Область техники
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые связываются с белком CD70 человека с высокой аффинностью и демонстрируют сильное ингибирование роста опухолевых клеток.
Предпосылки создания изобретения
Рецептор цитокинов CD27 является членом суперсемейства рецепторов факторов некроза опухолей (TFNR), которые играют роль в росте и дифференциации клеток, а также апоптозе. Лигандом CD27 является CD70, который относится к семейству факторов некроза опухолей - лигандов. CD70 представляет собой полипептид из 193 аминокислот, содержащий гидрофильный N-концевой домен из 20 аминокислот и C-концевой домен, содержащий 2 потенциальных сайта N-гликозилирования (присоединения сахара к атому N) (Goodwin, R.G. et al. (1993) Cell 73: 447-456; Bowman et al. (1994) Immunol 152: 1756-1761). На основе этих особенностей было установлено, что CD70 является трансмембранным белком типа II, содержащим экстраклеточную C-концевую часть.
CD70 кратковременно обнаруживается на активированных T- и B-лимфоцитах и дендритных клетках (Hintzen et al. (1994) J. Immunol. 152: 1762-1773; Oshima et al. (1998) Int. Immunol. 10: 517-26; Tesselaar et al. (2003) J. Immunol. 170: 33-40). Помимо экспрессии в нормальных клетках экспрессия CD70 была описана в различных типах раков, включая почечно-клеточные карциномы, метастатические раки молочной железы, опухоли головного мозга, лейкозы, лимфомы и карциномы носоглотки (Junker et al. (2005) J. Urol. 173: 2150-2153; Sloan et al. (2004) Am J Pathol. 164: 315-323; Held-Feindt and Mentlein (2002) Int J Cancer 98: 352-356; Hishima et al. (2000) Am J Surg Pathol. 24: 742-746; Lens et al. (1999) Br J Haematol. 106: 491-503). Также взаимодействие CD70 с CD27, как было предположено, играет роль в клеточноопосредованном аутоиммунном заболевании и ингибировании продукции TNF-альфа (Nakajima et al. (2000) J. Neuroimmunol. 109: 188-196).
Соответственно, CD70 представляет собой мишень в случае лечения рака, аутоиммунных нарушений и ряда других заболеваний, характеризующихся экспрессией CD70.
В WO 2006/0044643 описываются антитела против CD70, содержащие эффекторный домен антитела, который может опосредовать одну или более из ADCC, ADCP, CDC или ADC и или проявлять цитостатический или цитотоксический эффект по отношению к CD70-экспрессирующему раку, или проявлять имммуносупрессорный эффект по отношению к иммунологическому нарушению, характеризующемуся экспрессией CD70, в отсутствие конъюгации с цитостатическим или цитотоксическим агентом. Приводимые здесь в качестве примера антитела основаны на антигенсвязывающих областях двух моноклональных антител, обозначенных 1F6 и 2F2.
В WO 2007/038637 описываются полностью человеческие моноклональные антитела, которые связываются с CD70. Эти антитела характеризуются связыванием с CD70 человека с KD, составляющей 1×10-7 M или менее. Антитела также связываются с линиями опухолевых клеток почечноклеточной карциномы, которые экспрессируют CD70, такими как 786-О, и подвергаются интернализации в них.
Краткое изложение сущности изобретения
Здесь обеспечиваются антитела, или их антигенсвязывающие фрагменты, (называемые здесь антителами против CD70), которые связываются с белком CD70 человека и демонстрируют характеристики, отличные и, как правило, лучшие, (от) таковых антител против CD70, описанных в известном уровне техники. Лучшие характеристики этих антител являются преимущественными, что касается применения для лечения людей, особенно лечения CD70-экспресссирующих раков, а также иммунологических нарушений.
