Настоящее изобретение относится к продуцирующим L-лизин штаммам коринебактерий с усиленным геном lysE (ген-носитель, участвующий в экспорте лизина), в которых усиливают, прежде всего сверхэкспрессируют другие гены, выбранные из группы, включающей ген dapA (ген дигидродипиколинат-синтазы), ген lysC (ген аспартаткиназы), ген dapB (ген дигидродипиколинат-редуктазы) или ген рус (ген пируваткарбоксилазы), прежде всего ген dapA и ген lysC (ген аспартаткиназы), и к способу получения L-лизина.
Предпосылки создания изобретения
L-лизин представляет собой важную с коммерческой точки зрения L-аминокислоту, которая находит применение прежде всего в качестве добавки к кормам для животных. Потребность в этом продукте постоянно возрастает в кормам для животных. Потребность в этом продукте постоянно возрастает в последние годы.
L-лизин получают ферментативным путем с использованием продуцирующих L-лизин штаммов коринебактерий, прежде всего Corynebacterium glutamicum. Вследствие большого значения этой группы продуктов постоянно ведутся работы по совершенствованию способа их получения. Усовершенствования способа могут касаться технических и технологических сторон процесса ферментации, таких как, например, перемешивание и обеспечение кислородом, или состава питательных сред, например концентрации сахара в процессе ферментации, или обработки продукта, например с помощью ионообменной хроматографии, или присущих самому микроорганизму характеристик продуктивности.
Для улучшения характеристик продуктивности этих микроорганизмов применяют методы мутагенеза, селекции и отбора мутантов. Таким путем получают штаммы, которые обладают устойчивостью к антиметаболитам, таким как, например, S-(2-аминоэтил)цистеин, или которые являются ауксотрофами в отношении аминокислот, таких как, например, L-лейцин, и продуцируют L-лизин.
В течение ряда лет для улучшения продуцирующих L-лизин штаммов Corynebacterium glutamicum также применяют методы рекомбинантной ДНК, с помощью которых амплифицируют отдельные гены биосинтеза и исследуют влияние на продуцирование L-лизина.
Так, например, в ЕР-А 0088166 описано улучшение характеристик продуктивности в результате амплификации фрагмента ДНК, придающего устойчивость к аминоэтилцистеину. В ЕР-В 0387527 описано улучшение характеристик продуктивности в результате амплификации аллеля lysC, кодирующего устойчивую в отношении обратной связи ("feed back") аспартаткиназу. В ЕР-В 0197335 описано улучшение характеристик продуктивности в результате амплификации гена dapA, кодирующего дигидродипиколинат-синтазу. В ЕР-А 0219027 описано улучшение характеристик продуктивности в результате амплификации гена asd, кодирующего аспартатсемиальдегид-дегидрогеназу. У Pisabarro и др. (Journal of Bacteriology, 175 (9), 2743-2749, (1993)) описан ген dapB, кодирующий дигидродипиколинат-редуктазу.
Кроме того, проводились исследования воздействия амплификации генов первичного пути метаболизма на продуцирование L-лизина. Так, в частности, в ЕР-А 0219027 описано улучшение характеристик продуктивности в результате амплификации гена aspC, кодирующего аспартат-аминотрансферазу. В ЕР-В 0143195 и ЕР-В 0358940 описано улучшение характеристик продуктивности в результате амплификации гена ррс, кодирующего фосфоенолпируват-карбоксилазу. В заявке DE-A 19831609 описано улучшение характеристик продуктивности в результате амплификации гена рус, кодирующего пируваткарбоксилазу.
И, наконец, в заявке DE-A 19548222 говорится, что повышенная активность кодируемого геном-носителем lysE, участвующим в экспорте L-лизина, способствует продуцированию лизина.
Наряду с этими подходами, основанными на амплификации отдельного гена, также существуют подходы, основанные на одновременной амплификации двух или нескольких генов с целью увеличения продуцирования L-лизина коринебактериями. Так, например, в заявке DE-A 3823451 описано улучшение характеристик продуктивности в результате одновременной амплификации гена asd и гена dapA Escherichia coli. В заявке DE-A 3943117 описано улучшение характеристик продуктивности в результате одновременной амплификации аллеля lysC, кодирующего устойчивость в отношении обратной связи ("feed back"), и гена dapA с использованием плазмиды pJC50. В ЕР-А 0841395 описано улучшение характеристик продуктивности прежде всего в результате одновременной амплификации аллеля lysC, кодирующего устойчивость в отношении обратной связи ("feed back"), и гена dapB, при этом в результате дополнительной амплификации генов dapB, lysA и ddh удалось дополнительно улучшить характеристики продуктивности. В ЕР-А 0854189 описано улучшение характеристик продуктивности в результате одновременной амплификации аллеля lysC, кодирующего устойчивость в отношении обратной связи ("feed back"), гена dapA, гена dapB, гена lysA и гена aspC. В ЕР-А 0857784 описано улучшение характеристик продуктивности прежде всего в результате одновременной амплификации аллеля lysC, кодирующего устойчивость в отношении обратной связи ("feed back"), и гена lysA, при этом в результате дополнительной амплификации гена ррс удалось дополнительно улучшить характеристики продуктивности.
Несмотря на большое количество известных методов в данной области техники, сохраняется потребность в разработке новых подходов и в усовершенствовании существующих способов продуцирования лизина коринебактериями.
Задача изобретения
В основу изобретения была положена задача по созданию с помощью новых методов улучшенных продуцирующих L-лизин штаммов коринебактерий.
Описание изобретения
L-лизин представляет собой важную с коммерческой точки зрения L-аминокислоту, которая находит применение прежде всего в качестве добавки к кормам для животных.
Если в дальнейшем упоминается L-лизин или лизин, то при этом подразумеваются не только сами эти основания, но и их соответствующие соли, такие как гидрохлорид лизина или сульфат лизина.
Объектом изобретения являются продуцирующие L-лизин штаммы коринебактерий с усиленным геном lysE (ген-носитель, участвующий в экспорте лизина), отличающиеся тем, что они дополнительно содержат гены, выбранные из группы, включающей ген dapA (ген дигидродипиколинат-синтазы), ген lysC (ген аспартаткиназы), ген dapB (ген дигидродипиколинат-редуктазы) и ген рус (ген пируваткарбоксилазы), прежде всего ген dapA и ген lysC, которые по отдельности или в сочетании усиливают, прежде всего сверхэкспрессируют.
Кроме того, выявлена новая последовательность ДНК, которая расположена против хода транскрипции (5'-область) гена dapB и которая несет - 35-область промотора dapB и обладает преимуществом в отношении экспрессии гена dapB. Эта последовательность представлена в виде SEQ ID NO:1.
Объектом изобретения также является соответствующая реплицируемая ДНК, имеющая нуклеотидную последовательность, представленную в SEQ ID NO:1.
Следующим объектом изобретения являются мутанты МС20 и МА16 промотора dapA, представленные в виде SEQ ID NO:5 и SEQ ID NO:6, депонированные под номерами DSM 12868 и DSM 12867.
Еще одним объектом изобретения являются продуцирующие L-лизин штаммы коринебактерий с усиленным геном lysE, отличающиеся тем, что в них дополнительно одновременно усиливают, прежде всего сверхэкспрессируют, ген dapA и ген dapB.
И, наконец, объектом изобретения также являются продуцирующие L-лизин штаммы коринебактерий с усиленным геном lysE, отличающиеся тем, что в них дополнительно одновременно усиливают, прежде всего сверхэкспрессируют, ген dapA и ген lysC.
В контексте настоящего изобретения понятие "усиление" означает повышение в микроорганизме внутриклеточной активности одного или нескольких ферментов, кодируемых соответствующей ДНК, за счет, например, увеличения количества копий гена, соответственно генов, использования более сильного промотора или гена, кодирующего соответствующий фермент с высокой активностью, и при необходимости сочетания этих мер.
Объектом изобретения является далее способ получения L-лизина путем ферментации с использованием этих коринебактерий.
