Родственные заявки
Настоящая заявка заявляет приоритет согласно 35 Кодексу законов США §119 по предварительным заявкам на патент США №№61/324503, поданной 15 апреля 2010 года, и 61/381669, поданной 10 сентября 2010 года, полное содержание каждой из которых включено в настоящий документ с помощью ссылки.
Область техники
Настоящее изобретение, в целом, относится к композициям и методам лечения, которые могут быть применены для лечения инфекции Clostridium difficile (С.difficile), а также ассоциированных с С.difficile болезненных состояний и патологий, например, ассоциированная с С.diff. диарея (CDAD), которая может возникать в результате инфицирования бактериями С.difficile. Настоящее изобретение также относится к антителам или их антигенсвязывающим фрагментам, которые специфично связываются с эпитопами на токсине А и/или токсине В С.difficile, композициям, содержащим такие антитела, а также способам применения антител или композиций.
Уровень техники
С.difficile (или С.diff) представляет собой грамположительную образующую споры анаэробную бактерию, которая представляет ведущую причину госпитальной (приобретенной в больнице) диареи, связанной с применением антибиотиков, и псевдомембранозного колита. Инфекция С.difficile оценивается совокупно более чем 750000 случаев в год в США и является причиной большего количества смертей, чем все остальные кишечные инфекции вместе взятые (1). Во многих больницах С.difficile представляет больший риск для пациентов, чем устойчивая к метициллину Staphylococcus aureus (MRSA) или же любая другая бактерия (2). Ежегодные затраты на организацию профилактики и лечения ассоциированного с Clostridium difficile заболевания (CDAD) оцениваются как превышающие 3,2 миллиарда долларов в США (3). Недавние вспышки заболевания штаммами С.difficile с повышенной вирулентностью или устойчивостью к антибиотикам привели к безуспешности лечения, более частым рецидивам и повышенным коэффициентам смертности (4).
Как правило, CDAD вызывается нарушением микрофлоры толстой кишки вследствие применения антибиотиков, таких как клиндамицин, цефалоспорины и фторхинолоны (3,8). Это нарушение микроокружения толстой кишки вместе с воздействием спор С.difficile ведет к колонизации. Примерно у одной трети всех пациентов, которые подверглись колонизации, развивается CDAD (9), что может привести к тяжелой диарее, перфорации толстой кишки, колэктомии и смерти (10). CDAD возникает в результате приобретения и пролиферации С. difficile в кишечнике, где бактерия С.difficile продуцирует токсин А и токсин В, два важных фактора вирулентности CDAD. Токсины А и В С.difficile показывают значительную гомологию последовательности и структурную гомологию. Оба имеют С-концевой рецепторсвязывающий домен, содержащий многократно повторяющиеся последовательности, центральный гидрофобный домен и N-концевой глюкозилтрансферазный домен. Рецепторсвязывающий домен опосредует связывание токсинов с клетками кишечного эпителия через рецепторы распознавания хозяина, которые остаются недостаточно хорошо определенными для людей. После интернализации через эндосомальный путь центральный гидрофобный домен внедряется в мембрану эндосомы. Кислотный рН эндосомы запускает образование пор и перемещение аминоконцевых доменов токсинов в цитозоль. Гликозилирование целевых цитозольных Rho ОТРаз ведет к разрушению цитоскелета и клеточной смерти. Токсины А и В демонстрируют отличающиеся профили патологии с возможным синергизмом при вызываемом заболевании.
Недавние вспышки заболевания гипервирулентными штаммами С.difficile привели к повышенным уровням тяжелого заболевания, более частым рецидивам и повышенной смертности. Один гипервирулентный штамм, BI/NAP 1/027 токсинотип III, был в прошлом редко встречающимся, но сейчас он является эпидемическим. Гипервирулентные штаммы, такие как BI/NAP 1/027, продуцируют в несколько раз больше токсина А и токсина В, чем не являющиеся гипервирулентными штаммы С.difficile, делая эти штаммы более труднопреодолимыми для лечения после инфицирования. Так как резистентность гипервирулентных штаммов к широко применяемым противомикробным средствам и антибиотикам является растущей проблемой, которая делает эти штаммы более сложными для лечения и сдерживания, дополнительные подходы к лечению и более сильные терапевтические средства необходимы для борьбы с гипервирулентностью и рецидивами ассоциированных с гипервирулентными изолятами С.difficile заболеваний.
Современные способы лечения антибиотиками инфекции С.difficile включают применение ванкомицина и/или метронидазола; однако эти антибиотики ограничены неполными уровнями ответа и повышением частот повторного инфицирования и рецидивов. Сообщали о значительно более высоких частотах неудач для терапии метронидазолом с 2000 года (23-25). Высокие частоты рецидивов после лечения антибиотиками могут возникнуть в результате продолжительного нарушения нормальной микрофлоры толстой кишки, обеспечивающем возможность С.difficile получить преимущество при небольшой конкуренции (26-28). Риск рецидива повышен у пациентов, у которых уже был один рецидив, возрастая от около 20% после первого эпизода до более 60% после двух или более рецидивов (29, 30). Этот повышенный риск рецидива был связан с неспособностью обеспечить соответствующий гуморальный иммунный ответ антителами к токсину (31). Действительно, пациенты с самыми высокими титрами сывороточного IgG к токсину в конце антибактериальной терапии имели пониженный риск рецидива (32). В отдельном исследовании уровни антитела к токсину В в сыворотке коррелировали с защитой от рецидивирующего CDAD (33).
Распространенность инфекции С.difficile стабильно возрастала, особенно у пожилых людей, которые часто являются ослабленными. Приблизительно треть пациентов с инфекцией С.difficile имеют рецидивы их инфекции, обычно в пределах двух месяцев от первоначальной болезни. Повторные инфекции имеют тенденцию быть более тяжелыми, чем первоначальное заболевание; они чаще являются смертельными. Взрослые более старшего возраста и люди с ослабленными иммунными системами являются особо восприимчивыми к рецидивирующим инфекциям. При отсутствии незамедлительного и правильно проводимого лечения осложнения инфекции С.difficile включает обезвоживание, почечную недостаточность, перфорацию кишечника, токсический мегаколон, что может привести к разрыву толстой кишки и смерти.
Хотя в США инфекция С.difficile является наиболее часто встречающейся приобретенной госпитализированными пациентами инфекцией, она также может быть приобретена вне больниц в обществе. Оценивается, что ежегодно в США в обществе выявляют 20000 инфекций С.difficile. В международном масштабе заболеваемость является крайне изменчивой и зависит от множества факторов, включая частоту, с которой эндоскопию применяют для установления диагноза, схемы применения противомикробного средства и эпидемиологические особенности.
Таким образом, очевидно, что заболевание, вызываемое инфекцией С.difficile, ставит жизни людей всех возрастов под угрозу, как в условиях больницы, так и в обществе. В современном мире всегда присутствует риск инфицирования С. difficile для тех, кто сталкивается с госпитализацией, или для тех, кто находится на длительном стационарном лечении. Поскольку существует возможность заражения инфекцией С. difficile вне больничной среды, вероятность заражения инфекцией детей младшего возраста и младенцев велика. Дополнительно существует возможность, что схемы приема антибиотиков, применяемые в настоящее для лечения С. difficile, могут иметь эффективность менее оптимальной. Пациенты, которые страдают ассоциированным с С. difficile заболеванием, нуждаются в серьезном стационарном лечении и продолжительном пребывании в больнице. Затраты, связанные с высоким уровнем поддерживающего лечения в условиях стационара и лечением, которое необходимо для пациентов с ассоциированным с С. difficile заболеванием, велики и предполагают дорогостоящие ресурсы, как, например, большие количества персонала врачей и медсестер, лабораторные исследования и мониторинг, сопутствующая лекарственная терапия и дополнительные меры поддержки.
Следовательно, существует необходимость более эффективных лекарственных препаратов, лекарственных средств и лечебные средства, которые нацелены на угрожающие жизни заболевания, вызываемые С.difficile, и, в частности, на сильнодействующие токсины, продуцируемые С.difficile, для пользы в профилактике и лечении. Существует неудовлетворенная медицинская потребность в успешных и устойчивых средствах лечения ассоциированного с С.difficile заболевания, которые обеспечивают более низкую вероятность развития резистентности и более высокую вероятность успешного ответа пациента и устранения заболевания, приводящего к ликвидации заболевания.
Сущность изобретения
Настоящее изобретение обеспечивает, по меньшей мере частично, новые реактивы на основе антител и композиции, содержащие антитела к токсину А и/или токсину В С.difficile. Настоящие реактивы и композиции могут быть полезны для лечения все более возрастающих количеств субъектов, пораженных ассоциированной с С.difficile инфекцией и заболеванием, для обеспечения улучшенного качества жизни, для устранения CDAD и инфекции С.difficile и для содействия выживанию инфицированных индивидуумов.
В одном аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile и который перекрестно конкурирует за связывание с токсином А С. difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9692. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9694. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9888. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывается с эпитопом токсина А С. difficile, определяемым моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9692. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9694. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9888. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывается с токсином В С.difficile и который перекрестно конкурирует за связывание с токсином В С.difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9693. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9692. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывается с эпитопом токсина В С.difficile, определяемым моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9693. В варианте осуществления гибридомная клеточная линия депонирована под № доступа АТСС РТА-9692. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме. В варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина В С.difficile. В варианте осуществления антитело или его антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина В С.difficile в количестве от 0,1 до 1000 мкг.
В другом аспекте обеспечено моноклональное антитело РА-39 (№ доступа АТСС 9692) или его антигенсвязывающий фрагмент. В другом аспекте обеспечено моноклональное антитело РА-50 (№ доступа АТСС РТА-9694) или его антигенсвязывающий фрагмент. В другом аспекте обеспечено моноклональное антитело РА-38 (№ доступа АТСС РТА-9888) или его антигенсвязывающий фрагмент. В другом аспекте обеспечено моноклональное антитело РА-41 (№ доступа АТСС РТА-9693) или его антигенсвязывающий фрагмент. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В еще одном аспекте обеспечен вектор экспрессии, содержащий по меньшей мере одну молекулу нуклеиновой кислоты, кодирующую антитела или их антигенсвязывающие фрагменты, как описано выше и в настоящем документе. В еще одном аспекте обеспечен вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую тяжелую цепь или ее часть антител или их антигенсвязывающих фрагментов, как описано выше или в настоящем документе. В еще одном аспекте обеспечен вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую легкую цепь или ее часть антител или их антигенсвязывающих фрагментов, как описано выше или в настоящем документе. В еще одном аспекте обеспечен вектор экспрессии, содержащий по меньшей мере одну молекулу нуклеиновой кислоты, кодирующую тяжелую цепь или ее часть и легкую цепь или ее часть антител или их антигенсвязывающих фрагментов, как описано выше или в настоящем документе.
В другом аспекте обеспечены клетки-хозяева, трансформированные или трансфицированные любым из векторов экспрессии, описанных выше и в настоящем документе. В другом аспекте обеспечена плазмида, которая кодирует любое из антител или их антигенсвязывающие фрагменты, как описано выше и в настоящем документе.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина А С.difficile. В варианте осуществления антитело или антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина А С.difficile в количестве от 0,1 мкг до 1000 мкг или от 1 мкг/кг до 100000 мкг/кг. В другом варианте осуществления выделенное антитело или антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина А С.difficile в количестве, выбранном из от 0,5 мкг до 1000 мкг, или от 5 мкг до 250 мкг, или от 10 мг/кг до 50 мг/кг. В варианте осуществления антитело представляет собой моноклональное антитело РА-39 (№ доступа АТСС 9692) или его антигенсвязывающий фрагмент. В варианте осуществления антитело представляет собой моноклональное антитело РА-50 (№ доступа АТСС РТА-9694) или его антигенсвязывающий фрагмент. В варианте осуществления антитело представляет собой моноклональное антитело РА-38 (№ доступа АТСС РТА-9888) или его антигенсвязывающий фрагмент. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В другом аспекте обеспечено выделенное антитело к токсину В С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина В С.difficile. В этом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина В С.difficile в количестве, выбранном из от 0,5 мкг до 1000 мкг, 0,5 мкг, 5 мкг, 40 мкг, 50 мкг, 100 мкг, 200 мкг, 1000 мкг или от 10 мг/кг до 50 мг/кг. В варианте осуществления антитело представляет собой моноклональное антитело РА-39 (№ доступа АТСС 9692) или его антигенсвязывающий фрагмент. В варианте осуществления антитело представляет собой моноклональное антитело РА-41 (№ доступа АТСС РТА-9693) или его антигенсвязывающий фрагмент. В варианте осуществления моноклональное антитело или его антигенсвязывающий фрагмент находится в химерной или гуманизированной форме.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент при введении субъекту, инфицированному С.difficile, в комбинации с выделенным антителом или его антигенсвязывающим фрагментом, который специфично связывает и/или нейтрализует токсин В С.difficile, лечит CDAD и/или повышает выживаемость субъекта. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят одновременно. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят в разные моменты времени. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят последовательно. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина А. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин В С.difficile представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина В.
В другом аспекте обеспечено выделенное антитело к токсину В С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент при введении субъекту, инфицированному С.difficile, в комбинации с выделенным антителом или его антигенсвязывающим фрагментом, который специфично связывает и/или нейтрализует токсин А С.difficile, лечит CDAD и/или повышает выживаемость субъекта. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят одновременно. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят в разные моменты времени. В варианте осуществления антитела к токсину А и токсину В или их фрагменты вводят последовательно. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестие реагирует с ним при связывании токсина А. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин В С.difficile представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина В.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент при введении субъекту, инфицированному С.difficile, в комбинации с выделенным антителом или его антигенсвязывающим фрагментом, который специфично связывает токсин В С.difficile, лечит CDAD и/или повышает выживаемость субъекта. В варианте осуществления антитело к токсину А или его антигенсвязывающий фрагмент вводят в количестве от 1 мкг до 1000 мкг, или от 1 мкг до 250 мкг, или от 5 мкг до 100 мкг, и дозу антитела к токсину В или его антигенсвязывающего фрагмента вводят в количестве от 0,1 мкг до 1000 мкг, или от 1 мкг до 250 мкг, или от 5 мкг до 100 мкг. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile, представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина А. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин В С.difficile представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина В.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент при введении субъекту, инфицированному С.difficile, в комбинации с выделенным антителом или его антигенсвязывающим фрагментом, который специфично связывает токсин В С.difficile, лечит CDAD и/или повышает выживаемость субъекта. В варианте осуществления антитело к токсину А или его антигенсвязывающий фрагмент вводят в количестве 50 мг/кг, антитело к токсину В или его антигенсвязывающий фрагмент вводят в количестве 50 мг/кг. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile, представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина А. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин В С.difficile, представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно реагирует с ним при связывании токсина В.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile или антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или антигенсвязывающий фрагмент при введении субъекту, инфицированному С.difficile, в комбинации с выделенным антителом или его антигенсвязывающим фрагментом, который специфично связывает токсин В С.difficile, обеспечивает показатель эффективности лечения или выживаемости 50%, 60%, 70%, 80%, 90% или 100%. В варианте осуществления антитело или антигенсвязывающий фрагмент вводят q2d×4 в дозе 40-50 мг/кг. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин А С.difficile, представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно конкурирует за связывание токсина А с одним или несколькими моноклональными антителами, депонированными под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. В варианте осуществления выделенное антитело или антигенсвязывающий фрагмент, который специфично связывает токсин В С.difficile, представляет собой антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, его гуманизированную форму или моноклональное антитело, которое перекрестно конкурирует за связывание токсина В с одним или несколькими моноклональными антителами, депонированными под № доступа АТСС РТА-9692 или РТА-9693.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile, или антитело к токсину В С.difficile, или их антигенсвязывающий фрагмент, как описано в настоящем документе, где антитело или антигенсвязывающий фрагмент находится в форме или происходит от одного или нескольких из моноклонального антитела, гуманизированного антитела, антитела человека или химерного антитела.
В другом аспекте обеспечено выделенное антитело к токсину А С.difficile, или антитело к токсину В С.difficile, или их антигенсвязывающего фрагмента, как описано в настоящем документе, где антитело или его антигенсвязывающий фрагмент находится в форме или происходит от биспецифичного и бифункционального антитела.
В другом аспекте обеспечено биспецифичное антитело или его антигенсвязывающий фрагмент, который содержит (i) моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, его антигенсвязывающий фрагмент, гуманизированный варианта антитела или его антигенсвязывающего фрагмента, вариабельный домен тяжелой цепи антитела или его антигенсвязывающего фрагмента и/или вариабельный домен легкой цепи антитела или антигенсвязывающего фрагмента; и (ii) моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, гуманизированный вариант антитела или его антигенсвязывающего фрагмента, вариабельный домен тяжелой цепи антитела или его антигенсвязывающего фрагмента и/или вариабельный домен легкой цепи антитела или его антигенсвязывающего фрагмента.
В другом аспекте обеспечено биспецифичное антитело или его антигенсвязывающий фрагмент, где антитело содержит (1) моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, его антигенсвязывающий фрагмент, гуманизированный вариант антитела или его антигенсвязывающего фрагмента, вариабельный домен тяжелой цепи антитела или его антигенсвязывающего фрагмента и/или вариабельный домен легкой цепи антитела или антигенсвязывающего фрагмента; и (ii) выделенное моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692, его антигенсвязывающий фрагмент, гуманизированный вариант антитела или его антигенсвязывающего фрагмента, вариабельный домен тяжелой цепи антитела или антигенсвязывающего фрагмента и/или вариабельный домен легкой цепи антитела или его антигенсвязывающего фрагмента.
В различных вариантах осуществления обеспечено антитело или его антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антигенсвязывающий фрагмент выбран из Fab-фрагмента, Р(ab’)2-фрагмента и Fv-фрагмента. В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, как описано выше и в настоящем документе, где антитело или его антигенсвязывающий фрагмент представляет собой или содержит одноцепочечное антитело.
В другом аспекте обеспечена композиция, содержащая одно или несколько антител или их антигенсвязывающих фрагментов согласно настоящему изобретению, как описано выше и в настоящем документе, и фармацевтически приемлемый носитель, наполнитель, среду или разбавитель. В варианте осуществления композиция содержит по меньшей мере одно антитело к токсину А согласно настоящему изобретению, например, mAb РТА-9692, mAb PTA-9694, mAb PTA-9888, его антигенсвязывающий фрагмент или его гуманизированную форму, и по меньшей мере одно антитело к токсину В согласно настоящему изобретению, например, mAb РТА-9692, mAb РТА-9693, его антигенсвязывающий фрагмент или его гуманизированную форму. В варианте осуществления композиция содержит одно антитело к токсину А согласно настоящему изобретению, например mAb PTA-9888, его антигенсвязывающий фрагмент или его гуманизированную форму и одно антитело к токсину В согласно настоящему изобретению, например mAb 9693, его антигенсвязывающий фрагмент или его гуманизированную форму. В варианте осуществления композиция содержит одно антитело к токсину А согласно настоящему изобретению, например mAb PTA-9694, его антигенсвязывающий фрагмент или его гуманизированную форму и одно антитело к токсину В согласно настоящему изобретению, например mAb 9693, его антигенсвязывающий фрагмент или его гуманизированную форму. В варианте осуществления каждое mAb присутствует в композиции в одинаковом количестве. В варианте осуществления каждое mAb присутствует в композиции в соотношении 1:1 по весу относительно друг друга. В варианте осуществления каждое mAb присутствует в композиции в различных количествах. В варианте осуществления каждое mAb присутствует в композиции в соотношениях, отличных от 1:1, по весу относительно друг друга, причем соотношения являются такими, как обеспечено в настоящем документе. В варианте осуществления композиция дополнительно содержит дополнительное терапевтическое средство. В варианте осуществления дополнительное терапевтическое средство представляет собой антибиотик, противобактериальное, бактерицидное, бактериостатическое средство или их комбинацию. В варианте осуществления дополнительное терапевтическое средство представляет собой метронидазол, ванкомицин, фидаксомицин или их комбинацию.
В другом аспекте обеспечена композиция, содержащая вектор экспрессии согласно настоящему изобретению, как описано выше и в настоящем документе, и фармацевтически приемлемый носитель, наполнитель, среду или разбавитель. В другом аспекте обеспечена композиция, содержащая клетки-хозяева, несущие вектор экспрессии согласно настоящему изобретению, как описано выше и в настоящем документе, и фармацевтически приемлемый носитель, наполнитель, среду или разбавитель. В другом аспекте обеспечена композиция, содержащая плазмиду согласно настоящему изобретению, как описано выше и в настоящем документе, и фармацевтически приемлемый носитель, наполнитель, среду или разбавитель.
В другом аспекте обеспечен связывающий белок, содержащий по меньшей мере две полипептидные цепи, содержащие связывающие участки для связывания токсина А и токсина В С.difficile, где по меньшей мере одна полипептидная цепь содержит первый вариабельный домен тяжелой цепи, второй вариабельный домен тяжелой цепи и константный домен тяжелой цепи или его часть; и по меньшей мере одна полипептидная цепь содержит первый вариабельный домен легкой цепи, второй вариабельный домен легкой цепи и константный домен легкой цепи или его часть, где вариабельные домены, содержащие полипептидные цепи, образуют функциональные связывающие участки для токсина А и токсина В С.difficile. В варианте осуществления первый вариабельный домен тяжелой цепи и первый вариабельный домен легкой цепи связывающего белка образуют функциональный связывающий участок для токсина А С.difficile, и второй вариабельный домен тяжелой цепи и второй вариабельный домен легкой цепи связывающего белка образуют функциональный связывающий участок для токсина В С.difficile. В варианте осуществления первый вариабельный домен тяжелой цепи и первый вариабельный домен легкой цепи связывающего белка образуют функциональный связывающий участок для токсина В бактерии С.difficile, и второй вариабельный домен тяжелой цепи и второй вариабельный домен легкой цепи связывающего белка образуют функциональный связывающий участок для токсина А С. difficile. В варианте осуществления связывающий белок содержит Fc-область. В варианте осуществления связывающий белок нейтрализует токсичность токсина А и токсина В С.difficile. В различных вариантах осуществления связывающий белок имеет константу скорости ассоциации (Kon) с токсином А или токсином В, выбранную из по меньшей мере 102M-1c-1; по меньшей мере 103M-1c-1; по меньшей мере 104M-1с-1; по меньшей мере 105M-1c-1; по меньшей мере 106M-1с-1; или по меньшей мере 107М-1с-1, как измерено при помощи поверхностного плазменного резонанса. В различных вариантах осуществления связывающий белок имеет константу скорости диссоциации (Koff) с токсином А или токсином В, выбранную из не более 10-3c-1; не более 10-4c-1; не более 10-5с-1; или не более 10-6c-1, как измерено при помощи поверхностного плазменного резонанса. В различных вариантах осуществления связующий белок имеет константу диссоциации (KD) с токсином А или токсином В, выбранную из не более 10-7М; не более 10-8М; не более 10-9М; не более 10-10М; не более 10-11М; не более 10-12М; или не более 10-13М.
В другом аспекте обеспечена композиция, содержащая связывающий белок, как описано выше и в настоящем документе, фармацевтически приемлемый носитель, наполнитель, среду или разбавитель. В варианте осуществления композиция дополнительно содержит дополнительное терапевтическое средство. В варианте осуществления дополнительное терапевтическое средство композиции представляет собой антибиотик, противобактериальное, бактерицидное, бактериостатическое средство или их комбинацию. В варианте осуществления дополнительное терапевтическое средство композиции представляет собой метронидазол, ванкомицин, фидаксомицин, нитазоксанид, рифаксимин, рамопланин или их комбинацию.
В другом аспекте обеспечены гибридомные клеточные линии, депонированные под № доступа АТСС РТА-9692, РТА-9693, РТА-9494 или РТА-9888.
В другом аспекте обеспечен способ лечения субъекта с инфекцией С.difficile или ассоциированным с С.difficile заболеванием, включающий введение субъекту по меньшей мере одной композиции, как описано в настоящем документе. В варианте осуществления композиции включают одно или несколько антител согласно настоящему изобретению, предпочтительно в гуманизированной форме. В варианте осуществления композиции содержат по меньшей мере одно антитело к токсину А, приведенное в настоящем документе, в гуманизированной форме или его антигенсвязывающий фрагмент и по меньшей мере одно антитело к токсину В согласно настоящему изобретению в гуманизированной форме или его антигенсвязывающий фрагмент. В различных вариантах осуществления один или несколько терапевтических реактивов, лекарственных средств, соединений или ингредиентов могут быть включены в композиции. В варианте осуществления композиции дополнительно включают фармацевтически приемлемый носитель, разбавитель, среду или наполнитель. В варианте осуществления композиции вводят в количестве, эффективном для лечения инфекции С.difficile или ассоциированного с С.difficile заболевания, например ассоциированной с С.difficile диареи (CDAD). В варианте осуществления две композиции вводят субъекту в количестве, эффективном для лечения инфекции С.difficile или ассоциированного с С.difficile заболевания. В варианте осуществления две композиции вводят одновременно. В другом варианте осуществления две композиции вводят в разные моменты времени.
В другом аспекте обеспечен способ подавления или нейтрализации токсичности токсина А и токсина В С.difficile в отношении клетки, который включает воздействие на клетку эффективной подавляющей или нейтрализующей токсин А С.difficile дозой моноклонального антитела к токсину А согласно настоящему изобретению или его антигенсвязывающего фрагмента и эффективной подавляющей или нейтрализующей токсин В С.difficile дозой моноклонального антитела к токсину В согласно настоящему изобретению или его антигенсвязывающего фрагмента. В варианте осуществления антитело к токсину А и антитело к токсину В находятся в гуманизированной форме. В варианте осуществления антитело к токсину А и антитело к токсину В находятся в химерной форме. В варианте осуществления антитела или их антигенсвязывающие фрагменты вводят одновременно. В варианте осуществления антитела или их антигенсвязывающие фрагменты вводят в разные моменты времени. В варианте осуществления способа клетка присутствует у субъекта, и антитела или их антигенсвязывающие фрагменты вводят субъекту в количестве, эффективном для подавления или нейтрализации токсина А и токсина В С.difficile.
В другом аспекте обеспечен способ подавления и нейтрализации токсичности токсина С.difficile в отношении клетки, который включает воздействие на клетку эффективной подавляющей или нейтрализующей токсин С.difficile дозой по меньшей мере одной из композиций согласно настоящему изобретению, как описано в настоящем документе. В варианте осуществления способа на клетку воздействуют эффективной подавляющей или нейтрализующей токсин С.difficile дозой двух композиций, одна из которых содержит антитело к токсину А или его антигенсвязывающий фрагмент, и другая из которых содержит антитело к токсину В или его антигенсвязывающий фрагмент. В варианте осуществления антитела являются гуманизированными. В варианте осуществления антитела являются химерными. В вариантах осуществления две композиции вводят одновременно или в разные моменты времени. В варианте осуществления клетка присутствует у субъекта, и по меньшей мере одну композицию вводят субъекту в количестве, эффективном для подавления или нейтрализации токсина С.difficile.
В другом аспекте обеспечен способ подавления и нейтрализации токсинов, продуцируемых гипервирулентным штаммом С.difficile, который включает введение субъекту, нуждающемуся в этом, (i) антитела или его антигенсвязывающего фрагмента согласно настоящему изобретению, где антитело связывает и нейтрализует токсин А С.difficile, и (ii) антитела или его антигенсвязывающего фрагмента согласно настоящему изобретению, где антитело связывает и нейтрализует токсин В С.difficile, в количестве, эффективном для нейтрализации токсинов, продуцируемых гипервирулентным штаммом. В варианте осуществления антитела (i) и (ii) представляют собой гуманизированные антитела. В варианте осуществления антитела (i) и (ii) представляют собой химерные антитела. В вариантах осуществления антитела или их антигенсвязывающие фрагменты вводят одновременно или в разные моменты времени. В варианте осуществления токсины гипервирулентного штамма представляют собой токсин А и токсин В. В варианте осуществления гипервирулентный штамм С.difficile представляет собой один или несколько из BI/NAP1/027, CCL676, НМС553, Pitt45, CD196, montreal 5 или montreal 7.1. В варианте осуществления антитело к токсину А или его антигенсвязывающий фрагмент обладает нейтрализующей активностью в отношении токсина А, продуцируемого гипервирулентными штаммами С.difficile, как определено по значению ЕС50 от 2,6-12M до 7,7-11М или от 7,7-12M до 4,8-8M. В варианте осуществления антитело к токсину В или его антигенсвязывающий фрагмент обладает нейтрализующей активностью в отношении токсина В, продуцируемого гипервирулентными штаммами С.difficile, как определено по значению ЕС50 от 1,1-11М до 6,5-10М.
В другом аспекте обеспечен набор, содержащий антитело или его антигенсвязывающий фрагмент согласно настоящему изобретению, и как описано в настоящем документе, особенно в гуманизированной форме, и инструкции по применению. В варианте осуществления антитела или антигенсвязывающие фрагменты содержатся в том же самом контейнере в наборе. В варианте осуществления антитела или антигенсвязывающие фрагменты содержатся в отдельных контейнерах в наборе. В варианте осуществления набор содержит линкер для конъюгирования антител или их антигенсвязывающих фрагментов. В варианте осуществления набор содержит дополнительное терапевтическое средство, которое может быть антибиотиком, противобактериальным, бактерицидным или бактериостатическим средством. В варианте осуществления дополнительное терапевтическое средство представляет собой метронидазол, ванкомицин, фидаксомицин, нитазоксанид, рифаксимин, рамопланин или их комбинацию.
В другом аспекте обеспечено моноклональное антитело или его антигенсвязывающий фрагмент, особенно в гуманизированной форме, который нейтрализует токсин А или токсин В гипервирулентного штамма С.difficile. В варианте осуществления моноклональное антитело определено номером доступа АТСС РТА-9692, РТА-9694, РТА-9888 или РТА-9693 и продуцируется гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694, РТА-9888 или РТА-9693, соответственно. В варианте осуществления антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694, РТА-9888, РТА-9693 или РТА-9692, было гуманизировано или находится в химерной форме.
В варианте осуществления гипервирулентный штамм С.difficile представляет собой одно или несколько из BI/NAP1/027, CCL676, НМС553, Pitt45, CD196, montreal 5 и montreal 7.1.
В другом аспекте обеспечен способ лечения субъекта, который не проявляет симптомов, но который является чувствительным к или подверженным риску заражения инфекцией С.difficile, который включает: введение субъекту (i) антитела к токсину А С.difficile или его антигенсвязывающего фрагмента, приведенного и описанного в настоящем документе, и (ii) антитела к токсину В С.difficile или его антигенсвязывающего фрагмента, приведенного и описанного в настоящем документе, в количестве эффективном для лечения субъекта. В варианте осуществления способа субъекта госпитализируют.
В другом аспекте обеспечено гуманизированное моноклональное антитело, получаемое к токсину А С.difficile. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:1, и СН-область человека, и двух легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:3, и CL-область человека. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID N0:2, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:3, и CL-область человека. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:1, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:4, и CL-область человека. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:2, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:4, и CL-область человека. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:5, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:7, и CL-область человека. В варианте осуществления такое антитело к токсину А С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:6, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:7, и CL-область человека.
В другом аспекте обеспечено гуманизированное моноклональное антитело к токсину В С.difficile. В варианте осуществления такое антитело к токсину В С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:8, и СН-область человека, и двух полипептидов легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:10, и CL-область человека. В варианте осуществления такое антитело к токсину В С.difficile состоит из двух полипептидов тяжелых цепей, где каждая тяжелая цепь содержит VH-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:9, и СН-область человека, и двух легких цепей, где каждая легкая цепь содержит VL-область, содержащую аминокислотную последовательность, как изложено в SEQ ID NO:10, и CL-область человека.
В другом аспекте обеспечено моноклональное антитело или его фрагмент, получаемое к токсину А С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, причем каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, причем каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида тяжелой цепи антитела SEQ ID NO:14, изложена в SEQ ID NO:15 (Фиг.38В); последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида легкой цепи антитела SEQ ID NO:16, изложена в SEQ ID NO:17 (Фиг.38А).
В другом аспекте обеспечено моноклональное антитело или его фрагмент, получаемое к токсину А С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, причем каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, причем каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида тяжелой цепи антитела SEQ ID NO:18, изложена в SEQ ID NO:19 (Фиг.39В); последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида легкой цепи антитела SEQ ID NO:20, изложена в SEQ ID NO:21 (Фиг.39А).
В другом аспекте обеспечено моноклональное антитело или его фрагмент, получаемый к токсину В С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, причем каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, причем каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида тяжелой цепи антитела SEQ ID NO:22, изложена в SEQ ID NO:23 (Фиг.40В); последовательность нуклеиновой кислоты (или кДНК), кодирующей аминокислотную последовательность полипептида легкой цепи антитела SEQ ID NO:24, изложена в SEQ ID NO:25 (Фиг.40А).
В различных вариантах осуществления направленных на любое из вышеупомянутых гуманизированных моноклональных антител согласно настоящему изобретению, СН-область моноклонального антитела выбрана из IgGI, IgG2a, IgG2b, IgG3, IgG4, IgA, IgE или IgM. В варианте осуществления СН-область представляет собой IgGI. В варианте осуществления CL-область выбрана из изотипа к или λ. В варианте осуществления CL-область представляет собой изотип к. В других вариантах осуществления CDR, т.е. CDR1, CDR2 и/или CDR3 гуманизированных антител или их антигенсвязывающих фрагментов, как описано в настоящем документе, включены для связывания и/или нейтрализации токсина А и/или токсина В С.difficile в продукты и способы согласно настоящему изобретению.
В другом аспекте обеспечено антитело к токсину А С.difficile или его фрагмент, где V-область L-цепи содержит последовательность, выбранную из одной или нескольких из SEQ ID NO:3, SEQ ID NO:4 и SEQ ID NO:7. Также обеспечено антитело к токсину В С.difficile или его фрагмент, где V-область L-цепи содержит последовательность, как изложено в SEQ ID NO:10. Также обеспечено антитело к токсину А С.difficile или его фрагмент, где V-область Н-цепи содержит последовательность, выбранную из одной или нескольких из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:5 и SEQ ID NO:6. Также обеспечено антитело к токсину В С.difficile или его фрагмент, где V-область Н-цепи содержит последовательность, выбранную из одной или нескольких из SEQ ID NO:8 или SEQ ID NO:9.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который (i) специфично связывает токсин А С.difficile и который перекрестно конкурирует за связывание с токсином А С.difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, или который (ii) специфично связывается с эпитопом токсина А С.difficile, определяемым моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, где эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, содержит область за пределами рецепторсвязывающего домена, например транслокационный домен, токсина А С.difficile. В варианте осуществления антитело находится в гуманизированной форме. В варианте осуществления антитело находится в химерной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который (i) специфично связывает токсин А С.difficile и который перекрестно конкурирует за связывание с токсином А С.difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694, или который (ii) специфично связывается с эпитопом токсина А С.difficile, определяемым моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694, где эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694, содержит по меньшей мере два участка в рецепторсвязывающем домене, например С-концевые рецепторсвязывающие эпитопы, токсина А С.difficile. В варианте осуществления антитело находится в гуманизированной форме. В варианте осуществления антитело находится в химерной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который (i) специфично связывает токсин А С.difficile и который перекрестие конкурирует за связывание с токсином А С.difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888, или который (ii) специфично связывается с эпитопом токсина А С.difficile, определяемым моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888, где эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888, содержит С-концевые рецепторсвязывающие эпитопы токсина А С.difficile. В варианте осуществления антитело находится в гуманизированной форме. В варианте осуществления антитело находится в химерной форме.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который (i) специфично связывает токсин В С.difficile и который перекрестие конкурирует за связывание с токсином В С.difficile с моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693, или который (ii) специфично связывается с эпитопом токсина В С.difficile, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693, где эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693, содержит N-концевой ферментативный домен токсина В С.difficile. В варианте осуществления эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693, содержит 63 кДа фрагмент, получаемый обработкой токсина В каспазой 1, содержащий N-концевой ферментативный домен токсина В С.difficile. В варианте осуществления эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, содержит транслокационный домен токсина В С.difficile.
В варианте осуществления эпитоп, определяемый моноклональным антителом, продуцируемым гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, содержит 167 кДа фрагмент, получаемый обработкой токсина В каспазой 1, и 63 кДа белок, который содержит необработанный токсин В. В варианте осуществления антитело находится в гуманизированной форме. В варианте осуществления антитело находится в химерной форме.
В другом аспекте обеспечен способ получения моноклонального антитела, которое связывает и нейтрализует токсин А или токсин В С.difficile, включающий иммунизацию одного или нескольких животных-реципиентов неактивным анатоксином А с определенной периодичностью; бустер-иммунизацию животных увеличивающимися количествами активного токсина А или активного токсина В с определенной периодичностью; получение гибридомных клеток из иммунных клеток иммунизированного и подвергнутого бустер-иммунизации животного, слитых с подходящей иммортализованной клеточной линией, где гибридомные клетки продуцируют и секретируют антитела к токсину А, которые связывают и нейтрализуют токсин А С.difficile, или антитела к токсину В, которые связывают и нейтрализуют токсин В С.difficile. В варианте осуществления выделяют нейтрализующие моноклональные антитела к токсину А С.difficile и/или нейтрализующие моноклональные антитела к токсину В С.difficile. В вариантах осуществления способа этапы иммунизации и бустер-иммунизации включают применение адъюванта. В варианте осуществления адъювант представляет собой Quil А. В других вариантах осуществления способа этапы иммунизации и бустер-иммунизации осуществляют с периодичностью в три недели. В других вариантах осуществления животных-реципиентов иммунизируют двумя или тремя дозами анатоксина А с последующими от трех до пяти бустер-иммунизациями возрастающими дозами либо активного токсина А, либо активного токсина В.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который подавляет, блокирует или предотвращает токсичность токсина А С.difficile при помощи подавления, блокирования или предотвращения интернализации и цитотоксичности токсина А. В варианте осуществления антитело представляет собой моноклональное антитело. В варианте осуществления антитело представляет собой гуманизированное или химерное антитело. В варианте осуществления антитело представляет собой РА-39 (№ доступа АТСС РТА-9692) или гуманизированное РА-39. В варианте осуществления антитело представляет собой РА-50 (№ доступа АТСС РТА-964) или гуманизированное РА-50. В других вариантах осуществления антитело конкурирует с РА-39, гуманизированным PA-39, PA-50 или гуманизированным PA-50 за связывание с токсином А. В варианте осуществления антитело связывает один участок в области токсина А за пределами рецепторсвязывающего домена токсина А. В варианте осуществления антитело конкурирует с РА-39 или его гуманизированной формой путем связывания одного участка в области токсина А за пределами рецепторсвязывающего домена токсина А. В варианте осуществления антитело связывается с по меньшей мере двумя участками в рецепторсвязывающем домене токсина А. В варианте осуществления антитело конкурирует с PA-50 или его гуманизированной формой путем связывания с по меньшей мере с двумя участками в рецепторсвязывающем домене токсина А. В варианте осуществления антитело подавляет токсичность токсина А посредством смешанного конкурентного механизма действия. В варианте осуществления антитело подавляет токсичность токсина А посредством конкурентного механизма действия. Все из вышеупомянутых вариантов осуществления, как подразумевается, включают антигенсвязывающий фрагмент антитела.
В другом аспекте обеспечено выделенное антитело или его антигенсвязывающий фрагмент, который подавляет, блокирует или предотвращает токсичность токсина В С.difficile путем связывания с эпитопным участком в N-концевой ферментативной области токсина В. В варианте осуществления антитело представляет собой моноклональное антитело. В варианте осуществления антитело представляет собой гуманизированное или химерное антитело. В варианте осуществления антитело представляет собой РА-41 (№ доступа АТСС РТА-9693) или гуманизированную форму РА-41. В варианте осуществления антитело конкурирует с РА-41 или гуманизированным РА-41 за связывание с N-концевой ферментативной областью токсина В С.difficile. В варианте осуществления антитело конкурирует с РА-41 или гуманизированным РА-41 за связывание с одним участком в N-концевой ферментативной области токсина В С.difficile. В варианте осуществления антитело подавляет токсичность токсина В посредством смешанного конкурентного механизма действия.
В другом аспекте обеспечена вакцина или иммуноген, содержащий части, фрагменты или пептиды токсина А и/или токсина В С.difficile, содержащие этитопные области, распознаваемые и/или связываемые одним или несколькими из моноклонального антитела РА-39 (№ доступа АТСС РТА-9692), гуманизированной формы РА-39, моноклонального антитела PA-50 (№ доступа АТСС РТА-9694), гуманизированной формы РА-51, моноклонального антитела РА-41 (№ доступа АТСС РТА-9693), гуманизированной формы РА-41, антитела, которое конкурирует за связывание токсина А с моноклональным антителом РА-39 или его гуманизированной формой, антитела, которое конкурирует за связывание токсина А с моноклональным антителом РА-50 или его гуманизированной формой, или антитела, которое конкурирует за связывание токсина В с моноклональным антителом РА-41 или его гуманизированной формой. В варианте осуществления вакцина или иммуноген содержит части, фрагменты или пептиды токсина А и/или токсина В С.difficile, содержащие этитопные области, распознаваемые и/или связываемые одним или несколькими из моноклонального антитела РА-39 (№ доступа АТСС РТА-9692), гуманизированной формы РА-39 или антитела, которое конкурирует за связывание токсина А и токсина В с моноклональным антителом РА-39 или его гуманизированной формой. В варианте осуществления содержащие эпитоп части, фрагменты или пептиды токсина А и/или токсина В вакцины или иммуногена получены из белка токсина А или токсина В при помощи протеолитического расщепления. В варианте осуществления фрагменты, части или пептиды токсина А вакцины или иммуногена получены при помощи протеолитического расщепления энтерокиназой. В варианте осуществления фрагменты, части или пептиды токсина В вакцины или иммуногена получены при помощи протеолитического расщепления каспазой (каспаза 1). В варианте осуществления содержащие эпитоп части или фрагменты вакцины или иммуногена являются химически синтезированными или рекомбинантно созданные пептиды белка токсина А или токсина В. В варианте осуществления фрагменты, части или пептиды вакцины или иммуногена, содержащие одну или несколько этитопных областей токсина А и/или токсина В, которые распознаются и связываются антителом, получены из одного или нескольких амино-конца токсина А; амино-конца токсина В; карбокси-конца токсина А; карбокси-конца токсина В; рецепторсвязывающего домена токсина А; области за пределами рецепторсвязывающего домена токсина А; рецепторсвязывающего домена токсина В; N-концевой ферментативной области токсина В; глюкозилтрансферазного домена токсина А; глюкозилтрансферазного домена токсина В; протеолитического домена токсина А; протеолитического домена токсина В; гидрофобного порообразующего домена токсина А; гидрофобного порообразующего домена токсина В. В варианте осуществления размеры содержащих эпитоп фрагментов или частей токсина А или токсина В составляют <300 кДа, ~158-160 кДа, ~100-105 кДа, например 103 кДа, ~90-95 кДа, например 91 кДа, и/или ~63-68 кДа, например 63 кДа или 68 кДа. В варианте осуществления размеры содержащих эпитоп фрагментов или частей токсина А составляют ~158-160 кДа, ~90-95 кДа, например 91 кДа, и/или ~63-68 кДа, например 68 кДа. В варианте осуществления размеры содержащих эпитоп фрагментов или частей токсина В составляют ~100-105 кДа, например 103 кДа, и/или ~63-68 кДа, например 63 кДа. В любом из вариантов осуществления вакцины или иммуногена токсин А или токсин В или их фрагмент, часть или пептид является таковым любого из штаммов, приведенных в настоящем документе.
В другом аспекте обеспечен нейтрализации, подавления, блокирования, снижения, уменьшения интенсивности, излечивания или лечения инфекции С.difficile или ассоциированного с С.difficile заболевания у субъекта, нуждающегося в этом, включающий введение субъекту эффективного количества вышеописанных вакцины или иммуногена. В варианте осуществления способа у субъекта вызывают гуморальный ответ на токсин А и/или токсин В С.difficile после введения вакцины или иммуногена, таким образом продуцируя антитела к токсину А и/или токсину В, которые могут специфично нейтрализовать, подавлять, блокировать, снижать, уменьшать интенсивность, излечивать или лечить ассоциированное с С.difficile заболевание или CDAD, включая легкую-тяжелую диарею и в некоторых случаях ассоциированную с тяжелыми угрожающими жизни осложнениями, такими как псевдомембранозный колит, токсический мегаколон, перфорация кишечника, сепсис и смерть. В варианте осуществления способа антитела, образование которых вызывают посредством гуморального ответа, включают антитела, имеющие специфичности и механизмы действия, сходные с или идентичные mAb согласно настоящему изобретению или антителам, которые конкурируют с mAb согласно настоящему изобретению при нейтрализации токсина А и/или токсина В С.difficile или которые конкурируют с mAb согласно настоящему изобретению по механизму действия, вовлеченному в нейтрализацию токсина А и/или токсина В С.difficile.
В другом аспекте обеспечен способ нейтрализации, подавления или блокирования активности токсина А и/или токсина В в или по отношению к клетке, чувствительной к инфекции С.difficile, включающий контакт клетки в антителом или его антигенсвязывающим фрагментом в соответствии с настоящим изобретением, где антитело или его антигенсвязывающий фрагмент нейтрализует, подавляет или блокирует активность токсина А и/или токсина В в или по отношению к клетке при помощи конкурентного или смешанного конкурентного механизма действия. В варианте осуществления способа антитело представляет собой одно или несколько из моноклонального антитела, гуманизированного антитела или химерного антитела. В варианте осуществления способа клетка, например клетка кишечного эпителия, находится в субъекте, и антитело или его антигенсвязывающий фрагмент вводят субъекту в эффективном количестве. В варианте осуществления способа токсин представляет собой токсин А. В варианте осуществления способа токсин представляет собой токсин В. В варианте осуществления способа токсин представляет собой токсин А, и механизм действия представляет собой механизм действия конкурентного подавления. В варианте осуществления способа антителом или его антигенсвязывающий фрагмент представляет собой РА-50 (№ доступа АТСС РТА-9694), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-50 за нейтрализацию активности токсина А. В варианте осуществления способа токсин представляет собой токсин А, и механизм действия представляет собой механизм действия смешанного конкурентного подавления. В варианте осуществления способа антитело или его антигенсвязывающий фрагмент представляет собой РА-39 (№ доступа АТСС РТА-9692), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-39 за нейтрализацию активности токсина А. В варианте осуществления способа токсин представляет собой токсин В, и механизм действия представляет собой механизм действия смешанного конкурентного подавления. В варианте осуществления антитело или его антигенсвязывающий фрагмент представляет собой РА-41 (№ доступа АТСС РТА-9693), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-41 за нейтрализацию активности токсина В.
Эти и другие аспекты настоящего изобретения будут описаны более подробно в подробном описании настоящего изобретения.
Краткое описание графических материалов
На фиг.1А-1С демонстрируется специфичность mAb к токсину С.difficile по настоящему изобретению в отношении токсина А и/или токсина В посредством ELISA. Планшеты для ELISA покрывали токсином А (закрашенные кружки) или токсином В (пустые квадраты) на протяжении ночи при 4°С. Затем планшеты промывали и блокировали, мышиные mAb PA-38 (А), РА-39 (В) или РА-41 (С) титровали и вносили в планшеты. Связывание моноклональных антител определяли с помощью HRP-конъюгированных антител козы к IgG-Fc мыши. ОП измеряли на планшет-ридере SpectraMax M5 (Molecular Devices).
На фиг.2A-2D представлены результаты анализов характеристики связывания Biacore с помощью мышиных mAb PA-38, PA-39, PA-41 и РА-50. Специфичность связывания определяли с помощью устройства Biacore 3000 (GE Healthcare). mAb (PA-38 (2A), PA-39 (2B), PA-50 (2C), PA-41 (2D) или неспецифичные mAb в качестве контроля) ковалентно иммобилизовали на поверхности сенсорного чипа СМ5 (GE Healthcare) при приблизительно 10000 единицах резонанса (RU) в соответствии с инструкцией производителя для аминного присоединения. Эксперименты по связыванию проводили при 25°С в PBS. Очищенный токсин А или токсин В (List Biological Laboratories) при 30 нМ пропускали над контрольными (неспецифичные mAb) и тестовыми проточными ячейками при скорости потока 5 мкл/мин с фазой ассоциации (600 с для PA-38, PA-39 и PA-41; и 300 с для РА-50) и фазой диссоциации (300 с для РА-38, PA-39 и PA-41; и 600 с для РА-50). Графики представлены в RU относительно времени.
На фиг.3А-3Е и 3F-3H показаны результаты кинетики присоединения антитела к токсину, определяемого с помощью Biacore. Для фиг.3А-3Е, мышиные mAb захватывали с применением сенсорного чипа СМ5, подготовленного с набором для захвата антител мыши от Biacore. Токсин затем пропускали через проточные ячейки при различных концентрациях (0,4-100 нМ, двукратное возрастание) при скорости потока 30 мкл/мин. Все концентрации mAb тестировали в двух повторностях, и поверхность чипа регенерировали после каждого опыта с применением условий, указанных в наборе. Изменения в RU записывали и анализировали с использованием модели связывания 1:1 (Langmuir) в Bia Evaluation Software, которая рассчитывает kd mAb в отношении токсина. Фиг.3А: связывание РА-38 с токсином А; фиг.3В: связывание РА-50 с токсином А; фиг.3С: связывание PA-39 с токсином А; фиг.3D: связывание PA-39 с токсином В; и фиг.3Е: связывание PA-41 с токсином В. Для фиг.3F-3H, как и выше, мышиные mAb, например, mPA-50, mPA-41 или mPA-39, ковалентно связывали с сенсорным чипом СМ5 способом аминного присоединения. Токсин А (линия обозначенная "(красным)") или токсин В (линия обозначенная "(синим)") при 30 нМ пропускали над тестовыми проточными ячейками (mPA-50, mPA-41 или mPA-39) при скорости потока 5 мкл/мин. Результаты показывают, что mPA-50 избирательно связывает токсин А (фиг.3F), а mPA-41 избирательно связывает токсин В (фиг.3G). mPA-39 связывается предпочтительно с токсином А, но также проявляет кросс-реактивность к токсину В (фиг.3Н).
На фиг.4 демонстрируется активность нейтрализации in vitro активности токсина А с применением очищенных мышиных mAb PA-39 на клетках СНО-K1. Для измерений цитотоксичности токсин А инкубировали с различными концентрациями РА-39 в течение 1 часа при 37°С (пример 3А). Затем смеси mAb-токсин вносили к клеткам СНО-K1, выращенным в 96-луночных планшетах при 2000 клеток/лунку и инкубировали в течение 72 часов. Выживание клеток сравнивали в необработанных и обработанных культурах и вычисляли концентрацию mAb, требуемую для 50% нейтрализации цитотоксичности (ЕС50). Выживание клеток определяли посредством CellTiter-Blue; исходные данные нормализовали относительно необработанных контрольных лунок. Значения наносили на график с применением Prism, и кривые рассчитывали с применением модели сигмоидальной кривой «доза-эффект» (переменный наклон). Кривую затем применяли для определения ЕС50 mAb. Точки данных представляют собой среднее трех лунок на одном планшете.
На фиг.5 демонстрируется in vitro активность нейтрализации активности токсина В с применением очищенных мышиных mAb РА-41 на клетках СНО-K1. Для измерений цитотоксичности токсин В инкубировали с различными концентрациями РА-41 в течение 1 часа при 37°С (пример 3В). Затем смеси mAb-токсин вносили к клеткам СНО-K1, выращенным в 96-луночных планшетах при 2000 клеток/лунку и инкубировали в течение 72 часов. Выживание клеток сравнивали в необработанных и обработанных культурах и вычисляли концентрацию mAb, требуемую для 50% нейтрализации цитотоксичности (ЕС50). Выживаемость клеток определяли с помощью CellTiter-Blue; исходные данные нормализовали относительно необработанных контрольных лунок. Значения наносили на график с применением Prism, и кривые рассчитывали с применением модели сигмоидальной кривой «доза-эффект» (переменный наклон). Кривую затем применяли для определения ЕС50 mAb. Точки данных представляют собой среднее трех лунок на одном планшете.
На фиг.6 демонстрируется in vitro активность нейтрализации активности токсина А с применением очищенных мышиных mAb РА-38 и РА-50 на клетках Т-84 (пример 3С). Клетки Т-84 высевали (15000 клеток/лунку) в 96-луночные планшеты и обрабатывали комбинацией из протитрованного mAb (РА-38 (■) или РА-50 (▲)) и токсина А (60 нг/мл). После инкубации (72 часа) сравнивали выживание клеток в обработанных и необработанных культурах, и рассчитывали концентрацию mAb, требуемых для 50% нейтрализации цитотоксичности (ЕС50). Выживание клеток определяли посредством CellTiter-Blue; исходные данные нормализовали относительно необработанных контрольных лунок. Значения наносили на график с применением Prism, и кривые рассчитывали с применением модели сигмоидальной кривой «доза-эффект» (переменный наклон). Кривую затем применяли для определения ЕС50 mAb. Точки данных представляют собой среднее трех лунок на одном планшете.
На фиг.7 демонстрируются результаты тестирования мышиных mAb PA-38 (■) или РА-50 (▲) в отношении их способности блокировать или предотвращать индуцированную токсином А гемагглютинацию красных кровяных клеток кролика (RBC). Токсин А (2 мкг/мл) комбинировали с различными разведениями PA-38 или РА-50, и смесь вносили в планшеты, содержащие 50 мкл эритроцитов кролика. Планшеты инкубировали при 4°С в течение 4 часов. Гемагглютинацию определяли количественно как интенсивность цвета с применением анализа матрицы точек ImageQuant 400 (GE Healthcare). Данные представляли как % контроля, при этом 100% представляют собой отсутствие гемагглютинации. Точки данных представляют собой среднее трех лунок, проанализированных на одном планшете.
На фиг.8 демонстрируется активность mAb к токсину С.difficile по настоящему изобретению в предотвращении разрушения монослоя клеток Сасо-2 токсином А. Клетки Сасо-2 высевали (25000 клеток/лунку) в верхнюю камеру 96-луночного планшета Multiscreen Сасо-2 Assay (Millipore). После инкубации в течение 10-14 дней образование плотного монослоя подтверждали с помощью измерения трансэпителиального электрического сопротивления (TEER) с применением эпителиального вольтомметра (World Precision Instruments). После того, как выяснили и определили целостность монослоя, в верхнюю камеру планшета для анализа вносили токсин А (25 нг/мл) и серийно разведенное мышиное mAb (PA-38 (■) или РА-50 (▲)). Планшеты инкубировали в течение 18-24 часов, и измеряли значение TEER с применением вольтомметра. Целостность монослоя сравнивали в необработанных и обработанных токсином лунках. Данные подавления аппроксимировали с сигмоидальной кривой «доза-эффект» с нелинейной регрессией с применением программного обеспечения GraphPad Prism, чтобы определить концентрацию mAb, требуемую для 50% подавления токсина (ЕС50).
На фиг.9А-9С демонстрируется способность mAb к токсину A PA-38 (9A) и РА-50 (9В) нейтрализовать активность токсина A in vivo. Самкам мышей Swiss Webster (возраст 6-8-недель, 5 мышей/группа) инъецировали (i.p.) мышиное mAb PA-38 или мышиное mAb РА-50 в указанных количествах или PBS (200 мкл) в день 0. Нейтрализующую активность моноклонального антитела сравнения к токсину А, указанного в данном документе как CDA-1, оценивали при указанных количествах антитела (9С). mAb сравнения к токсину А, CDA-1, получали, синтезируя (DNA2.0) нуклеиновые кислоты, кодирующие вариабельные области тяжелой и легкой цепи 3D8 (W02006/121422 и US2005/0287150), которые клонировали в векторы экспрессии IgGI человека полной длины (pCON-гамма1 и pCON-каппа). mAb сравнения CDA-1 экспрессировали и продуцировали в клетках CHO-KSV1 и очищали, как описано в разделе Примеры в данном документе. Затем мышам инъецировали 100 нг токсина А (200 мкл) в день 1, и за ними наблюдали ежедневно в течение первых 72 часов и еженедельно после этого. Результаты показывают, что оба mAb PA-38 и РА-50 способны полностью подавлять ассоциированную с токсином А токсичность после однократной дозы 2 мкг mAb/животное, тогда как mAb сравнения CDA-1 (5 мкг/животное) не могло полностью подавить ассоциированную с токсином А С.difficile токсичность.
На фиг.10 демонстрируется способность mAb PA-41 нейтрализовать активность токсина В in vivo. Самкам мышей Swiss Webster (возраст 6-8-недель, 5 мышей/группа) инъецировали (i.p.) или мышиное mAb PA-41 в указанных количествах или PBS (200 мкл) в день 0. Затем мышам инъецировали 100 нг токсина В (200 мкл) в день 1, и за ними наблюдали ежедневно в течение первых 72 часов и еженедельно после этого. Результаты данного эксперимента показывают, что mAb PA-41 полностью подавляет ассоциированную с токсином В С.difficile токсичность после однократной дозы 5 мкг mAb/животное. Подобный эксперимент проводили с применением моноклонального антитела сравнения к токсину В, указанного в данном документе как mAb сравнения CDB-1. mAb сравнения к токсину В CDB-1 получали, синтезируя (DNA2.0) нуклеиновые кислоты, кодирующие вариабельные области тяжелой и легкой цепи 124 (WO2006/121422 и US2005/0287150), которые клонировали в векторы экспрессии IgGI человека полной длины (pCON-гамма1 и pCON-каппа). mAb сравнения CDB-1 экспрессировали и продуцировали в клетках CHO-KSV1 и очищали, как описано в разделе Примеры в данном документе. Результаты этих экспериментов показали отсутствие активности нейтрализации в отношении токсина В у mAb сравнения CDB-1, даже при количестве 250 мкг.
На фиг.11 демонстрируется возможность комбинации мышиных mAb РА-38 и РА-41 (РА-38+РА-41) настоящего изобретения для нейтрализации активности токсина А и токсина В in vivo. Самкам мышей Swiss Webster (возраст 6-8-недель, 5 мышей/группа) инъецировали (i.p.) комбинацию mAb РА-38+РА-41 или PBS (200 мкл) в день 0. Затем мышам инъецировали 100 нг комбинации токсина А и токсина В (200 мкл) в день 1, и за ними наблюдали ежедневно в течение первых 72 часов и еженедельно после этого. Графики для токсина, только РА-38 (пустые кружки) и только РА-41 (закрашенные ромбы) наложены на общий график. Результаты показывают, что только mAb РА-38 (пустые кружки), только mAb РА-41 (закрашенные ромбы) не было достаточно для подавления эффектов обоих токсинов, и они не защищали животных от инфекции С.difficile. В отличие от этого, комбинация РА-38 и РА-41 (РА-38+РА-41), при 50 мкг каждого (свободные, перевернутые треугольники), смогла защитить инфицированных животных и предотвратить смерть, связанную с токсином, у 4 из 5 тестовых животных. Комбинация РА-38 и РА-41 (РА-38+РА-41) по 5 мкг каждого (закрашенные кружки) обеспечила некоторую защиту от токсичности токсинов А и В С.difficile у инфицированных тестовых животных.