Описываемые здесь антитела против CD70 характеризуются очень высокой аффинностью к CD70 человека. Все предпочтительные варианты осуществления, описываемые здесь, демонстрируют аффинность к рекомбинантному CD70 человека (измеряемую с помощью поверхностного плазмонного резонанса с использованием BiacoreTM, описываемого здесь), которая значительно выше таковой самых сильных антител против CD70 известного уровня техники, предложенных для лечения людей, в том числе антител против CD70 «полностью человеческого происхождения» известного уровня техники. Кроме того, все предпочтительные варианты антител против CD70, описываемые здесь, демонстрируют лучшее связывание (т.е. с большей аффинностью) с CD70, представленным на поверхности линий клеток человека, особенно линий раковых клеток человека, по сравнению с антителами против CD70 известного уровня техники, предложенными для лечения людей. Это лучшее связывание с CD70 на клеточной поверхности особенно заметно относительно линий раковых клеток человека, которые экспрессируют CD70 в «низкой копийности», и имеет прямое отношение к применению антител для лечения людей. Более того, предпочтительные варианты антител против CD70, описываемые здесь, демонстрируют в значительной степени увеличенное связывание с раковыми клетками, изолированными от являющихся людьми пациентов, в частности, раковыми клетками, изолированными от пациентов с хроническим лимфолейкозом (CLL), по сравнению с антителами против CD70 известного уровня техники, предложенными для лечения людей.
Следовательно, в первом аспекте настоящего изобретения обеспечивается связывающееся с CD70 человека антитело или его антигенсвязывающий фрагмент, включающее(ий) по крайней мере один вариабельный домен тяжелой цепи (VH) и по крайней мере один вариабельный домен легкой цепи (VL), причем указанные VH- и VL-домены, при исследовании в виде Fab-фрагмента, демонстрируют скорость диссоциации (koff, измеряемую с использованием BiacoreTM) от CD70 человека, составляющую менее 7×10-4 сек-1, используя стандартный протокол BiacoreTM, описываемый здесь.
В предпочтительном варианте осуществления антитело или антигенсвязывающий фрагмент включает по крайней мере один вариабельный домен тяжелой цепи (VH) и по крайней мере один вариабельный домен легкой цепи (VL), причем указанные VH- и VL-домены демонстрируют скорость диссоциации от CD70 человека, составляющую 5×10-4 сек-1 или менее, или 2×10-4 сек-1 или менее, или 1×10-4 сек-1 или менее. Наиболее предпочтительно, когда антитело против CD70, при исследовании в виде Fab-фрагмента, будет демонстрировать скорость диссоциации от CD70 в диапазоне от 0,4×10-4 сек-1 до 4,8×10-4 сек-1.
Также обеспечивается связывающееся с CD70 человека антитело, включающее две Fab-области, причем каждая из Fab-областей связывается с CD70 человека и демонстрирует, при исследовании в виде Fab-фрагмента, скорость диссоциации от CD70 человека, составляющую 5×10-4 сек-1 или менее, или 2×10-4 сек-1 или менее, или 1×10-4 сек-1 или менее, а предпочтительно находящуюся в диапазоне от 0,4×10-4 сек-1 до 4,8×10-4 сек-1. Две Fab-области могут быть идентичными, или они могут отличаться по характеристикам связывания, например, аффинности к CD70 человека. Две Fab-области могут связываться с одним и тем же эпитопом или перекрывающимися эпитопами CD70 человека, или они могут связываться с отличными, не перекрывающимися эпитопами CD70 человека. Две Fab-области могут отличаться друг от друга по аминокислотной последовательности в одном или обоих VH- и VL-доменах.
Предпочтительные варианты антител против CD70, обеспечиваемых здесь, могут, помимо очень высокой аффинности к CD70, демонстрировать сильное блокирование или ингибирование взаимодействия между CD70 или его лигандом CD27. Предпочтительные антитела против CD70, которые демонстрируют как связывание с высокой аффинностью с CD70, так и сильное блокирование взаимодействия CD70/CD27, являются особенно преимущественными в качестве терапевтических средств для лечения показаний-заболеваний, в случае которых блокирование передачи сигнала от CD70/CD27 увеличивает терапевтическую эффективность (например, помимо уничтожения клеток, опосредуемого эффекторными функциями антитела против CD70), например, аутоиммунных заболеваний и раков, которые коэкспрессируют CD70 и CD27.
Не все из антител против CD70, описываемых здесь, демонстрируют сильное блокирование взаимодействия CD70/CD27 помимо связывания с высокой аффинностью с CD70. Здесь также описывается ряд антител против CD70, которые демонстрируют связывание с очень высокой аффинностью с CD70, но не демонстрируют значительное блокирование взаимодействия CD70/CD27. Характеристики этих антител описываются здесь в другом месте. Наличие не блокирующих антител против CD70 с высокой аффинностью может расширить диапазон возможностей терапевтического применения.
