I и FVIII-/- мышей анестезировали внутрибрюшинным введением смеси кетамина / ксилазина (100 мг/мл раствор) и помещали на платформу с подогревом (39°С) для недопущения падения температуры тела. В процедурной комнате поддерживали температуру 82°F. За десять минут до иссечения хвоста хвост погружали в 10 мл предварительно нагретого PBS (15 мл центрифужная пробирка, 39°С). Восьми-десяти мышам вводили рекомбинантный человеческий FVIIa (NovoSeven®, Novo Nordisk) или модифицированные FVII-полипептиды, растворенные в буфере, состоящем из 52 мМ хлорида натрия, 10,2 мМ дегидратированного хлористого кальция, 9,84 мМ глицилглицина, 0,01% полисорбата 80 и 165 мМ маннита, через хвостовую вену одной инъекцией. Группе мышей в качестве контроля вводили только носитель. Если инъекцию пропускали, животное исключали из исследования. Инъекции агента или носителя осуществляли за 5 минут до иссечения хвоста. Иссечение хвоста осуществляли с использованием лезвия бритвы в 5 мм от конца хвоста, и кровь собирали в PBS в течение 20 минут. В конце периода сбора крови определяли общую потерю крови. Собранную кровь перемешали и отобранную аликвоту в 1 мл каждого образца анализировали на содержание гемоглобина. Тритон Х-100 разбавили 1/4 в стерильной воде, и 100 мкл полученной смеси добавили к 1 мл образца, чтобы вызвать гемолиз. Абсорбцию образцов измеряли при длине волны 546 нм. Чтобы вычислить количество потерянной крови, абсорбцию сравнивали со стандартной кривой, полученной измерением оптической плотности при 546 нм известных объемов мышиной крови, разбавленной PBS и подвергнутой гемолизу, как описано выше с Тритоном Х-100.
Также были выполнены исследования зависимости реакции от дозы, в которых были исследованы дозы в 0,3, 1 или 3 мг/кг СВ553 (FVIIa дикого типа). Мыши, которые получили только носитель, потеряли 1002,3±60,71 мкл за 20 минут анализа. Потеря была значительно снижена у мышей, которым вводили 3 мг/кг СВ553, до 415,5±90,85 мкл (р<0,05 с помощью критерия Крускала-Уоллиса, а затем повторной проверки Данна). Снижение дозы до 1 мг/кг привело к потере крови 679,57±83,95 мкл, и более низкая доза 0,3 мг/кг привела к потере крови 852,42±94,46 мкл.
В отдельном исследовании СВ735 (K341D) и СВ945 (M298Q/K341D) оценивали в дозе 3 мг/кг. Инъекция носителя была использована в качестве контроля. Группа мышей, которые получили только носитель, потеряли 803±92,18 мкл и 813,1±82,66 мкл крови при 20 минутах анализа. Потеря крови аналогична потери при лечении СВ735 или СВ945, в результате которого потеря крови составила 746±110,5 мкл и 870,9±78,38 мкл соответственно.
В. Анализ FVIIa коагулянтной активности у FVIII-/- мышей
Мышиная модель гемофилии А с дефицитными по FVIII мышами (FVII-/- мышами) также была использована для оценки коагулянтной активности полипептидов FVIIa, с использованием тех же протоколов, как описано выше, за исключением того, что мышей не обрабатывали анти-FVIII антителами.
1. Исследование зависимости реакции от дозы для коагулянтной активности FVIIa дикого типа
Исследования зависимости реакции от дозы для коагулянтной активности белка NovoSeven® и СВ553 на FVII-/- мышах проводили при концентрациях 0,3, 1, 3 и 6 мг/кг. В эксперименте с белком NovoSeven® потеря крови в группе, которой вводили носитель, составляла 912,79±38,32 мкл, потеря крови при лечении дозами NovoSeven® 6 и 3 мг/кг значительно сократилась (до 361,74±55,28 мкл и 586,98±60,56 мкл, р 0,05 по критерию Крускала-Уоллиса и повторной проверке Данна). Снижение дозы до 1 мг/кг приводило к потере крови 674,84±46,88 мкл, и при самых низких протестированных дозах значение составляло 801,08±41,39 мкл. В эксперименте с СВ553 потеря крови в группе, которой вводили носитель, составляла 904,08±15,38 мкл. Потеря крови значительно снизилась (р<0,05 по критерию Крускала-Уоллиса и повторной проверке Данна) при введении СВ553 в дозе 6 мг/кг до 451,04±74,17 мкл. Снижение дозы до 3 мг/кг приводило к потере крови 695,75±60,50 мкл, а снижение дозы до 1 и 0,3 мг/кг приводило к потере крови, близкой потери при введении носителя (846,08±34,17 мкл и 936,43±31,39 мкл соответственно).
Пример 15. Определение связывания фактора VIIa с растворимым тканевым фактором
Способность вариантов FVIIa, экспрессированных в НЕК 293 или ВНК клетках, связывать растворимый тканевый фактор (рТФ) оценивали с помощью поверхностного плазмонного резонанса на приборе Biacore. Варианты FVIIa оценивали с помощью измерения профиля связывания при трех концентрациях протеазы в двух экспериментах, с использованием двух различных уровней рТФ, связанного с чипом Biacore CM5.
