ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет предварительной заявки на патент США 61/716605, поданной 21 октября 2012 г., которая включена посредством ссылки во всей своей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам, имеющим отношение к мутантным токсинам из Clostridium difficile.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Clostridium difficile (С. difficile) представляет собой грамположительную анаэробную бактерию, которую ассоциируют с желудочно-кишечным заболеванием у людей. Колонизация С. difficile обычно происходит в толстой кишке, если природная кишечная флора сокращается в результате лечения антибиотиками. Инфекция может приводить к антибиотикоассоциированной диарее и иногда к псевдомембранозному колиту вследствие выделения гликозилированных токсинов, токсина А и токсина В (308 и 270 кДа, соответственно), которые являются первичными факторами вирулентности С. difficile.
Токсин А и токсин В кодируются в пределах локуса патогенности (PaLoc) размером 19 тысяч оснований (т.о.) генами tcdA и tcdB, соответственно. В непатогенных штаммах С. difficile этот локус заменен альтернативной последовательностью, состоящей из 115 пар оснований.
Токсин А и токсин В оба являются мощными цитотоксинами. Эти белки гомологичны глюкозилтрансферазам, которые инактивируют малые ГТФазы (гуанозинтрифосфатазы) семейства Rho/Rac/Ras. Вытекающий из этого сбой в передаче сигналов вызывает утрату межклеточных контактов, нарушение регуляции актинового цитоскелета и/или апоптоз, что приводит к секреторной диарее в тяжелой форме, которая ассоциирована с инфекциями Clostridium difficile (CDI).
За последнее десятилетие число и тяжесть вспышек заболеваний, обусловленных С. difficile, в больницах, лечебно-реабилитационных центрах для престарелых и инвалидов и других лечебных учреждениях для хронических больных резко возросло. Ключевые факторы этого возрастания включают появление гипервирулентных патогенных штаммов, более широкое применение антибиотиков, усовершенствование методов обнаружения и увеличение случаев контакта с источниками спор, передающихся воздушно-капельным путем, в медицинских учреждениях.
Применение метронидазола и ванкомицина представляет собой принятый в настоящее время стандарт оказания медицинской помощи по лечению антибиотиками заболевания, ассоциированного с С. difficile (CDAD). Однако примерно у 20% пациентов, получающих такое лечение, случается рецидив инфекции после первого эпизода CDI, и примерно до 50% этих пациентов страдают от повторных рецидивов. Лечение рецидивов представляет собой очень серьезную проблему, и большинство рецидивов обычно происходит в течение одного месяца с момента предшествующего эпизода.
Соответственно, существует потребность в иммуногенных и/или терапевтических композициях и способах, которые направлены на С. difficile.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Эти и другие задачи решаются настоящим изобретением.
В одном из аспектов изобретение относится к мутантному токсину А из С. difficile. Мутантный токсин А содержит мутацию в остатках по положениям 285, 287, 700, 972 и 978 по сравнению с токсином А дикого типа. В одном из воплощений мутантный токсин А содержит SEQ ID NO: 183. В одном из воплощений мутантный токсин А является менее цитотоксичным, чем соответствующий токсин А дикого типа. В одном из воплощений мутантный токсин А содержит по меньшей мере один аминокислотный остаток, который химически модифицирован. В одном из аспектов изобретение относится к выделенному полипептиду, который содержит SEQ ID NO: 183.
В другом аспекте изобретение относится к мутантному токсину В из С. difficile. Мутантный токсин В содержит мутацию в остатках 286, 288, 698, 970 и 976 по сравнению с токсином В дикого типа. В одном из воплощений мутантный токсин В содержит SEQ ID NO: 184. В одном из воплощений мутантный токсин В является менее цитотоксичным, чем соответствующий токсин В дикого типа. В одном из воплощений мутантный токсин В содержит по меньшей мере один аминокислотный остаток, который химически модифицирован. В одном из аспектов изобретение относится к выделенному полипептиду, который содержит SEQ ID NO: 184.
Изобретение также относится к композициям и способам для применения при культивировании С. difficile и получении токсинов С. difficile. В одном из аспектов изобретение относится к культуральной среде, включающей гидролизат растительных белков и клетку С. difficile. В предпочтительном воплощении гидролизатом является гидролизат сои. Более предпочтительно, гидролизат сои представляет собой гидролизат сои SE50MK.
В другом аспекте изобретение относится к культуральной среде, включающей источник азота и клетку С. difficile. В одном из воплощений источником азота является дрожжевой экстракт. Предпочтительно, дрожжевой экстракт представляет собой HY YEST 412 (Kerry Biosciences).
В следующем аспекте изобретение относится к культуральной среде, включающей гидролизат растительных белков, дрожжевой экстракт и клетку С. difficile. В одном из воплощений среда не содержит источника углерода.
В предпочтительном воплощении среда дополнительно содержит источник углерода. Авторы изобретения открыли, что ферментация С. difficile в культуральной среде, содержащей по меньшей мере один источник углерода, обеспечивает высокие значения оптической плотности при 600 нм (OD600) и высокие выходы по продукту токсину по сравнению с ферментацией в отсутствие источника углерода. В одном из воплощений источником углерода является глюкоза, маннит, фруктоза и/или манноза.
В одном из воплощений клетка С. difficile не является генетически модифицированной. В другом воплощении клетка С. difficile является рекомбинантной клеткой С. difficile. В одном из воплощений в клетке С. difficile отсутствует эндогенный полинуклеотид, кодирующий токсин. В другом воплощении эта клетка содержит конститутивный промотор. В предпочтительном воплощении промотор представляет собой промотор фередоксина (fdx) Clostridium sporogenes. В дополнительном воплощении эта клетка не содержит нативного, регулируемого хромосомального промотора.
В другом аспекте изобретение относится к способу культивирования С. difficile. Способ включает культивирование клетки С. difficile в среде. В одном из воплощений среда содержит гидролизат сои и/или дрожжевой экстракт. В предпочтительном воплощении среда дополнительно содержит источник углерода. Предпочтительно, источником углерода является глюкоза.
В одном из воплощений стадию культивирования осуществляют в анаэробных условиях.
В одном из воплощений С. difficile выращивают в качестве посевной культуры. В одном из воплощений выращивание посевной культуры начинают путем инокуляции исходной культурой, которая была выращена в данной среде.
В одном из воплощений С. difficile выращивают в качестве ферментируемой культуры. В одном из воплощений ферментируемую культуру инокулируют посевной культурой, которая была выращена в данной среде. В альтернативном аспекте изобретение относится к способу культивирования С. difficile. Способ включает культивирование клетки С. difficile в среде для моноклональных антител (mAb).
В одном из аспектов изобретение относится к способу получения токсина С. difficile. Способ включает культивирование клетки С. difficile в среде. Способ дополнительно включает выделение токсина С. difficile из указанной среды.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1А-Н. Выравнивание последовательностей токсина А С. difficile дикого типа из штаммов 630, VPI10463, R20291, CD196 и мутантного токсина А, имеющего SEQ ID NO: 4, с использованием выравнивания CLUSTALW с параметрами по умолчанию.
Фиг. 2A-F. Выравнивание последовательностей токсина В дикого типа С. difficile из штаммов 630, VPI10463, R20291, CD196 и мутантного токсина В, имеющего SEQ ID NO: 6, с использованием выравнивания CLUSTALW с параметрами по умолчанию.
Фиг. 3. График, показывающий идентификацию токсин-негативных штаммов С. difficile дикого типа. Культуральные среды 13 штаммов С. difficile тестировали посредством ELISA (иммуноферментный твердофазный анализ) в отношении токсина А. Как проиллюстрировано, семь штаммов экспрессировали токсин А, а 6 штаммов нет (штаммы 1351, 3232, 7322, 5036, 4811 и VPI 11186).
Фиг. 4А и В. Результаты SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), иллюстрирующие, что тройной мутант A (SEQ ID NO: 4), двойной мутант В (SEQ ID NO: 5) и тройной мутант В (SEQ ID NO: 6) не глюкозилируют ГТФазы Rac1 или RhoA в анализах глюкозилирования in vitro с использованием уридиндифосфат(УДФ)-14С-глюкозы; тогда как токсин В дикого типа в количестве от 10 мкг до 1 нг глюкозилирует Rac1.
Фиг. 5. Вестерн-блот, показывающий устранение активности цистеиновой протеазы у мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно) по сравнению с наблюдаемыми фрагментами расщепления токсинов А и В дикого типа (SEQ ID NO: 1 и 2, соответственно). См. пример 13.
Фиг. 6. Графики, показывающие, что тройные мутантные токсины А и В (SEQ ID NO: 4 и 6, соответственно) демонстрируют остаточную цитотоксичность при тестировании в высоких концентрациях (например, приблизительно 100 мкг/мл) в анализе цитотоксичности in vitro на клетках IMR-90.
Фиг. 7. График, показывающий, что величины ЕС50 аналогичны для тройного мутантного токсина В (SEQ ID NO: 6) и мутантного по семи положениям токсина В (SEQ ID NO: 8).
Фиг. 8. График, представляющий результаты тестирований цитотоксичности in vitro, на котором изображена зависимость уровней АТФ (аденозинтрифосфат) (RLU) от возрастающих концентраций тройного мутантного TcdA (SEQ ID NO: 4) (верхняя панель) и тройного мутантного TcdB (SEQ ID NO: 6) (нижняя панель). Остаточную цитотоксичность мутантного токсина А и В можно полностью устранить, используя нейтрализующие антитела, специфичные к мутантному токсину А (верхняя панель - рАb (поликлональное антитело) А и mAb А3-25+А60-22) и к мутантному токсину В (нижняя панель - рАb В).
Фиг. 9. Изображения морфологического строения клеток IMR-90 через 72 часа после обработки. На панели А показаны ложнообработанные контрольные клетки. На панели В показано морфологическое строение клеток после обработки мутантным TcdB (SEQ ID NO: 6), инактивированным формалином. На панели С показано морфологическое строение клеток после обработки мутантным TcdB (SEQ ID NO: 6), инактивированным EDC (1-этил-3-(3-диметиламинопропил)-карбодиимид). На панели D показано морфологическое строение клеток после обработки токсином В дикого типа (SEQ ID NO: 2). На панели Ε показано морфологическое строение клеток после обработки тройным мутантным TcdB (SEQ ID NO: 6). Схожие результаты наблюдали для обработок с использованием TcdA.
Фиг. 10. График, показывающий титры нейтрализующих антител, как описано в примере 25 (исследование muCdiff2010-06).
Фиг. 11А-В. График, показывающий титры нейтрализующих антител, как описано в примере 26 (исследование muCdiff2010-07).
Фиг. 12. График, показывающий нейтрализующие антительные ответы на токсины А и В у хомяков после четырех иммунизаций, как описано в примере 27 (исследование hamС. difficile2010-02).
Фиг. 13А-В. График, показывающий нейтрализующие антительные ответы у хомяков после вакцинации химически инактивированными генетически мутантными токсинами и токсоидами от List Biologicals, как описано в примере 27 (исследование hamС. difficile e2010-02).
Фиг. 14. Кривые выживаемости для трех иммунизированных групп хомяков по сравнению с неиммунизированными контролями, как описано в примере 28 (исследование hamС. difficile 2010-02, продолжение).
Фиг. 15. График, показывающий относительный нейтрализующий антительный ответ на разные композиции мутантных токсинов из С. difficile у хомяков (исследование hamС. difficile 2010-03), как описано в примере 29.
Фиг. 16А-В. Графики, показывающие сильный относительный нейтрализующий антительный ответ на химически инактивированные генетически мутантные токсины А и В (SEQ ID NO: 4 и 6, соответственно) у яванских макак, как описано в примере 30.
Фиг. 17. Аминокислотные последовательности вариабельных областей легкой (VL) и тяжелой (VH) цепей из mAb А3-25 IgE. Сигнальный пептид выделен в рамку; CDR (определяющие комплементарность участки) выделены курсивным шрифтом и подчеркнуты; константный участок выделен жирным шрифтом и подчеркнут (полная последовательность не показана).
Фиг. 18. График, показывающий титрование индивидуальных моноклональных антител к токсину А в анализе нейтрализации токсинов с использованием уровней АТФ (количественно определенных посредством относительных световых единиц (RLU)) в качестве индикатора жизнеспособности клеток. В случае сравнения с токсином (4×EC50) в качестве в качестве контроля, моноклональные антитела (mAb) А80-29, А65-33, А60-22 и А3-25 показывали усиление нейтрализующего действия на токсин А при возрастании концентрации, но не достигали уровня кроличьего антитела к токсину А, взятого в качестве положительного контроля. MAb А50-10, А56-33 и А58-46 не оказывали нейтрализующего действия на токсин А. Для контроля, состоящего только из клеток, получали 1-1,5×106 RLU.
Фиг. 19. Картирование 8 групп эпитопов токсина В для mAb с использованием BiaCore.
Фиг. 20А-С. Синергетические нейтрализующие активности комбинаций mAb к токсину А: добавление нейтрализующих антител А60-22, А65-33 и А80-29 в различных разведениях к mAb А3-25 в возрастающих концентрациях синергетически усиливало нейтрализацию токсина А независимо от разведения. Проиллюстрированы значения RLU для контроля, состоящего только из токсина А, (4×ЕС50) (меньше 0,3×106) и для контролей, состоящих только из клеток, которые составляли 2-2,5×106 RLU, как изображено на графиках, показанных на Фиг. 20В и Фиг. 20С.
Фиг. 21. Синергетические нейтрализующие активности mAb к токсину В: нейтрализация токсина В под действием mAb 8-26, В60-2 и В59-3 проиллюстрирована на Фиг. 21 А. Нейтрализация токсина В синергетически возрастала после объединения В8-26 с В59-3 в разных разведениях (Фиг. 21В).
Фиг. 22. Вестерн-блот, показывающий, что экспрессия ГТФазы Rac1 ослаблена в экстрактах клеток, культивированных от 24 до 96 часов, обработанных генетически мутантным токсином В (SEQ ID NO: 6), но не в экстрактах клеток, обработанных токсином В дикого типа (SEQ ID NO: 2). Этот блот также показывает, что Rac1 глюкозилируется в обработанных токсином В экстрактах клеток, но не в экстрактах клеток, обработанных генетически мутантным токсином В.
Фиг. 23А-К. График, представляющий результаты тестирования цитотоксичности in vitro, на котором изображена зависимость уровней АТФ (RLU) от возрастающих концентраций: смеси культуральных сред для С. difficile и сывороток крови хомяков (■); смеси неочищенного токсина (сбора из культуральной среды) из соответствующего штамма и сывороток крови хомяков (●); смеси очищенного токсина (коммерческого препарата токсина, полученного от List Biologicals) и сывороток крови хомяков (▲); неочищенного токсина (▼) в качестве контроля и очищенного токсина (♦) в качестве контроля. Токсины из соответствующих штаммов добавляли к клеткам в концентрации 4×EC50. Фиг. 23 показывает, что иммуногенная композиция, содержащая мутантный TcdA (SEQ ID NO: 4) и мутантный TcdB (SEQ ID NO: 6), причем мутантные токсины были инактивированы с использованием EDC согласно, например, примеру 29, Таблице 15, как изложено в данном описании, индуцировала нейтрализующие антитела, которые демонстрировали нейтрализующую активность в отношении токсинов по меньшей мере из следующих 16 разных штаммов С. difficile, полученных из Центров по контролю и профилактике заболеваний (CDC), по сравнению с соответствующим контролем, состоящим только из токсина: 2007886 (Фиг. 23А), 2006017 (Фиг. 23 В), 2007070 (Фиг. 23С), 2007302 (Фиг. 23D), 2007838 (Фиг. 23Е), 2007886 (Фиг. 23F), 2009292 (Фиг. 23G), 2004013 (Фиг. 23Н), 2009141 (Фиг. 23I), 2005022 (Фиг. 23J), 2006376 (Фиг. 23К).
Фиг. 24. Иллюстрация типичной индуцированной EDC/NHS (N-гидроксисукцинимид) инактивации мутантных токсинов из С. difficile, в результате которой образуются по меньшей мере три возможных типа модификаций: сшивки, глициновые аддукты и бета-аланиновые аддукты. Панель А иллюстрирует образование сшивок. Карбоксильные группы тройных мутантных токсинов активируют путем добавления EDC и NHS. Активированные сложные эфиры взаимодействуют с первичными аминами с образованием стабильных амидных связей, в результате чего образуются внутри- и межмолекулярные сшивки. Панель В иллюстрирует образование глициновых аддуктов. После инактивации оставшиеся активированные эфиры гасят путем добавления глицина с образованием стабильных амидных связей. Панель С иллюстрирует образование бета-аланиновых аддуктов. Три моля NHS могут взаимодействовать с одним молем EDC с образованием активированного бета-аланина. Затем его приводят во взаимодействие с первичными аминами с образованием стабильных амидных связей.
Фиг. 25. Иллюстрация типичной EDC/NHS-индуцированной инактивации мутантных токсинов из С. difficile, в результате которой образуется по меньшей мере один из следующих типов модификаций: (А) сшивки, (В) глициновые аддукты и (С) бета-аланиновые аддукты.
Фиг. 26. График, представляющий результаты анализа цитотоксичности in vitro, на котором изображена зависимость уровней АТФ (RLU) (анализ АТФ, 72 ч) от возрастающих концентраций TcdB дикого типа, приобретенного у List Biologicals (□), тройного мутантного TcdB (SEQ ID NO: 86) (●) и мутантного по пяти положениям TcdB (SEQ ID NO: 184) (■). Клетки IMR-90 (*) использовали в качестве контроля.
Фиг. 27. График, показывающий конкурентное ингибирование опосредуемой тройным мутантным токсином В (SEQ ID NO: 86) цитотоксичности под действием мутантного по пяти положениям токсина В (SEQ ID NO: 184) на клетках IMR-90 (анализ АТФ, 72 ч). -●- представляет собой мутантный по пяти положениям токсин В (SEQ ID NO: 184) («РМ-В»); -▲- представляет собой тройной мутантный (ТМ) токсин в концентрации 200 нг/мл.
Фиг. 28. График, показывающий конечное значение OD и титр для тройного мутантного токсина В (мг/л) после перфузионной ферментации (CDF-5126). -●- представляет OD600 нм; -▲- представляет скорость потока перфузионной жидкости (количество объемов ферментера/2,0 ч); -■- представляет глюкозу (г/л); -●- представляет токсоид В (тройной мутантный, SEQ ID NO: 86).
Фиг. 29. График, показывающий конечное значение OD и титр для тройного мутантного токсина В (мг/л), полученные в результате другого перфузионного культивирования (CDF-5127). -●- представляет OD600 нм; -▲- скорость потока перфузионной жидкости (количество объемов ферментера/2,0 ч); -■- представляет глюкозу (г/л); -●- представляет токсоид В (тройной мутантный, SEQ ID NO: 86).
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В SEQ ID NO: 1 приведена аминокислотная последовательность для токсина А из С. difficile (TcdA) 630 дикого типа.
В SEQ ID NO: 2 приведена аминокислотная последовательность для токсина В из С. difficile (TcdB) 630 дикого типа.
В SEQ ID NO: 3 приведена аминокислотная последовательность для мутантного TcdA, имеющего мутацию в положениях 285 и 287, по сравнению с SEQ ID NO: 1.
В SEQ ID NO: 4 приведена аминокислотная последовательность для мутантного TcdA, имеющего мутацию в положениях 285, 287 и 700, по сравнению с SEQ ID NO: 1.
В SEQ ID NO: 5 приведена аминокислотная последовательность для мутантного TcdB, имеющего мутацию в положениях 286 и 288, по сравнению с SEQ ID NO: 2.
В SEQ ID NO: 6 приведена аминокислотная последовательность для мутантного TcdB, имеющего мутацию в положениях 286, 288 и 698, по сравнению с SEQ ID NO: 2.
В SEQ ID NO: 7 приведена аминокислотная последовательность для мутантного TcdA, имеющего мутацию в положениях 269, 272, 285, 287, 460, 462 и 700, по сравнению с SEQ ID NO: 1.
В SEQ ID NO: 8 приведена аминокислотная последовательность для мутантного TcdB, имеющего мутацию в положениях 270, 273, 286, 288, 461, 463 и 698, по сравнению с SEQ ID NO: 2.
В SEQ ID NO: 9 приведена последовательность ДНК, кодирующую токсин А из С. difficile (TcdA) 630 дикого типа.
В SEQ ID NO: 10 приведена последовательность ДНК, кодирующую токсин В из С. difficile (TcdB) 630 дикого типа.
В SEQ ID NO: 11 приведена последовательность ДНК, кодирующую SEQ ID NO: 3.
В SEQ ID NO: 12 приведена последовательность ДНК, кодирующую SEQ ID NO: 4.
В SEQ ID NO: 13 приведена последовательность ДНК, кодирующую SEQ ID NO: 5.
В SEQ ID NO: 14 приведена последовательность ДНК, кодирующую SEQ ID NO: 6.
В SEQ ID NO: 15 приведена аминокислотная последовательность для TcdA из С. difficile R20291 дикого типа.
В SEQ ID NO: 16 приведена последовательность ДНК, кодирующую SEQ ID NO: 15.
В SEQ ID NO: 17 приведена аминокислотная последовательность для TcdA из С. difficile CD196 дикого типа.
В SEQ ID NO: 18 приведена последовательность ДНК, кодирующую SEQ IDNO: 17.
В SEQ ID NO: 19 приведена аминокислотная последовательность для TcdA из С. difficile VP110463 дикого типа.
В SEQ ID NO: 20 приведена последовательность ДНК, кодирующую SEQ ID NO: 19.
В SEQ ID NO: 21 приведена аминокислотная последовательность для Tcd В из С. difficile R20291 дикого типа.
В SEQ ID NO: 22 приведена последовательность ДНК, кодирующую SEQ ID NO: 21.
В SEQ ID NO: 23 приведена аминокислотная последовательность для TcdB из С. difficile CD196 дикого типа.
В SEQ ID NO: 24 приведена последовательность ДНК, кодирующую SEQ ID NO: 23.
В SEQ ID NO: 25 приведена аминокислотная последовательность для TcdB С. difficile VP110463 дикого типа.
В SEQ ID NO: 26 приведена последовательность ДНК, кодирующую SEQ ID NO: 25.
В SEQ ID NO: 27 приведена последовательность ДНК для локуса патогенности С. difficile VPI10463 дикого типа.
В SEQ ID NO: 28 приведена аминокислотная последовательность для остатков 101-293 в SEQ ID NO: 1.
В SEQ ID NO: 29 приведена аминокислотная последовательность для остатков 1-542 в SEQ ID NO: 1.
В SEQ ID NO: 30 приведена аминокислотная последовательность для остатков 101-293 в SEQ ID NO: 2.
В SEQ ID NO: 31 приведена аминокислотная последовательность для остатков 1-543 в SEQ ID NO: 2.
В SEQ ID NO: 32 приведена аминокислотная последовательность для остатков 543-809 в SEQ ID NO: 1.
В SEQ ID NO: 33 приведена аминокислотная последовательность для остатков 544-767 в SEQ ID NO: 2.
В SEQ ID NO: 34 приведена аминокислотная последовательность для мутантного TcdA, где остатки 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 могут представлять собой остатки любой аминокислоты.
В SEQ ID NO: 35 приведена аминокислотная последовательность для мутантного Tcd В, где остатки 102, 270, 273, 286, 288, 384, 461, 463, 520, 543, 544, 587, 600, 653, 698 и 751 могут представлять собой остатки любой аминокислоты.
В SEQ ID NO: 36 приведена аминокислотная последовательность для вариабельной области легкой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 37 приведена аминокислотная последовательность для вариабельной области тяжелой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 38 приведена аминокислотная последовательность для CDR1 вариабельной области легкой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 39 приведена аминокислотная последовательность для CDR2 вариабельной области легкой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 40 приведена аминокислотная последовательность для CDR3 вариабельной области легкой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 41 приведена аминокислотная последовательность для CDR1 вариабельной области тяжелой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 42 приведена аминокислотная последовательность для CDR2 вариабельной области тяжелой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 43 приведена аминокислотная последовательность для CDR3 вариабельной области тяжелой цепи нейтрализующего антитела для TcdA из С. difficile (mAb А3-25).
В SEQ ID NO: 44 приведена последовательность ДНК, кодирующую SEQ ID NO: 3.
В SEQ ID NO: 45 приведена последовательность ДНК, кодирующую SEQ IDNO: 4.
В SEQ ID NO: 46 приведена последовательность ДНК, кодирующую SEQ ID NO: 5.
В SEQ ID NO: 47 приведена последовательность ДНК, кодирующую SEQ ID NO: 6.
В SEQ ID NO: 48 приведена нуклеотидная последовательность содержащего CpG (динуклеотид, состоящий из остатков цитозина и гуанидина, соединенных фосфатной группой) иммуностимулирующего олигонуклеотида ODN 24555.
В SEQ ID NO: 49 приведена аминокислотная последовательность для вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 50 приведена аминокислотная последовательность для сигнального пептида вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 51 приведена аминокислотная последовательность для CDR1 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 52 приведена аминокислотная последовательность для CDR2 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 53 приведена аминокислотная последовательность для CDR3 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 54 приведена аминокислотная последовательность для константного участка вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 55 приведена аминокислотная последовательность для вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 56 приведена аминокислотная последовательность для сигнального пептида вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 57 приведена аминокислотная последовательность для CDR1 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 58 приведена аминокислотная последовательность для CDR2 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 59 приведена аминокислотная последовательность для CDR3 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В8-26).
В SEQ ID NO: 60 приведена аминокислотная последовательность для вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 61 приведена аминокислотная последовательность для сигнального пептида вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 62 приведена аминокислотная последовательность для CDR1 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 63 приведена аминокислотная последовательность для CDR2 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 64 приведена аминокислотная последовательность для CDR3 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 65 приведена аминокислотная последовательность для константного участка вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 66 приведена аминокислотная последовательность для вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 67 приведена аминокислотная последовательность для сигнального пептида вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 68 приведена аминокислотная последовательность для CDR1 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 69 приведена аминокислотная последовательность для CDR2 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 70 приведена аминокислотная последовательность для CDR3 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В59-3).
В SEQ ID NO: 71 приведена аминокислотная последовательность для вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 72 приведена аминокислотная последовательность для сигнального пептида вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 73 приведена аминокислотная последовательность для CDR1 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 74 приведена аминокислотная последовательность для CDR2 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 75 приведена аминокислотная последовательность для CDR3 вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 76 приведена аминокислотная последовательность для константного участка вариабельной области тяжелой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 77 приведена аминокислотная последовательность для вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 78 приведена аминокислотная последовательность для сигнального пептида вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 79 приведена аминокислотная последовательность для CDR1 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 80 приведена аминокислотная последовательность для CDR2 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 81 приведена аминокислотная последовательность для CDR3 вариабельной области легкой цепи антитела, нейтрализующего TcdB из С. difficile, (mAb В9-30).
В SEQ ID NO: 82 приведена аминокислотная последовательность для мутантного TcdB, где остаток в положениях 102, 270, 273, 286, 288, 384, 461, 463, 520, 543, 544, 587, 600, 653, 698 и 751 может представлять собой остаток любой аминокислоты.
В SEQ ID NO: 83 приведена аминокислотная последовательность для мутантного TcdA, имеющего мутацию в положениях 269, 272, 285, 287, 460, 462 и 700, по сравнению с SEQ ID NO: 1, где метионин в положении 1 отсутствует.
В SEQ ID NO: 84 приведена аминокислотная последовательность для мутантного токсина А из С. difficile, имеющего мутацию в положениях 285, 287 и 700 по сравнению с SEQ ID NO: 1, где метионин в положении 1 отсутствует.
В SEQ ID NO: 85 приведена аминокислотная последовательность для мутантного токсина В из С. difficile, имеющего мутацию в положениях 270, 273, 286, 288, 461, 463 и 698, по сравнению с SEQ ID NO: 2, где метионин в положении 1 отсутствует.
В SEQ ID NO: 86 приведена аминокислотная последовательность для мутантного токсина В из С. difficile, имеющего мутацию в положениях 286, 288 и 698, по сравнению с SEQ ID NO: 2, где метионин в положении 1 отсутствует.
В SEQ ID NO: 87 приведена аминокислотная последовательность для TcdA из С. difficile 2004013 дикого типа.
В SEQ ID NO: 88 приведена аминокислотная последовательность для TcdA из С. difficile 2004111 дикого типа.
В SEQ ID NO: 89 приведена аминокислотная последовательность для TcdA из С. difficile 2004118 дикого типа.
В SEQ ID NO: 90 приведена аминокислотная последовательность для TcdA из С. difficile 2004205 дикого типа.
В SEQ ID NO: 91 приведена аминокислотная последовательность для TcdA из С. difficile 2004206 дикого типа.
В SEQ ID NO: 92 приведена аминокислотная последовательность для TcdA из С. difficile 2005022 дикого типа.
В SEQ ID NO: 93 приведена аминокислотная последовательность для TcdA из С. difficile 2005088 дикого типа.
В SEQ ID NO: 94 приведена аминокислотная последовательность для TcdA из С. difficile 2005283 дикого типа.
В SEQ ID NO: 95 приведена аминокислотная последовательность для TcdA из С. difficile 2005325 дикого типа.
В SEQ ID NO: 96 приведена аминокислотная последовательность для TcdA из С. difficile 2005359 дикого типа.
В SEQ ID NO: 97 приведена аминокислотная последовательность для TcdA из С. difficile 2006017 дикого типа.
В SEQ ID NO: 98 приведена аминокислотная последовательность для TcdA из С. difficile 2007070 дикого типа.
В SEQ ID NO: 99 приведена аминокислотная последовательность для TcdA из С. difficile 2007217 дикого типа.
В SEQ ID NO: 100 приведена аминокислотная последовательность для TcdA из С. difficile 2007302 дикого типа.
В SEQ ID NO: 101 приведена аминокислотная последовательность для TcdA из С. difficile 2007816 дикого типа.
В SEQ ID NO: 102 приведена аминокислотная последовательность для TcdA из С. difficile 2007838 дикого типа.
В SEQ ID NO: 103 приведена аминокислотная последовательность для TcdA из С. difficile 2007858 дикого типа.
В SEQ ID NO: 104 приведена аминокислотная последовательность для TcdA из С. difficile 2007886 дикого типа.
В SEQ ID NO: 105 приведена аминокислотная последовательность для TcdA из С. difficile 2008222 дикого типа.
В SEQ ID NO: 106 приведена аминокислотная последовательность для TcdA из С. difficile 2009078 дикого типа.
В SEQ ID NO: 107 приведена аминокислотная последовательность для TcdA из С. difficile 2009087 дикого типа.
В SEQ ID NO: 108 приведена аминокислотная последовательность для TcdA из С. difficile 2009141 дикого типа.
В SEQ ID NO: 109 приведена аминокислотная последовательность для TcdA из С. difficile 2009292 дикого типа.
В SEQ ID NO: 110 приведена аминокислотная последовательность для TcdB из С. difficile 2004013 дикого типа.
В SEQ ID NO: 111 приведена аминокислотная последовательность для TcdB из С. difficile 2004111 дикого типа.
В SEQ ID NO: 112 приведена аминокислотная последовательность для TcdB из С. difficile 2004118 дикого типа.
В SEQ ID NO: 113 приведена аминокислотная последовательность для TcdB из С. difficile 2004205 дикого типа.
В SEQ ID NO: 114 приведена аминокислотная последовательность для TcdB из С. difficile 2004206 дикого типа.
В SEQ ID NO: 115 приведена аминокислотная последовательность для TcdB из С. difficile 2005022 дикого типа.
В SEQ ID NO: 116 приведена аминокислотная последовательность для TcdB из С. difficile 2005088 дикого типа.
В SEQ ID NO: 117 приведена аминокислотная последовательность для TcdB из С. difficile 2005283 дикого типа.
В SEQ ID NO: 118 приведена аминокислотная последовательность для TcdB из С. difficile 2005325 дикого типа.
В SEQ ID NO: 119 приведена аминокислотная последовательность для TcdB из С. difficile 2005359 дикого типа.
В SEQ ID NO: 120 приведена аминокислотная последовательность для TcdB из С. difficile 2006017 дикого типа.
В SEQ ID NO: 121 приведена аминокислотная последовательность для TcdB из С. difficile 2006376 дикого типа.
В SEQ ID NO: 122 приведена аминокислотная последовательность для TcdB из С. difficile 2007070 дикого типа.
В SEQ ID NO: 123 приведена аминокислотная последовательность для TcdB из С. difficile 2007217 дикого типа.
В SEQ ID NO: 124 приведена аминокислотная последовательность для TcdB из С. difficile 2007302 дикого типа.
В SEQ ID NO: 125 приведена аминокислотная последовательность для TcdB из С. difficile 2007816 дикого типа.
В SEQ ID NO: 126 приведена аминокислотная последовательность для TcdB из С. difficile 2007838 дикого типа.
В SEQ ID NO: 127 приведена аминокислотная последовательность для TcdB из С. difficile 2007858 дикого типа.
В SEQ ID NO: 128 приведена аминокислотная последовательность для TcdB из С. difficile 2007886 дикого типа.
В SEQ ID NO: 129 приведена аминокислотная последовательность для TcdB из С. difficile 2008222 дикого типа.
В SEQ ID NO: 130 приведена аминокислотная последовательность для TcdB из С. difficile 2009078 дикого типа.
В SEQ ID NO: 131 приведена аминокислотная последовательность для TcdB из С. difficile 2009087 дикого типа.
В SEQ ID NO: 132 приведена аминокислотная последовательность для TcdB из С. difficile 2009141 дикого типа.
В SEQ ID NO: 133 приведена аминокислотная последовательность для TcdB из С. difficile 2009292 дикого типа.
В SEQ ID NO: 134 приведена аминокислотная последовательность для TcdA из С. difficile 014 дикого типа.
В SEQ ID NO: 135 приведена аминокислотная последовательность для TcdA из С. difficile 015 дикого типа.
В SEQ ID NO: 136 приведена аминокислотная последовательность для TcdA из С. difficile 020 дикого типа.
В SEQ ID NO: 137 приведена аминокислотная последовательность для TcdA из С. difficile 023 дикого типа.
В SEQ ID NO: 138 приведена аминокислотная последовательность для TcdA из С. difficile 027 дикого типа.
В SEQ ID NO: 139 приведена аминокислотная последовательность для TcdA из С. difficile 029 дикого типа.
В SEQ ID NO: 140 приведена аминокислотная последовательность для TcdA из С. difficile 046 дикого типа.
В SEQ ID NO: 141 приведена аминокислотная последовательность для TcdB из С. difficile 014 дикого типа.
В SEQ ID NO: 142 приведена аминокислотная последовательность для TcdB из С. difficile 015 дикого типа.
В SEQ ID NO: 143 приведена аминокислотная последовательность для TcdB из С. difficile 020 дикого типа.
В SEQ ID NO: 144 приведена аминокислотная последовательность для TcdB из С. difficile 023 дикого типа.
В SEQ ID NO: 145 приведена аминокислотная последовательность для TcdB из С. difficile 027 дикого типа.
В SEQ ID NO: 146 приведена аминокислотная последовательность для TcdB из С. difficile 029 дикого типа.
В SEQ ID NO: 147 приведена аминокислотная последовательность для TcdB из С. difficile 046 дикого типа.
В SEQ ID NO: 148 приведена аминокислотная последовательность для TcdA из С. difficile 001 дикого типа.
В SEQ ID NO: 149 приведена аминокислотная последовательность для TcdA из С. difficile 002 дикого типа.
В SEQ ID NO: 150 приведена аминокислотная последовательность для TcdA из С. difficile 003 дикого типа.
В SEQ ID NO: 151 приведена аминокислотная последовательность для TcdA из С. difficile 004 дикого типа.
В SEQ ID NO: 152 приведена аминокислотная последовательность для TcdA из С. difficile 070 дикого типа.
В SEQ ID NO: 153 приведена аминокислотная последовательность для Tc из С. difficile 075 дикого типа.
В SEQ ID NO: 154 приведена аминокислотная последовательность для TcdA из С. difficile 077 дикого типа.
В SEQ ID NO: 155 приведена аминокислотная последовательность для TcdA из С. difficile 081 дикого типа.
В SEQ ID NO: 156 приведена аминокислотная последовательность для TcdA из С. difficile 117 дикого типа.
В SEQ ID NO: 157 приведена аминокислотная последовательность для TcdA из С. difficile 131 дикого типа.
В SEQ ID NO: 158 приведена аминокислотная последовательность для TcdB из С. difficile 001 дикого типа.
В SEQ ID NO: 159 приведена аминокислотная последовательность для TcdB из С. difficile 002 дикого типа.
В SEQ ID NO: 160 приведена аминокислотная последовательность для TcdB из С. difficile 003 дикого типа.
В SEQ ID NO: 161 приведена аминокислотная последовательность для TcdB из С. difficile 004 дикого типа.
В SEQ ID NO: 162 приведена аминокислотная последовательность для TcdB из С. difficile 070 дикого типа.
В SEQ ID NO: 163 приведена аминокислотная последовательность для TcdB из С. difficile 075 дикого типа.
В SEQ ID NO: 164 приведена аминокислотная последовательность для TcdB из С. difficile 077 дикого типа.
В SEQ ID NO: 165 приведена аминокислотная последовательность для TcdB из С. difficile 081 дикого типа.
В SEQ ID NO: 166 приведена аминокислотная последовательность для TcdB из С. difficile 117 дикого типа.
В SEQ ID NO: 167 приведена аминокислотная последовательность для TcdB из С. difficile 131 дикого типа.
В SEQ ID NO: 168 приведена аминокислотная последовательность для TcdA из С. difficile 053 дикого типа.
В SEQ ID NO: 169 приведена аминокислотная последовательность для TcdA из С. difficile 078 дикого типа.
В SEQ ID NO: 170 приведена аминокислотная последовательность для TcdA из С. difficile 087 дикого типа.
В SEQ ID NO: 171 приведена аминокислотная последовательность для TcdA из С. difficile 095 дикого типа.
В SEQ ID NO: 172 приведена аминокислотная последовательность для TcdA из С. difficile 126 дикого типа.
В SEQ ID NO: 173 приведена аминокислотная последовательность для TcdB из С. difficile 053 дикого типа.
В SEQ ID NO: 174 приведена аминокислотная последовательность для TcdB из С. difficile 078 дикого типа.
В SEQ ID NO: 175 приведена аминокислотная последовательность для TcdB из С. difficile 087 дикого типа.
В SEQ ID NO: 176 приведена аминокислотная последовательность для TcdB из С. difficile 095 дикого типа.
В SEQ ID NO: 177 приведена аминокислотная последовательность для TcdB из С. difficile 126 дикого типа.
В SEQ ID NO: 178 приведена аминокислотная последовательность для TcdA из С. difficile 059 дикого типа.
В SEQ ID NO: 179 приведена аминокислотная последовательность для TcdB из С. difficile 059 дикого типа.
В SEQ ID NO: 180 приведена аминокислотная последовательность для TcdA из С. difficile 106 дикого типа.
В SEQ ID NO: 181 приведена аминокислотная последовательность для TcdB из С. difficile 106 дикого типа.
В SEQ ID NO: 182 приведена аминокислотная последовательность для TcdB из С. difficile 017 дикого типа.
В SEQ ID NO: 183 приведена аминокислотная последовательность для мутантного TcdA, имеющего мутацию в положениях 285, 287, 700, 972 и 978, по сравнению с SEQ ID NO: 1.
В SEQ ID NO: 184 приведена аминокислотная последовательность для мутантного TcdB, имеющего мутацию в положениях 286, 288, 698, 970 и 976, по сравнению с SEQ ID NO: 2.
В каждой из последовательностей от SEQ ID NO: 185 до SEQ ID NO: 195 приведена аминокислотная последовательность для типичного мутантного токсина.
В каждой из последовательностей от SEQ ID NO: 196 до SEQ ID NO: 212 приведена аминокислотная последовательность для типичного мутантного токсина А.
Каждая из последовательностей SEQ ID NO: 213 до SEQ ID NO: 222 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 223 до SEQ ID NO: 236 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 237 до SEQ ID NO: 243 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей SEQ ID NO: 244 и SEQ ID NO: 245 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 246 до SEQ ID NO: 249 приведена аминокислотная последовательность для типичного мутантного токсина В.
Каждая из последовательностей SEQ ID NO: 250 до SEQ ID NO: 253 приведена аминокислотная последовательность для типичного мутантного токсина А.
В SEQ ID NO: 254 приведена аминокислотная последовательность для типичного мутантного токсина.
В каждой из последовательностей от SEQ ID NO: 255 до SEQ ID NO: 263 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 264 до SEQ ID NO: 269 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 270 до SEQ ID NO: 275 приведена аминокислотная последовательность для типичного мутантного токсина.
В каждой из последовательностей от SEQ ID NO: 276 до SEQ ID NO: 323 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 324 до SEQ ID NO: 373 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 374 до SEQ ID NO: 421 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 422 до SEQ ID NO: 471 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 472 до SEQ ID NO: 519 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 568 до SEQ ID NO: 615 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 520 до SEQ ID NO: 567 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 616 до SEQ ID NO: 663 приведена аминокислотная последовательность для типичного мутантного токсина В.
В каждой из последовательностей от SEQ ID NO: 664 до SEQ ID NO: 711 приведена аминокислотная последовательность для типичного мутантного токсина А.
В каждой из последовательностей от SEQ ID NO: 712 до SEQ ID NO: 761 приведена аминокислотная последовательность для типичного мутантного токсина В.
ПОДРОБНОЕ ОПИСАНИЕ
Авторы изобретения неожиданным образом открыли, среди прочего, мутантный токсин А и мутантный токсин В из С. difficile и способы их получения. Эти мутанты частично характеризуются тем, что они являются иммуногенными и проявляют более низкую цитотоксичность по сравнению с соответствующим токсином в форме дикого типа. Настоящее изобретение также относится к их иммуногенным участкам, их биологическим эквивалентам и выделенным полинуклеотидам, которые включают нуклеиновокислотные
последовательности, кодирующие любое из перечисленного выше.
Иммуногенные композиции, описанные здесь, неожиданно демонстрировали способность вызывать образование новых нейтрализующих антител к токсинам С. difficile, и они могут обладать способностью обеспечивать активную и/или пассивную защиту от заражения С. difficile. Новые антитела направлены на различные эпитопы токсина А и токсина В. Кроме того, авторы изобретения открыли, что комбинация по меньшей мере двух нейтрализующих моноклональных антител неожиданно может демонстрировать синергетический эффект в отношении нейтрализации in vitro токсина А и токсина В.
Композиции по изобретению, описанные здесь, можно использовать для лечения, предупреждения, снижения риска, уменьшения числа случаев, ослабления тяжести и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания (CDAD), синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данная композиция не была введена.
Кроме того, авторы изобретения открыли рекомбинантную аспорогенную клетку С. difficile, которая может стабильно экспрессировать мутантный токсин А и мутантный токсин В С. difficile, и новые способы их получения.
Иммуногенные композиции
В одном из аспектов изобретение относится к иммуногенной композиции, содержащей мутантный токсин из С. difficile. Мутантный токсин из С. difficile содержит аминокислотную последовательность, имеющую по меньшей мере одну мутацию в глюкозилтрансферазном домене и по меньшей мере одну мутацию в домене цистеиновой протеазы по сравнению с соответствующим токсином из С. difficile дикого типа.
Термин «дикий тип», использованный в данном описании, относится к форме, обнаруживаемой в природе. Например, полипептидная или полинуклеотидная последовательность дикого типа представляет собой последовательность, имеющуюся у микроорганизма, который может быть выделен из природного источника и который не был целенаправленно модифицирован в результате человеческой деятельности. Настоящее изобретение также относится к выделенным полинуклеотидам, которые включают нуклеиновокислотные последовательности, кодирующие любое из перечисленного выше. Помимо этого, настоящее изобретение относится к применению любой из перечисленных выше композиций для лечения, предупреждения, снижения риска, ослабления тяжести, уменьшения числа случаев и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания (CDAD), синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данную композицию не вводят, а также к способам изготовления указанных композиций.
Как использовано в данном описании, «иммуногенная композиция» или «иммуноген» относится к композиции, которая вызывает иммунный ответ у млекопитающего, которому эту композицию вводят.
«Иммунный ответ» относится к развитию у реципиента-пациента полезного гуморального (антитело-опосредованного) и/или клеточного (опосредованного антиген-специфическими Т-клетками или продуктами их секреции) ответа на токсин из С. difficile. Иммунный ответ может быть гуморальным, клеточным или тем и другим.
Иммунный ответ может представлять собой активный ответ, индуцируемый в результате введения иммуногенной композиции, иммуногена. Альтернативно, иммунный ответ может представлять собой пассивный ответ, индуцируемый в результате введения антитела или примированных Т-клеток.
Наличие гуморального (антитело-опосредованного) иммунного ответа может быть определено, например, посредством анализов с применением клеток, известных в данной области техники, таких как анализ нейтрализующих антител, ELISA и т.д.
Клеточный иммунный ответ обычно вызывается в результате презентации эпитопов полипептидов вместе с молекулами класса I или класса II главного комплекса гистосовместимости (МНС) для активации антигенспецифических CD4+ хелперных Т-клеток и/или CD8+ цитотоксических Т-клеток. Ответ также может включать активацию моноцитов, макрофагов, естественных (NK) киллерных клеток, базофилов, дендритных клеток, астроцитов, клеток микроглии, эозинофилов или других компонентов врожденного иммунитета. Наличие клеточно-опосредованного иммунологического ответа может быть определено в анализах пролиферации (CD4+ Т-клеток) или анализов с использованием цитотоксических Т-лимфоцитов (CTL), известных в данной области техники.
В одном из воплощений иммуногенная композиция представляет собой вакцинную композицию. Как использовано в данном описании, «вакцинная композиция» представляет собой композицию, которая вызывает иммунный ответ у млекопитающего, которому вводят эту композицию. Вакцинная композиция может защитить иммунизированное млекопитающее от последующего заражения иммунизирующим агентом или агентом с иммунологической перекрестной реактивностью. Что касается ослабления симптомов или инфекции, то защита может быть полной или частичной при сравнении с невакцинированным млекопитающим, находящимся в тех же условия.
Иммуногенные композиции, описанные здесь, являются перекрестно-реактивными, то есть характеризуются способностью вызывать эффективный иммунный ответ (например, гуморальный иммунный ответ) на токсин, продуцируемый другим штаммом С. difficile, отличающимся от штамма, на основе которого изготовлена композиция. Например, иммуногенные композиции (например, на основе С. difficile 630), описанные здесь, могут способствовать образованию перекрестно-реактивных антител, которые могут связываться с токсинами, продуцируемыми многочисленными штаммами С. difficile (например, токсинами, продуцируемыми С. difficile R20291 и VPI10463). См., например, пример 37. Перекрестная реактивность указывает на возможность осуществления перекрестной защиты бактериальным иммуногеном, и наоборот.
Термин «оказывающий перекрестное защитное действие», использованный в данном описании, относится к способности иммунного ответа, индуцированного иммуногенной композицией, предотвращать или ослаблять инфекцию другим бактериальным штаммом или видом одного и того же рода. Например, иммуногенная композиция (например, на основе С. difficile 630), изложенная в данном описании, может индуцировать эффективный иммунный ответ у млекопитающего с целью ослабления у этого млекопитающего инфекции С. difficile и/или опосредованного С. difficile заболевания, вызванного штаммом, отличающимся от 630 (например, С. difficile R20291).
Типичные млекопитающие, у которых иммуногенная композиция или иммуноген вызывает иммунный ответ, включают любых млекопитающих, таких как, например, мыши, хомяки, приматы и люди. В предпочтительном воплощении иммуногенная композиция или иммуноген вызывает иммунный ответ у человека, которому вводят эту композицию.
Как описано выше, токсин A (TcdA) и токсин В (TcdB) гомологичны глюкозилтрансферазам, которые инактивируют малые ГТФазы семейства Rho/Rac/Ras. Действие TcdA и TcdB на клетки-мишени млекопитающих зависит от многостадийного механизма рецептор-опосредованного эндоцитоза, мембраной транслокации, аутопротеолитического процессинга и моноглюкозилирования ГТФаз. Многие из этих функциональных активностей были приписаны отдельным областям в первичной последовательности токсинов, и были получены изображения этих токсинов для демонстрации того, что данные молекулы имеют схожую структуру.
Ген дикого типа для TcdA имеет примерно 8130 нуклеотидов, которые кодируют белок, имеющий предсказанную молекулярную массу примерно 308 кДа и состоящий примерно из 2710 аминокислот. Как использовано в данном описании, TcdA из С. difficile дикого типа включает TcdA С. difficile из любого штамма С. difficile дикого типа. TcdA из С. difficile дикого типа может содержать аминокислотную последовательность TcdA из С.Difficile дикого типа, имеющую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с SEQ ID NO: 1 (полноразмерная последовательность) при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию.
В предпочтительном воплощении TcdA из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1, которая описывает аминокислотную последовательность дикого типа для TcdA из штамма С. difficile 630 (также описанного с номером доступа в Genbank YP_001087137.1 и/или CAJ67494.1). Штамм С. difficile 630 известен в данной области техники как являющийся штаммом ПЦР(полимеразная цепная реакция)-риботипа 012. SEQ ID NO: 9 описывает ген дикого типа для TcdA из штамма С. difficile 630, который также описан с номером доступа в Genbank NC_009089.1.
Другой типичный TcdA из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 15, которая описывает аминокислотную последовательность дикого типа для TcdA из штамма С. difficile R20291 (также описанного с номером доступа в Genbank YP_003217088.1). Штамм С. difficile R20291 известен в данной области техники как являющийся гипервирулентным штаммом и штаммом ПЦР-риботипа 027. Аминокислотная последовательность для TcdA из штамма С. difficile R20291 имеет примерно 98% идентичности с SEQ ID ΝΟ: 1. SEQ ID NO: 16 описывает ген дикого типа для TcdA из штамма С. difficile R20291, который также описан с номером доступа в Genbank NC_013316.1.
Еще один типичный TcdA из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 17, которая описывает аминокислотную последовательность дикого типа для TcdA из штамма С. difficile CD196 (также описанного с номером доступа в Genbank СВА61156.1). CD196 представляет собой штамм, выделенный при недавней вспышке заболевания в Канаде, и он известен в данной области техники как штамм ПЦР-риботипа 027. Аминокислотная последовательность для TcdA из штамма С. difficile CD196 имеет примерно 98% идентичности с SEQ ID NO: 1 и имеет примерно 100% идентичности с последовательностью для TcdA из штамма С. difficile R20291. SEQ ID NO: 18 описывает ген дикого типа для TcdA из штамма С. difficile CD196, который также описан с номером доступа в Genbank FN538970.1.
Другие примеры аминокислотной последовательности для TcdA из С. difficile дикого типа включают SEQ ID NO: 19, которая описывает аминокислотную последовательность дикого типа для TcdA из штамма С. difficile VP110463 (также описанного с номером доступа в Genbank САА63564.1). Аминокислотная последовательность для TcdA из штамма С. difficile VPI10463 имеет примерно 100% (99,8%) идентичности с SEQ ID NO: 1. SEQ ID NO: 20 описывает ген дикого типа для TcdA из штамма С. difficile VP110463, который также описан с номером доступа в Genbank Х92982.1.
Дополнительные примеры TcdA из С. difficile дикого типа включают TcdA из штаммов С. difficile дикого типа, полученных из Центров по контролю и профилактике заболеваний (CDC, Atlanta, GA). Авторы изобретения открыли, что аминокислотная последовательность TcdA из штаммов С. difficile дикого типа, полученных из CDC, имеет по меньшей мере примерно 99,3%-100% идентичности, при оптимальном выравнивании, с аминокислотными остатками 1-821 в SEQ ID NO: 1 (TcdA из С. difficile 630). См. Таблицу 1.
Авторы изобретения также открыли, что аминокислотная последовательность TcdA из штаммов С. difficile дикого типа может иметь по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, до примерно 100% идентичности, при оптимальном выравнивании (например, при оптимальном выравнивании полноразмерных последовательностей), с SEQ ID NO: 1.

