Область техники
Настоящее изобретение относится к модифицированным последовательностям нуклеиновых кислот, кодирующим фактор коагуляции VIII (FVIII) и фактор Виллебранда (VWF), а также их комплексы и их производные, рекомбинантным экспрессионным векторам, содержащим такие последовательности нуклеиновых кислот, клеткам-хозяевам, трансформированным такими рекомбинантными экспрессионными векторами, кодируемым такими последовательностями нуклеиновых кислот, рекомбинантным полипептидам и производным, которые действительно обладают биологическими активностями вместе с удлиненным полупериодом существования in vivo и/или увеличенным отношением фактической максимальной концентрации in vivo (в крови и т.п.) к ожидаемой по сравнению с немодифицированным белком дикого типа. Настоящее изобретение также относится к соответствующим последовательностям FVIII, которые дают в результате увеличенное количество продукта экспрессии. Настоящее изобретение, кроме того, относится к процессам производства таких рекомбинантных белков и их производных. Настоящее изобретение также относится к вектору переноса для применения в генной терапии людей, который включает такие модифицированные последовательности нуклеиновых кислот.
Предпосылки создания изобретения
Существуют различные нарушения свертываемости крови, вызываемые недостатками факторов свертывающей системы крови. Самыми часто встречающимися нарушениями являются гемофилии A и B, являющиеся следствием недостатков фактора свертывающей системы крови VIII и IX соответственно. Другим известным нарушением свертываемости крови является болезнь Виллебранда.
В плазме FVIII существует, главным образом, в виде нековалетного комплекса с VWF, и его коагулирующей функцией является ускорение зависимого от фактора IXa превращения фактора X в Xa. Вследствие образования комплекса из FVIII и VWF в течение длительного времени полагали, что функции FVIII и VWF представляют собой две функции одной и той же молекулы. Только в семидесятых годах стало ясно, что FVIII и VWF являются отдельными молекулами, которые образуют комплекс в физиологических условиях. Затем в восьмидесятых годах была определена константа диссоциации, составляющая приблизительно 0,2 нмоль/л (Leyte et al., Biochem J. 1989, 257: 679-683), и были изучены последовательности ДНК обеих молекул.
Классическая гемофилия или гемофилия A является наследственным нарушением свертываемости крови. Она является следствием сцепленного с X-хромосомой недостатка FVIII свертывающей системы крови и поражает почти исключительно мужчин с частотой, составляющей один-два индивидуума на 10000. Дефект X-хромосомы передается женщинами-носителями, которые сами не больны гемофилией. Клиническим проявлением гемофилии A является повышенная склонность к кровотечениям. До внедрения лечения концентратами FVIII средняя продолжительность жизни человека с тяжелой гемофилией составляла менее 20 лет. Использование концентратов FVIII из плазмы значительно улучшило ситуацию для страдающих гемофилией A пациентов, намного увеличив среднюю продолжительность жизни, дав большинству из них возможность жить более или менее нормальной жизнью. Однако существовали определенные проблемы с получаемыми из плазмы концентратами и их применением, самой серьезной из которых была передача вирусов. Пока серьезно поражали популяцию вирусы, вызывающие гепатит B, гепатит ни-A, ни-B и СПИД. С тех пор были разработаны недавно различные способы инактивации вирусов и новые в высокой степени очищенные концентраты FVIII, которые установили очень высокий стандарт безопасности также для получаемого из плазмы FVIII.
Клонирование кДНК для FVIII (Wood et al. 1984. Nature 312: 330-336; Vehar et al. 1984. Nature 312: 337-342) сделало возможной экспрессию FVIII рекомбинатно, что привело к разработке нескольких продуктов в виде рекомбинантного FVII, которые были разрешены регулирующими органами между 1992 и 2003. То обстоятельство, что центральный B-домен полипептидной цепи FVIII, находящийся между аминокислотами Arg-740 и Glu-1649, по-видимому, не является нужным для полной биологической активности, также привело к разработке FVIII с делецией В-домена.
Зрелая молекула FVIII состоит из 2332 аминокислот, которые можно сгруппировать в три гомологичных A-домена, два гомологичных C-домена и B-домен, которые располагаются в порядке A1-A2-B-A3-C1-C2. Полная аминокислотная последовательность зрелого FVIII человека представлена в SEQ ID NO: 15. Во время секреции в плазму FVIII процессируется внутриклеточно в ряд связанных ионом металла гетеродимеров, когда одноцепочечный FVIII расщепляется на границе B-A3 и в различных сайтах внутри B-домена. Этот процессинг приводит к образованию гетерогенных молекул тяжелых цепей, состоящих из A1, A2 и различных частей B-домена, которые имеют молекулярную массу, находящуюся в диапазоне от 90 кДа до 200 кДа. Тяжелые цепи связываются посредством иона металла с легкими цепями, которые состоят из A3, C1 и C2-домена (Saenko et al. 2002. Vox Sang. 83: 89-96). В плазме этот гетеродимерный FVIII связывается с высоким сродством с фактором Виллебранда (VWF), который защищает его от преждевременного катаболизма. Полупериод существования не активированного FVIII, связанного с VWF, составляет приблизительно 12 часов в плазме.
Фактор коагуляции FVIII активируется посредством протеолитического расщепления тромбином и FXa в положениях аминокислот Arg372 и Arg740 внутри тяжелой цепи и в Arg1689 в легкой цепи, что приводит к высвобождению фактора Виллебранда и образованию активированного гетеротримера FVIII, который будет образовывать теназный комплекс на фосфолипидных поверхностях с FIXa и FX при условии присутствия Ca2+. Гетеротример состоит из A1-домена, фрагмента массой 50 кДа, A2-домена, фрагмента массой 43 кДа, и легкой цепи (A3-C1-C2), фрагмента массой 73 кДа. Таким образом, активная форма FVIII (FVIIIa) состоит из A1-субъединицы, связанной через образуемую двухвалентным металлом ионную связь с расщепленной тромбином легкой цепью A3-C1-C2, и свободной A2-субъединицы, относительно слабо связанной с A1 и A3-доменом.
Во избежание чрезмерной коагуляции FVIIIa должен быть инактивирован вскоре после активации. Полагают, что инактивация FVIIIa посредством активированного белка C (APC) путем расщепления в Arg336 и Arg562 не является основной определяющей скорость стадией. Скорее ей является диссоциация нековалентно присоединенной A2-субъединицы от гетеротримера, которая, как полагают, является определяющей скорость стадией в инактивации FVIIIa после активации тромбином (Fay et al. 1991. J. Biol. Chem. 266: 8957, Fay & Smudzin 1992. J. Biol. Chem. 267: 13246-50). Она является быстрым процессом, что объясняет короткий полупериод существования FVIIIa в плазме, который составляет только 2,1 минуты (Saenko et al. 2002. Vox Sang. 83: 89-96).
Пациентам с тяжелой гемофилией A, подвергаемым профилактическому лечению, FVIII должен вводиться внутривенно приблизительно 3 раза в неделю из-за короткого полупериода существования FVIII в плазме, составляющего приблизительно 12-14 часов. Каждое внутривенное введение является трудоемким, сопровождающимся болью и влечет за собой риск инфицирования, особенно когда это делается в основном в домашних условиях самими пациентами или родителями детей, у которых диагностирована гемофилия A.
Поэтому было бы в высокой степени желательным создание FVIII с увеличенным полупериодом функционального существования, делающее возможным производство содержащих FVIII фармацевтических композиций, которые нужно вводить менее часто.
Было предпринято несколько попыток для продления полупериода существования не активированного FVIII или посредством уменьшения его взаимодействия с клеточными рецепторами (WO 03/093313A2, WO 02/060951A2), посредством ковалентного присоединения полимеров к FVIII (WO 94/15625, WO 97/11957 и US 4970300), посредством инкапсуляции FVIII (WO 99/55306), посредством введения новых сайтов связывания металла (WO 97/03193), посредством ковалентного присоединения A2-домена к A3-домену с помощью либо пептидной (WO 97/40145 и WO 03/087355), либо дисульфидной связи (WO 02/103024A2), или посредством ковалентного присоединения A1-домена к A2-домену (WO2006/108590).
Другой подход к увеличению полупериода функционального существования FVIII или VWF осуществляют посредством пегилирования FVIII (WO 2007/126808, WO 2006/053299, WO 2004/075923) или посредством пегилирования VWF (WO 2006/071801), который, имея в результате пегилирования увеличенный полупериод существования, мог бы опосредованно увеличить также полупериод существования FVIII, присутствующего в плазме.
Поскольку ни один из описанных выше подходов еще не привел к разрешению лекарственного средства FVIII и поскольку введение мутаций в последовательность FVIII дикого типа или введение химических модификаций влечет за собой, по крайней мере, теоретический риск создания иммуногенных вариантов FVIII, существует сохраняющаяся потребность в разработке модифицированных молекул фактора VIII коагуляции, которые демонстрируют удлиненный полупериод существования.
Ввиду потенциального риска тромбообразования более желательным является удлинение полупериода существования не активированной формы FVIII, чем FVIIIa.
VWF, который является отсутствующим, функционально дефектным или имеется в наличии только в уменьшенном количестве при различных формах болезни Виллебранда (VWD), представляет собой мультимерный адгезивный гликопротеин, присутствующий в плазме млекопитающих, который обладает множеством физиологических функций. Во время первичного гемостаза VWF функционирует в качестве медиатора между специфическими рецепторами на поверхности тромбоцита и компонентами экстраклеточного матрикса, такими как коллаген. Кроме того, VWF служит в качестве носителя и стабилизирующего белка для прокоагулянта FVIII. VWF синтезируется в эндотелиальных клетках и мегакариоцитах в виде молекулы-предшественника из 2813 аминокислот. Аминокислотная последовательность и последовательность кДНК VWF дикого типа сообщается в Collins et al. 1987, Proc Natl. Acad. Sci. USA 84: 4393-4397. Полипептид-предшественник, препро-VWF, состоит из сигнального пептида длиной 22 остатка, пропептида длиной 741 остаток и полипептида длиной 2050 остатков, обнаруживаемого в зрелом плазменном VWF (Fischer et al., FEBS Lett. 351: 345-348, 1994). После отщепления сигнального пептида в эндоплазматическом ретикулуме между двумя мономерами VWF образуется C-концевой дисульфидный мостик. Во время дальнейшего перемещения через секреторный путь добавляются 12 N-сочленяемых и 10 O-сочленяемых углеводных боковых цепей. Важнее, что димеры VWF подвергаются мультимеризации через N-концевые дисульфидные мостики, и пропептид длиной 741 аминокислот отщепляется ферментом PACE (расщепляющим спаренные основные аминокислоты ферментом)/фурином в позднем аппарате Гольджи. Пропептид, а также высокомолекулярные мультимеры VWF (VWF-HMWM) хранятся в тельцах Вейбеля-Паллада эндотелиальных клеток или в [альфа]-гранулах тромбоцитов.
После секреции в плазму протеаза ADAMTS 13 расщепляет VWF внутри A1-домена VWF. Поэтому VWF в плазме состоит из целого ряда мультимеров, простирающегося от единичных димеров с М.м. 500 кДа до мультимеров, состоящих из вплоть до более чем 20 димеров, с молекулярной массой, превышающей 10000 кДа. При этом VWF-HMWM обладают более сильной гемостатической активностью, которую можно определить через ристоцетин-кофакторную активность (VWF:RCo). Чем больше отношение VWF:RCo/VWF антиген, тем больше относительное количество высокомолекулярных мультимеров.
Дефекты в VWF являются причиной болезни Виллебранда (VWD), которая характеризуется более или менее отчетливым фенотипом нарушения свертываемости крови. VWD типа 3 является самой тяжелой формой, при которой VWF полностью отсутствует, VWD типа 1 связан с количественной потерей VWF, и ее фенотип может быть очень слабым. VWD типа 2 связан с качественными дефектами VWF и может быть настолько же тяжелой, как и VWD типа 3. VWD типа 2 имеет множество подформ, при этом некоторые из них связаны с потерей или уменьшением высокомолекулярных мультимеров. VWD типа 2a характеризуется потерей как средних, так и больших мультимеров. VWD типа 2B характеризуется потерей мультимеров с наибольшей молекулярной массой.
VWD является самым часто встречающимся нарушением свертываемости крови у людей и может подвергаться лечению с помощью заместительной терапии с использованием концентратов, содержащих VWF плазматического или рекомбинантного происхождения. VWF можно приготовить из плазмы человека, как, например, описано в EP 05503991. В EP 0784632 описывается способ выделения рекомбинантного VWF.
В плазме FVIII связывается с высоким сродством с VWF, который защищает его от преждевременного катаболизма и поэтому играет, помимо своей роли в первичном гемостазе, основную роль в регулировании уровней FVIII в плазме и, как следствие, также является центральным фактором для контролирования вторичного гемостаза. Полупериод существования не активированного FVIII, связанного с VWF, составляет приблизительно 12-14 часов в плазме. При болезни Виллебранда типа 3, при которой VWF не присутствует или почти не присутствует, полупериод существования FVIII составляет только приблизительно 6 часов, приводя к симптомам гемофилии А от небольшой до умеренной тяжести у таких пациентов вследствие уменьшенных концентраций FVIII. Стабилизирующий эффект VWF на FVIII был также использован для способствования рекомбинантной экспрессии FVIII в клетках CHO (Kaufman et al. 1989, Mol. Cell Biol.).
До настоящего времени стандартное лечение гемофилии A и VWD включает частые внутривенные инфузии препаратов FVIII и концентратов VWF или инфузии концентратов, включающих комплекс FVIII и VWF, получаемый из плазм людей-доноров, или в случае FVIII инфузии фармацевтических препаратов на основе рекомбинантного FVIII. Хотя эти заместительные терапии являются обычно эффективными, например, для пациентов с тяжелой гемофилией A, подвергаемых профилактическому лечению, FVIII должен вводиться внутривенно приблизительно 3 раза в неделю вследствие короткого полупериода существования FVIII в плазме, составляющего приблизительно 12 часов. Уже при уровнях, превышающих 1% от активности FVIII у не больных гемофилией, например, при повышении уровней FVIII на 0,01 Е/мл, тяжелая гемофилия A превращается в гемофилию A средней тяжести. При профилактической терапии схемы дозирования разрабатывают так, чтобы минимальные уровни активности FVIII не опускались ниже уровней, составляющих 2-3% от активности FVIII у не больных гемофилией. Каждое внутривенное введение является трудоемким, сопровождающимся болью и влечет за собой риск инфицирования, особенно когда это в основном делается при лечении в домашних условиях самими пациентами или родителями детей, у которых диагностирована гемофилия A. Кроме того, частые внутривенные инъекции неизбежно приводят к образованию шрамов, мешающему дальнейшим инфузиям. Поскольку профилактическое лечению при тяжелой гемофилии начинают рано в периоде жизни, при этом возраст детей часто составляет менее 2 лет, введение FVIII трижды в неделю в вены таких маленьких пациентов является даже более трудным. В течение ограниченного периода времени имплантация инфузионной системы может быть альтернативой. Несмотря на то обстоятельство, что могут иметь место многократные инфицирования, и инфузионные системы могут вызывать неудобство во время физической нагрузки, они, тем не менее, обычно считаются предпочтительными по сравнению с внутривенными инъекциями.
Полупериод in vivo существования VWF человека в кровотоке человека составляет приблизительно от 12 до 20 часов. При профилактическом лечении VWD, например, типа 3 также могло бы быть в высокой степени желательным нахождение путей для удлинения полупериода функционального существования VWF.
Другим подходом к увеличению полупериода функционального существования VWF является пегилирование VWF (WO 2006/071801), который, имея в результате пегилирования увеличенный полупериод существования, мог бы опосредованно увеличить также полупериод существования FVIII, присутствующего в плазме.
