Настоящее изобретение относится к биспецифическим, двухвалентным антителам против фактора роста эндотелия сосудов человека (VEGF/VEGF-A) и против ангиопоэтина-2 человека (ANG-2), к способам их получения, к фармацевтическим композициям, содержащим эти антитела, и к их применениям.
Предшествующий уровень техники
Ангиогенез вовлечен в патогенез ряда расстройств, которые включают солидные опухоли, внутриглазные неоваскулярные синдромы, такие как пролиферативные ретинопатии или возрастная макулярная дегенерация (ВМД), ревматоидный артрит и псориаз (Folkman, J., et al., J. Biol. Chem. 267 (1992) 10931-10934; Klagsbrun, M., et al., Annu. Rev. Physiol. 53 (1991) 217-239; и Garner, A., Vascular diseases, in: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G.K. (eds.), 2nd edition, Marcel Dekker, New York (1994), pp.1625-1710). В случае солидных опухолей неоваскуляризация дает возможность опухолевым клеткам приобретать преимущество роста и пролиферативную автономию по сравнению с нормальными клетками. Соответственно, наблюдается корреляция между плотностью микрососудов в срезах опухолей и выживанием пациента, как при раке молочной железы, так и при нескольких других опухолях (Weidner, N„ et al., N Engl J Med. 324 (1991) 1-8; Horak, E.R., et al., Lancet 340 (1992) 1120-1124; и Macchiarini, P., et al., Lancet 340 (1992) 145-146).
VEGF и антитела против VEGF
Фактор роста эндотелия сосудов человека (VEGF/VEGF-A) (SEQ ID No:105) описан, например, в Leung, D.W., et al., Science 246 (1989) 1306-9; Keck, P.J., et al., Science 246 (1989) 1309-12 и Connolly, D.T„ et al., J. Biol. Chem. 264 (1989) 20017-24. VEGF вовлечен в регуляцию нормального и аномального ангиогенеза и неоваскуляризацию, ассоциированную с опухолями и внутриглазными расстройствами (Ferrara, N., et al., Endocr. Rev. 18 (1997) 4-25; Berkman, R.A., et al., J. Clin. Invest. 91 (1993) 153-159; Brown, LF., et al., Human Pathol. 26 (1995) 86-91; Brown, L.F., et al., Cancer Res. 53 (1993) 4727-4735; Mattern, J., et al., Brit. J. Cancer. 73 (1996) 931-934; и Dvorak, H.F., et al., Am. J. Pathol. 146 (1995) 1029-1039). VEGF представляет собой гомодимерный гликопротеин, который выделен из нескольких источников. VEGF проявляет высокоспецифичную митогенную активность к эндотелиальным клеткам. VEGF обладает важными регуляторными функциями в формировании новых кровеносных сосудов в процессе эмбрионального васкулогенеза и в ангиогенезе в течение взрослой жизни (Carmeliet, P., et al., Nature, 380 (1996) 435-439; Ferrara, N, et al. Nature, 380 (1996) 439-442; обзор в Ferrara, N., et al, Endocr. Rev. 18 (1997) 4-25). Значимость роли, которую играет VEGF, продемонстрирована в исследованиях, показывающих, что инактивация единственного аллеля VEGF приводит в результате к эмбриональной летальности вследствие неспособности к развитию сосудистой сети (Carmeliet, P., et al., Nature, 380 (1996) 435-439; Ferrara, N, et al. Nature, 380 (1996) 439-442). Кроме того, VEGF обладает сильной хемоаттрактантной активностью в отношении моноцитов, могут индуцировать активатор плазминогена и ингибитор активатора плазминогена в эндотелиальных клетках, а также могут индуцировать проницаемость микрососудов. В связи с последней активностью иногда на него ссылаются как на фактор проницаемости сосудов (VPF). Сделан обзор выделения и свойств VEGF; см. Ferrara, N., et al., J. Cellular Biochem, 47 (1991) 211-218 и Connolly, D.T, J. Cellular Biochem., 47 (1991) 219-223. Альтернативный сплайсинг мРНК единственного гена VEGF образует пять изоформ VEGF.
Нейтрализующие антитела против VEGF подавляют рост ряда опухолевых клеточных линий человека у мышей (Kirn, K.J, et al., Nature 362 (1993) 841-844; Warren, S.R, et al., J. Clin. Invest. 95 (1995) 1789-1797; Borgstrom, P., et al., Cancer Res. 56 (1996) 4032-4039; и Melnyk, 0, et al., Cancer Res. 56 (1996) 921-924). WO 94/10202, WO 98/45332, WO 2005/00900 и WO 00/35956 относятся к антителам против VEGF. Гуманизированное моноклональное антитело бевацизумаб (имеющееся в продаже под торговым названием Авастин®) представляет собой антитело против VEGF, применяемое в противоопухолевой терапии (WO 98/45331).
Ранибизумаб (торговое название Луцентис®) представляет собой фрагмент моноклонального антитела, имеющий происхождение от того же родительского антитела, что и бевацизумаб (Авастин). Он намного меньше родительской молекулы и обладает созревшим сродством, обеспечивающим более сильное связывание с VEGF-A (WO 98/45331). Он является антиангиогенным, что одобрено для лечения "влажного" типа возрастной макулярной дегенерации (ВМД), распространенной формы возрастной потери зрения. Другим антителом против VEGF является, например, HuMab G6-31, описанное, например, в US 2007/0141065.
ANG-2 и антитела против ANG-2
Ангиопоэтин-2 человека (ANG-2) (альтернативно сокращенный как ANGPT2 или ANG2) (SEQ ID No:106) описан в Maisonpierre, P.C, et al, Science 277 (1997) 55-60 и Cheung.A.H, et al., Genomics 48 (1998) 389-91. Ангиопоэтины-1 и -2 (ANG-1 (SEQ ID No:107) и ANG-2 (SEQ ID No:106)) были открыты как лиганды к Tie, семейству тирозинкиназ, которые селективно экспрессируются внутри эндотелия сосудов. Yancopoulos, G.D., et al., Nature 407 (2000) 242-48. В настоящее время существует четыре точно определенных члена семейства ангиопоэтинов. Ангиопоэтин-3 и -4 (Апд-3 и Апд-4) могут представлять собой широко дивергировавшие копии одного и того же генного локуса у мыши и человека. Kirn, I., et al., FEBS Let, 443 (1999) 353-56; Kim, I., et at., J Biol Chem 274 (1999) 26523-28. ANG-1 и ANG-2 были исходно идентифицированы в экспериментах на тканевых культурах как агонист и антагонист, соответственно (см. для ANG-1: Davis, S., et al., Cell 87 (1996) 1161-69; и для ANG-2: Maisonpierre, P.C., et al., Science 277 (1997) 55-60). Все известные ангиопоэтины связываются, прежде всего, с Tie2, и оба Апд-1 и -2 связываются с Tie2 со сродством 3 нМ (Kd). Maisonpierre, P.C., et al., Science 277 (1997) 55-60. Было показано, что Апд-1 поддерживает выживание ЕС и стимулирует целостность эндотелия, Davis, S., et al., Cell 87 (1996) 1161-69; Kwak, H.J., et al., FEBS Lett 448 (1999) 249-53; Suri, С., et al., Science 282 (1998) 468-71; Thurston, G., et al., Science 286 (1999) 2511-2514; Thurston, G., et al., Nat. Med. 6 (2000) 460-63, тогда как ANG-2 обладал противоположным эффектом и стимулировал дестабилизацию и регрессию кровеносных сосудов в отсутствие факторов выживания VEGF или основного фактора роста фибробластов. Maisonpierre, P.C., et al., Science 277 (1997) 55-60. Однако многие исследования функции ANG-2 позволили предположить более сложную ситуацию. ANG-2 может быть комплексным регулятором ремоделирования сосудов, который играет роль как в спрутинге сосудов, так и в регрессии сосудов. В подтверждение таких ролей для ANG-2, экспрессионные анализы выявили, что ANG-2 быстро индуцируется, вместе с VEGF, в моделях ангиогенного спрутинга у взрослых, тогда как ANG-2 индуцируется в отсутствие VEGF в моделях васкулярной регрессии. Holash, J., et al., Science 284 (1999) 1994-98; Holash, J., et al., Oncogene 18 (1999) 5356-62. В соответствии с контекстно-зависимой ролью, ANG-2 специфично связывается с одним и тем же рецептором эндотелиальной специфичности, Tie-2, который активируется Апд-1, но обладает контекстно-зависимыми эффектами на его активацию. Maisonpierre, P.C., et al., Science 277 (1997) 55-60.
Анализы ангиогенеза роговицы показали, что и ANG-1, и ANG-2 обладают сходными эффектами, действуя синергично с VEGF, по стимуляции роста новых кровеносных сосудов. Asahara, Т., et al., Circ. Res. 83 (1998) 233-40. Вопрос о возможности того, что существует дозозависимый эндотелиальный ответ, был вызван в результате наблюдения, что in vitro при высокой концентрации ANG-2 может также быть проангиогенным. Kirn, I., et al., Oncogene 19 (2000) 4549-52. При высокой концентрации ANG-2 действует как фактор выживания апоптоза для эндотелиальных клеток в процессе апоптоза, обусловленного сывороточным голоданием, посредством активации Tie2 через биохимический путь киназы PI-3 и Akt. Kirn, I., et al., Oncogene 19 (2000) 4549-52.
Другие эксперименты in vitro позволили предположить, что во время пролонгированного воздействия эффекты ANG-2 могут претерпевать постепенный сдвиг от эффекта антагониста к эффекту агониста Tie2, и в более поздние моменты времени они могут вносить непосредственный вклад в формирование сосудистой трубки и стабилизации нового кровеносного сосуда. Teichert-Kuliszewska, К., et al., Cardiovasc. Res. 49 (2001) 659-70. Кроме того, если ЕС культивировали на фибриновом геле, активацию Tie2 за счет ANG-2 также наблюдали, что, возможно, предполагает, что действие ANG-2 может зависеть от стадии дифференциации ЕС.Teichert-Kuliszewska, К., et al., Cardiovasc. Res. 49 (2001) 659-70. В ЕС микрососудов, культивируемых в трехмерном коллагеновом геле, ANG-2 также может индуцировать активацию Tie2 и стимулировать образование капилляроподобных структур. Mochizuki, Y., et al., J. Cell. Sci. 115 (2002) 175-83. Использование 3-D сферической совместной культуры в качестве in-vitro модели созревания кровеносных сосудов продемонстрировало, что прямой контакт между ЕС и мезенхимными клетками уничтожает реактивность в отношении VEGF, тогда как присутствие VEGF и ANG-2 индуцирует спрутинг.Korff, Т., et al., Faseb J. 15 (2001) 447-57. Авторы Etoh, Т.Н. et al. продемонстрировали, что в ЕС, которые конститутивно экспрессируют Tie2, экспрессия ММР-1, -9 и и-РА претерпевала сильную понижающую регуляцию ANG-2 в присутствии VEGF. Etoh, Т., et al., Cancer Res. 61 (2001) 2145-53. На модели in vivo зрачковой перепонки Lobov, I.В. et al. показали, что ANG-2 в присутствии эндогенного VEGF стимулирует быстрое увеличение диаметра капилляра, ремоделирование базальной пластинки, пролиферацию и миграцию эндотелиальных клеток, а также стимулирует спрутинг новых кровеносных сосудов. Lobov, I.B., et al., Proc. Natl. Acad. Sci. USA 99 (2002) 11205-10. Напротив, ANG-2 стимулирует гибель эндотелиальных клеток и регрессию сосудов без эндогенного VEGF. Lobov, I.B., et al., Proc. Natl. Acad. Sci. USA 99 (2002) 11205-10. Подобным образом, в модели опухоли in vivo авторы Vajkoczy, P., et al. продемонстрировали, что многоклеточные агрегаты инициируют рост кровеносных сосудов путем ангиогенного спрутинга посредством одновременной экспрессии VEGFR-2 и ANG-2 эндотелием хозяина и опухоли. Vajkoczy, P., et al., J. Clin. Invest. 109 (2002) 777-85. Данная модель проиллюстрировала, что установившаяся сеть микрососудов растущих опухолей характеризуется непрерывным ремоделированием, предположительно опосредованным экспрессией VEGF и ANG-2 (Vajkoczy, P., et al., J. Clin. Invest. 109 (2002) 777-85).
Исследования Tie-2 и ангиопоэтина-1 на нокаут-мышах показывают сходные фенотипы и позволяют предположить, что стимулируемое ангиопоэтином-1 фосфорилирование Tie-2 опосредует ремоделирование и стабилизацию развивающегося сосуда, стимулируя созревание кровеносного сосуда в процессе ангиогенеза, и сохранение клеточной адгезии, поддерживаемой эндотелиальными клетками (Dumont, D.J., et al., Genes & Development, 8 (1994) 1897-1909; Sato, T.N., Nature, 376 (1995) 70-74; (Thurston, G., et al., Nature Medicine 6 (2000) 460-463). Считают, что роль ангиопоэтина-1 консервативна у взрослых, где он экспрессируется широко и конститутивно (Hanahan, D., Science, 277 (1997) 48-50; Zagzag, D., et al., Exp Neurology 159 (1999) 391-400). Напротив, экспрессия ангиопоэтина-2, в основном, ограничена сайтами ремоделирования сосудов, где, как считают, она блокирует конститутивную функцию ангиопоэтина-1 по стабилизации или созреванию, что дает возможность кровеносным сосудам вернуться в пластичное состояние, которое может быть более реактивным на сигналы спрутинга, или оставаться в нем (Hanahan, D., 1997; Holash, J., et al., Oncogene 18 (199) 5356-62; Maisonpierre, P.C., 1997). В исследованиях экспрессии ангиопоэтина-2 при патологическом ангиогенезе обнаружено, что многие типы опухолей проявляют экспрессию ангиопоэтина-2 кровеносных сосудов (Maisonpierre, P.C., et al., Science 277 (1997) 55-60). Функциональные исследования позволяют предположить, что ангиопоэтин-2 вовлечен в опухолевый ангиогенез, и связывают гиперэкспрессию ангиопоэтина-2 с повышенным опухолевым ростом в мышиной модели ксенотрансплантата (Ahmad, S.A., et al., Cancer Res., 61 (2001) 1255-1259). Другие исследования связывают гиперэкспрессию ангиопоэтина-2 с гиперваскулярностью опухоли (Eton, Т., et al., Cancer Res. 61 (2001) 2145-53; Tanaka, F., et al., Cancer Res. 62 (2002) 7124-7129).
В последние годы ангиопоэтин-1, ангиопоэтин-2 и/или Tie-2 предложены в качестве возможных противораковых терапевтических мишеней. Например, в каждом из US 6166185, US 5650490 и US 5814464 раскрыты антитела против лиганда и рецептора Tie-2. В исследованиях с использованием растворимого Tie-2 сообщали о снижении числа и размера опухолей у грызунов (Lin, 1997; Lin 1998). Авторы Siemeister, G., et al., Cancer Res. 59:3 (1999) 3185-91 создали линии клеток меланомы человека, экспрессирующие внеклеточный домен Tie-2, инъецировали их бестимусным мышам и сообщили, что растворимый Tie-2 приводит в результате к значимому ингибированию опухолевого роста и опухолевого ангиогенеза. С учетом того, что и ангиопоэтин-1, и ангиопоэтин-2 связывается с Tie-2, из этих исследований непонятно, будет ли ангиопоэтин-1, ангиопоэтин-2 или Tie-2 привлекательной мишенью для противораковой терапии. Однако считают, что эффективная терапия против ангиопоэтина-2 полезна при лечении заболеваний, таких как рак, при которых прогрессирование зависит от аберрантного ангиогенеза, где блокирование этого процесса может привести к предотвращению прогрессивного развития заболевания (Folkman, J., Nature Medicine. 1 (1995) 27-31).
Кроме того, некоторые группы сообщили о применении антител и пептидов, которые связываются с ангиопоэтином-2. См., например, US 6166185 и US 2003/10124129. WO 03/030833, WO 2006/068953, WO 03/057134 или US 2006/0122370.
Исследование эффекта очаговой экспрессии ангиопоэтина-2 показало, то антагонизация сигнала ангиопоэтина-1Л1е-2 ослабляет жесткую структуру сосудов, подвергая посредством этого ЕС воздействию активирующих сигналов от индукторов ангиогенеза, например, VEGF (Hanahan, D., Science, 277 (1997) 48-50). Этот проангиогенный эффект, являющийся результатом ингибирования ангиопоэтина-1, указывает на то, что терапия против ангиотензина-1 не будет эффективной противораковой терапией.
ANG-2 экспрессируется в процессе развития в сайтах, где происходит ремоделирование кровеносных сосудов. Maisonpierre, P.С., et al., Science 277 (1997) 55-60. У взрослых индивидуумов экспрессия ANG-2 ограничена сайтами ремоделирования кровеносных сосудов, а также в высоко васкуляризированных опухолях, включая глиому, Osada, H., et al., Int. J. Oncol. 18 (2001) 305-09); Кода, К., et al., Cancer Res. 61 (2001) 6248-54, печеночно-клеточный рак, Tanaka, S., et al., J. Clin. Invest. 103 (1999) 341-45, карциному желудка, Eton, Т., et al., Cancer Res. 61 (2001) 2145-53; Lee, J.H., et al., Int. J. Oncol. 18 (2001) 355-61, опухоль щитовидной железы, Bunone, G., et al., Am J Pathol 155 (1999) 1967-76, немелкоклеточный рак легкого, Wong, M.P., et al., Lung Cancer 29 (2000) 11-22, и рак ободочной кишки, Ahmad, S.A., et al., Cancer 92 (2001) 1138-43, и простаты, Wurmbach, J.H., et al., Anticancer Res. 20 (2000) 5217-20. Обнаружено, что некоторые опухолевые клетки экспрессируют ANG-2. Например, Tanaka, S., et al., J. Clin. Invest. 103 (1999) 341-45 обнаружили мРНК ANG-2 в 10 из 12 образцов печеночно-клеточного рака человека (НСС). Группа Ellis' сообщила, что ANG-2 экспрессируется повсеместно в эпителии опухоли. Ahmad, S.A., et al., Cancer 92 (2001) 1138-43. Другие исследователи сообщали о подобных открытиях. Chen, L, et al., J. Tongji Med. Univ. 21 (2001) 228-35. В результате определения уровней мРНК ANG-2 в архивированных образцах опухолей молочной железы человека Sfiligoi, С., et al., Int. J. Cancer 103 (2003) 466-74 сообщили, что мРНК ANG-2 в значительной степени связана с сопутствующей инвазией лимфоузлов, коротким периодом времени без заболевания и плохой общей выживаемостью. Tanaka, F., et al., Cancer Res. 62 (2002) 7124-29 проведен обзор суммарно 236 пациентов с немелкоклеточным раком легкого (НМКРЛ) с патологической стадией I-IIIA, соответственно. Используя иммуногистохимию, они обнаружили, что 16,9% из пациентов с НМКРЛ были положительными по ANG-2. Плотность микрососудов для опухоли, положительной по ANG-2, значительно выше, чем для отрицательных по ANG-2. Такой ангиогенный эффект ANG-2 наблюдали только тогда, когда экспрессия VEGF была высокой. Кроме того, положительная экспрессия ANG-2 была значимым фактором для предсказания плохой послеоперационной выживаемости. Tanaka, F., et al., Cancer Res. 62 (2002) 7124-7129. Однако они не обнаружили значимую корреляцию между экспрессией Ang-1 и плотностью микрососудов. Tanaka, F., et al., Cancer Res. 62 (2002) 7124-7129. Эти результаты позволяют предположить, что ANG-2 является индикатором пациентов с плохим прогнозом при нескольких типах рака.
Недавно, используя модель нокаут-мышей по ANG-2, группа Yancopoulos' сообщила, что ANG-2 требуется для постнатального ангиогенеза. Gale, N.W., et al., Dev. Cell 3 (2002) 411-23. Они показали, что программируемая в процессе развития регрессия сосудистой сети гиалоидной мембраны в глазу не происходит у нокаут-мышей по ANG-2, и кровеносные сосуды их сетчатки неспособны к спрутингу из центральной ретинальной артерии. Gale, N.W., et al., Dev. Cell 3 (2002) 411-23. Эти авторы также обнаружили, что делеция ANG-2 приводит в результате к глубоким дефектам в структурировании и функции сети лимфатических сосудов. Gale, N.W., et al., Dev. Cell 3 (2002) 411-23. Генетическое спасение Ang-1 корректирует лимфатические дефекты, но не дефекты ангиогенеза. Gale, N.W., et al., Dev. Cell 3 (2002) 411-23.
Peters с соавторами сообщили, что растворимый Tie2 при доставке либо в виде рекомбинантного белка, либо в вирусном экспрессионном векторе ингибирует in vivo рост карциномы молочной железы мыши и меланомы в мышиных моделях. Lin, P., et al., Proc. Natl. Acad. Sci. USA 95 (1998) 8829-34; Lin, P., et al., J. Clin. Invest. 100 (1997) 2072-78. Плотности кровеносных сосудов в опухолевых тканях, обработанных таким образом, были значительно снижены. Кроме того, растворимый Tie2 блокировал ангиогенез роговицы крыс, стимулированный средой, кондиционированной опухолевыми клетками. Lin, P., et al., J. Clin. Invest. 100 (1997) 2072-78. Кроме того, Isner и его группой продемонстрировано, что добавление ANG-2 к VEGF стимулировано значительно более длительную и более в большей степени кольцевую неоваскуляризацию, чем только VEGF. Asahara, Т., et al., Circ. Res. 83 (1998) 233-40. Избыток растворимого рецептора Tie2 препятствовал модулированию VEGF-индуцированной неоваскуляризации ANG-2. Asahara, Т., et al., Circ. Res. 83 (1998) 233-40. Siemeister, G., et al., Cancer Res. 59:3 (1999) 3185-91 с помощью ксенотрансплантатов бестимусных мышей показали, что гиперэкспрессия внеклеточных лиганд-связывающих доменов либо Flt-1, либо Tie2 в ксенотрансплантатах, приводящая в результате к значимому ингибированию биохимического пути, не может компенсироваться друг другом, что позволяет предположить, что биохимический путь рецептора VEGF и биохимический путь Tie2 следует считать двумя независимыми медиаторами, существенными для процесса ангиогенеза in vivo. Siemeister, G., et al., Cancer Res. 59:3 (1999) 3185-91. Это доказано недавней публикацией White, R., R., et al.,, Proc. Natl. Acad. Sci. USA 100 (2003) 5028-33. В их исследовании было продемонстрировано, что устойчивый к нуклеазе аптамер РНК, который специфично связывает и ингибирует ANG-2, значимо ингибирует неоваскуляризацию, индуцированную bFGF, в модели ангиогенеза микродивертикулов роговицы крыс.
Биспецифические антитела
В недавнем прошлом разработано широкое разнообразие форматов рекомбинантных антител, например, четырехвалентные биспецифические антитела, путем слияния, например, IgG-формата антитела и одноцепочечных доменов (см., например, Coloma, M.J., et al., Nature Biotech 15 (1997) 159-163; WO 2001/077342; и Morrison, S.L, Nature Biotech 25 (2007) 1233-1234).
Также разработано несколько других новых форматов, где сердцевинная структура антитела (IgA, IgD, IgE, IgG или IgM) больше не сохраняется, как, например, диа-, триа- или тетратела, минитела, несколько одноцепочечных форматов (scFv, Bis-scFv), которые способны к связыванию двух или более чем двух антигенов (Holliger, P., et al., Nature Biotech 23 (2005) 1126-1136; Fischer, N., Leger, O., Pathobiology 74 (2007) 3-14; Shen, J., et al., Journal of Immunological Methods 318 (2007) 65-74; Wu, С., et al., Nature Biotech. 25 (2007) 1290-1297).
Во всех таких форматах используют линкеры, либо для слияния сердцевины антитела (IgA, IgD, IgE, IgG или IgM) с другим связывающим белком (например, scFv), либо для слияния, например, двух Fab фрагментов или scFv (Fischer, N., Leger, О., Pathobiology 74 (2007) 3-14). Следует помнить, что в одном случае желательно сохранить эффекторные функции, такие как, например, комплемент-зависимая цитотоксичность (CDC) или антителозависимая клеточная цитотоксичность (ADCC), которые опосредованы связыванием рецептора Fc, путем сохранения высокой степени подобия встречающимся в природе антителам.
В WO 2007/024715 описаны иммуноглобулины с двойным вариабельным доменом, как сконструированные поливалентные и полиспецифические связывающие белки. Способ получения биологически активных димеров антитела описан в US 6897044. Конструкция Fy поливалентного антитела, имеющего по меньшей мере четыре вариабельных домена, которые связаны друг с другом посредством пептидных линкеров, описана в US 7129330. Димерные и мультимерные антигенсвязывающие структуры описаны в US 2005/0079170. Трех-или четырехвалентный моноспецифический антигенсвязывающий белок, содержащий три или четыре Fab фрагмента, связанные друг с другом ковалентно посредством соединительной структуры, где этот белок не является природным иммуноглобулином, описан в US 6511663. В WO 2006/020258 описаны четырехвалентные биспецифические антитела, которые могут эффективно экспрессироваться в прокариотических и эукариотических клетках, и полезны в терапевтических и диагностических методах. Способ разделения или преимущественного синтеза димеров, которые связаны посредством по меньшей мере одной межцепочечной дисульфидной связи, и димеров, которые не связаны посредством по меньшей мере одной межцепочечной дисульфидной связи, из смеси, содержащей эти два типа полипептидных димеров, описан в US 2005/0163782. Биспецифические тетравалентные рецепторы описаны в US 5959083. Сконструированные антитела с тремя или большим числом функциональных антигенсвязывающих сайтов описаны в WO 2001/077342.
