ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По данной заявке испрашивается приоритет по патентной заявке США № 61/258565, которая подана 5 ноября 2009 года, содержание которой включено в данный документ в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение в основном относится к области молекулярной биологии и белковой технологии. Более конкретно, изобретение относится к сигнальным последовательностям для секреции гетерологичных полипептидов бактериями. Изобретение также относится к продуцируемым прокариотами рекомбинантным полипептидам и их применению.
УРОВЕНЬ ТЕХНИКИ
Секреция гетерологичных полипептидов в периплазматическое пространство E. coli и других прокариотов или в их среды для культивирования зависит от различных параметров. Типично векторы для секреции полипептида, представляющего интерес, конструируют для того, чтобы поместить ДНК, кодирующую секреторную сигнальную последовательность, ближе к 5' относительно ДНК, кодирующей полипептид, представляющий интерес.
В последние годы расширились перспективы использования гетерологичных полипептидов, например, антител, в качестве диагностических и терапевтических средств для различных нарушений и заболеваний. Для многих исследовательских и клинических применений необходимы большие количества функционального полипептида, что, таким образом, требует увеличенных в масштабе и при этом экономичных систем для получения полипептида. В частности, можно использовать рекомбинантное получение антител с использованием различных экспрессирующих организмов-хозяев в диапазоне от прокариотов, таких как E. coli или B. subtilis, до клеток дрожжей, растений, насекомых и млекопитающих. Kipriyanov and Little (1999) Mol. Biotech. 12: 173-201.
По сравнению с другими системами получения полипептидов, бактерии, в частности, E. coli, обеспечивают многие уникальные преимущества. Используемое сырье (т.е. бактериальные клетки) недороги и просты в выращивании, вследствие чего снижается стоимость продуктов. Прокариотические организмы-хозяева растут значительно быстрее, чем, например, клетки млекопитающих, что допускает более быстрый анализ генетических манипуляций. Более короткое время жизни поколения и легкое масштабирование также делают бактериальную ферментацию более привлекательным средством для получения белка в больших количествах. Структура генома и биологическая активность многих бактериальных видов, включая E. coli, хорошо изучены, а также доступен широкий спектр подходящих векторов, что делает экспрессию желаемого антитела более удобной. По сравнению с эукариотами, процесс получения включает меньшее число стадий, включая манипуляцию рекомбинантными генами, стабильную трансформацию множественных копий в организм-хозяин, индукцию экспрессии и определение характеристик продуктов. Pluckthun and Pack (1997) Immunotech 3:83-105.
Для получения рекомбинантных полипептидов в бактериях использовались различные подходы. Рекомбинантные белки можно получить из бактерий или посредством повторной укладки телец включения, экспрессируемых в цитоплазме, или посредством экспрессии с последующей секрецией в бактериальную периплазму. Выбор между секрецией и повторной укладкой, как правило, обусловлен несколькими соображениями. Обычно секреция является более быстрой и более общеупотребительной стратегией получения антител. Kipriyanov and Little (1999), выше.
Экспрессию антител в прокариотических системах можно осуществлять в различных масштабах. Культуры во встряхиваемых колбах (в пределах 2-5 л) типично создают продукты в количестве менее чем 5 мг/л. Carter et al. (1992) Bio/Technology 10: 12-16 разработали систему ферментации при высокой плотности клеток, в которой достигали высокого уровня экспрессии фрагментов антител (до 2 г/л). Титры Fab' в граммах на литр, которых достигли Carter et al., в значительной степени обусловлены высокими плотностями клеток, что является результатом более точного контроля среды ферментатора, чем таковой в простой встряхиваемой колбе. Система содержит бицистронный оперон, сконструированный для совместной экспрессии фрагментов легких цепей и тяжелых цепей. Бицистронный оперон находится под управлением одного промотора phoA E. coli, который можно индуцировать посредством фосфатного голодания. Каждой цепи антитела предшествует сигнальная последовательность термостабильного энтеротоксина II (stII) E. coli, чтобы направить секрецию в периплазматическое пространство.
Общие обзоры получения антител в E. coli см. в Pluckthun and Pack (1997) Immunotech 3:83-105; Pluckthun et al. (1996) in ANTIBODY ENGINEERING: A PRACTICAL APPROACH, pp 203-252 (Oxford Press); Pluckthun (1994) in HANDBOOK OF EXP PHARMCOL VOL 3: THE PHARMCOL OF MONOCLONAL ANTIBODIES, pp. 269-315 (ed. M. Rosenberg and G.P. Moore; Springer-Verlag, Berlin).
Многие биологические анализы (такие как рентгенокристаллография) и клинические применения (такие как белковая терапия) требуют больших количеств белка. Соответственно, существует потребность в высокопродуктивных и при этом простых системах для получения правильно собранных, растворимых и функциональных гетерологичных полипептидов, таких как антитела.
Все цитируемые в данном документе ссылки, включая патентные заявки и публикации, включены в данный документ в качестве ссылки в полном объеме.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к новому средству для увеличения образования гетерологичных белков, которое содержит новые варианты области инициации трансляции (TIR), включая варианты TIR, содержащие котрансляционные сигнальные пептиды секреции (сигнальные пептиды, которые направляют транслокацию котрансляционным образом) и/или варианты TIR, содержащие посттрансляционные сигнальные пептиды секреции (сигнальные пептиды, которые направляют транслокацию посттрансляционным образом). Кроме того, в настоящем документе продемонстрировано увеличенное образование антител с использованием векторов, содержащих легкую цепь антитела, функционально связанную с TIR, содержащей ко- или посттрансляционный сигнальный пептид секреции, и тяжелую цепь антитела, функционально связанную с TIR, содержащей котрансляционный сигнальный пептид секреции для пиковой экспрессии. В настоящем документе предоставлены новые варианты TIR.
В одном из аспектов изобретение относится к вариантам областей инициации трансляции. В некоторых вариантах осуществления вариант содержит вариант области инициации трансляции (в некоторых вариантах осуществления прокариотическую посттрансляционную сигнальную последовательность секреции или прокариотическую котрансляционную сигнальную последовательность секреции). В некоторых вариантах осуществления вариант содержит варианты нуклеиновой кислоты сигнальной последовательности секреции, такой как PhoA, MalE, DsbA или STII. В некоторых вариантах осуществления вариант дополнительно содержит сайт рестрикции MlaI, BssHII или XbaI. В некоторых вариантах осуществления вариант содержит вариант области инициации трансляции, содержащий последовательность, представленную в таблице 3.
В одном из аспектов изобретение относится к вариантам сигнальных последовательностей секреции. В некоторых вариантах осуществления сигнальная последовательность секреции представляет собой прокариотическую посттрансляционную сигнальную последовательность секреции или прокариотическую котрансляционную сигнальную последовательность секреции. В некоторых вариантах осуществления сигнальная последовательность секреции представляет собой эукариотическую посттрансляционную сигнальную последовательность секреции или эукариотическую котрансляционную сигнальную последовательность секреции. В некоторых вариантах осуществления варианты представляют собой варианты нуклеиновой кислоты сигнальной последовательности секреции PhoA, MalE, DsbA или STII. В некоторых вариантах осуществления варианты содержат сигнальную последовательность секреции, представленную в таблице 3. Варианты сигнальных последовательностей секреции по изобретению пригодны для использования, например, в любом способе, описанном в настоящем документе.
В другом аспекте изобретение относится к полинуклеотиду, содержащему область инициации трансляции по изобретению. В некоторых вариантах осуществления область инициации трансляции содержит последовательность, представленную в таблице 3 (например, одну из SEQ ID NO: 1-42). В некоторых вариантах осуществления область инициации трансляции содержит одну из SEQ ID NO: 1-14, 16-24, 26-39, 41-42. Эти полинуклеотиды пригодны для использования, например, в любом из способов, описанных в настоящем документе.
В другом аспекте изобретение относится к полинуклеотиду, содержащему сигнальную последовательность секреции по изобретению. В некоторых вариантах осуществления сигнальная последовательность секреции содержит последовательность, представленную в таблице 3 (например, одну из SEQ ID NO: 1-42). В некоторых вариантах осуществления область инициации трансляции содержит одну из SEQ ID NO: 1-14, 16-24, 26-39, 41-42. Эти полинуклеотиды пригодны для использования, например, в любом из способов, описанных в настоящем документе.
В другом аспекте изобретение относится к полинуклеотиду, содержащему область инициации трансляции по изобретению, функционально связанную с полинуклеотидом, кодирующим гетерологичный полипептид, посредством чего при экспрессии гетерологичного полипептида в клетке-хозяине (например, прокариотической клетке-хозяине, например, клетке-хозяине E. coli) происходит укладка и сборка гетерологичного полипептида, чтобы сформировать биологически активный гетерологичный полипептид. Примеры гетерологичных полипептидов дополнительно описаны в настоящем документе. В некоторых вариантах осуществления гетерологичный полипептид представляет собой тяжелую цепь антитела. В некоторых вариантах осуществления гетерологичный полипептид представляет собой легкую цепь антитела. В некоторых вариантах осуществления гетерологичный полипептид представляет собой полипептид Fc. В некоторых вариантах осуществления гетерологичный полипептид представляет собой мультимерный полипептид. В некоторых вариантах осуществления гетерологичный полипептид представляет собой гетеромультимер. В некоторых вариантах осуществления область инициации трансляции представляет собой любую область инициации трансляции, описанную в настоящем документе, например, область инициации трансляции, которая содержит последовательность, представленную в таблице 3. В некоторых вариантах осуществления область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42. В некоторых вариантах осуществления область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-14, 36-39, 41-42. В некоторых вариантах осуществления область инициации трансляции содержит вариант сигнальной последовательности STII, DsbI, PhoA или MalE.
В другом аспекте изобретение относится к полинуклеотиду, содержащему (1) первую область инициации трансляции (TIR), функционально связанную с полинуклеотидом, кодирующим первый гетерологичный полипептид, где TIR содержит котрансляционную прокариотическую сигнальную последовательность секреции; и (2) второй TIR, функционально связанный с полинуклеотидом, кодирующим второй гетерологичный, где второй TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции, посредством чего при экспрессии антитела в клетке-хозяине происходит укладка и сборка первого и второго гетерологичных полипептидов, чтобы сформировать биологически активный полипептидный комплекс.
В другом аспекте изобретение относится к полинуклеотиду, кодирующему антитело, указанный полинуклеотид содержит (1) первую область инициации трансляции по изобретению, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, и (2) вторую область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, посредством чего при экспрессии антитела в клетке-хозяине (например, прокариотической клетке-хозяине, например, клетке-хозяине E. coli) происходит укладка и сборка тяжелой и легкой цепей, чтобы сформировать биологически активное антитело.
В некоторых вариантах осуществления первая область инициации трансляции содержит котрансляционную прокариотическую сигнальную последовательность секреции (например, сигнальную последовательность, которая направляет трансляцию через сигнал-узнающий пептид). В некоторых вариантах осуществления первая область инициации трансляции содержит сигнальную последовательность STII или DsbA. В некоторых вариантах осуществления первая область инициации трансляции содержит сигнальную последовательность DsbA. В некоторых вариантах осуществления первая область инициации трансляции содержит сигнальную последовательность PhoA или MalE. В некоторых вариантах осуществления первая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-10 и 36-42. В некоторых вариантах осуществления первая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-10 и 36-29 и 41 и 42. В некоторых вариантах осуществления первая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42. В некоторых вариантах осуществления первая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-14, 16-24, 26-39, 41-42.
В некоторых вариантах осуществления вторая область инициации трансляции содержит (i) котрансляционную прокариотическую сигнальную последовательность секреции или посттрансляционную прокариотическую сигнальную последовательность секреции (например, сигнальную последовательность, которая направляет трансляцию через путь sec). В некоторых вариантах осуществления вторая область инициации трансляции содержит сигнальную последовательность STII, DsbA, MalE или PhoA. В некоторых вариантах осуществления вторая область инициации трансляции содержит сигнальную последовательность PhoA или MalE. В некоторых вариантах осуществления вторая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42. В некоторых вариантах осуществления вторая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-14, 16-24, 26-39, 41-42.
В некоторых вариантах осуществления полинуклеотид, кодирующий антитело, дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность STII, PhoA или DsbA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность DsbA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность PhoA.
В другом аспекте изобретение относится к полинуклеотиду, содержащему (1) первую область инициации трансляции (TIR), функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где TIR содержит котрансляционную прокариотическую сигнальную последовательность секреции; и (2) вторую TIR, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции, посредством чего при экспрессии антитела в клетке-хозяине происходит укладка и сборка тяжелой и легкой цепей, чтобы сформировать биологически активное антитело.
В другом аспекте изобретение относится к полинуклеотиду, кодирующему фрагмент антитела (такой как моновалентный фрагмент антитела), указанный полинуклеотид содержит (1) первую область инициации трансляции по изобретению, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела; (2) вторую область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела; и (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc, посредством чего при экспрессии антитела в клетке-хозяине (например, прокариотической клетке-хозяине), происходит укладка и сборка тяжелой цепи, легкой цепи и полипептида Fc, чтобы сформировать биологически активное антитело (такой как антитело с одним Fab фрагментом). В некоторых вариантах осуществления третья область инициации трансляции содержит котрансляционную прокариотическую сигнальную последовательность секреции или посттрансляционную прокариотическую сигнальную последовательность секреции. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность STII, PhoA, MalE или DsbA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность DsbA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность PhoA. В некоторых вариантах осуществления третья область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42. В некоторых вариантах осуществления третья область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-14, 16-24, 26-39, 41-42.
В другом аспекте изобретение относится к полинуклеотиду, кодирующему антитело, указанный полинуклеотид содержит (1) первую область инициации трансляции по изобретению, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где первая область инициации трансляции содержит сигнальную последовательность STII или DsbA, и (2) вторую область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая область инициации трансляции содержит сигнальную последовательность STII, DsbA, MalE или PhoA, посредством чего при экспрессии антитела в клетке-хозяине (например, прокариотической клетке-хозяине) происходит укладка и сборка легкой и тяжелой цепей, чтобы сформировать биологически активное антитело. В некоторых вариантах осуществления первая область инициации трансляции содержит сигнальную последовательность DsbA, а вторая область инициации трансляции содержит сигнальную последовательность MalE или PhoA. В некоторых вариантах осуществления полинуклеотид, кодирующий антитело, дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность STII, PhoA или DsbA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность PhoA. В некоторых вариантах осуществления третья область инициации трансляции содержит сигнальную последовательность DsbA.
В некоторых вариантах осуществления интенсивность трансляции указанного варианта области инициации трансляции менее интенсивности трансляции области инициации трансляции дикого типа. В некоторых вариантах осуществления интенсивность трансляции указанного варианта области инициации трансляции превышает интенсивность трансляции область инициации трансляции дикого типа. В некоторых вариантах осуществления аминокислотная последовательность варианта области инициации трансляции не изменена относительно аминокислотной последовательности дикого типа. В некоторых вариантах осуществления аминокислотная последовательность варианта области инициации трансляции изменена относительно аминокислотной последовательности дикого типа. В некоторых вариантах осуществления область инициации трансляции содержит прокариотическую сигнальную последовательность секреции. В некоторых вариантах осуществления первая и вторая области инициации трансляции (и, в некоторых вариантах осуществления, третья область инициации трансляции) обеспечивают приблизительно одинаковые интенсивности трансляции. В некоторых вариантах осуществления относительная интенсивность трансляции составляет приблизительно один или два. В некоторых вариантах осуществления относительная интенсивность трансляции составляет приблизительно один. В некоторых вариантах осуществления относительная интенсивность трансляции составляет приблизительно два. В некоторых вариантах осуществления относительная интенсивность трансляции составляет один и/или два. В некоторых вариантах осуществления относительная интенсивность трансляции составляет приблизительно три или приблизительно четыре. В некоторых вариантах осуществления относительная интенсивность трансляции выбрана из одного или нескольких из 1, 2, 3, 4, 5 или более (например, 6 или 7 или более).
В некоторых вариантах осуществления полинуклеотид по изобретению дополнительно содержит промотор, функционально связанный с гетерологичным полипептидом. В некоторых вариантах осуществления промотор представляет собой прокариотический промотор, выбранный из группы, состоящей из промоторов phoA, tac, lpp, lac-lpp, lac, ara, trp и T7. В некоторых вариантах осуществления промотор представляет собой промотор phoA. В некоторых вариантах осуществления, включающих экспрессию тяжелой и легкой цепи антитела, полинуклеотид дополнительно содержит (a) первый промотор, где первый промотор функционально связан с легкой цепью и (b) второй промотор, где второй промотор функционально связан с тяжелой цепью. В некоторых вариантах осуществления первый и второй промоторы представляют собой промоторы phoA. В некоторых вариантах осуществления, включающих экспрессию тяжелой и легкой цепи антитела и полипептида Fc, полинуклеотид дополнительно содержит (c) третий промотор, где третий промотор функционально связан с полипептидом Fc. В некоторых вариантах осуществления третий промотор представляет собой полипептид Fc.
Когда экспрессируют полипептиды, которые содержат более чем один полипептид (например, антитело, содержащее тяжелую цепь и легкую цепь), полинуклеотид для экспрессии полипептида может представлять собой полицистронный полинуклеотид (т.е. один полинуклеотид, который содержит и экспрессирует несколько цистронов под регуляторным управлением одного промотора). Типичным примером полицистронного вектора является «бицистронный» вектор, который содержит и экспрессирует два различных полипептида под управлением одного промотора. При экспрессии бицистронного или полицистронного вектора сначала происходит транскрипция нескольких кодирующих областей (например, генов) в виде одной транскрипционной единицы, а затем раздельная трансляция. Цистрон относится к генетическому элементу, приблизительно эквивалентному трансляционной единице, содержащей нуклеотидную последовательность, кодирующую полипептидную цепь и смежные управляющие области (включая, например, TIR). В других вариантах осуществления полинуклеотид может содержать отдельные цистроны, которые относятся к одному полинуклеотиду, содержащему по меньшей мере две отдельные пары промотор-цистрон, где каждый цистрон находится под управлением своего собственного промотора. При экспрессии экспрессирующего вектора с отдельным цистроном, как процесс транскрипции, так и процесс трансляции различных генов происходит отдельно и независимо. В других вариантах осуществления полинуклеотид может содержать полицистронную часть и часть с отдельным цистроном.
В еще одном аспекте изобретение относится к векторам, содержащим полинуклеотид по изобретению. В некоторых вариантах осуществления векторы представляют собой экспрессирующие векторы.
В дополнительном аспекте изобретение относится к композициям, содержащим один или несколько полинуклеотидов по изобретению и носитель. В одном из вариантов осуществления носитель является фармацевтически приемлемым.
В одном из аспектов изобретение относится к клеткам-хозяевам, содержащим полинуклеотид или вектор по изобретению. В некоторых вариантах осуществления клетки-хозяева содержат полинуклеотид по изобретению, который кодирует антитело (в некоторых вариантах осуществления биспецифическое антитело или антитело с одним Fab фрагментом). Клетка-хозяин может содержать один или несколько полинуклеотидов, которые совместно кодируют антитело. Вектор может относиться к любому типу, например, к рекомбинантным векторам, таким как экспрессирующие векторы. Можно использовать любую из множества клеток-хозяев. В одном из вариантов осуществления клетка-хозяин представляет собой прокариотическую клетку, например, E. coli. В некоторых вариантах осуществления E. coli представляет собой штамм с дефицитом активностей эндогенных протеаз. В некоторых вариантах осуществления в генотипе E. coli отсутствуют гены degP и prc и присутствует мутантный ген spr.
В некоторых вариантах осуществления клетка-хозяин дополнительно содержит полинуклеотид, кодирующий прокариотический шаперон (такой как белки Dsb (DsbA, DsbB, DsbC, DsbD, FkpA и/или DsbG). В некоторых вариантах осуществления в клетке-хозяине происходит сверхэкспрессия шаперона. В некоторых вариантах осуществления шаперон представляет собой DsbA и/или DsbC.
В одном из аспектов клетка-хозяин содержит один или несколько полинуклеотидов, которые совместно кодируют антитело с одним Fab фрагментом. В одном из вариантов осуществления один полинуклеотид кодирует (a) компоненты легкой и тяжелой цепей антитела с одним Fab фрагментом, и (b) полипептид Fc. В одном из вариантов осуществления один полинуклеотид кодирует компоненты легкой цепи и полипептида Fc антитела с одним Fab фрагментом, и отдельный полинуклеотид кодирует полипептид тяжелой цепи. В одном из вариантов осуществления один полинуклеотид кодирует компоненты тяжелой цепи и полипептида Fc антитела с одним Fab фрагментом и отдельный полинуклеотид кодирует компонент легкой цепи антитела с одним Fab фрагментом. В одном из вариантов осуществления отдельные полинуклеотиды кодируют компонент легкой цепи антитела с одним Fab фрагментом, компонент тяжелой цепи антитела с одним Fab фрагментом и полипептид Fc, соответственно.
В настоящем документе описаны гетерологичные полипептиды. В некоторых вариантах осуществления гетерологичный полипептид представляет собой антитело. В некоторых вариантах осуществления антитело представляет собой моноклональное антитело. В других вариантах осуществления антитело представляет собой поликлональное антитело. В некоторых вариантах осуществления антитело выбирают из группы, состоящей из химерного антитела, аффинно зрелого антитела, гуманизированного антитела и антитела человека. В определенных вариантах осуществления антитело представляет собой биспецифическое антитело. В определенных вариантах осуществления антитело представляет собой фрагмент антитела. В некоторых вариантах осуществления антитело представляет собой моновалентное антитело. В некоторых вариантах осуществления антитело представляет собой Fab, Fab', Fab'-SH, F(ab')2 или scFv. В некоторых вариантах осуществления антитело представляет собой антитело с одним Fab фрагментом (т.е. вариабельный домен тяжелой цепи и вариабельный домен легкой цепи формируют один антигенсвязывающий фрагмент), содержащее Fc-область, где Fc-область содержит первый и второй полипептиды Fc, где первый и второй полипептиды Fc представлены в комплексе и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с молекулой Fab, содержащей указанный антигенсвязывающий фрагмент.
В некоторых вариантах осуществления антитело связывает (в некоторых вариантах осуществления специфически связывает) c-met. В некоторых вариантах осуществления антитело против c-met содержит (a) первый полипептид, содержащий вариабельный домен тяжелой цепи, который имеет последовательность:

последовательность CH1 и первый полипептид Fc; (b) второй полипептид, содержащий вариабельный домен легкой цепи, который имеет последовательность:
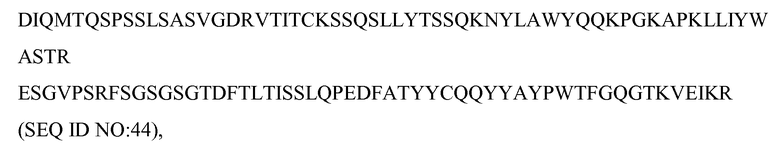
и последовательность CL1; и (c) третий полипептид, содержащий второй полипептид Fc, где вариабельный домен тяжелой цепи и вариабельный домен легкой цепи представлены в виде комплекса и формируют один антигенсвязывающий фрагмент, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с молекулой Fab, содержащей указанный антигенсвязывающий фрагмент. В некоторых вариантах осуществления первый полипептид содержит последовательность Fc, изображенную на фиг.7 (SEQ ID NO: 68), а второй полипептид содержит последовательность Fc, изображенную на фиг.8 (SEQ ID NO: 47). В некоторых вариантах осуществления первый полипептид содержит последовательность Fc, изображенную на фиг.8 (SEQ ID NO: 47), и второй полипептид содержит последовательность Fc, изображенную на фиг.7 (SEQ ID NO: 68).
В некоторых вариантах осуществления антитело против c-met содержит (a) первый полипептид, содержащий вариабельный домен тяжелой цепи, указанный полипептид содержит последовательность:

(b) второй полипептид, содержащий вариабельный домен легкой цепи, полипептид содержит последовательность

и третий полипептид, содержащий полипептид Fc, полипептид содержит последовательность

где вариабельный домен тяжелой цепи и вариабельный домен легкой цепи представлены в виде комплекса и формируют один антигенсвязывающий фрагмент, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с молекулой Fab, содержащей указанный антигенсвязывающий фрагмент.
В одном из вариантов осуществления антитело против c-met содержит вариабельный домен тяжелой цепи, который содержит одну или несколько из последовательностей CDR1-HC, CDR2-HC и CDR3-HC, изображенных на фиг.7 (SEQ ID NO: 52-53 и 66). В некоторых вариантах осуществления антитело содержит вариабельный домен легкой цепи, содержащий одну или несколько из последовательностей CDR1-LC, CDR2-LC и CDR3-LC, изображенных на фиг.7 (SEQ ID NO: 49-51). В некоторых вариантах осуществления вариабельный домен тяжелой цепи содержит последовательности FR1-HC, FR2-HC, FR3-HC и FR4-HC, изображенные на фиг.7 (SEQ ID NO: 62-65). В некоторых вариантах осуществления вариабельный домен легкой цепи содержит последовательности FR1-LC, FR2-LC, FR3-LC и FR4-LC, изображенные на фиг.7 (SEQ ID NO: 57-60).
В некоторых вариантах осуществления антитело содержит по меньшей мере одну характеристику, которая способствует гетеродимеризации, при этом минимизируя гомодимеризацию последовательностей Fc в фрагменте антитела. Такая характеристика(и) повышает выход и/или чистоту и/или гомогенность популяций иммуноглобулинов, получаемых способами по изобретению, как описано в настоящем документе. В одном из вариантов осуществления первый полипептид Fc и второй полипептид Fc встречаются/взаимодействуют в области контакта. В некоторых вариантах осуществления, где первый и второй полипептиды Fc встречаются в области контакта, область контакта второго полипептида (последовательности) Fc содержит выпуклость (также называемый «выступ»), которую можно разместить в полости (также обозначаемой «отверстием») в области контакта первого полипептида (последовательности) Fc. В одном из вариантов осуществления антитело содержит мутации Fc, образующие «выступы» и «отверстия», как описано в WO 2005/063816. Например, мутация отверстия может представлять собой одну или несколько из T366A, L368A и/или Y407V в полипептиде Fc, а мутация выступа может представлять собой T366W.
Изобретение также относится к способам, в которых используют вариант TIR и сигнальные последовательности по изобретению. Понятно, что любое из варианта TIR, сигнальных последовательностей и полинуклеотидов, описанных в настоящем документе, пригодно для использования в способах, например, способах по изобретению, описанных в настоящем документе. В дополнительном аспекте изобретение относится к способам получения гетерологичного полипептида по изобретению. Например, изобретение относится к способам получения гетерологичного полипептида (например, антитела, которое, как определено в настоящем документе, включает полноразмерное антитело и его фрагменты), указанный способ включает культивирование клетки-хозяина, содержащей полинуклеотид по изобретению (например, полинуклеотид, содержащий область инициации трансляции) с тем, чтобы экспрессировать полинуклеотид, посредством чего при экспрессии указанного полинуклеотида в клетке-хозяине (например, прокариотической клетке-хозяине) происходит укладка гетерологичного полипептида, чтобы сформировать биологически активный гетерологичный полипептид. В вариантах осуществления, включающих экспрессию антител, при экспрессии указанного полинуклеотида в клетке-хозяине происходит укладка и сборка легкой и тяжелой цепей, чтобы сформировать биологически активное антитело. В некоторых вариантах осуществления способ дополнительно включает извлечение гетерологичного полипептида (например, антитела) из культуры клетки-хозяина. В некоторых вариантах осуществления гетерологичный полипептид извлекают из среды для культивирования клетки-хозяина. В некоторых вариантах осуществления способ дополнительно включает объединение извлеченного гетерологичного полипептида (например, антитела) с фармацевтически приемлемым носителем, эксципиентом или носителем для получения фармацевтического состава, содержащего гетерологичный полипептид (например, антитело).
В одном из аспектов изобретение относится к способам секреции гетерологичного полипептида, представляющего интерес, из клетки, указанный способ включает культивирование клетки-хозяина, содержащей полинуклеотид по изобретению, с тем, чтобы экспрессировать полинуклеотид и секретировать гетерологичный полипептид.
В одном из аспектов изобретение относится к способам транслокации гетерологичного полипептида, представляющего интерес, из клетки, указанный способ включает культивирование клетки-хозяина, содержащей полинуклеотид по изобретению, с тем чтобы экспрессировать полинуклеотид и транслоцировать гетерологичный полипептид.
В другом аспекте изобретение относится к способу оптимизации секреции гетерологичного полипептида, представляющего интерес, в клетке, который содержит сравнение уровней экспрессии полипептида под управлением группы полинуклеотидных вариантов области инициации трансляции, где группа вариантов представляет диапазон интенсивностей трансляции, и определение оптимальной интенсивности трансляции для получения зрелого полипептида. В некоторых вариантах осуществления оптимальная интенсивность трансляции ниже интенсивности трансляции области инициации трансляции дикого типа. В некоторых вариантах осуществления оптимальная интенсивность трансляции выше интенсивности трансляции области инициации трансляции дикого типа. В некоторых вариантах осуществления варианты содержат полинуклеотидные варианты сигнальной последовательности секреции. В некоторых вариантах осуществления варианты сигнальных последовательностей секреции представляют собой сигнальные последовательности пути sec и/или сигнальные последовательности пути SRP. В некоторых вариантах осуществления варианты сигнальных последовательностей секреции представляют собой варианты сигнальных последовательностей PhoA, MalE, DsbA или STII. В некоторых вариантах осуществления вариант представляет собой один или несколько вариантов, представленных в таблице 3. В некоторых вариантах осуществления вариант содержит последовательность одной из SEQ ID NO: 1-14, 36-39, 41-42.
В одном из аспектов изобретение относится к гетерологичному полипептиду, полученному способом по изобретению, как описано в настоящем документе. В некоторых вариантах осуществления гетерологичный полипептид представляет собой антитело.
В одном из аспектов изобретение относится к использованию гетерологичного полипептида, созданного с использованием способов по изобретению, в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение. Гетерологичный полипептид может относиться к любой форме, описанной в настоящем документе, включая антитело, фрагмент антитела, полипептид (например, олигопептид) или их сочетание.
В одном из аспектов изобретение относится к использованию полинуклеотида по изобретению в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение.
В одном из аспектов изобретение относится к использованию экспрессирующего вектора по изобретению в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение.
В одном из аспектов изобретение относится к использованию клетки-хозяина по изобретению в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение.
В одном из аспектов изобретение относится к использованию промышленного изделия по изобретению в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение, иммунное (такое как аутоиммунное) нарушение и/или связанное с ангиогенезом нарушение (заживление ран).
В одном из аспектов изобретение относится к использованию набора по изобретению в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1: Транслокация указанных сигнальных пептидов через внутреннюю мембрану бактерий. Сигнальные пептиды связывающего мальтозу периплазматического белка (MalE) и щелочной фосфатазы (PhoA) направляют транслокацию из цитоплазмы в периплазму посттрансляционным образом с помощью молекулярного двигателя SecA. Сигнальные пептиды термостабильного энтеротоксина II (StII) и тиол:дисульфид обменивающего белка (DsbA) направляют транслокацию котрансляционным образом с помощью с помощью частицы узнавания сигнала (SRP).
Фиг.2: Относительная интенсивность области инициации транслокации для вариантов сигнального пептида. Нормализованная исходная активность щелочной фосфатазы клеток 27C7, несущих вектор со слиянием между любым сигнальным пептидом из STII, MalE, PhoA или DsbA и зрелым доменом гена щелочной фосфатазы (BAP) E. coli. Каждый столбик представляет отдельную культуру, которую инкубировали с хромогенным субстратом PNPP, и ферментативную активность определяли как поглощение это культуры при 410 нм ниже поглощения культуры, несущей пустой вектор (pBR322). Активности нормализовали к исходной активности клеток 27C7, несущих плазмиду pPho41. Белые столбцы представляют варианты сигнальных пептидов с сайтом рестрикции BssHII в положении -9 по отношению к первой паре оснований инициирующего кодона. Серые или заштрихованные столбцы представляют сайт MluI или XbaI в положении -9, соответственно. Все активности представляют собой среднее между семью и десятью повторными экспериментами. Границы ошибки приведены в виде неточности среднего с 95% доверительного интервала. Все различия в относительной интенсивности TIR между смежным столбцами являются статистически значимыми (P<<0,001). Столбцы представляют клоны SH1.2, SH2.41, SH3.38, SH4.60, SH5.34, SH6.52, SH8.36, SL1.2, SL2.74, SL3.72, MH1.92, MH2.100, ML1.97, ML2.123, MX1.wt, MX2.15, MX3.12, MX5.37, MX6.4, MX7.25, MX8.13, MX11.34, PH1.70, PH2.64, PH3.wt, PH4.67, PH5.71, PH6.77, PL1.104, PX2.41, PX3.wt, PX5.53, PX6.15, PX8.24, PX10.23, DH1.48, DH2.wt, DH3.79, DH7.72, DL1.wt, DL2.3, DL3.37 (в порядке слева направо).
Фиг.3: Установка для мониторинга частиц антител с манипуляциями с сигнальным пептидом тяжелой цепи. Клетки 64B4 растили в 25 мл фосфат-ограничивающей среды C.R.A.P. в течение 24 часов, и для анализа SDS-PAGE получали растворимые фракции, к тому же при нормализации осадка общего белка по оптической плотности (OD). (A) Образцы клеток, несущих плазмиду pBR-SS-5D5-1.1 (SS 1.1), pBR-MS-5D5-1.1 (MS 1.1), pBR-DS-5D5-1.1 (DS 1.1) или pBR-PS-5D5-1.1 (PS 1.1), получали посредством электрофореза в геле SDS-PAGE (масса в кДа обозначена слева), переносили на нитроцеллюлозу и анализировали присутствие частиц, содержащих тяжелые цепи, с использованием антитела, специфичного к α-Fc. Растворимые образцы (верхний блот) состояли из предположительно идентифицированных полос, соответствующих (сверху вниз): полноразмерному антителу, тяжелой-тяжелой-легкой (HHL), тяжелой-легкой (HL) или свободной тяжелой цепи (мономер тяжелой цепи). Нормализованные образцы общего белка (нижний блот) восстанавливали с использованием 0,2 M DTT, чтобы разрушить структуру дисульфидной связи и каждая отдельная дорожка мигрировала в одну полосу с кажущейся массой ~49 кДа. (B) Образцы из (A) прогоняли на отдельном геле SDS-PAGE (масса кДа обозначена справа), переносили на нитроцеллюлозу и анализировали комплексы, содержащие легкие цепи, с использованием антитела, специфичного к α-κLc. Растворимые образцы (верхний блот) состояли из предположительно идентифицированных полос, соответствующих (сверху вниз): полноразмерному антителу, HL, легкому-легкому (LL) димеру или свободной легкой цепи (мономер легкой цепи). Нормализованные образцы общего белка (нижний блот) восстанавливали с использованием 0,2 M DTT, и каждая отдельная дорожка мигрировала в отдельную полосу с кажущейся массой ~25 кДа. Сокращения: S = сигнальная последовательность STII, M = сигнальная последовательность MalE, D = сигнальная последовательность DsbA, P = сигнальная последовательность PhoA. XX#.# (например, DS1.1) относится к сигнальной последовательности тяжелой цепи, сигнальной последовательности легкой цепи, TIR тяжелой цепи, TIR легкой цепи, использованным в эксперименте.