Микроорганизмы, являющиеся объектом настоящего изобретения, могут продуцировать L-лизин из глюкозы, сахарозы, лактозы, фруктозы, мальтозы, мелассы, крахмала, целлюлозы или из глицерина и этанола, прежде всего из глюкозы или сахарозы. Они включают представителей коринебактерий, прежде всего из рода Corinebacterium. В роде Corynebacterium следует прежде всего отметить вид Corynebacterium glutamicum, который, как известно специалистам в данной области техники, обладает способностью продуцировать аминокислоты. К этому виду принадлежат штаммы дикого типа, такие как Corynebacterium glutamicum ATCC 13032, Brevibacterium flavum ATCC 14067, Corynebacterium melassecola ATCC 17965 и происходящие из них штаммы, соответственно мутанты. Примерами продуцирующих L-лизин мутантов коринебактерий являются Corynebacterium glutamicum FERM-P 1709, Brevibacterium flavum FERM-P 1708, Brevibacterium lactofermentum FERM-P 1712, Brevibacterium flavum FERM-P 6463, Brevibacterium flavum FERM-P 6464, Corynebacterium glutamicum DSM 5714, Corynebacterium glutamicum DSM 12866.
Позитивное влияние сверхэкспрессии гена lysE на продуцирование L-лизина известно из заявки DE-A 19548222.
Дополнительная усиленная экспрессия гена dapB или гена рус, прежде всего дополнительно усиленная экспрессия аллеля lysC, кодирующего устойчивую в отношении обратной связи аспартаткиназу, или прежде всего дополнительно усиленная экспрессия гена dapA, повышают продуцирование L-лизина.
При создании изобретения было установлено, что при заданной сверхэкспрессии гена lysE одновременная дополнительно усиленная экспрессия гена dapA и гена dapB позволяет достичь дополнительных преимуществ в отношении продуцирования L-лизина.
Объектом изобретения также является соответствующая реплицируемая ДНК, имеющая нуклеотидную последовательность, представленную в SEQ ID NO:1.
Было установлено, что при заданной сверхэкспрессии гена lysE предпочтительна также одновременная дополнительно усиленная экспрессия гена dapA и аллеля lysC.
Для достижения усиления (сверхэкспрессии) может быть увеличено количество копий соответствующего гена или могут быть вызваны мутации в промоторной и регуляторной областях либо в сайте связывания рибосомы, расположенном против хода транскрипции относительно структурного гена. Таким же образом действуют кассеты экспрессии, встраиваемые против хода транскрипции относительно структурного гена. Кроме того, экспрессию можно дополнительно повышать в процессе ферментативного получения L-лизина с помощью индуцибельных промоторов. Экспрессия также может быть усилена за счет увеличения продолжительности жизни мРНК. Кроме того, ферментативная активность также может быть усилена ингибированием разложения протеинов ферментов. При этом гены или генные конструкции либо могут находиться в различном количестве копий в плазмидах (бифункциональные векторы), либо их интегрируют в хромосомы и амплифицируют. В альтернативном варианте сверхэкспрессия рассматриваемых генов также может быть достигнута путем изменения состава среды и условий культивирования.
Соответствующие рекомендации можно найти у Martin и др. (Bio/Technology 5, 137-146 (1987)), у Guerrero и др. (Gene 138, 35-41 (1994)), Tsuchiya и Morinaga (Bio/Technology 6, 428-430 (1988)), у Eikmanns и др. (Gene 102, 93-98 (1991)), в ЕР 0472869, в US 4601893, у Schwarzer и  (Bio/Technology 9, 84-87 (1991)), у Reinscheid и др. (Applied and Environmental Microbiology 60, 126-132 (1994)), у LaBarre и др. (Journal of Bacteriology 175, 1001-1007 (1993), в заявке WO 96/15246, у Malumbres и др. (Gene 134, 15-24 (1993)), в JP-A 10-229891, у Jensen и Hammer (Biotechnology and Bioengineering 58, 191-195 (1998)) или в справочнике "Manual of Methods for General Bacteriology der American Society for Bacteriology" (Washington D.C., США, 1981) и в известных учебниках по генетике и молекулярной биологии.
(Bio/Technology 9, 84-87 (1991)), у Reinscheid и др. (Applied and Environmental Microbiology 60, 126-132 (1994)), у LaBarre и др. (Journal of Bacteriology 175, 1001-1007 (1993), в заявке WO 96/15246, у Malumbres и др. (Gene 134, 15-24 (1993)), в JP-A 10-229891, у Jensen и Hammer (Biotechnology and Bioengineering 58, 191-195 (1998)) или в справочнике "Manual of Methods for General Bacteriology der American Society for Bacteriology" (Washington D.C., США, 1981) и в известных учебниках по генетике и молекулярной биологии.
Применяемые согласно изобретению гены Corynebacterium glutamicum являются известными и могут быть выделены, соответственно сконструированы или синтезированы известными методами.
Методы сайтспецифического мутагенеза описаны среди прочего у Higuchi и др. (Nucleic Acids Research 16: 7351-7367 (1988)) или у Silver и др. в справочнике PCR Strategies, под ред. Innis, Glefand и Sninsky (Academic Press, London, Великобритания, 1995).
Для выделения рассматриваемого гена С.glutamicum прежде всего создают банк генов этого микроорганизма, например в Е.coli или при необходимости также в С.glutamicum. Создание банка генов описано в широко известных учебниках и справочниках. В качестве примера может служить учебник Winnacker: Gene und Klone, Eine  in die Gentechnology (изд-во Chemie, Weinheim, Германия, 1990) или справочник Sambrook и др.: Molecular Cloning, A Laboratory Manual (Cold Spring Harbor Laboratory Press, 1989). У Bathe и др. (Molecular and General Genetics, 252: 255-265, 1996) описан банк генов штамма С. glutamicum ATCC 13032, встроенный с помощью космидного вектора SuperCos I (Wahl и др., Proceedings of the National Academy of Sciences USA, 84: 2160-2164 (1987)) в штамм Е.coli K-12 NM554 (Raleigh и др., Nucleic Acids Research 16, 1563-1575 (1988)). В свою очередь у
in die Gentechnology (изд-во Chemie, Weinheim, Германия, 1990) или справочник Sambrook и др.: Molecular Cloning, A Laboratory Manual (Cold Spring Harbor Laboratory Press, 1989). У Bathe и др. (Molecular and General Genetics, 252: 255-265, 1996) описан банк генов штамма С. glutamicum ATCC 13032, встроенный с помощью космидного вектора SuperCos I (Wahl и др., Proceedings of the National Academy of Sciences USA, 84: 2160-2164 (1987)) в штамм Е.coli K-12 NM554 (Raleigh и др., Nucleic Acids Research 16, 1563-1575 (1988)). В свою очередь у  и др. (Molecular Microbiology 6 (3), 317-326) описан банк генов штамма С. glutamicum АТСС 13032, полученный с помощью космиды рНС79 (Hohn и Collins, Gene 11, 291-298 (1980)). Для создания банка генов С. glutamicum в Е. coli могут быть использованы также такие плазмиды, как pBR322 (Bolivar, Life Sciences, 25, 807-818 (1979)) или pUC19 (Norrander и др., Gene, 26: 101-106 (1983)). Равным образом также могут быть использованы бифункциональные векторы, такие как pJC1 (Cremer и др., Molecular and General Genetics, 220, 478-480 (1990)) или pEC5 (Eikmanns и др., Gene 102, 93-98 (1991)), которые реплицируются и в Е.coli, и в С.glutamicum. В качестве хозяев наиболее пригодны такие штаммы, которые имеют дефекты, полученные путем рестрикции и/или рекомбинации. Примером является штамм Е.coli DH5αmcr, описанный у Grant и др. (Proceedings of the National Academy of Sciences USA, 87: 4645-4649 (1990)). Другим примером являются описанные у
и др. (Molecular Microbiology 6 (3), 317-326) описан банк генов штамма С. glutamicum АТСС 13032, полученный с помощью космиды рНС79 (Hohn и Collins, Gene 11, 291-298 (1980)). Для создания банка генов С. glutamicum в Е. coli могут быть использованы также такие плазмиды, как pBR322 (Bolivar, Life Sciences, 25, 807-818 (1979)) или pUC19 (Norrander и др., Gene, 26: 101-106 (1983)). Равным образом также могут быть использованы бифункциональные векторы, такие как pJC1 (Cremer и др., Molecular and General Genetics, 220, 478-480 (1990)) или pEC5 (Eikmanns и др., Gene 102, 93-98 (1991)), которые реплицируются и в Е.coli, и в С.glutamicum. В качестве хозяев наиболее пригодны такие штаммы, которые имеют дефекты, полученные путем рестрикции и/или рекомбинации. Примером является штамм Е.coli DH5αmcr, описанный у Grant и др. (Proceedings of the National Academy of Sciences USA, 87: 4645-4649 (1990)). Другим примером являются описанные у  и др. (Applied and Environmental Microbiology, 60 (2), 756-759, (1994)) штаммы С.glutamicum RM3 и RM4, имеющие дефекты, полученные путем рестрикции.