Однако химическая конъюгация PEG или других молекул с терапевтическими белками всегда влечет за собой риск снижения специфической активности вследствие экранирования важных сайтов взаимодействия с другими белками, химическая конъюгация прибавляет дополнительную стадию при производстве таких белков, уменьшая конечные выходы продукта и делая производство более дорогостоящим. Также не известны долгосрочные эффекты на здоровье человека, поскольку известные в настоящее время пегилированные терапевтические белки не нужно вводить в течение всей жизни, как это могло бы быть случаем для VWF, вводимого для профилактики болезни Виллебранда, или для FVIII, вводимого при гемофилии A.
Поэтому могло бы быть в высокой степени желательным получение долгоживущего VWF, который не является химически модифицированным.
В известном уровне техники были описаны слияния факторов коагуляции с альбумином (WO 01/79271), альфа-фетопротеином (WO 2005/024044) и иммуноглобулином (WO 2004/101740) в качестве увеличивающих полупериод существования полипептидов. Указывалось, что они присоединены к карбоксильному концу или амино-концу или к обоим концам соответствующей терапевтической белковой составляющей, иногда связаны с помощью пептидных линкеров, предпочтительно с помощью линкеров, состоящих из глицина и серина.
Ballance и др. (WO 01/79271) описали полипептиды из множества различных терапевтических полипептидов, слитые в своих N- или C-концевых частях с сывороточным альбумином человека. Представлены длинные списки возможных партеров по слиянию без сообщения в отношении почти любого из этих белков экспериментальных данных, касающихся того, сохраняют ли соответствующие слитые с альбумином белки биологическую активность и имеют ли они улучшенные свойства. В указанном списке терапевтических полипептидов упомянуты также FVIII и VWF.
Невероятно, чтобы C-концевое слияние серьезно рассматривалось квалифицированным в данной области специалистом, поскольку C2-домен FVIII в весьма C-концевой части FVIII между аминокислотами 2303 и 2332 FVIII включает сайт связывания с мембранами тромбоцитов, который весьма важен для функционирования FVIII. Вот почему существует множество известных аминокислотных мутаций в этой области, которые вызывают гемофилию A. Поэтому вызывало удивление то, что относительно большой гетерологичный полипептид, вроде альбумина, можно слить с C-концевой частью FVIII без препятствования функционированию FVIII посредством препятствования связыванию с тромбоцитами. Кроме того, C2-домен также содержит сайт связывания с VWF. Этот сайт вместе с аминокислотной последовательностью 1649-1689 ответственен за связывание FVIII с высоким сродством с VWF. Поэтому квалифицированный в данной в области техники специалист не мог бы ожидать, что FVIII, слитый в своей C-концевой части с альбумином, будет сохранять свое связывание с VWF.
С удивлением было обнаружено, что в противоположность предсказанию Ballance и др. слияние альбумина с N-концом FVIII не секретируется в культуральную среду. Поэтому и по причинам, детализированным выше, с даже большим удивлением теперь было обнаружено, что FVIII, слитый в своей C-концевой части с альбумином, секретируется в культуральную среду и сохраняет свою биологическую функцию, в том числе связывание с мембранами активированных тромбоцитов и с VWF.
С удивлением было также обнаружено, что модифицированный FVIII настоящего изобретения демонстрирует увеличение отношения фактической максимальной концентрации in vivo к ожидаемой на приблизительно 20% по сравнению с FVIII дикого типа.
Квалифицированный в данной области специалист также не рассматривал бы слияние альбумина человека с N- или C-концом VWF. При N-концевом слиянии альбуминовая часть отщеплялась бы во время процессирования пропептида. Или, если бы пропептид был не включен, могла бы не происходить мультимеризация. Как обсуждалось выше, C-конец VWF весьма важен для первоначальной димеризации и секреции, что продемонстрировано Schneppenheim и др. (Schneppenheim R. et al. 1996. Defective dimerization of VWF subunits due to a Cys to Arg mutation in VWD type MD. Proc. Natl. Acad. Sci. USA 93: 3581-3586; Schneppenheim R. et al. 2001. Expression and characterization of VWF dimerization defects in different types of VWD. Blood 97: 2059-2066.), Baronciani et al. (Baronciani L. et al. 2000. Molecular characterization of a multiethnic group of 21 patients with VWD type 3. Thromb. Haemost 84: 536-540), Enayat et al. (Enayat MS et al. 2001. Aberrant dimerization of VWF as the result of mutations in the carboxy-terminal region: identification of 3 mutations in members of 3 different families with type 2A (phenotype MD) VWD. Blood 98: 674-680; и Tjernberg et al. 2006. Homozygous C2362F VWF induces intracellular retention of mutant VWF resulting in autosomal recessive severe VWD. Br J Haematol. 133: 409-418). Поэтому квалифицированный в данной области специалист не рассматривал бы слияние большого белка, вроде альбумина человека, с C- или N-концом VWF, так как он бы ожидал, что нормальная димеризация или мультимеризация VWF может быть нарушена. Так как мультимеры VWF с большей молекулярной массой являются самыми активными при первичном гемостазе, квалифицированный в данной области специалист искал бы другие пути для удлинения полупериода функционального существования VWF.
С удивлением теперь было обнаружено, что слияние гетерологичных полипептидов, таких как альбумин, с C-концевой частью VWF1 не только допускает экспрессию и секрецию химерных белков VWF из клеток млекопитающих, но также дает в результате модифицированные молекулы VWF, которые сохраняют значительную активность VWF и образуют высокомолекулярные мультимеры. Кроме того, такие модифицированные молекулы VWF демонстрируют удлиненный полупериод существования и/или увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой.
Краткое изложение сущности изобретения
Целью этого изобретения является обеспечение модифицированного FVIII или модифицированного VWF, а также комплексов модифицированного FVIII с немодифицированным VWF, комплексов немодифицированного FVIII с модифицированным VWF, а также комплексов модифицированного FVIII с модифицированным VWF с удлиненным полупериодом существования in vivo.
В используемом в настоящем изобретении смысле термин «модифицированный FVIII» или «модифицированный VWF» означает полипептиды FVIII или VWF, которые слиты с увеличивающими полупериод существования полипептидами, охватывающие также природные аллели, варианты, делеции и вставки FVIII или VWF.
Другой целью этого изобретения является обеспечение модифицированного FVIII или модифицированного VWF, а также комплексов модифицированного FVIII с немодифицированным VWF, комплексов немодифицированного FVIII с модифицированным VWF, а также комплексов модифицированного FVIII с модифицированным VWF с увеличенным отношением фактической максимальной концентрации in vivo к ожидаемой.
Другой целью настоящего изобретения является возможность экспрессии модифицированного FVIII или модифицированного VWF, а также комплексов модифицированного FVIII с немодифицированным VWF, комплексов немодифицированного FVIII с модифицированным VWF, а также комплексов модифицированного FVIII с модифицированным VWF клетками млекопитающих и сохранение у них соответствующих биологических активностей.
Итак, модифицированный FVIII или модифицированный VWF, а также комплексы модифицированного FVIII с немодифицированным VWF, комплексы немодифицированного FVIII с модифицированным VWF, а также комплексы модифицированного FVIII с модифицированным VWF настоящего изобретения имеют, как ни удивительно, сохраненную биологическую активность, увеличенный полупериод существования in vivo и увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой.
Дополнительным потенциальным преимуществом тех вариантов осуществления настоящего изобретения, в которых FVIII является модифицированным и в которых A2-домен остается присоединенным к A3-домену исключительно нековалентно после активации, является то, что увеличивается только полупериод существования неактивированной формы FVIII, тогда как полупериод существования активированной формы FVIII остается по существу тем же самым, что могло бы привести к уменьшенному риску тромбообразования по сравнению с вариантами FVIII, которые вызывают стабилизацию активированной формы FVIII.
Модифицированный FVIII или модифицированный VWF, а также комплексы модифицированного FVIII с немодифицированным VWF, комплексы немодифицированного FVIII с модифицированным VWF, а также комплексы модифицированного FVIII с модифицированным VWF настоящего изобретения можно создать посредством слияния составляющей в виде увеличивающего полупериод существования белка (HLEP) с C-концевой частью FVIII или C-концевой частью VWF.
HLEP в используемом в настоящем изобретении смысле выбирают из группы, состоящей из членов семейства альбуминов, которое включает альбумин, афамин, альфа-фетопротеин и связывающийся с витамином D белок, а также частей константной области иммуноглобулина и полипептидов, способных к связыванию в физиологических условиях с членами семейства альбуминов, а также частями константной области иммуноглобулина. Наиболее предпочтительным HLEP является альбумин человека.
Следовательно, настоящее изобретение относится к модифицированному FVIII или модифицированному VWF, а также комплексам модифицированного FVIII с немодифицированным VWF, комплексам немодифицированного FVIII с модифицированным VWF, а также комплексам модифицированного FVIII с модифицированным VWF, имеющему(им) в C-концевой части модифицированного FVIII и/или VWF слияние с HLEP, характеризующемуся(имся) тем, что модифицированный FVIII или модифицированный VWF, а также комплекс модифицированного FVIII с немодифицированным VWF, комплекс немодифицированного FVIII с модифицированным VWF или комплекс модифицированного FVIII с модифицированным VWF имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования FVIII дикого типа или VWF дикого типа или комплекса VWF дикого типа с FVIII дикого типа.
Настоящее изобретение также относится к C-концевым слияниям с более чем одним HLEP, причем HLEP, который подвергают слиянию несколько раз, может быть одним и тем же HLEP или может быть комбинацией различных HLEP.
Настоящее изобретение также относится к модифицированному FVIII, имеющему в C-концевой части слияние с HLEP, характеризующемуся тем, что модифицированный FVIII или модифицированный VWF, или комплекс модифицированного FVIII с немодифицированным VWF, комплекс немодифицированного FVIII с модифицированным VWF или комплекс модифицированного FVIII с модифицированным VWF имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для FVIII дикого типа или VWF дикого типа или комплекса VWF дикого типа с FVIII дикого типа.
Другим вариантом осуществления настоящего изобретения являются модифицированные полипептиды FVIII, имеющим в C-концевой части слияние с HLEP, характеризующиеся тем, что модифицированный FVIII секретируется в среду для ферментации в большем количестве, чем FVIII дикого типа.
Другим аспектом настоящего изобретения являются полинуклеотиды или комбинации полинуклеотидов, кодирующие модифицированный FVIII и/или модифицированный VWF.
Настоящее изобретение, кроме того, относится к плазмидам или векторам, включающим описанный здесь полинуклеотид, клеткам-хозяевам, включающим описанный здесь полинуклеотид, или плазмиду, или вектор.
Другим аспектом настоящего изобретения является способ продуцирования модифицированного FVIII или модифицированного VWF, или комплекса модифицированного FVIII с немодифицированным VWF, комплекса немодифицированного FVIII с модифицированным VWF, или комплекса модифицированного FVIII с модифицированным VWF, включающий:
(a) культивирование клеток-хозяев настоящего изобретения в таких условиях, что экспрессируется модифицированный фактор коагуляции; и
(b) необязательно выделение модифицированного фактора коагуляции из клеток-хозяев или из культуральной среды.
Настоящее изобретение, кроме того, имеет отношение к фармацевтическим композициям, включающим модифицированный FVIII или модифицированный VWF, или комплекс модифицированного FVIII с немодифицированным VWF, или комплекс немодифицированного FVIII с модифицированным VWF, или комплекс модифицированного FVIII с модифицированным VWF, полинуклеотид или плазмиду или вектор, описанные здесь.
Еще одним аспектом настоящего изобретения является применение модифицированного FVIII или модифицированного VWF, или комплекса модифицированного FVIII с немодифицированным VWF, или комплекса немодифицированного FVIII с модифицированным VWF, или комплекса модифицированного FVIII с модифицированным VWF, одного или нескольких полинуклеотидов или одной или нескольких плазмид или векторов или применение клеток-хозяев в соответствии с этим изобретением для производства лекарственного средства для лечения или профилактики нарушения свертываемости крови.
Подробное описание изобретения
Настоящее изобретение имеет отношение к комплексу, включающему FVIII и VWF, или одному из его индивидуальных полипептидных компонентов, причем по крайней мере один полипептидный компонент указанного комплекса слит в C-концевой части своего первичного продукта трансляции с N-концевой частью увеличивающего полупериод существования полипептида (HLEP).
Настоящее изобретение также имеет отношение к модифицированному FVIII или модифицированному VWF, или комплексу, включающему модифицированный FVIII и немодифицированный VWF, или комплексу, включающему немодифицированный FVIII и модифицированный VWF, или комплексу, включающему модифицированный FVIII и модифицированный VWF, причем модифицированный FVIII слит в C-концевой части первичного продукта трансляции FVIII с N-концевой частью HLEP, или модифицированный VWF слит в C-концевой части первичного продукта трансляции VWF с N-концевой частью HLEP.
В предпочтительных вариантах осуществления настоящее изобретение имеет отношение к модифицированному FVIII или модифицированному VWF, или комплексу, включающему модифицированный FVIII и немодифицированный VWF, или комплексу, включающему немодифицированный FVIII и модифицированный VWF, или комплексу, включающему модифицированный FVIII и модифицированный VWF, причем
a) модифицированный FVIII имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования FVIII дикого типа, или
b) модифицированный VWF имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования VWF дикого типа, или
c) комплекс, включающий модифицированный FVIII и немодифицированный VWF, имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа, или
d) комплекс, включающий немодифицированный FVIII и модифицированный VWF, имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа, или
e) комплекс модифицированного FVIII с модифицированным VWF имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа.
Предпочтительным вариантом осуществления настоящего изобретения является модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем модифицированный полипептид имеет полупериод функционального существования, увеличенный на по крайней мере 25% по сравнению с полупериодом функционального существования соответствующего полипептида дикого типа, или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, имеет полупериод функционального существования, увеличенный на по крайней мере 25% по сравнению с полупериодом функционального существования соответствующего комплекса FVIII дикого типа с VWF дикого типа.
Другим вариантом осуществления настоящего изобретения является модифицированный FVIII или модифицированный VWF, или комплекс, включающий модифицированный FVIII и немодифицированный VWF, или комплекс, включающий немодифицированный FVIII и модифицированный VWF, или комплекс, включающий модифицированный FVIII и модифицированный VWF, причем
a) модифицированный FVIII имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена FVIII дикого типа, или
b) модифицированный VWF имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена VWF дикого типа, или
c) комплекс, включающий модифицированный FVIII и немодифицированнный VWF, имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа, или
d) комплекс, включающий немодифицированный FVIII и модифицированный VWF, имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена соответствующего комплекса FVIII дикого типа с VWF дикого типа, или
e) комплекс, включающий модифицированный FVIII и модифицированный VWF, имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена соответствующего комплекса FVIII дикого типа с VWF дикого типа.
Предпочтительным вариантом осуществления настоящего изобретения является модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем модифицированный полипептид имеет полупериод существования в качестве антигена, увеличенный на по крайней мере 25% по сравнению с полупериодом существования в качестве антигена соответствующего полипептида дикого типа, или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, имеет полупериод существования в качестве антигена, увеличенный на по крайней мере 25% по сравнению с полупериодом существования в качестве антигена соответствующего комплекса FVIII дикого типа с VWF дикого типа.