Полиспецифические и поливалентные антигенсвязывающие полипептиды описаны в WO 1997/001580. В WO 1992/004053 описаны гомоконъюгаты, типично полученные из моноклональных антител класса IgG, которые связываются с одной и той же антигенной детерминантой, и ковалентно связанные посредством синтетического сшивания. Олигомерные моноклональные антитела с высоким авидитетом к антигену описаны в WO 1991/06305, где секретируются олигомеры, типично класса IgG, имеющие два или более чем два иммуноглобулиновых мономера, ассоциированные вместе с образованием четырехвалентных или шестивалентных молекул IgG. Антитела, полученные от овцы, и сконструированные конструкции антител описаны в US 6350860, где их можно применять при лечении заболеваний, где активность интерферона гамма является патогенной. В US 2005/0100543 описаны направляемые конструкции, которые являются поливалентными носителями биспецифических антител, то есть каждая молекула направляемой конструкции может служить в качестве носителя двух или более чем двух биспецифических антител. Генетически сконструированные биспецифические четырехвалентные антитела описаны в W01995/009917. В WO 2007/109254 описаны стабилизированные связывающие молекулы, которые состоят из стабилизированного scFv или включают его.
Комбинация ингибиторов VEGF и ANG-2
WO 2007/068895 относится к комбинации антагониста ANG-2 и VEGF, антагонистов KDR и/или FLTL. WO 2007/089445 относится к комбинациям ингибиторов ANG-2 и VEGF.
WO 2003/106501 относится к слитым белкам, связывающимся с ангиопоэтином и содержащим домен мультимеризации. WO 2008/132568 относится к слитым белкам, связывающимся с ангиопоэтином и VEGF. W0 2003/020906 относится к поливалентным белковым конъюгатам с множественными лиганд-связывающими доменами рецепторов.
WO 2009/136352 относится к антиангиогенным соединениям.
Краткое изложение сущности изобретения
Изобретение направлено на биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:1 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:2; и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:3 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:4.
В одном аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF; и
b) модифицированной тяжелой цепи и модифицированной легкой цепи первого полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:7 в качестве тяжелой цепи первого полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:5 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:8 в качестве модифицированной тяжелой цепи второго полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:6 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:11 в качестве тяжелой цепи первого полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:9 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:12 в качестве модифицированной тяжелой цепи второго полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:10 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:15 в качестве тяжелой цепи первого полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:13 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:16 в качестве модифицированной тяжелой цепи второго полноразмерного антитела, и аминокислотной последовательности SEQ ID NO:14 в качестве модифицированной легкой цепи второго полноразмерного антитела.
Следующими аспектами изобретения являются: фармацевтическая композиция, содержащая биспецифическое антитело, данная композиция для лечения рака, применение данного биспецифического антитела для получения лекарственного средства для лечения рака, способ лечения пациента, страдающего раком, путем введения данного биспецифического антитела пациенту, нуждающемуся в таком лечении.
Следующими аспектами изобретения являются: фармацевтическая композиция, содержащая биспецифическое антитело, данная композиция для лечения сосудистых заболеваний, применение данного биспецифического антитела для получения лекарственного средства для лечения сосудистых заболеваний, способ лечения пациента, страдающего сосудистым заболеванием, путем введения данного биспецифического антитела пациенту, нуждающемуся в таком лечении.
Следующим аспектом изобретения является молекула нуклеиновой кислоты, кодирующая цепь биспецифического антитела в соответствии с изобретением.
Далее в изобретении предложены экспрессионные векторы, содержащие нуклеиновую кислоту в соответствии с изобретением, способные экспрессировать эту нуклеиновую кислоту в прокариотической или эукариотической клетке-хозяине, и клетки-хозяева, содержащие такие векторы, для рекомбинантного продуцирования биспецифического антитела в соответствии с изобретением.
Далее изобретение включает прокариотическую или эукариотическую клетку-хозяина, содержащую вектор в соответствии с изобретением.
Далее изобретение включает способ получения биспецифического антитела в соответствии с изобретением, характеризующийся экспрессией нуклеиновой кислоты в соответствии с изобретением в прокариотической или эукариотической клетке-хозяине и выделение биспецифического антитела из клетки или из супернатанта клеточной культуры. Далее изобретение включает антитело, полученное таким способом получения биспецифического антитела.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 и SEQ ID NO:8.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:9, SEQ ID NO:10 SEQ ID NO:11 и SEQ ID NO:12.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15 и SEQ ID NO:16.
Биспецифические, двухвалентные антитела в соответствии с изобретением проявляют полезные эффекты для пациентов-людей, нуждающихся в терапии, направленной на VEGF и ANG-2. Антитела в соответствии с изобретением обладают в высокой степени ценными свойствами, дающими пользу пациенту, страдающему таким заболеванием, в частности, страдающему раком. Биспецифические антитела в соответствии с изобретением высокоэффективны при ингибировании опухолевого роста и/или ингибировании опухолевого ангиогенеза или сосудистых заболеваний. Биспецифические, двухвалентные антитела <VEGF-ANG-2> в соответствии с изобретением проявляют ценные фармакокинетические/-динамические свойства, такие как, например, стабильность, хороший (то есть медленный) клиренс (например, при низких дозах).
Биспецифические антитела в соответствии с изобретением высоко эффективны при
a) ингибировании опухолевого роста (например, посредством биспецифических антител в соответствии с изобретением стаз опухоли может быть уже достигнут при более низких концентрациях по сравнению с комбинацией двух моноспецифических антител (например, в моделях опухолей Colo205 и KPL-4 Примера 9 и 10 стаз опухоли был уже достигнут при 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина), и/или
b) ингибировании опухолевого ангиогенеза или сосудистых заболеваний (например, максимальные антиангиогенные эффекты посредством биспецифических антител в соответствии с изобретением могли быть уже достигнуты при более низких концентрациях по сравнению с комбинацией двух моноспецифических антител (например, в анализе ангиогенеза роговицы мыши Примера 8 максимальный антиангиогенный эффект был уже достигнут при 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина).
Описание графических материалов
Фиг.1. Примерный формат двухвалентного биспецифического антитела для примеров XMab, включающий домены СН3, модифицированные как выступы-во-впадины.
Фиг.2а. Примерный формат двухвалентного биспецифического антитела для примеров OAscFab, включающий домены СН3, модифицированные как выступы-во-впадины.
Фиг.2b. Примерный формат двухвалентного биспецифического антитела для примера OAscXFab1, включающий домены СН3, модифицированные как выступы-во-впадины.
Фиг.2с. Примерный формат двухвалентного биспецифического антитела для примеров OAscXFab2 и OAscXFab3, включающий домены СН3, модифицированные как выступы-во-впадины.
Фиг.3. Одновременное связывание <VEGF-Ang-2> XMab1 с VEGF (первая стадия) с последующим связыванием с hAng-2 (вторая стадия).
Фиг.4. Принцип ELISA для количественного определения связывания активных антител mAb <Ang2/VEGF>.
Фиг.5. Калибровочная кривая ELISA для количественного определения связывания активных антител <Ang2/VEGF>XMab1.
Фиг.6. Анализ ангиогенеза роговицы мыши - ингибирование прорастания сосудов из лимба роговицы в направлении градиента VEGF введением биспецифического антитела в соответствии с изобретением.
Фиг.7. Анализ ангиогенеза роговицы мыши - ингибирование ангиогенеза/прорастания сосудов из лимба роговицы в направлении градиента VEGF введением биспецифического антитела в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF>XMab1, <Ang2>Mab ANG2i-LC06 (LC06), <VEGF>Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF>Mab бевацизумаба (авастина).
Фиг.8. Ингибирование опухолевого роста in vivo в ксенотрансплантате мыши колоректального рака человека Colo205 (мелкие опухоли) биспецифическим антителом в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF>XMab1, <Ang2>Mab ANG2i-LC06 (LC06), <VEGF>Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF>Mab бевацизумаба (авастина).
Фиг.9. Ингибирование опухолевого роста in vivo в ксенотрансплантате мыши колоректального рака человека Colo205 (большие опухоли) биспецифическим антителом в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF>XMab1, <Ang2>Mab ANG2i-LC06 (LC06),<VEGF>Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF>Mab бевацизумаба (авастина).
Фиг. 10. Ингибирование опухолевого роста in vivo в ксенотрансплантате мыши рака молочной железы человека KPL-4 (мелкие опухоли) биспецифическим антителом в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF> XMab1, <Ang2> Mab ANG2i-LC06 (LC06), <VEGF> Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF> Mab бевацизумаба (авастина).
Фиг. 11. Ингибирование опухолевого роста in vivo в ксенотрансплантате мыши рака молочной железы человека KPL-4 (большие опухоли) биспецифическим антителом в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF> XMab1, <Ang2> Mab ANG2i-LC06 (LC06), <VEGF> Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF> Mab бевацизумаба (авастина).
Фиг. 12. Ингибирование опухолевого роста in vivo в ксенотрансплантате мыши рака желудка N87 биспецифическим антителом в соответствии с изобретением - сравнение биспецифического антитела <Ang2/VEGF> XMab1, <Ang2> Mab ANG2i-LC06 (LC06), <VEGF> Mab бевацизумаба (авастина) и комбинации ANG2i-LC06 и <VEGF> Mab бевацизумаба (авастина).
Фиг. 13. Ингибирование опухолевого роста in vivo в ксенотрансплантате колоректального рака Colo205 антителом XMab1 8 мг/кг и комбинацией антител LC06 и авастина (по 4 мг/кг).
Подробное описание изобретения
Изобретение направлено на биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека. Характеризующееся тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO: 1 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO: 2; и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO: 3 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO: 4.
В одном аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF;
b) модифицированной тяжелой цепи и модифицированной легкой цепи полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом.
Данный формат биспецифического, двухвалентного антитела для биспецифического антитела, специфично связывающегося с фактором роста эндотелия сосудов человека (VEGF) и ангиопоэтином-2 (ANG-2) человека, описан в WO 2009/080253 (см. примерную схему при включении доменов СН3, модифицированных как выступы-во-впадины, на Фиг.1). Антитела, основанные на данном формате биспецифического, двухвалентного антитела, названы ХМаЬ в примерах настоящего изобретения.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:7 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:5 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:8 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:6 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:11 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:9 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:12 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:10 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:15 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:13 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:16 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:14 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:19 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:17 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:20 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:18 в качестве модифицированной легкой цепи второго полноразмерного антитела.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:23 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:21 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:24 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:22 в качестве модифицированной легкой цепи второго полноразмерного антитела
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:27 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:25 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:28 в качестве модифицированной тяжелой цепи второго полноразмерного антитела и аминокислотной последовательности SEQ ID NO:26 в качестве модифицированной легкой цепи второго полноразмерного антитела.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 и SEQ ID NO:8.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11 и SEQ ID NO:12.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15 и SEQ ID NO:16.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:19 и SEQ ID NO:20.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:23 и SEQ ID NO:24.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:25, SEQ ID NO:26, SEQ ID NO:27 и SEQ ID NO:28.
В другом аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
а) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается с ANG-2, где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера.
Примерная схема данного формата биспецифического, двухвалентного антитела для данного биспецифического антитела, специфично связывающегося с фактором роста эндотелия сосудов человека (VEGF) и ангиопоэтином-2 (ANG-2) человека, показана на Фиг.2а, включающего домены СН3, модифицированные как выступы-во-впадины. Антитела на основе данного формата биспецифического, двухвалентного антитела названы OAscFab в примерах настоящего изобретения.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:30 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:31 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:29 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
В одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием
a) аминокислотной последовательности SEQ ID NO:33 в качестве тяжелой цепи первого полноразмерного антитела и аминокислотной последовательности SEQ ID NO:34 в качестве легкой цепи первого полноразмерного антитела, и
b) аминокислотной последовательности SEQ ID NO:32 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
В одной форме осуществления вариабельный домен тяжелой цепи (VH) антитела и вариабельный домен легкой цепи (VL) антитела тяжелой и легкой цепи второго полноразмерного антитела стабилизированы дисульфидом путем введения дисульфидной связи между нижеследующими положениями:
положением вариабельного домена тяжелой цепи 44 и положением вариабельного домена легкой цепи 100 (нумерация всегда в соответствии с индексом EU по Kabat (Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991))). Такая дополнительная дисульфидная стабилизация достигается введением дисульфидной связи между вариабельными доменами VH и VL тяжелой и легкой цепи второго полноразмерного антитела. Методы введения неприродных дисульфидных мостиков для стабилизации описаны, например, в WO 94/029350, Rajagopal, V., et al, Prot. Engin. 10 (1997) 1453-59; Kobayashi, et al., Nuclear Medicine & Biology, Vol.25 (1998) 387-393; или Schmidt, M, et al., Oncogene 18 (1999) 1711-1721.
Таким образом, в одной форме осуществления такое биспецифическое, двухвалентное антитело характеризуется содержанием дисульфидной связи между вариабельными доменами тяжелой и легкой цепи второго полноразмерного антитела, которая находится между положением 44 вариабельного домена тяжелой цепи и положением 100 вариабельного домена легкой цепи, и включает
a) аминокислотную последовательность SEQ ID NO:36 в качестве тяжелой цепи первого полноразмерного антитела, и аминокислотную последовательность SEQ ID NO:37 в качестве тяжелой цепи первого полноразмерного антитела, и
b) аминокислотную последовательность SEQ ID NO:35 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
В другом аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается с ANG-2, где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера, и где вариабельные домены VL и VH заменены друг другом.
Примерная схема данного формата биспецифического, двухвалентного антитела для данного биспецифического антитела, специфично связывающегося с фактором роста эндотелия сосудов человека (VEGF) и ангиопоэтином-2 (ANG-2) человека, показана на Фиг.2b, включая домены СН3, модифицированные как выступы-во-впадины. Антитела на основе данного формата биспецифического, двухвалентного антитела названы в примерах OAscXFab1.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) SEQ ID NO:39 в качестве тяжелой цепи первого полноразмерного антитела и SEQ ID NO:40 в качестве тяжелой цепи первого полноразмерного антитела, и
b) SEQ ID NO:38 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
В другом аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается с ANG-2,
где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера; и
где константные домены CL и СН1 заменены друг другом.
Примерная схема данного формата биспецифического, двухвалентного антитела для данного биспецифического антитела, специфично связывающегося с фактором роста эндотелия сосудов человека (VEGF) и ангиопоэтином-2 (ANG-2) человека, показана на Фиг.2с, включая домены СН3, модифицированные как выступы-во-впадины. Антитела на основе данного формата биспецифического, двухвалентного антитела названы в примерах OAscXFab2 и OAscXFab3.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) SEQ ID NO:42 в качестве тяжелой цепи первого полноразмерного антитела и SEQ ID NO:43 в качестве тяжелой цепи первого полноразмерного антитела, и
b) SEQ ID NO:41 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) SEQ ID NO:45 в качестве тяжелой цепи первого полноразмерного антитела и SEQ ID NO:46 в качестве тяжелой цепи первого полноразмерного антитела,и
b) SEQ ID NO:44 в качестве тяжелой цепи второго полноразмерного антитела, соединенной с легкой цепью второго полноразмерного антитела посредством пептидного линкера.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:29, SEQ ID NO:30 и SEQ ID NO:31.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:32, SEQ ID NO:33 и SEQ ID NO:34.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:35, SEQ ID NO:36 и SEQ ID NO:37.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:38, SEQ ID NO:39 и SEQ ID NO:40.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:41, SEQ ID NO:42 и SEQ ID NO:43.
Соответственно, одной формой осуществления изобретения является биспецифическое, двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся содержанием аминокислотных последовательностей SEQ ID NO:44, SEQ ID NO:45 и SEQ ID NO:46.
Предпочтительно домены СН3 биспецифического, двухвалентного антитела в соответствии с изобретением изменены с помощью технологии "выступы-во-впадины", которая подробно описана с несколькими примерами, например, в WO 96/027011, Ridgway J.B., et al., Protein Eng 9 (1996)617-621; и Merchant, A.M., et al., Nat Biotechnol 16 (1998) 677-681. При данном способе поверхности взаимодействия двух доменов СН3 изменены, чтобы повысить гетеродимеризацию обеих тяжелых цепей, содержащих эти два домена СН3. Каждый из двух доменов СН3 (двух тяжелых цепей) может представлять собой "выступ", тогда как другой представляет собой "впадину". Введение дисульфидного мостика стабилизирует гетеродимеры (Merchant, A.M, et al., Nature Biotech 16 (1998) 677-681; Atwell, S., et al. J. Mol. Biol. 270 (1997) 26-35) и повышает выход.
В предпочтительном аспекте изобретения все биспецифические антитела в соответствии с изобретением характеризуются тем, что
каждый домен СН3 тяжелой цепи и домен СН3 другой тяжелой цепи совпадают на границе, которая содержит исходную границу между доменами СН3 антитела;
где граница изменена таким образом, что способствует образованию биспецифического антитела, где это изменение характеризуется тем, что:
a) домен СН3 одной тяжелой цепи изменен,
таким образом, что в пределах исходной границы домена СН3 одной тяжелой цепи, которая совпадает с исходной границей домена СН3 другой тяжелой цепи в пределах биспецифического антитела,
аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, создавая посредством этого выступ в пределах границы домена СН3 одной тяжелой цепи, который может быть расположен в полости внутри границы домена СН3 другой тяжелой цепи,
и
b) домен СН3 другой тяжелой цепи изменен,
таким образом, что в пределах исходной границы домена СН3 одной тяжелой цепи, которая совпадает с исходной границей домена СН3 другой тяжелой цепи в пределах биспецифического антитела,
аминокислотный остаток заменен аминокислотным остатком, имеющим меньший объем боковой цепи, создавая посредством этого полость в пределах границы домена СН3, внутри которой может быть расположен выступ в пределах границы первого домена СН3.
Таким образом, антитело в соответствии с изобретением предпочтительно характеризуется тем, что
каждый домен СН3 тяжелой цепи полноразмерного антитела а) и домен СН3 тяжелой цепи полноразмерного антитела b) совпадает на границе, которая содержит изменение в исходной границе между доменами СН3 антитела;
где i) в домене СН3 одной тяжелой цепи
аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, создавая посредством этого выступ в пределах границы домена СН3 одной тяжелой цепи, который может быть расположен в полости внутри границы домена СН3 другой тяжелой цепи,
и где
ii) в домене СН3 другой тяжелой цепи
аминокислотный остаток заменен аминокислотным остатком, имеющим меньший объем боковой цепи, создавая посредством этого полость в пределах границы домена СН3, внутри которой может быть расположен выступ в пределах границы первого домена СН3.
Предпочтительно аминокислотный остаток, имеющий больший объем боковой цепи, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W).
Предпочтительно аминокислотный остаток, имеющий меньший объем боковой цепи, выбран из группы, состоящей из аланина (А), серина (S), треонина (Т), валина (V).
В одном аспекте изобретения оба домена СН3 дополнительно изменены путем введения цистеина (С) в качестве аминокислоты в соответствующих положениях каждого домена СН3, так что может образоваться дисульфидный мостик между обоими доменами СН3.
В одной форме осуществления биспецифическое антитело содержит мутацию T366W в домене СН3 "цепи выступа" и мутации T366S, L368A, Y407V в домене СН3 "цепи впадины". Дополнительный межцепочечный дисульфидный мостик между доменами СН3 можно также использовать (Merchant, A.M, et al., Nature Biotech 16 (1998) 677-681), например, путем введения мутации Y349C в домен СН3 "цепи выступа" и мутации Е356С или мутации S354C в домен СН3 "цепи впадины".
В другой форме осуществления биспецифическое антитело в соответствии с изобретением содержит мутации Y349C, T366W в одном из двух доменов СН3 и мутации Е356С, T366S, L368A, Y407VB другом из двух доменов СН3. В другой предпочтительной форме осуществления биспецифическое антитело содержит мутации Y349C, T366W в одном из двух доменов СН3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов СН3 (где дополнительная мутация Y349C в одном домене СН3 и дополнительная мутация Е356С или S354C в другом домене СН3 образуют межцепочечный дисульфидный мостик) (нумерация всегда соответствует индексу ED no Kabat; (Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991))). Но также альтернативно или дополнительно можно использовать другие технологии выступы-во-впадины, как описано в ЕР 1870459 А1. Таким образом, другим примером для биспецифического антитела являются мутации R409D; К370Е в домене СН3 "цепи выступа" и мутации D399K;
Е357К в домене СН3 "цепи впадины" (нумерация всегда соответствует индексу EU по Kabat (Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991))).
В другой форме осуществления биспецифическое антитело содержит мутацию T366W в домене СН3 "цепи выступа" и мутации T366S, L368A, Y407V в домене СН3 "цепи впадины" и дополнительно мутации R409D; К370Е в домене СН3 "цепи выступа" и мутации D399K; Е357К в домене СН3 "цепи впадины".
В другой форме осуществления биспецифическое антитело содержит мутации Y349C, T366W в дном из двух доменов СН3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов СН3, либо трехвалентное, биспецифическое антитело содержит мутации Y349C, T366W в одном из двух доменов СН3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов СН3 и дополнительно мутации R409D; К370Е в домене СН3 "цепи выступа" и мутации D399K; Е357К в домене СН3 "цепи впадины".
В одной форме осуществления изобретения биспецифическое антитело в соответствии с изобретением характеризуется тем, что обладает одним или более чем одним из нижеописанных свойств (определенных в анализах, которые описаны в Примерах 3-7):
- биспецифическое, двухвалентное антитело связывается с VEGF со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело связывается с ANG-2 со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело ингибирует ANG-2-индуцированное фосфорилирование Tie2 в клетках НЕК293, трансфицированных Tie2, с IC50 15нМ или менее (в одной форме осуществления с IC50 10 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание ANG-2 с Tie2 с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание VEGF с рецептором VEGF с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует VEGF-индуцированную пролиферацию клеток HUVEC с IC50 10 нМ или менее (в одной форме осуществления с IC50 5 нМ или менее).
В одной форме осуществления биспецифическое, двухвалентное антитело характеризуется содержанием
первого антигенсвязывающего сайта, который специфично связывается с VEGF человека, и второго антигенсвязывающего сайта, который специфично связывается с ANG-2 человека, характеризующихся тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:1 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:2; и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:3 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:4;
и обладает одним или более чем одним из нижеописанных свойств (определенных в анализах, которые описаны в Примерах 3-7):
- биспецифическое, двухвалентное антитело связывается с VEGF со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело связывается с ANG-2 со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело ингибирует ANG-2-индуцированное фосфорилирование Tie2 в клетках НЕК293, трансфицированных Tie2, с IC50 15нМ или менее (в одной форме осуществления с IC50 10 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание ANG-2 с Tie2 с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание VEGF с рецептором VEGF с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует VEGF-индуцированную пролиферацию клеток HUVEC с IC50 10 нМ или менее (в одной форме осуществления с IC50 5 нМ или менее).
В одном аспекте изобретения такое биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF;
b) модифицированной тяжелой цепи и модифицированной легкой цепи полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом;
и обладает одним или более чем одним из нижеописанных свойств (определенных в анализах, которые описаны в Примерах 3-7):
- биспецифическое, двухвалентное антитело связывается с VEGF со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело связывается с ANG-2 со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело ингибирует ANG-2-индуцированное фосфорилирование Tie2 в клетках НЕК293, трансфицированных Tie2, с IC50 15нМ или менее (в одной форме осуществления с IC50 10 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание ANG-2 с Tie2 с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание VEGF с рецептором VEGF с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует VEGF-индуцированную пролиферацию клеток HUVEC с IC50 10 нМ или менее (в одной форме осуществления с IC50 5 нМ или менее).
В одной форме осуществления биспецифическое, двухвалентное антитело характеризуется содержанием
первого антигенсвязывающего сайта, который специфично связывается с VEGF человека, и второго антигенсвязывающего сайта, который специфично связывается с ANG-2 человека, характеризующихся тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:1 с заменой не более чем одного аминокислотного остатка в CDR и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:2 с заменой не более чем одного аминокислотного остатка в CDR; и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:3 с заменой не более чем одного аминокислотного остатка в CDR и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:4 с заменой не более чем одного аминокислотного остатка в CDR.
В одной форме осуществления биспецифическое, двухвалентное антитело характеризуется содержанием
первого антигенсвязывающего сайта, который специфично связывается с VEGF человека, и второго антигенсвязывающего сайта, который специфично связывается с ANG-2 человека, характеризующихся тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:1 с заменой не более чем одного аминокислотного остатка в CDR и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:2 с заменой не более чем одного аминокислотного остатка в CDR; и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO:3 с заменой не более чем одного аминокислотного остатка в CDR и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO:4 с заменой не более чем одного аминокислотного остатка в CDR;
и обладает одним или более чем одним из нижеописанных свойств (определенных в анализах, которые описаны в Примерах 3-7):
- биспецифическое, двухвалентное антитело связывается с VEGF со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело связывается с ANG-2 со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело ингибирует ANG-2-индуцированное фосфорилирование Tie2 в клетках НЕК293, трансфицированных Tie2, с IC50 15 нМ или менее (в одной форме осуществления с IC50 10 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание ANG-2 с Tie2 с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание VEGF с рецептором VEGF с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует VEGF-индуцированную пролиферацию клеток HUVEC с IC50 10 нМ или менее (в одной форме осуществления с IC50 5 нМ или менее).
