Область техники, к которой относится изобретение
Настоящее изобретение относится к области аллергии. Более конкретно, изобретение относится к идентификации новых аллергенов млекопитающих и к диагностированию и лечению аллергии у млекопитающих.
Уровень техники
Перхоть собаки является основной причиной аллергии внутри помещений, и симптомы этой аллергии включают ринит, конъюнктивит, бронхит и астму. Аллергены собаки могут быть обнаружены не только в домах, где собак содержат в качестве домашних животных, но также и в других местах, таких как школы и детские сады, где собаки присутствуют непостоянно [1].
Аллергия на собак сопровождается и зависит от сенсибилизации к белкам шерсти и перхоти собаки. При подозрении на аллергию на собак клиническое обследование включает оценку сенсибилизации с помощью кожной пробы или измерения специфических IgE-антител, используя экстракт шерсти и/или перхоти собаки. Лабораторное иммунологическое исследование на специфические IgE, такое как Phadia ImmunoCAPTM, в большинстве случаев может обнаружить сенсибилизацию к аллергенам собаки, используя природный экстракт перхоти собаки, благодаря благоприятным условиям анализа и большой твердой фазе, доступной для прикрепления аллергена.
Экстракты шерсти и перхоти собаки содержат комплекс аллергенных и неаллергенных белков [2, 3]. К настоящему моменту было идентифицировано и изучено четыре аллергена собаки: Can f 1, Can f 2, Can f 3 и Can f 5 [4-6]. Два первых аллергена являются членами семейства белков липокалина, и они были очищены и экспрессированы как рекомбинантные белки [4, 7]. Can f 3, сывороточный альбумин собак, представляет собой относительно консервативный белок, проявляющий обширную перекрестную реактивность с другими альбуминами млекопитающих [8]. Can f 5, простатический калликреин собак, недавно был описан как главный аллерген собаки и было показано, что он перекрестно взаимодействует с простатспецифическим антигеном (PSA) человека [5].
Из аллергенов перхоти собаки, известных до настоящего времени, Can f 1 и Can f 5, по-видимому, являются наиболее важными, так как связывают IgE-антитела у приблизительно 50% и 70% индивидов, имеющих аллергию на собак, соответственно [5, 9]. Хотя у приблизительно 20-40% взрослых индивидов, имеющих аллергию на собак, было отмечено связывание IgE-антител с Can f 2 или Can f 3, по-видимому, незначительное количество индивидов реагирует только или предпочтительно на любой из этих аллергенов [5, 9]. В недавнем исследовании было обнаружено, что у небольшой части пациентов, страдающих аллергией на собак, отсутствует связывание IgE-антител с любым из Can f 1, Can f 2, Can f 3 и Can f 5, несмотря на то, что они сенсибилизированы к природному экстракту перхоти собаки [5].
Кроме аллергенов, описанных выше, был описан IgE-реактивный липокалин-подобный белок с молекулярной массой 18 кДа, отличный от Can f 1 и Can f 2, который был обозначен как Can f 4 [9]. Было показано, что у пятнадцати из 25 (60%) пациентов, страдающих аллергией на собак, была отмечена реактивность сывороточного IgE к полосе с молекулярной массой 18 кДа на иммуноблоттинге, что указывает на то, что этот белок возможно является важным аллергенным компонентом собаки. Однако аллерген был охарактеризован только как N-концевая последовательность, состоящая из 13 аминокислот, по результатам SDS PAGE геля, при этом последовательность не совпадает ни с одной известной белковой последовательностью собаки. Таким образом, полноразмерная белковая последовательность неизвестна, а клонирование рекомбинантного белка не было проведено. Ни кем не был выделен нативный белок, и не было выдвинуто предположений относительно того, каким образом можно осуществить клонирование Can f 4.
В реферате статьи Saarelainen et al., опубликованной в сборнике тезисов 3-его международного симпозиума по молекулярной аллергологии (Зальцбург, 18-20 апреля 2008 года), отмечается, что mAb, специфичное к Can f 4, распознавало белок с молекулярной массой 20 кДа в экстракте перхоти коровы.
В специальном издании научного журнала Allergy, в котором опубликованы рефераты XXVIII Конгресса европейской академии по аллергии и клинической иммунологии (EAACI) (Варшава, 6-10 июня 2009 года), присутствует реферат, в котором раскрыта информация о том, что была клонирована кДНК, кодирующая полноразмерный Can f 4 [Allergy 64 (Suppl. 90): 179-538]. Однако в реферате не дано указание, каким образом было проведено клонирование и не приведены ни нуклеотидная, ни аминокислотная последовательность.
Сущность изобретения
Авторы изобретения определили необходимость в данной области проведение дополнительной характеристики и проверки важности аллергена Can f 4.
Как указано выше, с помощью лабораторного иммунологического исследования для специфических IgE можно обнаружить большинство случаев сенсибилизации к аллергенам собаки, используя природный экстракт перхоти собаки, благодаря благоприятным условиям анализа и большой твердой фазы, доступной для присоединения аллергена. Однако было обнаружено, что при проведении мини-анализа или не лабораторного иммунологического исследования, например, на микрочипах, для определения аллергенов или при тестировании в кабинете врача, сочетание менее благоприятных условий исследования, более низкой емкости антитело-связывающего аллерген реагента и экстракта природного аллергена с ограниченной эффективностью, приводит к недостаточной диагностической чувствительности. Такая ситуация также может иметь место при проведении иммунологических исследований для специфических IgE к эпителию других животных. Таким образом, в некоторых случаях существует необходимость использовать чистые аллергенные белки, для того, чтобы получить достаточную чувствительность в диагностических тестах на специфические IgE. Другая необходимость в использовании рекомбинантных аллергенных компонентов связана со способом компонентной аллергодиагностики [24]. Согласно этому способу анализ IgE-ответа на индивидуальные аллергенные компоненты, а не на экстрактах целых аллергенов, позволяет лучше провести различие между сенсибилизацией к перекрестно-реактивному аллергену и сенсибилизацией к природному аллергену. Таким образом, существует необходимость идентифицировать и получить в виде рекомбинантного белка все возможные аллергенные компоненты из конкретного источника аллергена.
Авторы настоящего изобретения столкнулись с проблемой при попытке секвенировать и клонировать Can f 4 стандартными методами, известными в данной области техники. Проблемы были решены только при использовании нестандартных методов, как описано ниже в разделе «Результаты». Настоящее изобретение основано на неожиданном обнаружении, что IgE-антитело связывает как экстракт перхоти собаки, так и экстракт перхоти коровы, и на понимании того, что для того, чтобы определить последовательность нуклеиновой кислоты и осуществить клонирование Can f 4, авторы изобретения должны были сопоставить Can f 4 с белком другого организма.
В одном из аспектов изобретение относится к аллергену Can f 4, полученному рекомбинантными способами.
В другом аспекте изобретение относится к нуклеиновой кислоте, кодирующей указанный аллерген Can f 4, полученный рекомбинантными способами.
В других аспектах изобретение относится к вектору, содержащему указанную нуклеиновую кислоту, и клетке-хозяину, содержащей указанный вектор.
В дополнительных аспектах изобретение относится к аллергену Can f 4, полученному рекомбинантными способами, который может использоваться в in vitro диагностике аллергии I типа.
В другом аспекте изобретение относится к способу получения аллергенной композиции, предусматривающей стадию добавления аллергена Can f 4, полученного рекомбинантными способами, к композиции, содержащей аллергенный экстракт и/или по меньшей мере один очищенный аллергенный компонент.
В другом аспекте изобретение относится к аллергенной композиции, в которую добавлен аллерген Can f 4, полученный рекомбинантными способами. Такая аллергенная композиция может представлять экстракт аллергена или смесь очищенных и/или рекомбинантных аллергенных компонентов, не содержащая или содержащая пониженное количество аллергена Can f 4, в которую вводят аллерген Can f 4, полученный рекомбинантными способами, для связывания IgE пациентов, у которых IgE не связываются или имеют низкий уровень связывания с другими аллергенными компонентами композиции. В этом аспекте изобретение также относится к способу получения такой композиции, которая содержит стадию добавления аллергена Can f 4, полученного рекомбинантными способами, к аллергенной композиции, такой как экстракт аллергена (необязательно содержащей другие компоненты) или смесь очищенного природного или рекомбинантного аллергенных компонентов.
Кроме того, в другом аспекте изобретение относится к диагностическому тесту in vitro для диагностики аллергии I типа у пациента, где образцы биологических жидкостей, такие как образцы крови или сыворотки пациента, вступают в контакт с аллергеном Can f 4, полученным рекомбинантными способами, или композицией согласно предыдущему аспекту, и обнаруживается, содержит ли образец пациента IgE-антитела, которые специфически связываются с аллергеном Can f 4, полученным рекомбинантными способами или нет, где присутствие подобных IgE-антител, специфически связывающих с указанным аллергеном Can f 4, свидетельствует о наличии аллергии I типа. Такой диагностический метод может быть осуществлен любым способом, известным в данной области. Аллерген Can f 4, полученный рекомбинантными способами, можно, например, иммобилизировать на твердой подложке, например, стандартного лабораторного иммунологического исследования, на микрочипах или в горизонтальном проточном анализе.
В другом аспекте изобретение относится к диагностическому набору для осуществления метода согласно предыдущему аспекту, который включает аллерген Can f 4, полученный рекомбинантными способами.
Изобретение, кроме того, относится к способу лечения аллергии I типа у млекопитающих, предусматривающему введение индивиду, при необходимости, аллергена Can f 4, полученного рекомбинантными способами, или его формы, которая модифицирована, чтобы исключить или ослабить его IgE-ответ, как объяснено ниже. В одном из вариантов осуществления млекопитающее представляет собой собаку. В другом варианте осуществления млекопитающее может быть любым индивидом, имеющим аллерген млекопитающих, который имеет гомологию с аллергеном Can f 4, полученным рекомбинантными способами, при условии, что у индивида отмечена IgE-опосредованная перекрестная реактивность с указанным аллергеном млекопитающих, таким как Bos d 23k. Этот аспект изобретения также относится к применению аллергена Can f 4, полученного рекомбинантными способами, в иммунотерапии, в том числе, например, в компонентной иммунотерапии. Примеры модификаций включают, но ими не ограничены, фрагментацию, процессинг или тандемеризацию молекулы, делецию внутреннего сегмента(ов), замену аминокислотного остатка(ов), перегруппировку доменов или разрушение по меньшей мере части третичной структуры путем разрушения дисульфидных мостиков или путем связывания с другой макромолекулярной структурой или путем удаления способности белка связывать низкомолекулярные соединения.
В другом аспекте изобретение относится к аллергену Can f 4, полученному рекомбинантными способами, или к его форме, которая модифицирована так, чтобы исключить или ослабить его IgE-ответ, как объяснено ниже, для использования при лечении аллергии I типа.
В еще одном аспекте изобретение относится к фармацевтической композиции, содержащей аллерген Can f 4, полученный рекомбинантными способами, или к форме указанного аллергена Can f 4, которая модифицирована так, чтобы исключить или ослабить его IgE-ответ.
В указанных аспектах аллерген Can f 4, полученный рекомбинантными способами, может быть заменен его вариантом или фрагментом, имеющим общие антигенные детерминанты антител с аллергеном Can f 4 дикого типа, как определено ниже.
В одном из вариантов осуществления каждого указанного аспекта аллерген Can f 4, полученный рекомбинантными способами, имеет аминокислотную последовательность SEQ ID NO:2 и кодируется нуклеотидной последовательностью SEQ ID NO:1.
Кроме того, изобретение относится к аллергену Can f 4, полученному рекомбинантными способами, имеющему аминокислотную последовательность SEQ ID NO:2, который может быть использован в диагностике, а также в терапии.
Изобретение, кроме того, относится к аллергену Bos d 23k, полученному рекомбинантными способами, который используется в in vitro диагностике аллергии I типа.
