ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
1. Область изобретения
Настоящее изобретение относится к микроорганизму Corynebacterium sp., модифицированному для утилизации ксилозы, и способу получения L-лизина с применением такого микроорганизма.
2. Описание предшествующего уровня техники
Промышленные микроорганизмы утилизируют в качестве источника углерода сахар, такой как глюкоза, фруктоза и сахароза. В качестве сырья для получения этих источников углерода обычно используют сельскохозяйственные продукты, но они дороги и более ценны в качестве продуктов питания. Недавно вместо применения сельскохозяйственных продуктов в качестве традиционного сырья внимание было привлечено к целлюлозной биомассе, включая сельскохозяйственные отходы или отходы деревообрабатывающей промышленности, промышленные отходы и так далее, как к идеальному сахарному сырью для ферментации, поскольку она обладает преимуществами низкой стоимости и широкой распространенности.
Среди них ксилоза является вторым по распространенности лигноцеллюлозным углеводом в природе и представляет собой типичную целлюлозную биомассу. С использованием промышленных микроорганизмов из ксилозы были получены полезные вещества. Например, известен способ получения L-аминокислот посредством культивирования штамма Escherichia sp. в среде, содержащей смесь пентоз, включая глюкозу и ксилозу, где штамм модифицирован для увеличения экспрессии генного кластера xylABFGHR, кодирующего фермент (ксилозидазу), гидролизующий ксилозид, который представляет собой гликозид, имеющий происхождение от ксилозы, и затем выделения L-аминокислоты из среды (патент Японии №4665567).
С другой стороны, дифтериеподобная бактерия Corynebacterium glutamicum известна как грам-положительный штамм, используемый для получения различных L-аминокислот. Как описано выше, поскольку ксилоза является вторым по распространенности лигноцеллюлозным углеводом в природе, ожидают, что L-аминокислоты, такие как L-лизин, могут быть более экономично получены из Corynebacterium glutamicum с использованием ксилозы. Тем не менее, у Corynebacterium glutamicum отсутствуют важные гены, вовлеченные в метаболический путь ксилозы, которая является пентозой, и, таким образом, существует проблема, которая заключается том, что L-аминокислоты не могут быть получены из Corynebacterium glutamicum с использованием ксилозы. Для решения этой проблемы было сообщено о модификации Corynebacterium glutamicum таким образом, чтобы она была способна утилизировать ксилозу, посредством введения ксилозоизомеразы (XylA) и ксилулокиназы (XylB), имеющих происхождение из Escherichia coli (Kawaguchi et al., АЕМ 72:3418-3428, 2006).
Авторы настоящего изобретения приложили значительные усилия для получения L-аминокислот более экономичным способом, и в результате обнаружили, что при введении генов, имеющих происхождение из Erwinia carotovora, кодирующих XylA и XylB, в Corynebacterium glutamicum, такой вариант способен утилизировать ксилозу для продуцирования L-лизина и также демонстрирует улучшенную способность утилизировать ксилозу по сравнению с ранее известным дифтериеподобным микроорганизмом, в который введены xylA и xylB, имеющие происхождение из Escherichia coli, в чем и заключается настоящее изобретение.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задачей настоящего изобретения является обеспечение модифицированного микроорганизма Corynebacterium sp., способного продуцировать L-лизин посредством утилизации ксилозы.
Другой задачей настоящего изобретения является обеспечение способа получения L-лизина с применением модифицированного микроорганизма Corynebacterium sp.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 показана карта рестрикции вектора экспрессии pECCG122-pcj7-xylAB(Er) по настоящему изобретению.
На ФИГ. 2 показан график, на котором представлены характеристики роста родительского штамма и трансформанта с введенным вектором экспрессии, в соответствии с источником углерода, присутствующим в среде.
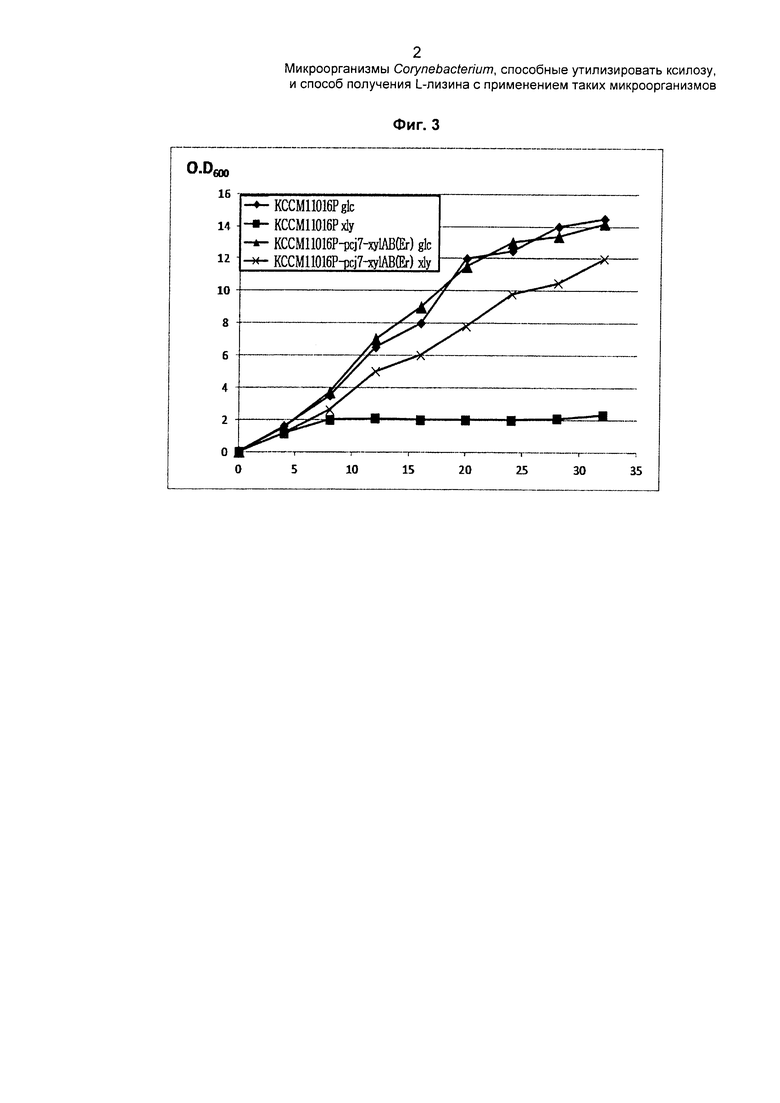
На ФИГ. 3 показан график, на котором представлены характеристики роста родительского штамма и трансформанта, где в хромосому введен pcj7-xylAB(Er), в соответствии с источником углерода, содержащимся в среде.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
В одном аспекте согласно настоящему изобретению предложен модифицированный микроорганизм Corynebacterium sp., способный продуцировать L-лизин посредством утилизации ксилозы, характеризующийся тем, что в него введены активности ксилозоизомеразы и ксилулокиназы, имеющие происхождение из Erwinia carotovora.
При использовании здесь термин «ксилозоизомераза (XylA)» означает фермент, катализирующий реакцию изомеризации ксилозы с образованием ксилулозы, которая вовлечена в метаболический путь ксилозы, и относительно задачи настоящего изобретения, этот фермент может представлять собой фермент, имеющий происхождение из Erwinia carotovora.