Предпочтительные варианты антител против CD70, описываемых здесь, демонстрирующие очень высокую аффинность к CD70 человека, также характеризуются сочетанием характеристик связывания, которое не демонстрируют антитела против CD70 известного уровня техники, предложенные для терапевтического применения для людей. Соответственно, предпочтительные антитела против CD70, описываемые здесь, характеризуются:
(a) связыванием в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO: 342) в CD70 человека;
(b) перекрестной реактивностью с гомологами CD70 у макака-резус (Macaca mulatta) и яванского макака (Macaca cynomolgus);
(c) связыванием как со встречающимся в природе CD70 человека, так и с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
Это сочетание характеристик связывания, которое не демонстрируют антитела известного уровня техники, предложенные для терапевтического применения для людей, указывает на связывание с новым эпитопом CD70, который отличен от эпитопов, с которыми связываются антитела против CD70 известного уровня техники.
Сочетание характеристик связывания, которое демонстрируют предпочтительные антитела против CD70, является преимущественным в рамках разработки лекарственных средств для людей. В частности, перекрестная реактивность с гомологами CD70 у обезьян дает возможность проводить токсикологические исследования антител против CD70, предложенных для терапевтического применения для людей, в моделях на приматах.
Предпочтительные антитела против CD70, описываемые здесь, кроме того, демонстрируют подходящие характеристики, которые имеют отношение к промышленному производству в виде терапевтического продукта в виде антитела. Как здесь описывается в другом месте, предпочтительные антитела против CD70, обеспечиваемых здесь, демонстрируют экспрессию на очень высоком уровне в рекомбинантных экспрессионных системах, используемых для промышленного производства терапевтических продуктов в виде антител клинической стадии. Уровни экспрессии, достигаемые в случае предпочтительных антител против CD70, в значительной степени превышают уровни, обычно достигаемые даже в случае терапевтических продуктов в виде «полностью человеческих» антител. Кроме того, продукты в виде предпочтительных антител против CD70 (продуцированных в результате рекомбинантной экспрессии в формате, подходящем для терапевтического применения для людей) демонстрируют исключительную термостабильность, которая превосходит таковую типичных терапевтических продуктов в виде антител.
Обеспечиваемые здесь антитела против CD70 с более высокой аффинностью к CD70 человека и другими преимущественными характеристиками, перечисленными выше, происходят от животных семейства Camelidae (верблюдовых) (например, происходят от лам). Происходящие от верблюдовых антитела против CD70 могут быть выделенными или рекомбинантно экспрессированными моноклональными антителами. Предпочтительные варианты осуществления могут быть гуманизированным (или приведенным к зародышевому типу) моноклональным антителом (например, гуманизированным вариантом происходящего от верблюдовых антитела против CD70), химерным антителом (например, животное семейства верблюдовых-человек химерным антителом) или гуманизированным химерным антителом (например, химерным антителом, включающим гуманизированные варианты VH- и VL-доменов верблюдовых и константные домена антитела человека).
Происходящие от верблюдовых антитела против CD70 могут включать по крайней мере один гипервариабельный петлевой участок или определяющий комплементарность участок, полученный из VH-домена или VL-домена вида семейства Camelidae. В конкретном варианте осуществления антитело против CD70, или его антигенсвязывающий фрагмент, может включать вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), причем VH- и VL-домены, или один или более их CDR, происходят от верблюдовых. В конкретных вариантах осуществления антитело или его антигенсвязывающий фрагмент могут включать VH- и VL-домены ламы, или приведенные к человеческому зародышевому типу варианты VH- и VL-доменов ламы. Это антитело, или антигенсвязывающий фрагмент, может проявлять «высокую степень гомологии с антителом человека», как здесь описывается.
Происходящие от верблюдовых антитела против CD70, описываемые здесь, обычно демонстрируют аминокислотные последовательности VH- и/или VL-областей, идентичные на по крайней мере 90% (например, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%) ближе всего соответствующей последовательности антитела человека зародышевого типа.
Дальнейшие предпочтительные варианты осуществления настоящего изобретения включают гуманизированные (или приведенные к человеческому зародышевому типу) варианты происходящих от верблюдовых антител против CD70. В частности, настоящим изобретением обеспечиваются гуманизированные или приведенные к человеческому зародышевому типу варианты происходящих от ламы антител против CD70, описываемых здесь.
