Область техники
Изобретение относится к области биотехнологии и ветеринарной вирусологии, и представляет собой экспрессирующий плазмидный вектор, обеспечивающий синтез в клетках Е. coli рекомбинантного белка, состоящего из фрагмента р30 вируса АЧС, тиоредоксина (Trx-Tag) и полигистидиновых участков (6xHis), и может быть использовано для изготовления вирусспецифических антигенов в тест-системы, предназначенные для исследования сывороток свиней методом иммуноблоттинга.
Уровень техники
АЧС - контагиозная, септическая болезнь свиней, характеризующаяся лихорадкой, признаками токсикоза, геморрагическим диатезом и высокой летальностью. К АЧС восприимчивы домашние свиньи и дикие кабаны независимо от породы и возраста, у которых болезнь может протекать сверхостро, остро, подостро, хронически и бессимптомно. Возбудитель АЧС передается от больных животных и выживших вирусоносителей контактным и алиментарным путем, а также трансплацентарно [1]. Поскольку вакцины против АЧС не разработаны, то для ликвидации и предупреждения распространения болезни применяется тотальный убой свиней в первой угрожаемой зоне радиусом до 20 км от очага инфекции, а также жесткие карантинные мероприятия во второй угрожаемой зоне радиусом до 100 км. С учетом этого лабораторная диагностика имеет решающее значение для принятия решений о ликвидации вспышек и контроля распространения болезни. Современные методы лабораторной диагностики АЧС, основанные на определении вирусных антигенов или ДНК, не всегда обеспечивают выявление хронически или бессимптомно инфицированных животных. В этих случаях наиболее информативными являются серологические методы, основанные на определении антител к вирусным белкам в сыворотках крови животных.
Согласно рекомендациям Международного эпизоотического бюро (МЭБ) для подтверждения сомнительных результатов лабораторной диагностики АЧС непрямыми вариантами иммуноферментного анализа (ИФА) или реакции иммунофлуоресценции следует использовать метод иммуноблоттинга. Также его применяют при нарушении условий хранения проб сывороток крови и для исключения ложноположительных результатов в случаях, когда антитела сывороток крови от привитых животных реагируют с антигенами клеток, использованных при изготовлении вакцин.
В рекомендованных МЭБ тест-системах для выявления АЧС методом иммуноблоттинга вирусные антигены происходят из живого вируса, что предполагает манипуляции с инфекционным агентом. Применение вместо них рекомбинантных белков гарантирует биологическую безопасность производства диагностических тест-систем и высокую специфичность входящих в их состав антигенов [2, 3].
Установлено, что в ранние сроки после заражения (3-6 суток) низко-, умеренно- и высоковирулентными штаммами вируса АЧС в сыворотках крови свиней преимущественно выявляются антитела к структурному белку вируса р30 [4]. Было показано, что чувствительность и специфичность ИФА при использовании рекомбинантного р30 для обнаружения антител к АЧС в сыворотках экспериментально инфицированных свиней и полевых сывороток от серопозитивных животных с бессимптомным течением болезни, составляют 96% и 99%, соответственно [5].
Таким образом, применение рекомбинантного белка р30 в качестве антигена для выявления вирусспецифических антител при АЧС методом иммуноблоттинга является предпочтительным решением.
Известен штамм клеток Е. coli M15 (Qiagen) с плазмидой рТТ9р30-9, экспрессирующий гибридный белок р30 вируса АЧС, выход которого при индукции IPTG составлял всего 1 мг с 1 дм3 бактериальной культуры [6], чего недостаточно для получения рекомбинантного р30 в препаративных количествах.
Патент RU 2463343 С1 раскрывает штамм клеток Е. coli BL21(DE3)pLysS, клон pTT9/ASFVp30 (клон 11), содержащий плазмиду, кодирующую, в том числе, рекомбинантный белок, включающий аминокислоты с 90-й по 190-ю белка р30 с молекулярной массой 14 кДа, в том числе, его конформационный эпитоп, а также целлюлозо-связывающий домен. Выход рекомбинантного белка после очистки на целлюлозном сорбенте достигает 2-4 мг с 1 дм3 бактериальной культуры [7].
В описании изобретения не были представлены электрофореграммы препаратов очищенного рекомбинантного белка, которые бы объективно свидетельствовали об отсутствии в нем примесей белков Е. coli, что принципиально важно для метода иммуноблоттинга. Кроме того, метод иммуноблоттинга предполагает электрофоретическое разделение полипептидов в денатурирующих условиях, в результате чего конформационные эпитопы теряют антигенную активность.
