ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к способам лечения заболевания, расстройства или патологического состояния, при которых положительный эффект оказывает повышение числа регуляторных Т-клеток (Трег-клеток), с использованием антагонистов дельта-подобного лиганда 4 (Dll4). Более конкретно, с помощью способов по изобретению можно лечить различные аутоиммунные заболевания, такие как рассеянный склероз и диабет, а также отторжение трансплантированных органов и болезнь «трансплантат против хозяина» (GVHD), путем блокирования связывания Dll4 с Notch-рецептором антагонистами Dll4, тем самым повышая число Трег-клеток. Изобретение дополнительно относится к способам предупреждения возникновения или рецидива таких заболеваний или расстройств у субъекта, который предрасположен или восприимчив к данным заболеваниям или расстройствам.
ОПИСАНИЕ ПРЕДШЕСТВУЮЩЕГО УРОВНЯ ТЕХНИКИ
Взаимодействие между Notch-рецепторами и их лигандами представляет собой эволюционно консервативный путь, важный не только для решения судьбы клеток, но также для регуляции направлений дифференцировки при гемопоэзе и в развивающемся тимусе (Artavanis-Tsakonas et al., 1999, Science 284:770-776; Skokos et al., 2007; J Exp Med 204:1525-1531; и Amsen et al., 2004, Cell 117:515-526). Недавно было показано, что ингибирование взаимодействия Dll4-Notch1 приводит к полному блокированию развития Т-клеток, сопровождаемому эктопическим появлением В-клеток и увеличением числа дендритных клеток (DC), которое может быть следствием конверсии дифференцировки предшественников Т-клеток в DC в тимусе (Hozumi et al., 2008, J Exp Med 205(11):2507-2513; Koch et al., 2008, J Exp Med 205(11):2515-2523; и Feyerabend et al., 2009, Immunity 30:1-13). Таким образом, накапливаются свидетельства того, что Notch-сигнальный путь критичен для определения направления дифференцировки клеток из гемопоэтических клеток-предшественников. Кроме того, был показан контроль по принципу обратной связи гомеостаза регуляторных Т-клеток (Трег-клеток) дендритными клетками in vivo (Darrasse-Jeze et al., 2009, J Exp Med 206(9):1853-1862). Однако роль Notch-сигнального пути в контроле происхождения и развития дендритных клеток и, соответственно, гомеостаза Трег-клеток еще неизвестен. Это является важным клиническим вопросом, поскольку найденные новые способы индукции размножения Трег-клеток можно было бы использовать в качестве лечения аутоиммунных заболеваний и расстройств.
Нуклеотидные и аминокислотные последовательности Dll4 человека (hDll4) показаны, соответственно, в SEQ ID NO:1 и 2. Антагонисты Dll4 и их применение раскрыты в WO 2007/143689, WO 2007/070671, WO 2008/076379, WO 2008/042236 и WO/2008/019144.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение частично основано на наблюдении автора изобретения, что антитело, которое специфично связывает Dll4 и блокирует связывание Dll4 с Notch-рецепторами, способно полностью предупреждать развитие экспериментального аутоиммунного энцефаломиелита (ЕАЕ) у мышей, животной модели рассеянного склероза человека, в то время как контрольное антитело не предупреждает ЕАЕ. Кроме того, автором настоящего изобретения было открыто, что этот эффект анти-Dll4 антитела ассоциирован с повышением числа Трег-клеток. В дополнение, также наблюдалось, что анти-Dll4 антитело предупреждает повышение уровня глюкозы в крови и сохраняет число и морфологию панкреатических островков у мышей NOD/ShiLtJ, животной модели диабета 1-го типа, и такие эффекты, по меньшей мере частично, опосредованы повышением числа Трег-клеток.
Поэтому, в первом аспекте, изобретение относится к способу увеличения числа Трег-клеток, включающему введение эффективного количества антагониста Dll4 нуждающемуся в этом субъекту, причем антагонист Dll4 блокирует взаимодействие между Dll4 и Notch-рецептором, и увеличивается число Трег-клеток.
Во втором аспекте изобретение относится к способу лечения или ослабления болезни, расстройства или патологического состояния, при которых положительный эффект оказывает повышение числа регуляторных Т-клеток (Трег-клеток), включающему введение терапевтически эффективного количества антагониста Dll4 нуждающемуся в этом субъекту. Заболеванием или расстройством, которое поддается лечению способами по изобретению, является любое заболевание, расстройство или состояние, на которое оказывает эффект, то есть улучшает, ослабляет, ингибирует или предупреждает, удаление, ингибирование или снижение активности Dll4, тем самым увеличивая число Трег-клеток у получающего лечение субъекта. Такие заболевания, расстройства или состояния включают, но не ограничены этим, ряд аутоиммунных заболеваний, таких как рассеянный склероз, сахарный диабет 1-го и 2-го типа, ревматоидный артрит, болезнь Крона, синдром Гудпасчера, болезнь Грейвса, синдром Гийена-Барре (GBS), болезнь Хашимото, идиопатическую тромбоцитопеническую пурпуру, синдром раздраженного кишечника, красную волчанку, смешанное заболевание соединительной ткани, миастению, нарколепсию, обыкновенную пузырчатку, пернициозную анемию, полимиозит, первичный билиарный цирроз, синдром Сьоргена, язвенный колит, васкулит, гранулематоз Вегенера и т.п., а также отторжение трансплантированных органов и GVHD.
В одном варианте осуществления изобретения антагонистом для использования в способах по изобретению является антитело к Dll4 или его фрагмент («анти-Dll4 АТ» или «Dll4-АТ»), которое специфично связывает Dll4 с высокой аффинностью и блокирует связывание Dll4 с Notch-рецепторами и/или блокирует сигнальные пути, активируемые взаимодействием Dll4-Notch. Антитело может быть поликлональным, моноклональным, химерным, гуманизированным или полностью антителом человека или его фрагментом. Фрагментом антитела может быть одноцепочечное антитело, Fab или (Fab')2.
В одном варианте осуществления изобретения Dll4-АТ или его антиген-связывающий фрагмент связывают эпитоп в N-концевом домене (остатки S27-R172) или в домене Дельта/Серрат/Лаг-2 (DSL-домене) (остатки V173-C217), или в N-концевом и DSL-домене (остатки S27-C217) белка hDll4 (SEQ ID NO:2). В другом варианте осуществления изобретения Dll4-АТ или его антиген-связывающий фрагмент связывают эпитоп в одном из EGF-доменов, то есть примерно аминокислотные остатки Q218-N251 (домен 1), E252-D282 (домен 2), D284-E322 (домен 3), E324-E360 (домен 4), S362-E400 (домен 5), K402-E438 (домен 6), H440-E476 (домен 7) или S480-E518 (домен 8) белка hDll4 (SEQ ID NO:2). В некоторых вариантах осуществления изобретения антитело или фрагмент антитела могут связывать конформационный эпитоп, включающий более одного эпитопа из перечисленных выше. Dll4-АТ или его фрагменты для использования в способах по изобретению способны связывать Dll4 человека с высокой аффинностью и имеют константу диссоциации (KD) примерно 1 нМ или меньше, примерно 500 пМ или меньше, примерно 300 пМ или меньше, примерно 200 пМ или меньше, примерно 100 или меньше или примерно 50 рМ или меньше, измеренную с помощью поверхностного плазмонного резонанса.
В одном варианте осуществления изобретения Dll4-АТ или его фрагмент включают в себя вариабельную область тяжелой цепи (HCVR), содержащую три определяющих комплементарность области тяжелой цепи, HCDR1, HCDR2 и HCDR3, имеющие аминокислотные последовательности, соответственно, SEQ ID NO: 22, 24 и 26. В другом варианте осуществления изобретения антитело или его фрагмент включают в себя вариабельную область легкой цепи (LCVR), содержащую три определяющих комплементарность области легкой цепи, LCDR1, LCDR2 и LCDR3, имеющие аминокислотные последовательности, соответственно, SEQ ID NO: 30, 32 и 34. В другом варианте осуществления изобретения Dll4-АТ или его фрагмент включают в себя последовательности CDR легкой и тяжелой цепей, содержащие комбинацию последовательностей CDR из SEQ ID NO: 22, 24, 26, 30, 32 и 34. В еще одном варианте осуществления изобретения Dll4-АТ включает в себя HCVR, содержащую аминокислотную последовательность SEQ ID NO:20 или 116, или LCVR, содержащую аминокислотную последовательность SEQ ID NO:28 или 118. В еще одном варианте осуществления изобретения Dll4-АТ включает в себя комбинацию HCVR/LCVR из SEQ ID NO:20/28 (REGN281) или 116/118 (REGN421).
В некоторых вариантах осуществления изобретения Dll4-АТ включает в себя комбинацию CDR1/CDR2/CDR3 тяжелой цепи и комбинацию CDR1/CDR2/CDR3 легкой цепи, выбранную из: SEQ ID NO:6/8/10 и SEQ ID NO:14/16/18, соответственно; SEQ ID NO:38/40/42 и SEQ ID NO:46/48/50, соответственно; SEQ ID NO:54/56/58 и SEQ ID NO:62/64/66, соответственно; SEQ ID NO:70/72/74 и SEQ ID NO:78/80/82, соответственно; SEQ ID NO:86/88/90 и SEQ ID NO:94/96/98, соответственно; и SEQ ID NO:102/104/106 и SEQ ID NO:110/112/114, соответственно. В другом варианте осуществления изобретения Dll4-АТ включает в себя HCVR, содержащую аминокислотную последовательность SEQ ID NO:4, 36, 52, 68, 84 или 100, или LCVR, содержащую аминокислотную последовательность SEQ ID NO:12, 44, 60, 76, 92 или 108. В еще одном варианте осуществления изобретения Dll4-АТ включает в себя комбинацию HCVR/LCVR, выбранную из: SEQ ID NO:4/12 (REGN279); SEQ ID NO:36/44 (REGN290); SEQ ID NO:52/60 (REGN306); SEQ ID NO:68/76 (REGN309); SEQ ID NO:84/92 (REGN310); и SEQ ID NO:100/108 (REGN289).
Нуклеотидные последовательности, кодирующие аминокислотные последовательности SEQ ID NO:4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 116 и 118, приведены в SEQ ID NO:3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101, 103, 105, 107, 109, 111, 113, 115 и 117, соответственно.
В другом варианте осуществления изобретения антагонистом Dll4, применимым в способах по изобретению, является слитый белок, содержащий по меньшей мере один растворимый Notch-рецептор или его фрагмент, способный связывать Dll4, слитый с мультимеризующим компонентом. В одном варианте осуществления изобретения растворимым Notch-рецептором является Notch1 или Notch4. В другом варианте осуществления изобретения антагонистом Dll4 является модифицированный белок Dll4, способный связывать Notch-рецептор(ы), но такое связывание не приводит к активации рецептора(ов). В некоторых вариантах осуществления изобретения антагонистом Dll4 по изобретению является слитый белок, содержащий внеклеточный домен Dll4, или его фрагмент, слитые с мультимеризующим компонентом, таким как иммуноглобулиновый домен, например, Fc-домен IgG человека. В некоторых вариантах осуществления изобретения антагонисты Dll4 включают низкомолекулярные соединения и другие агенты, которые могут блокировать взаимодействие Dll4-Notch.
В третьем аспекте изобретение относится к способу предупреждения или снижения частоты возникновения или рецидива аутоиммунного заболевания или расстройства, описанных выше, у субъекта, который предрасположен или восприимчив к развитию этого заболевания или расстройства, включающему введение субъекту профилактически эффективного количества антагониста Dll4. В родственном варианте осуществления изобретение дополнительно относится к способу предупреждения или снижения вероятности отторжения трансплантированных органов или GVHD у реципиента трансплантата, включающему введение профилактически эффективного количества антагониста Dll4 реципиенту до и/или после трансплантации.
В четвертом аспекте изобретение относится к способу предупреждения, лечения или ослабления аутоиммунного заболевания или расстройства, или отторжения трансплантированного органа, включающему введение субъекту, нуждающемуся в этом, эффективного количества антагониста Dll4 в комбинации с дополнительным терапевтическим агентом, например, иммуносупрессирующим агентом или иммуносупрессантом, противовоспалительным агентом, анальгетиком, агентом для снижения сахара в крови (например, инсулином, аналогами инсулина и т.п.), многие из которых могут иметь перекрывающиеся терапевтические эффекты друг с другом. Иммуносупрессанты, подходящие для применения в комбинации с антагонистом Dll4, включают, но не ограничены этим, глюкокортикостероиды, циклоспорин, метотрексат, интерферон β (IFN-β), такролимус, сиролимус, азатиоприн, меркаптопурин, опиоиды, микофенолят, TNF-связывающие белки, такие как инфликсимаб, этанерцепт, адалимумаб и т.п., цитотоксические антибиотики, такие как дактиномицин, антрациклины, митомицин С, блеомицин, митрамицин и т.п., антитела к клеткам иммунной системы, такие как анти-CD20 антитела, анти-CD3 антитела, и т.п. Противовоспалительные агенты и/или анальгетики, подходящие для комбинированных методов лечения с антагонистами Dll4, включают кортикостероиды, нестероидные противовоспалительные соединения (NSAID), такие как аспирин, ибупрофен, напроксен и т.п., антагонисты TNF-α, антагонисты IL-1, антагонисты IL-6, ацетаминофен, морфиномиметики, и т.п.
В пятом аспекте изобретение относится к фармацевтической композиции, содержащей антагонист Dll4, по меньшей мере один дополнительный терапевтический агент и фармацевтически приемлемый носитель. В одном варианте осуществления изобретения антагонистом Dll4 является Dll4-АТ или его фрагмент, которые специфично связывают Dll4 с высокой аффинностью и нейтрализуют функции Dll4, а по меньшей мере одним дополнительным терапевтическим агентом является любой из иммуносупрессантов, противовоспалительных агентов, анальгетиков, агентов для снижения сахара в крови и т.п., описанных выше.
В шестом аспекте изобретение относится к набору, включающему в себя контейнер, содержащий фармацевтическую композицию по настоящему изобретению и вкладыш в упаковку с инструкцией по применению. В одном варианте осуществления изобретения набор может включать в себя контейнер, содержащий антитело или его фрагмент, которые специфично связывают hDll4, другой контейнер, содержащий по меньшей мере один дополнительный терапевтический агент, описанный выше.
Другие задачи и преимущества изобретения будут очевидны при рассмотрении следующего подробного описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигурах 1A-1B показаны эффекты блокирования Dll4 на развитие Т-клеток и В-клеток. Мышам вводили анти-Dll4 антитело (REGN577) или контрольный Fc-фрагмент человека (hFc). Через четырнадцать дней собирали тимусы и анализировали Т-клеточные и В-клеточные субпопуляции проточной цитометрией. Фиг. 1А: на точечных диаграммах показано число CD4-CD8- (двойных негативных тимоцитарных предшественников или «DN»), CD4+CD8+ (двойных положительных тимоцитарных предшественников или «DP»), CD4+ или CD8+ (одиночных негативных тимоцитарных предшественников или «SP») и DN/CD44+CD25- (тимоцитарных предшественников на стадии DN1) T-клеток. Числа на точечных диаграммах представляют собой проценты (среднее ± SEM (стандартная ошибка среднего)) субпопуляций Т-клеток от общего количества клеток тимуса. Фиг. 1В: на гистограммах показаны проценты (среднее ± SD (среднеквадратичное отклонение)) В-клеток (B220+) от DN1-клеток (а именно, отбираемых как CD4-CD8-CD44+CD25-).
На фигурах 2А-2В показаны эффекты блокирования Dll4 на стадии развития В-клеток в костном мозге (фиг. 2А) и на гомеостаз В-клеток в селезенке (фиг. 2В). Числа на точечных диаграммах представляют собой проценты (среднее ± SEM) субпопуляций В-клеток от общего количества клеток в костном мозге или в селезенке. GC: В-клетки зародышевого центра; Т1 и Т2: субпопуляции В-клеток; М: краевые В-клетки; и Fo: фолликулярные В-клетки.
На фигурах 3A-3D показаны эффекты блокирования Dll4 на развитие дендритных клеток (DC). Фиг. 3A: на точечных графиках показано увеличение числа обычных DC («cDC»; B220-CD11C+) и плазмацитоидных DC («pDC»; PDCA1+B220+CD11C+) в тимусе после воздействия антител к Dll4. Числа на точечных диаграммах представляют среднее процентное содержание (среднее ± SEM) дендритных клеток относительно общего количества клеток на 14-й день. Фиг. 3В: на гистограмме показана кинетика размножения cDC и pDC в тимусе мышей, получавших воздействие Dll4-АТ (■), и мышей, получавших контрольное воздействие hFc (□). Фиг. 3C: на точечных диаграммах показаны эффекты Dll4-АТ на клетки-предшественники DC (MHCIIнизк.CD11cсредн.CD135+Sirp-αсредн.) и поздние клетки-предшественники DC (MHCIIнизк.CD11cсредн.) в тимусе. Числа на точечных диаграммах представляют среднее процентное содержание (среднее ± SEM) клеток-предшественников DC относительно общего количества клеток на 14-й день. Фиг. 3D: на точечных диаграммах показано присутствие MHCIIнизк.CD11cсредн. DC в DN1-популяции про-Т-клеток (CD4-CD8-CD44+CD25-) в тимусе мышей, получавших воздействие Dll4-АТ, но не в тимусе мышей, получавших воздействие контрольного антитела, hFc. Числа на точечных диаграммах представляют среднее число (среднее ± SEM) MHCIIнизк.CD11cсредн. DC в DN1-популяции про-Т-клеток на 3-й день.
На фигуре 4 показан эффект блокирования Dll4 на развитие в тимусе альтернативной дифференцировки дендритных клеток в незрелые DC (imDC), предшественниками которых является общий DN1-предшественник T/DC-клеток. CD45.1+Lin--отсортированные DN1-клетки переносили в тимус мышей-хозяев (CD45.2+), получавших воздействие Dll4-АТ (■)или контрольного антитела, hFc (□).
На фигуре 5 показан эффект блокирования Dll4 на сывороточный уровень CSF-1 (M-CSF), ключевого цитокина, вовлеченного в развитие DC. Уровень CSF-1 в сыворотке мышей, не получавших воздействие ( ), или получавших воздействие изотипного контрольного антитела (□), или Dll4-АТ (■) измеряли с помощью твердофазного иммуноферментного анализа (ELISA).