Еще одним вариантом осуществления настоящее изобретение является модифицированный FVIII или модифицированный VWF, или комплекс, включающий модифицированный FVIII и немодифицированный VWF, или комплекс, включающий немодифицированный FVIII и модифицированный VWF, или комплекс, включающий модифицированный FVIII и модифицированный VWF, причем
a) модифицированный FVIII имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для FVIII дикого типа, или
b) модифицированный VWF имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для VWF дикого типа, или
c) комплекс, включающий модифицированный FVIII и немодифицированный VWF, имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа, или
d) комплекс, включающий немодифицированный FVIII и модифицированный VWF, имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа, или
e) комплекс, включающий модифицированный FVIII и модифицированный VWF, имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для соответствующего комплекса, включающего FVIII дикого типа и VWF дикого типа.
Другим предпочтительным вариантом осуществления настоящего изобретения является модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем модифицированный полипептид имеет отношение фактической максимальной концентрации in vivo к ожидаемой, увеличенное на по крайней мере 10% по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для соответствующего полипептида дикого типа, или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, имеет отношение фактической максимальной концентрации in vivo к ожидаемой, увеличенное на по крайней мере 10% по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для соответствующего комплекса FVIII дикого типа с VWF дикого типа.
Другим предпочтительным вариантом осуществления настоящего изобретения является
а) модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем по крайней мере один полипептидный компонент указанного комплекса слит в C-концевой аминокислоте своего первичного продукта трансляции с N-концевой частью HLEP, или
b) модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем по крайней мере один полипептидный компонент указанного комплекса слит в C-концевой части своего первичного продукта трансляции с N-концевой аминокислотой HLEP, или
c) модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем по крайней мере один полипептидный компонент указанного комплекса слит в C-концевой аминокислоте своего первичного продукта трансляции с N-концевой аминокислотой HLEP.
Другим предпочтительным вариантом осуществления настоящего изобретения является модифицированный полипептид или комплекс, включающий указанный модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, описанные выше, причем модифицированный полипептид имеет по крайней мере 10% биологической активности полипептида дикого типа, или комплекс, включающий модифицированный полипептид, или комплекс, включающий указанные модифицированные полипептиды, имеет по крайней мере 10% биологической активности соответствующего комплекса FVIII дикого типа с VWF дикого типа.
В настоящее изобретение также включен способ приготовления модифицированного FVIII или модифицированного VWF, имеющего увеличенный полупериод функционального существования, включающий слияние N-концевой части увеличивающего полупериод существования полипептида с C-концевой частью первичного продукта трансляции FVIII или с C-концевой частью первичного продукта трансляции VWF, а также способ приготовления комплекса, включающего модифицированный FVIII и немодифицированный VWF, или комплекса, включающего немодифицированный FVIII и модифицированный VWF, или комплекса, включающего модифицированный FVIII и модифицированный VWF, посредством смешивания модифицированного FVIII, приготовленного описанным выше способом, с VWF дикого типа или посредством смешивания FVIII дикого типа с модифицированным VWF, приготовленным описанным выше способом, или посредством смешивания модифицированного FVIII и модифицированного VWF, приготовленных описанным выше способом.
В настоящее изобретение также включено применение
a) модифицированного FVIII, приготовленного описанным выше способом, и VWF дикого типа, или
b) FVIII дикого типа и модифицированного VWF, приготовленного описанным выше способом, или
c) модифицированного FVIII, приготовленного описанным выше способом, и модифицированного VWF, приготовленного описанным выше способом,
для производства комбинированного фармацевтического препарата для одновременного, раздельного или последовательного использования при лечении нарушений свертываемости крови, предпочтительно при лечении гемофилии A и/или болезни Виллебранда.
«Полупериодом функционального существования» в соответствии с настоящим изобретением является полупериод существования биологической активности модифицированного FVIII или модифицированного VWF, или комплекса модифицированного FVIII с немодифицированным VWF, или комплекса немодифицированного FVIII с модифицированным VWF, или комплекса модифицированного FVIII с модифицированным VWF после его введения млекопитающему и может быть измерен in vitro в образцах крови, взятых с различными интервалами времени у указанного млекопитающего после введения модифицированного FVIII или модифицированного VWF, или комплекса модифицированного FVIII с немодифицированным VWF, или комплекса немодифицированного FVIII с модифицированным VWF, или комплекса модифицированного FVIII с модифицированным VWF.
Выражения «слияние» или «слитый» относятся к добавлению аминокислот к C-концевой части FVIII и/или к C-концевой части VWF. При ссылке здесь на «слияние с C-концевой аминокислотой FVIII» или на «слияние с C-концевой аминокислотой VWF» подразумевается слияние как раз с C-концевой аминокислотой FVIII в положении 2332 зрелого FVIII дикого типа или как раз с C-концевой аминокислотой VWF в положении 2050 зрелого VWF дикого типа. Зрелый FVIII или зрелый VWF означает соответствующий полипептид после отщепления пропептида. Однако настоящим изобретением также охватывается «слияние с C-концевой частью FVIII» или «слияние с C-концевой частью VWF». Эти выражения в используемом в настоящем изобретении смысле могут также включать слияние с молекулой FVIII и/или VWF, соответственно, из которой делетирована одна или несколько аминокислот, вплоть до n аминокислот от C-концевой аминокислоты FVIII и/или VWF. Число n является целым числом, которое не должно превышать 5%, предпочтительно 1% от общего числа аминокислот FVIII и/или VWF. Обычно n равно 20, предпочтительно 15, более предпочтительно 10, еще предпочтительнее 5 или меньше (например, 1, 2, 3, 4 или 5).
В одном варианте осуществления модифицированный FVIII имеет следующую структуру:
N-FVIII-C-L1-H,
где
N представляет собой N-концевую часть FVIII,
L1 представляет собой химическую связь или линкерную последовательность,
H представляет собой HLEP, а
C представляет собой C-концевую часть FVIII.
В другом варианте осуществления модифицированный VWF имеет следующую структуру:
N-VWF-C-L1-H,
где
N представляет собой N-концевую часть VWF,
L1 представляет собой химическую связь или линкерную последовательность,
H представляет собой HLEP, а
C представляет собой С-концевую часть VWF.
L1 может быть химической связью или линкерной последовательностью, состоящей из одной или нескольких аминокислот, например 1-20, 1-15, 1-10, 1-5 или 1-3 (например, 1, 2 или 3) аминокислот, которые могут быть одинаковыми или отличными друг от друга. Обычно линкерные последовательности не присутствуют в соответствующем положении в факторе коагуляции дикого типа. Примеры подходящих аминокислот, присутствующих в L1, включают Gly и Ser.
Предпочтительные последовательности HLEP представлены ниже. Также настоящим изобретением охватываются слияния как раз с «N-концевой аминокислотой» соответствующего HLEP или слияния с «N-концевой частью» соответствующего HLEP, который включает N-концевые делеции одной или нескольких аминокислот HLEP.
Модифицированный FVIII или модифицированный VWF, или комплекс модифицированного FVIII с немодифицированным VWF, комплекс немодифицированного FVIII с модифицированным VWF, или комплекс модифицированного FVIII с модифицированным VWF настоящего изобретения может включать более одной последовательности HLEP, например две или три последовательности HLEP. Эти множественные последовательности HLEP могут быть слиты с C-концевой частью FVIII и/или C-концевой частью VWF в тандеме, например, в виде следующих друг за другом повторов.
FVIII может процессироваться протеолитически на различных стадиях. Например, как отмечены выше, во время своей секреции в плазму одноцепочечный FVIII расщепляется внутриклеточно на границе B-A3 и в различных сайтах внутри B-домена. Тяжелая цепь связывается посредством иона металла с легкой цепью, имеющей доменную структуру A3-C1-C2. FVIII активируется посредством протеолитического расщепления в положениях аминокислот Arg372 и Arg740 внутри тяжелой цепи и в Arg1689 в легкой цепи, создающего гетеротример активированного FVIII, состоящий из A1-домена, A2-домена и легкой цепи (A3-C1-C2), фрагмента массой 73 кДа. Таким образом, активная форма FVIII (FVIIIa) состоит из A1-субъединицы, связанной через образуемую двухвалентным металлом ионную связь с расщепленной тромбином легкой цепью A3-C1-C2, и свободной A2-субъединицы, относительно слабо связанной с A1 и A3-доменом.
Соответственно, настоящим изобретением также охватывается модифицированный FVIII, который представлен не в виде одноцепочечного полипептида, а состоит из нескольких полипептидов (например, одного или двух, или трех), которые связаны друг с другом посредством нековалетных связей.
Предпочтительно N-FVIII-C включает полноразмерную последовательность FVIII. Также включаются N-концевые, C-концевые или внутренние делеции FVIII при условии, что биологическая активность FVIII сохраняется. В используемом в настоящем изобретении смысле биологическая активность сохраняется, если FVIII с делециями сохраняет по крайней мере 10%, предпочтительно по крайней мере 25%, более предпочтительно по крайней мере 50%, наиболее предпочтительно по крайней мере 75% биологической активности FVIII дикого типа. Биологическая активность FVIII может быть определена специалистом, как описано ниже.
Подходящим анализом для определения биологической активности FVIII является, например, одноступенчатый или двухступенчатый анализ коагуляции (Rizza et al. 1982. Coagulation assay of FVIII:C and FIXa in Bloom ed. The Hemophilias. NY Churchchill Livingston 1992) или анализ FVIII:C с использованием хромогенного субстрата (S. Rosen, 1984. Scand J Haematol 33: 139-145, suppl.). Содержание этих ссылок включено сюда посредством ссылки.
Последовательность кДНК и аминокислотная последовательность зрелой формы дикого типа FVIII свертывающей системы крови человека представлены в SEQ ID NO: 14 и SEQ ID NO: 15 соответственно. Ссылка на положение аминокислоты конкретной последовательности означает положение указанной аминокислоты в белке FVIII дикого типа и не исключает наличие мутаций, например делеций, вставок и/или замен в других положениях в последовательности, на которую приводится ссылка. Например, мутация в «Glu2004» со ссылкой на SEQ ID NO: 15 не исключает, что в модифицированном гомологе отсутствует одна или несколько аминокислот в положениях с 1 по 2332 SEQ ID NO: 15.
Термины «фактор VIII свертывающей системы крови», «фактор VIII» и «FVIII» используются здесь взаимозаменяемо. «Фактор VIII свертывающей системы крови» включает фактор VIII дикого типа свертывающей системы крови, а также производные фактора VIII дикого типа свертывающей системы крови, обладающие активностью прокоагулянта фактора VIII дикого типа свертывающей системы крови. Производные могут иметь делеции, вставки и/или добавления по сравнению с аминокислотной последовательностью FVIII дикого типа. Термин «FVIII» включает протеолитически процессированные формы FVIII, например форму до активации, включающую тяжелую цепь и легкую цепь.
Термин «FVIII» включает любые варианты или мутанты FVIII, обладающие по крайней мере 25%, более предпочтительно по крайней мере 50%, наиболее предпочтительно по крайней мере 75% биологической активности фактора VIII дикого типа.
В качестве неограничивающих примеров молекулы FVIII включают мутанты FVIII, предотвращающие или уменьшающие расщепление под действием APC (Amano 1998. Thromb. Haemost. 79: 557-563), мутанты FVIII, дополнительно стабилизирующие A2-домен (WO 97/40145), мутанты FVIII, приводящие к увеличенной экспрессии (Swaroop et al. 1997. JBC 272: 24121-24124), мутанты FVIII, снижающие его иммуногенность (Lollar 1999. Thromb. Haemost. 82: 505-508), FVIII, воссозданный из отдельно экспрессионных тяжелых и легких цепей (Oh et al. 1999. Exp. Mol. Med. 31: 95-100), мутанты FVIII, уменьшающие вызывающее катаболизм FVIII связывание с рецепторами, вроде HSPG (гепарансульфат-протеогликанов) и/или LRP (белка, родственного рецепторам липопротеинов низкой плотности) (Ananyeva et al. 2001. TCM, 11: 251-257), варианты FVIII со стабилизированной дисульфидной связью (Gale et al., 2006. J. Thromb. Hemost. 4: 1315-1322), мутанты FVIII с улучшенными характеристиками секреции (Miao et al., 2004. Blood 103: 3412-3419), мутанты FVIII с увеличенной кофакторной специфической активностью (Wakabayashi et al., 2005. Biochemistry 44: 10298-10304), мутанты FVIII с улучшенным биосинтезом и секрецией, уменьшенным взаимодействием с шаперонами эндоплазматического ретикулума, улучшенным переносом из эндоплазматического ретикулума в аппарат Гольджи, увеличенной активацией или устойчивостью к инактивации и увеличенным полупериодом существования (резюмированные Pipe 2004. Sem. Thromb. Hemost. 30: 227-237). Все эти мутанты и варианты FVIII включены сюда в целом посредством ссылки.
VWF может процессироваться протеолитически на различных стадиях. Например, как отмечено выше, протеаза ADAMTS13 расщепляет VWF внутри A2-домена VWF. Соответственно, настоящим изобретением также охватывается модифицированный VWF, который был протеолитически расщеплен, например, ADAMTS13. Такое расщепление могло бы привести к мультимерным цепям VWF, которые включают на своих концах по крайней мере один или самое большее два мономера VWF, которые отщепились под действием ADAMTS 13.
Предпочтительно N-VWF-C включает полноразмерную последовательность VWF. Также включаются N-концевые, C-концевые или внутренние делеции VWF при условии, что биологическая активность VWF сохраняется. В используемом в настоящем изобретении смысле биологическая активность сохраняется, если VWF с делециями сохраняет по крайней мере 10%, предпочтительно по крайней мере 25%, более предпочтительно по крайней мере 50%, наиболее предпочтительно по крайней мере 75% биологической активности VWF дикого типа. Биологическая активность VWF дикого типа может быть определена специалистом, используя способы для определения ристоцетин-кофакторной активности (Federici AB et al. 2004. Haematologica 89: 77-85), связывания VWF с гликопротеином Ibα гликопротеинового комплекса Ib-V-IX тромбоцитов (Sucker et al. 2006. Clin Appl Thromb Hemost. 12: 305-310), или анализ связывания с коллагеном (Kallas & Talpsep. 2001. Annals of Hematology 80: 466-471).
«FVIII» и/или «VWF» в вышеприведенном определении также включают природные аллельные вариации, которые могут существовать или встречаться при переходе от одного индивидуума к другому. «FVIII» и/или «VWF» в вышеприведенном определении, кроме того, включает варианты FVIII и/или VWF. Такие варианты отличаются одним или несколькими аминокислотными остатками от последовательности дикого типа. Примеры таких отличий могут включать консервативные аминокислотные замены, т.е. замены в пределах групп аминокислот со схожими характеристиками, например, (1) небольших аминокислот, (2) кислотных аминокислот, (3) полярных аминокислот, (4) основных аминокислот, (5) гидрофобных аминокислот и (6) ароматических аминокислот. Примеры таких консервативных замен представлены в следующей таблице.
Один или несколько HLEP можно слить с C-концевой частью FVIII предпочтительно так, чтобы не вносились затруднения в способности FVIII к связыванию, например, с VWF, тромбоцитами или FIX.
Один или несколько HLEP можно слить с C-концевой частью VWF предпочтительно так, чтобы не вносились затруднения в способности VWF к связыванию, например, с FVIII, тромбоцитами, гепарином или коллагеном.