Новый Series S CM5 сенсорный чип (GE Healthcare Cat # BR 1006-68) соединили с бычим сывороточным альбумином и растворимым тканевым фактором с использованием прибора Biacore Т100. Соединение осуществляли с использованием буфера Biacore для соединения (30 мМ Na Hepes с рН 7,4,135 мМ NaCl, 1 мМ ЭДТА, 0,01% Твин-20), набора для соединения аминов (GE Healthcare Cat # BR-1000-50) и мастера протоколов программного обеспечения Biacore Т100. Для иммобилизации были использованы все четыре ячейки чипа. Ячейки 1 и 3 были использованы для связывания с 500 единицами ответа (RU) бычьего сывороточного альбумина, растворенного в ацетатном буфере с рН 4,0, и ячейки 2 и 4 были использованы для связывания с 500 и 250 RU рТФ (R & D Systems), растворенного в ацетатном буфере, рН 4,5.
Каждый вариант FVIIa и протеазу FVIIa дикого типа протестировали при трех концентрациях в двух повторностях. Протеазы разбавили до 60 нМ, 30 нМ и 15 нМ в 100 мкл буфера для анализа Biacore (200 мМ Na Hepes, рН 7,4,150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ 8000,0,1% БСА, 0,01% Твин-20) в 96-ячеечном планшете для анализа. Каждый образец исследовали на приборе Biacore T100 при времени контакта 120 секунд и времени диссоциации 180 секунд при скорости потока 10 мкл/мин. Буферный контроль также был исследован. Чип регенерировали 50 мМ ЭДТА, рН 7,0 в течение 60 секунд, а затем 30 секунд. Тест для оценки связывания FVIIa дикого типа с рТФ дал три набора кривых, которые дали К-d около 8 нМ.
Для анализа данных использовали программное обеспечение Biacore T100 Evaluation. В частности, был использован анализ на связывание кинетика / аффинность 1:1, который аппроксимирует данные изотермой Ленгмюра, и данные были индивидуально аппроксимированы для двух повторностей каждого варианта на двух ячейках с единицами ответа. Полученные аппроксимированном четыре значения Kd усреднили и представили в таблице 22. FVIIa полипептиды, содержащие мутацию M156Q, как правило, обладают более низким Кd, и тем самым более прочно связываются с рТФ.
Связывание вариантов FVIIa с растворимым ТФ
Пример 16
Скрининг вариантов FVIIa на устойчивость к TFPI с помощью поверхностного плазменного резонанса (SPR)
Относительную устойчивость различных вариантов FVIIa к ингибированию рекомбинантным растворимым человеческим TFPI оценивали с помощью высокопроизводительного поверхностного плазменного резонанса (SPR) на приборе Biacore T100. Относительную устойчивость вариантов FVIIa к ингибированию TFPI оценивали путем измерения относительного количества варианта FVIIa, связанного с растворимым TFPI, иммобилизованным на сенсоре Biacore CM5, по сравнению с количеством FVIIa дикого типа, связанного после стандартизированного времени введения и концентрации протеазы.
Для каждого эксперимента растворимый TFPI (R & D Systems) иммобилизовали на новом 4-ячеечном проточном Biacore CM5 Series S сенсорном чипе (GE Healthcare), используя протокол для связывания аминов, имеющийся в Biacore Т-100 Control Software (GE Healthcare), и реагенты, предусмотренные в наборе для связывания аминов (GE Healthcare). Все четыре доступных проточных ячейки были использовали для иммобилизации двух разных плотностей TFPI и бычьего сывороточного альбумина (БСА), который служил блокирующим агентом в указанных ячейках. БСА разбавили до 5 мкг/мл в ацетате натрия (рН 4,0) и иммобилизовали в проточных ячейках 1 и 3 в количестве 1000 и 2000 единиц ответа (RU) соответственно. Для иммобилизации TFPI, лиофилизированный растворимый TFPI (10 мкг) ресуспендировали в 100 мкл 1 × буфера для связывания (30 мМ HEPES, 135 мМ NaCl, 1 мМ ЭДТА, 0,01% Твин-20, рН 7,4) до концентрации 0,1 мг/мл. В общей сложности 20 мкл 0,1 мг / мл раствора TFPI разбавили до концентрации 10 мкг/мл ацетатом натрия с рН 4,0 для иммобилизации в проточных ячейках 2 и 4 до 1000 и 2000 RU соответственно. Буфер для соединения был использован в качестве рабочего буфера во время стадий иммобилизации.
Каждый образец FVIIa приготовили с окончательной концентрацией 320 нМ в 1 × рабочем буфере (20 мМ HEPES, 150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ 8000, 0,1% БСА, 0,01% Твин-20, рН 7,4), содержащем 620 нМ рТФ (человеческий фактор свертывания III, R & D Systems). Каждый вариант FVIIa разбавили в 10 раз в 1 × рабочем буфере до окончательного разведения 320 нМ. FVIIa / рТФ комплексы получили в окончательном объеме 120 мкл в двух повторностях, загружая 48 уникальных вариантов FVIIa на 96-ячеечный планшет для хранения, и оценивали с двумя инъекциями в одном анализе. FVIIa / рТФ комплекс инкубировали при комнатной температуре в течение 10-15 минут до первого введения образца.
Стандартизированный метод анализа связывания был осуществлен с помощью программного обеспечения Biacore Control (GE Healthcare), в каждом анализе при введении FVIIa время ассоциации составляло 180 секунд, время для диссоциации составляло 60 секунд при скорости потока 10 мкл/мин. Регенерацию сенсорного чипа после этапа диссоциации проводили в течение 30 секунд 10 мМ глицином, 500 мМ NaCl, pH 3,0, и затем в течение 60 секунд проводили период стабилизации 1 × рабочим буфером при скорости потока 10 мкл/мин. В каждом анализе для последующего анализа данных были записаны две точки, одна за 5 секунд до завершения фазы ассоциации (связывания), а вторая за 5 секунд завершения фазы диссоциации (диссоциации). Перед началом полного анализа сенсорный чип протестировали с помощью одного ввода 320 нМ FVIIa дикого типа / рТФ в течение 180 секунд, что должно дать ответ в, примерно, 400-450 RU и 750-850 RU после связывания с проточными ячейками 2 (1000 RU) и 4 (200 RU), соответственно.