Соответственно, в одном из воплощений аминокислотная последовательность TcdA из С. difficile дикого типа включает последовательность, содержащую по меньшей мере примерно 500, 600, 700 или 800 следующих друг за другом остатков, которая имеет по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с последовательностью равной длины, расположенной между остатками 1 и 900 в SEQ ID NO: 1, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию. Примеры включают штаммы, описанные выше (например, R20291, CD196 и т.д.), и штаммы, приведенные в Таблице 1.
В другом воплощении аминокислотная последовательность TcdA из С. difficile дикого типа включает последовательность, имеющую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой последовательностью, выбранной из SEQ ID NO: 87-109, при оптимальном выравнивании. См. Таблицу 1-а.

Ген дикого типа для TcdB имеет примерно 7098 нуклеотидов, которые кодируют белок, имеющий предсказанную молекулярную массу примерно 270 кДа и состоящий примерно из 2366 аминокислот. Как использовано в данном описании, TcdB из С. difficile дикого типа включает TcdB С. difficile из любого штамма С. difficile дикого типа. TcdB из С. difficile дикого типа может содержать аминокислотную последовательность дикого типа, имеющую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с SEQ ID NO: 2 при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию. В предпочтительном воплощении TcdB из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 2, которая описывает аминокислотную последовательность дикого типа для TcdB из штамма С. difficile 630 (также описанного с номером доступа в Genbank YP_001087135.1 и/или CAJ67492). SEQ ID NO: 10 описывает ген дикого типа для TcdB из штамма С. difficile 630, который также описан с номером доступа в Genbank NC_009089.1.
Другой типичный TcdB из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 21, которая описывает аминокислотную последовательность дикого типа для TcdB из штамма С. difficile R20291 (также описанного с номером доступа в Genbank YP_003217086.1 и/или СВЕ02479.1). Аминокислотная последовательность для TcdB из штамма С. difficile R20291 имеет примерно 92% идентичности с SEQ ID NO: 2. SEQ ID NO: 22 описывает ген дикого типа для TcdB из штамма С. difficile R20291, который также описан с номером доступа в Genbank NC_013316.1.
Дополнительный типичный TcdB из С. difficile дикого типа содержит аминокислотную последовательность, приведенную в SEQ ID NO: 23, которая описывает аминокислотную последовательность дикого типа для TcdB из штамма С. difficile CD196 (также описанного с номером доступа в Genbank YP_003213639.1 и/или СВА61153.1). SEQ ID NO: 24 описывает ген дикого типа для TcdB из штамма С. difficile CD196, который также описан с номером доступа в Genbank NC_013315.1. Аминокислотная последовательность для TcdB из штамма С. difficile CD196 имеет примерно 92% идентичности с SEQ ID NO: 2.
Другие примеры аминокислотной последовательности для TcdB из С. difficile дикого типа включают SEQ ID NO: 25, которая описывает аминокислотную последовательность дикого типа для TcdB из штамма С. difficile VP110463 (также описанного с номером доступа в Genbank Р18177 и/или САА37298). Аминокислотная последовательность для TcdB из штамма С. difficile VPI10463 имеет 100% идентичности с SEQ ID NO: 2. SEQ ID NO: 26 описывает ген дикого типа для TcdB из штамма С. difficile VPI10463, который также описан с номером доступа в Genbank Х53138.1.
Дополнительные примеры TcdB из С. difficile дикого типа включают TcdB из штаммов С. difficile дикого типа, полученных из Центров по контролю и профилактике заболеваний (CDC, Atlanta, GA). Авторы изобретения открыли, что аминокислотная последовательность TcdB из штаммов С. difficile дикого типа, полученных из CDC, имеет по меньшей мере примерно 96%-100% идентичности, при оптимальном выравнивании, с аминокислотными остатками 1-821 в SEQ ID NO: 2 (TcdB из С. difficile 630). См. Таблицу 2.
Таблица 2. Штаммы С. difficile дикого типа, полученные из CDC, и % идентичности аминокислотных остатков 1-821 TcdB из соответствующего штамма С. difficile дикого типа с аминокислотными остатками 1-821 в SEQ ID NO: 2 при оптимальном выравнивании.

Соответственно, в одном из воплощений аминокислотная последовательность TcdB из С. difficile дикого типа включает последовательность, содержащую по меньшей мере примерно 500, 600, 700 или 800 следующих друг за другом остатков, которая имеет по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с последовательностью равной длины, расположенной между остатками 1 и 900 в SEQ ID NO: 2, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию. Примеры включают штаммы, описанные выше (например, R20291, CD196 и т.д.), и штаммы, приведенные в Таблице 2.
В другом воплощении аминокислотная последовательность TcdB из С. difficile дикого типа включает последовательность, имеющую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, предпочтительно примерно 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой последовательностью, выбранной из SEQ ID NO: 110-133, при оптимальном выравнивании. См. Таблицу 2-а.
Гены токсинов А и В (tcdA и tcdB) составляют часть генетического локуса (локуса патогенности, PaLoc) размером 19,6 т.о., который включает 3 дополнительные небольшие открытые рамки считывания (ORF), tcdD, tcdE и tcdC, и могут быть полезными для проявления вирулентности. Известно, что PaLoc является стабильным и консервативным в токсикогенных штаммах. Он находится в одном и том же хромосомном сайте интеграции во всех токсикогенных штаммах, которые проанализированы на сегодняшний день. В нетоксикогенных штаммах локус патогенности (PaLoc) отсутствует. Соответственно, характерной чертой штаммов С. difficile дикого типа, изложенных в данном описании, является наличие локуса патогенности. Другой предпочтительной характерной чертой штаммов С. difficile дикого типа, изложенных в данном описании, является продуцирование как TcdA, так и TcdB.
В одном из воплощений штамм С. difficile дикого типа представляет собой штамм, имеющий локус патогенности, идентичный по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно на 100% локусу из С. difficile 630 или VP110463. Вся последовательность локуса патогенности С. difficile VP110463, зарегистрированная в базе данных EMBL (Европейская лаборатория по молекулярной биологии) как последовательность с номером доступа Х92982, также показана в SEQ ID NO: 26. Штаммы, у которых PaLoc идентичен таковому ссылочного штамма VP110463, обозначают как токсинотип 0. Штаммы токсинотипов I-VII, IX, XII-XV и XVIII-XXIV продуцируют как TcdA, так и TcdB, несмотря на вариации в имеющихся у них генах этихтоксинов.
На N-конце токсинов локализован глюкозилтрансферазный домен. Глюкозилтрансферазная активность токсинов связана с цитотоксической функцией токсинов. Без связи с механизмом или теорией считается, что глюкозилтрансферазная активность обоих токсинов катализирует моноглюкозилирование малых ГТФ-связывающих белков в суперсемействе Rho/Rac/Ras. После глюкозилирования этих ГТФ-связывающих белков физиология клетки резко меняется, что выражается в утрате структурной целостности и нарушении работы жизненно важных сигнальных путей в клетках хозяина, инфицированных токсинами. Мотив Asp-Xaa-Asp (DXD), который вовлечен в связывание марганца, уридин-дифосфата (УДФ) и глюкозы, представляет собой типичный характерный для глюкозилтрансфераз домен. Без связи с механизмом или теорией, считается, что остатки, играющие критическую роль для каталитической активности, такие как мотив DXD, являются одинаковыми у TcdB из известного «исторического» штамма, такого как 630, и у TcdB из гипервирулентного штамма, такого как R20291. Мотив DXD локализован в положениях остатков 285-287 TcdA из С. difficile дикого типа, соответствующих нумерации в SEQ ID NO: 1, и в положениях остатков 286-288 TcdB из С. difficile дикого типа, соответствующих нумерации в SEQ ID NO: 2.
Алгоритмы глобального выравнивания (например, программы анализа последовательностей) известны в данной области техники и могут быть использованы для оптимального выравнивания двух или более аминокислотных последовательностей токсинов с целью определения того, содержит ли данный токсин конкретный сигнатурный мотив (например, DXD в глюкозилтрансферазном домене, DHC в домене цистеиновой протеазы, описанном ниже, и т.д.). Оптимально выравненная(ые) последовательность(и) сравнивают с соответствующей ссылочной последовательностью (например, SEQ ID NO: 1 для TcdA или SEQ ID NO: 2 для TcdB) для определения присутствия данного сигнатурного мотива. «Оптимальное выравнивание» относится к выравниванию, дающему самый высокий балл для процента идентичности. Такое выравнивание можно выполнить, используя известные программы анализа последовательностей. В одном из воплощений для идентификации подходящих токсинов дикого типа используют выравнивание CLUSTAL (кластерный анализ попарных выравниваний) (такое как CLUSTALW) с параметрами по умолчанию, сравнивая запрашиваемую последовательность со ссылочной последовательностью. Относительную нумерацию консервативных остатков аминокислот основывают на нумерации остатков в ссылочной аминокислотной последовательности, чтобы учесть небольшие вставки или делеции (например, меньше пяти аминокислот) в пределах выравненной последовательности.
Использованный в данном описании термин "в соответствии с нумерацией в" относится к нумерации остатков в ссылочной последовательности, когда заданную аминокислотную или полинуклеотидную последовательность сравнивают со ссылочной последовательностью. Другими словами, номер или положение остатка в заданном полимере обозначается с отнесением к ссылочной последовательности, а не по фактическому выраженному числом положению этого остатка в пределах заданной аминокислотной или полинуклеотидной последовательности.
Например, заданная аминокислотная последовательность, как например, последовательность из гипервирулентного штамма С. difficile дикого типа, может быть выравнена относительно ссылочной последовательности (например, такой как последовательность исторического штамма С. difficile дикого типа, например, 630) посредством введения разрывов, при необходимости, для оптимизации совпадений остатков между этими двумя последовательностями. В этих случаях, несмотря на присутствие разрывов, присвоение номера остатку в заданной аминокислотной или полинуклеотидной последовательности выполняют по ссылочной последовательности, относительно которой она была выравнена. Как использовано в данном описании, "ссылочная последовательность" относится к определенной последовательности, используемой в качестве основы для сравнения последовательностей.
Если не указано иное, все ссылки в данном описании на положения аминокислот в TcdA относятся к нумерации в SEQ ID NO: 1. Если не указано иное, все ссылки в данном описании на положения аминокислот в TcdB относятся к нумерации в SEQ ID NO: 2.
Глюкозилтрансферазный домен TcdA, как использовано в данном описании, в типичном случае может начинаться от остатка 1, 101 или 102 и может заканчиваться на остатке 542, 516 или 293 в TcdA из С. difficile дикого типа, например, в SEQ ID NO: 1. Любое минимальное положение остатка можно комбинировать с максимальным положением остатка между остатками 1 и 542 в TcdA для определения последовательности глюкозилтрансферазного домена при условии включения участка мотива DXD. Например, в одном из воплощений глюкозилтрансферазный домен TcdA содержит SEQ ID NO: 27, что идентично остаткам 101-293 в SEQ ID NO: 1, и он содержит участок мотива DXD. В другом воплощении глюкозилтрансферазный домен TcdA содержит SEQ ID NO: 28, что идентично остаткам 1-542 в SEQ ID NO: 1.
Глюкозилтрансферазный домен TcdB, как использовано в данном описании, в типичном случае может начинаться от остатка 1, 101 или 102 и может заканчиваться на остатке 543, 516 или 293 в TcdB из С. difficile дикого типа, например, в SEQ ID NO: 2. Любое минимальное положение остатка можно комбинировать с максимальным положением остатка между остатками 1 и 543 в TcdB для определения последовательности глюкозилтрансферазного домена при условии включения участка мотива DXD. Например, в одном из воплощений глюкозилтрансферазный домен TcdB содержит SEQ ID NO: 29, что идентично остаткам 101-293 в SEQ ID NO: 2, и он содержит участок мотива DXD. В другом воплощении глюкозилтрансферазный домен TcdB содержит SEQ ID NO: 30, что идентично остаткам 1-543 в SEQ ID NO: 2.
Без связи с теорией или механизмом, считают, что N-конец TcdA и/или TcdB отщепляется в результате аутопротеолитического процесса с целью транслокации и высвобождения глюкозилтрансферазного домена в цитозоль клетки-хозяина, где он может взаимодействовать с ГТФазами Rac/Ras/Rho. Показано, что TcdA из С. difficile дикого типа расщепляется между L542 и S543. Показано, что TcdB из С. difficile дикого типа расщепляется между L543 и G544.
Домен цистеиновой протеазы связан с аутокаталитической протеолитической активностью токсина. Домен цистеиновой протеазы локализован в прямом направлении относительно глюкозилтрансферазного домена и может быть охарактеризован каталитической триадой аспартат, гистидин и цистеин (DHC), например, D589, Н655 и С700 в TcdA дикого типа и D587, Н653 и С698 в TcdB дикого типа. Без связи с механизмом или теорией считается, что для токсина из "исторического" штамма, такого как 630, и для TcdB из гипервирулентного штамма, такого как R20291, каталитическая триада является консервативной.
Домен цистеиновой протеазы в TcdA, как использовано в данном описании, в типичном случае может начинаться от остатка 543 и может заканчиваться на остатке 809, 769, 768 или 767 в TcdA дикого типа, например, в SEQ ID NO: 1. Любое минимальное положение остатка можно комбинировать с максимальным положением остатка между остатками 543 и 809 в TcdA дикого типа для определения последовательности домена цистеиновой протеазы при условии включения участка с мотивом DHC, представляющим собой каталитическую триаду. Например, в одном из воплощений домен цистеиновой протеазы в TcdA содержит последовательность SEQ ID NO: 32, имеющая участок с мотивом DHC, локализованным при остатках 47, 113 и 158 в SEQ ID NO: 32, которые в свою очередь соответствуют D589, Н655 и С700 в TcdA дикого типа согласно нумерации в SEQ ID NO: 1. Последовательность SEQ ID NO: 32 идентична остаткам 543-809 в SEQ ID NO: 1, TcdA.
Домен цистеиновой протеазы в TcdB, как использовано в данном описании, в типичном случае может начинаться от остатка 544 и может заканчиваться на остатке 801, 767, 755 или 700 в TcdB дикого типа, например, в SEQ ID NO: 2. Любое минимальное положение остатка можно комбинировать с максимальным положением остатка между остатками 544 и 801 в TcdB дикого типа для определения последовательности для домена цистеиновой протеазы при условии включения участка с мотивом DHC, представляющим собой каталитическую триаду. Например, в одном из воплощений домен цистеиновой протеазы в TcdB содержит последовательность SEQ ID NO: 33, которая имеет участок с мотивом DHC, локализованным при остатках 44, 110 и 115 в SEQ ID NO: 33, которые в свою очередь соответствуют D587, Н653 и С698 в TcdB дикого типа согласно нумерации в SEQ ID NO: 2. Последовательность SEQ ID NO: 33 идентична остаткам 544-767 в SEQ ID NO: 2, TcdB. В другом воплощении домен цистеиновой протеазы в TcdB включает остатки 544-801 в SEQ ID NO: 2, TcdB.
Мутантный токсин
Описанная в настоящем изобретении иммуногенная композиция включает мутантный токсин С. difficile. Термин «мутантный», использованный в данном описании, относится к молекуле, которая демонстрирует структуру или последовательность, отличающуюся от соответствующей структуры или последовательности дикого типа, например тем, что имеет сшивки по сравнению с соответствующей структурой дикого типа, и/или тем, что имеет по меньшей мере одну мутацию по сравнению с соответствующей последовательностью дикого типа, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию. Термин «мутантный», использованный в данном описании, также включает молекулу, которая демонстрирует функциональное свойство (например, отсутствующую глюкозилтрансферазную активность и/или отсутствующую активность цистеиновой протеазы), которое отличает ее от соответствующей молекулы дикого типа.
Токсин С. difficile из любых штаммов дикого типа, описанных выше, можно использовать в качестве источника, из которого получают мутантный токсин С. difficile. Предпочтительно, источником, из которого получают мутантный токсин С. difficile, является С. difficile 630.
Мутация может включать замену, делецию, укорочение или модификацию аминокислотного остатка дикого типа, обычно локализованного в этом положении. Предпочтительно, мутация представляет собой неконсервативную аминокислотную замену. Согласно настоящему изобретению также предложены выделенные полинуклеотиды, которые включают нуклеиновокислотные последовательности, кодирующие любой из мутантных токсинов, описанных в данном изобретении.
Термин «неконсервативная» аминокислотная замена, использованный в данном описании, относится к замене аминокислоты из одного класса на аминокислоту из другого класса согласно приведенной ниже Таблице 3.