В одном аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF,
и где тяжелая цепь первого полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:7 с заменой не более чем одного аминокислотного остатка в CDR, и легкая цепь первого полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:5 с заменой не более чем одного аминокислотного остатка в CDR, и
b) модифицированной тяжелой цепи и модифицированной легкой цепи полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом,
и где модифицированная тяжелая цепь второго полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:8 с заменой не более чем одного аминокислотного остатка в CDR, и модифицированная легкая цепь второго полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:6 с заменой не более чем одного аминокислотного остатка в CDR.
В одном аспекте изобретения биспецифическое антитело в соответствии с изобретением характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с VEGF,
и где тяжелая цепь первого полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:7 с заменой не более чем одного аминокислотного остатка в CDR, и легкая цепь первого полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:5 с заменой не более чем одного аминокислотного остатка в CDR, и
b) модифицированной тяжелой цепи и модифицированной легкой цепи полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом,
и где модифицированная тяжелая цепь второго полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:8 с заменой не более чем одного аминокислотного остатка в CDR, и модифицированная легкая цепь второго полноразмерного антитела содержит аминокислотную последовательность SEQ ID NO:6 с заменой не более чем одного аминокислотного остатка в CDR;
и обладает одним или более чем одним из нижеописанных свойств (определенных в анализах, которые описаны в Примерах 3-7):
- биспецифическое, двухвалентное антитело связывается с VEGF со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело связывается с ANG-2 со значением KD связывающего сродства 5 нМ или менее;
- биспецифическое, двухвалентное антитело ингибирует ANG-2-индуцированное фосфорилирование Tie2 в клетках НЕК293, трансфицированных Tie2, с IC50 15 нМ или менее (в одной форме осуществления с IC50 10 нМ или менее);
биспецифическое, двухвалентное антитело ингибирует связывание ANG-2 с Tie2 с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует связывание VEGF с рецептором VEGF с IC50 20 нМ или менее (в одной форме осуществления с IC50 15 нМ или менее);
- биспецифическое, двухвалентное антитело ингибирует VEGF-индуцированную пролиферацию клеток HUVEC с IC50 10 нМ или менее (в одной форме осуществления с IC50 5 нМ или менее).
Как используют в данной заявке, "антитело" относится к связывающему белку, который содержит антигенсвязывающие сайты. Термины "связывающий сайт" или "антигенсвязывающий сайт", как используют в данной заявке, обозначает участок (участки) молекулы антитела, с которым действительно связывается лиганд. Термин "антигенсвязывающий сайт" включает вариабельные домены тяжелой цепи антитела (VH) и вариабельные домены легкой цепи антитела (VL) (пару VH/VL)).
Специфичность антитела относится к селективному распознаванию антителом определенного эпитопа антигена. Природные антитела, например, являются моноспецифическими.
"Биспецифические антитела" в соответствии с изобретением представляют собой антитела, которые имеют две различные антигенсвязывающие специфичности. Антитела по настоящему изобретению специфичны к двум различным антигенам, VEGF в качестве первого антигена и ANG-2 в качестве второго антигена.
Термин "моноспецифическое" антитело, как используют в данной заявке, обозначает антитело, которое имеет один или более чем один связывающий сайт, каждый из которых связывается с одним и тем же эпитопом одного и того же антигена.
Термин "валентный", как используют в настоящей заявке, обозначает присутствие указанного числа связывающих сайтов в молекуле антитела. Как таковые, термины "двухвалентное", "четырехвалентное" и "шестивалентное" означают присутствие двух связывающих сайтов, четырех связывающих сайтов и шести связывающих сайтов, соответственно, в молекуле антитела. Биспецифические антитела в соответствии с изобретением являются "двухвалентными".
Термин "VEGF", как используют в данной заявке, относится к фактору роста эндотелия сосудов человека (VEGF/VEGF-A) (SEQ ID NO:47), который описан, например, в Leung, D.W., et al., Science 246 (1989) 1306-9; Keck, P.J., et al., Science 246 (1989) 1309-12 и Connolly, D.T., et al., J. Biol. Chem. 264 (1989) 20017-24. VEGF вовлечен в регуляцию нормального и аномального ангиогенеза и неоваскуляризации, ассоциированной с опухолями и внутриглазными расстройствами (Ferrara, N.. et al., Endocr. Rev. 18 (1997) 4-25; Berkman, R.A.,et al., J. Clin. Invest. 91 (1993) 153-159; Brown, L.F., et al., Human Pathol. 26 (1995) 86-91; Brown, L.F., et al., Cancer Res. 53 (1993) 4727-4735; Mattern, J., et al., Brit. J. Cancer. 73 (1996) 931-934; and Dvorak, H.F„ et al., Am. J. Pathol. 146 (1995) 1029-1039). VEGF представляет собой гомодимерный гликопротеин, который выделен из нескольких источников. VEGF проявляет высоко специфичную митогенную активность в отношении эндотелиальных клеток.
Термин "ANG-2", как используют в данной заявке, относится к ангиопоэтину-2 (ANG-2) человека (альтернативно сокращаемому как ANGPT2 или ANG2) (SEQ ID NO: 48), который описан, например, в Maisonpierre, P.C., et al, Science 277 (1997) 55-60 и Cheung, A.M., et al., Genomics 48 (1998) 389-91. Ангиопоэтины-1 и -2 были открыты как лиганды к Tie, семейству тирозинкиназ, которые селективно экспрессируются в пределах эндотелия кровеносных сосудов. Yancopoulos, G.D., et al., Nature 407 (2000) 242-48. Сейчас существует четыре отдельных члена семейства ангиопоэтинов. Ангиопоэтин-3 и -4 (Ang-3 и Ang-4) могут представлять собой широко дивергировавшие копии локуса одного и того же гена у мыши и человека. Kirn, I., et al., FEBS Let, 443 (1999) 353-56; Kirn, I., et al., J Biol Chem 274 (1999) 26523-28. ANG-1 и ANG-2 были исходно идентифицированы в экспериментах на тканевых культурах как агонист и антагонист, соответственно (см. для ANG-1: Davis, S., et al., Cell 87 (1996) 1161-69; и для ANG-2: Maisonpierre, P.С., et al., Science 277 (1997) 55-60). Все известные ангиопоэтины связываются, главным образом, с Tie2, и как Ang-1, так и -2 связываются с Tie2 со сродством 3 нМ (Kd). Maisonpierre, P.C., et al., Science 277 (1997) 55-60.
Антигенсвязывающие сайты биспецифического антитела по изобретению содержат шесть участков, определяющих комплементарность (CDR), которые вносят вклад в варьирующие степени сродства связывающего сайта к антигену. Существует три CDR вариабельного домена тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три CDR вариабельного домена легкой цепи (CDRL1, CDRL2 и CDRL3). Степень CDR и каркасных областей (FR) определяется путем сравнения со скомпилированной базой данных аминокислотных последовательностей, в которых эти области определены в соответствии с вариабельностью среди последовательностей. Также включенными в объем изобретения являются функциональные антигенсвязывающие сайты, состоящие из меньшего числа CDR (то есть, где связывающая специфичность определяется тремя, четырьмя или пятью CDR). Например, неполный набор 6 CDR может быть достаточен для связывания. В некоторых случаях достаточно домена VH или VL
Антитела по изобретению дополнительно содержат константные области иммуноглобулина одного или более чем одного класса иммуноглобулинов. Классы иммуноглобулинов включают изотипы IgG, IgM, IgA, IgD и IgE и, в случае IgG и IgA, их подтипы.
Термины "моноклональное антитело" или "композиция моноклонального антитела", как используют в данной заявке, относятся к препарату молекул антитела одной аминокислотной композиции.
Термин "химерное антитело" относится к антителу, содержащему вариабельный участок, то есть связывающий участок, из одного источника или вида и по меньшей мере участок константной области, имеющей происхождение из другого источника или вида, обычно полученному методами рекомбинантных ДНК. Химерные антитела, содержащие вариабельный участок мыши и константную область человека, являются предпочтительными. Другими предпочтительными формами "химерных антител", охваченными настоящим изобретением, являются формы, в которых константная область модифицирована или изменена по сравнению с исходным антителом для получения свойств в соответствии с изобретением, в частности, в отношении связывания C1q и/или связывания Fc рецептора (FcR). Такие химерные антитела также называют "антителами с переключенным классом". Химерные антитела являются продуктом экспрессируемых генов иммуноглобулинов, содержащих сегменты ДНК, кодирующие вариабельные участки иммуноглобулинов, и сегменты ДНК, кодирующие константные области иммуноглобулинов. Способы получения химерных антител, включающие общепринятые методы рекомбинантных ДНК и трансфекции генов, хорошо известны в данной области техники. См., например, Morrison, S.L, et al„ Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855; US 5202238 и US 5204244.
Термин "гуманизированное антитело" относится к антителам, в которых каркасные области или "участки, определяющие комплементарность" (CDR) модифицированы таким образом, что они содержат CDR иммуноглобулина другой специфичности по сравнению со специфичностью родительского иммуноглобулина. В предпочтительной форме осуществления CDR мыши прививают на каркасную область человеческого антитела с получением "гуманизированного антитела". См., например, Riechmann, L, et al., Nature 332 (1988) 323-327; и Neuberger, M.S., et al., Nature 314 (1985) 268-270. Особенно предпочтительные CDR соответствуют репрезентативным последовательностям, распознающим антигены, отмеченным выше для химерных антител. Другими формами "гуманизированных антител", охваченными настоящим изобретением, являются формы, в которых константная область дополнительно модифицирована или изменена по сравнению с исходным антителом для получения свойств в соответствии с изобретением, в частности, в отношении связывания C1q и/или связывания Fc рецептора (FcR).
Термин "человеческое антитело", как используют в данной заявке, подразумевают как включающий антитела, имеющие вариабельные и константные области, имеющие происхождение от последовательностей иммуноглобулинов зародышевой линии человека. Человеческие антитела хорошо известны на уровне техники (van Dijk, M.A., and van de Winkel, J.G., Curr. Opin. Chem. Biol. 5 (2001) 368-374). Человеческие антитела могут быть также получены в трансгенных животных (например, мышах), которые после иммунизации способны к продуцированию полного репертуара или к селекции человеческих антител в отсутствие продуцирования эндогенного иммуноглобулина. Перенос генетической информации иммуноглобулина зародышевой линии человека в таких мышей, мутантных по зародышевой линии, приведет в результате к продуцированию человеческих антител после антигенной стимуляции (см., например, Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 2551-2555; Jakobovits, A., et al., Nature 362 (1993) 255-258; Brueggemann, M., et al., Year Immunol. 7 (1993) 33-40). Человеческие антитела можно также продуцировать в библиотеках фагового дисплея (Hoogenboom, H.R., and Winter, G., J. Mol. Biol. 227 (1992) 381-388; Marks, J.D., et al., J. Mol. Biol. 222 (1991) 581-597). Методы Cole, A., et al. and Boerner, P., et al. также доступны для получения человеческих моноклональных антител (Cole, A., et al., Monoclonal Antibodies and Cancer Therapy, Liss, A.L, p.77 (1985); и Boerner, P., et al., J. Immunol. 147 (1991) 86-95). Как уже упомянуто для химерных и гуманизированных антител в соответствии с изобретением, термин "человеческое антитело", как используют в данной заявке, также включает такие антитела, которые модифицированы в константной области для получения свойств в соответствии с изобретением, в частности, в отношении связывания C1q и/или связывания FcR, например, путем "переключения класса", то есть изменения или мутации участков Fc (например, с IgG1 на IgG4 и/или мутации IgG1/IgG4).
Термин "рекомбинантное человеческое антитело", как используют в данной заявке, подразумевают как включающий все человеческие антитела, которые получают, экспрессируют, создают или выделяют рекомбинантными методами, как, например, антитела, выделенные из клетки-хозяина, такой как клетка NSO или СНО, или из животного (например, мыши), которое является трансгенным по генам иммуноглобулинов человека, или антитела, экспрессированные с использованием рекомбинантного экспрессионного вектора, трансфицированного в клетку-хозяина. Такие рекомбинантные человеческие антитела имеют вариабельные и константные области в перестроенной форме. Рекомбинантные человеческие антитела в соответствии с изобретением подвергнуты соматической сверхмутации in vivo. Таким образом, аминокислотные последовательности участков VH и VL рекомбинантных антител представляют собой последовательности, которые, хотя имеют происхождение от последовательностей VH и VL зародышевой линии человека и родственны им, могут не существовать в природе в пределах репертуара антител зародышевой линии человека in vivo.
"Вариабельный домен" (вариабельный домен легкой цепи (VL), вариабельный домен тяжелой цепи (VH), как используют в данной заявке, означает участок каждой из пары тяжелой и легкой цепей, который непосредственно вовлечен в связывание антитела с антигеном. Домены вариабельной легкой и тяжелой цепи имеют одну и ту же общую структуру, и каждый домен содержит четыре каркасных (FR) участка, последовательности которых являются широко консервативными, соединенные тремя "гипервариабельными участками" (или участками определения комплементарности, CDR). Каркасные участки принимают конформацию р-слоя, а CDR могут образовывать петли, соединяющие структуру р-слоя. CDR в каждой цепи удерживаются в своей трехмерной структуре каркасными участками и вместе образуют с CDR от другой цепи антигенсвязывающий сайт. Участки CDR3 тяжелой и легкой цепи антитела играют особенно важную роль в связывающей специфичности/сродстве антител в соответствии с изобретением и, следовательно, составляют следующий объект изобретения.
Термины "гипервариабельный участок" или "антигенсвязывающий участок антитела" при использовании в данной заявке относятся к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Гипервариабельная область содержит аминокислотные остатки от "участков, определяющих комплементарность" или "CDR". "Каркасные" или "FR" участки представляют собой те участки вариабельного домена, которые отличаются от каркасных участков, как определено в данной заявке. Таким образом, тяжелые и легкие цепи антитела содержат от N- к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. CDR на каждой цепи разделены такими каркасными аминокислотами. В частности, CDR3 тяжелой цепи является участком, который вносит наибольший вклад в связывание антигена. Участки CDR и FR определяют в соответствии со стандартным определением Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991) (включая нумерацию в соответствии с EU индексом по Kabat (сокращенно нумерацию по Kabat в данной заявке ниже)).
Как используют в данной заявке, термин "связывание" или "специфическое связывание" относится к связыванию антитела с эпитопом антигена (либо VEGF человека, либо ANG-2 человека) в анализе in vitro, предпочтительно в анализе плазменного резонанса (BIAcore, GE-Healthcare Uppsala, Sweden) (Пример 3) с очищенным антигеном дикого типа. Сродство связывания определяют терминами ка (константа скорости ассоциации антитела из комплекса антитело/антиген), KD (константа диссоциации) и KD (kD/ka). В одной форме осуществления связывание или специфичное связывание означает связывающее сродство (KD) 10-8 моль/л или менее, предпочтительно от 10-9 М до 10-13 моль/л.
Термин "эпитоп" включает любую полипептидную детерминанту, способную к специфичному связыванию с антителом. В некоторых формах осуществления эпитопная детерминанта включает химически активные поверхностные группировки молекул, таких как аминокислоты, сахарные боковые цепи, фосфорил или сульфонил, и в некоторых формах осуществления могут обладать специфичными трехмерными структурными характеристиками или специфичными характеристиками заряда. Эпитопом является участок антигена, который связывается антителом.
В некоторых формах осуществления указано, что антитело специфично связывает антиген, когда оно преимущественно распознает его антиген-мишень в комплексной смеси белков и/или макромолекул.
Термин "полноразмерное антитело" обозначает антитело, состоящее из двух "полноразмерных тяжелых цепей антитела" и двух "полноразмерных легких цепей антитела" (см. фиг.1). "Полноразмерная тяжелая цепь антитела" представляет собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 тяжелой цепи антитела (СН1), шарнирной области (HR) антитела, константного домена 2 тяжелой цепи антитела (СН2) и константного домена 3 тяжелой цепи антитела (СНЗ), сокращенно VH-CH1-HR-CH2-CH3; и необязательно константного домена 4 тяжелой цепи антитела (СН4) в случае антитела подкласса IgE. Предпочтительно "полноразмерная тяжелая цепь антитела" представляет собой полипептид, состоящий в направлении от N-конца к С-концу из VH, СН1, HR, CH2 и СН3. "Полноразмерная легкая цепь антитела" представляет собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL), сокращенно VL-CL. Константный домен легкой цепи антитела (CL) может представлять собой к (каппа) или ^ (лямбда). Две полноразмерные цепи антитела связаны вместе посредством межполипептидных дисульфидных связей между доменом CL и доменом СН1 и между шарнирными областями полноразмерных тяжелых цепей антитела. Примерами типичных полноразмерных антител являются природные антитела, такие как IgG (например, IgG1 и IgG2), IgM, IgA, IgD и IgE. Полноразмерные антитела в соответствии с изобретением могут быть от одного вида, например, человека, либо они могут представлять собой химеризованные или гуманизированные антитела. Полноразмерные антитела в соответствии с изобретением содержат два антигенсвязывающих сайта, каждый из которых образован парой VH и VL, которые оба специфично связываются с одним и тем же антигеном. С-конец тяжелой или легкой цепи данного полноразмерного антитела означает последнюю аминокислоту на С-конце данной тяжелой или легкой цепи. N-конец тяжелой или легкой цепи данного полноразмерного антитела означает последнюю аминокислоту на М-конце данной тяжелой или легкой цепи.
Термин "пептидный линкер", как используют в пределах изобретения, означает пептид с аминокислотными последовательностями, которые предпочтительно имеют синтетическое происхождение. Эти пептиды в соответствии с изобретением используют для соединения С-конца легкой цепи с N-концом тяжелой цепи второго полноразмерного антитела (которое специфично связывается со вторым антигеном) посредством пептидного линкера. Пептидный линкер в пределах тяжелой и легкой цепи второго полноразмерного антитела представляет собой пептид с аминокислотной последовательностью длиной по меньшей мере 30 аминокислот, предпочтительно длиной от 32 до 50 аминокислот. В одной форме осуществления пептидный линкер представляет собой пептид с аминокислотной последовательностью длиной от 32 до 40 аминокислот. В одной форме осуществления данный линкер представляет собой (GxS)n, где G=глицин, S=серин (х=3, n=8, 9 или 10 и m=0, 1, 2 или 3) или (х=4 и n=6, 7 или 8 и m=О, 1, 2 или 3), предпочтительно где х=4, n=6 или 7 и m=0, 1,2 или 3, более предпочтительно где х=4, n=7 и m=2. В одной форме осуществления данный линкер представляет собой (G4S)6G2.
Термин "константная область", как используют в пределах настоящей заявки, обозначает сумму доменов антитела, отличающихся от вариабельной области. Константная область не вовлечена непосредственно в связывание антигена, но проявляет различные эффекторные функции. В зависимости от аминокислотной последовательности константной области их тяжелых цепей, антитела делятся на классы: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на подклассы, такие как IgG1, IgG2, IgG3 и IgG4, IgA1 и lgA2. Константные области тяжелых цепей, которые соответствуют различным классам антител, называют α, δ, ε, γ и µ, соответственно. Константные области легкой цепи, которые можно найти во всех пяти классах антител, называют к (каппа) и А (лямбда).
Термин "константная область человеческого происхождения", как используют в настоящей заявке, обозначает константную область тяжелой цепи человеческого антитела подкласса IgG1, IgG2, IgG3 или IgG4 и/или константную область легкой цепи каппа или лямбда. Такие константные области хорошо известны на уровне техники и описаны, например, Kabat, E.A. (см., например, Johnson, G., and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218; Kabat, E.A., et al., Proc. Natl. Acad. Sci. USA 72 (1975) 2785-2788).
Предпочтительно биспецифические, двухвалентные антитела в соответствии с изобретением имеют константную область подкласса IgG1 человека.
Хотя антитела подкласса IgG4 проявляют сниженное связывание с Рс рецептором (FcγRIIIa), антитела других подклассов IgG проявляют связывание. Однако Pro238, Asp265, Asp270, Asn297 (потеря углевода Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435 являются остатками, которые при их изменении обеспечивают также сниженное связывание с Fc рецептором (Shields, R.L, et al., J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9 (1995) 115-119; Morgan, A., et al., Immunology 86 (1995) 319-324; EP 0307434).
В одной форме осуществления антитело в соответствии с изобретением обладает сниженным связыванием с FcR по сравнению с антителом IgG1, и биспецифическое, двухвалентное антитело относится в отношении связывания с FcR к подклассу IgG4 или к подклассу IgG1 с мутацией в S228, L234, L235 и/или D265, и/или содержит мутацию PVA236. В одной форме осуществления мутаций в биспецифическом, двухвалентном антителе находятся в IgG4 S228P и L235E и в IgG1 L234A и L235A.
В другом аспекте изобретения биспецифическое, двухвалентное антитело характеризуется содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с первым антигеном;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается со вторым антигеном,
где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера; и
где вариабельные домены VL и VH или константные домены CL и СН1 заменены друг другом.
Предпочтительно домены СН3 данного формата биспецифического, двухвалентного антитела изменены с помощью технологии "выступы-во-впадины", которая подробно описана с несколькими примерами, например, в WO 96/027011, Ridgway J.B., et al., Protein Eng 9 (1996) 617-621; и Merchant, A.M., et al., Nat Biotechnol 16 (1998) 677-681. При данном способе поверхности взаимодействия двух доменов СН3 изменены, чтобы повысить гетеродимеризацию обеих тяжелых цепей, содержащих эти два домена СН3. Каждый из двух доменов СН3 (двух тяжелых цепей) может представлять собой "выступ", тогда как другой представляет собой "впадину". Введение дисульфидного мостика стабилизирует гетеродимеры (Merchant, A.M, et al., Nature Biotech 16 (1998) 677-681; Atwell, S., et al. J. Mol. Biol. 270 (1997) 26-35) и повышает выход. Дополнительные подробности и формы осуществления см. выше.
Другой аспект изобретения составляет биспецифическое, двухвалентное антитело, характеризующееся содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с первым антигеном;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается со вторым антигеном,
где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера; и
где вариабельные домены VL и VH заменены друг другом.
Примерная схема данного формата биспецифического, двухвалентного антитела показана на Фиг.2b, включая домены СН3, модифицированные как выступы-во-впадины. Антитела на основе данного формата биспецифического, двухвалентного антитела названы в примерах OAscXFab1.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO:
39, и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO:40, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенного с легкой цепью второго полноразмерного антитела через пептидный линкер, SEQ ID NO:38.
Другой аспект изобретения составляет биспецифическое, двухвалентное антитело, характеризующееся содержанием
a) тяжелой цепи и легкой цепи первого полноразмерного антитела, которое специфично связывается с первым антигеном;
b) тяжелой цепи и легкой цепи второго полноразмерного антитела, которое специфично связывается со вторым антигеном,
где N-конец тяжелой цепи соединен с С-концом легкой цепи посредством пептидного линкера; и
где константные домены CL и СН1 заменены друг другом.
Примерная схема данного формата биспецифического, двухвалентного антитела показана на Фиг.2 с, включая домены СН3, модифицированные как выступы-во-впадины. Антитела на основе данного формата биспецифического, двухвалентного антитела названы в примерах OAscXFab2 и OAscXFab3.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO:42, и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO:43, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенного с легкой цепью второго полноразмерного антитела через пептидный линкер, SEQ ID NO:41.
В одной форме осуществления такое биспецифическое антитело характеризуется содержанием
a) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO:45, и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO:46, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенного с легкой цепью второго полноразмерного антитела через пептидный линкер, SEQ ID NO:44.
Антитело в соответствии с изобретением продуцируют рекомбинантными методами. Таким образом, один аспект настоящего изобретения составляет нуклеиновая кислота, кодирующая антитело в соответствии с изобретением, и следующий аспект составляет клетка, содержащая нуклеиновую кислоту, кодирующую антитело в соответствии с изобретением. Способы рекомбинантного n редуцирования широко известны на уровне техники и включают экспрессию белка в прокариотических и эукариотических клетках с последующим выделением антитела и обычно очисткой до фармацевтически приемлемой чистоты. Для экспрессии антител, как упомянуто выше, в клетке-хозяине нуклеиновые кислоты, кодирующие соответствующим образом модифицированные легкие и тяжелые цепи, встраивают в экспрессионные векторы стандартными способами. Экспрессию осуществляют в подходящих прокариотических и эукариотических клетках-хозяевах, таких как клетки СНО, клетки NSO, клетки SP2/0, клетки НЕК293, клетки COS, клетки PER.C6, клетки дрожжей или E. coli, и антитело выделяют из клеток (супернатанта или клеток после лизиса). Общие способы рекомбинантного продуцирования антител хорошо известны на уровне техники и описаны, например, в обзорных статьях Makrides, S.C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S., et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16 (2000) 151-160; Werner, R.G., Drug Res. 48 (1998) 870-880.