В другом аспекте изобретение относится к способу получения композиции аллергена, предусматривающему стадию добавления аллергена Bos d 23k, полученного рекомбинантными способами, к композиции, содержащей экстракт аллергена и/или по меньшей мере один очищенный аллергенный компонент.
В другом аспекте изобретение относится к аллергенной композиции, в которую добавлен аллерген Bos d 23k, полученный рекомбинантными способами. Такая аллергенная композиция может представлять собой экстракт аллергена или смесь очищенных и/или рекомбинантных аллергенных компонентов, не содержащую или содержащую незначительное количество аллергена Bos d 23k, где аллерген Bos d 23k, полученный рекомбинантными способами, добавляют для связывания IgE пациентов, у которых IgE не связывается или плохо связывается с другими аллергенными компонентами композиции. В этом аспекте изобретение также относится к способу получения такой композиции, который предусматривает стадию добавления аллергена Bos d 23k, полученного рекомбинантными способами, к аллергенной композиции, такой как экстракт аллергена (необязательно содержащий другие компоненты) или смесь очищенного нативного или рекомбинантного аллергенных компонентов.
Кроме того, в другом аспекте изобретение относится к диагностическому методу для диагностирования in vitro аллергии I типа у пациента, где образец биологической жидкости, такой как образец крови или сыворотки пациента, приводят в контакт с аллергеном Bos d 23k, полученным рекомбинантными способами, или композицией в соответствие с предыдущим аспектом, и обнаруживается, содержит ли образец пациента антитела IgE, которые специфически связывают аллерген Bos d 23k, полученный рекомбинантными способами, или нет, где присутствие подобных антител IgE, специфически связывающихся с вышеуказанным аллергеном Bos d 23k, свидетельствует о наличии аллергии I типа. Такой диагностический метод может быть осуществлен любым способом, известным в данной области. Аллерген Bos d 23k, полученный рекомбинантными способами, можно, например, иммобилизовать на твердой подложке, такой как подложка стандартного лабораторного иммунологического исследования, на микрочипах или в горизонтальном проточном анализе.
В другом аспекте изобретение относится к диагностическому набору для осуществления способа в соответствие с предыдущим аспектом, который содержит аллерген Bos d 23k, полученный рекомбинантными способами.
Изобретение, кроме того, относится к способу лечения аллергии I типа у млекопитающих, предусматривающему введение индивиду, при необходимости, аллергена Bos d 23k, полученного рекомбинантными способами, или его формы, которая модифицирована так, чтобы исключить или ослабить его IgE-связывающий ответ, как объяснено ниже. В одном из вариантов осуществления млекопитающее представляет собой корову. В другом варианте осуществления млекопитающее может быть любым индивидом, имеющим аллерген млекопитающих, который имеет гомологию с аллергеном Bos d 23k, полученным рекомбинантными способами, при условии, что у индивида отмечена IgE-опосредованная перекрестная реактивность с вышеуказанным аллергеном млекопитающих, таким как Can f 4. В данном аспекте изобретение также относится к применению аллергена Bos d 23k, полученного рекомбинантными способами, в иммунотерапии, в том числе, например, в компонентной иммунотерапии. Примеры модификаций включают, но ими не ограничиваются, фрагментацию, процессинг или тандемеризацию молекулы, делецию внутреннего сегмента(ов), замену аминокислотного остатка(ов), перегруппировку доменов или разрушение по меньшей мере части третичной структуры путем разрушения дисульфидных мостиков или путем связывания с другой макромолекулярной структурой, или путем удаления способности белка связывать низкомолекулярные соединения.
В другом аспекте изобретение относится к аллергену Bos d 23k, полученному рекомбинантными способами, или к форме, которая модифицирована так, чтобы исключить или ослабить его IgE-связывающий ответ, как объяснено ниже, для использования при лечении аллергии I типа.
В еще одном аспекте изобретение относится к фармацевтической композиции, содержащей аллерген Bos d 23k, полученный рекомбинантными способами, или к форме аллергена Bos d 23k, которая модифицирована так, чтобы исключить или ослабить его IgE-связывающий ответ.
В вышеуказанных аспектах аллерген Bos d 23k, полученный рекомбинантными способами, может быть заменен его вариантом или фрагментом, имеющим общие антигенные детерминанты антител для аллергена Bos d 23k дикого типа, как определено ниже.
В одном из вариантов осуществления каждого указанного аспекта, касающегося Bos d 23k, аллерген Bos d 23k, полученный рекомбинантными способами, имеет аминокислотную последовательность SEQ ID NO:4 и кодируется нуклеотидной последовательностью SEQ ID NO:3.
Определения
Can f 4 следует рассматривать как аллерген собаки, включенный в список International of Immunological Societies Allergen Nomenclature Sub-Committee (www.allergen.org).
Липокалины следует рассматривать как многочисленную и разнообразную группу белков, присутствующих в широком ряде организмов и вовлеченных во множество функций [25-27]. Эти белки характеризуются определенными консервативными структурными особенностями, но в остальном их аминокислотная последовательность не является высоко консервативной. Липокалины способны связывать небольшие, главным образом, гидрофобные молекулы, включая стероиды, жирные кислоты и феромоны. Одна подгруппа липокалинов, присутствующая в обонятельном аппарате млекопитающих, относится к одорант-связывающим белкам благодаря их способности обратимо связывать и высвобождать летучие соединения, вовлеченные в передачу обонятельных сигналов [28]. Несколько липокалинов млекопитающих были описаны как аллергены, вызывающие респираторные аллергические симптомы у сенсибилизированных людей [29].
Белок, обозначенный авторами изобретения Bos d 23k, описывается в «Примере» как белок с молекулярной массой 23 кДа, выделенный из перхоти коровы, содержащий аминокислотную последовательность в соответствии с гипотетическим бычьим генным продуктом, описанным в Acc No. XP_581277, выведенным из геномной последовательности Bos taurus.
Термин «Can f 4», не определенный в дальнейшем, следует рассматривать как полноразмерный, немодифицированный, интактный Can f 4.
Подобное определение также применяется с соответствующими изменениями в Bos d 23k.
Варианты и фрагменты аллергена Can f 4, имеющие общие антигенные детерминанты антител для аллергена Can f 4, следует рассматривать как фрагменты и варианты, для которых связывание IgE-антител из образца сыворотки типичного Can f 4-сенсибилизированного пациента может значительно ингибироваться аллергеном Can f 4. Подобный анализ ингибирования можно, например, провести согласно протоколу, описанному в примере 8 документа WO 2008/079095. Также указано, что варианты и фрагменты имеют сходные свойства связывания IgE с аллергеном как и Can f 4.
Подобное определение также применяется с соответствующими изменениями к Bos d 23k.
Форму вышеуказанного аллергена Can f 4, которая модифицирована так, чтобы исключить или уменьшить его IgE-ответ, следует рассматривать в контексте настоящего изобретения как значение аллергена Can f 4, который химически или генетически модифицирован так, чтобы исключить его иммунологические свойства, например, как проиллюстрировано выше, относительно иммунотерапевтического аспекта изобретения.
Подобное определение также применяется с соответствующими изменениями к Bos d 23k.
Гомолог аллергена следует рассматривать как значение аллергена, у которого последовательность нуклеиновой кислоты может гибридизоваться с последовательностью нуклеиновой кислоты другого аллергена. Результат гибридизации может зависеть от длины последовательностей, от того как гомология распространена по последовательностям и от экспериментальных условий, таких как концентрация соли, температура, жесткость промывки и т.д. Гомолог аллергена по настоящему изобретению может обладать более низкой идентичностью последовательности, чем идентичность, которую часто принимают для гомолога, то есть 65-70%. По определению в настоящем изобретении последовательность нуклеиновой кислоты гомолога аллергена может иметь более низкую идентичность последовательности к нуклеиновой кислоте другого аллергена, но все еще будет гибридизоваться с другим аллергеном благодаря сегментам последовательностей, показывающим более высокую идентичность.
Перекрестную реактивность к аллергену следует рассматривать как значение, при котором IgE-антитела индивида к первому аллергену также являются реактивными ко второму аллергену, который может или не может являться гомологом первого аллергена.
Краткое описание фигур
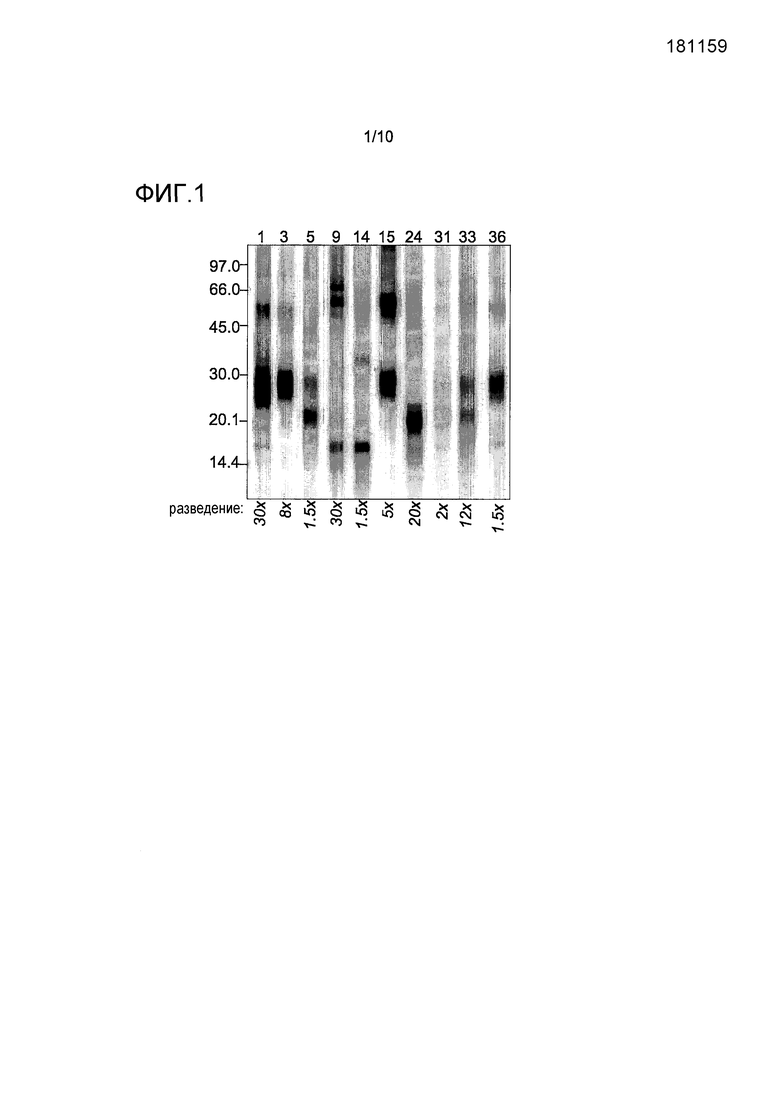
На фиг.1 показан иммуноблотинг связывания IgE-антител с белками экстракта перхоти собаки в невосстанавливающих условиях. Показаны десять из 37 проанализированных сывороток. Номер индивида указан выше и разведение сыворотки ниже каждой линии. Положения маркеров молекулярных масс белков указаны слева.