Фиг.4: Установка для мониторинга частиц антитела с манипуляциями сигнальным пептидом легкой цепи. Клетки 64B4 растили в 25 мл фосфат-ограничивающей среды C.R.A.P. в течение 24 ч и для анализа SDS-PAGE получали растворимые фракции, также при нормализации осадка общего белка по оптической плотности (OD). Образцы из клеток, несущих плазмиду pBR-DS-5D5-1.1 (DS1.1), pBR-DS-5D5-1.2 (DS1.2), pBR-DM-5D5-1.1 (DM1.1), pBR-DM-5D5-1.2 (DM1.2), pBR-DD-5D5-1.1 (DD1.1), pBR-DD-5D5-1.2 (DD1.2), pBR-DP-5D5-1.1 (DP1.1) или pBR-DP-5D5-1.2, получали посредством электрофореза в геле SDS-PAGE (масса в кДа обозначена слева), переносили на нитроцеллюлозу и анализировали присутствие частиц, содержащих тяжелую или легкую цепь, с использованием антитела, специфичного к α-Fc или α-κLc, соответственно, как указано вдоль правой стороны изображений. Растворимые образцы (верхний блот) состояли из предположительно идентифицированных полос, соответствующих (сверху вниз): полноразмерному антителу, тяжелому-тяжелому-легкому (HHL), димеру тяжелой-легкой цепи (HL) или свободной тяжелой цепи. Нормализованные образцы общего белка (средний и нижний блот) восстанавливали с использованием 0,2 M DTT, чтобы разрушить структуру дисульфидной связи, и каждая отдельная дорожка мигрировала в одну полосу с кажущейся массой ~49 кДа, когда анализировали с использованием антитела к α-Fc. Когда анализировали с использованием антитела, специфичного к α-κLc, все дорожки мигрировали в одинарную или двойную полосу с кажущейся массой или ~25 кДа или ~27 кДа и ~25 кДа. Сокращения: S = сигнальная последовательность STII, M = сигнальная последовательность MalE, D = сигнальная последовательность DsbA, P = сигнальная последовательность PhoA. XX#.# (например, DS1.1) относится к сигнальной последовательности тяжелой цепи, сигнальной последовательности легкой цепи, TIR тяжелой цепи, TIR легкой цепи, использованным в эксперименте.
ФИГ.5: Установка для мониторинга частиц антитела с течением времени из 10 л ферментаций. Клетки 64B4 растили до высокой плотности клеток в 10 л ферментации в течение трех суток, при этом через равные интервалы времени (время взятия указанных выше образцов на каждой дорожке составляло несколько часов после инокуляции) брали образцы, из которых для анализа SDS-PAGE получали растворимые фракции, также при нормализации осадка общего белка по оптической плотности (OD). Образцы из клеток, несущих плазмиду pBR-SS-5D5-1.1, совместно экспрессирующих несущую шаперон плазмиду pJJ247 (SS1.1+шапероны), pBR-DD-5D5-1.1 с pJJ247 (DD1.1+шапероны), pBR-DS-5D5-1.1 с pJJ247 (DM1.1+шапероны) или pBR-DP-5D5-1.1 с pJJ247 (DP1.1+шапероны), разделяли посредством электрофореза в геле SDS-PAGE (масса кДа обозначена слева), переносили на нитроцеллюлозу и анализировали присутствие частиц, содержащих тяжелую или легкую цепь, с использованием антитела, специфичного к α-Fc или α-κLc, соответственно, как указано вдоль правой стороны изображений. Растворимые образцы (верхний блот) состояли из предположительно идентифицированных полос, соответствующих (сверху вниз): полноразмерному антителу, тяжелой-тяжелой-легкой цепи (HHL), димеру тяжелой-легкой цепи (HL), димеру легкой-легкой цепи (LL) или свободной легкой цепи. Нормализованные образцы общего белка (средний и нижний блот) восстанавливали с использованием 0,2 мМ DTT, чтобы разрушить структуру дисульфидной связи, и каждая отдельная дорожка мигрировала в одну полосу с кажущейся массой ~49 кДа, когда анализировали с использованием α-Fc. Когда анализировали с использованием антитела, специфичного к α-κLc, все дорожки мигрировали в одну полосу с кажущейся массой также ~25 кДа. Сокращения: S = сигнальная последовательность STII, M = сигнальная последовательность MalE, D = сигнальная последовательность DsbA, P = сигнальная последовательность PhoA. XX#.# (например, DS1.1) относится к сигнальной последовательности тяжелой цепи, сигнальной последовательности легкой цепи, TIR тяжелой цепи, TIR легкой цепи, использованным в эксперименте.
Фиг.6: Накопление зрелого PhoA в индуцирующих условиях. Клетки 27C7 растили в 25 мл фосфат-ограничивающей среды C.R.A.P. в течение 24 часов, растворимые фракции нормализовали по оптической плотность (OD) и получали их для анализа SDS-PAGE. Зрелый домен гена phoA E. coli сливали с указанными вариантами TIR (верх) DsbA или STII (низ). Присутствие белка в геле визуализировали посредством окрашивания кумаси синим. Возникала предположительно идентифицированная полоса, соответствующая зрелому домену PhoA (справа), с массой ~47 кДа (масса указана слева).
Фиг.7: изображены аминокислотные последовательности каркасной области (FR), CDR, первого константного домена (CL или CH1) и Fc-области (Fc) MetMAb (OA5D5v2). На фигуре раскрыты последовательности легкой цепи согласно SEQ ID NO: 57-60, 49-51 и 61, соответственно, в порядке появления и последовательности тяжелой цепи согласно SEQ ID NO: 62-65, 52-53 и 66-68, соответственно, в порядке появления. Изображенная последовательность Fc содержит мутации «отверстия» (полости) T366S, L368A и Y407V, как описано в WO 2005/063816.
Фиг.8: изображена последовательность полипептида Fc (SEQ ID NO: 47), содержащего мутацию «выступа» (выпуклости) T366W, как описано в WO 2005/063816. В одном из вариантов осуществления полипептид Fc, содержащий эту последовательность, формирует комплекс с полипептидом Fc, содержащим последовательность Fc с фиг.7, чтобы создать Fc-область.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Основные способы
Способы и процедуры, описанные или упомянутые в настоящем документе, как правило, широко распространены, и специалисты в данной области обычно их применяют с использованием стандартных способов, таких как, например, широко используемые способы, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al. eds., (2003)); серии METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE (R. I. Freshney, ed. (1987)).
Определения
Термин «вектор», как применяют в настоящем документе, предназначен для обозначения молекулы нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. Одним типом вектора является «плазмида», которая обозначает замкнутую кольцевую двухцепочечную ДНК, в которой могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, где дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, содержащие бактериальный участок начала репликации и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) можно встраивать в геном клетки-хозяина после введения в клетку-хозяина и, тем самым, реплицировать наряду с геномом организма-хозяина. Кроме того, определенные векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы обозначают в настоящем документе как «рекомбинантные экспрессирующие векторы» (или просто «рекомбинантные векторы»). В целом, экспрессирующие векторы, пригодные для способов рекомбинантной ДНК, часто имеют форму плазмид. В настоящем описании «плазмида» и «вектор» могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее общеупотребительной формой вектора.
Термин «цистрон», как применяют в настоящем документе, предназначен для обозначения генетического элемента, во многом эквивалентного трансляционной единице, содержащей нуклеотидную последовательность, кодирующую полипептидную цепь, и смежные управляющие области. «Смежные управляющие области» включают, например, область инициации трансляции (TIR; как определено в настоящем документе ниже) и область терминации.
«Полицистронный» экспрессирующий вектор относится к одному вектору, который содержит и экспрессирует несколько цистронов под регулирующим управлением одного промотора. Типичным примером полицистронного вектора является «бицистронный» вектор, который содержит и экспрессирует два различных полипептида под управлением одного промотора. При экспрессии бицистронного или полицистронного вектора сначала происходит транскрипция нескольких генов в виде одной транскрипционной единицы, а затем раздельная трансляция.
Экспрессирующий вектор с «отдельным цистроном» в соответствии с настоящим изобретением относится к одному вектору, содержащему по меньшей мере две отдельных пары промотор-цистрон, где каждый цистрон находится под управлением своего собственного промотора. При экспрессии экспрессирующего вектора с отдельным цистроном процессы как транскрипции, так и трансляции различных генов происходят отдельно и независимо.
«Область инициации трансляции» или TIR или область трансляционной инициации или последовательность трансляционной инициации, как применяют в настоящем документе, относится к области нуклеиновой кислоты, обеспечивающей эффективность инициации трансляции гена, представляющего интерес. В целом, TIR в конкретном цистроне охватывает участок связывания рибосомы (RBS) и последовательности, расположенные ближе к 5'- и 3'-концам относительно RBS. Определено, что RBS как минимум содержит область Шайна-Дальгарно и инициирующий кодон (AUG). Соответственно, TIR также содержит по меньшей мере часть последовательности нуклеиновой кислоты, подлежащей трансляции. Предпочтительно TIR по изобретению содержит сигнальную последовательность секреции, кодирующая сигнальный пептид, которая предшествует последовательности, кодирующей легкую или тяжелую цепь в цистроне. Вариант TIR содержит варианты последовательностей (в частности, замены) в области TIR, которые изменяют свойства TIR, такие как его интенсивность трансляции, как определено в настоящем документе ниже. Предпочтительно, вариант TIR по изобретению содержит замены последовательности в первых от 2 до приблизительно 14, предпочтительно от приблизительно 4 до 12, более предпочтительно приблизительно в 6 кодонах сигнальной последовательности секреции, которая предшествует последовательности, кодирующей легкую или тяжелую цепь в цистроне.
Термин «интенсивность трансляции», как применяют в настоящем документе, относится к измерению секретируемого полипептида в системе контроля, где один или несколько вариантов TIR используют для того, чтобы направлять секрецию полипептида и результаты сравнивают с TIR дикого типа или каким-либо другим контролем при тех же условиях культивирования и анализа. Не ограничиваясь какой-либо теорией, «интенсивность трансляции», как применяют в настоящем документе, может содержать, например, меру стабильности мРНК, эффективности связывания рибосомы с участком связывания рибосомы и способ транслокации через мембрану.
«Сигнальная последовательность секреции» или «сигнальная последовательность» относится к последовательности нуклеиновой кислоты, которая кодирует короткий сигнальный пептид, который можно использовать для того, чтобы направлять новый синтезированный белок, представляющий интерес, через клеточную мембрану, обычно через внутреннюю мембрану или как через внутреннюю, так и через наружную мембраны прокариотов. По существу, происходит секреция белка, представляющего интерес, такого как полипептид легкой или тяжелой цепи иммуноглобулина, в периплазму прокариотических клеток-хозяев или в среду для культивирования. Сигнальный пептид, кодируемый сигнальной последовательностью секреции, может быть эндогенным для клеток-хозяев или он может быть экзогенным, включая сигнальные пептиды, нативные для полипептида, подлежащего экспрессии. Сигнальные последовательности секреции типично присутствуют на аминоконце полипептида, подлежащего экспрессии, и типично происходит их ферментативное удаление между биосинтезом и секрецией полипептида из цитоплазмы. Таким образом, сигнальный пептид обычно не присутствует на зрелом белковом продукте.
«Функционально связанный» относится к непосредственному соседству двух или более компонентов, где описанные таким образом компоненты находятся в связи, позволяющей им функционировать предполагаемым для них образом. Например, промотор функционально связан с кодирующей последовательностью, если в цис он управляет или модулирует транскрипцию связанной последовательности. В целом, но не обязательно, последовательности ДНК, которые «функционально связаны», являются смежными и, когда необходимо соединить две области, кодирующие белки, или в случае секреторной лидерной последовательности, смежны и у них совпадает рамка считывания. Однако, несмотря на то, что функционально связанный промотор, как правило, расположен выше по направлению считывания кодирующей последовательности, он не обязательно смежен с ней. Функционально связанные энхансеры могут быть расположены выше по направлению считывания, внутри или ниже по направлению считывания кодирующих последовательностей и на существенном расстоянии от промотор. Формирование связи осуществляют посредством рекомбинантных способов, известных в данной области, например, используя способ ПЦР, посредством ренатурации или посредством лигирования по удобным сайтам рестрикции. Если удобные сайты рестрикции не существуют, то синтетические олигонуклеотидные адапторы или линкеры используют в соответствии со стандартной практикой.
«Регуляторные элементы» как применяют в настоящем документе, относятся к нуклеотидным последовательностям, присутствующим в цис, которые необходимы для транскрипции и трансляции полинуклеотида, кодирующего гетерологичный полипептид, в полипептиды. Транскрипционные регуляторные элементы в норме содержат промотор, расположенный ближе к 5'-концу относительно последовательности гена, подлежащей экспрессии, сайты инициации и терминации транскрипции и сигнальную последовательность полиаденилирования. Термин «сайт инициации транскрипции» относится к нуклеиновой кислоте в конструкции, соответствующей первой нуклеиновой кислоте, встроенной в первичный транскрипт, т.е. предшественник мРНК; сайт инициации транскрипции может перекрываться с промоторными последовательностями.
«Промотор» относится к полинуклеотидной последовательности, которая управляет транскрипцией гена или последовательности, с которой он функционально связан. Промотор содержит сигналы для связывания РНК полимеразы и инициации транскрипции. Используемые промоторы должны быть функциональными в том типе клеток-хозяев, в которых планируют экспрессировать выбранную последовательность. Большое число промоторов, включая конститутивные, индуцибельные и репрессируемые промоторы из множества различных источников, хорошо известны в данной области (и идентифицированы в базах данных, таких как GenBank) и доступны в виде или внутри клонированных полинуклеотидов (например, из таких хранилищ, как ATCC, а также других коммерческих или индивидуальных источников). При использовании индуцибельных промоторов активность промотора повышается или снижается в ответ на сигнал.
Термин «клетка-хозяин» (или «рекомбинантная клетка-хозяин»), как применяют в настоящем документе, предназначен для обозначения клетки, которую генетически изменили или которая способна быть изменена генетически посредством введения экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. Следует понимать, что такие термины предназначены для того, чтобы указывать не только на конкретную рассматриваемую клетку, но также на потомство такой клетки. Поскольку определенные модификации могут возникнуть в последующих поколения вследствие или мутации или влияния окружающей среды, такое потомство фактически может не быть идентичным материнской клетке, но все еще включено в объем термина «клетка-хозяин», как применяют в настоящем документе.
«Выделенный» полипептид (например, антитело) представляет собой полипептид, который идентифицирован и отделен и/или извлечен из компонента его естественной среды. Загрязняющие компоненты его естественной среды представляют собой вещества, которые будут препятствовать диагностическому или терапевтическому использованию антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления полипептид должен быть очищен (1) более чем на 95% по массе полипептида, как определено посредством способа Лоури, и наиболее предпочтительно более чем на 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности при использовании секвенатора с вращающимся стаканом, или (3) до гомогенности посредством SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) при восстанавливающих или невосстанавливающих условиях с использованием кумаси синего или, предпочтительно, окрашивания серебром. Выделенный полипептид содержит полипептид in situ в рекомбинантных клетках, поскольку должен не присутствовать по меньшей мере один компонент естественной среды полипептида. Однако, обычно выделенный полипептид получают посредством по меньшей мере одной стадии очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена от по меньшей мере одной загрязняющей молекулы нуклеиновой кислоты, с которой она, как правило, ассоциирована в естественном источнике нуклеиновой кислоты. Выделенная молекула нуклеиновой кислоты отлична о той формы или окружения, в котором ее находят в природе. Следовательно, выделенные молекулы нуклеиновой кислоты отличают от молекулы нуклеиновой кислоты, как она существует в естественных клетках. Однако, выделенная молекула нуклеиновой кислоты содержит молекулу нуклеиновой кислоты, содержащуюся в клетках, которые, как правило, экспрессируют нуклеиновую кислоту (например, нуклеиновую кислоту, кодирующую антитело), где, например, молекула нуклеиновой кислоты находится в хромосомном положении, отличном от такового в естественных клетках.
«Полинуклеотид» или «нуклеиновая кислота», как взаимозаменяемо используют в настоящем документе, относится к полимерами нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги или любой субстрат, который может быть встроен в полимер посредством ДНК или РНК полимеразы или посредством реакции синтеза. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если имеет место, модификацию структуры нуклеотида можно внести до или после сборки полимера. Последовательность нуклеотидов может быть прервана ненуклеотидными компонентами. Полинуклеотид дополнительно можно модифицировать после синтеза, например, посредством конъюгации с меткой. Другие типы модификации включают, например, «кэпы», замену одного или нескольких встречающихся в природе нуклеотидов на аналог, межнуклеотидные модификации, такие как, например, модификации незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и заряженными связями (например, фосфотиоатами, фосфодитиоатами и т.д.), те, которые содержат боковые фрагменты, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, ply-L-лизин и т.д.), те, которые содержат интеркаляторы (например, акридин, псорален и т.д.), те, которые содержат хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), те, которые содержат алкилирующие средства, те, которые содержат модифицированные связи (например, α-аномерные нуклеиновые кислоты и т.д.), а также немодифицированные формы полинуклеотида(ов). Кроме того, любые гидроксильные группы, как правило, присутствующие в сахарах, можно заменить, например, фосфонатными группами, фосфатными группами, защищенными стандартными защитными группами или активированными для получения дополнительных связей с дополнительными нуклеотидами, или можно конъюгировать с твердыми или полутвердыми носителями. 5'- и 3'-концевую OH можно фосфорилировать или заместить аминами или фрагментами органических блокирующих групп от 1 до 20 углеродных атомов. Другие гидроксилы также можно дериватизировать в стандартные защитные группы. Полинуклеотиды также могут содержать аналогичные формы рибозных или дезоксирибозных сахаров, которые, как правило, известны в данной области, включая, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидорибозу, аналоги карбоциклических сахаров, α-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и основные нуклеозидные аналоги, такие как метилрибозид. Одну или несколько фосфодиэфирных связей можно заменить на альтернативные линкерные группы. Эти альтернативные линкерные группы включают в качестве неограничивающих примеров варианты осуществления, где фосфат замещают на P(O)S («тиоат»), P(S)S («дитиоат»), (O)NR2 («амидат»), P(O)R, P(O)OR', CO или CH2 («формацеталь»), в которых каждый R или R' независимо представляет собой H или замещенный или незамещенный алкил (C1-20), необязательно содержащий простую эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не все связи в полинуклеотиде должны быть идентичными. Предшествующее описание применимо ко всем полинуклеотидам, упомянутым в настоящем документе, включая РНК и ДНК.
«Олигонуклеотид», как применяют в настоящем документе, как правило, относится к коротким, как правило, одноцепочечным, как правило, синтетическим полинуклеотидам, которые, как правило, но не обязательно, имеют длину приблизительно менее 200 нуклеотидов. Термины «олигонуклеотид» и «полинуклеотид» не являются взаимоисключающими. Приведенное выше описание для полинуклеотидов в равной и полной мере применимо к олигонуклеотидам.
Как применяют в настоящем документе, «полипептид» относится, как правило, к пептидам и белкам из любого клеточного источника, которые содержат приблизительно более десяти аминокислот. «Гетерологичные» полипептиды представляют собой полипептиды, которые инородны для используемой клетки-хозяина, такие как белок человека, продуцируемый в E. coli. Несмотря на то, что гетерологичный полипептид может быть прокариотическим или эукариотическим, предпочтительно он является эукариотическим, более предпочтительно полипептидом млекопитающего и наиболее предпочтительно полипептидом человека. Предпочтительно, он является рекомбинантно получаемым или рекомбинантным полипептидом.
«Гетерологичные» полипептиды представляют собой полипептиды, инородные для используемой клетки-хозяина, такие как белок человека, продуцируемый в E. coli. Несмотря на то, что гетерологичный полипептид может быть прокариотическим или эукариотическим, предпочтительно он является эукариотическими, более предпочтительно полипептидом млекопитающего и наиболее предпочтительно полипептидом человека. Предпочтительно, он является рекомбинантно получаемым или рекомбинантным полипептидом.
Примеры полипептидов млекопитающих включают такие молекулы, как, например, ренин, гормон роста, включая гормон роста человека; бычий гормон роста; рилизинг-фактор гормона роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; 1-антитрипсин; A-цепь инсулина; B-цепь инсулина; проинсулин; тромбопоэтин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания, такие как фактор VIIIC, фактор IX, тканевой фактор и фактор фон Виллебранда; противосвертывающие факторы, такие как протеин C; предсердный натрийуретический фактор; сурфактант легких; активатор плазминогена, такой как урокиназа или активатор плазминогена человека тканевого типа или мочи (t-PA), и его варианты, такие как RETEVASE™ и ТНКАЗА™; бомбезин; тромбин; гематопоэтический фактор роста; фактор некроза опухоли α и β; антитела к домену(ам) ErbB2, такие как 2C4 (WO 01/00245; гибридома ATCC HB-12697), которые связываются с областью во внеклеточном домене ErbB2 (например, любой один или несколько остатков в области приблизительно от остатка 22 приблизительно до остатка 584 ErbB2, включительно), энкефалиназа; сывороточный альбумин, такой как сывороточный альбумин человека; ингибирующее вещество Мюллера; A-цепь релаксина; B-цепь релаксина; прорелаксин; гонадотропин-ассоциированный пептид мыши; микробный белок, такой как β-лактамаза; ДНКаза; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы гормонов или факторов роста; интегрин; белок A или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор, полученный из головного мозга (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов, такой как NGF; кардиотрофины (фактор гипертрофии сердца), такие как кардиотрофин-1 (CT-1); тромбоцитарный фактор роста (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-α и TGF-β, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); дез(1-3)-IGF-I (IGF-I головного мозга), белки, связывающие инсулиноподобный фактор роста; белки CD, такие как CD-3, CD-4, CD-8 и CD-19; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; морфогенетический белок кости (BMP); интерферон, такой как интерферон-α, -β и -γ; сывороточный альбумин, такой как сывороточный альбумин человека (HSA) или бычий сывороточный альбумин (BSA); колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-10; антитело против HER-2; лиганд Apo2; супероксиддисмутаза; T-клеточные рецепторы; поверхностные мембранные белки; стимулятор гемолиза; вирусный антиген, такой как, например, часть оболочки СПИД; транспортные белки; хоуминг-рецепторы; адрессины; регуляторные белки; антитела; и фрагменты любого из перечисленных выше полипептидов.
Предпочтительные полипептиды в настоящем документе включают сывороточный альбумин человека (HSA), 2C4, тканевой фактор, средство против тканевого фактора, средство против CD20, средство против HER-2, херегулин, средство против IgE, средство против CD11a, средство против CD18, VEGF и их рецепторы и антитела к ним, такие как rhuFab V2 и AVASTIN™, гормон роста и его варианты, такие как hGH, рецепторы гормона роста, рилизинг-белок гормона роста (GHRP), LIV-1 (EP 1263780), TRAIL, фактор некроза опухоли (TNF) и антитела к нему, рецептор TNF и связанные антитела, IgG к рецептору TNF, ассоциированные с рецептором TNF факторы (TRAF) и их ингибиторы, фактор VIII, домен фактора VIII B, интерфероны, такие как интерферон-γ, трансформирующий фактор роста (TGF), такой как TGF-β, средство против TGF, такое как средство против TGF-β, активин, ингибин, средство против активина, средство против ингибина, тканевые активаторы плазминогена и их варианты, такие как t-PA, RETEPLASE™ и ТНКаза, антитела против Fas, лиганд Apo-2; ингибитор лиганда Apo-2; рецептор Apo-2, Apo-3, факторы апоптоза, Ced-4, DcR3, рецептор смерти и агонистические антитела (DR4, DR5), лимфотоксин (LT), пролактин, рецептор пролактина, белки SOB, WISP (wnt-индуцируемые секретируемые белки), нейротоксин-3 (NT-3), фактор роста нервов (NGF) и средство против NGF, ДНКазу, антиген гепатита, антиген простого герпеса, лептин, инсулиноподобные факторы роста (IGF), такие как IGF-1 и IGF-2 и связывающие их белки и рецепторы, такие как IGFBP-1-IGFBP-6, инсулин, факторы роста фибробластов (FGF), такие как FGF-17, белок Toll, лиганды TIE, CD40 и средство против CD40, иммуноадгезины, субтилизин, фактор роста гепатоцитов (HGF), тромбопоэтин (TPO), интерлейкины, такие как IL-2, IL-12, IL-17, IL-22, IL-8, IL-9 и антитела к ним, и простатоспецифический антиген злокачественной опухоли (PSCA).
Особенно предпочтительными полипептидами являются рекомбинантные полипептиды, более предпочтительно антитела, которые включают моноклональные антитела и гуманизированные антитела. Такие антитела могут представлять собой полноразмерные антитела или фрагменты антител. Более предпочтительно, эти антитела представляют собой антитела человека или гуманизированные антитела. Еще более предпочтительно антитело представляет собой антитело против c-met, против IgE, против CD18, против VEGF, против тканевого фактора, 2C4, против Her-2, против CD20, против CD40 или против CD11a. Фрагменты антител, охватываемые определением полипептида, предпочтительно содержат легкую цепь, более предпочтительно легкую цепь κ. Такие предпочтительные фрагменты включают, например, Fab, Fab', F(ab')2 или слитый белок F(ab')2-лейциновая молния (LZ), и антитело с одним Fab фрагментом.
«Экспрессия» белка относится к превращению информации, кодируемой генов, в матричную РНК (мРНК) и затем в белок.
«Иммуноконъюгат» (взаимозаменяемо обозначаемый как «конъюгат антитело-лекарственное средство» или «КАЛС») обозначает антитело, конъюгированное с одним или несколькими цитотоксическими средствами, такими как химиотерапевтическое средство, лекарственное средство, ингибирующее рост средство, токсин (например, белковый токсин, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты), или радиоактивный изотоп (т.е. радиоконъюгат).
«Блокирующее» антитело или «антагонист» антитела представляет собой то, что ингибирует или снижает биологическую активность того антигена, который оно связывает. В некоторых вариантах осуществления блокирующие антитела или антагонистические антитела полностью ингибируют биологическую активность антигена.
«Агонистическое антитело», как применяют в настоящем документе, представляет собой антитело, которое имитирует по меньшей мере одну функциональную активность полипептида, представляющего интерес (например, HGF).
«Аффинность связывания», как правило, относится к силе совокупных нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнера по связыванию (например, антигена). Если не указано иначе, как применяют в настоящем документе, «аффинность связывания» относится к фактической аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающееся пары (например, антитело и антиген). Аффинность молекулы X к ее партнеру Y, как правило, можно представить посредством константы диссоциации (Kd). Желательно Kd составляет 1×10-7, 1×10-8, 5×10-8, 1×10-9, 3×10-9, 5×10-9 или даже 1×10-10 или выше. Аффинность можно измерить посредством общеизвестных в данной области способов, включая те, что описаны в настоящем документе. Антитела с низкой аффинностью, как правило, связывают антиген медленно и склонны к легкой диссоциации, тогда как антитела с высокой аффинностью как правило, связывают антиген быстрее и склонны дольше оставаться связанными. В данной области известны различные способы измерения аффинности связывания, среди которых можно использовать любые для целей по настоящему изобретению. Конкретные иллюстративные варианты осуществления описаны далее.
В одном из вариантов осуществления «Kd» или «значение Kd» в соответствии с данным изобретением измеряют посредством анализа связывания радиоактивно меченного антигена (RIA), который осуществляют с использованием Fab-версии антитела, представляющего интерес, и его антигена, как описано посредством следующего анализа, в котором измеряют аффинность связывания в растворе Fab с антигеном посредством уравновешивания Fab с минимальной концентрацией меченного [125I] антигена в присутствие серии титров немеченого антигена, и затем захватывая связанный антиген на планшете, покрытом антителом против Fab (Chen, et al., (1999) J. Mol. Biol. 293:865-881). Чтобы создать условия для анализа, микротитровальные планшеты (Dynex) покрывают в течение ночи с использованием 5 мкг/мл захватывающим антителом против Fab (Cappel Labs) в 50 мМ карбонате натрия (pH 9,6), а затем блокируют 2% (масс./об.) бычьим сывороточным альбумином в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В неадсорбирующем планшете (Nunc #269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями Fab, представляющего интерес (например, согласующегося с оценкой антитела против VEGF, Fab-12, в Presta et al., (1997) Cancer Res. 57:4593-4599). Затем Fab, представляющий интерес, инкубируют в течение ночи; однако инкубация может продолжаться в течение более длительного периода (например, 65 часов), чтобы гарантировать, что равновесие достигнуто. После этого смеси переносят в захватывающий планшет для инкубации при комнатной температуре (например, в течение одного часа). Затем раствор удаляют и планшет промывают восемь раз в 0,1% Tween-20 в PBS. Когда планшеты высохли, добавляют сцинтиллятор 150 мкл/лунка (Micro Scint-20; Packard) и выполняют подсчет планшетов на счетчике γ-излучения Topcount (Packard) в течение десяти минут. Концентрации каждого Fab, который дает 20% максимального связывания или менее выбирают для использования в анализе конкурентного связывания. Согласно другому варианту осуществления, Kd или значение Kd измеряют, используя анализ поверхностного плазмонного резонанса с использованием BIAcore™-2000 или a BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами с иммобилизованными антигенами CM5 при ~10 единицах ответа (RU). В кратком изложении, карбоксиметилированные декстрановые биосенсорные чипы (CM5, BIAcore Inc.) активировали N-этил-N'-(3-диметиламинопропил)-карбодиимидгидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией при скорости потока 5 мкл/мин, чтобы достичь приблизительно 10 единиц ответа (RU) спаренного белка. После инъекции антигена инъецируют 1 M этаноламин, чтобы блокировать непрореагировавшие группы. Для измерения кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) инъецируют в PBS с 0,05% Tween 20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. В некоторых вариантах осуществления последующие модификации используют для способа анализа поверхностного плазмонного резонанса: антитело иммобилизуют на биосенсорных чипах CM5, чтобы достичь приблизительно 400 RU, и для измерения кинетики инъецируют двукратные серийные разведения целевого белка в буфере PBST при 25°C со скоростью потока приблизительно 30 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания один-к-одному Ленгмюра (BIAcore Evaluation Software версии 3.2) посредством одновременной аппроксимации сенсограммы ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют в виде отношения koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. Если скорость образования превышает 106 М-1с-1 согласно описанному выше анализу поверхностного плазмонного резонанса, то скорость ассоциации можно определить, используя способ гашения флуоресценция, в котором измеряют повышение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 25°C 20 нМ антитела против антигена (Fab-формы) в PBS, pH 7,2, в присутствие увеличивающихся концентраций антигена, как измеряют в спектрометре, таком как оборудованным остановкой потока спектрофотометр (Aviv Instruments) или спектрофотометр SLM-Aminco серии 8000 (ThermoSpectronic) с красной перемешиваемой кюветой.
«Скорость образования» или «скорость ассоциации» или «kon» в соответствии с данным изобретением также можно определить, используя тот же способ поверхностного плазмонного резонанса, который описан выше, с помощью BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами с иммобилизованными антигенами CM5 при ~10 единицах ответа (RU). В кратком изложении, карбоксиметилированные декстрановые биосенсорные чипы (CM5, BIAcore Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимидгидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/мин, чтобы достичь приблизительно 10 единиц ответа (RU) спаренного белка. После инъекции антигена 1 M этаноламин инъецируют, чтобы блокировать непрореагировавшие группы. Для измерения кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) инъецируют в PBS с 0,05% Tween 20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. В некоторых вариантах осуществления последующие модификации используют для способа анализа поверхностного плазмонного резонанса: антитело иммобилизуют на биосенсорных чипах CM5, чтобы достичь приблизительно 400 RU, и для измерения кинетики двукратные серийные разведения целевого белка инъецируют в буфере PBST при 25°C со скоростью потока приблизительно 30 мкл/мин. Скорость ассоциации (kon) и скорость диссоциации (koff) вычисляют с использованием простой модели связывания один-к-одному Ленгмюра (BIAcore Evaluation Software версии 3.2) посредством одновременной аппроксимации сенсограммы ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляли в виде отношения koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. Однако, если скорость образования превышает 106 M-1с-1 согласно описанному выше анализу поверхностного плазмонного резонанса, то скорость образования предпочтительно определяют, используя способ гашения флуоресценции, в котором измеряют повышение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 25°C 20 нМ антитела против антигена (Fab-формы) в PBS, pH 7,2, в присутствие увеличивающиеся концентрации антигена, как измеряют в спектрометре, таком как оборудованным остановкой потока спектрофотометр (Aviv Instruments) или спектрофотометр SLM-Aminco серии 8000 (ThermoSpectronic) с красной перемешиваемой кюветой.
«Голое антитело» представляет собой антитело, которое не конъюгировано с гетерологичной молекулой, такой как цитотоксический фрагменты или радиоактивная метка.
Антитело, обладающее «биологической характеристикой» обозначенного антитела, представляет собой то, которое обладает одной или несколькими биологическими характеристиками этого антитела, которые отличают его от других антител, которые связываются с тем же антигеном.
Для того чтобы осуществлять скрининг антител, которые связываются с эпитопом на антигене, связанном антителом, представляющем интерес, можно осуществлять обычный перекрестный конкурентный анализ, такой как тот, который описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988).
Чтобы увеличить время полужизни антитела или полипептида, содержащего аминокислотные последовательности по данному изобретению, можно присоединить эпитоп связывания рецептора спасения к антителу (в частности, к фрагменту антитела), как описано, например, в патенте США 5739277. Например, молекулу нуклеиновой кислоты, кодирующую эпитоп связывания рецептора спасения, можно связать с сохранением рамки считывания с нуклеиновой кислотой, кодирующей полипептидную последовательность по данному изобретению, с тем, чтобы слитый белок, экспрессируемый посредством сконструированной молекулы нуклеиновой кислоты, содержал эпитоп связывания рецептора спасения и полипептидную последовательность по данному изобретению. Как применяют в настоящем документе, термин «эпитоп связывания рецептора спасения» относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за повышение времени полужизни молекулы IgG в сыворотке in vivo (например, Ghetie et al., Ann. Rev. Immunol. 18:739-766 (2000), таблица 1). Антитела с заменами в их Fc-области и увеличенным временем полужизни в сыворотке также описаны в WO 00/42072, WO 02/060919; Shields et al., J. Biol. Chem. 276:6591-6604 (2001); Hinton, J. Biol. Chem. 279:6213-6216 (2004)). В другом варианте осуществления время полужизни в сыворотке также можно увеличить, например, посредством присоединения другой полипептидной последовательности. Например, антитела или другие полипептиды, которые можно использовать в способах по изобретению, можно присоединять к сывороточному альбумину или части сывороточного альбумина, которая связывается с рецептором FcRn, или связывающему сывороточный альбумин пептиду с тем, чтобы сывороточный альбумин связывался с антителом или полипептидом, например, такие полипептидные последовательности раскрыты в WO 01/45746. В одном из предпочтительных вариантов осуществления прикрепляемый пептид сывороточного альбумина содержит аминокислотную последовательность DICLPRWGCLW (SEQ ID NO: 48). В другом варианте осуществления время полужизни Fab увеличивают этими способами. Последовательности связывающего сывороточный альбумин пептида также см. в Dennis et al. J. Biol. Chem. 277:35035-35043 (2002).