), или получавших воздействие изотипного контрольного антитела (□), или Dll4-АТ (■) измеряли с помощью твердофазного иммуноферментного анализа (ELISA).
На фиг. 6 показаны эффекты генетической делеции Dll4 после воздействия тамоксифена (ТАМ) на гомеостаз В-клеток и DC у мышей DLL4COIN, содержащих индуцируемую тамоксифеном конструкцию Cre-рекомбиназы, CreERT2. Числа на точечных диаграммах представляют средние процентные значения (среднее ± SEM) В-клеток, а также pDC и cDC, от общего количества клеток в тимусе.
На фигурах 7А-7С показаны эффекты блокирования/делеции Dll4 на гомеостаз Трег-клеток. Фиг. 7А: на точечной диаграмме показано увеличение числа Трег-клеток в тимусе мышей, получавших воздействие Dll4-АТ в течение двух недель, по сравнению с мышами, получавшими воздействие контрольного антитела, hFc. Числа на точечных диаграммах представляют средние процентные значения (среднее ± SEM) Трег-клеток от CD3+CD4+ T-клеток в тимусе. Фиг. 7В: на гистограмме показана кинетика развития Трег-клеток в тимусе (верхняя панель) и селезенке (нижняя панель), соответственно, мышей, получавших воздействие Dll4-АТ (■) и получавших воздействие контрольного антитела, hFc (□). Фиг. 7C: на точечной диаграмме показано увеличение числа Трег-клеток в тимусе мышей DLL4COIN, получавших воздействие тамоксифена, по сравнению с контрольными мышами DLL4COIN, получавшими воздействие кукурузного масла. Числа на точечных диаграммах представляют средние процентные значения (среднее ± SEM) Трег-клеток от CD3+CD4+ T-клеток в тимусе.
На фигурах 8A-8B показаны эффекты блокирования Dll4 на гомеостаз DC (фиг. 8A) и Трег-клетки (фиг. 8B) в тимусе мышей, экспрессирующих Dll4 человека (hDll4), наблюдаемые на 7-й и 14-й день после воздействия Dll4-АТ (REGN421) (1 мг/кг или 5 мг/кг) или воздействия hFc (5 мг/кг), два раза в неделю в течение 2 недель и на 28-й день после отмены воздействия. Числа на точечных диаграммах представляют средние процентные значения (среднее ± SEM) pDC и cDC (фиг. 8A) или Трег-клеток (фиг. 8B) от общего количества клеток в тимусе.
На фиг. 9А-9В показаны эффекты блокирования Dll4 на мышиную модель экспериментального аутоиммунного энцефаломиелита (ЕАЕ). Фиг. 9А: на графике показана частота возникновения заболевания (%) на группу воздействия. Фиг. 9В: на графике показано развитие ЕАЕ по средней оценке заболевания. Воздействие проводили антителами к Dll4 (REGN577) до индукции заболевания (▼); изотипным контрольными антителами до индукции заболевания (◆); антителами REGN577 после индукции заболевания (▲); или антителами к VLA-4 (PS/2) до индукции заболевания (■).
На фиг. 10 показаны эффекты блокирования Dll4 на продукцию IL-17 и IFN-γ в лимфатических узлах EAE-мышей. Уровень IL-17 (левая панель) и IFN-γ (правая панель) в лимфатических узлах EAE-мышей, получавших воздействие Dll4-АТ (■) или контрольных АТ, hFc (□), измеряли на 12-й и 18-й день с помощоью ELISA.
На фигурах 11А-11Е показаны эффекты антител к Dll4 на диабетическую модель мышей NOD. На фиг. 11А показан % возникновения диабета (два последовательных определения уровня глюкозы выше 250 мг/дл) у 9-недельных мышей, которые получали либо контрольное антитело, hFC (●), либо анти-Dll4 антитело (REGN577) (■). Также показан % возникновения диабета у пяти мышей, которые получали воздействие Dll4-АТ, и после получали инъекции антитела PC61 на 20-й неделе (◆). PC61 представляет собой анти-CD25 антитело и истощает Трег-клетки. На фигуре 11В показана измеренная с помощью ELISA продукция анти-инсулинового аутоантитела (□), аутоантитела к декарбоксилазе глутаминовой кислоты 65 (GAD65) (■) у мышей NOD, получавших воздействие Dll4-АТ или контроля, hFc, по сравнению с не получавшими воздействие мышами дикого типа (WT). На фигуре 11С показаны срезы поджелудочной железы, окрашенные гематоксилином и эозином (H&E), мышей NOD, получавших воздействие Dll4-АТ (левая панель) или контрольного антитела, hFc (правая панель). Черные стрелки указывают на отдельные панкреатические островки, а белые стрелки указывают на клеточные инфильтраты в островках (правая панель). На фигуре 11D показано число панкреатических островков (левая панель) или % инфильтрованных панкреатических островков (правая панель) в поджелудочной железе мышей, получавших воздействие контрольного антитела, hFc (□), или получавших воздействие Dll4-АТ (■). На фигуре 11Е показаны изменения уровня глюкозы в крови у мышей, получавших в начале заболевания Dll4-АТ (●) или контрольное антитело, hFc (□), через 42 дня после воздействия.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед описанием настоящих способов следует отметить, что настоящее изобретение не ограничено описанными конкретными способами и экспериментальными условиями, поскольку такие способы и условия могут изменяться. Следует также понимать, что используемая в настоящем документе терминология предназначена только для описания конкретных вариантов осуществления изобретения и не носит ограничительного характера, поскольку объем настоящего изобретения будет лимитирован только прилагаемой формулой изобретения.
Используемое в данном описании и приложенной формуле изобретения единственное число включает ссылку на множественное число, за исключением случаев, когда контекст явно указывает на обратное. Поэтому, например, ссылка на «способ» включает один или несколько способов и/или стадий описанного в настоящем документе типа, и/или которые будут очевидны специалистам в данной области при прочтении описания настоящего изобретения.
Если не указано иное, то все технические и научные термины, используемые в настоящем документе, имеют такое же значение, которое общеизвестно рядовым специалистам в области, к которой относится настоящее изобретение. Хотя на практике или при проверке настоящего изобретения можно использовать любые способы и материалы, аналогичные или идентичные описанным в настоящем документе, ниже описаны предпочтительные способы и материалы. Все указанные в настоящем документе публикации полностью включены в него путем ссылки.
ОПРЕДЕЛЕНИЯ
Используемый в настоящем документе термин «антагонисты Dll4» включает антитела к Dll4 и их фрагменты, способные блокировать связывание Dll4 с Notch-рецептором (таким как Notch1 и Notch4) и/или блокировать активируемые взаимодействием Dll4-Notch сигнальные пути (см., например, WO 2008/076379), слитые белки, содержащие внеклеточный домен Dll4, слитый с мультимеризующим компонентом, или их фрагменты (см., например, патентные публикации США №№ 2006/0134121 и 2008/0107648), пептиды и пептидоантитела (см., например, патент США № 7,138,370), и т.п., которые блокируют взаимодействие между Dll4 и Notch-рецептором. Так, в некоторых вариантах осуществления изобретения термин также охватывает антагонисты, такие как низкомолекулярные соединения, антитела или их антиген-связывающие фрагменты, и т.п., которые специфично связывают Notch-рецепторы (например, анти-Notch1 антитела, анти-Notch4 антитела и т.п.) и блокируют активируемые взаимодействием Dll4-Notch сигнальные пути.
Используемый в настоящем документе термин «антитело» предназначен для обозначения молекул иммуноглобулина, состоящих из четырех полипептидных цепей, двух тяжелых (Н) цепей и двух легких (L) цепей, связанных друг с другом дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно называемой в настоящем документе HCVR или VH) и константной области тяжелой цепи (СН). Константная область тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно называемой в настоящем документе LCVR или VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут дополнительно подразделяться на области гипервариабельности, которые называются определяющими комплементарность областями (CDR), между которыми находятся более консервативные области, называемыми каркасными областями (FR). Каждая область VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
Способы и методики для идентификации CDR в аминокислотных последовательностях HCVR и LCVR известны в данной области, и их можно применять для идентификации указанных аминокислотных последовательностей HCVR и/или LCVR, раскрытых в настоящем документе. Условия, которые можно использовать для идентификации границ CDR, включают определение по Кабату, определение по Чотии и AbM-определение. В общем определение по Кабату основано на вариабельности последовательности, определение по Чотии основано на расположении областей структурных петель, а AbM-определение является компромиссом между подходами по Кабату и Чотии. См., например, Kabat, "Sequences of Proteins of Immunological Interest," National Institutes of Health, Bethesda, Md. (1991); Al-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); и Martin et al., Proc. Natl. Acad. Sci. USA 86:9268-9272 (1989). Общедоступны также базы данных для идентификации последовательностей CDR в антителе.
Также возможна замена одного или нескольких остатков CDR или удаление одного или нескольких CDR. В научной литературе были описаны антитела, в которых один или два CDR не критичны для связывания. Padlan и др. (1995 FASEB J. 9:133-139) проанализировали области контакта между антителами и их антигенами, исходя из опубликованных кристаллических структур, и сделали вывод, что только примерно от одной пятой до одной третьей остатков CDR действительно контактируют с антигеном. Padlan также обнаружил множество антител, у которых один или два CDR не имели аминокислот в контакте с антигеном (см. также Vajdos et al. 2002 J Mol Biol 320:415-428).
Остатки CDR, не контактирующие с антигеном, можно идентифицировать, исходя из предшествующих исследований (например, остатки H60-H65 и CDRH2 часто не требуются), в областях CDR по Кабату, расположенных вне CDR по Чотии, с помощью молекулярного моделирования и/или эмпирически. Если CDR или его остаток(остатки) удаляют, то их обычно заменяют аминокислотной последовательностью, занимающей соответствующую позицию в последовательности другого антитела человека или консенсуса таких последовательностей. Позиции для замены внутри CDR и аминокислоты для замены также можно выбрать эмпирически. Эмпирическими заменами могут быть консервативные или неконсервативные замены.
Термин «антитело» также охватывает антитела с модифицированной схемой гликозилирования. В некоторых вариантах применения может оказаться полезным удалить нежелательные сайты гликозилирования, или, например, удалить остатки фукозы для усиления функции антителозависимой клеточной цитотоксичности (ADCC) (см. Shield et al. (2002) JBC 277:26733). В других вариантах применения удаление участка N-гликозилирования может снизить нежелательные иммунные реакции против терапевтических антител или увеличить аффинность антител. В других вариантах применения можно осуществить модификацию галактозилирования, чтобы изменить комплемент-зависимую цитотоксичность (CDC).
Используемый в настоящем документе термин «антигенсвязывающий фрагмент» антитела (или просто «фрагмент антитела») относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфично связывать hDll4 или любые другие предполагаемые белки-мишени. Фрагмент антитела может включать: Fab-фрагмент, F(ab')2-фрагмент, Fd-фрагмент, Fv-фрагмент, одноцепочечную молекулу Fv (scFv), dAb-фрагмент, минимальные распознающие элементы, состоящие из аминокислотных остатков, которые имитируют гипервариабельную область антитела (например, фрагмент, содержащий CDR, или выделенный CDR). Другие сконструированные молекулы, такие как диантитела, триантитела, тетраантитела и миниантитела также охвачены выражением «антигенсвязывающий фрагмент», используемым в настоящем документе. В некоторых вариантах осуществления изобретения антитело или фрагменты антитела по изобретению можно конъюгировать с терапевтической молекулой («иммуноконъюгат»), такой как цитотоксин, химиотерапевтическое соединение, иммуносупрессант или радиоактивный изотоп.
Антигенсвязывающий фрагмент антитела обычно будет содержать по меньшей мере один вариабельный домен. Вариабельный домен может иметь любой размер или аминокислотный состав и обычно будет содержать по меньшей мере одну CDR, смежную или с сохранением рамки считывания с одной или несколькими каркасными последовательностями. В антигенсвязывающих фрагментах, имеющих VH-домен, ассоциированный с VL-доменом, VH- и VL-домены могут быть расположены относительно друг друга в любом подходящем порядке. Например, вариабельная область может быть димерной и содержать димеры VH-VH, VH-VL или VL-VL. В альтернативном варианте, антигенсвязыывающий фрагмент антитела может содержать мономерный VH- или VL-домен.
В некоторых вариантах осуществления изобретения антигенсвязывающий фрагмент антитела может содержать по меньшей мере один вариабельный домен, ковалентно соединенный по меньшей мере с одним константным доменом. Неограничивающие примеры конфигураций вариабельных и константных доменов, которые можно найти среди антигенсвязывающих фрагментов антител по настоящему изобретению, включают: (i) VH-CH1; (ii) VH-CH2; (iii) VH-CH3; (iv) VH-CH1-CH2; (v) VH-CH1-CH2-CH3; (vi) VH-CH2-CH3; (vii) VH-CL; (viii) VL-CH1; (ix) VL-CH2; (x) VL-CH3; (xi) VL-CH1-CH2; (xii) VL-CH1-CH2-CH3; (xiii) VL-CH2-CH3 и (xiv) VL-CL. В любой конфигурации вариабельных и константных доменов, включая любые из примеров конфигураций, перечисленных выше, вариабельные и константные домены могут быть либо напрямую соединены друг с другом, либо могут быть соединены через полноразмерную или частичную шарнирную или линкерную область. Шарнирная область может состоять по меньшей мере из 2 (например, из 5, 10, 15, 20, 40, 60 или большего числа) аминокислот, что приводит к гибкому или среднегибкому соединению между соседними вариабельными и/или константными доменами в одной молекуле полипептида. Более того, антигенсвязывающий фрагмент антитела по настоящему изобретению может включать в себя гомодимер или гетеродимер (или другой мультимер) любых конфигураций вариабельных и константных доменов, перечисленных выше, в нековалентной ассоцииации друг с другом и/или с одним или несколькими VH- или VL-доменами (например, посредством дисульфидной(ых) связи(ей)).
Как и в случае с полноразмерными молекулами антител, антиген-связывающие фрагменты могут быть моноспецифичными или полиспецифичными (например, биспецифичными). Полиспецифичный антиген-связывающий фрагмент антитела обычно будет содержать по меньшей мере два различных вариабельных домена, причем каждый вариабельный домен способен специфично связывать отдельный антиген или другой эпитоп на одном антигене. Любой формат полиспецифичного антитела можно адаптировать для применения в контексте антигенсвязывающего фрагмента антитела по настоящему изобретению, используя рутинные методики, имеющиеся в данной области.
Используемый в настоящем документе термин «антитело человека» предназначен для включения антител с вариабельной и константной областями, полученными из иммуноглобулиновых последовательностей зародышевой линии человека. Моноклональные антитела человека по изобретению могут включать аминокислотные остатки, не кодируемые иммуноглобулиновыми последовательностями зародышевой линии человека (например, мутации, вызванные случайным или сайт-специфичным мутагенезом in vitro или соматическими мутациями in vivo), например, в CDR, особенно в CDR3. Однако используемый в настоящем документе термин «антитело человека» не подразумевает включение моноклональных антител, в которых в FR-последовательности человека встроены CDR последовательности, полученные из зародышевой линии млекопитающего другого вида (например, мыши).
Раскрытые в настоящем документе полностью антитела человека к Dll4 могут содержать одну или несколько аминокислотных замен, вставок и/или делеций в каркасных и/или CDR областях вариабельных доменов тяжелой и легкой цепей по сравнению с соответствующими последовательностями зародышевой линии. Такие мутации можно легко выявить, сравнивая аминокислотные последовательности, раскрытые в настоящем документе, с последовательностями зародышевой линии, доступными, например, в открытых базах данных последовательностей антител. Настоящее изобретение включает антитела и их антиген-связывающие фрагменты, которые получены из любых аминокислотных последовательностях, раскрытых в настоящем документе, в которых одна или несколько аминокислот в одной или нескольких каркасных и/или CDR областях мутированы относительно соответствующего(их) остатка(ов) зародышевой линии, из которой получено антитело, или относительно соответствующего(их) остатка(ов) другой зародышевой линии человека, или относительно консервативной аминокислотной замены соответствующего(их) остатка(ов) зародышевой линии (такие изменения последовательности собирательно именуются в настоящем документе «мутациями зародышевой линии»). Рядовой специалист в данной области, начиная с последовательностей вариабельных областей тяжелой и легкой цепей, раскрытых в настоящем документе, может легко получить множество антител и антиген-связывающих фрагментов, которые содержать одну или несколько реверсий относительно индивидуальной зародышевой линии или их комбинации. В некоторых вариантах осуществления изобретения все каркасные и/или CDR остатки в VH и/или VL-доменах мутированы обратно до остатков, найденных в исходной последовательности зародышевой линии, из которой было получено антитело. В других вариантах осуществления изобретения только некоторые остатки мутированы обратно до исходной последовательности зародышевой линии, например, только мутированные остатки, найденные в первых 8 аминокислотах FR1 или в последних 8 аминокислотах FR4, или только мутированные остатки, найденные в CDR1, CDR2 или CDR3. В других вариантах осуществления изобретения один или несколько каркасных и/или CDR-остатков мутированы в соответствующие остатки последовательности другой зародышевой линии (то есть, последовательности зародышевой линии, которая отличается от последовательности зародышевой линии, из которой исходно было получено антитело). Кроме того, антитела по настоящему изобретению могут содержать любую комбинацию двух или нескольких мутаций зародышевой линии в каркасной и/или CDR-областях, например, в которых некоторые индивидуальные остатки мутированы в соответствующие остатки в последовательности конкретной зародышевой линии, в то время как некоторые другие остатки, которые отличаются от последовательности исходной зародышевой линии, сохраняются или мутированы в соответствующие остатки другой зародышевой линии. После получения антитела и антигенсвязывающие фрагменты, которые содержат одну или несколько мутаций зародышевой линии, можно легко проверить по одному или нескольким желательным свойствам, таким как увеличенная специфичность связывания, увеличенная аффинность связывания, увеличенные или улучшенные антагонистические или агонистические биологические свойства (в зависимости от обстоятельств), сниженная иммуногенность и т.п. Антитела и антигенсвязывающие фрагменты, полученные по этому общему принципу, входят в объем настоящего изобретения.