Целью настоящего изобретения является усовершенствование генетической конструкции, кодирующей фрагмент белка р30 вируса АЧС и обеспечивающей охват максимального количества его гидрофильных участков и антигенных эпитопов, а также получение максимально растворимой формы рекомбинантного белка за счет лидирующей последовательности тиоредоксина, входящей в его структуру, и последовательностей, кодирующих полигистидиновые участки, обеспечивающие его эффективную очистку методом металлохелатной хроматографии в препаративных количествах.
Раскрытие изобретения
Технический результат от использования предлагаемого изобретения заключается в возможности поучения пригодного для иммуноблоттинга высокоочищенного модифицированного рекомбинантного р30 вируса АЧС с выходом не менее 15-20 мг с 1 дм3 бактериальной культуры.
Для достижения технического результата изобретения существенна совокупность следующих этапов:
1) дизайн конструкции гена TrxA-6xHis-p30e2-6xHis, предполагающий подбор олигонуклеотидных праймеров с сайтами рестрикции для фланкирования участков гена CP204L вируса АЧС с сохранением рамок считывания при клонировании;
2) выбор штамма Е. coli и реципиентной плазмиды для создания экспрессирующей конструкции;
3) оптимизация условий экспрессии и хроматографической очистки рекомбинантного белка;
4) оценка специфичности рекомбинантного белка методом иммуноблоттинга.
Лидирующая последовательность фрагмента тиоредоксина в составе экспрессирующего вектора в Е. coli обеспечивает сохранение рекомбинантного белка p30e2_TrxA_6xHis (Rec р30е2) в растворимой форме в цитоплазме продуцента, а наличие двух 6xHis участков позволяет достигать высокой степени очистки целевого продукта на металлохелатном сорбенте.
Предлагаются:
- нуклеотидная последовательность TrxA-6xHis-p30e2-6xHis (SEQ ID №1), представленная открытой рамкой считывания химерного белка Rec р30е2 (SEQ ID №3);
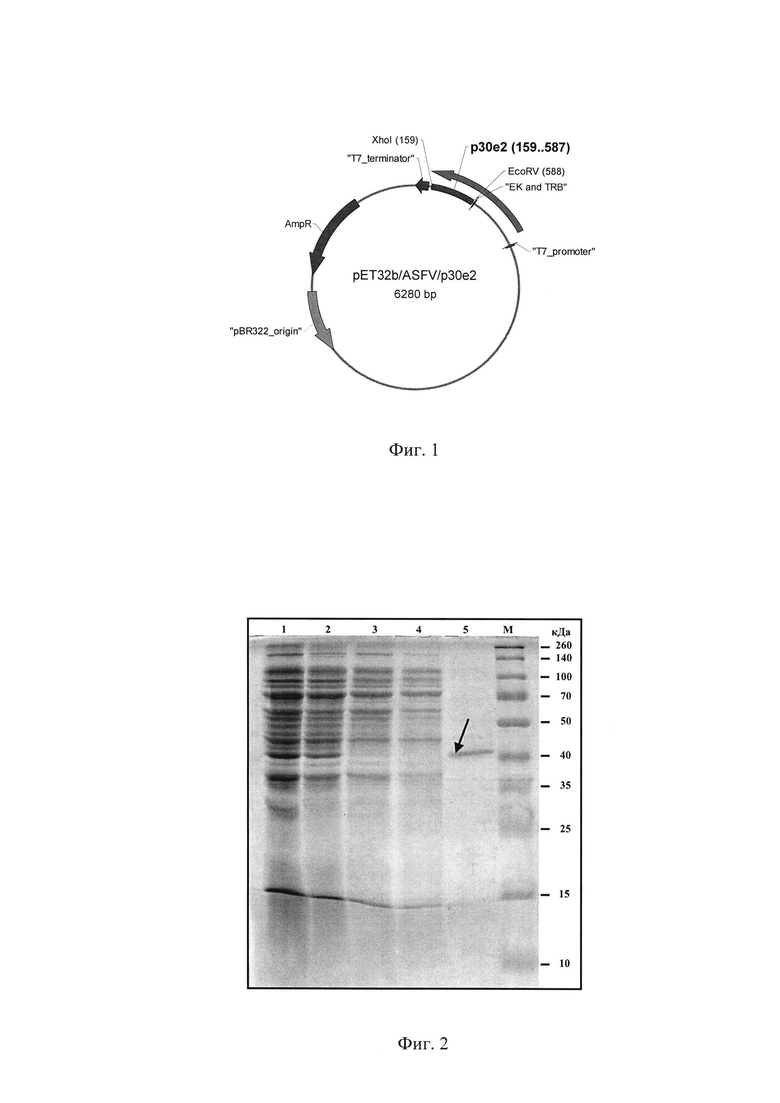
- рекомбинантная плазмида pET32b(+)ASFV/p30e2 (SEQ ID №2), обеспечивающая высокий уровень экспрессии Rec р30е2 в клетках Escherichia coli и содержащая ген TrxA-6xHis-p30e2-6xHis с последовательностью SEQ ID №1 под контролем промотора гена 10 бактериофага Т7 и терминатора Т7 (фиг. 1).