XylA представляет собой, без ограничения, ксилозоизомеразу, имеющую происхождение из Erwinia carotovora, и может включать последовательность, способную придавать способность утилизировать ксилозу при введении ее активности совместно с активностью ксилулокиназы, имеющей происхождение из Erwinia carotovora, в микроорганизм Corynebacterium sp., не имеющий ксилозоизомеразной активности. Кроме того, очевидно, что любая последовательность, имеющая активность, эквивалентную активности вышеуказанной последовательности, даже если она не имеет происхождения из Erwinia carotovora, включена в объем настоящего изобретения.
Например, возможно включение аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, содержащей консервативную последовательность аминокислотной последовательности SEQ ID NO: 1 и замену, делецию, вставку, присоединение или инверсию одной или нескольких аминокислот (может варьировать в зависимости от положений и типов аминокислотных остатков в трехмерной структуре белка, конкретно от 2 до 20, конкретно от 2 до 10, конкретнее от 2 до 5 аминокислот) в одном или более чем одном положении. При условии сохранения или усиления активности XylA возможно включение аминокислотной последовательности, на 80% или более, конкретнее на 90% или более, конкретнее на 95% или более, еще конкретнее на 97% или более гомологичной аминокислотной последовательности SEQ ID NO: 1, и замена, делеция, вставка, присоединение или инверсия аминокислот также включают видоизмененную последовательность, встречающуюся в природе у микроорганизма, имеющего активность XylA, или искусственно модифицированную последовательность.
При использовании здесь термин «гомология» относится к идентичности между двумя различными аминокислотными последовательностями или двумя различными нуклеотидными последовательностями, и она может быть определена способом, хорошо известным специалистам в данной области техники, например, без ограничения, BLAST 2.0, позволяющим рассчитывать такие параметры, как общий балльный показатель, идентичность и сходство.
При использовании здесь термин «ксилулокиназа» означает фермент, катализирующий реакцию образования ксилулозо-5-фосфата из ксилулозы, которая вовлечена в метаболический путь ксилозы, и относительно задачи настоящего изобретения, он может представлять собой фермент, имеющий происхождение из Erwinia carotovora.
XylB представляет собой, без ограничения, ксилулокиназу, имеющую происхождение из Erwinia carotovora, и может включать последовательность, способную придавать способность утилизировать ксилозу при введении ее активности совместно с активностью ксилозоизомеразы, имеющей происхождение из Erwinia carotovora, в микроорганизм Corynebacterium sp., не имеющий ксилулокиназной активности. Кроме того, очевидно, что любая последовательность, имеющая активность, эквивалентную активности указанной выше последовательности, даже если она не имеет происхождения из Erwinia carotovora, включена в объем настоящего изобретения.
Например, возможно включение аминокислотной последовательности SEQ ID NO: 2, или аминокислотной последовательности, содержащей консервативную последовательность аминокислотной последовательности SEQ ID NO: 2 и замену, делецию, вставку, присоединение или инверсию одной или нескольких аминокислот (может варьировать в зависимости от положений и типов аминокислотных остатков в трехмерной структуре белка, конкретно от 2 до 20, конкретно от 2 до 10, конкретнее от 2 до 5 аминокислот) в одном или более чем одном положении. При условии сохранения или усиления активности XylB возможно включение аминокислотной последовательности, на 80% или более, конкретнее на 90% или более, конкретнее на 95% или более, еще конкретнее на 97% или более гомологичной аминокислотной последовательности SEQ ID NO: 2, и замена, делеция, вставка, присоединение или инверсия аминокислот также включает видоизмененную последовательность, встречающуюся в природе у микроорганизма, имеющего активность XylB, или искусственно модифицированную последовательность.
При использовании здесь термин «ген, кодирующий ксилозоизомеразу (XylA)» (далее xylA) означает полинуклеотид, кодирующий описанную выше XylA.
Ген может включать нуклеотидную последовательность SEQ ID NO: 3, нуклеотидную последовательность, которая может гибридизоваться с зондом, имеющим происхождение от нуклеотидной последовательности SEQ ID NO: 3, в «строгих условиях», или нуклеотидную последовательность, в которой один или несколько нуклеотидов замещен(ы), удален(ы), вставлен(ы) или добавлен(ы) в одном или более чем одном положении нуклеотидной последовательности SEQ ID NO: 3. При условии сохранения или усиления активности XylA ген может содержать, без ограничения, нуклеотидную последовательность, на 80% или более, конкретнее на 90% или более, конкретнее на 95% или более, еще конкретнее на 97% или более гомологичную нуклеотидной последовательности SEQ ID NO: 3, нуклеотидную последовательность, замещенную кодонами, подходящими для клеток-хозяев, нуклеотидную последовательность, N-конец или С-конец которой удлинен или удален, или нуклеотидную последовательность, в которой старт-кодон модифицирован для контроля уровня экспрессии.
При использовании здесь термин «ген, кодирующий ксилулокиназу (XylB)» (далее xylB) означает полинуклеотид, кодирующий описанную выше XylB.
Ген может содержать нуклеотидную последовательность SEQ ID NO: 4, нуклеотидную последовательность, которая может гибридизоваться с зондом, имеющим происхождение от нуклеотидной последовательности SEQ ID NO: 4, в «строгих условиях», или нуклеотидную последовательность, в которой один или несколько нуклеотидов замещен(ы), удален(ы), вставлен(ы) или добавлен(ы) в одном или более чем одном положении нуклеотидной последовательности SEQ ID NO: 4. При условии сохранения или усиления активности XylB, ген может содержать, без ограничения, нуклеотидную последовательность, на 80% или более, конкретнее на 90% или более, конкретнее на 95% или более, еще конкретнее на 97% или более гомологичную нуклеотидной последовательности SEQ ID NO: 4, нуклеотидную последовательность, замещенную кодонами, подходящими для клеток-хозяев, нуклеотидную последовательность, N-конец или С-конец которой удлинен или удален, или нуклеотидную последовательность, в которой старт-кодон модифицирован для контроля уровня экспрессии.
При использовании здесь термин «строгие условия» означает условия, которые позволяют проводить специфическую гибридизацию полинуклеотидов, например гибридизацию в буфере для гибридизации при 65°С (3,5×SSC (0,15 M NaCl/0,15M цитрат натрия, рН 7,0), 0,02% Фиколл, 0,02% поливинилпирролидон, 0,02% бычий сывороточный альбумин, 0,5% SDS (додецилсульфат натрия), 2 мМ EDTA (этилендиаминтетрауксусная кислота), 2,5 мМ NaН2РO4, рН 7), и подробное описание приведено в предшествующем уровне техники (Molecular Cloning (A Laboratory Manual, J. Sambrook et al., Editors, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989) или в Current Protocols in Molecular Biology (F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York)).
Как описано выше, введение активностей XylA и XylB в микроорганизм Corynebacterium sp.можно осуществлять различными способами, хорошо известными в данной области техники. Например, существует способ введения полинуклеотида, содержащего нуклеотидные последовательности, кодирующие XylA и XylB, в хромосому, способ введения векторной системы, содержащей полинуклеотид, в микроорганизм, способ введения активного промотора перед нуклеотидными последовательностями, кодирующими XylA и XylB, способ введения xylA и xylB с модифицированным промотором, или способ введения модифицированных нуклеотидных последовательностей, кодирующих XylA и XylB, и тому подобное. Конкретнее, при введении нуклеотидных последовательностей, кодирующих XylA и XylB, для контроля их экспрессии может быть использован промотор pcj7, имеющий происхождение из Corynebacterium ammoniagenes (патент Кореи №10-0620092). В одном воплощении настоящего изобретения приобретение способности утилизировать ксилозу подтверждали как активность чужеродного гена, отсутствующего в родительском штамме, наблюдаемую после введения вектора экспрессии или хромосомной вставки.