и др. (Applied and Environmental Microbiology, 60 (2), 756-759, (1994)) штаммы С.glutamicum RM3 и RM4, имеющие дефекты, полученные путем рестрикции.
Затем банк генов может быть введен в индикаторный штамм путем трансформации (Hanahan, Journal of Molecular Biology 166, 557-580, 1983) или электропорации (Tauch и др., FEMS Microbiological Letters, 123: 343-347 (1994)). Индикаторный штамм отличается тем, что он имеет мутацию в рассматриваемом гене, которая обусловливает обнаруживаемый фенотип, например ауксотрофию. Индикаторные штаммы, соответственно мутанты могут быть получены из известных источников или коллекций штаммов, таких как, например, центр Genetic Stock Center Йельского университета (Нью-Хейвен, шт. Коннектикут, США), или при необходимости созданы самим исследователем. В качестве примера такого индикаторного штамма можно назвать нуждающийся в мезодиаминопимелиновой кислоте штамм Е.coli RDA8 (Richaud и др., C.R. Acad. Sci. Paris Ser. III 293: 507-512 (1981)), несущий мутацию (dap::Mu) в гене dapA.
После трансформации индикаторного штамма рекомбинантной плазмидой, несущей представляющий интерес ген, и экспрессии рассматриваемого гена индикаторный штамм приобретает соответствующие свойства, например становится прототрофом. Если клонированный фрагмент ДНК придает устойчивость, например к антиметаболитам, таким как S-(2-аминоэтил)цистеин, то несущий рекомбинантную плазмиду индикаторный штамм можно идентифицировать путем отбора на соответствующим образом дополненных питательных средах.
В случае известной, соответственно доступной из базы данных нуклеотидной последовательности представляющей интерес области гена хромосомная ДНК может быть выделена известными методами, такими как метод, описанный у Eikmanns и др. (Microbiology 140, 1817-1828, (1994)), и рассматриваемый ген может быть синтезирован с помощью полимеразной цепной реакции (ПЦР) с использованием соответствующего праймера и клонирован в соответствующем плазмидном векторе, таком как pCRIITOPO фирмы Invitrogen (Гронинген, Нидерланды). Обзор методов ПЦР можно найти в книге Newton и Graham: PCR (изд-во Spectrum Akademischer Verlag, Heidelberg, Германия, 1994).
Доступными для научной общественности базами данных нуклеотидных последовательностей являются, например, базы данных Европейских лабораторий молекулярной биологии (EMBL, Гейдельберг, Германия) или Национального центра информации в области биотехнологии (NCBI, Бетесда, шт. Мэриленд, США).
Выделение и клонирование гена lysE из штамма С.glutamicum ATCC 13032, а также нуклеотидная последовательность описаны в заявке DE-A 19548222.
Выделение, клонирование и секвенирование гена dapA из различных штаммов С.glutamicum описаны у Cremer и др. (Molecular and General Genetics 220: 478-480 (1990)), у Pisabarro и др. (Journal of Bacteriology 175: 2743-2749 (1993)) и у Bonassie и др. (Nucleic Acids Research 18: 6421 (1990)). Нуклеотидная последовательность гена dapA доступна под регистрационным номером Х53993.
Выделение, клонирование и секвенирование гена dapB из Brevibacterium lactofermentum описаны у Pisabarro и др. (Journal of Bacteriology 175: 2743-2749 (1993)). Нуклеотидная последовательность гена dapB доступна под регистрационным номером Х67737.
Выделение, клонирование и секвенирование гена lysC и аллелей гена lysC, кодирующих аспартаткиназу, устойчивую в отношении обратной связи, описано во многих публикациях. Так, например, у Kalinowski и др. (Molecular and General Genetics 224: 317-324 (1990)) описан аллель гена lysC из штамма С.glutamicum DM58-1. В DE-A 3943117 описано клонирование аллеля гена lysC из штамма С.glutamicum MH20. У Follettie и др. (Journal of Bacteriology 175: 4096-4103 (1993)) описан аллель гена lysC из штамма С.flavum N13, который в этой работе обозначен как ask. У Kalinowski и др. (Molecular Microbiology 5, 1197-1204 (1991)) описан ген lysC из штамма С. glutamicum ATCC 13032. Нуклеотидные последовательности гена lysC и различных аллелей гена lysC доступны среди прочего под регистрационными номерами Х57226 и Е06826.
Полученные таким образом гены могут быть, в частности, встроены по отдельности или в соответствующих комбинациях в плазмидные векторы, такие как pJC1 (Cremer и др., Molecular and General Genetics 220: 478-480 (1990)) или pEC5 (Eikmanns и др., Gene 102, 93-98 (1991)), и введены в требуемые штаммы коринебактерий, например в штамм МН20-22В (Schrumpf и др., Applied Microbiology and Biotechnology 37: 566-571 (1992)), путем трансформации, как это описано, например, у Tierbach и др. (Applied Microbiology and Biotechnology 29: 366-362 (1988)), или электропорации, как это описано, например, у Dunican и Shivnan (Bio/Technology 7, 1067-1070 (1989)), и экспрессированы в них. Выбранный штамм также можно трансформировать двумя плазмидными векторами, каждый из которых содержит рассматриваемый ген или рассматриваемые гены, и в результате этого достичь предпочтительно одновременно усиленной экспрессии второго или нескольких генов дополнительно к известному усилению гена lysE.
Примерами таких штаммов являются следующие:
- штамм MH20-22B/pJC33/pEC7lysE, в котором одновременно усиленно экспрессируют ген lysE и ген lysC, или
- штамм MH20-22B/pJC50/pEC7lysE, в котором одновременно усиленно экспрессируют ген lysE, ген lysC и ген dapA, или
- штамм MH20-22B/pJC23/pEC7lysE, в котором одновременно усиленно экспрессируют ген lysE и ген dapA, или
- штамм MH20-22B/pJC23/pEC7dapBlysE, в котором одновременно усиленно экспрессируют ген lysE, ген dapA и ген dapB.
Для продуцирования L-лизина созданные согласно изобретению микроорганизмы можно культивировать непрерывно или периодически с использованием периодического процесса (культивирование партий) или периодического процесса с подпиткой либо периодического процесса с повторяющейся подпиткой. Обзор известных методов культивирования представлен в учебнике Chmiel (Bioprozesstechnik 1. in die Bioverfahrenstechnik (изд-во Gustav Fischer, Stuttgart, 1991)) или в учебнике Storhas (Bioreaktoren und periphere Einrichtungen (изд-во Vieweg, Braunschweig/Wiesbaden, 1994)).
Используемая культуральная среда должна быть адаптирована к требованиям соответствующего штамма. Описания культуральных сред для различных микроорганизмов содержатся в справочнике "Manual of Methods for General Bacteriology" Американского общества бактериологии (Washington D.C., США, 1981). В качестве источника углерода могут быть использованы сахара и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, крахмал и целлюлоза, масла и жиры, такие как соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло, жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота. Эти вещества могут применяться по отдельности или в виде смеси. В качестве источника азота могут применяться органические азотсодержащие соединения, такие как пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкость, образующаяся после замачивания зерен кукурузы до разбухания, соевая мука и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота могут применяться по отдельности или в виде смеси. В качестве источника фосфора могут применяться кислый фосфат калия или дикалийгидрофосфат либо соответствующие содержащие натрий соли. Кроме того, культуральная среда должна содержать соли металлов, такие как сульфат магния или сульфат железа, которые необходимы для роста. И, наконец, в дополнение к вышеназванным соединениям могут использоваться такие важные для роста вещества, как аминокислоты и витамины. Вышеуказанные добавки могут вводиться в культуральную среду в виде одноразовой добавки или добавляться соответствующим образом в процессе культивирования.