- аминокислотная последовательность химерного белка Rec р30е2 (SEQ ID №3).
Дизайн гибридного гена TrxA-6xHis-p30e2-6xHis проводили в BioEdit 7.2.5 (http://www.mbio.ncsu.edu/bioedit) с учетом сохранения корректных рамок считывания при клонировании в вектор экспрессии. Нуклеотидные последовательности, кодирующие соответствующие участки гена CP204L вируса АЧС, получали методом ПЦР со специфичными праймерами на матрице ДНК вируса АЧС штамма Stavropol2008 (Ставрополь 01/08) [8].
Для получения вектора экспрессии была выбрана плазмида pET32b(+), которая содержала лидирующую последовательность фрагмента тиоредоксина Е. coli под контролем сильного промотора гена «10» фага Т7, обеспечивающего высокий уровень экспрессии в Е. coli, стабильно или индуцибельно экспрессирующую ДНК-зависимую РНК-полимеразу фага Т7 [9].
Экспрессирующая рекомбинантная плазмида pET32b(+)ASFV/p30e2 получена путем субклонирования полученной в ПЦР нуклеотидной последовательностей гена CP204L вируса АЧС с праймерами F-p30e2 и R-р30е2 (таблица 1) в вектор pET32b(+) по сайтам рестрикции EcoRV и XhoI (фиг. 1).
После трансформации клеток Е. coli штамма KRX (Promega) плазмидой pET32b(+)ASFV/p30e2 и последующего скрининга рекомбинантов с высоким уровнем экспрессии гибридного белка Rec р30е2, отобран штамм-продуцент Е. coli pET32b/ASFV/p30e2/1.
Экспрессия целевого белка в полученном штамме-продуценте осуществляется при культивировании на селективных средах с добавлением индуктора L-рамнозы, последующего сбора биомассы клеток, их разрушения, отделения нерастворимых белков и дебриса и очисткой белка Rec р30е2 методом металлохелатной хроматографии в нативных или денатурирующих условиях. Конечный выход целевого белка составляет не менее 15-20 мг с 1 дм3 бактериальной культуры.
Осуществление изобретения
При осуществлении изобретения, помимо методов, описанных в нижеследующих примерах, использовали ряд общепринятых методик, описанных в руководствах по молекулярной биологии [10].
Изобретение иллюстрируют нуклеотидные и аминокислотные последовательности:
Seq ID №1 Нуклеотидная последовательность TrxA-6xHis-p30e2-6xHis, кодирующая химерный белок Rec р30е2 (1…327 - лидирующая последовательность тиоредоксина (TrxA-Tag); 349…366, 916…933 -полигистидиновые участки (6xHis); 376…393 - нуклеотидная последовательность, кодирующая сайт расщепления тромбина (thrombin site); 460…474 - нуклеотидная последовательность, кодирующая сайт расщепления энтерокиназы (enterokinase site); 490…894 - нуклеотидная последовательность, кодирующая рекомбинантный р30е2).
Seq ID №2 Нуклеотидная последовательность рекомбинантной плазмиды pET32b(+)ASFV/p30e2 (746…1072 - лидирующая последовательность тиоредоксина (TrxA-Tag); 140…157, 707…724 - полигистидиновые участки (6xHis); 680…697 - нуклеотидная последовательность, кодирующая сайт расщепления тромбина (thrombin site); 599…613 - нуклеотидная последовательность, кодирующая сайт расщепления энтерокиназы (enterokinase site); 179…583 - нуклеотидная последовательность, кодирующая рекомбинантный р30е2).
Seq ID №3 Аминокислотная последовательность химерного белка Rec р30е2 (1…109 - лидирующая последовательность тиоредоксина (TrxA-Tag); 117…122, 306…311 - полигистидиновые участки (6xHis); 126…131 - сайт расщепления тромбина (thrombin site); 154…158 - сайт расщепления энтерокиназы (enterokinase site); 164…298 - последовательность рекомбинантного р30е2).