Анализ данных сначала проводили на Biacore Т100Evaluation Software (GE Healthcare) для проверки достоверности параметров анализа, что включало проверку, что связывание с указанной ячейкой является минимальным, проверку на смещение базового уровня и связывание при контрольном введении (рабочий буфер). Для этой цели были составлены таблицы с данными, в которых указаны количества вариантов FVIIa (в RU), связанных как в точке связывания, так и в точке диссоциации. Данные таблицы затем были экспортированы для дальнейшего анализа в таблицах Microsoft Excel. Сырые данные (связывание в RU) были скорректированы с помощью контрольного связывания с сенсорным чипом, а затем для каждого параметра вычисляли отношение количества связанного FVIIa дикого типа (в RU) к количеству связанного варианта FVIIa (в RU) и представляли как Связывание (дикий тип/ вариант) и Диссоциацию (дикий тип / вариант). Устойчивость к ингибированию TFPI отражается в увеличении соотношения для одного или обоих оцениваемых параметров. Например, значение Связывания (дикий тип / вариант) или Диссоциации (дикий тип / вариант) 20 для того или иного варианта FVIIa указывает, что этот вариант является в 20 раз более устойчивым к ингибированию TFPI, чем FVIIa дикого типа. Несколько вариантов проявили повышенную устойчивость к ингибированию TFPI. Например, СВ609, СВ637, СВ691, СВ856 и СВ857 относятся к группе, которая проявляла от 20 до 60 раз большую устойчивость, и отношение для вариантов, содержащих мутацию K341D по нумерации зрелого FVII (что соответствует химотрипсиновой нумерации), таких как СВ735, свидетельствует о существенной устойчивости к TFPI (более чем в 50-150 раз), и отношение для вариантов, содержащих мутацию K192D, например, 9СВ945 свидетельствует о существенной устойчивости к TFPI (более чем в 40-150 раз). В некоторых случаях скорость диссоциации была изменена больше, чем скорость ассоциации. Некоторые примеры вариантов, проявляющих такой профиль, включают СВ735, СВ854, СВ855, СВ856 и СВ857.
Устойчивость вариантов FVIIa к ингибированию TFPI
Пример 17. Определение концентрации каталитически активной протеазы с использованием титранта активного сайта 4-метилумбеллиферил р'-гуанидинобензоата (MUGB)
Концентрацию каталитически активного FVIIa в исходном растворе определяли титрованием комплекса FVIIa и растворимого тканевого фактора (рТФ) 4-метилумбеллиферил p'-гуанидинобензоатом (MUGB), флуорогенном эфирным субстратом, разработанным для активного цетра трипсин-подобных сериновых протеаз. Анализ проводили, как описано Раупе и др. (Biochemistry (1996) 35:7100-7106), с некоторыми незначительными модификациями. MUGB легко реагирует с FVIIa, но не с FVII или неактивной протеазой, с образованием эффективно стабильного ацил-ферментного интермедиата, в условиях насыщения концентрации MUGB деацилирование происходит особенно медленно и ограничивает скорость катализа. В этих условиях, протеаза FVIIa совершает один оборот каталитического цикла, высвобождая 4-метилумбеллифероновый флуорофор (4-MU). После того как первоначальный подъем флуоресценции откалибровали по внешней стандартной кривой флуоресценции 4-MU от концентрации, может быть рассчитана концентрация активных центров.
Анализы проводили с объемом 2 мл в кварцевых кюветах 1 см × 1 см при непрерывном перемешивании. Каждая реакционная смесь содержала 0,5 мкМ рТФ (R & D Systems Human) в буфере для анализа, содержащем 50 мМ HEPES, 100 мМ NaCl, 5 мМ CaCl2 и 0,1% ПЭГ 8000, рН 7,6. Был получен свежеприготовленный стандартный раствор 4-MU с исходной концентрацией 0,5 М в ДМСО, концентрация была подтверждена с помощью спектроскопии при абсорбции при 360 нм с использованием коэффициента экстинкции 19000 M-1см-1 в 50 мМ трис-буфере, рН 9,0. MUGB приготовили в исходной концентрации 0,04 М в ДМСО по сухому весу. Анализы начинали добавлением 4 мкл 4 мМ MUGB (конечная концентрация 8 мкМ) к 0,5 мкМ раствору рТФ (20,2 мкл 49,4 мкМ рТФ) в 1 × буфере для анализа, и сначала измеряли фоновый гидролиз MUGB за ~ 150-200 секунд перед добавлением FVIIa или варианта FVIIa до конечной концентрации 100-200 нМ по данным ELISA (Пример 2С.1) или титрования активного центра FFR-CMK (пример 6). Флуоресценцию высвобожденного 4-MU в фазе подъема реакции отслеживали в течение 1000-1200 секунд. Стандартную кривую свободного 4-MU получили титрованием калиброванного абсорбцией 4-MU в 1 × буфере для анализа, содержащем 0,5 мкМ рТФ, в шагах по 20 нМ до конечной концентрации 250 нМ.