Для контроля значения рН культуральной среды используют соответственно либо основания, такие как гидроксид натрия, гидроксид калия, аммиак, либо кислоты, такие как фосфорная кислота или серная кислота. Для контроля пенообразования добавляют антивспениватели, такие как полигликолевые эфиры жирных кислот. Для поддержания стабильности плазмид могут быть добавлены соответствующие конкретной среде вещества, обладающие избирательным действием, например антибиотики. Для поддержания аэробных условий в культуру вводят кислород или кислородсодержащие газовые смеси, такие как воздух. Температура культуральной среды в норме составляет от 20°С до 45°С, предпочтительно от 25°С до 40°С. Культивирование продолжают до тех пор, пока не образуется максимальное количество требуемого L-лизина. Как правило, эта цель достигается в течение 10-160 ч.
Концентрацию образовавшегося L-лизина можно определять с помощью анализаторов аминокислот путем ионообменной хроматографии и последующей реакции на колонках с обнаружением нингидрином по методу, описанному у Spackman и др. (Analytical Chemistry, 30, 1190 (1958)).
В соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ, Брауншвейг, Германия) были депонированы следующие штаммы:
- штамм Escherichia coli К-12 DH5α/pEC7lysE под регистрационным номером DSM 12871;
- штамм Escherichia coli K-12 DH5α/pEC7dapBlysE под регистрационным номером DSM 12875;
- штамм Corynebacterium glutamicum DSM5715/pJC23 под регистрационным номером DSM 12869;
- штамм Corynebacterium glutamicum DSM5715aecD::dapA(MA16) под регистрационным номером DSM 12867;
- штамм Corynebacterium glutamicum DSM5715aecD::dapA(MC20) под регистрационным номером DSM 12868.
Примеры
Пример 1
Получение кодирующей последовательности ДНК гена lysE
Хромосомную ДНК выделяли из штамма АТСС 13032 с помощью общепринятых методов (Eikmanns и др., Microbiology 140: 1817-1828 (1994)). С помощью полимеразной цепной реакции (ПЦР) амплифицировали фрагмент ДНК, несущий ген lysE. На основе известной последовательности гена lysE из С. glutamicum (Vrljic и др., Molecular Microbiology 22 (5), 815-826 (1996)) (регистрационный номер Х96471) выбирали следующие олигонуклеотиды в качестве праймеров для ПЦР:
LysBam1:
5'-СТС GAG AGC (GGA ТСС) GCG CTG ACT САС С-3';
LysBam2:
5'-GGA GAG TAC GGC (GGA ТСС) ACC GTG ACC-3'.
Представленные праймеры были синтезированы фирмой MWG Biotech (Эберсберг, Германия) и с их использованием проводили ПЦР согласно стандартному методу, описанному у Innis и др. (PCR protocols. A guide to methods and applications. Academic Press, 1990). С использованием этих праймеров оказалось возможным амплифицировать фрагмент ДНК длиной приблизительно 1,1 т.п.н., несущий ген lysE. Кроме того, праймеры содержали последовательность распознаваемого рестриктазой BamHI сайта, которая в вышеприведенных нуклеотидных последовательностях заключена в скобки.
Амплифицированный фрагмент ДНК длиной приблизительно 1,1 т.п.н., несущий ген lysE, идентифицировали с помощью электрофореза в 0,8%-ном агарозном геле, выделяли из геля и очищали с помощью набора QIAquick Gel Extraction Kit (каталожный номер 28704) фирмы Quiagen (Гильден, Германия).
Затем с помощью ДНК-лигазы фага Т4 фирмы Boehringer Mannheim (Маннгейм, Германия) фрагмент встраивали в вектор pUC18 (Norrander и др., Gene 26, 101-106 (1983)). Для этого вектор pUC18 полностью расщепляли рестриктазой Smal и обрабатывали щелочной фосфатазой (щелочная фосфатаза фирмы Boehringer Mannheim, Маннгейм, Германия). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α (Hanahan, DNA cloning. A practical approach, т.I, IRL-Press, Oxford, Washington DC, США). Отбор несущих плазмиду клеток осуществляли путем посева продуктов, полученных в результате трансформации, на LB-arap (Sambrook и др., Molecular Cloning a laboratory manual, 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.), дополненный 50 мг/л ампициллина. Плазмидную ДНК выделяли из одного трансформанта, проверяли с помощью обработки рестриктазой BamHI и последующего электрофореза на агарозном геле. Полученную плазмиду обозначили как pUC18lysE.
Пример 2
Получение гена dapB
Хромосомную ДНК выделяли из штамма Corynebacterium glutamicum ATCC 13032 аналогично примеру 1. Последовательность гена dapB из Corynebacterium glutamicum в принципе является известной (регистрационный номер Х67737). Однако опубликованная последовательность ДНК включает фрагмент, расположенный перед сайтом начала трансляции и состоящий только из 56 пар оснований. Поэтому проводили дополнительное секвенирование 5'-области, расположенной перед сайтом начала трансляции.
Для этого проводили секвенирование с помощью плазмиды pJC25 (EP-B 0435132) с использованием олигонуклеотидного праймера, связывающегося с областью известной последовательности гена dapB (регистрационный номер Х67737). Используемый для секвенирования праймер имел следующую последовательность:
5'-GAA CGC САА ССТ TGA ТТС С-3'.
Секвенирование проводили по методу, основанному на разрыве цепи, как это описано у Sanger и др., Proc. Natl. Acad. Sci. USA (74) 5463-5467 (1977). Реакцию секвенирования проводили с помощью набора AutoRead Sequencing Kit (фирма Pharmacia, Фрейбург, Германия). Электрофоретический анализ и обнаружение продукта секвенирования проводили с помощью ДНК-секвенатора типа A.L.F. фирмы Pharmacia (Фрейбург, Германия).
Полученную последовательность ДНК использовали для выбора второго праймера с целью получить дополнительные данные о последовательности области перед сайтом начала транскрипции. Для этой цели был выбран следующий праймер:
5'-СТТ TGC CGC CGT TGG GTT С-3'.
Реакцию секвенирования проводили по описанной выше методике. Новая последовательность, расположенная против хода транскрипции относительно гена dapB, представлена в виде SEQ ID NO:1. Последовательность, включающая нуклеотидную последовательность гена dapB, представлена в виде SEQ ID NO:2.
Ген dapB амплифицировали с помощью полимеразной цепной реакции. Для этого фирмой MWG Biotech были синтезированы два олигонуклеотидных праймера, выбранные на основе известной последовательности гена dapB:
P-dap:
5'-(AAG CTT) AGG TTG TAG GCG TTG AGC-3';
dapall:
5'-TTA ACT TGT TCG GCC АСА GC-3'.
5'-Праймер (праймер P-dap) содержит сайт рестрикции HindIII, заключенный в скобки в вышеприведенной последовательности. ПЦР проводили аналогично примеру 1. Таким путем амплифицировали фрагмент ДНК длиной приблизительно 1,1 т.п.н., который несет ген dapB и на одном конце имеет сайт, распознаваемый рестриктазой HindIII. Полученный в результате ПЦР фрагмент очищали на 0,8%-ном агарозном геле (набор QIAquick Gel Extraction Kit фирмы Qiagen, Гильден, Германия) и клонировали с помощью набора типа ТОРО ТА Cloning Kit (фирма Invitrogen, Леек, Нидерланды, каталожный номер К4550-01) в клонирующем векторе pCR2.1TOPO (фирма Invitrogen, Леек, Нидерланды). Полученным в результате лигирования продуктом трансформировали штамм Е.coli TOP10F' фирмы Invitrogen, продукт трансформации высевали на дополненный канамицином (50 мг/л) LB-агар, содержащий IPTG (0,16 мМ) и X-Gal (64 мг/л), и выделяли устойчивые к канамицину и окрашенные в белый цвет колонии. Плазмидную ДНК выделяли из трансформантов с помощью набора QIAprep Spin Miniprep Kit фирмы Qiagen и проверяли с помощью расщепления рестриктазой HindIII и последующего электрофореза в агарозном геле. Аминокислотную последовательность амплифицированного фрагмента ДНК проверяли путем секвенирования. Последовательность ПЦР-продукта согласуется с последовательностью, представленной в SEQ ID NO: 1. Полученную плазмиду обозначили как pCR2.1TOPOdapB.
