Область техники
Настоящее изобретение относится к области биотехнологии, а именно к антителам и применению данных антител. Более конкретно, настоящее изобретение относится к биспецифичным антителам, которые специфически связываются с HER2 и HER3. Изобретение также относится к нуклеиновой кислоте, кодирующей данное антитело, вектору экспрессии, способу получения антитела и применению антитела для лечения заболеваний или нарушений, связанных с HER2 и HER3.
Уровень техники
Группа рецепторов эпидермального фактора роста (human epidermal factor receptor, HER), также именуемая ErbB-рецепторами, относится к семейству трансмембранных рецепторных тирозинкиназ. Это семейство включает в себя рецептор эпидермального фактора роста (epidermal growth factor receptor, EGFR), также называемый ErbB-1 (или HER1), и гомологичные рецепторы ErbB-2 (HER2), ErbB-3 (HER3) и ErbB-4 (HER4). Эти рецепторы в большом количестве экспрессируются на поверхности эпителиальных клеток (обзор представлен в публикации Yarden and Pines 2012). Повышение уровня экспрессии HER рецепторов или их лигандов, таких как херегулин (HRG) или фактор эпидермального роста (EGF), часто наблюдается у пациентов с онкологическими заболеваниями (Wilson, Fridlyand et al. 2012).
Связывание лиганда с внеклеточным доменом тирозинкиназ вызывает процесс димеризации рецепторов, который может проходить как между двумя идентичными рецепторами (гомодимеризация), так и между разными рецепторами внутри одного семейства (гетеродимеризация). Димеризация активирует внутриклеточные домены тирозинкиназ и вызывает их аутофосфорилирование. Это, в свою очередь, запускает ряд нисходящих пролиферативных сигнальных каскадов, в том числе опосредованных митоген-активируемыми протеинкиназами, а также сигнальный путь AKT, направленный на выживание клетки (описано в работе Yarden and Pines, 2012). Для ErbB-2 не удалось обнаружить специфичных эндогенных лигандов, и предполагается, что в норме активация этого рецептора происходит путем гетеродимеризации (Sergina, Rausch et al. 2007), тогда как активация рецептора ErbB-3 может происходить с участием лигандов. К таким лигандам относятся, среди прочих, нейрегулин (NRG) или херегулин (HRG).
Нарушение регуляции ErbB-рецепторов приводит к образованию и росту опухолей (Yarden, Sliwkowski et al. 2001). Известным примером служит амплификация и гиперэкспрессия рецептора ErbB2, которая наблюдается в 20-30% случаев рака молочной железы и рака желудка. Несмотря на отсутствие высокоаффинных лигандов, ErbB2-рецепторы успешно участвуют в реализации сигнальных путей, направленных на выживание клеток. Это происходит посредством образования гетеродимеров с другими рецепторами ErbB-семейства, например, с ErbB-3-рецептором.
Лекарственные препараты, направленно действующие на ErbB2-рецепторы (Baselga, Swain et al. 2009), позволили значительно улучшить результаты лечения у многих пациентов с опухолями, характеризующимися повышенной экспрессией ErbB2. Однако у значительной части пациентов такое лечение оказывается неэффективным, а у тех, кто изначально отвечал на терапию, со временем развивается резистентность (Nahta, Yu et al. 2006). Активация ErbB3-рецептора его лигандом может обуславливать резистентность к ErbB-направленной терапии (Huang, Gao et al. 2010; Sergina, Rausch et al. 2007; Gijsen, King et al. 2011). Экспрессия ErbB3-рецептора повышается в опухолевых клетках, устойчивых к трастузумабу (huMAb4D5-8, rhuMAb HER2, HERCEPTIN®; патент США №5821337, Narayan, Wilken et al. 2009), и ассоциируется с неблагоприятным прогнозом у пациентов с раком молочной железы (Sassen, Rochon et al. 2008; Witton, Reeves et al. 2003). Несмотря на отсутствие активного киназного домена, ErbB3 образует гетеродимеры с другими представителями семейства ErbB рецепторов, что приводит к фосфорилированию фосфоинозитид-3-киназы (PI3K) и активации мощных внутриклеточных сигнальных каскадов. Рецептор ErbB3 является предпочтительным партнером для ErbB2 при димеризации (Graus-Porta, Beerli et al. 1997; Tzahar, Waterman et al. 1996). Было показано, что гетеродимер ErbB2/ErbB3 является наиболее мощным активатором каскада AKT по сравнению с другими димерами ErbB-рецепторов, при этом включение онкогенных сигнальных путей строго зависит от наличия ErbB3 (Holbro, Beerli et al. 2003; Lee-Hoeflich, Crocker et al. 2008). Однако, поскольку у ErbB3-рецептора отсутствует активный киназный домен, и в опухолевых клетках не происходит его амплификамии и значительной гиперэкспрессии, основной мишенью для лекарственных препаратов, атакующих данную гетеродимерную единицу, оставался рецептор ErbB2.
В патентном документе WO 2009068625 описано нанотело, специфичное в отношении HER2.
Другие HER2-aнтитeлa с различными свойствами описаны в Tagliabue et al. Int. J. Cancer 47:933-937 (1991); McKenzie et al. Oncogene 4:543-548 (1989); Maier et al. Cancer Res. 51:5361-5369 (1991); Bacus et al. Molecular Carcinogenesis 3:350-362 (1990); Stancovski et al. PNAS (USA) 88:8691-8695 (1991); Bacus et al. Cancer Research 52:2580-2589 (1992); Xu et al. Int. J. Cancer 53:401-408 (1993); WO 94/00136; Kasprzyk et al. Cancer Research 52:2771-2776 (1992); Hancock et al. Cancer Res. 51:4575-4580 (1991); Shawver et. al., Cancer Res. 54:1367-1373 (1994); Arteaga et al. Cancer Res. 54:3758-3765 (1994); Harwerth et al. J. Biol. Chem. 267:15160-15167 (1992); U.S. Patent No. 5,783,186; и Klapper et al. Oncogene 14:2099-2109 (1997); патент США №5783186; и Klapper et al. Oncogene 14: 2099-2109 (1997).
В международной заявке WO 97/35885 описаны HER3-антитела. В международной заявке WO 2003/013602 описаны ингибиторы HER-активности, включая HER-антитела. В международных публикациях WO 2007/077028 и WO 2008/100624 также описаны HER3-антитела.
В настоящее время активно ведутся разработки мультиспецифичных антител, которые специфично связываются с рецепторами семейства HER, с целью уменьшения нежелательных побочных эффектов и токсичности.
В патенте ЕА 022201 В1 описан агент, способный связываться с опухолевой клеткой и убивать опухолевую клетку или ингибировать пролиферацию опухолевых клеток, содержащий линкер на основе человеческого сывороточного альбумина (HSA), который может быть специфичен к HER2/HER3.
В патенте RU 2491294 C2 описано димерное антитело, которое специфично к HER2/HER3.
В патентном документе WO 2011143414 описано изолированное, рекомбинантное антитело, обладающее аффинностью связывания с сигнальной молекулой рецептора ErbB, выбранной из группы, состоящей из по меньшей мере одного из EGFR, ErbB2, ErbB3 и ErbB4. Вышеуказанное антитело может представлять собой моноклональное антитело, поликлональное антитело, одноцепочечное Fv антитело, димерное антитело, тримерное антитело, тетрамерное антитело, биспецифическое антитело, мини-антитело.
В патентном документе WO 2012158818 описано мультиспецифичное антитело, специфичное, в частности, к HER2 и HER3.
В патентных документах WO 2014182970, WO 2015066543, WO 2007084181 описаны биспецифические антитела к HER2/HER3.
В патентных документах WO 2008119353, WO 2015153765 и WO 2015173248 описаны способы получения биспецифического антитела к HER2/HER3.
В патентном документе ЕР 2727943 описаны триспецифические антитела к HER2/HER3/ EGFR.
Важным открытием явилось обнаружение в крови представителей семейства Camelidae (верблюды, ламы, викуньи) в значительном количестве особых неканонических антител с упрощенной структурой (Hamers Casterman С., Atarhouch Т., Muyldermans S., Robinson G., Hamers C, Bajyana Songa E., BendahmanN., Hamers R. 1993. Naturally occurring antibodies devoid of light chains. Nature. 363, 446-448). Такие антитела ("heavy chain antibody", HCAb) состоят из димера только одной укороченной тяжелой цепи (без СН1-домена), а легкая цепь при этом отсутствует. Антигенузнающий участок HCAb формируется лишь одним вариабельным доменом тяжелой цепи (VHH), который непосредственно связан через шарнирную область с Fc-доменом. Часто вместо VHH используют термин "однодоменное антитело", "nanobody" (нанотело), "мини-антитело" или "наноантитело". Как оказалось, такая монодоменная структура в изолированном виде, кроме малых размеров (12-15 кДа), обладает рядом преимуществ перед классическими IgG антителами, а именно - агрегационной, химической и термостабильностью. Антитела VHH можно эффективно клонировать и экспрессировать в бактериях и дрожжах. Обладая такими свойствами, они получили технологическое развитие как в терапевтическом направлении, развиваемом компанией "Ablynx", так и области лабораторной и промышленной хроматографии с линейкой носителей CaptureSelect.
Неканонические антитела (HCAb), состоящие из димера только тяжелой цепи иммуноглобулина, впервые обнаружены при электрофоретическом анализе иммуноглобулинов в сыворотке крови различных представителей семейства верблюдовых (Hamers Casterman С, Atarhouch Т., Muyldermans S., Robinson G., Hamers C, Bajyana Songa E., BendahmanN., Hamers R. 1993. Naturally occurring antibodies devoid of light chains. Nature. 363, 446-448). Относительная доля HCAb варьирует от примерно 15-25% (всех IgG) у лам и викуний до примерно 60-80% у верблюдов (С.В. Тиллиб "Верблюжьи наноантитела" - эффективный инструмент для исследований, диагностики и терапии Молекулярная биология, 2011, том 45, №1, с. 77-85).
Как предполагают, неканонические антитела (HCAb), по крайней мере, в случае верблюдовых, представляют собой результат относительно недавней эволюции генов канонических антител. Два константных домена тяжелых цепей, СН2 и СН3, в случае HCAb и классических антител высококонсервативны. В составе антител HCAb домен, соответствующий первому константному СН1-домену классических антител, отсутствует. Геном одногорбого верблюда (вид Dromedary) содержит кластер из примерно пятидесяти VH- и сорока VHH-генеративных генов, за которыми располагаются множественные гены D-сегментов, J-сегментов и гены константных участков (Сμ, Сγ, Сε, Сα). Очевидно, что некоторые из Сγ-генов предназначены для формирования HCAb (мутация приводит к потере СН1-домена), в то время как остальные - для формирования классических антител (с сохраняемым СН1-доменом). Одни и те же гены сегментов D и J могут случайным образом соединяться как с одним из VH-, так и с одним из VHH-генов. Это указывает на то, что гены VH и VHH находятся в одном и том же локусе (Nguyen V.K., Hamers R., Wyns L., Muyldermans S.1999. Loss of splice consensus signal is responsible for the removal of the entire CH1 domain of the functional camel IGG2A heavy-chain antibodies. Mol. Immunol. 36, 515-524; Woolven B.P., Frenken L., van der Logt P., Nicholls P.J. 1999. The structure of the llama heavy chain constant genes reveals a mechanism for heavy-chain antibody formation. Immunogenetics. 50, 98-101; Nguyen V.K., Hamers R., Wyns L., Muyldermans S.2000. Camel heavy-chain antibodies: diverse germline VHH and specific mechanisms enlarge the antigen-binding repertoire. EMBO J. 19, 921-931; De Genst E., Saerens D., Muyldermans S., Conrath K. 2006. Antibody repertoire development in camelids. Develop. Comp. Immunol. 30, 187-198).
Организация вариабельных доменов неканонических антител (VHH) и вариабельных доменов (VH) классических антител весьма сходна, т.к. у человека VH-домены подкласса IgG3 имеют особо выраженную гомологию с VH и VHH верблюдовых. В обоих случаях V-домены состоят из четырех консервативных каркасных участков FR, которые окружают три гипервариабельных участка, определяющих комплементарность, CDR. Также в обоих случаях формируется типичная для V-домена иммуноглобулина пространственная структура из двух β-слоев, один из которых состоит из четырех аминокислотных цепочек и второй - из пяти (Muyldermans S., Cambillau С., Wyns L. 2001. Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains. TIBS. 26, 230-235; De Genst E., Silence K., Decanniere K., Loris R., Kinne J., Muyldermans S. 2006. Molecular basis for the preferential cleft recognition by dromedary heavy chain antibodies. Proc. Natl. Acad. Sci. USA. 103, 4586-4591). В этой структуре все три гипервариабельных участка кластеризуются с одной стороны V-домена, где они участвуют в распознавании антигена и располагаются в петлях, соединяющих β-структуры. Однако имеются и важные отличия, связанные с функционированием VHH в формате одного домена. Так, гипервариабельные участки CDR1 и CDR3 VHH заметно увеличены. Часто в гипервариабельных участках VHH обнаруживаются цистеиновые остатки сразу в двух участках (чаще всего - в CDR1 и CDR3, реже - в CDR2 и CDR3). При исследовании кристаллических структур VHH показано, что эти цистеиновые остатки формируют дисульфидные связи, и это дополнительно стабилизирует структуру петель данного антитела (De Genst Е., Silence K., Decanniere K., Loris R., Kinne J., Muyldermans S. 2006. Molecular basis for the preferential cleft recognition by dromedary heavy chain antibodies. Proc. Natl. Acad. Sci. USA. 103, 4586-4591). Наиболее явный и воспроизводимый отличительный признак VHH - четыре замены гидрофобных аминокислотных остатков на гидрофильные во втором каркасном участке (Val37Phe, Gly44Glu, Leu45Arg, Trp47Gly, согласно нумерации Кабат (Kabat) и соавт. (Kabat Е., Wu Т.Т., Perry Н.М., Gottesman K.S., Foeller С. 1991. Sequence of proteins of immunological interest. US Public Health Services. NIH, Bethesda, MD, Publication no. 91-3242)). Этот каркасный участок у VH-домена высококонсервативен, обогащен гидрофобными аминокислотными остатками и особо важен при образовании связи с вариабельным доменом VL легкой цепи. В этом аспекте VHH-домен сильно отличается: указанные замены гидрофобных аминокислот на гидрофильные делают невозможной ассоциацию VHH и VL. Эти замены также объясняют обычно высокую растворимость VHH (наноантитела) при его получении в виде рекомбинантного белка (Nguyen V.K., Desmyter A., Muyldermans S. 2001. Functional heavy-chain antibodies in camelidae. Adv. Immunol. 79, 261-296).
Репертуары возможных паратопов (антигенсвязывающих структур антитела) HCAb и классических антител, по-видимому, могут заметно отличаться. Так как эти два типа антител сосуществуют в одном организме, то можно предполагать, что они не конкурируют, а взаимно дополняют друг друга. Действительно, не раз отмечалось, что оба типа антител могут возникать параллельно, взаимоисключающе или в разных соотношениях, по отношению к разным эпитопам антигенного материала при иммунизации одного и того же животного. Несмотря на предполагаемое меньшее разнообразие возможных паратопов у однодоменных антител по сравнению с классическими двухдоменными антителами, работы многих авторов убедительно продемонстрировали, что HCAb могут быть получены против самых разнообразных эпитопов весьма широкого спектра антигенов (Muyldermans S., Baral T.N., Retamozzo V.C., et al. 2009. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunopathol. 128(1-3), 178-183). Очевидно, этому способствуют заметно увеличенные гипервариабельные участки CDR1 и CDR3. Следует также отметить удивительно большое (в сравнении с V-доменами классических антител) число соматических гипермутаций в VHH, накапливающихся, по-видимому, в процессе аффинного созревания антител в ходе иммунизации (de Genst Е., Silence K., Decanniere K., Loris R., Kinne J., Wyns L., Muyldermans S. 2005. Strong in vivo maturation compensates for structurally restricted H3 loops in antibody repertoires. J. Biol. Chem. 280, 14114-14121). Рентгеноструктурный анализ показал, что антигенсвязывающие петлевые участки VHH способны образовывать необычные для классических V-доменов структуры (De Genst Е., Silence K., Decanniere K., Loris R., Kinne J., Muyldermans S. 2006. Molecular basis for the preferential cleft recognition by dromedary heavy chain antibodies. Proc. Natl. Acad. Sci. USA. 103, 4586-4591; de Genst E., Silence K., Decanniere K., Loris R., Kinne J., Wyns L., Muyldermans S. 2005. Strong in vivo maturation compensates for structurally restricted H3 loops in antibody repertoires. J. Biol. Chem. 280, 14114-14121). Если в случае VH- и VL-доменов классических антител все шесть гипервариабельных участков вносят более или менее одинаковый вклад в связывание антигена, то в случае VHH обычно для формирования паратопа более важен CDR3-участок. Показано, что CDR3-участок в VHH (но не в VH или VL) может образовывать необычные длинные пальцеобразные выступающие структуры, которые могут углубляться в структуру антигена, в частности, распознавать активные центры ферментов (De Genst Е., Silence K., Decanniere K., Loris R., Kinne J., Muyldermans S. 2006. Molecular basis for the preferential cleft recognition by dromedary heavy chain antibodies. Proc. Natl. Acad. Sci. USA. 103, 4586-4591). Малыми размерами антигенсвязывающего участка (VHH) и его способностью формировать необычные выступающие паратопы объясняется возможность получения HCAb, способных распознавать недоступные для классических антител эпитопы, например, при образовании антител, являющихся эффективными ингибиторами ферментов (Lauwereys М., Ghahroudi М., Desmyter A., Kinne J., Holzer W., De Genst E., Wyns L., Muyldermans S. 1998. Potent enzyme inhibitors derived from dromedary heavy chain antibodies. EMBO J. 17, 3512-3520).
При всем своем высоком потенциале уникальной, в сравнении с классическими IgG антителами, специфичности, использование монодоменных VHH для терапевтического применения в ряде случаев ограниченно из-за быстрого выведения этой малой молекулы из организма. Существует ряд решений по улучшению фармакокинетики VHH структур, которые включают использование химической конъюгации с ПЭГ (PEG) и ковалентное соединение с полипептидом, опосредующим уменьшенный клиренс из крови, такими как сывороточный альбумин человека (HSA) и Fc-фрагмент классического антитела человека, в виде гибридных белков, период полувыведения в крови которых составляет до трех недель (Kontermann RE. 2009 Strategies to extend plasma half-lives of recombinant antibodies. BioDrugs.; 23(2):93-109; Ken Coppieters, Torsten Dreier, Karen Silence, Hans de Haard, Marc Lauwereys, Peter Casteels, Els Beirnaert, Heidi Jonckheere, 2006 Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis Arthritis & Rheumatology.; 54(6): 1856-66; Gabrielle Richard, Ashley J Meyers, Michael D McLean, Mehdi Arbabi-Ghahroudi, Roger MacKenzie, J Christopher Hall 2013. In Vivo Neutralization of α-Cobratoxin with High-Affinity Llama Single-Domain Antibodies (VHHs) and a VHH-Fc Antibody. PLoS One 22; 8(7):e69495). Также показано успешное применение малых пептидов, присоединяемых с помощью методов генной инженерии к VHH и способных к высокоаффиному нековалентному взаимодействию с этими же компонентами (HSA и IgG) в крови человека (Jan Terje Andersen, Maria Gonzalez-Pajuelo, Stian Foss, Ole J.B. Landsverk, 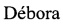 Pinto, Alexander Szyroki, Hans J. de Haard, Michael Saunders, Peter Vanlandschoot Inger Sandlie 2012, Selection of Nanobodies that Target Human Neonatal Fc Receptor Scientific Reports, 3, 1118). Однако технологичность и иммуногенность этих подходов остается пока неясной и находится только на стадии проверки применимости как в клинических исследованиях так и на более ранних стадиях исследований.
Pinto, Alexander Szyroki, Hans J. de Haard, Michael Saunders, Peter Vanlandschoot Inger Sandlie 2012, Selection of Nanobodies that Target Human Neonatal Fc Receptor Scientific Reports, 3, 1118). Однако технологичность и иммуногенность этих подходов остается пока неясной и находится только на стадии проверки применимости как в клинических исследованиях так и на более ранних стадиях исследований.
Кроме того, наибольшее ограничение при применении антител в качестве препаратов для лечения различных заболеваний накладывает агрегационная и химическая стабильность, аффинность и иммуногенность. Т.к. большинство моноклональных антител в настоящее время получают на основе мышиных, применение таких антител у пациентов приводит к развитию иммунного ответа на терапию антителами, например, аллергических реакций. Такие типы иммунного ответа могут повлечь за собой в конечном итоге потерю эффективности при лечении по меньшей мере и, в худшем случае, к возможным тяжелым анафилактическим реакциям. С другой стороны, агрегационно или химически нестабильные терапевтические антитела при хранении препарата ухудшают его терапевтические свойства и могут усиливать иммуногенность при введении в организм пациента.
В связи с вышесказанным, актуальным является создание биспецифических антител на основе антител VHH, которые эффективно воздействуют на сигнальную систему HER, в частности на HER2 и/или HER3, и при этом которые имели бы, по сравнению с известными ранее антителами, улучшенные функциональные и терапевтические свойства, в частности, повышенную агрегационную, химическую и термостабильность, и улучшенную аффинность, и в то же время отличались бы легкостью и простотой получения, в т.ч. и в промышленно значимых масштабах.
Биспецифическая молекула BCD090 селективно связывается с гетеродимером ErbB2/ErbB3, что приводит к образованию неактивного тримера. Эта молекула является более эффективным ингибитором активации ErbB3 и ErbB2 рецепторов, чем моноклональные антитела против ErbB2 или ErbB3.
Краткое описание изобретения
Настоящее изобретение относится к биспецифическим связывающим молекулам, в частности антителам, направленным для связывания с рецепторами HER2 и HER3. Такие антитела могут быть использованы для лечения заболевания или нарушения, опосредуемого HER2 и HER3, например, выбранного из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи. По сравнению с существующими в настоящее время способами лечения таких заболеваний, включая лечение антителами, предполагается, что биспецифичные связывающие молекулы, в частности антитела, направленные для связывания с рецепторами HER2 и HER3, по настоящему изобретению, в частности биспецифическая молекула BCD090, могут обеспечить наилучший клинический ответ, применяемые как отдельно, так и в сочетании с другой терапией.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, которое специфично связывается с HER2 и HER3. Данное антитело содержит вариабельный домен тяжелой цепи VHH, специфичный к HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2, связаны между собой пептидным линкером.
В некоторых вариантах биспецифическое антитело включает вариабельный домен тяжелой цепи VHH, содержащий аминокислотную последовательность по меньшей мере на 75% гомологичную последовательности SEQ ID NO: 3. В некоторых вариантах биспецифическое антитело включает вариабельный домен тяжелой цепи VHH, который содержит аминокислотную последовательность SEQ ID NO: 3.
В некоторых вариантах осуществления изобретения биспецифическое антитело включает вариабельный домен тяжелой цепи VHH, специфичный к HER3, который содержит аминокислотные последовательности, гомологичные по меньшей мере на 75% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, содержит аминокислотные последовательности, гомологичные по меньшей мере на 80% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, содержит аминокислотные последовательности, гомологичные по меньшей мере на 85% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, содержит аминокислотные последовательности, гомологичные по меньшей мере на 90% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, содержит аминокислотные последовательности, гомологичные по меньшей мере на 95% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH специфичный к HER3, содержит аминокислотные последовательности, гомологичные по меньшей мере на 98% последовательностям SEQ ID NO: 1-3. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, содержит аминокислотные последовательности, представленные последовательностями SEQ ID NO: 1-3.
В некоторых вариантах осуществления изобретения биспецифическое антитело включает вариабельный домен тяжелой цепи VHH, который конкурирует за связывание или связывается с тем же эпитопом, что связывающий домен, содержащий аминокислотную последовательность SEQ ID NO: 4.
В некоторых вариантах осуществления изобретения биспецифическое антитело включает последовательность вариабельного домена тяжелой цепи VHH, специфичного к HER3, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4. В некоторых вариантах осуществления изобретения последовательность вариабельного домена тяжелой цепи VHH, специфичного к HER3, является по меньшей мере на 95% гомологичной аминокислотной последовательности SEQ ID NO: 4. В некоторых вариантах осуществления изобретения последовательность вариабельного домена тяжелой цепи VHH, специфичного к HER3, является по меньшей мере на 98% гомологичной аминокислотной последовательности SEQ ID NO: 4. В отдельном варианте осуществления изобретения последовательность вариабельного домена тяжелой цепи VHH, специфичного к HER3, состоит из аминокислотной последовательности SEQ ID NO: 4.
В некоторых вариантах осуществления биспецифическое антитело включает антитело, специфичное к HER2, которое представляет собой полноразмерное антитело IgG. В некоторых вариантах осуществления изобретения полноразмерное антитело, специфичное к HER2, относится к изотипу IgG1, IgG2, IgG3, IgG4 человека. В некоторых вариантах осуществления изобретения полноразмерное антитело, специфичное к HER2, содержит тяжелую и легкую цепи, которые, соответственно, содержат аминокислотные последовательности, по меньшей мере на 80% гомологичные последовательностям SEQ ID NO: 5 и SEQ ID NO: 6. В некоторых вариантах осуществления изобретения полноразмерное антитело, специфичное к HER2, содержит тяжелую и легкую цепи, которые, соответственно, содержат аминокислотные последовательности, по меньшей мере на 90% гомологичные последовательностям SEQ ID NO: 5 и SEQ ID NO: 6. В некоторых вариантах осуществления изобретения полноразмерное антитело, специфичное к HER2, содержит тяжелую и легкую цепи, которые, соответственно, содержат аминокислотные последовательности, по меньшей мере на 95% гомологичные последовательностям SEQ ID NO: 5 и SEQ ID NO: 6. В некоторых вариантах осуществления изобретения полноразмерное антитело, специфичное к HER2, содержит тяжелую и легкую цепи, которые, соответственно, содержат аминокислотные последовательности, по меньшей мере на 98% гомологичные последовательностям SEQ ID NO: 5 и SEQ ID NO: 6. В отдельном варианте осуществления изобретения полноразмерное антитело, специфичное к HER2, содержит тяжелую и легкую цепи, которые, соответственно, содержат аминокислотные последовательности SEQ ID NO: 5 и SEQ ID NO: 6.
В некоторых вариантах осуществления изобретения биспецифическое антитело, которое специфично связывается с HER2 и HER3, включает антигенсвязывающий домен, который специфично связывается с HER2.
В некоторых вариантах осуществления изобретения биспецифическое антитело, которое специфично связывается с HER2 и HER3, представляет собой ассиметричное антитело.
В некоторых вариантах осуществления изобретения биспецифическое антитело включает вариабельный домен тяжелой цепи VHH, специфичный к HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2 которые связаны между собой пептидным линкером из более чем пяти аминокислот. В некоторых вариантах осуществления изобретения, аминокислотные остатки пептидного линкера выбраны из G, А, S, P, E, T, D и К. Вариабельный домен тяжелой цепи VHH присоединен к N-концу или С-концу легкой или тяжелой цепи антитела, специфичного против HER2, через пептидный линкер.
В некоторых вариантах осуществления изобретения биспецифическое антитело имеет константу диссоциации KD связывания с HER2 человека не более 10-10 М и обезьяны не более 10-7 М, KD связывания с HER3 человека не более 10-9 М и обезьяны не более 10-8 М.
В одном из аспектов настоящее изобретение относится к раствору для ингибирования биологической активности HER2 и/или HER3 у субъекта, который включает биспецифическое антитело, которое специфично связывается с HER2 и HER3, и один или несколько фармацевтически приемлемых эксципиентов.
В некоторых вариантах осуществления изобретения раствор для ингибирования биологической активности HER2 и/или HER3 у субъекта имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и хранении при t=4°С в течение более чем 6 месяцев содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
В некоторых вариантах осуществления изобретения раствор для ингибирования биологической активности HER2 и/или HER3 у субъекта имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и при повышении температуры до 37°С в течение более чем 2 недель содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
В некоторых вариантах осуществления изобретения раствор для ингибирования биологической активности HER2 и/или HER3 у субъекта имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и повышении температуры до 50°С в течение более чем 6 часов содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которое содержит нуклеотидную последовательность, кодирующую любую из биспецифичных связывающих молекул, описанных в настоящем документе.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая содержит нуклеотидную последовательность, которая кодирует любой вариабельный домен тяжелой цепи VHH, описанный в настоящем документе.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая содержит нуклеотидную последовательность, которая кодирует тяжелую цепь, легкую цепь или обе цепи антитела, специфичного к HER2, описанную в настоящем документе.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая содержит следующее:
a) любую нуклеотидную последовательность, которая кодирует вариабельный домен тяжелой цепи VHH, описанный в настоящем документе;
b) любую нуклеотидную последовательность, которая кодирует тяжелую цепь антитела, специфичного к HER2, описанную в настоящем документе;
c) любую нуклеотидную последовательность, которая кодирует легкую цепь антитела, специфичного к HER2, описанную в настоящем документе; или
d) любое сочетание а)-с).
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую любой вариабельный домен тяжелой цепи VHH, описанный в настоящем документе, и любую нуклеотидную последовательность, кодирующую тяжелую цепь, легкую цепь или обе цепи антитела, специфичного к HER2, описанные в настоящем документе, в некоторых случаях дополнительно содержит нуклеотидную последовательность, кодирующую пептидный линкер.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая кодирует любую из аминокислотных последовательностей SEQ ID NO: 1-6.
В одном из аспектов настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая содержит нуклеотидную последовательность, которая кодирует любое из биспецифических антител, которые специфично связываются с HER2 и HER3, описанных в настоящем документе. В некоторых вариантах осуществления изобретения выделенная молекула нуклеиновой кислоты содержит последовательность ДНК, которая кодирует любое из биспецифических антител, которые специфично связываются с HER2 и HER3, описанных в настоящем документе.
В одном из аспектов настоящее изобретение относится к вектору, содержащему любую из выделенных молекул нуклеиновой кислоты, описанных выше, при этом указанный вектор далее содержит последовательности, контролирующие экспрессию.
В одном из аспектов настоящее изобретение относится к способу получения клетки-хозяина для получения любого из вышеописанных антител, где способ включает трансформирование клетки любым вышеуказанным вектором.
Настоящее изобретение также относится к клетке-хозяину, содержащей любую из вышеописанных нуклеиновых кислот.
В одном из аспектов настоящее изобретение относится к способу получения любых биспецифических антител, которые специфично связывается с HER2 и HER3, описанные в настоящем документе, который включает получение клетки-хозяина, как описано выше, культивирование указанной клетки-хозяина в культуральной среде в условиях, достаточных для получения указанного антитела, а также, при необходимости, с последующим выделением и очисткой полученного антитела.
В одном из аспектов настоящее изобретение относится к фармацевтической композиции для профилактики или лечения заболевания или нарушения, опосредуемого HER2 и HER3, которая содержит любое биспецифическое антитело, которое специфично связывается с HER2 и HER3, описанное в настоящем документе, в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами.