После эндогенной активации FVIII во время коагуляции in vivo сохранение увеличенного полупериода функционального существования теперь активированного FVIII больше не может быть желательным, поскольку это могло бы привести к тромбозным осложнениям, что уже является случаем для такого активированного фактора дикого типа коагуляции, как FVIIa (Aledort 2004. J Thromb Haemost 2: 1700-1708), и что может быть более релевантным, если бы активированный фактор имел увеличенный полупериод функционального существования. Поэтому другой целью настоящего изобретения является обеспечение долгоживущих молекул FVIII, которые после эндогенной активации in vivo или при доступности кофактора действительно имеют полупериод функционального существования, сравнимый с таковым немодифицированого FVIII. В качестве неограничивающего примера этого можно достичь путем введения сайта расщепления, например, для фактора коагуляции между C-концевой частью FVIII и HLEP. При использовании таких соединяющих FVIII-HLEP последовательностей активация химерного белка FVIII настоящего изобретения будет вызывать сопутствующее полное отделение FVIIIa от составляющей в виде HLEP. Соответственно, в одном варианте осуществления полупериод функционального существования эндогенно активированного модифицированного FVIII является по существу одинаковым с полупериодом функционального существования активированного FVIII дикого типа (например, ±15%, предпочтительно ±10%).
В еще одном варианте осуществления настоящего изобретения, однако, один или несколько сайтов протеолитического расщепления, предпочтительно сайты расщепления тромбином в Arg740 и/или Arg372, мутированы или делетированы для того, чтобы предотвратить расщепление и иметь в результате инсерционный белок, который демонстрирует улучшенные свойства, вроде увеличенного полупериода функционального существования даже в виде активированной молекулы.
В другом варианте осуществления настоящего изобретения белки FVIII настоящего изобретения могут экспрессироваться в виде двух отдельных цепей (смотри ниже).
Модифицированный FVIII в соответствии с этим изобретением может быть одноцепочечным полипептидом или он может быть составлен из двух или трех полипептидных цепей, которые связаны посредством нековалентных связей, вследствие протеолитического процессирования.
В другом варианте осуществления настоящего изобретения мутированы или делетированы аминокислоты в сайте расщепления PACE/фуринон (Arg1648) или вблизи этого сайта для предотвращения расщепления PACE/фурином. Полагают, что это имеет следствием одноцепочечную слитую FVIII/HLEP молекулу с увеличенным полупериод существования.
В одном варианте осуществления настоящего изобретения модифицированный FVIII настоящего изобретения демонстрирует увеличенный полупериод функционального существования по сравнению с соответствующей формой FVIII, не содержащей присоединенного HLEP, и/или формой FVIII дикого типа. Полупериод функционального существования можно, например, определить in vivo в моделях гемофилии А на животных, вроде мышей с выключением FVIII, в которых можно было бы ожидать более длительное гемостатическое действие модифицированного FVIII по сравнению с FVIII дикого типа. Гемостатическое действие можно было бы проверить, например, с помощью определения периода времени до остановки кровотечения после клипирования хвоста.
В одном варианте осуществления настоящего изобретения полупериодом функционального существования является полупериод существования биологической активности FVIII после его введения млекопитающему и измеряется in vitro. Полупериод функционального существования модифицированного FVIII в соответствии с настоящим изобретением больше полупериода функционального существования FVIII, в котором отсутствует модификация, при проверке у одного и того же вида. Полупериод функционального существования увеличен предпочтительно на по крайней мере 10%, предпочтительно на по крайней мере 25%, более предпочтительно на по крайней мере 50% и даже предпочтительнее на по крайней мере 100% по сравнению с формой FVIII дикого типа.
Полупериод функционального существования модифицированного FVIII, включающего модификацию в виде HLEP, можно определить с помощью введения соответствующего модифицированного FVIII (и в сравнение с FVIII дикого типа) крысам, кроликам или другим видам экспериментальных животных внутривенно или подкожно и слежения за элиминацией биологической активности указанного модифицированного или соответственно немодифицированного фактора коагуляции в образцах крови, взятых с соответствующими интервалами после применения. Подходящими методами проверки являются описанные здесь анализы активности.
Полупериодом функционального существования в соответствии с другим вариантом осуществления настоящего изобретения является полупериод существования биологической активности VWF после его введения млекопитающему и измеряется in vitro. Полупериод функционального существования модифицированного VWF в соответствии с настоящим изобретением больше полупериода функционального существования VWF, в котором отсутствует модификация, при проверке у одного и того же вида. Полупериод функционального существования увеличен на по крайней мере 10%, предпочтительно на по крайней мере 25%, более предпочтительно на по крайней мере 50% и даже предпочтительнее на по крайней мере 100% по сравнению с VWF, в котором отсутствует модификация, и/или формой VWF дикого типа.
Полупериод функционального существования модифицированного VWF, включающего модификацию в виде HLEP, можно определить с помощью введения соответствующего модифицированного VWF (и в сравнении с таковым немодифицированного VWF) крысам, кроликам или другим видам экспериментальных животных внутривенно или подкожно и слежения за элиминацией биологической активности указанного модифицированного или соответственно немодифицированного VWF в образцах крови, взятых с соответствующими интервалами после применения. Подходящими методами проверки являются описанные здесь анализы активности.
В качестве суррогатного маркера для полупериода существования биологической активности можно измерить также уровни антигена в виде модифицированного FVIII или соответственно FVIII дикого типа или уровни антигена в виде модифицированного VWF или соответственно VWF дикого типа. Таким образом, настоящим изобретением также охватываются модифицированные молекулы FVIII и/или VWF, имеющие в C-концевой части FVIII и/или VWF слияние с HLEP, характеризующиеся тем, что модифицированный FVIII или модифицированный VWF, или комплекс модифицированного FVIII с немодифицированным VWF, или комплекс немодифицированного FVIII с модифицированным VWF, или комплекс модифицированного FVIII с модифицированным VWF имеет удлиненный полупериод существования FVIII и/или VWF антигена по сравнению с полупериодом существования FVIII и/или VWF антигена, в котором отсутствует указанная инсерция. «Полупериод существования FVIII антигена» в соответствии с настоящим изобретением является полупериодом существования антигена в виде FVIII после его введения млекопитающему и измеряется in vitro. «Полупериод существования VWF антигена» в соответствии с настоящим изобретением является полупериодом существования антигена в виде VWF после его введения млекопитающему и измеряется in vitro. Способы антигенного анализа основаны на специфических антителах в формате иммуноферментного анализа, известного специалисту и имеющегося в продаже (например, Dade Behring, Instrumentation Laboratory, Abbott Laboratories, Diagnostica Stago). Полупериод функционального существования и полупериод существования в качестве антигена можно рассчитать, используя моменты времени бета-фазы элиминации по формуле t1/2=ln2/k, принимая во внимание, что k является углом наклона линии регрессии.
В другом варианте осуществления полупериод функционального существования эндогенно активированного модифицированного FVIII удлинен по сравнению с таковым активированного FVIII дикого типа. Увеличение может быть больше 15%, например, составлять по крайней мере 20% или по крайней мере 50%. Снова значения таких полупериодов функционального существования можно рассчитать, как описано выше для полупериодов функционального существования. Увеличенные полупериоды существования эндогенного активированных модифицированных молекул FVIII могут быть полезны в ситуациях, в которых достижимы только очень низкие уровни FVIII, которые поэтому не являются тромбообразующими. Такие ситуации могут возникать, например, при лечении в виде генной терапии, при котором часто могут быть достигнуты только низкие скорости экспрессии. Следовательно, такие стабилизированные молекулы FVIII могли бы быть полезными, например, в генной терапии несмотря на риск тромбообразования, связанный с такими молекулами FVIII, если бы они вводились в виде белков в высоких или физиологических дозах.
В другом варианте осуществления настоящего изобретения модифицированный FVIII настоящего изобретения демонстрирует увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с таковым FVIII дикого типа, и модифицированный VWF настоящего изобретения демонстрирует увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с таковым VWF дикого типа. Отношение фактической максимальной концентрации in vivo к ожидаемой можно определить in vivo, например, у нормальных животных или в моделях гемофилии A на животных, вроде мышей с выключением FVIII, или в моделях VWD, вроде мышей с выключением VWF, в которых можно было бы ожидать, что увеличенный процент модифицированного FVIII или VWF настоящего изобретения будет выявляться с помощью анализов антигена или активности в кровотоке вскоре (от 5 до 10 мин) после внутривенного введения по сравнению с соответствующим FVIII дикого типа или VWF дикого типа.
Предпочтительно отношение фактической максимальной концентрации in vivo к ожидаемой увеличено на по крайней мере 10%, более предпочтительно на по крайней мере 20% и даже предпочтительнее на по крайней мере 40% по сравнению с формой FVIII дикого типа или VWF дикого типа.
В еще одном варианте осуществления настоящего изобретения константные области иммуноглобулинов или их части используются в качестве HLEP. Предпочтительно используют Fc-район, составленный из CH2- and CH3-доменов и шарнирной области IgG, более предпочтительно IgG1, или его фрагменты или варианты, включающие мутации, которые усиливают связывание с неонатальным рецептором для Fc-фрагмента (FcRn).
Другой целью настоящего изобретения является обеспечение долгоживущих молекул FVIII, которые после протеолитического процессирования in vivo действительно имеют полупериод функционального существования, сравнимый с таковым немодифицированного FVIII. Этого можно достичь с помощью сохранения определенных сайтов расщепления в модифицированном FVIII, приводящих к протеолитическому расщеплению, например, при контактировании с активированными факторами коагуляции, которое приводит к отделению FVIII от HLEP. Соответственно, в одном варианте осуществления полупериод функционального существования протеолитически процессированного модифицированного FVIII является по существу одинаковым с таковым немодифицированного FVIII, у которого отсутствует модификация, и/или он является по существу одинаковым с таковым FVIII дикого типа (например, ±15%, предпочтительно ±10%).
Еще одним вариантом осуществления настоящего изобретения являются модифицированные полипептиды FVIII, которые слиты с HLEP, например, альбумином, в C-концевой части молекулы FVIII, которые действительно имеют уменьшенное связывание с VWF или не связываются с VWF совсем.
Другой целью настоящего изобретения является обеспечение долгоживущих молекул VWF, которые после протеолитического процессирования in vivo действительно имеют функциональные свойства, сравнимые с таковыми немодифицированного VWF. Этого можно достичь с помощью сохранения или вставки определенных сайтов расщепления в модифицированном(ый) VWF (смотри ниже), приводящих к протеолитическому расщеплению, например, при контактировании с активированными факторами коагуляции, которое приводит к отделению VWF от HLEP. Соответственно, в одном варианте осуществления полупериод функционального существования протеолитически процессированного модифицированного VWF является по существу одинаковым с таковым немодифицированного VWF, у которого отсутствует модификация, и/или он является по существу одинаковым с таковым VWF дикого типа (например, ±15%, предпочтительно ±10%).
Другим предпочтительным вариантом осуществления настоящего изобретения является коэкспрессия VWF дикого типа и модифицированного VWF в соответствии с настоящим изобретением, имеющая следствием мультимеры VWF, включающие немодифицированные, а также модифицированные мономеры VWF.
Линкерные последовательности
В соответствии с этим изобретением составляющая в виде терапевтического полипептида может быть соединена с составляющей в виде HLEP с помощью пептидного линкера. Линкер должен быть неиммуногенным и может быть нерасщепляемым или расщепляемым линкером.
Нерасщепляемые линкеры могут быть составлены из чередующихся остатков глицина и серина, как показано в качестве примера в WO2007/090584.
В другом варианте осуществления настоящего изобретения пептидный линкер между составляющей в виде FVIII и/или VWF и составляющей в виде альбумина состоит из пептидных последовательностей, которые служат в качестве природных междоменных линкеров в белках человека. Предпочтительно такие пептидные последовательности в их природном окружении локализуются близко к поверхности белка и достижимы для иммунной системы, так что можно предположить наличие естественной толерантности к этой последовательности. Примеры даны в WO2007/090584.
Расщепляемые линкеры должны быть в достаточной степени гибкими для того, чтобы сделать возможным расщепление протеазами. В предпочтительном варианте осуществления расщепление линкера протекает сравнительно быстро по мере активации FVIII внутри слитого белка, если слитый белок является модифицированным FVIII.
Предпочтительно расщепляемый линкер включает последовательность, происходящую из
a) вводимого самого по себе терапевтического полипептида, если он содержит сайты протеолитического расщепления, которые протеолитически расщепляются во время активации терапевтического полипептида,
b) являющегося субстратом полипептида, расщепляемого протеазой, которая активируется или образуется при участии терапевтического полипептида,
c) полипептида, вовлеченного в коагуляцию или фибринолиз.
В более предпочтительном варианте осуществления линкерный район включает последовательность FVIII и/или VWF, которая должна приводить к уменьшенному риску неоантигенных свойств у экспрессированного слитого белка. Также в случае, когда терапевтическим белком является FVIII, который должен быть протеолитически активирован, кинетика расщепления пептидного линкера будет точнее отражать кинетику связанной с коагуляцией активации зимогена.
В предпочтительном варианте осуществления терапевтическим полипептидом является зимоген FVIII, а HLEP является альбумин. В этом случае линкерная последовательность либо происходит из последовательностей районов активации FVIII, из района расщепления любого субстрата FIX, например FX или FVII, либо из района расщепления любого являющегося субстратом полипептида, который расщепляется протеазой, в активацию которой вовлечен FIXa.
В предпочтительном в высокой степени варианте осуществления линкерный пептид происходит из самого FVIII и составлен из последовательностей, охватывающих сайты расщепления тромбином в положениях аминокислот 372, 740 и 1689 SEQ ID NO: 15, соответственно. В другом предпочтительном варианте осуществления линкерный пептид происходит из FX, FIX, FVII или FXI.
Предпочтительно линкерные пептиды являются расщепляемыми протеазами свертывающей системы крови, например, FIIa, FIXa, FXa, FXIa, FXIIa и FVIIa.
Указанные линкерные последовательности можно также использовать в модифицированном VWF настоящего изобретения.
Приводимые в качестве примеров комбинации из терапевтического полипептида, расщепляемого линкера и HLEP включают конструкции, перечисленные в WO2007/090584 (например, в таблице 2 и фиг. 4) и в WO2007/144173 (например, в таблицах 3a и 3b), но не ограничиваются ими.
Увеличивающие полупериод существования полипептиды (HLEP)
Как здесь используется, «увеличивающий полупериод существования полипептид» выбирают из группы, состоящей из альбумина, члена семейства альбуминов, константной области иммуноглобулина G и ее фрагментов и полипептидов, способных к связыванию в физиологических условиях с альбумином, членами семейства альбуминов, а также частями константной области иммуноглобулина. Он может быть полноразмерным увеличивающим полупериод существования белком, описанным здесь (например, альбумином, членом семейства альбуминов или константной областью иммуноглобулина G) или одним или несколькими его фрагментами, которые способны к стабилизации или удлинению по времени терапевтической активности или биологической активности фактора коагуляции. Длина таких фрагментов может составлять 10 или более аминокислот, или они могут включать по крайней мере приблизительно 15, по крайней мере приблизительно 20, по крайней мере приблизительно 25, по крайней мере приблизительно 30, по крайней мере приблизительно 50, по крайней мере приблизительно 100 или более следующих друг за другом аминокислот из последовательности HLEP или могут включать часть или все из специфических доменов соответствующего HLEP при условии, что фрагмент HLEP обеспечивает удлинение полупериода функционального существования, составляющее по крайней мере 25%, по сравнению с FVIII дикого типа или VWF дикого типа.
Часть HLEP предложенных конструкций с включением фактора коагуляции настоящего изобретения может быть вариантом нормального HLEP. Термин «варианты» включает вставки, делеции и замены, либо консервативные, либо неконсервативные, причем такие изменения не изменяют значительно активный сайт или активный домен, который обеспечивает биологические активности модифицированного FVIII или модифицированного VWF.
В частности, предложенные конструкции слияния FVIII с HLEP или VWF с HLEP настоящего изобретения могут включать встречающиеся в природе полиморфные варианты HLEP и фрагментов HLEP. HLEP может происходить из любого позвоночного, в частности любого млекопитающего, например человека, обезьяны, коровы, овцы или свиньи. HLEP не от млекопитающих включают, но без ограничения, HLEP курицы и лосося.