При использовании здесь термин «вектор» относится к ДНК-продукту, имеющему нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, функционально связанный с подходящей регуляторной последовательностью, для экспрессии целевого белка в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, произвольную последовательность оператора для регуляции транскрипции, последовательность, кодирующую сайт связывания рибосомы с подходящей мРНК, и последовательности, регулирующие терминацию транскрипции и трансляции. После трансформации подходящего хозяина возможны репликация или функционирование вектора независимо от генома хозяина или его интеграция в сам геном.
Вектор, используемый в настоящем изобретении, не ограничен конкретными примерами и может представлять собой любой вектор, известный в данной области техники, при условии, что возможна его репликация в хозяине. Примеры обычно используемых векторов представляют собой встречающиеся в природе или рекомбинантные плазмиды, косм иды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, λMBL3, λMBL4, λΙΧΙΙ, λASHII, λΑΡΙΙ, λt10, λt11, Charon4A, Charon21A или тому подобное. В качестве плазмидного вектора можно использовать вектор типа pBR, типа pUC, типа pBluescriptll, типа pGEM, типа pTZ, типа pCL, типа рЕТ или тому подобное.
Вектор, полезный в настоящем изобретении, не ограничен какими-либо конкретными примерами, и может быть использован любой известный вектор экспрессии. Конкретно, можно использовать векторы pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCCIBAC или тому подобное.
Кроме того, вектор, используемый в настоящем изобретении, может представлять собой вектор, позволяющий трансформировать клетки хозяина для вставки полинуклеотида, кодирующего целевой белок, в хромосому клетки-хозяина. Конкретными примерами таких векторов являются, без ограничения, шаттл-вектор pECCG112, который может самореплицироваться в обоих направлениях в Е. coli и/или в дифтериеподобных бактериях (патент Кореи №10-0057684).
При использовании здесь термин «трансформация» означает серию процедур для введения вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина для экспрессии кодируемого полинуклеотидом белка в клетке-хозяине. Полинуклеотид, вводимый в клетку-хозяина, может иметь любую форму, при условии, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме кассеты экспрессии, которая представляет собой структуру, включающую все элементы (промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосом, сигнал терминации трансляции и так далее), необходимые для самоэкспрессии. Кассета экспрессии может быть в форме самореплицирующегося вектора экспрессии. Кроме того, полинуклеотид может быть введен в клетку-хозяина сам по себе для функционального связывания с последовательностью, необходимой для экспрессии в клетке-хозяине.
Клетка-хозяин может представлять собой любой прокариотический микроорганизм, при условии, что он способен продуцировать L-лизин. Примеры клеток-хозяев могут включать Providencia sp., Corynebacterium sp. и микроорганизм Brevibacterium sp., конкретнее микроорганизм Corynebacterium sp. и еще конкретнее Corynebacterium glutamicum. В одном воплощении настоящего изобретения при введении XlyA и XlyB, имеющих происхождение из Erwinia carotovora, в штаммы КССМ11016Р, КССМ10770Р, KFCC10750 и CJ3P в качестве микроорганизма Corynebacteríum sp., не способные утилизировать ксилозу, им придают способность утилизировать ксилозу, и в результате они могут продуцировать L-аминокислоты, такие как L-лизин, посредством утилизации ксилозы в качестве источника углерода (Таблицы 1-6).
Микроорганизм Corynebacterium sp., способный продуцировать L-лизин, может представлять собой вариант, устойчивый к аналогу L-лизина. Аналог L-лизина ингибирует рост дифтериеподобных микроорганизмов, но такое ингибирование полностью или частично снижено, когда в среде также присутствует L-лизин. Примеры аналогов L-лизина включают, без ограничения, окса-L-лизин, гидроксамат L-лизина, S-(2-аминоэтил)-L-цистеин (АЕС), γ-метил L-лизин, α-хлоркапролактам и тому подобное. Вариант, обладающий устойчивостью к этим аналогам L-лизина, может быть получен традиционным искусственным мутагенезом дифтериеподобных организмов. Кроме того, при генетических манипуляциях, осуществляемых для получения микроорганизма, продуцирующего L-лизин, этот эффект может быть достигнут посредством улучшения экспрессии одного или более генов, кодирующих ферменты, вовлеченные в систему биосинтеза L-лизина. Примеры таких генов могут включать, без ограничения, ген дигидродипиколинатсинтазы (dapA), ген аспартокиназы (lysC), ген дигидродипиколинатредуктазы (dapB), ген диаминопимелатдекарбоксилазы (lysA), ген диаминопимелатдегидрогеназы (ddh), ген фосфоенолпируваткарбоксилазы (ррс), ген аспартатполуальдегиддегидрогеназы (asd) и ген аспартазы (aspA).
При использовании здесь термин «L-лизин» представляет собой одну из основных α-аминокислот, представляет собой незаменимую аминокислоту, не синтезирующуюся в организме, представляет собой одну из L-аминокислот и имеет химическую формулу NH2(CH2)4CH(NH2)COOH. L-лизин синтезируется из оксалоацетата в биосинтетическом пути L-лизина, и промежуточный процесс биосинтеза L-лизина катализирует NADPH-зависимая редуктаза. В процессе биосинтеза 1 молекулы L-лизина 3 молекулы NADPH непосредственно используются соответствующими ферментами и 1 молекула NADPH используется косвенно.
При использовании здесь термин «микроорганизм Corynebacterium sp., способный продуцировать L-лизин» означает микроорганизм Corynebacterium sp., модифицированный для получения L-лизина из ксилозы, что достигается введением генов, кодирующих ферменты, вовлеченные в метаболизм ксилозы и отсутствующие в микроорганизме Corynebacterium sp.Микроорганизм Corynebacterium sp. может представлять собой, без ограничения, Corynebacterium glutamicum, и ферменты, вовлеченные в метаболизм ксилозы, могут представлять собой, без ограничения, XylA и XylB.
В связи с этим, клетка-хозяин может представлять собой микроорганизм Corynebacterium sp., в котором улучшена экспрессия одного или более генов, кодирующих ферменты, вовлеченные в систему биосинтеза L-лизина, и гены, кодирующие ферменты, вовлеченные в систему биосинтеза L-лизина, могут представлять собой, без ограничения, ген дигидродипиколинатсинтазы (dapA), ген аспартокиназы (lysC), ген дигидродипиколинатредуктазы (dapB), ген диаминопимелатдекарбоксилазы (lysА), ген диаминопимелатдегидрогеназы (ddh), ген фосфоенолпируваткарбоксилазы (ррс), ген аспартатполуальдегиддегидрогеназы (asd), ген аспартазы (aspA) или тому подобное.
Кроме того, клетка-хозяин может представлять собой мутантный штамм, устойчивый к аналогу L-лизина. Мутантный штамм может быть получен мутацией микроорганизма Corynebacterium sp. Аналог L-лизина ингибирует рост дифтериеподобного микроорганизма, но такое ингибирование полностью или частично снижено, когда в среде также присутствует L-лизин. Примеры аналогов L-лизина включают, без ограничения, предпочтительно окса-L-лизин, гидроксамат L-лизина, S-(2-аминоэтил)-L-цистеин (АЕС), γ-метил L-лизин, α-хлоркапролактам и тому подобное.
