ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет заявки на патент США № 61/953629, поданной 14 марта 2014 г., и заявки на патент США № 62/107981, поданной 26 января 2015 г., и заявки на патент США № 62/108476, поданной 27 января 2015 г., содержание которых в полном объеме включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и в полном объеме включен в данный документ посредством ссылки. Указанная копия ASCII, созданная 12 марта 2015 г., называется P5804R1-WO_SL.txt, а ее размер составляет 21971 байт.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение в общем случае относится к областям молекулярной биологии и белковой технологии. В частности, изобретение относится к сигнальным пептидам для секреции гетерологичных полипептидов бактериями. Также изобретение относится к прокариотически полученным рекомбинантным полипептидам и их применениям.
УРОВЕНЬ ТЕХНИКИ
В последние годы появились растущие перспективы применения гетерологичных полипептидов, например, антител, в качестве диагностических и терапевтических агентов для различных нарушений и заболеваний. Для многих исследований и клинических применений необходимы большие количества функционального полипептида, следовательно, существует потребность в крупномасштабных, но при этом рентабельных системах для производства полипептидов. Исключительно удобным является рекомбинантное получение антител при помощи разнообразных экспрессионных хозяев, от прокариотов, таких как E. coli или B. subtilis, до дрожжей, растений, клеток насекомых и клеток млекопитающих. Kipriyanov and Little (1999) Mol. Biotech. 12:173-201.
По сравнению с другими системами для получения полипептидов бактерии, в частности, E. coli, обеспечивают множество уникальных преимуществ. Применяемые исходные материалы (т.е. бактериальные клетки) являются недорогими и легки в выращивании, что тем самым снижает стоимость продуктов. Прокариотические хозяева растут намного быстрее, чем, например, клетки млекопитающих, что позволяет проводить более быстрый анализ генетических манипуляций. Более короткий период воспроизведения и легкость увеличения масштаба также делают бактериальную ферментацию более привлекательным средством для получения белка в большом количестве. Геномная структура и биологическая активность многих бактериальных видов, включая E. coli, была хорошо изучена, при этом доступен широкий диапазон подходящих векторов, что делает экспрессию необходимого антитела более удобной. По сравнению с эукариотами в процесс производства входит меньше этапов, включая манипуляцию с рекомбинантными генами, стабильную трансформацию множественных копий в организм-хозяина, индукцию экспрессии и получение характеристик продуктов. Pluckthun and Pack (1997) Immunotech 3:83-105.
Для получения рекомбинантных полипептидов в бактериях применяли различные подходы. Рекомбинантные белки можно получать из бактерий как путем рефолдинга телец включения, экспрессируемых в цитоплазме, так и путем экспрессии с последующей секрецией в периплазму бактерии. Выбор между секрецией и рефолдингом в общем случае обусловлен несколькими соображениями. Секреция является более быстрой и чаще применяемой стратегией для получения антител. Kipriyanov and Little (1999), выше. При этом секреция и способность к рефолдингу E. coli часто ограничена более низким уровнем по сравнению с другими экспрессионными хозяевами.
Экспрессию антител в прокариотических системах можно осуществлять в разном масштабе. Общие обзоры получения антител в E. coli смотрите в Pluckthun and Pack (1997) Immunotech 3:83-105; Pluckthun et al. (1996) in Antibody Engineering: A Practical Approach, pp. 203-252 (Oxford Press); Pluckthun (1994) в Handbook of Exp Pharmcol vol 3: The Pharmcol of Monoclonal Antibodies, pp. 269-315 (ed. M. Rosenberg and G.P. Moore; Springer-Verlag, Berlin).
Для многих биологических методов анализа (таких как рентгеновская кристаллография) и клинических применений (таких как белковая терапия) необходимы большие количества белка. Соответственно, существует потребность в высокопроизводительных, но при этом простых системах для получения прошедших надлежащую сборку, растворимых и функциональных гетерологичных полипептидов, таких как антитела.
Все приведенные в данном документе ссылки, включая патентные заявки и публикации, в полном объеме включены посредством ссылки.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В изобретении предложены новые средства для повышения производства гетерологичных белков, таких как антитела. Применение периплазматической секреции в качестве средства для крупномасштабного производства гетерологичных белков (например, антител) может ограничиваться несколькими распространенными проблемами. Во-первых, эффективность секреции представляющего интерес белка может быть низкой. Во-вторых, во многих случаях предшественник не является полностью процессированным в зрелый белок. В-третьих, сверхэкспрессируемые гетерологичные белки часто неправильно сворачиваются, агрегируют в нерастворимые тельца включения или протеолизируются протеазами E. coli. В-четвертых, антитела являются сложными мультимерными белками, состоящими из двух разных полипептидов, тяжелых и легких цепей, которые должны экспортироваться в периплазму, правильно укладываться и образовывать соответствующие дисульфидные связи. Сложность сворачивания и секреции этого белка увеличивает трудности получения антител в E. coli. Секреция и способность к рефолдингу E. coli часто ограничена более низким уровнем по сравнению с другими экспрессионными хозяевами. Хотя было показано, что оптимизация TIR полезна для достижения более эффективной секреции белка, другие подходы, как правило, не улучшают секрецию гетерологичных белков в E. coli. Например, было показано, что оптимизация сигнального белка снижает экспрессию рекомбинантной циклодекстрин глюканотрансферазы (CGTазы) в периплазматическое пространство. Jonet et al., J Mol Microbiol Biotechnol (2012); 22:48-58.
В настоящей работе авторы изобретения демонстрируют, что повышение средней гидрофобности сигнального пептида повышает секрецию растворимого антитела в периплазму E. coli. Были разработаны вариантные сигнальные пептиды с повышенной средней гидрофобностью, а авторы изобретения продемонстрировали повышение периплазматической секреции растворимого антитела в случае применения вариантов сигнальных пептидов с повышенной средней гидрофобностью и снижение периплазматической секреции растворимого антитела в случае применения вариантов сигнальных пептидов со сниженной средней гидрофобностью.
В одном аспекте предложены способы повышения секреции тяжелой цепи антитела и/или легкой цепи антитела из клетки-хозяина E. coli, включающие культивирование клетки-хозяина E. coli, содержащей полинуклеотид, содержащий (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, при этом средняя гидрофобность сигнального пептида составляет более чем около 0,5; и/или (2) второй полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, при этом средняя гидрофобность второго сигнального пептида составляет более чем около 0,5, в результате чего после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи проходят фолдинг и сборку с образованием биологически активного антитела.
В одном аспекте предложены способы получения тяжелой цепи антитела и/или легкой цепи антитела из клетки-хозяина E. coli, включающие культивирование клетки-хозяина E. coli, содержащей полинуклеотид, содержащий (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, при этом средняя гидрофобность сигнального пептида составляет более чем около 0,5; и/или (2) второй полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, при этом средняя гидрофобность второго сигнального пептида составляет более чем около 0,5, в результате чего после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи проходят фолдинг и сборку с образованием биологически активного антитела. В некоторых вариантах реализации изобретения способ дополнительно включает выделение гетерологичного полипептида из культуры клеток-хозяев. В некоторых вариантах реализации изобретения гетерологичный полипептид выделяют из культуральной среды клеток-хозяев. В некоторых вариантах реализации изобретения способ дополнительно включает комбинирование выделенного гетерологичного полипептида с фармацевтически приемлемым носителем, вспомогательным веществом или носителем для получения фармацевтического препарата, содержащего гетерологичный полипептид.
В одном аспекте предложены способы получения тяжелой цепи антитела и/или легкой цепи антитела из клетки-хозяина E. coli, включающие культивирование клетки-хозяина E. coli, содержащей полинуклеотид, содержащий (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, при этом средняя гидрофобность сигнального пептида составляет более чем около 0,5; и/или (2) второй полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, при этом средняя гидрофобность второго сигнального пептида составляет более чем около 0,5, в результате чего после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи проходят фолдинг и сборку с образованием биологически активного антитела.
В одном аспекте предложены способы транслокации тяжелой цепи антитела и/или легкой цепи антитела из клетки-хозяина E. coli, включающие культивирование клетки-хозяина E. coli, содержащей полинуклеотид, содержащий (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, при этом средняя гидрофобность сигнального пептида составляет более чем около 0,5; и/или (2) второй полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, при этом средняя гидрофобность второго сигнального пептида составляет более чем около 0,5, в результате чего после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи проходят фолдинг и сборку с образованием биологически активного антитела.
В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,6. В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,7. В некоторых вариантах реализации изобретения средняя гидрофобность второго сигнального пептида составляет более чем около 0,6. В некоторых вариантах реализации изобретения средняя гидрофобность второго сигнального пептида составляет более чем около 0,7. В некоторых вариантах реализации изобретения средняя гидрофобность первого и второго сигнальных пептидов является сходной (например, приблизительно эквивалентной). В некоторых вариантах реализации изобретения средняя гидрофобность первого и второго сигнальных пептидов является разной.
В некоторых вариантах реализации изобретения первый и/или второй сигнальный пептид представляет собой вариантный котрансляционный сигнальный пептид. В некоторых вариантах реализации изобретения первый и/или второй сигнальный пептид представляет собой вариантный сигнальный пептид DsbA. В некоторых вариантах реализации изобретения вариантный сигнальный пептид DsbA содержит мутацию в остатке L11, при этом вариантный сигнальный пептид DsbA имеет большую среднюю гидрофобность, чем сигнальный пептид DsbA дикого типа с SEQ ID NO:3. В некоторых вариантах реализации изобретения мутация представляет собой L11I или S18Y. В некоторых вариантах реализации изобретения вариантный сигнальный пептид DsbA содержит последовательность SEQ ID NO:13 или 15.
В некоторых вариантах реализации изобретения сигнальный пептид представляет собой сигнальный пептид Sfmc. В некоторых вариантах реализации изобретения сигнальный пептид Sfmc обладает эффективностью TIR, которая отличается от эффективности TIR сигнального пептида Sfmc дикого типа. В некоторых вариантах реализации изобретения относительная эффективность трансляции (также называемая эффективностью TIR) сигнального пептида Sfmc составляет около 2, около 3, около 4, около 5, около 6, около 7 или более, например, около 8, около 9 или более. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида Sfmc составляет между 1 и 3, между 2 и 4, между 3 и 5, между 4 и 6, между 5 и 7, между 6 и 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида Sfmc составляет между 2 и 5, между 3 и 7 или между 4 и 8. В некоторых вариантах реализации изобретения сигнальный пептид представляет собой FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT. В некоторых вариантах реализации изобретения сигнальный пептид FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT обладает относительной эффективностью трансляции, которая отличается от относительной эффективности трансляции сигнального пептида FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT дикого типа. В некоторых вариантах реализации изобретения эффективность TIR сигнального пептида FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT составляет около 2, около 3, около 4, около 5, около 6, около 7 или более, например, около 8, около 9 или более. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT составляет между 1 и 3, между 2 и 4, между 3 и 5, между 4 и 6, между 5 и 7, между 6 и 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT составляет между 2 и 5, между 3 и 7 или между 4 и 8.
В некоторых вариантах реализации изобретения сигнальный пептид не представляет собой Sfmc. В некоторых вариантах реализации изобретения сигнальный пептид не представляет собой TorT. В некоторых вариантах реализации изобретения сигнальный пептид не представляет собой любой из FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL или TorT.
В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и/или второго сигнального пептида составляет около 1, около 2, около 3, около 4, около 5, около 6, около 7 или около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого сигнального пептида составляет около 5, а относительная эффективность трансляции второго сигнального пептида составляет около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого сигнального пептида составляет около 8, а относительная эффективность трансляции второго сигнального пептида составляет около 5. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида Sfmc составляет между 1 и 3, между 2 и 4, между 3 и 5, между 4 и 6, между 5 и 7, между 6 и 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции сигнального пептида Sfmc составляет между 2 и 5, между 3 и 7 или между 4 и 8.
В некоторых вариантах реализации изобретения полинуклеотид в клетке-хозяине дополнительно содержит промотор. В некоторых вариантах реализации изобретения промотор представляет собой прокариотический промотор, выбранный из группы, состоящей из промотора phoA, tac, lpp, lac-lpp, lac, ara и T7.
В некоторых вариантах реализации изобретения клетка-хозяин E. coli представляет собой штамм с дефицитом эндогенной протеазной активности. В некоторых вариантах реализации изобретения в генотипе E. coli отсутствуют гены degP и prc и присутствует мутантный ген spr. В некоторых вариантах реализации изобретения клетка-хозяин дополнительно содержит полинуклеотид, кодирующий по меньшей мере один прокариотический полипептид, выбранный из группы, состоящей из DsbA, DsbC, DsbG и FkpA. В некоторых вариантах реализации изобретения полинуклеотид кодирует как DsbA, так и DsbC.
В некоторых вариантах реализации изобретения способы дополнительно включают выделение антитела из культуры клеток-хозяев. В некоторых вариантах реализации изобретения антитело выделяют из культуральной среды клеток-хозяев. В некоторых вариантах реализации изобретения способы дополнительно включают комбинирование выделенного антитела с фармацевтически приемлемым носителем, вспомогательным веществом или носителем для получения фармацевтического препарата, содержащего антитело. В некоторых вариантах реализации изобретения по меньшей мере 50% образующихся полипептидных комплексов иммуноглобулина являются антителами. В некоторых вариантах реализации изобретения по меньшей мере 70% образующихся полипептидных комплексов иммуноглобулина являются антителами. В некоторых вариантах реализации изобретения по меньшей мере 80% образующихся полипептидных комплексов иммуноглобулина являются антителами. В некоторых вариантах реализации изобретения по меньшей мере 90% образующихся полипептидных комплексов иммуноглобулина являются антителами.
В некоторых вариантах реализации изобретения антитело представляет собой моноклональное антитело. В некоторых вариантах реализации изобретения антитело представляет собой химерное антитело, аффинно созревшее антитело, биспецифическое антитело, гуманизированное антитело или человеческое антитело. В некоторых вариантах реализации изобретения антитело представляет собой биспецифическое антитело.
В другом аспекте предложены вариантные сигнальные пептиды DsbA, причем вариант содержит H-область со средней гидрофобностью, составляющей более чем 0,5.
В другом аспекте предложены вариантные сигнальные пептиды DsbA, содержащие мутацию в остатке S11, при этом вариант имеет большую среднюю гидрофобность, чем сигнальный пептид DsbA с SEQ ID NO:3. В некоторых вариантах реализации изобретения мутация представляет собой L11I и/или S18Y.
В другом аспекте предложены вариантные сигнальные пептиды STII, содержащие мутацию в остатке S11, при этом вариантный сигнальный пептид STII имеет большую среднюю гидрофобность, чем сигнальный пептид STII с SEQ ID NO:1. В некоторых вариантах реализации изобретения мутация представляет собой S11A, S11I или S11L.
В другом аспекте предложены вариантные сигнальные пептиды, состоящие из, состоящие по существу из или содержащие последовательность SEQ ID NO:8, 11, 13, 15, 31 или 33.
В другом аспекте предложен любой из раскрытых в данном документе сигнальных пептидов, слитый с гетерологичным белком. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой тяжелую цепь антитела. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой легкую цепь антитела. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой легкую и тяжелую цепь антитела. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой мультимерный полипептид. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой иммуноадгезин.
В другом аспекте предложены полинуклеотидные последовательности, кодирующие любой из раскрытых в данном документе вариантных сигнальных пептидов.
В другом аспекте предложены полинуклеотидные последовательности, которые кодируют любой из раскрытых в данном документе вариантных сигнальных пептидов, функционально связанные с полинуклеотидом, кодирующим гетерологичный полипептид, причем после экспрессии гетерологичного полипептида в клетке-хозяине гетерологичный полипептид проходит фолдинг и сборку с образованием биологически активного гетерологичного полипептида.
В некоторых вариантах реализации изобретения клетка-хозяин представляет собой прокариотическую клетку-хозяина. В некоторых вариантах реализации изобретения клетка-хозяин представляет собой E. coli.
В другом аспекте предложены полинуклеотиды, кодирующие антитело, при этом указанные полинуклеотиды содержат (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, и (2) полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, причем после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи проходят фолдинг и сборку с образованием биологически активного антитела, при этом первый сигнальный пептид представляет собой вариантный сигнальный пептид любого из раскрытых в данном документе вариантных сигнальных пептидов. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит, состоит по существу из или содержит последовательность SEQ ID NO:8, 11, 13, 15, 31, 33 или 42. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой вариантный сигнальный пептид DsbA, причем вариант содержит H-область со средней гидрофобностью, составляющей более чем 0,5. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой вариантный сигнальный пептид DsbA, содержащий мутацию в остатке S11, при этом вариант имеет большую среднюю гидрофобность, чем сигнальный пептид DsbA с SEQ ID NO:3. В некоторых вариантах реализации изобретения мутация представляет собой L11I и/или S18Y. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой вариантный сигнальный пептид STII, содержащий мутацию в остатке S11, при этом вариантный сигнальный пептид STII имеет большую среднюю гидрофобность, чем сигнальный пептид STII с SEQ ID NO:1. В некоторых вариантах реализации изобретения мутация представляет собой S11A, S11I или S11L.
В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой сигнальный пептид. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит, состоит по существу из или содержит последовательность SEQ ID NO:8, 11, 13, 15, 31, 33 или 42. В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой вариантный сигнальный пептид DsbA, причем вариант содержит H-область со средней гидрофобностью, составляющей более чем 0,5. В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой вариантный сигнальный пептид DsbA, содержащий мутацию в остатке S11, при этом вариант имеет большую среднюю гидрофобность, чем сигнальный пептид DsbA с SEQ ID NO:3. В некоторых вариантах реализации изобретения мутация представляет собой L11I и/или S18Y. В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой вариантный сигнальный пептид STII, содержащий мутацию в остатке S11, при этом вариантный сигнальный пептид STII имеет большую среднюю гидрофобность, чем сигнальный пептид STII с SEQ ID NO:1. В некоторых вариантах реализации изобретения мутация представляет собой S11A, S11I или S11L.
В некоторых вариантах реализации изобретения полинуклеотид, кодирующий антитело, дополнительно содержит (3) полинуклеотид, кодирующий третий сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим Fc-полипептид. Третий сигнальный пептид может быть, например, любым из раскрытых в данном документе вариантных сигнальных пептидов. В некоторых вариантах реализации изобретения третий сигнальный пептид состоит, состоит по существу из или содержит последовательность SEQ ID NO:8, 11, 13, 15, 31, 33 или 42.
В некоторых вариантах реализации изобретения полинуклеотид дополнительно содержит промотор, функционально связанный с гетерологичным полипептидом. В некоторых вариантах реализации изобретения промотор представляет собой прокариотический промотор, выбранный из группы, состоящей из промотора phoA, tac, lpp, lac-lpp, lac, ara, trp и T7. В некоторых вариантах реализации изобретения промотор представляет собой промотор phoA.
В некоторых вариантах реализации изобретения полинуклеотид содержит (a) первый промотор, причем первый промотор функционально связан с легкой цепью, и (b) второй промотор, причем второй промотор функционально связан с тяжелой цепью. В некоторых вариантах реализации изобретения первый и второй промоторы оба являются промоторами phoA.
В некоторых вариантах реализации изобретения полинуклеотид дополнительно содержит (c) третий промотор, причем третий промотор функционально связан с Fc-полипептидом. В некоторых вариантах реализации изобретения промотор представляет собой промотор phoA.
В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой протеазу, иммуноадгезин, внеклеточный домен рецептора, гетеромультимерный белок или антитело.
В некоторых вариантах реализации изобретения антитело представляет собой моноклональное антитело. В некоторых вариантах реализации изобретения антитело представляет собой химерное антитело, биспецифическое антитело, гуманизированное антитело, фрагмент антитела или человеческое антитело. В некоторых вариантах реализации изобретения антитело представляет собой биспецифическое антитело.
Вектор, содержащий полинуклеотид любого из раскрытых в данном документе полинуклеотидов. В некоторых вариантах реализации изобретения вектор представляет собой экспрессионный вектор.
Композиция, содержащая любой из раскрытых в данном документе полинуклеотидов.
Клетка-хозяин, содержащая любой из раскрытых в данном документе полинуклеотидов. В некоторых вариантах реализации изобретения клетка-хозяин представляет собой прокариотическую клетку. В некоторых вариантах реализации изобретения прокариотическая клетка представляет собой E. coli. В некоторых вариантах реализации изобретения E. coli представляет собой штамм с дефицитом эндогенной протеазной активности. В некоторых вариантах реализации изобретения в генотипе E. coli отсутствуют гены degP и prc и присутствует мутантный ген spr. В некоторых вариантах реализации изобретения клетка-хозяин дополнительно содержит полинуклеотид, кодирующий прокариотический белок шаперона. В некоторых вариантах реализации изобретения прокариотический белок шаперона представляет собой DsbA и/или DsbC. В некоторых вариантах реализации изобретения клетка-хозяин сверхэкспрессирует прокариотический белок шаперона.
В другом аспекте предложены способы получения гетерологичного полипептида, включающие культивирование любой из описанных в данном документе клеток-хозяев так, чтобы происходила экспрессия нуклеиновой кислоты, причем после экспрессии указанного полинуклеотида в клетке-хозяине гетерологичный полипептид проходит фолдинг с образованием биологически активного гетерологичного полипептида. В некоторых вариантах реализации изобретения способ дополнительно включает выделение гетерологичного полипептида из культуры клеток-хозяев. В некоторых вариантах реализации изобретения гетерологичный полипептид выделяют из культуральной среды клеток-хозяев. В некоторых вариантах реализации изобретения способ дополнительно включает комбинирование выделенного гетерологичного полипептида с фармацевтически приемлемым носителем, вспомогательным веществом или носителем для получения фармацевтического препарата, содержащего гетерологичный полипептид.
В другом аспекте предложены способы секреции гетерологичного полипептида из клетки, включающие культивирование любой из предложенных в данном документе клеток-хозяев так, чтобы происходила экспрессия нуклеиновой кислоты и секреция гетерологичного полипептида.
В другом аспекте предложены способы транслокации гетерологичного полипептида из клетки, включающие культивирование любой из предложенных в данном документе клеток-хозяев так, чтобы происходила экспрессия нуклеиновой кислоты и транслокация гетерологичного полипептида.
Гетерологичный полипептид, полученный любым из предложенных в данном документе способов. В некоторых вариантах реализации изобретения полипептид представляет собой антитело.
В некоторых вариантах реализации изобретения применение вариантного сигнального пептида приводит, например, к повышению выработки гетерологичного полипептида (например, антитела, например, тяжелой цепи и/или легкой цепи), повышению секреции гетерологичного полипептида (например, антитела), повышению выработки зрелого гетерологичного полипептида (например, антитела), повышению секреции зрелого гетерологичного полипептида (например, антитела), повышению выработки растворимого гетерологичного полипептида (например, антитела), повышению секреции растворимого гетерологичного полипептида (например, антитела), повышению локализации телец включения на периплазматической стороне и/или повышению выработки гетерологичного полипептида, в результате чего происходит секреция, фолдинг и сборка гетерологичного полипептида в биологически активный полипептид (например, антитело), например, по сравнению с применением сигнального пептида дикого типа (невариантного). В некоторых вариантах реализации изобретения относительная эффективность трансляции (также называемая эффективностью TIR) вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 1. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 2, около 3, около 4, около 5, около 6, около 7 или более, например, около 8 или более. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 4. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 5. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 6. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и второго сигнальных пептидов является приблизительно одинаковой. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и второго сигнальных пептидов является разной.
В одном аспекте предложена полинуклеотидная последовательность, кодирующая вариантный сигнальный пептид согласно изобретению. В некоторых вариантах реализации изобретения варианты представляют собой варианты сигнального пептида PhoA, MalE, DsbA или STII. В некоторых вариантах реализации изобретения полинуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO:8, 11, 12, 13, 14, 15, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения полинуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения полинуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения полинуклеотид кодирует аминокислотную последовательность SEQ ID NO: 31 или 33. В некоторых вариантах реализации изобретения полинуклеотид кодирует аминокислотную последовательность SEQ ID NO: 32, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29 30, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 23, 24, 25, 26 или 28. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 29. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность одной из SEQ ID NO:27 или 30. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения полинуклеотид состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 37, 39 или 40.
В другом аспекте в изобретении предложен полинуклеотид, содержащий полинуклеотид, кодирующий вариантный сигнальный пептид согласно изобретению, функционально связанный с полинуклеотидом, кодирующим гетерологичный полипептид, причем после экспрессии гетерологичного полипептида в клетке-хозяине (например, прокариотической клетке-хозяине, например, клетке-хозяине E. coli) происходит фолдинг и сборка гетерологичного полипептида с образованием биологически активного гетерологичного полипептида. Примеры гетерологичных полипептидов дополнительно раскрыты в данном документе. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой тяжелую цепь антитела. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой легкую цепь антитела. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой Fc-полипептид. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой мультимерный полипептид. В некоторых вариантах реализации изобретения гетерологичный полипептид представляет собой гетеромультимер. В некоторых вариантах реализации изобретения сигнальный пептид представляет собой любой из раскрытых в данном документе вариантных сигнальных пептидов. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 31 или 33. В некоторых вариантах реализации изобретения сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 32, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 44, 45 или 46. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 26, 28, 30, 36, 37, 38, 45 или 46. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 27 или 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 37, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий вариантный сигнальный пептид, состоит из, состоит по существу из или содержит полинуклеотидную последовательность SEQ ID NO: 45 или 46.
В другом аспекте в изобретении предложен полинуклеотид, содержащий (1) полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим первый гетерологичный полипептид, и (2) полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим второй гетерологичный полипептид, причем после экспрессии антитела в клетке-хозяине происходит фолдинг и сборка первого и второго гетерологичных полипептидов с образованием биологически активного полипептидного комплекса.
В другом аспекте в изобретении предложен полинуклеотид, кодирующий антитело, при этом указанный полинуклеотид содержит (1) полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, и (2) полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, причем после экспрессии антитела в клетке-хозяине (например, прокариотической клетке-хозяине, например, клетке-хозяине E. coli) происходит фолдинг и сборка тяжелой и легкой цепей с образованием биологически активного антитела.
В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой сигнальный пептид (например, любой известный в данной области техники сигнальный пептид). В некоторых вариантах реализации изобретения сигнальный пептид представляет собой котрансляционный сигнальный пептид. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой сигнальный пептид DsbA. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой сигнальный пептид STII. В некоторых вариантах реализации изобретения первый сигнальный пептид представляет собой любой из раскрытых в данном документе вариантных сигнальных пептидов. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 31 или 33. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 32, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29 30, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 28, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 27 или 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 37, 39 или 40.
В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой сигнальный пептид (например, любой известный в данной области техники сигнальный пептид). В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой сигнальный пептид DsbA. В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой сигнальный пептид STII. В некоторых вариантах реализации изобретения второй сигнальный пептид представляет собой любой из раскрытых в данном документе вариантных сигнальных пептидов. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 31 или 33 В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 32, 35 или 35. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29 30, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26 или 28. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:27 или 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:37, 39 или 40.
В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид (например, любой известный в данной области техники сигнальный пептид), а второй сигнальный пептид состоит из, состоит по существу из или содержит раскрытый в данном документе сигнальный пептид, например, сигнальный пептид согласно любой из SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 и 35 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 36 и 38). В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO:8, 11, 12, 13, 14, 15, 31, 32, 33, 34 или 35. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 37, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29 30, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26 или 28. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 27 или 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий второй сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:37, 39 или 40.
В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид (например, любой известный в данной области техники сигнальный пептид), а первый сигнальный пептид состоит из, состоит по существу из или содержит раскрытый в данном документе сигнальный пептид, например, сигнальный пептид, содержащий, состоящий из или по существу состоящий из аминокислотной последовательности согласно любой из SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 и 35 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33). В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11, 12, 13, 14, 15, 31, 32, 33, 35 или 35. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 31 или 33. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 33, 34 или 35. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29. 30, 36, 37, 38 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 28, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 27 или 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий первый сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 37, 39 или 40.
В некоторых вариантах реализации изобретения полинуклеотид, кодирующий антитело, дополнительно содержит (3) полинуклеотид, кодирующий третий сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим Fc-полипептид. В некоторых вариантах реализации изобретения третий сигнальный пептид состоит из, состоит по существу из или содержит раскрытый в данном документе сигнальный пептид, например, сигнальный пептид, содержащий, состоящий из или по существу состоящий из аминокислотной последовательности согласно любой из 8, 11, 12, 13, 14, 15, 31, 32, 33, 34 и 35 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33). В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 8, 11 или 13. В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах реализации изобретения третий сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 12 или 15. В некоторых вариантах реализации изобретения третий сигнальный пептид состоит из, состоит по существу из или содержит аминокислотную последовательность SEQ ID NO: 31 или 33. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 27, 28, 29, 30, 36, 37, 38 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO: 23, 24, 25, 26, 28, 36, 37, 38, 39 или 40. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:29. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:27 и 30. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:36 или 38. В некоторых вариантах реализации изобретения полинуклеотид, кодирующий третий сигнальный пептид, состоит из, состоит по существу из или содержит последовательность SEQ ID NO:37, 39 или 40. В некоторых вариантах реализации изобретения третий сигнальный пептид представляет собой сигнальный пептид (например, любой известный в данной области техники сигнальный пептид).
В некоторых вариантах реализации, например, полинуклеотида, кодирующего антитело, который дополнительно содержит (3) полинуклеотид, кодирующий третий сигнальный пептид, первый сигнальный пептид содержит сигнальный пептид (например, любой известный в данной области техники сигнальный пептид), а второй сигнальный пептид содержит сигнальный пептид (например, любой известный в данной области техники сигнальный пептид). В некоторых вариантах реализации изобретения первый сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид, а второй сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид, раскрытый в данном документе, например, сигнальный пептид согласно любой из SEQ ID NO: 8, 11, 12, 13, 14, 15, 36, 37, 38, 39 и 40 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33). В некоторых вариантах реализации изобретения второй сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид, а первый сигнальный пептид состоит из, состоит по существу из или содержит сигнальный пептид, раскрытый в данном документе, например, сигнальный пептид согласно любой из SEQ ID NO: 8, 11, 12, 13, 14, 15, 36, 37, 38, 39 и 40 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33). В некоторых вариантах реализации, например, полинуклеотида, кодирующего антитело, который дополнительно содержит (3) полинуклеотид, кодирующий третий сигнальный пептид, первый сигнальный пептид состоит из, состоит по существу из или содержит раскрытый в данном документе сигнальный пептид, например, сигнальный пептид согласно любой из SEQ ID NO: 8, 11, 12, 13, 14, 15, 36, 3, 38, 39 и 40 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33), а второй сигнальный пептид состоит из, состоит по существу из или содержит раскрытый в данном документе сигнальный пептид, например, сигнальный пептид согласно любой из 8, 11, 12, 13, 14, 15, 36, 37, 38, 39 и 40 (в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 8, 11 и 13, а в некоторых вариантах реализации изобретения согласно любой из SEQ ID NO: 31 и 33).
В некоторых вариантах реализации изобретения применение вариантного сигнального пептида приводит, например, к повышению выработки гетерологичного полипептида (например, антитела), повышению секреции гетерологичного полипептида (например, антитела), повышению выработки зрелого гетерологичного полипептида (например, антитела), повышению секреции зрелого гетерологичного полипептида (например, антитела), повышению выработки растворимого гетерологичного полипептида (например, антитела), повышению секреции растворимого гетерологичного полипептида (например, антитела), повышению выработки гетерологичного полипептида, в результате чего происходит секреция, фолдинг и сборка гетерологичного полипептида в биологически активный полипептид (например, антитело), например, по сравнению с применением сигнального пептида дикого типа (невариантного). В некоторых вариантах реализации изобретения применение вариантного сигнального пептида приводит, например, к повышению выработки гетерологичного полипептида (например, антитела), повышению секреции гетерологичного полипептида (например, антитела), повышению выработки зрелого гетерологичного полипептида (например, антитела), повышению секреции зрелого гетерологичного полипептида (например, антитела), повышению выработки растворимого гетерологичного полипептида (например, антитела), повышению секреции растворимого гетерологичного полипептида (например, антитела), повышению выработки гетерологичного полипептида, в результате чего происходит секреция, фолдинг и сборка гетерологичного полипептида в биологически активный полипептид (например, антитело), например, по сравнению с применением сигнального пептида дикого типа (невариантного), при этом относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) является приблизительно эквивалентной. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 1. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 2, около 3, около 4, около 5, около 6, около 7 или более, например, около 8 или более. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 4. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 5. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 6. В некоторых вариантах реализации изобретения относительная эффективность трансляции вариантного сигнального пептида и сигнального пептида дикого типа (невариантного) составляет около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и второго сигнальных пептидов является приблизительно одинаковой. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и второго сигнальных пептидов является разной.
В одном аспекте в изобретении предложено применение гетерологичного полипептида, полученного способами согласно изобретению, в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение. Гетерологичный полипептид может принимать любую описанную в данном документе форму, включая антитело, фрагмент антитела, полипептид (например, олигопептид) или их комбинацию.
В одном аспекте в изобретении предложено применение раскрытого в данном документе сигнального пептида или раскрытого в данном документе полинуклеотида, кодирующего сигнальный пептид, в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение.
В одном аспекте в изобретении предложено применение раскрытого в данном документе экспрессионного вектора в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение.
В одном аспекте в изобретении предложено применение раскрытой в данном документе клетки-хозяина в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение.
В одном аспекте в изобретении предложено применение раскрытого в данном документе изделия в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное (например, аутоиммунное) нарушение и/или связанное с ангиогенезом нарушение (лечение ран).