Для анализа данных, данные о реакции импортировали в пакет программного обеспечения GraphPad Prism, и вклад фонового гидролиза вычитали из кривой путем экстраполяции первоначальной измеренной скорости самопроизвольного гидролиза MUGB, который, как правило, составлял менее 5% от общего подъема флуоресценции. Откорректированная кривая была аппроксимирована одним экспоненциальным уравнением с линейным компонентом (для учета медленных темпов деацилирования) вида Δ флуоресценции =Amp(1-e-kt)+Bt, где Amp=амплитуда фазы подъема в условиях анализа с насыщением, описанных выше, k представляет собой наблюдаемую константу скорости первого порядка для образования ацил-фермента, и В представляет собой константу скорости, связанную с полным оборотом MUGB. Концентрацию активных протеаз FVIIa рассчитывали путем сравнения параметров аппроксимирования для амплитуды с 4-MU стандартной кривой. Были измерены значения из нескольких проб, определены среднее и стандартное отклонения.
Если модификации очевидны для специалистов в данной области, предполагается, что это изобретение будет ограничено только границами формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ДОМЕНА GLA ФАКТОРА VII ИЛИ VIIa | 2004 |
|
RU2373282C2 |
| ПОЛИПЕПТИД ФАКТОРА VII СВЕРТЫВАНИЯ КРОВИ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2001 |
|
RU2326126C2 |
| ХИМЕРНЫЕ МОЛЕКУЛЫ ФАКТОРА VII | 2010 |
|
RU2563231C2 |
| ПОЛИПЕПТИД ФАКТОРА КОАГУЛЯЦИИ VII ЧЕЛОВЕКА, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2002 |
|
RU2325401C2 |
| МОДИФИЦИРОВАННЫЙ КОАГУЛИРУЮЩИЙ ФАКТОР VIIa С ПРОДЛЕННЫМ ВРЕМЕНЕМ ПОЛУЖИЗНИ | 2007 |
|
RU2466141C2 |
| АНТИТЕЛА ПРОТИВ ИНГИБИТОРА МЕТАБОЛИЧЕСКОГО ПУТИ ТКАНЕВОГО ФАКТОРА | 2009 |
|
RU2562114C2 |
| МОДИФИЦИРОВАННЫЕ ВИТАМИН К-ЗАВИСИМЫЕ ПОЛИПЕПТИДЫ | 2005 |
|
RU2396347C2 |
| НОВЫЕ АНТИТЕЛА К ТКАНЕВОМУ ФАКТОРУ В КАЧЕСТВЕ АНТИКОАГУЛЯНТОВ | 2003 |
|
RU2345789C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПОЛИПЕПТИДЫ ФАКТОРА VII И ПОЛИПЕПТИДЫ ФАКТОРА XI | 2002 |
|
RU2298416C2 |
| МИКРОВЕЗИКУЛЫ, ПОЛУЧЕННЫЕ ИЗ РЕКОМБИНАНТНЫХ ДРОЖЖЕЙ, ОБЛАДАЮЩИЕ ГЕМОСТАТИЧЕСКИМИ АКТИВНОСТЯМИ, И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2492184C2 |

























































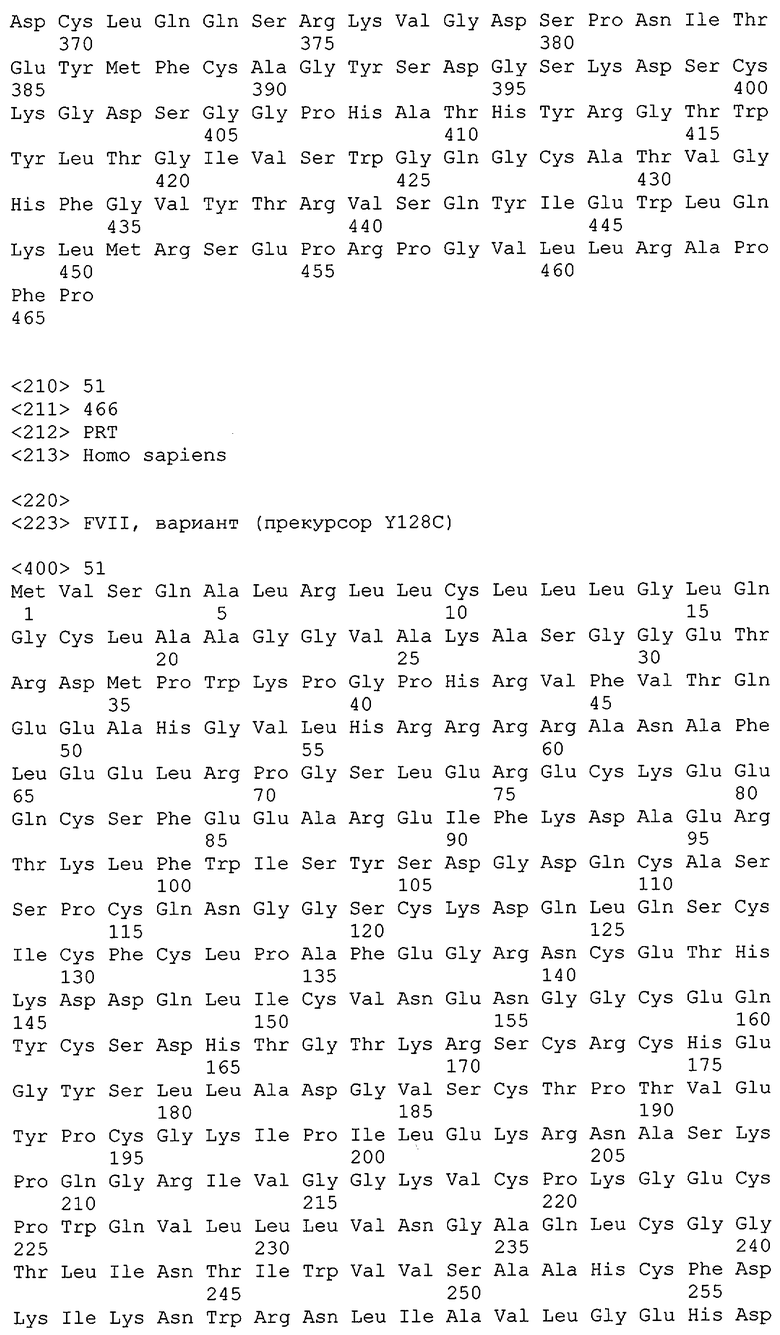










































































































































