Альбумин в качестве HLEP
Термины «сывороточный альбумин человека» (HSA) и «альбумин человека» (HA) и «альбумин» (ALB) используются в этой заявке взаимозаменяемо. Термины «альбумин» и «сывороточный альбумин» шире и охватывают сывороточный альбумин человека (и его фрагменты и варианты), а также альбумин из других видов (и его фрагменты и варианты).
Как здесь используется, «альбумин» относится в совокупности к полипептиду альбумину или аминокислотной последовательности альбумина, или фрагменту или варианту альбумина, обладающему одной или более функциональными активностями (например, биологическими активностями) альбумина. В частности, «альбумин» относится к альбумину человека или его фрагментам, особенно зрелой форме альбумина человека, представленной здесь в SEQ ID NO: 16, или альбумину из других позвоночных или его фрагментам, или аналогам или вариантам этих молекул или их фрагментов.
В частности, предложенные конструкции слияния с FVIII и/или VWF настоящего изобретения могут включать встречающиеся в природе полиморфные варианты альбумина человека и фрагментов альбумина человека. Вообще говоря, длина фрагмента или варианта альбумина будет составлять по крайней мере 10, предпочтительно по крайней мере 40, более предпочтительно более 70 аминокислот. Предпочтительно вариант альбумина может состоять или в альтернативном случае включать по крайней мере один цельный домен альбумина или фрагменты указанных доменов, например, домены 1 (аминокислоты 1-194 SEQ ID NO: 16), 2 (аминокислоты 195-387 SEQ ID NO: 16), 3 (аминокислоты 388-585 SEQ ID NO: 16), 1 + 2 (аминокислоты 1-387 SEQ ID NO: 16), 2 + 3 (аминокислоты 195-585 SEQ ID NO: 16) или 1 + 3 (аминокислоты 1-194 SEQ ID NO: 16 + аминокислоты 388-585 SEQ ID NO: 16). Каждый домен сам составлен из двух гомологичных субдоменов, а именно 1-105, 120-194, 195-291, 316-387, 388-491 и 512-585, с гибкими линкерными районами между субдоменами, включающими остатки с Lys106 до Glu119, с Glu292 до Val315 и с Glu492 до Ala511.
Часть альбумина предложенных конструкций слияния с FVIII и/или слияния с VWF настоящего изобретения может включать по крайней мере один субдомен или домен HA или его консервативные модификации.
Афамин, альфа-фетопротеин и связывающийся с витамином D белок в качестве HLEP
Помимо альбумина, альфа-фетопротеин, другой член семейства альбуминов, как утверждалось, увеличивает полупериод существования присоединенного терапевтического полипептида in vivo (WO 2005/024044). Семейство белков альбуминов, эволюционно родственных сывороточных белков-переносчиков, состоит из альбумина, альфа-фетопротеина (AFP; Beattie & Dugaiczyk 1982, Gene 20: 415-422), афамина (AFM; Lichenstein et al. 1994. J. Biol. Chem. 269: 18149-18154) и связывающегося в витамином D белка (DBP; Cooke & David 1985, J. Clin. Invest. 76: 2420-2424). Их гены представляют собой кластер множества генов со структурными и функциональными сходствами, картируемый в одной и той же хромосомной области у людей, мышей и крыс. Структурное сходство членов семейства альбуминов говорит об их пригодности в качестве HLEP. Поэтому другой целью настоящего изобретения является использование членов семейства альбуминов, их фрагментов и вариантов в качестве HLEP. Термин «варианты» включает вставки, делеции и замены, либо консервативные, либо неконсервативные, при условии, что требуемая функция все еще имеется.
Члены семейства альбуминов могут включать полную длину соответствующего белка AFP, AFM и DBP или могут включать один или несколько его фрагментов, которые способны к стабилизации или удлинению по времени терапевтической активности. Длина таких фрагментов может составлять 10 или более аминокислот, или они могут включать приблизительно 15, 20, 25, 30, 50 или более следующих друг за другом аминокислот из соответствующей белковой последовательности или могут включать часть или все из специфических доменов соответствующего белка при условии, что фрагменты HLEP обеспечивают удлинение полупериода функционального существования, составляющее по крайней мере 25%. Члены семейства альбуминов инсерционных белков настоящего изобретения могут включать встречающиеся в природе полиморфные варианты AFP, AFM и DBP.
Иммуноглобулины в качестве HLEP
В данной области техники известно, что константная область (Fc) иммуноглобулина G (IgG) увеличивает полупериод существования терапевтических белков (Dumont JA et al. 2006. BioDrugs 20: 151-160). Константная область тяжелой цепи IgG состоит из 3 доменов (CH1-CH3) и шарнирной области. Последовательность иммуноглобулина может происходить из любого млекопитающего или из субклассов IgG1, IgG2, IgG3 или IgG4, соответственно. В качестве HLEP могут также использоваться IgG и фрагменты IgG без антигенсвязывающего домена. Часть терапевтического полипептида соединяют с IgG или фрагментами IgG предпочтительно посредством шарнирной области антитела или пептидного линкера, который может быть даже расщепляемым. В нескольких патентах и заявках на патенты описывается слияние терапевтических белков с константными областями иммуноглобулина для увеличения полупериодов существования in vivo терапевтических белков. В US 2004/0087778 и WO 2005/001025 описываются белки слияния биологически активных пептидов с Fc-доменами или по крайней мере частями константных областей иммуноглобулинов, которые увеличивают полупериод существования пептида, который в противном случае быстро элиминировался бы in vivo. Были описаны слитые белки Fc-IFN-β, которые приобретают увеличенную биологическую активность, удлиненный полупериод существования в кровотоке и большую растворимость (WO 2006/000448). Были описаны белки Fc-EPO с удлиненным полупериодом существования в сыворотке и увеличенной in vivo активностью (WO 2005/063808), а также слияние Fc с G-CSF (WO 2003/076567), подобным глюкагону пептидом-1 (WO 2005/000892), факторами свертывающей системы крови (WO 2004/101740) и интерлейкином-10 (US 6,403,077), при этом все из них имели свойства увеличения полупериода существования.
Полинуклеотиды
Настоящее изобретение, кроме того, относится к полинуклеотиду, кодирующему модифицированный фактор коагуляции, предпочтительно модифицированный вариант FVIII и/или модифицированный вариант VWF, описанный в этой заявке. Термин «полинуклеотид(ы)» обычно относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, который может быть немодифицированной РНК или ДНК или модифицированной РНК или ДНК. Полинуклеотид может быть одно- или двухцепочечной ДНК, одно- или двухцепочечной РНК. Используемый здесь термин «полинуклеотид(ы)» также включает ДНК или РНК, которые включают одно или несколько модифицированных оснований и/или необычных оснований, таких как инозин. Будет понятно, что в ДНК и РНК можно ввести ряд модификаций, которые служат множеству полезных целей, известных квалифицированным в данной области техники специалистам. Термин «полинуклеотид(ы)», как он здесь применяется, охватывает такие химически, ферментативно или метаболически модифицированные формы полинуклеотидов, а также химические формы ДНК- и РНК-характеристки вирусов и клеток, включающих, например, простые и сложные клетки.
Квалифицированному специалисту будет понятно, что вследствие вырожденности генетического кода определенный полипептид может кодироваться различными полинуклеотидами. Эти «варианты» охватываются этим изобретением.
Предпочтительно полинуклеотид настоящего изобретения является выделенным полинуклеотидом. Термин «выделенный» полинуклеотид относится к полинуклеотиду, который по существу свободен от других последовательностей нуклеиновых кислот, например и без ограничения, других хромосомных и экстрахромосомных ДНК и РНК. Выделенные полинуклеотиды могут быть очищены из клетки-хозяина. Для получения выделенных полинуклеотидов могут использоваться общепринятые способы очистки нуклеиновых кислот, известные квалифицированным специалистам. Термин также включает рекомбинантные полинуклеотиды и химически синтезированные полинуклеотиды.
Настоящее изобретение, кроме того, относится к группе полинуклеотидов, которые вместе кодируют модифицированный FVIII и/или модифицированный VWF настоящего изобретения. Первый полинуклеотид в этой группе может кодировать N-концевую часть модифицированного FVIII и/или модифицированного VWF, а второй полинуклеотид может кодировать C-концевую часть модифицированного FVIII и/или модифицированного VWF.
Еще одним аспектом настоящего изобретения является плазмида или вектор, включающая полинуклеотид в соответствии с настоящим изобретением. Предпочтительно плазмида или вектор является экспрессионным вектором. В конкретном варианте осуществления вектором является вектор переноса для применения в генной терапии людей.
Настоящее изобретение также относится к группе плазмид или векторов, которые включают указанную выше группу полинуклеотидов. Первая плазмида или вектор может содержать указанный первый полинуклеотид, а вторая плазмида или вектор может содержать указанный второй полинуклеотид. В качестве примера и со ссылкой на фактор VIII коагуляции в первый экспрессионный вектор можно клонировать последовательности, кодирующие сигнальный пептид, A1- и A2-домены, остаток последовательности B-домена и HLEP, а во второй экспрессионный вектор можно клонировать последовательности, кодирующие A3, C1 и C2 с подходящей последовательностью сигнального пептида. Оба экспрессионных вектора котрансфецируют в подходящую клетку-хозяина, что будет вызывать экспрессию легких и тяжелых цепей молекулы FVIII настоящего изобретения и образование функционального белка.
Альтернативно, в первый экспрессионный вектор клонируют последовательности, кодирующие сигнальный пептид FVIII, A1- и A2-домены, а во второй экспрессионный вектор клонируют последовательности, кодирующие HLEP, A3, C1 и C2 FVIII с подходящей последовательностью сигнального пептида. Оба экспрессионных вектора котрансфецируют в подходящую клетку-хозяина, что будет вызывать экспрессию легких и тяжелых цепей молекулы FVIII настоящего изобретения и образование функционального белка.
Альтернативно, обе кодирующие последовательности клонируют в один экспрессионный вектор, используя либо две отдельные промоторные последовательности, либо один промотор и элемент в виде участка внутренней посадки рибосомы (IRES) для управления экспрессией обеих цепей FVIII.
Все еще другим аспектом настоящего изобретения является клетка-хозяин, включающая полинуклеотид, плазмиду или вектор настоящего изобретения, или группу полинуклеотидов, или группу плазмид или векторов, описанную здесь.
Клетки-хозяева настоящего изобретения могут использоваться в способе продуцирования модифицированного фактора коагуляции, предпочтительно модифицированной молекулы FVIII, который является частью этого изобретения. Этот способ включает:
(a) культивирование клеток-хозяев настоящего изобретения в таких условиях, что экспрессируется желаемый инсерционный белок; и
(b) необязательно выделение желаемого инсерционного белка из клеток-хозяев или из культуральной среды.
Предпочтительной является очистка модифицированного FVIII и/или модифицированного VWF настоящего изобретения до >80% чистоты, более предпочтительно >95% чистоты и особенно предпочтительным является фармацевтически чистое состояние, в котором чистота по отношению к загрязняющим макромолекулам, в частности другим белкам и нуклеиновым кислотам, превышает 99,9% и отсутствуют инфекционные и пирогенные агенты. Предпочтительно выделенный или очищенный модифицированный FVIII и/иди модифицированный VWF настоящего изобретения по существу не содержит другие, неродственные полипептиды.
Различные продукты настоящего изобретения применимы в качестве лекарственных средств. Соответственно, настоящее изобретение относится к фармацевтической композиции, включающей модифицированный FVIII и/или модифицированный VWF, описанный здесь, полинуклеотид настоящего изобретения, или плазмиду, или вектор настоящего изобретения.
Настоящее изобретение также относится к способу лечения индивидуума, страдающего нарушением свертываемости крови, таким как гемофилия A или B. Способ включает введение указанному индивидууму эффективного количества FVIII и/или модифицированного VWF, или модифицированного VWF, или комплекса модифицированного FVIII с немодифицированным VWF, или комплекса немодифицированного FVIII с модифицированным VWF, или комплекса модифицированного FVIII с модифицированным VWF, которые здесь описываются. В другом варианте осуществления способ включает введение индивидууму эффективного количества полинуклеотида настоящего изобретения, или плазмиды, или вектора настоящего изобретения. Альтернативно, способ может включать введение индивидууму эффективного количества клеток-хозяев настоящего изобретения, описанных здесь.
Экспрессия предложенных мутантов
Для продукции рекомбинантных мутантных белков на высоких уровнях в подходящих клетках-хозяевах необходима сборка вышеупомянутых модифицированных кДНК в эффективных транскрипционных единицах вместе с подходящими регуляторными элементами в рекомбинантном экспрессионном векторе, который может репродуцироваться в различных экспрессионных системах, в соответствии с методами, известными квалифицированным в данной области техники специалистам. Эффективные регулирующие транскрипцию элементы могли бы происходить из вирусов, имеющих клетки животных в качестве своих природных хозяев, или из хромосомной ДНК клеток животных. Предпочтительно могут использоваться комбинации промотор-энхансер, происходящие из вакуолизирующего обезьяньего вируса 40, аденовируса, полиомавируса BK, цитомегаловируса человека или длинного концевого повтора вируса саркомы Рауса, или комбинации промотор-энхансер, включающие гены, сильно конститутивно транскрибируемые в клетках животных, вроде бета-актина или GRP78. Для достижения устойчивых высоких уровней мРНК, транскрибируемой с кДНК, транскрипционная единица должна содержать в своей 3'-проксимальной части ДНК-район, кодирующий последовательность полиаденилирования, являющуюся терминатором транскрипции. Предпочтительно эта последовательность происходит из района ранней транскрипции вакуолизирующего обезьяньего вируса 40, гена бета-глобулина кролика или гена активатора тканевого плазминогена человека.
кДНК затем интегрируются в геном подходящей линии клеток-хозяев для экспрессии модифицированных белков FVIII и/или VWF. Предпочтительно эта линия клеток должна быть линией клеток позвоночных животных для обеспечения правильной укладки, образования дисульфидных связей, гликозилирования по аспарагину и других посттрансляционных модификаций, а также секреции в культуральную среду. Примерами других посттрасляционных модификаций являются О-сульфатирование тирозина и протеолитическое процессирование растущей полипептидной цепи. Примерами линий клеток, которые можно использовать, являются клетки COS обезьяны, L-клетки мыши, клетки C127 мыши, клетки BHK-21 хомяка, клетки 293 почки эмбриона человека и клетки CHO хомяка.
Рекомбинантный экспрессионный вектор, включающий соответствующие кДНК, можно ввести в линию клеток животных несколькими различными способами. Например, рекомбинантные экспрессионные векторы можно создать из векторов на основе различных вирусов животных. Их примерами являются векторы на основе бакуловируса, вируса коровьей оспы, аденовируса и предпочтительно бычьего папилломавируса.
Транскрипционные единицы, включающие соответствующие ДНК, можно также ввести в клетки животных вместе с другим рекомбинантным геном, который может функционировать в качестве доминантного селектируемого маркера в этих клетках, для облегчения выделения специфических клеточных клонов, в геном которых была интегрирована рекомбинантная ДНК. Примерами этого типа генов доминантных селектируемых маркеров являются аминогликозид-фосфотрансфераза Tn5, придающая устойчивость к генетицину (G418), гигромицин-фосфотрансфера, придающая устойчивость к гигромицину, и пуромицин-ацетилтрасфераза, придающая устойчивость к пуромицину. Рекомбинантный экспрессионный вектор, кодирующий такой селектируемый маркер, может либо быть тем же вектором, что и вектор, включающий кДНК желаемого белка, либо он может быть отдельным вектором, который вводится одновременно и интегрируется в геном клетки-хозяина, что часто является следствием прочной физической связи между различными транскрипционными единицами.