В одном аспекте в изобретении предложено применение раскрытого в данном документе набора в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигуры 1A, 1B и 1C: иллюстрируют влияние вариантов сигнальных пептидов на уровни полноразмерного антитела 5D5 и тяжелой цепи. Хозяйский штамм E. coli 64B4, содержащий экспрессионный вектор, выращивали в полной среде C.R.A.P. с ограниченным содержанием фосфатов в качалочной колбе в течение 24 ч, а образцы, соответствующие конечной точке, нормировали по ОП550. (1A) Верхняя панель: Вестерн-блоттинг для растворимых содержащих тяжелые цепи молекул. Лизировали клетки 64B4, несущие pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bMalE1-5D5 (mSTII1, bMalE1), pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1) или pBR-mSTII1-bPhoA1-5D5 (mSTII1, bPhoA1), и отделяли водорастворимую фракцию при помощи невосстанавливающего ДСН-ПААГ электрофореза, после чего проводили вестерн-блот-зондирование с конъюгированным с HRP антителом αFc. Полученный в результате блот демонстрирует, сверху вниз, наличие содержащих тяжелые цепи молекул, соответствующих полноразмерному антителу 5D5, тяжелой цепи-тяжелой цепи-легкой цепи и тяжелой цепи-легкой цепи. Нижняя панель: образцы, соответствующие конечной точке, нормировали к 1 ОП550 и осаждали. Все белки денатурировали и восстанавливали путем смешивания с трициновым буфером для образцов, содержащим 0,2 M ДТТ. Тяжелые цепи перемещались в виде одиночной полосы в ДСН-ПААГ при молекулярной массе около 49 кДа и были зондированы антителом αFc. Полосы тяжелых цепей, которые перемещались медленнее в геле, содержали предшественников, что подтверждалось белковым секвенированием по Эдману. (1B) Уровни растворимых тяжелых цепей в периплазме. Соответствующие конечной точке образцы клеток 64B4, несущие pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bMalE1-5D5 (mSTII1, bMalE1), pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1) или pBR-mSTII1-bPhoA1-5D5 (mSTII1, bPhoA1), подвергали осмотическому шоку. Собирали супернатант, денатурировали и восстанавливали трициновым буфером, содержащим 0,2 M ДТТ. Восстановленные тяжелые цепи перемещались в виде одиночной полосы при молекулярной массе около 49 кДа и были зондированы αFc. (1C) Верхняя панель: Невосстанавливающий вестерн-блоттинг для клеток 64B4, несущих pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1), pBR-mSTII1-mPhoA1-5D5 (mSTII1, mPhoA1) и pBR-mSTII1-mMalE1-5D5 (mSTII1, mMalE1). Средняя панель: Общее количество тяжелых цепей в 1 ОП550 осадке тех же образцов восстанавливали ДТТ и анализировали при помощи вестерн-блоттинга. Нижняя панель: Периплазматические экстракты из тех же образцов восстанавливали и анализировали при помощи вестерн-блоттинга с антителом αFc. Все экспрессионные плазмиды называли так, чтобы TIR-вариант первого сигнального пептида соответствовал легкой цепи, а TIR-вариант второго сигнального пептида соответствовал тяжелой цепи. Например, mSTII1-bSTII1-5D5 обозначает сигнальную последовательность STII с TIR 1 для легкой цепи и расположенным выше рестрикционным сайтом mluI; bSTII1 обозначает сигнальную последовательность STII с TIR 1 для HC с расположенной выше сигнальной последовательностью bst; 5D5 соответствует названию антитела.
Фигуры 2A и 2B: иллюстрируют влияние применения разных сигнальных пептидов на клеточную локализацию тяжелых цепей согласно данным электронной микроскопии с иммунным окрашиванием золотом. Клетки A64B4, несущие конструкции, экспрессирующие различные сигнальные пептиды, культивировали в качалочных колбах в течение 24 ч. Образцы, соответствующие конечной точке, фиксировали, заливали и делали криосрезы. Криосрезы зондировали при помощи HRP-конъюгированного антитела αFc и конъюгированного с золотом вторичного антитела αHRP. Иммуноокрашенные образцы визуализировали при помощи трансмиссионного электронного микроскопа (ТЭМ). (2A) иллюстрирует клетки A64B4, несущие pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1). (2B) иллюстрирует клетки A64B4, несущие pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1). Черные стрелки указывают на периплазматическое пространство. Маленькие черные стрелки указывают на сигналы от иммунного окрашивания золотом.
Фигуры 3A и 3B: иллюстрируют влияние гидрофобности сигнального пептида на накопление тяжелых цепей в периплазме. (3A) иллюстрирует уровни переплазматических растворимых тяжелых цепей в отсутствие легких цепей. Периплазматические экстракты из клеток 64B4, несущих pBR-bDsbA1-5D5HC (bDsbA1), pBR-bDsbA1 L11I-5D5HC (bDsbA1 L11I), pBR-bDsbA1 L11S-5D5HC (bDsbA1 L11S), pBR-bSTII1-5D5HC (bSTII1), pBR-bSTII1 S11L кодон1-5D5HC (bSTII1 кодон1) или pBR-bSTII1 S11L кодон2-5D5HC (bSTII1 кодон2), анализировали при помощи вестерн-блоттинга. Восстановленные тяжелые цепи перемещались при молекулярной массе около 49 кДа и были зондированы антителом αFc. (3B) иллюстрирует уровни переплазматических растворимых тяжелых цепей с коэкспрессией легких цепей. Периплазматические экстракты из 64B4, несущих pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bMalE1-5D5 (mSTII1, bMalE1), pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1), pBR-mSTII1-bPhoA1-5D5 (mSTII1, bPhoA1), pBR-mSTII1-bDsbA1 L11I-5D5 (mSTII1, bDsbA1 L11I), pBR-mSTII1-bDsbA1 L11S-5D5 (mSTII1, bDsbA1 L11S), pBR-mSTII1-bSTII1 S11L-5D5 (mSTII1, bSTII1 S11L) или pBR-mSTII1-bSTII1 S11I-5D5 (mSTII1, bSTII1 S11I), анализировали при помощи вестерн-блоттинга. Восстановленные тяжелые цепи выявляли при помощи антитела αFc.
Фигуры 4A и 4B: иллюстрируют влияние гидрофобности сигнального пептида на уровень полноразмерного 5D5. Верхние панели: полные клеточные лизаты из 64B4, содержащих (A) pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bDsbA1 L11I-5D5 (mSTII1, bDsbA1 L11I), pBR-mSTII1-bDsbA1 L11S-5D5 (mSTII1, bDsbA1 L11S), pBR-bDsbA1-5D5HC (bDsbA1) (B) pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-bDsbA1-5D5HC (bDsbA1), pBR-mSTII1-bSTII1 S11L-5D5 (mSTII1, bSTII1 S11L) или pBR-mSTII1-bSTII1 S11I-5D5 (mSTII1, bSTII1 S11I), анализировали при помощи невосстанавливающего ДСН-ПААГ геля, после чего проводили вестерн-блот-зондирование с антителом αFc. Указаны молекулы, содержащие тяжелые цепи. Нижние панели: общий белок тяжелых цепей в 1 ОП550 осадке из тех же образцов, что и на верхних панелях, анализировали при помощи невосстанавливающего ДСН-ПААГ геля и вестерн-блоттинга. Тяжелые цепи, перемещающиеся при 49 кДа, зондировали антителом αFc.
Фигура 5: иллюстрирует влияние сигнального пептида и его гидрофобности на клеточную локализацию телец включения. Соответствующие конечной точке образцы из культур 64B4, несущих pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bMalE1-5D5 (mSTII1, bMalE1), pBR-mSTII1-bPhoA1-5D5 (mSTII1, bPhoA1), pBR-mSTII1-bDsbA1-5D5 (mSTII1, bDsbA1), pBR-mSTII1-bDsbA1 L11S-5D5 (mSTII1, bDsbA1 L11S), pBR-mSTII1-bSTII1 S11L-5D5 (mSTII1, bSTII1 S11L), pBR-mSTII1-mMalE1-5D5 (mSTII1, mMalE1) или pBR-mSTII1-mPhoA1-5D5 (mSTII1, mPhoA1), фиксировали, заливали и делали ультратонкие срезы и визуализировали под ТЭМ-микроскопом. Черные стрелки указывают на периплазматическое пространство, а маленькие черные стрелки указывают на тельца включения.
Фигура 6: иллюстрирует вестерн-блоттинг-анализ для мутаций Ser/Tyr в С-концевой области. Верхняя панель: полные клеточные лизаты из клеток 64B4, содержащих pBR-mSTII1-bSTII1-5D5 (mSTII1, bSTII1), pBR-mSTII1-bSTII1 Y22S-5D5 (mSTII1, bSTII1 Y22S), pBR-bDsbA1-5D5HC (bDsbA1) или pBR-bDsbA1 S18Y-5D5HC (bDsbA1 S18Y), анализировали при помощи невосстанавливающего вестерн-блот-зондирования αFc. Указаны молекулы, содержащие тяжелые цепи, включая полноразмерное 5D5, тяжелая-тяжелая-легкая цепь и тяжелая-легкая цепь. Нижняя панель: общий белок в 1 ОП550 осадке восстанавливали при помощи ДТТ и анализировали при помощи вестерн-блоттинга. Восстановленные тяжелые цепи перемещались при ~49 кДа.
Фигуры 7A и 7B: иллюстрируют влияние гидрофобности сигнального пептида на уровни полноразмерного антитела и периплазматические уровни растворимых тяжелых цепей для mAb1 и mAb2. (7A) Верхняя панель иллюстрирует полные клеточные лизаты из 64B4, содержащих pBR-mSTII1-bSTII1-mAb1 (mSTII1, bSTII1), pBR-mSTII1-bSTII1 S11L-mAb1 (mSTII1, bSTII1 S11L), pBR-mSTII1-bDsbA1-mAb1 (mSTII1, bDsbA1) и pBR-mSTII1-bDsbA1 L11S-mAb1 (mSTII1, bDsbA1 L11S), которые анализировали при помощи невосстанавливающего ДСН-ПААГ геля, после чего проводили вестерн-блот-зондирование с антителом αFc. Содержащие тяжелые цепи молекулы указаны стрелками. Средняя панель: иллюстрирует периплазматические белки из тех же образцов, которые были выделены, восстановлены при помощи ДТТ и проанализированы при помощи ДСН-ПААГ геля с последующим вестерн-блот-зондированием с антителом αFc. Нижняя панель иллюстрирует общий белок тяжелой цепи в 1 ОП550 осадках из тех же образцов, которые были восстановлены при помощи ДТТ и проанализированы при помощи вестерн-блоттинга. (7B) Верхняя панель иллюстрирует полные клеточные лизаты из 64B4, содержащих pBR-mSTII1-bSTII1-mAb2 (mSTII1, bSTII1), pBR-mSTII1-bSTII1 S11L-mAb2 (mSTII1, bSTII1 S11L), pBR-mSTII1-bDsbA1-mAb2 (mSTII1, bDsbA1) и pBR-mSTII1-bDsbA1 L11S-mAb2 (mSTII1, bDsbA1 L11S), которые анализировали при помощи невосстанавливающего ДСН-ПААГ геля, после чего проводили вестерн-блот-зондирование с антителом αFc. Содержащие тяжелые цепи молекулы указаны стрелками. Средняя панель иллюстрирует периплазматические белки из тех же образцов, которые были выделены, восстановлены при помощи ДТТ и проанализированы при помощи ДСН-ПААГ геля с последующим вестерн-блот-зондированием с антителом αFc. Нижняя панель иллюстрирует общий белок тяжелой цепи в 1 ОП550 осадках из тех же образцов, которые были восстановлены при помощи ДТТ и проанализированы при помощи вестерн-блоттинга.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие методы
Методы и процедуры, которые описаны или на которые ссылается данный документ, в общем случае являются хорошо известными и обычно применяемыми с использованием традиционной методологии среди специалистов в данной области техники, например, это широко применяемые методологии, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al. eds., (2003)); серии METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, и ANIMAL CELL CULTURE (R. I. Freshney, ed. (1987)).
Определения
Подразумевается, что употребляемый в данном документе термин «вектор» относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. Одним из типов вектора является «плазмида», которая относится к кольцевой двухцепочечной ДНК-петле, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они внесены (например, бактериальные векторы, содержащие бактериальную точку начала репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут интегрироваться в геном клетки-хозяина после внесения в клетку-хозяина и, следовательно, реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называются в данном документе «рекомбинантными экспрессионными векторами» (или просто «рекомбинантными векторами»). В общем случае применяемые в технологии рекомбинантных ДНК экспрессионные векторы часто находятся в форме плазмид. В настоящем описании термины «плазмида» и «вектор» могут использоваться взаимозаменяемо, так как плазмида является наиболее часто применяемой формой вектора.
Подразумевается, что употребляемый в данном документе термин «цистрон» относится к генетическому элементу, в целом эквивалентному трансляционной единице, содержащей нуклеотидную последовательность, кодирующую полипептидную цепь, и смежные регуляторные области. «Смежные регуляторные области» включают, например, область инициации трансляции (TIR (от англ. «translational initiation region»); согласно приведенному ниже определению) и область терминации трансляции.
«Полицистронный» экспрессионный вектор относится к одиночному вектору, который содержит и экспрессирует несколько цистронов под управлением одного единственного промотора. Типовым примером полицистронного вектора является «дицистронный» вектор, который содержит и экспрессирует два разных полипептида под управлением одного промотора. После экспрессии дицистронного или полицистронного вектора несколько генов транскрибируются сначала в виде одной транскрипционной единицы, а затем транслируются по отдельности.
Экспрессионный вектор «с отдельными цистронами» в соответствии с настоящим изобретением относится к одиночному вектору, содержащему по меньшей мере две отдельных пары промотор-цистрон, при этом каждый цистрон находится под управлением своего промотора. После экспрессии экспрессионного вектора с отдельными цистронами процессы как транскрипции, так и трансляции разных генов происходят отдельно и независимо.
В контексте данного документа «область инициации трансляции» или TIR, или последовательность инициации трансляции относится к области нуклеиновой кислоты, обеспечивающей эффективность инициации трансляции представляющего интерес гена. В общем случае TIR в пределах конкретного цистрона содержит участок связывания рибосомы (УСР) и последовательности 5' и 3' к УСР. УСР содержит по определению, как минимум, область Шайна-Дальгарно и стартовый кодон (AUG). Соответственно, TIR также включает по меньшей мере часть нуклеотидной последовательности, предназначенной для трансляции. Предпочтительно TIR содержит секреторную сигнальную последовательность, кодирующую сигнальный пептид, которая предшествует последовательности, кодирующей легкую или тяжелую цепь, в цистроне. Вариант TIR включает варианты последовательностей (в частности, замены) с TIR-областью, которая изменяет свойства TIR, такие как эффективность трансляции, определенная ниже. Предпочтительно вариант TIR согласно изобретению содержит замещения последовательностей приблизительно в пределах первых 2-14, предпочтительно - приблизительно 4-12, более предпочтительно - приблизительно 6 кодонов секреторной сигнальной последовательности, которая предшествует последовательности, кодирующей легкую или тяжелую цепь, в цистроне.
Употребляемый в данном документе термин «эффективность трансляции» относится к измерению количества секретируемого полипептида в контрольной системе, в которой один или более вариантов TIR применяют для управления секрецией полипептида, а результаты сравнивают с TIR дикого типа или каким-либо другим контролем в тех же культуральных и экспериментальных условиях.
«Сигнальный пептид» (также называемый «сигнальной последовательностью») относится к короткому пептиду, который можно использовать для направления представляющего интерес свежесинтезированного белка через клеточную мембрану, обычно внутреннюю мембрану или как внутреннюю, так и внешнюю мембраны прокариотов. Сигнальный пептид, кодируемый секреторной сигнальной последовательностью, может быть эндогенным для клеток-хозяев или может быть экзогенным, включая сигнальные пептиды, нативные для предназначенного для экспрессии полипептида. Сигнальный пептид, как правило, находится в амино-конце предназначенного для экспрессии полипептида и, как правило, удаляется ферментативным путем на этапе между биосинтезом и секрецией полипептида из цитоплазмы. Таким образом, сигнальный пептид обычно не присутствует в зрелом белковом продукте. Сигнальные пептиды (например, прокариотические, например, сигнальные пептиды E. coli) обычно состоят из трех разных областей: N-концевой области, которая, как правило, содержит по меньшей мере 1 или 2 положительно заряженных аминокислотных остатка, гидрофобной коровой области, называемой Н-областью (также называемой Н-доменом), и С-концевой области, распознаваемой сигнальной пептидазой. Специалисту в данной области техники известно, как определить N-концевую область, Н-область и С-концевую область заданного сигнального пептида.
Под «средней гидрофобностью» пептида (или части пептида) подразумевается средняя гидрофобность, рассчитанная по формуле: средняя гидрофобность пептида (или части пептида)=общая (суммарная) гидрофобность пептида (или части пептида) /число аминокислот в пептиде (или части пептида). «Общую» или «суммарную» гидрофобность рассчитывают путем (a) приписывания каждой аминокислоте в пептиде (или части пептида) величины нормированной консенсусной гидрофобности в соответствии с Eisenberg, D. et al., J Mol Biol (1984) 179:125-142. Таблица I (страница 126), затем сложения величин нормированной консенсусной гидрофобности для аминокислот в пептиде (или части пептида). В некоторых вариантах реализации изобретения среднюю гидрофобность рассчитывают для Н-домена сигнального пептида.
«Функционально связанный» относится к контактному положению двух или более компонентов, причем описанные таким образом компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим образом. Например, промотор функционально связан с кодирующей последовательностью, если он действует в цис-положении, регулируя или модулируя транскрипцию связанной последовательности. В общем случае, но не обязательно, «функционально связанные» последовательности ДНК являются смежными и, в случае необходимости соединения двух кодирующих белок областей или в случае секреторной лидерной последовательности, смежными и находящимися в рамке считывания. При этом, хотя функционально связанный промотор в общем случае расположен выше кодирующей последовательности, он не обязательно является смежным с ней. Функционально связанные энхансеры могут быть расположены выше, в пределах или ниже кодирующих последовательностей и на значительном расстоянии от промотора. Связывание осуществляют известными в данной области техники рекомбинантными методами, например, при помощи методологии ПЦР, путем отжига или путем лигирования в удобные рестрикционные сайты. Если удобный рестрикционный сайт отсутствует, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с традиционной практикой.
В контексте данного документа «регуляторные элементы» относятся к нуклеотидным последовательностям, находящимся в цис-положении, необходимым для транскрипции и трансляции полинуклеотида, кодирующего гетерологичный полипептид, в полипептиды. Транскрипционные регуляторные элементы обычно включают промотор, расположенный 5' от предназначенной для экспрессии генной последовательности, участки инициации и терминации транскрипции и сигнальную последовательность полиаденилирования. Термин «сайт инициации транскрипции» относится к нуклеиновой кислоте в конструкции, соответствующей первой нуклеиновой кислоте, вставленной в первичный транскрипт, т.е. предшественник мРНК; сайт инициации транскрипции может перекрываться с промоторными последовательностями.
«Промотор» относится к полинуклеотидной последовательности, которая регулирует транскрипцию гена или последовательности, с которыми он функционально связан. Промотор включает сигналы для связывания РНК-полимеразы и инициации транскрипции. Применяемые промоторы функциональны в том клеточном типе клеток-хозяев, в котором предполагается экспрессия выбранной последовательности. В данной области техники хорошо известно (и определено в базах данных, таких как GenBank) большое число промоторов, включая конститутивные, индуцибельные и репрессируемые промоторы из множества разных источников, которые доступны в виде или в составе клонированных полинуклеотидов (например, из депозитариев, таких как ATCC, а также других коммерческих или частных источников). В случае индуцибельных промоторов активность промотора повышается или понижается в ответ на сигнал.
Подразумевается, что употребляемый в данном документе термин «клетка-хозяин» (или «рекомбинантная клетка-хозяин») относится к клетке, которая была генетически изменена или может быть генетически изменена путем внесения экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. Следует понимать, что такие термины относятся не только к конкретной рассматриваемой клетке, но и к потомству такой клетки. Так как у последующих поколений вследствие мутации или влияния окружающей среды могут возникать определенные модификации, такое потомство может по факту не быть идентичным родительской клетке, но при этом входит в понятие употребляемого в данном документе термина «клетка-хозяин».
«Выделенный» полипептид (например, антитело) представляет собой полипептид, который был выявлен и отделен и/или очищен от компонентов его естественного окружения. Контаминантные компоненты естественного окружения представляют собой материалы, которые препятствовали бы диагностическому или терапевтическому применению антитела, и которые могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах реализации изобретения полипептид будет очищен (1) более чем до 95% по массе полипептида согласно определению по методу Лоури, и, наиболее предпочтительно, более чем до 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности при помощи секвенатора с вращающимся стаканом, или (3) до гомогенности по ДСН-ПААГ (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) в восстанавливающих или невосстанавливающих условиях с применением голубого кумасси или, предпочтительно, серебряного красителя. Выделенный полипептид включает полипептид in situ в рекомбинантных клетках, если отсутствует по меньшей мере один компонент естественного окружения полипептида. Однако, как правило, выделенный полипептид получают при помощи по меньшей мере одного этапа очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая была выявлена и отделена по меньшей мере от одной контаминантной молекулы нуклеиновой кислоты, с которой она обычно связана в естественном источнике нуклеиновой кислоты. Выделенная молекула нуклеиновой кислоты отличается от той, что находится в естественной форме или окружении.
Взаимозаменяемо употребляемые в данном документе термины «полинуклеотид» или «нуклеиновая кислота» относятся к полимерам из нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги, или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой или путем синтетической реакции. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. В случае наличия, модификации в нуклеотидную структуру можно вносить до или после сборки полимера. Последовательность нуклеотидов может прерываться отличными от нуклеотидов компонентами. Полинуклеотид можно дополнительно модифицировать после синтеза, например, путем конъюгации с меткой. Другие типы модификаций включают, например, «кэпы», замещение одного или более нуклеотидов природного происхождения аналогом, межнуклеотидные модификации, такие как, например, модификации с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и с заряженными связями (например, тиофосфаты, дитиофосфаты и т.д.), модификации, включающие подвешенные компоненты, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), модификации интеркаляторами (например, акридином, псораленом и т.д.), модификации, включающие хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), модификации, включающие алкиляторы, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т.д.), а также немодифицированные формы полинуклеотида(ов). Кроме того, любая из гидроксильных групп, обычно присутствующих в сахарах, может быть замещена, например, фосфонатными группами, фосфатными группами, защищена стандартными защитными группами или активирована для получения дополнительных связей для дополнительных нуклеотидов, или может быть конъюгирована к твердым или полутвердым подложкам. 5' и 3' концевая OH-группа может быть фосфорилирована или замещена аминами или органическими кэпирующими группами компонентов из от 1 до 20 атомов углерода Другие гидроксилы также могут быть дериватизированы до стандартных защитных групп. Полинуклеотиды также могут содержать функционально аналогичные формы сахаров рибозы или дезоксирибозы, которые общеизвестны в данной области техники, включая, например 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидорибозу, карбоциклические сахарные аналоги, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и основные нуклеозидные агалоги, такие как метилрибозид. Одна или более фосфодиэфирных связей могут быть замещены альтернативными связующими группами. Эти альтернативные связующие группы включают, но не ограничиваются этим, варианты реализации изобретения, в которых фосфат замещен P(O)S («тиоатом»), P(S)S («дитиоатом»), (O)NR2 («амидатом»), P(O)R, P(O)OR', CO или CH2 («формацеталем»), где каждый R или R' независимо представляет собой Н или замещенный или незамещенный алкил (1-20 C), необязательно содержащий эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралкил. Не все связи в полинуклеотиде должны быть идентичными. Вышеприведенное описание применимо ко всем полинуклеотидам, перечисленным в данном документе, включая РНК и ДНК.
В контексте данного документа «олигонуклеотид» в общем случае относится к коротким, в общем случае одноцепочечным, в общем случае синтетическим полинуклеотидам, которые в общем случае, но не обязательно, имеют длину менее чем около 200 нуклеотидов. Термины «олигонуклеотид» и «полинуклеотид» не являются взаимоисключающими. Приведенное выше описание полинуклеотидов в равной степени и в полном объеме применимо к олигонуклеотидам.
В контексте данного документа «полипептид» в общем случае относится к полипептидам и белкам из любого источника клеток, содержащим более чем около десяти аминокислот. «Гетерологичные» полипептиды - это полипептиды, чужеродные для применяемой клетки-хозяина, такие как человеческий белок, вырабатываемый E. coli. Хотя гетерологичный полипептид может быть прокариотическим или эукариотическим, предпочтительно он является эукариотическим, более предпочтительно - полипептидом млекопитающего, и наиболее предпочтительно - полипептидом человека. Предпочтительно он является рекомбинантно полученным или рекомбинантным полипептидом. «Гетерологичные» полипептиды - это полипептиды, чужеродные для применяемой клетки-хозяина, такие как человеческий белок, вырабатываемый E. coli. Хотя гетерологичный полипептид может быть прокариотическим или эукариотическим, предпочтительно он является эукариотическим, более предпочтительно - полипептидом млекопитающего, и наиболее предпочтительно - полипептидом человека. Предпочтительно он является рекомбинантно полученным или рекомбинантным полипептидом.
Примеры полипептидов млекопитающих включают молекулы, такие как, например, ренин, гормон роста, включая человеческий гормон роста; бычий гормон роста; фактор, высвобождающий гормон роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; 1-антитрипсин; А-цепь инсулина; В-цепь инсулина; проинсулин; тромбопоэтин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VIIIC, фактор IX, тканевой фактор и фактор Виллебранда; противосвертывающие факторы, такие как протеин С; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или активатор плазминогена человеческой мочи или тканевой активатор плазминогена (t-PA) и его варианты, такие как RETEVASE™ и TNKASE™; бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухолей альфа и бета; антитела к домену(ам) ErbB2, такие как 2C4 (WO 01/00245; гибридома ATCC HB-12697), которое связывается с областью во внеклеточном домене ErbB2 (например, любым одним или более остатками в области приблизительно от остатка 22 приблизительно до остатка 584 ErbB2, включительно), энкефалиназа; сывороточный альбумин, такой как человеческий сывороточный альбумин; Мюллерова ингибирующая субстанция; А-цепь релаксина; В-цепь релаксина; прорелаксин; мышиный гонадотропин-ассоциированный пептид; микробный белок, такой как бета-лактамаза; ДНКаза; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы гормонов или факторов роста; интегрин; протеин A или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор головного мозга (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов, такой как NGF; кардиотрофины (факторы гипертрофии сердца), такие как кардиотрофин-1 (CT-1); тромбоцитарный фактор роста (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулин-подобный фактор роста-I и -II (IGF-I и IGF-II); des(I-3)-IGF-I (IGF-I головного мозга), белки, связывающие инсулин-подобный фактор роста; белки CD, такие как CD-3, CD-4, CD-8 и CD-19; эритропоэтин; остеиндуктивные факторы; иммунотоксины; костный морфогенетический белок (BMP); интерферон, такой как интерферон-альфа, -бета и -гамма; сывороточный альбумин, такой как человеческий сывороточный альбумин (ЧСА) или бычий сывороточный альбумин (БСА); колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-10; анти-HER-2 антитело; лиганд Apo2; супероксиддисмутаза; рецепторы Т-клеток; поверхностные белки мембраны; фактор ускорения распада; вирусный антиген, такой как, например, часть оболочки СПИД; транспортные белки; хоминг-рецепторы; аддрессины; регуляторные белки; антитела; и фрагменты любого из вышеперечисленных полипептидов.
В некоторых вариантах реализации изобретения полипептиды согласно данному документу включают человеческий сывороточный альбумин (ЧСА), 2C4, тканевой фактор, антитканевой фактор, анти-CD20, анти-HER-2, херегулин, анти-IgE, анти-CD11a, анти-CD18, VEGF, а также рецепторы и антитела к нему, такие как rhuFab V2 и AVASTIN™, гормон роста и его варианты, такие как hGH, рецепторы гормона роста, белок, высвобождающий гормон роста (GHRP), LIV-1 (EP 1,263,780), TRAIL, фактор некроза опухолей (ФНО) и антитела к нему, рецептор ФНО и родственные антитела, ФНО-рецептор-IgG, факторы, ассоциированные с рецептором ФНО (TRAF), и их ингибиторы, фактор VIII, В-домен фактора VIII, интерфероны, такие как интерферон-гамма, трансформирующие факторы роста (TGF), такие как TGF-бета, анти-TGF, такие как анти-TGF-бета, активин, ингибин, анти-активин, анти-ингибин, тканевые активаторы плазминогена и их варианты, такие как t-PA, RETEPLASE™ и TNKаза, анти-Fas антитела, лиганд Apo-2; ингибитор лиганда Apo-2; рецептор Apo-2, Apo-3, апоптотические факторы, Ced-4, DcR3, рецептор смерти и агонистические антитела (DR4, DR5), лимфотоксин (LT), пролактин, рецептор пролактина, белки SOB, WISP (wnt-индуцируемые секретируемые белки), нейротоксин-3 (NT-3), фактор роста нервов (NGF) и анти-NGF, ДНКаза, антиген гепатита, антиген простого герпеса, лептин, инсулин-подобные факторы роста (IGFs), такие как IGF-1 и IGF-2, и их связывающие белки и рецепторы, такие как IGFBP-1-IGFBP-6, инсулин, факторы роста фибробластов (FGF), такие как FGF-17, белок толл, лиганды TIE, CD40 и анти-CD40, иммуноадгезины, субтилизин, фактор роста гепатоцитов (HGF), тромбопоэтин (TPO), интерлейкины, такие как ИЛ-2, ИЛ-12, ИЛ-17, ИЛ-22, ИЛ-8, ИЛ-9, и антитела к ним и простат-специфический раковый антиген (PSCA).
В особенности предпочтительными полипептидами являются рекомбинантные полипептиды, в некоторых вариантах реализации изобретения - антитела, которые включают моноклональные антитела и гуманизированные антитела. Такие антитела могут быть полноразмерными антителами или фрагментами антител. В некоторых вариантах реализации изобретения эти антитела являются человеческими или гуманизированными антителами. В некоторых вариантах реализации изобретения антитело представляет собой анти-c-met, анти-IgE, анти-CD18, анти-VEGF, антитканевой фактор, 2C4, анти-Her-2, анти-CD20, анти-CD40 или анти-CD11a антитело. Фрагменты антител, включенные в определение полипептида, в некоторых вариантах реализации изобретения содержат легкую цепь, в некоторых вариантах реализации изобретения - легкую цепь каппа. Такие типовые фрагменты включают, например, Fab, Fab', F(ab')2 или продукт слияния F(ab')2-лейциновая молния (LZ) и антитело с одним плечом.
«Экспрессия» белка относится к конвертации информации, закодированной в гене, в матричную РНК (мРНК), а затем в белок.
«Иммуноконъюгат» (взаимозаменяемо называемый «конъюгатом антитело-лекарственный препарат» или «ADC») обозначает антитело, конъюгированное с одним или более цитотоксическими агентами, такими как химиотерапевтический агент, лекарственный препарат, агент, ингибирующий рост, токсин (например, белковый токсин, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат).
«Блокирующее» антитело или «антагонистическое» антитело - это такое антитело, которые ингибирует или снижает биологическую активность антигена, с которым оно связывается. В некоторых вариантах реализации изобретения блокирующие антитела или антагонистические антитела полностью блокируют биологическую активность антигена.
В контексте данного документа «агонистическое антитело» представляет собой антитело, которое имитирует по меньшей мере один вид функциональной активности представляющего интерес полипептида (например, HGF).
«Аффинность связывания» в общем случае относится к силе суммарных нековалентных взаимодействий между одним связывающим участком молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иное, в контексте данного документа «аффинность связывания» относится к характерной аффинности связывания, которая отображает 1:1 взаимодействие между участниками пары связывания (например, антителом и антигеном). Аффинность молекулы Х в отношении ее партнера Y можно в общем случае представить при помощи константы диссоциации (Kд). Желательно, чтобы Kд составляла 1×10-7, 1×10-8, 5×10-8, 1×10-9, 3×10-9, 5×10-9 или даже 1×10-10 или менее. Аффинность можно определить при помощи общих методов, известных в данной области техники, включая описанные в данном документе. Низкоаффинные антитела в общем случае медленно связывают антиген и имеют тенденцию к легкой диссоциации, в то время как высокоаффинные антитела в общем случае быстрее связывают антиген и имеют тенденцию оставаться дольше в связанном состоянии. В данной области техники известно множество способов определения аффинности, любой из которых можно использовать в целях настоящего изобретения. Далее описаны конкретные иллюстративные варианты реализации изобретения. В одном варианте реализации изобретения «Kд» или «значение Kд» в соответствии с данным изобретением определяют при помощи анализа связывания с радиомеченым антигеном (РИА), проводимого с Fab-версией представляющего интерес антитела и его антигеном, как описано для следующего метода анализа, в котором определяют аффинность связывания Fab с антигеном в растворе путем уравновешивания Fab минимальной концентрацией (125I)-меченого антигена в присутствии серии титров немеченого антигена, затем проводят захват связанного антигена на покрытом анти-Fab планшете (Chen, et al., (1999) J. Mol. Biol. 293:865-881). Чтобы создать условия для анализа, микротитровальные планшеты (Dynex) покрывают в течение ночи 5 мкг/мл захватывающего анти-Fab антитела (Cappel Labs) в 50 мM карбоната натрия (pH 9,6), после чего блокируют 2% (м/об) бычьим сывороточным альбумином в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В неадсорбирующем планшете (Nunc #269620), смешивают 100 пМ или 26 пМ [125I]-антигена с серийными разведениями представляющего интерес Fab (например, в соответствии с оценкой анти-VEGF антитела Fab-12 в Presta et al., (1997) Cancer Res. 57:4593-4599). Затем представляющий интерес Fab инкубируют в течение ночи; при этом инкубация может продолжаться и более долгий период (например, 65 часов), чтобы гарантировать достижение равновесия. После этого смеси переносят на захватывающий планшет для инкубации при комнатной температуре (например, в течение одного часа). Затем раствор удаляют, а планшет промывают восемь раз 0,1% Твин-20 в PBS. После высыхания планшетов добавляют 150 мкл/лунка сцинтиллятора (MicroScint-20; Packard), а планшеты считывают на гамма-счетчике Topcount (Packard) в течение десяти минут. Концентрации каждого Fab, которые дают менее чем или 20% от максимального связывания, отбирают для применения в конкурентном анализе связывания. В соответствии с другим вариантом реализации изобретения Kд или значение Kд определяют методом поверхностного плазмонного резонанса, используя BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами с иммобилизованным антигеном CM5 при ~10 единицах ответа (ЕО). Вкратце, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимид гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетата натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инжекцией при скорости потока 5 мкл/минута до достижения приблизительно 10 единиц ответа (ЕО) связанного белка. После инжекции антигена инжектируют 1 M этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) инжектируют в PBS с 0,05% Твин 20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. В некоторых вариантах реализации изобретения используют следующие модификации в методе поверхностного плазмонного резонанса: антитело иммобилизуют на биосенсорных чипах CM5 до достижения приблизительно 400 ЕО, а для кинетических измерений инжектируют двукратные серийные разведения белка-мишени в буфер PBST при 25°C при скорости потока приблизительно 30 мкл/минута. Скорости ассоциации (kасс) и скорости диссоциации (kдисс) рассчитывают, используя простую взаимно однозначную модель связывания Ленгмюра (программное обеспечение для оценки BIAcore, версия 3.2) путем одновременной подгонки сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kд) рассчитывают как соотношение kдисс/kасс. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. Если скорость ассоциации превышает 106 M-1 с-1 согласно данным вышеописанного метода поверхностного плазмонного резонанса, скорость ассоциации можно определить методом гашения флуоресценции, в котором определяют повышение или снижение интенсивности флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 25°C для 20 нМ анти-антиген антитела (в форме Fab) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена, при помощи спектрометра, такого как спектрофотометр с остановкой потока (Aviv Instruments) или спектрофотометр 8000-series SLM-Aminco (ThermoSpectronic) с перемешиваемой кюветой.