На фиг.12А и 12В демонстрируются фармакокинетические результаты (PK) на хомячках для мышиных mAb РА-38 и РА-41. Хомячкам i.p. давали дозы 2 мг/кг (○) или 10 мг/кг (■) mAb РА-38 (12А) или РА41 (12В). У животных брали кровь в определенные промежутки времени, и сыворотку анализировали, применяя для обнаружения ELISA с покрытыми токсином планшетами и антителами козы к IgG мыши, HPR-конъюгированными. Полученные в результате кривые иллюстрируют дозозависимый ответ у когорт 2 мг/кг и 10 мг/кг для каждого антитела. Анализ WinNonLin проводили на каждой кривой. Оба моноклональных антитела имеют конечный период полураспада больше 6 дней.
На фиг.13 иллюстрируются результаты выживания из исследования на хомячках, описанного в примере 5 В. В этом исследовании хомячков обрабатывали клиндамицином, заражали С. difficile (■ инфицированный контроль, группа 3), а затем обрабатывали ванкомицином (◆20 мг/кг, группа 4), комбинацией mAb РА-38+РА-41 (Δ 50, 50 мг/кг, группа 6) или комбинацией mAb PA-39+PA-41 (○ 50, 40 мг/кг, группа 7). Все животные в неинфицированном контроле (группа 1) и контроле, обработанном поликлональными Ab козы (группа 5), выжили. Животные, обработанные комбинацией mAb к токсину А и токсину В по настоящему изобретению, выжили и были защищены от токсичности С.difficile в течение срока исследования.
На фиг.14 показана средний вес тела (в граммах) хомячков из исследования, описанного в примере 5В. Экспериментальные группы животных следующие: неинфицированный контроль (◆, группа 1); контроль, обработанный ванкомицином (■, группа 4); группа, обработанная комбинацией мышиных mAb PA-38+PA-41 (х, группа 6), или группа, обработанная комбинацией мышиных mAb PA-39+PA-41 (●, группа 7). Животные в инфицированной контрольной группе (группа 3) не выжили в течение 5 дней, поэтому для группы 3 измерения веса тела после заражения произвести было невозможно.
На фиг.15A-15D изображены результаты посмертного вскрытия из исследования на хомячках, описанного в примере 5В. Оценивали типичных животные из каждой соответствующей группы из исследования: (А) группа 1, неинфицированный контроль; (B) группа 3, инфицированный контроль; (С) группа 6, группа, обработанная комбинацией мышиных mAb PA-38+PA-41, и (D) группа 7, группа, обработанная комбинацией мышиных mAb PA-39+PA-41. Стрелки указывают слепую кишку каждого хомячка. У инфицированной контрольной группы 3 (В) слепая кишка была заметно покрасневшей и воспаленной. В отличие от этого, слепые кишки хомячков в группе 6 (C) и группе 7 (D) были похожи на слепые кишки у здоровых, неинфицированных контрольных животных группы 1 (А).
Фиг.16А и 16В-1 и В-2. На фиг.16А иллюстрируются результаты выживания в исследовании на хомячках, описанном в примере 5С. В этом исследовании хомячков обрабатывали клиндамицином, заражали С.difficile (■ инфицированный контроль, группа 1), а затем различные группы животных обрабатывали ванкомицином (◆20 мг/кг, группа 2), комбинацией мышиных mAb PA-39+PA-41 (▲ 50+50 мг/кг, группа 3), только мышиным mAb РА-41 (○ 50 мг/кг, группа 4), только mAb PA-38 мыши (∇ 50 мг/кг, группа 5), только мышиным mAb PA-39 (□ 50 мг/кг, группа 6) или только мышиным mAb PA-50 (◇ 50 мг/кг, группа 7). Все животные в неинфицированном контроле (группа 1) выжили в продолжение хода исследования. На фиг.16 В-1 иллюстрируются результаты выживания животных в исследовании на хомячках, описанном в примере 5Е. Кривые выживания Каплана-Меера показаны для животных, обработанных клиндамицином, а затем зараженных С.difficile (■, инфицированный контроль, группа 2); обработанных ванкомицином (◇, 20 мг/кг, группа 3); обработанных комбинацией 1:1 гуманизированных mAb PA-50+PA-41 ((hPA-50+hPA-41), Δ, 50+50 мг/кг, группа 4); обработанных комбинацией 1:1 гуманизированных mAb PA-50+PA-41 ((hPA-50+hPA-41), ○, 20+20 мг/кг, группа 5); обработанных комбинацией 1:1 mAb сравнения CDA-1+CDB-1 препарата ((CDA-1+CDB-1), ∇, 50+50 мг/кг, группа 6) или обработанных комбинацией 1:1 mAb сравнения CDA-1+CDB-1 ((CDA-1+CDB-1), □, 20+20 мг/кг, группа 7). На фиг.16 В-2 иллюстрируются результаты изменения веса в исследовании на хомячках, описанном в примере 5Е. Среднее (±SD) веса тела животных по времени в группах с различной обработкой, описанных для фиг.16 В-1, сравнивали с неинфицированными контрольными животными.
На фиг.17А-17С показана обработка токсина В С.difficile каспазой 1 (А): токсин В С.difficile полной длины и его домены. (В): 3-8% Tris-ацетатный SDS-PAGE (восстанавливающий) анализ токсина В (TcdB) и обработанного каспазой 1 токсина В. Отмечали четыре фрагмента токсина: 193, 167, 103 и 63 кДа. (С): возможные фрагменты, образованные после обработки каспазой 1 токсина В.
На фиг.18А-18С показаны SDS-PAGE (А) и вестерн-блоттинг (В, С) обнаружение фрагментов токсина В С.difficile с применением mAb к токсину В. (А): SDS-PAGE анализ фрагментов токсина В, образованных при обработке каспазой 1 (так же, как на фиг.17В.); (В): вестерн-блоттинг обнаружение фрагментов токсина В с применением mAb РА-41. (С) Вестерн-блоттинг обнаружение фрагментов токсина В с применением мышиного mAb РА-39.
На фиг.19А-19Е показана характеристика мышиных mAb к токсину С.difficile с помощью связывания Biacore. Оценивали конкурентное связывание mAb к токсину. (А): mAb РА-41 связывает один эпитоп на токсине В. (В): mAb РА-39 связывает один эпитоп на токсине А. (С): mAb РА-39 и РА-41 связываются с различными эпитопами на токсине В. Связывание mAb РА-41 с токсином В является эпитопически отличным от связывания антитела сравнения к токсину В CDB-1. (D) mPA-41, иммобилизованное на чипе СМ5, захватывает токсин В, но не способно связывать дополнительное тА-41, что свидетельствует, что имеется только один эпитоп связывания mPA-41. Добавление mAb сравнения CDB-1 дало повышенный сигнал, демонстрируя, что mPA-41 и mAb сравнения CDB-1 связывают различные эпитопы на токсине В. (Е) Вестерн-блоттинг анализ с использованием токсина В, или необработанного, или обработанного каспазой 1, демонстрируют, что mPA-41 и mAb сравнения CDB-1 имеют различные паттерны связывания и связывают различные эпитопы на токсине В.
На фиг.20А-20С показано расщепление токсина А С.difficile с применением энтерокиназы (EK). (А): токсин А С.difficile полной длины и его домены. (В): 3-8% Tris-ацетатный SDS-PAGE (восстанавливающий) анализ токсина A (TcdA) и токсина А, обработанного EK. (С): возможные фрагменты, образованные после обработки токсина А EK при 25°С в течение 48 часов.
На фиг.21А-21С показаны окрашивание Кумасси синим (SDS-PAGE), (А), и вестерн-блоттинг (В, С) обнаружения фрагментов токсина А С.difficile с применением мышиных mAb к токсину А. (А): SDS-PAGE анализ фрагментов токсина А, образованных при обработке EK (так же, как на фиг.20В). (В): вестерн-блоттинг обнаружение фрагментов токсина А с применением mAb PA-50. (С): вестерн-блоттинг обнаружение фрагментов токсина А с применением mAb РА-39. На фиг.21В и 21C, полоса (кДа) составляет ~158 кДа и может рассматриваться как составляющая ~158-160 кДа.
Фиг.22А-1 и A-2-22F. На фиг.22А-1 и A-2-22D показана характеристика связывания мышиного mAb к токсину А с токсином А С.difficile с применением анализа связывания Biacore. Фиг.22А-1: отмечали, что mAb PA-50 связывает множественные сайты на токсине А. Фиг.22А-2: mAb PA-50 иммобилизовали на сенсорном чипе, а затем последовательно приводили в контакт с очищенным токсином А, дополнительным PA-50 и mAb сравнения CDA-1 (WO/2006/121422; US2005/0287150). Фиг.22В: токсин А, захваченный mAb сравнения CDA-1 на чипе Biacore, дополнительно связывает дополнительные CDA-1 и mAb PA-50, показывая различия в эпитопах токсина А, связанных антителами. Фиг.22С: эпитоп связывания mAb PA-39 на токсине А отличается от эпитопа связывания mAb сравнения CDA-1 на токсине А. Фиг.22D: конкурентное связывание мышиных mAb PA-50 и PA-39 с токсином А проводили с применением Biacore. mAb PA-50, иммобилизованное на чипе СМ5, захватывает токсин А, который может связывать дополнительное mPA-50 и также mPA-39, демонстрируя, что существуют множественные копии эпитопов для mPA-50 на токсине А, и что mPA-50 и mPA-39 связываются с отличными эпитопами токсина А. Фиг.22Е и 22F: результаты Biacore подтверждали вестерн-блоттинг анализом с использованием токсина А, который не обрабатывали и обрабатывали ферментом энтерокиназой (EK). (Е): mPA-39 и mAb сравнения CDA-1 показывают различные паттерны связывания с обработанным EK токсином А (полоска: TcdA/EK), таким образом отмечая различные домены и эпитопы связывания токсина A. (F): mPA-50 и mAb сравнения CDA-1 связываются с тем же доменом токсина А, но с разными эпитопами.
На фиг.23А и 23В демонстрируется нейтрализующая активность PA-41 in vitro в отношении панели из двадцати различных токсикогенных клинических изолятов С.difficile, включающих 6 BI/NAP 1/027 изолятов, 3 референс-штамма (VPI 10463, АТСС 43596 и 630), 2 токсин А-отрицательных/токсин В-положительных (toxA-/toxB+) изолята, 3 амбулаторных изолята и 6 других часто встречающихся клинических изолятов. На фиг.23А показана нейтрализующая активность мышиного mAb PA-41 на клетках СНО-K1 в отношении супернатантов, полученных от культур различных клинических изолятов С.difficile, представленных в примере 8, таблице 6. Очищенное mAb PA-41 последовательно разводили и затем смешивали с супернатантом с предварительно определенным коэффициентом разведения, который может вызывать >90% смерти клеток. Смеси инкубировали в течение 1 часа при 37°С и затем вносили к клеткам СНО-K1. Клетки инкубировали в течение 72 часов, и жизнеспособность клеток измеряли с применением Cell-Titer Blue. На фиг.23В иллюстрируется нейтрализующая активность гуманизированного mAb hPA-41, а также mAb сравнения CDB-1 в отношении 2 референс-штаммов С.difficile (VPI 10463 и АТСС 43596) и 6 BI/027/027 штаммов С.difficile (CCL678, НМС553, Pitt 45, CD196, Montreal 5.1 и Montreal 7.1). Показана нейтрализующая активность mAb hPA-41 (закрашенные квадраты) и mAb сравнения CDB-1 (закрашенные треугольники) на клетках СНО-K1 в отношении супернатанта от референс-штаммов (VPI 10483 и АТСС 43596) и BI/NAP 1/027 штаммов (CCL678, НМС553, Pitt 45, CD196, Montreal 5.1 и Montreal 7.1).
На фиг.24А и 24В демонстрируется нейтрализующая активность in vitro mAb PA-50 в отношении токсикогенных клинических изолятов С.difficile, описанных для фиг.23А и В. На фиг.24А показана нейтрализующая активность мышиного mAb PA-50 на клетках Т-84 в отношении супернатанта, полученного от культур различных клинических изолятов С.difficile, представленных в примере 8, таблице 6. Очищенные mAb PA-50 серийно разводили и затем смешивали с предварительно определенным коэффициентом разведения, который может вызывать >90% смерти клеток. Смеси инкубировали в течение 1 часа при 37°С и затем вносили к клеткам Т-84. Клетки инкубировали в течение 72 часов, и жизнеспособность клеток измеряли с применением Cell-Titer Blue. На фиг.24В показаны результаты сходных экспериментов, проведенных с применением гуманизированного mAb hPA-50 и mAb сравнения CDA-1 на клетках Т-84. Показана нейтрализующая активность hPA-50 (закрашенные квадраты) и сравнения CDA-1 (закрашенные треугольник) на клетках Т-84 в отношении супернатантов, полученных из шести BI/NAP1/027 штаммов (CCL678, НМС553, Pin 45, CD 196, Montreal 5.1 и Montreal 7.1). Для фиг.24А: *: N/A: Не применимо; токсин А не продуцировался токсин а-/токсин В+штаммами F1470, 8864, CCL13820 и CCL14402; ": Титр токсина А был очень низким; нет измеримой цитотоксичности на Т-84 с применением супернатанта; ^: Не применимо; токсин А не продуцировался токсин А-/токсин В+штаммами или концентрация была слишком низкой.
На фиг.25A-25D демонстрируется нейтрализация токсинов, продуцированных различными штаммами С. difficile. На фиг.25А и 25В показана нейтрализующая активность мышиного mAb PA-39 на клетках Т-84 в отношении супернатантов, полученных от культур различных клинических изолятов С. difficile, представленных в примере 8, таблице 6. mAb PA-39 (супернатант гибридомы) серийно разводили, и показан коэффициент разведения для каждого супернатанта. На фиг.25В показаны результаты подобных экспериментов, проведенных с применением мышиного mAb PA-39 и mAb сравнения CDA-1 на клетках Т-84. Показана нейтрализующая активность PA-39 (закрашенные квадраты) и mAb сравнения CDA-1 (закрашенные треугольники) на клетках Т-84 в отношении супернатантов, полученных от шести BI/NAP1/027 штаммов (CCL678, НМС553, Pitt45, CD196, Montreal 5.1 и Montreal 7.1). Для фиг.25А: *: N/A: Не применимо; токсин А не продуцировался токсин а-/токсин В+штаммами F1470, 8864, CCL13820 и CCL14402; ": титр токсина А был очень низким; нет измеримой цитотоксичности на Т-84 с применением супернатанта; ^: Не применимо; токсин А не продуцировался токсин A-/токсин В+штаммами или концентрация была слишком низкой. На фиг.25С, тестировали гуманизированное mAb к токсину А РА-50 и mAb сравнения к токсину А CDA-1 на нейтрализацию цитотоксичности культурального супернатанта С.difficile в отношении клеток Т-84. (пример 8, таблица 7). На фиг.25D, тестировали гуманизированное mAb к токсину В РА-41 и mAb сравнения к токсину А CDB-1 на нейтрализацию цитотоксичности культурального супернатанта С.difficile в отношении клеток Т-84 (пример 8, таблица 7).
На фиг.26 показано, что химерные mAb PA-41 (mAb сРА-41) эффективно нейтрализуют токсичность токсина В С. difficile на СНО-K1 в сравнении с аналогичными мышиными mAb PA-41. Получали два химерных mAb PA-41; у одного из которых сайт гликозилирования в VL-области удаляли, обозначено как cPA-41(NG), а у одного из которых сайт гликозилирования не удаляли, обозначено как cPA-41(G) на фигуре. Оба cPA-41(NG) и cPA-41(G) показали сходные уровни нейтрализации относительно токсина В (2 пг/мл, TechLab) на клетках СНО-K1, и оба химерных mAb нейтрализовали токсин В на уровне, сходном с таковым исходного мышиного mAb (mPA-41).
На фиг.27 показано, что химерное mAb PA-39 (сРА-39) эффективно нейтрализует токсичность токсина А С.difficile (1 пг/мл, Listlab) на клетках СНО-K1 в сравнении с исходным мышиным mAb PA-39 (mPA-39).
На фиг.28 показано, что химерное mAb PA-50 (сРА-50) эффективно нейтрализует токсичность токсина А С.difficile (60 пг/мл, TechLab) на клетках Т-84 в сравнении с исходным мышиным mAb PA-50 (mPA-50).
На фиг.29 показана in vitro нейтрализующая активность мышиного mAb PA-41 (тРА-41) и гуманизированного mAb PA-41 (hPA-41) в отношении токсина В С.difficile. Процент выживания клеток в сравнении с контролями измеряли с применением CellTiter Blue. mAb hPA-41 эффективно нейтрализует токсичность токсина В (2 пг/мл, Techlab) на клетках СНО-K1 с ЕС50 - 6 пМ, и был фактически равносильным с исходным мышиным моноклональным антителом (mPA-41).
На фиг.30 показана нейтрализующая активность in vitro мышиного mAb PA-39 (mPA-39) и гуманизированного mAb PA-39 (hPA-39) в отношении токсина А С.difficile. mAb hPA-39 эффективно нейтрализует токсичность токсина А (20 пг/мл, TechLab) на СНО-K1 клетках с ЕС50 - 50 пМ, и был фактически равносильным с исходным мышиным моноклональным антителом (mPA-39).
На фиг.31А-31Н показаны результаты исследований нейтрализующей активности in vitro и механизма действия (МОА) с применением описанных mAb. Основанные на клетках анализы с применением клеток СНО-K1 и клеток Т-84 клетки поводили, как описано в примерах 1, 3 и 7. На фиг.31А показано, что гуманизированное mAb PA-50 (hPA-50) эффективно нейтрализует токсичность токсина А (60 пг/мл, TechLab) на клетках Т-84 в сравнении с исходным мышиным mAb PA-50 (mPA-50). На фиг.31B-D показаны нейтрализующие активности mAb к токсину А PA-39 и PA-50 в сравнении с таковой mAb сравнения к токсину A CDA-1. Значения ЕС50 и максимального процента подавления таковые, как представлены в таблице А примера 7С. На фиг.31Е и F показана нейтрализующая активность mAb к токсину В РА-41 в сравнении с mAb сравнения к токсину В CDB-1. Значения ЕС5о и максимального процента подавления таковые, как представлены в таблице В примера 7С.На фиг.31G и Н показаны способ ELISA и результаты оценки способности mAb к токсину А блокировать интернализацию токсина А в клетки и оценки активностей этих mAb в предотвращении интернализации токсина А относительно контроля с поликлональным антителом козы к токсину А и контроля без антител.
На фиг.32А и 32В изображены аминокислотные последовательности VH-областей гуманизированного PA-39 (hPA-39), SEQ ID NO:1 и SEQ ID NO:2. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.33А и 33В изображены аминокислотные последовательности VL-областей гуманизированного PA-39 (hPA-39), SEQ ID NO:3 и SEQ ID NO:4. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.34А и 34В изображены аминокислотные последовательности VH-областей гуманизированного PA-50 (hPA-50), SEQ ID NO:5 и SEQ ID NO:6. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.35 изображена аминокислотная последовательность VL-области гуманизированного PA-50, SEQ ID NO:7. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.36А и 36В изображены аминокислотные последовательности VH-областей гуманизированного PA-41 (hPA-41), SEQ ID NO:8 и SEQ ID NO:9. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.37 изображена аминокислотная последовательность VL-области гуманизированного PA-41, SEQ ID NO:10. Аминокислотные остатки показаны при помощи однобуквенного кода. Номера над последовательностями указывают положения в соответствии с Kabat et al. Положения CDR подчеркнуты.
На фиг.38А и 38В показаны последовательность нуклеиновой кислоты и закодированная аминокислотная последовательность гуманизированного моноклонального антитела к токсину А С.difficile. На фиг.38А изображена аминокислотная последовательность легкой цепи гуманизированного моноклонального антитела к токсину А, изложенная в SEQ ID NO:16, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:17. На фиг.38В изображена аминокислотная последовательность тяжелой цепи гуманизированного моноклонального антитела, изложенная в SEQ ID NO:14, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:15.
На фиг.39А и 39В показаны последовательность нуклеиновой кислоты и закодированная аминокислотная последовательность гуманизированного моноклонального антитела к токсину А С.difficile. На фиг.39А изображена аминокислотная последовательность легкой цепи гуманизированного моноклонального антитела к токсину А, изложенная в SEQ ID NO:20, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:21. На фиг.39 В изображена аминокислотную последовательность тяжелой цепи гуманизированного моноклонального антитела к токсину А, изложенная в SEQ ID NO:18, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:19.
На фиг.40А и 40В показаны последовательность нуклеиновой кислоты и закодированная аминокислотная последовательность гуманизированного моноклонального антитела к токсину В С.difficile. На фиг.40А изображена аминокислотная последовательность легкой цепи гуманизированного моноклонального антитела к токсину В, изложенная в SEQ ID NO:24, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:25. На фиг.40 В изображена аминокислотная последовательность тяжелой цепи гуманизированного моноклонального антитела к токсину В, изложенная в SEQ ID NO:22, которая кодируется последовательностью нуклеиновой кислоты, изложенной в SEQ ID NO:23.
На фиг.41А-41С демонстрируется нейтрализующая активность in vitro Fab-фрагментов мышиных mAb в отношении токсина А и токсина В С.difficile в сравнении с эффективностью эквивалентного цельного антитела. (А): нейтрализующая активность Fab мышиных mAb РА-39 и РА-39 на клетках СНО-K1; токсин A (Techlab, 60 нг/мл); (В): нейтрализующая активность Fab мышиных mAb PA-41 и РА-41 на клетках СНО-K1; токсин В (Techlab, 2 пг/мл); (С): нейтрализующая активность Fab мышиных mAb РА-50 и РА-50 на клетках Т-84; токсин A (Techlab, 60 нг/мл).
На фиг.42А и 42В показаны профили концентрации антител, полученные в результате фармакокинетического (PK) исследования, описанного в примере 13 в данном документе. На фиг.42А изображены результаты PK (концентрация сывороточных антител в мг/мл) в течение 29 дней от животных, получающих однократную дозу очищенного, гуманизированного, mAb к токсину А РА-50 при концентрации 1 мг/кг (▲) или 5 мг/кг (■) в день 0. На фиг.42В изображены результаты PK (концентрация сывороточных антител в мг/мл) в течение 29 дней от животных, получающих однократную дозу очищенного, гуманизированного, mAb к токсину В PA-41 при концентрации 1 мг/кг (▲) или 5 мг/кг (■) в день 0.
Подробное описание изобретения
Настоящее изобретение охватывает антитела и их антигенсвязывающие фрагменты, которые обеспечивают терапевтические средства и лекарственные средства, не являющиеся антибиотиками, которые блокируют патогенные эффекты инфекции С.difficile и, предпочтительно, обеспечивают время для лечения толстой кишки и/или нормализации микрофлоры желудочно-кишечного тракта, например, толстой кишки, кишечника, кишечного тракта и т.д., до его восстановления. Моноклональные антитела, описываемые в настоящем документе, их антигенсвязывающие фрагменты, как, например, их гуманизированные или гибридные формы таковых, обеспечивают терапевтические средства и лекарственные препараты, не являющие антибиотиками, как для лечения активного заболевания, так и для предупреждения повторного заболевания, с тем, чтобы дать возможность пациенту, страдающему инфекцией С.difficile и CDAD, излечиться от своего заболевания без рецидива в дальнейшее заболевание или в более тяжелое заболевание или симптомы. В варианте осуществления антитела настоящего изобретения обладают терапевтической активностью к активному заболеванию, вызванному или ассоциированному с инфицированием субъектов С.difficile. В варианте осуществления антитела настоящего изобретения устраняют активное заболевание, вызванное или ассоциированное с инфицированном субъектов С.difficile. В варианте осуществления антитела настоящего изобретения обладают терапевтическим эффектом при уменьшении продолжительности и/или тяжести активного заболевания, вызванного или ассоциированного с инфицированном С.difficile у субъектов. В варианте осуществления антитела настоящего изобретения или части, или их фрагменты могут обеспечиваться в комбинации с антибиотическими терапевтическими средствами.
Моноклональные антитела (mAb) к токсинам А и В С.difficile были получены, как описано в настоящем документе. mAb к токсинам проявляют сильную активность как в исследованиях in vitro, так и в доклинических моделях на животных с инфекцией С.difficile in vivo. Более конкретно, mAb настоящего изобретения эффективно и долговременно защищают хомяков от смертности в релевантной или точной модели на хомяках с инфекцией С.difficile.
Антитела обеспечивают подходы к лечению CDAD без антибиотиков и могут предоставить возможность для прекращения приема антибиотиков и блокирования патогенных эффектов токсинов С.difficile, таким образом, обеспечивая время для исцеления кишечника и восстановления нормальной кишечной микрофлоры. mAb, описанные в настоящем документе, могут обеспечить терапевтическую полезность, благодаря их способности нейтрализовать токсины С.difficile и их можно использовать при пассивных или активных стратегиях лечения пациентов с многочисленными повторными проявлениями С.difficile. В частности, mAb могут быть использованы для предупреждения повторного проявления инфекции, для лечения тяжелых активных форм заболевания и для лечения пациентов с множественными рецидивами заболеваний, ассоциированных с С.difficile. mAb, описанные в настоящем документе, могут обеспечить эффективные средства нейтрализации токсинов А и В С. difficile с тем, чтобы предотвратить, блокировать или подавить повторное проявление инфекции, и/или тяжелые и активные формы заболевания и многочисленные рецидивы.
Применяемые в настоящем документе «токсин А» и «токсин В» относятся к цитотоксическим энтеротоксинам, продуцируемым микроорганизмом С.difficile. Токсины А и В являются главными детерминантами вирулентности С.difficile, a токсин-отрицательные штаммы являются непатогенными. Токсины А и В транскрибируются из локуса патогенности, который включает гены токсина tcdA (токсин А) и tcdB (токсин В), и три регуляторных гена, один из которых (tcdC) кодирует предполагаемый отрицательный регулятор транскрипции токсина. Оказывается, что белок TcdC ингибирует транскрипцию токсина в ходе ранней фазы экспоненциального роста бактериального жизненного цикла. Был описан участок автокаталитического расщепления между лейцином543 и глицином544 для токсина В. Расщепление происходит в результате активации домена аспартиловой протеазы цитозольным инозитол-фосфатом хозяина, и высвобождается активный гликозилтрансферазный домен.
В настоящем документе приводятся антитела и их антигенсвязывающие фрагменты, которые специфично связываются с токсином А и/или токсином В С.difficile, композиции, содержащие одно или несколько таких антител или их антигенсвязывающих фрагментов, векторы, содержащие последовательности нуклеиновых кислот, которые кодируют антитела или их антигенсвязывающие фрагменты, гибридомные клеточные линии, которые продуцируют антитела, и способы применения антител или их антигенсвязывающих фрагментов для лечения или предупреждения инфекции С.difficile или заболевания, ассоциированного с С.difficile.
Следует понимать, что при упоминании в настоящем документе выражения «антитела» или «иммуноглобулины» при описании настоящего изобретения и его различных аспектов и вариантов осуществления подразумевается, что это выражение в основном охватывает антигенсвязывающие фрагменты таких антител или иммуноглобулинов с тем, чтобы избежать чрезмерного повторения ассоциированной фразы «антигенсвязывающие фрагменты» каждый раз при упоминании выражение «антитела» или «иммуноглобулины». Таким образом, настоящее изобретение охватывает не только антитела, направленные против токсина А и токсина В С.difficile, то есть антигенов токсина А и токсина В, но также и фрагменты таких антител, которые связывают антигены токсина А и токсина В С.difficile, описанные далее в настоящем документе. В вариантах осуществления, такие антигенсвязывающие фрагменты способны нейтрализовать токсичность токсина А и/или токсина В подобно тому, как это происходит у интактного антитела.
Антитела, которые охватываются настоящим изобретением, включают выделенные антитела, которые специфично связывают токсин А С.difficile и которые конкурентно подавляют или перекрестие конкурируют за специфичное связывание с токсином А C. difficile выделенного моноклонального антитела, продуцируемого гибридомной клеточной линии, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. Эти антитела также включают выделенные антитела, которые специфично связываются с токсином А С. difficile, и которые специфично связываются с эпитопом на токсине А С. difficile, определяемым путем связывания выделенного моноклонального антитела, продуцируемого гибридомной клеточной линей, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. В некоторых вариантах осуществления эпитопы располагаются в С-концевом рецепторсвязывающем домене tcdA. В некоторых из этих вариантов осуществления антитела конкурентно подавляют или перекрестие конкурируют за связывание РА-50 с tcdA. В других вариантах осуществления эпитоп располагается в транслокационном домене tcdA. В некоторых из этих вариантов осуществления антитела конкурентно подавляют или перекрестие конкурируют за связывание РА-39 с tcdA. Такие выделенные антитела могут включать моноклональные антитела, поликлональные антитела, химерные антитела, антитела человека, гуманизированные антитела и антигенсвязывающие фрагменты или их части.
Эксперименты с использованием ферментативного (энтерокиназного) протеолиза токсина А для оценки специфичности моноклональных антител настоящего изобретения к токсину А С. difficile осуществляли, как показано в Примере 6. Эпитоп на токсине А, который распознается и связывается mAb PA-39, находится в пределах области, которая отличается от рецепторсвязывающего домена токсина А, то есть вне рецепторсвязывающего домена токсина А, и также отличается от эпитопа, связываемого антителом человека к токсину А, которое, как сообщают, связывает С-концевой рецепторсвязывающий домен токсина А (7). Как описано в Примерах 6 и 7 в настоящем документе и показано на фиг.22 и 31, анализы Biacore подтверждают одиночный участок связывания на токсине А для РА-39. Обнаружение с помощью вестерн-блоттинга расщепленного ферментативно токсина А демонстрирует, что РА-39 связывает область на токсине А, которая отделена от областей, связываемых РА-50 и препаратом сравнения mAb CDA-1. Активность PA-39 in vitro в исследовании эффективности токсина показывает сдвиги как в ЕС50, так и в максимальном проценте подавления по мере того, как больше токсина А добавляют в культуру, указывая на смешанный конкурентный механизм подавления РА-39. Обнаружение токсина А с помощью ELISA после защиты 100-кратным избытком РА-39 подтвердило, что подавление токсина с помощью РА-39 происходит путем предупреждения интернализации токсина и неблагоприятных эффектов токсина на клетку, например, цитотоксичности.
Кроме того, как описано в Примерах 6 и 7 в настоящем документе и показано на Фиг.22 и 31, анализ Biacore подтверждает по меньшей мере два участка связывания на токсине А для РА-50. Обнаружение с помощью вестерн-блоттинга расщепленного ферментативно токсина А демонстрирует, что РА-50 связывает область на токсине А, сходную с таковой, связываемой препаратом сравнения mAb CDA-1. Активность РА-50 in vitro в исследовании эффективности токсина показывает сдвиг в ЕС50 по мере того, как больше токсина А добавляют в культуру, указывая на конкурентный механизм подавления для РА-50. Обнаружение токсина А с помощью ELISA после защиты 100-кратным избытком РА-50 подтвердило, что подавление токсина с помощью РА-50 происходит путем предупреждения интернализации токсина и последующей цитотоксичности.
Антитела настоящего изобретения также включают выделенные антитела, которые специфично связывают токсин В С.difficile и которые конкурентно подавляют или перекрестие конкурируют за специфичное связывание с токсином В С.difficile выделенного моноклонального антитела, продуцируемого гибридомной клеточной линей, депонированной под № доступа АТСС РТА-9693 или РТА-9692. Антитела также включают выделенные антитела, которые специфично связываются с токсином В С.difficile и которые специфично связываются с эпитопом на токсине В С.difficile, определяемом путем связывания выделенного моноклонального антитела, продуцируемого гибридомной клеточной линии, депонированной под № доступа АТСС РТА-9693 или РТА-9692. В некоторых вариантах осуществления эпитоп находится в N-концевом ферментативном домене tcdB. В некоторых из этих вариантов осуществления антитела конкурентно подавляют или перекрестно конкурируют за связывание РА-41 с tcdB. В других вариантах осуществления эпитоп находится в транслокационном домене, например аминокислоты 850-1330 tcdB. В некоторых из этих вариантов осуществления антитела конкурентно подавляют или перекрестно конкурируют за связывание РА-39 с tcdB.
Эксперименты с использованием ферментативного (каспаза 1) протеолиза токсина В для оценки специфичности моноклональных антител к токсину В С.difficile настоящего изобретения осуществляли, как описано в Примере 6. mAb PA-41 (PTA-9693), как было показано, распознает фрагменты приблизительно по 103 и 63 кДа, которые происходят из N-концевого ферментативного домена токсина В. Анализ N-концевой последовательности большинства расщепленных фрагментов подтвердил этот анализ. mAb РА-41, как было показано, связывает уникальный эпитоп в пределах N-концевого ферментативного домена токсина В, отличного от С-концевого рецепторсвязывающего домена токсина В, связываемого антителом человека к токсину В (7). Как описано в примерах 6 и 7 в настоящем документе и показано на Фиг.19 и 31Е и F, анализы Biacore подтверждают одиночный участок связывания на токсине В для РА-41. Обнаружение с помощью вестерн-блоттинга ферментативно расщепленного токсина В показывает, что РА-41 связывает отличную область на токсине В, которая отличается от таковой, связываемой препаратом сравнения mAb CDB-1. Активность PA-41 in vitro в исследовании эффективности токсина показывает сдвиги как в ЕС5о, так и в максимальном проценте подавления по мере того, как больше токсина В добавляют в культуру, указывая на смешанный конкурентный механизм подавления РА-41.
Антитела, приведенные в настоящем документе, включают моноклональные антитела, продуцируемые гибридомами, которые были депонированы и которым были даны такие патентные обозначения для депонирования: РТА-9692 (для РА-39), PTA-9693 (для РА-41), РТА-9694 (для РА-50) и РТА-9888 (для РА-38). Эти гибридомы были депонированы в соответствии с и для соблюдения требований Будапештского договора о международном признании депонирования микроорганизмов для целей патентной методики Американской коллекцией типовых культур («АТСС»), P.O. Box 1549, Manassas, VA 20108 США в качестве Международного ведомства по депонированию, 6 января 2009 года (для РТА-9692, PTA-9693, РТА-9694) и 24 марта 2009 года (для РТА-9888), и им были даны вышеупомянутые патентные обозначения для депонирования. Как используется в настоящем документе, как на депонированные гибридомы, так и на моноклональные антитела, продуцируемые гибридомами, можно ссылаться с подобными обозначениями для депонирования АТСС или с номерами, обнаруживаемыми среди обозначений для депонирования АТСС. Например, РТА-9888 или 9888 можно использовать для ссылки на депонированную гибридому или на моноклональное антитело, продуцируемое гибридомой. Соответственно, названия моноклональных антител, описанных в данном документе, можно использовать взаимозаменяемо с названиями гибридом, которые их продуцируют. Специалисту в данной области будет понятно, когда название предназначено для ссылки на антитело или на гибридому, которая производит это антитело. Антигенсвязывающие участки, приводимые в настоящем документе, включают антигенсвязывающие участки вышеупомянутых депонированных антител.
Антитела настоящего изобретения проявляют ряд полезных свойств. Например, антитела к токсину А нейтрализуют или подавляют токсичность токсина А как in vitro, так и in vivo. В исследованиях нейтрализации in vitro с использованием клеток IMR-90, гуманизированных РА-39 и гуманизированных РА-41 были продемонстрированы эффективности нейтрализации (например, значения ЕС50) из расчета 46 пМ против токсина А на клетках и 5 пМ против токсина В на этих клетках, соответственно. По сравнению со значениями, описанными в литературе для нейтрализации моноклональными антителами человека к токсину А и токсину В (WO/2006/121422; US2005/0287150; Babcock et al., Infect. Immun., 2006), (7), EC50 значение нейтрализации 46 пМ для hPA-39, как оказалось, было выше, чем таковое, описанное для mAb человека к токсину А, и ЕС50 для значение нейтрализации 5 пМ hPA-41, как оказалось, было выше, чем таковое, описанное для mAb человека к токсину В. Соответственно, в исследованиях, описанных в настоящем документе, гуманизированные моноклональные антитела настоящего изобретения к токсину А С.difficile и токсину В С.difficile, в частности, гуманизированные формы этих моноклональных антител, показывают улучшенные характеристики нейтрализации против токсинов по сравнению с таковыми других антител к токсинам, о которых сообщалось.
В одном варианте осуществления антитело к токсину А настоящего изобретения нейтрализует или подавляет in vivo токсичность токсина А С.difficile в эффективной дозе. В другом варианте осуществления антитела к токсину В нейтрализуют или подавляют in vivo токсичность токсина В. В варианте осуществления эффективная доза одного или нескольких антител к токсину А настоящего изобретения обеспечивается для субъекта, инфицированного С.difficile. В варианте осуществления эффективная доза одного или нескольких антител к токсину А настоящего изобретения обеспечивается в комбинации с эффективной дозой одного или нескольких антител к токсину В настоящего изобретения для субъекта, инфицированного С.difficile. В варианте осуществления антитело к токсину А настоящего изобретения в комбинации 1:1 с антителом к токсину В настоящего изобретения обеспечиваются как эффективная доза для субъекта, инфицированного С. difficile. В варианте осуществления эффективная доза антитела к токсину А и антитела к токсину В настоящего изобретения может быть, например, ½:1, 1:1, 2:1, 3:1, 4:1 и т.д., комбинацией антител, обеспеченных для субъекта, инфицированного С.difficile. В варианте осуществления антитела являются гуманизированными. В варианте осуществления эти антитела включены в композицию. Для иллюстрации, эффективная доза антител к токсину А и/или токсину В может находиться в диапазоне от 0,1 мг до 1000 миллиграмм (мг). Антитела к токсину А и антитела к токсину В или их антигенсвязывающие участки могут вводиться субъекту в количестве, например, 0,1 мг/кг-150 мг/кг; в количестве 0,5 мг/кг-75 мг/кг; в количестве 1 мг/кг-100 мг/кг; в количестве 1 мг/кг-50 мг/кг; в количестве 2 мг/кг-40 мг/кг; в количестве 2 мг/кг-50 мг/кг; в количестве 5 мг/кг -50 мг/кг; в количестве 5 мг/кг-25 мг/кг; в количестве 10 мг/кг-40 мг/кг; в количестве 10 мг/кг-50 мг/кг; в количестве 10 мг/кг-25 мг/кг; или в количестве 15 мг/кг-50 мг/кг. В варианте осуществления вышеупомянутые количества могут включать различные соотношения антитела к токсину А и антител к токсину В, обеспечиваемых в комбинации.
Как используется в настоящем документе, выражение «нейтрализовать» относится к снижению, подавлению, блокированию, снижению интенсивности или элиминации неблагоприятного эффекта(эффектов) токсина(токсинов), которые антитело(антитела) специфично связывают. Нейтрализация неблагоприятного эффекта(эффектов) токсина(токсинов) включает 1) задержку, снижение, подавление или предупреждение появления или прогрессирования инфекции С.difficile или диареи, или заболевания, ассоциированного с С.difficile, 2) повышение выживаемости субъекта по сравнению со средней выживаемостью субъектов, которых не лечили антителом(антителами) и которые имеют инфекцию С.difficile или заболевание, ассоциированное с С.difficile, 3) элиминацию одного или нескольких симптомов или неблагоприятных эффектов или снижение тяжести одного или нескольких симптомов или неблагоприятных эффектов, ассоциированных с инфекцией С.difficile или диареей, или заболеванием, ассоциированным с С.difficile, 4) предоставление возможности для репопуляции нормальной микрофлоры желудочно-кишечного тракта субъектов, которые являются или были инфицированными С.difficile, 5) предупреждение повторного проявления инфекции С.difficile или заболевания, ассоциированного с С.difficile у субъектов, которые страдали от инфекции С.difficile или заболевания, ассоциированного с С.difficile, 6) выполнение показателя эффективности лечения по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% субъектов, которые имеют инфекцию С.difficile или заболевание, ассоциированное с С.difficile и/или 7) предупреждение смерти из-за CDAD или других неблагоприятных событий, ассоциированных с инфекцией С.difficile.
Антитела к токсину А и токсину В С.difficile настоящего изобретения могут быть использованы для лечения большого количества видов субъектов, включая человека (людей) и других отличных от человека (млекопитающих) животных. Субъекты, поддающиеся лечению в соответствии с настоящим изобретением, включают людей, приматов, кроме человека, собак, кошек, мышей, крыс, хомяков, морских свинок, коров, коз, овец, свиней, лошадей и других. Человеческие субъекты в настоящем документе могут также называться пациентами или индивидами. В частности, субъекты, включают пациентов-людей, имеющих инфекцию С.difficile или заболевание, ассоциированное с С.difficile. Такие люди-пациенты включают тех, которые являются пожилыми или с иммунодефицитом.
В соответствии с настоящим изобретением антитела к токсину А и токсину В С. difficile могут устранить заболевание С. difficile и повысить выживаемость субъекта. В одном варианте осуществления одно или несколько антител к токсину А и/или одно или несколько антител к токсину В при введении субъекту улучшает(улучшают) выживаемость субъекта по сравнению со средней выживаемостью субъектов, которых не лечили антителом(антителами) и которые имеют инфекцию С. difficile или заболевание, ассоциированное с С. difficile.
В некоторых вариантах осуществления доза или количество одного или нескольких антител к токсину А и к токсину В может находиться, например, в диапазоне от 0,2 мкг-250 мкг, или от 2 мкг-50 мкг, или от 5 мкг - 50 мкг, например, на основе исследований на мышах in vivo. В некоторых вариантах осуществления доза или количество одного или нескольких ее антител к токсину А и к токсину В и, в частности, комбинация антитела к токсину А и антитела к токсину В может находиться, например, в диапазоне от 2 мг/кг-40 мг/кг, 2 мг/кг-50 мг/кг, 5 мг/кг-40 мг/кг, 5 мг/кг-50 мг/кг, 10 мг/кг-40 мг/кг или 10 мг/кг-50 мг/кг, например, на основе исследований на хомяках in vivo.
В качестве другого примера антитела настоящего изобретения могут выполнить показатель эффективности лечения или выживаемости на по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 99%. В качестве другого примера эти антитела могут выполнить показатель эффективности лечения или выживаемости на 100%. В одном варианте осуществления одно или несколько антител к токсину А при введении субъекту вместе с одним или несколькими антителами к токсину В выполняют показатель эффективности лечения или выживаемости на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 99% или 100%. Как используется в настоящем документе,, «показатель эффективности лечения» относится к проценту субъектов, которые, по определению клинициста более не имеют инфекцию или заболевание, из популяции субъектов с инфекцией или заболеванием, которым вводили одно или несколько антител настоящего изобретения или одну или несколько их композиций.. «Показатель выживаемости», как используется в настоящем документе, относится к проценту субъектов, которые выживают за необходимый период времени, из популяции субъектов, которым вводили одно или несколько антител настоящего изобретения или одну или несколько их композиций. Примеры таких необходимых периодов времени приведены в настоящем документе в других местах.
Антитела к токсину А и токсину В, обеспечиваемые настоящим изобретением, могут дать возможность для восстановления нормальной кишечной флоры у субъекта, инфицированного С.difficile. Таким образом, такие антитела могут устранить заболевание у пациентов, которые подвергаются лечению. Антитела к токсину А и токсина В настоящего изобретения могут также продемонстрировать благоприятную фармакокинетику in vivo. Антитела к токсину А и токсину В настоящего изобретения могут также обеспечить пролонгированную или долго длящуюся терапию для субъекта, который был инфицирован С.difficile. Как используется в настоящем документе, выражение, «долго длящаяся» относится к терапии, которая приводит к отсутствию инфекции С.difficile или заболевания, ассоциированного с С.difficile, через один месяц или несколько после отмены лечения. Предпочтительно лечение приводит к отсутствию инфекции С.difficile или заболевания, ассоциированного с С.difficile, в течение двух или более месяцев. В некоторых вариантах осуществления терапия с использованием mAb настоящего изобретения приводит к лечению или снижению действия активной инфекции С.difficile и к снижению или уменьшению устойчивости инфекции. В других вариантах осуществления терапия, обеспечиваемая настоящим изобретением, приводит к отсутствию инфекции С. difficile или заболевания, ассоциированного с С. difficile, у субъектов в течение 1, 2, 3, 4, 5 или 6 месяцев. В других вариантах осуществления терапия, обеспечиваемая настоящим изобретением, приводит к отсутствию инфекции С.difficile или заболевания, ассоциированного с С.difficile у субъекта в течение больше, чем 6 месяцев. Антитела к токсину А и токсину В настоящего изобретения могут предотвратить повторное проявление инфекции С. difficile и/или заболевания, ассоциированного с С. difficile.
Природа повторного проявления CDAD усугубляется появлением гипервирулентных штаммов BI/NAP 1/027, которые, как было обнаружено, резистентны к двум из новейших антибиотиков последней надежды, левофлоксацину и моксифлоксацину. Такие штаммы вызывали вспышки с повышающейся частотой по всем США, Канаде и Западной Европе. Гипервирулентные штаммы включают группу близкородственных изолятов, характеризующихся как North American Pulsed Field Type 1 (NAP1), тип «BI», на основе рестрикционного картирования, и ПЦР-риботип 027, общеизвестных как BI/NAP1/027 (5). Гипервирулентность штаммов BI/NAP1/027 объясняли по меньшей мере частично возрастанием продукции токсинов А и В, двух вирулентных факторов CDAD (6). Изоляты BI/NAP1/027 продуцируют в 16-23 раза более высокие уровни токсинов А и В, чем другие штаммы (6). Очевидная приспособленность этих штаммов создает угрозу распространения по всему миру, подвергая риску возможность лечения антибиотиками других заболеваний, а также увеличения частоты повторных проявлений и тяжести CDAD. Хотя существуют антибиотики в стадии разработки для лечения CDAD, такие как нитазоксанид, рифаксимин, рамопланин, фидаксомицин, сообщалось о клинических изолятах С.difficile, которые устойчивы к рифаксимину. В недавно завершенной фазе 3 испытаний (91), фидаксомицин значительно снижал общую частоту повторного проявления CDAD по сравнению с ванкомицином, но не в отношении штаммов BI/NAP1/027. Вспышки гипервирулентных штаммов BI/NAP1/027 привели к увеличению времени госпитализации, неэффективности лечения, частот рецидивов и показателей смертности (3). Новые mAb, разработанные и описанные в настоящем документе, обеспечивают новые методы лечения для борьбы с возрастающей частотой возникновения и тяжестью CDAD.
В варианте осуществления mAb настоящего изобретения используются в лечении инфекции, вызванной различными штаммами С.difficile. В варианте осуществления штаммы С.difficile являются сильно инфекционными, а их токсины нейтрализуются mAb настоящего изобретения. В варианте осуществления токсины гипервирулентных штаммов С. difficile, включая BI/NAP1/027, нейтрализуются mAb настоящего изобретения. В варианте осуществления mAb настоящего изобретения обеспечивают терапевтический эффект при нейтрализации токсинов из широкого диапазона токсикогенных клинических изолятов, включая штаммы от амбулаторных изолятов. Предпочтительно mAb нейтрализуют токсины гипервирулентных изолятов, таких как BI/NAP 1/027, и по меньшей мере 90% или более других клинически значимых культур С.difficile. В частности, и как проиллюстрировано в примере 8 в настоящем документе, mAb настоящего изобретения, как было показано, могут нейтрализовать в значительной степени токсичность/активность девятнадцати различных клинических изолятов С.difficile, включая BI/NAP 1/027 и другие гипервирулентные штаммы С.difficile, например, CCL676, НМС553, Pitt45, CD196, montreal 5 и montreal 7.1. В соответствии с настоящим изобретением, обеспечиваются антитела, которые нейтрализуют токсин А и токсин В гипервирулентных штаммов С.difficile, например, без ограничения, как определено значением ЕС50, находящемся в диапазоне от 7,7-12М до 4,8-8M для mAb к токсину А и значением ЕС50, находящемся в диапазоне от 1,1-11M до 6,5-10М для mAb к токсину В. Кроме того, mAb настоящего изобретения обеспечиваются для использования при нейтрализации гипервирулентных штаммов С.difficile, включая изоляты госпитального и негоспитального происхождения, в качестве лечения от инфекции С.difficile и заболеваний, связанных с ними.
В других вариантах осуществления и в неограничивающем примере mAb настоящего изобретения проявляют значение нейтрализации EC50 в пределах 93 пМ-30 нМ или ЕС5о 46 пМ для нейтрализации токсина А и значение EC50 в пределах от 4 пМ -9,5 пМ, или ЕС50 5 пМ для нейтрализации токсина В в зависимости от использованного клеточного анализа in vitro. Как описано в настоящем документе в примере 3, в анализе, включающем клетки СНО-K1, в которых было использовано 8 мкг/мл токсина А, mAb к токсину А настоящего изобретения проявили значение ЕС50 93 пМ. В анализе, включающем клетки СНО-K1, в котором было использовано 8 пг/мл токсина В, mAb к токсину В настоящего изобретения проявило значение EC50 9,2 пМ. В анализе, включающем клетки Т-84, в котором было использовано 240 нг/мл токсина А, mAb к токсину А настоящего изобретения проявили значение EC50 146 пМ и 175 пМ. В анализе, основанном на клетках Сасо-2, mAb к токсину А настоящего изобретения нейтрализовали токсичность токсина А при уровнях ЕС50 196 пМ и 485 пМ. В красных кровяных клетках анализ геммаглютинации, в котором было использовано 8 мкг/мл токсина A, mAb к токсину А настоящего изобретения имели значение нейтрализации ЕС50 1,8 нМ и 30 нМ для предупреждения геммаглютинации RBC.
Антитела настоящего изобретения могут обладать любыми из, комбинациями, или всеми вышеупомянутые особенностями.
Как описано в примерах в настоящем документе, mAb настоящего изобретения демонстрируют превосходную эффективность нейтрализации токсинов как in vitro, так и in vivo в наиболее доступных доклинических моделях CDAD. Кроме того, mAb изобретения демонстрируют уникально обширную и эффективную нейтрализацию токсинов из многочисленных штаммов BI/NAP1/027. Более того, такие mAb продемонстрировали полную и длительную защиту от смертности в высокоточных моделях CDAD на хомяках. Эти результаты подтверждают способность mAb настоящего изобретения достаточно и эффективно блокировать патогенные эффекты токсинов С.difficile таким образом, что дают возможность заживления толстой кишки, восстановиться нормальной кишечной микрофлоре и устранить заболевание CDAD и/или инфекцию С. difficile.
В варианте осуществления антитела к токсину А включают те, которые конкурентно подавляют или перекрестие конкурируют за специфичное связывание с токсином А С. difficile выделенного моноклонального антитела, продуцированного гибридомной клеточной линией, депонированный под №№ доступа АТСС РТА-9692, РТА-9694 или РТА-9888. Предпочтительные антитела к токсину В включают те, которые конкурентно подавляют или перекрестно конкурируют за специфичное связывание с токсином В С. difficile выделенного моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692. В некоторых вариантах осуществления антитела включают таковые, которые конкурентно подавляют или перекрестно конкурируют за специфичное связывание с токсином А С. difficile выделенного моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888, и конкурентно подавляют или перекрестно конкурируют за специфичное связывание с токсином В С. difficile выделенного моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692. Все варианты осуществления дополнительно включают гуманизированные формы вышеописанных антител под №№ доступа АТСС РТА-9692, РТА-9694, РТА-9888 или РТА-9693.
Для определения конкурентного подавления или перекрестного конкурирования связывания могут быть использованы различные анализы, известные специалистам в данной области. Например, анализы перекрестного конкурирования могут быть использованы для определения, подавляет ли антитело конкурентно связывание с токсином А и/или токсином В других антител. Такие методы могут быть клеточными методами с использованием проточной цитометрии или твердофазного анализа связывания. Другие анализы, которые оценивают способность антител к перекрестному конкурированию за связывание токсина А и/или токсина В в твердой фазе или в жидкой фазе, также могут быть использованы. Примеры антител или их антигенсвязывающих фрагментов, которые охватываются настоящим изобретением, включают те, которые конкурентно подавляют специфичное связывание по меньшей мере на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 99%. Подавление можно оценить при различных молярных соотношениях или массовых соотношениях, например, эксперименты с конкурентным связыванием можно проводить с 2-кратным, 3-кратным, 4-кратным, 5-кратным, 7-кратным, 10-кратным или большим молярным избытком первого антитела по сравнению со вторым антителом.
Другие антитела, которые охватываются настоящим изобретением, включают те, которые специфично связываются с эпитопом на токсине А С.difficile, определяемом связыванием выделенного моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694 или РТА-9888. Тем не менее, другие антитела или антигенсвязывающие фрагменты, которые охватываются настоящим изобретением, включают те, которые специфично связываются с эпитопом на токсине В С.difficile, определяемом связыванием выделенного моноклонального антитела продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692. Тем не менее, другие антитела или антигенсвязывающие фрагменты, которые охватываются настоящим изобретением, включают те, которые специфично связываются с эпитопом на токсине А С.difficile, определяемом связыванием выделенного моноклонального антитела продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692, РТА-9694, или РТА-9888 и специфично связанного с эпитопом на токсине В С.difficile, определяемом связыванием выделенного моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 или РТА-9692.
Для определения эпитопа, можно использовать стандартные методы картирования эпитопов, известные в данном уровне техники. Например, фрагменты (пептиды) токсина (предпочтительно синтетические пептиды), которые связывают антитело, могут быть использованы, чтобы определить, связывает ли представляющее интерес антитело или его антигенсвязывающий фрагмент тот же эпитоп. Для линейных эпитопов синтезируются перекрывающиеся пептиды определенной длины (например, 8 или более аминокислот). Пептиды предпочтительно смещаются на 1 аминокислоту, так что получают серии пептидов, охватывающих каждые 8 аминокислотных фрагментов из последовательности токсина. Меньшее количество пептидов можно получить с использованием больших смещений, например, на 2 или 3 аминокислоты. Кроме того, могут быть синтезированы более длинные пептиды (например, 9-, 10- или 11-мерные). Связывание пептидов с антителами можно определить с использованием стандартных методик, включая поверхностный плазменный резонанс (например, Biacore) и ELISA анализы. Для исследования конформационных эпитопов, которые антитела, приводимые в настоящем документе, могут в некоторых вариантах осуществления связывать, могут использоваться большие фрагменты пептидов. Другие методы, в которых используется масс-спектрометрия для определения конформационных эпитопов, были описаны и могут быть использованы (см., например, Baerga-Ortiz et al., Protein Science 11:1300-1308, 2002 и ссылки, процитированные в настоящем документе). Тем не менее, другие методы для определения эпитопов приводятся в справочных работах лабораторий стандартов, таких как в разделе 6.8 ("Phage Display Selection and Analysis of B-cell Epitopes") и в разделе 9.8 ("Identification of Antigenic Determinants Using Synthetic Peptide Combinatorial Libraries") Current Protocols in Immunology, Coligan et al., eds., John Wiley & Sons. Эпитопы можно подтвердить путем введения одной или нескольких точечных мутаций или делеций в известный эпитоп, а затем тестирования связывания с одним или несколькими антителами, чтобы определить какие мутации снижают связывание антител.
Антитела или антигенсвязывающие фрагменты, обеспечиваемые в настоящем изобретении, могут специфично связывать токсин А и/или токсин В с субнаномолярной аффинностью. Антитела или антигенсвязывающие фрагменты могут иметь связывающую аффинность приблизительно 1×10-9 М или менее, приблизительно 1×10-10M или менее, или приблизительно 1×10-11 М или менее. В отдельном варианте осуществления связывающая аффинность составляет меньше, чем приблизительно 5×10-10М.
Антитела или антигенсвязывающие участки могут иметь константу скорости ассоциации (Kon) для токсина А или токсина В по меньшей мере 102M-1c-1; по меньшей мере 103M-1c-1; по меньшей мере 104M-1c-1; по меньшей мере 105M-1с-1; по меньшей мере 106M-1c-1; или по меньшей мере 107M-1c-1, измеренную с помощью поверхностного плазменного резонанса. Антитела или антигенсвязывающие фрагменты могут иметь константу скорости диссоциации (Koff) для токсина А или токсина В не больше, чем 10-3c-1; не больше, чем 10-4с-1; не больше, чем 10-5c-1; или не больше, чем 106с-1, измеренную с помощью поверхностного плазменного резонанса. Антитела или антигенсвязывающие фрагменты могут иметь константу диссоциации (KD) для токсина 7 А или токсина В не больше, чем 10-7М; не больше, чем 10-8М; не больше, чем 10-9М; не больше, чем 10-10М; не больше, чем 10-11М; не больше, чем 10-12М; или не больше, чем 10-13М.
Как используется в настоящем документе, выражения «антитело» или «иммуноглобулин» включают гликопротеины, содержащие по меньшей мере две тяжелых (Н) полипептидных цепи и две легких (L) полипептидных цепи, соединенных дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенная в настоящем документе как HCVR или VH) и константную область тяжелой цепи (CH). Константная область тяжелой цепи состоит из трех доменов СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенная в настоящем документе как LCVR или VL) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть далее подразделяться на области гипервариабельности, называемые гипервариабельными областями (CDR), чередующиеся с областями, которые являются более консервативными, называемые каркасными областями (FR). Каждая VH, и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В общей сложности вариабельные области полипептидов тяжелых и легких цепей содержат или формируют связывающий домен, который взаимодействует с/связывает антиген. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Кроме того, настоящее изобретение обеспечивает другие формы антител, такие как одноцепочечные антитела, рекомбинантно полученные антитела, биспецифичные, гетероспецифичные или мультимерные антитела, диатела и т.д., как описано далее в настоящем документе.
Выражение «антигенсвязывающий фрагмент» антитела, как используется в настоящем документе, относится к одной или нескольким частям антитела, которое сохраняет способность специфично связываться с антигеном (например, токсин А, токсин В, токсин А и токсин В и т.д.) или эпитопными областями антигена. Было показано, что антигенсвязывающая функция антитела может осуществляться фрагментами антитела полной длины. В варианте осуществления фрагменты моноклональных антител функционируют способом, сходным с интактными аналогичными моноклональными антителами. В варианте осуществления фрагменты моноклональных антител перекрестие реагируют с интактными аналогичными моноклональными антителами. В варианте осуществления фрагменты моноклональных антител могут быть использованы взаимозаменяемо с интактными аналогичными моноклональными антителами. Примеры связывающих фрагментов, охваченные выражением «антигенсвязывающий фрагмент» антитела, включают (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL, и CH1; (ii) фрагмент F(ab’)2, бивалентный фрагмент, включающий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и СН1; (iv) фрагмент Fv, состоящий из доменов VL и VH отдельного плеча антитела, (v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (vi) выделенную гипервариабельную область (CDR). Более того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут быть объединены с использованием рекомбинантных методов с помощью синтетического линкера, который позволяет им образовать одну белковую цепь, в которой области VL и VH располагаются парами для формирования моновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; and Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные антитела охватываются выражением "антигенсвязывающий фрагмент" антитела. Эти фрагменты антител получают с использованием обычных методик, таких как методики протеолитической фрагментации, как описано в J. Coding, Monoclonal Antibodies: Principles and Practice, pp 98-118 (N.Y. Academic Press 1983), которая включена в настоящем документе посредством ссылки, а также при помощи других техник, известных специалистам в данной области. Фрагменты подвергают скринингу на активность или практичность таким же образом, как и интактные антитела.