Примеры неконсервативных аминокислотных замен включают такую замену, когда остаток аспарагиновой кислоты (Asp, D) заменен на остаток аланина (Ala, А). Другие примеры включают замену остатка аспарагиновой кислоты (Asp, D) на остаток аспарагина (Asn, N); замену остатка аргинина (Arg, R), глутаминовой кислоты (Glu, Е), лизина (Lys, К) и/или гистидина (His, Н) на остаток аланина (Ala, А).
Консервативная замена относится к замене, производимой между аминокислотами одного и того же класса, например, в соответствии с Таблицей 3.
Мутантные токсины по изобретению могут быть получены методами, известными в данной области техники для внесения мутаций, такими как, например, сайтспецифический мутагенез, мутагенез с использованием мутагена (например, УФ излучения) и т.д. Предпочтительно используют сайт-специфический мутагенез. Альтернативно, молекулу нуклеиновой кислоты, имеющую целевую последовательность, можно синтезировать напрямую. Такие методы химического синтеза известны в данной области техники.
Мутантный токсин С. difficile по настоящему изобретению включает по меньшей мере одну мутацию в глюкозилтрансферазном домене по сравнению с соответствующим токсином С. difficile дикого типа. В одном из воплощений глюкозилтрансферазный домен включает по меньшей мере две мутации. Предпочтительно, мутация уменьшает или устраняет глюкозилтрансферазную ферментативную активность токсина по сравнению с глюкозилтрансферазной ферментативной активностью соответствующего токсина С. difficile дикого типа.
Типичные аминокислотные остатки в глюкозилтрансферазном домене TcdA, которые могут подвергаться мутации, включают по меньшей мере один остаток из следующих далее или любую их комбинацию: W101, D269, R272, D285, D287, Е460, R462, S541 и L542, по сравнению с TcdA из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 1. Другие типичные аминокислотные остатки, которые могут быть подвергнуты мутации, включают Е514, S517 и W519, по сравнению с TcdA из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 1.
Типичные мутации в глюкозилтрансферазном домене TcdA включают по меньшей мере одну мутацию из следующих далее или любую их комбинацию: W101A, D269A, R272A, D285A, D287A, Е460А, R462A, S541A и L542G, по сравнению с TcdA из С. difficile дикого типа. В предпочтительном воплощении глюкозилтрансферазный домен TcdA содержит мутацию L542G по сравнению с TcdA из С. difficile дикого типа. В другом предпочтительном воплощении глюкозилтрансферазный домен TcdA содержит мутации D285A и D287A по сравнению с TcdA из С. difficile дикого типа.
Типичные аминокислотные остатки в глюкозилтрансферазном домене TcdB, которые могут быть подвергнуты мутации, включают по меньшей мере один остаток из следующих далее или любую их комбинацию: W102, D270, R273, D286, D288, N384, D461, К463, W520 и L543, по сравнению с токсином В из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 2. Другие типичные аминокислотные остатки, которые могут быть подвергнуты мутации, включают Е515, S518 и W520, по сравнению с токсином В из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 2.
Типичные мутации в глюкозилтрансферазном домене TcdB включают по меньшей мере одну мутацию из следующих далее или любую их комбинацию: W102A, D270A, D270N, R273A, D286A, D288A, N384A, D461A, D461R, К463А, К463Е, W520A и L543A, по сравнению с TcdB из С. difficile дикого типа. В предпочтительном воплощении глюкозилтрансферазный домен TcdB содержит мутацию L543A по сравнению с TcdB из С. difficile дикого типа. В другом предпочтительном воплощении глюкозилтрансферазный домен TcdB содержит мутации D286A и D288A по сравнению с TcdB из С. difficile дикого типа.
Любую из мутаций, изложенных в данном описании выше, можно комбинировать с мутацией в домене цистеиновой протеазы. В настоящем изобретении мутантный токсин из С. difficile содержит по меньшей мере одну мутацию в домене цистеиновой протеазы относительно соответствующего токсина С. difficile дикого типа. Предпочтительно, мутация уменьшает или устраняет активность цистеиновой протеазы у токсина по сравнению с активностью цистеиновой протеазы у соответствующего токсина С. difficile дикого типа.
Типичные аминокислотные остатки в домене цистеиновой протеазы в TcdA, которые могут быть подвергнуты мутации, включают по меньшей мере один остаток из следующих далее или любую их комбинацию: S543, D589, Н655 и С700, по сравнению с TcdA из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 1. Типичные мутации в глюкозилтрансферазном домене TcdA включают по меньшей мере одну мутацию из следующих далее или любую их комбинацию: S543A, D589A, D589N, Н655А, С700А, по сравнению с TcdA из С. difficile дикого типа. В предпочтительном воплощении домен цистеиновой протеазы в TcdA содержит мутацию С700А по сравнению с TcdA из С. difficile дикого типа.
Типичные аминокислотные остатки в домене цистеиновой протеазы в TcdB, которые могут быть подвергнуты мутации, включают по меньшей мере один остаток из следующих далее или любую их комбинацию: G544, D587, Н653 и С698, по сравнению с TcdB из С. difficile дикого типа, в соответствии с нумерацией в SEQ ID NO: 2. Типичные мутации в глюкозилтрансферазном домене TcdB включают по меньшей мере одну мутацию из следующих далее или любую их комбинацию: G544A, D587A, D587N, Н653А, С698А, по сравнению с TcdB из С. difficile дикого типа. В предпочтительном воплощении домен цистеиновой протеазы в TcdB содержит мутацию С698А по сравнению с TcdB из С. difficile дикого типа. Дополнительные аминокислотные остатки в домене цистеиновой протеазы в TcdB, которые могут быть подвергнуты мутации, включают: К600 и/или R751, по сравнению с диким типом TcdB. Типичные мутации включают К600Е и/или R751E.
Соответственно, мутантный токсин из С. difficile по изобретению содержит глюкозилтрансферазный домен, имеющий мутацию, и домен цистеиновой протеазы, имеющий мутацию относительно соответствующего токсина С. difficile дикого типа. В одном из воплощений мутантный токсин содержит по меньшей мере одну мутацию в глюкозилтрансферазном домене и по меньшей мере одну мутацию в домене цистеиновой протеазы. В предпочтительном воплощении мутантный токсин А содержит по меньшей мере мутацию D285, D287 и С700. В предпочтительном воплощении мутантный токсин В содержит по меньшей мере мутацию D286, D288 и С698. Мутантные токсины могут содержать любую дополнительную мутацию, отдельно или в комбинации, как изложено в данном описании.
Типичный мутантный TcdA из С. difficile включает глюкозилтрансферазный домен, содержащий SEQ ID NO: 29, имеющую аминокислотную замену в положениях 285 и 287, и домен цистеиновой протеазы, содержащий SEQ ID NO: 32, имеющую аминокислотную замену в положении 158 относительно соответствующего токсина А из С. difficile дикого типа. Например, такой мутантный TcdA из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 4, где возможно не присутствует начальный метионин. В другом воплощении мутантный токсин А из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 84.
Другие типичные мутантные токсины А из С. difficile включают аминокислотную последовательность, приведенную в SEQ ID NO: 7, которая имеет мутацию D269A, R272A, D285A, D287A, Е460А, R462A и С700А по сравнению с SEQ ID NO: 1, где возможно не присутствует начальный метионин. В другом воплощении мутантный токсин А из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 83.
Другой типичный мутантный TcdA содержит SEQ ID NO: 34, при этом остаток в положениях 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 может представлять собой остаток любой аминокислоты.
В некоторых воплощениях для мутантного токсина из С. difficile наблюдается ослабление или устранение аутопротеолитического процессинга по сравнению с соответствующим токсином С. difficile дикого типа. Например, мутантный TcdA из С. difficile может содержать мутацию в одном из следующих далее остатков или любой их комбинации: S541, L542 и/или S543, по сравнению с соответствующим TcdA из С. difficile дикого типа. Предпочтительно, мутантный TcdA из С. difficile содержит по меньшей мере одну из следующих мутаций или любую их комбинацию: S541A, L542G и S543A, по сравнению с соответствующим TcdA из С. difficile дикого типа.
Другой типичный мутантный TcdA из С. difficile содержит мутацию S541A, L542, S543 и С700 по сравнению с соответствующим TcdA из С. difficile дикого типа.
Типичный мутантный токсин В из С. difficile включает глюкозилтрансферазный домен, содержащий SEQ ID NO: 31, имеющий аминокислотную замену в положениях 286 и 288, и домен цистеиновой протеазы, содержащий SEQ ID NO: 33, имеющий аминокислотную замену в положении 155 относительно соответствующего токсина В из С. difficile дикого типа. Например, такой мутантный TcdB из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 6, где возможно не присутствует начальный метионин. В другом воплощении мутантный токсин А из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 86.
Другие типичные мутантные TcdB из С. difficile включают аминокислотную последовательность, приведенную в SEQ ID NO: 8, которая имеет мутацию D270A, R273A, D286A, D288A, D461A, К463А и С698А по сравнению с SEQ ID NO: 2. В SEQ ID NO: 8 возможно не присутствует начальный метионин. В другом воплощении мутантный токсин А из С. difficile содержит аминокислотную последовательность, приведенную в SEQ ID NO: 85.
Другой типичный мутантный TcdB содержит SEQ ID NO: 35, причем остаток в положениях 101, 269, 272, 285, 287, 460, 462, 541, 542, 543, 589, 655 и 700 может представлять собой остаток любой аминокислоты.
В качестве другого примера, мутантный TcdB из С. difficile может содержать мутацию в положениях 543 и/или 544 по сравнению с соответствующим TcdB из С. difficile дикого типа. Предпочтительно, мутантный TcdB из С. difficile содержит мутацию L543 и/или G544 по сравнению с соответствующим TcdB из С. difficile дикого типа. Более предпочтительно, мутантный TcdB из С. difficile содержит мутацию L543G и/или G544A по сравнению с соответствующим TcdB из С. difficile дикого типа.
Другой типичный мутантный TcdB из С. difficile содержит мутацию L543G, G544A и С698 по сравнению с соответствующим TcdB из С. difficile дикого типа.
В одном из аспектов изобретение относится к выделенному полипептиду, имеющему мутацию по любому положению от аминокислотного остатка 1 до 1500 в соответствии с нумерацией в SEQ ID NO: 2, для определения типичного мутантного токсина В С. difficile. Например, в одном из воплощений выделенный полипептид содержит мутацию между аминокислотными остатками 830 и 990 в SEQ ID NO: 2. Типичные положения для мутаций включают положения 970 и 976 в соответствии с нумерацией в SEQ ID NO: 2. Предпочтительно, мутация между остатками 830 и 990 представляет собой замену. В одном из воплощений мутация представляет собой неконсервативную замену, где остаток аминокислот Asp (D) и/или Glu (Ε) заменен на остаток аминокислоты, которая не нейтрализуется при подкислении, такой как, например, лизин (К), аргинин (R) и гистидин (Н). Для определения мутантного токсина В из С. difficile типичные мутации включают: Е970К, E970R, Е970Н, Е976К, E976R, Е976Н в SEQ ID NO: 2.
В одном из воплощений выделенный полипептид содержит следующие замены D286A/D288A/C698A/E970K/E976K (SEQ ID NO: 184). Положения Е970 и Е976 являются консервативными в токсине В из всех рассмотренных штаммов С. difficile (см. Таблицу 2-а, SEQ ID NO: 110-133) за исключением штамма риботипа 078 и риботипа 126 (см. Таблицу 35 и Таблицу 37). В токсине В из штаммов риботипа 078 и риботипа 126 имеется глицин-970 (G970) вместо глутамата-970. Соответственно, в одном из воплощений выделенный полипептид содержит мутацию в G970 и Е976, такую как, например, G970K и Е976К. Мутантные токсины, изложенные выше и в данном описании, могут демонстрировать более низкую цитотоксичность по сравнению с соответствующим токсином дикого типа. См. примеры 8 и 15.
В другом аспекте изобретение относится к выделенному полипептиду, имеющему мутацию по любому положению от аминокислотного остатка 1 до 1500 в соответствии с нумерацией в SEQ ID NO: 1, для определения типичного мутантного токсина А из С. difficile. Например, в одном из воплощений выделенный полипептид содержит мутацию между аминокислотными остатками 832 и 992 в SEQ ID NO: 1. Типичные положения для мутаций включают положения 972 и 978 в соответствии с нумерацией в SEQ ID NO: 1. Предпочтительно, мутация между остатками 832 и 992 представляет собой замену. В одном из воплощений мутация представляет собой неконсервативную замену, где аминокислотный остаток Asp (D) и/или Glu (Ε) заменен на аминокислотный остаток, который не нейтрализуется при подкислении, такой как, например, лизин (К), аргинин (R) и гистидин (Н). Типичные мутации включают: D972K, D972R, D972H, D978K, D978R, D978H в SEQ ID NO: 1, для определения мутантного токсина А из С. difficile.
В одном из воплощений выделенный полипептид содержит следующие замены D285A/D287A/C700A/D972K/D978K (SEQ ID NO: 183). Остатки D972 и D978 являются консервативными в токсине А из всех проанализированных штаммов С. difficile (см. Таблицу 1-а, SEQ ID NO: 87-109). Мутантные токсины, изложенные выше и в данном описании, могут демонстрировать более низкую цитотоксичность по сравнению с соответствующим токсином дикого типа.
Далее описываются дополнительные типичные мутантные токсины. В одном из воплощений мутантный токсин TcdA включает (1) SEQ ID NO: 185; (2) полипептид из SEQ ID NO: 185, имеющий по меньшей мере 90%, 92%, 93%, 95%, 98%, 99% или 100% идентичности SEQ ID NO: 185; или (3) фрагмент по меньшей мере из 250, 280 или 300 аминокислот в SEQ ID NO: 185. В другом воплощении мутантный токсин TcdB включает (4) SEQ ID NO: 186; (5) полипептид из SEQ ID NO: 186, имеющий по меньшей мере 80%, 85%, 90%, 92%, 93%, 95%, 98%, 99% или 100% идентичности SEQ ID NO: 186; или (6) фрагмент по меньшей мере из 250, 280 или 300 аминокислот в SEQ ID NO: 186.
В одном из воплощений мутантный токсин TcdA состоит из менее чем 600, 675, 650, 625, 600, 575, 550, 525, 500, 475, 450, 425, 400, 375, 350, 325, 300, 275, 250 или 225 аминокислот. В одном из воплощений мутантный токсин состоит из менее чем 800, 775, 750, 725, 700, 675, 650, 625, 600, 575, 550 или 525 аминокислот. В одном из воплощений мутантный токсин содержит по меньшей мере 200, 225, 250, 270, 280, 300 или 310 аминокислот из SEQ ID NO: 185 или по меньшей мере 200, 225, 250, 270, 280, 300 или 310 аминокислот полипептида, имеющего по меньшей мере 80%, 85%, 90%, 92%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 185. В одном из воплощений мутантный токсин содержит по меньшей мере 400, 425, 450, 475, 500, 525, 550, 575, 600 или 610 аминокислот из SEQ ID NO: 186 или по меньшей мере 400, 425, 450, 475, 500, 525, 550, 575, 600 или 610 аминокислот полипептида, имеющего по меньшей мере 80%, 85%, 90%, 92%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 186.
В одном из воплощений мутантный токсин содержит слитый белок, который включает А) (1) SEQ ID NO: 185; (2) полипептид из SEQ ID NO: 185, имеющий по меньшей мере 90%, 92%, 93%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 185; или (3) фрагмент по меньшей мере из 250, 280 или 300 аминокислот из SEQ ID NO: 185, слитый с Б) (4) SEQ ID NO: 186; (5) полипептидом из SEQ ID NO: 186, имеющим по меньшей мере 80%, 85%, 90%, 92%, 93%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 186; или (6) фрагментом по меньшей мере из 250, 280 или 300 аминокислот из SEQ ID NO: 186. В следующем воплощении N-конец SEQ ID NO: 185 примыкает к С-концу SEQ ID NO: 186. В следующем воплощении N-конец SEQ ID NO: 185 примыкает к N-концу SEQ ID NO: 186. В следующем воплощении С-конец SEQ ID NO: 185 примыкает к С-концу SEQ ID NO: 186. Другие примеры мутантного токсина включают полипептид, имеющий любую из следующих далее аминокислотных последовательностей SEQ ID NO: 187, SEQ ID NO: 188, SEQ ID NO: 189, SEQ ID NO: 190, SEQ ID NO: 191, SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194 и SEQ ID NO: 195.
В другом воплощении мутантный токсин содержит слитый и/или гибридный полипептид, который содержит любую комбинацию аминокислотных последовательностей, выбранных из любой из следующих далее: SEQ ID NO: 223, SEQ ID NO: 224, SEQ ID NO: 225, SEQ ID NO: 226, SEQ ID NO: 227, SEQ ID NO: 228, SEQ ID NO: 229, SEQ ID NO: 230, SEQ ID NO: 231, SEQ ID NO: 232, SEQ ID NO: 233, SEQ ID NO: 234, SEQ ID NO: 235, SEQ ID NO: 236, SEQ ID NO: 237, SEQ ID NO: 238, SEQ ID NO: 239, SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242 и SEQ ID NO: 243. Например, в одном из воплощений мутантный токсин содержит аминокислотную последовательность, которая приведена в любой из следующих далее: SEQ ID NO: 254, SEQ ID NO: 270, SEQ ID NO: 271, SEQ ID NO: 272, SEQ ID NO: 273, SEQ ID NO: 274 или SEQ ID NO: 275.
Другой пример мутантного токсина включает фрагмент токсина дикого типа. Термин "фрагмент" мутантного токсина TcdA, использованный в данном описании, относится к пептидной последовательности, которая в целом имеет меньшее число следующих одна за другой аминокислот, чем соответствующая последовательность токсина TcdA из С. difficile дикого типа, например, последовательность, которая в целом содержит менее 2710 следующих одна за другой аминокислот. Фрагмент мутантного токсина TcdA также может содержать мутацию аминокислотного остатка, описанную в данной заявке. Термин "фрагмент" мутантного токсина TcdB, использованный в данном описании, относится к пептидной последовательности, которая в целом имеет меньшее число следующих одна за другой аминокислот, чем соответствующая последовательность токсина TcdB из С. difficile дикого типа, например, последовательность, которая в целом содержит менее 2366 следующих одна за другой аминокислот. Фрагмент мутантного токсина TcdB также может содержать мутацию аминокислотного остатка, описанную в данной заявке. Последовательности таких типичных мутантных токсинов включают SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33. В одном из воплощений мутантный токсин TcdA содержит по меньшей мере 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900 или 2000 следующих одна за другой аминокислот мутантного токсина А дикого типа, описанного в данном изобретении, например, SEQ ID NO: 1. В другом воплощении мутантный токсин содержит фрагмент генетически мутированного токсина А, описанного в данном изобретении. В одном из воплощений мутантный токсин TcdB содержит по меньшей мере 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900 или 2000 следующих одна за другой аминокислот мутантного токсина В дикого типа, описанного в данном изобретении, например, SEQ ID NO: 2. В другом воплощении мутантный токсин содержит фрагмент генетически мутированный токсина В, изложенного в данном описании. В одном из воплощений мутантный токсин содержит не более 3000, 2710, 2500, 2400, 2366, 2000, 1900, 1800, 1700, 1600, 1500, 1400, 1300, 1200, 1100, 1000, 900, 800, 700, 600, 500, 400 или 300 следующих одна за другой аминокислот. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В другом воплощении мутантный токсин слит по меньшей мере с одним другим пептидом или по меньшей мере с одним другим мутантным токсином таким образом, что это приводит к получению гибридной молекулы.
Еще один типичный фрагмент мутантного токсина TcdA включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 374 до SEQ ID NO: 421. Другой типичный фрагмент мутантного токсина TcdA включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 472 до SEQ ID NO: 519. Еще один типичный мутантный токсин TcdB включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 422 до SEQ ID NO: 471. Другой типичный фрагмент мутантного токсина TcdB включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 568 до SEQ ID NO: 615.
Далее описываются другие типичные мутантные токсины. В одном из воплощений мутантный токсин включает TcdA, который включает или состоит из по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 мутаций в аминокислотных остатках, выбранных из группы, состоящей из: W101, D287, Е514, D285, S517, W519 и С700, например, в соответствии с нумерацией в SEQ ID NO: 1. В дополнительных воплощениях мутанты TcdA содержат или состоят из по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 мутаций, выбранных из группы, состоящей из: замен W101A, D287A, E514Q, D285A, S517A, W519A и С700А и делеции W101.
Другой типичный мутантный токсин TcdA содержит аминокислотные замены W101, D287A и E514Q, например, в соответствии с нумерацией в SEQ ID NO: 1. В следующем воплощении, предложен белок TcdA, содержащий или состоящий из аминокислотных замен W101A, D287A, E514Q и W519A. Другое конкретное воплощение изобретения относится к белку TcdA, содержащему или состоящему из аминокислотных замен W101A, D287A, E514Q, W519A и С700А.
В другом воплощении мутантный токсин TcdA содержит мутации W101A, D287A, E514Q и D285A, например, в соответствии с нумерацией в SEQ ID NO: 1. В другом воплощении мутантный токсин TcdA содержит мутации W101A, D287A, E514QHS517A.
В другом воплощении в мутантный TcdA может быть добавлена дополнительная мутация, например мутация С700А. В дополнительных воплощениях в любое из описанных в данном изобретении воплощений мутантного токсина TcdA может быть добавлено до 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 дополнительных мутаций.
Другой типичный мутантный токсин TcdA включает аминокислотную последовательность, которая описана в SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209 или SEQ ID NO: 211. В другом воплощении мутантный токсин TcdA содержит аминокислотную последовательность SEQ ID NO: 210. В еще одном воплощении мутантный токсин TcdA содержит мутацию в положениях W101, D287, Е514, W519 и С700; при этом W101 заменен на любую аминокислоту, за исключением триптофана, D287 заменен на любую аминокислоту, кроме аспарагиновой кислоты, Е514 заменен на любую аминокислоту, кроме глутаминовой кислоты, W519 заменен на любую аминокислоту, кроме триптофана, и С700 заменен на любую аминокислоту, кроме цистеина, как приведено в SEQ ID NO: 212.
Дополнительные примеры мутантного токсина TcdA включают полипептид, имеющий аминокислотную последовательность, идентичную по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% первоначальной ссылочной последовательности (например, SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209, SEQ ID NO: 210, SEQ ID NO: 211 или SEQ ID NO: 212 либо SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85 или SEQ ID NO: 86).
В следующем воплощении мутантный токсин TcdA содержит мутации не более чем по 12 аминокислотным остаткам, не более чем по 11 аминокислотным остаткам, не более чем по 10 аминокислотным остаткам, не более чем по 9 аминокислотным остаткам, не более чем по 8 аминокислотным остаткам, не более чем по 7 аминокислотным остаткам, не более чем по 6 аминокислотным остаткам, не более чем по 5 аминокислотным остаткам, не более чем по 4 аминокислотным остаткам, не более чем по 3 аминокислотным остаткам, не более чем по 2 аминокислотным остаткам или по 1 аминокислотному остатку, например, в соответствии с SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209, SEQ ID NO: 210, SEQ ID NO: 211 и SEQ ID NO: 212 или SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85 или SEQ ID NO: 86.
Далее описываются другие типичные мутантные токсины. В одном из воплощений мутантный токсин представляет собой мутантный TcdB, который содержит по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 мутаций в аминокислотных остатках, выбранных из группы, состоящей из: W102, D288, Е515, D286, S518, W520 и С698, в соответствии с нумерацией в SEQ ID NO: 2. В другом воплощении мутантный токсин TcdB содержит по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 мутаций, выбранных из группы, состоящей из: замен W102A, D288A, E515Q, D286A, S518A, W520A и С698А и делеции W102. В одном из воплощений мутантный токсин TcdB содержит аминокислотную последовательность, приведенную в SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220 или SEQ ID NO: 221. В еще одном воплощении мутантный токсин TcdB содержит мутацию в положениях W102, D288, Е515, W520 и С698; при этом W102 заменен на любую аминокислоту за исключением триптофана, D288 заменен на любую аминокислоту кроме аспарагиновой кислоты, Е515 заменен на любую аминокислоту кроме глутаминовой кислоты, W520 заменен на любую аминокислоту кроме триптофана, и С698 заменен на любую аминокислоту кроме цистеина, как приведено в SEQ ID NO: 222.
Дополнительные примеры мутантного токсина TcdB включают полипептид, имеющий аминокислотную последовательность, идентичную по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% первоначальной ссылочной последовательности SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220, SEQ ID NO: 221 или SEQ ID NO: 222.
В следующем воплощении мутантный токсин TcdB содержит мутации не более чем по 12 аминокислотным остаткам, не более чем по 11 аминокислотным остаткам, не более чем по 10 аминокислотным остаткам, не более чем по 9 аминокислотным остаткам, не более чем по 8 аминокислотным остаткам, не более чем по 7 аминокислотным остаткам, не более чем по 6 аминокислотным остаткам, не более чем по 5 аминокислотным остаткам, не более чем по 4 аминокислотным остаткам, не более чем по 3 аминокислотным остаткам, не более чем по 2 аминокислотным остаткам или по 1 аминокислотному остатку, например, в соответствии с SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220, SEQ ID NO: 221 и/или SEQ ID NO: 222.
Еще один типичный мутантный токсин TcdA включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 276 до SEQ ID NO: 323. Еще один типичный мутантный токсин TcdB включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 324 до SEQ ID NO: 373.
Далее описываются другие типичные мутантные токсины. В одном из воплощений мутантный токсин TcdA содержит аминокислотную последовательность: (а) имеющую 50% идентичности или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98,5%, 99%, 99,5%, 99,8%, 99,9% или 100%) с SEQ ID NO: 224 или SEQ ID NO: 245; и/или (б) которая представляет собой фрагмент, состоящий по меньшей мере из «n» следующих одна за другой аминокислот в SEQ ID NO: 224 или SEQ ID NO: 245, при этом «n» равно 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 150, 250, 300, 400, 500, 540 или более), или фрагмент полипептида, имеющего 80% идентичности или больше с SEQ ID NO: 224 или SEQ ID NO: 245 и содержащего эпитоп из SEQ ID NO: 224 и/или SEQ ID NO: 245.
В одном из воплощений мутантный токсин TcdA содержит аминокислотную последовательность, приведенную в SEQ ID NO: 223, SEQ ID NO: 224, SEQ ID NO: 225, SEQ ID NO: 226, SEQ ID NO: 227, SEQ ID NO: 228, SEQ ID NO: 229, SEQ ID NO: 230, SEQ ID NO: 231, SEQ ID NO: 232, SEQ ID NO: 233, SEQ ID NO: 234, SEQ ID NO: 235 или SEQ ID NO: 236. Дополнительные типичные воплощения мутантного токсина TcdA содержат аминокислотную последовательность, приведенную в любом из следующих: SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253, SEQ ID NO: 254, SEQ ID NO: 255, SEQ ID NO: 256, SEQ ID NO: 257, SEQ ID NO: 258, SEQ ID NO: 259, SEQ ID NO: 260, SEQ ID NO: 261, SEQ ID NO: 262 и/или SEQ ID NO: 263.
В одном из воплощений мутантный токсин TcdB содержит аминокислотную последовательность (а), имеющую 80% идентичности или больше с SEQ ID NO: 238 или SEQ ID NO: 247; и/или (б) которая представляет собой фрагмент, состоящий по меньшей мере из 7 следующих одна за другой аминокислот в SEQ ID NO: 238 или SEQ ID NO: 247, или фрагмент полипептида, имеющего 80% идентичности или больше с SEQ ID NO: 238 или SEQ ID NO: 247 и содержащего эпитоп из SEQ ID NO: 238 и/или SEQ ID NO: 247.
В одном из воплощений мутантный токсин TcdB содержит аминокислотную последовательность: (а) имеющую 50% идентичности или больше (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 98,5%, 99%, 99,5%, 99,8%, 99,9% или более) с SEQ ID NO: 238 или SEQ ID NO: 247; и/или (б) которая представляет собой фрагмент, состоящий по меньшей мере из «n» следующих одна за другой аминокислот в SEQ ID NO: 238 или SEQ ID NO: 247, или фрагмент полипептида, имеющего 50% идентичности или больше с SEQ ID NO: 238 или SEQ ID NO: 247, при этом «n» равно 7 или больше (например, 8, 10, 12, 14, 16, 18, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 150, 250, 300, 400, 500, 540 или больше).
В одном из воплощений мутантный токсин TcdB содержит аминокислотную последовательность, приведенную в SEQ ID NO: 237, SEQ ID NO: 238, SEQ ID NO: 239, SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242 или SEQ ID NO: 243. Дополнительные типичные воплощения мутантного токсина TcdB содержат аминокислотную последовательность, приведенную в любом из следующих: SEQ ID NO: 246, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 249, SEQ ID NO: 264, SEQ ID NO: 265, SEQ ID NO: 266, SEQ ID NO: 267, SEQ ID NO: 268 и/или SEQ ID NO: 269.
Еще один типичный мутантный токсин TcdA включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 520 до SEQ ID NO: 567. Еще один типичный мутантный токсин TcdB включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 616 до SEQ ID NO: 663.
Далее описываются следующие дополнительные типичные мутантные токсины. В одном из воплощений мутантный токсин TcdA включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 664 до SEQ ID NO: 711. Еще один типичный мутантный токсин TcdB включает полипептид, имеющий любую из аминокислотных последовательностей от SEQ ID NO: 712 до SEQ ID NO: 761.
В некоторых случаях, как результат опосредованного клеткой хозяина процесса, полипептиды по изобретению могут включать начальный остаток метионина. В данной области техники известно, что в зависимости, например, от клетки хозяина, используемой в методике рекомбинантного получения, и/или условий ферментации или роста клетки хозяина N-концевой метионин, кодируемый кодоном инициации трансляции, может быть удален из полипептида после трансляции в клетках или N-концевой метионин может оставаться в выделенном полипептиде.
Соответственно, в одном из аспектов изобретение относится к выделенному полипептиду с аминокислотной последовательностью, приведенной в SEQ ID NO: 4, при этом начальный метионин (в положении 1) возможно не присутствует. В одном из воплощений начальный метионин в SEQ ID NO: 4 отсутствует. В одном из аспектов изобретение относится к выделенному полипептиду с аминокислотной последовательностью, приведенной в SEQ ID NO: 84, которая идентична SEQ ID NO: 4, но без начального метионина.
В другом аспекте выделенный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 6, при этом начальный метионин (в положении 1) возможно не присутствует. В одном из воплощений начальный метионин в SEQ ID NO: 6 отсутствует. В одном из аспектов изобретение относится к выделенному полипептиду с аминокислотной последовательностью, приведенной в SEQ ID NO: 86, которая идентична SEQ ID NO: 6, но без начального метионина.
В следующем аспекте выделенный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 7, при этом начальный метионин (в положении 1) возможно не присутствует. В одном из воплощений изобретение относится к выделенному полипептиду с аминокислотной последовательностью, приведенной в SEQ ID NO: 83, которая идентична SEQ ID NO: 7, но без начального метионина. В еще одном аспекте выделенный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 8, при этом начальный метионин (в положении 1) возможно не присутствует. В одном из воплощений выделенный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 85, которая идентична SEQ ID NO: 8, но без начального метионина.
В одном из аспектов изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 4, при этом начальный метионин (в положении 1) возможно не присутствует. В другом аспекте изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 6, при этом начальный метионин (в положении 1) возможно не присутствует. В следующем аспекте изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 7, при этом начальный метионин (в положении 1) возможно не присутствует. В еще одном аспекте изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 8, при этом начальный метионин (в положении 1) возможно не присутствует.
В другом аспекте изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 83. В одном из аспектов изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 84. В одном из аспектов изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 85. В другом аспекте изобретение относится к иммуногенной композиции, содержащей SEQ ID NO: 86.
Цитотоксичность
Кроме того, что они вызывают иммунный ответ у млекопитающего, иммуногенные композиции, описанные в данном изобретении, также имеют сниженную цитотоксичность по сравнению с соответствующим токсином С. difficile дикого типа. Предпочтительно, иммуногенные композиции являются безвредными и имеют цитотоксичность в диапазоне от минимальной (например, со снижением примерно 6-8 log10) цитотоксичности до ее полного отсутствия относительно цитотоксичности соответствующего токсина дикого типа при введении млекопитающим.
Использованный в данном описании термин "цитотоксичность" представляет собой установленный в данной области техники термин и относится к апоптотической гибели клеток и/или состоянию, при котором одна или более обычных биохимических или биологических функций клетки аномально нарушены по сравнению с идентичной клеткой, находящейся в одинаковых условиях, но в отсутствие цитотоксического агента. Количественно токсичность можно определить, например, в клетках или у млекопитающих, как количество агента, необходимое для индуцирования 50%-ной гибели клеток, (т.е. ЕС50 или ED50, соответственно), или другими методами, известными в данной области техники.
В данной области техники известны анализы для регистрации цитотоксичности, такие как анализы округления клеток (см., например, Kuehne et al. Nature. 2010, Oct 7, 467(7316): 711-3). Действие TcdA и TcdB вызывает округление клеток (например, утрату морфологической структуры) и гибель, и такое явление можно увидеть с использованием световой микроскопии. См., например, Фиг. 9.
Дополнительные типичные анализы цитотоксичности, известные в данной области техники, включают анализы глюкозилирования, относящиеся к анализам фосфовизуализации (phosphorimaging) Ras, меченой [14С]глюкозой (как описано в Busch et al., J. Biol. Chem., 1998, Jul 31, 273(31): 19566-72), и предпочтительно анализ цитотоксичности in vitro, описанный ниже в разделе Примеры, при этом ЕС50 может относиться к концентрации иммуногенной композиции, демонстрирующей по меньшей мере примерно 50% цитопатогенного эффекта (СРЕ) в клетке, предпочтительно человеческой диплоидной фибробластной клетке (например, клетке IMR90 (АТСС CCL-186™), по сравнению с идентичной клеткой, находящейся в идентичных условиях, но в отсутствие токсина. Анализ цитотоксичности in vitro также можно использовать для оценки концентрации композиции, которая ингибирует по меньшей мере примерно 50% индуцированного токсином из С. difficile дикого типа цитопатогенного эффекта (СРЕ) в клетке, предпочтительно человеческой диплоидной фибробластной клетке (например, клетке IMR90 (АТСС CCL-186™), по сравнению с идентичной клеткой, находящейся в идентичных условиях, но в отсутствие токсина. Дополнительные типичные анализы цитотоксичности включают анализы, описанные в Doern et al., J. Clin. Microbiol., 1992, Aug, 30(8): 2042-6. Цитотоксичность также может быть определена посредством измерения уровней АТФ в клетках, обработанных токсином. Например, можно использовать субстрат на основе люциферазы, такой как CELLTITERGLO® (Promega), который испускает люминесценцию, измеряемую в виде относительных световых единиц (RLU). В таком анализе жизнеспособность клеток может быть прямо пропорциональна количеству АТФ в клетках или величине RLU.
В одном из воплощений цитотоксичность иммуногенной композиции снижена по меньшей мере примерно в 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000 раз, 14000 раз, 15000 раз или больше по сравнению с соответствующим токсином С. difficile дикого типа. См., например, Таблицу 20.
В другом воплощении снижение цитотоксичности иммуногенной композиции составляет по меньшей мере примерно 2 log10, более предпочтительно примерно 3 log10 и наиболее предпочтительно примерно 4 log10 или больше относительно соответствующего токсина дикого типа, находящегося в идентичных условиях. Например, мутантный TcdB из С. difficile может иметь величину ЕС50 примерно 10-9 г/мл по результатам измерений в стандартном анализе цитопатического эффекта (СРЕ) по сравнению с типичным TcdB из С. difficile дикого типа, который может иметь величину ЕС50 по меньшей мере примерно 10-12 г/мл. См., например, Таблицы 7А, 7В, 8А и 8В в приведенном ниже разделе Примеры.
В еще одном воплощении цитотоксичность мутантного токсина из С. difficile имеет ЕС50 по меньшей мере примерно 50 мкг/мл, 100 мкг/мл, 200 мкг/мл, 300 мкг/мл, 400 мкг/мл, 500 мкг/мл, 600 мкг/мл, 700 мкг/мл, 800 мкг/мл, 900 мкг/мл, 1000 мкг/мл или больше по результатам измерений, например, с использованием анализа цитотоксичности in vitro, такого как анализ, изложенный в данном описании. Соответственно, в предпочтительном воплощении иммуногенные композиции и мутантные токсины являются биологически безвредными при введении млекопитающим.
Без связи с механизмом или теорией, ожидалось, что TcdA, имеющий мутацию в D285 и D287 по сравнению с TcdA дикого типа, и TcdB, имеющий мутацию в D286 и D288 по сравнению с TcdB дикого типа, будут дефектными по глюкозилтрансферазной активности и, следовательно, дефектными в отношении индуцирования цитопатического эффекта. Кроме того, ожидалось, что токсин, имеющий мутацию в мотиве DHC, будет дефектным в отношении аутокаталитического процессинга и, следовательно, не будет оказывать каких-либо цитотоксических эффектов.
Однако авторы изобретения неожиданным образом открыли, среди прочего, что типичный мутантный TcdA, имеющий SEQ ID NO: 4, и типичный мутантный TcdB, имеющий SEQ ID NO: 6, неожиданно демонстрировали цитотоксичность (хотя и существенно сниженную по сравнению с токсинами С. difficile 630 дикого типа), несмотря на выявление нарушенной глюкозилтрансферазной активности и нарушенной активности цистеиновой протеазы. Без связи с механизмом или теорией, полагают, что мутантные токсины оказывают влияние на цитотоксичность согласно новому механизму. Тем не менее, типичный мутантный TcdA, имеющий SEQ ID NO: 4, и типичный мутантный TcdB, имеющий SEQ ID NO: 6, неожиданно оказались иммуногенными. См. приведенные ниже примеры.
Образование сшивок
Хотя и существует возможность того, что образование химических сшивок в токсине дикого типа не приведет к инактивации токсина, авторы изобретения далее открыли, что образование химических сшивок с участием по меньшей мере одной аминокислоты мутантного токсина также снижает цитотоксичность мутантного токсина относительно идентичного мутантного токсина, у которого отсутствуют химические сшивки, и относительно соответствующего токсина дикого типа. Предпочтительно, чтобы мутантный токсин был подвергнут очистке до приведения в контакт с химическим сшивающим агентом.
Кроме того, несмотря на способность химических сшивающих агентов изменять подходящие эпитопы, авторы изобретения неожиданным образом открыли, что иммуногенные композиции, изготовленные на основе генетически модифицированного мутантного токсина из С. difficile, имеющего по меньшей мере одну вовлеченную в образование химических сшивок аминокислоту, вызывали образование многочисленных нейтрализующих антител или их связывающих фрагментов. Следовательно, после образования химических сшивок неожиданно оставались незатронутыми эпитопы, ассоциированные с молекулами нейтрализующих антител.
Образование сшивок (также называемое в данном описании как «химическая инактивация» или «инактивация») представляет собой процесс химического соединения двух или более молекул посредством ковалентной связи. Термины «реагенты для образования сшивок», «сшивающие агенты» и «кросслинкеры» относятся к молекулам, которые способны вступать во взаимодействие со специфическими функциональными группами (первичными аминами, сульфгидрилами, карбоксилами, карбонилами и т.д.) на пептидах, полипептидах и/или белках и/или химически присоединяться к ним. В одном из воплощений молекула может содержать два или более реакционно-способных конца, которые способны вступать во взаимодействие со специфическими функциональными группами (первичными аминами, сульфгидрилами, карбоксилами, карбонилами и т.д.) на пептидах, полипептидах и/или белках и/или химически присоединяться к ним. Предпочтительно, чтобы химический сшивающий агент был растворим в воде. В другом предпочтительном воплощении химический сшивающий агент представляет собой гетеробифункциональный сшивающий агент. В другом воплощении химический сшивающий агент не является бифункциональным сшивающим агентом. Химические сшивающие агенты известны в данной области техники.
В предпочтительном воплощении сшивающий агент представляет собой сшивающий агент нулевой длины. Сшивающий агент «нулевой длины» относится к сшивающему агенту, который будет опосредовать образование сшивки или напрямую сшивать между собой функциональные группы двух молекул. Например, при образовании сшивок между двумя полипептидами сшивающий агент нулевой длины будет приводить к образованию мостика или сшивки между карбоксильной группой из боковой цепи аминокислоты одного полипептида и аминогруппой другого полипептида без введения чего-либо извне. Сшивающие агенты нулевой длины могут катализировать, например, образование сложноэфирных связей между гидроксильными и карбоксильными группировками и/или образование амидных связей между карбоксильными и группировками и группировками первичных аминов.
Типичные подходящие химические сшивающие агенты включают формальдегид; формалин; ацетальдегид; пропиональдегид; водорастворимые карбодиимиды (RN=C=NR'), которые включают 1-этил-3-(3-диметиламинопропил)-карбодиимид (EDC), гидрохлорид 1-этил-3-(3-диметиламинопропил)-карбодиимида, мето-п-толуолсульфонат 1-циклогексил-3-(2-морфолинил-4-этил)карбодиимида (CMC), Ν,Ν'-дициклогексилкарбодиимид (DCC) и Ν,Ν'-диизопропилкарбодиимид (DIC) и их производные; и N-гидроксисукцинимид (NHS); фенилглиоксаль; и/или УДФ-диальдегид.
Предпочтительно, сшивающий агент представляет собой EDC. В том случае, когда полипептид мутантного токсина из С. difficile химически модифицирован с использованием EDC (например, путем приведения в контакт полипептида с EDC), в одном из воплощений полипептид содержит (а) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде. В одном из воплощений полипептид содержит (б) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде. В одном из воплощений полипептид содержит (в) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца полипептида. В одном из воплощений полипептид содержит (г) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина в полипептиде. В одном из воплощений полипептид содержит (д) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина во втором выделенном полипептиде. В одном из воплощений полипептид содержит (е) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина второго выделенного полипептида. В одном из воплощений полипептид содержит (ж) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца второго выделенного полипептида. В одном из воплощений полипептид содержит (з) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина во втором выделенном полипептиде. См., например, Фиг. 24 и Фиг. 25.
Термин «второй выделенный полипептид» относится к любому выделенному полипептиду, который присутствует в процессе взаимодействия с EDC. В одном из воплощений вторым выделенным полипептидом является полипептид мутантного токсина из С. difficile, имеющий последовательность, идентичную последовательности первого выделенного полипептида. В другом воплощении вторым выделенным полипептидом является полипептид мутантного токсина из С. difficile, имеющий последовательность, отличающуюся от последовательности первого выделенного полипептида.
В одном из воплощений полипептид содержит по меньшей мере две модификации, выбранные из модификаций (а)-(г). В типичном воплощении полипептид содержит (а) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде и (б) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде. В следующем воплощении полипептид содержит по меньшей мере три модификации, выбранные из модификаций (а)-(г). В еще одном воплощении полипептид содержит модификации (а), (б), (в) и (г).
Если в процессе химической модификации с использованием EDC присутствует более чем один мутантный полипептид, то в одном из воплощений полученная композиция содержит по меньшей мере одну из всех модификаций (а)-(з). В одном из воплощений композиция содержит по меньшей мере две модификации, выбранные из модификаций (а)-(з). В следующем воплощении композиция содержит по меньшей мере три модификации, выбранные из модификаций (а)-(з). В еще одном воплощении композиция содержит по меньшей мере четыре модификации, выбранные из модификаций (а)-(з). В другом воплощении композиция содержит по меньшей мере по одной из всех модификаций (а)-(з).
В типичном воплощении полученная композиция содержит (а) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде и (б) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде. В одном из воплощений композиция дополнительно содержит (в) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца полипептида и (г) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина в полипептиде.
В другом типичном воплощении полученная композиция содержит (д) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина во втором выделенном полипептиде; (е) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина во втором выделенном полипептиде; (ж) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и аминогруппой N-конца второго выделенного полипептида и (з) по меньшей мере одну сшивку между карбоксильной группой на С-конце полипептида и боковой цепью остатка лизина во втором выделенном полипептиде.
В другом типичном воплощении полученная композиция содержит (а) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде; (б) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде; (д) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина во втором выделенном полипептиде и (е) по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина во втором выделенном полипептиде.
В предпочтительном воплощении химический сшивающий агент включает формальдегид, более предпочтительно, агент, представляющий собой формальдегид в отсутствие лизина. В реакциях образования сшивок в качестве гасителя можно использовать глицин или другое соответствующее соединение с группой первичного амина. Соответственно, в другом предпочтительном воплощении химический агент включает формальдегид и применяется глицин.
В еще одном предпочтительном воплощении химический сшивающий агент включает EDC и NHS. Как известно в данной области техники, в протоколы реакций EDC-сочетания можно включать NHS. Тем не менее, авторы изобретения неожиданным образом открыли, что присутствие NHS может способствовать дальнейшему снижению цитотоксичности мутантного токсина из С. difficile по сравнению с соответствующим токсином дикого типа, по сравнению с генетически мутированным токсином и по сравнению с генетически мутированным токсином, который был подвергнут реакции образования химических сшивок с использованием EDC. См., например, пример 22. Соответственно, без связи с механизмом или теорией считается, что полипептид мутантного токсина, имеющий бета-аланиновую группировку, связанную с боковой цепью по меньшей мере одного остатка лизина в полипептиде (например, в результате взаимодействия полипептида мутантного токсина, EDC и NHS), может способствовать дальнейшему снижению цитотоксичности мутантного токсина по сравнению, например, с токсином из С. difficile (дикого типа или мутантным), у которого бета-аланиновая группировка отсутствует.
Использование EDC и/или NHS также может включать применение в качестве гасителя глицина или другого соответствующего соединения с группой первичного амина. В качестве гасителя можно использовать любое соединение, имеющее группу первичного амина, такое как, например метиловый эфир глицина и аланин. В предпочтительном воплощении соединение-гаситель представляет собой неполимерный гидрофильный первичный амин. Примеры неполимерного гидрофильного первичного амина включают, например, аминосахара, аминоспирты и аминополиолы. Конкретные примеры неполимерного гидрофильного первичного амина включают глицин, этаноламин, глюкамин, амин-функционализированный полиэтиленгликоль и олигомеры амин-функционализированного этиленгликоля.
В одном из аспектов изобретение относится к полипептиду мутантного токсина из С. difficile, в котором по меньшей мере одна боковая цепь аминокислоты химически модифицирована с использованием EDC и неполимерного гидрофильного первичного амина, предпочтительно глицина. Полученные глициновые аддукты (например, в результате обработки тройных мутантных токсинов с использованием EDC, NHS и гашения глицином) могут способствовать снижению цитотоксичности мутантного токсина по сравнению с соответствующим токсином дикого типа.
В одном из воплощений, когда полипептид мутантного токсина из С. difficile химически модифицирован EDC и глицином, полипептид содержит по меньшей мере одну модификацию, когда полипептид модифицирован с использованием EDC (например, по меньшей мере одну из любых модификаций (а)-(з), описанных выше), и по меньшей мере одну из следующих типичных модификаций: (и) глициновую группировку, связанную с карбоксильной группой на С-конце полипептида; (к) глициновую группировку, связанную с боковой цепью по меньшей мере одного остатка аспарагиновой кислоты в полипептиде; и (л) глициновую группировку, связанную с боковой цепью по меньшей мере одного остатка глутаминовой кислоты в полипептиде. См., например, Фиг. 24 и Фиг. 25.
В одном из воплощений по меньшей мере одна аминокислота мутантного TcdA из С. difficile вовлечена в образование химических сшивок и/или по меньшей мере одна аминокислота мутантного TcdB из С. difficile вовлечена в образование химических сшивок. Любой из мутантных токсинов, описанных в данном изобретении, может быть подвергнут реакции образования химических сшивок. В другом воплощении по меньшей мере одна аминокислота в SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 7 и/или SEQ ID NO: 8 вовлечена в образование химических сшивок. В одном из воплощений, по меньшей мере один аминокислотный остаток полипептида, имеющего аминокислотную последовательность, выбранную из любой последовательности от SEQ ID NO: 1 до SEQ ID NO: 761, вовлечен в образование химических сшивок. В другом воплощении, по меньшей мере один аминокислотный остаток полипептида, имеющего аминокислотную последовательность, выбранную из любой последовательности от SEQ ID NO: 183 до SEQ ID NO: 761, содержит модификацию, которая описана выше, например, любую из модификаций (а)-(л), как например, (а) по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде.
Например, по меньшей мере одна аминокислота может быть вовлечена в образование химических сшивок с использованием агента, который включает карбодиимид, такой как EDC. Карбодиимиды могут образовывать ковалентную связь между свободными карбоксильными группами (например, боковых цепей аспарагиновой кислоты и/или глутаминовой кислоты) и аминогруппами (например, в боковой цепи остатков лизина) с образованием стабильных амидных связей.
В качестве другого примера, по меньшей мере одна аминокислота может быть вовлечена в образование химических сшивок с использованием агента, который включает NHS. Сшивающие агенты на основе активированного сложного эфира NHS могут взаимодействовать с первичными аминами (например, на N-конце каждой полипептидной цепи и/или в боковой цепи остатков лизина) с получением амидной связи.
В другом воплощении по меньшей мере одна аминокислота может быть вовлечена в образование химических сшивок с использованием агента, который включает EDC и NHS. Например, в одном из воплощений изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, приведенную в SEQ ID NO: 4, где остаток метионина в положении 1 возможно не присутствует, при этом полипептид содержит по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC и NHS. В другом воплощении изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, приведенную в SEQ ID NO: 6, где остаток метионина в положении 1 возможно не присутствует, при этом полипептид содержит по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC и NHS. В еще одном воплощении изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, приведенную в SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 83, SEQ ID NO: 85, SEQ ID NO: 7 или SEQ ID NO: 8. Полипептид модифицируют путем приведения в контакт данного полипептида с EDC и NHS. См., например, Фиг. 24 и Фиг. 25.
Если в одном из воплощений полипептид мутантного токсина из С. difficile химически модифицирован посредством (например, в результате приведения в контакт с) EDC и NHS, то данный полипептид содержит по меньшей мере одну модификацию, когда полипептид модифицирован с использованием EDC (например, по меньшей мере одну из всех описанных выше модификаций (а)-(з)), и (I) бета-аланиновую группировку, связанную с боковой цепью по меньшей мере одного остатка лизина в полипептиде.
В другом аспекте изобретение относится к полипептиду мутантного токсина из С. difficile, при этом полипептид содержит по меньшей мере одну боковую цепь аминокислоты, химически модифицированную с использованием EDC, NHS и неполимерного гидрофильного первичного амина, предпочтительно глицина. В одном из воплощений полипептид содержит по меньшей мере одну модификацию, когда полипептид модифицирован с использованием EDC (например, по меньшей мере одну из всех описанных выше модификаций (а)-(з)), по меньшей мере одну модификацию, когда полипептид модифицирован глицином (например, по меньшей мере одну из всех описанных выше модификаций (и)-(л)), и (м) бета-аланиновую группировку, связанную с боковой цепью по меньшей мере одного остатка лизина в полипептиде. См., например, Фиг. 24 и Фиг. 25.
В одном из аспектов изобретение относится к полипептиду мутантного токсина из С. difficile, при этом боковая цепь по меньшей мере одного остатка лизина в полипептиде связана с бета-аланиновой группировкой. В одном из воплощений боковая цепь второго остатка лизина в полипептиде связана с боковой цепью остатка аспарагиновой кислоты и/или с боковой цепью остатка глутаминовой кислоты. «Второй» остаток лизина в полипептиде представляет собой остаток лизина в полипептиде, который не связан с бета-аланиновой группировкой. Боковая цепь аспарагиновой кислоты и/или боковая цепь глутаминовой кислоты, с которой связан второй остаток лизина, может представлять собой боковую цепь полипептида, использованную для образования внутримолекулярной сшивки, или боковую цепь второго полипептида, использованную для образования межмолекулярной сшивки. В другом воплощении боковая цепь по меньшей мере одного остатка аспарагиновой кислоты и/или боковая цепь по меньшей мере одного остатка глутаминовой кислоты в полипептиде связана с глициновой группировкой. Остаток аспарагиновой кислоты и/или остаток глутаминовой кислоты, который связан с глициновой группировкой, не является также связанным с остатком лизина.
В другом аспекте изобретение относится к мутантному токсину из С. difficile, при этом по меньшей мере одна боковая цепь аминокислоты токсина из С. difficile дикого типа химически модифицирована. В одном из воплощений по меньшей мере одна боковая цепь аминокислоты токсина А из С. difficile дикого типа и/или по меньшей мере одна боковая цепь аминокислоты токсина В из С. difficile дикого типа химически модифицированы с использованием EDC. Например, в одном из воплощений TcdA (SEQ ID NO: 1) и/или TcdB (SEQ ID NO: 2) химически модифицированы с использованием EDC. В другом воплощении токсин дикого типа химически модифицирован с использованием EDC и NHS. В одном из воплощений мутантный токсин включает химически модифицированный токсин А дикого типа, при этом токсин А дикого типа представляет собой любой токсин, описанный в Таблице 1-а. В другом воплощении мутантный токсин включает химически модифицированный токсин В дикого типа, при этом токсин В дикого типа представляет собой любой токсин, описанный в Таблице 2-а.
В качестве еще одного примера химически сшитого полипептида мутантного токсина из С. difficile рассматривается такой, в котором по меньшей мере одна аминокислота может быть вовлечена в образование химических сшивок с использованием агента, который включает формальдегид. Формальдегид может взаимодействовать с аминогруппой N-концевого аминокислотного остатка и боковыми цепями аргинина, цистеина, гистидина и лизина. Формальдегид и глицин могут образовывать аддукт шиффово основание, который может присоединяться к первичным N-концевым аминогруппам, остаткам аргинина и тирозина и в меньшей степени к остаткам аспарагина, глутамина, гистидина и триптофана.
Известно, что химический сшивающий агент снижает цитотоксичность токсина, если подвергнутый обработке токсин имеет более низкую токсичность (например, более низкую токсичность примерно на 100%, 99%, 95%, 90%, 80%, 75%, 60%, 50%, 25% или 10%), чем не подвергнутый обработке токсин, находящийся в идентичных условиях, по результатам измерений, например, с использованием анализа цитотоксичности in vitro или исследования токсичности на животных.
Предпочтительно, снижение цитотоксичности мутантного токсина из С. difficile под действием химического сшивающего агента составляет по меньшей мере примерно 2 log10, более предпочтительно примерно 3 log10 и наиболее предпочтительно примерно 4 log10 или больше относительно мутантного токсина, находящегося в идентичных условиях, но в отсутствие химического сшивающего агента. По сравнению с токсином дикого типа снижение цитотоксичности мутантного токсина под действием химического сшивающего агента предпочтительно составляет по меньшей мере примерно 5 log10, примерно 6 log10, примерно 7 log10, примерно 8 log10 или больше.
В другом предпочтительном воплощении химически инактивированный мутантный токсин из С. difficile демонстрирует значение ЕС50 больше или по меньшей мере примерно 50 мкг/мл, 100 мкг/мл, 200 мкг/мл, 300 мкг/мл, 400 мкг/мл, 500 мкг/мл, 600 мкг/мл, 700 мкг/мл, 800 мкг/мл, 900 мкг/мл, 1000 мкг/мл или более по результатам измерений, например, с использованием анализа цитотоксичности in vitro, такого как анализ, описанный в данном изобретении.
Реакционные условия для приведения в контакт мутантного токсина с химическим сшивающим агентом находятся в компетенции специалиста в данной области техники, и эти условия могут варьировать в зависимости от используемого агента. Однако авторы изобретения неожиданным образом открыли оптимальные реакционные условия для приведения в контакт полипептида мутантного токсина из С. difficile с химическим сшивающим агентом при сохранении функциональных эпитопов и снижении цитотоксичности мутантного токсина по сравнению с соответствующим токсином дикого типа.
Предпочтительно, реакционные условия выбирают с целью приведения в контакт мутантного токсина со сшивающим агентом, при этом мутантный токсин имеет минимальную концентрацию от примерно 0,5, 0,75, 1,0, 1,25, 1,5, 1,75, 2,0 мг/мл до максимальной, составляющей примерно 3,0, 2,5, 2,0, 1,5 или 1,25 мг/мл. Любое минимальное значение можно комбинировать с любым максимальным значением для определения диапазона подходящих для данной реакции концентраций мутантного токсина. Наиболее предпочтительно, для данной реакции мутантный токсин имеет концентрацию примерно 1,0-1,25 мг/мл.
В одном из воплощений агент, используемый в реакции, имеет минимальную концентрацию примерно 1 мМ, 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, 20 мМ, 30 мМ, 40 мМ или 50 мМ и максимальную концентрацию примерно 100 мМ, 90 мМ, 80 мМ, 70 мМ, 60 мМ или 50 мМ. Любое минимальное значение можно комбинировать с любым максимальным значением для определения диапазона подходящих для данной реакции концентраций химического агента.
В предпочтительном воплощении, где агент включает формальдегид, используемая концентрация предпочтительно представляет собой любую концентрацию от примерно 2 мМ до 80 мМ, наиболее предпочтительно примерно 40 мМ. В другом предпочтительном воплощении, где агент включает EDC, используемая концентрация предпочтительно представляет собой любую концентрацию от примерно 1,3 мМ до примерно 13 мМ, более предпочтительно от примерно 2 мМ до 3 мМ, наиболее предпочтительно примерно 2,6 мМ. В одном из воплощений концентрация EDC составляет не больше 5 г/л, 4 г/л, 3 г/л, 2,5 г/л, 2 г/л, 1,5 г/л, 1.0 г/л, 0,5 г/л из расчета на общий объем реакционной смеси, предпочтительно не больше 1 г/л, более предпочтительно не больше 0,5 г/л.
Типичные продолжительности реакции, в течение которых мутантный токсин взаимодействует с химическим сшивающим агентом, включают минимально примерно 0,5, 1, 2, 3, 4, 5, 6, 12, 24, 36, 48 или 60 часов и максимально примерно 14 суток, 12 суток, 10 суток, 7 суток, 5 суток, 3 суток, 2 суток, 1 сутки или 12, 11, 10, 9, 8, 7, 6, 5 часов, 4, 3, 2 часа или 1 час. Любое минимальное значение можно комбинировать с любым максимальным значением для определения диапазона подходящих продолжительностей реакции.
В предпочтительном воплощении стадия приведения в контакт мутантного токсина с химическим сшивающим агентом происходит в течение периода времени, достаточного для снижения цитотоксичности мутантного токсина из С. difficile до значения ЕС50, составляющего по меньшей мере примерно 1000 мкг/мл, в подходящей клетке человека, например, клетках IMR-90, в стандартном анализе цитотоксичности in vitro, по сравнению с идентичным мутантным токсином в отсутствие сшивающего агента. Более предпочтительно, эту стадию реакции осуществляют в течение периода времени, который по меньшей мере в два раза продолжительнее и наиболее предпочтительно по меньшей мере в три или больше раз продолжительнее периода времени, достаточного для снижения цитотоксичности мутантного токсина до значения ЕС50, составляющего по меньшей мере примерно 1000 мкг/мл, в подходящей клетке человека. В одном из воплощений продолжительность реакции не превышает примерно 168 часов (или 7 суток).
Например, в одном из воплощений, где агент включает формальдегид, мутантный токсин предпочтительно взаимодействует с агентом в течение примерно 12 часов, что, как было показано, представляет собой типичный период времени, которого было достаточно для снижения цитотоксичности мутантного токсина из С. difficile до значения ЕС50, составляющего по меньшей мере примерно 1000 мкг/мл, в подходящей клетке человека, например, клетках IMR-90, в стандартном анализе цитотоксичности in vitro, по сравнению с идентичным мутантным токсином в отсутствие сшивающего агента. В более предпочтительном воплощении реакцию осуществляют в течение примерно 48 часов, что по меньшей мере примерно в три раза продолжительнее периода времени, достаточного для данной реакции. В таком воплощении продолжительность реакции составляет предпочтительно не более чем примерно 72 часа.
В другом воплощении, где агент включает EDC, мутантный токсин предпочтительно взаимодействует с агентом в течение примерно 0,5 часа, более предпочтительно в течение по меньшей мере примерно 1 часа или наиболее предпочтительно в течение примерно 2 часов. В одном из воплощений мутантный токсин взаимодействует с EDC в течение примерно не более 5 часов, предпочтительно не более примерно 3 часов, более предпочтительно не более примерно 2 часов. В таком воплощении продолжительность реакции составляет предпочтительно не более чем примерно 6 часов.
Типичное значение рН, при котором мутантный токсин приводят в контакт с химическим сшивающим агентом, включает минимально примерно рН 5,5, 6,0, 6,5, 7,0 или 7,5 и максимально примерно рН 8,5, 8,0, 7,5, 7,0 или 6,5. Любое минимальное значение можно комбинировать с любым максимальным значением для определения диапазона подходящего рН. Предпочтительно, реакция протекает при рН 6,5-7,5, предпочтительно при рН 7,0.
Типичные температуры, при которых мутантный токсин приводят в контакт с химическим сшивающим агентом, включают минимальную температуру примерно 2°С, 4°С, 10°С, 20°С, 25°С или 37°С и максимальную температуру примерно 40°С, 37°С, 30°С, 27°С, 25°С или 20°С. Любое минимальное значение можно комбинировать с любым максимальным значением для определения диапазона подходящей температуры реакции. Предпочтительно, реакция протекает при примерно 20°С-30°С, наиболее предпочтительно при примерно 25°С.
Иммуногенные композиции, описанные выше, могут содержать один мутантный токсин из С. difficile (А или В). Таким образом, иммуногенные композиции могут быть помещены в отдельные флаконы (например, отдельный флакон для композиции, содержащей мутантный токсин А из С. difficile, и отдельный флакон для композиции, содержащей мутантный токсин В из С. difficile) в виде препарата или в наборе. Иммуногенные композиции могут быть предназначены для одновременного, последовательного или раздельного применения.
В другом воплощении иммуногенные композиции, описанные выше, могут содержать оба мутантных токсина из С. difficile (А и В). Мутантный токсин А из С. difficile и мутантный токсин В из С. difficile, описанные выше, могут быть объединены в любой комбинации для получения иммуногенной композиции. Соответственно, иммуногенные композиции могут быть объединены в одном флаконе (например, в одном флаконе, содержащем и композицию, включающую мутантный TcdA из С. difficile, и композицию, включающую мутантный TcdB из С. difficile). Предпочтительно, иммуногенные композиции содержат мутантный TcdA из С. difficile и мутантный TcdB из С. difficile.
Например, в одном из воплощений иммуногенная композиция включает SEQ ID NO: 4 и SEQ ID NO: 6, при этом по меньшей мере одна аминокислота каждой из SEQ ID NO: 4 и SEQ ID NO: 6 вовлечена в образование химических сшивок. В другом воплощении иммуногенная композиция включает мутантный токсин А из С. difficile, который содержит SEQ ID NO: 4 или SEQ ID NO: 7, и мутантный токсин В из С. difficile, который содержит SEQ ID NO: 6 или SEQ ID NO: 8, при этом по меньшей мере одна аминокислота каждого из мутантных токсинов С. difficile вовлечена в образование химических сшивок.
В другом воплощении иммуногенная композиция содержит любую последовательность, выбранную из SEQ ID NO: 4, SEQ ID NO: 84 и SEQ ID NO: 83, и любую последовательность, выбранную из SEQ ID NO: 6, SEQ ID NO: 86 и SEQ ID NO: 85. В другом воплощении иммуногенная композиция содержит SEQ ID NO: 84 и иммуногенную композицию, содержащую SEQ ID NO: 86. В другом воплощении иммуногенная композиция содержит SEQ ID NO: 83 и иммуногенную композицию, содержащую SEQ ID NO: 85. В другом воплощении иммуногенная композиция содержит SEQ ID NO: 84, SEQ ID NO: 83, SEQ ID NO: 86 и SEQ ID NO: 85.
В другом воплощении иммуногенная композиция содержит полипептид, имеющий любую последовательность, выбранную из последовательностей от SEQ ID NO: 1 до SEQ ID NO: 761, и второй полипептид, имеющий любую последовательность, выбранную из последовательностей от SEQ ID NO: 1 до SEQ ID NO: 761.
Очевидно, что любую из композиций по изобретению, например, иммуногенных композиций, содержащих мутантный токсин А и/или мутантный токсин В, можно комбинировать в разных соотношениях или количествах для оказания терапевтического эффекта. Например, мутантный TcdA из С. difficile и мутантный TcdB из С. difficile могут присутствовать в иммуногенной композиции в соотношении для А:В, изменяющемся в диапазоне от 0,1:10 до 10:0,1. В другом воплощении, например, мутантный TcdB из С. difficile и мутантный TcdA из С. difficile могут присутствовать в иммуногенной композиции в соотношении для В:А, изменяющемся в диапазоне от 0,1:10 до 10:0,1. В одном из предпочтительных воплощений это соотношение является таким, что общее количество мутантного TcdB, включенного в композицию, превышает общее количество мутантного TcdA.
В одном из аспектов иммуногенная композиция способна связываться с нейтрализующим антителом или его связывающим фрагментом. Предпочтительно, нейтрализующее антитело или его связывающий фрагмент представляет собой антитело или его связывающий фрагмент, описанные здесь ниже. В одном из типичных воплощений иммуногенная композиция способна связываться с антителом к токсину А или его связывающим фрагментом, при этом антитело к токсину А или его связывающий фрагмент содержит вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 36, и вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 37. Например, иммуногенная композиция может содержать мутантный TcdA из С. difficile, SEQ ID NO: 4 или SEQ ID NO: 7. В качестве другого примера, иммуногенная композиция может содержать SEQ ID NO: 84 или SEQ ID NO: 83.
В другом типичном воплощении иммуногенная композиция способна связываться с антителом к токсину В или его связывающим фрагментом, при этом антитело к токсину В или его связывающий фрагмент содержит вариабельную область легкой цепи от В8-26 и вариабельную область тяжелой цепи от В8-26. Например, иммуногенная композиция может содержать мутантный TcdB из С. difficile, SEQ ID NO: 6 или SEQ ID NO: 8. В качестве другого примера иммуногенная композиция может содержать SEQ ID NO: 86 или SEQ ID NO: 85.
Рекомбинантная клетка
В другом аспекте изобретение относится к рекомбинантной клетке или ее потомству. В одном из воплощений клетка или ее потомство содержит полинуклеотид, кодирующий мутантный TcdA из С. difficile и/или мутантный TcdB из С. difficile.
В другом воплощении рекомбинантная клетка или ее потомство содержит нуклеиновокислотную последовательность, которая кодирует полипептид, имеющий по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой из SEQ ID N0: 4, SEQ ID N0: 6, SEQ ID NO: 7 или SEQ ID NO: 8, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию. В одном из воплощений рекомбинантная клетка или ее потомство содержит нуклеиновокислотную последовательность, которая кодирует полипептид, имеющий по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой из последовательностей от SEQ ID NO: 1 до SEQ ID NO: 761.
В другом воплощении рекомбинантная клетка или ее потомство содержит нуклеиновокислотную последовательность, которая кодирует полипептид, имеющий по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой из SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 83 или SEQ ID NO: 85, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию.
В дополнительном воплощении рекомбинантная клетка или ее потомство содержит нуклеиновокислотную последовательность, имеющую по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% идентичности с любой последовательностью из SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 44, SEQ ID NO: 45, SEQ ID NO: 46 или SEQ ID NO: 47, при оптимальном выравнивании, например, с применением программ GAP или BESTFIT, использующих значения веса разрыва по умолчанию.
Рекомбинантная клетка может происходить из любой клетки, полезной для рекомбинантного получения полипептида по настоящему изобретению, например, из прокариота или эукариота. Предпочтительно, рекомбинантная клетка происходит из любой клетки, которая подходит для экспрессии гетерологических нуклеиновокислотных последовательностей размером более чем примерно 5000, 6000, предпочтительно примерно 7000 и более предпочтительно примерно 8000 нуклеотидов или более. Прокариотическая клетка-хозяин может представлять собой любую грамотрицательную или грамположительную бактерию. В типичных воплощениях в прокариотической клетке хозяина отсутствует эндогенный полинуклеотид, кодирующий токсин, и/или спора.
Грамотрицательные бактерии включают, но не ограничиваются этим, Campylobacter, Ε. coli, Flavobacteríum, Fusobacterium, Helicobacter, llyobacter, Neisseria, Pseudomonas, Salmonella и Ureaplasma. Например, рекомбинантная клетка может происходить из клетки Pseudomonas fluorescens, как описано в публикации патентной заявки США 2010013762, абзацах [0201]-[0230], которая включена в данное описание посредством ссылки.
Грамположительные бактерии включают, но не ограничиваются этим, Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus и Streptomyces. Предпочтительно, клетка происходит из клетки С. difficile.
Авторы изобретения идентифицировали штаммы С. difficile дикого типа, в которых отсутствует эндогенный полинуклеотид, кодирующий токсин из С. difficile. Штаммы, в которых отсутствуют гены эндогенных токсинов А и В, включают следующие штаммы, доступные через Американскую коллекцию типовых культур (АТСС) (Manassas, VA): С. difficile 1351 (АТСС 43593™), С. difficile 3232 (АТСС ВАА-1801™), С. difficile 7322 (АТСС 43601™), С. difficile 5036 (АТСС 43603™), С. difficile 4811 (АТСС 43602™) и С. difficile VPI 11186 (АТСС 700057™).
Соответственно, в одном из воплощений рекомбинантая клетка С. difficile происходит из штамма, описанного в данном изобретении. Предпочтительно, рекомбинантая клетка С. difficile или ее потомство происходят из группы, состоящей из С. difficile 1351, С. difficile 5036 и С. difficile VPI 11186. Более предпочтительно, рекомбинантая клетка С. difficile или ее потомство происходит из клетки С. difficile VPI 11186.
В предпочтительном воплощении ген спорообразования рекомбинантной клетки С. difficile или ее потомства является инактивированным. Споры могут быть инфекционными, высоко резистентными и способствовать сохранению С. difficile в аэробных условиях вне хозяина. Споры также могут вносить вклад в выживаемость С. difficile внутри хозяина при проведении антимикробной терапии. Соответственно, клетка С. difficile, у которой отсутствует ген спорообразования, полезна при получении безвредной иммуногенной композиции для введения млекопитающим. Помимо этого, применение таких клеток способствует безопасности в процессе производства, например, безопасность в отношении защиты предприятия, будущей продукции и персонала.
Примеры генов спорообразования для направленной инактивации включают, среди прочих, spo0A, spollE, σE, σG и σK. Предпочтительно, ген spo0A является инактивированным.
Способы инактивации гена спорообразования в С. difficile известны в данной области техники. Например, ген спорообразования может быть инактивирован посредством целенаправленной вставки селектируемого маркера, такого как, маркер устойчивости к антибиотику. См., например, Heap et al., J. Microbiol. Methods. 2010 Jan, 80(1): 49-55; Heap et al., J. Microbiol. Methods, 2007 Sept, 70(3): 452-464; и Underwood et al., J. Bacteriol. 2009 Dec, 191(23): 7296-305. Кроме этого см., например, заявку Minton и др. WO 2007/148091 под названием «Молекулы ДНК и способы» ("DNA Molecules and Methods"), включенную в данное описание посредством ссылки во всей своей полноте, на страницах 33-66 или соответствующую публикацию США US 20110124109 А1, абзацы [00137]-[0227].
Композиции, относящиеся к культивированию клетки С. difficile
Изобретение также относится к композициям и способам, применяемым при культивировании С. difficile и получении токсинов С. difficile. В одном из аспектов изобретение относится к культуральной среде, содержащей источник азота и клетку С. difficile.
Подходящие источники азота для культуральных сред включают: HY-SOY (Quest), AMI-SOY (Quest), NZ-SOY (Quest), NZ-SOY BL4 (Quest), NZ-SOY BL7 (Quest), SHEFTONE D (Sheffield), SE50M (DMV), SE50 (DMV), SE50 MK (DMV), соевый пептон (Gibco), BACTO-SOYTON (Difco), NUTRISOY 2207 (ADM), BAKES NUTRISOY (ADM), NUTRISOY FLOUR, соевую муку, бакто-дрожжевой экстракт (Difco), дрожжевой экстракт (Gibco), дрожжевой экстракт HY-YEST 412 (Quest), дрожжевой экстракт HY-YEST 441 (Quest), дрожжевой экстракт HY-YEST 444 (Quest), дрожжевой экстракт HY-YEST 455 (Quest), бакто-мальт-экстракт (Difco), кукурузный экстракт и PROFLO (Traders).
В одном из аспектов изобретение относится к культуральной среде, включающей гидролизат растительных белков и клетку С. difficile. Может быть приемлемым любой гидролизат растительных белков. Примерами подходящих гидролизатов растительных белков являются гидролизат семян хлопка, гидролизат гороха и гидролизат сои.
В предпочтительном воплощении гидролизатом растительных белков является гидролизат сои. Предпочтительно, гидролизат сои представляет собой SE50MK (Friesland-Campaigna). Другие примеры соевых продуктов, которые можно использовать в данном изобретении, и их источники, включают: Tekniscience: соевый пептон А1, соевый пептон А2, соевый пептон A3, растительный пептон Е1, растительный пептон ЕТ1 и пшеничный пептон Е1; Quest: HY-Soy, HY-Soy Τ, AMI-Soy, NZ-Soy, NZ-Soy BLA и NZ-Soy BL7; DMV: SE50M, SE70M, SE50MK, SE50MK-NK (Friesland-Campaigna), WGE80BT, WGE80M, CNE50M и SE70BT; Marcor: соевый пептон, тип AB, соевый пептон, тип АС, соевый пептон, тип SL, соевый пептон, тип II, и соевый пептон, тип F; Oxoid: пептон растительного происхождения и пептон растительного происхождения №1; Gibco: соевый пептон; и Difco: Bacsoytone.
Концентрации гидролизата растительных белков в культуральной среде могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация гидролизата растительных белков в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 30 г/л. Концентрация гидролизата растительных белков в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В другом аспекте изобретение относится к культуральной среде, включающей дрожжевой экстракт (например, в качестве источника азота) и клетку Clostridium difficile. Наиболее предпочтительно, дрожжевой экстракт представляет собой HY YEST 412 (Kerry Biosciences).
Концентрации дрожжевого экстракта в культуральной среде могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация дрожжевого экстракта в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 20 г/л. Концентрация дрожжевого экстракта в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
Авторы изобретения обнаружили, что рост С. difficile может поддерживаться в культуральной среде, включающей гидролизат растительных белков без дрожжевого экстракта, и в культуральной среде, которая включает дрожжевой экстракт без гидролизата растительных белков. Однако, как было замечено, при культивировании клеток и/или продуцировании токсина с применением культуральной среды, включающей как гидролизат растительных белков, так и дрожжевой экстракт, были отмечены более высокие выходы, чем при применении культуральной среды, включающей гидролизат растительных белков в отсутствие дрожжевого экстракта, и более высокие выходы, чем при применении культуральной среды, включающей дрожжевой экстракт в отсутствие гидролизата растительных белков.
Соответственно, авторы изобретения открыли, что комбинация гидролизата растительных белков и дрожжевого экстракта способствует поддержанию максимального роста С. difficile и/или продуцированию токсина. В одном из аспектов изобретение относится к культуральной среде, включающей гидролизат растительных белков, дрожжевой экстракт и клетку С. difficile. Гидролизатом растительных белков может быть любой подходящий гидролизат растительных белков, известный в данной области техники, как описано выше. Предпочтительно, гидролизатом является гидролизат сои. Более предпочтительно, гидролизат сои представляет собой SE50MK (Friesland-Campaigna). В предпочтительном воплощении дрожжевой экстракт представляет собой HY YEST 412.
В одном из воплощений среда не содержит источника углерода. Авторы изобретения обнаружили, что использование культуральной среды, включающей гидролизат сои/дрожжевой экстракт в отсутствие источника углерода, позволяет достичь значений 2-3 для OD600 и выходов по продукту токсину 10-15 мг/л.
Тем не менее, неожиданно было обнаружено, что культуральная среда, содержащая источник углерода, усиливает эффективность культивирования клеток С. difficile и продуцирования токсина по сравнению со средой в отсутствие источника углерода. Кроме того, авторы изобретения неожиданным образом обнаружили, что не происходило заметного ингибирования продуцирования мутантного токсина в результате культивирования клетки С. difficile в анаэробных условиях, включающих использование источника углерода в присутствии среды, содержащей гидролизат сои. Соответственно, в предпочтительном воплощении среда дополнительно содержит источник углерода. Более предпочтительно, в одном из воплощений культуральная среда включает источник углерода, причем данная среда дополнительно содержит рекомбинантную клетку С. difficile. Еще более предпочтительно, рекомбинантная клетка С. difficile содержит конститутивный промотор. В предпочтительном воплощении промотор представляет собой промотор фередоксина (fdx) Clostridium sporogenes. В другом предпочтительном воплощении рекомбинантная клетка С. difficile не содержит регулируемого хромосомального промотора, который может отрицательно регулироваться посредством, например, глюкозной репрессии. В наиболее предпочтительном воплощении клетка С. difficile представляет собой рекомбинантную клетку или ее потомство, как описано выше. В культуральной среде может быть использован любой источник углерода. Подходящие источники углерода включают глюкозу, декстрозу, маннит, фруктозу и/или маннозу. Предпочтительно, источником углерода в культуральной среде является глюкоза. В предпочтительном воплощении культуральная среда не содержит арабинозы, ксилозы, сахарозы, лактозы, мальтозы, глицерина, рамнозы и/или галактозы.
Концентрации источника углерода (например, глюкозы, маннита, фруктозы и/или маннозы) в культуральной среде могут изменяться, например, от минимального значения 1, 5, 10, 15, 20, 30, 40, 50 или 60 г/л до максимального значения 150, 100, 90, 80, 75, 70, 50, 40 или 30 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация источника углерода в культуральной среде составляет 5-70 г/л. В одном из воплощений концентрация источника углерода в культуральной среде составляет примерно 10 г/л. В другом воплощении концентрация источника углерода в культуральной среде составляет более 10 г/л. В другом воплощении концентрация источника углерода (например, глюкозы) в культуральной среде составляет 45-75 г/л, предпочтительно 55-65 г/л, более предпочтительно 55-65 г/л. В другом воплощении концентрация источника углерода в культуральной среде составляет примерно 60 г/л. Концентрация источника углерода в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В одном из воплощений культуральная среда дополнительно включает производное хлорамфеникола. Типичные производные хлорамфеникола включают любое из тиамфеникола, флорфеникола, хлорамфеникола сукцината и фторамфеникола (fluoramphenicol). Предпочтительно, культуральная среда включает тиамфеникол. Без связи с механизмом или теорией считается, что производное хлорамфеникола помогает предотвратить потерю плазмиды в процессе ферментации и повысить эффективность продуцирования клеток и/или токсина по сравнению с культуральной средой в отсутствие производного хлорамфеникола.
Концентрации производного хлорамфеникола в культуральной среде могут изменяться, например, от минимального значения 5, 10, 15, 20 или 30 мг/л до максимального значения 100, 75, 50 или 40 мг/л. В другом воплощении концентрация производного хлорамфеникола в культуральной среде может изменяться, например, от минимального значения 0,5 г/л, 1 г/л, 1,5 г/л, 2 г/л, 2,5 г/л, 3 г/л, 3,5 г/л, 4 г/л, 4,5 г/л или 5 г/л до максимального значения 10 г/л, 9,5 г/л, 9 г/л, 8,5 г/л, 8 г/л, 7,5 г/л, 7 г/л, 6,5 г/л, 6 г/л, 5,5 г/л, 5 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация производного хлорамфеникола в культуральной среде составляет 5-20 мг/л, наиболее предпочтительно примерно 15 мг/л. В другом предпочтительном воплощении концентрация производного хлорамфеникола в культуральной среде составляет от 1 г/л до 10 г/л, предпочтительно от 1 г/л до 5 г/л, наиболее предпочтительно примерно 3 г/л. Концентрация производного хлорамфеникола в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В одном из воплощений культуральная среда дополнительно содержит защищающее клетки вещество, такое как полиэтиленгликоль, поливиниловый спирт или полиол плуроник. В одном из воплощений культуральная среда содержит полиэтиленгликоль, такой как полиэтиленгликоль 2000 (PPG 2000).
Концентрации защищающего клетки вещества, такого как полиэтиленгликоль, в культуральной среде могут изменяться, например, от минимального значения 0,01, 0,05, 0,10, 0,15, 0,20, 0,25, 0,30, 0,35, 0,40, 0,45, 0,50, 0,55, 0,60, 0,65, 0,70, 0,75, 0,80, 0,85, 0,90, 0,95 или 1 мл/л до максимального значения 2, 1, 0,90, 0,80, 0,70, 0,60, 0,50, 0,40, 0,30, 0,20 мл/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация защищающего клетки вещества, такого как полиэтиленгликоль, в культуральной среде составляет от 0,01 до 0,50 мл/л, наиболее предпочтительно примерно 0,25 мл/л. Концентрация защищающего клетки вещества в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В предпочтительном воплощении культуральная среда содержит гидролизат растительных белков, дрожжевой экстракт и источник углерода, при этом гидролизат растительных белков представляет собой гидролизат сои, дрожжевой экстракт представляет собой HY YEST 412 (Kerry Biosciences), а источник углерода представляет собой глюкозу, маннит, фруктозу и/или маннозу. В наиболее предпочтительном воплощении культуральная среда содержит гидролизат сои SE50MK, дрожжевой экстракт HY YEST 412, глюкозу и тиамфеникол, при рН 7.
В одном из предпочтительных воплощений культуральная среда содержит гидролизат сои SE50MK в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкозу в концентрации 10 г/л и тиамфеникол в концентрации 15 мг/л, при рН 7. В одном из предпочтительных воплощений культуральная среда содержит гидролизат сои SE50MK в концентрации примерно 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации примерно 20 г/л, глюкозу примерно 10 г/л и тиамфеникол в концентрации примерно 15 мг/л, при рН 7. Культуральная среда также может содержать PPG 2000 в концентрации примерно 0,25 мл/л. В другом воплощении культуральная среда также может содержать декстрозу.
В другом предпочтительном воплощении культуральная среда содержит гидролизат сои SE50MK в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкозу в концентрации 60 г/л и тиамфеникол в концентрации 15 мг/л, при рН 7. В другом предпочтительном воплощении культуральная среда содержит гидролизат сои SE50MK в концентрации примерно 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации примерно 20 г/л, глюкозу в концентрации примерно 60 г/л и тиамфеникол в концентрации примерно 15 мг/л, при рН 7. Культуральная среда также может содержать PPG 2000 в концентрации примерно 0,25 мл/л. В другом воплощении культуральная среда также может содержать декстрозу.
В одном из воплощений культуральная среда дополнительно содержит себя полипропиленгликоль 2000 (PPG 2000). Культуральная среда может содержать от примерно 0,05 мл PPG 2000 до примерно 1 мл PPG 2000 на один литр среды, предпочтительно культуральная среда содержит примерно 0,25 мл PPG 2000/л среды.
Культуральная среда, описанная в данном изобретении, может быть приемлемой для культивирования любой клетки С. difficile. В одном из воплощений клетка не является генетически модифицированной. В другом воплощении клетка является генетически модифицированной, такой как рекомбинантная клетка или ее потомство, как описано выше. В одном из воплощений в клетке С. difficile отсутствует эндогенный полинуклеотид, кодирующий токсин. В предпочтительном воплощении клетка С. difficile происходит из VPI 11186.
В другом воплощении клетка содержит конститутивный промотор. В предпочтительном воплощении промотор представляет собой промотор фередоксина (fdx) Clostridium sporogenes. В другом предпочтительном воплощении клетка не содержит регулируемого хромосомального промотора, который может отрицательно регулироваться посредством, например, глюкозной репрессии. В наиболее предпочтительном воплощении клетка С. difficile представляет собой рекомбинантную клетку или ее потомство, как описано выше.
Авторы изобретения также открыли, что среда для моноклональных антител поддерживает рост С. difficile. Соответственно, в одном из аспектов изобретение относится к культуральной среде, которая включает среду для моноклональных антител. В одном из воплощений среда представляет собой среду SFM4MAb™ (Thermo Scientific). Для этой среды было продемонстрировано достижение величин OD600 примерно 10 и выхода по продукту токсину примерно 40 мг/л.
В одном из воплощений рН культуральной среды может изменяться, например, от минимального значения 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 или 7,0 до максимального значения 8,0, 7,9, 7,8, 7,7, 7,6, 7,5, 7,4, 7,3, 7,2 или 7,1. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении рН культуральной среды составляет от 6,0 до 8,0, более предпочтительно от 6,5 до 7,5. Наиболее предпочтительно рН составляет 7,0.
Подходящие титрующие вещества для поддержания рН известны в данной области техники, в том числе, например, NH4OH, Nа2СО3 и NaOH. Предпочтительно, титрующим веществом для поддержания рН является NaOH.
Культуральная среда может включать фосфатсодержащие ингредиенты, такие как Nа2НРО4, КН2РО4. Однако, в предпочтительном воплощении культуральная среда не включает фосфатсодержащего ингредиента.
Способы, относящиеся к культивированию клетки С. difficile
В другом аспекте изобретение относится к способу культивирования С. difficile. Способ включает культивирование клетки С. difficile в среде, которая описана выше.
В одном из воплощений выращивание С. difficile в соответствии со способами по изобретению протекает по меньшей мере за две фазы: выращивание посевного материала культуры и ферментация. Сначала выращивают посевную культуру путем инокуляции исходной культурой, например, из рабочего банка клеток. Посевной материал используют либо для инокуляции второй посевной культуры, либо для инокуляции относительно большой ферментируемой культуры. Как очевидно в данной области техники, число используемых посевных культур может зависеть, например, от размера и объема на стадии ферментации.
Соответственно, в одном из аспектов изобретение относится к способу культивирования С. difficile. Способ включает культивирование клетки С. difficile в первой культуральной среде в условиях, способствующих росту клетки; инокулирование второй культуральной среды всей указанной первой средой или ее частью после указанного первого культивирования; культивирование указанной инокулированной второй среды в условиях, способствующих росту клетки. Способ также может включать выделение токсина С. difficile из указанной второй среды. В одном из воплощений С. difficile выращивают в первой культуральной среде, обозначаемой как посевная культура. В одном из воплощений посевная культура включает культуральную среду, описанную выше, и инокулят из исходной культуры, которая была выращена в данной среде.
Фазу (или фазы) выращивания посевного материала обычно выполняют для количественного масштабирования микроорганизма из сохраняемой культуры с тем, чтобы его можно было использовать в качестве инокулята для фазы ферментации. Фаза выращивания посевного материала также может быть выполнена с целью обеспечения обновления микроорганизмов, находящихся в состоянии относительного покоя в сохраняемых культурах, и для их роста в активно растущих культурах.
Объем и количество жизнеспособных клеток, используемых для инокуляции ферментируемой культуры, можно регулировать с большей точностью, если они взяты из активно растущей культуры (например, посевной культуры), нежели из сохраняемой культуры.
Кроме того, для количественного масштабирования С. difficile с целью инокуляции среды для ферментации можно использовать больше одной фазы (например, две или три фазы) выращивания посевного материала. Альтернативно, при желании, рост С. difficile в фазе ферментации можно осуществлять непосредственно из сохраняемой культуры путем проведения прямой инокуляции.
Первая культуральная среда включает культуральную среду, описанную выше. Например, в одном из воплощений первая культуральная среда содержит гидролизат растительных белков и клетку С. difficile. Предпочтительно, гидролизатом растительных белков является гидролизат сои, наиболее предпочтительно гидролизат сои SE50MK. В другом воплощении первая культуральная среда содержит дрожжевой экстракт и клетку С. difficile. Предпочтительно, дрожжевой экстракт представляет собой HY YEST 412. В следующем воплощении первая культуральная среда содержит гидролизат растительных белков и дрожжевой экстракт. В еще одном воплощении первая культуральная среда дополнительно содержит источник углерода. Предпочтительно, источником углерода является глюкоза. В одном из воплощений культуральная среда содержит рекомбинантную клетку С. difficile, когда источник углерода включен, при этом рекомбинантная клетка содержит конститутивный промотор. В другом воплощении первая культуральная среда дополнительно содержит производное хлорамфеникола. Предпочтительно, культуральная среда содержит тиамфеникол.
Концентрации гидролизата растительных белков в первой культуральной среде (например, среде с посевной культурой) могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация гидролизата растительных белков в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 30 г/л. Концентрация гидролизата растительных белков в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
Концентрации дрожжевого экстракта в первой культуральной среде (например, среде с посевной культурой) могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация дрожжевого экстракта в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 20 г/л. Концентрация дрожжевого экстракта в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
Концентрации источника углерода в первой культуральной среде (например, среде с посевной культурой) могут изменяться, например, от минимального значения 1, 5, 10, 15 или 20 г/л до максимального значения 100, 75, 50, 40 или 30 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация источника углерода в культуральной среде составляет 5-20 г/л, наиболее предпочтительно примерно 10 г/л.
В одном из воплощений культуральная среда дополнительно включает производное хлорамфеникола, выбранное из группы, состоящей из тиамфеникола, флорфеникола, хлорамфеникола сукцината и фторамфеникола. Предпочтительно, культуральная среда включает тиамфеникол. Концентрации производного хлорамфеникола в первой культуральной среде (например, среде с посевной культурой) могут изменяться, например, от минимального значения 5, 10, 15, 20 или 30 мг/л до максимального значения 100, 75, 50 или 40 мг/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация производного хлорамфеникола в культуральной среде составляет 5-20 мг/л, наиболее предпочтительно примерно 15 мг/л.
В одном из воплощений рН первой культуральной среды может изменяться, например, от минимального значения 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 или 7,0 до максимального значения 8,0, 7,9, 7,8, 7,7, 7,6, 7,5, 7,4, 7,3, 7,2 или 7,1. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении рН первой культуральной среды составляет от 6,0 до 8,0, более предпочтительно от 6,5 до 7,5. Наиболее предпочтительно, рН составляет 7,0.
В предпочтительном воплощении первая культуральная среда содержит гидролизат сои, дрожжевой экстракт HY YEST 412, глюкозу, тиамфеникол, при рН 7. Более предпочтительно, культуральная среда содержит гидролизат сои SE50MK в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкозу в концентрации 10 г/л, тиамфеникол в концентрации 15 мг/л, при рН 7.
Чтобы начать фазу ферментации, часть посевной культуры или всю посевную культуру, содержащую С. difficile, можно использовать для инокуляции ферментируемой культуральной среды. Соответствующая концентрация посевной культуры для использования с целью инокуляции ферментируемой среды может быть определена специалистами в данной области техники и может изменяться, например, в диапазоне 0,1-10%. В конкретных примерах могут быть использованы концентрации 0,5, 1, 5, 5,5, 6, 6,25, 6,5, 7, 8, 9, 10%.
Ферментацию можно использовать для получения максимального количества клеток в анаэробных условиях крупномасштабного производства. В одном из воплощений С. difficile выращивают в качестве ферментируемой культуры. В одном из воплощений ферментируемую культуру инокулировали посевной культурой, которая была выращена в данной среде.
Вторая культуральная среда включает культуральную среду, описанную выше. Например, в одном из воплощений вторая культуральная среда содержит гидролизат растительных белков и клетку С. difficile. Предпочтительно, гидролизатом растительных белков является гидролизат сои, наиболее предпочтительно гидролизат сои SE50MK. В другом воплощении вторая культуральная среда содержит дрожжевой экстракт и клетку С. difficile. Предпочтительно, дрожжевой экстракт представляет собой HY YEST 412. В следующем воплощении вторая культуральная среда содержит гидролизат растительных белков и дрожжевой экстракт. В еще одном воплощении вторая культуральная среда дополнительно содержит источник углерода. Предпочтительно, источником углерода является глюкоза. В одном из воплощений культуральная среда содержит рекомбинантную клетку С. difficile, когда источник углерода включен, при этом рекомбинантная клетка содержит конститутивный промотор. В другом воплощении вторая культуральная среда дополнительно содержит производное хлорамфеникола. Предпочтительно, культуральная среда содержит тиамфеникол.
Концентрации гидролизата растительных белков во второй культуральной среде могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация гидролизата растительных белков в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 30 г/л. Концентрация гидролизата растительных белков в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
Концентрации дрожжевого экстракта во второй культуральной среде могут изменяться, например, от минимального значения 5, 10, 20, 30, 40 или 50 г/л до максимального значения 200, 150, 100 или 75 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация дрожжевого экстракта в культуральной среде составляет 10-50 г/л, наиболее предпочтительно примерно 20 г/л.
Концентрации источника углерода во второй культуральной среде могут изменяться, например, от минимального значения 10, 20, 30, 40, 50 или 60 г/л до максимального значения 150, 100, 90, 80 или 70 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация источника углерода в культуральной среде составляет 50-70 г/л, наиболее предпочтительно примерно 60 г/л. В одном из воплощений концентрация источника углерода во второй культуральной среде оказывается больше концентрации источника углерода в первой культуральной среде. Концентрация источника углерода в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В одном из воплощений культуральная среда дополнительно содержит производное хлорамфеникола, выбранное из группы, состоящей из тиамфеникола, флорфеникола, хлорамфеникола сукцината и фторамфеникола. Предпочтительно, культуральная среда содержит тиамфеникол. Концентрации производного хлорамфеникола во второй культуральной среде могут изменяться, например, от минимального значения 5, 10, 15, 20 или 30 мг/л до максимального значения 100, 75, 50 или 40 мг/л. В другом воплощении концентрация производного хлорамфеникола в культуральной среде может изменяться, например от минимального значения 0,5 г/л, 1 г/л, 1,5 г/л, 2 г/л, 2,5 г/л, 3 г/л, 3,5 г/л, 4 г/л, 4,5 г/л или 5 г/л до максимального значения 10 г/л, 9,5 г/л, 9 г/л, 8,5 г/л, 8 г/л, 7,5 г/л, 7 г/л, 6,5 г/л, 6 г/л, 5,5 г/л, 5 г/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация производного хлорамфеникола в культуральной среде составляет 5-20 мг/л, наиболее предпочтительно примерно 15 мг/л. В другом предпочтительном воплощении концентрация производного хлорамфеникола в культуральной среде составляет от 1 г/л до 10 г/л, предпочтительно от 1 г/л до 5 г/л, наиболее предпочтительно примерно 3 г/л. Концентрация производного хлорамфеникола в культуральной среде, описанной в данной изобретении, может быть рассчитана на общий объем культуральной среды.
В одном из воплощений культуральная среда дополнительно содержит защищающее клетки вещество, такое как полиэтиленгликоль, поливиниловый спирт или полиол плуроник. В одном из воплощений культуральная среда содержит полиэтиленгликоль, такой как полиэтиленгликоль 2000 (PPG 2000).
Концентрации защищающего клетки вещества, такого как полиэтиленгликоль, в культуральной среде могут изменяться, например, от минимального значения 0,01, 0,05, 0,10, 0,15, 0,20, 0,25, 0,30, 0,35, 0,40, 0,45, 0,50, 0,55, 0,60, 0,65, 0,70, 0,75, 0,80, 0,85, 0,90, 0,95 или 1 мл/л до максимального значения 2, 1, 0,90, 0,80, 0,70, 0,60, 0,50, 0,40, 0,30, 0,20 мл/л. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении концентрация защищающего клетки вещества, такого как полиэтиленгликоль, в культуральной среде составляет от 0,01 мл/л до 0,50, наиболее предпочтительно примерно 0,25 мл/л. Концентрация защищающего клетки вещества в культуральной среде, описанной в данном изобретении, может быть рассчитана на общий объем культуральной среды.
В одном из воплощений рН второй культуральной среды может изменяться, например, от минимального значения 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 или 7,0 до максимального значения 8,0, 7,9, 7,8, 7,7, 7,6, 7,5, 7,4, 7,3, 7,2 или 7,1. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В предпочтительном воплощении рН второй культуральной среды составляет от 6,0 до 8,0, более предпочтительно от 6,5 до 7,5. Наиболее предпочтительно, рН составляет 7,0.
В предпочтительном воплощении вторая культуральная среда содержит гидролизат сои, дрожжевой экстракт HY YEST 412, глюкозу, тиамфеникол, при рН 7. Более предпочтительно, культуральная среда содержит гидролизат сои в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкозу в концентрации 60 г/л, тиамфеникол в концентрации 15 мг/л, при рН 7.
В одном из воплощений культивирование осуществляют в анаэробных условиях. В одном из воплощений стадии культивирования способов по изобретению (и внесение посевного материала, и ферментацию) осуществляют в анаэробных условиях, хотя для любой из этих фаз также можно использовать аэробные условия.
Методики анаэробного культивирования таких бактерий, как С. difficile, известны в данной области техники, и могут использовать, например, газообразный азот или смесь газообразных азота и водорода. Этот газ можно барботировать через среду (например, путем разбрызгивания) в процессе ферментации или пропускать через зону над жидкостью в культуральной камере (например, через свободное пространство в камере над жидкостью).
Культивирование клетки С. difficile может быть проведено в анаэробной камере при температуре приблизительно 30±1°С, 31±1°С, 32±1°С, 33±1°С, 34±1°С, 35±1°С, 36±1°С, 37±1°С, 38±1°С или 39±1°С, предпочтительно при примерно 37±1°С. Культивирование может быть проведено в течение периодов времени, изменяющихся, например, от минимального значения 1, 2, 3, 4, 7, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23 или 24 часа до максимального значения 9 суток, 8 суток, 7 суток, 6, суток, 5 суток, 4 суток, 3 суток, 48 часов, 36 часов или 24 часа. Любое минимальное значение можно комбинировать с любым максимальным значением для определения подходящего диапазона. В предпочтительном воплощении культивирование этой клетки протекает в течение 11-48 часов, наиболее предпочтительно в течение примерно 24 часов. В другом воплощении культивирование этой клетки протекает в течение 5-25 часов, 10-20 часов, предпочтительно в течение примерно 15 часов. Мониторинг роста можно осуществлять, измеряя оптическую плотность (OD) среды.
В одном из воплощений способ дополнительно включает выделение токсинов С. difficile из указанной среды. Токсины С. difficile можно выделить и/или очистить из культур, используя известные методы очистки белков. Затем очищенные токсины могут быть, например, инактивированы с использованием химического метода инактивации.
В альтернативном аспекте изобретение относится к способу культивирования С. difficile. Способ включает культивирование клетки С. difficile в среде для моноклональных антител.
В другом аспекте изобретение относится к способу продуцирования токсина С. difficile. Способ включает культивирование бактерии С. difficile в среде, которая описана выше, в условиях, способствующих продуцированию данного токсина. Способ дополнительно включает выделение токсина С. difficile из среды.
В одном из воплощений способ включает культивирование клетки С. difficile в первой среде в условиях, способствующих росту С. difficile; инокулирование второй среды всей указанной первой средой или ее частью после указанного культивирования; культивирование указанной инокулированной второй среды в условиях, способствующих росту С. difficile и продуцированию токсина; и выделение токсина С. difficile из указанной второй среды. В одном из воплощений культивирование первой или второй сред, включающих С. difficile, осуществляют в анаэробных условиях.
В одном из воплощений клетку С. difficile культивируют в системе непрерывного культивирования. В другом воплощении изобретение относится к способу культивирования С. difficile в перфузионной культуре. Неожиданным образом, авторы изобретения обнаружили, что рекомбинантную клетку С. difficile, экспрессирующую мутантный токсин, можно с успехом культивировать в анаэробной непрерывной культуре и в анаэробной перфузионной культуре. Преимущество непрерывной системы и/или перфузионной системы заключается в том, что порции свежей среды можно добавлять непрерывно. К тому же, побочные продукты, образующиеся при получении токсина, могут быть удалены в процессе производства при сохранении жизнеспособности клеток в системе.
Система для непрерывного культивирования может включать обеспечение клеток свежей порцией среды при одновременном удалении отработанной среды и клеток из биореактора. Непрерывное культивирование может включать перфузионное культивирование, при котором выводимая жидкость содержит культуральную среду, которая по существу не содержит клеток или содержит клетки в концентрации, значительно ниже концентрации в биореакторе. При перфузионном культивировании клетки могут быть сохранены, например, посредством фильтрации, ультразвуковой фильтрации, центрифугирования или седиментации.
В одном из воплощений отработанные среды извлекают и фильтруют, что предотвращает удаление клеток из биореактора. Фильтр может представлять собой фильтр поперечного потока (cross-flow) и/или фильтр тангенциального потока. В одном из воплощений указанная система фильтрации содержит фильтр с полыми волокнами. В другом воплощении удаление клеток из биореактора предотвращают посредством стадии центрифугирования. В другом воплощении удаление клеток из биореактора предотвращают посредством стадии ультразвуковой фильтрации. В другом воплощении удаление клеток из биореактора предотвращают посредством системы седиментации. В другом воплощении указанная система фильтрации содержит составленную из плоских пластин кассету.
В еще одном воплощении система для перфузии содержит фильтр с полыми волокнами, которые удерживают клетки, но не желаемый продукт. Клетки возвращают в биореактор, а отработанную среду, содержащую желаемый продукт, пропускают через фильтр с желаемым отсечением по молекулярной массе. На таком фильтре желаемый продукт будет задерживаться. Не задержанные на фильтре отходы могут быть удалены или использованы повторно.
Способ получения мутантного токсина С. difficile
В одном из аспектов изобретение относится к способу получения мутантного токсина С. difficile. В одном из воплощений способ включает культивирование любой рекомбинантной клетки или ее потомства, описанных выше, в условиях, подходящих для экспрессии полипептида. Способ дополнительно включает стадию выделения токсина из среды.
В другом воплощении способ включает культивирование рекомбинантной клетки или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С. difficile, где указанная клетка содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, и где мутантный токсин содержит глюкозилтрансферазный домен, имеющий по меньшей мере одну мутацию, и домен цистеиновой протеазы, имеющий по меньшей мере одну мутацию по сравнению с соответствующим токсином Clostridium difficile дикого типа. В одном из воплощений в клетке отсутствует эндогенный полинуклеотид, кодирующий токсин.
В следующем воплощении способ включает культивирование рекомбинантной клетки С. difficile или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С. difficile, при этом указанная клетка содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, и в этой клетке отсутствует эндогенный полинуклеотид, кодирующий токсин С. difficile.
В другом аспекте изобретение относится к способу получения мутантного токсина из С. difficile. Способ включает стадии: (а) приведения клетки С. difficile в контакт с рекомбинантной клеткой Escherichia coli, при этом у клетки С. difficile отсутствует эндогенный полинуклеотид, кодирующий токсин из С. difficile, и клетка Е. coli содержит полинуклеотид, кодирующий мутантный токсин из С. difficile; (б) культивирования клетки С. difficile и клетки £. coli в условиях, подходящих для переноса полинуклеотида из клетки Е. coli в клетку С. difficile; (в) отбора клетки С. difficile, содержащей полинуклеотид, кодирующий мутантный токсин из С. difficile; (г) культивирования клетки С. difficile со стадии (в) в условиях, подходящих для экспрессии полинуклеотида; и (д) выделения мутантного токсина С. difficile.
В способе по изобретению рекомбинантная клетка Е. coli содержит гетерологический полинуклеотид, кодирующий мутантный токсин из С. difficile, изложенный в данном описании. Полинуклеотид может представлять собой ДНК или РНК. В одном из типичных воплощений полинуклеотид, кодирующий мутантный токсин из С. difficile, является кодон-оптимизированным для использования кодонов в Е. coli. Методы получения кодон-оптимизированного полинуклеотида известны в данной области техники.
В одном из воплощений полинуклеотид содержит нуклеиновокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% последовательности полинуклеотида, кодирующего мутантный TcdA из С. difficile, который описан выше. В одном из воплощений полинуклеотид содержит нуклеиновокислотную
последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% последовательности полинуклеотида, кодирующего полипептид, имеющий любую последовательность, выбранную из последовательностей от SEQ ID NO: 1 до SEQ ID NO: 761. Типичный полинуклеотид, кодирующий мутантный токсин А из С. difficile, содержит SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 44 и SEQ ID NO: 45.
В другом воплощении полинуклеотид содержит нуклеиновокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% последовательности полинуклеотида, кодирующего мутантный TcdB из С. difficile, который описан выше. Типичный полинуклеотид, кодирующий мутантный токсин В из С. difficile, содержит SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 46 и SEQ ID NO: 47. В другом воплощении полинуклеотид кодирует SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85 или SEQ ID NO: 86.
В одном из воплощений клетка Ε. coli, включающая гетерологический полинуклеотид, представляет собой клетку Е. coli, которая является стабильным хозяином гетерологического полинуклеотида, кодирующего мутантный токсин из С. difficile. Типичные клетки Е. coli включают клетку, выбранную из группы, состоящей из компетентных клеток Е. coli MAX Efficiency® Stbl2™ (Invitrogen, Carlsbad, CA), химически компетентных Ε. coli Stbl3™ в формате One Shot® (Invitrogen, Carlsbad, CA), компетентных клеток Ε. coli ElectroMAX™ Stbl4™ (Invitrogen) и Ε. coli CA434. В предпочтительном воплощении клонирующая клетка-хозяин Е. coli не представляет собой DH5α. Более предпочтительно, клонирующая клетка-хозяин Е. coli представляет собой компетентную клетку Е. coli MAX Efficiency® Stbl2™.
Способ по изобретению дополнительно включает стадию культивирования клетки С. difficile и клетки Е. coli в условиях, подходящих для переноса полинуклеотида из клетки Е. coli в клетку С. difficile с образованием в результате рекомбинантной клетки С. difficile. В предпочтительном воплощении условия культивирования являются приемлемыми для переноса полинуклеотида из клетки Е. coli (донорной клетки) в клетку С. difficile (реципиентную клетку) с получением в результате генетически стабильного наследования.
Наиболее предпочтительно, чтобы условия культивирования подходили для конъюгирования бактерий и были известны в данной области техники. «Конъюгирование» относится к конкретному процессу переноса полинуклеотида, при котором однонаправленный перенос полинуклеотида (например, из бактериальной плазмиды) осуществляется от одной бактериальной клетки (т.е. «донора») к другой (т.е. «реципиенту»). Процесс конъюгирования включает приведение в контакт донорной клетки и реципиентной клетки. Предпочтительно, донорная клетка Е. coli представляет собой клетку Е. coli СА434.
Типичные подходящие условия (конъюгирования) для переноса полинуклеотида из клетки Е. coli в клетку С. difficile включают выращивание жидких культур С. difficile в бульоне с сердечно-мозговым экстрактом (BHI; Oxoid) или бульоне Шедлера для анаэробов (SAB; Oxoid). В другом воплощении твердые культуры С. difficile можно выращивать на свежем кровяном агаре (FBA) или BHI-агаре. Предпочтительно, С. difficile выращивают при 37°С в анаэробных условиях (например, 80% N2, 10% СО2 и 10% Н2 (об./об.)). В одном из воплощений подходящие условия включают выращивание Е. coli в аэробных условиях в бульоне Лурия-Бертани (LB) или на LB-агаре при 37°С. Типичные условия, подходящие для конъюгативного переноса в С. difficile, включают выращивание Е. coli в анаэробных условиях на FBA. Как известно в данной области техники, в жидкие и твердые среды могут быть включены антибиотики. Примеры таких антибиотиков включают циклосерин (250 мкг/мл), цефокситин (8 мкг/мл), хлорамфеникол (12,5 мкг/мл), тиамфеникол (15 мкг/мл) и эритромицин (5 мкг/мл).
Кроме того, способ по изобретению включает стадию отбора полученной рекомбинантной клетки С. difficile, которая содержит полинуклеотид, кодирующий мутантный токсин из С. difficile. В типичном воплощении рекомбинантная клетка С. difficile является реципиентом кодирующего мутантный токсин С. difficile полинуклеотида из рекомбинантной клетки Е. coli, перенос которого осуществляется посредством конъюгирования.
Способ по изобретению включает стадию культивирования рекомбинантной клетки или ее потомства в условиях, подходящих для экспрессии полинуклеотида, кодирующего мутантный токсин С. difficile, в результате чего осуществляется продуцирование мутантного токсина С. difficile. Подходящие условия для экспрессии рекомбинантной клеткой данного полинуклеотида включают условия культивирования, подходящие для выращивания клетки С. difficile, которые известны в данной области техники. Например, подходящие условия могут включать культивирование трансформантов С. difficile в бульоне с сердечно-мозговым экстрактом (BHI; Oxoid) или бульоне Шедлера для анаэробов (SAB; Oxoid). В другом воплощении твердые культуры С. difficile можно выращивать на FBA- или BHI-arape. Предпочтительно, С. difficile выращивают при 37°С в анаэробных условиях (например, 80% N2, 10% СO2 и 10% Н2 (об./об.)).
В одном из воплощений способ по изобретению включает стадию выделения полученного мутантного токсина С. difficile. Способы выделения белка С. difficile известны в данной области техники.
В другом воплощении способ включает стадию очистки полученного мутантного токсина С. difficile. Способы очистки полипептида, такие как хроматография, известны в данной области техники.
В типичном воплощении способ дополнительно включает стадию приведения в контакт выделенного мутантного токсина Clostridium difficile с химическим сшивающим агентом, описанным выше. Предпочтительно, агент включает формальдегид, этил-3-(3-диметиламинопропил)карбодиимид или комбинацию EDC и NHS. Типичные реакционные условия описаны выше и в приведенном ниже разделе Примеры.
В другом аспекте изобретение относится к иммуногенной композиции, содержащей мутантный токсин из С. difficile, изложенный в данном описании, полученный любым способом, предпочтительно любым из способов, описанных выше.
Антитела
Неожиданно оказалось, что иммуногенные композиции по изобретению, описанные выше, вызывали образование новых антител in vivo, и это свидетельствует о том, что нативная структура соответствующего токсина С. difficile дикого типа в иммуногенных композициях сохранена (например, сохранен антигенный эпитоп) и что иммуногенные композиции содержат эпитоп. Антитела, вырабатываемые на токсин из одного штамма С. difficile, могут быть способны связываться с соответствующим токсином, продуцируемым другим штаммом С. difficile. То есть, антитела и их связывающие фрагменты могут быть «перекрестно-реактивными», что относится к способности взаимодействовать с похожими антигенными сайтами на токсинах, получаемых из многочисленных штаммов С. difficile. Перекрестная реактивность также включает способность антитела взаимодействовать или связываться с антигеном, который не стимулирует его образование, т.е. реакцию между антигеном и антителом, которое вырабатывалось на другой, но похожий антиген.
В одном из аспектов авторы изобретения неожиданным образом обнаружили моноклональные антитела, обладающие нейтрализующим действием в отношении токсинов С. difficile, и способы их получения. Антитела по изобретению могут нейтрализовать цитотоксичность токсина С. difficile in vitro, ингибировать связывание токсина С. difficile с клетками млекопитающих и/или могут нейтрализовать энтеротоксичность токсина С. difficile in vivo. Настоящее изобретение также относится к выделенным полинуклеотидам, которые включают нуклеиновокислотные последовательности, кодирующие любое из перечисленного выше. Помимо этого, настоящее изобретение относится к применению любой из перечисленных выше композиций для лечения, предупреждения, снижения риска, ослабления тяжести, уменьшения числа случаев и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данную композицию не вводят, а также к способам изготовления указанных композиций.
Кроме того, авторы изобретения открыли, что комбинация по меньшей мере двух нейтрализующих моноклональных антител неожиданно может демонстрировать синергетический эффект в отношении нейтрализации TcdA или TcdB. Антитела к токсину или их связывающие фрагменты могут быть полезны в подавлении инфекции С. difficile.
«Антитело» представляет собой белок, содержащий по меньшей мере одну или две вариабельные области тяжелой (Н) цепи (в данном описании имеющие сокращение VH) и по меньшей мере одну или две вариабельные области легкой (L) цепи (в данном описании имеющие сокращение VL). Кроме того, VH- и VL-области можно подразделить на участки гипервариабельности, обозначаемые термином «определяющие комплементарность участки» («CDR»), чередующиеся с областями, которые являются более консервативными и обозначаются термином «каркасные области» (FR). Протяженность каркасной области и CDR точно определена (см. Kabat Ε. Α., et al. Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication №91-3242, 1991 и Chothia C. et al., J. Mol. Biol., 196: 901-917, 1987). Термин «антитело» включает интактные иммуноглобулины типов IgA, IgG, IgE, IgD, IgM (а также их подтипов), где легкие цепи иммуноглобулина могут быть цепями типов каппа или лямбда.
Молекулы антител могут быть полноразмерными (например, антитело lgG1 или lgG4). Антитела могут быть различных изотипов, включая: IgG (например, lgG1, lgG2, lgG3, lgG4), IgM, lgA1, lgA2, IgD или IgE. В одном из предпочтительных воплощений антитело представляет собой антитело IgG-изотипа, например, lgG1. В другом предпочтительном воплощении антитело представляет собой антитело IgE.
В другом воплощении молекула антитела включает «антигенсвязывающий фрагмент» или «связывающий фрагмент», как использовано в данном описании, что относится к части антитела, которая специфически связывается с токсином из С. difficile (например, токсином А). Связывающим фрагментом является, например, молекула, в которой одна или более иммуноглобулиновых цепей не являются полноразмерными, но которая специфически связывается сданным токсином.
Примеры связывающих частей, охватываемых термином «связывающий фрагмент» антитела, включают (1) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL (константная область легкой цепи) и СН1 (константная область 1 тяжелой цепи); (2) F(аb’)2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (3) Fd-фрагмент, состоящий из доменов VH и СН1; (4) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (5) фрагмент dAb (однодоменное антитело) (Ward et al., Nature, 341: 544-546, 1989), который состоит из домена VH; и (6) выделенный определяющий комплементарность участок (CDR), имеющий каркас, достаточный для осуществления специфического связывания, например, антигенсвязывающую часть вариабельной области.
Связывающий фрагмент вариабельной области легкой цепи и связывающий фрагмент вариабельной области тяжелой цепи, например, два домена Fv-фрагмента, VL и VH, могут быть соединены друг с другом методами рекомбинантного синтеза посредством синтетического линкера, что позволяет им находиться в составе одной белковой цепи, в которой VL- и VH-области объединяются в пару с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science, 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA, 85: 5879-5883). Такие одноцепочечные антитела также охватываются термином «связывающий фрагмент» антитела. Такие части антитела получают, используя методы, известные в данной области техники, и скрининг данных частей на предмет их применимости осуществляют аналогично тому, как если бы они представляли собой интактные антитела.
Как использовано в данном описании, антитело, которое «специфически связывается» с или является «специфичным» к конкретному полипептиду или эпитопу на конкретном полипептиде, представляет собой антитело, которое связывается с этим конкретным полипептидом или эпитопом на конкретном полипептиде без существенного связывания с любым другим полипептидом или эпитопом данного полипептида. Например, при ссылке на биологическую молекулу (например, белок, нуклеиновую кислоту, антитело и т.д.), которая «специфически связывается» с мишенью, понимают, что данная биологическая молекула связывается со своей молекулой-мишенью и не связывается в значительной степени с другими молекулами в гетерогенной популяции молекул, которые включают данную мишень, по результатам измерений в указанных условиях (например, условиях иммуноанализа в случае антитела). Реакция связывания между антителом и его мишенью является определяющим признаком присутствия данной мишени в гетерогенной популяции молекул. Например, термин «специфическое связывание» или «специфически связывается» относится к способности антитела или его связывающего фрагмента связываться с токсином С. difficile дикого типа и/или мутантным токсином С. difficile с аффинностью, по меньшей мере в два раза превышающей его аффинность в отношении неспецифического антигена.
В типичном воплощении антитело представляет собой химерное антитело. Химерное антитело может быть получено методами рекомбинантной ДНК, известными в данной области техники. Например, ген, кодирующий константную область Fc молекулы мышиного (или другого вида) моноклонального антитела может быть отщеплен ферментами рестрикции для удаления области, кодирующей Fc антитела мыши, и заменен на эквивалентную часть гена, кодирующего константную область Fc антитела человека. Химерное антитело может быть создано такими методами рекомбинантной ДНК, при которых ДНК, кодирующая вариабельные области мышиного антитела, может быть лигирована с ДНК, кодирующей константные области человеческого антитела.
В другом типичном воплощении антитело или его связывающий фрагмент гуманизируют методами, известными в данной области техники. Например, после получения мышиных антител, CDR данного антитела может быть заменено по меньшей мере частью CDR антитела человека. Гуманизированные антитела также можно создавать, заменяя последовательности вариабельной области Fv мышиного антитела, которые напрямую не участвуют в связывании антигена, на эквивалентные последовательности из вариабельных областей Fv антитела человека. Общие способы создания гуманизированных антител известны в данной области техники.
Например, моноклональные антитела, направленные на TcdA из С. difficile или TcdB из С. difficile, также могут быть получены стандартными методами, такими как метод гибридом (см., например, Kohler and Milstein, 1975, Nature, 256: 495-497). Кратко, бессмертную клеточную линию сливают с лимфоцитами от млекопитающего, иммунизированного TcdA из С. difficile, TcdB из С. difficile или мутантным токсином из С. difficile, описанным в данном изобретении, и проводят скрининг культуральных супернатантов полученных гибридомных клеток на предмет идентификации гибридомы, продуцирующей моноклональное антитело, которое связывается с TcdA из С. difficile или TcdB из С. difficile. Обычно бессмертная клеточная линия происходит из того же вида млекопитающих, что и лимфоциты. Обнаружение гибридомных клеток, продуцирующих моноклональное антитело по изобретению, осуществляют, проводя скрининг супернатантов гибридомных культур на предмет антител, которые связываются с TcdA из С. difficile или TcdB из С. difficile, с использованием такого анализа, как ELISA. Человеческие гибридомы могут быть получены аналогичным образом.
В качестве альтернативы получению антител посредством иммунизации и отбора, антитела по изобретению также могут быть идентифицированы путем скрининга комбинаторной библиотеки рекомбинантных иммуноглобулинов с использованием TcdA из С. difficile, TcdB из С. difficile или мутантного токсина из С. difficile, описанных в данном изобретении. Библиотека рекомбинантных антител может представлять собой scFv-библиотеку или Fab-библиотеку, например. Кроме того, антитела по изобретению, описанные здесь, могут быть использованы в исследованиях конкурентного связывания для идентификации дополнительных антител к TcdA или TcdB и их связывающих фрагментов. Например, дополнительные антитела к TcdA или TcdB и их связывающие фрагменты могут быть идентифицированы путем скрининга библиотеки человеческих антител и идентификации в этой библиотеке молекул, которые конкурируют с антителами по изобретению, изложенными в данном описании, в анализе конкурентного связывания.
Помимо этого, антитела, охватываемые настоящим изобретением, включают рекомбинантные антитела, которые могут быть образованы с использованием методов фагового дисплея, известных в данной области техники. В методах фагового дисплея фаг можно использовать для выявления антигенсвязывающих доменов, экспрессированных из репертуара или библиотеки антител (например, человеческих или мышиных). Фаг, экспрессирующий антигенсвязывающий домен, который связывается с иммуногеном, описанным в данном изобретении (например, мутантным токсином из С. difficile), может быть отобран или идентифицирован с помощью антигена, например, с использованием меченого антигена.
Также в объем изобретения включены антитела и их связывающие фрагменты, в которых выполнены замены, делеции или вставки по конкретным аминокислотам. В частности, предпочтительные антитела имеют аминокислотные замены в каркасной области, например, для улучшения связывания с антигеном. Например, отобранные в небольшом количестве остатки в акцепторном каркасе иммуноглобулиновой цепи могут быть заменены соответствующими донорными аминокислотами. Предпочтительные расположения замен включают аминокислотные остатки, расположенные вблизи CDR, или те, которые способны взаимодействовать с CDR. Критерии выбора аминокислот от донора описаны в патенте США №5585089 (например, колонки 12-16). Акцепторный каркас может представлять собой каркасную последовательность или консенсусную последовательность зрелого человеческого антитела.
Как использовано в данном описании, «нейтрализующее антитело или его связывающий фрагмент» относится к соответствующему антителу или его связывающему фрагменту, которые связываются с патогеном (например, TcdA или TcdB из С. difficile) и ослабляют инфекционность и/или активность этого патогена (например, снижают цитотоксичность) у млекопитающего и/или в клеточной культуре по сравнению с патогеном, находящимся в идентичных условиях, но в отсутствие нейтрализующего антитела или его связывающего фрагмента. В одном из воплощений нейтрализующее антитело или его связывающий фрагмент способны нейтрализовать по меньшей мере примерно 70%, 75%, 80%, 85%, 90%, 95%, 99% или более биологической активности патогена по сравнению с биологической активностью патогена, находящегося в идентичных условиях, но в отсутствие нейтрализующего антитела или его связывающего фрагмента.
Использованный в данном описании термин «антитело к токсину или его связывающий фрагмент» относится к антителу или его связывающему фрагменту, которые связываются с соответствующим токсином из С. difficile (например, токсином А или токсином В из С. difficile). Например, антитело к токсину А или его связывающий фрагмент относится к антителу или его связывающему фрагменту, которые связываются с TcdA.
Антитела или их связывающие фрагменты, описанные в данном изобретении, могут быть индуцированы у любого млекопитающего дикого типа и/или трансгенного млекопитающего, включая, например, мышей, людей, кроликов и коз.
В том случае, когда иммуногенная композиция, описанная выше, представляет собой композицию, которая ранее была введена популяции, как например, для вакцинации, антительный ответ, индуцированный у субъектов, можно использовать для нейтрализации токсинов из того же штамма и из штамма, который не стимулировал продуцирование этого антитела. См., например, пример 37, в котором показаны результаты исследований, относящихся к перекрестной реактивности антител, индуцированных под действием иммуногенной композиции, при сравнении штамма 630 и токсинов из различных штаммов С. difficile дикого типа.
В одном из аспектов изобретение относится к антителу или его связывающему фрагменту, специфичным к TcdA из С. difficile. Моноклональные антитела, которые специфически связываются с TcdA, включают А65-33, А60-22, А80-29 и/или, предпочтительно, А3-25.
В одном из аспектов изобретение относится к антителу или его связывающему фрагменту, специфичным к TcdA из любого штамма С. difficile дикого типа, как например, описанным выше, например, к SEQ ID NO: 1. В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к иммуногенной композиции, описанной выше. Например, в одном из воплощений антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 4 или SEQ ID NO: 7. В другом воплощении антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 4 или SEQ ID NO: 7, при этом по меньшей мере одна аминокислота в SEQ ID NO: 4 или SEQ ID NO: 7 вовлечена в образование химических сшивок посредством формальдегида, EDC, NHS или комбинации EDC и NHS. В другом воплощении антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 84 или SEQ ID NO: 83.
Антитела или их связывающие фрагменты, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, которые по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно на 98%, более предпочтительно примерно на 99% или наиболее предпочтительно примерно на 100% идентичны вариабельным областям тяжелой и легкой цепей А65-33, А60-22, А80-29 и/или, предпочтительно, А3-25, также могут связываться с TcdA.
В одном из воплощений антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи А3-25, которая описана в SEQ ID NO: 37.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи А3-25, которая описано в SEQ ID NO: 36.
Еще в одном аспекте антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области тяжелой цепи, описанной в SEQ ID NO: 37, и вариабельную область легкой цепи, имеющую аминокислотную последовательность, по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности вариабельной области легкой цепи, описанной в SEQ ID NO: 36.
В другом воплощении антитела или их связывающие фрагменты, имеющие определяющие комплементарность участки (CDR) вариабельных областей тяжелых цепей и/или вариабельных областей легких цепей А65-33, А60-22, А80-29 и/или, предпочтительно, А3-25, также могут связываться с TcdA. CDR вариабельной области тяжелой цепи А3-25 показаны ниже в Таблице 4.

