ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка претендует на приоритет предварительной заявки на патент US 61/478474, поданной 22 апреля 2011, предварительной заявки на патент US 61/478899, поданной 25 апреля 2011. Содержание упомянутых выше заявок полностью включено в настоящее описание посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям, содержащим мутантные токсины Clostridium difficile, и способам их применения.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Clostridium difficile (С.difficile) представляет собой грамположительную анаэробную бактерию, связанную с желудочно-кишечным заболеванием у людей. Колонизация С.difficile обычно происходит в толстой кишке, если естественная кишечная флора ослаблена в результате лечения антибиотиками. Инфекция может привести к антибиотикоассоциированной диарее и иногда к псевдомембранозному колиту вследствие секреции гликозилирующих токсинов, токсина А и токсина В (308 и 270 кДа, соответственно), которые являются основными факторами вирулентности С.difficile.
Токсин А и токсин В кодируются в пределах локуса патогенности (PaLoc), длина которого составляет 19 т.п.н., генами tcdA и tcdB соответственно. У непатогенных штаммов С.difficile данный локус заменен альтернативной последовательностью, длина которой составляет 115 пар оснований.
Оба токсина, токсин А и токсин В, являются сильными цитотоксинами. Эти белки представляют собой гомологичные глюкозилтрансферазы, которые инактивируют малые ГТФазы семейства Rho/Rac/Ras. В результате прерывание в передаче сигнала вызывает утрату межклеточных контактов, нарушение регуляции актинового цитоскелета и/или апоптоз, приводя в результате к выраженной секреторной диарее, обусловленной инфекциями Clostridium difficile (CDI).
За последнее десятилетие резко возросли число случаев и тяжесть вспышек эпидемии С.difficile в больницах, центрах сестринского ухода и других лечебных учреждениях для хронических больных. Ключевые факторы этого возрастания включают возникновение гипервирулентных патогенных штаммов, усиленное применение антибиотиков, улучшенные способы обнаружения и повышенное воздействие воздушно-капельных спор в лечебных учреждениях.
Метронидазол и ванкомицин представляют принятый в настоящее время стандарт лечения антибиотиками диареи, вызванной С.difficile (CDAD; С.difficile associated disease). Однако приблизительно у 20% пациентов, получающих такое лечение, происходит рецидив инфекции после первого эпизода CDI, и вплоть до приблизительно 50% этих пациентов страдают дополнительными рецидивами. Лечение этих рецидивов является достаточно большой проблемой, и большинство рецидивов обычно встречается в течение месяца после предыдущего эпизода.
Соответственно, существует необходимость в иммуногенных и/или терапевтических композициях и способах их применения, направленных на С.difficile.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Эти и другие цели достигнуты настоящим изобретениям.
В одном аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин А С.difficile. Этот мутантный токсин А С.difficile включает глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию, по сравнению с соответствующим токсином А С.difficile дикого типа. В одном воплощении, по меньшей мере одна аминокислота мутантного токсина А С.difficile поперечно-сшита химическим образом.
В одном аспекте изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, описанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и где данный полипептид включает по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS).
В одном воплощении, по меньшей мере одна аминокислота мутантного токсина С.difficile поперечно-сшита химическим образом.
В одном воплощении, по меньшей мере одна аминокислота поперечно-сшита химическим образом с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC), N-гидроксисукцината или комбинации EDC и NHS.
В одном воплощении, иммуногенная композиция распознается соответствующим нейтрализующим антителом против токсина или его связывающим фрагментом.
В одном воплощении иммуногенная композиция проявляет пониженную цитотоксичность по сравнению с соответствующим токсином С.difficile дикого типа.
В другом аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает глюкозилтрансферазный домен, имеющий последовательность SEQ ID NO:29, которая имеет аминокислотные замены в положении 285 и 287, и цистеинпротеазный домен, имеющий последовательность SEQ ID NO:32, которая имеет аминокислотную замену в положении 158, по сравнению с соответствующим токсином А С.difficile дикого типа, где по меньшей мере одна аминокислота мутантного токсина А С.difficile поперечно-сшита химическим образом.
В следующем аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, где по меньшей мере одна аминокислота мутантного токсина А С.difficile поперечно-сшита химическим образом.
Еще в одном другом аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 или SEQ ID NO:8.
В одном аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин В С.difficile. Этот мутантный токсин В С.difficile включает глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен; имеющий по меньшей мере одну мутацию по сравнению с соответствующим токсином В С.difficile дикого типа.
В другом аспекте изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, изложенную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и где данный полипептид включает боковую цепь аминокислоты, химически модифицированную 1-этил-3-(3-диметиламинопропил)карбодиимидом (EDC) и N-гидроксисукцинимидом (NHS).
В другом аспекте, изобретение относится к иммуногенной композиции, включающей мутантный токсин В С.difficile, который включает глюкозилтрансферазный домен, имеющий последовательность SEQ ID NO:31, которая имеет аминокислотные замены в положении 286 и 288, и цистеинпротеазный домен, имеющий последовательность SEQ ID NO:33, которая имеет аминокислотную замену в положении 155, по сравнению с соответствующим токсином В С.difficile дикого типа, где по меньшей мере одна аминокислота мутантного токсина В С.difficile поперечно-сшита химическим образом.
В следующем аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота мутантного токсина В С.difficile поперечно-сшита химическим образом.
В одном аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, и мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита химическим образом.
В следующих аспектах изобретение относится к рекомбинантной клетке или потомству этой клетки, которая включает полинуклеотид, кодирующий любой из вышеуказанных мутантных токсинов С.difficile, причем в этой клетке отсутствует эндогенный полинуклеотид, кодирующий токсин.
В другом аспекте изобретение относится к антителу или к связывающему фрагменту антитела, который связывается с иммуногенной композицией, включающей мутантный токсин С.difficile.
В одном аспекте изобретение относится к способу лечения инфекции С.difficile у млекопитающего. Указанный способ включает введение млекопитающему иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, и мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита с использованием формальдегида.
В другом аспекте способ лечения С.difficile инфекции С.difficile у млекопитающего включает введение млекопитающему иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, и мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)-карбодиимида и/или N-гидроксисукцинимида (NHS).
В одном аспекте изобретение относится к способу индукции иммунного ответа на инфекцию С.difficile у млекопитающего. Данный способ включает введение млекопитающему иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, и мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита с использованием формальдегида.
В другом аспекте способ индукции иммунного ответа на инфекцию С.difficile у млекопитающего включает введение млекопитающему иммуногенной композиции, включающей мутантный токсин А С.difficile, который включает SEQ ID NO:4, и мутантный токсин В С.difficile, который включает SEQ ID NO:6, где по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)-карбодиимида и/или N-гидроксисукцинимида (NHS).
В одном воплощении способы лечения или способы индукции иммунного ответа осуществляют у млекопитающего, нуждающегося в этом.
В одном воплощении способы лечения или способы индукции иммунного ответа включают млекопитающее, имеющее рецидивирующую инфекцию С.difficile.
В одном воплощении способы лечения или способы индукции иммунного ответа включают парентеральное введение композиции.
В одном воплощении способы лечения или способы индукции иммунного ответа включают иммуногенную композицию, дополнительно включающую адъювант.
В одном воплощении адъювант включает гель гидроксида алюминия и CpG олигонуклеотид. В другом воплощении адъювант включает ISCOMATRIX.
В одном воплощении выделенный полипептид включает по меньшей мере одну боковую цепь остатка аспарагиновой кислоты полипептида или по меньшей мере одну боковую цепь остатка глутаминовой кислоты полипептида, химически модифицированную глицином.
В одном воплощении выделенный полипептид включает по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; и по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида.
В одном воплощении выделенный полипептид включает бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида.
В одном воплощении выделенный полипептид включает глициновую группировку, сшитую с боковой цепью остатка аспарагиновой кислоты полипептида или с боковой цепью остатка глутаминовой кислоты полипептида.
В одном воплощении выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:4, в которой остаток метионина в положении 1 возможно отсутствует, и боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой.
В одном воплощении выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:6, в которой остаток метионина в положении 1 возможно отсутствует, и боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой.
В одном воплощении выделенный полипептид включает боковую цепь второго остатка лизина полипептида, сшитую с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
В одном воплощении выделенный полипептид включает боковую цепь остатка аспарагиновой кислоты или боковую цепь остатка глутаминовой кислоты полипептида, сшитую с глициновой группировкой.
В одном воплощении выделенный полипептид имеет значение EC50 по меньшей мере приблизительно 100 мкг/мл.
В одном аспекте иммуногенная композиция включает выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:4, в которой остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и где эти полипептиды имеют по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием 1-этил-3-(3-диметиламинопропил) карбодиимида (EDC) и N-гидроксисукцинимида (NHS).
В одном воплощении полипептид включает по меньшей мере одно из: а) по меньшей мере одной бета-аланиновой группировки, сшитой с боковой цепью остатка лизина полипептида; б) по меньшей мере одной поперечной сшивки между боковой цепью остатка лизина полипептида и боковой цепью остатка аспарагиновой кислоты; и в) по меньшей мере одной поперечной сшивки между боковой цепью остатка лизина полипептида и боковой цепью остатка глутаминовой кислоты.
В одном воплощении выделенный полипептид имеет значение ЕСво по меньшей мере приблизительно 100 мкг/мл.
В одном аспекте иммуногенная композиция включает выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и а) где боковая цепь по меньшей мере одного остатка лизина SEQ ID NO:4 сшита с бета-аланиновой группировкой, и б) где боковая цепь по меньшей мере одного остатка лизина SEQ ID NO:6 сшита с бета-аланиновой группировкой.
В одном воплощении иммуногенная композиция включает боковую цепь второго остатка лизина SEQ ID NO:4, сшитую с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты, и где второй остаток лизина SEQ ID NO:6 сшит с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
В одном воплощении иммуногенная композиция включает боковую цепь остатка аспарагиновой кислоты или боковую цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, описанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, сшитую с глициновой группировкой.
В одном воплощении иммуногенная композиция включает боковую цепь остатка аспарагиновой кислоты или боковую цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, описанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, сшитую с глициновой группировкой.
В одном воплощении выделенный полипептид имеет значение ЕСбо по меньшей мере приблизительно 100 мкг/мл.
В одном аспекте иммуногенная композиция включает выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:84, и выделенный полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID NO:86, где каждый полипептид включает: а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; б) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида; в) бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида; и г) глициновую группировку, сшитую с боковой цепью по меньшей мере одного остатка аспарагиновой кислоты полипептида или с боковой цепью по меньшей мере одного остатка глутаминовой кислоты полипептида.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1А-Н представлено выравнивание последовательности токсина А С.difficile дикого типа из штаммов 630, VPI10463, R20291, CD196, и мутантного токсина А, имеющего последовательность SEQ ID NO:4, с использованием выравнивания CLUSTALW, параметры по умолчанию.
На Фиг.2A-F представлено выравнивание последовательности токсина В С.difficile дикого типа из штаммов 630, VPI10463, R20291, CD196, и мутантного токсина В, имеющего последовательность SEQ ID NO:6, с использованием выравнивания CLUSTALW, параметры по умолчанию.
На Фиг.3 представлен график, показывающий идентификацию штаммов, отрицательных по токсину С.difficile дикого типа. Культуральную среду 13 штаммов С.difficile тестировали с помощью ELISA на токсин А. Как проиллюстрировано, семь штаммов экспрессировало токсин А, и 6 штаммов не экспрессировало его (штаммы 1351, 3232, 7322, 5036, 4811 и VPI11186).
На Фиг.4А и В представлены результаты электрофореза в ДСН-ПААГ, иллюстрирующие, что тройной мутант A (SEQ ID NO:4), двойной мутант В (SEQ ID NO:5) и тройной мутант В (SEQ ID NO:6) не гликозилируют ГТФазы Rac1 или RhoA в анализах гликозилирования in vitro с UDP-14C-глюкозой; тогда как от 10 мкг до 1 нг токсина В дикого типа гликозилирует Rac1.
На Фиг.5 представлены результаты Вестерн-блота, показывающие нейтрализацию активности цистеиновой протеазы в мутантных токсинах А и В (SEQ ID NO:4 и 6, соответственно) по сравнению с наблюдением расщепленных фрагментов токсинов А и В дикого типа (SEQ ID NO:1 и 2, соответственно). См. Пример 13.
На Фиг.6 представлены графики, показывающие, что тройные мутантные токсины А и В (SEQ ID NO:4 и 6, соответственно) проявляют остаточную цитотоксичность при тестировании при высоких концентрациях (например, приблизительно 100 мкг/мл) в результате анализа цитотоксичности in vitro в клетках IMR-90.
На Фиг.7 представлен график, показывающий, что значения ЕСво аналогичны для тройного мутантного токсина В (SEQ ID NO:6) и гепта-мутантного токсина В (SEQ ID NO:8).
На Фиг.8 изображен график, на котором представлены результаты тестов цитотоксичности in vitro, где уровни АТФ (RLU) нанесены на график против возрастающих концентраций тройного мутантного TcdA (SEQ ID NO:4) (верхняя панель) и тройного мутантного TcdB (SEQ ID NO:6) (нижняя панель). Остаточная цитотоксичность мутантных токсинов А и В может быть полностью нейтрализована нейтрализующими антителами, специфичными к мутантному токсину А (верхняя панель - pAb А и mAb А3-25+А60-22) и к мутантному токсину В (нижняя панель - pAb В).
На Фиг.9 представлены изображения морфологии клеток IMR-90 через 72 часа после обработки. На панели А показаны контрольные клетки, обработанные имитатором. На панели В показана морфология клеток после обработки мутантным TcdB (SEQ ID NO:6), инактивированным формалином. На панели С показана морфология клеток после обработки мутантным TcdB (SEQ ID NO:6), инактивированным EDC. На панели D показана морфология клеток после обработки токсином В дикого типа (SEQ ID NO:2). На панели Е показана морфология клеток после обработки тройным мутантным TcdB (SEQ ID NO:6). Подобные результаты наблюдали для обработок TcdA.
На Фиг.10 представлен график, показывающий титры нейтрализующего антитела, описанного в Примере 25 (исследование muCdiff2010-06).
На Фиг.11 представлен график, показывающий титры нейтрализующего антитела, описанного в Примере 26 (исследование muCdiff2010-07).
На Фиг.12 представлен график, показывающий ответы нейтрализующего антитела против токсинов А и В у хомячков после четырех иммунизации, как описано в Примере 27 (исследование ham C.difficile 2010-02)
На Фиг.13 представлен график, показывающий ответы нейтрализующего антитела у хомячков после вакцинации химически инактивированными, генетически мутантными токсинами и токсоидами List Biological, как описано в Примере 27 (исследование ham C.difficile 2010-02).
На Фиг.14 представлены кривые выживаемости трех иммунизированных групп хомячков по сравнению с не иммунизированными контролями, как описано в Примере 28 (исследование ham C.difficile 2010-02, продолжение).
На Фиг.15 представлен график, показывающий относительный ответ нейтрализующего антитела против различных препаратов мутантных токсинов С.difficile у хомячков (исследование ham C.difficile 2010-03), как описано в Примере 29.
На Фиг.16А-В представлены графики, показывающие сильный ответ нейтрализующего антитела против химически инактивированных генетических мутантных токсинов А и В (SEQ ID NO:4 и 6, соответственно) у яванских макак, как описано в Примере 30.
На Фиг.17 представлены аминокислотные последовательности вариабельных областей легких (VL) и тяжелых (HL) цепей А3-25 mAb IgE. Сигнальный пептид - выделен цветом; CDR - выделены курсивом и подчеркнуты; константная область - выделена полужирным шрифтом и подчеркнута (полноразмерная последовательность не показана).
На Фиг.18 представлен график, показывающий титрование индивидуальных антител к токсину А в анализе нейтрализации токсина с использованием уровней АТФ (количественно определяемый в относительных световых единицах - RLU) как индикатора выживаемости клеток. По сравнению с контролем токсина (4×ЕС50) mAb A80-29, А65-33, А60-22 и А3-25 обладали возрастающими нейтрализующими действиями на токсин А с возрастанием концентрации, но не действовали на уровень положительных контрольных антител кролика против токсина A. mAb A50-10, А56-33 и А58-46 не нейтрализовали токсин А. Контроль, представляющий собой только клетки, имел 1-1,5×106 RLU.
На Фиг.19 представлено картирование 8 групп эпитопов mAb к токсину В с помощью BiaCore.
На Фиг.20А-С представлены синергетические нейтрализующие активности комбинаций mAb к токсину А: Добавление различных разведении нейтрализующих антител А60-22, А65-33 и A80-29 к возрастающим концентрациям А3-25 mAb синергетическим путем повышало нейтрализацию токсина А независимо от разведения. Проиллюстрирован контроль RLU только токсина А (4х EC50) (менее 0,3×106), а контроли, представляющие собой только клетки, имели 2-2,5×106 RLU, как изображено на графиках, показанных на Фиг.20 В и Фиг.20С.
На Фиг.21 представлены синергетические нейтрализующие активности mAb к токсину В: Нейтрализация токсина В антителами mAb 8-26, В60-2 и В59-3 проиллюстрирована на Фиг.21А. Нейтрализация токсина В возрастала синергетическим путем после объединения В8-26 с разведениями В59-3 (Фиг.21В).
На Фиг.22 представлены результаты Вестерн-блота, показывающие, что экспрессия ГТФазы Rac1 снижена в экстрактах, обработанных генетическим мутантным токсином В (SEQ ID NO:6) с 24 до 96 часов, но не снижена в экстрактах, обработанных токсином В (SEQ ID NO:2) дикого типа. Результаты блота также показывают, что Rac1 гликозилирован в экстрактах, обработанных токсином В, но не гликозилирован в экстрактах, обработанных генетическим мутантным токсином В.
На Фиг.23А-К представлены графики, на которых показаны результаты тестов цитотоксичности in vitro, в которых уровни АТФ (RLU) нанесены на график против возрастающих концентраций культуральной среды С.difficile и пула сывороток хомячков (■); необработанный токсин (собранная культуральная среда) от соответствующего штамма и пул сывороток хомячка (●); очищенный токсин (коммерческий токсин, полученный от List Biologicals) и пул сывороток хомячка (▲); необработанный токсин (▼), контроль; и очищенный токсин (♦), контроль. Токсины из соответствующих штаммов добавляли к клеткам при значениях 4×ЕС50. На фиг.23 показано, что иммуногенная композиция, включающая мутантный TcdA (SEQ ID NO:4) и мутантный TcdB (SEQ ID NO:6), где мутантные токсины были инактивированы EDC в соответствии, например, с Примером 29, таблицей 15, описанной здесь, индуцировала нейтрализующие антитела, которые проявляли нейтрализующую активность против токсинов по меньшей мере из следующих 16 различных штаммов CDC С.difficile по сравнению с соответствующим контролем, представляющим собой только токсин: 2007886 (Фиг.23А); 2006017 (Фиг.23В); 2007070 (Фиг.23С); 2007302 (Фиг.23D); 2007838 (Фиг.23Е); 2007886 (Фиг.23F); 2009292 (Фиг.23G); 2004013 (Фиг.23Н); 2009141 (Фиг.231); 2005022 (Фиг.23J); 2006376 (Фиг.23К).
На Фиг.24 представлена иллюстрация примерной инактивации EDC/NHS мутантных токсинов С.difficile, приводящей в результате по меньшей мере к трем возможным типам модификаций: поперечным сшивкам, образованию аддуктов глицина и аддуктов бета-аланина. Панель А иллюстрирует поперечное сшивание. Карбоксильные остатки тройных мутантных токсинов инактивируют добавлением EDC и NHS. Активированные эфиры взаимодействуют с первичными аминами с образованием стабильных амидных связей, приводящих в результате к внутри- и межмолекулярным поперечным сшивкам. Панель В иллюстрирует образование аддуктов глицина. После инактивации остаточные активированные сложные эфиры гасят добавлением глицина с образованием стабильных амидных связей. Панель С иллюстрирует образование аддуктов бета-аланина. Три моль NHS может взаимодействовать с одним моль EDC с образованием активированного бета-аланина. Затем он взаимодействует с первичными аминами с образованием стабильных амидных связей.
На Фиг.25 представлена иллюстрация примерной инактивации EDC/NHS мутантных токсинов С.difficile, приводящей в результате по меньшей мере к одному из следующих типов модификаций: поперечные сшивки, аддукты глицина и аддукты бета-аланина.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1 описывает аминокислотную последовательность токсина А С.difficile 630 (TcdA) дикого типа.
SEQ ID NO:2 описывает аминокислотную последовательность токсина В С.difficile 630 (TcdB) дикого типа.
SEQ ID NO:3 описывает аминокислотную последовательность мутантного TcdA, имеющего мутацию в положениях 285 и 287 по сравнению с SEQ ID NO:1.
SEQ ID NO:4 описывает аминокислотную последовательность мутантного TcdA, имеющего мутацию в положениях 285, 287 и 700 по сравнению с SEQ ID NO:1.
SEQ ID NO:5 описывает аминокислотную последовательность мутантного TcdB, имеющего мутацию в положениях 286 и 288 по сравнению с SEQ ID NO:2.
SEQ ID NO:6 описывает аминокислотную последовательность мутантного TcdB, имеющего мутацию в положениях 286, 288 и 698 по сравнению с SEQ ID NO:2.
SEQ ID NO:7 описывает аминокислотную последовательность мутантного TcdA, имеющего мутацию в положениях 269, 272, 285, 287, 460, 462 и 700 по сравнению с SEQ ID NO:1
SEQ ID NO:8 описывает аминокислотную последовательность мутантного TcdB, имеющего мутацию в положениях 270, 273, 286, 288, 461, 463 и 698 по сравнению с SEQ ID NO:2
SEQ ID NO:9 описывает последовательность ДНК, кодирующую токсин А С.difficile 630 (TcdA) дикого типа.
SEQ ID NO:10 описывает последовательность ДНК, кодирующую токсин В С.difficile 630 (TcdB) дикого типа.
SEQ ID NO:11 описывает последовательность ДНК, кодирующую SEQ ID NO:3.
SEQ ID NO:12 описывает последовательность ДНК, кодирующую SEQ ID NO:4.
SEQ ID NO:13 описывает последовательность ДНК, кодирующую SEQ ID NO:5.
SEQ ID NO:14 описывает последовательность ДНК, кодирующую SEQ ID NO:6.
SEQ ID NO:15 описывает аминокислотную последовательность TcdA С.difficile R20291 дикого типа.
SEQ ID NO:16 описывает последовательность ДНК, кодирующую SEQ ID NO: 15.
SEQ ID NO:17 описывает аминокислотную последовательность TcdA С.difficile CD196 дикого типа.
SEQ ID NO:18 описывает последовательность ДНК, кодирующую SEQ ID NO: 17.
SEQ ID NO:19 описывает аминокислотную последовательность TcdA С.difficile VPI10463 дикого типа.
SEQ ID NO:20 описывает последовательность ДНК, кодирующую SEQ ID NO: 19.
SEQ ID NO:21 описывает аминокислотную последовательность TcdB С.difficile R20291 дикого типа.
SEQ ID NO:22 описывает последовательность ДНК, кодирующую SEQ ID NO:21.
SEQ ID NO:23 описывает аминокислотную последовательность TcdB С.difficile CD196 дикого типа.
SEQ ID NO:24 описывает последовательность ДНК, кодирующую SEQ ID NO: 23.
SEQ ID NO:25 описывает аминокислотную последовательность TcdB С.difficile VPI10463 дикого типа.
SEQ ID NO:26 описывает последовательность ДНК, кодирующую SEQ ID NO: 25.
SEQ ID NO:27 описывает последовательность ДНК локуса патогенности С.difficile VPI10463 дикого типа.
SEQ ID NO:28 описывает аминокислотную последовательность для остатков 101-293 SEQ ID NO:1.
SEQ ID NO:29 описывает аминокислотную последовательность для остатков 1-542 SEQ ID NO:1.
SEQ ID NO:30 описывает аминокислотную последовательность для остатков 101-293 SEQ ID NO:2.
SEQ ID NO:31 описывает аминокислотную последовательность для остатков 1-543 SEQ ID NO:2.
SEQ ID NO:32 описывает аминокислотную последовательность для остатков 543-809 SEQ ID NO:1.
SEQ ID NO:33 описывает аминокислотную последовательность для остатков 544-767 SEQ ID NO; 2.
SEQ ID NO:34 описывает аминокислотную последовательность мутантного TcdA, в которой остатки 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 могут представлять собой любую аминокислоту.
SEQ ID NO:35 описывает аминокислотную последовательность мутантного TcdB, в которой остатки 102, 270, 273, 286, 288, 384, 461, 463, 520, 543, 544, 587, 600, 653, 698 и 751 могут представлять собой любую аминокислоту.
SEQ ID NO:36 описывает аминокислотную последовательность вариабельной легкой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:37 описывает аминокислотную последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:38 описывает аминокислотную последовательность CDR1 вариабельной легкой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:39 описывает аминокислотную последовательность CDR2 вариабельной легкой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:40 описывает аминокислотную последовательность CDR3 вариабельной легкой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:41 описывает аминокислотную последовательность CDR1 вариабельной тяжелой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:42 описывает аминокислотную последовательность CDR2 вариабельной тяжелой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:43 описывает аминокислотную последовательность CDR3 вариабельной тяжелой цепи нейтрализующего антитела к TcdA С.difficile (A3-25 mAb).
SEQ ID NO:44 описывает последовательность ДНК, кодирующую SEQ ID NO:3.
SEQ ID NO:45 описывает последовательность ДНК, кодирующую SEQ ID NO:4.
SEQ ID NO:46 описывает последовательность ДНК, кодирующую SEQ ID NO: 5.
SEQ ID NO:47 описывает последовательность ДНК, кодирующую SEQ ID NO:6.
SEQ ID NO:48 описывает нуклеотидную последовательность иммуностимуляторного олигонуклеотида ODN CpG 24555.
SEQ ID NO:49 описывает аминокислотную последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В8-26 mAb).
SEQ ID NO:50 описывает аминокислотную последовательность сигнального пептида вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:51 описывает аминокислотную последовательность CDR1 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:52 описывает аминокислотную последовательность CDR2 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:53 описывает аминокислотную последовательность CDR3 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:54 описывает аминокислотную последовательность константной области вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:55 описывает аминокислотную последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:56 описывает аминокислотную последовательность сигнального пептида вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:57 описывает аминокислотную последовательность CDR1 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:58 описывает аминокислотную последовательность CDR2 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:59 описывает аминокислотную последовательность CDR3 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb).
SEQ ID NO:60 описывает аминокислотную последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В59-3 mAb).
SEQ ID NO:61 описывает аминокислотную последовательность сигнального пептида вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:62 описывает аминокислотную последовательность CDR1 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В59-3 mAb).
SEQ ID NO:63 описывает аминокислотную последовательность CDR2 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В59-3 mAb).
SEQ ID NO:64 описывает аминокислотную последовательность CDR3 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В59-3 mAb).
SEQ ID NO:65 описывает аминокислотную последовательность константной области вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:66 описывает аминокислотную последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:67 описывает аминокислотную последовательность сигнального пептида вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:68 описывает аминокислотную последовательность CDR1 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:69 описывает аминокислотную последовательность CDR2 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:70 описывает аминокислотную последовательность CDR3 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb).
SEQ ID NO:71 описывает аминокислотную последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (В9-30 mAb).
SEQ ID NO:72 описывает аминокислотную последовательность сигнального пептида вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:73 описывает аминокислотную последовательность CDR1 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:74 описывает аминокислотную последовательность CDR2 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:75 описывает аминокислотную последовательность CDR3 вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:76 описывает аминокислотную последовательность константной области вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:77 описывает аминокислотную последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:78 описывает аминокислотную последовательность сигнального пептида вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:79 описывает аминокислотную последовательность CDR1 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:80 описывает аминокислотную последовательность CDR2 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:81 описывает аминокислотную последовательность CDR3 вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb).
SEQ ID NO:82 описывает аминокислотную последовательность мутантного TcdB, где остаток в положении 102, 270, 273, 286, 288, 384, 461, 463, 520, 543, 544, 587, 600, 653, 698 и 751 может представлять собой любую аминокислоту.
SEQ ID NO:83 описывает аминокислотную последовательность мутантного TcdA, имеющего мутацию в положениях 269, 272, 285, 287, 460, 462 и 700 по сравнению с SEQ ID NO:1, в которой метионин в положении 1 отсутствует.
SEQ ID NO:84 описывает аминокислотную последовательность мутантного токсина А С.difficile, имеющего мутацию в положениях 285, 287 и 700 по сравнению с SEQ ID NO:1, в которой метионин в положении 1 отсутствует.
SEQ ID NO:85 описывает аминокислотную последовательность мутантного токсина В С.difficile, имеющего мутацию в положениях 270, 273, 286, 288, 461, 463 и 698 по сравнению с SEQ ID NO:2, в которой метионин в положении 1 отсутствует.
SEQ ID NO:86 описывает аминокислотную последовательность мутантного токсина В С.difficile, имеющего мутацию в положениях 286, 288 и 698 по сравнению с SEQ ID NO:2, в которой метионин в положении 1 отсутствует.
SEQ ID NO:87 описывает аминокислотную последовательность TcdA С.difficile 2004013 дикого типа.
SEQ ID NO:88 описывает аминокислотную последовательность TcdA С.difficile 2004111 дикого типа.
SEQ ID NO:89 описывает аминокислотную последовательность TcdA С.difficile 2004118 дикого типа.
SEQ ID NO:90 описывает аминокислотную последовательность TcdA С.difficile 2004205 дикого типа.
SEQ ID NO:91 описывает аминокислотную последовательность TcdA С.difficile 2004206 дикого типа.
SEQ ID NO:92 описывает аминокислотную последовательность TcdA С.difficile 2005022 дикого типа.
SEQ ID NO:93 описывает аминокислотную последовательность TcdA С.difficile 2005088 дикого типа.
SEQ ID NO:94 описывает аминокислотную последовательность TcdA С.difficile 2005283 дикого типа.
SEQ ID NO:95 описывает аминокислотную последовательность TcdA С.difficile 2005325 дикого типа.
SEQ ID NO:96 описывает аминокислотную последовательность TcdA С.difficile 2005359 дикого типа.
SEQ ID NO:97 описывает аминокислотную последовательность TcdA С.difficile 2006017 дикого типа.
SEQ ID NO:98 описывает аминокислотную последовательность TcdA С.difficile 2007070 дикого типа.
SEQ ID NO:99 описывает аминокислотную последовательность TcdA С.difficile 2007217 дикого типа.
SEQ ID NO:100 описывает аминокислотную последовательность TcdA С.difficile 2007302 дикого типа.
SEQ ID NO:101 описывает аминокислотную последовательность TcdA С.difficile 2007816 дикого типа.
SEQ ID NO:102 описывает аминокислотную последовательность TcdA С.difficile 2007838 дикого типа.
SEQ ID NO:103 описывает аминокислотную последовательность TcdA С.difficile 2007858 дикого типа.
SEQ ID NO:104 описывает аминокислотную последовательность TcdA С.difficile 2007886 дикого типа.
SEQ ID NO:105 описывает аминокислотную последовательность TcdA С.difficile 2008222 дикого типа.
SEQ ID NO:106 описывает аминокислотную последовательность TcdA С.difficile 2009078 дикого типа.
SEQ ID NO:107 описывает аминокислотную последовательность TcdA С.difficile 2009087 дикого типа.
SEQ ID NO:108 описывает аминокислотную последовательность TcdA С.difficile 2009141 дикого типа.
SEQ ID NO:109 описывает аминокислотную последовательность TcdA С.difficile 2009292 дикого типа.
SEQ ID NO:110 описывает аминокислотную последовательность TcdB С.difficile 2004013 дикого типа.
SEQ ID NO:111 описывает аминокислотную последовательность TcdB С.difficile 2004111 дикого типа.
SEQ ID NO:112 описывает аминокислотную последовательность TcdB С.difficile 2004118 дикого типа.
SEQ ID NO:113 описывает аминокислотную последовательность TcdB С.difficile 2004205 дикого типа.
SEQ ID NO:114 описывает аминокислотную последовательность TcdB С.difficile 2004206 дикого типа.
SEQ ID NO:115 описывает аминокислотную последовательность TcdB С.difficile 2005022 дикого типа.
SEQ ID NO:116 описывает аминокислотную последовательность TcdB С.difficile 2005088 дикого типа.
SEQ ID NO:117 описывает аминокислотную последовательность TcdB С.difficile 2005283 дикого типа.
SEQ ID NO:118 описывает аминокислотную последовательность TcdB С.difficile 2005325 дикого типа.
SEQ ID NO:119 описывает аминокислотную последовательность TcdB С.difficile 2005359 дикого типа.
SEQ ID NO:120 описывает аминокислотную последовательность TcdB С.difficile 2006017 дикого типа.
SEQ ID NO:121 описывает аминокислотную последовательность TcdB С.difficile 2006376 дикого типа.
SEQ ID NO:122 описывает аминокислотную последовательность TcdB С.difficile 2007070 дикого типа.
SEQ ID NO:123 описывает аминокислотную последовательность TcdB С.difficile 2007217 дикого типа.
SEQ ID NO:124 описывает аминокислотную последовательность TcdB С.difficile 2007302 дикого типа.
SEQ ID NO:125 описывает аминокислотную последовательность TcdB С.difficile 2007816 дикого типа.
SEQ ID NO:126 описывает аминокислотную последовательность TcdB С.difficile 2007838 дикого типа.
SEQ ID NO:127 описывает аминокислотную последовательность TcdB С.difficile 2007858 дикого типа.
SEQ ID NO:128 описывает аминокислотную последовательность TcdB С.difficile 2007886 дикого типа.
SEQ ID NO:129 описывает аминокислотную последовательность TcdB С.difficile 2008222 дикого типа.
SEQ ID NO:130 описывает аминокислотную последовательность TcdB С.difficile 2009078 дикого типа.
SEQ ID NO:131 описывает аминокислотную последовательность TcdB С.difficile 2009087 дикого типа.
SEQ ID NO:132 описывает аминокислотную последовательность TcdB С.difficile 2009141 дикого типа.
SEQ ID NO:133 описывает аминокислотную последовательность TcdB С.difficile 2009292 дикого типа.
SEQ ID NO:134 описывает аминокислотную последовательность TcdA С.difficile 014 дикого типа.
SEQ ID NO:135 описывает аминокислотную последовательность TcdA С.difficile 015 дикого типа.
SEQ ID NO:136 описывает аминокислотную последовательность TcdA С.difficile 020 дикого типа.
SEQ ID NO:137 описывает аминокислотную последовательность TcdA С.difficile 023 дикого типа.
SEQ ID NO:138 описывает аминокислотную последовательность TcdA С.difficile 027 дикого типа.
SEQ ID NO:139 описывает аминокислотную последовательность TcdA С.difficile 029 дикого типа.
SEQ ID NO:140 описывает аминокислотную последовательность TcdA С.difficile 046 дикого типа.
SEQ ID NO:141 описывает аминокислотную последовательность TcdB С.difficile 014 дикого типа.
SEQ ID NO:142 описывает аминокислотную последовательность TcdB С.difficile 015 дикого типа.
SEQ ID NO:143 описывает аминокислотную последовательность TcdB С.difficile 020 дикого типа.
SEQ ID NO:144 описывает аминокислотную последовательность TcdB С.difficile 023 дикого типа.
SEQ ID NO:145 описывает аминокислотную последовательность TcdB С.difficile 027 дикого типа.
SEQ ID NO:146 описывает аминокислотную последовательность TcdB С.difficile 029 дикого типа.
SEQ ID NO:147 описывает аминокислотную последовательность TcdB С.difficile 046 дикого типа.
SEQ ID NO:148 описывает аминокислотную последовательность TcdA С.difficile 001 дикого типа.
SEQ ID NO:149 описывает аминокислотную последовательность TcdA С.difficile 002 дикого типа.
SEQ ID NO:150 описывает аминокислотную последовательность TcdA С.difficile 003 дикого типа.
SEQ ID NO:151 описывает аминокислотную последовательность TcdA С.difficile 004 дикого типа.
SEQ ID NO:152 описывает аминокислотную последовательность TcdA С.difficile 070 дикого типа.
SEQ ID NO:153 описывает аминокислотную последовательность TcdA С.difficile 075 дикого типа.
SEQ ID NO:154 описывает аминокислотную последовательность TcdA С.difficile 077 дикого типа.
SEQ ID NO:155 описывает аминокислотную последовательность TcdA С.difficile 081 дикого типа.
SEQ ID NO:156 описывает аминокислотную последовательность TcdA С.difficile 117 дикого типа.
SEQ ID NO:157 описывает аминокислотную последовательность TcdA С.difficile 131 дикого типа.
SEQ ID NO:158 описывает аминокислотную последовательность TcdB С.difficile 001 дикого типа.
SEQ ID NO:159 описывает аминокислотную последовательность TcdB С.difficile 002 дикого типа.
SEQ ID NO:160 описывает аминокислотную последовательность TcdB С.difficile 003 дикого типа.
SEQ ID NO:161 описывает аминокислотную последовательность TcdB С.difficile 004 дикого типа.
SEQ ID NO:162 описывает аминокислотную последовательность TcdB С.difficile 070 дикого типа.
SEQ ID NO:163 описывает аминокислотную последовательность TcdB С.difficile 075 дикого типа.
SEQ ID NO:164 описывает аминокислотную последовательность TcdB С.difficile 077 дикого типа.
SEQ ID NO:165 описывает аминокислотную последовательность TcdB С.difficile 081 дикого типа.
SEQ ID NO:166 описывает аминокислотную последовательность TcdB С.difficile 117 дикого типа.
SEQ ID NO:167 описывает аминокислотную последовательность TcdB С.difficile 131 дикого типа.
SEQ ID NO:168 описывает аминокислотную последовательность TcdA С.difficile 053 дикого типа.
SEQ ID NO:169 описывает аминокислотную последовательность TcdA С.difficile 078 дикого типа.
SEQ ID NO:170 описывает аминокислотную последовательность TcdA С.difficile 087 дикого типа.
SEQ ID NO:171 описывает аминокислотную последовательность TcdA С.difficile 095 дикого типа.
SEQ ID NO:172 описывает аминокислотную последовательность TcdA С.difficile 126 дикого типа.
SEQ ID NO:173 описывает аминокислотную последовательность TcdB С.difficile 053 дикого типа.
SEQ ID NO:174 описывает аминокислотную последовательность TcdB С.difficile 078 дикого типа.
SEQ ID NO:175 описывает аминокислотную последовательность TcdB С.difficile 087 дикого типа.
SEQ ID NO:176 описывает аминокислотную последовательность TcdB С.difficile 095 дикого типа.
SEQ ID NO:177 описывает аминокислотную последовательность TcdB С.difficile 126 дикого типа.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы изобретения неожиданно обнаружили мутантный токсин А и токсин В С.difficile и способы их применения. Эти мутанты частично охарактеризованы как обладающие иммуногенностью и проявляющие пониженную цитотоксичность по сравнению с формой дикого типа соответствующего токсина. Настоящее изобретение также относится к их иммуногенным участкам, к их биологическим эквивалентам и к выделенным полинуклеотидам, которые включают нуклеиново-кислотные последовательности, кодирующие любое из указанного выше.
Иммуногенные композиции, описанные здесь, неожиданно показали способность вызывать образование новых нейтрализующих антител против токсинов С.difficile, и они могут быть способны придавать активную и/или пассивную защиту против провокации С.difficile. Новые антитела направлены против различных эпитопов токсина А и токсина В. Кроме того, авторы изобретения обнаружили, что комбинация по меньшей мере двух из нейтрализующих моноклональных антител может неожиданно проявлять синергетический эффект при соответствующей нейтрализации in vitro токсина А и токсина В.
Композиции по изобретению, описанные здесь, можно применять для лечения, предупреждения, снижения риска, снижения частоты встречаемости, снижения тяжести и/или замедления начала инфекции С.difficile, заболевания, обусловленного С.difficile (CDAD), его синдрома, состояния, симптома и/или осложнения у млекопитающего по сравнению с млекопитающим, которому не вводили данную композицию.
Кроме того, авторы изобретения открыли рекомбинантную аспорогенную клетку С.difficile, которая может стабильно экспрессировать мутантный токсин А и токсин В С.difficile, и новые способы ее получения.
Иммуногенные композиции
В одном аспекте, изобретение относится к иммуногенной композиции, включающей мутантный токсин С.difficile. Этот мутантный токсин С.difficile включает аминокислотную последовательность, имеющую по меньшей мере одну мутацию в глюкозилтрансферазном домене и по меньшей мере одну мутацию в цистеинпротеазном домене относительно соответствующего токсина С.difficile дикого типа.
Термин "дикий тип", как используют здесь, относится к форме, обнаруживаемой в природе. Например, полипептидная или полинуклеотидная последовательность дикого типа представляет собой последовательность, находящуюся в организме, которая может быть выделена из источника в природе, и которая целенаправленно не модифицирована с помощью манипуляций человека. Настоящее изобретение также относится к выделенным полинуклеотидам, которые включают нуклеиново-кислотные последовательности, кодирующие любое из указанного выше. Кроме того, настоящее изобретение относится к применению любой из вышеуказанных композиций для лечения, предупреждения, снижения риска, снижения частоты встречаемости, снижения тяжести и/или замедления начала инфекции С.difficile, заболевания, обусловленного С.difficile (CDAD), его синдрома, состояния, симптома и/или осложнения у млекопитающего по сравнению с млекопитающим, которому не вводят данную композицию, а также к способам получения таких композиций.
При использовании здесь, "иммуногенная композиция" или "иммуноген" относятся к композиции, которая вызывает иммунный ответ у млекопитающего, которому вводят эту композицию.
"Иммунный ответ" относится к развитию полезного гуморального (опосредованного антителами) и/или клеточного (опосредованного антигенспецифичными Т клетками или продуктами их секреции) ответа, направленного против токсина С.difficile у пациента-реципиента. Иммунный ответ может представлять собой гуморальный, клеточный или оба типа ответа.
Иммунный ответ может представлять собой активный ответ, индуцированный введением иммуногенной композиции, иммуногена. Альтернативно, иммунный ответ может представлять собой пассивный ответ, индуцированный введением антитела или примированных Т-клеток.
Наличие гуморального (антитело-опосредованного) иммунного ответа можно определить, например, с помощью анализов на основе клеток, известных в данной области техники, таких как анализ нейтрализующего антитела, ELISA (твердофазный иммуноферментный анализ) и т.д.
Клеточный иммунный ответ в характерном случае вызван презентированием полипептидных эпитопов в ассоциации с молекулами ГКГ (главного комплекса гистосовместимости) класса 1 или класса II, активирующим антигенспецифичные CD4+ Т-хелперные клетки и/или CD8+ цитотоксические Т-клетки. В этот ответ может быть также вовлечена активация моноцитов, макрофагов, клеток NK (естественных киллеров), базофилов, дендритных клеток, астроцитов, клеток микроглии, эозинофилов или других компонентов врожденного иммунитета. Наличие клеточно-опосредованного иммунологического ответа можно определить с помощью анализов пролиферации (CD4+ Т клеток) или анализов CTL (цитотоксических Т-лимфоцитов), известных в данной области техники.