Настоящее изобретение также включает анти-Dll4 антитела, содержащие варианты любых аминокислотных последовательностей HCVR, LCVR и/или CDR, раскрытых в настоящем документе, имеющие одну или несколько консервативных замен. Например, настоящее изобретение включает анти-Dll4 антитела, имеющие аминокислотные последовательности HCVR, LCVR и/или CDR, например, с 10 или меньше, 8 или меньше, 6 или меньше, 4 или меньше, 2 или 1 консервативными аминокислотными заменами относительно любых аминокислотных последовательностей HCVR, LCVR и/или CDR, раскрытых в настоящем изобретении. В одном варианте осуществления изобретения HCVR содержит аминокислотную последовательность SEQ ID NO:116 с 10 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения HCVR содержит аминокислотную последовательность SEQ ID NO:116 с 8 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения HCVR содержит аминокислотную последовательность SEQ ID NO:116 с 6 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения HCVR содержит аминокислотную последовательность SEQ ID NO:116 с 4 или меньше консервативными заменами в ней. В еще одном варианте осуществления изобретения HCVR содержит аминокислотную последовательность SEQ ID NO:116 с 2 или 1 консервативными заменами в ней. В одном варианте осуществления изобретения LCVR содержит аминокислотную последовательность SEQ ID NO:118 с 10 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения LCVR содержит аминокислотную последовательность SEQ ID NO:118 с 8 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения LCVR содержит аминокислотную последовательность SEQ ID NO:118 с 6 или меньше консервативными заменами в ней. В другом варианте осуществления изобретения LCVR содержит аминокислотную последовательность SEQ ID NO:118 с 4 или меньше консервативными заменами в ней. В еще одном варианте осуществления изобретения LCVR содержит аминокислотную последовательность SEQ ID NO:118 с 2 или 1 консервативными заменами в ней.
Предполагается, что «нейтрализующее», или «блокирующее» антитело относится антителу, связывание которого с Dll4 приводит к ингибированию биологической активности Dll4. Такое ингибирование биологической активности Dll4 можно оценить, измеряя один или нескольких показателей биологической активности Dll4. Данные показатели биологической активности Dll4 можно оценить с помощью одного или нескольких методов анализа in vitro или in vivo, известных в данной области. Например, способность антитела нейтрализовать активность Dll4 оценивают по ингибированию связывания Dll4 с Notch-рецептором. Аналогичным образом термин также применим к антителам против других мишеней, таких как Notch1 и Notch4; такие антитела ингибируют биологическую активность мишеней, тем самым ингибируя взаимодействие Dll4 и Notch или сигнальные пути, активируемые этим взаимодействием.
Термин «специфично связывается» и т.п. означает, что антитело или его антиген-связывающий фрагмент образуют комплекс с антигеном, относительно стабильный при физиологических условиях. Специфичное связывание можно описать константной диссоциации по меньшей мере примерно 1×10-6 M или меньше (например, меньшая KD означает более сильное связывание). Способы для определения специфичности связывания двух молекул хорошо известны в данной области и включают, например, равновесный диализ, поверхностный плазмонный резонанс и т.п. Однако выделенное антитело, которое специфично связывает hDll4, может проявлять перекрестную реактивность с другими антигенами, такими как молекулы Dll4 из других биологических видов. Более того, в контексте настоящего изобретения полиспецифичные антитела (например, биспецифичные), которые связывают hDll4 и один или несколько дополнительных антигенов, тем не менее считаются антителами, которые «специфично связывают» hDll4.
Предполагается, что используемый в настоящем документе термин «KD» относится к константе диссоциации конкретного взаимодействия антитела с антигеном.
Термин «высокоаффинное антитело» относится к антителам, которые связывают hDll4 с KD примерно 1 нМ или меньше, примерно 500 пМ или меньше, примерно 400 пМ или меньше, примерно 300 пМ или меньше, примерно 200 пМ или меньше, или примерно 100 пМ или меньше, или примерно 50 пМ или меньше, измеренной с помощью поверхностного плазмонного резонанса, например BIACORE™, или ELISA для измерения аффинности раствора.
Используемый в настоящем документе термин «поверхностный плазмонный резонанс» относится к оптическому явлению, которое позволяет проводить анализ биоспецифичных взаимодействий в режиме реального времени путем детектирования изменений концентраций белка в биосенсорной матрице, например, с использованием системы BIACORE™ (Pharmacia Biosensor AB, Uppsala, Sweden и Piscataway, N.J.).
Термин «эпитоп» представляет собой область антигена, которая связывается с антителом. Эпитопы можно определить, как структурные или функциональные. Функциональные эпитопы представляют собой субпопуляцию структурных эпитопов и включают остатки, которые вносят непосредственный вклад в аффинность взаимодействия. Эпитопы также могут быть конформационными, то есть могут состоять из несвязанных между собой аминокислот. В некоторых вариантах осуществления изобретения эпитопы могут включать детерминанты, которые являются химически активными группировками молекул на поверхности, таких как аминокислоты, гликозидные боковые цепи, фосфорильные группы или сульфонильные группы, и в некоторых вариантах осуществления изобретения могут иметь определенные трехмерные структурные характеристики и/или определенные характеристики заряда. Эпитоп обычно включает по меньшей мере 3, более типично, по меньше мере 5 или 8-10 аминокислот в уникальной пространственной конформации.
Предполагается, что используемый в настоящем документе термин «воздействие» или «лечение» означает как профилактические (или предупреждающие), так и терапевтические методики, если не указано иное. Нуждающиеся в лечении субъекты включают не только субъектов с уже существующим конкретным состоянием, расстройством или заболеванием, но также и субъектов, предрасположенных или восприимчивых к развитию такого состояния, расстройства или заболевания, и которым профилактические методики принесут пользу в том, что снизят возникновение или рецидив, или развитие при возникновении, такого состояния, расстройства или заболевания по сравнению с отсутствием лечения.
Фраза «терапевтически эффективное количество», «профилактически эффективное количество» или «эффективное количество» означает количество, которое обеспечивает желаемый эффект, для которого его вводят. Точное количество будет зависеть от цели лечения, возраста и размеров субъекта, получающего лечение, пути введения и т.п. и может быть определено специалистом в данной области с использованием известных методик (см., например, Loyd (1999) The Art, Science and Technology of Pharmaceutical Compounding).
ОБЩЕЕ ОПИСАНИЕ
Настоящее изобретение основано частично на открытии, что блокирование Dll4 с помощью Dll4-специфичного антитела приводит к повышению числа Трег-клеток, что, в свою очередь, предупреждает, снижает или замедляет развитие EAE или диабета у мышей. Для описания полностью антитела человека к Dll4, включая рекомбинантное антитело Dll4 человека, см. международную патентную публикацию № WO 2008/076379.
ТЕРАПЕВТИЧЕСКОЕ ВВЕДЕНИЕ И СОСТАВЫ
Настоящее изобретение относится к способам предупреждения, лечения или ослабления заболевания или расстройства, при котором положительный эффект оказывает повышение числа Трег-клеток, включающим введение терапевтически эффективного количества фармацевтической композиции, содержащей антагонист Dll4, такой как антитело к Dll4. Фармацевтическая композиция, содержащая антагонист Dll4, может дополнительно содержать один или несколько дополнительных терапевтических агентов, таких как иммуносупрессанты, противовоспалительные агенты, анальгетики, агенты для снижения уровня глюкозы в крови и т.п. (см. следующий раздел). Терапевтические композиции по изобретению можно вводить с подходящими носителями, вспомогательными веществами и другими агентами, которые включают в составы, для того чтобы улучшить перенос, доставку, переносимость и т.п. Множество соответствующих составов можно найти в фармацевтическом справочнике, известном всем специалистам в области фармацевтической химии: Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, PA. Эти составы включают, например, порошки, пасты, мази, гели, воски, масла, липиды, липидсодержащие (катионные или анионные) везикулы (например, LIPOFECTIN™), конъюгаты ДНК, безводные абсорбционные пасты, эмульсии масло-в-воде и вода-в-масле, эмульсии карбовакса (полиэтиленгликолей разного молекулярного веса), полутвердые гели и полутвердые смеси, содержащие карбовакс. См. также Powell et al. "Compendium of excipients for parenteral formulations" PDA (1998) J Pharm Sci Technol 52:238-311.
Для системного введения терапевтически эффективную дозу можно сначала определить из анализов in vitro. Например, доза может быть разработана в моделях на животных, чтобы достичь диапазона концентрации в кровотоке, который включают IC50, определенную на культуре клеток. Такую информацию можно использовать для более точного определения доз, приемлемых для человека. Исходные дозы также можно оценить на основе данных in vivo, например, в моделях на животных, используя методики, хорошо известные в данной области. Рядовой специалист в данной области без труда может оптимизировать введение человеку на основе данных, полученных на животных.
Дозировка может варьировать в зависимости от возраста и размера (например, веса тела или площади поверхности тела) субъекта, которому планируется вводить соединение, целевого заболевания, условий, пути введения и т.п. Для системного введения антагонистов Dll4, в частности, антител к Dll4, типичными диапазонами дозировок для внутривенного введения, являются дневная доза от примерно 0,01 до примерно 100 мг/кг веса тела, от примерно 0,1 до примерно 50 мг/кг, или от примерно 0,2 до примерно 10 мг/кг. Для подкожного введения антитела можно вводить от примерно 1 мг до примерно 800 мг, от примерно 10 мг до примерно 500 мг, от примерно 20 мг до примерно 400 мг, от примерно 30 мг до примерно 300 мг или от примерно 50 мг до примерно 200 мг, при концентрации антител по меньшей мере примерно 25 мг/мл, примерно 50 мг/мл, примерно 75 мг/мл, примерно 100 мг/мл, примерно 125 мг/мл, примерно 150 мг/мл, примерно 175 мг/мл, примерно 200 мг/мл или примерно 250 мг/мл, по меньшей мере от 1 до 5 раз в день, от 1 до 5 раз в неделю, от 1 до 5 раз в месяц. В альтернативном варианте, антитела можно первоначально вводить внутривенной инъекцией с последующим подкожным введением.
Известны различные системы доставки, которые можно использовать для введения фармацевтической композиции по изобретению, например инкапсулирование в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать мутантные вирусы, опосредованный рецепторами эндоцитоз (см., например, Wu et al. 1987, J. Biol. Chem. 262: 4429-4432). Способы введения включают без ограничения интрадермальный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный и пероральный пути. Композицию можно вводить любым удобным путем, например инфузией или болюсной инъекцией, путем всасывания через эпителиальные или кожно-слизистые выстилки (например, слизистую оболочку ротовой полости, слизистую оболочку прямой кишки и кишечника и т.д.), и можно вводить вместе с другими биологически активными агентами. Введение может быть системным или местным.
Фармацевтическую композицию также можно доставить в везикулах, в частности, липосомах (см. Langer (1990) Science 249:1527-1533; Treat et al. (1989) в Liposomes in the Therapy of Infectious Disease and Cancer, Lopez Berestein and Fidler (eds.), Liss, New York, pp. 353-365; Lopez-Berestein, ibid., pp. 317-327; см. в общем тот же источник).
В некоторых ситуациях фармацевтическую композицию можно доставить в системе контролируемого высвобождения. В одном варианте осуществления можно использовать насос (см., Langer, supra; Sefton (1987) CRC Crit. Ref. Biomed. Eng. 14:201). В другом варианте осуществления изобретения можно использовать полимерные материалы; см. Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida (1974). В еще одном варианте осуществления изобретения систему контролируемого высвобождения можно поместить близко от мишени композиции, таким образом, необходима будет только часть системной дозы (см., например, Goodson, в Medical Applications of Controlled Release, supra, vol. 2, pp. 115-138, 1984).
Препараты для инъекций могут включать лекарственные формы для внутривенных, подкожных, внутрикожных и внутримышечных инъекций, капельных инфузий и т.п. Эти препараты для инъекций можно приготовить общеизвестными способами. Например, препараты для инъекций можно приготовить, растворяя, ресуспендируя или эмульгируя антитело или его соль, описанные выше, в стерильной водной среде или масляной среде, обычно используемой для инъекций. В качестве водной среды для инъекций существует, например, физиологический раствор, изотонический раствор, содержащий глюкозу и другие вспомогательные агенты и т.п., которые можно использовать в комбинации с соответствующим солюбилизирующим агентом, таким как спирт (например, этанол), многоатомный спирт (например, полипропиленгликоль, полиэтиленгликоль), неионное поверхностно-активное вещество (например, полисорбат 80, НСО-50 (аддукт гидрированного касторового масла с полиоксиэтиленом (50 моль))), и т.п. В качестве масляной среды используют, например, кунжутное масло, соевое масло и т.п., которые можно использовать в комбинации с солюбилизирующим агентом, таким как бензилбензоат, бензиловый спирт и т.п. Приготовленным таким образом раствором для инъекции предпочтительно заполнить подходящие ампулы. Фармацевтическую композицию по настоящему изобретению можно доставить подкожно или внутривенно с использованием стандартных иглы и шприца. В дополнение, при подкожном введении часто применяют шприц-ручку для доставки фармацевтической композиции по настоящему изобретению. Такая шприц-ручка может быть для многократного использования или одноразовой. В шприце-ручке для многократного использования обычно применяют заменяемый картридж, который содержит фармацевтическую композицию. После введения всей фармацевтической композиции в картридже и опустошения картриджа, пустой картридж легко удаляется и заменяется новым картриджем, который содержит фармацевтическую композицию. Затем шприц-ручку можно использовать повторно. В одноразовых шприц-ручках нет заменяемого картриджа. Вместо этого, одноразовая шприц-ручка продается заполненной фармацевтической композицией, содержащейся в резервуаре устройства. После полного израсходования фармацевтической композиции в резервуаре выбрасывают все устройство.
Различные шприцы-ручки и автоинжекторные устройства доставки для многократного применения используют для подкожной доставки фармацевтической композиции по настоящему изобретению. Примеры включают, но несомненно не ограничены этим, AUTOPEN™ (Owen Mumford, Inc., Woodstock, UK), шприц-ручку DISETRONIC™ (Disetronic Medical Systems, Burghdorf, Switzerland), шприц-ручку HUMALOG MIX 75/25™, шприц-ручку HUMALOG™, шприц-ручку HUMALIN 70/30™ (Eli Lilly and Co., Indianapolis, IN), NOVOPEN™ I, II и III (Novo Nordisk, Copenhagen, Denmark), NOVOPEN JUNIOR™ (Novo Nordisk, Copenhagen, Denmark), шприц-ручку BD™ pen (Becton Dickinson, Franklin Lakes, NJ), OPTIPEN™, OPTIPEN PRO™, OPTIPEN STARLET™ и OPTICLIK™ (Sanofi-Aventis, Frankfurt, Germany), не говоря о других. Примеры одноразовых шприц-ручек, применяемых для подкожной доставки фармацевтической композиции по настоящему изобретению, включают, но несомненно не ограничены этим, шприц-ручку SOLOSTAR™ (Sanofi-Aventis), FLEXPEN™ (Novo Nordisk) и KWIKPEN™ (Eli Lilly).
Предпочтительно, чтобы фармацевтические композиции для перорального или парентерального применения, описанные выше, изготавливали в лекарственных формах в виде стандартной дозы, соответствующей дозе активных ингредиентов. Такие лекарственные формы в виде стандартной дозы включают, например, таблетки, пилюли, капсулы, инъекции (ампулы), суппозитории и т.п. Содержащееся количество антагониста Dll4, такого как антитело к Dll4, в общем составляет от примерно 0,1 до примерно 800 мг на лекарственную форму в виде стандартной дозы; особенно в случае инъекций, предпочтительно, чтобы антитело содержалось в количестве от примерно 5 до примерно 100 мг, и в количестве от примерно 10 до примерно 250 мг для других лекарственных форм.
В некотором варианте осуществления изобретения может быть желательно вводить фармацевтические композиции по изобретению местно в область, которой необходимо воздействие; этого можно достичь, например (а не в качестве ограничения), с помощью местной инфузии в течение хирургического вмешательства, местного нанесения, например, с помощью инъекции, через катетер, или посредством имплантата, причем имплантат является пористым, непористым или гелеобразным материалом, включая мембраны, такие как силиконовые мембраны, волокна или доступные в продаже заменители кожи.
КОМБИНИРОВАННЫЕ МЕТОДЫ ЛЕЧЕНИЯ
В терапевтических способах по изобретению антагонист Dll4 можно вводить один или в комбинации с одним или несколькими дополнительными терапевтическими агентами, такими как иммуносупрессирующие агенты или иммуносупрессанты, противовоспалительные агенты, анальгетики, прямые или непрямые агенты для снижения уровня глюкозы в крови и т.п. Подходящие иммуносупрессанты включают, но не ограничены этим, глюкокортикостероиды, циклоспорин, метотрексат, интерферон β (IFN-β), такролимус, сиролимус, азатиоприн, меркаптопурин, опиоиды, микофенолят, TNF-связывающие белки, такие как инфликсимаб, этанерцепт, адалимумаб и т.п., цитотоксические антибиотики, такие как дактиномицин, антрациклины, митомицин С, блеомицин, митрамицин и т.п., антитела к клеткам иммунной системы, такие как анти-CD20 антитела, анти-CD3 антитела, и т.п. Противовоспалительные агенты и/или анальгетики, подходящие для комбинированных методов лечения с антагонистами Dll4, включают кортикостероиды, нестероидные противовоспалительные соединения (NSAID), такие как аспирин, ибупрофен, напроксен и т.п., антагонисты TNF-α (например, Инфликсимаб или РЕМИКАД® от Centocor Inc.; голимумаб от Centocor Inc.; этанерцепт или ЭНБРЕЛ® от Amgen/Wyeth; адалимумаб или ХУМИРА® от Abbott Laboratories и т.п.), антагонисты IL-1 (например, связывающие IL-1 слитые белки, например, АРКАЛИСТ® от Regeneron Pharmaceuticals, Inc., см. патент США № 6927044; КИНЕРЕТ® от Amgen и т.п.), антагонисты IL-6 (например, антитела к рецептору IL-6, раскрытые в патенте США № 7582298, и АКТЕМРА® от Roche), ацетаминофен, морфиномиметики, и т.п. Подходящие агенты для снижения уровня глюкозы в крови включают, но не ограничены этим, инсулин и его аналоги, бигуаниды, сульфонамиды и их производные с мочевиной, ингибиторы альфа-глюкозидазы, тиазолидиндион и его производные, ингибиторы дипептидил-пептидазы 4, гуаровую камедь, репаглинид, натеглинид, эксенатид, прамлинтид, бенфлуорекс, лираглутид, митиглинид, ингибиторы альдозоредуктазы и т.п.