В варианте осуществления были созданы фрагменты Fab mAb настоящего изобретения и протестированы на активность нейтрализации в клеточных анализах, как описано в примере 10 в настоящем документе. Таким образом, фрагменты антител, такие как фрагменты Fab mAb настоящего изобретения могут также быть использованы для связывания и нейтрализации токсина А и/или токсина В С.difficile.
«Выделенное антитело», как используется в настоящем документе, предназначено для обозначения антитела, которое по существу не содержит других антител, имеющих различные антигенные специфичности (например, выделенное антитело, которое специфично связывается с токсином А по существу не содержит антитела, которые специфично связывают антигены, отличные от токсина А). Выделенное антитело, которое специфично связывается с эпитопом, изоформой или вариантом токсина А или токсина В, однако в общем имеет перекрестную реактивность к другим родственным антигенам, например, из других штаммов С.difficile. Кроме того, выделенное антитело, которое специфично связывается с эпитопом, изоформой или вариантом токсина А также может специфично связывать токсин В, а выделенное антитело, которое специфично связывается с эпитопом, изоформой или вариантом токсина В также может специфично связывать токсин А. В некоторых вариантах осуществления, однако, выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывается с эпитопом, изоформой или вариантом токсина А, также не связывает специфично токсин В. В других еще вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент, который специфично связывается с эпитопом, изоформой или вариантом токсина В также не связывает специфично токсин А. Более того, выделенное антитело может по существу не содержать другой клеточный материал и/или химические вещества. Антитела, которые по существу не содержат другие антитела, имеющие различные антигенные специфичности, или другие материалы и/или химические вещества, и/или белки, могут быть выделенными и/или очищенными антителами. Антитела могут быть очищены способами, которые обычно осуществляются специалистами в данной области, например, аффинная хроматография, хроматография на белке А и тому подобное. Как используется в настоящем документе, «специфичное связывание» относится к связыванию антител с предопределенным или родственным антигеном. Типично антитело связывается с аффинностью, которая по меньшей мере в два раза больше, чем его аффинность для связывания с неспецифичном антигеном (например, BSA, казеин), отличным от предопределенного антигена или близкородственного антигена. В варианте осуществления антитело настоящего изобретения может связывать линейный эпитоп целевого антигена, например, токсина А и/или токсина В. В варианте осуществления антитело настоящего изобретения может связывать конформационный эпитоп целевого антигена, например, токсина А и/или токсина В.
Выделенные антитела настоящего изобретения охватывают различные изотипы тяжелых и легких цепей антител (иммуноглобулинов), такие как классы тяжелых цепей или изотипов IgGI, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgAsec, IgD, IgE и их подтипов, например, IgG2a, IgG2b; и изотипы легких цепей к и Х и их подтипы. В одном варианте осуществления выделенные антитела состоят из изотипа IgG2a или IgG1k. Как используется в настоящем документе, «изотип» относится к классу антител (например, IgM или IgGI, или λ1), которые кодируются генами константных областей тяжелых и легких цепей. Антитела или их антигенсвязывающие области могут быть полной длины или могут включать только антигенсвязывающий фрагмент, такой как константный и/или вариабельный домен антител IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgAsec, IgD или IgE, или могут состоять из фрагмента Fab, фрагмента F(ab’)2 и фрагмента Fv.
Антитела настоящего изобретения могут быть поликлональными, моноклональными или смесью поликлональных и моноклональных антител. Антитела могут быть получены различными методами, хорошо известными в данном уровне техники. Методики для индукции поликлональных антител хорошо известны. В качестве неограничивающего примера поликлональные антитела индуцируют путем введения белка токсина А и/или токсина В подкожно новозеландским белым кроликам, у которых сначала берут кровь для получения преиммунной сыворотки. Токсин А и/или токсин В можно инъецировать в общем объеме 100 мкл на участок в шесть различных участков типично с одним или несколькими адъювантами. Затем у кроликов берут кровь через две недели после первой инъекции и периодически бустер-иммунизируют тем же антигеном три раза каждые шесть недель. Образец сыворотки собирается через 10 дней после каждой бустер-иммунизации. Поликлональные антитела получают из сыворотки, предпочтительно путем аффинной хроматографии с использованием токсина А и/или токсина В для захвата антитела. Эта и другие методики индукции поликлональных антител описаны в Harlow, E. and Lane, D., Eds., Antibodies: A Laboratory Manual (1988), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, содержания которых включены в настоящем документе посредством ссылки.
Производство моноклональных антител может осуществляться с помощью методов, которые также хорошо известны в данном уровне техники. Выражение «моноклональное антитело», как используется в настоящем документе, относится к получению молекул антител одного молекулярного состава. Моноклональное антитело проявляет единственную связывающую специфичность и аффинность к отдельному эпитопу данного антигена или иммуногена. Процесс производства моноклональных антител включает получение иммунных соматических клеток с потенциалом для продукции антитела, в частности лимфоцитов В, которые были ранее иммунизированы представляющим интерес антигеном либо in vivo, либо in vitro, или и то, и другое, которые являются подходящими для слияния с В-клетками миеломной линии. Моноклональные антитела могут быть получены с использованием иммунных клеток и миеломных клеток от различных видов, таких как мышиные и человеческие клетки и клеточные линии или, например, в мышиных штаммах, которые были генетически модифицированы, чтобы поддерживать иммунную систему человека, как описано ниже.
Хотя моноклональные антитела, направленные против токсина А и токсина В, типично получали путем иммунизации животных анатоксинами (неактивные формы токсина А и токсина В) и/или неактивными фрагментами этих токсинов, mAb настоящего изобретения были созданы путем разработки и использования новой стратегии иммунизации. В соответствии с настоящим изобретением mAb, описанные в настоящем документе и депонированные, были получены путем иммунизации животных анатоксинами с последующей бустер-иммунизацией животных активной (неанатоксиновой) формой токсина А и/или токсина В (см. пример 1 в настоящем документе). Бустер-иммунизация активной формой токсина А и токсина В способствовала идентификации тех иммунизированных животных, которые вырабатывали уникальные защитные антитела в силу новой схемы иммунизации. Не желая быть связанными теорией, режим бустер-иммунизации с активными токсином А и/или токсином В был более высокоиммуногенным у реципиентных животных. Те животные, которые переносили возрастающие дозы бустер-иммунизации активного токсина А и токсина В, которые типично детальны для животных, не использовавшихся ранее в опытах, продуцировали высокоэффективные нейтрализующие антитела, которые защищали этих животных и вносили вклад в их выживаемость, несмотря на то, что они получали активный токсин. Получение гибридом от животных, которые проявили эффективный иммунный ответ против токсина А или токсина В, дало высокоэффективные моноклональные антитела к токсину А и токсину В, которые обеспечивают высокий уровень защиты как in vitro, так и in vivo. При получении антител, включая поликлональные и моноклональные антитела, могут использоваться адъюванты. Неограничивающие примеры адъювантов, которые подходят для использования, включают неполный адъювант Фрейнда фосфат алюминия, гидроксид алюминия, Ribi (например, монофосфорил-липид А, димиколат трегалозы, скелет клеточной стенки микобактерии и Tween® 80, с 2% скваленом), сапонины, Quil А или алюм. Ответ цитотоксических Т-лимфоцитов (CTL) может быть примирован конъюгацией токсинов (или фрагментов, неактивных производных или их аналогов) с липидами, таким как, например, трипальмитоил-8-глицерилцистеинилсерилсерин.
В других вариантах осуществления дополнительные методы иммунизации могут быть использованы для создания моноклональных антител, направленных против токсина А и/или токсина В. Например, иммунизация животных in vivo (например, мышей) может проводиться с необходимым типом и количеством белка или полипептида, например, анатоксина или токсина. Такие иммунизации при необходимости повторяются с интервалами в несколько недель, чтобы получить достаточный титр антител. После иммунизации животные могут быть использованы в качестве источника антителопродуцирующих лимфоцитов. После последней бустер-иммунизации антигенами, животных умерщвляют и клетки селезенки удаляют. Мышиные лимфоциты дают более высокий процент стабильных слияний с описанными в настоящем документе мышиными миеломными линиями. Среди них подходящей является линия мышей BALB/c. Однако другие линии мышей, кролики, хомяки, овцы, козы и лягушки могут быть также использованы в качестве реципиентов для получения антителопродуцирующих клеток. См.; Coding (в Monoclonal Antibodies: Principles and Practice, 2d ed., pp.60-61, Orlando, Fla., Academic Press, 1986). В частности, могут быть использованы линии мышей, которые имеют гены иммуноглобулинов человека, встроенные в геном (и которые не могут продуцировать мышиные иммуноглобулины). Примеры включают в линии мышей HumAb, полученных Medarex (сейчас Bristol Myers Squibb)/GenPharm International, и линии XenoMouse, полученные Abgenix. Такие мыши продуцируют в полной мере молекулы иммуноглобулинов человека в ответ на иммунизацию.
Те антителопродуцирующие клетки, которые находятся в стадии делящегося плазмобласта, сливаются предпочтительно. Соматические клетки могут быть получены, например, из лимфатических узлов, селезенок и периферической крови антиген-примированных животных, а предпочтительные лимфатические клетки зависят в большой степени от их эмпирической применимости в определенной системе слияния. Лимфоциты, секретирующие антитела, затем сливают с (мышиными) миеломными В-клетками или трансформированными клетками, которые способны к репликации на неопределенный срок в культуре клеток, тем самым создавая иммортализованную секретирующую иммуноглобулины клеточную линию. Полученные в результате слитые клетки или гибридомы культивируют, а полученные в результате колонии подвергают скринингу на продукцию необходимых моноклональных антител. Колонии, продуцирующие такие антитела, клонируются, субклонируются и выращиваются как in vivo (как асциты), так и in vitro для продукции больших количеств антитела. Описания методологии получения гибридом и технологии можно найти в Kohler and Milstein, Nature 256:495 (1975) или Harlow, E. and Lane, D., Eds., Antibodies: A Laboratory Manual (1988), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY„ которые включены в настоящем документе посредством ссылки.
В качестве альтернативы соматические клетки человека, способные продуцировать антитело, в особенности В-лимфоциты, подходят для слияния с миеломными клеточными линиями. В то время как В-лимфоциты из подвергнутых биопсии селезенок, миндалин или лимфатических узлов индивидуума могут быть использованы, более легко доступные В-лимфоциты (PBL) периферической крови являются предпочтительными. Кроме того, В-клетки человека могут быть непосредственно иммортализованы вирусом Эпштейна-Барра (Cole et al., 1995, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77-96). Хотя методики гибридизации соматических клеток являются предпочтительными, в принципе, могут быть использованы и другие методы получения моноклональных антител, такие как вирусная или онкогенная трансформация В-лимфоцитов.
Миеломные клеточные линии, подходящие для использования в методиках слияния с получением гибридом, предпочтительно не являются антителопродуцирующими, имеют высокую эффективность слияния и недостатки ферментов, которые делают их неспособными к росту в определенных селективных средах, которые поддерживают рост необходимых гибридом. Примеры таких миеломных клеточных линий, которые могут быть использованы для производства слитых клеточных линий, включают Р3-X63/Ag8, X63-Ag8.653, NSl/l.Ag 4.1, Sp2/0-Agl4, FO, NSO/U, MPC-11, МРСП-Х45-GTG 1.7, S194/5XX0 Bul, полученные от мышей; R210.RCY3, Y3-Ag 1.2.3, IR983F и 4 В210 полученные от крыс; и U-266, GM1500-GRG2, LICR-LON-HMy2, UC729-6, полученные от людей (Goding, в Monoclonal Antibodies: Principles and Practice, 2d ed., pp.65-66, Orlando, Fla., Academic Press, 1986; Campbell, в Monoclonal Antibody Technology, Laboratory Techniques in Biochemistry and Molecular Biology Vol.13, Burden and Von Knippenberg, eds. pp.75-83, Amsterdam, Elseview, 1984).
Слияние с миеломными клетками млекопитающих или других партнеров слияния, способных реплицироваться неограниченно в культуре клеток, осуществляется путем стандартных и хорошо известных методов, например, с использованием полиэтиленгликоля («PEG») или других агентов слияния (см. Milstein and Kohler, Eur. J. Immunol. 6:511 (1976), которая включена посредством ссылки).
В других вариантах осуществления антитела могут быть рекомбинантными антителами. Выражение «рекомбинантные антитела», как используется в настоящем документе, предназначено для включения антител, которые получают, экспрессируют, создают или выделяют рекомбинантными способами, таких как антитела, выделенные из животных (например, мышей), которые являются трансгенными по генам иммуноглобулинов других видов, антитела, экспрессированные с использованием рекомбинантного вектора экспрессии, трансфицированного в клетку-хозяина, антитела, выделенные из рекомбинанта, комбинаторной библиотеки антител или антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, которые включают сплайсинг последовательностей генов иммуноглобулинов с другими последовательностями ДНК.
В других еще вариантах осуществления антитела могут быть химерными или гуманизированными антителами. Как используется в настоящем документе выражение, выражение «химерное антитело» относится к антителу, в котором объединяются вариабельные или гипервариабельные области иммуноглобулина (Ig) мыши с константной областью Ig человека, или константными и вариабельным каркасными областями. В некоторых вариантах осуществления химерное антитело включает вариабельную область любого из депонированных антител, приведенных в настоящем документе, и константную область человека. В некоторых вариантах осуществления константная область человека является константной областью IgG человека, такой как константная область IgGI человека. Химерные антитела могут быть получены способом, который приводится ниже в примерах, или любым способом, известным специалистам в данном уровне техники. Как используется в настоящем документе, выражение «гуманизированное антитело» относится к антителу, которое сохраняет по существу только антигенсвязывающие CDR от исходного антитела, например, мышиное моноклональное антитело в ассоциации с каркасными областями человека (см., например, Waldmaim, 1991, Science 252:1657). Такие химерные или гуманизированные антитела, которые сохраняют специфичность связывания мышиного антитела, но имеют константные/каркасные области Ig человека, как ожидается, имеют сниженную иммуногенность при введении in vivo. Таким образом, химерные и гуманизированные антитела предпочтительно сохраняют нейтрализующие токсин активности обеспеченных моноклональных антител и подходят для повторного дозирования (например, у людей). Специалист в данной области может использовать известные способы (например, клеточные анализы in vitro) для сравнения активности гуманизированных антител с депонированными моноклональными антителами, приведенными в настоящем документе, и для определения того, лечат ли гуманизированные антитела и/или предотвращают рецидив установленной инфекции С.difficile. Специалист в данной области может также использовать способы, описанные в настоящем документе, включая модель инфекции С.difficile на хомяках, описанную ниже.
Последовательности гуманизированных mAb могут быть сконструированы с помощью следующего иллюстративного, неограничивающего способа. Во-первых, идентифицируются каркасные аминокислотные остатки, важные для структуры CDR. Параллельно последовательности VH и VL человека, имеющие высокую гомологию по отношению к VH и VL мыши соответственно, выбираются из последовательностей известных иммуноглобулинов (зародышевая линия) человека. Последовательности CDR из mAb мыши вместе с каркасными аминокислотными остатками, важными для поддержания структуры CDR, пересаживают в выбранные каркасные последовательности человека. Кроме того, каркасные аминокислотные остатки человека, которые, как было обнаружено, являются атипичными в соответствующей подгруппе V области, заменяются на типичные остатки для снижения потенциальной иммуногенности полученного в результате гуманизированного mAb. Эти гуманизированные области VH и VL клонируют в векторы экспрессии, например, pCON Gammal и pCON kappa (Lonza Biologies, Berkshire, UK) соответственно. Эти векторы кодируют константную область (области) генов тяжелых и легких цепей иммуноглобулина человека. Клетки 293Т можно временно трансфицировать этими векторами экспрессии с использованием системы Effectene (Qiagen, Valencia, CA). Клеточные супернатанты, содержащие секретированное химерное mAb, можно собрать после трансфекции, например, через три дня, и очистить с использованием хроматографии на белке А. Другие векторы экспрессии и клетки-хозяина можно использовать для рекомбинантного получения описанных антител, что будет понятно специалистам в данной области.
Другие способы гуманизации антител или антигенсвязывающих фрагментов хорошо известны в данном уровне техники и включают способы, которые приводятся, например, в патентах США №№5585089; 5693761; 5693762; и 6180370. Методы осуществления гуманизации, которые приводятся в этих патентах, включены в настоящем документе посредством ссылки в полном их объеме. Антитела или антигенсвязывающие фрагменты, гуманизированные в соответствии со способами, которые приводятся в этих патентах, также приводятся в настоящем документе.
В варианте осуществления гуманизированное mAb (hmAb) к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок или его фрагмент, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:1, и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:3, и CL-область, человека, например, С-область κ-цепи. В варианте осуществления гуманизированное mAb к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок или его фрагмент, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:2 и СН-область человека, например, С-область IgG1, и (ii), двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:3 и CL-область человека, например, С-область κ-цепи. В варианте осуществления гуманизированное mAb к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, состоящий из (i), двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:1 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:4 и CL-область человека, например, С-область κ-цепи. В варианте осуществления гуманизированное mAb к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:2 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:4, и CL-область человека, например, С-область κ-цепи. Такие гуманизированные mAb к токсину А С.difficile охватывают mAb hPA-39 настоящего изобретения.
В варианте осуществления гуманизированное mAb к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:5 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:7 и CL-область человека, например, С-область κ-цепи. В варианте осуществления гуманизированное mAb к токсину А С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность как которая изложена в SEQ ID NO:6 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:7 и CL-область человека, например, С-область κ-цепи. Такие гуманизированные mAb к токсину А С.difficile охватывают mAb hPA-50 настоящего изобретения.
В варианте осуществления гуманизированные mAb к токсину В С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:8 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:10 и CL-область человека, например, С-область κ-цепи. В варианте осуществления гуманизированные mAb к токсину В С.difficile настоящего изобретения охватывает иммуноглобулиновый белок, который состоит из (i) двух полипептидов тяжелых (Н) цепей, где каждая Н-цепь содержит VH-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:9 и СН-область человека, например, С-область IgG1, и (ii) двух полипептидов легких (L) цепей, где каждая L-цепь содержит VL-область, включающую аминокислотную последовательность, которая изложена в SEQ ID NO:10 и CL-область человека, например, С-область κ-цепи. Такие гуманизированные mAb к токсину А С.difficile охватывают mAb bPA-41 настоящего изобретения.
С-области L-цепи и Н-цепи описанных выше гуманизированных антител настоящего изобретения могут включать С-область (CL) к L-цепи человека и С-область (CH) Н-цепи IgGI человека, имеющие последовательности, содержащиеся в Genbank под № доступа NW_001838785 и в Genbank под № доступа MW_001838121 соответственно. В других вариантах осуществления гуманизированные антитела включают С-область Н-цепи человека, выбранную из изотипов IgG2a, IgG2b, IgG3 или IgG4.
В иллюстративном варианте осуществления настоящее изобретение охватывает моноклональное антитело или его фрагмент, созданное к токсину А С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновых кислот (или кДНК), кодирующая последующую аминокислотную последовательность полипептида тяжелой цепи SEQ ID NO:14 изложена в SEQ ID NO:15 (Фиг.38В); последовательность нуклеиновых кислот (или cDNA), кодирующая последующую аминокислотную последовательность полипептида легкой цепи SEQ ID NO:16 изложена в SEQ ID NO:17 (Фиг.38А).
В иллюстративном варианте осуществления настоящее изобретение охватывает моноклональное антитело или его фрагмент, созданное к токсину А С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновых кислот (или кДНК), кодирующая последующую аминокислотную последовательность полипептида тяжелой цепи SEQ ID NO:18 изложена в SEQ ID NO:19 (Фиг.39В); последовательность нуклеиновых кислот (или кДНК), кодирующая последующую аминокислотную последовательность полипептида легкой цепи SEQ ID NO:20 изложена в SEQ ID NO:21 (Фиг.39А).
В иллюстративном варианте осуществления настоящее изобретение охватывает моноклональное антитело или его фрагмент, созданное к токсину В С.difficile, где антитело состоит из двух полипептидов тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и двух полипептидов легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновых кислот (или кДНК), кодирующая последующую аминокислотную последовательность полипептида тяжелой цепи SEQ ID NO:22 изложена в SEQ ID NO:23 (Фиг.40В); последовательность нуклеиновых кислот (или cDNA), кодирующая последующую аминокислотную последовательность полипептида легкой цепи SEQ ID NO:24 изложена в SEQ ID NO:25 (Фиг.40А).
Также настоящим изобретением охвачены части или фрагменты описанных выше гуманизированных антител к токсину А и токсину В С.difficile. Такие части или фрагменты включают гипервариабельные области (CDR) V-областей полипептидов как Н- цепей, так и L-цепей, что специалисты в данной области могут определить обычным способом; Р(ab)-фрагменты, F(ab’)-фрагменты, Р(ab’)2-фрагменты, Fc-фрагменты, Fd-фрагменты и т.п. В варианте осуществления части или фрагменты гуманизированных антител, содержащих V-области или их функциональные части, в оптимальном случае будут связываться с соответствующим токсином и нейтрализовать активность токсина. В варианте осуществления такие функциональные части или фрагменты гуманизированных антител в оптимальном случае нейтрализуют активность токсина на уровне, подобном уровню полного гуманизированного антитела, если не лучше.
В соответствии с настоящим изобретением предоставлены молекулярно клонированные, гуманизированные mAb, направленные против токсинов А или В С.difficile. Такие гуманизированные mAb были выделены и характеризованы как описано в Примере 9, Разделах D и Е ниже в настоящем документе. В варианте осуществления константная область (CL) легкой цепи каждого из гуманизированных антител относится к классу каппа (к). В варианте осуществления константная область (СН) тяжелой цепи каждого из гуманизированных антител относится к изотипу IgG1. В других вариантах осуществления СН-область гуманизированных антител относится к изотипу IgG2a, IgG2b, IgG3, IgG4, IgA, IgE, IgA или IgM. Было обнаружено, что гуманизированные mAb, содержащие уникальные вариабельные (V) области, связывают и нейтрализуют активность либо токсина А, либо токсина В С.difficile. VL-и VH-области гуманизированных mAb могут образовывать часть полной молекулы иммуноглобулина (Ig) или антитела, или их можно применять в качестве частей или фрагментов антитела, в частности, частей или фрагментов, обладающих связывающей и/или нейтрализующей активностью. Неограничивающие примеры фрагментов антител включают фрагменты Fab, F(ab)2 и F(ab') или F(ab')2. Варианты осуществления настоящего изобретения направлены на гуманизированные mAb к токсину А С.difficile, или их фрагменты, обладающие активностью против токсина А С.difficile, где V-область L-цепи выбрана из одной или нескольких из SEQ ID NO:3, SEQ ID NO:4 и SEQ ID NO:7. В варианте осуществления настоящее изобретение направлено на CDR, а именно CDR1, CDR2, и/или CDR3, в VH- и VL-областях описанных антител. Варианты осуществления настоящего изобретения направлены на гуманизированные mAb к токсину В С.difficile, или их фрагменты, обладающие активностью против токсина В С.difficile, где V-область L-цепи изложена в SEQ ID NO:10. Варианты осуществления настоящего изобретения направлены на гуманизированные mAb к токсину А С.difficile, или их фрагменты, обладающие активностью против токсина А С.difficile, где V-область Н-цепи выбрана из одной или нескольких из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:5 и SEQ ID NO:6. Варианты осуществления настоящего изобретения направлены на гуманизированные mAb к токсину В С.difficile, или их фрагменты, обладающие активностью против токсина В С.difficile, где V-область Н-цепи выбрана из одной или нескольких из SEQ ID NO:8 или SEQ ID NO:9. В варианте осуществления изобретения направлен на CDR, а именно CDR1, CDR2, и/или CDR3, в VH- и VL-областях описанных антител.
В другом варианте осуществления настоящее изобретение охватывает нуклеиновые кислоты, которые кодируют антигенсвязывающие части, CDR или вариабельные (V) области антител к токсину А и/или токсина В С. difficile настоящего изобретения. В различных вариантах осуществления части CDR или V-области получают из РА-38, РА-39, РА-41, или РА-50, или их гуманизированных вариантов, описанных в настоящем документе. В дополнительных вариантах осуществления настоящее изобретение охватывает аминокислотные последовательности антигенсвязывающих частей, CDR или V-областей, которые кодируются соответствующими нуклеиновыми кислотами.
В соответствии с другим вариантом осуществления моноклональные антитела настоящего изобретения можно модифицировать так, они были в форме биспецифичного антитела, бифункционального антитела, мультиспецифичного антитела или гетерофункционального антитела. Неограничивающие примеры биспецифичных и гетероспецифичных антител и процедур получения таких антител можно найти в ряде иллюстрирующих подобные примеры публикаций, например, UA20090060910, WO 2009/058383, WO 2009/030734, WO 2007/093630, USP 6071517, WO 2008/024188, UA 20070071675, USP 7442778, USP 7235641, USP 5932448 и USP 5292668. Термин «биспецифичное антитело» понимают как включающий любое средство, например, белок, пептид, или белковый или пептидный комплекс, которое имеет две различных специфичности связывания, и которое связывается с или взаимодействует с (а) токсином А С.difficile и (b) токсином В С.difficile. В одном варианте осуществления биспецифичное антитело включает РА-39 или РА-50, или их антигенсвязывающий фрагмент, и РА-41, или его антигенсвязывающий фрагмент. В варианте осуществления биспецифичное антитело включает химерную или гуманизированную форму РА-39 или РА-50, или их антигенсвязывающего фрагмента, и РА-41, или его антигенсвязывающего фрагмента. Соответственно, биспецифичное антитело, включающее РА-39 и РА-41, или их химерные или гуманизированные формы, или его антигенсвязывающий фрагмент, будет связывать как токсин А, так и токсин В С.difficile. Точно так же, биспецифичное антитело, включающее РА-50 и РА-41, или их химерные или гуманизированные формы, или его антигенсвязывающий фрагмент, будет связывать как токсин А, так и токсин В С.difficile. Термин «мультиспецифичное антитело» понимают как включающий любое средство, например, белок, пептид, или белковый или пептидный комплекс, которое имеет более двух различных специфичностей связывания и которое связывается с или взаимодействует с (а) токсином А С.difficile, (b) токсином В С.difficile и (с) по меньшей мере одним другим компонентом. Соответственно, настоящее изобретение включает, но без ограничений, биспецифичные, триспецифичные, тетраспецифичные и другие мультиспецифичные антитела. В одном варианте осуществления антитела или антигенсвязывающие фрагменты биспецифичных или мультиспецифичных антител являются гуманизированными.
Термин «биспецифичные антитела» дополнительно включает в себя диатела. Диатела обеспечивают терапевтические антитела, имеющие двойную специфичность и способные нацеливаться на множество различных эпитопов в одной молекуле. Диатела являются бивалентными, биспецифичными антителами, у которых VH и VL домены экспрессированы на одной полипептидной цепи, но использующими линкер, который является слишком коротким для того, чтобы сделать возможным спаривание двух доменов на одной цепи, тем самым вынуждая домены спариваться с комплементарными доменами другой цепи и создавая два антигенсвязывающих участка (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poijak, R.J., et al. (1994) Structure 2:1121-1123). Две антигенсвязывающие области биспецифичного антитела либо химически связаны, либо экспрессируются клеткой, созданной методами генной инженерии для продуцирования биспецифичного антитела. (см., в целом, Fanger et al., 1995 Drug News & Perspec. 8(3): 133-137). В варианте осуществления эффективное количество биспецифичного антитела можно вводить субъекту с инфекцией С.difficile и/или заболеванием, ассоциированным с С.difficile, и биспецифичное антитело нейтрализует токсичность токсина А и токсина В у субъекта.
В определенных вариантах осуществления антитела могут быть антителами человека. Термин «антитело человека», применяемый в настоящем документе, понимают как включающий антитела, имеющие вариабельные и константные области Ig, полученные от последовательностей иммуноглобулина зародышевой линии клеток человека. Антитела человека могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии клеток человека (например, мутации, введенные случайным образом или сайт-специфическим мутагенезом in vitro или соматической мутацией in vivo). Однако, термин «антитело человека», применяемый в настоящем документе, не понимают как включающий антитела, у которых последовательности CDR, полученные из зародышевой линии другого вида млекопитающих, такого как мыши, были привиты на каркасные последовательности человека (называемые в настоящем документе «гуманизированные антитела»). Антитела человека, направленные против токсина А и/или токсина В, можно получить с применением трансгенных мышей, генетически модифицированных и разводимых для экспрессии компоненты иммунной системы человека, а не мышиной системы.
Полностью человеческие моноклональные антитела также можно получить путем иммунизации мышей, трансгенных по большим частям локусов тяжелых и легких цепей иммуноглобулина человека. См., например, патенты США 5591669, 5598369, 5545806, 5545807, 6150584 и цитируемые в них ссылки, содержание которых включено в данный документ посредством ссылки. Эти животные были генетически модифицированы таким образом, что имеет место функциональная делеция при продуцировании эндогенных (например, мышиных) антител. Животные дополнительно модифицируют с тем, чтобы они содержали весь или часть локуса гена иммуноглобулина зародышевой линии человека, так что иммунизация этих животных приводит к продуцированию полностью человеческих антител к представляющему интерес антигену. После иммунизации этих мышей (например, мыши XenoMouse (Abgenix), HumAb (Medarex/GenPharm)) получают моноклональные антитела в соответствии со стандартной гибридомной технологией. Эти моноклональные антитела имеют аминокислотные последовательности иммуноглобулина человека и, следовательно, не будут провоцировать гуморальный иммунный ответ у человека на мышиное антитело (НАМА) при введении людям.
Специалистам в данной области будет понятно, что в настоящем документе также приведены нуклеиновые кислоты и полинуклеотиды, которые кодируют описанные антитела или их антигенсвязывающие фрагменты. Также будет понятно, что в настоящем документе приведены нуклеиновые кислоты и полинуклеотиды, которые содержат последовательность, кодирующую антитела или их антигенсвязывающий фрагменты. Следовательно, настоящим изобретением обеспечиваются векторы и плазмиды, сконструированные так, чтобы содержать и/или экспрессировать антитело-кодирующие нуклеиновые кислоты и полинуклеотиды. Применяемая в настоящем документе «кодирующая область» относится к области нуклеотидной последовательности, которая кодирует полипептидную последовательность;
кодирующая область может включать в себя область, кодирующую часть белка, которая позже отщепляется, как, например, сигнальный пептид. В некоторых случаях нуклеотидные и аминокислотные последовательности могут включать в себя последовательности, которые кодируют сигнальные пептиды или которые являются сигнальными пептидами. Настоящее изобретение включает в себя каждую из этих последовательностей с или без части последовательности, которая кодирует сигнальный пептид или является сигнальным пептидом.
Антитела, приведенные в настоящем документе, можно клонировать с помощью следующего способа, а также других способов, известных рядовым специалистам в данной области. В качестве неограничивающего примера, от гибридомных клеток получают общую РНК и обратно транскрибируют кДНК с применением олиго-dT праймера. РНКазу Н можно применять для удаления РНК с получением одноцепочечной кДНК. Для удаления свободных нуклеотидов можно применять очистку с помощью центрифужной колонки. Затем с помощью терминальной трансферазы можно добавить 3’ поли-dG-хвост. ПЦР-амплификацию можно провести с применением олиго-dC-праймера с добавлением вырожденного праймера к константной области. Можно провести примерно 40 циклов для надежной амплификации тяжелой цепи. Затем можно провести прямое секвенирование ПЦР-продуктов.
В определенных вариантах осуществления антитело или его антигенсвязывающий фрагмент кодируется молекулой нуклеиновой кислоты, которая является высоко гомологичной к вышеупомянутым молекулам нуклеиновых кислот. Гомологичная молекула нуклеиновой кислоты может включать нуклеотидную последовательность, которая по меньшей мере на 90% идентична приведенной в настоящем документе нуклеотидной последовательности. Гомологичная молекула нуклеиновой кислоты может включать нуклеотидную последовательность, которая по меньшей мере на 95% идентична, по меньшей мере на 97% идентична, по меньшей мере на 98% идентична или по меньшей мере на 99% идентична приведенной в настоящем документе нуклеотидной последовательности. Гомологию можно рассчитать с помощью различных, общедоступных программных средств, хорошо известных рядовому специалисту в данной области. Иллюстративные инструменты включают в себя систему BLAST, доступную на веб-сайте Национального центра биотехнологической информации (NCBI) при Национальном институте здравоохранения.
Одним способом выявления высоко гомологичных нуклеотидных последовательностей является с помощью гибридизации нуклеиновых кислот. Также в настоящем документе приведены антитела, обладающие свойствами связывания токсина А и/или токсина В и другими функциональными свойствами, описанными в настоящем документе, которые кодируются молекулами нуклеиновой кислоты, которые гибридизуются в условиях высокой жесткости с молекулами нуклеиновых кислот, кодирующими антитела настоящего изобретения. Выявление родственных последовательностей также можно осуществить с помощью полимеразной цепной реакции (ПЦР) и других амплификационных методик, подходящих для клонирования родственных последовательностей нуклеиновых кислот. Для таких методик ПЦР-праймеры обычно выбирают для амплификации таких частей представляющих интерес последовательностей нуклеиновых кислот, как CDR.
Термин «условия высокой жесткости», применяемый в настоящем документе, относится к параметрам, с которыми знакома данная область техники. Параметры для гибридизации нуклеиновых кислот можно найти в справочниках, в которых собраны такие способы, например, Molecular Cloning: A Laboratory Manual, J. Sambrook, et al., eds. Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, или Current Protocols in Molecular Biology, F.M. Ausubel, et al., eds., John Wiley & Sons, Inc., New York. Одним неограничивающим примером условий высокой жесткости является гибридизация при 65°С в гибридизационном буфере (3,5Х SSC, 0,02% фикола, 0,02% поливинилпирролидона, 0,02% бычьего сывороточного альбумина, 2,5 мМ NaH2PO4 (рН 7), 0,5% SDS, 2 мМ EDTA). SSC представляет собой 0.15М хлорид натрия/0,015М цитрат натрия, рН 7; SDS означает додецилсульфат натрия; и ЭДТА означает этилендиаминтетрауксусную кислоту. После гибридизации мембрану, на которую перенесена нуклеиновая кислота, промывают, например, в 2Х SSC при комнатной температуре, а затем в 0,1-0,5Х SSC/0,1X SDS при температурах до 68°С.
В настоящем документе также приведены векторы (например, векторы экспрессии) или плазмиды, содержащие описанные молекулы нуклеиновых кислот, помимо других последовательностей нуклеиновых кислот, например, точку начала репликации, промотор, энхансер, терминаторные последовательности, необходимые для экспрессии белка, полипептида или пептида. Данные векторы можно применять для трансформации или трансфекции клеток-хозяев для продуцирования антител или их антигенсвязывающих фрагментов со специфичностью связывания и/или характеристиками связывания описываемых в настоящем документе антител или антигенсвязывающих фрагментов. В одном варианте осуществления векторы могут содержать выделенную молекулу нуклеиновой кислоты, кодирующую тяжелую цепь или ее часть приведенных антител и антигенсвязывающие фрагментов. В другом варианте осуществления векторы могут содержать последовательности нуклеиновой кислоты, кодирующие легкую цепь или ее часть. В следующем варианте осуществления векторы настоящего изобретения могут содержать последовательность для тяжелой цепи или ее части и последовательность легкой цепи или ее части. В следующем варианте осуществления приведены плазмиды, которые дают описываемые в настоящем документе антитела или антигенсвязывающие фрагменты.
Также приведены модифицированные версии антител настоящего изобретения. Модификации в антителе или его антигенсвязывающем фрагменте, как правило, производят в нуклеиновой кислоты, которая кодирует антитело или его антигенсвязывающий фрагмент, и они могут включать делеции, точечные мутации, укорочения, аминокислотные замены и добавления аминокислот или неаминокислотных фрагментов. В альтернативном варианте модификации можно произвести непосредственно в полипептиде, как, например, путем расщепления, добавления линкерной молекулы, добавления детектируемого фрагмента, такого как биотин, добавление жирной кислоты и тому подобное. Модификации также охватывают гибридные белки, содержащие всю или часть аминокислотной последовательности антитела или антигенсвязывающего фрагмента. Модификации дополнительно охватывают присоединение или совмещение антитела с другим средством, таким как цитотоксическое средство, лекарственное средство или терапевтическое средство.
Модифицированные полипептиды включают полипептиды, которые специально модифицированы так, чтобы изменить признак полипептида, не имеющий отношения к его физиологической активности. Например, цистеиновые остатки можно заменить или удалить для предупреждения нежелательных дисульфидных связей. Аналогичным образом, некоторые аминокислоты можно изменить для усиления экспрессии полипептида путем устранения протеолиза протеазами в системе экспрессии (например, двухосновных аминокислотных остатков в дрожжевых системах экспрессии, в которых присутствует КЕХ2-протеазная активность). Кроме того, можно изменить одну или несколько аминокислот, в частности, в константной области Ig, для предупреждения протеолитического распада антитела посредством ферментов после определенного пути введения, например, перорального введения, как описано, например, в WO 2006/071877, опубликованной 6 июля 2006 года.
Модификации удобно получить путем изменения молекулы нуклеиновой кислоты, которая кодирует полипептид. Мутации нуклеиновой кислоты, которая кодирует полипептид, предпочтительно сохраняют аминокислотные рамки считывания кодирующей последовательности, и, предпочтительно, не создают областей в нуклеиновой кислоте, которые могут гибридизироваться с образованием вторичных структур, таких как шпильки или петли, которые могут быть вредны для экспрессии модифицированного полипептида.
Модификации можно осуществить в любых антителах или их антигенсвязывающих фрагментов, путем выбора аминокислотной замены или с помощью случайного мутагенеза выбранного сайта в нуклеиновой кислоте, которая кодирует полипептид. Модифицированные полипептиды затем можно экспрессировать и протестировать на одну или более видов активности, чтобы определить, какая мутация дает модифицированный полипептид с требуемыми свойствами. Дополнительные мутации могут быть внесены в модифицированные полипептиды (или немодифицированные полипептиды), которые являются молчащими в отношении аминокислотной последовательности полипептида, но которые обеспечивают предпочтительные кодоны для трансляции в конкретном хозяине. Предпочтительные кодоны для трансляции нуклеиновой кислоты, например, в Е. coli, хорошо известны рядовым специалистам в данной области. Тем не менее, в некодирующие последовательности последовательности-клона или кДНК-клона могут быть внесены другие мутации для усиления экспрессии полипептида. Активность модифицированных полипептидов можно протестировать путем клонирования гена, кодирующего модифицированный полипептид, в вектор экспрессии, введения вектора в подходящую клетку-хозяина, экспрессии модифицированного полипептида и тестирования на функциональную способность полипептидов, описываемых в настоящем документе. Приведенные выше процедуры хорошо известны рядовым специалистам в данной области.
Специалисту также будет понятно, что в полипептидах можно осуществить консервативные аминокислотные замены для получения функционально эквивалентных полипептидов. Применяемая в настоящем документе «консервативная аминокислотная замена» относится к аминокислотной замене, которая не изменяет относительные характеристики заряда или размера белка, в котором произведена аминокислотная замена. Модифицированные полипептиды могут быть получены в соответствии со способами изменения полипептидной последовательности, которые известны рядовому специалисту в данной области, как, например, те, которые можно найти в справочниках, в которых собраны такие способы, например, Molecular Cloning:
A Laboratory Manual, J. Sambrook, et al., eds., Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, или Current Protocols in Molecular Biology, F.M. Ausubel, et al., eds., John Wiley & Sons, Inc., New York. Консервативные замены аминокислот включают замены, сделанные среди аминокислот в пределах следующих иллюстративных групп: (а) М, I, L, V; (b) F, Y, W; (с) K, R, H; (d) A, G; (е) S, T; (f) Q, N и (g) E, D.
Консервативные аминокислотные замены в полипептидах, как правило, осуществляют путем изменения нуклеиновой кислоты, кодирующей полипептид. Такие замены могут быть произведены при помощи ряда способов, известных рядовому специалисту в данной области. Например, аминокислотные замены можно осуществить с помощью ПЦР-направленного мутирования, сайт-направленного мутагенеза или путем химического синтеза гена, кодирующего полипептид. Если аминокислотные замены осуществляют в небольшом фрагменте полипептида, замены можно осуществить путем непосредственного синтеза пептида. Активность функционально эквивалентных фрагментов полипептидов можно протестировать путем клонирования гена, кодирующего измененный полипептид, в бактериальный вектор экспрессии или вектор экспрессии у млекопитающего, введения вектора в подходящую клетку-хозяина, экспрессии измененного полипептида, и тестирования на функциональную возможность полипептидов, раскрываемых в настоящем документе.
Антитело к токсину или его антигенсвязывающую часть настоящего изобретения можно модифицировать или связать другой функциональной молекулой, например, другим пептидом или белком. Кроме того, антитело или часть антитела можно функционально связать, например, с помощью химического связывания, генетического слияния, нековалентной ассоциации и т.д., с одной или несколькими другими молекулярными структурами, такими как другое антитело, обнаруживаемое средство, цитотоксическое средство, терапевтическое средство, фармацевтическое средство, и/или белок, или пептид, который может выступить посредником ассоциации с другой молекулой, например, коровой областью стрептавидина или полигистидиновым хвостом.
Производный белок или антитело могут быть получены путем сшивания или связывания двух или более белков или антител одного и того же или разных типов. Подходящие сшивающие или связующие средства включают те, которые гетеробифункциональны, при этом имеют две различные реакционно-способные группы, разделенные соответствующим спейсером (например, сложный м-малеимидобензоил-N-гидроксисукцинимидиновый эфир), или гомобифункциональны (например, дисукцинимидилсуберат), и коммерчески доступны (Pierce Chemical Company, Rockford, IL). Антитело к токсину или его антигенсвязывающий фрагмент настоящего изобретения можно конъюгировать с другой молекулярной структурой, такой как метка. Обнаруживаемые средства или метки, с помощью которых белок может быть дериватизирован или помечен, включают флуоресцентные соединения, ферменты, простетические группы (например, стрептавидин/биотин и авидин/биотин), хемилюминесцентные материалы, биолюминесцентные материалы, химические структуры и радиоактивные материалы. Примеры обнаруживаемых флуоресцентных соединений включают флуоресцеин, флуоресцеинизотиоцианат (FITC), родамин и фикоэритрин. Белок или антитело также можно дериватизировать с определяемыми ферментами, такими как щелочная фосфатаза (АР), пероксидаза хрена, бета-галактозидаза, ацетилхолинэстераза, глюкозооксидаза и т.д. Такие ферментативно дериватизированные белки или антитела становятся обнаруживаемыми при добавлении специфического субстрата или фермента с получением обнаруживаемым продуктом реакции. Белки, дериватизированные простетической группой, такой как биотин, могут быть обнаружены путем косвенного измерения связывания авидина или стрептавидина.
Меченые белки и антитела можно применять в качестве диагностических и/или экспериментальных средств или реагентов для выделения известного или предопределенного антигена с помощью стандартных методик, таких как аффинная хроматография или иммунопреципитация, или для обнаружения известного или предопределенного антигена с тем, чтобы определить уровни белка в тканях, как часть клинической процедуры тестирования, например, для мониторинга эффективности данной схемы лечения. В варианте осуществления антигеном, подлежащим определению, может быть токсин в клеточном лизате или в образце, взятом у пациента.
В частном варианте осуществления антитела или антигенсвязывающие фрагменты настоящего изобретения применяют в сочетании, например, в виде фармацевтической композиции, содержащей два или более различных антител или их антигенсвязывающих фрагментов (например, один или несколько, направленных к токсину А, и один или несколько, направленных к токсину В, два или более, направленных к токсину А, или два или более направленных к токсину В, и т.д.). Комбинации антител или их антигенсвязывающих фрагментов можно объединить в одной терапии (т.е. вводить одновременно) для достижения желаемого терапевтического эффекта. В другом варианте антитела или их антигенсвязывающие фрагменты можно вводить по отдельности (то есть в разные моменты времени). Отсюда следует, что антитела или их антигенсвязывающие фрагменты можно хранить вместе или по отдельности. Антитела или их антигенсвязывающие фрагменты можно хранить в водной среде или в лиофилизированной форме, которую можно ресуспендировать перед применением.
В другом варианте осуществления предоставлены композиции, содержащие одно или несколько выделенных антител или их антигенсвязывающий фрагмент. Также предоставлены композиции, содержащие комбинацию из одного или нескольких из вышеупомянутых антител или их антигенсвязывающих фрагментов. Также предоставлены композиции, каждая из которых содержит одно или несколько из вышеупомянутых антител или их антигенсвязывающих фрагментов, каковые композиции предназначены для применения в комбинации. Такие композиции могут включать физиологически или фармацевтически приемлемый носитель, наполнитель, среду или разбавитель. Физиологически или фармацевтически приемлемый носитель, наполнитель, среду или разбавитель может быть смешан с выделенным антителом или его антигенсвязывающим фрагментом. В варианте осуществления композиции включают комбинацию множества (например, двух или более) выделенных антител или их антигенсвязывающих фрагментов. В варианте осуществления одно или несколько антител или их антигенсвязывающих фрагментов композиции специфично связывают токсин А С.difficile и нейтрализуют его токсические эффекты, в то время как одно или несколько антител или их антигенсвязывающих фрагментов специфично связывают токсин В С.difficile и нейтрализуют его токсические эффекты, В одном варианте осуществления как одно или несколько антител или их антигенсвязывающих фрагментов, которые специфично связывают токсин А С.difficile и нейтрализуют его токсические эффекты, так и одно или несколько, которые специфично связывают токсин В С.difficile и нейтрализуют его токсические эффекты, являются гуманизированными.
В частном варианте осуществления композиция содержит комбинацию антитела к токсину А или его антигенсвязывающего фрагмента, описываемого в настоящем документе, и одного антитела к токсину В или его антигенсвязывающего фрагмента, описываемого в настоящем документе. В такой композиции антитело к токсину А и антитело к токсину В могут присутствовать в равных количествах или в соотношениях, например, 1:1. В другом варианте, в такой композиции антитело к токсину А и антитело к токсину В могут присутствовать в различных количествах и соотношениях, как, например, 1/2:1,2:1,3:1;4:1 и т.д. В варианте осуществления антитела композиции являются гуманизированными. В одном варианте осуществления композиция содержит комбинацию mAb PTA-9888, его антигенсвязывающего фрагмента или его гуманизированной формы и mAb 9693, его антигенсвязывающего фрагмента или его гуманизированной формы. В варианте осуществления композиция содержит комбинацию mAb PTA-9694, его антигенсвязывающего фрагмента или его гуманизированной формы и mAb 9693, его антигенсвязывающего фрагмента или его гуманизированной формы. В варианте осуществления композиция содержит комбинацию любого из следующих: mAb РТА-9692, его антигенсвязывающий фрагмент или его гуманизированная форма и mAb 9693, его антигенсвязывающий фрагмент или его гуманизированная форма; или mAb PTA-9888, его антигенсвязывающий фрагмент или его гуманизированная форма и mAb 9692, его антигенсвязывающий фрагмент или его гуманизированная форма; или mAb PTA-9694, его антигенсвязывающий фрагмент или его гуманизированная форма и mAb 9692, его антигенсвязывающий фрагмент или его гуманизированная форма.
Фармацевтические композиции также могут быть введены В комбинированной терапии, т.е. в сочетании с другими терапевтическими средствами. Например, комбинированная терапия может включать в себя композицию, содержащую одно или несколько антител или их антигенсвязывающих фрагментов, приведенных в настоящем документе, по меньшей мере с одной другой традиционной терапией. Такие дополнительные терапевтические средства включают терапевтические средства, являющиеся антибиотикам, и терапевтические средства, не являющиеся антибиотиками. Дополнительные терапевтические средства включают вакцину на основе анатоксина С.difficile, ампициллин/амоксициллин, ванкомицин, метронидазол, фидаксомицин, линезолид, нитазоксанид, рифаксимин, рамопланин, дифимицин (также называемый PAR-101 или ОРТ-80), клиндамицин, цефалоспорины (такие как, цефалоспорины второго и третьего поколения), фторхинолоны (такие как, гатифлоксацин или моксифлоксацин), макролиды (например, эритромицин, кларитромицин, азитромицин), пенициллины, аминогликозиды, триметоприм-сульфаметоксазол, хлорамфеникол, тетрациклин, имипенем и меропенем. Дополнительные терапевтические средства также включают антибиотики, противобактериальные средства, бактерицидные средства или бактериостатические средства. В варианте осуществления дополнительное терапевтическое средство может быть малой молекулой или химическим соединением с низкой молекулярной весом, которая нацелена на С.difficile и/или его токсины. В варианте осуществления дополнительным терапевтическим средством является ОРТ-80. Терапевтические средства, не являющиеся антибиотиками, включают толевамер, высокомолекулярный анионный полимер, который связывает токсины А и В посредством неспецифических механизмов на основе заряда.
В качестве альтернативы, предполагается, что антитела или их антигенсвязывающие фрагменты по настоящему изобретению можно применять в комбинации с другими антителами или их антигенсвязывающими фрагментами. Другие дополнительные терапевтические средства включают нормальный объединенный иммуноглобулин, внутривенный иммуноглобулин или поликлональные иммуноглобулины к токсину А и токсину В в сыворотке крови. Другие антитела включают mAb человека, направленные к токсину А или токсина В С.difficile, которые описаны и сообщаются в опубликованной литературе (например, WO/2006/121422; US2005/0287150).
В настоящем документе также охвачен способ, который включает применение антител или их антигенсвязывающих фрагментов по настоящему изобретению для лечения или профилактики, т.е. для лечения, устранения, ослабления, ликвидации, предупреждения или замедление инфекции С.difficile или ассоциированного с С.difficile заболевания, патологии, или их развития или прогрессирования. CDAD, как правило, спровоцирован путем нарушением кишечной флоры посредством применения антибиотиков, таких как клиндамицин, цефалоспорины и фторхинолоны. Это расстройство микросреды кишечника наряду с воздействием спор С.difficile приводит к колонизации пострадавших лиц. Примерно у одной трети всех пациентов, которые подверглись колонизации, развивается CDAD, которое может привести к тяжелой диарее, перфорации толстой кишки, колэктомии и смерти. Поэтому, приведены способы, в соответствии с которыми пациенту вводят одно или несколько антител настоящего изобретения или композицию, описанную в настоящем документе, для лечения инфекции С.difficile или CDAD.
Применяемое в настоящем документе «лечить» относится к любому благоприятному действию на пациента с инфекцией С.difficile или ассоциированным с С.difficile заболеванием, сообщаемому путем введения антител или композиции или комбинации композиций, приведенных в настоящем документе. Например, и без ограничения, такими благоприятными действиями могут быть устранение одного или нескольких симптомов или неблагоприятных эффектов или снижение или ослабление тяжести одного или нескольких симптомов или побочных эффектов, которые являются результатом инфекции или заболевания; замедление, остановка или купирование прогрессирования инфекции или заболевания; повторная колонизация, возрождение или вторичное заселение нормальной и естественной микрофлоры желудочно-кишечного тракта, толстой кишки, кишечника и т.д., или излечения от инфекции или заболевания (т.е. врач должен оценить субъекта и определить, что у субъекта более нет инфекции или заболевания). Симптомы или неблагоприятные эффекты, ассоциированные с инфекцией С.difficile, включают обезвоживание, диарею, судороги, почечную недостаточность, перфорацию кишечника, токсический мегаколон, что может привести к разрыву толстой кишки и смерти. Приведенные композиции можно применять для снижения, уменьшения, ослабления или устранения любого или всех симптомов или неблагоприятных эффектов, приведенных в настоящем документе.
Применяемая в настоящем документе «инфекция С.difficile» относится к инфекции, причиной которой является присутствие С.difficile в кишечной флоре, где она ранее не присутствовала, или изменение присутствия С.difficile в кишечной флоре (например, увеличение общего количества С.difficile по отношению к одному или нескольким другим бактериям и т.д.), что приводит или может привести к неблагоприятному эффекту(ам) и/или повышению уровня токсинов А и/или В в кишечнике или других органах и тканях, входящих в состав в желудочно-кишечного тракта. Как правило, причиной CDAD является приобретение и размножения С.difficile в кишечнике. In vivo токсины А и В проявляют различные патологические профили с сильной синергией, служащей причиной заболевания. У кроликов и мышей, например, токсин А является энтеротоксином, что вызывает диарею, тогда как токсин В не вызывает жидкостный ответ у этих видов. Тем не менее, токсин В сильнее цитотоксичен in vitro. Все чаще сообщают о токсин-А-отрицательных, токсин-В-положительных (А-/В+) штаммах С.difficile. А-/В+-штаммы не производят токсин А из-за делеций повторяющихся доменов гена tcdA, но все же способны вызвать клиническое заболевание. В противоположность этому, на сегодняшний день не существует каких-либо сообщений о токсин-А-положительных, токсин-В-отрицательных (А+/В-) штаммах у людей.
Инфекция С.difficile обычно проявляется в виде легкой-умеренной диареи, иногда со спазмами в животе. Иногда наблюдаются псевдомембраны, которые представляют собой приросшие желтовато-белые бляшки на слизистой оболочке кишечника. В редких случаях у пациентов с инфекцией С.difficile может быть острый брюшной и скоротечный, угрожающий жизни колит, который возникает в результате нарушения нормальной бактериальной флоры толстой кишки, колонизации С.difficile и выделения токсинов, которые вызывают воспаление и повреждение слизистой оболочки. Лечение антибиотиками является ключевым фактором, меняющим кишечную флору. В то время, как нормальная кишечная флора сопротивляется колонизации и повышенному росту С.difficile, применение антибиотика, который подавляет нормальную флору, позволяет размножаться бактериям С.difficile. С.difficile присутствует у 2-3% здоровых взрослых и до 70% здоровых детей. В одном из его аспектов, mAb настоящего изобретения используют для лечения субъектов, у которых не обнаруживают симптомов, но которые восприимчивы или подвергаются риску приобретения инфекции С.difficile и тому, чтобы стать пораженными ассоциированными с ней заболеваниями. Таких субъектов можно госпитализировать или можно оставить вне больничной обстановки.
Главным фактором риска для заболевания, связанного с С.difficile, является предшествующее воздействие антибиотиков. Наиболее распространенные антибиотики, подразумеваемые при колите, вызванном С.difficile, включают цефалоспорины (особенно второго и третьего поколения), ампициллин/амоксициллин и клиндамицин. Редко подразумеваемыми антибиотиками являются макролиды (например, эритромицин, кларитромицин, азитромицин) и другие пенициллины. Соединения или другие средства, которые иногда описаны в литературе как вызывающие заболевание, включают аминогликозиды, фторхинолоны, триметоприм-сульфаметоксазол, метронидазол, хлорамфеникол, тетрациклин, имипенем и меропенем. Даже кратковременное воздействие любого отдельного антибиотика может вызвать колит С.difficile, особенно, если нормальная кишечная флора подвергается неблагоприятному воздействию или уничтожена. Длительный курс антибиотиков или применение двух или более антибиотиков увеличивает риск заболевания. Было показано, что антибиотики, традиционно применяемые для лечения колита С.difficile, вызывают заболевание. Другие факторы риска, ассоциированные с инфицированном С. difficile, включают пожилой возраст (>65 лет); ослабленную иммунную систему; недавнюю госпитализацию (особенно совместное использование больничной палаты с инфицированным пациентом, интенсивная терапия, пребывание в отделении и длительное пребывание в стационаре); проживание в доме престарелых, хосписе или другом заведении длительного лечения; брюшную хирургию; хронические заболевания толстой кишки (например, воспалительные заболевания кишечника (IBD) или колоректальный рак); прием по рецепту или без рецепта антацидных средств, которые могут снизить желудочную кислотность и сделать возможным более легкое прохождение С. difficile в кишечный тракт, и предшествующая инфекция С. difficile. Другие факторы, ассоциированные с заболеванием С. difficile, включают противоопухолевые средства, прежде всего метотрексат, гемолитико-уремический синдром, злокачественные новообразования, кишечную ишемию, почечную недостаточность, некротический энтероколит, болезнь Гиршпрунга, IBD и нехирургические процедуры желудочно-кишечного тракта, в том числе назогастральные зонды. Субъекты, которым можно вводить приведенные в настоящем документе композиции, включают любой из описанных субъектов, подверженных риску инфицирования С.difficile.
Несмотря на то, что большинство пациентов с колитом С.difficile выздоравливают без специальной терапии, симптомы могут быть длительными и изнурительными. Ассоциированная с С.difficile диарея может быть тяжелым состоянием с уровнем смертности до 25% у пожилых пациентов, которые являются болезненными. Обзоры, которые сосредоточены на более серьезно больных пациентах, указывают уровни смертности 10-30%. Инфекция С.difficile чаще встречается у пожилых людей, и пожилой возраст может способствовать восприимчивости к колонизации и заболеванию. Несмотря на то, что младенцы и маленькие дети часто несут С.difficile и ее токсины, манифестная инфекция является редкостью. Перекрестное инфицирования С.difficile распространена в отделениях для новорожденных, но похоже, что у новорожденных не развивается диарея, ассоциированная с С.difficile.
В настоящем документе приведен ряд способов с применением гуманизированных антител настоящего изобретения и/или композиций, приведенных в настоящем документе. Например, приведен способ лечения субъекта, который имеет инфекцию С.difficile или заболевание, проявляет любой из приведенных в настоящем документе симптомов или неблагоприятных эффектов или имеет какое-либо из приведенных в настоящем документе заболеваний. В одном варианте осуществления способ снижает, уменьшает или ослабляет тяжесть заболевания, ассоциированного с инфекцией С. difficile, или С. difficile-ассоциированного заболевания у субъекта. В качестве другого примера приведен способ лечения субъекта, который страдает от С. difficile-ассоциированной диареи.
Также предложен способ нейтрализации токсина А и/или токсина В С. difficile у субъекта. В качестве примера приведен способ нейтрализации комбинированных системных токсина А и токсина В С. difficile. В одном варианте осуществления комбинированные системные токсин А и токсина В С. difficile нейтрализуют путем введения как гуманизированного антитела к токсину А или его антигенсвязывающего фрагмента, так и гуманизированного антитела к токсину В или его антигенсвязывающего фрагмента, или композиции, содержащей эти антитела. В другом аспекте комбинированные системные токсин А и токсин В С. difficile нейтрализуют путем введения антитела или его антигенсвязывающего фрагмента, который специфично связывает как токсин А, так и токсин В, или композиции, содержащей антитела (например, в гуманизированной форме). В некоторых вариантах осуществления гуманизированные антитела или композиции вводят в сочетании с другим терапевтическим средством, который нацелен на С. difficile.
В качестве другого варианта осуществления также предоставлен способ восстановления нормальной флоры желудочно-кишечного тракта у субъекта, инфицированного С. difficile, тем самым эффективно излечивая инфекцию, вызванную С. difficile и/или и ее токсинами. В качестве еще одного варианта осуществления также предоставлен способ уменьшения восприимчивости субъекта к инфекции С.difficile или заболевания, ассоциированного с С.difficile. В качестве другого варианта осуществления представлен способ предупреждения инфекции С.difficile или заболевания, ассоциированного с С.difficile, у субъекта.
В вышеупомянутых способах субъекту вводят одно или несколько антител или композиций, приведенных в настоящем документе (например, композицию, содержащую моноклональное антитело или его антигенсвязывающий фрагмент, направленный против токсина А С.difficile, и композицию, содержащую моноклональное антитело или его антигенсвязывающий фрагмент, направленный против токсина В С.difficile). Композиции можно вводить субъекту в одно время или в разные моменты времени. Композиции можно вводить субъекту в виде смеси в композиции, содержащей фармацевтически приемлемый носитель, среду или наполнитель, и, необязательно, другой антибиотик, средство, не являющееся антибиотиком, лекарственное средство или терапевтическое эффективное средство против С.difficile и/или ее энтеротоксинов.