CDR вариабельной области легкой цепи А3-25 показаны ниже в Таблице 5.

В одном из воплощений антитело или его связывающий фрагмент содержит аминокислотные последовательности определяющих комплементарность участков (CDR) тяжелой цепи, которые показаны в SEQ ID NO: 41 (CDR Н1), 42 (CDR Н2) и 43 (CDR Н3), и/или аминокислотные последовательности CDR легкой цепи, которые показаны в SEQ ID NO: 38 (CDR L1), 39 (CDR L2) и 40 (CDR L3).
В одном из типичных воплощений антитело или его связывающий фрагмент, специфичные к токсину А из С. difficile, специфически связываются с эпитопом в N-концевом участке TcdA, например, эпитопом между аминокислотами 1-1256 в TcdA, в соответствии с нумерацией в SEQ ID NO: 1.
В предпочтительном воплощении антитело или его связывающий фрагмент, специфичные к токсину А из С. difficile, специфически связываются с эпитопом в С-концевом участке токсина А, например, эпитопом между аминокислотами от 1832 до 2710 в TcdA, в соответствии с нумерацией в SEQ ID NO: 1. Примеры включают А3-25, А65-33, А60-22, А80-29.
В еще одном воплощении антитело или его связывающий фрагмент, специфичные к токсину А из С. difficile, специфически связываются с эпитопом в пределах области «транслокации» токсина А С. difficile, например, эпитопом, который предпочтительно включает остатки 956-1128 в TcdA, в соответствии с нумерацией в SEQ ID NO: 1, таким как эпитоп между аминокислотами 659-1832 в TcdA, в соответствии с нумерацией в SEQ ID NO: 1.
В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к TcdB из С. difficile. Например, антитело или его связывающий фрагмент могут быть специфичны к TcdB из любого штамма С. difficile дикого типа, такому, как описанный выше, например, к SEQ ID NO: 2. В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к иммуногенной композиции, описанной выше. Например, в одном из воплощений антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 6 или SEQ ID NO: 8.
В другом воплощении антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 6 или SEQ ID NO: 8, при этом по меньшей мере одна аминокислота в SEQ ID NO: 6 или SEQ ID NO: 8 вовлечена в образование химических сшивок посредством формальдегида, EDC, NHS или комбинации EDC и NHS. В другом воплощении антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая содержит SEQ ID NO: 86 или SEQ ID NO: 85.
Моноклональные антитела, которые специфически связываются с TcdB, включают антитела, продуцируемые клонами В2-31, В5-40, В70-2, В6-30, В9-30, В59-3, В60-2, В56-6 и/или, предпочтительно, В8-26, описанными в данном изобретении.
Антитела или их связывающие фрагменты, которые также могут связываться с TcdB, включают таковые, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, которые идентичны по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% вариабельным областям тяжелой и легкой цепей В2-31, В5-40, В70-2, В6-30, В9-30, В59-3, В60-2, В56-6, предпочтительно В8-26, В59-3 и/или В9-30.
В одном из воплощений антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области тяжелой цепи А3-25, которая приведена в SEQ ID NO: 49.
В одном из воплощений антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области тяжелой цепи А3-25, которая приведена в SEQ ID NO: 60.
В одном из воплощений антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области тяжелой цепи А3-25, которая приведена в SEQ ID NO: 71.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области легкой цепи А3-25, которая приведена в SEQ ID NO: 55.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области легкой цепи А3-25, которая приведена в SEQ ID NO: 66.
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области легкой цепи А3-25, которая приведена в SEQ ID NO: 77.
Аминокислотная последовательность вариабельной области тяжелой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В8-26) приведена в SEQ ID NO: 49. См. Таблицу 25-а.
Аминокислотная последовательность вариабельной области легкой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В8-26) приведена в SEQ ID NO: 55. См. Таблицу 25-b.

В одном из воплощений антитело или его связывающий фрагмент включает аминокислотные последовательности CDR тяжелой цепи, которые показаны в SEQ ID NO: 51 (CDR Н1), 52 (CDR Н2) и 53 (CDR НЗ), и/или аминокислотные последовательности CDR легкой цепи, которые показаны в SEQ ID NO: 57 (CDR L1), 58 (CDR L2) и 59 (CDR L3).
Аминокислотная последовательность вариабельной области тяжелой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В59-3) приведена в SEQ ID NO: 60. См. Таблицу 26-а.


Аминокислотная последовательность вариабельной области легкой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В59-3) приведена в SEQ ID NO: 66. См. Таблицу 26-b.

В одном из воплощений антитело или его связывающий фрагмент включает аминокислотные последовательности CDR тяжелой цепи, которые показаны в SEQ ID NO: 62 (CDR Н1), 63 (CDR Н2) и 64 (CDR Н3), и/или аминокислотные последовательности CDR легкой цепи, которые показаны в I SEQ ID NO: 68 (CDR L1), 69 (CDR L2) и 70 (CDR L3).
Аминокислотная последовательность вариабельной области тяжелой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В9-30) приведена в SEQ ID NO: 71. См. Таблицу 27-а.


Аминокислотная последовательность вариабельной области легкой цепи нейтрализующего антитела для TcdB из С. difficile (mAb В9-30) приведена в SEQ ID NO: 77. См. Таблицу 27-b.