В то же время, в настоящем изобретении активности известных ферментов, вовлеченных в биосинтез L-лизина, можно дополнительно контролировать, чтобы дополнительно улучшить продуцирование L-лизина. Конкретно, в настоящем изобретении гены asd, dapB, и ddh, каждый из которых кодирует аспартатполуальдегиддегидрогеназу, дигидродипиколинатредуктазу и диаминопимелатдегидрогеназу, сверхэкспрессированы для дополнительного контроля активностей ферментов, таким образом улучшая продуцирование L-лизина.
Согласно одному воплощению настоящего изобретения, авторы настоящего изобретения отобрали гены ЕСА0097(ху/А) (SEQ ID NO: 1), имеющий происхождение из Erwinia carotovora (SCRI1043), и ECA0096(xylB) (SEQ ID NO: 2) как подходящие гены, кодирующие XylA и XylB, для введения в микроорганизм Corynebacterium sp.(Пример 1), и клонировали отобранные гены, кодирующие XylA и XylB, чтобы сконструировать вектор экспрессии ECCG122-pcj7-xylA-xylB (далее pECCG122-pcj7-xylAB(Er)), одновременно экспрессирующий xylA и xylB (далее хуlAB(Еr)). Вектор экспрессии вводили в Corynebacterium glutamicum КССМ11016Р (этот микроорганизм раскрыт как KFCC10881, и повторно депонирован в Международном органе по депонированию согласно Будапештскому договору под регистрационным номером КССМ11016Р, патенты Кореи №№10-0159812 и 10-0397322) для получения трансформанта, сверхэкспрессирующего хуlAB(Еr). Было подтверждено, что полученный трансформант растет, утилизируя ксилозу в качестве источника углерода (ФИГ. 2), и продуцирует L-лизин посредством утилизации как глюкозы, так и ксилозы по отдельности или посредством утилизации глюкозы и ксилозы одновременно (Таблица 1). Кроме того, для экспрессии хуlАВ(Еr) на хромосоме, конструировали рекомбинантный вектор pDZTn-pcj7-xylAB(Er) для вставки в хромосому, и трансформировали им КССМ11016Р, и посредством второго кроссинговера конструировали трансформант KCCM11016P-pcj7-xylAB(Er), несущий хуlАВ(Еr), функционально связанный с промотором pcj7 внутри транспозона на хромосоме. Также было подтверждено, что трансформант растет, утилизируя ксилозу в качестве источника углерода (ФИГ. 3) и продуцирует L-лизин посредством утилизации как глюкозы, так и ксилозы по отдельности, или посредством утилизации глюкозы и ксилозы одновременно (Таблица 2). Кроме того, для сравнения эффектов улучшения способности утилизировать ксилозу между введением xylAB (далее хуlАВ(Ес)), имеющих происхождение из Е. Coli, о котором сообщалось ранее, и введением xylAB(Er), имеющем происхождение из Erwinia carotovora, по настоящему изобретению, штамм (KCCM11016P-pcj7-xylAB(Ec)) получали введением хуlAВ(Ес) в КССМ11016Р, и его характеристики - способность утилизировать ксилозу и продуцировать L-лизин - сравнивали с таковыми для полученного KCCM11016P-pcj7-xylAB(Er). В результате было обнаружено, что уровень потребления ксилозы KCCM11016P-pcj7-xylAB(Er) значительно возрастал по сравнению с таковым для KCCM11016P-pcj7-xylAB(Ec), что указывает на улучшение ферментативного получения L-лизина (Таблица 3). Кроме того, для подтверждения того, что различные микроорганизмы Corynebacterium sp. демонстрируют одинаковые результаты, в штамм КССМ10770Р, продуцирующий L-лизин, вводили pDZTn-pcj7-xylAB(Er) для получения трансформанта KCCM10770P-pcj7-xylAB(Er), и подтверждали, что трансформант способен продуцировать L-лизин посредством утилизации как глюкозы, так и ксилозы по отдельности, или посредством утилизации глюкозы и ксилозы одновременно (Таблица 4). pDZTn-pcj7-xy/A8(Er) также вводили в другой штамм KFCC10750, продуцирующий L-лизин (этот микроорганизм раскрыт как KFCC10750, и повторно депонирован в Международном органе по депонированию согласно Будапештскому договору под регистрационным номером КССМ11347Р, патент Кореи №10-0073610) для получения трансформанта KFCC10750-pcj7-xylAB(Er). Также подтверждали, что этот трансформант способен продуцировать L-лизин посредством утилизации как глюкозы, так и ксилозы по отдельности, или посредством утилизации глюкозы и ксилозы одновременно (Таблица 5). Кроме того, pDZTn-pcj7-xylAB(Er) также вводили в другой штамм CJ3P, продуцирующий L-лизин, для получения трансформанта CJ3P-pcj7-xylAB(Er). Также подтверждали, что этот трансформант способен продуцировать L-лизин посредством утилизации как глюкозы, так и ксилозы по отдельности, или посредством утилизации глюкозы и ксилозы одновременно (Таблица 6).
Соответственно, авторы настоящего изобретения обозначили трансформант, который растет, утилизируя ксилозу в среде, и также продуцирует L-лизин посредством утилизации ксилозы и глюкозы в среде, как «СА01-2195» и депонировали его согласно Будапештскому договору в Корейском центре культур микроорганизмов (КССМ, расположен в Hongjae 1-Dong, Seodaemun-Gu, Сеул, Корея) 29 декабря 2011, под регистрационным номером № КССМ11242Р. Таким образом, этот депонированный организм признан Международным органом по депонированию согласно Будапештскому договору.
В другом аспекте настоящего изобретения предложен способ получения L-лизина, включающий стадии (1) культивирования модифицированного микроорганизма Corynebacterium sp., способного продуцировать L-лизин посредством утилизации ксилозы в культуральной среде, содержащей ксилозу в качестве источника углерода, с получением культуральной жидкости; и (2) выделения L-лизина из культуральной жидкости.
При использовании здесь термин «культивирование» означает, что микроорганизм культивируют в искусственно контролируемых условиях окружающей среды. В настоящем изобретении способ культивирования микроорганизма Corynebacterium sp. может быть осуществлен с применением способа, широко известного в области техники. Конкретно, примеры способов культивирования включают, без ограничения, периодическое культвирование, культивирование с подпиткой или повторяющееся непрерывное культивирование с подпиткой.
Среда, применяемая для культивирования, должна надлежащим образом соответствовать требованиям конкретного микроорганизма, одновременно с контролем температуры, рН и так далее, в аэробных условиях в типичной среде, содержащей надлежащий источник углерода, источник азота, аминокислоты, витамины и так далее. Культуральные среды для штамма Corynebacterium раскрыты, например, в Manual of Methods for General Bacteriology, American Society for Bacteriology, Washington D.C., USA, 1981. Возможные источники углерода могут включать сахара и углеводы, такие как сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза, в дополнение к смеси глюкозы и ксилозы в качестве основного источника углерода, масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло, жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота. Эти вещества могут быть использованы по отдельности или в смеси. Возможные источники азота могут включать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; аминокислоты, такие как глутаминовая кислота, метионин и глутамин; и органические источники азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизаты казеина, рыба и продукты ее распада, и обезжиренный соевый жмых или продукты его распада. Эти источники азота могут быть использованы по отдельности или в комбинации. Среда может содержать дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли в качестве источников фосфора. Возможные источники фосфора могут включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли. Кроме того, могут быть использованы неорганические соединения, такие как хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца и карбонат кальция. В дополнение к вышеперечисленным веществам, могут быть включены важные для роста вещества, такие как аминокислоты и витамины.