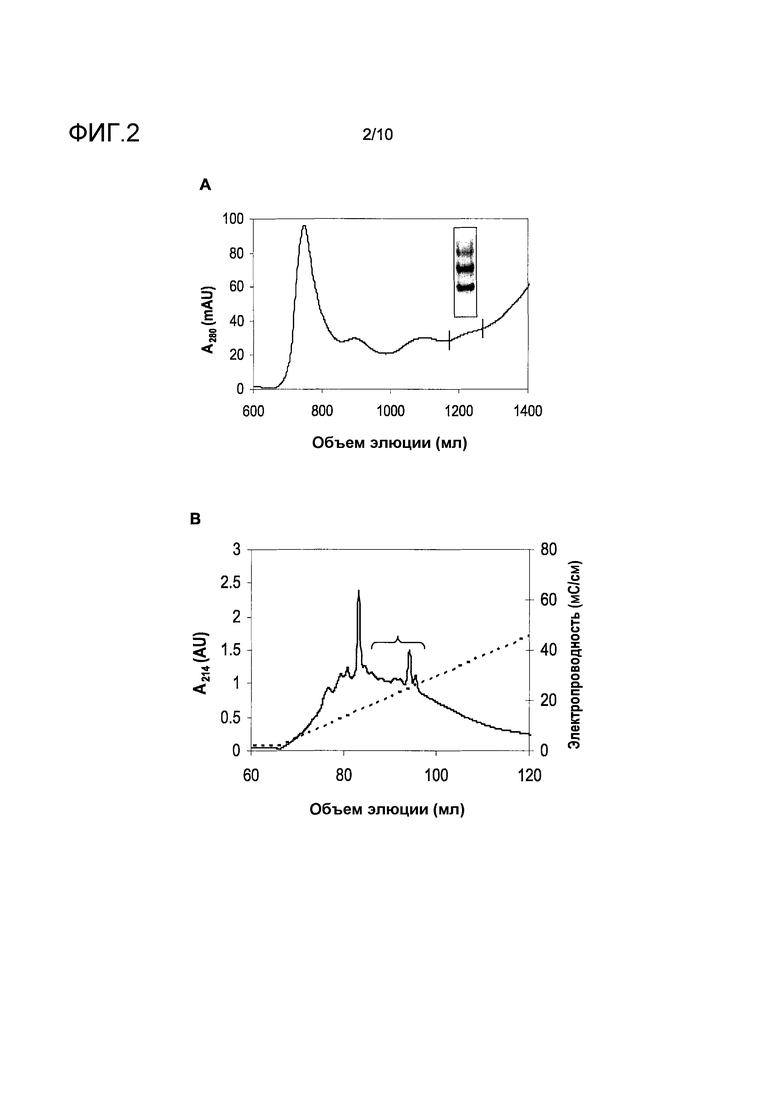
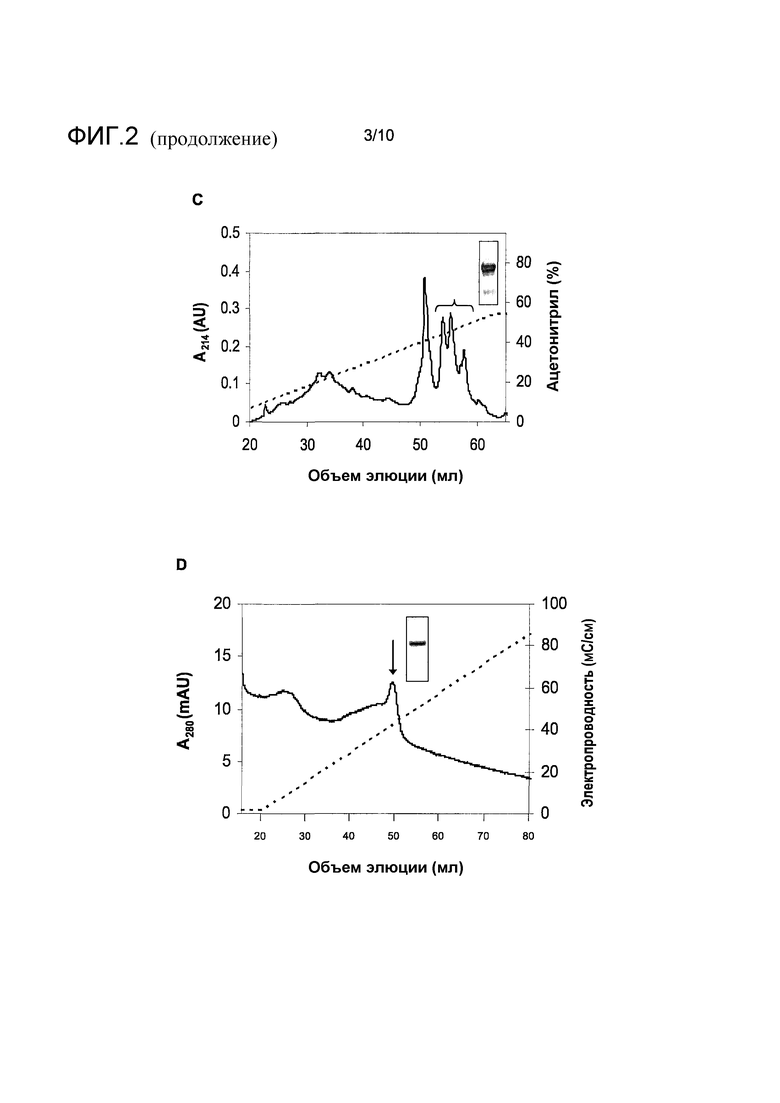
На фиг.2 показана очистка белка перхоти собаки с молекулярной массой 16 кДа, идентифицированного иммуноблоттингом в четыре последовательные стадии. A, препаративная эксклюзионная хроматография. Объединенные фракции обозначены вертикальными столбиками. B, анион-обменная хроматография. Заштрихованная линия обозначает электропроводность (правая ось y). Фракции, объединенные для дальнейшей очистки, обозначены горизонтальной скобкой. C, обращенно-фазная хроматография. Заштрихованная линия обозначает концентрацию ацетонитрила (правая ось y). Фракции, объединенные для дальнейшей очистки, обозначены горизонтальной скобкой. D, анион-обменная хроматография. Заштрихованная линия обозначает электропроводность (правая ось y). Пик, содержащий чистый мишеневый белок, обозначен стрелкой.
На фиг.3 показаны пептидные последовательности, полученные из триптических фрагментов очищенного белка перхоти собаки с молекулярной массой 16 кДа.
На фиг.4 показана последовательность кДНК (как SEQ ID NO:1 из перечня последовательностей) и установленная аминокислотная последовательность (SEQ ID NO:2) Can f 4. Предполагаемый сайт расщепления сигнального пептида обозначен стрелкой.
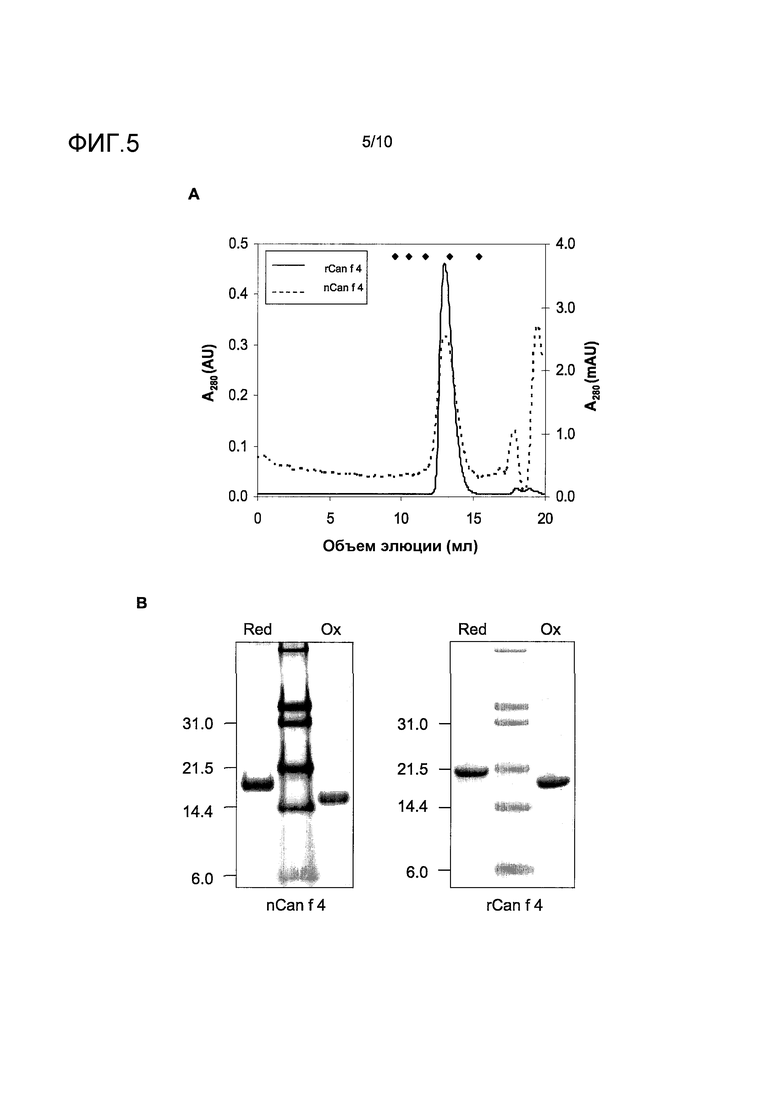
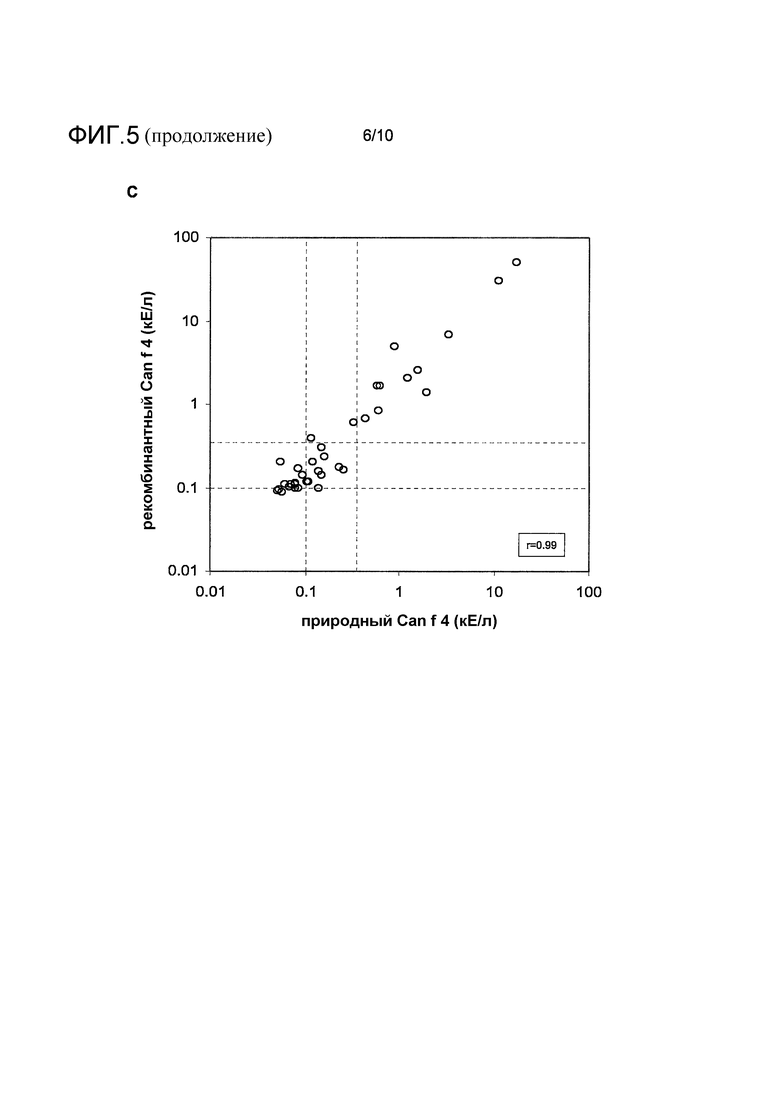
На фиг.5 показан сравнительный биохимический и иммунологический анализ очищенного nCan f 4 и rCan f 4. A, аналитическая гель-фильтрация. Заштрихованная линия: nCan f 4 (правая ось y); сплошная линия: rCan f 4 (левая ось y). Объемы элюции калибровочных белков с молекулярной массой 6,5, 13,7, 29, 43 и 75 кДа отмечены ромбами. B, SDS электрофорез в полиакриламидном геле образцов nCan f 4 и rCan f 4 при невосстанавливающих (ox) и восстанавливающих (red) условиях. Размеры соответствующих маркеров молекулярных масс белков указаны слева. C, анализ активности связывания IgE-антител (кЕ/л) при использовании ImmunoCAP. Проанализированы сыворотки 37 индивидов с аллергией на собаку. Заштрихованные линии указывают на уровень 0,10 и 0,35 кЕ/л.