В соответствии с одной формой осуществления находится способ получения биспецифического антитела в соответствии с изобретением, включающий стадии
a) трансформации клетки-хозяина векторами, содержащими молекулы нуклеиновой кислоты, кодирующие данное антитело;
b) культивирования клетки-хозяина в условиях, которые дают возможность синтеза данной молекулы антитела; и
c) выделения данной молекулы антитела из культуры.
Биспецифические антитела соответствующим образом отделяют от культуральной среды общепринятыми методами очистки иммуноглобулинов, такими как, например, белок А-сефароза, хроматография на гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография. ДНК и РНК, кодирующую Моноклональные антитела, легко выделяют и секвенируют с использованием общепринятых методов. Клетки гибридомы могут служить в качестве источника такой ДНК и РНК. Как только ДНК выделена, ее можно встраивать в экспрессионные векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки НЕК 293, клетки СНО или клетки миеломы, которые иначе не продуцируют белок иммуноглобулин, чтобы получить синтез рекомбинантных моноклональных антител в клетках-хозяевах.
Варианты (или мутанты) аминокислотной последовательности биспецифического антитела получают путем введения соответствующих изменений нуклеотидов в ДНК антитела или путем нуклеотидного синтеза. Такие модификации можно осуществлять, однако, только в очень ограниченном диапазоне. Например, модификации не изменяют вышеупомянутые характеристики антитела, такие как изотип IgG и связывание антигена, но могут улучшать выход рекомбинантного продуцирования, стабильность белка, либо облегчать очистку.
Термин "клетка-хозяин", как используют в настоящей заявке, означает любой вид клеточной системы, которая может быть сконструирована для получения антител в соответствии с настоящим изобретением. В одной форме осуществления в качестве клеток-хозяев используют клетки НЕК293 и клетки СНО. Как используют в данной заявке, выражения "клетка", "клеточная линия" и "клеточная культура" используют взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, выражения "трансформанты" и "трансформированные клетки" включают первичные клетки-субъекты и культуры, полученные от них, независимо от числа пересевов. Также понятно, что все потомство может не быть точно идентичным по содержанию ДНК за счет преднамеренных или случайных мутаций. Включены варианты потомства, которые обладают такой же функцией или биологической активностью на основании скрининга, что и исходно трансформированная клетка.
Экспрессия в клетках NSO описана, например, Barnes, L.M., et al., Cytotechnology 32 (2000) 109-123; Barnes, L.M., et al., Biotech. Bioeng. 73 (2001) 261-270. Транзитная экспрессия описана, например, Durocher, Y., et al., Nucl. Acids. Res. 30 (2002) E9. Клонирование вариабельных доменов описано Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; и Norderhaug, L, et al., J. Immunol. Methods 204 (1997) 77-87. Предпочтительная транзитная экспрессионная система (НЕК 293) описана Schlaeger, E.-J. и Christensen, К. в Cytotechnology 30 (1999) 71-83 и Schlaeger, E.-J. в J. Immunol. Methods 194 (1996) 191-199.
Контрольные последовательности, которые пригодны для прокариот, например, включают промотор, необязательно операторную последовательность и сайт связывания рибосомы. Известно, что эукариотические клетки используют промоторы, энхансеры и сигналы полиаденилирования.
Нуклеиновая кислота "оперативно сцеплена", когда она помещена в функциональные взаимоотношения с другой нуклеиново-кислотной последовательностью. Например, ДНК для препоследовательности или секреторного лидера оперативно сцеплена с ДНК для полипептида, если она экспрессируется в виде пребелка, который участвует в секреции полипептида; промотор или энхансер оперативно сцеплен с кодирующей последовательностью, если он влияет на транскрипцию последовательности; либо сайт связывания рибосомы оперативно сцеплен с кодирующей последовательностью, если он находится в таком положении, чтобы способствовать трансляции. В целом, "оперативно сцепленный" означает, что последовательности ДНК, являющиеся сцепленными, являются непрерывными, а в случае секреторного лидера непрерывными и находящимися в одной рамке считывания. Однако энхансеры необязательно должны быть непрерывными. Сцепление осуществляют путем лигирования в удобных сайтах рестрикции. Если таких сайтов не существует, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
Очистку антител осуществляют в целях удаления клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков, стандартными методами, включая обработку щелочью/ДСН, Бэндинг CsCI, колоночная хроматография, электрофорез в агарозном геле и другое хорошо известны в данной области техники. См. Ausubel, F., et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987). Хорошо разработаны и широко распространены различные методы, используемые для очистки белка, такие как аффинная хроматография с белками микроорганизмов (например, аффинная хроматография с белком А или белком G), ионообменная хроматография (например, катионообменная (карбоксиметиловые смолы), анионообменная (аминоэтиловые смолы) и комбинированная обменная), тиофильное поглощение (например, с бета-меркаптоэтанолом и другими SH лигандами), хроматография гидрофобного взаимодействия или ароматическая адсорбционная хроматография (например, с фенилсефарозой, аза-аренофильными смолами или мета-аминофенилбороновой кислотой), аффинная хроматография с-хелатами металлов (например, с Ni(II)- и Cu(II)-аффинным материалом), эксклюзионная хроматография и электрофоретические методы (такие как гель-электрофорез, капиллярный электрофорез) (Vijayalakshmi, M.A., Appl. Biochem. Biotech. 75 (1998) 93-102).
Теперь обнаружено, что биспецифические антитела против VEGF человека и ANG-2 человека в соответствии с настоящим изобретением обладают ценными характеристиками, такими как высокая стабильность, и ценными фармакокинетическими/фармакодинамическими свойствами, такие как, например, хороший (то есть медленный) клиренс (например, в низких дозах).
Биспецифические, двухвалентные антитела в соответствии с изобретением проявляют пользу для пациентов-людей, нуждающихся в терапии, направленной на VEGF и ANG-2.
Кроме того, они обладают биологической или фармакологической активностью и проявляют ингибирование опухолевого роста и/или ингибирование опухолевого ангиогенеза in vivo.
Биспецифические антитела в соответствии с изобретением высокоэффективны при
а) ингибировании опухолевого роста (например, с биспецифическими антителами в соответствии с изобретением стаз опухоли может быть уже достигнут при более низких концентрациях по сравнению с комбинацией двух моноспецифических антител (например, в моделях опухолей COL0205 и KPL4 Примера 9 и 10 стаз опухоли был уже достигнут с 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина), и/или b) ингибировании опухолевого ангиогенеза или сосудистых заболеваний (например, максимальные антиангиогенные эффекты с биспецифическими антителами в соответствии с изобретением могут быть уже достигнуты при более низких концентрациях по сравнению с комбинацией двух моноспецифических антител (например, в анализе ангиогенеза роговицы мыши Примера 8 максимальный антиангиогенный эффект был уже достигнут с 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина).
Наконец, двухвалентное биспецифическое антитело против VEGF человека и ANG-2 человека в соответствии с настоящим изобретением может обладать ценным профилем эффективности/токсичности может обеспечивать полезные эффекты для пациента, нуждающегося в терапии против VEGF и против ANG-2.
Один аспект изобретения составляет фармацевтическая композиция, содержащая антитело в соответствии с изобретением. Другой аспект изобретения составляет применение антитела в соответствии с изобретением для получения фармацевтической композиции. Следующий аспект изобретения составляет способ получения фармацевтической композиции, содержащей антитело в соответствии с изобретением. В другом аспекте в настоящем изобретении предложена композиция, например, фармацевтическая композиция, содержащая антитело в соответствии с настоящим изобретением, приготовленная вместе с фармацевтически приемлемым носителем.
Одну форму осуществления изобретения составляет биспецифическое антитело в соответствии с изобретением для лечения рака.
Другой аспект изобретения составляет фармацевтическая композиция для лечения рака.
Другой аспект изобретения составляет применение антитела в соответствии с изобретением для получения лекарственного средства для лечения рака.
Другой аспект изобретения составляет способ лечения пациента, страдающего раком, путем введения антитела в соответствии с изобретением пациенту, нуждающемуся в таком лечении.
Другой аспект изобретения составляет фармацевтическая композиция для предупреждения метастазов.
Изобретение включает биспецифическое антитело в соответствии с изобретением для предупреждения метастазов.
Другой аспект изобретения составляет применение антитела в соответствии с изобретением для получения лекарственного средства для предупреждения метастазов.
Другой аспект изобретения составляет способ предупреждения метастазов у пациента, страдающего первичным раком, путем введения биспецифического антитела в соответствии с изобретением пациенту, нуждающемуся в таком превентивном лечении.
Авторы изобретения смогли показать высокоэффективное предупреждение спонтанных метастазов/вторичных опухолей in vivo в ортотопической и в подкожной модели рака (см. Пример 9) (в противоположность экспериментальной модели, где раковые клетки инъецируют i.v. Данная модель подобна клинической ситуации, где клетки диссеминируют из первичной опухоли и метастазируют во вторичный орган, такой как легкое или печень (где вторичные опухоли).
Термин "метастаз" в соответствии с изобретением относится к перемещению раковых клеток из первичной опухоли в один или более чем один сайт в другом месте у пациента, где затем развиваются вторичные опухоли. Способы определения того, метастазировал ли рак, известны в данной области техники и включают сканирование костей, рентген грудной клетки, КГ сканирование, МРТ сканирование и тесты на опухолевые маркеры.
Термин "предупреждение метастазов" или "предупреждение вторичных опухолей", как используют в данной заявке, имеет такое же значение и относится к профилактическому агенту против метастазов у пациента, страдающего раком, ингибируя или уменьшая таким путем дальнейшее перемещение раковых клеток из первичной опухоли в один или более чем один сайт в другом месте у пациента. Это означает, что метастаз первичной опухоли или рака предупреждается, замедляется или уменьшается, и, следовательно, развитие вторичных опухолей предупреждается, замедляется или уменьшается. Предпочтительно метастаз, то есть вторичные опухоли, легкого предупреждаются или уменьшаются за счет того, что метастатическое перемещение раковых клеток из первичной опухоли в легкое предупреждается или уменьшается.
Как используют в данной заявке, "фармацевтический носитель" включает любое и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические агенты и агенты, замедляющие всасывание, и тому подобное, которые являются физиологически совместимыми. Предпочтительно носитель пригоден для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии).
Композицию по настоящему изобретению можно вводить разнообразными способами, известными в данной области техники. Как понятно специалистам в данной области техники, путь и/или режим введения варьирует в зависимости от желаемых результатов. Чтобы ввести соединение по изобретению определенными путями введения, может быть необходимо покрыть соединение веществом, которое предотвращает его инактивацию, или вводить совместно с ним. Например, соединение можно вводить субъекту в подходящем носителе, например, в липосомах, или разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Фармацевтические носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий непосредственно перед применением. Использование таких сред и агентов для фармацевтически активных веществ известно в данной области техники.
Выражения "парентеральное введение" и "вводимый парентерально", как используют в данной заявке, означает режимы введения, иные, чем энтеральное и местное введение, обычно путем инъекции, и включает без ограничения внутривенную, внутримышечную, внутриартериальную, подоболочечную, интракапсулярную, внутриглазную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, интраспинальную, эпидуральную и интрастернальную инъекцию и инфузию.
Термин рак, как используют в данной заявке, относится к пролиферативным заболеваниям, таким как лимфомы, лимфоцитарные лейкозы, рак легкого, немелкоклеточный рак легкого (НМКРЛ), бронхиолоальвеолярный рак легкого, рак кости, рак поджелудочной железы, рак кожи, рак головы или шеи, подкожная или внутриглазная меланома, рак матки, рак яичника, рак прямой кишки, рак анальной области, рак желудка, рак ЖКТ, рак ободочной кишки, рак молочной железы, рак шейки матки, карцинома фаллопиевых труб, карциномы эндометрия, карцинома шейки матки, карцинома влагалища, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркома мягкой ткани, рак уретры, рак пениса, рак простаты, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточный рак, карцинома почечной лоханки, мезотелиома, печеночно-клеточный рак, рак желчного пузыря, новообразования центральной нервной системы (ЦНС), опухоли позвоночного столба, глиома ствола головного мозга, полиморфная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, чешуйчато-клеточные карциномы, аденома гипофиза и саркома Юинга, включая трудно поддающиеся лечению варианты любого из вышеуказанных раков или комбинацию одного или более чем одного из вышеуказанных раков.
Другой аспект изобретения составляет биспецифическое антитело в соответствии с изобретением или фармацевтическая композиция в качестве антиангиогенного агента. Такой антиангиогенный агент можно применять для лечения рака, в частности, солидных опухолей, и других сосудистых заболеваний.
Одну форму осуществления изобретения составляет биспецифическое антитело в соответствии с изобретением для лечения сосудистых заболеваний.
Другой аспект изобретения составляет фармацевтическая композиция для лечения сосудистых заболеваний.
Другой аспект изобретения составляет применение антитела в соответствии с изобретением для получения лекарственного средства для лечения сосудистых заболеваний.
Другой аспект изобретения составляет способ лечения пациента, страдающего сосудистыми заболеваниями, путем введения антитела в соответствии с изобретением пациенту, нуждающемуся в таком лечении.
Термин "сосудистые заболевания" включает рак, воспалительные заболевания, атеросклероз, ишемию, травму, сепсис, ХОБЛ, астму, диабет, ВМД, ретинопатию, удар, ожирение, острое легочное повреждение, кровотечение, сосудистую утечку, например, индуцированную цитокинами, аллергию, болезнь Грейвса, аутоиммунный тиреоидит Хасимото, идиопатическую тромбоцитопеническую пурпуру, гигантоклеточный артериит, ревматоидный артрит, системную красную волчанку (СКВ), волчаночный нефрит, болезнь Крона, рассеянный склероз, неспецифический язвенный колит, особенно для солидных опухолей, внутриглазные неоваскулярные синдромы, такие как пролиферативные ретинопатии или возрастная макулярная дегенерация (ВМД), ревматоидный артрит и псориаз (Folkman, J., et al., J. Biol. Chem. 267 (1992) 10931-10934; Klagsbrun, M., et al., Annu. Rev. Physiol. 53 (1991) 217-239; и Garner, A., Vascular diseases, In: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G.K., (eds.), 2nd edition, Marcel Dekker, New York (1994), pp 1625-1710).
Данные композиции могут также содержать адъюванты, такие как консерванты, увлажняющие агенты, эмульгирующие агенты и диспергирующие агенты. Предотвращение присутствия микроорганизмов можно обеспечить как методами стерилизации, см. выше, так и путем включения различных антибактериальных и противогрибковых агентов, например, парабена, хлорбутанола, фенола, сорбиновой кислоты и тому подобного. Может быть также желательным включать в композиции изотонические агенты, такие как сахара, хлорид натрия и тому подобное. Кроме того, пролонгированное всасывание инъекционной фармацевтической формы может быть вызвано путем включения агентов, которые замедляют всасывание, таких как моностеарат алюминия и желатин.
Независимо от выбранного пути введения, соединения по настоящему изобретению, которые можно использовать в подходящей гидратированной форме, и/или фармацевтические композиции по настоящему изобретению включают в фармацевтически приемлемые лекарственные формы общепринятыми способами, известными специалистам в данной области техники.
Фактические уровни дозы активных ингредиентов в фармацевтических композициях по настоящему изобретению могут варьировать таким образом, чтобы получить количество активного ингредиента, эффективное для достижения желаемого терапевтического ответа для конкретного пациента, композиции и режима введения, не являющееся токсичным для пациента. Выбранный уровень дозы зависит от ряда фармакокинетических факторов, включающих активность конкретных применяемых композиций по настоящему изобретению, путь введения, время введения, скорость выделения конкретного применяемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или вещества, используемые в комбинации с конкретными применяемыми композициями, возраст, пол, массу, состояние, общее состояние здоровья и анамнез пациента, подлежащего лечению, и подобные факторы, хорошо известные в медицинских областях.
Композиция должна быть стерильной и текучей до такой степени, чтобы эту композицию можно было доставлять через шприц. Кроме воды, носитель предпочтительно представляет собой изотонический забуференный физиологический раствор.
Соответствующую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, за счет сохранения необходимого размера частиц в случае дисперсии и путем использования сурфактантов. Во многих случаях предпочтительно включать в композицию изотонические агенты, например, сахара, многоатомные спирты, такие как маннит или сорбит, и хлорид натрия.
Как используют в данной заявке, выражения "клетка", "клеточная линия" и "клеточная культура" используют взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова "трансформанты" и "трансофрмированные клетки" включают первичную клетку-субъект и культуры, имеющие происхождение от нее, независимо от числа пересевов. Также понятно, что все потомство может не быть точно идентичным по содержанию ДНК за счет преднамеренных или случайных мутаций. Включены варианты потомства, которые обладают такой же функцией или биологической активностью на основании скрининга, что и исходно трансформированная клетка. Где подразумевают отдельные обозначения, будет понятно из контекста.
Термин "трансформация", как используют в данной заявке, относится к процессу переноса векторов/нуклеиновой кислоты в клетку-хозяина. Если в качестве клеток-хозяев используют клетки без труднопреодолимых барьеров клеточной стенки, трансфекцию осуществляют, например, методом кальцийфосфатной преципитации, как описано Graham, F.L, van der Eb, A.J., Virology 52 (1973) 546-467. Однако можно также использовать другие способы введения ДНК в клетки, такие как ядерная инъекция или слияние протопластов. Если используют прокариотические клетки или клетки, которые содержат существенные конструкции клеточной стенки, одним из методов трансфекции является, например, обработка кальцием с использованием хлорида кальция, как описано Cohen, S.N., etal., PNAS. 69 (1972) 2110-2114.
Как используют в данной заявке, "экспрессия" относится к процессу, посредством которого нуклеиновая кислота транскрибируется в мРНК и/или к процессу, посредством которого транскрибированная мРНК (также называемая транскриптом) впоследствии транслируется в пептиды, полипептиды или белки. Транскрипты и кодируемые полипептиды все вместе называют генным продуктом. Если полинуклеотид имеет происхождение из геномной ДНК, экспрессия в эукариотической клетке может включать сплайсинг мРНК.
"Вектор" представляет собой молекулу нуклеиновой кислоты, в частности, самореплицирующуюся, которая переносит встроенную молекулу нуклеиновой кислоты в клетки-хозяева и/или между ними. Этот термин включают векторы, которые действуют, в основном, для встраивания ДНК или РНК в клетку (например, хромосомной интеграции), реплицирующиеся векторы, которые действуют, в основном, для репликации ДНК или РНК, и экспрессионные векторы, которые действуют для транскрипции и/или трансляции ДНК или РНК. Включены также векторы, которые обеспечивают более чем одну из описанных функций.
"Экспрессионный вектор" представляет собой полинуклеотид, который при введении в соответствующую клетку-хозяина может транскрибироваться и транслироваться в полипептид. "Экспрессионная система" обычно относится к подходящей клетке-хозяину, содержащей экспрессионный вектор, который может действовать для выхода желаемого продукта экспрессии.
Приведенные ниже примеры, перечень последовательностей и графические материалы предложены, чтобы способствовать пониманию настоящего изобретения, действительный объем которого представлен в прилагаемой формуле изобретения. Понятно, что в вышеизложенных способах можно производить модификации без отклонения от сущности изобретения.
Описание перечня последовательностей (аминокислотные последовательности)
SEQ ID NO:1 вариабельный домен тяжелой цепи VH of<VEGF>бевацизумаб
SEQ ID NO:2 вариабельный домен легкой цепи VL<VEGF>бевацизумаб
SEQ ID NO:3 вариабельный домен тяжелой цепи VH of<ANG-2>E6Q
SEQ ID NO:4 вариабельный домен легкой цепи VL of<ANG-2>E6Q
SEQ ID NO:5 XMab1-<VEGF> легкая цепь
SEQ ID NO:6 XMab1-<ANG2> легкая цепь
SEQ ID NO:7 XMab1-<VEGF> тяжелая цепь
SEQ ID NO:8 XMab1-<ANG2> тяжелая цепь
SEQ ID NO:9 XMab2-<VEGF> легкая цепь
SEQ ID NO:10 XMab2-<ANG2> легкая цепь
SEQ ID NO:11 XMab2-<VEGF> тяжелая цепь
SEQ ID NO:12 XMab2-<ANG2> тяжелая цепь
SEQ ID NO:13 XMab3-<VEGF> легкая цепь
SEQ ID NO:14 XMab3-<ANG2> легкая цепь
SEQ ID NO:15 XMab3-<VEGF> тяжелая цепь
SEQ ID NO:16 XMab3-<ANG2> тяжелая цепь
SEQ ID NO:17 XMab4-<VEGF> легкая цепь
SEQ ID NO:18 XMab4-<ANG2> легкая цепь
SEQ ID NO:19 XMab4-<VEGF> тяжелая цепь
SEQ ID NO:20 XMab4-<ANG2> тяжелая цепь
SEQ ID NO:21 XMab5-<VEGF> легкая цепь
SEQ ID NO:22 XMab5-<ANG2> легкая цепь
SEQ ID NO:23 XMab5-<VEGF> тяжелая цепь
SEQ ID NO:24 XMab5-<ANG2> тяжелая цепь
SEQ ID NO:25 XMab6-<VEGF> легкая цепь
SEQ ID NO:26 XMab6-<ANG2> легкая цепь
SEQ ID NO:27 XMab6-<VEGF> тяжелая цепь
SEQ ID NO:28 XMab6-<ANG2> тяжелая цепь
SEQ ID NO:29 OAscFab1-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:30 OAscFab1-<VEGF> тяжелая цепь
SEQ ID NO:31 OAscFab1-<VEGF> легкая цепь
SEQ ID NO:32 OAscFab2-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:33 OAscFab2-<VEGF> тяжелая цепь
SEQ ID NO:34 OAscFab2-<VEGF> легкая цепь
SEQ ID NO:35 OAscFab3-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:36 OAscFab3-<VEGF> тяжелая цепь
SEQ ID NO:37 OAscFab3-<VEGF> легкая цепь
SEQ ID NO:38 OAscXFab1-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:39 OAscXFab1-<VEGF> тяжелая цепь
SEQ ID NO:40 OAscXFab1-<VEGF> легкая цепь
SEQ ID NO:41 OAscXFab2-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:42 OAscXFab2-<VEGF> тяжелая цепь
SEQ ID NO:43 OAscXFab2-<VEGF> легкая цепь
SEQ ID NO:44 OAscXFab3-<ANG2> тяжелая цепь и легкая цепь, соединенные пептидом
SEQ ID NO:45 OAscXFab3-<VEGF> тяжелая цепь
SEQ ID NO:46 OAscXFab3-<VEGF> легкая цепь
SEQ ID NO:47 Фактор роста эндотелия сосудов человека (VEGF)
SEQ ID NO:48 Ангиопоэтин-2 человека (ANG-2)
Экспериментальные методы
Примеры
Материалы и общие методы
Общая информация относительно нуклеотидных последовательностей легких и тяжелых цепей иммуноглобулинов человека приведена в: Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991). Аминокислоты цепей антитела пронумерованы и названы в соответствии с нумерацией EU (Edelman, G.M., et al., Proc. Natl. Acad. Sci. USA 63 (1969) 78-85; Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD, (1991)).
Методы рекомбинантных ДНК
Для манипуляций с ДНК использованы стандартные методы, как описано в Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реагенты использовали в соответствии с инструкциями изготовителя.
Синтез генов
Желаемые генные сегменты могут быть получены из олигонуклеотидов, полученных путем химического синтеза. Генные сегменты, которые фланкированы уникальными сайтами эндонуклеаз рестрикции, собирали путем отжига и лигирования нуклеотидов, включая ПЦР амплификацию, а затем клонировали посредством указанных сайтов рестрикции, например, KpnI/SacI или Ascl/PacI, в клонирующем векторе pGA4 на основе pPCRScript (Stratagene). Последовательности ДНК субклонированных генных фрагментов подтверждали с помощью секвенирования ДНК.
Фрагменты для генного синтеза заказывали в соответствии с приведенными описаниями в Geneart (Regensburg, Germany). Все генные сегменты, кодирующие легкие и тяжелые цепи биспецифических антител Ang-2/VEGF, синтезировали с 5'-концевой последовательностью ДНК, кодирующей лидерный пептид (MGWSCIILFLVATATGVHS), который направляет белки на секрецию в эукариотических клетках, и с уникальными сайтами рестрикции на 5' и 3' концах синтезированного гена. Последовательности ДНК, несущие тяжелые цепи, стабилизированные дисульфидами и модифицированные как "выступы-во-впадины", были сконструированы с мутациями S354C и T366W в "выступах" тяжелой цепи и мутациями Y349C, T366S, L368A и Y407V во "впадинах" тяжелой цепи.
Определение последовательности ДНК
Последовательности ДНК определяли с помощью двунитевого секвенирования, проведенного в MediGenomix GmbH (Martinsried, Germany) или Sequiserve GmbH (Vaterstetten, Germany).
Анализ последовательности ДНК и белка и управление данными последовательностей
Для создания, картирования, анализа, аннотации и иллюстрации последовательностей использовали программное обеспечение GCG (Genetics Computer Group, Madison, Wisconsin) версия 10.2, и Infomax's Vector NT1 Advance suite, версия 8.0.
Экспрессионные векторы
Для экспрессии желаемых вариантов антител использовали экспрессионные плазмиды для транзитной экспрессии (например, в клетках НЕК293 EBNA или HEK293-F) или для стабильной экспрессии (например, в клетках СНО) на основе либо организации кДНК с промотором CMV-интрон А, либо геномной организации с промотором CMV (например, Фиг.2В).