В одном из воплощений антитело или его связывающий фрагмент содержит аминокислотные последовательности CDR тяжелой цепи, которые показаны в SEQ ID NO: 73 (CDR Н1), 74 (CDR Н2) и 75 (CDR Н3), и/или аминокислотные последовательности CDR легкой цепи, которые показаны в SEQ ID NO: 79 (CDR L1), 80 (CDR L2) и 81 (CDR L3).
В одном из аспектов изобретение относится к антителу или его связывающему фрагменту, специфичным к TcdB С. difficile дикого типа из любого штамма С. difficile, такому, как описанный выше, например, к SEQ ID NO: 2. В другом аспекте изобретение относится к антителу или его связывающему фрагменту, специфичному к иммуногенной композиции, описанной выше. Например, в одном из воплощений антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая включает SEQ ID NO: 6 или SEQ ID NO: 8. В другом воплощении антитело или его связывающий фрагмент специфичны к иммуногенной композиции, которая включает SEQ ID NO: 6 или SEQ ID NO: 8, при этом по меньшей мере одна аминокислота в SEQ ID NO: 6 или SEQ ID NO: 8 вовлечена в образование химических сшивок посредством формальдегида, EDC, NHS или комбинации EDC и NHS.
Антитела или их связывающие фрагменты, имеющие вариабельные области тяжелой цепи и вариабельные области легкой цепи, которые идентичны по меньшей мере примерно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, предпочтительно примерно 98%, более предпочтительно примерно 99% или наиболее предпочтительно примерно 100% вариабельным областям тяжелой и легкой цепей В2-31, В5-40, В70-2, В6-30, В9-30, В59-3, В60-2, В56-6 и/или, предпочтительно, В8-26, также могут связываться с TcdB.
В одном из воплощений антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области тяжелой цепи В8-26 (SEQ ID NO: 49).
В другом воплощении антитело или его антигенсвязывающий фрагмент включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области легкой цепи В8-26 (SEQ ID NO: 55).
В еще одном аспекте антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области тяжелой цепи В8-26 (SEQ ID NO: 49), и вариабельную область легкой цепи, имеющую аминокислотную последовательность, идентичную по меньшей мере примерно на 60%, 65%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности вариабельной области легкой цепи В8-26 (SEQ ID NO: 55).
В другом воплощении антитела или их связывающие фрагменты, имеющие CDR вариабельной области тяжелых цепей и/или вариабельной области легких цепей В2-31, В5-40, В70-2, В6-30, В9-30, В59-3, В60-2, В56-6 и/или, предпочтительно, В8-26, также могут связываться с TcdB.
В одном из воплощений антитело или его связывающий фрагмент включает аминокислотные последовательности определяющих комплементарность участков тяжелой цепи (CDR) В8-26 и/или аминокислотные последовательности CDR легкой цепи В8-26.
В предпочтительном воплощении антитело или его связывающий фрагмент, специфичные к токсину В из С. difficile, специфически связываются с эпитопом в N-концевом участке токсина В, например, эпитопом, расположенным между аминокислотами 1 и 1256 в TcdB, в соответствии с нумерацией в SEQ ID NO: 2. Примеры включают В2-31, В5-40, В8-26, В70-2, В6-30 и В9-30.
В типичном воплощении антитело или его связывающий фрагмент, специфичные к токсину В из С. difficile, специфически связываются с эпитопом в С-концевом участке токсина В, например, эпитопом между аминокислотами от 1832 до 2710 в TcdB, в соответствии с нумерацией в SEQ ID NO: 2.
В еще одном воплощении антитело или его связывающий фрагмент, специфичные к токсину В из С. difficile, специфически связываются с эпитопом в пределах области «транслокации» токсина В из С. difficile, например, эпитопом, который предпочтительно включает остатки 956-1128 в TcdB, в соответствии с нумерацией в SEQ ID NO: 2, таким как эпитоп между аминокислотами 659-1832 в TcdB. Примеры включают В59-3, В60-2 и В56-6.
Комбинации антител
Антитело к токсину или его связывающий фрагмент можно вводить в комбинации с другими антителами к токсину из С. difficile (например, другими моноклональными антителами, поликлональным гамма-глобулином) или их связывающим фрагментом. Комбинации, которые можно использовать, включают антитело к токсину А или его связывающий фрагмент и антитело к токсину В или его связывающий фрагмент.
В другом воплощении комбинация включает антитело к токсину А или его связывающий фрагмент и другое антитело к токсину А или его связывающий фрагмент. Предпочтительно, комбинация включает моноклональное нейтрализующее антитело для токсина или его связывающий фрагмент и другое моноклональное нейтрализующее антитело для токсина или его связывающий фрагмент. Неожиданным образом авторы изобретения обнаружили, что такая комбинация приводила к синергетическому эффекту в отношении нейтрализации цитотоксичности токсина А. Например, комбинация включает комбинацию по меньшей мере двух из следующих моноклональных нейтрализующих антител для токсина А: А3-25, А65-33, А60-22 и А80-29. Более предпочтительно, комбинация включает антитело А3-25 и по меньшей мере одно из следующих моноклональных нейтрализующих антител для токсина А: А65-33, А60-22 и А80-29. Наиболее предпочтительно, комбинация включает все четыре антитела: А3-25, А65-33, А60-22 и А80-29.
В следующем воплощении комбинация включает антитело к токсину В или его связывающий фрагмент и другое антитело к токсину В или его связывающий фрагмент. Предпочтительно, комбинация включает моноклональное нейтрализующее антитело для токсина В или его связывающий фрагмент и другое моноклональное нейтрализующее антитело для токсина В или его связывающий фрагмент. Неожиданным образом авторы изобретения обнаружили, что такая комбинация приводила к синергетическому эффекту в отношении нейтрализации цитотоксичности токсина В. Более предпочтительно, комбинация включает комбинацию по меньшей мере двух из следующих моноклональных нейтрализующих антител для токсина В: В8-26, В9-30 и В59-3. Наиболее предпочтительно, комбинация включает все три антитела: В8-26, В9-30 и В59-3.
В еще одном воплощении комбинация включает антитело к токсину В или его связывающий фрагмент и другое антитело к токсину В или его связывающий фрагмент. Как указано ранее, авторы изобретения открыли, что комбинация по меньшей мере двух нейтрализующих моноклональных антител неожиданно может демонстрировать синергетический эффект в отношении нейтрализации токсина А и токсина В.
В другом воплощении на основе агентов по изобретению могут быть изготовлены композиции в виде смеси, или агенты по изобретению могут быть подвергнуты химическому или генетическому связыванию с использованием признанных в данной области техники методов с получением тем самым ковалентно связанных антител (или ковалентно связанных фрагментов антител), обладающих связывающими свойствами как антитела к токсину А, так и антитела к токсину В. Для объединенной композиции можно руководствоваться определением одного или более параметров, таких как аффинность, авидность или биологическая эффективность агента по отдельности или в комбинации с другим агентом.
Такие комбинированные терапии предпочтительно являются аддитивными и/или синергетическими в проявлении своей терапевтической активности, например, в ингибировании, предупреждении (например, рецидива) и/или лечении связанных с С. difficile заболеваний или расстройств. Применение таких комбинированных терапий может снизить дозировку терапевтического агента (например, смеси антитела или фрагмента антитела либо сшитого или генетически слитого биспецифического антитела или фрагмента антитела), необходимую для достижения желаемого эффекта.
Очевидно, что в любой из композиций по изобретению, например, антитело к токсину А и/или токсину В или их связывающие фрагменты могут быть скомбинированы в разных соотношениях или количествах для оказания терапевтического эффекта. Например, антитело к токсину А и токсину В или его соответствующий связывающий фрагмент могут присутствовать в композиции в соотношении для А:В, изменяющемся в диапазоне от 0,1:10 до 10:0,1. В другом воплощении антитело к токсину А и токсину В или его соответствующий связывающий фрагмент могут присутствовать в композиции в соотношении для В:А, изменяющемся в диапазоне от 0,1:10 до 10:0,1.
В другом аспекте изобретение относится к способу получения нейтрализующего антитела к TcdA из С. difficile. Способ включает введение иммуногенной композиции, описанной выше, млекопитающему и извлечение антитела из этого млекопитающего. В предпочтительном воплощении иммуногенная композиция включает мутантный TcdA из С. difficile, имеющий SEQ ID NO: 4, при этом по меньшей мере одна аминокислота мутантного TcdA из С. difficile вовлечена в образование химических сшивок, предпочтительно с использованием формальдегида или 1-этил-3-(3-диметиламинопропил)карбодиимида. Типичные нейтрализующие антитела к TcdA, которые могут быть получены, включают А65-33, А60-22, А80-29 и/или А3-25.
В еще одном аспекте изобретение относится к способу получения нейтрализующего антитела к TcdB из С. difficile. Способ включает введение иммуногенной композиции, описанной выше, млекопитающему и извлечение антитела из этого млекопитающего. В предпочтительном воплощении иммуногенная композиция включает мутантный TcdB из С. difficile, имеющий SEQ ID NO: 6, при этом по меньшей мере одна аминокислота мутантного TcdB из С. difficile вовлечена в образование химических сшивок, предпочтительно с использованием формальдегида или 1-этил-3-(3-диметиламинопропил)карбодиимида. Типичные нейтрализующие антитела к TcdB, которые могут быть получены, включают В2-31, В5-40, В70-2, В6-30, В9-30, В59-3, В60-2, В56-6 и/или В8-26.
Композиции
Композиции по настоящему изобретению (такие как, например, композиции, содержащие мутантный токсин из С. difficile, иммуногенные композиции, антитела и/или их связывающие фрагменты, описанные в данном изобретении) могут существовать в разнообразных формах. Они включают, например, полутвердые и твердые лекарственные формы, суппозитории, жидкие формы, такие как жидкие растворы (например, инъекционные и инфузионные растворы), дисперсии или суспензии, липосомы и/или высушенную форму, такую как, например, лиофилизированная порошковая форма, высушенная сублимационной сушкой форма, высушенная распылительной сушкой форма и/или высушенная в виде пены форма. Что касается суппозиториев, то связующие вещества и носители включают, например, полиалкиленгликоли или триглицериды; такие суппозитории могут быть изготовлены из смесей, содержащих композиции по изобретению. В типичном воплощении композиция существует в форме, которая подходит для растворения или суспендирования в жидких разбавителях перед инъекцией. В другом типичном воплощении на основе композиции готовят эмульсию, или ее инкапсулируют в липосомы или микрочастицы, как например из полилактида, полигликолида или сополимера.
В предпочтительном воплощении композицию лиофилизируют и повторно растворяют непосредственно перед применением.
В одном из аспектов настоящее изобретение относится к фармацевтическим композициям, которые включают любую из композиций, описанных в данном изобретении (таких как, например, композиции, содержащие мутантный токсин из С. difficile, иммуногенные композиции, антитела и/или их связывающие фрагменты, описанные в данном изобретении), изготовленных вместе с фармацевтически приемлемым носителем. «Фармацевтически приемлемые носители» включают любые растворители, диспергирующие среды, стабилизаторы, разбавители и/или буферы, которые являются физиологически приемлемыми.
Типичные стабилизаторы включают углеводы, такие как сорбит, маннит, крахмал, декстран, сахароза, трегалоза, лактоза и/или глюкоза; инертные белки, такие как альбумин и/или казеин; и/или другие большие, медленно метаболизируемые макромолекулы, такие как полисахариды, например хитозан, полимолочные кислоты, полигликолевые кислоты и сополимеры (такие как латексная функционализированная агароза SEPHAROSE™, агароза, целлюлоза и т.д.), аминокислоты, полимерные аминокислоты, сополимеры аминокислот и липидные агрегаты (такие как капельки масла или липосомы). Кроме того, эти носители могут действовать в качестве иммуностимулирующих агентов (т.е. адъювантов).
Предпочтительно, композиция содержит трегалозу. Предпочтительные количества трегалозы (% по массе) составляют минимально от примерно 1%, 2%, 3% или 4% до максимально примерно 10%, 9%, 8%, 7%, 6% или 5%. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений композиция содержит примерно 3%-6% трегалозы, наиболее предпочтительно 4,5% трегалозы, например, на дозу 0,5 мл.
Примеры подходящих разбавителей включают дистиллированную воду, физиологический раствор, забуференный фосфатом физиологический раствор, глицерин, спирт (такой как этанол), растворы Рингера, раствор декстрозы, сбалансированные солевые растворы Хенкса и/или эксципиент для лиофилизации.
Типичные буферы включают буферы на основе фосфата (такого как фосфат калия, фосфат натрия); ацетата (такого как ацетат натрия); сукцината (такого как сукцинат натрия); глицина; гистидина; карбоната, триса (трис(гидроксиметил)аминометан) и/или бикарбоната (такого как бикарбонат аммония). Предпочтительно, композиция содержит трис-буфер. Предпочтительные количества трис-буфера значения от минимального, составляющего примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ до максимального, составляющего примерно 100 мМ, 50 мМ, 20 мМ, 19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений композиция содержит примерно 8 мМ-12 мМ трис-буфер, наиболее предпочтительно, 10 мМ трис-буфер, например, на дозу 0,5 мл.
В другом предпочтительном воплощении композиция содержит буфер на основе гистидина. Предпочтительные количества буфера на основе гистидина включают значения от минимального, составляющего примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ, до максимального, составляющего примерно 100 мМ, 50 мМ, 20 мМ, 19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений композиция содержит примерно 8 мМ-12 мМ буфер на основе гистидина, наиболее предпочтительно, 10 мМ буфер на основе гистидина, например, на дозу 0,5 мл.
В еще одном предпочтительном воплощении композиция содержит фосфатный буфер. Предпочтительный количества фосфатного буфера включают значения от минимального, составляющего примерно 1 мМ, 5 мМ, 6 мМ, 7 мМ, 8 мМ, 9 мМ, 10 мМ, до максимального, составляющего примерно 100 мМ, 50 мМ, 20 мМ, 19 мМ, 18 мМ, 17 мМ, 16 мМ, 15 мМ, 14 мМ, 13 мМ, 12 мМ или 11 мМ. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений композиция содержит примерно 8 мМ-12 мМ фосфатный буфер, наиболее предпочтительно, 10 мМ фосфатный буфер, например, на дозу 0,5 мл.
Значения рН буфера будут выбраны предпочтительно с целью стабилизации активного вещества и могут быть установлены специалистами в данной области техники известными методами. Предпочтительно, рН буфера будет находиться в диапазоне физиологического рН. Таким образом, предпочтительные диапазоны рН составляют от примерно 3 до примерно 8; более предпочтительно от примерно 6,0 до примерно 8,0; еще более предпочтительно от примерно 6,5 до примерно 7,5; и наиболее предпочтительно при значениях от примерно 7,0 до примерно 7,2.
В некоторых воплощениях фармацевтические композиции могут включать поверхностно-активное вещество. Подходящим является любое поверхностно-активное вещество независимо от того, является ли оно амфотерным, неионным, катионным или анионным. Типичные поверхностно-активные вещества включают поверхностно-активные вещества на основе сложных эфиров полиоксиэтиленсорбитана (например, TWEEN®), такие как полисорбат 20 и/или полисорбат 80; простые эфиры на основе жирных спиртов и полиоксиэтилена, происходящие из лаурилового, цетилового, стеарилового и олеилового спиртов (известные как поверхностно-активные вещества под названием Brij), такие как простой монолауриловый эфир триэтиленгликоля (Brij 30); тритон Х-100 или трет-октилфеноксиполиэтоксиэтанол; и сложные эфиры сорбитана (широко известные как сланы (SPAN)), такие как триолеат сорбитана (спан 85) и монолаурат сорбитана, и их комбинации. Предпочтительные поверхностно-активные вещества включают полисорбат 80 (моноолеат полиоксиэтиленсорбитана).
Предпочтительные количества полисорбата 80 (% по массе) включают значения от минимального, составляющего примерно 0,001%, 0,005% или 0,01%, до максимального, составляющего примерно 0,010%, 0,015%, 0,025% или 1,0%. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений композиция содержит примерно 0,005%-0,015% полисорбата 80, наиболее предпочтительно 0,01% полисорбата 80.
В типичном воплощении иммуногенная композиция содержит трегалозу и фосфат 80. В другом типичном воплощении иммуногенная композиция содержит трис-буфер и полисорбат 80. В другом типичном воплощении иммуногенная композиция содержит буфер на основе гистидина и полисорбат 80. В еще одном типичном воплощении иммуногенная композиция содержит фосфатный буфер и полисорбат 80.
В одном из типичных воплощений иммуногенная композиция содержит трегалозу, трис-буфер и полисорбат 80. В другом типичном воплощении иммуногенная композиция содержит трегалозу, буфер на основе гистидина и полисорбат 80. В еще одном типичном воплощении иммуногенная композиция содержит трегалозу, фосфатный буфер и полисорбат 80.
Композиции, описанные в данном изобретении, также могут содержать компоненты нефти, компоненты животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло и/или минеральное масло. Примеры включают гликоли, такие как пропиленгликоль или полиэтиленгликоль.
В некоторых воплощениях фармацевтическая композиция дополнительно содержит формальдегид. Например, в предпочтительном воплощении фармацевтическая композиция, которая дополнительно содержит формальдегид, включает иммуногенную композицию, при этом мутантный токсин из С. difficile в этой иммуногенной композиции был приведен в контакт с химическим сшивающим агентом, который представляет собой формальдегид. Количество формальдегида, присутствующего в фармацевтической композиции, может изменяться от минимального значения примерно 0,001%, 0,002%, 0,003%, 0,004%, 0,005%, 0,006%, 0,007%, 0,008%, 0,009%, 0,010%, 0,013% или 0,015%, до максимального значения примерно 0,020%, 0,019%, 0,018%, 0,017% 0,016%, 0,015%, 0,014%, 0,013%, 0,012% 0,011% или 0,010%. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. В одном из воплощений фармацевтическая композиция содержит примерно 0,010% формальдегида.
В некоторых альтернативных воплощениях фармацевтические композиции, описанные здесь, не содержат формальдегида. Например, в предпочтительном воплощении фармацевтическая композиция, не содержащая формальдегида, включает иммуногенную композицию, при этом по меньшей мере одна аминокислота мутантного токсина С. difficile вовлечена в образование химических сшивок с использованием агента, который представляет собой EDC. Более предпочтительно, в таком воплощении мутантный токсин из С. difficile не был приведен в контакт с химическим сшивающим агентом, который представляет собой формальдегид. В качестве другого типичного воплощения рассматривается фармацевтическая композиция, находящаяся в лиофилизированной форме, которая не содержит формальдегида.
В другом воплощении композиции, описанные здесь, могут содержать адъювант, описанный ниже. Предпочтительные адъюванты усиливают врожденный иммунный ответ на иммуноген, не вызывая конформационных изменений в иммуногене, которые могут оказывать влияние на качественную форму иммунного ответа.
Типичные адъюванты включают 3-де-О-ацилированный монофосфориллипид A (MPL™) (см. GB 2220211 (GSK)); гель гидроксида алюминия, такой как Alhydrogel™ (Brenntag Biosector, Denmark); соли алюминия (такие как гидроксид алюминия, фосфат алюминия, сульфат алюминия), которые можно использовать вместе с иммуностимулирующим агентом, таким как MPL или 3D-MPL, QS-21, полимерные или мономерные аминокислоты, такие как полиглутаминовая кислота или полилизин, или без него.
Еще одним типичным адъювантом является иммуностимулирующий олигонуклеотид, такой как CpG-содержащий олигонуклеотид (см. например, WO 1998/040100, WO 2010/067262) или сапонин, и такой иммуностимулирующий олигонуклеотид, как CpG-содержащий олигонуклеотид (см. например, WO 00/062800). В предпочтительном воплощении адъювант представляет собой CpG-содержащий олигонуклеотид, наиболее предпочтительно CpG-содержащие олигодезоксинуклеотиды (CpG ODN). Предпочтительные CpG ODN относятся к классу В, которые активируют предпочтительно В-клетки. В аспектах изобретения CpG ODN имеет нуклеиновокислотную последовательность 5' T*C*G*T*C*G*T*T*T*T*T*C*G*G*T*G*C*T*T*T*T 3' (SEQ ID NO: 48), где * указывает на фосфотиоатную связь. CpG ODN с такой последовательностью известен как CpG 24555, который описан в WO 2010/067262. В предпочтительном воплощении CpG 24555 используют вместе с солью гидроксида алюминия, такой как Alhydrogel.
Другой класс типичных адъювантов включает сапониновые адъюванты, такие как Stimulon™ (QS-21, представляющий собой тритерпеновый гликозид или сапонин, Aquila, Framingham, Mass.) или изготовленные из них частицы, такие как ISCOM (иммуностимулирующие комплексы) и адъювант ISCOMATRIX®. Соответственно, композиции по настоящему изобретению могут быть доставлены в форме ISCOM, ISCOM, содержащих В-субъединицу холерного токсина (СТВ), липосом или могут быть инкапсулированы в такие соединения, как акрилаты или поли(DL-лактид-ко-гликозид), с образованием микросфер, размер которых подходит для всасывания. Обычно термин «ISCOM» относится к иммуногенным комплексам, образуемым между гликозидами, такими как тритерпеноидные сапонины (в частности, Quil А), и антигенами, содержащими гидрофобный участок. В предпочтительном воплощении адъювантом является адъювант ISCOMATRIX.
Другие типичные адъюванты включают RC-529, гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) и полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA).
Еще одним классом типичных адъювантов являются гликолипидные аналоги, включая N-гликозиламиды, N-гликозилмочевины и N-гликозилкарбаматы, каждый из которых замещен в остатке сахара аминокислотой.
Возможно, фармацевтическая композиция содержит два или более различных адъювантов. Предпочтительные комбинации адъювантов включают любую комбинацию адъювантов, в том числе, например, по меньшей мере два адъюванта из следующих: квасцов, MPL, QS-21, ISCOMATRIX, CpG и Alhydrogel. Типичная комбинация адъювантов включает комбинацию CpG и Alhydrogel.
Альтернативно, в одном из воплощений композицию вводят млекопитающему без адъюванта.
Композиции, описанные здесь, можно вводить любым способом введения, таким как, например, парентеральный, местный, внутривенный, мукозальный, пероральный, подкожный, интраартериальный, интракраниальный, интратекальный, внутрибрюшинный, интраназальный, внутримышечный, интрадермальный, инфузионный, ректальный и/или трансдермальный способы, для профилактического и/или терапевтического применений. В предпочтительном воплощении способом введения композиции является парентеральное, более предпочтительно, внутримышечное введение. Типичное внутримышечное введение выполняют в мышцы рук и ног.
Композиции, описанные здесь, можно вводить в комбинации с такими терапиями, которые по меньшей мере частично эффективны в предотвращении и/или лечении инфекции С. difficile. Например, композицию по изобретению можно вводить до, одновременно с или после лечения биопрепаратами, терапии пробиотиками, фекальных трансплантаций, иммунотерапии (например, внутривенным иммуноглобулином); и/или принятого стандарта оказания медицинской помощи по лечению антибиотиками заболевания, ассоциированного с С. difficile (CDAD), такого как применение метронидазола и/или ванкомицина.
Композицию по настоящему изобретению, имеющую отношение к токсину А и токсину В, можно вводить млекопитающему в любой комбинации. Например, иммуногенную композицию, содержащую мутантный TcdA из С. difficile, можно вводить млекопитающему до, одновременно с или после введения иммуногенной композиции, содержащей мутантный TcdB из С. difficile. И наоборот, иммуногенную композицию, содержащую мутантный TcdB из С. difficile, можно вводить млекопитающему до, одновременно с или после введения иммуногенной композиции, содержащей мутантный TcdA из С. difficile.
В другом воплощении композицию, содержащую антитело к токсину А или его связывающий фрагмент, можно вводить млекопитающему до, одновременно с или после введения композиции, содержащей антитело к токсину В или его связывающий фрагмент. И наоборот, композицию, содержащую антитело к токсину В или его связывающий фрагмент, можно вводить млекопитающему до, одновременно с или после введения композиции, содержащей антитело к токсину А или его связывающий фрагмент.
В следующем воплощении композицию по настоящему изобретению можно вводить млекопитающему до, одновременно с или после введения фармацевтически приемлемого носителя. Например, адъювант можно вводить до, одновременно с или после введения композиции, содержащей мутантный токсин из С. difficile. Соответственно, композиция по настоящему изобретению и фармацевтически приемлемый носитель могут быть упакованы в одном флаконе или они могут быть упакованы в отдельных флаконах и смешаны перед применением. Композиции могут быть изготовлены для введения разовых доз и/или введения многократных доз.
Способы защиты от инфекции и/или лечения инфекции С. difficile у млекопитающего
В одном из аспектов изобретение относится к способу индукции иммунного ответа на токсин из С. difficile у млекопитающего. Способ включает введение эффективного количества композиции, описанной в данном изобретении, млекопитающему. Например, способ может включать введение количества, эффективного для вызывания у млекопитающего иммунного ответа на соответствующий токсин из С. difficile.
В типичном воплощении изобретение относится к способу индукции иммунного ответа на TcdA из С. difficile у млекопитающего. Способ включает введение млекопитающему эффективного количества иммуногенной композиции, содержащей мутантный TcdA из С. difficile. В другом типичном воплощении изобретение относится к способу индукции иммунного ответа на TcdB из С. difficile у млекопитающего. Способ включает введение млекопитающему эффективного количества иммуногенной композиции, содержащей мутантный TcdB из С. difficile.
В следующем воплощении способ включает введение млекопитающему эффективного количества иммуногенной композиции, содержащей мутантный TcdA из С. difficile, и эффективного количества иммуногенной композиции, содержащей мутантный TcdB из С. difficile. В дополнительных аспектах, композиции, описанные в данном изобретении, можно использовать для лечения, предупреждения, снижения риска, ослабления тяжести, уменьшения числа случаев и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данную композицию не вводят. Способ включает введение эффективного количества композиции млекопитающему.
С учетом тяжести инфекции общепризнаны три клинических синдрома, вызываемые инфекцией С. difficile. Самой тяжелой формой является псевдомембранозный колит (РМС), который характеризуется профузной диареей, абдоминальной болью, системными признаками болезни и отличительным, выявленным при эндоскопическом обследовании видом толстой кишки.
Антибиотикоассоциированный колит (ААС) также характеризуется профузной диареей, абдоминальной болью и болезненной чувствительностью, системными признаками (например, лихорадкой) и лейкоцитозом. Поражение кишечника при ААС оказывается менее тяжелым, чем при РМС, отсутствует выявляемый при эндоскопическом обследовании характерный при РМС вид толстой кишки, и смертность является низкой.
И наконец, антибиотикоассоциированная диарея (AAD), также известная как ассоциированная с С. difficile диарея (CDAD), представляет собой относительно мягкий синдром и характеризуется как диарея от легкой до умеренной степени тяжести, при которой отсутствуют как воспаление толстой кишки (которое характеризуется, например, абдоминальной болью и болезненной чувствительностью), так и системные признаки инфекции (например, лихорадка).
Эти три различных синдрома, как правило, возникают в порядке возрастания частоты их возникновения. То есть, обычно РМС возникает реже, чем ААС, и обычно AAD считается наиболее частым клиническим проявлением вызванного С. difficile заболевания.
Частым осложнением инфекции С. difficile является рецидивирующее или повторно проявляющееся заболевание, которое возникает у 20% включительно всех субъектов, которые выздоравливают от вызванного С. difficile заболевания. Клинически рецидив может быть охарактеризован как AAD, ААС или РМС. Пациенты, которые один раз переносили рецидив, с большей вероятностью перенесут его снова.
Как использовано в данном описании, условия протекания инфекции С. difficile включают, например, слабую, от слабой до умеренной, умеренную и тяжелую инфекцию С. difficile. Условие протекания инфекции С. difficile может меняться в зависимости от наличия и/или тяжести симптомов инфекции.
Симптомы инфекции С. difficile могут включать физиологические, биохимические, гистологические и/или поведенческие симптомы, такие как, например, диарея; колит; колит со спазмами, лихорадка, лейкоциты в кале и воспаление по результатам биопсии толстой кишки; псевдомембранозный колит; гипоальбуминемия; анасарка; лейкоцитоз; сепсис; абдоминальная боль; бессимптомное носительство; и/или осложнения и промежуточные патологические фенотипические признаки, присутствующие в процессе развития инфекции, и их комбинации и т.д. Соответственно, например, введение эффективного количества композиций, описанных в данном изобретении, может, например, лечить, предупреждать, диарею, абдоминальную боль, спазмы, лихорадку, воспаление по результатам биопсии толстой кишки, гипоальбуминемию, анасарку, лейкоцитоз, сепсис и/или бессимптомное носительство и т.д., а также снижать риск, ослаблять тяжесть, уменьшать число случаев и/или замедлять начало указанных заболеваний, по сравнению с млекопитающим, которому данная композиция не была введена.
Факторы риска инфекции С. difficile могут включать, например, применение в настоящее время или в ближайшем будущем антимикробного средства (при этом учитывается любой антимикробный агент с антибактериальным спектром и/или активностью, направленной на анаэробные бактерии, в том числе, например, антибиотики, которые вызывают уничтожение нормальной микробиоты толстой кишки, например, клиндамицин, цефалоспорины, метронидазол, ванкомицин, фторхинолоны (включая левофлоксацин, моксифлоксацин, гатифлоксацин и ципрофлоксацин), линезолид и т.д.); отмену в настоящее время или в ближайшем будущем назначенного метронидазола или ванкомицина; госпитализацию в настоящее время или в ближайшем будущем в медицинское учреждение (такое как больница, лечебное учреждение для хронических больных и т.д.) и медработников; лечение в настоящее время или в ближайшем будущем ингибиторами протонного насоса, Н2-антагонистами и/или метотрексатом или их комбинациями; наличие в настоящее время или риск желудочно-кишечных заболеваний, таких как воспалительное заболевание кишечника; осуществление у млекопитающего в прошлом, в настоящее время или в ближайшем будущем хирургической операции в области желудочно-кишечного тракта или процедуры в области желудочно-кишечного тракта; рецидив инфекции С. difficile и/или CDAD в прошлом или в настоящее время, например, у пациентов, у которых инфекция С. difficile и/или CDAD была один раз или более одного раза; и людей в возрасте по меньшей мере примерно 65 и выше.
В способах, изложенных в данном описании, млекопитающим может быть любое млекопитающее, такое как, например, мыши, хомяки, приматы и люди. В предпочтительном воплощении млекопитающим является человек. Согласно настоящему изобретению, человек может представлять собой индивидуумов, у которых уже была обнаружена инфекция С. difficile, ассоциированные с С. difficile заболевание, синдром, состояние, симптом и/или их осложнение; индивидуумов, у которых в настоящий момент имеется инфекция С. difficile, ассоциированные с С. difficile заболевание, синдром, состояние, симптом и/или их осложнение; и индивидуумов с риском инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения.
Типичные индивидуумы, которые уже демонстрировали симптомы инфекции С. difficile, включают индивидуумов, которые уже демонстрировали или демонстрируют в настоящий момент описанные выше симптомы; индивидуумов, которые имели или имеют в данный момент инфекцию С. difficile и/или ассоциированное с С. difficile заболевание (CDAD); и индивидуумов, которые имеют рецидив инфекции С. difficile и/или CDAD.
Примеры пациентов с риском инфекции С. difficile включают индивидуумов с риском прохождения планового применения антимикробных средств или проходящих его в данный момент; индивидуумов с риском подвергания или подвергающихся в данный момент синдрому отмены назначенного метронидазола или ванкомицина; индивидуумов с риском прохождения плановой госпитализации в медицинское учреждение (такое как больница, лечебное учреждение для хронических больных и т.д.) или проходящих ее в данный момент и медработников; и/или индивидуумов с риском прохождения планового лечения или проходящих в данный момент плановое лечение ингибиторами протонного насоса, Н2-антагонистами и/или метотрексатом или их комбинацией; индивидуумов, которые подвергались или подвергаются в данный момент желудочно-кишечным заболеваниям, таким как воспалительное заболевание кишечника; индивидуумов, которые подвергались или подвергаются в данный момент хирургической операции в области желудочно-кишечного тракта или процедурам в области желудочно-кишечного тракта; и индивидуумов, которые подвергались или подвергаются в данный момент рецидиву инфекции С. difficile и/или CDAD, например, пациентов, у которых инфекция С. difficile и/или CDAD была один раз или более одного раза; индивидуумов, возраст которых составляет примерно 65 и выше. Имеющие такой риск пациенты могут вскоре продемонстрировать симптомы инфекции С. difficile или могут не продемонстрировать данные симптомы.
В случае бессимптомных пациентов профилактику и/или лечение можно начинать в любом возрасте (например, в возрасте примерно 10, 20 или 30 лет). Тем не менее, согласно одному из воплощений, нет необходимости начинать лечение до тех пор, пока пациент не достигнет возраста по меньшей мере примерно 45, 55, 65, 75 или 85 лет. Например, композиции, описанные в данном изобретении, можно вводить бессимптомному пациенту-человеку, возраст которого составляет 50-85 лет.
В одном из воплощений способ предупреждения, снижения риска, ослабления тяжести, уменьшения числа случаев и/или задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего включает введение эффективного количества композиции, описанной в данном изобретении, млекопитающему, нуждающемуся в этом, млекопитающему с риском инфекции С. difficile и/или млекопитающему, восприимчивому к инфекции С. difficile. Эффективное количество включает, например, количество, достаточное для предупреждения, снижения риска, ослабления тяжести, уменьшения числа случаев и/или задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данную композицию не вводят. Введение эффективного количества композиций, описанных в данном изобретении, может, например, предотвращать, снижать риск, ослаблять тяжесть, уменьшать число случаев и/или задерживать начало диареи; абдоминальной боли, спазмов, лихорадки, воспаления по результатам биопсии толстой кишки, гипоальбуминемии, анасарки, лейкоцитоза, сепсиса и/или бессимптомного носительства и т.д. по сравнению с млекопитающим, которому данная композиция не была введена. В предпочтительном воплощении способ включает введение эффективного количества иммуногенной композиции, описанной в данном изобретении, млекопитающему, нуждающемуся в этом, млекопитающему с риском инфекции С. difficile и/или млекопитающему, восприимчивому к инфекции С. difficile.
В дополнительном воплощении способ лечения, ослабления тяжести и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего включает введение эффективного количества композиции, описанной в данном изобретении, млекопитающему, предположительно имеющему инфекцию С. difficile или страдающему в настоящее время от инфекции С. difficile. Эффективное количество включает, например, количество, достаточное для лечения, ослабления тяжести и/или для задержки начала инфекции С. difficile, ассоциированного с С. difficile заболевания, синдрома, состояния, симптома и/или их осложнения у млекопитающего по сравнению с млекопитающим, которому данную композицию не вводят.
Введение эффективного количества композиции может улучшить по меньшей мере один признак или симптом инфекции С. difficile у субъекта, например, у такого, который описан ниже. Введение эффективного количества композиций, описанных в данном изобретении, может, например, ослаблять тяжесть и/или уменьшать число случаев диареи; ослаблять тяжесть и/или уменьшать число случаев абдоминальной боли, спазмов, лихорадки, воспаления по результатам биопсии толстой кишки, гипоальбуминемии, анасарки, лейкоцитоза, сепсиса и/или бессимптомного носительства и т.д. по сравнению с млекопитающим, которому данная композиция не была введена. Возможно, что наличие симптомов, признаков и/или факторов риска инфекции определяют до начала лечения. В предпочтительном воплощении способ включает введение эффективного количества антитела и/или его связывающего фрагмента, описанного в данном изобретении, млекопитающему, предположительно имеющему инфекцию С. difficile или страдающему в настоящее время от инфекции С. difficile.
Соответственно, эффективное количество композиции относится к количеству, достаточному для достижения желаемого эффекта (например, профилактического и/или терапевтического эффекта) в способах по настоящему изобретению. Например, количество иммуногена для введения может изменяться от минимального значения примерно 1 мкг, 5 мкг, 25 мкг, 50 мкг, 75 мкг, 100 мкг, 200 мкг, 500 мкг или 1 мг до максимального значения примерно 2 мг, 1 мг, 500 мкг, 200 мкг на одну инъекцию. Любое минимальное значение можно комбинировать с любым максимальным значением для выявления подходящего диапазона. Обычно, для каждой инъекции человеку используют примерно 10, 20, 50 или 100 мкг иммуногена.
Количество композиции по изобретению, вводимой субъекту, может зависеть от типа и тяжести инфекции и/или от характеристик индивидуума, таких как общее состояние здоровья, возраст, пол, масса тела и переносимость лекарственных средств. Оно также может зависеть от степени, тяжести и типа заболевания. Эффективное количество также может варьировать в зависимости от таких факторов, как путь введения, место целевого воздействия, физиологическое состояние пациента, возраст пациента, является ли пациент человеком или животным, другие применяемые терапии и является ли лечение профилактическим или терапевтическим. Специалист в данной области будет способен определить соответствующие количества в зависимости от этих и других факторов.
Для применения в способах, изложенных в данном описании, эффективное количество может включать одну эффективную дозу или несколько эффективных доз (как, например, 2, 3, 4 дозы или более). Для оптимизации безопасности и эффективности может потребоваться титрование эффективных дозировок.
Сочетание количества и частоты введения дозы, адекватное для осуществления профилактического и/или терапевтического применений, определяется как профилактически или терапевтически эффективный режим лечения. В случае профилактического и/или терапевтического режима лечения обычно вводят более чем в одну дозировку композиции до тех пор, пока не будет достигнут надлежащий иммунный ответ. Обычно выполняют мониторинг иммунного ответа и, если иммунный ответ начинает ослабевать, вводят повторные дозировки.
Можно вводить нескольких дозировок композиции в течение некоторого периода времени. Можно выполнять мониторинг лечения, анализируя во времени антительные ответы или ответы активированных Т-клеток или В-клеток на терапевтический агент (например, иммуногенную композицию, содержащую мутантный токсин из С. difficile). Ослабление ответа является показанием к введению бустер-дозировки.
ПРИМЕРЫ
Пример 1. Идентификация токсин-негативных штаммов С. difficile
Для идентификации штаммов С. difficile, не содержащих гены токсинов (А и В) и не демонстрирующих экспрессию токсинов, тестировали 13 штаммов С. difficile. Культуральные среды 13 штаммов С. difficile тестировали посредством ELISA в отношении токсина А. Семь штаммов экспрессировали токсин А: С. difficile 14797-2, С. difficile 630, С. difficile BDMS, С. difficile W1194, С. difficile 870, С. difficile 1253 и С. difficile 2149. См. Фиг. 3.
Шесть штаммов не экспрессировали токсин А и не содержали целый локус патогенности: С. difficile 1351 (АТСС 43593™), С. difficile 3232 (АТСС ВАА-1801™), С. difficile 7322 (АТСС 43601™), С. difficile 5036 (АТСС 43603™), С. difficile 4811 (4 АТСС 3602™) и С. difficile VPI 11186 (АТСС 700057™). VPI 11186 был выбран на основании его эффективности принимать плазмидную ДНК путем конъюгации.
Те же 13 штаммов тестировали в анализе с применением множественной ПЦР с использованием праймеров за пределами локуса патогенности (PaLoc; Braun et al., Gene, 1996, Nov 28, 181(1-2): 29-38). Результаты ПЦР для этих 6 штаммов, негативных в отношении токсина А по данным ELISA, демонстрировали отсутствие амплификации ДНК каких-либо генов из PaLoc (tcdA-tcdE). Присутствовали последовательности, фланкирующие PaLoc (cdd3 и cdu2) (данные не показаны).
Пример 2. Инактивация пути спорообразования у С. difficile VP111186
Инактивация спорообразующей функции промышленного штамма С. difficile облегчает крупномасштабную ферментацию в безопасных условиях производства. Для создания аспорогенного штамма С. difficile использовали систему ClosTron. См. Heap et al., J. Microbiol. Methods, 2009 Jul, 78(1): 79-85. Система ClosTron позволяет осуществлять направленную инактивацию генов с использованием интрона группы II в случае сайт-специфической инсерционной инактивации гена spo0Al у клостридий. Для токсин-негативного промышленного штамма VPI 11186 проводили инактивацию спорообразования, используя технологию ClosTron. Отбирали мутанты, устойчивые к эритромицину, и наличие инсерционной кассеты подтверждали с использованием ПЦР (данные не показаны). Была подтверждена неспособность двух независимых клонов образовывать споры.
Пример 3. Генетическая модификация генов токсинов А и В с целью инактивации цитотоксической функции
Открытые рамки считывания (ORF) генов полноразмерных мутантных токсинов А и В на основе геномных последовательностей штамма 630Δ были сконструированы и синтезированы под заказ в Blue Heron Biotech. См., например, SEQ ID NO: 9-14. Активный центр, связанный с проявлением глюкозилтрансферазной активности, обуславливающей клеточную токсичность, был изменен в результате двух аллельных замен: D285A/D287A (см. SEQ ID NO: 3) для токсина А и D286A/D288A (см. SEQ ID NO: 5) для токсина В. Были внесены мутации в два нуклеотида в каждом кодоне аспарагиновой кислоты (D) с целью создания кодона для аланина (А). См., например, SEQ ID NO: 9-14. Помимо этого, под заказ в Blue Heron Biotech была сконструирована пара векторов, экспрессирующих мутантные токсины, не содержащие остатков цистеина. Семь остатков цистеина в мутантном токсине А и 9 остатков цистеина в мутантном токсине В были заменены на аланин. Такие замены включают замены каталитических остатков цистеина токсинов А и В, осуществляющих аутокаталитический протеолиз. Кроме того, там, где это необходимо, были введены «молчащие» мутации для устранения сайтов узнавания ферментами рестрикции, использованных при конструировании векторов.
Пример 4. Экспрессирующий вектор рМТL84121fdx
Плазмидный «челночный» вектор, использованный для экспрессии антигена мутантного токсина С. difficile, был выбран из модульной системы серии pMTL8000, разработанной в лаборатории проф. Минтона (Minton lab) (см. Heap et al., J. Microbiol. Methods. 2009, Jul, 78(1): 79-85). Выбранный вектор pMTL84121fdx содержит Gram+ репликон плазмиды С. difficile pCD6, селектируемый маркер catP (хлорамфеникол/тиамфеникол), Gram- репликон р15а и осуществляющий функцию переноса ген tra (tra function) и промотор фередоксина (fdx) С.sporogenes и отдаленный сайт множественного клонирования (MCS). Экспериментальные данные подтвердили, что низкокопийный репликон р15а обеспечивает большую стабильность в Е. coli, чем альтернативный Со1Е1. Промотор fdx был выбран в силу того, что он дает более высокий уровень экспрессии, чем другие промоторы, протестированные в экспериментах с репортерными конструкциями на основе генов CAT (хлорамфениколацетилтрансфераза) (например, tcdA, tcdB; или гетерологическими tetR или xylR) (данные не показаны).
Пример 5. Клонирование модифицированных ORF токсинов в pMTL.84121 fdx
Полноразмерные открытые рамки считывания (ORF) мутантных токсинов А и В на основе геномных последовательностей штамма 630Δ субклонировали, используя сайты множественного клонирования NdeI и BglII вектора pMTL84121fdx, с применением стандартных методов молекулярной биологии. Для облегчения клонирования ORF были фланкированы расположенным в непосредственной близости сайтом NdeI, содержащим стартовый кодон, и сайтом BglII непосредственно вниз по течению относительно стоп-кодона.
Пример 6. Сайт-специфический мутагенез TcdA для создания тройного мутанта
Выполняли замену каталитического остатка цистеина домена протеазы, осуществляющей аутокаталитический протеолиз (т.е. С700А для TcdA и С698А для TcdB) в SEQ ID NO: 3 и 5, т.е. в каждом из «двойных мутантов». Чтобы провести мутагенез мутантного токсина А, фрагмент NdeI-HindIII размером 2,48 т.о. из экспрессирующей плазмиды TcdA D285A/D287A субклонировали в pUC19 (разрезанный (рестриктазами) таким же образом) и сайт-специфический мутагенез проводили на этой матрице. После подтверждения наличия новых аллелей в анализе последовательности ДНК модифицированные фрагменты NdeI-HindIII повторно вводили в экспрессирующий вектор pMTL84121fdx для создания «тройных мутантов», т.е. SEQ ID NO: 4 и SEQ ID NO: 6.
Пример 7. Сайт-специфический мутагенез TcdB для создания тройного мутанта
Чтобы провести мутагенез мутантного токсина В, фрагмент NdeI-EcoHI размером 3,29 т.о. из плазмиды для мутантного токсина В модифицировали и вводили повторно. Поскольку в имеющихся в наличии клонирующих векторах сайт EcoNI отсутствует, в pUC19 (полученном с использованием NdeI-SmaI) субклонировали фрагмент NdeI-EcoRV чуть большего размера 3,5 т.о. После проведения мутагенеза модифицированный внутренний фрагмент NdeI-EcoNI размером 3,3 т.о. «вырезали» и использовали для замены соответствующего фрагмента экспрессирующего вектора рМТL84121fdх для мутантного токсина В. Поскольку было обнаружено, что эффективность клонирования с использованием этой направленной стратегии была достаточно низкой, параллельно была предпринята попытка применения альтернативной стратегии для введения аллеля С698А, включающей замену на фрагмент размером 1,5 т.о., получаемый под действием DraIII. В результате применения обеих стратегий независимо были получены желаемые рекомбинанты.
Пример 8. Создание дополнительных вариантов мутантного токсина с использованием сайт-специфического мутагенеза
Было сконструировано по меньшей мере двенадцать разных вариантов мутантных токсинов С. difficile. С использованием сайт-специфического мутагенеза (набор Quickchange®) в участки генов, кодирующих N-концевые фрагменты мутантных токсинов, были введены аллельные замены. Кроме того, были разработаны рекомбинантные токсины в качестве референсных контролей сравнения для оценки способности этой основанной на использовании плазмид системы вырабатывать белок, эквивалентный в количественном выражении по биологической активности природным токсинам, очищенным из штаммов С. difficile дикого типа. В этом случае, аллельные замены вводили для восстановления исходной глюкозилтрансферазной активности. Помимо этого, под заказ в Blue Heron Biotech была сконструирована пара векторов, экспрессирующих мутантные токсины, не содержащие остатков цистеина.
Эти двенадцать вариантов токсинов включают (1) мутантный токсин А из С. difficile, имеющий мутацию D285A/D287A, (SEQ ID NO: 3); (2) мутантный токсин В из С. difficile, имеющий мутацию D286A/D288A, (SEQ ID NO: 5); (3) мутантный токсин А из С. difficile, имеющий мутацию D285A/D287A С700А, (SEQ ID NO: 4); (4) мутантный токсин В из С. difficile, имеющий мутацию D286A/D288A С698А, (SEQ ID NO: 6); (5) рекомбинантный токсин А, имеющий SEQ ID NO: 1; (6) рекомбинантный токсин В, имеющий SEQ ID NO: 2; (7) мутантный токсин А из С. difficile, имеющий мутацию С700А; (8) мутантный токсин В из С. difficile, имеющий мутацию С698А; (9) мутантный токсин А из С. difficile, имеющий мутацию С700А, C597S, C1169S, C1407S, C1623S, C2023S и C2236S; (10) мутантный токсин В из С. difficile, имеющий мутацию С698А, C395S, C595S, C824S, C870S, C1167S, C1625S, C1687S и C2232S; (11) мутантный токсин А из С. difficile, имеющий мутацию D285A, D287A, С700А, D269A, R272A, Е460А и R462A, (SEQ ID NO: 7); и (12) мутантный токсин В из С. difficile, имеющий мутацию D270A, R273A, D286A, D288A, D461A, К463А и С698А, (SEQ ID NO: 8).
Мутантные по пяти положениям токсины также конструировали сайт-специфическим мутагенезом и с использованием тех же материалов и методов, которые описаны выше, например, в примерах 1-7. Мутантный по пяти положениям токсин В включал следующие замены D286A/D288A/C698A/E970K/E976K (SEQ ID NO: 184). Мутантный по пяти положениям токсин В экспрессировали в VPI 11186 spoOA-негативных клетках, как описано выше. Для подтверждения экспрессии мутантного по пяти положениям токсина В проводили вестерн-блоттинг с использованием mAb №В8-26 (SEQ ID NO: 49), специфичного к N-концевому эпитопу токсина В. На второй, третьей и четвертой дорожках, начиная с левого края, в качестве белка сравнения использовали 50 нг, 30 нг и 10 нг очищенного тройного мутантного токсина В (SEQ ID NO: 86), соответственно. На пятой и шестой дорожках, начиная с левого края, анализировали концентрат клеточного лизата, содержащего мутантный по пяти положениям токсин В, в разведении 1:100 и в разведении 1:1000, соответственно. По оценкам, количество белка в концентрированном растворе мутантного по пяти положениям токсина В составляет примерно 1000 мкг/мл. Как показано на блоте, мутантный по пяти положениям токсин В (концентрированный) демонстрирует белковую зону ожидаемого размера при примерно 270 кДа.
Пример 9. Стабильность трансформантов
Реаранжированные плазмиды получали с применением широко используемого лабораторного штамма Е. coli DH5α. Для сравнения, в результате трансформации с использованием в качестве хозяина Е. coli Stbl2™ от Invitrogen получали медленно растущие рекомбинанты полноразмерного мутантного токсина после трех суток роста при 30°С на чашках Петри на LB-среде с хлорамфениколом (25 мкг/мл). Более низкую эффективность клонирования получали при использовании родственных штаммов Е. coli Stbl3™ и Stbl4™, несмотря на то, что было установлено, что эти линии стабильны в плане поддержания плазмид. Далее трансформанты выращивали в агаре или в жидкой культуре при 30°С, используя для селекции хлорамфеникол. Также было обнаружено, что применение LB-сред (по Миллеру) улучшает выход и рост трансформантов в сравнении со средами на основе сои, не содержащими триптона животного происхождения.
Пример 10. Трансформация С. difficile плазмидой pMTL84121fdx, кодирующей гены токсинов дикого типа или генетически мутантных токсинов
Трансформацию С. difficile с использованием конъюгационного переноса в Е. coli проводили по существу так, как описано в Heap et al., Journal of Microbiological Methods, 2009. 78(1): p.79-85. Клетки хозяина Ε. coli CA434 трансформировали плазмидой pMTL84121 fdx, кодирующей гены токсинов дикого типа или вариантов мутантного токсина. Клетка-хозяин Е. coli СА434 представляет собой промежуточное звено, необходимое для перенесения экспрессирующих плазмид в промышленный штамм С. difficile VPI 11186 spo0AI. СА434 представляет собой производное Е. coli НВ101. Этот штамм несет конъюгативную плазмиду Tra+Mob+R702, которая придает устойчивость к канамицину (Km), тетрациклину (Тс), сульфаниламидам (Su), стрептомицину/спектиномицину (Sm/Spe) и гигромицину (Нg) (обусловленную Тn1831). Готовили химически компетентные или электрокомпетентные клетки СА434 и отбирали трансформанты, содержащие экспрессирующий вектор, на чашках со средой LB по Миллеру с хлорамфениколом (САМ) при 30°С. Отбирали медленно растущие колонии, появляющиеся через 3 суток, и накапливали в 3 мл культуральных сред LB с хлорамфениколом до середины логарифмической фазы роста (приблизительно 24 ч, 225 об./мин, орбитальный шейкер при 30°С). Культуры Е. coli собирали посредством низкоскоростного (5000×g) центрифугирования, чтобы избежать повреждения пилей, и клеточный осадок после центрифугирования осторожно ресуспендировали в 1 мл забуференного фосфатом физиологического раствора (PBS), используя пипетку для переноса материала с широким отверстием. Клетки концентрировали центрифугированием на малых оборотах. Большую часть PBS удаляли, выливая через край, и подсушенные осадки после центрифугирования переносили в камеру для культивирования анаэробов и ресуспендировали в 0,2 мл культуры С. difficile, наносили в виде пятен на чашки с агаром BHIS (агар с сердечно мозговым экстрактом, дополненным 1-цистеином, гемином и NaHCO3) и оставляли расти в течение 8 ч или в течение ночи. В случае трансформантов мутантного токсина А более хороших результатов достигали с применением конъюгирования в течение ночи. Клеточные бляшки соскабливали в 0,5 мл PBS и 0,1 мл высевали на чашки с селективной средой BHIS, дополненной тиамфениколом в концентрации 15 мкг/мл (более мощным аналогом хлорамфеникола) и смесью D-циклосерина/цефокситина для уничтожения донорных клеток Е. coli. Трансформанты, появляющиеся через 16-24 ч, очищали посредством повторного посева штрихом на новую чашку с BHIS (вместе с добавками) и после этого культуры тестировали на предмет экспрессии рекомбинантных токсинов или мутантных токсинов. Из клонов, демонстрирующих хорошую экспрессию, готовили образцы, устойчивые при хранении в глицерине, и концентрированные образцы для затравки. Также готовили минипрепараты (minipreps) плазмид из 2 мл культур, используя модифицированную методику с применением набора Qiagen, в которой клетки предварительно обрабатывали лизоцимом (не обязательно). ДНК минипрепаратов С. difficile использовали в качестве матрицы для ПЦР-секвенирования с целью подтверждения целостности клонов. Альтернативно, из трансформантов Stbl2™ Ε. coli готовили максипрепарат (maxiprep) плазмидной ДНК и проводили секвенирование.
Пример 11. Анализ экспрессии в С. difficile тройного мутантного токсина А и В (SEQ ID NO: 4 и 6, соответственно) и мутантного по семи положениям токсина В (SEQ ID NO: 8)
Трансформанты выращивали или в культурах объемом 2 мл (для рутинного анализа), или в колбах с закрытым отводным отверстием (для экспериментов по определению зависимости от времени). Образцы (2 мл) центрифугировали в течение непродолжительного времени (10000 об./мин, 30 с) для концентрирования клеток; супернатанты декантировали и концентрировали 10х (Amicon-ultra, 30 кДа); осадки после центрифугирования подсушивали и замораживали при -80°С. Осадки клеток после центрифугирования размораживали на льду, ресуспендировали в 1 мл лизирующего буфера (трис-HCl, рН 7,5; 1 мМ EDTA (этилендиаминтетраукусная кислота, 15% глицерина) и обрабатывали ультразвуком (импульс 1×20 с при использовании микронаконечника). Лизат центрифугировали при 4°С и супернатант концентрировали в 5 раз. Образцы супернатанта и лизата объединяли с образцом буфера и прогревали (10 мин, 80°С), затем наносили на два одинаковых, приготовленных на трис-ацетате 3-8%-ных геля для SDS-PAGE (Invitrogen). Один гель окрашивали Кумасси, второй подвергали электроблоттингу для выполнения вестерн-анализа. Чтобы обнаружить варианты мутантных токсинов А и В, использовали образцы специфичной к токсину А и специфичной к токсину В кроличьей антисыворотки (Fitgerald; Biodesign). Экспрессию мутантного по семи положениям токсина В (SEQ ID NO: 8) также подтверждали вестерн-блот-гибридизацией.
Пример 12. Устранение глюкозилтрансферазной активности мутантных токсинов
Двойные генетически мутантные (DM) токсины А и В (SEQ ID NO: 3 и 5, соответственно) и тройные генетически мутантные (ТМ) токсины А и В (SEQ ID NO: 4 и 6, соответственно) не переносили 14С-глюкозу на 10 мкг ГТФаз RhoA, Rac1 и Cdc42 в анализах глюкозилирования in vitro в присутствии УДФ-14С-глюкозы [30 мкМ], 50 мМ HEPES (N-2-гидроксиэтил-пиперазин-N-2-этансульфоновая кислота), рН 7,2, 100 мМ KCl, 4 мМ МgСl2, 2 мМ МnСl2, 1 мМ DTT (дитиотреит) и BSA (бычий сывороточный альбумин) в концентрации 0,1 мкг/мкл. Однако, контрольные токсины А и В дикого типа (имеющие SEQ ID NO: 1 и 2, соответственно) эффективно переносили 14С-глюкозу на ГТФазы в низкой дозе 10 и 1 нг каждый (и ниже - данные не показаны) (Фиг. 4А и 4В) даже в присутствии 100 мкг мутантного токсина (Фиг. 4 В), что указывало по меньшей мере на 100000-кратное уменьшение в сравнении с соответствующими токсинами дикого типа. Похожие результаты были обнаружены для ГТФазы Cdc42 (данные не показаны).
Конкретно, на Фиг. 4В, токсин А и токсин В дикого типа (1 нг) или тройной мутантный токсин А и тройной мутантный токсин В (100 мкг) инкубировали с ГТФазой RhoA в присутствии УДФ-14С-глюкозы в течение 2 ч при 30°С. Как проиллюстрировано, 1 нг TcdA и TcdB дикого типа переносили 14С-глюкозу на RhoA, однако 100 мкг тройного мутантного токсина А и тройного мутантного токсина В нет. В том случае, когда в реакционную смесь вносили 1 нг TcdA или TcdB дикого типа вместе с 100 мкг соответствующего тройного мутантного токсина А или тройного мутантного токсина В, обнаруживали глюкозилирование RhoA, что указывало на отсутствие ингибиторов глюкозилирования. Была установлена чувствительность обнаружения для глюкозилирующей активности, составляющая 1 нг токсина дикого типа на фоне 100 мкг мутантного токсина (соотношение 1:100000). Данные результаты показывают, что мутации в активном центре глюкозилтрансферазы в тройном мутантном токсине А и тройном мутантном токсине В снижали любую измеряемую (менее чем в 100000 раз активность по сравнению с активностью соответствующих токсинов дикого типа) глюкозилтрансферазную активность. Кроме того, для количественного определения глюкозилтрансферазной активности был разработан похожий анализ с использованием осаждения трихлоруксусной кислотой (ТСА) глюкозилированных ГТФаз.
Пример 13. Устранение аутокаталитической активности цистеиновой протеазы
Функцию аутокаталитического расщепления у тройных генетических мутантов (ТМ) А и В (SEQ ID NO: 4 и 6, соответственно) устраняли, когда подвергали мутациям сайт расщепления для цистеиновой протеазы. Как проиллюстрировано на Фиг. 5, токсины А и В дикого типа (wt) (SEQ ID NO: 3 и 5, соответственно) расщепляются в присутствии инозит-6-фосфата. Двойные мутантные токсины А и В (SEQ ID NO: 3 и 5, соответственно) также расщепляются в присутствии инозит-6-фосфата (данные не показаны) аналогично токсинам дикого типа. Токсин A (SEQ ID NO: 3; 308 кДа) расщепляется на 2 фрагмента 245 и 60 «Да. Токсин В (SEQ ID NO: 5; 270 кДа) расщепляется на два фрагмента 207 и 63 кДа. Тройные генетические мутантные (ТМ) токсины А и В (SEQ ID NO: 4 и 6, соответственно) остаются неизменными с массой 308 и 270 кДа, соответственно, даже в присутствии инозит-6-фосфата. См. Фиг. 5. Таким образом, активность цистеиновой протеазы была устранена в результате генетической модификации.
Более конкретно, как показано на Фиг. 5, по одному мкг тройного мутанта А и тройного мутанта В инкубировали в течение 90 минут при комнатной температуре (21±5°С) параллельно с TcdA и TcdB дикого типа от List Biologicals. Реакцию расщепления проводили в пробе объемом 20 мкл, содержащей трис-HCI, рН 7,5, 2 мМ DTT в присутствии или в отсутствие инозит-6-фосфата (10 мМ для TcdA и 0,1 мМ для Tcd В). Затем реакционную смесь в полном объеме наносили на 3-8%-ный гель для проведения SDS/PAGE; белковые зоны визуализировали окрашиванием серебром. Как проиллюстрировано, Tcd А и TcdB wt в присутствии инозит-6-фосфата расщеплялись на две белковые зоны 245 кДа и 60 кДа и, соответственно, 207 кДа и 63 кДа. Тройной мутантный токсин А и тройной мутантный токсин В не расщеплялись, подтверждая тем самым, что мутация С700А в тройном мутантном токсине А и мутация С698А в тройном мутантном токсине В блокировали расщепление.
Пример 14. Остаточная цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно)
Генетически мутантные токсины оценивали в отношении их цитотоксичности в анализе цитотоксичности in vitro на клетках IMR90, диплоидной клеточной линии фибробластов легкого человека. Эти клетки чувствительны к обоим токсинам А и В. В качестве альтернативного предпочтительного воплощения в анализе цитотоксичности могут быть использованы обычные клетки Vero из почки, взятой у Cercopithecus aethiops, так как для них наблюдали надлежащую чувствительность к токсину А и В. Предпочтительно, в анализе цитотоксичности не используют клетки колоректальной аденокарциномы человека НТ-29, поскольку они демонстрировали значительно сниженную чувствительность к токсинам по сравнению с клеточными линиями Vero и IMR90. См., например, приведенную ниже Таблицу 6.