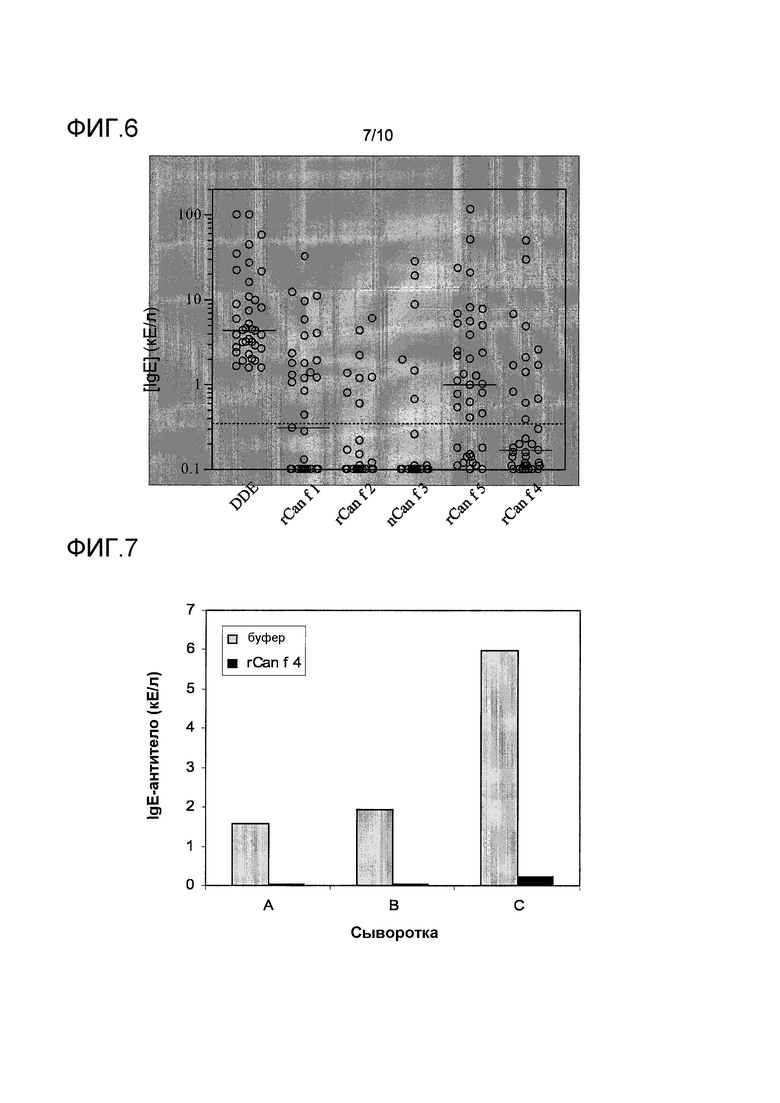
На фиг.6 показана концентрация IgE-антител в отношении экстракта перхоти собаки (DDE), rCan f 1, rCan f 2, nCan f 3, rCan f 5 и rCan f 4 среди 37 индивидов с аллергией на собаку. Горизонтальные столбики указывают средние значения. Пунктирная линия указывает на уровень 0,35 кЕ/л. Значения ниже 0,1 кЕ/л приравнивали к 0,1 кЕ/л.
На фиг.7 показано ингибирование связывания IgE-антител с Bos d 23k, вызванного rCan f 4. Три Bos d 23k-реактивные сыворотки (A-C) преинкубировали с 100 мкг/мл rCan f 4 (черные столбики) или буфером (отрицательный контроль, серые столбики), до момента измерения связывания IgE с иммобилизованным Bos d 23k.
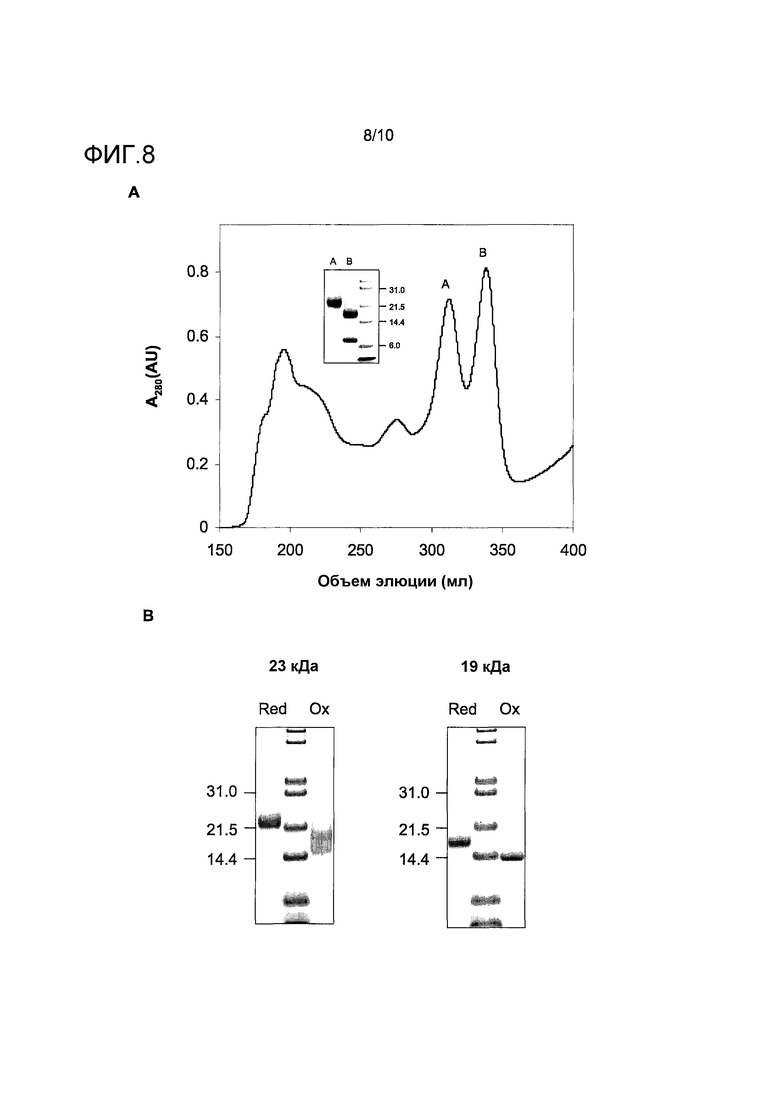
На фиг.8 показана очистка двух белков, связывающих IgE-антитела, Bos d 23k и Bos d 2, из перхоти коровы. A, фракционирование путем эксклюзионной хроматографии. Главные фракции от двух пиков, обозначенных A и B, объединяли и подвергали SDS-электрофоретическому анализу при восстанавливающих условиях (вставка). Размеры соответствующих маркеров молекулярных масс белков указаны справа. B, SDS электрофорез в полиакриламидном геле образцов очищенного Bos d 23k и Bos d 2 после дальнейшей очистки путем гидрофобного взаимодействия и анион-обменной хроматографии при невосстанавливающих (ox) и восстанавливающих (red) условиях. Размеры соответствующих маркеров молекулярных масс белков указаны слева от каждого геля.
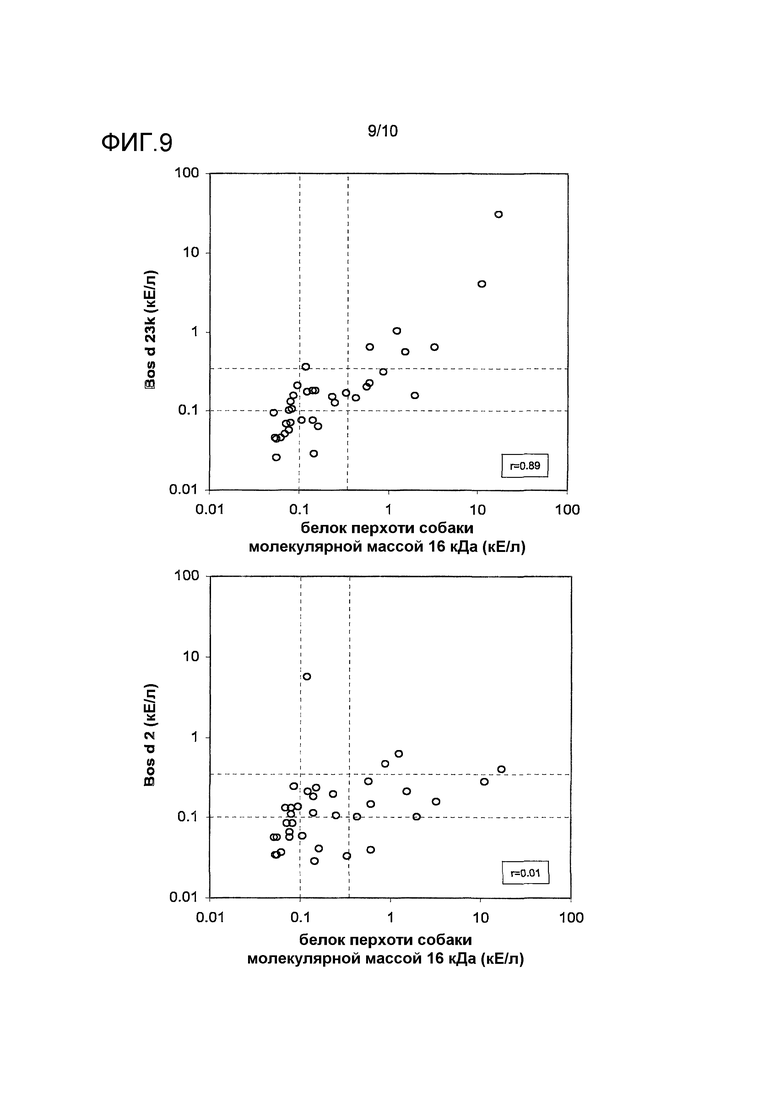
На фиг.9 показано сравнение связывания IgE-антител с Can f 4, Bos d 23k (верхний график) и Bos d 2 (нижний график). Указан коэффициент корреляции (r) для каждого сравнения. Проанализированы сыворотки 37 индивидов с аллергией на собаку. Заштрихованные линии указывают уровень 0,10 и 0,35 кЕ/л.
На фиг.10 показано выравнивание аминокислотной последовательности Can f 4 (как SEQ ID NO:2) и Bos d 23k (как SEQ ID NO:4).
Подробное описание изобретения
Пример ниже иллюстрирует настоящее изобретение с выделением и использованием липокалин-подобного белка Can f 4 от собаки. Также представлена часть, иллюстрирующая перекрестную реактивность между Can f 4 и аллергеном коровы. Пример является только иллюстративным и его не следует рассматривать как ограничивающий изобретение, которое определено объемом приложенной формулы изобретения.
ПРИМЕР
МАТЕРИАЛЫ И МЕТОДЫ
Иммуноблот-анализ IgE
Осуществляли иммуноблот-анализ экстракта перхоти собаки, разделенного посредством SDS электрофореза в полиакриламидном геле при невосстанавливающих условиях, используя гомогенный 12,5% ExcelGel (GE Healthcare Life Sciences, Uppsala, Sweden) и нанесенного электроблоттингом на нитроцеллюлозную мембрану Hybond ECL (GE Healthcare Life Sciences). В качестве маркеров молекулярной массы использовали набор LMW (GE Healthcare Life Sciences). Белковые блоты блокировали в течение 1 ч при комнатной температуре, используя блокирующий буфер (50 мМ фосфат pH 7,4, 0,1% (об./об.) Tween-20, 0,9% (масс./об.) NaCl, 0,3% (масс./об.) декстран T10) и затем инкубировали в течение ночи с сывороткой каждого пациента, разводили в 1,5-30 раз в блокирующем буфере. После промывания в блокирующем буфере с 0,5% (об./об.) Tween-20 мембрану инкубировали в течение 4 ч при комнатной температуре с 125I-меченными человеческими анти-IgE антителами в блокирующем буфере и после промывания связанные IgE детектировали рентгенографически, используя фосфоимиджер и сканер Typhoon 9410 Variable Mode Imager (GE Healthcare Life Sciences).