В дальнейшем аспекте настоящего изобретения обеспечивается химерное животное семейства верблюдовых-человек антитело, которое связывается с CD70 человека, причем антигенсвязывающие части антитела (например, VH- и/или VL-домены или их CDR) происходят от верблюдовых, а константные области антитела происходят из антитела человека. В частности, настоящим изобретением обеспечивается химерное лама-человек антитело, которое связывается с CD70 человека.
В дальнейшем аспекте настоящего изобретения обеспечивается гуманизированный вариант химерного животное семейства верблюдовых-человек антитела, которое связывается с CD70 человека, причем антигенсвязывающие части антитела (например, VH- и/или VL-домены или их CDR) являются гуманизированными вариантами происходящих от верблюдовых последовательностей, а константные области антитела происходят из антитела человека. В частности, настоящим изобретением обеспечивается гуманизированный вариант химерного лама-человек антитела, которое связывается с CD70 человека.
Предпочтительные (но не ограничивающие) варианты антител против CD70, или их антигенсвязывающие фрагменты, определены ниже в соответствии с конкретными структурными характеристиками, т.е. точно определенными аминокислотными последовательностями либо CDR (одной или более из SEQ ID NO: 49-59, 262 или 263 (CDR3 тяжелой цепи), или SEQ ID NO: 26-37, 249, 258 или 259 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20, 248, 256 или 257 (CDR1 тяжелой цепи), или одной из последовательностей CDR, представленных в виде SEQ ID NO: 148-168, 271 или 273 (CDR3 легкой цепи), или SEQ ID NO: 109-128 или 270 (CDR2 легкой цепи), или SEQ ID NO: 77-95 или 250-253, 267 или 268 (CDR1 легкой цепи), либо полноразмерных вариабельных доменов (одной или более из SEQ ID NO: 177-188, 212-223, 274 или 275 (VH) или SEQ ID NO: 189-211, 230-245, 276 или 277 (VL)). Все из этих антител связываются с CD70 человека с высокой аффинностью, демонстрируя, при исследовании в виде Fab-фрагмента, скорость диссоциации от CD70 человека, составляющую 5×10-4 сек-1 или менее, и типично находящуюся в диапазоне от 0,4×10-4 сек-1 до 4,8×10-4 сек-1.
Настоящим изобретением также обеспечиваются гуманизированные/приведенные к зародышевому типу варианты этих антител, плюс варианты аффинности и варианты, содержащие консервативные аминокислотные замены, определенные здесь. В частности, обеспечиваются химерные антитела, содержащие VH- и VL-домены, происходящие от верблюдовых, или их приведенные к человеческому зародышевому типу варианты, слитые с константными доменами антител человека, в частности, IgG1, IgG2, IgG3 или IgG4. Вариабельные домены тяжелой цепи, определенные выше, могут использоваться в качестве однодоменных антител, или могут быть включены в обычное четырехцепочечное антитело или другие антигенсвязывающие белки, такие как, например, Fab-, Fab′-, F(ab′)2-, биспецифические Fab′- и Fv-фрагменты, диатела, линейные антитела, одноцепочечные молекулы антител, одноцепочечный вариабельный фрагмент (scFv) и полиспецифические антитела.