В одном воплощении иммуногенная композиция представляет собой вакцинную композицию. При использовании здесь, "вакцинная композиция" представляет собой композицию, которая вызывает иммунный ответ у млекопитающего, которому вводят эту композицию. Вакцинная композиция может защищать иммунизированное млекопитающее против последующей провокации иммунизирующим агентом или иммунологически перекрестно-реактивным агентом. Эта защита может быть полной или частичной в отношении снижения симптомов или инфекции по сравнению невакцинированным млекопитающим в тех же условиях.
Иммуногенные композиции, описанные здесь, являются перекрестно-реактивными, что означает, что они обладают такими характеристиками, что способны вызывать эффективный иммунный ответ (например, гуморальный иммунный ответ) против токсина, продуцируемого другим штаммом С.difficile, отличающимся от штамма, от которого имеет происхождение композиция. Например, иммуногенные композиции (например, имеющие происхождение от С.difficile 630), описанные здесь, могут вызывать перекрестно-реактивные антитела, которые могут связываться с токсинами, продуцируемыми множественными штаммами С.difficile (например, с токсинами, продуцируемыми С.difficile R20291 и VPI10463). См., например, Пример 37. Перекрестная реактивность является показателем потенциала перекрестной защиты бактериального иммуногена, и наоборот.
Термин "перекрестно-реактивный", при использовании здесь, относится к способности иммунного ответа, индуцированного иммуногенной композицией, предотвращать или ослаблять инфекцию различными штаммами или видами бактерий одного и того же рода. Например, иммуногенная композиция (например, имеющая происхождение от С.difficile 630), описанная здесь, может индуцировать у млекопитающего эффективный иммунный ответ, ослабляющий инфекцию С.difficile и/или ослабляющий заболевание, обусловленное С.difficile, вызванное штаммом, отличающимся от 630 (например, С.difficile R20291).
Иллюстративные млекопитающие, у которых иммуногенная композиция или иммуноген вызывает иммунный ответ, включают любых млекопитающих, таких как, например, мыши, хомячки, приматы и люди. В предпочтительном воплощении иммуногенная композиция или иммуноген вызывает иммунный ответ у человека, которому вводят эту композицию.
Как описано выше, токсин A (TcdA) и токсин В (TcdB) представляют собой гомологичные глюкозилтрансферазы, которые инактивируют малые ГТФазы семейства Rho/Rac/Ras. Действие TcdA и TcdB на клетки-мишени млекопитающих зависит от многоступенчатого механизма рецептор-опосредованного эндоцитоза, мембранной транслокации, аутопротеолитического процессинга и моногликозилирования ГТФаз. Многие из этих функциональных активностей приписаны дискретным участкам в пределах первичной последовательности токсинов, и токсины визуализированы, чтобы показать, что их молекулы подобны по структуре.
Ген TcdA дикого типа имеет приблизительно 8130 нуклеотидов, которые кодируют белок, имеющий выведенную молекулярную массу, составляющую примерно 308 кДа, и включающий приблизительно 2710 аминокислот. При использовании здесь, TcdA С.difficile дикого типа включает TcdA С.difficile из любого штамма С.difficile дикого типа. TcdA С.difficile дикого типа может включать аминокислотную последовательность TcdA С.difficile дикого типа, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью SEQ ID NO:1 (полноразмерной) при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT при использовании параметров массы гэпов по умолчанию.
В предпочтительном воплощении TcdA С.difficile дикого типа включает аминокислотную последовательность, описанную в SEQ ID NO:1, которая описывает аминокислотную последовательность TcdA дикого типа из штамма 630 С.difficile (также раскрытую в GenBank, номер доступа YP_001087137.1 и/или CAJ67494.1). Штамм 630 С.difficile известен в данной области техники как представляющий собой штамм ПЦР-риботипа 012. SEQ ID NO:9 описывает ген TcdA дикого типа из штамма 630 С.difficile, который также раскрыт в GenBank, номер доступа МС_009089.1.
Другой пример TcdA С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:15, которая описывает аминокислотную последовательность TcdA дикого типа из штамма R20291 С.difficile (также раскрытую в GenBank, номер доступа YP_003217088.1). Штамм R20291 С.difficile известен в данной области техники как гипервирулентный штамм и штамм ПЦР-риботипа 027. Аминокислотная последовательность TcdA из штамма R20291 С.difficile обладает примерно 98% идентичностью с SEQ ID NO:1. SEQ ID NO:16 описывает ген TcdA дикого типа из штамма R20291 С.difficile, который также раскрыт в GenBank, номер доступа NC_013316.1.
Следующий пример TcdA С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:17, которая описывает аминокислотную последовательность TcdA дикого типа из штамма CD196 С.difficile (также раскрытую в GenBank, номер доступа СВА61156.1). CD196 представляет собой штамм из недавней вспышки эпидемии в Канаде, и известен в данной области техники как штамм ПЦР-риботипа 027. Аминокислотная последовательность TcdA из штамма CD196 С. difficile обладает примерно 98% идентичностью SEQ ID NO:1 и обладает примерно 100% идентичностью TcdA из штамма R20291 С.difficile. SEQ ID NO:18 описывает ген TcdA дикого типа из штамма CD196 С.difficile, который также раскрыт в GenBank, номер доступа FN538970.1.
Следующий пример TcdA С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:19, которая описывает аминокислотную последовательность TcdA дикого типа из штамма VP110463 С.difficile (также раскрытую в GenBank, номер доступа САА63564.1). Аминокислотная последовательность TcdA из штамма VP110463 С.difficile обладает примерно 100% (99,8%) идентичностью SEQ ID NO:1. SEQ ID NO:20 описывает ген TcdA дикого типа из штамма VP110463 С.difficile, который также раскрыт в GenBank, номер доступа Х92982.1.
Дополнительные примеры TcdA С.difficile дикого типа включают TcdA из штаммов С.difficile дикого типа, которые могут быть получены из Центров контроля и профилактики заболеваний (CDC (Centers for Disease Control and Prevention) Atlanta, GA). Авторы изобретения обнаружили, что аминокислотная последовательность TcdA из штаммов С.difficile дикого типа, которые могут быть получены из CDC, включает по меньшей мере примерно 99,3%-100% идентичность при оптимальном выравнивании аминокислотным остаткам 1-821 SEQ ID NO:1 (TcdA из С.difficile 630). См. таблицу 1.
Авторы изобретения также обнаружили, что аминокислотная последовательность TcdA из штаммов С.difficile дикого типа может включать по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% до примерно 100% идентичность при оптимальном выравнивании (например, при оптимальном выравнивании полноразмерных последовательностей) SEQ ID NO:1.
Таблица 1: Штаммы С.difficile дикого типа, полученные из CDC, и процент идентичности аминокислотных остатков 1-821 TcdA из соответствующего штамма С.difficile дикого типа аминокислотным остаткам 1-821 SEQ ID NO:1 при оптимальном выравнивании.
Соответственно, в одном воплощении аминокислотная последовательность TcdA С.difficile дикого типа включает последовательность по меньшей мере из примерно 500, 600, 700 или 800 непрерывных остатков, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью последовательности равной длины между остатками 1-900 SEQ ID NO:1 при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT, используя параметры массы гэпов по умолчанию. Примеры включают штаммы, описанные выше (например, R20291, CD196 и т.д.), и штаммы, перечисленные в таблице 1.
В другом воплощении аминокислотная последовательность TcdA С.difficile дикого типа включает последовательность, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью любой последовательности, выбранной из SEQ ID NO:87-109 при оптимальном выравнивании. См. таблицу 1-а.
Ген TcdB дикого типа имеет приблизительно 7098 нуклеотидов, которые кодируют белок, имеющий выведенную молекулярную массу, составляющую примерно 270 кДа, и включающий приблизительно 2366 аминокислот. Как используют здесь, TcdB С.difficile дикого типа включает TcdB С.difficile из любого штамма С.difficile дикого типа. TcdB С.difficile дикого типа может включать аминокислотную последовательность дикого типа, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью SEQ ID NO:2 при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT при использовании параметров массы гэпов по умолчанию. В предпочтительном воплощении TcdB С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:2, которая описывает аминокислотную последовательность TcdB из штамма 630 С.difficile (также раскрытую в GenBank, номер доступа YP_001087135.1 и/или CAJ67492). SEQ ID NO:10 описывает ген TcdB дикого типа из штамма 630 С.difficile, который также раскрыт в GenBank, номер доступа NC_009089.1.
Другой пример TcdB С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:21, которая описывает аминокислотную последовательность TcdB дикого типа из штамма R20291 С.difficile (также раскрытую в GenBank, номер доступа YP_003217086.1 и/или СВЕ02479.1). Аминокислотная последовательность TcdB из штамма R20291 С.difficile обладает примерно 92% идентичностью SEQ ID NO:2. SEQ ID NO:22 описывает ген TcdB дикого типа из штамма R20291 С.difficile, который также раскрыт в GenBank, номер доступа NC_013316.1.
Дополнительный пример TcdB С.difficile дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO:23, которая описывает аминокислотную последовательность TcdB дикого типа из штамма С.difficile CD196 (также раскрытую в GenBank, номер доступа YP_003213639.1 и/или СВА61153.1). SEQ ID NO:24 описывает ген TcdB дикого типа из штамма CD196 С.difficile, который также раскрыт в GenBank, номер доступа NC_013315.1. Аминокислотная последовательность TcdB из штамма CD196 С.difficile обладает примерно 92% идентичностью SEQ ID NO:2.
Следующий пример аминокислотной последовательности TcdB С.difficile дикого типа включает SEQ ID NO:25, которая описывает аминокислотную последовательность TcdB дикого типа из штамма VPI10463 С.difficile (также раскрытую в GenBank, номер доступа Р18177 и/или САА37298). Аминокислотная последовательность TcdB из штамма VPI10463 С.difficile обладает 100% идентичностью SEQ ID NO:2. SEQ ID NO:26 описывает ген TcdB дикого типа из штамма VPI10463 С.difficile, который также раскрыт в GenBank, номер доступа Х53138.1.
Дополнительные примеры TcdB С.difficile дикого типа включают TcdB из штаммов С.difficile дикого типа, которые могут быть получены из Центров контроля и профилактики заболеваний (CDC, Atlanta, GA). Авторы изобретения обнаружили, что аминокислотная последовательность TcdB из штаммов С.difficile дикого типа, которые могут быть получены из CDC, включает по меньшей мере примерно 96%-100% идентичность при оптимальном выравнивании аминокислотным остаткам 1-821 SEQ ID NO:2 (TcdB из штамма 630 С.difficile). См. таблицу 2.
Таблица 2: Штаммы С.difficile дикого типа, полученные из CDC, и % идентичности аминокислотных остатков 1-821 TcdB из соответствующего штамма С.difficile дикого типа аминокислотным остаткам 1-821 SEQ ID NO:2 при оптимальном выравнивании.
Соответственно, в одном воплощении аминокислотная последовательность TcdB С.difficile дикого типа включает последовательность по меньшей мере из примерно 500, 600, 700 или 800 непрерывных остатков, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью последовательности равной длины между остатками 1-900 SEQ ID NO:2 при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT, используя параметры массы гэпов по умолчанию. Примеры включают штаммы, описанные выше (например, R20291, CD196 и т.д.), и штаммы, перечисленные в таблице 2.
В другом воплощении аминокислотная последовательность TcdB С.difficile дикого типа включает последовательность, обладающую примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью любой последовательности, выбранной из SEQ ID NO:110-133 при оптимальном выравнивании. См. таблицу 2-а.
Гены токсинов А и В (tcdA и tcdB) составляют часть генетического локуса длиной 19,6 т.п.н. (локуса патогенности, PaLoc), включающего 3 дополнительные малые открытые рамки считывания (OPC): tcdD, tcdE и tcdC, и могут считаться полезными для вирулентности. Известно, что PaLoc стабилен и консервативен в токсикогенных штаммах. Он присутствует в одном и том же сайте хромосомной интеграции во всех токсикогенных штаммах, проанализированных к настоящему времени. В нетоксикогенных штаммах локус патогенности (PaLoc) отсутствует.Соответственно, характерным для штаммов С.difficile дикого типа, описанных здесь, является присутствие локуса патогенности. Другой предпочтительной характеристикой штаммов С.difficile дикого типа, описанных здесь, является продуцирование обоих токсинов, TcdA и TcdB.
В одном воплощении штамм С.difficile дикого типа представляет собой штамм, имеющий локус патогенности, который по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно на 98%, более предпочтительно примерно на 99% или наиболее предпочтительно примерно на 100% локусу патогенности штаммов С.difficile 630 или VP110463. Полноразмерная последовательность локуса патогенности С.difficile VP110463 зарегистрирована в базе данных EMBL под номером доступа последовательности Х92982, также представленной в SEQ ID NO:26. Штаммы, в которых PaLoc идентичен данному локусу эталонного штамма VP110463, называют штаммами токсинотипа 0. Штаммы токсинотипов I-VII, IX, XII-XV и XVIII-XXIV продуцируют оба токсина, TcdA и TcdB, несмотря на вариации в их генах токсинов.
На N-конце токсинов находится глюкозилтрансферазный домен. Глюкозилтрансферазная активность токсинов связана с цитотоксической функцией токсинов. Без связи с каким-либо механизмом или теорией, считают, что Глюкозилтрансферазная активность обоих токсинов катализирует моногликозилирование малых ГТФ-связывающих белков в надсемействе Rho/Rac/Ras. После гликозилирования этих ГТФ-связывающих белков клеточная физиология резко модифицируется, что приводит в результате к утрате структурной целостности и нарушению важных путей передачи сигнала клеток-хозяев, инфицированных токсинами. Мотив Asp-Xaa-Asp (DXD), вовлеченный в связывание марганца, уридиндифосфата (УДФ) и глюкозы, является типичным характеристическим мотивом для глюкозилтрансферазного домена. Без связи с каким-либо механизмом или теорией, считают, что остатки, принципиально важные для каталитической активности, такие как мотив DXD, не варьируют между TcdB из известного "исторического" штамма, такого как 630, и TcdB из гипервирулентного штамма, такого как R20291. Мотив DXD локализован в остатках 285-287 TcdA С.difficile дикого типа в соответствии с нумерацией SEQ ID NO:1 и в остатках 286-288 TcdB С.difficile дикого типа в соответствии с нумерацией SEQ ID NO:2.
Глобальные алгоритмы выравнивания (например, программы анализа последовательностей) известны в данной области техники и могут быть использованы для оптимального выравнивания двух или более чем двух аминокислотных последовательностей токсинов, чтобы определить, включает ли токсин определенный опознавательный мотив (например, DXD в глюкозилтрансферазном домене, DHC в домене цистеиновой протеазы, описанном ниже, и т.д.). Оптимально выровненную последовательность(и) сравнивают с соответствующей эталонной последовательностью (например, SEQ ID NO:1 для TcdA или SEQ ID NO:2 для TcdB), чтобы определить существование опознавательного мотива. "Оптимальное выравнивание" относится к выравниванию, дающему самый высокий балл процента идентичности. Такой анализ можно проводить, используя известные программы анализа последовательностей. В одном воплощении для идентификации подходящих токсинов дикого типа используют выравнивание CLUSTAL (например, CLUSTALW) с параметрами по умолчанию путем сравнения запрашиваемой последовательности с эталонной последовательностью. Относительная нумерация консервативных аминокислотных остатков основана на нумерации остатков эталонной аминокислотной последовательности, чтобы учитывать малые инсерции или делеции (например, пяти аминокислот или менее) внутри выровненной последовательности.
При использовании здесь, термин "в соответствии с нумерацией" относится к нумерации остатков эталонной последовательности при сравнении данной аминокислотной или полинуклеотидной последовательности с эталонной последовательностью. Иными словами, номер положения остатка данного полимера обозначают по отношению к эталонной последовательности, а не к действительному номеру положения остатка внутри данной аминокислотной или полинуклеотидной последовательности.
Например, данная аминокислотная последовательность, например, последовательность гипервирулентного штамма С.difficile дикого типа, может выть выровнена с эталонной последовательностью (например, с такой, как последовательность исторического штамма С.difficile, например, 630) путем введения гэпов при необходимости, чтобы оптимизировать совпадения остатков между двумя последовательностями. В этих случаях, несмотря на присутствие гэпов, нумерацию остатка в данной аминокислотной или полинуклеотидной последовательности делают по отношению к эталонной последовательности, с которой выравнивают данную последовательность. Как используют здесь, "эталонная последовательность" относится к определенной последовательности, используемой в качестве основы для сравнения последовательностей.
Если не указано иное, все ссылки здесь на положения аминокислот TcdA относятся к нумерации SEQ ID NO:1. Если не указано иное, все ссылки здесь на положения аминокислот TcdB относятся к нумерации SEQ ID NO:2.
Глюкозилтрансферазный домен TcdA, как используют здесь, может начинаться в иллюстративном остатке 1, 101 или 102 и может заканчиваться в иллюстративном остатке 542, 516 или 293 TcdA С.difficile дикого типа, например, SEQ ID NO:1. Любое минимальное положение остатка может быть объединено с максимальным положением остатка между остатками 1 и 542 TcdA, чтобы определить, включен ли участок мотива DXD в последовательность глюкозилтрансферазного домена. Например, в одном воплощении глюкозилтрансферазный домен TcdA включает SEQ ID NO:27, идентичную остаткам 101-293 SEQ ID NO:1, и этот домен включает участок мотива DXD. В другом воплощении глюкозилтрансферазный домен TcdA включает SEQ ID NO:28, идентичную остаткам 1-542 SEQ ID NO:1.
Глюкозилтрансферазный домен TcdB, как используют здесь, может начинаться в иллюстративном остатке 1, 101 или 102 и может заканчиваться в иллюстративном остатке 543, 516 или 293 TcdB С.difficile дикого типа, например, SEQ ID NO:2. Любое минимальное положение остатка может быть объединено с максимальным положением остатка между остатками 1 и 543 TcdB, чтобы определить, включен ли участок мотива DXD в последовательность глюкозилтрансферазного домена. Например, в одном воплощении глюкозилтрансферазный домен TcdB включает SEQ ID NO:29, идентичную остаткам 101-293 SEQ ID NO:2, и он включает участок мотива DXD. В другом воплощении глюкозилтрансферазный домен TcdB включает SEQ ID NO:30, идентичную остаткам 1-543 SEQ ID NO:2.
Без связи с какой-либо теорией или механизмом, считают, что N-конец TcdA и/или TcdB расщепляется посредством процесса аутопротеолиза, в результате которого происходит транслокация глюкозилтрансферазного домена и его высвобождение в цитозоль клетки-хозяина, где он может взаимодействовать с ГТФазами Rac/Ras/Rho. Показано, что TcdA С.difficile дикого типа расщепляется между L542 и S543. Показано, что TcdB С.difficile дикого типа расщепляется между L543 and G544.
Цистеинпротеазный домен связан с аутокаталитической протеолитической активностью токсина. Цистеинпротеазный домен локализован ниже глюкозилтрансферазного домена и может характеризоваться каталитической триадой аспартат, гистидин и цистеин (DHC), например, D589, Н655 и С700 TcdA дикого типа и D587, Н653 и С698 TcdB дикого типа. Без связи с каким-либо механизмом или теорией, считают, что каталитическая триада консервативна между токсином из "исторического" штамма, такого как 630, и TcdB из гипервирулентного штамма, такого как R20291.
Цистеинпротеазный домен TcdA, как используют здесь, может начинаться в иллюстративном остатке 543 и может заканчиваться в иллюстративном остатке 809 769, 768, or 767 of a TcdA дикого типа, e.g., SEQ ID NO:1. Любое минимальное положение остатка может быть объединено с максимальным положением остатка между остатками 543 и 809 TcdA дикого типа, чтобы определить, включен ли участок каталитической триады DHC в цистеинпротеазный домен. Например, в одном воплощении цистеинпротеазный домен TcdA включает SEQ ID NO:32, которая имеет участок мотива DHC, локализованного в остатках 47, 113 и 158 SEQ ID NO:32, которые, соответственно, совпадают с D589, Н655 и С700 TcdA дикого типа в соответствии с нумерацией SEQ ID NO:1. SEQ ID NO:32 идентична остаткам 543-809 SEQ ID NO:1 TcdA.
Цистеинпротеазный домен TcdB, как используют здесь, может начинаться в иллюстративном остатке 544 и может заканчиваться в иллюстративном остатке 801, 767, 755 или 700 TcdB дикого типа, например, SEQ ID NO:2. Любое минимальное положение остатка может быть объединено с максимальным положением остатка между остатками 544 и 801 TcdB дикого типа, чтобы определить, включен ли участок каталитической триады DHC в цистеинпротеазный домен. Например, в одном воплощении цистеинпротеазный домен TcdB включает SEQ ID NO:33, которая включает участок мотива DHC, локализованного в остатках 44, 110 и 115 SEQ ID NO:33, которые, соответственно, совпадают с D587, Н653 и С698 TcdB дикого типа в соответствии с нумерацией SEQ ID NO:2. SEQ ID NO:33 идентична остаткам 544-767 SEQ ID NO:2 TcdB. В другом воплощении цистеинпротеазный домен TcdB включает остатки 544-801 of SEQ ID NO:2 TcdB.
В настоящем изобретении иммуногенная композиция включает мутантный токсин С.difficile. Термин "мутантный", как используют здесь, относится к молекуле, проявляющей структуру или последовательность, отличающуюся от соответствующей структуры или последовательности дикого типа, например, тем, что имеет поперечные сшивки по сравнению с соответствующей структурой дикого типа и/или имеет по меньшей мере одну мутацию по сравнению с соответствующей последовательностью дикого типа при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT при использовании значений массы гэпа по умолчанию. Термин "мутантный", как используют здесь, кроме того, включает молекулу, проявляющую функциональное свойство (например, элиминированную активность глюкозилтрансферазы и/или элиминированную активность цистеиновой протеазы), отличающееся от соответствующего свойства молекулы дикого типа.
Токсин С.difficile из любого из штаммов дикого типа, описанных выше, можно использовать в качестве источника, из которого получают мутантный токсин С.difficile. Предпочтительно источником, из которого получают мутантный токсин С.difficile, является С.difficile 630.
Мутация может включать замену, делецию, укорочение или модификацию аминокислотного остатка дикого типа, обычно локализованного в данном положении. Предпочтительно мутация представляет собой неконсервативную аминокислотную замену. В настоящем изобретении также рассмотрены выделенные полинуклеотиды, включающие нуклеиново-кислотные последовательности, кодирующие любой из мутантных токсинов, описанных здесь.
"Неконсервативная" аминокислотная замена, как используют здесь, относится к замене аминокислоты из одного класса аминокислотой из другого класса в соответствии с приведенной ниже таблицей 3:
Примеры неконсервативной аминокислотной замены включают замену, при которой остаток аспарагиновой кислоты (Asp, D) заменен остатком аланина (Ala, А). Другие примеры включают замену остатка аспарагиновой кислоты (Asp, D) остатком аспарагина (Asn, N); замену остатка аргинина (Arg, R), глутаминовой кислоты (Glu, E), лизина (Lys, K) и/или гистидина (His, H) остатком аланина (Ala, А).
Консервативная замена относится к замене аминокислот из одного и того же класса, например, в соответствии с таблицей 3.
Мутантные токсины по изобретению могут быть получены методами, известными в данной области техники для получения мутаций, такими как, например, следующие методы: сайт-направленный мутагенез, мутагенез с использованием мутагена (например, УФ света) и т.д. Предпочтительно используют сайт-направленный мутагенез. Альтернативно можно синтезировать молекулу нуклеиновой кислоты, имеющую целевую последовательность. Такие способы химического синтеза известны в данной области техники.
В настоящем изобретении мутантный токсин С.difficile включает по меньшей мере одну мутацию в глюкозилтрансферазном домене по сравнению с соответствующим токсином С.difficile дикого типа. В одном воплощении глюкозилтрансферазный домен включает по меньшей мере две мутации. Предпочтительно мутация снижает или элиминирует ферментативную активность глюкозилтрансферазы токсина по сравнению с ферментативной активностью глюкозилтрансферазы соответствующего токсина С.difficile дикого типа.
Примерные аминокислотные остатки в глюкозилтрансферазном домене TcdA, которые могут претерпевать мутацию, включают по меньшей мере один из следующих остатков: W101, D269, R272, D285, D287, Е460, R462, S541 и L542 или любую их комбинацию по сравнению с TcdA С.difficile дикого типа, в соответствии с нумерацией SEQ ID NO:1.
Примерные мутации в глюкозилтрансферазном домене TcdA включают по меньшей мере одну из следующих мутаций: W101A, D269A, R272A, D285A, D287A, Е460А, R462A, S541A и L542G или любую их комбинацию по сравнению с TcdA С.difficile дикого типа. В предпочтительном воплощении глюкозилтрансферазный домен TcdA включает мутацию L542G по сравнению с TcdA С.difficile дикого типа. В другом предпочтительном воплощении глюкозилтрансферазный домен TcdA включает мутацию D285A и мутацию D287A по сравнению с TcdA С.difficile дикого типа.
Примерные аминокислотные остатки в глюкозилтрансферазном домене TcdB, которые могут претерпевать мутацию, включают по меньшей мере один из следующих остатков: W102, D270, R273, D286, D288, N384, D461, K463, W520 и L543 или любую их комбинацию по сравнению с TcdA С.difficile дикого типа в соответствии с нумерацией SEQ ID NO:2.
Примерные мутации в глюкозилтрансферазном домене TcdB включают по меньшей мере одну из следующих мутаций: W102A, D270A, D270N, R273A, D286A, D288A, N384A, D461A, D461R, K463A, K463E, W520A и L543A или любую их комбинацию по сравнению с TcdB С.difficile дикого типа. В предпочтительном воплощении глюкозилтрансферазный домен TcdB включает L543A по сравнению с TcdB С.difficile дикого типа. В другом предпочтительном воплощении глюкозилтрансферазный домен TcdB включает мутации D286A и D288A по сравнению с TcdB С.difficile дикого типа.
Любую из мутаций, описанных выше здесь, можно комбинировать с мутацией в домене цистеиновой протеазы. В настоящем изобретении мутантный токсин С.difficile включает по меньшей мере одну мутацию в цистеинпротеазном домене относительно соответствующего токсина С.difficile дикого типа. Предпочтительно мутация снижает или элиминирует активность цистеиновой протеазы токсина по сравнению активностью цистеиновой протеазы соответствующего токсина С.difficile дикого типа.
Примерные аминокислотные остатки в цистеинпротеазном домене TcdA, которые могут претерпевать мутацию, включают по меньшей мере один из следующих остатков: S543, D589, Н655 и С700 или любую их комбинацию по сравнению с TcdA С.difficile дикого типа в соответствии с нумерацией SEQ ID NO:1. Примерные мутации в цистеинпротеазном домене TcdA включают по меньшей мере одну из следующих мутаций: S543A, D589A, D589N, Н655А, С700А или любую их комбинацию по сравнению с TcdA С.difficile дикого типа. В предпочтительном воплощении цистеинпротеазный домен TcdA включает мутацию С700А по сравнению с TcdA С.difficile дикого типа.
Примерные аминокислотные остатки в цистеинпротеазном домене TcdB, которые могут претерпевать мутацию, включают по меньшей мере один из следующих остатков: G544, D587, Н653 и С698 по сравнению с TcdB С.difficile дикого типа в соответствии с нумерацией SEQ ID NO:2. Примерные мутации в цистеинпротеазном домене TcdA включают по меньшей мере одну из следующих мутаций: G544A, D587A, D587N, Н653А, С698А или любую их комбинацию по сравнению с TcdB С.difficile дикого типа. В предпочтительном воплощении цистеинпротеазный домен TcdB включает мутацию С698А по сравнению с TcdB С.difficile дикого типа. Дополнительные аминокислотные остатки в цистеинпротеазном домене TcdB, которые могут претерпевать мутацию, включают следующие остатки: К600 и/или R751 по сравнению с TcdB дикого типа. Примерные мутации включают К600Е и/или R751E.
Соответственно, мутантный токсин С.difficile по изобретению включает глюкозилтрансферазный домен, имеющий мутацию, и цистеинпротеазный домен, имеющий мутацию относительно соответствующего токсина С.difficile дикого типа.
Примерный мутантный TcdA С.difficile включает глюкозилтрансферазный домен, включающий SEQ ID NO:29, имеющую аминокислотную замену в положениях 285 и 287, и цистеинпротеазный домен, включающий SEQ ID NO:
32, имеющую аминокислотную замену в положении 158, по сравнению с соответствующим токсином А С.difficile дикого типа. Например, такой мутантный TcdA С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO:4, в которой начальный метионин возможно отсутствует.В другом воплощении мутантный токсин А С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO:84.
Дополнительные примеры мутантного токсина А С.difficile включают аминокислотную последовательность, описанную в SEQ ID NO:7, которая имеет мутацию D269A, R272A, D285A, D287A, Е460А, R462A и С700А по сравнению с SEQ ID NO:1, в которой начальный метионин возможно отсутствует. В другом воплощении мутантный токсин А С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO:83.
Другой примерный мутантный TcdA включает SEQ ID NO:34, в которой остаток в положениях 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 может представлять собой любую аминокислоту.
В некоторых воплощениях мутантный токсин С.difficile проявляет сниженный или элиминированный аутопротеолитический процессинг по сравнению с соответствующим токсином С.difficile дикого типа. Например, мутантный TcdA С.difficile может включать мутацию в одном из следующих остатков: S541, L542 и/или S543 или любую их комбинацию по сравнению с соответствующим TcdA С.difficile дикого типа. Предпочтительно мутантный TcdA C.difficile включает по меньшей мере одну из следующих мутаций: S541A, L542G и S543A или любую их комбинацию по сравнению с соответствующим TcdA С.difficile дикого типа.
Другой примерный мутантный TcdA С.difficile включает мутацию S541A, L542, S543 и С700 по сравнению с соответствующим TcdA С.difficile дикого типа.
Примерный мутантный токсин В С.difficile включает глюкозилтрансферазный домен, включающий SEQ ID NO:31, имеющую аминокислотную замену в положениях 286 и 288, и цистеинпротеазный домен, включающий SEQ ID NO:33, имеющую аминокислотную замену в положении 155 по сравнению с соответствующим токсином В С.difficile дикого типа. Например, такой мутантный TcdB С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO:6, в которой начальный метионин возможно отсутствует.В другом воплощении мутантный токсин В С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO: 86.
Дополнительные примеры мутантного TcdB С.difficile включают аминокислотную последовательность, описанную в SEQ ID NO:8, которая имеет мутацию D270A, R273A, D286A, D288A, D461A, K463A и С698А по сравнению с SEQ ID NO:2, SEQ ID NO:8, в которой начальный метионин возможно отсутствует.В другом воплощении мутантный токсин В С.difficile включает аминокислотную последовательность, описанную в SEQ ID NO:85.
Другой примерный мутантный TcdB включает SEQ ID NO:35, в которой остаток в положениях 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 может представлять собой любую аминокислоту.
В качестве другого примера, мутантный TcdB С.difficile может включать мутацию в положениях 543 и/или 544 по сравнению с соответствующим TcdB С.difficile дикого типа. Предпочтительно мутантный TcdB С.difficile включает мутацию L543 и/или G544 по сравнению с соответствующим TcdB С.difficile дикого типа. Более предпочтительно мутантный TcdB С.difficile включает мутацию L543G и/или G544A по сравнению с соответствующим TcdB С.difficile дикого типа.
Другой примерный мутантный TcdB С.difficile включает мутацию L543G, G544A и С698 по сравнению с соответствующим TcdB С.difficile дикого типа.
В одном аспекте изобретение относится к выделенному полипептиду, имеющему мутацию в любом положении от аминокислотного остатка 1 до 1500 в соответствии с нумерацией SEQ ID NO:2, для определения иллюстративного мутантного токсина В С.difficile. Например, в одном воплощении выделенный полипептид включает мутацию между остатками аминокислот 830 и 990 SEQ ID NO:2. Иллюстративные положения для мутаций включают положения 970 и 976 в соответствии с нумерацией SEQ ID NO:2. Предпочтительно мутация между остатками 830 и 990 представляет собой замену. В одном воплощении мутация представляет собой неконсервативную замену, при которой аминокислотный остаток Asp (D) и/или a Glu (E) заменен аминокислотным остатком, который не нейтрализуется при подкислении, таким как, например, лизин (К), аргинин (R) и гистидин (Н). Примеры мутаций включают: E970K, E970R, Е970Н, E976K, E976R, Е976Н SEQ ID NO:2 для определения мутантного токсина В С.difficile.
В другом аспекте изобретение относится к выделенному полипептиду, имеющему мутацию в любом положении от аминокислотного остатка 1 до 1500 в соответствии с нумерацией SEQ ID NO:1, для определения примерного мутантного токсина А С.difficile. Например, в одном воплощении выделенный полипептид включает мутацию между остатками аминокислот 832 и 992 в SEQ ID NO:1. Иллюстративные положения для мутаций включают положения 972 и 978 в соответствии с нумерацией SEQ ID NO:1. Предпочтительно мутация между остатками 832 и 992 представляет собой замену. В одном воплощении мутация представляет собой неконсервативную замену, при которой аминокислотный остаток Asp (D) и/или Glu (E) заменен аминокислотным остатком, который не нейтрализуется при подкислении, таким как, например, лизин (К), аргинин (R) и гистидин (Н). Примеры мутаций включают: D972K, D972R, D972H, D978K, D978R, D978H SEQ ID NO:1 для определения мутантного токсина А С.difficile.
Полипептиды по изобретению могут включать начальный остаток метионина, в некоторых случаях в результате процесса, опосредованного клеткой-хозяином. В зависимости, например, от клетки-хозяина, используемой в методе рекомбинантного продуцирования, и/или от условий ферментации или роста клетки-хозяина, в данной области техники известно, что N-концевой метионин, кодируемый кодоном инициации трансляции, может быть удален из полипептида после трансляции в клетках, либо N-концевой метионин может оставаться присутствующим в выделенном полипептиде.
Соответственно, в одном аспекте изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, описанную в SEQ ID NO:4, в которой начальный метионин (в положении 1) возможно отсутствует.В одном воплощении начальный метионин SEQ ID NO:4 отсутствует.В одном аспекте изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, описанную в SEQ ID NO:84, которая идентична SEQ ID NO:4, за исключением отсутствия начального метионина.
В другом аспекте выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:6, в которой начальный метионин (в положении 1) возможно отсутствует.В одном воплощении начальный метионин в SEQ ID NO:6 отсутствует.В одном аспекте изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, описанную в SEQ ID NO:86, которая идентичная SEQ ID NO:6, за исключением отсутствия начального метионина.
В следующем аспекте выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:7, в которой начальный метионин (в положении 1) возможно отсутствует.В одном воплощении, изобретение относится к выделенному полипептиду, включающему аминокислотную последовательность, описанную в SEQ ID NO:83, идентичную SEQ ID NO:7, за исключением отсутствия начального метионина. Еще в одном аспекте выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:8, в которой начальный метионин (в положении 1) возможно отсутствует.В одном воплощении выделенный полипептид включает аминокислотную последовательность, описанную в SEQ ID NO:85, идентичную SEQ ID NO:8, за исключением отсутствия начального метионина.
В одном аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:4, в которой начальный метионин (в положении 1) возможно отсутствует.В другом аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:6, в которой начальный метионин (в положении 1) возможно отсутствует.В следующем аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:7, в которой начальный метионин (в положении 1) возможно отсутствует. Еще в одном аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:8, в которой начальный метионин (в положении 1) возможно отсутствует.
В другом аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:83. В одном аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:84. В одном аспекте изобретение относится к иммуногенной композиции, включающей SEQ ID NO:85. В другом аспекте, изобретение относится к иммуногенной композиции, включающей SEQ ID NO:86.
Кроме того, для получения иммунного ответа у млекопитающего иммуногенные композиции для введения млекопитающим, описанные здесь, также обладают пониженной цитотоксичностью по сравнению с соответствующим токсином С.difficile дикого типа. Предпочтительно иммуногенные композиции безопасны и обладают цитотоксичностью от минимальной (например, снижение цитотоксичности составляет примерно 6-8 log10) до нулевой относительно цитотоксичности соответствующего токсина дикого типа.
Как используют здесь, термин "цитотоксичность" представляет собой термин, как его понимают в данной области техники, и относится к апоптической клеточной гибели и/или состоянию, при котором одна или более чем одна обычная биохимическая или биологическая функция клетки аберрантно нарушена по сравнению с идентичной клеткой в идентичных условиях, но в отсутствие цитотоксического агента. Токсичность можно определить количественно, например, в клетках или в млекопитающих, как количество агента, необходимое для индукции 50% клеточной гибели (то есть ЕС50 или ED50, соответственно), или другими способами, известными в данной области техники.
Анализы для определения цитотоксичности известны в данной области техники, например, анализы округления клеток (см., например, статью Kuehne et al. Nature. 2010 Oct 7; 467(7316):711-3). Действие TcdA и TcdB вызывает округление клеток (например, утрату морфологии) и их гибель, и такое явление видимо с помощью световой микроскопии. См., например, фиг.9.
Дополнительные примеры анализов цитотоксичности, известные в данной области техники, включают анализы гликозилирования, относящиеся к анализам формирования изображения на люминесцентном фосфорном покрытии Ras, меченого [14С]глюкозой (как описано в статье Busch et al., J Biol Chem. 1998 Jul 31;273(31):19566-72), и предпочтительно анализ цитотоксичности in vitro, описаны в Примерах ниже, при котором EC50 может относиться к концентрации иммуногенной композиции, проявляющей по меньшей мере примерно 50% цитопатогенного действия (СРЕ) в клетке, предпочтительно в диплоидном фибробласте человека (например, в клетке IMR90 cell (ATCC CCL-186™)), по сравнению с идентичной клеткой в идентичных условиях в отсутствие токсина. Анализ цитотоксичности in vitro можно также использовать, чтобы оценивать концентрацию композиции, ингибирующую примерно 50% цитопатогенного действия (СРЕ), индуцированного токсином С.difficile дикого типа, предпочтительно в клетке, предпочтительно в диплоидном фибробласте человека (например, в клетке IMR90 cell (ATCC CCL-186™)), по сравнению с идентичной клеткой в идентичных условиях в отсутствие токсина. Дополнительные примерные анализы цитотоксичности включают анализы, описанные в статье Doern et al., J Clin Microbiol. 1992 Aug; 30(8):2042-6. Цитотоксичность можно также определять путем измерения уровней АТФ в клетках, обработанных токсином. Например, можно использовать субстрат на основе люциферазы, такой как CELLTITERGLO® (Promega), испускающий люминесценцию, измеряемую в относительных световых единицах (ОСЕ). При таком анализе выживаемость клеток может быть прямо пропорциональна количеству АТФ в клетках или значениям ОСЕ.
В одном воплощении цитотоксичность иммуногенной композиции снижена по меньшей мере примерно в 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000 раз или более по сравнению с соответствующим токсином С.difficile дикого типа. См., например, таблицу 20.
В другом воплощении цитотоксичность иммуногенной композиции снижена по меньшей мере примерно на 2-log-io, более предпочтительно примерно на 3-log10 и наиболее предпочтительно примерно на 4-log10 или более по сравнению с соответствующим токсином дикого типа в идентичных условиях. Например, мутантный TcdB С.difficile может иметь значение ЕС50, составляющее примерно 10-9 г/мл, измеренное в стандартном анализе цитопатического действия (СРЕ), по сравнению с примерным TcdB С.difficile дикого типа, который может иметь значение EC50, составляющее по меньшей мере примерно 10-12 г/мл. См., например, таблицы 7А, 7В, 8А и 8В в разделе Примеров ниже.
Еще в одном воплощении цитотоксичность мутантного токсина С.difficile имеет значение EC50, составляющее по меньшей мере примерно 50 мкг/мл, 100 мкг/мл, 200 мкг/мл, 300 мкг/мл, 400 мкг/мл, 500 мкг/мл, 600 мкг/мл, 700 мкг/мл, 800 мкг/мл, 900 мкг/мл, 1000 мкг/мл или более, измеренное, например, с помощью анализа цитотоксичности in vitro, такого как анализ, описанный здесь. Соответственно, в предпочтительном воплощении иммуногенные композиции и мутантные токсины биологически безопасны для введения млекопитающим.
Без связи с каким-либо механизмом или теорией, ожидали, что TcdA, имеющий мутацию D285 и D287 по сравнению с TcdA дикого типа, и TcdB, имеющий мутацию D286 и D288 по сравнению с TcdB дикого типа, дефекты по активности глюкозилтрансферазы и, следовательно, дефектны при индукции цитопатического эффекта. Кроме того, ожидали, что токсин, имеющий мутацию в мотиве DHC, дефектен по аутокаталитическому процессингу, и, следовательно, не обладает какими-либо цитотоксическими действиями.
Тем не менее, авторы изобретения неожиданно обнаружили, среди прочего, что иллюстративный мутантный TcdA, имеющий SEQ ID NO:4, и иллюстративный мутантный TcdB, имеющий SEQ ID NO:6, неожиданно проявляли цитотоксичность (хотя и значительно пониженную по сравнению с токсинами С.difficile 630 дикого типа), несмотря на то, что они проявляли дисфункциональную активность глюкозилтрансферазы и дисфункциональную активность цистеиновой протеазы. Без связи с каким-либо механизмом или теорией, считают, что мутантные токсины проявляют цитотоксичность посредством нового механизма. Тем не менее, иллюстративный мутантный TcdA, имеющий SEQ ID NO:4, и иллюстративный мутантный TcdB, имеющий SEQ ID NO:6, неожиданно обладали иммуногенностью. См. Примеры ниже.
Хотя химическое поперечное сшивание токсина дикого типа имеет потенциальный недостаток в инактивировании токсина, авторы изобретения дополнительно обнаружили, что химическое поперечное сшивание по меньшей мере одной аминокислоты мутантного токсина дополнительно снижало цитотоксичность мутантного токсина относительно идентичного мутантного токсина без химических поперечных связей, и по сравнению с соответствующим токсином дикого типа. Предпочтительно мутантный токсин очищают перед приведением в контакт с химическим сшивающим агентом.
Кроме того, несмотря на потенциал агентов химического сшивания к изменению полезных эпитопов, авторы изобретения неожиданно обнаружили, что генетически модифицированный мутантный токсин С.difficile, имеющий по меньшей мере одну поперечно-сшитую химическим образом аминокислоту, приводит в результате к иммуногенной композиции, вызывающей образование множественных нейтрализующих антител или их связывающих фрагментов. Соответственно, эпитопы, связывающиеся с молекулами нейтрализующих антител, неожиданно сохранялись после химического сшивания.
Поперечное сшивание (также называемое здесь "химической инактивацией" или "инактивацией") представляет собой процесс химического соединения двух или более чем двух молекул ковалентной связью. Термины "сшивающие реагенты", "сшивающие агенты" и "сшиватели" относятся к молекулам, способным взаимодействовать с определенными функциональными группами и/или химически атаковать эти группы (первичные амины, сульфгидрилы, карбоксилы, карбонилы и т.д.) на пептидах, полипептидах и/или белках. В одном воплощении эта молекула может содержать два или более чем два реакционных конца, способных взаимодействовать с определенными функциональными группами и/или химически атаковать эти группы (первичные амины, сульфгидрилы, карбоксилы, карбонилы и т.д.) на пептидах, полипептидах и/или белках. Предпочтительно химический сшивающий агент растворим в воде. В другом предпочтительном воплощении химический сшивающий агент представляет собой гетеробифункциональный сшиватель. В другом воплощении химический сшивающий агент не является бифункциональным сшивателем. Химические сшивающие агенты известны в данной области техники.