В некоторых вариантах осуществления изобретения фармацевтическая композиция, которая содержит любое биспецифическое антитело, которые специфично связывается с HER2 и/или HER3, описанное в настоящем документе, предназначена для профилактики или лечения, заболевания или нарушения, опосредуемого HER2 и HER3, выбранного из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
В одном из аспектов настоящее изобретение относится к способу ингибирования биологической активности HER2 и/или HER3 у субъекта, нуждающемуся в таком ингибировании, который включает введение субъекту эффективного количества любого биспецифического антитела, которое специфично связывается с HER2 и HER3, описанного в настоящем документе.
В одном из аспектов настоящее изобретение относится к способу лечения заболевания или нарушения, опосредованного HER2 и/или HER3, который включает введение субъекту, нуждающемуся в таком лечении, любого биспецифического антитела, которые специфично связывается с HER2 и HER3, описанного в настоящем документе, или любого раствора, описанного в настоящем документе, который содержит любое биспецифическое антитело, которое специфично связывается с HER2 и HER3, описанного в настоящем документе, или любой фармацевтической композиции, описанной в настоящем документе, которая содержит любое биспецифическое антитело, которое специфично связывается с HER2 и HER3, описанное в настоящем документе, в терапевтически эффективном количестве.
В некоторых вариантах осуществления изобретения способ лечения предназначен для лечения заболевания или нарушения, где заболевание или нарушение выбрано из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
В одном из аспектов настоящее изобретение относится к применению любого биспецифического антитела, которое специфично связывается с HER2 и HER3, описанного в настоящем документе, или любого раствора, описанного в настоящем документе, который содержит любое биспецифическое антитело, которое специфично связывается с HER2 и HER3, описанного в настоящем документе, или любой фармацевтической композиции, описанной в настоящем документе, которая содержит любое биспецифическое антитело, которое специфично связывается с HER2 и HER3, описанное в настоящем документе, для лечения у субъекта, нуждающегося в таком лечении, заболевания или нарушения, опосредуемого HER2 и/или HER3.
В некоторых вариантах осуществления изобретения применение для лечения у субъекта, нуждающегося в таком лечении, заболевания или нарушения, опосредуемого HER2 и/или HER3, где заболевание или нарушение выбрано из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
Краткое описание чертежей
Фиг. 1. Схема получения биспецифического антитела anti-HER2/anti-HER3.
Фиг. 2. Последовательность белка HER3-His6-FLAG (HER3HF).
Фиг. 3. Последовательность белка 3DHER3-His6-EPEA (3DHER3HE).
Фиг. 4. Последовательность белка HER3-Fc-His6 (HER3-Fc).
Фиг. 5. Последовательность контрольного антитела ahti-HER3 AV203 с опубликованной последовательностью (от AVEO, Biogen, WO 2011/136911 А2)
Фиг. 6. Очищенные препараты белков HER3HF, 3DHER3HE, HER3-Fc.
Фиг. 7. Очищенный препарат контрольного антитела против HER3.
Фиг. 8. Титр антител против HER3 человека, полученный путем иммунизации Lama glama.
Фиг. 9. Карта плазмиды для фагового дисплея Fab фрагментов антител.
Фиг. 10. Карта плазмиды для секреционной экспрессии Fab фрагментов антител в E. coli.
Фиг. 11. График зависимости сигнала связывания поликлонального фага со специфическими и неспецифическими антигенами после селекции иммунной фаговой библиотеки.
Фиг. 12. Иммуноферментный анализ взаимодействия моноспецифического aнти-HER3 антитела с HER3 и другими антигенами.
Фиг. 13. ИФА связывания BCD-090 антитела с различными ортологичными белками HER2.
Фиг. 14. ИФА связывания BCD-090 антитела с различными ортологичными белками HER3.
Фиг. 15. Кривые ассоциации/диссоциации для комплекса HER3 человека/ BCD090, полученные для двух различных концентраций аналита.
Фиг.16. Кривые ассоциации/диссоциации для комплекса HER3 макаки резус/ BCD090, полученные для двух различных концентраций аналита.
Фиг. 17. Кривые ассоциации/диссоциации для комплекса HER2 макаки циномолгус/ BCD090, полученные для семи различных концентраций аналита.
Фиг. 18. Кривые ассоциации/диссоциации для комплекса HER2 человека/ВСD090.
Фиг. 19. Сенсограмма измерения одновременного связывания BCD090 антителом обоих лигандов HER2 и HER3 человека на приборе OctetRED96.
Фиг. 20. График зависимости процента живых клеток линии MCF7 в зависимости от концентрации антител в среде через 72 часа в СO2 инкубаторе. Планки погрешностей отражают стандартное отклонение, рассчитанное по трем повторам для каждой концентрации антитела.
Фиг. 21. График зависимости процента живых клеток линии ВТ474 в зависимости от концентрации антител в среде через 72 часа в СO2 инкубаторе. Планки погрешностей отражают стандартное отклонение, рассчитанное по трем повторам для каждой концентрации антитела.
Фиг. 22. График зависимости процента живых клеток линии A431NS в зависимости от концентрации антител в среде через 72 часа в СO2 инкубаторе. Планки погрешностей отражают стандартное отклонение, рассчитанное по трем повторам для каждой концентрации антитела.
Фиг. 23. График зависимости процента живых клеток линии SKBR-3 в зависимости от концентрации антител в среде через 72 часа в СO2 инкубаторе. Планки погрешностей отражают стандартное отклонение, рассчитанное по трем повторам для каждой концентрации антитела.
Фиг. 24. Определение термоколлоидной стабильности по точке агрегации белка методом динамического светорассеяния (DLS) для BCD-090 антитела. График зависимости среднего размера частиц (Z-average) от температуры в четырех буферных растворах.
Фиг.25. Определение термической стабильности при длительном воздействии методом «Термостресс 50°С» для BCD090 антитела в четырех различных буферных растворах.
Фиг. 26. Хроматограммы, отражающие результат анализа термостресса биспецифического антитела BCD090: красный - интактный, синий - 72 ч инкубации при 50°С. Длина волны 220 nm.
Фиг.27. Схемы методов выявления BCD090 в сыворотке крови человека.
Фиг. 28. Открываемость различных концентраций BCD090 в сыворотке человека до и после хранения при 37°С.
Фиг.29. График зависимости количества погибших клеток линии ВТ474 от концентрации антител в среде.
Фиг. 30. Показатели торможения роста опухоли линии MCF7 в экспериментальных группах, отражающие противоопухолевую активность препарата.
Фиг. 31. Динамика роста опухоли линии MCF7 в экспериментальных группах.
Фиг. 32. Показатели торможения роста опухоли линии SK-BR-3 в экспериментальных группах, отражающие противоопухолевую активность препарата.
Фиг. 33. Динамика роста опухоли линии SK-BR-3 в экспериментальных группах.
Фиг. 34. Показатели торможения роста опухоли линии ВТ474 в экспериментальных группах, отражающие противоопухолевую активность препарата.
Фиг. 35. Динамика роста опухоли линии ВТ474 в экспериментальных группах.
Фиг. 36. Модель комплекса HER3 (4LEO) и VHH-aHER3.
Подробное описание изобретения
Определения и общие методы
Если иное не определено в настоящем документе, научные и технические термины, используемые в связи с настоящим изобретением, будут иметь значения, которые обычно понятны специалистам в данной области. Примерные способы и материалы описаны ниже, хотя на практике или при тестировании настоящего изобретения могут также использоваться способы и материалы, подобные или эквивалентные тем, что описаны в настоящем документе. Все публикации и другие ссылочные материалы, упомянутые в настоящем документе, включены во всей их полноте путем отсылки. В случае противоречий настоящее описание, включая определения, будет превалировать. Хотя в настоящем документе цитируется ряд документов, такое цитирование не является признанием того, что любой из этих документов образует часть общеизвестных знаний в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы клеточной и тканевой культуры, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
В этом описании и вариантах осуществления изобретения слова «иметь» и «содержать» или их вариации, такие как «имеет», «имеющий», «содержит» или «содержащий», следует понимать как включение указанного целого или группы целых, но не исключение любого другого целого или группы целых.
Определения, связанные с антителом
"HER-рецептор" является рецепторной тирозиновой протеинкиназой, которая относится к семейству HER-рецепторов и включает рецепторы EGFR, HER2, HER3 и HER4 и других представителей такого семейства, которые будут идентифицированы в будущем. HER-рецептор, как правило, может содержать внеклеточный домен, который может связывать лиганд HER; липофильный трансмембранный домен; консервативный внутриклеточный тирозинкиназный домен; и находящийся на карбоксильном конце домен передачи сигнала, несущий несколько остатков тирозина, которые могут быть фосфорилированы. Предпочтительно HER-рецептор представляет собой HER-рецептор человека с нативной последовательностью.
Внеклеточный домен HER2 содержит четыре домена: домен I (аминокислотные остатки примерно от 1 до 195), домен II (аминокислотные остатки примерно от 196 до 319), домен III (аминокислотные остатки примерно от 320 до 488) и домен IV (аминокислотные остатки примерно от 489 до 630) (нумерация остатков без сигнального пептида). См. Garrett et al. Mol. Cell. 11: 495-505 (2003), Cho et al. Nature 421: 756-760 (2003), Franklin et al. Cancer Cell 5: 317-328 (2004) и Plowman et al. Proc. Natl. Acad. Sci. 90: 1746-1750 (1993). Также см. фиг. 1 в данном описании.
Термины "ErbB1", "HER1", "рецептор эпидермального фактора роста" и "EGFR" используют в настоящем описании взаимозаменяемо, и они относятся к EGFR, который описан, например, в Carpenter et al. Ann. Rev. Biochem. 56: 881-914 (1987), включая его встречающиеся в природе мутантные формы (например, делеционный мутант EGFR, который описан в Humphrey et al. PNAS (USA) 87: 4207-4211 (1990)). ErbB1 относится к гену, кодирующему белковый продукт EGFR.
Выражения "ErbB2" и "HER2" используют в настоящем описании взаимозаменяемо, и они относятся к белку HER2 человека, описанному, например, в Semba et al., PNAS (USA) 82: 6497-6501 (1985) и Yamamoto et al. Nature 319: 230-234 (1986) (номер доступа в Genebank X03363). Термин "erbB2" относится к гену, кодирующему ErbB2 человека, и "neu" относится к гену, кодирующему p185neu крысы. Предпочтительный HER2 представляет собой HER2 человека с нативной последовательностью.
"ErbB3" и "HER3" относятся к полипептиду рецептора, который описан, например, в патентах США №5183884 и 5480968, а также в Kraus et al. PNAS (USA) 86: 9193-9197 (1989).
Термины "ErbB4" и "HER4" в настоящем описании относятся к полипептиду рецептора, который описан, например, в заявке на выдачу патента ЕР №599274; Plowman et al., Proc. Natl. Acad. Sci. USA, 90: 1746-1750 (1993); и Plowman et al., Nature, 366: 473-475 (1993), включая его изоформы, которые описаны, например, в WO 99/19488, опубликованной 22 апреля 1999.
Под "лигандом HER" подразумевают полипептид, который связывается и/или активирует рецептор HER. Лиганд HER, представляющий особый интерес согласно настоящему изобретению, является лигандом HER человека с нативной последовательностью, таким как эпидермальный фактор роста (EGF) (Savage et al., J. Biol. Chem. 247: 7612-7621 (1972)); трансформирующий фактор роста альфа (TGF-α) (Marquardt et al., Science 223: 1079-1082 (1984)); амфирегулин, также известный как аутокринный фактор роста шванномы и кератиноцитов (Shoyab et al. Science 243: 1074-1076 (1989); Kimura et al. Nature 348: 257-260 (1990); и Cook et al. Mol. Cell. Biol. 11: 2547-2557 (1991)); бетацеллюлин (Shing et al., Science 259: 1604-1607 (1993); и Sasada et al. Biochem. Biophys. Res. Commun. 190: 1173 (1993)); гепарин-связывающий эпидермальный фактор роста (HB-EGF) (Higashiyama et al., Science 251: 936-939 (1991)); эпирегулин (Toyoda et al., J. Biol. Chem. 270: 7495-7500 (1995); и Komurasaki et al. Oncogene 15: 2841-2848 (1997)); херегулин (См. ниже); нейрегулин-2 (NRG-2) (Carraway et al., Nature 387: 512-516 (1997)); нейрегулин-3 (NRG-3) (Zhang et al., Proc. Natl. Acad. Sci. 94: 9562-9567 (1997)); нейрегулин-4 (NRG-4) (Harari et al. Oncogene 18: 2681-89 (1999)); и cripto-(CR-1) (Kannan et al. J. Biol. Chem. 272(6): 3330-3335 (1997)). Лиганды HER, которые связывают EGFR, включают EGF, TGF-α, амфирегулин, бетацеллюлин, HB-EGF и эпирегулин. Лиганды HER, которые связывают HER3, включают херегулины. Лиганды HER, способные связывать HER4, включают бетацеллюлин, эпирегулин, HB-EGF, NRG-2, NRG-3, NRG-4 и херегулины.
Используемые в данном документе термины «связывающийся с человеческим HER2», или «специфически связывающийся с человеческим HER2», или «анти-HER2-антитeлo» являются взаимозаменяемыми и относятся к антителу, специфически связывающемуся с человеческим HER2-антигеном с аффинностью связывания, которая имеет значение KD 1×10-8 моль/л или ниже при 25°С, в одном воплощении имеет значение KD 1×10-9 моль/л или ниже при 25°С. Аффинность связывания определяют в стандартном анализе связывания при 25°С, таком как методика поверхностного плазменного резонанса (BIAcore®, GE-Healthcare, Упсала, Швеция). Способ определения значения KD аффинности связывания описан в примере 2b). Таким образом, термин «антитело, связывающееся с человеческим HER2», используемый в данном документе, относится к антителу, специфически связывающемуся с человеческим HER2-антигеном с аффинностью связывания со значением KD 1×10-8 моль/л или ниже (предпочтительно 1×10-8 моль/л - 1,0×10-12 моль/л) при 25°С.
Используемые в данном документе термины «связывающийся с человеческим HER3», или «специфически связывающийся с человеческим HER3», или «aнти-HER3-антитело» являются взаимозаменяемыми и относятся к антителу, специфически связывающемуся с человеческим HER3-антигеном с аффинностью связывания, которая имеет значение KD 1×10-8 моль/л или ниже при 25°С, в одном воплощении имеет значение KD 1×10-9 моль/л или ниже при 25°С. Аффинность связывания определяют в стандартном анализе связывания при 25°С, таком как методика поверхностного плазменного резонанса (BIAcore®, GE-Healthcare, Упсала, Швеция). Способ определения значения KD аффинности связывания описан в примере 2b). Таким образом, термин «антитело, связывающееся с человеческим HER3», используемый в данном документе, относится к антителу, специфически связывающемуся с человеческим HER3-антигеном с аффинностью связывания со значением KD 1×10-8 моль/л или ниже (предпочтительно 1×10-8 моль/л - 1,0×10-12 моль/л) при 25°С.
Человеческий HER3 (ErbB-3, ERBB3, c-erbB-3, c-erbB3, тирозин-киназный рецептор erbB-3) кодирует рецептор эпидермального фактора роста (EGFR), член семейства тирозинкиназных рецепторов, которое также включает HER1 (также известный как EGFR), HER2 и HER4 (Kraus, М.Н. et al, PNAS 86 (1989) 9193-9197; Plowman, G.D. et al, PNAS 87 (1990) 4905-4909; Kraus, M.H. et al, PNAS 90 (1993) 2900-2904). Как и прототипичный рецептор эпидермального фактора роста, трансмембранный рецептор HER3 состоит из внеклеточного лигандсвязывающего домена (ECD), домена димеризации с ECD, трансмембранного домена, внутриклеточного домена тирозинкиназного белка (TKD) и С-концевого фосфорилированного домена. Этот связанный с мембраной белок имеет херегулин (HRG)-связывающий домен HER3 во внеклеточном домене, но не имеет активного киназного домена. Таким образом, он может связывать этот лиганд, но не передает сигнал внутрь клетки через фосфорилирование белка. Тем не менее, он формирует гетеродимеры с другими членами HER-семейства, которые обладают киназной активностью. Гетеродимеризация приводит к активации рецептор-опосредованного сигнального пути и трансфосфорилированию его внутриклеточного домена. Образование димеров между членами HER-семейства расширяет возможности передачи сигнала HER3 и является средством не только распределения сигнала, но и усиления сигнала. Например, гетеродимер HER2/HER3 индуцирует один из наиболее важных митогенных сигналов через путь PI3K и AKT среди членов семейства HER (Sliwkowski М.Х., et at, J. Biol. Chem. 269 (1994) 14661-14665; Alimandi M, et al, Oncogene. 10 (1995) 1813-1821; Hellyer, N.J., J. Biol. Chem. 276 (2001) 42153-4261; Singer, E., J. Biol. Chem. 276 (2001) 44266-44274; Schaefer, K.L, Neoplasia 8 (2006) 613-622).
Амплификация этого гена и/или сверхэкспрессия его белка были обнаружены при многих раковых заболеваниях, в том числе при опухолях предстательной железы, мочевого пузыря и молочной железы. Были охарактеризованы транскрипционные варианты альтернативного сплайсинга, кодирующие различные изоформы. У одной изоформы отсутствует межмембранная область, и эта изоформа секретируется за пределы клетки. Эта форма действует путем модуляции активности формы, связанной с мембраной. Также сообщалось о дополнительных вариантах сплайсинга, но они не были детально описаны.
Термин «антитело», как использовано в данном описании, включает целые антитела и любой антигенсвязывающий фрагмент (т.е. «антигенсвязывающую часть») или его отдельные цепи. Термин "антитело" относится к гликопротеину, содержащему по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно называемую в данном описании как VH) и константную область тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, СН2 и СН 3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно называемой в данном описании как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), разбросанные между областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включая различные клетки иммунной системы (например, эффекторными клетками), и первый компонент (Clq) классической системы комплемента.
«Биспецифичное антитело» представляет собой антитело, содержащее антигенсвязывающий домен или антигенсвязывающие домены, которые способны к специфическому связыванию с двумя различными эпитопами на одной биологической молекуле или способны к специфическому связыванию с эпитопами на двух различных биологических молекулах. Биспецифичное антитело также упоминается в настоящем документе, как обладающее «двойной специфичностью» или как являющееся антителом с «двойной специфичностью».
Под биспецифическим антителом BCD-090 понимают биспецифическое антитело, которое специфично связывается с HER2 и HER3. Данное антитело содержит вариабельный домен тяжелой цепи VHH, специфичный к HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2. В некоторых вариантах осуществления изобретения вариабельный домен тяжелой цепи VHH, специфичный к HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2, связаны между собой пептидным линкером.
Термин «пептидный линкер» в настоящем документе означает любой пептид с возможностью соединения доменов с длиной в зависимости от доменов, которые он связывает между собой, содержащий любую аминокислотную последовательность. Предпочтительно пептидный линкер имеет длину более 5 аминокислот и состоит из любого набора аминокислот, выбранного из G, A, S, Р, Е, Т, D, K.
HER2-HER3-связывающие антитела включают также антитела тяжелых цепей (HCAb). Исключения в отношении структуры Н2L2 общепринятых антител встречаются в некоторых изотипах иммуноглобулинов, обнаруженных в семействе верблюдовых (Camelidae) (верблюдах, дромедарах (одногорбых верблюдах и ламах; Hamers-Casterman et al., 1993 Nature 363: 446; Nguyen et al., 1998 J. Mol. Biol. 275: 413), ковровых акулах (воббегонгах) (Nuttal et al., Mol Immunol. 38:313-26, 2001), усатых акулах-няньках (Greenberg et al., Nature 374:164-73, 1995; Roux et al., Proc. Natl. Acad. Sci. USA 95: 11804) и пятнистых химерах (Nguyen et al., "Heavy-chain antibodies in Camelidae; a case of evolutionary innovation," 2002 Immunogenetics 54(1): 39-47)). Такие антитела, очевидно, могут образовывать антигенсвязывающие области с использованием только вариабельной области тяжелой цепи, так что функциональные антитела являются димерами только тяжелых цепей (называемыми «антителами тяжелых цепей» или обозначенными как "HCAb"). Таким образом, некоторые варианты данных HER2-HER3-связывающих антител могут быть антителами тяжелых цепей (HCAb), которые специфически связываются с HER2 и HER3. Например, антитела тяжелых цепей, которые являются антителами класса IgG и лишены легких цепей, продуцируются животными семейства Camelidae, которое включает в себя верблюдов, дромедаров и лам (Hamers-Casterman et al., Nature 363: 446-448 (1993)). HCAb имеют молекулярную массу приблизительно 95 кДа вместо молекулярной массы приблизительно 160 кДа общепринятых IgG-антител. Их связывающие домены состоят только из вариабельных доменов тяжелой цепи, часто обозначаемые в настоящем описании как VHH, чтобы отличить их от общепринятых VH. Muyldermans et al., J. Mol. Recognit. 12:131-140 (1999). Вариабельный домен антител тяжелой цепи иногда называют нанотелом (Cortez-Retamozo et al., Cancer Research 64:2853-57, 2004). Библиотека нанотел может быть генерирована из иммунизированного дромедара, как описано Conrath et al., Antimicrob Agents Chemother 45: 2807-12, 2001, или с использованием рекомбинантных способов.
Поскольку первый константный домен (CH1) отсутствует (сплайсирован во время процессинга мРНК вследствие утраты сигнала сплайсингового консенсуса), за вариабельным доменом VHH следует непосредственно шарнирная область, домены СH2 и СH3 (Nguyen et al., Mol. Immunol. 36:515-524 (1999); Woolven et al., Immunogenetics 50:98-101 (1999)). VHH верблюдовых рекомбинируется с константными областями IgG2 и IgG3, которые содержат шарнирную область, домены СН2 и СН3 и лишены домена CH1 (Hamers-Casterman et al., supra). Например, IgG1 ламы является общепринятым изотипом антитела (Н2L2), в котором VH рекомбинируется с константной областью, которая содержит шарнирную область, домены CH1, СН2 и СН3, тогда как IgG2 и IgG3 ламы являются изотипами только тяжелой цепи, которая лишена доменов СН1 и которая не содержит легких цепей.
Хотя HCAb лишены легких цепей, они имеют антигенсвязывающий репертуар. Механизм генетического генерирования HCAb обсуждается в Nguyen et al., Adv. Immunol 79:261-296 (2001) и Nguyen et al., Immunogenetics 54:39-47 (2002). Акулы, включая усатую акулу-няньку, обнаруживают сходные содержащие рецептор антигена отдельные мономерные V-домены. Irving et al., J. Immunol. Methods 248:31-45 (2001); Roux et al., Proc. Natl. Acad. Sci. USA 95:11804 (1998).
VHH содержат малые интактные антигенсвязывающие фрагменты (например, фрагменты, которые имеют молекулярную массу 15 кДа, 118-136 остатков). Было обнаружено, что VHH-домены верблюдовых связываются с антигеном с высокой аффинностью (Desmyter et al., J. Biol. Chem. 276:26285-90, 2001), с VHH-аффинностью обычно в наномолярном диапазоне и сравнимыми с аффинностью фрагментов Fab и scFv. VHH являются высокорастворимыми и более стабильными, чем соответствующие производные фрагментов scFv и Fab. VH-фрагменты было относительно трудно получить в растворимой форме, но могут быть получены улучшения в растворимости и специфическом связывании, когда каркасные остатки изменяют таким образом, чтобы они были более VHH-подобными. (См., например, Reichman et al., J Immunol Methods 1999, 231:25-38). VHH несут аминокислотные замены, которые делают их более гидрофильными и предотвращают пролонгированное взаимодействие с BiP (связывающий белок тяжелой цепи иммуноглобулина), который обычно связывается с Н-цепью в эндоплазматическом ретикулуме (ER) во время укладки и сборки, пока он не вытесняется L-цепью. Вследствие увеличенной гидрофильности VHH секреция из ER улучшается.
Функциональные VHH могут быть получены протеолитическим расщеплением HCAb иммунизированного животного семейства верблюдовых, прямым клонированием генов VHH из В-клеток иммунизированного верблюда с получением рекомбинантных VHH, или из наивных («необученных») или синтетических библиотек. VHH с желаемой антигенной специфичностью могут быть также получены с использованием метода фагового дисплея. Использование VHH в фаговом дисплее является гораздо более простым и более эффективным по сравнению с Fab или scFv, так как только один домен должен быть клонирован и экспрессирован для получения функционального антигенсвязывающего фрагмента. Muyldermans, Biotechnol. 74:277-302 (2001); Chahroudi et al., FEBS Lett. 441:521-526 (1997) и van der Linden et al., J. Biotechnol. 80:261-270 (2000). Способы получения антител, имеющих тяжелые цепи верблюдовых, описаны также в публикациях патентов США No. 20050136049 и 20050037421.
Способы рибосомного дисплея могут быть использованы для идентификации и выделения молекул scFv и/или VHH, имеющих желаемые активность и аффинность связывания. Irving et al., J. Immunol. Methods 248:31-45 (2001). Рибосомный дисплей и отбор имеет потенциал для генерирования больших библиотек дисплея (1014).
В других вариантах осуществления предоставлены УНн-подобные молекулы, генерируемые посредством способа «верблюдизации», модификацией неверблюдовых VH, таких как VHH человека, для улучшения их растворимости и предотвращения неспецифического связывания. Это достигается заменой остатков на VL-стороне VH VHH-подобными остатками, имитируя, таким образом, более растворимые VHH-фрагменты. Ожидается, что «верблюдизированные» VH-фрагменты, в частности фрагменты, основанные на каркасной области антитела человека, должны проявлять в значительной степени уменьшенную иммунную реакцию при введении in vivo пациенту, и, следовательно, ожидается, что они будут иметь значимые преимущества для терапевтических применений. Davies et al., FEBS Lett. 339:285-290 (1994); Davies et al., Protein Eng. 9:531-537 (1996); Tanha et al., J. Biol. Chem. 276:24774-24780 (2001) и Riechmann et al., Immunol. Methods 231:25-38 (1999).
Термин "идентичность" или "гомологичность" следует толковать как означающее процентное содержание остатков аминокислот в кандидатной последовательности, которые идентичны остаткам соответствующей последовательности, с которой ее сравнивают, после сравнения последовательностей и введения "брешей", если необходимо достичь максимального процента идентичности для полной последовательности и не учитывая любые консервативные замещения как часть идентичности последовательности. Ни N- или С-концевой удлиняющей, ни инсерционные сегменты не следует толковать как уменьшающие идентичность или гомологичность. Методы и компьютерные программы для сравнения хорошо известны. Идентичность последовательности можно определить, используя программное обеспечение для анализа последовательности (например, Sequence Analysis Software Package, Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Ave., Madison, WI 53705). Данное программное обеспечение подходит для подобных последовательностей путем определения степени гомологичности для разнообразных замещений, делеций (элиминирований) и других модификаций.
Термин "идентичность" или "гомологичность" в контексте последовательностей нуклеиновых кислот или аминокислот, относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия. Существует целый ряд различных алгоритмов, известных в данной области, которые могут быть использованы для измерения идентичности нуклеотидной или аминокислотной последовательностей. Например, полинуклеотидные последовательности можно сравнить с использованием FASTA, Gap или BESTFIT, которые являются программами в Wisconsin Package версии 10.0, Genetics Computer Group (GCG), Мэдисон, штат Висконсин. FASTA, которая включает, например, программы FASTA2 и FASTA3, обеспечивает выравнивание и процентную идентичность последовательности в областях наилучшего покрытия между запрашиваемой и искомой последовательностями (Pearson, Methods Enzymol. 183:63 98 (1990); Pearson, Methods Mol. Biol. 132: 185-219 (2000); Pearson, Methods Enzymol. 266: 227-258 (1996); Pearson, J. Mol. Biol. 276: 71-84 (1998)). Если не указано иное, используются параметры по умолчанию для конкретной программы или алгоритма. Например, процент идентичности последовательности между последовательностями нуклеиновых кислот может быть определен с использованием FASTA с параметрами по умолчанию (размер слова 6 и фактора NOPAM для балльной матрицы) или с использованием Gap с параметрами по умолчанию, как это предусмотрено в GCG версии 6.1.
Фразу "гомологичный", что касается полипептидной последовательности антитела, следует толковать как антитело, проявляющее по крайней мере, 70%-ную, предпочтительно 80%-ную, более предпочтительно 90%-ную и наиболее предпочтительно 95%-ную идентичность последовательности относительно полипептидной последовательности. Термин в отношении последовательности нуклеиновой кислоты следует толковать как последовательность нуклеотидов, проявляющих, по крайней мере, 85%-ную, предпочтительно 90%-ную, более предпочтительно 95%-ную и наиболее предпочтительно 97%-ную идентичность последовательности относительно последовательности нуклеиновой кислоты.
Суммарную гомологию определяют следующим образом:
(1) вначале отыскивают последовательность антитела или его фрагмента с минимальным расстоянием редактирования по Дамерау-Левенштейну по отношению к последовательности, применительно к которой определяется степень гомологии, затем
(2) указанное расстояние Дамерау-Левенштейна нормируют таким образом, что расстояние между идентичными последовательностями принимается за 100% гомологии, а расстояние до рандомной последовательности такой же длины - за 0%,
(3) повторяют процедуру с остальными отрезками и
(4) определяют суммарную гомологию как средневзвешенное по длине отрезков число.
Предлагается модификация(и) аминокислотных последовательностей HER2-HER3-антител, описанных в настоящей публикации. Например, может быть желательным улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности HER2-HER3-антитела получают введением соответствующих изменений нуклеотидов в нуклеиновую кислоту HER2-HER3-антитела или пептидным синтезом. Такие модификации включают, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях HER2-HER3-антитела. Осуществляют любое сочетание делеции, инсерции и замены, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает требуемыми характеристиками. Изменения аминокислот также могут изменять посттрансляционные процессы в HER2-HER3-антителе, такие как изменение количества или положения сайтов гликозилирования.
Применимый способ идентификации некоторых остатков или областей HER2-HER3-антитела, которые являются предпочтительными участками для мутагенеза, называют "мутагенезом с использованием сканирования аланином", который описан Cunningham and Wells Science, 244: 1081-1085 (1989). В данном случае идентифицируют остаток или группу остатков-мишеней (например, заряженных остатков, таких как arg, asp, his, lys и glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтительно аланином или полиаланином), чтобы повлиять на взаимодействие аминокислот с антигеном HER2 и/или HER3. Те участки аминокислот, которые проявляют функциональную чувствительность к заменам, затем улучшают введением дополнительных или других вариантов в участки или вместо участков замены. Таким образом, хотя участок введения варианта аминокислотной последовательности предварительно определяют, нет необходимости в предварительном определении природы мутации как таковой. Например, чтобы проанализировать эффективность мутации в данном участке проводят ala-сканирование или случайный мутагенез в кодоне-мишени или области-мишени и экспрессированные варианты HER2-HER3-антител подвергают скринингу в отношении требуемой активности.