Предпочтительными вариантами антител против CD70 являются антитела, или их антигенсвязывающие фрагменты, включающие вариабельный домен тяжелой цепи, включающий CDR3 вариабельного домена тяжелой цепи, CDR2 вариабельного домена тяжелой цепи и CDR1 вариабельного домена тяжелой цепи, причем указанный CDR3 вариабельного домена тяжелой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 262 и SEQ ID NO: 263, и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности;
указанный CDR2 вариабельного домена тяжелой цепи необязательно включает аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 306 [X1X2X3X4X5X6X7X8X9YYADSVKX10], где
X1 представляет собой любую аминокислоту, предпочтительно D, T, S или E,
X2 представляет собой любую аминокислоту, предпочтительно I,
X3 представляет собой любую аминокислоту, предпочтительно N, S, T или Y,
X4 представляет собой любую аминокислоту, предпочтительно N, M, S или T,
X5 представляет собой любую аминокислоту, предпочтительно E, D, Y или H,
X6 представляет собой любую аминокислоту, предпочтительно G, D, S или N,
X7 представляет собой любую аминокислоту, предпочтительно G, Y, S, D или M,
X8 представляет собой любую аминокислоту, предпочтительно T, E, S, N, Y или R,
X9 представляет собой любую аминокислоту, предпочтительно T, A или R, и
X10 представляет собой любую аминокислоту, предпочтительно G или S,
и аминокислотных последовательностей SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, SEQ ID NO: 249, SEQ ID NO: 258 и SEQ ID NO: 259 и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности; и
указанный CDR1 вариабельного домена тяжелой цепи необязательно включает аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 307 [X1YYMN], где
X1 представляет собой любую аминокислоту, предпочтительно V, G or A,
аминокислотных последовательностей SEQ ID NO: 308 [X1X2AMS], где
X1 представляет собой любую аминокислоту, предпочтительно D, T, S, N или G, и
X2 представляет собой любую аминокислоту, предпочтительно Y, S или P,
и аминокислотных последовательностей SEQ ID NO: 10, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 248, SEQ ID NO: 256 и SEQ ID NO: 257 и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности.
Вариабельный домен тяжелой цепи может включать любую из перечисленных последовательностей CDR3 вариабельного домена тяжелой цепи (HCDR3) в сочетании с любой из последовательностей CDR2 вариабельного домена тяжелой цепи (HCDR2) и любой из последовательностей CDR1 вариабельного домена тяжелой цепи (HCDR1). Однако особенно предпочтительными являются некоторые сочетания HCDR3 и HCDR2 и HCDR1, которые являются встречающимися в природе сочетаниями, которые происходят из одного общего VH-домена. Эти предпочтительные сочетания перечислены в таблице 6 и таблице 14A.
Антитело или его антигенсвязывающий фрагмент может, кроме того, включать вариабельный домен легкой цепи (VL), который спаривается с VH-доменом с образованием антигенсвязывающего домена. Предпочтительными вариабельными доменами легкой цепи являются те, которые включают CDR3 вариабельного домена легкой цепи, CDR2 вариабельного домена легкой цепи и CDR1 вариабельного домена легкой цепи, причем указанный CDR3 вариабельного домена легкой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO: 159, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 162, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 165, SEQ ID NO: 166, SEQ ID NO: 166, SEQ ID NO: 168, SEQ ID NO: 271 и SEQ ID NO: 273, и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности;
указанный CDR2 вариабельного домена легкой цепи необязательно включает аминокислотную последовательность, выбираемую из группы, состоящей из
(а) аминокислотных последовательностей SEQ ID NO: 310 [X1TX2X3RHS], где
X1 представляет собой любую аминокислоту, предпочтительно N или S,
X2 представляет собой любую аминокислоту, предпочтительно N, S или A, и
X3 представляет собой любую аминокислоту, предпочтительно S, N или T,
(b) аминокислотных последовательностей SEQ ID NO: 311 [YYSDSX1X2X3QX4S], где
X1 представляет собой любую аминокислоту, предпочтительно Y, V или L,
X2 представляет собой любую аминокислоту, предпочтительно K или S,
X3 представляет собой любую аминокислоту, предпочтительно H или N, и
X4 представляет собой любую аминокислоту, предпочтительно G или S,
(c) аминокислотных последовательностей SEQ ID NO: 312 [X1NX2NRPS], где