Серийно разведенные образцы генетически мутантного токсина или токсина wt добавляли к монослоям клеток, выращенных в 96-луночных планшетах для тканевых культур. После инкубации при 37°С в течение 72 ч планшеты оценивали на предмет метаболически активных клеток, измеряя уровни АТФ в клетках посредством добавления люциферазного реагента CellTiterGlo® (Promega, Madison, WI), генерирующего люминесценцию, выражаемую в виде относительных единиц люминесценции (RLU). Большие значения RLU указывают на то, что клетки являются жизнеспособными, низкие значения RLU указывают на то, что клетки не являются метаболически активными и находятся в фазе отмирания. Уровень цитотоксичности, выраженный в виде ЕС50, определяют как количество токсина wt или генетически мутантного токсина, которое вызывает уменьшение RLU на 50% по сравнению с уровнями в контрольных клеточных культурах (подробности этого анализа приводятся ниже). Как показано на Фиг. 6, в Таблице 7А и Таблице 8А, величины ЕС50 для TcdA и TcdB составляли примерно 0,92 нг/мл и 0,009 нг/мл, соответственно. Величины ЕС50 для тройного мутантного токсина А и тройного мутантного токсина В составляли примерно 8600 нг/мл и 74 нг/мл, соответственно. Несмотря на приблизительно 10000-кратное снижение цитотоксичности относительно токсинов wt, оба генетически мутантных токсина все еще демонстрировали низкие остаточные уровни цитотоксичности. Эту остаточную цитотоксичность можно было блокировать нейтрализующими моноклональными антителами к токсинам, что указывало на то, что она была специфической для тройных мутантных токсинов, но по всей вероятности не была связана с известными ферментативными активностями токсинов wt (глюкозилированием или аутопротеолизом).
Оба токсина wt демонстрируют сильную цитотоксичность in vitro, при этом небольших количеств токсинов достаточно, чтобы вызвать различные эффекты в клетках млекопитающих, такие как округление клеток (цитопатический эффект или СРЕ) и потерю метаболической активности (по результатам измерений уровней АТФ). В связи с этим были разработаны два анализа in vitro (анализ СРЕ или округления клеток и анализ АТФ) для подтверждения того, что у мутантных токсинов как лекарственных субстанций не сохраняется никакой остаточной цитотоксичности. Результаты выражают в виде величины ЕС5о, которая представляет собой количество токсина или мутантного токсина, вызывающее 1) развитие СРЕ у 50% клеток или 2) уменьшение на 50% уровней АТФ по результатам измерений в относительных световых единицах.
В анализе СРЕ готовят серийные разведения образца лекарственной субстанции и осуществляют инкубацию с клетками IMR90, которые исследуют с точки зрения возможного цитопатического эффекта. Результаты анализа СРЕ оценивают в баллах по шкале от 0 (нормальные клетки) до 4 (округление приблизительно 100% клеток) и балл 2 (округление приблизительно 50% клеток) определяют как величину ЕС50 тестируемого образца. Этот способ используют для тестирования мутантного токсина как лекарственной субстанции в концентрации 1000 мкг/мл, являющейся максимальной переносимой концентрацией, которую можно протестировать этом анализе без (учета) влияния матрикса. Соответственно, никакой детектируемой цитотоксичности не приводится, если ЕС50 превышает 1000 мкг/мл.
Анализ АТФ основан на измерении величины сигнала люминесценции, генерированного с использованием АТФ, который пропорционален числу метаболически активных клеток. Максимальная переносимая концентрация, которую можно протестировать в этом анализе без искажения результатов анализа, составляет примерно 200 мкг/мл. Поэтому никакой детектируемой цитотоксичности в этом анализе не приводится, если ЕС50 превышает 200 мкг/мл.
Мутантные токсины А и В в разных концентрациях добавляли к клеткам параллельно с контрольными токсинами. Конечные результаты данного анализа состояли в определении жизнеспособности клеток по уровням АТФ в клетках с использованием CellTiter-Glo® (Promega). Степень люминесценции пропорциональна уровням АТФ или количеству жизнеспособных клеток.
Цитотоксичность in vitro (ЕС50) для токсина А дикого типа (wt) составляла 920 пкг/мл и 9 пкг/мл для токсина В. Цитотоксичность in vitro (ЕС50) для мутантного токсина A (SEQ ID NO: 4) составляла 8600 нг/мл и 74 нг/мл для мутантного токсина В (SEQ ID NO: 6). Несмотря на то, что эти величины соответствуют снижению в 9348 раз и 8222 раза, соответственно, остаточная цитотоксичность была обнаружена для обоих мутантных токсинов.
Другими словами, цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно) в анализе цитотоксичности in vitro на клетках IMR-90 оказалась значительно меньше цитотоксичности токсинов А и В wt (SEQ ID NO: 1 и 2, соответственно). Как проиллюстрировано на Фиг. 6, несмотря на то, что оба тройных мутантных токсина демонстрировали значительное снижение цитотоксичности (в 104 раз) относительно токсина wt, остаточную цитотоксичность для обоих тройных мутантных токсинов наблюдали в более высоких концентрациях.
Кроме того, остаточную цитотоксичность каждого тройного мутантного токсина можно было полностью нейтрализовать (например, по меньшей мере со снижением токсичности, составляющим 6-8 lоg10 относительно токсичности токсина дикого типа) специфичными к данному токсину антителами. См. приведенный ниже пример 16.
Анализы с применением клеточных культур более чувствительны в отношении обнаружения цитотоксичности, чем животные модели in vivo. При доставке либо внутрибрюшинным (в./б.), либо внутривенным (в./в.) путями в мышиной модели летального заражения TcdA wt имеет LD50 приблизительно 50 нг на одну мышь, тогда как TcdB wt оказывает более сильное действие с LD50 приблизительно 5 нг на одну мышь. В противоположность этому, в описанных выше анализах in vitro, основанных на применении клеточных культур, получают значения ЕС50 100 пг на одну лунку для TcdA wt и 2 пг на одну лунку для TcdB wt.
Пример 15. Остаточная цитотоксичность генетически мутантного по семи положениям токсина В (SEQ ID NO: 8) и цитотоксичность мутантного по пяти положениям токсина В (SEQ ID NO: 184)
Как проиллюстрировано на Фиг. 7, величины ЕС50 оказываются примерно одинаковыми в случае тройного мутантного токсина В (SEQ ID NO: 6) (20,78 нг/мл) и мутантного по семи положениям токсина В (SEQ ID NO: 8) (35,9 нг/мл), и это указывает на то, что четыре дополнительные мутации с целью дальнейшей модификации активного центра глюкозилтрансферазы и субстратсвязывающего сайта ГТФазы не привели к дальнейшему снижению цитотоксичности генетически мутантных токсинов. В случае двойного мутантного токсина В (SEQ ID NO: 5) величины ЕС50 также были схожи с таковыми для токсинов, мутантных по трем и по семи положениям (данные не показаны). Это наблюдение подтверждает, что механизм, обуславливающий цитотоксичность мутантных токсинов, неожиданным образом не зависит от активности глюкозилтрансферазы и механизма распознавания субстрата.
Мутантный по пяти положениям токсин В (SEQ ID NO: 184) оценивали в отношении его цитотоксичности в анализе цитотоксичности in vitro по уровню АТФ в клетках IMR-90, как описано выше. См., например, пример 14. Как показано на Фиг. 26, цитотоксичность мутантного по пяти положениям токсина В оказалась существенно меньше таковой токсина В дикого типа (например, SEQ ID NO: 2), который получали из коммерческих источников от List Biologicals, и тройного мутантного токсина В (SEQ ID NO: 86). Контроль по типу «только клетки» относится к клеткам IMR-90. См. приведенную ниже Таблицу 62, показывающую соответствующие величины ЕС50.

Кроме того, мутантный по пяти положениям токсин В («РМ-В») (SEQ ID NO: 184) оценивали в отношении конкурентного ингибирования цитотоксичности, опосредуемой тройным мутантным токсином В (SEQ ID NO: 86), на клетках IMR-90. См. Фиг. 27. Для приготовления образцов во все лунки вносили тройной мутантный токсин В (ТМ В) (SEQ ID NO: 86) в постоянной концентрации 200 нг/мл (10×ЕС50)- В лунки, содержащие тройной мутантный токсин В, добавляли мутантный по пяти положениям токсин В в 2-кратных серийных разведениях, начиная с 5 мкг/мл. Затем образцы переносили в 96-луночный планшет, содержащий клетки IMR-90, далее инкубировали в течение 72 часов. Анализ АТФ завершали, как описано выше, например, в примере 14. Как можно видеть на Фиг. 27, мутантный по пяти положениям токсин В конкурентно ингибировал цитотоксичность тройного мутантного токсина В.
Пример 16. Остаточная цитотоксичность тройных мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно)
Для дальнейшей оценки природы остаточной цитотоксичности мутантные токсины (SEQ ID NO: 4 и 6) смешивали, инкубировали с соответствующими упомянутыми ранее нейтрализующими их антителами и смесь добавляли к монослою клеток IMR90.
Результаты (Фиг. 8) показали, что остаточную цитотоксичность мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно) можно полностью устранить, используя нейтрализующие антитела, специфичные к мутантному токсину А (верхняя панель, Фиг. 8) и мутантному токсину В (нижняя панель, Фиг. 8). Мутантные токсины А (верхняя панель) и В (нижняя панель) в возрастающих концентрациях инкубировали с кроличьими поликлональными антителами (рАb, разведение 1:10) или мышиными моноклональными антителами к токсину (разведение 1:50 из концентрированного раствора, содержащего IgG в концентрации 3,0 мг/мл), после чего добавляли к клеткам IMR90. После обработки путем инкубации в течение 72 ч с клетками IMR90 при 37°С добавляли субстрат CellTiter-Glo® и относительные световые единицы (RLU) измеряли на спектрофотометре с использованием программы для измерения люминесценции с целью определения уровней АТФ. Чем ниже уровень АТФ, тем выше токсичность. Контроли включали TcdA и TcdB, добавленные в концентрациях, в 4 раза превышающих соответствующие им величины ЕС50.
В опубликованных сообщениях говорится, что мутации в глюкозилтрансферазном или домене аутокаталитического протеолиза у токсинов приводят к полной инактивации токсичности. Однако данные авторов данного изобретения не согласуются с такими опубликованными сообщениями, и это можно было бы отнести к повышению концентраций высокоочищенных мутантных токсинов, протестированных в исследованиях авторов изобретения, в отличие от неочищенных культуральных лизатов, использованных в опубликованных сообщениях; к увеличению промежутков времени, для которых наблюдали округление обработанных мутантными токсинами клеток (например, 24 часа, 48 часов, 72 часа или 96 часов), в отличие от наблюдений, сделанных в течение менее чем 12 часов; к применению клеточных линий, демонстрирующих значительно более высокую чувствительность к токсинам в анализах цитотоксичности по настоящему изобретению, в отличие от клеток колоректальной аденокарциномы человека НТ-29 в анализах цитотоксичности, описанных в опубликованных сообщениях; и/или к неизвестной активности или процессу, отличающимся от гликозилирования, которые могли бы быть движущей силой для проявления остаточной токсичности мутантных токсинов.
Пример 17. Новый механизм цитотоксичности генетически мутантных токсинов
Чтобы исследовать механизм остаточной цитотоксичности генетически мутантных токсинов, клетки IMR-90 обрабатывали токсином В wt (SEQ ID NO: 2) или генетически мутантным токсином В (SEQ ID NO: 6), и глюкозилирование ГТФазы Rac1 изучали в зависимости от времени обработки. Собирали образцы в интервале от 24 до 96 часов и готовили клеточные экстракты. Чтобы отличить глюкозилированную Rac1 от неглюкозилированной Rac1, использовали вестерн-блоттинг с применением двух антител к Rac1. Одно антитело распознает обе формы Rac1 (23А8), а другое (102) распознает только неглюкозилированную Rac1. Как проиллюстрировано на Фиг. 22, в случае токсина В общие уровни Rac1 оставались неизменными во времени, при этом по большей части Rac1 является глюкозилированной. С другой стороны, обработка генетически мутантным токсином В (SEQ ID NO: 6) приводила к значительному снижению общих уровней Rac1, однако для всех моментов времени Rac1 была неглюкозилированной. Это указывает на то, что на уровень Rac1 отрицательно влияет обработка генетически мутантным токсином, а не токсином wt. Как проиллюстрировано на Фиг. 22, в указанные моменты времени уровень актина был одинаков в клетках, обработанных токсином и генетически мутантным токсином В, и одинаков в случае ложнообработанных клеток. Это является демонстрацией того, что механизм цитотоксичности, проявляемый генетически мутантными токсинами, отличается от механизма, представляющего собой путь глюкозилирования, приводимого в действие токсином дикого типа.
Пример 18. Химическая обработка генетически мутантных токсинов
Несмотря на то, что генетически модифицированные мутантные токсины, демонстрировавшие снижение цитотоксической активности, составляющее 4 log, являются предпочтительными, обсуждалась возможность дальнейшего снижения (2-4 log) цитотоксической активности. Проводили оценку двух стратегий химической инактивации.
В первом способе для инактивации мутантных токсинов используют формальдегид и глицин. Инактивация формальдегидом осуществляется в результате образования шиффова основания (имина) между формальдегидом и первичными аминами на белке. Затем шиффовы основания приводят во взаимодействие с рядом аминокислотных остатков (Arg, His, Тrр, Туr, Gin, Asn) с образованием либо внутри-, либо межмолекулярных сшивок. Введение таких сшивок фиксирует структуру белка, делая его неактивным. Кроме того, формальдегид может взаимодействовать с глицином с образованием шиффова основания. Затем это глицил-шиффово основание может взаимодействовать с аминокислотными остатками с образованием межмолекулярных сшивок белок-глицин. Применение формальдегида приводило к снижению цитотоксической активности генетически мутантных токсинов до значений ниже пределов обнаружения (снижение цитотоксичности для тройного мутантного токсина В (SEQ ID NO: 6) составляло более 8 lоg10, а снижение для тройного мутантного токсина A (SEQ ID NO: 4) составляло более 6 lоg10). Однако со временем наблюдали реверсию, когда инактивированные формальдегидом (FI) тройные мутантные токсины инкубировали при 25°С. Цитотоксическую реверсию можно предотвратить путем добавления небольшого количества формальдегида (0,01-0,02%) в растворы FI тройных мутантных токсинов при хранении. См. пример 23.
В другом способе для получения инактивированных мутантных токсинов используют обработку 1-этил-3-(3-диметиламинопропил)карбодиимидом (ЕDС)/N-гидроксисукцинимидом (NHS). В этом способе EDC/NHS взаимодействует с карбоксильными группами на белке с образованием активированных сложных эфиров. Затем активированные эфиры приводят во взаимодействие с первичными аминами на белке с образованием стабильных амидных связей. Как и в случае реакции с использованием формальдегида, в результате этой реакции образуются внутри- и межмолекулярные сшивки. Амидная связь, образованная в результате обработки EDC/NHS, является более стабильной и не подверженной реверсии по сравнению с лабильной иминной связью, образованной в результате инактивации формалином. Помимо сшивок, образованных в результате взаимодействия активированных эфиров с первичными аминами на полипептиде, могут быть получены как глициновые, так и бета-аланиновые аддукты. Без связи с механизмом или теорией считается, что глициновые аддукты получаются тогда, когда глицин добавляют для гашения непрореагировавших активированных эфиров. Аминогруппа глицина взаимодействует с активированным эфиром на полипептиде с образованием стабильных амидных связей. Без связи с механизмом или теорией считается, что бета-аланиновые аддукты образуются в результате взаимодействия активированного бета-аланина с первичными аминами на полипептиде. В результате этой реакции образуются стабильные амидные связи. Активированный бета-аланин получается при взаимодействии трех молей NHS с одним молем EDC.
Чтобы достичь снижения цитотоксической активности 2-4 log относительно генетически модифицированных мутантных токсинов (6-8 log относительно нативных токсинов), химически инактивированные мутантные токсины должны иметь значения ЕС50 не меньше 1000 мкг/мл. Помимо снижения цитотоксической активности было бы выгодно сохранить ключевые эпитопы, определенные с использованием дот-блот-анализа. На сегодняшний день установлен ряд реакционных условий, отвечающих как критериям сниженной цитотоксичности, так и критериям распознавания эпитопов. Было приготовлено несколько партий инактивированных мутантных токсинов для исследований на животных, и данные анализа некоторых репрезентативных партий приведены в Таблицах 7А и 7В, Таблицах 8А и 8В.



Пример 19. Ферментация и очистка
Ферментации начинали, исходя из замороженного источника рекомбинантной Clostridium difficile, содержащей fdx-промотор, описанный выше. Замороженные концентраты представляли собой клеточную суспензию, подведенную до СD600 2,0 и содержащую 20% глицерина. Стартовую культуру инокулировали из расчета 0,2 мл культурального концентрата на 500 мл среды SHYG10 (гидролизат сои SE50MK в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкоза в концентрации 10 г/л, тиамфеникол в концентрации 15 мг/л, рН 7). Среду помещали в имеющую отверстия для сообщения с атмосферой бутыль емкостью 500 мл. Инокулирование выполняли в стандартном боксе биологической безопасности, бутыль продували сильной струей азота и затем содержимое бутыли инкубировали в статичном состоянии (с закрытыми отверстиями) в течение приблизительно 16-18 часов при 37°С в стандартном инкубаторе.
Для фазы ферментации использовали биореактор емкостью десять литров, содержащий 8 л среды SHYG60 (гидролизат сои SE50MK в концентрации 30 г/л, дрожжевой экстракт HY YEST 412 в концентрации 20 г/л, глюкоза в концентрации 60 г/л, тиамфеникол в концентрации 15 мг/л, рН 7). Ферментер, который работал при 37°С, 400 об./мин (1,47 м/с) с барботированием азотом с интенсивностью 0,1 об./об./мин, инокулировали содержимым бутыли с инокулятом емкостью 500 мл. Значение рН регулировали на уровне 7,0, автоматически добавляя 5 н. NaOH. Обычно ферментацию проводили в течение приблизительно 24 часов до достижения максимальной эффективности. Рост в ходе ферментации регистрировали по измерениям СD600. Образцы, отобранные для количественной оценки мутантного токсина, центрифугировали при приблизительно 5000×g и полученные осадки после центрифугирования замораживали при -70°С. Затем осадок клеток после центрифугирования размораживали и ресуспендировали в буфере, содержащем 20 мМ трис, 3 мМ NaCl, 0,5 мМ EDTA, рН 6,5, и обрабатывали ультразвуком с амплитудой 40 в течение 20 секунд. Полученный клеточный лизат центрифугировали при 5000×g в течение 10 мин. Супернатант объединяли с загрузочным буфером и восстанавливающим агентом и подвергали PAGE в 3,8%-ном геле в трис-ацетатном буфере при 150 вольт в течение 50-55 мин параллельно с достоверными стандартами токсинов. Гель окрашивали в течение ночи, затем отмывали от красителя и проводили количественный анализ на сканирующем денситометре. Обычно наблюдали значения OD600 приблизительно 10-12 для концентраций токсинов 80-120 мг/л.
Следующая далее таблица представляет собой пример данных по ферментации мутантного токсина А.

Модификации композиции и способы культивирования рекомбинантной клетки С. difficile и/или получения мутантных токсинов были протестированы и включены в объем данного изобретения. Например, хотя в качестве предпочтительного источника азота был выбран SE50MK (и в отдельных случаях SE50MK-NK, при этом оба поставляются Friesland-Campaigna), был выявлен ряд других гидролизатов сои от других производителей, которые хорошо себя показали в данном процессе. Применение культуральной среды, содержащей гидролизат сои в отсутствие дрожжевого экстракта, обеспечивало 30-40% от ожидаемого выхода.
Было показано, что дрожжевые экстракты от альтернативных производителей способствуют получению эквивалентных выходов. Применение культуральной среды, содержащей дрожжевой экстракт в отсутствие гидролизата сои, обеспечивало 60-70% от ожидаемого выхода.
В результате использования культуральной среды на основе гидролизата сои/дрожжевого экстракта, в отсутствие источника углерода, получали СD600 примерно 2-3 и токсин в концентрации примерно 10-15 мг/л. Несмотря на то, что глюкоза была предпочтительным источником углерода для культивирования, эквивалентные результаты были получены при использовании маннита. Скрининг источников углерода в бутылях показал, что С. difficile также может утилизировать фруктозу и маннозу, что, как ожидается, также будет способствовать высоким значениям OD/выхода по сравнению с культуральной средой в отсутствие источника углерода. Как оказалось, оптимальному росту не способствовали следующие источники углерода: арабиноза, ксилоза, сахароза, лактоза, мальтоза, глицерин, рамноза и галактоза.
Кроме того, оказалось, что увеличение продолжительности ферментации до 48 часов (требующее добавления большего количества глюкозы) по существу не улучшало выхода.
Далее, после проведения ферментации при рН 6,5 и 7,5 получали выходы в ожидаемом диапазоне 80-120 мг/л. После проведения ферментации при более экстремальных значениях рН (рН 6,0 или 8,0) все же получали ожидаемые значения OD600, но выходы токсина (40-60 мг/л) были снижены.
Ферментация с применением культуральной среды в отсутствие тиамфеникола приводила к утрате плазмиды, например, примерно на 10-20% для плазмиды, кодирующей мутантный токсин В, и примерно на 30-40% для плазмиды, кодирующей мутантный токсин А. Соответственно, ферментация с применением культуральной среды в отсутствие производного хлорамфеникола была допустима.
Также были протестированы альтернативные режимы производственной ферментации и включены в объем данного изобретения. Например, ферментация может протекать при 400 об./мин или меньше и может быть использована подача слоя азота, оба эти метода были опробованы и успешно использованы.
Кроме того, была протестирована среда для моноклональных антител SFM4MAb и было показано, что на ней можно получить количество клеток, соответствующее значению OD600 примерно 10, и концентрацию мутантного токсина примерно 40 мг/л.
Наконец, добавление в ферментируемую среду фосфатсодержащего ингредиента по-видимому снижает выработку токсина по сравнению с культуральной средой в отсутствие фосфатсодержащего ингредиента.
По окончании ферментации ферментер охлаждают. Клеточную суспензию извлекают посредством непрерывного центрифугирования и ресуспендируют в соответствующем буфере. Лизиса клеточной суспензии достигают посредством гомогенизации при высоком давлении. В случае мутантного токсина А гомогенат подвергают флокуляции и проводят непрерывное центрифугирование флокулированного раствора. Этот раствор фильтруют и затем переносят для последующей обработки. В случае мутантного токсина В гомогенат осветляют посредством непрерывного центрифугирования и затем переносят для последующей обработки.
Мутантный токсин A (SEQ ID NO: 4) очищают, используя две хроматографические стадии с последующей заменой на конечный буфер. Осветленный лизат загружают на колонку для гидрофобной хроматографии (HIC), и связавшийся мутантный токсин элюируют, используя градиент цитрата натрия. Пул продукта с колонки для HIC затем загружают на катионообменную (СЕХ) колонку, и связавшийся мутантный токсин А элюируют, используя градиент хлорида натрия. В пуле после СЕХ-колонки, содержащем очищенный мутантный токсин А, производят замену на конечный буфер, используя диафильтрацию. В растворе очищенного мутантного токсина А производят замену на конечный буфер для промежуточной лекарственной субстанции, используя диафильтрацию. После диафильтрации ретентат фильтруют через 0,2 микронный фильтр, после чего проводят химическую инактивацию с получением конечной лекарственной субстанции. Концентрацию белка подводят до целевого значения 1-3 мг/мл.
Мутантный токсин В (SEQ ID NO: 6) очищают, используя две хроматографические стадии с последующей заменой на конечный буфер. Осветленный лизат загружают на анионообменную (АЕХ) колонку и связавшийся мутантный токсин элюируют, используя градиент хлорида натрия. К пулу продукта с АЕХ-колонки добавляют цитрат натрия и загружают на колонку для гидрофобной хроматографии (HIC). Связавшийся мутантный токсин элюируют, используя градиент цитрата натрия. В пуле после HIC, содержащем очищенный полипептид мутантного токсина (SEQ ID NO: 6), производят замену на конечный буфер, используя диафильтрацию. В растворе очищенного мутантного токсина В производят замену на конечный буфер для промежуточной лекарственной субстанции, используя диафильтрацию. После диафильтрации ретентат фильтруют через 0,2 микронный фильтр, после чего проводят химическую инактивацию с получением конечной лекарственной субстанции. Концентрацию белка подводят до целевого значения 1-3 мг/мл.
Пример 20. Инактивация формальдегидом/глицином
После очистки генетически мутантные токсины А и В (SEQ ID NO: 4 и 6, соответственно) инактивируют в течение 48 часов при 25°С, используя 40 мМ (1,2 мг/мл) формальдегид. Инактивацию проводят при рН 7,0±0,5 в буфере на основе 10 мМ фосфата, 150 мМ хлорида натрия, содержащем 40 мМ (3 мг/мл) глицин. Продолжительность периода инактивации устанавливают такой, чтобы она в три раза превышала продолжительность периода, необходимого для снижения ЕС50 в клетках IMR90 до значения более чем 1000 мкг/мл. Через 48 часов уменьшение биологической активности составляет 7-8 log10 относительно природного токсина. После 48 часов инкубации, в растворе инактивированного мутантного токсина производят замену на буфер для конечной лекарственной субстанции посредством диафильтрации. Например, используя ультрафильтрационную кассету из регенерированной (из ацетата целлюлозы) целлюлозы с отсечением 100 кДа, инактивированный токсин концентрируют до 1-2 мг/мл и производят замену буфера.
Пример 21. Инактивация N-(3-диметиламинопропил)-N-этилкарбодиимидом (ЕDС)/N-гидроксисукцинимидом (NHS)
После очистки генетически мутантные токсины (SEQ ID NO: 4 и SEQ ID NO: 6, соответственно) инактивируют в течение 2 часов при 25°С, используя 0,5 мг EDC и 0,5 мг NHS на один мг очищенного генетически мутантного токсина А и В (приблизительно 2,6 мМ и 4,4 мМ, соответственно). Реакцию гасят, добавляя глицин до конечной концентрации 100 мМ, и реакционные смеси инкубируют в течение еще 2 часов при 25°С. Инактивацию проводят при рН 7,0±0,5 в буфере на основе 10 мМ фосфата, 150 мМ хлорида натрия. Продолжительность периода инактивации устанавливают такой, чтобы она в три раза превышала продолжительность периода, необходимого для снижения ЕС50 в клетках IMR90 до значения более чем 1000 мкг/мл. Через 2 часа уменьшение биологической активности составляет 7-8 log10 относительно природного токсина. После 4 часов инкубации, в растворе инактивированного мутантного токсина производят замену на буфер для конечной лекарственной субстанции посредством диафильтрации. Например, используя ультрафильтрационную кассету из регенерированной (из ацетата целлюлозы) целлюлозы с отсечением 100 кДа, инактивированный токсин концентрируют до 1-2 мг/мл и производят замену буфера.
Если не указано иное, приведенные ниже термины, использованные в разделе Примеры, относятся к композиции, изготовленной в соответствии с описанием настоящего изобретения в примере 21: «EDC/NHS-обработанный тройной мутантный токсин», «EDC-инактивированный мутантный токсин», «мутантный токсин [А/В] как лекарственная субстанция», «EI мутантный токсин», «EDC/NHS-инактивированный тройной мутантный токсин». Например, приведенные ниже термины являются синонимами: «EDC/NHS-обработанный тройной мутантный токсин А», «EDC-инактивированный мутантный токсин А», «мутантный токсин А как лекарственная субстанция», «EI мутантный токсин А», «EDC/NHS-инактивированный тройной мутантный токсин А». В качестве другого примера, синонимами являются приведенные ниже термины: «EDC/NHS-обработанный тройной мутантный токсин В», «EDC-инактивированный мутантный токсин В», «мутантный токсин В как лекарственная субстанция», «EI мутантный токсин В», «EDC/NHS-инактивированный тройной мутантный токсин В».
Мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию получают, используя периодический процесс, который включает (1) ферментацию токсин-негативного штамма С. difficile (VPI 11186), содержащего плазмиду, кодирующую соответствующий полипептид тройного генетически мутантного токсина (в среде, содержащей гидролизат сои, дрожжевой экстракт HY YEST™ 412 (Sheffield Bioscience), глюкозу и тиамфеникол), (2) очистку генетически мутантного токсина («промежуточной лекарственной субстанции») из освобожденного от клеток лизата с использованием методик ионообменной и гидрофобной хроматографии до чистоты по меньшей мере более 95%, (3) химическую инактивацию посредством обработки EDC/NHS с последующим гашением/блокированием глицином и (4) переведение в матрицу конечного буфера.
Пример 22. Исследования, обосновывающие условия инактивации и изготовления композиций
Для оптимизации химической инактивации генетически мутантных токсинов выполняли статистический расчет эксперимента (DOE). Факторы, рассматриваемые при DOE, включали температуру, концентрацию формальдегида/глицина, концентрацию EDC/NHS и время (Таблицы 9 и 10). Для мониторинга потери биологической активности определяли значения ЕС50 в клетках IMR-90. Кроме того, также наблюдали за морфологией клеток IMR-90 в различные моменты времени после обработки. См. Фиг. 9, демонстрирующую морфологию через 72 часа после обработки. Для определения влияния на структуру белка проводили мониторинг эпитопного распознавания, используя дот-блот-анализ с применением панели моноклональных антител, выработанных в отношении разных доменов токсина.

При инактивации мутантных токсинов из С. difficile формальдегидом/глицином конечные реакционные условия выбирали таким образом, чтобы достигался желаемый уровень снижения цитотоксической активности (7-8 log10) при одновременном улучшении распознавания эпитопов до максимального значения. См. приведенный выше пример 20.
При инактивации мутантных токсинов из С. difficile с использованием EDC/NHS конечные реакционные условия выбирали таким образом, чтобы достигался желаемый уровень снижения цитотоксической активности (7-8 log10) при одновременном улучшении распознавания эпитопов до максимального значения. См. приведенный выше пример 21.
В альтернативном воплощении реакцию EDC-NHS гасили, добавляя аланин, который в достаточной степени гасил данную реакцию. Применение аланина может приводить к модификации белка мутантного токсина, похожей на модификацию в случае гашения реакции глицином. Например, гашение посредством добавления аланина может приводить к присоединению аланиновой группировки к боковой цепи остатка глутаминовой кислоты и/или аспарагиновой кислоты мутантного токсина. В другом альтернативном воплощении реакцию EDC-NHS гасили, добавляя сложный глицинметиловый эфир, который в достаточной степени гасил данную реакцию.
Получение химически инактивированных тройных мутантных токсина А и токсина В из С. difficile в оптимизированных условиях приводило к дальнейшему снижению остаточной цитотоксичности до недетектируемого уровня (больше 1000 мкг/мл - самой высокой концентрации, которая может быть протестирована в анализе СРЕ) при сохранении антигенности по результатам измерений их реакционной способности в отношении токсин-специфических нейтрализующих антител. Результаты, приведенные в Таблице 28, демонстрируют постадийное снижение цитотоксичности от токсина wt к EDC/NHS-обработанным тройным мутантным токсинам. Иммунофлуоресцентное мечение подтверждало, что тройные мутантные токсины (SEQ ID NO: 4 и 6) и мутантные токсины как лекарственные субстанции демонстрировали сравнимое связывание с клетками IMR-90, подтверждая тем самым, что потеря цитотоксичности не обуславливалась ухудшением связывания с этими клетками (данные не показаны). В сравнении с мутантным токсином А как лекарственной субстанцией показано, что для мутантного токсина В как лекарственной субстанции достигали более высокой кратности снижения цитотоксичности, которое соответствует приблизительно в 600 раз более высокой наблюдаемой эффективности TcdB в сравнении с TcdA.
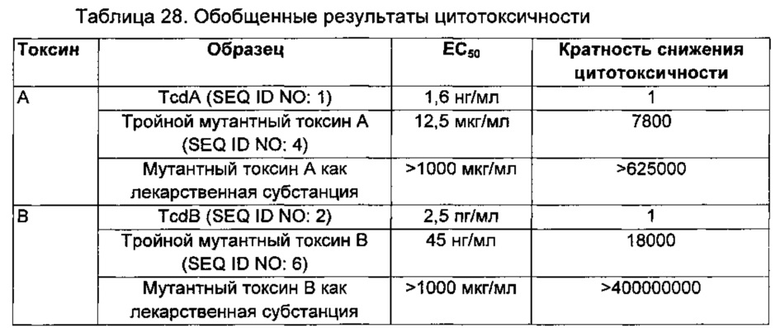
Также оценивали результаты анализа цитотоксичности для мутантного токсина В, модифицированного с использованием только EDC или с использованием EDC и сульфо-NHS. См. Таблицу 29.

Условия. Тройной мутантный токсин В («ТМ TcdB») (SEQ ID NO: 6) модифицировали при массовом соотношении мутантный токсин В:ЕОС:сульфо-NHS, составляющем 1:0,5:0,94. Это соотношение является эквимолярным (скорректированным на более высокое значение молекулярной массы (MW) сульфо-NHS) для стандартной EDC/NHS-реакции, которая описана в примере 21. Чтобы определить влияние сульфо-NHS, соотношение сульфо-NHS варьировали от 0,5х до 2х стандартного соотношения. Реакции в двух повторах проводили в 1х PBS, рН 7,0, при 25°С и инициировали путем добавления раствора EDC. Через 2 часа реакции гасили добавлением 1 М раствора глицина с рН 7,0 (конечная концентрация 0,1 М) и реакционную смесь инкубировали в течение еще 2 часов. Погашенные реакционные смеси подвергали обессоливанию и раствор мутантного токсина В как лекарственной субстанции («ТМ TcdB-EDC») концентрировали с использованием устройств Vivaspin 20, осуществляли стерильное фильтрование в стерильные флаконы и передавали для проведения оценки посредством анализа цитотоксичности.
Применение сульфо-NHS при таком же молярном соотношении уменьшало ЕС50 до примерно 250 мкг/мл по сравнению со значением для NHS, превышающим 1000 мкг/мл. Даже при двукратном молярном соотношении сульфо-NHS, по-видимому, не является столь же эффективным, как NHS в плане снижения цитотоксичности. См. Таблицу 30.
Для определения числа и типа модификаций выполняли пептидное картирование на образцах тройного мутантного токсина В, инактивированного как с использованием EDC/NHS, так и ЕDС/сульфо-NHS. В обоих образцах были обнаружены схожие количества глициновых аддуктов, сшивок и дегидроаланиновых модификаций. Однако в сульфо-NHS-инактивированном образце никакого бета-аланина не было обнаружено.
Токсин В дикого типа (SEQ ID NO: 2) инактивировали, используя стандартный протокол (см. пример 21): соотношение токсин B:EDC:NHS 1:0,5:0,5; 25°С в течение 2 часов в 1х PBS, рН 7,0, затем гашение 1 М глицином (конечная концентрация 0,1 М) и инкубирование в течение еще 2 часов. Образец подвергали обессоливанию, концентрировали и передавали для проведения анализа цитотоксичности. Значения ЕС50 для этих образцов составляли меньше 244 нг/мл.
Пример 23. Исследования реверсии
Для определения того, имеет ли место реверсия у мутантных токсинов С. difficile, инактивированных либо формальдегидом/глицином, либо EDC/NHS, образцы инактивированных мутантных токсинов (1 мг/мл) инкубировали при 25°С в течение пяти-шести недель. Каждую неделю отбирали аликвоты и определяли величины ЕС50 на клетках IMR90. Один образец, инактивированный формальдегидом/глицином, не содержал формальдегид, а один образец содержал 0,01% формальдегида. Величины ЕС50 измеряли посредством анализа СРЕ.

При 25°С в отсутствие остаточного формальдегида наблюдается частичная реверсия (Таблица 11). Через пять недель цитотоксическая активность увеличивалась приблизительно в 3 раза. Несмотря на увеличение цитотоксической активности, через пять недель снижение все еще составляло 7 log10 относительно нативного токсина. Реверсия была полностью предотвращена посредством введения формалина в концентрации 0,010%. Никакой реверсии не наблюдали в образце, инактивированном EDC/NHS. В продолжение всей 6-недельной инкубации значения ЕС50 оставались на исходном уровне, составляющем больше 1000 мкг/мл, для всех четырех партий как EDC/NHS-обработанного тройного мутантного токсина A (SEQ ID NO: 4), так и EDC/NHS-обработанного тройного мутантного токсина В (SEQ ID NO: 6). В противоположность этому, величины ЕС50 для FI-обработанного тройного мутантного токсина A (SEQ ID NO: 4) и FI-обработанного тройного мутантного токсина В (SEQ ID NO: 6) не были стабильными и снижались до неприемлемо низких величин ЕС50, указывая на увеличение цитотоксичности или реверсию инактивации. См. Таблицу 11.
В дополнение к стабильному снижению цитотоксичности до недетектируемого уровня (более 1000 мкг/мл по результатам измерений в анализе СРЕ) мутантные токсины, инактивированные с использованием EDC/NHS, сохраняли важные эпитопы, которые являются мишенями для токсин-нейтрализующих mAb. См. Таблицу 31. Для FI мутантных токсинов была продемонстрирована потеря этих же антигенных детерминант.

Пример 24. Доклинические исследования иммуногенности
Ключевые задачи доклинических исследований включают тестирование композиций, содержащих мутантные токсины А и В из С. difficile, на мелких животных и на отличных от людей приматах (NHP). Мышей и хомяков иммунизировали для определения, среди других параметров, того, могут ли композиции на основе С. difficile способствовать выработке нейтрализующих антител к мутантным токсинам А и В. Антигены тестировали на предмет индуцирования нейтрализующих антительных ответов в сыворотке крови после серии иммунизаций, проведенных мышам, хомякам и яванским макакам. Композиции на основе генетически и/или химически инактивированных мутантных токсинов готовили либо в нейтральном буфере, либо в алюминий-фосфатном буфере, либо в некоторых воплощениях в буфере, содержащем ISCOMATRIX в качестве адъюванта. Обычно нейтрализующие антительные ответы тестировали примерно через две-четыре недели после каждого введения бустерной или конечной дозы.
Анализ нейтрализации токсинов демонстрирует способность антисыворотки нейтрализовать цитотоксический эффект, опосредуемый TcdA или TcdB из С. difficile, и ввиду этого позволяет измерить функциональную активность антител, присутствующих в образце. Анализ нейтрализации токсинов проводили на клеточной линии фибробластов легких человека, IMR-90, которая чувствительна как к TcdA, так и к TcdB. Кратко, в 96-луночный титрационный микропланшет рассевали клетки IMR-90, служащие в качестве мишени при определении токсин-опосредуемой цитотоксичности. Каждый тестируемый образец сыворотки анализировали отдельно по способности нейтрализовать TcdA и TcdB. Тестируемые образцы антисыворотки в соответствующих серийных разведениях смешивали с TcdA или TcdB в фиксированных концентрациях и инкубировали при 37°С в течение 90 минут в инкубаторе с увлажнением (37°С/5% СO2), чтобы дать возможность осуществиться нейтрализации токсинов. Для качественного контроля все планшеты содержали стандарт сравнения и контроли, включающие антитела к токсину с известным титром. Через 90 минут к монослою клеток IMR-90 добавляли смесь токсина с образцами антисыворотки и планшеты инкубировали в течение еще 72 часов. После этого в используемый в анализе планшет добавляли субстрат CellTiter-Glo® для определения уровней аденозинтрифосфата (АТФ), присутствующего в метаболически активных клетках, и измеряли в виде относительных единиц люминесценции (RLU). Высокий уровень АТФ указывает на высокую жизнеспособность клеток, и уровни прямо пропорциональны степени нейтрализации токсина антителом, присутствующим в образце. Для обработки доклинических данных строили график зависимости данных RLU от величины разведения тестируемого образца антисыворотки с получением четырехпараметрической логистической (4-PL) регрессионной аппроксимирующей кривой для ответа. Титры нейтрализующих антител выражали в виде величины разведения образца, при которой наблюдали 50%-ное снижение цитотоксичности.
Пример 25. Исследование иммуногенности на мышах: muC. difficile2010-06
Цель данного исследования состояла в оценке иммуногенности двух форм мутантного токсина В из С. difficile (SEQ ID NO: 6), каждая из которых была химически инактивирована разными методами. В этом исследовании в качестве контроля использовали необработанный мутантный токсин В (SEQ ID NO: 6) (инактивированный генетически, но не инактивированный химически) с адъювантом и без него.
Группы из 10 мышей иммунизировали внутримышечно (в./м.), используя 10 мкг иммуногена в соответствии с Таблицей 12.
Результаты. Не наблюдали никаких неблагоприятных явлений у мышей после каждого введения вакцин-кандидатов. Как проиллюстрировано на Фиг. 10, у мышей в каждой группе вырабатывались очень сильные нейтрализующие антитела к токсину В после введения третьей дозы соответствующих мутантных токсинов.
По-видимому, на основании титров на 12-й неделе, у мышей вырабатывались эффективные нейтрализующие ответы на EDC-инактивированный мутантный токсин В (группа 2) и инактивированные формалином мутантные токсины (группа 1).
В случае отсутствия химической инактивации нейтрализующие ответы на генетически мутантный токсин В (SEQ ID NO: 6) вырабатывались после двух введений дозы (группы 3-4, 8-я неделя), которые усиливались после введения третьей дозы (группы 3-4, 12-я неделя).
Пример 26. Исследование иммуногенности на мышах: muC. difficile 2010-07
Цель данного исследования состояла в оценке иммуногенности химически инактивированных мутантных токсинов А и В из С. difficile (SEQ ID NO: 4 и 6, соответственно) либо по отдельности, либо в комбинации. Для всех групп готовили композиции на основе иммуногенов с использованием фосфата алюминия в качестве адъюванта.
Группы из 5 мышей иммунизировали внутримышечно, используя 10 мкг иммуногена в соответствии с Таблицей 13.