Инсерции в аминокислотных последовательностях включают слияния с амино- и/или карбоксильным концом длиной в диапазоне от одного остатка до полипептидов, содержащих сто или более остатков, а также инсерции внутри последовательности одного или нескольких аминокислотных остатков. Примеры концевых инсерции включают HER2-aнтитeлo с N-концевым остатком метионила или антитело, слитое с цитотоксическим полипептидом. Другие инсерционные варианты молекулы HER2-HER3-антитела включают слияние N- или С-конца HER2-HER3-антитела с ферментом (например, в случае ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
Вариантом другого типа является вариант с аминокислотными заменами. Такие варианты имеют, по меньшей мере, один аминокислотный остаток в молекуле HER2-HER3-антитела, замененный другим остатком. Места, представляющие наибольший интерес для мутагенеза путем замен, включают гипервариабельные области или CDR, но также предполагаются изменения и в области FR или Fc. Консервативные замены показаны в таблице 1 под заголовком "предпочтительные замены". Если такие замены приводят к изменению биологической активности, то могут быть введены дополнительные существенные изменения, названные "примерами заменам" в таблице А, или изменения, дополнительно описанные ниже при описании классов аминокислот, и может быть проведен скрининг продуктов.
Существенные модификации биологических свойств антитела осуществляют отбором замен, которые существенно отличаются по
своему влиянию на поддержание (а) структуры полипептидного остова в области замены, например, в виде слоистой или спиральной конформации, (b) заряда или гидрофобности молекулы в сайте-мишени, или (с) величины боковой цепи. Аминокислоты могут быть сгруппированы в соответствии со сходствами в свойствах их боковых цепей (в A.L. Lehninger, Biochemistry, second ed., pp. 73-75, Worth Publishers, New York (1975)):
(1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M),
(2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (С), Tyr (Y), Asn (N), Gln (Q),
(3) кислые: Asp (D), Glu (E),
(4) основные: Lys (K), Arg (R), His(H).
Альтернативно встречающиеся в природе остатки могут быть разделены на группы на основе общих свойств боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены влекут за собой замену представителя одного из указанных классов представителем другого класса.
Любой остаток цистеина, не вовлеченный в поддержание правильной конформации HER2-HER3-антитела, также может быть заменен, обычно серином, чтобы повысить стабильность молекулы к окислению и предотвратить аномальное поперечное сшивание. Наоборот, может быть добавлена цистеиновая связь (связи) к антителу, чтобы повысить его стабильность (особенно когда антитело представляет собой фрагмент антитела, такой как Fv-фрагмент).
Особенно предпочтительный тип варианта с заменами содержит замену одного или нескольких остатков гипервариабельной области исходного антитела (например, гуманизированного антитела или антитела человека). В общем, полученные в результате вариант(ты), отобранные для дальнейшей разработки, будут иметь улучшенные биологические свойства по сравнению с исходным антителом, из которого они созданы. Подходящий способ создания таких вариантов с заменами заключается в созревании аффинности с использованием фагового дисплея. Коротко, несколько сайтов гипервариабельной области (например 6-7 сайтов) подвергают мутациям, чтобы создать все возможные аминокислотные замены в каждом сайте. Полученные таким образом варианты антитела представляют в виде дисплея в моновалентной форме на частицах нитчатого фага в виде слияний с продуктом гена III М13, упакованного в каждой частице. Представленные в фаговом дисплее варианты затем подвергают скринингу в отношении их биологической активности (например, аффинности связывания), как описано в данной публикации. Чтобы идентифицировать подходящие для модификации сайты гипервариабельной области можно осуществить мутагенез посредством сканирования аланином, идентифицируя остатки гипервариабельной области, вносящие существенный вклад в связывание антигена. Альтернативно или дополнительно может быть полезным анализ кристаллической структуры комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и HER2 и/или HER3 человека. Такие остатки в области контакта и соседние остатки являются кандидатами на замену способами, разработанными согласно изобретению. После создания таких вариантов панель вариантов подвергают скринингу, как описано в данной публикации, и антитела с превосходящими свойствами в одном или нескольких подходящих анализах можно отобрать для дальнейшей разработки.
В другом типе аминокислотного варианта антитела изменена исходная картина гликозилирования антитела. Под изменением подразумевают делецию одного или нескольких углеводных остатков, обнаруженных в антителе, и/или добавление одного или нескольких сайтов гликозилирования, которые не присутствуют в антителе.
Гликозилирование антител обычно является либо N-связанным, либо О-связанным. N-связанный относится к связыванию углеводного остатка с боковой цепью остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где X означает любую аминокислоту, за исключением пролина, являются последовательностями узнавания для ферментативного связывания углеводного остатка с боковой цепью аспарагина. Таким образом, присутствие любой из указанных трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. О-связанное гликозилирование относится к связыванию одного из сахаров N-ацетилгалактозамина, галактозы или ксилозы с гидроксиаминокислотой, чаще всего серином или треонином, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление сайтов гликозилирования к антителу обычно осуществляют посредством изменения аминокислотной последовательности, так чтобы она содержала одну или несколько из описанных выше трипептидных последовательностей (для сайтов N-связанного гликозилирования). Изменение также можно осуществить добавлением или заменой одного или нескольких остатков серина или треонина к последовательности исходного антитела (для сайтов О-связанного гликозилирования).
В том случае, когда антитело содержит Fc-область, может быть изменен связанный с ним углевод. Например, антитела со зрелой углеводной структурой, в которой утрачена фукоза, связанная с Fc-областью антитела, описаны в заявке на выдачу патента США № US 2003/0157108 A1, Presta, L. См. также US 2004/0093621 A1 (Kyowa Hakko Kogyo Co., Ltd). Антитела с разветвляющимся N-ацетилглюкозамином (GlcNAc) в олигосахаридной структуре, связанной с Fc-областью антитела, описаны в WO 03/011878, Jean-Mairet et al. и в патенте США No. 6602684, Umana et al. Антитела, по меньшей мере, с одним остатком галактозы в олигосахаридной структуре, связанной с Fc-областью антитела, описаны в WO 97/30087, Patel et al. См., также заявки на выдачу патента WO 98/58964 (Raju, S.) и WO 99/22764 (Raju, S.), касающиеся антител с измененным углеводом, связанным с их Fc-областью. В настоящем изобретении предполагаются композиции антител, содержащие антитело главного типа с такими углеводными структурами, связанными с одной или двумя тяжелыми цепями Fc-области.
Термин «антигенсвязывающая часть» антитела (или просто «часть антитела»), как использовано в данном описании, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин "антигенсвязывающая часть" антитела включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и СН 1; (ii) Р(аb')2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и СН 1; (iv) Fv-фрагмент, состоящий из доменов VL и VH в едином плече антитела, (v) dAb-фрагмент (Ward et al. (1989) Nature 341:544-546), который состоит из домена VH/VHH; и (vi) выделенная определяющая комплементарность область (CDR). Кроме того, две области Fv-фрагмента, VL и VH, кодируются разными генами, они могут быть соединены при помощи рекомбинантных способов с использованием синтетического линкера, который дает возможность получать их в виде единой белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные молекулы также включены в термин "антигенсвязывающая часть" антитела. Такие фрагменты антител получают с использованием общепринятых способов, известных специалистам в данной области, и эти фрагменты подвергают скринингу таким же образом, как и интактные антитела.
Предпочтительно CDR антигенсвязывающего участка или весь антигенсвязывающий участок антител по изобретению имеет происхождение из мыши, ламы или донорской человеческой библиотеки или по существу человеческое происхождение с определенными аминокислотными остатками, измененными, например, замещенными разными аминокислотными остатками с тем, чтобы оптимизировать конкретные свойства антитела, например KD, koff, IC50, ЕС50, ED50. Предпочтительно каркасные участки антитела по изобретению имеют человеческое происхождение или по существу человеческое происхождение (по крайней мере на 80, 85, 90, 95, 96, 97, 98 или 99% человеческое происхождение).
В других вариантах осуществления антигенсвязывающий участок антитела по изобретению может происходить из других нечеловеческих видов, включая мыши, ламы, кролика, крысу или хомяка, но не ограничиваясь ими. Альтернативно, антигенсвязывающий участок может происходить из человеческих видов.
Термин «вариабельный» относится к тому факту, что определенные сегменты вариабельных доменов широко отличаются в последовательности среди антител. Домен V опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность неравномерно распределяется на участке вариабельных доменов из 110 аминокислот. Напротив, V области состоят из инвариантных фрагментов, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемых "гипервариабельными областями" или CDR. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в основном принимающих конфигурацию бета-листов, связанных тремя гипервариабельными областями, которые образуют петли, связывающие, и в некоторых случаях являющиеся частью бета-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в тесной близости с помощью FR и с гипервариабельными областями другой цепи вносят вклад в образование антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest. 5 th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (АЗКЦ, ADCC).
Термин "гипервариабельная область" по данному описанию относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Обычно гипервариабельная область содержит аминокислотные остатки из "области, определяющей комплементарность" или "CDR", и/или такие остатки из "гипервариабельной петли".
"Каркасные области" (FR) представляют собой остатки вариабельного домена, отличные от CDR остатков. Обычно каждый вариабельный домен имеет четыре FR, определяемые как FR1, FR2, FR3 и FR4. Если CDR определяются согласно Kabat, FR остатки легкой цепи локализуются, приблизительно, в области остатков 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4), а остатки FR тяжелой цепи локализуются, приблизительно, в области остатков 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103-113 (HCFR4) в тяжелой цепи. Если участки CDR содержат аминокислотные остатки из гипервариабельных петель, FR остатки легкой цепи локализуются, приблизительно, в остатках 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4) в легкой цепи, a FR остатки тяжелой цепи локализуются, примерно, в остатках 1-25 (HCFRI), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4) в остатках тяжелой цепи. В некоторых примерах, когда CDR содержит аминокислоты как из CDR по Kabat, так и аминокислоты из гипервариабельной петли, FR cooтветствующим образом корректируются. Например, когда CDRH1 включает аминокислоты Н26-Н35, остатки FR1 тяжелой цепи находятся в положениях 1-25, а остатки FR2 находятся в положениях 36-49.
Антитело по данному изобретению, "которое связывает" целевой антиген, представляет собой антитело, которое связывает антиген с достаточной аффинностью так, что антитело можно применять в качестве диагностического и/или терапевтического агента при нацеливании на белок или клетку или ткань, экспрессирующую антиген, и в незначительной степени перекрестно реагирует с другими белками. По данным аналитических методов: сортинга флуоресцентно-активированных клеток (FACS), радиоиммунопреципитации (RIA) или ИФА (ELISA), в таких вариантах изобретения степень связывания антитела с белком, не являющимся "мишенью" (с "нецелевым белком"), составляет менее 10% от связывания антитела с конкретным белком-мишенью. По отношению к связыванию антитела с молекулой-мишенью термин "специфическое связывание" или выражения "специфически связывается с" или "специфический к" конкретному полипептиду или эпитопу на конкретном полипептиде-мишени означает связывание, которое заметно (измеримо) отличается от неспецифического взаимодействия (например, в случае bH1-44 или bH1-81 неспецифическое взаимодействие представляет собой связывание с бычьим сывороточным альбумином, казеином, фетальной бычьей сывороткой или нейтравидином). Специфическое связывание можно определять количественно, например, определяя связывание молекулы по сравнению со связыванием контрольной молекулы. Например, специфическое связывание можно определять конкурентной реакцией с другой молекулой, аналогичной мишени, например, с избытком немеченой мишени. В этом случае специфическое связывание указывается, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. В данном описании термин "специфическое связывание" или выражения "специфически связывается с" или "специфический к" конкретному полипептиду или эпитопу на конкретном полипептиде-мишени можно характеризовать на примере молекулы, имеющей Kd к мишени по меньшей мере около 200 нМ, или же по меньшей мере около 150 нМ, или же по меньшей мере около 100 нМ, или же по меньшей мере около 60 нМ, или же по меньшей мере около 50 нМ, или же по меньшей мере около 40 нМ, или же по меньшей мере около 30 нМ, или же по меньшей мере около 20 нМ, или же по меньшей мере около 10 нМ, или же по меньшей мере около 8 нМ, или же по меньшей мере около 6 нМ, или же по меньшей мере около 4 нМ, или же по меньшей мере около 2 нМ, или же по меньшей мере около 1 нМ или выше. В одном варианте изобретения термин "специфическое связывание" относится к связыванию, при котором молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде, практически не связываясь с каким-либо другим полипептидом или эпитопом на полипептиде.
Термин «Ка», как использовано в данном описании, относится к скорости ассоциации конкретного взаимодействия антитело-антиген, тогда как термин или «Kd» относится к скорости диссоциации конкретного взаимодействия антитело-антиген.
"Аффинность связывания" обычно относится к силе совокупных нековалентных взаимодействий между единичным сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иначе, "аффинность связывания" относится к внутренней (характерной, истинной) аффинности связывания, которая отражает 1:1 взаимодействие между членами пары связывания (например, антителом и антигеном). Аффинность молекулы X к своему партнеру Y обычно можно представить константной диссоциации (Kd). Желательно, чтобы величина Kd составляла, примерно, 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерять обычными методами, известными в уровне техники, включая методы по данному описанию. Низкоаффинные антитела обычно медленно связываются с антигеном и имеют тенденцию легко диссоциировать, тогда как высокоаффинные антитела обычно быстрее связывают антиген и имеют тенденцию дольше оставаться в связанном состоянии. В уровне техники известны различные методы измерения аффинности связывания, любой из этих методов можно использовать для целей настоящего изобретения.
В одном варианте изобретения "Kd" или "величину Kd" по данному изобретению измеряют методами поверхностного плазменного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (например, от 0.78 нМ до 500 нМ) вводят в PBS с 0.05% Tween 20 (PBST) при 25°С при скорости потока, примерно, 25 мкл/мин. Величины скорости ассоциации (kon) и скорости ассоциации (koff) рассчитывают, применяя простую модель Ленгмюра для связывания один-плюс-один (BIAcore Evaluation Software версия 3.2), с помощью одновременного получения сенсограммы ассоциации и диссоциации. Константу равновесной диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen, Y., et al. (1999) J. Mol. Biol. 293: 865-881. Если по данным вышеуказанного метода поверхностного плазмонного резонанса скорость ассоциации превышает 106 М-1 сек-1, тогда ее можно определять методом тушения флуоресценции, который измеряет увеличение или уменьшение интенсивности флуоресцентной эмиссии (возбуждение = 295 нм; эмиссия (излучение) = 340 нм, полоса 16 нм) при 25°С раствора антитела против антигена (Fab форма) с концентрацией 20 нМ в PBS, рН 7.2, в присутствии увеличивающихся концентраций антигена, измеряемых с помощью спектрометра, такого как спектрофотометр остановленного потока (Aviv Instruments) или спектрофотометр SLM-Aminco (ThermoSpectronie) серии 8000 с кюветой с перемешиванием.
"Скорость ассоциации" ("on-rate") или "kon" по данному изобретению можно также определять тем же самым описанным выше методом поверхностного плазменного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламино пропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (например, от 0.78 нМ до 500 нМ) вводят в PBS с 0.05% Tween 20 (PBST) при 25°С при скорости потока, примерно, 25 мкл/мин. Величины скорости ассоциации (kon) и скорости ассоциации (koff) рассчитывают, применяя простую модель Ленгмюра для связывания один-плюс-один (BIAcore Evaluation Software версия 3.2), одновременно получая сенсограммы ассоциации и диссоциации. Константу равновесной диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen, Y., et al. (1999) J. Mol. Biol. 293: 865-881. Однако, если по данным вышеуказанного метода поверхностного плазменного резонанса скорость ассоциации превышает 106 М-1 сек-1, тогда ее можно определять методом тушения флуоресценции, который измеряет увеличение или уменьшение интенсивности флуоресцентной эмиссии (возбуждение = 295 нм; эмиссия (излучение) = 340 нм, полоса 16 нм) при 25°С раствора антитела против антигена (Fab форма) с концентрацией 20 нМ в PBS, рН 7.2, в присутствии увеличивающихся концентраций антигена, измеряемых с помощью спектрометра, такого как спектрофотометр остановленного потока (Aviv Instruments) или спектрофотометр SLM-Aminco (ThermoSpectronic) серии 8000 с кюветой с перемешиванием.
Если специально не указано иначе, выражения "биологически активный", и "биологическая активность", и "биологические характеристики", по отношению к полипептиду по данному изобретению, означают обладание способностью связываться с биологической молекулой.
Выражение "биологическая молекула" относится к нуклеиновой кислоте, белку, углеводу, липиду и их комбинации. В одном варианте изобретения биологическая молекула существует в природе.
Участки антител, такие как Fab- и F(ab')2-фрагменты, могут быть получены из целых антител с использованием традиционных методов, таких как папаиновый или пепсиновый гидролиз целых антител. Более того, антитела, части антител и молекулы иммуноадгезии могут быть получены с использованием стандартных методов рекомбинантной ДТПС, например, как описано в настоящем документе.
Термин «рекомбинантное антитело» означает антитело, которое экспрессируется из клетки или клеточной линии, содержащей нуклеотидную последовательность (нуклеотидные последовательности), которая кодирует антитела, при этом указанная нуклеотидная последовательность (нуклеотидные последовательности) не ассоциирована с клеткой в природе.
Термин «вариантное» антитело, используемый в данном документе, относится к антителу, имеющему аминокислотную последовательность, которая отличается от аминокислотной последовательности его «родительского» антитела путем добавления, удаления и/или замены одного или более аминокислотных остатков относительно последовательности родительского антитела. В предпочтительном варианте осуществления изобретения вариантное антитело содержит по меньшей мере одно или более (например, от одного до двенадцати, например, два, три, четыре, пять, шесть, семь, восемь или девять, десять, одиннадцать или двенадцать; и в некоторых вариантах осуществления изобретения от одного до примерно десяти) добавлений, делеций и/или замен аминокислот относительно родительского антитела. В некоторых вариантах осуществления изобретение добавления, делеций и/или замены осуществляются на CDR-участках вариантного антитела. Идентичность или гомология по отношению к последовательности вариантного антитела определяется в настоящем документе как процент аминокислотных остатков в последовательности вариантного антитела, которые идентичны остаткам родительского антитела, после выравнивания последовательностей и введения гэпов, если это необходимо, для достижения максимального процента идентичности последовательности. Вариантное антитело сохраняет способность связываться с тем же антигеном, и предпочтительно эпитопом, с которым связывается родительское антитело, и в некоторых вариантах осуществления изобретения по меньшей мере одно свойство или биологическая активность превосходит аналогичные свойства родительского антитела. Например, вариантное антитело может иметь, например, более выраженную аффинность связывания, более длительный период полувыведения, более низкое значение ИК50 или повышенную способность подавлять биологическую активность антигена по сравнению с родительским антителом. Особый интерес в настоящем документе представляет вариантное антитело, показывающее биологическую активность, превышающую по меньшей мере в 2 раза (предпочтительно, по меньшей мере в 5 раз, 10 раз или 20 раз) биологическую активность родительского антитела.
Термин «химерное антитело» относится в широком смысле к антителу, которое содержит одну или более областей из одного антитела, и одну или более областей из одного или нескольких других антител, как правило, антитело, частично человеческого происхождения и частично нечеловеческого происхождения, то есть полученное частично из не относящегося к человеку животного, например, мыши, крысы или другого грызуна или верблюдовых, таких как лама или альпака. Химерные антитела являются предпочтительными по сравнению с нечеловеческими антителами для того, чтобы снизить риск иммунного ответа, направленного против антител у человека, например, ответа, направленного против мышиных тел у человека в случае мышиного антитела. Примером типичного химерного антитела является то, в котором последовательности вариабельного участка являются мышиными, в то время как последовательности константного участка являются человеческими. В случае химерного антитела нечеловеческие части могут быть подвергнуты дальнейшему изменению с целью гуманизации антитела.
Термин «гуманизация» относится к факту, что когда антитело имеет полностью или частично нечеловеческое происхождение, например, антитело мыши или ламы, полученное при иммунизации мышей или лам, соответственно, с представляющим интерес антигеном, или является химерным антителом на основе такого антитела мыши или ламы, можно заменить некоторые аминокислоты, в частности, в каркасных областях и константных доменах тяжелой и легкой цепей, с тем чтобы избежать или свести к минимуму иммунный ответ у человека. Специфичность взаимодействия антитела с антигеном-мишенью присуща главным образом аминокислотным остаткам, расположенных в шести CDR-участках тяжелой и легкой цепи. Поэтому аминокислотные последовательности внутри CDR-участков, являются гораздо более вариабельными между отдельными антителами, по сравнению с последовательностями вне CDR-участков. Поскольку последовательности CDR-участков отвечают за большинство антитело-антиген взаимодействий, можно экспрессировать рекомбинантные антитела, которые имитируют свойства специфического природного антитела, или в более общем плане какого-либо специфического антитела с данной аминокислотной последовательностью, например, путем конструирования экспрессионных векторов, которые экспрессируют последовательности CDR-участков из специфического антитела в каркасные последовательности другого антитела. В результате, можно «гуманизировать» нечеловеческое антитело и в значительной степени сохранить специфичность связывания и аффинность исходного антитела. Несмотря на то, что невозможно точно предсказать иммуногенность и тем самым иммунный ответ, направленный против антитела у человека, на конкретное антитело, нечеловеческие антитела, как правило, более иммуногенны, чем человеческие антитела. Химерные антитела, у которых инородные (например, грызуна или верблюда) константные участки были заменены последовательностями человеческого происхождения, показали в целом более низкую иммуногенность, чем антитела полностью инородного происхождения, и существует тенденция использовать в терапевтических антителах гуманизированные или полностью человеческие антитела. Химерные антитела или другие антитела нечеловеческого происхождения, таким образом, могут быть гуманизированы, чтобы снизить риск иммунного ответа, направленного против антитела, у человека.
Для химерных антител, гуманизация обычно включает в себя модификацию каркасных участков последовательностей вариабельного участка. Аминокислотные остатки, которые являются частью участков, определяющих комплементарность (CDR-участков), чаще всего не будут изменяться в связи с гуманизацией, хотя в некоторых случаях это может быть желательным, чтобы изменить отдельные аминокислотные остатки CDR-участка, например, чтобы удалить участок гликозилирования, участок дезамидирования, участок изомеризации аспартата или нежелательный остаток цистеина или метионина. N-связанное гликозилирование происходит путем присоединения олигосахаридной цепи к остатку аспарагина в трипептидной последовательности Asn-X-Ser или Asn-X-Thr, где X может быть любой аминокислотой, кроме Pro. Удаление участка N-гликозилирования может быть достигнуто путем мутирования Asn или Ser/Thr остатка другим остатком, предпочтительно путем консервативной замены. Дезамидирование остатков аспарагина и глутамина может происходить в зависимости от таких факторов, как рН и обнажение поверхности. Остатки аспарагина особенно восприимчивы к дезамидированию, прежде всего, если они присутствуют в последовательности Asn-Gly, и в меньшей степени в других дипептидных последовательностях, таких как Asn-Ala. При наличии такого дезамидированного участка, в частности, Asn-Gly в последовательности CDR-участка, может быть предпочтительным удалить этот участок, как правило, путем консервативной замены для удаления одного из вовлеченных остатков.
В данной области техники известны многочисленные способы гуманизации последовательности антитела; см., например, обзор Almagro & Fransson, Front Biosci. 13:1619-1633 (2008). Одним из наиболее часто используемых методов является трансплантация CDR-участков, например, когда химерные антитела мышиного происхождения задействуют идентификацию человеческих зародышевых генных эквивалентов к мышиным генам вариабельного участка и трансплантацию последовательностей мышиных CDR-участков в этот каркас. Трансплантация CDR-участка может быть основана на определениях CDR-участков по Kabat, хотя в более поздней публикации (Magdelaine-Beuzelin et al., Crit Rev. Oncol Hematol. 64:210-225 (2007)) предполагается, что определение по IMGT® (the international ImMunoGeneTics information system®, www.imgt.org) может улучшить результат гуманизации (см Lefranc et al., Dev. Comp Immunol. 27:55-77 (2003)). В некоторых случаях, трансплантация CDR-участкка может уменьшить специфичность и аффинность связывания, и, следовательно, биологическую активность, в CDR-трансплантированном нечеловеческом антителе, по сравнению с родительским антителом, из которого получены CDR-участки. Обратные мутации (иногда именуемые «ремонт каркасного участка»), могут применяться в выбранных положениях CDR-трансплантированного антитела, как правило, в каркасных участках, для того, чтобы восстановить специфичность и аффинность связывания родительского антитела. Определение позиций для возможных обратных мутаций может быть выполнено с использованием информации, имеющейся в литературе и в базах данных антител. Аминокислотные остатки, которые являются кандидатами для обратных мутаций, как правило, расположены на поверхности молекулы антитела, в то время как остатки, которые углублены или имеют низкую степень обнажения поверхности обычно не будут подвержены изменениям. Метод гуманизации, альтернативный трансплантации CDR-участка и обратной мутации, представляет собой изменение поверхности, при котором неэкспонированные на поверхности остатки нечеловеческого происхождения, сохраняются, в то время как экспонированные на поверхности остатки изменяются в человеческие остатки.
В некоторых случаях может также быть предпочтительным изменение одного или более остатков аминокислот CDR-участков с целью повышения аффинности связывания с целевым эпитопом. Это известно как «созревание аффинности» и в некоторых случаях может выполняться в связи с гуманизацией, например, в ситуациях, когда гуманизация антитела приводит к снижению специфичности или аффинности связывания, и не представляется возможным в достаточной степени улучшить специфичность или аффинность связывания с помощью только обратных мутаций. Различные методы созревания аффинности известны в данной области техники, например, способ in vitro сканирующего насыщающего мутагенеза, описанный Burks et al., Proc Natl Acad Sci USA, 94:412-417 (1997), и способ пошагового in vitro созревания аффинности, предложенный Wu et al., Proc Natl Acad Sci USA 95:6037-6042 (1998).
Определение "выделенный" ("изолированный"), применяемое для описания различных антител по данному описанию, означает антитело, идентифицированное и выделенное и/или регенерированное из клетки или клеточной культуры, в которой оно экспрессируется. Примеси (загрязняющие компоненты) из природной среды представляют собой материалы, которые, как правило, мешают диагностическому или терапевтическому применению полипептида, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах изобретения антитело очищают (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности, при использовании секвенатора с вращающейся стеклянной чашечкой (секвенатора Эдмана), или (2) до гомогенности методом SDS-PAGE в невосстанавливающих или восстанавливающих условиях с применением окрашивания Кумасси бриллиантовым голубым или, предпочтительно, серебром. Выделенное антитело включает антитела in situ внутри рекомбинантных клеток, так как по меньшей мере один компонент природной среды полипептида отсутствует. Обычно выделенный полипептид получают в результате по меньшей мере одной стадии очистки.
Термин «эпитоп» при использовании в данном документе относится к части (детерминанте) антигена, который специфически связывается со связывающей молекулой (например, и антитело или родственная молекула, такие как биспецифичная связывающая молекула). Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или углеводы, или боковые цепи Сахаров, и, как правило, имеют специфические трехмерные структурные характеристики, а также специфические характеристики зарядов. Эпитоп может быть «линейным» или «конформационным.» В линейном эпитопе, все точки взаимодействия между белком (например, антиген) и взаимодействующей молекулой (такой как антитело) происходит линейно вдоль первичной аминокислотной последовательности белка. В конформационном эпитопе, точки взаимодействия происходят через аминокислотные остатки на белке, отделенные друг от друга в первичной аминокислотной последовательности. Когда желаемый эпитоп антигена определен, можно генерировать антитела к этому эпитопу с использованием методик, хорошо известных в данной области техники. Кроме того, генерация и характеристика антител или других связывающих молекул могут пролить свет на информацию о желательных эпитопах. Основываясь на этой информации, можно затем конкурентно скринировать связывающие молекулы для связывания с теми же или аналогичными эпитопами, например, путем проведения исследований конкуренции, чтобы найти связывающие молекулы, которые конкурируют за связывание с антигеном.
Можно определить, связывается ли антитело или другая связывающая молекула с тем же эпитопом или перекрестно конкурирует за связывание с Her2 и Her3 с биспецифичной связывающей молекулой по данному изобретению, используя хорошо известные в данной области техники методы. В одном варианте осуществления изобретения возможно связывание биспецифичной связывающей молекуле, предложенной в данном изобретении, с Her2 и Her3 в условиях насыщения с последующим измерением способности испытуемого антитела связываться с указанным антигеном-мишенью. Если испытуемое антитело способно связываться с антигеном-мишенью в одно время с референсной биспецифичной связывающей молекулой, то испытуемое антитело связывается с эпитопом, отличным от эпитопа референсной биспецифичной связывающей молекулы. Тем не менее, если испытуемое антитело не способно связываться с антигеном-мишенью одновременно, то испытуемое антитело связывается с тем же эпитопом, перекрывающимся эпитопом или эпитопом, который находится в непосредственной близости от эпитопа, связанного с биспецифичной связывающей молекулой. Этот эксперимент может быть выполнен с использованием ИФА, РИА, анализа межмолекулярных взаимодействий на BIACORE™, биослойной интерферометрии или проточной цитометрии. Для того, чтобы проверить, кросс-конкурирует ли биспецифичная связывающая молекула по данному изобретению с другой связывающей молекулой, можно использовать описанный выше способ конкурентного анализа в двух направлениях, то есть определить, блокирует ли известная связывающая молекула испытуемую связывающую молекулу, и наоборот. Такие эксперименты по перекрестной конкуренции могут быть выполнены, например, с помощью прибора IBIS МХ96 SPR или системы Octet™.
В одном варианте осуществления изобретения связывающая молекула, предложенная в данном изобретении, представляет собой моноклональное антитело. Используемое в данном документе сокращение «mAb» относится к моноклональному антителу, то есть антителу, которые синтезировано и выделено отдельной клональной популяцией клеток. Клональная популяция может быть клональной популяцией иммортализованных клеток. В некоторых вариантах осуществления изобретения иммортализованные клетки в клональной популяции являются гибридными клетками, гибридомами, которые обычно получают путем слияния отдельных В-лимфоцитов от иммунизированных животных с отдельными клетками лимфоцитарной опухоли. Гибридомы представляют собой тип сконструированных клеток и не встречаются в природе.
Класс (изотип) и подкласс антител может быть определен любым способом, известным в данной области техники. В целом, класс и подкласс антитела может быть определен с помощью антител, специфичных к конкретному классу и подклассу антител. Такие антитела коммерчески доступны. Класс и подкласс может быть определен с помощью ИФА, вестерн-блот анализа, а также другими методами. В другом варианте класс и подкласс может быть определен посредством секвенирования всех или части константных доменов тяжелой и/или легкой цепей антител, сравнения их аминокислотных последовательностей с известными аминокислотными последовательностями различных классов и подклассов иммуноглобулинов и определения класса и подкласса антител.
"Зависимая от антител опосредованная клетками цитотоксичность" и "ADCC" относятся к опосредованному клетками ответу, при котором неспецифичные цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), узнают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках суммирована в таблице 3 на странице 464 в публикации Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991). Чтобы оценить активность в ADCC представляющей интерес молекулы можно осуществить анализы ADCC in vitro, такие как анализы, описанные в патентах США №5500362 или 5821337. Применимые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и природные клетки-киллеры (NK). Альтернативно или дополнительно ADCC-активность представляющей интерес молекулы можно оценить in vivo, например, в животной модели, такой как модель, описанная в Clynes et al. PNAS (USA) 95: 652-656 (1998).
"Эффекторными клетками человека" являются лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Предпочтительно клетки экспрессируют, по меньшей мере, FcγRIII и осуществляют ADCC-эффекторную функцию. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (РВМС), природные клетки-киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы; при этом предпочтительны РВМС и NK-клетки. Эффекторные клетки могут быть выделены из их природного источника, например из крови или РВМС, как описано в настоящей публикации.