Изобретение иллюстрируют графические материалы.
Фиг. 1 - Схема рекомбинантной плазмиды pET32b(+)ASFV/p30e2. Примечания: обозначено положение нуклеотидной вставки «р30е2»; стрелками указаны открытые рамки считывания гена TrxA-6xHis-p30e2-6xHis; сайты энтерокиназы и тромбина «ЕК and Trb»; сайтов рестрикции; «T7_promoter» - промотер фага Т7; «T7_terminator» - последовательность терминального участка фага Т7; «AmpR» - bla (β-lactamase) ген, кодирующий резистентность к ампициллину, карбенициллину и другим производным антибиотикам; «pBR 322_origin» - инициация репликации ColE1/pMB1/pBR322/pUC.
Фиг. 2 - Электрофореграммы исходного лизата клеток Е. coli (штамм-продуцент KRX pET32b/ASFV/p30e2/1) (1), фракций в процессе отмывки сорбента (2-4) и фракции очищенного рекомбинантного белка (5). М - окрашенные маркеры молекулярной массы белков Spectra™ Multicolor 10 - 260 кДа.
Фиг. 3 - Специфическое окрашивание иммунострипа (указано стрелкой) в результате взаимодействия рекомбинантного белка р30 вируса АЧС со специфическими антителами положительной контрольной сыворотки крови свиньи, иммунной к вирусу АЧС (К+), и отсутствие окрашивания иммунострипа после инкубирования его с сывороткой крови от интактной свиньи (К-). Примечание: К+ - положительная инактивированная контрольная сыворотка, К- - отрицательная контрольная сывороток.
Изобретение иллюстрируют следующие примеры.
Пример 1. Получение специфической нуклеотидной вставки, кодирующей участок гена CP204L (р30) вируса АЧС
Аминокислотная последовательность структурного белка р30 вируса АЧС составляет 201 аминокислоту (ген CP204L). На первом этапе был проведен биоинформатический анализ с целью поиска трансмембранных регионов и/или гидрофобных участков, которые могут негативно повлиять на экспрессию белка или последующую очистку. Используя алгоритм по поиску протяженных гидрофобных участков «Ноор and Woods» (BioEdit 2.7.5), а также сервер «ТМНММ Server 2.0» (http://www.cbs.dtu.dk/services/TMHMM/) локализованы 2 перекрывающихся участка для клонирования.
В качестве объекта для расчета праймеров была выбрана нуклеотидная последовательность генома вируса АЧС штамма Georgia/2007 (GeneBank FR682468.1). При подборе праймеров соблюдали минимальное содержание димерных структур и самокомплементарных регионов, которые могут приводить к образованию шпилек с высокой температурой плавления. В праймеры F-p30e2 и R-p30e2 включены сайты рестрикции EcoRV и XhoI. Данные сайты рестрикции позволяют переклонировать нуклеотидную вставку в необходимый прокариотический и эукариотический экспрессирующий вектор. Последовательности рассчитанных олигонуклеотидных праймеров указаны в таблице 1.

ПЦР проводили на приборе Терцик МС2 («ДНК-технология», Россия). В качестве матрицы использовали геномную ДНК вируса АЧС штамма Ставрополь 01/08 (Stavropol2008), циркулирующего на территории РФ [8]. Препаративные реакции проводили в объеме 25 мкл. Реакционная смесь состояла из: 1х буфер для термостабильной pfu ДНК-полимеразы; 10 пмоль каждого праймера; 2,5 мМ смеси дезоксирибонуклеотидтрифосфатов; 2,5 ед. pfu-полимеразы. Поверх этой смеси наслаивали 20 мкл минерального масла.
Режим амплификации участка гена CP204L (р30): горячий старт: 95°С - 3 мин × 1 цикл; 24 цикла: денатурация 95°С - 30 сек, отжиг 54°С - 30 сек, элонгация 72°С - 30 сек; заключительная элонгация: 72°С - 3 мин × 1 цикл.
ПЦР-продукты (амплификаты) выделяли из 1% агарозного геля набором GeneJET Plasmid Miniprep Kit (Thermo Scietific, США) по протоколу производителя.
Очищенные ПЦР-продукты рестрицировали соответствующими эндонуклеазами 1 час при 37°С: EcoRV+XhoI в буфере Red (Thermo Scietific, США) по рекомендованному протоколу производителя. Продукты рестрикции подвергали фракционированию в 1% агарозном геле с последующим выделением набором GeneJET Gel Extraction Kit (Thermo Scietific, США).