«Скорость ассоциации» или «kасс» в соответствии с данным изобретением также можно определить тем же описанным выше методом поверхностного плазмонного резонанса, используя BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами с иммобилизованным антигеном CM5 при ~10 единицах ответа (ЕО). Вкратце, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимид гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетата натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инжекцией при скорости потока 5 мкл/минута до достижения приблизительно 10 единиц ответа (ЕО) связанного белка. После инжекции антигена инжектируют 1 M этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) инжектируют в PBS с 0,05% Твин 20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. В некоторых вариантах реализации изобретения используют следующие модификации в методе поверхностного плазмонного резонанса: антитело иммобилизуют на биосенсорных чипах CM5 до достижения приблизительно 400 ЕО, а для кинетических измерений инжектируют двукратные серийные разведения белка-мишени в буфер PBST при 25°C при скорости потока приблизительно 30 мкл/минута. Скорости ассоциации (kасс) и скорости диссоциации (kдисс) рассчитывают, используя простую взаимно однозначную модель связывания Ленгмюра (программное обеспечение для оценки BIAcore, версия 3.2) путем одновременной подгонки сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kд) рассчитывают как соотношение kдисс/kасс. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293:865-881. При этом, если скорость ассоциации превышает 106 M-1 с-1 согласно данным вышеописанного метода поверхностного плазмонного резонанса, скорость ассоциации предпочтительно определяют методом гашения флуоресценции, в котором определяют повышение или снижение интенсивности флуоресценции (возбуждение=295 нм; испускание=340 нм, полоса пропускания 16 нм) при 25°C для 20 нМ анти-антиген антитела (в форме Fab) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена, при помощи спектрометра, такого как спектрофотометр с остановкой потока (Aviv Instruments) или спектрофотометр 8000-series SLM-Aminco (ThermoSpectronic) с перемешиваемой кюветой.
«Оголенное антитело» представляет собой антитело, которое не конъюгировано с гетерологичной молекулой, такой как цитотоксический компонент или радиометка.
Антитело, имеющее «биологическую характеристику» указанного антитела, является антителом, которое обладает одной или более биологическими характеристиками того антитела, которые отличают его от других антител, которые связываются с тем же антигеном.
С целью проведения скрининга в отношении антител, которые связываются с эпитопом на антигене, связываемым представляющим интерес антителом, можно проводить стандартный анализ перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988).
Чтобы повысить время полужизни антител или полипептида, содержащего аминокислотные последовательности согласно данному изобретению, можно присоединить к антителу (в особенности к фрагменту антитела) эпитоп связывания рецептора реутилизации, как описано, например, в патенте США 5739277. Например, молекулу нуклеиновой кислоты, кодирующую эпитоп связывания рецептора реутилизации, можно связать в рамке с нуклеиновой кислотой, кодирующей полипептидную последовательность согласно данному изобретению, так, чтобы слитый белок, экспрессируемый сконструированной молекулой нуклеиновой кислоты, содержал эпитоп связывания рецептора реутилизации и полипептидную последовательность согласно данному изобретению. Употребляемый в данном документе термин «эпитоп связывания рецептора реутилизации» относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за повышение in vivo сывороточного времени полужизни молекулы IgG (например, Ghetie et al., Ann. Rev. Immunol. 18:739-766 (2000), Таблица 1). Антитела с заменами в Fc-области и повышенным сывороточным временем полужизни также описаны в WO00/42072, WO 02/060919; Shields et al., J. Biol. Chem. 276:6591-6604 (2001); Hinton, J. Biol. Chem. 279:6213-6216 (2004)). В другом варианте реализации изобретения сывороточное время полужизни также можно повысить путем, например, присоединения других полипептидных последовательностей. Например, антитела или другие полипептиды, применимые в способах согласно изобретению, можно присоединять к сывороточному альбумину или части сывороточного альбумина, которая связывается с рецептором FcRn, или связывающему сывороточный альбумин белку так, чтобы сывороточный альбумин связывался с антителом или полипептидом, например, такие полипептидные последовательности раскрыты в WO01/45746. В одном предпочтительном варианте реализации изобретения предназначенный для присоединения пептид сывороточного альбумина содержит аминокислотную последовательность DICLPRWGCLW (SEQ ID NO: 48). В другом варианте реализации изобретения время полужизни Fab повышают этими способами. См. также Dennis et al. J. Biol. Chem. 277:35035-35043 (2002) в отношении связывающих сывороточный альбумин пептидных последовательностей.
Под «фрагментом» подразумевается часть полипептида или молекулы нуклеиновой кислоты, которая предпочтительно содержит по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или более полной длины стандартной молекулы нуклеиновой кислоты или полипептида. Фрагмент может содержать 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100, 200, 300, 400, 500, 600 или более нуклеотидов или 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 120, 140, 160, 180, 190, 200 аминокислот или более.
Термин «антитело» употребляется в данном документе в самом широком смысле и включает различные структуры антител, включая, но не ограничиваясь этим, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител в той мере, пока они демонстрируют необходимую антигенсвязывающую активность.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются этим, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела; линейные антитела; одноцепочечные молекулы антител (например, scFv); и мультиспецифические антитела, образуемые фрагментами антител.
«Антитело, которое связывается с тем же эпитопом», что и стандартное антитело, относится к антителу, которое блокирует связывание стандартного антитела с антигеном в конкурентном анализе на 50% или более, и, наоборот, стандартное антитело блокирует связывание антитела с антигеном в конкурентном анализе на 50% или более. В данном документе предложен типовой конкурентный анализ.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остаток тяжелой и/или легкой цепи получен из другого источника или вида.
«Класс» антитела относится к типу константного домена или константной области его тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них могут быть дополнительно поделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Термин «Fc-область» употребляется в данном документе для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Данный термин включает нативные последовательности Fc-области и вариантные Fc-области. В одном варианте реализации изобретения Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбокси-конца тяжелой цепи. При этом С-концевой лизин (Lys447) Fc-области может присутствовать или отсутствовать. Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой индексом EU, описанной в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена в общем случае состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в общем случае расположены в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактное антитело» и «цельное антитело» взаимозаменяемо употребляются в данном документе для обозначения антитела, имеющего структуру, в значительной степени сходную со структурой нативного антитела, или имеющего тяжелые цепи, которые содержат Fc-область согласно определению данного документа.
«Человеческое антитело» - это антитело, которое имеет аминокислотную последовательность, которая соответствует последовательности антитела, вырабатываемого человеком или клеткой человека, или полученного из нечеловеческого источника, в котором используется репертуар человеческих антител или других кодирующих человеческое антитело последовательностей. Из этого определения человеческого антитела специальным образом исключено гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
«Человеческая консенсусная каркасная область» -это каркасная область, в которой представлены наиболее часто встречающиеся аминокислотные остатки из набора каркасных последовательностей VL или VH человеческого иммуноглобулина. В общем случае набор последовательностей VL или VH человеческого иммуноглобулина относится к подгруппе последовательностей вариабельного домена. В общем случае подгруппа последовательностей представляет собой подгруппу согласно Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3. В одном варианте реализации изобретения в случае VL подгруппа представляет собой подгруппу каппа I согласно Kabat et al., выше. В одном варианте реализации изобретения в случае VH подгруппа представляет собой подгруппу III согласно Kabat et al., выше.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В определенных вариантах реализации изобретения гуманизированное антитело содержит практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют областям нечеловеческого антитела, а все или практически все FR соответствуют областям человеческого антитела. Гуманизированное антитело, необязательно, может содержать по меньшей мере часть константной области антитела, полученную из человеческого антитела. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подверглось гуманизации.
Употребляемый в данном документе термин «гипервариабельная область» или «HVR» относится к каждой из областей вариабельного домена антитела, последовательности которых являются гипервариабельными («определяющие комплементарность области» или «CDR») и/или образуют структурно определенные петли («гипервариабельные петли»), и/или содержат антиген-контактирующие остатки («антигенные контакты»). В общем случае антитела содержат шесть HVR: три в VH (H1, H2, H3) и три в VL (L1, L2, L3). Типовые HVR согласно данному документу включают:
(a) гипервариабельные петли, соответствующие аминокислотным остаткам 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2) и 96-101 (H3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(b) CDR, соответствующие аминокислотным остаткам 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (H3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991));
(c) антигенные контакты, соответствующие аминокислотным остаткам 27c-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (H2) и 93-101 (H3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(d) комбинации (a), (b) и/или (c), включая аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (H2), 93-102 (H3) и 94-102 (H3).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) нумеруются в данном документе в соответствии с Kabat et al., выше.
Термин «мультиспецифическое антитело» употребляется в самом широком смысле и, в частности, включает антитело, которое имеет полиэпитопную специфичность. Такие мультиспецифические антитела включают, но не ограничиваются этим, антитело, содержащее вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), при этом единица VHVL имеет полиэпитопную специфичность, антитела, содержащие два или более домена VL и VH, где каждая единица VHVL связывается с отдельным эпитопом, антитела, содержащие два или более одиночных вариабельных домена, где каждый одиночный вариабельный домен связывается с отдельным эпитопом, полноразмерные антитела, фрагменты антител, такие как Fab, Fv, dsFv, scFv, диатела, биспецифические диатела и триатела, фрагменты антител, которые были ковалентно или нековалентно связаны. «Полиэпитопная специфичность» относится к способности специфически связываться с двумя или более разными эпитопами на одной мишени или на разных мишенях. «Моноспецифичность» относится к способности связывать только один эпитоп. В соответствии с одним вариантом реализации изобретения мультиспецифическое антитело представляет собой антитело IgG, которое связывается с каждым эпитопом с аффинностью, составляющей от 5 мкМ до 0,001 пМ, от 3 мкМ до 0,001 пМ, от 1 мкМ до 0,001 пМ, от 0,5 мкМ до 0,001 пМ или от 0,1 мкМ до 0,001 пМ.
Термин «антитело с одним плечом» или «антитела с одним плечом» относится к антителу, которое содержит (1) вариабельный домен, соединенный пептидной связью с полипептидом, содержащим домен CH2, домен СН3 или домен CH2-CH3, и (2) второй домен CH2, CH3 или CH2-CH3, при этом вариабельный домен не соединен пептидной связью с полипептидом, содержащим второй домен CH2, CH3 или CH2-CH3. В одном варианте реализации изобретения антитело с одним плечом содержит три полипептида (1) первый полипептид, содержащий вариабельный домен (например, VH), CH1, CH2 и CH3, (2) второй полипептид, содержащий вариабельный домен (например, VL) и домен CL, и (3) третий полипептид, содержащий домен CH2 и CH3. В одном варианте реализации изобретения третий полипептид не содержит вариабельный домен. В другом варианте реализации изобретения антитело с одним плечом частично содержит шарнирную область, содержащую два остатка цистеина, которые образуют дисульфидные связи, соединяющие константные тяжелые цепи. В одном варианте реализации изобретения вариабельные домены антитела с одним плечом образуют антигенсвязывающую область. В другом варианте реализации изобретения вариабельный домен антитела с одним плечом представляет собой одиночный вариабельный домен, при этом каждый одиночный вариабельный домен представляет собой антигенсвязывающую область.
Упоминаемый в данном документе термин технология «выступ-во-впадину» или «KnH» относится к технологии управления спариванием двух полипептидов in vitro или in vivo путем внесения выпуклости (выступа) в один полипептид и углубления (впадины) в другой полипептид на поверхности контакта, где происходит их взаимодействие. Например, KnH вносили в поверхность связывания Fc:Fc, поверхности контакта CL:CH1 или поверхности контакта VH/VL антител (например, US2007/0178552, WO 96/027011, WO 98/050431 и Zhu et al. (1997) Protein Science 6:781-788). В особенности это целесообразно при управлении спариванием двух разных тяжелых цепей во время получения мультиспецифических антител. Например, мультиспецифические антитела, содержащие KnH в Fc-областях, могут дополнительно содержать одиночные вариабельные домены, связанные с каждой Fc-областью, или дополнительно содержать разные вариабельные домены тяжелой цепи, которые спариваются со сходными или отличными вариабельными доменами легкой цепи. Технологию KnH также можно применять для спаривания внеклеточных доменов двух разных рецепторов или каких-либо других полипептидных последовательностей, которые содержат разные распознающие мишень последовательности (например, включая аффитела, пептитела и другие продукты слияния Fc).
Расщепление антител папаином приводит к получению двух идентичных антигенсвязывающих фрагментов, называемых фрагментами «Fab», и остаточного «Fc»-фрагмента, обозначение которых отражает способность легко кристаллизироваться. Фрагмент Fab состоит из полной L-цепи вместе с вариабельным доменом Н-цепи (VH) и первым константным доменом тяжелой цепи (CH1). Обработка антитела пепсином приводит к получению крупного одиночного фрагмента F(ab')2, который приблизительно соответствует двум дисульфидно связанным фрагментам Fab, обладает дивалентной антигенсвязывающей активностью и способен перекрестно связывать антиген. Фрагменты Fab' отличаются от фрагментов Fab содержанием нескольких дополнительных остатков в карбокси-конце домена CH1, включая один или более цистеинов из шарнирной области антитела. Fab'-SH в данном документе обозначает Fab', в котором остаток(ки) цистеина константного домена несут свободные тиольные группы. Фрагменты антител F(ab')2 изначально были получены в виде пары фрагментов Fab' с шарнирными цистеинами между ними. Также известны другие химические сопряжения фрагментов антител.
Употребляемый в данном документе термин «иммуноадгезин» обозначает молекулы, в которых скомбинированы специфичность связывания гетерологичного белка («адгезин») и эффекторные функции константных доменов иммуноглобулина. Структурно иммуноадгезины содержат продукт слияния аминокислотной последовательности с необходимой специфичностью, при этом аминокислотная последовательность отличается от участка распознавания и связывания антигена антитела (т.е. является «гетерологичной» по сравнению с константной областью антитела), и последовательность константного домена иммуноглобулина (например, последовательность CH2 и/или CH3 IgG). Типовые последовательности адгезинов включают смежные аминокислотные последовательности, которые содержат часть рецептора или лиганда, которая связывается с представляющим интерес белком. Последовательности адгезинов также могут представлять собой последовательности, которые связывают представляющий интерес белок, но не являются последовательностями рецепторов или лигандов (например, последовательности адгезинов в пептидотелах). Такие полипептидные последовательности можно отбирать или выявлять различными способами, включая технологию фагового дисплея и высокопроизводительные методы сортинга. Последовательность константного домена иммуноглобулина в иммуноадгезине может быть получена из любого иммуноглобулина, такого как подтипы IgG-1, IgG-2, IgG-3 или IgG-4, IgA (включая IgA-1 и IgA-2), IgE, IgD и IgM.
«Шарнирная область» в контексте антитела или полуантитела в общем случае простирается по определению от Glu216 до Pro230 человеческого IgG1 (Burton, Molec. Immunol.22:161-206 (1985)). Шарнирные области других изотипов IgG можно выравнивать с последовательностями IgG1 путем размещения первого и последнего остатков цистеина, образующих S-S связи между тяжелыми цепями, в тех же позициях.
«Нижняя шарнирная область» Fc-области обычно определяется как группа остатков, расположенных непосредственно С-терминально по отношению к шарнирной области, т.е. остатки от 233 до 239 Fc-области. До настоящего изобретения связывание Fc-гамма-R в общем случае связывали с аминокислотными остатками в нижней шарнирной области Fc-области IgG.
«Домен СН2» Fc-области человеческого IgG обычно простирается приблизительно от остатка 231 приблизительно до 340 IgG. Уникальность домена СН2 состоит в том, что он не является близко спаренным с другим доменом. Напротив, две N-связанные разветвленные углеводные цепи расположены между двумя доменами СН2 интактной молекулы нативного IgG. Предполагалось, что углевод может обеспечивать замещение для спаривания домен-домен и помогать стабилизировать домен СН2. Burton, Molec. Immunol.22:161-206 (1985).
«Домен СН3» содержит группу остатков, расположенных С-терминально по отношению к домену СН2 в Fc-области (т.е. приблизительно от аминокислотного остатка 341 приблизительно до аминокислотного остатка 447 IgG).
Функциональная «Fc-область» обладает «эффекторной функцией» нативной последовательности Fc-области. Типовые «эффекторные функции» включают связывание C1q; КЗЦ; связывание Fc-рецептора; АЗКЦ; фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, рецептор В-клеток; РВК) и т.д. Для таких эффекторных функций в общем случае необходимо, чтобы Fc-область была скомбинирована со связывающим доменом (например, вариабельным доменом антитела), что может быть достигнуто при помощи различных раскрытых методов анализа, например, приведенных в определениях в данном документе.
«Нативная последовательность Fc-области» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области природного происхождения. Нативная последовательность человеческих Fc-областей содержит нативную последовательность Fc-области человеческого IgG1 (аллотипы А и отличные от А); нативную последовательность Fc-области человеческого IgG2; нативную последовательность Fc-области человеческого IgG3; и нативную последовательность Fc-области человеческого IgG4, а также их варианты природного происхождения.
«Вариантная Fc-область» содержит аминокислотную последовательность, которая отличается от нативной последовательности Fc-области за счет по меньшей мере одной аминокислотной модификации, предпочтительно одной или более аминокислотных замен. Предпочтительно вариантная Fc-область содержит по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или Fc-областью родительского полипептида, например, от около одной до около десяти аминокислотных замен, и предпочтительно от около одной до около пяти аминокислотных замен в нативной последовательности Fc-области или в Fc-области родительского полипептида. Вариантная Fc-область согласно данному документу предпочтительно обладает по меньшей мере около 80% гомологии с нативной последовательностью Fc-области и/или с Fc-областью родительского полипептида, и наиболее предпочтительно - по меньшей мере около 90% гомологии с ними, более предпочтительно - по меньшей мере около 95% гомологии с ними.
В контексте данного документа «Fc-комплекс» относится к двум доменам СН2 Fc-области, взаимодействующим друг с другом, и/или двум доменам СН3 Fc-области, взаимодействующим друг с другом, при этом домены СН2 и/или домены СН3 взаимодействуют посредством связей и/или сил (например, ван-дер-ваальсовых, гидрофобных, гидрофильных сил), которые относятся к пептидным связям.
В контексте данного документа «Fc-компонент» относится к шарнирной области, домену СН2 или домену СН3 Fc-области.
В контексте данного документа «Fc-СН-компонент» или «FcCH» относится к полипептиду, содержащему домен СН2, домен СН3 или домены СН2 и СН3 Fc-области.
В контексте данного документа «мутант антитела» или «вариант антитела» относится к варианту аминокислотной последовательности антитела, в которой были модифицированы один или более аминокислотных остатков видозависимого антитела. Такие мутанты обязательно обладают менее чем 100% идентичности или сходства последовательностей с видозависимым антителом. В одном варианте реализации изобретения мутант антитела имеет аминокислотную последовательность, имеющую по меньшей мере 75% идентичности или сходства последовательностей с аминокислотной последовательностью вариабельного домена тяжелой или легкой цепи видозависимого антитела, более предпочтительно - по меньшей мере 80%, более предпочтительно - по меньшей мере 85%, более предпочтительно - по меньшей мере 90% и наиболее предпочтительно - по меньшей мере 95%. Идентичность или сходство по отношению к этой последовательности определяют в данном документе как процент аминокислотных остатков в кандидатной последовательности, которые являются идентичными (т.е. такой же самый остаток) или сходными (т.е. аминокислотный остаток, принадлежащий такой же группе на основании общих свойств боковых цепей, См. ниже) с остатками видозависимого антитела после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимального процента идентичности последовательностей. В некоторых вариантах реализации изобретения N-концевые, C-концевые или внутренние продления, делеции или вставки в последовательность антитела за пределами вариабельного домена не должны учитываться так такие, которые влияют на идентичность или сходство последовательностей.
«Нарушение» или «заболевание» представляет собой любое патологическое состояние, которое можно улучшить посредством лечения веществом/молекулой или способом согласно данному изобретению. Они включают хронические и острые нарушения или заболевания, включая те патологические состояния, которые могут предрасполагать млекопитающее к рассматриваемому нарушению. Неограничивающие примеры нарушений, которые можно лечить согласно данному документу, включают злокачественные и доброкачественные опухоли; карциному, бластому и саркому.
«Лечение» относится как к терапевтическим, так и к профилактическим или превентивным мерам. Нуждающиеся в лечении включают тех, у кого уже имеется доброкачественная, предраковая или неметастатическая опухоль, а также тех, у кого необходимо предотвратить появления или повторное появление рака.
Термин «терапевтически эффективное количество» относится к количеству терапевтического агента для лечения или предотвращения заболевания или нарушения у млекопитающего. В случае рака терапевтически эффективное количество терапевтического агента может снижать число раковых клеток; снижать первоначальный объем опухоли; ингибировать (т.е. в некоторой степени замедлять и, предпочтительно, останавливать) инфильтрацию раковых клеток в периферические органы; ингибировать (т.е. в некоторой степени замедлять и, предпочтительно, останавливать) опухолевые метастазы; ингибировать в некоторой степени рост опухоли; и/или облегчать в некоторой степени один или более симптомов, связанных с нарушением. В той степени, в которой лекарственный препарат может предотвращать рост и/или уничтожать раковые клетки, он может быть цитостатическим и/или цитотоксическим. В случае раковой терапии эффективность in vivo можно, например, определять путем оценки продолжительности жизни, времени до прогрессирования заболевания (ВДП), скорости ответа (СО), продолжительности ответа и/или качества жизни.
В контексте данного документа «аутоиммунное заболевание» представляет собой незлокачественное заболевание или нарушение, возникающее из и направленное против собственных тканей индивида. В контексте данного документа из аутоиммунных заболеваний исключены, в частности, злокачественные или раковые заболевания или патологические состояния, в частности, В-клеточная лимфома, острый лимфобластный лейкоз (ОЛЛ), хронический лимфоцитарный лейкоз (ХЛЛ), лейкоз ворсистых клеток и хронический миелобластный лейкоз. Примеры аутоиммунных заболеваний или нарушений включают, но не ограничиваются этим, воспалительные ответы, такие как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); системную склеродермию и склероз; ответы, связанные с воспалительным заболеванием кишечника (таким как болезнь Крона и язвенный колит); синдром острой дыхательной недостаточности (включая синдром острой дыхательной недостаточности у взрослых; СОДНВ); дерматит; менингит; энцефалит; увеит; колит; гломерулонефрит; аллергические состояния, такие как экзема и астма, и другие состояния, включая инфильтрацию Т-клеток и хронические воспалительные ответы; атеросклероз; нарушение адгезии лейкоцитов; ревматоидный артрит; системную красную волчанку (СКВ); сахарный диабет (например, сахарный диабет I типа или инсулин-зависимый сахарный диабет); множественный склероз; синдром Рейно; аутоиммунный тиреоидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные ответы, связанные с острой и замедленной гиперчувствительностью, опосредуемые цитокинами и Т-лимфоцитами, как правило, обнаруживаемыми при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; пернициозную анемию (болезнь Аддисона); заболевания, сопровождающиеся диапедезом лейкоцитов; воспалительное расстройство центральной нервной системы (ЦНС); синдром полиорганной недостаточности; гемолитическую анемию (включая, но не ограничиваясь этим, криоглобулинемию или Кумбс-положительную анемию); миастению гравис; заболевания, опосредуемые комплексом антиген-антитело; синдром Гудпасчера; антифосфолипидный синдром; аллергический неврит; болезнь Грейвса; миастенический синдром Ламберта-Итона; буллезный пемфигоид; пемфигус; аутоиммунные полиэндокринопатии; болезнь Рейтера; синдром мышечной скованности; болезнь Бехчета; гигантоклеточный артериит; иммунный комплексный нефрит; IgA-нефропатию; IgM-полинефропатии; иммунную тромбоцитопеническую пурпуру (ИТП) или аутоиммунную тромбоцитопению и т.д.
Термины «рак» и «раковый» относятся к или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется неконтролируемым клеточным ростом. В это определение входит доброкачественный и злокачественный рак. Под «ранней стадией рака» или «ранней стадией опухоли» подразумевается рак, которые не является инвазивным или метастатическим или классифицируется как рак 0, I или II стадии. Примеры рака включают, но не ограничиваются этим, карциному, лимфому, бластому (включая медуллобластому и ретинобластому), саркому (включая липосаркому и саркому синовиальных клеток), нейроэндокринные опухоли (включая карциноидные опухоли, гастриному и рак островковых клеток), мезотелиому, шванному (включая нейрому слухового нерва), менингиому, аденокарциному, меланому и лейкоз или лимфолейкоз. Более конкретные примеры таких типов рака включают плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, включая мелкоклеточный рак легкого (МКРЛ), немелкоклеточный рак легкого (НМКРЛ), аденокарциному легкого и плоскоклеточную карциному легкого, рак брюшной полости, гепатоцеллюлярный рак, рак желудка, включая рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы (включая метастатический рак молочной железы), рак толстой кишки, рак прямой кишки, рак толстой и прямой кишки, карциному эндометрия или матки, карциному слюнной железы, рак почек, рак простаты, рак вульвы, рак щитовидной железы, гепатокарциному, анальную карциному, карциному полового члена, рак яичка, рак пищевода, опухоли желчных протоков, а также рак головы и шеи и множественную миелому.
Термин «предраковый» относится к заболеванию или росту, который, как правило, предшествует раку или развивается в рак. «Предраковый рост» включает клетки, которые характеризуются аномальной регуляцией клеточного цикла, пролиферацией или дифференцировкой, что можно определить по маркерам регуляции цикла, клеточной пролиферации или дифференцировки.
Под «дисплазией» подразумевается любой аномальный рост или развитие ткани, органа или клеток. Предпочтительно дисплазия является дисплазией высокой степени или предраковой.
Под «метастазом» подразумевается распространение рака от его изначального участка в другие участки тела. Раковые клетки могут отрываться от первичной опухоли, проникать в лимфатические и кровеносные сосуды, циркулировать в кровотоке и расти в отдаленных очагах (метастазировать) в нормальных тканях в любом месте организма. Метастаз может быть локальным или отдаленным. Метастаз представляет собой многостадийный процесс, возможный при условии отрыва опухолевых клеток от первичной опухоли, перемещения через кровоток и остановки в отдаленном участке. В новом участке клетки налаживают кровоснабжение и могут расти, образуя массу, несущую угрозу для жизни.
Это поведение регулируется как стимуляторными, так и ингибиторными молекулярными путями в раковой клетке, и также имеют значение взаимодействия между опухолевой клеткой и клетками-хозяевами в отдаленном участке.
Под «неметастатическим» подразумевается рак, который является доброкачественным или который остается в первичном участке и не проникает в лимфатические или кровеносные сосуды или ткани, отличные от первичного участка. В общем случае неметастатический рак представляет собой любой рак 0, I или II стадии и, в некоторых случаях, рак III стадии.
Под «первичной опухолью» или «первичным раком» подразумевается исходный рак, а не метастатическое поражение, локализованное в другой ткани, органе или локации в организме субъекта.
Под «доброкачественной опухолью» или «доброкачественным раком» подразумевается рак, который остается локализованным в исходном участке и не обладает способность инфильтрировать, инвазировать или метастазировать в отдаленный участок.
Под «опухолевой нагрузкой» подразумевается число раковых клеток, размер опухоли или объем рака в организме. Опухолевая нагрузка также называется опухолевой массой.
Под «числом опухолей» подразумевается число опухолей.
Под «субъектом» подразумевается млекопитающее, включая, но не ограничиваясь этим, человека или отличное от человека млекопитающее, такое как крупный рогатый скот, лошадиные, собачьи, овечьи или кошачьи. Предпочтительно субъект является человеком.
Термин «противораковая терапия» относится к терапии, применяемой при лечении рака. Примеры противораковых терапевтических агентов включают, но не ограничиваются этим, например, химиотерапевтические агенты, агенты, ингибирующие рост, цитотоксические агенты, агенты, применяемые в лучевой терапии, антиангиогенные агенты, апоптотические агенты, антитубулиновые агенты и другие агенты для лечения рака, анти-CD20 антитела, ингибиторы фактора роста тромбоцитов (например, Гливек™ (иматиниба мезилат)), ингибитор COX-2 (например, целекоксиб), интерфероны, цитокины, антагонисты (например, нейтрализующие антитела), которые связываются с одной или более следующих мишеней: ErbB2, ErbB3, ErbB4, PDGFR-бета, BlyS, APRIL, BCMA или рецептором(ами) VEGF, TRAIL/Apo2, и другие биоактивные и органические химические агенты и т.д. В изобретение также включены их комбинации.
Употребляемый в данном документе термин «цитотоксический агент» относится к веществу, которое ингибирует или препятствует функции клеток и/или приводит к разрушению клеток. Данный термин включает в себя радиоактивные изотопы (например, I131, I125, Y90 и Re186), химиотерапевтические агенты и токсины, такие как ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты.
«Химиотерапевтический агент» представляет собой химическое соединение, применяемое в лечение рака. Примеры химиотерапевтического агента включают химическое соединение, применяемое в лечение рака. Примеры химиотерапевтического агента включают алкилирующие агенты, такие как тиотепа и ЦИТОКСАН® циклофосфамид; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметиломеламин; ацетогенины (в особенности буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитромочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в особенности калихеамицин гамма 1I и калихеамицин омега I1 (См., например, Agnew, Chem Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин A; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарциностатиновый хромофор и родственные хромофоры хромопротеиновых энедииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин АДРИАМИЦИН® (включая морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, порфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолона пропионат, эпитиостанол, мепитиостан, тестолактон; антиадреналины, такие как аминоглютетимид, митотан, трилостан; восполнитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамидгликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; эллиптиния ацетат; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2"-трихлортриэтиламин; трихотецены (в особенности токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-C»); циклофосфамид; тиотепу; таксоиды, например, паклитаксел ТАКСОЛ® (Bristol- Myers Squibb Oncology, Princeton, N.J.), не содержащий кремофор АБРАКСАНТМ, препараты из наночастиц паклитаксела, сконструированные на основе альбумина (American Pharmaceutical Partners, Schaumberg, Illinois) и доксетаксел ТАКСОТЕР® (Rhône- Poulenc Rorer, Antony, France); хлоранбуцил; GEMZAR® (гемцитабин); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин НАВЕЛБИН®; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселоду; ибандронат; иринотекан (камптозар, СРТ-11) (включая схему лечения иринотеканом с 5-FU и лейковорином); ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин; бортезомиб ВЕЛКАД; леналидомид РЕВЛИМИД; лейковорин (ЛВ); оксалиплатин, включая схему лечения оксалиплатином (ФОЛФОКС); ингибиторы PKC-альфа, Raf, H-Ras, EGFR (например, эрлотиниб (ТарцеваТМ)) и VEGF-A, которые снижают клеточную пролиферацию, и фармацевтически приемлемые соли, кислоты или производные любого из приведенных выше соединений.
В данное определение также включены антигормональные агенты, действие которых состоит в регулировании или ингибировании действия гормонов на опухоли, такие как антиэстрогены и селективные модуляторы рецепторов эстрогенов (SERM), включая, например, тамоксифен (включая тамоксифен НОЛВАДЕКС®), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен ФАРЕСТОН; ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует выработку эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, мегестрола ацетат МЕГАЗ®, экземестан АРОМАЗИН®, форместан, фадрозол, ворозол РИВИЗОР®, летрозол ФЕМАРА® и анастрозол АРМИДЕКС®; и антиандрогены, такие как флутамид, нилутамид, бикалутамид, леупролид и госерелин; а также троксацитабин (1,3-диоксолановый аналог нуклеозида цитозина); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, которые ингибируют экспрессию генов в сигнальных путях, участвующих в аберрантной пролиферации клеток, таких как, например PKC-альфа, Ralf и H-Ras; рибозимы, такие как ингибиторы экспрессии VEGF (например, рибозим АНГИОЗИМ®) и ингибиторы экспрессии HER2; вакцины, такие как вакцины для генной терапии, например вакцина АЛЛОВЕКТИН®, вакцина ЛЕУВЕКТИН® и вакцина ВАКСИД®; ПРОЛЕЙКИН® rIL-2; ингибитор топоизомеразы 1 ЛУРТОТЕКАН®; АБАРЕЛИКС® rmRH; винорелбин и эсперамицины (См. патент США № 4675187), и фармацевтически приемлемые соли, кислоты или производные любого из приведенных выше соединений.
Употребляемый в данной заявке термин «пролекарство» относится к форме предшественника или производного фармацевтически активного вещества, которая является менее цитотоксической по отношению к опухолевым клеткам по сравнению с родительским лекарством и может быть ферментативно активирована или преобразована в более активную родительскую форму. См., например, Wilman, ʺProdrugs in Cancer Chemotherapyʺ Biochemical Society Transactions, 14, pp. 375-382, 615th Meeting Belfast (1986) и Stella et al., ʺProdrugs: A Chemical Approach to Targeted Drug Delivery,ʺ Directed Drug Delivery, Borchardt et al., (ed.), pp. 247-267, Humana Press (1985). Пролекарства согласно данному изобретению включают, но не ограничиваются этим, фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, модифицированные D-аминокислотами пролекарства, гликозилированные пролекарства, β-лактамсодержащие пролекарства, необязательно замещенные феноксиацетамидсодержащие пролекарства или необязательно замещенные фенилацетамидсодержащие пролекарства, 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые могут быть преобразованы в более активный цитотоксический свободный лекарственный препарат. Примеры цитотоксических лекарственных препаратов, которые могут быть дериватизированы в форму пролекарства для применения в данном изобретении, включают, но не ограничиваются этим, описанные выше химиотерапевтические агенты.
Под «лучевой терапией» подразумевается применение направленных гамма-лучей или бета-лучей, чтобы индуцировать достаточное повреждение клетки так, чтобы ограничить ее способность к нормальному функционированию или полностью разрушить клетку. Следует понимать, что в данной области техники существует много способов определения дозы и продолжительности облучения. Типовое облучение проводят в виде одного сеанса, а типовые дозы облучения соответствуют диапазону от 10 до 200 единиц (грей) в день.
«Биологически активный» или «функциональный» полипептид (такой как гетерологичный полипептид) - это полипептид, способный демонстрировать один или более из своих природных видов активности в структурных, регуляторных, биохимических или биофизических явлениях.
«Биологически активное» или «функциональное» антитело - это антитело, способное демонстрировать один или более из своих природных видов активности в структурных, регуляторных, биохимических или биофизических явлениях. Например, биологически активное антитело может обладать способностью специфически связывать антиген, а связывание может, в свою очередь, устранять или изменять клеточное или молекулярное явление, такое как сигнальная трансдукция или ферментативная активность. Биологически активное антитело также может блокировать активацию лиганда рецептора или действовать как агонистическое антитело. Способность антитела демонстрировать один или более из своих природных видов активности зависит от нескольких факторов, включая правильный фолдинг и сборку полипептидных цепей.