Антагонист Dll4, такой как hDll4-АТ или его фрагменты, и дополнительные терапевтические агенты, описанные выше, можно вводить совместно или по отдельности. При использовании отдельных лекарственных составов антитело или его фрагмент по изобретению и дополнительные агенты можно вводить одновременно или отдельно через интервалы времени, например, последовательно, в соответствующем порядке.
НАБОРЫ
Изобретение дополнительно относится к промышленному изделию или набору, содержащим упаковочный материал, контейнер и фармацевтический агент, содержащийся в контейнере, причем фармацевтический агент включает в себя по меньшей мере один антагонист Dll4, такой как антитело к Dll4, и по меньшей мере один дополнительный терапевтический агент, и в котором упаковочный материл включает в себя этикетку или упаковочный вкладыш, в которых приведены показания и инструкция по применению. В одном варианте осуществления изобретения антагонист Dll4 и дополнительный терапевтический агент могут содержаться в отдельных контейнерах.
ПРИМЕРЫ
Следующие примеры приведены, для того чтобы предоставить средним специалистам в данной области полное содержание и описание того, как осуществить и применить способы и композиции по изобретению, и не подразумевает ограничения объема того, что авторы рассматривают в качестве своего изобретения. Были приняты меры, для того чтобы обеспечить точность в отношении используемых чисел (например, количеств, температуры и т.п.), но следует принимать во внимание некоторые экспериментальные ошибки и отклонения. Если не указано иное, то части представляют собой массовые части, температура выражена по шкале Цельсия, а давление представляет собой атмосферное или близкое к нему, и указанные погрешности являются средним ± SEM.
В примерах ниже для окрашивания клеток для проточной цитометрии использовали следующие антитела в Dulbecco's PBS 1 X (GIBCO® INVITROGEN™) с добавленными 3% FCS: для DC - антитела к сигнально-регуляторному белку α (Sirp-α; кат. № P84; BD Biosciences), антитела к B220 (кат. № RA3-6B2), антитела к PDCA-1 (кат. № eBio927), антитела к CD8 (кат. № 53-6.7), антитела к CD11b (кат № M1/70), антитела к MHCII (кат № M5/114.15.2), антитела к CD11c (кат № N418) и антитела к CD135 (кат № A2F10), соответственно; для T, B и NK-клеток, - антитела к CD4 (кат № GK1.5 или L3T4), антитела к CD3 (кат № 145-2C11), антитела к CD25 (кат № PC61 или 7D4), антитела к CD44 (кат № IM7), антитела к FoxP3 (кат № FJK16s); и антитела к F4/80 (кат № BM8), антитела к NK1.1 (кат. № PK136), антитела к IgM (кат № II/41), антитела к IgD (кат № 26-11c), антитела к CD43 (кат № S7), антитела к CD21 (кат № eBio4E3), антитела к HSA (кат № M1/69) и антитела к CD23 (кат № B3B4), соответственно (все от eBioscience).
Пример 1: эффект блокирования Dll4 на развитие В-клеток, дендритных клеток и Т-клеток
Было показано, что ингибирование взаимодействия Dll4-Notch1 приводит к полному блоку Т-клеточного развития, сопровождаемому эктопическим появлением В-клеток и увеличением числа дендритных клеток (DC), которое может быть следствием конверсии дифференцировки предшественников Т-клеток в DC в тимусе (Hozumi et al., 2008, J Exp Med 205(11):2507-2513; Koch et al., 2008, J Exp Med 205(11):2515-2523; и Feyerabend et al., 2009, Immunity 30:1-13). Однако все еще неизвестно, на какую конкретную стадию развития DC влияет блокирование Dll4.
Для ответа на этот вопрос 6-недельным мышам C57BL/6 (Jackson Labs) вводили с помощью подкожных инъекций 5 или 25 мг/кг анти-Dll4 антитела (REGN577) (n=5) или Fc-фрагмента человека (контроль) (n=5) два раза в неделю в течение четырех недель. REGN577 было получено внутри компании на основе опубликованной последовательности (WO 2007/143689). REGN 577 связывает Dll4 человека и мыши, но не связывает на детектируемом уровне Dll1 и JAG1 человека. Через четырнадцать (14) дней после инъекции собирали тимусы и селезенки и расщепляли при 37°C с течение 30 мин в полной среде RPMI 1640 (Invitrogen) с 10% фетальной сыворотки теленка (FCS) и содержащей коллагеназу D (Sigma Aldrich). Для остановки реакции добавляли 2 мМ EDTA, и суспензию органов пропускали через 70-мм клеточное сито. Костный мозг (ВМ) собирали у каждой мыши, промывая бедренную и большую берцовую кости в полной среде RPMI 1640 с 10% FCS, и клетки ресуспендировали в среде RPMI. Субпопуляции Т-клеток, В-клеток и DC оценивали проточной цитометрией после окрашивания клеток антителами к специфичным маркерам, описанным выше. Окрашенные клетки анализировали на BD™ LSR II Flow Cytometer (BD Biosciences), и данные анализировали, используя программное обеспечение FlowJo (версия 8.8.6; Tree Star Inc.).
На фигурах 1A и 1B показаны популяции Т-клеток и В-клеток в тимусе. Как показано на фиг. 1А, блокирование Dll4 вызывало значительное увеличение числа двойных негативных («DN»; CD4-CD8-) Т-клеток и снижение числа двойных положительных («DP»; CD4+CD8+) Т-клеток в тимусе. В дополнение, это же воздействие вызывало эктопическое появление В-клеток в тимусе, которые возникают из предшественников Т-клеток (то есть клеток CD44+CD25-CD4-CD8- на стадии DN1) (см. фиг. 1В). В отличие от этого, блокирование Dll4 не оказывало никакого эффекта на развитие В-клеток в костном мозге (фиг. 2А) или на периферические субпопуляции В-клеток в селезенке (фиг. 2В). Кроме того, блокирование Dll4 вызывало увеличение числа обычных DC («cDC»; B220-CD11C+) и плазмацитоидных DC («pDC»; PDCA1+B220+CD11C+) в тимусе (фиг. 3А) с значительным увеличением количества начиная с 7-го дня (p<0,001) по 14-й день (p<0,001) и длилось до 21-го дня (p<0,01) (фиг. 3B) после первичной инъекции антител к Dll4. Числа на точечных диаграммах представляют среднее процентное содержание DC относительно общего количества клеток на 14-й день. Кроме того, DC размножались в конечностях мышей, получавших воздействие антител к Dll4 (REGN577). В таблице 1 показана кратность увеличения процентного содержания и абсолютного числа DC в селезенке после воздействия Dll4-АТ по сравнению с контрольными мышами (получавшими hFc).
Известно, что cDC, pDC и моноциты лимфоидной ткани имеют общего предшественника, называемого «предшественник макрофагов и DC» или «MDP», который можно идентифицировать по его поверхностному фенотипу «Lin-cKitвысок.CD115+FLT3+», в то время как фактический предшественник, называемый «общий предшественник DC» или «CDP» с «Lin-cKitнизк.CD115+FLT3+», ограничен тем, что он продуцирует только cDC и pDC. Хотя у моноцитов могут развиться многие из фенотипических признаков DC в воспалительных условиях, линии cDC, pDC и моноцитов различаются по времени, за которое они достигают тканей, и никогда ни моноциты, ни pDC не развиваются в cDC при стационарных условиях. Полагают, что в отличие от моноцитов и pDC, cDC в лимфоидной ткани появляются из костного мозга в качестве незрелых клеток, которые должны дополнительно дифференцироваться и делиться в лимфоидных органах. Предшественники DC (MHCIIнизк.CD11cсредн.CD135+Sirp-αсредн.) и поздние клетки-предшественники DC (MHCIIнизк.CD11cсредн.) являются предшественниками преимущественно для cDC, которые появляются в костном мозге (Liu et al., 2009, Science 324:392-397).
Для того чтобы идентифицировать какой-либо эффект антител к Dll4 на гомеостаз предшественников DC, содержание MDP и CDP в тимусе, костном мозге и селезенке оценивали проточной цитометрией. MDP и CDP детектировались только в костном мозге, но ни в тимусе, ни в селезенке (данные не показаны). Кроме того, блокирование Dll4 не вызывало размножения ранних предшественников в костном мозге по сравнению с мышами, получавшими контрольное воздействие. Таким образом, результат позволяет предположить, что антитела к Dll4 могут действовать на более поздней стадии развития DC чем MDP и CDP, например, на стадии предшественников DC.
Соответственно, исследовали предшественников DC и поздних предшественников DC в тимусе и костном мозге, используя проточную цитометрию. Как показано на фиг. 3С, DC (MHCIIнизк.CD11cсредн.), которые в норме присутствуют в костном мозге, размножались только в тимусе через 14 дней после воздействия антител к Dll4 (p<0,001), в то время как в костном мозге этих же мышей не детектировалось никакого размножения DC (MHCIIнизк.CD11cсредн.) (данные не показаны). Таким образом, увеличение числа DC, началом которого является стадия предшественников DC, было ограничена тимусом. Для того чтобы определить источник DC (MHCIIнизк.CD11cсредн.) в тимусе, проводили проточную цитометрию для идентификации DC (MHCIIнизк.CD11cсредн.) в DN1-популяции про-Т-клеток (CD4-CD8-CD44+CD25-). Как показано на фиг. 3D, DC (MHCIIнизк.CD11cсредн.) детектировались в DN1-популяции про-Т-клеток на 3-й день после блокирования Dll4. DC (MHCIIнизк.CD11cсредн.) не детектировались при отсутствии воздействия антител к Dll4, а также в DN2, DN3 и DN4-попуяциях Т-клеток после воздействия антител к Dll4 (данные не показаны). После воздействия антител к Dll4 не наблюдалось никаких изменений гомеостаза периферических DC (данные не показаны). Таким образом, блокирование Dll4 индуцировало значительное увеличение количества DC (MHCIIнизк.CD11cсредн.) в DN1-популяции про-Т-клеток в тимусе на 3-й день (p<0,01) (фиг. 3D) с пиком размножения, приходящимся на 14-й день (p<0,001) (данные не показаны). В то время как субпопуляции зрелых DC размножались с 7-го (p<0,001) по 21-й день (p<0,01) в тимусе, как было указано выше (см. фиг. 3В).
Для того чтобы проверить, могут ли источником размножения DC быть некоммитированные предшественники Т-клеток, DN1-клетки, отсортированные по CD45.1+Lin-, переносили в тимус мышей-хозяев (CD45.2+), получавших воздействие Dll4-АТ. Было найдено, что клетки CD45.1+ накапливались на стадии DN1 (данные не показаны), и незрелые DC (imDC) детектировались и размножались в тимусе (фиг. 4) (p<0,01). Никакие клетки не детектировались у мышей, получавших воздействие контрольного антитела, возможно, вследствие того что большинство DN1-перенесенных клеток элиминировались негативной селекцией Т-клеток. Был сделан вывод о том, что блокирование Dll4 стимулирует развитие альтернативной линии DC в тимусе, источником которой является общий DN1-предшественник Т- и дендритных клеток.
Лиганд Fms-подобной тирозинкиназы 3 (Flt3-L) является достаточным и необходимым для дифференцировки клеток-предшественников костного мозга в DC и развития периферических DC. Уровень Flt3-L в сыворотке не изменялся у мышей дикого типа, получавших воздействие антител к Dll4 (данные не показаны). Кроме того, как показано в таблицах 2 и 3 ниже, процентное содержание DC в тимусе увеличивалось у мышей дикого типа (WT) (таблицы 2 и 3), у мышей, нокаутных по FLt3-L (Flt3-L-/-) (p<0,05) (таблица 2), и у мышей, нокаутных по FLt3-R (Flt3-R-/-) (p<0,001) (таблица 3), всех получавших Dll4-АТ, по сравнению с мышами, получавшими контрольное антитело. Таким образом, блокирование Dll4 вызывает Flt3-независимое увеличение числа DC в тимусе.
Ранее было показана способность ранних предшественников Т-клеток изменять направление дифференцировки в направлении не-Т-клеточного фенотипа (James P. Di Santo, 2010, Science 329:44-45). Для того чтобы определить эффект воздействия антител к Dll4 на гены, участвующие в дифференцировке линий Т-клеток относительно В-клеток и DC, проводили матричный анализ экспрессии генов на тимоцитах и про-Т-клетках. Было найдено, что экспрессия генов, необходимых для дифференцировки в Т-клетки (например, Tcf7, Gata3 и Ets1), была снижена, в то время как экспрессия генов (Lyl1, SfpH), которые каждый могут блокировать развитие Т-клеток, была повышена (данные не показаны; см. Di Santo, 2010, supra). Наиболее интересно то, что экспрессия генов, контролирующих развитие DC (PU.1 и Spi-B) и B-клеток также была повышена (данные не показаны; см. M. Merad et al., 2009, Blood 113:3418-3427). В дополнение, экспрессия RelB и Id2, а также интерферон-регуляторных факторов (IRF) 2, 4 и 8 - ключевых транскрипционных факторов, участвующих в развитии субпопуляции DC, была повышена (данные не показаны; см. Merad et al., 2009, supra). Наконец, было найдено, что экспрессия гена CSF-1 (M-CSF), ключевого цитокина, участвующего в развитии DC, повышена при воздействии антител к Dll4 (p<0,05; данные не показаны). Кроме того, уровень CSF-1 в сыворотке был повышен при воздействии антител к Dll4 (фиг. 5; p<0,05) (см. B. Francke, et al., 2008, Blood 111:150-159). Таким образом, можно сделать вывод о том, что блокирование сигнального пути Dll4-Notch снижает экспрессию транскрипционных факторов, специфичных для Т-клеточной дифференцировки, одновременно повышая экспрессию других факторов, критичных для развития DC.
Пример 2: эффект удаления Dll4 на развитие Т-клеток
Для того чтобы оценить, является ли наблюдаемый в примере 1 эффект Dll4 на развитие DC характерным для Dll4, были получены мыши DLL4COIN, у которых Dll4 кондиционно инактивирован. «Кондиционные в результате инверсии (COIN)» аллели представляют собой аллели, в основе которых лежит обратимый элемент («COIN-элемент»), обеспечивающий опосредованные рекомбиназой кондиционные мутации. Мыши DLL4COIN содержат индуцируемую тамоксифеном конструкцию Cre-рекомбиназы, CreERT2, которая кодирует Cre-рекомбиназу, слитую с мутантным доменом, связывающим эстрогеновые лиганды (ERT2). CreERT2 по существу неактивна в отсутствие тамоксифена и также не активируется эндогенными эстрогенами. Действие тамоксифена у мышей будет активировать CreERT2 и будет вызывать инверсию COIN-элемента, который блокирует транскрипцию всех экзонов, расположенных ниже от места встраивания COIN-элемента, таким образом блокируя экспрессию Dll4. Подробное описание рекомбиназной системы CreERT2 см. в Feil et al., 1997, Biochemical and Biophysical Research Communications 237:752-757.
Мышам DLL4COIN (n=6) вводили внутрибрюшинно (i.p.) тамоксифен (TAM) (кат. № T-5648, Sigma) в количестве 3 мг/150 мкл кукурузного масла на мышь три (3) раза в неделю в течение 2 недель. Контрольным мышам DLL4COIN (n=6) вводили кукурузное масло без тамоксифена. Аналогичным образом мыши дикого типа, C5B1/6, получали воздействие тамоксифена (n=6) или только кукурузного масла (n=6). Мышей контролировали на наличие недомоганий (например, по состоянию меха, низкой активности и т.п.), инфекций и избыточной потери веса тела. Мышей взвешивали приблизительно три раза в неделю. Любую мышь, которая теряла более 20% веса тела, выводили из эксперимента. После 2 недель воздействия собирали тимусы, и проводили анализ клеток тимуса проточной цитометрией.
Как показано на фиг. 6, в отсутствие Dll4 (то есть у мышей, получавших тамоксифен) В-клетки, а также pDC и cDC, размножались в тимусе по сравнению с мышами, получавшими кукурузное масло, что указывает на то, что эффекты Dll4 на развитие и гомеостаз DC, наблюдаемые в примере 1, были несомненно характерны для Dll4. Таким образом, сигнальный путь Dll4-Notch, по-видимому, поддерживает Т-клеточную дифференцировку в популяции про-Т-клеток, подавляя возможность дифференцировки не по Т-клеточному пути.
Пример 3: эффект блокирования или удаления Dll4 на гомеостаз Т рег -клеток
Недавно было показано, что Трег-клетки необходимы для поддержания нормального числа DC. При истощении Трег-клеток наблюдается компенсаторное зависимое от Fms-подобной тирозинкиназы 3 (Flt3) повышение числа DC (Liu et al., 2009, supra). Кроме того, двумя независимыми группами был показан контроль по принципу обратной связи гомеостаза регуляторных Т-клеток дендритными клетками in vivo; то есть увеличение числа DC приводит к увеличению деления и накоплению Трег-клеток, что может предупреждать развитие аутоиммунных заболеваний (Darrasse-Jeze G. et al., 2009, J Exp. Med. 206(9):1853-1862; и Swee LK et al., 2009, Blood 113(25):6277-6287).
Для того чтобы определить, может ли блокирование Dll4 влиять на гомеостаз Трег-клеток, число Трег-клеток в тимусе мышей, получавших воздействие либо антител к Dll4, либо Fc человека (контроль) в примере 1, измеряли проточной цитометрией. Как показано на фиг. 7А, блокирование Dll4 приводило к сильному увеличению числа Трег-клеток в тимусе на 14-й день после первичной инъекции. Увеличение числа Трег-клеток начиналось на 7-й день (p<0,001) и достигало максимума на 14-й день в тимусе (p<0,001) после первичной инъекции (см. фиг. 7В), в то время как на периферии (то есть, в селезенке) Трег-клетки начали появляться только в интервале между 14-м и 21-м днями (p<0,05) (фиг. 7B и таблица 4). В таблице 4 показана кратность увеличения процентного содержания и абсолютного числа Трег-клеток в селезенке после воздействия антител к Dll4 по сравнению с контрольными мышами (получавшими hFc).