Пример 3
Клонирование гена IvsE в векторе рЕС7
Фрагмент, содержащий ген lysE из плазмиды pUC18lysE (пример 1), встраивали по описанной ниже методике в вектор рЕС7. Основой вектора рЕС7 является бифункциональный вектор рЕС5, способный реплицироваться в Е.coli и С.glutamicum (Eikmanns и др., 102: 93-98 (1991)). Из плазмиды рЕС5 удаляли те сайты рестрикции BamHI, которые не находятся в полилинкере, частично расщепляя плазмиду рЕС5 рестриктазой BamHI. Из агарозного геля выделяли фрагмент ДНК длиной приблизительно 7,2 т.п.н. и выступающие концы обрабатывали полимеразой Кленова (фирма Boehringer Mannheim). Полученный таким путем фрагмент ДНК лигировали (лигаза фага Т4, фирма Boehringer Mannheim). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α и выделяли устойчивые к канамицину колонии на LB-агаре, содержащем канамицин (50 мг/л). Из одного трансформанта выделяли плазмидную ДНК (набор типа QIAprep Spin Miniprep Kit фирмы Qiagen) и проверяли путем расщепления рестриктазами BamHI и PstI. Полученную таким путем плазмиду обозначили как рЕС6.
Плазмиду рЕС6 полностью расщепляли рестриктазой XhoI. Фрагмент ДНК, несущий терминатор trp, лигировали с фрагментом ДНК вектора (лигаза фага Т4, фирма Boehringer Mannheim). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α и выделяли устойчивые к хлорамфениколу колонии на LB-агаре, содержащем хлорамфеникол (50 мг/л). Из одного трансформанта выделяли плазмидную ДНК (набор типа QIAprep Spin Miniprep Kit фирмы Qiagen) и проверяли путем расщепления рестриктазами BamHI и Xhol. Полученную таким путем плазмиду обозначили как рЕС7.
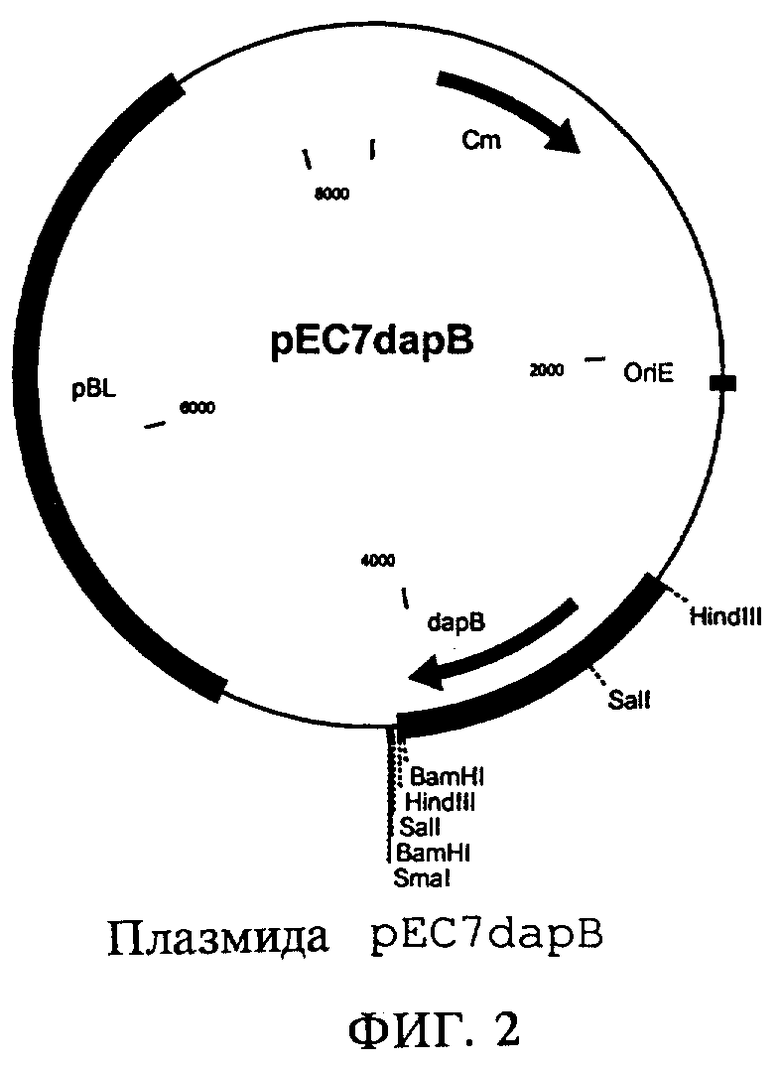
Описанную в примере 1 плазмиду pUC18lysE полностью расщепляли рестриктазой BamHI и аналогично примеру 1 выделяли BamHI-фрагмент длиной 1,1 т.п.н., содержащий ген lysE. Вектор рЕС7 также полностью расщепляли рестриктазой BamHI и обрабатывали щелочной фосфатазой. Лигировали BamHI-фрагмент вектора и BamHI-фрагмент гена lysE (Rapid DNA Ligation Kit, фирма Boehringer Mannheim) и полученным продуктом трансформировали штамм Е.coli DH5α. Трансформанты, несущие плазмиду, отбирали на LB-агаре, содержащем хлорамфеникол (10 мг/л). Выделяли плазмидную ДНК (набор типа QIAprep Spin Miniprep Kit фирмы Qiagen) и проверяли путем расщепления рестриктазой BamHI. Полученную таким образом плазмиду обозначили как pEC7lysE (фиг.1). Штамм, полученный путем трансформации штамма Е.coli DH5α плазмидой pEC7lysE, обозначили как DH5α/pEC7lysE.