Термины "Fc-рецептор" и "FcR" используют для описания рецептора, который связывается с Fc-областью антитела. Предпочтительным FcR является FcR человека с нативной последовательностью. Кроме того, предпочтительным FcR является FcR, который связывает IgG-антитело (гамма-рецептор), и к предпочтительным рецепторам относятся рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсируемые формы указанных рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене основанный на тирозине мотив активации иммунорецептора (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене основанный на тирозине мотив ингибирования иммунорецептора (ITIM) (см. обзор в  , Annu. Rev. Immunol. 15: 203-234 (1997)). Обзор, посвященный FcR, представлен в Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995). Другие FcR, включая FcR, которые будут идентифицированы в будущем, включены в настоящем описании в термин "FcR". Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)).
, Annu. Rev. Immunol. 15: 203-234 (1997)). Обзор, посвященный FcR, представлен в Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995). Другие FcR, включая FcR, которые будут идентифицированы в будущем, включены в настоящем описании в термин "FcR". Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)).
"Комплемент-зависимая цитотоксичность" и "CDC" относятся к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе со своим антигеном. Чтобы оценить активацию комплемента можно осуществить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
"Нативные антитела" обычно являются гетеротетрамерными гликопротеидами с молекулярной массой примерно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, тогда как количество дисульфидных связей между тяжелыми цепями варьирует в разных изотипах иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет равномерно расположенные внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следует несколько константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на другом конце. Константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Полагают, что конкретные аминокислотные остатки образуют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи.
Термин «вектор» при использовании в настоящем документе означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она соединена. В некоторых вариантах осуществления изобретения вектор представляет собой плазмиду, т.е. кольцевую двухцепочечную часть ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. В некоторых вариантах осуществления изобретения вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. В некоторых вариантах осуществления изобретения векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). В других вариантах осуществления изобретения векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, и таким образом реплицируются вместе с геном хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально соединены. Такие векторы упоминаются в данном документе как «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы»).
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») при использовании в данном документе означает клетку, в которую введен рекомбинантный экспрессионный вектор. Настоящее изобретение относится к клеткам-хозяевам, которые могут включать, например, вектор в соответствии с настоящим изобретением, описанным выше. Настоящее изобретение относится также к клеткам-хозяевам, которые включают, например, нуклеотидную последовательность, кодирующую тяжелую цепь или ее антигенсвязывающие части, нуклеотидную последовательность, кодирующую легкую цепь или ее антигенсвязывающие части, или обе из них, полноразмерного антитела и/или нуклеотидную последовательность, кодирующую вариабельный домен тяжелой цепи VHH, биспецифичного антитела по данному изобретению. Следует понимать, что «рекомбинантная клетка-хозяин» и «клетка-хозяин» означают не только конкретную заявленную клетку, но также и потомство такой клетки. Поскольку модификации могут проходить в последующих поколениях вследствие мутации или воздействий окружающей среды, такое потомство не может, на самом деле, быть идентичным родительской клетке, но такие клетки по-прежнему включены в объем термина «клетка-хозяин» при использовании в настоящем документе.
Молекулы нуклеиновых кислот и векторы
Настоящее изобретение также относится к молекулам нуклеиновых кислот, и последовательностям, кодирующим биспецифичные связывающие молекулы по данному изобретению, описанным в данном документе. В некоторых вариантах осуществления изобретения различные молекулы нуклеиновых кислот кодируют первый домен и антитело или второй домен аминокислотной последовательности биспецифичной связывающей молекулы. Там, где первый домен представляет собой VHH, а второй домен содержит тяжелую цепь и/или легкую цепь, в некоторых вариантах осуществления изобретения различные нуклеиновые кислоты кодируют тяжелую цепь и аминокислотные последовательности легкой цепи. В других вариантах осуществления изобретения та же молекула нуклеиновой кислоты кодирует последовательность тяжелой цепи и аминокислот легкой цепи. В конкретной варианте осуществления изобретения молекула нуклеиновой кислоты может кодировать аминокислотную последовательность первого связывающего домена VHH и аминокислотную последовательность легкой цепи второго связывающего домена, необязательно включающей любую последовательность соединяющего их пептидного линкера.
Ссылка на нуклеотидную последовательность, охватывает его комплимент, если не указано иное. Таким образом, ссылка на нуклеиновую кислоту, имеющую определенную последовательность следует понимать как охватывающие ее комплементарную цепь с ее комплементарной последовательностью. Термин «полинуклеотид», упоминаемое в данном документе, означает полимерную форму нуклеотидов, по меньшей мере 10 оснований в длину, либо рибонуклеотидов, либо дезоксинуклеотидов или модифицированную форму любого типа нуклеотида. Термин включает в себя одно- и двухцепочечные формы.
Настоящее изобретение также относится к нуклеотидным последовательностям, которые по меньшей мере на 75%, 80%>, 85%), 90%>, 95%, 97%, 98% или 99% гомологичны или идентичны одной или нескольким из вышеупомянутых нуклеотидных последовательностей или нуклеотидной последовательности, кодирующей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-3. В определенных вариантах осуществления нуклеотидные последовательности по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологичны или идентичны нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO: 4. Настоящее изобретение также относится к нуклеотидным последовательностям, которые по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичны одной или нескольким из вышеупомянутых нуклеотидных последовательностей или нуклеотидной последовательности, кодирующей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5-6.
В одном из аспектов настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, выбранную из SEQ ID NO: 1-6. Молекула нуклеиновой кислоты может также содержать любую комбинацию указанных нуклеотидных последовательностей. В одном варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO: 3. В другом варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 1-3. В одном варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую SEQ ID NO: 4. В другом варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 4 и 5. В другом варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 4 и 6. В другом варианте осуществления изобретения молекула нуклеиновой кислоты содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 4, 5 и 6.
В любом из указанных выше вариантах осуществления изобретения молекулы нуклеиновых кислот могут быть выделенными.
В еще одном аспекте настоящее изобретение относится к вектору, подходящему для экспрессии любой из нуклеотидных последовательностей, описанных в настоящем документе. Термин «вектор» при использовании в настоящем документе означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она соединена. В некоторых вариантах осуществления изобретения вектор представляет собой плазмиду, т.е. кольцевую двухцепочечную часть ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. В некоторых вариантах осуществления изобретения вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. В некоторых вариантах осуществления изобретения векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). В других вариантах осуществления изобретения векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, и таким образом реплицируются вместе с геном хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально соединены. Такие векторы упоминаются в данном документе как «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы»).
Настоящее изобретение относится к векторам, содержащим молекулы нуклеиновых кислот, которые кодируют любую из аминокислотных последовательностей биспецифичных связывающих молекул или их частей (например, последовательностей тяжелой цепи первого и/или тяжелой и/или легкой цепи второго связывающих доменов) как описано в настоящем документе. Настоящее изобретение далее относится к векторам, содержащим молекулы нуклеиновых кислот, кодирующих слитые белки, модифицированные антитела, фрагменты антител.
Молекула нуклеиновой кислоты по данному изобретению может быть выделена из любого источника, который продуцирует биспецифичную связывающую молекулу или ее часть. В определенных вариантах осуществления изобретения молекула нуклеиновой кислоты данному изобретению может быть синтезирована, а не выделена.
В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты по данному изобретению может содержать нуклеотидную последовательность, кодирующую VHH первого домена и/или VH второго домена биспецифичной связывающей молекулы по данному изобретению, соединенного в рамках считывания с нуклеотидной последовательности, кодирующей константный домен тяжелой цепи из любого источника. Аналогичным образом, молекула нуклеиновой кислоты по данному изобретению может содержать нуклеотидную последовательность, кодирующую VL домен второй области биспецифичной связывающей молекулы по данному изобретению, объединенной в рамках считывания с нуклеотидной последовательностью, кодирующей константный домен легкой цепи из любого источника.
В еще одном аспекте настоящего изобретения молекулы нуклеиновой кислоты, кодирующие вариабельный домен тяжелой цепи (VHH) первого связывающего домена и/или вариабельный домен тяжелой (VH) и/или легкой цепи (VL) второго связывающего домена, могут «преобразовываться» по всей длине генов антитела. В одном варианте осуществления изобретения молекулы нуклеиновых кислот, кодирующие домены VHH и/или VH и/или VL преобразуются в гены антитела по всей длине путем вставки в экспрессионный вектор, уже кодирующей константные домены тяжелой цепи (СН) или легкой цепи (CL), соответственно, так что VHH/VH сегмент функционально соединен с СН сегментом(-ами) в векторе и/или VL сегмент оперативно соединен с CL сегментом в векторе. В другом варианте осуществления изобретения молекулы нуклеиновых кислот, кодирующие VHH/VH и/или VL домены преобразуются в гены по всей длине антитела путем соединения, например, лигирования, молекулы нуклеиновой кислоты, кодирующей VHH/VH и/или VL домены, к молекуле нуклеиновой кислоты, кодирующей СН и/или CL домены с использованием стандартных молекулярно-биологических методов. Молекулы нуклеиновых кислот, кодирующие по всей длине тяжелую и/или легкую цепи, могут затем экспрессироваться из клетки, в которую они были введены.
Молекулы нуклеиновой кислоты могут использоваться для экспрессии большого количества рекомбинантных биспецифичных связывающих молекул. Молекулы нуклеиновой кислоты могут использоваться для получения человеческих антител, гуманизированных антител, химерных антител, биспецифических антител, одноцепочечных антител, иммуноадгезинов, диател, мутировавших антител и производных антител, как описано в настоящем документе.
В другом варианте осуществления изобретения молекулы нуклеиновых кислот и векторы могут быть использованы для изготовления мутировавших биспецифичных связывающих молекул. Антитела могут мутировать в вариабельных доменах тяжелой цепи первого и/или тяжелой и/или легкой цепей второго связывающего домена, например, для изменения связывающей способности биспецифичной связывающей молекулы. Например, мутация может произойти в одном или нескольких CDR-участках, чтобы увеличить или уменьшить KD биспецифичной связывающей молекулы, чтобы увеличить или уменьшить koff или изменить специфичность связывания антитела в отношении HER2 и/или HER3. В другом варианте осуществления изобретения одной или более мутациям подвергнут аминокислотный остаток, который, как известно, изменен по сравнению с зародышевой линией в антителе, соответствующем первому или второму связывающему домену биспецифичной связывающей молекулы по данному изобретению. Эти мутации могут быть сделаны в CDR-участке или каркасном участке вариабельного домена, или в константном домене. В предпочтительном варианте осуществления изобретения мутации произведены в вариабельном домене. В другой варианте осуществления изобретения одной или нескольким мутациям подвергнут аминокислотный остаток, который, как известно, изменен по сравнению с зародышевой линией в CDR-участке или каркасном участке вариабельного домена биспецифичной связывающей молекулы по данному изобретению.
В другой варианте осуществления изобретения каркасный участок(-и) мутирует таким образом, что полученный каркасный участок(-и) имеет аминокислотную последовательность соответствующего гена зародышевой линии. Мутации могут осуществляться в каркасном участке или константном домене, чтобы увеличить период полувыведения биспецифичной связывающей молекулы. См., например, WO 00/09560. Мутация в каркасном участке или константном домене также может быть произведена для изменения иммуногенности биспецифичной связывающей молекулы и/или обеспечить участок для ковалентного или нековалентного связывания с другой молекулой. В соответствии с изобретением биспецифичная связывающая молекула может иметь мутации в любом одном или нескольких из CDR-участков или каркасных участков вариабельного домена или в константном домене.
В некоторых вариантах осуществления изобретения биспецифичные связывающие молекулы по данному изобретению экспрессируются путем вставки ДНК, кодирующей частично или полностью последовательность первого и второго связывающего домена (например, последовательности тяжелой/VHH или тяжелой и легкой цепи, где связывающий домен содержит последовательности тяжелой/VHH или тяжелой и легкой цепи), полученный, как описано выше, в экспрессионных векторах таким образом, что гены функционально соединены с необходимыми последовательностями, контролирующими экспрессию, такими как транскрипционные и трансляционные контрольные последовательности. Экспрессионные векторы включают плазмиды, ретровирусы, аденовирусы, адено-ассоциированные вирусы (AAV), вирусы растений, такие как вирус мозаики цветной капусты, вирусы табачной мозаики, космиды, YAC, EBV полученные эписомы и тому подобное. Молекулы ДНК могут быть лигированы в вектор таким образом, что последовательности, контролирующие транскрипцию и трансляцию в векторе, выполняют предусмотренную функцию регуляции транскрипции и трансляции ДНК. Экспрессионный вектор и последовательности контроля экспрессии могут быть выбраны таким образом, чтобы быть совместимыми с используемой экспрессирующей клеткой-хозяином. Молекулы ДНК, кодирующие частично или по всей длине последовательности первого и второго связывающих доменов (например, последовательности тяжелой и легкой цепи, где связывающий домен содержит последовательность тяжелой и легкой цепи) могут быть введены в отдельные векторы. В одном варианте осуществления изобретения любая комбинация указанных выше молекул ДНК вводится в тот же экспрессионный вектор. Молекулы ДНК могут быть введены в экспрессионный вектор стандартными способами (например, лигированием комплементарных сайтов рестрикции на фрагменте гена антитела и вектора или лигированием тупых концов, если сайты рестрикции отсутствуют).
Подходящим вектором является тот, который кодирует функционально законченные последовательности СН или CL человеческого иммуноглобулина с конструированием соответствующего места рестрикции так, что любая последовательность VHH и/или VH и/или VL может быть легко включена и экспрессирована, как описано выше. НС - и LC-кодирование генов в таких векторах может содержать интронные последовательности, что приводит к общему увеличению белковых продуктов антитела путем стабилизации соответствующей мРНК. Интронные последовательности находится в окружении сплайс-донора и сплайс-акцептора сайтов, которые определяют, где будет происходить сплайсинг РНК. Расположение интронных последовательностей может быть либо в вариабельных или константных участках цепей антитела, или как в вариабельных, так и константных участках, когда используются несколько интронов. Прекращение полиаденилирования и транскрипции может произойти вниз по ходу сайта нативной хромосомы кодируемых участков. Рекомбинантный экспрессионный вектор также может кодировать сигнальный пептид, который облегчает выработку цепочки антитела клеткой-хозяином. Ген цепочки антитела может быть клонирован в вектор таким образом, что сигнальный пептид соединен с рамкой считывания аминоконца цепи иммуноглобулина. Сигнальным пептидом может быть сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть, сигнальный пептид белка не иммуноглобулиновой природы).
Помимо цепочки генов антител, рекомбинантная экспрессия векторов по данному изобретению может нести регулирующие последовательности, которые контролируют экспрессию генов цепи антител в клетке-хозяине. Специалистам в этой области будет понятно, что дизайн экспрессионного вектора, включая выбор регулирующих последовательностей, может зависеть от таких факторов, как селекция клетки-хозяина для трансформации, уровень экспрессии желаемого белка, и т.д. Предпочтительные регулирующие последовательности для экспрессирующей клетки-хозяина млекопитающих включают вирусные элементы обеспечивающие высокий уровень экспрессии белков в клетках млекопитающих, таких как промотеры и/или энхансеры полученные из ретровирусной LTR, цитомегаловируса (CMV) (например, CMV промотера/энхансера), обезьяньего вируса 40 (SV40) (например, SV40 промотера/энхансера), аденовируса (например, аденовирус большого позднего промотора (AdMLP)), вирус полиомы, а также сильных промотеров млекопитающих, таких как нативный иммуноглобулин и актин промотеров. Для дальнейшего описания вирусных регулирующих элементов и их последовательностей см., например, патенты США 5,168,062, 4,510,245 and 4,968,615. Способы экспрессии связывающих молекул, таких как антитела растений, в том числе описание промотеров и векторов, а также трансформация растений, известны в данной области техники. См., например, патент США 6,517,529. Методы экспрессии полипептидов в бактериальных клетках или клетках грибов, например, дрожжевых клетках, также хорошо известны в данной области техники.
В дополнение к генам цепи антитела и регулирующим последовательностям, рекомбинантные векторы экспрессии изобретения могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селектируемого маркера. Ген селектируемого маркера облегчает селекцию клеток-хозяев, в которые был введен вектор (см., например, патенты США 4,399,216, 4,634,665 и 5,179,017). Например, обычно ген селектируемого маркера придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую вектор введен. Например, гены селектируемого маркера включают ген дигидрофолат редуктазы (DHFR) (для использования в dhfr-клетках-хозяевах при селекции/амплификации метотрексата), ген нео (для селекции G418) и ген синтетазы глутамата.
Термин «последовательность контроля экспрессии», используемый в данном описании, означает полинуклеотидные последовательности, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, к которым они лигированы. Контролирующие экспрессию последовательности включают соответствующие последовательности инициации транскрипции, терминации, промотора и энхансера; эффективные сигналы процессинга РНК, такие как сплайсинг и сигналы полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые повышают эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, которые повышают стабильность белка; и, при желании, последовательности, которые усиливают секрецию белка. Характер таких контролирующих последовательностей различается в зависимости от организма-хозяина; в прокариотах такие контролирующие последовательности, как правило, включают промотер рибосомального сайта связывания, а также последовательности терминации транскрипции; в эукариотах, как правило, такие контролирующие последовательности включают промотеры и последовательности терминации транскрипции. Термин «контролирующие последовательности» включает, как минимум, все компоненты, наличие которых имеет важное значение для экспрессии и процессинга, и может также включать дополнительные компоненты, чье присутствие является полезным, например, лидирующие последовательности и последовательности слившихся клеток.
Клетки-хозяева и способы получения биспецифичной связывающей молекулы
Дополнительный аспект настоящего изобретения относится к способам получения биспецифичных связывающих молекул по данному изобретению. Один вариант осуществления изобретения относится к способу получения биспецифичных связывающих молекул, как определено в настоящем документе, содержащему введение рекомбинантной клетки-хозяина, способной экспрессировать биспецифичную связывающую молекулу, культивацию указанной клетки-хозяина в условиях, подходящих для экспрессии биспецифичной связывающей молекулы, и выделение полученной биспецифичной связывающей молекулы. Биспецифичные связывающие молекулы, полученные такой экспрессией в таких рекомбинантных клетках-хозяевах упоминаются в данном документе как «рекомбинантные биспецифичные связывающие молекулы». Там, где биспецифичные связывающие молекулы являются антителами, они называются «рекомбинантными антителами». Изобретение также относится к потомству клеток таких клеток-хозяев и биспецифичных связывающих молекул получаемых аналогично.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») при использовании в данном документе означает клетку, в которую введен рекомбинантный экспрессионный вектор. Настоящее изобретение относится к клеткам-хозяевам, которые могут включать, например, вектор в соответствии с настоящим изобретением, описанным выше. Настоящее изобретение относится также к клеткам-хозяевам, которые включают, например, нуклеотидную последовательность, кодирующую тяжелую цепь или ее антигенсвязывающие части или VHH, нуклеотидную последовательность, кодирующую легкую цепь или ее антигенсвязывающие части, или обе из них, первого связывающего домена или второго связывающего домена или полноразмерного антитела, входящих в состав биспецифичной связывающей молекулы по данному изобретению. Следует понимать, что «рекомбинантная клетка-хозяин» и «клетка-хозяин» означают не только конкретную заявленную клетку, но также и потомство такой клетки. Поскольку модификации могут проходить в последующих поколениях вследствие мутации или воздействий окружающей среды, такое потомство не может, на самом деле, быть идентичным родительской клетке, но такие клетки по-прежнему включены в объем термина «клетка-хозяин» при использовании в настоящем документе.
Молекулы нуклеиновой кислоты, кодирующие биспецифичные связывающие молекулы по изобретению и векторы, содержащие эти молекулы нуклеиновой кислоты, могут быть использованы для трансфекции подходящего млекопитающего или его клетки, растения или его клетки, бактериальной или дрожжевой клетки-хозяина. Преобразование может происходить любым известным способом для введения полинуклеотидов в клетку-хозяина. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают декстран-опосредованную трансфекцию, осаждение фосфата кальция, полибрен-опосредованную трансфекцию, слияние протопластов, инкапсуляцию полинуклеотида(-ов) в липосомы и прямую микроинъекцию ДНК в ядра. В дополнение, молекулы нуклеиновых кислот могут быть введены в клетки млекопитающих вирусными векторами. Способы трансформации клеток хорошо известны в данной области техники. См., например, патенты США, 4,399,216, 4,912,040, 4,740,461 и 4,959,455. Способы трансформации клеток растений хорошо известны в данной области, включая, например, Agrobacterium-опосредованную трансформацию, биолистическую трансформацию, прямую инъекцию, электропорацию и вирусную трансформацию. Методы трансформации клеток бактерий и дрожжей также хорошо известны в данной области.
Клеточные линии млекопитающих используемые в качестве хозяев для трансформации, хорошо известны в данной области и включают множество иммортализованных доступных клеточных линий. К ним относятся, в частности, клетки яичников китайского хомячка (CHO), NS0 клетки, клетки SP2, HEK-293Т клетки, 293 Фристайл клетки (Invitrogen), NIH-3T3 клетки, клетки HeLa, клетки почек хомячка (BHK), клетки почек африканских зеленых мартышек (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), А549 клетки и ряд других клеточных линий. Клеточные линии особого предпочтения выбираются путем определения, какие клеточные линии имеют высокие уровни экспрессии. Другими клеточными линиями, которые могут быть использованы, являются клеточные линии насекомых, такие как Sf9 или Sf21 клетки. Когда векторы рекомбинантной экспрессии, кодирующих биспецифичные связывающие молекулы, вводятся в клетки-хозяева млекопитающих, связывающие молекулы продуцируются путем культивирования клеток-хозяев в течение времени, достаточного для экспрессии связывающей молекулы в клетках-хозяевах или, предпочтительнее, выделения связывающей молекулы в питательной среде, в которой выращиваются клетки-хозяева. Биспецифичные связывающие молекулы могут быть восстановлены из питательной среды с использованием стандартных методов очистки белка. Клетки-хозяева растений, например, включают Nicotiana, Arabidopsis, ряску, кукурузу, пшеницу, картофель и т.д. Клетки бактерий хозяина включают виды Е. coli и Streptomyces. Дрожжевые клетки-хозяева включают Schizosaccharomyces pombe, Saccharomyces cerevisiae и Pichia pastoris.
Кроме того, экспрессию биспецифичных связывающих молекул по данному изобретению из продуцирующей клеточной линии можно усилить с помощью ряда известных методов. Например, система экспрессии гена глутамин синтетазы (система GS) является достаточно распространенной для усиления экспрессии при определенных условиях. Система GS обсуждается в целом или частично в связи с патентами ЕР 0216846, 0256055, 0323997 и 0338841.
Вполне вероятно, что биспецифичные связывающие молекулы различных клеточных линий или трансгенные животные будут отличаться друг от друга профилем гликозилирования. Однако все биспецифичные связывающие молекулы, кодируемыми молекулами нуклеиновой кислоты, описанными в данном документе, или содержащими аминокислотные последовательности, приведенные в настоящем документе, являются частью данного изобретения, независимо от состояния гликозилирования связывающих молекул и в целом, независимо от наличия или отсутствия посттрансляционных модификаций.
В некоторых вариантах осуществления изобретения связывающие домены биспецифичной связывающей молекулы могут быть спарены вместе посредством инновационной молекулой линкера, предназначенной для защиты от протеолитической деградации связывающей молекулы. Такой линкер обычно лишен сайта протеолитического расщепления.
Способ получения
Способы получения биспецифичных антител известны в данной области. Традиционное получение полноразмерных биспецифичных антител основано на коэкспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина, при этом две цепи имеют разные специфичности (Millstein et al., Nature, 305: 537-539 (1983)). Вследствие случайного выбора тяжелых и легких цепей иммуноглобулина такие гибридомы (квадромы) продуцируют возможную смесь 10 разных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистка правильной молекулы, которую обычно осуществляют, используя стадии аффинной хроматографии, является довольно громоздкой, а выходы продукта низкими. Сходные способы описаны в WO 93/08829 и в Traunecker et al., EMBO J., 10: 3655-3659 (1991).
Согласно другому способу вариабельные домены антител с требуемыми специфичностями связывания (участки связывания антитело-антиген) сливают с последовательностями константных доменов иммуноглобулина. Слияние предпочтительно осуществляют с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнирной области, областей СН2 и СНЗ. Предпочтительно иметь первую константную область тяжелой цепи (СH1), содержащую сайт, необходимый для связывания легкой цепи, присутствующий, по меньшей мере, в одном из слияний. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и в случае необходимости легкой цепи иммуноглобулина, встраивают в отдельные экспрессирующие векторы и котрансфицируют в подходящий организм хозяина. Это дает большую гибкость при корректировке взаимных соотношений трех полипептидных фрагментов в таких вариантах, когда неравные соотношения трех полипептидных цепей, используемых в конструкции, дают оптимальные выходы. Однако можно встраивать кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессирующий вектор в том случае, когда экспрессия, по меньшей мере, двух полипептидных цепей в равных соотношениях дает высокие выходы или когда соотношения не имеют особого значения.
В предпочтительном варианте указанного способа вариабельные домены тяжелой цепи VHH, который специфично связывается с HER3, сливают с полноразмерным антителом или его антигенсвязывающим доменом, которые специфично связываются с HER2.
В предпочтительном варианте указанного способа биспецифичные антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном плече, например, VHH, и гибридной пары тяжелой цепи-легкой цепи иммуноглобулина (обеспечивающей вторую специфичность связывания) в другом плече. Обнаружено, что такая асимметричная структура облегчает отделение требуемого биспецифичного соединения от нежелательных сочетаний цепей иммуноглобулина, так как наличие легкой цепи иммуноглобулина только в одной половине биспецифичной молекулы обеспечивает простой способ разделения. Такой способ описан в WO 94/04690. Более подробное описание создания биспецифичных антител см., например, в Suresh et al., Methods in Enzymology, 121: 210 (1986).
Биспецифичные антитела включают поперечно сшитые антитела или "гетероконъюгаты" антител. Например, одно из антител в гетероконъюгате может быть связано с авидином, другое с биотином. Такие антитела, например, были предложены для направления клеток иммунной системы к нежелательным клеткам (патент США №4676980) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и ЕР 03089). Гетероконъюгированные антитела могут быть получены с использованием любого удобного способа образования поперечных сшивок. Подходящие поперечно сшивающие агенты хорошо известны в данной области и описаны в патенте США №4676980 наряду с описанием ряда способов образования поперечных сшивок.
Способы создания биспецифичных антител из фрагментов антител также описаны в литературе. Например, биспецифичные антитела могут быть получены с использованием химического связывания. Brennan с соавторами (Science, 229: 81 (1985)) описывают способ, при котором интактные антитела протеолитически расщепляют, чтобы создать F(ab')2-фрагменты. Полученные фрагменты восстанавливают в присутствии агента, образующего комплексы дитиола, арсенита натрия, чтобы стабилизировать соседние дитиолы и предотвратить образование межмолекулярного дисульфида. Затем созданные Fab'-фрагменты превращают в производные тионитробензоата (TNB). Один из Fab'-TNB-производных затем снова превращают в Fab'-тиол восстановлением с использованием меркаптоэтиламина и смешивают с эквимолярным количеством другого Fab'-TNB-производного с образованием биспецифичного антитела. Полученные биспецифичные антитела можно использовать в качестве агентов для избирательной иммобилизации ферментов.
Недавние достижения облегчили прямое извлечение Fab'-SH-фрагментов из Е. coli, которые могут быть химически связаны с образованием биспецифичных антител. В публикации Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описано получение полностью гуманизированной молекулы биспецифичного антитела на основе F(ab')2. Каждый Fab'-фрагмент отдельно секретировался из клеток Е. coli, и его подвергали химическому связыванию in vitro с образованием биспецифичного антитела. Образованное таким образом биспецифичное антитело способно связываться с клетками, сверхэкспрессирующими рецептор HER2 и/или HER3, и нормальными Т-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против опухолевых мишеней молочной железы человека.
Предполагаются антитела с двумя валентностями, а также с более чем двумя валентностями. Например, могут быть получены триспецифичные антитела. Tutt et al. J. Immunol. 147: 60 (1991), а также четырехспецифичные антитела.
Фармацевтические композиции
Другим аспектом изобретения является фармацевтическая композиция, содержащая в качестве активного ингредиента (или в качестве единственного активного ингредиента) биспецифичную связывающую молекулу по данному изобретению, которая специфична к HER2 и HER3. Фармацевтическая композиция может включать любую из биспецифичных связывающих молекул, как описано в данном документе. В некоторых вариантах осуществления изобретения композиции предназначены для улучшения, профилактики или лечения нарушений, которые могут быть связаны с активностью HER2 и/или HER3.
Как правило, биспецифичные связывающие молекулы по данному изобретению пригодны для применения в виде лекарственных форм в сочетании с одним или более фармацевтически приемлемым эксципиентом, например, как описано далее.
Фармацевтические композиции по данному изобретению могут содержать по меньшей мере одну биспецифичную связывающую молекулу и одну или более дополнительных связывающих молекул (например, антител), которые нацелены на один или более соответствующих поверхностных рецепторов.
Фармацевтическая композиция является «стерильной», если она асептична, т.е. свободная от микроорганизмов и их спор.
Фармацевтическая композиция является «стабильной» если активный агент сохраняет свою физическую стабильность и/или химическую стабильность и/или биологическую активность в течении всего срока годности при температуре хранения, например, при температуре 2-8°С. Предпочтительно, чтобы активные агент сохранял и физическую и химическую стабильность, а также биологическую активность. Период хранения выбирается на основании результатов исследования стабильность при ускоренном и естественном хранении.
Термин «эксципиент» используется в данном документе для описания любого ингредиента, отличающегося от соединения(-ий) по данному изобретению. Выбор инертного эксципиента будет в значительной степени зависеть от таких факторов, как конкретный способ введения, действие эксципиента на растворимость и стабильность и характер лекарственной формы. При использовании в данном документе «фармацевтически приемлемый эксципиент» включает все без исключения растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и задерживающие абсорбцию агенты и подобные физиологически совместимые вещества. Некоторыми примерами фармацевтически приемлемых эксципиентов являются вода, физиологический раствор, фосфатный буфер, декстроза, глицерин, этанол и т.п., а также их комбинации. Во многих случаях будет предпочтительным включить в композицию изотонические агенты, например, сахара, полиспирты, такие как маннитол, сорбит, или хлорид натрия. Дополнительными примерами фармацевтически приемлемых веществ являются увлажняющие агенты или небольшое количество вспомогательных веществ, таких как увлажняющие или эмульгирующие вещества, консерванты или буферы, которые увеличивают продолжительность хранения или эффективность антитела.
Под «буферным агентом» понимается раствор способный сохранять значение рН благодаря взаимодействию кислотных и щелочных компонентов, входящих в его состав. В общем случае, преимущественными являются значения рН фармацевтической композиции от 4,5 до 7,0. В качестве буферных агентов могут быть использованы, например, ацетатный, фосфатный, цитратный, гистидиновый, сукцинатный и т.п. буферные растворы, но, не ограничиваясь ими.