X1 представляет собой любую аминокислоту, предпочтительно V, I или Y, и
X2 представляет собой любую аминокислоту, предпочтительно N или T,
(d) аминокислотных последовательностей SEQ ID NO: 313 [GDNX1X2PL], где
X1 представляет собой любую аминокислоту, предпочтительно Y, и
X2 представляет собой любую аминокислоту, предпочтительно R или M,
(e) аминокислотных последовательностей SEQ ID NO: 314 [X1DDX2RPS], где
X1 представляет собой любую аминокислоту, предпочтительно D или G, и
X2 представляет собой любую аминокислоту, предпочтительно S или I,
и аминокислотных последовательностей SEQ ID NO: 113, SEQ ID NO: 116, SEQ ID NO: 120 и SEQ ID NO: 270, и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности; и
указанный CDR1 вариабельного домена легкой цепи необязательно включает аминокислотную последовательность, выбираемую из группы, состоящей из
(а) аминокислотных последовательностей SEQ ID NO: 315 [GLX1SGSX2TX3X4X5YPX6], где
X1 представляет собой любую аминокислоту, предпочтительно S или T,
X2 представляет собой любую аминокислоту, предпочтительно V или A,
X3 представляет собой любую аминокислоту, предпочтительно S или T,
X4 представляет собой любую аминокислоту, предпочтительно S, T или G,
X5 представляет собой любую аминокислоту, предпочтительно N или H,
X6 представляет собой любую аминокислоту, предпочтительно G, D или E,
(b) аминокислотных последовательностей SEQ ID NO: 316 [TLX1SX2X3X4X5GX6YDIS], где
X1 представляет собой любую аминокислоту, предпочтительно S, N или I,
X2 представляет собой любую аминокислоту, предпочтительно G или A,
X3 представляет собой любую аминокислоту, предпочтительно N или D,
X4 представляет собой любую аминокислоту, предпочтительно N или S,
X5 представляет собой любую аминокислоту, предпочтительно V или I,
X6 представляет собой любую аминокислоту, предпочтительно N или S,
(c) аминокислотных последовательностей SEQ ID NO: 317 [QGGNLX1LYGAN], где
X1 представляет собой любую аминокислоту, предпочтительно G или W,
(d) аминокислотных последовательностей SEQ ID NO: 318 [RGDX1LX2X3YX4X5N], где
X1 представляет собой любую аминокислоту, предпочтительно S или T,
X2 представляет собой любую аминокислоту, предпочтительно E или R,
X3 представляет собой любую аминокислоту, предпочтительно R или N,
X4 представляет собой любую аминокислоту, предпочтительно G или H,
X5 представляет собой любую аминокислоту, предпочтительно T или A,
(e) аминокислотных последовательностей SEQ ID NO: 319 [GX1X2SGSVTSX3NFPT], где
X1 представляет собой любую аминокислоту, предпочтительно V или L,
X2 представляет собой любую аминокислоту, предпочтительно K или T,
X3 представляет собой любую аминокислоту, предпочтительно T или D,
и аминокислотных последовательностей SEQ ID NO: 82, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253, SEQ ID NO: 267 и SEQ ID NO: 268, и вариантов любой из приведенных последовательностей, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности.
Вариабельный домен легкой цепи может включать любую из перечисленных последовательностей CDR3 вариабельного домена легкой цепи (LCDR3) в сочетании с любой из последовательностей CDR2 вариабельного домена легкой цепи (HCDR2) и любой из последовательностей CDR1 вариабельного домена легкой цепи (LCDR1). Однако особенно предпочтительными являются некоторые сочетания LCDR3 и LCDR2 и LCDR1, которые являются встречающимися в природе сочетаниями, которые происходят из одного общего VL-домена. Эти предпочтительные сочетания перечислены в таблице 7 и таблице 15A.
Любое конкретное антитело против CD70 или его антигенсвязывающий фрагмент, включающее(ий) VH-домен, спаренный с VL-доменом с образованием центра связывания с антигеном CD70, будет включать сочетание 6 CDR: CDR3 вариабельного домена тяжелой цепи (HCDR3), CDR2 вариабельного домена тяжелой цепи (HCDR2), CDR1 вариабельного домена тяжелой цепи (HCDR1), CDR3 вариабельного домена легкой цепи (LCDR3), CDR2 вариабельного домена легкой цепи (LCDR2) и CDR1 вариабельного домена легкой цепи (LCDR1). Хотя все сочетания 6 CDR, выбираемых из групп последовательностей CDR, перечисленных выше, являются допустимыми и находятся в объеме настоящего изобретения, особенно предпочтительными являются некоторые сочетания 6 CDR, которые являются встречающимися в природе сочетаниями в одном Fab, демонстрирующем связывание с высокой аффинностью с CD70 человека.