Пример 2. Конструирование экспрессирующего плазмидного вектора pET32b(+)ASFV/p30e2
Акцепторную плазмиду pET32b(+) (Novagen) рестрицировали эндонуклеазами EcoRV и XhoI в буфере Red в течение 4 часов при 37°С. По завершению рестрикции эндонуклеазы инактивировали прогреванием 80°С 20 минут. Весь объем вносили в лунки 1% агарозного геля для последующего электрофореза. Линеаризованную плазмиду размером ~5900 п. н. выделяли из геля набором GeneJET Gel Extraction Kit.
Для получения плазмиды pET32b(+)ASFV/p30e2 очищенный рестрицированный ПЦР-продукт, полученный с использованием праймеров F-p30e2 и R-p30e2, лигировали с линеаризованной плазмидой pET32b(+), рестрицированной EcoRV/XhoI. Лигирование проводили с использованием Т4 ДНК-лигазы в рекомендованном буфере («α фермент», Россия) при комнатной температуре (25±2)°С в течение часа, соотношение вставки к вектору составляло 2:1.
Полученной лигазной смесью трансформировали клетки Е. coli штамма KRX с генотипом [F′, traD36, ΔompP, proA+В+, lacIq, Δ(lacZ)M15] ΔompT, endA1, recA1, gyrA96 (Nalr), thi-1, hsdR17 (rk -, mk +), e14-(McrA), relA1, supE44, Δ(lac-proAB), Δ(rhaBAD): T7 RNA polymerase.
Для этого к 50 мкл суспензии клеток Е. coli вносили 3 мкл лигазной смеси, инкубировали на льду в течение 10 минут, подвергали «термошоку» (42°С - 20 сек), охлаждали, а затем переносили суспензию в чашку Петри с твердой агаризованной питательной средой, содержащей ампициллин в концентрации 100 мкг/см3, с последующим инкубированием в термостате при 37°С в течение 18 часов. Единичные колонии анализировали методом ПЦР с использованием матрицы ДНК клонов и праймеров, использованных при получении ПЦР-продукта. Положительные в ПЦР клоны наращивали в 3 см3 питательного бульона SOB (ампициллин 100 мкг/см3) и проводили выделение плазмидной ДНК набором реактивов GeneJET Plasmid Miniprep Kit по протоколу производителя (ThermoScientific). Секвенирование для верифицирования конструкций проводили по методу Сэнгера. В результате секвенирования установили, что в области вставки плазмиды pET32b(+)ASFV/p30e2 штамма-продуцента pET32b/ASFV/p30e2/1, не содержатся мутации, то есть кодируется корректная последовательность гена TrxA-6xHis-p30e2-6xHis. Карта экспрессирующей плазмидной конструкции приведена на фиг. 1, ее нуклеотидная последовательность - SEQ ID №2. Аминокислотная последовательность продукта экспрессии приведена в SEQ ID №3.
Полученный рекомбинантный белок р30 штамма Ставрополь 01/08 (Stavropol2008) Rec р30е2 кодирует последовательность с 56 по 190 аминокислоту и соответствуют таковому других изолятов вируса АЧС, циркулирующего на территории РФ.
Пример 3. Индукция экспрессии и хроматографическая очистка белка Rec р30е2
Штамм продуцент pET32b/ASFV/p30e2/1 высевали из аликвот в 20 см3 жидкой питательной среды SOB, содержащей 100 мкг/см3 ампициллина, культивировали на термошейкере 14 часов при 37°С («ночная культура») M.R. Green & J. Sambrook (2012) [10].
На следующий день проводили пересев «ночной культуры» на питательную среду в соотношении среды к расплодке 25:1, соответственно, и культивирование по методу F.W. Studier и соавт.(2005) [11] в среде SOB-5052 с 50 мкг/см3 ампициллина и с использованием шейкера в течение 2-3 часов при температуре (37,0±0,5)°С и умеренном встряхивании (110-120 об/мин) до концентрации (6-7)×107 жизнеспособных клеток в см3, при этом оптическая плотность бактериальной культуры при длине волны 600 нм (OD600) составляла 0,4-0,5 (процедура подращивания пересеянной на свежеприготовленную среду «ночной» бактериальной культуры). Далее инкубировали бактериальную культуру в течение 1-2 часов до концентрации (7-9)×107 жизнеспособных клеток в см3 (OD600 0,5-0,6) при температуре (28,0±0,5)°С и также при постоянном встряхивании, затем, вносили индуктор экспрессии рекомбинантного белка - 20% раствор L-рамнозы, в отношении 1:200 (5 см3 индуктора / 1000 см3 бактериальной культуры) [9, 11, 12], и продолжали инкубировать при тех же параметрах еще 16-18 часов («автоиндукция» экспрессии рекомбинантного белка Rec р30е2).