Под «изотоническими агентами» понимается вспомогательное вещество или смесь двух и более вспомогательных веществ, которые обеспечивают изотоническое осмотическое давление раствора. «Изотоническим» считается раствор создающий осмотическое давление от примерно 250 до 350 мОсм/кг. В качестве изотоноческих агентов могут быть использованы полиолы, моно- и дисахара, аминокислоты, соли металлов, например, хлорид натрия, и т.п., но, не ограничиваясь ими.
Под «стабилизатором» понимается вспомогательное вещество или смесь двух и более вспомогательных веществ, которые обеспечивают физическую и/или химическую стабильность активного агента. В качестве стабилизаторов могут быть использованы аминокислоты, например, аргинин, гистидин, глицин, лизин, глутамин, пролин, но, не ограничиваясь ими; поверхностно-активные вещества, например, полисорбат 20 (торговое наименование Tween 20), полисорбат 80 (торговое наименование Tween 80), полиэтилен-полипропилен гликоль и его кополимеры (торговые наименования Полоксамер (Poloxaner), Плуроник (Pluronic)), натрия додецилсульфат (SDS), но, не ограничиваясь ими; антиоксиданты, например, метионин, ацетилцистеин, аскорбиновая кислота, монотиоглицерол, соли серистых кислот, и т.п., но не ограничиваясь ими; хелатирующие агенты, например, ЭДТА, ДТПА, цитрат натрия и т.п., но не ограничиваясь ими.
Фармацевтические композиции по настоящему изобретению и способы их изготовления будут бесспорно очевидными для специалистов в этой области. Такие композиции и способы их изготовления можно найти, например, в Remington, The Science and Practice of Pharmacy, 21th Edition, Troy, Beringer., Lippincott Williams and Wilkins., Philadelphia, PA 2006. Производство фармацевтических композиций предпочтительно должно соответствовать требованиям GMP (надлежащей производственной практики).
Фармацевтическая композиция по данному изобретению может изготавливаться, упаковываться или широко продаваться, в виде единичной стандартной дозы или множества единичных стандартных доз. Используемый в данном документе термин «единичная стандартная доза» означает дискретное количество фармацевтической композиции, содержащего заранее определенное количество активного ингредиента. Количество активного ингредиента обычно равно дозировке активного ингредиента, который будет вводиться субъекту, или удобной части такой дозировки, например, половине или трети такой дозировки.
Любой способ введения пептидов, белков или антител, принятый в данной области, может соответствующим образом использоваться для биспецифичной связывающей молекулы по данному изобретению.
Фармацевтические композиции по настоящему изобретению, как правило, пригодны для парентерального введения. Используемый в данном документе термин «парентеральное введение» фармацевтической композиции включает любой способ введения, для которого характерно физическое нарушение целостности ткани субъекта и введение фармацевтической композиции через нарушение в ткани, что обычно приводит к прямому попаданию в кровоток, в мышцу или во внутренний орган. Таким образом, парентеральное введение включает, помимо прочего, введение фармацевтической композиции путем инъекции композиции посредством введения композиции через хирургический разрез, путем нанесения композиции с помощью проникающей в ткани нехирургической раны и т.п. В частности, предполагается, что парентеральное введение включает, помимо прочего, подкожную, внутрибрюшинную, внутримышечную, внутривенную, внутриартериальную, интратекальную, внутрижелудочковую, интрауретральную, внутричерепную, внутрисуставнную инъекцию или инфузии; и почечные диализные инфузионные методики. Внутриопухолевая доставка, например, внутриопухолевая инъекция, также может оказаться полезной. Также предусмотрена региональная перфузия. Предпочтительные варианты осуществления изобретения включают внутривенный и подкожный пути.
Лекарственные формы фармацевтических композиций, подходящие для парентерального введения, обычно содержат активный ингредиент в сочетании с фармацевтически приемлемым носителем/эксципиентом, например, стерильной водой или стерильным изотоническим раствором. Такие лекарственные формы могут быть изготовлены, упакованы или проданы в форме, подходящей для болюсного введения или для непрерывного введения. Инъекционные лекарственные формы могут быть изготовлены, упакованы или проданы в стандартной лекарственной форме, например, в ампулах, или в многодозовых контейнерах, содержащих консервант. Лекарственные формы для парентерального введения включают, помимо прочего, суспензии, растворы, эмульсии в масляных или водных основах, пасты и тому подобное. Такие лекарственные формы могут также содержать один или более дополнительных ингредиентов, включая, помимо прочего, суспендирующие, стабилизирующие или диспергирующие агенты. В одном варианте осуществления изобретения композиции для парентерального введения активный ингредиент предоставляется в сухой форме (то есть, порошок или гранулы) для растворения с подходящей основой (например, стерильная апирогенная вода) до парентерального введения восстановленного состава. Парентеральные лекарственные формы и также включают водные растворы, которые могут содержать наполнители, такие как соли, углеводы и буферные агенты (предпочтительно рН от 3 до 9, еще более предпочтительно рН от 4,5 до 7), но для некоторых видов применения более подходящей лекарственной формой может являться стерильный неводный раствор или сухая форма для использования в сочетании с подходящей основой, такой как стерильная апирогенная вода. Примером формы для парентерального введения являются растворы или взвеси в стерильных водных растворах, например, водные растворы пропиленгликоля или декстрозы. Такие лекарственные формы при необходимости могут быть буферными. Другие подходящие лекарственные формы для парентерального введения могут включать те, которые содержат активный ингредиент в микрокристаллической форме или в липосомальном препарате. Лекарственные формы для парентерального введения могут быть выполнены для немедленного и/или модифицированного высвобождения. Лекарственные формы с модифицированным высвобождением включают отсроченное, замедленное, пульсирующее, контролируемое, нацеленное и программируемое высвобождение.
Например, в одном из аспектов стерильные растворы для инъекций могут быть получены путем включения биспецифичной связывающей молекулы в необходимом количестве в соответствующий растворитель с одним или комбинацией ингредиентов, перечисленных выше, по мере необходимости, с последующей фильтрационной стерилизацией. Обычно дисперсии получают введением активного соединения в стерильный растворитель, который содержит основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов, способы получения представляют собой сушку вымораживанием (лиофилизацию), которая дает порошок активного ингредиента плюс любой дополнительный желательный ингредиент из его предварительно стерилизованного фильтрацией раствора. Надлежащая текучесть раствора может поддерживаться, например, применением материалов покрытия, таких как лецитин, поддержанием необходимого размера частиц в случае дисперсий и применением поверхностно-активных веществ. Пролонгированное всасывание инъекционных композиций может быть осуществлено путем включения в композицию агента, который задерживает абсорбцию, например, моностеараты и желатин, и/или путем покрытий с модифицированным высвобождением (например, покрытий с медленным высвобождением).
Биспецифичные связывающие молекулы по данному изобретению могут также вводиться интраназально или ингаляционно, обычно в форме сухого порошка (самостоятельно, в виде смеси или в виде частиц со смешанными компонентами, например, смешанными с подходящим фармацевтически приемлемым наполнителем) из ингалятора с сухим порошком, такого как аэрозольный контейнер под давлением, помпа, спрей, распылитель (предпочтительно распылитель, в котором использован принцип электрогидродинамики для получения мелкодисперсного тумана) или небулайзер, в котором используется или не используется подходящий пропеллент, или в виде назальных капель.
Контейнер под давлением, помпа, спрей, распылитель или небулайзер обычно содержит раствор или суспензию связывающей молекулы по данному изобретению, включая, например, подходящее вещество для диспергирования, растворения или продления высвобождения активного вещества, пропеллент в качестве растворителя.
До использования в виде сухого порошка или суспензии, лекарственный препарат обычно микронизируют до размера подходящего для доставки путем ингаляции (обычно менее 5 микрон). Этого можно достичь любым подходящим способом измельчения, таким как размол на спиральной струйной мельнице, размол на струйной мельнице с кипящим слоем, сверхкритическая очистка жидкости для формирования наночастиц, гомогенизация высоким давлением или распылительная сушка.
Капсулы, блистеры и картриджи для использования в ингаляторе или инсуфляторе могут выполнены так, чтобы содержать порошковую смесь соединения по данному изобретению, подходящей порошковой основы и модификатор активности.
Подходящая формула раствора для использования в распылителе, в котором использован принцип электрогидродинамики для получения мелкодисперсного тумана, может содержать подходящую дозу биспецифичной связывающей молекулы по данному изобретению на одно нажатие, и объем на нажатие может варьироваться, например, от 1 мкл до 200 мкл, более предпочтительно от 1 мкл до 100 мкл.
В лекарственные формы по данному изобретения, предназначенные для ингаляции/интраназального введения, могут быть добавлены подходящие ароматизаторы, такие как ментол и левоментол, или подсластители, такие как сахарин или натрия сахарин.
Лекарственные формы для парентерального введения могут быть выполнены для немедленного и/или модифицированного высвобождения. Лекарственные формы с модифицированным высвобождением включают отсроченное, замедленное, пульсирующее, контролируемое, нацеленное и программируемое высвобождение.
В случае порошковых ингаляторов и аэрозолей, единица дозирования устанавливается посредством клапана, который доставляет отмеренное количество. Единицы в соответствии с данным изобретением обычно устанавливают для введения отмеренной дозы или «впрыска» связывающей молекулы по данному изобретению. Общая суточная доза будет обычно вводиться в виде однократной дозы или еще чаще - в виде разделенных доз в течение суток.
Биспецифичные молекулы по данному изобретению также могут быть выполнены в лекарственной форме для перорального введения. Пероральное введение может включать глотание, так что соединение поступает в желудочно-кишечный тракт и/или буккально, лингвально или сублингвально поступает в кровоток непосредственно из полости рта.
Лекарственные формы, пригодные для перорального введения, включают твердые, полутвердые и жидкие системы, такие как таблетки; мягкие или твердые капсулы, содержащие мульти- или наночастицы, жидкости или порошки; пастилки (включая заполненные жидкостью); жевательные формы; гели; быстро растворимые лекарственные формы; пленки; суппозитории; спреи; и щечные/мукоадгезивные пластыри.
Жидкие лекарственные формы включают суспензии, растворы, сиропы и эликсиры. Такие лекарственные формы могут быть использованы как наполнители в мягких или жестких капсулах (например, из желатина или гидроксипропилметилцеллюлозы) и обычно содержат носитель, например, воду, этанол, полиэтиленгликоль, пропиленгликоль, метилцеллюлозу или подходящее масло и один или более эмульгаторов и/или суспендирующих агентов. Жидкие лекарственные формы могут быть также изготовлены путем восстановления твердого вещества, например, из саше.
В одном варианте осуществления изобретения биспецифичная связывающая молекула по данному изобретению является частью композиции, содержащей гистидин и ацетат в концентрации от 1 до 100 мМ в буфере с рН от 5,0 до 6,5.
Терапевтическое применение биспецифичной связывающей молекулы по данному изобретению
В одном аспекте биспецифичная связывающая молекула по данному изобретению применяется в лечении нарушений, которые связаны с активностью HER2 и HER3.
Термин "нарушение" означает любое состояние, которое можно улучшить в результате лечения по настоящему изобретению. В определение данного термина входят хронические и острые нарушения или заболевания, включающие в себя патологические состояния, которые вызывают предрасположенность млекопитающего к возникновению данного нарушения. Неограничивающие примеры подлежащих лечению заболеваний включают в себя доброкачественные и злокачественные опухоли; лейкозы и лимфоидные злокачественные новообразования, в частности рак молочной железы, яичника, желудка, эндометрия, слюнной железы, легкого, почки, ободочной кишки, щитовидной железы, поджелудочной железы, предстательной железы или мочевого пузыря; нейронные, глиальные, астроцитальные, гипоталамусные и другие гландулярные, макрофаговые, эпителиальные, стромальные и бластоцельные нарушения; воспалительные, ангиогенные и иммунологические нарушения. Предпочтительным подлежащим лечению нарушением согласно изобретению является злокачественная опухоль.
Термины "рак" или "раковый" относятся к физиологическому состоянию или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым(/-ой) ростом/пролиферацией клеток. Это определение охватывает доброкачественные и злокачественные раковые заболевания. Примеры раковых заболеваний включают, но без ограничения, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры таких раковых заболеваний включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, печеночноклеточный рак, рак желудка, включая рак ЖКТ, рак поджелудочной железы, глиобластому, глиому, цервикальный рак, рак яичника, рак печени, рак мочевого пузыря, рак молочной железы, рак толстой кишки, колоректальный рак, карциному эндометрия и матки, карциному слюнных желез, рак почки (почечноклеточную карциному), рак предстательной железы (рак простаты), рак наружных женских половых органов, рак щитовидной железы, печеночную карциному, карциному анального канала, карциному пениса, меланому и различные типы рака головы и шеи.
«Лечить», «лечение» и «терапия» относятся к методу смягчения или устранения биологического расстройства и/или по меньшей мере одного из сопутствующих ему симптомов. Используемый в данном документе, чтобы «облегчить» болезнь, заболевание или состояние, означает уменьшение тяжести и/или частоты возникновения симптомов заболевания, расстройства или состояния. Кроме того, содержащиеся в данном документе ссылки в на «лечение» включает ссылки на лечебную, паллиативную и профилактическую терапию.
Под выражением "снижает или ингибирует" понимают способность вызывать общее снижение, предпочтительно, на 20% или более, более предпочтительно, на 50% или более и, наиболее предпочтительно, на 75%, 85%, 90%), 95% или более. "Снижает или ингибирует" может относиться к наличию или размеру метастазов, размеру первичной опухоли или к размеру или числу кровеносных сосудов при ангиогенных расстройствах.
В одном аспекте субъект лечения или пациент является млекопитающим, предпочтительно человеческим субъектом. Вышеупомянутый субъект может быть мужского или женского пола и любого возраста.
Термин "терапевтически эффективное количество" относится к количеству антитела или фрагмента антитела для лечения заболевания или расстройства у субъекта. В случае опухоли (например, раковой опухоли) терапевтически эффективное количество антитела или фрагмента антитела (например, антитела или фрагмента антитела, которое специфически связывает HER2 и HER3) может уменьшать число раковых клеток; уменьшать начальный размер опухоли; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) инфильтрацию раковыми клетками периферических органов; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) метастазирование опухоли; ингибировать, до некоторой степени, рост опухоли; и/или облегчать, до некоторой степени, один или более симптомов, обусловленных расстройством. Антитело или фрагмент антитела может, до некоторой степени, предупреждать рост и/или убивать имеющиеся раковые клетки, оно может вызывать цитостатический и/или цитотоксический эффект. При терапии рака эффективность in vivo можно определять, например, оценивая продолжительность жизни, время до прогрессирования заболевания (ТТР), частоту ответа опухоли на лечение (RR), продолжительность ответа и/или качество жизни.
Используемые в данном документе термины «совместное назначение», «совместно назначенный» и «в сочетании с» относящиеся к биспецифичным связывающим молекулам с одним или более другими терапевтическими агентами, как предполагается, означают, сслылаются или включают:
1) одновременное введение такой комбинации биспецифичной связывающей молекулы по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы вместе в одной лекарственной форме, из которой указанные компоненты высвобождаются практически одновременно указанному пациенту,
2) одновременное введение такой комбинации биспецифичной связывающей молекулы по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы отдельно в разных лекарственных формах, введение которых происходит практически в одно и то же время указанному пациенту, после чего указанные компоненты высвобождаются практически одновременно указанному пациенту,
3) последовательное введение такой комбинации биспецифичной связывающей молекулы по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы отдельно друг от друга в отдельных лекарственных формах, которые принимаются в последовательно по времени указанным пациентом со значимым временным интервалом между каждым введением, после чего указанные компоненты высвобождаются в практически разное время указанному пациенту; а также
4) последовательное введение такой комбинации биспецифичной связывающей молекулы по данному изобретению и терапевтического агента пациенту, который нуждается в лечении, когда такие компоненты сформулированы вместе в единый лекарственной форме, из которой высвобождение указанных компонентов происходит контролируемым образом, после чего они одновременно, последовательно или совместно высвобождаются в одно и то же время и/или разное время указанному пациенту, где каждая часть может быть введена одним или разными путями.
Биспецифичные связывающие молекулы по данному изобретению могут назначаться без дополнительного терапевтического лечения, т.е. в качестве самостоятельной терапии. Кроме того, лечение биспецифичными связывающими молекулами по данному изобретению может включать по меньшей мере одно дополнительное терапевтическое лечение (комбинированная терапия). В некоторых вариантах осуществления изобретения биспецифичная связывающая молекула может вводиться совместно или быть сформулирована с другим медикаментом/препаратом для лечения рака.
Термин "цитотоксическое средство" в используемом в настоящем описании смысле относится к веществу, которое ингибирует или предотвращает функционирование клеток и/или вызывает разрушение клеток. Подразумевается, что термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические средства и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины, имеющие происхождение из бактерий, грибов, растений или животных, включая их фрагменты и/или варианты.
"Химиотерапевтическим средством" является химическое соединение, применимое для лечения злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN®); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбохинон, метуредопа и уредопа; этиленимины и метилмеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилмеламин; ацетогенины (в частности, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включая синтетический аналог топотекан (HYCAMTIN®), СРТ-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности калихеамицин гамма II и калихеамицин омега II (см., например, Agnew, Chem. Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин А; эсперамицин; а также хромофор неокарциностатина и родственные хромофоры хромопротеинов энедииновые антибиотики, аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая ADRIAMICIN®, морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин, доксорубицино HCl в инъецируемых липосомах (DOXOL®), липосомный доксорубицин TLC D-99 (MYOCET®), пегилированный липосомный доксорубицин (CAELYX®) и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (GEMZAR®), тегафур (UFTORAL®), капецитабин (XELODA®), эпотилон и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; средства, подавляющие функции надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазихон; элфорнитин; ацетат эллиптиния; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (в частности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("ara-С"); тиотепа; таксоид, например паклитаксел (TAXOL®), препарат паклитаксела на основе сконструированных связанных с альбумином наночастиц (ABRAXANETM) и доцетаксел (TAXOTERE®); хлорамбуцил; 6-тиогуанин; меркаптопурин; метотрексат; средства на основе платины, такие как цисплатин, оксалиплатин и карбоплатин; алкалоиды барвинка, которые предотвращают полимеризацию тубулина из образующихся микротрубочек, включая винбластин (VELBAN®), винкристин (ONCOVIN®) виндезин (ELDISINE®, FILDESIN®) и винорелбин (NAVELBINE®); этопозид (VP-16); ифосфамид; митоксантрон; лейковорин; новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота, включая бексаротен (TARGRETIN®); бифосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат (DIDROCAL®), NE-58095, золедроновую кислоту/золедронат (ZOMETA®), алендронат (FOSAMAX®), памидронат (AREDIA®), тилудронат (SKELID®) или ризендронат (ACTONEL®); троксацитабин (1,3-диоксолановый нуклеозидный аналог цитозина); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, которые ингибируют экспрессию генов в путях передачи сигналов, вовлеченных в пролиферацию аберрантных клеток, таких как, например, PKC-альфа, Raf, H-Ras и рецептора эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 (например, LURTOTECAN®); rmRH (например, ABARELIX®); BAY439006 (сорафениб; Bayer); SU-11248 (Pfizer); перифосин, ингибитор ЦОГ-2 (например, целекоксиб или эторикоксиб), ингибитор протеосом (например, PS341); бортезомиб (VELCADE®); CCI-779; типифарниб (811577); орафениб, АВТ510; ингибитор Всl-2, такой как облимерсен натрия (GENASENSE®); пиксантрон; ингибиторы EGFR (см. определение ниже); ингибиторы тирозинкиназ (см. определение ниже); и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше средств; а также сочетания двух или более указанных выше средств, такие как CHOP, сокращенное название комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращенное название схемы лечения оксалиплатином (ELOXATINTM) в сочетании с 5-FU и лейковорином.
Также в указанное определение включены противогормональные средства, которые действуют, регулируя или ингибируя действие гормонов на опухоли, такие как антиэстрогены со смешанным профилем агонист/антагонист, включая тамоксифен (NOLVADEX®), 4-гидрокситамоксифен, триоксифен, торемифен (FARESTON®); идоксифен, дролоксифен, ралоксифен (EVISTA®), триоксифен, кеоксифен и избирательные модуляторы рецепторов эстрогена (SERM), такие как SERM3; чистые антиэстрогены без агонистических свойств, такие как фулвестрант (FASLODEX®) и ЕМ800 (такие средства могут блокировать димеризацию рецепторов эстрогена (ER), ингибировать связывание ДНК, усиливать метаболизм ER и/или снижать уровни ER); ингибиторы ароматазы, включая стероидные ингибиторы ароматазы, такие как форместан и эксеместан (AROMASIN®), и нестероидные ингибиторы ароматазы, такие как анастразол (ARIMIDEX®), летрозол (FEMARA®) и аминоглутетимид и другие ингибиторы ароматазы, включая ворозол (RIVISOR®), ацетат мегестрола (MEGASE®), фадрозол, имидазол; агонисты рилизинг-гормона лютеинизирующего гормона, включая лейпролид (LUPRON® и ELIGARD®), гозерелин, бузерелин и триптерелин; половые стероиды, включая прогестины, такие как ацетат мегестрола и ацетат медроксипрогестерона, эстрогены, такие как диэтилстилбестрол и премарин и андрогены/ретиноиды, такие как флуоксиместерон, полностью трансретиноевая кислота и фенретинид; онапристон; антипрогестероны; понижающие регуляторы рецепторов эстрогенов (ERD); антиандрогены, такие как флутамид, нилутамид и бикалутамид; тестолактон; и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше средств; а также сочетания двух или более указанных выше средств.
Подразумевается, что биспецифичные связывающие молекулы по данному изобретению могут использоваться в способах лечения, как описано выше, могут использоваться в лечении, как описано выше, и/или могут использоваться в производстве медикаментов для лечения, как описано выше.
Дозы и пути введения
Биспецифичные связывающие молекулы по данному изобретению будут вводиться в количестве, эффективном для лечения состояния, о котором идет речь, т.е. в дозах и в течение периодов времени, необходимых для достижения желаемого результата. Терапевтически эффективное количество может изменяться в зависимости от таких факторов, как конкретное состояние, по поводу которого проводится лечение, возраста, пола и веса пациента, а также является ли введение биспецифичных связывающих молекул самостоятельным лечением или оно проводится в комбинации с одним или более дополнительных анти-аутоиммунных или противовоспалительных методов лечения.
Схемы приема лекарственных средств можно регулировать, чтобы обеспечить оптимальный желаемый ответ. Например, может быть введен один болюс, несколько разделенных доз могут быть введены в течение некоторого времени, или доза могут быть пропорционально уменьшена или увеличена в зависимости от остроты терапевтической ситуации. Особенно полезным является изготовление парентеральных композиций в стандартной лекарственной форме для простоты введения и однородности дозирования. Стандартная лекарственная форма при использовании в данном документе, относится к физически дискретным единицам, пригодным в качестве единичных доз для пациентов/субъектов, подлежащих лечению; каждая единица содержит заданное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта в сочетании с требуемым фармацевтическим носителем. Спецификация для стандартных лекарственных форм по настоящему изобретению, как правило, диктуется и непосредственно зависит от (а) уникальных характеристик химиотерапевтического агента и конкретного терапевтического или профилактического эффекта, которые должны быть достигнуты, и (b) ограничений, присущих в технике компаундирования такого активного соединения для лечения чувствительности у субъектов.
Таким образом, квалифицированным специалистам понятно, исходя из раскрытия, представленного в данном документе, что дозы и режим дозирования корректируются в соответствии со способами, хорошо известными в терапевтической области. Это означает, что может быть легко установлена максимально переносимая доза и может быть также определено эффективное количество, обеспечивающее обнаруживаемый терапевтический эффект для пациента, так же как и требования к времени введения каждого агента для достижения видимого терапевтического эффекта для пациента. Таким образом, хотя некоторые дозы и схемы режима введения приведены в качестве примеров в данном документе, эти примеры никоим образом не ограничивают дозы и режимы введения, которые могут понадобиться для пациента в практике применения настоящего изобретения.
Следует отметить, что значения дозировки могут изменяться, в зависимости от типа и тяжести состояния, которое следует облегчить, и может включать одну или более доз. Кроме того, необходимо понимать, что для любого конкретного пациента, конкретные схемы введения должны быть скорректированы через некоторое время согласно индивидуальной потребности и на усмотрение медицинского работника, который осуществляет введение или контролирует введение композиций, и что диапазоны концентрации, приведенные в данном описании, приведены только в качестве примера и не предназначены для ограничения объема или практики заявленных композиций. Кроме того, режим дозирования с композициями по данному изобретению может быть основан на различных факторах, включая тип заболевания, возраст, вес, пол, состояния здоровья пациента, тяжесть состояния, путь введения и конкретную используемую биспецифичную связывающую молекулу. Таким образом, режим дозирования может широко варьироваться, но может определяться регулярно с помощью стандартных методов. Например, дозы могут быть скорректированы на основе фармакокинетических и фармакодинамических параметров, которые могут включать клинические эффекты, такие как токсические эффекты или лабораторные значения. Таким образом, настоящее изобретение охватывает индивидуальное повышение дозы, которое определяется квалифицированным специалистом. Определение необходимой дозы и режимы хорошо известны в соответствующей области техники и будут понятны специалисту в данной области после ознакомления с идеями, раскрытыми в данном документе.
Примеры подходящих способов введения предусмотрены выше.
Предполагается, что подходящая доза биспецифичной связывающей молекулы по данному изобретению будет в диапазоне от 0,1-200 мг/кг, предпочтительно 0,1-100 мг/кг, в том числе около 0,5-50 мг/кг, например, около 1-20 мг/кг. Биспецифичная связывающая молекула может быть введена, например, в дозе по меньшей мере 0,25 мг/кг, например, по меньшей мере 0,5 мг/кг, в том числе не менее 1 мг/кг, например, по меньшей мере 1,5 мг/кг, например, также как не менее 2 мг/кг, например, по меньшей мере 3 мг/кг, в том числе по меньшей мере 4 мг/кг, например, по меньшей мере 5 мг/кг; и например вплоть до максимально 50 мг/кг, в том числе вплоть до максимально 30 мг/кг, например, вплоть до максимально 20 мг/кг, в том числе вплоть до максимально 15 мг/кг. Введение будет повторяться обычно в подходящие промежутки времени, например, раз в неделю, раз в две недели, один раз каждые три недели или один раз каждые четыре недели, и так долго, как будет сочтено целесообразным ответственным врачом, который может в некоторых случаях увеличить или уменьшить дозу при необходимости.
Диагностическое использование и композиции
Биспецифичные связывающие молекулы по настоящему изобретению также используются в диагностических процессах (например, in vitro, ex vivo). Например, биспецифичные связывающие молекулы могут использоваться для обнаружения или измерения уровня HER2 и/или HER3 в образцах, полученных от пациента (например, образец ткани или образец жидкости тела, такой как воспалительный экссудат, кровь, сыворотка крови, жидкость кишечника, слюна или моча). Подходящие методы обнаружения и измерения включают иммунологические методы, такие как проточная цитометрия, твердофазный иммуноферментный анализ (ИФА), хемилюминесцентный анализ, радиоиммуноанализ и иммуногистология. Изобретение далее включает наборы (например, диагностические наборы), содержащие биспецифичные связывающие молекулы, описанные в данном документе.
Для наилучшего понимания изобретения приводятся следующие примеры. Эти примеры приведены только в иллюстративных целях и не должны толковаться как ограничивающие сферу применения изобретения в любой форме.
Все публикации, патенты и патентные заявки, указанные в этой спецификации включены в данный документ путем отсылки. Хотя вышеупомянутое изобретение было довольно подробно описано путем иллюстрации и примера в целях исключения двусмысленного толкования, специалистам в данной области на основе идей, раскрытых в данном изобретении, будет вполне понятно, что могут быть внесены определенные изменения и модификации без отклонения от сущности и объема прилагаемых вариантов осуществления изобретения.
Примеры
Пример 1
Схема получения биспецифического антитела antiHER2/antiHER3 (BCD090) антитела
На Фиг. 1 представлена принципиальная схема создания и оптимизации биспецифического антитела antiHER2/antiHER3.
Провели иммунизацию ламы рекомбинантным антигеном HER3HF. Далее выделяли фракцию монунуклеарных клеток, из которых впоследствии выделили суммарную РНК. После этого провели ряд генноинженерных процедур, направленных на создание фаговой Fab библиотеки. Все вышеописанные процедуры более подробно описаны в Примере 3.
Далее провели несколько раундов селекции полученной библиотеки (Пример 4) и скринингов (Примеры 5, 6, 7). После этого секвенировали последовательности выбранных клонов. Для проведения функционального анализа выбранных кандидатов при помощи генноинженерных процедур перевели их в следующий формат: полноразмерный IgG, где вместо VH домена присутствует VHH домен. После выбора наиболее аффинного и функционально-активного кандадата создали генноинженерную конструкцию, в которой на N-конец легкой цепи aHER2 IgG HER2 был клонирован ген анти-HER3 VHH Fab с разделенный от легкой цепи специальным гибким амикислотным линкером, необходимым для корректного независимого фолдинга обоих полипептидов в составе фьюжн молекулы и независимого связывания с антигенами HER2 и HER3 (Пример 17).
Пример 2
Получение рекомбинантных антигенов и антител в суспензионной культуре клеток млекопитающих
Антитела и антигены продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия CHO-K1), согласно опубликованным протоколам [Biotechnol Bioeng. 2005 Sep 20; 91(6):670-677, Liao Metal., 2004; Biotechnol Lett. 2006 Jun; 28(11):843-848; Biotechnol Bioeng. 2003 Nov 5; 84(3):332-342]. Использовали клетки, конститутивно экспрессирующие ген белка EBNA1 (Epstein-Barrvirus nuclear antigen 1). Суспензионное культивирование осуществляли в колбах на орбитальном шейкере, с использованием бессывороточных сред производства компании Life Technologies Corporation, согласно инструкциям производителя. Для транзиентной экспрессии клетки в концентрации 2Е+6 кл/мл трансфецировали с помощью линейного полиэтиленимина (ПЭИ «МАХ», «Polysciences»). Соотношение ДНК/ПЭИ составляло 1:3-1:10. Через 5-7 дней после трансфекции культуральную среду центрифугировали при 2000 g в течение 20 минут и фильтровали через фильтр с размером пор 0,22 мкм. Целевые белки выделяли из культуральной жидкости с помощью аффинной хроматографии.
Рекомбинантные белки HER3, содержащие на С-конце белка шесть аминокислот гистидина (Фиг. 2, 3), выделяли и очищали из культуральной жидкости, используя смолу Profinity IMAC Ni-charged (компании Bio-Rad). До очистки в культуральную жидкость добавляли NiCl2 до концентрации 1 мМ. Далее в культуральную жидкость добавляли Profinity IMAC Ni-charged в соотношении 1 к 200 и перемешивали на качалке 1 час при комнатной температуре. Переносили сорбент на колонку Thermo scientific Polypropylene columns объемом 5 мл и промывали его фосфатно-солевым буфером (PBS). Связавшийся белок элюировали раствором 0,3М имидазола рН8 с 0,3M NaCl. Далее белок переводили в PBS (рН 7,4) с помощью диализа, используя SnakeSkin Dialysis Tubing, фильтровали (0,22 мкм), переносили в пробирки и хранили при -70°С. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (Фиг. 6).