Другие типы генов селектируемых маркеров, которые можно использовать вместе с кДНК желаемого белка, основаны на различных транскрипционных единицах, кодирующих дигидрофолатредуктазу (dhfr). После введения этого типа гена в клетки, в которых отсутствует эндогенная dhfr-активность, предпочтительно клетки CHO (DUKX-B11, DG-44), он сделает возможным рост этих клеток в средах, в которых отсутствуют нуклеозиды. Примером такой среды является F12 Хэма без гипоксантина, тимидина и глицина. Эти гены dhfr можно вводить вместе с включающими кДНК FVIII транскрипционными единицами в клетки CHO вышеотмеченного типа, либо соединенные в одном и том векторе, либо в различных векторах, создавая тем самым линии клеток dhfr+, продуцирующие рекомбинантный белок.
При выращивании указанных выше линий клеток в присутствии цитотоксического dhfr-ингибитора метотрексата будут появляться новые линии клеток, устойчивые к метотрексату. Эти линии клеток могут продуцировать рекомбинантный белок с увеличенной скоростью из-за увеличенного числа связанных dhfr и транскрипционных единиц для желаемого белка. При размножении этих линий клеток в увеличивающихся концентрациях метотрексата (1-10000 нМ) можно получить новые линии клеток, которые продуцируют желаемый белок с очень высокой скоростью.
Указанные выше линии клеток, продуцирующие желаемый белок, можно выращивать в большом масштабе, либо в суспензионной культуре, либо на различных твердых подложках. Примерами этих подложек являются микроподложки на основе декстрановых или коллагеновых матриц, или твердые подложки в форме полых волокон или различных керамических материалов. При выращивании в суспензионной клеточной культуре или на микроподложках культивирование указанных выше линий клеток можно проводить либо в виде культивирования в замкнутом объеме, либо в виде перфузионного культивирования с непрерывной подачей кондиционированной среды в течение длительных периодов времени. Таким образом, в соответствии с настоящим изобретением указанные выше линии клеток полностью подходят для разработки промышленного процесса для продукции желаемых рекомбинантных мутантных белков.
Очистка и препарат
Рекомбинантный модифицированный белок FVIII и/или рекомбинантный модифицированный белок VWF, который накапливается в среде секретирующих клеток указанных выше типов, можно сконцентрировать и очистить с помощью ряда биохимических и хроматографических способов, включающих способы, в которых используются различия по размеру, заряду, гидрофобности, растворимости, специфическому сродству и т.д. между желаемым белком и другими веществами в среде для культивирования клеток.
Примером такой очистки является адсорбция рекомбинантного мутантного белка моноклональным антителом, направленным против, например, HLEP, предпочтительно альбумина человека, или направленным против соответствующего фактора коагуляции, которое иммобилизовано на твердой подложке. После адсорбции модифицированного FVIII и/или модифицированного VWF подложкой, промывки и десорбции белок можно далее очистить с помощью ряда хроматографических методов, основанных на указанных выше свойствах. Порядок стадий очистки выбирают, например, в соответствии с пропускной способностью и избирательностью стадий, устойчивостью подложки или других аспектов. Например, предпочтительными стадиями очистки являются, но без ограничения, стадии ионообменной хроматографии, стадии иммуноаффинной хроматографии, стадии аффинной хроматографии, стадии хроматографии на основе гидрофобных взаимодействий, стадии хроматографии с использованием красителя, стадии хроматографии на гидроксиапатите, стадии мультимодальной хроматографии и стадии гель-фильтрации.
Для минимизации теоретического риска вирусных заражений в процесс могут быть включены дополнительные стадии, позволяющие эффективно инактивировать или устранять вирусы. Например, такими стадиями являются термообработка в жидком или твердом состоянии, обработка растворителями и/или детергентами, излучение в видимой или ультрафиолетовой области спектра, гамма-излучение или нанофильтрация.
Модифицированные полинуклеотиды (например, ДНК) этого изобретения можно также интегрировать в вектор переноса для применения в генной терапии людей.
Различные варианты осуществления, описанные здесь, можно комбинировать друг с другом. Настоящее изобретение будет далее описано подробнее в следующих его примерах. Это описание конкретных вариантов осуществления настоящего изобретения будет сделано в соединении с прилагаемыми фигурами.
Модифицированный FVIII и/или модифицированный VWF, описанный в этом изобретении, можно составить в фармацевтические препараты для терапевтического применения. Очищенный белок можно растворить в общепринятых, физиологически совместимых, водных буферных растворах, к которым могут быть необязательно добавлены фармацевтические наполнители, для обеспечения фармацевтических препаратов.
Такие фармацевтические носители и наполнители, а также подходящие фармацевтические препараты хорошо известны в данной области техники (смотри, например, "Pharmaceutical Formulation Development of Peptides and Proteins", Frokjaer et al., Taylor & Francis (2000) или "Handbook of Pharmaceutical Excipients", 3rd edition, Kibbe et al., Pharmaceutical Press (2000)). В частности, фармацевтическую композицию, включающую вариант полипептида настоящего изобретения, можно приготовить в лиофилизированной или стабильной жидкой форме. Вариант полипептида можно лиофилизировать с помощью ряда известных в данной области техники процедур. Лиофилизированные препараты воссоздают перед использованием путем добавления одного или нескольких фармацевтически приемлемых разбавителей, таких как стерильная вода для инъекции или стерильный физиологический солевой раствор.
Препараты композиции доставляют индивидууму любым фармацевтически подходящим способом введения. Различные системы доставки известны и могут использоваться для введения композиции с использованием любого подходящего пути. Предпочтительно композиции настоящего изобретения вводят системно. Для системного применения инсерционные белки настоящего изобретения составляют для парентеральной (например, внутривенной, подкожной, внутримышечной, внутрибрюшинной, интрацеребральной, внутрилегочной, интраназальной или чрескожной) или энтеральной (например, оральной, вагинальной или ректальной) доставки в соответствии с общепринятыми способами. Самыми предпочтительными путями введения являются внутривенное и подкожное введение. Препараты можно вводить безостановочно с помощью инфузии или с помощью болюсной инъекции. Некоторые препараты включают системы замедленного высвобождения.
Инсерционные белки настоящего изобретения вводят пациентам в терапевтически эффективной дозе, означающей дозу, которая является достаточной для вызова желаемых эффектов, предупреждения или уменьшения тяжести или распространения заболевания или симптома, подвергаемого лечению, без достижения дозы, которая вызывает недопустимые неблагоприятные побочные эффекты. Точная доза зависит от многих факторов, например симптома, препарата, способа введения, и должна быть определена в преклинических и клинических испытаниях для каждого соответствующего симптома.
Фармацевтические композиции настоящего изобретения можно вводить отдельно или в соединении с другими терапевтическими средствами. Эти средства могут быть включены в виде части одного и того же лекарственного средства. Одним примером такого средства является комбинация модифицированного FVIII с немодифицированным VWF или комбинация немодифицированного FVIII с модифицированным VWF, или комбинация модифицированного FVIII с модифицированным VWF.
Краткое описание чертежей
Фиг. 1: Уровни антигена в виде FVIII дикого типа и полипептидов С-концевого слияния FVIII с альбумином и уровни их активности.
Фиг. 2: Сравнение фармакокинетики FVIII:Ag (антигена) человека у мышей, у которых выключен VWF, после внутривенной инъекции 100 Е (FVIII:Ag)/кг FVIII дикого типа и слитого с FVIII белка 1656 (среднее значение; n=4/момент времени).
Фиг. 3: Отношения VWF:RCo/VWF:Ag для супернатантов клеточных культур, содержащих рекомбинантный VWF дикого типа (1570/1212), рекомбинантный слитый c VWF белок (1572/1212), содержащий на своем C-конце связанный альбумин, или клеточной культуры для комбинированной экспрессии, содержащей смесь рекомбинантного VWF дикого типа (1570/1212) и рекомбинантного слитого с VWF белка (1572/1212), трансфецированных в соотношении 5:1. В каждом случае были получены значения, составляющие приблизительно 0,8, которые близки к 1, что является теоретическим отношением NHP в соответствии с определениями единиц.
Фиг. 4: Электрофорез в SDS-агарозных гелях рекомбинантного VWF дикого типа (1570/1212), экспрессированного в клетках HEK, (B) и рекомбинантного слитого с VWF белка (1572/1212), также экспрессированного в клетках HEK, (A). Полосы выявляли, используя либо антитела против VWF, либо антитела против альбумина (HSA).
Фиг. 5: Сравнение фармакокинетики рекомбинантного VWF человека дикого типа и рекомбинантного слитого с VWF белка после внутривенной инъекции 100 МЕ VWF:Ag крысам (среднее значение; n=2-3/момент времени).
Примеры
Пример 1: Создание векторов для экспрессии молекул C-концевого слияния FVIII с альбумином
Экспрессионную плазмиду на основе pIRESpuro3 (BD Biosciences), содержащую последовательность кДНК полноразмерного FVIII в ее сайте множественного клонирования, (pF8-FL) использовали сначала для создания FVIII с делецией В-домена. Для этого использовали олигонуклеотиды F8-1 и F8-2 (SEQ ID NO: 1 и 2) в эксперименте сайт-направленного мутагенеза в соответствии со стандартными протоколами (с использованием набора для сайт-направленного мутагенеза QuickChange XL, Stratagene, La Jolla, CA, США), используя pF8-FL в качестве матрицы для делетирования B-домена. На второй стадии встраивали последовательность, кодирующую аминокислотную последовательность RRGR, для соединения R740 A2-домена с R1648 А3-домена. Это выполняли в другом цикле сайт-направленного мутагенеза, используя праймеры F8-3 и F8-4 (SEQ ID NO 3: and 4). Результирующую плазмиду назвали pF8-457.
Конструкцию для слияния FVIII с альбумином создавали поэтапно. Сначала сайт расщепления PinAI вставляли в 3'-конец FVIII. Для этого создавали с использованием pF8-457 в качестве матрицы, используя праймеры для ПЦР We2827 и We2828 (SEQ ID NO: 5 и 6), ПЦР-фрагмент, который впоследствии очищали из геля, разрезали рестрикционными эндонуклеазами BspE1 и Notl и лигировали с pF8-457, предварительно расщепленной ферментами BspE1 и Notl. Затем результирующую плазмиду (pF8-1433) разрезали ферментами PinAI и Notl и встраивали фрагмент, полученный с помощью ПЦР на содержащей кДНК альбумина человека плазмиде, используя праймеры We 2829 и We 2830 (SEQ ID NO: 7 и 8), и впоследствии расщепленный ферментами PinAI и Notl. Результирующая экспрессионная плазмида (pF8-1434) содержала последовательность, кодирующую FVIII с делецией В-домена, за которой следовал сайт для PinAI для включения линкеров (кодирующих аминокислотную последовательность ThrGly) и последовательность, кодирующая альбумин человека. Аминокислотная последовательность, кодируемая pF8-1434, представлена в виде SEQ ID NO: 9.
Линкерные последовательности, разделяющие FVIII и альбумин в качестве составляющих, можно было легко встроить во вновь созданный сайт PinAI, описанный выше. Встраивание двух линкерных последовательностей описывается далее. Кроме того, исходя из pF8-1434, в довершение можно было бы делетировать линкер TG, и можно ввести даже делеции внутри C-конца FVIII или N-конца альбумина, используя сайт-направленный мутагенез.
Встраивание расщепляемого линкера, происходящего из сайта расщепления тромбином FVIII: Сначала создавали ПЦР-фрагмент, содержащий последовательность, кодирующую сайт расщепления тромбином в положении 372, с помощью ПЦР, используя праймеры We2979 и We2980 (SEQ ID NO: 10 и 11) и pF8-457 в качестве матрицы. Этот фрагмент очищали, расщепляли ферментом PinAI и лигировали с расщепленной ферментом PinAI pF8-1434. Секвенирование подтвердило встраивание фрагмента в правильной ориентации, результирующую плазмиду назвали pF8-1563.
Встраивание гибкого линкера из глицинов/серинов: ПЦР-фрагмент, содержащий последовательность, кодирующую линкер из 31 аминокислот - глицинов/серинов, амплифицировали с помощью ПЦР с pFVII-937, описанной в WO2007/090584, используя праймеры We2991 и We2992 (SEQ ID NO: 12 и 13). Этот фрагмент затем очищали, расщепляли рестрикционной эндонуклеазой PinAI и лигировали с расщепленной PinAI pF8-1434. Секвенирование подтвердило встраивание фрагмента в правильной ориентации, результирующую плазмиду назвали pF8-1568.
Используя протоколы и плазмиды, описанные выше, и применяя методы молекулярной биологии, известные квалифицированным в данной области техники специалистам (и описанные, например, в Current Protocols in Molecular Biology, Ausubel FM et al. (eds.) John Wiley & Sons, Inc.; http://www.currentprotocols.com/WileyCDA/), специалист может создать другие конструкции для замены альбумина другим HLEP или встраивания любого другого линкера в описанный сайт для PinAI. Перенос кДНК для FVIII/альбумина в подходящие векторы, подобные pIRESneo3 (Invitrogen) и pEE12.4 (Lonza), позволил экспрессировать соответствующий белок слияния FVIII с альбумином в клетках CHO и отобрать экспрессирующие его клоны.
Пример 2: Трансфекция и экспрессия белков FVIII и VWF
Экспрессионные плазмиды репродуцировали в E.coli TOP10 (Invitrogen, Carlsbad, CA, США) и очищали, используя стандартные протоколы (Qiagen, Hilden, Германия). Клетки HEK-293 (Invitrogen) трансфецировали, используя реагент липофектамин 2000 (Invitrogen), и выращивали в бессывороточной среде (Invitrogen 293 Express) в присутствии 4 мкг/мл пуромицина и необязательно 0,5 МЕ/мл VWF. Клетки CHO (CHO-S, Invitrogen; CHOK1SV, Lonza) трансфецировали, используя реагент липофектамин 2000 (Invitrogen), и выращивали в бессывороточной среде (Invitrogen CD CHO, 6 мМ глютамин в случае CHO-S и CD-CHO в случае CHOK1SV) в присутствии 500-1000 мкг/мл генетицина (только в случае CHO-S). Для экспрессии FVIII необязательно добавляли 0,5 МЕ/мл VWF. Для экспрессии VWF котрасфецировали экспрессионную плазмиду, кодирующую PACE/фурин, (pFu-797), описанную в WO2007/144173. В другом эксперименте две плазмиды, кодирующие VWF дикого типа и VWF, слитый на C-конце с альбумином, котрансфецировали с pFu-797, что давало в результате мультимеры VWF, включающие мономеры в виде VWF дикого типа и мономеры в виде слитого с альбумином VWF (смотри фиг. 3). Популяции трансфецированных клеток высевали из T-флаконов во вращающиеся флаконы или малогабаритные ферментеры, из которых супернатанты собирали для очистки.
В таблице 2 перечислены данные по экспрессии в HEK-293 при использовании конструкций, описанных в примере 1.
Пример 3: Увеличенная скорость экспрессии белка слияния FVIII с альбумином
На фиг. 1 суммированы результаты исследования экспрессии белка слияния FVIII с альбумином в бессывороточной культуре клеток. Клетки HEK-293 трансфецировали в трех повторах с использованием pF8-1434 (С-концевое слияние FVIII с альбумином) и pF8-457 (FVIII дикого типа), соответственно, засевали в T80-флаконы равным количеством клеток и выращивали в отсутствие стабилизирующего VWF. Супернатанты культур затем собирали через 96, 120 и 144 часов и проверяли на активность FVIII.