«Фрагментом» обозначают часть полипептида или молекулы нуклеиновой кислоты, которая содержит предпочтительно - по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или более полноразмерной эталонной молекулы нуклеиновой кислоты или полипептида. Фрагмент может содержать 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100, 200, 300, 400, 500, 600 или более нуклеотидов или 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 120, 140, 160, 180, 190, 200 аминокислот или более.
Термины «антитело» и «иммуноглобулин» используют взаимозаменяемо в самом широком смысле, и они включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, поливалентные антитела, полиспецифические антитела (например, биспецифические антитела при условии, что они проявляют желательную биологическую активность), а также могут включать определенные фрагменты антител (как более подробно описано в настоящем документе). Антитело может представлять собой антитело человека, гуманизированное и/или аффинно зрелое антитело.
Термин «вариабельный» относится к тому факту, что определенные части вариабельных доменов сильно различаются по последовательности среди антител, и они используются в связывании и специфичности каждого конкретного антитела в отношении его конкретного антигена. Однако, вариабельность не равномерно распределена по всем вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых определяющим комплементарность областями (CDR) или гипервариабельными областями вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высоко консервативные части вариабельных доменов называют каркасной областью (FR). Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре области FR, в значительной степени принимающие конфигурацию β-листа, которые соединены тремя CDR, которые формируют петли, соединяющие структуру β-листа и в некоторых случаях формирующие ее часть. CDR в каждой цепи удерживаются вместе в непосредственной близости посредством областей FR и вместе с CDR из другой цепи участвуют в формировании антигенсвязывающего участка антитела (см. Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности.
Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых «Fab» фрагментами, каждый с одним антигенсвязывающим участком, и остаточный «Fc» фрагмент, в названии которого отражена его способность легко кристаллизоваться. Обработка пепсином дает фрагмент F(ab')2, который содержит два антигенсвязывающих участка и все еще способен к перекрестному связыванию антигена.
«Fv» представляет собой минимальный фрагмент антитела, который содержит полный участок распознавания и связывания антигена. В двухцепочечных частицах Fv эта область состоит из димера вариабельных доменов одной тяжелой и одной легкой цепи, связанных крепкой нековалентной связью. В одноцепочечных частицах Fv вариабельные домены одной тяжелой и одной легкой цепи могут быть ковалентно связаны посредством гибкого пептидного линкера так, что легкая и тяжелая цепи могут объединяться в «димерную» структуру, аналогичную таковой в двухцепочечных частицах Fv. В этой конфигурации три CDR каждого вариабельного домена взаимодействуют, чтобы определить антигенсвязывающий участок на поверхности димера VH-VL. Совместно шесть CDR дают антителу антигенсвязывающую специфичность. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичных для определенного антигена) способен распознавать и связывать антиген, хотя и с меньшей аффинностью, чем целый участок связывания.
Fab фрагмент также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Фрагменты Fab' отличаются от Fab фрагментов добавкой из нескольких остатков на карбоксильном конце домена CH1 тяжелой цепи, содержащей один или несколько цистеинов из шарнирной области антитела. В настоящем документе Fab'-SH обозначает Fab', в котором остаток(и) цистеина из константных доменов несут свободную тиоловую группу. F(ab')2 фрагменты антител первоначально получали в виде пар фрагментов Fab', которые содержат шарнирные цистеины между ними. Также известны другие химические способы связывания фрагментов антител.
«Легкие цепи» антител (иммуноглобулинов) позвоночного любого вида можно отнести к одному из двух четко различаемых типов, называемых каппа (κ) и лямбда (λ), на основе аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей, иммуноглобулины можно отнести к различным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM и некоторые из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называют α, Δ, ε, γ и μ, соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. «Фрагменты антител» содержат только часть интактного антитела, где часть предпочтительно сохраняет по меньшей мере одну, предпочтительно большинство или все функции, в норме связанные с этой частью, когда она находится в интактном антителе. Примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv фрагменты; диатела; линейные антитела; одноцепочечные молекулы антител; и полиспецифические антитела, сформированные из фрагментов антител. В одном из вариантов осуществления фрагмент антитела содержит антигенсвязывающий участок интактного антитела и, таким образом, сохраняет способность связывать антиген. В другом варианте осуществления фрагмент антитела, например, содержащий Fc-область, сохраняет по меньшей мере одну биологическую функцию, в норме связанную с Fc-областью, когда она находится в интактном антителе, такую как связывание FcRn, модуляция времени полужизни антитела, функция АЗКЦ и связывание комплемента. В одном из вариантов осуществления фрагмент антитела представляет собой моновалентное антитело, которое имеет время полужизни in vivo по существу схожее с интактным антителом. Например, такой фрагмент антитела может содержать на антигенсвязывающем фрагменте, соединенном с Fc, последовательность, способную придавать фрагменту стабильность in vivo.
Термин «гипервариабельная область», «HVR» или «HV», когда используется в настоящем документе, относится к областям вариабельных доменов антитела, которые гипервариабельны по последовательности и/или формируют петли, определяемые структурой. В целом, антитела содержат шесть гипервариабельных областей; три в VH (H1, H2, H3) и три в VL (L1, L2, L3). В настоящем документе используется и охвачено множество описаний гипервариабельных областей. Определяющие комплементарность области по Kabat (CDR) основаны на вариабельности последовательностей и являются наиболее общеупотребительными (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Chothia вместо этого опирается на расположение структурных петель (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)). Гипервариабельные области AbM представляют компромисс между CDR по Kabat и структурными петлями по Chothia, и их используют в Oxford Molecular's AbM antibody modeling software. «Контактные» гипервариабельные области основаны на анализе доступных комплексных кристаллических структур. Остатки из каждой из этих гипервариабельных областей указаны ниже.
Гипервариабельные области могут содержать следующие «протяженные гипервариабельные области»: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (H2) и 93-102, 94-102 или 95-102 (H3) в VH. Остатки вариабельного домена пронумерованы в соответствии с Kabat et al., выше, для каждого из этих определений.
Остатки «каркасной области» или «FR» представляют собой те остатки вариабельного домена, которые отличны от остатков гипервариабельной области, как определено в настоящем документе.
«Гуманизированные» формы антител, не принадлежащих человеку, (например, антител мыши) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не принадлежащему человеку. Преимущественно гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиентного антитела заменяют на остатки из гипервариабельной области вида, не являющегося человеком (донорное антитело), такого как мышь, крыса, кролик или примат, не являющийся человеком, которая обладает желаемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют на соответствующие не принадлежащие человеку остатки. Кроме того, гуманизированные антитела могут содержать остатки, которые не найдены в реципиентном антителе или в донорном антителе. Эти модификации выполняют для того, чтобы дополнительно улучшить характеристики антитела. В целом, гуманизированное антитело должно содержать по существу все из по меньшей мере одного и типично из двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют таковым иммуноглобулина, не принадлежащего человеку, и все или по существу все из FR из последовательности иммуноглобулина человека. Гуманизированное антитело необязательно должно содержать по меньшей мере часть константной области иммуноглобулина (Fc), типично из иммуноглобулина человека. Более подробно, см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). Также см. Следующие обзорные статьи и цитируемые в них ссылки: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1: 105-115 (1998); Harris, Biochem. Soc. Transactions 23: 1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
«Химерные» антитела (иммуноглобулины) имеют часть тяжелой и/или легкой цепи, идентичную или гомологичную соответствующим последовательностям в антителах, полученных от конкретных видов или принадлежащих к конкретному классу или подклассу антител, тогда как остальная цепь(и) идентична или гомологична соответствующим последовательностям антител, полученным из других видов или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител при условии, что они проявляют желательную биологическую активность (патент США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)). Гуманизированное антитело, как применяют в настоящем документе, является подмножеством химерных антител.
«Одноцепочечные Fv» или «scFv» фрагменты антител содержат домены VH и VL антитела, где эти домены присутствуют в одной полипептидной цепи. В целом, scFv полипептид дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет scFv формировать желаемую структуру для связывания антигена. Обзор по scFv см. в Pluckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
«Антиген» представляет собой предварительно определяемый антиген, с котором антитело может избирательно связываться. Антиген-мишень может представлять собой полипептид, углевод, нуклеиновую кислоту, липид, гаптен или другое встречающееся в природе или синтетическое соединение. Предпочтительно, антиген-мишень представляет собой полипептид.
Термин «диатела» относится к небольшим фрагментам антител с двумя антигенсвязывающими участками, эти фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одну и ту же полипептидную цепь (VH-VL). Используя линкер, который слишком короток для того, чтобы сделать возможным образование пары между двумя доменами в одной и той же цепи, домены заставляют образовывать пару с комплементарными доменами другой цепи и создавать два антигенсвязывающих участка. Более полно диатела описаны, например, в EP 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993).
«Антитело человека» представляет собой то, которое обладает аминокислотной последовательностью, которая соответствует таковой антитела, продуцируемого человеком и/или полученного с использованием любых способов получения антител человека, как описано в настоящем документе. Определение антитела человека конкретно исключает гуманизированное антитело, содержащее принадлежащие человеку антигенсвязывающие остатки.
«Аффинно зрелое» антитело представляет собой антитело с один или несколькими изменениями в его одном или нескольких CDR, которые ведут к улучшению аффинности антитела по отношению антигену в сравнении с родительским антителом, которое не обладает этим изменением(ями). Предпочтительно аффинно зрелые антитела должны обладать наномолярными или даже пикомолярными аффинностями по отношению к антигену-мишени. Аффинно зрелые антитела получают известными в данной области способами. Marks et al. Bio/Technology 10:779-783 (1992) описывают созревание аффинности посредством перетасовки доменов VH и VL. Случайный мутагенез CDR и/или каркасных остатков описан авторами Barbas et al., Proc Nat. Acad. Sci, USA 91:3809-3813 (1994); Schier et al., Gene 169: 147-155 (1995); Yelton et al., J. Immunol. 155: 1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995); и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
«Рецептор Fc» или «FcR» описывает рецептор, который связывается с Fc-областью антитела. Предпочтительный FcR представляет собой нативную последовательность FcR человека. Кроме того, предпочтительный FcR представляет собой тот, который связывает антитело IgG (γ-рецептор) и включает подклассы рецепторов FcγRI, FcγRII и FcγRIII, включая аллельные варианты этих рецепторов и формы с альтернативным сплайсингом. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), который имеет схожие аминокислотные последовательности, которые отличаются в первую очередь их цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит иммунорецепторный основанный на тирозине активирующий мотив (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный основанный на тирозине ингибирующий мотив (ITIM) в своем цитоплазматическом домене, (см. обзор M. in Daëron, Annu. Rev. Immunol. 15:203-234 (1997)). Обзор FcR приведен у Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). В настоящем документе другие FcR, включая те, которые будут идентифицированы в будущем, охватывает термин «FcR». Термин также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регулирует гомеостаз иммуноглобулинов. В WO 00/42072 (Presta) описаны варианты антител с улучшенным или уменьшенным связыванием с FcR. Содержание этой публикации патента конкретно включено в настоящий документ в качестве ссылки. Также см. Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001).
Известны способы измерения связывания с FcRn (см., например, Ghetie 1997, Hinton 2004). Связывание с FcRn человека in vivo и время полужизни в сыворотке для полипептидов, связывающих FcRn человека с высокой аффинностью, можно анализировать, например, в трансгенных мышах или трансфицированных клеточных линиях человека, экспрессирующих FcRn человека, или в приматах, которым вводят полипептиды вариантов Fc.
Полипептидные варианты с измененными аминокислотными последовательностями Fc-области и повышенной или пониженной способностью к связыванию C1q, описаны в патенте США № 6194551B1 и WO 99/51642. Содержание этих публикаций патентов конкретно включены в настоящий документ в качестве ссылки. Также см. Idusogie et al., J. Immunol. 164:4178-4184 (2000).
Термин «Fc-область», как применяют в настоящем документе, как правило, относится к димерному комплексу, содержащему C-концевые полипептидные последовательности тяжелой цепи иммуноглобулина, где C-концевая полипептидная последовательность представляет собой ту, которую можно получить посредством расщепления интактного антитела папаином. Fc-область может содержать нативные последовательности Fc или варианты последовательностей Fc. Несмотря на то, что границы последовательности Fc тяжелой цепи иммуноглобулина могут варьировать, обычно последовательность Fc тяжелой цепи IgG человека определяют как простирающуюся от аминокислотного остатка приблизительно в положении Cys226, или приблизительно от положения Pro230, до карбоксильного конца последовательности Fc. Последовательность Fc иммуноглобулина в целом содержит два константных домена, домен CH2 и домен CH3, и необязательно содержит домен CH4. Под «полипептидом Fc» в настоящем документе понимают один из полипептидов, который составляет Fc-область. Полипептид Fc можно получить из любого подходящего иммуноглобулина, такого как субтипы IgG1, IgG2, IgG3 или IgG4, IgA, IgE, IgD или IgM. В некоторых вариантах осуществления полипептид Fc содержит частично или полностью шарнирную последовательность дикого типа (как правило, на своем N-конце). В некоторых вариантах осуществления полипептид Fc не содержит функциональную шарнирную последовательность или шарнирную последовательность дикого типа.
Как применяют в настоящем документе, «мутант антитела» или «вариант антитела» относится к варианту аминокислотной последовательности антитела, где модифицированы один или несколько аминокислотных остатков антитела, зависящего от вида. Такие мутанты неизбежно имеют менее чем 100% идентичность или сходство последовательности с антителом, зависящим от вида. В одном из вариантов осуществления мутант антитела должен иметь аминокислотную последовательность, обладающую по меньшей мере 75% идентичностью или сходством аминокислотных последовательностей с аминокислотной последовательностью вариабельного домена или тяжелой или легкой цепи антитела, зависящего от вида, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%. Идентичность или сходство по отношению к этой последовательности определяют в настоящем документе как процентную долю аминокислотных остатков в последовательности-кандидате, которые идентичны (т.е. такие же остатки) или схожи (т.е. аминокислотный остаток из той же группы, основанной на общих свойствах боковой цепи, см. ниже) с остатками антитела, зависящего от вида, после выравнивания последовательностей и, в случае необходимости, вставления пропусков, чтобы достичь максимальный процент идентичности последовательностей. Ни одно из N-концевого, C-концевого или внутреннего удлинений, делеций или вставок в последовательность антитела за пределами вариабельного домена не следует рассматривать в качестве влияющего на идентичность или сходство последовательностей.
«Нарушение» или «заболевание» представляет собой любое состояние, на которое оказывает благоприятный эффект лечение с использованием вещества/молекулы или способа по изобретению. Оно включает хронические и острые нарушения или заболевания, включая те патологические состояния, которые предрасполагают млекопитающее к обсуждаемому нарушению. Неограничивающие примеры нарушений, подлежащих лечению в настоящем документе, включают злокачественные и доброкачественные опухоли; карциному, бластому и саркому.
«Лечение» относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. Нуждающиеся в лечении включают тех, кто уже имеет доброкачественную, предзлокачественную или неметастатическую опухоль, а также тех, у которых следует предотвратить возникновение или повторное возникновение злокачественной опухоли.
Термин «терапевтически эффективное количество» относится к количеству терапевтического средства для лечения или предотвращения заболевания или нарушения у млекопитающего. В случае злокачественных опухолей, терапевтически эффективное количество терапевтического средства может снижать число клеток злокачественных; снижать размер первичной опухоли; ингибировать (т.е. до некоторой степени замедлять и предпочтительно останавливать) инфильтрацию периферических органов злокачественными клетками; ингибировать (т.е. до некоторой степени замедлять и предпочтительно останавливать) метастазы опухоли; ингибировать, до некоторой степени, рост опухоли; и/или облегчать до некоторой степени один или несколько симптомов, ассоциированных с нарушением. В тех случаях, когда лекарственное средство может предотвращать рост и/или убивать существующие злокачественные клетки, оно может быть цитостатическим и/или цитотоксическим. Для терапии злокачественной опухоли эффективность in vivo можно измерить, например, посредством оценки длительности выживания, времени до развития заболевания (TTP), скоростей ответа (RR), длительности ответа и/или качества жизни.
«Аутоиммунное заболевание» в настоящем документе представляет собой незлокачественное заболевание или нарушение, возникающее из и направленное против собственных тканей индивидуума. Аутоиммунные заболевания в настоящем документе специально исключают озлокачествленные или злокачественные заболевания или состояния, в частности, исключая B-клеточную лимфому, острый лимфобластный лейкоз (ALL), хронический лимфоцитарный лейкоз (CLL), волосатоклеточный лейкоз и хронический миелобластный лейкоз. Примеры аутоиммунных заболеваний или нарушений включают в качестве неограничивающих примеров воспалительные реакции, такие как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); системную склеродермию и склероз; реакции, ассоциированные с воспалительным заболеванием кишечника (таким как болезнь Крона и язвенный колит); респираторный дистресс синдром (включая респираторный дистресс синдром взрослых; РДСВ); дерматит; менингит; энцефалит; увеит; колит; гломерулонефрит; аллергические состоянии, такие как экзема и астма и другие состояния, вовлекающие инфильтрацию T-клетками и хронические воспалительные реакции; атеросклероз; дефицит адгезии лимфоцитов; ревматоидный артрит; системную красную волчанку (СКВ); сахарный диабет (например, сахарный диабет I типа или инсулинзависимый сахарный диабет); рассеянный склероз; синдром Рейно; аутоиммунный тиреоидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные ответы, ассоциированные с острой и замедленной гиперчувствительностью, опосредованной цитокинами и T-лимфоцитами, которые типично находят при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; пернициозной анемии (болезнь Аддисона); заболевания, в которые вовлечен диапедез лейкоцитов; воспалительные нарушения центральной нервной системы (ЦНС); синдром полиорганного повреждения; гемолитическая анемия (включая в качестве неограничивающих примеров криоглобинемию или Кумбс-положительную анемию); миастения гравис; заболевания, опосредованные комплексами антиген-антитело; синдром Гудпасчера; антифосфолипидный синдром; аллергический неврит; болезнь Грейвса; миастенический синдром Ламберта-Итона; буллезный пемфигоид; пемфигус; аутоиммунные полиэндокраниопатии; болезнь Рейтера; синдром скованного человека; болезнь Бехчета; гигантоклеточный артериит; нефрит иммунных комплексов; IgA-нефропатия; IgM-полинейропатии; иммунная тромбоцитопеническая пурпура (ИТП) или аутоиммунная тромбоцитопения и т.д.
Термины «злокачественная опухоль» и «злокачественный» относятся к или описывают физиологическое состояние у млекопитающих, которое типично характеризуется неконтролируемым клеточным ростом. В это определение включены доброкачественные и злокачественные опухоли. Под «ранней стадией злокачественной опухоли» или «ранней стадией опухоли» понимают злокачественную опухоль, которая не инвазивна или не метастазирует или классифицирована как стадия 0, I или II злокачественной опухоли. Примеры злокачественной опухоли включают в качестве неограничивающих примеров карциному, лимфому, бластому (включая медуллобластому и ретинобластому), саркому (включая липосаркому и синовиальноклеточную саркому), нейроэндокринные опухоли (включая карциноидные опухоли, гастриному и злокачественную опухоль островковых клеток), мезотелиому, шванному (включая нейрому слухового нерва), менингиому, аденокарциному, меланому и лейкоз или злокачественные новообразования лимфоидной ткани. Более конкретные примеры таких злокачественных опухолей включают плоскоклеточную злокачественную опухоль (например, эпителиальную плоскоклеточную злокачественную опухоль), злокачественную опухоль легких включая мелкоклеточную злокачественную опухоль легких (SCLC), немелкоклеточную злокачественную опухоль легких (NSCLC), аденокарциному легкого и сквамозную карциному легкого, злокачественную опухоль брюшины, гепатоцеллюлярную злокачественную опухоль, злокачественную опухоль желудка, включая злокачественную опухоль желудочно-кишечного тракта, злокачественную опухоль поджелудочной железы, глиобластому, злокачественную опухоль шейки матки, злокачественную опухоль яичников, злокачественную опухоль печени, злокачественную опухоль мочевого пузыря, гепатому, злокачественную опухоль молочной железы (включая метастатическую злокачественную опухоль молочной железы), злокачественную опухоль ободочной кишки, злокачественную опухоль прямой кишки, злокачественную опухоль толстой кишки, карциному эндометрия или матки, карциному слюнной железы, злокачественную опухоль почки, злокачественную опухоль предстательной железы, злокачественную опухоль женских наружных половых органов, злокачественную опухоль щитовидной железы, печеночную карциному, анальную карциному, пенильную карциному, злокачественную опухоль яичка, злокачественную опухоль пищевода, опухоли желчных путей, а также злокачественную опухоль головы и шеи и множественную миелому.
Термин «предзлокачественный» относится к состоянию или росту, который типично предшествует или развивается в злокачественную опухоль. «Предзлокачественный» рост должен иметь клетки, которые характеризуются ненормальной регуляцией клеточного цикла, пролиферации или дифференциации, которые можно определить посредством маркеров регуляции клеточного цикла, клеточной пролиферации или дифференциации.
Под «дисплазией» понимают любой ненормальный рост или развитие ткани, органа или клеток. Предпочтительно, дисплазия имеет высокую степень или является предзлокачественной.
Под «метастазами» понимают распространение злокачественной опухоли из ее первичной локализации в другие места в организме. Злокачественные клетки могут отрываться от первичной опухоли, проникать в лимфатические и кровеносные сосуды, циркулировать с током крови и расти в удаленном очаге (метастазировать) в нормальных тканях в другом месте в организме. Метастазы могут быть локализованными или отдаленными. Метастаз представляет собой последовательный процесс, зависящий от отрыва опухолевых клеток от первичной опухоли, переноса по кровотоку и остановки в удаленном месте. В новом месте клетки обеспечивают кровоснабжение и могут расти до формирования массы, опасной для жизни.
Как стимулирующие, так и ингибирующие молекулярные пути в опухолевой клетке регулируют ее поведение, и взаимодействия между опухолевой клеткой и клетками организма-хозяина в удаленном месте также имеют значение.
Под «неметастатической» понимают злокачественную опухоль, которая является доброкачественной или которая остается в первичной локализации и не проникла в лимфатический или кровеносный сосудистую систему или ткани, отличные от первичной локализации. В целом, неметастатическая злокачественная опухоль представляет собой любую злокачественную опухоль, которая представляет собой стадию 0, I или II злокачественной опухоли и иногда стадию III злокачественной опухоли.
Под «первичной опухолью» или «первичной злокачественной опухолью» понимают исходную злокачественную опухоль, а не метастатическое повреждение, локализованное в другой ткани, органе или локализации в организме субъекта.
Под «доброкачественной опухолью» понимают опухоль, которая сохраняет локализацию в месте возникновения и не обладает способностью к инфильтрации, инвазии или метастазированию в удаленное место.
Под «опухолевой массой» понимают множество злокачественных клеток, размер опухоли или количество злокачественной опухоли в организме. Опухолевую массу также обозначают как опухолевая нагрузка.
Под «опухолевым числом» понимают число опухолей.
Под «субъектом» понимают млекопитающее, включая в качестве неограничивающих примеров человека или млекопитающее, не относящееся к человеку, такое как корова, лошадь, собака, овца или кошка. Предпочтительно, субъектом является человек.
Термин «противоопухолевая терапия» относится к терапии, которую можно использовать в лечении опухоли. Примеры противоопухолевых терапевтических средств включают, но без ограничения, например, химиотерапевтические средства, ингибирующие рост средства, цитотоксические средства, средства, используемые в лучевой терапии, антиангиогенные средства, апоптотические средства, антитубулиновые средства и другие средства для лечения опухоли, антитела против CD20, ингибиторы тромбоцитарного фактора роста (например, гливек™ (иматиниба мезилат)), ингибитор ЦОГ-2 (например, целекоксиб), интерфероны, цитокины, антагонисты (например, нейтрализующие антитела), которые связываются с одним или несколькими из следующих рецепторов-мишеней ErbB2, ErbB3, ErbB4, PDGFR-β, BlyS, APRIL, BCMA или VEGF, TRAIL/Apo2 и другие биологически активные и органические химические средства и т.д. Их сочетания также включены в изобретение.
Термин «цитотоксическое средство», как применяют в настоящем документе, относится к веществу, которое ингибирует или препятствует функции клеток и/или вызывает разрушение клеток. Предусмотрено, что этот термин включает радиоактивные изотопы (например, I131, 1125, Y90 и Re186), химиотерапевтические средства и токсины, такие как ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты.
«Химиотерапевтическое средство» представляет собой химическое соединение, которое можно использовать в лечении злокачественной опухоли. Примеры химиотерапевтических средств включают химическое соединение, которое можно использовать в лечении злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие средства, такие как тиотепа и цитоксан® циклофосфамид; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в частности, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекана); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлоретамин, мехлоретаминоксид гидрохлорид, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихимицин, в частности, калихимицин γ1I и калихимицин ωI1 (см., например, Agnew, Chem Intl. Ed. Engl, 33: 183-186 (1994)); динемицин, включая динемицин A; бисфосфонаты, такие как клодронат; эсперамицин; а также хромофор неокарциностатина и родственные ему хромофоры хромопротеинового энедиинового антибиотика), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицин, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин адриамицин® (включая морфолинодоксорубицин, цианморфолинодоксорубицин, 2-пирролиндоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; противоадреналовые средства, такие как аминоглутетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатрексат; дефофамин; демеколцин; диазиквон; элфорнитин; ацетат эллиптиния; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (в частности, токсин T-2, верракурин A, роридин A и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-C»); циклофосфамид; тиотепу; таксоиды, например паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), не содержащий кремофор, сконструированный с альбумином препарат наночастиц паклитаксела абраксан™ (American Pharmaceutical Partners, Schaumberg, Illinois), и и доксетаксел таксотер® (Rhone-Poulenc Rorer, Antony, France); хлоранбуцил; гемцитабин гемзар®; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин, оксалиплатин и карбоплатин; винбластин; платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин навельбин®; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселоду; ибандронат; иринотекан (камптозар, CPT-11) (включая схему лечения иринотеканом вместе с 5-FU и лейковорином); ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин; бортезомиб велкад; леналидомид ревлимид; лейковорин (LV); оксалиплатин, включая схему лечения оксалиплатином (фолфокс); ингибиторы PKC-α, Raf, H-Ras, EGFR (например, эрлотиниб (тарцева™)) и VEGF-A, которые снижают клеточную пролиферацию и фармацевтически приемлемые соли, кислоты или производные любого указанного выше.
Также в это определение включены противогормональные средства, которые действуют, регулируя или ингибируя воздействие гормона на опухоли, такие как антиэстрогены и избирательные модуляторы рецепторов эстрогена (SERM), включая, например, тамоксифен (включая нолвадекс® тамоксифен), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен фарестон; ингибиторы ароматазы, которые ингибируют фермент ароматазу, регулирующую продукцию эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, ацетат мегестрола мегаза®, экземестан аромазин®, форместан, фадрозол, ворозол ривисор®, летрозол фемара® и анастрозол аримидекс®; и противоандрогенные средства, такие как флутамид, нилутамид, бикалутамид, леупролид и гозерелин; а также троксацитабин (аналог 1,3-диоксолана нуклеозида цитозина); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, ингибирующие экспрессию генов в путях передачи сигнала, которые вовлечены в патологическую пролиферацию клеток, такие как, например, ингибитор экспрессии PKC-α, Raf и H-Ras; рибозимы, такие как ингибитор экпсрессии VEGF (например, рибозим ангиозим®) и ингибитор экпсрессии HER2; вакцины, такие как генотерапевтические вакцины, например, вакцина алловектин®, вакцина лейвектин® и вакцина ваксид®; rIL-2 пролейкин®; луртотекан® ингибитор топоизомеразы 1; rmRH абареликс®; винорелбин и эсперамицины (см. патент США № 4675187) и фармацевтически приемлемые соли, кислоты или производные любого их указанных выше.
Термин «пролекарство», как используют в этой заявке, относится к предшественнику или производному фармацевтически активного вещества, которое менее цитотоксично в отношении опухолевых клеток по сравнению с исходным лекарственным средством и способно к ферментативной активации или превращению в более активную исходную форму. См., например, Wilman, «Prodrugs in Cancer Chemotherapy» Biochemical Society Transactions, 14, pp. 375-382, 615th Meeting Belfast (1986) и Stella et al., «Prodrugs: A Chemical Approach to Targeted Drug Delivery», Directed Drug Delivery, Borchardt et al., (ed.), pp. 247-267, Humana Press (1985). Пролекарства по данному изобретению включают в качестве неограничивающих примеров фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, модифицированные D-аминокислотами пролекарства, гликозилированные пролекарства, β-лактамсодержащие пролекарства, необязательно замещенные феноксиацетамидсодержащие пролекарства или необязательно замещенные фенилацетамидсодержащие пролекарства, 5-фторцитозин и другие 5-фторуридиновые пролекарства, которые можно превратить в более активное цитотоксическое свободное лекарственное средство. Примеры цитотоксических лекарственных средств, которые можно превратить в пролекарственную форму для использования в этом изобретении включают в качестве неограничивающих примеров те химиотерапевтические средства, которые описаны выше.
Под «лучевой терапией» понимают использование направленных γ-лучей или β-лучей, чтобы вызывать достаточное повреждение клетки с тем, чтобы ограничить ее способность нормально функционировать или совсем уничтожить клетку. Следует принимать во внимание, что в данной области известно много путей для определения дозы и длительности лечения. Типичное лечение доставляют в виде однократного введения и типичные дозы варьируют от 10 до 200 единиц (Грэй) в сутки.
«Биологически активный» или «функциональный» полипептид (такой как гетерологичный полипептид) представляет собой полипептид, который способен проявлять одну или несколько своих естественных активностей в структурных, регуляторных, биохимических или биофизических актах.
«Биологически активное» или «функциональное» антитело представляет собой антитело, которое способно проявлять одну или несколько своих естественных активностей в структурных, регуляторных, биохимических или биофизических актах. Например, биологически активное антитело может обладать способностью специфически связывать антиген, и это связывание может в свою очередь вызывать или изменять клеточный или молекулярный акт, такой как передача сигнала или ферментативная активность. Биологически активное антитело также может блокировать активацию рецептора лигандом или действовать в качестве агонистического антитела. Способность антитела проявлять одну или несколько своих естественных активностей зависит от нескольких факторов, включая правильную укладку и сборку полипептидных цепей. Как применяют в настоящем документе, биологически активное антитело, созданное посредством раскрытых способов, типично содержит гетеротетрамеры, содержащие две идентичные цепи L и две идентичные цепи H, которые соединены посредством нескольких дисульфидных связей и правильно уложены.
Композиции по изобретению и способы их использования
В одном из аспектов настоящее изобретение относится к вариантам TIR. Таким образом, для заданного TIR можно создать группу вариантов последовательностей аминокислот или нуклеиновых кислот с диапазоном интенсивностей трансляции, тем самым предоставляя удобное средство, посредством которого корректируют этот фактор для оптимальной секреции многих различных полипептидов. Использование репортерного гена, экспрессируемого под управлением этих вариантов, такого как PhoA, предоставляет способ количественного определения относительных интенсивностей трансляции различных областей инициации трансляции. Варианты или мутанты TIR можно предоставить на фоне плазмидного вектора, тем самым предоставляя набор плазмид, в которые можно встроить ген, представляющий интерес, и измерить его экспрессию с тем, чтобы установить оптимальный диапазон интенсивности трансляции для максимальной экспрессии зрелого полипептида.
Мутагенез TIR выполняют посредством общепринятых способов, которые ведут к изменениям кодонов, которые могут изменять аминокислотную последовательность, хотя молчащие изменения в нуклеотидной последовательности предпочтительны. Измерения в TIR могут содержать, например, изменения числа или расстояния последовательностей Шайна-Дальгарно, наряду с изменениями в сигнальной последовательности. Один способ генерации мутантных сигнальных последовательностей состоит в генерации «банка кодонов» в начале кодирующей последовательности, который не меняет аминокислотную последовательность сигнальной последовательности (т.е. все изменения являются молчащими). Это можно осуществить посредством изменения положения третьего нуклеотида каждого кодона; дополнительно, некоторые аминокислоты, такие как лейцин, серин и аргинин, имеют множество первых и вторых положений, которые могут увеличивать сложность создания банка. Этот способ мутагенеза подробно описан в Yansura et al. (Methods: A Companion to Methods in Enzymol. 4: 151-158 (1992)). По существу, фрагмент ДНК, кодирующий сигнальную последовательность и начало зрелого полипептида, синтезируют так, что изменяют третье (и, возможно, первое и второе, как описано выше) положение каждого из первых 6-12 кодонов. Дополнительные нуклеотиды, расположенные ниже по направлению транскрипции относительно этих кодонов, предоставляют сайт для связывания комплементарного праймера, используемого в создании нижней цепи. Обработка верхней кодирующей цепи и праймера нижней цепи ДНК полимеразой I (Кленова) приведет к группе фрагментов двухцепочечной ДНК, содержащих случайные кодоны. Праймеры конструируют так, чтобы они содержали полезные сайты клонирования, которые затем можно использовать для встраивания фрагментов ДНК в подходящий вектор, тем самым делая возможной амплификацию банка кодонов. Альтернативные способы включают, например, замену всего rbs случайными нуклеотидами (Wilson et al., BioTechniquesl7:944-952 (1994)), и использование библиотек фагового дисплея (см., например, Barbas et al., Proc. Natl. Acad. Sci. U.S.A.89:4457-4461 (1992); Garrard et al., Gene128: 103-109 (1993)).
Бактериальная транслоказа Sec облегчает экспорт белков у прокариот. Секреторные белки могут быть нацелены на транслоказу Sec посредством двух различных механизмов, т.е. посредством котрансляционного и посттрансляционного нацеливания. Потом происходит высвобождение секреторного белка, содержащего сигнальную последовательность, из рибосомы в его состоянии завершенного синтеза и направление к транслоказе Sec. У различных грамотрицательных бактерий секреторные белки направляются к транслоказе Sec посредством специфичного для секреции шаперона SecB, который поддерживает эти белки в надлежащем неуложенном состоянии для транслокации. Во время котрансляционного нацеливания частица узнавания сигнала (SRP) связывается с сигнальной последовательностью секреторного белка, в то время как она выходит из рибосомы, и происходит нацеливание полного тройного комплекса SRP/рабосомы/возникающей цепи секреторного белка на транслоказу Sec.
Например, сигнальные пептиды связывающего мальтозу периплазматического белка (MalE) и щелочной фосфатазы (PhoA) направляют транслокацию из цитоплазмы в периплазму посттрансляционным образом с помощью молекулярного двигателя SecA. Другими образцовыми сигнальными пептидами, которые направляют транслокацию посттрансляционным образом, являются dsbC, lolA, ompA, lamb и lpp. Сигнальные пептиды термостабильного энтеротоксина II (stII) и тиол:дисульфид обменивающего белка (dsbA) направляют транслокацию котрансляционным образом с помощью частицы узнавания сигнала (SRP). Другими образцовыми сигнальными пептидами, которые направляют транслокацию котрансляционным образом, являются yral, tort, tolB, sfmC, nikA и sfmC. Также см. обзор по Sec- и Tat-опосредованной секреции белка через бактериальную цитоплазматическую мембрану авторов Natale et al. (Natale et al. (2008) Biochemica et Biophysica Acta 1778: 1735-56.)