В культуральную среду также могут быть добавлены подходящие предшественники. Вышеперечисленные вещества могут подходящим образом быть добавлены в культуральную среду, без ограничения, партией, партией с подпиткой или непрерывно в течение культивирования. рН культуры может быть скорректирован подходящим добавлением основных соединений, таких как гидроксид натрия, гидроксид калия и аммиак, или кислотных соединений, таких как фосфорная кислота или серная кислота.
Пеногаситель, такой как сложный эфир полигликоля и жирной кислоты, можно применять для подавления образования пены. Для поддержания аэробных условий в культуральную жидкость можно вводить кислород или кислородосодержащий газ (например, воздух). Температура культуральной жидкости в норме составляет от 27°С до 37°С, конкретно от 30°С до 35°С. Культивирование можно продолжать до тех пор, пока продуцирование L-лизина не достигнет желаемого уровня. Это обычно достигается за период от 10 до 100 часов. L-лизин может выделяться в культуральную среду или быть включен в клетки.
Более того, стадию выделения L-лизина из культуральной жидкости можно осуществлять способами, известными в области техники. Конкретно, известный способ выделения L-лизина представляет собой, без ограничения, центрифугирование, фильтрацию, экстракцию, распыление, сушку, испарение, осаждение, кристаллизацию, электрофорез, дифференциальную растворимость (например, осаждение сульфатом аммония), хроматографию (например, ионообменную, аффинную, гидрофобную и гель-хроматографию) или тому подобное.
Далее сущность и эффекты настоящего изобретения будут описаны более подробно в Примерах. Тем не менее, эти Примеры приведены только в иллюстративных целях, и подразумевают, что эти Примеры не ограничивают объем изобретения.
Пример 1: Выбор чужеродного гена
ЕСА0097(хуlА) (аминокислотная последовательность: SEQIDNO:1, нуклеотидная: SEQ ID NO: 3) и ECA0096(xylB) (аминокислотная последовательность: SEQ ID NO: 2, нуклеотидная: SEQ ID NO: 4), имеющие происхождение из Erwinia carotovora (SCRI1043), были выбраны в качестве чужеродных генов для получения модифицированного микроорганизма Corynebacterium sp., способного утилизировать ксилозу.
Пример 2: Конструирование вектора экспрессии xvlAB, имеющего происхождение из Erwinia carotovora
Гены, кодирующие XylA и XylB, имеющие происхождение из Erwinia carotovora, отобранные в Примере 1, расположены рядом друг с другом. Информацию (Номер доступа № ВХ 950851) о хуlA и хуlВ(Еr) и окружающей нуклеотидной последовательности получали из US NIH GenBank и на основании полученной нуклеотидной последовательности синтезировали праймеры для амплификации хуlАв(Еr), имеющего происхождение из Erwinia carotovora.
SEQ ID NO: 5: 5′-ACACATATGCAAGCCTATTTTGAACAGATC-3′
SEQ ID NO: 6: 5′-AGAACTAGTGCCTTTTGGTGGTGTTTAAGT-3′
Для получения фрагмента хуlАB(Еr) проводили ПЦР (полимеразная цепная реакция), используя хромосомную ДНК штамма SCRI1043 Erwinia carotovora в качестве матрицы и пару праймеров (SEQ ID NO: 5 и 6). В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene) и ПЦР проводили с денатурацией при 94°С в течение 5 минут с последующим повторяющимся 30 раз циклом, включающим денатурацию при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и полимеризацию при 72°С в течение 3 минут, и затем с полимеризацией при 72°С в течение 7 минут. В результате получали генный фрагмент 3122 п.о., содержащий хуlАB(Еr) (SEQ ID NO: 17) размером 2844 п.о. (SEQ ID NO: 18). Для получения промотора pcj7, имеющего происхождение из Corynebacterium ammoniagenes (KR0620092), проводили ПЦР, используя геномную ДНК Corynebacterium ammoniagenes CJHB100 (KR0620092) в качестве матрицы и пару праймеров (SEQ ID NO: 15 и 16). В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene), и ПЦР проводили с денатурацией при 94°С в течение 5 минут, с последующим повторяющимся 30 раз циклом, включающим денатурацию при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд с полимеризацией при 72°С в течение 1 минуты, и затем с полимеризацией при 72°С в течение 7 минут. В результате получали полинуклеотид размером 318 п.о. (SEQ ID NO: 14).

pcj7 клонировали в шаттл-вектор pECCG122 в Е. coli-Corynebacterium, используя ферменты рестрикции Xbal и Spel, и затем фрагмент хуlАВ(Еr) клонировали, используя Ndel и Spel, получая таким образом вектор pECCG122-pcj7-xylAB(Er) (ФИГ. 1). На ФИГ. 1 показана карта рестрикции экспрессионного вектора pECCG122-pcj7-xylAB(Er) по настоящему изобретению.
Пример 3: Разработка штамма, продуцирующего L-лизин, с введенным xvlAB, имеющим происхождение из Erwinia carotovora, и оценка способности утилизировать ксилозу
Каждый вектор экспрессии рЕСС0122-рсj7-хуlАB(Еr), полученный в Примере 2, вводили в Corynebacterium glutamicum КССМ11016Р (патенты Кореи №№10-0159812 и 10-0397322) для получения трансформанта Corynebacterium glutamicum СА01-2195, экспрессирующего xylAB(Er).
Для сравнения способности утилизировать ксилозу между КССМ11016Р и СА01-2195, штаммы культивировали в среде для посева, содержащей глюкозу или ксилозу в качестве источника углерода, и сравнивали их характеристики роста. Их также культивировали в среде для получения продукта, содержащей глюкозу или ксилозу в качестве источника углерода, и сравнивали их характеристики продуцирования L-лизина.
Сначала, для сравнения характеристик роста, штаммы высевали в 25 мл среды для посева [источник углерода (глюкоза или ксилоза) 10 г/л, пептон 10 г/л, дрожжевой экстракт 10 г/л, мочевина 5 г/л, КН2РO4 4 г/л, К2НРO4 8 г/л, MgSO47H2O 0,5 г/л, биотин 100 мкг/л, тиамин-HCl 1 мг/л, рН 7.0], соответственно. Поглощение (OD600) культуральной жидкости измеряли в течение культивирования штаммов при 30°С в течение 32 часов и сравнивали друг с другом (ФИГ. 2). ФИГ.2 представляет собой график, на котором показаны характеристики роста КССМ11016Р и СА01-2195, в соответствии с источником углерода, содержащимся в среде, где (♦) обозначает КССМ11016Р, культивированный в среде, содержащей глюкозу, (■) обозначает КССМ11016Р, культивированный в среде, содержащей ксилозу, (▲) обозначает СА01-2195, культивированный в среде, содержащей глюкозу, и (×) обозначает СА01-2195, культивированный в среде, содержащей ксилозу.
Как показано на ФИГ. 2, никаких различий в характеристиках роста между КССМ11016Р и СА01-2195 в среде для посева, содержащей глюкозу в качестве источника углерода, не было. Однако в среде для посева, содержащей ксилозу в качестве источника углерода, СА01-2195 рос до определенного уровня, в то время как рост КССМ11016Р был затруднен. Таким образом, видно, что СА01-2195 способен расти, утилизируя ксилозу, содержащуюся в среде в качестве единственного источника углерода.