Пример 4
Клонирование гена dapB в векторе рЕС7
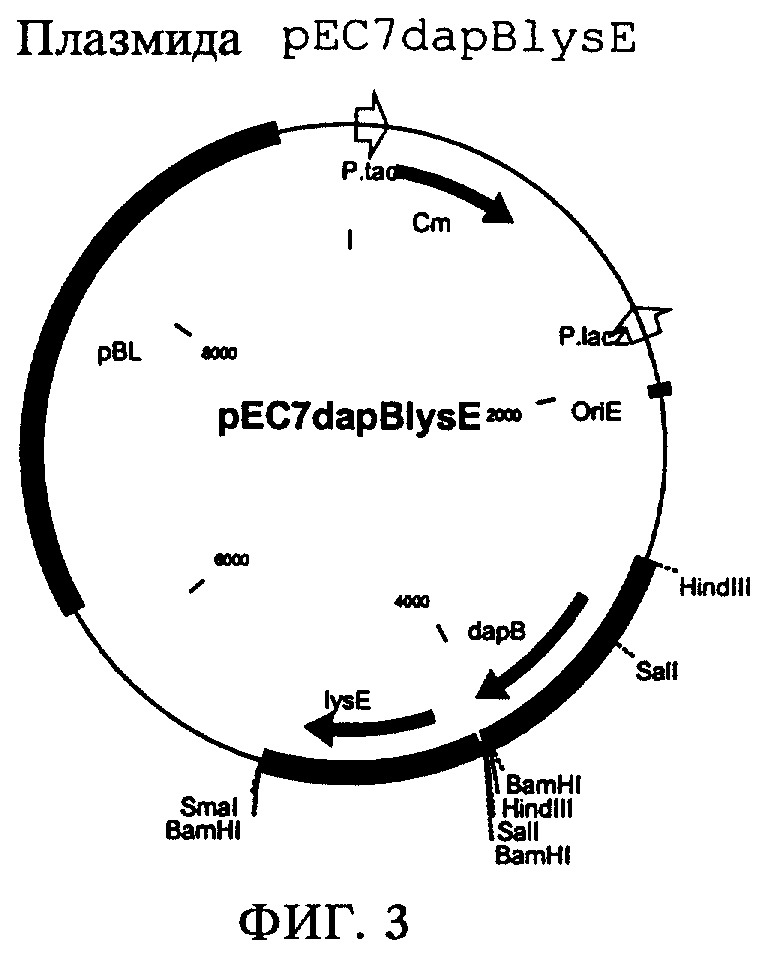
Из плазмиды pCR2.1TOPOdapB (из примера 2) выделяли фрагмент ДНК длиной приблизительно 1,1 т.п.н., несущий ген dapB. Для этого плазмиду pCR2.1TOPOdapB полностью расщепляли рестриктазой HindIII и выделяли фрагмент ДНК длиной приблизительно 1,1 т.п.н., несущий ген dapB. Полученный таким образом фрагмент ДНК, несущий ген dapB, лигировали с помощью ДНК-лигазы фага Т4 (фирма Boehringer Mannheim) с вектором рЕС7 (пример 3), также полностью расщепленным рестриктазой HindIII и обработанным щелочной фосфатазой (щелочная фосфатаза, фирма Boehringer Mannheim). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α и выделяли устойчивые к канамицину колонии на LB-агаре, дополненном канамицином (50 мг/л). Из одного трансформанта выделяли плазмидную ДНК (набор типа QIAprep Spin Miniprep Kit, фирма Qiagen) и проверяли путем расщепления рестриктазой HindIII. Полученную таким образом плазмиду обозначили как pEC7dapB (фиг.2). Полученный штамм Escherichia coli обозначили как DH5α/pEC7dapB.
Пример 5
Получение плазмиды, содержащей одновременно ген dapB и ген IvsE
Из плазмиды pCR2.1TOPOdapB, содержащей ген dapB из штамма С. glutamicum ATCC 13032, выделяли ген dapB в виде HindIII-фрагмента. Для этого плазмиду полностью расщепляли рестриктазой HindIII и фрагмент ДНК, несущий ген dapB, выделяли из 0,8%-ного агарозного геля (набор QIAquick Gel Extraction Kit, фирма Qiagen). Кроме того, вектор рЕС7 (пример 3) полностью расщепляли рестриктазой HindIII и обрабатывали щелочной фосфатазой.
Полученный таким образом линеаризованный фрагмент вектора лигировали с фрагментом длиной 1,1 т.п.н., содержащим ген dapB, с помощью лигазы фага Т4 (фирма Boehringer Mannheim) и полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α. Трансформанты, несущие плазмиду, отбирали на LB-агаре, содержащем хлорамфеникол (10 мг/л). Выделяли плазмидную ДНК (набор типа QIAprep Spin Miniprep Kit, фирма Qiagen, Гильден, Германия) и проверяли путем расщепления рестриктазой HindIII. Полученную таким образом плазмиду обозначили как pEC7lysEdapB. Эта плазмида может автономно реплицироваться как в Escherichia coli, так и в микроорганизмах рода Corinebacterium и придает своему хозяину устойчивость к антибиотику хлорамфениколу.
Плазмида pEC7lysEdapB, как показано на фиг.3, содержит одновременно ген dapB, кодирующий дигидродипиколинат-редуктазу, и ген lysE, участвующий в экспорте лизина. Штамм, полученный в результате трансформации штамма E.coli DH5a плазмидой pEC7lysEdapB, обозначили как DH5α/pEC7lysEdapB.
Пример 6
Трансформация штамма МН20-22В плазмидами pJC1, pJC33 и pJC50
Плазмида pJC1 представляет собой плазмиду, реплицируемую и в Escherichia coli, и в Corynebacterium glutamicum (Cremer и др.. Molecular and General Genetics 220: 478-480 (1990)). Плазмида pJC33 (Cremer и др., Applied and Environmental Microbiology 57 (6), 1746-1752 (1991)) является производной указанной плазмиды и содержит ген lysC (Fbr) из штамма С.glutamicum МН20-22В.
Основой плазмиды pJC50 также является вектор pJC1, и она несет ген lysC (Fbr) из штамма С.glutamicum МН20-22В и ген dapA из штамма С.glutamicum АТСС 13032 (DE-A 3943117).
В штамм МН20-22В методом электропорации (Haynes и Britz, FEMS Microbiology Letters (61), 329-334 (1989)) вводили плазмиды pJC1, pJC33 и pJC50. Штамм С.glutamicum МН20-22В представляет собой устойчивый к АЕС и продуцирующий лизин штамм, который депонирован под регистрационным номером DSM 5715.
Полученные путем электропорации трансформанты выделяли на агаре, предназначенном для селекции [LBHIS-агар (18,5 г/л бульона, используемого для инфузии в мозг и сердце, 0,5М сорбит, 5 г/л бакто-триптона, 2,5 г/л экстракта бакто-дрожжей, 5 г/л NaCl, 18 г/л бакто-агара)] и дополненном 15 мг/л канамицина. Плазмидную ДНК выделяли общепринятыми методами (Peters-Wendisch и др., Microbiology, 144, 915-927 (1998)), расщепляли соответствующими рестриктазами и проверяли. Полученные штаммы обозначили как MH20-22B/pJC1, MH20-22B/pJC33 и MH20-22B/pJC50.
Пример 7
Трансформация с использованием плазмид pEC7lysE и pEC7dapBlysE
В дальнейшем для введения второй плазмиды использовали штаммы, полученные в соответствии с примером 6.
В описанные выше штаммы MH20-22B/pJC1, MH20-22B/pJC33 и MH20-22B/pJC50 путем электропорации вводили плазмиды pEC7lysE и pEC7dapBlysE.
Трансформированные бактерии отбирали по признаку устойчивости содержащихся в них плазмид к антибиотикам. Полученные с помощью электропорации трансформанты выделяли на агаре, предназначенном для селекции (LHBIS-агар с добавлением 15 мг/л канамицина и 7,5 мг/л хлорамфеникола). Плазмидную ДНК выделяли, расщепляли соответствующими рестриктазами и проверяли.
Пример 8
Получение L-лизина
Полученные в примере 7 различные штаммы С.glutamicum культивировали в пригодной для получения лизина питательной среде и определяли содержание лизина в надосадочной жидкости культуры.
Для этого различные штаммы сначала инкубировали в течение 24 ч при 33°С на агаровых пластинах, содержащих соответствующие антибиотики (агар со средой, используемой для инфузии в мозг и сердце, дополненный канамицином (25 мг/л), хлорамфениколом (10 мг/л)). Эти высеянные на агаровые пластины культуры вносили в среду для предварительной культуры (10 мл среды в колбах Эрленмейера объемом 100 мл). В качестве среды для предварительной культуры использовали полную среду CgIII. В нее добавляли канамицин (25 мг/л) и хлорамфеникол (10 мг/л). Предварительную культуру инкубировали в течение 24 ч и при 33°С на шейкере при 240 об/мин. Эту предварительную культуру вносили в основную культуру в таком количестве, чтобы начальная оптическая плотность (ОП) при 660 нм основной культуры составляла 0,2. Для основной культуры использовали среду ММ следующего состава:
Состав среды ММ:
соли:
Значения рН для ЖЗК (жидкость, образующаяся при замачивании зерен кукурузы до разбухания), МПС (3-морфолинпропансульфоновая кислота) и раствора солей доводили до 7 с помощью аммиачной воды и автоклавировали. Затем добавляли стерильный субстрат и растворы витаминов, а также сухой автоклавированный СаСО3.
Культивирование осуществляли в среде объемом 10 мл, находящейся в колбе Эрленмейера объемом 100 мл с дефлекторами. Добавляли канамицин (25 мг/л) и хлорамфеникол (10 мг/л). Культивирование осуществляли при 33°С и 80%-ной влажности воздуха.