Предпочтительные сочетания 6 CDR включают, но без ограничения, сочетания CDR3 вариабельного домена тяжелой цепи (HCDR3), CDR2 вариабельного домена тяжелой цепи (HCDR2), CDR1 вариабельного домена тяжелой цепи (HCDR1), CDR3 вариабельного домена легкой цепи (LCDR3), CDR2 вариабельного домена легкой цепи (LCDR2) и CDR1 вариабельного домена легкой цепи (LCDR1), выбираемые из группы, состоящей из:
(i) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 27, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 160, LCDR2, включающего SEQ ID NO: 119, и LCDR1, включающего SEQ ID NO: 250;
(ii) HCDR3, включающего SEQ ID NO: 49, HCDR2, включающего SEQ ID NO: 26, HCDR1, включающего SEQ ID NO: 10, LCDR3, включающего SEQ ID NO: 148, LCDR2, включающего SEQ ID NO: 109, и LCDR1, включающего SEQ ID NO: 77;
(iii) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 27, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 149, LCDR2, включающего SEQ ID NO: 110, и LCDR1, включающего SEQ ID NO: 78;
(iv) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 28, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 150, LCDR2, включающего SEQ ID NO: 111, и LCDR1, включающего SEQ ID NO: 79;
(v) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 28, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 151, LCDR2, включающего SEQ ID NO: 110, и LCDR1, включающего SEQ ID NO: 80;
(vi) HCDR3, включающего SEQ ID NO: 51, HCDR2, включающего SEQ ID NO: 29, HCDR1, включающего SEQ ID NO: 12, LCDR3, включающего SEQ ID NO: 152, LCDR2, включающего SEQ ID NO: 110, и LCDR1, включающего SEQ ID NO: 80;
(vii) HCDR3, включающего SEQ ID NO: 52, HCDR2, включающего SEQ ID NO: 30, HCDR1, включающего SEQ ID NO: 13, LCDR3, включающего SEQ ID NO: 153, LCDR2, включающего SEQ ID NO: 112, и LCDR1, включающего SEQ ID NO: 81;
(viii) HCDR3, включающего SEQ ID NO: 53, HCDR2, включающего SEQ ID NO: 31, HCDR1, включающего SEQ ID NO: 14, LCDR3, включающего SEQ ID NO: 154, LCDR2, включающего SEQ ID NO: 113, и LCDR1, включающего SEQ ID NO: 82;
(ix) HCDR3, включающего SEQ ID NO: 54, HCDR2, включающего SEQ ID NO: 32, HCDR1, включающего SEQ ID NO: 15, LCDR3, включающего SEQ ID NO: 155, LCDR2, включающего SEQ ID NO: 114, и LCDR1, включающего SEQ ID NO: 83;
(x) HCDR3, включающего SEQ ID NO: 55, HCDR2, включающего SEQ ID NO: 33, HCDR1, включающего SEQ ID NO: 16, LCDR3, включающего SEQ ID NO: 156, LCDR2, включающего SEQ ID NO: 115, и LCDR1, включающего SEQ ID NO: 84;
(xi) HCDR3, включающего SEQ ID NO: 56, HCDR2, включающего SEQ ID NO: 34, HCDR1, включающего SEQ ID NO: 17, LCDR3, включающего SEQ ID NO: 157, LCDR2, включающего SEQ ID NO: 116, и LCDR1, включающего SEQ ID NO: 85;
(xii) HCDR3, включающего SEQ ID NO: 57, HCDR2, включающего SEQ ID NO: 35, HCDR1, включающего SEQ ID NO: 18, LCDR3, включающего SEQ ID NO: 158, LCDR2, включающего SEQ ID NO: 117, и LCDR1, включающего SEQ ID NO: 84;
(xiii) HCDR3, включающего SEQ ID NO: 58, HCDR2, включающего SEQ ID NO: 36, HCDR1, включающего SEQ ID NO: 19, LCDR3, включающего SEQ ID NO: 159, LCDR2, включающего SEQ ID NO: 118, и LCDR1, включающего SEQ ID NO: 86;
(xiv) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 27, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 161, LCDR2, включающего SEQ ID NO: 120, и LCDR1, включающего SEQ ID NO: 88;
(xv) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 27, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 162, LCDR2, включающего SEQ ID NO: 121, и LCDR1, включающего SEQ ID NO: 89;