Кроме экспрессионной кассеты антитела, векторы содержали:
- точку начала репликации, которая дает возможность репликации этой плазмиды в Е. coli, и
- ген β-лактамазы, который придает устойчивость к ампциллину в Е. coli.
Транскрипционная единица гена антитела состоит из следующих элементов:
- уникальный сайт(ы) рестрикции на 5' конце,
- немедленно-ранний энхансер и промотор от цитомегаловируса человека,
- с последующей последовательностью интрона А в случае организации кДНК,
- 5'-нетранслируемую область гена антитела человека,
- сигнальную последовательность тяжелой цепи иммуноглобулина,
- цепь антитела человека (тяжелую цепь, модифицированную тяжелую цепь или легкую цепь) либо в виде кДНК, либо в виде геномной организации, с экзон-интронной организацией иммуноглобулина,
- 3'-нетранслируемую область с последовательностью сигнала полиаденилирования,
- уникальный сайт(ы) рестрикции на 3' конце.
Для транзитных и стабильных трансфекций большие количества плазмид получали путем получения плазмидных препаратов из трансформированных культур Е. coli (Nucleobond AX, Macherey-Nagel). Методы клеточных культур
Стандартные методы клеточных культур использовали, как описано в Current Protocols in Cell Biology (2000), Bonifacino, J.S., Dasso, M., Harford, J.B., Lippincott-Schwartz, J. and Yamada, K.M. (eds.), John Wiley & Sons, Inc.
Транзитные трансфекции в системе HEK293-F
Рекомбинантные варианты иммуноглобулинов экспрессировали путем транзитной трансфекции клеток эмбриональных почек человека 293-F, используя экспрессионную систему FreeStyle™ 293 в соответствии с инструкцией изготовителя (Invitrogen, USA). Кратко, суспензию клеток FreeStyle™ 293-F культивировали в экспрессионной среде FreeStyle™ 293 при 37°С/8% CO2, и клетки высевали в свежую среду при плотности 1-2×106 жизнеспособных клеток/мл на сутки трансфекций. Комплексы DNA-293fectin™ готовили в среде Opti-MEM® I (Invitrogen, USA), используя 325 мкл 293fectin™ (Invitrogen, Germany) и 250 мкг плазмидной ДНК тяжелой и легкой цепи в молярном отношении 1:1 для конечного объема трансфекции 250 мл для моноспецифических родительских антител. Комплексы "выступы-во-впадины" ДНК-293fectin с двумя тяжелыми цепями и одной легкой цепью готовили в среде Opti-MEM® I (Invitrogen, USA), используя 325 мкл 293fectin™ (Invitrogen, Germany) и 250 мкг плазмидной ДНК тяжелой цепи "выступы-во-впадины" 1 и 2 и легкой цепи, обычно в молярном отношении 1:1:1, для конечного объема трансфекции 250 мл (OAscFab и OAscXFab). Для оптимизации выхода экспрессии это отношение можно варьировать. Комплексы XMab ДНК-293fectin готовили в среде Opti-MEM® I (Invitrogen, USA), используя 325 мкл 293fectin™ (Invitrogen, Germany) и 250 мкг плазмидной ДНК тяжелой цепи "выступы-во-впадины" 1 и 2 и легкой цепи в молярном отношении 1:1:1, для конечного объема трансфекции 250 мл. Для оптимизации выхода экспрессии это отношение можно варьировать. Супернатанты клеточной культуры, содержащие антитело, собирали через 7 суток после трансфекции путем центрифугирования при 14000 g в течение 30 минут и фильтровали через стерильный фильтр (0,22 мкм). Супернатанты хранили при -20°С до очистки.
Определение белка
Концентрацию белка очищенных антител и их производных определяли путем определения оптической плотности (OD) при 280 нм, используя коэффициент молярной экстинкции, вычисленный на основании аминокислотной последовательности в соответствии с Расе, C.N., et. al., Protein Science 4 (1995) 2411-1423.
Определение концентрации антитела в супернатантах
Концентрацию антител и их производных в супернатантах клеточных культур определяли путем иммунопреципитации с гранулами агарозы с белком А (Roche). 60 мкл агарозных гранул с белком А промывают три раза в TBS-NP40 (50 мМ Трис, рН 7,5, 150 мМ NaCI, 1% Нонидет-Р40). Затем 1-15 мл супернатанта клеточной культуры наносят на агарозные гранулы с белком А, уравновешенные в TBS-NP40. После инкубации в течение 1 ч при комнатной температуре гранулы промывают на фильтрационной колонке Ultrafree-MC (Amicon) один раз 0,5 мл TBS-NP40, дважды 0,5 мл 2х фосфатно-солевого буфера (2×ФСБ, Roche) и кратковременно четыре раза 0,5 мл 100 мМ Na-цитратом рН 5,0. Связанное антитело элюируют добавлением 35 мкл буфера для образцов NuPAGE® LDS (Invitrogen). Половину образца объединяют с восстанавливающим агентом для образца NuPAGE® или оставляют невосстановленным, соответственно, и нагревают в течение 10 мин при 70°С. Затем 20 мкл наносят на 4-12% Бис-Трис ДСН-ПААГ NuPAGE® (Invitrogen) (с буфером MOPS для невосстановленного ДСН-ПААГ и с буфером MES с антиоксидантной добавкой для буфера пробега NuPAGE® (Invitrogen) для восстановленного ДСН-ПААГ) и окрашивают Кумасси синим.
Концентрацию антител и их производных в супернатантах клеточных культур измеряли путем хроматографии белок А-ВЭЖХ. Кратко, супернатанты клеточных культур, содержащие антитела и их производные, которые связываются с белком А, наносили на колонку HiTrap белок A (GE Healthcare) в 50 мМ K2HPO4, 300 мМ NaCl, pH 7,3 и элюировали из матрикса 550 мМ уксусной кислотой, рН 2,5 на системе ВЭЖХ Dionex. Элюированный белок определяли количественно с помощью поглощения УФ света и интегрирования площадей пиков. Очищенное стандартное антитело IgG1 служило в качестве стандарта.
Альтернативно концентрацию антител и их производных в супернатантах клеточных культур измеряли путем сэндвич-анализа IgG ELISA. Кратко, 96-луночные микротитрационные планшеты StreptaWell с высоким связыванием стрептавидина A (Roche) покрывали 100 мкл/лунка биотинилированной иммобилизованной молекулой IgG против человека F(ab')2<h-Fcгамма>BI (Dianova) при 0,1 мкг/мл в течение 1 ч при комнатной температуре или альтернативно в течение ночи при 4°С и последовательно промывали три раза 200 мкл/лунка ФСБ, 0,05% Твин (ФСБТ, Sigma). 100 мкл/лунка серийных разведении в ФСБ (Sigma) соответствующих супернатантов клеточных культур, содержащих антитело, добавляли в лунки и инкубировали в течение 1-2 ч на встряхивателе для микротитрационных планшетов при комнатной температуре. Лунки промывали три раза 200 мкл/лунка ФСБТ, и связанное антитело обнаруживали с помощью 100 мкл F(ab')2<hFcгамма>POD (Dianova) при 0,1 мкг/мл в качестве идентифицирующего антитела в течение 1-2 ч на встряхивателе для микротитрационных планшетов при комнатной температуре. Несвязанное идентифицирующее антитело отмывали три раза 200 мкл/лунка ФСБТ, и связанное идентифицирующее антитело обнаруживали путем добавления 100 мкл ABTS/лунка. Определение поглощения осуществляли на спектрометре Tecan Fluor при длине волны измерения 405 нм (эталонная длина волны 492 нм).
Очистка биспецифических антител
Биспецифические антитела очищали из супернатантов клеточных культур путем аффинной хроматографии, используя белок А-сефарозу™ (GE Healthcare, Sweden) и экскпюзионную хроматографию Superdex200. Кратко, стерильно профильтрованные супернатанты клеточных культур наносили на колонку HiTrap ProteinA HP (5 мл), уравновешенную буфером ФСБ (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, pH 7,4). Несвязанные белки отмывали буфером для уравновешивания. Антитело и варианты антитела элюировали 0,1 М цитратным буфером, pH 2,8, и фракции, содержащие белок, нейтрализовали 0,1 мл 1 М Трис, pH 8,5. Затем элюированные белковые фракции объединяли, концентрировали с помощью центрифужного фильтровального устройства Amicon Ultra (MWCO: 30 К, Millipore) до объема 3 мл и наносили на колонку для гель-фильтрации Superdex200 HiLoad 120 мл 16/60 (GE Healthcare, Sweden), уравновешенную 20 мМ гистидином, 140 мМ NaCI, pH 6,0. Фракции, содержащие очищенные биспецифические антитела менее чем с 5% высокомолекулярных агрегатов, объединяли и хранили в виде аликвот по 1,0 мг/мл при -80°С.
ДСН-ПААГ электрофорез
Систему готового геля NuPAGE® (Invitrogen) использовали в соответствии с инструкцией изготовителя. В частности, использовали готовые 4-20% Трис-глициновые гели NuPAGE® Novex® и Трис-глициновый буфер пробега с ДСН Novex® (см., например, Фиг.3). Восстановление образцов было достигнуто путем добавления агента для восстановления образцов NuPAGE® перед проведением гель-электрофореза.
Аналитическая эксклюзионная хроматография
Эксклюзионную хроматографию для определения агрегации и олигомерного состояния антител проводили с помощью хроматографии ВЭЖХ. Кратко, антитела, очищенные белком А, наносили на колонку Tosoh TSKgel G3000SW в 300 мМ NaCl, 50 мМ KH2PO4/K2HPO4, pH 7,5 на системе ВЭЖХ Agilent 1100 или на колонку Superdex 200 (GE Healthcare) в 2×ФСБ на системе ВЭЖХ Dionex. Элюированный белок определяли количественно путем поглощения в УФ свете и интегрирования площадей пиков. Стандарт для гель-фильтрации BioRad 151-1901 служил в качестве стандарта (см., например, Фиг.4).
Масс-спектрометрия
Суммарную дегликозилированную массу кроссоверных антител определяли и подтверждали посредством масс-спектрометрии с ионизацией электрораспылением (ИЭР-МС). Кратко, 100 мкг очищенных антител дегликозилировали 50 мЕ N-гликозидазы F (PNGaseF, ProZyme) в 100 мМ KH2PO4/K2HPO4, pH 7 при 37°С в течение 12-24 ч при концентрации белка вплоть до 2 мг/мл, а затем обессоливали посредством ВЭЖХ на колонке Sephadex G25 (GE Healthcare). Массу соответствующих тяжелых и легких цепей определяли с помощью ИЭР-МС после дегликозилирования и восстановления. Кратко, 50 мкг антитела в 115 мкл инкубировали с 60 мкл 1М ТСЕР и 50 мкл 8 М гуанидина гидрохлорида с последующим обессоливанием. Суммарную массу и массу восстановленных тяжелых и легких цепей определяли посредством ИЭР-МС на системе МС Q-Star Elite, оборудованной источником NanoMate.
Получение клеточной линии HEK293-Tie2
С целью определения взаимодействия антител к ангиопоэтину-2 со стимулированным ANGPT2 фосфорилированием Tie2 и связывания ANGPT2 с Tie2 на клетках была получена рекомбинантная клеточная линия HEK293-Tie. Кратко, плазмиду на основе pcDNA3 (RB22-pcDNA3 Topo hTie2), кодирующую полноразмерный Tie2 человека (SEQ ID 108) под контролем промотора CMV и маркер устойчивости к неомицину, трансфицировали, используя FuGENE (Roche Applied Science) в качестве реагента трансфекции, в клетки НЕК293 (АТСС), и устойчивые клетки отбирали в DMEM с 10% FCS, 500 мкг/мл G418. Индивидуальные клоны выделяли посредством цилиндра для клонирования, а затем анализировали на экспрессию Tie2 с помощью FACS (сортировки флуоресцентно-активированных клеток). Клон 22 был идентифицирован как клон с высокой и стабильной экспрессией Tie2 даже в отсутствие G418 (HEK293-Tie2 клон 22). Затем HEK293-Tie2 клон 22 использовали для клеточных анализов: индуцированного ANGPT2 фосфорилирования Tie2 и клеточного анализа связывания лиганда ANGPT2.
Анализ индуцированного ANGPT2 фосфорилирования Tie2
Ингибирование индуцированного ANGPT2 фосфорилирования Tie2 антителами к ANGPT2 измеряли в соответствии с описанным ниже принципом анализа. HEK293-Tie2 клон 22 стимулировали ANGPT2 в течение 5 минут в отсутствие или в присутствии ANGPT2, и P-Tie2 определяли количественно с помощью сэндвич-анализа ELISA. Кратко, 2×105 клеток HEK293-Tie2 клон 22 на лунку выращивали в течение ночи на покрытом поли-D-лизином 96 луночном микротитрационном планшете в 100 мкл DMEM, 10% FCS, 500 мкг/мл генетицина. На следующие сутки готовили титрационный ряд антител ANGPT2 в микротитрационном планшете (4-кратно концентрированные, 75 мкл конечный объем/лунка, два повтора) и смешивали с 75 мкл разведения ANGPT2 (R&D systems # 623-AN) (3,2 мкг/мл в качестве раствора 4-кратной концентрации). Антитела и ANGPT2 предварительно инкубировали в течение 15 мин при комнатной температуре. 100 мкп смеси добавляли к клеткам HEK293-Tie2 клон 22, (предварительно инкубированным в течение 5 мин с 1 мМ NaV3O4, Sigma #S6508) и инкубировали в течение 5 мин при 37°С. Затем клетки промывали 200 мкл охлажденного во льду ФСБ+1 мМ NaV3O4 на лунку и подвергали лизису путем добавления 120 мкл буфера для лизиса (20 мМ Трис, рН 8,0, 137 мМ NaCl, 1% NP-40, 10% глицерин, 2 мМ ЭДТА, 1 мМ NaV3O4, 1 мМ ФМСФ и 10 мкг/мл апротинина) на лунку на льду. Клетки подвергали лизису в течение 30 мин при 4°С на встряхивателе для микротитрационных планшетов, и 100 мкл лизата переносили непосредственно на микротитрационный планшет для p-Tie2 ELISA (R&D Systems, R&D #DY990) без предварительного центрифугирования и без определения суммарного белка. Количества P-Tie2 определяли количественно в соответствии с инструкциями изготовителя, и значения IC50 для ингибирования определяли, используя плагин анализа XLfit4 для Excel (одноцентровая модель доза-ответ 205). Значения ICso можно сравнивать внутри эксперимента, но они могут варьировать от одного эксперимента к другому.
Анализ индуцированной VEGF пролиферации HUVEC
Индуцированная VEGF пролиферация HUVEC (эндотелиальные клетки пупочной вены человека, Promocell #C-12200) была выбрана для измерения клеточной функции антител к VEGF. Кратко, 5000 клеток HUVEC (низкое число пассажей, не более 5 пассажей) на 96 лунок инкубировали в 100 мкл истощенной среды (ЕВМ-2 эндотелиальная базовая среда 2, Promocell #С-22211, 0,5% FCS, пенициллин/стрептомицин) в покрытом коллагеном I 96-луночном микротитрационном планшете BD Biocoat Collagen (BD #354407/35640) в течение ночи. Варьирующие концентрации антитела смешивали с rhVEGF (конечная концентрация 30 нг/мл, BD #354107) и предварительно инкубировали в течение 15 минут при комнатной температуре. Затем смесь добавляли к клеткам HUVEC и инкубировали их в течение 72 ч при 37°С, 5% CO2. На сутки анализа планшет уравновешивали до комнатной температуры в течение 30 мин и определяли жизнеспособность/пролиферацию клеток, используя набор для люминесцентного анализа жизнеспособности клеток CellTiter-GloTM в соответствии с руководством (Promega, #G7571/2/3). Люминесценцию определяли на спектрофотометре.
Пример 1а
Экспрессия и очистка молекул биспецифического, двухвалентного антитела с обменом доменов <VEGF-ANG-2>XMab
В соответствии с методами, описанными выше в материалах и методах, молекулы биспецифического, двухвалентного антитела с обменом доменов <VEGF-ANG-2>XMab1, XMab1 и XMab3 экспрессировали и очищали. Участки VH и VL<VEGF>(SEQ ID NO:1 и SEQ ID NO:2) основаны на бевацизумабе. Участок VH<ANG2>(SEQ ID NO:3) был получен путем мутации E6Q (исходная аминокислота глутаминовая кислота (Е) в положении 6 была заменена глутамином (Q)) последовательности VH ANG2i-LC06 (которая описана в заявке РСТ №РСТ/ЕР2009/007182 (W02010/040508), и которая представляет собой фрагмент, подвергнутый дальнейшему созреванию, последовательности, полученной посредством фагового дисплея). Участок VL<ANG2>(SEQ ID NO:4) был получен из последовательностей VL ANG2i-LC06 (см. заявку РСТ №РСТ/ЕР2009/007182 (W02010/040508)). Молекулы биспецифического, двухвалентного антитела с обменом доменов <VEGF-ANG-2>XMab1, XMab2 и ХМаЬЗ экспрессировали и очищали. Релевантные аминокислотные последовательности легкой и тяжелой цепи этих биспецифических, двухвалентных антител представлены в SEQ ID NO:5-8 (XMab1), в SEQ ID NO:9-12 (XMab2) и в SEQ ID NO:13-16 (ХМаЬЗ). Примерную структуру см. на Фиг.1.
Биспецифические, двухвалентные антитела <VEGF-ANG-2>XMab4, XMab5 и ХМаЬб (с релевантными аминокислотными последовательностями легкой и тяжелой цепи, представленными в SEQ ID NO:17-20 (XMab4), в SEQ ID NO:21-24 (XMab5) и в SEQ ID NO:25-28 (XMab6)) экспрессировали и очищали аналогично.
Связывающее сродство и другие свойства определяли или определяют, как описано.
Пример 1b
Экспрессия и очистка молекул биспецифического, двухвалентного антитела <VEGF-ANG-2>OAscFab
В соответствии с методами, описанными выше в материалах и методах, молекулы биспецифического, двухвалентного антитела <VEGF-ANG-2>OAscFab1, OAscFab2, OAscFab3 экспрессировали и очищали. Участки VH и VL<VEGF> (SEQ ID NO:1 и SEQ ID NO:2) основаны на бевацизумабе. Участок VH<ANG2> (SEQ ID NO:3) был получен путем мутации E6Q (исходная аминокислота глутаминовая кислота (Е) в положении 6 была заменена глутамином (Q)) последовательности VH ANG2i-LC06 (которая описана в заявке РСТ №РСТ/ЕР2009/007182 (W02010/040508), и которая представляет собой фрагмент, подвергнутый дальнейшему созреванию, последовательности, полученной посредством фагового дисплея). Участок VL<ANG2>E6Q (SEQ ID NO:4) был получен из последовательности VL ANG2i-LC06 (см. заявку РСТ №РСТ/ЕР2009/007182 (WO 2010/040508) (WO 2010/040508)). Релевантные аминокислотные последовательности легкой и тяжелой цепи этих биспецифических, двухвалентных антител представлены в SEQ ID NO:29-31 (OAscFabI), в SEQ ID NO:32-34 (OAscFab2) и в SEQ ID NO:35-37 (OAscFab3). Примерную структуру см. на Фиг.2а. Экспрессию OAscFabI, OAscFabI, OAscFab2 и OAscFab3 подтверждали с помощью Вестерн-блоттинга. Очистка OAscFab2 и OAscFab3 привела к описанным ниже выходам.
Связывающее сродство и другие свойства определяют, как описано.
Пример 1с
Экспрессия и очистка молекул биспецифического, двухвалентного антитела с обменом доменов <VEGF-ANG-2>OAscXFab
В соответствии с методами, описанными выше в материалах и методах, молекулы биспецифического, двухвалентного антитела с обменом доменов <VEGF-ANG-2>OAscXFab1, OAscXFab2, OAscXFab3 экспрессировали и очищали. Участки VH и VL<VEGF> (SEQ ID NO:1 и SEQ ID NO:2) основаны на бевацизумабе. Участок VH<ANG2>E6Q (SEQ ID NO:3) был получен путем мутации E6Q (исходная аминокислота глутаминовая кислота (Е) в положении 6 была заменена глутамином (Q)) последовательности VH ANG2i-LC06 (которая описана в заявке РСТ №РСТ/ЕР2009/007182 (W02010/040508), и которая представляет собой фрагмент, подвергнутый дальнейшему созреванию, последовательности, полученной посредством фагового дисплея). Участок VL<ANG2>E6Q (SEQ ID NO:4) был получен из последовательности VL ANG2i-LC06 (см. заявку РСТ №РСТ/ЕР2009/007182 (W02010/040508) (W02010/040508)). Релевантные аминокислотные последовательности легкой и тяжелой цепи этих биспецифических, двухвалентных антител представлены в SEQ ID NO:38-40 (OAscXFabI), в SEQ ID NO:41-43 (OAscXFab2) и в SEQ ID NO:44-46 (OAscXFab3). Примерную структуру см. на Фиг.2b (OAscXFabI) и Фиг.2 с (OAscXFab2, OAscXFab3). Экспрессию подтверждали с помощью Вестерн-блоттинга.
Связывающее сродство и другие свойства определяют, как описано.
Пример 2
Стабильность биспецифических антител
Температура денатурации (метод SYPRO оранжевого)
Чтобы определить температуру, при которой происходит денатурация белка (то есть индуцированная температурой утрата структуры белка), использовали способ, основанный на гидрофобном флуоресцентном красителе (SYPRO оранжевый, Invitrogen), который проявляет сильную флуоресценцию в гидрофобных средах. При денатурации белка гидрофобные пэтчи становятся доступными для растворителя, что приводит к повышенной флуоресценции. При температурах выше температуры денатурации интенсивности флуоресценции снова снижаются, следовательно, температуру, при которой достигается максимальная интенсивность, определяют как температуру денатурации. Этот метод описан Ericsson, U.B., et al., Anal Biochem 357 (2006) 289-298 и He, F., et al., Journal of Pharmaceutical Sciences 99 (2010) 1707-1720.
Образцы белка при концентрации примерно 1 мг/мл в 20 мМ His/HisCI, 140 мМ NaCI, pH 6,0 смешивали с SYPRO оранжевым (5000х исходный раствор, до достижения конечного разведения 1:5000. Объем 20 мкл переносили в 384-луночный планшет, и зависимую от температуры флуоресценцию регистрировали в системе ПЦР в реальном времени LightCycler® 480 (Roche Applied Sciences) при скорости нагревания 0,36°С/мин.
Температура агрегации по динамическому светорассеянию (ДСР)
Температуру, при которой происходит термически индуцированная агрегация белка, определяли по динамическому светорассеянию (ДСР). ДСР дает информацию о распределении макромолекул по размеру в растворе, выведенную на основании колебаний интенсивностей рассеянного света в микросекундном масштабе. Когда образцы постепенно нагревают, агрегация начинается при определенной температуре, вызывая увеличение размера частиц. Температуру, при которой размеры частиц начинают увеличиваться, определяют как температуру агрегации. Температуры агрегации и денатурации необязательно должны быть идентичными, поскольку денатурация необязательно является необходимым условием агрегации.
Для измерений температуры агрегации использовали считывающее устройство для планшетов DynaPro DLS (Wyatt technologies). Перед измерением образцы фильтровали через 384-луночные фильтрационные планшеты (384-луночная фильтрационная система Millipore MultiScreen, 0,45 мкм) в оптические 384-луночные планшеты (Corning #3540). Объем образца 35 мкл использовали при концентрации белка примерно 1 мг/мл в буфере для препарата (20 мМ цитрат, 180 мМ сахароза, 20 мМ аргинин, 0,02% полисорбат 20). Каждую лунку покрывали 20 мкл вазелинового масла (Sigma) во избежание испарения. Образцы нагревали от 25°С до 80°С со скоростью 0,05°С/мин, и данные ДСР снимали непрерывно для максимального числа 15 образцов на пробег.
Скорость агрегации по ДСР
ДСР является чувствительным методом для обнаружения агрегатов макромолекул в растворе, поскольку агрегаты вызывают сильные сигналы светорассеяния. Следовательно, склонность молекулы к агрегации можно прослеживать по времени путем повторного снятия данных ДСР. Для ускорения потенциальной агрегации до практических скоростей измерения проводили при 50°С.
Приготовление образца осуществляли, как описано выше. Данные ДСР записывали в течение вплоть до 100 часов. Скорости агрегации (нм/сутки) вычисляли как наклон кривой линейного соответствия средних диаметров по времени.
Стабильность в буфере для препарата
Чтобы оценить биспецифические молекулы на их стабильность в отношении агрегации/фрагментации, образцы инкубировали в течение 3 недель при 40°С в буфере для препарата (20 мМ цитрат, 180 мМ сахароза, 20 мМ аргинин, 0,02% полисорбат 20) при концентрации белка примерно 1 мг/мл. Контрольный образец хранили в течение 3 недель при -80°С.
Эксклюзионную хроматографию для количественного определения агрегатов и низкомолекулярных (LMW) молекул проводили с помощью ВЭЖХ. Количество 25-100 мкг белка наносили на колонку Tosoh TSKgel G3000SWXL в 300 мМ NaCl, 50 мМ фосфат калия, рН 7,5 на системе ВЭЖХ Ultimate3000 (Dionex). Элюированный белок определяли количественно на основании поглощения УФ света при 280 нм.