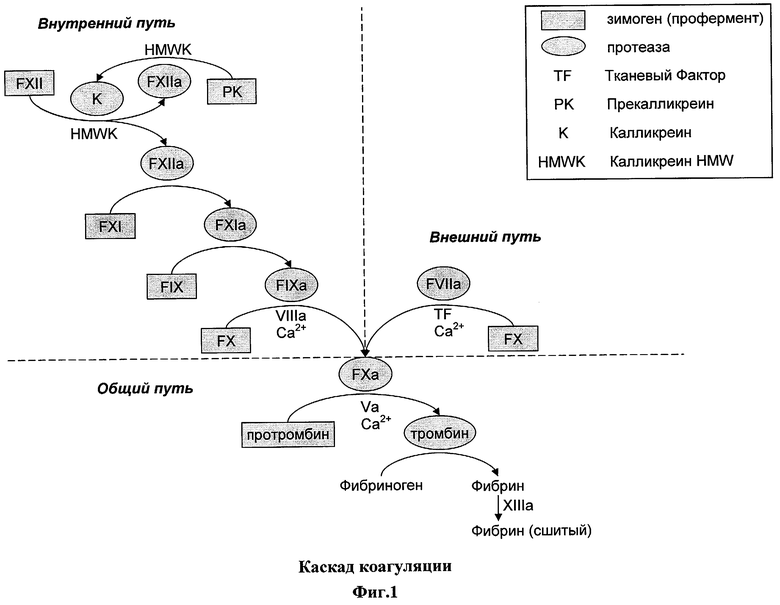
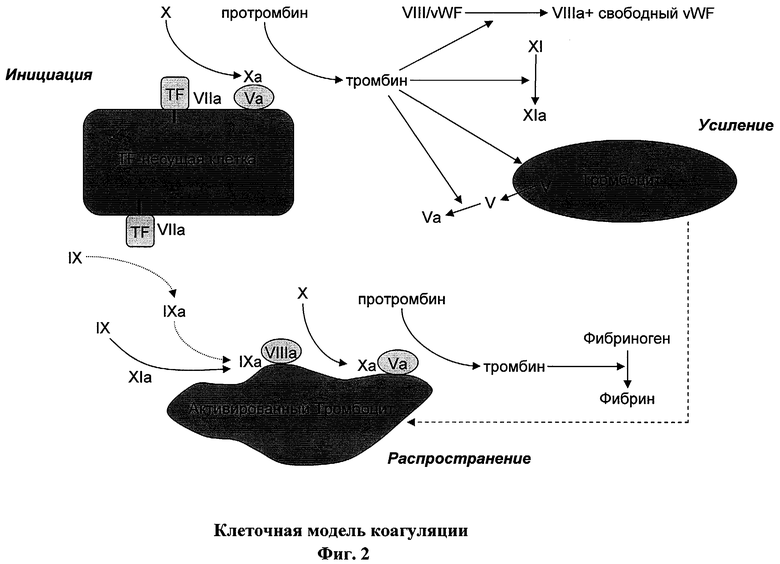
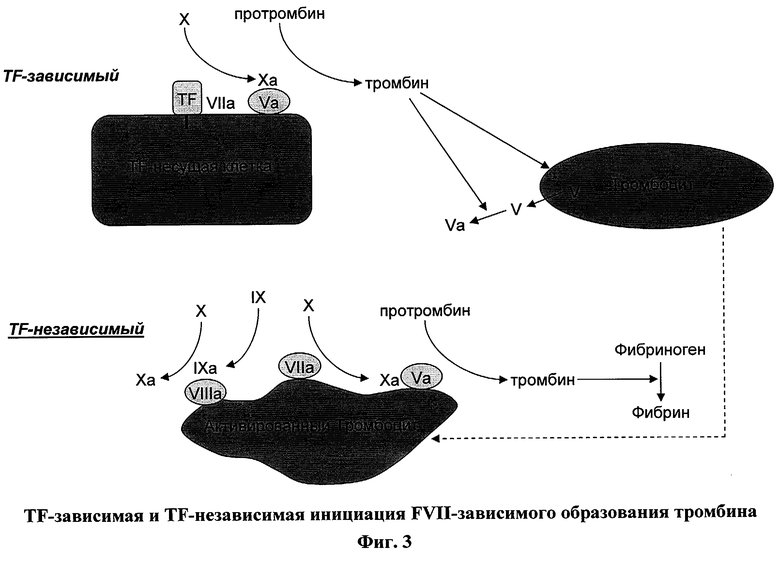
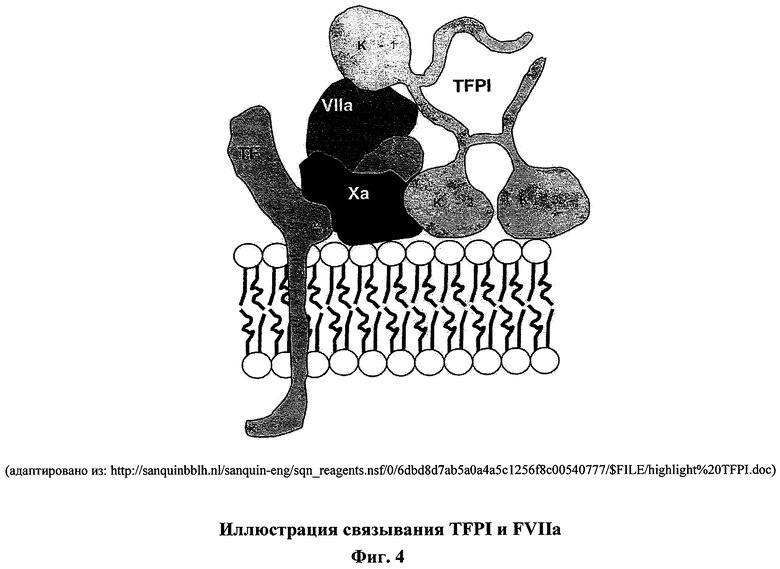

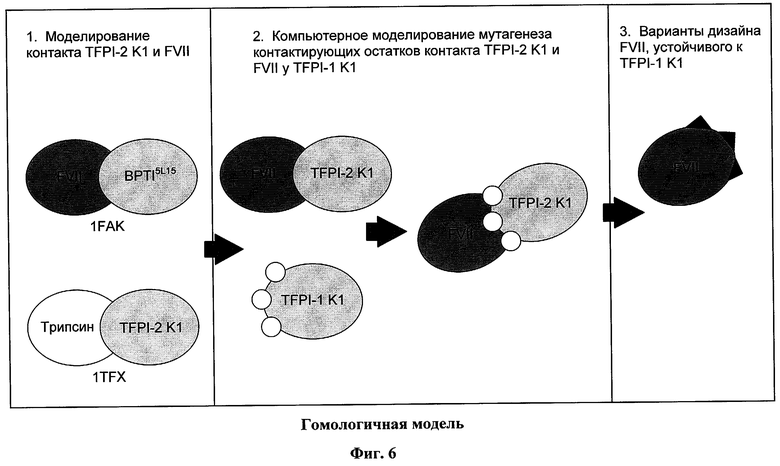
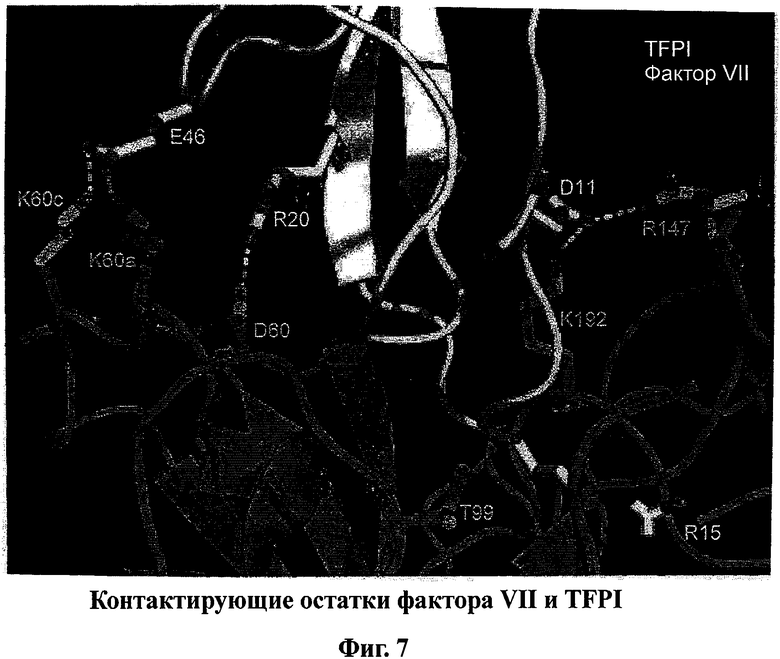
Изобретение относится к области биотехнологии, конкретно к получению модифицированного фактора FVII, и может быть использовано в медицине. Полипептид FVII модифицируют путем замены аминокислотного остатка, выбранной из D196L, D196I, D196Y, К197Е, K197D, K197L, К197М, K197I, K197V, K197F, K197W, K197Y, K199D и K199E, в аминокислотной последовательности FVII, представленной в виде SEQ ID NO: 3, или в соответствующих остатках его предшественника с SEQ ID NO: 1 или SEQ ID NO: 2. Полипептид может содержать дополнительные модификации аминокислот, улучшающие его функциональные свойства. Изобретение позволяет получить модифицированный полипептид FVII, который при активации обладает увеличенной коагулянтной активностью и/или повышенной устойчивостью к ингибиторам по сравнению с немодифицированным полипептидом FVII с последовательностью, приведенной в SEQ ID NO: 3. 9 н. и 39 з.п. ф-лы, 7 ил., 23 табл., 17 пр.