В предпочтительном воплощении сшивающий агент представляет собой сшивающий агент нулевой длины. Сшиватель "нулевой длины" относится к сшивающему агенту, опосредующему или осуществляющему прямую поперечную сшивку между функциональными группами двух молекул. Например, при сшивании двух полипептидов сшиватель нулевой длины приведет в результате к образованию мостиковой связи или поперечной сшивки между карбоксильной группой от боковой цепи аминокислоты одного полипептида и аминогруппой другого полипептида без интеграции с внешней стороной. Сшивающие агенты нулевой длины могут катализировать, например, образование сложноэфирных связей между гидроксильной и карбоксильной группировками и/или образование амидных связей между карбоксильной и первичной аминогруппировкой.
Примерные подходящие химические сшивающие агенты включают формальдегид; формалин; ацетальдегид; проиональдегид; водорастворимые карбодиимиды (RN=C=NR′), которые включают 1-этил-3-(3-диметиламинопропил)-карбодиимид (EDC), 1-этил-3-(3-диметиламинопропил)-карбодиимида гидрохлорид, 1-циклогексил-3-(2-морфолинил-(4-этил)карбодиимида мето-пара-толуолсульфонат (CMC), N,N′-дициклогексилкарбодиимид (DCC) и N,N′-диизопропилкарбодиимид (DIC), и их производные, а также N-гидроксисукцинимид (NHS); фенилглиоксаль; и/или УДФ-диальдегид.
Предпочтительно сшивающий агент представляет собой EDC. При химической модификации мутантного токсина С.difficile с помощью EDC (например, путем приведения в контакт полипептида с EDC) в одном воплощении полипептид включает (а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида. В одном воплощении полипептид включает (b) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида. В одном воплощении полипептид включает (с) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца полипептида. В одном воплощении полипептид включает (d) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина полипептида. В одном воплощении, полипептид включает (е) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида. В одном воплощении полипептид включает (f) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида. В одном воплощении, полипептид включает (g) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца второго выделенного полипептида. В одном воплощении полипептид включает (h) по меньшей мере одну поперечную сшивку между карбоксильной группой С-конца полипептида и боковой цепью остатка лизина второго выделенного полипептида. См., например, фиг.24 и фиг.25.
"Второй выделенный полипептид" относится к любому выделенному полипептиду, присутствующему при взаимодействии с EDC. В одном воплощении второй выделенный полипептид представляет собой полипептид мутантного токсина С.difficile, имеющий последовательность, идентичную первому выделенному полипептиду. В другом воплощении, второй выделенный полипептид представляет собой полипептид мутантного токсина С.difficile, имеющий последовательность, отличающуюся от первого выделенного полипептида.
В одном воплощении полипептид включает по меньшей мере две модификации, выбранные из модификаций (a)-(d). В примерном воплощении полипептид включает (а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида и (b) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида. В следующем воплощении полипептид включает по меньшей мере три модификации, выбранные из модификаций (а)-(d). Еще в одном воплощении полипептид включает модификации (а), (b), (с) и (d).
Если при химической модификации с помощью EDC присутствует более чем один мутантный полипептид, в одном воплощении полученная в результате композиция включает по меньшей мере одну из любых модификаций (a)-(h). В одном воплощении композиция включает по меньшей мере две модификации, выбранные из модификаций (a)-(h). В следующем воплощении композиция включает по меньшей мере три модификации, выбранные из модификаций (а)-(h). Еще в одном воплощении композиция включает по меньшей мере четыре модификации, выбранные из модификаций (a)-(h). В другом воплощении композиция включает по меньшей мере одну каждую модификацию (a)-(h).
В примерном воплощении полученная в результате композиция включает (а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; и (b) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида. В одном воплощении композиция дополнительно включает (с) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца полипептида; и (d) по меньшей мере одну поперечную сшивку между карбоксильной группой С-конца полипептида и боковой цепью остатка лизина полипептида.
В другом примерном воплощении полученная в результате композиция включает (е) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида; (f) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида; (g) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца второго выделенного полипептида; и (h) по меньшей мере одну поперечную сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина второго выделенного полипептида.
В следующем примерном воплощении полученная в результате композиция включает (а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; (b) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида; (е) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида; и (f) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина второго выделенного полипептида.
В предпочтительном воплощении химический сшивающий агент включает формальдегид, более предпочтительно агент включает формальдегид при отсутствии лизина. Глицин или другое подходящее соединение с первичным амином можно использовать в качестве гасителя в реакциях сшивания. Соответственно, в другом предпочтительном воплощении химический агент включает формальдегид и использование глицина.
Еще в одном предпочтительном воплощении химический сшивающий агент включает EDC и NHS. Как известно в данной области техники, NHS можно включать в протоколы сочетания посредством EDC. Тем не менее, авторы изобретения неожиданно обнаружили, что NHS может способствовать дополнительному снижению цитотоксичности мутантного токсина С.difficile по сравнению с соответствующим токсином дикого типа, по сравнению с генетически мутированным токсином и по сравнению с генетически мутированным токсином, поперечно-сшитым химическим образом с использованием EDC. См., например, Пример 22. Соответственно, без связи с каким-либо механизмом или теорией, наличие у полипептида мутантного токсина бета-аланиновой группировки, сшитой с боковой цепью по меньшей мере одного остатка лизина полипептида (например, в результате взаимодействия полипептида мутантного токсина, EDC и NHS), может способствовать дополнительному снижению цитотоксичности мутантного токсина по сравнению, например, с токсином С.difficile (дикого типа или мутантным), у которого отсутствует группировка бета-аланина.
Использование EDC и/или NHS может также включать использование глицина или другого подходящего соединения с первичным амином в качестве гасителя. В качестве гасителя можно использовать любое соединение, включающее первичный амин, такое как, например, метиловый эфир глицина и аланин. В предпочтительном воплощении соединение-гаситель представляет собой неполимерный гидрофильный первичный амин. Примеры неполимерного гидрофильного первичного амина включают, например, аминосахара, аминоспирты и аминополиолы. Конкретные примеры неполимерного гидрофильного первичного амина включают глицин, этаноламин, глюкамин, полиэтиленгликоль, функционализированный амином, и олигомеры этиленгликоля, функционализированные амином.
В одном аспекте изобретение относится к полипептиду мутантного токсина С.difficile, в котором по меньшей мере одна боковая цепь аминокислоты химически модифицирована с помощью EDC и неполимерного гидрофильного первичного амина, предпочтительно глицина. Полученные в результате аддукты глицина (например, в результате взаимодействия тройных мутантных токсинов, обработанных EDC, NHS и погашенных глицином) могут способствовать снижению цитотоксичности мутантного токсина по сравнению с соответствующим токсином дикого типа.
В одном воплощении, при химической модификации полипептида мутантного токсина С.difficile с помощью EDC и глицина, полипептид включает по меньшей мере одну модификацию в результате его модификации с помощью EDC (например, по меньшей мере одну из любых модификаций (а)-(h), описанных выше) и по меньшей мере одну из следующих примерных модификаций: (i) глициновую группировку, сшитую с карбоксильной группой С-конца полипептида; G) глициновую группировку, сшитую с боковой цепью по меньшей мере одного остатка аспарагиновой кислоты полипептида; и (k) глициновую группировку, сшитую с боковой цепью по меньшей мере одного остатка глутаминовой кислоты полипептида. См., например, фиг.24 и фиг.25.
В одном воплощении по меньшей мере одна аминокислота мутантного TcdA С.difficile поперечно-сшита химическим образом, и/или по меньшей мере одна аминокислота мутантного TcdB С.difficile поперечно-сшита химическим образом. В другом воплощении по меньшей мере одна аминокислота SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:7 и/или SEQ ID NO:8 поперечно-сшита химическим образом. Например, по меньшей мере одна аминокислота может быть поперечно-сшитой химическим образом с использованием агента, включающего карбодиимид, такой как EDC. Карбодиимиды могут образовать ковалентную связь между свободной карбоксильной (например, от боковых цепей аспарагиновой кислоты и/или глутаминовой кислоты) и аминогруппой (например, в боковой цепи остатков лизина) с образованием стабильных амидных связей.
В качестве другого примера, по меньшей мере одна аминокислота может быть поперечно-сшитой химическим образом с использованием агента, включающего NHS. Сшиватели, представляющие собой активированные сложные эфиры NHS, могут взаимодействовать с первичными аминами (например, на N-конце каждой полипептидной цепи и/или в боковой цепи остатков лизина) с образованием амидной связи.
В другом воплощении по меньшей мере одна аминокислота может быть поперечно-сшитой химическим образом с использованием агента, включающего EDC и NHS. Например, в одном воплощении изобретение относится к выделенному полипептиду имеющему аминокислотную последовательность, описанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, где полипептид включает по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC и NHS. В другом воплощении изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, описанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где полипептид включает по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC и NHS. Еще в одном воплощении изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, описанную в SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:7 или SEQ ID NO:8. Полипептид модифицируют путем приведения полипептида в контакт с EDC и NHS. См., например, фиг.24 и фиг.25.
При химической модификации полипептида мутантного токсина С.difficile с использованием EDC и NHS (например, путем приведения в контакт с этими агентами), в одном воплощении полипептид включает по меньшей мере одну модификацию в результате модификации полипептида с использованием EDC (например, по меньшей мере одну из любых модификаций (а)-(h), описанных выше) и (l) бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида.
В другом аспекте изобретение относится к полипептиду мутантного токсина С.difficile, включающему по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC, NHS и неполимерного гидрофильного первичного амина, предпочтительно глицина. В одном воплощении полипептид включает по меньшей мере одну модификацию, когда он модифицирован с использованием EDC (например, по меньшей мере одну из любых модификаций (а)-(h), описанных выше), по меньшей мере одну модификацию, когда он модифицирован глицином (например, по меньшей мере одну из любых модификаций (i)-(k), описанных выше) и (l) бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида. См., например, фиг.24 и фиг.25.
В одном аспекте изобретение относится к полипептиду мутантного токсина С.difficile, в котором боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой. В одном воплощении боковая цепь второго остатка лизина полипептида сшита с боковой цепью остатка аспарагиновой кислоты и/или с боковой цепью остатка глутаминовой кислоты. "Второй" остаток лизина полипептида включает остаток лизина полипептида, который не сшит с бета-аланиновой группировкой. Боковая цепь аспарагиновой кислоты и/или боковая цепь глутаминовой кислоты, с которой сшит второй остаток лизина, может представлять собой боковую цепь полипептида, образующую внутримолекулярную поперечную сшивку, или боковую цепь второго полипептида, образующую межмолекулярную поперечную сшивку. В другом воплощении боковая цепь по меньшей мере одного остатка аспарагиновой кислоты и/или боковая цепь по меньшей мере одного остатка глутаминовой кислоты полипептида сшита с глициновой группировкой. Остаток аспарагиновой кислоты и/или остаток глутаминовой кислоты, который сшит с глициновой группировкой, также не сшит с остатком лизина.
В качестве еще одного примера поперечно-сшитого химическим образом полипептида мутантного токсина С.difficile, по меньшей мере одна аминокислота может быть поперечно-сшитой химическим образом с использованием агента, включающего формальдегид. Формальдегид может взаимодействовать с аминогруппой N-концевого аминокислотного остатка и боковыми цепями аргинина, цистеина, гистидина и лизина. Формальдегид и глицин могут образовать аддукт, представляющий собой шиффово основание, который может присоединяться к первичным N-концевым аминогруппам, остаткам аргинина и тирозина, и в меньшей степени к остаткам аспарагина, глутамина, гистидина и триптофана.
Указано, что химический сшивающий агент снижает цитотоксичность токсина, если обработанный токсин обладает меньшей токсичностью (например, примерно на 100%, 99%, 95%, 90%, 80%, 75%, 60%, 50%, 25% или на 10% меньшей токсичностью), чем необработанный токсин в идентичных условиях, при измерении, например, с использованием анализа цитотоксичности in vitro или токсичности животных.
Предпочтительно химический сшивающий агент снижает цитотоксичность мутантного токсина С.difficile, где снижение составляет по меньшей мере примерно 2-log10, более предпочтительно примерно 3-log10 и наиболее предпочтительно примерно 4-log10 или более относительно мутантного токсина в идентичных условиях, но в отсутствие химического сшивающего агента. По сравнению с токсином дикого типа химический сшивающий агент предпочтительно снижает цитотоксичность мутантного токсина, где снижение составляет по меньшей мере примерно 5-log10, примерно 6-log10, примерно 7-log10, примерно 8-log10 или более.
В другом предпочтительном воплощении химически инактивированный мутантный токсин С.difficile проявляет значение EC50, большее или по меньшей мере примерно равное 50 мкг/мл, 100 мкг/мл, 200 мкг/мл, 300 мкг/мл, 400 мкг/мл, 500 мкг/мл, 600 мкг/мл, 700 мкг/мл, 800 мкг/мл, 900 мкг/мл, 1000 мкг/мл или более при измерении, например, с использованием анализа цитотоксичности in vitro, такого как описано здесь.
Условия реакции для приведения в контакт мутантного токсина с химическим сшивающим агентом находятся в пределах компетенции специалистов в данной области техники, и эти условия можно варьировать в зависимости от используемого агента. Тем не менее, авторы изобретения неожиданно обнаружили оптимальные условия реакции для приведения в контакт полипептида мутантного токсина С.difficile с химическим сшивающим агентом, сохраняя при этом функциональные эпитопы и снижая цитотоксичность мутантного токсина по сравнению с соответствующим токсином дикого типа.
Предпочтительно для приведения в контакт мутантного токсина со сшивающим агентом выбраны таким образом, что мутантный токсин имеет минимальную концентрацию, составляющую примерно 0,5, 0,75, 1,0, 1,25, 1,5, 1,75, 2,0 мг/мл, до максимума, составляющего примерно 3,0, 2,5, 2,0, 1,5 или 1,25 мг/мл. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить диапазон подходящих концентраций мутантного токсина для взаимодействия. Наиболее предпочтительно мутантный токсин имеет концентрацию, составляющую примерно 1,0-1,25 мг/мл на реакцию.
В одном воплощении агент, используемый в реакции, имеет минимальную концентрацию, составляющую примерно 1 мМ, 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, 20 мМ, 30 мМ, 40 мМ или 50 мМ, и максимальную концентрацию, составляющую примерно 100 мМ, 90 мМ, 80 мМ, 70 мМ, 60 мМ или 50 мМ. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить диапазон подходящих концентраций химического агента для реакции.
В предпочтительном воплощении, где агент включает формальдегид, используемая концентрация предпочтительно представляет собой любую концентрацию, составляющую примерно от 2 мМ до 80 мМ, наиболее предпочтительно примерно 40 мМ. В другом предпочтительном воплощении, где агент включает EDC, используемая концентрация предпочтительно представляет собой любую концентрацию, составляющую от примерно 1,3 мМ до примерно 13 мМ, более предпочтительно примерно от 2 мМ до 3 мМ, наиболее предпочтительно примерно 2,6 мМ.
Примерные периоды времени реакции, при которых мутантный токсин контактирует с химическим сшивающим агентом, включают минимум, составляющий примерно 0,5, 1, 2, 3, 4, 5, 6, 12, 24, 36, 48 или 60 часов, и максимум, составляющий примерно 14 суток, 12 суток, 10 суток, 7 суток, 5 суток, 3 суток, 2 суток, 1 сутки или 12 часов. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить диапазон подходящих периодов времени реакции.
В предпочтительном воплощении стадия осуществления контакта мутантного токсина с химическим сшивающим агентом происходит в течение периода времени, достаточного для снижения цитотоксичности мутантного токсина С.difficile до значения ЕС50, составляющего по меньшей мере примерно 1000 мкг/мл в подходящей клетке человека, например, в клетках IMR-90, в стандартном анализе цитотоксичности in vitro по сравнению с идентичным мутантным токсином в отсутствие сшивающего агента. Более предпочтительно стадию взаимодействия проводят в течение периода времени, по меньшей мере в два раза более длительного, и наиболее предпочтительно по меньшей мере в три или более раза более длительного, чем период времени, достаточный для снижения цитотоксичности мутантного токсина до значения ЕС50, составляющего по меньшей мере примерно 1000 мкг/мл в подходящей клетке человека. В одном воплощении время реакции не превышает примерно 168 часов (или 7 суток).
Например, в одном воплощении, где агент включает формальдегид, мутантный токсин предпочтительно контактирует с агентом в течение примерно 12 часов, что, как было показано, составляет достаточный период времени для снижения цитотоксичности мутантного токсина С.difficile до значения EC50, составляющего по меньшей мере примерно 1000 мкг/мл в подходящей клетке человека, например, в клетках IMR-90, в стандартном анализе цитотоксичности in vitro по сравнению с идентичным мутантным токсином в отсутствие сшивающего агента. В более предпочтительном воплощении эту реакцию проводят в течение примерно 48 часов, что по меньшей мере примерно в три раза дольше, чем достаточный период для взаимодействия. В таком воплощении время реакции предпочтительно не превышает примерно 72 часа.
В другом воплощении, где агент включает EDC, мутантный токсин предпочтительно контактирует с агентом в течение примерно 0,5 часов, более предпочтительно по меньшей мере примерно 1 часа или наиболее предпочтительно примерно 2 часов. В таком воплощении время реакции предпочтительно не превышает примерно 6 часов.
Примерное значение рН, при котором мутантный токсин контактирует с химическим сшивающим агентом, включает минимум примерно рН 5,5, 6,0, 6,5, 7,0 или 7,5 и максимум примерно рН 8,5, 8,0, 7,5, 7,0 или 6,5. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить диапазон подходящих значений рН. Предпочтительно реакцию осуществляют при рН 6,5-7,5, предпочтительно при рН 7,0.
Примерные температуры, при которых мутантный токсин контактирует с химическим сшивающим агентом, включает минимум, составляющий примерно 2°С, 4°С, 10°С, 20°С, 25°С или 37°С, и максимальную температуру, составляющую примерно 40°С, 37°С, 30°С, 27°С, 25°С или 20°С. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить диапазон подходящей температуры реакции. Предпочтительно реакцию осуществляют при температуре, составляющей примерно от 20°С до 30°С, наиболее предпочтительно примерно при 25°С.
Иммуногенные композиции, описанные выше, могут включать один мутантный токсин С.difficile (А или В). Соответственно, иммуногенные композиции могут занимать отдельные флаконы (например, отдельный флакон для композиции, включающей мутантный токсин А С.difficile, и отдельный флакон для композиции, включающей мутантный токсин В С.difficile) в препарате или в наборе. Иммуногенные композиции могут быть предназначены для одновременного, последовательного или раздельного применения.
В другом воплощении иммуногенные композиции, описанные выше, могут включать оба мутантных токсина С.difficile (А и В). Любую описанную комбинацию мутантного токсина А С.difficile и мутантного токсина В С.difficile можно объединять для получения иммуногенной композиции. Соответственно, иммуногенные композиции можно объединять в одном флаконе (например, во флаконе, содержащем обе композиции: композицию, включающую мутантный TcdA С.difficile, и композицию, включающую мутантный TcdB С.difficile). Предпочтительно иммуногенные композиции включают мутантный TcdA С.difficile и мутантный TcdB С.difficile.
Например, в одном воплощении иммуногенная композиция включает SEQ ID NO:4 и SEQ ID NO:6, причем, по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и SEQ ID NO:6 поперечно-сшита химическим образом. В другом воплощении иммуногенная композиция включает мутантный токсин А С.difficile, включающий SEQ ID NO:4 или SEQ ID NO:7, и мутантный токсин В С.difficile, включающий SEQ ID NO:6 или SEQ ID NO:8, причем, по меньшей мере одна аминокислота каждого из мутантных токсинов С.difficile поперечно-сшита химическим образом.
В другом воплощении иммуногенная композиция включает любую последовательность, выбранную из SEQ ID NO:4, SEQ ID NO:84 и SEQ ID NO:83, и любую последовательность, выбранную из SEQ ID NO:6, SEQ ID NO:86 и SEQ ID NO:85. В другом воплощении иммуногенная композиция включает SEQ ID NO:84, и иммуногенная композиция включает SEQ ID NO:86. В другом воплощении иммуногенная композиция включает SEQ ID NO:83, и иммуногенная композиция включает SEQ ID NO:85. В другом воплощении иммуногенная композиция включает SEQ ID NO:84, SEQ ID NO:83, SEQ ID NO:86 и SEQ ID NO:85.
Понятно, что любые из композиций по изобретению, например, иммуногенные композиции, включающие мутантный токсин А и/или мутантный токсин В, можно объединять в различных соотношениях или количествах для получения терапевтического эффекта. Например, мутантный TcdA С.difficile и мутантный TcdB С.difficile могут находиться в иммуногенной композиции в соотношении, находящемся в диапазоне от 0,1:10 до 10:0,1 А:В. В другом воплощении, например, мутантный TcdB С.difficile и мутантный TcdA С.difficile могут находиться в иммуногенной композиции в соотношении, находящемся в диапазоне от 0,1:10 до 10:0,1 В:А. В одном предпочтительном воплощении это соотношение является таким, что композиция включает большее суммарное количество мутантного TcdB, чем суммарное количество мутантного TcdA.
В одном аспекте иммуногенная композиция способна к связыванию с нейтрализующим антителом или его связывающим фрагментом. Предпочтительно нейтрализующее антитело или его связывающий фрагмент являются такими, как описано здесь ниже. В одном примерном воплощении иммуногенная композиция способна к связыванию с антителом против токсина А или его связывающим фрагментом, где антитело против токсина А или его связывающий фрагмент включает вариабельную легкую цепь, имеющую аминокислотную последовательность SEQ ID NO:36, и вариабельную тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO:37. Например, иммуногенная композиция может включать мутантный TcdA С.difficile SEQ ID NO:4 или SEQ ID NO:7. В качестве другого примера иммуногенная композиция может включать SEQ ID NO:84 или SEQ ID NO:83.
В другом примерном воплощении иммуногенная композиция способна к связыванию с антителом против токсина В или его связывающим фрагментом, где антитело против токсина В или его связывающий фрагмент включает вариабельную легкую цепь В8-26 и вариабельную тяжелую цепь В8-26. Например, иммуногенная композиция может включать мутантный TcdB С.difficile SEQ ID NO:6 или SEQ ID NO:8. В качестве другого примера иммуногенная композиция может включать SEQ ID NO:86 или SEQ ID NO:85.
Рекомбинантная клетка
В другом аспекте изобретение относится к рекомбинантной клетке или к ее потомству. В одном воплощении клетка или ее потомство включают полинуклеотид, кодирующий мутантный TcdA С.difficile и/или мутантный TcdB С.difficile.
В другом воплощении рекомбинантная клетка или ее потомство включают нуклеиново-кислотную последовательность, кодирующую полипептид, обладающий по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичностью любой из SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:7 или SEQ ID NO:8 при оптимальном выравнивании, например, с использованием программ GAP или BESTFIT при использовании параметров массы гэпов по умолчанию.
В другом воплощении рекомбинантная клетка или ее потомство включают нуклеиново-кислотную последовательность, кодирующую полипептид, обладающий по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% or наиболее предпочтительно примерно 100% идентичностью любой из SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:83 или SEQ ID NO:85 при оптимальном выравнивании, например, с использованием программ GAP или BESTFIT при использовании параметров массы гэпов по умолчанию.
В дополнительном воплощении рекомбинантная клетка или ее потомство включают нуклеиново-кислотную последовательность, обладающую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% or наиболее предпочтительно примерно 100% идентичностью любой из SEQ ID NO: 11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:46 или SEQ ID NO:47 при оптимальном выравнивании, например, с использованием программ GAP или BESTFIT при использовании параметров массы гэпов по умолчанию.
Рекомбинантная клетка может иметь происхождение от любой клетки, полезной при рекомбинантном продуцировании полипептида по настоящему изобретению, например, прокариота или эукариота. Предпочтительно рекомбинантная клетка имеет происхождение от любой клетки, пригодной для экспрессии гетерологичных нуклеиново-кислотных последовательностей, имеющих размер, больший, чем примерно 5000, 6000, предпочтительно примерно 7000 и более предпочтительно примерно 8000 нуклеотидов или более. Прокариотическая клетка-хозяин может представлять собой любую грамотрицательную или грамположительную бактерию. В примерных воплощениях Прокариотическая клетка-хозяин не имеет эндогенного полинуклеотида, кодирующего токсин и/или спору.
Грамотрицательные бактерии включают, но не ограничены ими, следующие бактерии: Campylobacter, Е.coli, Flavobacterium, Fusobacterium, Helicobacter, llyobacter, Neisseria, Pseudomonas, Salmonella и Ureaplasma. Например, рекомбинантная клетка может иметь происхождение от клетки Pseudomonas fluorescens, как описано в публикации заявки на патент US 2010013762, параграфы [0201]-[0230], которая включена в настоящее описание посредством ссылки.
Грамположительные бактерии включают, но не ограничены ими, следующие бактерии: Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus и Streptomyces. Предпочтительно клетка имеет происхождение от клетки С.difficile.
Авторы изобретения идентифицировали штаммы С.difficile дикого типа, не имеющие эндогенного полинуклеотида, кодирующего токсин С.difficile. Штаммы, не имеющие эндогенных генов токсина А и В, включают следующие штаммы, которые доступны из Американской коллекции типовых клеточных культур (АТСС) (Manassas, VA): С.difficile 1351 (АТСС 43593™), С.difficile 3232 (АТСС ВАА-1801™), С.difficile 7322 (АТСС 43601™), С.difficile 5036 (АТСС 43603™), С.difficile 4811 (АТСС 43602™) и С.difficile VPI 11186 (АТСС 700057™).
Соответственно, в одном воплощении рекомбинантная клетка С.difficile имеет происхождение от штамма, описанного здесь. Предпочтительно рекомбинантная клетка С.difficile или ее потомство имеет происхождение от группы, состоящей из С.difficile 1351, С.difficile 5036 и С.difficile VPI 11186. Более предпочтительно рекомбинантная клетка С.difficile или ее потомство имеет происхождение от клетки С.difficile VPI 11186.
В предпочтительном воплощении ген споруляции рекомбинантной клетки С.difficile или ее потомства инактивирован. Споры могут быть инфекционными, высоко устойчивыми и способствовать персистенции С.difficile в аэробных окружающих средах вне хозяина. Споры могут также вносить вклад в выживание С.difficile внутри хозяина в процессе антибактериальной терапии. Соответственно, клетка С.difficile, не имеющая гена споруляции, полезна для продуцирования безопасной иммуногенной композиции для введения млекопитающим. Кроме того, применение таких клеток способствует безопасности в процессе получения, например, безопасности для защиты оборудования, будущих препаратов и сотрудников.
Примеры генов споруляции для направленной инактивации включают среди прочего spo0A, spoIIE, σE, σG и σK. Предпочтительно инактивируют ген spo0A.
Способы инактивации гена споруляции С.difficile известны в данной области техники. Например, ген споруляции можно инактивировать путем направленной инсерции селективного маркера, такого как маркер устойчивости к антибиотику. См., например, Heap et al., J Microbiol Methods. 2010 Jan; 80(1):49-55; Heap et al., J. Microbiol. Methods, 2007 Sept; 70(3):452-464; и Underwood et al., J Bacteriol. 2009 Dec; 191(23):7296-305. См. также, например, Minton et al., WO 2007/148091, "DNA Molecules and Methods", полностью включенный в настоящее описание посредством ссылки, с.33-66, или соответствующую публикацию US 20110124109 А1, параграфы [00137]-[0227].
Способ продуцирования мутантного токсина С.difficile
В одном аспекте изобретение относится к способу продуцирования мутантного токсина С.difficile. В одном воплощении этот способ включает культивирование любой рекомбинантной клетки или ее потомства, раскрытых выше, в условиях, подходящих для экспрессии полипептида.
В другом воплощении этот способ включает культивирование рекомбинантной клетки или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С.difficile, причем, клетка содержит полинуклеотид, кодирующий мутантный токсин С.difficile, a мутант содержит глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию, по сравнению с соответствующим токсином Clostridium difficile дикого типа. В одном воплощении, в клетке отсутствует эндогенный полинуклеотид, кодирующий токсин.
В следующем воплощении, этот способ включает культивирование рекомбинантной клетки С.difficile или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С.difficile, причем, клетка содержит полинуклеотид, кодирующий мутантный токсин С.difficile, и в клетке отсутствует эндогенный полинуклеотид, кодирующий токсин С.difficile.
В другом аспекте, изобретение относится к способу продуцирования мутантного токсина С.difficile. Этот способ включает следующие стадии: (а) приведение клетки С.difficile в контакт с рекомбинантной клеткой Escherichia coii, причем в клетке С.difficile отсутствует эндогенный полинуклеотид, кодирующий токсин С.difficile, а клетка Е. coii содержит полинуклеотид, кодирующий мутантный токсин С.difficile, (б) культивирование клетки С.difficile и клетки Е. coii в условиях, подходящих для переноса полинуклеотида из клетки Е. coii в клетку С.difficile; (в) отбор клетки С.difficile, содержащей полинуклеотид, кодирующий мутантный токсин С.difficile; (г) культивирование клетки С.difficile со стадии (в) в условиях, подходящих для экспрессии полинуклеотида; и (д) выделение мутантного токсина С.difficile.
В способе по изобретению рекомбинантная клетка Е. coli содержит гетерологичный полинуклеотид, кодирующий мутантный токсин С.difficile, раскрытый здесь. Полинуклеотид может представлять собой ДНК или РНК. В одном иллюстративном воплощении полинуклеотид, кодирующий мутантный токсин С.difficile, оптимизирован по кодонам для частоты использования кодонов Е. coli. Способы оптимизации кодонов полинуклеотида известны в данной области техники.
В одном воплощении полинуклеотид включает последовательность нуклеиновой кислоты, которая по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична полинуклеотиду, кодирующему мутантный TcdA С.difficile, как раскрыто выше. Иллюстративный полинуклеотид, кодирующий мутантный токсин А С.difficile, включает SEQ ID NO: 11, SEQ ID NO:12, SEQ ID NO:44 и SEQ ID NO:45.
В другом воплощении, полинуклеотид включает последовательность нуклеиновой кислоты, которая по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична полинуклеотиду, кодирующему мутантный TcdB С.difficile, как раскрыто выше. Иллюстративный полинуклеотид, кодирующий мутантный токсин В С.difficile, включает SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:46 и SEQ ID NO:47. В другом воплощении полинуклеотид кодирует SEQ ID NO:83, SEQ ID NO:84, SEQ ID NO:85 или SEQ ID NO:86.
В одном воплощении клетка Е. coli, содержащая гетерологичный полинуклеотид, представляет собой клетку E. coli, которая является стабильным хозяином гетерологичного полинуклеотида, кодирующего мутантный токсин С.difficile. Иллюстративные клетки Е. coli включают клетки, выбранные из группы, состоящей из следующих клеток: компетентных клеток Е. coli MAX Efficiency® Stbl2™ (Invitrogen, Carlsbad, CA), химически компетентных клеток E. coli One Shot® Stbl3™ (Invitrogen, Carlsbad, CA), компетентных клеток Е. coil ElectroMAX™ Stbl4™ (Invitrogen) и E. coli СА434. В предпочтительном воплощении клонирующая клетка-хозяин Е. coli не относится к штамму DH5α.
Более предпочтительно, клонирующая клетка-хозяин Е. coli представляет собой компетентную клетку Е. coli MAX Efficiency® Stbl2™.
Способ по изобретению дополнительно включает стадию культивирования клетки С.difficile и клетки Е. coli в условиях, подходящих для переноса полинуклеотида из клетки Е. coli в клетку С.difficile, результатом которого является рекомбинантная клетка С.difficile. В предпочтительном воплощении условия культивирования подходят для переноса полинуклеотида из клетки Е. coli (клетки-донора) в клетку С.difficile (клетку-реципиента), приводящего в результате к генетически стабильному наследованию.
Наиболее предпочтительно, условия культивирования подходят для бактериальной конъюгации, которая известна в данной области техники. "Конъюгация" относится к определенному процессу переноса полинуклеотида, при котором происходит однонаправленный перенос полинуклеотида (например, из бактериальной плазмиды) от одной клетки бактерий (то есть "донора") к другой (то есть "реципиенту"). В процесс конъюгации вовлечен контакт клетки-донора с клеткой-реципиентом. Предпочтительно клетка-донор Е. со//' представляет собой клетку Е. coli СА434.
Иллюстративные подходящие условия (конъюгации) для переноса полинуклеотида из клетки Е. coli в клетку С.difficile включают выращивание жидких культур С.difficile в бульоне с сердечно-мозговым экстрактом (ВН1;
Oxoid) или в бульоне Шедлера для анаэробов (SAB; Oxoid). В другом воплощении можно выращивать твердые культуры С.difficile на свежем кровяном агаре (FBA) или на агаре ВН1. Предпочтительно С.difficile выращивают при 37°С в анаэробной среде (например, 80% N2, 10% СО2 и 10% Н2 [об./об.]). В одном воплощении подходящие условия включают выращивание Е. coli аэробно в среде Лурия-Бертани (Luria-Bertani; LB) или на агаре LB при 37°С. Для конъюгативного переноса в С.difficile иллюстративные подходящие условия включают выращивание Е. coli анаэробно на FBA. В жидкие и твердые среды можно включать антибиотики, как известно в данной области техники. Примеры таких антибиотиков включают циклосерин (250 мкг/мл), цефокситин (8 мкг/мл), хлорамфеникол (12,5 мкг/мл), тиамфеникол (15 мкг/мл) и эритромицин (5 мкг/мл).
Способ по изобретению дополнительно включает стадию отбора полученной в результате рекомбинантной клетки С.difficile, содержащей полинуклеотид, кодирующий мутантный токсин С.difficile. В иллюстративном воплощении рекомбинантная клетка С.difficile является реципиентом полинуклеотида, кодирующего мутантный токсин С.difficile, из рекомбинантной клетки Е. coli посредством конъюгации.
Способ по изобретению включает стадию культивирования рекомбинантной клетки или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С.difficile, приводящего в результате к продуцированию мутантного токсина С.difficile. Подходящие условия для экспрессии рекомбинантной клеткой полинуклеотида включают условия культивирования, подходящие для выращивания клетки С.difficile, известные в данной области техники. Например, подходящие условия могут включать культивирование трансформантов С.difficile в бульоне с сердечно-мозговым экстрактом (BHI; Oxoid) или в бульоне Шедлера для анаэробов (SAB; Oxoid). В другом воплощении можно выращивать твердые культуры С.difficile на агаре FBA или BHI. Предпочтительно С.difficile выращивают при 37°С в анаэробной среде (например, 80% N2, 10% CO2 и 10% Н2 [об/об]).
В одном воплощении способ по изобретению включает стадию выделения полученного в результате мутантного токсина С.difficile. Способы выделения белка из С.difficile известны в данной области техники.
В другом воплощении способ включает стадию очистки полученного в результате мутантного токсина С.difficile. Способы очистки полипептида, такие как хроматография, известны в данной области техники.
В иллюстративном воплощении способ дополнительно включает стадию приведения выделенного мутантного токсина Clostridium difficile в контакт с химическим сшивающим агентом, описанным выше. Предпочтительно, этот агент включает формальдегид, этил-3-(3-диметиламинопропил)карбодиимид или комбинацию EDC и NHS. Иллюстративные условия взаимодействия описаны выше и в разделе Примеров ниже.
В другом аспекте изобретение относится к иммуногенной композиции, включающей мутантный токсин С.difficile, раскрытый здесь, полученный любым способом, предпочтительно любым из способов, раскрытых выше.
Антитела
Неожиданно, иммуногенные композиции по изобретению, раскрытые выше, вызывали образование новых антител in vivo, что позволяет предположить, что иммуногенные композиции включают сохраненную нативную структуру (например, сохраненный антигенный эпитоп) соответствующего токсина С.difficile дикого типа, и что иммуногенные композиции включают эпитоп.Антитела, продуцируемые против токсина из одного штамма С.difficile, могут обладать способностью к связыванию с соответствующим токсином, продуцируемым другим штаммом С.difficile. Таким образом, антитела и их связывающие фрагменты могут быть "перекрестно-реагирующими", что относится к способности к взаимодействию с аналогичными антигенными сайтами на токсинах, продуцируемых множественными штаммами С.difficile. Перекрестная реактивность также включает способность антитела к взаимодействию или к связыванию с антигеном, не стимулировавшим его продуцирование, то есть к взаимодействию между антигеном и антителом, образованным против другого, но аналогичного антигена.
В одном аспекте, авторы изобретения неожиданно обнаружили моноклональные антитела, обладающие нейтрализующим действием на токсины С.difficile, и способы получения этих антител. Антитела по изобретению могут нейтрализовать цитотоксичность токсина С.difficile in vitro, ингибировать связывание токсина С.difficile с клетками млекопитающих и/или могут нейтрализовать энтеротоксичность токсина С.difficile in vivo. Настоящее изобретение также относится к выделенным полинуклеотидам, включающим нуклеиново-кислотные последовательности, кодирующие любое из раскрытого выше. Кроме того, настоящее изобретение относится к применению любой из раскрытых выше композиций для лечения, предупреждения, снижения риска, уменьшения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому не вводят эту композицию, а также к способам получения таких композиций.
Кроме того, авторы изобретения обнаружили, что комбинация по меньшей мере двух нейтрализующих моноклональных антител может проявлять неожиданный синергетический эффект при соответствующей нейтрализации TcdA или TcdB. Антитела против токсина или их связывающие фрагменты могут быть полезны при ингибировании инфекции С.difficile.
"Антитело" представляет собой белок, включающий по меньшей мере одну или две вариабельные области тяжелой цепи (Н) (сокращенные здесь как VH) и по меньшей мере одну или две вариабельные области легкой цепи (L) (сокращенные здесь как VL). Области VH и VL можно дополнительно подразделить на гипервариабельные участки, также называемые "участками, определяющими комплементарность" (CDR), чередующиеся с участками, которые являются более консервативными, называемыми "каркасными областями" (FR). Протяженность каркасной области и CDR точно определена (см. Kabat, E. A., et al., Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No.91-3242, 1991, и Chothia, С. Et al., J. Mol. Biol. 196:901-917, 1987). Термин «антитело» включает интактные иммуноглобулины, относящиеся к типам IgA, IgG, IgE, IgD, IgM (а также к их подтипам), в которых легкие цепи иммуноглобулинов могут относиться к типам каппа или лямбда.
Молекулы антитела могут быть полноразмерными (например, антитело IgG1 или IgG4). Антитела могут относиться к различным изотипам, включающим: IgG (например, IgG1, IgG2, IgG3, IgG4), IgM, IgA1, IgA2, IgD или IgE. В одном предпочтительном воплощении антитело относится к изотипу IgG, например, IgG1. В другом предпочтительном воплощении антитело представляет собой антитело IgE.
В другом воплощении молекула антитела включает «антигенсвязывающий фрагмент» или «связывающий фрагмент», как используют здесь, который относится к участку антитела, специфично связывающемуся с токсином С.difficile (например, с токсином А). Связывающий фрагмент представляет собой, например, молекулу, в которой одна или более чем одна цепь иммуноглобулина не является полноразмерной, но которая специфично связывается с токсином.
Примеры связывающих участков, включенных в термин «связывающий фрагмент» антитела, включают следующие участки: (1) Fab фрагмент, моновалентный фрагмент, состоящий из следующих доменов: VL, VH, CL и СН1; (2) F(ab′)2 фрагмент, бивалентный фрагмент, включающий два Fab фрагмента, связанных дисульфидным мостиком в шарнирной области; (3) Fd фрагмент, состоящий из доменов VH и СН1; (4) Fv фрагмент, состоящий из доменов VL и VH одного плеча антитела, (5) dAb фрагмент (Ward et al., Nature 341:544-546, 1989), состоящий из домена VH; и (6) выделенный гипервариабельный участок (CDR), имеющий достаточный каркасный участок, чтобы специфично связывать, например, антигенсвязывающий участок вариабельной области.
Связывающий фрагмент вариабельной области легкой цепи и связывающий фрагмент вариабельной области тяжелой цепи, например, два домена фрагмента Fv, VL и VH, могут быть соединены с использованием рекомбинантных методов, синтетическим линкером, обеспечивающим их получение в виде единой цепи белка, в которой участки VL и VH спарены с образованием моновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела также включены в термин «связывающий фрагмент» антитела. Эти участки антитела получают, используя методы, известные в данной области техники, и скрининг этих участков на их применимость проводят так же, как для интактных антител.
При использовании здесь, антитело, которое «специфично связывается» или является «специфичным» к определенному полипептиду или к эпитопу определенного полипептида, представляет собой антитело, которое связывается с определенным полипептидом или эпитопом на определенном полипептиде, но по существу не связывается с каким-либо другим полипептидом или эпитопом полипептида. Например, что касается биомолекулы (например, белка, нуклеиновой кислоты, антитела и т.д.), которая «специфично связывается» с мишенью, эта биомолекула связывается со своей молекулой-мишенью и не связывается в значимом количестве с другими молекулами в гетерогенной популяции молекул, включающих эту мишень, что измеряют в заданных условиях (например, в условиях иммунологического анализа в случае антитела). Связывающее взаимодействие между антителом и его мишенью является определителем присутствия мишени в гетерогенной популяции молекул. Например, «специфичное связывание» или «специфично связывается» относится к способности антитела или его связывающего фрагмента к связыванию с токсином дикого типа и/или с мутантным токсином С.difficile со сродством, по меньшей мере в два раза более высоким, чем сродство к неспецифическому антигену.
В иллюстративном воплощении, антитело представляет собой химерное антитело. Химерное антитело может быть получено методами рекомбинантных ДНК, известными в данной области техники. Например, ген, кодирующий константную область Fc молекулы моноклонального антитела мыши (или другого вида) можно расщеплять ферментами рестрикции, чтобы удалить участок, кодирующий Fc мыши, и заменить эквивалентным участком гена, кодирующего константную область Fc человека. Химерное антитело может быть также создано методами рекомбинантных ДНК, при которых ДНК, кодирующую вариабельные области мыши, можно лигировать с ДНК, кодирующей константные области человека.
В другом иллюстративном воплощении антитело или его связывающий фрагмент гуманизируют способами, известными в данной области техники. Например, когда получены мышиные антитела, CDR антитела может быть заменен по меньшей мере частью CDR человека. Гуманизированные антитела могут быть также созданы путем замены последовательности вариабельной области Fv мыши, которые непосредственно не вовлечены в связывание антигена, эквивалентными последовательностями из вариабельных областей Fv человека. Общие способы создания гуманизированных антител известны в данной области техники.
Например, моноклональные антитела, направленные против TcdA С.difficile или TcdB С.difficile, могут быть также получены стандартными методами, такими как метод гибридом (см., например, статью Kohler and Milstein, 1975, Nature, 256: 495-497). Кратко, иммортализованную линию клеток подвергают слиянию с лимфоцитом от млекопитающего, иммунизированного TcdA C.difficile, TcdB C.difficile или мутантным токсином С.difficile, раскрытым здесь, и культуральные супернатанты полученных в результате клеток гибридомы подвергают скринингу, чтобы идентифицировать гибридому, продуцирующую моноклональное антитело, которое связывается с TcdA С.difficile или TcdB C.difficile. В характерном случае иммортализованная линия клеток имеет происхождение от того же вида млекопитающих, что и лимфоциты. Клетки гибридомы, продуцирующие моноклональное антитело по изобретению, обнаруживают путем скрининга культуральных супернатантов гибридом на антитела, связывающие TcdA C.difficile или TcdB C.difficile, с использованием анализа, такого как ELISA. Гибридомы человека могут быть получены аналогично.