Затем, для сравнения характеристик продуцирования L-лизина КССМ11016Р и СА01-2195, 1 мл культуральной жидкости для посева высевали в 24 мл среды для получения продукта [источник углерода, (NH4)2SO4 40 г/л, соевый белок 2,5 г/л, твердый кукурузный экстракт 5 г/л, мочевина 3 г/л, КН2РО4 1 г/л, MgSO47H2O 0,5 г/л, биотин 100 мкг/л, тиамин-HCl 1 мг/л, СаСO3 30 г/л, рН 7,0], и культивировали при 35°С и 200 об/мин в течение 72 часов. В это время использовали глюкозу 100 г/л, глюкозу 50 г/л плюс ксилозу 50 г/л, и глюкозу 70 г/л плюс ксилозу 30 г/л качестве источника углерода. Далее, измеряли и сравнивали концентрацию L-лизина, концентрацию оставшейся ксилозы и концентрацию оставшейся глюкозы в каждой культуральной жидкости (Таблица 1).

прекращения реакции)
Как показано в Таблице 1, при использовании среды, не содержащей ксилозу (глюкоза 100 г/л), концентрация L-лизина, продуцированного родительским штаммом, была эквивалентна концентрации L-лизина, продуцированного штаммом СА01-2195. Однако когда использовали среду, содержащую ксилозу (глюкоза 50 г/л плюс ксилоза 50 г/л, и глюкоза 70 г/л плюс ксилоза 30 г/л), СА01-2195 продуцировал L-лизин, потребляя как глюкозу, так и ксилозу, в то время как родительский штамм продуцировал L-лизин, потребляя только глюкозу, но не ксилозу.
Данный результат указывает на то, что микроорганизм Corynebacterium sp., не способный утилизировать ксилозу, способен потреблять ксилозу после введения xylAB, имеющего происхождение из Erwinia carotovora.
Пример 4: Конструирование рекомбинантного вектора (pDZTn-pcj7-xylAB(Er)) для вставки в хромосому xylAB, имеющего происхождение из Erwinia carotovora, и рекомбинантного вектора (pDZTn-pcj7-xylAB(Ec)) для вставки в хромосому xylAB, имеющего происхождение из Е. соli
Для экспрессии хуlАB(Еr) на хромосоме конструировали рекомбинантный вектор pDZTn-pcj7-xylAB(Er) для вставки в хромосому. Для получения фрагмента pcj7-xylAB(Er), проводили ПЦР, используя pECCG122-pcj7-xylAB(Er), полученный в Примере, в качестве матрицы и пару праймеров (SEQ ID NO: 7 и 8). В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene), и ПЦР проводили с денатурацией при 94°С в течение 5 минут, с последующим повторяющимся 30 раз циклом, включающим денатурацию при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и полимеризацию при 72°С в течение 3 минут, и затем с полимеризацией при 72°С в течение 7 минут. В результате получали генный фрагмент 3440 п.о. (SEQ ID NO: 9). Впоследствии, pcj7-xylAB(Er) 3440 п.о. клонировали в вектор pDZTn (патент Кореи №10-1126041), обработанный ферментом рестрикции Spel, используя набор BD In-Fusion, таким образом получая рекомбинантный вектор pDZTn-pcj7-xylAB(Er).

Далее, для экспрессии xylAB, имеющего происхождение из Е. coli, на хромосоме, конструировали рекомбинантный pDZTn-pcj7-xylAB(Ec) для вставки в хромосому. Для получения промотора pcj7, проводили ПЦР, используя фрагмент pcj7, полученный в Примере 2, в качестве матрицы и пару праймеров (SEQ ID NO: 7 и 10). В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene), и ПЦР проводили с денатурацией при 94°С в течение 5 минут, с последующим повторяющимся 30 раз циклом, включающим денатурацию при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и полимеризацию при 72°С в течение 1 минуты, и затем с полимеризацией при 72°С в течение 7 минут. В результате получали генный фрагмент 318 п.о. Для получения фрагмента хуlАВ(Ес), проводили ПЦР, используя хромосомную ДНК Е. coli К12 в качестве матрицы и пару праймеров (SEQ ID NO: 11 и 12). В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene), и ПЦР проводили с денатурацией при 94°С в течение 5 минут, с последующим повторяющимся 30 раз циклом, включающим денатурацию при 94°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и полимеризацию при 72°С в течение 3 минут, и затем с полимеризацией при 72°С в течение 7 минут. В результате получали фрагмент хуlАВ(Ес) 3145 п.о. (SEQ ID NO: 13). Впоследствии, продукты ПЦР области pcj7 размером 318 п.о. и хуlАВ(Ес) размером 3145 п.о. клонировали в вектор pDZTn, обработанный ферментом рестрикции Spel, используя набор BD In-Fusion, таким образом получая итоговый рекомбинантный вектор pDZTn-pcj7-xylAB(Ec).
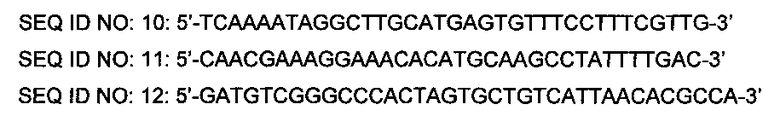
Пример 5: Разработка штамма, продуцирующего L-лизин. с введенным xylAB, имеющим происхождение из Erwinia, и оценка способности утилизировать ксилозу
Полученным вектором pDZTn-pcj7-xylAB(Er) трансформировали КССМ11016Р, и посредством второго кроссинговера получали КССМ11016Р-pcj7-xylAB(Er), несущий хуlАВ(Еr) с заменой одной копии промотора pcj7 внутри транспозона на хромосоме.
Для сравнения способности утилизировать ксилозу между КССМ11016Р-pcj7-xylAB(Er) и КССМ11016Р, штаммы культивировали в среде для посева, содержащей глюкозу или ксилозу в качестве источника углерода, и сравнивали их характеристики роста, и штаммы культивировали в среде для получения продукта, содержащей глюкозу или ксилозу в качестве источника углерода, и сравнивали их характеристики продуцирования L-лизина так же, как в Примере 3.
ФИГ. 3 представляет собой график, на котором показаны характеристики роста штаммов, в соответствии с источником углерода, содержащимся в среде, где (♦) обозначает КССМ11016Р, культивированный в среде, содержащей глюкозу, (■) обозначает КССМ11016Р, культивированный в середе, содержащей ксилозу, (▲) обозначает KCCM11016P-pcj7-xylAB(Er), культивированный в среде, содержащей глюкозу, и (×) обозначает KCCM11016P-pcj7-xylAB(Er), культивированный в среде, содержащей ксилозу.
Различий в характеристиках роста между KCCM11016P-pcj7-xylAB(Er) и КССМ11016Р в среде для посева, содержащей глюкозу в качестве источника углерода, не было. Однако, в среде для посева, содержащей ксилозу в качестве источника углерода, KCCM11016P-pcj7-xylAB(Er) рос до определенного уровня, тогда как рост КССМ11016Р был затруднен. Таким образом, можно увидеть, что при вставке хуlАВ(Еr) в хромосому штамм был способен расти, утилизируя ксилозу, содержащуюся в среде.