Через 48 ч измеряли ОП при длине волны 660 нм с помощью прибора типа Biomek 1000 (фирма Beckmann Instruments GmbH, Мюнхен). Количество образовавшегося лизина определяли с помощью анализатора аминокислот фирмы Eppendorf-BioTronik (Гамбург, Германия) с использованием ионообменной хроматографии и последующей дериватизации на колонках с обнаружением нингидрином. Содержание глюкозы определяли с помощью анализатора сахара фирмы Skalar Analytik GmbH (Эркеленц, Германия).
В таблице 1 представлены результаты опыта.
Пример 9
Клонирование гена aecD в векторе pUC18
Плазмиду pSIR21 (Rossol, диссертация, Университет Билефельда, 1992) полностью расщепляли ферментами BglII и EcoRV и выделяли фрагмент ДНК длиной 1,4 т.п.н., содержащий ген aecD (регистрационный номер М89931) (Rossol и , Journal of Bacteriology 174(9), 2968-2977 (1992)) из штамма С.glutamicum ATCC 13032. Выделенный фрагмент ДНК встраивали с помощью ДНК-лигазы фага Т4 по методу, описанному у Sambrook и др. (Molecular Cloning: A Laboratory Manual (1989), Cold Spring Harbor Laboratory Press), лигировали с плазмидой pUC18, полностью расщепленной с помощью ферментов BamHI и SmaI. Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α. Отбор трансформантов осуществляли на планшетах со средой, используемой для инфузии в мозг и сердце, дополненной 100 мг/л ампициллина. Из одной колонии выделяли плазмидную ДНК. Полученную плазмиду обозначили как pUC18::aecD.
Пример 10
Клонирование гена dapA в плазмиде pSP72
Фрагмент, содержащий ген dapA, выделяли из плазмиды pJC20 (Cremer J., диссертация, Университет Дюссельдорфа, 1989) в виде SphI-BamHI-фрагмента. Вектор pSP72 (фирма Promega Corporation, США) полностью расщепляли ферментами SphI и BamHI и обрабатывали щелочной фосфатазой. В этот вектор лигированием с помощью ДНК-лигазы фага Т4 встраивали фрагмент, несущий ген dapA. Затем фрагментом ДНК трансформировали штамм Е. coli XL1 Blue (Bullock, Fernandez и Short, BioTechniques (5) 376-379 (1987)). Отбор трансформантов осуществляли на LB-среде, содержащей 100 мг/л ампициллина. Из одного трансформанта выделяли плазмидную ДНК, которую обозначили как pSP72::dapA.
Пример 11
Мутагенез промотора dapA и получение плазмид pSP72::dapA(MC20) и pSP72::dapA(MA16)
Для мутагенеза промоторной области использовали набор для сайтнаправленного мутагенеза типа Quickchange фирмы Stratagene. На основе имеющейся последовательности гена dapA создавали представленные ниже праймеры, которые использовали для введения мутаций.
Для получения плазмиды pSP72::dapA(MC20) использовали:
праймер dap1 для МС20:
ССА ААТ GAG AGA TGG TAA CCT TGA ACT СТА TGA GCA
праймер dap2 для МС20:
GTG СТС АТА GAG TTC AAG GTT АСС АТС ТТС CCT CAT TTG G
Для получения плазмиды pSP72::dapA (MA16) использовали:
праймер dap3 для МА16
ССА ААТ GAG GGA AGA AGG TAT ААТ TGA ACT СТА TGA GCA
праймер dap4 для МА16
GTG СТС АТА GAG TTC ААТ TAT АСС TTC TTC CCT CAT TTG G
ПЦР проводили согласно инструкциям производителя (фирма Stratagene) набора для сайтнаправленного мутагенеза Quickchange с использованием плазмиды pSP72::dapA (из примера 10) в качестве матрицы.
Продуктами, полученными в результате мутагенеза, трансформировали штамм Е.coli XL1-Blue. Отбор трансформантов проводили на LB-среде, содержащей 100 мг/л карбенициллина. Из одного трансформанта выделяли плазмидную ДНК, при этом путем расщепления с помощью BstEII контролировали отсутствие сайта BstEII. Плазмида, в которой отсутствовал сайт BstEII, содержала требуемую мутацию.
Полученной плазмидой трансформировали мутантный штамм Е.coli RDA8 с дефектным геном dapA. Полученные в результате трансформации продукты высевали на LB-среду, содержащую 100 мг/л карбенициллина, с целью проверить комплементацию мутации в гене dapA. Из одного трансформанта каждой группы выделяли ДНК и полученные плазмиды обозначили как pSP72::dapA(MC20) и pSP72::dapA(MA16) соответственно. Секвенирование плазмид осуществляли с использованием обратного и универсального праймера для секвенирования по методу, основанному на разрыве цепи, который описан у Sanger и др., Proceedings of National Academy of Sciences of the USA, (74) 5463-5467 (1977). Реакцию секвенирования проводили с помощью набора типа Auto Read Sequencing Kit (фирма Pharmacia, Фрейбург, Германия). Электрофоретический анализ и обнаружение продукта секвенирования осуществляли с помощью ДНК-секвенатора типа A.L.F. (фирма Pharmacia, Фрейбург, Германия).
Пример 12
Получение плазмид pK19mobsacBaecD::dapA(MC20) и pK19mobsacBaecD::dapA(MA16) (обратное клонирование содержащих мутации фрагментов)
Плазмиды pSP72::dapA(MC20) и pSP72::dapA(MA16) (из примера 11) полностью расщепляли рестриктазами PvuII и SmaI. PvuII-SmaI-фрагмент длиной 1450 пар оснований, несущий ген dapA, который имеет мутантный промотор МС20, соответственно МА16, встраивали лигированием с помощью ДНК-лигазы фага Т4 в расщепленный с помощью StuI вектор pUC18::aecD (из примера 9). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α. Отбор трансформантов осуществляли на LB-среде, содержащей 100 мг/л ампициллина. Из одного трансформанта каждой группы выделяли плазмидную ДНК. Таким путем получали плазмиды pUC18aecD::dapA (MC20) и pUC18aecD::dapA (MA16).
Плазмиды pUC18aecD::dapA(MC20) и pUC18aecD::dapA(MA16) подвергали частичному расщеплению рестриктазой EcoRI и полному расщеплению рестриктазой SalI с целью получить фрагмент длиной 3,0 т.п.н., несущий aecD::dapA(MA16), соответственно aecD::dapA(MC20). Этот фрагмент встраивали лигированием с помощью ДНК-лигазы фага Т4 в расщепленный и обработанный щелочной фосфатазой вектор pK19mobsacB ( и др., Gene (145) 69-73 (1994)). Полученным в результате лигирования продуктом трансформировали штамм Е.coli DH5α (Hanahan, DNA cloning. A practical approach, т.I, IRL-Press, Oxford, Washington DC, США (1985)). Отбор трансформантов осуществляли на LB-среде, содержащей 50 мг/л канамицина. Из одного трансформанта каждой группы выделяли плазмидную ДНК. Таким путем получали плазмиды pK19mobsacBaecD::dapA (MC20) и pK19mobsacBaecD::dapA (MA16).
Плазмидной ДНК трансформировали штамм Е.coli S17-1 (Simon, Priefer и , Bio/Technology (1) 784-791 (1983)). Отбор трансформантов осуществляли на LB-среде, содержащей 50 мг/л канамицина. Из одного трансформанта каждой группы выделяли плазмидную ДНК и проверяли. Полученные таким образом штаммы обозначили как S17-1/pK19mobsacBaecD::dapA (MC20) и S17-l/pK19mobsacBaecD::dapA (MA16).