Результаты выявили эффект увеличения экспрессии, вызываемый составляющей в виде альбумина при ее присутствии в качестве интегральной части молекулы FVIII, в клеточной культуре. Следовательно, продуктивность была несомненно увеличена в случае слитого белка по сравнению с FVIII дикого типа (фиг. 1).
Пример 4: Очистка белков FVIII
К содержащему молекулу FVIII супернатанту от экспрессирующей культуры добавляли количество иммуноаффинной смолы, достаточное для почти полного связывания активности FVIII. Иммуноаффинная смола была приготовлена путем ковалентного связывания подходящего мАт против FVIII со смолой Sephacryl S1000, используемой в качестве носителя. После промывки смолы ею наполняли хроматографическую колонку, и снова осуществляли промывку. Элюирование осуществляли, используя буфер, содержащий 250 мМ CaCl2 и 50% этиленгликоля.
Полученные при иммуноаффинной хроматографии (IAC) фракции, содержащие активность FVIII:C, объединяли, подвергали диализу против буфера для препарата (включающего наполнители: натрия хлорид, сахарозу, гистидин, кальция хлорид и Tween 80) и концентрировали. Образцы либо хранили замороженными, либо лиофилизировали, используя подходящий цикл лиофилизации.
Альтернативно, содержащий FVIII супернатант от клеточной культуры концентрируют/очищают сначала с помощью ионообменной хроматографии, а затем с помощью дальнейшей очистки, используя иммуноаффинную хроматографию (IAC). В этом случае элюат, полученный при ионообменной хроматографии, загружают на колонку для IAC, используя вышеотмеченную смолу.
Пример 5: Анализ активности FVIII и FVIII антигена
Для определения активности FVIII:C in vitro использовали либо анализ коагуляции (например, с использованием реагента Pathromtin SL и плазмы с недостатком FVIII, поставляемой Dade Behring, Германия), либо анализ с использованием хромогенного субстрата (например, анализ FVIII:C Coamatic, созданный Haemochrom). Эти анализы выполняли в соответствии с инструкциями производителей.
FVIII антиген (FVIII:Ag) определяли с помощью ELISA, исполнение которого известно квалифицированным в данной области техники специалистам. Вкратце, микропланшеты инкубировали со 100 мкл на каждую лунку антитела для захвата (овечьего IgG против FVIII человека, Cedarlane CL20035K-C, разведенного 1:200 в буфере A [Sigma C3041]) в течение 2 часов при температуре окружающей среды. После промывки планшетов три раза буфером B (Sigma P3563) последовательные разведения проверяемого образца в буфере для разведения образца (Cedarlane), а также последовательные разведения препарата FVIII (CSL Behring; 200 - 2 МЕ/мл) в буфере для разведения образца (в объемах на лунку: 100 мкл) инкубировали в течение двух часов при температуре окружающей среды. После трех стадий промывки буфером B в каждую лунку добавляли 100 мкл антитела для выявления (овечьего IgG против FVIII человека, Cedarlane CL20035K-D, меченного пероксидазой) в разведении 1:2 в буфере В и инкубировали в течение еще одного часа при температуре окружающей среды. После трех стадий промывки буфером B в каждую лунку добавляли 100 мкл раствора субстрата (1:10 (в объемном отношении) TMB OUVF:буфер для TMB OUVG, Dade Behring) и инкубировали в течение 30 минут при температуре окружающей среды в темноте. Добавление 100 мкл стоп-реагента (Dade Behring, OSFA) делало образцы готовыми к считыванию в подходящем для микропланшетов считывающем устройстве при длине волны 450 нм. Затем рассчитывали концентрации в проверяемых образцах, используя стандартную калибровочную кривую, построенную с использованием препарата FVIII в качестве эталона.
Пример 6: Оценка фармакокинетики слитого с FVIII белка у мышей, у которых выключен VWF, после однократной внутривенной инъекции
Для сравнения фармакокинетики FVIII дикого типа (ДНК 457) и белка слияния с С-концом FVIII (ДНК 1656) оба варианта FVIII вводили мышам внутривенно. Линия мышей, у которых выключен VWF (Denis C. et al, Proc. Natl. Acad. Sci. USA, 1998, Vol. 95, 9524-9529), была выбрана, поскольку среди прочих функций VWF служит в качестве носителя и стабилизирующего белка для FVIII, защищая, тем самым, FVIII от преждевременного расщепления, например, протеазами и от преждевременной элиминации из кровотока. Для немодифицированного FVIII весьма важным является ненарушенное взаимодействие с VWF, что иллюстрируется случаями гемофилии А, вызванной мутацией в C-концевом районе, приводящей к уменьшению связывания с VWF. В случае модифицированного FVIII такое связывание может, однако, быть даже нежелательным, для исследования или достижения улучшенной фармакокинетики. Соответственно, оба продукта вводились внутривенно в дозе 100 Е (FVIII:Ag)/кг в виде болюса двум группам мышей (таблица 3). Образцы крови отбирали ретроорбитально с соответствующими интервалами с началом через 5 минут после применения проверяемых веществ и вплоть до 24 часов. Один образец крови/мышь отбирали, превращали в плазму и хранили замороженным при -20°C до анализа. Концентрацию FVIII:Ag человека определяли, используя анализ ELISA, специфичный для FVIII человека, или комплексный ELISA, специфичный для альбумина человека и FVIII, соответственно. Для расчета фармакокинетических параметров использовали среднюю концентрацию в плазме образцов, для каждого момента времени группы. Полупериод существования рассчитывали, используя моменты времени бета-фазы элиминации по формуле t1/2=ln2/k, принимая во внимание, что k является углом наклона линии регрессии. Результат представлен на фиг. 2. Удивительно, что слитый с FVIII белок 1656 (t1/2=3,06 ч, между 5 и 960 мин) имел в приблизительно 3-4 раза более длинный конечный полупериод существования по сравнению с FVIII дикого типа (t1/2=0,8 ч, между 5 и 240 мин). Кроме того, отношение фактической максимальной концентрации in vivo к ожидаемой для слитого с FVIII белка 1656 было увеличено на приблизительно 20% по сравнению с FVIII дикого типа (таблица 4).
Подвергаемые лечению группы для сравнения фармакокинетики FVIII у мышей, у которых выключен VWF
Биодоступность (%) FVIII дикого типа и модифициованнного FVIII, слитого с FVIII белка 1656, после внутривенной инъекции мышам, у которых выключен VWF
Пример 7: Создание векторов для экспрессии VWF дикого типа и белков слияния VWF с альбумином
Сначала создавали экспрессионную плазмиду, содержащую последовательность кДНК полноразмерного VWF в ее сайте множественного клонирования. Для этого амплифицировали кодирующую VWF последовательность с помощью полимеразной цепной реакции (ПЦР), используя набор праймеров VWF+ и VWF- (SEQ ID NO: 17 и 18), в стандартных условиях, известных квалифицированным в данной области техники специалистам (и описанным в Current Protocols in Molecular Biology, Ausubel FM et al. (eds.) John Wiley & Sons, Inc.; http://www.currentprotocols.com/WileyCDA/), с плазмиды, содержащей кДНК VWF (имеющейся в продаже, например, pMT2-VWF от ATCC, № 67122). Результирующий ПЦР-фрагмент расщепляли рестрикционной эндонуклеазой EcoRI и лигировали с экспрессионным вектором pIRESpuro3 (BD Biosciences, Franklin Lakes, NJ, США), который был линеаризован с помощью EcoRI. Результирующую экспрессионную плазмиду, содержащую кДНК VWF дикого типа 3' от промотора CMV, назвали pVWF-1570.
ПЦР-фрагмент, содержащий последовательность, кодирующую линкер из 31 аминокислот - глицинов/серинов, и кДНК альбумина человека, амплифицировали с pFVII-937, описанной в WO2007/090584, используя праймеры We2994 и We1335 (SEQ ID NO: 19 и 20). Этот ПЦР-фрагмент затем расщепляли рестрикционной эндонуклеазой Notl и лигировали с расщепленной Notl pVWF-1570. Результирующую плазмиду, содержащую последовательности, кодирующие VWF дикого типа, линкерную последовательность и альбумин человека, назвали pVWF-1574.
Для достижения экспрессии слитого белка требовалось делетирование нескольких оснований между VWF и линкерной последовательностью. Это осуществляли с помощью сайт-направленного мутагенеза в соответствии со стандартными протоколами (с использованием набора для сайт-направленного мутагенеза QuickChange XL, Stratagene, La Jolla, CA, США), используя олигонуклеотиды We2995 и We2996 (SEQ ID NO: 21 и 22). Результирующая экспрессионная плазмида, названная pVWF-1572, содержала кодирующую VWF последовательность в рамке считывания с последовательностями, кодирующими линкер из 31 аминокислот - глицинов/серинов и альбумин человека. Аминокислотная последовательность экспрессируемого рекомбинантного слитого с VWF белка представлена в виде SEQ ID NO: 25. Аминокислотная последовательность препробелка VWF человека представлена в виде SEQ ID NO: 24.
Используя протоколы и плазмиды, описанные выше, и применяя методы молекулярной биологии, известные квалифицированным в данной области техники специалистам (и описанные, например, в Current Protocols in Molecular Biology, там же), специалист может создать другие конструкции для замены последовательности альбумина последовательностью другого HLEP или последовательности линкера другой линкерной последовательностью.
Пример 8: Очистка VWF и белков слияния VWF с альбумином
Супернатанты клеточных культур, содержащие рекомбинантный VWF дикого типа (rVWF дикого типа) или рекомбинантные белки слияния VWF с альбумином (rVWF-FP), стерилизовали фильтрованием через 0,2 мкм фильтр и подвергали диализу против равновесного буфера (EB; 10 мМ Трис-HCl, 10 мМ CaCl2, pH 7,0). Этот материал затем вносили в колонку Heparin Fractogel, уравновешенную буфером EB. Колонку промывали буфером EB, и белки VWF элюировали с использованием 500 мМ NaCl в EB. Пик элюирования концентрировали и подвергали диализу против буфера FB (3 г/л натрия хлорида, 20 г/л глицина, 5,5 г/л тринатриевого цитрата дигидрата, pH 7,0). Наконец, материал стерилизовали фильтрованием и замораживали в аликвотах. При необходимости применяли дополнительные стадии очистки, включающие анионо- и/или катионообменную хроматографию, HIC и SEC.
Пример 9: Анализ активности VWF и VWF антигена
Образцы анализировали с помощью иммунотурбидиметрического определения VWF:Ag (OPAB03, Siemens Healthcare Diagnostics, Marburg, Германия) и связывания с коллагеном (Technozym VWF:CBA ELISA, Исх. № 5450301 с набором-калибратором 5450310 и контрольным набором 5450312, Technoclone, Vienna, Австрия), как описано производителями.
Проверку VWF:RCo осуществляли, используя реагент BC VWF Siemens Healthcare Diagnostics, Marburg, Германия, в соответствии с описанием производителей. Международный концентрированный стандарт использовали в качестве основного стандартного препарата для калибровки собственного стандартного препарата для каждодневного применения.
Отношения VWF:RCo к VWF:Ag рассчитывали для сравнения этого параметра для различных проверяемых конструкций. Как показано на фиг. 3, отношение VWF:RCo/VWF:Ag было соизмеримо для rVWF дикого типа и рекомбинантного белка C-концевого слияния VWF с альбумином.
Для фармакокинетических анализов VWF антиген определяли с помощью ELISA, исполнение которого известно квалифицированным в данной области техники специалистам. Вкратце, микропланшеты инкубировали со 100 мкл на каждую лунку антитела для захвата (кроличьего IgG против VWF человека, Dako A0082 [Dako, Hamburg, Германия], разведенного 1:200 в буфере A [Sigma C3041, Sigma-Aldrich, Munich, Германия]) в течение ночи при температуре окружающей среды. После промывки планшетов три раза буфером B (Sigma P3563) каждую лунку инкубировали с 200 мкл буфера С Sigma P3688) в течение 1,5 часов при температуре окружающей среды (для блокирования). После трех дополнительных стадий промывки буфером В последовательные разведения проверяемого образца в буфере В, а также последовательные разведения стандартной плазмы человека (ORKL21; 20-0,2 мЕ/мл; Siemens Healthcare Diagnostics, Marburg, Германия) в буфере В (в объемах на лунку: 100 мкл) инкубировали в течение 1,5 часов при температуре окружающей среды. После трех стадий промывки буфером B в каждую лунку добавляли 100 мкл антитела для выявления (кроличьего IgG против VWF человека, Dako P0226, меченного пероксидазой) в разведении 1:16000 в буфере В и инкубировали в течение 1 часа при температуре окружающей среды. После трех стадий промывки буфером B в каждую лунку добавляли 100 мкл раствора субстрата (OUVF, Siemens Healthcare Diagnostics) и инкубировали в течение 30 минут при температуре окружающей среды в темноте. Добавление 100 мкл стоп-реагента (OSFA, Siemens Healthcare Diagnostics) делало образцы готовыми к считыванию в подходящем для микропланшетов считывающем устройстве при длине волны 450 нм. Затем рассчитывали концентрации в проверяемых образцах, используя стандартную калибровочную кривую, построенную с использованием стандартной плазмы человека в качестве эталона.
Пример 10: Анализ мультимеров VWF и белков слияния VWF с альбумином
Анализ мультимеров VWF проводили с помощью электрофореза в SDS-агарозных гелях, как ранее описывалось (Tatewaki et al., Thromb. Res. 52: 23-32 (1988), и Metzner et al., Haemophilia 4 (Suppl. 3): 25-32 (1998)) с небольшими изменениями. Вкратце, после уравновешивания в подвижном буфере готовые к применению 1% агарозные минигели (BioRad) использовали для стандартизации способа в максимальной степени. Соизмеримые количества VWF антигена подвергали электрофорезу в SDS-агарозных гелях. После Вестерн-блоттинга VWF-белковые полосы выявляли, используя антитела против VWF (DAKO, № прод. 0854) или альбумина, а затем меченные щелочной фосфатазой антитела против IgG (SIGMA, № прод. 1305), и цветную реакцию определяли количественно с помощью денситометрии.
Используя рекомбинантный VWF дикого типа (1570/797) и рекомбинантный слитый с VWF белок (1572/797) было можно продемонстрировать с помощью Вестерн-блоттинга и выявления с использованием антител против альбумина или VWF, что рекомбинантный слитый с VWF белок образует нормальное распределение мультимеров, выявляемых с помощью как антител против альбумина, так и антител против VWF (фиг. 4). Это служит подтверждением того, что, хотя каждая субъединица мультимерного VWF содержит альбумин, формируется обычная картина мультимеров VWF. Очевидно, что составляющая в виде альбумина не ингибирует ни N-концевую димеризацию, ни C-концевую мультимеризацию молекул VWF.
Пример 11: Оценка фармакокинетики VWF и белка слияния VWF с альбумином у крыс после однократной внутривенной инъекции
Рекомбинантный слитый с VWF белок и рекомбинантный VWF дикого типа вводили внутривенно каждой из 4 крыс CD в общем. Доза составляла 100 Е (VWF:Ag)/кг веса тела, в объеме инъекции 4 мл/кг.
Образцы крови отбирали ретроорбитально с соответствующими интервалами с началом через 5 минут после применения проверяемых веществ, используя меняющиеся схемы взятия образцов, что давало в результате образцы от 2 животных/момент времени (t=0, 5, 30, 90 мин, 4 ч, 1 день для подмножества № 1 и 0, 15 мин, 1, 2, 8 ч и 2 дня для подмножества № 2). Схема была разработана так, чтобы минимизировать возможные эффекты взятия образцов на концентрацию в плазме, которую определяли. Кровь превращали в плазму и хранили глубоко замороженной до анализа. Впоследствии определяли уровень VWF:Ag в плазме с помощью ELISA, описанного в примере 9. Для расчета фармакокинетических параметров использовали среднюю концентрацию в плазме. Полупериод существования рассчитывали, используя моменты времени бета-фазы элиминации по формуле t1/2=ln2/k, принимая во внимание, что k является углом наклона линии регрессии.