Очистка белка с молекулярной массой 16 кДа из перхоти собаки
Перхоть собаки (Allergon, Välinge, Швеция) экстрагировали в 20 мМ MOPS pH 7,6, 0,5 М NaCl (MBS), осветляли центрифугированием, отфильтровывали через 0,45-мкм смешанный фильтр сложного эфира целлюлозы (Millipore, Billerica, MA) и наносили на колонку Superdex 75 (GE Healthcare Life Sciences) для проведения эксклюзионной хроматографии (SEC). Фракции, содержащие полосу с молекулярной массой 16 кДа, наблюдаемую в иммуноблот-анализе, концентрировали в перемешиваемой ячейке Amicon (Millipore), используя фильтр YM-3, обессоливали на высокодисперсной колонке Sephadex G25 (GE Healthcare Life Sciences), используя 20 мМ Tris-HCl pH 8,0. Обессоленный препарат затем наносили на колонку Source Q (GE Healthcare Life Sciences) для проведения анион-обменной хроматографии (AIEC) и элюировали линейным градиентом NaCl 0-0,5 М. Дальнейшую очистку проводили путем обращенно-фазной хроматографии (RPC), используя колонку Source 15 RPC (GE Healthcare Life Sciences) и элюировали линейным градиентом ацетонитрила 0-54% в воде, содержащей 0,05% трифторуксусной кислоты (TFA). Фракции, содержащие целевой белок, идентифицировали путем SDS электрофореза в полиакриламидном геле и объединяли. После восстановления, алкилирования и расщепления трипсином, пептиды очищенного белка выделяли путем RPC и анализировали, определяя аминокислотную последовательность. Для оценки связывания IgE-антител с использованием ImmunoCAP белок с молекулярной массой 16 кДа подвергали заключительному этапу доочистки путем катион-обменной хроматографии (CIEC), используя колонку SP Sepharose FF (GE Healthcare Life Sciences), уравновешенную в 20 мМ цитрате pH 4,0 и элюировали линейным градиентом NaCl 0-1 М.
Очистка IgE-связывающих белков из перхоти коровы
Перхоть коровы (Allergon) экстрагировали и фракционировали путем SEC, как описано выше. Фракции, содержащие доминантную полосу с молекулярной массой 23 кДа, объединяли, обрабатывали NH4SO4 до конечной концентрации 1 М и далее очищали путем гидрофобной хроматографии (HIC), используя колонку с фенил-Сефарозой HP (GE Healthcare Life Sciences). Полосу с молекулярной массой 23 кДа элюировали в потоке по фракциям и обессоливали в 20 мМ Bis-Tris пропане pH 8,5 на колонке с высокодисперсной Sephadex G25 (GE Healthcare Life Sciences) и впоследствии наносили на колонку Source 15Q (GE Healthcare Life Sciences), уравновешенную тем же самым буфером. Элюцию проводили линейным градиентом NaCl 0-0,4 М, и фракции, содержащие полосу с молекулярной массой 23 кДа, объединяли. Концентрацию белка конечного препарата определяли по спектральной поглощательной способности при 280 нм, используя расчетный коэффициент экстинкции 1,04 на мг/мл.
Фракции, содержащие доминантную полосу с молекулярной массой 19 кДа, объединяли и далее очищали посредством HIC, как описано выше. Белок с молекулярной массой 19 кДа элюировали линейным градиентом 0-1 М NH4SO4 в 20 мМ Tris-HCl pH 8,0 и объединяли пики фракций. Обессоливание и хроматографию AIEC на колонке Source 15Q проводили, как описано выше. Концентрацию белка конечного препарата определяли по спектральной поглощательной способности при 280 нм, используя расчетный коэффициент экстинкции 1,04 на мг/мл.
Анализ белка
Если иначе не определено, SDS электрофорез в полиакриламидном геле образцов белка при восстанавливающих (4% β-меркаптоэтанол) и невосстанавливающих условиях проводили, используя 10% гель NuPAGE (Invitrogen, Carlsbad, CA) и Mark 12 (Invitrogen) в качестве маркеров молекулярной массы. После электрофоретического разделения белки визуализировали путем окрашивания Coomassie Brilliant Blue. Анализ N-концевой последовательности выделенных полос белка проводили, используя Hewlett-Packard G1000A (HewlettPackard, Palo Alto, CA). Аналитическую хроматографию SEC проводили на колонке Superdex 75 HR 10/30 (GE Healthcare Life Sciences), уравновешенной MBS. Калибровку молекулярных масс колонки проводили, используя набор для калибровки гель-фильтрационных колонок LMW (GE Healthcare Life Sciences).
Для проведения анализа масс пептидов (PMF) путем матричной лазерной десорбционной времяпролетной (MALDI-TOF) масс-спектрометрии (MS), подготовку образца RPC-очищенного белка в растворе, включая восстановление, алкилирование и расщепление трипсином, проводили по существу как описано [10], используя Bruker Daltonics Autoflex 2 (Bruker Daltonics, Bremen, Germany). Анализ проводили путем тандемной масс-спектрометрии (MS/MS) для того, чтобы идентифицировать отобранные пептиды. Для того, чтобы идентифицировать белки, соответствующие результатам, полученным при PMF и MS/MS анализе, исследовали базу данных MSDB, используя сервер Mascot (Matrixscience, Landon, UK).
Расщепление трипсином в геле отдельных белковых полос из SDS электрофореза в полиакриламидном геле проводили по существу согласно Shevchenko et al. [11]. Подготовку образца и анализ масс пептидов проводили, как описано выше.
Клонирование, экспрессия и очистка рекомбинантного Can f 4
Тотальную РНК получали из бокового сегмента языка собаки, используя набор RNAqueous Kit (Ambion, Austin, Texas). Полиаденилированную РНК получали из тотальной РНК, используя набор mRNA Purification Kit (GE Healthcare Life Sciences), и первую цепочку кДНК получали, используя набор First-Strand cDNA Synthesis kit (GE Healthcare Life Sciences). ПЦР с быстрой амплификацией концов кДНК (3'RACE) проводили согласно Frohman [12], используя внутренние («nested») обратные олигонуклеотидные праймеры 5'-ATGAAGATCCTACTGTTGTGTC-3' и 5'-CAGCTACCCCTTCCTAATG-3', оба несущие терминальный сайт рестрикции NdeI для клонирования. Семь независимых 3'RACE-клонов выделяли и полностью секвенировали, посредством чего можно было идентифицировать кодирующую последовательность Can f 4. Секвенирование ДНК проводили, используя генетический анализатор Applied Biosystems 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA). Анализы и оценки последовательности ДНК и аминокислотной последовательности проводили, используя программы GCG Wisconsin Package (Accelrys, San Diego, CA, USA). Предсказание сигнального пептида проводили, используя SignalP (www.cbs.dtu.dk/services/SignalP). С целью экспрессии белка кодирующую последовательность Can f 4 амплифицировали, используя праймеры 5'-GTCAGCATATGCAGCTACCCCTTCCTAATG-3' и 5'-ACTGACTCGAGTTCATGGTTGGGACAGTTGTC-3', и клонировали между сайтами NdeI и XhoI вектора pET-23a(+) (Novagen, Madison, WI, USA). Рекомбинантный Can f 4 получали как белок, меченный шестью аминокислотными остатками гистидина с C-конца, в E. coli BL21, используя биореактор 3-L (Belach Bioteknik, Solna, Sweden).
Для очистки rCan f 4 собранные клетки повторно суспендировали в 20 мМ Tris-HCl pH 8,0 и лизировали путем пропускания суспензии через гомогенизатор Emulsiflex C5 (Avestin Inc., Canada) при 15000-17000 кПа. После осветления центрифугированием и фильтрацией, супернатант наносили на колонку Chelating Sepharose FF (GE Healthcare Life Sciences), насыщенную NiSO4. Промывку колонки проводили 20 мМ имидазолом в 20 мМ Tris-HCl pH 8,0, 0,15 М NaCl и рекомбинантный белок элюировали в линейном градиенте имидазола 20-500 мМ в том же самом буфере. Дальнейшую очистку рекомбинантного белка проводили путем хроматографии AIEC в 20 мМ Tris-HCl pH 8,0 используя колонку Q Sepharose FF (GE Healthcare Life Sciences). Белок элюировали, используя линейный градиент NaCl 0-0,5 М, и фракции объединяли в соответствии с результатами SDS электрофореза в полиакриламидном геле. Концентрацию белка конечного препарата определяли по спектральной поглощательной способности при 280 нм, используя расчетный коэффициент экстинкции 0,78 на мг/мл. Интактность рекомбинантного белка подтверждали N-концевым секвенированием.
Индивиды с аллергией на собаку и пыльцевые аллергические контроли
В исследовании использовались сыворотки 37 индивидов с аллергией на собаку из Испании (n=23), Швеции (n=10) и Америки (n=4) (таблица 2). Все пациенты имели установленный врачом диагноз аллергии на собаку, с такими симптомами, как астма, риноконъюктивит и крапивница, положительную кожную пробу и положительный тест ImmunoCAP (Phadia, Uppsala, Sweden) для специфического IgE к экстракту перхоти собаки. В целях контроля использовали сыворотки 44 индивидов с аллергией на пыльцу, не имеющих диагноз или описанные симптомы аллергии на собаку. Все образцы и клинические данные собирали с одобрения локального этического комитета в каждом центре.
Измерения специфических IgE-антител
IgE-связывающая активность очищенных и рекомбинантных аллергенов проверяли, используя стандартные и экспериментальные тесты ImmunoCAPTM (Phadia). Экспериментальные тесты ImmunoCAP подготавливали, как описано [13]. Специфичность исследования экспериментальных тестов оценивали, используя отрицательную контрольную сыворотку, с добавленным IgE-миелома в конечной концентрации 0, 1000 или 3000 кЕ/л. Эксперимент по ингибированию IgE проводили путем преинкубации образцов сыворотки с рекомбинантным Can f 4 в конечной концентрации 100 мкг/мл до измерения связывания IgE-антител с белком перхоти коровы с молекулярной массой 23 кДа, иммобилизованным на твердой фазе ImmunoCAP. Результаты вычисляли как средние значения повторных определений.
Результаты
Иммуноблот-анализ сывороток индивидов с аллергией на собаку
Образцы сывороток 10 индивидов с аллергией на собаку подвергали IgE-иммуноблот-анализу, используя экстракт перхоти собаки при невосстанавливающих условиях (фиг.1). Для сравнения для тех же сывороток были получены данные анализа ImmunoCAP для специфических IgE к rCan f 1, rCan f 2, nCan f 3 и rCan f 5. Иммуноблот-анализ выявлял связывание IgE с по меньшей мере 8 различными белковыми полосами. Из данных 8 полос полосы в области участка, соответствующего молекулярной массе 23 кДа, коррелировали с выявленным ImmunoCAP анализом связыванием IgE с rCan f 1 и rCan f 2 (индивиды 5, 24, 31 и 33), полосы в области участка, соответствующего молекулярной массе 28 кДа, с реактивностью rCan f 5 (индивиды 1, 3, 5, 15, 31, 33 и 36) и полосы в области участка, соответствующего молекулярной массе приблизительно 60 кДа, с реактивностью nCan f 3 (индивиды 9, 15 и 33).
Связывание IgE с полосой с молекулярной массой 16 кДа на иммуноблоте обнаруживали для сывороток 4 индивидов (1, 9, 14 и 36), из которых один индивид (индивид 14) вызывал особенно интенсивный сигнал. Тот факт, что в случае данной сыворотки не наблюдалось связывание IgE с любым из rCan f 1, rCan f 2, nCan f 3 или rCan f 5 в анализе ImmunoCAP, позволял предпологать, что полоса с молекулярной массой 16 кДа представляет собой новый аллерген.