Результаты:
площади ALMW: 0,5%
площади Д мономер: -1,2% площади
Пример 3
Связывающие свойства биспецифического антитела <VEGF-Ang-2>
А) Связывающие свойства, охарактеризованные с помощью анализа поверхностного плазмонного резонанса (SPR)
Одновременное связывание обоих антигенов было подтверждено путем применения поверхностного плазменного резонанса (SPR) с использованием прибора BIAcore Т100 (GE Healthcare Biosciences AB, Uppsala, Sweden). VEGF был иммобилизован на сенсорном чипе СМ5 с использованием стандартной химии аминного сочетания. На первой стадии <VEGF-Ang-2>XMAb впрыскивали при концентрации 10 мкг/мл в буфере HBS (10 мМ ГЭПЭС, 150 мМ NaCI, 0,05% Твин 20, рН 7,4) при 25°С.После связывания антитела с иммобилизованным VEGF на второй стадии впрыскивали hAng-2 при 10 мкг/мл (Фиг.3).
В следующем эксперименте определяли сродство и кинетику связывания <VEGF-Ang-2>XMab. Кратко, Поликлональные антитела козы <hIgG-Fгамма> иммобилизовали на чипе СМ4 посредством аминного сочетания для презентирования биспецифического антитела против Ang-2 и VEGF. Связывание измеряли в буфере HBS при 25°С или 37°С. Очищенный Ang-2-His (фирмы R&D или очищенный в лаборатории) или VEGF (R&D systems или очищенный в лаборатории) добавляли при различных концентрациях от 0,37 нМ до 30 нМ или от 3,7 нМ до 200 нМ в растворе. Ассоциацию измеряли путем впрыскивания в течение 3 минут; диссоциацию измеряли путем промывания поверхности чипа буфером HBS в течение 10 минут, и значение KD оценивали, используя модель связывания Ленгмюра 1:1. В связи с гетерогенностью препарата Ang-2 связывание 1:1 невозможно было наблюдать. Поэтому значения KD являются кажущимися значениями. Сродство <VEGF-Ang-2> XMab к VEGF было определено как крайне высокое, вычисленная скорость диссоциации находилась за пределами описания к Biacore даже при 37°С. В таблице 1 суммированы константы связывания для обоих антигенов.
В) Анализ на количественное определение связывания активного биспецифического <Ang2/VEGF>XMab1
В дополнение к анализу SPR был разработан анализ ELISA (твердофазный иммуноферментный анализ) с целью количественного определения связывания биспецифических антител mAb<Ang2/VEGF>. В данном анализе на первой стадии hAng2 непосредственно наносили в качестве покрытия в лунки микротитрационного планшета Maxisorp (MTP). В то же время, образцы/эталонные стандарты (mAb<Ang2/VEGF>) предварительно инкубировали в лунках другого MTP с дигоксигенилированным VEGF. После предварительной инкубации и покрытия избыток несвязанного Ang2 удаляли путем промывания MTP, покрытого Ang2. Затем предварительно проинкубированную смесь <Ang2/VEGF> и VEGF-Dig переносили в MTP, покрытый hAng2, и инкубировали. После инкубации избыток раствора для предварительной инкубации удаляли путем промывания с последующей инкубацией с антителом против дигоксигенина, меченым пероксидазой хрена. Конъюгат антитело-фермент катализирует цветную реакцию с субстратом ABTS®. Сигнал измеряли с помощью считывающего устройства для ELISA при длине волны 405 нм (эталонная длина волны: 490 нм ([405/490] нм)). Значения поглощения каждого образца определяли в двух повторах. (Схема, на которой приведен пример данной системы тестирования, представлена на Фиг.4 и калибровочная кривая ELISA для количественного определения представлена на Фиг.5).
Пример 4
Фосфорилирование Tie2
С целью подтверждения, что активности, связанные с антителом против ANGPT2, сохраняются в биспецифическом двухвалентном антителе <VEGF-ANGPT2>XMAb1, был проведен анализ фосфорилирования Tie2. Эффективность XMAb1 определяли в анализе ANGPT2-стимулированного фосфорилирования Tie2, как описано выше.
Было показано, что XMAb1 препятствует АМСРТ2-стимулированному фосфорилированию Tie2 в анализе АМОРТ2-стимулированного фосфорилирования Tie2, как описано выше. IC50 для XMAb1 составляла 7,4 нМ +/-2,3.
Пример 5
Ингибирование связывания huANG-2 с Tie-2 (ELISA)
Взаимодействие в анализе ELISA проводили на 384-луночных микротитрационных планшетах (MicroCoat, DE, №по каталогу 464718) при КГ. После каждой стадии инкубации планшеты промывали 3 раза ФСБТ. Планшеты ELISA покрывали 5 мкг/мл белка Tie-2 в течение 1 часа (ч). Затем лунки блокировали ФСБ с добавлением 0,2% Твин-20 и 2% БСА (Roche Diagnostics GmbH, DE) в течение 1 ч. Разведение очищенных биспецифических антител ХМаЬ в ФСБ инкубировали вместе с 0,2 мкг/мл ангиопоэтина-2 человека (R&D Systems, UK, №по каталогу 623-AN) в течение 1 ч при КГ. После промывания к смеси добавляли 0,5 мкг/мл биотинилированного антитела к ангиопоэтину-2 клон ВАМ0981 (R&D Systems, UK) и разведенный 1:3000 стрептавидин-HRP (Roche Diagnostics GmbH, DE, №по каталогу11089153001) в течение 1 ч. Затем планшеты промывали 3 раза ФСБТ. Планшеты проявляли свежеприготовленным реагентом ABTS (Roche Diagnostics GmbH, DE, буфер №204530001, таблетки №11112422001) в течение 30 минут при КГ. Поглощение измеряли при 405 нм и определяли IC50. ХМаМ показало ингибирование связывания ANG-2 с Tie-2 с IC50 12 нМ.
Пример 6
Ингибирование связывания hVEGF с рецептором hVEGF (ELISA)
Тест проводили на 384-луночных микротитрационных планшетах (MicroCoat, DE, №по каталогу 464718) при КГ. После каждой стадии инкубации планшеты промывали 3 раза ФСБТ. Сначала планшеты покрывали 1 мкг/мл белка hVEGF-R (R&D Systems, UK, №по KaTanory321-FL) в течение 1 часа (ч). Затем лунки блокировали ФСБ с добавлением 0,2% Твин-20 и 2% БСА (Roche Diagnostics GmbH, DE) в течение 1 ч. Разведения очищенных биспецифических антител ХМаЬ в ФСБ инкубировали вместе с 0,15 мкг/мл huVEGF121 (R&D Systems, UK, №по каталогу 298-VS) в течение 1 ч при КГ. После промывания к смеси добавляли 0,5 мкг/мл антитела против VEGF клон Mab923 (R&D Systems, UK) и 1:2000 F(ab')2 против IgG мыши, конъюгированного с пероксидазой хрена (HRP) (GE Healthcare, UK, №по каталогу NA9310 V), в течение 1 ч. Затем планшеты промывали 6 раз ФСБТ. Планшеты проявляли свежеприготовленным реагентом ABTS (Roche Diagnostics GmbH, DE, буфер №204530001, таблетки №11112422001) в течение 30 минут при КГ. Поглощение измеряли при 405 нм и определяли IC50. ХМаЫ показало ингибирование связывания VEGF с рецептором VEGF с IC50 10 нМ.
Пример 7
Пролиферация HUVEC
С целью подтверждения того, что активности, связанные с антителом против VEGF, сохранены в биспецифическом двухвалентном антителе <VEGF-ANG2>XMAb1, проводили анализ VEGF-индуцированной пролиферации HUVEC. Было показано, что XMAb1 препятствует VEGF-индуцированной пролиферации HUVEC в степени, сравнимой с бевацизумабом, в анализе VEGF-индуцированной пролиферации HUVEC, как описано выше. XMAb1 зависимо от концентрации препятствует VEGF-индуцированной пролиферации HUVEC сравнимо с родительским антителом бевацизумабом (авастином). Юбо составляла 1,1 нМ для бевацизумаба и 2,3 нМ для XMAn.
Пример 8
Анализ ангиогенеза микродивертикулов роговицы мыши
Самок мышей Balb/c в возрасте от 8 до 10 недель покупали у Charles River, Suizfeld, Germany. Протокол был модифицирован в соответствии со способом, описанным Rogers, M.S., et al., Nat Protoc 2 (2007) 2545-2550. Кратко, микродивертикулы шириной примерно 500 мкм препарировали под микроскопом у анестезированной мыши примерно на 1 мм от лимба роговицы к верху роговицы, используя лезвие скальпеля и острый микропинцет.Имплантировали диск (Nylaflo®, Pall Corporation, Michigan) диаметром 0,6 мм, и поверхность области имплантации выправляли. Диски инкубировали в соответствующем факторе роста или в носителе в течение по меньшей мере 30 мин. После 3, 5 и 7 суток (или альтернативно только после 3 суток) глаза фотографировали и измеряли сосудистый ответ.Анализ обсчитывали путем вычисления процента площади новых сосудов против общей площади роговицы.
На диски наносили 300 нг VEGF или ФСБ в качестве контроля и имплантировали на 7 суток. Мониторинг прорастания сосудов из лимба в диск осуществляли по времени на сутки 3, 5 и 7. За одни сутки до имплантации диска антитела (<Ang-2/VEGF>XMAb1, <hVEGF> авастин (бевацизумаб)) вводили внутривенно в дозе 10 мг/кг для авастина и XMAb1. Животные в контрольной группе получали носитель. Объем введения составлял 10 мл/кг.
Для тестирования эффекта XMAb1 на VEGF-индуцированный ангиогенез in vivo авторы изобретения проводили анализ ангиогенеза роговицы мыши. В данном анализе диск Nylaflo, пропитанный VEGF, имплантировали в дивертикул бессосудистой роговицы на фиксированном расстоянии до лимбических сосудов. Сосуды сразу прорастают в роговицу в направлении увеличения градиента VEGF. Результаты авторов изобретения демонстрируют, что системное введение XMAb1 (10 мг/кг) почти полностью ингибирует прорастание сосудов из лимба в направлении градиента VEGF на сутки исследования от 3 до 5 (Фиг.6). В следующем эксперименте проводили непосредственные сравнительные исследования. На диски наносили 300 нг VEGF или ФСБ в качестве контроля и имплантировали на 3 суток. Мониторинг прорастания сосудов из лимба в диск осуществляли по времени на сутки 3. За одни сутки до имплантации диска антитела (биспецифическое антитело <Ang-2/VEGF>XMAb1, родительское антитело <VEGF> бевацизумаб (авастин), родительское антитело <Ang-2>ANG2i-LC06 и комбинацию антитела<VEGF>бевацизумаба (авастина) и антитела <Ang-2>ANG2i-LC06) вводили внутривенно в дозе 10 мг/кг для бевацизумаба (авастина), 10 мг/кг для XMAb1, 10 мг/кг для бевацизумаба (авастина) и 10 мг/кг для ANG2i-LC06. Комбинацию бевацизумаба (авастина) и ANG2i-LC06 вводили в дозе 10 мг/кг для бевацизумаба (авастина) и 10 мг/кг для ANG2i-LC06. Животные в контрольной группе получали носитель. Объем введения составлял 10 мл/кг.
Результаты авторов изобретения (см. фиг.7 и приведенную ниже таблицу) демонстрируют, что системное введение XMAb1 (10 мг/кг) почти полностью ингибирует прорастание сосудов из лимба в направлении градиента VEGF на сутки исследования 3 сравнимо с комбинацией бевацизумаба и ANG2i-LC06. Монотерапия антителом против Ang-2, напротив, только слабо ингибировала VEGF-индуцированный ангиогенез (Фиг.7). Максимальный эффект мог быть уже достигнут при более низких концентрациях 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг бевацизумаба (авастина).
Пример 9
Эффективность in vivo биспецифического антитела <VEGF-ANG-2> в модели ксенотрансплантата Colo205 у бежевых мышей SCID Клеточные линии и условия культивирования
Клетки колоректального рака человека Colo205 (АТСС No. CCL-222). Опухолевую клеточную линию культивировали обычным путем в среде RPMI 1640 (РАА, Laboratories, Austria) с добавлением 10% фетальной бычьей сыворотки (РАА Laboratories, Austria) и 2 мМ L-глутамина при 37°С в атмосфере, насыщенной водой, при 5% CO-. Для трансплантации использовали пассаж 2-5.
Животные:
Самок бежевых мышей SCID в возрасте 4-5 недель (приобретенных у Charles River, Germany) по прибытии поддерживали в условиях, свободных от специфических патогенов, при суточных циклах 12 ч свет/12 ч темнота в соответствии с действующим руководством (GV-Solas; Felasa; TierschG). Протокол экспериментального исследования был проверен и одобрен местным правительством. По прибытии животных поддерживали в карантинном отделе вивария в течение в течение одной недели для привыкания к новому окружению и наблюдения. Постоянный мониторинг состояния здоровья проводили на регулярной основе. Диетический корм (Provimi Kliba 3337) и воду (подкисленную до рН 2,5-3) обеспечивали свободно. Возраст мышей в начале исследования составляет примерно 10 недель.
Инъекция опухолевых клеток:
На сутки инъекции опухолевые клетки собирали (трипсин-ЭДТА) из культуральных флаконов (Greiner) и переносили в 50 мл культуральной среды, промывали один раз и ресуспендировали в ФСБ. После дополнительной стадии отмывки в ФСБ и фильтрования (клеточное сито; Falcon диаметр 100 мкм) конечный титр клеток доводили до 2,5×107/мл. Суспензию опухолевых клеток осторожно смешивали переносящей пипеткой во избежание агрегации клеток. После этого клеточную суспензию заполняли в 1,0 мл туберкулиновый шприц (Braun Melsungen), используя широкую иглу (1,10×40 мм); для инъекции размер иглы меняли (0,45×25 мм), и для каждой инъекции использовали новую иглу. Анестезию осуществляли, используя ингаляционное устройство Stephens для мелких животных с камерой предварительной инкубации (плексиглас), индивидуальной маской для носа для мышей (силикон) и не воспламеняемое и не взрывчатое соединение для анестезии изофлуран (CH-Pharma) в закрытой системе циркуляции. За двое суток до инъекции шерсть животных сбривали, и для инъекции клеток кожу анестезированных животных осторожно поднимали анатомическими ножницами, и 100 мкл клеточной суспензии (=2,5×106 клеток) инъецировали подкожно в правый бок животных.
Обработка животных
Обработку животных начинали на сутки рандомизации при среднем объеме опухоли ~100 мм3, соответственно. Мышей обрабатывали один раз в неделю i.p. различными соединениями, как указано в приведенной ниже таблице.
Мониторинг:
Животных контролировали 2 раза в неделю на их состояние здоровья. Массу тела документировали 2 раза в неделю после инъекции клеток. Размеры опухоли измеряли с помощью циркуля, начиная на сутки постановки, а затем 2 раза в неделю в течение всего периода обработки. Объем опухоли вычисляли в соответствии с протоколом NCI (масса опухоли=1/2ab2, где "а" и "b" представляют собой длинный и короткий диаметры опухоли соответственно). Критерием окончания была критическая масса опухоли (вплоть до 1,7 г или диаметр более 1,5 см), потеря массы тела более 20% от базовой, изъязвление опухоли или плохое общее состояние животных.
Результаты (см. Фиг.8) показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMAb1 проявляет высокое ингибирование опухолевого роста в модели ксенотрансплантата опухоли Colo205 у бежевых мышей SCID по сравнению с обработкой моноспецифическими антителами. Эффективность комбинации ANG2i-LC06 и бевацизумаба показала сравнимые результаты с XMAb1. Максимальная эффективность XMAb1 была достигнута уже при 10 мг/кг.
Во втором эксперименте анализировали эффект XMAb1 на опухоли большего размера.
Обработка животных
Обработку животных начинали на сутки рандомизации при среднем объеме опухоли ~400 мм3, соответственно. Мышей обрабатывали один раз в неделю i.p. различными соединениями, как указано в приведенной ниже таблице.
Мониторинг:
Животных контролировали 2 раза в неделю на их состояние здоровья. Массу тела документировали 2 раза в неделю после инъекции клеток. Размеры опухоли измеряли с помощью циркуля, начиная на сутки постановки, а затем 2 раза в неделю в течение всего периода обработки. Объем опухоли вычисляли в соответствии с протоколом NCI (масса опухоли=1/2ab2, где "а" и "b" представляют собой длинный и короткий диаметры опухоли соответственно). Критерием окончания была критическая масса опухоли (вплоть до 1,7 г или диаметр более 1,5 см), потеря массы тела более 20% от базовой, изъязвление опухоли или плохое общее состояние животных.
Результаты (см. Фиг.9) показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMAb1 проявляет высокое ингибирование опухолевого роста в модели ксенотрансплантата опухоли Colo205 у бежевых мышей SCID по сравнению с обработкой моноспецифическими антителами, которые не проявляют эффективность в больших опухолях по сравнению с контролем. Эффективность комбинации ANG2J-LC06 и бевацизумаба показала сравнимые результаты с XMAb1. Максимальная эффективность XMAb1 была достигнута уже при 10 мг/кг.
Взятые вместе, эти результаты демонстрируют, что независимо от размера опухоли XMAb1 проявляет лучшую эффективность по сравнению с обработкой моноспецифическими антителами.
Стаз опухоли в этих моделях мог быть уже достигнут при более низких концентрациях 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина.
Пример 10
Эффективность in vivo биспецифического антитело <VEGF-ANG-2> в ортотопической модели ксенотрансплантата KPL-4 у бежевых мышей SCID
Опухолевая клеточная линия
Линия клеток рака молочной железы человека KPL-4 ((Kurebayashi, J., et al., Br. J. Cancer 79 (1999) 707-17)) основана из злокачественного плеврального выпота пациента с раком молочной железы с воспалительным метастазом кожи. Опухолевую клеточную линию культивировали обычным путем в среде RPMI 1640 (РАА, Laboratories, Austria) с добавлением 10% фетальной бычьей сыворотки (РАА Laboratories, Austria) и 2 мМ L-глутамина при 37°С в атмосфере, насыщенной водой, при 5% CO2. Пересев культуры осуществляли с расщеплением трипсином/ЭДТА 1× (PAN) три раза в неделю.
Мыши
По прибытии самок бежевых мышей SCID (возраст 10-12 недель; масса тела 18-20 г) Charles River, Suizfeld, Germany) поддерживали в карантинном отделе вивария, одобренном AALAAC, в течение одной недели для привыкания к новому окружению и для наблюдения. Проводили постоянный мониторинг состояния здоровья. Мышей держали в SPF-условиях в соответствии с международным руководством (GV-Solas; Felasa; TierschG) при суточных циклах 12 ч свет/12 ч темнота. Диетический корм (Kliba Provimi 3347) и воду (фильтрованную) обеспечивали свободно. Протокол экспериментального исследования был проверен и одобрен местным правительством (Regierung von Oberbayern; регистрационный №211.2531.2-22/2003).
Инъекция опухолевых клеток
На сутки инъекции опухолевые клетки собирали (трипсин-ЭДТА) из культуральных флаконов (Greiner TriFlask) и переносили в 50 мл культуральной среды, промывали один раз и ресуспендировали в ФСБ. После дополнительной стадии отмывки в ФСБ и фильтрования (клеточное сито; Falcon диаметр 100 мкм) конечный титр клеток доводили до 1,5×108/мл. Суспензию опухолевых клеток осторожно смешивали переносящей пипеткой во избежание агрегации клеток. Анестезию осуществляли, используя ингаляционное устройство Stephens для мелких животных с камерой предварительной инкубации (плексиглас), индивидуальной маской для носа для мышей (силикон) и не воспламеняемое и не взрывчатое соединение для анестезии изофлуран (Pharmacia-Upjohn, Germany) в закрытой системе циркуляции. За двое суток до инъекции шерсть животных сбривали. Для инъекции i.m.f.p. клетки инъецировали ортотопически в объеме 20 мкл в жировое тело правой предпоследней паховой молочной железы каждой анестезированной мыши. Для ортотопической имплантации клеточную суспензию инъецировали через кожу под сосок, используя микролитровый шприц Гамильтона и иглу 30G×1/2".
Обработка животных
Обработку животных начинали на сутки рандомизации при среднем объеме опухоли ~80 мм3, соответственно. Мышей обрабатывали один раз в неделю i.p. различными соединениями, как указано в приведенной ниже таблице.
Мониторинг опухолевого роста
Животных контролировали 2 раза в неделю на их состояние здоровья. Массу тела документировали 2 раза в неделю после инъекции клеток. Размеры опухоли измеряли с помощью циркуля, начиная на сутки постановки, с начала периода обработки 2 раза в неделю. Объем опухоли вычисляли в соответствии с протоколом NCl (В. Teicher; Anticancer drug development guide, Humana Press, 1997, Chapter 5, page 92) (масса опухоли=1/2ab2, где "а" и "b" представляют собой длинный и короткий диаметры опухоли соответственно).
Критерием окончания была критическая масса опухоли (вплоть до 1,7 г или диаметр более 1,5 см), потеря массы тела более 20% от базовой, изъязвление опухоли или плохое общее состояние животных.
Результаты (см. Фиг.10) показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMAb1 проявляет высокое ингибирование опухолевого роста в модели ксенотрансплантата опухоли KPL-4 у бежевых мышей SCID по сравнению с обработкой моноспецифическими антителами. Эффективность комбинации ANG2i-LC06 и бевацизумаба показала сравнимые результаты с XMAb1. Максимальная эффективность XMAb1 была достигнута уже при 10 мг/кг.
Во втором эксперименте анализировали эффект XMAb1 на опухоли большего размера.
Обработка животных
Обработку животных начинали на сутки рандомизации при среднем объеме опухоли ~160 мм3, соответственно. Мышей обрабатывали один раз в неделю i.p. различными соединениями, как указано в приведенной ниже таблице.
Мониторинг:
Животных контролировали 2 раза в неделю на их состояние здоровья. Массу тела документировали 2 раза в неделю после инъекции клеток. Размеры опухоли измеряли с помощью циркуля, начиная на сутки постановки, с начала периода обработки 2 раза в неделю. Объем опухоли вычисляли в соответствии с протоколом NCl (В. Teicher; Anticancer drug development guide, Humana Press, 1997, Chapter 5, page 92) (масса опухоли=1/2ab2, где "а" и "b" представляют собой длинный и короткий диаметры опухоли соответственно).
Критерием окончания была критическая масса опухоли (вплоть до 1,7 г или диаметр более 1,5 см), потеря массы тела более 20% от базовой, изъязвление опухоли или плохое общее состояние животных.
Результаты (см. Фиг.11) показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMAb1 проявляет высокое ингибирование опухолевого роста в модели ксенотрансплантата опухоли KPL-4 у бежевых мышей SCID по сравнению с обработкой моноспецифическими антителами. Эффективность комбинации ANG2i-LC06 и бевацизумаба показала сравнимые результаты с XMAb1. Максимальная эффективность XMAb1 была достигнута уже при 10 мг/кг.
Взятые вместе, эти результаты демонстрируют, что независимо от размера опухоли XMAb1 проявляет лучшую эффективность по сравнению с обработкой моноспецифическими антителами.
Стаз опухоли в этих моделях мог быть уже достигнут при более низких концентрациях 10 мг/кг XMAb1 по сравнению с комбинацией 10 мг/кг ANG2i-LC06+10 мг/кг авастина.
Пример 11
Эффект обработки XMAb1 на плотность микрососудов в подкожном ксенотрансплантате Colo205
Плотность кровеносных сосудов оценивали путем подсчета всех сосудов среза опухоли. Сосуды метили флуоресцентным антителом против мыши CD34 (клон МЕС14.7) на срезах, заключенных в парафин. Сосуды подсчитывали, и плотность микрососудов вычисляли как число сосудов на мм2. Все результаты выражали в виде среднего ±СКО. Для определения значимых различий экспериментальных групп использовали метод Дуннетта. р<0,05 считали статистически значимой. Результаты показывают, что суммарная внутриопухолевая MVD была снижена в обработанных опухолях. Обработка ANG2i-LC06 снижала MVD на 29%, бевацизумаб на число, меньшее или равное 0%, бевацизумаб+ANG2i-LC06 на 15% и XMAb1 на 28%.
Пример 12
Эффективность in vivo биспецифического антитела <VEGF-ANG-2> в модели подкожного ксенотрансплантата N87 у бежевых мышей SCID
Опухолевая клеточная линия
Линия клеток рака ЖКТ человека N87 (NCI-N87 (АТСС No.CRL 5822)). Опухолевые клетки культивировали обычным путем в среде RPMI 1640 (РАА, Laboratories, Austria) с добавлением 10% фетальной бычьей сыворотки (РАА Laboratories, Austria) и 2 мМ L-глутамина при 37°С в атмосфере, насыщенной водой, при 5% CO2. Пересев культуры осуществляли с расщеплением трипсином/ЭДТА 1× (PAN) три раза в неделю.