1. Модифицированный полипептид фактора VII (FVII), включающий аминокислотную модификацию в полипептиде FVII или его активных фрагментах, где
эта модификация представляет собой замену аминокислотного остатка D196, K197 или K199 в полипептиде FVII, имеющем последовательность аминокислот, приведенную в SEQ ID NO: 3, или в соответствующих остатках предшественника в полипептиде FVII, имеющего последовательность аминокислот, приведенную в SEQ ID NO: 1 или SEQ ID NO: 2;
модифицированный полипептид FVII содержит 1, 2, 3, 4, 5, 6 или 7 аминокислотных замен в полипептиде с последовательностью, приведенной в SEQ ID NO: 3; и
одну или более модификаций остатков, в дополнение к заменам остатков D196, K197 или K199, приводящую(их) к дальнейшему улучшению функциональных свойств модифицированного полипептида FVII; и
аминокислотная замена остатка D196, K197 или K199 выбрана из D196L, D196I, D196Y, K197Е, K197D, K197L, K197М, K197I, K197V, K197F, K197W, K197Y, K199D и K199E,
за счет чего модифицированный полипептид FVII, когда он активирован, обладает увеличенной коагулянтной активностью и/или повышенной устойчивостью к ингибиторам по сравнению с немодифицированным полипептидом FVII с последовательностью, приведенной в SEQ ID NO: 3.
2. Модифицированный полипептид FVII по п. 1, в котором модификация полипептида FVII выбрана из K197Е, K197D, K197L, K197М, K197I, K197V, K197F, K197W, K199D и K199Е.
3. Модифицированный полипептид FVII по п. 1, в котором модификация полипептида FVII представляет собой замену аминокислоты в положении 197 на Y.
4. Модифицированный полипептид FVII по п. 1, в котором произведена дополнительная модификация, представляющая собой замену аминокислоты по позиции, выбранной из D196, K197, K199, G237, Т239, R290 и K341, и
где первая модификация и дополнительная модификация затрагивают различные аминокислоты.
5. Модифицированный полипептид FVII по п. 4, в котором дополнительные модификации аминокислот выбраны из D196K, D196R, D196Y, D196F, D196W, D196L, D196I, K197Y, K197А, K197Е, K197D, K197L, K197М, K197I, K197V, K197F, K197W, K199А, K199D, K199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, K341Е, K341R, K341N, K341М, K341D и K341Q.
6. Модифицированный полипептид FVII по любому из пп. 1-5, включающий модификации, выбранные из D196R/K197E/K199E, D196K/K197E/K199E, D196R/K197E/K199E/R290E, D196R/K197M/K199E, D196R/K197M/K199E/R290E, D196K/K197L, D196F/K197L, D196L/K197L, D196M/K197L, D196W/K197L, D196F/K197E, D196W/K197E, K196V/K197E, K197E/K341Q, K197L/K341Q, K197E/G237V/K341Q, K197Е/ K199Е, K197E/G237V, K199E/K341Q и K197E/K199E/K341Q.
7. Модифицированный полипептид фактора VII (FVII) по любому из пп. 1-6, включающий две аминокислотные замены, выбранные из D196K, D196R, D196A, D196Y, D196F, D196M, D196W, D196L, D196I, K197Y, K197А, K197Е, K197D, K197L, K197М, K197I, K197V, K197F, K197W, K199А, K199D, K199Е, G237W, G237T, G237I, G237V, Т239А, R290A, R290E, R290D, R290N, R290Q, R290K, K341Е, K341R, K341N, K341М, K341D и K341Q, где, по крайней мере, одна замена выбрана из D196L, D196I, D196Y, K197Е, K197D, K197L, K197М, K197I, K197V, K197F, K197W, K197Y, K199D и K199Е, и вторая модификация отлична от первой.
8. Модифицированный полипептид FVII по п. 7, включающий замены, выбранные из D196R/K197E/K199E, D196K/K197E/K199E, D196R/K197E/K199E/R290E, D196R/K197M/K199E, D196R/K197M/K199E/R290E, D196K/K197L, D196F/K197L, D196L/K197L, D196M/K197L, D196W/K197L, D196F/K197E, D196W/K197E, D196V/K197E, K197E/K341Q, K197L/K341Q, K197E/G237V/K341Q, K197Е/ K199Е, K197E/G237V, K199E/K341Q, K197E/K199E/K341Q и K197E/G237V/M298Q.
9. Модифицированный полипептид FVII по любому из пп. 1-8, где ингибитором является ингибитор пути тканевого фактора (TFPI).
10. Модифицированный полипептид FVII по любому из пп. 1-9, включающий одну или более дополнительных модификаций аминокислоты (аминокислот) в позициях, соответствующих позиции Q176, М298 или Е296 в полипептиде FVII с последовательностью аминокислот, приведенной в SEQ ID NO 3, или в соответствующих остатках в полипептиде FVII.
11. Модифицированный полипептид FVII по п. 10, в котором одна или более модификаций аминокислоты (аминокислот) в позициях Q176, М298 или Е296 выбраны из Q176A, M298Q, E296V и Е296А.
12. Модифицированный полипептид FVII по п. 11, включающий модификации аминокислот, выбранные из K197E/G237V/M298Q, K197E/G237V/M298Q/K341Q, K197E/K199E/G237V/M298Q/K341Q и K197E/M298Q.
13. Модифицированный полипептид FVII по любому из пп. 1-12, в котором немодифицированный полипептид FVII состоит из последовательности аминокислотных остатков, приведенной в SEQ ID NO: 3.
14. Модифицированный полипептид FVII по п. 1, включающий последовательность аминокислот, приведенную в любой из SEQ ID NO: 21, 23-26, 28, 29, 39-43, 125, 128-133, 139-146, 229, 230, 232-235, 242-244, 248 и 250.
15. Модифицированный полипептид FVII по п. 1 или 2, состоящий из последовательности аминокислот, приведенной в любой из SEQ ID NO: 21, 23-26, 28, 29, 39-43, 125, 128-133, 139-146, 229, 230, 232-235, 242-244, 248 и 250.
16. Модифицированный полипептид FVII по любому из пп. 1-12 и 14, который является активным или зрелым полипептидом.
17. Модифицированный полипептид FVII по любому из пп. 1-16, содержащий химическую модификацию или посттрансляционную модификацию.