Авторы настоящего изобретения разработали библиотеки новых вариантов сигнальных пептидов области инициации трансляции (TIR) (фиг.2, таблица 2) для сигнальных пептидов, представляющих два основных пути секреции для транспортировки через внутреннюю мембрану в E. coli: sec (PhoA, MalE) и SRP (DsbA, STII). Каждая библиотека содержит панель векторов, которые содержат варианты TIR с различными интенсивностями трансляции, которые предоставляют средство, с помощью которого легко корректировать уровень трансляции для заданного белка, представляющего интерес.
Типично, варианты TIR должны быть представлены в плазмидном векторе с подходящими элементами для экспрессии гена, представляющего интерес. Например, типичная конструкция должна содержать промотор ближе к 5'-концу относительно сигнальной последовательности, сайт распознавания рестрикционного фермента ближе к 3'-концу относительно сигнальной последовательности для встраивания гена, представляющего интерес, или репортерного гена и селективный маркер, такой как маркер устойчивости к лекарственному средству, для отбора и/или поддержания бактерий, трансформированных полученными плазмидами. Плазмидные векторы дополнительно рассмотрены и проиллюстрированы в настоящем документе. Промоторы, пригодные для использования с прокариотическими организмами-хозяевами, известны в данной области и некоторые проиллюстрированы и описаны в настоящем документе.
Можно использовать любой репортерный ген, который можно некоторым образом определять количественно. Таким образом, например, образование щелочной фосфатазы можно определять количественно в виде меры секретируемого уровня продукта гена phoA. Другие примеры включают, например, β-лактамазные гены.
В целом, можно создать группу векторов с диапазоном интенсивностей TIR для каждого цистрона вектора этой группы. Эта ограниченная группа обеспечивает сравнение уровней экспрессии каждой цепи, а также выхода полноразмерных продуктов при различных комбинациях интенсивностей TIR. Интенсивности TIR можно определить посредством количественного определения уровня экспрессии репортерного гена, как описано в деталях в Simmons et al. патент США № 5840523. Для цели по данному изобретению, комбинация интенсивностей трансляции для конкретной пары TIR в векторе представлена посредством (N-легкая, M-тяжелая), где N представляет собой относительную интенсивность TIR легкой цепи, а M представляет собой относительную интенсивность TIR тяжелой цепи. Например, (3-легкая, 7-тяжелая) обозначает, что вектор обеспечивает относительную интенсивность TIR приблизительно 3 для экспрессии легкой цепи и относительную интенсивность TIR приблизительно 7 для экспрессии тяжелой цепи. Основываясь на сравнении интенсивностей трансляции, желаемые индивидуальные TIR выбирают для комбинирования в конструкциях экспрессирующих векторов по изобретению. Сконструированные таким образом векторы можно использовать для трансформации подходящего организма-хозяина. Предпочтительно, организмом-хозяином является прокариотический организм-хозяин. Более предпочтительно, организмом-хозяином является E. coli.
Секретируемый уровень полипептидов можно определить, например, посредством функциональных анализов на полипептид, представляющий интерес, если они доступны, радиоиммунного анализа (RIA), иммуноферментного анализа (ELISA) или посредством PAGE и визуализации правильной молекулярной массы полипептида, представляющего интерес. Способы определения уровня секретируемого полипептида хорошо известны в данной области и некоторые из них приведены в качестве примера в настоящем документе.
Антитела
Антитела по изобретению предпочтительно являются моноклональными. Объем изобретения также охватывает фрагменты Fab, Fab', Fab'-SH и F(ab')2 антител, предоставленных в настоящем документе. Эти фрагменты антител можно создавать с помощью традиционных средств, таких как ферментативное расщепление, или можно создавать рекомбинантными способами. Такие фрагменты антител могут быть химерными или гуманизированными. Эти фрагменты можно использовать для диагностических и терапевтических целей, изложенных ниже.
Соответственно, в некотором варианте осуществления антитело против c-met представляет собой антитело с одним Fab фрагментом (т.е. вариабельный домен тяжелой цепи и вариабельный домен легкой цепи формируют один антигенсвязывающий фрагмент), содержащее Fc-область, где Fc-область содержит первый и второй полипептиды Fc, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с Fab молекулой, содержащей указанный антигенсвязывающий фрагмент. Для лечения патологических состояний, требующего антагонистической функции, и когда бивалентность антитела ведет к нежелательному агонистическому эффекту, моновалентное свойство антитела с одним Fab фрагментом (т.е. антитела, содержащего один антигенсвязывающий фрагмент) ведет к и/или обеспечивает антагонистическую функцию при связывании антитела с молекулой-мишенью. Кроме того, антитело с одним Fab фрагментом, содержащее Fc-область, отличается превосходными фармакокинетическими показателями (такими как увеличенное время полужизни и/или сниженная скорость клиренса in vivo) по сравнению с Fab-формами, обладающими схожими/по существу идентичными характеристиками связывания антигенов, таким образом преодолевая основной недостаток использования стандартных моновалентных Fab антител. Антитела с одним Fab фрагментом раскрыты, например, в WO2005/063816; Martens et al., Clin Cancer Res (2006), 12: 6144. В некоторых вариантах осуществления антитело с одним Fab фрагментом представляет собой фрагмент моновалентного антитела, где фрагмент антитела содержит первый полипептид, содержащий вариабельный домен легкой цепи, второй полипептид, содержащий вариабельный домен тяжелой цепи и указанный первый полипептид Fc, и третий полипептид, содержащий указанный второй полипептид Fc, посредством чего вариабельный домен тяжелой цепи и вариабельный домен легкой цепи формируют один антигенсвязывающий фрагмент и посредством чего первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с Fab-молекулой, содержащей указанный антигенсвязывающий фрагмент.
В некоторых вариантах осуществления антитело связывает (в некоторых вариантах осуществления специфично связывает) c-met. В некоторых вариантах осуществления антитело против c-met содержит (a) первый полипептид, содержащий вариабельный домен тяжелой цепи, обладающий последовательностью:

последовательность CH1 и первый полипептид Fc; (b) второй полипептид, содержащий вариабельный домен легкой цепи, обладающий последовательностью:
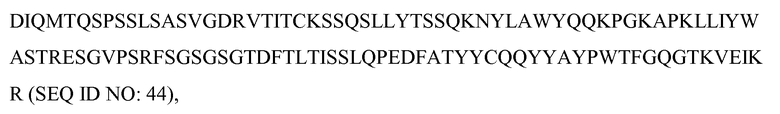
и последовательность CL1; и (c) третий полипептид, содержащий второй полипептид Fc, где вариабельный домен тяжелой цепи и вариабельный домен легкой цепи представлены в виде комплекса и формируют один антигенсвязывающий фрагмент, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с Fab-молекулой, содержащей указанный антигенсвязывающий фрагмент. В некоторых вариантах осуществления первый полипептид содержит последовательность Fc, изображенную на фиг.7 (SEQ ID NO: 68), и второй полипептид содержит последовательность Fc, изображенную на фиг.8 (SEQ ID NO: 47). В некоторых вариантах осуществления первый полипептид содержит последовательность Fc, изображенную на фиг.8 (SEQ ID NO: 47), и второй полипептид содержит последовательность Fc, изображенную на фиг.7 (SEQ ID NO: 68).
В некоторых вариантах осуществления антитело против c-met содержит (a) первый полипептид, содержащий вариабельный домен тяжелой цепи, указанный полипептид содержит последовательность:

(b) второй полипептид, содержащий вариабельный домен легкой цепи, полипептид содержит последовательность

и третий полипептид, содержащий последовательность Fc, полипептид содержит последовательность

где вариабельный домен тяжелой цепи и вариабельный домен легкой цепи представлены в виде комплекса и формируют один антигенсвязывающий фрагмент, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с Fab-молекулой, содержащей указанный антигенсвязывающий фрагмент.
В одном из аспектов антитело против c-met содержит:
(a) по меньшей мере одну, две, три, четыре или пять последовательностей гипервариабельных областей (CDR), выбранные из группы, состоящей из:
(i) CDR-L1, содержащей последовательность A1-A17, где A1-A17 предсталяет собой KSSQSLLYTSSQKNYLA (SEQ ID NO: 49)
(ii) CDR-L2, содержащей последовательность B1-B7, где B1-B7 представляет собой WASTRES (SEQ ID NO: 50)
(iii) CDR-L3, содержащей последовательность C1-C9, где C1-C9 представляет собой QQYYAYPWT (SEQ ID NO: 51)
(iv) CDR-H1, содержащей последовательность D1-D10, где D1-D10 представляет собой GYTFTSYWLH (SEQ ID NO: 52)
(v) CDR-H2, содержащей последовательность E1-E18, где E1-E18 представляет собой GMIDPSNSDTRFNPNFKD (SEQ ID NO: 53) и
(vi) CDR-H3, содержащей последовательность F1-F11, где F1-F11 представляет собой T/SYGSYVSPLDY (SEQ ID NO: 54);
и (b) по меньшей мере один вариант CDR, где последовательность варианта CDR содержит модификации по меньшей мере одного остатка последовательности, представленной в (i)-(vi). В одном из вариантов осуществления CDR-H3 содержит TYGSYVSPLDY (SEQ ID NO: 55). В одном из вариантов осуществления CDR-H3 содержит SYGSYVSPLDY (SEQ ID NO: 56). В одном из вариантов осуществления антитело по изобретению, содержащее эти последовательности (в комбинации, как описано в настоящем документе), представляет собой гуманизированное антитело или антитело человека.
В одном из вариантов осуществления антитело против c-met содержит вариабельный домен тяжелой цепи, содержащий одну или несколько из последовательностей CDR1-HC, CDR2-HC и CDR3-HC, изображенных на фиг.7 (SEQ ID NO: 52-53 и 66). В некоторых вариантах осуществления антитело содержит вариабельный домен легкой цепи, содержащий одну или несколько из последовательностей CDR1-LC, CDR2-LC и CDR3-LC, изображенных на фиг.7 (SEQ ID NO: 49-51). В некоторых вариантах осуществления вариабельный домен тяжелой цепи содержит последовательность FR1-HC, FR2-HC, FR3-HC и FR4-HC, изображенную на фиг.7 (SEQ ID NO: 62-65). В некоторых вариантах осуществления вариабельный домен легкой цепи содержит последовательность FR1-LC, FR2-LC, FR3-LC и FR4-LC, изображенную на фиг.7 (SEQ ID NO: 57-60).
Варианты HVR в антителе против c-met по изобретению могут содержать модификации одного или нескольких остатков в HVR. В одном из вариантов осуществления вариант HVR-L2 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации из следующих положений: B1 (M или L), B2 (P, T, G или S), B3 (N, G, R или T), B4 (I, N или F), B5 (P, I, L или G), B6 (A, D, T или V) и B7 (R, I, M или G). В одном из вариантов осуществления вариант HVR-H1 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации из следующих положений: D3 (N, P, L, S, A, I), D5 (I, S или Y), D6 (G, D, T, K, R), D7 (F, H, R, S, T или V) и D9 (M или V). В одном из вариантов осуществления вариант HVR-H2 содержит 1-4 (1, 2, 3 или 4) замен в любой комбинации из следующих положений: E7 (Y), E9 (I), E10 (I), E14 (T или Q), E15 (D, K, S, T или V), E16 (L), E17 (E, H, N или D) и E18 (Y, E или H). В одном из вариантов осуществления вариант HVR-H3 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации из следующих положений: F1 (T, S), F3 (R, S, H, T, A, K), F4 (G), F6 (R, F, M, T, E, K, A, L, W), F7 (L, I, T, R, K, V), F8 (S, A), F10 (Y, N) и F11 (Q, S, H, F). Буква(ы) в круглых скобках после каждого положения обозначает иллюстративную замену (т.е. замещение) аминокислоты; специалисту в данной области должно быть очевидно, что применимость других аминокислот в качестве заменяющих аминокислот в контексте, описанном в настоящем документе, можно оценить обычным путем с применением способов, известных в данной области и/или описанных в настоящем документе. В одном из вариантов осуществления HVR-L1 содержит последовательность SEQ ID NO: 49. В одном из вариантов осуществления F1 в варианте HVR-H3 представляет собой T. В одном из вариантов осуществления F1 в варианте HVR-H3 представляет собой S. В одном из вариантов осуществления F3 в варианте HVR-H3 представляет собой R. В одном из вариантов осуществления F3 в варианте HVR-H3 представляет собой S. В одном из вариантов осуществления F7 в варианте HVR-H3 представляет собой T. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой T или S, F3 представляет собой R или S и F7 представляет собой T.
В одном из вариантов осуществления антитело против c-met по изобретению содержит вариант HVR-H3, где F1 представляет собой T, F3 представляет собой R и F7 представляет собой T. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой S. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой T и F3 представляет собой R. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой S, F3 представляет собой R и F7 представляет собой T. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой T, F3 представляет собой S, F7 представляет собой T и F8 представляет собой S. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H3, где F1 представляет собой T, F3 представляет собой S, F7 представляет собой T и F8 представляет собой A. В некоторых вариантах осуществления указанное антитело с вариант HVR-H3 дополнительно содержит HVR-L1, HVR-L2, HVR-L3, HVR-H1 и HVR-H2, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 50, 51, 52 и 53. В некоторых вариантах осуществления эти антитела дополнительно содержат консенсусную последовательность каркасной области тяжелой цепи подгруппы III человека. В одном из вариантов осуществления этих антител консенсусная последовательность каркасной области содержит замену в положении 71, 73 и/или 78. В некоторых вариантах осуществления этих антител положение 71 представляет собой A, 73 представляет собой T и/или 78 представляет собой A. В одном из вариантов осуществления этих антител эти антитела дополнительно содержат консенсусную последовательность каркасной области легкой цепи κI человека.
В одном из вариантов осуществления антитело против c-met по изобретению содержит вариант HVR-L2, где B6 представляет собой V. В некоторых вариантах осуществления указанное антитело с вариантом HVR-L2 дополнительно содержит HVR-L1, HVR-L3, HVR-H1, HVR-H2 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 51, 52, 53, 54. В некоторых вариантах осуществления указанное антитело с вариантом HVR-L2 дополнительно содержит HVR-L1, HVR-L3, HVR-H1, HVR-H2 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 51, 52, 53, 55. В некоторых вариантах осуществления указанное антитело с вариантом HVR-L2 дополнительно содержит HVR-L1, HVR-L3, HVR-H1, HVR-H2 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 51, 52, 53, 56. В некоторых вариантах осуществления эти антитела дополнительно содержат консенсусную последовательность каркасной области тяжелой цепи подгруппы III человека. В одном из вариантов осуществления этих антител консенсусная последовательность каркасной области содержит замену в положении 71, 73 и/или 78. В некоторых вариантах осуществления этих антител положение 71 представляет собой A, 73 представляет собой T и/или 78 представляет собой A. В одном из вариантов осуществления этих антител эти антитела дополнительно содержат консенсусную последовательность каркасной области легкой цепи κI человека.
В одном из вариантов осуществления антитело против c-met по изобретению содержит вариант HVR-H2, где E14 представляет собой T, E15 представляет собой K и E17 представляет собой E. В одном из вариантов осуществления антитело по изобретению содержит вариант HVR-H2, где E17 представляет собой E. В некоторых вариантах осуществления указанное антитело с вариантом HVR-H3 дополнительно содержит HVR-L1, HVR-L2, HVR-L3, HVR-H1 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 50, 51, 52, 54. В некоторых вариантах осуществления указанное антитело с вариантом HVR-H2 дополнительно содержит HVR-L1, HVR-L2, HVR-L3, HVR-H1 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 50, 51, 52, 55. В некоторых вариантах осуществления указанное антитело с вариантом HVR-H2 дополнительно содержит HVR-L1, HVR-L2, HVR-L3, HVR-H1 и HVR-H3, где каждая по порядку содержит последовательность, представленную в SEQ ID NO: 49, 50, 51, 52, 55. В некоторых вариантах осуществления эти антитела дополнительно содержат консенсусную последовательность каркасной области тяжелой цепи подгруппы III человека. В одном из вариантов осуществления этих антител консенсусная последовательность каркасной области содержит замену в положении 71, 73 и/или 78. В некоторых вариантах осуществления этих антител положение 71 представляет собой A, 73 представляет собой T и/или 78 представляет собой A. В одном из вариантов осуществления этих антител эти антитела дополнительно содержат консенсусную последовательность каркасной области легкой цепи κI человека.
В данной области известны другие антитела против c-met, пригодные для использования в способах по изобретению.
В одном из аспектов антитело против c-met содержит по меньшей мере одну характеристику, которая способствует гетеродимеризации, при этом минимизируя гомодимеризацию, последовательностей Fc в фрагменте антитела. Такая характеристика(и) улучшает выход и/или чистоту и/или гомогенность популяций иммуноглобулинов. В одном из вариантов осуществления антитело содержит мутации Fc, образующие «выступы» и «отверстия», как описано в WO2005/063816. Например, мутация отверстия может представлять собой одну или несколько из T366A, L368A и/или Y407V в полипептиде Fc, а мутация выступ может представлять собой T366W. Мутации выступа и отверстия в Fc дополнительно описаны в настоящем документе.
Моноклональные антитела получают из популяции по существу гомогенных антител, т.е. индивидуальные антитела, содержащиеся в популяции, идентичны за исключением возможных встречающихся в природе мутаций, которые могут быть представлены в незначительных количествах. Таким образом, определение «моноклональный» указывает на характер антитела как не являющегося смесью отдельных антител.
Моноклональные антитела по изобретению можно получить с использованием гибридомного способа, впервые описанного авторами Kohler et al., Nature, 256:495 (1975), или можно получить способами рекомбинантной ДНК (патент США № 4816567).
В гибридомном способе мышь или другое подходящее животное-хозяина, такое как хомяк, иммунизируют для того, чтобы вызвать образование лимфоцитов, которые продуцируют или способны продуцировать антитела, которые должны специфично связываться с белком, используемым для иммунизации. Антитела к антигену можно индуцировать у животных посредством множественных подкожных (sc) или интраперитонеальных (ip) инъекций антигена и адъюванта. Антиген можно получать, используя способы, хорошо известные в данной области, некоторые из которых дополнительно описаны в настоящем документе. Например, рекомбинантное получение антигена человека и мыши описано ниже. В одном из вариантов осуществления животных иммунизируют антигеном, слитым с Fc-частью тяжелой цепи иммуноглобулина. В предпочтительном варианте осуществления животных иммунизируют слитым белком антиген-IgG1. Животных, как правило, иммунизируют против иммуногенных конъюгатов или производных антигена, полученных с использованием монофосфориллипида A (MPL)/дикриномиколата трегалозы (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT), и раствор инъецируют внутрикожно в нескольких местах. Через две недели проводят повторную иммунизацию животных. Через 7-14 суток у животных берут кровь и анализируют титр антител в сыворотке. Повторную иммунизацию животных проводят до тех пор, пока титр не выйдет на плато.
Альтернативно, лимфоциты можно иммунизировать in vitro. Затем лимфоциты сливают с миеломными клетками, использующими подходящее средство для слияния, такое как полиэтиленгликоль, чтобы сформировать гибридомную клетку (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Полученные таким образом гибридомные клетки высевают и растят в подходящей среде для культивирования, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживание неслитых материнских миеломных клеток. Например, если у материнских миеломных клеток отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), среда для культивирования для гибридом типично должна содержать гипоксантин, аминоптерин и тимидин (среда HAT), эти вещества предотвращают рост клеток с дефицитом HGPRT.
Предпочтительные миеломные клетки представляют собой те, которые эффективно сливаются, поддерживают стабильно высокий уровень образования антитела посредством выбранных антителопродуцирующих клеток и чувствительны к среде, такой как среда HAT. Среди них предпочтительными линиями миеломных клеток являются линии миеломы мыши, такие как те, что получены из опухолей мыши MOPC-21 и MPC-11, доступные в Salk Institute Cell Distribution Center, San Diego, California USA, и клетки SP-2 или X63-Ag8-653, доступные в American Type Culture Collection, Rockville, Maryland USA. Линии миеломы человека и гетеромиеломных клеток мыши-человека также описаны в отношении получения моноклональных антител человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Образование моноклональных антител, направленных против антигена, анализируют в среде для культивирования, в которой выращивают гибридомные клетки. Предпочтительно, специфичность связывания моноклональных антител, продуцируемых гибридомными клетками определяют посредством иммунопреципитации или анализа связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA).
Аффинность связывания моноклонального антитела можно определить, например, посредством анализа Скетчарда согласно Munson et al., Anal. Biochem., 107:220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с желаемой специфичностью, аффинностью и/или активностью, можно осуществлять субклонирование клонов посредством серийных разведений и выращивание стандартными способами (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Подходящие среды для культивирования с этой целью включают, например, среду D-MEM или RPMI-1640. Кроме того, гибридомные клетки можно растить in vivo в виде асцитных опухолей у животного.
Моноклональные антитела, секретируемые субклонами, надлежаще отделяют от среды для культивирования, асцитной жидкости или сыворотки посредством стандартных процедур очистки иммуноглобулинов, таких как, например, белок A-сефароза, хроматография на гидроксиапатите, электрофорез в геле, диализ или аффинная хроматография.
Антитела по изобретению можно получать, используя комбинаторные библиотеки для скрининга клонов синтетических антител с желаемой активностью или активностями. По существу, клоны синтетических антител отбирают посредством скрининга фаговых библиотек, содержащих фаг, в котором представлены различные фрагменты вариабельной области (Fv) антитела, слитой с белком оболочки фага. Пэннинг таких фаговых библиотек проводят посредством аффинной хроматографии против желаемого антигена. Клоны, экспрессирующие фрагменты Fv, способные связываться с желаемым антигеном, адсорбируют на антигене и, таким образом, отделяют от несвязывающих клонов в библиотеке. Затем элюируют связывающие клоны из антигена и их можно дополнительно обогатить посредством дополнительных циклов адсорбции на антигене/элюирования. Любые антитела по изобретению можно получить посредством разработки подходящей процедуры антигенного скрининга для отбора клона фага, представляющего интерес, с последующим конструированием клона антитела с использованием последовательностей Fv из клона фага, представляющего интерес, и подходящих последовательностей константной области (Fc), описанных в Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3. Образцовый способ создания антител раскрыт в примерах.
Антиген-связывающий домен антитела формируют из двух вариабельных (V) областей приблизительно из 110 аминокислоты, по одной из легкой (VL) и тяжелой (VH) цепей, каждая из которых представляет три гипервариабельные петли или определяющих комплементарность области (CDR). Вариабельные домены можно представить функционально на фаге, или в виде одноцепочечных Fv-фрагментов (scFv), в которых VH и VL ковалентно связаны через короткий гибкий пептид, или в виде Fab-фрагментов, в которых каждый из них слит с константным доменом и взаимодействует нековалентно, как описано в Winter et al., αnn. Rev. Immunol, 12: 433-455 (1994). Как применяют в настоящем документе, кодирующие scFv клоны фага и кодирующие Fab клоны фага совместно обозначают как «Fv клоны фага» или «Fv клоны».
Репертуары VH и VL генов можно раздельно клонировать посредством полимеразной цепной реакции (ПЦР) и выполнять их случайную рекобминацию в фаговых библиотеках, в которых затем можно проводить поиск антигенсвязывающих клонов, как описано в Winter et al., Ann. Rev. Immunol, 12: 433-455 (1994). Библиотеки из иммунизированных источников обеспечивают антитела с высокой аффинностью к иммуногену без необходимости создавать гибридомы. Альтернативно, наивный репертуар можно клонировать для того, чтобы предоставить один источник антител человека к широкому диапазону не ауто-, а также аутоантигенов без какой-либо иммунизации, как описано авторами Griffiths et al., EMBO J 12: 725-734 (1993). Наконец, наивные библиотеки также можно создать синтетическим путем посредством клонирования нереаранжированных сегментов V-генов из стволовых клеток и использования праймеров для ПЦР, содержащих случайную последовательность, чтобы кодировать высоко вариабельные области CDR3 и выполнить реаранжировку in vitro, как описано авторами Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992).
Нитчатые фаги используют для представления фрагментов антител посредством слияния с минорным белком оболочки pIII. Фрагменты антител можно представлять в виде одноцепочечных Fv-фрагментов, в которых домены VH и VL соединены в одну и ту же полипептидную цепь посредством гибкого полипептидного спейсера, например, как описано авторами Marks et al., J. Mol. Biol., 222: 581-597 (1991), или в виде Fab-фрагментов, в котором одна цепь слита с pIII, а другая секретируется в периплазму бактериальной клетки-хозяина, где сборка структуры Fab-белок оболочки, которая представлена на поверхности фага посредством вытеснения некоторых белков оболочки дикого типа, например, как описано в Hoogenboom et al., Nucl. Acids Res., 19: 4133-4137 (1991).
В целом, нуклеиновые кислоты, кодирующие фрагменты генов антител получают из иммунных клеток, полученных у человека или животных. Если желательно получить библиотеку, которая смещена в пользу клонов, нацеленных на конкретных антиген, то индивидуума иммунизируют антигеном для создания антительного ответа, и для конструирования библиотеки собирают клетки селезенки и/или циркулирующие B-клетки другие лимфоциты периферической крови (PBL). В предпочтительном варианте осуществления библиотеку фрагментов генов антител человека, смещенную в пользу антиген-реактивных клонов, получают посредством создания антительного ответа у трансгенных мышей, несущих массив генов функциональных иммуноглобулинов человека (и у которых отсутствует система образования функциональных эндогенных антител) так, что иммунизация антигеном вызывает в B-клетках образование антител человека против антигена. Создание трансгенных мышей, продуцирующих антитела человека, описано ниже.
Дополнительного обогащения популяций антиген-реактивных клеток можно добиться посредством использования подходящей процедуры скрининга для выделения B-клеток, экспрессирующих антиген-специфическое связанное с мембраной антитело, например, посредством разделения клеток аффинной хроматографией на антигене или адсорбции клеток на антигене, меченном флуорохромом, с последующей флуоресцентной сортировкой клеток (FACS).
Альтернативно, использование клеток селезенки и/или B-клеток или других PBL от неиммунизированного донора обеспечивает более полное представление возможного репертуара антител, а также позволяет конструировать библиотеку антител, используя любые виды животных (человека или животных, не относящихся к человеку), у которых антиген не является антигенным. Для библиотек, включающих конструирование генов антител in vitro, стволовые клетки собирают у индивидуумов, чтобы предоставить нуклеиновые кислоты, кодирующие нереаранжированные сегменты генов антител. Иммунные клетки, представляющие интерес, можно получить от различных видов животных, таких видов как человек, мышь, крыса, зайцеобразные, волчьи, собачьи, кошачьи, свиньи, коровы, лошадиные и птицы и т.д.
Нуклеиновую кислоту, кодирующую сегменты вариабельных генов антител (включая сегменты VH и VL) получают из клеток, представляющих интерес, и амплифицируют. В случае библиотек реаранжированных VH и VL генов, желаемую ДНК можно получить посредством выделения геномной ДНК или мРНК из лимфоцитов с последующей полимеразной цепной реакцией (ПЦР) с праймерами, совпадающими с 5'- и 3'-концами реаранжироваанных VH и VL генов, как описано в Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86: 3833-3837 (1989), тем самым получая разнообразные репертуары генов V для экспрессии. V гены можно амплифицировать из кДНК и геномной ДНК, с обратными праймерами на 5'-конце экзона, кодирующего зрелый V-домен, и прямыми праймерами, размещенными внутри J-сегмента, как описано в Orlandi et al. (1989) и в Ward et al., Nature, 341: 544-546 (1989). Однако для амплификации из кДНК обратные праймеры также могут быть размещены в лидерном экзоне, описано в Jones et al., Biotechnol, 9: 88-89 (1991), а прямые праймеры в константной области, как описано в Sastry et al., Proc. Natl. Acad. Sci. (USA), 86: 5728-5732 (1989). Чтобы максимизировать комплементарность, вырожденность можно встроить в праймеры, как описано в Orlandi et al. (1989) или Sastry et al. (1989). Предпочтительно, разнообразие библиотеки максимизируют посредством использования праймеров для ПЦР, нацеленных на каждое семейство V-генов, чтобы амплифицировать все доступные аранжировки VH и VL, присутствующие в образце нуклеиновой кислоты иммунной клетки, например, как описано в способе по Marks et al., J. Mol. Biol, 222: 581-597 (1991) или как описано в способе по Orum et al., Nucleic Acids Res., 21: 4491-4498 (1993). Для клонирования амплифицированного ДНК в экспрессирующие векторы, внутри праймера для ПЦР можно ввести редкие сайты рестрикции в качестве метки на одном конце, как описано в Orlandi et al. (1989), или посредством дополнительной ПЦР амплификации с меченным праймером, как описано в Clackson et al., Nature, 352: 624-628 (1991).
Репертуары синтетически реаранжированных V-генов можно получить in vitro из сегментов V-генов. Большинство сегментов VH-генов человека клонировано и секвенировано (представлено в Tomlinson et al., J. Mol. Biol, 227: 776-798 (1992)) и картировано (представлено в Matsuda et al., Nature Genet., 3: 88-94 (1993); эти клонированные сегменты (включая все основные конформации петли H1 и H2) можно использовать для создания разнообразных репертуаров VH-генов с использованием праймеров для ПЦР, кодирующих петли H3 с разнообразными последовательностями и длинами, как описано в Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992). Также можно создавать репертуары VH, в которых все разнообразие последовательностей сфокусировано в длинной петле H3 одной длины, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 89: 4457-4461 (1992). Сегменты Vκ и Vλ человека клонированы и секвенированы (представлены в Williams and Winter, Eur. J. Immunol, 23: 1456-1461 (1993)), и их можно использовать для получения синтетических репертуаров легкой цепи. Репертуары синтетических V-генов, основанные на спектре укладок VH и VL и длин L3 и H3, должны кодировать антитела со значительным структурным разнообразием. После амплификации ДНК, кодирующей V-гены, сегменты V-генов зародышевой линии можно реаранжировать in vitro в соответствии со способами по Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992).
Репертуары фрагментов антител можно конструировать посредством комбинирования репертуаров VH и VL генов вместе несколькими путями. Каждый репертуар можно создавать в различных векторах, и векторы рекомбинированы in vitro, например, как описано в Hogrefe et al., Gene, 128: 119-126 (1993), или in vivo посредством комбинаторной инфекции, например, системы loxP, описанной в Waterhouse et al., Nucl. Acids Res., 21:2265-2266 (1993). В подходе с рекомбинацией in vivo используют двухцепочечные свойства Fab-фрагментов, чтобы преодолеть предел для размера библиотеки, налагаемый эффективностью трансформации E. coli. Наивные репертуары VH и VL клонируют раздельно, один в фагемиду и другой в фаговый вектор. Затем две библиотеки комбинируют посредством фаговой инфекции бактерий, содержащих фагемиды, с тем, чтобы каждая клетка содержала отличающуюся комбинацию, а размер библиотеки ограничен только числом присутствующих клеток (приблизительно 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo с тем, чтобы происходила рекомбинация VH и VL генов в одном репликоне и их совместная упаковка в фаговые вирионы. Эти огромные библиотеки обеспечивают большие количества разнообразных антител с хорошей аффинностью (Kd -1 приблизительно 10-8 M).
Альтернативно, репертуары можно клонировать последовательно в один и тот же вектор, например, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или собирать вместе посредством ПЦР и затем клонировать, например, как описано в Clackson et al., Nature, 352: 624-628 (1991). ПЦР сборку также можно использовать для соединения ДНК VH и VL с ДНК, кодирующей гибкий пептидный спейсер для формирования репертуаров одноцепочечных Fv (scFv). В еще одном другом способе, «ПЦР сборку в клетке» используют для комбинирования VH и VL генов внутри лимфоцитов посредством ПЦР и затем клонировать репертуары связанных генов, как описано в Embleton et al., Nucl. Acids Res., 20:3831-3837 (1992).
Антитела, продуцируемые посредством наивных библиотек (или естественных или синтетических), могут обладать умеренной аффинностью (Kd -1 от приблизительно 106 до 107 M-1), но созревание аффинности также можно имитировать in vitro посредством конструирования и повторного отбора из вторичных библиотек, как описано в Winter et al. (1994), выше. Например, мутации можно вводить случайным образом in vitro с использованием полимеразы, допускающей ошибки (представлено в Leung et al., Technique, 1: 11-15 (1989)), в способе по Hawkins et al., J. Mol. Biol, 226: 889-896 (1992) или в способе по Gram et al., Proc. Natl. Acad. Sci USA, 89: 3576-3580 (1992). Дополнительно, созревание аффинности можно осуществлять посредством случайного мутирования одного или нескольких CDR, например, используя ПЦР с праймерами, несущими случайную последовательность, перекрывающими CDR, представляющую интерес, в отобранных индивидуальных Fv клонах и скрининга клонов с более высокой аффинностью. В WO 96/07754 (опубликовано 14 марта 1996 года) описан способ индуцировать мутагенез в определяющей комплементарность области легкой цепи иммуноглобулина для создания библиотеки генов легкой цепи. Другой эффективный подход состоит в том, чтобы рекомбинировать домены VH или VL, отобранные посредством фагового дисплея с репертуарами встречающихся в природе вариантов V-домена, полученными от неиммунизированных доноров, и проводить скрининг на предмет более высокой аффинности в нескольких раундах повторной перетасовки цепей, как описано в Marks et al., Biotechnol, 10:779-783 (1992). Этот способ делает возможным получение антител и фрагментов антител с аффинностями в диапазоне 10-9 M.
Последовательность нуклеиновой кислоты, кодирующую желаемый антиген-мишень, можно сконструировать с использованием аминокислотной последовательности желаемой области антигена.
Нуклеиновые кислоты, кодирующие антиген-мишень, можно получать различными известными в данной области способами. Эти способы включают в качестве неограничивающих примеров химический синтез любыми способами, описанными в Engels et al., Agnew. Chem. Int. Ed. Engl, 28: 716-734 (1989), такими как триэфирный, фосфитный, фосфорамидитный и H-фосфонатный способы. В одном из вариантов осуществления в конструировании ДНК, кодирующей антиген, используют кодоны, предпочитаемые экспрессирующей клеткой-хозяином. Альтернативно, ДНК, кодирующую антиген, можно выделить из геномной или кДНК библиотеки.
После конструирования молекулы ДНК, кодирующей антиген, молекулу ДНК функционально связывают с последовательностью, управляющей экспрессией, в экспрессирующем векторе, таком как плазмида, где клетка-хозяин, трансформированная вектором, распознает управляющую последовательность. В целом, плазмидные векторы содержат последовательности репликации и управляющие последовательности, которые получены от видов, совместимых с клеткой-хозяином. Вектор, как правило, несет сайт репликации, а также последовательности, которые кодируют белки, которые способны обеспечить фенотипический отбор в трансформированных клетках. Подходящие векторы для экспрессии в прокариотических и эукариотических клетках-хозяевах известны в данной области и некоторые дополнительно описаны в настоящем документе. Можно использовать эукариотические организмы, такие как дрожжи, или клетки, полученные от многоклеточных организмов, таких как млекопитающие.