Гуманизированное моноклональное антитело к токсину А или его антигенсвязывающий фрагмент и/или гуманизированное моноклональное антитело к токсину В или его антигенсвязывающий фрагмент, или фармацевтически приемлемую композицию, содержащую гуманизированные антитела или их антигенсвязывающие фрагменты, по отдельности или вместе, можно применять в любом из описанных способов по настоящему изобретению.
Применяемый в настоящем документе «фармацевтически приемлемый носитель» или «физиологически приемлемый носитель» включает в себя всевозможные соли, растворители, дисперсионные среды, покрытия, противобактериальные и противогрибковые средства, изотонические и задерживающие абсорбцию средства и т.п., которые являются физиологически совместимыми. Предпочтительно, носитель подходит для перорального, внутривенного, внутрибрюшинного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения, активное соединение, то есть антитело, может быть покрыто материалом для защиты соединения от воздействия кислот и других природных условий, которые могут инактивировать соединение.
При введении фармацевтические препараты настоящего изобретения применяют в фармацевтически приемлемых количествах и в фармацевтически приемлемых композициях. Термин «фармацевтически приемлемый» означает нетоксичный, физиологически приемлемый материал, который не затрагивает эффективность биологической активности активных ингредиентов. Такие препараты обычно могут содержать соли, буферные средства, консерванты, совместимые носители и, необязательно, другие терапевтические средства, такие как дополнительные иммуностимулирующие средства, включая адъюванты, хемокины и цитокины. При применении в медицине соли должны быть фармацевтически приемлемыми, но фармацевтически неприемлемые соли обычно можно применять для получения фармацевтически приемлемых солей и не исключаются из объема настоящего изобретения.
Соль сохраняет требуемую биологическую активность исходного соединения и не вызывает каких-либо нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J. Pharm. Sci. 66: 1-19). Примеры таких солей включают соли присоединения кислоты и соли присоединения основания. Соли присоединения кислоты включают соли, полученные из нетоксичных неорганических кислот, таких как соляная, азотная, фосфорная, сернокислая, бромоводородная, йодоводородная, фосфорная и им подобных, а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенилзамещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Соли присоединения основания включают соли, полученные из щелочно-земельных металлов, таких как натрий, калий, магний, кальций и т.п., а также нетоксичные органические амины, такие как N,N’-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, этилендиаминацетат (EDTA), с или без противоиона, такого как натрий или кальций, прокаин и т.п.
Любую из композиций настоящего изобретения, при желании, можно скомбинировать с фармацевтически приемлемым носителем. Термин «фармацевтически приемлемый носитель», применяемый в настоящем документе, означает один или несколько совместимых твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые пригодны для введения человеку. Термин «носитель» обозначает органический или неорганический ингредиент, природный или синтетический, с которым комбинируют активный ингредиент для облегчения применения. Компоненты фармацевтических композиций также смешиваемы с молекулам приведенных композиций и друг с другом таким образом, чтобы не было взаимодействия, которое существенно ухудшило бы требуемую фармацевтическую эффективность.
Фармацевтические композиции могут содержать подходящие буферные средства, включая: уксусную кислоту в соли, лимонную кислоту в соли, борную кислоту в соли и фосфорную кислоты в соли.
Фармацевтические композиции также могут содержать при необходимости подходящие консерванты, такие как: бензалкония хлорид; хлорбутанол и парабены.
Фармацевтические композиции традиционно могут быть представлены в виде стандартной лекарственной формы и могут быть получены любым из способов, хорошо известных в области фармацевтики. Все способы включают этап приведения активного средства в ассоциацию с носителем, который составляет один или несколько вспомогательных ингредиентов. В целом, композиции получают путем равномерного и тщательного приведения активного соединения в ассоциацию с жидким носителем, мелкодисперсным твердым носителем, или и тем, и другим, а затем, если необходимо, формования продукта.
Антитела к токсину А и токсину В настоящего изобретения или их части можно давать в соответствии со схемами приема, которые можно скорректировать для получения оптимального желаемого ответа, такого как терапевтический или профилактический ответа, у отдельного субъекта. Например, можно вводить однократную болюсную дозу, можно вводить несколько раздельных доз с течением времени или дозу можно пропорционально уменьшать или увеличивать, как может требоваться конкретной терапевтической ситуацией. Парентеральные композиции можно упаковать или приготовить в стандартной лекарственной форме для удобства введения и единообразия дозировки. Стандартная лекарственная форма относится к физически дискретным единицам, предоставленным в виде однократных доз для субъектов, подлежащих лечению, где каждая единица содержит предопределенное количество активного соединения, рассчитанное для получения требуемого терапевтического эффекта, в сочетании с требуемым фармацевтическим носителем, средой, наполнителем или разбавителем. Требования к стандартным лекарственным формам настоящего изобретения продиктованы и напрямую зависят от (а) уникальных характеристик активного соединения и конкретного терапевтического эффекта, который необходимо достигнуть, и (b) ограничений, присущих области техники, относящейся к получению составов такого активного соединения для лечения чувствительности у индивидуумов.
Композиции, подходящие для парентерального введения обычно включают стерильный водный или неводный препарат, который предпочтительно является изотоничным с кровью реципиента. Этот препарат можно составить в соответствии с известными методами с применением подходящих диспергирующих или увлажняющих средств и суспендирующих средств. Стерильный инъекционный препарат также может быть стерильным инъекционным раствором или суспензий в нетоксичном парентерально приемлемом разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. К числу приемлемых сред и растворителей, которые могут быть использованы, относятся вода, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителя или суспензионной среды традиционно используют стерильные жирные масла. Для этой цели можно использовать любое мягкое жирное масло, в том числе синтетические моно- или диглицериды. Кроме того, при приготовлении лекарственных форм для инъекций можно использовать жирные кислоты, такие как олеиновая кислота. Составы-носители, пригодные для перорального, подкожного, внутрибрюшинного, внутривенного, внутримышечного и т.д. введения, можно найти в Remington’s Pharmaceutical Sciences, Mack Publishing Co., Easton, PA.
Активные компоненты можно приготовить с носителями, которые будут защищать компоненты от быстрого высвобождения, как, например, состав с контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Можно применять биоразлагаемые, биосовместимые полимеры, например, этиленвинилацетат, полиангидриды, полигликолевую кислоту, коллаген, сложные полиортоэфиры и полимолочную кислоту. Многие способы получения таких составов запатентованы или общеизвестны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Антитело и композиции настоящего изобретения в качестве терапевтических средств можно вводить любым традиционным путем, в том числе инъекцией или постепенным вливанием в течение долгого времени. Путь введения может представлять собой, в качестве неограничивающих примеров, пероральный, внутривенный, подкожный, внутрибрюшинный, внутримышечный, интратекальный, внутриполостной, ретроорбитальным, вагинальный, ректальный, ингаляционный, аспирация, кожный, с помощью суппозитория или трансдермальный.
Антитела и композиции настоящего изобретения вводят в эффективных количествах или дозах. «Эффективное количество» представляет собой такое количество антитела или антигенсвязывающего фрагмента или композиции(ий), приведенных в настоящем документе, которое отдельно или вместе с дополнительными дозами или другим терапевтическим средством(ами), производит требуемый ответ, например, лечит, ослабляет, ликвидирует, устраняет или предупреждает инфекцию С. difficile, диарею или заболевание, ассоциированное с С. difficile, у субъекта. Это может включать только замедление прогрессирования инфекции, диареи или заболевания в течение длительного периода, например, дольше одной недели, двух недель, трех недель, месяца, двух месяцев, трех месяцев или более трех месяцев. Однако такие эффективные количества в оптимальном случае бессрочно лечат или останавливают прогрессирование инфекции, диареи или заболевания. Это можно отслеживать с помощью стандартных способов. Требуемым ответом на лечение заболевания или состояния также может быть замедление начала или даже предупреждение начала инфекции или заболевания.
Конечно эффективные количества будут зависеть от конкретной инфекции или заболевания, которую лечат, тяжести инфекции или заболевания, индивидуальных показателей пациента, включая возраст, физическое состояние, размер и вес, продолжительность лечения, характер сопутствующей терапии (если таковая имеется), конкретные пути введения, и подобных факторов в рамках знания и компетентности практикующего врача. Эти факторы хорошо известны специалистам в данной области и могут быть изучены посредством всего лишь общепринятого тестирования или экспериментирования. Как правило, предпочтительно, чтобы была использована максимальная доза отдельных компонентов или их комбинации, а именно самая высокая безопасная доза в соответствии с результатами медицинской оценки. Однако, специалистам в данной области будет понятно, что пациент может настаивать на более низкой дозе или предельно допустимой дозе по медицинским причинам, психологическим причинам или практически по любой Другой причине.
Фармацевтические композиции, используемые в упомянутых ранее методах, предпочтительно являются стерильными и содержат эффективное количество одного или более антител или антигенсвязывающих фрагментов, приведенных в данном документе, для получения желаемого ответа, в единице массы или объема пригодной для введения пациенту. Например, ответ может быть измерен путем определения физиологических эффектов композиции, таких как ослабление симптомов заболевания. Другие анализы будут известны специалистам в данной области и могут быть использованы для измерения уровня ответа.
Дозы или количества композиций, вводимых субъекту, могут быть выбраны в соответствии с различными показателями, в частности, в соответствии с использованным способом введения и состоянием субъекта. Другие факторы включают желаемый период лечения. В случае, если ответ у субъекта недостаточен, при применении начальных доз, могут применяться более высокие дозы (или эффективно более высокие дозы по другому, более локализованному пути доставки) в той мере, в какой позволяет переносимость пациента.
В целом дозы или количества могут варьироваться в пределах от приблизительно 1 мкг/кг до приблизительно 100000 мкг/кг. Неограничивающие примеры диапазонов доз составляют терапевтически или профилактически эффективное количество антитела, части антитела или композиции настоящего изобретения включают от 0,1 мг/кг до 100 мг/кг; от 0,1 мг/кг до 60 мг/кг; от 0,5 мг/кг до 75 мг/кг; от 0,5 мг/кг до 25 мг/кг; от 0,75 мг/кг до 40 мг/кг; от 1 мг/кг до 50 мг/кг; или от 1 мг/кг до 5 мг/кг. Следует иметь в виду, что для любого определенного человека, пациента или субъекта, конкретные дозы и схемы приема, должны быть скорректированы с течением времени в соответствии с индивидуальной потребностью и профессиональным суждением квалифицированного практикующего врача, который управляет или координирует введение антител и/или композиций. Такие диапазоны доз являются только примерными и не предназначены ограничить объем или применение данного изобретения. Основываясь на композиции, доза может быть доставлена непрерывно, например, путем непрерывной накачки или через определенные периоды времени. Желаемые интервалы времени многократных доз определенной композиции могут быть определены без чрезмерного экспериментирования специалистами в данной области. Специалистам в данной области будут известны другие протоколы для введения композиций, в которых количество дозы, график введения, места введения, способ введения и т.п. отличаются от упомянутых ранее.
Введение композиции млекопитающим, отличных от человека, например, для испытательных целей или ветеринарных лечебных целей, осуществляют, по существу, в тех же условиях, как описано выше.
Также, приведенными в данном документе являются наборы, содержащие антитела по настоящему изобретению или композиции, содержащие антитела по настоящему изобретению и инструкции по применению. Наборы могут дополнительно содержать по крайней мере один дополнительный реактив, такой как дополнительное терапевтическое средство, или одно или более дополнительных антител или антигенсвязывающих фрагментов, как приведено в данном документе (например, антитело или его антигенсвязывающий фрагмент к токсину А, если первое антитело или его антигенсвязывающий фрагмент в наборе представляет собой антитело или его антигенсвязывающий фрагмент к токсину В, и наоборот).
Компоненты наборов могут быть упакованы, или в водное средство, или в лиофилизированную форму. Если антитела или их антигенсвязывающие фрагменты используются в наборах в виде конъюгатов (например, биспецифичного конъюгата антител), то компоненты таких конъюгатов могут быть поставлены, или в полностью конъюгированной форме, в форме промежуточных продуктов, или в виде отдельных частиц, чтобы быть сконъюгированными пользователем набора в соответствии с приведенными инструкциями по применению.
Набор может включать носитель, который является разделенным на отсеки, с получением в нем в строгой изоляции одного или более контейнерных устройств или серии контейнерных устройств, таких как пробирки, ампулы, сосуды, бутылки, шприцы и т.п.Первое контейнерное устройство или серия контейнерных устройств может содержать одно или более антител или их антигенсвязывающих фрагментов. Второе контейнерное устройство или серия контейнерных устройств может содержать одно или более антител или их антигенсвязывающих фрагментов, в которых антитела или их антигенсвязывающие фрагменты являются отличными от тех в первых контейнерных устройствах или некоторого другого дополнительного терапевтического средства. Наборы, приведенные в данном документе, могут дополнительно включать третий контейнер, который содержит молекулу для связывания антител или антигенсвязывающих фрагментов, содержащиеся в первом и втором контейнерах.
Как используется в данном документе по отношению к полипептидам, белкам или их фрагментам, «изолированный» означает отделенный от его естественной окружающей среды и присутствующий в количестве, достаточном, чтобы позволить его определение или применение. Выделенный, если имеют в виду белок или полипептид, означает, например: (i) получаемый выборочно экспрессионным клонированием или (ii) очищенные, как с помощью хроматографии или электрофореза. Изолированные белки или полипептиды могут быть, но могут и не быть, практически чистыми. Термин «практически чистый» означает, что белки или полипептиды являются в основном свободными от других веществ, с которыми они могут быть найдены в природе, или в т vivo системах, в некоторой степени практические и подходят для их использования по назначению. Практически чистые полипептиды могут быть получены способами, хорошо известными в данной области. Так как выделенные белки могут быть смешаны с фармацевтически приемлемым носителем в фармацевтическом препарате, белок может составлять лишь небольшой процент по массе препарата. Белок, тем не менее, выделяют в том, что он был отделен от веществ, с которыми он может быть естественно связан в живых системах, т.е. выделены из других природных белков.
Также предоставлены способы оценки средства-кандидата для эффективности в лечении инфекции С.difficile или ассоциированного с С.difficile заболевания. Такие способы могут включать этапы лечения субъекта средством, которое повышает риск инфекции С. difficile или ассоциированного с С. difficile заболевания у субъекта, инокуляцию субъекта С. difficile, лечение субъекта с помощью средства-кандидата и оценку эффективности лечения средством-кандидатом. Как используется в данном документе, «средство, которое повышает риск инфекции С. difficile или ассоциированного с С. difficile заболевания» является любьм средством, которое, как полагают, способствует началу или прогрессированию инфекции С.difficile или ассоциированного с С. difficile заболевания. Таким средством может быть антибиотическое или неантибиотическое средство. Например, средство может быть любым из антибиотиков, описанных в данном документе. Иллюстративно, таким антибиотиком может быть клиндамицин, метронидазол, ванкомицин, фидаксомицин, нитазоксанид, рифаксимин, рамопланин или их комбинации.
В этих способах, средство-кандидат может быть введено субъекту до или после инокуляции С.difficile. Средством-кандидатом может быть любое средство, который, как полагают, имеет потенциал для лечения или профилактики инфекции С.difficile или ассоциированного с С.difficile заболевания. Средства-кандидаты, которые могут быть антибиотиками или неантибиотиками, включают антитела или их антигенсвязывающие фрагменты, которые специфически связывают токсин А и/или токсин В С.difficile.
Эти способы включают любые из in vitro и in vivo способов описанных в примерах ниже.
Конечной целью лечения CDAD является прекращение всех антибиотиков и восстановление нормальной микрофлоры кишечника. В соответствии с настоящим изобретением, mAb к токсину А и В, описанные в данном документе, могут обеспечить неантибиотические терапии, направленные на блокирование патогенных эффектов токсинов С.difficile, позволить прекратить антибиотики и тем самым обеспечить время для исцеления толстой кишки и восстановления нормальной микрофлоры кишечника. Моноклональные антитела данного изобретения продемонстрировали полную и долговременную (>37 дней) защиту в жесткой модели CDAD на хомяках. Основываясь на их исключительных характеристиках и свойствах, mAb к токсину А и В С.difficile настоящего изобретения предоставляют новые возможности лечения и лекарственные препараты для пациентов, которым плохо оказывали помощь существующие терапии, включая тех индивидуумов, которые страдают наиболее тяжелыми случаями заболевания.
Настоящее изобретение дополнительно включает вакцину или иммуноген, содержащий части, фрагменты или пептиды токсина А и/или токсина В С.difficile, содержащие эпитопные области, узнаваемые и/или связываемые одним или более из моноклонального антитела РА-39 (АТСС № доступа РТА-9692), гуманизированной формы РА-39, моноклонального антитела РА-50 (АТСС № доступа РТА-9694), гуманизированной формы РА-50, моноклонального антитела РА-41 (АТСС № доступа РТА-9693), гуманизированной формы РА-41, антитела, которое конкурирует за связывание токсина А с моноклональным антителом РА-39 или его гуманизированной формой, антитела, которое конкурирует за связывание токсина А с моноклональным антителом РА-50 или его гуманизированной формой или антитела, которое конкурирует за связывание токсина В с моноклональным антителом РА-41 или его гуманизированной формой. В варианте осуществления вакцина или иммуноген включают части, фрагменты или пептиды токсина А и токсина В С.difficile, содержащие эпитопные области, распознаваемые и/или связываемые одним или более из моноклонального антитела РА-39 (АТСС № доступа РТА-9692), гуманизированной формы РА-39 или антитела, которое конкурирует за связывание токсина А и токсина В с моноклональным антителом РА-39 или его гуманизированной формой. В варианте осуществления эпитоп-содержащие части, фрагменты или пептиды токсина А и/или токсина В данной вакцины или иммуногена, получают из белка токсина А или токсина В протеолитическим расщеплением. В варианте осуществления фрагменты, части или пептиды токсина А данной вакцины или иммуногена получают протеолитическим расщеплением энтерокиназой. В варианте осуществления фрагменты, части или пептиды токсина В данной вакцины или иммуногена получают протеолитическим расщеплением каспазой (каспазой 1). В варианте осуществления эпитоп-содержащие части или фрагменты вакцины или иммуногена, химически или рекомбинантно синтезировали пептиды белка токсина А и токсина В. В варианте осуществления фрагменты, части или пептиды вакцины или иммуногена, содержащие одну или более эпитопных областей токсина А и/или токсина В, распознаваемые и связываемые антителом, получают из одного или более из амино-конца токсина А; амино-конца токсина В; карбокси-конца токсина А; карбокси-конца токсина В; рецепторсвязывающего домена токсина А; области вне рецепторсвязывающего домена токсина А; N-терминальной ферментативной области токсина В; гликозилтрансферазного домена токсина А; гликозилтрансферазного домена токсина В; протеолитического домена токсина А; протеолитического домена токсина В; гидрофобного, порообразующего домена токсина А; или гидрофобного, порообразующего домена токсина В.
В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из амино-конца токсина А или токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из С-конца токсина А или токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из гликозилтрасферазного домена токсина А или токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из протеолитического домена токсина А или токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из гидрофобного, порообразующего домена токсина А или токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из рецепторсвязывающего домена токсина А. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей распознаваемые и связываемые антителами, получают из рецепторсвязывающего домена токсина В. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из области вне рецепторсвязывающего домена токсина А. В некоторых вариантах осуществления фрагменты, содержащие одну или более эпитопных областей, распознаваемые и связываемые антителами, получают из N-концевой ферментативной области токсина В. В варианте осуществления, эпитоп-содержащие фрагменты или части токсина А и/или токсина В имеют размер <300 кДа. Эпитоп-содержащие фрагменты или части токсина А и/или токсина В имеют размер ~158-160 кДа, ~100-105 кДа, например, 103 кДа, ~90-95 кДа, например, 91 кДа, и/или ~63-68 кДа, например, 63 кДа или 68 кДа. В других вариантах осуществления эпитоп-содержащие фрагменты или части токсина А имеют размер ~158-160 кДа; ~90-95 кДа, например, 91 кДа, и/или ~63-68 кДа, например, 68 кДа. В других вариантах осуществления эпитоп-содержащие фрагменты или части токсина В имеют размер ~100-105 кДа, например, 103 кДа, и/или ~63-68 кДа, например, 63 кДа.
Такие части, фрагменты или пептиды токсинов при введении в форме вакцины или иммуногена субъекту, зараженному С.difficile или пораженному ассоциированным с С.difficile заболеванием, могут вызвать гуморальный ответ у субъекта, то есть антитела, имеющие специфичности к токсину А и/или токсину В, таким образом позволяя субъекту обеспечить иммунный ответ по отношению к токсинам и нейтрализовать, блокировать, снизить, уменьшить интенсивность, излечить или лечить ассоциированное с С. difficile заболевание, инфекцию или CDAD у субъекта. Соответственно, в другом варианте осуществления обеспечен способ нейтрализации, блокирования, снижения, уменьшения интенсивности, излечения или лечения инфекции С. difficile или ассоциированного с С. difficile заболевания у субъекта, нуждающегося в этом, включающий введение субъекту эффективного количества вышеописанных вакцины или иммуногена. В варианте осуществления субъект проявляет гуморальный ответ на токсин А и/или токсин В С.difficile, таким образом нейтрализуя, блокируя, снижая, уменьшая интенсивность, излечивая или осуществляя лечение ассоциированного с С.difficile заболевания, инфекции или CDAD у субъекта. В другом варианте осуществления субъект проявляет клеточный иммунный ответ на токсин А и/или токсин В С.difficile. В другом варианте осуществления субъект проявляет как гуморальный, так и клеточный иммунный ответ на токсин А и/или токсин В С.difficile.
В другом варианте осуществления настоящее изобретение включает способ нейтрализации, подавления или блокирования активности токсина А и/или токсина В в или по отношению к клетке, чувствительной к инфекции С.difficile, включающий контакт клетки с антителом или его антигенсвязывающим фрагментом в соответствии с настоящим изобретением, где антитело или его антигенсвязывающий фрагмент нейтрализует, подавляет или блокирует активность токсина А и/или токсина В в или по отношению к клетке при помощи конкурентного или смешанного конкурентного механизма действия. В варианте осуществления антитело представляет собой одно или несколько из моноклонального антитела, гуманизированного антитела или химерного антитела. В варианте осуществления клетка находится в субъекте, и антитело или его антигенсвязывающий фрагмент вводят субъекту в эффективном количестве. В варианте осуществления клетка находится в пределах желудочно-кишечного тракта субъекта, например, клетка кишечного эпителия. В варианте осуществления токсин представляет собой токсин А. В варианте осуществления токсин представляет собой токсин В. В варианте осуществления токсин представляет собой токсин А, и механизм действия антитела представляет собой механизма действия конкурентного подавления. В варианте осуществления токсин представляет собой токсин А, и антитело или его антигенсвязывающий фрагмент представляет собой РА-50 (№ доступа АТСС РТА-9694), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-50 за нейтрализацию активности токсина А. В варианте осуществления токсин представляет собой токсин А, и механизм действия антитела представляет собой механизм действия смешанного конкурентного подавления. В варианте осуществления токсин представляет собой токсин А, и антитело или его антигенсвязывающий фрагмент представляет собой РА-39 (№ доступа АТСС РТА-9692), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-39 за нейтрализацию активности токсина А. В варианте осуществления токсин представляет собой токсин В, и механизм действия антитела представляет собой механизм действия смешанного конкурентного подавления. В варианте осуществления токсин представляет собой токсин В, и антитело или его антигенсвязывающий фрагмент представляет собой РА-41 (№ доступа АТСС РТА-9693), его гуманизированную форму или антитело или его фрагмент, который конкурирует с РА-41 за нейтрализацию активности токсина В.
Как применяется в настоящем документе, выражение «конкурентный ингибитор» токсина относится к нейтрализующему токсин ингибитору, например, антителу, средству или малой молекуле или химической структуре, которая проявляет правосторонний сдвиг ЕС50 на кривой нейтрализации без изменения максимального процента нейтрализации по мере того, как концентрация токсина увеличивается в культуре. Таким образом, конкурентный ингибитор, как правило, способен преодолеть цитотоксический эффект токсина при добавлении большего количества ингибитора. Выражение «неконкурентный ингибитор» токсина относится к нейтрализующему токсин ингибитору, который проявляет снижение максимального процента нейтрализации без изменения концентрации, давая полумаксимальное значение ответа (EC50) по мере того, как концентрация токсина увеличивается в культуре. Таким образом, неконкурентный ингибитор, как правило, неспособен полностью преодолеть цитотоксический эффект токсина при добавлении большего количества ингибитора. Выражение «смешанный конкурентный ингибитор» токсина относится к нейтрализующему токсин ингибитору, который проявляет некоторую степень как конкурентного, так и неконкурентного ингибирования по мере того, как концентрация токсина увеличивается в культуре. Например, смешанный конкурентный ингибитор токсина может связывать токсин и обнаруживает свой эффект, блокируя связывание токсина с клеткой, а также блокируя другие цитотоксические эффекты токсина; таким образом обнаруживая смешанный конкурентный механизм действия.
Примеры
Пример 1
Получение нейтрализующих моноклональных антител против токсина А и/или токсину В С.difficile
А. Подготовка иммуногена
Нейтрализующие моноклональные антитела, направленные против токсина А и/или токсина В С.difficile получали, иммунизируя мышей анатоксином токсина А С.difficile (неактивная форма токсина) и активными формами токсина А и/или токсина В. Мышиные mAb (PA-38: mAb к токсину А, АТСС №РТА-9888, PA-39: mAb к токсинам А и В, АТСС №РТА-9692, РА-41: mAb к токсину В АТСС №РТА-9693 и РА-50: mAb к токсину А, АТСС №РТА-9694) получали, иммунизируя животных анатоксином А и последующими иммунизациями активной формой токсина А и/или токсина В. Анатоксин токсина А, токсин А, токсин В (List Biological Laboratories Inc., Campbell, CA) и токсин A (TechLab Inc, Blacksburg, VA) хранили при 4°С до применения. Токсины и анатоксин были получены из штамма VPI 10463, широко используемого референс-штамма С.difficile. Адъювант Quil A (Accurate Chemical, Westbury, NY) добавляли в требуемый объем анатоксина или токсина и смешивали. Смесь готовили за 60 минут до иммунизации и хранили на льду до момента иммунизации. Для заключительной бустер-дозы перед слиянием, требуемый токсин разводили в PBS и хранили на льду до применения в иммунизации.
В. Иммунизация и слияние
Тридцать самок мышей BALB/c (Charles River Labs, Wilmigton, MA) получали две иммунизирующие дозы (для РА-50) или три иммунизирующие дозы (для PA-38, PA-39 и РА-41) анатоксина токсина А (10 мкг) подкожно с интервалами в три недели перед получением бустерных иммунизации с повышающимися дозами активного токсина А или активного токсина В, также с интервалами в три недели. Для РА-38, одна мышь получала бустерную иммунизацию каждые три недели при в сумме трех бустер-дозах с токсином A (List Biological Laboratories Inc.), при этом каждая бустер-доза повышается в дозе от 500 нг до 2 мкг, а заключительная бустер-доза токсина А (8 мкг) за три дня до спленэктомии. Для PA-39 и РА-41, две мыши получали три или пять бустерных иммунизации, соответственно, каждые три недели токсином В, при этом каждая бустер-доза повышается в дозе от 2 мкг до 12,5 мкг, а заключительная бустер-доза токсина В (20 мкг) за три дня до спленэктомии. Для РА-50 одна мышь получала бустерную иммунизацию каждые три недели при в сумме четырех бустер-дозах с токсином А (TechLab Inc), при этом каждая бустер-доза повышается в дозе от 20 нг до 2,5 мкг, а заключительная бустер-доза (10 мкг) за три дня до спленэктомии.
Иммунизирующие и бустерные дозы анатоксина и токсина, соответственно, вводили в сочетании с адъювантом, например Quil A (10 мкг). Бустер-иммунизация активной формой токсина А или токсина В служила для идентификации животных, которые могут иметь выработанные защитные антитела. Сыворотки от иммунизированных животных серийно разводили и тестировали в отношении нейтрализации цитотоксического эффекта токсина А на клетках СНО-K1, как описано ниже. Животных с самым высоким титром нейтрализующих антител выбирали для слияний и бустер-иммунизации токсином без адъюванта.
После бустер-иммунизации животных умерщвляли, и изолированные спленоциты сливали с клеточной линией Sp2/0 с применением стандартных способов. Гибридомы суспендировали в селективной среде, RPMI-1640, 10% FBS, 10% ВМ Condimed-Hl (Roche Applied Science, Indianapolis, IN) и бета-меркаптоэтанол (для PA-38 и PA-39) или Hybridoma-SFM и 10% FBS (для РА-41 и РА-50), которая содержала 100 мкМ гипоксантин, 1 мкг/мл азасерин и 16 мкМ тимидин для селективного давления. Гибридомы высевали в 96-луночные плоскодонные планшеты для культуры тканей (BD Biosciences, San Jose, CA). Планшеты инкубировали при 37°С в течение 3 дней с последующим добавлением ростовой среды НТ (селективная среда без азасерина). Инкубация продолжалась еще в течение 4-7 дней до скрининга гибридомных супернатантов в отношении нейтрализующей активности.
В первичном скрининге на РА-38, 608 гибридомных супернатантов тестировали на способность нейтрализовать цитотоксический эффект токсина A (List Laboratories) на клетках СНО-K1 (АТСС №CCL-61, Manassas, VA). В первичном скрининге на РА-39 и РА-41, 2416 гибридомных супернатантов тестировали на способность нейтрализовать цитотоксический эффект токсина В (List Laboratories) на клетках СНО-K1. В первичном скрининге на РА-50, 1440 гибридомных супернатантов тестировали на способность нейтрализовать цитотоксический эффект токсина A (Techlab Inc.) на клетках Т-84. Во втором анализе исследовали подавление опосредованной токсином агглютинации эритроцитов кролика. На основании процедуры скрининга выделили четыре mAb, которые обозначили РА-38 (к токсину А), РА-39 (к токсину А/В), РА-50 (к токсину А) и РА-41 (к токсину В) и который эффективно подавляли или нейтрализовали токсины С.difficile в анализах скрининга. Гибридомные клеточные линии, которые продуцировали данные mAb, клонировали дважды с помощью предельного разведения для получения клональных клеточных линий. С применением IsoStrip Mouse Monoclonal Antibody Isotyping Kit (Roche Applied Science, Indianapolis, IN) определили, что mAb PA-38, PA-39, PA-41 и PA-50 представляют собой изотипы IgG2a,K, IgG1.K, IgG1.K и IgG1.K, соответственно. Гибридомные клеточные линии, продуцирующие mAb, обозначаются тем же названием, что и mAb, которые они продуцируют.
С.Скрининг: нейтрализация цитотоксического эффекта токсина А или В на клетках
Гибридомные супернатанты подвергали скринингу в отношении способности нейтрализовать цитотоксический эффект токсина А или токсина В на клетках. Разработали высокопроизводительный способ для обработки тысяч гибридомных супернатантов одновременно. Для анализа на цитотоксичность использовали или клетки СНО-K1 (для РА-38, РА-39 и РА-41) или клетки Т-84 (для РА-50). Клетки вносили в планшеты для анализа (96-луночные планшеты, белая непрозрачная стенка, прозрачное плоское дно; Perkin Elmer, Waltham, МА) с применением роботизированной системы Biomek FX (Beckman Coulter, Brea, CA). Планшеты для анализа инкубировали в течение 4 часов при 37°С, чтобы дать клеткам возможность прикрепиться к лункам. Для анализа на Т-84, токсин А разводили до 240 нг/мл. Для анализа на СНО-K1, токсин А разводили до 2 мкг/мл или токсин В разводили до 6 нг/мл. Разведенный токсин вносили в планшеты для разведения реактивов (96-луночные круглодонные планшеты; BD, Franklin Lakes, NJ) вручную в боксе (BSC). Гибридомные супернатанты собирали вручную и вносили в лунки планшетов для разведения реактивов с применением системы Biomek FX. Смесь супернатанта и токсина инкубировали при 37°С в течение 1 часа и вносили в планшеты для анализа, содержащие клетки с применением системы Biomek FX. После инкубации при 37°С в течение 72 часов в каждую лунку вносили 20 мкл/лунку CellTiter-Blue (Promega, Madison, WI). Планшеты инкубировали в течение дополнительных 4 часов, а затем считывали на планшет-ридере SpectraMax M5 (Molecular Devices, Sunnyvale, CA) с применением длины волны возбуждения 560 нм и длины волны излучения 590 нм. Выживание клеток сравнивали в необработанных и обработанных токсином культурах. Процент выживания клеток размещали на графике относительно концентрации.
D. Получение мышиных mAb по настоящему изобретению Для получения выделенных и/или очищенных mAb настоящего изобретения применяли in vivo и in vitro способы продукции. Для in vivo продукции мышиных mAb получали асцитную жидкость путем инъецирования соответствующей гибридомной клеточной линии в брюшную полость пристан-примированным мышам BALB/c. mAb очищали до >95% гомогенности с помощью осаждения сульфатом аммония с последующей хроматографией с белком А. Очищенные антитела ресуспендировали в фосфатно-солевом буфере (PBS).
Для in vitro продукции в малом масштабе (<100 мг) мышиные mAb антитела очищали от гибридомных супернатантов, выращенных в культуре. Гибридомы культивировали в Hybridoma-SFM (Invitrogen) и 10% FBS. Клеточные линии пересевали и размножали в Т-150 матрасах три раза в неделю, чтобы обеспечить то, что концентрация клеток не превышает 2×106 клеток/мл. Супернатанты, содержащие PA-39 (IgG1, κ), PA-41 (IgG1, κ) и РА-50 (IgG1, κ) очищали от примесей путем центрифугирования при 2000 оборотах в минуту в течение 10 минут и фильтрации. Очищенный от примесей материал разбавляли до конечной концентрации подвижным буфером (60 мМ глицин/3 М NaCl, рН 8,5) и загружали на колонку с белком А, уравновешенную подвижным буфером. После отмывки колонки, mAb PA-39 или РА-41 элюировали 0,1 ацетатом натрия, рН 5,5, и нейтрализовали до рН 7,0. Супернатанты, содержащие PA-38 (IgG2a, κ) очищали от примесей путем центрифугирования при 2000 оборотах в минуту в течение 10 минут и фильтрации. Очищенный от примесей материал доводили до конечной концентрации 25 мМ натрийфосфатным буфером/100 мМ NaCl, рН 7,0 и загружали на колонку с белком А, уравновешенную 50 мМ натрийфосфатным буфером /0,5 М NaCl. Колонку отмывали, mAb PA-38 элюировали 0,1 М ацетатом натрия, рН 3,0, и затем элюированное антитело нейтрализовали до рН 7,0.
Для in vitro продукции в больших количеств mAb (>100 мг), гибридомы инокулировали в биореактор WAVE (GE Healthcare, Piscataway, NJ) при начальной плотности 2×105 клеток/мл с Hybridoma-SFM с 5% Ultra Low IgG FBS. Подсчет клеток производили, и жизнеспособность клеток отслеживали ежедневно. Примерно на день 6 или 7, когда продукция антител замедлилась, культивирование прекращали. Культуру очищали и затем концентрировали в 10-20 раз с помощью тангенциальной поточной фильтрации. Антитело загружали на колонку с белком А, уравновешенную 60 мМ глицином 3 М NaCl при рН 8,5. Колонку отмывали тем же буфером, и антитело элюировали 50 мМ ацетатом, рН 3,5. Объединенное антитело нейтрализовали до рН 7,4 с помощью 1 М Tris, концентрировали до 10 мг/мл, и диафильтровали в PBS. Очищенные mAb стерилизовали фильтрованием и хранили при -80°С.
Пример 2
Специфичность и аффинность mAb к токсину А и/или токсину В С. difficile по настоящему изобретению токсину А и/или токсину В
A. ELISA для определения специфичности mAb к токсину А и/или токсину В
ELISA-планшеты (BD Biosciences) покрывали 50 нг/лунку токсина A (List Laboratories) или 25 нг/лунку токсина В (List Laboratories) на протяжении ночи при 4°С. После отмывки планшетов в PBS-(PBS без кальция или магния, 0,05% Tween 20) лунки блокировали с помощью 200 мкл блокирующего буфера (PBS без кальция или магния, 0,1% Tween 20, 2,5% обезжиренного молока) в течение одного часа при 37°С. Этап отмывки повторяли, вносили гибридомные супернатанты или очищенные mAb на один час при 37°С. Планшет отмывали и инкубировали в течение одного часа при 37°С с конъюгированным с пероксидазой хрена (HPR) антителом козы к IgG-Fc мыши (Jackson Immunoresearch, West Grove, PA). Планшет обработали ABTS peroxidase substrate system (KPL, Gaithersburg, MD), ABTS peroxidase stop solution (KPL) и считывали на планшет-ридере SpectraMax (Molecular Devices) при 405 нм.
Данные титрации показаны на фиг.1А-С. На фиг.1А демонстрируется, что РА-38 связывается с токсином А, но не с токсином В. На фиг.1В демонстрируется, что РА-39 может связываться как с токсином А, так и с токсином В. На фиг.1C демонстрируется, что РА-41 связывается с токсином В, но не с токсином А. В. Реактивность mAb к токсину А и В в Biacore Устройство Biacore 3000 (GE Healthcare) применяли для определения специфичности связывания mAb настоящего изобретения с токсином А и/или В. mAb иммобилизовали при приблизительно 10000 единиц резонанса (RU) в сенсорных чипах СМ5 (GE Healthcare) в соответствии с инструкциями производителя для аминного присоединения. Контрольную поверхность с изотипно-совпадающими антителами несоответствующей специфичности (Southern Biotech) применяли в качестве контроля. Эксперименты по связыванию проводили при 25°С в HPS-EP буфере на основе HEPES (GE Healthcare). Очищенный токсин А или токсин В (30 нМ; List Biological Laboratories) пропускали над контрольной и тестовой проточными ячейками при скорости 5 мкл/мин. Если требовалось, дополнительные mAb (100 нМ) пропускали над проточной ячейкой при 5 мкл/мин, чтобы проверить мультивалентное или конкурентное связывание.
Как показано на фиг.2A-D, mAb РА-38 (фиг.2А) и mAb PA-50 (фиг.2С) специфично связывается с токсином А; mAb PA-41 (фиг.2D) специфично связывается с токсином В и mAb РА-39 (фиг.2В) связывается преимущественно с токсином А, но также демонстрирует связывание с токсином В. Результаты из этих данных согласуются с данными ELISA (фиг.1А-С) и демонстрируют специфичности связывания mAb настоящего изобретения к токсину А и/или токсину В.
С. Аффинность связывания
Анализ Biacore также применяли для определения авидности связывания mAb настоящего изобретения со своим соответствующим токсином. mAb захватывали с использованием сенсорного чипа СМ5, подготовленного с помощью набора для захвата антител мыши от Biacore. Затем токсин пропускали через проточные ячейки при различных концентрациях (0,4-100 нМ, двукратное возрастание). Все концентрации токсинов тестировали в двух повторностях, и поверхность чипа регенерировали после каждого опыта с применением условий, указанные в наборах. Изменения в RU записывали и анализировали с применением модели связывания 1:1 (Langmuir) в Bia Evaluation Software, которая рассчитывает kd mAb в отношении токсина. Данные ассоциации и диссоциации и аппроксимация иллюстрируются на фиг.3А-Е.
С помощью анализа Biacore определили, что kd mAb в отношении токсина А составляет 1,0 нМ для РА-38, 0,16 нМ для РА-39 и 0,16 нМ для РА-50. Определили, что kd mAb в отношении токсина В составляет 2,4 нМ для РА-39 и 0,59 нМ для PA-41. Эти результаты показали, что mAb настоящего изобретения связывали токсин А и/или токсин В с наномолярной и субнаномолярной аффинностью.
Пример 3
In vitro анализы нейтрализации на основе клеток
Анализы цитотоксичности на основе клеток, использующие клетки СНО-K1 или клетки Т-84, применяли для оценки нейтрализующих активностей описанных mAb к токсину А и токсину В.
А. Нейтрализация цитотоксического эффекта токсина А на клетках СНО-K1
Клетки СНО-K1 высевали (2000 клеток в 50 мкл/лунку) на планшеты для анализа (96-луночные планшеты, белая непрозрачная стенка, прозрачное плоское дно (Perkin Elmer)). Клеткам дали возможность прикрепляться в течение 4 часов перед обработкой. Равные объемы (35 мкл) 2 мкг/мл токсина A (List Biological Laboratories) и серийно разведенных mAb смешивали на планшетах для разбавления реактивов (96-луночные круглодонные планшеты (Falcon)) в течение 1 часа при температуре 37°С, а затем 50 мкл смеси вносили в каждую лунку планшетов. После инкубации в течение 72 часов в каждую лунку вносили 20 мкг/мл CellTiter-Blue (Promega). Планшеты инкубировали в течение дополнительных 4 часов, затем считывали на планшет-ридере SpectraMax M5 (Molecular Devices) с применением длины волны возбуждения 560 нм и длины волны излучения 590 нм. Выживание клеток сравнивали в необработанных и обработанных токсином культурах. Процент выживания клеток размещали на графике относительно концентрации mAb. Данные подавления аппроксимировали с сигмоидальной кривой доза-эффект с нелинейной регрессией с применением программного обеспечения GraphPad Prism, и рассчитывали концентрацию mAb, требуемую для 50% нейтрализации цитотоксичности (ЕС50). Как показано на фиг.4, mAb PA-39 полностью нейтрализовало активность токсина А на клетках СНО-K1 с ЕС50 - 93 пМ. В. Нейтрализация цитотоксического эффекта токсина В на клетках СНО-K1 Анализ цитотоксичности на СНО-K1 применяли для оценки нейтрализующей активности специфичных mAb к токсину В. Как и в случае оценки mAb к токсину А, токсин В (8 пг/мл, TechLab) инкубировали в течение 1 часа при 37°С с серийно разведенными mAb перед внесением к СНО-K1 (2000 клеток/лунку) в 96-луночном планшете. После 72 часов, в каждую лунку вносили 20 мкл/лунку CellTiter-Blue (Promega). Планшеты инкубировали в течение дополнительных 4 часов, а затем считывали на планшет-ридере SpectraMax M5 (Molecular Devices) с применением длины волны возбуждения 560 нм и длины волны излучения 590 нм. Жизнеспособность клеток определяли с помощью CellTiter-Blue; выживание клеток сравнивали между обработанной и необработанной культурами. Данные подавления аппроксимировали с сигмоидальной кривой доза-эффект с нелинейной регрессией с применением программного обеспечения GraphPad Prism, и рассчитывали концентрацию mAb, требуемую для 50% нейтрализации цитотоксичности (ЕС50). Как показано на фиг.5, РА-41 показал высокую степень активности (EC50 - 9,2 пМ) в нейтрализации цитотоксичности токсина В на клетках СНО-K1.
Хотя mAb PA-39 продемонстрировали связывание с токсином В, исходя из анализов ELISA и Biacore, это mAb не имело активности in vitro в отношении токсина В в анализах на основе клеток СНО-K1 и других клеток. Уже сообщалось об антителах, которые связываются как с токсином А, так и с токсином В, но не имеют функциональной активности в нейтрализации токсина А или токсина В w vitro анализах на основе клеток (46, 92 и 93). Настоящее изобретение включает новое mAb с двойной способностью связывать как токсин А, так и токсин В, а также нейтрализовать цитотоксичность токсина С.difficile, например, токсина А.
С.Нейтрализация цитотоксического эффекта токсина А на клетках Т-84
Анализ цитотоксичности на Т-84 применяли для оценки нейтрализующей активности описанных mAb к токсину А. Клетки Т-84 высевали (15000 клеток в 50 мкл/лунку) в планшеты для анализа (96-луночные планшеты, белая непрозрачная стенка, прозрачное плоское дно (Perkin Elmer)). Клеткам дали возможность прикрепляться в течение 4 часов перед обработкой. Равные объемы (35 мкл) 240 нг/мл токсина A (TechLab) и серийно разведенные mAb смешивали в планшетах для разведения реактивов (96-луночные круглодонные планшеты (Falcon)) в течение 1 часа при 37°С, а затем в каждую лунку планшетов анализа вносили по 50 мкл смеси. После инкубации в течение 72 часов, в каждую лунку вносили по 20 мкл/лунку CellTiter-Blue (Promega). Планшеты инкубировали в течение дополнительных 4 часов, затем считывали на планшет-ридере SpectraMax M5 (Molecular Devices) с применением длины волны возбуждения 560 нм и длины волны излучения 590 нм. Выживание клеток сравнивали в необработанных и обработанных токсином культурах. Данные подавления аппроксимировали с сигмоидальной кривой доза-эффект с нелинейной регрессией с применением программного обеспечения GraphPad Prism, и рассчитывали концентрацию mAb, требуемую для 50% нейтрализации цитотоксичности (ЕС50). Как показано на фиг.6, mAb РА-38 и РА-50 полностью нейтрализовали активность токсина А на клетках Т-84 с ЕС50 - 175 пМ и 146 пМ, соответственно. В анализе на клетках Т-84 mAb РА-39 показало минимальную активность в отношении токсина А, а РА-41 не было активным.
D. Гемагглютинация красных кровяных клеток кролика (RBC)
Способность mAb настоящего изобретения блокировать связывание токсина А с клеточными рецепторами оценивали с помощью анализа гемагглютинации. Для этого анализа равные объемы (30 мкл/лунку) токсина А (8 мкг/мл; TechLab) и серийно разведенных mAb смешивали в планшетах для разведения реактивов (96-луночные круглодонные планшеты (Falcon)) в течение 1 часа при температуре 4°С. Красные кровяные клетки кролика (RBC) (Colorado Serum Co., Denver, CO) три раза промывали PBS и ресуспендировали в PBS. 60 мкл 1% RBC суспензии вносили в лунки 96-луночных планшетов, содержащих смесь токсин A-mAb, планшеты инкубировали при 4°С в течение 4 часов. Свободный токсин А вызывает гемагглютинацию RBC. Соответственно, добавление mAb к токсину А, которое связывается с токсином А, как ожидается, предотвращает гемагглютинацию. Степень гемагглютинации определяли с помощью устройства ImageQuant 400; полная гемагглютинация давала более сильный сигнал по сравнению с RBC в суспензии. Значения ЕС5о рассчитывали из данных подавления с применением аппроксимации сигмоидальной кривой доза-эффект с нелинейной регрессией от GraphPad Prism. Как показано на фиг.7, mAb PA-38 (закрашенные квадраты) и mAb PA-50 (закрашенные треугольники) полностью нейтрализовали активность токсина А на RBC с ЕС50 - 30 нМ и 1,8 нМ, соответственно. Оказалось, что РА-38 и PA-50 нейтрализуют токсин А, блокируя связывание токсина А с его рецептором. Обнаружили, что mAb PA-39 и РА-41 оказались неактивными в данном анализе.
Е. Анализ монослоя на Сасо-2
Клетки Сасо-2 высевали (25000 клеток в 75 мкл/лунку) в верхнюю камеру 96-луночных планшетов Multiscreen Сасо-2 (Millipore Billerica, MA) с 250 мкл среды, внесенной в нижнюю камеру. Клеткам дали возможность расти в течение 10 дней с регулярными заменами среды каждые 3-4 дня. После инкубации в течение 10-14 дней образование плотного монослоя подтверждали с помощью измерения трансэпителиального электрического сопротивления (TEER) с использованием эпителиального вольтомметра (модель: EVOMX, World Precision Instruments, Sarasota, FL). После того, как выяснили и определили целостность монослоя, равные объемы (60 мкл) токсина А (при 50 нг/мл) и серийно разведенного mAb смешали в течение 1 часа при 37°С, а затем вносили в верхнюю камеру планшета анализа. Планшеты инкубировали в течение 18-24 часов, а затем измеряли значение TEER с помощью вольтомметра. Целостность монослоя сравнивали в необработанных и обработанных токсином лунок. Как показано на фиг.8, данные подавления аппроксимировали с сигмоидальной кривой доза-эффект с нелинейной регрессией с применением программного обеспечения GraphPad Prism, чтобы определить концентрацию mAb, требуемую для 50% подавления токсина (EC50). mAb PA-38 и PA-50 нейтрализовали нарушение монослоев Сасо-2 токсином А с ЕС50 -485 пМ и 196 пМ, соответственно. Обнаружили, что другие mAb оказались неактивными в этом анализе.
Без желания быть связанными теорией, in vitro результаты на основе клеток показывают, что РА-38 и PA-50, по-видимому, представляют собой один класс mAb к токсину А, в то время как РА-39 представляет собой другой класс mAb к токсину А. mAb PA-38 и РА-50, по-видимому, связывают эпитоп токсина А, важный для связывания с рецептором; mAb PA-39, по-видимому, связывает токсин таким способом, который более прямо блокирует цитотоксические эффекты токсина A in vitro.
Пример 4
Оценка in vivo эффективности mAb к токсину А и к токсину В С. difficile по настоящему изобретению на мышах
Мышиную модель in vivo использовали для измерения способности описанных в данном документе mAb нейтрализовать циркулирующие токсины С. difficile у животного. In vivo нейтрализующие активности mAb PA-38, PA-39, PA-41 или РА-50, вводимых по отдельности или в комбинации, тестировали в отношении эффектов комбинированных системных токсинов А и В С. difficile (Techlab) на тестовых животных.
В экспериментах использовали самок Swiss Webster, 4-6 мышей/группа (возраст: ~6-8 недель в начале исследования; Charles River Laboratories). Мышей акклиматизировали в лаборатории, как минимум в течение 4 дней, и перед использованием животных проверяли их здоровье. Эксперименты на животных проводили в соответствии с утвержденным IACUC протоколом.
Первоначальные эксперименты проводили для определения токсичности токсинов А и В на мышах. Животным вводили 0, 20, 100, 500, 2500 нг/токсина/животное интраперитонеально (i.p.) и дозу, которая была летальной для животных, выбирали для применения в последующих экспериментах по нейтрализации антителами. Контрольные мыши, которым инъецировали PBS, были незатронутыми. Дозу токсина A (TechLab) 100 нг выбрали для экспериментов по нейтрализации, так как обнаружили, что это был самый низкий уровень дозы, являющийся летальным для 100% мышей в течение 24 часов после инъекции. Аналогично, дозу токсина В (TechLab) 100 нг выбрали для экспериментов по нейтрализации, так как это был самый низкий уровень дозы, являющийся летальным для 100% мышей в течение 24 часов после инъекции.
Для оценки нейтрализующей активности mAb к токсину, мышам (5 на группу) интраперитонеально вводили одну инъекцию каждого mAb при различных уровнях дозы в день 0, с последующей интраперитонеальной инъекцией 100 нг/200 мкл токсина А или токсина В в день 1. Животных наблюдали ежедневно в течение 3 дней и, затем, еженедельно до 21 дня после инъекции токсина. Выживание животных было главным ожидаемым результатом исследования.
Для экспериментов по нейтрализации все дозы антитела составляли в PBS без кальция или магния (PBS-, Invitrogen, Carlsbad, СА). В день 0 мышам интраперитонеально вводили одну инъекцию mAb PA-38, PA-39, PA-41 или РА-50 при разных уровнях дозы (200 мкл/доза/животное), с последующей инъекцией токсина (интраперитонеально в место, отличное от места инъекции антител) в день 1. Состояние здоровья животных отслеживали ежедневно в течение первых 3-4 дней и, затем, дважды в неделю в течение 21 дня после введения токсина. Наблюдения за животными, находящимися в клетках, (например, согнутая поза, спутанная шерсть, малоподвижные) записывали так же, как и выживание.
Оценили отличающиеся уровни дозы РА-38 (0,2 мкг - 250 мкг/животное) и РА-50 (0,2 мкг - 100 мкг/животное). На этой модели обнаружили, что РА-38 и РА-50 нейтрализуют дозу токсина А 100 нг и обеспечивают 100% выживание при таких низких уровнях дозы, как 2 мкг mAb/животное, как показано на фиг.9А и В. В отличие от них, человеческое моноклональное антитело сравнения к токсину А (WO/2006/121422 и US2005/0287150), упомянутое в данном документе как mAb сравнения CDA-1, при 5 мкг/животное, не защищало животных от связанной с токсином смерти, в отличие от mAb по настоящему изобретению (фиг.9С). Оценили дозы PA-41 в диапазоне от 0,5 мкг до 250 мкг, и обнаружили, что одна доза 5 мкг mAb/животное полностью нейтрализует токсичность дозы токсина В 100 нг, как показано на фиг.10. Не наблюдали, чтобы mAb РА-39 (100 мкг/животное) обеспечивало замедление связанной с токсином смерти мышей ни от токсина А, ни от токсина В.
После того как нейтрализующая активность индивидуальных антител к токсину А (РА-38, РА-50) или к токсину В (PA-41) была явно показана т vivo, проводили эксперимент для тестирования комбинации mAb (РА-38+PA-41) при уровнях дозы 5 и 50 мкг каждого mAb по отношению к комбинированной летальной дозе токсинов (100 нг токсина А и 100 нг токсина В) на такой же мышиной модели w vivo. Кроме того, индивидуальные моноклональные антитела включали в качестве контролей. Как показано на фиг.11, комбинация mAb РА-38 и PA-41 показала защиту от комбинации токсинов как при 50 мкг/животное (4 из 5 выжили), так и при 5 мкг/животное (1 из 5 выжили) по сравнению с активностью каждого из mAb по отдельности (все животные погибли в течение 24 часов после введения токсина).
Пример 5
Оценка mAb к токсину А и к токсину В С.difficile по настоящему изобретению на модели ассоциированной с С.difficile диареи (CDAD) у золотистых сирийских хомячков
Модель CDAD на хомячках воспроизводит ключевые аспекты заболевания CDAD у людей. В результате лечения антибиотиками уничтожается нормальная флора толстой кишки, и хомячки становятся легко восприимчивыми к инфекции С.difficile. Инфекция приводит к тяжелой диарее, псевдомембранозному колиту и смерти. Модель CDAD на хомячках использовали для оценки потенциальной эффективности mAb по настоящему изобретению для предотвращения заболевания и смерти животных, связанных с заражением живыми бактериями С.difficile. Эти эксперименты проводили в соответствии с утвержденными IACUC протоколами.
А. Фармакокинетический анализ
Перед проведением исследования эффективности на модели хомячков с использованием хомячков, зараженных живыми микроорганизмами С.difficile, осуществили фармакокинетические исследования на нормальных неинфицированных хомячках. Золотистым сирийским хомячкам (Harlan) интраперитонеально инъецировали 0,2 мг/животное или 1 мг/животное очищенных mAb РА-38 или mAb РА-41. Образцы крови собирали при помощи методик ретро-орбитального или с пункцией сердца (терминального) забора крови через 0,125, 0,25, 1, 2, 4, 7, 10, 14 и 21 дней. Образцы крови центрифугировали при 8000 оборотов в минуту в течение 10 минут для получения сыворотки.
Концентрацию mAb в сыворотке определяли при помощи ELISA. Девяносто шестилуночные планшеты для ELISA (BD Biosciences) покрывали в течение ночи токсином А (TechLab) или токсином В (TechLab) по 250 нг/лунку при 4°С. Планшеты три раза промывали PBS/0,05% Tween-20® (PBS-T) и блокировали 200 мкл блокирующего буфера (PBS без кальция или магния, 0,1% Tween 20®, 2,5% обезжиренного молока) в течение одного часа при комнатной температуре. Стандартный образец антител (очищенные mAb PA-38 или mAb PA-41) разводили в 1% объединенной сыворотке от не подвергавшихся воздействию хомячков для получения стандартной кривой с диапазоном 0,3-1000 нг/мл. Разведенные тестовые и стандартные образцы инкубировали в течение одного часа при комнатной температуре.
Планшеты промывали (как описано выше) и инкубировали в течение одного часа при комнатной температуре с HRP-конъюгированными антителами козы, специфичными к Fey IgG мыши (Jackson Immunoresearch). Планшеты обрабатывали ABTS peroxidase substrate system (KPL), остановили ABTS peroxidase stop solution (KPL) и считывали на планшет-ридере SpectraMax (Molecular Devices) при 405 нм. Концентрацию mAb у каждого хомячка в различные моменты времени рассчитали с использованием стандартных кривых. Приблизительно 10% образцов не имели титра антител, вероятно, из-за пропуска инъекции или отсутствия абсорбции, эти образцы не включили в расчет параметров PK. Некомпартментный фармакокинетический анализ проводили с использованием WinNonLin, версия 4.0 (Pharsight Corp, Mountain View, CA), и данные показаны в таблице 1 и на фиг.12А и В. Как указано, Cmax и площадь под кривой (AUC) были зависимыми от дозы. Каждое из антител продемонстрировало время полужизни больше, чем 6 дней, что обеспечило сохранение антител в исследованиях эффективности, описанных ниже.
В. Оценка mAb к токсину А и к токсину В С.difficile по настоящему изобретению, в комбинации, на модели, ассоциированной с С.difficile диареи (CDAD) у золотистых сирийских хомячков
Эксперимент по эффективности осуществили для оценки способности мышиных mAb к токсину А и токсину В по настоящему изобретению влиять на выживаемость инфицированных животных в in vivo модели ассоциированной с С.difficile диареи у хомячков. Самцов золотистых сирийских хомячков (~90 г) (Crl:LVG(SYR)) (Charles River Laboratories, Inc., Kingston, NY) предварительно обработали однократной подкожной дозой клиндамицина (Sigma, St. Louis, составили в PBS при 5 мг/мл) при 50 мг/кг для нарушения нормальной флоры толстой кишки. На следующий день хомячки в соответствующих тестовых группах получили пероральную дозу (1×10 CFU в 0,5 мл) суспензии С.difficile (штамм АТСС 43596). Штамм 43596 ранее использовали в моделях на хомячках для оценки нейтрализующих антител. Животных еженедельно взвешивали и отслеживали состояние здоровья и выживание.
Тестовые антитела включали комбинации мышиных mAb по настоящему изобретению, т.е. комбинацию mAb РА-38 и РА-41 или комбинацию mAb РА-39 и РА-41. Поликлональные антитела козы к токсину А и токсину В С.difficile (Techlab) включали в качестве положительного контроля. mAb и контрольные реактивы вводили как описано в таблице 2.
Хомячки из группы 1 не получали никакой обработки на протяжении всего исследования. Хомячков из групп 2-7 предварительно обработали путем однократной подкожной дозы клиндамицин фосфата при 50 мг/кг (день -1). Хомячкам из групп 5-7 вводили Поликлональные антитела козы (группа 5) или комбинации mAb (группа 6 и 7), как указано в таблице 2, посредством интраперитонеального введения непосредственно после обработки клиндамицином. Через 24 часа каждого хомячка из групп 3-7 инокулировали 0,5 мл соответствующей суспензии С.difficile АТСС 43596 (106-107 CFU/мл) через желудочный зонд (день 0). После первоначальной обработки антителами в день -1 проводили последующие три обработки этих групп каждые два дня, один раз в день, в дни 1, 3 и 5. Ванкомицин (20 мг/кг BID) вводили животным из группы 4 через желудочный зонд дважды в день с интервалом примерно 6 часов в дни 1-5. Введение ванкомицина (животные из группы 4) начинали примерно через 20-24 часа после того, как животных инокулировали С.difficile.