Очистка белка перхоти собаки с молекулярной массой 16 кДа, идентифицированного путем иммуноблоттинга
Три этапа процесса очистки, включающие SEC, с последующей AIEC и RPC, приводили к получению белка с молекулярной массой 16 кДа с чистотой 90-95% (фиг.2). Анализ N-концевой последовательности данного белка не давал никакого результата, что позволяло предположить, что данный N-конец заблокирован. Для того чтобы преодолеть данную проблему, белок восстанавливали, алкилировали и расщепляли трипсином для того, чтобы вызвать образование внутренних пептидов. Четыре подобных триптических пептида разделяли путем RPC и анализировали N-концевым секвенированием, получив последовательности, показанные на фиг.3. Поиск в базе данных BLAST по данным пептидным последовательностям не идентифицировал точное совпадение с любым известным белком собаки. Однако несовершенные совпадения пептидов 1 и 4 с записью базы данных, представляющей одорант-связывающие белки коровы и свиньи, позволяли предположить, что очищенный белок собаки мог бы быть связан с данным семейством белков. Более того, поиск TBLASTN базы данных генома собаки для пептида 4, используя сокращенные параметры точности, приводил к совпадениям с концептуальными трансляциями трех смежных сегментов в терминальном участке X хромосомы. PMF анализ очищенного белка не приводил к какому-либо существенному совпадению с известной белковой последовательностью.
IgE-связывающую активность белка перхоти собаки с молекулярной массой 16 кДа оценивали путем иммунологического исследования ImmunoCAP, вслед за заключительным этапом дополнительной очистки посредством CIEC, чтобы в дальнейшем увеличить чистоту препарата. Анализировали сыворотки 37 индивидов с аллергией на собаку и показывали, что связывание IgE с очищенным белком в ImmunoCAP хорошо коррелирует с детекцией полосы с молекулярной массой 16 кДа в иммуноблот-анализе экстракта перхоти собаки, показывая, что очищенный белок представлял собой полосу с молекулярной массой 16 кДа, наблюдаемую на иммуноблоттинге.
Связь между эпителиальными аллергенами собаки и коровы
Неожиданная помощь в идентификации и клонировании аллергена собаки с молекулярной массой 16 кДа пришла в результате проведения отдельного ряда экспериментов в лаборатории заявителей изобретения на эпителиальных аллергенах других видов животных. Одна сыворотка, которая была разделена между исследованиями аллергенов собаки и коровы, показала существенное связывание IgE-антител и с экстрактом перхоти собаки, и с экстрактом перхоти коровы, тогда как была нереактивной в ImmunoCAP ко всем rCan f 1, rCan f 2, nCan f 3 и rCan f 5, что позволяло поставить вопрос о возможности того, что данная сыворотка может определять новый аллерген собаки.
Обнаружено, что белок с молекулярной массой 23 кДа в экстракте перхоти связывает IgE из данной сыворотки и может быть высоко обогащен посредством SEC (фиг.8, A). Обнаружено, что смежный пик в SEC фракционировании содержит главный аллерген Bos d 2, липокалин с молекулярной массой 19 кДа, как подтверждено PMF анализом (данные не приведены). Дальнейшая очистка данных белков посредством HIC и AIEC (данные не приведены) приводила к высокочистым препаратам (фиг.8, B), которые могут быть использованы как реактивы для иммунологического исследования.
Белок с молекулярной массой 23 кДа идентифицировали путем значительного совпадения (p<0,05) PMF-данных с записью базы данных XP_581277 (как описано SEQ ID NO:4), представляющей бычий липокалин, аннотируемый как "подобный одорант-связывающему белку". Далее, анализ N-концевой последовательности белка с молекулярной массой 23 кДа показал последовательность EAQGDASQFT, соответствующую остаткам 19-28 той же самой записи базы данных, и таким образом подтверждая PMF-совпадение. Данный белок в дальнейшем именуется как Bos d 23k.
Экспериментальные тесты ImmunoCAP, проводимые и с Bos d 23k и с Bos d 2, использовали для того, чтобы оценить корреляцию в связывании IgE с белком перхоти собаки с молекулярной массой 16 кДа. Сыворотки 37 индивидов с аллергией на собаку анализировали, и результаты показаны на фиг.9. В то время как белок перхоти собаки с молекулярной массой 16 кДа и Bos d 2 не показывали корреляции в связывании IgE (r=0,01), существенная корреляция наблюдалась к Bos d 23k (r=0,89), что позволяло предположить перекрестную реактивность и структурное подобие между аллергеном собаки и Bos d 23k.
Клонирование и анализ последовательности белка перхоти собаки с молекулярной массой 16 кДа
Возможное подобие последовательности между белком перхоти собаки с молекулярной массой 16 кДа и Bos d 23k натолкнуло на поиск в базе данных, нацеленный на идентификацию белка собаки или последовательности его гена. Поиск BLASTN базы данных генома собаки по последовательности бычьего белка (XP_581277) приводил к совпадению в трансляции нуклеотидных позиций 338441-338307 (Acc. No. AAEX02025758), сегмент размером 431454 п.о. последовательности генома собаки [14]. Интересно, что теоретическая трансляция данной геномной области содержала точное совпадение 5 остатков из аминокислотных остатков 9-13 представленной N-концевой последовательности Can f 4 [9]. В более высоком (upstream) положении (339006-338926) была обнаружена нуклеотидная последовательность, кодирующая аминокислоты, соответствующие остаткам 1-8 последовательности Can f 4, а также предполагаемый сигнальный пептид.
С целью клонирования кДНК, соответствующей идентифицированному сегменту AAEX02025758, который, как предполагают, кодирует Can f 4, проводили амплификацию путем 3'RACE. Использовали олигонуклеотидные праймеры, основанные на геномной последовательности, кодирующей первую часть предполагаемого сигнального пептида и участок после предполагаемого сайта расщепления, вместе с первой цепочкой кДНК, полученной от poly-A РНК языка собаки в качестве матрицы. Получали отдельный продукт амплификации, который клонировали, анализировали и впоследствии использовали для экспериментов для экспрессии белка.
Полная последовательность ДНК и аминокислотная трансляция клонированной кДНК показаны на фиг.4. Идентифицировали открытую рамку считывания, кодирующую 174 аминокислотных остатка, из которых первые 16 остатков были предсказаны SignalP для сформирования сигнального пептида. Зрелый белок, выведенный из клонированной кДНК, состоял из 158 аминокислотных остатков, включая два цистеина, и обладал предсказанной молекулярной массой 17,6 кДа и изоэлектрической точкой 6,53. Для того чтобы оценить наличие разнообразия последовательности, выделяли и секвенировали семь независимых 3'RACE-клонов. Среди клонов обнаруживали несколько нуклеотидных замен (данные не приведены), но ни одна не вызывала изменения в аминокислотной последовательности белка. Во всех проанализированных клонах стоп-кодон, терминирующий открытую рамку считывания, следовал за нетранслированным сегментом нуклеотида 213, предшествующим poly-A участку.
Предсказанный сайт расщепления сигнального пептида между остатками 16 и 17 сделало бы остаток глутамина в положении 17 первым в зрелом белке. N-концевой остаток глутамина может подвергаться циклизации в пироглутамат, что, как известно, вызывает блокировку N-концевого секвенирования деградацией по Эдману, что согласуется с неудачей заявителей изобретения определить последовательность интактного белка (15). Далее, при повторной проверке результатов PMF для нативного белка, идентифицировали, что триптический фрагмент массой 1759,98 Да точно соответствует предполагаемому фрагменту, предсказанному для остатка 17-32 предполагаемого первичного продукта трансляции, модифицированного циклизацией в пироглутамат. Кроме того, идентифицировали триптические фрагменты, соответствующие аминокислотным остаткам 33-51, 56-63, 66-73 и 95-104.
Кроме того, аминокислотная последовательность, закодированная клонированной кДНК, содержала все четыре триптических пептидных последовательности, полученных из очищенного природного белка молекулярной массой 16 кДа (фиг.3) с точным соответствием для пептидов 2-4 и двух консервативных аминокислотных замен в пептиде 1. В целом, фрагменты белка, идентифицированные PMF и пептидным секвенированием, представляли степень покрытия 58% установленной аминокислотной последовательности Can f 4 (таблица 1). В своей совокупности, результаты обеспечивают убедительное доказательство того, что очищенный аллерген молекулярной массой 16 кДа и белок, закодированный клонированной кДНК, оба представляют аллерген собаки Can f 4.
Путем выравнивания последовательности клонированной кДНК с геномной последовательностью Acc. No. AAEX02025758, обнаруживали, что кодирующая часть гена Can f 4 охватывает в общей сложности 5916 п.о. и включает 6 экзонов: 339006-338929 (экзон 1), 338447-338307 (экзон 2), 335150-335082 (экзон 3), 334581-334468 (экзон 4), 333609-333511 (экзон 5) и 333114-333091 (экзон 6). Точное место сплайсинга между экзонами 3 и 4 не может быть однозначно выведено из последовательности, и границы данных экзонов могут вместо этого быть 335150-335079 и 334578-334468, соответственно.
Аминокислотные последовательности, выведенные из кДНК Can f 4 и сегментов гена, отклонялись друг от друга в 5 положениях. В положении 20 последовательности зрелого Can f 4 остаток изолейцина в трансляции кДНК соответствовал остатку валина в последовательности, полученной из гена, в положении 30 остаток аспарагиновой кислоты соответствовал остатку глутаминовой кислоты, в положении 38 остаток метионина соответствовал остатку лейцина, в положении 51 остаток серина соответствовал остатку лейцина, и в положении 81 остаток тирозина соответствовал или остатку аспарагиновой кислоты, или остатку цистеина, в зависимости от точного места сплайсинга между экзонами 3 и 4.
В дополнение к последовательности гена Can f 4, описанной выше, обнаружено, что запись базы данных AAEX02025758 содержит два других сегмента, относящихся к Can f 4, объясняя три несовпадения, полученных в геноме собаки путем поиска TBLASTN по последовательности триптического пептида 4. Три сегмента, относящихся к Can f 4, были организованы в тандем на расстоянии 5,4 и 6,7 т.п.о. друг от друга, с одним сегментом, описанным выше, расположенным в самом нижнем (downstream) положении. Сегмент, относящийся к Can f 4, расположенный в середине, обладал структурой экзон/интрон подобной структуре гена Can f 4, описанной выше, и его выведенная аминокислотная последовательность отличалась по 24 положениям по сравнению с геном Can f 4 и по 28 положениям по сравнению с кДНК Can f 4. Интересно, что это полностью соответствовало последовательности триптического пептида 1, которая отклонялась по двум положениям от аминокислотной последовательности, кодированной кДНК. В сегменте, относящимся к Can f 4, локализованном в самом верхнем (upstream) положении, последовательность, соответствующая экзону 4, не могла быть идентифицирована.
Can f 4 принадлежит к суперсемейству липокалинов и проявляет 38-39% идентичности последовательности к бычьему (XP_581277) и свиному [16] (NP_998961) одорант-связывающим белкам. Выравнивание аминокислотной последовательности Can f A и Bos d 23k показано на фиг.10. Нужно отметить, что кроме разрозненной группы подобных или идентичных остатков, C-концевой участок белков является высоко консервативным, так же как и оба остатка цистеина (положение 62 и 154 в Can f 4). Другие консервативные элементы, особенность липокалинов [17], включают мотив GxW около N-конца (положение 13-15), остаток глицина (положение 118). Другой мотив, типичный для липокалинов [17], YxxxYxG, присутствует в Bos d 23k (положение 74-80), но только частично консервативен в Can f 4. Интересно, что, несмотря на принадлежность к сходному семейству белков, Can f 4 продемонстрировал только 24% и 26% идентичности аминокислотной последовательности к Can f 1 и Can f 2, соответственно.