Композиции согласно изобретению и способы их применения
В данном документе предложены способы, в которых применяют сигнальные пептиды и вариантные сигнальные пептиды, подходящие, например, для способов получения гетерологичных полипептидов (например, антител, например, полноразмерных антител). Способы получения характеристик сигнальных пептидов известны в данной области техники. Согласно одной схеме, сигнальные пептиды обычно состоят из трех разных областей: N-концевой области, которая содержит 1 или 2 положительно заряженных аминокислотных остатка, гидрофобной коровой области, часто называемой Н-областью (также называемой Н-доменом), и С-концевой области, распознаваемой сигнальной пептидазой. В некоторых вариантах реализации изобретения длина Н-области составляет около 10-16 остатков. Гидрофобность сигнального пептида можно рассчитать, используя шкалу Айзенберга. См. Eisenberg, D. et al., J Mol Biol (1984) 179:125-142. Вкратце, каждой аминокислоте приписывают значение нормированной консенсусной гидрофобности (См. Eisenberg, et al., выше, в Таблице I (страница 126). Суммарную гидрофобность (также называемую общей гидрофобностью) рассчитывают путем суммирования значений консенсусной гидрофобности для каждой аминокислоты сигнального пептида (или, например, чтобы рассчитать общее значение гидрофобности для Н-области, суммируют значения консенсусной гидрофобности для каждой аминокислоты в Н-области). Среднюю гидрофобность рассчитывают по следующей формуле: Средняя гидрофобность = общая (суммарная) гидрофобность /число аминокислот. В некоторых вариантах реализации изобретения рассчитывают среднюю гидрофобность всего сигнального пептида. В некоторых вариантах реализации изобретения рассчитывают среднюю гидрофобность Н-области (также называемой Н-доменом).
Мутации в последовательности сигнального пептида можно проводить известными в данной области техники способами. Обзор методов генетической модификации ДНК подобным образом приведен в Mutagenesis: a Practical Approach, M.J. McPherson, Ed., (IRL Press, Oxford, UK. (1991), и включает, например, сайт-направленный мутагенез, кассетный мутагенез и мутагенез с применением полимеразной цепной реакции (ПЦР). Другие способы мутагенеза включают сайт-направленный мутагенез QuickChange и ПЦР с перекрывающимися праймерами.
В другом аспекте предложено применение сигнального пептида, обладающего большей средней гидрофобностью (например, средней гидрофобностью Н-области или средней гидрофобностью всего сигнального пептида), чем сигнальный пептид DsbA, например, для применения в любом из способов согласно изобретению, например, получения гетерологичного полипептида (например, антитела), секреции гетерологичного полипептида из клетки, получения растворимого гетерологичного полипептида, секреции в периплазму растворимого гетерологичного полипептида, получения зрелого гетерологичного полипептида, секреции в периплазму зрелого гетерологичного полипептида, транслокации гетерологичного полипептида, оптимизации секреции гетерологичного полипептида. Типовые сигнальные пептиды, которые обладают большей средней гидрофобностью чем DsbA, включают: FlgI, NikA, AsmA, TolB, YraI, FecB, CemH, TreA, FocC, TraU, SfmL, TorT, SfmC.
В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,5. В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,6. В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,7. В некоторых вариантах реализации изобретения средняя гидрофобность первого сигнального пептида составляет более чем около 0,52, 0,53, 0,54, 0,55, 0,56, 0,57, 0,58, 0,59, 0,6, 0,61, 0,62, 0,63, 0,64, 0,65, 0,66, 0,67, 0,68, 0,69, 0,7 или больше. В некоторых вариантах реализации изобретения средняя гидрофобность второго сигнального пептида составляет более чем около 0,6. В некоторых вариантах реализации изобретения средняя гидрофобность второго сигнального пептида составляет более чем около 0,7. В некоторых вариантах реализации изобретения средняя гидрофобность второго сигнального пептида составляет более чем около 0,52, 0,53, 0,54, 0,55, 0,56, 0,57, 0,58, 0,59, 0,6, 0,61, 0,62, 0,63, 0,64, 0,65, 0,66, 0,67, 0,68, 0,69, 0,7 или больше. В некоторых вариантах реализации изобретения средняя гидрофобность первого и второго сигнальных пептидов является сходной (например, приблизительно эквивалентной). В некоторых вариантах реализации изобретения средняя гидрофобность первого и второго сигнальных пептидов является разной.
Сигнальные пептиды DsbA и STII хорошо известны в данной области техники. Последовательности сигнальных пептидов DsbA и STII приведены в Таблицах 7 и 8. Последовательности N-концевой области, Н-области и С-концевой области DsbA и STII приведены в Таблицах 7 и 8.
Способы получения характеристик выработки и секреции гетерологичного полипептида известны в данной области техники, а некоторые способы описаны и проиллюстрированы в данном документе. Например, культивируют хозяйский штамм, несущий экспрессионный(ые) вектор(ы), кодирующий(ие) вариантные сигнальные пептиды, функционально связанные с гетерологичным белком (например, антителом), и выделяют полипептид. Растворимую фракцию отделяют при помощи невосстанавливающего электрофореза ДСН-ПААГ с последующим анализом методом вестерн-блоттинга для определения уровня вырабатываемого полноразмерного гетерологичного белка. Можно определить наличие или отсутствие уровня зрелого полипептида и полипептида-предшественника, например, методом секвенирования по Эдману белка, выделенного с полос геля вестерн-блоттинга, и охарактеризовать выделенный полипептид, как содержащий или не содержащий сигнальный пептид, как хорошо известно в данной области техники. Выработку полного антитела (или, например, других гетеромультимерных белков) можно определить путем проведения вестрен-блоттинга в денатурирующих условиях. Активность гетерологичного полипептида (например, антитела) можно определить, применяя рутинные функциональные методы анализа, хорошо известные в данной области техники. Функцию белка можно определить, применяя подходящий функциональный анализ. Например, активность связывания можно исследовать методом ELISA, Biacore и другими методами, хорошо известными в данной области техники. Другие функции можно исследовать, применяя методы анализа, хорошо известные в данной области техники как такие, которые подходят для специфического гетерологичного полипептида.
Область инициации трансляции (TIR) является основным определяющим фактором общего уровня трансляции белка. TIR включает полинуклеотид, который кодирует сигнальную последовательность и простирается от участка непосредственно выше последовательности Шайна-Дальгарно до участка приблизительно на двадцать нуклеотидов ниже стартового кодона. Модификации этой полинуклеотидной последовательности (в некоторых вариантах реализации изобретения - модификация в пределах первых от около 2 до около 14, от около 4 до 12, около 6 кодонов последовательности, кодирующей сигнальный пептид) могут изменять эффективность инициации трансляции, тем самым корректируя уровень трансляции нижерасположенного белка. TIR имеют разную трансляционную эффективность. В некоторых вариантах реализации изобретения первая и вторая области инициации трансляции (функционально связанные, например, с первым и вторым гетерологичным полипептидом) (и, в некоторых вариантах реализации изобретения, третья область инициации трансляции, функционально связанная, например, с третьим гетерологичным полипептидом) обеспечивают приблизительно одинаковую эффективность трансляции. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет около одного или двух. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет около одного. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет около двух. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет один и/или два. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет около трех или около четырех. В некоторых вариантах реализации изобретения относительная эффективность трансляции выбрана из одного или более из одного, двух, трех, четырех, пяти или более (например, шести или семи или более). В некоторых вариантах реализации изобретения относительная эффективность трансляции первой и второй TIR составляет около одного. В некоторых вариантах реализации изобретения относительная эффективность трансляции (также называемая эффективностью TIR) первой и второй TIR составляет около двух. В некоторых вариантах реализации изобретения относительная эффективность трансляции первой, второй и третьей TIR составляет около одного. В некоторых вариантах реализации изобретения относительная эффективность трансляции первой, второй и третьей TIR составляет около двух. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет около 2, около 3, приблизительно 4, около 5, около 6, около 7 или более, например, около 8, около 9 или более. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет между 1 и 3, между 2 и 4, между 3 и 5, между 4 и 6, между 5 и 7 или между 6 и 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции составляет между 2 и 5, между 3 и 7 или между 4 и 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого и/или второго сигнального пептида составляет около 1, около 2, около 3, около 4, около 5, приблизительно 6, около 7, или около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первого сигнального пептида составляет около 5, а относительная эффективность трансляции второго сигнального пептида составляет около 8. В некоторых вариантах реализации изобретения относительная эффективность трансляции первой и второй TIR является приблизительно одинаковой. В некоторых вариантах реализации изобретения относительная эффективность трансляции первой и второй TIR является разной.
В некоторых вариантах реализации изобретения полинуклеотид, кодирующий сигнальный пептид (такой как вариантный сигнальный пептид), предложен в виде вектора с соответствующими элементами для экспрессии представляющего интерес гена. В некоторых вариантах реализации изобретения вектор содержит промотор 5' к сигнальной последовательности, сайт распознавания рестрикционных ферментов 3' к сигнальной последовательности для вставки представляющего интерес гена или репортерного гена и селективный маркер, такой как маркер устойчивости к лекарственному препарату для отбора и/или сохранения бактерий, трансформированных результирующими плазмидами. Плазмидные векторы дополнительно обсуждаются и иллюстрируются в данном документе. Промоторы, подходящие для применения с прокариотическими хозяевами, известны в данной области техники, а некоторые проиллюстрированы и описаны в данном документе.
Можно применять любой репортерный ген, который поддается какой-либо количественной оценке. Таким образом, например, выработку щелочной фосфатазы можно количественно оценить, измеряя секретируемый уровень генного продукта phoA. Другие примеры включают, например, ген β-лактамазы.
Секретируемый уровень полипептидов можно определять, например, путем проведения функционального анализа представляющего интерес полипептида, например, радиоиммуноанализа (РИА), ферментного иммуноанализа (ELISA), или при помощи ПААГ и визуализации правильной молекулярной массы представляющего интерес полипептида. Способы определения уровня секретируемого полипептида хорошо известны в данной области техники, а некоторые проиллюстрированы в данном документе.
Полипептиды
Типовые гетерологичные полипептиды включают трансмембранную молекулу (например, рецептор, такой как рецептор тирозинкиназы) или лиганд, такой как фактор роста. Типовые гетерологичные полипептиды включают молекулы, такие как ренин; гормон роста, включая человеческий гормон роста и бычий гормон роста; фактор, высвобождающий гормон роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; альфа-1-антитрипсин; А-цепь инсулина; В-цепь инсулина; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VIIIC, фактор IX, тканевой фактор (TF) и фактор Виллебранда; противосвертывающие факторы, такие как протеин С; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или активатор плазминогена человеческой мочи или тканевой активатор плазминогена (t-PA); бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухолей альфа и бета; энкефалиназа; RANTES (регулятор активации экспрессии и секреции нормальных T-клеток); человеческий воспалительный белок макрофагов (MIP-1-альфа); сывороточный альбумин, такой как человеческий сывороточный альбумин; Мюллерова ингибирующая субстанция; А-цепь релаксина; В-цепь релаксина; прорелаксин; мышиный гонадотропин-ассоциированный пептид; микробный белок, такой как бета-лактамаза; ДНКаза; IgE; антиген цитотоксических Т-лимфоцитов (CTLA), такой как CTLA-4; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы гормонов или факторов роста; протеин A или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор головного мозга (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов, такой как NGF-β; тромбоцитарный фактор роста (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-β1, TGF-β2, TGF-β3, TGF-β4 или TGF-β5; инсулин-подобный фактор роста-I и -II (IGF-I и IGF-II); des(1-3)-IGF-I (IGF-I головного мозга), белки, связывающие инсулин-подобный фактор роста; белки CD, такие как CD3, CD4, CD8, CD19, CD20 и CD40; эритропоэтин; остеиндуктивные факторы; иммунотоксины; костный морфогенетический белок (BMP); интерферон, такой как интерферон-альфа, -бета и -гамма; колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-10; супероксиддисмутаза; рецепторы Т-клеток; поверхностные белки мембраны; фактор ускорения распада; вирусный антиген, такой как, например, часть оболочки СПИД; транспортные белки; хоминг-рецепторы; аддрессины; регуляторные белки; интегрины, такие как CD11a, CD11b, CD11c, CD18, ICAM, VLA-4 и VCAM; опухоль-ассоциированный антиген, такой как рецептор HER2, HER3 или HER4; и фрагменты любого из вышеперечисленных полипептидов.
Иммуноадгезины однозначно рассматриваются как гетерологичные полипептиды в соответствии с изобретением.
Антитела
Типовые мишени для антител или гетеромультимерных полипептидов, или полипептидов, или иммуноадгезинов включают, но не ограничиваются этим, следующий список:
BMPI, BMP2, BMP3B (GDFIO), BMP4, BMP6, BMP8, CSFI (M-CSF), CSF2 (GM-CSF), CSF3 (G-CSF), EPO, FGFI (aFGF), FGF2 (bFGF), FGF3 (int-2), FGF4 (HST), FGF5, FGF6 (HST-2), FGF7 (KGF), FGF9, FGF10, FGF1 1, FGF12, FGF12B, FGF14, FGF16, FGF17, FGF19, FGF20, FGF21, FGF23, IGF1, IGF2, IFNAI, IFNA2, IFNA4, IFNA5, IFNA6, IFNA7, IFNBI, IFNG, IFNWI, FELI, FELI (EPSELON), FELI (ZETA), IL1A, IL1 B, IL2, IL3, IL4, IL5, IL6, IL7, IL8, IL9, IL10, IL1 1, IL12A, I L12B, IL13, IL14, IL15, I L16, IL17, IL17B, IL18, IL19, IL20, IL22, IL23, IL24, IL25, IL26, IL27, IL28A, IL28B, I L29, IL30, PDGFA, PDGFB, TGFA, TGFB1, TGFB2, TGFB3, LTA (ФНО-b), LTB, ФНО (ФНО-a), ФHOSF4 (OX40 лиганд), ФНОSF5 (CD40 лиганд), ФНОSF6 (FasL), ФНОSF7 (CD27 лиганд), ФНОSF8 (CD30 лиганд), ФНОSF9 (4-1 BB лиганд), ФНОSFIO (TRAIL), ФНОSF1 I (TRANCE), ФНОSF12 (AP03L), ФНОSF13 (April), ФНОSF13B, ФНОSF14 (HVEM- L), ФНОSF15 (VEGI), ФНОSF18, HGF (VEGFD), VEGF, VEGFB, VEGFC, ILIR1, IL1 R2, IL1 RL1, IL1 RL2, IL2RA, IL2RB, IL2RG, IL3RA, IL4R, IL5RA, IL6R, IL7R, IL8RA, IL8RB, IL9R, ILIORA, ILIORB, IL1 1 RA, IL12RB1, IL12RB2, IL13RA1, IL13RA2, IL15RA, IL17R, IL18R1, IL20RA, IL21 R, IL22R, IL1 HY1, IL1 RAP, IL1 RAPL1, IL1 RAPL2, IL1 RN, IL6ST, IL18BP, IL18RAP, IL22RA2, AIFI, HGF, LEP (лептин), PTN, THPO, CCLI (I-309), CCL2 (MCP - 1/MCAF), CCL3 (MIP-la), CCL4 (MIP-lb), CCL5 (RANTES), CCL7 (MCP-3), CCL8 (mcp-2), CCLH (эотаксин), CCL13 (MCP-4), CCL15 (MIP-ld), CCL16 (HCC-4), CCL17 (TARC), CCL18 (PARC), CCL19 (MDP-3b), CCL20 (MIP- 3a), CCL21 (SLC/экзодус-2), CCL22 (MDC/STC-I), CCL23 (MPIF-I), CCL24 (MPIF-2/эотаксин-2), CCL25 (TECK), CCL26 (эотаксин-3), CCL27 (CTACK/ILC), CCL28, CXCLI (GROI), CXCL2 (GR02), CXCL3 (GR03), CXCL5 (ENA-78), CXCL6 (GCP-2), CXCL9 (MIG), CXCL10 (IP 10), CXCL1 1 (I- TAC), CXCL12 (SDFI), CXCL13, CXCL14, CXCL16, PF4 (CXCL4), PPBP (CXCL7), CX3CL1 (SCYDI), SCYEI, XCLI (лимфотактин), XCL2 (SCM-lb), BLRI (MDR15), CCBP2 (D6/JAB61), CCR1 (CKRI/HM145), CCR2 (mcp-IRB/RA), CCR3 (CKR3/CMKBR3), CCR4, CCR5 (CMKBR5/ChemR13), CCR6 (CMKBR6/CKR-L3/STRL22/DRY6), CCR7 (CKR7/EBII), CCR8 (CMKBR8/TERI/CKR- LI), CCR9 (GPR-9-6), CCRLI (VSHKI), CCRL2 (L-CCR), XCRI (GPR5/CCXCRI), CMKLRI, CMKORI (RDCI), CX3CR1 (V28), CXCR4, GPR2 (CCRIO), GPR31, GPR81 (FKSG80), CXCR3 (GPR9/CKR-L2), CXCR6 (TYMSTR /STRL33/Bonzo), HM74, IL8RA (IL8Ra), IL8RB (IL8Rb), LTB4R (GPR16), TCPIO, CKLFSF2, CKLFSF3, CKLFSF4, CKLFSF5, CKLFSF6, CKLFSF7, CKLFSF8, BDNF, C5R1, CSF3, GRCCIO (CIO), EPO, FY (DARC), GDF5, HDFIA, DL8, PRL, RGS3, RGS13, SDF2, SLIT2, TLR2, TLR4, TREMI, TREM2, VHL, ABCFI; ACVRI; ACVRIB; ACVR2; ACVR2B; ACVRLI; AD0RA2A; аггрекан; AGR2; AICDA; AIFI; AIGI; AKAPI; AKAP2; AMH; AMHR2; ANGPTI; ANGPT2; ANGPTL3; ANGPTL4; ANPEP; APC; APOCI; AR; AZGPI (цинк-a-гликопротеин); B7.1; B7.2; BAD; BAFF (BLys); BAGI; BAH; BCL2; BCL6; BDNF; BLNK; BLRI (MDR15); BMPI; BMP2; BMP3B (GDFIO); BMP4; BMP6; BMP8; BMPRIA; BMPRIB; BMPR2; BPAGI (плектин); BRCAI; C19orflO (IL27w); C3; C4A; C5; C5R1; CANTI; CASP1; CASP4; CAVI; CCBP2 (D6/JAB61 ); CCLI (1-309); CCLII (эотаксин); CCL13 (MCP-4); CCL15 (MlP-ld); CCL16 (HCC-4); CCL17 (TARC); CCL18 (PARC); CCL19 (MIP-3b); CCL2 (MCP -1 ); MCAF; CCL20 (MIP-3a); CCL21 (MTP-2); SLC; экзодус-2; CCL22 (MDC/STC-I); CCL23 (MPIF-1 ); CCL24 (MPIF-2/эотаксин-2); CCL25 (TECK); CCL26 (эотаксин-3); CCL27 (CTACK/ILC); CCL28; CCL3 (MTP-la); CCL4 (MDP-lb); CCL5 (RANTES); CCL7 (MCP-3); CCL8 (mcp-2); CCNAI; CCNA2; CCNDI; CCNEI; CCNE2; CCRI (CKRI/HM145); CCR2 (mcp-IRB/RA);CCR3 (CKR3/CMKBR3); CCR4; CCR5 (CMKBR5/ChemR13); CCR6 (CMKBR6/CKR-L3/STRL22/DRY6); CCR7 (CKR7/EBII); CCR8 (CMKBR8/TERI/CKR-LI); CCR9 (GPR-9-6); CCRLI (VSHKI); CCRL2 (L-CCR); CD164; CD19; CDIC; CD20; CD200; CD22; CD24; CD28; CD3; CD37; CD38; CD3E; CD3G; CD3Z; CD4; CD40; CD40L; CD44; CD45RB; CD52; CD69; CD72; CD74; CD79A; CD79B; CD8; CD80; CD81; CD83; CD86; CDHI (E-кадгерин); CDH10; CDH12; CDH13; CDH18; CDH19; CDH20; CDH5; CDH7; CDH8; CDH9; CDK2; CDK3; CDK4; CDK5; CDK6; CDK7; CDK9; CDKNIA (p21Wapl/Cipl); CDKNIB (p27Kipl); CDKNIC; CDKN2A (P16INK4a); CDKN2B; CDKN2C; CDKN3; CEBPB; CERI; CHGA; CHGB; хитиназа; CHST10; CKLFSF2; CKLFSF3; CKLFSF4; CKLFSF5; CKLFSF6; CKLFSF7; CKLFSF8; CLDN3; CLDN7 (клаудин-7); CLN3; CLU (кластерин); CMKLRI; CMKORI (RDCI); CNRI; COL18A1; COLIAI; COL4A3; COL6A1; CR2; CRP; CSFI (M-CSF); CSF2 (GM-CSF); CSF3 (GCSF); CTLA4; CTNNBI (b-катенин); CTSB (катепсин B); CX3CL1 (SCYDI); CX3CR1 (V28); CXCLI (GROI); CXCL10 (IP-10); CXCLII (l-TAC/IP-9); CXCL12 (SDFI); CXCL13; CXCL14; CXCL16; CXCL2, (GR02); CXCL3 (GR03); CXCL5 (ENA-78/LIX); CXCL6 (GCP-2); CXCL9 (MIG); CXCR3 (GPR9/CKR-L2); CXCR4; CXCR6 (TYMSTR /STRL33/Bonzo); CYB5; CYCI; CYSLTRI; DAB2IP; DES; DKFZp451 J01 18; DNCLI; DPP4; E2F1; ECGFI; EDGI; EFNAI; EFNA3; EFNB2; EGF; EGFR; ELAC2; ENG; EN01; EN02; EN03; EPHB4; EPO; ERBB2 (Her-2); EREG; ERK8; ESRI; ESR2; F3 (TF); FADD; FasL; FASN; FCERIA; FCER2; FCGR3A; FGF; FGFI (aFGF); FGF10; FGF11; FGF12; FGF12B; FGF13; FGF14; FGF16; FGF17; FGF18; FGF19; FGF2 (bFGF); FGF20; FGF21; FGF22; FGF23; FGF3 (int-2); FGF4 (HST); FGF5; FGF6 (HST-2); FGF7 (KGF); FGF8; FGF9; FGFR3; FIGF (VEGFD); FELI (EPSILON); FILI (ZETA); FLJ12584; FLJ25530; FLRTI (фибронектин); FLTI; FOS; FOSLI (FRA-I); FY (DARC); GABRP (GABAa); GAGEBI; GAGECI; GALNAC4S-6ST; GAT A3; GDF5; GFI 1; GGT1; GM- CSF; GNASI; GNRHI; GPR2 (CCRIO); GPR31; GPR44; GPR81 (FKSG80); GRCCIO (CIO); GRP; GSN (гельзолин); GSTPI; HAVCR2; HDAC4; HDAC5; HDAC7A; HDAC9; HGF; HIFIA; HDPI; гистамин и рецепторы гистамина; HLA-A; HLA-DRA; HM74; HMOXI; HUMCYT2A; ICEBERG; ICOSL; ID2; IFN-a; IFNAI; IFNA2; IFNA4; IFNA5; IFNA6; IFNA7; IFNB1; IFNгамма; DFNWI; IGBPI; IGFI; IGFIR; IGF2; IGFBP2; IGFBP3; IGFBP6; IL-I; IL10; MORA; IL10RB; IL1 1; IL1 1 RA; IL-12; IL12A; IL12B; IL12RB1; IL12RB2; IL13; IL13RA1; IL13RA2; IL14; IL15; IL15RA; IL16; IL17; IL17B; IL17C; IL17R; IL18; IL18BP; IL18R1; IL18RAP; IL19; IL1A; IL1 B; ILIF10; IL1 F5; IL1 F6; IL1 F7; IL1 F8; IL1 F9; IL1 HYI; IL1 Rl; IL1 R2; IL1 RAP; IL1 RAPL1; IL1 RAPL2; IL1 RL1; IL1 RL2, ILIRN; IL2; IL20; IL20RA; IL21 R; IL22; IL22R; IL22RA2; IL23; IL24; IL25; IL26; IL27; I L28A; IL28B; IL29; IL2RA; IL2RB; IL2RG; IL3; IL30; IL3RA; IL4; IL4R; IL5; IL5RA; IL6; IL6R; IL6ST (гликопротеин 130); EL7; EL7R; EL8; IL8RA; DL8RB; IL8RB; DL9; DL9R; DLK; INHA; INHBA;INSL3; INSL4; IRAKI; ERAK2; ITGAI; ITGA2; ITGA3; ITGA6 (a6 интегрин); ITGAV; ITGB3; ITGB4 (b4 интегрин); JAGI; JAKI; JAK3; JUN; K6HF; KAN; KDR; KITLG; KLF5 (GC Box BP); KLF6; KLKIO; KLK12; KLK13; KLK14; KLK15; KLK3; KLK4; KLK5; KLK6; KLK9; KRT1; KRT19 (кератин 19); KRT2A; KHTHB6 (специфический для волос кератин типа Н); LAMAS; LEP (лептин); Lingo-p75; Lingo-Troy; LPS; LTA (ФНО-b); LTB; LTB4R (GPR16); LTB4R2; LTBR; MACMARCKS; MAG или Omgp; MAP2K7 (c-Jun); MDK; MIBI; мидкин; MEF; MIP-2; MKI67; (Ki-67); MMP2; MMP9; MS4A1; MSMB; MT3 (металлотионеин-lll); MTSSI; MUCI (муцин); MYC; MYD88; NCK2; нейрокан; NFKBI; NFKB2; NGFB (NGF); NGFR; NgR-Lingo; NgR-Nogo66 (Nogo); NgR- p75; NgR-Troy; NMEI (NM23A); N0X5; NPPB; NROBI; NR0B2; NRIDI; NR1 D2; NR1 H2; NR1 H3; NR1 H4; NR1 I2; NR1 I3; NR2C1; NR2C2; NR2E1; NR2E3; NR2F1; NR2F2; NR2F6; NR3C1; NR3C2; NR4A1; NR4A2; NR4A3; NR5A1; NR5A2; NR6A1; NRPI; NRP2; NT5E; NTN4; ODZI; OPRDI; P2RX7; PAP; PARTI; PATE; PAWR; PCA3; PCNA; PDGFA; PDGFB; PECAMI; PF4 (CXCL4); PGF; PGR; фосфакан; PIAS2; PIK3CG; PLAU (uPA); PLG; PLXDCI; PPBP (CXCL7); PPID; PRI; PRKCQ; PRKDI; PRL; PROC; PROK2; PSAP; PSCA; PTAFR; PTEN; PTGS2 (COX-2); PTN; RAC2 (p21 Rac2); RARB; RGSI; RGS13; RGS3; RNFIIO (ZNF144); ROB02; S100A2; SCGB1 D2 (липофилин B); SCGB2A1 (маммаглобин 2); SCGB2A2 (маммаглобин 1); SCYEI (эндотелиальный моноцит-активирующий цитокин); SDF2; SERPINAI; SERPINA3; SERP1 NB5 (маспин); SERPINEI (PAI-I); SERPDMF1; SHBG; SLA2; SLC2A2; SLC33A1; SLC43A1; SLIT2; SPPI; SPRRIB (Sprl); ST6GAL1; STABI; STAT6; STEAP; STEAP2; TB4R2; TBX21; TCPIO; TDGFI; TEK; TGFA; TGFBI; TGFBIII; TGFB2; TGFB3; TGFBI; TGFBRI; TGFBR2; TGFBR3; THIL; THBSI (тромбоспондин-1); THBS2; THBS4; THPO; TIE (Tie-1); TMP3; тканевой фактор; TLRIO; TLR2; TLR3; TLR4; TLR5; TLR6; TLR7; TLR8; TLR9; ФНО; ФНО-a; ФНОAEP2 (B94); ФНОAIP3; ФНОRSFIIA; ФНОRSFIA; ФНОRSFIB; ФНОRSF21; ФНОRSF5; ФНОRSF6 (Fas); ФНОRSF7; ФНОRSF8; ФНОRSF9; ФНОSFIO (TRAIL); ФНОSFI 1 (TRANCE); ФНОSF12 (AP03L); ФНОSF13 (April); ФНОSF13B; ФНОSF14 (HVEM-L); ФНОSF15 (VEGI); ФНОSF18; ФHOSF4 (OX40 лиганд); ФНОSF5 (CD40 лиганд); ФНОSF6 (FasL); ФНОSF7 (CD27 лиганд); ФНОSF8 (CD30 лиганд); ФНОSF9 (4-1 BB лиганд); TOLLIP; Толл-подобные рецепторы; TOP2A (топоизомераза Ea); TP53; TPMI; TPM2; TRADD; TRAFI; TRAF2; TRAF3; TRAF4; TRAF5; TRAF6; TREMI; TREM2; TRPC6; TSLP; TWEAK; VEGF; VEGFB; VEGFC; версикан; VHL C5; VLA-4; XCLI (лимфотактин); XCL2 (SCM-lb); XCRI(GPR5/CCXCRI); YYI; и ZFPM2.
В некоторых вариантах реализации изобретения мишени включают белки CD, такие как CD3, CD4, CD8, CD16, CD19, CD20, CD34; CD64, CD200, представителей семейства рецепторов ErbB, таких как рецептор EGF, рецептор HER2, HER3 или HER4; молекулы клеточной адгезии, такие как LFA-1, Mad, p150.95, VLA-4, ICAM-1, VCAM, альфа4/бета7 интегрин и альфа(v)/бета3 интегрин, включая любые их альфа- или бета-субъединицы (например, анти-CD1 1 a, анти-CD18 или анти-CD1 1 b антитела); факторы роста, такие как VEGF-A, VEGF-C; тканевой фактор (TF); альфа-интерферон (альфаIFN); ФНОальфа, интерлейкин, такой как IL-1 бета, IL-3, IL-4, IL-5, IL-8, IL-9, IL-13, IL17A/F, IL-18, IL-13Rальфа1, IL13Rальфа2, IL-4R, IL-5R, IL-9R, IgE; антигены группы крови; рецептор flk2/flt3; рецептор ожирения (OB); рецептор mpl; CTLA-4; RANKL, RANK, белок RSV F, протеин C и т.д., белок подобный рецептору липопротеинов низкой плотности (LRP)-1 или LRP-8 или рецептор трансферрина, и по меньшей мере одну мишень, выбранную из группы, состоящей из 1) бета-секретазы (BACE1 или BACE2), 2) альфа-секретазы, 3) гамма-секретазы, 4) тау-секретазы, 5) белка-предшественника амилоида (APP), 6) рецептора смерти 6 (DR6), 7) пептида бета-амилоида, 8) альфа-синуклеина, 9) паркина, 10) хантингтина, 11) p75 NTR и 12) каспазы-6.
Понятно, что антитело (например, мультиспецифическое антитело, например, биспецифическое антитело) может связывать по меньшей мере две молекулы-мишени, например, по меньшей мере две молекулы-мишени, выбранные из группы, состоящей из: IL-1 альфа и IL-1 бета, IL-12 и IL-18; IL-13 и IL-9; IL-13 и IL-4; IL-13 и IL-5; IL-5 и IL-4; IL-13 и IL-1 бета; IL-13 и IL-25; IL-13 и TARC; IL-13 и MDC; IL-13 и MEF; IL-13 и TGF-β; IL-13 и агониста LHR; IL-12 и TWEAK, IL-13 и CL25; IL-13 и SPRR2a; IL-13 и SPRR2b; IL-13 и ADAM8, IL-13 и PED2, IL17A и IL17F, CD3 и CD19, CD138 и CD20; CD138 и CD40; CD19 и CD20; CD20 и CD3; CD38 и CD138; CD38 и CD20; CD38 и CD40; CD40 и CD20; CD-8 и IL-6; CD20 и BR3, ФНОальфа и TGF-бета, ФНОальфа и IL-1 бета; ФНОальфа и IL-2, ФНО альфа и IL-3, ФНОальфа и IL-4, ФНОальфа и IL-5, ФНОальфа и IL6, ФНОальфа и IL8, ФНОальфа и IL-9, ФНОальфа и IL-10, ФНОальфа и IL-11, ФНОальфа и IL-12, ФНОальфа и IL-13, ФНОальфа и IL-14, ФНОальфа и IL-15, ФНОальфа и IL-16, ФНОальфа и IL-17, ФНОальфа и IL-18, ФНОальфа и IL-19, ФНОальфа и IL-20, ФНОальфа и IL-23, ФНОальфа и IFNальфа, ФНОальфа и CD4, ФНОальфа и VEGF, ФНОальфа и MIF, ФНОальфа и ICAM-1, ФНОальфа и PGE4, ФНОальфа и PEG2, ФНОальфа и лиганда RANK, ФНОальфа и Te38; ФНОальфа и BAFF; ФНОальфа и CD22; ФНОальфа и CTLA-4; ФНОальфа и GP130; ФНОa и IL-12p40; VEGF и HER2, VEGF-A и HER2, VEGF-A и PDGF, HER1 и HER2, VEGF-A и VEGF-C, VEGF-C и VEGF-D, HER2 и DR5, VEGF и IL-8, VEGF и MET, VEGFR и рецептора MET, VEGFR и EGFR, HER2 и CD64, HER2 и CD3, HER2 и CD16, HER2 и HER3; EGFR(HERI) и HER2, EGFR и HER3, EGFR и HER4, IL-13 и CD40L, IL4 и CD40L, ФНОR1 и IL-1 R, ФНОR1 и IL-6R, и ФНОR1, и IL-18R, EpCAM и CD3, MAPG и CD28, EGFR и CD64, CSPG и RGM A; CTLA-4 и BTN02; IGF1 и IGF2; IGF1/2 и Erb2B; MAG и RGM A; NgR и RGM A; NogoA и RGM A; OMGp и RGM A; PDL-I и CTLA-4; и RGM A и RGM B.
В некоторых вариантах реализации изобретения мишенью является анти-VEGF, анти-c-met, анти-IgE, анти-CD11, анти-CD18, анти-CD40, анти-тканевой фактор (TF), анти-HER2 и анти-TrkC антитела. Также рассматриваются антитела, направленные против неполипептидных антигенов (таких как опухолеассоциированные гликолипидные антигены). Дополнительные типовые мишени для антител, входящие в настоящее изобретение, включают белки CD, такие как CD3, CD4, CD8, CD19, CD20, CD34 и CD46; представителей семейства рецепторов ErbB, таких как рецептор EGF, рецептор HER2, HER3 или HER4; молекулы клеточной адгезии, такие как LFA-1, Mac1, p150.95, VLA-4, ICAM-1, VCAM, α4/β7 интегрин и αv/β3 интегрин, включая их субъединицы α или β (например, анти-CD11a, анти-CD18 или анти-CD11b антитела); факторы роста, такие как VEGF; тканевой фактор (TF); TGF-β, интерферон альфа (α-IFN); интерлейкин, такой как IL-8; IgE; антигены группы крови Apo2, рецептор смерти; рецептор flk2/flt3; рецептор ожирения (OB); рецептор mpl; CTLA-4; протеин С и т.д. Биспецифические антитела в явной форме предусмотрены изобретением.