Мыши
По прибытии самок бежевых мышей SCID (возраст 10-12 недель; масса тела 18-20 г) Charles River, Suizfeld, Germany) поддерживали в карантинном отделе вивария, одобренном AALAAC, в течение одной недели для привыкания к новому окружению и для наблюдения. Проводили постоянный мониторинг состояния здоровья. Мышей держали в SPF-условиях в соответствии с международным руководством (GV-Solas; Felasa; TierschG) при суточных циклах 12 ч свет/12 ч темнота. Диетический корм (Kliba Provimi 3347) и воду (фильтрованную) обеспечивали свободно. Протокол экспериментального исследования был проверен и одобрен местным правительством (Regierung von Oberbayern; регистрационный №211.2531.2-22/2003).
Инъекция опухолевых клеток
На сутки инъекции клеток клетки собирали из культуральных флаконов (Greiner Т 75), переносили в 50 мл культуральной среды, промывали один раз и ресуспендировали в ФСБ. После дополнительного промывания ФСБ концентрацию клеток измеряли Vi-Cell™ (анализатор жизнеспособности клеток, Beckman Coulter, Madison, Wisconsin, USA). Суспензию опухолевых клеток (ФСБ) осторожно смешивали (для уменьшения агрегации клеток) и держали на льду. Клеточную суспензию заполняли в 1,0 мл шприц. Для инъекции использовали размер иглы 0,45×25 мм. Для образования первичных опухолей 5×106 опухолевых клеток N87 в объеме 100 мкл ФСБ инъецировали подкожно в правый бок каждой мыши.
Обработка животных
Обработку животных начинали на сутки рандомизации при среднем объеме опухоли ~130 мм3, соответственно. Мышей обрабатывали один раз в неделю i.p. различными соединениями, как указано в приведенной ниже таблице.
Мониторинг опухолевого роста
Животных контролировали 1 раз в неделю на их состояние здоровья. Массу тела документировали один раз в неделю после инъекции клеток. Размеры опухолей измеряли с помощью циркуля на сутки постановки, с начала периода обработки один раз в неделю. Объем опухоли вычисляли в соответствии с протоколом NCI (В. Teicher; Anticancer drug development guide, Humana Press, 1997, Chapter 5, page 92) (масса опухоли=1/2ab2, где "а" и "b" представляют собой длинный и короткий диаметры опухоли соответственно).
Критерием окончания была критическая масса опухоли (вплоть до 1,7 г или диаметр более 1,5 см), потеря массы тела более 20% от базовой, изъязвление опухоли или плохое общее состояние животных.
Результаты показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMAb1 показало высокое ингибирование опухолевого роста в модели ксенотрансплантата опухоли N87 у бежевых мышей SCID по сравнению с обработкой моноспецифическими антителами (Фиг. 12).
Пример 13
Эффективность in vivo биспецифического антитела XMab1 в модели ксенотрансплантата рака Colo205 у мышей
В данном примере использовалась модель, описанная в Примере 9.
Проводилось сравнение ингибирования опухолевого роста in vivo в ксенотрансплантате колоректального рака Colo205 антителом <VEGF-ANG-2> XMab1 в дозе 8 мг/кг и комбинацией антител LC06 и авастина (каждое в дозе по 4 мг/кг).
Результаты показывают, что биспецифическое двухвалентное антитело <VEGF-ANG-2> XMab1 оказывает более сильный эффект ингибирования опухолевого роста в модели ксенотрансплантата колоректального рака Colo205 по сравнению с обработкой комбинацией моноспецифических антител при введении антитела XMab1 и комбинации LC06 и авастина в эквимолярном количестве (Фиг. 13): ΔTGI - разница в ингибировании опухолевого роста.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИ-VEGF/АНТИ-ANG-2 АНТИТЕЛА | 2009 |
|
RU2542382C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИ-VEGF/АНТИ-ANG-2 АНТИТЕЛА | 2014 |
|
RU2640253C2 |
| АНТИТЕЛА ПРОТИВ АНГИОПОЭТИНА-2 ЧЕЛОВЕКА | 2009 |
|
RU2569461C2 |
| АНТИТЕЛА ПРОТИВ АНГИОПОЭТИНА-2 ЧЕЛОВЕКА | 2009 |
|
RU2569107C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| БИСПЕЦИФИЧЕСКИЕ ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2604189C2 |
| СОСТАВ, СОДЕРЖАЩИЙ АНТИТЕЛО | 2019 |
|
RU2806628C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2573914C2 |
| БИСПЕЦИФИЧНЫЕ АНТИ-HER2/АНТИ-HER3 АНТИТЕЛА | 2016 |
|
RU2653443C2 |



















































































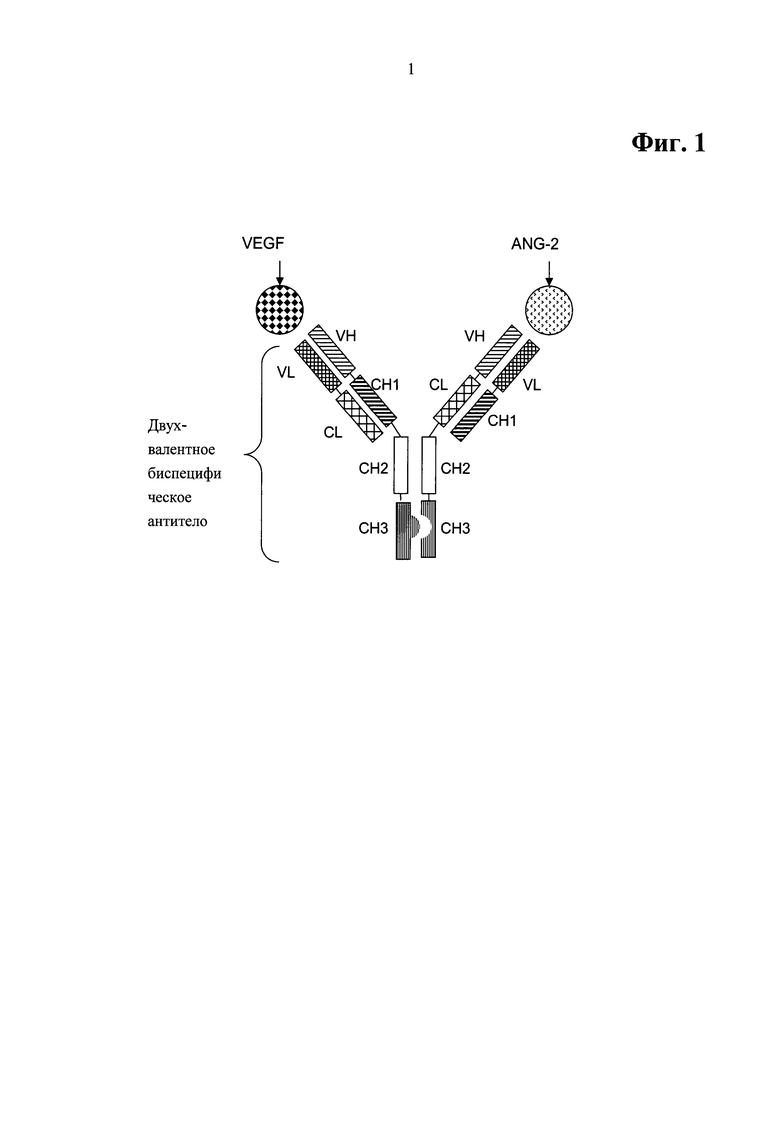
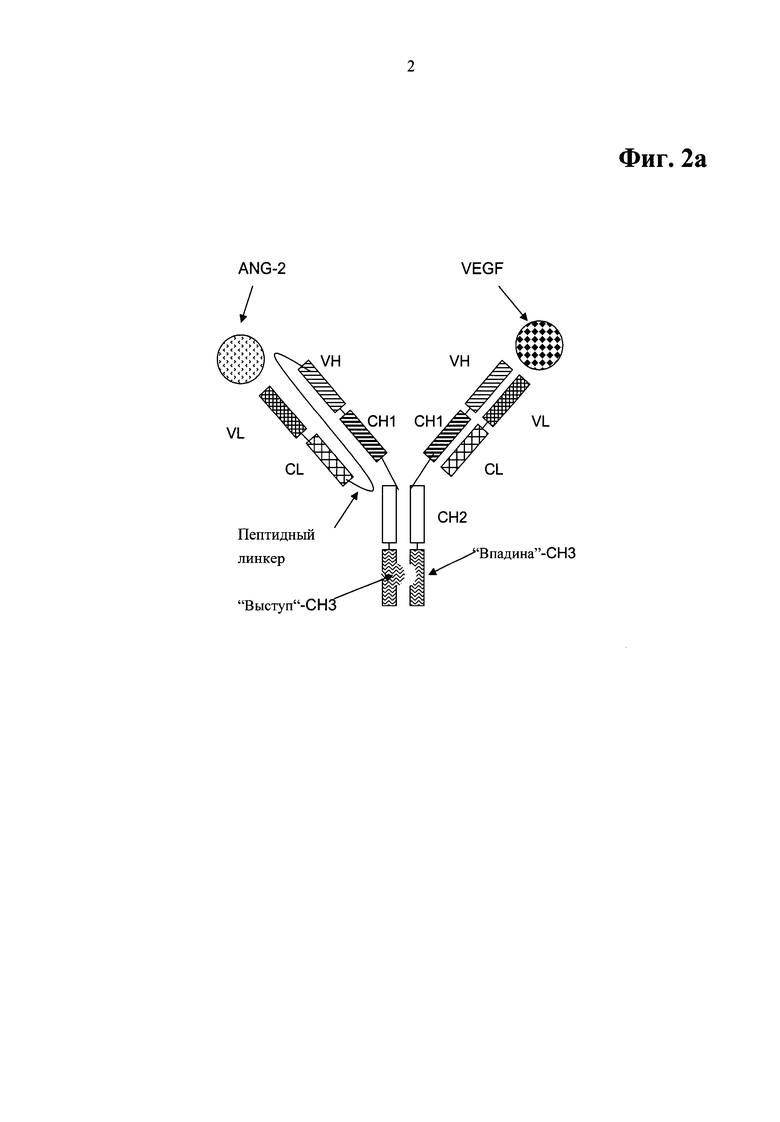
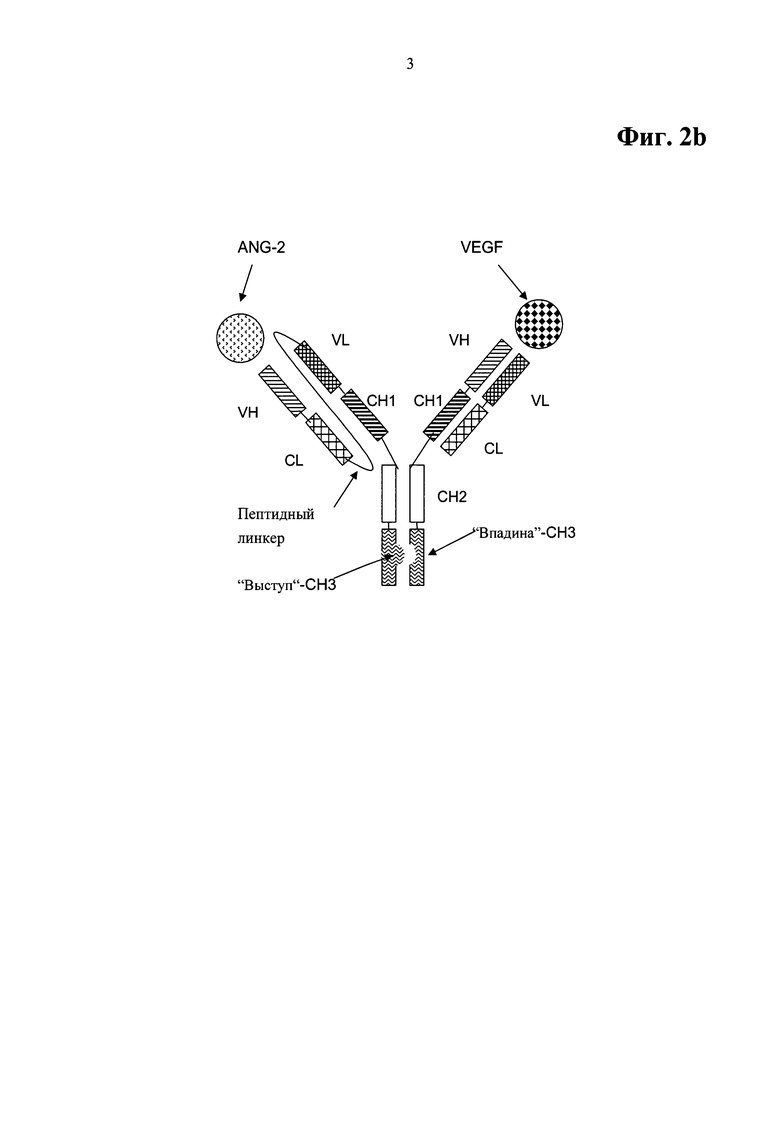
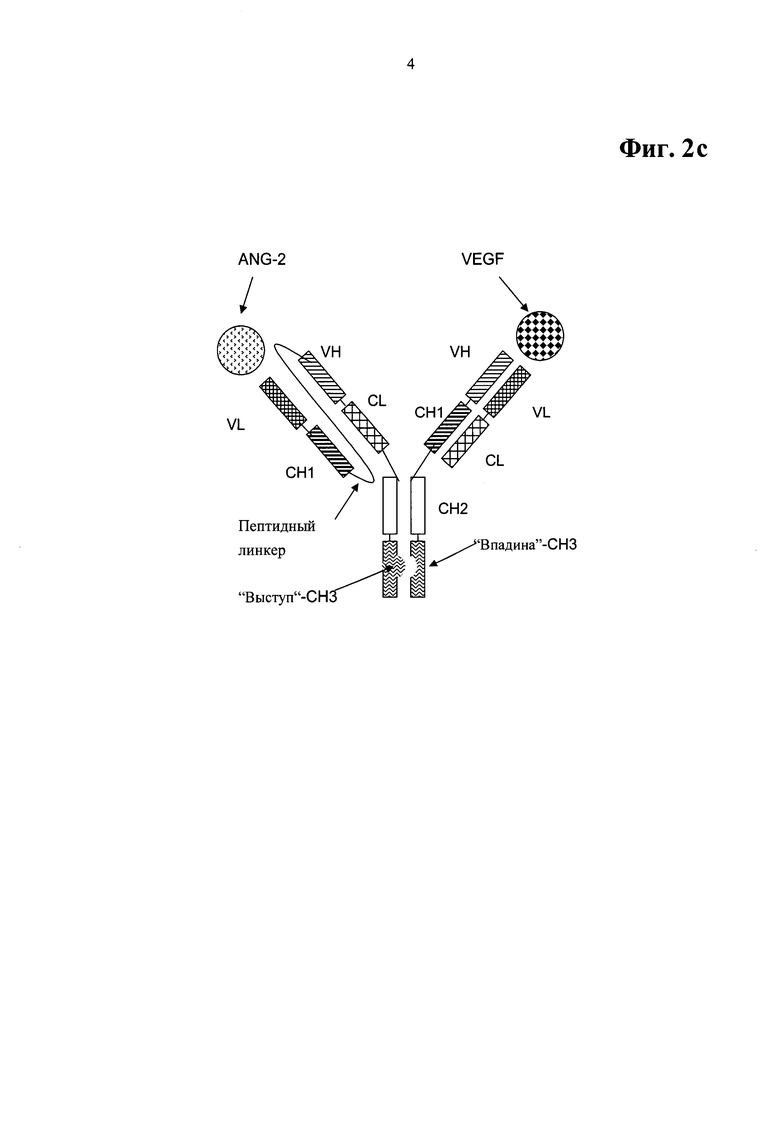
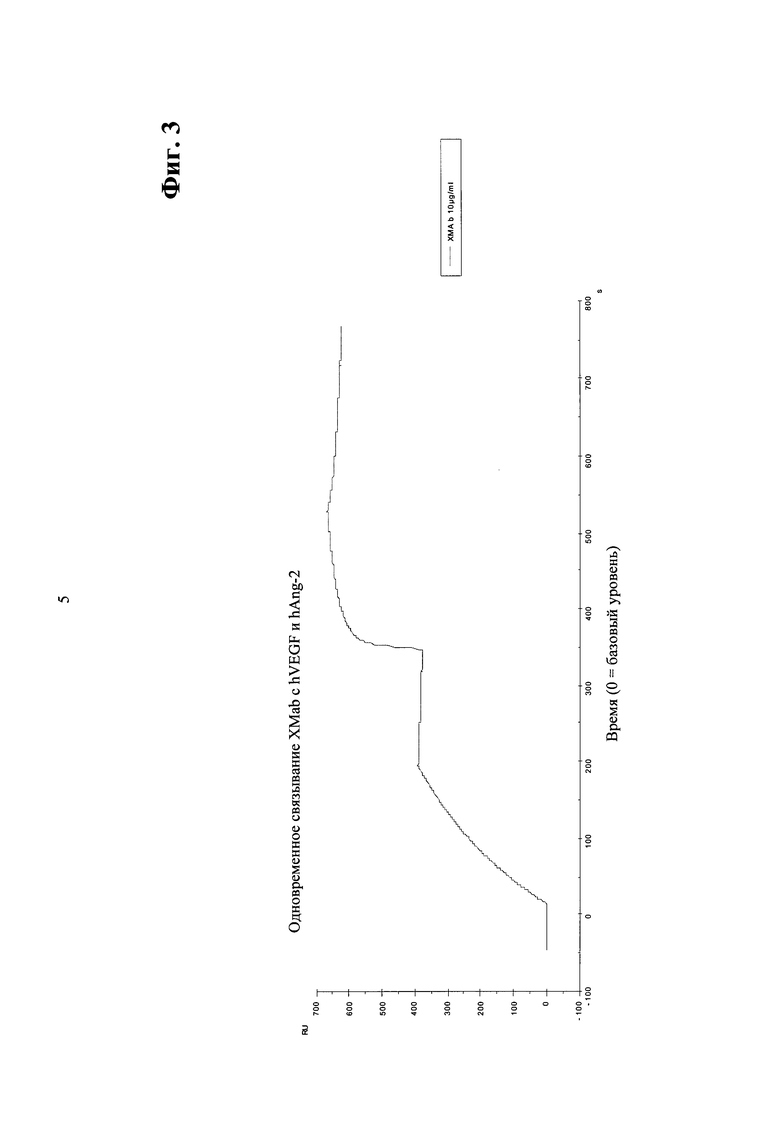
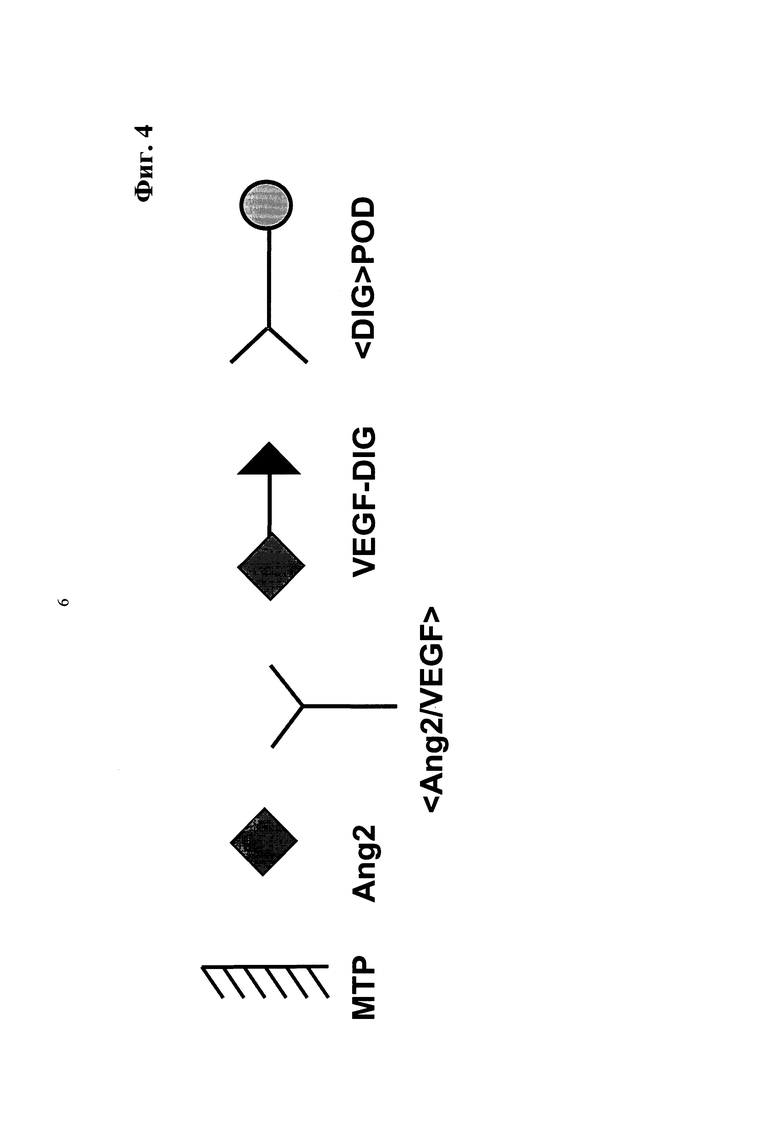
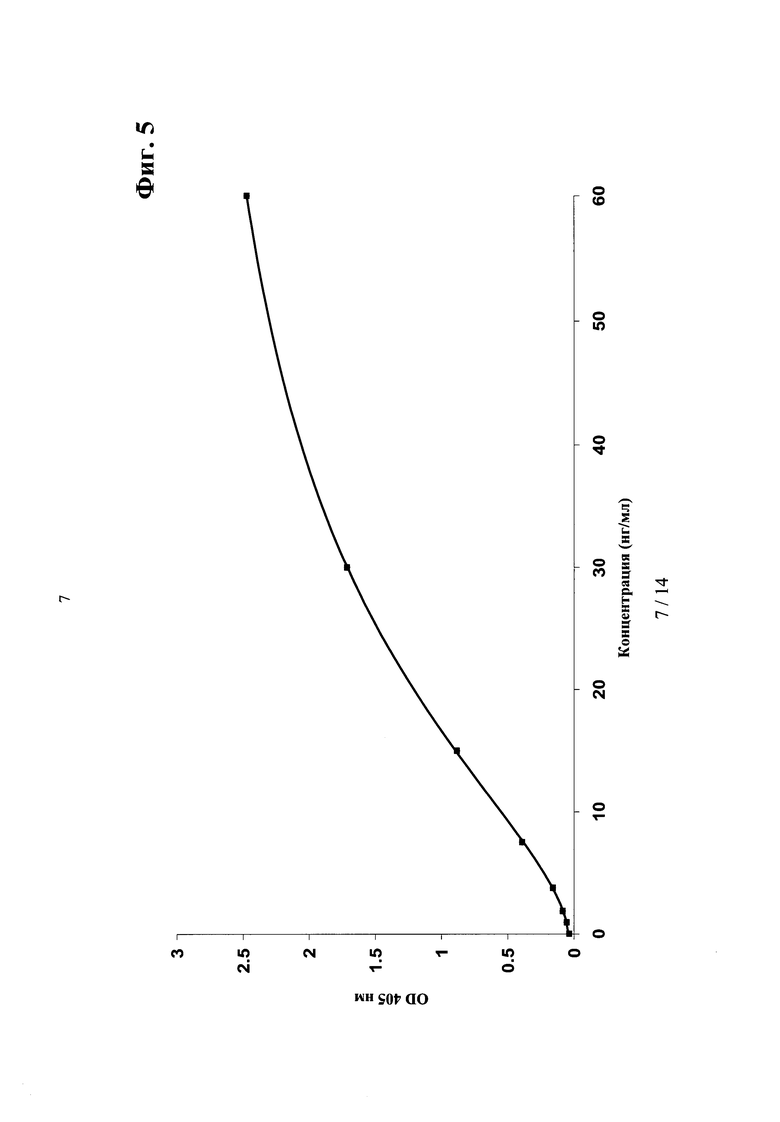
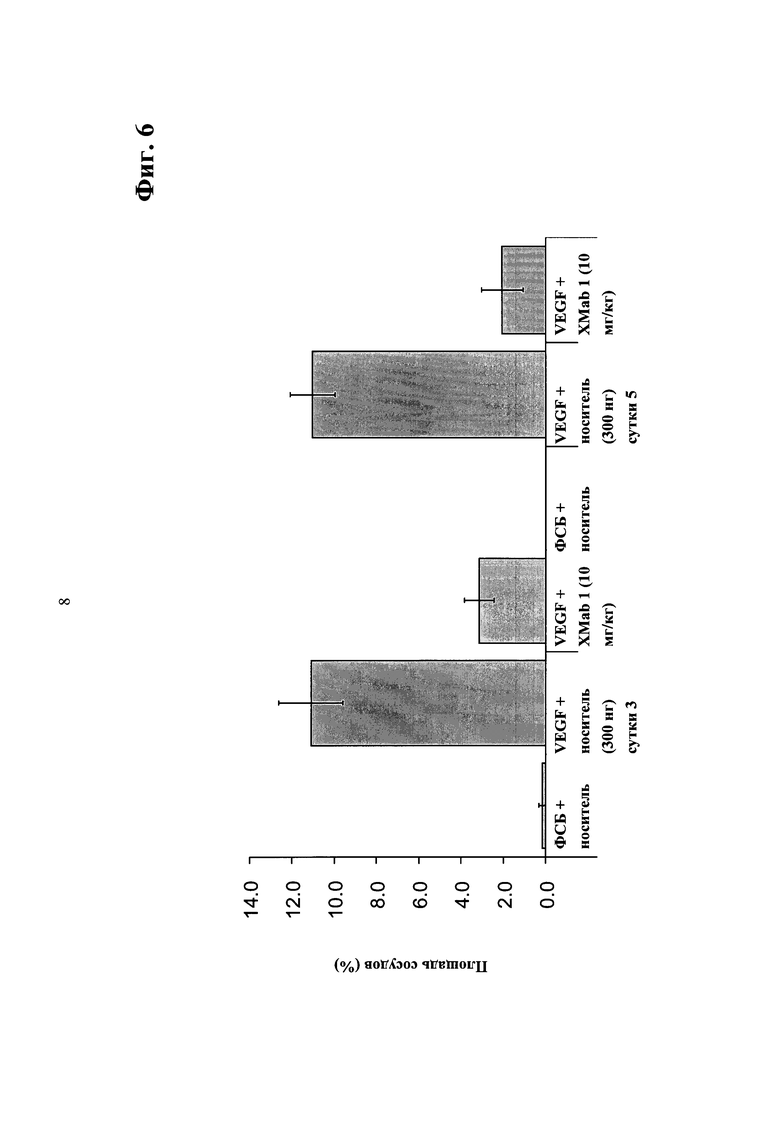
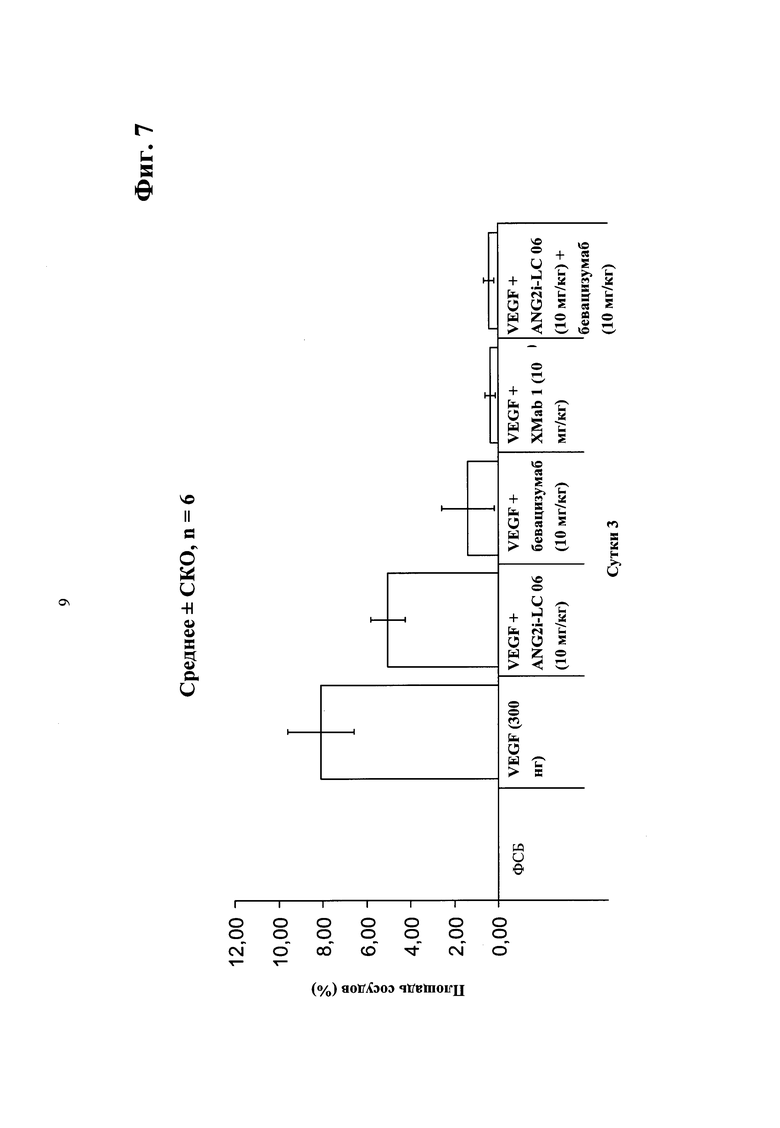
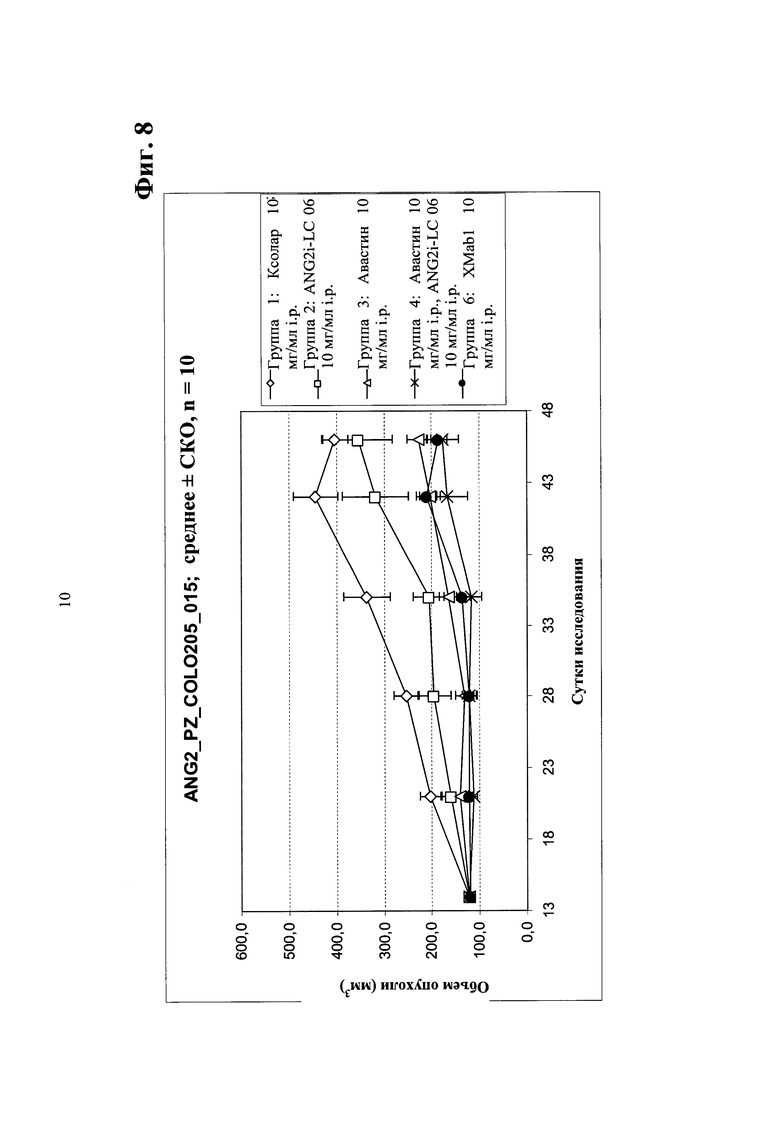
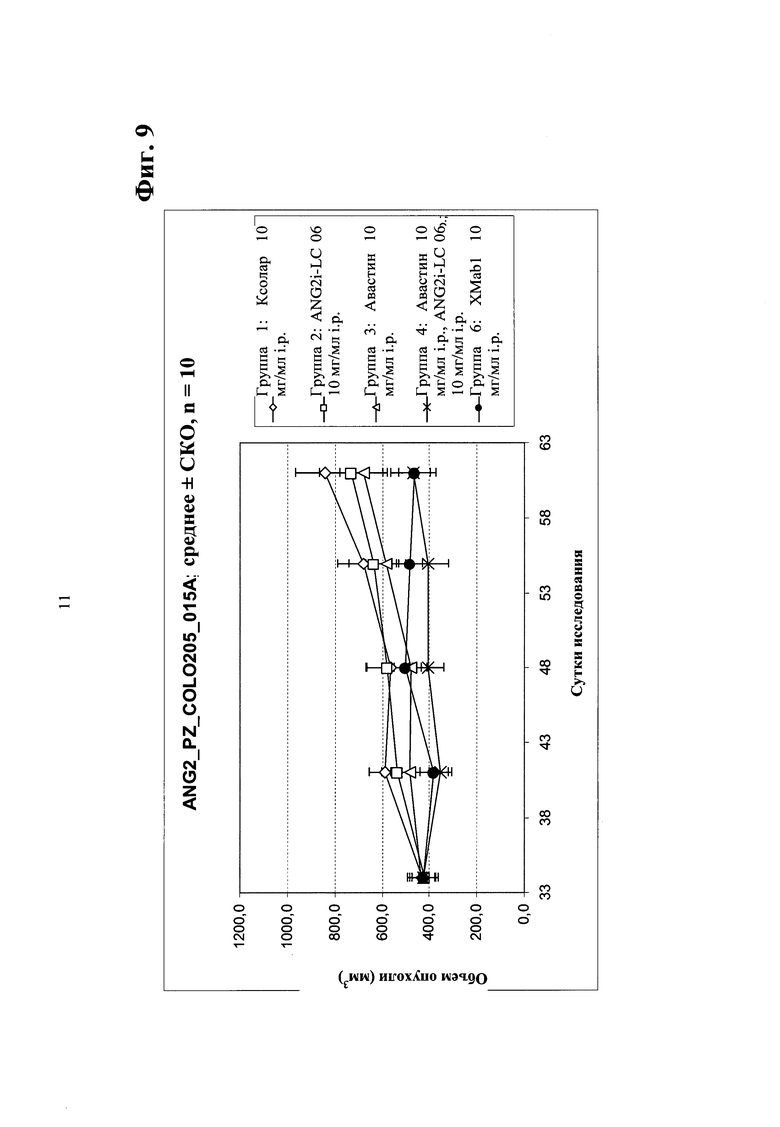