Получение рекомбинантного Can f 4
Рекомбинантный Can f 4, исключая сигнальный пептид, экспрессировали в E. coli в виде белка, меченного с С-конца шестью аминокислотными остатками гистидина. Рекомбинантный белок очищали от растворимой клеточной фракции посредством IMAC и AIEC. Для того чтобы оценить уровень агрегации рекомбинантного белка, образец препарата подвергали аналитической SEC, параллельно с очищенным природным Can f 4. Как показано на фиг.5, A, в обоих случаях на хроматограмме преобладал одиночный симметричный пик, соответствуя молекулярной массе приблизительно 16,5 кДа, как определено при использовании калибраторов. В анализе путем SDS электрофореза в полиакриламидном геле (фиг.5, B) и природный и рекомбинантный Can f 4 показали сдвиг в уровне миграции при восстанавливающих условиях, позволяя предположить, что два остатка цистеина, присутствующих в белке, были включены в формирование дисульфидного мостика. Следует отметить, что на значение кажущейся молекулярной массы Can f 4, показанной предварительным иммуноблоттингом (фиг.1), влиял тот факт, что экстракт перхоти собаки, используемый для анализа, наносили на разделяющий гель при невосстанавливающих условиях, что приводило к заниженной оценке относительно размера белка, указанного путем последующих исследований образцов очищенного белка при восстанавливающих условиях. Увеличение размера, продемонстрированное rCan f 4 по сравнению с природным белком, согласовывалось с добавлением C-концевой метки из шести остатков гистидина в рекомбинантном белке и почти полному сохранению остатка метионина, который является инициатором биосинтеза полипептидной цепи.
Анализ специфического связывания антитела IgE с rCan f 4 у индивидов с аллергией на собаку
Иммунологическую активность рекомбинантного Can f 4 оценивали по сравнению с природным белком, очищенным из перхоти собаки. Каждый белок иммобилизовывали на подложку ImmunoCAPTM и проверяли связывание белков с IgE-антителами, используя образцы сыворотки 37 индивидов с аллергией на собаку (фиг.5, C). Два набора данных показывали очень строгую корреляцию (r=0,99), демонстрируя, что рекомбинантный аллерген имеет близкое сходство со своим природным аналогом в отношении IgE-связывающих детерминант.
Частота и величина реактивности IgE-антител к Can f 4 по сравнению с другими аллергенами перхоти собаки проиллюстрированы на фиг.6. Более полный набор данных по индивидам с аллергией на собаку и их примеры сенсибилизации представлены в таблице 2. Среди 37 проанализированных сывороток, 13 (35%) демонстрировали связывание IgE-антител с Can f 4. Одна из 13 сывороток, реактивная к Can f 4, не показывала связывание IgE ни с одним из других тестируемых аллергенов собаки. Отсутствие корреляции в связывании IgE можно было наблюдать между Can f 4 и Can f 1 или Can f 2, в соответствии с их высоко неоднородной первичной структурой. Специфичность исследования экспериментального теста ImmunoCAP Can f 4 демонстрировалась путем низких результатов, полученных при тестировании отрицательной контрольной сыворотки с добавленной IgE-миелома в пределах 3000 кЕ/л (таблица 3).
Для того чтобы исследовать появление Can f 4-специфических IgE-антител у атопических индивидов, не имеющих аллергии на собаку, сыворотки 44 индивидов с аллергией на пыльцу, не имеющих поставленный диагноз или описанные симптомы аллергии на собаку, тестировали со всеми доступными компонентами аллергена собаки и спектром пыльцевых экстрактов (таблица 4). Семь из данных сывороток (16%) показывали положительный ответ на экстракт перхоти собаки с одним исключением при уровне 2 кЕ/л или ниже, один из которых также показывал связывание IgE-антител с rCan f 4. Ни одна сыворотка, отрицательная к перхоти собаки, не показывала положительный ответ на rCan f 4.
Перекрестная реактивность между rCan f 4 и Bos d 23k
Для того чтобы изучить степень перекрестной реактивности между Can f 4 и Bos d 23k, показанную путем их корреляции в связывание IgE-антител, эксперимент по ингибированию IgE проводили с тремя сыворотками, реактивными к бычьему белку. Очищенный бычий белок присоединяли к подложке ImmunoCAP и измеряли связывание IgE из сывороток, преинкубированных или с rCan f 4 или с буфером. Как можно увидеть на фиг.7, rCan f 4 почти полностью (≥96%) ингибировал связывание IgE с белком перхоти коровы во всех трех сыворотках, демонстрируя, что все детерминанты связывания IgE с бычьим белком, распознаваемые сыворотками, используемыми в эксперименте, были общими с rCan f 4.
Обсуждение
Как описано в данной заявке, заявители изобретения выделяли, клонировали и охарактеризовывали IgE-связывающий белок из перхоти собаки, идентифицированный как Can f 4. До данной работы была известна только N-концевая последовательность данного аллергена, состоящая из 13 остатков. Обнаружено, что Can f 4 принадлежит к разнообразному суперсемейству липокалинов и показывает сходные характеристики с одорант-связывающими белками других видов, включая корову и свинью. Обнаружено, что Can f 4 вступает в реакцию перекрестной реактивности с одорант-связывающим белком с молекулярной массой 23 кДа, очищенным из перхоти коровы.
Очистка природного Can f 4 привела к очень низкому выходу и из-за дефицита белка в экстракте перхоти, и из-за низкого хроматографического разрешения. Наличие Can f 4 в нескольких пиках в RPC и широкий пик Can f 4 на этапе CIEC показало некоторую степень гетерогенности белка. В то время как возможны несколько объяснений данного поведения, включая модификацию белка, частичную деградацию и наличие изоформ, N-связанное гликозилирование маловероятно, так как последовательность Can f 4 не содержала потенциального сайта для присоединения N-гликана. Среди четырех полученных триптических пептидных последовательностей одна показала отклонение в двух положениях по сравнению с аминокислотной последовательностью, выведенной из кДНК клонов и идентифицированного геномного сегмента. Таким образом, возможно, что варианты изоформ способствовали гетерогенности, наблюдаемой в течение очистки природного белка, даже при том, что доказательства вариабельности последовательности в нуклеотидной последовательности кДНК семи независимых клонов не были получены. Дополнительно, секвенирование триптического фрагмента и анализ MALDI-TOF совместно подтвердили 58% аминокислотной последовательности, выведенной от кДНК клонов, позволяя предложить ограниченный полиморфизм. Даже если нельзя исключить, что, по меньшей мере, один из идентифицированных дополнительных геномных сегментов, связанных с Can f 4, экспрессировался и может дать начало варьирующей форме Can f 4, единственное указание заявителей изобретения на данный аспект - это совпадение двух аминокислотных остатков триптического фрагмента 1, что отклонялось от последовательности кДНК. Никакая другая последовательность пептида, предпочтительно совпадающая с другим сегментом, связанным с Can f 4, и всеми кДНК клонов, не были ясно получены из гена Can f 4, для которого положения нуклеотидов детализированы в настоящей заявке.
Независимо от разновидности изоформ рекомбинантная форма Can f 4 приводила к представленному исключительному биохимическому и иммунологическому соответствию с очищенным природным белком. Два белка, элюированные в точно таком же объеме в аналитической SEC и их связывание с IgE-антителами показали очень высокую корреляцию. Что наиболее важно, ни при каких случаях не наблюдалось связывание IgE с природным, но не с рекомбинантным аллергеном. Фактически, наблюдалось несколько более высокое связывание IgE в экспериментальных тестах, несущих рекомбинантный белок, но это происходило, наиболее вероятно, из-за более низкого, чем оптимальная концентрация, взаимодействия природного белка, вызванного низким выходом очистки.
Важность Can f 4 как аллергена перхоти собаки оценивали путем ImmunoCAP тестирования сывороток 37 индивидов с аллергией на собаку. В недавнем исследовании, включающем ту же самую группу индивидов, заявители изобретения сообщали, что 49% показывали связывание IgE-антител с Can f 1, 22% c Can f 2, 16% c Can f 3 и 70% c Can f 5. В данной работе заявители изобретения обнаружили, что 13 из 37 индивидов (35%) были сенсибилизированы к Can f 4. Таким образом, Can f 4 распознавался в большинстве случаев, чем и rCan f 2, и nCan f 3, которые выступали как минорные аллергены в данной исследуемой группе индивидов. Из 13 Can f 4-реактивных сывороток, одна не показывала связывание IgE ни с одним из других протестированных аллергенов собаки, что позволяет предположить, что Can f 4 может быть релевантным как независимый сенсибилизатор при аллергии на собаку и важным дополнением к компонентной аллергодиагностике. Это понятие подкреплено уникальностью Can f 4 и в последовательности и в связывании IgE по сравнению с Can f 1 и Can f 2. Тот факт, что только один из 44 контролей пыльцевых аллергенов, не имеющих чувствительность к аллергенам собаки, показывал слабый ответ IgE-связывания c Can f 4, позволяет предположить, что распознавание IgE данного аллергена в результате других аэроаллергенных сенсибилизаций происходит нечасто.
По сравнению с экстрактом перхоти собаки экстракт перхоти коровы, как оказалось, содержит намного более высокое количество аллергенов и поэтому дает более удовлетворительные результаты очистки. Судя по анализу экстракта перхоти коровы путем SDS электрофореза в полиакриламидном геле, доминирующие белки в диапазоне молекулярной массы 10-40 кДа были представлены Bos d 2 и представленным здесь белком с молекулярной массой 23 кДа (Bos d 23k), по-видимому, присутствующих в сходном количестве. Bos d 2 был хорошо известен как главный (мажорный) аллерген в перхоти коровы, получен как рекомбинантный аллерген и структурно охарактеризован [18-22]. Напротив, о Bos d 23k намного менее известно, хотя он может быть идентичен белковой полосе с молекулярной массой 22 кДа, показанной иммуноблот-анализом экстракта перхоти коровы, о чем было сообщено ранее [21-23].
Аминокислотная последовательность Bos d 23k, состоящая из 154 остатков, предсказывала молекулярную массу 17,8 кДа, почти точно совпадающую с молекулярной массой Can f 4. Несмотря на данный факт, Bos d 23k показал значительно более медленный уровень миграции, чем Can f 4, при проведении SDS электрофореза в полиакриламидном геле. Поскольку два белка имеют сходство последовательностей и, как можно предположить, обладают сходной третичной структурой, вероятным объяснением наблюдаемого электрофоретического различия могло бы быть различие в гликозилировании. Действительно, анализ последовательностей показывает, что Bos d 23k содержит потенциальный сайт N-гликозилирования в положении остатка аспарагина 45 зрелого белка, тогда как в последовательности Can f 4 не присутствует ни одного подобного сайта.
Среди известных аллергенов перхоти собаки и коровы сывороточный альбумин (Can f 3 и Bos d 6, соответственно) показывает единственную хорошо распознаваемую перекрестную реактивность к аллергенам других видов. Перекрестная реактивность между Can f 4 и Bos d 23k, таким образом, обуславливает новую иммунологическую ассоциацию между аллергенами перхоти собаки и коровы. Учитывая их относительно низкий уровень полной идентичности последовательности, 37%, широкая перекрестная реактивность между двумя белками кажется несколько неожиданной. Однако возможно, что наблюдаемая перекрестная реактивность происходит из-за части белков, имеющих более высокое сходство последовательности, чем белки в целом. В частности, C-концевая часть двух белков значительно консервативна по сегменту из 20 аминокислотных остатков между положением 136/133 и 155/152, показывая 75% идентичности. Несмотря на демонстрируемую перекрестную реактивность между двумя белками, IgE-связывающий ответ был выше к Can f 4, чем к Bos d 23k, для всех протестированных сывороток, подразумевая, что Can f 4, а не Bos d 23k, являлся основным сенсибилизатором в группе индивидов, исследуемой в настоящем описании.