Результаты. Не наблюдали никаких неблагоприятных явлений у мышей после каждого введения вакцин-кандидатов. Как проиллюстрировано на Фиг. 11, после двух введений дозы химически инактивированных генетически мутантных токсинов уровень нейтрализующих антител к токсину А (группы 3-5) повышался до титров 3-4 log10, тогда как уровень нейтрализующих антител к токсину В (группы 1-2, 5) оставался в диапазоне от низкого до не подлежащего обнаружению, что согласуется с данными исследования на мышах, описанными выше (Фиг. 10). Уровень нейтрализующих токсин В антител возрастал до 2-3 1одю в группах 1, 2 и 5 после введения третьей дозы (титры на 12-й неделе) и достигал своего пика через две недели после введения четвертой дозы (титры на 14-й неделе). Титры нейтрализующих токсин А антител в группах 3-5 практически не увеличивались после третьей (титры на 12-й неделе) и четвертой иммунизаций (титры на 14-й неделе).
Пример 27. Исследование иммуногенности на хомяках: ham С. difficile 2010-02
Цель данного исследования состояла в оценке иммуногенности и защитного потенциала тройных мутантных и химически инактивированных мутантных токсинов А и В из С. difficile в модели сирийского золотого хомяка. Модель сирийского золотого хомяка представляет собой самую доступную модель заражения для имитации CDAD у человека. В этом исследовании использовали те же партии мутантных токсинов А и В, которые использовали в исследовании на мышах muC. difficile2010-07. В качестве контроля одна группа получала мутантные токсины без алюминийсодержащего адъюванта.
Группы из 5 сирийских золотых хомяков иммунизировали внутримышечно, используя 10 мкг иммуногена, в соответствии с Таблицей 14.
Результаты. Не наблюдали никаких неблагоприятных явлений после иммунизации мутантными токсинами. Как проиллюстрировано на Фиг. 12, после введения одной дозы мутантных токсинов нейтрализующие токсин А антительные ответы составляли 2-3 log10 для инактивированных формалином мутантных токсинов (группы 1-2) и 3-4 log10 для EDC-инактивированных мутантных токсинов (группа 3). Введение второй дозы усиливало образование антител к токсину А во всех трех группах. Как оказалось, после введения третьей дозы количество антител к токсину А во всех трех группах не увеличивалось. Аналогичный результат наблюдали после четвертой иммунизации, когда увеличение титра наблюдали в группе с инактивацией формалином, которая не содержала адъюванта на основе алюминия (группе 2).
Нейтрализующие токсин В антительные ответы не были обнаружены в группах с инактивированными формалином мутантными токсинами (группах 1-2) и составляли чуть больше 2 log10 для EDC-инактивированных мутантных токсинов (группа 3) после введения одной дозы. После введения второй дозы титры нейтрализующих токсин В антител в двух группах с инактивацией формалином (группах 1-2) возрастали до 3-4 log10, в то время как в группе с EDC-инактивацией (группе 3) они возрастали до 4-5 log10. Для всех трех групп возрастание титров нейтрализующих токсин В антител наблюдали после введения третьей и/или четвертой доз, при этом во всех группах максимального значения титра достигали на 16-й неделе (после введения последней дозы). См. Фиг. 12.
На Фиг. 13 приведено сравнение уровня нейтрализующих антительных ответов на химически инактивированные генетически мутантные токсины (Фиг. 12) с ответами, вызванными токсоидами от компании List Biological Laboratories, Inc. (Campbell, California) (также обозначенной в данном описании как «List Βiο» или «List Biologicals») (т.е. токсоидами, приобретенными у List Biological Laboratories, которые были получены посредством инактивации формалином токсинов дикого типа; контрольным реагентом, использованным для обоснования модели заражения хомяков).
Как использовано в данном описании, «FI» на рисунках и в таблицах относится к обработке токсинов формалином/глицином в течение 2 суток при 25°С, если не указано иное. Как использовано в данном описании, «EI» на рисунках и в таблицах относится к обработке с использованием EDC/NHS в течение 4 часов при 30°С, если не указано иное. На Фиг. 13: 5 животных хомяков обрабатывали соответствующей композицией на основе мутантного токсина, тогда как 11 животных хомяков обрабатывали токсоидом, приобретенным у List Biologicals.
Данные, приведенные на Фиг. 13, показывают, что соответствующие титры нейтрализующих токсин А (Фиг. 13А) и токсин В (Фиг. 13В) антител, индуцированных у хомяков при введении согласно Таблице 14 иммуногенной композиции, содержащей EDC-инактивированные мутантные токсины, после двух введений дозы оказываются выше соответствующих титры нейтрализующих антител, вырабатываемых при введении токсоидов от List Biologicals.
Пример 28. Исследование иммуногенности на хомяках: С. difficile ham2010-02 (продолжение)
Для оценки защитной эффективности мутантных токсинов иммунизированным хомякам, наряду с одной контрольной группой неиммунизированных животных, сначала вводили перорально дозу антибиотика клиндамицина (30 мг/кг) для уничтожения нормальной кишечной флоры. Через пять суток осуществляли контрольное заражение хомяков, используя пероральное введение спор С. difficile дикого типа (штамм 630, по 100 колониеобразующих единиц (КОЕ) на одно животное). Мониторинг состояния животных проводили один раз в сутки в течение одиннадцати суток после контрольного заражения на предмет обнаружения признаков CDAD, которая у хомяков известна под названием «мокрый хвост». По результатам применения системы клинической балльной оценки ряда разных параметров, животных, у которых обнаружена тяжелая форма CDAD, подвергали эвтаназии. Параметры включали активность после стимуляции, обезвоживание, состав фекалий, температуру и массу и т.д., которые известны в данной области техники.
На 11-е сутки исследование завершали, и всех выживших животных подвергали эвтаназии. На Фиг. 14 показаны кривые выживаемости для каждой из трех иммунизированных групп (групп 1-3, в соответствии с Таблицей 14) в сравнении с неиммунизированными контролями. Как можно видеть, у всех неиммунизированных животных развивалась тяжелая форма CDAD, и потребовалась эвтаназия в интервале от 1 до 3 суток после контрольного заражения (0% выживаемости). Для обеих групп, которым вводили инактивированный формалином мутантный токсин, получали кривые 60%-ной выживаемости, при этом не было необходимости в эвтаназии животных до 3-х суток (группа 1) или 4-х суток (группа 2). Для группы, которой вводили EDC-инактивированный мутантный токсин, получали кривую 80%-ной выживаемости, при этом потребовалась эвтаназия 1 животного (из 5) на 7-е сутки. Таким образом, хомяки были защищены от летального заражения спорами С. difficile.
Пример 29. Исследование иммуногенности на хомяках: hamС. difficile2010-03: иммуногенность генетически и химически инактивированных мутантных токсинов из С. difficile
Цель данного исследования состояла в оценке иммуногенности безадъювантных тройных мутантных и химически инактивированных мутантных токсинов А и В из С. difficile (SEQ ID NO: 4 и 6, соответственно) в модели сирийского золотого хомяка. В этом исследовании использовали те же партии мутантных токсинов А и В (SEQ ID NO: 4 и 6, соответственно), которые использовали в исследовании на мышах muC. difficile2010-07. В качестве контроля одной группе (группе 1) давали забуференный фосфатом физиологический раствор в виде плацебо.
Группы из пяти или десяти сирийских золотых хомяков иммунизировали иммуногеном в соответствии с Таблицей 15. Животным вводили три дозы. Кроме того, введение животным доз выполняли каждые две недели.

Результаты. См. Фиг. 15. Никаких антител к токсину А или В не обнаруживали в контрольной группе, получавшей плацебо. После введения одной дозы наблюдали титры нейтрализующих токсин А антител с 2-3 log10 для группы с инактивацией формалином (группы 2) и группы с генетически мутантным токсином (группы 4) и с 3-4 log10 для группы с EDC-инактивацией (группы 3). Уровень нейтрализующих токсин А антител возрастал в каждой из этих групп (2-4) после второй иммунизации релевантными мутантными токсинами (см. сравнение титров на 2-й неделе относительно 3-й недели на Фиг. 15). После третьей дозы мутантных токсинов (введенной на 4-й неделе) титры нейтрализующих токсин А антител в группах 2-4 возрастали в сравнении с соответствующими титрами на 4-й неделе.
Нейтрализующие токсин В антитела обнаруживали после введения второй дозы, при этом уровень антител, нейтрализующих инактивированный формалином (группа 2) и EDC-инактивированный (группа 3) токсин В, возрастал до 3-4 log10 и в случае генетического тройного мутанта (группа 4) до 2-3 log10. После третьей иммунизации (4-я неделя) титры нейтрализующих токсин В антител возрастали до 3-4 log10 в случае инактивированных формалином мутантных токсинов (группа 2) и генетически мутантных токсинов (группа 4) и до 4-5 log10 в случае EDC-инактивированных мутантных токсинов (группа 3).
В случае нейтрализующих антител как к токсину А, так и к токсину В максимальные титры наблюдали на 6-й неделе (после введения 3-й дозы) у всех вакцинированных групп (групп 2-4).
Оценка иммуногенных композиций, дополненных адъювантом Alhydrogel/CpG или ISCOMATRIX
У хомяков, иммунизированных иммуногенной композицией, включающей химически инактивированный мутантный токсин вместе с Alhydrogel, ISCOMATRIX или Alhydrogel/CpG24555 (Alh/CpG), вырабатывалась сильная нейтрализующая токсины антисыворотка. Было обнаружено, что максимальные антительные ответы на токсин А и токсин В были в 2-3-раза выше и были статистически значимыми в группах, иммунизированных мутантными токсинами в составе композиций вместе с Alh/CpG или ISCOMATRIX, в сравнении с вакциной, изготовленной только с Alhydrogel. См. Таблицу 32, показывающую титры нейтрализующих на 50% антител. Хомяков (n=10/группа) иммунизировали в./м. в 0-ю, 2-ю и 4-ю недели, используя, по 10 мкг каждого, мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию в составе композиций вместе с 100 мкг Alhydrogel или 200 мкг CpG 24555+100 мкг Alhydrogel или 10 единицами ISCOMATRIX. В каждый тестируемый момент времени отбирали кровь для приготовления сыворотки и ее исследовали в анализе нейтрализации токсинов с точки зрения функциональной антитоксинной активности. Средние геометрические титры (GMT) приведены в Таблице 32. Звездочки (*) указывают на статистическую значимость (р<0,05) при сравнении с титрами в группе с Alhydrogel.

Тестировали защитную эффективность иммуногенной композиции, содержащей в своем составе мутантные токсины как лекарственные субстанции вместе с этими адъювантами. Хомяков иммунизировали и давали перорально клиндамицин (30 мг/кг) на 5-й неделе и проводили контрольное заражение в соответствии со способом, описанным выше. Одна группа неиммунизированных хомяков (n=5) была включена в качестве контроля. Более высокую эффективность наблюдали у хомяков, иммунизированных мутантными токсинами как лекарственными субстанциями с добавлением адъювантов либо Alh/CpG, либо ISCOMATRIX (выживаемость 100%), в сравнении только с Alhydrogel (выживаемость 70%). Таким образом, хомяки были защищены от летального заражения спорами С. difficile.
Пример 30. Вакцинация Clostridium difficile на яванских макаках
Цель этого исследования заключалась в тестировании иммуногенности EDC-инактивированных и инактивированных формалином мутантных токсинов из С. difficile в низких и высоких дозах на яванских макаках. На основе всех мутантных токсинов готовили композиции с использованием ISCOMATRIX® в качестве адъюванта за исключением одной группы, которая служила в качестве контроля без адъюванта (группа 5).

Результаты. На Фиг. 16 показаны нейтрализующие токсин антительные ответы для этих животных на 0-й, 2-й, 3-й, 4-й, 6-й, 8-й и 12-й неделях. Титры антител к токсину А составляли 2-3 log10 для всех пяти групп после введения одной дозы (титры на 2-й неделе). Эти титры возрастали после каждого последующего введения дозы для каждой группы. У этих животных не наблюдали снижения титра в интервале от 3-й до 4-й недель. Для всех групп максимальные титры находились в диапазоне 4-5 log10. Во все тестируемые моменты времени для группы без адъюванта ISCOMATRIX (группы 5) получали самые низкие титры, что указывало на применимость ISCOMATRIX при усилении иммунных ответов. В безадъювантной контрольной группе (группе 5) максимальных титров достигали на 12-й неделе, как и в группе, иммунизированной EDC-инактивированными мутантными токсинами в высокой дозе (группе 4); во всех других группах максимальных титров достигали на 6-й неделе, через две недели после введения последней дозы. Титры во всех группах возрастали после введения второй дозы (временная точка - 3-я неделя). И антительные ответы на токсин А, и антительные ответы на токсин В не снижались в интервале от 3-й недели до 4-й недели. После введения третьей дозы (временная точка - 6-я неделя) титры нейтрализующих токсин В антител во всех группах составляли 3-4 log10, за исключением группы с инактивацией формалином и с низкой дозой (группы 1) и группы с инактивацией EDC и с высокой дозой (группы 4), титры в которых составляли чуть больше 4 log10. Максимальные титры наблюдали на 12-й неделе для всех групп за исключением группы с инактивацией EDC и с низкой дозой (группы 3), для которой получали максимальные титры на 8-й неделе. Для всех групп получали максимальные титры, превышающие 4 log10.
Пример 31. Получение моноклональных антител
Несмотря на то, что токсины А и В обладают значительной структурной гомологией, было обнаружено, что нейтрализующие активности антител являются токсин-специфичными. В данном изобретении были идентифицированы различные антитела, которые специфичны к индивидуальному токсину и направлены на различные эпитопы и функциональные домены и которые характеризуются высокой аффинностью и сильной нейтрализующей активностью по отношению к нативным токсинам. Антитела выделяли из мышей, которых иммунизировали либо имеющимся в продаже инактивированным формалином (FI) мутантным токсином, либо рекомбинантным мутантным голотоксином (SEQ ID NO: 4 и 6), сделанным нетоксичным посредством введения специфических мутаций в его каталитический центр с целью получения mAb к токсинам А и В, соответственно. Эпитопное картирование антител показало, что значительное большинство mAb к токсину А (49 из 52) было направлено на некаталитический С-концевой домен токсина.
Моноклональные антитела к токсину В были направлены на три домена этого белка. Из всех 17 mAb, специфичных к токсину В, 6 обладали специфичностью к N-концевому (например, аминокислотам 1-543 TcdB из С. difficile дикого типа, как например, из штамма 630), 6 к С-концевому (например, аминокислотам 1834-2366 TcdB из С. difficile дикого типа, как например, из штамма 630) и 5 к расположенному в срединной части домену транслокации (например, аминокислотам 799-1833 TcdB из С. difficile дикого типа, как например, из штамма 630). Таким образом, подход с использованием мутантных токсинов С. difficile (например, SEQ ID NO: 4 и 6) в качестве иммунизирующих антигенов, дает ключевое преимущество, заключающееся в презентации большинства, если не всех, антигенных эпитопов по сравнению со способом с применением инактивации формалином, который имеет тенденцию неблагоприятно воздействовать на антигенную структуру мутантного токсина.
Пример 32. Определение характеристик mAb к токсину А, А3-25, которое содержит вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 36, и вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 37
MAb А3-25 представляло особый интерес вследствие того, что все попытки определения иммуноглобулинового (lg) изотипа этого антитела с использованием общедоступных наборов для изотипирования на IgG, IgM и IgA терпели неудачу. Дальнейший анализ вестерн-блоттингом с использованием антисыворотки, специфичной к Η-цепи lg, показал, что А3-25 имеет lgE-изотип, что является редким событием при получении mAb. Этот факт был дополнительно подтвержден секвенированием нуклеотидной последовательности мРНК, выделенной из гибридомных клеток А3-25. Аминокислотные последовательности, предсказанные на основании нуклеотидных последовательностей вариабельных областей Н- и L-цепи А3-25, показаны на Фиг. 17.
Для проведения дальнейших оценок mAb А3-25 в животной модели инфекции С. difficile и заболевания, вызываемого С. difficile, его lg-изотип был изменен на мышиный lgG1 посредством молекулярного привития вариабельной области от ε-Н-цепи на мышиную тяжелую цепь у в соответствии с опубликованными в литературе методами.
Пример 33. Нейтрализующая способность и эпитопное картирование токсин-специфических антител
Далее, в попытке идентификации функциональных/нейтрализующих антител все моноклональные антитела оценивали по способности нейтрализовать токсины дикого типа в стандартном анализе цитопатического эффекта (СРЕ) или в более точном и количественном анализе, основанном на измерении АТФ как индикатора жизнеспособности клеток.
Из всех 52 специфичных к токсину А антител четыре mAb (А3-25, А65-33, А60-22 и А80-29; Таблица 17 и Фиг. 18) демонстрировали различные уровни нейтрализующей активности. С целью картирования эпитопов данных антител были проведены анализ конкурентного связывания с применением BiaCore и анализы ингибирования гемагглютинации (HI). Результаты показали, что эти антитела могут быть направлены на разные эпитопы белка токсина А (Таблица 17). Чтобы далее провести идентификацию расположения сайтов связывания на белке, антитела индивидуально оценивали в вестерн-блот- или дот-блот-анализах с использованием фрагментов токсинов с известными последовательностями. Было обнаружено, что все 4 нейтрализующих mAb направлены на С-концевой участок данного токсина.
Было обнаружено, что из всех 17 специфичных к токсину В антител 9 являются нейтрализующими. Из девяти нейтрализующих mAb шесть были направлены на N-концевой домен, а остальные три на домен транслокации в токсине В (Таблица 18). Основываясь на результатах анализа конкурентного связывания с применением Biacore, эти девять нейтрализующих моноклональных антител можно свести к четырем эпитопным группам, как показано на Фиг. 19.

Пример 34. Идентификация новых комбинаций на основе антител к токсину А со значительно повышенной нейтрализующей активностью
Четыре mAb к токсину А (А3-25, А65-33, А60-22 и А80-29) демонстрировали неполную или частичную нейтрализацию токсина А при индивидуальном тестировании в анализе нейтрализации, основанном на измерении уровней АТФ. MAb А3-25 было самым сильнодействующим антителом, а остальные три оказывали меньшее нейтрализующее действие, при этом активность А80-29 едва превышала фоновую (Фиг. 18). Тем не менее, когда А3-25 комбинировали с каким-либо одним из трех остальных mAb, синергетический эффект в отношении нейтрализации наблюдали для всех трех комбинаций, который был гораздо больше общей суммы нейтрализующих активностей отдельных антител, что показано на Фиг. 20А-С. Помимо этого, все три комбинации демонстрировали способность к полной нейтрализации, обычно наблюдаемую при использовании поликлональных антител к токсину А.
Пример 35. Идентификация новых комбинаций на основе антител к токсину В, демонстрирующих значительно повышенную нейтрализующую активность
Авторы изобретения также наблюдали синергетический эффект при нейтрализации с использованием mAb к токсину В из разных эпитопных групп, идентифицированных посредством BiaCore-анализа. MAb к токсину В В8-26, наиболее доминирующее mAb из группы 1, комбинировали со многими mAb из группы 3. Комбинации были оценены в анализе специфичной в отношении токсина В нейтрализации и результаты показаны на Фиг. 21 и в Таблице 19.

Наблюдали синергетический эффект при нейтрализации в случае, когда В8-26 комбинировали с mAb из эпитопной группы 3 (Фиг. 21В), но не с любым другим mAb (данные не показаны).
Пример 36. Скрининг in vitro безвредных и эффективных композиций на основе мутантных токсинов с помощью mAb
Генетически мутантные токсины А и В из С difficile (например, SEQ ID NO: 4 и 6), созданные генно-инженерным методом, демонстрировали остаточную цитотоксичность в анализе цитотоксичности in vitro. Несмотря на то, что для каждого мутантного токсина из С. difficile авторами изобретения было достигнуто снижение цитотоксичности, составляющее приблизительно 4 log (Таблица 20), предпочтительной оказалась дальнейшая химическая инактивация мутантных токсинов, такая как обработка формалином. Однако, приводящие к инактивации химические воздействия могут быть жесткими и могут неблагоприятно влиять на ключевые антигенные эпитопы этих токсинов или мутантных токсинов.
Для оптимизации биологического процесса выполняли статистический расчет эксперимента (DOE) по химической инактивации тройных мутантных Tcd А и В (1 мг/мл) с использованием обработки формалином и EDC/NHS. Для оптимизации инактивации формалином тройного мутантного TcdA авторы изобретения варьировали концентрации формалина/глицина (20-40 мМ), рН (6,5-7,5) и температуру (25-40°С). Для тройного мутантного TcdB авторы изобретения варьировали концентрацию формалина/глицина от 2 до 80 мМ, а температура и рН составляли 25°С и 7,0 соответственно. Продолжительность инкубации для всех обработок формалином составляла 24 часа. Что касается инактивации формалином, то «40/40» в Таблицах 21 и 23 представляет собой концентрацию формалина и глицина, использованную в реакции. В случае обработки с использованием EDC/NHS авторы изобретения варьировали количества EDC/NHS от 0,25 до 2,5 мг/мг тройного мутантного TcdA и от 0,125 до 2,5 мг/мг тройного мутантного TcdB и инкубировали в течение четырех часов при 25°С. По завершении реакций все образцы обессоливали, переводя в 10 мМ фосфат, рН 7,0. После очистки подвергнутые обработке Tcd анализировали на предмет остаточной цитотоксичности и распознавания эпитопов моноклональными антителами (mAb) в дот-блот-анализе. Задача состояла в определении условий обработки, которые снижают цитотоксичность до желаемого уровня (ЕС50 больше 1000 мкг/мл) без отрицательного воздействия на эпитопы, распознаваемые панелью нейтрализующих mAb (++++ или +++). В таких условиях обработки (отмеченных галочкой «√» в Таблицах 21-24) получали потенциально безвредные и эффективные иммуногенные композиции которые сохраняли реакционную способность по меньшей мере четырех нейтрализующих mAb, демонстрируя при этом снижение цитотоксичности 6-8 log10 в сравнении с соответствующей цитотоксичностью токсина дикого типа. Отдельные результаты проиллюстрированы в Таблицах 21-24. Дополнительные данные, полученные для различных условий обработки тройных мутантных токсинов, и данные анализов цитотоксичности in vitro и нейтрализации токсинов показаны в Таблице 33 и Таблице 34. Кроме этого см., например, приведенные выше примеры 20 и 21, в которых приводятся дополнительные подробности, касающиеся предпочтительных условий обработки мутантных токсинов посредством введения сшивок.



Условия реакции образования химических сшивок для образцов тройного мутантного токсина A (SEQ ID NO: 4) согласно Таблице 33
Образцы 1-4 подвергали модификации, используя EDC/NHS. Условия: 30°С, 20 мМ MES (2-(N-морфолино)этансульфоновая кислота)/150 мМ NaCl, рН 6,5. Реакции инициировали путем добавления EDO Через 2 часа протекания реакции в образцы А, В и С добавляли 1 М глицин до конечной концентрации глицина 50 мМ. В образец D глицина не добавляли. Реакции проводили при разных массовых соотношениях мутантный токсин A (SEQ ID NO: 4):EDC:NHS, которые указаны ниже.
1 - L44166-157А 1:0,25:0,25 (масс.:масс.:масс).
2- L44166-157B 1:1,25:1,25.
3- L44166-157C 1:2,5:2,5.
4- L44166-157D 1:2,5:2,5.
Образец 5 - L44905-160A, 80 мМ НСНО, 80 мМ глицин, 80 мМ Na3P04, рН 7, мутантный токсин A (SEQ ID NO: 4) в концентрации по белку 1 мг/мл, продолжительность реакции 48 ч при 25°С.
Образец 6 - L44166-166, EDC/NHS-модификация мутантного токсина А (SEQ ID NO: 4) при 25°С в 20 мМ MES/150 мМ NaCl, рН 6,5. Соотношение мутантный токсин A (SEQ ID NO: 4):EDC:NHS составляет 1:0,5:0,5. Реакцию инициировали добавлением EDC. Через 2 часа протекания реакции добавляли 1 М глицин до конечной концентрации глицина 0,1 М и инкубацию продолжали в течение еще 2 часов. По прошествии этого времени реакционный буфер заменяли на 1х PBS на Сефадексе G25.
Образец 7 - L44905-170A, 80 мМ НСНО, 80 мМ глицин, 80 мМ Na3PO4, рН 7, мутантный токсин A (SEQ ID NO: 4) в концентрации по белку 1 мг/мл, продолжительность реакции 48 ч при 35°С. Эта реакция с формалином была направлена на получение избыточного сшивания, поэтому связывание антигена будет сильно ослаблено.
Образец 8 - L44897-61, 32 мМ НСНО/80 мМ глицин, продолжительность реакции 72 ч при 25°С.
Образец 9 - L44897-63, 80 мМ НСНО/80 мМ глицин, продолжительность реакции 72 ч при 25°С.
Продолжительность всех приведенных ниже реакций составляла 24 ч.
Образец 10 - L44897-72, пробирка №1, 25°С, 80 мМ NaPi, рН 6,5, 20 мМ НСНО/20 мМ глицин.
Образец 11 - L44897-72, пробирка №2, 25°С, 80 мМ NaPi, рН 6,5, 40 мМ НСНО/40 мМ глицин.
Образец 12 - L44897-72, пробирка №3, 32,5°С, 80 мМ NaPi, рН 7,0, 30 мМ НСНО/30 мМ глицин.
Образец 13 - L44897-72, пробирка №4, 32,5°С, 80 мМ NaPi рН 7,0, 30 мМ НСНО/30 мМ глицин.
Образец 14 - L44897-72, пробирка №5, 32,5°С, 80 мМ NaPi, рН 7,0, 30 мМ НСНО/30 мМ глицин.
Образец 15 - L44897-75, пробирка №6, 25°С, 80 мМ NaPi, рН 7,5, 20 мМ НСНО/20 мМ глицин.
Образец 16 - L44897-75, пробирка №7, 25°С, 80 мМ NaPi, рН 7,5, 40 мМ НСНО/40 мМ глицин.
Образец 17 - L44897-75, пробирка №8, 40°С, 80 мМ NaPi, рН 6,5, 20 мМ НСНО/20 мМ глицин.
Образец 18 - L44897-75, пробирка №9, 40°С, 80 мМ NaPi, рН 6,5, 40 мМ НСНО/40 мМ глицин.
Образец 19 - L44897-75, пробирка №10, 40°С, 80 мМ NaPi, рН 7,5, 20 мМ НСНО/20 мМ глицин.
Образец 20 - L44897-75, пробирка №11, 40°С, 80 мМ NaPi, рН 7,5, 40 мМ НСНО/40 мМ глицин.
Реакции с участием следующих 8 образцов проводили при 25°С в течение указанных периодов времени в 80 мМ NaPi, рН 7,0, содержащем 78 мМ НСНО и 76 мМ глицин.
Образец 21 - L44897-101 (до модификации) контрольный образец ТхА-контроль на нулевой момент времени, не подвергнутый модификации или воздействию НСНО/глицина.
Образец 22 - L44897-101, 2 ч.
Образец 23 - L44897-101, 4 ч.
Образец 24 - L44897-101, 6 ч.
Образец 25 - L44897 102, 24 ч.
Образец 26 - L44897-103, 51 ч.
Образец 27 - L44897-104, 74 ч.
Образец 28 - L44897-105, 120 ч.
Образец 29 (L44980-004) представлял собой EDC/NHS-модифицированный мутантный токсин A (SEQ ID NO: 4) (тройной мутантный токсин A (SEQ ID NO: 4)-EDC). Используют следующие реакционные условия: 25°С, буфер - 20 мМ MES/150 мМ NaCl, рН 6,6. Соотношение тройной мутантный токсин A (SEQ ID NO: 4):EDC:NHS составляло 1:0,5:0,5 (масс.:масс.:масс). Реакцию инициировали добавлением EDC. Через 2 часа протекания реакции добавляли глицин до конечной концентрации 0,1 М и взаимодействие продолжали в течение еще 2 часов при 25°С. Реакцию заканчивали путем обессоливания на Сефадексе G25.
Следующие 12 образцов и 2 контроля относились к экспериментам по реверсии, при этом образцы инкубировали при 25°С и 37°С.
Реакция 1 - 25°С, 80 мМ NaPi, рН 7,0, только 40 мМ НСНО (без глицина), продолжительность реакции 24 часа.
Реакция 2 - 25°С, 80 мМ NaPi, рН 7,0, 40 мМ НСНО/40 мМ глицин, продолжительность реакции 24 часа.
Образец Реакция
30 Реакция №1, 0-я неделя, 25°С
31 Реакция №1, 1-я неделя, 25°С
32 Реакция №1, 2-я неделя, 25°С
33 Реакция №1, 3-я неделя, 25°С
34 Реакция №1, 4-я неделя, 25°С
35 Реакция №1, 3-я неделя, 37°С
36 Реакция №2, 0-я неделя, 25°С
37 Реакция №2, 1-я неделя, 25°С
38 Реакция №2, 2-я неделя, 25°С
39 Реакция №2, 3-я неделя, 25°С
40 Реакция №2, 4-я неделя, 25°С
41 Реакция №2, 3-я неделя, 37°С
42 ТхА-контроль, 3-я неделя, 25°С
43 ТхА-контроль, 3-я неделя, 37°С
Следующие 4 образца получали в результате проведения реакции в течение указанных промежутков времени при 25°С в 80 мМ NaPi, рН 7,04; 40 мМ НСНО/40 мМ глицин.
44 L44897-116-6; 29,5 ч
45 L44897-116-7; 57,5 ч
46 L44897-116-8; 79,5 ч
47 L44897-116-9; 123,5 ч
Образец 48 - L44897-139, продолжительность реакции 48 ч при 25°С, 80 мМ NaPi, рН 7,0; 40 мМ НСНО/40 мМ глицин.
Образец 49 - L44166-204, EDC/NHS-модификация мутантного токсина А (SEQ ID NO: 4), 25°С, буфер 1х PBS, рН 7,0. Соотношение мутантный токсин А (SEQ ID NO: 4):EDC:NHS составляло 1:0,5:0,5 (масс.:масс.:масс). Продолжительность реакции с EDC/NHS 2 часа, затем добавляли 1 М глицин до конечной концентрации 0,1 М и реакцию проводили в течение еще 2 часов. Используя Сефадекс G25, проводили замену буфера на 20 мМ L-гистидин/100 мМ NaCl, рН 6,5.

Условия реакции образования химических сшивок для образцов мутантного токсина В со ссылкой на Таблицу 34
Тройной мутантный токсин В (SEQ ID NO: 6) подвергали реакции образования химических сшивок и тестировали в соответствии с приведенными ниже реакционными условиями. Образцы L44905-86 тестировали в эксперименте, включающем три варианта реакции с участием формалина и две температуры инкубации. Один раз в указанные сутки отбирали 6 образцов, что в общей сложности составило 18 образцов. Первый образец в данном списке представляет собой необработанный контроль (с учетом которого получается в целом 19 образцов). Необработанный контроль включал необработанный полипептид тройного мутантного токсина В (SEQ ID NO: 6).
Реакция 1 («Rxn1») - 80 мМ НСНО, 80 мМ глицин, 80 мМ NaPO4, рН 7, тройной мутантный токсин В (SEQ ID NO: 6) в концентрации по белку 1 мг/мл.
Реакция 2 («Rxn2») - 80 мМ НСНО, без глицина, 80 мМ NaPO4, рН 7, тройной мутантный токсин В (SEQ ID NO: 6) в концентрации по белку 1 мг/мл.
Реакция 3 («Rxn3») - 80 мМ НСНО, без глицина, 80 мМ NaPO4, рН 7, тройной мутантный токсин В (SEQ ID NO: 6) в концентрации по белку 1 мг/мл + блокирование цианоборгидридом. Блокирование цианоборгидридом включало добавление 80 мМ CNBrH4 к обессоленной конечной реакционной смеси и инкубирование в течение 24 ч при 36°С.
Для образца L34346-30A - 0,5 г EDC и NHS на один грамм тройного мутантного токсина В (SEQ ID NO: 6), 4 часа при 30°С в 20 мМ MES, 150 мМ NaCl, рН 6,5.
Для образца L34346-30B - 0,5 г EDC и NHS на один грамм тройного мутантного токсина В (SEQ ID NO: 6), 2 часа при 30°С, затем добавление глицина (конечная концентрация г/л) и инкубирование в течение еще 2 часов при 30°С в 20 мМ MES, 150 мМ NaCl, рН 6,5. Единственным различием между двумя этими реакциями для L34346-30A и L34346-30B является добавление глицина в реакционную смесь с L34346-30B.
Пример 37. Антитела, индуцированные иммуногенными композициями, способны нейтрализовать токсины различных штаммов С. difficile
Чтобы оценить, могут ли антитела, индуцированные иммуногенными композициями, содержащими мутантные токсины как лекарственные субстанции, нейтрализовать широкий спектр токсинов с различными последовательностями, проводили секвенирование для штаммов, представляющих различные риботипы и токсинотипы, с целью идентификации степени генетического разнообразия среди различных штаммов в сравнении с мутантными токсинами как лекарственными субстанциями. Культуральные супернатанты, содержащие секретированные токсины из различных штаммов, далее тестировали в анализе нейтрализации in vitro, используя образцы сыворотки от иммунизированных хомяков с целью определения степени охвата иммуногенной композицией и определения способности иммуногенной композиции защищать от различных токсинов из циркулирующих клинических штаммов.
Для тестирования нейтрализации токсинов, экспрессированных штаммами, полученными из CDC, использовали как клетки НТ-29 (линию раковых клеток толстой кишки), так и клетки IMR-90. Клетки НТ-29 более чувствительны к TcdA; ЕС50 для очищенного TcdA при использовании этих клеток составляет 100 пг/мл по сравнению с 3,3 нг/мл для TcdB. С другой стороны, клетки IMR-90 более чувствительны к TcdB; ЕС50 для очищенного TcdB при использовании этих клеток лежит в диапазоне 9-30 пг/мл по сравнению с 0,92-1,5 нг/мл для TcdA. Полученная в анализе специфичность для обоих TcdA и TcdB в этих клеточных линиях была подтверждена с использованием как поликлональных, так и моноклональных специфичных к токсинам антител. Для нормирования данных анализа проводили тестирование фильтратов культур 24 полученных из CDC изолятов в концентрации, в четыре раза превышающей соответствующую величину ЕС50. Уровни токсинов для трех из этих штаммов были слишком низкими для тестирования в анализе нейтрализации.
Двадцать четыре штамма, представляющих различные риботипы/токсинотипы, охватывающие более 95% циркулирующих штаммов С. difficile в США и Канаде, были получены из CDC. Среди этих изолятов были штаммы, представляющие риботипы 027, 001 и 078, три эпидемических штамма, вызывающих CDAD в Соединенных Штатах, Канаде и Соединенном Королевстве (UK). Штаммы 2004013 и 2004118 представляли риботип 027; штамм 2004111 представлял риботип 001, и штаммы 2005088, 2005325 и 2007816 представляли риботип 078. Для идентификации степени генетического различия между вызывающими данное заболевание клиническими изолятами и штаммом 630 было проведено полное секвенирование генов токсинов (tcdA и tcdB) из этих клинических штаммов. См. Таблицу 35. Аминокислотные последовательности токсинов выравнивали, используя ClustalW в программном обеспечении Megalign™ (DNASTAR® Lasergene®), и анализировали на предмет идентичности последовательностей. В случае TcdA анализ геномного выравнивания показал, что все клинические изоляты и штамм 630 имели в целом примерно 98-100% идентичности аминокислотных последовательностей. С-концевая часть гена tcdA отличалась не намного. Этот же анализ проводили для гена tcdB, который демонстрировал большее отличие в последовательности. В частности, штаммы 2007838/NAP7/126 и 2007858/NAP1/unk5 демонстрировали наиболее отличающиеся от штамма 630 картины в N-концевом (79-100%) и С-концевом доменах (88-100%; данные не показаны).
Смесь сывороток крови хомяков (HS) собирали от сирийских золотых хомяков, которых иммунизировали иммуногеном, включающим мутантный TcdA (SEQ ID NO: 4) и мутантный TcdB (SEQ ID NO: 6), при этом мутантные токсины были инактивированы EDC, например, в соответствии с примером 29, Таблица 15, описанным выше, и на их основе были изготовлены композиции вместе с фосфатом алюминия. Результаты, приведенные в Таблице 35, показывают, что в анализе нейтрализации in vitro под действием сыворотки крови от иммунизированных хомяков был нейтрализован по меньшей мере токсин В из соответствующих культуральных супернатантов.
На Фиг. 23 изображены результаты анализа нейтрализации с использованием препаратов токсинов из различных штаммов С. difficile на клетках IMR-90. Данные показывают, что нейтрализующие TcdB антитела в образцах антисыворотки хомяков были способны нейтрализовать токсины из всех протестированных 21 изолята, включая гипервирулентные штаммы и TcdA-негативный, TcdB-позитивный штамм. По меньшей мере 16 разных штаммов С. difficile получали из CDC (Atlanta, GA) (как описано ранее) и культивировали в средах для культивирования С. difficile в подходящих условиях, которые известны в данной области техники и которые описаны выше. Культуральные супернатанты, содержащие секретированные токсины, анализировали в отношении определения их цитотоксичности (ЕС50) на монослоях IMR-90 и после тестировали в стандартном анализе нейтрализации in vitro в концентрации, в 4 раза превышающей ЕС50, используя различные разведения образцов сыворотки от хомяков, иммунизированных мутантным токсином А как лекарственной субстанцией и мутантным токсином В как лекарственной субстанцией, на основе которых готовят композиции вместе с фосфатом алюминия. Неочищенный токсин получали из культуральных супернатантов каждого штамма и очищенный токсин (коммерческий препарат токсина, полученный от List Biologicals, а не очищенный из соответствующих супернатантов) тестировали на предмет цитотоксичности в отношении клеток IMR-90, используя описанный выше анализ цитотоксичности in vitro.
На приведенных на Фиг. 23А-К графиках показаны результаты тестирований цитотоксичности in vitro (как описано ранее), на которых изображена зависимость уровней АТФ (RLU) от возрастающих концентраций: сред для культивирования С. difficile и смеси сывороток крови хомяков (■); неочищенного токсина и смеси сывороток крови хомяков (●); очищенного токсина и смеси сывороток крови хомяков (▼); неочищенного токсина (♦), контроля; и очищенного токсина (♦), контроля. Токсины из соответствующих штаммов добавляли к клеткам в концентрации, соответствующей величинам 4хЕС50.
Как показано на Фиг. 23А-К, у хомяков, которые получали описанный иммуноген, неожиданным образом вырабатывались нейтрализующие антитела, которые демонстрировали нейтрализующую активность в отношении токсинов по меньшей мере из следующих 16 разных полученных из CDC штаммов С. difficile в сравнении с соответствующим контролем, состоящим только из токсина: 2007886 (Фиг. 23А); 2006017 (Фиг. 23 В); 2007070 (Фиг. 23С); 2007302 (Фиг. 23D); 2007838 (Фиг. 23Е); 2007886 (Фиг. 23F); 2009292 (Фиг. 23G); 2004013 (Фиг. 23Н); 2009141 (Фиг. 23I); 2005022 (Фиг. 23J); 2006376 (Фиг. 23К). Также см. Таблицу 35 для дополнительных штаммов С. difficile, выделенные из которых токсины тестировали и нейтрализовали, используя иммуногенную композицию, содержащую мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию вместе с фосфатом алюминия.
В другом исследовании культуральные супернатанты, содержащие токсины, секретированные из различных штаммов С. difficile (полученных из CDC и из больницы г. Лидса, Соединенное Королевство), тестировали анализе нейтрализации in vitro, используя образцы сыворотки от хомяков, которым вводили мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию, на основе которых готовили композиции с Alhydrogel. См. Таблицу 36 для плана эксперимента. Результаты показаны в Таблице 37 и Таблице 38.

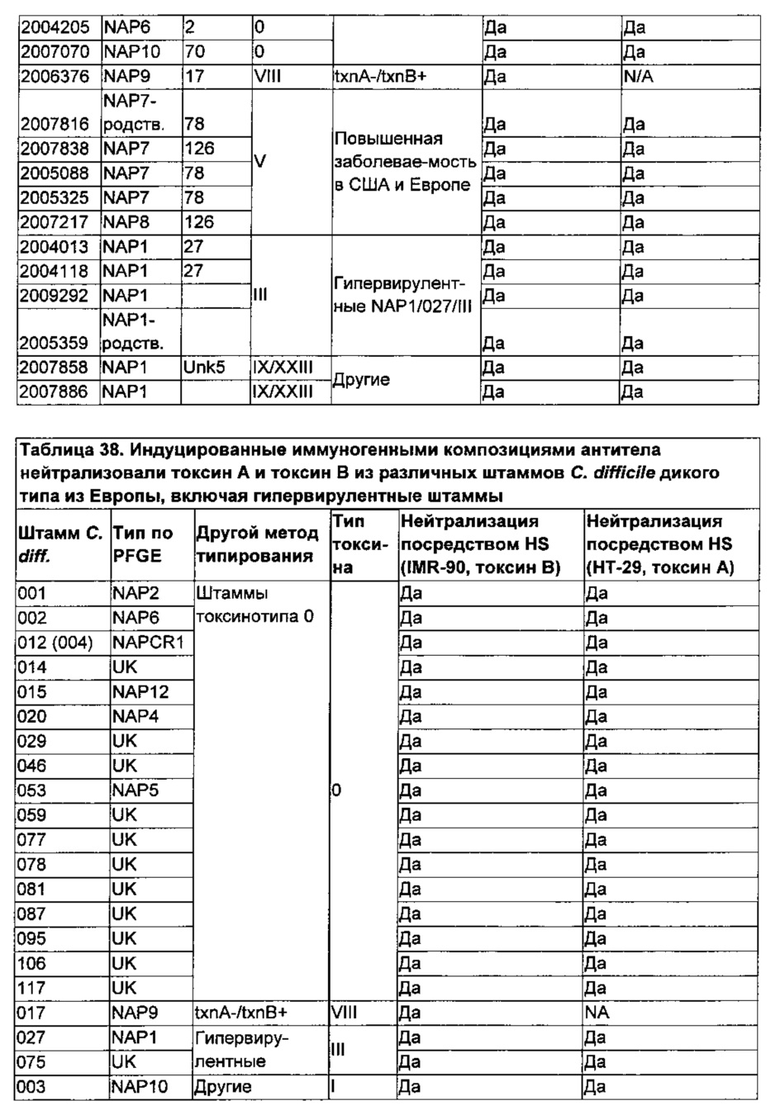

Пример 38. Пептидное картирование EDC/NHS тройных мутантных токсинов
Чтобы охарактеризовать EDC/NHS-инактивированные тройные мутантные токсины, проводили эксперименты по пептидному картированию на четырех партиях обработанного EDC/NHS тройного мутантного токсина A (SEQ ID NO: 4) и на четырех партиях обработанного EDC/NHS тройного мутантного токсина В (SEQ ID NO: 6). После расщепления мутантных токсинов трипсином полученные пептидные фрагменты разделяли, используя обращенно-фазовую высокоэффективную жидкостную хроматографию (HPLC). Масс-спектральный анализ использовали для идентификации модификаций, которые являются результатом процесса инактивации. И для мутантного токсина А как лекарственной субстанции, и для мутантного токсина В как лекарственной субстанции были идентифицированы более 95% от теоретических триптических пептидов. Были идентифицированы сшивки и глициновые аддукты (глицин использовали в качестве блокирующего агента). Кроме того, и в мутантном токсине А как лекарственной субстанции, и в мутантном токсине В как лекарственной субстанции были обнаружены бета-аланиновые аддукты. Без связи с механизмом или теорией считается, что бета-аланиновые аддукты образуются в результате взаимодействия трех молей NHS с одним молем EDC, при этом образуется NHS-активированный бета-аланин. Затем эта молекула может взаимодействовать с группами лизина с образованием бета-аланиновых аддуктов (+70 Да). В образцах EDC/NHS-обработанного тройного мутантного токсина В также обнаружено присутствие дегидроаланина (-34 Да) в низких уровнях (0,07 моль/моль белка). Дегидроаланин образуется в результате десульфонирования остатка цистеина. Такие же тип и степень модификации наблюдали для всех четырех партий каждого мутантного токсина, что указывает на то, что при использовании данного способа образуется сходный продукт. Пептидное картирование (с охватом более чем 95% последовательностей) подтверждает наличие модификаций. Обобщенные результаты модификаций показаны в Таблице 39. См. также Фиг. 24-25. Кроме этого, увеличилась гетерогенность размера и заряда тройного мутантного токсина А как лекарственной субстанции и тройного мутантного токсина В как лекарственной субстанции в сравнении с гетерогенностью размера и заряда соответствующего тройного мутантного токсина А и тройного мутантного токсина В в отсутствие химической инактивации. В результате этого, профили, полученные при проведении гельпроникающей хроматографии (SEC) и анионообменной (АЕХ) хроматографии, содержали относительно широкие пики (данные не показаны).

Пример 39. Изготовление готового лекарственного продукта
Иммуногенная композиция на основе С. difficile (готовый лекарственный продукт) содержит два активных фармацевтических ингредиента (мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию). Типичный готовый лекарственный продукт представляет собой лиофилизированную композицию, содержащую 10 мМ трис-буфер, рН 7,4, 4,5% (масс./масс.) дигидрата трегалозы и 0,01% (масс./об.) полисорбата 80, в том числе любой мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию. См. Таблицу 40. Иммуногенную композицию для введения посредством инъекции готовят путем ресуспендирования лиофилизированной вакцины с использованием либо разбавителя, либо разбавителя, содержащего Alhydrogel. Плацебо будет включать в себя стерильный обычный физиологический раствор для инъекций (0,9%-ный раствор хлорида натрия).
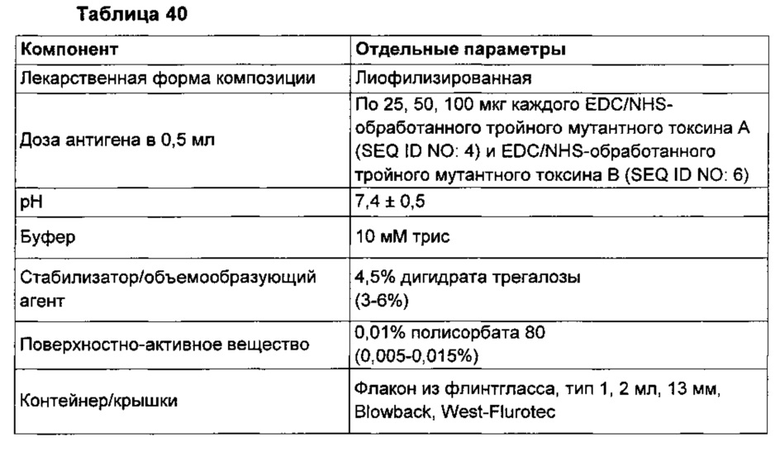
Подготовка буферных растворов
Воду для инъекций (WFI) добавляют в сосуд с перемешиванием. Непрерывно перемешивая, добавляют эксципиенты и растворяют до полного перехода их в раствор. Измеряют рН. При необходимости рН подводят до 7,4±0,1, используя HCl. Раствор разбавляют до конечной массы с использованием WFI, затем фильтруют, используя 0,22 мкм стерилизующий фильтр высокой емкости (SHC) Express XL150 от Millipore. Перед фильтрацией отбирают нефильтрованный образец для оценки уменьшения микрофлоры. Отбирают образец профильтрованного буфера для измерения осмоляльности и рН.
Приготовление композиций
Размороженные образцы мутантных токсинов как лекарственных субстанций объединяют в сосуде для приготовления композиций с учетом предварительно рассчитанных количеств в следующем порядке выполнения операций: сначала в сосуд добавляют 50% от объема целевого буфера для разведения, чтобы достичь концентрации 0,6 мг/мл, затем добавляют мутантный токсин А как лекарственную субстанцию и перемешивают в течение 5 минут при 100 об./мин. Затем в этот сосуд добавляют мутантный токсин В как лекарственную субстанцию, раствор также разбавляют до достижения разведения 0,6 мг/мл и затем перемешивают в течение еще 5 минут при 100 об./мин. Отбирают образец и тестируют общую концентрацию мутантных токсинов. Раствор разбавляют до 100 процентов объема с учетом получения необходимой в способе величины концентрации мутантных токсинов, затем перемешивают в течение 15 минут при 100 об./мин. Из приготовленного раствора лекарственного продукта отбирают образец для определения рН и микрофлоры до фильтрации. Приготовленный лекарственный продукт затем фильтруют, используя SHC-фильтр Express XL150 от Millipore, с хранением в течение ночи или направляют в линию розлива для проведения стерилизующей фильтрации.
Приготовленный нерасфасованный продукт направляют на участок розлива, отбирают образец для контроля микрофлоры и затем осуществляют стерилизующую фильтрацию, используя два соединенных последовательно SHC-фильтра Express XL150 от Millipore. Приготовленным нерасфасованным продуктом наполняют апирогенные стеклянные флаконы до намеченного объема заполнения 0,73 мл. Наполненные флаконы неплотно закрывают пробками и затем загружают в сублимационную сушилку. Выполняют цикл лиофилизации так, как показано в Таблице 41. По завершении цикла камеру для лиофильной сушки наполняют азотом до 0,8 ат и затем пробки устанавливают полностью. Камеру разгружают и флаконы закрывают, используя колпачки «флип-офф».

Данные по стабильности лекарственного продукта суммированы в Таблице 42. Данные свидетельствуют о том, что лекарственный продукт физически и химически стабилен при хранении при 2-8°С в течение по меньшей мере 3 месяцев или по меньшей мере 1 месяца при 25° или 40°С. Для обоих условий хранения уровень примесей, обнаруженных посредством гельпроникающей хроматографии (SEC), не изменялся, также не было изменений в антигенности in vitro на всем протяжении тестирования до самых последних моментов времени.

Пример 40. Разбавители для вакцин
Что касается физиологического раствора, то для лиофилизированного лекарственного продукта без какого-либо адъюванта в качестве разбавителя используют 60 мМ NaCl, чтобы гарантировать получение изотонического раствора при повторном разведении.
Alhydrogel: Alhydrogel «85» 2% (Brenntag) представляет собой имеющийся в продаже продукт качества «для надлежащей производственной практики» (GMP), состоящий из октаэдрических кристаллических пластинок гидроксида алюминия. Типичная композиция разбавителя Alhydrogel показана в Таблице 43. Такую типичную композицию можно использовать в комбинации с лекарственным продуктом, описанным выше.