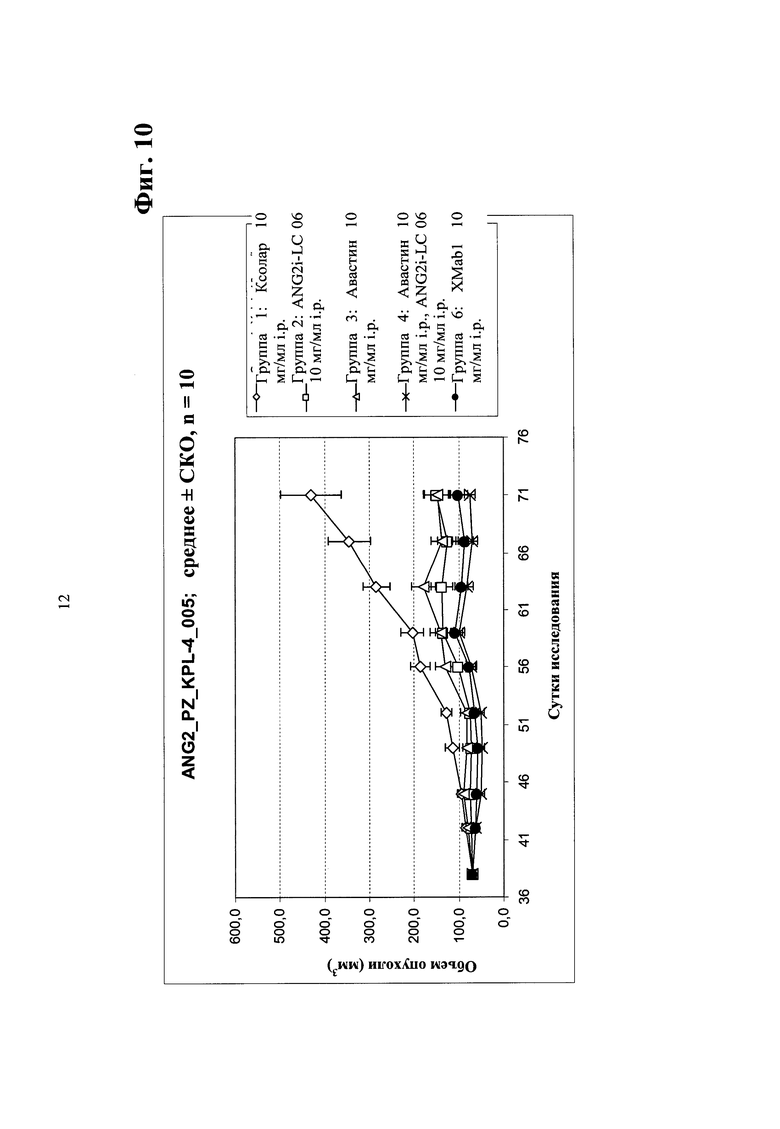
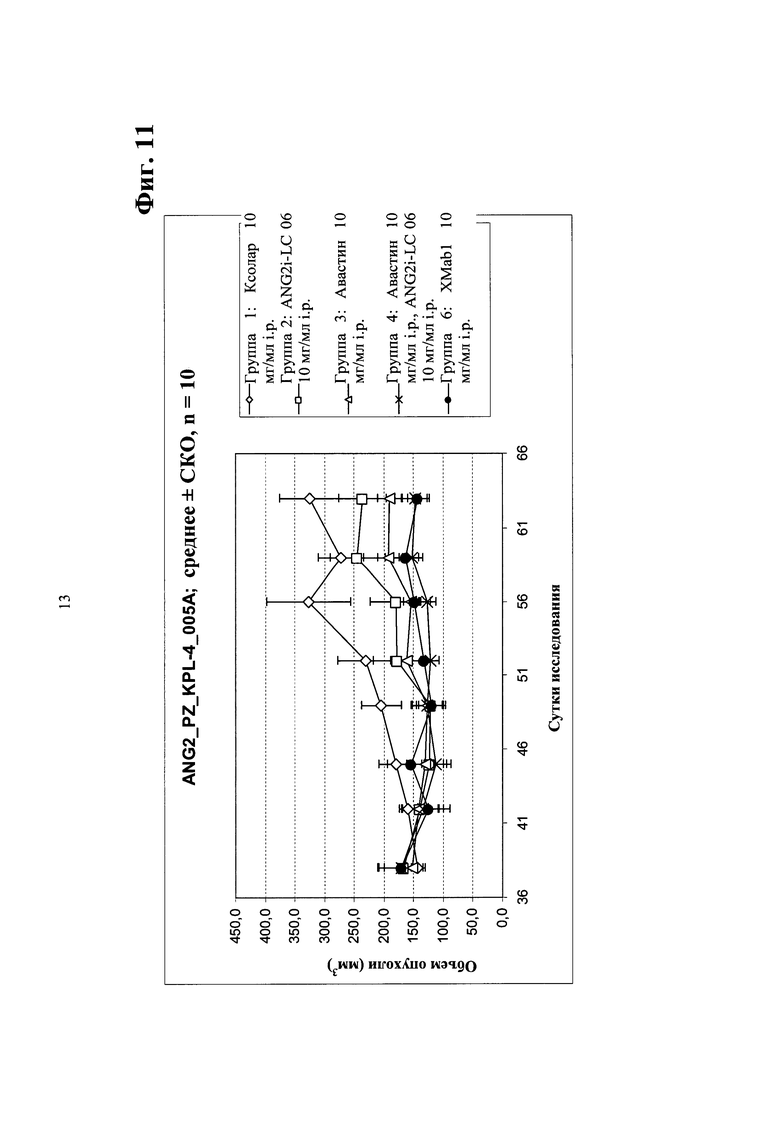
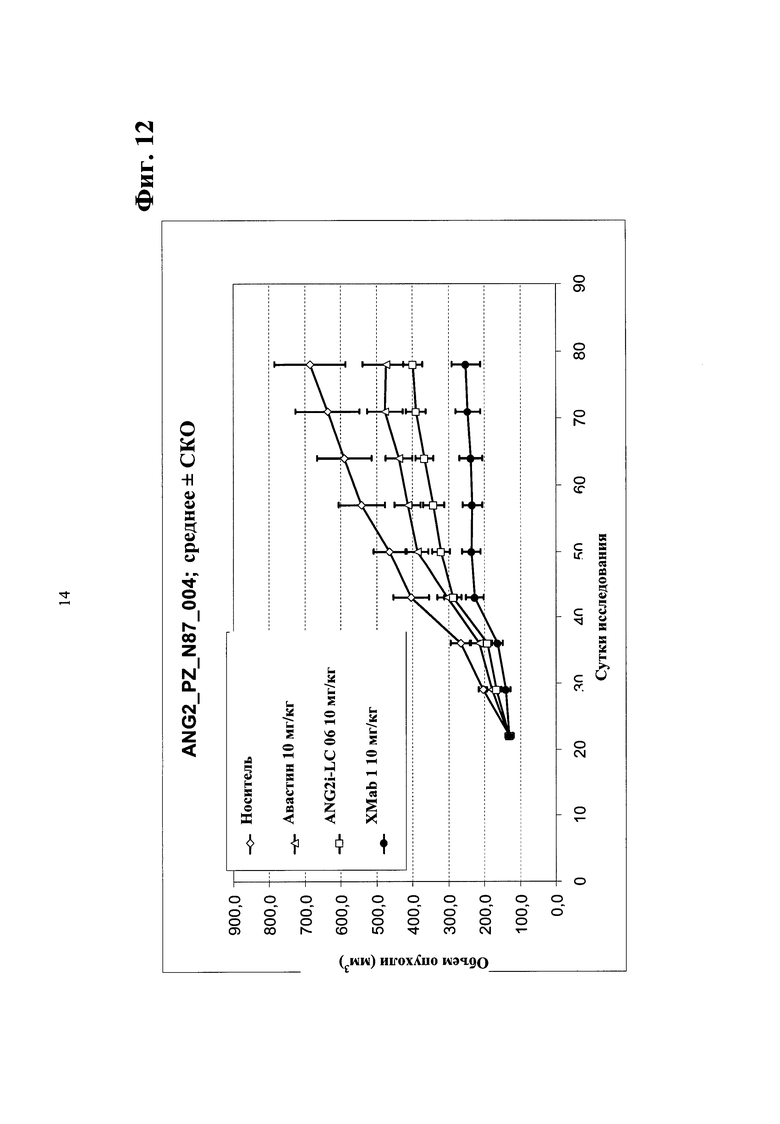
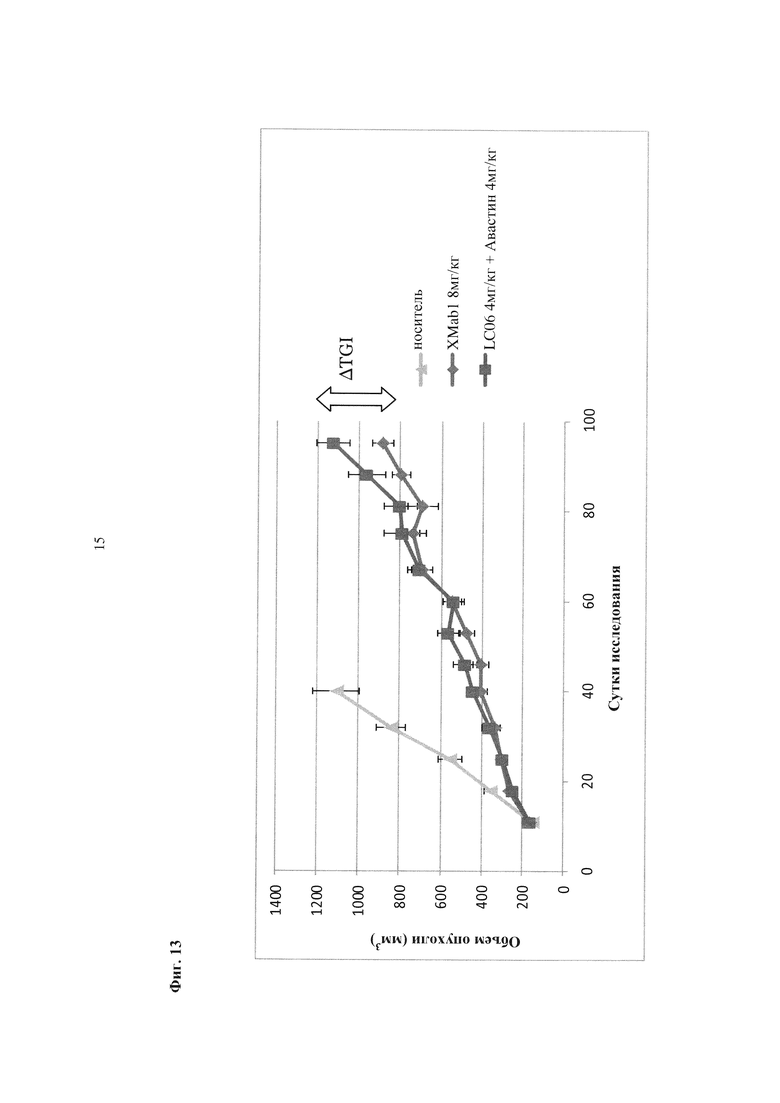
Изобретение относится к области биохимии, в частности к биспецифическому двухвалентному антителу, которое специфично связывается с фактором роста эндотелия сосудов человека (VEGF) и с ангиопоэтина-2 человека (ANG-2). Также раскрыты: фармацевтическая композиция для лечения рака, связанного с VEGF и ANG-2, содержащая терапевтически эффективное количество вышеуказанного биспецифического антитела, нуклеиновая кислота, кодирующая указанное антитело, экспрессионный вектор, содержащий указанную нуклеиновую кислоту, прокариотическая или эукариотическая клетка-хозяин, содержащая указанный вектор. Раскрыт способ получения указанного биспецифического антитела. Изобретение позволяет эффективно лечить VEGF- и ANG-2-опосредуемый рак. 6 н. и 3 з.п. ф-лы, 15 ил., 11 табл., 13 пр.
1. Биспецифическое двухвалентное антитело, которое специфично связывается с VEGF и ANG-2 человека, содержащее первый антигенсвязывающий сайт, который специфично связывается с VEGF человека, и второй антигенсвязывающий сайт, который специфично связывается с ANG-2 человека, характеризующееся тем, что указанное антитело содержит:
i) тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфично связывается с VEGF, с первым антигенсвязывающим сайтом, содержащим в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO: 1 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO: 2; и
ii) модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфично связывается с ANG-2, где константные домены CL и СН1 заменены друг другом, со вторым антигенсвязывающим сайтом, содержащим в качестве вариабельного домена тяжелой цепи (VH) SEQ ID NO: 3 и в качестве вариабельного домена легкой цепи (VL) SEQ ID NO: 4,
где указанное биспецифическое двухвалентное антитело имеет константную область подкласса lgG1 человека и где указанное биспецифическое двухвалентное антитело содержит мутацию T366W в домене СН3 одной тяжелой цепи и мутации T366S, L368A, Y407V в домене СН3 другой тяжелой цепи и дополнительно содержит мутацию Y349C в СН3 одной тяжелой цепи и мутацию S354C в домене СН3 другой тяжелой цепи.
2. Биспецифическое антитело по п. 1, характеризующееся тем, что содержит:
a) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO: 7 и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO: 5 и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела SEQ ID NO: 8 и в качестве модифицированной легкой цепи второго полноразмерного антитела SEQ ID NO: 6.
3. Биспецифическое антитело по п. 1, характеризующееся тем, что содержит:
а) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO: 11 и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO: 9 и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела SEQ ID NO: 12 и в качестве модифицированной легкой цепи второго полноразмерного антитела SEQ ID NO: 10.
4. Биспецифическое антитело по п. 1, характеризующееся тем, что содержит:
a) в качестве тяжелой цепи первого полноразмерного антитела SEQ ID NO: 15 и в качестве легкой цепи первого полноразмерного антитела SEQ ID NO: 13 и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела SEQ ID NO: 16 и в качестве модифицированной легкой цепи второго полноразмерного антитела SEQ ID NO: 14.
5. Фармацевтическая композиция, содержащая терапевтически эффективное количество антитела по пп. 1-4, для применения в лечении рака, ассоциированного с VEGF и ANG-2.
6. Нуклеиновая кислота, кодирующая биспецифическое антитело по любому из пп. 1-4.
7. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 6, способный к экспрессии указанной нуклеиновой кислоты в прокариотической или эукариотической клетке-хозяине.
8. Прокариотическая или эукариотическая клетка-хозяин, содержащая вектор по п. 7, для продуцирования антитела по любому из пп. 1-4.
9. Способ получения биспецифического антитела по пп. 1-4, включающий стадии
a) трансформации клетки-хозяина экспрессионным вектором по п. 7;
b) культивирования клетки-хозяина в условиях, которые дают возможность синтеза молекулы антитела; и
c) выделения молекулы антитела из культуры.
| WO 2009142460 A2 от 26.11.2009 | |||
| US 0007504379 B2 от 17.03.2009 | |||
| US 20090304694 A1 от 10.12.2009 | |||
| Asahara | |||
| T, and et al., Tie2 Receptor Ligands, Angiopoietin-1 and Angiopoietin-2, Modulate VEGF-Induced Postnatal Neovascularization, Circ Res., 1998; Vol 83, pp.233-240 | |||
| КОМПОЗИЦИЯ АНТАГОНИСТА VEGF И АНТИПРОЛИФЕРАТИВНОГО СРЕДСТВА | 2004 |
|
RU2353353C2 |