Затем оценивали характеристики продуцирования L-лизина KCCM11016P-pcj7-xylAB(Er) и КССМ11016Р и сравнивали их друг с другом (Таблица 2).
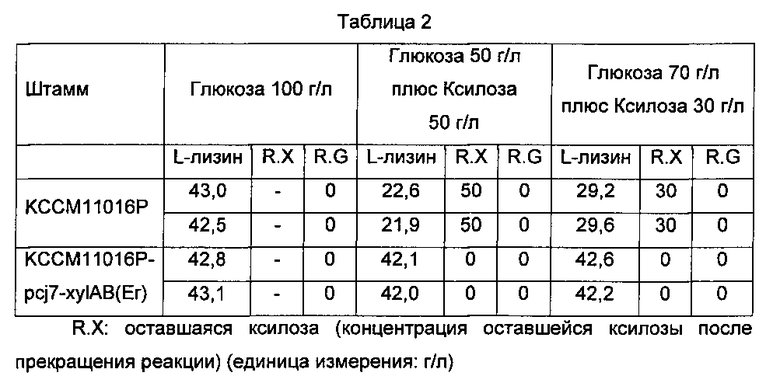
R.G: оставшаяся глюкоза (концентрация оставшейся глюкозы после прекращения реакции)
Как показано в Таблице 2, при использовании среды, не содержащей ксилозу (глюкоза, 100 г/л), концентрация L-лизина, продуцированного КССМ11016Р, была эквивалентна концентрации L-лизина, продуцированного KCCM11016P-pcj7-xylAB(Er). Однако, при использовании среды, содержащей ксилозу (глюкоза 50 г/л плюс ксилоза 50 г/л и глюкоза 70 г/л плюс ксилоза 30 г/л), штамм КССМ11016Р продуцировал L-лизин, потребляя только глюкозу, тогда как KCCM11016P-pcj7-xylAB(Er) продуцировал L-лизин, потребляя как глюкозу, так и ксилозу.
Пример 6: Получение штамма, продуцирующего L-лизин, с введенным xvlAB, имеющим происхождение из Е. соli, и сравнение его способности утилизировать ксилозу со способностью утилизировать ксилозу штамма KCCM11016P-pcj7-xylAB(Ec)
Для сравнения эффектов улучшения способности утилизировать ксилозу между введением ранее известного xylAB, имеющего происхождение из Е. coli, и введением XylAB, имеющего происхождение из Erwinia carotovora, по настоящему изобретению, получали штамм посредством введения xylAB(Ec) в КССМ11016Р, и сравнивали его способность утилизировать ксилозу и характеристики продуцирования L-лизина с таковыми для полученного КССМ11016P-pcj7-xylAB(Er).
Для получения штамма, в который введен хуlAВ(Ес), рекомбинантным вектором pDZTn-pcj7-xylAB(Ec), полученным в Примере 4, трансформировали КССМ11016Р, и посредством второго кроссинговера получали штамм KCCM11016P-pcj7-xylAB(Ec), имеющий хуlАВ(Ес), функционально связанный с промотором pcj7 внутри транспозона на хромосоме.
Так же, как в Примере 3, для сравнения способности использовать ксилозу между KCCM11016P-pcj7-xylAB(Er) и KCCM11016P-pcj7-xylAB(Ec) их культивировали в среде для образования продукта, содержащей 50 г/л глюкозы и 50 г/л ксилозы в качестве источника углерода, и сравнивали их характеристики продуцирования L-лизина и оценивали способность утилизировать ксилозу, измеряли концентрации оставшейся ксилозы в культуральной жидкости через 15 часов (Таблица 3).

Как показано в Таблице 3, два штамма демонстрировали эквивалентную способность продуцировать L-лизин. Скорость потребления ксилозы штаммом KCCM11016P-pcj7-xylAB(Er) была выше, чем штаммом KCCM11016P-pcj7-xylAB(Ec), что указывает на улучшение эффективности продуцирования при ферментации. То есть, данный результат указывает на то, что введение xylAB(Еr), имеющего происхождение из Erwinia carotovora, по настоящему изобретению, демонстрирует более выраженный эффект улучшения эффективности продуцирования L-лизина при ферментации чем введение ранее известного хуlАВ(Ес), имеющего происхождение из Е. coli.
Пример 7: Разработка штамма, имеющего происхождение от КССМ10770Р, с введенным xylAB, имеющим происхождение из Erwinia carotovora, и оценка способности утилизировать ксилозу
Для оценки того, демонстрирует ли введение хуlАВ(Еr) тот же эффект в других штаммах, отличных от КССМ11016Р, продуцирующих L-лизин, pDZTn-pcj7-xylAB(Er) вводили в штамм КССМ 10770Р, продуцирующий L-лизин (патент Кореи №10-0924065), депонированный в Международном органе по депонированию согласно Будапештскому договору. После введения способом электропорации посредством второго кроссинговера был получен штамм, имеющий хуlАВ(Еr) с заменой одной копии промотора pcj7 внутри транспозона на хромосоме, и штамм назвали Corynebacterium glutamicum KCCM10770P-pcj7-xylAB(Er).
Способность Corynebacterium glutamicum KCCM10770P и Corynebacterium glutamicum KCCM10770P-pcj7-xylAB(Er) по настоящему изобретению утилизировать ксилозу и продуцировать L-лизин измеряли таким же образом, как в Примере 3 (Таблица 4).
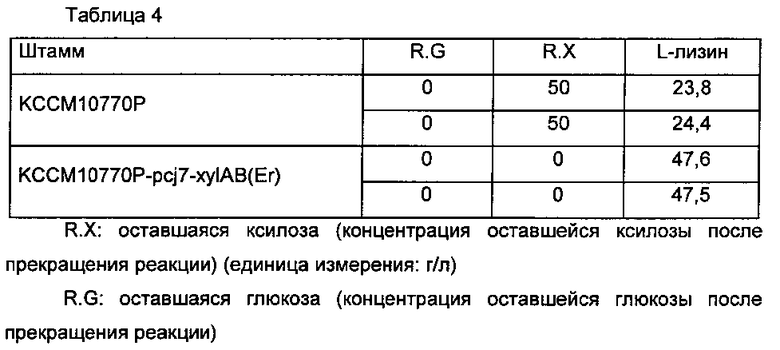
Как показано в Таблице 4, при введении хуlАВ(Еr) в штамм КССМ10770Р, продуцирующий L-лизин, штамм полностью потреблял ксилозу, в отличие от родительского штамма, не способного утилизировать ксилозу, и его способность продуцировать L-лизин также возрастала.
Данный результат демонстрирует, что при введении xylAB, имеющего происхождение из Erwinia carotovora, в различные микроорганизмы Corynebacterium sp., а также в микроорганизмы Corynebacterium sp., определенные конкретным номером доступа, микроорганизмы полностью потребляют ксилозу в качестве источника углерода, тем самым эффективно продуцируя аминокислоты, такие как L-лизин.
Пример 8: Разработка штамма, имеющего происхождение от KFCC10750, с введенным xylAB, имеющим происхождение из Erwinia carotovora, и оценка способности утилизировать ксилозу
Для оценки того, демонстрирует ли введение хуlАВ(Еr) тот же самый эффект в других штаммах, отличных от КССМ11016Р, продуцирующих L-лизин, pDZTn-pcj7-xylAB(Er) вводили в штамм KFCC10750, продуцирующий L-лизин (патент Кореи №10-0073610). После введения способом электропорации посредством второго кроссинговера был получен штамм, имеющий хуlАВ(Ех) с заменой одной копии промотора pcj7 внутри транспозона на хромосоме, и штамм назвали Corynebacterium glutamicum KFCC10750-pcj7-xylAB(Er).