Химерный рекомбинантный белок HER3, содержащий 2 домена Fc иммуноглобулина на С-конце белка (Фиг. 4), выделили и очистили из культуральной жидкости, с помощью аффинной хроматографии на колонке HiTrap rProtein A Sepharose FF объемом 5 мл (GE Healthcare). Осветленную культуральную жидкость пропускали через колонку, которая была уравновешена фосфатно-солевым буфером (PBS, рН 7,4). Затем колонку промыли PBS, чтобы вымыть неспецифическисвязывающие компоненты. Связанный антиген элюировали 0,1 М глициновым буфером рН3. Главный протеиновый пик элюции собрали и довели его рН до нейтрального с помощью 1М TrisHCl (рН 8). Все стадии очистки проводили на хроматографической системе AKTA prime plus при скорости потока 110 см/ч. Далее белок перевели в PBS с помощью диализа, используя SnakeSkin Dialysis Tubing, профильтровали (0,22 мкм), перенесли в пробирки и хранили при -70°С. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (Фиг. 6).
Исследуемые и контрольное антитело IgG1 (Фиг. 5.) очищали на колонке HiTrapr ProteinAFF объемом 1 мл (GE Healthcare) согласно методике, описанной выше для антигена HER3-Fc. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (Фиг. 7).
Пример 3
Иммунизация ламы HER3 человека и создание Fab библиотеки фаговых антител ламы
Животное Lama Glama последовательно иммунизировали 5 раз путем подкожного введения антигенного материала, смешанного с равным объемом полного (при первой инъекции) или неполного (при остальных инъекциях) адъюванта Фрейнда. В качестве антигена использовали смесь рекомбинантных белков (по 0,1 мг каждого из белков на 1 инъекцию), одним из которых был человеческий HER3HF. Инъекции антигенов проводили на следующих сроках: 0, 2, 4, 5, 8 недели. Взятие крови (50 мл) проводили через 5 дней после каждой инъекции, начиная с третьей. В качестве антикоагулянта применяли 3,8% цитрат натрия в соотношении 1:9. Отобранную кровь ламы разводили в 2 раза стерильным физиологическим раствором. 30 мл разбавленного раствора крови наслаивали на среду Lymphoprep™ (Axis-Shield, Norway) с плотностью 1,077 г/мл объемом 15 мл и проводили центрифугирование в течение 20 мин при 800 g. Мононуклеарные клетки (лимфоциты и моноциты) отбирали из интерфазной зоны плазма/среда Lymphoprep, после чего промывали стерильным раствором PBS.
Полученный титр сывороточного иммуноглобулина против HER3, оцененный согласно стандартному протоколу, оказался не менее 1/8000 (Фиг. 8.), что является удовлетворительным для приготовления библиотеки антител.
Суммарную РНК мононуклеарных клеток ламы выделяли с помощью набора RNeasy Mini Kit согласно предлагаемому протоколу (QIAGEN). Концентрацию РНК определяли с помощью Nanovue (GE Healthcare) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле.
Реакцию обратной транскрипции проводили с использованием набора MMLV RT kit (от Evrogen) согласно рекомендованному протоколу с использованием обратной транскриптазы MMuLV и рандом-гексамерных олигонуклеотидов в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции для получения генов вариабельных доменов, фланкированных сайтами рестрикции, с использованием набора олигонуклеотидов и протоколам авторов [J Biol Chem. 1999 Jun 25; 274(26): 18218-30]. Далее, соединение генов вариабельных доменов легких и тяжелых цепей в один фрагмент проводили путем последовательных реакций рестрикции, лигирования и амплификации. Гены тяжелых цепей соединяли с генами каппа и отдельно с генами лямбда легких цепей. При этом расчетное количество молекул матрицы во всех реакциях составляло не менее 1011. Полученный ДНК препарат VL-CK-VH обрабатывали рестриктазами NheI /Есо91I и лигировали в оригинальную фагмиду рН5. Строение полученной фагмиды показано на (Фиг. 9). Продукты лигирования трансформировали в электрокомпетентные клетки штамма SS320, полученные согласно протоколам [Methods Enzymol. 2000; 328: 333-63.]. Репертуар производной каппа Fab библиотеки составил 1,3Е+7 и лямбда Fab библиотеки - 8,4Е+7, соответственно. Препарат фага библиотек был приготовлен согласно процедуре, описанной ранее [J Mol Biol. 1991 Dec 5; 222(3): 581-97].
Пример 4
Селекция Fab-библиотек фаговых антител
Специфичные фаговые Fab-антитела против HER3 были получены из фаговой Fab-дисплейной библиотеки, полученной путем иммунизации ламы. Селекцию проводили на рекомбинантном белке 3DHER3HE, как описано ранее [Nat Biotechnol. 1996 Mar; 14(3):309-14; J Mol Biol. 1991 Dec 5; 222(3): 581-97]. Для осуществления селекции методом пэннинга, белок 3DHER3HE (в 50 мМ карбонатном буфере (рН 9,5) адсорбировали в течение ночи при 4°С на поверхности сорбирующих пробирок EIA/RIA high binding (Greineer bio-one). Пробирки промывали несколько раз PBST (PBS рН 7,4 и 0,1% Твина 20 v/V), а затем блокировали вакантные центры связывания белка на поверхности пробирок раствором обезжиренного молока (0,5% m/V в PBST рН 7,4). Инкубировали раствор обезжиренного молока в пробирках в течение 1 ч. Далее отмывали пробирки раствором PBST. Затем развели фаговую библиотеку в 2-4 мл раствора PBST с 0,5% обезжиренным молоком до конечной концентрации 1013 фаговых частиц на мл. Добавляли подготовленную библиотеку в пробирки со связанным на поверхности антигеном и инкубировали в течение 1 ч при перемешивании. Не связавшиеся фаги удаляли в ходе серий отмывок пробирок раствором PBST. Количество отмывок увеличивали от первого раунда к третьему раунду: 20-30-40 раз, соответственно. Оставшиеся связанными фаговые частицы элюировали с пластиковой пробирки 0,1 М раствором Gly-HCl (рН 2,2) в течение 15 мин при перемешивании. Затем переносили раствор в чистые пластиковые пробирки и нейтрализовали 1 М Tris-HCl (рН 8) 5:1. Бактерии штамма Е. coli TG1 инфицировали полученными фагами, фаги амплифицировали и использовали в следующем цикле селекции.
После второго и третьего раунда селекции на 3DHER3HE, проведенный ИФА анализ препарата поликлонального фага показал значительное обогащение (Фиг. 11). Полученный пул клонов, обогащенных Fab человека, специфичных против 3DHER3HE, был реклонирован в экспрессионную плазмиду pLL (Фиг. 10), содержащую myc-tag и His6 tag на С- конце гена СН1 тяжелой цепи. Клетки экспрессионного штамма E. coli Bl21Gold трансформировали библиотекой клонов и высевали на твердую среду, содержащую ампициллин (100 мкг/мл) и канамицин (30 мкг/мл). Культивировали в течение 15 часов при 30°С. Одиночные колонии переносили стерильным наконечником для автоматической пипетки в лунки 96-луночных планшетов, содержащие по 100 мкл среды 2TY с ампициллином (100 мкг/мл), канамицином (30 мкг/мл) и глюкозой (0,4%). Планшеты заклеивали воздухопроницаемой пленкой Breathseal и культивировали в течение 16 часов на орбитальном шейкере при 30°С, 900 об/мин. Затем клетки осаждали центрифугированием планшетов при 2500 об/мин в течение 20 минут, а супернатанты использовали для анализа содержащихся в них Fab-фрагментов антител.
Пример 5
Анализ специфического связывания Fab с HER3 человека
ИФА использовали для измерения связывания исследуемых Fab-фрагментов с HER3 человека. В качестве позитивного контроля использовали Fab антитела AV203 с опубликованной последовательностью (WO 2011/136911 А2). Для анализа специфического связывания лунки планшетов (от Nunc ImmunoMaxisorp) покрывали 50 мкл (0,5 мкг/мл в 1X покрывающем карбонатном буфере) 3DHER3HE, герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному ИФА протоколу с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем GenetixQ-pix2xt (от Molecular Device) и Tecan Freedom EVO 200 (от Tecan). Для блокирования неспецифического связывания в лунки планшетов добавляли блокирующий буфер ВВ (200 мкл 0,5% нежирного молока в PBS). Планшеты инкубировали на шейкере в течение часа при комнатной температуре. После отмывок фосфатно-солевым буфером, содержащим Твин-20 (PBST), добавляли по 50 мкл на лунку тестируемого клеточного супернатанта, содержащего исследуемый Fab. Планшеты снова инкубировали, встряхивая, один час при комнатной температуре, после чего каждую лунку планшетов пять раз промывали PBST. После промывания добавляли (50 мкл/лунку) античеловеческого Fab HRP-конъюгированного вторичного антитела (от Pierce-ThermoScientific) в соотношении 1:7500 в PBS-Твин. Планшеты встряхивали на ротационном шейкере (50 мин, комнатная температура) и промывали пять раз PBST. Колориметрический сигнал развивали добавлением ТМВ (50 мкл/лунку) до насыщения (в среднем 10-12 мин), затем дальнейшее развитие останавливали добавлением стопового раствора (25 мкл/лунку, 1% серная кислота). Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала. Клоны, у которых цветовой сигнал превышал фоновый более чем в 5 раз, были проверены в конкурентном ИФА анализе для выявления антагонистических Fab, блокирующих взаимодействие лиганда и рецептора.
Пример 6
Конкурентный ИФА анализ блокирования взаимодействия rhHRG1-β1ECD лиганда и Her3 рецептора
Конкурентный ИФА был использован для проверки антагонистической способности отобранных ранее специфичных Fab против HER3 человека. В качестве позитивного контроля антагониста использовали Fab антитела AV203 (WO 2011/136911 А2). Лиганд rhHRG1-β1ECD (от R&D Systems) иммобилизовали в лунки планшетов (от Nunc Immuno Maxisorp) по 50 мкл с концентрацией 1 мкг/мл в 1X карбонатном буфере и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартным ИФА протоколам с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем GenetixQ-pix2xt (от Molecular Device) и Tecan Freedom EVO 200 (от Tecan). Для блокирования неспецифического связывания в лунки планшетов добавляли блокирующий буфер ВВ (200 мкл 0,5% нежирного молока в PBS). Планшеты инкубировали на шейкере в течение часа при комнатной температуре.
Параллельно 40 мкл тестируемого клеточного супернатанта, содержащего исследуемый Fab, смешивали в несвязывающем 96-луночном планшете с 40 мкл HER3-Fc с концентрацией 2 мкг/мл в 0,5%молоке на PBS-Твин. Инкубировали 1 час при комнатной температуре с перемешиванием при 500 об/мин.
После отмывок от блокирующего буфера в планшет, содержащий лиганд rhHRG1-β1ECD, переносили описанную выше реакционную смесь Fab и HER3-Fc рецептора в расчете 80 мкл на лунку. Планшеты снова инкубировали, встряхивая, 45 мин при комнатной температуре, после чего каждую лунку планшетов пять раз промывали PBST. Добавляли 50 мкл/лунку античеловеческого Fab HRP-конъюгированного вторичного антитела (от Pierce-ThermoScientific) в соотношении 1:7500 в PBST. Планшеты встряхивали на ротационном шейкере 45 мин при комнатной температуре и промывали 5 раз PBST. Колориметрический сигнал развивали добавлением ТМВ (50 мкл/лунку) до насыщения (в среднем 10-12 мин), затем дальнейшее развитие останавливали добавлением стопового раствора (25 мкл/лунку, 1% серная кислота). Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала.
Клоны, показавшие блокирование на уровне контрольного Fab антитела AV203, были отмечены как позитивные и использовались в дальнейших анализах. Гены вариабельных доменов позитивных клонов были секвенированы, согласно стандартным протоколам на аппарате Applied Biosystems 3130 Genetic Analyzer (Applied Biosystems).
Пример 7
Сравнительный скрининг анти-Her3 Fab кандидатов человека по koff
Koff-скрининг производился с использованием прибора Pall Forte Bio Octet QK 96. Анти FABCH1-биосенсоры в течение 30 минут регидратировались в рабочем буфере, содержащем 10 мМ PBS, рН 7.2-7.4, 0.1% Твин-20, 0,1% BSA. В исследуемые образцы супернатантов Е. coli добавляли 10× рабочий буфер до конечной концентрации 1×. Затем анти FABCH1-биосенсоры погружались в супернатанты Е. coli, содержащие Fab-фрагменты кандидатов антител, на 12 часов при 4°С. Сенсоры с иммобилизованными на поверхности Fab - фрагментами переносились в лунки с рабочим буфером, где прописывалась базовая линия (в течение 60 сек). Далее сенсоры переносились в лунки с раствором аналита (3DHER3HE, 30 мкг/мл) для ассоциации комплекса антиген-Fab (в течение 300 сек). Затем сенсоры возвращали в лунки, содержащие рабочий буфер, для последующей стадии диссоциации (в течение 600 сек). После каждого эксперимента использованные сенсоры регенерировались путем трехкратного помещения их в буфер для регенерации (Gly-HCl, рН 1,7) после чего могли быть использованы в следующем эксперименте. Анализ полученных кривых проводился с помощью программного обеспечения Octet Data Analysis (версия 7.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. В качестве контроля использовался Fab воспроизведенного антитела AV203 (WO 2011/136911 А2). Результаты представлены в Таблице 1. Кандидаты МР3:Н3(1), МР3:А4(2), МР3:В4(3) имеют сходный либо лучший показатель Koff по сравнению с показателем Koff Fab фрагмента воспроизведенного антитела AV203.
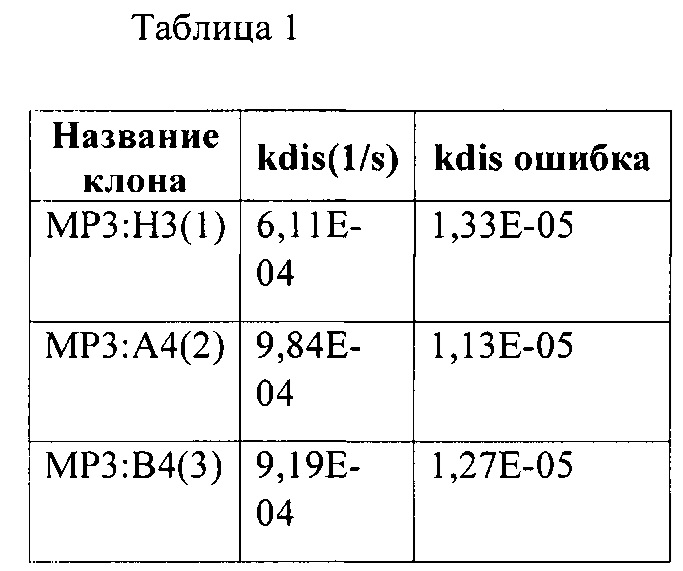
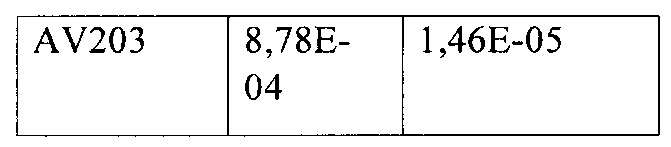
Пример 8
Конструирование библиотеки VHH-aHER3 мутантных монодоменных антител, специфичных против HER3
Для создания библиотек VHH-aHER3 мутантных монодоменных антител, специфичных против HER3, был синтезирован пул генов, кодирующих последовательность VHH-aHER3, имеющих по четыре соседних рандомизированнных триплета NNK в области каждого CDR участка (Таблица 2). Полученная ДНК рандомизированного гена была клонирована в фаговую плазмиду рН6, описанную в работе [FEBS Lett. 1997 Sep 15; 414(3):521-6], согласно описанному в работе протоколу, для дисплея в составе белка капсида gpIII. Трансформация, указанных конструкций в штамм SS320 дала на выходе от 2Е+8 независимых трансформантов для библиотеки, согласно процедуре [Methods Enzymol. 2000; 328: 333-63]. Препараты фага мутантных VHH-aHER3 библиотек были приготовлены согласно процедуре, описанной ранее [Mol Biol. 1991 Dec 5; 222(3): 581-97].
Селекции полученных фаговых мутантных VHH-aHER3 библиотек проводили в условиях аналогичных, описанных выше (Пример 4).
После третьего раунда селекции на препарате рекомбинантного HER3 человека, проведенный ИФА анализ препаратов поликлонального фага показал значительное обогащение и превышение более 10 раз над фоном неспецифического связывания. Пулы генов из обогащенных фаговых библиотек VHH-aHER3 мутантных монодоменных антител, специфичных против HER3 человека, были реклонированы в экспрессионную плазмиду рЕТ22b(+) компании Novagen, содержащую myc-tag пептид на С-конце для ELISA детекции.
Пример 9
Выделение VHH-aHER3 мутантных монодоменных антител, специфичных против HER3
Для анализа связывания исследуемых мутантных монодоменных антител с HER3 человека использовали ИФА по методике, описанной в Примере 5. Количество тестируемых клонов-продуцентов VHH-aHER3 мутантных монодоменных антител, составило 1200 штук. В качестве позитивного контроля использовали исходный вариант VHH-aHER3 монодоменного антитела с последовательностью SEQ ID NO:4.
В результате анализа было отобрано 37 позитивных клонов, давших сигнал выше или аналогичный контрольному исходному VHH-aHER3 монодоменному антителу (значения в интервале 0,7-1,0 отн.ед.).
Пример 10
Характеризация VHH-aHER3 мутантных монодоменных антител, специфичных против HER3
Тридцать семь отобранных в результате ИФА скрининга (Пример 9) кандидатов позитивных клонов, давших сигнал выше или аналогичный контрольному VHH-aHER3 монодоменному антителу были секвенированы на секвенаторе Applied Biosystems 3130, согласно протоколам рекомендуемым компанией производителем. В результате было получено 26 уникальных по последовательности клона. Полученные уникальные кандидаты далее были проанализированы по количественной характеристике кинетической константы диссоциации на приборе OctetRED96 согласно методике, описанной в Примере 11.
Полученные количественные кинетические данные специфического взаимодействия VHH-aHER3 мутантных монодоменных антител и их последовательности приведены в Таблице 2.
В результате получены мутанты VHH-аНЕR3 монодоменных антител, имеющие до девяти замен во всех трех CDR и, при этом, сохраняющие или имеющие усиленную аффинность к HER3 рецептору. Кроме того, аминокислотный состав таких замен совершенно разнообразный. Таким образом, показана возможность создания высокоаффинных, специфичных мутантных форм VHH-aHER3 полипептида, имеющих до 25% замен по совокупным последовательностям CDR (Подсчитали варианты с максимальным количеством замен - до 9 замен, к общему количеству позиций CDRs- около 40).
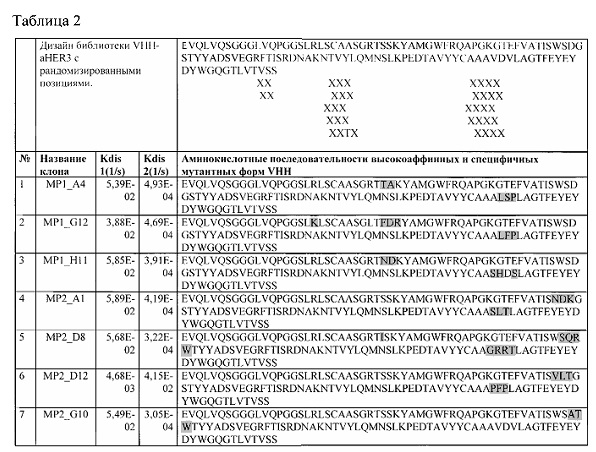
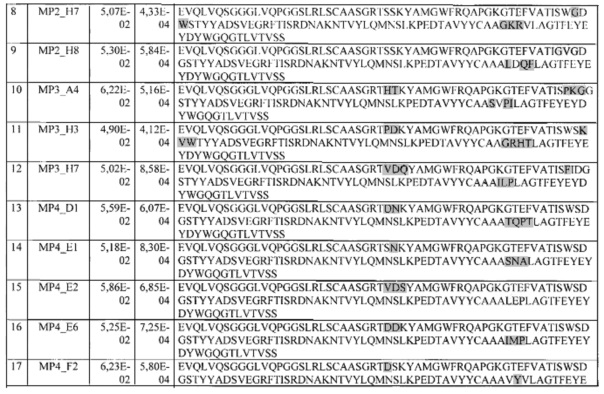
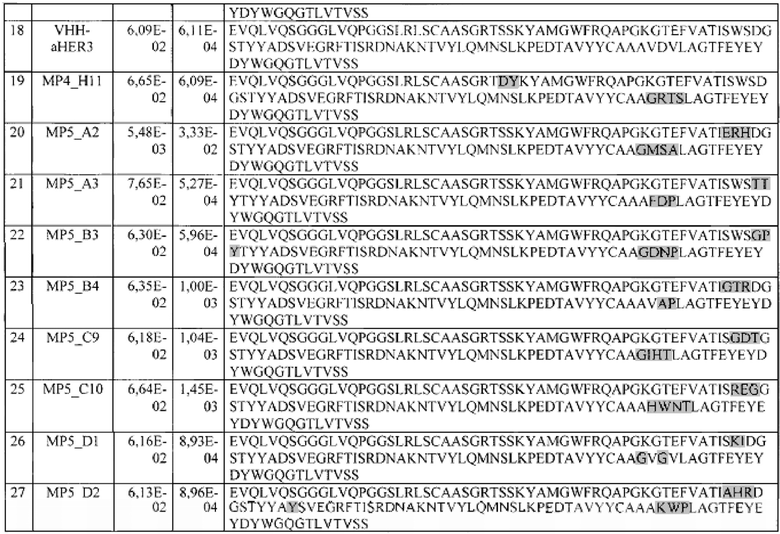
Пример 11
Сравнительный скрининг VHH-aHER3 мутантных монодоменных антител по koff
Koff-скрининг производился с использованием прибора Pall Forte Bio Octet QK 96. Анти Streptavidin-биосенсоры (SA) в течение 30 минут регидратировались в рабочем буфере, содержащем 10 мМ PBS, рН 7.2-7.4, 0.1% Твин-20, 0,1% BSA. Далее происходила иммобилизация на биосенсоры биотинилированного антигена HER3HF (20 мкг/мл). Сенсоры с иммобилизованным биотинилированным антигеном переносились в лунки с рабочим буфером, где прописывалась базовая линия (в течение 60 сек.). В исследуемые образцы супернатантов Е. coli добавляли 10× рабочий буфер до конечной концентрации 1×. Затем биосенсоры погружались в супернатанты Е. coli, содержащие Fab-фрагменты кандидатов антител для ассоциации комплекса антиген-антитело (в течение 300 сек). Затем сенсоры возвращали в лунки, содержащие рабочий буфер, для последующей стадии диссоциации (в течение 600 сек). Анализ полученных кривых проводился с помощью программного обеспечения Octet Data Analysis (версия 7.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. В качестве контроля использовался Fab воспроизведенного антитела AV203 (WO 2011/136911 А2).
Пример 12
Конструирование и наработка биспецифических антител VHH-IgG1 против HER2 и HER3
Гены вариабельных доменов легких и тяжелых цепей антитела против HER2 человека [WO/1999/031140] были синтезированы в оригинальном кодоновом составе и клонированы в плазмиды, содержащие гены константных доменов легких и тяжелых цепей соответственно, для транзиентной экспрессии в клетках CHO-K1, как описано в Примере 2. Полученные конструкции были валидированы секвенированием (SEQ ID NO: 5 и SEQ ID NO: 6 аминокислотные последовательности вариабельных доменов тяжелой и легкой цепей соответственно).
Кроме того, на базе последовательности обнаруженного высокоаффиного VHH Fab против HER3 (Пример 7) был синтезирован ген, кодирующий аминокислотную последовательность SEQ ID NO 4. При этом были введены мутации E44G/R45T на участке межсубъединичного контакта вариабельных доменов VH и VL, которые, как было показано ранее, [заявка RU 2014138740], обуславливают стабильность VHH производного в комплексе с легкой цепью антитела.
Далее, по стандартной генно-инженерной процедуре на N-конец легкой цепи в вышеописанную конструкцию с легкой цепью антитела против HER2 был клонирован ген анти-HER3, кодирующий аминокислотную последовательность SEQ ID NO 4, разделенный от легкой цепи гибким аминокислотным линкером.
Пример 13
Иммуноферментный анализ взаимодействия анти-HER3 антитела с HER3 и другими антигенами
ИФА использовали для измерения сравнительной аффинности антитела к HER3 и другим антигенам. Для анализа связывания лунки планшетов (medium binding от Greiner bio one) покрывали 50 мкл HER3HF, 3DHER3HE, HER2-Fc, EGFR-Fc, IGFR-Fc, IL12-Fc, CD3ED-Fc, GCSF, cMET, IL17-Fc, IL6R-Fc (5 мкг/мл в 1× карбонатном буфере), герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному ИФА протоколу, описанному в Примере 5. Тестируемые антитела использовались в концентрации 5 мкг/мл в PBST.
Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала (Фиг. 12). Анти-HER3 антитело, представляющее собой полноразмерный IgG, где вместо VH домена присутствует VHH домен, специфически связывается с полноразмерным HER3 и его третьим доменом и не связывается с другими исследуемыми антигенами.
Пример 14
Иммуноферментный анализ взаимодействий биспецифического anti- HER2/antiHER3 (BCD090) антитела с HER2, HER3 рецепторами разных организмов
ИФА использовали для измерения сравнительной аффинности биспецифического antiHER2/antiHER3 антитела (далее BCD090) к рецепторам HER2, HER3 разных организмов. Для анализа связывания лунки планшетов (medium binding от Greiner bio one) заполняли 50 мкл HER2-Fc человека, HER2His яванского макака, мыши, крысы, HER2FE кролика, морской свинки, HER3-Fc человека, макаки, мыши, HER3His крысы, HER3FE морской свинки (1 мкг/мл в 1× покрывающем карбонатном буфере), герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному ИФА протоколу, описанному в Примере 5. Биспецифическое антитело BCD090 аффинно связывается с HER2 человека, HER2 яванского макака, HER3 человека и со всеми антигенами ортологами HER3: макаки, мыши, крысы, морской свинки. BCD090 в 2 раза слабее связывается с HER2 морской свинки и не связывается с другими исследуемыми рецепторами (Фиг. 13, Фиг. 14).
Пример 15
Анализ взаимодействий antiHER2/antiHER3 антитела (BCD090) с рецепторами HER3 человека и макаки резус и HER2 макаки циномолгус на приборе Octet RED 96
Константы аффинности связывания антитела с рецепторами HER3 человека и макаки резус и HER2 макаки циномолгус получили с помощью прибора OctetRed 96 (от ForteBio). Антитела были неспецифически иммобилизованы на поверхность аминреактивных сенсоров второго поколения (ForteBio, Pall) по стандартному протоколу согласно инструкции производителя для подготовки и иммобилизации AR2G сенсоров. Анализ был произведен при 30°С с использованием PBS, содержащего 0,1% Твин-20 и 0,1% BSA в качестве рабочего буфера. Титрование рецепторов производили с использованием рабочего буфера от различных концентраций в зависимости от предполагаемой аффинности.
Кривые связывания с вычетом базового сигнала анализировали с помощью программы Octet Data Analysis (версия 7.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. BCD090 антитело специфически связывается с антигенами HER3 человека и макаки резус и HER2 макаки циномолгус (Фиг. 15, 16, 17). Анализ показал, что BCD090 антитело с высокой аффинностью связывается с HER3 человека, аффинность к HER3 макаки резус уступает в 6 раз. BCD090 антитело также связывается с HER2 макаки циномолгус, но константа аффинности для HER2 циномолгус уступает константе аффинности для HER2 человека примерно на 4 порядка. Полученные константы аффинности представлены в Таблице 3.
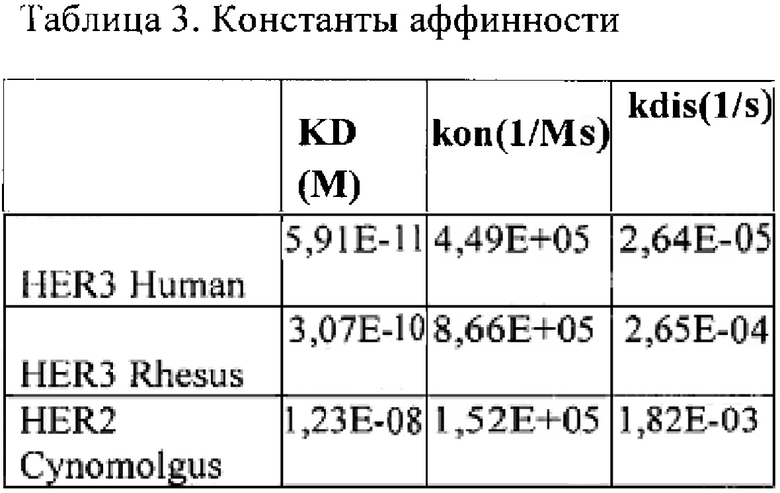
Пример 16
Анализ взаимодействий antiHER2/antiHER3 (BCD090) антитела с HER2 человека с помощью прибора Biacore Т200
Эксперименты по исследованию аффинности связывания BCD090 антитела к антигену HER2 человека проводили на оптическом анализаторе Biacore Т200 (Ge Healthcare Life Sciences), принцип метода которого основан на эффекте поверхностного плазмонного резонанса для регистрации межмолекулярных взаимодействий. В работе использовали стандартные оптические чипы СМ5 с карбоксиметилдекстрановым покрытием. Антитело BCD090 было неспецифически (по NH2-группам) иммобилизовано на поверхность чипа в концентрации 20 мкг/мл согласно протоколу, соответствующему инструкции производителя. После чего проводилась инжекция антигена в концентрациях 1-15 мкг/мл в течение 5 минут при скорости потока 10 мкл/мин в рабочем буфере (HBS-EP, 0.01 М HEPES рН 7.4, 0.15 М NaCl, 3 mM EDTA, 0.005% v/v Surfactant Р20). Диссоциацию комплекса детектировали в реальном времени при температуре 37°С.
Кривые специфических взаимодействий антигенов с иммобилизованными на оптическим чипе антителами получали в виде разности сигналов между опытным и контрольным каналом. Анализ полученных данных производился с использованием программы BIA Evalution 3.1 и модели Heterogeneous Ligand. Константа аффинности рассчитывалась, как среднее значение из полученных в эксперименте результатов.
Биспецифическое антитело BCD090 обладает высокой аффинностью к HER2 человека.
Аффинность BCD090 к HER2 составила KD1/KD2 -0,55 pMol. Кривые ассоциации/диссоциации представлены на Фиг. 18.