Необязательно ДНК, кодирующая антиген, функционально связана с секреторной лидерной последовательностью, которая ведет к секреции продукта экспрессии клеткой-хозяином в среду для культивирования. Примеры секреторных лидерных последовательностей включают stII, экотин, lamB, GD герпеса, lpp, щелочную фосфатазу, инвертазу и α-фактор. Также подходящей для применения по настоящему документу является лидерная последовательность белка A из 36 аминокислот (Abrahmsen et al., EMBO J, 4: 3901 (1985)).
Клетки-хозяева трансфицируют и предпочтительно трансформируют описанными выше экспрессирующими или клонирующими векторами по данному изобретению и культивируют в стандартных питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих желаемые последовательности.
Трансфекция относится к захвату экспрессирующего вектора клеткой-хозяином не зависимо от того, фактически экспрессируются какие-либо кодирующие последовательности или нет. Как правило, специалистам в данной области известно множество способов трансфекции, например, преципитация с CaPO4 и электропорация. Трансфекцию считают в целом успешной, когда в клетке-хозяине возникает какой-либо признак работы этого вектора. Способы трансфекции хорошо известны в данной области, и некоторые дополнительно описаны в настоящем документе.
Трансформация обозначает введение ДНК в организм с тем, чтобы была возможна репликация ДНК, в виде или внехромосомного элемента или состанвой части хромосомы. В зависимости от используемой клетки-хозяина, трансформацию выполняют с использованием стандартных способов, подходящих для таких клеток. Способы трансформации хорошо известны в данной области, и некоторые дополнительно описаны в настоящем документе.
Прокариотические клетки-хозяева, используемые для получения антигена, можно культивировать, как в целом описано в Sambrook et al., выше.
Клетки-хозяева млекопитающих, используемые для получения антигена, можно культивировать в различных средах, которые хорошо известны в данной области, и некоторые из них описаны в настоящем документе.
Клетки-хозяева, указанные в этом раскрытии, охватывают клетки в культуре in vitro, а также клетки, которые находятся внутри животного-хозяина.
Очистку антигена можно выполнять, используя принятые в данной области способы, некоторые из которых описаны в настоящем документе.
Очищенный антиген можно прикрепить к подходящей матрице, такой как агарозные гранулы, акриламидные гранулы, стеклянные гранулы, целлюлоза, различные акриловые сополимеры, гидроксилметакрилатные гели, полиакриловые и полиметакриловые сополимеры, нейлон, нейтральные и ионные носители и т.п. для использования в разделении клонов фагового диспля аффинной хроматографией. Прикрепление антигенного белка к матрице можно выполнять способами, которые описаны в Methods in Enzymology, vol. 44 (1976). Обычно используемый способ прикрепления белковых лигандов к полисахаридным матрицам, например, к агарозе, декстрану или целлюлозе, включает активацию носителя галогенцианами и последующее образование связи первичных алифатических или ароматических аминов пептидного лиганда с активированной матрицей.
Альтернативно, антиген можно использовать для того, чтобы покрыть лунки адсорбирующих планшетов, экспрессировать на клетках-хозяевах, фиксированных на адсорбирующих планшетах, или использовать в клеточной сортировке, или конъюгировать с биотином для захвата с использованием гранул, покрытых стрептавидином, или использовать в любом другом известном в данной области способе пэннинга библиотек фагового дисплея.
Образцы фаговой библиотеки приводят в контакт с иммобилизованным антигеном в условиях, подходящих для связывания по меньшей мере части фаговых частиц с адсорбентом. Обычно условия, включая pH, ионную силу, температуру и т.п. выбирают так, чтобы имитировать физиологические условия. Фаги, связанные с твердой фазой, промывают и затем элюируют кислотой, например, как описано в Barbas et al., Proc. Natl. Acad. Sci USA, 88: 7978-7982 (1991), или щелочью, например, как описано в Marks et al., J. Mol. Biol, 222: 581-597 (1991), или посредством межантигенной конкуренции, например, в процедуре, похожей на способ антигенной конкуренции по Clackson et al., Nature, 352: 624-628 (1991). Фаги можно обогатить 20-1000-кратно в одном раунде отбора. Кроме того, обогащенные фаги можно растить в бактериальной культуре и подвергать дополнительным раундам отбора.
Эффективность отбора зависит от многих факторов, включая показатели кинетики диссоциации во время промывания и возможность одновременного захвата антигеном нескольких фрагментов антител на одном фаге. Антитела с высокими показателями кинетики диссоциации (и низкими аффинностями связывания) можно удерживать, используя короткие промывания, поливалентный фаговый дисплей и высокую плотность покрытия антигеном в твердой фазе. Высокая плотность не только стабилизирует фаг через поливалентные взаимодействия, но и способствует повторному связыванию фага, который диссоциировал. Отбору антител с низкими показателями кинетики диссоциации (и высокими аффинностями связывания) может способствовать использование длительных промываний и моновалентного фагового дисплея, как описано в Bass et al., Proteins, 8: 309-314 (1990) и в WO 92/09690, и низкой плотности покрытия антигеном, как описано в Marks et al., Biotechnol, 10: 779-783 (1992).
Возможно проведение отбора фаговых антител с различными аффинностями к антигену, даже с аффинностями, которые различаются незначительно. Однако, случайная мутация отобранного антитела (например, как выполняют в некоторых способах созревания аффинности, описанных выше) вероятно может дать начало многим мутантам, в большинстве связывающимся с антигеном, и некоторые из которых обладают более высокой аффинностью. При ограничении по антигену можно отобрать редкий фаг с высокой аффинностью. Чтобы удержать все мутанты с более высокой аффинностью, фаги можно инкубировать с избытком биотинилированного антигена, но при концентрации биотинилированного антигена более низкой молярности, чем целевая молярная константа аффинности к антигену. Затем связывающиеся с высокой аффинностью фаги можно захватить с помощью парамагнитных бус, покрытых стрептавидином. Такой «равновесный захват» позволяет отбирать антитела в соответствии с их аффинностями связывания с чувствительностью, которая делает возможным выделение мутантных клонов со всего лишь в два раза более высокой аффинностью из большого избытка фагов с более низкой аффинностью. Условиями, используемыми при промывании фагов, связанных с твердой фазой, также можно манипулировать для того, чтобы проводить различия на основе показателей кинетики диссоциации.
Антигенные клоны могут быть отобраны по активности. Fv клоны, соответствующие таким антигенным антителам, могут быть отобраны посредством (1) выделения антигенных клонов из фаговой библиотеки, как описано выше, и необязательно, амплификации выделенной популяции клонов фага посредством выращивания популяции в подходящем бактериальном организме-хозяине; (2) выбор антигена и второго белка, против которых желательна блокирующая и неблокирующая активность, соответственно; (3) адсорбирования антигенсвязывающих клонов фага с иммобилизованным антигеном; (4) использования избытка второго белка, чтобы элюировать любые нежелательные клоны, которые распознают антигенсвязывающие детерминанты, которые перекрывают или являются частью связывающих детерминант второго белка; и (5) элюирования клонов, которые остались адсорбированными после стадии (4). Необязательно, клоны с желаемыми блокирующим/неблокирующими свойствами можно дополнительно обогатить посредством повторения процедур отбора, описанных в настоящем документе, один или несколько раз.
ДНК, кодирующую полученные из гибридомы моноклональные антитела или Fv клоны фагового дисплея по изобретению, легко выделяют и секвенируют, используя стандартные процедуры (например, используя олигонуклеотидные праймеры, сконструированные для специфической амплификации кодирующих областей тяжелой и легкой цепи, представляющих интерес, из гибридомной или фаговой ДНК матрицы). После выделения, ДНК можно поместить в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьяны, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые иначе не продуцируют белок иммуноглобулина, чтобы добиться синтеза желаемых моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии ДНК, кодирующей антитело, в бактериях включают Skerra et al., Curr. Opinion in Immunol, 5: 256 (1993) и Pluckthun, Immunol. Revs, 130: 151 (1992).
ДНК, кодирующую Fv клоны по изобретению, можно комбинировать с известными ДНК последовательностями, кодирующими константные области тяжелой цепи и/или легкой цепи (например, подходящие ДНК последовательности можно получить из Kabat et al., выше), чтобы сформировать клоны, кодирующие полноразмерные или неполноразмерные тяжелые и/или легкие цепи. Следует принимать во внимание, что с этой целью можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получить от человека или животного любого вида. Fv клон, полученный из ДНК вариабельного домена животного одного вида (такого как человек) и затем слитый с ДНК константной области животного другого вида, чтобы сформировать кодирующую последовательность(и) для «гибридной» полноразмерной тяжелой цепи и/или легкой цепи, включен в определение «химерного» и «гибридного» антитела, как применяют в настоящем документе. В предпочтительном варианте осуществления Fv клон, полученный из вариабельной ДНК человека, сливают с ДНК константной области человека, чтобы сформировать кодирующую последовательность(и) для всех полноразмерных или неполноразмерных тяжелых и/или легких цепей человека.
ДНК, кодирующую антитело, полученное из гибридомы по изобретению, также можно модифицировать также можно модифицировать, например, посредством замены кодирующей последовательности константных доменов тяжелой и легкой цепи человека вместо гомологичных последовательностей мыши, полученных из гибридомного клона (например, как в способе по Morrison et al., Proc. Natl Acad. Sci. USA, 81:6851-6855 (1984)). ДНК, кодирующую полученное из гибридомного или Fv клона антитело или фрагмент, можно дополнительно модифицировать посредством ковалентного связывания с кодирующей последовательностью иммуноглобулина всей или части кодирующей последовательности для полипептида не иммуноглобулина. Таким образом, получают «химерные» или «гибридные» антитела, которые обладают специфичностью связывания полученных из Fv клона или гибридомного клона антител по изобретению.
Антигенная специфичность
Настоящее изобретение применимо к антителам с любой подходящей антигенсвязывающей специфичностью. Предпочтительно, антитела по изобретению специфичны к антигенам, которые представляют собой биологически важные полипептиды. Более предпочтительно, антитела по изобретению можно использовать для терапии или диагностики заболеваний или нарушений у млекопитающего. Неограничивающие примеры терапевтических антител включают антитела против VEGF, против c-met, против IgE, против CD11, против CD18, против CD40, против тканевого фактора (TF), против HER2 и против TrkC. Также предусмотрены антитела, направленные против неполипептидных антигенов (таких как ассоциированные с опухолями гликолипидные антигены).
Когда антиген представляет собой полипептид, он может представлять собой трансмембраную молекулу (например, рецептор, такой как рецепторная тирозинкиназа) или лиганд, такой как фактор роста. Типичные антигены, включюат молекулы, такие как ренин; гормон роста, включая гормон роста человека и гормон роста коровы; релизинг-фактор гормона роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; α-1-антитрипсин; A-цепь инсулина; B-цепь инсулина; проинсулин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания, такие как фактор VIIIC, фактор IX, тканевой фактор (TF) и фактор Виллебранда; противосвертывающие факторы, такие как белок C; предсердный натрийуретический фактор; сурфактант легких; активатор плазминогена, такой как урокиназа или активатор плазминогена мочи или тканевого типа (t-PA) человека; бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухоли-α и -β; энкефалиназа; RANTES (белок, экспрессируемый и секретируемый нормальными Т-клетками при активации); макрофагальный воспалительный белок человека (MIP-1-α); сывороточный альбумин, такой как сывороточный альбумин человека; ингибирующее вещество Мюллера; A-цепь релаксина; B-цепь релаксина; прорелаксин; гонадотропин-ассоциированный пептид мыши; микробный белок, такой как β-лактамаза; ДНКаза; IgE; ассоциированный с цитотоксическими T-лимфоцитами антиген (CTLA), такой как CTLA-4; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы для гормонов или факторов роста; белок A или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор кости (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6), или фактор роста нервов, такой как NGF-β; фактор роста тромбоцитов (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-α и TGF-β, включая TGF-β1, TGF-β2, TGF-β3, TGF-β4 или TGF-β5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); дез(1-3)-IGF-I (IGF-I головного мозга), белки, связывающие инсулиноподобный фактор роста; белки CD, такие как CD3, CD4, CD8, CD19, CD20 и CD40; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; морфогенетический белок кости (BMP); интерферон, такой как интерферон-α, -β и -γ; колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-10; супероксиддисмутаза; T-клеточные рецепторы; поверхностные мембранные белки; стимулятор гемолиза; вирусный антиген, такой как, например, часть оболочки СПИД; транспортные белки; рецепторы хоуминга; адрессины; регуляторные белки; интегрины, такие как CD11a, CD11b, CD11c, CD18, ICAM, VLA-4 и VCAM; ассоциированный с опухолью антиген, такой как рецептор HER2, HER3 или HER4; и фрагменты любых перечисленных выше полипептидов.
Типичные антигены для антител, охватываемые настоящим изобретением, включают белки CD, такие как CD3, CD4, CD8, CD19, CD20, CD34 и CD46; члены семейства рецепторов ErbB, такие как рецептор EGF, рецептор HER2, HER3 или HER4; молекулы клеточной адгезии, такие как LFA-1, Mac1, p150.95, VLA-4, ICAM-1, VCAM, α4/β7 интегрин и αv/β3 интегрин, включая их α или β субъединицы (например, антитела против CD11a, против CD18 или против CD11b); факторы роста, такие как VEGF; тканевой фактор (TF); TGF-

Фрагменты антител
Настоящее изобретение относится к фрагментам антител. В определенных случаях полезно использовать фрагменты антител, а не целые антитела. Меньший размер фрагментов допускает быстрый клиренс и может вести к улучшенному доступу к солидным опухолям.
Разработаны различные способы получения фрагментов антител. Традиционно эти фрагменты получали посредством протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24: 107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако, теперь эти фрагменты можно получать непосредственно с помощью рекомбинантных клеток-хозяев. E. coli могут экспрессировать и секретировать все фрагменты антител Fab, Fv и ScFv, таким образом допуская гибкое получение больших количеств этих фрагментов. Фрагменты антител можно выделить из фаговых библиотек антител, рассмотренных выше. Альтернативно, Fab'-SH фрагменты можно непосредственно извлекать из E. coli и химически связывать, чтобы сформировать F(ab')2 фрагменты (Carter et al., Bio/Technology 10: 163-167 (1992)). Согласно другому подходу, F(ab')2 фрагменты можно выделять непосредственно из культуры рекомбинантных клеток-хозяев. Fab и F(ab')2 фрагменты с увеличенным временем полужизни in vivo, содержащий остатки эпитопа, связывающего рецептор спасения, описаны в патенте США № 5869046. Другие способы получения фрагментов антител очевидны практикующим специалистам. В других вариантах осуществления выбранное антитело представляет собой одноцепочечный Fv фрагмент (scFv) (см., например, WO 93/16185; патенты США №№ 5571894 и 5587458). Fv и sFv представляют собой единственные частицы с интактными антигенсвязывающими участками, у которых отсутствуют константные области; таким образом, они подходят для ослабленного неспецифического связывания во время использования in vivo. Можно конструировать sFv слитые белки, чтобы произвести слияние эффекторного белка на N- или C-конце sFv. См. Antibody Engineering, ed. Borrebaeck, выше. Фрагмент антитела также может представлять собой «линейное антитело», например, как описано, например, в патенте США № 5641870. Такие линейные фрагменты антител могут быть моноспецифическими или биспецифическими.
Соответственно, в некотором варианте осуществления антитело против c-met представляет собой антитело с одним Fab фрагментом (т.е. вариабельный домен тяжелой цепи и вариабельный домен легкой цепи формируют один антигенсвязывающий фрагмент), содержащее Fc-область, где Fc-область содержит первый и второй полипептид Fc, где первый и второй полипептиды Fc представлены в виде комплекса и формируют Fc-область, которая повышает стабильность указанного фрагмента антитела по сравнению с Fab молекулой, содержащей указанный антигенсвязывающий фрагмент. Антитела с одним Fab фрагментом дополнительно описаны в настоящем документе.
Гуманизированные антитела
Настоящее изобретение относится к гуманизированным антителам. В данной области известны различные способы гуманизации не принадлежащих человеку антител. Например, гуманизированное антитело может содержать один или несколько аминокислотных остатков, введенных в него из источника, который не принадлежит человеку. Эти не принадлежащие человеку аминокислотные остатки часто обозначают как «импортные» остатки, которые типично взяты из «импортного» вариабельного домена. Гуманизацию можно выполнять по существу после способа согласно Winter'у и коллегам (Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-327; Verhoeyen et al. (1988) Science 239: 1534-1536), посредством замены последовательностями гипервариабельных областей соответствующих последовательностей антитела человека. Соответственно, такие «гуманизированные» антитела представляют собой химерные антитела (патент США № 4816567), где по существу менее чем интактный вариабельный домен человека заменен соответствующей последовательностью из видов, не являющихся человеком. На практике, гуманизированные антитела типично представляют собой антитела человека, в которых некоторые остатки гипервариабельной области и возможно некоторые FR остатки заменены остатками из аналогичных сайтов в антителах грызунов.
Для снижения антигенности очень важно выбрать вариабельные домены человека, как легкие и тяжелые, для использования в получении гуманизированных антител. Согласно так называемому способу «наилучшего соответствия», проводят скрининг последовательности вариабельного домена антитела грызуна против целой библиотеки известных последовательностей вариабельного домена человека. Затем ту последовательность человека, которая ближе всего к таковой грызуна, принимают в качестве каркасной области человека для гуманизированного антитела (Sims et al. (1993) J. Immunol. 151:2296; Chothia et al. (1987) J. Mol. Biol. 196:901. В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека конкретной подгруппы легкой или тяжелой цепи. Ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al. (1992) Proc. Natl. Acad. Sci. USA, 89:4285; Presta et al. (1993) J. Immunol, 151:2623.
Кроме того, важно гуманизировать антитела с охранением высокой аффинности к антигену и других благоприятных биологических свойств. Чтобы достичь этой цели, согласно одному способу, гуманизированные антитела получают посредством процесса анализа родительских последовательностей и различных умозрительных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов обычно доступны и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и визуализируют возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Изучение этих визуализаций дает возможность анализа возможной роли остатков в функционировании последовательности иммуноглобулина-кандидата, т.е. анализа остатков, которые влияют на способность иммуноглобулина-кандидата связывать свой антиген. Таким образом, остатки FR из последовательности-реципиента и импортируемой последовательности можно выбирать и комбинировать с тем, чтобы добиться желаемой характеристики антитела, такой как повышенная аффинность в отношении антигена-мишени(ей). В целом, остатки гипервариабельной области непосредственно и наиболее существенно вовлечены во вленияние на связывание антигена.
Антитела человека
Антитела человека по изобретению можно конструировать, комбинируя последовательность(и) вариабельных доменов Fv клона, выбранную из библиотек фагового дисплея, полученных от человека, с известной последовательностью(ями) константных доменов человека как описано выше. Альтернативно, моноклональные антитела человека по изобретению можно создать гибридомным способом. Клеточные линии миеломы человека и гетеромиеломная клеточная линия мыши-человека для получения моноклональных антител человека описаны, например, в Kozbor J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol, 147:86 (1991).
Можно создавать трансгенных животных (например, мышей), которые после иммунизации способны продуцировать полный репертуар антител человека в отсутствие эндогенного образования иммуноглобулинов. Например, описано, что гомозиготная делеция гена соединительной области (JH) тяжелой цепи антитела у химерных мышей и мышей с мутацией в зародышевой линии ведет к полному ингибированию эндогенного образования антител. Перенос массива генов иммуноглобулинов зародышевой линии человека в таких мышей с мутацией в зародышевой линии должен привести к образованию антител человека после антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255 (1993); Bruggermann et al., Year in Immunol, 7:33 (1993).
Также можно использовать перетасовку генов для получения антител человека из антител, не принадлежащих человеку, например, антител грызуна, где антитело человека обладает схожими аффинностями и специфичностями с не принадлежащим человеку антителом. В соответствии с этим способом, который также называют «импринтингом эпитопа», фрагмент вариабельной области тяжелой или легкой цепи не принадлежащего человеку антитела, полученный с помощью способов фагового дисплея, как описано выше, заменяют на репертуар генов V-домена человека, создавая популяцию scFv или Fab химер из не принадлежащей человеку цепи/цепи человека. Отбор с использованием антигена ведет к выделению химерных scFv или Fab из не принадлежащей человеку цепи/цепи человека, где цепь человека восстанавливает антигенсвязывающий участок, разрушенный при удалении соответствующей не принадлежащей человеку цепи в первичном клоне фагового дисплея, т.е. эпитоп определяет (запечатлевает) выбор цепи-партнера человека. Когда процесс повторяют для того, чтобы заменить оставшуюся не принадлежащую человеку цепь, получают антитело человека (см. PCT WO 93/06213, опубликовано 1 апреля 1993 года). В отличие от традиционной гуманизации не принадлежащих человеку антител посредством пересадки CDR, этот способ предоставляет антитела, полностью принадлежащие человеку, которые не содержат остатки FR или CDR, которые получены не от человека.
Биспецифические антитела
Биспецифические антитела представляют собой моноклональные антитела, предпочтительно антитела человека или гуманизированные антитела, которые обладают специфичностями связывания для по меньшей мере двух различных антигенов. В настоящем случае одна из специфичностей связывания предназначена для антигена, а другая для любого другого антигена. Типичные биспецифические антитела могут связываться с двумя различными эпитопами антигена. Биспецифические антитела также можно использовать для локализации цитотоксических средств в клетках, которые экспрессируют антиген. Эти антитела обладают антигенсвязывающим Fab фрагментом и Fab фрагментом, который связывает цитотоксическое средство (например, сапорин, средство против интерферона-α, алкалоид барвинка, A-цепь рицина, метотрексат или гаптен с радиоактивным изотопом). Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител (например, F(ab')2 биспецифические антитела).
В данной области известны способы создания биспецифических антител. Традиционно рекомбинантное получение биспецифических антител основано на совместной экспрессии двух пар тяжелой цепи-легкой цепи иммуноглобулина, где две тяжелые цепи обладают различными специфичностями (Milstein and Cuello, Nature, 305: 537 (1983)). Вследствие случайного ассортимента тяжелых и легких цепей иммуноглобулинов, эти гибридомы (квадрогибридомы) продуцируют потенциальную смесь из 10 различных молекул антител, из которых только одна имеет правильную биспецифическуюю структуру. Очистка правильной молекулы, которую обычно выполняют с помощью стадий аффинной хроматографии, является довольно тяжелой, и выход продукта низок. Схожие процедуры раскрыты в WO 93/08829, опубликованной 13 мая 1993 года, и в Traunecker et al., EMBO J., 10: 3655 (1991).
Согласно другому и более предпочтительному подходу, вариабельные домены антител с желаемыми специфичностями связывания (участки связывания антитело-антиген) сливают с последовательностями константных доменов иммуноглобулинов. Слияние предпочтительно проводят с константным доменом тяжелой цепи иммуноглобулина, содержащей по меньшей мере часть шарнирной области, областей CH2 и CH3. Предпочтительно иметь первую константную область тяжелой цепи (CH1), содержащую сайт, необходимый для связывания легкой цепи, присутствующей в по меньшей мере одном из слияний. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и, при желании, легкую цепь иммуноглобулина, встраивают в отдельные экспрессирующие векторы, и совместно трансфицируют в подходящий организм-хозяин. Это обеспечивает большую гибкость в корректировании взаимных пропорций трех полипептидных фрагментов в вариантах осуществления, когда неравные соотношения трех полипептидных цепей, использованные в конструировании, обеспечивают оптимальный выход. Однако можно встроить кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессирующий вектор, когда экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях ведет к высокому выходу или когда отношения не имеют конкретного значения.
В предпочтительном варианте осуществления этого подхода биспецифические антитела создают из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном Fab фрагменте и гибридной пары тяжелой цепи-легкой цепи иммуноглобулина (обеспечивающей вторую специфичность связывания) в другом Fab фрагменте. Обнаружено, что эта асимметричная структура облегчает отделение желаемого биспецифического соединения от нежелательных комбинаций цепей иммуноглобулинов, поскольку присутствие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий путь разделения. Этот подход раскрыт в WO 94/04690. Более подробно о создании биспецифических антител см., например, в Suresh et al., Methods in Enzymology, 121:210 (1986).
Согласно другому подходу, область контакта между парой молекул антител можно сконструировать для того, чтобы максимизировать процентную долю гетеродимеров, которые извлекают из культуры рекомбинантных клеток. Предпочтительная область контакта содержит по меньшей мере часть домена CH3 константного домена антитела. В этом способе одну или несколько маленьких боковых цепей аминокислот из области контакта первой молекулы антитела заменяют на более крупные боковые цепи (например, тирозин или триптофан) (выступы или выпуклости). Компенсирующие «пустоты» (отверстия) размера, идентичного или схожего с большой боковой цепью(ями), создают на области контакта второй молекулы антитела посредством замены больших боковых цепей аминокислота на меньшие (например, аланин или треонин). Это предоставляет механизм для повышения выхода гетеродимера относительно других нежелательных конечных продуктов, таких как гомодимеры. Выступы и отверстия дополнительно описаны в настоящем документе.
Биспецифические антитела включают сшитые антитела или «гетероконъюгаты». Например, одно из антител в гетероконъюгате может быть связано с авидином, другое с биотином. Такие антитела предложены, например, для нацеливания клеток иммунной системы на нежелательные клетки (патент США № 4676980) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/00373 и EP 03089). Гетероконъюгированные антитела можно создавать, используя любые удобные способы сшивания. Подходящие сшивающие средства хорошо известны в данной области и раскрыты в патенте США № 4676980, вместе с множеством способов сшивания.
Способы создания биспецифических антител из фрагментов антител также описаны в литературе. Например, можно получать биспецифические антитела, используя химическую связь. Brennan et al., Science, 229: 81 (1985) описывают процедуру, где интактные антитела протеолитически расщепляют для создания F(ab')2 фрагментов. Эти фрагменты восстанавливают в присутствие дитиолового комплексообразователя арсенита натрия, чтобы стабилизировать винициальные дитиолы и предотвратить межмолекулярное образование дисульфидов. Созданные Fab' фрагменты затем превращают в производные тионитробензоата (TNB). Затем одно Fab'-TNB производных повторно превращают в Fab'-тиол посредством восстановления меркаптоэтиламином и смешивают с эквимолярным количеством другого Fab'-TNB производного, чтобы сформировать биспецифическое антитело. Полученные биспецифические антитела можно использовать в качестве средств для избирательной иммобилизации ферментов.
Недавние успехи облегчили непосредственное извлечение Fab'-SH фрагментов из E. coli, которые можно химически связывать для формирования биспецифических антител. Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описывают получение полностью F(ab')2 молекулы гуманизированного биспецифического антитела. Каждый фрагмент Fab' раздельно секретировали из E. coli и подвергали непосредственному химическому связыванию in vitro, чтобы сформировать биспецифическое антитело. Сформированное таким образом биспецифическое антитело способно связываться с клетками, сверхэекспрессирующими рецептор HER2, и нормальными T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против мишени опухоли молочной железы человека.
Также описаны различные способы получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получали с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992). Пептиды лейциновых молний из белков Fos и Jun связывали с Fab' частями двух различных антитела посредством слияния генов. Гомодимеры антител восстанавливали в шарнирной области, чтобы сформировать мономеры и затем повторно окисляли, чтобы сформировать гетеродимеры антител. Этот способ также можно использовать для получения гомодимеров антител. Технология «диател», описанная авторами Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993), предоставляет альтернативный механизм для создания фрагментов биспецифических антител. Фрагменты содержат вариабельный домен (VH) тяжелой цепи, соединенный с вариабельным доменом (VL) легкой цепи посредством линкера, который слишком короток, чтобы сделать возможным спаривание между двумя доменами в одной и той же цепи. Соответственно, домены VH и VL одного фрагмента вынуждены образовывать пары с комплементарными доменами VL и VH другого фрагмента, тем самым формируя два антигенсвязывающих участка. Также сообщалось о другой стратегии создания фрагментов биспецифических антител с помощью одноцепочечных Fv (sFv) димеров. См. Gruber et al., J. Immunol, 152:5368 (1994).
Предусмотрены антитела более чем с двумя валентностями. Например, можно получать триспецифические антитела. Tutt et al. J. Immunol. 147: 60 (1991).
Поливалентные антитела
Интернализация (и/или катаболизм) поливалентного антитела может происходить быстрее, чем у бивалентного антитела, посредством экспрессии клеткой антигена, с которым связываются антитела. Антитела по настоящему изобретению могут представлять собой поливалентные антитела (которые отличны от класса IgM) с тремя или более антигенсвязывающими участками (например, тетравалентные антитела), которые можно легко получать посредством рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Поливалентное антитело может содержать домен димеризации и три или более антигенсвязывающих участка. Предпочтительный домен димеризации содержит (или состоит из) Fc-области или шарнирной области. В этом сценарии антитело должно содержать Fc-область и три или более антигенсвязывающих участка ближе к N-концу относительно Fc-области. Предпочтительное поливалентное антитело в настоящем документе содержит (или состоит из) от трех до приблизительно восьми, но предпочтительно четыре антигенсвязывающих участка. Поливалентное антитело содержит по меньшей мере одну полипептидную цепь (и предпочтительно две полипептидные цепи), где полипептидная цепь(и) содержит два или более вариабельных домена. Например, полипептидная цепь(и) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидную цепь Fc-области, X1 и X2 представляют аминокислоту или полипептид, и n равно 0 или 1. Например, полипептидная цепь(и) может содержать: цепь VH-CH1-гибкий линкер-VH-CH1-Fc-область; или цепь VH-CH1-VH-CH1-Fc-область. Поливалентное антитело в настоящем документ предпочтительно дополнительно содержит по меньшей мере два (и предпочтительно четыре) полипептида вариабельного домена легкой цепи. Поливалентное антитело в настоящем документе может содержать, например, от приблизительно двух до приблизительно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, предусмотренные здесь, содержат вариабельный домен легкой цепи и, необязательно, дополнительно содержат домен CL.
Варианты антител
В некоторых вариантах осуществления предусмотрена модификация(и) аминокислотных последовательностей антител, описанных в настоящем документе. Например, может быть желательно улучшить аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотных последовательностей антитела получают посредством введения подходящих изменений нуклеотидов в нуклеиновую кислоту антитела или посредством синтеза пептидов. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Любое сочетание делеции, вставки и замены выполняют для того, чтобы прийти к конечной конструкции при условии, что конечная конструкция обладает желаемыми характеристиками. Изменения аминокислот можно вводить в рассматриваемую аминокислотную последовательность антитела во время создания этой последовательности.
Эффективный способ идентификации определенных остатков или областей антитела, которые представляют собой предпочтительные положения для мутагенеза, называется «аланин-сканирующим мутагенезом», как описано авторами Cunningham and Wells (1989) Science, 244: 1081-1085. Здесь остаток или группу целевых остатков идентифицируют (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют нейтральными или отрицательно заряженными аминокислотами (наиболее предпочтительно аланин или полиаланин), чтобы оказать влияние на взаимодействие аминокислот с антигеном. Затем эти положения аминокислот, проявляющие функциональную чувствительность к заменам, уточняют посредством введения дополнительных или других вариантов в сайты замены или для них. Таким образом, несмотря на то, что сайт для введения разнообразия аминокислотных последовательностей определяют предварительно, свойства мутации per se не нужно определять предварительно. Например, чтобы проанализировать эффективность мутации в заданном сайте, в целевом сайте или области проводят аланин-сканирующий или случайный мутагенез и проводят скрининг экспрессированных иммуноглобулинов на желаемую активность.
Вставки аминокислотной последовательности включают амино- и/или карбокси-концевые слияния, длина которых варьирует от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности из одного или нескольких аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым метиониловым остатком или антитело, слитое с цитотоксическим полипептидом. Другие варианты вставок в молекуле антитела включают слияние N- или C-конца антитела с ферментом (например, для ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
Гликозилирование полипептидов типично является или N-связанным или O-связанным. N-связанный относится к присоединению молекулы углевода к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту за исключением пролина, представляют собой последовательности распознавания для ферментативного присоединения молекулы углевода к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров N-ацетилгалактозамин, галактоза или ксилоза к гидроксиаминокислоте, чаще всего к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление сайтов гликозилирования к антителу удобно выполнять посредством изменения аминокислотной последовательности так, чтобы она содержала одну или несколько описанных выше трипептидных последовательностей (для N-связанных сайтов гликозилирования). Также изменение можно выполнять посредством добавления одного или нескольких остатков серина или треонина или замены на них в последовательности исходного антитела (для O-связанных сайтов гликозилирования).
Когда антитело содержит Fc-область, присоединенный к ней углевод может быть изменен. Например, антитела со зрелой углеводной структурой, в которой отсутствует фукоза, присоединенная к Fc-области антител, описаны в заявке на патент США № US 2003/0157108 (Presta, L.). Также см. US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). На антитела с разветвляющим N-ацетилглюкозамином (GlcNAc) в углеводе, присоединенном к Fc-области антитела, приведены ссылки в WO 2003/011878, Jean-Mairet et al. и патенте США № 6602684, Umana et al. Антитела с по меньшей мере одним остатком галактозы в олигосахариде, присоединенном к Fc-области антитела, описаны в WO 1997/30087, Patel et al. Также см. WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.) в отношении антител с измененным углеводом, прикрепленным к их Fc-областям. Об антигенсвязывающих молекулах с модифицированным гликозилированием также см. в US 2005/0123546 (Umana et al.).
Предпочтительный вариант гликозилирования в настоящем документ содержит Fc-область, где в углеводной структуре, присоединенной к Fc-области, отсутствует фукоза. Такие варианты имеют улучшенную функцию АЗКЦ. Необязательно Fc-область дополнительно содержит в себе одну или несколько замен аминокислот, которые дополнительно улучшают АЗКЦ, например, замены в положениях 298, 333 и/или 334 Fc-области (нумерация остатков по Eu). Примеры публикаций, связанных с «дефукозилированными» или «дефицитными по фукозе» антителами, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; Okazaki et al. J. Mol. Biol. 336: 1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры клеточных линий, продуцирующих дефукозилированные антитела, включают клетки Led13 CHO, дефицитные по фукозилированию белка (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); заявка на патент США № US 2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., в частности, в примере 11), и клеточные линии с нокаутом, такие как клетки CHO с нокаутом гена α-1,6-фукозилтрансферазы, FUT8 (Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004)).