Способность Corynebacterium glutamicum KFCC10750 и Corynebacterium glutamicum KFCC10750-pcj7-xylAB(Er) по настоящему изобретению утилизировать ксилозу и продуцировать L-лизин измеряли таким же образом, как в Примере 3 (Таблица 5).

Как показано в Таблице 5, при введении хуlAВ(Еr) в штамм KFCC10750, образующий L-лизин, штамм полностью потреблял ксилозу, в отличие от родительского штамма, не способного утилизировать ксилозу, и его способность продуцировать L-лизин также возрастала.
Данный результат демонстрирует, что при введении xylAB, имеющего происхождение из Erwinia carotovora, в различные микроорганизмы Corynebacterium sp., а также в микроорганизмы Corynebacterium sp, определенные конкретным номером доступа, они полностью потребляют ксилозу в качестве источника углерода, тем самым эффективно продуцируя аминокислоты, такие как L-лизин.
Пример 9: Разработка штамма, имеющего происхождение от CJ3P, с введенным xylAB, имеющим происхождение из Erwinia carotovora, и оценка способности утилизировать ксилозу
Для оценки того, демонстрирует ли введение хуlАВ(Еr) тот же самый эффект в других штаммах, продуцирующих L-лизин, отличных от КССМ11016Р, pDZTn-pcj7-xylAB(Er) вводили в штамм CJ3P, продуцирующий L-лизин. Штамм CJ3P представляет собой штамм Corynebacterium glutamicum, который обладает способностью продуцировать L-лизин вследствие введения мутаций P458S, V59A, и T311I в 3 типа генов, рус, horn, и lysC штамма дикого типа, на основе методики, описанной Binder et al. (Genome Biology 2102, 13:R40). После введения способом электропорации посредством второго кроссинговера был получен штамм, имеющий хуlАВ(Еr) с заменой одной копии промотора pcj7 внутри транспозона на хромосоме, и штамм назвали Corynebacterium glutamicum CJ3P-pcj7-xylAB(Er).
Способность утилизировать ксилозу и продуцировать L-лизин Corynebacterium glutamicum CJ3P и Corynebacterium glutamicum CJ3P-pcj7-xylAB(Er) по настоящему изобретению измеряли таким же образом, как в Примере 3 (Таблица 6).
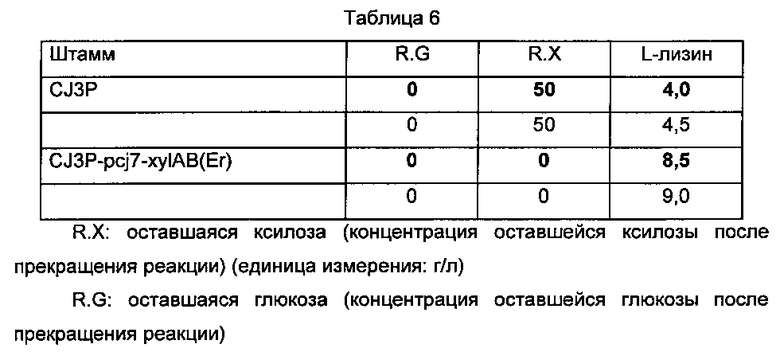
Как показано в Таблице 6, при введении хуlАВ(Еr) в штамм CJ3P, продуцирующий L-лизин, штамм полностью потреблял ксилозу, в отличие от родительского штамма, не способного утилизировать ксилозу, и его способность продуцировать L-лизин также возрастала.
Данный результат демонстрирует, что при введении xylAB, имеющего происхождение из Erwinia carotovora, в различные микроорганизмы Corynebacterium sp., а также в микроорганизмы Corynebacterium sp, определенные конкретным номером доступа, они полностью потребляют ксилозу в качестве источника углерода, тем самым эффективно продуцируя аминокислоты, такие как L-лизин.
Таким образом, микроорганизм Corynebacterium sp., экспрессирующий хуlАВ(Еr), способен расти, утилизируя ксилозу, находящуюся в среде, а также способен продуцировать L-лизин посредством утилизации ксилозы и глюкозы, находящихся в среде.
Эффект изобретения
При использовании микроорганизма Corynebacterium sp по настоящему изобретению, способного продуцировать L-лизин посредством утилизации ксилозы, L-лизин может быть получен посредством утилизации ксилозы, как второго по распространенности в природе лигноцеллюлозного углевода. Таким образом, данный микроорганизм может быть широко применен для эффективного и экономичного получения L-лизина.















Изобретение относится к биотехнологии и представляет собой модифицированный Corynebacterium glutamicum, способный продуцировать L-лизин посредством утилизации ксилозы, в который введены активности ксилозоизомеразы (XylA) и ксилулокиназы (XylB), имеющие происхождение из Erwinia carotovora. Настоящее изобретение также относится к способу получения L-лизина, включающему стадию культивирования указанного модифицированного микроорганизма, использующего ксилозу в качестве источника углерода, и выделения L-лизина из культуры. Изобретение позволяет получать L-лизин с высокой степенью эффективности. 2 н. и 3 з.п. ф-лы, 3 ил., 6 табл., 9 пр.
1. Модифицированный Corynebacterium glutamicum, способный продуцировать L-лизин посредством утилизации ксилозы, в который введены активности ксилозоизомеразы (XylA) и ксилулокиназы (XylB), имеющие происхождение из Erwinia carotovora.
2. Микроорганизм по п. 1, где XylA содержит аминокислотную последовательность SEQ ID NO: 1 и XylB содержит аминокислотную последовательность SEQ ID NO: 2.
3. Микроорганизм по п. 1, где XylA кодируется полинуклеотидной последовательностью SEQ ID NO: 3 и XylB кодируется полинуклеотидной последовательностью SEQ ID NO: 4.
4. Микроорганизм по п. 1, где введение активностей XylA и XylB осуществляют посредством вставки полинуклеотида, содержащего нуклеотидные последовательности, кодирующие XylA и XylB, в хромосому, введения векторной системы, содержащей этот полинуклеотид, в микроорганизм, введения активного промотора перед нуклеотидными последовательностями, кодирующими XylA и XylB, введения XylA и XylB с модифицированным промотором или введения модифицированных нуклеотидных последовательностей, кодирующих XylA и XylB.
5. Способ получения L-лизина, включающий стадии:
(1) культивирования модифицированного Corynebacterium glutamicum, способного продуцировать L-лизин посредством утилизации ксилозы, по любому из пп. 1-4 в культуральной среде, содержащей ксилозу в качестве источника углерода, с получением культуральной жидкости; и
(2) выделения L-лизина из культуральной жидкости.
| KAWAGUCHI H | |||
| Engineering of a xylose metabolic pathway in Corynebacterium glutamicum, Appl Environ Microbiol, 2006 May, 72(5):3418-28 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНОВ УТИЛИЗАЦИИ КСИЛОЗЫ | 2005 |
|
RU2283346C1 |
| BLOMBACH B | |||
| et.al | |||
| Carbohydrate metabolism in Corynebacterium glutamicum and applications for the metabolic engineering of L-lysine production strains, Appl Microbiol Biotechnol | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |

