Область техники
Настоящее изобретение относится к способам получения и применения антител, которые связываются с человеческим белком альфа-энолазы (ENO1).
Уровень техники
Опухоли возникают в результате аберрантной, неограниченной пролиферации единственной клетки, генерирующей клон трансформированных клеток. Опухолевые клетки могут экспрессировать уникальные антигены, которые могут распознаваться иммунной системой. Опухолеспецифические антигены включают мутированные онкогены, мутированные нормальные клеточные белки, аберрантно экспрессированные клеточные белки, аномальные белки клеточной поверхности и онкогенные вирусные белки. Иммунная система рассматривает эти опухолеспецифические антигены, как не свои, и может вырабатывать антитела для устранения этих чужеродных антиген-несущих опухолевых клеток, не затрагивая при этом здоровые клетки. Таким образом, идентификация иммуногенных опухолеспецифических антигенов может использоваться для клинических, прогностических или терапевтических применений при лечении злокачественного новообразования.
Некоторые злокачественные опухоли могут быть идентифицированы с помощью плеврального выпота, который представляет собой избыточную жидкость в пространстве между легкими и грудной клеткой. Карцинома легкого, карцинома молочной железы и лимфома обусловливают примерно 75% всех злокачественных плевральных выпотов. Злокачественный плевральный выпот может обогащаться лимфоцитарными инфильтратами и опухолевыми клетками. Опухолеспецифические иммунные комплексы или аутоантитела, такие как антитела к p53, антитела к белкам ядра, и антитела к L-Myc обнаружены в жидкостях выпота и ассоциируются с неблагоприятным прогнозом. Некоторые опухолеспецифические антигены также были идентифицированы в злокачественных выпотах, включая фрагменты цитокератина 19, нейрон-специфическую энолазу (ENO2), антиген плоскоклеточной карциномы и растворимый HLA-I, и т.д.
Альфа-энолаза (энолаза-1, ENO1) представляет собой многофункциональный белок, который был впервые обнаружен в качестве ключевого фермента пути гликолиза. При нормальных условиях ENO1 экспрессируется в цитозоли. Однако также было обнаружено, что ENO1 экспрессируется на клеточной поверхности множества опухолевых клеток в виде рецептора плазминогена и на активированных гематопоэтических клетках, таких как нейтрофилы, лимфоциты и моноциты. Известно, что положительная регуляция белков рецептора плазминогена может индуцировать каскадный ответ урокиназной системы активации плазминогена (uPAS).
Система урокиназной активации плазминогена (uPAS) состоит из урокиназного активатора плазминогена (uPA), распознающего его рецептор (uPAR), и двух специфических ингибиторов, ингибитора 1 активатора плазминогена (PAI-1) и ингибитора 2 активатора плазминогена (PAI-2). Урокиназный активатор плазминогена превращает профермент плазминогена в активную сериновую протеазу, плазмин. Плазмин вовлечен в ряд процессов ремоделирования ткани, таких как ремоделирование базальной мембраны (BM) и внеклеточного матрикса (ECM), которые требуются при опухолевой прогрессии и метастазировании. Кроме того, было представлено, что uPAS может быть вовлечен в неопластическую эволюцию, воздействуя на опухолевый ангиогенез, пролиферацию злокачественных клеток и миграцию, интраваскуляризацию и рост в месте метастазирования.
Конкретно, активация плазминогена может приводить к деградациям внеклеточного матрикса, которые, в свою очередь, могут приводить к повышенному метастазированию опухолевых клеток и инфильтрации иммунных клеток. Другими словами, экспрессия ENO1 на поверхности опухолевых клеток в виде рецептора плазминогена может повышать активность инвазии опухолевых клеток. Таким образом, ENO1 представляет собой потенциальную мишень для противоопухолевой терапии.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Воплощения изобретения относится к агентам направленного связывания, которые специфично связываются с человеческим ENO1, ингибируя таким образом связывание лиганда (например, плазминогена) с ENO1. Путем ингибирования связывания плазминогена с ENO1 агенты направленного связывания по изобретению могут ингибировать активацию плазминогена, приводя к уменьшенной деградации внеклеточного матрикса, что, в свою очередь, предотвращает или уменьшает диссоциацию опухолевых клеток из внеклеточного матрикса. Таким образом, агенты направленного связывания согласно воплощениям изобретения могут использоваться для ингибирования опухолевого роста и метастазирования. Механизмы, с помощью которых этого можно достичь, могут включать, но не ограничиваются этим, ингибирование связывания лиганда (такого как плазминоген) с его рецептором ENO1, или устранение интер-реакций между рецептором ENO1 и его лигандами, уменьшая посредством этого эффективную концентрацию ENO1.
Согласно одному воплощению изобретения агент направленного связывания представляет собой антитело, которое может связываться с человеческим ENO1 для предотвращения связывания его лигандов (например, плазминогена) с ENO1. Предотвращение связывания плазминогена с рецептором может предотвращать активацию плазминогена. Это приводит к ингибированию урокиназной системы активации плазминогена (uPAS) во внеклеточном матриксе опухолевых клеток.
Согласно некоторым воплощениям изобретения, антитело может связываться с ENO1 с высокой аффинностью, такой как Kd менее чем 0,3нМ. Такие агенты тесного связывания могут ингибировать ENO1 с высокой эффективностью.
Согласно некоторым воплощениям изобретения, агент направленного связывания представляет собой антитело, которое может связываться с человеческим ENO1 и ингибировать индуцированную активность плазмина или опухолевых клеток с высокой эффективностью, как например, ингибирование на 80%, 90%, или на 100%. Анализы ингибирования могут осуществляться путем индуцирования экспрессии ENO1 (следовательно, активации плазминогена) в опухолевой клетке (такой как человеческие клетки лимфомы U937) с помощью обработки с использованием 10 микрограмм/мл липополисахарида (LPS) в течение 5 часов. Ингибирование такой индуцированной активности плазмина может быть проанализировано с использованием антитела при походящей концентрации. Используя антитело по изобретению, такое ингибирование может быть детектировано при таких низких концентрациях антитела, как 20 микрограмм/мл или менее.
Согласно некоторым воплощениям изобретения, агент направленного связывания представляет собой антитело, которое может связываться с человеческим ENO1. Такое антитело может использоваться для ингибирования инвазивной активности опухолевой клетки. Например, антитела по изобретению могут ингибировать более чем на 50%, 60%, или на 70% инвазивную активность клеток CL1-5 немелкоклеточного рака легкого при настолько низких концентрациях антитела, как 50 микрограмм/мл или менее.
Согласно некоторым воплощениям изобретения, агент направленного связывания представляет собой антитело, которое может связываться с человеческим ENO1 для ингибирования деградации внеклеточного матрикса, ингибируя посредством этого диссоциацию опухолевых клеток из внеклеточного матрикса. Например, антитело по изобретению может ингибировать более чем на 40%, 50%, или 60% опосредованную плазминогеном диссоциацию клеток CL1-5 из коллагена или фибронектина при настолько низких концентрациях антитела, как 50 микрограмм/мл или менее.
Согласно воплощениям изобретения, агент направленного связывания (т.е. антитело) может иметь аминокислотную последовательность тяжелой цепи, содержащую область, определяющую комплементарность (CDR), содержащую одну из последовательностей CDR, включенных в последовательность 1.
Согласно воплощениям изобретения, агент направленного связывания (т.е. антитело) может иметь аминокислотную последовательность легкой цепи, содержащую область, определяющую комплементарность (CDR), содержащую одну из последовательностей CDR, включенных в последовательность 2.
Согласно некоторым воплощениям изобретения, агент направленного связывания представляет собой антитело, которое может представлять собой моноклональное антитело или поликлональное антитело.
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность легкой цепи, содержащую любую из последовательностей LCDR1, LCDR2 или LCDR3, включенных в последовательность 2.
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность легкой цепи, содержащую любые две из последовательностей LCDR1, LCDR2 или LCDR3, включенных в последовательность 2 (то есть,LCDR1 и LCDR2, LCDR1 и LCDR3 или LCDR2 и LCDR3).
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность легкой цепи, которая содержит последовательности LCDR1, LCDR2 или LCDR3, включенные в последовательность 2. Согласно некоторым воплощениям изобретения, антитело может представлять собой гуманизированное антитело или полностью человеческое моноклональное антитело.
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность тяжелой цепи, содержащую любую из последовательностей HCDR1, HCDR2 или hCDR3, включенных в последовательность 1.
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность тяжелой цепи, содержащую любые две из последовательностей HCDR1, HCDR2 или HCDR3, включенных в последовательность 1 (то есть, HCDR1 и HCDR2, HCDR1 и HCDR3 или HCDR2 и HCDR3).
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит аминокислотную последовательность тяжелой цепи, которая содержит последовательности HCDR1, HCDR2 и HCDR3, включенные в последовательность 1. Согласно некоторым воплощениям изобретения, антитело может представлять собой гуманизированное антитело или полностью человеческое моноклональное антитело.
Согласно некоторым воплощениям изобретения, антитело, которое может связываться с человеческим белком ENO1, содержит последовательность легкой цепи, содержащую CDR, включающую одну из последовательностей CDR, включенных в последовательность 2. Согласно некоторым воплощениям изобретения, антитело может связываться с человеческим белком ENO1 и содержит аминокислотную последовательность тяжелой цепи, содержащую CDR, включающую последовательность, представленную в последовательности 1. Согласно некоторым воплощениям изобретения, антитело может представлять собой гуманизированное или полностью человеческое моноклональное антитело.
Согласно некоторым воплощениям изобретения, антитело может связываться с человеческим белком ENO1 и содержит аминокислотную последовательность тяжелой цепи, содержащую одну из последовательностей CDR, включенных в последовательность 1, и аминокислотную последовательность легкой цепи, содержащую одну из последовательностей CDR, включенных в последовательность 2. Согласно некоторым воплощениям изобретения, антитело может представлять собой гуманизированное или полностью человеческое моноклональное антитело.
Согласно некоторым воплощениям изобретения, агент направленного связывания (т.е. антитело) может конкурировать за связывание плазминогена с человеческим белком ENO1. Согласно некоторым воплощениям изобретения, указанный агент направленного связывания содержит аминокислотную последовательность тяжелой цепи, содержащую, по меньшей мере, одну из последовательностей CDR, включенных в последовательность 1, и аминокислотную последовательность легкой цепи, содержащую по меньшей мере, одну из последовательностей CDR, включенных в последовательность 2.
Согласно некоторым воплощениям изобретения, агент направленного связывания может связываться с эпитопом, содержащим аминокислотную последовательность 296FD Q D D W G A W Q K F TA309 (SEQ ID NO:9) или 326K R I A K A V N EK S336 (SEQ ID NO:10) человеческого белка ENO1 (GenBank: AAH50642.1). Согласно некоторым воплощениям изобретения, указанный агент направленного связывания содержит аминокислотную последовательность тяжелой цепи, содержащую одну из последовательностей CDR, включенных в последовательность 1, и аминокислотную последовательность легкой цепи, содержащую одну из последовательностей CDR, включенных в последовательность 2.
Согласно некоторым воплощениям изобретения, агент связывания по изобретению содержит антиген-связывающий сайт внутри молекулы, не являющейся антителом. Например, такой агент связывания может содержать одну или несколько CDR, например, набор CDR в каркасе белка, не являющегося антителом, как обсуждается дополнительно ниже.
Некоторые воплощения изобретения относятся к способам для анализа уровня человеческого белка ENO1 у пациента или в образце пациента. Способ по изобретению включает контакт антитела к ENO1 с биологическим образцом пациента и детектирование уровня связывания между указанным антителом и человеческим белком ENO1 в указанном образце. В более конкретных воплощениях, биологическим образцом является кровь или плазма.
Другие воплощения изобретения относятся к композициям, содержащим агент направленного связывания, который может включать антитело или его функциональный фрагмент и фармацевтически приемлемый носитель.
Еще другие воплощения изобретения относятся к способам эффективного лечения объекта (например, человека или животного), страдающего заболеванием или расстройством, связанным с ENO1. Способ может включать отбор объекта, нуждающегося в лечении неопластического или не неопластического заболевания, и введение объекту терапевтически эффективной дозы антитела (которое может быть гуманизированным или полностью человеческим моноклональным антителом), которое специфично связывается с белком ENO1.
Антитело по изобретению может использоваться для лечения заболевания или расстройства, связанного с человеческим белком ENO1. Заболевание или расстройство, связанное с человеческим белком ENO1, может представлять собой любое патологическое состояние, возникающее в результате аберрантной активации или экспрессии человеческого белка ENO1. Примеры таких заболеваний включают такие, где человеческий белок ENO1 аберрантно взаимодействует со своими лигандами, изменяя посредством этого свойства клеточной адгезии или передачи клеточных сигналов. Это изменение в свойствах клеточной адгезии или передачи клеточных сигналов может приводить к возникновению неопластических заболеваний или некоторых иммунных заболеваний.
Например, заболевание, связанное с человеческим белком ENO1, может представлять собой неопластическое заболевание, такое как злокачественное новообразование легкого, молочной железы, поджелудочной железы, печени, ободочной и прямой кишки и предстательной железы.
Согласно некоторым воплощениям изобретения, агент направленного связывания (например, антитело) может связываться с пептидом, содержащим аминокислотную последовательность 296FD Q D D W G A W Q K F T308 (SEQ ID NO:9) или 326K R I A K A V N EK S336 (SEQ ID NO:10) человеческого белка ENO1 (GenBank: AAH50642.1) и может использоваться для лечения заболевания или расстройства, связанного с человеческим белком ENO1, как отмечено выше.
Дополнительные воплощения изобретения относятся к способам ингибирования ENO1-индуцированной диссоциации опухолевых клеток объекта из внеклеточного матрикса. Эти способы могут включать отбор объекта (например, человека или животного), нуждающегося в лечении ENO1-индуцированной клеточной диссоциации, и введение указанному объекту терапевтически эффективного количества антитела, где указанное антитело специфично связывается с ENO1. Антитело может быть гуманизированным или полностью человеческим моноклональным антителом.
Дополнительные воплощения изобретения относятся к применениям антитела при получении лекарственного средства для лечения у объекта (например, человека или животного) заболевания или расстройства, связанного с ENO1, где указанное антитело специфично связывается с ENO1. Антитело может быть гуманизированным или полностью человеческим моноклональным антителом.
Согласно некоторым воплощениям изобретения, агенты направленного связывания, описанные в данном документе, могут использоваться при получении лекарственного средства для лечения у животного клеточной диссоциации из внеклеточного матрикса, индуцированной белком ENO1, где указанное антитело специфично связывается с ENO1. Антитело может быть гуманизированным или полностью человеческим моноклональным антителом.
Некоторые воплощения изобретения, описанные в данном документе, относятся к моноклональным антителам, которые связываются с человеческим ENO1 и воздействуют на функции человеческого ENO1. Другие воплощения изобретения относятся к препаратам антител к ENO1 с целевыми свойствами для лечебных применений. Такие свойства могут включать высокую аффинность связывания с ENO1, способность нейтрализации активности ENO1 in vitro и in vivo, и способность ингибировать ENO1-индуцированную клеточную диссоциацию, рост и метастазирование опухолей.
В некоторых воплощениях, изобретение относится к антителу, которое может связываться с человеческим ENO1 с очень высокой аффинностью (т.е., с низкой Kd). Например, человеческое, кроличье, мышиное, химерное или гуманизированное антитело, которое способно связываться с ENO1 с Kd менее, чем примерно 10-5, 10-6, 10-7, 10-8, 10-9, 10-10 или примерно 10-11M, или соответствует любому интервалу или значению между ними. Измерения аффинности и/или авидности может осуществляться с использованием тИФА и/или BIACORE, как описано в данном документе, или согласно методам, известным в данной области.
Понятно, что воплощения изобретения не ограничены любой конкретной формой антитела или способом генерирования или получения. Например, антитело к ENO1 может представлять собой полноразмерное антитело (например, содержи интактный человеческий Fc-участок) или фрагмент антитела (например, Fab, Fab' или F(ab')2, FV или Dab (Dab представляют собой кратчайшие функциональные связывающие элементы человеческих антител). Кроме того, антитело может быть получено из гибридомы, которая секретирует антитело, или из рекомбинантно полученной клетки, которая трансформирована или трансфецирована геном или генами, кодирующими антитело.
Другие воплощения изобретения относятся к выделенным молекулам нуклеиновых кислот, кодирующим любые из антител, описанных в данном документе, к векторам, содержащим выделенные молекулы нуклеиновых кислот, кодирующих антитело к ENO1, или к клетке-хозяину, трансформированной любой из таких молекул нуклеиновых кислот.
Кроме того, некоторые воплощения изобретения относятся к способу получения антитела к ENO1 с помощью культивирования клеток-хозяев при условиях, где молекула нуклеиновой кислоты экспрессируется с получением антитела, с последующим извлечением антитела. Следует понимать, что воплощения изобретения также могут включать любую молекулу нуклеиновой кислоты, которая кодирует антитело или фрагмент антитела по изобретению, включая последовательности нуклеиновых кислот, оптимизированные для повышения выхода антител или их фрагментов при трансфекции в клетки-хозяева для продуцирования антитела.
Следующие воплощения изобретения могут относиться к способам получения антител с высокой аффинностью к человеческому ENO1 путем иммунизации млекопитающего с использованием человеческого белка ENO1, его фрагмента и одной или нескольких ортологических последовательностей или их фрагментов.
Другие воплощения относятся к генерации и идентификации выделенных антител, которые могут специфично связываться с человеческим ENO1. Ингибирование биологической активности ENO1 может осуществляться этими антителами для предотвращения ENO1-индуцированной клеточной диссоциации, инвазии и других целевых эффектов злокачественных новообразований.
Другие воплощения изобретения относятся к фармацевтическим композициям, содержащим эффективное количество антитела к ENO1. Композиция может дополнительно содержать фармацевтически приемлемый носитель или разбавитель. Еще в других воплощениях, антитело к ENO1 или его фрагмент конъюгировано с терапевтическим агентом. Терапевтический агент может представлять собой, например, токсин или радиоизотоп.
Еще другие воплощения изобретения относятся к способам лечения заболеваний или патологических состояний, ассоциированных с экспрессией ENO1 у пациента. Способы могут включать введение пациенту эффективного количества антитела к ENO1. Антитело к ENO1 может вводиться индивидуально или в комбинации с дополнительными антителами или химиотерапевтическим лекарственным средством или с лучевой терапией. Например, моноклональная, олигоклональная или поликлональная смесь антитела к ENO1, которая блокирует клеточную диссоциацию, может вводиться в комбинации с лекарственным средством, которое, как было продемонстрировано, прямо ингибирует пролиферацию опухолевых клеток. Способ может осуществляться in vivo, и пациентом предпочтительно является человек. В предпочтительном воплощении, способ касается лечения заболевания или расстройства, связанного с ENO1, включающего, в частности, непластическое заболевание, такое как злокачественное новообразование легкого, молочной железы, поджелудочной железы, печени, ободочной и прямой кишки и предстательной железы и/или солидные опухоли.
Некоторые воплощения изобретения относятся к способу мониторинга развития злокачественного новообразования. Способ может включать определение представленности белков альфа-энолазы (ENO1) в образце (например, в опухолевых клетках), где повышенный уровень ENO1 коррелирует с тяжестью злокачественного новообразования. Согласно воплощениям изобретения, представленность может быть определена путем измерения связывания ENO1-специфичного антитела с белками ENO1.
Некоторые воплощения изобретения относятся к способу детектирования злокачественного новообразования. Такой способ может включать определение представленности ENO1-специфичных антиел в образце сыворотки, где низкий уровень ENO1-специфичных антител указывает на присутствие злокачественной опухоли.
Краткое Описание Чертежей
ФИГ. 1A и 1B демонстрируют результаты исследований локализации ENO1 на клеточной поверхности в злокачественных клетках с использованием проточной цитометрии (ФИГ. 1A) и иммуноокрашивания (ФИГ. 1B), соответственно. Подробные процедуры осуществляли, как описано в Примере 1. Данные продемонстрировали, что ENO1 локализован на клеточной поверхности в злокачественных клетках (NHRI-L89 & CA926), но не в нормальных клетках эпителия легкого (NHBE & SAEC).
ФИГ. 2 демонстрирует ингибирование инвазивной активности высокоинвазивных клеток аденокарциномы легкого CL1-5 с помощью поликлонального антитела против ENO1 клеточной поверхности. Подробные процедуры осуществляли, как описано в Примере 2. Данные демонстрируют, что введение поликлонального антитела к ENO1 ослабляет инвазию CL1-5.
ФИГ. 3 демонстрирует, что поликлональное антитело против человеческого ENO1 ослабляет инвазивную способность клеток CL1-5F4 в легком. Введение антитела животному и образование метастатической колонии в легком отслеживали через 12 и 19 дней после опухолевой инъекции с использованием системы IVIS. Подробные процедуры осуществляли, как описано в Примере 3. Данные демонстрируют, что поликлональное антитело к ENO1 ингибирует миграцию CL1-5F4 в легкое in vivo.
ФИГ. 4 демонстрирует генерирование гибридом антитела к ENO1 и подтверждение каждого клона моноклонального антитела с помощью проточной цитометрии. Процедуры иммунизации мышей и генерирование гибридомы и продуцирование каждого антитела и подтверждение антитела с помощью проточной цитометрии описаны в Примере 4. Данные показали, что все 5 гибридомных антител распознают поверхностный ENO1 на клетках CL1-5F4 аденокарциномы легкого.
ФИГ. 5A демонстрирует связывание ENO1 в тИФА 5 антител, выделенных из асцитов индивидуальных гибридом. Очистку с сульфатом аммония, очистку на колонке с протеином A и очистку SDS-PAGE осуществляли, как описано в Примере 5. Данные на ФИГ. 5B демонстрируют значения Kd 5 различных антител к человеческому ENO1.
ФИГ. 6 демонстрирует результаты фибринолитического анализа для U937 5 антител, выделенных из асцитов индивидуальных гибридом, соответственно. Индукцию экспрессии ENO1 с помощью LPS в клетках U937 клеточной линии лимфомы человека и анализ активности плазмина осуществляли, как описано в Примере 6. Эти данные демонстрируют, что 5 различных антител к ENO1 обладают различными ингибирующими активностями против рецептора плазминогена ENO1 и что ингибирующие активности коррелируют со значениями Kd.
ФИГ. 7 демонстрирует результаты ингибирования инвазивных активностей клеток CL1-5, обработанных 5 различными антителами, выделенными из асцитов индивидуальных гибридом, соответственно. Процедуры подробно описаны в Примере 7. Эти данные демонстрируют, что все 5 различных антител к ENO1 могут ингибировать инвазивную активность клеток CL1-5. Интересно, что даже если клон 8 имеет Kd примерно в 7,2-раза выше, чем у клона 10, ингибирование инвазивной активности против клеток CL1-5 с помощью клона 8 аналогично эффекту клона 10.
ФИГ. 8A демонстрирует результаты инвазивных активностей клеток CL1-5, обработанных различными концентрациями антитела EN10 mAb, выделенного из гибридомы. Процедуры осуществляли, как подробно описано в Примере 8. Эти данные демонстрируют, что антитело EN10mAb ингибирует инвазивную активность CL1-5 в дозозависимой манере.
ФИГ. 8B демонстрирует результаты инвазивных активностей клеток U937, обработанных различными концентрациями EN10 mAb, выделенного из гибридомы, после того как индуцировали экспрессию поверхностного ENO1 в клетках с помощью LPS. Процедуры осуществляли, как подробно описано в Примере 8. Эти данные демонстрируют, что EN10 mAb ингибирует инвазивную активность клеток U937 в дозозависимой манере.
ФИГ. 9 демонстрирует, что EN10 mAb распознает ENO1 клеточной поверхности на клетках U937, обработанных с помощью LPS. Процедуры осуществляли, как подробно описано в Примере 9.
ФИГ. 10A демонстрирует адгезивную активность клеток CL1-5 карциномы легкого по отношению к матриксным белкам. Анализ адгезии осуществляли, как описано в Примере 10. Эти данные демонстрируют, что клетки CL1-5 обладают более высокими адгезивными активностями по отношению к коллагену и фибриногену.
ФИГ. 10B демонстрирует результаты ингибирования диссоциации клеток CL1-5 из фибронектина, обработанного EN10 mAb. Анализ ассоциированной с клетками адгезии осуществляли, как описано в Примере 10. Эти данные демонстрируют, что EN10 mAb ингибирует активность диссоциации клеток CL1-5 от фибронектина в дозозависимой манере.
ФИГ. 10C демонстрирует результаты ингибирования диссоциации клеток CL1-5 из коллагена, обработанного EN10 mAb. Анализ ассоциированной с клетками адгезии осуществляли, как описано в Примере 10. Эти данные демонстрируют, что EN10 mAb ингибирует активность диссоциации клеток CL1-5 от коллагена в дозозависимой манере.
На ФИГ. 11A изображена аминокислотная последовательность вариабельной области тяжелой цепи EN10 mAb (SEQ ID NO: 1). Показаны каркасные области (FR1, FR2, FR3 и FR4) и CDR (HCDR1, HCDR2 и HCDR3). Клонирование EN10 mAb осуществляли, как описано в Примере 11.
На ФИГ. 11B изображены аминокислотные последовательности вариабельной области легкой цепи EN10 mAb(SEQ ID NO: 2). Показаны каркасные области (FR1, FR2, FR3 и FR4) и CDR (LCDR1, LCDR2 и LCDR3). Клонирование EN10 mAb осуществляли, как описано в Примере 11.
ФИГ. 12A демонстрирует активности EN10 mAb связывания делеционных мутантов ENO1. Эпитоп связывания EN10 mAb локализован между аминокислотными остатками номер 293 и 434 человеческого белка ENO1. Большую часть делеций ENO1 для определения области связывания EN10 mAb осуществляли, как описано в Примере 12.
ФИГ. 12B демонстрирует 12% SDS PAGE 6 C-концевых делеционных мутантов белка ENO1, очищенных из E.coli. Процедуры очистки делеционных мутантов ENO1 подробно описаны в Примере 12.
ФИГ. 12C демонстрирует активности связывания EN10 mAb и 6 C-концевых делеционных мутантов ENO1. Эпитоп связывания EN10 mAb локализован между аминокислотными остатками номер 296 и 336 человеческого белка ENO1. Большую часть делеций ENO1 для определения области связывания EN10 mAb осуществляли, как описано в Примере 12.
На ФИГ. 13A изображена кристаллическая структура и экспонированные на поверхности аминокислотные остатки между аминокислотными остатками номер 296 и 336 человеческого ENO1. Прогнозирование структуры описано в Примере 13.
ФИГ. 13B демонстрирует 12% SDS PAGE 11 мутантных белков ENO1, полученных мутагенезом в виде сканирования аланином, очищенных из E.coli. Процедуры для очистки мутантных белков ENO1 подробно описаны в Примере 13.
ФИГ. 13C демонстрирует тИФА связывания ENO1 и значения Kd 11 мутантов сканирования аланином против EN10 mAb. Результат предполагает, что последовательности пептида 1 ENO1, FD Q D D W G A W Q K F TA (SEQ ID NO: 9), и пептида 2, K R I A K A V N EK S (SEQ ID NO:10), локализованные между аминокислотными остатками номер 296 и 336 человеческого ENO1, вовлечены в связывание EN10 mAb. Сканирование аланином осуществляли, как описано в Примере 13.
ФИГ. 13D демонстрирует последовательности пептида 1 ENO1 (FD Q D D W G A W Q K F TA (SEQ ID NO: 9)) и пептида 2 (K R I A K A V N EK S (SEQ ID NO:10)) между аминокислотными остатками номер 296 и 336 человеческого ENO1(SEQ ID NO:49), которые участвуют в связывании человеческого ENO1 и EN10 mAb.
ФИГ. 14A демонстрируют 3D-структуру человеческого белка ENO1. На ФИГ.14B изображена предполагаемая структура EN10 mAb Fab (представленного в виде ленточной спирали, легкая цепь - серая, тяжелая цепь - черная) в комплексе с областью связывания плазминогена человеческого белка ENO1. Прогнозирование кристаллической структуры белка ENO1 и EN10 mAb осуществляли, как описано в Примере 14.
ФИГ. 15A демонстрирует экспрессирующий вектор для генерирования химерного EN10 mAb мышь-человек. Процедуры для очистки химерного антитела EN10 mAb описаны в Примере 15.
На ФИГ. 15B изображены результаты, полученные при применении химерного антитела для определения аффинности связывания и констант кинетики EN10 mAb. Процедуры экспрессии, очистки и анализа Kd химерного антитела осуществляли, как подробно описано в Примере 15.
ФИГ. 16 демонстрирует ингибирующие эффекты EN10 mAb в отношении роста опухоли легкого в присутствии компонента комплемента. Введение EN10 mAb и замедление опухолевого роста осуществляли, как описано в Примере 16. Данные демонстрируют, что введение EN10 mAb вместе с компонентом комплемента дважды в неделю обладало эффективностью, аналогичной эффективности обработки такой же дозой коммерческого лекарственного средства ErbituxTM в мышиной модели ксенотрансплантата CL1-5.
ФИГ. 17A, 17B и 17C демонстрируют результаты, полученные при блокаде опухолевого распространения злокачественного новообразования поджелудочной железы в анализе метастатического колониеобразования селезенки-печени с помощью EN10 mAb. Введение EN10 mAb и ингибирование метастатического опухолевого роста с помощью обработки антителом осуществляли, как описано в Примере 17. Результаты исследования выявили, что введение 10 мг/кг EN10 mAb дважды в неделю уменьшало количество метастатических опухолевых узлов, размер опухоли и массу опухоли мышей по сравнению с аналогичными параметрами от такой же дозы контрольного IgG в мышиной метастатической модели селезенки-печени.
ОПРЕДЕЛЕНИЯ
Если не определено иное, то научные и технические термины, использованные в данном документе, имеют тот же смысл, который вкладывается в них обычным специалистом в данной области. Кроме того, до тех пор, пока иное не требуется по контексту, термины в единственном числе будут включать значения во множественном числе, и термины во множественном числе будут включать значения в единственном числе. Как правило, номенклатура и методы, использованные в отношении клеточной и тканевой культуры, молекулярной биологии, и белковой и олиго- или полинуклеотидной химии и гибридизации, описанные в данном документе, хорошо известны и широко используются в данной области.
Стандартные методы используются для рекомбинантных ДНК, синтеза олигонуклеотидов, тканевой культуры и трансформации (например, электропорации, липофекции). Ферментативные реакции и методы очистки осуществляются согласно описаниям производителя или так, как они обычно осуществляются в данной области или так, как они описаны в данном документе. Вышеописанные методы и процедуры, как правило, осуществляют согласно стандартным методам, хорошо известным в данной области и как описано в различных общих и в более конкретных публикациях, которые цитируются и обсуждаются на протяжении настоящего описания. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual (3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001)), которая включена в данный документ ссылкой. Номенклатура, применяемая в этой связи, а также лабораторные процедуры и методы аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, описанные в данном документе, хорошо известны и широко применяются в данной области. Стандартные методы используются для химического синтеза, химического анализа, фармацевтических препаратов, составов и для доставки, и для лечения пациентов.
Применяемые согласно настоящему описанию следующие термины до тех пор, пока не указано иное, следует понимать, как имеющие следующие значения: используемый в настоящей заявке термин «и/или» следует понимать как конкретное описание каждого из двух конкретных признаков или компонентов вместе с другим или без него. Например, «A и/или B» следует понимать, как описание каждого из (i) A, (ii) B и (iii) A и B, как если бы каждый был представлен в данном документе индивидуально.
Антагонист может представлять собой полипептид, нуклеиновую кислоту, углевод, липид, низкомолекулярное соединение, оигонуклеотид, олигопептид, интерферирующую РНК (РНКи), антисмысловую РНК, рекомбинантный белок, антитело или их конъюгаты или сшитые белки. Для обзора РНКи см. Milhavet O, Gary D S, Mattson M P. (Pharmacol Rev. 2003 December; 55(4):629-48. Review.) и способов с использованием антисмысловой последовательности см. Opalinska J B, Gewirtz A M. (Sci STKE. 2003 Oct. 28; 2003 (206): pe47).
Аберрантная активация или экспрессия «ENO1», связанная с заболеванием, может быть любой аномальной, нежелательной или патологической клеточной адгезией, например, опухолеспецифическая клеточная адгезия. Заболевания, связанные с клеточной адгезией, включают, в частности, не солидные опухоли, такие как лейкоз или лимфома, а также солидные опухоли, такие как меланома, немелкоклеточный рак легкого, гепатоклеточная (печеночная) карцинома, рак желудочно-кишечного тракта, рак головы и шеи, рак системы печени, желудка, молочной железы, яичников, легкого, матки, вульвы, ободочной и прямой кишки и поджелудочной железы.
Термин ENO1 относится к гетеродимерной молекуле энолазы, состоящей из ENO1 и ENO2 или ENO3.
Используемый в настоящей заявке термин «антитело» относится, как правило и в основном, к иммуноглобулинам, аутоантителам, моноклональным антителам и поликлональным антителам, а также к их активным фрагментам. Фрагмент может быть активным по части его связывания с распознаваемым антигеном или он может быть активным по части его биологической функции. Антитела по изобретению могут быть химерными, гуманизированными или человеческими, полученными с использованием методов, хорошо известных в данной области.
Используемый в настоящей заявке термин «моноклональное антитело» относится к антителам, которые являются химически или иммунологически гомогенными, как правило, полученными с помощью гибридом. См. A Laboratory Manual, Harlow and Lane, eds., Cold Spring Harbor, N.Y. (1988).
Используемый в настоящей заявке термин «поликлональное антитело» относится к антителам, которые получают с помощью более чем одного клона плазматических клеток, синтезирующих антитела (B-лимфоциты) в ответ на один антиген. Они, как правило, вырабатываются животными после их иммунизации антигеном.
Используемый в настоящей заявке термин «химерное антитело» относится к антителам, которые содержат последовательности из более чем одного источника. Например, такие антитела могут содержать последовательности из источников, не относящихся к человеку, которые затем модифицируют путем введения человеческих последовательностей. Используемый в настоящей заявке термин «гуманизированное антитело» относится к антителу, в котором минимальные участки антитела, не относящегося к человеку, вводят в другое человеческое антитело. Используемый в настоящей заявке термин «человеческое антитело» относится к антителу, в котором по существу каждая часть белка является по существу неимуногенной у людей и содержит только минорные изменения или вариации последовательности.
Используемый в настоящей заявке термин «антитело, специфичное к альфа-энолазе» относится к антителу, которое обладает высокой специфичностью к ENO1 млекопитающих, но не к ENO2 или ENO3. Аналогично, термин «ENO1-специфичное антитело» относится к антителу, которое связывается с белком альфа-энолазы.
Термин «нейтрализующий», когда он относится к агенту направленного связывания, такому как антитело, относится к способности указанного агента направленного связывания устранять или существенно уменьшать активность антигена-мишени. Соответственно, «нейтрализующее» ENO1 антитело способно устранять или значительно уменьшать активность ENO1. Нейтрализующее ENO1 антитело может, например, действовать путем блокировки связывания ENO1 с плазминогеном. Путем блокировки данного связывания опосредованная плазминогеном диссоциация клеток значительно или полностью устраняется. В идеале, нейтрализующее антитело против ENO1 усиливает клеточную адгезию.
Термин «полипептид» используется в настоящей заявке в качестве общего наименования, которое относится к нативному белку, его фрагментам или аналогам полипептидной последовательности. Следовательно, нативный белок, его фрагменты и аналоги представляют собой компоненты полипептидного происхождения. Предпочтительные полипептиды согласно изобретению содержат молекулы тяжелых цепей иммуноглобулина человека и молекулы легкой цепи каппа иммуноглобулина человека, а также молекулы антитела, образованные комбинациями, содержащими молекулы тяжелой цепи иммуноглобулина с молекулами легкой цепи иммуноглобулина, такими как молекулы легкой цепи каппа иммуноглобулина или легкой цепи лямбда и наоборот, а также их фрагментов и аналогов. Предпочтительные полипептиды согласно изобретению могут также содержать только молекулы тяжелой цепи иммуноглобулина человека или их фрагменты.
Используемый в настоящей заявке термин «полинуклеотид» означает полимерную форму длиной по меньшей мере 10 оснований, состоящих либо из рибонуклеотидов, либо дезоксинуклеотидов или модифицированных форм любого типа нуклеотидов, или РНК или РНК-ДНК гетеродуплексов. Термин включает одноцепочечную и двухцепочечную форму ДНК.
Термин «CDR-область» или «CDR» предназначен для обозначения гипервариабельных областей тяжелой и легкой цепей иммуноглобулина, как определено у Kabat et al., 1991 (Kabat, E. A. et al., (1991) Sequences of Proteins of Immunological Interest, 5th Edition. US Department of Health and Human Services, Public Service, NIH, Washington), и в более поздних изданиях. Антитело, как правило, содержит 3 CDR тяжелой цепи и 3 CDR легкой цепи. Термин CDR или множество CDR используется здесь с целью обозначения, соответственно ситуации, одной из этих областей или нескольких или даже всех этих областей, которые содержат большинство аминокислотных остатков, ответственных за связывание с помощью аффинности антитела к антигену или эпитопа, который его распознает.
Среди шести коротких последовательностей CDR, третья CDR тяжелой цепи (HCDR3) обладает вариабельностью большего размера (большее разнообразие по существу обусловлено механизмом реаранжировки генов, который его вызывает). Последовательность может быть настолько короткой, как 2 аминокислоты, хотя самый длинный известный размер соответствует 26. Длина CDR также может варьироваться согласно длине, которая может обеспечиваться конкретным исходным каркасом. Функционально, HCDR3 частично играет роль в определении специфичности антитела (Segal et al., PNAS, 71:4298-4302, 1974).
Термин «набор CDR», на который ссылаются в настоящей заявке, включает CDR1, CDR2 и CDR3. Таким образом, набор HCDR относится к HCDR1, HCDR2 и HCDR3 (HCDR относится к вариабельной области CDR тяжелой цепи), и набор LCDR относится к LCDR1, LCDR2 и LCDR3 (LCDR относится к вариабельной области CDR легкой цепи). До тех пор, пока не установлено иное, «набор CDR» включает HCDR и LCDR.
Термин «соответствует» используется в настоящей заявке для обозначения того, что полинуклеотидная последовательность является гомологом (т.е., идентична, связана не строго эволюционно) всей или части эталонной полинуклеотидной последовательности, или что полипептидная последовательность идентична эталонной полипептидной последовательности.
Напротив, термин «комплементарен» используется в настоящей заявке для обозначения того, что комплементарная последовательность гомологичная всей или части эталонной полинуклеотидной последовательности. Для иллюстрации, нуклеотидная последовательность «TATAC» соответствует эталонной последовательности «TATAC» и комплементарна эталонной последовательности «GTATA».
Термин «идентичность последовательности» означает, что две полинуклеотидные или аминокислотные последовательности идентичны (т.е., на основе сравнения нуклеотид-нуклеотид или остаток-остаток) в области сравнения. Термин «процент идентичности последовательности» означает процент, который рассчитывают путем сравнения двух оптимально выровненных последовательностей в области сравнения, определяя количество положений, в которых присутствуют идентичные нуклеотидные основания (например, A, T, C, G, U, или I) или аминокислотные остатки в обеих последовательностях с получением количества совпадающих положений, и путем деления количества совпадающих положений на суммарное количество положений в области сравнения (т.е., размер области), и путем умножения результата на 100 с получением процента идентичности последовательностей.
Используемые в настоящей заявке термины «существенная идентичности» или «по существу идентичен» обозначают характеристику полинуклеотидной или аминокислотной последовательности, которые содержат последовательность, которая имеет, по меньшей мере, 85% идентичности последовательности, предпочтительно, по меньшей мере, 90% - 95% идентичности последовательности, более предпочтительно, по меньшей мере, 99% идентичности последовательности по сравнению с эталонной последовательностью в области сравнения, составляющей, по меньшей мере, 18 положений нуклеотидов (6 аминокислот), часто в области, по меньшей мере, 24-48 положений нуклеотидов (8-16 аминокислот), где процент идентичности последовательности рассчитывают путем сравнения эталонной последовательности с последовательностью, которая может включать делеции или вставки, которые составляют суммарно 20 процентов или менее эталонной последовательности в области сравнения. Эталонная последовательность может представлять собой часть более крупной последовательности.
Используемые в настоящей заявке двадцать стандартных аминокислот и их аббревиатуры используются согласно общепринятому употреблению. Стереоизомеры (например, D-аминокислоты) двадцати стандартных аминокислот, синтетических аминокислот, таких как α,α-дизамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нестандартные аминокислоты также могут быть подходящими компонентами для полипептидов согласно настоящему изобретению. Примеры нестандартных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин, и другие сходные аминокислоты и иминокислоты (например, 4-гидроксипролин). В полипептидном обозначении, используемом в настоящей заявке, левое направление представляет собой N-концевое направление, и правое направление представляет собой С-концевое направление согласно стандартным употреблениям и условным обозначениям.
Аналогично, до тех пор, пока не указано иное, левый конец одноцепочечной полинуклеотидной последовательности представляет собой 5'-конец; левое направление двухцепочечной полинуклеотидной последовательности относится к 5ʹ-направлению. Направление от 5ʹ к 3ʹ прироста образующихся РНК-транскриптов относится к направлению транскрипции; области последовательности на ДНК-цепи, содержащие такую же последовательность, как РНК и которые располагаются в 5ʹ-области по отношению к 5ʹ-концу РНК-транскрипта, относятся к «вышерасположенным последовательностям»; области последовательности на ДНК-цепи, содержащие такую же последовательность, что и РНК и которая располагается в 3ʹ-области по отношению к 3ʹ-концу РНК-транскрипта, относятся к «нижерасположенным последовательностям».
Применительно к полипептидам термин «существенная идентичность» означает, что две пептидные последовательности при оптимальном выравнивании, таком как с помощью программ GAP или BESTFIT с использованием параметров пропуска масс по умолчанию, имеют, по меньшей мере, 80% идентичности последовательности, предпочтительно, по меньшей мере, 90% идентичности последовательности, более предпочтительно, по меньшей мере, 95% идентичности последовательности, и наиболее предпочтительно, по меньшей мере, 99% идентичности последовательности. Предпочтительно, положения остатков, которые не идентичны, отличаются консервативными аминокислотными заменами.
Как обсуждалось в настоящей заявке, предполагается, что минорные вариации в аминокислотных последовательностях антител или иммуноглобулиновых молекул охвачены настоящим изобретением, при условии что вариации в аминокислотной последовательности сохраняют, по меньшей мере, примерно 75%, более предпочтительно, по меньшей мере, 80%, 90%, 95%, и наиболее предпочтительно, примерно 99% идентичности последовательности с антителами или молекулами иммуноглобулина, описанными в настоящей заявке. Конкретно, рассматриваются консервативные аминокислотные замены.
Приведет ли аминокислотная замена к получению функционального пептида, может быть легко определено оценкой специфической активности полипептидного производного. Анализы подробно описаны в настоящей заявке. Фрагменты или аналоги антител или иммуноглобулиновых молекул могут быть легко получены специалистами в данной области. Предпочтительные N- и C-концы фрагментов или аналогов присутствуют около границ функциональных доменов. Структурные и функциональные домены могут быть идентифицированы путем сравнения данных нуклеотидной и/или аминокислотной последовательности с публичными или частными базами данных. Известны методы идентификации белковых последовательностей, которые укладываются в известные трехмерные структуры. Bowie et al., (1991) Science 253:164. Таким образом, вышеописанные примеры демонстрируют, что специалист в данной области может распознать мотивы последовательностей и структурные конформации, которые могут использоваться для определения структурных и функциональных доменов, соответствующих антителам, описанным в настоящей заявке.
Следующий аспект изобретения представляет собой агент направленного связывания или молекулу антитела, содержащую домен VH, который имеет, по меньшей мере, примерно 60, 70, 80, 85, 90, 95, 98 или примерно 99% идентичности аминокислотной последовательности с доменом VH любого из антител, представленных в последовательностях 1, в прилагаемом списке последовательностей, антитела, описанного в настоящей заявке, или с HCDR (например, HCDR1, HCDR2 или HCDR3), представленными в последовательностях 1. Агент направленного связывания или молекула антитела также необязательно может содержать домен VL, который имеет, по меньшей мере, примерно 60, 70, 80, 85, 90, 95, 98 или примерно 99% идентичности аминокислотной последовательности с доменом VL любого из антител, представленных в последовательностях 2, в прилагаемом списке последовательностей, антитела, описанного в настоящей заявке, или с LCDR (например, LCDR1, LCDR2, или LCDR3), представленными в последовательностях 2. Алгоритмы, которые могут использоваться для расчета % идентичности двух аминокислотных последовательностей, включают, например, BLAST (Altschul et al., (1990) J. Mol. Biol. 215: 405-410), FASTA (Pearson and Lipman (1988) PNAS USA 85: 2444-2448), или алгоритм Смита-Ватермана (Smith and Waterman (1981) J. Mol. Biol. 147: 195-197), например, применяя параметры по умолчанию. В некоторых воплощениях, агент направленного связывания или антитело, которое имеет такую же идентичность аминокислотной последовательности, как описано выше, демонстрирует по существу, такую же активность, как и упомянутые антитела. Например, по существу такая же активность включает по меньшей мере одну активность, которая отличается от активности эталонных антител не более чем примерно на 50%, 40%, 30%, 20%, 10%, 5%, 2%, 1% или менее.
Антиген-связывающий сайт, как правило, образуется с помощью вариабельного домена тяжелой цепи иммуноглобулина (VH) и вариабельного домена легкой цепи иммуноглобулина (VL) с антиген-связывающим интерфейсом, образованным с помощью шести полипептидных петель, называемых областями, определяющими комплементарность (CDR). Существуют три CDR в каждом VH (HCDR1, HCDR2, HCDR3) и в каждом VL (LCDR1, LCDR2, LCDR3), вместе с каркасными областями (FR).
Как правило, домен VH образует пару с доменом VL с получением антиген-связывающего сайта антитела, хотя индивидуальный домен VH или VL может использоваться для связывания антигена. Домен VH (например, из последовательности 1) может образовывать пару с доменом VL (например, из последовательности 2), так чтобы образовался антиген-связывающий сайт антитела, содержащий оба домена, VH и VL. Аналогичные воплощения представлены для других доменов VH и VL, раскрытых в настоящей заявке. В других воплощениях, VH-цепи в последовательностях 1 образуют пары с гетерологичным доменом VL. Также из уровня техники известна разнородность легкой цепи. И также, аналогичные воплощения представлены изобретением для других доменов VH и VL, раскрытых в настоящей заявке. Таким образом, VH родительской или любой из цепей антитела на последовательностях 2 может образовывать пару с VL родительского или любого из антител на последовательностях 1, и 2 или другого антитела.
Антиген-связывающий сайт может содержать набор H и/или L CDR родительского антитела или любого из антител в последовательностях 1 и 2, содержащих настолько много, как двадцать, шестнадцать, десять, девять или менее, например, одну, три, четыре или пять аминокислотных дополнений, замен, делеций и/или вставок внутри раскрытого набора H и/или L CDR. такие модификации могут потенциально осуществляться по любому остатку внутри набора CDR.
Предпочтительные аминокислотные замены представляют собой такие, которые: (1) уменьшают чувствительность к протеолизу, (2) уменьшают чувствительность к окислению, (3) изменяют аффинность связывания образующихся белковых комплексов, (4) изменяют аффинности связывания и (4) придают или модифицируют другие физико-химические или функциональные свойства таких аналогов. Аналоги могут включать различные мутантные белки последовательности, отличной от природной пептидной последовательности. Например, могут быть сделаны одиночные или множество аминокислотных замен (предпочтительно консервативных аминокислотных замен) в природной последовательности (предпочтительно в области полипептида вне домен-образующих межмолекулярных контактов). Консервативная аминокислотная замена не должна существенно изменять структурные характеристики родительской последовательности (например, замена аминокислоты не должна приводить к тенденции разрушения спирали, которая присутствует в родительской последовательности, или к нарушению других типов вторичной структуры, которые характеризуют родительскую последовательность).
Следующий аспект изобретения представляет собой молекулу антитела, содержащую домен VH, который имеет, по меньшей мере, примерно 60, 70, 80, 85, 90, 95, 98 или примерно 99% идентичности аминокислотной последовательности с доменом VH любого из антител, представленных в последовательности 1, в прилагаемом списке последовательностей, антитела, описанного в настоящей заявке, или с HCDR (например, HCDR1, HCDR2 или HCDR3), представленными в последовательностях 1. Молекула антитела также необязательно может содержать домен VL, который имеет, по меньшей мере, 60, 70, 80, 85, 90, 95, 98 или 99% идентичности аминокислотной последовательности с доменом VL любого из антител, представленных в последовательностях 2, в прилагаемом списке последовательностей, антитела, описанного в настоящей заявке, или с LCDR (например, LCDR1, LCDR2, или LCDR3), представленными в последовательностях 2. Алгоритмы, которые могут использоваться для расчета % идентичности двух аминокислотных последовательностей, включают, например, BLAST, FASTA или алгоритм Смита-Ватермана.
Варианты доменов VH и VL и CDR согласно настоящему изобретению, включающие такие, для которых аминокислотные последовательности представлены в настоящей заявке и которые могут применяться в агентах направленного связывания и в антителах к человеческому белку ENO1, могут быть получены посредством методов изменения последовательности или мутации и скрининга на предмет поиска свойства направленного воздействия на антиген с целевыми характеристиками. Примеры целевых характеристик включают, в частности: повышенную аффинность связывания с антигеном относительно известных антител, которые специфичны к антигену; повышенная нейтрализация антигенной активности относительно известных антител, которые специфичны к антигену, если эта активность известна; определенная конкурентная способность с известным антителом или лигандом антигена в конкретном молярном соотношении; способность к иммунопреципитации комплекса; способность связываться с конкретным эпитопом; линейный эпитоп, например, пептидная последовательность, идентифицированная с использованием сканирования пептидного связывания, как описано в настоящей заявке, например с использованием пептидов, скринированных в линейной и/или ограниченной конформации; конформационный эпитоп, образованный разомкнутыми остатками; способность модулировать новую биологическую активность человеческого белка ENO1 или молекулы, располагающейся ниже по сигнальному пути. Такие методы также представлены в настоящей заявке.
Следующий аспект настоящего изобретения относится к агентам направленного связывания (т.е. к антителу), включающим такие агенты, для которых аминокислотные последовательности связываются с пептидом эпитопа, содержащим аминокислотную последовательность, которая имеет, по меньшей мере, примерно 60, 70, 80, 85, 90, или примерно 92% идентичности аминокислотной последовательности, представленной в последовательностях 9 или 10 на белке ENO1, и которые могут использоваться для лечения заболевания или расстройства, связанного с человеческим белком ENO1. Заболевание или расстройство, связанное с человеческим белком ENO1, может представлять собой любое патологическое состояние, возникающее в результате аберрантной активации или экспрессии человеческого белка ENO1. В одном примере, заболевание, связанное с человеческим белком ENO1, представляет собой неопластическое заболевание, такое как немелкоклеточный рак легкого, гепатоклеточная (печени) карцинома, рак желудочно-кишечного тракта (желудка), рак молочной железы, аденокарцинома протока поджелудочной железы.
Антиген-связывающий сайт антитела, состоящий из домена VH и домена VL, как правило, образуется с помощью шести петель полипептида: три из вариабельного домена легкой цепи (VL) и три из вариабельного домена тяжелой цепи (VH). Анализ антител известной атомной структуры выявил связь между последовательностью и трехмерной структурой антитело-комбинирующих сайтов. Эти связи предполагают, что за исключением третьей области (петли) в доменах VH петли связывающих сайтов имеют одну из небольшого числа основных конформаций цепей: канонические структуры. Каноническая структура, образованная в конкретной петле, как было представлено, определяется ее размером и присутствием определенных остатков в ключевых сайтах в обеих областях, в области петли и в каркасной области.
Из области техники, как правило, доступны методы, которые требуются для создания замен внутри аминокислотных последовательностей CDR, доменов VH или VL антитела и/или агентов связывания. Вариантные последовательности могут быть получены путем замен, по которым может или не может быть сделан прогноз о том, могут ли они иметь минимальный или полезный эффект в отношении активности, и могут тестироваться на предмет способности связывания и/или нейтрализации и/или любого другого свойства. Варианты аминокислотной последовательности вариабельного домена любого из доменов, VH и VL, чьи последовательности конкретно раскрыты в настоящей заявке, могут применяться согласно настоящему изобретению, как обсуждалось.
Используемый в настоящей заявке термин «полипептидный фрагмент» относится к полипептиду, который содержит N-концевую и/или C-концевую делецию, но где оставшаяся аминокислотная последовательность идентична соответствующим положениям в природной последовательности, выведенной, например, из последовательности полноразмерной кДНК. Длина фрагментов, как правило, составляет, по меньшей мере, примерно 5, 6, 8 или 10 аминокислот, предпочтительно, по меньшей мере, примерно 14 аминокислот, более предпочтительно, по меньшей мере, примерно 20 аминокислот, обычно, по меньшей мере, примерно 50 аминокислот и еще более предпочтительно, по меньшей мере, примерно 70 аминокислот. Используемый в настоящей заявке термин «аналог» относится к полипептидам, которые состоят из сегмента, составляющего по меньшей мере, примерно 25 аминокислот, которые обладают существенной идентичностью с участком предполагаемой аминокислотной последовательности и которые обладают, по меньшей мере, одним из следующих свойств: (1) специфичное связывание с человеческим белком ENO1 при подходящих условиях связывания, (2) способность блокировать соответствующее связывание лиганда/ белка ENO1, или (3) способность ингибировать активность белка ENO1. Как правило, полипептидные аналоги содержат консервативную аминокислотную замену (или вставку или делецию) по отношению к природной последовательности. Длина аналогов, как правило, составляет по меньшей мере 20 аминокислот, предпочтительно, по меньшей мере, 50 аминокислот, и часто могут соответствовать по длине полноразмерному природному полипептиду.
Используемый в настоящей заявке «агент направленного связывания» представляет собой агент, например, антитело или его связывающий фрагмент, который предпочтительно связывается с сайтом-мишенью. В одном воплощении, агент направленного связывания специфичен только для одного сайта-мишени. В других воплощениях, агент направленного связывания специфичен для более чем одного сайта-мишени. В одном воплощении, агент направленного связывания может представлять собой моноклональное антитело, и сайт-мишень может представлять собой эпитоп. Как описано ниже, агент направленного связывания может содержать, по меньшей мере, один антиген-связывающий домен антитела, где указанный домен сшит или содержится внутри гетерологичного белка.
Антитело может быть олигоклональным, поликлональным антителом, химерным антителом, антителом с CDR-трансплантатом, полиспецифичным антителом, биспецифичным антителом, каталитическим антителом, химерным антителом, гуманизированным антителом, полностью человеческим антителом, анти-идиотипическим антителом и антителом, которое может быть меченным в растворимой или связанной форме, а также к их фрагментам или производным или индивидуально, или в комбинации с другими аминокислотными последовательностями, полученными известными методами. Антитело может быть из любых видов. Термин антитело также включает связывающие фрагменты антител согласно изобретению; примерные фрагменты включают Fv, Fab, Fabʹ, одноцепочечное антитело (svFC), димерную вариабельную область (диатело) вариабельную область, стабилизированную дисульфилными связями (dsFv). «Связывающие фрагменты» антитела могут быть получены с помощью технологий рекомбинантной ДНК или с помощью ферментативного или химического расщепления интактных антител. Связывающие фрагменты включают Fab, Fabʹ, F(abʹ)2, Fv, и одноцепочечные антитела.
Гидролиз антител ферментом папаином приводит к получению двух идентичных антиген-связывающих фрагментов, известных как «Fab»-фрагменты, а также «Fc»-фрагмент, обладающий антиген-связывающей активностью, но имеющий способность кристаллизоваться. Гидролиз антител ферментом пепсин приводит к получению F(abʹ)2-фрагмента, в котором две руки молекулы антитела остаются связанными и содержат два антиген-связывающих сайта. F(abʹ)2-фрагмент обладает способностью сшивать антиген. Используемый в настоящей заявке термин «Fv» относится к минимальному фрагменту антитела, который сохраняет оба сайта - распознавания антигена и связывания антигена. Используемый в настоящей заявке термин «Fab» относится к фрагменту антитела, который содержит константный домен легкой цепи и CH1 домен тяжелой цепи. Термин «mAb» относится к моноклональному антителу.
Термин «эпитоп» включает любую белковую детерминанту, способную специфично связываться с иммуноглобулином или с T-клеточным рецептором. Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или боковые цепи сахаров и могут, но не всегда, иметь специфические характеристики трехмерной структуры, а также специфические характеристики заряда. Говорят, что антитело специфично связывается с антигеном, когда константа диссоциации составляет≤1 мкM, предпочтительно≤100 нM, и наиболее предпочтительно≤10 нM.
«Активный» или «активность» касательно полипептида ENO1 относится к области полипептида ENO1, которая обладает биологической или иммунологической активностью нативного полипептида ENO1. Используемый в настоящей заявке термин «биологический» относится к биологической функции, которая происходит из активности нативного полипептида ENO1. Предпочтительная биологическая активность ENO1 включает, например, ENO1-индуцированную активность плазминогена.
Используемый в настоящей заявке термин «млекопитающее» относится к любому животному, которое считается млекопитающим. Предпочтительно, млекопитающее представляет собой человека. Термин «объект» включает человека и ветеринарные объекты.
Используемый в настоящей заявке термин «фармацевтический агент или лекарственное средство» относится к химическому соединению или к композиции, способной индуцировать целевой терапевтический эффект при правильном введении пациенту. Другие химические термины используются в настоящей заявке согласно обычному употреблению в данной области, как описано в качестве примера в The McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)).
Используемый в настоящей заявке термин «по существу чистый» означает целевой компонент, который присутствует с преобладанием (т.е., на молярной основе более представлен, чем любой другой индивидуальный компонент в композиции), и предпочтительно, по существу чистая фракция представляет собой композицию, где целевой компонент содержит по меньшей мере примерно 50 процентов (на молярной основе) от всех присутствующих макромолекулярных веществ. Как правило, по существу чистая композиция будет содержать более чем примерно 80 процентов всех макромолекулярных компонентов, присутствующих в композиции, более предпочтительно, более чем примерно 85%, 90%, 95%, и 99%. Наиболее предпочтительно, целевой компонент очищен до существенной гомогенности (загрязняющий компонент не может быть детектирован в композиции с помощью стандартных методов детектирования), где композиция состоит по существу из единственного макромолекулярного компонента.
Используемый в настоящей заявке термин «мониторинг» относится к способу детектирования и/или наблюдения за развитием злокачественного новообразования путем определения представленности белка ENO1 в опухолевых клетках.
Методы определения представленности ENO1 включают в частности измерение связывания белков ENO1 и ENO1-специфичных антител, вестерн-блот анализ, проточную цитометрию, иммуногистохимию (IHC), ОТ-ПЦР, и/или микроэррей-анализ.
ПРИМЕРЫ
Практическое осуществление настоящего изобретения будет применять технологии, включающие стандартные методы клеточной биологии, клеточной культуры, технологии антител и генетической инженерии, которые известны специалисту в данной области. Такие методы в полной мере описаны в научной литературе.
Следующие примеры иллюстрируют разработку и применение ENO1-специфичных антител для подавления опухолевого роста путем индукции иммунного ответа против ENO1.
Пример 1
Локализация ENO1 на клеточной поверхности злокачественных клеток
Для анализа клеточной локализации ENO1 на злокачественных клетках осуществляли проточную цитометрию для подтверждения локализации на клеточной поверхности ENO1. Опухолевые клетки получали из плевральных выпотов 17 пациентов с раком легкого. Среди них опухолевые клетки, полученные от 2 пациентов со злокачественным новообразованием, опухолевые клетки выпота NHRI-L89 и CA926 являются репрезентативными случаями, представленным в данном документе. Клетки NHBE и SAEC представляют собой нормальные человеческие первичные клетки легкого. Интактные цельные клетки окрашивали с использованием антисыворотки к ENO1 или без нее и анализировали поверхностное распределение ENO1 с использованием антитела, специфичного к белку ENO1 в анализе проточной цитометрии (как представлено на ФИГ. 1A). Клеточную локализацию ENO1 клеток клеточной линии NHRI-L89 рака легкого дополнительно подтверждали с помощью иммуногистохимии (ФИГ. 1B). Вкратце, 1×104 клеток NHRI-L89 растили в среде DMEM, содержащей 10% FCS на предметном стекле, покрытом фибронектином. После культивирования в течение ночи, препарат промывали с помощью PBS и блокировали с использованием козьей сыворотки в течение 1 часа. Интактные цельные клетки NHRI-L89 окрашивали с использованием контроля (CTL) или антитела к ENO1 для поверхностного ENO1, с последующим окрашиванием с помощью PI для ядерного детектирования. Окрашивания визуализировали под конфокальным микроскопом (400х; ФИГ. 1B).
Результаты данного исследования представлены на ФИГ. 1A (проточная цитометрия) и на 1B (иммуноокрашивание). Инкубирование NHRI-L89 и CA926 с антителом к ENO1 сдвигает гистограмму вправо по сравнению с инкубированием клеток с контрольным антителом. Нормальные первичные клетки легкого человека NHBE и SAEC демонстрируют значительные сдвиги (ФИГ. 1A). Этот результат выявляет, что белок ENO1 экспрессируется на клетках рака легкого, но не экспрессируется на нормальных первичных клетках (ФИГ. 1A). Когда дополнительно анализировали клеточную локализацию ENO1 с помощью иммуногистохимии, интактные клетки NHRI-L89, окрашенные с помощью антитела к ENO1 демонстрирует сигналы на клеточной поверхности (ФИГ.1B). Этот результат предполагает, что ENO1 экспрессируется на поверхности клеток рака легкого и клеток некоторых злокачественных новообразований и, таким образом, может быть потенциальной мишенью для иммунотерапии.
Пример 2
Антитело к ENO1 ингибирует инвазивную способность клеток рака легкого человека
Для исследования эффектов антитела к ENO1 в отношении клеточной инвазии, оценивали инвазивную активность клеток CL1-5 с использованием анализа Transwell путем добавления антитела к ENO1 к клеткам CL1-5, выращенным на микропористых фильтрах (Becton Dickinson, Franklin Lakes, N.J.), покрытых внеклеточным матриксом (матригель) (Becton Dickinson). После смешивания с разведенным 1:20, 1:100 и 1:500 поликлональным антителом к ENO1, соответственно, высевали 2×104 клеток CL1-5 в верхнюю камеру со средой, содержащей 2% FBS, двухкамерной аналитической системы и инкубировали в течение 24 часов в присутствии среды 10% FBS в нижней камере. Мышиное поликлональное антитело к GST использовали в качестве группы отрицательного контроля. Две камеры разделяли с помощью микропористого фильтра (размер пор 12 мкм), покрытого матригелем. После периода инкубирования фильтры, покрытые матригелем, окрашивали и подсчитывали под микроскопом количество клеток, проникших в фильтр, покрытый матригелем. Исследование повторяли три раза. Данные представлены в виде среднего±SD. T-тест использовали для сравнения активности между группами. P значения <0,05 рассматривали, как статистически значимые.
Результаты представлены на ФИГ. 2. Не было значительных различий в количестве клеток между не обработанной (160±8; N=3) группой и группой, обработанной поликлональным антителом к GST(168±12; N=3). Однако когда клетки CL1-5 вводили с различными разведениями поликлонального антитела к ENO1, количество клеток CL1-5 уменьшалось в интервале от 52±12(N=3) для разведения 1:20 156±30 (N=3) для разведения 1:500; уменьшение пропорционально концентрациям поликлонального антитела к ENO1. Это статистическое различие между группами разведения 1:20 поликлонального антитела ENO1 и разведения 1:20 поликлонального антитела к GST (P<0,05). Эти результаты предполагают, что введение антитела ENO1 в опухолевые клетки CL1-5 ингибирует активность клеточной инвазии in vitro.
Пример 3
Поликлональное антитело к ENO1 ослабляет способность тканевой инвазии клеток CL1-5F4 в легкое.
Для исследования эффекта введения антитела ENO1 в отношении метастазирования клеток рака легкого и оценки потенциала антитела к ENO1 в противораковой терапии, высокоинвазивные клетки CL1-5F4 с репортерным геном люциферазы (1×106 клеток/на мышь) внутривенно инъецировали мышам NOD-SCID вместе с антителом к ENO1 и контрольной антисывороткой (CTL) дважды в неделю. Метастатическое колониеобразование в легком отслеживали через 12 и 19 дней после инъекции опухоли с использованием системы IVIS. Мышей, инъецированных клетками CL1-5F4 с нокдауном гена ENO1, использовали в качестве отрицательного контроля. После того, как мышей умерщвляли в дни 12 и 19, метастатические колонии и размер опухолей визуализировали и подсчитывали.
Результаты представлены на ФИГ.3. Когда мышам, несущим клетки CL1-5F4 с репортерным геном люциферазы, вводили контрольное антитело, то легкое мыши начинало демонстрировать флуоресцентные сигналы на 12 день. С другой стороны, сигналы не детектировали у мышей, обработанных поликлональным антителом к ENO1, или у мышей, несущих клетки CL1-5F4 и с нокдауном гена ENO1. На 19 день сигнал мышей, обработанных контрольным антителом, становился сильнее, указывая на присутствие большей клеточной инвазии или опухолевого роста. В тот же день сигналы еще не детектировали у мышей, обработанных поликлональным антителом к ENO1 или у мышей, несущих клетки CL1-5F4 с нокдауном гена ENO1. Эти результаты выявляют, что метастазирование клеток CL1-5F4 нарушалось с помощью антитела к ENO1 или при нокдауне гена ENO1 (ФИГ.3; верхняя панель). После дней 12 и 19, легкие каждой группы обработки удаляли и определяли опухолевые узлы. Результаты представлены в нижней панели на ФИГ.3. Было мало опухолевых узлов, появившихся у мышей в группе обработки контрольным антителом, наблюдался четкий размер опухоли. Однако мыши, обработанные поликлональным антителом ENO1, или мыши, несущие нокдаун CL1-5F4ENO1, демонстрировали нечеткие размеры опухоли, указывая на задержку опухолевого метастазирования в этих группах мышей. Результаты данного исследования предполагают, что введение антитела к ENO1 обладало анти-метастатическими эффектами или анти-пролиферирующими эффектами в отношении опухолевых клеток in vivo, и антитело к ENO1 обладало потенциалом для применения в противоопухолевой терапии.
Пример 4
Получение моноклонального антитела к ENO1 человека.
Из примеров 2 и 3 ясно, что антитело к ENO1 может ослаблять инвазию и метастазирование опухолевых клеток. Таким образом, антитело к ENO1 потенциально может быть разработано в качестве терапевтического антитела. Для получения моноклонального антитела против человеческого ENO1, мышей BALB/c прививали с использованием очищенного рекомбинантного человеческого антигена ENO1 (50 мкг/на мышь), с последующим стимулированием вызванного ENO1 гуморального ответа с использованием эмульгированного CpG-адюванта. Спленоциты сбирали, сливали с клетками Fo и затем разводили согласно стандартному протоколу. Гибридомные клетки, секретирующие моноклональные антитела, которые распознают антиген ENO1, отбирали с помощью тИФА на основе антигена. Отобранные клоны затем подтверждали с помощью активности связывания с ENO1 клеточной поверхности на клетках аденокарциномы легкого CL1-5F4 с использованием окрашивания цельных интактных клеток в анализе проточной цитометрии, как описано в Примере 1.
Результаты представлены на ФИГ.4. Все из выделенных клонов антител распознают ENO1 на клеточной поверхности CL1-5F4, что наблюдали с помощью сдвига гистограмм вправо по сравнению с контрольным антителом.
Пример 5
тИФА связывания ENO1 с использованием 5 выделенных клонов антител
Для исследования аффинности связывания ENO1 с помощью 5 клонов антител, выделенных выше, индивидуальные гибридомы выращивали в среде RPMI, содержащей 10% FCS. Через одну неделю после культивирования 1×106 клеток собирали, промывали с помощью PBS, ресуспендировали в 200мкл среды RPMI и инъецировали мышам SCID с помощью IP-инъекции (внутрибрюшинно). Через 3 недели асциты мышей собирали и разводили до 15 мл. Антитело дополнительно очищали с помощью преципитации 40% сульфатом аммония и хроматографии на колонке с протеином A (набор «Montage antibody purification kit» Millipore). Очищенное антитело концентрировали с использованием устройства центрифуги с фильтром Amicon Ultra-15, следуя протоколам, представленным производителем (Millpore). Степень чистоты антител анализировали с помощью 12% SDS PAGE. Человеческий белок ENO1 (400нг) наносили в виде покрытия на 96-луночный тИФА-планшет и планшет дважды промывали с помощью PBS. Серии разведений от 1×10-11 до 1×10-7 M добавляли к планшету, и планшет инкубировали при 37°C в течение 1 часа. Козий IgG к мышиному белку, конъюгированный с HPRT, добавляли через 1 час инкубирования и затем добавляли TMB. OD405 считывали для расчета активности. Каждое исследование повторяли три раза, и данные представляли в виде среднего±SD. OD-значения и концентрации антител использовали для создания графика множественного рассеяния с использованием SigmaplotTM. Значения Kd клонов прогнозировали с помощью 4-параметрического логистического анализа.
Результаты данного эксперимента представлены на ФИГ.5. Все из гибридом антител имеют продуктивность о 20,4 мг до 4,6 мг на мышь. Значения Kd антител находятся в интервалеот 1,77×10-10 ± 0,12 до 1,6×10-5±0,2 M (N=3). Эти результаты предполагают, что эти 5 клонов антител распознают человеческий белок ENO1, и что клон 10 обладает лучшей аффинностью с Kd примерно 1,77×10-10±0,12M (N=3).
Пример 6
Антагонистические активности 5 выделенных клонов антител по отношению к рецептору плазминогена
Для исследования антагонистической активности по отношению к ENO1, проявляемой выделенным антителом к человеческому ENO1, клетки U937 клеточной линии лимфомы человека растили в RPMI, содержащей 10% FCS. Клетки обрабатывали с использованием 10 микрограмм/мл LPS в течение 6 часов для индукции экспрессии белка ENO1 на клеточной поверхности. 1,5×106клеток/мл в PBS пре-инкубировали вместе с 1микрограмм/мл человеческого Lys-плазминогена и 10 микрограмм/мл других клонов антител к ENO1 в течение одного часа, соответственно. Образцы промывали дважды с помощью PBS и затем добавляли 3нM тканеспецифического активатора плазминогена и 0,5мM хромогенного субстрата S-2251. Через один час инкубации при 37°C считывали OD405 для расчета активностей. Каждое исследование повторяли три раза и анализировали антагонистические активности. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
Результаты данного исследования представлены на ФИГ.6. Каждый из 5 клонов антител обладает антагонистической активностью по отношению к рецептора плазминогена с ингибированием в интервале от 52% до 100% LSP-индуцированной ENO1-специфичной активности. Эти результаты предполагают, что антагонистическая активность рецептора плазминогена пропорциональна значению Kd каждого клона, и что клон 10 обладает лучшей ингибирующей активностью, близкой к 100% ингибирования ENO1-специфичной активности, индуцированной LPS.
Пример 7
Моноклональное антитело к ENO1 ингибирует инвазивную способность клеток рака легкого человека
Ингибирование инвазивной способности клеток CL1-5 с помощью каждого клона антитела оценивали с использованием анализа Transwell с использованием микропористых фильтров (Becton Dickinson, Franklin Lakes, N.J.), покрытых внеклеточным матриксом (матригель) (Becton Dickinson). После смешивания с 10 микрограмм/мл каждого из 5 клонов антител, 2±104 клеток высевали в верхнюю камеру двухкамерной аналитической системы и инкубировали в течение 24 часов со средой, содержащей 10% FBS в нижней камере. IgG к мышиному белку и 5-аза-2-дезоксицитидин (5ADC) использовали в качестве отрицательного контроля и положительного контроля, соответственно. Две камеры разделяли с помощью микропористого фильтра (размер пор 12 мкм), покрытого матригелем. После периода инкубирования фильтры, покрытые матригелем, окрашивали и подсчитывали под микроскопом количество клеток, проникших в фильтр, покрытый матригелем. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. Значения P<0,05 рассматривали как статистически значимые.
Результаты представлены на ФИГ.7. Эти результаты демонстрируют, что все клоны антител к человеческому ENO1 обладают ингибирующими активностями против инвазивной способности CL1-5, и что клон 10 обладает лучшей ингибирующей активностью, примерно 65,5±0,3% (N=3) от группы контрольного IgG.
На основе вместе взятых результатов из Примеров 5, 6 и 7 клон 10 антитела к человеческому ENO1 выбрали для дальнейшей разработки и обозначили его в данном описании как EN10 mAb.
Пример 8
EN10 mAb ингибирует инвазивную способность клеток рака легкого человека и клеток лимфомы
Ингибирование инвазивной способности клеток CL1-5 с помощью EN10 mAb оценивали с использованием анализа Transwell с использованием микропористых фильтров (Becton Dickinson, Franklin Lakes, N.J.), покрытых внеклеточным матриксом (матригель) (Becton Dickinson). После смешивания с 10, 50 и 100 микрограмм/мл EN10 mAb, соответственно, 2±104 клеток высевали в верхнюю камеру со средой, содержащей 2% FBS в двухкамерной аналитической системе и инкубировали в течение 24 часов в присутствии среды 10% FBS в нижней камере. IgG к мышиному белку использовали в качестве отрицательного контроля. Две камеры разделяли с помощью микропористого фильтра (размер пор 12 мкм), покрытого матригелем. После периода инкубирования фильтры, покрытые матригелем, окрашивали и подсчитывали под микроскопом количество клеток, проникших в фильтр, покрытый матригелем. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активностей между группами. P-значения <0,05 рассматривали, как статистически значимые. Результаты представлены на ФИГ.8A.
Аналогично, ингибирование инвазивной способности клеток U937 с помощью EN10 mAb оценивали с использованием анализа Transwell с использованием микропористых фильтров (Becton Dickinson, Franklin Lakes, N.J.), покрытых внеклеточным матриксом (матригель) (Becton Dickinson). Клетки U937 клеточной линии лимфомы человека растили в RPMI, содержащей 10% FCS. Клетки обрабатывали с помощью 10 микрограмм/мл LPS в течение 6 часов для индукции экспрессии белка ENO1 на клеточной поверхности. После смешивания с 10, 50 и 100 микрограмм/мл EN10 mAb, соответственно, 2±104 клеток высевали в верхнюю камеру двухкамерной аналитической системы и инкубировали в течение 24 часов со средой, содержащей 10% FBS и 10нM MCP-1 в нижней камере. IgG к мышиному белку использовали в качестве отрицательного контроля. Две камеры разделяли с помощью микропористого фильтра (размер пор 8 мкм), покрытого матригелем. После периода инкубирования клетки в нижней камере считали с помощью гемоцитометра под микроскопом. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
Результаты представлены на ФИГ. 8A и 8B. Инвазивная активность CL1-5 ингибировалась до степени в интервале от 31±2 до 77±1% (N=3) контрольного IgG, когда клетки вводили вместе с EN10 mAb от 10 микрограмм/мл до 100 микрограмм/мл (ФИГ. 8A). Эти результаты аналогичны результатам Примера 4, и ингибирование пропорционально концентрациям EN10 mAb с EC50, которая по оценкам составила примерно 30 микрограмм/мл.
Для оценки того, применимо ли ингибирование инвазии к другим опухолевым клеткам, клетки U937 обрабатывали с использованием EN10 mAb в интервале от 5 микрограмм/мл до 50 микрограмм/мл аналогичным образом. Инвазивная активность U937 ингибировалась до степени в интервале от 91,2±2 до 49,1±1% (N=3) контрольного IgG (ФИГ. 8B). Эти результаты выявляют, что инвазивная активность клеток CL1-5 и U937 ингибируется EN10 mAb в дозозависимой манере, в основном путем ингибирования активности рецептора плазминогена ENO1.
Пример 9
EN10 mAb распознает LSP-индуцированный поверхностный ENO1 в клетках лимфомы
Клетки U937 клеточной линии лимфомы человека растили в RPMI, содержащей 10% FCS. Клетки обрабатывали с помощью 1 микрограмм/мл LPS в течение 6 часов для индукции экспрессии белка ENO1 на клеточной поверхности. Для анализа проточной цитометрии интактные цельные клетки окрашивали или нет с помощью EN10 mAb (1:300 разведение), визуализировали с использованием FITC-конъюгированного козьего антитела к IgG мыши (Jackson Lab) и анализировали с помощью проточного цитометра FACScan (Becton Dickinson). Экспрессию ENO1 измеряли с помощью полученной интенсивности флуоресценции.
Результаты из этих экспериментов представлены на ФИГ. 9. Инкубирование U937 с LPS и обработка с помощью EN10 mAb сдвигает гистограмму вправо по сравнению с инкубированием клеток без обработки LPS, нос обработкой EN10 mAb, выявляя, что клетки U937 экспрессируют ENO1 на своей поверхности. Эти данные поддерживают, что EN10 mAb распознает LSP-индуцированный поверхностный ENO1 на клетках лимфомы.
Пример 10
EN10 mAb ингибирует диссоциацию клеток CL-5 из коллагена и фибронектина
Для оценки пути сигнальной трансдукции между рецептором плазминогена ENO1-плазмином и внеклеточными субстратами, наносили на ночь на необработанные тИФА-планшеты покрытие в виде 1 мг/мл желатина, 100 мкг/мл фибриногена, 10 мкг/мл коллагена и 10 мкг/мл фибронектина, соответственно. Клетки CL1-5 (4×104 клеток) высевали на планшет и добавляли 50 мкг/мл EN10 mAb к 200мкл DMEM, содержащей 10% FCS. Клетки инкубировали при 37°С в течение 24 часов и затем промывали дважды с помощью PBS. 10% WST добавляли и реакционные смеси инкубировали при 37°C в течение 4 часов. Относительное количество клеток оценивали с помощью считывания OD450. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
Результаты из этих экспериментов представлены на ФИГ. 10. Эти данные выявляют, что значения OD450 на планшетах, покрытых с помощью фибронектина и коллагена составили, 2,45±0,37(N=3) и 1,83±0,44 (N=3). Значения оказались гораздо выше, чем их аналоги для планшетов с желатином и фибронектином, которые не имели значительных отличий от фона. Существует значительное различие между группой, обработанной EN10 mAb, и необработанной группой. Эти результаты предполагают, что клетки CL1-5 благоприятствуют связыванию с фибронектином и с коллагеном, и EN10 mAb не вовлечено в путь клеточной ассоциации, когда клетки инкубировали в среде без протеаз нижних сигнальных путей, например, плазминогена и tPA. Данные на ФИГ. 10A предполагают, что активность рецептора плазминогена ENO1 не вовлечена в путь клеточной ассоциации во внеклеточном маткриксе.
Мы дополнительно тестировали, принимает ли ENO1 участие в пути клеточной диссоциации во внеклеточном матриксе. На необработанный тИФА-планшет наносили в течение ночи покрытие в виде 1 микрограмм/мл фибронектина и 10 микрограмм/мл коллагена. Клетки CL1-5 (4×104 клеток) высевали на планшет, и добавляли 0, 6,25, 12,5, 25, и 50 микрограмм/мл EN10 mAb, соответственно, к 200 мкл DMEM, содержащей 10% FCS. Кроме того, добавляли 10 микрограмм/мл Glu-плазминогена и 2нM tPA. Клетки инкубировали при 37°C в течение 24 часов и затем промывали дважды с помощью PBS. 10% WST добавляли и реакционные смеси инкубировали при 37°C в течение 4 часов. Относительное количество клеток в планшете оценивали с помощью считывания OD450. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
Результаты из этих экспериментов представлены на ФИГ. 10B и ФИГ. 10C. Данные указывают, что количество клеток прямо пропорционально концентрациям обработок EN10 mAb в обоих внеклеточных матриксах, когда среда содержит нижние по сигнальным путям протеазы рецептора ENO1, плазминоген и tPA. Существует значительное различие между группой обработки 50 микрограмм EN10 mAb и группой обработки контрольным IgG (P<0,05) в обоих исследованиях внеклеточного матрикса. Эти результаты предполагают, что ENO1 вовлечен в путь диссоциации клеток CL1-5 от внеклеточного матрикса, в основном за счет усиления протеазной активности плазминогена и tPA. EN10 mAb, функционируя в качестве антагониста ENO1, блокирует рецепторную активность ENO1, приводя к ингибированию активации плазмина и tPA, и, таким образом, ингибирует активность диссоциации клеток CL1-5 из внеклеточных матриксов и инвазию.
Пример 11
Получение генов, кодирующих моноклональные антитела
Клонирование гена, кодирующего антитело EN10 mAb, осуществляли согласно процедурам, оипсанным ниже.
(1) клонирование кДНК генов антитела и получение
Гибридому культивировали в среде RPMI (производитель Gibco), содержащей 10% FCS. После того, как количество клеток достигало примерно 10×106/мл, клетки собирали с помощью центрифугирования и затем добавляли TRIzol® (производитель Invitrogen) к экстракту суммарной РНК согласно инструкции производителя. Клонирование вариабельной области кДНК антитела осуществляли с использованием набора мышиных праймеров для Ig (производство Novagen) согласно прилагаемой инструкции производителя.
Синтез 1-ой цепи кДНК осуществляли согласно руководству SuperScript® III "First-Strand Synthesis System" (производство Invitrogen).
1-ую цепь кДНК получали с использованием 5 мкг суммарной РНК в качестве матрицы. Пять микрограммов суммарной гибридомной РНК, 1 мкл 50нг/мкл случайных праймеров и 1мкл 10мM dNTP смешивали и добавляли воду, обработанную DEPC до объема 10мкл в 200мкл ПЦР-пробирке. Реакционную смесь инкубировали при 65°C в течение 5 мин, и затем помещали на лед в течение по меньшей мере 1 минуты. Десять (10)мкл смеси синтеза кДНК, содержащей 2мкл 10× RT буфера, 4 мкл 25мM MgCl2, 2мкл DTT, 1мкл 4 единицы RNaseOUT™, и 1мкл 200 единиц недолгим центрифугированием. Реакционную пробирку инкубировали в течение 10 мин при 25°C с последующим инкубированием в течение 50 мини при 50°C. Реакцию останавливали при 85°C в течение 5 мин и охлаждали на льду. Пробирку недолго центрифугировали, чтобы собрать реакционную смесь, и добавляли 1 мкл РНК-азы H и инкубировали в течение 20 мин при 37°C.
Амплификация с помощью ПЦР генов тяжелой цепи и генов легкой цепи
Готовили реакционный раствор, имеющий состав 5 мкл кДНК, 5 мкл 10× реакционного буфера, 1 мкл 10мM dNTP-микс, 1мкл 2,5 ед. Taqполимеразы и 1 мкл прямого праймера 1 и 1мкл обратного праймера 2, обеспеченных набором праймеров, в конечном объеме 50 мкл с дважды дистиллированной водой и подвергали ПЦР.
Для амплификации легкой цепи и тяжелой цепи антитела использовали цикл 94°C в течение 10 минут, затем цикл 94°C в течение 1 мин, 52°C в течение 1 мин, и 72°C в течение 1 мин повторяли 35 раз и и реакцию инкубировали при 72°C в течение еще 10 минут. Реакционный раствор подвергали электрофорезу на 2%-агарозном геле для анализа продуктов реакции. Продукты с корректной молекулярной массой, примерно 463 п.о. для тяжелой цепи и 451 п.о. для легкой цепи лигировали в вектор pCR 2.1-TOPO (производство Invitrogen) для субклонирования согласно прилагаемому руководству. Затем использовали праймеры M13прямой (5ʹ-GTAAACAAC GACGGCGAG--3ʹ(SEQ ID NO:11) и M13 обратный (5ʹ-CAG GAA ACA GCT ATG AC--3ʹ (SEQ ID NO:12) ) для определения нуклеотидной последовательности. На основе информации о последовательности последовательность антитела транслировали в белковые последовательности с помощью инструмента трансляции ExPASY-Translation Tool. Полученные последовательности EN10 mAb содержат аминокислотную последовательность тяжелой цепи, содержащую области, определяющие комплементарность (CDR), которые определяли с помощью метода, опубликованного у Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda Md. (1991), vols. 1-3.
На ФИГ. 11A изображена аминокислотная последовательность вариабельной области тяжелой цепи EN10 mAb (SEQ ID NO: 1). Каркасные области (FR1, FR2, FR3, и FR4) и CDR (HCDR1(SEQ ID NO: 3), HCDR2(SEQ ID NO: 4), и HCDR3(SEQ ID NO: 5) указаны.
На ФИГ. 11B изображены аминокислотные последовательности вариабельной области легкой цепи EN10 mAb (SEQ ID NO: 2). Каркасные области (FR1, FR2, FR3, и FR4) и CDR (LCDR1(SEQ ID NO: 3), LCDR2(SEQ ID NO: 4), и LCDR3(SEQ ID NO: 5) указаны.
Пример 12
Картирование эпитопов
Картирование эпитопов антитела
Для определения эпитопа EN10Mab на человеческом белке ENO1, конструировали два прямых праймера с нуклеотидные последовательностями 5ʹ-GGATCCGCAGCAAACTTCAGGGAAGCCATG-3ʹ (SEQ ID NO:13), и 5ʹ-GGATCCTCGAAGATCCCTTTGACCAGGATG-3ʹ (SEQ ID NO:14),и обратный праймер (5ʹ-TCAGGCTGAAAATCTCTCATCCGC-3ʹ (SEQ ID NO:15). Экспрессирующую плазмиду E.coli pTRC-HIS ENO1, содержащую кДНК человеческого гена ENO1, использовали в качестве матрицы для амплификации делеционных мутантов ENO1. Праймеры с SEQ NO:13 и SEQ NO:14 использовали в качестве прямых праймеров, а с SEQ ID NO:15 в качестве обратногопраймера для амплификации мутантов Δ1-189(Фиг. 12A) и Δ1-297 (Фиг. 12A), соответственно. Другой набор праймеров, имеющих последовательности 5ʹ-GGATCCTATCTATTCTCAAGATCCATGCC-3ʹʹ (SEQ ID NO:16) и 5ʹ-CTCGAGGTCATGGTGTCTCATCGTTCGCTCGAG-3ʹ(SEQ ID NO:17), использовали для амплификации делеционного мутанта Δ297-434. Для амплификации каждого мутанта готовили реакционный раствор, имеющий состав 1мкл 1:1000 разведенной ДНК-матрицы примерно 0,1нг, 5 мкл 10х реакционного буфера, 1мкл 10мM dNTP-микса, 1мкл 2,5 ед. Taq-полимеразы, 1мкл прямого праймера и 1мкл обратного праймера, в конечном объеме 50 мкл с дважды дистиллированной водой и подвергали ПЦР. Использовали цикл 94°C в течение 10 мин., затем цикл 94°C в течение 1 мин, 52°C в течение 1 мин и 72°C в течение 1 мин, повторяли 35 раз, и реакцию инкубировали при 72°C в течение еще 10 минут. Данный реакционный раствор подвергали электрофорезу на 2%-агарозном геле для анализа продуктов реакции. Реакционные продукты с корректными молекулярными массами лигировали в вектор pCR 2.1-TOPO (производство Invitrogen) для субклонирования согласно прилагаемому руководству. Прайемры M13 прямой (5ʹ-GTAAACAACGACGGCGAG--3ʹ (SEQ ID NO: 11) и M13 обратный (5ʹ- CAG GAAACA GCT ATG AC--3ʹ (SEQ ID NO: 12)) затем использовали для определения нуклеотидной последовательности. Каждый мутантный клон с корректной последовательностью гидролизовали ферментами рестрикции BamHI и XhoI, и продукты гидролиза подвергали электрофорезу на 2%-агарозном геле. Фрагмент вставки каждого мутанта вырезали из агарозного геля и очищали с использованием набора «Gene Clean Kit» согласно прилагаемому руководству, которое обеспечивает производитель(BIO101). ДНК-фрагмент BamHI и XhoI каждого мутанта лигировали по сайтам BamHI и XhoI экспрессирующего вектора E.coli pTRC His A (Invitrogen). Полученную плазмиду трансформирвали в E.coliBL21 Rosseta. Мутантный белок ENO1 экспрессировали в E.coli с помощью индукции IPTG и очищали согласно прилагаемому руководству, предложенному производителем (Qiagen), после чего осадок бактерий подвергали воздействию ультразвука. Степень чистоты каждого мутанта анализировали с помощью 12% SDS PAGE. Для определения активности связывания каждого мутантного белка, 400нг человеческого белка наносили в виде покрытия на 96-луночный тИФА-планшет, и планшет промывали с помощью PBS. 10 мкг EN10 mAb добавляли к планшету и планшет инкубировали при 37°C в течение 1 часа. После промывки комплекса связывания дважды с помощью PBS добавляли козьи IgG к мышиному белку, конъюгированные с HPRT. Через 1 час после инкубирования добавляли TMB. Аффинность связывания определяли путем считывания OD 405. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
Результаты представлены на ФИГ. 12A. ENO1 мутанты Δ1-189и Δ1-297 имеют значения OD405, составляющие примерно 1,43±0,18 и 1,56±0,08 (N=3)(это 1,43 и 1,56 на ФИГ. 12A), которые составляют примерно 42% и 39% от значений дикого типа ENO1 (2,87±0,08) (N=3), соответственно. Однако когда детектировали аминокислотные остатки от 297до 434, то активность связывания данного мутантного ENO1 с EN10 mAb терялась по сравнению с BSA-фоном. эти результаты предполагают, что аминокислотные остатки от 297 до 434 требуются для связывания белка ENO1 с EN10 mAb и уменьшение активности связывания мутантов Δ1-182 и Δ1-297 может быть обусловлено нестабильностью или конформационными изменениями мутантных белков.