В одном из аспектов изобретение относится к фрагменту антитела, содержащему по меньшей мере одну характеристику, которая способствует гетеродимеризации, при этом минимизируя гомодимеризацию, Fc последовательностей в фрагменте антитела. Такая характеристика(и) улучшает выход и/или чистоту и/или гомогенность популяций иммуноглобулинов, получаемых посредством способов по изобретению, как описано в настоящем документе. В одном из вариантов осуществления первый полипептид Fc и второй полипептид Fc встречаются/взаимодействуют в области контакта. В некоторых вариантах осуществления, где первый и второй полипептиды Fc встречаются в области контакта, область контакта второго полипептида Fc (последовательность) содержит выпуклость (также обозначаемую «выступ»), который может быть размещен в полости (также обозначаемой «отверстие») в области контакта первого полипептида Fc (последовательности). В одном из вариантов осуществления первый полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать полость, или второй полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать выпуклость, или и то и другое. В одном из вариантов осуществления первый полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать полость, и второй полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать выпуклость. В одном из вариантов осуществления область контакта второго полипептида Fc содержит выпуклость, которая может быть размещена в полости в области контакта первого полипептида Fc, где полость или выпуклость, или и то и другое, введена в область контакта первого и второго полипептидов Fc, соответственно. В некоторых вариантах осуществления, где первый и второй полипептиды Fc встречаются в области контакта, область контакта первого полипептида Fc (последовательность) содержит выпуклость, которая может быть размещена в полости в области контакта второго полипептида Fc (последовательности). В одном из вариантов осуществления второй полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать полость, или первый полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать выпуклость, или и то и другое. В одном из вариантов осуществления второй полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать полость, и первый полипептид Fc изменен относительно шаблонного/исходного полипептида, чтобы кодировать выпуклость. В одном из вариантов осуществления область контакта первого полипептида Fc содержит выпуклость, которая может быть размещена в полости в области контакта второго полипептида Fc, где выпуклость или полость, или и то и другое, введена в область контакта первого и второго полипептидов Fc, соответственно.
В одном из вариантов осуществления и выпуклость, и полость содержит встречающийся в природе аминокислотный остаток. В одном из вариантов осуществления полипептид Fc, содержащий выпуклость, создают посредством замены исходного остатка из области контакта шаблонного/исходного полипептида на импортный остаток, который имеет больший объем боковой цепи, чем исходный остаток. В одном из вариантов осуществления полипептид Fc, содержащий выпуклость, создают посредством способа, содержащего стадию, где полинуклеотид, кодирующий исходный остаток из области контакта указанного полипептида заменяют на полинуклеотид, кодирующий импортный остаток, который имеет больший объем боковой цепи, чем исходный. В одном из вариантов осуществления исходный остаток представляет собой треонин. В одном из вариантов осуществления исходный остаток представляет собой T366. В одном из вариантов осуществления импортный остаток представляет собой аргинин (R). В одном из вариантов осуществления импортный остаток представляет собой фенилаланин (F). В одном из вариантов осуществления импортный остаток представляет собой тирозин (Y). В одном из вариантов осуществления импортный остаток представляет собой триптофан (W). В одном из вариантов осуществления импортный остаток представляет собой R, F, Y или W. В одном из вариантов осуществления выпуклость создают посредством замены двух или более остатков в шаблонном/исходном полипептиде. В одном из вариантов осуществления полипептид Fc, содержащий выпуклость, содержит замену треонина в положении 366 на триптофан, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al. (pp. 688-696 in Sequences of proteins of immunological interest, 5th ed., Vol. 1 (1991; N1H, Bethesda, MD)).
В некоторых вариантах осуществления полипептид Fc, содержащий полость, создают посредством замены исходного остатка в области контакта шаблонного/исходного полипептида на импортный остаток, который имеет меньший объем боковой цепи, чем исходный остаток. Например, полипептид Fc, содержащий полость, можно создать посредством способа, содержащего стадию, где полинуклеотид, кодирующий исходный остаток из области контакта указанного полипептида, заменяют на полинуклеотид, кодирующий импортный остаток, который имеет меньший объем боковой цепи, чем исходный. В одном из вариантов осуществления исходный остаток представляет собой треонин. В одном из вариантов осуществления исходный остаток представляет собой лейцин. В одном из вариантов осуществления исходный остаток представляет собой тирозин. В одном из вариантов осуществления импортный остаток представляет собой не цистеин (C). В одном из вариантов осуществления импортный остаток представляет собой аланин (A). В одном из вариантов осуществления импортный остаток представляет собой серин (S). В одном из вариантов осуществления импортный остаток представляет собой треонин (T). В одном из вариантов осуществления импортный остаток представляет собой валин (V). Полость можно создать посредством замены одного или нескольких исходных остатков шаблонного/исходного полипептида. Например, в одном из вариантов осуществления полипептид Fc, содержащий полость, содержит замену двух или более исходных аминокислот, выбранных из группы, состоящей из треонина, лейцина и тирозина. В одном из вариантов осуществления полипептид Fc, содержащий полость, содержит два или более импортных остатка, выбранных из группы, состоящей из аланина, серина, треонина и валина. В некоторых вариантах осуществления полипептид Fc, содержащий полость, содержит замену двух или более исходных аминокислот, выбранных из группы, состоящей из треонина, лейцина и тирозина, и где указанные исходные аминокислоты заменяют на импортные остатки, выбранные из группы, состоящей из аланина, серина, треонина и валина. В некоторых вариантах осуществления исходная аминокислота, которую заменяют, представляет собой T366, L368 и/или Y407. В одном из вариантов осуществления полипептид Fc, содержащий полость, содержит замену треонина в положении 366 на серин, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al. выше. В одном из вариантов осуществления полипептид Fc, содержащий полость, содержит замену лейцина в положении 368 на аланин, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al. выше. В одном из вариантов осуществления полипептид Fc, содержащий полость, содержит замену тирозина в положении 407 на валин, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al. выше. В одном из вариантов осуществления полипептид Fc, содержащий полость, содержит две или более аминокислотных замены, выбранных из группы, состоящей из T366S, L368A и Y407V, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al. выше. В некоторых вариантах осуществления этих фрагментов антител, полипептид Fc, содержащий выпуклость, содержит замену треонина в положении 366 на триптофан, нумерация аминокислот соответствует схеме нумерации по EU согласно Kabat et al., выше.
В одном из вариантов осуществления антитело содержит Fc мутации, составляющие «выступы» и «отверстия», как описано в WO2005/063816. Например, мутация отверстия может представлять собой одну или несколько из T366A, L368A и/или Y407V в полипептиде Fc, и мутация выступа может представлять собой T366W.
Другой тип варианта представляет собой вариант с заменой аминокислоты. Эти варианты имеют по меньшей мере один аминокислотный (по меньшей мере два, по меньшей мере три, по меньшей мере 4 или более) остаток в молекуле антитела, замененный на другой остаток. Участки наибольшего интереса для мутагенеза замен включают гипервариабельные области, но также предусмотрены изменения FR. Консервативные замены приведены в таблице A под заголовком «предпочтительные замены». Если такие замены ведут к изменению биологической активности, тогда можно вводить более значительные замены, обозначенные как «типичные замены» в таблице A, или как дополнительно описано ниже в отношении классов аминокислот, и проводить скрининг продуктов.

Значительные модификации биологических свойств антитела осуществляют посредством выбора замен, которые значительно отличаются по эффекту, оказываемому ими на сохранение (a) структуры полипептидного остова в области замены, например, в виде конформации листа или спирали, (b) заряда или гидрофобности молекулы в целевом участке или (c) объема боковой цепи. Встречающиеся в природе остатки делятся на группы на основе общих свойств боковой цепи:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: asp, glu;
(4) основные: his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены должны влечь за собой обмен члена одного из этих классов на другой класс.
Один тип варианта с заменой включает замену одного или нескольких остатков гипервариабельной области родительского антитела (например, гуманизированного антитела или антитела человека). В целом, полученный вариант(ы), отобранный для дальнейшей разработки, должен обладать улучшенными биологическими свойствами по отношению к родительскому антителу, из которого он создан. Удобный путь создания таких вариантов с заменой включает созревание аффинности с использованием фагового дисплея. В кратком изложении, в несколько участков гипервариабельной области (например, 6-7 участков) вводят мутации для того, чтобы создать все возможные замены аминокислот в каждом участке. Созданные таким образом антитела представляют на частицах нитчатого фага в виде слияний с продуктом гена III M13, упакованных в каждую частицу. Затем проводят скрининг представленных на фагах вариантов по их биологической активности (например, аффинности связывания) как раскрыто в настоящем документе. Для того чтобы идентифицировать участки-кандидаты гипервариабельной области для модификации, можно осуществлять аланин-сканирующий мутагенез, чтобы идентифицировать остатки гипервариабельной области, вносящие значительный вклад в связывание антигена. Альтернативно, или дополнительно, может быть полезным анализировать кристаллическую структуру комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактные остатки и соседние остатки являются кандидатами для замены в соответствии со способами, выработанными в настоящем документе. После создания таких вариантов, панель вариантов подвергают скринингу, как описано в настоящем документе, и антитела с превосходными свойствами в одном или нескольких релевантных анализах можно выбрать для дальнейшей разработки.
Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности антитела, получают посредством различных известных в данной области способов. Эти способы включают в качестве неограничивающих примеров выделение из природного источника (в случае встречающихся в природе вариантов аминокислотной последовательности) или получение посредством опосредованного олигонуклеотидами (или сайт-направленного) мутагенеза, ПЦР мутагенеза и кассетного мутагенеза ранее полученного варианта или невариантной версии антитела.
Может быть желательно ввести одну или несколько аминокислотных модификаций в Fc-области полипептидов иммуноглобулинов по изобретению, тем самым создавая вариант Fc-области. Вариант Fc-области может содержать последовательность Fc-области человека (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях, включая таковые цистеина шарнирной области.
В соответствии с этим описанием и представлениями в данной области предусмотрено, что в некоторых вариантах осуществления антитело, используемое в способах по изобретению, может содержать одно или несколько изменений по сравнению с соответствующим антителом дикого типа, например, в Fc-области. Тем не менее, эти антитела должны сохранять по существу те же характеристики, необходимые для терапевтической эффективности по сравнению с соответствующим им диким типом. Например, полагают, что в Fc-области можно создать определенные изменения, которые должны привести к измененным (т.е. или улучшенным или ухудшенным) связыванию C1q и/или комплемент-зависимой цитотоксичности (КЗЦ), например, как описано в WO 99/51642. Другие примеры вариантов Fc-области также см. в Duncan & Winter Nature 322:738-40 (1988); патенте США № 5648260; патенте США № 5624821; и WO 94/29351. В WO 00/42072 (Presta) и WO 2004/056312 (Lowman) описаны варианты антител с улучшенным или ухудшенным связыванием с FcR. Содержание этих публикаций патентов прямо включено в настоящий документ в качестве ссылки. Также см. Shields et al. J. Biol. Chem. 9(2): 6591-6604 (2001). Антитела с увеличенным временем полужизни и улучшенным связыванием с неонатальным рецептором Fc (FcRn), которые отвечают за перено материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US 2005/0014934A1 (Hinton et al.). Эти антитела содержат в себе Fc-область с одной или несколькими заменами, которые улучшают связывание Fc-области с FcRn. Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной способностью к связыванию C1q описаны в патенте США № 6194551B1, WO 99/51642. Содержание этих публикаций патентов явно включено в настоящий документ в качестве ссылки. Также см. Idusogie et al., J. Immunol. 164: 4178-4184 (2000).
Производные антител
Антитела по настоящему изобретению можно дополнительно модифицировать, чтобы они содержали дополнительные небелковые фрагменты, которые известны в данной области и легко доступны. Предпочтительно, фрагменты, подходящие для получения производных антитела, представляют собой водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают в качестве неограничивающих примеров полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (или гомополимеры или статистические сополимеры) и декстран или поли(n-винил пирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры полипропиленоккида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоль пропиональдегид может обладать преимуществами в производстве вследствие его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может варьировать, и если присоединено более одного полимера, они могут представлять собой одни и те же или различные молекулы. В целом, число и/или тип полимеров, используемых для получения производных, можно определить, основываясь на соображениях, включающих в качестве неограничивающих примеров конкретные свойства или функции антитела, подлежащие улучшению, возможность использования производного антитела в терапии при определенных условиях и т.д.
Скрининг антител с желаемыми свойствами
Антитела по настоящему изобретению можно охарактеризовать по их физическим/химическим свойствам и биологическим функциям посредством различных анализов, известных в данной области (некоторые из которых описаны в настоящем документе). Например, антитела можно дополнительно охарактеризовать посредством серии анализов, включающих в качестве неограничивающих примеров N-концевое секвенирование, анализ аминокислот, неденатутрирующую эксклюзионную жидкостную хроматографию высокого давления (ЖХВД), масс-спектрометрию, ионообменную хроматографию и расщепление папаином.
В определенных вариантах осуществления анализируют биологическую активность антител, получаемых в настоящем документе. В некоторых вариантах осуществления тестируют активность связывания антигена антителами по настоящему изобретению. Анализы связывания антигена, которые известны в данной области и могут быть использованы в настоящем документе, включают без ограничения любые анализы прямого или конкурентного связывания с использованием таких способов, как вестерн-блоттинг, радиоиммунный анализ, ELISA (твердофазный иммуноферментный анализ), иммунологические «сэндвич»-анализы, анализы иммунопреципитации, флуоресцентные иммунологические анализы, и иммунологические анализы с белком A. Иллюстративные анализы предоставлены ниже в разделе с примерами.
Векторы, клетки-хозяева и рекомбинантные способы
Для рекомбинантного получения гетерологичного полипептида (например, антитела), кодирующую его нуклеиновую кислоту выделяют и встраивают в реплицируемый вектор для дальнейшего клонирования (амплификация ДНК) или для экспрессии. ДНК, кодирующую полипептид (например, антитело), легко выделить и секвенировать, используя стандартные процедуры (например, используя олигонуклеотидные зонды, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Доступно множество векторов. Выбор вектора частично зависит от клетки-хозяина, подлежащей использованию. В целом, предпочтительные клетки-хозяева имеют любое прокариотическое происхождение. Следует принимать во внимание, что с этой целью можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получить от человека или любых видов животных.
a. Создание антител с использованием прокариотических клеток-хозяев:
i. Конструкция вектора
Полинуклеотидные последовательности, кодирующие полипептидные компоненты полипептида (например, антитела) по изобретению можно получить, используя стандартные рекомбинантные способы. Желаемые полинуклеотидные последовательности можно выделить и секвенировать из антителопродуцирующих клеток, таких как гибридомные клетки. Альтернативно, полинуклеотиды можно синтезировать, используя синтезатор нуклеотидов или ПЦР способы. После получения, последовательности, кодирующие полипептиды, встраивают в рекомбинантный вектор, способный к репликации и экспрессии гетерологичных полинуклеотидов в прокариотических организмах-хозяевах. Многие векторы, которые доступны и известны в данной области, можно использовать для целей по настоящему изобретению. Выбор подходящего вектора зависит в основном от размера нуклеиновых кислот, подлежащих встраиванию в вектор, и конкретной клетки-хозяина, подлежащей трансформации вектором. Каждый вектор содержит различные компоненты, в зависимости от его функции (амплификация или экспрессия гетерологичного полинуклеотида или и то и другое) и его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора, как правило, включают в качестве неограничивающих примеров: участок начала репликации, селекционный маркерный ген, промотор, участок связывания рибосомы (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
В целом, плазмидные векторы, содержащие репликон и управляющие последовательности, которые получают от видов, совместимых с клеткой-хозяином, используют применительно к этим организмам-хозяевам. Вектор, как правило, несет сайт репликации, а также маркерные последовательности, которые способны обеспечить фенотипический отбор в трансформированных клетках. Например, E. coli типично трансформируют с использованием плазмиды pBR322, полученной из вида E. coli. pBR322 содержит гены, кодирующие устойчивость к ампициллину (Amp) и тетрациклину (Tet) и, таким образом, предоставляет удобное средство для идентификации трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаги также могут содержать или могут быть модифицированы так, чтобы содержать промоторы, которые может использовать микробный организм для экспрессии эндогенных белков. Примеры производных pBR322, используемых для экспрессии конкретных антител, подробно описаны в Carter et al., патенте США № 5648237.
Кроме того, фаговые векторы, содержащие репликон и управляющие последовательности, которые совместимы с микроорганизмом-хозяином, можно использовать в качестве трансформирующих векторов применительно к этим организмам-хозяевам. Например, бактериофаг, такой как λGΕΜ.ΤΜ.-11, можно использовать в создании рекомбинантного вектора, который можно использовать для трансформации восприимчивых клеток-хозяев, таких как E. coli LE392.
Экспрессирующий вектор по изобретению может содержать две или более пары промотор-цистрон, каждая кодирует полипептидные компоненты. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную ближе к 5'-концу относительно цистрона, которая модулирует его экспрессию. Прокариотические промоторы типично делят в два класса, индуцибельные и конститутивные. Индуцибельный промотор представляет собой промотор, который инициирует повышенные уровни транскрипции цистрона, находящегося под его управлением, в ответ на изменения в условиях культивирования, например, на присутствие или отсутствие питательного вещества или изменение температуры.
Широко известно большое количество промоторов, узнаваемых различными потенциальными клетками-хозяевами. Выбранный промотор может быть функционально связан с ДНК цистрона, кодирующей легкую или тяжелую цепь, посредством удаления промотора из ДНК источника путем расщепления рестрикционными ферментами и встраивания выделенной последовательности промотора в вектор по изобретению. Как нативную последовательность промотора, так и многие гетерологичные промоторы можно использовать, чтобы направлять амплификацию и/или экспрессию целевых генов. В некоторых вариантах осуществления используют гетерологичные промоторы, поскольку они, как правило, делают возможной более интенсивную транскрипцию и более высокий выход экспрессируемого целевого гена по сравнению с нативным промотором целевого полипептида.
Промоторы, пригодные для использования с прокариотическими организмами-хозяевами, включают промотор PhoA, промоторные системы β-лактамазы и лактозы, триптофановую (trp) промоторную систему и гибридные промоторы, такие как промотор tac или trc. Однако, также подходят другие промоторы, которые могут функционировать в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, тем самым позволяя опытному работнику оперативно лигировать их с цистронами, кодирующими целевые легкие и тяжелые цепи (Siebenlist et al., (1980) Cell 20: 269), с использованием линкеров или адапторов, чтобы обеспечить любые необходимые сайты рестрикции.
В одном из аспектов изобретения каждый цистрон в рекомбинантном векторе содержит компонент сигнальной последовательности секреции, который направляет транслокацию экспрессируемых полипептидов через мембрану. В целом, сигнальная последовательность может представлять собой компонент вектора или она может представлять собой часть ДНК целевого полипептида, которую встраивают в вектор. Сигнальная последовательность, выбранная для целей данного изобретения, не должна быть той, которую распознает и процессирует (т.е. расщепляет сигнальной пептидазой) клетка-хозяин. Для прокариотических клеток-хозяев, которые не распознают и не процессируют сигнальные последовательности, нативные для гетерологичных полипептидов, сигнальную последовательность заменят на выбранную прокариотическую сигнальную последовательность, например, из сигнальных полипептидов по настоящему изобретению. Кроме того, вектор может содержать сигнальную последовательность, выбранную из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Lpp или термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP.
В одном из аспектов изобретения один или несколько полинуклеотидов (например, экспрессирующие векторы) совместно кодируют антитело с одним Fab фрагментом. В одном из вариантов осуществления один полинуклеотид кодирует (a) компоненты легкой и тяжелой цепей антитела с одним Fab фрагментом и (b) полипептид Fc. В одном из вариантов осуществления один полинуклеотид кодирует компоненты легкой и тяжелой цепей антитела с одним Fab фрагментом, а отдельный полинуклеотид кодирует полипептид Fc. В одном из вариантов осуществления отдельные полинуклеотиды кодируют компонент легкой цепи антитела с одним Fab фрагментом, компонент тяжелой цепи антитела с одним Fab фрагментом и полипептид Fc, соответственно. Получение антитела с одним Fab фрагментом описано, например, в WO 2005063816.
Прокариотические клетки-хозяева, подходящие для экспрессии антител по изобретению, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры бактерий, которые можно использовать, включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, виды Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления используют грамотрицательные клетки. В одном из вариантов осуществления клетки E. coli используют в качестве организмов-хозяев по изобретению. Примеры штаммов E. coli вклчают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219; ATCC депозит № 27325) и их производные, включая штамм 33D3, обладающий генотипом W3110 ΔfhuA (Δtonα) ptr3 lac Iq lacL8 ΔompTA(nmpc-fepE) degP41 kanR (патент США № 5639635) и штаммы 63C1 и 64B4. Также подходят другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coliλ 1776 (ATCC 31537) и E. coli RV308(ATCC 31608). Эти примеры являются иллюстративными, а не ограничивающим. Способы конструирования производных любой из указанных выше бактерий, обладающих определенными генотипами, известны в данной области и описаны, например, в Bass et al., Proteins, 8:309-314 (1990). В целом, необходимо выбрать подходящие бактерии, принимая во внимание возможность репликации репликона в клетках бактерии. Например, E. coli, виды Serratia или Salmonella можно надлежаще использовать в качестве организма-хозяина, когда широко известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410 используют для того, чтобы снабдить репликон. Типично клетка-хозяин должна секретировать минимальные количества протеолитических ферментов и дополнительные ингибиторы протеазы можно по желанию встраивать в клеточную культуру.
ii. Получение антител
Клетки-хозяева трансформируют описанными выше экспрессирующими векторами и культивируют в стандартных питательных средах, модифицированных соответствующим образом, чтобы индуцировать промоторы, отобрать трансформанты или амплифицировать гены, кодирующие желаемые последовательности.
Трансформация обозначает введение ДНК в прокариотический организм-хозяин с тем, чтобы ДНК была реплицируемой, или в качестве внехромосомного элемента или составной части хромосомы. В зависимости от используемой клетки-хозяина, трансформацию выполняют с использованием стандартных способов, подходящих для таких клеток. Обработку кальцием с использованием хлорида кальция, как правило, используют для бактериальных клеток, которые содержат значительные барьеры клеточной стенки. В другом способе трансформации используют полиэтиленгликоль/ДМСО. Еще один используемый способ представляет собой электропорацию.
Прокариотические клетки, используемые для получения полипептидов по изобретению, выращивают в средах, известных в данной области и подходящих для культуры выбранных клеток-хозяев. Примеры подходящих сред включают среду Лурия-Бертани (LB) плюс необходимые питательные добавки. В некоторых вариантах осуществления среды также содержат селекционное средство, выбранное на основе конструкции экспрессирующего вектора, чтобы избирательно разрешать рост прокариотических клеток, содержащих экспрессирующий вектор. Например, ампициллин добавляют в среды для роста клеток, экспрессирующих ген устойчивости к ампициллину.
Любые необходимые добавки, помимо углерода, азота и неорганических источников фосфата, также можно включать в подходящих концентрациях отдельно или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно среда для культивирования может содержать одно или несколько восстанавливающих средств, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при подходящих температурах. Для роста E. coli предпочтительная температура находится в диапазоне, например, от приблизительно 20°C до приблизительно 39°C, более предпочтительно от приблизительно 25°C до приблизительно 37°C, даже более предпочтительно приблизительно 30°C. pH среды может представлять собой любой pH в диапазоне от приблизительно 5 до приблизительно 9, в основном в зависимости от организма-хозяина. Для E. coli pH предпочтительно составляет от приблизительно 6,8 до приблизительно 7,4 и более предпочтительно приблизительно 7,0.
Если в экспрессирующем векторе по изобретению используют индуцибельный промотор, экспрессию белка индуцируют в условиях, подходящих для активации промотора. В одном из аспектов изобретения промоторы PhoA используют для управления транскрипцией полипептидов. Соответственно, трансформированные клетки-хозяева культивируют в ограниченной по фосфату среде для индукции. Предпочтительно, ограниченная по фосфату среда представляет собой среду C.R.A.P. (см., например, Simmons et al., J. Immunol. Methods (2002), 263: 133-147) или среды, описанные в WO 2002/061090. Можно использовать многие другие индукторы, соответствующие конструкции используемого вектора, как известно в данной области.
В одном из вариантов осуществления экспрессируемые полипептиды по настоящему изобретению секретируются в и извлекаются из периплазмы клеток-хозяев. Извлечение белка типично включает разрушение микроорганизма, как правило, таким средствами, как осмотический шок, разрушение ультразвуком или лизин. После разрушения клеток, обломки клеток или цельные клетки можно удалять посредством центрифугирования или фильтрования. Белки можно дополнительно очистить, например, посредством аффинной хроматографии на смоле. Альтернативно, белки могут быть транспортированы в среды для культивирования и выделены в них. Клетки можно удалить из культуры, а культуральный супернатант фильтровать и концентрировать для дальнейшей очистки полученных белков. Кроме того, экспрессируемые полипептиды можно выделить и идентифицировать с использованием широко известных способов, таких как электрофорез в полиакриламидном геле (PAGE) и анализ вестерн-блоттинга.
В одном из аспектов изобретения получение антитела проводят в большом количестве посредством процесса ферментации. Для получения рекомбинантных белков доступны различные крупномасштабные процедуры ферментации с подпиткой. Крупномасштабная ферментация имеет емкость по меньшей мере 1000 литров, предпочтительно приблизительно емкость от 1000 до 100000 литров. Эти ферментаторы используют перемешивающие импеллеры, чтобы распределять кислород и питательные вещества, в частности, глюкозу (предпочтительный источник углерода/энергии). Мелкомасштабная ферментация, как правило, относится к ферментации в ферментаторе, который обладает объемной емкостью не более чем приблизительно 100 литров и может варьировать от приблизительно 1 литра до приблизительно 100 литров.
В процессе ферментации индукцию экспрессии белка типично инициируют после того, как клетки выросли в подходящих условиях до желаемой плотности, например, OD550 приблизительно 180-220, на этой стадии клетки находятся в ранней стационарной фазе. Можно использовать различные индукторы, соответствующие конструкции используемого вектора, как известно в данной области и описано выше. Клетки можно выращивать в течение более коротких периодов перед индукцией. Обычно клетки индуцируют в течение приблизительно 12-50 часов, хотя можно использовать более длительное или более короткое время индукции.
Чтобы улучшить выход и качество образования полипептидов по изобретению, можно модифицировать различные условия ферментации. Например, чтобы улучшить правильную сборку и укладку секретируемых полипептидов антител, дополнительные векторы, сверхэкспрессирующие шапероны, такие как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпропил-цис,транс-изомераза с шаперонной активностью) можно использовать для совместной трансформации клеток прокариотического организма-хозяина. Показано, что шапероны способствуют правильной укладке и растворимости гетерологичных белков, продуцируемых в бактериальных клетках-хозяевах. Chen et al., (1999) J. Biol. Chem. 274: 19601-19605; Georgiou et al., патент США № 6083715; Georgiou et al., патент США № 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275: 17100-17105; Ramm and Pluckthun, (2000) J. Biol. Chem. 275: 17106-17113; Arie et al., (2001) Mol. Microbiol. 39: 199-210. В некоторых вариантах осуществления осуществляют экспрессию DsbA и C в бактериальной клетке-хозяине.
Чтобы минимизировать протеолиз экспрессируемых гетерологичных белков (в частности, тех, которые чувствительны к протеолизу), для настоящего изобретения можно использовать определенные штаммы организма-хозяина с дефицитом протеолитических ферментов. Например, штаммы клетки-хозяина можно модифицировать, чтобы осуществить генетическую мутацию(и) в генах, кодирующих известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их сочетания. Некоторые штаммы E. coli с дефицитом протеаз доступны и описаны, например, в Joly et al., (1998), выше; Georgiou et al., патенте США № 5264365; Georgiou et al., патенте США № 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном из вариантов осуществления штаммы E. coli с дефицитом протеолитических ферментов и трансформированные плазмидами, сверхэкспрессирующими один или несколько шаперонов, используют в качестве клеток-хозяев в экспрессирующей системе по изобретению.
iii. Очистка антител
Можно использовать стандартные известные в данной области способы очистки белка. Следующие процедуры являются образцами подходящих процедур очистки: фракционирование на иммуноаффинной или ионообменной колонке, преципитация этанолом, ВЭЖХ с обращенной фазой, хроматография на диоксиде кремния или катионообменной смоле, такой как DEAE, хроматофокусирование, SDS-PAGE, преципитация сульфатом аммония и гель-фильтрация с использованием, например, Sephadex G-75.
В одном из аспектов белок A, иммобилизованный на твердой фазе, используют для иммуноаффинной очистки антительных продуктов по изобретению. Белок A представляет собой 41 кДа белок клеточной стенки из Staphylococcus aureus, который с высокой аффинностью связывается с Fc-областью антитела. Lindmark et al., (1983) J. Immunol. Meth. 62: 1-13. Твердая фаза, на которой иммобилизуют белок A, предпочтительно представляет собой колонку, содержащую поверхность из стекла или диоксида кремния, более предпочтительно колонку со стеклом с контролируемым размером пор или колонку с кремниевой кислотой. В некоторых применениях колонка покрыта реактивом, таким как глицерин, в попытке предотвратить неспецифическое прилипание загрязнителей.
В качестве первой стадии очистки, препарат, полученный из клеточной культуры, как описано выше, наносят на твердую фазу с иммобилизованным белком A, чтобы сделать возможным специфическое связывание антитела, представляющей интерес, с белком A. Затем твердую фазу промывают для того, чтобы удалить загрязнители, неспецифически связанные с твердой фазой. Наконец, антитело, представляющее интерес, извлекают из твердой фаза посредством элюирования.
Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам (взаимозаменяемо обозначаемым «конъюгаты антитело-лекарственное средство» или «КАЛС»), содержащим любые антитела, описанные в настоящем документе, которые конъюгированы с цитотоксическим средством, таким как химиотерапевтическое средство, лекарственное средство, ингибирующее рост средство, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или их фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат).
Использование конъюгатов антитело-лекарственное средство для локальной доставки цитотоксических или цитостатических средств, т.е. лекарственных средств для того, чтобы убить или подавить опухолевые клетки при лечении злокачественной опухоли (Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drg. Del. Rev. 26: 151-172; патент США № 4975278), делает возможной нацеленную доставку фрагмента лекарственного средства в опухоли и внутриклеточное накопление в ней, где системное введение этих неконъюгированных лекарственных средств может привести к неприемлемым уровням токсичности для нормальных клеток, а также для опухолевых клеток, которые желательно элиминировать (Baldwin et al., (1986) Lancet pp. (15 марта 1986 года): 603-05; Thorpe, (1985) «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review» in Monoclonal Antibodies'84: Biological And Clinical Applications, A. Pinchera et al. (eds), pp. 475-506). Таким образом, стремятся к максимальной эффективности при минимальной токсичности. Сообщалось, что в этих стратегиях эффективны как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother., 21: 183-87). Лекарственные средства, используемые в этих способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986) выше). Токсины, используемые в конъюгатах антитело-токсин, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al (2000) Jour. of the Nat. Cancer Inst. 92(19): 1573-1581; Mandler et al., (2000) Bioorganic & Med. Chem. Letters 10: 1025-1028; Mandler et al., (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (EP 1391213; Liu et al., (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихимицин (Lode et al., (1998) Cancer Res. 58:2928; Hinman et al., (1993) Cancer Res. 53:3336-3342). Токсины могут проявлять свои цитотоксические и цитостатические эффекты посредством механизмов, включая связывание с тубулином, связывание с ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства обычно неактивны или менее активны, когда они конъюгированы с большими антителами или лигандами белкового рецептора.
Зевалин® (ибритутомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело-радиоизотоп, состоящий из IgG1 κ моноклонального антитела мыши, направленного против антигена CD20, найденного на поверхности нормальных и злокачественных B-лимфоцитов, и радиоизотопа 111In или 90Y, связанного посредством линкера-хелатора из тиомочевины (Wiseman et al., (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al., (2002) Blood 99(12):4336-42; Witzig et al., (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al., (2002) J. Clin. Oncol. 20(15):3262-69). Несмотря на то, что зевалин обладает активностью против B-клеточной неходжкинской лимфомы (НХЛ), введение ведет к тяжелым и длительным цитопениям у большинства пациентов. MYLOTARG™ (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитело-лекарственное средство, состоящий из антитела hu CD33, связанного с калихимицином, одобрен в 2000 для лечения острого миелолейкоза посредством инъекции (Drugs of the Future (2000) 25(7):686; патенты США №№ 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Кантузумаб мертанзин (Immunogen, Inc.), конъюгат антитело-лекарственное средство, состоящий из антитела huC242, связанного через дисульфидный линкер SPP с майтанзиноидным фрагментом лекарственного средства, DM1, достиг испытаний фазы II для лечения злокачественных опухолей, которые экспрессируют CanAg, таких как опухоли ободочной кишки, поджелудочной железы, желудка и другие. MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen Inc.), конъюгат антитело-лекарственное средство, состоящий из моноклонального антитела против простатоспецифического мембраного антигена (PSMA), связанного с майтанзиноидным фрагментом лекарственного средства, DM1, находится в разработке для потенциального лечения опухолей простаты. Пептиды ауристатин, ауристатин E (AE) и монометилауристатин (MMAE), синтетические аналоги доластатина, конъюгированы с химерными моноклональными антителами cBR96 (специфичны к Lewis Y в карциномах) и cAC10 (специфичны к CD30 в гематологических злокачественных новообразованиях) (Doronina et al., (2003) Nature Biotechnology 21 (7):778-784) и находятся в разработке терапевтического средства.
В настоящем документе описаны химиотерапевтические средства, которые можно использовать в создании иммуноконъюгатов (например, выше). Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A цепь дифтерии, несвязывающие активные фрагменты дифтерийного токсина, A цепь экзотоксина (из Pseudomonas aeruginosa), A цепь рицина, A цепь арбина, A цепь модессина, α-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецины. См., например, WO 93/21232, публикация 28 октября 1993 года. Для получения радиоконъюгированных антител доступны различные радионуклиды. Примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического средства получают с использованием различных бифункциональных связывающих белки средств, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производны имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат), и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в Vitetta et al., Science, 238: 1098 (1987). Меченная 14C 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является типичным хелатирующим средством для конъюгации радионуклеотида с антителом. См. WO 94/11026.
Конъюгаты антитела и одного или нескольких низкомолекулярных токсинов, таких как калихимицин, майтанзиноиды, доластатины, ауростатины, трихотецен и CC1065 и производные этих токсинов, которые обладают активностью токсина, также предусмотрены в настоящем документе.
i. Майтанзин и майтанзиноиды
В некоторых вариантах осуществления иммуноконъюгат содержит антитело (полноразмерное или фрагменты) по изобретению, конъюгированное с одной или несколькими молекулами майтанзиноида.
Майтанзиноиды представляют собой ингибиторы митоза, которые действуют посредством ингибирования полимеризации тубулина. Майтанзин впервые выделен из восточно-африканского кустарника Maytenus serrata (патент США № 3896111). Впоследствии обнаружили, что определенные микробы также продуцируют майтанзиноиды, такие как майтанзинол и C-3 сложные эфиры майтанзинола (патент США № 4151042). Синтетический майтанзинол и его производные и аналоги раскрыты, например, в патентах США №№ 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533.