В одном варианте реализации изобретения антитело(а), например, антитело(а), применяемое(ые) в способах согласно данному документу, могут характеризоваться любыми из признаков, по отдельности или в комбинации, как описано ниже в разделах 1-6:
1. Фрагменты антител
В определенных вариантах реализации изобретения предложенное в данном документе антитело представляет собой фрагмент антитела. Фрагменты антител включают, но не ограничиваются этим, фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv, антитело с одним плечом и другие фрагменты, описанные ниже. Обзор некоторых фрагментов антител см. в Hudson et al. Nat. Med. 9:129-134 (2003). Обзор фрагментов scFv См., например, в Pluckthün, в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), pp. 269-315 (1994); также См. WO 93/16185; и патенты США № 5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное in vivo время полужизни, См. в патенте США № 5869046.
Диатела представляют собой фрагменты антител с двумя антигенсвязывающими участками, которые могут бать бивалентными или биспецифическими. См., например, EP 404,097; WO 1993/01161; Hudson et al., Nat. Med. 9:129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах реализации изобретения однодоменное антитело является человеческим однодоменным антителом (Domantis, Inc., Waltham, MA; См., например, патент США № 6248516 B1).
Антитела с одним плечом (т.е. вариабельный домен тяжелой цепи и вариабельный домен легкой цепи образуют одно антигенсвязывающее плечо) раскрыты, например, в WO 2005/063816; Martens et al., Clin Cancer Res (2006), 12: 6144. В случае лечения патологических состояний, для которых необходима антагонистическая функция, а бивалентность антитела приводит к нежелательному агонистическому эффекту, моновалентность, характерная для антитела с одним плечом (т.е. антитела, содержащего одно антигенсвязывающее плечо) приводит к и/или гарантирует наличие антагонистической функции после связывания антитела с молекулой-мишенью. Кроме того, антитело с одним плечом, содержащее Fc-область, имеет превосходящие фармакокинетические характеристики (такие как увеличенное время полужизни и/или пониженная скорость выведения in vivo) по сравнению с Fab-формами, имеющими сходные/практически идентичные антигенсвязывающие характеристики, и тем самым преодолеваются основные недостатки применения традиционных моновалентных Fab-антител. Методы получения антител с одним плечом включают, но не ограничиваются этим, метод конструирования «выступ-во-впадину» (См., например, патент США № 5731168). Примером антитела с одним плечом является онартузумаб.
Фрагменты антител можно получать различными методами, включая, но не ограничиваясь этим, протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, E. coli или фагом), как описано в данном документе.
2. Химерные и гуманизированные антитела
В определенных вариантах реализации изобретения предложенное в данном документе антитело представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в патенте США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В одном примере химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область, полученную от мыши, крысы, хомяка, кролика или отличного от человека примата, такого как обезьяна) и человеческую константную область. В дополнительном примере химерное антитело представляет собой антитело «с переключенным классом», класс или подкласс которого был изменен относительно родительского антитела. Химерные антитела включают их антигенсвязывающие фрагменты.
В определенных вариантах реализации изобретения химерное антитело является гуманизированным антителом. Как правило, нечеловеческое антитело гуманизируют, чтобы снизить его иммуногенность для людей, в то же время сохраняя специфичность и аффинность родительского нечеловеческого антитела. В общем случае гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR, например, CDR (или их части), получены из нечеловеческого антитела, а FR (или их части) получены из последовательностей человеческого антитела. Необязательно, гуманизированное антитело также содержит по меньшей мере часть человеческой константной области. В некоторых вариантах реализации изобретения некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками из нечеловеческого антитела (например, антитела, из которого получены остатки HVR), например, для сохранения или улучшения специфичности или аффинности антитела.
Обзор гуманизированных антител и способов их получения приведен, например, в Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008), а их дополнительное описание приведено, например, в Riechmann et al., Nature 332:323-329 (1988); Queen et al., Proc. Nat'l Acad. Sci. USA 86:10029-10033 (1989); патентах США № 5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36:25-34 (2005) (описывающей приживление SDR (a-CDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (описывающей «изменение поверхности»); Dall'Acqua et al., Methods 36:43-60 (2005) (описывающей «перетасовку FR»); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (описывающей подход «управляемого отбора» для перетасовки FR).
Человеческие каркасные области, которые можно использовать для гуманизации, включают, но не ограничиваются этим: каркасные области, выбранные методом «наилучшего соответствия» (См., например, Sims et al. J. Immunol. 151:2296 (1993)); каркасные области, полученные из консенсусной последовательности человеческих антител из конкретной подгруппы вариабельных областей легкой или тяжелой цепи (См., например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285 (1992); и Presta et al. J. Immunol., 151:2623 (1993)); человеческие зрелые (соматически мутированные) каркасные области или каркасные области человеческой зародышевой линии (См., например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)); и каркасные области, полученные после скрининга библиотек FR (См., например, Baca et al., J. Biol. Chem. 272:10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
3. Человеческие антитела
В определенных вариантах реализации изобретения предложенное в данном документе антитело представляет собой человеческое антитело. Человеческие антитела можно получать при помощи различных методов, известных в данной области техники. Общее описание человеческих антител приведено в van Dijk and van de Winkel, Opin. Pharmacol. 5: 368-74 (2001) и Lonberg, Curr. Opin. Immunol. 20:450-459 (2008).
Человеческие антитела можно получать путем введения иммуногена трансгенному животному, которое было модифицировано так, чтобы вырабатывать интактные человеческие антитела или интактные антитела с человеческими вариабельными областями в ответ на стимуляцию антигеном. Такие животные, как правило, содержат все или часть локусов человеческого иммуноглобулина, которые замещают эндогенные локусы иммуноглобулина, или которые присутствуют внехромосомно или случайным образом интегрированы в хромосомы животного. У таких трансгенных мышей эндогенные локусы иммуноглобулина в общем случае инактивированы. Обзор способов получения человеческих антител от трансгенных животных См. в Lonberg, Nat. Biotech. 23:1117-1125 (2005). См. также, например, патенты США № 6075181 и 6150584, описывающие технологию XENOMOUSETM; патент США № 5770429, описывающий технологию HuMab®; патент США № 7041870, описывающий технологию K-M MOUSE®, и заявку на патент США № US 2007/0061900, описывающую технологию VelociMouse®). Человеческие вариабельные области из интактных антител, вырабатываемых такими животными, можно дополнительно модифицировать, например, комбинируя с разными человеческими константными областями.
Человеческие антитела также можно получать способами на основе гибридом. Были описаны клеточные линии человеческой миеломы и мышино-человеческой гетеромиеломы для получения человеческих моноклональных антител. (См., например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147: 86 (1991).) Человеческие антитела, полученные при помощи технологии человеческой В-клеточной гибридомы, также описаны в Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные способы включают описанные, например, в патенте США № 7189826 (описывающем получение моноклональных человеческих антител IgM из клеточных линий гибридомы) и Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (описывающей человек-человеческие гибридомы). Технология человеческой гибридомы (технология триомы) также описана в Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3):185-91 (2005).
Человеческие антитела также можно получать путем выделения последовательностей вариабельных доменов Fv-клонов, отобранных из человеческих библиотек фагового дисплея. Затем такие последовательности вариабельных доменов можно комбинировать с необходимым человеческим константным доменом. Методы отбора человеческих антител из библиотек антител описаны ниже.
4. Полученные из библиотек антитела
Антитела согласно изобретению можно выделять методом скрининга комбинаторных библиотек для получения антител с необходимой активностью или активностями. Например, в данной области техники известно множество способов для генерации библиотек фагового дисплея и скрининга таких библиотек в отношении антител, обладающих необходимыми характеристиками связывания. Обзор таких способов приведен, например, в Hoogenboom et al. in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001), а дополнительное описание приведено, например, в McCafferty et al., Nature 348:552-554; Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, в Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2): 119-132(2004).
В определенных методах фагового дисплея репертуары генов VH и VL клонируют отдельно при помощи полимеразной цепной реакции (ПЦР) и повторно комбинируют случайным образом в фаговых библиотеках, после чего можно проводить их скрининг в отношении антигенсвязывающего фага, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Фаговый дисплей, как правило, представляет фрагменты антител, как в виде одноцепочечных фрагментов Fv (scFv), так и в виде фрагментов Fab. Библиотеки из иммунизированных источников обеспечивают высокоаффинные антитела к иммуногену, не требуя конструирования гибридом. В альтернативном варианте можно клонировать наивный репертуар (например, от человека), чтобы обеспечить единый источник антител для широкого диапазона чужих, а также своих антигенов без какой-либо иммунизации, как описано в Griffiths et al., EMBO J, 12: 725-734 (1993). И наконец, наивные библиотеки также можно получать синтетически - путем клонирования неперестроенных генных V-сегментов из стволовых клеток и применения ПЦР-праймеров, содержащих случайную последовательность, для кодирования высокавариабельных областей CDR3 и осуществления перестройки in vitro, как описано в Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992). Патентные публикации, описывающие фаговые библиотеки человеческих антител, включают, например: патент США № 5750373 и публикации патентов США № 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
5. Мультиспецифические антитела
В определенных вариантах реализации изобретения предложенное в данном документе антитело представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностью связывания в отношении по меньшей мере двух разных участков. В определенных вариантах реализации изобретения одна специфичность связывания направлена на c-met, а другая на любой другой антиген. В определенных вариантах реализации изобретения биспецифические антитела могут связываться с двумя разными эпитопами на c-met. Биспецифические антитела также можно применять для локализации цитотоксических агентов в клетках, которые экспрессируют c-met. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методы получения таких мультиспецифических антител включают, но не ограничиваются этим, рекомбинантную коэкспрессию двух пар тяжелая цепь-легкая цепь иммуноглобулина, имеющих разные специфичности (См. Milstein and Cuello, Nature 305: 537 (1983), WO 93/08829, и Traunecker et al., EMBO J. 10: 3655 (1991)), и метод конструирования «выступ-во-впадину» (См., например, патент США № 5731168). Мультиспецифические антитела также можно получать путем конструирования с использованием эффекта электростатического взаимодействия для получения Fc-гетеродимерных молекул антитела (WO 2009/089004A1); перекрестного связывания двух или более антител или фрагментов (См., например, патент США № 4676980 и Brennan et al., Science, 229: 81 (1985)); применения лейциновых молний для получения биспецифических антител (См., например, Kostelny et al., J. Immunol., 148(5):1547-1553 (1992)); применения технологии «диател» для получения фрагментов биспецифических антител (См., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)); и применения одноцепочечных димеров Fv (sFv) (См., например, Gruber et al., J. Immunol., 152:5368 (1994)); и получения триспецифических антител, как описано, например, в Tutt et al. J. Immunol. 147: 60 (1991).
Также в данный документ включены сконструированные антитела с тремя или более антигенсвязывающими участками, включая антитела-осьминоги (См., например, US 2006/0025576A1).
Антитело или фрагмент согласно данному документу также включает «FAb двойного действия» или «DAF», содержащее антигенсвязывающий участок, который связывается с c-met, а также другим, отличным антигеном, таким как EGFR (См., например, US 2008/0069820).
Способы получения биспецифических антител известны в данной области техники. Традиционно, рекомбинантное получение биспецифических антител основано на коэкспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина, где две тяжелые цепи имеют разные специфичности (Milstein and Cuello, Nature, 305: 537 (1983)). Вследствие случайного подбора тяжелых и легких цепей иммуноглобулина эти гибридомы (квадромы) вырабатывают потенциальную смесь из 10 разных молекул антител, из которых одна имеет правильную биспецифическую структуру. Очистка правильной молекулы, которую обычно проводят при помощи этапов аффинной хроматографии, является достаточной трудоемкой, а выход продукта низким. Похожие процедуры раскрыты в WO 93/08829, опубликованной 13 мая 1993 г., и в Traunecker et al., EMBO J., 10: 3655 (1991).
В соответствии с другим и более предпочтительным подходом, вариабельные домены антитела с необходимой специфичностью связывания (участками связывания антитело-антиген) сливают с последовательностями константного домена иммуноглобулина. Слияние предпочтительно проводят с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнирной области, области CH2 и CH3. Предпочтительно, чтобы первая константная область тяжелой цепи (CH1), содержащая участок, необходимый для связывания легкой цепи, присутствовала по меньшей мере в одном из продуктов слияния. ДНК, кодирующие продукты слияния тяжелой цепи иммуноглобулина и, при желании, легкой цепи иммуноглобулина, вставляют в отдельные экспрессионные векторы и котрансфицируют в подходящий организм-хозяина. Это обеспечивает большую гибкость в коррекции взаимного соотношения трех полипептидных фрагментов в вариантах реализации изобретения, в которых неэквивалентные соотношения трех полипептидных цепей, применяемые в конструкции, обеспечивают оптимальный выход. При этом можно вставлять кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессионный вектор, когда экспрессия по меньшей мере двух полипептидных цепей в эквивалентном соотношении приводит к высокому выходу или когда соотношение не имеет особого значения.
В предпочтительном варианте реализации этого подхода биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном плече и гибридной пары тяжелая цепь-легкая цепь иммуноглобулина (обеспечивающей вторую специфичность связывания) во втором плече. Было обнаружено, что эта ассиметричная структура облегчает отделение необходимого биспецифического соединения от нежелательных комбинаций цепей иммуноглобулина, так как присутствие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ отделения. Этот подход раскрыт в WO 94/04690. Дополнительные подробности генерации биспецифических антител см., например, в Suresh et al., Methods in Enzymology, 121:210 (1986).
В соответствии с другим подходом поверхность раздела между парой молекул антител можно сконструировать так, чтобы максимизировать процентное содержание гетеродимеров, которые выделяют из культуры рекомбинантных клеток. Предпочтительная поверхность раздела содержит по меньшей мере часть домена CH3 константного домена антитела. В этом способе одну или более небольших аминокислотных боковых цепей с поверхности раздела первой молекулы антитела замещают более крупными боковыми цепями (например, тирозин или триптофан) (выступы или выпуклости). На поверхности раздела второй молекулы антитела создают компенсирующие «углубления» (впадины) идентичного или сходного размера с крупной(ыми) боковой(ыми) цепью(ями) (например, аланин или треонин). Это обеспечивает механизм для повышения выхода гетеродимеров по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры. Выступы и впадины дополнительно описаны в данном документе.
6. Варианты антител
В определенных вариантах реализации изобретения предусмотрены варианты аминокислотных последовательностей предложенных в данном документе антител. Например, может существовать необходимость в улучшении аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотных последовательностей антител можно получать, внося соответствующие модификации в нуклеотидную последовательность, кодирующую антитело, или при помощи пептидного синтеза. Такие модификации включают, например, делеции из и/или вставки в, и/или замены остатков в аминокислотных последовательностях антител. Для получения конечной конструкции можно осуществлять любые комбинации делеций, вставок и замен, при условии, что конечная конструкция обладает необходимыми характеристиками, например, связывает антиген.
В определенных вариантах реализации изобретения предложены варианты антител, содержащие одну или более аминокислотных замен. Представляющие интерес для замещающего мутагенеза участки включают HVR и FR. Аминокислотные замены можно вносить в представляющее интерес антитело, и проводить скрининг продуктов в отношении необходимой активности, например, сохранения/улучшения связывания антигена, снижения иммуногенности или улучшения АЗКЦ или КЗЦ.
Один из типов заместительных вариантов подразумевает замещение одного или более остатков гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). В общем случае полученный(ые) в результате вариант(ы), отобранный(ые) для дополнительного исследования, будет(ут) иметь модификации (например, улучшения) определенных биологических свойств (например, повышенную аффинность, сниженную иммуногенность) по сравнению с родительским антителом и/или будет(ут) в значительной степени сохранять определенные биологические свойства родительского антитела. Типовой заместительный вариант представляет собой аффинно созревшее антитело, которое можно легко получить, например, используя методы созревания аффинности на основе фагового дисплея, такие как описаны в данном документе. Вкратце, мутируют один или более остатков HVR, а вариантные антитела представляют на фаге и проводят их скрининг в отношении конкретной биологической активности (например, аффинности связывания).
Вставки в аминокислотные последовательности включают амино- и/или карбокси-конечные слияния длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или множества аминокислотных остатков внутри последовательности. Примеры концевых вставок включают антитело с N-концевым остатком метионила. Другие инсерционные варианты антител включают слияние N- или C-конца антитела с ферментом (например, для ADEPT) или полипептид, который увеличивает сывороточное время полужизни антитела.
В определенных вариантах реализации изобретения предложенное в данном документе антитело изменяют, чтобы повысить или понизить степень гликозилирования антитела. Добавление или удаление участков гликозилирования в антителе удобно осуществлять путем изменения аминокислотной последовательности так, чтобы создать или удалить один или более участков гликозилирования.
Если антитело содержит Fc-область, можно изменять присоединенный к ней углевод. Нативные тела, вырабатываемые клетками млекопитающих, как правило, содержат разветвленный 2-антенарный олигосахарид, который в общем случае присоединен посредством N-связи к Asn297 домена CH2 Fc-области. См., например, Wright et al. TIBTECH 15:26-32 (1997). Олигосахарид может содержать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» 2-антенарной олигосахаридной структуры. В некоторых вариантах реализации изобретения модификации олигосахарида в антителе согласно изобретению можно осуществлять с целью создания вариантов антител с определенными улучшенными свойствами.
В одном варианте реализации изобретения предложены варианты антител, содержащие углеводную структуру, в которой отсутствует фукоза, присоединенная (прямым или непрямым образом) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют, рассчитывая среднее количество фукозы в составе сахарной цепи в Asn297 по отношению к сумме всех гликоструктур, присоединенных к Asn 297 (например, комплексных, гибридных и структур с высоким содержанием маннозы), по данным масс-спектрометрии MALDI-TOF, как описано, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному приблизительно в позиции 297 в Fc-области (нумерация EU остатков Fc-области); при этом Asn297 также может бать расположен приблизительно на ± 3 аминокислоты выше или ниже позиции 297, т.е. между позициями 294 и 300, вследствие незначительных вариаций последовательностей в антителах. Такие фукозилированные варианты могут иметь улучшенную функцию АЗКЦ. См., например, публикации патентов США № US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, связанных с «дефукозозилированными» или «фукозодефицитными» вариантами антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры клеточных линий, способных вырабатывать дефукозозилированные антитела, включают клетки Lec13 CHO с дефицитом фукозилирования белка (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); заявка на патент США US 2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., в особенности Пример 11) и клетки с генным нокаутом, такие как клетки СНО с нокаутом гена альфа-1,6-фукозилтрансферазы, FUT8, (См., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94(4):680-688 (2006); and WO 2003/085107).
Дополнительно предложены варианты антител с разделенными пополам олигосахаридами, например, в которых 2-антенарный олигосахарид, присоединенный к Fc-области антитела, разделен пополам GlcNAc. Такие варианты антител могут иметь пониженную степень фукозилирования и/или улучшенную функцию АЗКЦ. Примеры таких вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США № 6602684 (Umana et al.); и US 2005/0123546 (Umana et al.). Также предложены варианты антител с по меньшей мере одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Такие варианты антител могут иметь улучшенную функцию КЗЦ. Такие варианты антител описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
В определенных вариантах реализации изобретения в Fc-область предложенного в данном документе антитела можно вносить одну или более аминокислотных модификаций, тем самым получая вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-область человеческого IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одной или более аминокислотных позициях.
В определенных вариантах реализации изобретения в изобретении предусмотрен вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для применений, в которых важным является in vivo время полужизни антитела, а некоторые эффекторные функции (такие как комплемент-зависимая и АЗКЦ) являются необязательными или вредными.
Антитела со сниженной эффекторной функцией включают антитела с заменой одного или более остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 ( патент США № 6737056). Такие Fc-мутанты включают Fc-мутантов с заменами в двух или более аминокислотных позициях 265, 269, 270, 297 и 327, включая так называемый Fc-мутант «DANA» с заменой остатков 265 и 297 на аланин (патент США № 7332581).
Описаны некоторые варианты антител с улучшенным или пониженным связыванием с FcR. (См., например, патент США № 6737056; WO 2004/056312, и Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001).)
В определенных вариантах реализации изобретения вариант антитела содержит Fc-область с одной или более аминокислотными заменами, которые улучшают АЗКЦ, например, заменами в позициях 298, 333 и/или 334 Fc-области (нумерация остатков EU).
В некоторых вариантах реализации изобретения проводят замены в Fc-области, которые приводят к изменению (т.е. улучшению или снижению) связывания C1q и/или комплемент-зависимой цитотоксичности (КЗЦ), например, как описано в патенте США № 6194551, WO 99/51642, и Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Антитела с повышенным временем полужизни и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Эти антитела содержат Fc-область с одной или более заменами в ней, которые улучшают связывание Fc-области с FcRn. Такие Fc-варианты включают варианты с заменами в одном или более остатках Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, заменой остатка 434 Fc-области (патент США № 7371826).
Также См. Duncan & Winter, Nature 322:738-40 (1988); патент США № 5648260; патент США № 5624821; и WO 94/29351 в отношении других примеров вариантов Fc-области.
В определенных вариантах реализации изобретения может возникать необходимость в создании цистеин-сконструированных антител, например, «thioMAb», в которых один или более остатков антитела замещены остатками цистеина. В конкретных вариантах реализации изобретения замещаемые остатки находятся в доступных участках антитела. Путем замещения этих остатков цистеином в доступные участки антитела помещают реактивные тиольные группы, которые можно использовать для конъюгации антитела с другими компонентами, такими как лекарственные компоненты или линкерно-лекарственные компоненты, чтобы создать иммуноконъюгат, как дополнительно описано в данном документе. В определенных вариантах реализации изобретения любой один или более из следующих остатков можно заменить цистеином: V205 (нумерация Кабата) легкой цепи; A118 (нумерация EU) тяжелой цепи; и S400 (нумерация EU) Fc-области тяжелой цепи. Цистеин-сконструированные антитела можно получать, как описано, например, в патенте США № 7521541.
Выступ-во-впадину
Применение технологии выступ-во-впадину как метода получения мультиспецифических антител и/или антител с одним плечом, и/или иммуноадгезинов хорошо известно в данной области техники. См. патент США № 5731168, выданный 24 марта 1998 г., закрепленный за Genentech, заявку согласно РСТ № WO2009089004, опубликованную 16 июля 2009 г., и закрепленную за Amgen, и публикацию патента США № 20090182127, опубликованную 16 июля 2009 г., и закрепленную за Novo Nordisk A/S. Также См. Marvin and Zhu, Acta Pharmacologica Sincia (2005) 26(6):649-658 и Kontermann (2005) Acta Pharacol. Sin., 26:1-9. Краткое обсуждение приведено в данном документе.
«Выпуклостью» называется по меньшей мере одна аминокислотная боковая цепь, которая выступает с поверхности раздела первого полипептида и, следовательно, может размещаться в компенсирующем углублении смежной поверхности раздела (т.е. поверхности раздела второго полипептида) так, чтобы стабилизировать гетеромультимер и тем самым способствовать, например, гетеромультимерной формации по сравнению с гомомультимерной формацией. Выпуклость может находиться на исходной поверхности раздела или может быть привнесена синтетически (например, путем изменения нуклеиновой кислоты, кодирующей поверхность раздела). Обычно, чтобы кодировать выпуклость, изменяют нуклеиновую кислоту, кодирующую поверхность раздела первого полипептида. Для достижения этого нуклеиновую кислоту, кодирующую по меньшей мере один «исходный» аминокислотный остаток на поверхности раздела первого полипептида, заменяют нуклеиновой кислотой, кодирующей по меньшей мере один «вносимый» аминокислотный остаток, который имеет больший объем боковой цепи, чем исходный аминокислотный остаток. Следует понимать, что может быть более одного исходного и соответствующего вносимого остатка. Верхняя граница для числа заменяемых исходных остатков соответствует общему числу остатков на поверхности раздела первого полипептида. Объем боковых цепей различных аминокислотных остатков приведен в следующей таблице.
ТАБЛИЦА B
Свойства аминокислотных остатков
a Молекулярная масса аминокислоты минус масса воды. Значения из Handbook of Chemistry and Physics, 43rd ed. Cleveland, Chemical Rubber Publishing Co., 1961.
b Значения из A.A. Zamyatnin, Prog. Biophys. Mol. Biol. 24:107-123, 1972.
c Значения из C. Chothia, J. Mol. Biol. 105:1-14, 1975. Доступная площадь поверхности определена на Фигурах 6-20 по этой ссылке.
Предпочтительными вносимыми остатками для образования выпуклости являются в общем случае аминокислотные остатки природного происхождения, которые предпочтительно выбраны из аргинина (R), фенилаланина (F), тирозина (Y) и триптофана (W). Наиболее предпочтительными являются триптофан и тирозин. В одном варианте реализации изобретения исходный остаток для образования выпуклости имеет небольшой объем боковой цепи, такой как аланин, аспарагин, аспарагиновая кислота, глицин, серин, треонин или валин. Типовые аминокислотные замены в домене СН3 для образования выпуклости включают, без ограничений, замену T366W.
«Углубление» относится по меньшей мере к одной аминокислотной боковой цепи, которая утоплена в поверхность раздела второго полипептида и, следовательно, в ней размещается соответствующая выпуклость смежной поверхности раздела первого полипептида. Углубление может находиться на исходной поверхности раздела или может быть привнесено синтетически (например, путем изменения нуклеиновой кислоты, кодирующей поверхность раздела). Обычно, чтобы кодировать углубление, изменяют нуклеиновую кислоту, кодирующую поверхность раздела второго полипептида. Для достижения этого нуклеиновую кислоту, кодирующую по меньшей мере один «исходный» аминокислотный остаток на поверхности раздела второго полипептида, заменяют ДНК, кодирующей по меньшей мере один «вносимый» аминокислотный остаток, который имеет меньший объем боковой цепи, чем исходный аминокислотный остаток. Следует понимать, что может быть более одного исходного и соответствующего вносимого остатка. Верхняя граница для числа заменяемых исходных остатков соответствует общему числу остатков на поверхности раздела второго полипептида. Объем боковых цепей ля различных аминокислотных остатков приведен выше в Таблице В. Предпочтительными вносимыми остатками для образования углубления обычно являются аминокислотные остатки природного происхождения, которые предпочтительно выбраны из аланина (A), серина (S), треонина (T) и валина (V). Наиболее предпочтительными являются серин, аланин или треонин. В одном варианте реализации изобретения исходный остаток для образования углубления имеет большой объем боковой цепи, такой как тирозин, аргинин, фенилаланин или триптофан. Типовые аминокислотные замены в домене СН3 для образования углубления включают, без ограничений, замены T366S, L368A и Y407A.
«Исходный» аминокислотный остаток представляет собой остаток, который замещают «вносимым» остатком, который может иметь больший или меньший объем боковой цепи по сравнению с исходным остатком. Вносимый аминокислотный остаток может быть аминокислотным остатком природного или неприродного происхождения, но предпочтительно является первым. Аминокислотные остатки «природного происхождения» - это остатки, кодируемые генетическом кодом и приведенные выше в Таблице В. Под аминокислотным остатком «неприродного происхождения» подразумевается остаток, который не кодируется генетическим кодом, но который способен ковалентно связывать смежный(ые) аминокислотный(ые) остаток(ки) в полипептидной цепи. Примерами аминокислотных остатков неприродного происхождения являются норлейцин, орнитин, норвалин, гомосерин и другие аналоги аминокислотных остатков, такие как описанные, например, в Ellman et al., Meth. Enzym. 202:301-336 (1991). Для получения таких аминокислотных остатков неприродного происхождения можно использовать способы согласно Noren et al. Science 244: 182 (1989) и Ellman et al., выше. Вкратце, они включают химическую активацию супрессорной тРНК с аминокислотным остатком неприродного происхождения с последующей in vitro транскрипцией и трансляцией РНК. Способ согласно данному изобретению включает замену по меньшей мере одного исходного аминокислотного остатка, но можно проводить замену и более одного исходного остатка. Обычно не более чем все остатки на поверхности раздела первого или второго полипептида будут составлять замещаемые исходные аминокислотные остатки. Как правило, исходные остатки для замещения являются «углубленными». Под «углубленным» подразумевается, что остаток практически недоступен для растворителя. В общем случае вносимый остаток не является цистеином для предотвращения возможного окисления или неправильного спаривания дисульфидных связей.
«Выпуклость» может быть размещена в углублении, что означает, что пространственное расположение выпуклости и углубления на поверхности раздела первого полипептида и второго полипептида, соответственно, совпадает, а размеры выпуклости и углубления являются такими, что выпуклость может расположиться в углублении без существенного нарушения нормальной связи на поверхности раздела первого и второго полипептидов. Так как выпуклости, такие как Tyr, Phe и Trp, как правило, не перпендикулярны оси поверхности раздела и имеют предпочтительные конформации, выравнивание выпуклости с соответствующим углублением основано на моделировании пары выпуклость/углубление на основании трехмерной структуры, которую получают методами рентгеновской кристаллографии или ядерного магнитного резонанса (ЯМР). Этого можно достичь, используя широко применяемые в данной области техники методы.
Под «исходной или матричной» нуклеиновой кислотой подразумевается нуклеиновая кислота, кодирующая представляющий интерес полипептид, который может быть «изменен» (т.е. генетически сконструирован или мутирован) так, чтобы кодировать выпуклость или углубление. Исходная или стартовая нуклеиновая кислота может быть нуклеиновой кислотой природного происхождения или может содержать нуклеиновую кислоту, которая ранее подвергалась изменению (например, фрагмент гуманизированного антитела). Под «изменением» нуклеиновой кислоты подразумевается, что исходная нуклеиновая кислота была мутирована путем вставки, удаления или замены по меньшей мере одного кодона, кодирующего представляющий интерес аминокислотный остаток. Обычно кодон, кодирующий исходный остаток, замещают кодоном, кодирующим вносимый остаток. Обзор методов генетической модификации ДНК подобным образом приведен в Mutagenesis: a Practical Approach, M.J. McPherson, Ed., (IRL Press, Oxford, UK. (1991), и включает, например, сайт-направленный мутагенез, кассетный мутагенез и мутагенез с применением полимеразной цепной реакции (ПЦР). Путем мутирования исходной/матричной нуклеиновой кислоты осуществляют, соответственно, замену исходного/матричного полипептида, кодируемого исходной/матричной нуклеиновой кислотой.
Выпуклость или углубление можно «привносить» в поверхность раздела первого или второго полипептида синтетическим образом, например, при помощи рекомбинантных технологий, in vitro пептидного синтеза, описанных ранее методов внесения аминокислотных остатков неприродного происхождения, путем ферментативного или химического сопряжения пептидов или некоторым образом комбинируя эти методы. Соответственно, «привнесенные» выпуклость или углубление являются «неприродными по происхождению» или «ненативными», что означает, что они не существуют в природе или в исходном полипептиде (например, гуманизированном моноклональном антителе).
В общем случае вносимый аминокислотный остаток для образования выпуклости имеет относительно небольшое число «ротамеров» (например, около 3-6). «Ротамер» представляет собой наиболее энергетически выгодную конформацию аминокислотной боковой цепи. Количество ротамеров в различных аминокислотных остатках обсуждается в Ponders and Richards, J. Mol. Biol. 193: 775-791 (1987).
В одном варианте реализации изобретения первый Fc-полипептид и второй Fc-полипептид стыкуются/взаимодействуют на поверхности раздела. В некоторых вариантах реализации изобретения, в которых первый и второй полипептиды стыкуются на поверхности раздела, поверхность раздела второго Fc-полипептида (последовательность) содержит выпуклость (также называемую «выступом»), которая может размещаться в углублении (также называемом «впадиной») на поверхности раздела первого Fc-полипептида (последовательности). В одном варианте реализации изобретения первый Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать углубление, или второй Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать выпуклость, или имеют место оба варианта. В одном варианте реализации изобретения первый Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать углубление, а второй Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать выпуклость. В одном варианте реализации изобретения поверхность раздела второго Fc-полипептида содержит выпуклость, которая может размещаться в углублении на поверхности раздела первого Fc-полипептида, при этом углубление или выпуклость, или оба эти элемента были привнесены в поверхности раздела первого и второго Fc-полипептидов, соответственно. В некоторых вариантах реализации изобретения, в которых первый и второй полипептиды стыкуются на поверхности раздела, поверхность раздела первого Fc-полипептида (последовательность) содержит выпуклость, которая может размещаться в углублении на поверхности раздела второго Fc-полипептида (последовательности). В одном варианте реализации изобретения второй Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать углубление, или первый Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать выпуклость, или имеют место оба варианта. В одном варианте реализации изобретения второй Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать углубление, а первый Fc-полипептид был изменен относительно матричного/исходного полипептида так, чтобы кодировать выпуклость. В одном варианте реализации изобретения поверхность раздела первого Fc-полипептида содержит выпуклость, которая может размещаться в углублении на поверхности раздела второго Fc-полипептида, при этом выпуклость или углубление, или оба эти элемента были привнесены в поверхности раздела первого и второго Fc-полипептидов, соответственно.
В одном варианте реализации изобретения и выпуклость, и углубление содержат аминокислотный остаток природного происхождения. В одном варианте реализации изобретения Fc-полипептид, содержащий выпуклость, получают путем замещения исходного остатка на поверхности раздела матричного/исходного полипептида вносимым остатком, имеющим больший объем боковой цепи, чем исходный остаток. В одном варианте реализации изобретения Fc-полипептид, содержащий выпуклость, получают способом, включающим этап, на котором полинуклеотид, кодирующий исходный остаток на поверхности раздела указанного полипептида, замещают полинуклеотидом, кодирующим вносимый остаток, имеющий больший объем боковой цепи, чем исходный. В одном варианте реализации изобретения исходным остатком является треонин. В одном варианте реализации изобретения исходным остатком является T366. В одном варианте реализации изобретения вносимым остатком является аргинин (R). В одном варианте реализации изобретения вносимым остатком является фенилаланин (F). В одном варианте реализации изобретения вносимым остатком является тирозин (Y). В одном варианте реализации изобретения вносимым остатком является триптофан (W). В одном варианте реализации изобретения вносимым остатком является R, F, Y или W. В одном варианте реализации изобретения выпуклость получают путем замещения двух или более остатков в матричном/исходном полипептиде. В одном варианте реализации изобретения Fc-полипептид, содержащий выпуклость, содержит замещение треонина в позиции 366 триптофаном, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. (pp. 688-696 в Sequences of proteins of immunological interest, 5th ed., Vol. 1 (1991; NIH, Bethesda, MD)).