Для того чтобы оценить, является ли наблюдаемое увеличение числа Трег-клеток характерным для молекулы Dll4, также определяли с помощью проточной цитометрии число Трег-клеток в тимусе мышей DLL4COIN из примера 2. Как и в случае блокирования Dll4 антителами к Dll4, кондиционная инактивация Dll4 в результате действия тамоксифена также приводила к увеличению числа Трег-клеток в тимусе по сравнению с мышами, получавшими кукурузное масло (см. фиг. 7С), а также мышами дикого типа, получавшими тамоксифен (данные не показаны). Таким образом, сигнальный путь Dll4-Notch поддерживает гомеостаз DC и, следовательно, Трег-клеток, и дифференцировку по Т-клеточному пути.
Аналогичный эксперимент был проведен на мышах, экспрессирущих Dll4 человека («Dll4-гуманизированных мышах») с использованием антитела к Dll4 (REGN421, имеющего последовательности HCVR и LCVR-SEQ ID NO:116 и 118, соответственно), которое, как известно, связывают N-концевой и DSL-домен Dll4 человека. Dll4-гуманизированных мышей получали замещением всего внеклеточного домена мышиного гена Dll4 соответствующей внеклеточной областью гена Dll4 человека (7 т.п.о.) в эмбриональных стволовых (ES) клетках мышей F1 C57BL/6/129. Получали гомозиготных по hDll4 мышей, и проводили их обратное скрещивание с мышами C57BL/6. Dll4-гуманизированным мышам вводили по 5 мг/кг hFc (контроль; n=6), или 1 мг/кг (n=6) или 5 мг/кг (n=6) антитела REGN421 дважды в неделю в течение двух недель. По две мыши из каждой группы воздействия умерщвляли на 7-й день, и по 2 и больше мышей на группу умерщвляли на 14-й день. Собирали тимусы и клетки окрашивали и исследовали проточной цитометрией. Остальных мышей оставляли восстанавливаться на дополнительные 4 недели без воздействия, и на 28-й день после отмены воздействия их умерщвляли, и анализировали клетки тимуса, используя проточную цитометрию. После двух недель воздействия в тимусе мышей, получавших антитела к Dll4, наблюдалось увеличение числа cDC и pDC (фиг. 8A), а также значительное увеличение популяции Трег-клеток (фиг. 8В) (p<0,01). В тимусе мышей, которые получали антитела к Dll4 в течение 2 недель с последующими 4 неделями без воздействия, число как DC, так и Трег-клеток вернулось к нормальному уровню в конце (фиг. 8А и 8В). В то же время увеличение количества DC и Трег-клеток также наблюдалось в периферических органах мышей, получавших антитела к Dll4, по сравнению с мышами, получавшими hFc (данные не показаны).
Пример 4: эффект блокирования Notch-рецептора на Т рег -клетки
Было показано, что увеличение числа DC приводит к увеличению числа Трег-клеток (Darrasse-Jeze G. et al., 2009). Как указано выше, наблюдалось, что при блокировании Dll4 в тимусе возрастало число как DC, так и Трег-клеток (фиг 3A и фиг. 7A). В дополнение, возрастание как процентного содержания, так и абсолютного числа DC и Трег-клеток, также было найдено в периферических органах мышей, получавших антитела к Dll4 (таблицы 1 и 4). Для того чтобы определить, приведет ли блокирование Notch-рецепторов к такому же фенотипу как и удаление Dll4, исследовали мышей, нокаутных (КО) по никастрину (Nic-/-). Никастрин представляет собой молекулу, участвующую в Notch-сигнальном пути, и генетическое удаление никастрина приводит у никастрин-дефицитных мышей к блокированию передачи сигнала от Notch-рецепторов 1, 2, 3 и 4 (Aifantis et al., неопубликованные данные). Было показано, что нокаутные по никастрину мыши имеют фенотип, аналогичный мышам с удаленным или блокированным Dll4 с повышением числа Трег-клеток, как в процентном содержании, так и в абсолютных значениях, в тимусе, а также в селезенке (см. таблицу 5).
(p<0,1)
(p<0,0001)
(p<0,01)
(p<0,0001)
Наконец, когда клетки костного мозга из мышей Nic-/- перенесли летально облученным мышам дикого типа (WT), у химер Nic-/-→WT наблюдалось размножение Трег-клеток в тимусе, позволяя предположить, что такое размножение являлось автономным клеточным эффектом; и блокирование Dll4 в мышах-реципиентах антителами к Dll4 не давало дополнительного эффекта (см. таблицу 6).
Эти результаты позволяют предположить, что нарушение сигнального пути Dll4-Notch в результате блокирования либо Dll4, либо Notch-рецепторов, приводит к аналогичным фенотипам в отношении размножения числа Трег-клеток.
Для того чтобы определить, коррелирует ли размножение Трег-клеток при блокировании Dll4 с числом DC (Darrasse-Jeze G. et al., 2009, supra), получали мышей, у которых отсутствуют DC, и тестировали с антителами к Dll4, как в примере 1. Трансгенные мыши, которые экспрессируют рецептор дифтерийного токсина приматов (DTR), получают чувствительность своих клеток к дифтерийному токсину (DT), которые в противном случае не чувствительны к DT. DT проникает в клетки путем взаимодействия своей В-субъединицы с клеточным DTR, и после эндоцитоза А-субъединица DT высвобождается и катализирует АДФ-рибизилирование фактора элонгации 2, приводя к ингибированию синтеза белков с последующим быстрым апоптозом как в делящихся, так и в терминально дифференцированных клетках. Специфичность и временную регуляцию удаления клеток можно регулировать промоторными/энхансерными элементами, специфичными к типу клеток, и схемой введения токсина, соответственно. Для придания чувствительности к DT дендритным клеткам Jung et al. (2002, Immunity 17:211-220) создали мышей (мыши CD11cre-DTR), которые несут трансген, кодирующий DTR обезьян, слитый с зеленым флуоресцентным белком (DTR-GFP) под контролем промотора гена CD11c мыши. Так как CD11c экспрессируется во всех дендритных клетках, то все субпопуляции дендритных клеток мыши, экспрессирующие CD11c, удалялись при введении дифтерийного токсина.
На полученных таким образом трансгенных мышей без DC воздействовали антителами к Dll4 или контрольными антителами, hFc, в соответствии с протоколом, описанным в примере 1. Через 14 дней после начала воздействия собирали тимусы и селезенку и подготавливали для анализа. Уровень экспрессии Dll4 на поверхности определенных субпопуляций DC или Т-клеток оценивали проточной цитометрией, для того чтобы определить, с какой определенной субпопуляцией связываются антитела к Dll4. Результаты показали, что дендритные клетки и Т-клетки не экспрессируют детектируемый уровень Dll4 на своей поверхности (данные не показаны). Это наблюдение подтверждено публикацией, что Dll4 экспрессируется на поверхности эпителиальных клеток тимуса (TEC) (Koch et at. 2008, supra). Однако наиболее важно то, что было найдено, что воздействие антител к Dll4 на мышей без DC не вызывало увеличения количества Трег-клеток, в то время как у мышей дикого типа (то есть, мышей, у которых DC не были удалены), получавших антитела к Dll4, значительно повышался процент Трег-клеток среди клеток CD3+CD4+ (p<0,001), позволяя предположить, что увеличение числа Трег-клеток при воздействии антител к Dll4 по меньшей мере частично опосредовано размножением DC.
Пример 5: эффект блокирования Dll4 на экспериментальный аутоиммунный энцефаломиелит (ЕАЕ)
Природные регуляторные Т-клетки CD4+CD25+FoxP3+ (то есть, Трег-клетки) играют важную роль в поддержании аутотолерантности и подавлении аутоиммунных заболеваний, таких как диабет 1-го типа, аутоиммунный энцефаломиелит, GVHD и воспалительная болезнь кишечника (IBD) (Darrasse-Jeze G. et al., 2009, supra; Swee LK et al., 2009, supra; и McGreachy et al., 2005, 175(5):3025-3032).
Для того чтобы посмотреть, будет ли повышенное число Трег-клеток в результате блокирования Dll4 предупреждать аутоиммунные заболевания, на мышиной модели исследовали влияние блокирования Dll4 на EAE. Мышиную модель ЕАЕ получали инъекциями в подушечку стопы мышей C57BL/6 пептида гликопротеина миелина олигодендроцитов (MOG), эмульгированного в полном адъюванте Фрейнда (CFA) с последующей (через 24 часа) инъекцией столбнячного токсина (РТХ) для индукции заболевания. Оценку заболевания определяли, исходя из следующих симптомов: (0) нет симптомов; (1) слабо двигающийся хвост; (2) слабо двигающийся хвост со слабостью задних конечностей; (3) частичный паралич задних конечностей; (4) полный паралич задних конечностей; (5) паралич всех конечностей; и (6) агония. За 12-24 часа до иммунизации мышам в группе введения «до индукции» (n=10) также вводили подкожной инъекцией по 25 мг/кг либо анти-Dll4 антитела (REGN577), либо изотипного контрольного антитела (антитела человека, специфичного к CD20, полученного внутри компании в соответствии с описанием в US 2008/0260641), либо PS/2 (крысиного/мышиного IgG2b против мышиной интегриноподобной молекулы клеточной адгезии, VLA-4; ATCC № CRL-1911), в то время как мыши из группы введения «после индукции» (n=10) получали тоже самое в день возникновения симптомов. Известно, что антитело PS/2 обостряет рецидивы заболевания и увеличивает накопление CD4+ T-клеток в центральной нервной системе в мышиной модели рецидивирующего экспериментального аутоиммунного энцефаломиелита (R-EAE) (Theien BE et al., 2001, J Clin Invest 107(8):995-1006). Инъекции антител проводили два раза в неделю в течение двух недель. В конце эксперимента осторожно удаляли спинной мозг у мышей, измельчали и инкубировали в среде RPMI 1640, содержащей коллагеназу D (Sigma Aldrich). Для остановки реакции добавляли 2 мМ EDTA, смесь пропускали через 70-мм клеточное сито и содержание клеток анализировали проточной цитометрией.
Как показано на фиг. 9А и 9В, после инъекций MOG у мышей, получавших изотипное контрольное антитело, развивались симптомы (то есть, они имели оценки заболевания выше «0»), начиная примерно с 10-14 дня с пиком между 15-м и 21-м днем. В отличие от этого, у мышей, получавших антитела к Dll4, полностью предупреждалось развитие заболевания по сравнению с мышами, получавшими контрольные антитела. В таблице 7 показана кратность увеличения процентного содержания и абсолютного числа Трег-клеток в тимусе и селезенке мышей, получавших антитела к Dll4, по сравнению с мышами, получавшими контрольные антитела.
Трег-клетки, по-видимому, размножаются преимущественно в тимусе примерно в районе 18-го дня, и значительное увеличение количества наблюдалось в периферических органах (то есть в селезенке) только после 21-го дня.
В этих конкретных экспериментальных условиях воздействие антител к Dll4 после стадии индукции не показывало значительного улучшения в развитии заболевания. Дозировки и/или частоту введения антител к Dll4 можно дополнительно регулировать, что входит в компетенцию специалиста в данной области. Однако важно, что у мышей, которым вводили антитела к Dll4 до индукции, наблюдалось значительное снижение инфильтрации клеток в спинной мозг на 18-й день по сравнению с мышами, которые получали контрольные антитела (см. таблицу 8 ниже). Инфильтрация клеток, наблюдаемая в спинном мозге мышей, получавших контрольные антитела, может быть главным действующим фактором развития заболевания у этих мышей.
Как показано в таблице 8, наблюдается 8-кратное снижение (p<0,0001) количества макрофагов (F4/80+), 2,7-кратное снижение количества NK-клеток (p<0,0001), 1,7-кратное снижение CD11b-клеток (p<0,001) и 2,5-кратное снижение В-клеток (p<0,001) в спинном мозге мышей, получавших антитела к Dll4, по сравнению со спинным мозгом мышей, получавших контрольные антитела, на 21-й день.
Кроме того, продукция IL-17 и IFN-γ в лимфатических узлах мышей, получавших антитела к Dll4, была значительно снижена (p<0,001) (фиг. 10). Таким образом, Dll4 может участвовать в патогенезе ЕАЕ, опосредуя развитие Th1, и воздействие антител к Dll4 может предупредит развитие заболевания, блокируя секрецию цитокинов Th1 и Th17.
Пример 6: эффект блокирования Dll4 на диабет
Эффект блокирования Dll4 на диабет также проверяли на мышах NOD/ShiLtJ («мышах NOD»), полигенной модели диабета 1-го типа (Makino S et al., 1980, Jikken Dobutsu 29 (1):1-13; Serreze DV et al., 1997, J Immunol 158 (8):3978-86). Диабет у мышей NOD/ShiLtJ отличается инсулитом и инфильтрацией лейкоцитов в панкреатические островки. Заметное снижение содержания панкреатического инсулина возникает спонтанно у самок в возрасте около 12 недель и несколько недель спустя у самцов. Соответственно, уровень глюкозы в плазме поднимается выше 250 мг/дл. Уровень глюкозы в крови мышей NOD проверяли дважды в неделю, используя ONETOUCH® mini (LifeScan, Inc.). Считали, что мыши имеют диабет, после двух последовательных определений уровня глюкозы в крови выше 250 мг/дл. Начало диабета датировали по первому из последовательных измерений, показывающих диабет. Мышам вводили с помощью инъекций hFc (n=5) или анти-Dll4 антитело (Regn577) (n=10) в количестве 25 мг/кг дважды в неделю в течение 7 недель, начиная с 9-недельного возраста. Уровень глюкозы в крови контролировали раз в неделю по образцам из хвоста.
Как показано на фиг. 11А, у мышей, получавших hFc, начинал развиваться спонтанный диабет с уровнем глюкозы в крови выше 250 мг/дл после 13-недели жизни (●). В отличие от этого, у мышей, получавших анти-Dll4 антитело (REGN577), не наблюдалось признаков повышения уровня глюкозы по 25-ю неделю жизни (■) и измерения продолжали еще в течение дополнительных 10 недель. Введение антител к Dll4 до начала диабета предупреждало его развитие, и у животных, получавших эти антитела, по-видимому, так и не развивался диабет. Интересно отметить, что когда 5 из 10 мышей, получавших антитела к Dll4, вводили с помощью инъекций анти-CD25 моноклональное антитело (PC61) в возрасте 20 недель для истощения Трег-клеток (◆), их уровень глюкозы в крови начинал повышаться через 1-2 недели, и у мышей развивался диабет. Это указывало на то, что профилактический эффекта антител к Dll4 на диабет 1-го типа опосредован, по меньшей мере частично, Трег-клетками (фиг. 11А).
Антитела к инсулину и GAD65 являются двумя стандартными аутоантителами, которые находят в сыворотке диабетических мышей NOD, а также у индивидуумов с диабетом. Соответственно, уровень аутоантител в сыворотке мышей, получавших контроль hFc или Dll4-АТ, измеряли с помощью ELISA. Как показано на фиг. 11В, воздействие Dll4-АТ блокировало продукцию анти-инсулиновых (□) и анти-GAD65 (■) аутоантител на уровне, аналогичном уровню мышей C57BL/6 дикого типа, не получавших воздействие (то есть, не-NOD мышей; животных отрицательного контроля). В отличие от этого мыши NOD (диабетические), которые получали контроль hFc, имели высокий уровень аутоантител в своей сыворотке. В дополнение, когда срезы поджелудочной железы 23-недельных мышей, которые получали Dll4-АТ и не проявляли никаких симптомов диабета, окрашивали H&E (гематоксилином и эозином), то наблюдалось нормальное число панкреатических островков (клеток, которые продуцируют инсулин или глюкагон, а их разрушение прямо коррелирует с частотой возникновения диабета) с сохраненной морфологией (фиг. 11С, левая панель и фиг. 11D, левая панель). Кроме того, у мышей, получавших DLL4-АТ, не наблюдалось никакой инфильтрации клеток в островки (фиг. 11С, левая панель и фиг. 11D, правая панель). В отличие от этого, диабетические животные, которые получали контрольное антитело, hFc, имели значительно меньшее число панкреатических островков (фиг. 11С, правая панель, и фиг. 11D, левая панель) в их поджелудочной железе относительно мышей, получавших Dll4-АТ, а оставшееся очень маленькое число островков содержало высокий уровень клеточных инфильтратов (фиг. 11С, правая панель, и фиг. 11D, правая панель). Таким образом, Dll4-АТ было способно предупреждать диабет полностью в течение продолжительного периода времени, и его эффект, по-видимому, по меньше мере частично, опосредован размножением Трег-клеток; однако возможно, что в защитный эффект Dll4-АТ на панкреатические островки и/или инсулин могут быть вовлечены дополнительный(ые) механизм(ы).
Фактические значения уровня глюкозы в крови диабетических мышей, получавших Dll4-АТ, определяли и сравнивали со значениями у мышей, получавших контроль hFc. Мышам с диабетом вводили по 25 мг/кг Dll4-АТ (n=3) или контроля, hFc (n=4), в начале заболевания (день 0). После введения Dll4-АТ у мышей с диабетом значительно снизился уровень глюкозы с 350 мг/дл до нормального уровня (примерно 120-130 мг/дл) (фиг. 11Е). Этот эффект продолжался в среднем от 4 до 5 недель. В общем, дополнительно было замечено, что когда диабетические мыши, имеющие уровень глюкозы в крови ниже 350 мг/дл, получали Dll4-АТ, их уровень глюкозы падал до нормального уровня, и этот эффект сохранялся в течение большего интервала времени относительно мышей, имеющих уровень глюкозы в крови выше 350 мг/дл на момент воздействия. Это указывает на то, что существует некоторое окно возможности длительного и эффективного воздействия для контроля уровня глюкозы в крови с помощью антител к Dll4. Таким образом, не касаясь каких-либо конкретных механизмов, описанных в настоящем документе, эти наблюдения позволяют предположить, что антитела к Dll4 имеют огромный терапевтический потенциал в отношении диабета 1-го типа.