Результаты выживания в обработанных mAb и контрольных группах проиллюстрированы на фиг.13. Обобщение смертности хомячков во всех группах приведено в таблице 3. Всех хомячков, инфицированных С.difficile без какой-либо обработки (инфицированный контроль, группа 3) нашли погибшими в день 2 или день 3 исследования. В группе, обработанной ванкомицином, (группа 4), семь из восьми хомячков нашли погибшими между днями 15 и 19. Как обычно наблюдается в этой модели, большинство (88%) хомячков, обработанных ванкомицином, повторно заболели и погибли от инфекции С.difficile через две недели после прекращения терапии. Напротив, все хомячки, обработанные комбинацией mAb РА-39+РА-41 (группа 7), и 7 из 8 хомячков, обработанных комбинацией mAb PA-38+РА-41 (группа 6), выжили до конца исследования (37 дней после инфекции). Кроме того, все животные из группы, обработанной поликлональными антителами козы (группа 5) были живы до конца исследования. У всех выживших хомячков были нормальные ЖКТ при посмертном вскрытии (см. фиг.15А, С и D).
Эти результаты показывают, что комбинация mAb PA-39 и РА-41 и комбинация mAb РА-38 и РА-41 эффективно и длительно защищали хомячков от тяжелого заболевания, как первоначально, так и от последующих рецидивов заболевания. Продолжительность эффекта обработки mAb (37 дней) значительно превысила интервал (две недели) для возникновения инфекции С.difficile после обработки клиндомицином в модели на хомячках.
Вес тела животных в группах, обработанных поликлональными антителами и mAb, a также в контрольных группах проиллюстрирован на фиг.14. Хомячки в неинфицированной контрольной группе (группа 1) постоянно набирали вес, в диапазоне от 13 до 29 г, на протяжении исследования. Все животные инфицированного контроля умерли до первого, после инокуляции, измерения веса. Средний вес тела животных, обработанных ванкомицином, поликлональными антителами козы, комбинацией mAb PA-38+РА-41 и комбинацией mAb PA-39+РА-41 значительно снизился в течение первой недели после инфицирования. После этого средний вес тела в группах, обработанных mAb, а также группах, обработанных поликлональными антителами, постоянно увеличивался, и был подобен таковому в неинфицированном контроле до конца исследования, указывая на то, что не было явной токсичности.
В общем, в этом исследовании на хомячках было показано, что комбинации mAb по настоящему изобретению эффективно и длительно защищали хомячков от смертности в соответствующих и строгих моделях на хомячках инфекции С.difficile. Эти полученные результаты подтверждают механизм, при помощи которого комбинации mAb защищали животных от заболевания С.difficile в течение времени, достаточно продолжительного для обеспечения естественного развития и репопуляции нормальной флоры кишечника у обработанных mAb животных по сравнению с неинфицированными животными (фиг.15A-D). Таким образом, mAb по настоящему изобретению обеспечивали терапевтическую защиту инфицированных животных и влияли на устранение ассоциированного с С.difficile заболевания, восстановление здоровья ЖКТ и выживание.
С. Оценка индивидуальных mAb к токсину А и/или токсину В С.difficile по настоящему изобретению на модели ассоциированной с С.difficile диареи (CDAD) у золотистых сирийских хомячков
Дополнительное исследование на хомячках провели для оценки эффективности индивидуальных мышиных mAb по настоящему изобретению, вводимых инфицированным животным, по сравнению с mAb, вводимыми в комбинации. Обрабатываемые группы данного исследования представлены в таблице 4.
В этом исследовании хомячков в группах 1-7 предварительно обработали однократной подкожной дозой клиндамицин фосфата при 50 мг/кг (день -1). Животным в группах 3-7 интраперитонеально вводили mAb непосредственно после обработки клиндамицином. Через 24 часа каждому хомячку из групп 1-7 инокулировали 0,5 мл суспензии С.difficile через желудочный зонд (день 0), как описано в разделе В выше. Обработки тестовым mAb вводили животным в группах 3-7 в однократной дозе в дни 1, 3 и 5. Ванкомицин вводили животным дважды в день в дни 1-5 (группа 2). Хомячков обследовали на жизнеспособность дважды в день. Вес тела отмечали один раз в неделю. Проводили вскрытие животных, которых обнаружили мертвыми или которых подвергли эвтаназии в ходе исследования. В конце исследования (40 дней после инокуляции) проводили конечное вскрытие всех остальных хомячков.
Выживание обработанных mAb и контрольных групп животных изображено на фиг.16А, а день смерти хомячков во всех группах обобщен в таблице 5.
В инфицированной контрольной группе (группа 1) всех семь хомячков нашли погибшими в день 2. У всех этих хомячков при посмертном обследовании были воспаленные ЖКТ. В обработанной ванкомицином группе (группа 2), все семь хомячков погибли между днем 12 и 19 исследования. Сроки этих смертей были подобны срокам, которые наблюдались ранее на этой модели при обработке ванкомицином. Посмертное обследование показало, что у всех хомячков были воспаленные ЖКТ, что свидетельствовало об инфекции С.difficile.
Обработка комбинацией mAb PA-39+РА-41 (группа 3) в данном исследовании была очень эффективна в отношении защиты инфицированных хомячков. Шесть из семи хомячков в группе 3 выжили до конца исследования. Одного хомячка нашли погибшим в день 12 исследования. Посмертное обследование показало, что у этого хомячка был воспаленный ЖКТ, что типично для инфекции С.difficile.
Среди обработок одними антителами (группы 4-7), только mAb РА-39 (группа 6) показало некоторую защитную активность у обработанных животных. В этой группе хомячков находили погибшими в продолжение дней 2-12. В группах, обработанных индивидуальными mAb, РА-41 (группа 4), РА-38 (группа 5) или РА-50 (группа 7), хомячков находили погибшими в дни 2 и 3. При конечном вскрытии у всех хомячков в этих группах были воспаленные ЖКТ, что свидетельствует об инфекции С.difficile. В отличие от этого, у всех обработанных хомячков, которые выжили, были нормальные ЖКТ.
Результаты этого исследования показывают, что обработка комбинацией mAb РА-39+РА-41 успешно защищала хомячков от развития болезни в течение более одного месяца после прекращения обработки, и они сходны с таковыми, полученными в описанном выше исследовании из примера 5 В, в котором восемь из восьми хомячков, обработанных комбинацией РА-39+РА-41, выжили при инфекции С.difficile. В этом исследовании mAb РА-39, в качестве обработки одними mAb, проявило некоторую активность в отношении защиты инфицированных С.difficile хомячков от заболевания С.difficile.
D. Определение концентрации антител в конечных образцах крови и оценка наличия С.difficile в конечных образцах слепой кишки хомячков
Кровь собирали от животных, которых в ходе исследования обнаружили умирающими. В конце исследования также собрали кровь от всех животных, которые остались живы. Образцы крови обработали для сбора сыворотки, если ниже не указано иное. Обработанные образцы заморозили при <-70°С для возможного дальнейшего анализа.
После in vivo исследований эффективности на хомячках, описанных выше, исследовали наличие mAb в конечных образцах крови, взятых у животных в исследованиях. Для исследования комбинаций mAb, описанных в примере 5В, получили конечные образцы крови в день 37 исследования от восьми животных из группы 7, которые получили дозу (Q2d×4) комбинации mAb РА-39 (50 мг/кг)+mAb PA-41 (40 мг/кг). В сыворотке, собранной из этих конечных образцов крови, обнаружили РА-39 на уровне 3,3±3,4 мкг/мл, и обнаружили PA-41 на уровне 2,4±1,7 мкг/мл. В случае исследования индивидуальных mAb, описанного в примере 5С, были получены конечные образцы крови в день 40 исследования от шести животных из группы 3, которые получили дозу (Q2d×4) комбинации mAb РА-39 (50 мг/кг)+mAb PA-41 (50 мг/кг), и образцы крови обработали для получения плазмы. В плазме, собранной из этих конечных образцов крови, обнаружили РА-39 на уровне 1,8±1,4 мкг/мл, и обнаружили PA-41 на уровне 3,4±3,2 мкг/мл. Предел обнаружения антител в этих анализах составил 1,6 нг/мл. Таким образом, определяемые уровни mAb измеряли у животных в течение нескольких недель. Это поддерживает способ действия, при котором эти mAb обеспечивают терапевтический эффект на протяжении режима обработки и после введения последних доз mAb.
При конечном вскрытии каждого хомячка из группы 3 в примере 5С подвергли осмотру слепую кишку, и каждая оказалась нормальной. Не наблюдалось воспаление или покраснение, и содержимое слепых кишок было относительно твердым по консистенции. Стенку каждой слепой кишки разрезали стерильным одноразовым скальпелем. Для каждого хомячка использовали новый скальпель, чтобы избегнуть перекрестного загрязнения. Небольшое количество кала удалили из слепой кишки стерильным тампоном и поместили в стерильную пробирку для тестирования. 10 мкл инокуляционную петлю использовали для сбора образца кала из пробирки и посева образца штрихом на чашку Петри с агаром, содержащую CCFA со средой из лошадиной крови (Remel, Lot 735065), которая является селективной для С.difficile.
Чашки Петри поместили в анаэробный бокс и инкубировали в течение 48 часов при температуре 37°С. Одну чашку Петри засевали штрихом с С.difficile ATCC 43596 из исходной культуры и инкубировали совместно со штриховыми культурами из фекалий для сравнения колоний. Колонии, напоминающие С.difficile, обнаружили на чашках Петри от всех шести хомячков. Результаты этих экспериментов показывают, что хотя выжившие животные, обработанные mAb по настоящему изобретению, все еще являлись хозяевами С.difficile, их нормальная флора репопулировала с восстановлением нормального микробного равновесия в кишечнике, которое способствовало их общей выживаемости.
Е. Оценка гуманизированных mAb к токсину А и/или токсину В С.difficile по настоящему изобретению и mAb сравнения к токсину А и/или токсину В на модели ассоциированной с С.difficile диареи (CDAD) у золотистых сирийских хомячков
Дополнительные исследования на хомячках проводили с целью оценки in vivo эффективности комбинации гуманизированных mAb к токсину А и токсину В по настоящему изобретению по сравнению с комбинацией человеческого mAb сравнения к токсину A CDA-1 и человеческого mAb сравнения к токсину В CDB-1 при введении соответствующих комбинаций антител инфицированным С.difficile животным. Группы обработки данного исследования представлены в таблице 5А.
Тестовые антитела включали комбинации гуманизированных mAb по настоящему изобретению, то есть комбинацию гуманизированного mAb PA-50 к токсину А и гуманизированного mAb PA-41 к токсину В (hPA-41+hPA-50), или комбинацию человеческого mAb сравнения к токсину А, именуемого mAb сравнения CDA-1, и человеческого mAb сравнения к токсину В, именуемого mAb сравнения CDB-1 (CDA-1+CDB-1), в количествах, указанных в таблице 5А. mAb сравнения синтезировали (DNA 2.0) на основе опубликованных областей 3D8 и 124 тяжелой и легкой цепи (WO2006/121422 и US2005/0287150), клонированных в полноразмерных векторах экспрессии IgGI (pCON-гамма и pCON-каппа), экспрессированных в клетках СНО-KSV1 и очищенных с использованием способов, описанных в данном документе. Сочетания mAb и контрольную обработку применяли, как описано в таблице 5А.
Способы обработки были, в основном, такими, как описано в части В данного примера выше. Вкратце, в примере 5Е данного исследования использовали золотистых сирийских хомячков (Charles River Laboratories, Stone Ridge, NY, возраст 50 дней). Хомячки в контрольной группе 1 были неинфицированными (и необработанными). Животных в группах 2-7 предварительно обрабатывали однократной подкожной дозой клиндамицин фосфата при 50 мг/кг, чтобы нарушить нормальную флору толстого кишечника (день -1). Животным групп 4-7 интраперитонеально вводили дозы непосредственно после обработки клиндамицином. Через 24 часа каждому животному из групп 2-7 инокулировали 0,5 мл суспензии С.difficile (ATCC 43596, штамм 545) через желудочный зонд (день 0), (то есть пероральная доза). Дополнительные введения тестовых обработок применили к животным в группах 4-7 в однократной дозе в день 1, 3 и 5. Ванкомицин вводили животным группы 3 дважды в день, с разницей примерно в 6 часов, в день 1-5. Животных еженедельно взвешивали и ежедневно отслеживали состояние здоровья и выживаемость в течение 39 дней. По окончании осуществили вскрытие, и после анаэробной культивации при 37°С в течение 48 часов на селективной среде определили титры микроорганизмов С.difficile в слепой кишке. Предел обнаружения составил 20 CFU/г содержимого слепой кишки. Это исследование и описанные выше исследования на хомячках были проведены в Ricerca Biosciences (Concord, ОН) в соответствии с директивами Институционального комитета по уходу за животными и их использованию.
Результаты выживаемости для обработанных mAb и контрольных групп показаны на фиг.16В-1. Данные исследования о смертности представлены в таблице 5В ниже. Краткое описание выживаемости хомячков представлено в таблице 5С ниже.
30
Как отмечено в таблице 5В, все 4 неинфицированных контрольных хомячка (группа 1) выжили до конца исследования. Все инфицированные контрольные (группа 2) животные погибли в дни 2 и 3. В обработанной ванкомицином группе (группа 3) все исследуемые животные погибли между днем 13 и 22. В группе 4 с hPA-50+hPA-41 (50, 50 мг/кг) девять из десяти животных выжили до конца исследования. Одного хомячка из этой группы обнаружили погибающим в день 8, с изменением цвета ЖКТ на красный, тогда как у остальных девяти выживших животных в этой группе был нормальный ЖКТ. Все десять хомячков в группе 5 выжили до конца исследования и имели нормальные ЖКТ. В группах с mAb сравнения девять из десяти животных, которым давали дозу 50 мг/кг (группа 6), погибли между днями 5 и 14; одно животное погибло в день 28 исследования. Все десять хомячков, обработанных 20 мг/кг комбинации mAb сравнения, (группа 7) погибли между днем 5 и 14 исследования.
Как отмечено в таблице 5С, все четыре животных в неинфицированной контрольной группе 1 выжили до конца исследования (40 дней). У всех хомячков, инфицированных С.difficile без какой-либо обработки (инфицированные контрольные, группа 2), была средняя выживаемость 2 дня; животные в этой группе не выжили до конца исследования. В обработанной ванкомицином группе (группа 3) средняя выживаемость составила 20 суток, животные не выжили до дня 40. Обе обработки комбинациями mAb hPA-50+hPA-41 были эффективными в отношении защиты инфицированных животных в этом исследовании (группы 4 и 5). Все (100%) хомячков, обработанных комбинацией гуманизированных mAb PA-50+РА-41 (20 мг/кг каждого, группа 5) и 90% хомячков, обработанных комбинацией гуманизированных mAb PA-50+РА-41 (50 мг/кг каждого; группа 4), выжили до конца исследования (40 дней после инфицирования). При посмертном вскрытии у всех выживших хомячков были, в основном, нормальные ЖКТ. В отличие от этого, средняя выживаемость животных, получавших комбинацию mAb сравнения к токсину А и к токсину В, была сходной для обеих доз mAb сравнения; в двух группах, обработанных комбинациями mAb сравнения, все животные погибли. В частности, для животных, получавших комбинацию CDA-1+CDB-1 (50 мг/кг каждого; группа 6), средняя выживаемость составила 14 дней, а для животных, получавших комбинацию CDA-1+CDB-1 (20 мг/кг каждого; группа 7), средняя выживаемость составила 11 дней.
В исследовании дополнительные оценки включали измерения веса тела, макроскопическое вскрытие и определение титра микроорганизмов С.difficile в слепой кишке по окончании исследования. Средний вес тела животных, обработанных ванкомицином или комбинацией РА-50/РА-41, снизился в течение первой недели после инфицирования, а затем восстановился (фиг.16В-2). В день 39 средний вес тела животных, обработанных комбинацией РА-50/РА-41, был сходным с таковым у здоровых неинфицированных животных, которых содержали параллельно (Р>0,05). Средний вес тела животных, обработанных комбинацией антител сравнения CDA1/CDB1, неуклонно снижался в ходе исследования.
В день 39 ЖКТ 19 выживших животных, обработанных комбинацией РА-50/РА-41, оказались сходными с таковыми у неинфицированных животных; и титры С. difficile в слепой кишке были либо необнаруживаемыми (<1,3 log10 CFU, n=11), либо низкими (4,15±0,76 log10 CFU, т=8). В отличие от этого, наблюдали воспаленные ЖКТ у некоторых или всех животных в других обработанных группах в момент смерти. С. difficile обнаружили у 4 из 4 необработанных животных (среднее CFU=8,96±0,59 log10, P<0,0001 по отношению к РА-50/РА-41) и у 4 из 4 обработанных ванкомицином животных (среднее CFU=6,01±0,93 log10, P<0,017 по отношению к РА-50/РА-41), в отношении которых осуществили анализы слепой кишки. У большинства хомячков, обработанных комбинацией антител сравнения CDA1/CDB1, практически не было содержимого слепой кишки, что исключает проведение количественного анализа титров С. difficile.
Для статистического анализа в данном исследовании и вышеупомянутых исследованиях данные нейтрализации были пригодны для логистического уравнения с четырьмя параметрами с использованием GraphPad Prism (v. 4,0 GraphPad Software, San Diego, CA). Для сравнения данных средней и общей выживаемости использовали двусторонние t-критерии или логарифмические ранговые критерии, соответственно.
Результаты исследования показывают, что обработка инфицированных С.difficile животных комбинацией гуманизированных mAb PA-50 и РА-41 при обоих уровнях дозы способствует эффективной и длительной защите хомячков от тяжелых заболеваний, как первоначально, так и при последующих рецидивах заболевания. Продолжительность эффекта обработки комбинацией гуманизированных mAb (40 дней) значительно улучшила долговременную выживаемость животных по сравнению с обработкой ванкомицином или mAb сравнения к токсину А и к токсину В в качестве контроля.
Как свидетельствуют in vivo исследования на животных, комбинированная обработка комбинацией гуманизированных mAb PA-50/PA-41 была высокоэффективной по отношению к инфекции С.difficile на хорошо организованной модели золотистых сирийских хомячков в отношении CDI и терапии. Короткий курс обработки PA-50/PA-41 привел к 95% выживаемости к дню 39 после инфицирования по сравнению с 0% выживаемости животных, которых не обработали, получили стандартную антибактериальную терапию или mAb сравнения. Через 39 дней после инфицирования у животных, обработанных PA-50/PA-41, был нормальный вес и не было никаких очевидных повреждений ЖКТ. С.difficile нельзя выделить у большинства животных, что отражает >7-log10 клиренса по отношению к необработанным животным. Одно из возможных объяснений этих результатов является то, что mAb-опосредованная нейтрализация токсинов в отсутствие антибиотиков обеспечила восстановление защитной микрофлоры в ЖКТ животных.
Сообщалось, что либо токсин А, либо токсин В, по отдельности, могут вызвать смертельное заболевание в модели CDI на хомячках, и, как правило, для максимальной эффективности лечения требуются mAb к обоим токсинам. В исследованиях, описанных выше, мышиные mAb PA-50 и РА-41 не показали благоприятного воздействия на выживание при применении индивидуально при дозах 50 мг/кг в модели на хомячках, что подчеркивает необходимость в комбинированной обработке.
В целом в этом исследовании на хомячках, подобно результатам описанного выше использования комбинации mAb мыши, продемонстрировали, что комбинации гуманизированных mAb по настоящему изобретению способствуют эффективной и длительной защите хомячков от смертности в строгой модели на хомячках инфекции С.difficile. He будучи ограниченными теорией, эти данные подтверждают механизм действия, с помощью которого комбинации гуманизированных mAb защищают животных от заболевания С.difficile и/или обеспечивают осуществление у животных реакции против инфекции С.difficile, в течение времени, достаточно длительного, чтобы обеспечить естественное развитие и репопуляцию нормальной флоры кишечника у обработанных гуманизированными mAb животных, таким образом обеспечивая терапевтическую защиту в отношении инфицированных животных, эффективное устранение ассоциированного с С.difficile заболевания и восстановление здоровья ЖКТ и выживание.
Пример 6
Связывание mAb с областями токсина А и токсина В С.difficile
Эксперименты проводили для определения областей эпитопа токсина А и токсина В С.difficile, с которым связываются mAb настоящего изобретения. Как токсин А, так и токсин В, продуцируемые С.difficile, составляют приблизительно 300 кДа и имеют значительную гомологию последовательности и структуры. Оба имеют С-концевой рецепторсвязывающий домен, который содержит клостридиальные повторяющиеся олигопептиды (CROP), центральный гидрофобный домен, который, как полагают, вызывает образование пор и опосредует включение токсина в мембрану эндосомы, и протеолитический домен, который расщепляет N-концевой ферментативный домен, таким образом, позволяя глюкозилтрансферазе проникнуть в цитозоль. Последовательности нуклеиновых кислот, кодирующие токсины С.difficile, а также другие белки С.difficile, были опубликованы и также доступны в базе данных Национального центра биотехнологической информации (NCBI) (т.е. www.ncbi.nlm.nih.gov). Например, для С.difficile штамма VPI 10463, последовательности ДНК, кодирующие токсин А и токсин В, можно найти под № доступа NCBI х92982; к тому же № доступа NCBI NC_009089, область 795842-803975, предоставляет последовательность ДНК для токсина А из полной геномной последовательности хромосомы С.difficile 630, в то время как № доступа NCBI NC_009089, область 787393-794493, предоставляет последовательность ДНК, кодирующую токсин В из последовательности хромосомы С.difficile 630.
А. Картирование антителосвязывающего домена токсина В С.difficile
Токсин В С.difficile полной длины состоит из трех основных доменов: N-концевого ферментативного домена (63 кДа), проявляющего глюкозилтрансферазную (GT) активность, и С-концевого (59 кДа), связывающего клеточный рецептор, которые находятся на обоих концах предполагаемого транслокационного домена (148 кДа) (фиг.17А и С). Некоторые фрагменты токсина В получали при помощи ферментативного расщепления токсина В полной длины с использованием фермента каспазы 1 (фиг.17С). Вслед за обработкой токсина В каспазой 1 (соотношение фермент/токсин ~1 U/мкг токсина) при 37°С в течение 96 часов получили четыре основных фрагмента, в том числе два фрагмента, которые содержат С-конец (193 и 167 кДа), и два фрагмента, которые содержат N-конец (103 и 63 кДа), как обнаружено посредством SDS-PAGE (фиг.17В). Другие меньшие фрагменты, такие как 26 и 14 кДа, по-видимому, также получили, но не смогли обнаружить в анализе с 3-8% Tris-ацетатным гелем.
SDS-PAGE и вестерн-блоттинг анализы осуществляли на токсине В, который не обрабатывали или обрабатывали каспазой 1 (фиг.18А-С). mAb PA-41 распознавало как фрагменты 103 кДа, так и 63 кДа обработанного каспазой 1 токсина В (правый ряд на фиг.18В), таким образом указывая на то, что РА-41 связывается с N-концевым ферментативным доменом токсина В. Анализ N-концевой последовательности подтвердил, что РА-41 связывается с 63 кДа N-концевым ферментативным доменом токсина В. Интересно отметить, что необработанный токсин В с полосой 63 кДа (левый ряд, фиг.18 В) не распознавался РА-41, что позволяет предположить, что в полосах два фрагменты с таким же молекулярным весом (63 кДа), по-видимому, являются разными белками.
mAb PA-39 связывало 167 кДа фрагмент токсина В, обработанного каспазой 1 (фиг.18С, правый ряд), а также 63 кДа белок в препарате необработанного токсина В (фиг.18С, левый ряд), что указывает таким образом на то, что PA-39 связывает эпитоп в транслокационном домене токсина В. Таким образом, на основе результатов SDS-PAGE/вестерн-блоттинг анализов токсина В С.difficile, обработанного каспазой 1, наблюдали, что mAb РА-41 и PA-39 по-разному взаимодействовали с токсином В. В то время, как обнаружили, что mAb РА-41 связывает эпитоп в N-концевом ферментативном домене токсина В, обнаружили, что mAb PA-39 связывает эпитоп в транслокационном домене токсина (аминокислоты 850-1330). Эти полученные результаты также подтвердили SDS-PAGE/вестерн-блоттинг анализами фрагментов токсина В с применением расщепления энтерокиназой.
Конкурентное связывание mAb к токсину В с токсином В также осуществили с использованием Biacore. Как видно из фиг.19А-Е, mAb PA-39 и РА-41 связывают разные эпитопные области токсина В. mAb PA-39 и РА-41 мыши, как обнаружили, связываются с единственным участком или эпитопом на токсине В; эти mAb, как обнаружили, не связываются с С-концевым связывающим клеточный рецептор (СКВ) доменом. Для РА-41 мыши аффинность связывания токсина В составляла 0,59 мМ. Дополнительно, участок на токсине В, связанный РА-41, не блокировал связывание mAb к токсину В CDB-1 (WO/2006/121422; US2005/0287150), (фиг.19D). Эти полученные результаты согласуются с результатами из вестерн-блоттинг анализов. Согласно наблюдениям из фиг.19С и Е, mAb сравнения к токсину В CDB-1 связывает токсин В по эпитопам, отличным от таковых для mAb PA-39 и PA-41.
В. Картирование антителосвязывающего домена токсина А С.difficile
Токсин А С.difficile полной длины имеет молекулярный вес 310 кДа (фиг.20А) и содержит три главных домена: N-концевой функциональный домен с глюкозилтрансферазной (GT) активностью (~63кДа) и С-концевой СКВ домен (~101 кДа), которые находится на обоих концах гидрофобного домена (~144 кДа).
Некоторые фрагменты токсина А получили при помощи ферментативного расщепления токсина полной длины с применением фермента энтерокиназы (ЕК). Вслед за обработкой токсина А энтерокиназой (соотношение фермент/токсин: приблизительно 3 мЕ/мкг токсина) при 25°С в течение 48 часов получили девять основных фрагментов, в том числе четыре С-концевых фрагмента (~223, ~158-160, ~91 и ~68 кДа) и три N-концевых фрагмента (~195, ~181 и ~127 кДа). Также наблюдали меньшие фрагменты (~53 и ~42 кДа) (фиг.20В и С).
SDS-PAGE и вестерн-блоттинг анализы осуществляли на токсине А, который обрабатывали или не обрабатывали энтерокиназой (фиг.21А-С). Токсин А полной длины и его фрагменты с молекулярным весом ~223, ~158-160, ~91 и ~68 кДа распознавались mAb PA-50 (фиг.21В); Определение N-концевой последовательности подтвердило, что 68 кДа фрагмент содержит часть С-концевого рецепторсвязывающего (CRB) домена. Паттерн связывания mAb PA-50 показывает, что mAb связывается с фрагментами токсина А, содержащими С-конец. Взятые вместе результаты указывают на то, что mAb PA-50 связывается с С-концевыми рецепторсвязывающими эпитопами на токсине А. mAb PA-39 связывало фрагменты, содержащие С-конец (-223 и -158-160 кДа), а также -181 кДа фрагменты, содержащие N-конец (фиг.21C), таким образом указывая на то, что mAb PA-39 связывает эпитоп в области за пределами рецепторсвязывающего домена токсина А. Множественные участки связывания (по меньшей мере два участка связывания) также идентифицировали в исследовании взаимодействия mAb PA-50 и токсина А с применением Biacore анализа (фиг.22А-1). В сравнительных анализах с применением Biacore анализа, иммобилизованное PA-50 мыши, в частности, связывало токсин А с аффинностью 0,16 нМ. Также обнаружили, что после захвата на сенсорном чипе mAb PA-50 мыши, токсин А был способен связывать дополнительный РА-50 и, впоследствии, mAb сравнения к токсину A CDA-1 (WO/2006/121422; US2005/0287150), (фиг.22А-2). Дополнительно, токсин А, захваченный mAb сравнения к токсину А CDA-1 на чипе Biacore, далее связывал дополнительные mAb CDA-1 и РА-50, указывая на то, что mAb сравнения CDA-1 связывается с множественными повторами на токсине А, которые являются отличными от связывающих эпитопов для mAb РА-50 на токсине А (фиг.22В). Таким образом, как определено из этих результатов, эпитоп mAb РА-50 присутствует во множественных копиях на токсине А и не перекрывается с эпитопом для CDA-1. Кроме того, mAb РА-39 связывало эпитоп(ы) на токсине А, отличные от эпитопа(ов) токсина А, связывающихся mAb сравнения CDA-1 (фиг.22С). mAb РА-39 и РА-50, как обнаружили, связывают разные эпитопы на токсине А (фиг.22D). Вестерн-блоттинг анализ показал, что РА-39 и mAb сравнения CDA-1 имеют разные паттерны связывания в отношении обработанного EK токсина А, таким образом указывая на разные связывающие домены и разные эпитопы на токсине А (фиг.22Е). Вестерн-блоттинг анализ показал, что РА-50 и mAb сравнения CDA-1 связываются с одним и тем же доменом обработанного EK токсина А (фиг.22F), но с разными эпитопами (фиг.22В).
Как описано в А и В этого примера, связывающие участки для мышиных mAb РА-50 и РА-41 были локализованы в определенных областях токсинов ограниченным протеолизом токсинов с последующим вестерн-блоттингом. Мышиное mAb РА-50 распознавало токсин А полной длины и некоторые продукты расщепления энтерокиназой, включая большой 223 кДа фрагмент и карбоксиконцевые фрагменты размером 68, 91 и 160 кДа (фиг.22F). Определение N-концевой последовательности подтвердило, что фрагмент 68 кДа соответствует карбоксиконцевому домену токсина А. Такие же фрагменты распознавались mAb сравнения CDA-1. Мышиное mAb РА-41 связывало токсин В полной длины, а также аминоконцевые фрагменты 63 и 103 кДа, полученные расщеплением каспазой-1 (фиг.18 В), в то время как mAb сравнения CDB-1 распознавало неодинаковый набор продуктов расщепления каспазой-1. Определение N-концевой последовательности подтвердило, что фрагмент 63 кДа соответствует аминоконцевому домену токсина В. Совместно, данные указывают на то, что mPA-50 связывает множественные участки в пределах рецепторсвязывающего домена токсина А, а mPA-41 связывает один участок в пределах ферментативного домена токсина В.
Пример 7
mAb к токсину А и токсину В С.difficile - исследования механизма действия
А. Клеточные анализы in vitro, применяемые для исследования механизма действия
Чтобы оценить механизм действия mAb к токсину, осуществляли анализы in vitro с применением разных концентраций токсина А или токсина В. В этих анализах использовали либо клетки СНО-K1, либо Т-84, как описано в примере 3 выше.
Вкратце, применяли анализ на СНО-K1, чтобы оценить нейтрализующую эффективность mAb к токсину А и к токсину В (РА-39 и РА-41). Производили посев клеток СНО-K1 (2000 клеток/лунка) в 96-луночные планшеты. Клеткам давали возможность прикрепиться в течение 4 часов до обработки. Разные концентрации (60, 30, 15 или 6 нг/мл) токсина С.difficile (штамм VPI 10463) инкубировали с серийно разведенными mAb, смешивали в 96-луночных круглодонных планшетах в течение 1 часа при 37°С, и смеси затем добавили в планшеты для культивирования клеток. После инкубации в течение 72 часов добавили 20 мкл/лунка CellTiter-Blue; смеси инкубировали в течение дополнительных 4 часов; и измерили процент клеточного выживания по сравнению с контролями.
Анализ цитотоксичности на Т-84 также применяли, чтобы оценить нейтрализующую эффективность mAb к токсину А. Производили посев клеток Т-84 (клеточная линия карциномы толстой кишки человека) (15000 клеток/лунка) в 96-луночные планшеты. Клеткам давали возможность прикрепиться в течение 4 часов до обработки. Разные концентрации (240, 120, 60 или 30 нг/мл) токсина С.difficile (штамм VPI 10463) инкубировали с серийно разведенными mAb, смешивали в 96-луночных круглодонных планшетах в течение 1 часа при 37°С, и смеси затем добавили в планшеты для культивирования клеток. После инкубации в течение 72 часов добавили 20 мкл/лунка CellTiter-Blue; смеси инкубировали в течение дополнительных 4 часов; и измерили процент клеточного выживания по сравнению с контролями.
В. ELISA, показывающий, что mAb к токсину А предотвращают интернализацию токсина А в клетки
В эксперименте, предназначенном для дальнейшей оценки механизма действия mAb к токсину С.difficile, каждое тестовое антитело (PA-39, PA-50, mAb сравнения к токсину A CDA-1 и контроль - поликлональные антитела козы к токсину А) перемешивали и инкубировали в течение 1 часа при 100х его ЕС90 значения с СС90 концентрацией токсина А для клеток Vero, чтобы обеспечить полную нейтрализацию при высокой цитотоксической концентрации токсина А. Смесь затем инкубировали с клетками Vero при 37°С в течение 15 минут. Клетки затем отмывали PBS, фиксировали и пермеабилизировали. Антитело (РА-38) к токсину А, меченное пероксидазой хрена (HRP), которое не конкурирует за связывание с тестируемыми антителами, применяли для исследования в отношении интернализованного токсина А и обнаруживали с применением хемилюминесценции (фиг.31G). В этом анализе только токсин А, который связался и интернализован в клетку, будет обнаруживаться зондом, таким образом, с выходом хемилюминесцентного сигнала на основе хемилюминесцентной реакции HRP. При хемилюминесцентном обнаружении применяется фермент, чтобы катализировать реакцию (т.е. катализируемое окисление люминола пероксидом) между ферментом HRP и его субстратом в присутствии пероксида, что приводит к возникновению видимого света. Окисленный люминол излучает свет по мере того, как он распадается до его основного состояния. Как только субстрат катализируется HRP, световой сигнал определяют количественно при помощи люминометра (Analyst GT).
С. Результаты нейтрализующей активности и исследования МОА
mAb к токсину А
В анализе клеточной цитотоксичности, который применяли, чтобы оценить нейтрализующую активность и механизм действия антител к токсину А, токсин А добавляли в увеличивающихся концентрациях к клеткам, как описано выше в части А этого примера. Результаты этих экспериментов, в которых оценивали нейтрализацию токсина А mAb к токсину A PA-39, PA-50 и mAb сравнения CDA-1, показаны на фиг.31B-D и ниже в таблице А.
(нМ)
Как видно из данных, представленных в таблице А, активность in vitro mAb PA-39 в анализе эффективности по отношению к токсину показывает сдвиги как в ЕС50, так и максимальном проценте подавления по мере того, как добавляют больше токсина А в культуру, что указывает на смешанный конкурентный механизм подавления для РА-39. Обнаружение с помощью ELISA токсина А после защиты 100-кратным избытком РА-39 подтвердило, что подавление токсина посредством РА-39 происходит путем предотвращения интернализации токсина и цитотоксического эффекта токсина. Активность in vitro mAb PA-50 в анализе эффективности по отношению к токсину показывает сдвиг в ЕС5о по мере того, как добавляют больше токсина А в культуру, что указывает на конкурентный механизм подавления для РА-50. Обнаружение с помощью ELISA токсина А после защиты 100-кратным избытком РА-50 подтвердило, что подавление токсина посредством РА-50 происходит путем предотвращения интернализации токсина.
mAb к токсину В
В анализе клеточной цитотоксичности, который применяли, чтобы оценить нейтрализующую активность и механизм действия антитела к токсину В, токсин В добавляли в увеличивающихся концентрациях к клеткам, как описано выше в части А этого примера. Результаты экспериментов по эффективности, в которых оценивали нейтрализацию токсина В mAb к токсину В РА-41 и mAb сравнения CDB-1, показаны на фиг.31Е и F и в таблице В ниже. Как видно из данных, представленных в таблице, активность in vitro PA-41 в анализе эффективности по отношению к токсину показывает сдвиги как в ЕС50, так и максимальном проценте подавления по мере того, как добавляют больше токсина В в культуру, что указывает на смешанный конкурентный механизм подавления для РА-41.
На основе вышеописанных анализов в этом примере, ингибитор, который обладает способностью полностью нейтрализовать увеличивающиеся концентрации токсина А или токсина В просто посредством добавления больших концентраций ингибитора, будет проявлять сдвиг в ЕС50 по мере того, как антитело связывает и нейтрализует более высокие концентрации токсина, и таким образом, будет считаться конкурентным ингибитором. Ингибитор, который не способен ослабить токсические эффекты увеличивающихся концентраций токсина, будет проявлять сниженный максимальный процент эффекта при более высоких концентрациях ингибитора, но не будет проявлять сдвиг в EC50 и, таким образом, будет считаться неконкурентным ингибитором. Дополнительно, ингибитор, который проявляет как сдвиг в ЕС50, так и сниженный максимальный процент эффекта в результате увеличивающихся концентраций ингибитора при более высоких концентрациях токсина, будет считаться смешанным конкурентным ингибитором. В некоторой степени оценивание механизма действия с применением анализов клеточной цитотоксичности следует осуществлять с учетом ошибки, связанной с воспроизводимостью анализа и фоновой цитотоксичностью наблюдаемой в контрольных лунках, не содержащих ингибитор, который воздействует на плато максимального процента подавления. Следовательно, незначительный сдвиг вправо в значениях ЕС50 можно наблюдать по мере того, как концентрации токсина увеличиваются вследствие этих эффектов.
В соответствии с вышеизложенным в отношении нейтрализации токсина А и МОА, РА-39 mAb считается смешанным конкурентным ингибитором вследствие сдвига в ЕС50 и сниженного плато, наблюдаемого в отношении кривых цитотоксичности РА-39 (фиг.31В), а также сниженного максимального процента эффекта, рассчитанного в таблице А. Эти данные подтверждаются результатами ELISA (фиг.31Н), которые показывают степень цитотоксического эффекта при высоких концентрациях РА-39, таким образом, указывая на то, что по меньшей мере некоторый из МОА РА-39 происходит внутриклеточно. Наблюдали, что mAb PA-50 является конкурентным ингибитором, о чем свидетельствуют сдвиг вправо в значениях ЕС50, наблюдаемый в кривых анализа цитотоксичности (фиг.31С), и данные, представленные в таблице А, показывающие минимальное изменение максимального процента подавления. Эти данные подтверждаются результатами ELISA (фиг.31Н), которые показывают полное подавление связывания токсина А и интернализации при высоких концентрациях PA-50.
Согласно наблюдениям из фиг.31D, mAb сравнения CDA-1 показывает минимальный сдвиг в ЕС50, но значительное снижение максимального процента эффекта по мере того, как количество токсина увеличивается. Если результаты, показанные на фиг.31D, рассматривать с данными, представленными в таблице А, mAb сравнения CDA-1 демонстрирует неконкурентный механизм действия, поскольку вся его активность наблюдается за пределами клетки. Стерическое несоответствие связывания токсина и клеточной интернализации, по-видимому, отвечает за выход графиков с данными на фиг.31D, таким образом подтверждая неконкурентный МОА для mAb сравнения CDA-1.
В соответствии с вышеизложенным в отношении нейтрализации и МОА токсина В, считается, что mAb PA-41 проявляет смешанный конкурентный механизм действия вследствие сдвига вправо значений EC50 и сниженного максимального процента эффекта, наблюдаемого на фиг.31Е и в данных, представленных в таблице В.
Результаты, наблюдаемые в отношении mAb PA-41, противоречат таковым, наблюдаемым в отношении mAb сравнения CDB-1, которое проявляет сниженный максимальный процент эффекта, но более низкую степень сдвига ЕС50. В данном случае механизм действия для активности mAb сравнения по отношению к токсину В менее понятен, особенно принимая во внимание рассмотрения ошибки, упомянутой выше.
Пример 8
Тестирование mAb настоящего изобретения относительно панели изолятов или штаммов С.difficile, включая гипервирулентные изоляты или штаммы
Чтобы оценить способность mAb к токсину А и В С.difficile настоящего изобретения нейтрализовать токсины из широкого диапазона соответствующих изолятов С.difficile, нейтрализующую активность mAb тестировали относительно набора из двадцати токсикогенных клинических изолятов или штаммов С.difficile, включая гипервирулентные изоляты BI/NAP1/027. Поскольку С.difficile проявляет значительную межштаммовую гетерогенность по генам, кодирующим токсины А и В, эти исследования выполняли для изучения степени нейтрализации токсина описанными mAb, и в частности, mAb РА-50 и PA-41.
Панель токсикогенных клинических изолятов С.difficile (таблица 6) выбрали в отношении географического и генетического разнообразия на основе типа токсина, риботипа и анализов с рестрикционными эндонуклеазами из международной коллекции изолятов или штаммов С.difficile, поддерживаемых в TechLab (Blacksburg, VA).
Как классифицировано в таблице 6, штаммы С.difficile включают 3 референс-штамма (VPI 10463 (АТСС 43255), 630 (АТСС ВАА-1382) и 545 (АТСС 43596), шесть полученных в больницах штаммов BI/NAP1/027 (CCL678, НМС553, Pitt45, CD196, 5, 7.1), два токсин А-/токсин В+(tcdA-tcdB+) штамма (F1470, 8864), три амбулаторных изолята (МН5, CCL13820 и CCL 14402) и другие часто встречающиеся клинические изоляты (Pitt2, CCL14137, UVA17, UVA30/TL42, Pittl02, Pitt7). Дополнительно изоляты 13 (CCL13820) и 19 (Pitt102), которые классифицируют как «амбулаторный изолят» и «часто встречающийся клинический изолят (отличный от риботипа 027)» соответственно, в таблице 6 также представляют собой штаммы toxA-/toxB+. Культуральные супернатанты, содержащие эти токсины С.difficile получили в TechLab, стерилизовали посредством фильтрации и хранили при 4°С. Присутствие токсинов в культуральных супернатантах подтверждали с применением набора токсин С.difficilelsamtOKcvoi (TechLab) и анализа цитотоксичности.
Цитотоксическую активность каждого содержащего токсин культурального супернатанта оценивали на клетках СНО-K1 (использовали для определения активности токсина В культуральных супернатантов) и клетках Т-84 (использовали для определения активности токсина А культуральных супернатантов) путем обработки клеток титрованными культуральными супернатантами.
Чтобы протестировать нейтрализующие активности mAb настоящего изобретения, культуральные супернатанты, содержащие токсин, применяли при максимальном разведении, что приводило к >95% потери жизнеспособности клеток. Супернатанты токсинов предварительно смешивали с различными концентрациями mAb в течение 1 часа и затем добавили к клеткам для инкубации при 37°С в течение 72 часов. Жизнеспособность клеток измеряли с применением Cell-Titer Blue (Promega). Процент выживаемости в обработанных лунках сравнивали с необработанными контрольными лунками и строили график, чтобы рассчитать in vitro нейтрализующую активность mAb (ЕС50). В первой серии экспериментов, на фиг.23А показана активность mAb РА-41 при нейтрализации супернатантов, содержащих токсин, с использованием клеток СНО-K1. РА-41 эффективно (диапазон ЕС50 от 1,1-11M до 6,5-10М) нейтрализовало супернатанты всех токсикогенных штаммов С.difficile, включая все гипервирулентные штаммы, за исключением трех штаммов токсин A-/токсин В+. Сообщалось, что существуют значительные отличия в последовательностях ферментативных доменов токсин A-/токсин В+ от стандартных штаммов С.difficile. Поскольку РА-41 связывается с ферментативным доменом токсина В, разнообразие последовательностей этого домена может объяснить менее эффективную нейтрализующую активность РА-41 по отношению к токсину В из двух штаммов токсин A-/токсин В+.
Эксперименты также проводили, чтобы оценить активность как hPA-41, так и человеческого mAb сравнения CDB-1 по отношению к токсину В (WO/2006/121422; US2005/0287150) к гипервирулентным штаммам С.difficile на клеточной линии СНО-K1. В этих исследованиях наблюдали, что hPA-41 показало значительную нейтрализующую активность к супернатантам от всех 6 штаммов BI/NAP 1/027, в то время как mAb сравнения CDB-1 показало минимальную активность (фиг.23В). Также в этих исследованиях наблюдали, что нейтрализующая активность mAb hPA-41 была в >1000 раз больше, чем таковая mAb сравнения CDB-1 при нейтрализации токсичности штаммов BI/NAP 1/027. Нейтрализующая активность hPA-41 и mAb сравнения CDB-1 по отношению к 2 референс-штаммам (VPI 10463 и АТСС 43596) и 6 штаммам BI/027/027 (CCL678, НМС553, Pitt 45, CD196, Montreal 5.1 и Montreal 7.1) проиллюстрирована на фиг 23 В. В этих исследованиях обнаружили, что hPA-41 является неактивным по отношению к трем изолятам риботипа 017 (таблица 6), которые являются токсин А-/В+, хотя mAb hPA-41 к токсину В проявляло в значительной степени большую нейтрализующую активность, чем mAb сравнения, по отношению к другим штаммам в панели.
Активность mAb РА-50 при нейтрализации супернатантов, содержащих токсин А, культур С.difficile, с использованием клеток Т-84, находилась в диапазоне от 2,6-12М до 7,7-11M, как показано на фиг.24А. РА-50 полностью нейтрализовало супернатанты от всех доступных штаммов, которые продуцируют токсин А, включая гипервирулентные штаммы. РА-50 не нейтрализовало четыре штамма токсин A-/токсин В+(F1470, 8864, CCL 13820, CCL 14402), поскольку эти штаммы не продуцируют какой-либо токсин А. hPA-50 также было в значительной степени более эффективным при нейтрализации активности оставшихся штаммов в панели. В других сравнительных исследованиях обнаружили, что нейтрализующая активность hPA-50 по отношению ко всем 6 штаммам BI/NAP 1/027 С.difficile, продуцирующим токсин А, была в>100 раз больше, чем таковая mAb сравнения CDA-1 (WO/2006/121422; US2005/0287150) (фиг.24В). hPA-50 также показало большую нейтрализующую активность, чем mAb сравнения CDA-1, по отношению к продуцирующим токсин А референс-штаммам VPI 10463 и 545 С.difficile. Подобным образом, mAb РА-39 нейтрализовало токсин А в супернатантах культур С.difficile, со значениями ЕС50, находящимися в диапазоне от 7,7-12M до 4,8-8M, как показано на (фиг.25А). Как можно заметить в отношении mAb РА-50, четыре штамма токсин A-/токсин В+ не были нейтрализованы РА-39. Более того, результаты сравнительных исследований продемонстрировали, что нейтрализующая активность mAb РА-39 по отношению ко всем 6 штаммам BI/NAP 1/027 была в>100 раз больше, чем таковая человеческого mAb сравнения к токсину А CDA-1; РА-39 также показало в значительной степени большую нейтрализующую активность по отношению к оставшимся штаммам в панели (фиг.25В). Отметили, что один штамм с токсином, CCL 14402, не снижал жизнеспособность клеток Т-84 достаточно, чтобы дать возможность точно измерить нейтрализующую активность mAb в данном анализе.
В этих исследованиях на клеточной линии СНО-K1, hPA-41 показало высокие уровни нейтрализующей активности по отношению к супернатантам из всех 6 штаммов BI/NAP 1/027, в то время как mAb сравнения CDB-1 показало минимальную активность. Наблюдали, что гуманизированное РА-41 имело нейтрализующую активность в >1000 раз больше, чем таковая mAb сравнения CDB-1, при нейтрализации гипервирулентных штаммов BI/NAP 1/027 С.difficile. Нейтрализующие активности hPA-41 и CDB-1 в этих исследованиях по отношению к 2 референс-штаммам (VPI 10463 и АТСС 43596) и 6 штаммам BI/027/027 (CCL678, НМС553, Pin 45, CD196, Montreal 5.1 и Montreal 7.1) проиллюстрированы на фиг.23В. Подобным образом, hPA-41 показало в этих исследованиях в значительной степени более высокую нейтрализующую активность по сравнению с mAb сравнения CDB-1 по отношению к другим штаммам в панели, за исключением трех изолятов риботипа 017, которые являются А-/В+. Подобные эксперименты осуществляли, чтобы оценить нейтрализующую активность hPA-39 и таковую человеческого mAb сравнения к токсину A CDA-1 на клетках Т-84. Результаты показали, что нейтрализация всех 6 штаммов BI/NAP 1/027 hPA-39 была в >100 раз больше, чем таковая mAb сравнения CDA-1 (фиг.25В). Гуманизированное РА-39 также показало в значительной степени более высокую нейтрализующую активность в исследованиях, чем mAb сравнения CDA-1, по отношению к оставшимся штаммам в панели. Таким образом, mAb hPA-41 и hPA-39 проявили высокие уровни нейтрализующих активностей к гипервирулентным штаммам С.difficile по отношению ко всем тестируемым штаммам. Это указывает на то, что эпитопы, распознаваемые этими гуманизированными mAb, являются высоко консервативными для совокупности различных штаммов.
Подобно таблице 6, в таблице 7 представлены результаты вышеописанных экспериментов in vitro по нейтрализации токсина С.difficile, показывающие панель токсикогенных штаммов С.difficile, изолированных из Северной Америки и Европы, и полученные в результате значения EC50, полученные при помощи гуманизированных mAb к токсину и mAb CDA-1 и CDB-1 препаратов сравнения. Панель включает риботипы 001, 002, 003, 012, 014, 017, 027 и 078 приблизительно пропорционально частотам, наблюдаемым клинически (94, 95), за исключением риботипа 017 штаммов tcdA-tcdB+, которые представлены в чрезмерном количестве в панели. Супернатанты от штаммов tcdA-tcdB+ применяли в качестве средств для идентификации клеток, которые не поддавались цитолизу супернатантами, содержащими только токсин В, и таким образом были бы подходящими для изучения цитотоксичности, опосредованной токсином A. VPI 10463 включили в панель, и это позволило сравнить результаты, полученные с очищенными и неочищенными токсинами.
В этих исследованиях гуманизированное mAb PA-50 нейтрализовало токсин А независимым от штаммов образом. Значение медианы EC50 составило 32 пМ (диапазон: от 20 до 127 пМ, таблица 7), и наблюдали крутые кривые доза-эффект с наклонами Хилла, которых типично было больше, чем два (фиг.25С). РА-50 было более активным, чем CDA1, по отношению к каждому тестируемому изоляту. Наибольшую разницу в эффективности наблюдали в отношении гипервирулентных штаммов 027, в отношении которых РА-50 было приблизительно в 1000 раз более эффективным, чем CDA1 (Р=0,0002), и в отношении штамма риботипа 078, включенного в панель.
Гуманизированное mAb PA-41 эффективно подавляло каждый из штаммов tcdA-tcdB+ со значением медианы EC50 23 пМ (диапазон: от 7,7 до 129 пМ, таблица 7), и, в сущности, полную нейтрализацию наблюдали при более высоких концентрациях (фиг.250). PA-41 было в общем более эффективным, чем CDB1, по отношению к штаммам tcdA-tcdB+ и было приблизительно в 500 раз более эффективным против гипервирулентных штаммов 027 (Р=0,003). CDB1, но не PA-41, было эффективным при нейтрализации токсина В из штаммов tcdA-tcdB+ риботипа 017. В конечном счете, PA-41 и РА-50 проявляли подобные активности по отношению к неочищенным и очищенным формам токсина из референс-штамма VPI 10463 (таблица 7 и фиг.25С и 25D).
Как показано в этом примере, гуманизированные mAb PA-50 и РА-41 показали высокий уровень нейтрализующей активности по отношению к токсинам А и В С.difficile, соответственно, из генетически различных штаммов, иллюстративных для наблюдающихся в настоящее время вспышек CDI. Степень активности этих mAb является значительной в свете генотипической и фенотипической вариации в пределах токсинов. В частности, РА-50 и РА-41 с пикомолярной активностью нейтрализовали токсины, продуцируемые гипервирулентными антибиотикоустойчивыми штаммами 027, в то время как mAb сравнения, как обнаружили, имели наномолярную активность. Этот результат может отражать ослабленное связывание CDA-1 с токсином А из штаммов 027, как сообщалось ранее (90). Токсин В из штаммов 027 проявляет заметное расхождение в последовательностях по отношению к другим штаммам С.difficile. Различия в последовательностях сконцентрированы в пределах карбоксиконцевого рецепторсвязывающего домена и ассоциированы с усиленной цитотоксичностью in vitro. Однако такое расхождение в последовательностях не воздействует на нейтрализующую активность РА-41, которое связывает эпитоп в пределах аминоконцевого домена токсина В. РА-50 и РА-41 с пикомолярными эффективностями нейтрализовали все шесть штаммов 027 в панели, включая штамм CD 196, который предшествует недавнему возрастанию преобладания 027. В общем, полученные результаты указывают на то, что эпитопы для РА-50 и РА-41 являются широко консервативными в происхождении 027.
CDI типично вызывается штаммами С.difficile, которые продуцируют оба токсина А и В. Однако штаммы tcdA-tcdB+ также связывали с заболеванием. Клинически релевантные штаммы tcdA-tcdB+ представляют собой преимущественно риботип 017. Штаммы риботипа 017, как сообщалось, проявляли сниженную патогенность в отношении хомячков и кодировали атипичный tcdB, аминоконцевая область которого имеет 70-80% идентичности последовательностей как с tcdB из VPI 10463, так и с летальным токсином (tcsL) из С.sordellii. Фенотиически, tcdB риботипа 017 имеет характеристики гибрида и проявляет рецепторсвязывающие свойства и специфичности гликозилирования, типичные для токсинов tcdB и tcsL, соответственно. Атипичная аминоконцевая область tcdB 017 предоставляет вероятное объяснение, по какой причине в этом исследовании этот токсин не был нейтрализован РА-41. Хотя штаммы 017 могут регионально преобладать и вызывать локальные вспышки CDI, в общем, как определили, они включают <2% штаммов, встречающихся в недавних клинических исследованиях международной фазы 3 исследовательских терапий для лечения CDI (94, 95).
Пример 9
Получение химерных mАb
Химерные моноклональные антитела, которые включают вариабельную область исходного mAb мыши и константную область IgG1 человека, получили и охарактеризовали. Химерные mAb получили, чтобы подтвердить, что клонировали вариабельные области мыши с подходящей активностью по отношению к токсину и специфичностью связывания, и что конструирование вариабельных областей мыши с константной областью человека незначительно изменяет связывающие и нейтрализующие свойства каждого клонированного mAb. Химерные mAb типично проявляют такую же связывающую активность, как и исходные mAb мыши.
Все из mAb PA-38, PA-39, РА-41 и РА-50 содержат легкие каппа-цепи. Для получения химерных mAb последовательности нуклеиновых кислот, кодирующие вариабельные области тяжелых и легких цепей, помещали внутрь подходящих векторов экспрессии, таких как, но без ограничений, pCON гамма1 и pCON каппа, соответственно (Lonza Biologies, Berkshire, UK). Подходящие плазмиды кодируют либо константную область легкой каппа-цепи человека, либо константную область тяжелой цепи IgG1 человека.
Для получения химерного mAb вариабельную область тяжелой цепи каждого mAb клонировали в плазмиду pCON гамма1. Ген полной тяжелой цепи субклонировали в плазмиду, содержащую ген легкой цепи для образования одной плазмиды, которая кодировала гены как тяжелой, так и легкой цепи. Клетки 293F временно трансфицировали этим вектором экспрессии с применением Effectene (Qiagen, Valencia, СА) согласно предложенному протоколу изготовителя. Клеточный супернатант, содержащий секретируемое химерное mAb, собирали через семь дней после трансфекции и очищали с применением хроматографии с белком А. Эффективности и активности химерного(химерных) mAb сравнивали с таковыми mAb мыши в анализах цитотоксичности и гемагглютинации.
В соответствии с вышеприведенной методикой, химерные mAb (сРА-39 и сРА-41) образовали на основе исходных mAb PA-39 и РА-41 мыши. Концентрации этих химерных mAb в неочищенных клеточных супернатантах находились в диапазоне приблизительно 2-11 мкг/мл. В частности, неочищенный супернатант из трансфицированной клеточной культуры 293F, продуцирующей сРА-39, содержал 10,6 мкг/мл химерного mAb, в то время как неочищенные супернатанты из нескольких трансфицированных клеточных культур 293F, продуцирующих сРА-41, содержали 9,6-10,9 мкг/мл химерного mAb.
Химерные mAb сравнивали с их соответствующими исходными mAb мыши в отношении способности нейтрализовать токсины С.difficile in vitro путем проведения анализов цитотоксичности (сРА-39: клетки СНО-K1, сРА-41: клетки СНО-K1 и сРА-50: клетки Т-84), как описано ранее (пример 3). Все химерные mAb, как обнаружили, были в равной степени эффективными при сравнении с их исходным mAb мыши, как показано на фиг.26 (РА-41), фиг.27 (PA-39) и фиг.28 (РА-50). Эти результаты демонстрируют успешность химеризации и продукцию функциональных химерных mAb с эффективностями нейтрализации токсинов, эквивалентными таковым исходных mAb мыши.
Пример 10
Гуманизация mAb мыши и тестирование гуманизированных mAb настоящего изобретения на эффективность нейтрализации токсинов in vitro
Гуманизированные mAb были получены способами, известными в настоящем уровне техники. Примеры и описания различных гуманизированных mAb включают, например, зенапакс (65, 66) синагис (67-69), герцептин (70-72), милотарг (73, 74), ксолар (75-77), раптива (78-80), авастин (81, 82) и тисабри (83). Можно получать гуманизированные моноклональные антитела, которые эффективны в минимизации иммуногенности, таким образом, что активность mAb в людях не подвергается нежелательным воздействиям (84-87). Предпочтительно гуманизированные mAb демонстрируют нейтрализующую токсин активность, которая находится в пределах двух порядков от исходных mAb мыши. Кроме того, гуманизированные mAb оптимально демонстрируют сильную эффективность в модели инфекции С.difficile на хомячках.
Для получения гуманизированных форм mAb мыши к токсину А и/или к токсину В применяли общепринятые способы пересадки гипервариабельного участка (CDR), как описано в настоящем документе. Гуманизированное(ые) mAb сравнивали с исходным(и) mAb мыши в отношении нейтрализующей токсин активности in vitro и in vivo. В соответствии с настоящим изобретением гуманизированное(ые) mAb могут сохранять активность против токсина исходного(ых) mAb мыши и могут подходить для повторного дозирования у людей.
А. Молекулярное клонирование генов тяжелых и легких цепей mAb мыши
Для клонирования генов антител применяли общепринятые способы (88). Вкратце, общую РНК очищали из 1×107 гибридомных клеток с применением реагента TRIzol (Invitrogen) согласно рекомендуемому протоколу изготовителя. 5 мкг общей РНК обратно транскрибировали с применением Superscript II Reverse Transcriptase (Invitrogen), с использованием праймера oligo-dT. Полученную в результате кДНК обрабатывали РНКазой Н для удаления РНК-матрицы и затем очищали с применением QIAquick PCR purification kit (Qiagen) для удаления свободных нуклеотидов и праймеров. Следующим шагом к 3'-концу кДНК добавляли хвост из гуанидиновых нуклеотидов с применением фермента терминальной трансферазы (NEB) в присутствии dGTP согласно рекомендуемому протоколу изготовителя. Полученную в результате кДНК с хвостом затем подвергали ПЦР с применением одного праймера, который отжигается с константной областью либо тяжелой, либо легкой цепи, и одного универсального праймера, который отжигается с гуанозиновым хвостом на кДНК. Как для тяжелых, так и для легких цепей применяли универсальный праймер 5TATATCTAGAATTCCCCCCCCCCCCCCCCC3’ SEQ ID NO:11. Для амплификации легкой цепи, применяли праймер 5’TATAGAGCTCAAGCTTGGATGGTGGGAAGATGGATACAGTTGGTGC3’ (SEQ ID NO:12), тогда как тяжелую цепь амплифицировали с применением праймера 5TATAGAGCTCAAGCTTCCAGTGGATAGAC(CAT)GATGGGG(GC)TGT(TC)GTTTT GGC3’ (SEQ ID NO:13), где последовательности в скобках указывают на вырожденности оснований. Полученную в результате амплифицированную с помощью ПЦР ДНК очищали с применением QIAquick PCR purification kit (Qiagen) и секвенировали. ПЦР-реакции осуществляли и секвенировали в трех повторностях, чтобы убедиться, что никаких ошибок не было внесено во время амплификации ДНК-фрагментов из приблизительно 500 пар оснований.