В заключение данная заявка сообщает о клонировании и характеризации аллергена собаки Can f 4. Рекомбинантный Can f 4 будет важным в компонентной аллергодиагностике аллергии на собаку, и его перекрестная реактивность с высоко мажорным белком бычьей перхоти поднимает вопрос о возможности взаимосвязи между аллергией к перхоти собаки и коровы.
Идентификация белкового фрагмента Can f 4
Индивиды с аллергией на собаку: демографические показатели, симптомы аллергии и сенсибилизация к аллергенам собаки*
NA, Северная Америка;
SE, Швеция;
e5, экстракт перхоти собаки;
RC, риноконъюктивит;
A, астма;
U, крапивница
*Аллерген-специфические IgE-антитела в международных килоединицах на литр: менее 0,35 кЕ/л
Специфичность исследования rCan f 4
посредством экспериментального ImmunoCAP теста
Индивиды с аллергией на пыльцу,
не имеющие диагностируемых или описанных симптомов аллергии на собаку:
демографические показатели и сенсибилизация к аллергенам*
DE, Германия;
e5, экстракт перхоти собаки;
t3, экстракт пыльцы березы;
t9, экстракт пыльцы оливы;
g6, экстракт пыльцы тимофеевки луговой;
w21, экстракт пыльцы Parietaria judaica;
w6, экстракт пыльцы рыжика посевного
*Аллерген-специфические IgE-антитела в международных килоединицах на литр; отрицательное: меньше 0,35 кЕ/л
Содержание перечня последовательностей
Ссылки
1. Custovic A, Green R, Taggart SCO, Smith A, Pickering CAC, Chapman MD, et al. Domestic allergens in public places II: dog (Can f 1) and cockroach (Bla g 2) allergens in dust and mite, cat, dog and cockroach allergens in the air in public buildings. Clinical and Experimental Allergy 1996; 26: 1246-52.
2. Spitzauer S. Allergy to mammalian proteins: At the borderline between foreign and self? International Archives of Allergy and Immunology 1999; 120: 259-69.
3. Spitzauer S, Schweiger C, Anrather J, Ebner C, Scheiner O, Kraft D, et al. Characterisation of dog allergens by means of immunoblotting. International Archives of Allergy and Immunology 1993; 100: 60-7.
4. Konieczny A, Morgenstern JP, Bizinkauskas CB, Lilley CH, Brauer AW, Bond JF, et al. The major dog allergens, Can f 1 and Can f 2, are salivary lipocalin proteins: cloning and immunological characterization of the recombinant forms. Immunology 1997; 92: 577-86.
5. Mattsson L, Lundgren T, Everberg H, Larsson H, Lidholm J. Prostatic kallikrein: a new major dog allergen. J Allergy Clin Immunol 2009; 123: 362-8.
6. Yman L, Brandt R, Ponterius G. Serum albumin-an important allergen in dog epithelia extracts. Int Arch Allergy Appl Immunol 1973; 44: 358-68.
7. de Groot H, Goei KGH, van Swieten P, Aalberse RC. Affinity purification of a major and a minor allergen from dog extract: Serologic activity of affinity-purified Can f I and of Can f I-depleted extract. Journal of Allergy and Clinical Immunology 1991; 87: 1056-65.
8. Boutin Y, Hebert H, Vrancken ER, Mourad W. Allergenicity and cross-reactivity of cat and dog allergenic extracts. Clinical Allergy 1988; 18: 287-93.
9. Saarelainen S, Taivainen A, Rytkonen-Nissinen M, Auriola S, Immonen A, Mantyjarvi R, et al. Assessment of recombinant dog allergens Can f 1 and Can f 2 for the diagnosis of dog allergy. Clinical and Experimental Allergy 2004; 34: 1576-82.
10. Moverare R, Everberg H, Carlsson R, Holtz A, Thunberg R, Olsson P, et al. Purification and characterization of the major oak pollen allergen Que a 1 for component-resolved diagnostics using ImmunoCAP(R). International Archives of Allergy and Immunology 2008; 146: 203-11.
11. Shevchenko A, Wilm M, Vorm O, Mann M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Analytical Chemistry 1996; 68: 850-8.
12. Frohman MA. Rapid amplification of complementary DNA ends for generation of full-length complementary DNAs: thermal RACE. Methods in Enzymology 1993; 218: 340-56.
13. Marknell DeWitt A, Niederberger V, Lehtonen P, Spitzauer S, Sperr WR, Valent P, et al. Molecular and immunological characterization of a novel timothy grass (Phleum pratense) pollen allergen, Phl p 11. Clinical and Experimental Allergy 2002; 32: 1329-40.
14. Lindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, Kamal M, et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005; 438: 803-19.
15. Allen G. Sequencing of proteins and peptides. 1 ed. Amsterdam: Elsevier Science Publishers B.V.; 1981.
16. Spinelli S, Ramoni R, Grolli S, Bonicel J, Cambillau C, Tegoni M. The structure of the monomeric porcine odorant binding protein sheds light on the domain swapping mechanism. Biochemistry 1998; 37: 7913-8.
17. Tegoni M, Pelosi P, Vincent F, Spinelli S, Campanacci V, Grolli S, et al. Mammalian odorant binding proteins. Biochim Biophys Acta 2000; 1482: 229-40.
18. Mantyjarvi. Complementary DNA cloning of the predominant allergen of bovine dander: a new member in the lipocalin family. Journal of Allergy and Clinical Immunology 1996; 97: 1297-303.
19. Rouvinen J, Rautiainen J, Virtanen T, Zeiler T, Kauppinen J, Taivainen A, et al. Probing the molecular basis of allergy, three-dimensional structure of the bovine lipocalin allergen Bos d 2. Journal of Biological Chemistry 1999; 274: 2337-43.
20. Virtanen T, Zeiler T, Rautiainen. J, Taivainen A, Pentikaineri J, Rytkonen M, et al. Immune reactivity of cow-asthmatic dairy farmers to the major allergen of cow (BDA20) and to other cow-derived proteins. The use of purified BDA20 increases the performance of diagnostic tests in respiratory cow allergy. Clin Exp Allergy 1996; 26: 188-96.
21. Ylonen J, Mantyjarvi R, Taivainen A, Virtanen T. Comparison of the antigenic and allergenic properties of three types of bovine epithelial material. International Archives of Allergy and Immunology 1992; 99: 112-7.
22. Ylonen J, Mantyjarvi R, Taivainen A, Virtanen T. IgG and IgE antibody responses to cow dander and urine in farmers with cow-induced asthma. Clinical and Experimental Allergy 1992; 22: 83-90.
23. Rautiainen J, Pentikainen J, Rytkonen M, Linnala-Kankkunen A, Pelkonen J, Virtanen T, et al. Molecular analysis of allergenic proteins in bovine dander. Allergy 1996; 51: 378-82.
24. Valenta R, Lidholm J, Niederberger V, Hayek B, Kraft D, Gronlund H. The recombinant allergen-based concept of component-resolved diagnostics and immunotherapy (CRD and CRIT). Clinical & Experimental Allergy 1999; 29: 896-904.
25. Akerstrom B, Flower DR, Salier JP. Lipocalins: unity in diversity. Biochim Biophys Acta. 2000; 1482(1-2): 1-8.
26. Flower DR, North AC, Sansom CE. The lipocalin protein family: structural and sequence overview. Biochim Biophys Acta. 2000; 1482(1-2): 9-24.
27. Gutierrez G, Ganfornina MD, Sanchez D. Evolution of the lipocalin family as inferred from a protein sequence phylogeny. Biochim Biophys Acta. 2000; 1482(1-2): 35-45.
28. Tegoni M, Pelosi P, Vincent F, Spinelli S, Campanacci V, Grolli S, Ramoni R, Cambillau C. Mammalian odorant binding proteins. Biochim Biophys Acta. 2000; 1482(1-2): 229-40.
29. Mantyjarvi R, Rautiainen J, Virtanen T. Lipocalins as allergens. Biochim Biophys Acta. 2000; 1482(1-2): 308-17.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ АЛЛЕРГЕН-ПРОСТАТИЧЕСКИЙ КАЛЛИКРЕИН | 2007 |
|
RU2502074C2 |
| НОВЫЙ АЛЛЕРГЕН | 2011 |
|
RU2608494C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ АЛЛЕРГЕННЫХ БЕЛКОВ И ПЕПТИДОВ | 2009 |
|
RU2519674C2 |
| КАЛИБРОВОЧНЫЙ РЕАГЕНТ И СПОСОБ | 2012 |
|
RU2629310C2 |
| НОВЫЕ АЛЛЕРГЕНЫ ПШЕНИЦЫ | 2008 |
|
RU2502742C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЭКСТРАКТА АЛЛЕРГЕНА | 2011 |
|
RU2572230C2 |
| Способ получения экстракта аллергена | 2011 |
|
RU2706685C2 |
| Ветеринарный продукт | 2018 |
|
RU2819906C2 |
| НОВЫЕ МУТАНТНЫЕ АЛЛЕРГЕНЫ | 2001 |
|
RU2285042C2 |
| СПОСОБЫ ОЧИСТКИ ЭКСТРАКТА АЛЛЕРГЕНА | 2019 |
|
RU2809823C2 |





Настоящее изобретение относится к биотехнологии, конкретно к новым аллергенам собаки, и может быть использовано в медицине. Рекомбинантным путем получают аллерген Can f 4. Полученный аллерген может быть использован в in vitro диагностике или лечении аллергии, вызванной перхотью или шерстью собак. Настоящее изобретение позволяет получить рекомбинантным путем аллерген Can f 4 с полностью установленной аминокислотной последовательностью. 10 н. и 1 з.п. ф-лы, 10 ил., 5 табл., 1 пр.
1. Аллерген Can f 4 с аминокислотной последовательностью SEQ ID NO: 2, полученный рекомбинантными способами.
2. Молекула нуклеиновой кислоты, кодирующая аллерген Can f 4, по п. 1.
3. Молекула нуклеиновой кислоты по п. 2 с последовательностью SEQ ID NO: 1.
4. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 2 или 3.
5. Клетка-хозяин, содержащая вектор по п. 4, продуцирующая аллерген по п. 1.
6. Применение аллергена Can f 4, полученного рекомбинантными способами, по п. 1 в in vitro диагностике аллергии I типа.
7. Способ in vitro диагностики аллергии I типа, предусматривающий стадии
- приведения в контакт образца биологической жидкости пациента с подозрением на наличие аллергии I типа с аллергеном Can f 4 по п. 1; и
- обнаружение наличия в образце IgE-антител, специфически связывающихся с аллергеном Can f 4 по п. 1,
где наличие подобных IgE-антител, специфически связывающихся с вышеуказанным аллергеном Can f 4 по п. 1, свидетельствует о наличии аллергии I типа.
8. Диагностический набор для in vitro диагностики аллергии I типа, отличающийся тем, что содержит аллерген Can f 4 по п. 1.
9. Применение аллергена Can f 4, полученного рекомбинантными способами, по п. 1 в in vitro диагностике аллергии, вызванной перхотью или шерстью собак.
10. Применение аллергена Can f 4, полученного рекомбинантными способами, по п. 1 в терапии аллергии, вызванной перхотью или шерстью собак.
11. Применение аллергена Can f 4, имеющего аминокислотную последовательность SEQ ID NO: 2, для получения варианта или фрагмента аллергена Can f 4, которые имеют общие антигенные детерминанты антител и аллергена Can f 4, или формы вышеуказанного аллергена Can f 4 или ее варианта или фрагмента, которые имеют общие антигенные детерминанты антител и аллергена Can f 4.
| LUNDGREN T | |||
| et al., Cloning and characterization of dog allergen Can f 4, 28th Congress of the European Academy of Allergy and Clinical Immunology, Poland, Warzawa, 2009, p | |||
| Вагонетка для движения по одной колее в обоих направлениях | 1920 |
|
SU179A1 |
| EP 2000531 A1, 10.12.2008 | |||
| SAARLAINEN S | |||
| et al., | |||