Результаты приведенных выше экспериментов выявили существование ранее неизвестной регуляторной петли, которая контролирует число Трег-клеток и DC in vivo. Этот регуляторный контур, вероятно, необходим для баланса между иммунным ответом и толерантностью, но, наиболее важно, в первый раз выявляет связь между тремя важными составляющими иммунной системы, то есть Dll4-DC-Трег-клетками. Таким образом, терапия антагонистами Dll4 представляет эффективную методику контроля числа Трег-клеток in vivo, и, соответственно, контроля развития аутоиммунных заболеваний и сопутствующих состояний.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ЛЕЧЕНИЯ ДИАБЕТА АНТАГОНИСТАМИ DLL4 | 2011 |
|
RU2587624C2 |
| АНТИТЕЛА ЧЕЛОВЕКА К ДЕЛЬТА-ПОДОБНОМУ ЛИГАНДУ-4 ЧЕЛОВЕКА | 2007 |
|
RU2448979C2 |
| СПОСОБ ЛЕЧЕНИЯ РАКА АНТАГОНИСТОМ DLL4 И ХИМИОТЕРАПЕВТИЧЕСКИМ СРЕДСТВОМ | 2010 |
|
RU2571220C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА ЧЕЛОВЕКА К РЕЦЕПТОРУ IL-4 ЧЕЛОВЕКА | 2007 |
|
RU2445318C2 |
| АНТИТЕЛА ЧЕЛОВЕКА С ВЫСОКОЙ АФФИННОСТЬЮ К ФАКТОРУ РОСТА НЕРВОВ ЧЕЛОВЕКА | 2008 |
|
RU2473564C2 |
| ВЫСОКОАФФИННЫЕ ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ АНГИОПОЭТИНУ-2 | 2010 |
|
RU2545399C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА С ВЫСОКОЙ АФФИННОСТЬЮ К РЕЦЕПТОРАМ IL-4 ЧЕЛОВЕКА | 2009 |
|
RU2539774C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА С ВЫСОКОЙ АФФИННОСТЬЮ К РЕЦЕПТОРАМ IL-4 ЧЕЛОВЕКА | 2014 |
|
RU2663106C2 |
| ТЕРАПЕВТИЧЕСКИЕ DLL4-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2011 |
|
RU2605928C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ RET, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2821548C2 |





























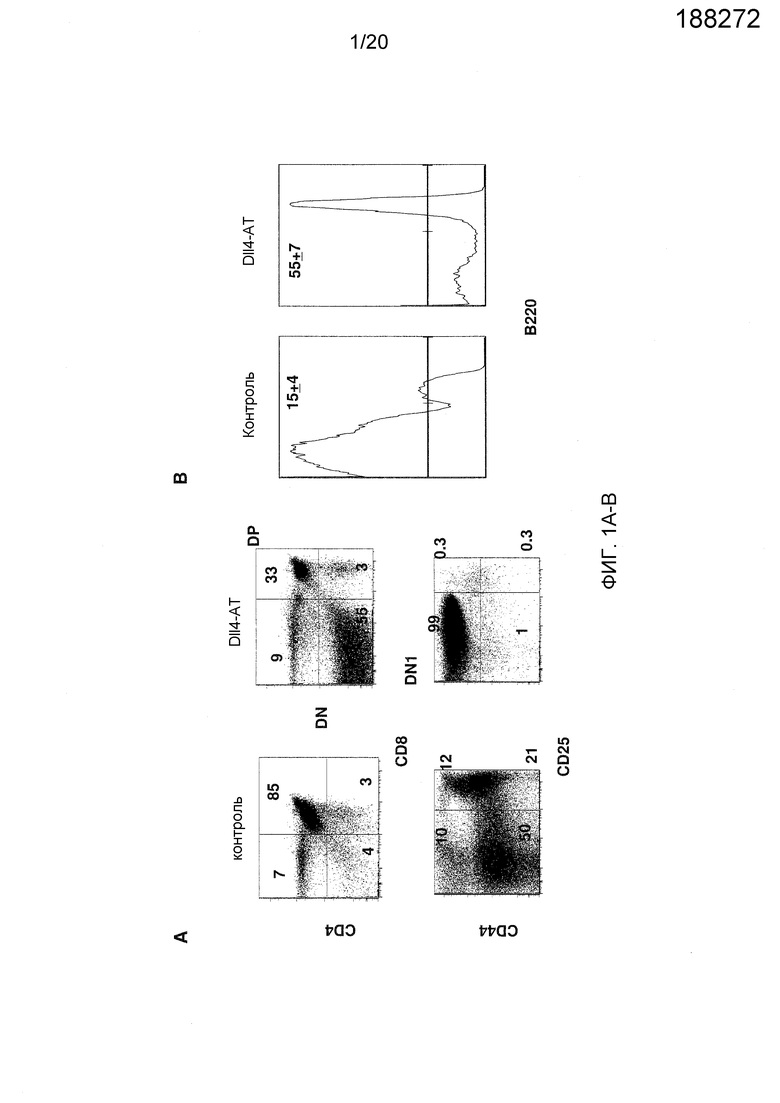
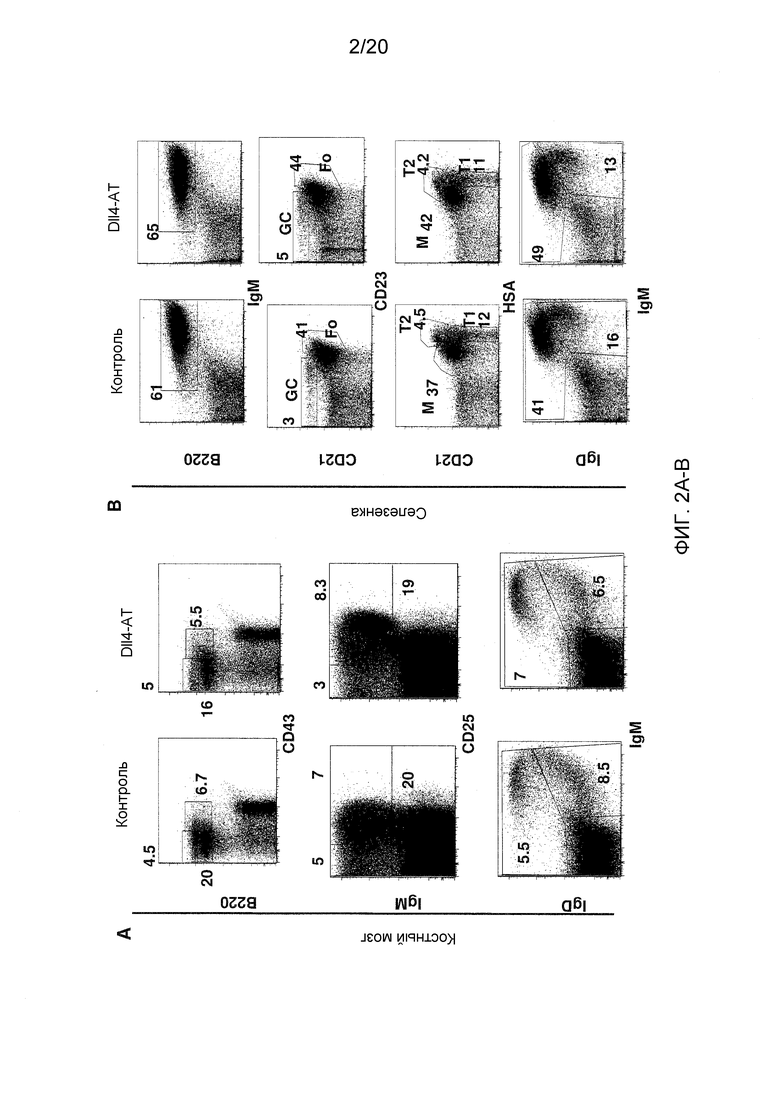
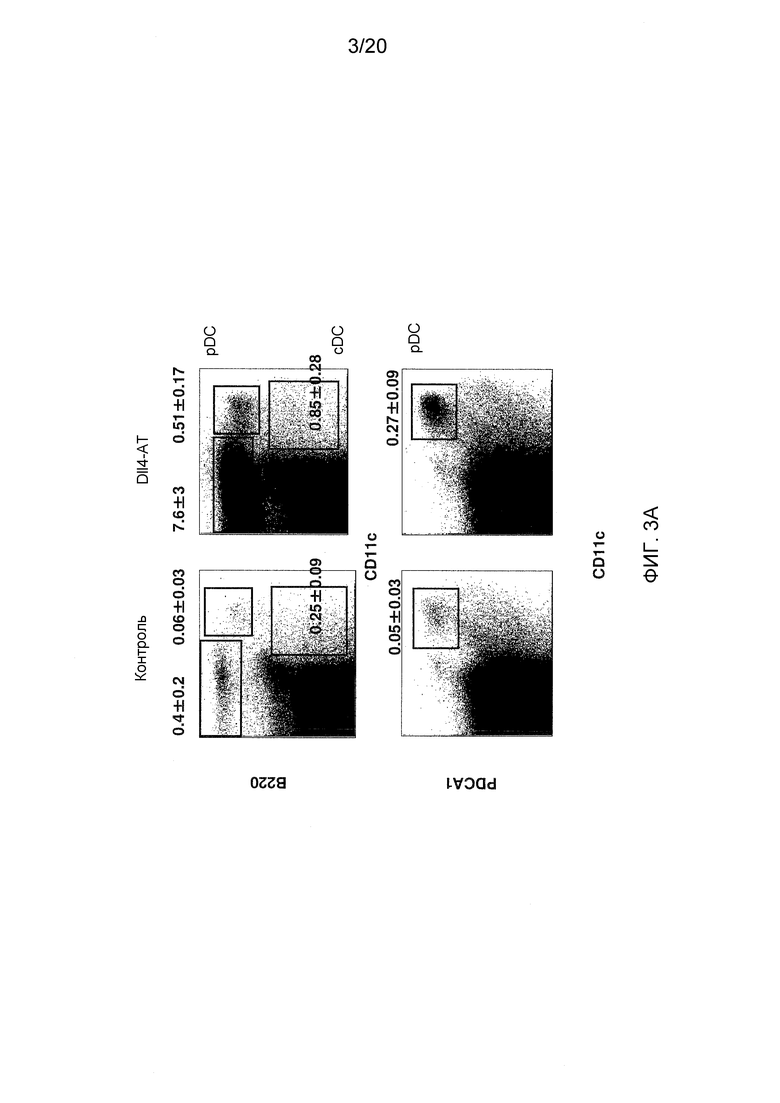
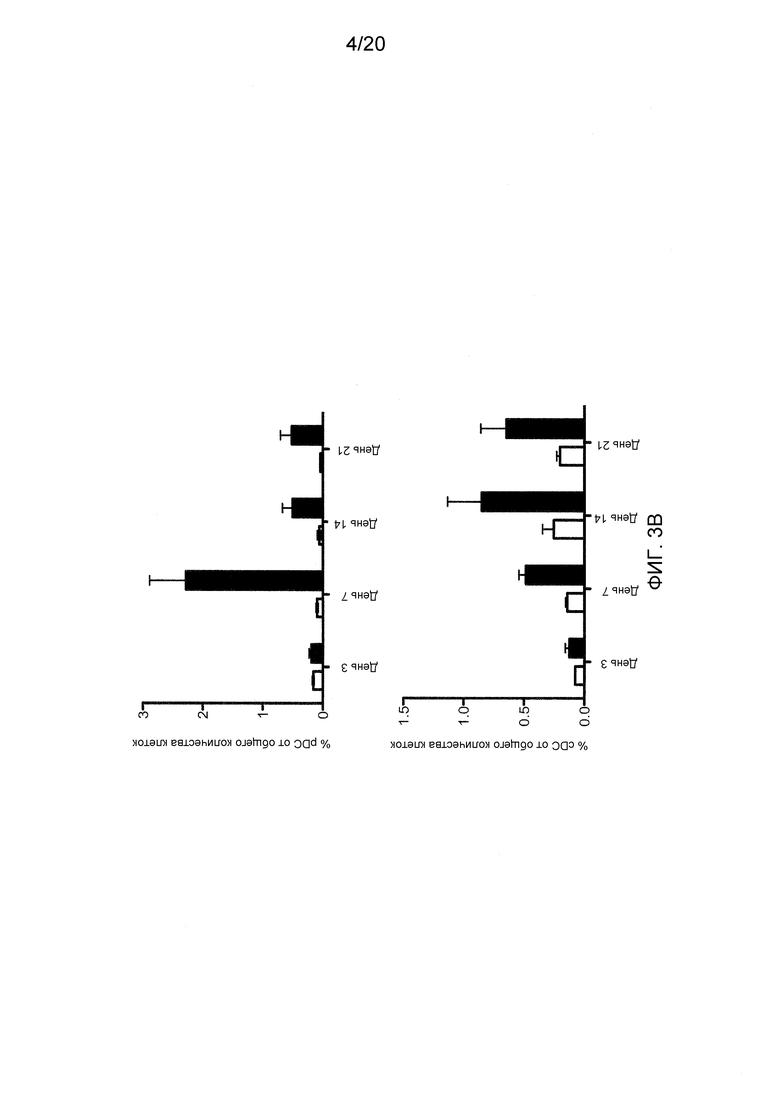
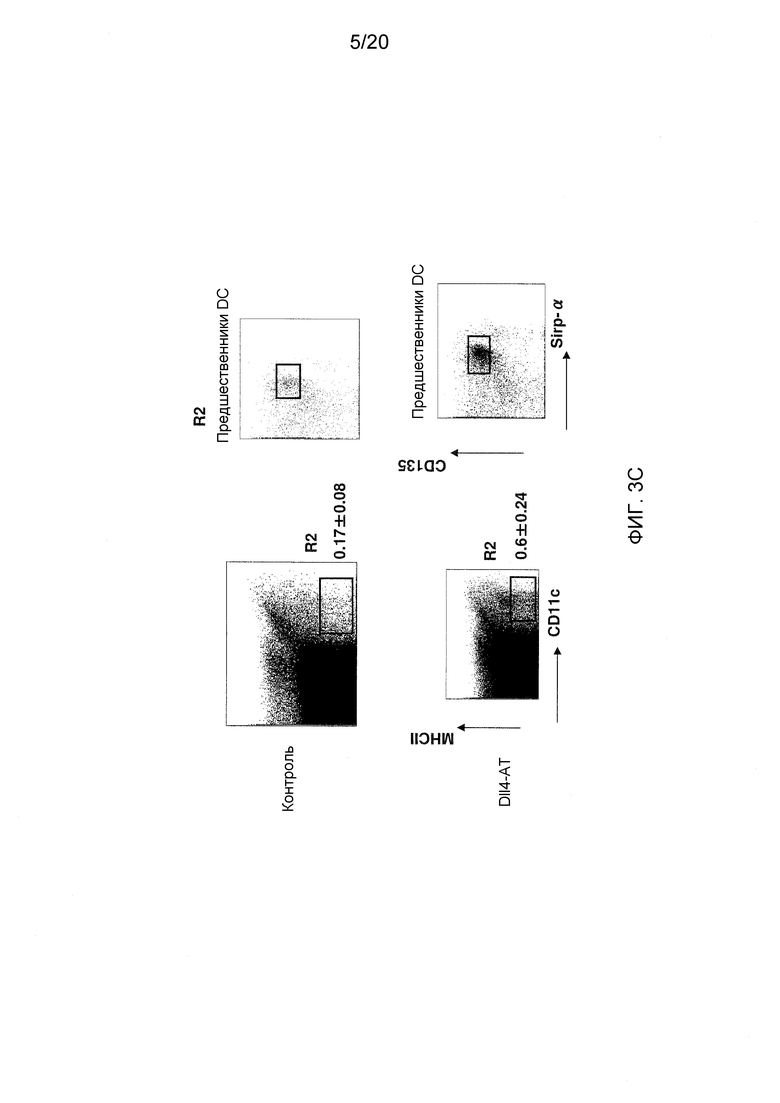
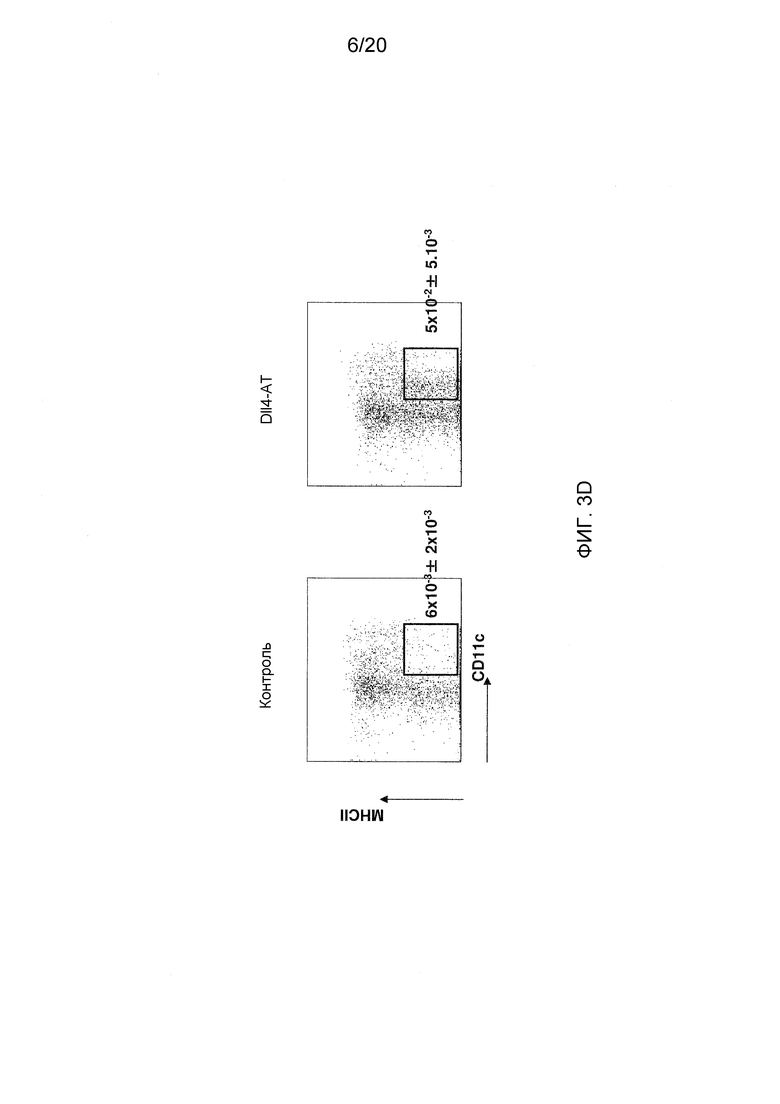
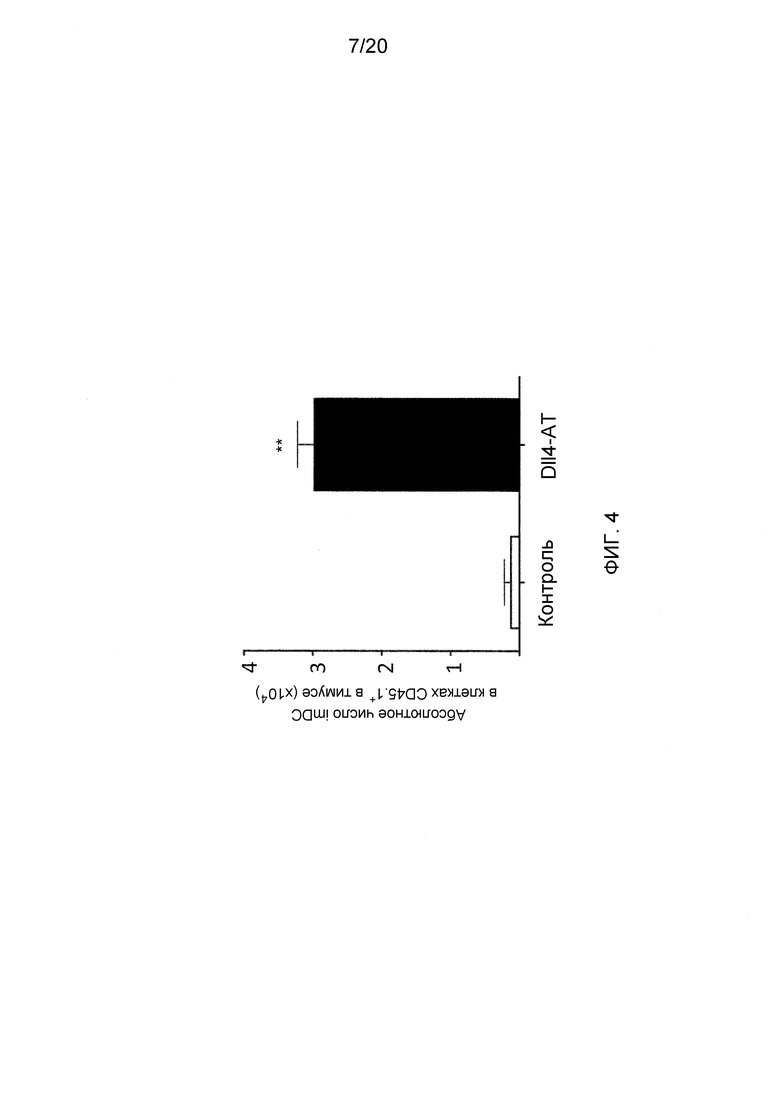
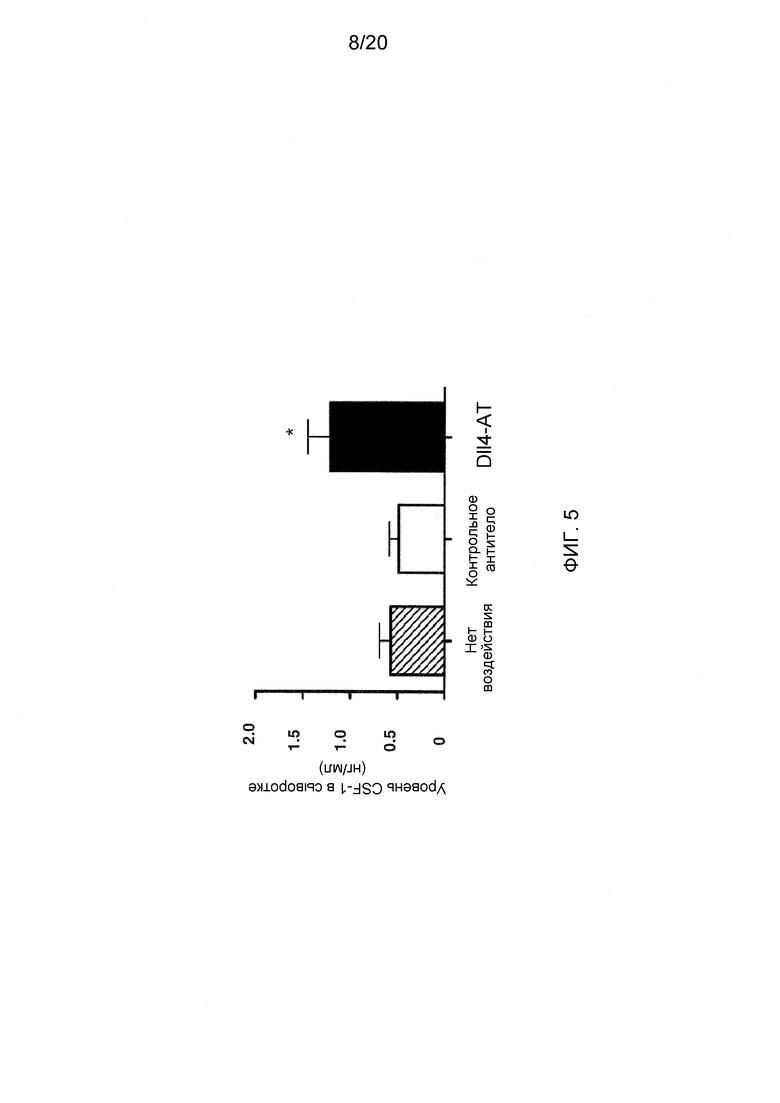

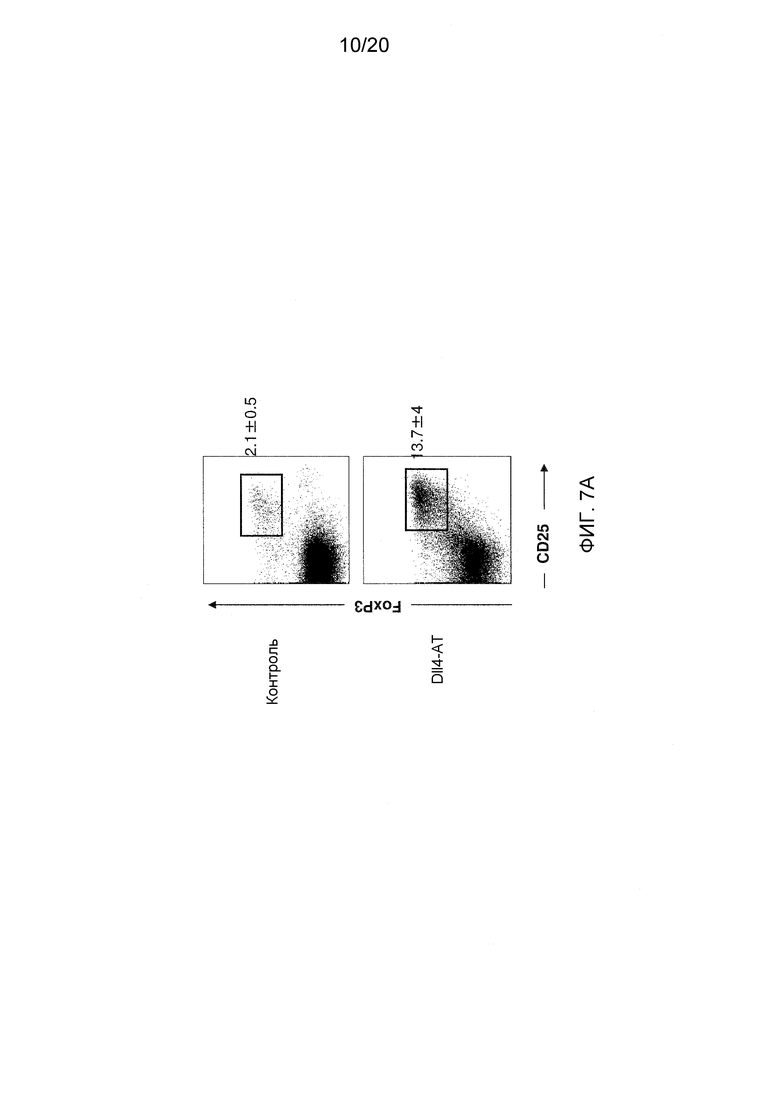
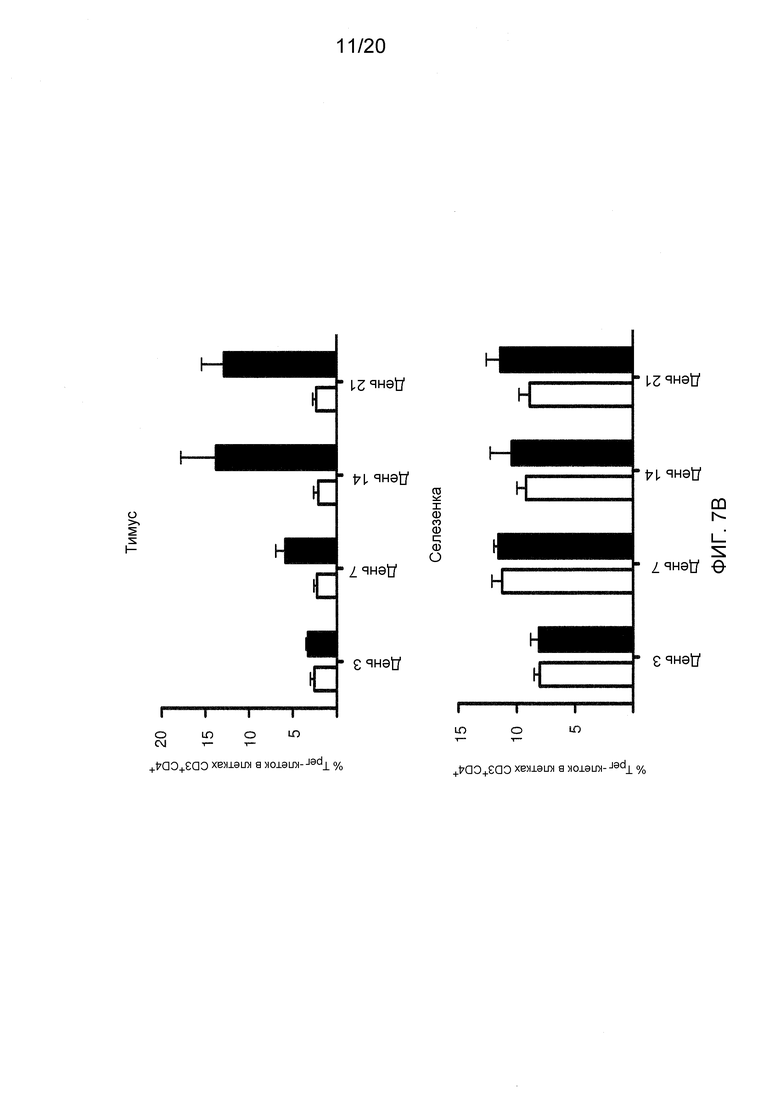