В качестве альтернативы получению антител путем иммунизации и отбора, антитела по изобретению могут быть также идентифицированы путем скрининга рекомбинантной комбинаторной библиотеки иммуноглобулинов с TcdA C.difficile, TcdB C.difficile или мутантным токсином С.difficile, раскрытым здесь. Рекомбинантная библиотека антител может представлять собой, например, библиотеку scFv или библиотеку Fab. Кроме того, антитела по изобретению, раскрытые здесь, можно использовать в анализах конкурентного связывания для идентификации дополнительных антител анти-TcdA или анти-TcdB и их связывающих фрагментов. Например, дополнительные антитела анти-TcdA или анти-TcdB и их связывающие фрагменты могут быть идентифицированы путем скрининга библиотеки антител человека и идентифицирующих молекул в пределах библиотеки, конкурирующих с антителами по изобретению, раскрытыми здесь, в анализе конкурентного связывания.
Кроме того, антитела, включенные в настоящее изобретение, включают рекомбинантные антитела, которые могут быть получены путем использования способов фагового дисплея, известных в данной области техники. При способах фагового дисплея фаг можно использовать для представления антигенсвязывающих доменов, экспрессируемых из репертуара или библиотеки антител (например, человека или мыши). Фаг, экспрессирующий антигенсвязывающий домен, связывающийся с иммуногеном, раскрытым здесь (например, с мутантным токсином С.difficile), можно отобрать или идентифицировать с использованием антигена, например, используя меченый антиген.
В пределах объема изобретения также находятся антитела и их связывающие фрагменты, в которых конкретные аминокислоты заменены, делегированы или добавлены. В частности, предпочтительные антитела имеют аминокислотные замены в каркасной области, например, для улучшения связывания с антигеном. Например, выбранное небольшое число акцепторных остатков каркасной области цепи иммуноглобулина может быть заменено соответствующими донорными аминокислотами. Предпочтительные положения замен включают аминокислотные остатки, примыкающие к CDR, или остатки, способные к взаимодействию с CDR. Критерии для отбора аминокислот из донора раскрыты в патенте US №5585089 (например, колонки 12-16). Акцепторная каркасная область может представлять собой последовательность каркасной области зрелого антитела человека или согласованную последовательность.
При использовании здесь, «нейтрализующее антитело или его связывающий фрагмент» относится к соответствующему антителу или его связывающему фрагменту, который связывается с _атогенна_ (например, с TcdA или TcdB С.difficile) и снижает инфекционность и/или активность _атогенна (например, снижает цитотоксичность) у млекопитающего и/или в культуре клеток по сравнению с _атогенна_ в идентичных условиях в отсутствие нейтрализующего антитела или его связывающего фрагмента. В одном воплощении, нейтрализующее антитело или его связывающий фрагмент способны к нейтрализации биологической активности _атогенна по меньшей мере примерно на 70%, 75%, 80%, 85%, 90%, 95%, 99% или более по сравнению с биологической активностью _атогенна в идентичных условиях в отсутствие нейтрализующего антитела или его связывающего фрагмента.
При использовании здесь, термин «антитело против токсина или его связывающий фрагмент" относится к антителу или его связывающему фрагменту, которые связываются с соответствующим токсином С.difficile (например, с токсином А или токсином В С.difficile). Например, антитело против токсина А или его связывающий фрагмент относится к антителу или к его связывающему фрагменту, которые связываются с TcdA.
Антитела или их связывающие фрагменты, раскрытые здесь, можно индуцировать у любого млекопитающего, дикого типа и/или трансгенного, включая, например, мышей, людей, кроликов и коз.
Когда раскрытая выше иммуногенная композиция представляет собой композицию, которая ранее была введена популяции, например, для вакцинации, ответ антител, полученный у субъектов, можно применять для нейтрализации токсинов из того же штамма и из штамма, не стимулировавшего продуцирование антител. См., например, Пример 37, в котором показаны исследования, относящиеся к перекрестной реактивности, созданной иммуногенной композицией, между штаммом 630 и токсинами из различных штаммов С.difficile дикого типа.
В одном аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к TcdA С.difficile. Моноклональные антитела, специфично связывающиеся с TcdA, включают А65-33; А60-22; А80-29 и/или предпочтительно А3-25.
В одном аспекте изобретение относится к антителу или к его связывающему фрагменту, специфичному к TcdA из любого штамма С.difficile дикого типа, такого как штаммы, раскрытые выше, например, к SEQ ID NO:1. В другом аспекте изобретение относится к антителу или к его связывающему фрагменту, специфичному к раскрытой выше иммуногенной композиции. Например, в одном воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:4 или SEQ ID NO:7. В другом воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:4 или SEQ ID NO:7, в которой по меньшей мере одна аминокислота SEQ ID NO:4 или SEQ ID NO:7 поперечно-сшита с использованием формальдегида, EDC, NHS или комбинацией EDC и NHS. В другом воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:84 или SEQ ID NO:83.
Антитела или их связывающие фрагменты, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно на 98%, более предпочтительно примерно на 99% или наиболее предпочтительно примерно на 100% идентичные вариабельным областям тяжелой и легкой цепи А65-33; А60-22; А80-29 и/или, предпочтительно, А3-25, также могут связываться с TcdA.
В одном воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи А3-25, как описано в SEQ ID NO:37.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи А3-25, как описано в SEQ ID NO:36.
Еще в одном дополнительном аспекте антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи, описанной в SEQ ID NO:37, и вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи, описанной в SEQ ID NO: 36.
В другом воплощении антитела или их связывающие фрагменты, имеющие гипервариабельные участки (CDR) вариабельных тяжелых цепей и/или вариабельных легких цепей А65-33; А60-22; А80-29 и/или предпочтительно А3-25, могут также связываться с TcdA. Участки CDR вариабельной тяжелой цепи А3-25 показаны в таблице 4 ниже.
Участки CDR вариабельной области легкой цепи А3-25 показаны в таблице 5 ниже.
В одном воплощении антитело или его связывающий фрагмент включает аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи, как показано в SEQ ID NO: 41 (CDR H1), 42 (CDR H2) и 43 (CDR Н3), и/или аминокислотные последовательности CDR легкой цепи, как показано в SEQ ID NO:38 (CDR L1), 39 (CDR L2) и 40 (CDR L3).
В одном иллюстративном воплощении антитело или его связывающий фрагмент, специфичное к токсину А С.difficile, специфично связывается с эпитопом в пределах N-концевого участка TcdA, например, с эпитопом между аминокислотами 1-1256 TcdA, в соответствии с нумерацией SEQ ID NO:1.
В предпочтительном воплощении антитело или его связывающий фрагмент, специфичное к токсину А С.difficile, специфично связывается с эпитопом в пределах С-концевого участка токсина А, например, с эпитопом между аминокислотами 1832-2710 TcdA, в соответствии с нумерацией SEQ ID NO: 1. Примеры включают А3-25; А65-33; А60-22; А80-29.
Еще в одном другом воплощении антитело или связывающий фрагмент, специфичное к токсину А С.difficile, специфично связывается с эпитопом внутри участка "транслокации" токсина А С.difficile, например, с эпитопом, предпочтительно включающим остатки 956-1128 TcdA, в соответствии с нумерацией SEQ ID NO:1, например, с эпитопом между аминокислотами 659-1832 TcdA, в соответствии с нумерацией SEQ ID NO:1.
В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к TcdB С.difficile. Например, антитело или его связывающий фрагмент может обладать специфичностью к TcdB из любого штамма С.difficile дикого типа, такого как штаммы, раскрытые выше, например, к SEQ ID NO:2. В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к раскрытой выше иммуногенной композиции. Например, в одном воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:6 или SEQ ID NO:8.
В другом воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:6 или SEQ ID NO:8, в которой по меньшей мере одна аминокислота SEQ ID NO:6 или SEQ ID NO:8 является перекрестно-сшитой формальдегидом, EDC, NHS или комбинацией EDC и NHS. В другом воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:86 или SEQ ID NO:85.
Моноклональные антитела, специфично связывающиеся с TcdB, включают антитела, продуцируемые клонами В2-31; В5-40, В70-2; В6-30; В9-30; В59-3; В60-2; В56-6; и/или, предпочтительно, В8-26, раскрытыми здесь.
Антитела или их связывающие фрагменты, которые также могут связываться с TcdB, включают антитела, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно на 99% или наиболее предпочтительно примерно на 100% идентичные вариабельным областям тяжелой и легкой цепи В2-31; В5-40, В70-2; В6-30; В9-30; В59-3; В60-2; В56-6, предпочтительно В8-26, В59-3 и/или В9-30.
В одном воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи А3-25, как описано в SEQ ID NO:49.
В одном воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи А3-25, как описано в SEQ ID NO:60.
В одном воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи А3-25, как описано в SEQ ID NO:71.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи А3-25, как описано в SEQ ID NO:55.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую аминокислотная последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи А3-25, как описано в SEQ ID NO:66.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи А3-25, как описано в SEQ ID NO:77.
Аминокислотная последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb) описана в SEQ ID NO:49. См. таблицу 25-а.
Аминокислотная последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B8-26 mAb) описана в SEQ ID NO:55. См. таблицу 25-b.
В одном воплощении антитело или его связывающий фрагмент включает аминокислотные последовательности CDR тяжелой цепи, как показано в SEQ ID NO:51 (CDR H1), 52 (CDR H2) и 53 (CDR H3), и/или аминокислотные последовательности CDR легкой цепи, как показано в SEQ ID NO:57 (CDR L1), 58 (CDR L2) и 59 (CDR L3).
Аминокислотная последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B59-3 mAb) описана в SEQ ID NO:60. См. таблицу 26-а.
Аминокислотная последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (В59-3 mAb) описана в SEQ ID NO:66. См. таблицу 26-b.
В одном воплощении антитело или его связывающий фрагмент включает аминокислотные последовательности CDR тяжелой цепи, как показано в SEQ ID NO:62 (CDR H1), 63 (CDR H2) и 64 (CDR H3), и/или аминокислотные последовательности CDR легкой цепи, как показано в SEQ ID NO:68 (CDR L1), 69 (CDR L2) и 70 (CDR L3).
Аминокислотная последовательность вариабельной тяжелой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb) описана в SEQ ID NO:71. См. таблицу 27-а.
Аминокислотная последовательность вариабельной легкой цепи нейтрализующего антитела к TcdB С.difficile (B9-30 mAb) описана в SEQ ID NO:77. См. таблицу 27-b.
В одном воплощении антитело или его связывающий фрагмент включает аминокислотные последовательности CDR тяжелой цепи, как показано в SEQ ID NO:73 (CDR H1), 74 (CDR H2) и 75 (CDR H3), и/или аминокислотные последовательности CDR легкой цепи, как показано в SEQ ID NO:79 (CDR L1), 80 (CDR L2) и 81 (CDR L3).
В одном аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к TcdB С.difficile из любого штамма С.difficile дикого типа, такого как штаммы, раскрытые выше, например, к SEQ ID NO: 2. В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к раскрытой выше иммуногенной композиции. Например, в одном воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:6 или SEQ ID NO:8. В другом воплощении антитело или его связывающий фрагмент являются специфичными к иммуногенной композиции, включающей SEQ ID NO:6 или SEQ ID NO:8, в которой по меньшей мере одна аминокислота SEQ ID NO:6 или SEQ ID NO:8 поперечно-сшита с использованием формальдегида, EDC, NHS или комбинацией EDC и NHS.
Антитела или их связывающие фрагменты, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно на 98%, более предпочтительно примерно на 99% или наиболее предпочтительно примерно на 100% идентичные вариабельным областям тяжелой и легкой цепи В2-31; В5-40, В70-2; В6-30; В9-30; В59-3; В60-2; В56-6 и/или предпочтительно В8-26, также могут связываться с TcdB.
В одном воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи В8-26 (SEQ ID NO:49).
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи В8-26 (SEQ ID NO:55).
Еще в одном дополнительном аспекте антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи В8-26 (SEQ ID NO:49), и вариабельную область легкой цепи, включающую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи В8-26 (SEQ ID NO:55).
В другом воплощении антитела или их связывающие фрагменты, имеющие CDR вариабельных тяжелых цепей и/или вариабельных легкий цепей В2-31; В5-40, В70-2; В6-30; В9-30; В59-3; В60-2; В56-6 и/или предпочтительно В8-26, могут также связываться с TcdB.
В одном воплощении антитело или его связывающий фрагмент включает аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи В8-26 и/или аминокислотные последовательности CDR легкой цепи В8-26.
В предпочтительном воплощении антитело или его связывающий фрагмент, специфичный к токсину В С.difficile, специфично связывается с эпитопом в пределах N-концевого участка токсина В, например, с эпитопом между аминокислотами 1-1256 TcdB, в соответствии с нумерацией SEQ ID NO:2. Примеры включают В2-31; В5-40; В8-26; В70-2; В6-30 и В9-30.
В иллюстративном воплощении антитело или его связывающий фрагмент, специфичное к токсину В С.difficile, специфично связывается с эпитопом в пределах С-концевого участка токсина В, например, с эпитопом между аминокислотами 1832-2710 TcdB, в соответствии с нумерацией SEQ ID NO:2.
Еще в одном другом воплощении антитело или его связывающий фрагмент, специфичное к токсину В С.difficile, специфично связывается с эпитопом в пределах участка "транслокации" токсина В С.difficile, например, с эпитопом, предпочтительно включающим остатки 956-1128 TcdB, в соответствии с нумерацией SEQ ID NO:2, например, с эпитопом между аминокислотами 659-1832 TcdB. Примеры включают В59-3; В60-2 и В56-6.
Комбинации антител
Антитело против токсина или его связывающий фрагмент можно вводить в комбинации с другими антителами против токсинов С.difficile (например, с другими моноклональными антителами, поликлональным гамма-глобулином) или их связывающими фрагментами. Комбинации, которые можно применять, включают антитело против токсина А или его связывающий фрагмент и антитело против токсина В или его связывающий фрагмент.
В другом воплощении, комбинация включает антитело против токсина А или его связывающий фрагмент и другое антитело против токсина А или его связывающий фрагмент.Предпочтительно комбинация включает нейтрализующее моноклональное антитело против токсина А или его связывающий фрагмент и другое нейтрализующее моноклональное антитело против токсина А или его связывающий фрагмент.Неожиданно авторы изобретения обнаружили, что такая комбинация приводит в результате к синергетическому эффекту при нейтрализации цитотоксичности токсина А. Например, комбинация включает комбинацию по меньшей мере двух из следующих нейтрализующих моноклональных антител против токсина А: А3-25; А65-33; А60-22 и А80-29. Более предпочтительно, комбинация включает антитело А3-25 и по меньшей мере одно из следующих нейтрализующих моноклональных антител против токсина А: А65-33; А60-22 и А80-29. Наиболее предпочтительно, комбинация включает все четыре антитела: А3-25; А65-33; А60-22 и А80-29.
В следующем воплощении, комбинация включает антитело против токсина В или его связывающий фрагмент и другое антитело против токсина В или его связывающий фрагмент.Предпочтительно комбинация включает нейтрализующее моноклональное антитело против токсина В или его связывающий фрагмент и другое нейтрализующее моноклональное антитело против токсина В или его связывающий фрагмент.Неожиданно авторы изобретения обнаружили, что такая комбинация приводит в результате к синергетическому эффекту при нейтрализации цитотоксичности токсина В. Более предпочтительно, комбинация включает комбинацию по меньшей мере двух из следующих нейтрализующих моноклональных антител против токсина В: В8-26; В9-30 и В59-3. Наиболее предпочтительно, комбинация включает все три антитела: В8-26; В9-30 и В59-3.
Еще в одном другом воплощении, комбинация включает антитело против токсина В или его связывающий фрагмент и другое антитело против токсина В или его связывающий фрагмент. Как указано выше, авторы изобретения обнаружили, что комбинация по меньшей мере двух из нейтрализующих моноклинальных антител может проявлять неожиданный синергетический эффект при соответствующей нейтрализации токсина А и токсина В.
В другом воплощении, агенты по изобретению можно включать в препарат в виде смеси, либо химически или генетически сшитыми с использованием методов, известных в данной области техники, с получением в результате ковалентно сшитых антител (или ковалентно сшитых фрагментов антител), обладающих связывающими свойства как с токсином А, так и с токсином В. При приготовлении комбинированного препарата можно руководствоваться определением одного или более чем одного из параметров, таких как сродство, авидность или биологическая эффективность одного агента или в комбинации с другим агентом.
Такие комбинированные терапии предпочтительно являются аддитивными и/или синергетическими по своей терапевтической активности, например, при ингибировании, предупреждении (например, рецидива) и/или лечении заболеваний или расстройств, обусловленных С.difficile. Введение таких комбинированных терапий может снизить дозировку терапевтического агента (например, смеси антител или фрагментов антител, либо поперечно-сшитого или генетически слитого биспецифического антитела или фрагмента антитела), необходимую для достижения желаемого эффекта.
Понятно, что любую из композиций по изобретению, например, антитело против токсина А и/или против токсина В или его связывающий фрагмент, можно комбинировать в различных соотношениях или количествах для терапевтического эффекта. Например, антитела против токсина А и/или против токсина В или их соответствующие связывающие фрагменты могут присутствовать в композиции в соотношении, находящемся в диапазоне от 0,1:10 до 10:0,1 А:В. В другом воплощении, антитела против токсина А и/или против токсина В или их соответствующие связывающие фрагменты могут присутствовать в композиции в соотношении, находящемся в диапазоне от 0,1:10 до 10:0,1 В:А.
В другом аспекте изобретение относится к способу получения нейтрализующего антитела против TcdA С.difficile. Этот способ включает введение иммуногенной композиции, как раскрыто выше, млекопитающему и выделение антитела из млекопитающего. В предпочтительном воплощении иммуногенная композиция включает мутантный TcdA С.difficile, имеющий SEQ ID NO:4, где по меньшей мере одна аминокислота мутантного TcdA С.difficile поперечно-сшита химическим образом, предпочтительно с использованием формальдегида или 1-этил-3-(3-диметиламинопропил)карбодиимида. Иллюстративные нейтрализующие антитела против TcdA, которые могут быть получены, включают следующие антитела: А65-33; А60-22; А80-29 и/или А3-25.
Еще в одном другом аспекте изобретение относится к способу получения нейтрализующего антитела против TcdB С.difficile. Этот способ включает введение иммуногенной композиции, как раскрыто выше, млекопитающему и выделение антитела из млекопитающего. В предпочтительном воплощении, иммуногенная композиция включает мутантный TcdB С.difficile, имеющий SEQ ID NO:6, где по меньшей мере одна аминокислота мутантного TcdB С.difficile поперечно-сшита химическим образом, предпочтительно с использованием формальдегида или 1-этил-3-(3-диметиламинопропил)карбодиимида. Иллюстративные нейтрализующие антитела против TcdB, которые могут быть получены, включают следующие антитела: В2-31; В5-40, В70-2; В6-30; В9-30; В59-3; В60-2; В56-6 и/или В8-26.
Препараты
Композиции по настоящему изобретению (такие как, например, композиции, включающие мутантный токсин С.difficile, иммуногенные композиции, антитела и/или связывающие фрагменты антител, раскрытые здесь) могут находиться в разнообразных формах. Эти формы включают, например, полужидкие и твердые лекарственные формы, суппозитории, жидкие формы, такие как жидкие растворы (например, инъекционные и инфузионные растворы), дисперсии или суспензии, липосомы, и/или сухую форму, такую как, например, лиофилизированная порошкообразная форма, форма, полученная сублимационной сушкой, форма, полученная распылительной сушкой, и/или форма, высушенная во вспененном состоянии. Для суппозиториев связующие агенты и носители включают, например, полиалкиленгликоли или триглицериды; такие суппозитории можно формовать из смесей, содержащих композиции по изобретению. В иллюстративном воплощении композиция находится в форме, пригодной для растворения или для суспендирования в жидких носителях перед инъекцией. В другом иллюстративном воплощении композицию эмульгируют или инкапсулируют в липосомы или микрочастицы, например, из полилактида, полигликолида или их сополимера.
В предпочтительном воплощении композицию лиофилизируют и восстанавливают непосредственно перед применением.
В одном аспекте настоящее изобретение относится к фармацевтическим композициям, включающим любую из композиций, раскрытых здесь (такую как, например, композиции, включающие мутантный токсин С.difficile, иммуногенные композиции, антитела и/или связывающие фрагменты антител, раскрытые здесь), включенную в препарат вместе с фармацевтически приемлемым носителем. "Фармацевтически приемлемые носители" включают любые растворители, дисперсионные среды, стабилизаторы, разбавители и/или буферы, которые являются физиологически пригодными.
Примеры стабилизаторов включают углеводы, такие как сорбит, маннит, крахмал, декстран, сахароза, трегалоза, лактоза и/или глюкоза; инертные белки, такие как альбумин и/или казеин; и/или другие крупные, медленно метаболизируемые макромолекулы, такие как полисахариды, например, хитозан, полимеры молочной кислоты, полимеры гликолевой кислоты и их сополимеры (такие как агароза SEPHAROSE™, функционализированная латексом, агароза, целлюлоза и т.д.), аминокислоты, полимерные аминокислоты, сополимеры аминокислот и липидные агрегаты (такие как капельки масла или липосомы). Дополнительно эти носители могут действовать как иммуностимулирующие агенты (то есть адъюванты).
Предпочтительно композиция включает трегалозу. Предпочтительные количества трегалозы (масс.%) составляют от минимального значения примерно 1%, 2%, 3% или 4% до максимального значения примерно 10%, 9%, 8%, 7%, 6% или 5%. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении композиция включает примерно 3%-6% трегалозы, наиболее предпочтительно 4,5% трегалозы, например, на дозу, составляющую 0,5 мл.
Примеры подходящих разбавителей включают дистиллированную воду, физиологический раствор, фосфатно-солевой буферный раствор, глицерин, спирт (такой как этанол), растворы Рингера, раствор декстрозы, сбалансированные солевые растворы Хэнкса и/или эксципиент для лиофилизации.
Иллюстративные буферы включают фосфатные (такие как фосфат калия, фосфат натрия); ацетатные (такие как ацетат натрия); сукцинатные (такие как сукцинат натрия); глициновые; гистидиновые; карбонатные, Трис (трис(гидроксиметил)аминометан) и/или бикарбонатные (такие как бикарбонат аммония) буферы. Предпочтительно композиция включает Трис-буфер. Предпочтительные количества Трис-буфера составляют от минимального значения примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ до максимального значения примерно 100 мМ, 50 мМ, 20 мМ,19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении композиция включает примерно 8 мМ-12 мМ Трис-буфера, наиболее предпочтительно 10 мМ Трис-буфера, например, на дозу, составляющую 0,5 мл.
В другом предпочтительном воплощении композиция включает гистидиновый буфер. Предпочтительные количества гистидинового буфера составляют от минимального значения примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ до максимального значения примерно 100 мМ, 50 мМ, 20 мМ,19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении композиция включает примерно 8 мМ-12 мМ гистидинового буфера, наиболее предпочтительно 10 мМ гистидинового буфера, например, на дозу, составляющую 0,5 мл.
Еще в одном другом предпочтительном воплощении композиция включает фосфатный буфер. Предпочтительные количества фосфатного буфера составляют от минимального значения примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ до максимального значения примерно 100 мМ, 50 мМ, 20 мМ, 19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении композиция включает примерно 8 мМ-12 мМ фосфатного буфера, наиболее предпочтительно, 10 мМ фосфатного буфера, например, на дозу, составляющую 0,5 мл.
Значение рН буфера, как правило, выбрано таким образом, чтобы стабилизировать выбранное активное вещество, и может быть установлено специалистами в данной области техники известными способами. Предпочтительно рН буфера находится в интервале физиологического значения рН. Таким образом, предпочтительные интервалы рН составляют от примерно 3 до примерно 8; более предпочтительно от примерно 6,0 до примерно 8,0; еще более предпочтительно от примерно 6,5 до примерно 7,5; и наиболее предпочтительно от примерно 7,0 до примерно 7,2.
В некоторых воплощениях фармацевтические композиции могут включать сурфактант. Подходит любой сурфактант независимо от того, является ли он амфотерным, неионным, катионным или анионным. Иллюстративные сурфактанты включают сурфактанты, представляющие собой полиоксиэтиленовые сложные эфиры сорбитана (Например, TWEEN®), такие как полисорбат 20 и/или полисорбат 80; простые эфиры полиоксиэтилена и жирных кислот, образованные из лаурилового, цетилового, стеарилового и олеилового спиртов (известные как сурфактанты Бридж), такие как монолауриловый эфир триэтиленгликоля (Бридж 30); Тритон Х 100 или трет-октилфеноксиполиэтоксиэтанол; и сложные эфиры сорбитана (общеизвестные как SPAN), такие как сорбитантриолеат (Span 85) и сорбитанмонолаурат, а также их комбинации. Предпочтительные сурфактанты включают полисорбат 80 (полиоксиэтиленсорбитанмоноолеат).
Предпочтительные количества полисорбата 80 (масс.%) составляют от минимального значения примерно 0,001%, 0,005% или 0,01% до максимального значения примерно 0,010%, 0,015%, 0,025% или 1,0%. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении композиция включает примерно 0,005%-0,015% полисорбата 80, наиболее предпочтительно 0,01% полисорбата 80.
В иллюстративном воплощении иммуногенная композиция включает трегалозу и фосфат 80. В другом иллюстративном воплощении иммуногенная композиция включает Трис-буфер и полисорбат 80. В другом иллюстративном воплощении иммуногенная композиция включает гистидиновый буфер и полисорбат 80. Еще в одном другом иллюстративном воплощении иммуногенная композиция включает фосфатный буфер и полисорбат 80.
В одном иллюстративном воплощении иммуногенная композиция включает трегалозу, Трис-буфер и полисорбат 80. В другом иллюстративном воплощении иммуногенная композиция включает трегалозу, гистидиновый буфер и полисорбат 80. Еще в одном другом иллюстративном воплощении иммуногенная композиция включает трегалозу, фосфатный буфер и полисорбат 80.
Композиции, раскрытые здесь, могут дополнительно включать ингредиенты нефтяного, животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло и/или минеральное масло. Примеры включают гликоли, такие как пропиленгликоль или полиэтиленгликоль.
В некоторых воплощениях фармацевтическая композиция дополнительно включает формальдегид. Например, в предпочтительном воплощении фармацевтическая композиция, дополнительно включающая формальдегид, имеет иммуногенную композицию, где мутантный токсин С.difficile иммуногенной композиции приведен в контакт с химическим поперечно-сшивающим агентом, включающим формальдегид. Количество формальдегида, присутствующее в фармацевтической композиции, может варьировать от минимального значения примерно 0,001%, 0,002%, 0,003%, 0,004%, 0,005%, 0,006%, 0,007%, 0,008%, 0,009%, 0,010%, 0,013% или 0,015% до максимального значения примерно 0,020%, 0,019%, 0,018%, 0,017% 0,016%, 0,015%, 0,014%, 0,013%, 0,012% 0,011% или 0,010%. Любое минимальное значение можно объединять с любым максимальным значением для определения подходящего диапазона. В одном воплощении фармацевтическая композиция включает примерно 0,010% формальдегида.
В некоторых альтернативных воплощениях фармацевтические композиции, раскрытые здесь, не включают формальдегид. Например, в предпочтительном воплощении фармацевтическая композиция, не включающая формальдегид, имеет иммуногенную композицию, где по меньшей мере одна аминокислота мутантного токсина С.difficile поперечно-сшита химическим образом агентом, включающим EDC. Более предпочтительно, в таком воплощении мутантный токсин С.difficile не был приведен в контакт с химическим поперечно-сшивающим агентом, включающим формальдегид. В качестве другого иллюстративного воплощения, фармацевтическая композиция, находящаяся в лиофилизированной форме, не включает формальдегид.
В другом воплощении, композиции, раскрытые здесь, могут включать адъювант, как описано ниже. Предпочтительные адъюванты усиливают собственный иммунный ответ на иммуноген, не вызывая конформационных изменений в иммуногене, которые могут повлиять на качественную форму иммунного ответа.
Примеры адъювантов включают 3-де-O-ацилированный монофосфориллипид A (MPL™) (см. GB 2220211 (GSK)); гель гидроксида алюминия, такой как Alhydrogel™ (Brenntag Biosector, Denmark); соли алюминия (такие как гидроксид алюминия, фосфат алюминия, сульфат алюминия), которые можно применять в присутствии или в отсутствие иммуностимулирующего агента, такого как MPL или 3-DMP, QS-21, полимерные или мономерные аминокислоты, такие как полиглутаминовая кислота или полилизин.
Еще одним другим иллюстративным адъювантом является иммуностимулирующий олигонуклеотид, такой как CpG олигонуклеотид (см., например, WO 1998/040100, WO 2010/067262), или сапонин и иммуностимулирующий олигонуклеотид, такой как CpG олигонуклеотид (см., например, WO 00/062800). В предпочтительном воплощении адъювант представляет собой CpG олигонуклеотид, наиболее предпочтительно CpG олигодезоксинуклеотиды (CpG ODN). Предпочтительные CpG ODN принадлежат к классу В, который преимущественно активирует В клетки. В аспектах изобретения CpG ODN имеет следующую нуклеиново-кислотную последовательность: 5′ T*C*G*T*C*G*T*T*T*T*T*C*G*G*T*G*C*T*T*T*T 3′ (SEQ ID NO:48), где * указывает на фосфотиоатную связь. CpG ODN, имеющий эту последовательность, известен как CpG 24555, который описан в документе WO 2010/067262. В предпочтительном воплощении CpG 24555 применяют вместе с солью гидроксидом алюминия, такой как Alhydrogel.
Следующий класс иллюстративных адъювантов включает сапониновые адъюванты, такие как Stimulon™ (QS-21, который представляет собой тритерпенгликозид или сапонин, Aquila, Framingham, Mass.) или полученные из него частицы, такие как адъювант ИСКОМ (иммуностимулирующие комплексы) и ISCOMATRIX®. Соответственно, композиции по настоящему изобретению могут быть доставлены в форме ИСКОМ, ИСКОМ, содержащих СТВ, липосом, либо инкапсулированными в соединениях, таких как акрилаты или сополимер поли-ОЬ-лактида и гликозида, с образованием микросфер, имеющих размер, подходящий для адсорбции. В характерном случае термин "ИСКОМ" относится к иммуногенным комплексам, образованным между гликозидами, такими как тритерпеновые сапонины (в частности, Quil А), и антигенами, содержащими гидрофобный участок. В предпочтительном воплощении адъювант представляет собой адъювант ISCOMATRIX.
Другие иллюстративные адъюванты включают RC-529, GM-CSF и полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA).
Еще одним классом иллюстративных адъювантов являются аналоги гликолипидов, включающие N-гликозиламины, N-гликозилмочевины и N-гликозилкарбаматы, каждый из которых замещен в остатке сахара аминокислотой.
Возможно, фармацевтическая композиция включает два или более чем два различных адъюванта. Предпочтительные комбинации адъювантов включают любую комбинацию адъювантов, включающую, например, по меньшей мере два из следующих адъювантов: квасцы, MPL, QS-21, ISCOMATRIX, CpG и Alhydrogel. Иллюстративная комбинация адъювантов включает комбинацию CpG и Alhydrogel.
Альтернативно, в одном воплощении композицию вводят млекопитающему в отсутствие адъюванта.
Композиции, раскрытые здесь, можно вводить посредством любого пути введения, такого как, например, парентеральный, местный, внутривенный, введение в слизистую оболочку, пероральный, подкожный, внутриартериальный, интракраниальный, подоболочечный, интраперитонеальный, интраназальный, внутримышечный, внутрикожный, инфузионный, ректальный и/или чрескожный путь, для профилактических и/или терапевтических применений. В предпочтительном воплощении путь введения композиции представляет собой парентеральный путь, более предпочтительно внутримышечное введение. Типично внутримышечное введение осуществляют в мышцы руки или ноги.
Композиции, раскрытые здесь, можно вводить в комбинации с терапиями, по меньшей мере частично эффективными при предупреждении и/или лечении инфекции С.difficile. Например, композицию по изобретению можно вводить до, во время или после биотерапии; пробиотической терапии; кишечных имплантатов (stool implants); иммунотерапии (такой как внутривенный иммуноглобулин) и/или общепринятого стандарта лечения заболевания, обусловленного С.difficile (CDAD), антибиотиками, такими как метронидазол и/или ванкомицин.
Композицию по настоящему изобретению, относящуюся к токсину А и токсину В, можно вводить млекопитающему в любой комбинации. Например, иммуногенную композицию, включающую мутантный TcdA С.difficile, можно вводить млекопитающему до, во время или после введения иммуногенной композиции, включающей мутантный TcdB С.difficile. Наоборот, иммуногенную композицию, включающую мутантный TcdB С.difficile, можно вводить млекопитающему до, во время или после введения иммуногенной композиции, включающей мутантный TcdA С.difficile.
В другом воплощении композицию, включающую антитело против токсина А или его связывающий фрагмент, можно вводить млекопитающему до, во время или после введения композиции, включающей антитело против токсина В или его связывающий фрагмент. Наоборот, композицию, включающую антитело против токсина В или его связывающий фрагмент, можно вводить млекопитающему до, во время или после введения композиции, включающей антитело против токсина А или его связывающий фрагмент.
В следующем воплощении композицию по настоящему изобретению можно вводить млекопитающему до, во время или после введения фармацевтически приемлемого носителя. Например, адъювант можно вводить до, во время или после введения композиции, включающей мутантный токсин С.difficile. Соответственно, композиция по настоящему изобретению и фармацевтически приемлемый носитель могут быть упакованы в один флакон, применением. Препараты композиций можно готовить для введения однократной дозы и/или для введения многократных доз.
Способы защиты от инфекции С.difficile и/или лечения инфекции С.difficile у млекопитающего
В одном аспекте изобретение относится к способу индукции иммунного ответа на токсин С.difficile у млекопитающего. Этот способ включает введение млекопитающему эффективного количества композиции, раскрытой здесь. Например, этот способ может включать введение количества, эффективного для получения у млекопитающего иммунного ответа на соответствующий токсин С.difficile.
В иллюстративном воплощении изобретение относится к способу индукции у млекопитающего иммунного ответа на TcdA С.difficile. Этот способ включает введение млекопитающему эффективного количества иммуногенной композиции, включающей мутантный TcdA С.difficile. В другом иллюстративном воплощении изобретение относится к способу индукции у млекопитающего иммунного ответа на TcdB С.difficile. Этот способ включает введение млекопитающему эффективного количества иммуногенной композиции, включающей мутантный TcdB С.difficile.
В следующем воплощении способ включает введение млекопитающему эффективного количества иммуногенной композиции, включающей мутантный TcdA С.difficile, и эффективного количества иммуногенной композиции, включающей мутантный TcdB С.difficile. В дополнительных аспектах композиции, раскрытые здесь, можно применять для лечения, предупреждения, снижения риска, снижения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения у млекопитающего по сравнению с млекопитающим, которому не вводят эту композицию. Способ включает введение млекопитающему эффективного количества композиции.
На основании тяжести инфекции распознают три клинических симптома, обусловленных инфекцией С.difficile. Самой тяжелой формой является псевдомембранозный колит (ПМК), характеризующийся профузной диареей, болью в животе, системными признаками заболевания и отчетливой эндоскопической картиной ободочной кишки.
Колит, связанный с применением антибиотиков (ААС) также характеризуется профузной диареей, болью в животе и болезненностью при пальпации, системными признаками (например, лихорадкой) и лейкоцитозом. Повреждение кишечника при ААС является менее тяжелым, чем при ПМК, характерная эндоскопическая картина ободочной кишки при ПМК отсутствует, и смертность является низкой.
Наконец, диарея, связанная с применением антибиотиков (AAD, также известная как диарея, обусловленная С.difficile (CDAD)) представляет собой относительно легкия синдром и характеризуется диареей, от легкой до среднетяжелой, при отсутствии как обширного воспаления кишечника (которое характеризуется, например, болью в животе и болезненностью при пальпации), так и системных признаков инфекции (например, лихорадки).
Эти три отдельных синдрома в характерном случае встречаются в возрастающем порядке частоты. То есть, ПМК в характерном случае встречается реже, чем ААС, а AAD в характерном случае является наиболее частым клиническим проявлением заболевания, обусловленного С.difficile.
Частым осложнением инфекции С.difficile является повторное или рецидивирующее заболевание, которое встречается у пациентов в количестве вплоть до 20% всех субъектов, выздоровевших после заболевания С.difficile. Рецидив может характеризоваться клинически как AAD, ААС или ПМК. У пациентов, у которых рецидив возник один раз, более вероятно снова возникает рецидив.
При использовании здесь, состояния инфекции С.difficile включают, например, легкую, от легкой до среднетяжелой, среднетяжелую и тяжелую инфекцию С.difficile. Состояние инфекции С.difficile может варьировать в зависимости от присутствия и/или тяжести симптомов инфекции.
Симптомы инфекции С.difficile могут включать физиологические, биохимические, гистологические и/или поведенческие симптомы, такие как, например, диарея; колит; колит, сопровождающийся спазмами, лихорадкой, лейкоцитами в фекалиях и воспалением, выявляемом при биопсии ободочной кишки; псевдомембранозный колит; гипоальбуминемия; анасарка; лейкоцитоз; сепсис; боль в животе; бессимптомное носительство и/или осложнения и промежуточные патологические фенотипы, присутствующие во время развития инфекции, а также их комбинации и т.д. Соответственно, например, введение эффективного количества композиций, раскрытых здесь, может, например, лечить, снижать риск, снижать тяжесть, снижать частоту встречаемости и/или замедлять возникновение следующих симптомов: диареи; боли в животе, спазмов, лихорадки, воспаления, выявляемого при биопсии ободочной кишки, гипоальбуминемии, анасарки; лейкоцитоза; сепсиса и/или бессимптомного носительства и т.д. по сравнению с млекопитающим, которому не вводили композицию.
Факторы риска инфекции С.difficile могут включать, например, применение в настоящем или в ближайшем будущем антибактериального средства (включая любое антибактериальное средство, обладающее антибактериальным спектром действия и/или активностью против анаэробных бактерий, включая, например, антибиотики, которые нарушают нормальную кишечную микрофлору, например, клиндамицин, цефалоспорины, метронидазол, ванкомицин, фторхинолоны (включающие левофлоксацин, моксифлоксацин, гатифлоксацин и ципрофлоксацин), линезолид и т.д.); отмена в настоящем или в ближайшем будущем назначенного метронидазола или ванкомицина; госпитализация в настоящем или в ближайшем будущем в лечебное учреждение (такое как больница, учреждение по уходу за хроническими больными и т.д.) и медицинские работники; лечение в настоящем или в ближайшем будущем ингибиторами протонного насоса, антагонистами Н2 и/или метотрексатом, либо их комбинацией; наличие или риск желудочно-кишечных заболеваний, таких как воспалительное кишечное заболевание;
желудочно-кишечная операция или процедура на желудочно-кишечном тракте у млекопитающего в прошлом, настоящем или ближайшем будущем; рецидив инфекции С.difficile и/или CDAD в прошлом или в настоящем, например, пациенты, страдавшие инфекцией С.difficile и/или CDAD один или более чем один раз; а также люди в возрасте по меньшей мере примерно 65 лет и старше.
В способах, раскрытых здесь, млекопитающее может представлять собой любое млекопитающее, такое как, например, мыши, хомячки, приматы и люди. В предпочтительном воплощении млекопитающее представляет собой человека. Согласно настоящему изобретению, человек может включать индивидуумов, у которых проявлялась инфекция С.difficile, заболевание, обусловленное С.difficile, его синдром, состояние, симптом и/или осложнение; индивидуумов, у которых в настоящее время проявляется инфекция С.difficile, заболевание, обусловленное С.difficile, его синдром, состояние, симптом и/или осложнение; и индивидуумов, имеющих риск инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения.
Примеры индивидуумов, у которых проявились симптомы инфекции С.difficile, включают индивидуумов, у которых проявились или проявляются описанные выше симптомы; индивидуумов, страдавших или страдающих инфекцией С.difficile и/или заболеванием, обусловленным С.difficile (CDAD); и индивидуумов, имевших рецидив инфекции С.difficile и/или CDAD.
Примеры пациентов, имеющих риск инфекции С.difficile, включают индивидуумов, имеющих риск планового применения или проходящих в настоящее время плановое применение антибактериальных средств; индивидуумов, имеющих риск отмены или проходящих в настоящее время отмену назначенного метронидазола или ванкомицина; индивидуумов, имеющих риск госпитализации или в настоящее время госпитализированных в медицинское учреждение (такое как больница, учреждение по уходу за хроническими больными и т.д.), и медицинских работников; и/или индивидуумов, имеющих риск планового лечения или проходящих в настоящее время плановое лечение ингибиторами протонного насоса, антагонистами Н2 и/или метотрексатом, либо их комбинацией; индивидуумов, страдавших или страдающих желудочно-кишечными заболеваниями, такими как воспалительное кишечное заболевание; индивидуумов, прошедших или проходящих желудочно-кишечную операцию или процедуры на желудочно-кишечном тракте; и индивидуумов, страдавших или страдающих рецидивом инфекции С.difficile и/или а CDAD, например, пациентов, страдавших инфекцией С.difficile и/или CDAD один или более чем один раз; а также людей в возрасте по меньшей мере примерно 65 лет и старше. У таких пациентов, имеющих риск, в настоящее время могут проявляться или не проявляться симптомы инфекции С.difficile.
У бессимптомных пациентов профилактику и/или лечение можно начинать в любом возрасте (например, примерно в возрасте 10, 20 или 30 лет). В одном воплощении, тем не менее, нет необходимости начинать лечение до тех пор, пока пациент не достигнет возраста по меньшей мере примерно 45, 55, 65, 75 или 85 лет. Например, композиции, раскрытые здесь, можно вводить человеку, не имеющему симптомов, в возрасте 50-85 лет.
В одном воплощении способ предупреждения, снижения риска, снижения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения у млекопитающего включает введение эффективного количества композиции, раскрытой здесь, млекопитающему, нуждающемуся в этом, млекопитающему, имеющему риск инфекции и/или млекопитающему, склонному к инфекции С.difficile. Эффективное количество включает, например, количество, достаточное для предупреждения, снижения риска, снижения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения у млекопитающего по сравнению с млекопитающим, которому не вводят композицию. Введение эффективного количества композиций, раскрытых здесь, может, например, предупреждать, снижать риск, снижать тяжесть, снижать частоту встречаемости и/или замедлять возникновение следующих симптомов: диареи; боли в животе, спазмов, лихорадки, воспаления, выявляемого при биопсии ободочной кишки, гипоальбуминемии, анасарки; лейкоцитоза; сепсиса и/или бессимптомного носительства и т.д. по сравнению с млекопитающим, которому не вводили композицию. В предпочтительном воплощении способ включает введение эффективного количества иммуногенной композиции, раскрытой здесь, млекопитающему, нуждающемуся в этом, млекопитающему, обладающему риском инфекции и/или млекопитающему, склонному к инфекции С.difficile.
В дополнительном воплощении способ лечения, снижения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения у млекопитающего включает введение эффективного количества композиции, раскрытой здесь, млекопитающему с подозрением на инфекцию или страдающему в настоящее время инфекцией С.difficile. Эффективное количество включает, например, количество, достаточное для лечения, снижения тяжести, снижения частоты встречаемости и/или замедления возникновения инфекции С.difficile, заболевания, обусловленного С.difficile, его синдрома, состояния, симптома и/или осложнения у млекопитающего по сравнению с млекопитающим, которому не вводят композицию.