В некоторых вариантах реализации изобретения Fc-полипептид, содержащий углубление, получают путем замещения исходного остатка на поверхности раздела матричного/исходного полипептида вносимым остатком, имеющим меньший объем боковой цепи, чем исходный остаток. Например, Fc-полипептид, содержащий углубление, можно получить способом, включающим этап, на котором полинуклеотид, кодирующий исходный остаток на поверхности раздела указанного полипептида, замещают полинуклеотидом, кодирующим вносимый остаток, имеющий меньший объем боковой цепи, чем исходный. В одном варианте реализации изобретения исходным остатком является треонин. В одном варианте реализации изобретения исходным остатком является лейцин. В одном варианте реализации изобретения исходным остатком является тирозин. В одном варианте реализации изобретения вносимым остатком является цистеин (C). В одном варианте реализации изобретения вносимым остатком является аланин (A). В одном варианте реализации изобретения вносимым остатком является серин (S). В одном варианте реализации изобретения вносимым остатком является треонин (T). В одном варианте реализации изобретения вносимым остатком является валин (V). Углубление можно получить путем замещения одного или более исходных остатков матричного/исходного полипептида. Например, в одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит замещение двух или более исходных аминокислот, выбранных из группы, состоящей из треонина, лейцина и тирозина. В одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит два или более вносимых остатка, выбранных из группы, состоящей из аланина, серина, треонина и валина. В некоторых вариантах реализации изобретения Fc-полипептид, содержащий углубление, содержит замещение двух или более исходных аминокислот, выбранных из группы, состоящей из треонина, лейцина и тирозина, а указанные исходные аминокислоты замещены вносимыми остатками, выбранными из группы, состоящей из аланина, серина, треонина и валина. В некоторых вариантах реализации изобретения замещаемой исходной аминокислотой является T366, L368 и/или Y407. В одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит замещение треонина в позиции 366 серином, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. выше. В одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит замещение лейцина в позиции 368 аланином, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. выше. В одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит замещение тирозина в позиции 407 валином, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. выше. В одном варианте реализации изобретения Fc-полипептид, содержащий углубление, содержит две или более аминокислотные замены, выбранные из группы, состоящей из T366S, L368A и Y407V, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. выше. В некоторых вариантах реализации этих фрагментов антител, Fc-полипептид, содержащий выпуклость, содержит замещение треонина в позиции 366 триптофаном, нумерация аминокислот соответствует схеме нумерации EU по Kabat et al. выше.
В одном варианте реализации изобретения антитело содержит Fc-мутации, составляющие «выступы» и «впадины», как описано в WO2005/063816. Например, мутацией, приводящей к образованию впадины, может быть одна или более из T366A, L368A и/или Y407V в Fc-полипептиде, а мутацией, приводящей к образованию выступа, может быть T366W.
В определенных вариантах реализации изобретения предложенное в данном документе антитело можно дополнительно модифицировать так, чтобы оно содержало дополнительные небелковые компоненты, которые известны в данной области техники и легкодоступны. Компоненты, подходящие для дериватизации антитела, включают, но не ограничиваются этим, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, но не ограничиваются этим, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоль/пропиленгликоль, карбоксиметилцеллюлозу, декстрин, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилен/малеиновый ангидрид, полиаминокислоты (как гомополимеры, так и случайные сополимеры) и декстран или поли(n-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксид/этиленоксид, полиоксиэтилированные полиолы (например, глицерол), поливиниловый спирт и их смеси. Полиэтиленгликоль пропиональдегид может иметь преимущество при производстве благодаря своей стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным и неразветвленным. Число полимеров, присоединенных к антителу, может варьироваться, а если присоединено более одного полимера, они могут представлять собой одинаковые или разные молекулы. В общем случае число и/или тип полимеров, применяемых в дериватизации, можно определить на основании факторов, включая, но не ограничиваясь этим, конкретные свойства или функции антитела, нуждающиеся в улучшении, того, будет ли антитело применяться в терапии в определенных условиях, и т.д.
В другом варианте реализации изобретения предложены конъюгаты антитела и небелкового компонента, который может избирательно нагреваться под воздействием излучения. В одном варианте реализации изобретения небелковый компонент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Излучение может быть любой длины волны и включает, но не ограничивается этим, длины волн, которые не наносят вред обычным клеткам, но которые нагревают небелковый компонент до температуры, при которой клетки, близко расположенные к конъюгату антитело-небелковый компонент, погибают.
В одном варианте реализации изобретения медикамент представляет собой иммуноконъюгат, содержащий антитело (такое как антитело c-met), конъюгированное с одним или более цитотоксическими агентами, такими как химиотерапевтические агенты или лекарственные препараты, агенты, ингибирующие рост, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты), или радиоактивные изотопы.
В одном варианте реализации изобретения иммуноконъюгат представляет собой конъюгат антитело-лекарственный препарат (ADC), в котором антитело конъюгировано с одним или более лекарственными препаратами, включая, но не ограничиваясь этим, майтанзиноид (См. патенты США № 5208020, 5416064 и Европейский патент EP 0 425 235 B1); ауристатин, такой как лекарственные препараты монометилауристатина DE и DF (MMAE и MMAF) (См. патенты США № 5635483 и 5780588, и 7498298); доластатин; калихеамицин или его производное (См. патенты США № 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman et al., Cancer Res. 53:3336-3342 (1993); и Lode et al., Cancer Res. 58:2925-2928 (1998)); антрациклин, такой как дауномицин или доксорубицин (См. Kratz et al. Current Med. Chem. 13:477-523 (2006); Jeffrey et al., Bioorganic & Med. Chem. Letters 16:358-362 (2006); Torgov et al., Bioconj. Chem. 16:717-721 (2005); Nagy et al., Proc. Natl. Acad. Sci. USA 97:829-834 (2000); Dubowchik et al., Bioorg. & Med. Chem. Letters 12:1529-1532 (2002); King et al., J. Med. Chem. 45:4336-4343 (2002); и патент США № 6630579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и CC1065.
В другом варианте реализации изобретения иммуноконъюгат содержит описанное в данном документе антитело, конъюгированное с ферментативно активным токсином или его фрагментом, включая, но не ограничиваясь этим, А-цепь дифтерии, несвязывающие активные фрагменты токсина дифтерии, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модессина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор момордики харантской, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом варианте реализации изобретения иммуноконъюгат содержит описанное в данном документе антитело, конъюгированное с радиоактивным атомом, для получения радиоканъюгата. Для получения радиоканъюгатов доступно множество радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если радиоканъюгат применяют для выявления, он может содержать радиоактивный атом для сцинтиграфических исследований, например, tc99m или I123, или спиновую метку для визуализации методом ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента можно получать, применяя множество бифункциональных агентов для белкового сопряжения, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (СПДП), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (СМЦК), иминотиолан (ИТ), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидо соединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензен). Например, иммунотоксин рицин можно получить так, как описано в Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминпентауксусная кислота (MX-ДТПА) является типовым хелатирующим агентом для конъюгации радионуклеотида к антителу. См. WO94/11026. Линкер может представлять собой «отщепляемый линкер», облегчающий высвобождение цитотоксического препарата в клетку. Например, можно использовать кислотно-лабильный линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); патент США № 5208020).
Иммуноконъюгаты или ADC согласно данному документу явным образом включают, но не ограничиваются конъюгатами, полученными при помощи перекрестно-сшивающих реагентов, включая, но не ограничиваясь этим, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A).
Векторы, клетки-хозяева и рекомбинантные методы
Для рекомбинантного получения гетерологичного полипептида (например, антитела) выделяют кодирующую его нуклеиновую кислоту и вставляют в реплицируемый вектор для дополнительного клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую полипептид (например, антитело), нетрудно выделить и секвенировать, применяя традиционные процедуры (например, применяя олигонуклеотидные зонды, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Многие векторы являются доступными. Выбор вектора частично зависит от предназначенной для применения клетки-хозяина. В общем случае предпочтительные клетки-хозяева имеют любое прокариотическое происхождение. Следует понимать, что в этих целях можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области могут быть получены от любого вида человека или животного.
a. Получение антител при помощи прокариотических клеток-хозяев:
i. Конструирование векторов
Полинуклеотидные последовательности, кодирующие полипептидные компоненты полипептида (например, антитела) согласно изобретению, можно получить при помощи стандартных рекомбинантных технологий. Необходимые полинуклеотидные последовательности можно выделять и секвенировать из вырабатывающих антитела клеток, таких как клетки гибридомы. В альтернативном варианте полинуклеотиды можно синтезировать при помощи нуклеотидного синтезатора или ПЦР-технологий. После получения последовательности, кодирующие полипептиды вставляют в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. Многие доступные и известные в данной области техники векторы можно применять в целях настоящего изобретения. Выбор подходящего вектора зависит, главным образом, от размера нуклеиновых кислот, предназначенных для вставки в вектор, и конкретной клетки-хозяина, предназначенной для трансформации вектором. Каждый вектор содержит различные компоненты в зависимости от его функции (амплификации или экспрессии гетерологичного полинуклеотида, или обоих вариантов) и его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора в общем случае включают, но не ограничиваются этим: точку начала репликации, ген селективного маркера, промотор, участок связывания рибосомы (УСР), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
В общем случае плазмидные векторы, содержащие репликон и регуляторные последовательности, которые получены от вида, совместимого с клеткой-хозяином, применяют в отношении этих хозяев. Вектор обычно несет сайт репликации, а также маркерные последовательности, которые способны обеспечить фенотипический отбор в трансформированных клетках. Например, E. coli, как правило, трансформируют pBR322 - плазмидой, полученной от вида E. coli. pBR322 содержит гены, кодирующие устойчивость к ампициллину (Amp) и тетрациклину (Tet), и тем самым обеспечивает простое средство для выявления трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаги также могут содержать или быть модифицированы так, чтобы содержать промоторы, которые могут использоваться микроорганизмом для экспрессии эндогенных белков. Примеры производных pBR322, применяемых для экспрессии конкретных антител, подробно описаны в Carter et al., патенте США № 5648237.
Кроме того, фаговые векторы, содержащие репликон и регуляторные последовательности, которые совместимы с микроорганизмом-хозяином, можно применять в качестве трансформирующих векторов в отношении этих хозяев. Например, бактериофаг, такой как λGEM.TM.-11, можно использовать при создании рекомбинантного вектора, который можно использовать для трансформации восприимчивых клеток-хозяев, таких как E. coli LE392.
Экспрессионный вектор согласно изобретению может содержать две или более пары промотор-цистрон, кодирующие каждый из полипептидных компонентов. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5') цистрона, которая модулирует его экспрессию. Прокариотические промоторы, как правило, делятся на два класса - индуцибельные и конститутивные. Индуцибельный промотор представляет собой промотор, который инициирует повышение уровней транскрипции цистрона, находящегося под его управлением, в ответ на изменения культуральных условий, например, присутствие или отсутствие питательных веществ или изменение температуры.
Хорошо известно большое число промоторов, распознаваемых множеством потенциальных клеток-хозяев. Выбранный промотор может быть функционально связан с цистронной ДНК, кодирующей легкую или тяжелую цепь, путем удаления промотора из источника ДНК при помощи расщепления рестрикционными ферментами и вставки выделенной промоторной последовательности в вектор согласно изобретению. Для управления амплификацией и/или экспрессией генов-мишеней можно использовать как нативную промоторную последовательность, так и множество гетерологичных промоторов. В некоторых вариантах реализации изобретения используют гетерологичные промоторы, так как они в общем случае позволяют получить более высокий уровень транскрипции и выход экспрессируемого гена-мишени по сравнению с нативным промотором полипептида-мишени.
Промоторы, подходящие для применения с прокариотическими хозяевами, включают промотор PhoA, системы промоторов β-лактамазы и лактозы, систему промотора триптофана (trp) и гибридные промоторы, такие как промотор tac или trc. При этом подходят также другие промоторы, которые функциональны в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности были опубликованы, следовательно, у специалиста в данной области есть возможность функционально лигировать их в цистроны, кодирующие легкие и тяжелые цепи-мишени (Siebenlist et al., (1980) Cell 20: 269), используя линкеры или адаптеры, чтобы обеспечить необходимые рестрикционные сайты.
Область инициации трансляции (TIR) является основным определяющим фактором общего уровня трансляции белка. TIR включает полинуклеотид, который кодирует сигнальную последовательность и простирается от участка непосредственно выше последовательности Шайна-Дальгарно до участка приблизительно на двадцать нуклеотидов ниже стартового кодона. В общем случае вектор содержит TIR. TIR и вариантные TIR известны в данной области техники, и способы получения TIR известны в данной области техники. Можно создать ряд вариантов нуклеотидных последовательностей с некоторым диапазоном трансляционной эффективности, тем самым обеспечивая удобное средство для корректировки этого фактора для оптимальной секреции многих разных полипептидов. Применение репортерного гена, слитого с этими вариантами, такого как PhoA, обеспечивает способ количественной оценки относительной эффективности трансляции разных областей инициации трансляции. Вариантные или мутантные TIR могут принадлежать фоновому генотипу плазмидного вектора, тем самым обеспечивая наличие группы плазмид, в которые можно вставить представляющий интерес ген и измерить его экспрессию так, чтобы установить оптимальный диапазон эффективности трансляции для максимальной экспрессии зрелого полипептида. Вариантные TIR раскрыты в патенте США 8241901.
В одном аспекте изобретения каждый цистрон в рекомбинантном векторе содержит компонент секреторной сигнальной последовательности, который управляет транслокацией экспрессируемых полипептидов через мембрану. В общем случае сигнальная последовательность может быть компонентом вектора или она может быть частью ДНК полипептида-мишени, которая вставлена в вектор. Сигнальная последовательность, выбранная в целях данного изобретения, должна распознаваться и процессироваться (т.е. отщепляться сигнальной пептидазой) клеткой-хозяином. В случае прокариотических клеток-хозяев, которые не распознают и не процессируют сигнальные последовательности, нативные по отношению к гетерологичным полипептидам, сигнальную последовательность замещают прокариотической сигнальной последовательностью, выбранной, например, из сигнальных полипептидов согласно настоящему изобретению. Кроме того, вектор может содержать сигнальную последовательность, выбранную из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Lpp или термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP.
В одном аспекте один или более полинуклеотидов (например, экспрессионных векторов) вместе кодируют антитело. В одном варианте реализации изобретения один полинуклеотид кодирует легкую цепь антитела, а отдельный полинуклеотид кодирует тяжелую цепь антитела. В одном варианте реализации изобретения один полинуклеотид кодирует легкую цепь и тяжелую цепь антитела. В некоторых вариантах реализации изобретения один или более полинуклеотидов (например, экспрессионных векторов) вместе кодируют антитело с одним плечом. В одном варианте реализации изобретения один полинуклеотид кодирует (a) легкую и тяжелую цепь антитела с одним плечом и (b) Fc-полипептид. В одном варианте реализации изобретения один полинуклеотид кодирует легкую и тяжелую цепь антитела с одним плечом, а отдельный полинуклеотид кодирует Fc-полипептид. В одном варианте реализации изобретения отдельные полинуклеотиды кодируют компонент легкой цепи антитела с одним плечом, компонент тяжелой цепи антитела с одним плечом и Fc-полипептид, соответственно. Получение антитела с одним плечом описано, например, в WO 2005063816.
Прокариотические клетки-хозяева, подходящие для экспрессии антител согласно изобретению, включают архебактерии и эубактерии, такие как грамотрицательные и грамположительные микроорганизмы. Примеры подходящих бактерий включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, вид Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном варианте реализации изобретения применяют грамотрицательные клетки. В одном варианте реализации изобретения клетки E. coli применяют в качестве хозяев в целях изобретения. Примеры штаммов E. coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219; ATCC Deposit № 27325) и его производные, включая штамм 33D3, имеющий генотип W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTΔ(nmpc-fepE) degP41 kanR (патент США № 5639635), и штаммы 63C1, 66F8 и 64B4. Также подходят другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coliλ 1776 (ATCC 31537) и E. coli RV308(ATCC 31608). Эти примеры являются иллюстративными, а не ограничивающими. Способы конструирования производных любых из вышеупомянутых бактерий, имеющих определенные генотипы, известны в данной области техники и описаны, например, в Bass et al., Proteins, 8:309-314 (1990). В общем случае необходимо выбрать подходящие бактерии, принимая во внимание способность репликона к репликации в клетках бактерий. Например, E. coli, виды Serratia или Salmonella можно использовать в качестве хозяев, когда для поддержания репликона применяют хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Как правило, клетка-хозяин должна секретировать минимальное количество протеолитических ферментов, а в клеточную культуру желательно включать дополнительные ингибиторы протеаз.
ii. Получение антител
Клетки-хозяев трансформируют вышеописанными экспрессионными векторами и культивируют в традиционной питательной среде, модифицированной надлежащим образом для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих необходимые последовательности.
Трансформация означает внесение ДНК в прокариотические организмы-хозяев так, чтобы ДНК могла реплицироваться как в качестве внехромосомного элемента, так и в качестве хромосомного интегранта. В зависимости от применяемой клетки-хозяина трансформацию проводят, применяя стандартные методы, подходящие для таких клеток. Обработку кальцием, в которой применяют хлорид кальция, в общем случае используют для бактериальных клеток, которые имеют прочную клеточную оболочку. В другом способе трансформации применяют полиэтиленгликоль/ДМСО. Еще одним применяемым методом является электропорация.
Прокариотические клетки, применяемые для получения полипептидов согласно изобретению, выращивают в известных в данной области техники средах, подходящих для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают бульон Луриа (LB) плюс необходимые питательные добавки. В некоторых вариантах реализации изобретения среда также содержит селекционный агент, выбранный на основании конструкции экспрессионного вектора, чтобы избирательно создавать возможность для роста прокариотических клеток, содержащих экспрессионный вектор. Например, в среду для роста клеток, экспрессирующих ген устойчивости к ампициллину, добавляют ампициллин.
Любые необходимые добавки, кроме источников углерода, азота и неорганических фосфатов, также могут быть включены в подходящих концентрациях, внесены отдельно или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно, культуральная среда может содержать один или более восстанавливающих агентов, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликоля, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяев культивируют при подходящих температурах. Для роста E. coli, например, предпочтительная температура соответствует диапазону от около 20°C до около 39°C, более предпочтительно - от около 25°C до около 37°C, еще более предпочтительно - около 30°C. pH среды может соответствовать любому рН в диапазоне от около 5 до около 9 в зависимости, главным образом, от организма-хозяина. Для E. coli pH предпочтительно составляет от около 6,8 до около 7,4 и более предпочтительно - около 7,0.
Если в экспрессионном векторе согласно изобретению применяют индуцибельный промотор, экспрессия белка индуцируется в условиях, подходящих для активации промотора. В одном аспекте изобретения для регуляции транскрипции полипептидов используют промоторы PhoA. Соответственно, для индукции трансформированные клетки-хозяев культивируют в среде с ограниченным содержанием фосфатов. Предпочтительно среда с ограниченным содержанием фосфатов представляет собой среду C.R.A.P (См., например, Simmons et al., J. Immunol. Methods (2002), 263:133-147) или среды, описанные в WO2002/061090. Как известно в данной области техники, в соответствии с применяемой векторной конструкцией можно использовать множество других индукторов.
В одном варианте реализации изобретения экспрессируемые полипептиды согласно изобретению секретируются в и выделяются из периплазмы клеток-хозяев. Выделение белков, как правило, включает разрушение микроорганизма, в общем случае вследствие осмотического шока, обработки ультразвуком или лизиса. После разрушения клеток клеточный дебрис или цельные клетки можно удалять путем центрифугирования или фильтрации. Белки можно дополнительно очищать, например, при помощи аффинной хроматографии на смоле. В альтернативном варианте белки можно переносить в культуральную среду и выделять там. Клетки можно удалять из культуры и получать отфильтрованный и концентрированный клеточный супернатант для дополнительной очистки белков. Экспрессируемые полипептиды можно дополнительно выделять и идентифицировать, используя широко известные способы, такие как электрофорез в полиакриламидном геле (ПААГ) и метод вестерн-блоттинга.
В одном аспекте изобретения получение антител осуществляют в большом количестве посредством процесса ферментации. Для получения рекомбинантных белков доступны различные крупномасштабные ферментационные процедуры с подпиткой. Емкость крупномасштабной ферментации составляет по меньшей мере 1000 литров, предпочтительно - от около 1000 до 100000 литров. В этих ферментаторах используются лопастные мешалки для распределения кислорода и питательных веществ, в особенности глюкозы (предпочтительный источник углерода/энергии). Мелкомасштабная ферментация в общем случае относится к ферментации в ферментаторе, объемная емкость которого составляет не более 100 литров и может соответствовать диапазону от около 1 литра до около 100 литров.
В ходе ферментационного процесса индукция экспрессии белка, как правило, инициируется после того, как клетки выросли в подходящих условиях до необходимой плотности, например, ОП550, составляющей около 180-220, на стадии которой клетки находятся в ранней стационарной фазе. Как известно в данной области техники и описано выше, в соответствии с применяемой векторной конструкцией можно использовать множество индукторов. Перед индукцией клетки можно выращивать в течение более коротких периодов. Обычно клетки индуцируют в течение около 12-50 часов, хотя можно применять и большее или меньшее время индукции.
Чтобы улучшить выход и качество полипептидов согласно изобретению, можно модифицировать различные условия ферментации. Например, чтобы улучшить правильную сборку и фолдинг секретируемых полипептидов антител можно применять дополнительные векторы, сверхэкспрессирующие белки шаперонов, такие как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил цис, транс-изомераза с активностью шаперона), для котрансфомации прокариотических клеток-хозяев. Было продемонстрировано, что белки шаперонов облегчают правильный фолдинг и растворимость гетерологичных белков, вырабатываемых в бактериальных клетках-хозяевах. Chen et al., (1999) J. Biol. Chem. 274:19601-19605; Georgiou et al., патент США №6083715; Georgiou et al., патент США №6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275:17100-17105; Ramm and Pluckthun, (2000) J. Biol. Chem. 275:17106-17113; Arie et al., (2001) Mol. Microbiol. 39:199-210. В некоторых вариантах реализации изобретения DsbA и C экспрессируются (например, сверхэкспрессируются) в бактериальной клетке-хозяине. В некоторых вариантах реализации изобретения DsbA, DsbC и FkpA экспрессируются (например, сверхэкспрессируются) в бактериальной клетке-хозяине.
Чтобы минимизировать протеолиз экспрессируемых гетерологичных белков (в особенности тех, которые являются протеолитически чувствительными), в целях настоящего изобретения можно использовать определенных штаммы клеток-хозяев с дефицитом протеолитических ферментов. Например, штаммы клеток-хозяев можно модифицировать так, чтобы привнести генетическую(ие) мутацию(ии) в гены, кодирующие известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их комбинации. Доступны некоторые штаммы E. coli с дефицитом протеаз, описанные, например, в Joly et al., (1998), выше; Georgiou et al., патент США № 5264365; Georgiou et al., патент США № 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном варианте реализации изобретения в качестве клеток-хозяев в экспрессионной системе согласно изобретению применяют штаммы E. coli с дефицитом протеолитических ферментов, трансформированные плазмидами, сверхэкспрессирующими один или более белков шаперонов и/или FkpA.
iii. Очистка антител
Можно применять стандартные методы очистки белка, известные в данной области техники. Следующие процедуры являются типовыми подходящими процедурами очистки: фракционирование на иммуноаффинных или ионообменных колонках, преципитация этанолом, обратно-фазовая ВЭЖХ, хроматография на силикагеле или на катион-обменной смоле, такой как ДЭАЭ, хроматофокусирование, ДСН-ПААГ, преципитация сульфатом аммония и гель-фильтрация с применением, например, Сефадекс G-75.
В одном аспекте для иммуноаффинной очистки продуктов антител согласно изобретению используют протеин А, иммобилизованный на твердой подложке. Протеин А представляет собой 41 кДа белок клеточной оболочки из Staphylococcus aureas, который связывается с высокой аффинностью с Fc-областью антител. Lindmark et al., (1983) J. Immunol. Meth. 62:1-13. Твердая подложка, на которой иммобилизуют протеин А, предпочтительно является колонкой, содержащей стеклянную или кварцевую поверхность, более предпочтительно - колонку со стеклом с контролируемым размером пор или колонку с кремниевой кислотой. В некоторых аспектах колонку покрывают реагентом, таким как глицерол, пытаясь предотвратить неспецифическую адгезию контаминантных веществ.
На первом этапе очистки препарат, полученный из описанной выше клеточной культуры наносят на протеин А, иммобилизованный на твердой подложке, чтобы сделать возможным специфическое связывание представляющего интерес антитела с протеином А. Затем твердую подложку промывают для удаления контаминантных веществ, неспецифически связанных с твердой подложкой. В конце представляющее интерес антитело отделяют от твердой подложки путем элюирования.
Также в изобретении предложены иммуноконъюгаты (взаимозаменяемо называемые «конъюгатами антитело-лекарственный препарат» или «ADC»), содержащие любое из описанных в данном документе антител, конъюгированное, например, с цитотоксическим агентом, таким как химиотерапевтический агент, лекарственный препарат, агент, ингибирующий рост, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат).
Применения
Гетерологичный полипептид можно применять, например, для очистки, выявления и нацеливания на специфический полипептид, который он распознает, включая как in vitro и in vivo диагностические и терапевтические методы.
В одном аспекте антитело согласно изобретению можно применять в методах иммуноанализа для качественного и количественного определения специфических антигенов в биологических образцах. Традиционные способы выявления связывания антиген-антитело включают, например, ферментный иммуносорбентный анализ (ELISA), радиоиммуноанализ (РИА) или тканевую иммуногистохимию. Во многих методах в целях выявления применяют метку, связанную с антителом. Метка, применяемая с антителом, представляет собой любой функциональный элемент, который не препятствует связыванию с антителом. Известно большое количество меток, включая радиоизотопы 32P, 32S, 14C, 125I, 3H и 131I, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люцифераза, например, люцифераза светляков и бактериальная люцифераза ( патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидаза хрена (HRP), щелочная фосфатаза, бета-галактозидаза, глюкоамилаза, лизозим, сахаридоксидазы, например, глюкооксидаза, галактозооксидаза и глюкозо-6-фосфат дегидрогеназа, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, лактопероксидаза, биотин/авидин, спиновые метки, бактериофагальные метки, стабильные свободные радикалы, визуализирующие радионуклиды (такие как технеций) и тому подобное.
Для ковалентного связывания этих меток с гетерологичными полипептидами существуют традиционные способы. Например, для мечения антител вышеописанными флуоресцентными, хемилюминесцентными и ферментными метками можно использовать связующие агенты, такие как диальдегиды, карбодиимиды, дималеимиды, бис-имидаты, бис-диазотизированный бензидин и тому подобное. См., например, патент США № 3940475 (флуориметрия) и патент США № 3645090 (ферменты); Hunter et al. Nature 144: 945 (1962); David et al. Biochemistry 13:1014-1021 (1974); Pain et al. J. Immunol. Methods 40:219-230 (1981); и Nygren Histochem. and Cytochem 30:407-412 (1982). Предпочтительными метками в контексте данного документа являются ферменты, такие как пероксидаза хрена и щелочная фосфатаза. Конъюгация такой метки, включая ферменты, с полипептидом антитела является стандартной процедурой для специалиста в области техники иммуноанализа. См., например, O'Sullivan et al., "Methods for the Preparation of Enzyme-antibody Conjugates for Use in Enzyme Immunoassay," в Methods in Enzymology, ed. J. J. Langone и H. Van Vunakis, Vol. 73 (Academic Press, New York, N.Y., 1981), pp. 147-166. Такие способы связывания подходят для применения с гетерологичными полипептидами согласно настоящему изобретению.
В качестве альтернативы мечению гетерологичного полипептида можно проводить анализ антигена в биологических жидкостях методами конкурентного иммуноанализа с применением конкурентного стандартного антигена, меченного выявляемым веществом, и немеченого гетерологичного полипептида. В этом методе анализа смешивают биологический образец, меченые стандартные антигены и гетерологичный полипептид и определяют количество меченого стандартного антигена, связанного с немеченым гетерологичным полипептидом. Количество исследуемого антигена в биологическом образце обратно пропорционально количеству меченого стандартного антигена, связанного с гетерологичным полипептидом.
В одном аспекте гетерологичный полипептид (такой как антитело) исключительно удобен для выявления и определения профилей экспрессии специфических поверхностных антигенов in vitro или in vivo. Как обсуждалось ранее, в общем случае агликозилированное антитело не проявляет эффекторные функции (т.е. активность АЗКЦ или КЗЦ). Следовательно, когда антитело связывается с антигеном клеточной поверхности, оно не будет инициировать нежелательные цитотоксические эффекты. Поверхностный антиген может быть специфическим по отношению к конкретному типу клеток или тканей и, следовательно, служить маркером этого типа клеток или тканей. Предпочтительно поверхностный антигенный маркер дифференциально экспрессируется на различных стадиях дифференцировки конкретных типов клеток или тканей. Таким образом, антитело, направленное против такого поверхностного антигена, можно применять для скрининга клеточных или тканевых популяций, экспрессирующих маркер. Например, антитело можно применять для скрининга и выделения стволовых клеток, таких как эмбриональные стволовые клетки, гемопоэтические стволовые клетки и мезенхимальные стволовые клетки. Также антитело можно применять для выявления опухолевых клеток, экспрессирующих опухоль-ассоциированные поверхностные антигены, такие как c-met, рецепторы HER2, HER3 или HER4.
Антитело или другой гетерологичный полипептид можно применять в качестве агента для аффинной очистки. В этом способе полипептид иммобилизуют на твердой подложке, такой как смола Сефадекс или фильтровальная бумага, используя хорошо известные в данной области техники методы. Иммобилизованный полипептид приводят в контакт с образцом, содержащим предназначенный для очистки антиген, после чего подложку промывают подходящим растворителем, который удаляет практически весь материал в образце за исключением предназначенного для очистки антигена, который связан с иммобилизованным полипептидом. В конце подложку промывают другим подходящим растворителем, таким как глициновый буфер, pH 5,0, который высвобождает антиген из полипептида.
В одном аспекте в изобретении предложено применение гетерологичного полипептида, полученного способами согласно изобретению, в приготовлении медикамента для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение и/или иммунное (например, аутоиммунное) нарушение. Гетерологичный полипептид может принимать любую описанную в данном документе форму, включая антитело, фрагмент антитела, полипептид (например, олигопептид) или их комбинацию. В некоторых вариантах реализации изобретения антиген является человеческой белковой молекулой, а субъект является человеком.
Полипептиды можно применять для диагностирования, лечения, ингибирования или предотвращения заболеваний, нарушений или патологических состояний, связанных с аномальной экспрессией или активностью одной или более молекул антигенов, включая, но не ограничиваясь этим, злокачественные и доброкачественные опухоли; нелейкозные и лимфоидные злокачественные опухоли; нейрональные, глиальные, астроцитальные, гипоталамичесике и другие железистые, макрофагальные, эпителиальные, стромальные и бластоцельные нарушения; и воспалительные, ангиогенные и иммунологические нарушения.
В определенных вариантах реализации изобретения субъекту вводят белок (например, антитело). В определенных вариантах реализации изобретения субъекту вводят иммуноконъюгат, содержащий антитело. Предпочтительно иммуноконъюгат и/или антиген, с которым он связан, интернализируется клеткой.
Гетерологичные полипептиды можно применять в терапии как одни, так и в комбинации с другими композициями. Например, гетерологичный полипептид можно совместно вводить с антителом, химиотерапевтическими агентами (включая коктейли химиотерапевтических агентов), другими цитотоксическими агентами, антиангиогенными агентами, цитокинами и/или агентами, ингибирующими рост. В случае, когда гетеролигичный полипептид ингибирует рост опухоли, может быть целесообразно комбинировать гетерологичный полипептид с одним или более другими терапевтическим агентами, которые также ингибируют рост опухоли. В альтернативном или дополнительном варианте пациент может проходить комбинированную лучевую терапию (например, внешнюю лучевую терапию или терапию радиоактивно меченым агентом, таким как антитело). Вышеуказанная комбинированная терапия включает комбинированное введение (когда два или более агентов включены в состав одного или отдельных препаратов) и отдельное введение, в случае которого введение антитела может происходить до и/или после применения дополнительной терапии.
Гетерологичный полипептид (и, необязательно, дополнительный терапевтический агент) вводят любым подходящим путем, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и интраназальное и, в случае необходимости местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, антитело удобно вводить путем импульсной инфузии, в частности, с убывающими дозами антитела. Предпочтительно дозу вводят путем инъекции, наиболее предпочтительно - внутривенной или подкожной инъекции, в зависимости, частично, от того, является ли введение краткосрочным или постоянным.
Приготовление, дозирование и введение гетерологичного полипептида проводят в соответствии с принятой медицинской практикой. Учитываемые в этом контексте факторы включают конкретное заболевание, лечение которого проводят, конкретное млекопитающее, лечение которого проводят, место доставки агента, способ введения, схему введения и другие факторы, известные практикующим врачам. Антитело, необязательно, приготовлено с одним или более агентами, применяемыми на сегодняшний день для лечения данного заболевания. Эффективное количество других агентов зависит от количества антитела, присутствующего в препарате, типа нарушения или лечения и других факторов, обсуждаемых выше. Их обычно применяют в таких же дозировках и такими же путями введения, которые были ранее описаны в данном документе, или в дозировках, составляющих от около 1 до 99% ранее приведенных дозировок.
Для предотвращения или лечения заболевания соответствующая дозировка антитела (применяемого одного или в комбинации с другими агентами, такими как химиотерапевтические агенты) будет зависеть от типа заболевания, лечение которого проводят, типа антитела, тяжести и курса заболевания, от того, вводят ли антитело в превентивных или терапевтических целях, предыдущей терапии, клинической истории пациента и ответа на антитело, а также решения лечащего врача. Антитело удобно вводить пациенту за один раз или в течение ряда сеансов лечения. В зависимости от типа и тяжести заболевания начальная кандидатная дозировка для введения пациенту составляет от около 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг - 10 мг/кг) антитела, будь то, например, путем одного или более отдельных введений или путем непрерывной инфузии. В случае повторных введений в течение нескольких дней или более, в зависимости от состояния, лечение продолжают до проявления необходимой супрессии симптомов заболевания. Предпочтительная дозировка антитела соответствует диапазону от около 0,05 мг/кг до около 10 мг/кг. Можно вводить более высокую начальную дозу с последующей одной или более меньшими дозами. При этом можно применять другие режимы дозирования. Прогресс этой терапии легко отслеживать традиционными способами и методами анализа.
Изделия
В другом варианте реализации изобретения предложено изделие, содержащее описанные выше материалы, применяемые для лечения нарушений. Изделие содержит емкость и этикетку или вкладыш, находящиеся на емкости или прикрепленные к емкости. Подходящие емкости включают, например, бутылки, флаконы, шприцы и т.д. Емкости могут быть выполнены из множества материалов, включая стекло или пластик. Емкость содержит композицию, эффективную в случае лечения патологического состояния, и может иметь стерильное входное отверстие (например, емкость может представлять собой пакет для внутривенного раствора или флакон с пробкой, прокалываемой гиподермической иглой для инъекций). По меньшей мере один активный агент в композиции представляет собой антитело согласно изобретению. На этикетке или вкладыше указано, что композицию применяют для лечения выбранного патологического состояния, такого как рак. Кроме того, изделие может содержать (a) первую емкость с содержащейся в ней композицией, при этом композиция содержит антитело; и (b) вторую емкость с содержащейся в ней композицией, при этом композиция содержит дополнительный цитотоксический агент. Изделие в этом варианте реализации изобретения может дополнительно содержать вкладыш с указанием того, что первую и вторую антитело-содержащие композиции можно применять для лечения рака. В альтернативном или дополнительном варианте изделие может дополнительно содержать вторую (или третью) емкость, содержащую фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БВДИ), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Оно может дополнительно содержать другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Следующие примеры приведены исключительно для иллюстрации практической реализации настоящего изобретения, а не в качестве ограничения. Содержание всех патентов и научной литературы, перечисленных в данном документе, явным образом и в полном объеме включено посредством ссылки.