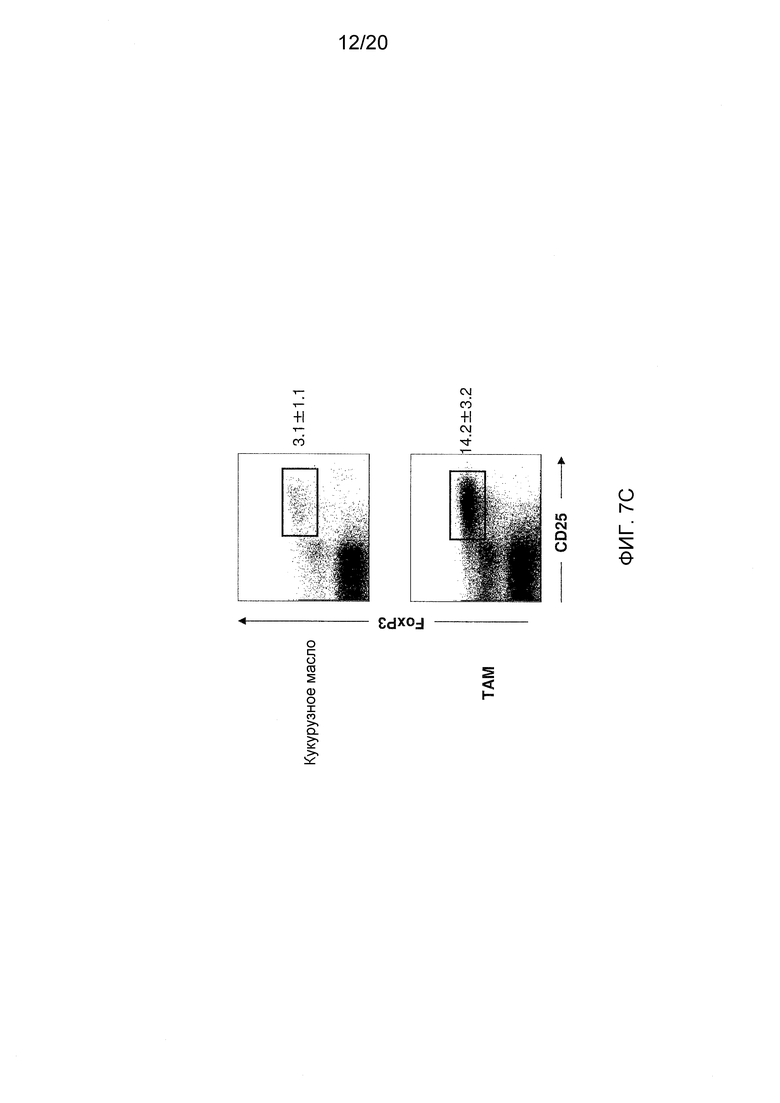
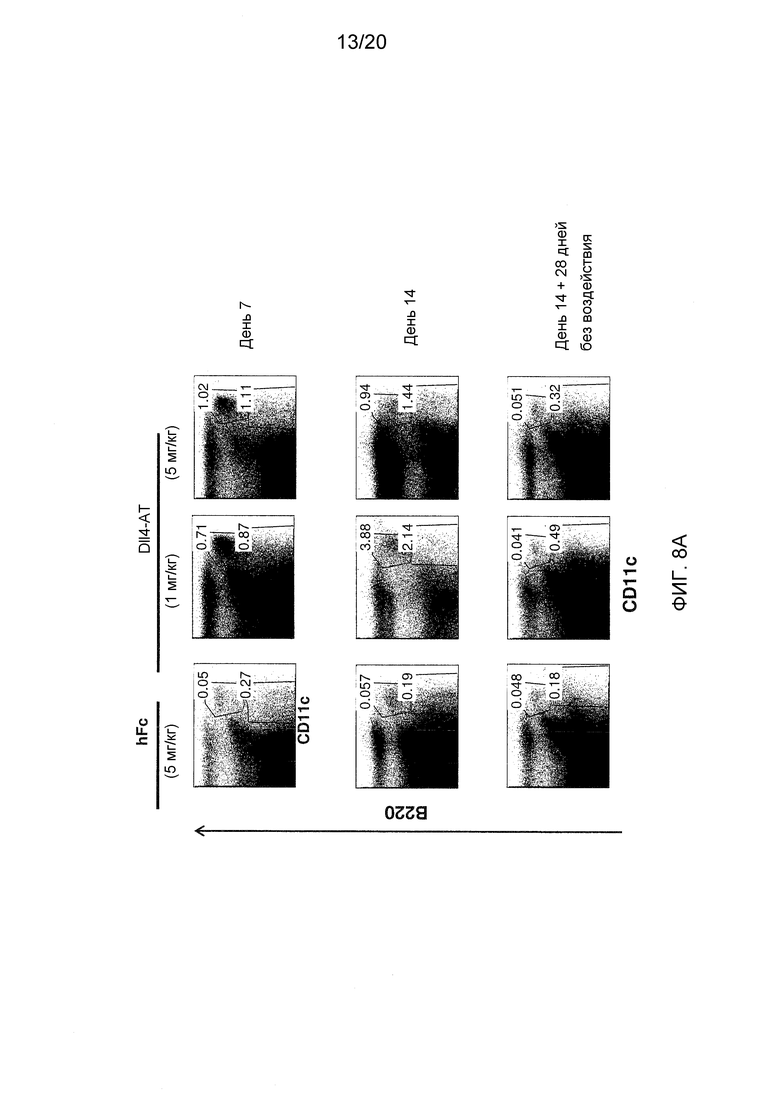

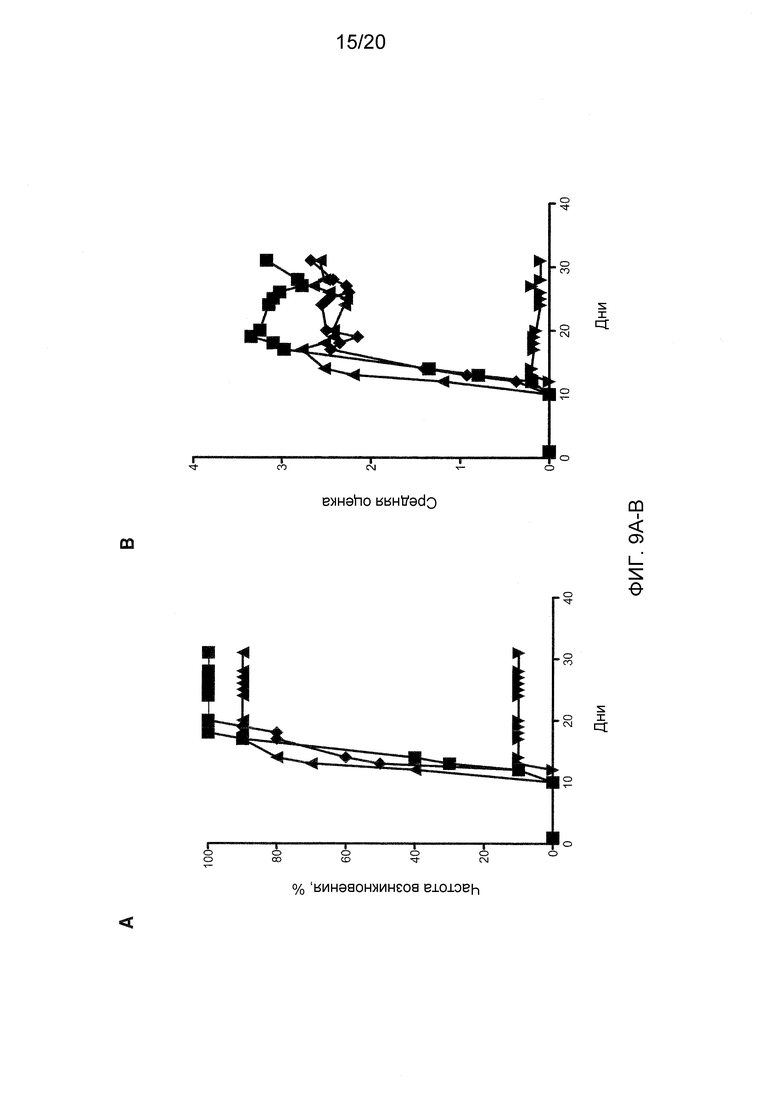
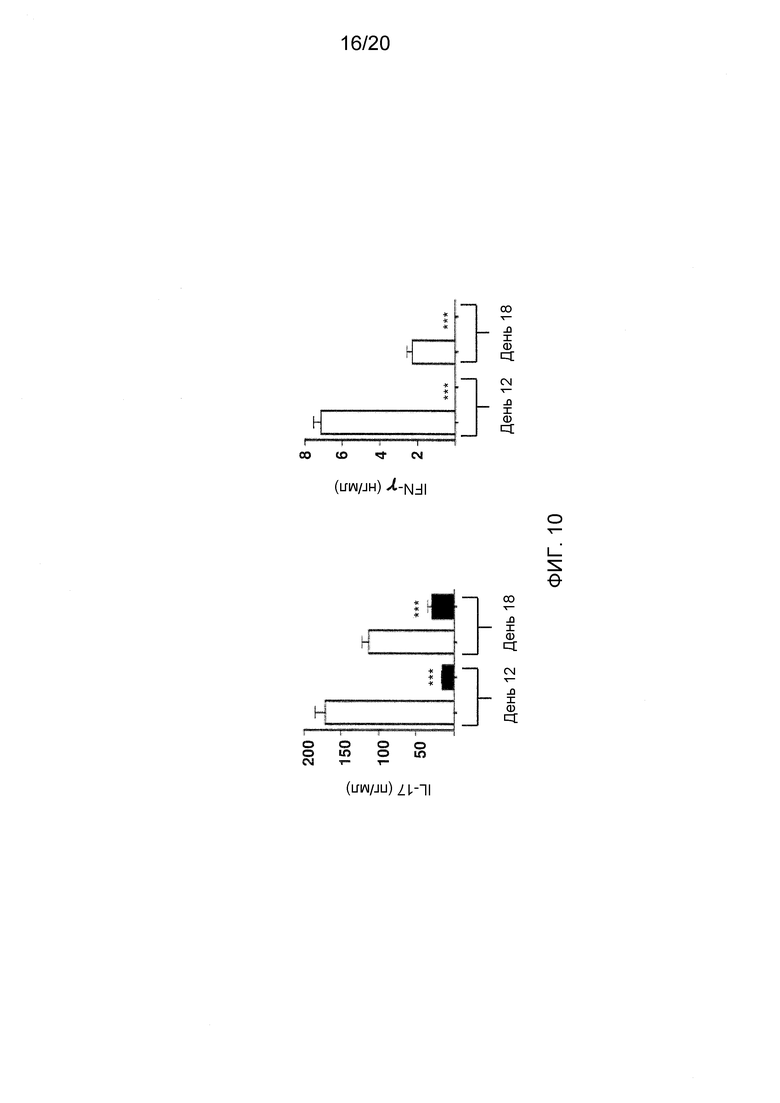
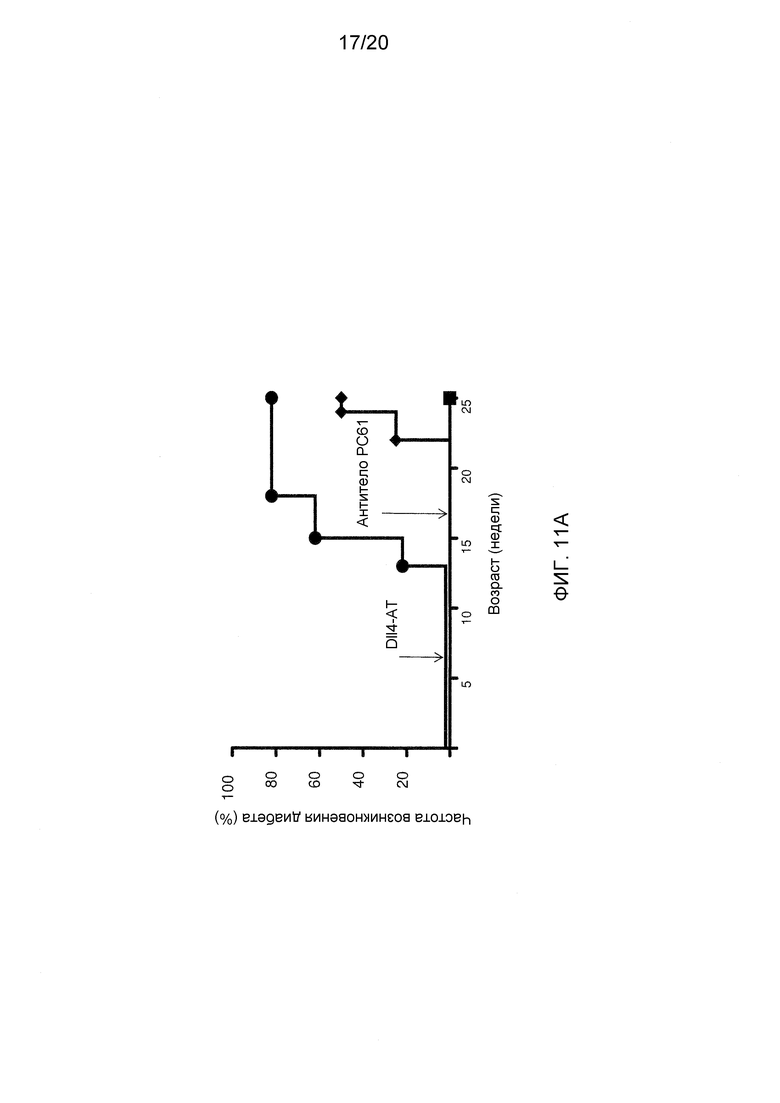
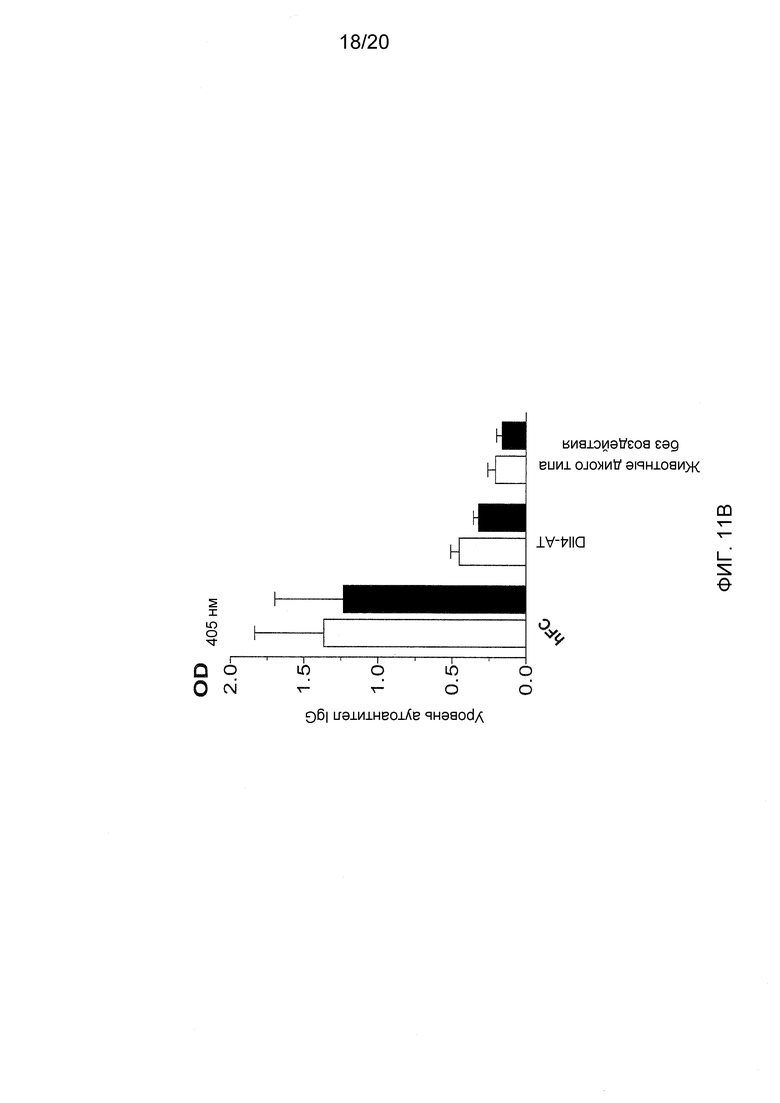
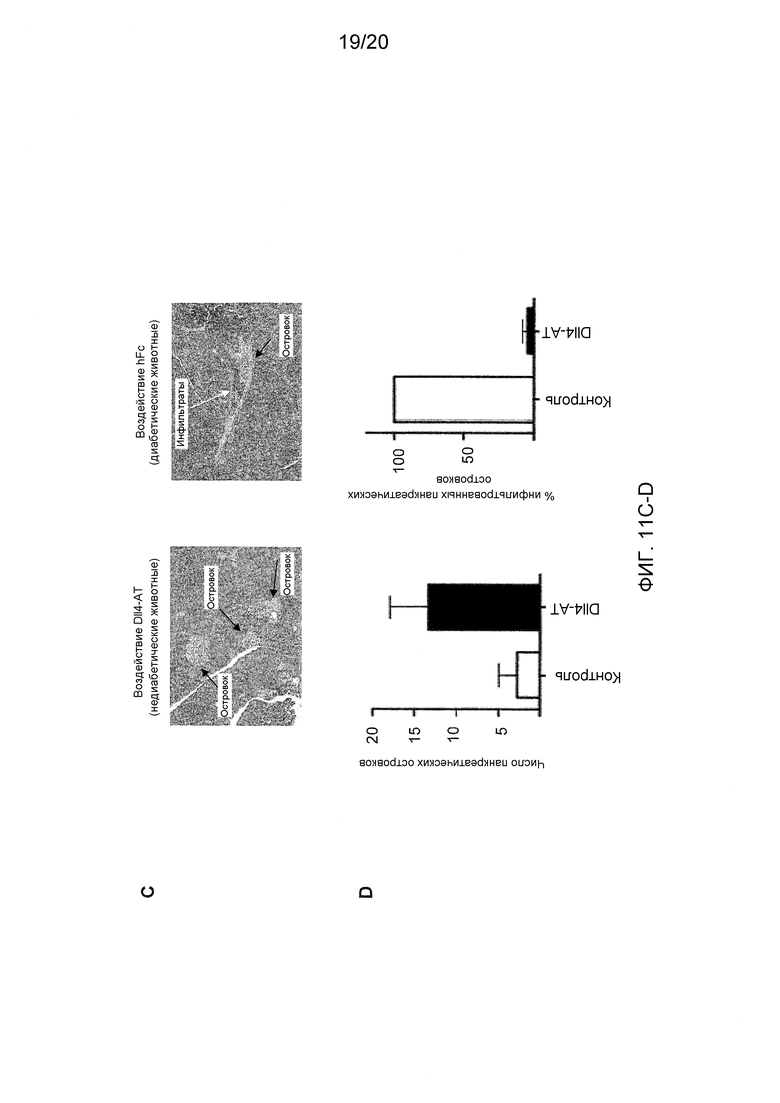
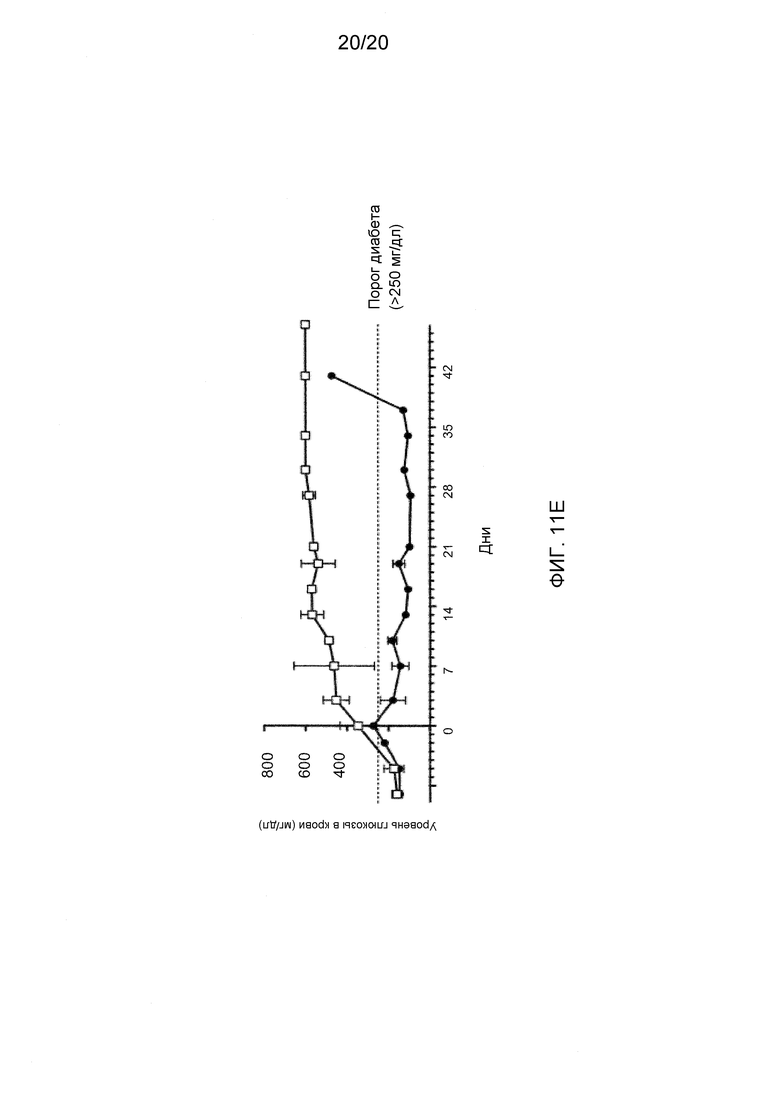
Изобретение относится к области иммунологии и медицины. Предложено применение антитела или его антиген-связывающего фрагмента, которые специфически связываются с дельта-подобным лигандом 4 (Dll4) человека, в предупреждении рецидивирования рассеянного склероза или снижении развития данного заболевания при его рецидивировании у субъекта. Также рассмотрено применение указанного антитела или его антиген-связывающего фрагмента в изготовлении соответствующего лекарственного средства. Применение антитела или его антиген-связывающего фрагмента по настоящему изобретению обеспечивает блокирование сигнальных путей от Dll4-Notch и повышение числа регуляторных Т-клеток. 2 н. и 6 з.п. ф-лы, 11 ил., 8 табл., 6 пр.
1. Применение антитела или его антиген-связывающего фрагмента, которые специфически связываются с дельта-подобным лигандом 4 (Dll4) человека, в предупреждении рецидивирования рассеянного склероза или снижении развития данного заболевания при его рецидивировании у субъекта, у которого ранее диагностирован рассеянный склероз, причем указанное антитело или его антиген-связывающий фрагмент содержат вариабельную область тяжелой цепи (HCVR), содержащую последовательности CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 22, 24 и 26, соответственно, и вариабельную область легкой цепи (LCVR), содержащую последовательности CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 30, 32 и 34, соответственно, и повышают число регуляторных Т-клеток.