Введение эффективного количества композиции может улучшить по меньшей мере один признак или симптом инфекции С.difficile у субъекта, такого как описанные выше субъекты. Введение эффективного количества композиций, раскрытых здесь, может, например, снизить тяжесть и/или снизить частоту встречаемости следующих симптомов: диареи; боли в животе, спазмов, лихорадки, воспаления, выявляемого при биопсии ободочной кишки, гипоальбуминемии, анасарки; лейкоцитоза; сепсиса и/или бессимптомного носительства и т.д. по сравнению с млекопитающим, которому не вводили композицию. Возможно, наличие симптомов, признаков и/или факторов риска инфекции определяют перед началом лечения. В предпочтительном воплощении способ включает введение эффективного количества антитела и/или его связывающего фрагмента, раскрытого здесь, млекопитающему с подозрением на инфекцию или страдающему в настоящее время инфекцией С.difficile.
Соответственно, эффективное количество композиции относится к количеству, достаточному для достижения желаемого эффекта (например, профилактического и/или терапевтического эффекта) при способах по настоящему изобретению. Например, количество иммуногена для введения может варьировать от минимального значения примерно 1 мкг, 5 мкг, 25 мкг, 50 мкг, 75 мкг, 100 мкг, 200 мкг, 500 мкг или 1 мг до максимального значения примерно 2 мг, 1 мг, 500 мкг, 200 мкг на инъекцию. Любое минимальное значение можно объединять с любым максимальным значением, чтобы определить подходящий диапазон. В характерном случае применяют примерно 10, 20, 50 или 100 мкг на иммуноген на каждую инъекцию человеку.
Количество композиции по изобретению, вводимое субъекту, может зависеть от типа и тяжести инфекции и/или от характеристик индивидуума, таких как общее состояние здоровья, возраст, пол, масса тела и переносимость лекарственных средств. Оно может также зависеть от степени, тяжести и типа заболевания. Эффективное количество может также варьировать в зависимости от таких факторов, как путь введения, сайт-мишень, физиологическое состояние пациента, возраст пациента, представляет ли собой пациент человека или животное, другие вводимые терапии, и является ли лечение профилактическим или терапевтическим. Специалист в данной области техники способен определить соответствующие количества в зависимости от этих и других факторов.
Эффективное количество может включать одну эффективную дозу или множество эффективных доз (такое как, например, 2, 3, 4 дозы или более) для применения в способах, раскрытых здесь. Эффективные дозировки может быть необходимо титровать для оптимизации безопасности и эффективности.
Сочетание количества и частоты дозы, адекватное для осуществления профилактических и/или терапевтических применений, определяют как профилактически или терапевтически эффективную схему. При профилактической и/или терапевтической схеме композицию в характерном случае вводят более чем в одной дозе до достижения достаточного иммунного ответа. В характерном случае проводят мониторинг иммунного ответа, и повторные дозы вводят, если иммунный ответ начинает ослабевать.
Композиции можно вводить в многократных дозах в течение периода времени. Мониторинг лечения можно осуществлять путем анализа антитела или активированных Т-клеточных или В-клеточных ответов на терапевтический агент (например, на иммуногенную композицию, включающую мутантный токсин С.difficile) со временем. Если ответ снижается, назначают бустерную дозу.
ПРИМЕРЫ
Пример 1: Идентификация токсин-отрицательных штаммов С.difficile
Чтобы идентифицировать штаммы С.difficile, в которых отсутствуют гены токсинов (А и В) и экспрессия токсина, было протестировано 13 штаммов С.difficile. Культуральную среду 13 штаммов С.difficile тестировали с использованием твердофазного иммуноферментного анализа ELISA на токсин А. Токсин А экспрессировался в следующих семи штаммах: С.difficile 14797-2, С.difficile 630, С.difficile BDMS, С.difficile W1194, С.difficile 870, С.difficile 1253 и С.difficile 2149. См. фиг.3.
Токсин А не экспрессировался, и отсутствовал полноразмерный локус патогенности в следующих шести штаммах: С.difficile 1351 (АТСС 43593™), С.difficile 3232 (АТСС ВАА-1801™), С.difficile 7322 (АТСС 43601™), С.difficile 5036 (АТСС 43603™), С.difficile 4811 (АТСС 3602™) и С.difficile VPI 11186 (АТСС 700057™). Штамм VPI 11186 был отобран на основании его эффективности при захвате плазмидной ДНК посредством конъюгации.
Те же 13 штаммов были протестированы в мультиплексном анализе ПЦР с использованием праймеров, расположенных снаружи локуса патогенности (PaLoc; Braun et al., Gene. 1996 Nov 28;181(1-2):29-38.). Результаты ПЦР показали, что ДНК из 6 штаммов, отрицательных по токсину А на основании ELISA, не амплифицировали какие-либо гены из PaLoc (tcdA-tcdE). Присутствовали последовательности, фланкирующие PaLoc (cdd3 и cdu2) (данные не представлены).
Пример 2: Инактивация пути споруляции в штамме С.difficile VPI 11186
Нокаутирование спорообразующей функции штамма-продуцента С.difficile способствует крупномасштабной ферментации в безопасных рабочих условиях. Для создания аспорогенного штамма С.difficile использовали систему ClosTron. См. Heap et al., J Microbiol Methods. 2009 Jul;78(1):79-85. Система ClosTron дает возможность направленной инактивации гена интроном группы II для сайт-направленной инсерционной инактивации клостридиального гена spo0A1. Токсин-отрицательный штамм-продуцент VPI11186 подвергали инактивации споруляции с использованием технологии ClosTron. Были отобраны мутанты, устойчивые к эритромицину, и присутствие инсерционной кассеты подтверждали с использованием ПЦР (не показано). Была подтверждена неспособность к образованию спор для двух клонов.
Пример 3: Генетическая модификация генов токсинов А и В для инактивации цитотоксической функции
Полноразмерные открытые рамки считывания (ОРС) мутантных токсинов А и В, основанные на геномных последовательностях штамма 630Д, были сконструированы для синтеза на заказ в фирме Blue Heron Biotech. См., например, SEQ ID NO:9-14. Активный центр для глюкозилтрансферазной активности, ответственный за клеточную цитотоксичность, был изменен двумя аллельными заменами: D285A/D287A (см. SEQ ID NO:3) для токсина А и D286A/D288A (см. SEQ ID NO:5) для токсина В. Два нуклеотида были мутированы в каждом кодоне аспартата (D) с образованием кодона для аланина (А). См., например, SEQ ID NO:9-14. Кроме того, была сконструирована пара векторов, экспрессирующих мутантные токсины, в которых отсутствуют остатки цистеина, путем выполнения синтеза на заказ в фирме Blue Heron Biotech. Семь остатков цистеина из мутантного токсина А и 9 остатков цистеина из мутантного токсина В были заменены аланином. Эти замены включали каталитические цистеины аутокаталитической протеазы токсинов А и В. При необходимости также были введены молчащие мутации, чтобы удалить сайты ферментов рестрикции, используемые для конструирования векторов.
Пример 4: Экспрессионный вектор pMTL84121fdx
Плазмидный челночный вектор, используемый для экспрессии антигена, представляющего собой мутантный токсин С.difficile, был выбран из модульной системы серии pMTL8000, разработанной лабораторией Minton (см. Heap et al., J Microbiol Methods. 2009 Jul;78(1):79-85). Выбранный вектор pMTL84121fcfx содержит репликон плазмиды pCD6 Грам+ С.difficile, селективный маркер catP (хлорамфеникол/триамфеникол), репликон р15а Грам- и функцию tra, а также промотор ферредоксина С.sporogenes (fdx) и дистальный множественный сайт клонирования (МСК). Эмпирические данные позволяют предположить, что низкокопийный репликон р15а придает более высокую стабильность в Е. coli, чем альтернативный репликон ColE1. Промотор fdx был выбран, поскольку он обеспечивает более высокую экспрессию, чем другие промоторы, протестированные в экспериментах с репортерной конструкцией CAT (например, tcdA, tcdB или гетерологичные гены tetR или xyIR) (данные не представлены).
Пример 5: Клонирование модифицированных ОРС токсинов в векторе pMTL84121fdx
Полноразмерные открытые рамки считывания (ОРС) мутантных токсинов А и В, основанные на геномных последовательностях штамма 630Δ, были субклонированы с использованием сайтов NdeI и BglII множественного сайта клонирования вектора pMTI-84121fdx, используя стандартные методы молекулярной биологии. Чтобы облегчить клонирование, ОРС были фланкированы проксимальным сайтом NdeI, содержащим стартовый кодон, и сайтом BgIII, расположенным непосредственно ниже стоп-кодона.
Пример 6: Свит-направленный мутагенез TcdA с получением тройного мутанта
Каталитический остаток цистеина домена аутокаталитической протеазы был заменен (то есть С700А для TcdA и С698А для TcdB) в SEQ ID NO:3 и 5, то есть в каждом из "двойных мутантов". Для мутагенеза мутантного токсина А фрагмент NdeI-HindIII длиной 2,48 т.п.н. из экспрессионной плазмиды TcdA D285A/D287A субклонировали в векторе pUC19 (рестрицированном теми же ферментами рестрикции), и на этой матрице проводили сайт-направленный мутагенез. Как только новые аллели были подтверждены анализом последовательности ДНК, модифицированные фрагменты NdeI-HindIII повторно вводили в экспрессионный вектор pMTI-84121fdx для создания "тройных мутантов", то есть SEQ ID NO:4 и SEQ ID NO:6.
Пример 7: Сайт-направленный мутагенез TcdB с получением тройного мутанта
Для мутагенеза мутантного токсина В фрагмент NdeI-EcoNI длиной 3,29 т.п.н. из плазмиды, несущей мутантный токсин В, модифицировали и вводили повторно. Поскольку сайт EcoNI отсутствует в доступных клонирующих векторах, фрагмент NdeI-EcoRV несколько большего размера, составляющего 3,5 т.п.н., субклонировали в векторе pUC19 (подготовленном путем рестрикции NdeI-SmaI). После мутагенеза модифицированный внутренний фрагмент NdeI-EcoNI размером 3,3 т.п.н. вырезали и использовали для замены соответствующего фрагмента экспрессионного вектора мутантного токсина В pMTL84121fdx. Поскольку было обнаружено, что эффективность клонирования данной направленной стратегии достаточно низка, параллельно была предпринята альтернативная стратегия введения аллеля С698А, включающая замену фрагмента DraIII размером 1,5 т.п.н. Обе стратегии независимо привели к получению желаемых рекомбинантов.
Пример 8: Создание дополнительных вариантов мутантных токсинов с использованием сайт-направленного мутагенеза
Было сконструировано по меньшей мере двенадцать различных вариантов мутантных токсинов С.difficile. Аллельные замены были введены в N-концевые фрагменты генов мутантных токсинов путем сайт-направленного мутагенеза (набор Quickchange®). Были также сконструированы мутантные токсины в качестве сравнительных контролей, чтобы оценить способность данной системы на основе плазмиды образовывать белок, количественно эквивалентный по биологической активности нативным токсинам, очищенным из штаммов С.difficile дикого типа. В данном случае вводили аллельные замены для реверсии исходных замен глюкозилтрансферазы. Кроме того, была сконструирована пара векторов, несущих мутантный токсин, выполняя синтез на заказ в фирме Blue Heron Biotech.
Двенадцать вариантов токсина включают следующие варианты: (1) мутантный токсин А С.difficile, имеющий мутацию D285A/D287A (SEQ ID NO:3); (2) мутантный токсин В С.difficile, имеющий мутацию D286A/D288A (SEQ ID NO:5); (3) мутантный токсин С.difficile А, имеющий мутацию D285A/D287A С700А (SEQ ID NO:4); (4) мутантный токсин В С.difficile, имеющий мутацию D286A/D288A С698А (SEQ ID NO:6); (5) рекомбинантный токсин А, имеющий последовательность SEQ ID NO:1; (6) рекомбинантный токсин В, имеющий последовательность SEQ ID NO:2; (7) мутантный токсин А С.difficile, имеющий мутацию С700А; (8) мутантный токсин В С.difficile, имеющий мутацию С698А; (9) мутантный токсин А С.difficile, имеющий мутацию С700А C597S, C1169S, C1407S, C1623S, C2023S и C2236S; (10) мутантный токсин В С.difficile, имеющий мутацию С698А C395S, C595S, C824S, C870S, C1167S, C1625S, C1687S и C2232S; (11) мутантный токсин А С.difficile, имеющий мутацию D285A, D287A, С700А, D269A, R272A, Е460А и R462A (SEQ ID NO:7); и (12) мутантный токсин В С.difficile, имеющий мутацию D270A, R273A, D286A, D288A, D461A, K463A и С698А (SEQ ID NO:8).
Пример 9: Стабильность трансформантов
Перегруппированные плазмиды были получены с использованием традиционно используемого лабораторного штамма DH5α E. coli. Напротив, в результате трансформаций с использованием штамма-хозяина Stbl2™ E. coli от фирмы Invitrogen получили медленно растущие рекомбинанты полноразмерных мутантных токсинов после трех суток роста при 30°С на чашках со средой LB, содержащей хлорамфеникол (25 мкг/мл). Более низкие эффективности клонирования были получены с использованием родственных штаммов Stbl3™ и Stbl4™ E. coli, хотя было обнаружено, что эти линии стабильны при сохранении плазмиды. Затем трансформанты размножали на агаре или в жидкой культуре с отбором на основе хлорамфеникола при 30°С. Было также обнаружено, что использование среды LB (Миллера) улучшает выход и рост трансформантов по сравнению со стерильной триптоново-соевой средой.
Пример 10: Трансформация С.difficile вектором pMTL84121fdx, кодирующим гены дикого типа или генетических мутантных токсинов
Трансформацию С.difficile путем конъюгативного переноса из Е. coli проводили по существу, как описано у Heap et al., Journal of Microbiological Methods, 2009. 78(1): p.79-85. Штамм-хозяин CA434 Е. coli трансформировали вектором pMTL84121fdx, кодирующим гены дикого типа или вариантов мутантных токсинов. Штамм-хозяин CA434 Е. coli является промежуточным хозяином для мобилизации экспрессионных плазмид в штамме-продуценте VPI 11186 spoOAl С.difficile. Штамм CA434 представляет собой производное штамма НВ101 Е. coli. Этот штамм несет конъюгативную плазмиду Tra+Mob+R702, которая придает устойчивость к Km, Tc, Su, Sm/Spe и Hg (за счет Tn1831). Готовили химически компетентные или электрокомпетентные клетки CA434, и трансформанты экспрессионного вектора отбирали на чашках со средой Миллера LB САМ при 30°С. Медленно растущие колонии, появляющиеся через 3 суток, собирали и амплифицировали в 3 мл культурах LB с хлорамфениколом до среднелогарифмической фазы (примерно 24 ч, 225 об/мин, орбитальный встряхиватель, 30°С). Культуры Е. coli собирали с использованием низкоскоростного (5000 g) центрифугирования во избежание ломки пилей, и осадки клеток мягко ресуспендировали пипеткой с широким отверстием в 1 мл фосфатно-солевого буфера (ФСБ). Клетки концентрировали с использованием низкоскоростного центрифугирования. Большую часть ФСБ удаляли путем переворачивания, и осушенные осадки переносили в анаэробную камеру и ресуспендировали в 0,2 мл культуры С.difficile, наносили пятнами на чашки с агаром с сердечно-мозговой вытяжкой (BHIS) и оставляли расти на 8 ч или в течение ночи. В случае трансформантов мутантного токсина А лучшие результаты были достигнуты при ночной конъюгации. Клеточные пэтчи соскребали в 0,5 мл ФСБ, и 0,1 мл высевали на селективную среду BHIS с добавлением 15 мкг/мл триамфеникола (более эффективного аналога хлорамфеникола) и D-циклосерина/цефокситина, чтобы уничтожить клетки-доноры Е. coli. Трансформанты, появляющиеся спустя 16-24 ч, очищали путем повторного рассева на новую чашку BHIS (с добавками), и последующие культуры тестировали на экспрессию рекомбинантных токсинов или мутантных токсинов. Глицериновые постоянные культуры и посевной материал готовили из клонов, показывающих хорошую экспрессию. Мини-препараты плазмид также готовили из 2 мл культур, используя модифицированный метод набора Qiagen, при котором клетки предварительно обрабатывали лизоцимом (необязательно). Мини-препарат ДНК С.difficile использовали в качестве матрицы для секвенирования с использованием ПЦР для проверки целостности клонов. Альтернативно готовили макси-препарат ДНК из трансформантов Stbl2™ E. coli и секвенировали.
Пример 11: Анализ экспрессии тройных мутантов токсина А и В С.difficile (SEQ ID NO:4 и 6 соответственно) и гепта-мутанта токсина В (SEQ ID NO:8)
Трансформанты выращивали либо в культурах объемом 2 мл (для рутинного анализа), либо в колбах, закрытых крышками с отверстием (для экспериментов по определению зависимости от времени). Образцы (2 мл) центрифугировали в течение короткого периода (10000 об/мин, 30 с), чтобы сконцентрировать клетки: супернатанты декантрировали и концентрировали в 10 раз (Amicon-ultra 30k); осадки подсушивали и замораживали при -80°С. Осадки клеток оттаивали на льду, ресуспендировали в 1 мл буфера для лизиса (Трис-HCl рН 7,5; 1 мМ ЭДТА, 15% глицерин) и обрабатывали ультразвуком (1×20 с импульс микронаконечником). Лизат центрифугировали при 4°С, и супернатант концентрировали в 5 раз. Образцы супернатанта и лизата объединяли с буфером для образца и обрабатывали нагреванием (10 мин, 80°С), после чего наносили на дубликаты 3-8% Трис-ацетатных полиакриламидных гелей с додецилсульфатом натрия (ДСН-ПААГ) (Invitrogen). Один гель окрашивали Кумасси, второй подвергали электропереносу для анализа с использованием Вестерн-блоттинга. Антисыворотку кролика, специфичную к токсину А и специфичную к токсину В (Fitgerald; Biodesign), использовали для обнаружения вариантов мутантного токсина А и В. Экспрессию гепта-мутантного токсина В (SEQ ID NO:8) также подтверждали с использованием Вестерн-блот-гибридизации.
Пример 12: Нейтрализация глюкозилтрансферазной активности мутантных токсинов
Генетические двойные мутантные (DM) токсины А и В (SEQ ID NO:3 и 5, соответственно) и тройные мутантные (ТМ) токсины А и В (SEQ ID NO:4 и 6, соответственно) не осуществляли перенос 14С-глюкозы в 10 мкг ГТФаз RhoA, Rad и Cdc42 в анализах гликозилирования in vitro в присутствии UDP-14C-глюкозы [30 мкМ], 50 мМ ГЭПЭС, рН 7,2, 100 мМ KCI, 4 мМ MgCl2, 2 мМ MnCl2 1 мМ дитиотрейтола ДТТ и 0,1 мкг/мкл бычьего сывороточного альбумина БСА. Тем не менее, контрольные токсины А и В дикого типа (имеющие SEQ ID NO:1 и 2, соответственно) эффективно переносили 14С-глюкозу на ГТФазы при низкой дозе, составляющей 10 и 1 нг каждая (и более низкие данные не представлены) (Фиг.4А и 4В), даже в присутствии 100 мкг мутантного токсина (Фиг.4 В), что показывает по меньшей мере 100000-кратное снижение по сравнению с соответствующими токсинами дикого типа. Подобные результаты были определены для ГТФазы Cdc42 (данные не представлены).
В частности, на фиг.4В токсин А и токсин В дикого типа (1 нг) или тройной мутантный токсин А и тройной мутантный токсин В (100 мкг) инкубировали с ГТФазой RhoA в присутствии UDP-14C-глюкозы в течение 2 ч при 30°С. Как проиллюстрировано, 1 нг TcdA и TcdB дикого типа переносили 14С-глюкозу на RhoA, но 100 мкг тройного мутантного токсина А и мутантный токсин В не переносили глюкозу. Когда в реакционную смесь с соответствующими 100 мкг тройного мутантного токсина А или тройного мутантного токсина В вносили 1 нг TcdA или TcdB дикого типа, обнаруживали гликозилирование RhoA, что указывает на отсутствие ингибиторов гликозилирования. Было установлено, что чувствительность обнаружения активности гликозилирования составляет 1 нг токсина дикого типа на фоне 100 мкг мутантного токсина (отношение 1:100000). Эти результаты показывают, что при мутациях в активном центре глюкозилтрансферазы в тройном мутантном токсине А и тройном мутантном токсине В снижена, если вообще измерима (менее чем в 100000 раз меньшая активность по сравнению с активностью соответствующих токсинов дикого типа) глюкозилтрансферазная активность. Аналогичный анализ был также разработан для количественного определения глюкозилтрансферазной активности путем осаждения трихлоруксусной кислотой (ТХУ) гликозилированных ГТФаз.
Пример 13: Нейтрализация аутокаталитической активности цистеиновой протеазы
Функция аутокаталитического расщепления в тройных генетически мутантных токсинах А и В (ТМ) (SEQ ID NO:4 и 6, соответственно) была нейтрализована в том случае, когда был мутирован сайт расщепления цистеиновой протеазы. Как проиллюстрировано на фиг.5, токсины А и В дикого типа (wt) (SEQ ID NO:3 и 5, соответственно) расщепляются в присутствии инозит-6-фосфата. Двойные мутантные токсины А и В (SEQ ID NO:3 и 5, соответственно) также расщепляются в присутствии инозит-6-фосфата (данные не представлены) аналогично токсинам дикого типа. Токсин A (SEQ ID NO:3) массой 308 кДа расщепляется на 2 фрагмента массой 245 и 60 кДа. Токсин В (SEQ ID NO:5) массой 270 кДа расщепляется на 2 фрагмента массой 207 и 63 кДа. Тройные генетически мутантные токсины А и В (ТМ) (SEQ ID NO:4 и 6, соответственно) остаются неизменными, имеющими массу 308 и 270 кДа соответственно, даже в присутствии инозит-6-фосфата. См. фиг.5. Следовательно, активность цистеиновой протеазы была инактивирована в результате генетической модификации.
Более конкретно, на фиг.5, один микрограмм тройного мутантного токсина А и тройного мутантного токсина В инкубировали в течение 90 минут при комнатной температуре (21±5°С) параллельно с TcdA и TcdB дикого типа от фирмы List Biologicals. Реакцию расщепления проводили в объеме 20 мкл в Трис-HCl, рН 7,5, 2 мМ дитиотрейтола (ДТТ), в присутствии или в отсутствие инозит-6-фосфата (10 мМ для TcdA и 0,1 мМ для Ted В). Затем весь объем реакционной смеси наносили на 3-8% ДСН-ПААГ; полосы белка визуализировали окрашиванием серебром. Как проиллюстрировано, wt Ted А и TcdB расщеплялись на две полосы белка массой 245 кДа и 60 кДа, и 207 кДа и 63 кДа, соответственно, в присутствии инозит-6-фосфата. Тройной мутантный токсин А и тройной мутантный токсин В не расщеплялись, что подтверждает, что мутация С700А в тройном мутантном токсине А и мутация С698А в тройном мутантном токсине В блокирует расщепление.
Пример 14: Остаточная цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO:4 и 6, соответственно)
Генетически мутантные токсины оценивали на их цитотоксичность с использованием анализа цитотоксичности in vitro в клетках IMR90, линии диплоидных фибробластов легкого человека. Эти клетки чувствительны как к токсину А, так и к токсину В. В качестве альтернативного предпочтительного воплощения в анализе цитотоксичности можно использовать нормальные клетки почки Vero от Cercopithecus aethiops, поскольку наблюдали, что они обладают умеренной чувствительностью к токсинам А и В. Предпочтительно в анализе цитотоксичности не используют клетки аденокарциномы прямой и ободочной кишки человека НТ-29, поскольку они показали значительно сниженную чувствительность к токсинам по сравнению с линиями клеток Vero и IMR90. См., например, таблицу 6 ниже.
Серийно разведенные образцы генетически мутантных токсинов или wt токсинов добавляли к монослоям клеток, выращенных в 96-луночных планшетах для тканевых культур. После инкубации при 37°С в течение 72 ч планшеты оценивали на метаболически активные клетки путем измерения клеточных уровней АТФ с использованием добавления реагента на основе люциферазы CellTiterGlo® (Promega, Madison, WI), образующего люминесценцию, выраженную в относительных единицах люминесценции (RLU). Высокое число RLU показывает, что клетки жизнеспособны, низкое число RLU показывает, что клетки метаболически неактивны и погибают. Уровень цитотоксичности, выраженный в виде EC50, определяют как количество wt токсина или генетически мутантного токсина, вызывающее 50% снижение RLU по сравнению с уровнями в контролях культур клеток (подробности этого анализа приведены ниже). Как показано на фиг.6, в таблице 7А и таблице 8А, значения EC50 TcdA и TcdB составляли примерно 0,92 нг/мл и 0,009 нг/мл, соответственно. Значения EC50 тройного мутантного токсина А и тройного мутантного токсина В составляли примерно 8600 нг/мл и 74 нг/мл, соответственно. Несмотря на приблизительно 10000-кратное снижение цитотоксичности по сравнению с wt токсинами, оба генетически мутантных токсина все же показали низкие остаточные уровни цитотоксичности. Эта остаточная цитотоксичность может быть блокирована путем нейтрализации моноклональных антител против токсинов, что указывает на то, что она специфична для мутантных токсинов, но, вероятно, не связана с известными ферментативными активностями wt токсинов (гликозилированием или аутопротеолизом).
Оба wt токсина проявляют сильную цитотоксичность in vitro, причем, небольших количеств токсинов достаточно, чтобы вызвать различные действия на клетки млекопитающих, такие как округление клеток (цитопатический эффект или СРЕ) и отсутствие метаболической активности (измеренной на основании уровней АТФ). Следовательно, было разработано два анализа in vitro (анализ СРЕ или округления клеток и анализ АТФ), чтобы подтвердить, что остаточная цитотоксичность лекарственных средств на основе мутантных токсинов сохраняется. Эти результаты выражены в значении ЕС50, которое представляет собой количество токсина или мутантного токсина, вызывающее: 1) развитие СРЕ в 50% клеток или 2) 50% снижение уровней АТФ, измеренных в относительных световых единицах.
При анализе СРЕ образец лекарственного вещества серийно разводят и инкубируют с клетками IMR90, которые наблюдают на потенциальный цитопатический эффект.Анализ СРЕ оценивают по шкале от 0 (нормальные клетки) до 4 (приблизительно 100% округления клеток), и балл 2 (приблизительно 50% округления клеток) определяют как значение EC50 тестируемого образца. Этот способ используют для тестирования лекарственного вещества на основе мутантных токсинов при концентрации, составляющей 1000 мкг /мл, которая представляет собой максимальную переносимую концентрацию, которую можно тестировать в данном анализе без интерференции матрикса. Следовательно, отсутствие обнаруживаемой цитотоксичности считается как ЕС50, составляющее более 1000 мкг/мл.
Анализ АТФ основан на измерении количества сигнала люминесценции, образующегося от АТФ, которое пропорционально числу метаболически активных клеток. Максимальная переносимая концентрация, которую можно тестировать в данном анализе без интерференции анализа, составляет примерно 200 мкг/мл. Следовательно, отсутствие обнаруживаемой цитотоксичности считается как EC50, составляющее более 200 мкг/мл.
Различные концентрации мутантных токсинов А и В добавляли к клеткам параллельно с контролями токсинов. Конечные показатели данного анализа представляли собой жизнеспособность клеток, измеренную на основании клеточных уровней АТФ с использованием CellTiter-Glo® (Promega). Степень люминесценции пропорциональна уровням АТФ или числу жизнеспособных клеток.
Цитотоксичность in vitro (EC50) токсина А дикого типа (wt) составляла 920 пг/мл, и 9 пг/мл для токсина В. Цитотоксичность in vitro (EC50) мутантного токсина A (SEQ ID NO:4) составляла 8600 нг/мл, и 74 нг/мл для мутантного токсина В (SEQ ID NO:6). Хотя эти значения представляют снижения в 9348 и в 8222 раза соответственно, остаточная Цитотоксичность была обнаружена в обоих мутантных токсинах.
Иными словами, Цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO:4 и 6, соответственно) была значительно снижена в анализах цитотоксичности in vitro в клетках IMR-90 относительно цитотоксичности wt токсинов А и В (SEQ ID NO:1 и 2 соответственно). Как проиллюстрировано на фиг.6, хотя оба тройных мутантных токсина проявляли значительное снижение в цитотоксичности (104 кратное) относительно wt токсина, остаточную Цитотоксичность наблюдали при более высоких концентрациях обоих тройных мутантных токсинов.
Кроме того, остаточная Цитотоксичность каждого тройного мутантного токсина может быть полностью нейтрализована (например, по меньшей мере 6-8 log10 снижение токсичности относительно токсичности токсина дикого типа) антителами, специфичными к токсину. См. Пример 16 ниже.
Анализы клеточных культур более чувствительны для определения цитотоксичности, чем модели на животных in vivo. При доставке либо интраперитонеальным i.p., либо внутривенным i.v путем в мышиной модели летального инфицирования, wt ТсаА имеет LD50, составляющую примерно 50 нг на мышь, тогда как wt TcdB более сильным, причем LD50 составляет примерно 5 нг на мышь. Напротив, анализы in vitro на основе клеточных культур, описанные выше, имеют значения EC50, составляющие 100 пг на лунку для wt TcdA и 2 пг на лунку для wt TcdB.
Пример 15: Остаточная цитотоксичность генетически гепта-мутантного токсина В (SEQ ID NO:8)
Как проиллюстрировано на фиг.7, значения EC50 аналогичны для мутантов, включающих тройной мутантный токсин В (SEQ ID NO:6) (20,78 нг/мл) и гепта-мутантный токсин В (SEQ ID NO:8) (35,9 нг/мл), что указывает на то, что четыре дополнительных мутации, дополнительно модифицирующие активный центр глюкозилтрансферазы и центр связывания субстрата ГТФазы, не приводят к дальнейшему снижению цитотоксичности генетически мутантных токсинов. Значения ЕС50 также были аналогичны для двойного мутантного токсина В (SEQ ID NO:5) и для тройного и гепта-мутантного токсина (данные не представлены). Это наблюдение позволяет предположить, что механизм цитотоксичности мутантных токсинов неожиданно не зависит от механизма глюкозилтрансферазы и распознавания субстрата.
Пример 16: Остаточная цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO:4 и 6, соответственно)
Для дополнительной оценки природы остаточной цитотоксичности, мутантные токсины (SEQ ID NO:4 и 6) смешивали и инкубировали с их соответствующими нейтрализующими антителами, после чего смесь добавляли к монослою клеток IMR90.
Результаты (фиг.8) показали, что остаточная цитотоксичность мутантных токсинов А и В (SEQ ID NO:4 и 6 соответственно) может быть полностью нейтрализована нейтрализующими антителами, специфичными к мутантному токсину А (верхняя панель, фиг.8) и к мутантному токсину В (нижняя панель, фиг.8). Возрастающие концентрации мутантного токсина А (верхняя панель) и В (нижняя панель) инкубировали с поликлональными антителами кролика против токсина (pAb, разведение 1:10) или с моноклональными антителами мыши (разведение 1:50 из исходного раствора, содержащего 3,0 мг IgG/мл), после чего добавляли к клеткам IMR90. После 72-часовой обработки инкубацией с клетками IMR90 при 37°С добавляли субстрат CellTiter-Glo®, и относительные световые единицы (RLU) измеряли в спектрофотометре с программой люминесценции для измерения уровней АТФ. Чем ниже уровень АТФ, тем выше токсичность. Контроли включали TcdA и TcdB, добавленные при их 4-кратных соответствующих значениях ЕС50.
Опубликованные сведения позволяют предположить, что мутации в домене глюкозилтрансферазы или аутокаталитической протеазы токсинов приводят в результате к полной инактивации токсичности. Тем не менее, данные авторов изобретения не согласуются с этими опубликованными сведениями, что может быть связано с повышенными концентрациями высокоочищенных мутантных токсинов, тестируемыми в исследованиях авторов изобретения, в противоположность сырым лизатам культуры в опубликованных сведениях; увеличенными периодами времени, в течение которых наблюдали округление клеток, обработанных мутантными токсинами (например, 24 часа, 48 часов, 72 часа или 96 часов), в противоположность наблюдениям, проведенным в течение менее 12 часов; с использованием клеточных линий, проявляющих значительно более высокую чувствительность к токсинам в настоящих анализах цитотоксичности, в противоположность клеткам аденокарциномы прямой и ободочной кишки человека НТ-29 в анализах цитотоксичности, раскрытых в опубликованных сведениях; и/или с неизвестной активностью или процессом, отличающимся от гликозилирования, который может управлять остаточной цитотоксичностью мутантных токсинов.
Пример 17: Новый механизм цитотоксичности генетически мутантных токсинов
Чтобы исследовать механизм остаточной цитотоксичности генетически мутантных токсинов, клетки IMR-90 обрабатывали wt токсином В (SEQ ID NO:2) или генетически мутантным токсином В (SEQ ID NO:6), и исследовали гликозилирование ГТФазы Rac1 в зависимости от времени обработки. Образцы собирали от 24 до 96 часов и готовили клеточные экстракты. Гликозилированный белок Rac1 отличают от не гликозилированного Rac1 с использованием Вестерн-блоттингов с двумя антителами к Rac1. Одно антитело распознает обе формы Rac1 (23А8), а другое (102) распознает только не гликозилированный Rac1. Как проиллюстрировано на фиг.22, для токсина В общие уровни Rac1 оставались неизменными со временем, причем, большая часть Rac1 гликозилирована. С другой стороны, обработка генетически мутантным токсином В (SEQ ID NO:6) приводила в результате к значительному снижению общего количества Rac1, тем не менее, Rac1 был не гликозилирован во все моменты времени. Это показывает, что обработка генетически мутантным токсином, но не wt токсином, отрицательно влияла на общий уровень Rac1. Как проиллюстрировано на фиг.22, в указанные моменты времени уровень актина был аналогичным в клетках, обработанных токсином и генетически мутантным токсином В, и аналогичным в клетках, обработанных имитатором. Это показывает, что генетически мутантные токсины проявляют цитотоксичность посредством механизма, отличающегося от биохимического пути гликозилирования, управляемого токсином дикого типа.
Пример 18: Химическая обработка генетически мутантных токсинов
Хотя предпочтительны генетически модифицированные мутантные токсины, показывающие 4-log снижение цитотоксической активности, было рассмотрено дальнейшее снижение (от 2 до 4 log) цитотоксической активности. Оценили две стратегии химической инактивации.
При первом способе для инактивации мутантных токсинов используют формальдегид и глицин. Инактивация формальдегидом происходит за счет образования шиффова основания (имина) между формальдегидом и первичными аминами на белке. Затем эти шиффовы основания могут взаимодействовать с рядом аминокислотных остатков (Arg, His, Trp, Tyr, Gln, Asn) с образованием либо внутри-, либо межмолекулярных поперечных сшивок. Это поперечное сшивание фиксирует структуру белка, делая ее неактивной. Кроме того, формальдегид может взаимодействовать с глицином с образованием шиффова основания. Затем глицильное шиффово основание может взаимодействовать с аминокислотными остатками с образованием межмолекулярных белок-глициновых поперечных сшивок. Формальдегид снижает цитотоксическую активность генетически мутантных токсинов до значений, находящихся ниже обнаружимых пределов (снижение циттоксичности более чем на 8 log10 для тройного мутанта В (SEQ ID NO:6) и более чем на 6 log10 для тройного мутанта A (SEQ ID NO:4). Тем не менее, наблюдали реверсию со временем при инкубации тройных мутантных токсинов, инактивированных формальдегидом (Fl, formaldehyde-inactivated) при 25°С.Цитотоксическую реверсию можно предотвратить путем добавления низкого количества формальдегида (0,01-0,02%) в раствор для хранения FI-тройных мутантных токсинов. См. Пример 23.
При другом способе для получения инактивированных мутантных токсинов используют обработку 1-этил-3-(3-диметиламинопропил)-карбодиимидом (EDC)/N-гидроксисукцинимидом (NHS). При этом способе EDC/NHS взаимодействуют с карбоксильными группами на белке с образованием активированных сложных эфиров. Затем эти активированные сложные эфиры могут взаимодействовать с первичными аминами на белке с образованием стабильных амидных связей. Как и при реакции с формальдегидом, эта реакция приводит в результате к внутри- и межмолекулярным поперечным сшивкам. Амидная связь, образующаяся в результате обработки EDC/NHS, более стабильна и необратима, чем лабильная имидная связь, образующаяся в результате инактивации формалином. Кроме поперечных сшивок, образующихся в результате взаимодействия активированных сложных эфиров с первичными аминами на полипептиде, могут образоваться аддукты как с глицином, так и с бета-аланином. Без связи с каким-либо механизмом или теорией, аддукты глицина получают при добавлении глицина для гашения не прореагировавших активированных сложных эфиров. Амин глицина взаимодействует с активированным сложным эфиром на полипептиде с образованием стабильных амидных связей. Без связи с каким-либо механизмом или теорией, аддукты бета-аланина образуются в результате взаимодействия активированного бета-аланина с первичными аминами на полипептиде. Эта реакция приводит в результате к стабильным амидным связям. Активированный бета-аланин получают путем взаимодействия трех моль NHS с одним моль EDC.
Для достижения 2-4 log снижения цитотоксической активности относительно генетически модифицированных мутантных токсинов (6-8 log относительно нативных токсинов) химически инактивированные мутантные токсины должны иметь значения ЕС50, составляющие не менее 1000 мкг/мл. Кроме снижения цитотоксической активности, было бы предпочтительно сохранить ключевые эпитопы, которые определяют с использованием дот-блот-анализа. К настоящему времени идентифицирован ряд условий реакции, удовлетворяющих критериям, как снижения цитотоксичности, так и распознавания эпитопов. Несколько партий инактивированных мутантных токсинов получено для исследований на животных, и аналитические данные для нескольких репрезентативных партий представлены в таблицах 7А и 7В и в таблицах 8А и 8В.
условия)
Пример 19: Очистка
По окончании ферментации ферментер охлаждают. Клеточную суспензию выделяют путем непрерывного центрифугирования и ресуспендируют в подходящем буфере. Лизис клеточной суспензии достигается путем гомогенизации высокого давления. Для мутантного токсина А гомогенат _локкулирова, и _локкулированный раствор претерпевает непрерывное центрифугирование. Этот раствор фильтруют, а затем переносят на последовательную переработку. Для мутантного токсина В гомогенат осветляют путем непрерывного центрифугирования, а затем переносят на последовательную переработку.
Мутантный токсин A (SEQ ID NO:4) очищают, используя две хроматографические стадии с последующей заменой конечного буфера. Осветленный лизат загружают на колонку для хроматографии с гидрофобным взаимодействием (ХГВ), и связанный мутантный токсин элюируют, используя градиент цитрата натрия. Затем объединенный продукт с колонки ХГВ загружают на катионообменную колонку (СЕХ), и связанный мутантный токсин А элюируют, используя градиент хлорида натрия. Пул СЕХ, содержащий очищенный мутантный токсин А, подвергают замене буфера на конечный буфер путем осветления. Очищенный мутантный токсин А подвергают замене буфера на конечный промежуточный буфер лекарственного вещества путем диафильтрации. После диафильтрации ретентат фильтруют через фильтр с размером пор 0,2 микрон, после чего подвергают химической инактивации с получением конечного лекарственного вещества. Целевая концентрация белка составляет 1-3 мг/мл.
Мутантный токсин В (SEQ ID NO:6) очищают, используя две хроматографические стадии с последующей заменой конечного буфера. Осветленный лизат загружают на анионообменную колонку (АЕХ), и связанный мутантный токсин элюируют, используя градиент хлорида натрия. К объединенному продукту с колонки АЕХ добавляют цитрат натрия и загружают на колонку для хроматографии с гидрофобным взаимодействием (ХГВ). Связанный мутантный токсин элюируют, используя градиент цитрата натрия. Пул ХГВ, содержащий очищенный полипептид мутантного токсина (SEQ ID NO:6), подвергают замене буфера на конечный буфер путем диафильтрации. Очищенный мутантный токсин В подвергают замене буфера на конечный промежуточный буфер лекарственного вещества путем диафильтрации. После диафильтрации ретентат фильтруют через фильтр с размером пор 0,2 микрон, после чего подвергают химической инактивации с получением конечного лекарственного вещества. Целевая концентрация белка составляет 1-3 мг/мл.
Пример 20: Инактивация формальдегидом/глицином
После очистки генетически мутантные токсины А и В (SEQ ID NO:4 и 6, соответственно) инактивируют в течение 48 часов при 25°С, используя 40 мМ (1,2 мг/мл) формальдегида. Инактивацию проводят при рН 7,0±0,5 в буфере, состоящем из 10 мМ фосфата, 150 мМ хлорида натрия, содержащем 40 мМ (3 мг/мл) глицина. Период инактивации устанавливают таким образом, чтобы он в три раза превышал период, необходимый для снижения ЕС50 в клетках IMR90 более чем до 1000 мкг/мл. После 48 часов биологическая активность снижается на 7-8 log10 относительно нативного токсина. После 48-часовой инкубации инактивированный мутантный токсин подвергают замене буфера на конечный буфер лекарственного вещества путем диафильтрации. Например, используя кассету для ультрафильтрации на 100 кДа, состоящую из регенерированного ацетата целлюлозы, инактивированный токсин концентрируют до 1-2 мг/мл и заменяют буфер.
Пример 21: Инактивация N-(3-диметиламинопропил)-N′-этилкарбодиимидом (EDC)/N-гидроксисукцинимидом (NHS)
После очистки генетически мутантные токсины (SEQ ID NO:4 и SEQ ID NO:6) инактивируют в течение 2 часов при 25°С, используя 0,5 мг EDC и 0,5 мг NHS на мг очищенного генетически мутантного токсина А и В (приблизительно 2,6 мМ и 4,4 мМ соответственно). Реакцию гасят добавлением глицина до конечной концентрации, составляющей 100 мМ, и реакционные смеси инкубируют дополнительно в течение 2 часов при 25°С. Инактивацию проводят при рН 7,0±0,5 в буфере, состоящем из 10 мМ фосфата, 150 мМ хлорида натрия. Период инактивации устанавливают таким образом, чтобы он в три раза превышал период, необходимый для снижения EC50 в клетках IMR90 более чем до 1000 мкг/мл. После 2 часов биологическая активность снижается на 7-8 lig10 относительно нативного токсина. После 4-часовой инкубации инактивированный мутантный токсин подвергают замене буфера на конечный буфер лекарственного вещества путем диафильтрации. Например, используя кассету для ультрафильтрации на 100 кДа, состоящую из регенерированного ацетата целлюлозы, инактивированный токсин концентрируют до 1-2 мг/мл и заменяют буфер.