Для работы использовали хелатообразующий сорбент из гидрофильной матрицы - Ni2+-сефароза (HIS-Select Nickel Affinity Gel, Sigma), pH буферных растворов в хроматографической системе 7,8±0,2.
Для приготовления лизата клеток бактериальную культуру рекомбинантного клона переносили в предварительно взвешенные полипропиленовые пробирки объемом 50 см3, охлаждали бактериальную культуру в течение 1 часа в бытовом холодильнике при 4±0,5°С, осаждали клетки бактериальной культуры центрифугированием при 3400 g в течение 30 мин и температуре 4±0,5°С. Супернатант удаляли, а осадок замораживали. После замораживания готовили лизат, для чего клетки ресуспендировали из расчета на 1 г осадка 10 см3 лизирующего буферного раствора и переносили в стеклянные флаконы по 4 см3. Биомассу разрушали тремя сериями ультразвуковой обработки с постоянным охлаждением смеси при частоте 18 кГц, каждая серия - три раза по 40 сек с перерывом в 1 мин, с перерывом между сериями - 15 минут. Процесс озвучивания контролировали микроскопически, окрашивая аликвоты лизата фуксином Пфейфера. Обломки клеток удаляли центрифугированием на настольной центрифуге при 13000 g в течение 5 мин. Супернатант отбирали и смешивали с двукратным буферным раствором для нанесения в соотношении 1:1, подводя рН пробы до значения 7,8±0,2. Содержание общего белка в лизате определяли по О.Н. Lowry et al. (1972) [13]. Пробу фильтровали через мембранные фильтры Millipore 0,45 мкм в стеклянные флаконы, объемом 10 см3.
Хроматографическую очистку рекомбинантного белка проводили при постоянной скорости подачи растворов перистальтическим насосом 0,5 см3/мин. Для элюции рекомбинантного белка использовали буферный раствор (рН 7,8±0,2): ФБР с 300 мМ NaCl; 500 мМ Imidazol; 0,25 мг/см3 Pefablock SC (Roche). Фракцию с очищенным рекомбинантным белком диализовали в ФБР с 300 мМ NaCl и 0,25 мг/см3 Pefablock SC (Roche) (рН 7,8±0,2) в течение 16-18 часов при температуре (4,0±2,0)°С.
Раствор рекомбинантного белка делили на аликвоты и хранили в замороженном виде в бытовом или низкотемпературном холодильнике.
Эффективность очистки рекомбинантного белка оценивали в 10% ДСН-ПААГ (SDS-PAAG) электрофорезе по U.K. Laemmle (1970) [14] (фиг. 2). Выход очищенного рекомбинантного Rec р30е2 составил не менее 15,0-20,0 мг с 1 дм3 бактериальной культуры, что в пять раз выше, чем при использовании прототипа.
Пример 4. Оценка специфичности рекомбинантного белка Rec р30е2 методом иммуноблоттинга
Разведения хроматографически очищенного белка Rec р30е2 подвергали фракцированию в 10% ДСН-ПААГ (SDS-PAAG) электрофорезе по U.K. Laemmle (1970) [14] с последующим электропереносом полипептидов из полиакриламидного геля на нитроцеллюлозную мембрану в полусухой буферной системе по методам J. Kyhse-Andersen (1984) [15]. Мембрану после электропереноса блокировали 2,0%-ным раствором обезжиренного сухого молока на фосфатном буферном растворе с 0,1% твина-20 (ФБР-т) (рН 7,8±0,2) при 4±0,5°С в течение 16-18 часов, после чего мембрану нарезали на единичные треки.
Постановку реакции иммуноблоттинга осуществляли, основываясь на методиках МЭБ [2, 3].