Пример 17
Анализ взаимодействия биспецифического антитела BCD090 с обоими антигенами HER2 и HER3 человека на приборе OctetRED96
Анализ одновременного взаимодействий BCD090 антитела с антигенами HER2 и HER3 человека проводили с помощью классического «сэндвич» метода анализа на приборе OctetRed 96 (Pall-ForteBio). Анализ был произведен при 30°С с использованием PBS, содержащим 0,1% Твин-20 и 0,1% БСА в качестве рабочего буфера. В ходе эксперимента первый антиген (HER2) в концентрации 5 мкг/мл и объемом 200 мкл/лунку был иммобилизован на аминореактивных сенсорах. Далее сенсоры погружались в раствор антител 200 мкл/лунку антител с концентрацией 20 мкг/мл. После этого сенсоры погружались или в раствор второго антигена HER3 200 мкл/лунку с концентрацией 10 мкг/мл и параллельно в физиологический раствор, не содержащий белка.
Кривые связывания с вычетом базового сигнала анализировали с помощью программы Octet Data Analysis (версия 8.2) согласно стандартной процедуре.
Сенсограмма, приведенная на Фиг. 19, свидетельствует о том, что молекула BCD090 антитела способна одновременно связывать два антигена: HER2 и HER3 человека.
Пример 18
Анализ взаимодействий BCD090 антител с FcRn and Fcγ-рецепторами при помощи Octet RED 96
Для анализа взаимодействия антител с FcgRIIIaV, FcgRIa, FcRn использовался прибор Forte bio Octet RED96 и биосенсоры, покрытые стрептавидином (SA-Streptavidin). В качестве контроля использовалось антитело Трастузумаб.
В ходе эксперимента проводили ориентированную иммобилизацию биотин-меченных рецепторов на поверхность, покрытых стрептавидином, сенсоров. Готовили серию разведений антител от концентрации 1000 (100 мкг/мл в случае FcgRIa) мкг/мл с шагом 2 на 7 точек и помещали в лунки 96-луночного планшета. Далее проводилась стадия ассоциации путем погружения сенсоров в растворы антител с различной концентрацией, после этого проводилась стадия диссоциации путем погружения сенсоров в рабочий буфер.
Для анализа аффинности антител к FcgRIIIaV, FcgRIa использовался рабочий буфер РН7.4 (PBS, 0,1% Tween, 0,1% BSA), для анализа аффинности к FcRn - рабочий буфер РН6.0.
Проводили SteadyState анализ полученных кривых с вычетом референсного сигнала с использованием модели 2:1 (гетерогенный лиганд) согласно стандартной процедуре. Анализ показал, что биспецифическое антитело специфически связывается с рецепторами FcRn, FcgRIIIaV, FcgRIa.
Анализ аффинности к Fcg-рецепторам свидетельствует о том, что эффекторные функции BCD090 предположительно будут сопоставимы с эффекторными функциями Трастузумаба. Анализ аффинности к FcRn-рецептору позволяет предположить, что фармакокинетика биспецифического антитела сходна с фармакокинетикой антитела Трастузумаб. Результаты представлены в Таблице 4.
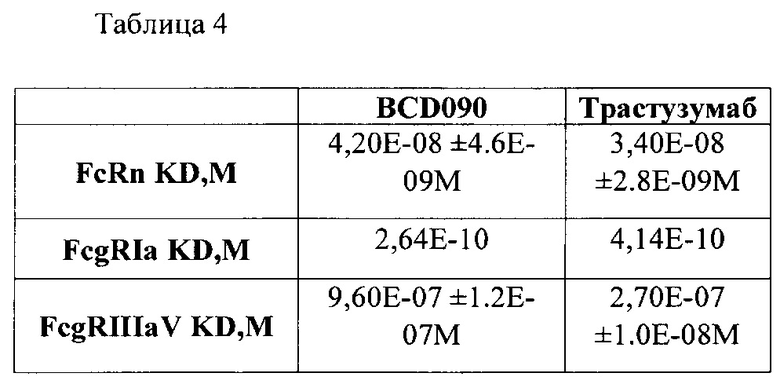
Пример 19
Анализ ингибирование пролиферации опухолевых клеток биспецифическим антителом BCD090
Для оценки способности антитела BCD090 ингибировать херегулинзависимую пролиферацию опухолевых клеток были использованы клеточные линии MCF7, ВТ474, A431NS, SKBR-3.
Состав среды для анализа на клеточной линии MCF7: DMEM, 2 mM Glutamine, 0,3% FBS, 10 мкг/мл, 25 нг/мл HRG (rhHRG1-β1).
Состав среды для анализа на клеточной линии ВТ474: DMEM, 2 mM Glutamine, 5% FBS, 10 мкг/мл, 2,5 нг/мл HRG.
Готовили растворы анализируемых антител в среде для анализа (RPMI, 2 mM Glutamine, 0,3% FBS, 10 мкг/мл инсулина в случае MCF7 и ВТ474, 5 нг/мл HRG, в случае SKBR-3) в концентрациях: 400 мкг/мл, 80 мкг/мл, 16 мкг/мл, 3,2 мкг/мл, 640 нг/мл, 128 нг/мл, 25.6 нг/мл, 5.1 нг/мл, 1.02 нг/мл. Полученные растворы перенесли в лунки 96-луночного в трех повторах.
К растворам антител добавили клетки в количестве 4000 клеток на лунку в среде для анализа.
Планшеты инкубировали при 37°С, 5% СO2 в течение 72 часов. Затем измеряли количество живых пролиферирующих клеток путем добавления витального красителя AlamarBlue и анализа уровня флюоресценции на приборе Fluoroskan Ascent.
Как показано на Фиг. 20, 21, 22, 23 биспецифическое моноклональное антитело BCD090 значительно сильнее ингибирует пролиферацию клеточных линий MCF7, ВТ474, A431NS, SKBR-3, чем контрольные антитела AV203 и Трастузумаб.
В Таблице 5 указаны показатели ЕС50 для различных антител для всех клеточных линий.
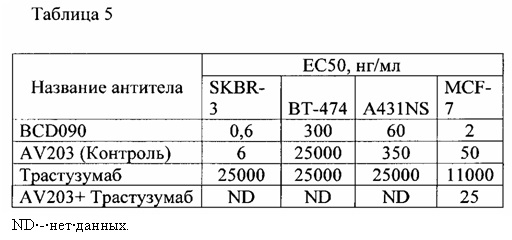
Пример 20
Определение термической стабильности по точке агрегации белка методом динамического светорассеяния (DLS).
Определение точки агрегации исследуемых образцов осуществляли на приборе Zetasizer Nano ZSP. Для этого раствор помещали в кварцевую обеспыленную кювету, которую постепенно нагревали в установке от 50 до 85°С. Шаг нагрева 1.5°С при постоянном измерении интенсивности рассеянного света. Интенсивность рассеянного света детектировали при угле θ=173°. Данные представлены на Фиг. 24 и в Таблице 6.
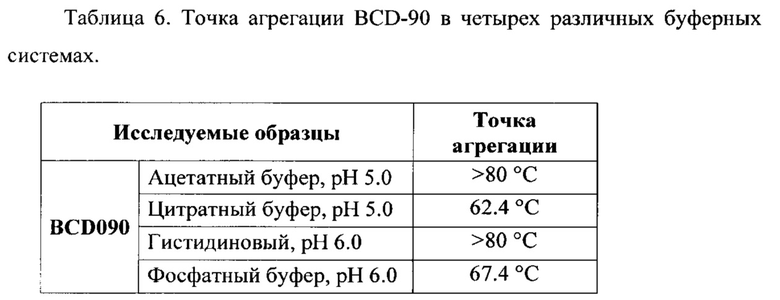
По результатам данного теста можно заключить, что молекула BCD090 имеет хорошую термоколлоидную стабильность и может быть пригодна для технологического и фармацевтического использования.
Пример 21
Определение термической стабильности при длительном воздействии методом Термостресс 50°С.
Исследуемые образцы в концентрации белка разделяли на 3 части по 1 пробирке на каждый состав откладывали в холодильник на хранение при +4°С, остальные устанавливали в термостат для пробирок и термостатировали при 50°С в течение 24 и 72 часа. После окончания прогрева выдерживали при комнатной температуре около 15 мин, центрифугировали при 8000 rpm в течение 5 минут. Супернатанты передавали на анализ методом эксклюзионной ВЭЖХ (SEC HPLC) с УФ-детектором. Хроматографию проводили на системе ВЭЖХ Agilent 1100 на колонке Tosoh TSK-Gel G3000SWXL, 7.8 мм × 30 см, кат. №08541 с предколонкой Tosoh TSKgel Guard SWXL, 6.0 мм × 4.0 см, с диаметром частиц 7 мкм, кат. №08543. Детектирование проводилось на длинах волн 220 и 280 нм.
Результирующие данные стабильности BCD090 в различных буферах при длительном прогреве при 50°С на Фиг. 25. На Фиг. 26 представлены совмещенные хроматограммы: красный - интактный, синий - 72 ч инкубации при 50°С.
По результатам данного теста можно заключить, что молекула BCD090 имеет хорошую термическую стабильность (разница между содержанием агрегатов в растворе до термостресса и после составила не более 5%) и может быть пригодна для технологического и фармацевтического использования.
Пример 22
Изучение стабильности моноклонального биспецифического антитела BCD090 в сыворотке крови человека
Для оценки стабильности BCD090 в сыворотку крови человека, полученную от 7 здоровых доноров и стабилизированную 0,01% мертиолятом, добавляли антитела до концентраций 4, 20 и 100 мкг/мл. Полученную смесь инкубировали в течение 7 дней при 37°С. Через 7 дней проводили определение концентрации BCD090 двумя методами (Фиг. 27).
При использовании первого метода на планшете иммобилизировали рекомбинантный HER2 человека, детекция уровня связанного antiHER2/antiHER3 антитела проводилась 3DHER3HE человека, меченным биотином, и конъюгатом стрептавидина с пероксидазой хрена. При использовании второго метода на планшете также иммобилизировали рекомбинантный HER2 человека, а детекция связанного antiHER2/antiHER3 проводилась с помощью конъюгата поликлональных антител козы против Fc фрагмента IgG человека с пероксидазой хрена. В качестве контроля 100% стабильности использовали образы сыворотки крови человека в которые были добавлены antiHER2/antiHER3 в аналогичных концентрациях непосредственно перед проведением анализа.
Измерение концентрации antiHER2/antiHER3 проводилось следующим образом. Контрольные и хранившиеся в течение 7 суток при 37°С образцы сыворотки крови с добавленными antiHER2/antiHER3 разводились до концентрации 100 нг/мл, соответственно в 40, 200 и 1000 раз в PBST. Концентрация определялась по калибровочной кривой (значения калибровочных точек от 6,25 до 200 нг/мл), калибровочные стандарты также разводились PBST. Далее анализ проводился по стандартной методике. 100 мкл предварительно разведенных исследуемых образцов и калибровочных стандартов вносились в лунки планшета с предварительно иммобилизованным рекомбинантным HER2 человека. Иммобилизация проводилась при концентрации HER2 2 мкг/мл из 20 мМ карбонатного буфера, рН 9,5, свободные участки связывания блокировались с помощью 200 мкл 0,5% сухого молока крупного рогатого скота в PBS. Планшеты инкубировались при постоянном встряхивании 300 об/мин при комнатной температуре. После 3-кратного промывания лунок 250 мкл PBST в лунки добавляли по 100 мкл биотинилированного 3DHER3HE, 3 мкг/мл (первый метод) или 100 мкл поликлональных антител козы к Fc фрагменту IgG человека в рабочем разведении 1:25000 в PBST (второй метод). Планшеты инкубировались при постоянном встряхивании (300 об/мин) при комнатной температуре. После 3-кратного промывания лунок 250 мкл PBST в те лунки, в которые добавляли биотинилированный 3DHER3HE, добавили по 100 мкл конъюгата стрептавидина с пероксидазой в рабочем разведении 1:20000. После инкубации и отмывок активность пероксидазы проявлялась с помощью 100 мкл субстрата ТМБ в течение 10-15 минут, реакция останавливалась добавлением 50 мкл 10% серной кислоты. Оптическая плотность в лунках определялась с помощью планшетного спектрофотометра Tecan Sunrise (Tecan) при 450 нм. Количество связавшегося antiHER2/antiHER3 антитела было прямо пропорционально оптической плотности. Концентрация antiHER2/antiHER3, рассчитанная по калибровочной кривой, умножалась на фактор разведения образца и выражалась в % от теоретически добавленной в сыворотку концентрации (Открываемость, %).
По результатам обоих экспериментов (Фиг. 28) не было обнаружено достоверных отличий между концентрацией antiHER2/antiHER3 определенной в сыворотке крови человека после инкубации в течение 7 суток при 37°С и концентрацией, определенной в сыворотке без инкубации. Таким образом, можно сделать вывод о том, что моноклональное биспецифическое антитело antiHER2/antiHER3 BCD090 стабильно в сыворотке крови человека.
Пример 23
Определение антителозависимой цитотоксичности (ADCC)
Для анализа ADCC использовались клетки-мишени линии ВТ474 (АТСС, НТВ-20) и мононуклеарные клетки периферической крови (РВМС) здорового донора.
Клетки линии культивировались в среде DMEM с 10% эмбриональной бычьей сыворотки (FBS) при 37°С и 5% СO2. Клетки были флюоресцентно мечены с помощью кальцеина AM (Santa Cruz Biotechnology, SC-203865) и ресуспендированы в среде RPMI1640 до плотности 2Е+5 кл/мл.
Мононуклеарные клетки периферической крови (РВМС) выделяли из венозной крови здорового донора с помощью разделения клеток в градиенте плотности. После этого клетки были дважды отмыты в PBS и ресуспендированы в RPMI1640 с 10% FBS до плотности 5Е+6 кл/мл.
Для анализа ADCC в лунки 96-луночного планшета внесли по 100 мкл антител с концентрациями от 20 пг/мл до 20 мкг/мл. К ним добавили по 50 мкл целевых клеток и по 50 мкл эффекторных клеток. Планшет инкубировался в течение 4 часов при 37°С и 5% СO2. После планшет был центрифугирован, далее по 100 мл супернатанта перенесено в новый планшет. Определена флюоресценция при длине волны возбуждения/испускания 485/538 нм. Флюоресценция спонтанно высвобождаемого кальцеина определялось в образцах, содержащих по 50 мкл целевых клеток и 150 мкл RPMI1640 с 10% FBS. Флюоресценция кальцеина при полном лизисе целевых клеток определялась в образцах, содержащих 50 мкл целевых клеток и 150 мкл RPMI1640 с 10% FBS и 1,4% Тритона X100. Эффективность ADCC определялась по формуле:
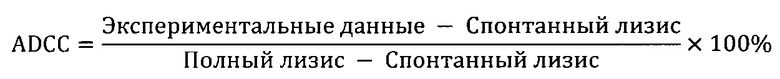
Результаты показаны на Фиг. 29.
Пример 24
Оценка противоопухолевой активности BCD090 на модели подкожных ксенографтов с использованием опухолевых линий человека MCF-7, ВТ474, SK-BR-3 на иммунодефицитных мышах
Взвесь опухолевых клеток вводили подкожно в правый бок иммунодефицитным мышам. За 3 дня до и еженедельно после введения опухолевых линий MCF-7 и ВТ474 (эстрогензависимые опухолевые линии) мышам вводили гормон эстрадиол. Объем вводимой взвеси опухолевых клеток составлял 0,5 мл.
Концентрация клеток MCF-7 составила 10Е+6 кл/мл в бессывороточной среде RPMI1640. Концентрация клеток ВТ474 составила 10Е+6 кл/мл в бессывороточной среде RPMI1640. Концентрация клеток SK-BR-3 составила 4Е+6 кл/мл в бессывороточной среде RPMI1640. Вместе с опухолевыми клетками вводили Матригель (сокращенный фактор роста) в соотношении 1:1.
Введение препаратов антител начинали с момента достижения объема опухоли 100 мм3. Исследуемый препарат и раствор плацебо вводили животным внутрибрюшинно два раза в неделю. В ходе эксперимента два раза в неделю измеряли размеры опухолевого узла и рассчитывали его объем по формуле:
V=π/6*L*W*H, где L, W, H - линейные размеры опухоли.
Критерием эффективности применения препарата служил индекс торможения роста опухоли (ТРО), расчет которого проводили по формуле:
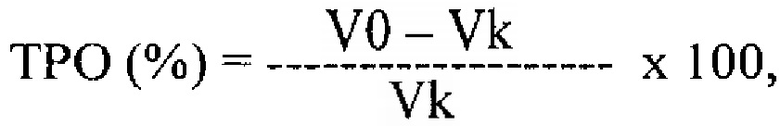
где Vk и V0 - средний объем опухоли (мм3) в контрольной и опытной группах, соответственно.
Также рассчитывали индекс прироста опухоли, характеризующий увеличение объема опухоли на каждой временной точке, в сравнении с размерами на момент начала лечения. Расчет показателя проводили по формуле:
Ii=Vi/V0,
где, I - индекс прироста опухоли, i - сутки эксперимента, V0 - объем опухоли в день начала лечения.