Если не указано иное, приведенные ниже термины, используемые в разделе Примеров, относятся к композиции, полученной в соответствии с настоящим описанием в Примере 21: «обработанный EDC/NHS тройной мутантный токсин»; «инактивированный EDC мутантный токсин»; «лекарственное вещество на основе мутантного токсина [А/В]»; «EI-мутантный токсин»; «EDC/NHS-тройной мутантный токсин». Например, следующие термины являются синонимами: «обработанный EDC/NHS тройной мутантный токсин А»; «инактивированный EDC мутантный токсин А»; «лекарственное вещество на основе мутантного токсина А»; «El-мутантный токсин А»; «EDC/NHS-тройной мутантный токсин А». В качестве другого примера, следующие термины являются синонимами: «обработанный EDC/NHS тройной мутантный токсин В»; «инактивированный EDC мутантный токсин В»; «лекарственное вещество на основе мутантного токсина В»; «EI-мутантный токсин В»; «EDC/NHS-тройной мутантный токсин В».
Каждое лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В получают, используя периодический процесс, включающий следующие стадии: (1) ферментацию токсин-отрицательного штамма С.difficile (VPI 11186), содержащего плазмиду, кодирующую соответствующий генетический тройной мутантный полипептид токсина (в среде, включающей гидролизат сои, дрожжевой экстракт HY YEST™ 412 (Sheffield Bioscience), глюкозу и триамфеникол), (2) очистку генетически мутантного токсина («промежуточного лекарственного вещества») из бесклеточного лизата с использованием методов ионообменной и гидрофобной хроматографии, по меньшей мере более чем до 95% чистоты, (3) химическую инактивацию обработкой EDC/NHS с последующим гашением/кэпированием глицином и (4) замену буфера на конечную буферную среду.
Пример 22: Исследования, подтверждающие условия инактивации и препарат
Чтобы оптимизировать химическую инактивацию генетически мутантных токсинов, было разработано статистическое планирование эксперимента (DOE). Факторы, исследованные в DOE, включали температуру, концентрацию формальдегида/глицина, концентрацию EDC/NHS и время (таблица 9 и 10). Для мониторинга потери биологической активности определяли значения ЕС50 в клетках IMR90. Кроме того, также наблюдали клеточную морфологию клеток IMR-90 в различные моменты времени после обработки. См. фиг.9, где показана морфология через 72 часа после обработки. Чтобы определить действие на структуру белка, мониторинг распознавания эпитопов проводили, используя дот-блот анализ с использованием панели моноклональных антител, индуцированных против различных доменов токсина.
При инактивации мутантных токсинов С.difficile формальдегидом/глицином конечные условия реакции были выбраны таким образом, что желаемый уровень снижения цитотоксической активности (7-8 log10) был достигнут при максимизации распознавания эпитопов. См. Пример 20 выше.
При инактивации мутантных токсинов С.difficile EDC/NHS конечные условия реакции были выбраны таким образом, что желаемый уровень снижения цитотоксической активности (7-8 log10) был достигнут при максимизации распознавания эпитопов. См. Пример 21 выше.
В альтернативном воплощении реакцию EDC-NHS гасили добавлением аланина, который в достаточной степени гасил реакцию. Использование аланина может привести в результате к модификации на белке мутантного токсина, подобной модификации при гашении реакции глицином. Например, гашение добавлением аланина может привести в результате к аланиновой группировке на боковой цепи остатка глутаминовой кислоты и/или аспарагиновой кислоты мутантного токсина. В другом альтернативном воплощении реакцию EDC-NHS гасили добавлением метилового эфира глицина, который в достаточной степени гасил реакцию.
Получение химически неактивного тройного мутантного токсина А и токсина В С.difficile в оптимизированных условиях привело в результате к дальнейшему снижению остаточной цитотоксичности до необнаружимого уровня (более 1000 мкг/мл, то есть самой высокой концентрации, протестированной посредством анализа СРЕ) при сохранении антигенности, измеренной по их реактивности на токсин-специфичные нейтрализующие антитела. Результаты, представленные в таблице 28, демонстрируют ступенчатое снижение цитотоксичности по сравнению с wt токсином посредством тройных мутантных токсинов, обработанных EDC/NHS. Мечение иммунофлуоресцентной меткой подтвердило, что тройные мутантные токсины (SEQ ID NO:4 и 6) и лекарственные вещества на основе мутантных токсинов проявляли сравнимое связывание с клетками IMR-90, что позволяет предположить, что утрата цитотоксичности не является следствием сниженного связывания с клетками (данные не представлены). По сравнению с лекарственным веществом на основе мутантного токсина А, лекарственное вещество на основе мутантного токсина В достигало более высокой кратности снижения цитотоксичности, что согласуется с приблизительно в 600 раз более высокой эффективностью TcdB по сравнению с TcdA.
Также оценивали результаты анализа цитотоксичности для мутантного токсина В, модифицированного только EDC или EDC и сульфо-NHS. См. таблицу 29.
Условия: Тройной мутантный токсин В ("TM TcdB") (SEQ ID NO:6) модифицировали при массовых отношениях мутантный токсин В:ЕОС:сульфо-NHS, составляющих 1:0,5:0,94. Это отношение представляет собой молярный эквивалент (с поправкой на более высокую молекулярную массу сульфо-NHS) стандартной реакции EDC/NHS, как описано в Примере 21. Чтобы определить влияние сульфо-NHS, долю сульфо-NHS изменяли от 0,5× до 2× стандартной доли. Дубликаты реакций ставили в 1×ФСБ, рН 7,0 при 25°С и инициировали добавлением раствора EDC. Через 2 часа реакционные смеси гасили добавлением 1 М глицина, рН 7,0 (0,1 М конечная концентрация) и инкубировали еще в течение 2 часов. Гашеные реакционные смеси обессоливали, и лекарственное вещество на основе мутантного токсина В ("TM TcdB-EDC") концентрировали с использованием устройств Vivaspin 20 и стерильно фильтровали в стерильные флаконы и подвергали оценке в анализе цитотоксичности.
При такой же молярной доле сульфо-NHS снижал ЕС50 примерно до 250 мкг/мл по сравнению с более чем 1000 мкг/мл для NHS. Даже при двукратной молярной доле сульфо-NHS оказывается не настолько эффективным, как NHS, при снижении цитотоксичности. См. таблицу 30.
Чтобы определить число и тип модификаций, было проведено пептидное картирование на обоих образцах тройного мутантного токсина В, инактивированных EDC/NHS и ЕОС/сульфо-NHS. В обоих образцах наблюдали сходные количества аддуктов глицина, поперечных сшивок и модификаций дегидроаланином. Тем не менее, в образце сульфо-NHS не наблюдали бета-аланина.
Токсин В дикого типа (SEQ ID NO:2) инактивировали, используя стандартный протокол (см. Пример 21); токсин В:EDC:NHS1:0,5:0,5, 25°С в течение 2 часов в 1×ФСБ рН 7,0, затем гашение 1 М глицином (конечная концентрация 0,1 М) и дополнительная инкубация в течение 2 часов. Образец обессоливали, концентрировали и подвергали анализу цитотоксичности. Значение ЕС50 Для данных образцов составляли менее 244 нг/мл.
Пример 23: Исследования реверсии
Чтобы определить, происходит ли реверсия мутантных токсинов С.difficile, инактивированных либо формальдегидом/глицином, либо EDC/NHS, образцы инактивированных мутантных токсинов (1 мг/мл) инкубировали при 25°С в течение пяти-шести недель. Аликвоты отбирали каждую неделю, и определяли значения ЕС50 в клетках IMR90. Один образец, инактивированный формальдегидом/глицином, не содержал формальдегид, и один образец содержал 0,01% формальдегид. Значения EC50 измеряли с использованием анализа СРЕ.
При 25°С в отсутствие остаточного формальдегида наблюдали частичную реверсию (Таблица 11). После пяти недель цитотоксическая активность возрастала приблизительно в 3 раза. Хотя цитотоксическая активность возрастала, после пяти недельь ее снижение все еще составляло 7 log10 относительно нативного токсина. Реверсия была полностью предотвращена в результате включения формалина при концентрации, составляющей 0,010%. В образце, инактивированном EDC/NHS, реверсию не наблюдали. На протяжении всей 6-недельной инкубации значения ЕС50 оставались на исходном уровне, составляющем более 1000 мкг/мл, для всех четырех партий, как тройного мутантного токсина A (SEQ ID NO:4), обработанного EDC/NHS, так и для тройного мутантного токсина В (SEQ ID NO:6), обработанного EDC/NHS. Напротив, значения ЕС50 тройного мутантного токсина A (SEQ ID NO:4), обработанного FI, и тройного мутантного токсина В (SEQ ID NO:6), обработанного FI, были нестабильными и снижались до неприемлемо низких значений ЕС50, то указывает на возрастание цитотоксичности или реверсии инактивации. См. таблицу 11.
В дополнение к стабильному снижению цитотоксичности до необнаружимого уровня (более 1000 мкг/мл, как измерено с использованием анализа СРЕ), в мутантных токсинах, инактивированных с использованием EDC/NHS, сохранялись важные эпитопы, являющиеся мишенями токсин-нейтрализующих mAb. См. таблицу 31. В мутантных токсинах, инактивированных FI, показана потеря тех же антигенных детерминант.
b Значения, определенные с использованием анализа Biacore™, используя множественные нейтрализующие mAb, направленные на различные не перекрывающиеся эпитопы токсина
с Значения представляют собой средние значения двух экспериментов
d Для первых трех строк нейтрализующие mAb "1," "2," "3" относятся к mAb А60-22, А80-29 и А65-33 к токсину А, соответственно. Для нижних трех строк нейтрализующие mAb "1," "2," "3" относятся к mAb B8-26, В59-3 и В-56-15 к токсину В, соответственно.
Пример 24: Доклинические исследования иммуногенности
Ключевые цели доклинических исследований включают тестирование композиций, включающих мутантные токсины А и В С.difficile, у небольших животных и нечеловекообразных приматов (НЧП). Мышей и хомячков иммунизировали, чтобы определить среди прочего, способны ли композиции С.difficile вызывать нейтрализующие антитела против мутантных токсинов А и В. Антигены тестировали на индукцию нейтрализующих ответов сывороточных антител после серии иммунизации у мышей, хомячков и яванских макак. Генетически и/или химически инактивированные мутантные токсины включали либо в нейтральный буфер, либо в буфер, представляющий собой фосфат алюминия, либо в буфер, содержащий ISCOMATRIX в качестве адъюванта в некоторых воплощениях. Нейтрализующие ответы антител обычно тестировали в течение примерно от двух до четырех недель после каждой бустерной или конечной дозы.
Анализ нейтрализации токсинов демонстрирует способность антисыворотки к нейтрализации цитотоксического действия, опосредованного TcdA или TcdB С.difficile, и, следовательно, способен измерять функциональную активность антител, присутствующих в образце. Анализ нейтрализации токсина проводили на линии клеток фибробластов легкого человека IMR-90, чувствительной как к TcdA, так и к TcdB. Кратко, 96-луночный микротитрационный планшет засевали клетками IMR-90, служащими в качестве мишени цитотоксичности, опосредованной токсином. Каждый тестируемый образец сыворотки анализировали отдельно на способность к нейтрализации TcdA и TcdB. Соответствующие серийные разведения тестируемых антисывороток смешивали с фиксированными концентрациями TcdA или TcdB и инкубировали при 37°С в течение 90 минут во влажной камере (37°С/5% CO2), чтобы дать возможность осуществления нейтрализации токсинов. Для контроля качества все планшеты включали сравнительные стандартные контроли, включающие антитела против токсинов, имеющих известные титры. После 90 минут смесь токсина и антисыворотки добавляли к монослою клеток IMR-90, и планшеты инкубировали дополнительно в течение 72 часов. Затем в аналитический планшет добавляли субстрат CellTiter-Glo®, чтобы определить уровни аденозинтрифосфата (АТФ), присутствующего в метаболически активных клетках, и измеряли в относительных единицах люминесценции (RLU). Высокий уровень АТФ указывает на высокую жизнеспособность клеток, и уровни прямо пропорциональны количеству нейтрализации токсина антителом, присутствующим в образце. Для получения доклинических данных, данные RLU наносили на график против значения разведения тестируемого образца антисыворотки с получением соответствия кривой четырехпараметрической логарифмической регрессии (4-PL). Титры нейтрализации выражали в виде значения разведения образца, проявляющего 50% снижение цитотоксичности.
Пример 25: Исследование иммуногенности мыши: mu C.difficile 2010-06
Цель данного исследования состояла в оценке иммуногенности двух форм мутантного токсина В С.difficile (SEQ ID NO:6), каждая из которых химически инактивирована различными способами. В данном исследовании в качестве контроля использовали необработанный мутантный токсин В (SEQ ID NO: 6) (генетически инактивированный, но химически не инактивированный) с адъювантом или без адъюванта.
Группы из 10 мышей иммунизировали внутримышечно (IM) 10 мкг иммуногена согласно таблице 12.
bхимическая инактивация = Обработка EDC/NHS, 30°С в течение 2 часов
cконцентрация иона алюминия = 0,5 мг/мл
Результаты: После каждого введения вакцин-кандидатов вредные эффекты у мышей отсутствовали. Как проиллюстрировано на фиг.10, у мышей в каждой группе развивались стабильные нейтрализующие антитела против токсина В после третьей дозы соответствующих мутантных токсинов.
На основании титров недели 12 оказалось, что у мышей, которым вводили EDC-инактивированный мутантный токсин В (Группа 2) и формалин-инактивированных мутантных токсинов (Группа 1), развивались сильные нейтрализующие ответы.
В отсутствие химической инактивации, генетически мутантный токсин В (SEQ ID NO:6) генерировал нейтрализующие ответы после двух доз (Группы 3-4, неделя 8), которые претерпевали бустер-эффект после третьей дозы (Группы 3-4, неделя 12).
Пример 26: Исследование иммуногенности мышей: mu C.difficile 2010-07:
Цель данного исследования состояла в оценке иммуногенности химически инактивированных мутантных токсинов А и В С.difficile (SEQ ID NO:4 и 6, соответственно), либо отдельно, либо в комбинации. Иммуногены для всех групп включали в препарат, содержащий фосфат алюминия в качестве адъюванта.
Группы из 5 мышей иммунизировали внутримышечно 10 мкг иммуногена согласно таблице 13.
bОбработка EDC = Обработка EDC/NHS в течение 4 ч при 30°С; мутантный токсин не обладал цитотоксичностью и сохранял связывание со всеми протестированными антителами, специфичными к мутантному токсину
сконцентрация иона алюминия = 0,5 мг/мл
Результаты: После каждого введения вакцин-кандидатов вредные эффекты у мышей отсутствовали. Как проиллюстрировано на фиг.11, после двух доз химически инактивированных генетически мутантных токсинов титры нейтрализующих антител против токсина А (Группы 3-5) повышались до 3-41одю, тогда как нейтрализующие антитела против токсина В (Группы 1-2, 5) оставались на уровне от низкого до необнаружимого, что согласуется с данными исследования на мышах, описанного выше (фиг.10). Титры нейтрализующих антител против токсина В повышались до 2-3 log10 в группах 1, 2 и 5 после третьей дозы (титры недели 12) и достигали пика через две недели после четвертой дозы (титры недели 14). Титры нейтрализующих антител против токсина А в группах 3-5 незначительно повышались после третьей (титры недели 12) и четвертой иммунизации (титры недели 14).
Пример 27: Исследование иммуногенности хомячков: ham C. difficile 2010-02:
Цель данного исследования состояла в оценке иммуногенности и защитного потенциала тройных мутантных и химически инактивированных мутантных токсинов А и В С.difficile в модели сирийского золотистого хомячка. Модель сирийского золотистого хомячка представляет собой лучшую доступную модель провокации для стимуляции CDAD человека. В данном исследовании использовали те же партии мутантных токсинов А и В, которые были использованы в исследовании на мышах mu C.difficile 2010-07. В качестве контроля одна группа получала мутантные токсины без адъюванта, содержащего алюминий.
Группы из 5 сирийских золотистых хомячков иммунизировали внутримышечно 10 мкг иммуногена согласно таблице 14.
bОбработка EDC=Обработка EDC/NHS в течение 4 ч при 30°С; мутантный токсин не обладал цитотоксичностью и сохранял связывание со всеми протестированными антителами, специфичными к мутантному токсину
сконцентрация иона алюминия = 0,5 мг/мл
1. Животные; 15 сирийских золотистых хомячков, самки, возраст 6-8 нед./100-130 г каждая.
2. Вакцинация: IM, 0,05 мл каждая, согласно описанному выше режиму. Токсоиды предоставлены Process Development и включены в разбавитель, представляющий собой AIP04, в группах препаратов. Группа 2 служит в качестве контрольной группы без адъюванта.
3. Забор крови: У всех хомячков берут кровь на неделе 0, 4, 8 и 12, непосредственно перед каждой иммунизацией.
4. Анализ образцов сыворотки: Анализ нейтрализации
Результаты: После каждого введения вакцин-кандидатов вредные эффекты отсутствовали. Как проиллюстрировано на фиг.12, после однократной дозы мутантных токсинов нейтрализующие антитела против токсина А составляли 2-3 log10 для формалин-инактивированных мутантных токсинов (Группы 1-2) и 3-4 log10 для EDC-инактивированных мутантных токсинов (Группа 3). После второй дозы антитела против токсина А повышались во всех трех группах. Антитела против токсина А во всех трех группах, как оказалось, не повышались после третьей дозы. Подобный результат наблюдали после четвертой иммунизации, где возрастание титра наблюдали в формалин-инактивированной группе, не содержащей алюминиевый адъювант (Группа 2).
Нейтрализующие ответы против токсина В не обнаруживались в группах формалин-инактивированных мутантных токсинов (Группы 1-2) и были только чуть выше 2 log10 для EDC-инактивированных мутантных токсинов (Группа 3) после однократной дозы. После второй дозы титры нейтрализующих антител против токсина В в двух формалин-инактивированных группах (Группы 1-2) повышались до 3-4 log10, при этом титры в EDC-инактивированных группах (Группа 3) повышались до 4-5 log10. Для всех трех групп возрастание титров нейтрализующих антител против токсина В наблюдали после третьей и/или четвертой дозы, причем, во всех группах титры достигали пика на неделе 16 (после последней дозы). См. фиг.12.
На фиг.13 уровень ответов нейтрализующих антител против химически инактивированных генетически мутантных токсинов (фиг.12) по сравнению с ответами, вызванными токсоидами от List Biological Laboratories, Inc. (Campbell, California) (также обозначаемыми как "List Bio" или "List Biologicals") (то есть токсоиды, приобретенные от List Biological Laboratories, готовили путем инактивации токсинов дикого типа формалином; этот контрольный реагент использовали для установления модели провокации на хомячках).
При использовании здесь, "FI" в графических материалах и таблицах относится к обработке токсинов формалином/глицином в течение 2 суток при 25°С, если не указано иное. При использовании здесь, "Е1" в графических материалах и таблицах относится к обработке EDC/NHS в течение 4 часов при 30°С, если не указано иное. На фиг.135 животных-хомячков обрабатывали соответствующей композицией мутантных токсинов, тогда как 11 животных-хомячков обрабатывали токсоидом, приобретенным от List Biological.
Данные на фиг.13 показывают, что у хомячков, получивших инъекции в соответствии с таблицей 14, титры соответствующих нейтрализующих антител против токсина А (Фиг.13А) и токсина В (Фиг.13В), индуцированных иммуногенной композицией, включающей EDC-инактивированные мутантные токсины после двух доз выше, чем титры соответствующих нейтрализующих антител, вызванных токсоидами List Biologicals.
Пример 28: Исследование иммуногенности хомячков: С.difficile ham 2010-02 (продолжение)
Для оценки защитной эффективности мутантных токсинов, иммунизированные хомячки параллельно с одной контрольной группой неиммунизированных животных сначала получали пероральную дозу антибиотика клиндамицина (30 мг/кг) для нарушения нормальной кишечной флоры. Через пять суток хомячков заражали пероральной дозой спор С.difficile дикого типа (штамм 630, 100 колониеобразующих единиц (кое) на животное). Мониторинг животных проводили ежесуточно в течение семи суток после провокации на признаки CDAD, которые у хомячков известны как "мокрый хвост". Используя систему клинической оценки ряда различных параметров, животных, у которых была определена тяжелая CDAD, подвергали эвтаназии. Эти параметры включали следующие параметры: активность после стимуляции, обезвоживание, экскременты, температуру и массу тела и т.д., известные в данной области техники.
На сутки 11 исследование заканчивали, и всех выживших животных подвергали эвтаназии. На фиг.14 показаны кривые выживаемости для каждой из трех иммунизированных групп (Группы 1-3 согласно таблице 14) по сравнению с неиммунизированными контролями. Как видно, у всех неиммунизированных животных развивалась тяжелая CDAD, и требовалась эвтаназия на сутки 1-3 после провокации (выживаемость 0%). Кривые выживаемости групп, которым вводили формалин-инактивированный мутантный токсин, составляли 60%, причем, животным не требовалась эвтаназия вплоть до суток 3 (Группа 1) или суток 4 (Группа 2). Кривые выживаемости группы, которой вводили EDC-инактивированный мутантный токсин, составляли 80%, причем, одному животному (из 5) требовалась эвтаназия на сутки 7. Соответственно, хомячки были защищены от летального заражения спорами С.difficile.
Пример 29: Исследование иммуногенности хомячков: ham C.difficile 2010-03: Иммуногенность генетически и химически инактивированных мутантных токсинов С.difficile
Цель данного исследования состояла в оценке иммуногенности тройных мутантных и химически инактивированных мутантных токсинов А и В С.difficile (SEQ ID NO:4 и 6, соответственно) в модели сирийских золотистых хомячков. В данном исследовании использовали те же партии мутантных токсинов А и В, которые были использованы в исследовании на мышах mu C.difficile 2010-07. В качестве контроля одна группа получала фосфатно-солевой буферный раствор в качестве плацебо.
Группы, состоящие из пяти или десяти сирийских золотистых хомячков, иммунизировали иммуногеном согласно таблице 15. Животные получали три дозы. Кроме того, животных дозировали каждые две недели.
Результаты: См. фиг.15. В контрольное группе плацебо не наблюдали антител против токсина А или В. После одной дозы наблюдали нейтрализующие антитела против токсина А, титр которых составлял 2-3 log10 для групп формалин-инактивированного (Группа 2) и генетически мутантного токсина (Группа 4) и 3-4 log10 для EDC-инактивированной группы (Группа 3). Титр нейтрализующих антител против токсина А повышался в каждой из этих групп (2-4) после второй иммунизации соответствующими мутантными токсинами (сравнительные титры недели 2-недели 3 на фиг.15). После третьей дозы мутантных токсинов (вводимой на неделе 4) титры нейтрализующих антител против токсина А в группах 2-4 возрастали по сравнению с их титрами на неделе 4.
Нейтрализующие антитела против токсина В были обнаружимы после второй дозы, причем, титры нейтрализующих антител против формалин-инактивированного (группа 2) и EDC-инактивированного (группа 3) токсина В возрастали до 3-4 log10 и до 2-3 log10 для генетических тройных мутантов (Группа 4). После третьей иммунизации (неделя 4) титры нейтрализующих антител против токсина В повышались до 3-4 log10 для формалин-инактивированных мутантных токсинов (Группа 2) и генетически мутантных токсинов (Группа 4) и до 4-5 log10 для EDC-инактивированных мутантных токсинов (Группа 3).
Для нейтрализующих антител, как против токсина А, так и против токсина В пиковые титры наблюдали на неделе 6 (3 недели после дозирования) для всех вакцинированных групп (Группы 2-4).
Оценка иммуногенных композиций, содержащих адъюванты Alhydrogel/CpG или ISCOMATRIX
У хомячков, иммунизированных иммуногенной композицией, включающей химически инактивированный мутантный токсин, включенный в препарат, содержащий Alhydrogel, ISCOMATRIX или Alhydrogel/CpG24555 (Alh/CpG), развивались стабильные антитоксинные антисыворотки. Наблюдали, что пиковые ответы против токсина А и против токсина В были в 2-3-раза выше, и были статистически значимыми в группах, иммунизированных мутантными токсинами, включенными в Alh/CpG или ISCOMATRIX, по сравнению с вакциной, в которую был включен только Alhydrogel. См. таблицу 32, в которой показаны 50% титры нейтрализации. Хомячков (n=10/группа) иммунизировали IM на 0, 2 и 4 неделе 10 мкг каждого лекарственного вещества, на основе мутантного токсина А и на основе мутантного токсина В, включенного в препарат, содержащий 100 мкг Alhydrogel, либо 200 мкг CpG 24555 и 100 мкг Alhydrogel, либо 10 ед. ISCOMATRIX. Сыворотки собирали в каждый момент времени и анализировали в анализе нейтрализации токсина на функциональную антитоксинную активность. Геометрические средние титров представлены в таблице 32. Звездочкой (*) указана статистическая значимость (р<0,05) по сравнению с титрами в группе Alhydrogel.
Была протестирована защитная активность иммуногенных композиций, включающих лекарственные вещества на основе мутантных токсинов, включенных в препараты, содержащие данные адъюванты. Хомячков иммунизировали и давали перорально клиндамицин (30 мг/кг) на неделе 5, и заражали согласно описанному выше способу. Одну группу иммунизированных хомячков (n=5) включали в качестве контроля. Повышенную эффективность наблюдали у хомячков, иммунизированных лекарственными веществами на основе мутантных токсинов, содержащими адъюванты Alh/CpG или ISCOMATRIX (выживаемость 100%), по сравнению с одним Alhydrogel (выживаемость 70%). Соответственно, хомячки были защищены от летального заражения спорами С.difficile.
Пример 30: Вакцинация Clostridium difficile у яванских макак
Цель данного исследования состояла в тестировании иммуногенности низких и высоких доз EDC-инактивированных и формалин-инактивированных мутантных токсинов С.difficile у яванских макак. Все мутантные токсины включали в препарат, содержащий ISCOMATRIX® в качестве адъюванта, за исключением одной группы, служащей в качестве контроля без адъюванта (Группа 5).
Звездочка "*" на фиг.16 относится к наличию только 4 макак в группе на неделе 12, у одного макака в группе был произведен терминальный забор крови на неделе 8
Вакцинация: IM, 0,5 мл на дозу, на неделе 0, 2 и 4. Композиции мутантных токсинов готовили, как описано выше. Композиции мутантных токсинов включали в препараты, содержащие ISCOMATRIX, за исключением группы 5, получавшей препарат в буфере без адъюванта.
Забор крови: Недели -2, 0, 2, 3, 4, 6, 8 и 12. Эвтаназия и терминальные заборы крови у животных с самыми высокими титрами С.difficile на неделе 8.
Анализ образцов сыворотки: ELISA белка и анализы нейтрализации
Результаты: На фиг.16 показаны ответы нейтрализующих антител против токсинов у этих животных на неделях 0, 2, 3, 4, 6, 8 и 12. Титры против токсина А составляли 2-3 log10 для всех пяти групп после однократной дозы (титры недели 2). Эти титры повышались после каждой последующей дозы для каждой группы. У этих животных падение титра между неделями 3 и 4 отсутствовало. Для всех групп пиковые титры составляли 4-5 log10. Во все моменты времени в группе без адъюванта ISCOMATRIX (Группа 5) были самые низкие титры, что показывает пользу ISCOMATRIX при усилении иммунных ответов. Контрольная группа без адъюванта (Группа 5) достигала пиковых титров на неделе 12, как и группа, иммунизированная высокой дозой EDC-инактивированных мутантных токсинов (Группа 4); все остальные группы достигали пиковых титров на неделе 6, через две недели после последней дозы. Титры во всех группах повышались после второй дозы (момент времени неделя 3). Как и для ответов против токсина А, ответы против токсина В не снижались с недели 3 до недели 4. После третьей дозы (момент времени неделя 6) титры нейтрализующих антител пртив токсина В во всех группах составляли 3-4 log10, за исключением группы низкой дозы формалин-инактивированного токсина (Группа 1) и высокой дозы EDC-инактивированного токсина (Группа 4), где в обеих группах титры составляли чуть более 4 log10. пиковые титры наблюдали на неделе 12 для всех групп за исключением группы низкой дозы EDC-инактивированного токсина (Группа 3), которая имела пиковые титры на неделе 8. Во всех группах пиковые титры составляли более 4 log10.
Пример 31: Получение моноклональных антител
Хотя токсины А и В обладают высокой структурной гомологией, было обнаружено, что нейтрализующие активности антител являются токсиноспецифичными. В данном изобретении было идентифицировано несколько антител, специфичных к индивидуальному токсину и направленных на различные эпитопы и функциональные домены, и данные антитела обладают высоким сродством и сильной нейтрализующей активностью по отношению к нативным токсинам. Антитела были выделены из мышей, иммунизированных либо имеющимся в продаже формалин-инактивированным (FI)-мутантным токсином, либо рекомбинантным полноразмерным мутантным токсином (SEQ ID NO:4 и 6), который делали нетоксичным путем введения определенных мутаций в его каталитический сайт для получения mAb к токсину А и В, соответственно. Картирование эпитопов антител показало, что подавляющее большинство mAb против токсина А (49 из 52) были направлены на некаталитический С-концевой домен токсина.
Моноклональные антитела против токсина В были направлены на три домена этого белка. Из 17 mAb, специфичных к токсину В, 6 были специфичны к N-концу (например, аминокислотам 1-543 TcdB С.difficile дикого типа, такого как 630), 6 к С-концу (например, к аминокислотам 1834-2366 TcdB С.difficile дикого типа, такого как 630) и 5 к среднему домену транслокации (например, к аминокислотам 799-1833 TcdB С.difficile дикого типа, такого как 630). Таким образом, метод использования мутантных токсинов С.difficiles (например, SEQ ID NO:4 и 6) в качестве иммунизирующих антигенов обеспечивает ключевое преимущество, состоящее в презентировании большинства, если не всех, антигенных эпитопов по сравнению со способом инактивации формалином, который склонен к вредному влиянию на антигенную структуру мутантного токсина.
Пример 32: Характеризация mAb к токсину А А3-25, включающего вариабельную легкую цепь, имеющую аминокислотную последовательность SEQ ID NO:36, и вариабельную тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO:37
Антитело mAb А3-25 представляло особый интерес, поскольку все попытки определения изотипирования иммуноглобулина (Ig) данного антитела с использованием общедоступных наборов для изотипирования для IgG, IgM и IgA были безуспешны. Дальнейший анализ с использованием вестерн-блоттинга с использованием антисывороток, специфичных к Ig Н-цепи, показал, что А3-25 имеет изотип IgE, что является редким событием при получении mAb. Это было дополнительно подтверждено секвенированием нуклеотидов мРНК, выделенной из клеток гибридомы А3-25. Аминокислотные последовательности, выведенные на основании нуклеотидных последовательностей вариабельных областей Н- и L-цепи А3-25, представлены на фиг.17.
Чтобы дополнительно оценить А3-25 mAb на инфекцию и заболевание, обусловленное С difficile, в модели на животных, изотип Ig этого антитела был изменен на IgG1 мыши путем молекулярного прививания участка ε Н цепи на тяжелую цепь γ мыши согласно опубликованным способам.
Пример 33: Нейтрализующая способность и картирование эпитопов токсиноспецифичных антител
Далее, при попытке идентифицировать функциональные/нейтрализующие антитела, все моноклональные антитела оценивали на способность к нейтрализации токсинов дикого типа в стандартном анализе цитопатического эффекта (СРЕ) или в более строгом и количественном анализе, основанном на измерении АТФ в качестве показателя жизнеспособности клеток.
Из суммарного количества, составляющего 52 антитела, специфичных к токсину А, четыре mAb (А3-25, А65-33, А60-22 и А80-29 (таблица 17 и фиг.18) проявляли варьирующие уровни нейтрализующей активности. Для картирования эпитопов антитела был проведен анализ конкурентного связывания BiaCore и анализ ингибирования гемагглютинации (HI). Результаты показали, что эти антитела могут быть направлены на различные эпитопы белка токсина А (таблица 17). Чтобы дополнительно идентифицировать положение сайтов связывания на белке, антитела индивидуально оценивали в вестерн-блот-анализе или в дот-блот-анализе, используя фрагменты токсина, имеющие известные последовательности. Было обнаружено, что все 4 нейтрализующих mAb направлены на С-концевой участок токсина.
Из суммарного количества, составляющего 17 антител, специфичных к токсину В, было обнаружено, что 9 являются нейтрализующими. Из девяти нейтрализующих mAb шесть было направлено на N-конец, а другие три на домен транслокации токсина В (таблица 18). На основании анализа конкурентного связывания Biacore девять нейтрализующих моноклональных антител можно сгруппировать на четыре группы эпитопов, как показано на фиг.19.
Пример 34: Идентификация новых комбинаций антител к токсину А, обладающих значительно усиленной нейтрализующей активностью
Четыре mAb к токсину А (А3-25, А65-33, А60-22 и А80-29) показали неполную или частичную нейтрализацию токсина А при индивидуальном тестировании в анализе нейтрализации на основе АТФ. mAb А3-25 представляло собой наиболее эффективное антитело, а три другие обладали меньшей нейтрализующей способностью, причем А80-29 лишь немного выше фоновой (фиг.18). Тем не менее, при комбинировании А3-25 с любым из трех других mAb во всех трех комбинациях наблюдали синергетический эффект нейтрализации, который был значительно выше, чем суммарная нейтрализация индивидуальных антител, как показано на фиг.20А-С. Кроме того, все три комбинации проявляли полную нейтрализующую способность, обычно наблюдаемую для поликлональных антител против токсина А.
Пример 35: Идентификация новых комбинаций антител к токсину В, проявляющих значительно усиленную нейтрализующую активность
Авторы изобретения также наблюдали синергетическую нейтрализацию для mAb к токсину В из различных групп эпитопов, идентифицированных с использованием анализа BiaCore. mAb B8-26 к токсину В, наиболее преобладающее mAb из группы 1, комбинировали с множественными mAb из группы 3. Эти комбинации оценивали в анализе нейтрализации, специфичной к токсину В, и его результаты представлены на фиг.21 и в таблице 19.
Синергетический нейтрализующий эффект наблюдали при комбинировании B8-26 с mAb группы 3 эпитопов (фиг.21В), но не наблюдали с какими-либо другими mAb (данные не представлены).
Пример 36: Скрининг in vitro с использованием mAb на безопасные и эффективные композиции мутантных токсинов
Генетически мутантные токсины А и В С.difficile (например, SEQ ID NO:4 и 6), полученные методами генной инженерии, проявляли остаточную цитотоксичность при использовании анализа цитотоксичности in vitro. Хотя авторы изобретения достигли снижения цитотоксичности, составляющего приблизительно 4 log для каждого мутантного токсина С.difficile (таблица 20), дополнительная химическая инактивация мутантных токсинов, например, обработкой формалином, была предпочтительна. Тем не менее, обработки при химической инактивации могут быть жесткими, и могут оказывать вредное влияние на ключевые антигенные эпитопы этих токсинов или мутантных токсинов.
Для оптимизации биопроцесса была разработана статистическая схема эксперимента (DOE) для химической инактивации тройного мутантного Ted А и В (1 мг/мл) с использованием обработки формалином и EDC/NHS. Чтобы оптимизировать инактивацию формалином тройного мутантного TcdA, авторы изобретения варьировали концентрацию формалина/глицина (20-40 мМ), рН (6,5-7,5) и температуру (25-40°С). Для тройного мутантного TcdB авторы изобретения варьировали концентрацию формалина/глицина от 2 до 80 мМ, а температура и рН составляли 25°С и 7,0 соответственно. Время инкубации для всех обработок формалином составляло 24 часа. Для инактивации формалином "40/40" в таблицах 21 и 23 представляет концентрацию формалина и глицина, используемую в реакции. Для обработки EDC/NHS авторы изобретения варьировали концентрации EDC/NHS от 0,25 до 2,5 мг/мг тройного мутантного TcdA и от 0,125 до 2,5 мг/мг тройного мутантного TcdB и инкубировали в течение четырех часов при 25°С. По окончании реакций все образцы обессоливали в 10 мМ фосфате, рН 7,0. После очистки обработанные Ted анализировали на остаточную цитотоксичность и распознавание mAb эпитопов с использованием дот-блот-анализов. Цель состояла в идентификации условий обработки, снижающих цитотоксичность до желаемого уровня (ЕСво более 1000 мкг/мл) без отрицательного воздействия на эпитопы, распознаваемые панелью нейтрализующих mAb (++++ или +++). Условия обработки (отмеченные пометкой " " в таблицах 21-24) давали потенциально безопасные и эффективные иммуногенные композиции, сохраняющие реактивность по меньшей мере к четырем нейтрализующим mAb, проявляя при этом снижение цитотоксичности, составляющее 6-8 log10, относительно цитотоксичности соответствующих токсинов дикого типа. Избранные результаты представлены в таблицах 21-24. Дополнительные данные по варьированию условий обработки тройных мутантных токсинов и данные анализов цитотоксичности in vitro и нейтрализации токсинов представлены в таблице 33 и таблице 34. См. также, например, Примеры 20 и 21 выше, в которых приведены дополнительные подробности, касающиеся предпочтительных условий сшивающей обработки мутантных токсинов.
" в таблицах 21-24) давали потенциально безопасные и эффективные иммуногенные композиции, сохраняющие реактивность по меньшей мере к четырем нейтрализующим mAb, проявляя при этом снижение цитотоксичности, составляющее 6-8 log10, относительно цитотоксичности соответствующих токсинов дикого типа. Избранные результаты представлены в таблицах 21-24. Дополнительные данные по варьированию условий обработки тройных мутантных токсинов и данные анализов цитотоксичности in vitro и нейтрализации токсинов представлены в таблице 33 и таблице 34. См. также, например, Примеры 20 и 21 выше, в которых приведены дополнительные подробности, касающиеся предпочтительных условий сшивающей обработки мутантных токсинов.
Условия реакции химического сшивания для образцов тройного мутантного токсина A (SEQ ID NO:4), представленных в таблице 33
Образцы 1-4 модифицировали EDC/NHS. Условия: 30°С, 20 мМ морфолиноэтансульфоновая кислота МЭС/150 мМ NaCl pH 6,5. Реакции инициировали добавлением EDC. После 2 часов реакции в образцы А, В и С добавляли 1 М глицин до конечной концентрации глицина, составляющей 50 мМ. В образец D глицин не добавляли. Реакции ставили с различными массовыми отношениями мутантный токсин A (SEQ ID NO:4):EDC:NHS, как указано ниже.
1 L44166-157A 1:0,25:0,25 масс: масс: масс
2 L44166-157 В 1:1,25:1,25
3 L44166-157C 1:2,5:2,5
4 L44166-157D 1:2,5:2,5
Образец 5 L44905-160A 80 мМ НСНО, 80 мМ глицин, 80 мМ NaPO4 pH 7, 1 мг/мл белка мутантного токсина A (SEQ ID NO:4), реакция 48 ч при 25°С.
Образец 6 L44166-166 модификация EDC/NHS мутантного токсина А (SEQ ID NO:4) при 25°С в 20 мМ МЭС/150 мМ NaCl pH 6,5. Мутантный токсин А (SEQ ID NO:4):EDC:NHS=1:0,5:0,5. Реакцию инициировали добавлением EDC. После 2 часов реакции добавляли 1 М глицин до конечной концентрации глицина, составляющей 0,1 М, и дополнительно инкубировали в течение 2 часов. После этого времени буфер реакции заменяли 1×ФСБ на Сефадексе G25.
Образец 7 L44905-170A 80 мМ НСНО, 80 мМ глицин, 80 мМ NaPO4 pH 7, 1 мг/мл белка мутантного токсина A (SEQ ID NO:4), реакция 48 ч при 35°С. Эта реакция с формалином была направлена на получение избыточного сшивания, поэтому связывание антигена должно было быть резко уменьшено.
Образец 8 L44897-61 32 мМ НСНО/80 мМ глицин, реакция 72 ч при 25°С.
Образец 9 L44897-63 80 мМ НСНО/80 мМ глицин, реакция 72 ч при 25°С.
Все следующие реакции проводили в течение периода реакции, составляющего 24 ч.
Образец 10 L44897-72 Пробирка №1 25°С, 80 мМ NaPi pH 6,5, 20 мМ НСНО/20 мМ глицин
Образец 11 L44897-72 Пробирка №2 25°С, 80 мМ NaPi pH 6,5, 40 мМ НСНО/40 мМ глицин
Образец 12 L44897-72 Пробирка №3 32,5°С, 80 мМ NaPi pH 7,0, 30 мМ НСНО/30 мМ глицин
Образец 13 L44897-72 Пробирка №4 32,5°С, 80 мМ NaPi pH 7,0, 30 мМ НСНО/30 мМ глицин
Образец 14 L44897-72 Пробирка №5 32,5°С, 80 мМ NaPi pH 7,0, 30 мМ НСНО/30 мМ глицин
Образец 15 L44897-75 Пробирка №6 25°С, 80 мМ NaPi pH 7,5, 20 мМ НСНО/20 мМ глицин
Образец 16 L44897-75 Пробирка №7 25°С, 80 мМ NaPi pH 7.5, 40 мМ НСНО/40 мМ глицин
Образец 17 L44897-75 Пробирка №8 40°С, 80 мМ NaPi pH 6,5, 20 мМ НСНО/20 мМ глицин
Образец 18 L44897-75 Пробирка №9 40°С, 80 мМ NaPi pH 6,5, 40 мМ НСНО/40 мМ глицин
Образец 19 L44897-75 Пробирка №10 40°С, 80 мМ NaPi pH 7,5, 20 мМ НСНО/20 мМ глицин
Образец 20 L44897-75 Пробирка №11 40°С, 80 мМ NaPi pH 7,5, 40 мМ НСНО/40 мМ глицин
Следующие 8 образцов подвергали взаимодействию при 25°С в течение указанных периодов в 80 мМ NaPi pH 7,0, содержащем 78 мМ НСНО и 76 мМ глицина:
Образец 21 L44897-101 (премодификация) ТхА контроль: контрольный образец, время 0, не модифицирован или не подвергнут воздействию НСНО/глицина
Образец 22 L44897-101,2 ч
Образец 23 L44897-101,4 ч
Образец 24 L44897-101,6 ч
Образец 25 L44897 102, 24 ч
Образец 26 L44897-103, 51 ч
Образец 27 L44897-104, 74 ч
Образец 28 L44897-105, 120 ч
Образец 29 (L44980-004) представлял собой модифицированный EDC/NHS мутантный токсин A (SEQ ID NO:4) (тройной мутантный токсин А (SEQ ID NO:4)-EDC). Условия реакции представляют собой: 25°С, буфер представлял собой 20 мМ МЭС/150 мМ NaCl pH 6,6. Тройной мутантный токсин A (SEQ ID NO:4):EDC:NHS=1:0,5:0,5 масс:масс:масс. Реакцию инициировали добавлением EDC. После 2 часов реакции добавляли 1 М глицин до конечной концентрации глицина, составляющей 0,1 М, и дополнительно инкубировали в течение 2 часов при 25°С. Реакцию останавливали обессоливанием на Сефадексе G25.
Следующие 12 образцов и 2 контроля представляли собой эксперименты по реверсии, где образцы инкубировали при 25°С и 37°С.
Реакция 1 = 25°С, 80 мМ NaPi pH 7,0, только 40 мМ НСНО (без глицина), реакция 24 часа.
Реакция 2 = 25°С, 80 мМ NaPi pH 7,0, 40 мМ НСНО/40 мМ глицин, реакция 24 часа Образец Реакция
30 Реакция №1 Неделя 0, 25°С
31 Реакция №1 Неделя 1, 25°С
32 Реакция №1 Неделя 2, 25°С
33 Реакция №1 Неделя 3, 25°С
34 Реакция №1 Неделя 4, 25°С
35 Реакция №1 Неделя 3, 37°С
36 Реакция №2 Неделя 0, 25°С
37 Реакция №2 Неделя 1, 25°С
38 Реакция №2 Неделя 2, 25°С
39 Реакция №2 Неделя 3, 25°С
40 Реакция №2 Неделя 4, 25°С
41 Реакция №2 Неделя 3, 37°С
42 Т×А контроль Неделя 3, 25°С
43 Т×А контроль Неделя 3, 37°С
Следующие 4 образца были получены путем взаимодействия в течение указанного времени при 25°С в 80 мМ NaPi pH 7,0, 40 мМ НСНО/40 мМ глицин.