ПРИМЕРЫ
Материалы и методы
Штаммы и плазмиды. Применяемые в этом исследовании штаммы и плазмиды перечислены в Таблице 1. Чтобы сконструировать векторы, кодирующие только тяжелые цепи, экспрессионные векторы для полноразмерного антитела расщепляли при помощи SpeI и NsiI (New England Biolabs). Полученный в результате фрагмент, содержащий последовательность сигнального пептида и последовательность тяжелой цепи, клонировали в сайты SpeI и NsiI полученного из pBR322 клонирующего вектора, содержащего усеченный фрагмент легкой цепи (122-237), промотор phoA, слитый с последовательностью тяжелой цепи, и терминатор транскрипции t0 лямбда. Зондирование методом вестерн-блоттинга с антителом ακLc подтвердило, что с вектором, кодирующим только тяжелую цепь, легкая цепь не экспрессировалась. Чтобы сконструировать PhoA-репортерные плазмиды для измерения эффективности трансляции, представляющий интерес вариант сигнального пептида сливали с геном phoA посредством обычной ПЦР или ПЦР со сращиванием перекрывающимися расширениями (SOE). Полученный в результате фрагмент клонировали в сайты SpeI и NotI (New England Biolabs) в pPhoA51 [1]. Сайт-специфические мутации в последовательности сигнальных пептидов вводили при помощи набора для сайт-направленного мутагенеза QuickChange (Stratagene).
Таблица 1. Применяемые в этом исследовании штаммы и плазмиды
Рост бактерий. Бактерии выращивали в среде Луриа-Бертани (LB) или полной среде C.R.A.P. (Simmons et al., Journal of immunological methods, 2002) в качалочных колбах с перемешиванием при 37°C или при 30°C, как указано. Антибиотики добавляли в следующих концентрациях: карбенициллин 50 мкг/мл, тетрациклин 20 мкг/мл. Чтобы индуцировать экспрессию белка, штамм-хозяин 64B4, несущий экспрессионный вектор для полноразмерного антитела или экспрессионный вектор только для тяжелой цепи, инокулировали в 5 мл LB, дополненной 20 мкл/мл тетрациклина и 5 мM фосфата натрия, pH 7, и инкубировали со встряхиванием при 30°C в течение ночи. 0,5 мл ночной культуры инокулировали в 25 мл полной среды C.R.A.P. с ограниченным содержанием фосфатов [2] и выращивали бактерии при 30°C со встряхиванием в течение 24 ч. Для конечной точки культивирования определяли оптическую плотность на 550 нм. Бактериальные образцы собирали для проведения вестерн-блоттинга или N-терминального секвенирования.
Измерение эффективности трансляции. Эффективность трансляции вариантов сигнальных пептидов определяли методом анализа со щелочной фосфатазой, адаптированного относительно ранее опубликованного [1,3]. Штамм 27C7, несущий PhoA-репортерный вектор, инокулировали в 5 мл селективной LB и выращивали при 30°C со встряхиванием в течение ночи. Ночную культуру 100-кратно разводили в 5 мл селективной LB и инкубировали при 30°C со встряхиванием в течение еще 4 часов. Бактерии нормировали к 1 OП на длине волны 600 нм и осаждали. Осадок немедленно суспендировали в 1 мл жесткой среды AP (Simmons et al., Nature Biotechnology, 1996) и хранили в течение ночи при -20°C. На следующий день бактерии размораживали и инкубировали с 20 мкл толуола со встряхиванием при 37°C в течение по меньшей мере 1 часа. Затем 40 мкл каждого образца добавляли в 1 мл 1 M Трис-HCL, pH 8, содержащей 1 мM нитрофенил фосфат динатриевой соли гексагидрата (PNPP, Sigma-Aldrich), и инкубировали при комнатной температуре в течение 1 часа. Затем, чтобы остановить реакцию, добавляли 100 мкл 1 M фосфата натрия, pH 6,5. 200 мкл каждого образца немедленно переносили в лунки 96-луночного планшета и измеряли ОП410 при помощи планшета-ридера (Molecular Technologies). Относительную эффективность трансляции рассчитывали, вычитая A410 для 27C7/pBR322 (пустой вектор) из A410 для каждого образца и затем деля полученное число на A410 для 27C7/pPhoA86. Относительная эффективность TIR для pPhoA86 была определена как равная 1.
Выделение антител и вестерн-блоттинг. Из культур в качалочных колбах собирали образцы, соответствующие конечной точке. Чтобы определить общий уровень тяжелых цепей, бактерии нормировали к 1 ОП на 550 нм и собирали путем центрифугирования при 16000 xg и 4°C в течение 3 мин. Осадок пересуспендировали в 200 мкл трицинового буфера с 0,2 M дитиотреитола (ДТТ, Sigma) и грели при 95°C в течение 5 мин, чтобы разрушить дисульфидные связи и денатурировать белки.
Чтобы выделить из бактерий растворимые белки, бульон с цельными клетками разводили в охлажденном лизисном буфере (10 мM Трис, pH 6,8, 5 мM ЭДТУ, 0,2 мг/мл лизозима и 5 мM иодуксусной кислоты) до конечной ОП550, составляющей 3, и инкубировали на льду в течение 10 мин. Затем образцы дважды подвергали действию ультразвука при 10×1 импульсах в секунду и центрифугировали в течение 15 мин при 16000 xg и 4°C. Аккуратно собирали супернатант. 100 мкл супернатанта смешивали с 100 мкл трицинового буфера и кипятили при 95°C в течение 5 мин.
Периплазматические белки выделяли, как описано в [4, 5]. Вкратце, 10 ОП550 клеток собирали путем центрифугирования при 3000 xg и 4°C в течение 20 мин. Осадок аккуратно пересуспендировали в 1 мл холодного буфера ТСБ (200 мM Трис, pH 8,0, 0,5 мM сахарозы, 1 мM ЭДТУ) с 1 таблеткой коктейля ингибиторов протеаз (Roche). Образцы инкубировали на льду в течение 30 мин и центрифугировали при 16000 xg и 4°C в течение 30 мин. Аккуратно собирали супернатант. 100 мкл супернатанта смешивали с 100 мкл трицинового буфера с 0,2 M ДТТ и грели при 95°C в течение 5 мин.
Образцы белков в трициновом буфере с или без ДТТ загружали в 10% Бис-Трис ДСН-ПААГ гели (Life Technologies) и разделяли методом электрофореза. Чтобы гарантировать загрузку одинакового количества белка, гели с 1 ОП лизатами окрашивали голубым кумасси. Белки из неокрашенных гелей переносили на целлюлозные мембраны (Biorad) методом полусухого переноса iBlot (Life Technologies) или методом влажного переноса (Biorad) с буфером CAPS (10 мM N-циклогексил-3-аминопропансульфоновая кислота, 3% метанол, pH 11). Молекулы, содержащие тяжелые цепи, зондировали козьим, направленным против человеческой Fc вторичным антителом, конъюгированным с HRP (Pierce). Молекулы, содержащие легкие цепи, зондировали козьим, направленным против человеческой kLc антителом, конъюгированным с HRP (Bethyl Laboratories). Белки-мишени на иммуноблотах выявляли методом усиленной хемилюминесценции (GE Healthcare).
Секвенирование по Эдману N-конца тяжелой цепи. Образцы из культур в качалочных колбах, соответствующие конечной точке, нормировали к 1 ОП550 и собирали центрифугированием при 16000 xg в течение 3 мин. Осадок пересуспендировали в 200 мкл трицинового буфера с 0,2 M ДТТ и грели при 95°C в течение 5 мин. Белки разделяли методом электрофореза в 8% или 10% Бис-Трис ДСН-ПААГ гелях (Life Technologies). Также в качестве контроля загружали 1 ОП550 или 4 ОП550 клеточного лизата из 64B4, несущего пустой вектор pBR322. После электрофореза белки переносили на мембраны PVDF методом влажного переноса (Biorad) в буфере CAPS. Вырезали полосу тяжелой цепи на ~49 кДа на мембране и анализировали методом секвенирования по Эдману с применением Applied Biosystems Procise Sequencer Model 494HT. Соотношение между зрелым белком и предшественником оценивали на основании пиковой интенсивности. Для полуколичественного анализа рассчитывали пикомолярные величины для каждой аминокислоты при помощи программы для анализа последовательностей SEQX против нескорректированных фенилтиогидантоиновых аминокислотных стандартов (Henzel et al., Journal of Chromatography, 1987). В среднем использовали 10 циклов для получения повторяемого графика выхода для расчета линейной регрессии, а начальный выход основных и второстепенных последовательностей определяли по пересечению графиков с осью у. Эффективность процессинга тяжелых цепей рассчитывали как процентную долю секретируемых тяжелых цепей (начальный выход зрелых тяжелых цепей/(начальный выход предшественников + начальный выход зрелых тяжелых цепей)).
Трансмиссионная электронная микроскопия (ТЭМ). Образцы из культур в качалочных колбах, соответствующие конечной точке, сначала фиксировали в модифицированном фиксаторе Карновского (2% параформальдегид и 2,5% глутаральдегид в 0,1 M буфере какодилата натрия, рН 7,2), а затем пост-фиксировали в 1% водном тетроксиде осмия (EM Sciences, Hatfield, PA) в течение 1 ч с последующей инкубацией в течение ночи в 0,5% уранилацетате при 4°C. Затем образцы дегидратировали в ряде концентраций этанола (50%, 70%, 90%, 100%) с последующей обработкой пропиленоксидом (каждый этап занимал 15 мин) и заливали в Eponate 12 (Ted Pella, Redding, CA). Ультратонкие срезы (80 нм) получали при помощи микротома Ultracut (Leica), окрашивали 0,2% цитратом свинца и исследовали на трансмиссионном электронном микроскопе (ТЭМ) JEOL JEM-1400 (TEM) при 120 кВ. Цифровые изображения получали при помощи камеры GATAN Ultrascan 1000 CCD.
Электронная микроскопия с иммунным окрашиванием золотом (иммуноЭМ). В ЭМ-экспериментах с иммунным окрашиванием золотом готовили образцы для получения криосрезов. Для получения криосрезов клетки фиксировали в 4% параформальдегиде с 0,1% глутаральдегидом в фосфатном буфере (0,1 M; pH 7,2), несколько раз промывали в PBS, заливали в 12% желатин и инфильтрировали в 2,3 М сахарозы в течение ночи при 4°C. Затем образцы размещали на стержнях для криоультрамикротомии и замораживали в камере для получения криосрезов (с подачей жидкого азота). Ультратонкие криосрезы (100 нм) получали при помощи алмазного ножа (Diatome) при -80°C при помощи ультрамикротома (Ultracut; Leica), оборудованного камерой для получения криосрезов. Размороженные криосрезы переносили на покрытые формваром и углеродом EM-решетки (Nickel) с каплей 2,3 M сахарозы и проводили иммуномечение (См. ниже) и контрастное окрашивание в отношении EM 0,5% уранилацетатом в 2% метилцеллюлозе в течение 1 мин при КТ. Для иммуномечения золотом: Размороженные криосрезы на решетках блокировали в блокирующем агенте (Aurion Inc) в течение 30 мин и инкубировали с HRP-конъюгированным козьим антителом против человеческой Fc (Pierce) в течение 45 мин при комнатной температуре, после чего инкубировали с козьим анти-HRP конъюгированным с золотом антителом (Jackson ImmunoResearch) в течение 30 мин. Затем проводили контрастное окрашивание срезов, как описано выше. Меченые иммунозолотом срезы визуализировали и исследовали на трансмиссионном электронном микроскопе (ТЭМ) JEOL JEM-1400 при 120 кВ. Цифровые изображения получали при помощи камеры GATAN Ultrascan 1000 CCD.
Результаты и обсуждение
Выработку антител (например, полноразмерных антител) в E. coli можно достичь путем секреции тяжелой и легкой цепи антитела из цитоплазмы в периплазму [2]. Секреция опосредуется сигнальным пептидом E. coli, который слит с N-концом тяжелой цепи и легкой цепи. Окислительное окружение и ферменты в периплазме облегчают сборку тяжелой цепи и легкой цепи в антитело. Применение периплазматической секреции в качестве средства для высокоуровневой выработки гетерологичных белков (например, антител) может ограничиваться несколькими часто встречающимися проблемами. Во-первых, эффективность секреции представляющего интерес белка (например, антитела) может быть низкой. Во-вторых, во многих случаях предшественник не является полностью процессированным в зрелый белок. В-третьих, сверхэкспрессируемые гетерологичные белки часто неправильно сворачиваются, агрегируют в нерастворимые тельца включения или протеолизируются протеазами E. coli. В-четвертых, антитела представляю собой сложные мультимерные белки, состоящие из двух разных полипептидов - тяжелой (HC) и легкой цепей (LC), которые должны экспортироваться в периплазму, надлежащим образом сворачиваться и образовывать соответствующие дисульфидные связи. Сложность фолдинга этого белка плюс секреция усложняют задачу получения антител в E. coli.
Хотя было показано, что оптимизация TIR полезна для достижения более эффективной секреции белка, другие подходы, как правило, не улучшают секрецию гетерологичных белков в E. coli. Например, было показано, что оптимизация сигнального белка снижает экспрессию рекомбинантной циклодекстрин глюканотрансферазы (CGTазы) в периплазматическое пространство. Jonet et al., J Mol Microbiol Biotechnol (2012); 22:48-58.
Два аспекта, связанные с сигнальным пептидом, влияют на накопление белка в периплазме: эффективность трансляции и эффективность транслокации. Область инициации трансляции (TIR) является основным определяющим фактором общего уровня трансляции белка. TIR включает полинуклеотид, который кодирует сигнальную последовательность и простирается от участка непосредственно выше последовательности Шайна-Дальгарно до участка приблизительно на двадцать нуклеотидов ниже стартового кодона. Модификации этой полинуклеотидной последовательности могут изменять эффективность инициации трансляции, тем самым корректируя уровень трансляции нижерасположенного белка. В предыдущих исследованиях, в которых изучали эффект мутаций в последовательности сигнального пептида, в общем случае не рассматривали потенциальное влияние на эффективность TIR вследствие изменений в нуклеотидной последовательности; однако в нашем предыдущем исследовании изучали применение разных сигнальных пептидов для управления выработкой полного антитела, учитывая при этом эффективность TIR. При относительной эффективности TIR, составляющей около 1, слияние сигнального пептида DsbA с тяжелой цепью в общем случае приводило к более высокому уровню выработки полноразмерного антитела, чем слияние сигнальных пептидов STII, MalE или PhoA (сигнальным пептидом легкой цепи в каждом исследовании был STII) (Фигура 1A). В настоящем исследовании мы предполагаем, что сигнальный пептид DsbA является более гидрофобным, чем другие исследуемые сигнальные пептиды, и демонстрируем, что гидрофобность сигнального пептида является важной для транслокации тяжелой цепи в периплазму и выработки полноразмерного антитела.
Сигнальные пептиды влияют на периплазматическое накопление и процессинг тяжелой цепи антитела. Мы исследовали влияние вариантов сигнальных пептидов на накопление тяжелых цепей в периплазме. Для всех вариантов относительная эффективность TIR составляла ~1, чтобы можно было учесть возможное влияние изменения гидрофобности сигнального пептида на эффективность транскрипции. В предыдущих исследованиях обычно не учитывали эффективность трансляции модифицированных сигнальных пептидов. Если тщательно не контролировать этот фактор как в настоящем исследовании, модуляция гидрофобности сигнального пептида может привести к изменению эффективности трансляции TIR (например, посредством изменений в полинуклеотидной последовательности TIR). Изменения эффективности трансляции TIR могут влиять на эффективность секреции и уровни выработки белка.
N-конец тяжелой цепи антитела 5D5 сливали с сигнальным пептидом STII (bSTII1), сигнальным пептидом DsbA (bDsbA1), сигнальным пептидом MalE (bMalE1) или сигнальным пептидом PhoA (bPhoA1), каждый со сходным уровнем трансляции (Таблица 3). В каждом случае легкую цепь антитела 5D5 сливали с таким же сигнальным пептидом STII (mSTII1). Экспрессию как тяжелой цепи, так и легкой цепи индуцировали после ограничения содержания фосфатов в культурах в качалочных колбах. Периплазматические белковые экстракты выделяли методом осмотического шока или центрифугированием. Вестерн-блоттинг супернатанта показал, что применение сигнального пептида DsbA приводило к значительному повышению уровней растворимой периплазматической тяжелой цепи по сравнению с применением сигнальных пептидов STII, MalE или PhoA (Фиг. 1B). Так как эффективность TIR для bMalE1 и bPhoA1 выше, чем для bSTII1 и bDsbA1, чтобы исключить возможность того, что более высокая TIR может приводить к неэффективной секреции, мы сконструировали дополнительные сигнальные пептиды mMalE1 и mPhoA1 с эффективностью TIR немного ниже, чем у bSTII1 и bDsbA1 (Таблица 3). Слияние как mMalE1, так и mPhoA1 с тяжелой цепью приводило к значительно меньшим уровням полноразмерного 5D5 и растворимой периплазматической тяжелой цепи по сравнению с bDsbA1 (Фиг. 1C).
Чтобы определить влияние сигнальных пептидов на секрецию тяжелой цепи из цитоплазмы в периплазму, мы отслеживали присутствие зрелой тяжелой цепи и тяжелой цепи-предшественника. Несекретированный периплазматический белок в цитоплазме или во внутренней мембране будет сохранять последовательность сигнального пептида. Во время транслокации в периплазму сигнальный пептид отщепляется пептидазами [6,7]. Следовательно, сигнальный белок, который высвобождается в периплазму, не будет содержать сигнальный пептид. Разница в молекулярной массе между предшественником и зрелой тяжелой цепью составляет только 1-2 кДа, что трудно разрешить при помощи ДСН-ПААГ. Соответственно, проводили секвенирование N-концевого белка, чтобы различить предшественник и зрелую тяжелую цепь. Восстановленные тяжелые цепи из образцов цельных клеток перемещались в виде одной полосы в ДСН-ПААГ. Белки из ДСН-ПААГ переносят на мембрану PVDF. Вырезали полосы тяжелых цепей с мембраны PVDF и проводили секвенирование по Эдману. Когда тяжелые цепи были слиты с сигнальными пептидами STII, MalE или PhoA, выявляли последовательности, соответствующие как предшественнику, так и зрелой тяжелой цепи (Таблица 4), что позволяет предположить, что некоторое количество тяжелых цепей удерживалось в цитоплазме или во внутренней мембране. Когда тяжелая цепь была слита с сигнальным пептидом DsbA, последовательности, соответствующие предшественнику, присутствовали в очень малом или невыявляемом количестве. Мы оценили соотношение между зрелой тяжелой цепью и тяжелой цепью-предшественником при помощи пиковой интенсивности и провели полуколичественный анализ процентного содержания секретируемой тяжелой цепи, используя начальный выход. Оба метода показали, что в случае слияния с сигнальным пептидом DsbA, большая часть секвенированного пептида соответствовала зрелой тяжелой цепи (Таблица 4), что позволяет предположить, что тяжелые цепи эффективно секретировались в периплазматическое пространство (Таблица 4). Следовательно, сигнальный пептид DsbA опосредует более эффективный процессинг тяжелой цепи (из тяжелой цепи-предшественника в зрелую тяжелую цепь), чем сигнальный пептид STII, MalE или PhoA.
Также мы использовали электронную микроскопию с иммунным окрашиванием золотом (иммуноЭМ), чтобы напрямую визуализировать клеточную локализацию тяжелой цепи в случае ее слияния с разными сигнальными пептидами. Для бактерий, собранных из культур в качалочных колбах, проводили получение криосрезов, зондирование αFc-HRP антителом, а потом вторичным антителом αHRP-18 нм золотые частицы, и анализировали под трансмиссионным электронным микроскопом (ТЭМ). Для бактерий, экспрессирующих STII-легкую цепь и STII-тяжелую цепь, в периплазме фиксировали очень мало сигналов иммунозолота. Большую часть золотых меток наблюдали со стороны цитоплазмы, что указывает на то, что тяжелая цепь находилась по существу в цитоплазме (Фигура 2A). В противоположность этому, для бактерий, экспрессирующих STII-легкую цепь и DsbA-тяжелую цепь, золотые метки выявляли по существу со стороны периплазмы (Фигура 2B). Вместе эти результаты демонстрируют, что при сходном уровне трансляции применение сигнального пептида DsbA приводило к более эффективной транслокации тяжелой цепи и большему накоплению тяжелых цепей в периплазме по сравнению с тремя другими сигнальными пептидами.
Гидрофобность сигнального пептида важна для секреции тяжелой цепи антитела в периплазму и накопления в ней. Чтобы исследовать механизм, приводящий к большей эффективности сигнального пептида DsbA для секреции тяжелой цепи, мы сравнивали аминокислотные последовательности сигнального пептида DsbA и сигнальных пептидов STII, MalE и PhoA. Несмотря на разнообразие в первичных последовательностях, сигнальные пептиды обычно состоят из трех характерных областей: N-концевой области, которая содержит 1 или 2 положительно заряженных аминокислотных остатка, гидрофобной коровой области, часто называемой Н-областью, и С-концевой области, распознаваемой сигнальной пептидазой [8]. Как Н-область, так и полноразмерная последовательность сигнального пептида DsbA являются более гидрофобными, чем у сигнальных пептидов STII, MalE и PhoA (Таблица 2). Нас заинтересовало, играет ли гидрофобность сигнального пептида важную роль в транслокации тяжелой цепи антитела в периплазму.
Гидрофобность сигнальных пептидов DsbA и STII модулировали методом сайт-направленного мутагенеза. Замена лейцина 11 (L11) на серин (S) в сигнальном пептиде DsbA снижает общую и среднюю гидрофобность сигнального пептида DsbA (Таблица 7 и Таблица 2). В качестве контроля создавали мутацию с заменой лейцина 11 на изолейцин (I) в сигнальном пептиде DsbA. В последовательности сигнального пептида STII серин 11 был мутирован на лейцин или изолейцин для повышения гидрофобности (Таблица 7 и Таблица 2). Каждый из мутированных сигнальных пептидов сливали со зрелым белком PhoA для измерения эффективности TIR. Варианты сигнальных пептидов с разной гидрофобностью со сходной эффективность TIR с TIR1 сигнального пептида STII или TIR1 сигнального пептида DsbA (Таблица 3) сливали с тяжелой цепью антитела 5D5. Чтобы исключить возможные эффекты вследствие потенциального взаимодействия легкой цепи с тяжелой цепью и/или конкуренции за один и тот же механизм секреции, мы сконструировали плазмиды, которые экспрессировали только тяжелую цепь, но не легкую цепь.
Влияние вариантов сигнальных пептидов с разной гидрофобностью на накопление тяжелых цепей в периплазме исследовали методом вестерн-блоттинга периплазматических экстрактов (Фигура 3B). В отсутствие экспрессии легкой цепи применение сигнального пептида DsbA приводило к большей растворимости тяжелой цепи в периплазме, чем применение сигнального пептида STII. Снижение гидрофобности сигнального пептида DsbA вследствие мутации L11S сильно снижало периплазматический уровень растворимой тяжелой цепи, в то время как применение сигнального пептида DsbA с контрольной мутацией L11I не показало больших изменений уровня тяжелых цепей (Фигура 3A). С другой стороны, повышение гидрофобности сигнального пептида STII повышало уровень растворимой тяжелой цепи в периплазме (Фигура 3A).
Чтобы дополнительно подтвердить влияние гидрофобности сигнального пептида на секрецию тяжелой цепи 5D5, мы создавали варианты сигнальных пептидов (Таблица 2; Таблица 8) с диапазоном разных значений гидрофобности и сливали их с тяжелой цепью 5D5. В сигнальной последовательности STII серин 11 был мутирован на аланин (A), который повышает общую и среднюю гидрофобность, тирозин (Y), который только немного повышает гидрофобность, или на глутамин (Q), который снижает гидрофобность STII. Также мы создали высокогидрофобный вариант сигнального пептида STII путем введения тройной мутации с заменой аланина 6 (A6) на лейцин, аланина 10 (A10) на лейцин и серина 11 (S11) на лейцин (STII A6L, A10L, S11L). В последовательности DsbA лейцин 11 был мутирован на аланин (A) или глутамин (Q), чтобы снизить общую и среднюю гидрофобность. Все варианты сигнальных пептидов имели сходную эффективность TIR с TIR1 сигнального пептида STII или TIR1 сигнального пептида DsbA (Таблица 3).
Эффективность секреции тяжелой цепи 5D5 в периплазму в отсутствие легкой цепи определяли методом N-концевого секвенирования, как описано выше (Таблица 5). Применение сигнального пептида DsbA привело к тому, что большинство тяжелых цепей представляло собой зрелые тяжелые цепи, а тяжелые цепи-предшественники присутствовали в очень малом количестве, что указывает на то, что секреция тяжелой цепи 5D5, опосредуемая сигнальным пептидом DsbA, является эффективной. Сходные результаты наблюдали для контроля - варианта DsbA с L11I. Снижение гидрофобности сигнального пептида DsbA вследствие L11A, L11S или L11Q снижало эффективность секреции, так как было выявлено больше предшественников, чем зрелых тяжелых цепей 5D5. С другой стороны, ни STII, ни STII S11Q не опосредовали эффективную секрецию тяжелой цепи 5D5 согласно данным неколичественного секвенирования по Эдману, в то время как STII S11L улучшал секрецию. Полуколичественное секвенирование по Эдману показало, что STII и STII S11Y опосредовали неэффективную секрецию тяжелой цепи 5D5; при этом STII с мутацией S11Q повышал процентное содержание секретируемой тяжелой цепи 5D5. Повышение гидрофобности сигнального пептида STII вследствие мутации S11A или S11L повышало эффективность секреции. Кроме того, мутации в одном остатке S11L было достаточно для эффективной секреции тяжелой цепи 5D5, так как высокогидрофобный вариант сигнального пептида STII со сходной эффективностью трансляции (STII A6L, A10L, S11L) дополнительно не повышал эффективность секреции. Вместе, модулирование гидрофобности сигнального пептида и учет эффективности TIR сильно влияют на эффективность секреции тяжелой цепи 5D5 в периплазму.
Сходные результаты наблюдали в случае коэкспрессии легкой цепи с тяжелой цепью. Когда легкую цепь антитела 5D5 сливали с сигнальным пептидом STII (mSTII1), применение сигнального пептида DsbA L11S приводило к значительно меньшему количеству водорастворимой тяжелой цепи в периплазме, чем применение сигнальных пептидов DsbA или DsbA L11I. Аналогично, применение сигнального пептида STII S11L или STII S11I опосредовало большее накопление растворимых тяжелых цепей в периплазме, чем применение сигнального пептида STII (Фиг. 3B). Вместе, модулирование гидрофобности сигнального пептида значительно изменяло уровень накопления тяжелой цепи 5D5 в периплазме. Повышение гидрофобности путем одной аминокислотной замены значительно улучшало периплазматический уровень растворимых тяжелых цепей, в то время как снижение гидрофобности имело противоположный эффект.
Мы дополнительно анализировали влияние гидрофобности сигнального пептида на процессинг тяжелой цепи методом N-концевого секвенирования (Таблица 4). В каждом случае легкую цепь 5D5 сливали с сигнальным пептидом STII (mSTII1). Методом N-концевого секвенирования были зафиксированы в основном сигналы зрелой тяжелой цепи и очень мало сигналов предшественников, когда тяжелая цепь была слита с сигнальными пептидами DsbA и DsbA L11I. Рассчитанное на основании пиковой интенсивности соотношение зрелой тяжелой цепи и тяжелой цепи-предшественника для конструкции с сигнальным пептидом DsbA составило более чем 10:1. Мутация L11S в сигнальном пептиде DsbA снижала соотношение зрелая цепь/предшественник до 1:3, так как было зафиксировано больше сигналов предшественника, чем зрелой тяжелой цепи. Сигнальный пептид STII демонстрировал меньший сигнал зрелой тяжелой цепи по сравнению с сигналом предшественника с соотношением зрелая цепь/предшественник 1:3. Улучшение гидрофобности сигнального пептида STII путем введения мутации S11L или S11I приводило к большему количеству зрелой тяжелой цепи по сравнению с тяжелой цепью-предшественником (3:1 и 4:1). Полуколичественная оценка с использованием начального выхода по данным секвенирования по Эдману подтвердила влияние гидрофобности сигнального пептида на секрецию тяжелой цепи 5D5. Применение сигнального пептида DsbA приводило к ~86% секретируемой тяжелой цепи; мутация L11S снижала процент секретируемой тяжелой цепи до 39%. STII опосредовал 39% секретируемой тяжелой цепи; мутации S11L и S11I повышали секрецию тяжелой цепи до более чем 60%. Вместе, гидрофобность сигнального пептида была важна для процессинга и секреции в периплазму тяжелой цепи антитела.
Влияние гидрофобности сигнального пептида на секрецию 5D5 также подтверждалось исследованиями высокогидрофобного сигнального пептида из SfmC. SfmC представляет собой прогнозируемый белок пилин, локализованный в периплазме. Секреция SfmC зависит от частицы узнавания сигнала (ЧУС) в котрансляционном пути секреции (Huber et al., J Bacteriol. 2005. 2983-2991; Zhou et al., 2014. PLOS One). Сигнальный пептид SfmC является более гидрофобным, чем сигнальные пептиды DsbA или STII (Таблица 3). Мы создали варианты TIR1 и TIR2 сигнального пептида SfmC и слили каждый из них с тяжелой цепью 5D5. Оба варианта опосредовали очень эффективную секрецию тяжелой цепи 5D5 (100% секретируемой тяжелой цепи).
Влияние гидрофобности сигнального пептида на уровни полноразмерного антитела. Далее мы анализировали влияние гидрофобности сигнального пептида на выработку полноразмерного антитела. Собирали лизаты из клеток-хозяев, экспрессирующих тяжелую цепь и легкую цепь 5D5 и анализировали уровни прошедшего полную сборку антитела 5D5 методом невосстанавливающего ДСН-ПААГ электрофореза с последующим вестерн-блоттингом. В каждом случае легкую цепь 5D5 сливали с сигнальным пептидом STII (используя нуклеотидную последовательность mSTII 1). Слияние сигнального пептида DsbA L11S с тяжелой цепью приводило к существенному снижению уровня полноразмерного 5D5 по сравнению с применением сигнальных пептидов DsbA или DsbA L11I, слитых с тяжелой цепью (Фигура 4A). С другой стороны, обе мутации - S11L и L11I - в сигнальном пептиде STII повышали уровень полноразмерного антитела 5D5 по сравнению с применением сигнального пептида STII (Фигура 4B). Следовательно, повышение гидрофобности сигнального пептида стимулировало выработку полноразмерного антитела 5D5.
Мы хотели узнать, распространяется ли влияние гидрофобности сигнального пептида на секрецию тяжелых цепей других антител. С этой целью мы сливали сигнальные пептиды STII, STII S11L, DsbA или DsbA L11S TIR1 с тяжелой цепью mAb1 или mAb2 и определяли эффективность секреции тяжелой цепи в отсутствие легкой цепи методом секвенирования по Эдману (Таблица 6). Для тяжелых цепей обоих антител DsbA опосредовал большую секрецию тяжелой цепи, чем STII. Снижение гидрофобности DsbA вследствие мутации L11S снижало процент секретируемой тяжелой цепи, а повышение гидрофобности STII вследствие мутации S11L оказывало обратный эффект.
Мы дополнительно исследовали влияние гидрофобности сигнального пептида на выработку полноразмерного антитела для двух других моноклональных антител - mab 1 и mab 2 (Фигура 7). Mab 1 и mab 2 являются полноразмерными антителами IgG1. В обоих случаях легкую цепь сливали с mSTII1, а тяжелую цепь сливали с STII, DsbA, STII S11L или DsbA L11S со сходной эффективностью TIR. Уровни полноразмерного антитела, периплазматические уровни растворимой тяжелой цепи и общие уровни тяжелой цепи определяли, как описано ранее. В случае mAb1 слияние сигнальных пептидов STII или DsbA с тяжелой цепью приводило к сходным уровням полноразмерного антитела и периплазматической растворимой тяжелой цепи; при этом применение варианта сигнального пептида DsbA L11S со сниженной гидрофобностью значительно снижало уровни полноразмерного mAb1 и периплазматической растворимой тяжелой цепи, что позволяет предположить, что гидрофобность сигнального пептида является важным фактором для выработки mAb1 и секреции в периплазму. В случае mAb2 применение сигнального пептида STII приводило к меньшему количеству полноразмерного антитела и меньшему количеству периплазматической растворимой тяжелой цепи, чем применение сигнального пептида DsbA. Повышение гидрофобности сигнального пептида STII вследствие мутации S11L приводило к повышению уровня полноразмерного mAb2, а также накопления периплазматической растворимой тяжелой цепи, в то время как снижение гидрофобности сигнального пептида DsbA вследствие мутации L11S оказывало обратный эффект. Следовательно, гидрофобность сигнального пептида является важной для выработки и секреции mAb2 в периплазму E. coli.
Модуляция гидрофобности сигнального пептида изменяла клеточную локализацию телец включения. Сверхэкспрессируемые рекомбинантные белки в E. coli часто находятся в составе крупных нерастворимых агрегатов, известных как тельца включения, в цитоплазме или периплазме. Образование телец включения, содержащих представляющие интерес белки, часто связывают с низкими уровнями растворимых белков-мишеней. Кроме того, образование телец включения в цитоплазме указывает на неэффективную секрецию белка в периплазму. При помощи трансмиссионной электронной микроскопии (ТЭМ) мы наблюдали тельца включения непосредственно в цитоплазме клеток-хозяев, экспрессирующих легкую цепь, слитую с mSTII1, и тяжелую цепь, слитую с сигнальным пептидом STII, MalE или PhoA (Фигура 5), что позволяет предположить неэффективную секрецию белка в этих клетках. В противоположность этому, в случае клеток-хозяев, экспрессирующих STII-LC и DsbA-HC, тельца включения наблюдали обычно реже и они были локализированы в основном в периплазме (Фигура 5), что указывает на эффективную секрецию белка. Интересно, что применение сигнального пептида STII S11L (с повышенной гидрофобностью) продемонстрировало фенотип, сходный с сигнальным пептидом DsbA: тельца включения были локализированы в основном со стороны периплазмы (Фигура 5). Применение сигнального пептида DsbA L11S (со сниженной гидрофобностью) приводило к наблюдению телец включения по существу в цитоплазме (Фигура 5). В общей сложности модуляция гидрофобности сигнального пептида изменяла клеточную локализацию телец включения.