Для дальнейшего исследования эпитопа EN10 mAb в белке ENO1 конструировали 5 обратных праймеров, имеющих структуру 5ʹ-CTCGAGAGGGATCTTCGATAGACACCACTGGG -3ʹ (SEQ ID NO:18), 5ʹ-CTCGAGCTACCTGGATTCCTGCACTGGCTG-3ʹ (SEQ ID NO:19),5ʹ-CTCGAGACTTCTCGTTCACGGCCTTGGCGATC-3ʹ (SEQ ID NO:20), 5ʹ-CTCGAGACTTCTCGTTCACGGCCTTGGCGATCC-3ʹ (SEQ ID NO:21), 5ʹ-CTCGAGCAGTCTCCCCCGAACGATGAGACACC-3ʹ (SEQ ID NO:22), и 5ʹ- CTCGAG CACCAGTCTTGATCTGCCCAGTGCAC-3ʹ (SEQ ID NO:23). Экспрессирующую плазмиду E.coli pTRC-HIS ENO1, содержащую кДНК человеческого гена ENO1, использовали в качестве матрицы для амплификации делеционных мутантов ENO1. SEQ ID NO:16 использовали в качестве прямого праймера для амплификации делеционных мутантов 296-434, 316-434, 336-434, 376-434 и 396-434 с последовательностями праймеров SEQ ID NO:18, SEQ ID:19, SEQ ID:20, SEQ ID:21, SEQ ID:22, и SEQ ID:23, соответственно. Для амплификации каждого мутанта готовили реакционный раствор, имеющий состав 1 мкл разведенной 1:1000 матрицы ДНК примерно 0,1нг, 5 мкл 10× реакционного буфера, 1 мкл 10мM dNTP-микс, 1мкл 2,5 ед. Taq-полимеразы, 1 мкл прямого праймера и 1мкл обратного праймера, в конечном объеме 50 мкл с дважды дистиллированной водой и подвергали ПЦР. Использовали цикл 94°C в течение 10 минут. Затем цикл 94°C в течение 1 мин, 52°C в течение 1 мин, и 72°C в течение 1 мин повторяли 35 раз, и реакцию инкубировали при 72°C в течение еще 10 минут. Данный реакционный раствор подвергали электрофорезу на 2%-агарозном геле для анализа продуктов реакции. Реакционные продукты с корректными молекулярными массами лигировали в вектор pCR 2.1-TOPO (производство Invitrogen) для субклонирования согласно прилагаемому руководству. Праймеры M13 прямой (5ʹ-GTAAACAACGACGGCGAG--3ʹ (SEQ ID NO:11) и M13 обратный (5ʹ-CAGGAA ACAGCTATGAC--3ʹ (SEQ ID NO:12) затем использовали для определения нуклеотидной последовательности. Каждый мутационный клон с корректной последовательностью гидролизовали с использованием ферментов рестрикции BamHI и XhoI и продукт гидролиза подвергали электрофорезу на 2%-агарозном геле. ДНК-фрагмент вставки каждого мутанта выделяли из агарозного геля и очищали с использованием набора «Gene Clean Kit» согласно прилагаемому руководству, которое обеспечивает производитель(BIO101). ДНК-фрагмент BamHI и XhoI каждого мутанта лигировали по сайтам BamHI и XhoI экспрессирующего вектора E.coli pTRC His A (Invitrogen). Полученную плазмиду трансформирвали в E.coli BL21 Rosseta. Мутантный белок ENO1 экспрессировали в E.coli с помощью индукции IPTG и очищали согласно прилагаемому руководству, предложенному производителем (Qiagen), после чего осадок бактерий подвергали воздействию ультразвука. Чистоту каждого мутанта анализировали с помощью 12% SDS-PAGE. Для определения активности связывания каждого мутантного белка 400нг человеческого белка ENO1 или мутантного белка наносили в виде покрытия на 96-луночный тИФА-планшет, и планшет промывали с помощью PBS. 10 мкг EN10 mAb добавляли и инкубировали при 37°C в течение 1 часа. После промывки комплекса связывания дважды с помощью PBS добавляли козьи IgG к мышиному белку, конъюгированные с HPRT. Через 1 час инкубации добавляли TMB. Аффинность связывания определяли путем считывания OD 405. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. T-тест использовали для сравнения активности между группами. P-значения <0,05 рассматривали, как статистически значимые.
12% SDS PAGE каждого мутанта и белка дикого типа представлен на ФИГ.12B. Молекулярная масса каждого мутанта увеличилась от 296-343 до дикого типа. Данные результат предполагает, что мы можем получать цельный белок из каждого мутанта, даже если можно наблюдать некоторую деградацию мутантов 336-434 и 376-434. Как представлено на ФИГ.12C, нет значительных различий между аффинностью связывания EN10 mAb белка ENO1 дикого типа и делеционных мутантов 336-434, 376-434 и 369-343. Однако, когда делетируют аминокислотные остатки от 296 до 316 и от 317 до 336, активность связывания EN10 mAb этих двух мутантов ENO1 теряется по сравнению с уровнем клеточного лизата E.coli. Эти результаты предполагает, что аминокислотные остатки от 296 до 336(FDQDDWGAWQKFTASAGIQVVGDDLTVTNPKRIAKAVNEKS, SEQ ID NO.49) важны для связывания белка ENO1 с EN10 mAb.
Пример 13
Сканирование аланином
Для дополнительного исследования того, какие остатки от 296 до 336 ENO1 человека важны для связывания EN10 mAb, кристаллическую структуру ENO1 загружали из банка белковой базы данных (pdb-entry: 2PSN). После структурного анализа прогнозировали, что аминокислотные остатки D300, W301, G302, Q305, K306, A309, K326, K330, N333, E334, и K335 экспонированы на поверхности белка и являются кандидатами для мутаций для анализа того, действительно ли они важны для связывания EN10 mAb. 10 из этих 11 остатков выбирали, чтобы подвергнуть их мутации за исключением A309,который подвергали мутации до глицина с использованием набора сайт-направленного мутагенеза QuickChange II согласно руководству, прилагаемому производителем (Agilent Technology). Генерировали следующие мутагенные олигонуклеотиды для сканирования аланином (Таблица 1) с помощью Genomics BioScience and Technology Co., Ltd.
Таблица1: Олигонуклеотидные последовательности 5ʹ-GATCCCTTTGACCAGGATGCCTGGGGAGCTTGGCAG-3ʹ (SEQ ID NO:24) 5ʹ-CTGCCAAGCTCCCCAGGCATCCTGGTCAAAGGGATC-3ʹ (SEQ ID NO: 25) 5ʹ-CCCTTTGACCAGGATGACGCGGGAGCTTGGCAGAAG-3ʹ (SEQ ID NO: 26) 5ʹ-CTTCTGCCAAGCTCCCGCGTCATCCTGGTCAAAGGG-3ʹ (SEQ ID NO:27 ) 5ʹ-CTTTGACCAGGATGACTGGGCAGCTTGGCAGAAGTTC-3ʹ (SEQ ID NO: 28) 5ʹ-GAACTTCTGCCAAGCTGCCCAGTCATCCTGGTCAAAG-3ʹ (SEQ ID NO:29)
5ʹ-GACTGGGGAGCTTGGGCGAAGTTCACAGCCAGTGCA-3ʹ (SEQ ID NO:30)
5ʹ-TGCACTGGCTGTGAACTTCGCCCAAGCTCCCCAGTC-3ʹ (SEQ ID NO:31)
5ʹ-GGGGAGCTTGGCAGGCGTTCACAGCCAGTGCAGG-3ʹ (SEQ ID NO:32)
5ʹ-CCTGCACTGGCTGTGAACGCCTGCCAAGCTCCCC-3ʹ (SEQ ID NO:33)
5ʹ-GGCAGAAGTTCACAGGCAGTGCAGGAATCCAGGTAG-3ʹ (SEQ ID NO:34)
5ʹ-CTACCTGGATTCCTGCACTGCCTGTGAACTTCTGCC-3ʹ (SEQ ID NO:35)
C5ʹ-TCACAGTGACCAACCCAGCGAGGATCGCCAAGGCC-3ʹ (SEQ ID NO:36)
5ʹ-GCCTTGGCGATCCTCGCTGGGTTGGTCACTGTGAG-3ʹ (SEQ ID NO:37)
5ʹ-CAACCCAAAGAGGATCGCCGCGGCCGTGAACGAGAAG-3ʹ (SEQ ID O:38)
5ʹ-CTTCTCGTTCACGGCCGCGGCGATCCTCTTTGGGTTG-3ʹ (SEQ ID NO:39) 5ʹ-GAGGATCGCCAAGGCCGTGGCCGAGAAGTCCTGCAAC-3ʹ (SEQ ID O:40)
5ʹ-GTTGCAGGACTTCTCGGCCACGGCCTTGGCGATCCTC-3ʹ (SEQ ID NO:41)
5ʹ-GATCGCCAAGGCCGTGAACGCGAAGTCCTGCAACTG-3ʹ C(SEQ ID NO:42)
5ʹ-GCAGTTGCAGGACTTCGCGTTCACGGCCTTGGCGATC-3ʹ(SEQ ID NO:43)
5ʹ-GCCAAGGCCGTGAACGAGGCGTCCTGCAACTGCCTC-3ʹ (SEQ ID NO:44)
5ʹ-GAGGCAGTTGCAGGACGCCTCGTTCACGGCCTTGGC-3ʹ (SEQ ID NO:45)
5ʹ-CAAGGCCGTGAACGCGGCGTCCTGCAACTGCCTCCTG-3ʹ (SEQ ID NO:46)
5ʹ-CAGGAGGCAGTTGCAGGACGCCGCGTTCACGGCCTTG-3ʹ (SEQ ID NO:47)
Для амплификации каждого мутанта готовили реакционный раствор, имеющий состав 3 мкл матрицы ДНК примерно 30нг, 5 мкл 10× реакционного буфера, 1 мкл 10мM dNTP-микс, 1мкл 2,5 ед. pfu-полимеразы, 12,5 мкл 125нг прямого праймера и 12,5 мкл 125нг обратного праймера, в конечном объеме 50 мкл с дважды дистиллированной водой и подвергали ПЦР. Использовали цикл 95°C в течение 10 минут. Затем цикл 95°C в течение 30 секунд, 55°C в течение 30 секунд, и 68°C в течение 6 мин повторяли 16 раз. После реакции ПЦР добавляли 1 мкл DpnI в каждую ПЦР-пробирку и инкубировали при 37°C в течение 1 часа и затем DpnI нагревали для инактивации при 80°C в течение 20 минут. Продукты реакции трансформировали в 50 мкл компетентных клеток XL1-Blue согласно руководству, прилагаемому производителем (производство Invitrogen). Праймер ENO1 R400-420 (5ʹ-GCAAGGGGCACCAGTCTTGATCTG--3ʹ (SEQ ID NO:48)) использовали для определения нуклеотидной последовательности. Каждую плазмиду с мутантным клоном с корректной последовательностью трансформировали в E.coli BL21 Rosseta. Мутантный белок ENO1 экспрессировали в E.coli с помощью индукции IPTG и очищали согласно прилагаемому руководству, предложенному производителем (Qiagen), после чего осадок бактерий подвергали воздействию ультразвука. Чистоту каждого мутанта анализировали с помощью 12% SDS-PAGE.
Для определения активности связывания каждого мутантного белка 400нг/100мкл человеческого белка ENO1 или мутантного белка ENO1 наносили в виде покрытия на 96-луночный тИФА-планшет на ночь при 4°C, и планшет промывали с помощью PBS. Планшет блокировали с помощью 1% BSA (масс./об.) в PBS при комнатной температуре в течение 1 часа, затем промывали снова с помощью 1x PBS. Первичное антитело (EN10 mAb), разведенное с помощью последовательных 2-кратных разведений с получением 15 различных концентраций, добавляли к планшету и инкубировали при 37°C в течение 1 часа. После проведения реакции планшет промывали 3 раза с помощью 1× PBS. Разведенное 1/8000 козьего антитела к мышиному белку-HRP добавляли и инкубировали при 37°C в течение 1 часа, затем планшет промывали 3 раза с помощью 1× PBS. Затем добавляли TMB субстрат и реакции давали пройти при комнатной температуре в течение 30 минут. Реакцию останавливали путем добавления 1N HCl, и значения OD 450 считывали для определения активности. Каждое исследование повторяли три раза. Данные представлены в виде среднего ± SD. OD-значения и концентрации антител использовали для получения графика множественного рассеяния с использованием SigmaplotTM. Значения Kd прогнозировали с помощью 4-параметрического логистического анализа.
Согласно результатам большей части делеционного исследования ENO1, представленных в Примере 9, пептидная последовательность FDQDDWGAWQKFTASAGIQVVGDDLTVTNPKRIAKAVNEKS(SEQ ID NO:49) от остатка номер 296 до 336 требуется для тесного связывания белка ENO1 с EN10 mAb. Используемый в настоящей заявке термин «тесное связывание» относится к связыванию между специфическим агентом связывания (например, антителом, scFv- или Fab-фрагментом) и лиганд/мишень (например, пептидом, белком или клеткой) с константой диссоциации(Kd) 10 нM или менее, предпочтительно, 1 нM или менее.
Вышеописанные делеционные эксперименты идентифицируют остатки 296-336 на ENO1 как область связывания антитела. Для дополнительной характеристики фактических сайтов связывания (например, эпитопов), из банка белковой базы данных загружали кристаллическую структуру ENO1 (pdb-entry: 2PSN) для анализа положений остатков из этой области. Есть одиннадцать аминокислотных остатков, включающих D300, W301, G302, Q305, K306, A309, K326, K330, N333, E334, и K335, экспонированных на белковой поверхности (ФИГ.13A, предполагаемый эпитоп). С помощью сайт-направленного мутагенеза эти 11 аминокислот подвергали мутации и полученные мутантные белки экспрессировали вE.coli и очищали, соответственно (ФИГ.13B). Каждый очищенный мутантный белок ENO1 анализировали на предмет любого изменения Kd (по сравнению со связыванием ENO1) с использованием тИФА.
Эти результаты выявили, что есть три функциональных класса аминокислотных остатков в этих мутантах. Аминокислотные остатки W301 и K330 важны для связывания между белком ENO1 и EN10 mAb. Если эти два аминокислотных остатка подвергают мутации до аланина, соответственно, то активности связывания этих двух мутантов ENO1 с EN10 mAb значительно нарушаются. Второй класс аминокислотных остатков включает A309, E334, K335 и D300. Если E334, K335 и D300 подвергают мутации до аланина или A309 подвергают мутации до глицина, то активности связывания этих мутантов ENO1 с EN10 mAb нарушаются. Оставшиеся аминокислотные остатки, включающие G302, Q305, K306, N333, и K326, принадлежат к группе аминокислотных остатков, которые не обладают значительными эффектами связывания в отношении связывания белка ENO1 с EN10 mAb (ФИГ.13C и Таблица 2). Эти результаты предполагают, что W301, K330A309, E334, K335 и D300 важны для белок-белкового связывания между ENO1 и EN10 mAb. Эти аминокислотные остатки принадлежат к последовательностям пептида 1 ENO1,296FD Q D D W G A W Q K F TA309(ФИГ.13D,SEQ ID NO:9) и пептида 2,326 K R I A K A V N EK S336 (ФИГ.13D,SEQ ID NO:10), которые могут представлять собой эпитопы связывания EN10 mAb в аминокислотных остатках от 296 до 335(ФИГ.13D; SEQ ID NO:49) человеческого ENO1.
Таблица 2: Значения Kd мутантов
Пример 14
Структурный анализ ENO1 и EN10 mAb
Кристаллическую структуру Энолазы альфа загружали из банка белковой базы данных (pdb-entry: 2PSN). Структуру Fab-фрагмента EN10 mAb строили с помощью гомологичного моделирования. Первая стадия состояла в обнаружении структуры матрицы с высокой гомологией последовательности с EN10 mAb. По базе данных NCBI (www.ncbi.nlm.nih.gov.) проводили поиск, и было обнаружено множество высоко гомологичных вариантов. Выбрали каркас тяжелой цепи (pdb-entry: 3DGG) (идентичность 89%) и каркас легкой цепи (pdb-entry: 1F6L) (идентичность 95%). Гомологичную модель антитела EN10 mAb конструировали с использованием программ Accelrys Discovery Studio. Структуры ENO1 и антитела использовали для стыковки (докинга). С целью облегчения компьютерной загрузки только CDR-петли вариабельного домена антител стыкуются с ENO1. Каркас антитела фиксирован. Так как с помощью процедуры докинга исследуют относительно большие области около стартового положения, очень аккуратное исходное положение партнеров по докингу не требуется. При использовании ZDock, структуры отклоняются с движением оси x-y-z (отклонение вдоль линии центров, в ангстремах отклонение в плоскости перпендикулярной линии центров, ангстрем-вращательное отклонение, в градусах) и приблизительно 5000 положений генерируют для каждой процедуры докинга. Анализ кластеризации результатов ZDock проводили с обоими пороговыми значениями 5Å и 10Å. Остатки взаимодействия ENO1 выбирали для картирования эпитопов.
3D-структура человеческого белка ENO1 представлена на ФИГ.14A. Аминокислотные остатки W301, K330,300D, E334, AND K335 помечены. Критические остатки сайта связывания плазминогена на белке ENO1, опубликованные Wang et al. указаны (250-FFRSGKY 256). На ФИГ.14B представлено, что комплекс связывания ENO1 и EN10 mAb прогнозируется примерно на 5Å - 15 Å поблизости от сайта связывания плазминогена ENO1. Пробел между сайтом связывания плазминогена и EN10 mAb составляет немногими меньше, чем 34 Å, что соответствует молекулярному размеру человеческого плазминогена. Данное исследование предполагает, что связывание ENO1 и EN10 mAb защищает сайт связывания плазминогена ENO1, предотвращает избыток плазминогена по отношению к ENO1, приводит к нарушению активности рецептора плазминогена ENO1, и предотвращает диссоциацию клеток из внеклеточного матрикса.
Пример 15
Анализ «Biacore»
Анализ «Biacore» с использованием растворимого белка ENO1 для связывания антител, иммобилизованных на чипах CM5, осуществляли для EN10 mAb для оценки их аффинности к растворимому антигену.
Экспрессирующий вектор pTCAE8-ENO1, содержащий химерное антитело из мышиной вариабельной области и человеческой Fc, как представлено на ФИГ.15A, вводили в клетки-хозяева для получения клеток, экспрессирующих рекомбинатное антитело. В качестве клеток-хозяев для экспрессии использовали клетки FreeStyle293 (производство Invitrogen). Вектор вводили в клетки-хозяева с помощью липофектамина 2000 согласно прилагаемому руководству производителя (производство Invitrogen.). Примерно 2,5 микрограмм вектора, экспрессирующего антитело, линеаризовали с помощью ферментов рестрикции, ген вводили в 4×106 клеток, и клетки инокулировали в 6-луночный культуральный планшет. Добавляли агент, соответствующий маркеру селекции экспрессирующего вектора, и клетки непрерывно культивировали с образованием стабильного пула.
Надосадочную жидкость культуры, содержащую человеческое антитело IgG, получали с помощью метода, описанного ниже. Антитело-продуцирующие клетки акклиматизировали в среде Free styleTM293 Expression Medium (GIBCO). Эти клетки культивировали во флаконах для культивирования тканевой клеточной культуры, и надосадочную жидкость культуры собирали, когда степень выживаемости клеток составляла 90%. Собранную надосадочную жидкость фильтровали через фильтры 10 мкм и 0,2 мкм (производство Millpore) для удаления загрязнений. Надосадочную жидкость, содержащую антитело, очищали на аффинной колонке с протеином A (производство Millipore), PBS в качестве буфера адсорбции и 20 мM натрий цитратного буфера (pH 3) в качестве элюирующего буфера. Элюированные фракции доводили до pH около 6 путем добавления 50 мM натрий фосфатного буфера (pH 7). Для получения антитела раствор заменяли на PBS с использованием диализной мембраны (10000 порог молекулярной массы, производитель Spectrum Laboratories) и стерилизовали фильтрацией через мембранный фильтр (производство Millpore), имеющий размер пор 0,22 мкм с получением очищенного антитела. Концентрацию очищенного антитела определяли с помощью измерения поглощения при 280 нм и путем превращения измеренного значения на основе приравнивания оптимальной плотности 1,45 к 1 мг/мл.
Для кинетики связывания использовали измерение поверхностного плазмонного резонанса (SPR) с использованием BIAcore 2000 (BIAcore, Inc., Piscataway, N.J.), как описано ранее (Karlsson&Falt, (1997) J. Immunol Methods 200:121-133). Биосенсорные чипы из карбоксиметилированного декстрана (CM5, BIAcore Inc.) активировали с использованием N-этил-Nʹ-(3-диметиламинопропил)-карбодиимид гидрохлорида (EDC) и N-гидроксисукцинимида (NHS) согласно инструкциям поставщика. Химерное антитело EN10 mAb разводили с использованием 10 мM ацетата натрия, pH 4,8, в 5 мкг/мл перед инъекцией со скоростью потока 20 мкл/мин для достижения приблизительно 100 единиц ответа (RU) конденсированного белка с последующей инъекцией 1M этаноламина для блокировки непрореагировавших групп. Для измерений кинетики двухкратные последовательные разведения ENO1 (0,3125 нM - 40 нM) инъецировали в HBS-PBiacore рабочий буфер, предоставленный производителем (BIAcore, Inc., Piscataway, N.J.) при 25°C со скоростью потока 25 мкл/мин, и корректировали ответы связывания на EN10 mAb с помощью вычитания ответов пустого потока клеток. Использовали скорости ассоциации (kon или ka) и скорости диссоциации (koff или kd) рассчитывали с использованием простой модели связывания Ленгмюра 1:1 с отдельными приспособлениями kon и koff. (BIAcore.TM. Evaluation Software version 3.2).
Таблица 3
Результаты представлены на ФИГ.15B и в Таблице 3. kon и koff связывания EN10 mAb с ENO1 составили 3,57×105 и 8,271×10-5, соответственно, и Kd равно 2. 311±0,003×10-10моль/л. Kd, анализированное с помощью SPR, оказалось очень близким к значению для связывания тИФА в Примере 2.
Пример 16
Ингибирующий эффект антитела EN10 в отношении опухолевого роста в присутствии компонента комплемента.
EN10 mAb обладает приемлемой аффинностью с Kd примерно 2,311±0,003×10-10моль/л и потенциалом для дальнейшего развития. Для оценки терапевтических эффектов EN10 mAb, осуществляли мышиную модель CL1-5 ксенотрансплантатов. Клетки аденокарциномы легкого CL1-5F4 (1×106клеток/на мышь; 5 мышей/на группу) инокулировали подкожно в день 0. Терапевтическую процедуру осуществляли через 14 дней после инокуляции опухоли путем введения 10 (мг/кг) изотипического контроля (CTL), EN10 mAb, или антитело ErbituxTM дважды в неделю. Сыворотку крольчат давали через 2ч после инъекции. Размер опухоли и массу тела каждой мыши измеряли еженедельно. Данные представлены в виде среднего ± SD для индивидуальных групп.
Результаты представлены на ФИГ.16. Через 14 дней размер опухоли мышей, несущих CL1-5, составил примерно 30-100 мм3, и отсутствовали значительные различия в размере опухоли среди контрольной группы, EN10 mAb, и группы обработки Erbitux после 25 дня. После 29 дня опухоль мышей в контрольной группе начала расти экспоненциально и отсутствовали значительные различия опухолевого роста у мышей в группах обработки EribituxTM и EN10 mAb. После дня 36, средний размер опухоли контрольной группы мышей составил 4330±990 мм3 (N=5), и для мышей, обработанных 10 мг/кг EN10 mAb и Erbitux, средний размер опухоли составил 358,6±240мм3 (N=5) и 219±98мм3 (N=5),соответственно. Средний размер опухоли в обеих группах обработки EN10 mAb и ErbituxTM был значительно меньше по сравнению с размером контрольной группы с значением P 0,004 и 0,003, соответственно. Нет значительных различий в размерах опухоли между группами обработки EN10 mAb и ErbituxTM (P=0,41). Этот результат выявляет, что EN10 mAb и Erbitux обладают ингибирующей активностью опухолевого роста в отношении клеток CL1-5 при объединении с компонентом комплемента C3 в мышиной модели ксенотрансплантатов, и EN10 mAb обладает хорошей эффективностью в качестве реагента для противоопухолевой терапии.
Пример 17
Блокада распространения опухоли злокачественных новообразований поджелудочной железы в анализе метастатического колониеобразования в печени-селезенке.
Результаты Примеров 2 и 3 предполагают, что введение антитела к ENO1 может ослаблять инвазию и метастазирование опухолевых клеток и результаты Примера 16 выявили, что ENO1 mAb обладает эффективностью ингибирования опухолевого роста при объединении с компонентом комплемента. Для оценки антиметастатической активности EN10 mAb в отношении опухолевых клеток in vivo, осуществляли анализ метастазирования в печени-селезенке. Клетки аденокарциномы поджелудочной железы PDAC1 (1×106/на мышь; 5 мышей/на группу) инъецировали в селезенку. Затем 10 мг/мл EN10 mAb и контрольное IgG вводили мышам каждой группе IP (внутрибрюшинно), соответственно. Мышей умерщвляли через 6 недель после инокуляции опухоли. Количество опухолевых узлов в печени и массу органов, включающих легкие, селезенку, печень, поджелудочную железу и почки считали и измеряли, соответственно. Данные представлены в виде среднего ± SD для индивидуальных групп.