Пример 13
Получение штаммов С.glutamicum DSM5715aecD::dapA (MC20) и DSM5715aecD::dapA (MA16)
Плазмиды pK19mobsacBaecD::dapA (MC20) и pK19mobsacBaecD::dapA (MA16) методом конъюгации ( и др., Journal of Bacteriology (172) 1663-1666 (1990)) с S17-1/pK19mobsacBaecD::dapA (MC20) и S17-1/pK19mobsacBaecD::dapA (MA16) (из примера 12) переносили в штамм С. glutamicum DSM5715. Для отбора трансконъюгантов полученные в результате конъюгации продукты высевали на среду, используемую для инфузии в мозг и сердце, содержащую налидиксиновую кислоту и канамицин. Полученные трансконъюганты инкубировали в течение ночи в 10 мл среды, используемой для инфузии в мозг и сердце. Затем аликвотные количества высевали на пластины, содержащие сахарозу (агар со средой, используемой для инфузии в мозг и сердце, содержащий 10% сахарозы) с целью произвести отбор по признаку потери чувствительности к сахарозе. Выделяли устойчивые к сахарозе клоны и повторно проверяли на содержащих хлорамфеникол и канамицин агаровых пластинах (среда, используемая для инфузии в мозг и сердце, содержащая 15 мг/л канамицина, и среда, используемая для инфузии в мозг и сердце, содержащая 10 мг/л хлорамфеникола).
Выделяли колонии, имеющие следующий фенотип:
устойчивые к сахарозе,
чувствительные к канамицину,
чувствительные к хлорамфениколу.
Встраивание фрагментов гена dapA в ген aecD проверяли методом Саузерн-блоттинга (Sambrook и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989)).
Пример 14
Получение штаммов С. glutamicum DSM5715aecD::dapA (MC20)/pEC7lysE, DSM5715aecD::dapA (MA16)/pEC7lysE, DSM5715aecD::dapA (MC20)/pEC7, DSM5715aecD::dapA (MA16)/pEC7 и DSM5715/pEC7
Как описано в примере 3, ген lysE находится в векторе рЕС7. Эту плазмиду pEC7lysE, а также плазмиду рЕС7 вводили путем электропорации (Haynes, FEMS Microbiology Letters 61: 329-334 (1989)) в штаммы DSM5715aecD::dapA (MC20), DSM5715aecD::dapA (MA16) и DSM5715 (из примера 13) с целью получить штаммы С. glutamicum DSM5715aecD::dapA (MC20)/pEC7lysE, DSM5715aecD::dapA (MA16)/pEC7lysE, DSM5715aecD::dapA (MC20)/pEC7, DSM5715aecD::dapA (MA16)/pEC7 и DSM5715/pEC7. Отбор трансформантов осуществляли на агаре со средой, используемой для инфузии в мозг и сердце, содержащей 25 мг/л канамицина. Из одного трансформанта каждой группы выделяли плазмидную ДНК и проверяли.
Таким путем получали следующие штаммы:
DSM5715aecD::dapA (MC20)/pEC7lysE,
DSM5715aecD::dapA (MA16)/pEC7lysE,
DSM5715aecD::dapA (MC20)/pEC7,
DSM5715aecD::dapA (MA16)/pEC7 и
DSM5715/pEC7.
Пример 15
Получение лизина с помощью полученных в примере 14 штаммов
Штаммы DSM5715aecD::dapA (MC20)/pEC7lysE, DSM5715aecD::dapA (MA16)/pEC7lysE, DSM5715aecD::dapA (MC20)/pEC7, DSM5715aecD::dapA (MA16)/pEC7 и DSM5715/pEC7 культивировали аналогично примеру 8 в среде ММ для получения продукта после предварительного культивирования в среде CgIII (Kase и Nakayama, Agricultural and Biological Chemistry 36(9) 1611-1621 (1972)). После инкубации в течение 48 ч определяли оптическую плотность при 660 нм и концентрацию образовавшегося L-лизина.
В таблице 2 представлены результаты опыта.
Описание чертежей
На приложенных к описанию чертежах показано:
на фиг.1 - плазмида pEC7lysE,
на фиг.2 - плазмида pEC7dapB и
на фиг.3 - плазмида pEC7dapBlysE.
Используемые на чертежах сокращения имеют следующие значения:
Cm: ген, обусловливающий устойчивость к хлорамфениколу,
dapB: ген dapB из С.glutamicum,
lysE: ген lysE из С.glutamicum,
рус: ген рус из С.glutamicum,
OriE: кодируемое плазмидой начало репликации Е. coli,
pBL: фрагмент ДНК плазмиды pBL1,
EcoRI: сайт рестрикции рестриктазы EcoRI,
EcoRV: сайт рестрикции рестриктазы EcoRV,
HincII: сайт рестрикции рестриктазы HincII,
HindIII: сайт рестрикции рестриктазы HindIII,
KpnI: сайт рестрикции рестриктазы KpnI,
SalI: сайт рестрикции рестриктазы SalI,
SmaI: сайт рестрикции рестриктазы Smal,
SphI: сайт рестрикции рестриктазы SphI,
PvuII: сайт рестрикции рестриктазы PvuII,
BamHI: сайт рестрикции рестриктазы BamHI.




Изобретение относится к биотехнологии. L-лизин получают путем ферментации коринебактерий, в которых по отдельности или вместе усиливают нуклеотидные последовательности генов биосинтеза L-лизина, причем коринебактерии имеют мутации МС20 или МА16 в промоторе гена dapA и усиленный ген lysE. Данное изобретение позволяет получать L-лизин с высокой степенью эффективности. 7 з.п. ф-лы, 3 ил., 2 табл.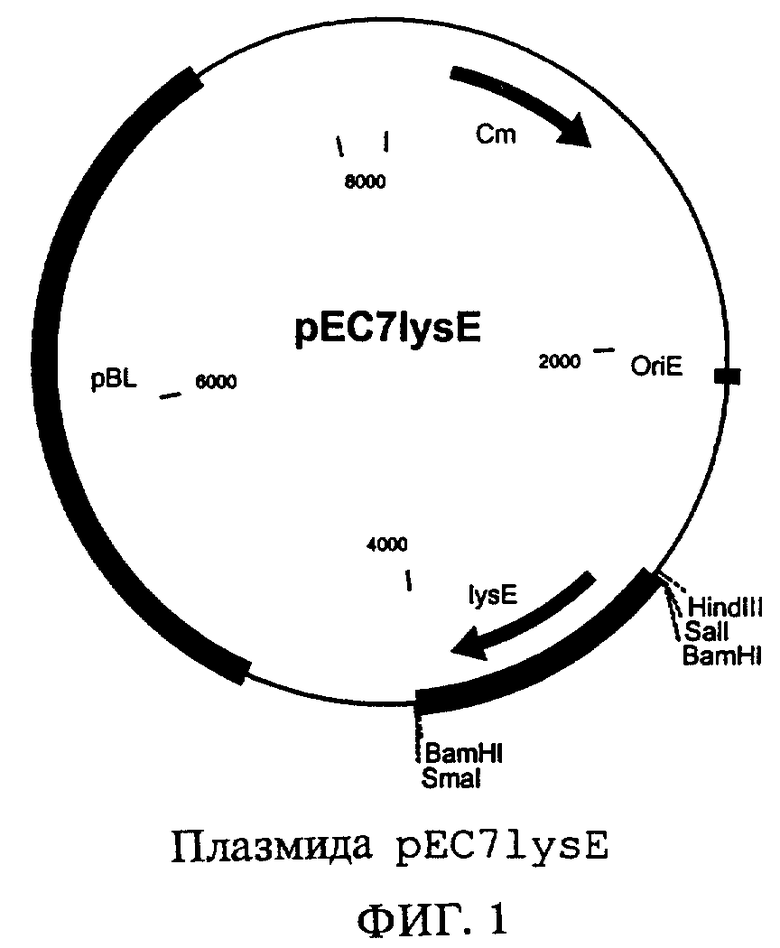
а) ферментацию с использованием продуцирующих L-лизин бактерий, в которых усиливают описанные гены,
б) накопление L-лизина в среде или в клетках бактерий и
в) выделение L-лизина.
| DE 19548222, 26.06.1997 | |||
| DE 3823451, 18.01.1990 | |||
| EP 0841395, 13.03.1998 | |||
| DE 3943117, 04.07.1991 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА ПОСРЕДСТВОМ КУЛЬТИВИРОВАНИЯ ШТАММА БАКТЕРИЙ | 1992 |
|
RU2099424C1 |