Исследования, выполненные с адъювантом Alhydrogel, демонстрируют 100%-ное связывание мутантного токсина А как лекарственной субстанции и мутантного токсина В как лекарственной субстанции с 1 мг Al/мл Alhydrogel при рН 6,0-7,5. Максимальное связывание обеих лекарственных субстанций отмечали при самой высокой тестируемой концентрации белка (300 мкг/мл для каждого).
Связывание белков с Alhydrogel также тестировали, используя композицию лиофилизированного лекарственного продукта, содержащую каждую лекарственную субстанцию в концентрации 200 мкг/мл и Alhydrogel в диапазоне от 0,25 до 1,5 мг/мл. Лекарственный продукт повторно разводили, используя разбавители, содержащие разные концентрации Alhydrogel, и измеряли процентную долю каждого связавшегося мутантного токсина. Для всех протестированных концентраций Alhydrogel было продемонстрировано 100%-ное связывание антигенов.
Также оценивали кинетику связывания белков с Alhydrogel при целевой дозе мутантного токсина А как лекарственной субстанции и мутантного токсина В как лекарственной субстанции (по 200 мкг/мл каждого). Результаты показывают, что за весь 24-часовой период времени при комнатной температуре (КТ) с Alhydrogel связалось 100% мутантных токсинов как лекарственных субстанций.
CpG 24555 и Alhydrogel: CpG 24555 представляет собой синтетический 21-мерный олигодезоксинуклеотид (ODN), имеющий последовательность 5-TCG TCG ТТТТТС GGT GCT ТТТ-3 (SEQ ID NO: 48). Типичная композиция для комбинации разбавителей CpG 24555 и Alhydrogel показана в Таблице 44. Такую типичную композицию можно использовать в комбинации с лекарственным продуктом, описанным выше.

ISCOMATRIX®: адъювант ISCOMATRIX® представляет собой известный в данной области техники адъювант на основе сапонина. Типичная композиция адъюванта ISCOMATRIX® показана в Таблице 45. Такую типичную композицию можно использовать в комбинации с лекарственным продуктом, описанным выше.
Пример 41. Иммуногенность композиций на основе мутантных токсинов как лекарственных субстанций, дополненных адъювантом Alhydrogel, в NHP-модели и доклиническая проверка концепции
Иммуногенность композиций на основе мутантного токсина А как лекарственной субстанции и мутантного токсина В как лекарственной субстанции, дополненных адъювантом Alhydrogel, оценивали на NHP, в частности на яванских макаках. У NHP, иммунизированных с двухнедельными интервалами (в 0-ю, 2-ю, 4-ю недели) композициями на основе мутантного токсина А как лекарственной субстанции и мутантного токсина В как лекарственной субстанции, по 10 мкг каждого на одну дозу (приготовленных вместе с Alhydrogel), развивались сильные нейтрализующие антительные ответы на токсины. См. Таблицу 46. И нейтрализующие антительные ответы на токсин А, и нейтрализующие антительные ответы на токсин В достигали диапазона защитного действия после третьей иммунизации и сохранялись в пределах или выше этого диапазона защитного действия по меньшей мере до 33-й недели (последняя исследуемая временная точка).
Яванских макак (n=8) иммунизировали в./м. в 0-ю, 2-ю и 4-ю недели, используя мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию, по 10 мкг каждого, в составе композиции вместе с 250 мкг Alhydrogel. В каждый тестируемый момент времени отбирали кровь для приготовления сыворотки и ее исследовали в анализе нейтрализации токсинов с точки зрения функциональной антитоксинной активности. GMT приведены в Таблице 46. Диапазон защитных титров, приведенный в этой таблице, описывает диапазон титров нейтрализующих антител, который коррелирует со значительным снижением рецидива инфекции С. difficile в исследовании терапии моноклональными антителами от Merck.
Корреляция титров защитных антител человека в исследовании терапии с использованием mAb от Merck с титрами, индуцированными у NHP под действием вакцины-кандидата от Pfizer
По всей видимости, фаза 2 исследования эффективности использования mAb от Merck/Medarex (Lowy et al., N. Engl. J. Med., 2010, Jan, 21, 362(3): 197-205) демонстрирует корреляцию между уровнем нейтрализующих токсины mAb в сыворотке крови и предупреждением рецидива CDAD. Оказалось, что после введения токсин-специфических mAb людям, присутствующие в сыворотке крови реципиентов-людей антитела с уровнями в диапазоне 10-100 мкг/мл, защищают от рецидивов (уменьшение на 70% рецидива CDAD).
Иммуногенные композиции, содержащие мутантные токсины как лекарственные субстанции, тестировали для проверки того, способны ли такие иммуногенные композиции индуцировать потенциально эффективные нейтрализующие антительные ответы у людей, сравнивая опубликованные данные фазы 2 исследования от Merck/Medarex с уровнями антител, индуцированными иммуногенными композициями в NHP-модели. Сравнение осуществляли, используя ранее опубликованные характеристики mAb от Merck/Medarex для преобразования диапазона этих mAb в сыворотке крови, полученной от субъектов, которые не проявляли никаких признаков рецидивов (10-100 мкг/мл), в титры нейтрализующих на 50% антител, и сравнивая эти титры («диапазон защитных титров») с титрами, наблюдавшимися в доклинических моделях, изложенных в данном описании. Как показано в Таблице 46, иммуногенные композиции, содержащие мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию, дополненные адъювантом Alhydrogel, генерировали иммунные ответы у NHP, которые достигали «диапазона защитного действия» после введения третьей дозы и оставались в пределах или выше этого диапазона до 33-й недели. Уровень токсин-нейтрализующих антител, индуцированных у NHP под действием иммуногенной композиции на основе С. difficile по изобретению, сравним с уровнями антител в сыворотке крови у субъектов в исследовании от Merck/Medarex, которые, как оказалось, были защищены от рецидивов CDAD.
Пример 42. Иммуногенность композиций на основе мутантных токсинов как лекарственных субстанций, дополненных адъювантом ISCOMATRIX или Alhydrogel/CpG 24555 (Alh/CpG), в NHP-модели
Присутствие в композиции как ISCOMATRIX, так и Alh/CpG статистически значимо повышало у NHP уровни титров нейтрализующих токсины А и В антител в сравнении с вакциной, введенной только с Alhydrogel (Таблица 47). Превышающие фон антительные ответы к токсинам вырабатывались в ранние тестируемые моменты времени под действием вакцины, введенной либо с Alh/CpG, либо с ISCOMATRIX (2-4-я неделя) в сравнении с введением только с Alhydrogel (4-6-я неделя), что может оказывать важное влияние на защиту от рецидива CDAD у людей. В сравнении с Alhydrogel, введение иммуногенной композиции, дополненной адъювантом Alh/CpG или ISCOMATRIX, вызывало образование нейтрализующих токсины антител с титрами, которые быстрее достигали диапазона защитного действия (см. также пример 41) и оставались в пределах или выше этого диапазона до 33-й недели.
Как показано в Таблице 47, яванских макак иммунизировали в./м. в 0-ю, 2-ю и 4-ю недели, используя мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию, по 10 мкг каждого, в составе композиции вместе с 250 мкг Alhydrogel (n=8) или 500 мкг CpG+250 мкг Alhydrogel (n=10) или 45 единицами ISCOMATRIX (n=10). В каждый тестируемый момент времени отбирали кровь для приготовления сыворотки и ее исследовали в описанном выше анализе нейтрализации токсинов с точки зрения функциональной антитоксинной активности. GMT приведены в таблицах. Звездочки (*) указывают на статистическую значимость (р<0,05) при сравнении с титрами в группе с Alhydrogel. Диапазон защитных титров представляет диапазон титров нейтрализующих антител, который коррелирует со значительным снижением рецидива инфекции С. difficile в исследовании терапии моноклональными антителами от Merck.

Также оценивали влияние дозы мутантного токсина А как лекарственной субстанции и мутантного токсина В как лекарственной субстанции, введенной в присутствии адъювантов ISCOMATRIX или Alh/CpG, на титры нейтрализующих токсины антител, генерированных у NHP. В одном исследовании NHP вводили низкую (10 мкг) или высокую (100 мкг) дозу каждого мутантного токсина как лекарственной субстанции в составе композиции вместе с ISCOMATRIX. Ответы сравнивали в каждый тестируемый момент времени после иммунизации. Как показано в Таблице 48, титры нейтрализующих токсины антител были высокими в обеих подвергнутых обработке группах. Титры антител к токсину А были примерно одинаковы для большинства тестируемых моментов времени в группах, получавших низкую дозу и высокую дозу, в то время как для титров антител к токсину В наблюдалась тенденция к более высоким значениям в группе, получавшей высокую дозу.

Как показано в Таблице 48, яванских макак (n=5) иммунизировали в./м. в 0-ю, 2-ю и 4-ю недели, используя мутантный токсин А как лекарственную субстанцию и мутантный токсин В как лекарственную субстанцию, по 10 мкг или 100 мкг каждого, в составе композиции вместе с 45 единицами ISCOMATRIX. В каждый тестируемый момент времени отбирали кровь для приготовления сыворотки и ее исследовали в анализе нейтрализации токсинов с точки зрения функциональной антитоксинной активности. GMT приведены в указанной таблице. Диапазон защитных титров представляет диапазон титров нейтрализующих антител, который коррелирует со значительным снижением рецидива инфекции С. difficile в исследовании терапии моноклональными антителами от Merck.
В целях улучшения кинетики возникновения антительных ответов на токсин В NHP иммунизировали постоянной дозой мутантного токсина А как лекарственной субстанции (10 мкг), которую смешивали с возрастающей дозой мутантного токсина В как лекарственной субстанции (10, 50 или 100 мкг) в присутствии адъювантов ISCOMATRIX или Alh/CpG. Независимо от адъюванта, у групп, получавших более высокие дозы мутантного токсина В как лекарственной субстанции (или 50, или 100 мкг), наблюдалась тенденция индуцировать более сильные нейтрализующие антительные ответы к токсину В в сравнении с дозой 10 мкг мутантного токсина В как лекарственной субстанции (Таблица 50, отмечено * для указания на статистически значимые увеличения). Эта тенденция наблюдалась для большинства тестируемых моментов времени после окончательной иммунизации. Однако, в некоторых случаях для нейтрализующих антительных ответов к токсину А было показано статистически значимое уменьшение (отмечено ^ в Таблице 49) при увеличении количества мутантного токсина В.
Как показано в Таблице 49 и Таблице 50, NHP (10 в одной группе) иммунизировали в./м. в 0-ю, 2-ю и 4-ю недели мутантным токсином А как лекарственной субстанцией и мутантным токсином В как лекарственной субстанцией в разных соотношениях (10 мкг мутантного токсина А как лекарственной субстанции плюс что-либо одно из 10, 50 или 100 мкг мутантного токсина В как лекарственной субстанции; что обозначено в Таблице 49 и Таблице 50 как 10A:10В, 10А:50В и 10A:100В, соответственно) в составе композиции вместе с ISCOMATRIX (45 единиц на одну дозу) или с Alh/CpG/ (250 мкг/500 мкг на одну дозу). В Таблице 49 показаны титры антител к токсину А. В Таблице 50 показаны титры антител к токсину В. В этих таблицах приведены GMT. Диапазон защитных титров представляет диапазон титров нейтрализующих антител, который коррелирует со значительным снижением рецидива инфекции С. difficile в исследовании терапии моноклональными антителами от Merck. Символ ^ представляет статистически значимое уменьшение титров нейтрализующих антител (р<0,05) в сравнении с группой 10A:10В. Символ звездочка, *, представляет статистически значимое увеличение титров нейтрализующих антител (р<0,05) в сравнении с группой 10A:10В.
Пример 43. Проводимое на яванских макаках в течение пяти недель исследование токсичности с повторными в./м. введениями иммуногенной композиции, с 4-х недельным периодом восстановления
Проводимое на яванских макаках в течение пяти недель исследование токсичности с повторными в./м. введениями PF-06425095 (иммуногенной композиции, содержащей тройной мутантный токсин А как лекарственную субстанцию и тройной мутантный токсин В как лекарственную субстанцию в комбинации с адъювантами гидроксидом алюминия и CpG 24555) выполняли для оценки возможной токсичности и иммуногенности тройного мутантного токсина А как лекарственной субстанции из С. difficile и тройного мутантного токсина В как лекарственной субстанции из С. difficile в комбинации с адъювантами гидроксидом алюминия и CpG 24555 (PF-06425095). PF-06425095 с содержанием тройного мутантного токсина А как лекарственной субстанции и тройного мутантного токсина В как лекарственной субстанции 0,2 или 0,4 мг/доза (группы с иммуногенной композицией в низкой и высокой дозе, соответственно), 0,5 мг алюминия в виде гидроксида алюминия и 1 мг CpG 24555 и одну лишь комбинацию адъювантов (гидроксид алюминия + CpG 24555; PF-06376915) вводили в./м. яванским макакам (6 животных разных полов/группа) в качестве примирующей дозы, после чего вводили 3 бустер-дозы (1-е, 8-е, 22-е и 36-е сутки). Отдельная группа животных (6 животных разных полов) получала 0,9%-ный изотонический физиологический раствор с рН приблизительно 7,0. В состав разбавителя для иммуногенной композиции входили 10 мМ трис-буфер с рН 7,4, 4,5% дигидрата трегалозы и 0,1% полисорбата 80. Контрольный разбавитель для адъюванта представлял собой 10 мМ буфер на основе гистидина с 60 нМ NaCl с рН 6,5. Общий вводимый объем дозы составлял 0,5 мл на одну инъекцию. Все дозы вводили в левую и/или правую четырехглавую мышцу. Отобранные животные прошли 4-х недельный период наблюдения без введения доз для оценки обратимости любого рода эффектов, наблюдаемых во время фазы введения доз в этом исследовании.
В этом исследовании не получено никаких данных о неблагоприятных эффектах. Композиция PF-06425095 хорошо переносилась и вызывала только местную воспалительную реакцию без признаков системной токсичности. Во время фазы введения доз наблюдали дозозависимые повышения, по сравнению с уровнями, отмеченными перед тестированием, уровней фибриногена (до значений от 23,1% до 2,3х) на 4-е и 38-е сутки, С-реактивного белка на 4-е (до значений от 2,1x до 27,5х) и 38-е сутки (до значений от 2,3х до 101,5х) и глобулина (до значений от 11,1% до 24,1%) на 36-е и/или 38-е сутки в группах, обработанных иммуногенной композицией, что находилось в соответствии с ожидаемым воспалительным ответом на введение адъювантной иммуногенной композиции.
Эти повышенные уровни фибриногена и С-реактивного белка, отмеченные на 4-е сутки, частично возвращались к прежним значениям на 8-е сутки, при этом повышенные уровни фибриногена (до значений от 25,6% до 65,5%) и С-реактивного белка (до значений 4,5х и 5,6х) оставались только в группе, получавшей иммуногенную композицию в высокой дозе. Повышение уровня интерлейкина(IL)-6 наблюдали в группах, получавших иммуногенную композицию в низкой и высокой дозах, на момент времени 1-е сутки, 3 часа (до значений в отдельных случаях от 8,3х до 127,2х на 1-е сутки, 0 часов, дозозависимым образом), и 36-е сутки, 3 часа (до значений в отдельных случаях от 9,4х до 39,5х на 36-е сутки, 0 часов). Не наблюдали никаких изменений в случае других цитокинов (IL-10, IL-12, интерферон-индуцибельного белка (ΙΡ-10) и фактора некроза опухолей α (TNF-α)). Повышение уровней таких белков острой фазы воспаления и цитокина были частью ожидаемого нормального физиологического ответа на введение чужеродного антигена. Не наблюдали никаких связанных с PF-06425095 или связанных с адъювантом изменений в данных параметрах клинической патологии в фазе восстановления (во время фазы восстановления уровень цитокинов не оценивали). Кроме того, имелись локальные изменения в местах инъекции, которые были сопоставимы по частоте возникновения и тяжести в случаях контрольной группы, получавшей адъювант, и групп, получавших иммуногенную композицию в низкой и высокой дозах; следовательно, они не имели непосредственного отношения к PF-06425095. Во время фазы введения доз эти изменения включали хроническое/активное воспаление от минимального до умеренного, которое характеризовалось расслоением мышечных волокон под действием инфильтратов, состоящих из макрофагов, которые часто содержали окрашиваемый основными красителями гранулированный материал (интерпретируемый как алюминийсодержащий адъювант), лимфоцитов, плазматических клеток, нейтрофилов, эозинофилов и некротического дебриса, и отеком. Окрашиваемый основными красителями гранулированный материал также присутствовал во внеклеточном пространстве в таких очагах хронического/активного воспаления. В конце фазы восстановления наблюдали хроническое воспаление от минимального до умеренного и наличие инфильтрата мононуклеарных клеток и минимального фиброз. Эти данные, касающиеся места инъекции, описывают местный воспалительный ответ на введение адъюванта. Другие микроскопические изменения включали увеличение от минимального до умеренного насыщенности лимфоидными клетками подвздошных (дренирующих) лимфатических узлов и минимальное увеличение насыщенности клетками зародышевых центров в селезенке, что было отмечено во время фазы введения доз в контрольной группе, получавшей адъювант, и в группах, получавших иммуногенную композицию в низкой и высокой дозах. В конце фазы восстановления данные этих микроскопических исследований демонстрировали низкую степень тяжести. Такие эффекты представляют собой иммунологический ответ на антигенную стимуляцию, и были фармакологическим ответом на введение адъюванта или PF-06425095. Не было зарегистрировано никакого связанного с тестируемым продуктом повышения уровня антител к ДНК.
С учетом отсутствия данных о неблагоприятных эффектах уровень, не вызывающий видимых неблагоприятных эффектов (NOAEL), в этом исследовании относится к группе, получавшей иммуногенную композицию в высокой дозе (0,4 мг тройного мутантного токсина А как лекарственной субстанции и тройного мутантного токсина В как лекарственной субстанции на одну дозу в составе PF-06425095), введенной в виде двух инъекций по 0,5 мл для четырех доз.
Пример 44. Эффективность серопозитивных сывороток крови NHP, пассивно перенесенных хомякам
Группам из 5 сирийских золотых хомяков вводили пероральную дозу антибиотика клиндамицина (30 мг/кг) для уничтожения нормальной кишечной флоры. Через пять суток осуществляли контрольное заражение хомяков, используя пероральное введение спор С. difficile дикого типа (штамм 630, по 100 КОЕ на одно животное), и вводили внутрибрюшинно (в./б.) образцы сыворотки крови NHP согласно Таблице 51. Без связи с механизмом или теорией считается, что симптомы заболевания после контрольного заражения спорами обычно проявляются примерно через 30-48 часов после контрольного заражения.
Образцы сыворотки крови NHP, которые вводили хомякам, представляли собой пул образцов сыворотки крови NHP, демонстрирующих самый высокий титр (антисыворотка к токсину А и антисыворотка к токсину В) после трех иммунизаций мутантным токсином А как лекарственной субстанцией и мутантным токсином В как лекарственной субстанцией (соотношения А:В составляли 10:10, 10:50 и 10:100) в составе композиций вместе с ISCOMATRIX (см. пример 42, Таблицу 49 и Таблицу 50). У NHP отбирали кровь для приготовления образцов сыворотки в моменты времени, соответствующие 5-й, 6-й и 8-й неделям (иммунизации осуществляли в 0-ю, 2-ю и 4-ю недели), как описано в примере 42. Результаты показаны в приведенных ниже Таблицах 52-54. Символ «+» указывает среднее геометрическое значение (GM) в скобках, которое не включает животного №3, не являющегося респондером. «*ТВ» представляет собой заключительный отбор крови, день подвергания животного эвтаназии, который не является одним и тем же для всех животных. 


В другом исследовании сирийским золотым хомякам вводили пероральную дозу антибиотика клиндамицина (30 мг/кг) для уничтожения нормальной кишечной флоры. Через пять суток осуществляли контрольное заражение хомяков, используя пероральное введение спор С. difficile дикого типа (штамм 630, по 100 КОЕ на одно животное), и вводили внутрибрюшинно (в./б.) образцы сыворотки крови NHP согласно Таблице 55. Без связи с механизмом или теорией считается, что симптомы заболевания после контрольного заражения спорами обычно проявляются примерно через 30-48 часов после контрольного заражения.
Образцы сыворотки крови NHP, которые вводили хомякам, представляли собой пул образцов, собранных от NHP после трех иммунизаций мутантным токсином А как лекарственной субстанцией и мутантным токсином В как лекарственной субстанцией (соотношения А:В составляли 10:10, 10:50 и 10:100) в составе композиций вместе с Alhydrogel и CpG 24555 (см. пример 42, Таблицу 49 и Таблицу 50). У NHP отбирали кровь для приготовления образцов сыворотки в моменты времени, соответствующие 5-й, 6-й, 8-й и 12-й неделям, как описано в примере 42 (NHP иммунизировали в 0-ю, 2-ю и 4-ю недели). Результаты показаны в приведенных ниже Таблицах 56-59. Образцы сыворотки от хомяков исследовали далее для определения величины ингибирующей концентрации (IC50), которую определяли с использованием описанного выше анализа нейтрализации токсинов. Уровень нейтрализующих токсины антител, индуцированных у хомяков под действием иммуногенной композиции на основе С. difficile по изобретению, сравним с уровнями антител в сыворотке крови у субъектов в исследовании от Merck/Medarex, которые, как оказалось, были защищены от рецидивов CDAD.


Пример 45. Определение характеристик мутантных токсинов как лекарственных субстанций
Первичная структура тройного мутантного токсина А показана в SEQ ID NO: 4. NH2-κoнцeвoй остаток Met в положении 1 в SEQ ID NO: 4 происходит из стартового кодона в SEQ ID NO: 12 и отсутствует в выделенном белке (например, см. SEQ ID NO: 84). Таким образом, в примерах 12-45 «SEQ ID NO: 4» относится к SEQ ID NO: 4, где начальный метионин (в положении 1) отсутствует. Оба, и очищенный тройной мутантный токсин A (SEQ ID NO: 4) (промежуточная лекарственная субстанция - партия L44993-132) и EDC/NHS-обработанный тройной мутантный токсин A (SEQ ID NO: 4) («мутантный токсин А как лекарственная субстанция» - партия L44898-012) обнаруживают одинаковую NH2-концевую последовательность, начинающуюся с SLISKEELIKLAYSI (положения 2-16 в SEQ ID NO: 4).
Первичная структура тройного мутантного токсина В представлена в SEQ ID NO: 6. NH2-κoнцeвoй остаток Met в положении 1 в SEQ ID NO: 6 происходит из стартового кодона и отсутствует в выделенном белке например, см. SEQ ID NO: 86). Таким образом, в примерах 12-45 «SEQ ID NO: 6» относится к SEQ ID NO: 6, где начальный метионин (в положении 1) отсутствует. Оба, и очищенный тройной мутантный токсин В (SEQ ID NO: 6) (промежуточная лекарственная субстанция - партия 010) и EDC/NHS-обработанный тройной мутантный токсин В (SEQ ID NO: 6) («мутантный токсин В как лекарственная субстанция» - партия L44906-153) обнаруживают одинаковую NH2-концевую последовательность, начинающуюся с SLVNRKQLEKMANVR (положения 2-16 в SEQ ID NO: 6).
Для оценки вторичной и третичной структуры тройного мутантного токсина A (SEQ ID NO: 4) и мутантного токсина А как лекарственной субстанции использовали спектроскопию кругового дихроизма (КД). КД-спектроскопию также использовали для оценки вторичной и третичной структуры тройного мутантного токсина В (SEQ ID NO: 6) и мутантного токсина В как лекарственной субстанции. КД-спектроскопию также использовали для оценки возможного влияния рН на структуру. Влияние обработки с использованием EDC на тройной мутантный токсин А анализировали, сравнивая данные КД, полученные для мутантного токсина А как лекарственной субстанции, с данными, полученными для тройного мутантного токсина А. Влияние обработки с использованием EDC на тройной мутантный токсин В (SEQ ID NO: 6) анализировали, сравнивая данные КД, полученные для мутантного токсина В как лекарственной субстанции, с данными, полученными для тройного мутантного токсина В.
Данные КД в дальней ультрафиолетовой (УФ) области для мутантного токсина А как лекарственной субстанции получали при разных значениях рН. Спектры, зарегистрированные при рН 5,0-7,0, свидетельствуют о высоком относительном содержании α-спиралей во вторичной структуре, позволяя предположить, что полипептидный остов белка принимает четко выраженную конформацию с преобладанием α-спиралей.
Кроме того, для мутантного токсина А как лекарственной субстанции получали спектры КД в ближней УФ-области. Сильная отрицательная эллиптичность между 260 и 300 нм является указанием на то, что боковые цепи ароматических аминокислот находятся в строго фиксированном жестком окружении, т.е. мутантный токсин А как лекарственная субстанция обладает третичной структурой. В самом деле, в спектре можно различить характерные признаки, обусловленные индивидуальными типами боковых цепей ароматических аминокислот: плечо приблизительно при 290 нм и самый большой отрицательный пик приблизительно при 283 нм обусловлены поглощением поляризованного света упорядоченными боковыми цепями триптофана, отрицательный пик при 276 нм происходит от боковых цепей тирозина, а минорные плечи при 262 и 268 нм указывают на остатки фенилаланина, участвующие в контактах, формирующих третичную структуру. Результаты измерений в дальней и ближней УФ-областях свидетельствуют о том, что мутантный токсин А как лекарственная субстанция сохраняет компактно уложенную структуру при физиологических значениях рН. Почти идентичные спектры КД в дальней и ближней УФ-областях, наблюдаемые при рН 5,0-7,0, указывают на то, что в пределах этого диапазона рН отсутствуют какие-либо детектируемые структурные изменения. Данные КД невозможно было получить при рН 3,0 и 4,0, поскольку данный белок при этих значениях рН не растворяется. Сравнение спектров КД в дальней и ближней УФ-областях для мутантного токсина А как лекарственной субстанции с таковыми для тройного мутантного токсина А говорит о том, что спектры обоих белков по-существу идентичны во всех исследуемых экспериментальных условиях, указывая на то, что обработка с использованием EDC не оказывала никакого детектируемого влияния на вторичную и третичную структуру тройного мутантного токсина А. Эти данные согласуются с результатами гель-фильтрации и аналитического ультрацентрифугирования, которые показывают полное отсутствие каких-либо детектируемых изменений в радиусах Стокса и коэффициентах седиментации/трения, соответственно.
Мутантный токсин А как лекарственная субстанция (а также тройной мутантный токсин А) содержит 25 остатков триптофана, которые распределены по всей первичной последовательности и могут служить в качестве подходящих внутренних флуоресцентных зондов. Для мутантного токсина А как лекарственной субстанции были получены спектры испускания флуоресценции в диапазоне 300-400 нм как функция от температуры. При 6,8°С мутантный токсин А как лекарственная субстанция демонстрирует характерный для триптофана спектр испускания флуоресценции после возбуждения при 280 нм. Максимальное испускание флуоресценции наблюдается приблизительно при 335 нм, указывая на то, что остатки триптофана в большей степени находятся в неполярном окружении, типичном для внутренней части белков, чем в полярном водном окружении. Результаты, полученные из спектров испускания флуоресценции, вместе с представленными в этом сообщении результатами экспериментов по КД подтверждают, что в мутантном токсине А как лекарственной субстанции сохраняется компактно уложенная структура.
Флуоресценцию внешнего зонда 8-анилино-1-нафталинсульфоновой кислоты (ANS) использовали для того, чтобы охарактеризовать возможные конформационные изменения в мутантном токсине А как лекарственной субстанции и тройном мутантном токсине А при изменении рН. Из этих результатов видно, что по-существу не наблюдается никакого увеличения интенсивности флуоресценции ANS, когда или мутантный токсин А как лекарственную субстанцию, или тройной мутантный токсин А титруют данным зондом при рН 7,0, что указывает на отсутствие какого-либо экспонирования гидрофобных поверхностей на этих белках в данных условиях. Смещение рН в сторону 2,6 приводит к резкому увеличению квантового выхода флуоресценции ANS при увеличении концентрации зонда, до тех пор, пока квантовый выход флуоресценции не достигнет кажущегося насыщения. Это увеличение квантового выхода флуоресценции ANS указывает на то, что при низких значениях рН (2,6) и мутантный токсин А как лекарственная субстанция, и тройной мутантный токсин А претерпевают рН-индуцированное конформационное изменение, приводящее к экспонированию гидрофобных поверхностей. Такие конформационные изменения указывают на то, что EDC-индуцированная модификация и инактивация тройного мутантного токсина А не ограничивали конформационной подвижности мутантного токсина А как лекарственной субстанции (DS).
Влияние EDC-обработки на гидродинамические свойства тройного мутантного токсина А оценивали с использованием гельпроникающей хроматографии на колонке G4000 SWXL. Мутантный токсин А как лекарственную субстанцию и тройной мутантный токсин А наносили на колонку G4000 SWXL, уравновешенную при рН 7,0, 6,0 и 5,0. Данные указывают на то, что с использованием гельпроникающей хроматографии не удается обнаружить каких-либо различий в радиусе Стокса у мутантного токсина А как лекарственной субстанции и тройного мутантного токсина А. Таким образом, обработка с использованием EDC не сказывается резко на гидродинамических свойствах и, соответственно, в целом на форме молекулы тройного мутантного токсина А.
Дальнейший анализ тройного мутантного токсина А и мутантного токсина А как лекарственной субстанции проводили, используя метод многоуглового лазерного светорассеяния (MALLS). Обработка тройного мутантного токсина А с использованием EDC приводила к образованию гетерогенной смеси, состоящей из различных многомерных и мономерных компонентов. Такая гетерогенность является отражением введения большого числа EDC-индуцированных меж- и внутримолекулярных ковалентных связей между карбоксильными группами и первичными аминогруппами белка.
Полученные данные определяют физические и химические характеристики тройного мутантного токсина А и мутантного токсина А как лекарственной субстанции (тройного мутантного токсина А, обработанного EDC) и описывают ключевые признаки их первичной, вторичной и третичной структуры. Полученные данные демонстрируют, что обработка тройного мутантного токсина А с использованием EDC приводила к ковалентной модификации его полипептидной цепи, но не влияла на вторичную и третичную структуру данного белка. Обработка EDC приводит к образованию внутри- и межмолекулярных сшивок. Биохимические и биофизические параметры, полученные для мутантного токсина А как лекарственной субстанции (а также тройного мутантного токсина А) представлены в Таблице 60.

Данные КД в дальней УФ-области для мутантного токсина В как лекарственной субстанции получали при разных значениях рН. Спектры, зарегистрированные при рН 5,0-7,0, свидетельствуют о высоком относительном содержании α-спиралей во вторичной структуре, позволяя предположить, что полипептидный остов белка принимает четко выраженную конформацию с преобладанием α-спиралей.
Кроме того, для мутантного токсина В как лекарственной субстанции получали спектры КД в ближней УФ-области. Сильная отрицательная эллиптичность между 260 и 300 нм является указанием на то, что боковые цепи ароматических аминокислот находятся в строго фиксированном жестком окружении, т.е. мутантный токсин В как лекарственная субстанция обладает третичной структурой. В самом деле, в спектре можно различить характерные признаки, обусловленные индивидуальными типами боковых цепей ароматических аминокислот: плечо приблизительно при 290 нм и самый большой отрицательный пик приблизительно при 283 нм обусловлены поглощением поляризованного света упорядоченными боковыми цепями триптофана, отрицательный пик при 276 нм происходит от боковых цепей тирозина, а минорные плечи при 262 и 268 нм указывают на остатки фенилаланина, участвующие в контактах, формирующих третичную структуру. Результаты измерений в дальней и ближней УФ-областях свидетельствуют о том, что мутантный токсин В как лекарственная субстанция сохраняет компактно уложенную структуру при физиологических значениях рН. Почти идентичные спектры КД в дальней и ближней УФ-областях, наблюдаемые при рН 5,0-7,0, указывают на то, что в пределах этого диапазона рН отсутствуют какие-либо детектируемые изменения вторичной или третичной структур. Данные КД невозможно было получить при рН 3,0 и 4,0, поскольку данный белок при этих значениях рН не растворяется.
Сравнение спектров КД в дальней и ближней УФ-областях для мутантного токсина В как лекарственной субстанции с таковыми для тройного мутантного токсина В говорит о том, что спектры обоих белков очень похожи в диапазоне рН 5,0-7,0, указывая на то, что обработка с использованием EDC не оказывала никакого детектируемого влияния на вторичную и третичную структуру белка.
Тройной мутантный токсин В содержит 16 остатков триптофана, которые распределены по всей первичной последовательности и могут служить в качестве подходящих внутренних флуоресцентных зондов. Для мутантного токсина В как лекарственной субстанции были получены спектры испускания флуоресценции в диапазоне 300-400 нм как функция от температуры. При 7°С мутантный токсин В как лекарственная субстанция демонстрирует характерный для триптофана спектр испускания флуоресценции после возбуждении при 280 нм. Максимальное испускание флуоресценции наблюдается приблизительно при 335 нм, указывая на то, что остатки триптофана в большей степени находятся в неполярном окружении, типичном для внутренней части белков, чем в полярном водном окружении. Эти результаты вместе с результатами экспериментов по КД (см. выше) подтверждают, что в мутантном токсине В как лекарственной субстанции сохраняется компактно уложенная структура.
Флуоресценцию внешнего зонда 8-анилино-1-нафталинсульфоновой кислоты (ANS) использовали для того, чтобы охарактеризовать возможные конформационные изменения в мутантном токсине В как лекарственной субстанции и тройном мутантном токсине В при изменении рН. Из этих результатов видно, что по-существу не наблюдается никакого увеличения интенсивности флуоресценции ANS, когда или мутантный токсин В как лекарственную субстанцию, или тройной мутантный токсин В титруют данным зондом при рН 7,0, что указывает на отсутствие какого-либо экспонирования гидрофобных поверхностей на этих белках в данных условиях. Смещение рН в сторону 2,6 приводит к резкому увеличению квантового выхода флуоресценции ANS при увеличении концентрации зонда, до тех пор, пока квантовый выход флуоресценции не достигнет кажущегося насыщения. Это увеличение квантового выхода флуоресценции ANS указывает на то, что при низких значениях рН (2,6) мутантный токсин В как лекарственная субстанция претерпевает рН-индуцированное конформационное изменение, приводящее к экспонированию гидрофобных поверхностей. Такие конформационные изменения указывают на то, что EDC-индуцированная модификация и инактивация тройного мутантного токсина В не ограничивали конформационной подвижности мутантного токсина В как лекарственной субстанции (DS).
Влияние EDC-обработки на гидродинамические свойства тройного мутантного токсина В оценивали с использованием гельпроникающей хроматографии на колонке G4000 SWXL. Мутантный токсин В как лекарственную субстанцию и тройной мутантный токсин В наносили на колонку G4000 SWXL, уравновешенную при рН 7,0, 6,0 и 5,0. Данные указывают на то, что с использованием гельпроникающей хроматографии не удается обнаружить каких-либо различий в радиусе Стокса у мутантного токсина В как лекарственной субстанции и тройного мутантного токсина В, и таким образом, обработка с использованием EDC не сказывается резко на гидродинамических свойствах и, соответственно, в целом на форме молекулы данного белка.
Дальнейший анализ тройного мутантного токсина В и мутантного токсина В как лекарственной субстанции проводили, используя метод многоуглового лазерного светорассеяния (MALLS). Обработка тройного мутантного токсина В с использованием EDC приводила к образованию более гетерогенной смеси, состоящей из различных многомерных и мономерных компонентов. Такая гетерогенность является отражением введения большого числа EDC-индуцированных меж- и внутримолекулярных ковалентных связей между карбоксильными группами и первичными аминогруппами белка.
Полученные данные определяют физические и химические характеристики тройного мутантного токсина В и мутантного токсина В как лекарственной субстанции (тройного мутантного токсина В, обработанного EDC) и описывают ключевые признаки их первичной, вторичной и третичной структуры. Полученные данные демонстрируют, что обработка тройного мутантного токсина В с использованием EDC приводила к ковалентной модификации его полипептидной цепи, но не влияла на вторичную и третичную структуру данного белка. Обработка EDC приводит к образованию внутри- и межмолекулярных сшивок. Основные биохимические и биофизические параметры, полученные для мутантного токсина В как лекарственной субстанции (а также тройного мутантного токсина В) представлены в Таблице 61.

Пример 46. Перфузионная ферментация для токсоида В (тройного мутанта)
Посевные культуры для токсоида В (тройного мутанта): каждую бутыль емкостью 1 л, содержащую 400 мл среды, инокулировали 1 мл посевного материала, инкубировали при 37°С, стационарно в течение ночи (приблизительно 15 ч). Конечная OD500 должна составлять 3,0-4,0. Рабочий объем: 3 л (2,7 л среды + 300 мл инокулята). Каждый ферментер был оснащен 1 крыльчаткой типа Rushton и трубчатым барботером. Начальные условия: температура: 37°С, поток N2: барботировали со скоростью приблизительно 0,5 vvm (объем/объем в минуту). Контроллеры: рН поддерживали при 7,0, используя 5 н. NaOH. Пенообразование регулировали путем автоматического добавления PPG-2000, который добавляли перед стерилизацией в количестве
0. 25 мл/л в находящуюся в ферментере среду.
Перфузионное культивирование С. difficile осуществляли с использованием батареи из 2 секционных кассет SARTOCON, с фильтрующим материалом HYDROSART с размером пор 0,2 мкм, с площадью поверхности одной кассеты 0,1 квадратный метр. По 3 л добавляли в стеклянные ферментеры емкостью 10 л. Перфузию начинали, когда OD достигает целевого значения, равного приблизительно 4, в течение 2-часовых интервалов времени при увеличении скорости по схеме 0,75 л/ч, 1,5 л/ч, 2,25 л/ч, 3 л/ч для примера 1, Фиг. 28. Перфузию будут начинать, когда в среде для ферментации OD будет достигнуто целевое значение, приблизительно равное 4, в течение 2-часовых интервалов времени при увеличении скорости по схеме 0,75 л/ч, 1,5 л/ч, 3 л/ч и 6 л/ч для примера 2, Фиг. 29. При получении стартового сигнала для начала перфузии запускали рециркуляционный насос с желаемой скоростью перекачки 1,3 л/мин.
Пример 1, Фиг. 28: окончательно получали OD 50 и титр для токсоида В 243 мг/л.
Пример 2, Фиг. 29: окончательно получали OD 59 и титр для токсоида В 306 мг/л.
Согласно изобретению также предложены следующие воплощения, которые определены в приведенных ниже пунктах.
Пункт 1. Выделенный полипептид, содержащий SEQ ID NO: 183.
Пункт 2. Выделенный полипептид, содержащий SEQ ID NO: 184.
Пункт 3. Иммуногенная композиция, содержащая выделенный полипептид, содержащий SEQ ID NO: 183.
Пункт 4. Иммуногенная композиция, содержащая выделенный полипептид, содержащий SEQ ID NO: 184.
Пункт 5. Культуральная среда, содержащая гидролизат сои, дрожжевой экстракт и глюкозу, где среда содержит бактерию Clostridium difficile, происходящую из VPI 11186, при этом у бактерии отсутствует эндогенный полинуклеотид, кодирующий токсин, при этом бактерия содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, и дополнительно содержит промотор фередоксина (fdx) Clostridium sporogenes.
Пункт 6. Способ культивирования Clostridium difficile, включающий культивирование С. difficile в среде, содержащей гидролизат сои, дрожжевой экстракт и глюкозу, где среда содержит бактерию Clostridium difficile, происходящую из VPI 11186, при этом у бактерии отсутствует эндогенный полинуклеотид, кодирующий токсин, при этом бактерия содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, и дополнительно содержит промотор фередоксина (fdx) Clostridium sporogenes.
Пункт 7. Способ получения токсина Clostridium difficile, включающий культивирование С. difficile в среде в условиях, подходящих для продуцирования токсина и выделения токсина из среды; при этом среда содержит гидролизат сои, дрожжевой экстракт и глюкозу и при этом содержит бактерию Clostridium difficile, происходящую из VPI 11186, при этом у бактерии отсутствует эндогенный полинуклеотид, кодирующий токсин, при этом бактерия содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, и дополнительно содержит промотор фередоксина (fdx) Clostridium sporogenes.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ, ОТНОСЯЩИЕСЯ К МУТАНТНОМУ ТОКСИНУ CLOSTRIDIUM DIFFICILE, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2592686C2 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2017 |
|
RU2773880C1 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2012 |
|
RU2754446C2 |
| Способ получения аминокислотных последовательностей, кодирующих наноантитела, нейтрализующие токсин А Clostridioides difficile | 2023 |
|
RU2833930C1 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ОТ ОТРАВЛЕНИЙ | 2007 |
|
RU2465893C2 |
| АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ АССОЦИИРОВАННЫХ С CLOSTRIDIUM DIFFICILE ИНФЕКЦИИ И ЗАБОЛЕВАНИЙ | 2011 |
|
RU2630663C9 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К ТОКСИНАМ CLOSTRIDIUM DIFFICILE | 2013 |
|
RU2628305C2 |
| СПОСОБ АМПЛИФИКАЦИИ ДНК НА ОСНОВЕ ВНЕДРЕНИЯ В ЦЕПЬ | 2014 |
|
RU2673733C2 |
| Новые гибридные ActRIIB белки-ловушки лигандов для лечения заболеваний, связанных с мышечной атрофией | 2016 |
|
RU2733492C2 |
| СПОСОБ ДЕТЕКТИРОВАНИЯ И ХАРАКТЕРИЗАЦИИ ТОКСИНОГЕННОГО ШТАММА CLOSTRIDIUM DIFFICILE | 2010 |
|
RU2612789C2 |

































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к выделенному полипептиду, вызывающему образование нейтрализующих антител к токсинам Clostridium difficile. Указанный полипептид характеризуется аминокислотной последовательностью, представленной в любой из SEQ ID NO: 4 или SEQ ID NO: 6. Настоящее изобретение также относится к иммуногенной композиции, содержащей выделенный полипептид. Изобретение также относится к рекомбинантной клетке Clostridium difficile. Указанная рекомбинантная клетка предназначена для экспрессии выделенного полипептида. Изобретение относится к способам культивирования Clostridium difficile и получения указанного выделенного полипептида. Настоящее изобретение позволяет получать выделенный полипептид и иммуногенную композицию, вызывающие образование нейтрализующих антител к токсинам Clostridium difficile. 7 н. и 32 з.п. ф-лы, 59 ил., 71 табл., 46 пр.
1. Применение среды, содержащей гидролизат сои, дрожжевой экстракт и глюкозу, для культивирования бактерии, представляющей собой рекомбинантную бактерию Clostridium difficile, в которой отсутствует эндогенный полинуклеотид, кодирующий токсин, где данная бактерия содержит полинуклеотид, кодирующий мутантный токсин С. difficile, который содержит аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 4 или SEQ ID NO: 6, под контролем промотора фередоксина (fdx) Clostridium sporogenes.
2. Применение по п. 1, где среда дополнительно содержит полиэтиленгликоль 2000.
3. Применение по п. 1, где среда дополнительно содержит тиамфеникол.
4. Применение по п. 1, где бактерия происходит из С. difficile VPI 11186.
5. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile VPI 11186 (АТСС 700057™).
6. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile 1351 (АТСС 43593™).
7. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile 3232 (АТСС ВАА-1801™).
8. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile 7322 (АТСС 43601™).
9. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile 5036 (АТСС 43603™).
10. Применение по п. 1, где бактерия представляет собой рекомбинантную С. difficile 4811 (АТСС 43602™).
11. Способ культивирования Clostridium difficile, включающий культивирование С. difficile в среде, содержащей гидролизат сои, дрожжевой экстракт и глюкозу, где бактерия представляет собой бактерию Clostridium difficile, происходящую из Clostridium difficile VPI 11186, где у данной бактерии отсутствуют эндогенные полинуклеотиды, кодирующие токсин А и токсин В, где данная бактерия содержит полинуклеотид, кодирующий мутантный токсин С. difficile, который содержит аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 4 или SEQ ID NO: 6, под контролем промотора фередоксина (fdx) Clostridium sporogenes.
12. Способ получения мутантного токсина Clostridium difficile, включающий культивирование С. difficile в среде в условиях, подходящих для продуцирования этого токсина, и выделение токсина из среды, где среда содержит гидролизат сои, дрожжевой экстракт и глюкозу и где бактерия представляет собой бактерию Clostridium difficile, происходящую из Clostridium difficile VPI 11186, где у данной бактерии отсутствуют эндогенные полинуклеотиды, кодирующие токсин А и токсин В, где данная бактерия содержит полинуклеотид, кодирующий мутантный токсин из С. difficile, который содержит аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 4 или SEQ ID NO: 6, под контролем промотора фередоксина (fdx) Clostridium sporogenes.
13. Выделенный полипептид, вызывающий образование нейтрализующих антител к токсинам С. difficile, содержащий аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 4 или SEQ ID NO: 6, где полипептид содержит по меньшей мере одну аминокислотную боковую цепь, которая химически модифицирована.
14. Выделенный полипептид по п. 13, содержащий по меньшей мере одну аминокислотную боковую цепь, которая химически модифицирована 1-этил-3-(3-диметиламинопропил)карбодиимидом (EDC).
15. Выделенный полипептид по п. 13, содержащий по меньшей мере одну аминокислотную боковую цепь, которая химически модифицирована N-гидроксисукцинимидом (NHS).
16. Выделенный полипептид по п. 13, где по меньшей мере одна боковая цепь остатка аспарагиновой кислоты в полипептиде или по меньшей мере одна боковая цепь остатка глутаминовой кислоты в полипептиде химически модифицирована глицином.
17. Выделенный полипептид по п. 13, содержащий по меньшей мере одну сшивку между боковой цепью остатка аспарагиновой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде.
18. Выделенный полипептид по п. 13, содержащий по меньшей мере одну сшивку между боковой цепью остатка глутаминовой кислоты в полипептиде и боковой цепью остатка лизина в полипептиде.
19. Выделенный полипептид по п. 13, содержащий бета-аланиновую группировку, связанную с боковой цепью по меньшей мере одного остатка лизина в полипептиде.
20. Выделенный полипептид по п. 13, содержащий глициновую группировку, связанную с боковой цепью остатка аспарагиновой кислоты в полипептиде или с боковой цепью остатка глутаминовой кислоты в полипептиде.
21. Выделенный полипептид по п. 13, имеющий значение ЕС50 по меньшей мере примерно 100 г/мл.
22. Иммуногенная композиция, вызывающая образование нейтрализующих антител к токсинам С. difficile и содержащая выделенный полипептид по любому из пп. 13-21 в эффективном количестве.
23. Иммуногенная композиция по п. 22, дополнительно содержащая второй выделенный полипептид по любому из пп. 13-21.
24. Рекомбинантная клетка С. difficile, в которой отсутствует эндогенные полинуклеотиды, кодирующие токсин А и токсин В, содержащая нуклеиновокислотную последовательность, кодирующую выделенный полипептид, который содержит аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 4 или SEQ ID NO: 6, предназначенная для экспрессии указанного выделенного полипептида.
25. Рекомбинантная клетка по п. 24, происходящая из клетки Clostridium difficile, выбранной из группы, состоящей из Clostridium difficile 1351, Clostridium difficile 3232, Clostridium difficile 7322, Clostridium difficile 5036, Clostridium difficile 4811 и Clostridium difficile VPI 11186.
26. Рекомбинантная клетка по п. 24, представляющая собой клетку Clostridium difficile VPI 11186.
27. Рекомбинантная клетка по п. 24, где ген спорообразования в клетке Clostridium difficile является инактивированным.
28. Способ получения выделенного полипептида по любому из пп. 13-21, включающий культивирование рекомбинантной клетки С. difficile по любому из пп. 24-27 в условиях, подходящих для экспрессии содержащейся в ней нуклеиновокислотной последовательности.
29. Способ по п. 28, дополнительно включающий выделение указанного полипептида.
30. Способ по п. 28, дополнительно включающий приведение в контакт выделенного полипептида с этил-3-(3-диметиламинопропил)карбодиимидом.
31. Способ по п. 30, где приведение в контакт осуществляют в течение не более 4 часов.
32. Способ по п. 30, где приведение в контакт осуществляют при примерно 25°С.
33. Способ по п. 30, дополнительно включающий приведение в контакт выделенного полипептида с N-гидроксисукцинимидом.
34. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile VPI 11186 (АТСС 700057™).
35. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile 1351 (АТСС 43593™).
36. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile 3232 (АТСС ВАА-1801™).
37. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile 7322 (АТСС 43601™).
38. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile 5036 (АТСС 43603™).
39. Способ по п. 28, где рекомбинантная клетка представляет собой клетку рекомбинантной С. difficile 4811 (АТСС 43602™).
| FANG A | |||
| et al | |||
| Production of Clostridium difficile toxin in a medium totally free of both animal and dairy proteins or digests, Proceedings of the National Academy of Sciences, 2009 | |||
| MICHAELS J.D | |||
| et al | |||
| Способ предохранения металлов и сплавов от окисления | 1926 |
|
SU8018A1 |
| HEAP J.T | |||
| et al | |||
| A modular system for Clostridium shuttle plasmids, Journal of Microbiological Methods, 2009 | |||
| MOCRIEF J.S | |||
| et al | |||
| Genetic characterization of toxin A-negative, toxin B-positive Clostridium difficile isolates by PCR, Journal of Clinical Microbiology, 2000 | |||
| HEAP J.T | |||
| et al | |||
| The ClosTron: Mutagenesis in Clostridium refined and streamlined, Journal of Microbiological Methods, 2010 | |||
| ВАРФОЛОМЕЕВА Н.А | |||
| и др | |||
| Генно-инженерный подход к получению фрагментов токсина А и В для диагностики и иммунотерапии Clostridium difficile инфекции | |||
| Молекулярная генетика, микробиология и вирусология, 2003. | |||