Результаты представлены на ФИГ. 17 A, 17B и 17C. Когда опухолевые узлы и размеры сравнивали, то средний размер опухолевых узлов и размер группы обработки контрольным IgG составили 34±10 (N=3) и 8500±3500 (N=5), соответственно. Однако когда те же мыши такого же типа обрабатывали с использованием 10 мг/кг EN10 mAb, то средний размер опухолевых узлов и размеров обработанных мышей составил 10±4 (N=3) и 500±120 (N=5), соответственно. Есть значительное количество опухолевых узлов и различи в размерах между группами обработки 10 мг/кг ENO1 и обработки контрольным IgG (P<0,05)(ФИГ 17A и 17B). Когда средние массы органов сравнивали между группами обработки 10 мг/кг ENO1 и обработки контрольным IgG, то не было существенных различий в массах органов, включающих легкие, селезенку, поджелудочную железу и почки. Однако мыши, обработанные 10 мг/кг IgG, имели средний размер печени примерно 7,2±3г (N=5), что статистически отличалось по сравнению с размером печени мышей, обработанных 10 мг/кг EN10 mAb (1,6±0,2г(N=5), P<0,05). Эти результаты предполагают, что введение EN10 mAb ингибирует метастазирование клеток PDAC1 из первичного места опухоли в печень.
Суммируя, EN10 mAb использует его антагонистическую активность рецептора плазминогена ENO1 для ингибирования активации плазминогена, индуцируя посредством этого отрицательную регуляцию протеазной активности на клеточной поверхности, которая, в свою очередь, приводит к ингибированию диссоциации опухолевых клеток из внеклеточного матрикса. В результате антитела к ENO1 могут ингибировать инвазивную способность опухолевых клеток. Эти данные поддерживают тот факт, что антитела к ENO1 (например, EN10 mAb) обладают подходящей аффинностью, эффективностью и потенциалом в качестве терапевтического антитела для лечения злокачественных новообразований.
Дополнительно к ингибированию диссоциации опухолевых клеток из внеклеточного матрикса и, таким образом, ингибирования инвазии опухолевых клеток, антитело по изобретению также может использоваться для диагностики злокачественных новообразований. Как отмечено выше и представлено на ФИГ. 1A и ФИГ. 1B, опухолевые клетки экспрессируют ENO1 на клеточной поверхности, в то время как нормальные нет. Таким образом, можно детектировать связывание антитела с ENO1 клеточной поверхности в качестве индикатора злокачественных новообразований.
Например, согласно воплощениям изобретения, тестируемый образец может быть реактивным по отношению к антителу к ENO1 (например, EN10 mAb) и связывание антитела с образцом может быть определено, как например, с помощью тИФА, сортинга флуоресцентных клеток, флуоресцентной микроскопии и любого подходящего метода, известного в данной области. Если связывание детектируют, или оно выше порогового значения (например, эталонное значение для нормального образца), то специалист может заключить, что образец содержит опухолевые клетки.