Параметры постановки иммуноблоттинга подобраны следующие:
- инкубирование иммунострипов в течение 1,5 ч при (25,0±2,5)°С с исследуемыми сыворотками в разведениях 1:20 или с исследуемыми 20% суспензиями проб органов на ФБР-т;
- трехкратная отмывка тест-полосок в ФБР-т по 5 мин при (25,0±2,5)°С (общее время отмывки составляет 15 мин);
- инкубирование иммунострипов в течение 45 мин при (25,0±2,5)°С в рабочем разведении пероксидазного конъюгата с протеином А (рабочее разведение конъюгата 1:2000);
- четырехкратная отмывка тест-полосок в ФБР-т по 5 мин при (25,0±2,5)°С (общее время отмывки составляет 20 мин);
- окрашивание иммунострипов в растворе хромогенного субстрата в течение 10-15 с при (25,0±2,5)°С;
- остановка реакции погружением иммунострипов в дистиллированную воду;
- визуальный учет результатов реакции.
При учете результатов проводят сравнение каждого иммунострипа с отрицательным (К-) и положительным (К+) контрольными образцами. Реакцию считают положительной, если выявляют окрашенную полосу различной интенсивности и ширины на уровне положительного контроля по центру иммунострипа, отрицательной - при отсутствии специфического окрашивания (фиг. 3).
Данные результаты показали, что наличие последовательности тиоредоксина в рекомбинантном белке Rec р30е2, не влияет на специфичность серологических реакций при использовании указанного белка в качестве антигена, также это позволило упростить процедуру выделения и хроматографической очистки.
Источники информации
1. Sanchez-Vizcaino, J.M. (2006). African swine fever. In Diseases of Swine, 9 Edition, pp 93-102. Ed A.D. Leman, B.E. Straw, W.L. Mengeling, S Dallaire and D.J. Taylor. Iowa State Univesity Press, 2000. - P. 159-165.
2. Immunoblotting OIE for Serological Diagnosis of African Swine Fever (SOP/CISA/ASF/IB/1/2008) [Электронный ресурс]. - Режим доступа: http://asf-referencelab.info/asf/images/files/SOPs/SOP-AFSIB12008.pdf. - Загл. с экрана.
3. Manual of diagnostic tests and vaccines for terrestrial animals (mammals, birds and bees). Capter 2.8.1. African swine fever/ Office International des Epizooties [Электронный ресурс]. - Paris, France, 2008. - 6th ed. - Режим доступа: http://www.oie.int/fileadmin/Home/fr/Health_standards/tahm/2.08.01_ASF.pdf. - Загл. с экрана.
4. Середа, А.Д. Белки вируса африканской чумы свиней / А.Д. Середа, Д.В. Колбасов // Научный журнал КубГАУ. - 2012. - №77(03). - С. 21-37. - 0421200012\0181. http://ej.kubagro.ru/2012/03/pdf/48.pdf.
5. Serodiagnosis of African swine fever using the recombinant protein p30 expressed in insect larvae / M.G. Barderas, A. Wigdorovit, F. Merelo, F. Beitia, C. Alonso, M.V. Borca, J.M. Escribano // Journal of Virological Methods. - 2000. - Vol. 89, N 1-2. - P. 129-136.
6. Копытов, B.O. Клонирование и экспрессия генов структурных белков р30 и р72 вируса африканской чумы свиней: дис.... канд. биол. наук: 03.00.06: защищена 23.12.04 / Копытов Валерий Олегович. - Покров, 2004. - 140 с.
7. Пат. RU 2463343 С1. Штамм клеток E.coli BL21(DE3)pLysS, клон pTT9/ASFVp30, содержащий рекомбинантную плазмиду со встройкой участка гена CP204L вируса африканской чумы свиней, кодирующего конформационный эпитоп белка р30, для изготовления диагностических препаратов / В.О. Копытов, С.Ж. Цыбанов, А.С. Казакова, Т.Э. Южук, С.А. Белянин, А.Г. Гузалова, Н.Н. Власова, Д.В. Колбасов (RU). - №2011141517/10; заявл. 13.10.2011; опубл. 10.10.2012, Бюл. №28. - 8 с. Режим доступа http:www.freepatent.ru/images/patents/54/2463343/patent-2463343.pdf.
8. Пат. RU 2439152 С1. Штамм «Ставрополь 01/08» вируса АЧС для вирусологических, молекулярно-генетических и мониторинговых исследований / В.М. Балышев, Д.В. Колбасов, В.В. Куриннов, Ю.Ф. Калантаенко, С.Ж. Цыбанов, А.Н. Жуков, А.П. Васильев (RU). - №2010126641/10; заявл. 30.06.2010; опубл. 10.01.2012, Бюл. №1. - 6 с. Режим доступа: http://www.freepatent.ru/images/patents/19/2439152/patent-2439152.pdf.