Наблюдение за животными проводили в течение 35 суток, с момента начала лечения.
Препарат BCD090 показал более выраженную противоопухолевую активность в сравнении с моноспецифическими антителами против HER2 и HER3 на всех опухолевых линиях по индексу ТРО. Данные представлены на Фиг. 30, 31, 32, 33, 34, 35.
Пример 25
Создание стабильной клеточной линии для BCD090
Стабильная клеточная линия - продуцент моноклонального антитела BCD090 была получена путем трансфекции методом электропорации с использованием прибора Neon Transfection System (Life Technologies) родительской суспензионной клеточной линии CHO-S векторными конструкциями, содержащими легкие и тяжелые цепи антитела в оптимизированном соотношении. Клональные линии с высоким уровнем продуктивности (более 1000 мг/л) были получены с использованием роботизированной платформы ClonePix (Molecular Devices) и предварительных этапов селекции минипулов с использованием антибиотиков в разных форматах культивирования. Анализ продуктивности проводился с использованием аналитической системы Octet RED96 (Pall Life Sciences). Дизайн эксперимента по подбору базовой среды и схемы культивирования осуществлялся на базе автоматизированной системы Biomek FX robotics (Beckman Coulter). Для культивирования продуцента использовали бессывороточные среды и фидинги, не содержащие белков животного происхождения. Наработку препарата BCD090 для преклинических исследований проводили в ферментере HyClone single-use bioreactor (Thermo Fisher Scientific) с рабочим объемом 50 л.
Пример 26
Очистка BCD090 из продукционной среды стабильной клеточной линии.
Осветление культуральной жидкости, содержащей биспецифическое антитело BCD090 проводили на глубинном фильтре Millistak С0НС (Merck-Millipore). Первичную очистку антитела проводили на аффинном сорбенте с белком А, используя специфическую элюцию целевого белка при рН 3.3-3.8 в глициновом буфере. Полученный элюат инкубировали в кислом рН в течение 30-60 минут для вирусной инактивации, а затем нейтрализовали 1М раствором Tris-OH. Финальную хроматографическую очистку проводили на сорбенте CHT™ Ceramic Hydroxyapatite (Bio-Rad) для удаления остаточных ДНК, белков клеток продуцента, отщепленного лиганда аффинного сорбента, агрегатов и фрагментов антител. Для этого раствор белка наносили на подготовленный сорбент при рН 7.5, с последующей специфической элюцией биспецифического антитела в градиенте хлорида натрия. Очищенный белок подвергали противовирусной фильтрации с использованием набора фильтров Viresolve PRO (Merck Millipore). Концентрирование выполняли в тангенциальном потоке на кассетах с пределом отсечения 50 кДа с последующей диафильтрацией против гистидинового буфера (рН 6.0). Концентрация белка в финальном растворе составляла не менее 20 мг/мл.
Пример 27
Гомологичное моделирование комплекса HER3 и antiHER3 VHH
Структура комплекса взаимодействия была получена с помощью программного обеспечения Schrodinger Suite 2016-1. В качестве структуры мишени была взята кристаллическая структура PDB 4LEO [Cancer Res. 2013 73: 5183-5194]. Для моделирования структуры антитела был применен протокол гомологичного фолдинга антитела с использованием шаблонной структуры PDB 2Х10 [Cell.Mol.Life Sci. 2010 67: 1519-1535] для построения каркасных регионов. Петля HCDR3 была промоделирована de novo с помощью алгоритма Prime [Proteins 2014 82(2): 1646-1655]. Полученные десять конформаций этой петли с наилучшими характеристиками (энергия, схожесть структуры петли с имеющимися в базе данных и проч.) были отранжированы по значению свободной энергии, после чего была выбрана наилучшая структура по этому параметру и произведен докинг со структурой мишени с помощью алгоритма Piper [Proteins 2006 65(2): 392-406]. В результате был получен репертуар поз, из которых была выбрана поза с наилучшей энергией связывания. Финальная структура была подвержена 5ns молекулярной динамики с использованием пакета Desmond [Bowers K.J., et al (2006) Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters, Proceedings of the ACM/IEEE Conference on Supercomputing (SC06), Tampa, Florida, November 11-17, 2006] и энергетического поля OPLS_2005 [Banks, J.L., et al (2005) Integrated Modeling Program, Applied Chemical Theory (IMPACT), J Comput Chem. 26(16): 1752-1780]. Из траектории симуляции был получен промежуток времени, на котором комплекс малоподвижен, и итоговая структура была взята из него.
Изображение (Фиг. 36) строилось с помощью программы для просмотра структур PyMol [The PyMOL Molecular Graphics System, Version 1.8  , LLC].
, LLC].


| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С ДОМЕНОМ II HER2, И ЕГО КИСЛЫЕ ВАРИАНТЫ | 2009 |
|
RU2543664C2 |
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| АНТИ-PD-1-АНТИТЕЛА, СПОСОБ ИХ ПОЛУЧЕНИЯ И СПОСОБ ПРИМЕНЕНИЯ | 2016 |
|
RU2656181C1 |
| ТЕРАПИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, РЕЗИСТЕНТНОЙ К ПРЕПАРАТАМ НА ОСНОВЕ ПЛАТИНЫ | 2005 |
|
RU2403065C2 |
| ВВЕДЕНИЕ ФИКСИРОВАННЫХ ДОЗ HER-АНТИТЕЛ | 2005 |
|
RU2438705C2 |
| АНТИ-LRP6 АНТИТЕЛА | 2011 |
|
RU2587625C2 |
| СПОСОБЫ ИДЕНТИФИКАЦИИ ОПУХОЛЕЙ, ВОСПРИИМЧИВЫХ К ЛЕЧЕНИЮ АНТИТЕЛАМИ ПРОТИВ ErbB2 | 2003 |
|
RU2338751C2 |
| НЕКОНКУРЕНТНЫЕ В ОТНОШЕНИИ НЕЙРЕГУЛИНА АЛЛОСТЕРИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО HER3 И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2704228C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К PD-L1 | 2017 |
|
RU2665790C1 |
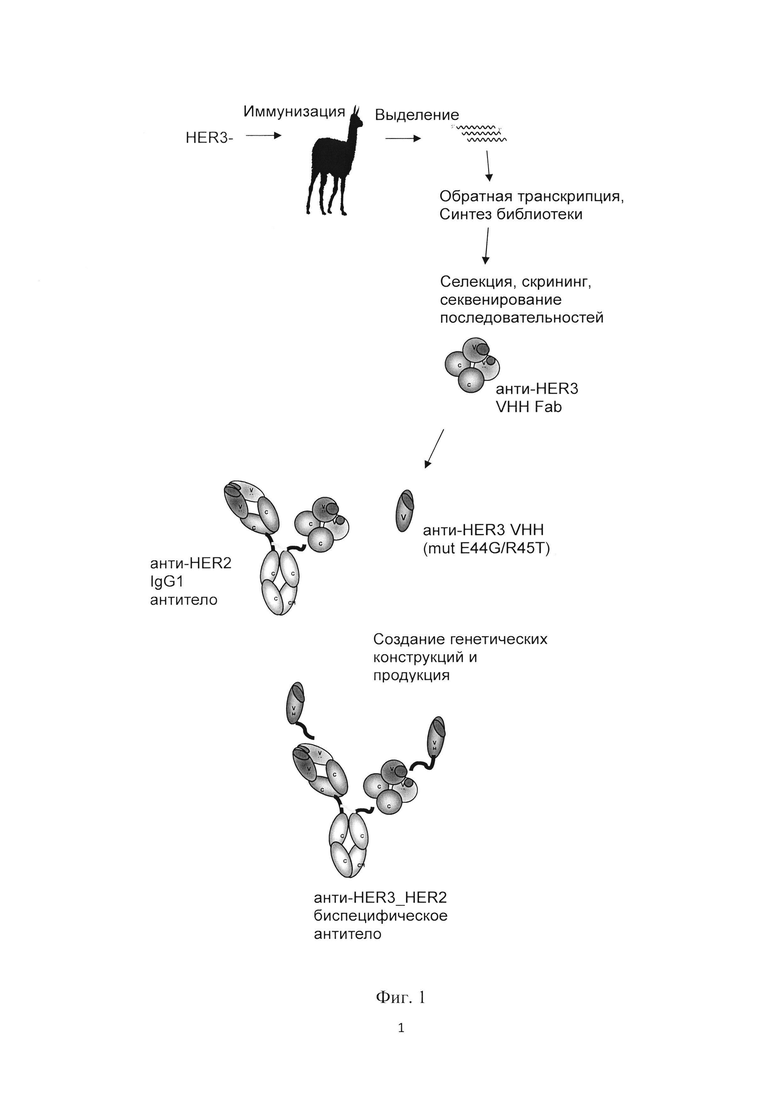




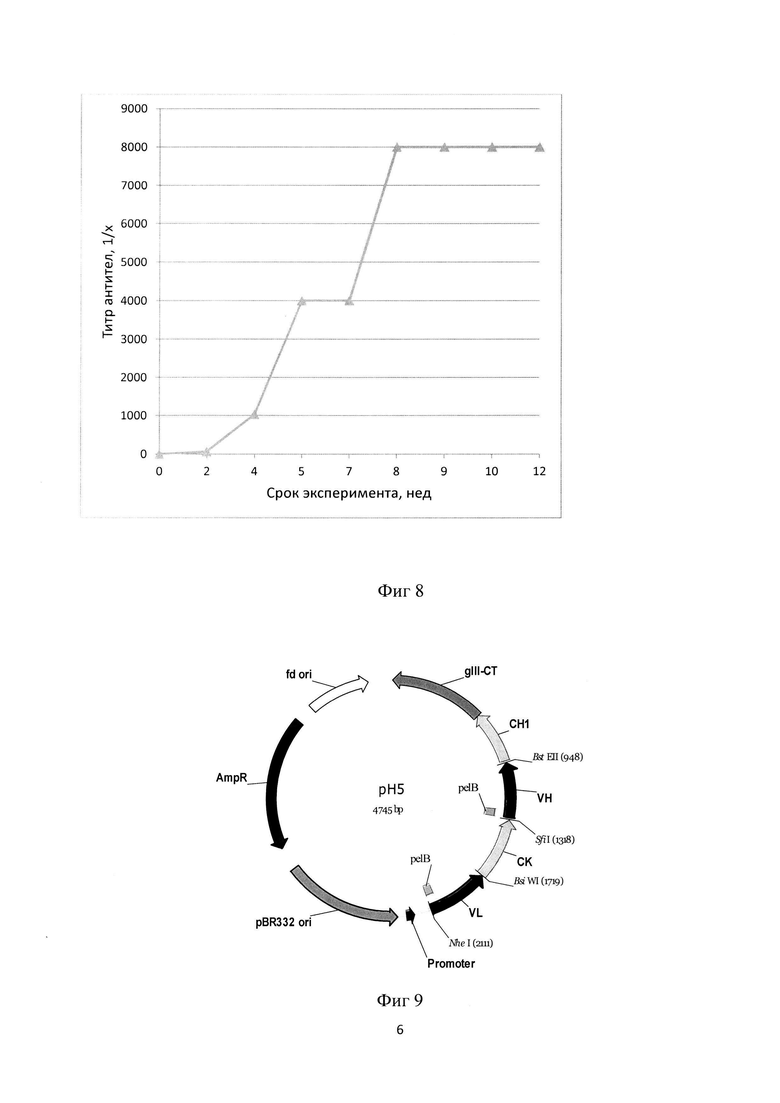
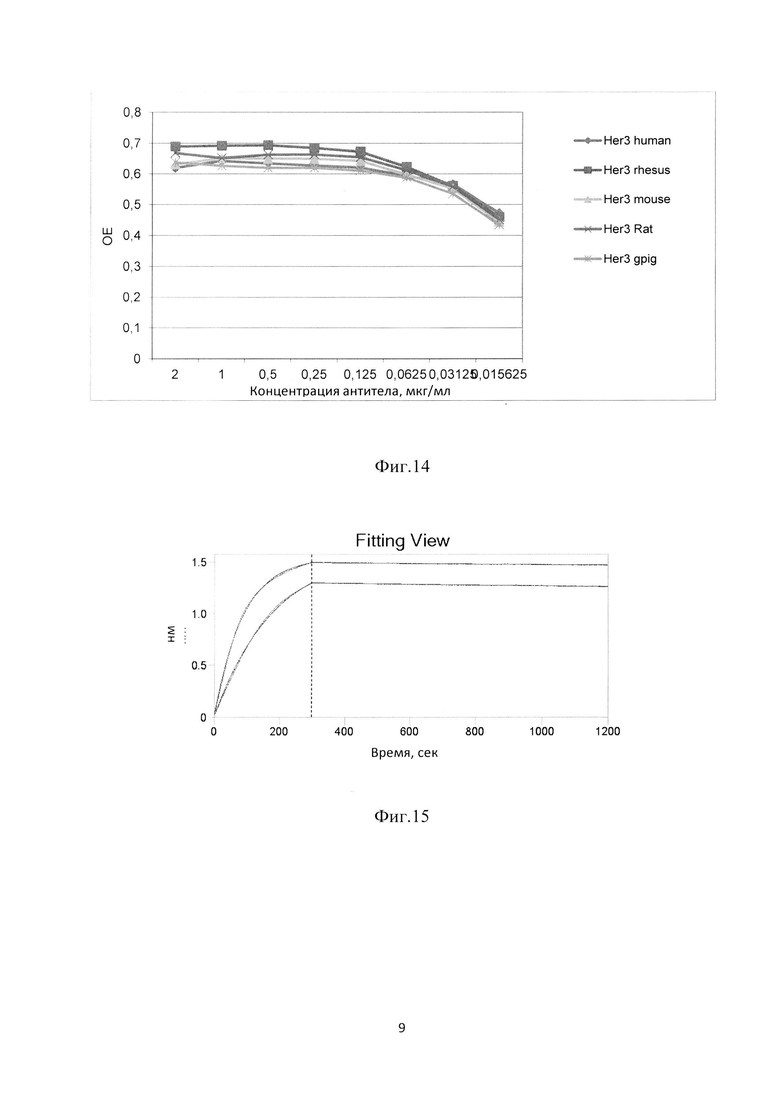
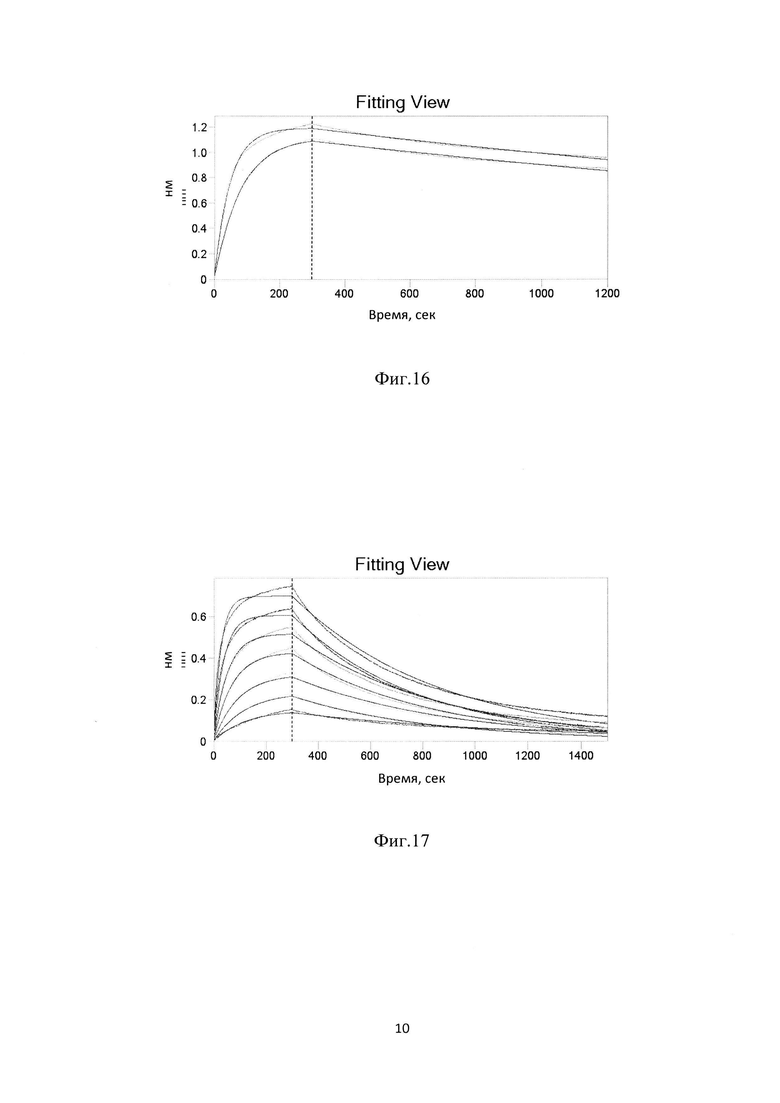
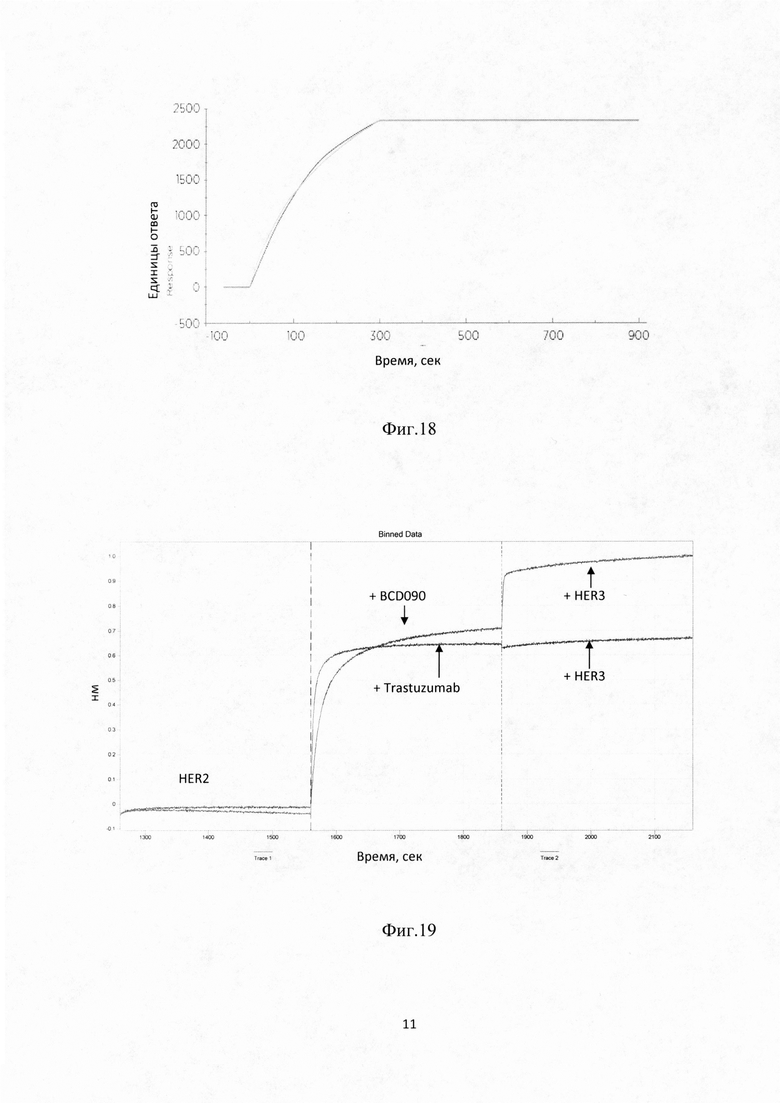
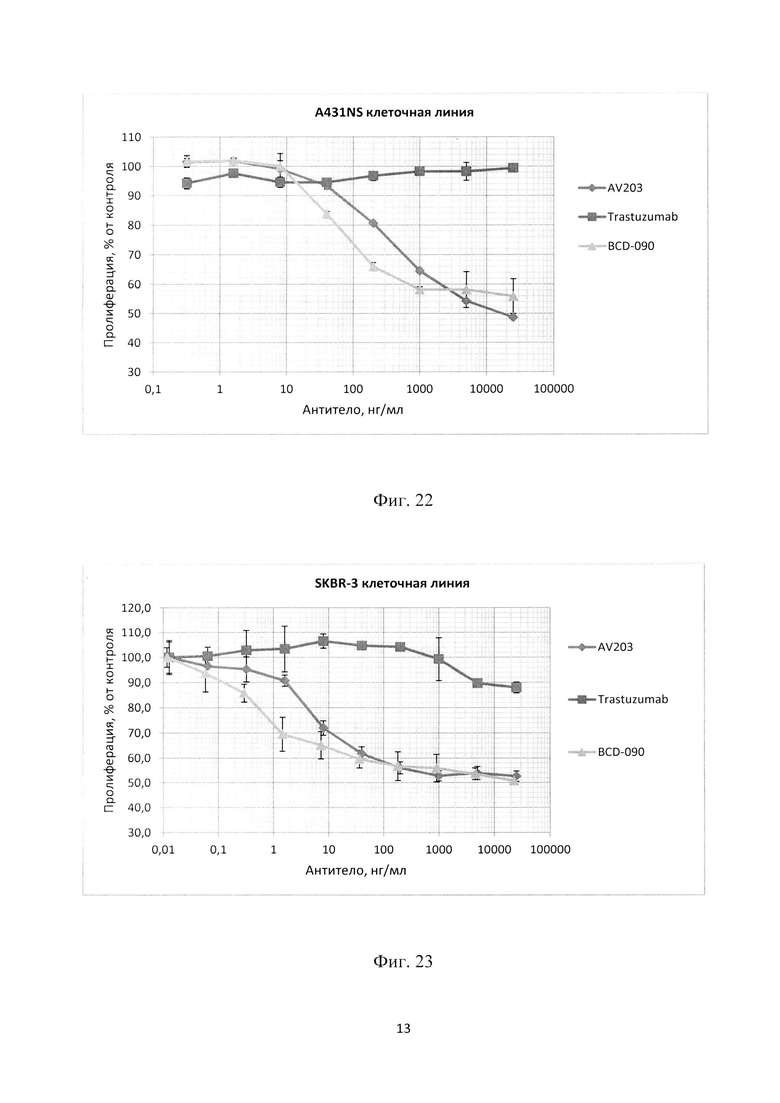
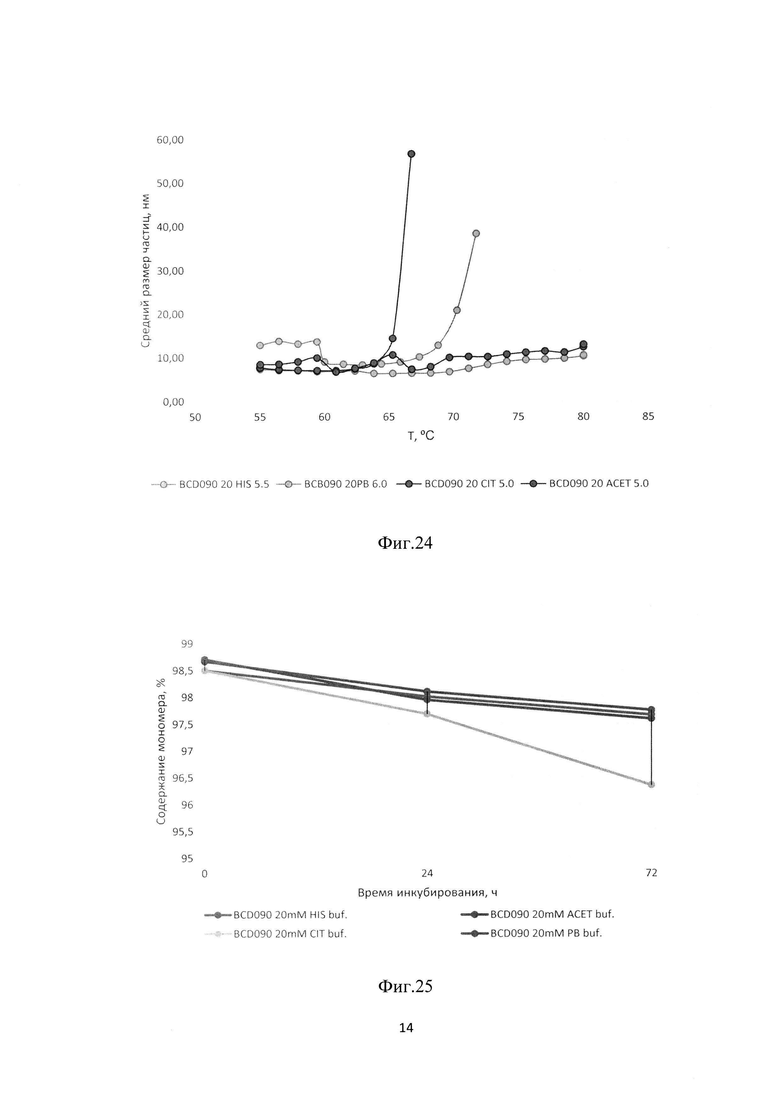
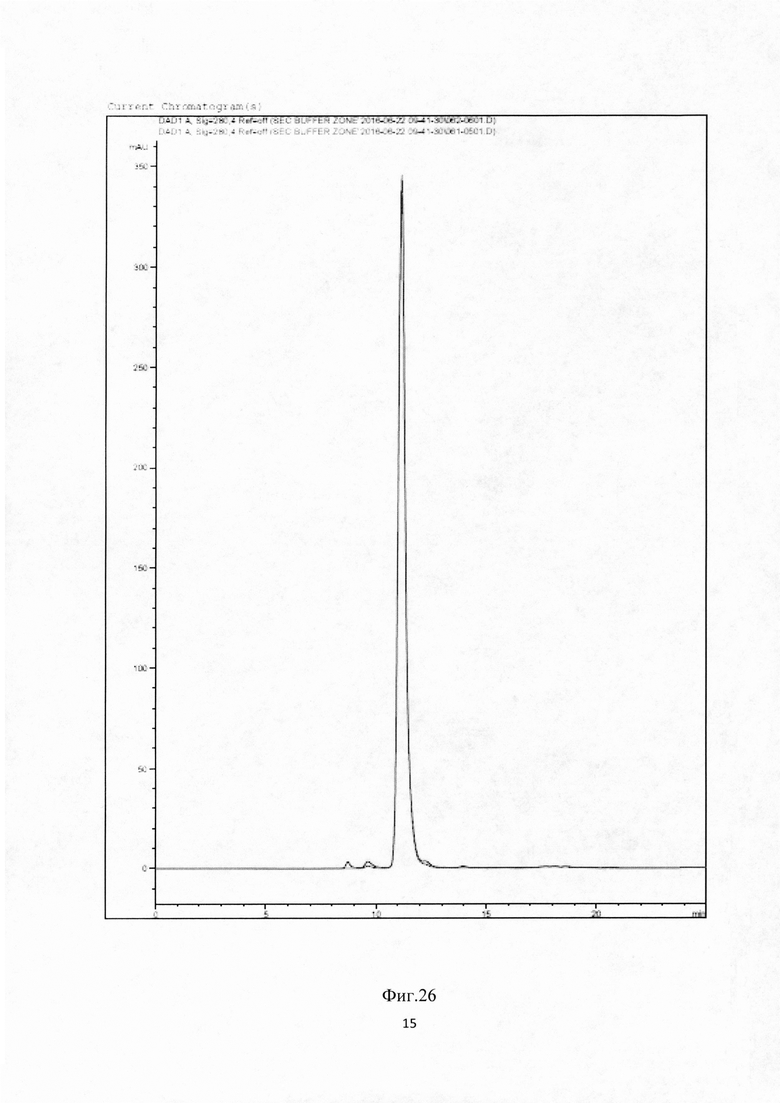
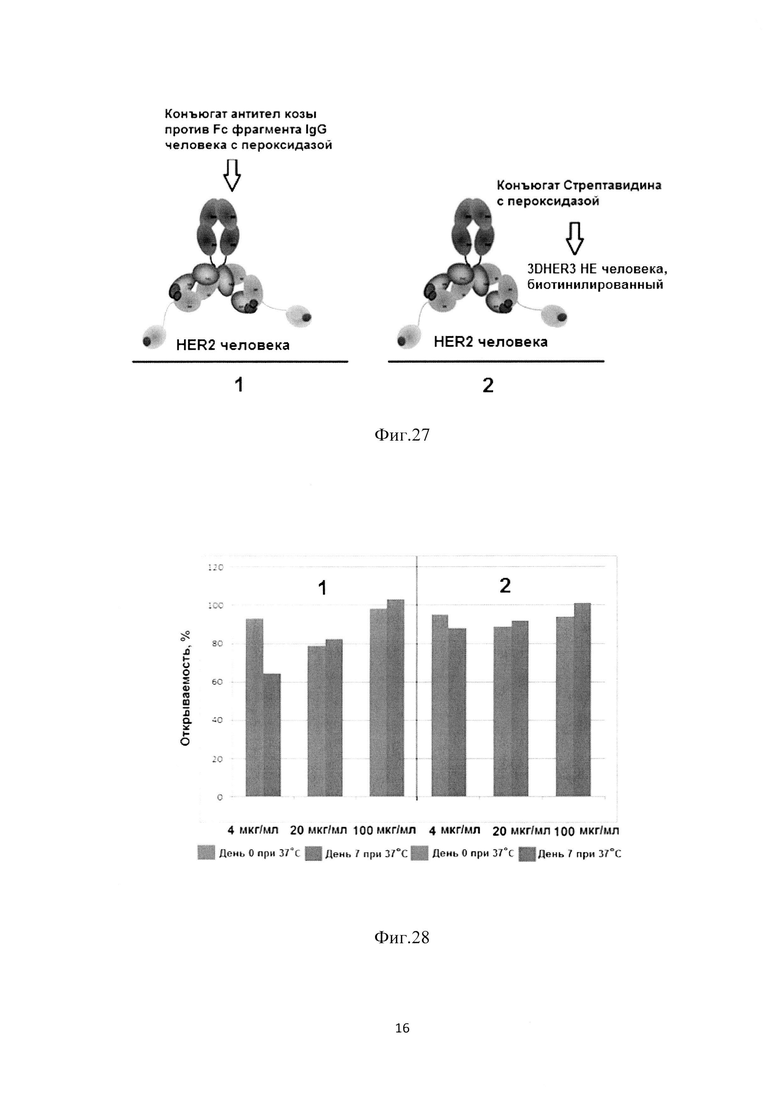
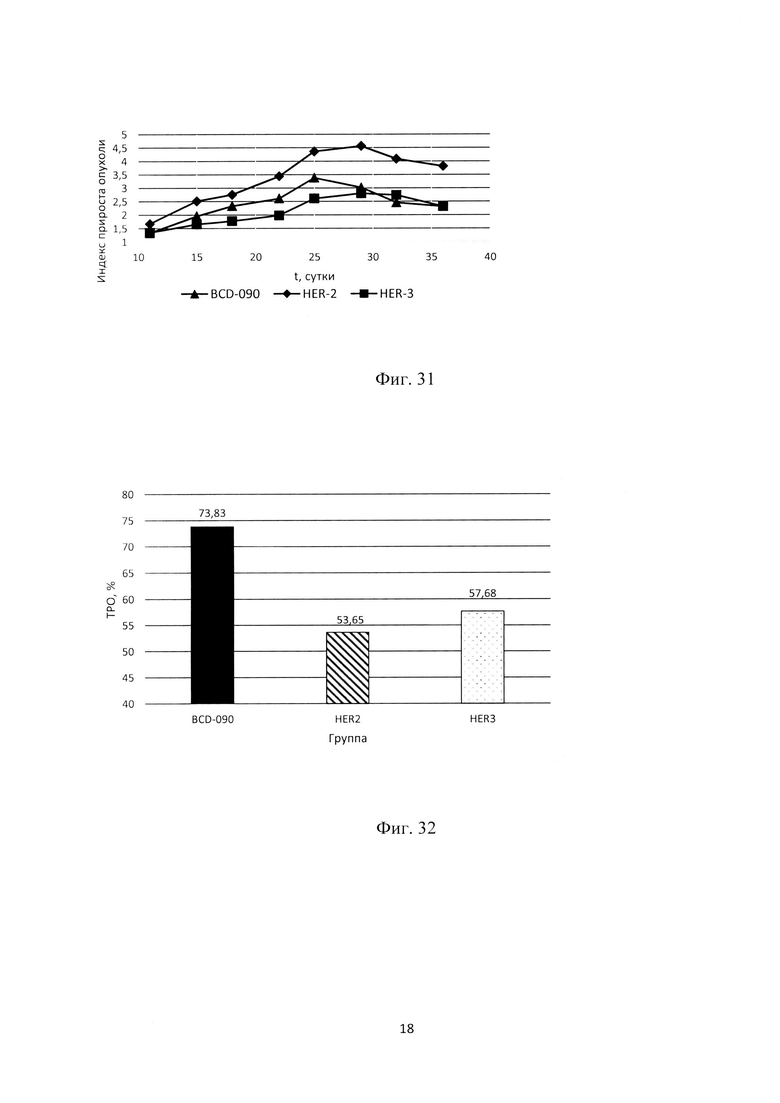
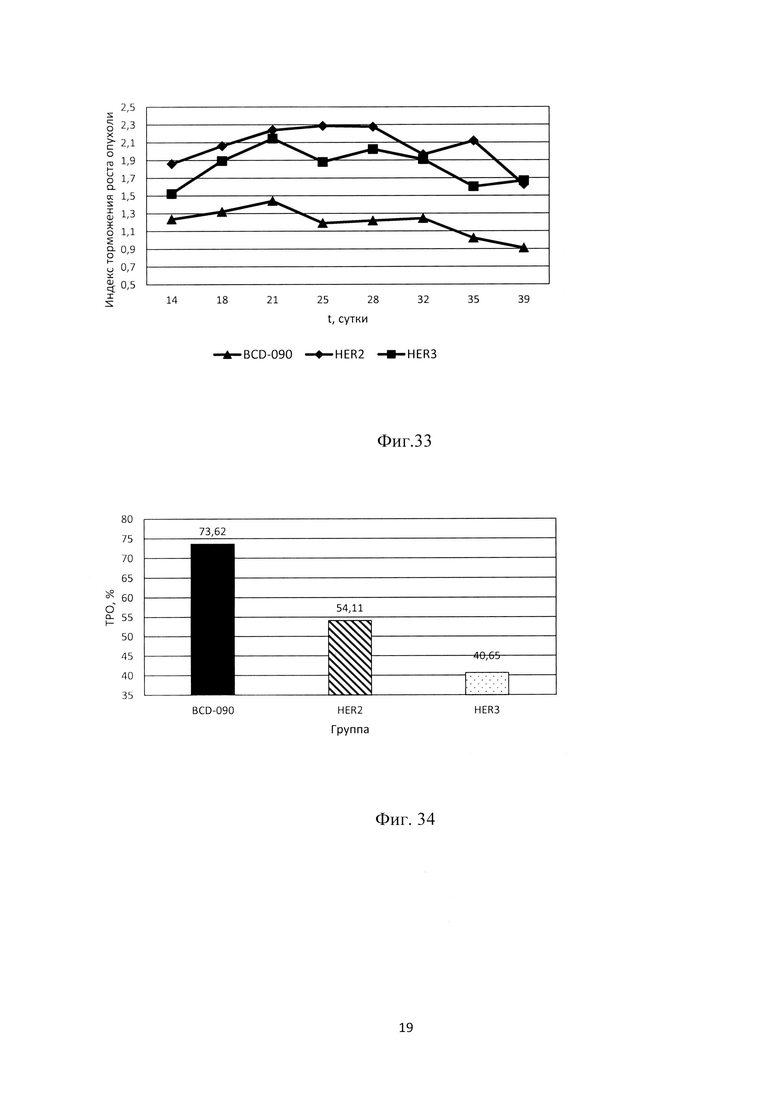
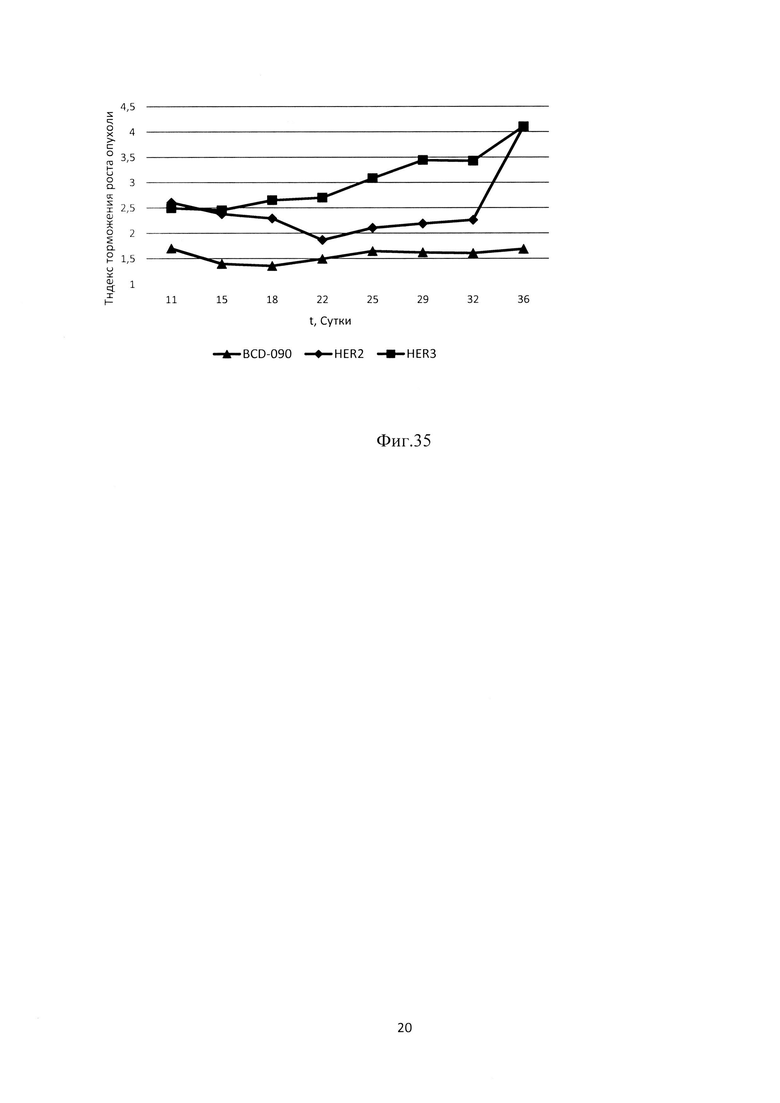

Изобретение относится к биотехнологии. Описано биспецифическое антитело, которое специфично связывается с HER2 человека или яванского макака и HER3, включающее: вариабельный домен тяжелой цепи VHH, который специфично связывается с HER3, где вариабельный домен тяжелой цепи VHH содержит аминокислотные последовательности, по меньшей мере на 75% гомологичные последовательностям SEQ ID NO: 1-3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2 человека или яванского макака. Изобретение также относится к ДНК, кодирующей указанное антитело, соответствующему экспрессионному вектору и способу продукции, а также к способу лечения с использованием указанного биспецифичного антитела. 11 н. и 20 з.п. ф-лы, 36 ил., 7 табл., 27 пр.
1. Биспецифическое антитело, которое специфично связывается с HER2 человека или яванского макака и HER3, включающее:
1) вариабельный домен тяжелой цепи VHH, который специфично связывается с HER3, где вариабельный домен тяжелой цепи VHH содержит аминокислотные последовательности по меньшей мере на 75% гомологичные последовательностям SEQ ID NO: 1-3;
2) антитело или его антигенсвязывающий домен, которые специфично связываются с HER2 человека или яванского макака.
2. Биспецифическое антитело по п. 1, где вариабельный домен тяжелой цепи VHH, который специфично связывается с HER3, и антитело или его антигенсвязывающий домен, которые специфично связываются с HER2 человека или яванского макака, соединены пептидным линкером.
3. Биспецифическое антитело по пп. 1, 2, где вариабельный домен тяжелой цепи VHH содержит аминокислотные последовательности, по меньшей мере на 90% гомологичные последовательностям SEQ ID NO: 1-3.
4. Биспецифическое антитело по п. 3, где вариабельный домен тяжелой цепи VHH содержит аминокислотные последовательности SEQ ID NO: 1-3.
5. Биспецифическое антитело по любому из пп. 1, 2, где вариабельный домен тяжелой цепи VHH содержит аминокислотную последовательность, по меньшей мере на 90% гомологичную аминокислотной последовательности SEQ ID NO: 4.
6. Биспецифическое антитело по п. 5, где вариабельный домен тяжелой цепи VHH содержит аминокислотную последовательность SEQ ID NO: 4.
7. Биспецифическое антитело по любому из пп. 1, 2, где антитело, специфичное против HER2, представляет собой полноразмерное антитело IgG.
8. Биспецифическое антитело по п. 7, где полноразмерное антитело IgG относится к изотипу IgG1, IgG2, IgG3, IgG4 человека.
9. Биспецифическое антитело по любому из пп. 1, 2, включающее антигенсвязывающий домен, который специфично связывается с HER2.
10. Биспецифическое антитело по п. 8, где полноразмерное антитело или антигенсвязывающий домен, специфичные к HER2 человека или яванского макака, содержат тяжелую и легкую цепи, которые включают аминокислотные последовательности, по меньшей мере на 80% гомологичные последовательностям SEQ ID NO: 5 и SEQ ID NO: 6 соответственно.
11. Биспецифическое антитело по п. 10, где полноразмерное антитело или антигенсвязывающий домен, специфичные к HER2 человека или яванского макака, содержат тяжелую и легкую цепи, которые включают аминокислотные последовательности SEQ ID NO: 5 и SEQ ID NO: 6, соответственно.
12. Биспецифическое антитело по п. 2, где пептидный линкер имеет длину более 5 аминокислот и состоит из любого набора аминокислот, выбранных из G, A, S, Р, Е, Т, D, K.
13. Биспецифическое антитело по пп. 2 и 12, где часть VHH присоединена к N-концу или С-концу легкой или тяжелой цепи антитела или антигенсвязывающего домена, специфичных к HER2 человека или яванского макака.
14. Биспецифическое антитело по любому из пп. 1, 2, имеющее константу диссоциации Kd связывания с HER2 человека не более 10-10 М и обезьяны не более 10-7 М, KD связывания с HER3 человека не более 10-9 М и обезьяны не более 10-8 М.
15. Раствор для ингибирования биологической активности HER2 человека или яванского макака и HER3 у субъекта, включающий биспецифическое антитело по любому из пп. 1-14 и один или несколько фармацевтически приемлемых эксципиентов.
16. Раствор по п. 15, который имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и хранении при t=4°С в течение более чем 6 месяцев содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
17. Раствор по п. 15, который имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и при повышении температуры до 37°С в течение более чем 2 недель содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
18. Раствор по п. 15, который имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и повышении температуры до 50°С в течение более чем 6 часов содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
19. Выделенная, молекула нуклеиновой кислоты, которая кодирует антитело по любому из пп. 1-14.
20. Выделенная молекула нуклеиновой кислоты по п. 19, где нуклеиновая кислота представляет собой ДНК.
21. Экспрессионный вектор, содержащий нуклеиновую кислоту по любому из пп. 19, 20.
22. Способ получения клетки-хозяина для получения антитела по любому из пп. 1-14, включающий трансформирование клетки вектором по п. 21.
23. Клетка-хозяин для получения антитела по любому из пп. 1-14, содержащая нуклеиновую кислоту по любому из пп. 19, 20.
24. Способ получения антитела по любому из пп. 1-14, заключающийся в культивировании клетки-хозяина по п. 23 в культуральной среде в условиях, достаточных для получения указанного антитела, при необходимости, с последующим выделением и очисткой полученного антитела.
25. Фармацевтическая композиция для профилактики или лечения заболевания или нарушения, опосредуемого HER2 человека или яванского макака и HER3, содержащая антитело по любому из пп. 1-14, в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами.
26. Фармацевтическая композиция по п. 25, предназначенная для профилактики или лечения заболевания или нарушения, опосредуемого HER2 человека или яванского макака и HER3, выбранного из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
27. Способ ингибирования биологической активности HER2 человека или яванского макака и HER3 у субъекта, нуждающемуся в таком ингибировании, включающий введение субъекту эффективного количества антитела по любому из пп. 1-14.
28. Способ лечения заболевания или нарушения, опосредованного HER2 человека или яванского макака и HER3, включающий введение субъекту антитела по любому из пп. 1-14, или раствора по п. 15, или фармацевтической композиции по пп. 25, 26, нуждающемуся в таком лечении, в терапевтически эффективном количестве.
29. Способ лечения заболевания или нарушения по п. 28, где заболевание или нарушение выбрано из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
30. Применение антитела по любому из пп. 1-14, раствора по п. 15 или фармацевтической композиции по пп. 25, 26 для лечения у субъекта, нуждающегося в таком лечении, заболевания или нарушения, опосредуемого HER2 человека или яванского макака и HER3.
31. Применение по п. 30, где заболевание или нарушение выбрано из группы: рака молочной железы (РМЖ), злокачественных новообразований желудка, немелкоклеточного рака легкого, злокачественных новообразований головы и/или шеи.
| WO2015173248 A1, 19.11.2015 | |||
| СПОСОБ РАЗГРУЗКИ ДАВЛЕНИЯ В ТОПЛИВНОЙ РАМПЕ НЕПОСРЕДСТВЕННОГО ВПРЫСКА (ВАРИАНТЫ) И ТОПЛИВНАЯ СИСТЕМА | 2017 |
|
RU2727942C2 |
| ВЫСОКОЭФФЕКТИВНЫЕ КОНЪЮГАТЫ И ГИДРОФИЛЬНЫЕ СШИВАЮЩИЕ АГЕНТЫ (ЛИНКЕРЫ) | 2009 |
|
RU2487877C2 |