В. Гуманизация вариабельных областей mАb
Чтобы получить последовательности гуманизированных mAb, вначале идентифицировали каркасные аминокислотные остатки, важные для структуры CDR. Параллельно из числа известных последовательностей иммуноглобулина человека выбирали последовательности VH и VL человека с высокой гомологией по отношению к VH и VL мыши, соответственно. Последовательности CDR из mAb мыши вместе с любыми каркасными аминокислотными остатками, важными для поддержания структуры CDR, при необходимости, пересаживали в отобранные каркасные последовательности человека. К тому же, каркасные аминокислотные остатки человека, которые, как было обнаружено, были атипичными в соответствующей подгруппе V-области, замещали типичными остатками в попытке снизить потенциальную иммуногенность полученного в результате гуманизированного mAb. Эти гуманизированные области VH и VL клонировали в векторы экспрессии, такие как, но без ограничений, pCON Гамма1 и pCON каппа (Lonza Biologies, Berkshire, UK) соответственно. Эти векторы кодируют гены константной области(ей) тяжелых и легких цепей иммуноглобулина человека. Клетки 293F временно трансфицировали этими векторами экспрессии с применением Effectene system (Qiagen, Valencia, CA). Клеточные супернатанты, содержащие секретированное гуманизированное mAb, собирали через семь дней после трансфекции и очищали с применением хроматографии с белком А.
С. Получение клональных стабильных клеток СНО, экспрессирующих гуманизированные mАb
Получение стабильных клеток/клеточных линий СНО дает возможность для продукции достаточных количеств mAb для тестирования как в клеточных анализах in vitro, так и в модели ассоциированной с С.difficile диареи (CDAD) на золотистых сирийских хомячках. В качестве всего одного примера, можно применять клетки СНО K1 SV от Lonza Biologies (Berkshire, UK) с применением системы селекции с глутамин-синтетазой и амплификации (GS) для получения стабильно трансфицированных клеток СНО. Система Lonza GS типично дает высокопродуктивные клеточные линии СНО, которые могут продуцировать значительные количества гуманизированных mAb.
Клетки СНО K1 SV выращивали в культуральной среде CD для клеток СНО (Invitrogen), дополненной IX глутамином (Invitrogen) и IX Н/Т Supplement (Invitrogen). 1×107 жизнеспособных клеток электропорировали при 290 Вт, бесконечном сопротивлении и 960 мкФ с использованием 40 мкг линеаризованной плазмидной ДНК, ресуспендированной в 100 мкл стерильного буфера ТЕ. Клетки переносили в Т-150 матрас, содержащий 50 мл полной среды CD для СНО, и инкубировали в течение приблизительно 48 часов при 37°С и 8,0% СО2. Клетки центрифугировали и ресуспендировали до конечной плотности 3,3×105 клеток/мл в селективной среде GS (CD для СНО+IX GS Supplement (JRH Biosciences)+IX Н/Т Supplement), содержащей 100 мкМ MSX (Sigma), высевали 5000 жизнеспособных клеток/лунку в 96-луночные планшеты (Coming) и инкубировали в течение приблизительно 3-4 недель пока не стали появляться колонии первичных клеток (клоны трансфицированных клеток). Приблизительно 300 клеточных колоний (клонов) отбирали для продукции рекомбинантного mAb путем осторожного удаления 20 мкл супернатанта и осуществления анализа ELISA в 96-луночном формате. Вкратце, 96-луночные планшеты покрывали иммобилизованным антителом (антитело козы к антителу человека), и затем добавляли супернатанты от клонированных трансфектантов СНО (разведенные 1:800), чтобы дать возможность связаться с иммобилизованным антителом, связанным с лунками планшета. После отмывки в планшет добавляли вторичное антитело (антитело козы к антителу человека, конъюгированное со щелочной фосфатазой) и давали возможность связаться с антителом человека в образце перед тем, как проводится отмывка для удаления неспецифичного связывания. Планшет затем анализировали в отношении активности щелочной фосфатазы с применением набора 1-Step PNPP (Thermo, Rockford, IL), чтобы идентифицировать клоны, продуцирующие наибольшее количество секретируемого антитела. Клоны, продуцирующие высокие количества mAb, выращивали в культуральной среде CD для клеток СНО, дополненной IX глутамином и IX Н/Т Supplement. Клеточные супернатанты, содержащие секретированное гуманизированное mAb, собирали и очищали с применением белка А. Клоны, проявляющие наилучшую продуктивность, субклонировали путем предельного разведения и постепенно увеличивали, чтобы продуцировать граммовые количества рекомбинантного, гуманизированного, моноклонального антитела.
D. Гуманизированные mAb hPA-39, hPA-41 и hPA-50
Молекулярно клонированные, гуманизированные mAb выделяли, как описано выше, и характеризовали (смотри раздел Е ниже). Константная (CL) область легкой (L) цепи каждого гуманизированного антитела относится к классу каппа (κ); константная область (СН) тяжелой (Н) цепи каждого гуманизированного антитела относится к изотипу IgG1. Гуманизированные mAb, содержащие уникальные вариабельные (V) области, как было обнаружено, связывают и нейтрализуют активность либо токсина А, либо токсина В С.difficile. V-области L- и Н-цепей гуманизированных mAb могут образовывать часть полного иммуноглобулина (Ig) или молекулы антитела, составленной из двух полипептидов Н-цепи и двух полипептидов L-цепи, типично связанных путем образования дисульфидных связей, или они могут представлять собой отдельные части или фрагменты антитела, в частности, части или фрагменты антитела, которые связывают токсин А и/или токсин В и/или которые нейтрализуют активность токсинов. Неограничивающие примеры подходящих фрагментов или частей иммуноглобулина, содержащих V-область, включают F(ab), F(ab’) или F(ab’)2 фрагменты.
Гуманизированные mAb к токсину А и токсину В С.difficile получали согласно вышеописанным методикам. Процесс гуманизации давал несколько гуманизированных mAb (hmAb) к токсину А С.difficile, которые связывали токсин А и нейтрализовали активность токсина А на восприимчивых клетках. Примеры таких hmAb включают гуманизированное mAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:1 (Фиг.32А) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:3 (Фиг.33А) и к С-область человека; hmAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:2 (Фиг.32В) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:3 (Фиг.33А) и к С-область человека; hmAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:1 (Фиг.32А) и С-область IgG1 человека, и полипептид L-цепи, включающий VL-область SEQ ID NO:4 (Фиг.33В) и к С-область человека; и hmAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:2 (Фиг.32В) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:4 (Фиг.33В) и к С-область человека. Такие гуманизированные mAb к токсину А С.difficile охватывают hmAb PA-39 (hPA-39) настоящего изобретения. Полный иммуноглобулин hPA-39 с двумя L-цепями и двумя Н-цепями можно продуцировать в клетке-хозяине, которая коэкспрессирует и секретирует полипептид Н-цепи hPA-39, составленный из VH-области настоящего изобретения (например, SEQ ID NO:1; SEQ ID NO:2) и подходящей СН-области, например изотипа IgG1, такой которая содержится под № доступа Genbank NW_001838121, и полипептид L-цепи hPA-39, составленный из VL-области hPA-39 настоящего изобретения (например, SEQ ID NO:3; SEQ ID NO:4) и подходящей CL-области, например подтипа к, такой которая содержится под № доступа Genbank NW_001838785.
Другие примеры гуманизированных mAb к токсину А С.difficile настоящего изобретения включают гуманизированное mAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:5 (Фиг.34А) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:7 (Фиг.35) и к С-область человека; и гуманизированное mAb к токсину А С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:6 (Фиг.34В) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:7 (Фиг.35) и к С-область человека. Такие гуманизированные mAb к токсину А С.difficile охватывают гуманизированное mAb PA-50 (hPA-50) настоящего изобретения. Полный иммуноглобулин hPA-50 с двумя L-цепями и двумя Н-цепями можно продуцировать в клетке-хозяине, которая коэкспрессирует и секретирует полипептид Н-цепи hPA-50, состоящий из VH-области настоящего изобретения (например, SEQ ID NO:5; SEQ ID NO:6) и подходящей СН-области, например изотипа IgG1, такой которая содержится под № доступа Genbank NW_001838121, и полипептид L-цепи hPA-50, состоящий из VL-области настоящего изобретения (SEQ ID NO:7) и CL-области, например подтипа к, такой которая содержится в пределах № доступа Genbank NW_001838785.
Процесс гуманизации дополнительно давал гуманизированные mAb к токсину В С.difficile, которые связывали токсин В и нейтрализовали активность токсина В на восприимчивых клетках in vitro. Примеры таких hmAb включают гуманизированное mAb к токсину В С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:8 (Фиг.36А) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:10 (Фиг.37) и к С-область человека; и гуманизированное mAb к токсину В С.difficile, включающее полипептидную последовательность Н-цепи, включающей VH-область SEQ ID NO:9 (Фиг.36В) и С-область IgG1 человека, и полипептидную последовательность L-цепи, включающей VL-область SEQ ID NO:10 (Фиг.37) и к С-область человека. Такие гуманизированные mAb к токсину В С.difficile охватывают гуманизированное mAb PA-41 (hPA-41) настоящего изобретения. Полный иммуноглобулин hPA-41 с двумя L-цепями и двумя Н-цепями можно продуцировать в подходящей клетке-хозяине, которая коэкспрессирует и секретирует полипептид Н-цепи hPA-41, состоящий из VH-области hPA-41 настоящего изобретения (например, SEQ ID NO:8; SEQ ID NO:9) и подходящей СН-области, например изотипа IgG1, такой которая содержится под № доступа Genbank NW_001838121, и полипептид L-цепи hPA-41, состоящий из VL-области hPA-41 настоящего изобретения (например, SEQ ID NO:10) и CL-области, например подтипа κ, такой которая содержится под № доступа Genbank NW_00183 8785.
Кроме того, получали гуманизированные, клонированные mAb (hmAb), которые связывают токсин А или токсин В С.difficile и сильно нейтрализуют активность токсинов. hmAb PA-50 к токсину А С.difficile состоит из полипептидов двух тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и полипептидов двух легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида тяжелой цепи hPA-50 SEQ ID NO:14 изложена в SEQ ID NO:15 (Фиг.38 В); последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида легкой цепи hPA-50 SEQ ID NO:16 изложена в SEQ ID NO:17. (Фиг.38А). hmAb PA-39 к токсину А С.difficile состоит из полипептидов двух тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и полипептидов двух легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида тяжелой цепи hPA-39 SEQ ID NO:18 изложена в SEQ ID NO:19 (Фиг.39В); последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида легкой цепи hPA-39 SEQ ID NO:20 изложена в SEQ ID NO:21 (Фиг.39А). hmAb PA-41 к токсину В С.difficile состоит из полипептидов двух тяжелых цепей, при этом каждая тяжелая цепь содержит VH-область и СН-область человека, и полипептидов двух легких цепей, при этом каждая легкая цепь содержит VL-область и CL-область человека. Последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида тяжелой цепи hPA-41 SEQ ID NO:22 изложена в SEQ ID NO:23 (Фиг.40 В); последовательность нуклеиновой кислоты (или кДНК), кодирующая аминокислотную последовательность полипептида легкой цепи hPA-41 SEQ ID NO:24 изложена в SEQ ID NO:25 (Фиг.40А).
Для применения в mAb сравнения получали моноклональные антитела CDA-1 и CDB1 (7, 89). Последовательности ДНК, кодирующие вариабельные области тяжелых и легких цепей Ig 3D8 и 124 (WO 2006/121422 и US 2005/0287150), синтезировали (ДНК 2.0) и клонировали в векторы pCON-гамма1 и pCON-каппа. mAb IgG1, κ полной длины экспрессировали в стабильно трансфицированных клетках CHO-K1SV и очищали, как описано выше. При тестировании на связывающую аффинность относительно токсинов А и В при помощи Biacore, подавление опосредуемых токсином цитопатических эффектов и гемагглютинацию согласно опубликованным способам (7), препараты CDA1 и CDB1 проявили ожидаемые уровни активности.
Е. In Vitro характеристика гуманизированных mAb
Эксперименты по нейтрализации токсина С.difficile проводили in vitro, чтобы сравнить функциональную активность гуманизированных mAb с таковой исходных mAb мыши. Как показано на Фиг.29, гуманизированное mAb PA-41 (hPA-41) эффективно нейтрализовало цитотоксичность токсина В (ЕС50 - 6 пМ) по сравнению с ЕС50 - 9 пМ в случае с mAb PA-41 (mPA-41) мыши. Подобным образом гуманизированное mAb РА-39 (hPA-39) и гуманизированное mAb PA-50 (hPA-50), как было обнаружено, были в равной степени эффективными по сравнению с их исходными mAb мыши в нейтрализации токсина А с применением клеток СНО-K1 или клеток Т-84 соответственно, как показано на Фиг.30 для hPA-39 и на Фиг.31 для hPA-50. Эти результаты демонстрируют, что исходные mAb мыши были успешно гуманизированы, и что гуманизированные mAb были функциональными и эффективными.
Из mAb к токсину С.difficile, изученных в этих исследованиях, РА-50 проявило отличающуюся кривую нейтрализации доза-эффект с коэффициентами Хилла, которые типично составляли больше двух, указывая на кооперативное подавление. Кооперативные взаимодействия часто встречаются в природе и часто характеризуются крутыми кривыми доза-эффект и коэффициентами Хилла >1. Лекарственные средства, которые проявляют кооперативную активность, были ассоциированы с усиленной клинической активностью при лечении вирусных инфекций. К тому же, РА-50 связывает токсин А мультивалентным способом, условие, которое часто является необходимым, но недостаточным для кооперативности.
Пример 11
Получение Fab-фрагментов из мышиных mAb к токсину по настоящему изобретению
А. Приготовление Fab-фрагментов
Fab-фрагментацию осуществляли с применением Mouse IgG1 Fab and F(ab’)2 Preparation Kit (Pierce) в соответствии с инструкциями изготовителя и реагентами, укомплектованными в наборе. Одинаковый протокол фрагментации применяли для всех mAb; PA-39, PA-41 и РА-50. Вкратце, иммобилизованную взвесь фицина (750 мкл) промывали буфером для расщепления (75 мМ цистеина, рН 5,6) перед тем, как добавляли приблизительно 3 мг mAb, и смесь инкубировали при 37°С в течение четырех часов с постоянным вращением с переворачиванием. После того, как расщепление завершилось, взвесь центрифугировали, и собирали продукт расщепления. Взвесь промывали три раза связывающим буфером с белком А и промытый материал добавляли к продукту завершенного расщепления. Колонку NAb с белком А уравновешивали связывающим буфером с белком А и добавляли образец расщепленного антитела. Колонку и образец инкубировали при комнатной температуре в течение 10 минут. Колонку центрифугировали при 1000g в течение одной минуты, чтобы собрать элюат, который содержал Fab-фрагменты. Колонку промывали три раза связывающим буфером с белком А. Элюат собирали, производили замену буфера на PBS и концентрировали.
B. SDS-PAGE Fab-фрагментов
Образцы анализировали посредством SDS-PAGE с применением Novex gel system (Invitrogen), и все реагенты, перечисленные ниже, были от Invitrogen, если не указано иное. Образцы смешивали с буфером для образцов NuPage и восстанавливали с использованием DTT. Восстановленные и невосстановленные образцы инкубировали при 100°С в течение 10 минут. После загрузки образцов (4 мкг) в 4-12% Bis Tris NuPage gel осуществляли электрофорез с использованием подвижного буфера MOPS при 180V в течение 60 минут. После электрофореза гель инкубировали с фиксатором (40% метанол, 10% уксусная кислота) в течение 20 минут, ополаскивали водой и окрашивали красителем Simply Blue на протяжении ночи при постоянном вращении. С.In vitro характеристика Fab
Эксперименты по in vitro нейтрализации токсина С.difficile проводили, чтобы сравнить функциональную активность Fab (▲) с таковой цельного mAb (■), исходя из количества связывающих участков. Fab PA-39 сильно нейтрализовал цитотоксичность токсина А на клетках СНО-K1 по сравнению с цельным PA-39 (EC50 - 880 пМ и EC50 -200 пМ, соответственно) (Фиг.41А). Было обнаружено, что Fab PA-41 было одинаково эффективным с цельным РА-41 при нейтрализации активности токсина В на клетках СНО-K1 (EC50 - 88 пМ и ЕС50 - 80 пМ, соответственно) (Фиг.41В). Fab PA-50 имел значение EC50 1,8 нМ по сравнению со значением ЕС50 100 пМ цельного mAb РА-50 при нейтрализации токсина А на клетках Т-84 (Фиг.41С).
Пример 12
Иммуногистохимический анализ гуманизированных mAb к токсину С.difficile на образцах человеческой ткани
Ценность иммуногистохимии (IHC) при исследовании экспрессии данного антигена в том, что она позволяет оценить детали микроанатомии и гетерогенность в нормальных и опухолевых тканях. IHC имеет преимущество над другими способами анализа, поскольку она может непосредственно определить местонахождение белков в индивидуальных типах клеток. Можно обнаруживать отличия генной экспрессии в нормальной и опухолевой ткани, в то же время отмечая изменения в числе и составе клеток. Ограничения этой техники включают возможные ложноотрицательные результаты вследствие низких уровней экспрессии исследуемой молекулы, а также ложноположительные результаты (кросс-реактивность) вследствие связывания антител с похожими эпитопами или эпитопами, которые являются общими с другими антигенами. Чтобы устранить эти ограничения, данное исследование проводили при самой низкой возможной концентрации каждого из антител, которые показывали сильное специфичное окрашивание на положительных специфичных к токсину контрольных образцах от инъецированных мышей.
Гуманизированные mAb PA-41 и РА-50 биотинилировали, чтобы определить иммуногистохимический паттерн связывания в выборке из замороженных тканей человека, включающих надпочечник, мочевой пузырь, костный мозг, молочную железу, мозжечок, кору головного мозга, шейку матки, толстую кишку, пищевод, глаз, фаллопиеву трубу, сердце, подвздошную кишку, тощую кишку, почку, печень, легкое, лимфатический узел, мышцу, яичник, периферический нерв, поджелудочную железу, паращитовидную железу, гипофиз, плаценту, простату, кожу, тонкую кишку, спинной мозг, селезенку, желудок, семенник, тимус, щитовидную железу, мочеточник, матку и белые клетки крови. Каждую ткань из вышеприведенных 37 разных типов тканей человека окрашивали каждым антителом. Работающие IHC-анализы разработали для обоих антител. Несоответствующее контрольное антитело изотипа IgG1, κ человека включали для всех образцов.
Для препаратов ткани из замороженных образцов, залитых в ОСТ Compound (Optimal Cutting Temperature embedding compound; Sakura, Torrance, CA), делали срезы по 5 микрон и помещали их на положительно заряженные предметные стекла. Способы и условия IHC-окрашивания для каждого антитела и тканевого образца разрабатывали, тестировали и оптимизировали. Процедуру IHC с непосредственным биотинилированием осуществляли с применением только нарезанных замороженных нефиксированных срезов ткани. Предметные стекла вытаскивали из криостата, давали возможность высохнуть на воздухе в течение 10 минут при комнатной температуре, фиксировали в 95% этаноле в течение 5 минут при комнатной температуре и затем промывали в трех последовательных кюветах с промывочным буфером Tris-забуференный физиологический раствор/0,1% Tween-20 (TBST; DakoCytomation) в течение 3 минут. Все последующие промывки осуществляли таким способом. Активность эндогенной пероксидазы блокировали при помощи 5-минутной инкубации при комнатной температуре с готовым к применению Peroxidase Block. После промывки буфером кроме того блокировали активность эндогенного биотина с помощью 15-минутных инкубаций, каждая с авидином, за которым следовал биотин, причем за каждым этапом следовали промывки буфером. В случае с РА-41, предметные стекла кроме того инкубировали с блокирующим белки реактивом Background Sniper в течение 10 минут при комнатной температуре без последующей промывки буфером. Предметные стекла инкубировали с тестируемым продуктом или реактивом для отрицательного контроля (1,25 мкг/мл для РА-41 и 10 мкг/мл для РА-50) в течение 30 минут при комнатной температуре. Первичное антитело РА-50 разводили 1:350 в разбавителе Dako, в то время как первичное антитело РА-41 разводили 1:3520 в разбавителе Dako с пролином (250 мМ, 0,576 г, Genzyme, CA) и гистидином (15 мМ, 0,046 г, Genzyme, CA), добавленными к 20 мл разбавителя (рН 7,7). Вслед за промывками в TBST на срезы тканей наносили реактив для обнаружения АВС (1:50 в TBST) для анализов обоих антител и инкубировали в течение 30 минут при комнатной температуре с последующими промывками буфером. Иммунную реакцию визуализировали с помощью инкубации с раствором 3,3’-диаминобензидин тетрагидрохлорида (DAB) в течение 5 минут при комнатной температуре. Предметные стекла ополаскивали деионизированной (DI) водой 3 раза, каждый в течение 30-60 секунд, контрастно окрашивали модифицированным гематоксилином Майера (DakoCytomation), подсинивали в 0,2% аммиаке, дегидратировали при помощи набора спиртов повышающейся концентрации, просветляли в ксилоле и накрывали покровным стеклом. Интерпретацию окрашенных предметных стекол осуществляли при помощи микроскопического исследования. Как правило, морфологически анализ ткани на предметном стекле после окрашивания антителом определял, присутствует ли адекватное количество ткани и представлены ли соответствующим образом обозначенные элементы нормальной ткани. Образцы, не соответствующие вышеуказанным стандартам, отбрасывались из анализа патологом, проводящим исследование.
Система оценок включала полуколичественный анализ интенсивности окрашивания. Интенсивность окрашивания тестируемого продукта оценивали по сравнению с интенсивностью предметного стекла с контрольной тканью, содержащего соседний срез, окрашенный антителом отрицательного контроля. Окрашивание среза, меченного реагентом отрицательного контроля, рассматривали как «фоновое» окрашивание. Бал «0» указывает на отсутствие окрашивания по сравнению с фоном; «1+» указывает на слабое окрашивание; «2+» указывает на умеренное окрашивание и «3+» указывает на сильное окрашивание. В соответствии со стандартной патологической практикой, интенсивность окрашивания описывалась при самом высоком уровне интенсивности, наблюдаемой во всех элементах ткани.
Результаты IHC-анализа для обоих гуманизированных mAb PA-50 и РА-41 были таковы, положительного окрашивание (0%) не проявлялось в любых из протестированных образцов тканей человека. Устойчивое сильное окрашивание (например, 3+) отмечалось в контрольных тканях мышцы ноги мышей, инъецированных токсином (токсин А для РА-50 и токсин В для РА-41) на всем протяжении исследования. В случае с РА-50, истинно положительно окрашивания не наблюдалось ни для какого образца ткани (т.е. 100% клеток показали 0% окрашивание). В случае с РА-41 истинно позитивное окрашивание не проявлялось в 37 протестированных тканях человека, однако, слабое (1+) позитивное окрашивание наблюдали как самую высокую интенсивность окрашивания в нормальной печени (из-за липохромного пигмента), нормальном легком (легочные макрофаги с инородным телом) и нормальной мышце (реакция, соответствующая артефактному окрашиванию). Такие значения слабого окрашивания для РА-41 рассматривались как несущественные относительно всех контролей, и в силу минимального изменения окрашивания на протяжении анализа.
Пример 13
Фармакокинетический анализ гуманизированных mAb к токсину С.difficile на приматах, кроме человека
Фармакокинетическое (PK) исследование проводили на ранее подвергавшихся воздействию яванских макаках с использованием очищенных, гуманизированных mAb РА-41 или mAb РА-50. В этом исследовании самцам яванских макак (Масаса fascicularis), ранее получавшим препарат, инъецировали внутривенно 1 мг/кг/животное или 5 мг/кг/животное очищенное гуманизированное mAb РА-41 или mAb РА-50. Исследование осуществляли в соответствии с принципами и методиками Комитета по содержанию и использованию лабораторных животных (IACUC).
В таблице 8 представлены параметры PK-исследования, показывающие, что каждое mAb (в концентрации 10 мг/кг) вводили внутривенно при двух уровнях доз животным, ранее подвергавшимся воздействию.
Животные получали однократную внутривенную инъекцию исследуемого антитела в начале исследования. После этого получали образцы крови при помощи венопункции периферических сосудов от каждого животного в 14 отдельных моментов времени в течение 29 дней (т.е. до введения препарата; через 0,5, 2, 6, 12 и 24 часа в день 1 (после введения); и в дни 3, 4, 7, 9, 12, 15, 22 и 29). Образцы крови собирали в пробирки для отделения сыворотки и хранили на водном льду до коагуляции. После коагуляции, образцы крови центрифугировали при 1800 g в течение 15 минут при 4°С для получения сывороток. Образцы сывороток хранили при -70°С до применения.
Концентрацию mAb в сыворотках определяли посредством ELISA. Девяностошестилуночные планшеты для ELISA (Thermo Fisher Scientific, Rochester, NY) покрывали в течение ночи при 4°С токсином A (Techlab) или токсином В (Techlab) при 100 нг/лунку. Планшеты отмывали три раза PBS/0,05% Tween-20® (PBS-T) и блокировали 200 мкл блокирующего буфера (PBS без кальция или магния, 0,1% Tween 20®, 1% казеин) в течение одного часа при комнатной температуре. Эталонное стандартное антитело (очищенное mAb РА-41 или mAb PA-50) разводили в 1% объединенной сыворотке ранее не подвергавшихся воздействию яванских макак (Bioreclamation) для получения стандартной кривой в диапазоне 0,3-4000 нг/мл. Разведенные тестируемые образцы и стандарты тестировали в трех повторностях и инкубировали в течение одного часа при комнатной температуре.
Планшеты отмывали шесть раз PBS-T и инкубировали в течение одного часа при комнатной температуре с HRP-конъюгированными антителами козы к IgG1 человека (The Binding Site, San Diego, CA). Планшеты проявляли с использованием 1 компонентного пероксидазного субстрата SureBlue TMB (KPL), блокировали 1н. соляной кислотой (Thermo Fisher Scientific) и считывали на планшет-ридере SpectraMax (Molecular Devices) при 450 нм. Концентрацию mAb у каждой обезьяны в разные моменты времени рассчитывали с применением стандартных кривых. Некомпартментный фармакокинетический анализ проводили с применением WinNonLin, версия 4.0 (Pharsight Corp., Mountain View, CA). Результаты РК для гуманизированного mAb PA-50 показаны на Фиг.42А; результаты для гуманизированного mAb РА-41 показаны на Фиг.42В. Для РА-50 при дозах 1 мг/кг и 5 мг/кг среднее T1/2 (дни) составляло 14,5±0,3 и 12,3±1,5 соответственно. Для РА-41 при дозах 1 мг/кг и 5 мг/кг среднее Т1/2 (дни) составляло 8,9±1,3 и 9,2±3,3 соответственно.
Список литературы
1. Bartlett, J. G., T. W. Chang, M. Gurwith, S. L. Gorbach, and A. B. Onderdonk. 1978. Antibiotic-associated pseudomembranous colitis due to toxin-producing clostridia. The New England Journal of Medicine 298:531-534.
2. Kyne, L., R. J. Farrell, and C. P. Kelly. 2001. Clostridium difficile. Gastroenterol. Clin North Am 30:753-777.
3. Kelly, С.P. and J. T. LaMont. 2008. Clostridium difficile-more difficult than ever. The New England Journal of Medicine 359:1932-1940.
4. MacCannell, D. R., T. J. Louie, D. B. Gregson, M. Laverdiere, A. C. Labbe, F. Laing, and S. Henwick. 2006. Molecular analysis of Clostridium difficile PCR ribotype 027 isolates from Eastern and Western Canada. J Clin Microbiol 44:2147-2152.
5. McDonald, L. C., G. E. Killgore, A. Thompson, R. C. Owens, Jr., S. V. Kazakova, S. P. Sambol, S. Johnson, and D. N. Gerding. 2005. An epidemic, toxin gene-variant strain of Clostridium difficile. The New England Journal of Medicine 353:2433-2441.
6. Wamy, M., J. Pepin, A. Fang, G. Killgore, A. Thompson, J. Brazier, E. Frost, and L. C. McDonald. 2005. Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 366:1079-1084.
7. Babcock, G. J., T. J. Broering, H. J. Hemandez, R. B. Mandell, K. Donahue, N. Boatright, A. M. Stack, I. Lowy, R. Graziano, D. Molrine, D. M. Ambrosino, and W. D. Thomas, Jr. 2006. Human monoclonal antibodies directed against toxins A and B prevent Clostridium difficile-induced mortality in hamsters. Infect Immun 74:6339-6347.
8. Bartlett, J. G. 1981. Antimicrobial agents implicated in Clostridium difficile toxin-associated diarrhea of colitis. Johns Hopkins Med J 149:6-9.
9. McFarland, L. V., M. E. Mulligan, R. Y. Kwok, and W. E. Stamm. 1989. Nosocomial acquisition of Clostridium difficile infection. The New England Journal of Medicine 320:204-210.
10. Zilberberg, M. D., A. F. Shorr, and M. H. Kollef. 2008. Increase in adult Clostridium difficile-related hospitalizations and case-fatality rate. United States, 2000-2005. Emerg. Infect Dis 14:929-931.
11. Riley, Т. V., Codde, J. P., and Rous, I. L. Increased length of hospital stay due to Clostridium difficile-associated diarrhoea. Lancet 345, 455-456. 2-18-1995.
12. O’Brien, J. А., В. J. Lahue, J. J. Caro, and D. M. Davidson. 2007. The emerging infectious challenge of clostridium difficile-associated disease in Massachusetts hospitals: clinical and economic consequences. Infect Control Hosp Epidemiol. 28:1219-1227.
13. Redelings, M. D., F. Sorvillo, and L. Mascola. 2007. Increase in Clostridium difficile-related mortality rates, United States, 1999-2004. Emerg. Infect Dis 13:1417-1419.
14. Loo, V. G., L. Poirier, M. A. Miller, M. Oughton, M. D. Libman, S. Michaud, A. M. Bourgault, T. Nguyen, C. Frenette, M. Kelly, A. Vibien, P. Brassard, S. Fenn, K. Dewar, T. J. Hudson, R. Horn, P. Rene, Y. Monczak, and A. Dascal. 2005. A predominantly clonal multi-institutional outbreak of Clostridium difficile-associated diarrhea with high morbidity and mortality. The New England Journal of Medicine 353:2442-2449.
15. Warny, M. K. C. 2003. Pathogenicity of Clostridium difficile toxins. Washington DC:
ASM Press 366:503-524.
16. Kelly, C. P., C. Pothoulakis, and J. T. LaMont. 1994. Clostridium difficile colitis. The New England Journal of Medicine 330:257-262.
17. Lyerly, D. M., D. E. Lockwood, S. H. Richardson, and T. D. Wilkins. 1982. Biological activities of toxins A and В of Clostridium difficile. Infect Immun. 35:1147-1150.
18. McFarland, L. V., H. W. Beneda, J. E. Clamdge, and G. J. Raugi. 2007. Implications of the changing face of Clostridium difficile disease for health care practitioners. Am J Infect Control 35:237-253.
19. Drudy, D., T. Quinn, R. O'Mahony, L. Kyne, P. O’Gaora, and S. Fanning. 2006. High-level resistance to moxifloxacin and gatifloxacin associated with a novel mutation in gyrB in toxin-A-negative, toxin-B-positive Clostridium difficile. J Antimicrob Chemother 58:1264-1267.
20. Rupnik, M., N. Kato, M. Grabnar, and H. Kato. 2003. New types of toxin A-negative, toxin B-positive strains among Clostridium difficile isolates from Asia. J Clin Microbiol 41:1118-1125.
21. Voth, D. E. and J. D. Ballard. 2005. Clostridium difficile toxins: mechanism of action and role in disease. Clin Microbiol Rev 18:247-263.
22. Reineke, J., S. Tenzer, M. Rupnik, A. Koschinski, O. Hasselmayer, A. Schrattenholz, H. Schild, and C. Eichel-Streiber. 2007. Autocatalytic cleavage of Clostridium difficile toxin B. Nature 446:415-419.
23. Aslam, S., R. J. Hamill, and D. M. Musher. 2005. Treatment of Clostridium difficile-associated disease: old therapies and new strategies. Lancet Infect Dis 5:549-557.
24. Musher, D. M., S. Aslam, N. Logan, S. Nallacheru, I. Bhaila, F. Borchert, and R. J. Hamill. 2005. Relatively poor outcome after treatment of Clostridium difficile colitis with metronidazole. Clin Infect Dis 40:1586-1590.
25. Pepin, J., M. E. Alary, L. Valiquette, E. Raiche, J. Ruel, K. Fulop, D. Godin, and C. Bourassa. 2005. Increasing risk of relapse after treatment of Clostridium difficile colitis in Quebec, Canada. Clin Infect Dis 40:1591-1597.
26. Fekety, R. and A. B. Shah. 1993. Diagnosis and treatment of Clostridium difficile colitis. JAMA 269:71-75.
27. Fekety, R., L. V. McFarland, C. M. Surawicz, R. N. Greenberg, G. W. Elmer, and M. E. Mulligan. 1997. Recurrent Clostridium difficile diarrhea: characteristics of and risk factors for patients enrolled in a prospective, randomized, double-blinded trial. Clin Infect Dis 24:324-333.
28. Pothoulakis, C. and J. T. LaMont. 1993. Clostridium difficile colitis and diarrhea. Gastroenterol. Clin North Am 22:623-637.
29. McFarland, L. V., C. M. Surawicz, R. N. Greenberg, R. Fekety, G. W. Elmer, K. A. Moyer, S. A. Melcher, K. E. Bowen, J. L. Cox, Z. Noorani, and. 1994. A randomized placebo-controlled trial of Saccharomyces boulardii in combination with standard antibiotics for Clostridium difficile disease. JAMA 271:1913-1918.
30. McFarland, L. V., G. W. Elmer, and C. M. Surawicz. 2002. Breaking the cycle: treatment strategies for 163 cases of recurrent Clostridium difficile disease. Am J Gastroenterol. 97:1769-1775.
31. Kyne, L., M. Wamy, A. Qamar, and C. P. Kelly. 2000. Asymptomatic carriage of Clostridium difficile and serum levels of IgG antibody against toxin A. The New England Journal of Medicine 342:390-397.
32. Kyne, L., M. Wamy, A. Qamar, and C. P. Kelly. 2001. Association between antibody response to toxin A and protection against recurrent Clostridium difficile diarrhoea. Lancet 357:189-193.
33. Leav, В., Blair, В., Leney, M., Knauber, M., Reilly, С., Lowy, I., Kohberger, R., Gerding, D. N., Kelly, C., Katchar, K., Baxter, R., and Ambrosino, D. 2008. Serum anti-toxin В antibody correlates with protection from recurrent Clostridium difficile associated diarrhea (CDAD). ICAAC/IDSA, Poster B-1295. 2008.
34. Wilcox, M. H., W. N. Fawley, C. D. Settle, and A. Davidson. 1998. Recurrence of symptoms in Clostridium difficile infection-relapse or reinfection? J Hosp Infect 38:93-100.
35. Jodlowski, Т. Z., R. Oehler, L. W. Kam, and I. Melnychuk. 2006. Emerging therapies in the treatment of Clostridium difficile-associated disease. Ann. Pharmacother 40:2164-2169.
36. Missaghi, В., А. J. Valenti, and R. C. Owens, Jr. 2008. Clostridium difficile Infection: A Critical Overview. Curr. Infect Dis Rep 10:165-173.
37. Optimer Pharmaceuticals, News Release, November 10. 2008.
38. Louie, Т. J., J. Peppe, C. K. Watt, D. Johnson, R. Mohammed, G. Dow, K. Weiss, S. Simon, J. F. John, Jr., G. Garber, S. Chasan-Taber, and D. M. Davidson. 2006. Tolevamer, a novel nonantibiotic polymer, compared with vancomycin in the treatment of mild to moderately severe Clostridium difficile-associated diarrhea. Clin Infect Dis 43:411-420.
39. Louie, Т. G. M. G. D. e. al. 2007. Results of a phase III trial comparing tolevamer, vancomycin and metronidazole in patients with Clostridium difficile-associated diarrhea (CDI). 47th Interscience Conference on Antimicrobial Agents and Chemotherapy.
40. Kelly, C. P., C. Pothoulakis, J. Orellana, and J. T. LaMont. 1992. Human colonic aspirates containing immunoglobulin A antibody to Clostridium difficile toxin A inhibit toxin A-receptor binding. Gastroenterology 102:35-40.
41. Salcedo, J., S. Keates, C. Pothoulakis, M. Wamy, I. Castagliuolo, J. T. LaMont, and C. P. Kelly. 1997. Intravenous immunoglobulin therapy for severe Clostridium difficile colitis. Gut 41:366-370.
42. Leung, D. Y., C. P. Kelly, M. Boguniewicz, C. Pothoulakis, J. T. LaMont, and A. Flores. 1991. Treatment with intravenously administered gamma globulin of chronic relapsing colitis induced by Clostridium difficile toxin. J Pediatr 118:633-637.
43. Medarex and Massachusetts Biologic Laboratories, News Release, November 3. 2008.
44. Kamiya, S., K. Yamakawa, X. Q. Meng, H. Ogura, and S. Nakamura. 1991. Production of monoclonal antibody to Clostridium difficile toxin A which neutralizes enterotoxieity but not haemagglutination activity. FEMS Microbiol Lett 65:311-315.
45. Kink, J. A. and J. A. Williams. 1998. Antibodies to recombinant Clostridium difficile toxins A and В are an effective treatment and prevent relapse of C. difficile-associated disease in a hamster model of infection. Infect Immun 66:2018-2025.
46. Lyerly, D. M., C. J. Phelps, J. Toth, and T. D. Wilkins. 1986. Characterization of toxins A and В of Clostridium difficile with monoclonal antibodies. Infect Immun 54:70-76.
47. Harlow, E. and Lane, D. Antibodies, a laboratory manual. Cold Spring harbor Laboratory Press, Cold Spring harbor, New York. 1988.
48. dark, G. F., H. C. Krivan, T. D. Wilkins, and D. F. Smith. 1987. Toxin A from Clostridium difficile binds to rabbit erythrocyte glycolipids with terminal Gal alpha 1-3 Gal beta l-4GlcNAc sequences. Arch Biochem Biophys 257:217-229.
49. Anton, P. M., M. O'Brien, E. Kokkotou, B. Eisenstein, A. Michaelis, D. Rothstein, S. Paraschos, C. P. Kelly, and C. Pothoulakis. 2004. Rifalazil treats and prevents relapse of clostridium difficile-associated diarrhea in hamsters. Antimicrob Agents Chemother 48:3975-3979.
50. Boss, S. M., C. L. Gries, B. K. Kirchner, G. D. Smith, and P. C. Francis. 1994. Use of vancomycin hydrochloride for treatment of Clostridium difficile enteritis in Syrian hamsters. Lab AnimSci 44:31-37.
51. Freeman, J., S. D. Baines, D. Jabes, and M. H. Wilcox. 2005. Comparison of the efficacy of ramoplanin and vancomycin in both in vitro and in vivo models of clindamycin-induced Clostridium difficile infection. J Antimicrob Chemother 56:717-725.
52. Kink, J. A. and J. A. Williams. 1998. Antibodies to recombinant Clostridium difficile toxins A and B are an effective treatment and prevent relapse of C. difficile-associated disease in a hamster model of infection. Infect Immun 66:2018-2025.
53. Kokkotou, E., A. C. Moss, A. Michos, D. Espinoza, J. W. Cloud, N. Mustafa, M. O'Brien, C. Pothoulakis, and C. P. Kelly. 2008. Comparative efficacies of rifaximin and vancomycin for treatment of Clostridium difficile-associated diarrhea and prevention of disease recurrence in hamsters. Antimicrob Agents Chemother 52:1121-1126.
54. Kurtz, С. В., E. P. Cannon, A. Brezzani, M. Pitruzzello, С.Dinardo, E. Rinard, D. W. Acheson, R. Fitzpatrick, P. Kelly, K. Shackett, A. T. Papoulis, P. J. Goddard, R. H. Barker, Jr., G. P. Palace, and J. D. Klinger. 2001. GT160-246, a toxin binding polymer for treatment of Clostridium difficile colitis. Antimicrob Agents Chemother 45:2340-2347.
55. McVay, C. S. and R. D. Rolfe. 2000. In vitro and in vivo activities of nitazoxanide against Clostridium difficile. Antimicrob Agents Chemother 44:2254-2258.
56. Razaq, N., S. Sambol, К. Nagaro, W. Zukowski, A. Cheknis, S. Johnson, and D. N. Gerding. 2007. Infection of hamsters with historical and epidemic BI types of Clostridium difficile. J Infect Dis 196:1813-1819.
57. Femie, D. S., R. O. Thomson, I. Batty, and P. D. Walker. 1983. Active and passive immunization to protect against antibiotic associated caecitis in hamsters. Developmental Biology Standard 53:325-332.
58. Giannasca, P. J., Z. X. Zhang, W. D. Lei, J. A. Boden, M. A. Giel, T. P. Monath, and W. D. Thomas, Jr. 1999. Serum antitoxin antibodies mediate systemic and mucosal protection from Clostridium difficile disease in hamsters. Infect Immun 67:527-538.
59. Kirn, P. H., J. P. laconis, and R. D. Rolfe. 1987. Immunization of adult hamsters against Clostridium difficile-associated ileocecitis and transfer of protection to infant hamsters. Infect Immun 55:2984-2992.
60. Lyerly, D. M., E. F. Bostwick, S. B. Binion, and T. D. Wilkins. 1991. Passive immunization of hamsters against disease caused by Clostridium difficile by use of bovine immunoglobulin G concentrate. Infect Immun 59:2215-2218.
61. Kelly, С.P., С.Pothoulakis, F. Vawa, I. Castagliuolo, E. F. Bostwick, J. C. O'Keane, S. Keates, and J. T. LaMont. 1996. Anti-Clostridium difficile bovine immunoglobulin concentrate inhibits cytotoxicity and enterotoxicity of C. difficile toxins. Antimicrob Agents Chemother 40:373-379.
62. Delmee, M., M. Homel, and G. Wauters. 1985. Serogrouping of Clostridium difficile strains by slide agglutination. J Clin Microbiol 21:323-327.
63. Delmee, M., V. Avesani, N. Delferriere, and G. Burtonboy. 1990. Characterization of flagella of Clostridium difficile and their role in serogrouping reactions. J Clin Microbiol 28:2210-2214.
64. Barbut, F., A. Richard, K. Hamadi, V. Chomette, B. Burghoffer, and J. C. Petit. 2000. Epidemiology of recurrences or reinfections of Clostridium difficile-associated diarrhea. J Clin Microbiol 38:2386-2388.
65. Carswell, C. I., G. L. Plosker, and A. J. Wagstaff. 2001. Daclizumab: a review of its use in the management of organ transplantation. BioDrugs 15:745-773.
66. Wiland, A. M. and B. Philosophe. 2004. Daclizumab induction in solid organ transplantation. Expert Opin Biol Ther 4:729-740.
67. Fenton, C., L. J. Scott, and G. L. Plosker. 2004. Palivizumab: a review of its use as prophylaxis for serious respiratory syncytial virus infection. Paediatr. Drugs 6:177-197.
68. Wu, H„ D. S. Pfarr, Y. Tang, L. L. An, N. K. Patel, J. D. Watkins, W. D. Huse, P. A. Kiener, and J. F. Young. 2005. Ultra-potent antibodies against respiratory syncytial virus: effects of binding kinetics and binding valence on viral neutralization. J Mol Biol 350:126-144.
69. Romero, J. R. Palivizumab prophylaxis of respiratory syncytial virus disease from 1998 to 2002:results from four years of palivizumab usage. Pediatr.Infect.Dis 22, S46-S54. 2003.
70. Carter, P., L. Presta, C. M. German, J. B. Ridgway, D. Henner, W. L. Wong, A. M. Rowland, C. Kotts, M. E. Carver, and H. M. Shepard. 1992. Humanization of an anti-pi 85HER2 antibody for human cancer therapy. Proc Nati Acad Sci U S A 89:4285-4289.
71. Emens, L. A. 2005. Trastuzumab: targeted therapy for the management of HER-2/neu-overexpressing metastatic breast cancer. Am J Ther 12:243-253.
72. Finn, R. S. and D. J. Slamon. 2003. Monoclonal antibody therapy for breast cancer:
herceptin. Cancer Chemother Biol Response Modif. 21:223-233.
73. Giles, F., E. Estey, and S. O’Brien. 2003. Gemtuzumab ozogamicin in the treatment of acute myeloid leukemia. Cancer 982095-2104.
74. Siemoneit, К., S. Cardoso Mda, K. Koemer, A. Wolpi, and B. Kubanek. 1995. Human monoclonal antibodies for the immunological characterization of a highly conserved protein domain of the hepatitis C virus glycoprotein El. Clin Exp Immunol 101:278-83.
75. Casale, Т. В. 2004. Omalizumab: an effective anti-IgE treatment for allergic asthma and rhinitis. Drugs Today (Bare.) 40:367-376.
76. Holgate, S. Т., R. Djukanovic, T. Casale, and J. Bousquet. 2005. Anti-immunoglobulin E treatment with omalizumab in allergic diseases: an update on anti-inflammatory activity and clinical efficacy. Clin Exp Allergy 35:408-416.
77. Presta, L. G., S. J. Lahr, R. L. Shields, J. P. Porter, C. M. Gorman, B. M. Fendly, and P. M. Jardieu. 1993. Humanization of an antibody directed against IgE. The Journal of Immunology 151:2623-2632.
78. Jordan, J. K. 2005. Efalizumab for the treatment of moderate to severe plaque psoriasis. Ann. Pharmacother 39:1476-1482.
79. Werther, W. A., T. N. Gonzalez, S. J. O'Connor, S. McCabe, B. Chan, T. Hotaling, M. Champe, J. A. Fox, P. M. Jardieu, P. W. Berman, and L. G. Presta. 1996. Humanization of an anti-lymphocyte function-associated antigen (LFA)-1 monoclonal antibody and reengineering of the humanized antibody for binding to rhesus LFA-1. The Journal of Immunology 157:4986-4995.
80. Leonardi, C. L. 2004. Current concepts and review of efalizumab in the treatment of psoriasis. Dermatol. Clin 22:427-435.
81. Ferrara, N., K. J. Hillan, H. P. Gerber, and W. Novotny. 2004. Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nat Rev Drug Discov. 3:391-400.
82. Presta, L. G., H. Chen, S. J. O'Connor, V. Chisholm, Y. G. Meng, L. Krummen, M. Winkler, and N. Ferrara. 1997. Humanization of an anti-vascular endothelial growth factor monoclonal antibody for the therapy of solid tumors and other disorders. Cancer Res 57:4593-4599.
83. Steinman, L. 2005. Blocking adhesion molecules as therapy for multiple sclerosis: natalizumab. Nat Rev Drug Discov. 4:510-518.
84. Fagnani, R. 1994. The immunogenicity of foreign monoclonal antibodies in human disease applications: problems and current approaches. Immunol Ser. 61:3-22.
85. Mateo, С., E. Moreno, K. Amour, J. Lombardero, W. Harris, and R. Perez. 1997. Humanization of a mouse monoclonal antibody that blocks the epidermal growth factor receptor: recovery of antagonistic activity. Immunotechnology. 3:71-81.
86. Reichert, J. M., C. J. Rosensweig, L. B. Faden, and M. C. Dewitz. 2005. Monoclonal antibody successes in the clinic. Nat Biotechnol 23:1073-1078.
87. Stephens, S., S. Emtage, 0. Vetterlein, L. Chaplin, C. Bebbington, A. Nesbitt, M. Sopwith, D. Athwal, C. Novak, and M. Bodmer. 1995. Comprehensive pharmacokinetics of a humanized antibody and analysis of residual anti-idiotypic responses. Immunology 85:668-674.
88. Co, M. S., N. M. Avdalovic, P. C. Caron, M. V. Avdalovic, D. A. Scheinberg, and C. Queen. 1992. Chimeric and humanized antibodies with specificity for the CD33 antigen. J Immunol. 148:1149-1154.
89. LowyJ. et al. 2010. Treatment with monoclonal antibodies against Clostridium difficile toxins. N. Engl. J Med., 362, 197-205.
90. Babcock,G.J. et al. Human Monoclonal Antibodies Neutralize Toxins Produced by Epidemic Strains of Clostridium difficile, the Infectious Diseases Society of America 43rd Annual Meeting, October 6-9,2005; San Francisco, California.
91. Optimer Pharmaceuticals Reports Positive Data from its North American Phase 3 CDI Study ofOPT-80. Optimer Pharmaceuticals, Press Release (2008).
92. Rothman SW, Toxicon. 1988; 26(6):583-97.
93. W02005US0047100 to Diversa.
94. Cheknis, AK. et al. Distribution of Clostridium difficile strains from a North American, European and Australian trial of treatment for C. difficile infections: 2005-2007. Anaerobe 15: 230-233 (2009).
95. Gerding D, et al. Restriction endonuclease analysis (REA) typing of Clostridium difficile in a phase 3 treatment trial of fidaxomicin vs vancomycin: Decreased cure rate for epidemic BI/NAP 1/027 strain. 49th Interscience Conference on Antimicrobial Agents and Chemotherapy, Abstract, LI-1642, San Francisco, CA, September 12-15, 2009.
Хотя настоящее изобретение было описано в деталях с целью иллюстрации, следует понимать, что такая детальность предназначена исключительно для этой цели и специалисты в данной области могут сделать изменения без отклонения от сущности и объема настоящего изобретения, которые определяются следующей формулой изобретения.
Содержания всех ссылок, патентов и опубликованных заявок на патенты, цитируемые на всем протяжении этой заявки, включаются в данный документ посредством ссылки.