В то время как изобретение описано, применительно к неограниченному количеству воплощений, при этом специалистам в данной области, которым полезно настоящее описание, будет понятно, что могут быть разработаны другие воплощения, которые не выходят за рамки объема изобретения, описанного в данном документе. Соответственно, объем изобретения следует ограничивать только приложенной формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПЕЦИФИЧЕСКИЕ АНТИТЕЛА К АЛЬФА-ЕНОЛАЗЕ И СПОСОБ ПРИМЕНЕНИЯ ПРИ ИММУННЫХ ЗАБОЛЕВАНИЯХ | 2014 |
|
RU2707812C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АЛЬФА-ЭНОЛАЗЕ, И СПОСОБЫ ПРИМЕНЕНИЯ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2014 |
|
RU2761662C2 |
| АНТИ-PD-1 АНТИТЕЛА | 2017 |
|
RU2752832C2 |
| АНТИТЕЛА ПРОТИВ PD-L1 И ИХ ВАРИАНТЫ | 2017 |
|
RU2770590C2 |
| АНТИТЕЛО, НАЦЕЛИВАЮЩЕЕСЯ НА CD3, БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2808138C1 |
| СПОСОБЫ И КОМПОЗИЦИИ НА ОСНОВЕ АНТИТЕЛ К CD73 И ИХ ВАРИАНТОВ | 2020 |
|
RU2841372C2 |
| КАНИНИЗИРОВАННЫЕ МЫШИНЫЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ PD-1 | 2014 |
|
RU2676158C1 |
| АНТИТЕЛА ПРОТИВ IL2 | 2006 |
|
RU2425054C2 |
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ ПРОДУКТА ДЛЯ АДРЕСНОЙ ДОСТАВКИ ПРИ ЛЕЧЕНИИ РАКА | 2012 |
|
RU2659094C2 |











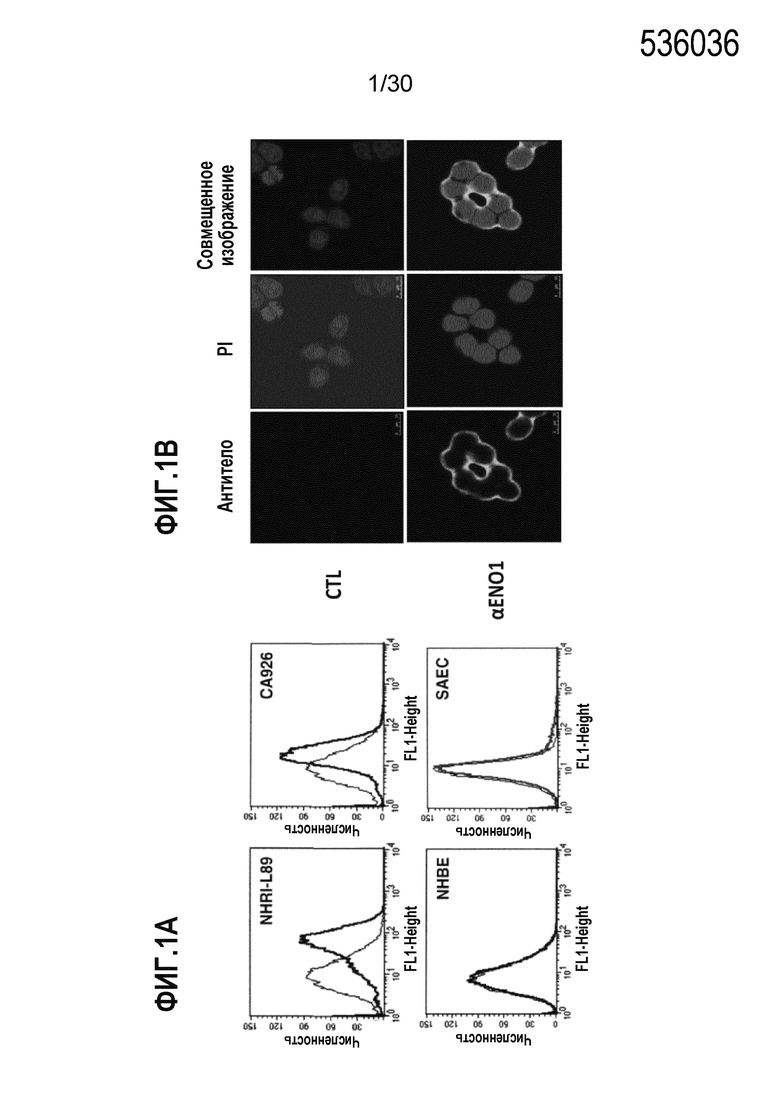
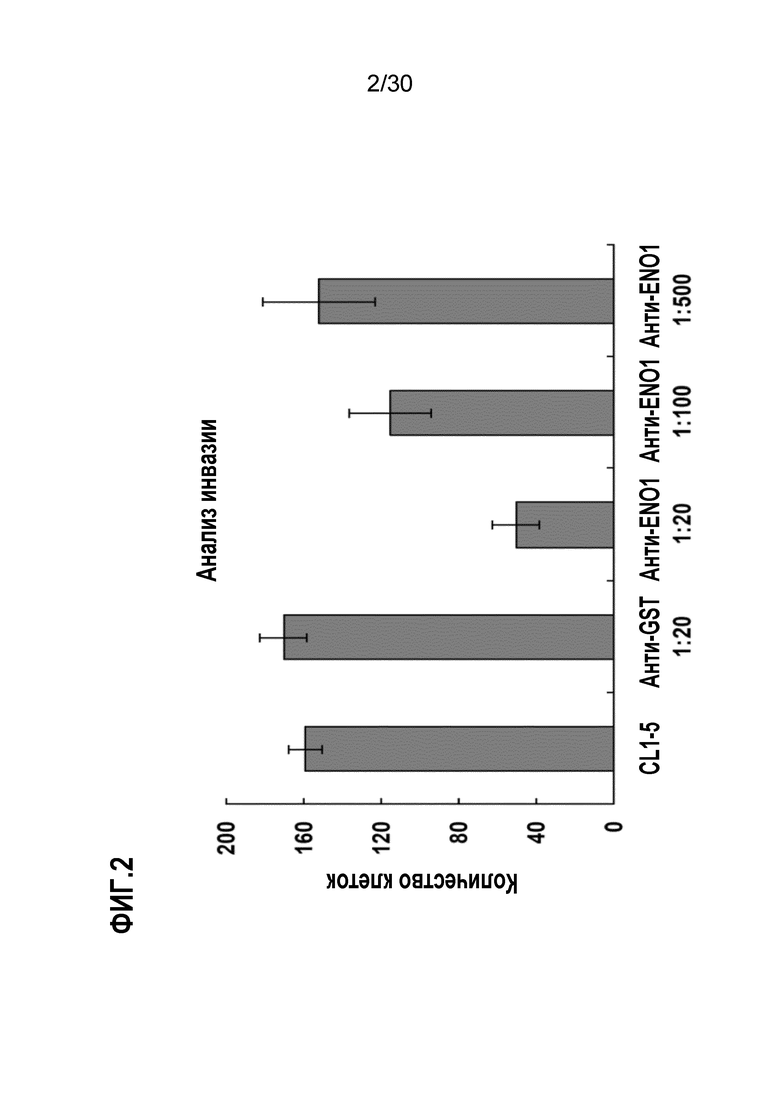


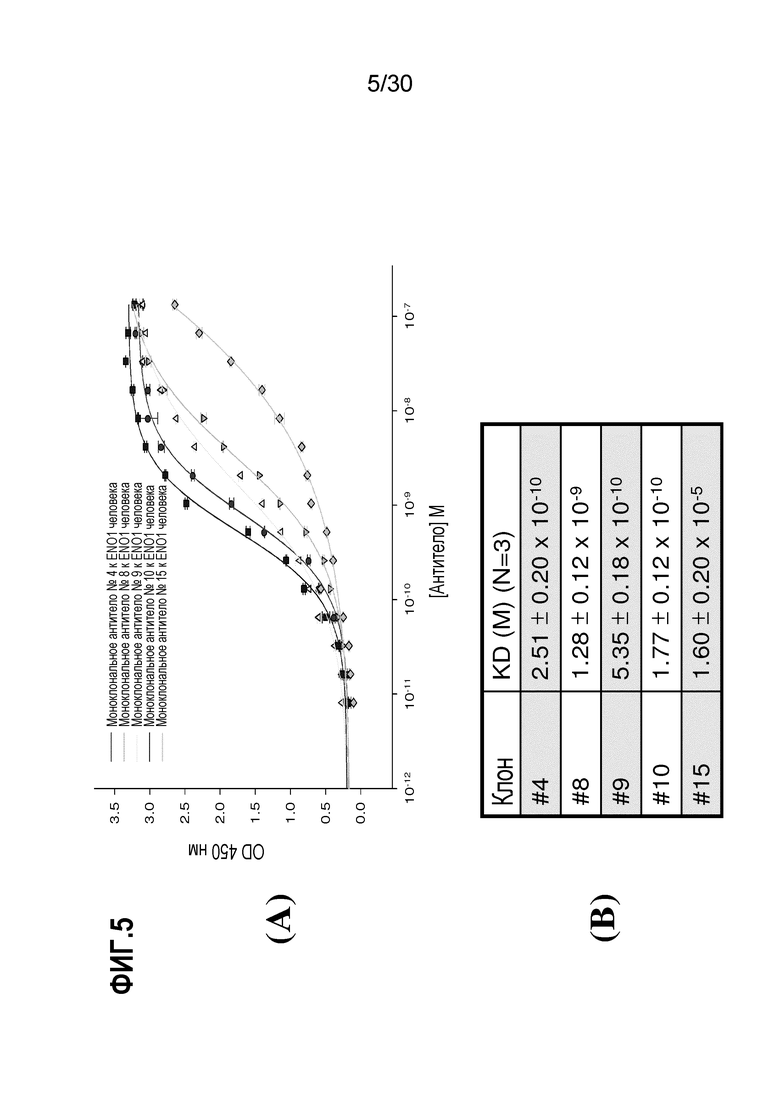
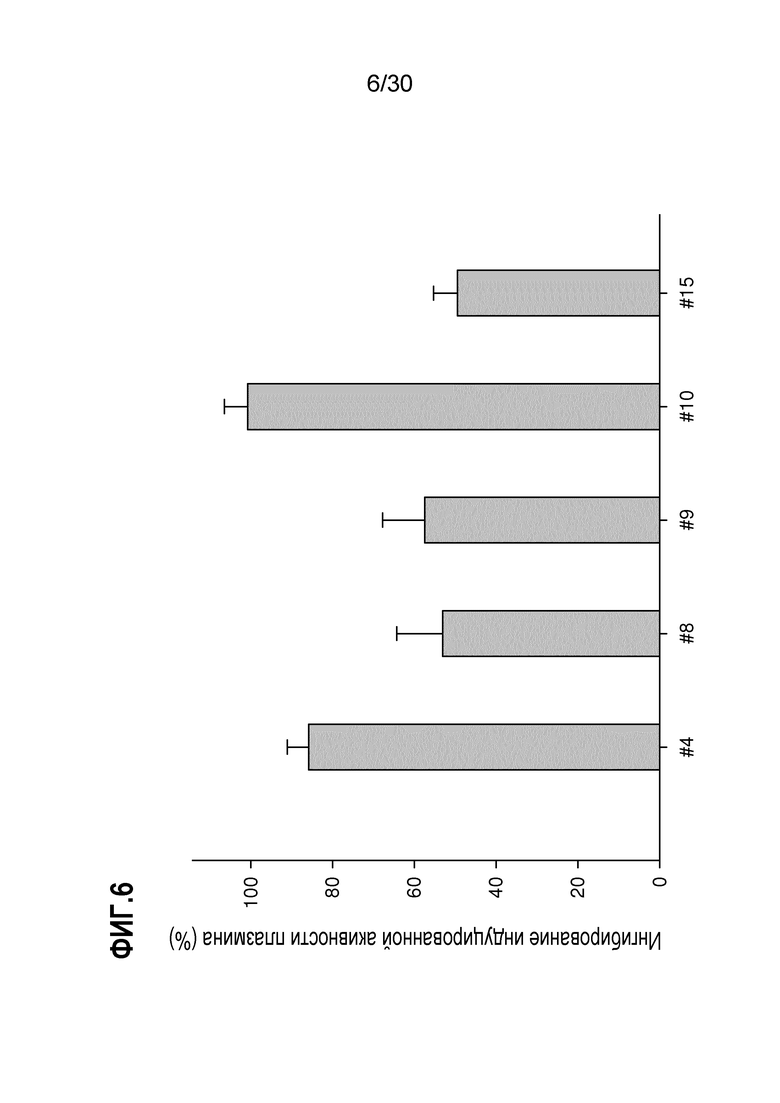
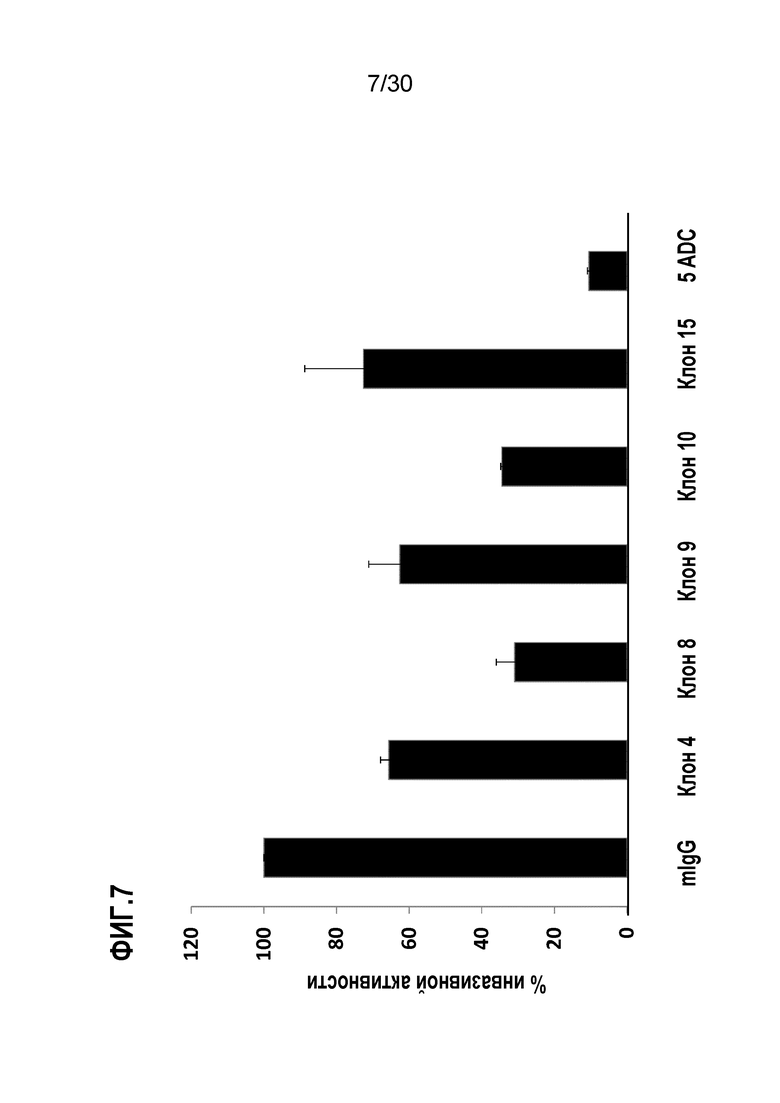
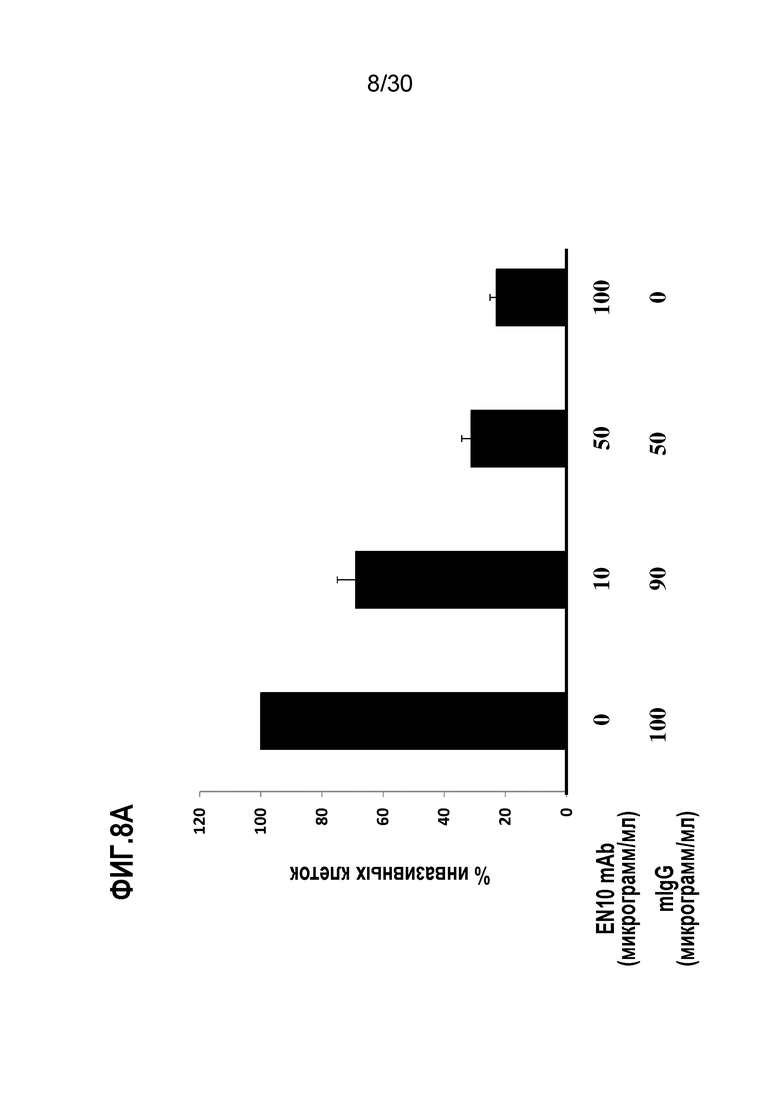
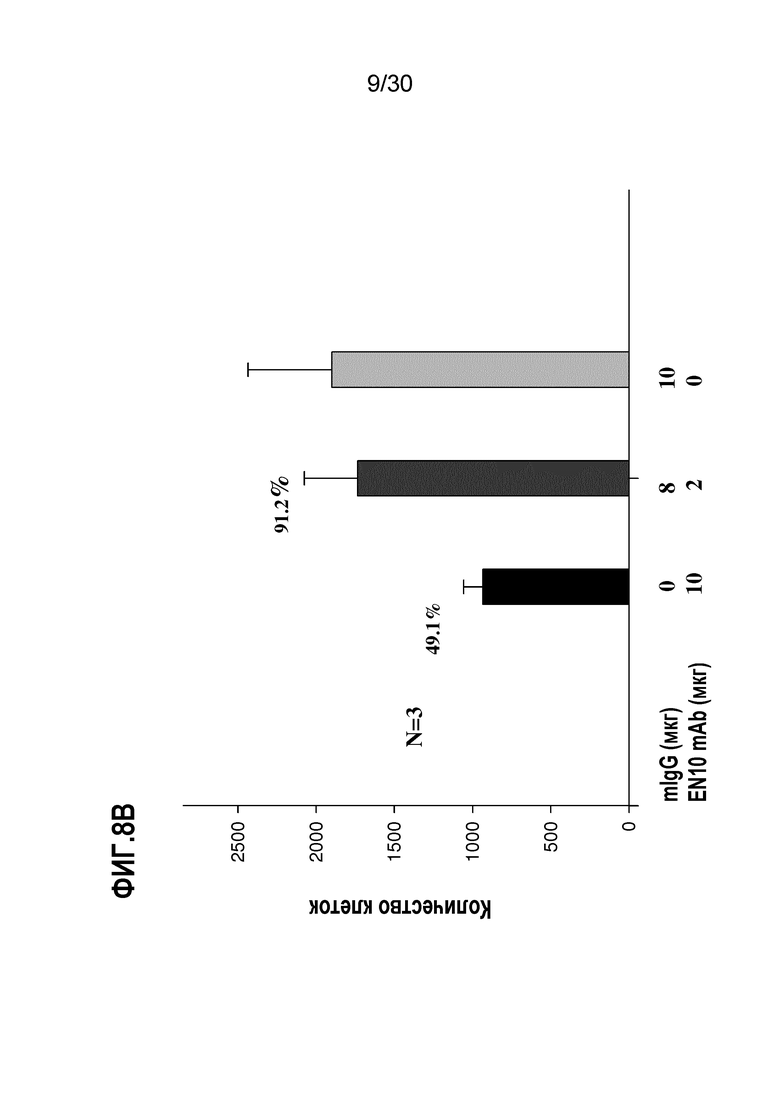
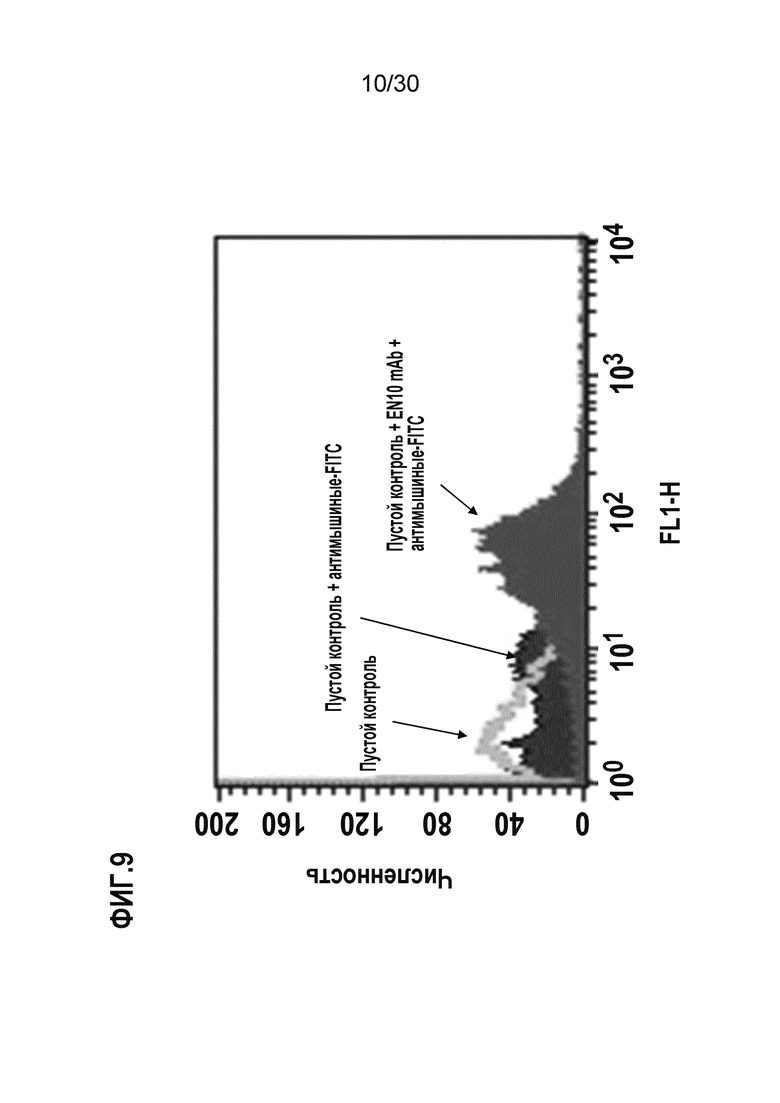
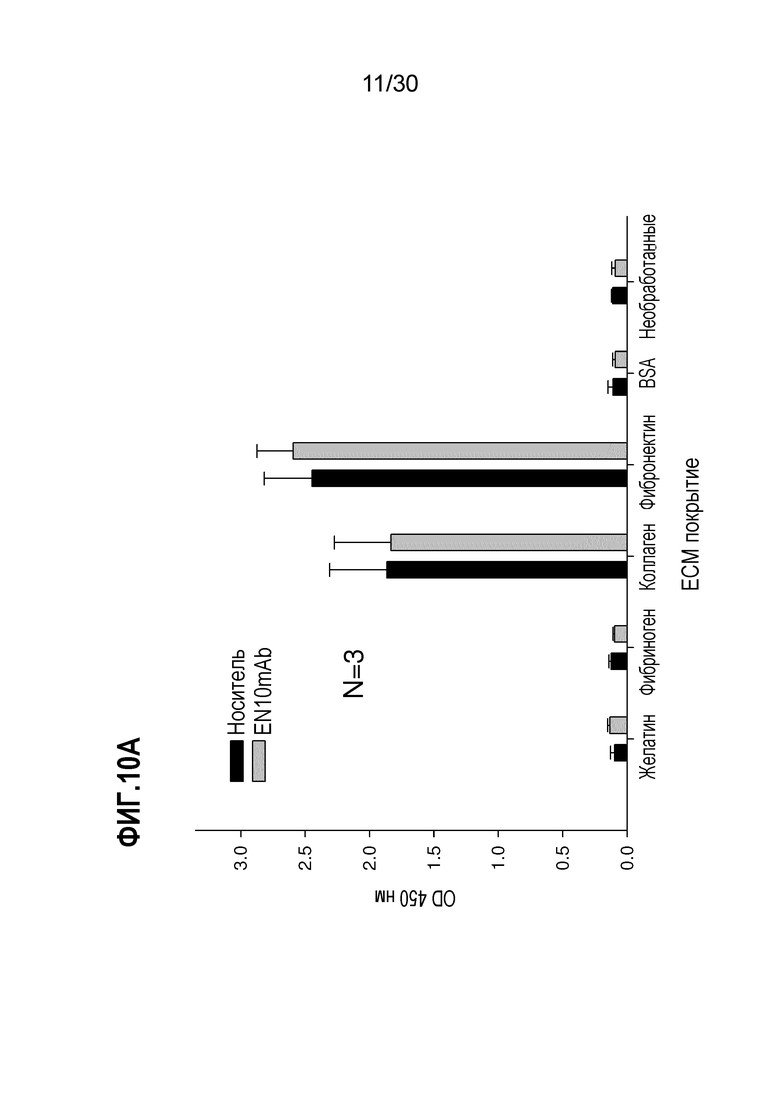

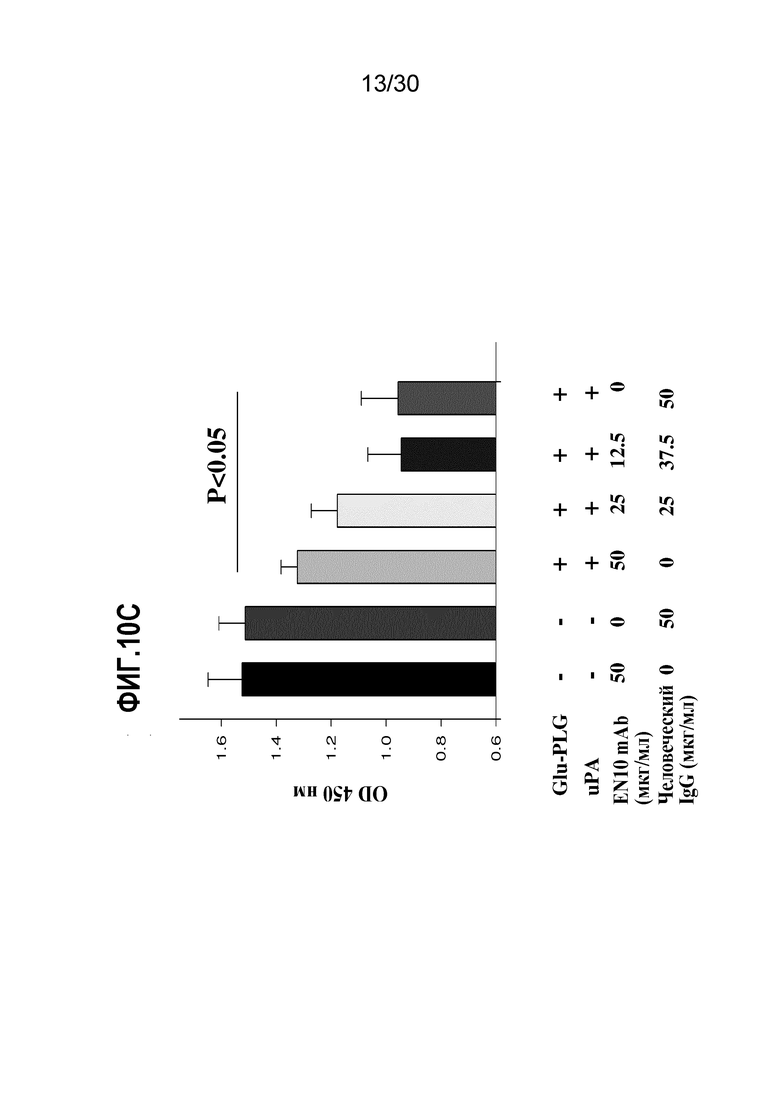


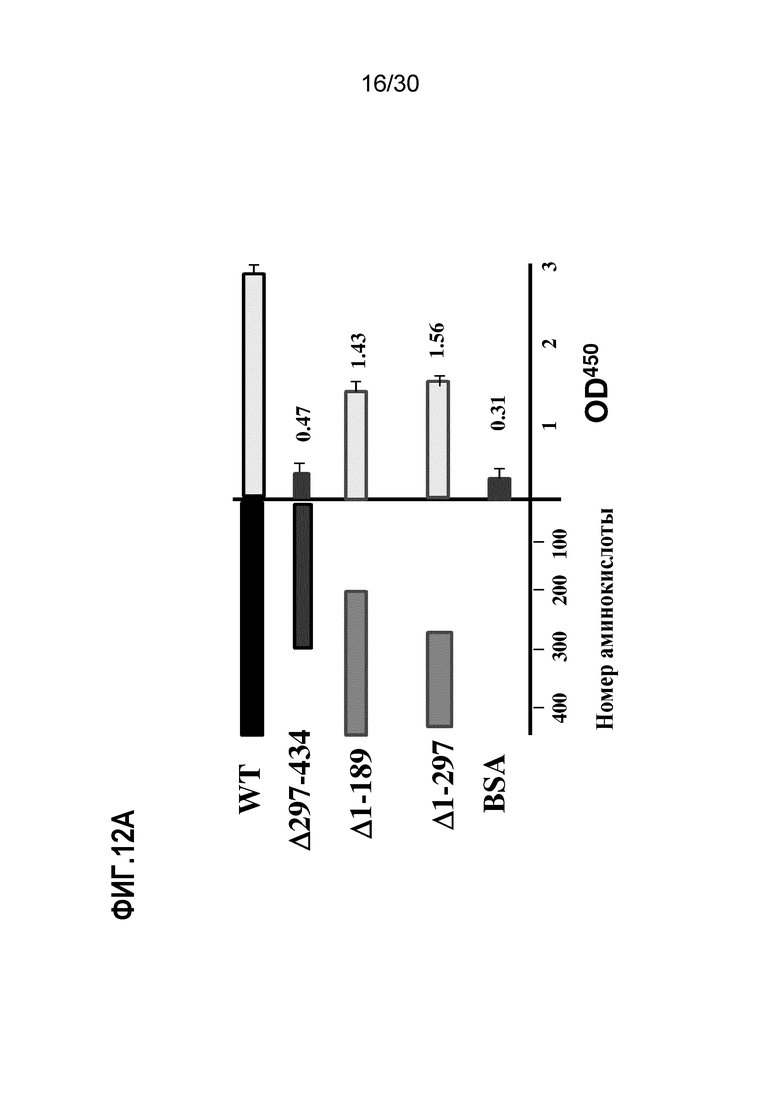
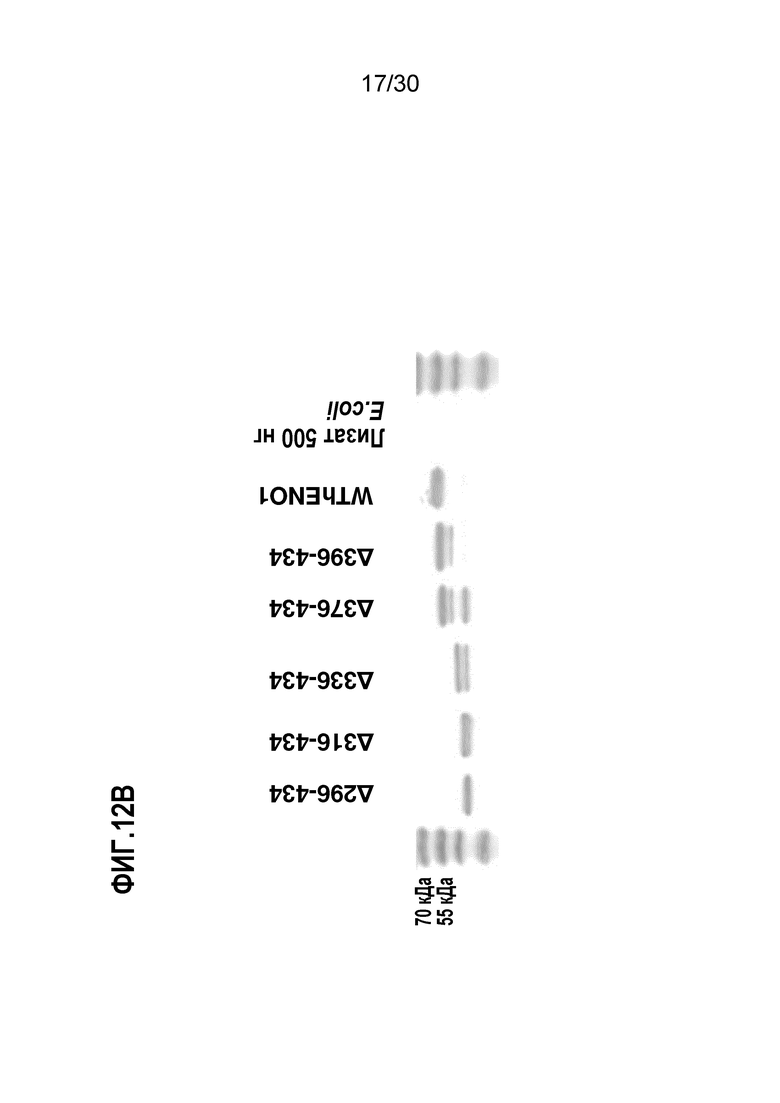
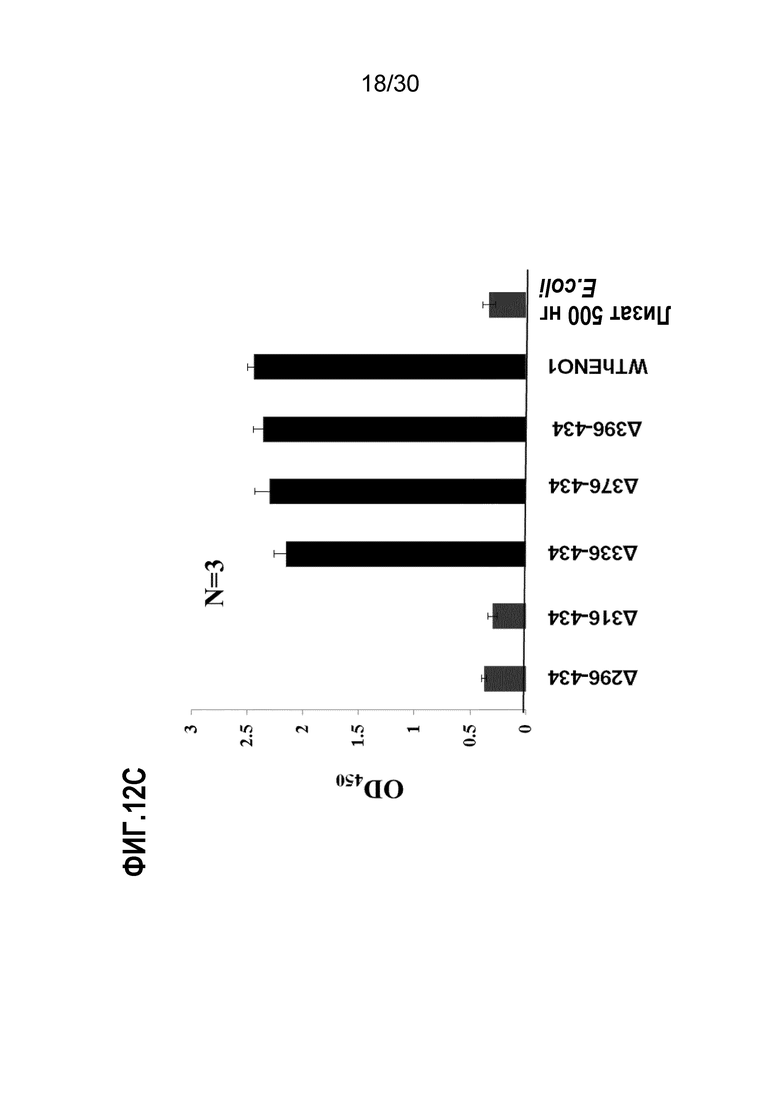
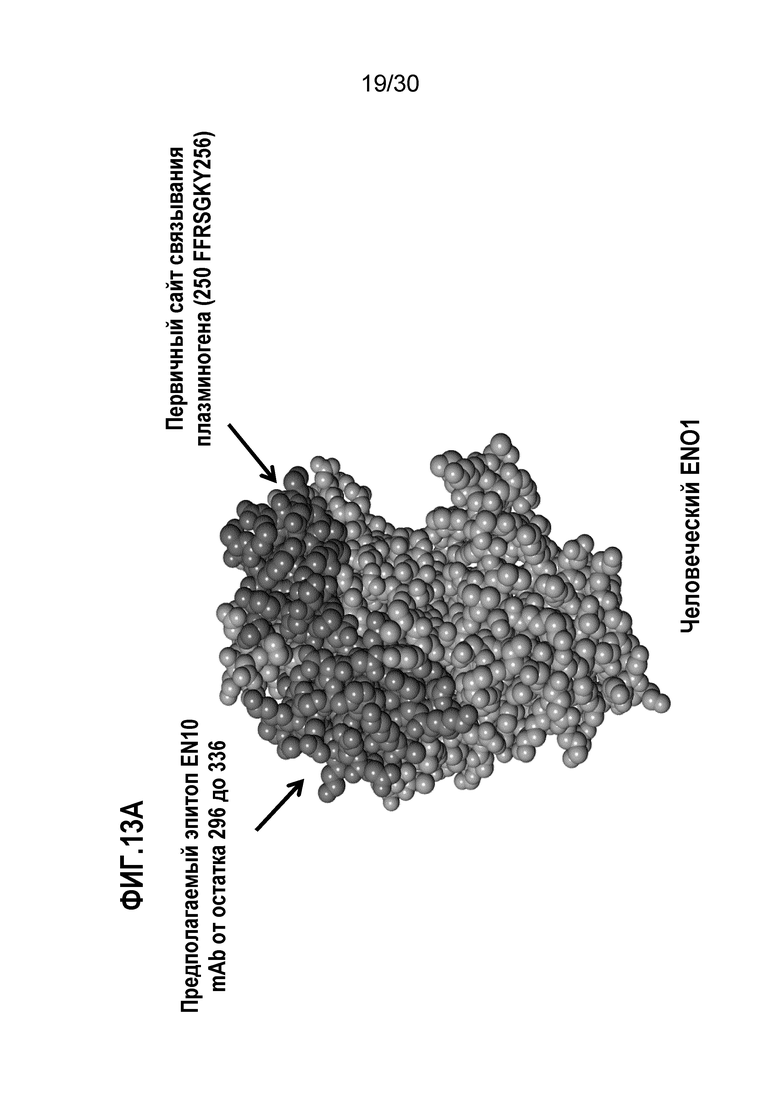


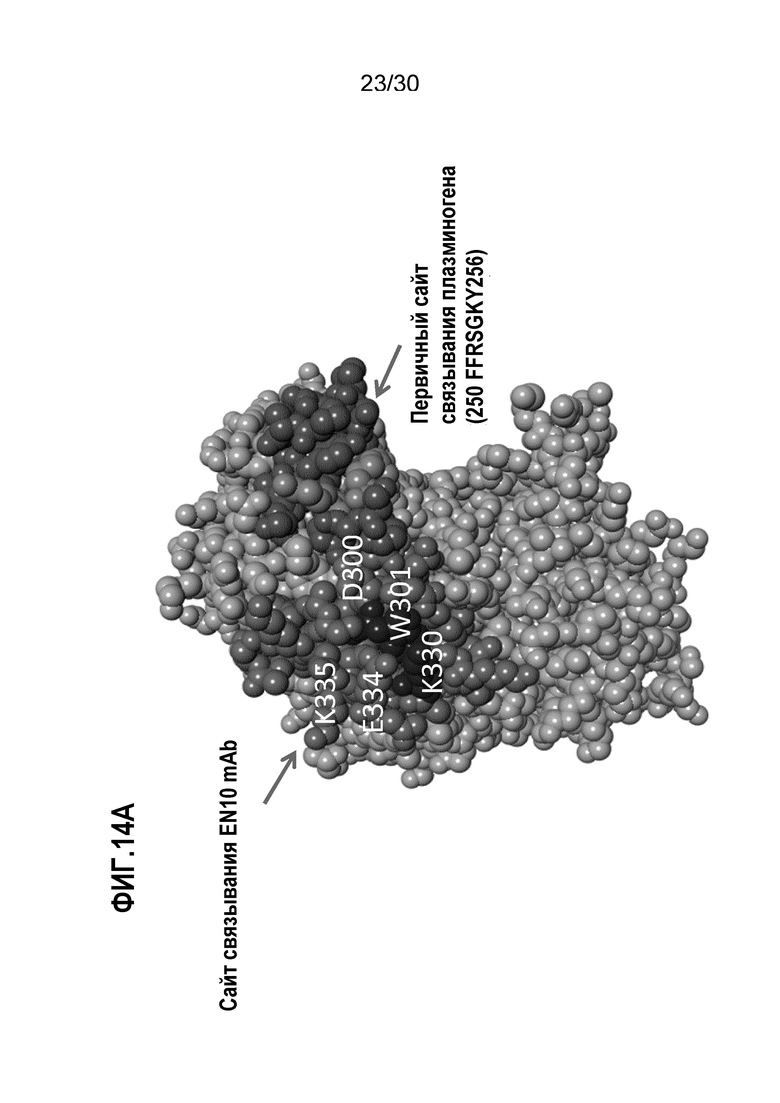
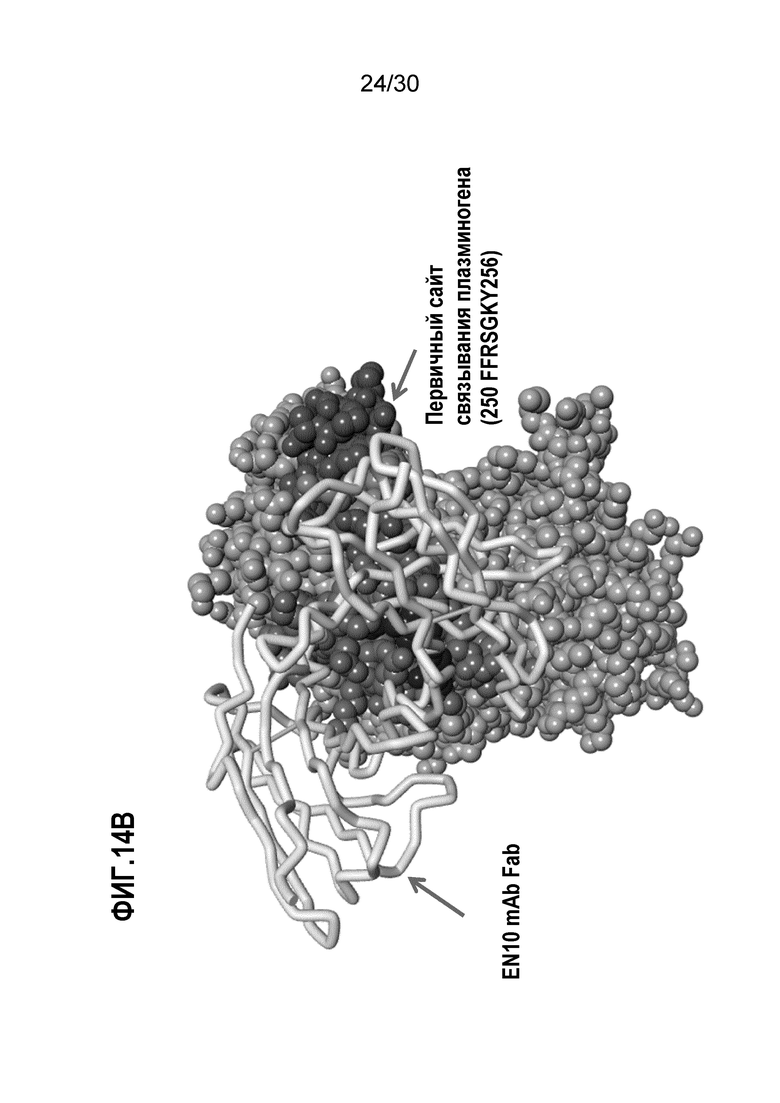
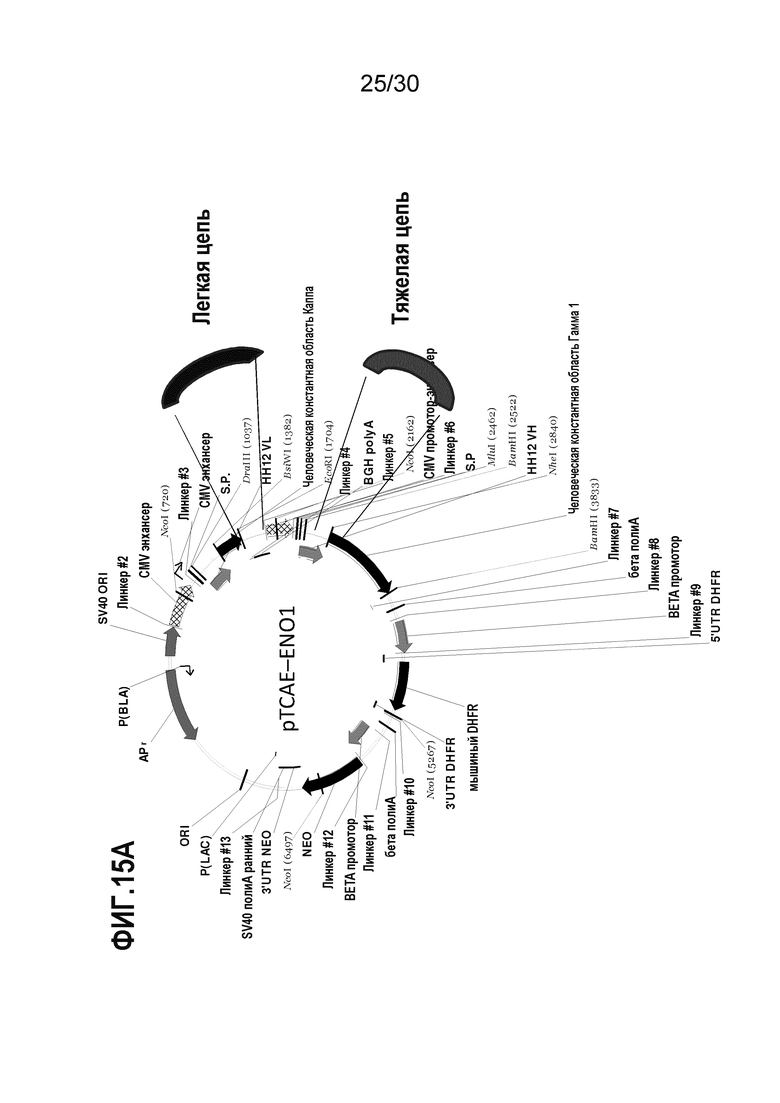
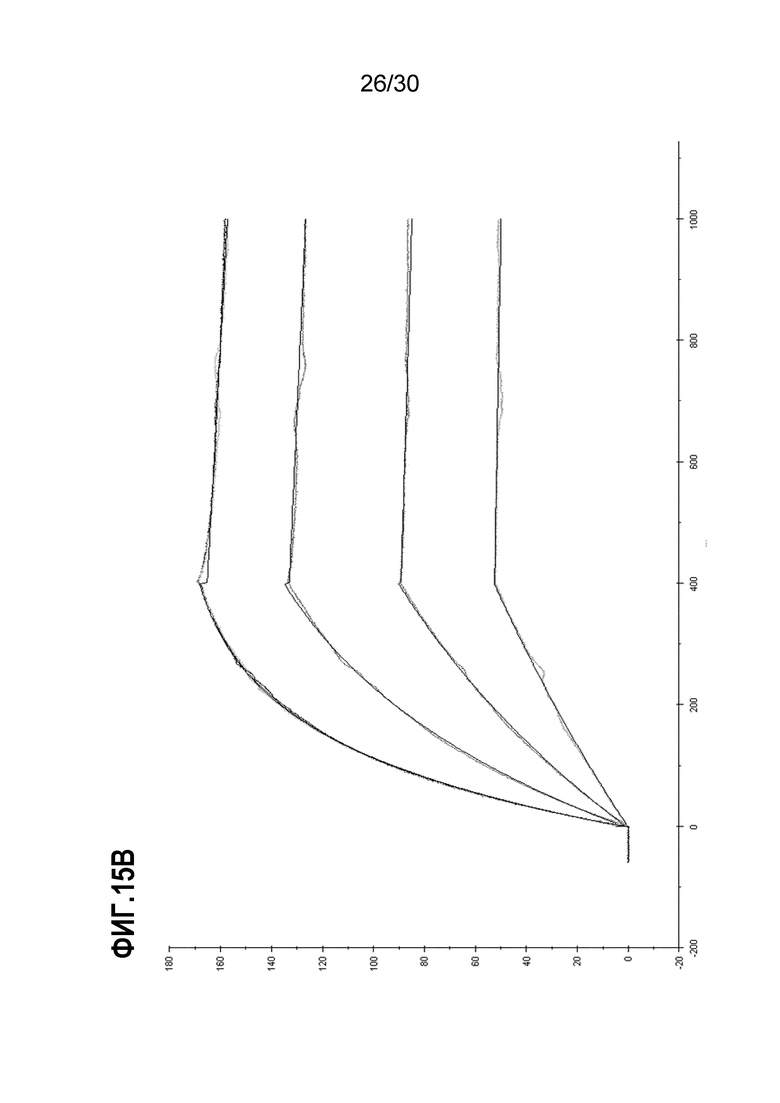
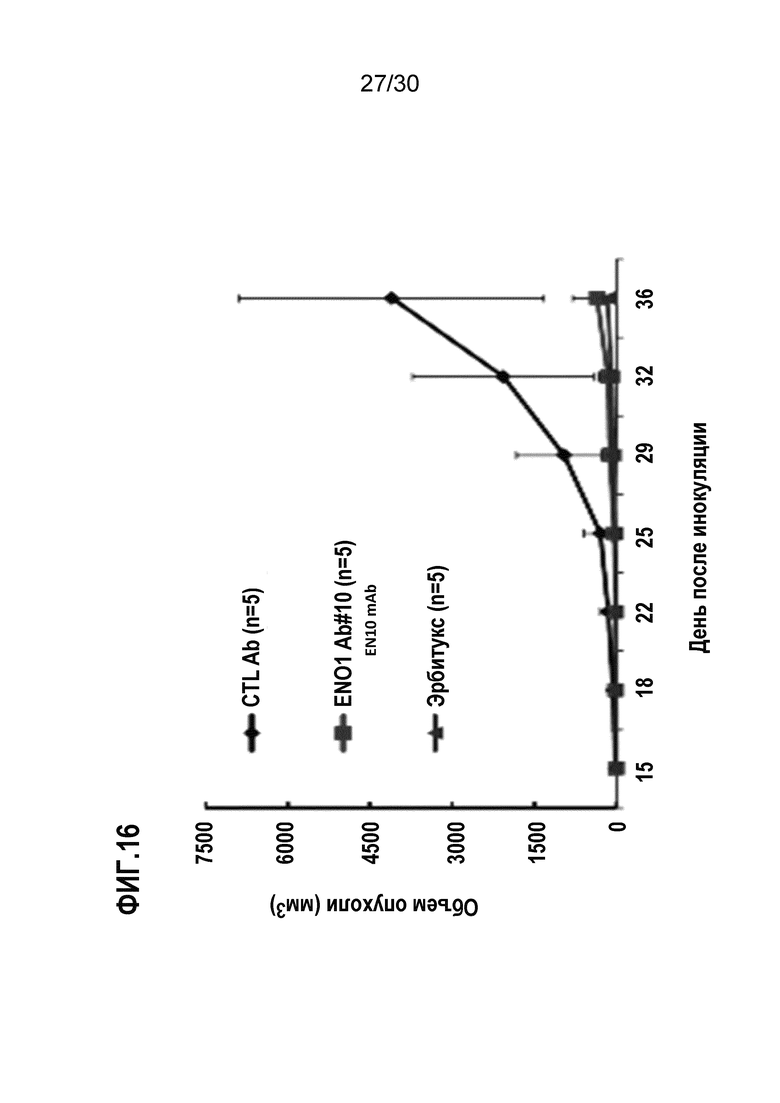
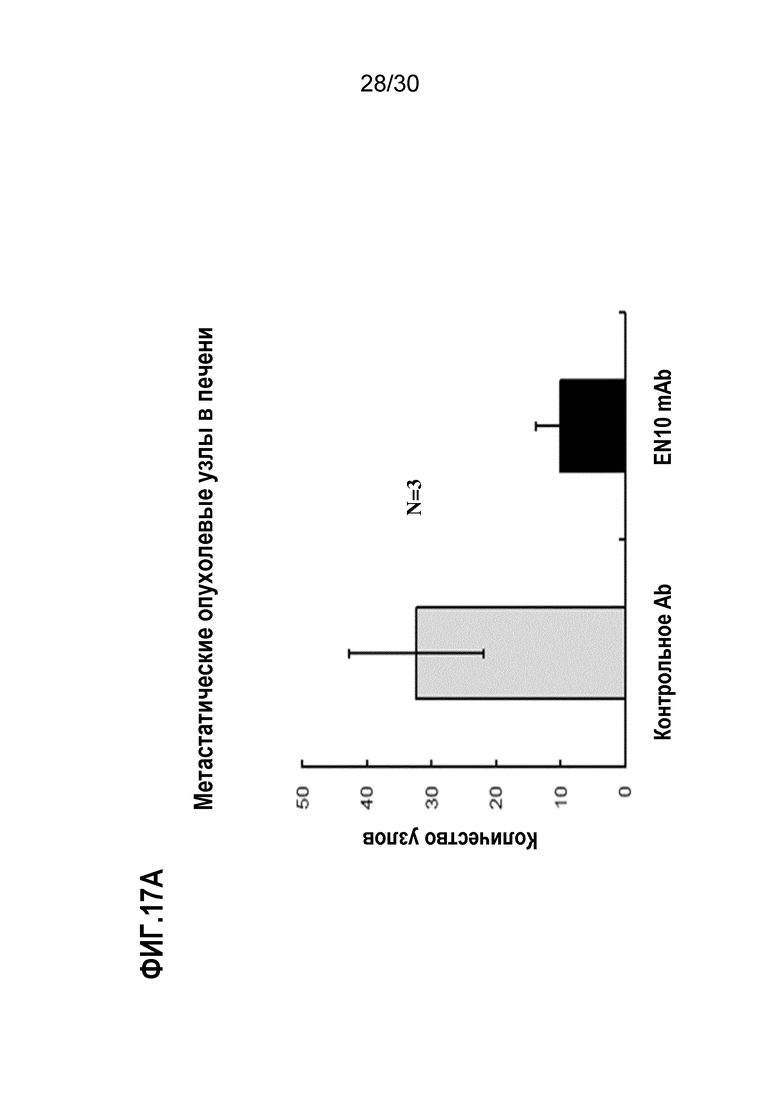
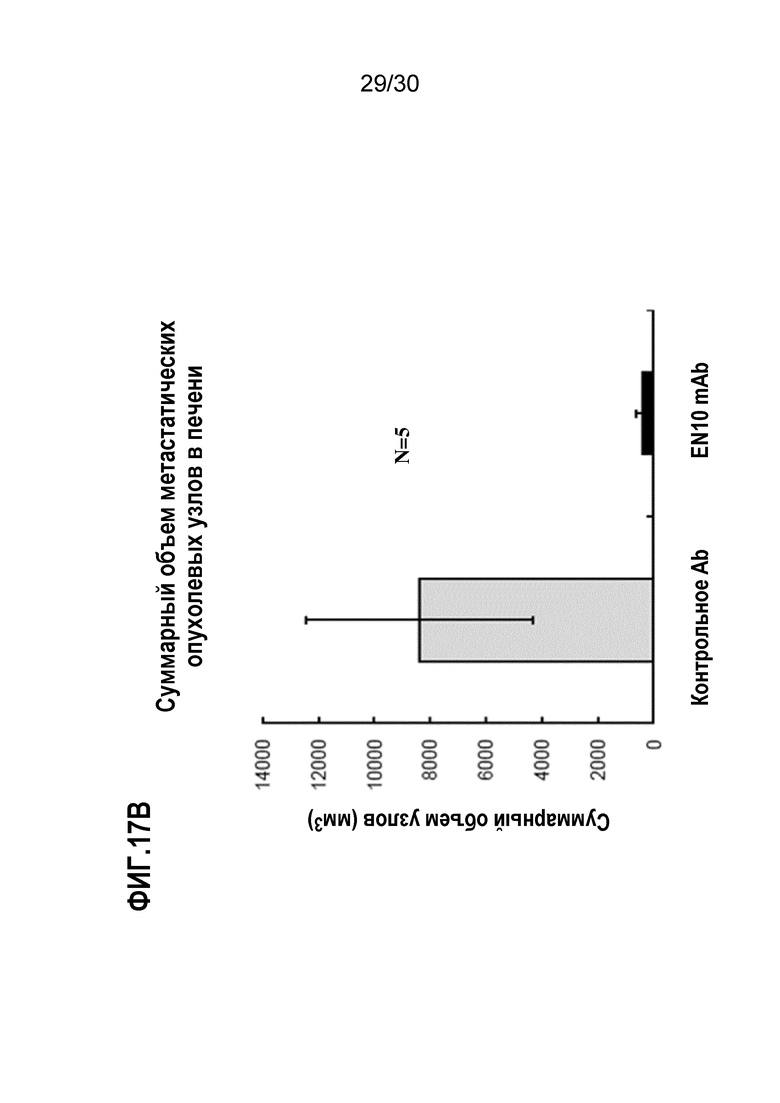
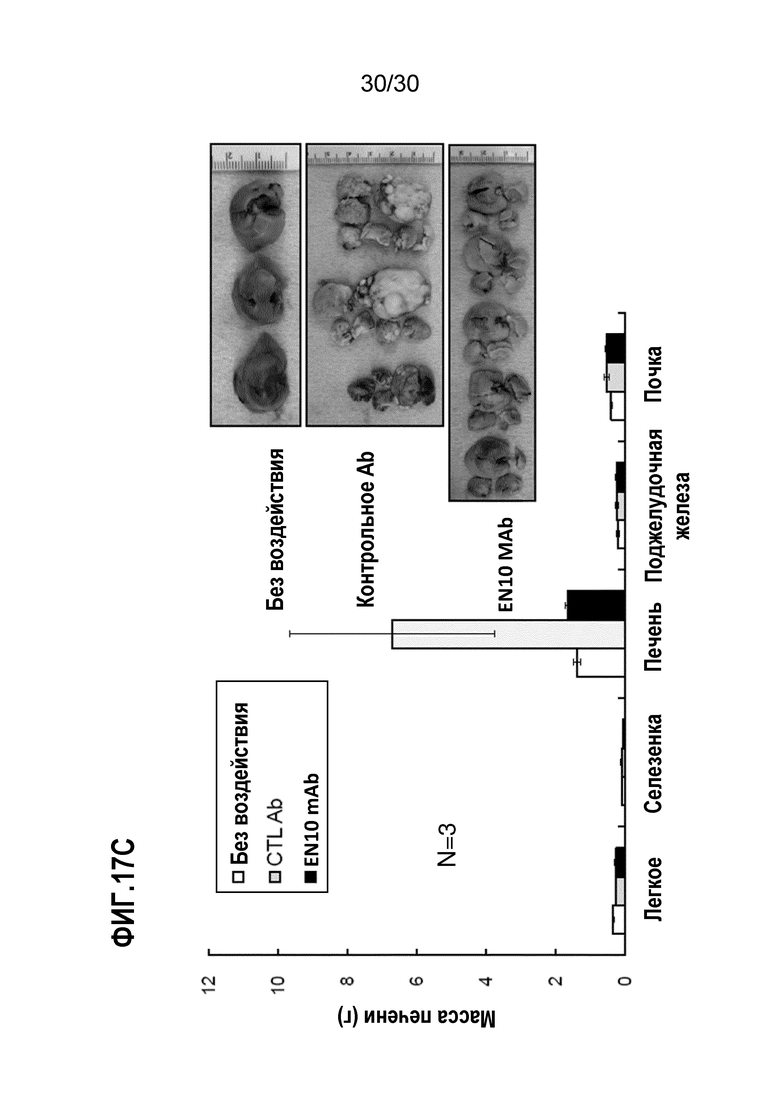
Настоящее изобретение относится к области иммунологии. Предложено антитело против альфа-энолазы (ENO1), а также его scFv- и Fab-фрагмент. Кроме того, описана фармацевтическая композиция для лечения злокачественного новообразования; а также способ диагностики злокачественного новообразования и способ ингибирования инвазии злокачественного новообразования. Предложенное антитело способно связываться с пептидами, включающими аминокислотную последовательность FDQDDWGAWQKFTA и/или KRIAKAVNEKS человеческого белка ENO1, и обладает подходящей активностью связывания (аффинность связывания составляет около 10-10 моль/л). Таким образом, данное изобретение может найти дальнейшее применение в терапии и диагностике заболеваний, связанных с ENO1. 4 н. и 4 з.п. ф-лы, 17 ил., 3 табл., 17 пр.
1. Антитело или его scFv- или Fab-фрагмент против альфа-энолазы, содержащее HCDR1 GYTFTSCVMN (SEQ ID NO:3), HCDR2 YINPYNDGTKYNEKFKG (SEQ ID NO:4), HCDR3 GFYYGNFDN (SEQ ID NO:5), LCDR1 RASENIYSYLT (SEQ ID NO:6), LCDR2 NAKTLPE (SEQ ID NO:7) и LCDR3 QHHYGTPYT (SEQ ID NO:8).