Майтанзиноидные фрагменты лекарственных средств представляют собой привлекательные лекарственные фрагменты в конъюгатах антитело-лекарственное средство, поскольку они: (i) относительно доступны для получения посредством ферментации или химической модификации, получения производных продуктов ферментации, (ii) поддаются получению производных с использованием функциональных групп, подходящих для конъюгации через не дисульфидные линкеры с антителами, (iii) стабильны в плазме и (iv) эффективны против различных линий опухолевых клеток.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения и их терапевтическое использование раскрыты, например, в патентах США №№ 5208020, 5416064 и европейском патенте EP 0425235 B1, описания которых явно включены в настоящий документ в качестве ссылки. В Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описаны иммуноконъюгаты, содержащие майтанзиноид, обозначенный DM1, связанный с моноклональным антителом C242, направленным против злокачественной опухоли толстой кишки человека. Обнаружено, что конъюгат обладает высокой цитотоксичностью против культивируемых клеток злокачественной опухоли ободочной кишки и проявляет противоопухолевую активность в анализе роста опухоли in vivo. В Chari et al., Cancer Research 52: 127-131 (1992) описаны иммуноконъюгаты, в которых майтанзиноид конъюгировали через дисульфидный линкер с антителом A7 мыши, связывающимся с антигеном на линиях клеток опухоли ободочной кишки человека, или с другим моноклональным антителом TA.1 мыши, которое связывает онкоген HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид тестировали in vitro на клеточной линии злокачественной опухоли молочной железы человека SK-BR-3, которая экспрессирует 3×105 поверхностных антигенов HER-2 на клетку. Конъюгат лекарственного средства достигал степени цитотоксичности, схожей с лекарственным средством, не содержащим майтанзиноид, которую можно повысить посредством увеличения числа молекул майтанзиноида на молекулу антитела. Конъюгат A7-майтанзиноид показывал низкую системную цитотоксичность у мышей.
Конъюгаты антитело-майтанзиноид получают посредством химического связывания антитела с молекулой майтанзиноида без значительного снижения биологической активности антитела или молекулы майтанзиноида. См., например, патент США № 5208020 (раскрытие которого, таким образом, явно включено в качестве ссылки). В среднем 3-4 конъюгированные молекулы майтанзиноида на молекулу антитела показали эффективность в усилении цитотоксичности целевых клеток без отрицательного влияния на функцию или растворимость антитела, хотя даже от одной молекулы токсина/антитела следует ожидать усиления цитотоксичности по сравнению с использованием голого антитела. Майтанзиноиды хорошо известны в данной области и их можно синтезировать известными способами или выделить из естественных источников. Подходящие майтанзиноиды раскрыты, например, в патенте США № 5208020 и в других патентах и непатентых публикациях, указанных выше в настоящем документе. Предпочтительные майтанзиноиды представляют собой майтанзинол и аналоги майтанзинола, модифицированные по ароматическому кольцу или в других положениях молекулы майтанзинол, такие как различные сложные эфиры майтанзинола.
В данной области известно множество линкерных групп для создания конъюгатов антитело-майтанзиноид, включая, например, те, которые раскрыты в патенте США № 5208020 или патенте EP 0425235 B1, Chari et al., Cancer Research 52: 127-131 (1992) и патентной заявке США № 10/960602, которая подана 8 октября 2004 года, описания которых явно включены в настоящий документ в качестве ссылки. Конъюгаты антитело-майтанзиноид, содержащие линкерный компонент SMCC, можно получать, как раскрыто в патентной заявке США № 10/960602, которая подана 8 октября 2004 года. Линкерные группы включают дисульфидные группы, тиоэфирные группы, кислотно-лабильные группы, фотолабильные группы, пептидазо-лабильные группы или эстеразо-лабильные группы, как раскрыто в идентифицированных выше родителях, дисульфидные и тиоэфирные группы являются предпочтительными. Дополнительные линкерные группы описаны и проиллюстрированы примерами в настоящем документе.
Конъюгаты антитела и майтанзиноида можно создавать с использованием различных бифункциональных связывающих белок средств, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат), и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Особенно предпочтительные связывающие средства включают N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP) для обеспечения дисульфидной связи.
Линкер можно присоединять к молекуле майтанзиноида в различных положениях в зависимости от типа связи. Например, сложноэфирную связь можно сформировать посредством реакции с гидроксильной группой, используя стандартные способы связывания. Реакция может проходить в положении C-3, содержащем гидроксильную группу, положении C-14, модифицированном гидроксиметилом, положении C-15, модифицированном гидроксильной группой, и положении C-20, имеющем гидроксильную группу. В предпочтительном варианте осуществления связь формируют в положении C-3 майтанзинола или аналога майтанзинола.
ii. Ауристатины и доластатины
В некоторых вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с доластатинами или пептидными аналогами и производными доластатинов, ауристатинами (патенты США №№ 5635483 и 5780588). Показано, что доластатины и ауристатины вмешиваются в динамику микротрубочек, GTP гидролиз и деление ядер и клеток (Woyke et al (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают противоопухолевой (патент США № 5663149) и противогрибковой активностью (Pettit et al., (1998) Antimicrob. Agents Chemother. 42:2961-2965). Доластатиновый или ауристатиновый фрагмент лекарственного средства можно присоединить к антителу через N(амино)-конец или C(карбокси)-конец пептидного фрагмента лекарственного средства (WO 02/088172).
Типичные ауристатиновые варианты осуществления включают связанные с N-концом монометилауристатиновые фрагменты лекарственных средств DE и DF, раскрытые в «Monomethylvaline Compounds Capable of Conjugation to Ligands», США с серийным № 10/983340, подано 5 ноября 2004 года, раскрытие которого явно включено в качестве ссылки в полном объеме.
Типично, фрагменты лекарственных средств на пептидной основе можно получать посредством формирования пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получать, например, в соответствии со способом синтеза в жидкой фазе (см. E. Schroder and K. Liibke, «The Peptides», volume 1, pp. 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Ауристатиновые/доластатиновые фрагменты лекарственных средств можно получать в соответствии со способами из: патентов США №№ 5635483 и 5780588; Pettit et al., (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al., (1998) Anti-Cancer Drug Design 13:243-277; Pettit, G.R., et al., Synthesis, 1996, 719-725; и Pettit et al., (1996) J. Chem. Soc. Perkin Trans. 1 5:859-863. Также см. Doronina (2003) Nat. Biotechnol. 21(7):778-784; «Monomethylvaline Compounds Capable of Conjugation to Ligands», США с серийным № 10/983340, подано 5 ноября 2004 года, включено, таким образом, в качестве ссылки в полном объеме (раскрыты, например, линкеры и способы получения соединений монометилвалина, таких как MMAE и MMAF, конъюгированных с линкерами).
iii. Калихимицин
В других вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с одной или несколькими молекулами калихимицина. Антибиотики семейства калихимицинов способны создавать двухцепочечные разрывы ДНК в субпикомолярных концентрациях. О получении конъюгатов семейства калихимицинов см. в патентах США №№ 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296 (все выданы American Cyanamid Company). Структурные аналоги калихимицина, которые можно использовать, включают в качестве неограничивающих примеров, γ1 1, α2 1, α3 1, N-ацетил-γ1, PSAG и θ1 1 (Hinman et al., Cancer Research 53:3336-3342 (1993), Lode et al., Cancer Research 58:2925-2928 (1998) и указанн выше патенты США, выданные American Cyanamid). Другое противоопухолевое лекарственное средство, в которым антитело может быть конъюгировано, представляет собой QFA, который представляет собой антифолат. Как калихимицин, так и QFA имеет внутриклеточные сайты действия и не без труда пересекают плазматическую мембрану. Следовательно, клеточный захват этих средство через опосредованную антителами интернализацию значительно усиливает их цитотоксические эффекты.
iv. Другие цитотоксические средства
Другие противоопухолевые средства, которые можно конъюгировать с антителами по изобретению, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство средств, которые совместно известны как LL-E33288, комплекс, описанный в патентах США №№ 5053394 и 5770710, а также эсперамицины (патент США № 5877296).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A цепь дифтерии, несвязывающие активные фрагменты дифтерийного токсина, A цепь экзотоксина (из Pseudomonas aeruginosa), A цепь рицина, A цепь абрина, A цепь модессина, α-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецен. См., например, WO 93/21232, которая опубликована 28 октября 1993 года.
Кроме того, в настоящем изобретении рассмотрен иммуноконъюгат, сформированный между антителом и соединением с нуклеолитической активностью (например, рибонуклеаза или ДНК эндонуклеаза, такая как дезоксирибонуклеаза; ДНКаза).
Для избирательного разрушения опухоли, антитело может содержать высоко радиоактивный атом. Различные радиоактивные изотопы доступны для получения радиоконъюгированных антител. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Когда конъюгат используют для обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, Tc99m или I123, или спиновую метку для визуализации ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как опять йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки можно встраивать в конъюгат известными способами. Например, пептид можно биосинтезировать или можно синтезировать посредством химического аминокислотного синтеза с использованием подходящих предшественников аминокислот, содержащих, например, фтор-19 вместо водорода. Такие метки, как Tc99m или I123, Re186, Re188 и In111 можно присоединять через остаток цистеина в пептиде. Иттрий-90 можно присоединять через остаток лизина. Способ IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80: 49-57 можно использовать для встраивания йода-123. В «Monoclonal Antibodies in Immunoscintigraphy» (Chatal, CRC Press 1989) подробно описаны другие способы.
Конъюгаты антитела и цитотоксического средства можно создать, используя различные бифункциональные связывающие белки средства, такие как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в Vitetta et al., Science 238: 1098 (1987). Меченная 14С 1-изотиосианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является типичным хелатирующим средством для конъюгации радионуклеотида с антителом. См. WO94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотно-лабильный линкер, пептидазо-чувствительный линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Research 52: 127-131 (1992); патент США № 5208020).
Соединения по изобретению явно предполагают, но не ограничены КАЛС, полученными с использованием кросс-линкерных реактивов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые коммерчески доступны (например, в Pierce Biotechnology, Inc., Rockford, IL., U.S.A). См. Страницы 467-498, 2003-2004 Applications Handbook and Catalog.
v. Получение конъюгатов антитело-лекарственное средство
В конъюгатах антитело-лекарственное средство (КАЛС) по изобретению антитело (Ab) конъюгируют с одним или несколькими фрагментами лекарственных средств (D), например, от приблизительно 1 до приблизительно 20 фрагментов лекарственного средства на антитело, через линкер (L). КАЛС формулы I можно получать различными путями, использующими реакции органической химии, условия и реактивы, известные специалистам в данной области, включая: (1) реакцию нуклеофильной группы антитела с бивалентным линкерным реактивом, чтобы сформировать Ab-L, через ковалентную связь, с последующей реакцией с фрагментом лекарственного средства D; и (2) реакцию нуклеофильной группы фрагмента лекарственного средства с бивалентным линкерным реактивом, чтобы сформировать D-L, через ковалентную связь, с последующей реакцией с нуклеофильной группой антитела. Дополнительные способы получения КАЛС описаны в настоящем документе.
Линкер может состоять из одного или нескольких линкерных компонентов. Типичные линкерные компоненты содержат 6-малеимидокапроил («MC»), малеимидопропаноил («MP»), валин-цитруллин («val-cit»), аланин-фенилаланин («ala-phe»), п-аминобензилоксикарбонил («PAB»), N-сукцинимидил-4-(2-пиридилтио)пентаноат («SPP»), N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат («SMCC»), и N-сукцинимидил-(4-йодацетил)аминобензоат («SIAB»). Дополнительные линкерные компоненты известны в данной области и некоторые описаны в настоящем документе. Также см. «Monomethylvaline Compounds Capable of Conjugation to Ligands», США серийный № 10/983340, подано 5 ноября 2004 года, содержание которого включено, таким образом, в качестве ссылки в полном объеме.
В некоторых вариантах осуществления линкер может содержать аминокислотные остатки. Типичные аминокислотные линкерные компоненты включают дипептиды, трипептиды, тетрапептиды или пентапептиды. Типичные дипептиды включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Типичные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, которые входят в аминокислотный линкерный компонент, включают те, что встречаются в природе, а также минорные аминокислоты и не встречающиеся в природе аналоги аминокислот, такие как цитруллин. Аминокислотные линкерные компоненты можно конструировать и оптимизировать их чувствительность для ферментативного расщепления конкретными ферментами, например, опухоль-ассоциированной протеазой, катантигеном B, C и D или протеазой плазмином.
Нуклеофильные группы на антителах включают в качестве неограничивающих примеров: (i) N-концевые аминогруппы, (ii) аминогруппы боковой цепи, например лизин, (iii) тиоловые группы боковой цепи, например, цистеин, и (iv) гидрокси- или аминогруппы сахара, когда антитело гликозилировано. Амино-, тиоловые и гидроксильные группы являются нуклеофильными и способны вступать в реакцию с образованием ковалентных связей с электрофильными группами на линкерных фрагментах и линкерных реактивах, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Определенные антитела содержат восстанавливаемые межцепные дисульфиды, т.е. цистеиновые мостики. Можно создать реакционноспособные антитела для конъюгации с линкерными реактивами посредством обработки восстанавливающим средством, таким как DTT (дитиотреитол). Таким образом, каждый цистеиновый мостик теоретически должен формировать два реакционноспособных нуклеофильных тиола. Дополнительные нуклеофильные группы можно вводить в антитела через реакцию лизинов с 2-иминотиоланом (реактив Трота), которая ведет к превращению амина в тиол. Реакционноспособные тиоловые группы можно вводить в антитело посредством введения одного, двух, трех, четырех или более остатков цистеина (например, получение мутантных антител, содержащих один или несколько ненативных цистеиновых аминокислотных остатка).
Конъюгаты антитело-лекарственное средство по изобретению также можно получать посредством модификации антитела для того, чтобы ввести электрофильные фрагменты, которые могут реагировать с нуклеофильными заместителями на линкерном реактиве или лекарственном средстве. Сахара гликозилированных антител могут быть окислены, например, с помощью окисляющих реактивов перйодатов, чтобы сформировать альдегидные или кетоновые группы, которые могут реагировать с аминогруппой линкерных реактивов или фрагментов лекарственных средств. Получаемые иминовые группы основний Шиффа могут формировать стабильную связь или могут быть восстановлены, например, посредством боргидридных реактивов, чтобы сформировать стабильные аминные связи. В одном из вариантов осуществления реакция углеводной части гликозилированного антитела или с галактозоксидазой или метаперйодатом натрия может давать карбонильные (альдегидные и кетоновые) группы в белке, которые могут реагировать с соответствующими группами на лекарственном средстве (Hermanson, Bioconjugate Techniques). В другом варианте осуществления белки, содержащие N-концевые остатки серина или треонина, могут реагировать с метаперйодатом натрия, что ведет к образованию альдегида вместо первой аминокислоты (Geoghegan & Stroh, (1992) Bioconjugate Chem. 3: 138-146; патент США № 5362852). Такой альдегид может реагировать с нуклеофильным фрагментом лекарственного средства или линкером.
Аналогичным образом, нуклеофильные группы на фрагменте лекарственного средства включают в качестве неограничивающих примеров: аминовые, тиольные, гидроксильные, гидразидные, оксимные, гидразинные, тиосемикарбазонные, гидразинкарбоксилатные и арилгидразидные группы, способные вступать в реакцию для образования ковалентных связей с электрофильными группами на линкерных фрагментах и линкерных реактивах, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы.
Альтернативно, слитый белок, содержащий антитело и цитотоксическое средство, можно создать, например, рекомбинантными способами или пептидным синтезом. Длина ДНК может содержать соответствующие области, кодирующие две части конъюгата, или смежно друг с другом или разделенные областью, кодирующей линкерный пептид, который не разрушает желаемые свойства конъюгата.
В еще одном другом варианте осуществления антитело можно конъюгировать с «рецептором» (таким как стрептавидин) для использовании в предварительном нацеливании на опухоль, где конъюгат антитело-рецептор вводят индивидууму, с последующим удалением несвязанного конъюгата из циркуляции с использованием очищающего средства и затем введением «лиганда» (например, авидина), который конъюгирован с цитотоксическим средством (например, радионуклеотидом).
Фармацевтические составы
Терапевтические составы гетерологичного полипептида получают для хранения посредством смешивания гетерологичного полипептида, обладающего желаемой степенью чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме водных растворов, лиофилизированных или других высушенных составов. Приемлемые носители, эксципиенты или стабилизаторы не токсичны для реципиентов в используемых дозах и концентрациях и содержат буферы, такие как фосфатный, цитратный, гистидиновый и из других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу, или декстрины; хелатирующие средства, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексные соединения с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные средства, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Состав в настоящем документе также может содержать более чем одно активное соединение по мере необходимости для конкретного показания, подлежащего лечению, предпочтительно соединения с комплементарными активностями, которые не оказывают нежелательного влияния друг на друга. Такие молекулы надлежаще присутствуют в сочетании в количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты также могут быть заключены в микрокапсуле, полученной, например, способами коацервации или интерфазной полимеризацией, например, в гидроксиметилцеллюлозной или желатиновой микрокапсуле и поли-(метилметилметакрилатной) микрокапсуле, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы раскрыты в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Составы, подлежащие использованию для введения in vivo, должны быть стерильными. Это легко выполнить посредством фильтрования через мембраны для стерилизующего фильтрования.
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, форма этих матриц представляет собой профилированные изделия, например, пленки или микрокапсулы. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли(винилалкоголь)), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагающийся этиленвинилацетат, разрушаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и лейпролид ацетата), и поли-D-(-)-3-гидроксимасляную кислоту. Тогда как полимеры, такие как этиленвинилацетат и молочная кислота-гликолевая кислота, делают возможным высвобождение молекулы в течение более чем 100 суток, определенные гидрогели высвобождают белки в течение более короткого периода времени. Когда инкапсулированные антитела остаются в организме в течение длительного времени, они могут денатурировать или образовывать агрегаты в результате воздействия влаги при 37°C, что ведет к потере биологической активности и возможным изменениям иммуногенности. Можно разработать рациональные стратегии для стабилизации в зависимости от задействованного механизма. Например, если обнаружено, что механизм агрегации представляет собой образование межмолекулярных S-S связей через тио-дисульфидный обмен, стабилизации можно добиться посредством модификации сульфгидрильных остатков, лиофилизации из кислых растворов, управления содержанием влаги, использования подходящих добавок и разработки конкретных композиций полимерных матриц.
Использование
Гетерологичный полипептид по настоящему изобретению можно использовать, например, для очистки, обнаружения и нацеливания конкретного полипептида, который он распознает, включая диагностические и терапевтические способы как in vitro, так и in vivo.
В одном из аспектов антитело по изобретению можно использовать в иммунологических анализах для качественного или количественного измерения конкретных антигенов в биологических образцах. Стандартные способы обнаружения связывания антиген-антитело включают, например, твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) или иммуногистохимия ткани. Во многих способах можно использовать метку, связанную с антителом с целью обнаружения. Метка, используемая с антителом, представляет собой любую поддающуюся обнаружению функциональность, которая не препятствует его связыванию с антителом. Известно множество меток, включая радиоизотопы 32P, 32S, 14C, 125I, 3H и 131I, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люцифераза светлячка и бактериальная люцифераза (патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, лактопероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы, радионуклиды для визуализации (такие как технеций) и т.п.
Доступны стандартные способы для ковалентного связывания этих меток с гетерологичными полипептидами. Например, связывающие средства, такие как диальдегиды, карбодиимиды, дималеимиды, бис-имидаты, бис-диазотированный бензидин и т.п., можно использовать для мечения антител описанными выше флуоресцентными, хемилюминесцентными и ферментативными метками. См., например, патент США № 3940475 (флуорометрия) и патент США № 3645090 (ферменты); Hunter et al. Nature 144: 945 (1962); David et al. Biochemistry 13: 1014-1021 (1974); Pain et al. J. Immunol. Methods 40:219-230 (1981); и Nygren Histochem. and Cytochem 30:407-412 (1982). Предпочтительными метками в настоящем документе являются ферменты, такие как пероксидаза хрена и щелочная фосфатаза. Конъюгация такой метки, включая ферменты, с полипептидом антитела является стандартной подручной процедурой для среднего специалиста в способах иммунологического анализа. См., например, O'Sullivan et al., «Methods for the Preparation of Enzyme-Antibody Conjugates for Use in Enzyme Immonoassay» в Methods in Enzymology, ed. J. J. Langone and H. Van Vunakis, Vol. 73 (Academic Press, New York, N.Y., 1981), pp. 147-166. Такие способы связывания пригодны для использования с гетерологичными полипептидами по данному изобретению.
Альтернативно мечению гетерологичного полипептида, антиген можно анализировать в биологических жидкостях посредством конкурентного иммунологического анализа с использованием стандарта конкурирующего антигена, меченого поддающимся обнаружению веществом, и немеченого гетерологичного полипептида. В этом анализе объединяют биологический образец, меченные стандарты антигена и гетерологичный полипептид и определяют количество меченного стандарта антигена, связанного с немеченым гетерологичным полипептидом. Количество тестируемого антигена в биологическом образце обратно пропорционально количеству меченого стандарта антигена, связанного с гетерологичным полипептидом.
В одном из аспектов, гетерологичный полипептид (такой как антитело) по изобретению в частности можно использовать для обнаружения и определения профиля экспрессии конкретных поверхностных антигенов in vitro или in vivo. Как рассмотрено выше, как правило, дегликозилированное антитело не проявляет эффекторные функции (т.е. АЗКЦ или КЗЦ активность). Следовательно, когда антитело связывается с антигеном клеточной поверхности, оно не должно инициировать нежелательные цитотоксические события. Поверхностный антиген может быть специфичен для конкретного типа клеток или тканей, следовательно, выступая в качестве маркера типа клеток или тканей. Предпочтительно, на различных стадиях дифференциации конкретных типов клеток или тканей происходит дифференциальная экспрессия поверхностного антигенного маркера. Антитело, направленное против такого поверхностного антигена, таким образом, можно использовать для скрининга популяций клеток или тканей, экспрессирующих маркер. Например, антитело по изобретению можно использовать для скрининга и выделения стволовых клеток, таких как эмбриональные стволовые клетки, гематопоэтические стволовые клетки и мезенхимальные стволовые клетки. Антитело по изобретению также можно использовать для обнаружения опухолевых клеток, экспрессирующих опухоль-ассоциированные поверхностные антигенные, такие как c-met, рецепторы HER2, HER3 или HER4.
Антитело или другой гетерологичный полипептид по изобретению можно использовать в качестве средства для аффинной очистки. В этом процессе полипептид иммобилизуют на твердой фазе, такой как смола Sephadex или фильтровальная бумага, используя способы, широко известные в данной области. Иммобилизованный полипептид приводят в контакт с образцом, содержащим антиген, подлежащий очистке, и после этого подложку промывают подходящим растворителем, который должен удалять по существу все вещество в образце, за исключением антигена, подлежащего очистке, который связан с иммобилизованным полипептидом. Наконец, подложку промывают другим подходящим растворителем, таким как глициновый буфер, pH 5,0, который должен высвобождать антиген из полипептида.
В одном из аспектов изобретение относится к использованию гетерологичного полипептида, созданного с использованием способов по изобретению, в получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как злокачественная опухоль, опухоль, клеточное пролиферативное нарушение и/или иммунное (такое как аутоиммунное) нарушение. Гетерологичный полипептид может иметь любую форму, описанную в настоящем документе, включая антитело, фрагмент антитела, полипептид (например, олигопептид) или их сочетание. В некоторых вариантах осуществления антиген представляет собой молекулу белка человека, а субъект представляет собой человеческий субъект.
Гетерологичные полипептиды по изобретению можно использовать для диагностирования, лечения, подавления или предотвращения заболеваний, нарушений или состояний, ассоциированы с ненормальной экспрессией и/или активностью одной или нескольких молекул антигенов, включая в качестве неограничивающих примеров злокачественные и доброкачественные опухоли; не лейкемии и лимфоидные злокачественные новообразования; нейрональные, глиальные, астроцитарные, гипоталамические и другие железистые, макрофагальные, эпителиальные, стромальные и бластоцельные нарушения; и воспалительные, ангиогенные и иммунологические нарушения.
В определенных вариантах осуществления иммуноконъюгат, содержащий антитело, вводят субъекту. Предпочтительно, клетка интернализует иммуноконъюгат и/или антиген, с которым он связывается.
В терапии гетерологичные полипептиды по настоящему изобретению можно использовать или отдельно или в сочетании с другими композициями. Например, гетерологичный полипептид можно совместно вводить с антителом, химиотерапевтическим средством(ами) (включая комбинации химиотерапевтических средств), другое цитотоксическое средство(а), антиангиогенное средство(а), цитокины и/или ингибирующее рост средство(а). Когда гетерологичный полипептид ингибирует рост опухоли, в частности, может быть желательно комбинировать гетерологичный полипептид с одним или несколькими другими терапевтическими средствами, которые также ингибируют рост опухоли. Альтернативно или дополнительно, пациент может получать комбинированную лучевую терапию (например, внешнее облучение или терапию радиоактивно меченным средством, таким как антитело). Такие комбинированные терапии, отмеченные выше, включают комбинированное введение (где два или более средства включены в один или в раздельные составы) и раздельное введение, в случае которого введение антитела может происходить перед и/или после введения вспомогательной терапии или терапий.
Гетерологичный полипептид (и необязательно, вспомогательное терапевтическое средство) вводят с помощью любого подходящего средства, включая парентеральное, подкожное, интрапетиронеальное, внутрилегочное и интраназальное введение и, при желании введение в поражение для местного лечения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, интраперитонеальное или подкожное введение. Кроме того, антитело надлежаще вводят посредством импульсных инфузий, в частности, со снижающимися дозами антитела. Предпочтительно дозирование дают посредством инъекций, наиболее предпочтительно внутривенных или подкожных инъекций, в зависимости, от части, от того, является ли введение коротким или хроническим.
Композицию гетерологичного полипептида по изобретению следует формулировать, дозировать и вводить таким образом, который согласуется с надлежащей медицинской практикой. Факторы для принятия решения в этом контексте включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки средства, способ введения, режим введения и другие факторы, известные медицинским практикам. Антитело не необходимо, а необязательно формулируют с одним или несколькими средствами, используемыми в настоящее время для того, чтобы предотвратить или лечить рассматриваемое нарушение. Эффективное количество таких других средств зависит от количества антитела, присутствующего в составе, типа нарушения или лечения и других факторов, рассмотренных выше. Их, как правило, используют в тех же дозах и с использованием путей введении, как применяют выше в настоящем документе, или от приблизительно 1 до 99% от использованных выше доз.
Для профилактики или лечения заболевания подходящая доза антитела (когда используют отдельно или в сочетании с другими средствами, такими как химиотерапевтические средства) должна зависеть от типа заболевания, подлежащего лечению, типа антитела, тяжести и течения заболевания, вводят ли антитело для профилактических или терапевтических целей, предшествующей терапии, истории болезни пациента и ответа на антитело и усмотрения лечащего врача. Антитело надлежаще вводят пациенту за один раз или в серии воздействий. В зависимости от типа и тяжести заболевания, от приблизительно 1 мкг/кг до 15 мг/кг (например, от 0,1 мг/кг до 10 мг/кг) антитела представляет собой начальную дозу-кандидата для введения пациенту, например, посредством одного или нескольких раздельных введений или посредством непрерывной инфузии. Для повторных введений в течение нескольких суток или более, в зависимости от состояния, лечение продолжают до тех пор, пока не наступит желаемое подавление симптомов заболевания. Предпочтительная доза антитела должна находиться в диапазоне от приблизительно 0,05 мг/кг до приблизительно 10 мг/кг. Можно вводить начальную более высокую загрузочную дозу с последующей одной или несколькими более низкими дозами. Однако могут быть эффективны другие схемы дозирования. Мониторинг прогресса этой терапии легко осуществлять посредством общепринятых способов и анализов.
Промышленные изделия
В другом варианте осуществления изобретения предусмотрено промышленное изделие, содержащее вещества, которые можно использовать для лечения нарушений, описанных выше. Промышленное изделие содержит контейнер и этикетку или вкладыш в упаковку на контейнере или связанные с ним. Подходящие контейнеры включают, например, бутылки, сосуды, шприцы и т.д. Контейнеры можно сформировать из различных материалов, таких как стекло или пластмасса. Контейнер содержи композицию, которая эффективна для лечения состояния, и может иметь стерильный порт доступа (например, контейнер может представлять собой пакет или сосуд с внутривенным раствором, который имеет пробку, прокалываемую иглой для подкожных инъекций). По меньшей мере одно активное средство в композиции представляет собой антитело по изобретению. Этикетка или вкладыш в упаковку указывает на то, что композицию используют для лечения выбранного состояния, такого как злокачественная опухоль. Кроме того, промышленное изделие может содержать (a) первый контейнер с композицией, содержащейся в нем, где композиция содержит антитело; и (b) второй контейнер с композицией, содержащейся в нем, где композиция содержит a дополнительное цитотоксическое средство. Промышленное изделие в этом варианте осуществления изобретения может дополнительно содержать вкладыш в упаковку, указывающий на то, что композиции первого и второго антитела можно использовать для лечения злокачественной опухоли. Альтернативно или дополнительно, промышленное изделие может дополнительно содержат второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекции (BWFI), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Он может дополнительно содержать другие вещества, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Следующие примеры предназначены для того, чтобы только иллюстрировать практическое осуществление настоящего изобретения, и предоставлены для ограничения. Раскрытия всех патентных и научных литературных источников, процитированных в настоящем документе, явно включены в полном объеме в качестве ссылки.
ПРИМЕРЫ
Вещества и способы
Бактериальные штаммы и среды - Штаммы и плазмиды, использованные в этом исследовании, перечислены в таблице 1. Для культур во встряхиваемых колбах все штаммы растили в среде Лурия-Бертани (LB) или фосфат-ограничивающей среде C.R.A.P. (1) при 30 или 37°C, где указано. Среда ферментатора по существу представляла собой то, что описано в литературном источнике 1. Антибиотики добавляли в следующих концентрациях: карбенициллин 50 мкг/мл, канамицин 50 мкг/мл, хлорамфеникол 12,5 мкг/мл или тетрациклин 20 мкг/мл.
Конструирование и оценка относительных библиотек TIR - Сигнальные пептиды термостабильного энтеротоксина II (stII), связывающего мальтозу периплазматического белка (malE), щелочной фосфатазы (phoA) или белка тиол-дисульфидного обмена (dsbA) ПЦР-амплифицировали и сливали со зрелым доменом гена phoA, используя вырожденные праймеры, которые вводили молчащие мутации кодонов на основе неоднозначности (2) в первые шесть аминокислот после инициирующего кодона родительского гена с сайтом рестрикции BssHII, MluI или XbaI на девять пар оснований (п.о.) выше по направлению транскрипции от указанного инициирующего кодона (см. таблицу 2). Также создавали ДНК, кодирующую кодоны каждой сигнальной последовательности дикого типа. Затем эти вставки обычным путем клонировали в сайты SpeI/NotI (New England Biolabs) плазмиды pPho41 (3) и трансформировали в компетентные клетки JM109 (Promega), которые извлекали в течение одного часа и субклонировали в 200 мл LB с добавлением карбенициллина при 37°C в течение шестнадцати часов и затем выделяли с помощью Maxi Prep (Qiagen). Аликвоту извлеченных клеток из каждой библиотеки высевали на чашки с селективным LB-агаром для того, чтобы определить размер библиотеки; все полученные библиотеки обладают ~10-100× покрытием относительно теоретических размеров библиотек. Затем очищенную ДНК трансформировали в компетентные клетки 27C7 и высевали на чашки с LB-агаром с добавлением карбенициллина и 100 мкг/мл 5-бром-4-хлор-3-индолилфосфата (BCIP; Sigma) и растили при 37°C в течение шестнадцати часов. Колонии, которые выглядели светло-синие, предположительно указывали на то, что они несли вариант TIR, отражая по меньшей мере низкий уровень активности PhoA, тогда как темно-синие колонии указывали на то, что клетки несли мощные варианты TIR (4), и белые колонии подразумевали, что клетки несли варианты TIR с низкой экспрессией PhoA или вовсе без нее; процентная доля синих колоний на данной чашке с агаром для каждой библиотеки варьировала ~ от 2 до 70%. ДНК из отдельных колоний, показывающих различные оттенки синего, выделяли с помощью Mini Prep (Qiagen), секвенировали посредством SRS Analysis (Genentech, Inc.), повторно трансформировали в компетентные клетки 27C7 и затем тестировали на их исходные активности PhoA, как предварительно описано (3). В кратком изложении, колонии растили в селективной LB при 30°C в течение шестнадцати часов и разбавляли 1:100 в свежих средах и растили в течение дополнительных четырех часов при 30°C. Затем культуры нормализовали, основываясь на оптической плотности (OD550), и ресуспендировали в строгих AP средах (3), затем хранили при -20°C в течение ночи. Затем клетки размораживали, частично пермеабилизировали с использованием обработки толуолом (Sigma) (5) и аэрировали при 37°C в течение одного часа. Сорок микролитров каждой культуры затем добавляли в раствор, содержащий 1 мМ 4-нитрофенилфосфатгексагидрата динатрия (PNPP; Promega) в 1 M буфере Трис-HCl (pH 8,0) и инкубировали в темноте при комнатной температуре в течение одного часа. Реакции останавливали добавлением 100 мкл натрий-фосфатного буфера (pH 6,5) и считывали поглощение при 410 нм (A410) в течение 20 минут. Относительные интенсивности TIR вычисляли посредством сначала вычитания фонового поглощения из A410 каждого образца из культуры, содержащей пустой вектор (pBR322), а затем делили на скорректированное поглощение из культуры, несущей плазмиду pPho41. Все приведенные значения TIR являются результатом по меньшей мере семи повторных экспериментов.
Конструирование экспрессирующих антитела векторов - Сигнальные пептиды обычным способом клонировали в предварительно описанную двухцистронную систему (1). Варианты сигнальных пептидов тяжелых цепей создавали посредством слияния сигнального пептида, представляющего интерес, с помощью ПЦР сплайсинга расширением перекрытия (SOE-PCR) с тяжелой цепью, представляющей интерес, и клонировали в сайты BssHII/HpaI (New England Biolabs). Варианты сигнальных пептидов легких цепей создавали аналогичным образом с использованием SOE-ПЦР и клонировали в сайты MluI/PacI (New England Biolabs) или XbaI/PacI (New England Biolabs), как точно определено нуклеотидной последовательностью индивидуального варианта TIR (таблица 2). Все последовательности конструкций подтверждали посредством SRS Analysis (Genentech, Inc).
Мелкомасштабная индукция и анализ - Клетки растили в 5 мл селективной LB с добавлением 5 мМ фосфата натрия (pH 7,0) при 30°C в течение 16 часов. Затем аликвоту клеток 500 мкл использовали для инокуляции 25 мл селективной фосфат-ограничивающей среды C.R.A.P. и растили в течение 24 часа при 30°C. Где указано, клетки, несущие плазмиду pJJ247, индуцировали изопропил-β-D-тиогалактозидом (IPTG) до конечной концентрации 1,0 мМ, когда клетки достигали OD600~2,0. Образцы цельного бульона в конечной точке брали и разбавляли до OD600~3,0 лизирующим буфером (10 мМ Трис pH 6,8, 5 мМ ЭДТА, 0,2 мг/мл лизоцим (Sigma), 5 мМ йодуксусная кислота (Sigma)) и инкубировали на льду в течение 10 минут. Образцы разрушали ультразвуком, центрифугировали для удаления обломков клеток и затем анализировали с использованием анализа SDS-PAGE (10% Бис-Трис, Invitrogen). Образцы цельноклеточного лизата нормализовали до эквивалентных оптических плотностей, восстанавливали 0,2 M дитиотреитолом (DTT, Sigma) и анализировали с использованием анализа SDS-PAGE. На все дорожки загружали эквивалентные объемы образцов и анализировали с использованием или антитела человека против Fc (Southern Biotech) в разведении 1:200000 или антитела мыши против κLc (Southern Biotech) в разведении 1:200000. Все антитела конъюгировали с HRP и иммуноблоты визуализировали с использованием Western Lightning-ECL (PerkinElmer) и экспонированием мембраны Biomax XAR Film (Kodak). Образцы белка также анализировали посредством окрашивания кумаси синим, следуя стандартным способам.