Мутации в С-концевом домене сигнального пептида не изменяют выработку полноразмерного антитела. С-концевая область сигнального пептида является ключевой для расщепления сигнального пептида пептидазами. Предыдущие исследования показали, что в этой области, как правило, содержатся аминокислотные остатки с небольшими боковыми цепями в позиции -1 и -3 в участке расщепления [6]. Оба сигнальных пептида STII и DsbA содержат небольшой остаток (Ala) в позиции -1 и -3 (Фигура 3A). При этом в позиции -2 в последовательности DsbA находится небольшой остаток Ser, в то время как в такой же позиции в последовательности STII находится объемный остаток (Tyr (Y)). Фигуга 6 демонстрирует, что изменения в размере боковой цепи в аминокислотной позиции -2 в участке расщепления не влияет на выработку антитела 5D5. Мутация с заменой Tyr22 на Ser в сигнальном пептиде STII не меняет уровень 5D5. Аналогично, мутация с заменой Ser18 на Tyr в сигнальном пептиде DsbA не оказывает влияние на уровень 5D5. Следовательно, объем боковой цепи в аминокислотной позиции -2 не влияет на уровни 5D5 в периплазме.
Таблица 2: Аминокислотные последовательности вариантов сигнальных пептидов
Подчеркнуто = мутации в аминокислотных последовательностях
Суммарную гидрофобность (сум. гидро) рассчитывают на основании нормированной консенсусной шкалы, разработанной Eisenberg et al. [20].
Средн. гидро = суммарная гидрофобность сигнальной последовательности, деленная на число аминокислотных остатков
Сум. гидро (H) = суммарная гидрофобность Н-области
Средн. гидро (H) = средняя гидрофобность Н-области.
Таблица 3. Последовательности ДНК вариантов сигнальных пептидов и относительная эффективность TIR
Жирный шрифт = сайт рестрикции BssHII, MluI или XbaI
Подчеркнуто = мутации, введенные для изменения аминокислотной последовательности.
Таблица 4. Секреция тяжелой цепи 5D5 с коэкспрессией легкой цепи 5D5
* Оценочное соотношение зрелая цепь/предшественник основано на грубой оценке пиковой интенсивности.
** % секретированной HC определяют при помощи полуколичественной оценки начального выхода зрелых тяжелых цепей и предшественников, как описано в разделе Материалы и методы.
Таблица 5. Секреция тяжелой цепи 5D5 в отсутствие легкой цепи 5D5
* % секретированной HC определяют при помощи полуколичественной оценки начального выхода зрелых тяжелых цепей и предшественников, как описано в разделе Материалы и методы.
Таблица 6. Секреция тяжелой цепи mAb1 и тяжелой цепи mAb2 в отсутствие легкой цепи.
* % секретированной HC определяют при помощи полуколичественной оценки начального выхода зрелых тяжелых цепей и предшественников, как описано в разделе Материалы и методы.
Таблица 7. Аминокислотные последовательности сигнальной последовательности DsbA, сигнальной последовательности STII и вариантных сигнальных последовательностей с измененной гидрофобностью. N-концевая область выделена жирным, Н-область выделена курсивом, С-концевая область обозначена обычным шрифтом. Мутированные остатки подчеркнуты.
Таблица 8. Аминокислотная последовательность сигнальной последовательности DsbA, сигнальной последовательности STII, вариантных сигнальных последовательностей с измененной гидрофобностью и сигнальной последовательности SfmC. N-концевая область выделена жирным, Н-область выделена курсивом, С-концевая область обозначена обычным шрифтом. Мутированные остатки подчеркнуты.
Частичный список литературы
1. Simmons, L.C. and D.G. Yansura, NatBiotechnol, 1996.14(5): p. 629-34.
2. Simmons, L.C., et al., J Immunol Methods, 2002. 263(1-2): p. 133-47.
3. US 8361744.
4. Quan, S., et al., MethodsMolBiol, 2013. 966: p. 359-66.
5. Oliver, D.B. and J. Beckwith, Cell, 1982.30(1): p. 311-9.
6. Auclair, S.M., M.K. Bhanu, and D.A. Kendall, ProteinSci, 2012. 21(1): p. 13-25.
7. Josefsson, L.G. and L.L. Randall, Cell, 1981. 25(1): p. 151-7.
8. Izard JW, K.D., MolMicrobiol, 1994. 13(5): p. 765-73.
Хотя вышеприведенное изобретение было довольно подробно описано при помощи иллюстраций и примеров в целях ясности понимания, описание и примеры не следует воспринимать как ограничивающие объем изобретения. Содержание всей патентной и научной литературы, на которую ссылается данный документ, явным образом в полном объеме включено посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GENENTECH, INC. ET AL.
<120> СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ
<130> P5804R1-WO
<140>
<141>
<150> 62/108476
<151> 2015-01-27
<150> 62/107981
<151> 2015-01-26
<150> 61/953629
<151> 2014-03-14
<160> 62
<170> PatentIn версии 3.5
<210> 1
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 1
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 2
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 2
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 3
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 3
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Leu Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 4
<211> 21
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 4
Met Lys Gln Ser Thr Ile Ala Leu Ala Leu Leu Pro Leu Leu Phe Thr
1 5 10 15
Pro Val Thr Lys Ala
20
<210> 5
<211> 21
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 5
Met Lys Gln Ser Thr Ile Ala Leu Ala Leu Leu Pro Leu Leu Phe Thr
1 5 10 15
Pro Val Thr Lys Ala
20
<210> 6
<211> 26
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 6
Met Lys Ile Lys Thr Gly Ala Arg Ile Leu Ala Leu Ser Ala Leu Thr
1 5 10 15
Thr Met Met Phe Ser Ala Ser Ala Leu Ala
20 25
<210> 7
<211> 26
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 7
Met Lys Ile Lys Thr Gly Ala Arg Ile Leu Ala Leu Ser Ala Leu Thr
1 5 10 15
Thr Met Met Phe Ser Ala Ser Ala Leu Ala
20 25
<210> 8
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 8
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 9
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 9
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 10
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 10
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 11
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 11
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ile Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 12
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 12
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Ser Ala
20
<210> 13
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 13
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ile Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 14
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 14
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ser Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 15
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 15
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Leu Val Leu Ala Phe Ser
1 5 10 15
Ala Tyr Ala
<210> 16
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 16
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcat ctatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 17
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 17
acgcgtatta tgaagaaaaa catcgctttt cttcttgcat ctatgttcgt tttttctatt 60
gctacaaacg cttac 75
<210> 18
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 18
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggtt tagttttagc gtttagcgca 60
tcggcg 66
<210> 19
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 19
gcgcgcatta tgaaacaatc cacgattgcc ctggcactct taccgttact gtttacccct 60
gtgacaaaag cc 72
<210> 20
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 20
acgcgtatta tgaaacagtc tactatcgct ctggcactct taccgttact gtttacccct 60
gtgacaaaag cc 72
<210> 21
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 21
gcgcgcatta tgaaaattaa gactggagca cgcatcctcg cattatccgc attaacgacg 60
atgatgtttt ccgcctcggc tctcgcc 87
<210> 22
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 22
acgcgtatta tgaagatcaa gacaggcgcg cgcatcctcg cattatccgc attaacgacg 60
atgatgtttt ccgcctcggc tctcgcc 87
<210> 23
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 23
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcat taatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 24
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 24
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcat tgatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 25
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 25
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcac tcatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 26
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 26
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcaa ttatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 27
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 27
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcat ctatgttcgt tttttctatt 60
gctacaaacg cttcggct 78
<210> 28
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 28
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggta ttgttttagc gtttagcgca 60
tcggcg 66
<210> 29
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 29
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggtt ctgttttagc gtttagcgca 60
tcggcg 66
<210> 30
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 30
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggtt tagttttagc gtttagcgca 60
tacgcg 66
<210> 31
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 31
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ala Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 32
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 32
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Gln Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 33
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 33
Met Lys Lys Asn Ile Leu Phe Leu Leu Leu Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 34
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 34
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ala Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 35
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 35
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Gln Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 36
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 36
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcag caatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 37
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 37
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcac aaatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 38
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 38
gcgcgcatta tgaagaaaaa catcctcttt cttcttctac taatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 39
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 39
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggtg ctgttttagc gtttagcgca 60
tcggcg 66
<210> 40
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 40
gcgcgcatta tgaaaaaaat ttggctcgcc ctggctggtc aggttttagc gtttagcgca 60
tcggcg 66
<210> 41
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 41
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Tyr Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 42
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 42
Met Met Thr Lys Ile Lys Leu Leu Met Leu Ile Ile Phe Tyr Leu Ile
1 5 10 15
Ile Ser Ala Ser Ala His Ala
20
<210> 43
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 43
Met Met Thr Lys Ile Lys Leu Leu Met Leu Ile Ile Phe Tyr Leu Ile
1 5 10 15
Ile Ser Ala Ser Ala His Ala
20
<210> 44
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 44
gcgcgcatta tgaagaaaaa catcgctttt cttcttgcat acatgttcgt tttttctatt 60
gctacaaacg cttacgct 78
<210> 45
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 45
tctagaatta tgatgactaa aatcaagctt ctaatgctca ttatatttta tttaatcatt 60
tcggccagcg cccatgct 78
<210> 46
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
олигонуклеотид»
<400> 46
tctagaatta tgatgacgaa aatcaagcta ctgatgctca ttatatttta tttaatcatt 60
tcggccagcg cccatgct 78
<210> 47
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 47
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Leu Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 48
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 48
Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10
<210> 49
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 49
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ile Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 50
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 50
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ser Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 51
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 51
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 52
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 52
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 53
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 53
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ile Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 54
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 54
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Leu Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 55
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 55
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Ala Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 56
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 56
Met Lys Lys Ile Trp Leu Ala Leu Ala Gly Gln Val Leu Ala Phe Ser
1 5 10 15
Ala Ser Ala
<210> 57
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 57
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 58
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 58
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ala Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 59
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 59
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Gln Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 60
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 60
Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Tyr Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 61
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 61
Met Lys Lys Asn Ile Leu Phe Leu Leu Leu Leu Met Phe Val Phe Ser
1 5 10 15
Ile Ala Thr Asn Ala Tyr Ala
20
<210> 62
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: Синтетический
пептид»
<400> 62
Met Met Thr Lys Ile Lys Leu Leu Met Leu Ile Ile Phe Tyr Leu Ile
1 5 10 15
Ile Ser Ala Ser Ala His Ala
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИЯ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2010 |
|
RU2585488C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2828630C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2739500C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2705274C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| СИСТЕМА ЭКСПРЕССИИ И СЕКРЕЦИИ | 2013 |
|
RU2670491C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| АНТИТЕЛО К GIPR И ЕГО СЛИТЫЙ С GLP-1 БЕЛОК, А ТАКЖЕ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800370C2 |
| ТРАНСГЕННЫЕ ГЕНЕТИЧЕСКИЕ МЕТКИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2729112C2 |
| ФАГОВЫЙ ДИСПЛЕЙ С ПРИМЕНЕНИЕМ КОТРАНСЛЯЦИОННОЙ ТРАНСЛОКАЦИИ СЛИТЫХ ПОЛИПЕПТИДОВ | 2006 |
|
RU2412249C2 |
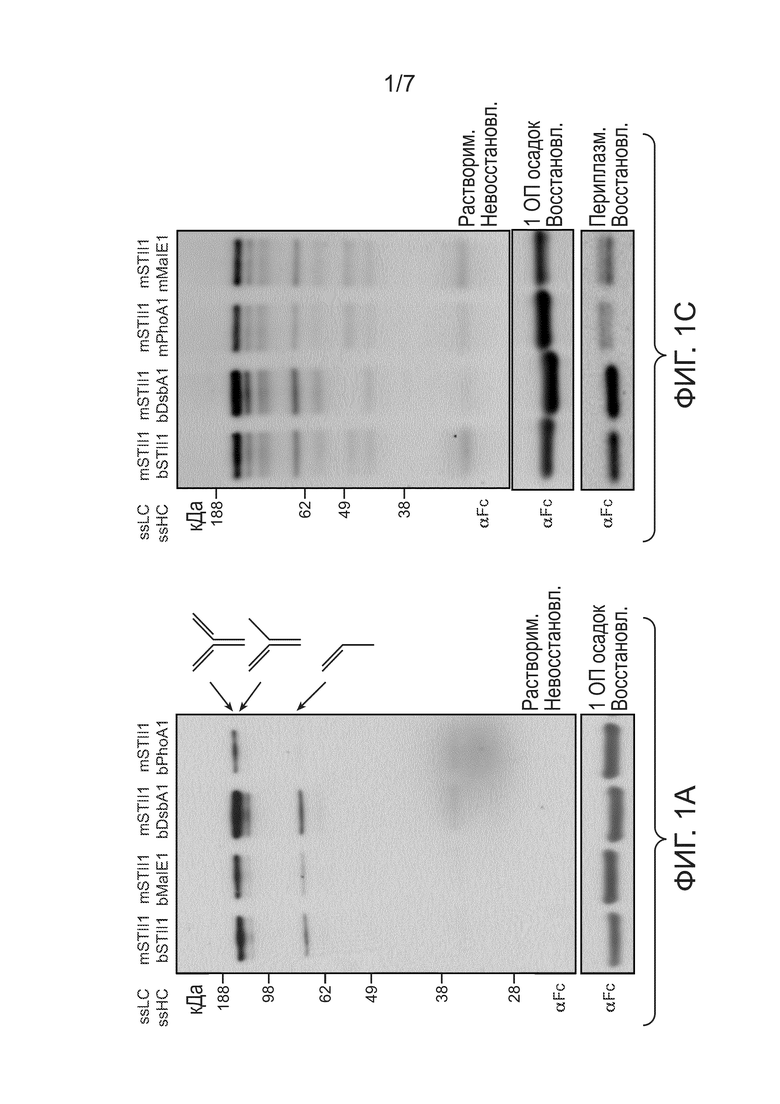
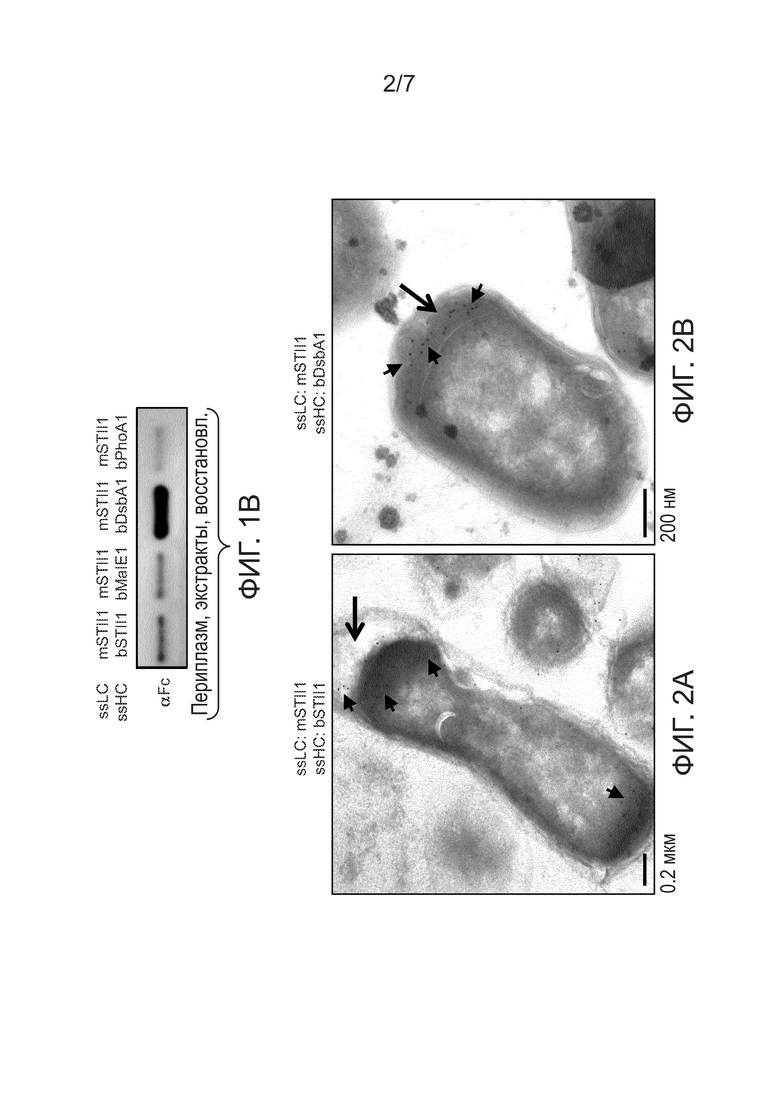
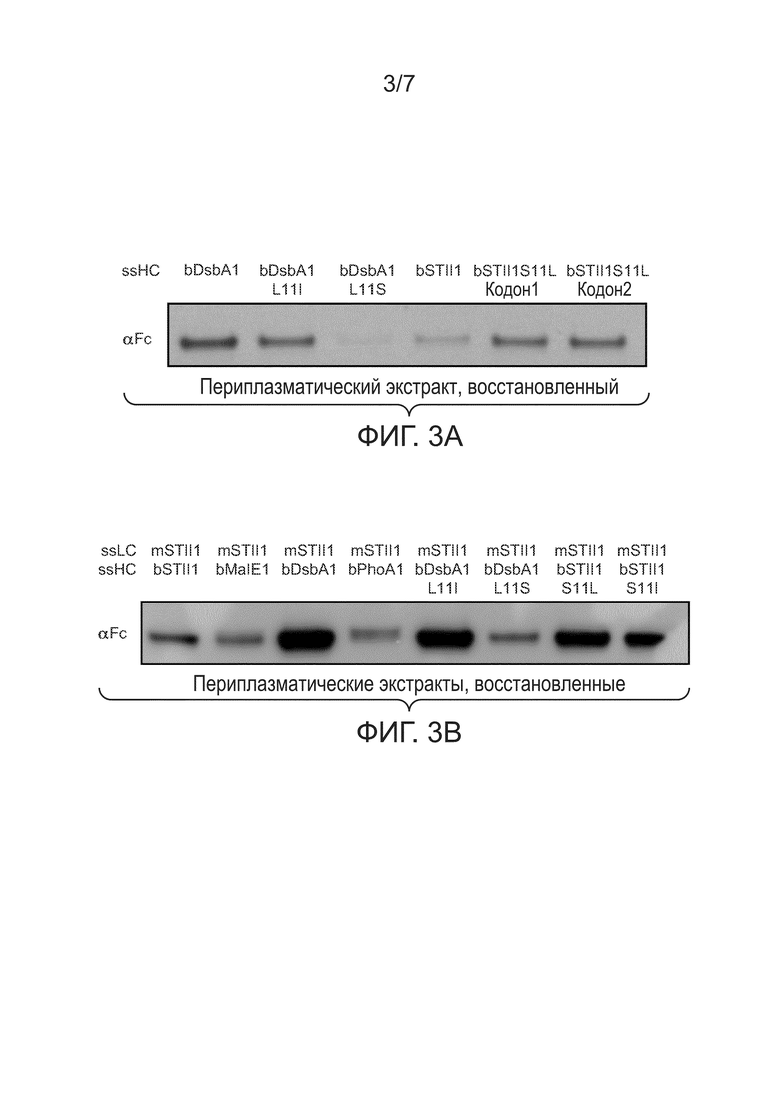
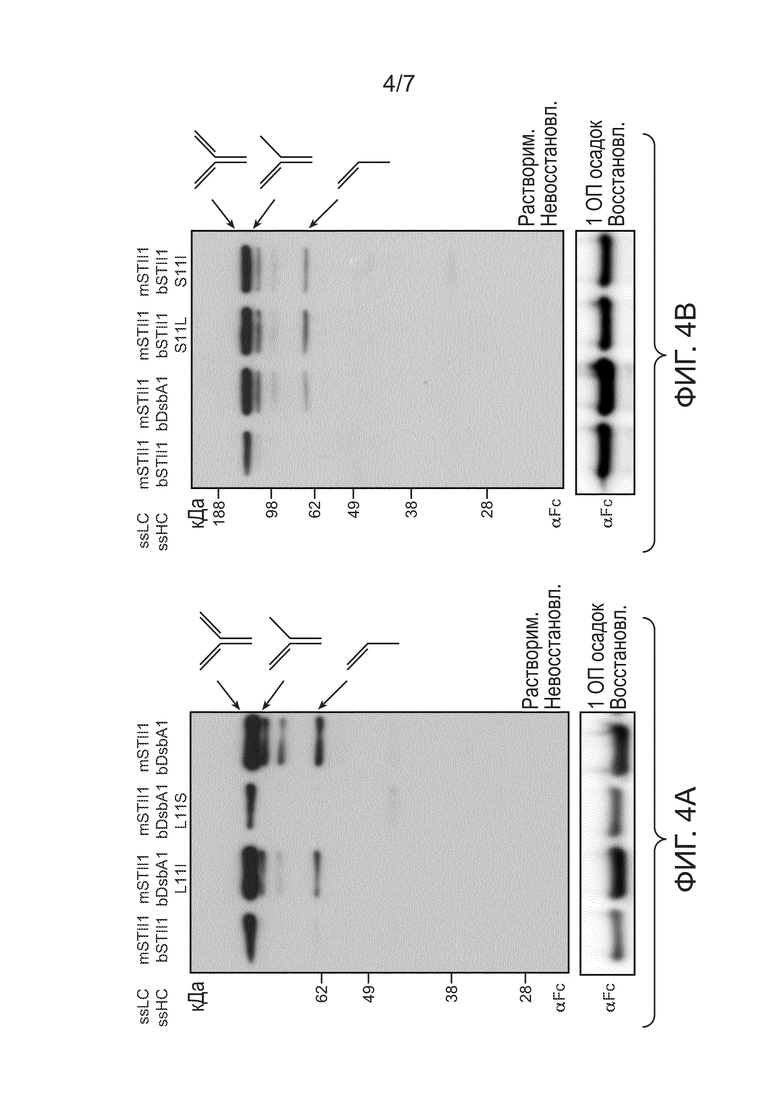
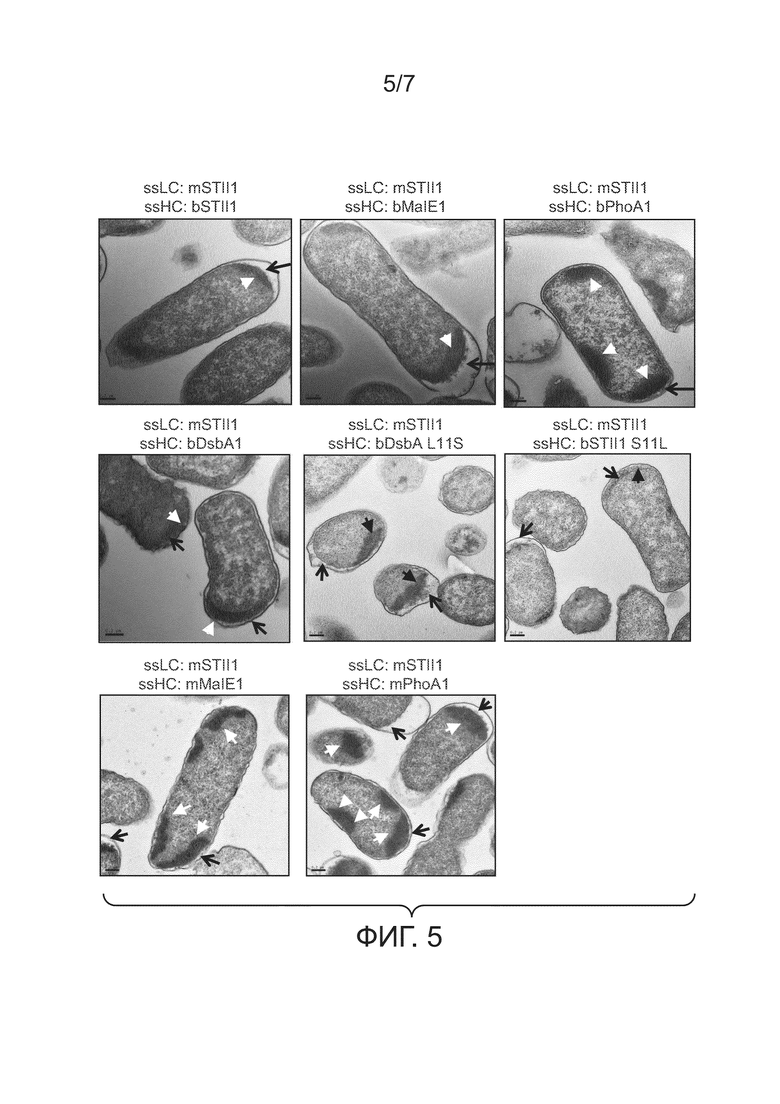
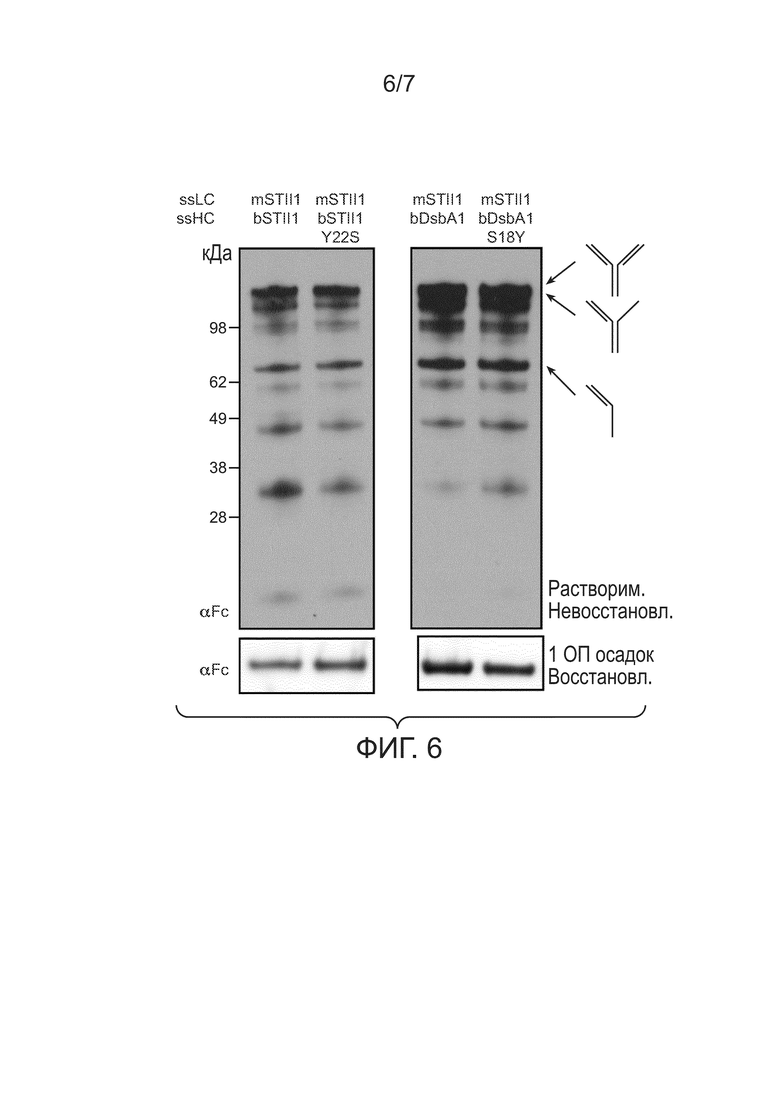

Настоящее изобретение относится к области биотехнологии. Предложен полинуклеотид, кодирующий антитело, вектор для получения антитела, клетка-хозяин. Кроме того, рассмотрен способ получения гетерологичного полипептида, способ секреции гетерологичного полипептида, способ транслокации гетерологичного полипептида. Данное изобретение может найти дальнейшее применение в получении антител с повышенной периплазматической секрецией. 6 н. и 22 з.п. ф-лы, 9 табл., 13 ил.
1. Полинуклеотид, кодирующий антитело, причем указанный полинуклеотид содержит (1) первый полинуклеотид, кодирующий первый сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим тяжелую цепь антитела, и (2) полинуклеотид, кодирующий второй сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим легкую цепь антитела, причем после экспрессии антитела в клетке-хозяине тяжелая и легкая цепи уложены и собраны с образованием биологически активного антитела, причем каждый из первого сигнального пептида и второго сигнального пептида содержит вариантный сигнальный пептид, выбранный из группы, состоящей из последовательности SEQ ID NO:8, 11, 13, 15, 31, 33 и 41, и
где полинуклеотид дополнительно содержит (i) первый промотор, где первый промотор функционально связан с полинуклеотидом, кодирующим тяжелую цепь, и (ii) второй промотор, где второй промотор функционально связан с полинуклеотидом, кодирующим легкую цепь.
2. Полинуклеотид по п. 1, отличающийся тем, что полинуклеотид, кодирующий антитело, дополнительно содержит (3) полинуклеотид, кодирующий третий сигнальный пептид, функционально связанный с полинуклеотидом, кодирующим Fc-полипептид.
3. Полинуклеотид по п. 2, отличающийся тем, что третий сигнальный пептид состоит из, состоит по существу из или содержит вариантный сигнальный пептид, выбранный из группы, состоящей из:
a) вариантного сигнального пептида DsbA, отличающегося тем, что вариант содержит H-область со средней гидрофобностью, составляющей более чем 0,5;
b) вариантного сигнального пептида DsbA, имеющего большую среднюю гидрофобность, чем сигнальный пептид DsbA дикого типа с SEQ ID NO:3;
c) вариантного сигнального пептида DsbA, имеющего большую среднюю гидрофобность, чем сигнальный пептид DsbA дикого типа с SEQ ID NO:3, где вариантный сигнальный пептид DsbA включает мутацию в L11 и/или S18;
d) вариантного сигнального пептида DsbA, имеющего большую среднюю гидрофобность, чем сигнальный пептид DsbA дикого типа с SEQ ID NO:3, где вариантный сигнальный пептид DsbA включает мутацию L11I и/или S18Y;
e) вариантного сигнального пептида STII, содержащего мутацию в остатке S11, причем вариантный сигнальный пептид STII имеет большую среднюю гидрофобность, чем сигнальный пептид STII с SEQ ID NO:1;
f) вариантного сигнального пептида STII, содержащего мутацию в остатке S11, причем вариантный сигнальный пептид STII имеет большую среднюю гидрофобность, чем сигнальный пептид STII с SEQ ID NO:1, где мутация представляет собой S11A, S11I или S11L; и
g) последовательности SEQ ID NO:8, 11, 13, 15, 31, 33 или 41.
4. Полинуклеотид по любому из пп. 1-3, отличающийся тем, что первый и второй промоторы представляют собой прокариотические промоторы, независимо выбранные из группы, состоящей из промотора phoA, tac, lpp, lac-lpp, lac, ara, trp и T7.
5. Полинуклеотид по п. 4, отличающийся тем, что новый промотор представляет собой промотор phoA.
6. Полинуклеотид по п. 4, отличающийся тем, что первый и второй промоторы оба являются промоторами phoA.
7. Полинуклеотид по любому из пп. 1-6, дополнительно содержащий (c) третий промотор, причем третий промотор функционально связан с Fc-полипептидом.
8. Полинуклеотид по п. 7, отличающийся тем, что промотор представляет собой промотор phoA.
9. Полинуклеотид по любому из пп. 1-8, отличающийся тем, что антитело представляет собой моноклональное антитело.
10. Полинуклеотид по п. 9, отличающийся тем, что антитело представляет собой химерное антитело, биспецифическое антитело, гуманизированное антитело, фрагмент антитела или человеческое антитело.
11. Полинуклеотид по п. 9, отличающийся тем, что фрагмент антитела представляет собой антитело с одним плечом.
12. Полинуклеотид по п. 9, отличающийся тем, что антитело представляет собой биспецифическое антитело.
13. Вектор для получения антитела, содержащий полинуклеотид по любому из пп. 1-12.
14. Вектор по п. 13, отличающийся тем, что вектор представляет собой экспрессионный вектор.
15. Клетка-хозяин для получения антитела, содержащая вектор по п. 14.
16. Клетка-хозяин по п. 15, отличающаяся тем, что клетка-хозяин представляет собой прокариотическую клетку.
17. Клетка-хозяин по п. 16, отличающаяся тем, что прокариотическая клетка представляет собой E. coli.
18. Клетка-хозяин по п. 17, отличающаяся тем, что E. coli представляет собой штамм с дефицитом эндогенной протеазной активности.
19. Клетка-хозяин по п. 17 или 18, отличающаяся тем, что в генотипе E. coli отсутствуют гены degP и prc и присутствует мутантный ген spr.
20. Клетка-хозяин по любому из пп. 15-19, отличающаяся тем, что клетка-хозяин дополнительно содержит полинуклеотид, кодирующий прокариотический белок шаперона.
21. Клетка-хозяин по п. 20, отличающаяся тем, что прокариотический белок шаперона представляет собой DsbA и/или DsbC.
22. Клетка-хозяин по п. 20 или 21, отличающаяся тем, что клетка-хозяин сверхэкспрессирует прокариотический белок шаперона.
23. Способ получения гетерологичного полипептида (такого как тяжелая цепь и/или легкая цепь антитела), причем указанный способ включает культивирование клетки-хозяина по любому из пп. 15-22 таким образом, чтобы происходила экспрессия нуклеиновой кислоты, причем после экспрессии указанного полинуклеотида в клетке-хозяине гетерологичный полипептид уложен с образованием биологически активного гетерологичного полипептида.
24. Способ по п. 23, отличающийся тем, что способ дополнительно включает выделение гетерологичного полипептида из культуры клеток-хозяев.
25. Способ по п. 24, отличающийся тем, что гетерологичный полипептид выделяют из культуральной среды клеток-хозяев.
26. Способ по п. 24 или 25, отличающийся тем, что способ дополнительно включает комбинирование выделенного гетерологичного полипептида с фармацевтически приемлемым носителем, вспомогательным веществом или носителем с получением фармацевтического препарата, содержащего гетерологичный полипептид.
27. Способ секреции гетерологичного полипептида (такого как тяжелая цепь и/или легкая цепь антитела) из клетки, причем указанный способ включает культивирование клетки-хозяина по любому из пп. 15-22 таким образом, чтобы происходила экспрессия нуклеиновой кислоты и секреция гетерологичного полипептида.
28. Способ транслокации гетерологичного полипептида (такого как тяжелая цепь и/или легкая цепь антитела) из клетки, причем указанный способ включает культивирование клетки-хозяина по любому из пп. 15-22 так, чтобы происходила экспрессия нуклеиновой кислоты и транслокация гетерологичного полипептида.
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| EISENBERG D et al | |||
| "Analysis of membrane and surface protein sequence with the hydrophobic moment plot", J | |||
| Mol | |||
| Biol., 1984, 179, pp | |||
| Плуг с фрезерным барабаном для рыхления пласта | 1922 |
|
SU125A1 |
| I на стр | |||
| Ударно-вращательная врубовая машина | 1922 |
|
SU126A1 |
| GION WR et al | |||
| "Expression of antibodies using single open | |||