18. Модифицированный полипептид FVII по п. 18, где полипептид FVII гликозилирован, карбоксилирован, гидроксилирован, сульфатирован, фосфорилирован, альбумирован, либо соединен с полиэтиленгликольным фрагментом (PEG).
19. Модифицированный полипептид FVII по любому из пп. 1-18, который является одноцепочечным полипептидом.
20. Модифицированный полипептид FVII по любому из пп. 1-18, который является двухцепочечным полипептидом.
21. Модифицированный полипептид FVII по любому из пп. 1-18, представляющий собой активированную двуцепочечную форму полипептида фактора VII (FVIIa).
22. Модифицированный полипептид FVII по любому из пп. 1-18, представляющий собой активированную двуцепочечную форму полипептида фактора VII (FVIIa), который расщеплен между остатками, соответствующими аргинину в положении 152 и изолейцину в положении 153 в SEQ ID NO:3.
23. Молекула нуклеиновой кислоты, кодирующая модифицированный полипептид FVII по любому из пп. 1-22.
24. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по п. 23, оперативно связанную с промотором экспрессии молекулы нуклеиновой кислоты.
25. Вектор по п. 24, где вектор представляет собой прокариотический вектор, вирусный вектор или эукариотический вектор.
26. Вектор по п. 24, где вектор представляет собой вектор для млекопитающих.
27. Вектор по п. 24, являющийся вирусным вектором, выбранным из аденовируса, аденосвязанного вируса, ретровируса, вируса герпеса, лентивируса, поксвируса и цитомегаловируса.
28. Не человеческая клетка для экспрессии модифицированного полипептида FVII по любому из пп. 1-22, включающая вектор по любому из пп. 24-27.
29. Клетка по п. 28, являющаяся эукариотической клеткой.
30. Изолированная клетка для экспрессии модифицированного полипептида FVII по любому из пп. 1-22, включающая вектор по любому из пп. 24-27, где клетка является эукариотической клеткой; и клетка не является клеткой эмбриона человека.
31. Клетка по п. 29 или 30, где эукариотическая клетка является клеткой млекопитающих.
32. Клетка по п. 31, выбранная из клеток почек детенышей хомяков (ВНК-21) и клеток СНО.
33. Клетка по п. 32, которая является дрожжевой клеткой.
34. Клетка по п. 33, которая является клеткой Pichia sp.
35. Клетка по п. 34, которая является клеткой Pichia pastoris.
36. Способ получения модифицированного полипептида FVII, включающий культурирование клетки по любому из пп. 28-35 в условиях, обеспечивающих экспрессию модифицированного полипептида FVII; и выделение экспрессированного полипептида FVII.
37. Способ лечения субъекта, включающий введение субъекту полипептида по любому из пп. 1-22, в котором субъект страдает заболеванием или состоянием, выбранным из группы, включающей нарушения коагуляции крови, гематологические нарушения, геморрагические нарушения, гемофилия, недостаток фактора VII, кровотечение, хирургическое кровотечение, или кровотечение по причине травмы.
38. Способ по п. 37, в котором лечение заболевания или состояния производится введением зимогенной или активной формы FVII.
39. Способ по п. 37, в котором заболевание или состояние представляет собой гемофилию А, гемофилию В или гемофилию С.
40. Способ по п. 37, в котором заболевание или состояние представляет собой гемофилию, которая является врожденной или приобретенной.
41. Способ по п. 37, в котором заболевание или состояние вызвано кровотечением, которое является осложнением после операции или травмы.
42. Способ по п. 41, в котором кровотечение проявляется как острый гемартроз, хроническая гемофильная артропатия, гематомы, гематурия, кровотечения в центральной нервной системе, желудочно-кишечные кровотечения или кровоизлияние в мозг.
43. Способ по п. 41, в котором кровотечение происходит из-за удаления зубов.
44. Способ по п. 41, в котором хирургия является хирургией на сердце, ангиопластикой, легочной хирургией, абдоминальной хирургией, хирургией позвоночника, сосудистой хирургией, стоматологической хирургией или трансплантацией органов.
45. Способ по п. 44, в котором хирургия является трансплантационной хирургией, выбранной из трансплантации костного мозга, сердца, легких, поджелудочной железы и печени.
46. Способ по п. 37, в котором у субъекта вырабатываются аутоантитела к фактору VIII или фактору IX.
47. Использование модифицированного полипептида FVII по любому из пп. 1-22 для лечения заболеваний или состояний, выбранных из группы, включающей нарушения коагуляции крови, гематологические нарушения, геморрагические нарушения, гемофилию, недостаток фактора VII, нарушения, связанные с кровотечением, хирургические кровотечения или кровотечения в результате травмы.
48. Применение модифицированного полипептида FVII по любому из пп. 1-22 для приготовления лекарственного средства для лечения заболеваний или состояний, которые выбраны из группы, включающей нарушения коагуляции крови, гематологические нарушения, геморрагические нарушения, гемофилию, недостаток фактора VII, нарушения, связанные с кровотечением, хирургические кровотечения или кровотечения в результате травмы.
| WO 2004083361 A2, 30.09.2004 | |||
| WO 2007031559 A2, 22.03.2007 | |||
| WO 2006114105 A2, 02.11.2006 | |||
| МОЛЕКУЛЫ, ПОДОБНЫЕ ФАКТОРУ VII ИЛИ VIIA | 2001 |
|
RU2278123C2 |
| GENG J | |||
| P | |||
| ET AL., Properties of a recombinant chimeric protein in which the gamma-carboxyglutamic acid and helical stack domains of human anticoagulant protein C are replaced by those of human coagulation factor VII, THROMBOSIS AND | |||