Крупномасштабная индукция - Ферментацию выполняли, как предварительно описано (1). В кратком изложении, аликвоту криосохраненных клеток 500 мкл из 5 мл культуры в избирательной LB использовали для инокуляции 500 мл избирательной LB и растили при 30°C в течение 16 часов. Затем инокулировали 10 л ферментатор (по существу, как описано в литературном источнике 1) и растили клетки до высокой плотности с использованием алгоритма на компьютерной основе для подачи концентрированного раствора глюкозы, основываясь на потребностях ферментации. Где указано, клетки, несущие плазмиду pJJ247, индуцировали изопропил-β-D-тиогалактозидом (IPTG) до конечной концентрации 1,0 M, когда клетки достигали OD550~2,00. Образцы цельного бульона и нормализованные OD550 образцы брали через равные интервалы времени и всю ферментацию заканчивали после 2-3 суток. Мониторинг приспособленности культуры осуществляли обычным способом, используя параметры, измеренные онлайн и оффлайн. Образцы анализировали с использованием анализа SDS-PAGE, как описано выше.
ВЭЖХ анализ образцов - Образцы из экспериментов по мелкомасштабной или крупномасштабной индукции анализировали на общую концентрацию (нерастворимых и растворимых) тяжелой или легкой цепей посредством ранее разработанного способа ВЭЖХ анализа с обращенной фазой (Lisa Wong, личный контакт). Образцы анализировали на частицы антител, содержащие легкую цепь, посредством двухколоночного ВЭЖХ анализа с обращенной фазой и белком L (Analytical Operations, Genentech, Inc.). Титры антител получали посредством сравнения площадей пиков на хроматограмме с таковыми стандартной кривой, созданной посредством внесения пустых образцов с известными количествами молекулы, представляющей интерес.
Штаммы и плазмиды, использованные в этом исследовании
Lc = легкая цепь
5D5 = клон 5D5.v2 моноклонального антитела против c-met. Последовательности тяжелой и легкой цепей 5D5.v2 представлены на фиг.7 и также описаны, например, в WO 2006/015371; Jin et al., Cancer Res (2008) 68:4360.
Варианты сигнальной последовательности
Жирный курсив = последовательность, которую варьировали (т.е. первые шесть аминокислот после инициирующего кодона)
Курсив = сайт рестрикции BssHII, MluI или XbaI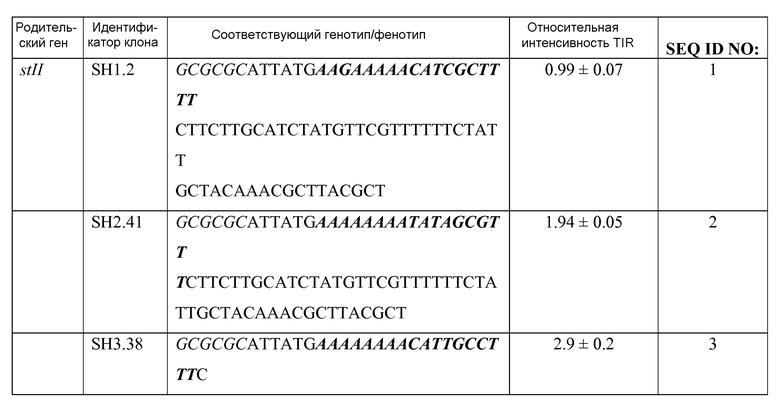




Результаты/обсуждение
Авторы настоящего изобретения разработали библиотеки новых вариантов сигнальных пептидов области инициации трансляции (TIR) (фиг.2, таблица 2) для сигнальных пептидов, представляющих два основных секреции пути для транспортировки через внутреннюю мембрану в E. coli: sec (PhoA, MalE) и SRP (DsbA, STII). Каждая библиотека содержит панель векторов, которая содержит варианты TIR с различными интенсивностями трансляции, предоставляющими средство, с помощью которого легко корректировать уровень трансляции для заданного белка, представляющего интерес. Сигнальные пептиды связывающего мальтозу периплазматического белка (MalE) и щелочной фосфатазы (PhoA) направляют транслокацию из цитоплазмы в периплазму посттрансляционным образом с помощью молекулярного двигателя SecA. Сигнальные пептиды термостабильного энтеротоксина II (stII) и белка тиол:дисульфидного обмена (dsbA) направляют транслокацию котрансляционным образом с помощью частицы узнавания сигнала (SRP) (фиг.1).
Во время конструирования библиотеки, сайт рестрикции BssHII, MluI или XbaI встраивали на девять пар оснований (п. о.) выше по напралению транскрипции относительно инициирующего кодона родительского гена. В зависимости от типа присутствующего сайта рестрикции, наблюдали различные диапазоны интенсивностей TIR (фиг.2). В целом, последовательности, несущие сайт MluI, проявляли наименьший диапазон интенсивностей TIR (~1-3), тогда как расположенный выше по направлению транскрипции сайт BssHII делал возможным умеренный диапазон интенсивностей TIR (~1-8), а сайт XbaI наибольший диапазон (~1-11). Эти сайты рестрикции, присутствующие в нетранслируемой области, включены в TIR (3). Несмотря на то, что нельзя исключать того, что могут существовать более мощные варианты TIR для каких-либо исследованных сочетаний сигнального пептида/сайта рестрикции, эти результаты, по-видимому, характерны для средних интенсивностей TIR каждой исследованной библиотеки сигнальных пептидов.
Серию плазмид конструировали для того, чтобы проиллюстрировать эффект уровня трансляции и сигнального пептида на секрецию. В каждом случае ген, представляющий интерес, встраивали ниже по направлению транскрипции относительно промотора phoA, trp Шайна-Дальгарно и сигнальной последовательности, обладающей различными относительными интенсивностями TIR. После трансформации и индукции промотора phoA в масштабе встряхиваемой колбы, лизаты из цельных клеток, экспрессирующих гетерологичный белок, клон 5D5.v2 антитела против c-met, анализировали посредством SDS-PAGE. В этих экспериментах варьриовала TIR или тяжелой цепи или легкой цепи, причем соотвествующую легкую цепь или тяжелую цепь, соответственно, оставляли неизменной.
На фиг.3 показаны результаты манипуляции с сигнальным пептидом тяжелой цепи. Когда анализировали с использованием антитела, специфичного к α-Fc, один вариант TIR ssDsbA-тяжелой цепи давал четкое увеличение полноразмерного антитела (FL-Ab), а также димера тяжелой-легкой цепи (HL) и частицы из тяжелой-тяжелой-легкой цепи (HHL) относительно других вариантов сигнального пептида (фиг.3A, верхний блот). Исследование общей тяжелой цепи из этих образцов выявило относительно схожие уровни между всеми исследованными слияними сигнальных пептидов (фиг.3A, нижний блот). Когда легкую цепь визуализировали с использованием антитела к α-κLc, получали схожие результаты, причем один вариант TIR ssDsbA-тяжелой цепи снова проявлял самый высокий уровень FL-Ab (фиг.3B, верхний блот). К удивлению, у образца слияния тяжелой цепи TIR один DsbA отсутствовали частицы с более низкой молекулярной массой - практически осуществленный димер легкой-легкой цепи (LL) и свободную легкую цепь наблюдали в других образцах. В целом, в случае, когда посттрансляционные сигналы (MalE, PhoA) сливали с тяжелой цепью, общая легкая цепь, по-видимому, экспрессировалась больше, чем в случаях котрансляционных (STII, DsbA) слияний сигнальных пептидов (фиг.3B, нижний блот). В целом, наблюдали следующую иерархию в отношении образования сигнального пептида, слитого с тяжелой цепью, и полноразмерного антитела: DsbA>STII>MalE>PhoA. В частности, вариант TIR DsbA приводил к повышенной экспрессии (например, полноразмерного антитела) по сравнению с вариантом TIR STII, несмотря на то, что относительная интенсивность TIR не менялась (т.е. обе TIR мощные).
На фиг.4 показаны результаты манипуляции с сигнальным пептидом легкой цепи. Изменение сигнального пептида легкой цепи с одного варианта TIR STII на один или два варианта TIR PhoA создавало заметное повышение титра FL-Ab (фиг.4, верхний блот). Модификация сигнального пептида, слитого с легкой цепью, по-видимому, не оказывала эффект на общее количество экспрессируемой тяжелой цепи (фиг.4, средний блот), но значительно изменяла общее количество присутствующей легкой цепи, причем наибольшее накопление процессированной легкой цепи возникало в образцах с вариантом два TIR STII или DsbA, слитым с легкой цепью (фиг.4, нижний блот). Когда выполняли слияние с посттрансляционными сигнальными пептидами, наблюдали две полосы в образцах общей легкой цепи, указывающие на непроцессированную легкую цепь. В целом, наблюдали следующую иерархию в отношении образования слияний сигнального пептида с легкой цепью и полноразмерного антитела: PhoA>MalE>STII>DsbA.
Установка для мониторинга частиц антитела с течением времени из 10 л ферментации выявляла результаты, схожие с экспериментами во встряхиваемых колбах, представленные на фиг.3 и 4. Наибольшее количество FL-Ab наблюдали в образцах с вариантом TIR, полученным из DsbA, который слит с тяжелой цепью, и сигнальным пептидом, полученным из DsbA или PhoA, который слит с легкой цепью (фиг.5, верхний блот). Эти образцы также показывали больше частиц димеров HHL и HL, чем слияние TIR один STII с тяжелой цепью. Дополнительно, димер LL и свободную легкую цепь было легко видно в образцах с сигнальным пептидом TIR один PhoA, слитым с легкой цепью. Исследование образцов со сниженным общим белком выявило, что слияние сигнального пептида DsbA приводило к большему количеству общей тяжелой цепи, чем слияние STII в индуцирующих экспрессию условиях ферментации (фиг.5, средний блот). Аналогичным образом, для слияний сигнального пептида легкой цепи более высокое накопление легкой цепи наблюдали со слияниями сигнального пептида TIR один DsbA, чем TIR один STII (фиг.5, нижний блот). Однако, самое высокое накопление легкой цепи наблюдали со слиянием сигнального пептида TIR один PhoA. Две полосы, которые наблюдали в образцах общей легкой цепи, взятых из встряхиваемых колб, выглядели только как одна полоса в образцах ферментации, что указывает на то, что процессин легкой цепи проходил более эффективно во время 10 л ферментации.
Авторы настоящего изобретения сливали сигнальные пептиды со зрелым доменом гена phoA E. coli (mPhoA), чтобы дополнительно исследовать различия в уровнях экспрессии общей легкой или тяжелой цепи, когда выполняли слияние с вариантами TIR, полученными из STII или DsbA, и осуществляли экспрессию в культурах в встряхиваемых колбах в индуцирующих условиях. Наблюдали схожие эффекты, оказываемые на уровни экспрессии общей легкой и тяжелой цепей (фиг.6). Когда индуцировали экспрессию белка, экспрессия mPhoA показывала одновременный подъем с повышенной интенсивностью TIR для слияний сигнального пептида DsbA, вплоть до интенсивности TIR семь (наивысшая интенсивность TIR, используемая в данном исследовании). Наблюдали схожее повышение экспрессии mPhoAc повышением интенсивности TIR для слияний сигнального пептида STII достигали интенсивности TIR вплоть до шести или восьми, тем самым казалось, что количество mPhoA снижено по сравнению с mPhoA, присутствующим в образце TIR три STII. К удивлению, образование большего количества тяжелой и легкой цепи происходило с использованием TIR DsbA с интенсивностью один, чем с использованием TIR STII с интенсивностью один, и направляемая STII транслокация PhoA достигала максимального количества при более никзой концентрации общего белка, чем направляемая Dba транслокация PhoA. Кроме того, изменение последовательности TIR с STII на DsbA увеличивало динамический диапазон эффекта TIR.
В образцах из 10 л ферментации анализировали титры антител с использованием ВЭЖХ анализа на основе белка L (таблица 3). Данные ВЭЖХ хорошо соответствовали качественным уровням титров, выявленным посредством анализа вестерн-блоттинга (фиг.3, 4). Когда сигнальную последовательность тяжелой цепи меняли с варианта TIR, полученного из STII, на вариант TIR, полученный из Dsb, титры FL-Ab возрастали на ~40-90%. Наивысшие титры получали, когда слияние тяжелой цепи DsbA один спаривали с легкой цепью слитой с сигнальными пептидами TIR один DsbA, MalE или PhoA. Наивысшие титры создавали, когда легкую цепь сливали с сигнальными пептидами TIR MalE или PhoA.
В отличие от этого, титр FL-Ab падал, когда посттрансляционный сигнальный пептид сливали с тяжелой цепью, причем слияние сигнального пептида TIR один PhoA и TIR один MalE покаывало 70% и 60% падение титра, соответственно. Авторы настоящего изобретения пришли к выводу о том, что экспрессия тяжелой цепи была оптимизирована, когда для того, чтобы управлять трансляцией, использовали котрансляционный сигнальный пептид (например, DsbA).
Авторы настоящего изобртеения тестировали эффект сверхэкспрессии шаперонов. Сверхэкспрессия шаперонов DsbA и DsbC (иногда обозначаемых DsbA/C в настоящем документе) усиливала эффекты сигнального пептида DsbA, слитого с тяжелой цепью, и сигналов DsbA, PhoA или MalE, слитых с легкой цепью. Когда сравнивали экспрессию FL-Ab с помощью сигнала TIR один STII, слитого с тяжелой и легкой цепями (SS1.1+шапероны), от приблизительно 2- до 2,5-кратное повышение титра FL-Ab наблюдали при использовании слияния TIR один DsbA-тяжелой цепи в паре со слиянием легкой цепи TIR один MalE, PhoA или DsbA.
Авторы настоящего изобретения исследовали зависимость между сигнальным пептидом, слитым с легкой цепью и тяжелой цепью антитела, и конечными титрами антитела. Титры полностью собранного антитела (FL-Ab) были наивысшими, когда котрансляционный (например, DsbA или STII) сигнальный пептид сливали с N-концом тяжелой цепи, при этом варианты TIR, полученые из DsbA, приводили к максимальным наблюдавшимся выходам FL-Ab. Таким образом, варианты TIR DsbA могут делать возможными более высокие уровни трансляции белка-пассажира, чем варианты TIR STII, в индуцирующих условиях, тем самым приводя к более высоким уровням экспрессии процессированного белка-пассажира. В отличие от этого, титры антител резко падали, когда любой посттрансляционный сигнальный пептид (т.е. MalE или PhoA) сливали с тяжелой цепью. Этот эффект может быть обусловлен протеолизом или может быть обусловлен отличающимся путем укладки, по которому следует тяжелая цепь (6). Исследование уровней общей тяжелой цепи из образцов, экспрессирующих сигнальный пептид TIR один PhoA или MalE, слитый с тяжелой цепью, выявило легкий сдвиг кажущейся массы, возможно, вследствие присутствия непроцессированной тяжелой цепи (фиг.3A, нижний блот).
Слияние посттрансляционных вариантов TIR, направляемых MalE или направляемых PhoA, сигнального пептида с легкой цепь приводило к высокому накоплению процессированной легкой цепи и повышенным титрам антител относительно STII-опосредованной транслокации во время 10 л ферментации (фиг.5, нижний блот). Повышенные выходы легкой цепи, а также FL-Ab также наблюдали, когда легкую цепь транслоцировали посредством вариантов TIR DsbA по сравнению с легкой цепью, транслоцированной посредством вариантов TIR STII. Однако, количество общей легкой цепи, экспрессируемой с варианта TIR один DsbA, не было высоким при этом с вариантов TIR один PhoA или MalE. Что интересно, анализ образцов, взятых в динамике из 10 л ферментации, показывает, что титры FL-Ab из экспериментов с легкой цепью, слитой с вариантами TIR MalE, DsbA или PhoA, продолжали расти с течением времени, тогда как слияния с вариантами TIR STII достигали не только более низкого максимального титра, но достигали этого уровня титра в значительно более ранний момент времени (фиг.5, верхний блот). Таким образом, эти данные приводят к выводу о том, что легкую цепь можно эффективно транслоцировать ко- или посттрансляционным образом, тогда как тяжелая цепь требует котрансляционной транслокации для пиковой экспрессии.
Экспрессия антитела против c-met с одним Fab фрагментом: авторы настоящего изобретения оценивали зависимость между сигнальным пептидом, слитым с легкой цепью, тяжелой цепью и Fc антитела с одним Fab фрагментом, и конечными титрами антитела. Плазмиды конструировали с использованием STII сигнальных последовательностей с TIR 1 для легкой цепи, тяжелой цепи и полипептида Fc, используя сигнальную последовательность PhoA с TIR 1 (SEQ ID NO: 29) для легкой цепи и сигнальную последовательность DsbA с TIR 1 (SEQ ID NO: 40) для тяжелой цепи и Fc фрагмента; и с использованием сигнальной последовательности PhoA с TIR 1 для легкой цепи и Fc фрагмента и сигнальной последовательности DsbA с TIR 1 для HC. Относительные значения титров брали из образцов из конца эксперимента и измеряли с использованием описанного выше ВЭЖХ анализа с обращенной фазой и белком L. Значения относительных титров нормализовали к титру для случая в первом ряду таблицы 4 - сигнальные последовательности STII и TIR=1 для LC, HC и Fc без совместной экспрессии DsbA/C.
Результаты представлены в таблице 4. Относительные титры антител с одним Fab фрагментом были наивысшими, когда котрансляционный (например, DsbA) сигнальный пептид сливали с N-концом тяжелой цепи, посттрансляционный (например, PhoA) сигнальный пептид сливали с N-концом легкой цепи и посттрансляционный (например, PhoA) сигнальный пептид сливали с Fc-областью, а экспрессия происходила в присутствие DsbA/C. В целом, в отношении экспрессии сигнального пептида, слитого с легкой цепью, тяжелой цепью и Fc фрагментом, и антитела с одним Fab фрагментом в присутствие DsbA/C наблюдали следующую иерархию: P.D.D>P.D.P>S.S.S. Уровни экспрессии в отсутствие DsbA/C были схожими во всех протестированных образцах, в которых наибольшее количество антител, секретированных в периплазму, образовывало агрегаты. Совместная экспрессия шаперонов дисульфидной связи увеличивала образуемое уложенное антитело, таким образом, выявляя повышенную экспрессию антитела, реализованную посредством оптимизации TIR.
Итак, данная технология предлагает новое средство для повышения выхода уложенного антитела, например, в E. coli, посредством манипуляции экспрессией легкой цепи и тяжелой цепи путем отбора из нового массива вариантов TIR и, кроме того, с помощью ко- или посттрансляционных сигнальных последовательностей для легкой цепи и котрансляционной сигнальной последовательности для тяжелой цепи. Улучшенную экспрессию антител с одним Fab фрагментом, содержащих тяжелую цепь, легкую цепь и Fc-область, также осуществляли с использованием полученных новых вариантов TIR, описанных в настоящем документе, и, кроме того, с помощью ко- или посттрансляционных сигнальных последовательностей для легкой цепи, котрансляционной сигнальной последовательности для тяжелой цепи и ко- или посттрансляционной сигнальной последовательности для полипептида Fc. Полезность этого способа, по-видимому, широко применима к широкому спектру антител (например, к биспецифическим антителам, содержащим мутации выступа и отверстия), производным антител и в целом к продуцированию рекомбинантных белков на основе бактерий.
Частичный список цитируемой литературы
1. Simmons, L. C, Reilly, D., Klimowski, L., Raju, T. S., Meng, G., Sims, P., Hong, K., Shields, R. L., Damico, L. A., Rancatore, P., and Yansura, D. G. (2002) Journal of immunological methods 263(1-2), 133-147
2. Stemmer, W. P., Morris, S. K., Kautzer, C. R., and Wilson, B. S. (1993) Gene 123(1), 1-7
3. Simmons, L. C, and Yansura, D. G. (1996) Nature biotechnology 14(5), 629-634
4. Le Calvez, H., Green, J. M., and Baty, D. (1996) Gene 170(1), 51-55
5. Jackson, R. W., and DeMoss, J. A. (1965) Journal of bacteriology 90(5), 1420-1425
6. Kadokura, H., and Beckwith, J. (2009) Cell 138(6), 1164-1173
Несмотря на то что указанное выше относится к конкретным вариантам осуществления, понятно, что настоящее изобретение не ограничено этим. Средним специалистам в данной области должно быть ясно, что различные модификации раскрытых вариантов осуществления можно выполнять без отклонения от общей идеи изобретения. Подразумевается, что все такие модификации входят в объем настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2015 |
|
RU2748026C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2828630C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2739500C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2705274C2 |
| АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА FcγRIIB И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2404991C2 |
| СИСТЕМА ЭКСПРЕССИИ И СЕКРЕЦИИ | 2013 |
|
RU2670491C2 |
| БАКТЕРИЯ Е.coli, ПРОДУЦИРУЮЩАЯ ГЕТЕРОЛОГИЧНЫЙ ПОЛИПЕПТИД, И СПОСОБ ПРОДУЦИРОВАНИЯ ПОЛИПЕПТИДА | 2001 |
|
RU2287574C2 |
| ГУМАНИЗИРОВАННЫЕ АНТАГОНИСТЫ, НАПРАВЛЕННЫЕ ПРОТИВ c-met | 2005 |
|
RU2398777C2 |
| СПОСОБ ОЧИСТКИ ГЕТЕРОЛОГИЧНОГО ПОЛИПЕПТИДА | 2004 |
|
RU2337968C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2012 |
|
RU2650775C2 |
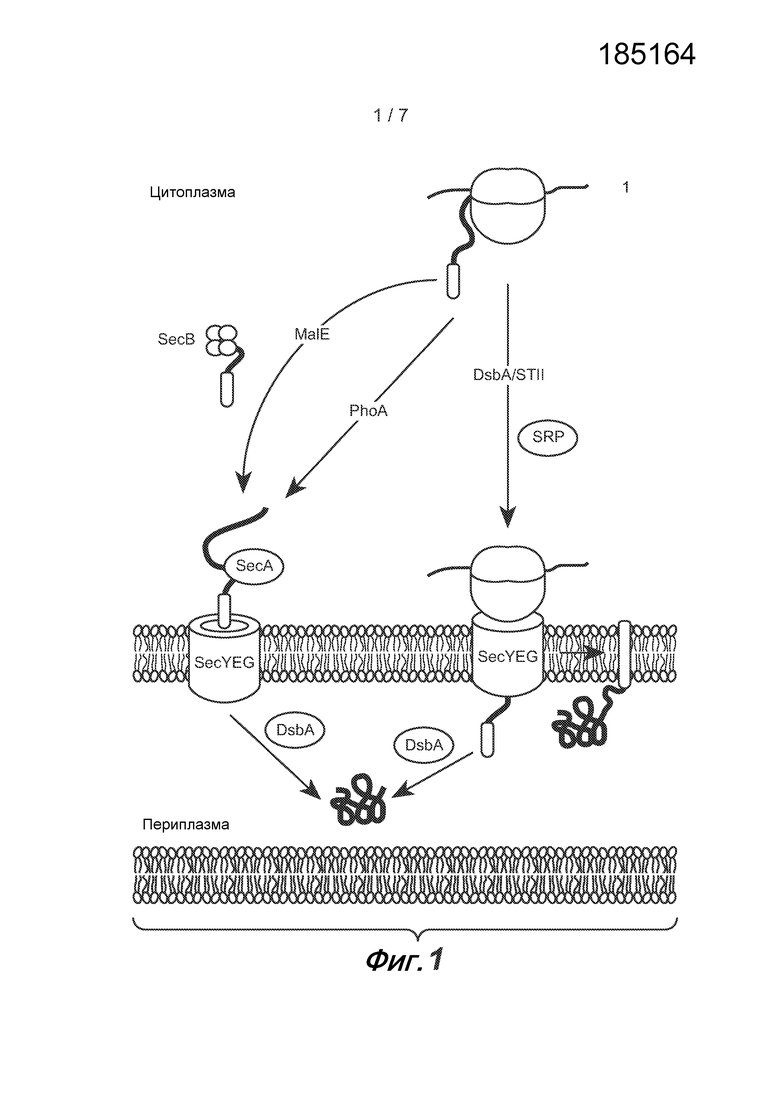






























Настоящее изобретение к биохимии, в частности к способу получения антитела. Указанный способ предусматривает культивирование клетки-хозяина, содержащей полинуклеотид, содержащий первую область инициации трансляции (TIR), функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, и вторую TIR, функционально связанную с полинуклеотидом, кодирующим лёгкую цепь антитела, и извлечение антитела из культуры клетки-хозяина. Первая TIR содержит вариант котрансляционной прокариотической сигнальной последовательности секреции DsbA и вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции. Благодаря наличию указанного полинуклеотида в клетке-хозяине происходит укладка и сборка тяжёлой и лёгкой цепей с формированием биологически активного антитела. Настоящее изобретение также раскрывает полинуклеотид и клетки-хозяева для получения антитела, т.е. для осуществления указанного выше способа. Настоящее изобретение позволяет увеличить выход получаемого антитела. 5 н. и 64 з.п. ф-лы, 7 ил., 4 табл., 1 пр.
1. Способ получения антитела, причем указанный способ включает культивирование клетки-хозяина, содержащей полинуклеотид, который содержит (1) первую область инициации трансляции (TIR), функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где TIR содержит вариант котрансляционной прокариотической сигнальной последовательности секреции DsbA; и (2) вторую TIR, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции, посредством чего при экспрессии антитела в клетке-хозяине происходит укладка и сборка тяжелой и легкой цепей, чтобы сформировать биологически активное антитело, и извлечение антитела из культуры клетки-хозяина.
2. Способ по п. 1, где первая область инициации трансляции содержит последовательность одной из SEQ ID NO: 36-42.
3. Способ по п. 1, где вторая область инициации трансляции содержит вариант сигнальной последовательности STII, DsbA, MalE или PhoA.
4. Способ по п. 3, где вторая область инициации трансляции содержит полинуклеотид, кодирующий вариант сигнальной последовательности PhoA или MalE.
5. Способ по п. 3, где вторая область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42.
6. Способ по п. 1, где клетка-хозяин дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc.
7. Способ по п. 6, где третья область инициации трансляции содержит вариант сигнальной последовательности STII, Phol, MalE или DsbA.
8. Способ по п. 6, где третья область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42.
9. Способ по п. 3, где клетка-хозяин дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc.
10. Способ по п. 9, где третья область инициации трансляции содержит последовательность варианта сигнальной последовательности STII, Phol, MalE или DsbA.
11. Способ по п. 9, где третья область инициации трансляции содержит последовательность одной из SEQ ID NO: 1-42.
12. Способ по любому из пп. 1, 2-8, 10 или 11, где первая и вторая области инициации трансляции обеспечивают приблизительно равные интенсивности трансляции.
13. Способ по п. 12, где относительная интенсивность трансляции составляет приблизительно один или два.
14. Способ по любому из пп. 6-8, 10 или 11, где первая, вторая и третья области инициации трансляции обеспечивают приблизительно равные интенсивности трансляции.
15. Способ по п. 14, где относительная интенсивность трансляции составляет приблизительно один или два.
16. Способ по любому из пп. 1, 2 или 5, где полинуклеотид в клетке-хозяине дополнительно содержит промотор.
17. Способ по п. 16, где промотор представляет собой прокариотический промотор, выбранный из группы, состоящей из промотора phoA, tac, lpp, lac-lpp, lac, ara и T7.
18. Способ по любому из пп. 1, 2 или 5, где клетка-хозяин представляет собой прокариотическую клетку.
19. Способ по п. 18, где прокариотическая клетка представляет собой Е. coli.
20. Способ по п. 19, где Е. coli представляет собой штамм с дефицитом активностей эндогенных протеаз.
21. Способ по п. 19, где генотип Е. coli не содержит гены degP и pre и содержит мутантный ген spc.
22. Способ по любому из пп. 1, 2 или 5, где клетка-хозяин дополнительно содержит полинуклеотид, кодирующий по меньшей мере один прокариотический полипептид, выбранный из группы, состоящей из DsbA, DsbC, DsbG и FkpA.
23. Способ по п. 22, где полинуклеотид кодирует как DsbA, так и DsbC.
24. Способ по любому из пп. 1, 2 и 5, где клетка-хозяин содержит один или несколько полинуклеотидов, совместно кодирующих антитело.
25. Способ по любому из пп. 1, 2 или 5, где антитело извлекают из среды для культивирования клетки-хозяина.
26. Способ по п. 25, где способ дополнительно содержит объединение извлеченного антитела с фармацевтически приемлемым носителем, эксципиентом или носителем для того, чтобы получить фармацевтический состав, содержащий антитело.
27. Способ по п. 25, где по меньшей мере 50% тяжелых и легких цепей образуют антитело.
28. Способ по п. 25, где по меньшей мере 70% тяжелых и легких цепей образуют антитело.
29. Способ по любому из пп. 1, 2 и 5, где антитело представляет собой моноклональное антитело.
30. Способ по п. 29, где антитело представляет собой химерное антитело, аффинно зрелое антитело, биспецифическое антитело, гуманизированное антитело, фрагмент антитела или антитело человека.
31. Способ по п. 30, где фрагмент антитела представляет собой антитело с одним Fab фрагментом.
32. Способ по п. 31, где антитело связывает c-met.
33. Способ по п. 29, где антитело представляет собой биспецифическое антитело.
34. Полинуклеотид для получения антитела, содержащий (1) TIR, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где TIR содержит вариант котрансляционной прокариотической сигнальной последовательности секреции DsbA; и (2) вторую TIR, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции.
35. Полинуклеотид по п. 34, где первая TIR содержит последовательность одной из SEQ ID NO: 36-42.
36. Полинуклеотид по п. 34, где вторая TIR содержит вариант сигнальной последовательности STII, DsbA, MalE или PhoA.
37. Полинуклеотид по п. 36, где вторая TIR содержит последовательность одной из SEQ ID NO: 1-42.
38. Полинуклеотид по п. 36, где вторая TIR содержит последовательность одной из SEQ ID NO: 1, 2, 8, 9, 11, 13, 29, 36, 37 и 40.
39. Полинуклеотид по п. 34, где полинуклеотид дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc.
40. Способ по п. 39, где третья TIR содержит вариант сигнальной последовательности STII, Phol, MalE или DsbA.
41. Полинуклеотид по п. 39, где третья TIR содержит последовательность одной из SEQ ID NO: 1-42.
42. Полинуклеотид по п. 36, где полинуклеотид дополнительно содержит (3) третью область инициации трансляции, функционально связанную с полинуклеотидом, кодирующим полипептид Fc.
43. Полинуклеотид по п. 42, где третья TIR содержит последовательность варианта сигнальной последовательности STII, Phol, MalE или DsbA.
44. Полинуклеотид по п. 43, где третья TIR содержит последовательность одной из SEQ ID NO: 1-42.
45. Полинуклеотид по любому из пп. 34-44, где первая и вторая TIR обеспечивают приблизительно равные интенсивности трансляции.
46. Полинуклеотид по п. 45, где относительная интенсивность трансляции составляет приблизительно один или два.
47. Полинуклеотид по любому из пп. 39-44, где первая, вторая и третья области инициации трансляции обеспечивают приблизительно равные интенсивности трансляции.
48. Полинуклеотид по п. 47, где относительная интенсивность трансляции составляет приблизительно один или два.
49. Полинуклеотид по любому из пп. 34, 35 или 38, где полинуклеотид дополнительно содержит промотор.
50. Полинуклеотид по п. 49, где промотор представляет собой прокариотический промотор, выбранный из группы, состоящей из промотора phoA, tac, lpp, lac-lpp, lac, ara и Т7.
51. Полинуклеотид по любому из пп. 34, 35 или 38, где антитело представляет собой моноклональное антитело.
52. Полинуклеотид по п. 51, где антитело представляет собой химерное антитело, аффинно зрелое антитело, биспецифическое антитело, гуманизированное антитело, фрагмент антитела или антитело человека.
53. Полинуклеотид по п. 52, где фрагмент антитела представляет собой антитело с одним Fab фрагментом.
54. Полинуклеотид по п. 53, где антитело связывает c-met.
55. Полинуклеотид по п. 51, где антитело представляет собой биспецифическое антитело.
56. Клетка-хозяин для получения антитела, содержащая полинуклеотид по любому из пп. 34, 35 или 38.
57. Клетка-хозяин по п. 56, где клетка-хозяин представляет собой прокариотическую клетку.
58. Клетка-хозяин для получения антитела, содержащая полинуклеотид для получения антитела, содержащий (1) TIR, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где TIR содержит вариант котрансляционной прокариотической сигнальной последовательности секреции DsbA; и (2) вторую TIR, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции, где клетка-хозяин представляет собой прокариотическую клетку, причем прокариотическая клетка представляет собой Е. coli.
59. Клетка-хозяин по п. 58, где первая TIR содержит последовательность одной из SEQ ID NO: 36-42.
60. Клетка-хозяин по п. 58, где вторая TIR содержит последовательность одной из SEQ ID NO: 1, 2, 8, 9, 11, 13, 29, 36, 37 и 40.
61. Клетка-хозяин по п. 58, где Е. coli представляет собой штамм с дефицитом активностей эндогенных протеаз.
62. Клетка-хозяин по п. 58, где генотип Е. coli не содержит гены degP и prc и содержит мутантный ген spr.
63. Полинуклеотид по п. 58, где клетка-хозяин дополнительно содержит полинуклеотид, кодирующий по меньшей мере один прокариотический полипептид, выбранный из группы, состоящей из DsbA, DsbC, DsbG и FkpA.
64. Полинуклеотид по п. 63, где полинуклеотид кодирует как DsbA, так и DsbC.
65. Клетка-хозяин для получения антитела, содержащая полинуклеотид для получения антитела, содержащий (1) TIR, функционально связанную с полинуклеотидом, кодирующим тяжелую цепь антитела, где TIR содержит вариант котрансляционной прокариотической сигнальной последовательности секреции DsbA; и (2) вторую TIR, функционально связанную с полинуклеотидом, кодирующим легкую цепь антитела, где вторая TIR содержит котрансляционную или посттрансляционную прокариотическую сигнальную последовательность секреции, где клетка-хозяин представляет собой прокариотическую клетку, причем прокариотическая клетка представляет собой Е. coli, где клетка-хозяин дополнительно содержит полинуклеотид, кодирующий прокариотический белок шаперон.
66. Клетка-хозяин по п. 65, где первая TIR содержит последовательность одной из SEQ ID NO: 36-42.
67. Клетка-хозяин по п. 65, где вторая TIR содержит последовательность одной из SEQ ID NO: 1, 2, 8, 9, 11, 13, 29, 36, 37 и 40.
68. Клетка-хозяин по п. 65, где Е. coli представляет собой штамм с дефицитом активностей эндогенных протеаз.
69. Клетка-хозяин по любому из пп. 65-68, где прокариотический белок шаперон представляет собой DsbA и/или DsbC.