Включение в список какой-либо ссылки не является признанием того, что данная ссылка является известным уровнем техники.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ, ОТНОСЯЩИЕСЯ К МУТАНТНОМУ ТОКСИНУ CLOSTRIDIUM DIFFICILE, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2592686C2 |
| Способ получения аминокислотных последовательностей, кодирующих наноантитела, нейтрализующие токсин А Clostridioides difficile | 2023 |
|
RU2833930C1 |
| АНТИКИНОВЫЕ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С НЕСКОЛЬКИМИ СС-ХЕМОКИНАМИ | 2010 |
|
RU2574786C2 |
| МОНОКЛОНАЛЬНЫЕ АНТЕТЕЛА К CD44, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ ПЛОСКОКЛЕТОЧНОЙ КАРЦИНОМЫ ГОЛОВЫ И ШЕИ | 2011 |
|
RU2599447C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ, ВЫЗВАННЫХ Staphylococcus aureus | 2015 |
|
RU2636780C1 |
| Композиции и способы, имеющие отношение к мутантному токсину из Clostridium Difficile | 2013 |
|
RU2630671C2 |
| АНТИТЕЛА, КОТОРЫЕ СПЕЦИФИЧНО СВЯЗЫВАЮТ АЛЬФА-ТОКСИН STAPHYLOCOCCUS AUREUS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620065C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-31 И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2444528C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2671481C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2431638C2 |



















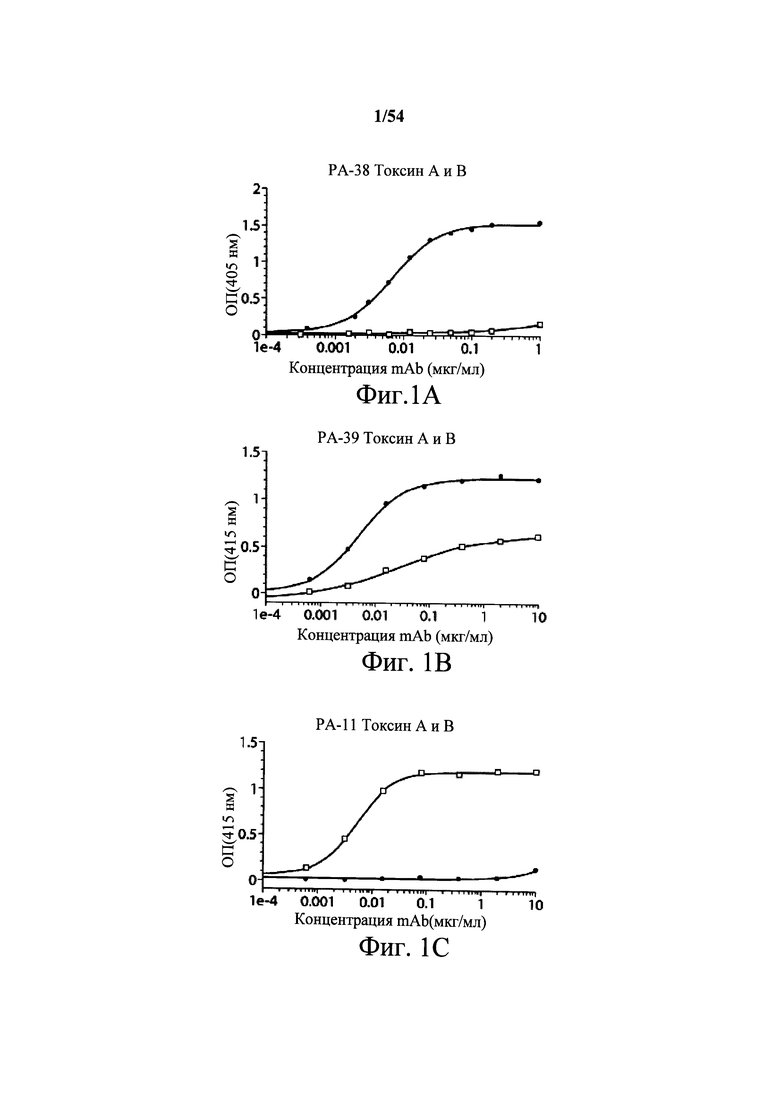
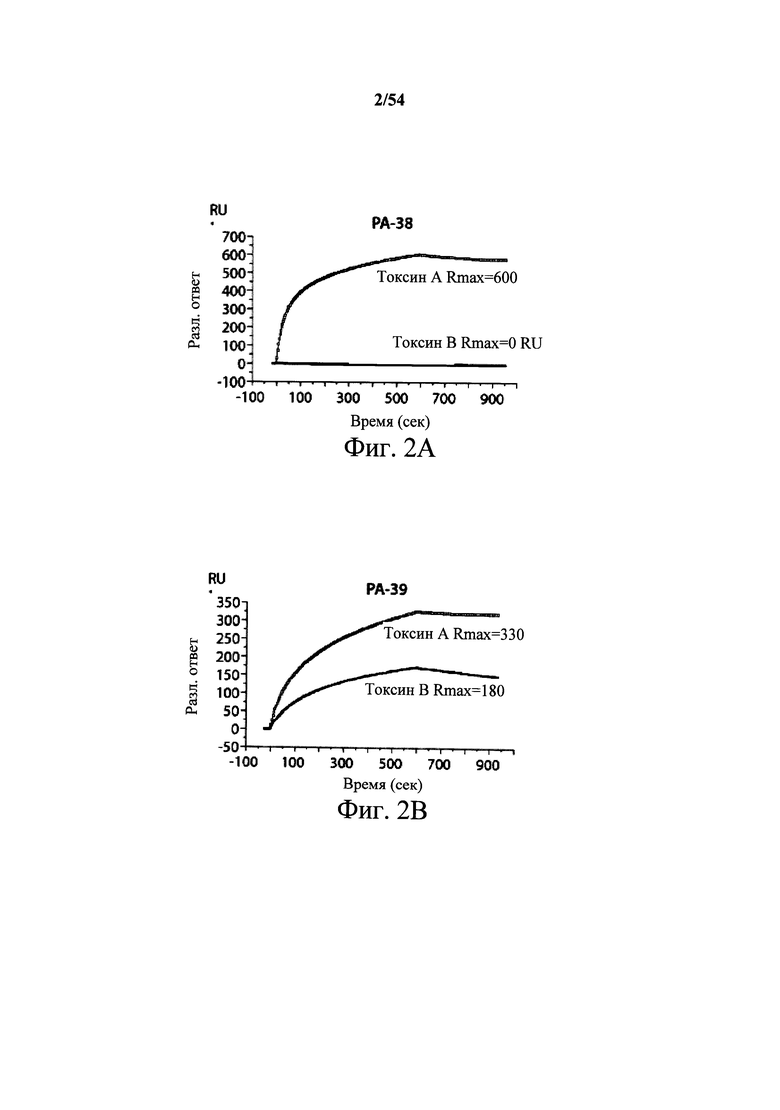
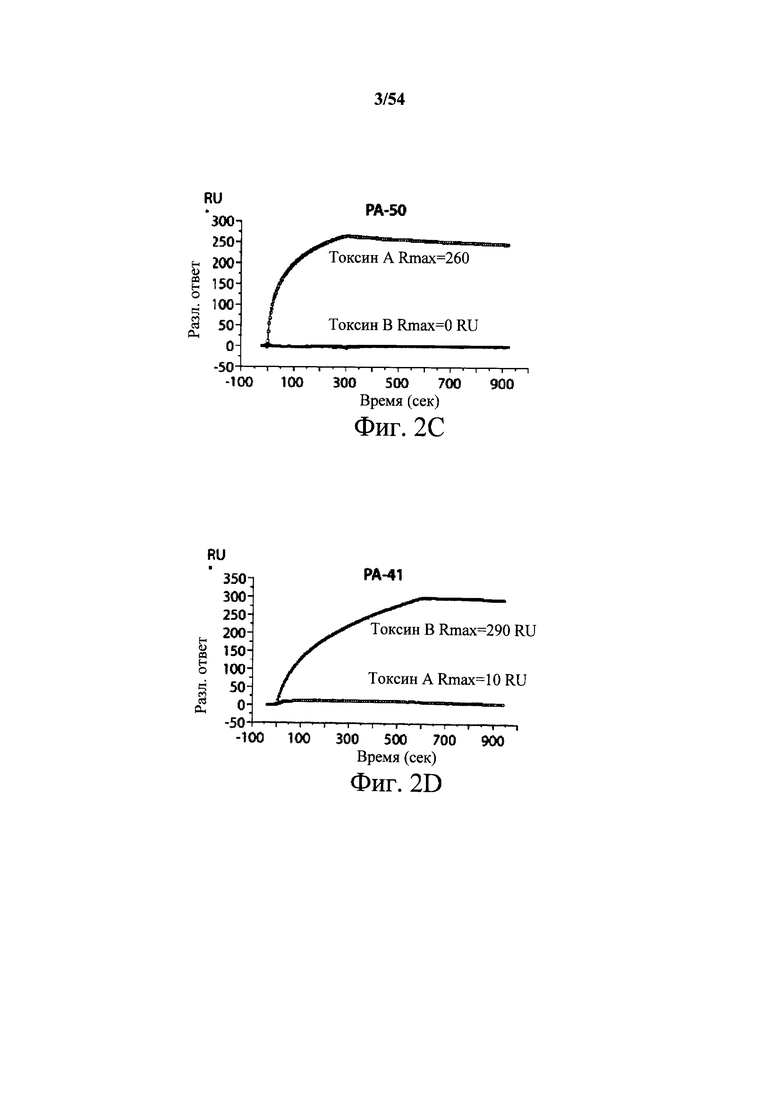
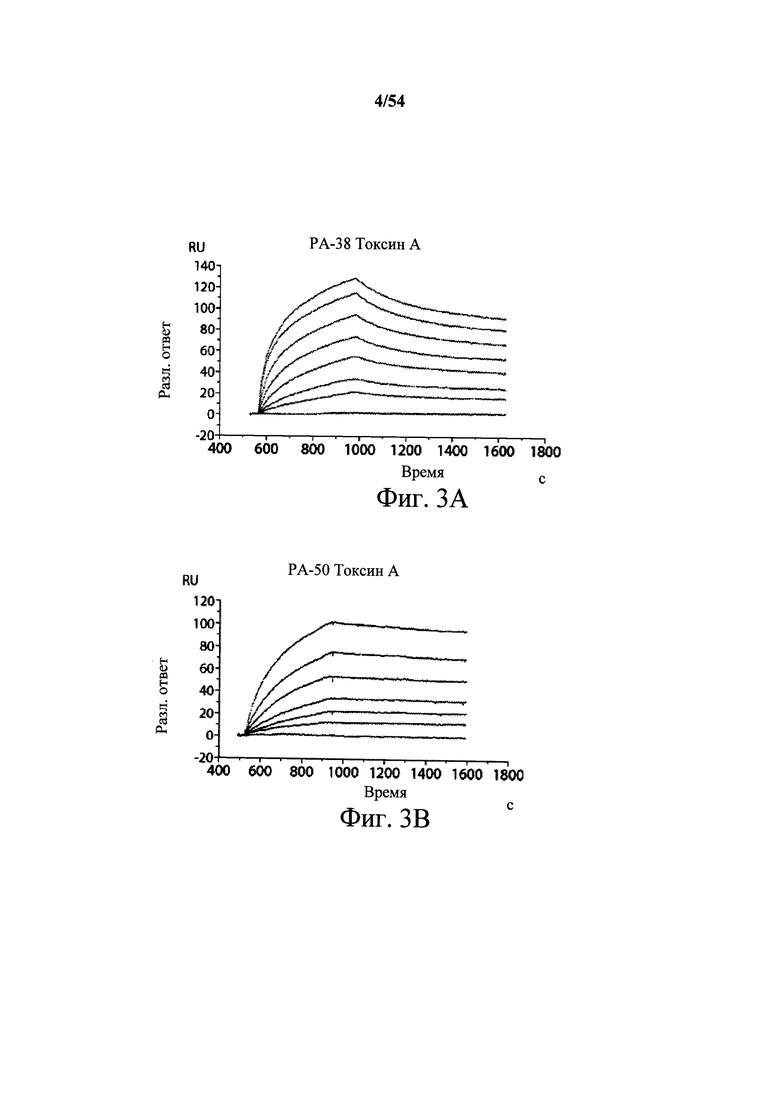
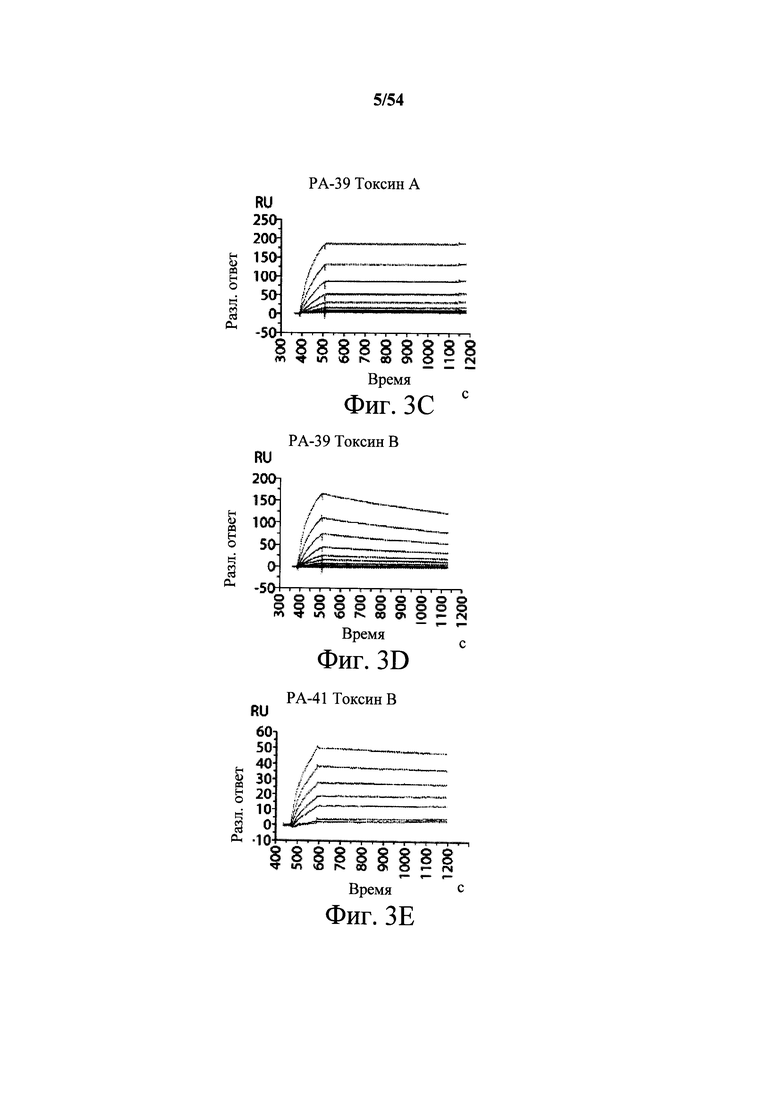
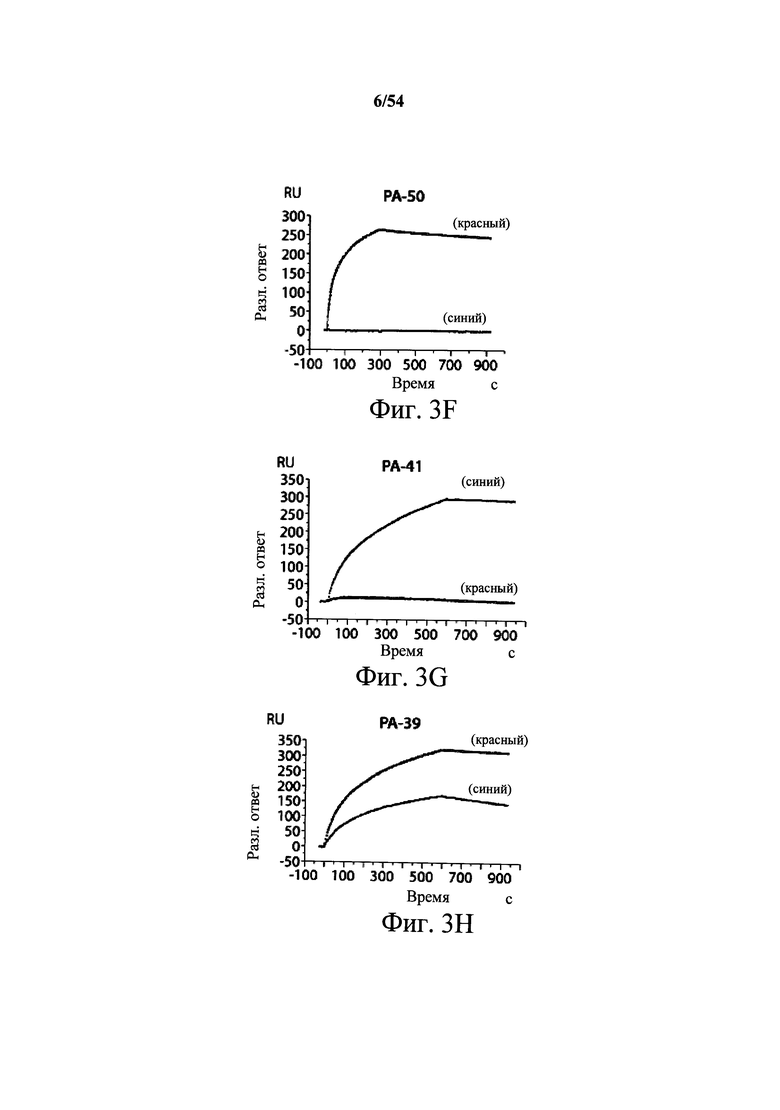
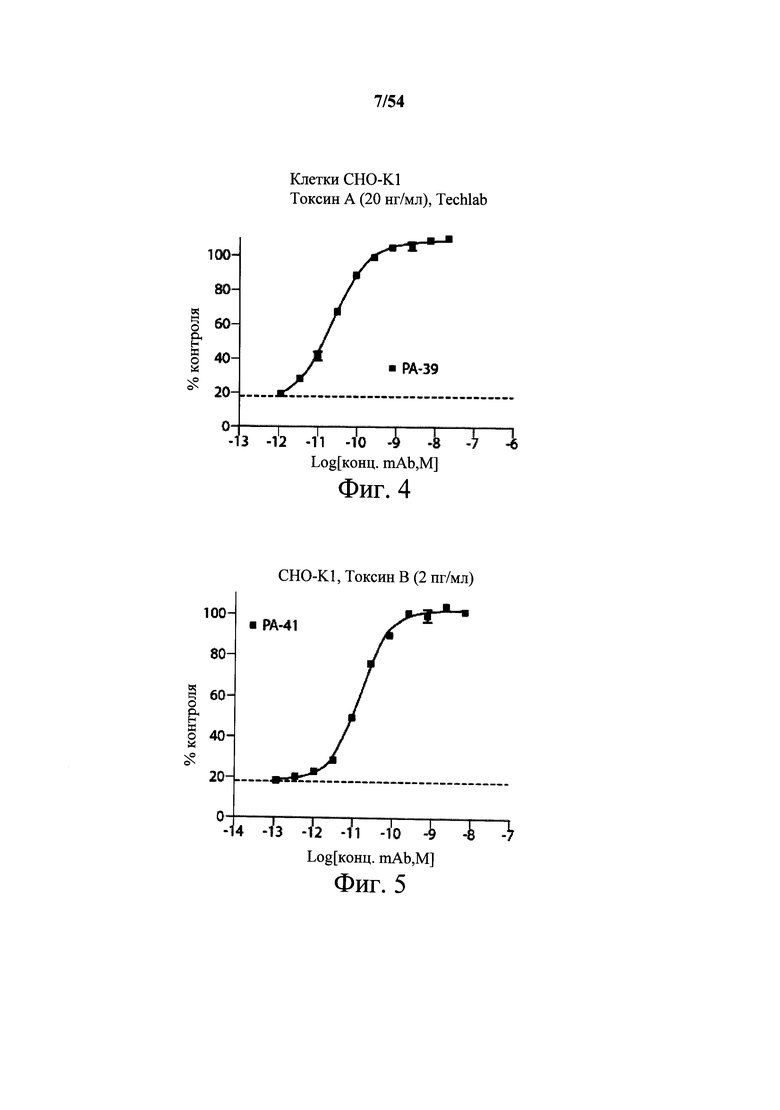
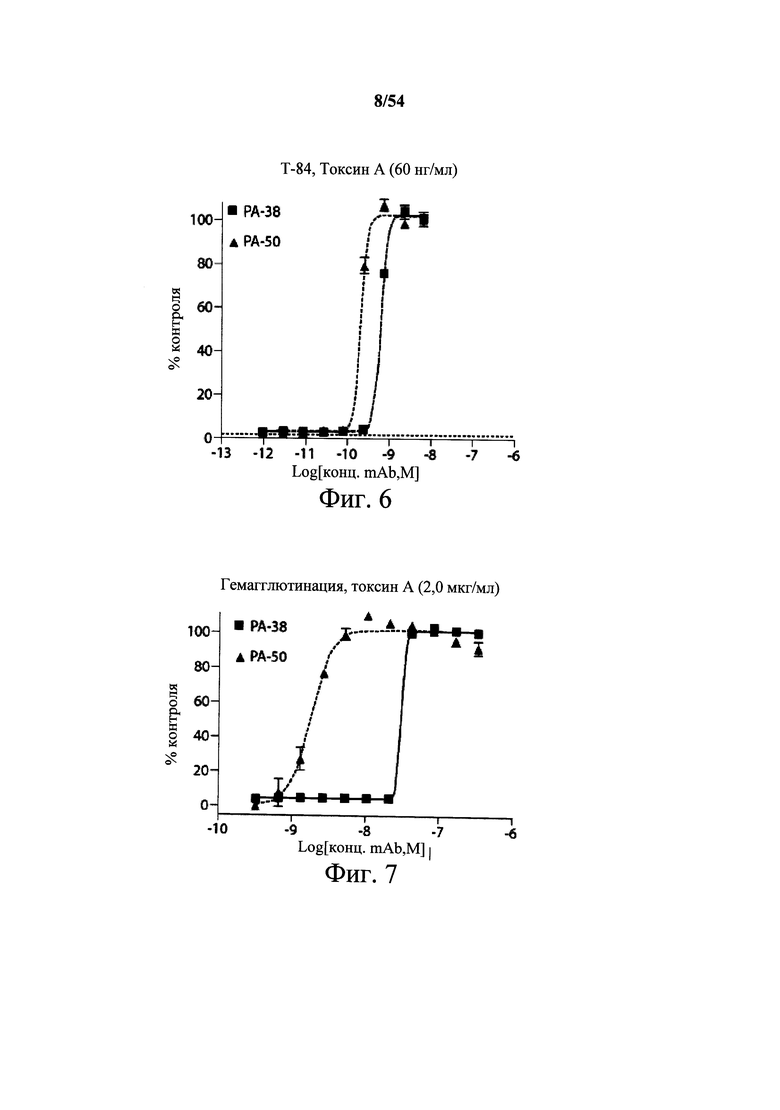
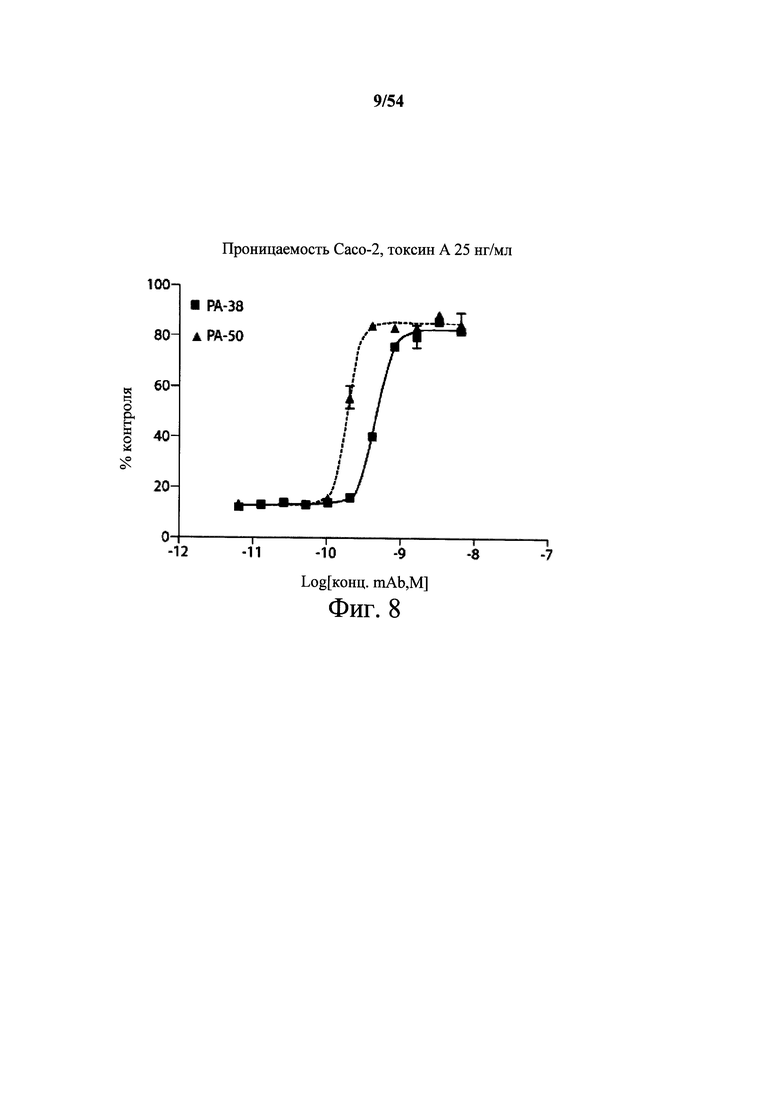
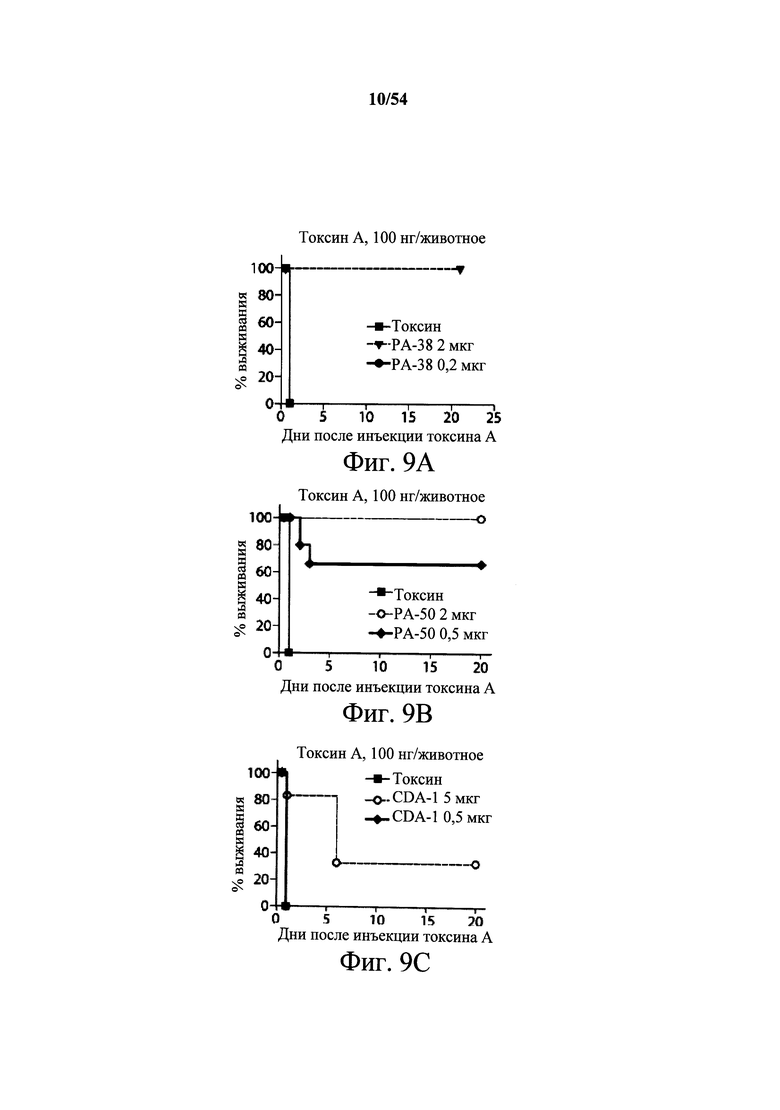
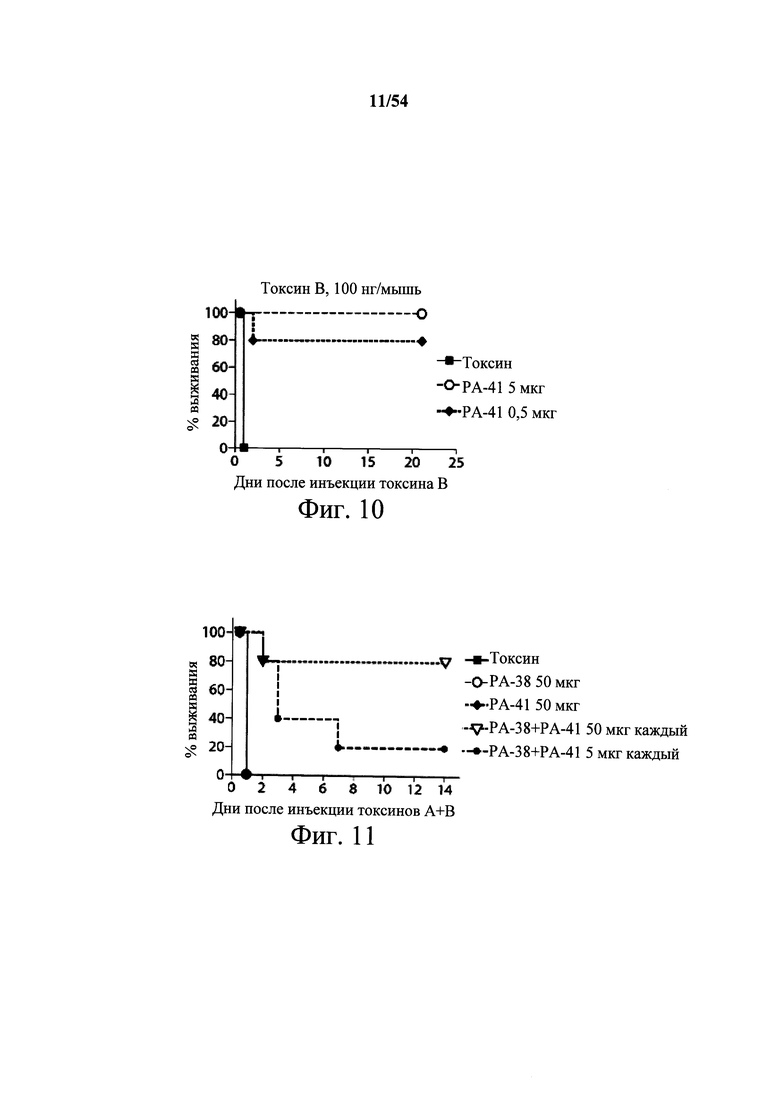
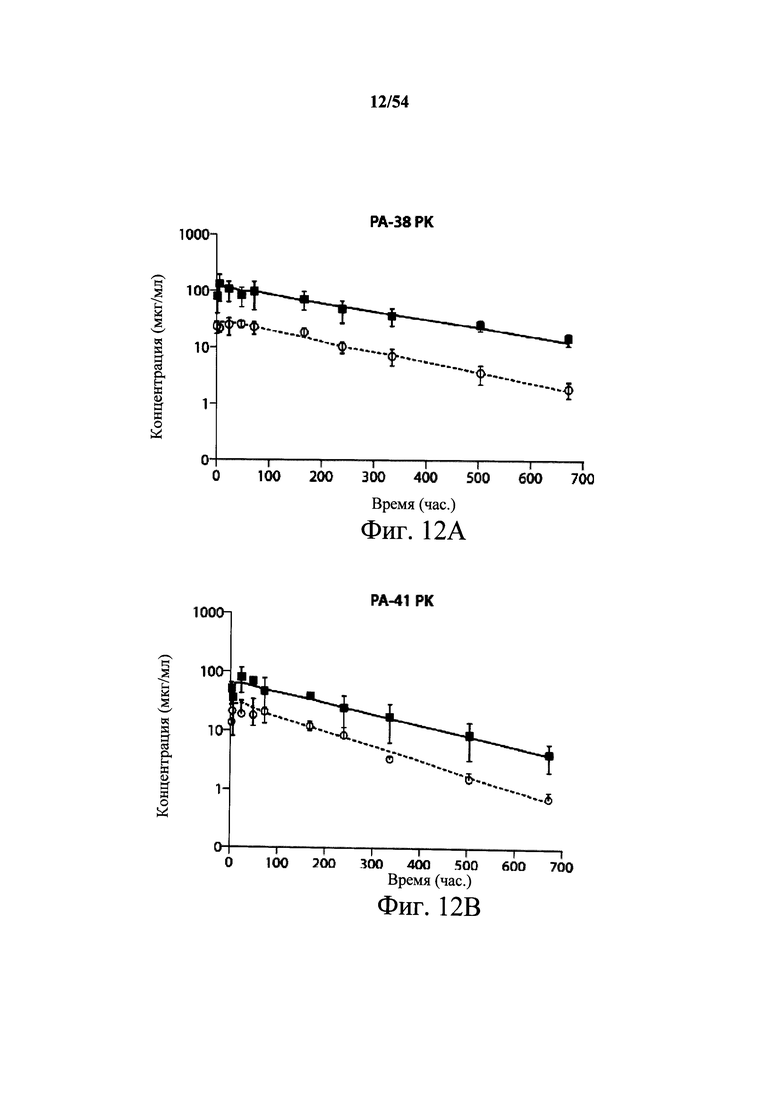
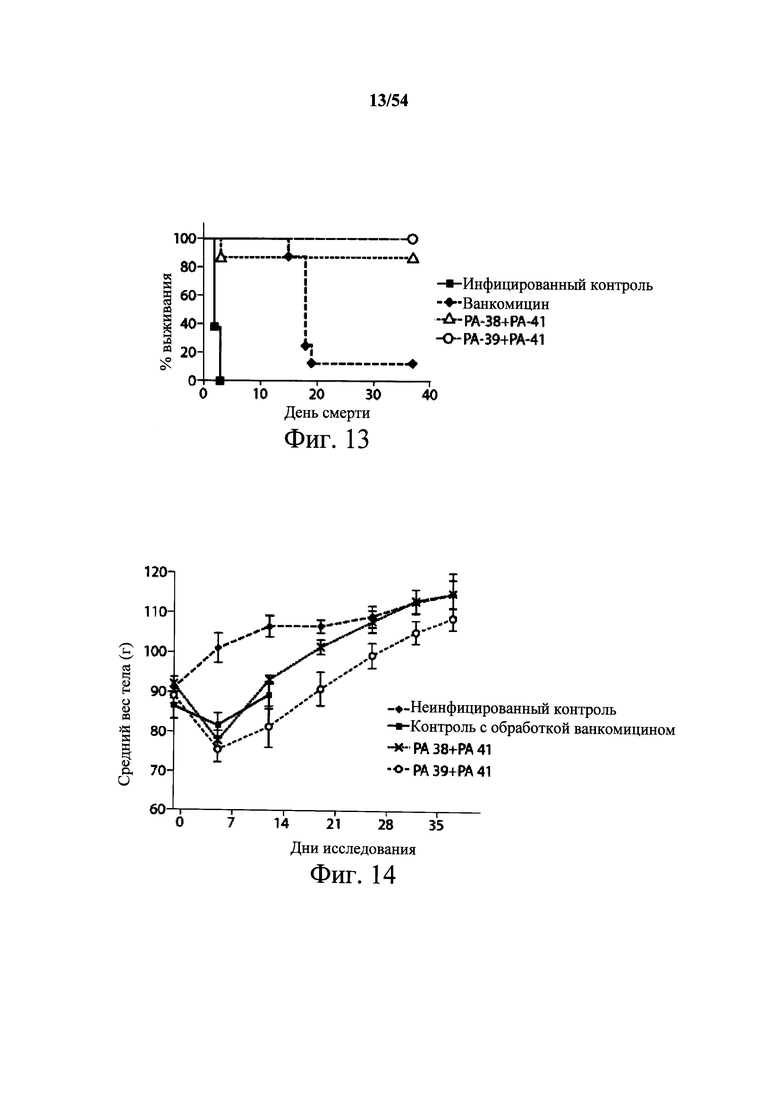

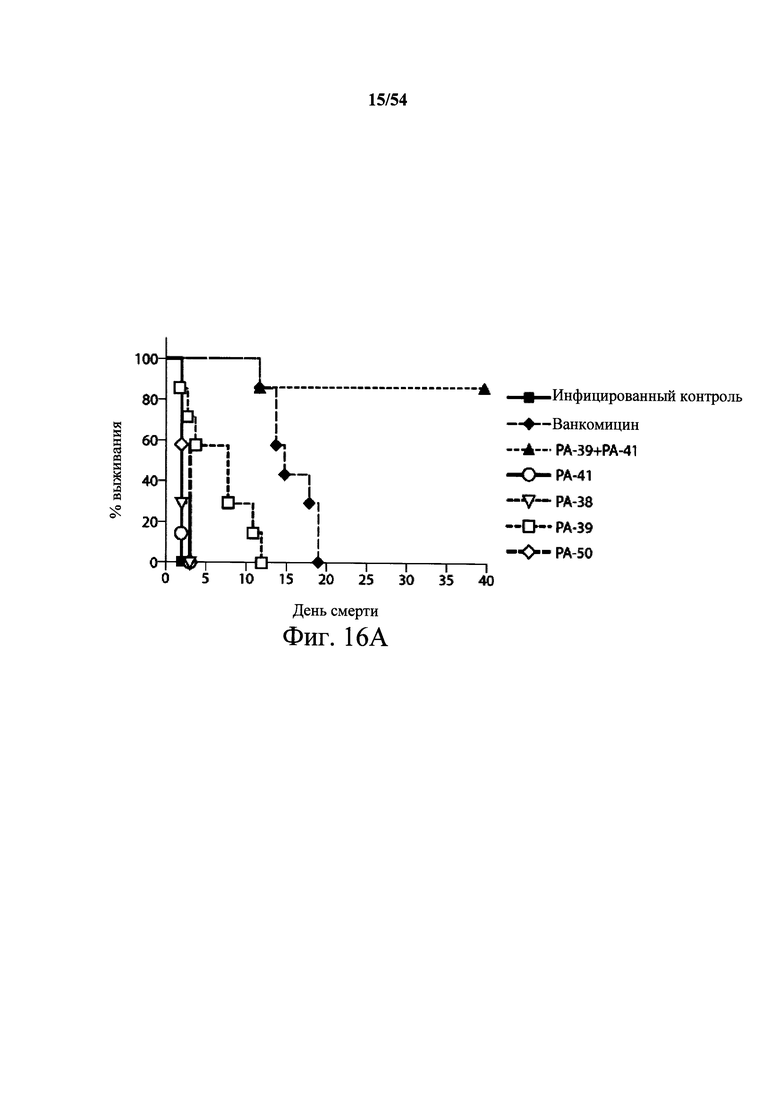
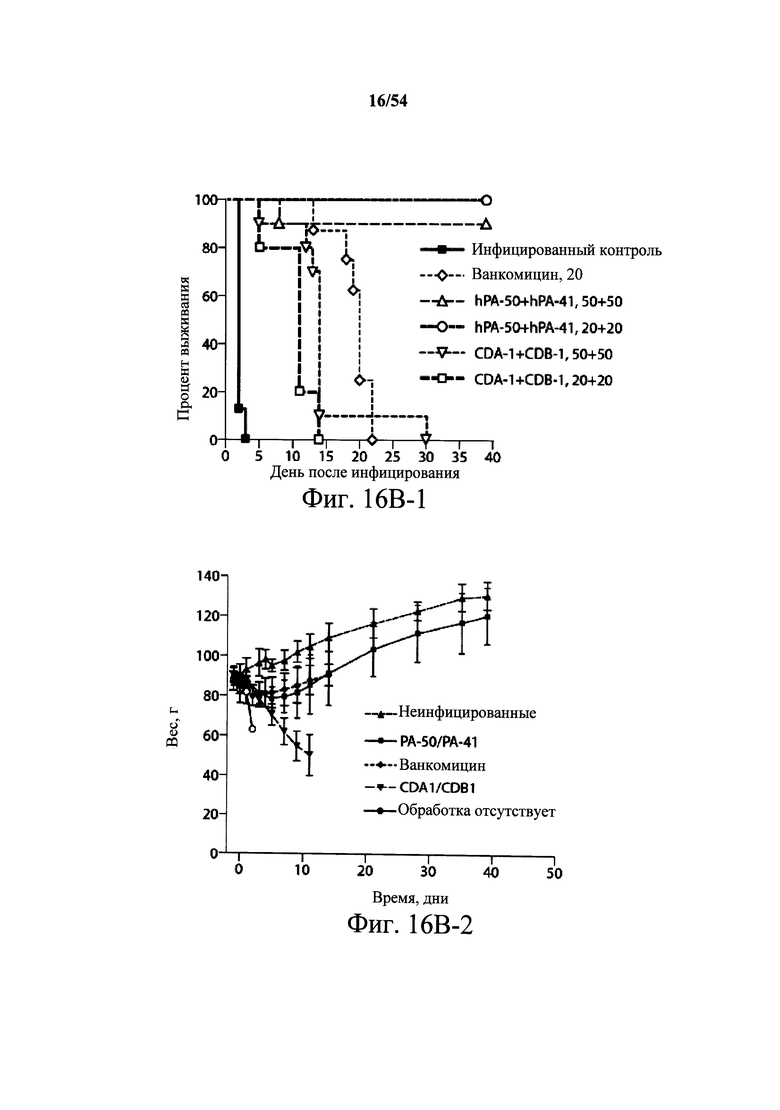
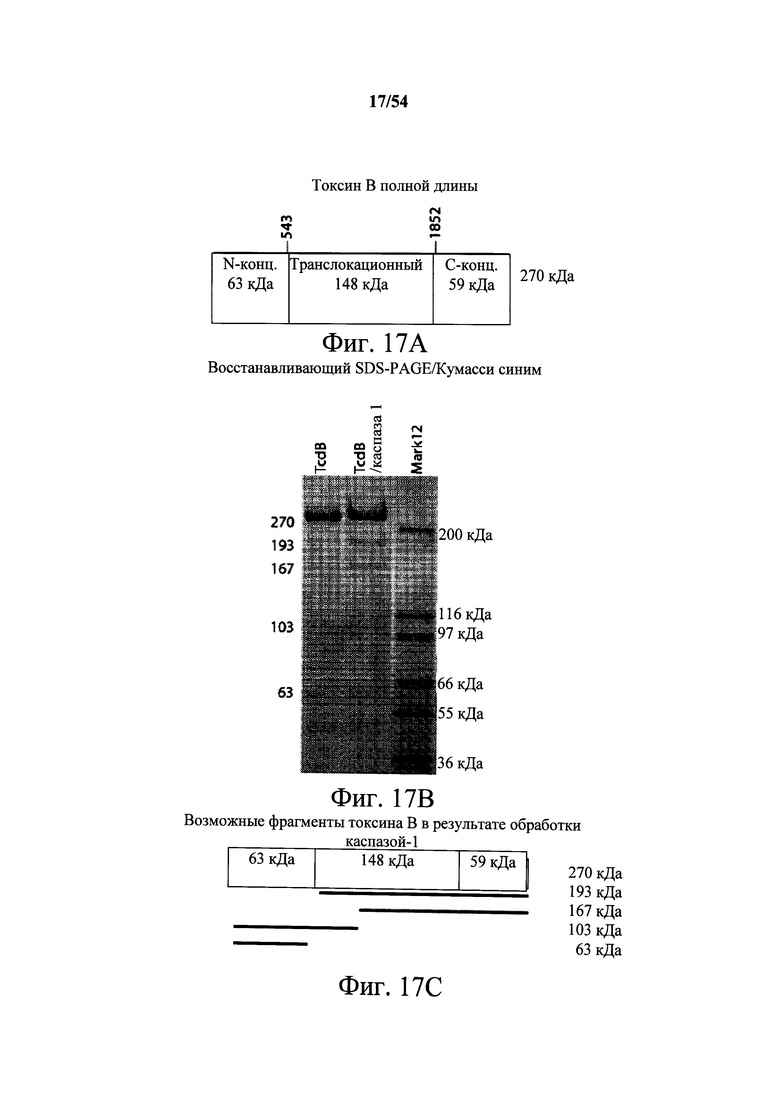
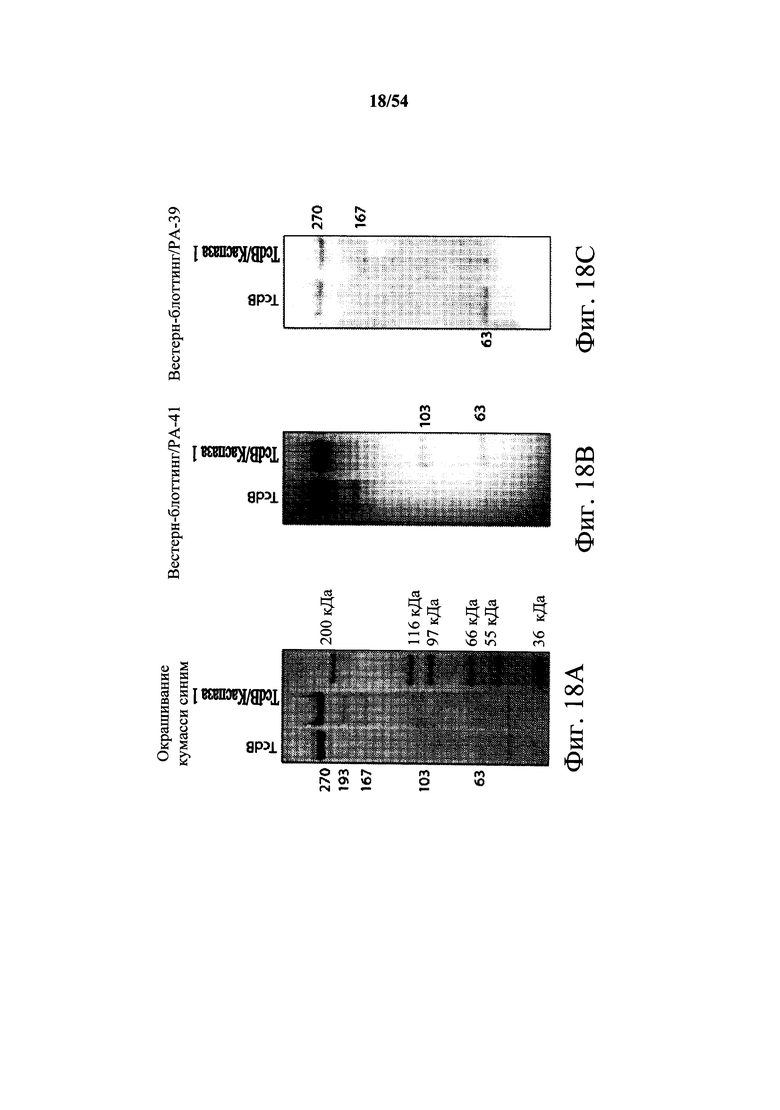
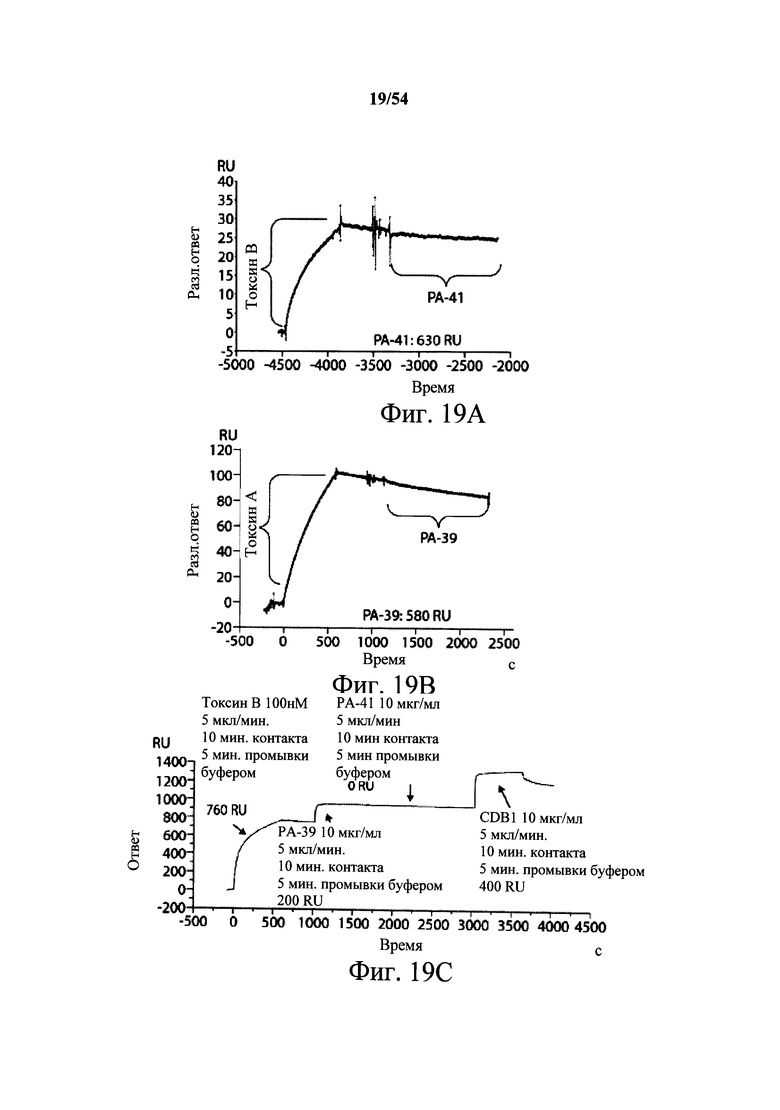
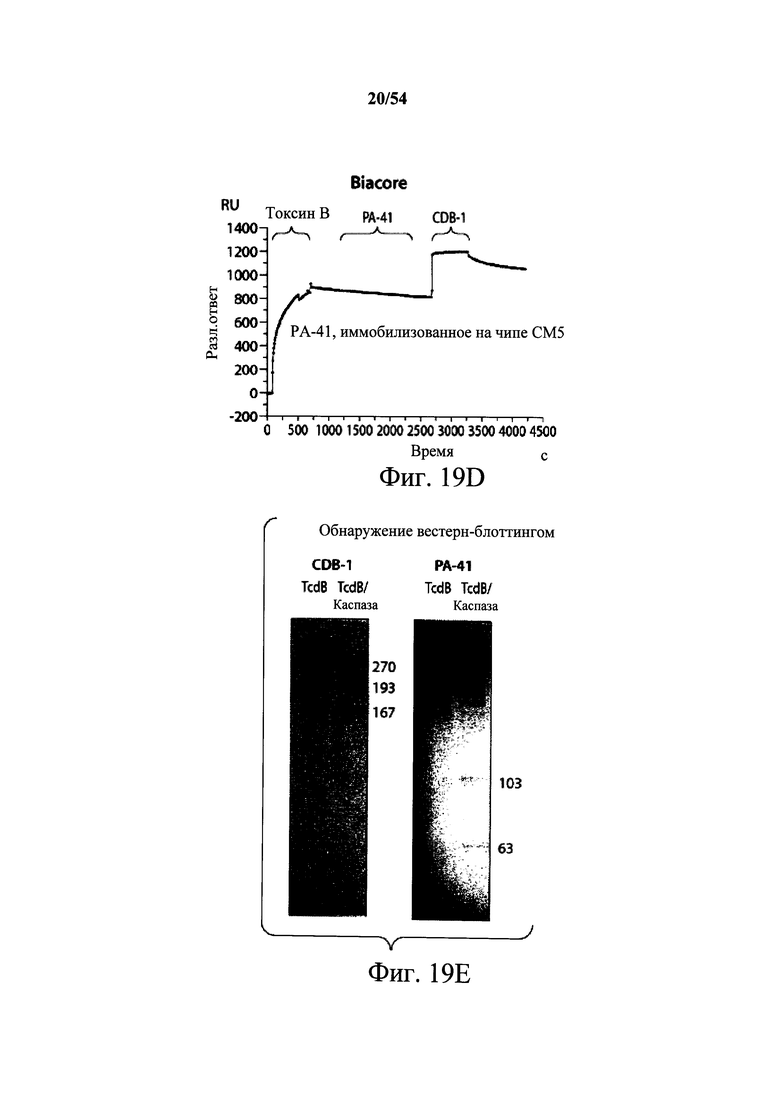
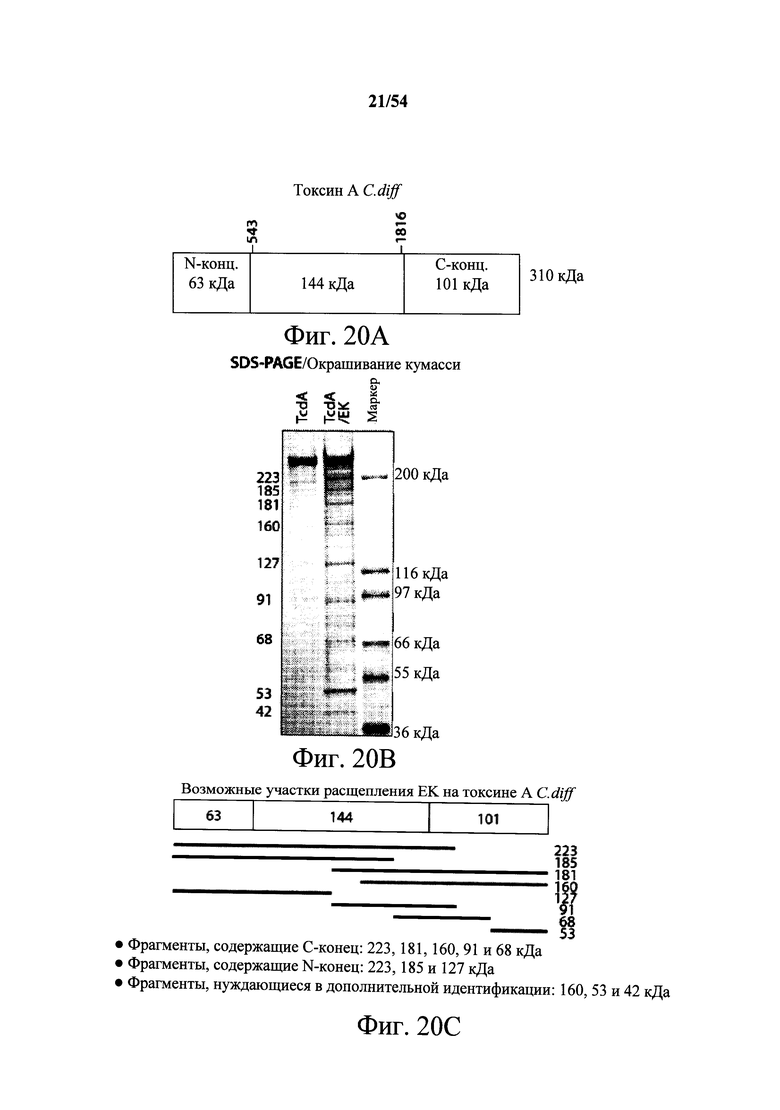
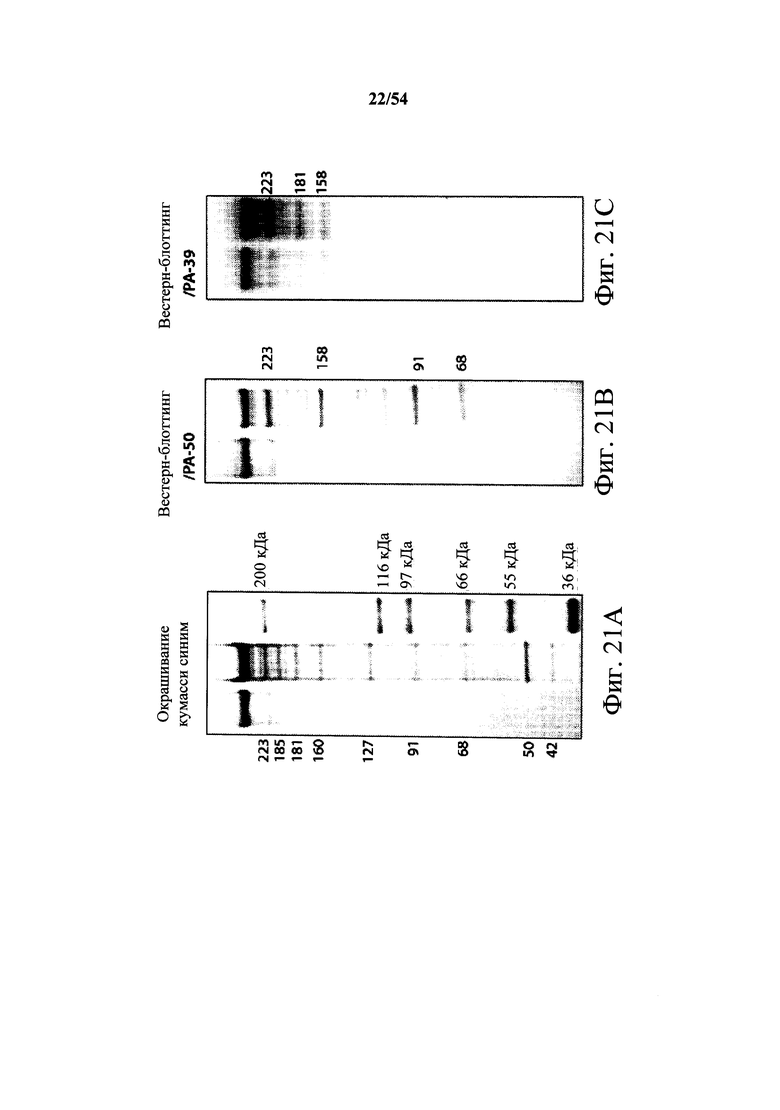
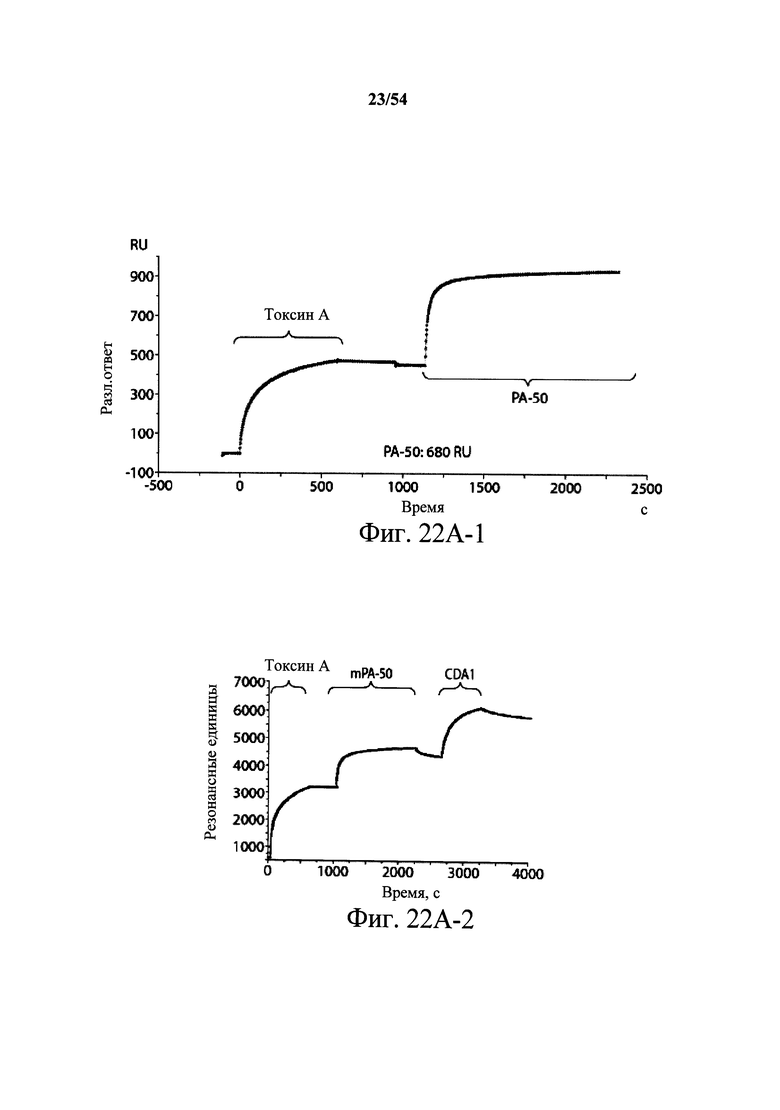
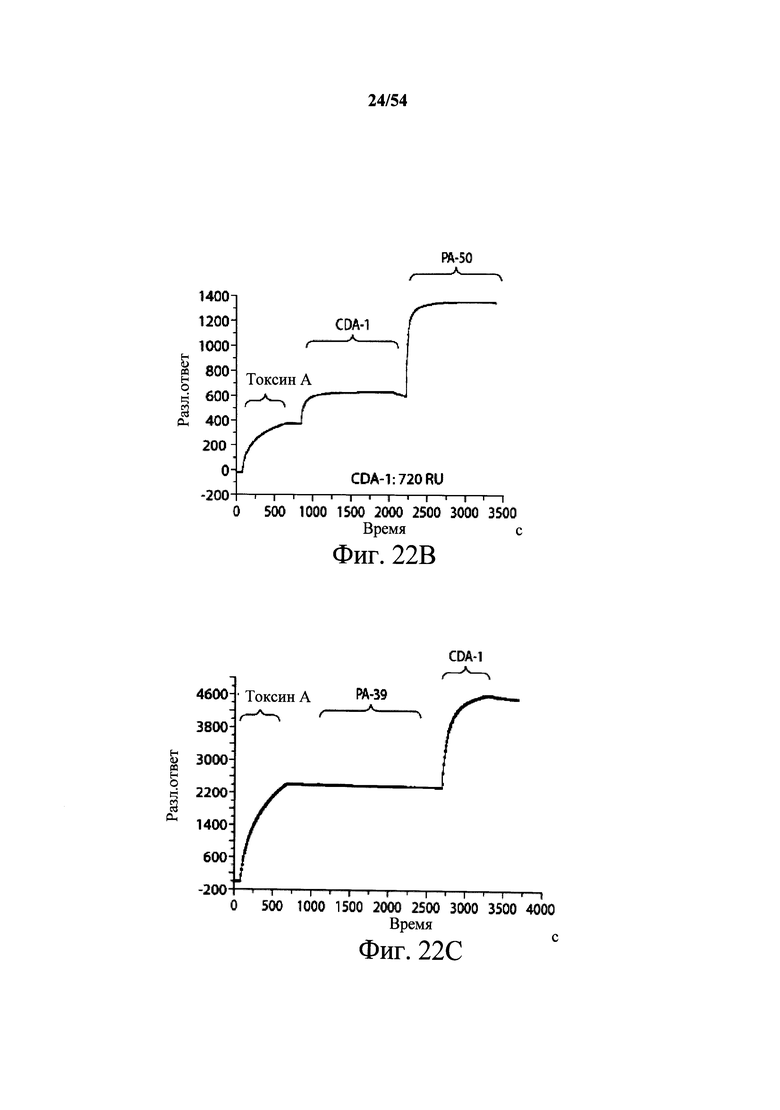
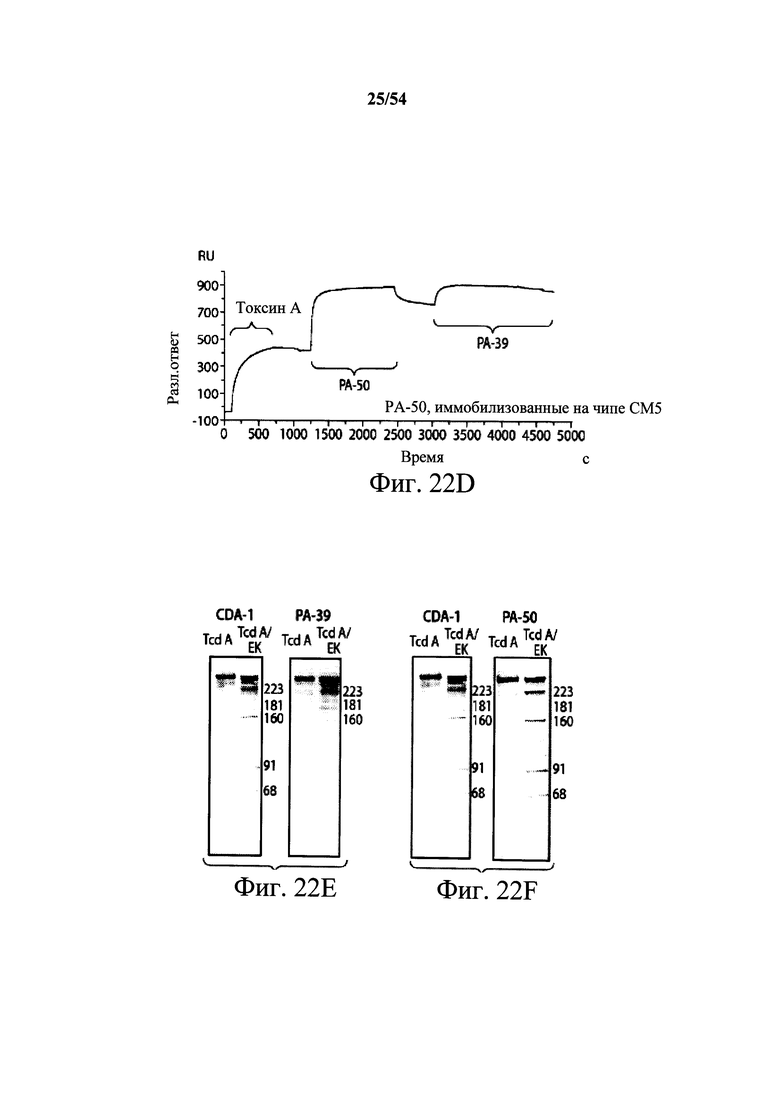
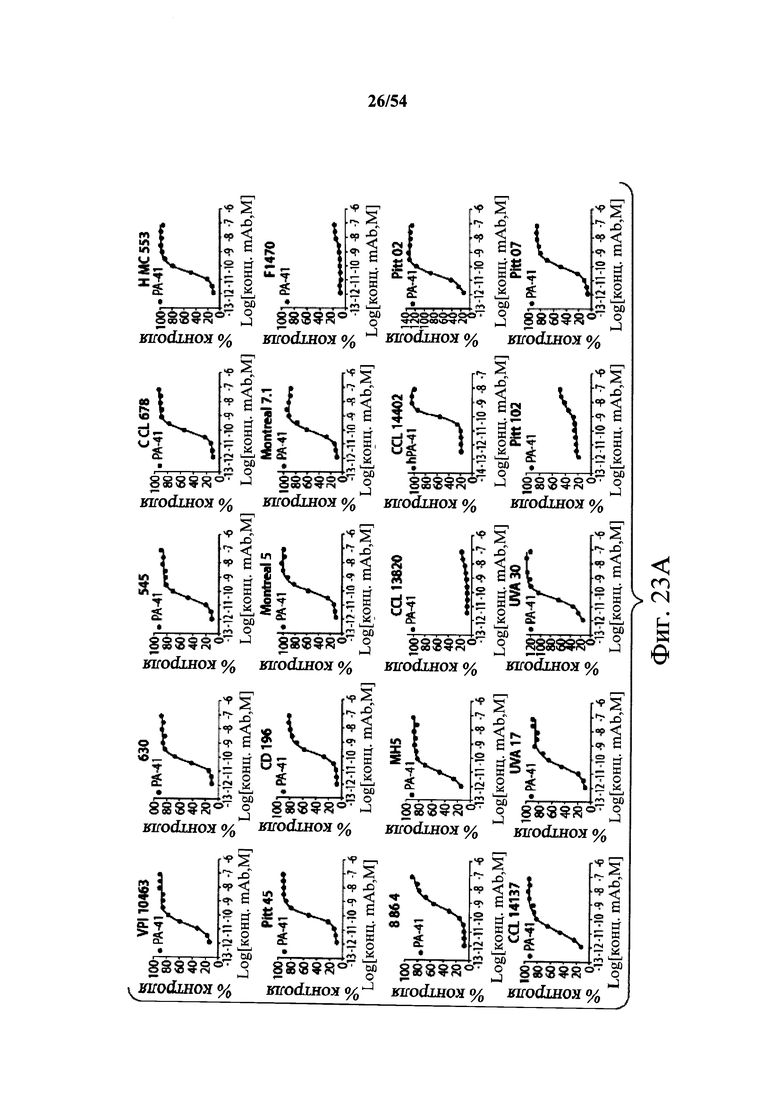
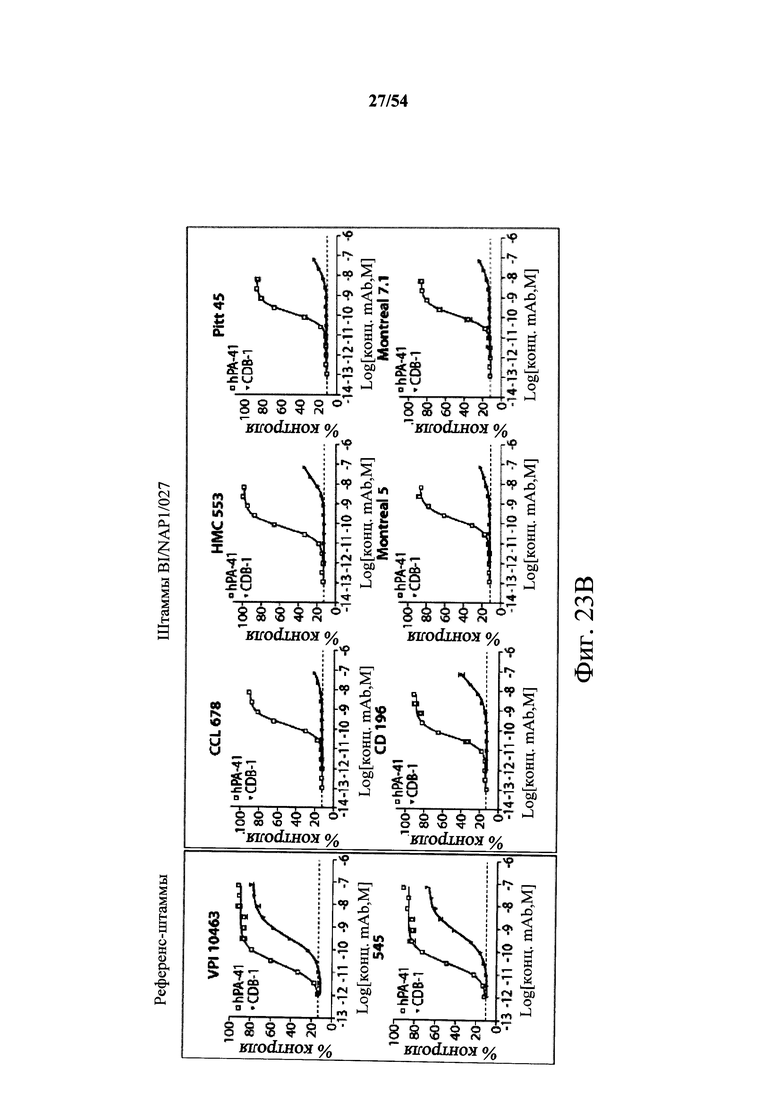
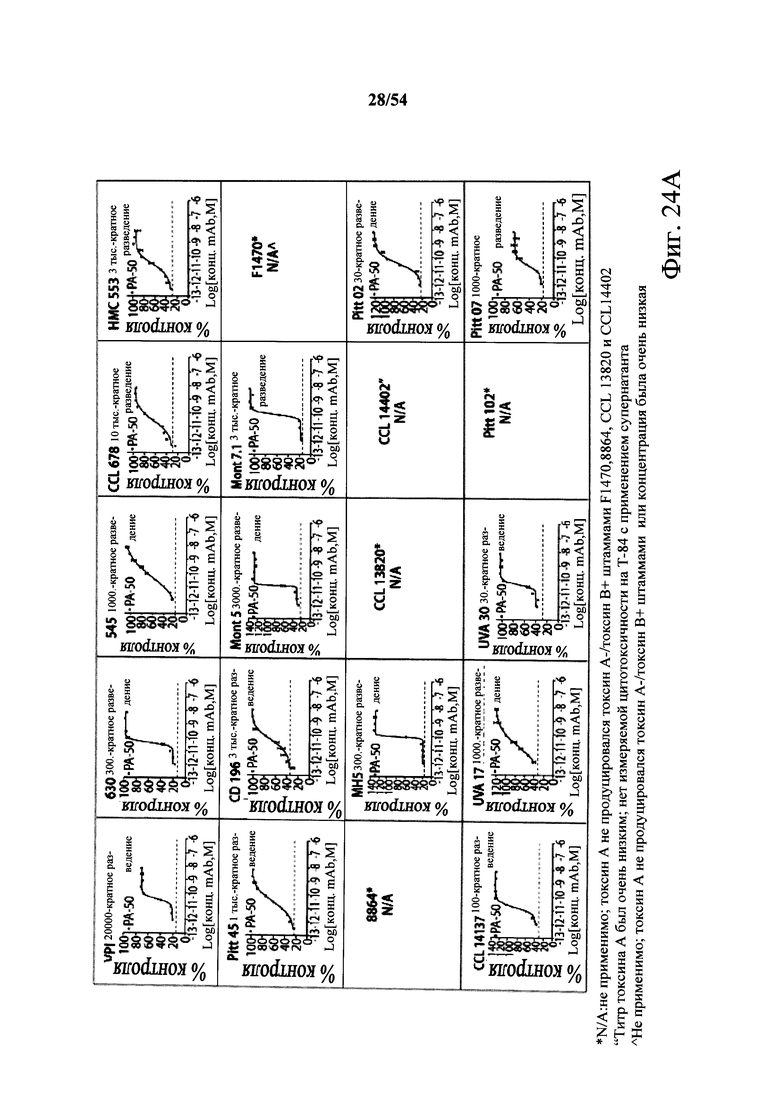
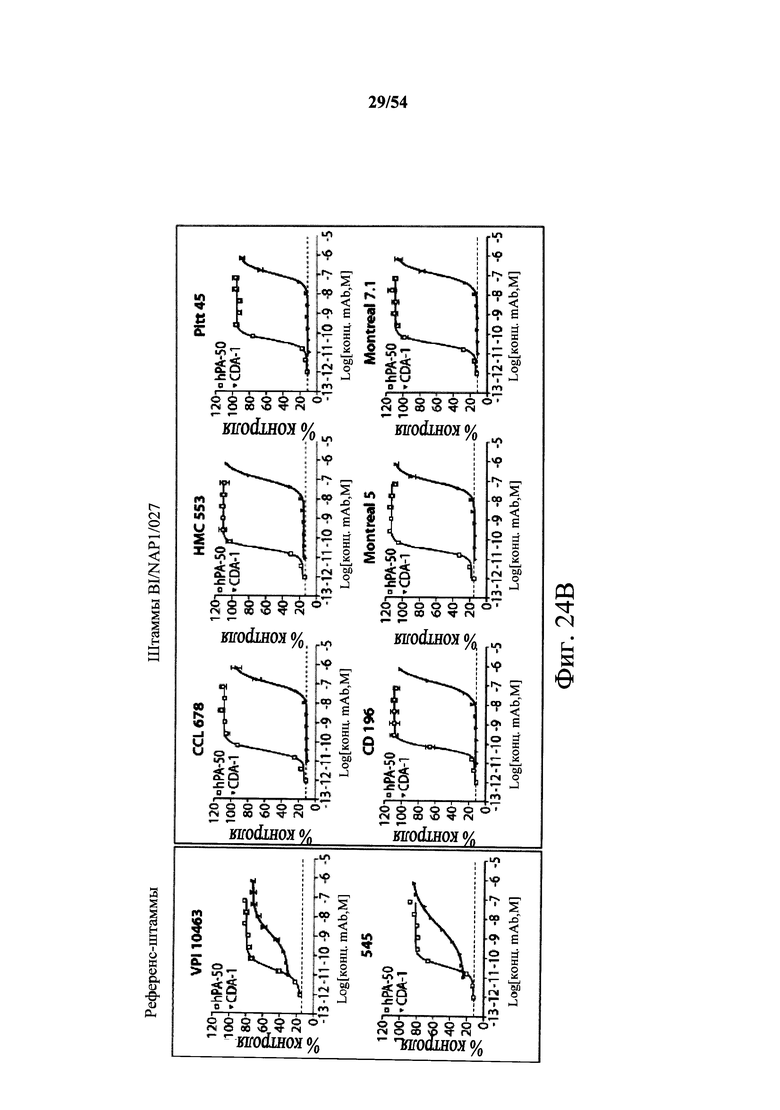
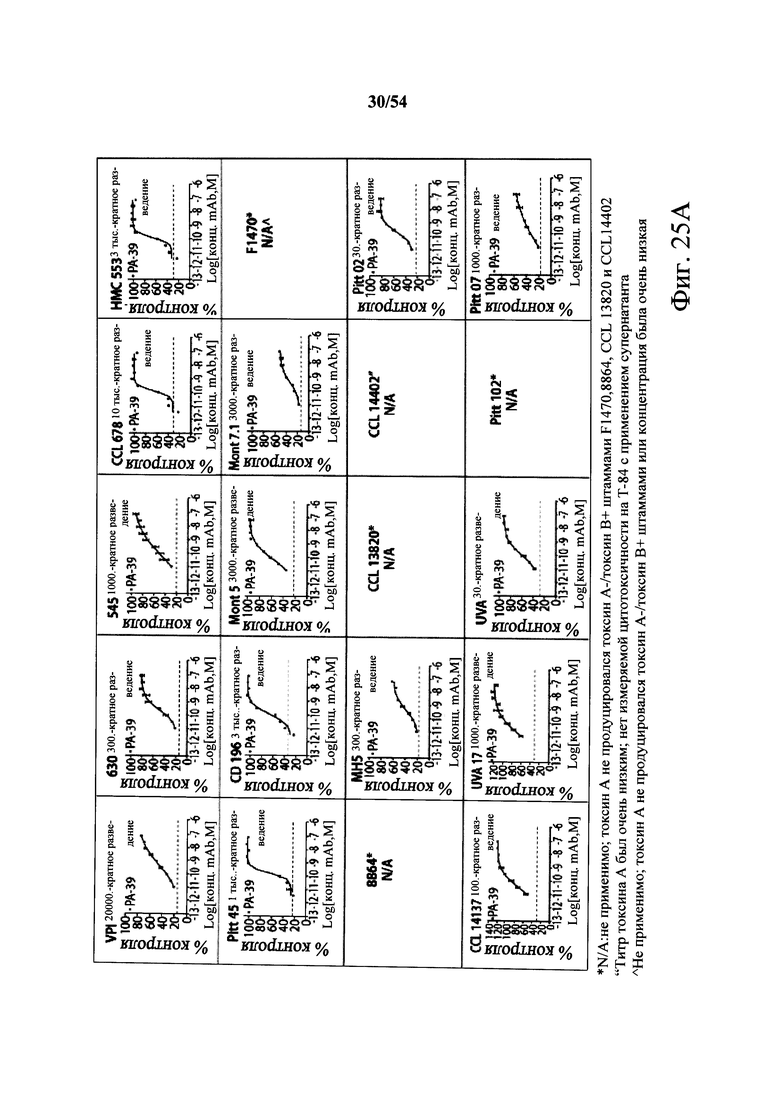
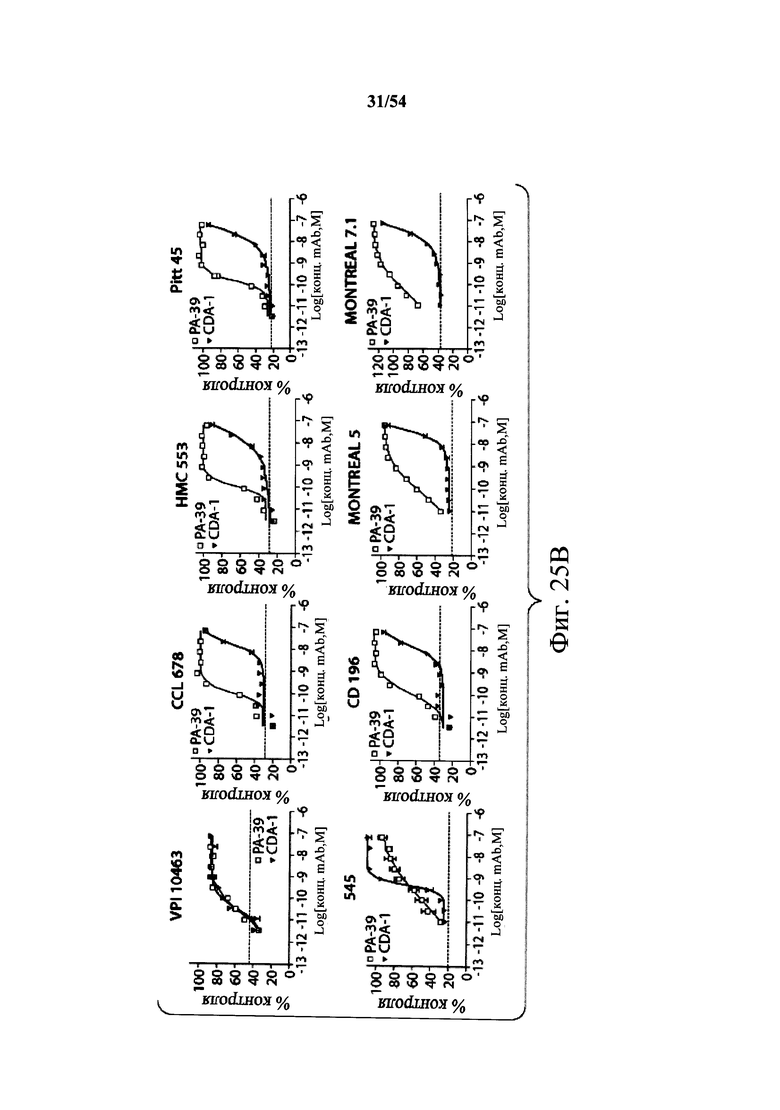
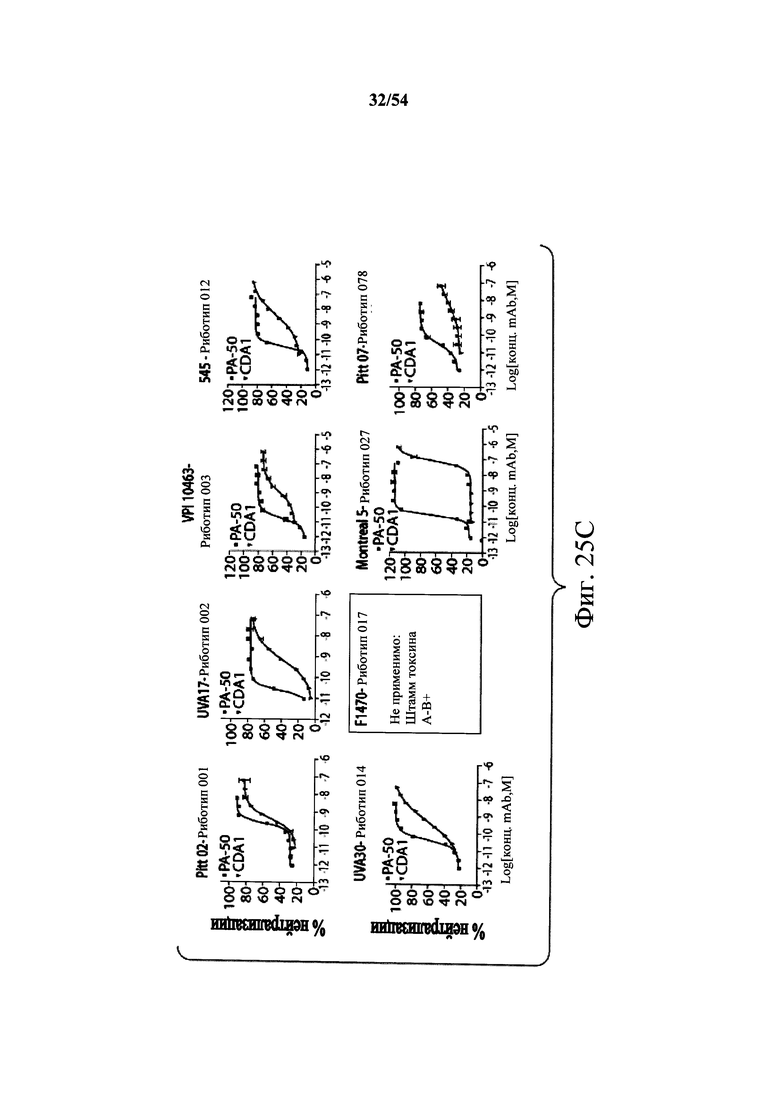
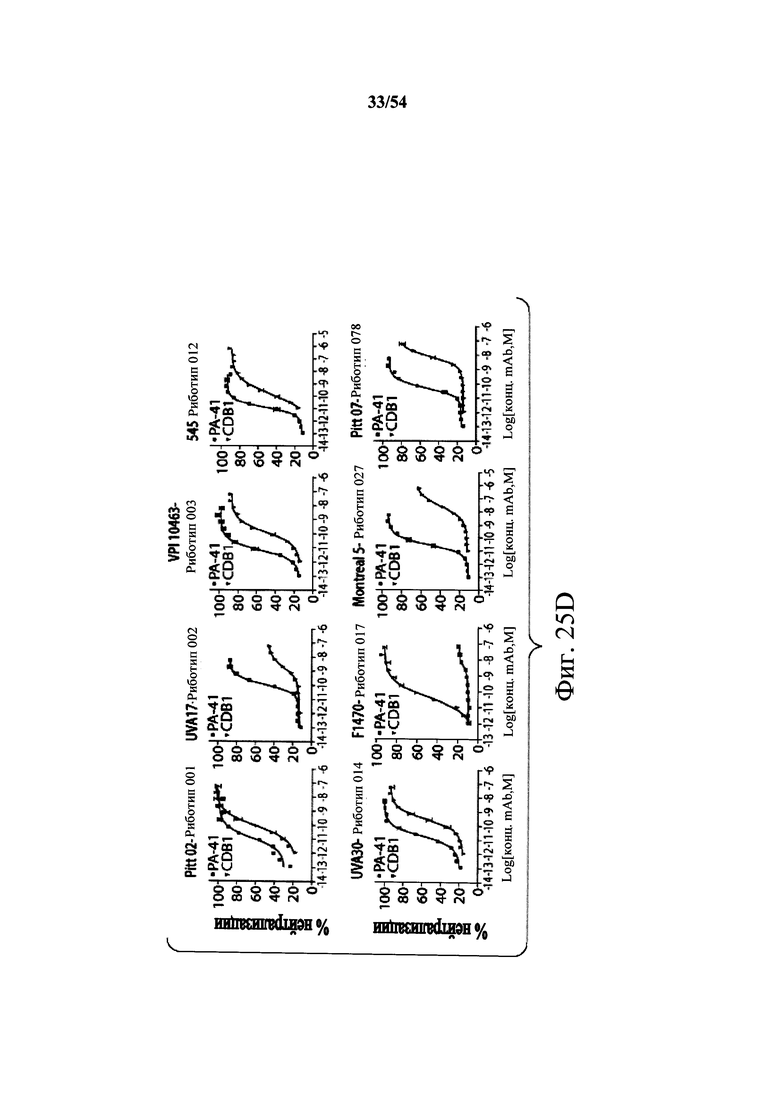
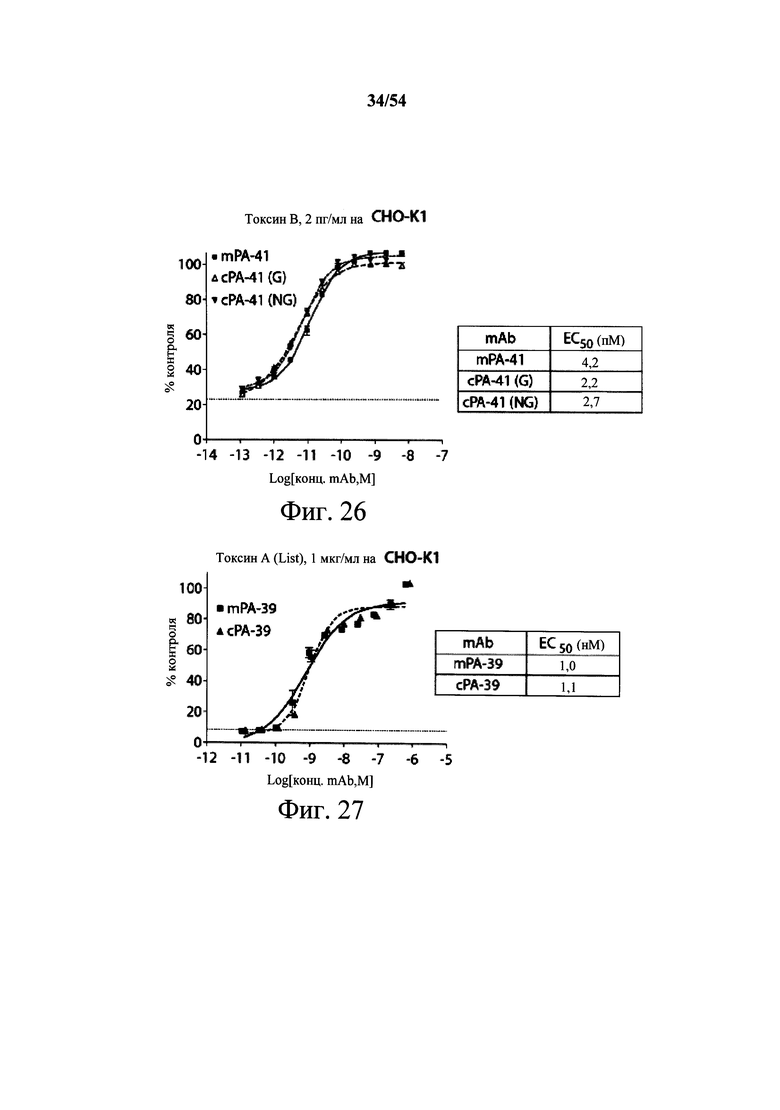
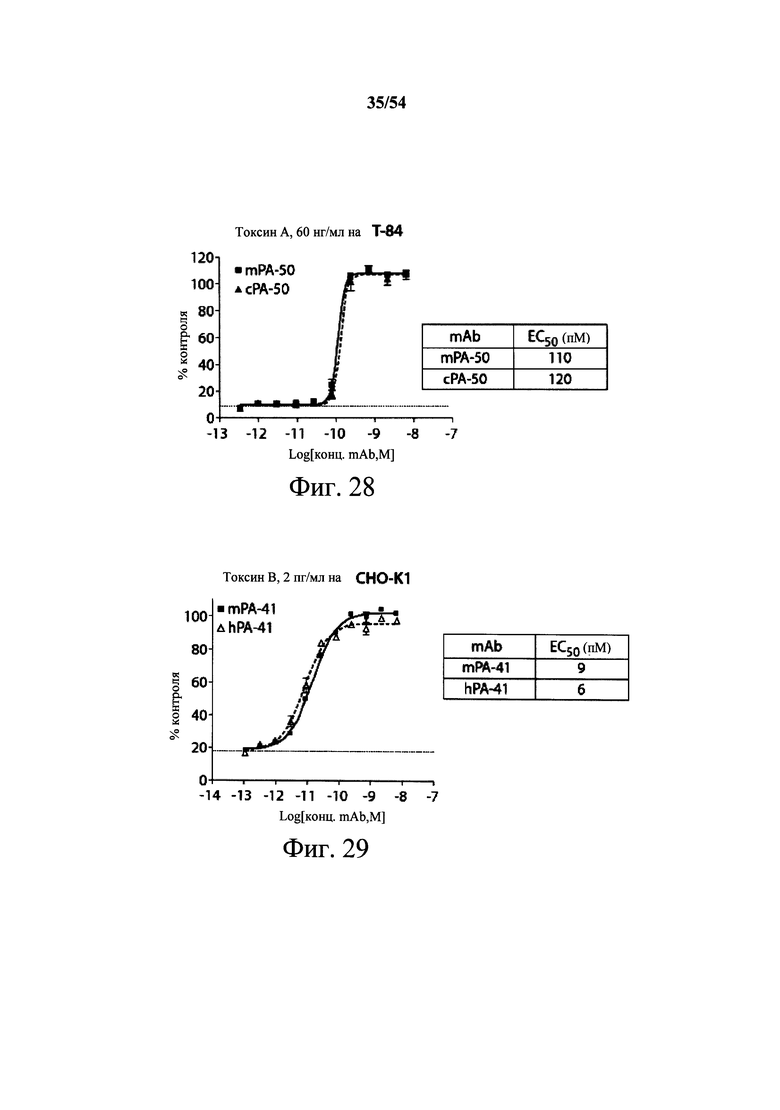
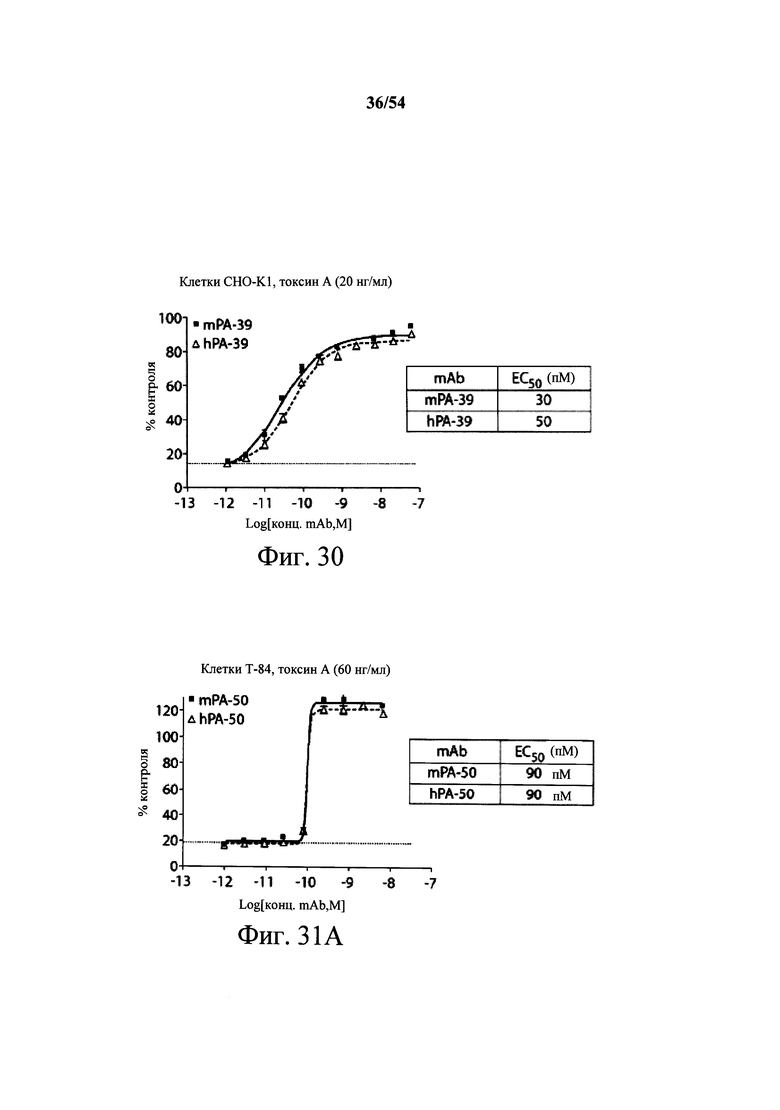
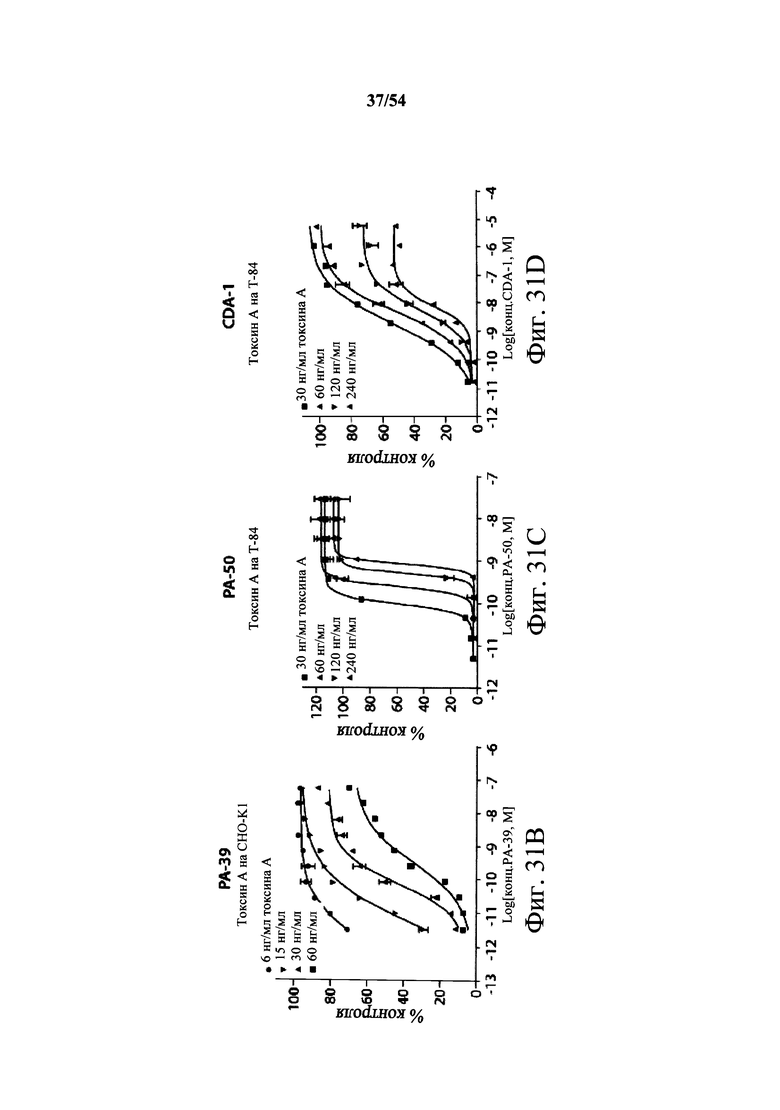
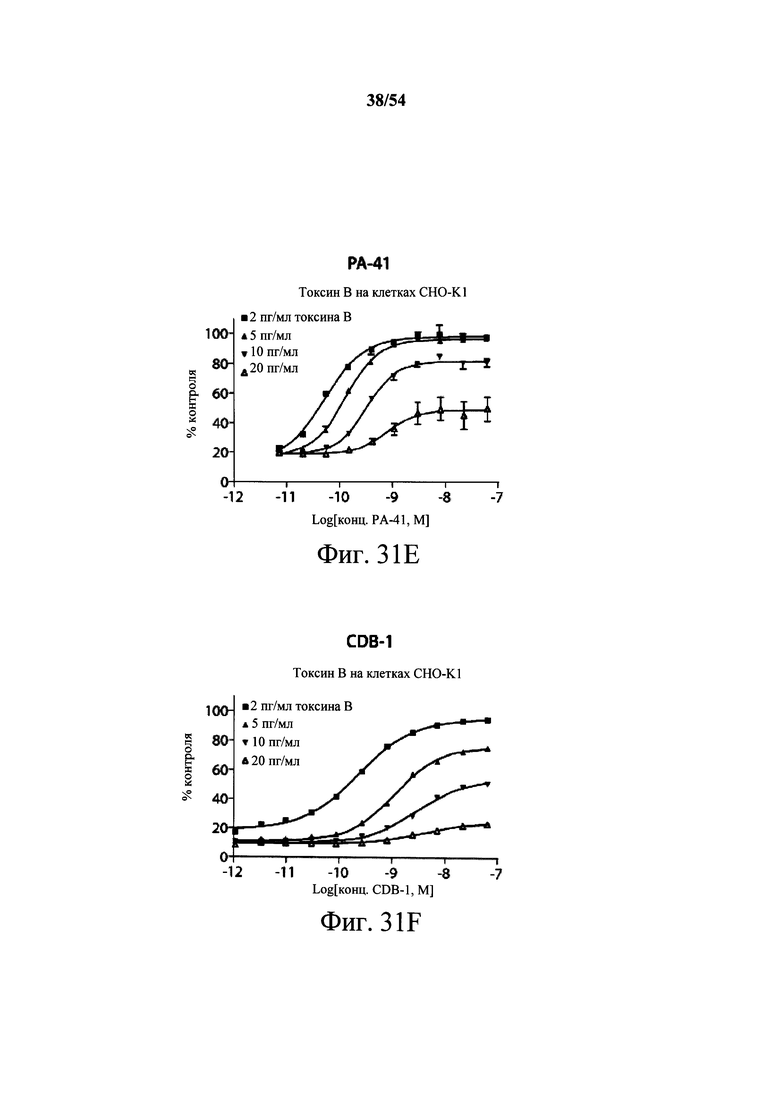
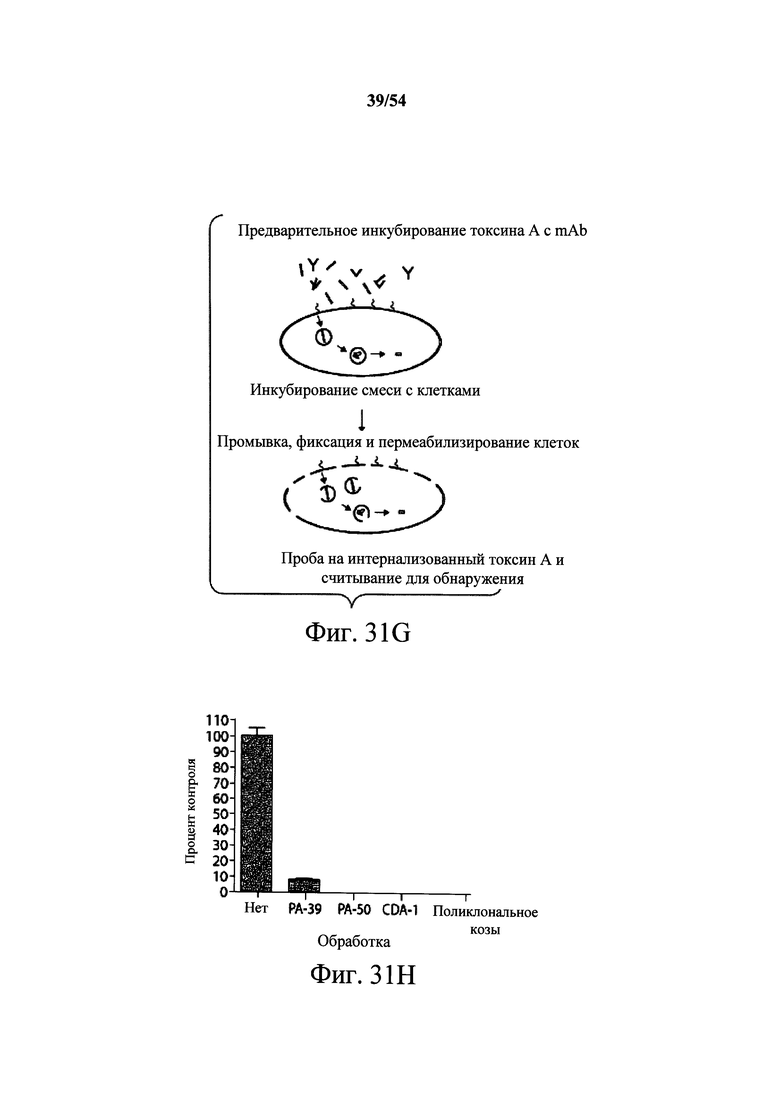













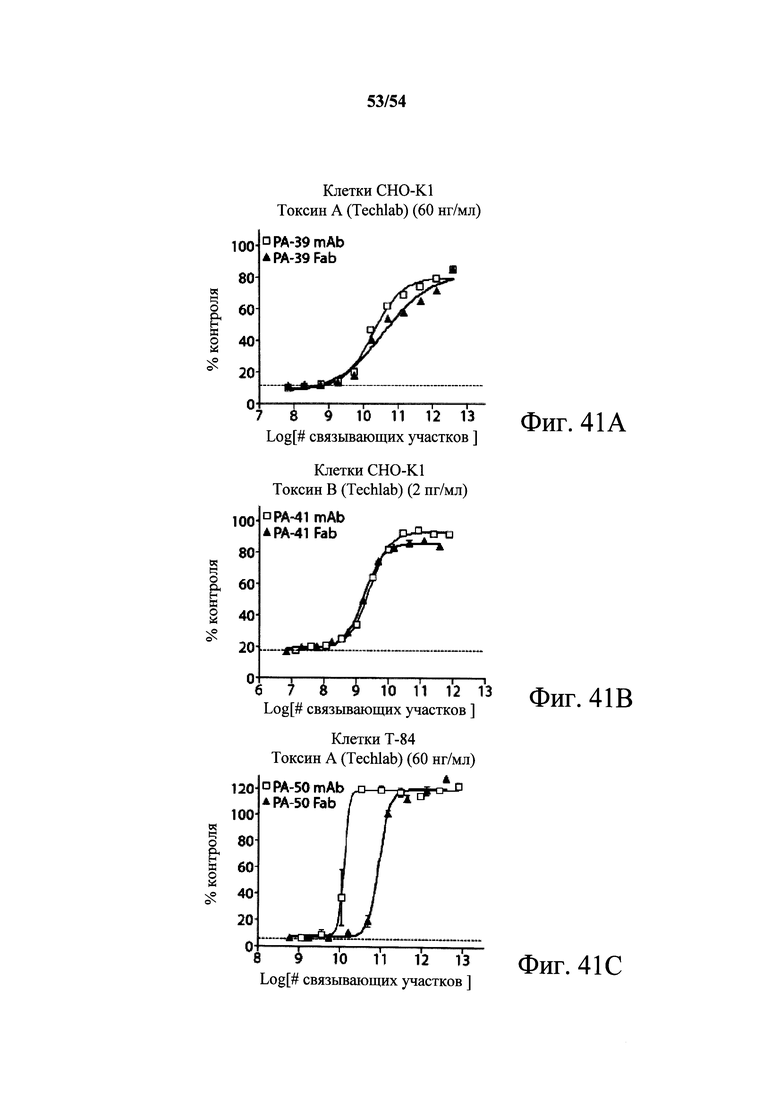
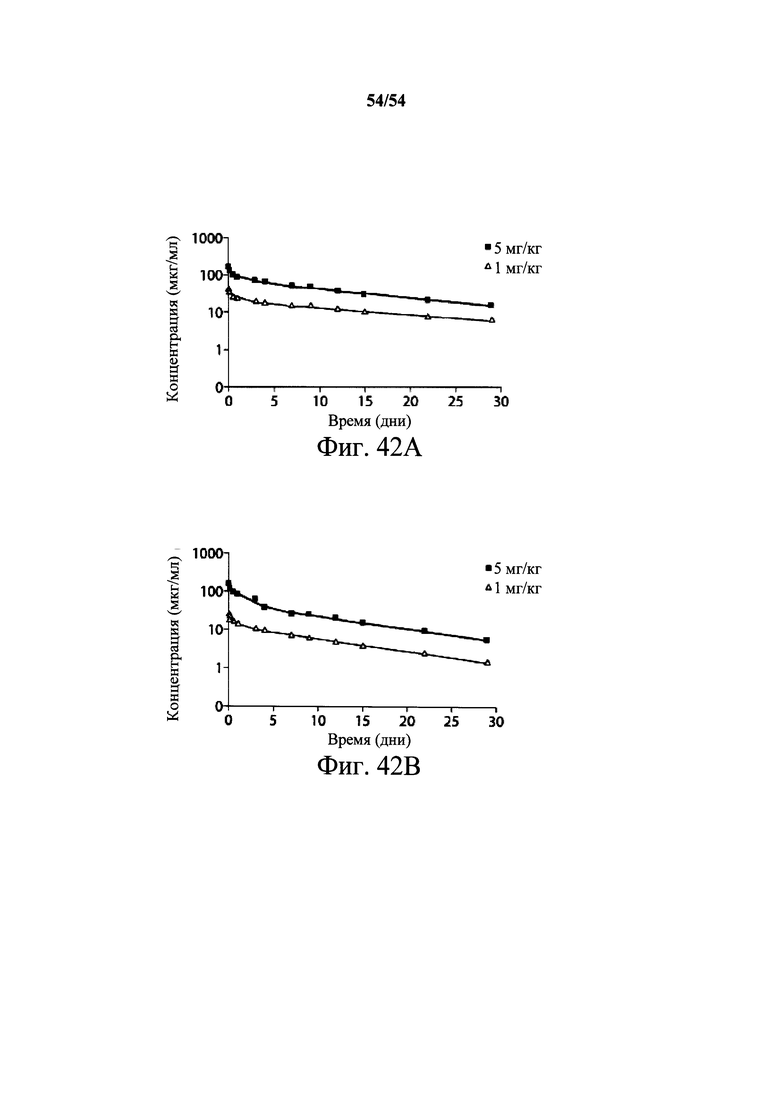
Изобретение относится к биотехнологии. Описаны выделенные антитела или их антигенсвязывающие фрагменты, которые специфично связывают токсин В и/или А С. difficile. Также представлены композиции и наборы, содержащие такие антитела. Представлены гибридомные клеточные линии, депонированные под номерами доступа АТСС РТА-9693, АТСС РТА-9692, АТСС РТА-9694, АТСС РТА-9888, где гибридома продуцирует моноклональное антитело, которое специфично связывает токсин С. difficile. Изобретение расширяет арсенал средств для ингибирования или нейтрализации токсичности, вызванной токсином С. difficile. 31 н. и 19 з.п. ф-лы, 42 ил., 8 табл., 13 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент специфично связывает токсин В С. difficile и представляет собой моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 (РА-41), его химерную или гуманизированную форму или его антигенсвязывающий фрагмент и где антитело или его антигенсвязывающий фрагмент специфично связывает токсин В С. difficile с KD около 0,59 нМ, определенной с помощью анализа Biacore, нейтрализует цитотоксичность токсина В на клетках СНО-K1 с EC50 около 9,2 пМ и/или специфично связывается с 63 кДа N-концевым ферментативным доменом обработанного каспазой-1 токсина В.
2. Выделенное антитело или его антигенсвязывающий фрагмент, которое:
(a) специфично связывает токсин А С. difficile и представляет собой моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694 (РА-50), его химерную или гуманизированную форму или его антигенсвязывающий фрагмент и где антитело или его антигенсвязывающий фрагмент специфично связывает токсин А С. difficile с KD около 0,16 нМ, определенной с помощью анализа Biacore, нейтрализует цитотоксичность токсина А на клетках Т-84 с EC50 около 146 пМ и/или специфично связывается с С-концевым связывающим клеточный рецептор (CRB) доменом токсина А; или
(b) специфично связывает токсин А С. difficile и представляет собой моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692 (РА-39), его химерную или гуманизированную форму или его антигенсвязывающий фрагмент и где антитело или его антигенсвязывающий фрагмент специфично связывает токсин А С. difficile с KD около 0,16 нМ, определенной с помощью анализа Biacore, нейтрализует цитотоксичность токсина А на клетках СНО-K1 с EC50 около 93 пМ и/или специфично связывается с С-концевым связывающим клеточный рецептор (CRB) доменом токсина А; или
(c) специфично связывает токсин А С. difficile и представляет собой моноклональное антитело, продуцируемое гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888 (РА-38), его химерную или гуманизированную форму или его антигенсвязывающий фрагмент и где антитело или его антигенсвязывающий фрагмент специфично связывает токсин А С. difficile с KD около 1,0 нМ, определенной с помощью анализа Biacore, и/или нейтрализует цитотоксичность токсина А на клетках Т-84 с EC50 около 175 пМ.
3. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент
специфично связывает токсин В С. difficile, и VH включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 8 или SEQ ID NO: 9, и VL включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 10.
4. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент:
(a) специфично связывает токсин А С. difficile, и VH включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 1 или SEQ ID NO: 2, и VL включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 3 или SEQ ID NO: 4; или
(b) специфично связывает токсин А С. difficile, и VH включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 5 или SEQ ID NO: 6, и VL включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 7.
5. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент
специфично связывает токсин В С. difficile, и VH включает CDR1, включающий аминокислотные остатки 31-36 последовательности SEQ ID NO: 8, CDR2, включающий аминокислотные остатки 50-66 последовательности SEQ ID NO: 8, и CDR3, включающий аминокислотные остатки 99-108 последовательности SEQ ID NO: 8, и VL включает CDR1, включающий аминокислотные остатки 24-34 последовательности SEQ ID NO: 10, CDR2, включающий аминокислотные остатки 50-56 последовательности SEQ ID NO: 10, и CDR3, включающий аминокислотные остатки 89-97 последовательности SEQ ID NO: 10.
6. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент:
(a) специфично связывает токсин А С. difficile, и VH включает CDR1, включающий аминокислотные остатки 31-35 последовательности SEQ ID NO: 1, CDR2, включающий аминокислотные остатки 50-65 последовательности SEQ ID NO: 1, и CDR3, включающий аминокислотные остатки 95-102 последовательности SEQ ID NO: 1, и VL включает CDR1, включающий аминокислотные остатки 24-34 последовательности SEQ ID NO: 3, CDR2, включающий аминокислотные остатки 50-56 последовательности SEQ ID NO: 3, и CDR3, включающий аминокислотные остатки 89-97 последовательности SEQ ID NO: 3; или
(b) специфично связывает токсин А С. difficile, и VH включает CDR1, включающий аминокислотные остатки 31-35 последовательности SEQ ID NO: 5, CDR2, включающий аминокислотные остатки 50-65 последовательности SEQ ID NO: 5, и CDR3, включающий аминокислотные остатки 95-102 последовательности SEQ ID NO: 5, и VL включает CDR1, включающий аминокислотные остатки 24-33 последовательности SEQ ID NO: 7, CDR2, включающий аминокислотные остатки 49-55 последовательности SEQ ID NO: 7, и CDR3, включающий аминокислотные остатки 88-94 последовательности SEQ ID NO: 7.
7. Выделенное антитело, которое представляет собой антитело к токсину В С. difficile, или его антигенсвязывающий фрагмент по любому из пп. 1, 3 или 5, где антитело или его антигенсвязывающий фрагмент специфично связывает токсин В С. difficile и:
V-область L-цепи содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 10; и/или
V-область Н-цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 8 или SEQ ID NO: 9.
8. Выделенное антитело, которое представляет собой антитело к токсину А С. difficile, или его антигенсвязывающий фрагмент по любому из пп. 2, 4 или 6, где антитело или его антигенсвязывающий фрагмент специфично связывает токсин А С. difficile и:
(а) V-область L-цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 3 или SEQ ID NO: 4, и/или V-область Н-цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1 или SEQ ID NO: 2; или
(b) V-область L-цепи содержит аминокислотную последовательность SEQ ID NO: 7 и/или V-область Н-цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 5 или SEQ ID NO: 6.
9. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антитело или его антигенсвязывающий фрагмент ингибирует, блокирует или предотвращает токсичность токсина А С. difficile при помощи ингибирования, блокирования или предотвращения интернализации и цитотоксичности токсина А и/или ингибирует, блокирует или предотвращает токсичность токсина В С. difficile путем связывания с эпитопным участком в N-концевой ферментативной области токсина В.
10. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антитело или антигенсвязывающий фрагмент нейтрализует токсичность in vivo токсина А или токсина В С. difficile или обоих.
11. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 3-6, где антитело или его антигенсвязывающий фрагмент:
(i) представляет собой моноклональное антитело;
(ii) находится в гуманизированной форме;
(iii) является человеческим; или
(iv) находится в химерной форме.
12. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, включающее СН-область, выбранную из IgG1, IgG2a, IgG2b, IgG3, IgG4, IgA, IgE или IgM.
13. Антитело или его антигенсвязывающий фрагмент по п. 12, где СН-область включает IgG1.
14. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, 13, включающее CL-область человека, выбранную из изотипа κ или λ.
15. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, 13, где антитело или его антигенсвязывающий фрагмент содержит одноцепочечное антитело.
16. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, 13, где антигенсвязывающий фрагмент выбран из Fab-фрагмента, F(ab')2-фрагмента или Fv-фрагмента.
17. Набор для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащий антитело к токсину В или его антигенсвязывающий фрагмент по любому из пп. 1, 3, 5, 7 и 9-16 и инструкции по применению.