44 L44897-116-6 29,5 ч
45 L44897-116-7 57,5 ч
46 L44897-116-8 79,5 ч
47 L44897-116-9 123,5 ч
Образец 48 L44897-139 48 ч взаимодействия при 25°С, 80 мМ NaPi pH 7,0, 40 мМ НСНО/40 мМ глицин.
Образец 49 L44166-204 модификация EDC/NHS мутантного токсина А (SEQ ID NO:4). 25°С, буфер 1×ФСБ pH 7,0. Мутантный токсин A (SEQ ID NO:4):EDC:NHS=1:0,5:0,5 масс:масс:масс.2 часа взаимодействие с EDC/NHS, затем добавляли 1 М глицин до конечной концентрации 0,1 М и дополнительно подвергали взаимодействию в течение 2 часов. Буфер заменяли на Сефадексе G25 20 мМ L-гистидин/100 мМ NaCl pH 6,5.
Условия реакции химического поперечного сшивания для образцов тройного мутантного токсина В, представленных в таблице 34
Тройной мутантный токсин В (SEQ ID NO:6) подвергали химическому поперечному сшиванию и тестировали согласно следующим условиям реакции. Образцы L44905-86 тестировали в эксперименте, включающем три варианта взаимодействия с формалином и две температуры инкубации. Каждые сутки отбирали 6 образцов из суммарного количества, составляющего 18 образцов. Первый образец в перечне представляет собой необработанный контроль (который составляет в сумме 19 образцов). Необработанный контроль включал необработанный полипептид тройного мутантного токсина В (SEQ ID NO:6).
Реакция 1 («Rxn1»)=80 мМ НСНО, 80 мМ глицин, 80 мМ NaPO4 рН 7, 1 мг/мл белка тройного мутантного токсина В (SEQ ID NO:6)
Реакция 2 («Rxn2»)=80 мМ НСНО, без глицина, 80 мМ NaPO4 pH 7, 1 мг/мл белка тройного мутантного токсина В (SEQ ID NO:6)
Реакция 3 («Rxn3»)=80 мМ НСНО, без глицина, 80 мМ NaPO4 pH 7, 1 мг/мл белка тройного мутантного токсина В (SEQ ID NO:6)+кэпирование цианоборгидридом. Кэпирование цианоборгидридом включало добавление 80 мМ CNBrH4 в конечную обессоленную реакционную смесь и инкубацию в течение 24 ч при 36°С.
Для образца L34346-30A 0,5 г EDC и NHS на грамм тройного мутантного токсина В (SEQ ID NO:6), 4 часа при 30°С в 20 мМ МЭС, 150 мМ NaCl, pH 6,5.
Для образца L34346-30B 0,5 г EDC и NHS NHS на грамм тройного мутантного токсина В (SEQ ID NO:6), 2 часа при 30°С с последующим добавлением глицина (конечная концентрация в г/л) и инкубацией еще в течение 2 часов при 30°С в 20 мМ МЭС, 150 мМ NaCl, pH 6,5. Единственное различие между этими двумя реакциями для L34346-30A и L34346-30B состоит в добавлении глицина в реакционную смесь L34346-30B.
Пример 37: Антитела, индуцированные иммуногенными композициями, способны к нейтрализации токсинов из различных штаммов С.difficile
Чтобы установить, могут ли антитела, индуцированные иммуногенными композициями, включающими лекарственные вещества на основе мутантных токсинов, нейтрализовать широкий спектр разнообразных последовательностей токсинов, штаммы, представляющие разнообразные риботипы и токсинотипы, были секвенированы для идентификации степени генетического разнообразия среди различных штаммов по сравнению с лекарственными веществами на основе мутантных токсинов. Затем супернатанты культур, содержащие секретированные токсины из различных штаммов, тестировали в анализе нейтрализации in vitro с использование сывороток от иммунизированных хомячков на определение покрытия иммуногенной композиции и на определение способности иммуногенной композиции к защите против разнообразных токсинов от циркулирующих клинических штаммов.
Оба типа клеток, НТ-29 (линия клеток карциномы ободочной кишки) и IMR-90, использовали для тестирования нейтрализации токсинов, экспрессируемых из штаммов CDC. Клетки НТ-29 обладают большей чувствительностью к TcdA; где EC50 очищенного TcdA в этих клетках составляет 100 пг/мл по сравнению с 3,3 нг/мл для TcdB. С другой стороны, клетки IMR-90 обладают большей чувствительностью к TcdB; где ЕС50 очищенного TcdB в этих клетках находится в интервале 9-30 пг/мл по сравнению с 0,92-1,5 нг/мл для TcdA. Специфичность анализа как для TcdA, так и для TcdB в этих линиях клеток была подтверждена путем использования как поликлональных, так и моноклональных токсиноспецифичных антител. Для нормализации анализа фильтраты культур 24 изолятов CDC тестировали при концентрации, составляющей их четырехкратное соответствующее значение ЕС50. Три из штаммов имели слишком низкие уровни токсина для тестирования в анализе нейтрализации.
Двадцать четыре штамма, представляющих разнообразные риботипы/токсинотипы, охватывающие более 95% циркулирующих штаммов С.difficile в США и Канаде, были получены из CDC. Среди этих изолятов находились штаммы, представляющие риботипы 027, 001 и 078, три эпидемических штамма CDAD в США, Канаде и Великобритании. Штаммы 2004013 и 2004118 представляли риботип 027; штамм 2004111 представлял риботип 001, и штаммы 2005088, 2005325 и 2007816 представляли риботип 078. Чтобы идентифицировать степень генетического разнообразия между клиническими изолятами возбудителей заболевания и штаммом 630, гены токсинов (tcdA и tcdB) из этих клинических штаммов были полностью секвенированы. См. таблицу 35. Аминокислотные последовательности токсинов выравнивали, используя программу ClustalW в пакете программ Megalign™ (DNASTAR® Lasergene®), и анализировали на идентичность последовательности. Для tcdA анализ геномного выравнивания показал, что все клинические изоляты и штамм 630 в целом обладают примерно 98-100% идентичностью аминокислотной последовательности. С-концевой участок гена tcdA обладал несколько большим разнообразием. Такой же анализ был проведен для гена tcdB, который проявлял большее разнообразие последовательности. В частности, штаммы 2007838/NAP7/126 и 2007858/NAP1/unk5 проявляли паттерны, наиболее отличающиеся от штамма 630, в N-концевом (79-100%) и С-концевом доменах (88-100%; данные не представлены).
Пул сывороток хомячков (HS) был собран от сирийских золотистых хомячков, которые были иммунизированы иммуногеном, включающим мутантный TcdA (SEQ ID NO:4) и мутантный TcdB (SEQ ID NO:6), причем, мутантные токсины были инактивированы EDC согласно, например, Примеру 29, таблице 15, как описано выше, и включены в препарат, содержащий фосфат алюминия. Результаты в таблице 35 показывают, что по меньшей мере токсин В из соответствующих супернатантов культур был нейтрализован в анализе нейтрализации in vitro сыворотками от иммунизированных хомячков.
На фиг.23 изображены результаты анализа нейтрализации с использованием препаратов токсинов из различных штаммов С.difficile на клетках IMR-90. Данные показывают, что нейтрализующие антитела к TcdB в антисыворотках хомячков способны к нейтрализации токсинов из всех протестированных изолятов в количестве 21, включающих гипервирулентные штаммы и TcdA-отрицательный, TcdB-положительный штамм. По меньшей мере 16 различных штаммов С.difficile было получено из CDC (Atlanta, GA) (описаны ранее), и их культивировали в культуральной среде для С.difficile в подходящих условиях, как известно в данной области техники и как описано выше. Супернатанты культур, содержащие секретированные токсины, анализировали на определение их цитотоксичности (EC50) на монослоях IMR-90, а затем тестировали в стандартном анализе нейтрализации in vitro при 4-кратных ЕС50, используя различные разведения сывороток от хомячков, иммунизированных лекарственным веществом на основе мутантного токсина А и лекарственным веществом на основе мутантного токсина В, включенным в препарат, содержащий фосфат алюминия. Необработанный токсин, полученный из супернатантов культуры каждого штамма, и очищенный токсин (имеющийся в продаже токсин, полученный от List Biologicals) (не очищенный из соответствующих супернатантов) тестировали на цитотоксичность на клетках IMR-90, используя описанный выше анализ цитотоксичности in vitro.
На фиг.23А-K графики представляют результаты тестов цитотоксичности in vitro (описанных ранее), на которых уровни АТФ (RLU) нанесены на график против возрастающих концентраций следующих препаратов: культуральной среды С.difficile и пула сывороток хомячков (■); необработанного токсина и пула сывороток хомячков (●); очищенного токсина и пула сывороток хомячков (▲); необработанного токсина (▼), контроль; и очищенного токсина (♦), контроль. Токсины из соответствующих штаммов добавляли к клеткам при 4х значениях EC50.
Как показано на фиг.23А-K, у хомячков, получавших описанный иммуноген, неожиданно развивались нейтрализующие антитела, проявляющие нейтрализующую активность против токсинов по меньшей мере из следующих 16 различных штаммов CDC С.difficile по сравнению с соответствующим контролем, представляющим собой только токсин: 2007886 (Фиг.23А); 2006017 (Фиг.23В); 2007070 (Фиг.23С); 2007302 (Фиг.23D); 2007838 (Фиг.23Е); 2007886 (Фиг.23F); 2009292 (Фиг.23G); 2004013 (Фиг.23Н); 2009141 (Фиг.231); 2005022 (Фиг.23J); 2006376 (Фиг.23K). Дополнительные штаммы С.difficile, токсины которых были протестированы и нейтрализовались иммуногенной композицией, включающей лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В, включенное в препарат, содержащий фосфат алюминия, см. также в таблице 35.
В другом исследовании супернатанты культур, содержащие секретированные токсины из различных штаммов С.difficile (полученных из CDC и из Leeds Hospital, UK), были протестированы в анализе нейтрализации in vitro с использованием сывороток от хомячков, которым вводили лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В, включенное в препарат, содержащий Alhydrogel. Схему эксперимента см. в таблице 36. Результаты представлены в таблице 37 и в таблице 38.
«НС» = неизвестный статус
НП, неприменимо; штамм не продуцирует токсин А; не был протестирован в анализе нейтрализации токсина А
Пример 38: Пептидное картирование тройных мутантных токсинов EDC/NHS
Чтобы охарактеризовать инактивированные EDC/NHS тройные мутантные токсины, были проведены эксперименты по пептидному картированию на четырех партиях обработанного EDC/NHS тройного мутантного токсина A (SEQ ID NO:4) и на четырех партиях обработанного EDC/NHS тройного мутантного токсина В (SEQ ID NO:6). После ферментативного расщепления мутантных токсинов трипсином полученные в результате пептидные фрагменты разделяли, используя ВЭЖХ с обращенной фазой. Для идентификации модификаций, возникающих в результате процесса инактивации, использовали масс-спектральный анализ. Для обоих лекарственных веществ, на основе мутантного токсина А и на основе мутантного токсина В, было идентифицировано более 95% теоретических триптических пептидов. Были идентифицированы поперечные сшивки и аддукты глицина (глицин использовали в качестве кэпирующего агента). В обоих лекарственных веществах, на основе мутантного токсина А и на основе мутантного токсина В, также наблюдали аддукты бета-аланина. Без связи с каким-либо механизмом или теорией, по-видимому, аддукты бета-аланина являются результатом взаимодействия трех моль NHS с одним моль EDC, в результате которого образуется бета-аланин, активированный NHS. Затем эта молекула может взаимодействовать с группами лизина с образованием аддуктов бета-аланина (+70 Да). В образцах обработанного EDC/NHS тройного мутантного токсина В также наблюдали низкие уровни (0,07 моль/моль белка) дегидроаланина (-34 Да). Дегидроаланин является результатом десульфонилирования остатка цистеина. Модификации одного и того же типа и одной и той же степени наблюдали во всех четырех партиях мутантного токсина, что указывает на то, что данным способом получают постоянный продукт. Пептидное картирование (при покрытии более 95% последовательности) подтверждает, что модификации присутствуют. Краткое изложение модификаций представлено в таблице 39. См. также фиг.24-25. Кроме того, гетерогенность заряда и размера лекарственного вещества на основе тройного мутантного токсина А и лекарственного вещества на основе тройного мутантного токсина повышается по сравнению с гетерогенностью заряда и размера соответствующего тройного мутантного токсина А и тройного мутантного токсина В в отсутствие химической инактивации. В результате, профили эксклюзионной хроматографии (ЭХ) и анионообменной хроматографии (АОХ) имели относительно широкие пики (данные не представлены).
Пример 39: Получение лекарственного препарата
Иммуногенная композиция (лекарственный продукт) С.difficile содержит два активных фармацевтических ингредиента (лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В). Иллюстративный лекарственный продукт представляет собой лиофилизированный препарат, содержащий 10 мМ Трис буфер рН 7,4, 4,5% (масс./масс.) дигидрата трегалозы и 0,01% (масс./об.) полисорбата 80, и включает каждое из лекарственных веществ, на основе мутантного токсина А и мутантного токсина В. См. таблицу 40. Иммуногенную композицию готовят для инъекции путем ресуспендирования лиофилизированной вакцины либо разбавителем, либо разбавителем, содержащим Alhydrogel. Плацебо включает стерильный нормальный физиологический раствор для инъекций (0,9% хлорид натрия).
Приготовление буфера
Воду для инъекций (ВДИ) добавляют в сосуд для смешивания. При перемешивании добавляют эксципиенты и растворяют до получения раствора. Измеряют рН. При необходимости рН доводят до 7,4±0,1 с использованием HCl. Раствор разбавляют ВДИ до конечной массы, затем фильтруют, используя 0,22 мкм фильтр Millipore Express SHC XL150. Перед фильтрованием берут образец на снижение биологической нагрузки перед фильтрованием. Берут образец фильтрованного буфера на осмолярность и рН.
Приготовление препарата
Размороженные лекарственные вещества на основе мутантных токсинов объединяют в сосуде для приготовления лекарственного препарата на основании предварительно рассчитанных количеств в следующем порядке действий: сначала в сосуд добавляют 50% целевого объема буфера для разведения до достижения 0,6 мг/мл, затем добавляют лекарственное вещество на основе мутантного токсина А и перемешивают в течение 5 минут при 100 об/мин. Затем в сосуд добавляют лекарственное вещество на основе мутантного токсина В, и раствор дополнительно разводят до точки разведения, составляющей 0,6 мг/мл, а затем перемешивают еще в течение 5 минут при 100 об/мин. Отбирают образец и тестируют на суммарную концентрацию мутантных токсинов. Раствор разводят до 100% объема на основании рабочего значения концентрации мутантных токсинов, затем перемешивают в течение 15 минут при 100 об/мин. Берут образец лекарственного продукта в виде препарата на рН и биологическую нагрузку перед фильтрованием. Затем лекарственный продукт в виде препарата фильтруют, используя фильтр Millipore Express SHC XL150, для хранения в течение ночи, или переносят на линию заполнения для стерильного фильтрования.
Нерасфасованный лекарственный продукт в виде препарата переносят в зону заполнения, берут образец на биологическую нагрузку, а затем стерильно фильтруют через два последовательно соединенных фильтра Millipore Express SHC XL150. Нерасфасованный лекарственный продукт в виде препарата заполняют в апирогенные стеклянные флаконы при целевом объеме заполнения, составляющем 0,73 мл. Заполненные флаконы частично закрывают пробками, а затем загружают в лиофилизатор. Цикл лиофилизации выполняют, как показано в таблице 41. После завершения цикла камеру лиофилизации снова заполняют азотом до 0,8 атм, а затем полностью устанавливают пробки. Камеру разгружают, и флаконы закрывают, используя предохранительные колпачки.
Данные стабильности лекарственного продукта суммированы в таблице 42. Эти данные позволяют предположить, что лекарственный продукт является физически и химически стабильным во время хранения при 2-8°С в течение по меньшей мере 3 месяцев или по меньшей мере 1 месяца при 25° или при 40°С. При обоих условиях хранения уровень примесей, обнаруженных с использованием эксклюзионной хроматографии (ЭХ), не меняется, как отсутствуют и изменения в антигенности in vitro на протяжении последних протестированных моментов времени.
Пример 40: Разбавители вакцины
Для физиологического раствора в качестве разбавителя используют 60 мМ NaCl для лиофилизированного лекарственного препарата без какого-либо адъюванта, чтобы обеспечить изотонический раствор после восстановления.
Alhydrogel: Alhydrogel «85» 2% (Brenntag) представляет собой имеющийся в продаже препарат марки надлежащей практики организации производства (Good Manufacturing Practice; GMP), состоящий из октаэдрических кристаллических слоев гидроксида алюминия. Иллюстративный препарат разбавителя Alhydrogel показан в таблице 43. Иллюстративный препарат можно применять в комбинации с описанным выше лекарственным препаратом.
Исследования, проведенные с адъювантом Alhydrogel, показывают 100% связывание лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В с 1 мг Al/мл Alhydrogel, pH 6,0-7,5. Максимальное связывание обоих лекарственных веществ наблюдали при самой высокой тестируемой концентрации белка (300 мкг/мл каждого).
Связывание белков с Alhydrogel также тестировали с лиофилизированным лекарственным препаратом, содержащим 200 мкг/мл каждого лекарственного вещества и Alhydrogel в интервале от 0,25 до 1,5 мг/мл. Лекарственный препарат восстанавливали разбавителями, содержащими различные концентрации Alhydrogel, и измеряли процент связывания каждого мутантного токсина. Для всех протестированных концентраций Alhydrogel продемонстрировано 100% связывание антигенов.
Также оценивали кинетику связывания белков с Alhydrogel при целевой дозе лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В (200 мкг/мл каждый). Результаты показывают, что 100% лекарственных веществ на основе мутантных токсинов связывается с Alhydrogel на протяжении всего времени удерживания, составляющего 24 часа.
CpG 24555 и Alhydrogel: CpG 24555 представляет собой синтетический 21-мерный олигодезоксинуклеотид (ОДН), имеющий следующую последовательность: 5-TCG TCG TTTTTC GGT GCT TTT-3 (SEQ ID NO:48). Иллюстративный препарат для комбинации разбавителей CpG 24555 и Alhydrogel показан в таблице 44. Иллюстративный препарат можно применять в комбинации с описанным выше лекарственным препаратом.
ISCOMATRIX®: Адъювант ISCOMATRIX® представляет собой адъювант на основе сапонина, известный в данной области техники. Иллюстративный препарат для адъюванта ISCOMATRIX® представлен в таблице 45. Иллюстративный препарат можно применять в комбинации с описанным выше лекарственным препаратом.
Пример 41: Иммуногенность композиций, содержащих лекарственные вещества на основе мутантных токсинов и адъювант Alhydrogel, в модели нечеловекообразных приматов (НЧП) и доклиническая проверка концепции
Оценивали иммуногенность композиций, содержащих лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В и адъювант Alhydrogel, у НЧП, в частности, у яванских макак. У НЧП, иммунизированных через двухнедельные интервалы (недели 0, 2, 4) композициями, содержащими 10 мкг каждого из лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В (включенными в препарат, содержащий Alhydrogel) на дозу, развивались стабильные нейтрализующие антитоксинные ответы. См. таблицу 46. Оба нейтрализующих ответа, против токсина А и против токсина В, достигали защитного диапазона после третьей иммунизации и оставались в пределах защитного диапазона или выше по меньшей мере на протяжении недели 33 (последний исследованный момент времени).
Яванских макак (n=8) иммунизировали IM на неделе 0, 2 и 4 10 мкг каждого из лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В, включенных в препарат, содержащий 250 мкг Alhydrogel. Сыворотки собирали в каждый момент времени и анализировали в анализе нейтрализации токсина на функциональную антитоксинную активность. Средние геометрические значения титра GMT приведены в таблице 46. Защитные диапазоны титра, приведенные в таблице, отражают титры нейтрализующих антител, которые коррелируют со значительным снижением рецидива инфекции С.difficile в клиническом испытании терапии моноклональным антителом Merck.
Корреляция защитных титров антитела человека из клинического испытания терапии Merck mAb с титрами, вызванными вакциной-кандидатом Pfizer, у НЧП
Исследование эффективности фазы 2, проведенное с mAb Merck/Medarex (Lowy et al., N Engl J Med. 2010 Jan 21;362(3):197-205), по-видимому, показывает корреляцию между уровнями нейтрализующих антитоксинных mAb в сыворотке и предотвращением рецидива CDAD. После введения токсин-специфичных mAb людям уровни сывороточного антитела у людей-реципиентов в диапазоне от 10 до 100 мкг/мл, по-видимому, защищают против рецидивов (70% снижение рецидива CDAD).
Иммуногенные композиции, включающие лекарственные вещества на основе мутантных токсинов, тестировали, чтобы измерить, способны ли иммуногенные композиции индуцировать потенциально эффективные ответы нейтрализующих антител у людей, путем сравнения опубликованных данных исследования Merck/Medarex фазы 2 с уровнями антител, индуцированных иммуногенными композициями в модели НЧП. Это было выполнено путем использования опубликованных ранее характеристик mAb Merck/Medarex для преобразования диапазона этих mAb в сыворотке, полученной от субъектов, не проявляющих признаков рецидивов (10-100 мкг/мл), в 50% титры нейтрализации и сравнения этих титров («защитный диапазон титров») с титрами, наблюдаемыми в доклинических моделях, раскрытых в данной работе. Как показано в таблице 46, иммуногенные композиции, включающие лекарственное вещество на основе мутантного токсина А и лекарственное вещество на основе мутантного токсина В, включенные в препарат, содержащий Alhydrogel, вызывали иммунные ответы у НЧП, которые достигали «защитного диапазона» после третьей дозы и оставались в пределах этого диапазона или выше на протяжении недели 33. Уровень токсин-нейтрализующих антител, индуцированных у НЧП иммуногенными композициями на основе С.difficile, сравнимы с сывороточными уровнями антител у субъектов испытания Merck/Medarex, которые оказались защищенными от рецидивов CDAD.
Пример 42: Иммуногенность композиций, содержащих лекарственные вещества на основе мутантных токсинов и адъювант ISCOMATR1X или Alhydrogel/CpG 24555 (Alh/CpG) в модели НЧП
У НЧП как ISCOMATR1X, так и Alh/CpG статистически значимо повышали титры нейтрализации против токсинов А и В по сравнению с вакциной, введенной только с Alhydrogel (Таблица 47). Антитоксинные ответы выше фонового уровня были индуцированы в более ранние моменты времени вакциной, введенной либо с Alh/CpG, либо с ISCOMATR1X (неделя 2-4), по сравнению с одним Alhydrogel (неделя 4-6), что может обладать важным действием на защиту от рецидива CDAD у людей. По сравнению с Alhydrogel иммуногенная композиция в сочетании с адъювантом Alh/CpG или ISCOMATRIX, вызывала титры нейтрализации против токсинов, достигающие защитные диапазоны (см. также Пример 41) быстрее, и они оставались в пределах этого диапазона или выше на протяжении недели 33.
Как показано в таблице 47, яванских макак иммунизировали IM на неделе 0, 2 и 4 10 мкг каждого из лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В, включенных в препарат, содержащий 250 мкг Alhydrogel (n=8), либо 500 мкг CpG и 250 мкг Alhydrogel (n=10), либо 45 ед. ISCOMATR1X (n=10). Сыворотки собирали в каждый момент времени и анализировали в описанном выше анализе нейтрализации токсина на функциональную антитоксинную активность. GMT перечислены в таблицах. Звездочками (*) указана статистическая значимость (р<0,05) по сравнению с титрами в группе Alhydrogel. Защитный диапазон титра представляет собой диапазон титра нейтрализующих антител, который коррелирует со значимым снижением рецидива инфекции С.difficile согласно испытанию терапии mAb Merck/Medarex.
Также оценивали влияние дозы лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В, введенного в присутствии адъювантов ISCOMATRIX или Alh/CpG, на титры нейтрализующих антител против токсинов, индуцируемые у НЧП. В одном исследовании НЧП вводили низкую (10 мкг) или высокую (100 мкг) дозу каждого лекарственного вещества на основе мутантного токсина, включенного в препарат, содержащий ISCOMATRIX. Ответы сравнивали в каждый момент времени после иммунизации. Как показано в таблице 48, титры нейтрализации против токсинов были стабильны в обеих группах обработки. Титры против токсина А были почти эквивалентны в большинстве моментов времени между группами низкой дозы и высокой дозы, тогда как для титров против токсина В существовала тенденция к повышению в группе высокой дозы.
Как показано в таблице 48, яванских макак (п=5) иммунизировали IM на неделе 0, 2 и 4 10 мкг или 100 мкг каждого из лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В, включенных в препарат, содержащий 45 ед. ISCOMATRIX. Сыворотки собирали в каждый момент времени и анализировали в описанном выше анализе нейтрализации токсина на функциональную антитоксинную активность. GMT перечислены в таблицах. Защитный диапазон титра представляет собой диапазон титра нейтрализующих антител, который коррелирует со значимым снижением рецидива инфекции С.difficile согласно испытанию терапии mAb Merck/Medarex.
При попытке усилить кинетику ответов против токсина В НЧП иммунизировали постоянной дозой лекарственного вещества на основе мутантного токсина А (10 мкг), которую смешивали с возрастающей дозой лекарственного вещества на основе мутантного токсина В (10, 50 или 100 мкг) в присутствии адъювантов ISCOMATRIX или Alh/CpG. Независимо от адъюванта для групп, получавших более высокие дозы лекарственного вещества на основе мутантного токсина В (либо 50, либо 100 мкг), существовала тенденция к индукции более высоких нейтрализующих ответов против токсина В по сравнению с дозой, составляющей 10 мкг лекарства на основе мутантного токсина В (таблица 50, отметкой * указаны статистически значимые возрастания). Эту тенденцию наблюдали в большинстве моментов времени после конечной иммунизации. Тем не менее, в некоторых случаях нейтрализующие ответы против токсина А проявляли статистически значимое снижение (отмечено ^ в таблице 49) при увеличении количества мутантного токсина В.
Как показано в таблице 49 и в таблице 50, НЧП (10 на группу) иммунизировали IM на неделе 0, 2 и 4 различными соотношениями лекарственного вещества на основе мутантного токсина А и лекарственного вещества на основе мутантного токсина В (10 мкг лекарственного вещества на основе мутантного токсина А и либо 10, либо 50, либо 100 мкг лекарственного вещества на основе мутантного токсина В; обозначены 10А:10 В, 10А:50 В и 10А:100 В, соответственно, в таблице 49 и в таблице 50), включенных в препарат, содержащий ISCOMATRIX (45 ед. на дозу) или Alh/CpG/ (250 мкг/500 мкг на дозу). В таблице 49 показаны титры против токсина А. В таблице 50 показаны титры против токсина В. GMT перечислены в таблицах. Защитный диапазон титра представляет собой диапазон титра нейтрализующих антител, который коррелирует со значимым снижением рецидива инфекции С.difficile согласно испытанию терапии mAb Merck/Medarex. Символ А представляет собой статистически значимое снижение титров нейтрализации (р<0,05) по сравнению с группой 10А:10 В. Символ звездочка * представляет собой статистически значимое повышение нейтрализующих титров (р<0,05) по сравнению с группой 10А:10В.
Пример 43: Пятинедельное исследование токсичности повторяющихся доз 1М иммуногенных композиций у яванских макак с 4-недельным восстановительным периодом
Пятинедельное исследование токсичности повторяющейся дозы IM с PF-06425095 (иммуногенной композицией, включающей лекарственное вещество на основе тройного мутантного токсина А и лекарственное вещество на основе тройного мутантного токсина В в комбинации с адъювантами, представляющими собой гидроксид алюминия и CpG 24555) у яванских макак было проведено для оценки потенциальной токсичности и иммуногенности лекарственного вещества на основе тройного мутантного токсина А и тройной лекарственного вещества на основе тройного мутантного токсина В С.difficile в комбинации с адъювантами, представляющими собой гидроксид алюминия и CpG 24555 (PF-06425095). PF-06425095 в количестве, составляющем 0,2 или 0,4 мг/доза лекарственного вещества на основе тройного мутантного токсина А и лекарственного вещества на основе тройного мутантного токсина В (группы иммуногенной композиции низкой и высокой дозы соответственно), 0,5 мг алюминия в виде гидроксида алюминия и 1 мг CpG 24555 и одну комбинацию адъювантов (гидроксида алюминия и CpG 24555; PF-06376915) вводили IM яванским макакам (б/пол/группа) в виде примирующей дозы с последующими 3 бустерными дозами (сутки 1, 8, 22 и 36). Отдельная группа животных (6/пол) получала 0,9% изотонический физиологический раствор, имеющий рН приблизительно 7,0. Носитель иммуногенной композиции состоял из 10 мМ Трис буфера при рН 7,4, 4,5% тригидрата трегалозы и 0,1% полисорбата 80. Контроль носителя адъюванта состоял из 10 мМ гистидинового буфера, содержащего 60 нМ NaCl, при рН 6,5. Суммарный объем дозы составлял 0,5 мл на инъекцию. Все дозы вводили в левую и/или правую четырехглавую мышцу. Выбранных животных подвергали 4-недельному периоду наблюдения без введения дозы для оценки обратимости каких-либо эффектов, наблюдаемых во время фазы дозирования в исследовании.
В данном исследовании никаких вредных эффектов не было обнаружено. PF-06425095 хорошо переносился и приводил только к местной воспалительной реакции при отсутствии данных системной токсичности. Во время фазы дозирования в группах, обработанных иммуногенной композицией, наблюдали дозозависимые повышения следующих показателей по сравнению с данными до тестирования: фибриногена (от 23,1% до 2,3х) на сутки 4 и 38, С-реактивного белка на сутки 4 (от 2,1х до 27,5х) и 38 (от 2,3х до 101,5х) и глобулина (от 11,1% до 24,1%) на сутки 36 и/или 38, что соответствовало ожидаемому воспалительному ответу на введение иммуногенной композиции, содержащей адъюванты.
Повышения фибриногена и С-реактивного белка, отмеченные на сутки 4, частично восстановились к суткам 8, причем повышения фибриногена (от 25,6% до 65,5%) и С-реактивного белка (от 4,5х до 5,6х) наблюдали только в группе высокой дозы иммуногенной композиции. Повышения интерлейкина (IL)-6 наблюдали в группах низкой и высокой дозы иммуногенной композиции на сутки 1, час 3 (от 8,3х до 127,2х индивидуальных значений суток 1, час О, дозозависимо) и на сутки 36, час 3 (9,4х до 39,5х индивидуальных значений суток 36, час 0). Изменений в других цитокинах (IL-10, IL-12, интерферон-индуцибельном белке (IP-10) и факторе некроза опухоли α (TNF-α) не наблюдали. Повышения этих белков острой фазы и цитокина составляли часть ожидаемого нормального физиологического ответа на введение чужеродного антигена. В фазе восстановления изменения этих параметров клинической патологии, связанные с PF 06425095 или связанные с адъювантом, отсутствовали (во время фазы восстановления цитокины не оценивали). Кроме того, были локализованные изменения в сайтах инъекции, которые имели сходную встречаемость и тяжесть в контрольной группе адъюванта и в группах низкой и высокой дозы иммуногенной композиции; следовательно, они напрямую не связаны с PF-06425095. Во время фазы дозирования эти изменения включали хронически-активное воспаление от минимального до умеренного, характеризующееся разделением мышечных волокон инфильтратами макрофагов, которые часто содержали базофильный гранулярный материал (интерпретируемый как адъювант, содержащий алюминий), лимфоцитов, плазматических клеток, нейтрофилов, эозинофилов, некротической тканью и отеком. Базофильный гранулярный материал также присутствовал внеклеточно в пределах этих очагов хронического активного воспаления. По окончании фазы восстановления хроническое воспаление и инфильтраты мононуклеарных клеток были от минимальных до умеренных, и фиброз был минимальным. Эти явления в сайте инъекции представляют собой местный воспалительный ответ на адъювант. Другие микроскопические изменения включали от минимальной до умеренной повышенной насыщенности лимфоидными клетками в подвздошном (дренирующем) лимфатическом узле и минимальную повышенную насыщенность зародышевыми центрами в селезенке, которые были отмечены во время фазы дозирования в контрольной группе адъюванта и в группах низкой и высокой дозы иммуногенной композиции. По окончании фазы восстановления эти микроскопические явления имели меньшую тяжесть. Эти явления представляют собой иммунологический ответ на антигенную стимуляцию, и представляют собой фармакологический ответ на адъювант или PF-06425095. Повышение антител против ДНК, связанное с тестируемым препаратом, отсутствовало.
На основании отсутствия вредных явлений максимальная доза, не вызывающая обнаруживаемого вредного воздействия (NOAEL; no observed adverse effect level), в данном исследовании представляет собой группу высокой дозы иммуногенной композиции (0,4 мг лекарственного вещества на основе тройного мутантного токсина А и лекарственного вещества на основе тройного мутантного токсина В/доза в виде препарата PF-06425095), вводимой в виде двух инъекций по 0,5 мл на четыре дозы.
Пример 44: Эффективность серопозитивных сывороток НЧП, пассивно переносимых хомячкам
Группам из 5 сирийских золотистых хомячков вводили перорально дозу антибиотика клиндамицина (30 мг/кг) для нарушения нормальной кишечной флоры. После пяти суток хомячков заражали пероральной дозой спор С.difficile дикого типа (штамм 630, 100 кое на животное), и вводили интраперитонеально (IP) сыворотки НЧП согласно таблице 51. Без связи с каким-либо механизмом или теорией, симптомы заболевания после заражения спорами обычно проявляются, начиная примерно с 30-48 часов после заражения.
Сыворотки НЧП, вводимые хомячкам, объединяли с образцами сыворотки НЧП, проявляющими самый высокий титр (сыворотки против токсина А и сыворотки против токсина В), после трех иммунизации лекарственным веществом на основе мутантного токсина А и лекарственным веществом на основе мутантного токсина В (соотношения 10:10, 10:50 и 10:100 А:В), включенным в препарат, содержащий ISCOMATRIX (см. Пример 42, таблицу 49 и таблицу 50). Сыворотки НЧП собирали в моменты времени на неделе 5, 6 и 8 (иммунизации осуществляли на неделе 0, 2 и 4), как описано в Примере 42. Результаты представлены в таблицах 52-54 ниже. Символом "+" указано геометрическое среднее (GM) в (), в которое не включено животное №3, нереспондер. "*ТВ" представляет собой терминальный забор крови на сутки, когда животных подвергали эвтаназии, которые неодинаковы для всех животных.
В другом исследовании сирийским золотистым хомячкам вводили перорально дозу антибиотика клиндамицина (30 мг/кг) для нарушения нормальной кишечной флоры. После пяти суток хомячков заражали пероральной дозой спор С.difficile дикого типа (штамм 630, 100 кое на животное), и вводили интраперитонеально (IP) сыворотки НЧП согласно таблице 55. Без связи с каким-либо механизмом или теорией, симптомы заболевания после заражения спорами обычно проявляются, начиная примерно с 30-48 часов после заражения.
Сыворотки НЧП, вводимые хомячкам, объединяли с образцами сыворотки НЧП, проявляющими самый высокий титр (сыворотки против токсина А и сыворотки против токсина В), после трех иммунизации лекарственным веществом на основе мутантного токсина А и лекарственным веществом на основе мутантного токсина В (соотношения 10:10, 10:50 и 10:100 А:В), включенным в препарат, содержащий Alhydrogel и CpG 24555 (см. Пример 42, таблицу 49 и таблицу 50). Сыворотки НЧП собирали в моменты времени на неделе 5, 6 и 8 (иммунизации осуществляли на неделе 0, 2 и 4), как описано в Примере 42. Результаты представлены в таблицах 56-59 ниже. Сыворотки от хомячков дополнительно исследовали, чтобы определить значение ингибиторной концентрации (IC50), которое определяли, используя описанный выше анализ нейтрализации токсина. Уровень токсин-нейтрализующих антител, индуцированных у хомячков, иммунизированных иммуногенной композицией С.difficile по изобретению, сравним с уровнями сывороточных антител у субъектов клинического испытания Merck/Medarex, которые оказались защищенными от рецидивов CDAD.
Пример 45: Характеризация лекарственных веществ на основе мутантных токсинов
Первичная структура тройного мутантного токсина А представлена в SEQ ID NO:4. NH2-концевой остаток Met в положении 1 SEQ ID NO:4 имеет происхождение от кодона инициации SEQ ID NO:12 и отсутствует в выделенном белке (например, см. SEQ ID NO:84). Соответственно, в Примере 12 - Примере 45 "SEQ ID NO:4" относится к SEQ ID NO:4, в которой начальный метионин (в положении 1) отсутствует. Как для очищенного тройного мутантного токсина A (SEQ ID NO:4) (Промежуточное лекарственное вещество - партия L44993-132), так и для EDC/NHS-обработанного тройного мутантного токсина A (SEQ ID NO:4) ("лекарственное вещество на основе мутантного токсина А" - партия L44898-012) показана единая МН2-концевая последовательность, начинающаяся с SLISKEELIKLAYSI (положения 2-16 SEQ ID NO:4).
Первичная структура тройного мутантного токсина В представлена в SEQ ID NO:6. NH2-концевой остаток Met в положении 1 SEQ ID NO:6 имеет происхождение от кодона инициации SEQ ID NO:12 и отсутствует в выделенном белке (например, см. SEQ ID NO:86 Соответственно, в Примере 12 - Примере 45 "SEQ ID NO:6" относится к SEQ ID NO:6, в которой начальный метионин (в положении 1) отсутствует. Как для очищенного тройного мутантного токсина В (SEQ ID NO:6) (Промежуточное лекарственное вещество - партия 010), так и для EDC/NHS-обработанного тройного мутантного токсина В (SEQ ID NO:6) ("лекарственное вещество на основе мутантного токсина В" -партия L44906-153) показана единая NH2-концевая последовательность, начинающаяся с SLVNRKQLEKMANVR (положения 2-16 SEQ ID NO:6).
Чтобы установить вторичную и третичную структуру тройного мутанта А (SEQ ID NO:4) и лекарственного вещества на основе мутантного токсина А, использовали спектроскопию кругового дихроизма (КД). Спектроскопию КД также использовали для установления вторичной и третичной структуры тройного мутантного токсина В (SEQ ID NO:6) и лекарственного вещества на основе мутантного токсина В. Спектроскопию КД также использовали, чтобы установить потенциальные воздействия рН на структуру. Воздействие обработки EDC на тройной мутантный токсин А анализировали путем сравнения данных КД, полученных для лекарственного вещества на основе мутантного токсина А, с данными, полученными для тройного мутантного токсина А. Воздействие обработки EDC на тройной мутантный токсин В анализировали путем сравнения данных КД, полученных для лекарственного вещества на основе мутантного токсина В, с данными, полученными для тройного мутантного токсина В.
Данные КД в далеком ультрафиолетовом (УФ) свете для лекарственного вещества на основе мутантного токсина А были получены при различных значениях рН. Спектры, снятые при рН 5,0-7,0, являются показателем высокой доли α-спиралей во вторичной структуре, что позволяет предположить, что полипептидный каркас белка принимает четко определенную конформацию с преобладанием α-спиралей.
Были также получены спектры лекарственного вещества на основе мутантного токсина А в ближнем ультрафиолетовом (УФ) свете. Сильная отрицательная эллиптичность между 260 и 300 нм указывает на то, что ароматические боковые цепи находятся в уникальной прочной окружающей среде, то есть лекарственное вещество на основе мутантного токсина А обладает третичной структурой. Действительно, в пределах спектра можно отличить следующие характеристические признаки, вызванные индивидуальными типами ароматических боковых цепей: плечо приблизительно при 290 нм и самый высокий отрицательный пик приблизительно при 283 нм являются следствием поглощения поляризованного света упорядоченными боковыми цепями триптофана, отрицательный пик при 276 нм образуется за счет боковых цепей тирозина, и минорные плечи при 262 и 268 нм являются показателем остатков фенилаланина, участвующих в третичных контактах. Результаты спектров далекого и ближнего УФ свидетельствуют о том, что лекарственное вещество на основе мутантного токсина А сохраняет компактную складчатую структуру при физиологических значениях рН. Почти идентичные спектры КД, наблюдаемые в далеком и ближнем УФ свете, при значениях рН 5,0-7,0 указывают на то, что в данном диапазоне рН обнаружимых структурных изменений не происходит. Данные КД не могут быть сняты при значениях рН 3,0 и 4,0, поскольку при данных показателях рН белок был нерастворим. При сравнении спектров КД лекарственного вещества на основе мутантного токсина А в далеком и ближнем УФ со спектрами тройного мутантного токсина А спектры обоих белков по существу идентичны при всех изученных условиях эксперимента, что указывает на то, что обработка EDC не имеет обнаружимых воздействий на вторичную и третичную структуру тройного мутантного токсина А. Данное открытие согласуется с результатами гель-фильтрации и аналитического ультрацентрифугирования, которые не показывает обнаружимых изменений в радиусах Стокса и коэффициентах седиментации/трения, соответственно.
Лекарственное вещество на основе мутантного токсина А (как и тройной мутантный токсин А) содержит 25 остатков триптофана, распределенных на протяжении всей первичной последовательности, которые могут служить в качестве удобных внутренних флуоресцентных зондов. Были получены спектры испускания флуоресценции лекарственного вещества на основе мутантного токсина А между 300 и 400 нм в зависимости от температуры. При 6,8°С лекарственное вещество на основе мутантного токсина А проявляет характеристический спектр испускания флуоресценции триптофана при возбуждении при 280 нм. Максимум испускания флуоресценции наблюдают приблизительно при 335 нм, что указывает на то, что остатки триптофана находятся в неполярном окружении, более характерном для внутренней структуры белков, чем полярное водное окружение. Результаты спектров испускания флуоресценции вместе с результатами экспериментов по КД, представленные в данной работе, подтверждают, что лекарственное вещество на основе мутантного токсина А сохраняет компактную складчатую структуру.
Флуоресценцию внешнего зонда, представляющего собой 8-анилино-1-нафталинсульфоновую кислоту (АНС), использовали для характеризации возможных конформационных изменений лекарственного вещества на основе мутантного токсина А и тройного мутантного токсина А при изменениях рН. Как видно из результатов, при титровании как лекарственного вещества на основе мутантного токсина А, так и тройного мутантного токсина этим зондом при рН 7,0 повышение интенсивности флуоресценции АНС по существу отсутствовало, что позволяет предположить, что гидрофобные поверхности на белках в данных условиях не открываются. Сдвиг рН до 2,6 приводит к резкому повышению квантового выхода флуоресценции АНС при увеличении концентрации зонда до тех пор, пока квантовый выход флуоресценции не достигнет кажущегося насыщения. Это повышение квантового выхода флуоресценции АНС указывает на то, что при низком рН (2,6) как лекарственное вещество на основе мутантного токсина А, так и тройной мутантный токсин А претерпевают индуцированные рН конформационные изменения, открывающие гидрофобные поверхности. Такие конформационные изменения указывают на то, что EDC-индуцированная модификация и инактивация тройного мутантного токсина А не ограничивает конформационную пластичность лекарственного вещества на основе мутантного токсина A (DS).