9. Novagen рЕТ System Manual. - 11th Edition. User Protocol TB055 Rev. С 0611 JN 1-63Х[Электронный ресурс]. - Режим доступа: http://www.emdmillipore.com/chemdat/en_CA/Merck-US-Site/USD/ViewProductDocuments-File?ProductSKU=EMD_BIO-71867&DocumentType=USP&DocumentId=/emd/biosciences/userprotocols/en-US/TB055.pdf&DocumentSource=GDS. - Загл. с экрана.
10. Green, M.R. Molecular Cloning: A Laboratory Manual. - 4th ed. / M.R. Green and J. Sambrook. - New York: Cold Spring Harbor Laboratory Press, 2012. - 2096 p.
11. Studier, F.W. Protein production by auto-induction in high density shaking cultures/ F.W. Studier // Protein Expr Purif. - 2005. - Vol. 41, №1. - P. 207-34. - Режим доступа http://www.biology.bnl.gov/cellbio/studier.html. - Загл. с экрана.
12. Single Step (KRX) Competent Cells. Technical Bulletin. - Promega, 2009. - Part TB352. - 11 p.
13. Protein masurement with the folin phenol reagents/ O.H. Lowry, M.G. Rosebrough, A.L. Farr [et al.] // J. Gen. Virol. - 1972. - Vol.14. - №1. - P. 111-114.
14. Laemmle, U.K. Clevage of structural proteins during the assembly of the head of bacteriophage Т4 / U.K. Laemmli // Nature. - 1970. - Vol. 227. - P. 680-685.
15. Kyhse-Andersen, J. Electroblotting of multiple gels:a simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose / J. Kyhse-Andersen // J. of Biochem. and Biophys. Methods. - 1984. - Vol. 10. - №3/4. - P. 203-209.











Изобретение относится к биотехнологии и ветеринарной вирусологии. Предложен экспрессирующий плазмидный вектор, обеспечивающий синтез в клетках Е. coli рекомбинантного белка, состоящего из фрагмента р30 вируса африканской чумы свиней (АЧС), тиоредоксина и двух полигистидиновых участков. Изобретение может быть использовано для изготовления вирусспецифических антигенов в интересах исследования сывороток свиней методом иммуноблоттинга. 3 ил., 1 табл., 4 пр.
Экспрессирующий плазмидный вектор pET32b(+)ASFV/p30e2 (SEQ ID №2), конструктивное выполнение которого раскрыто на фиг. 1, состоящий из плазмиды pET32b, фрагмента гена CP204L белка вируса африканской чумы свиней р30 и двух полигистидиновых последовательностей, обеспечивающий синтез в клетках Escherichia coli кодируемого геном TrxA-6xHis-p30e2-6xHis (SEQ ID №1) рекомбинантного белка Rec р30е2 (SEQ ID №3), используемого в качестве вирусспецифического антигена для диагностического исследования сывороток свиней методом иммуноблоттинга.
| ШТАММ КЛЕТОК E.coli BL21(DE3)pLysS, КЛОН pTT9/ASFVp30, СОДЕРЖАЩИЙ РЕКОМБИНАНТНУЮ ПЛАЗМИДУ СО ВСТРОЙКОЙ УЧАСТКА ГЕНА СР204L ВИРУСА АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ, КОДИРУЮЩЕГО КОНФОРМАЦИОННЫЙ ЭПИТОП БЕЛКА p30, ДЛЯ ИЗГОТОВЛЕНИЯ ДИАГНОСТИЧЕСКИХ ПРЕПАРАТОВ | 2011 |
|
RU2463343C1 |
| СПОСОБ ПОЛУЧЕНИЯ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ К БЕЛКУ Р30 ВИРУСА АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ С ИСПОЛЬЗОВАНИЕМ РЕКОМБИНАНТНЫХ КОНСТРУКЦИЙ | 2013 |
|
RU2534343C1 |
| СПОСОБ И ТЕСТ-СИСТЕМА ДЛЯ ОБНАРУЖЕНИЯ ДНК ВИРУСА АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ С ПОМОЩЬЮ СПЕЦИФИЧЕСКИХ ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ В ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2007 |
|
RU2360971C1 |
| СТАТОР ДЛЯ ЭЛЕКТРИЧЕСКОЙ МАШИНЫ | 2015 |
|
RU2674438C1 |
| АГРЕГАТ ПОДЗЕМНОГО РЕМОНТА СКВАЖИН С НЕПРЕРЫВНОЙ КОЛОННОЙ ГИБКИХ ТРУБ | 1996 |
|
RU2154146C2 |