2. Применение антитела или его антиген-связывающего фрагмента, которые специфически связываются с дельта-подобным лигандом 4 (Dll4) человека, в изготовлении лекарственного средства для применения в предупреждении рецидивирования рассеянного склероза или снижении развития данного заболевания при его рецидивировании у субъекта, у которого ранее диагностирован рассеянный склероз, причем указанное антитело или его антиген-связывающий фрагмент содержат вариабельную область тяжелой цепи (HCVR), содержащую последовательности CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 22, 24 и 26, соответственно, и вариабельную область легкой цепи (LCVR), содержащую последовательности CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 30, 32 и 34, соответственно, и повышают число регуляторных Т-клеток.
3. Применение по п.1 или 2, в котором антитело или его антиген-связывающий фрагмент содержат последовательность HCVR из SEQ ID NO: 20 или SEQ ID NO: 116.
4. Применение по п.1 или 2, в котором антитело или его антиген-связывающий фрагмент содержат последовательность LCVR из SEQ ID NO: 28 или SEQ ID NO: 118.
5. Применение по п.1 или 2, в котором антитело или его антиген-связывающий фрагмент содержат комбинацию HCVR/LCVR из SEQ ID NO: 20/28.
6. Применение по п.1 или 2, в котором антитело или его антиген-связывающий фрагмент содержат комбинацию HCVR/LCVR из SEQ ID NO: 116/118.
7. Применение по п.1 или 2, в котором лекарственное средство предназначено для применения в лечении или улучшении состояния при рассеянном склерозе в комбинации с по меньшей мере одним дополнительным терапевтическим агентом, выбранным из группы, состоящей из иммуносупрессанта, противовоспалительного агента и анальгетика.
8. Применение по п.7, в котором дополнительным терапевтическим агентом является по меньшей мере один, выбранный из группы, состоящей из глюкокортикостероидов, циклоспорина, метотрексата, нестероидных противовоспалительных соединений (NSAID), антагонистов TNF-α, антагонистов IL-1, антагонистов IL-6 и опиоидов.
| Колосоуборка | 1923 |
|
SU2009A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| JURYNCZYK MACIEJ et al | |||
| "Inhibition of Notch signaling enhances tissue repair in an animal model of multiple sclerosis", Journal of neuroimmunology, 2005, 170(1): 3-10 | |||
| FEYERABEND T.B | |||
| et al., "Deletion of Notch1 Converts Pro-T Cells to Dendritic Cells and | |||