2. Антитело или его scFv- или Fab-фрагмент по п. 1, отличающееся тем, что тяжелая цепь содержит последовательность SEQ ID NO:1.
3. Антитело или его scFv- или Fab-фрагмент по п. 1, отличающееся тем, что легкая цепь содержит последовательность SEQ ID NO:2.
4. Антитело или его scFv- или Fab-фрагмент по п. 1, отличающееся тем, что тяжелая цепь содержит последовательность SEQ ID NO:1 и легкая цепь содержит последовательность SEQ ID NO:2.
5. Антитело или его scFv- или Fab-фрагмент по любому из пп. 1-4, отличающееся тем, что антитело представляет собой моноклональное антитело.
6. Фармацевтическая композиция для лечения злокачественного новообразования, связанного с повышенной активностью или экспрессии альфа-энолазы, содержащая антитело или его scFv- или Fab-фрагмент по любому из пп. 1-5.
7. Способ диагностики злокачественного новообразования, связанного с повышенной активностью или экспрессии альфа-энолазы, включающий
реакцию образца от тестируемого объекта с антителом или с его scFv- или Fab-фрагментом по любому из пп. 1-5 для определения степени связывания, где степень связывания, которая составляет выше, чем эталонное значение, выявляет злокачественное новообразование.
8. Способ ингибирования инвазии злокачественного новообразования, связанного с повышенной активностью или экспрессией альфа-энолазы, включающий введение объекту, нуждающемуся в этом, антитела или его scFv- или Fab-фрагмента по любому из пп. 1-5.
| EP 1832605 A1, 12.09.2007 | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| GILKESON GS et al., "Molecular characterization of anti-DNA antibodies induced in normal mice by immunization with bacterial DNA | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| ROARK J.H | |||
| et al., "Breakdown of B cell tolerance in a mouse model of systemic lupus erythematosus." J Exp Med., 1995, 181(3):1157-67 | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| ADAMUS G | |||
| et al., "Anti-enolase-alpha autoantibodies in cancer-associated retinopathy: epitope mapping and cytotoxicity on retinal cells." J Autoimmun., 1998; 11(6):671-7 | |||
| LUNDBERG K | |||
| et al., "Antibodies to citrullinated alpha-enolase peptide 1 are specific for rheumatoid arthritis and cross-react with bacterial enolase." Arthritis Rheum., 2008; 58(10):3009-19 | |||
| WALTER M | |||
| et al | |||
| "Autoreactive epitopes within the human α-enolase and their recognition by sera from patients with endometriosis." Journal of autoimmunity, 1995, 8(6): 931-945 | |||
| LOPEZ-ALEMANY R | |||
| et al., "Inhibition of cell surface mediated plasminogen activation by a monoclonal antibody against α-Enolase." American journal of hematology, 2003, 72(4): 234-242 | |||
| LEU SY-JYE et al., "Generation and characterization of anti-α-enolase single-chain antibodies in chicken." Veterinary immunology and immunopathology, 2010, 137(3): 251-260 | |||
| МОИСЕЕНКО В.М | |||
| "Моноклональные антитела в лечении злокачественных опухолей", Практическая онкология, 2003, Т | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