Результат представлен на фиг. 5 (n=2/момент времени; среднее значение). Было рассчитано, что конечные полупериоды существования составляют 32,4 мин для рекомбинантного слитого с VWF белка и 2,6 мин для рекомбинантного VWF дикого типа. Отношение фактической максимальной концентрации in vivo к ожидаемой было также увеличено для рекомбинантного слитого с VWF белка, составляя 42,1%, по сравнению с 16,1% для рекомбинантного VWF дикого типа.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ КОАГУЛИРУЮЩИЙ ФАКТОР VIIa С ПРОДЛЕННЫМ ВРЕМЕНЕМ ПОЛУЖИЗНИ | 2007 |
|
RU2466141C2 |
| СОДЕРЖАЩИЕ ФАКТОР ФОН ВИЛЛЕБРАНДА (vWF) ПРЕПАРАТЫ И СПОСОБЫ, НАБОРЫ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2010 |
|
RU2579977C2 |
| ПОЛИПЕПТИДЫ, МОДУЛИРУЮЩИЕ SIGLEC-ЗАВИСИМЫЕ ИММУННЫЕ ОТВЕТЫ | 2017 |
|
RU2776807C2 |
| Химерный улучшенный рекомбинантный фактор VIII | 2015 |
|
RU2647769C2 |
| ПОЛИПЕПТИДЫ НА ОСНОВЕ МОДИФИЦИРОВАННОГО ФАКТОРА VII И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571931C2 |
| УЛУЧШЕНИЕ КЛИНИЧЕСКИХ ПАРАМЕТРОВ ПОСРЕДСТВОМ ЭКСПРЕССИИ ФАКТОРА VIII | 2019 |
|
RU2799048C2 |
| ПЕЧЕНЬ-СПЕЦИФИЧНЫЕ КОНСТРУКЦИИ, ЭКСПРЕССИОННЫЕ КАССЕТЫ ФАКТОРА VIII И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774874C2 |
| ГЛИКОЗИЛИРОВАННЫЕ СЛИТЫЕ БЕЛКИ VWF С УЛУЧШЕННОЙ ФАРМАКОКИНЕТИКОЙ | 2017 |
|
RU2782212C2 |
| КОНЪЮГИРОВАННЫЕ МОЛЕКУЛЫ ФАКТОРА VIII | 2009 |
|
RU2573587C2 |
| СПОСОБ ОТДЕЛЕНИЯ ФАКТОРА VIII ОТ ПРОДУКТОВ КРОВИ | 2017 |
|
RU2734783C2 |






















































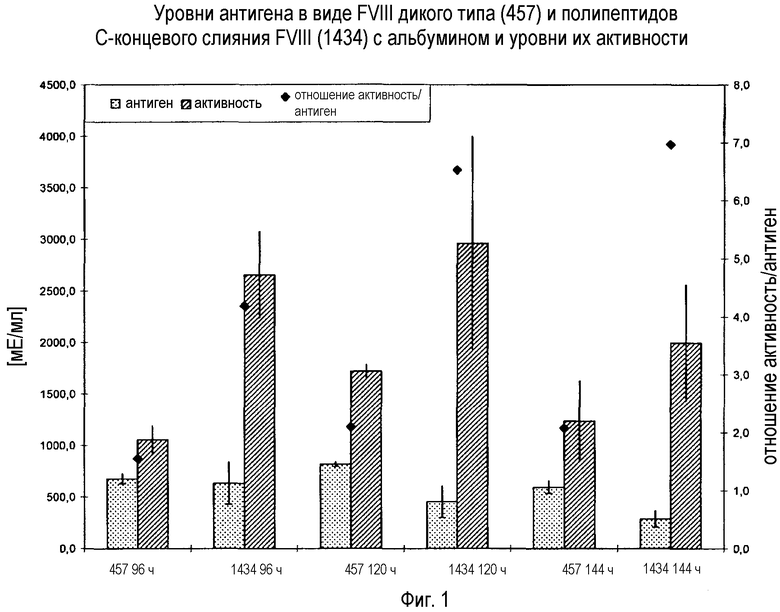
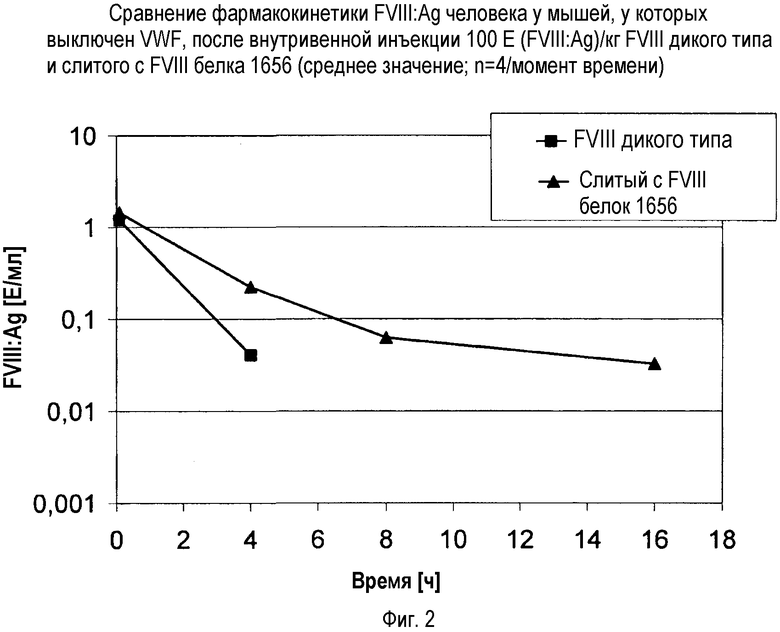
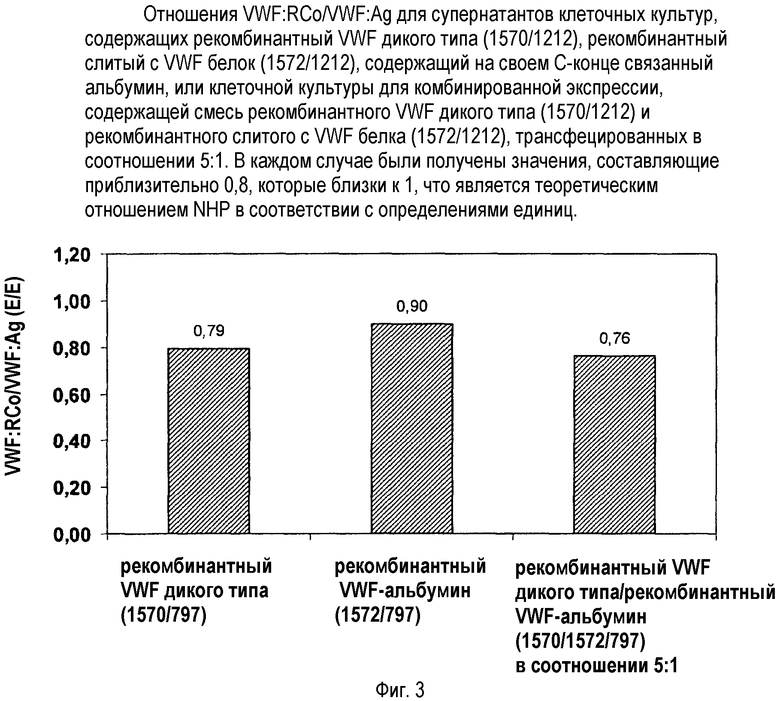
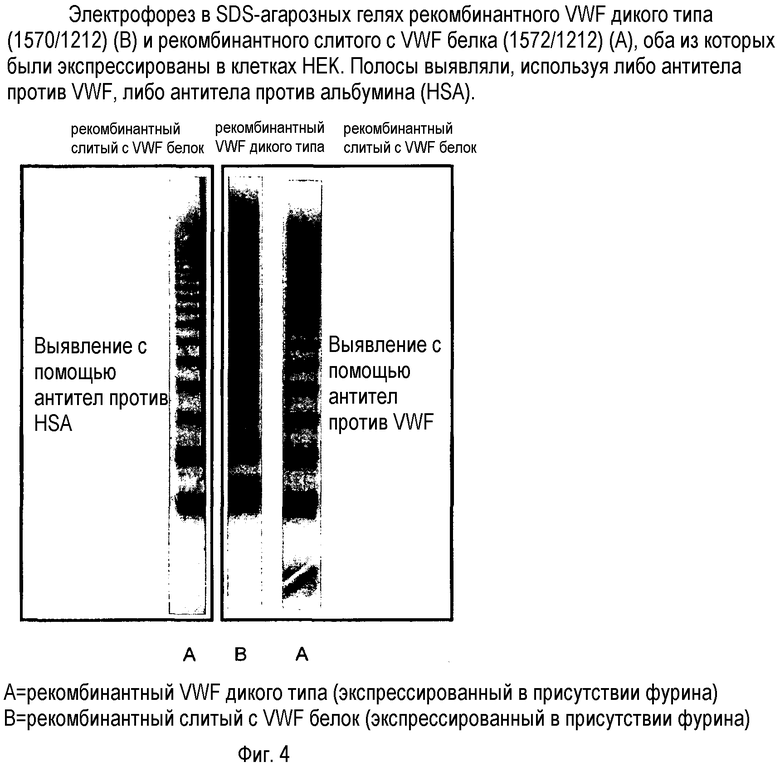
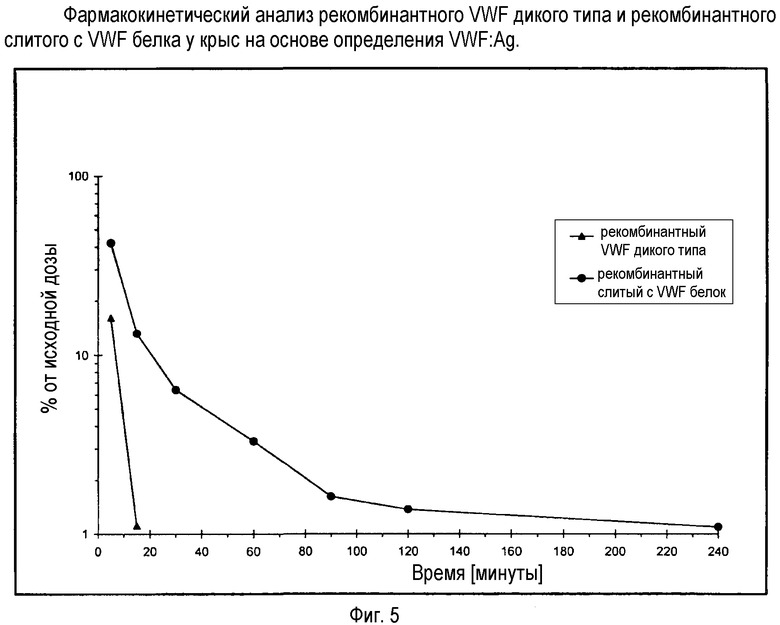
Изобретение относится к области биотехнологии, конкретно к модифицированному фактору Виллебранда (VWF), и может быть использовано в медицине. Рекомбинантным путем получают модифицированный VWF, слитый в С-концевой части своего первичного продукта трансляции с N-концевой частью альбумина посредством линкера SSGGSGGSGGSGGSGGSGGSGGSGGSGGSGS. Полученный модифицированный VWF используют в составе фармацевтической композиции для лечения или профилактики нарушения свертываемости крови. Изобретение позволяет получить модифицированный VWF, который сохраняет способность к N-концевой димеризации и С-концевой мультимеризации, имея при этом удлиненный полупериод функционального существования в плазме крови по сравнению с полупериодом функционального существования VWF. 8 н. и 9 з.п. ф-лы, 5 ил., 4 табл., 11 пр.
1. Модифицированный фактор Виллебранда (VWF), включающий VWF, слитый в С-концевой части своего первичного продукта трансляции с N-концевой частью альбумина посредством линкера SSGGSGGSGGSGGSGGSGGSGGSGGSGGSGS, где модифицированный VWF имеет удлиненный полупериод функционального существования по сравнению с полупериодом функционального существования VWF.
2. Модифицированный VWF по п.1, где модифицированный VWF имеет полупериод функционального существования, увеличенный по крайней мере на 25% по сравнению с полупериодом функционального существования VWF.
3. Модифицированный VWF по п.1, где модифицированный VWF имеет удлиненный полупериод существования в качестве антигена по сравнению с полупериодом существования в качестве антигена VWF.
4. Модифицированный VWF по п.3, где модифицированный VWF имеет полупериод существования в качестве антигена, увеличенный по крайней мере на 25% по сравнению с полупериодом существования в качестве антигена VWF.
5. Модифицированный VWF по п.1, где модифицированный VWF имеет увеличенное отношение фактической максимальной концентрации in vivo к ожидаемой по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для VWF.
6. Модифицированный VWF по п.5, где модифицированный VWF имеет отношение фактической максимальной концентрации in vivo к ожидаемой, увеличенное по крайней мере на 10% по сравнению с отношением фактической максимальной концентрации in vivo к ожидаемой для VWF.
7. Модифицированный VWF по п.1, где модифицированный VWF имеет по крайней мере 10% биологической активности VWF.
8. Полинуклеотид, кодирующий модифицированный VWF по любому из пп.1-7.
9. Экспрессионный вектор, содержащий полинуклеотид по п.8.
10. Клетка-хозяин, способная экспрессировать экзогенный генетический материал, содержащая полинуклеотид по п.8 или вектор по п.9.
11. Способ получения модифицированного VWF, включающий:
(a) культивирование клетки-хозяина по п.10 в таких условиях, что экспрессируется модифицированный VWF; и
(b) выделение модифицированного VWF из клетки-хозяина или из культуральной среды.
12. Фармацевтическая композиция для лечения или профилактики нарушения свертываемости крови, включающая модифицированный VWF по любому из пп.1-7, полинуклеотид по п.8 или вектор по п.9.
13. Применение модифицированного VWF по любому из пп.1-7, полинуклеотида по п.8, вектора по п.9 или клетки-хозяина по п.10 для производства лекарственного средства для лечения или профилактики нарушения свертываемости крови.
14. Применение по п.13, где нарушение свертываемости крови представляет собой гемофилию А.
15. Применение по п.13, где нарушение свертываемости крови представляет собой болезнь Виллебранда.
16. Применение по любому из пп.13-15, где лечение включает генную терапию людей.
15. Способ приготовления модифицированного VWF, имеющего увеличенный полупериод функционального существования, включающий слияние N-концевой альбумина с С-концевой частью первичного продукта трансляции VWF посредством линкера SSGGSGGSGGSGGSGGSGGSGGSGGSGGSGS.
| EP 1867660 A1, 19.12.2007 | |||
| CHUANG V.T | |||
| ET AL., Pharmaceutical strategies utilizing recombinant human serum albumin, PHARM | |||
| RESEARCH, NY, 2002, v.19, n.5, p.569-577 | |||
| СПОСОБ УПАРИВАНИЯ ЩЕЛОЧНО-АЛЮМИНАТНЫХРАСТВОРОВ | 0 |
|
SU179271A1 |
| US 5364771 A, 15.11.1994 | |||
| GENSANA M | |||
| et al., Influence of von Willebrand factor on the reactivity of human factor VIII inhibitors with factor VIII, | |||