18. Набор для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащий антитело к токсину А или его антигенсвязывающий фрагмент по любому из пп. 2, 4, 6, 8 и 9-16 и инструкции по применению.
19. Набор для лечения инфекции Clostridium difficile (С difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащий антитело к токсину В или его антигенсвязывающий фрагмент по любому из пп. 1, 3, 5, 7 и 9-16 и антитело к токсину А или его антигенсвязывающий фрагмент по любому из пп. 2, 4, 6, 8 и 9-16 и инструкции по применению.
20. Композиция для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащая терапевтически эффективное количество антитела к токсину В или его антигенсвязывающего фрагмента по любому из пп. 1, 3, 5, 7 и 9-16 и фармацевтически приемлемый носитель, наполнитель или разбавитель.
21. Композиция для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащая терапевтически эффективное количество антитела к токсину А или его антигенсвязывающего фрагмента по любому из пп. 2, 4, 6, 8 и 9-16 и фармацевтически приемлемый носитель, наполнитель или разбавитель.
22. Композиция для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащая терапевтически эффективное количество антитела к токсину В или его антигенсвязывающего фрагмента по любому из пп. 1, 3, 5, 7 и 9-16 и антитела к токсину А или его антигенсвязывающего фрагмента по любому из пп. 2, 4, 6, 8 и 9-16 и фармацевтически приемлемый носитель, наполнитель или разбавитель.
23. Композиция по п. 20, дополнительно содержащая по меньшей мере одно дополнительное терапевтическое средство.
24. Композиция по п. 23, где по меньшей мере одно дополнительное терапевтическое средство представляет собой метронидазол, ванкомицин, фидаксомицин, нитазоксанид, рифаксимин, рамопланин или их комбинацию.
25. Композиция для лечения инфекции Clostridium difficile (С. difficile) и ассоциированных с С. difficile болезненных состояний и патологий, содержащая (i) антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин В С. difficile; и (ii) антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С. difficile;
где антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин В С. difficile, выбрано из:
(а) моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693 (РА-41), его химерной или гуманизированной формы или его антигенсвязывающего фрагмента и где антитело или его антигенсвязывающий фрагмент специфично связывает токсин В С. difficile с KD около 0,59 нМ, определенной с помощью анализа Biacore, нейтрализует цитотоксичность токсина В на клетках СНО-K1 с EC50 около 9,2 пМ и/или специфично связывается с 63 кДа N-концевым ферментативным доменом обработанного каспазой-1 токсина В;
(b) антитела или его антигенсвязывающего фрагмента, в котором вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 VH-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693, и вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 VL-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9693;
(c) антитела или его антигенсвязывающего фрагмента, в котором VH включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 8 или SEQ ID NO: 9 и VL включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 10;
(d) антитела или его антигенсвязывающего фрагмента, в котором VH включает CDR1, включающий аминокислотные остатки 31-36 последовательности SEQ ID NO: 8, CDR2, включающий аминокислотные остатки 50-66 последовательности SEQ ID NO: 8, и CDR3, включающий аминокислотные остатки 99-108 последовательности SEQ ID NO: 8, и VL включает CDR1, включающий аминокислотные остатки 24-34 последовательности SEQ ID NO: 10, CDR2, включающий аминокислотные остатки 50-56 последовательности SEQ ID NO: 10, и CDR3, включающий аминокислотные остатки 89-97 последовательности SEQ ID NO: 10;
(e) антитела или его антигенсвязывающего фрагмента, в котором V-область L-цепи содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 10, и V-область Н-цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 8 или SEQ ID NO: 9.
26. Композиция по п. 25, где антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С. difficile, включает:
(a) CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи (VH) и CDR1, CDR2 и CDR3 вариабельной области легкой цепи (VL) моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692; или
(b) CDR1, CDR2 и CDR3 VH-области и CDR1, CDR2 и CDR3 VL-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694; или
(с) CDR1, CDR2 и CDR3 VH-области и CDR1, CDR2 и CDR3 VL-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888.
27. Композиция по любому из пп. 20-26 для ингибирования или нейтрализации токсичности, вызванной токсином А или токсином В С. difficile, для клетки.
28. Композиция по любому из пп. 20-26 для нейтрализации токсинов, продуцируемых гипервирулентным штаммом С. difficile.
29. Композиция по любому из пп. 20-26 для лечения субъекта, который не проявляет симптомов, но который является чувствительным к или имеет повышенный риск заражения инфекцией С. difficile, ассоциированным с С. difficile заболеванием или ассоциированной с С. difficile диареей (CDAD).
30. Гибридомная клеточная линия, депонированная под № доступа АТСС РТА-9693, где гибридома продуцирует моноклональное антитело, которое специфично связывает токсин В С. difficile.
31. Гибридомная клеточная линия, депонированная под № доступа АТСС РТА-9692, где гибридома продуцирует моноклональное антитело, которое специфично связывает токсин А С. difficile.
32. Гибридомная клеточная линия, депонированная под № доступа АТСС РТА-9694, где гибридома продуцирует моноклональное антитело, которое специфично связывает токсин А С. difficile.
33. Гибридомная клеточная линия, депонированная под № доступа АТСС РТА-9888, где гибридома продуцирует моноклональное антитело, которое специфично связывает токсин А С. difficile.
34. Биспецифичное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с токсином В и токсином A Clostridium difficile (С. difficile) и содержит:
(i) первую антигенсвязывающую область, включающую антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин В С. difficile по любому из пп. 1, 3, 5 и 7; и
(ii) вторую антигенсвязывающую область, включающую антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С. difficile по любому из пп. 2, 4, 6 и 8.
35. Биспецифичное антитело или его антигенсвязывающий фрагмент по п. 34, где вторая антигенсвязывающая область включает:
(a) антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С. difficile и включает CDR1, CDR2 и CDR3 в вариабельной области тяжелой цепи (VH) и CDR1, CDR2 и CDR3 в вариабельной области легкой цепи (VL) моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9692; или
(b) антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С. difficile и включает CDR1, CDR2 и CDR3 в VH-области и CDR1, CDR2 и CDR3 в VL-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9694; или
(c) антитело или его антигенсвязывающий фрагмент, которое специфично связывает токсин А С difficile и включает CDR1, CDR2 и CDR3 в VH-области и CDR1, CDR2 и CDR3 в VL-области моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной под № доступа АТСС РТА-9888.
36. Композиция, содержащая терапевтически эффективное количество биспецифичного антитела или его антигенсвязывающего фрагмента по п. 34 или 35 и фармацевтически приемлемый носитель, наполнитель или разбавитель, для лечения инфекции С. difficile, ассоциированного с С. difficile заболевания или ассоциированной с С. difficile диареи (CDAD).
37. Композиция, содержащая биспецифичное антитело или его антигенсвязывающий фрагмент по п. 34 или 35 и фармацевтически приемлемый носитель, наполнитель или разбавитель, для ингибирования или нейтрализации токсичности, вызванной токсином А С. difficile, для клетки.
38. Композиция, содержащая биспецифичное антитело или его антигенсвязывающий фрагмент по п. 34 или 35 и фармацевтически приемлемый носитель, наполнитель или разбавитель, для ингибирования или нейтрализации токсичности, вызванной токсином В С. difficile, для клетки.
39. Композиция, содержащая биспецифичное антитело или его антигенсвязывающий фрагмент по п. 34 или 35 и фармацевтически приемлемый носитель, наполнитель или разбавитель, для нейтрализации токсинов, продуцируемых гипервирулентным штаммом С. difficile.
40. Композиция, содержащая биспецифичное антитело или его антигенсвязывающий фрагмент по п. 34 или 35 и фармацевтически приемлемый носитель, наполнитель или разбавитель, для лечения субъекта, который не проявляет симптомов, но который является чувствительным к или имеет повышенный риск заражения инфекцией С. difficile, ассоциированным с С. difficile заболеванием или ассоциированной с С. difficile диареей (CDAD).
41. Композиция по любому из пп. 36-40, дополнительно содержащая дополнительное терапевтическое средство.
42. Композиция по любому из пп. 36-40, где гипервирулентный штамм С. difficile выбирают из одного или более из BI/NAP1/027, CCL678, НМС553, Pitt45, CD196, montreal 5, montreal 7.1, MH5, Pitt2, CCL14137, UVA17, UVA30/TL42 и Pitt7.
43. Применение композиции по любому из пп. 20-26 для лечения инфекции С. difficile, ассоциированного с С. difficile заболевания или ассоциированной с С. difficile диареи (CDAD).
44. Выделенная нуклеиновая кислота, которая кодирует антитело к токсину В или его антигенсвязывающий фрагмент по любому из пп. 1, 3, 5, 7, 9-16.
45. Выделенная нуклеиновая кислота, которая кодирует антитело к токсину А или его антигенсвязывающий фрагмент по любому из пп. 2, 4, 6, 8-16.
46. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 44 или 45.
47. Клетка-хозяин, трансформированная или трансфицированная вектором экспрессии по п. 46, где клетка-хозяин экспрессирует нуклеиновую кислоту.
48. Способ ex vivo ингибирования или нейтрализации токсичности, вызванной токсином А С. difficile, для клетки, который включает воздействие на клетку эффективной ингибирующей или нейтрализующей токсин А С. difficile дозой антитела или его антигенсвязывающего фрагмента по любому из пп. 2, 4, 6 или 8.
49. Способ ex vivo ингибирования или нейтрализации токсичности, вызванной токсином В С. difficile, для клетки, который включает воздействие на клетку эффективной ингибирующей или нейтрализующей токсин В С. difficile дозой антитела или его антигенсвязывающего фрагмента по любому из пп. 1, 3, 5 или 7.
50. Способ ex vivo ингибирования или нейтрализации токсичности, вызванной токсином А и токсином В С. difficile, для клетки, который включает воздействие на клетку эффективной ингибирующей или нейтрализующей токсин А С. difficile дозой антитела или его антигенсвязывающего фрагмента по любому из пп. 2, 4, 6 или 8 и эффективной ингибирующей или нейтрализующей токсин В С. difficile дозой антитела или его антигенсвязывающего фрагмента по любому из пп. 1, 3, 5 или 7.
| WO 2007024715 A2, 01.03.2007 | |||
| EP 1766093 А2, 28.03.2007 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Пер | |||
| с англ | |||
| / под ред | |||
| У | |||
| Пола | |||
| М.: Мир, 1987-1989, 360 с., Т.1, стр.270 | |||
| РОЙТ и др | |||
| "Иммунология", Пер | |||
| с англ | |||
| - М.: Мир, 2000, стр.110-111, 151. | |||