Воздействие обработки EDC на гидродинамические свойства тройного мутантного токсина А оценивали, используя эксклюзионную хроматографию на колонке G4000 SWXL. Лекарственное вещество на основе мутантного токсина А и тройной мутантный токсин А впрыскивали на колонку G4000 SWXL, уравновешенную при рН 7,0, 6,0 и 5,0. Данные указывают на отсутствие различий в радиусе Стокса лекарственного вещества на основе мутантного токсина А и тройного мутантного токсина А, которые могут быть обнаружены при использовании эксклюзионной хроматографии. Таким образом, обработка EDC не оказывает резкого влияния на гидродинамические свойства и, соответственно, на общую молекулярную форму тройного мутантного токсина А.
Дальнейший анализ тройного мутантного токсина А и лекарственного вещества на основе мутантного токсина А был проведен с использованием метода многоугольного рассеяния лазерного излучения (MALLS). Обработка тройного мутантного токсина А EDC привела в результате к образованию гетерогенной смеси, состоящей из различных мультимерных и мономерных соединений. Такая гетерогенность отражает введение большого числа EDC-индуцированных межмолекулярных и внутримолекулярных ковалентных связей между карбоксильными и первичными аминными группами белка.
Полученные данные обеспечивают физические и химические характеристики тройного мутантного токсина А и лекарственного вещества на основе мутантного токсина А (тройного мутантного токсина А, обработанного EDC) и описывают ключевые признаки их первичной, вторичной и третичной структуры. Полученные данные показывают, что обработка тройного мутантного токсина А EDC приводит в результате к ковалентной модификации его полипептидной цепи, но не влияет на вторичную и третичную структуру белка. Обработка EDC приводит к внутри- и межмолекулярному поперечному сшиванию. Биохимические и биофизические параметры, полученные для лекарственного вещества на основе мутантного токсина А (а также для тройного мутантного токсина А) представлены в таблице 60.
Данные КД в далеком ультрафиолетовом (УФ) свете для лекарственного вещества на основе мутантного токсина В были получены при различных значениях рН. Спектры, снятые при рН 5,0-7,0, являются показателем высокой доли α-спиралей во вторичной структуре, что позволяет предположить, что полипептидный каркас белка принимает четко определенную конформацию с преобладанием α-спиралей.
Были также получены спектры лекарственного вещества на основе мутантного токсина В в ближнем ультрафиолетовом (УФ) свете. Сильная отрицательная эллиптичность между 260 и 300 нм указывает на то, что ароматические боковые цепи находятся в уникальной прочной окружающей среде, то есть лекарственное вещество на основе мутантного токсина В обладает третичной структурой. Действительно, в пределах спектра можно отличить следующие характеристические признаки, вызванные индивидуальными типами ароматических боковых цепей: плечо приблизительно при 290 нм и самый высокий отрицательный пик приблизительно при 283 нм являются следствием поглощения поляризованного света упорядоченными боковыми цепями триптофана, отрицательный пик при 276 нм образуется за счет боковых цепей тирозина, и минорные плечи при 262 и 268 нм являются показателем остатков фенилаланина, участвующих в третичных контактах. Результаты спектров далекого и ближнего УФ свидетельствуют о том, что лекарственное вещество на основе мутантного токсина В сохраняет компактную складчатую структуру при физиологических значениях рН. Очень сходные спектры КД, наблюдаемые в далеком и ближнем УФ свете, при значениях рН 5,0-7,0 указывают на то, что в данном диапазоне рН обнаружимых структурных изменений не происходит. Данные КД не могут быть сняты при значениях рН 3,0 и 4,0, поскольку при данных показателях рН белок был нерастворим.
При сравнении спектров КД лекарственного вещества на основе мутантного токсина В в далеком и ближнем УФ со спектрами тройного мутантного токсина В спектры обоих белков являются очень сходными между значениями рН 5,0 и 7,0, что указывает на то, что обработка EDC не имеет обнаружимых воздействий на вторичную и третичную структуру белка.
Тройной мутантный токсин В содержит 16 остатков триптофана, распределенных на протяжении всей первичной последовательности, которые могут служить в качестве удобных внутренних флуоресцентных зондов. Были получены спектры испускания флуоресценции лекарственного вещества на основе мутантного токсина В между 300 и 400 нм в зависимости от температуры. При 7°С лекарственное вещество на основе мутантного токсина В проявляет характеристический спектр испускания флуоресценции триптофана при возбуждении при 280 нм. Максимум испускания флуоресценции наблюдают приблизительно при 335 нм, что указывает на то, что остатки триптофана находятся в неполярном окружении, более характерном для внутренней структуры белков, чем полярное водное окружение. Результаты спектров испускания флуоресценции вместе с результатами экспериментов по КД (см. выше) подтверждают, что лекарственное вещество на основе мутантного токсина В сохраняет компактную складчатую структуру.
Флуоресценцию внешнего зонда, представляющего собой 8-анилино-1-нафталинсульфоновую кислоту (АНС), использовали для характеризации возможных конформационных изменений лекарственного вещества на основе мутантного токсина В и тройного мутантного токсина В при изменениях рН. Как видно из результатов, при титровании как лекарственного вещества на основе мутантного токсина В, так и тройного мутантного токсина В этим зондом при рН 7,0 повышение интенсивности флуоресценции АНС по существу отсутствовало, что позволяет предположить, что гидрофобные поверхности на белках в данных условиях не открываются. Сдвиг рН до 2,6 приводит к резкому повышению квантового выхода флуоресценции АНС при увеличении концентрации зонда в присутствии лекарственного вещества на основе мутантного токсина В до тех пор, пока квантовый выход флуоресценции не достигнет кажущегося насыщения. Это повышение квантового выхода флуоресценции АНС указывает на то, что при низком рН (2,6) лекарственное вещество на основе мутантного токсина В претерпевают индуцированные рН конформационные изменения, открывающие гидрофобные поверхности. Такие конформационные изменения указывают на то, что EDC-индуцированная модификация и инактивация тройного мутантного токсина В не ограничивает конформационную пластичность лекарственного вещества на основе мутантного токсина В (DS).
Воздействие обработки EDC на гидродинамические свойства тройного мутантного токсина В оценивали, используя эксклюзионную хроматографию на колонке G4000 SWXL. Лекарственное вещество на основе мутантного токсина В и тройной мутантный токсин В впрыскивали на колонку G4000 SWXL, уравновешенную при рН 7,0, 6,0 и 5,0. Данные указывают на отсутствие различий в радиусе Стокса лекарственного вещества на основе мутантного токсина В и тройного мутантного токсина В, которые могут быть обнаружены при использовании эксклюзионной хроматографии. Таким образом, обработка EDC не оказывает резкого влияния на гидродинамические свойства и, соответственно, на общую молекулярную форму белка.
Дальнейший анализ тройного мутантного токсина В и лекарственного вещества на основе мутантного токсина В был проведен с использованием метода многоугольного рассеяния лазерного излучения (MALLS). Обработка тройного мутантного токсина В EDC привела в результате к образованию более гетерогенной смеси, состоящей из различных мультимерных и мономерных соединений. Такая гетерогенность отражает введение большого числа EDC-индуцированных межмолекулярных и внутримолекулярных ковалентных связей между карбоксильными и первичными аминными группами белка.
Полученные данные обеспечивают физические и химические характеристики тройного мутантного токсина В и лекарственного вещества на основе мутантного токсина В (тройного мутантного токсина В, обработанного EDC) и описывают ключевые признаки их первичной, вторичной и третичной структуры. Полученные данные показывают, что обработка тройного мутантного токсина В EDC приводит в результате к ковалентной модификации его полипептидной цепи, но не влияет на вторичную и третичную структуру белка. Обработка EDC приводит к внутри- и межмолекулярному поперечному сшиванию. Основные биохимические и биофизические параметры, полученные для лекарственного вещества на основе мутантного токсина В (а также для тройного мутантного токсина В) представлены в таблице 61.
Аспекты изобретения
В следующих пунктах (П) описаны дополнительные воплощения изобретения:
П1. Выделенный полипептид, включающий аминокислотную последовательность, указанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и где полипептид включает по меньшей мере одну боковую цепь аминокислоты, химически модифицированную 1-этил-3-(3-диметиламинопропил)карбодиимидом (EDC) и N-гидроксисукцинимидом (NHS).
П2. Выделенный полипептид, включающий аминокислотную последовательность, указанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и где полипептид включает по меньшей мере одну боковую цепь аминокислоты, химически модифицированную 1-этил-3-(3-диметиламинопропил)карбодиимидом (EDC) и N-гидроксисукцинимидом (NHS).
П3. Выделенный полипептид по пункту П1 или П2, где по меньшей мере одна боковая цепь остатка аспарагиновой кислоты полипептида или по меньшей мере одна боковая цепь остатка глутаминовой кислоты полипептида химически модифицирована глицином.
П4. Выделенный полипептид по любому из пунктов П1-ПЗ, включающий:
а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; и
б) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида.
П5. Выделенный полипептид по любому из пунктов П1-П4, включающий бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида.
П6. Выделенный полипептид по пункту П4, включающий глициновую группировку, сшитую с боковой цепью остатка аспарагиновой кислоты полипептида или с боковой цепью остатка глутаминовой кислоты полипептида.
П7. Выделенный полипептид, включающий аминокислотную последовательность, указанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и где боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой.
П8. Выделенный полипептид, включающий аминокислотную последовательность, указанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и где боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой.
П9. Выделенный полипептид по пункту П7 или П8, где боковая цепь второго остатка лизина полипептида сшита с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
П10. Выделенный полипептид по любому из пунктов П7-П9, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида сшита с глициновой группировкой.
П11. Выделенный полипептид по любому из пунктов П1-П10, где полипептид имеет значение EC50, составляющее по меньшей мере примерно 100 мкг/мл.
П12. Иммуногенная композиция, содержащая выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и где полипептиды имеют по меньшей мере одну боковую цепь аминокислоты, химически модифицированную 1-этил-3-(3-диметиламинопропил)карбодиимидом (EDC) и N-гидроксисукцинимидом (NHS).
П13. Иммуногенная композиция по пункту П12, где полипептид включает по меньшей мере любое из следующего:
а) по меньшей мере одну бета-аланиновую группировку, сшитую с боковой цепью остатка лизина полипептида;
б) по меньшей мере одну поперечную сшивку между боковой цепью остатка лизина полипептида и боковой цепью остатка аспарагиновой кислоты; и
в) по меньшей мере одну поперечную сшивку между боковой цепью остатка лизина полипептида и боковой цепью остатка глутаминовой кислоты.
П14. Иммуногенная композиция по пункту П12, где полипептиды имеют значение EC50, составляющее по меньшей мере примерно 100 мкг/мл.
П15. Иммуногенная композиция, содержащая выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, и
а) где боковая цепь по меньшей мере одного остатка лизина SEQ ID NO:4 сшита с бета-аланиновой группировкой, и
б) где боковая цепь по меньшей мере одного остатка лизина SEQ ID NO:6 сшита с бета-аланиновой группировкой.
П16. Иммуногенная композиция по пункту П15, где боковая цепь второго остатка лизина SEQ ID NO:4 сшита с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты, и где второй остаток лизина SEQ ID NO:6 сшит с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
П17. Иммуногенная композиция по любому из пунктов П12-П16, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, указанную в SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, сшита с глициновой группировкой.
П18. Иммуногенная композиция по любому из пунктов П12-П16, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, указанную в SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, сшита с глициновой группировкой.
П19. Иммуногенная композиция по любому из пунктов П12-П18, где полипептид имеет значение ЕС50, составляющее по меньшей мере примерно 100 мкг/мл.
П20. Иммуногенная композиция, содержащая выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:84, и выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO:86, где каждый полипептид включает:
а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида;
б) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида;
в) бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида; и
г) глициновую группировку, сшитую с боковой цепью по меньшей мере одного остатка аспарагиновой кислоты полипептида или с боковой цепью по меньшей мере одного остатка глутаминовой кислоты полипептида.
П21. Иммуногенная композиция, содержащая мутантный токсин А Clostridium difficile, включающий глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию, по сравнению с соответствующим токсином А Clostridium difficile дикого типа.
П22. Композиция по пункту П21, где мутация представляет собой неконсервативную аминокислотную замену.
П23. Композиция по пункту П22, где замена включает замену аланина.
П24. Композиция по любому из пунктов П21-П23, где токсин A Clostridium difficile дикого типа включает последовательность, обладающую по меньшей мере 95% идентичностью SEQ ID NO:1.
П25. Композиция по пункту П24, где токсин A Clostridium difficile дикого типа включает последовательность, обладающую по меньшей мере 98% идентичностью SEQ ID NO:1.
П26. Композиция по пункту П25, где токсин A Clostridium difficile дикого типа включает SEQ ID NO:1.
П27. Композиция по любому из пунктов П21-П26, где глюкозилтрансферазный домен включает по меньшей мере две мутации.
П28. Композиция по пункту П27, где по меньшей мере две мутации находятся в положениях аминокислот 101, 269, 272, 285, 287, 269, 272, 460, 462, 541 или 542, в соответствии с нумерацией SEQ ID NO:1.
П29. Композиция по любому из пунктов П21-П26, где глюкозилтрансферазный домен включает SEQ ID NO:29.
ПЗО. Композиция по пункту П29, где глюкозилтрансферазный домен включает по меньшей мере две неконсервативные мутации, находящиеся в положениях 101, 269, 272, 285, 287, 269, 272, 460, 462, 541 или 542 аминокислот SEQ ID NO:29 или в любой их комбинации.
П31. Композиция по любому из пунктов П21-П26, где цистеинпротеазный домен включает мутацию, находящуюся в положениях 700, 589, 655, 543, или в любой их комбинации, в соответствии с нумерацией SEQ ID NO:1.
П32. Композиция по любому из пунктов П21-П26, где цистеинпротеазный домен включает SEQ ID NO:32.
ПЗЗ. Композиция по пункту П32, где цистеинпротеазный домен включает неконсервативную мутацию, находящуюся в положениях 1, 47, 113, 158 SEQ ID NO:32 или в любой их комбинации.
П34. Композиция по пункту П21, где мутантный токсин A Clostridium difficile включает SEQ ID NO:4.
П35. Композиция по пункту П21, где мутантный токсин A Clostridium difficile включает SEQ ID NO:84.
П36. Композиция по пункту П21, где мутантный токсин A Clostridium difficile включает SEQ ID NO:7.
П37. Композиция по пункту П21, где мутантный токсин A Clostridium difficile включает SEQ ID NO:83.
П38. Композиция по любому из пунктов П21-П33, где по меньшей мере одна аминокислота мутантного токсина A Clostridium difficile поперечно-сшита химическим образом.
П39. Композиция по пункту П38, где аминокислота поперечно-сшита химическим образом с использованием формальдегида.
П40. Композиция по пункту П38, где аминокислота поперечно-сшита химическим образом с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида.
П41. Композиция по пункту П38 или П40, где аминокислота поперечно-сшита химическим образом с использованием N-гидроксисукцинимида.
П42. Композиция по любому из пунктов П21-П41, где композиция распознается нейтрализующим антителом против токсина А или его связывающим фрагментом.
П43. Иммуногенная композиция, содержащая мутантный токсин А Clostridium difficile, включающий глюкозилтрансферазный домен, включающий SEQ ID NO:29, имеющую аминокислотную замену в положениях 285 и 287, и цистеинпротеазный домен, включающий SEQ ID NO:32, имеющую аминокислотную замену в положении 158, по сравнению с соответствующим токсином A Clostridium difficile дикого типа, где по меньшей мере одна аминокислота мутантного токсина A Clostridium difficile поперечно-сшита химическим образом.
П44. Иммуногенная композиция, содержащая SEQ ID NO:4 или SEQ ID NO:7, где по меньшей мере одна аминокислота SEQ ID NO:4 или SEQ ID NO:7 поперечно-сшита химическим образом.
П45. Композиция по пункту П43 или П44, где по меньшей мере одна аминокислота поперечно-сшита с использованием формальдегида.
П46. Композиция по пункту П43 или П44, где по меньшей мере одна аминокислота поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)-карбодиимида.
П47. Композиция по пункту П43, П44 или П46, где по меньшей мере одна аминокислота поперечно-сшита с использованием N-гидроксисукцинимида.
П48. Композиция по пункту П43 или П44, где композиция распознается нейтрализующим антителом против токсина А или его связывающим фрагментом.
П49. Иммуногенная композиция, содержащая SEQ ID NO:4.
П50. Иммуногенная композиция, содержащая SEQ ID NO:84.
П51. Иммуногенная композиция, содержащая SEQ ID NO:7.
П52. Иммуногенная композиция, содержащая SEQ ID NO:83.
П53. Композиция по любому из пунктов П49-П52, где по меньшей мере одна аминокислота поперечно-сшита химическим образом.
П54. Композиция по любому из пунктов П21-П51, где композиция проявляет пониженную цитотоксичность по сравнению с соответствующим токсином A Clostridium difficile дикого типа.
П55. Выделенный полипептид, включающий SEQ ID NO:84.
П56. Выделенный полипептид, включающий SEQ ID NO:86.
П57. Выделенный полипептид, включающий SEQ ID NO:83.
П58. Выделенный полипептид, включающий SEQ ID NO:85.
П59. Иммуногенная композиция, содержащая мутантный токсин В Clostridium difficile, включающий глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию, по сравнению с соответствующим токсином В Clostridium difficile дикого типа.
П60. Композиция по пункту П59, где мутация представляет собой неконсервативную аминокислотную замену.
П61. Композиция по пункту П60, где замена включает замену аланина. П62. Композиция по любому из пунктов П59-П61, где токсин В Clostridium difficile дикого типа включает последовательность, обладающую по меньшей мере 95% идентичностью SEQ ID NO:2.
П63. Композиция по пункту П62, где токсин В Clostridium difficile дикого типа включает последовательность, обладающую по меньшей мере 98% идентичностью SEQ ID NO:2.
П64. Композиция по пункту П63, где токсин В Clostridium difficile дикого типа включает SEQ ID NO:2.
П65. Композиция по любому из пунктов П59-П64, где глюкозилтрансферазный домен включает по меньшей мере две мутации.
П66. Композиция по пункту П65, где по меньшей мере две мутации находятся в положениях аминокислот 102, 286, 288, 270, 273, 384, 461, 463, 520 или 543 в соответствии с нумерацией SEQ ID NO:2.
П67. Композиция по любому из пунктов П59-П64, где глюкозилтрансферазный домен включает SEQ ID NO:31.
П68. Композиция по пункту П67, где глюкозилтрансферазный домен включает по меньшей мере две неконсервативные мутации, находящиеся в положениях аминокислот 102, 286, 288, 270, 273, 384, 461, 463, 520 или 543 SEQ ID NO:31.
П69. Композиция по любому из пунктов П59-П64, где цистеинпротеазный домен включает мутацию, находящуюся в положениях 698, 653, 587, 544 или в любой их комбинации, в соответствии с нумерацией SEQ ID NO:2.
П70. Композиция по любому из пунктов П59-П64, где цистеинпротеазный домен включает SEQ ID NO:33.
П71. Композиция по пункту П70, где цистеинпротеазный домен включает неконсервативную мутацию, находящуюся в положениях 1, 44, 110, 155 SEQ ID NO:33 или в любой их комбинации.
П72. Композиция по пункту П59, где мутантный токсин В Clostridium difficile включает SEQ ID NO:6.
П73. Композиция по пункту П59, где мутантный токсин В Clostridium difficile включает SEQ ID NO:86.
П74. Композиция по пункту П59, где мутантный токсин В Clostridium difficile включает SEQ ID NO:8.
П75. Композиция по пункту П59, где мутантный токсин В Clostridium difficile включает SEQ ID NO:85.
П76. Композиция по любому из пунктов П59-П71, где по меньшей мере одна аминокислота мутантного токсина В Clostridium difficile поперечно-сшита химическим образом.
П77. Композиция по пункту П76, где аминокислота поперечно-сшита химическим образом с использованием формальдегида.
П78. Композиция по пункту П76, где аминокислота поперечно-сшита химическим образом с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида.
П79. Композиция по пункту П76 или П78, где по меньшей мере одна аминокислота поперечно-сшита с использованием N-гидроксисукцинимида.
П80. Композиция по любому из пунктов П59-П79, где композиция распознается нейтрализующим антителом против токсина В или его связывающим фрагментом.
П81. Иммуногенная композиция, содержащая мутантный токсин В Clostridium difficile, включающий глюкозилтрансферазный домен, включающий SEQ ID NO:31, имеющую аминокислотную замену в положениях 286 и 288, и цистеинпротеазный домен, включающий SEQ ID NO:33, имеющую аминокислотную замену в положении 155, по сравнению с соответствующим токсином В Clostridium difficile дикого типа, где по меньшей мере одна аминокислота мутантного токсина В Clostridium difficile поперечно-сшита химическим образом.
П82. Иммуногенная композиция, содержащая SEQ ID NO:6 или SEQ ID NO:8, где по меньшей мере одна аминокислота SEQ ID NO:6 или SEQ ID NO:8 поперечно-сшита химическим образом.
П83. Композиция по пункту П81 или П82, где по меньшей мере одна аминокислота поперечно-сшита с использованием формальдегида.
П84. Композиция по пункту П81 или П82, где по меньшей мере одна аминокислота поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида.
П85. Композиция по пункту П81, П82 или П84, где по меньшей мере одна аминокислота поперечно-сшита с использованием N-гидроксисукцинимида.
П86. Композиция по пункту П81 или П82, где композиция распознается нейтрализующим антителом против токсина В или его связывающим фрагментом.
П87. Иммуногенная композиция, содержащая SEQ ID NO:6.
П88. Иммуногенная композиция, содержащая SEQ ID NO:86.
П89. Иммуногенная композиция, содержащая SEQ ID NO:8.
П90. Иммуногенная композиция, содержащая SEQ ID NO:85.
П91. Композиция по любому из пунктов П59-П89, где композиция проявляет пониженную цитотоксичность по сравнению с соответствующим токсином В Clostridium difficile дикого типа.
П92. Иммуногенная композиция, содержащая SEQ ID NO:4, и иммуногенная композиция, содержащая SEQ ID NO:6, где по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и 6 поперечно-сшита химическим образом.
П93. Иммуногенная композиция, содержащая SEQ ID NO; 84, и иммуногенная композиция, содержащая SEQ ID NO:86, где по меньшей мере одна аминокислота каждой из SEQ ID NO:84 и 86 поперечно-сшита химическим образом.
П94. Композиция по пункту П92 или П93, где по меньшей мере одна аминокислота поперечно-сшита с использованием формальдегида.
П95. Композиция по пункту П92 или П93, где по меньшей мере одна аминокислота поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида.
П96. Композиция по любому из пунктов П92, П93 или П95, где по меньшей мере одна аминокислота поперечно-сшита с использованием N-гидроксисукцинимида.
П97. Рекомбинантная клетка или ее потомство, содержащая SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO; 13, SEQ ID NO:14, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:46 или SEQ ID NO:47.
П98. Рекомбинантная клетка или ее потомство, содержащая нуклеиново-кислотную последовательность, кодирующую SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:7 или SEQ ID NO:8.
П99. Рекомбинантная клетка или ее потомство, содержащая нуклеиново-кислотную последовательность, кодирующую SEQ ID NO:84.
П100. Рекомбинантная клетка или ее потомство, содержащая нуклеиново-кислотную последовательность, кодирующую SEQ ID NO:86.
П101. Рекомбинантная клетка или ее потомство, содержащая нуклеиново-кислотную последовательность, кодирующую SEQ ID NO:83.
П102. Рекомбинантная клетка или ее потомство, содержащая нуклеиново-кислотную последовательность, кодирующую SEQ ID NO:85.
П103. Рекомбинантная клетка по пункту П97 или П98, имеющая происхождение от грамположительной бактериальной клетки.
П104. Рекомбинантная клетка по любому из пунктов П97, П98 или П99, имеющая происхождение от клетки Clostridium difficile.
П105. Рекомбинантная клетка по любому из пунктов П97-П104, где отсутствует эндогенный полинуклеотид, кодирующий токсин.
П106. Клетка по любому из пунктов П104 или П105, имеющая происхождение от клетки Clostridium difficile, выбранной из группы, состоящей из Clostridium difficile 1351, Clostridium difficile 3232, Clostridium difficile 7322, Clostridium difficile 5036, Clostridium difficile 4811 и Clostridium difficile VPI 11186.
П107. Клетка по пункту П106, представляющая собой клетку Clostridium difficile VPI 11186.
П108. Клетка по пункту П106 или П107, где ген споруляции клетки Clostridium difficile инактивирован.
П109. Клетка по пункту П108, где ген споруляции включает ген spoOA или ген spollE.
П110. Способ продуцирования мутантного токсина Clostridium difficile, включающий культивирование рекомбинантной клетки или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин Clostridium difficile, где клетка содержит полинуклеотид, кодирующий мутантный токсин Clostridium difficile, и где мутант включает глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию, по сравнению с соответствующим токсином Clostridium difficile дикого типа.
П111. Способ по пункту П110, где в клетке отсутствует эндогенный полинуклеотид, кодирующий токсин.
П112. Способ по пункту П110, где рекомбинантная клетка или ее потомство включает клетку по любому из пунктов П97-П111.
П113. Способ по пункту П110, дополнительно включающий выделение мутантного токсина Clostridium difficile.
П114. Способ по пункту П113, дополнительно включающий приведение выделенного мутантного токсина Clostridium difficile в контакт с формальдегидом.
П115. Способ по пункту П114, где контакт осуществляют в течение не более чем 14 суток.
П116. Способ по пункту П115, где контакт осуществляют в течение не более чем 48 часов.
П117. Способ по пункту П114, где контакт осуществляют примерно при 25°С.
П118. Способ по пункту П113, дополнительно включающий приведение выделенного мутантного токсина Clostridium difficile в контакт с 1-этил-3-(3-диметиламинопропил)карбодиимидом.
П119. Способ по пункту П118, где контакт осуществляют в течение не более чем 24 часов.
П120. Способ по пункту П120, где контакт осуществляют в течение не более чем 4 часов.
П121. Способ по пункту П118, где контакт осуществляют примерно при 25°С.
П122. Способ по пункту П118, дополнительно включающий приведение выделенного мутантного токсина Clostridium difficile в контакт с N-гидроксисукцинимидом.
П123. Иммуногенная композиция, полученная способом по любому из пунктов П110-П122.
П124. Способ получения нейтрализующего антитела против токсина А Clostridium difficile, включающий введение иммуногенной композиции млекопитающему, причем указанная иммуногенная композиция включает SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота SEQ ID NO:4 поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида, и выделение антитела из млекопитающего.
П125. Способ получения нейтрализующего антитела против токсина А Clostridium difficile, включающий введение иммуногенной композиции млекопитающему, причем указанная иммуногенная композиция включает SEQ ID NO:84, где по меньшей мере одна аминокислота в SEQ ID NO:84 поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида, и выделение антитела из млекопитающего.
П126. Способ получения нейтрализующего антитела против токсина В Clostridium difficile, включающий введение иммуногенной композиции млекопитающему, причем указанная иммуногенная композиция включает SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота в SEQ ID NO:6 поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида, и выделение антитела из млекопитающего.
П127. Способ получения нейтрализующего антитела против токсина В Clostridium difficile, включающий введение иммуногенной композиции млекопитающему, причем указанная иммуногенная композиция включает SEQ ID NO:86, где по меньшей мере одна аминокислота в SEQ ID NO:86 поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)-карбодиимида, N-гидроксисукцинимида или комбинацией 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида, и выделение антитела из млекопитающего.
П128. Антитело или связывающий фрагмент антитела, специфичное к иммуногенной композиции, причем указанная иммуногенная композиция включает SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, или SEQ ID NO:7, где остаток метионина в положении 1 возможно отсутствует.
П129. Антитело или связывающий фрагмент антитела по пункту П128, где по меньшей мере одна аминокислота SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, или SEQ ID NO:7, где остаток метионина в положении 1 возможно отсутствует, поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П130. Антитело или связывающий фрагмент антитела, включающее аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи, указанные в SEQ ID NO: 41 (CDR H1), SEQ ID NO:42 (CDR H2) и SEQ ID NO:43 (CDR НЗ), и аминокислотные последовательности CDR легкой цепи, указанные в SEQ ID NO:38 (CDR L1), SEQ ID NO:39 (CDR L2) и SEQ ID NO:40 (CDR L3).
П131. Антитело или связывающий фрагмент антитела по любому из пунктов П128, П129 или П130, где антитело или связывающий фрагмент антитела содержит тяжелую цепь, включающую аминокислотную последовательность, представленную в SEQ ID NO:37, и легкую цепь, включающую аминокислотную последовательность, представленную в SEQ ID NO: 36.
П132. Композиция, содержащая комбинацию двух или более чем двух антител или связывающих фрагментов антител, выбранных из любого антитела или связывающего фрагмента антитела по любому из пунктов П128-П131.
П133. Антитело или связывающий фрагмент антитела, специфичное к иммуногенной композиции, причем указанная иммуногенная композиция включает SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, или SEQ ID NO:8, где остаток метионина в положении 1 возможно отсутствует.
П134. Антитело или связывающий фрагмент антитела по пункту П133, где по меньшей мере одна аминокислота SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, или SEQ ID NO:8, где остаток метионина в положении 1 возможно отсутствует, поперечно-сшита с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П135. Антитело или связывающий фрагмент антитела, включающее аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи, указанные в SEQ ID NO:51 (CDR H1), SEQ ID NO:52 (CDR H2) и SEQ ID NO:53 (CDR НЗ), и аминокислотные последовательности CDR легкой цепи, указанные в SEQ ID NO:57 (CDR L1), SEQ ID NO:58 (CDR L2) и SEQ ID NO:59 (CDR L3).
П136. Антитело или связывающий фрагмент антитела, включающее аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи, указанные в SEQ ID NO: 61 (CDR H1), SEQ ID NO:62 (CDR H2) и SEQ ID NO:63 (CDR НЗ), и аминокислотные последовательности CDR легкой цепи, указанные в SEQ ID NO:68 (CDR L1), SEQ ID NO:69 (CDR L2) и SEQ ID NO:70 (CDR L3).
П137. Антитело или связывающий фрагмент антитела, включающее аминокислотные последовательности гипервариабельных участков (CDR) тяжелой цепи, указанные в SEQ ID NO:73 (CDR H1), SEQ ID NO:74 (CDR H2) и SEQ ID NO:75 (CDR НЗ), и аминокислотные последовательности CDR легкой цепи, указанные в SEQ ID NO:79 (CDR L1), SEQ ID NO:80 (CDR L2) и SEQ ID NO:81 (CDR L3).
П138. Композиция, содержащая комбинацию двух или более чем двух антител или связывающих фрагментов антител, выбранных из любого антитела или связывающего фрагмента антитела по любому из пунктов П133-П137.
П139. Способ лечения инфекции Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и 6 поперечно-сшита с использованием формальдегида.
П140. Способ лечения инфекции Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и SEQ ID NO:6 поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П141. Способ лечения инфекции Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO:84, и иммуногенной композиции, включающей SEQ ID NO:86, где по меньшей мере одна аминокислота каждой из SEQ ID NO:84 и SEQ ID NO:86 поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П142. Способ индукции иммунного ответа к Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и SEQ ID NO:6 поперечно-сшита с использованием формальдегида.
П143. Способ индукции иммунного ответа к Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO:4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO:6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере одна аминокислота каждой из SEQ ID NO:4 и SEQ ID NO:6 поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П144. Способ индукции иммунного ответа к Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO; 84, и иммуногенной композиции, включающей SEQ ID NO:86, где по меньшей мере одна аминокислота каждой из SEQ ID NO:84 и SEQ ID NO:86 поперечно-сшита с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
П145. Способ по любому из пунктов П139-П144, где млекопитающее представляет собой млекопитающее, нуждающееся в этом.
П146. Способ по любому из пунктов П139-П144, где млекопитающее имеет рецидивирующую инфекцию Clostridium difficile.
П147. Способ по любому из пунктов П139-П144, при котором композицию вводят парентерально.
П148. Способ по любому из пунктов П139-П144, где композиция дополнительно включает адъювант.
П149. Способ по пункту П148, где адъювант включает алюминий.
П150. Способ по пункту П148, где адъювант включает гель гидроксида алюминия и CpG-олигонуклеотид.
П151. Способ по пункту П148, где адъювант включает ISCOMATRIX®.
| название | год | авторы | номер документа |
|---|---|---|---|
| Композиции и способы, имеющие отношение к мутантному токсину из Clostridium Difficile | 2013 |
|
RU2630671C2 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2012 |
|
RU2754446C2 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2017 |
|
RU2773880C1 |
| АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ АССОЦИИРОВАННЫХ С CLOSTRIDIUM DIFFICILE ИНФЕКЦИИ И ЗАБОЛЕВАНИЙ | 2011 |
|
RU2630663C9 |
| Способ получения аминокислотных последовательностей, кодирующих наноантитела, нейтрализующие токсин А Clostridioides difficile | 2023 |
|
RU2833930C1 |
| АНТИТЕЛА ПРОТИВ G-CSFR И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2605595C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ОТ ОТРАВЛЕНИЙ | 2007 |
|
RU2465893C2 |
| КОМПОЗИЦИИ ДЛЯ ПРЕДУПРЕЖДЕНИЯ И/ИЛИ ЛЕЧЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ ВИРУСОМ ВИЧ-1 | 2012 |
|
RU2603262C2 |
| МУТАНТНЫЙ ВИРУС БЫЧЬЕЙ ДИАРЕИ | 2006 |
|
RU2394594C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АЛЬФА-ЭНОЛАЗЕ, И СПОСОБЫ ПРИМЕНЕНИЯ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2656153C1 |






























































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































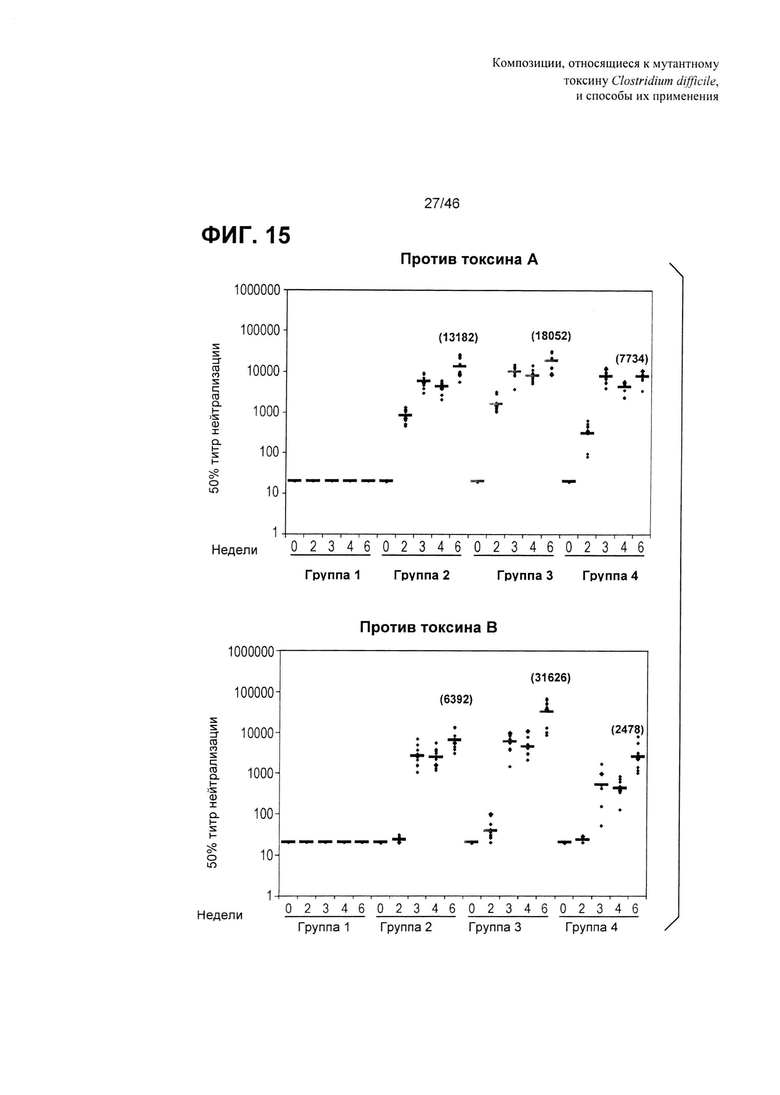




Изобретение относится к области биотехнологии, конкретно к получению иммуногенных мутантных токсинов Clostridium difficile, и может быть использовано в медицине для лечения инфекции C. difficile у млекопитающего. Мутантный токсин включает глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и цистеинпротеазный домен, имеющий по меньшей мере одну мутацию по сравнению с соответствующим токсином С. difficile дикого типа. Эти мутантные токсины дополнительно включают по меньшей мере одну аминокислоту, выбранную из лизина, аспарагиновой кислоты или глутаминовой кислоты, сшитую химическим путем с использованием формальдегида, 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC), N-гидроксисукцинимида (NHS) или комбинации EDC/NHS. Изобретение позволяет эффективно индуцировать нейтрализующие антитела против токсина C. difficile. 8 н. и 13 з.п. ф-лы, 25 ил., 61 табл., 45 пр.
1. Выделенный полипептид для индукции нейтрализующих антител против токсина A Clostridium difficile, включающий аминокислотную последовательность, указанную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и где полипептид содержит по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты данного полипептида, химически модифицированных с помощью 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS).
2. Выделенный полипептид по п. 1, где по меньшей мере одна боковая цепь остатка аспарагиновой кислоты полипептида или по меньшей мере одна боковая цепь остатка глутаминовой кислоты полипептида химически модифицирована глицином.
3. Выделенный полипептид по п. 1, включающий:
а) по меньшей мере одну поперечную сшивку между боковой цепью остатка аспарагиновой кислоты полипептида и боковой цепью остатка лизина полипептида; и
б) по меньшей мере одну поперечную сшивку между боковой цепью остатка глутаминовой кислоты полипептида и боковой цепью остатка лизина полипептида.
4. Выделенный полипептид по п. 1, включающий бета-аланиновую группировку, сшитую с боковой цепью по меньшей мере одного остатка лизина полипептида.
5. Выделенный полипептид по п. 3, включающий глициновую группировку, сшитую с боковой цепью остатка аспарагиновой кислоты полипептида или с боковой цепью остатка глутаминовой кислоты полипептида.
6. Выделенный полипептид для индукции нейтрализующих антител против токсина A Clostridium difficile, включающий аминокислотную последовательность, указанную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и где боковая цепь по меньшей мере одного остатка лизина полипептида сшита с бета-аланиновой группировкой с помощью сшивающего агента, выбранного из формальдегида, EDC, NHS или комбинации EDC/NHS.
7. Выделенный полипептид по п. 6, где боковая цепь второго остатка лизина полипептида сшита с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
8. Выделенный полипептид по п. 6, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида сшита с глициновой группировкой.
9. Выделенный полипептид по любому из пп. 1 или 6, где полипептид имеет значение ЕС50, составляющее по меньшей мере примерно 100 мкг/мл.
10. Иммуногенная композиция для индукции нейтрализующих антител против токсинов Clostridium difficile, содержащая выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, где указанные полипептиды содержат по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты, химически модифицированных с помощью 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS), и где указанные выделенные полипептиды содержатся в композиции в эффективном количестве.
11. Иммуногенная композиция по п. 10, где полипептид включает по меньшей мере любое из следующего:
а) по меньшей мере одну бета-аланиновую группировку, сшитую с боковой цепью остатка лизина полипептида;
б) по меньшей мере одну поперечную сшивку между боковой цепью остатка лизина полипептида и боковой цепью остатка аспарагиновой кислоты; и
в) по меньшей мере одну поперечную сшивку между боковой цепью остатка лизина полипептида и боковой цепью остатка глутаминовой кислоты.
12. Иммуногенная композиция по п. 10, где полипептиды имеют значение ЕС50, составляющее по меньшей мере примерно 100 мкг/мл.
13. Иммуногенная композиция для индукции нейтрализующих антител против токсинов Clostridium difficile, содержащая выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и выделенный полипептид, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, и
а) где боковая цепь по меньшей мере одного остатка лизина в SEQ ID NO: 4 сшита с бета-аланиновой группировкой, и
б) где боковая цепь по меньшей мере одного остатка лизина в SEQ ID NO: 6 сшита с бета-аланиновой группировкой,
с помощью сшивающего агента, выбранного из формальдегида, EDC, NHS или комбинации EDC/NHS, и где указанные выделенные полипептиды содержатся в композиции в эффективном количестве.
14. Иммуногенная композиция по п. 13, где боковая цепь второго остатка лизина SEQ ID NO: 4 сшита с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты, и где второй остаток лизина SEQ ID NO: 6 сшит с боковой цепью остатка аспарагиновой кислоты или с боковой цепью остатка глутаминовой кислоты.
15. Иммуногенная композиция по любому из пп. 10-14, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, указанную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, сшита с глициновой группировкой.
16. Иммуногенная композиция по любому из пп. 10-14, где боковая цепь остатка аспарагиновой кислоты или боковая цепь остатка глутаминовой кислоты полипептида, имеющего аминокислотную последовательность, указанную в SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, сшита с глициновой группировкой.
17. Иммуногенная композиция по п. 10 или 13, где полипептид имеет значение ЕС50. составляющее по меньшей мере примерно 100 мкг/мл.
18. Способ лечения инфекции Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты каждой из SEQ ID NO: 4 и 6 поперечно-сшит с использованием формальдегида.
19. Способ лечения инфекции Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты каждой из SEQ ID NO: 4 и SEQ ID NO: 6 поперечно-сшит с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
20. Способ индукции иммунного ответа к Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты каждой из SEQ ID NO: 4 и SEQ ID NO: 6 поперечно-сшит с использованием формальдегида.
21. Способ индукции иммунного ответа к Clostridium difficile у млекопитающего, включающий введение млекопитающему иммуногенной композиции, включающей SEQ ID NO: 4, где остаток метионина в положении 1 возможно отсутствует, и иммуногенной композиции, включающей SEQ ID NO: 6, где остаток метионина в положении 1 возможно отсутствует, где по меньшей мере один из остатков лизина, аспарагиновой кислоты или глутаминовой кислоты каждой из SEQ ID NO: 4 и SEQ ID NO: 6 поперечно-сшиты с использованием 1-этил-3-(3-диметиламинопропил)карбодиимида, N-гидроксисукцинимида или комбинации 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимида.
| WO 2010094970, 26.08.2010 | |||
| TEICHERT M | |||
| ET AL, Application of Mutated Clostridium difficile Toxin A for Determination of Glucosyltransferase-Dependent Effects, INFECTION AND IMMUNITY, 2006, v.74, n.10, p.6006 - 6010 | |||
| СПОСОБ ИММУНИЗАЦИИ ПТИЦЫ ПРОТИВ ИНФЕКЦИИ, ВЫЗВАННОЙ БАКТЕРИЯМИ CLOSTRIDIUM | 2007 |
|
RU2409385C2 |