(xvi) HCDR3, включающего SEQ ID NO: 50, HCDR2, включающего SEQ ID NO: 27, HCDR1, включающего SEQ ID NO: 11, LCDR3, включающего SEQ ID NO: 163, LCDR2, включающего SEQ ID NO: 122, и LCDR1, включающего SEQ ID NO: 90;
(xvii) HCDR3, включающего SEQ ID NO: 51, HCDR2, включающего SEQ ID NO: 29, HCDR1, включающего SEQ ID NO: 12, LCDR3, включающего SEQ ID NO: 164, LCDR2, включающего SEQ ID NO: 123, и LCDR1, включающего SEQ ID NO: 91;
(xviii) HCDR3, включающего SEQ ID NO: 51, HCDR2, включающего SEQ ID NO: 29, HCDR1, включающего SEQ ID NO: 12, LCDR3, включающего SEQ ID NO: 164, LCDR2, включающего SEQ ID NO: 124, и LCDR1, включающего SEQ ID NO: 91;
(xix) HCDR3, включающего SEQ ID NO: 59, HCDR2, включающего SEQ ID NO: 37, HCDR1, включающего SEQ ID NO: 12, LCDR3, включающего SEQ ID NO: 165, LCDR2, включающего SEQ ID NO: 125, и LCDR1, включающего SEQ ID NO: 92;
(xx) HCDR3, включающего SEQ ID NO: 59, HCDR2, включающего SEQ ID NO: 37, HCDR1, включающего SEQ ID NO: 20, LCDR3, включающего SEQ ID NO: 165, LCDR2, включающего SEQ ID NO: 126, и LCDR1, включающего SEQ ID NO: 93;
(xxi) HCDR3, включающего SEQ ID NO: 59, HCDR2, включающего SEQ ID NO: 37, HCDR1, включающего SEQ ID NO: 20, LCDR3, включающего SEQ ID NO: 166, LCDR2, включающего SEQ ID NO: 127, и LCDR1, включающего SEQ ID NO: 92;
(xxii) HCDR3, включающего SEQ ID NO: 59, HCDR2, включающего SEQ ID NO: 37, HCDR1, включающего SEQ ID NO: 20, LCDR3, включающего SEQ ID NO: 167, LCDR2, включающего SEQ ID NO: 128, и LCDR1, включающего SEQ ID NO: 94;
(xxiii) HCDR3, включающего SEQ ID NO: 59, HCDR2, включающего SEQ ID NO: 37, HCDR1, включающего SEQ ID NO: 20, LCDR3, включающего SEQ ID NO: 168, LCDR2, включающего SEQ ID NO: 110, и LCDR1, включающего SEQ ID NO: 95;
(xxiv) HCDR3, включающего SEQ ID NO: 262, HCDR2, включающего SEQ ID NO: 258, HCDR1, включающего SEQ ID NO: 256, LCDR3, включающего SEQ ID NO: 271, LCDR2, включающего SEQ ID NO: 110, и LCDR1, включающего SEQ ID NO: 267;
(xxv) HCDR3, включающего SEQ ID NO: 263, HCDR2, включающего SEQ ID NO: 259, HCDR1, включающего SEQ ID NO: 257, LCDR3, включающего SEQ ID NO: 273, LCDR2, включающего SEQ ID NO: 270, и LCDR1, включающего SEQ ID NO: 268.
Дополнительные предпочтительные антитела против CD70, демонстрирующие связывание с высокой аффинностью с CD70 человека, включают выделенные антитела или их антигенсвязывающие фрагменты, включающие вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 177, SEQ ID NO: 178, SEQ ID NO: 179, SEQ ID NO: 180, SEQ ID NO: 181, SEQ ID NO: 182, SEQ ID NO: 183, SEQ ID NO: 184, SEQ ID NO: 185, SEQ ID NO: 186, SEQ ID NO: 187, SEQ ID NO: 188, SEQ ID NO: 212, SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220, SEQ ID NO: 221, SEQ ID NO: 222, SEQ ID NO: 223, SEQ ID NO: 224, SEQ ID NO: 225, SEQ ID NO: 226, SEQ ID NO: 227, SEQ ID NO: 228, SEQ ID NO: 229, SEQ ID NO: 274 и SEQ ID NO: 275, и аминокислотных последовательностей, идентичных на по крайней мере 90%, 95%, 97%, 98% или 99% одной из приведенных последовательностей, и необязательно включающие вариабельный домен легкой цепи, имеющий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 189, SEQ ID NO: 190, SEQ ID NO: 191, SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196, SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201, SEQ ID NO: 202, SEQ ID NO: 203, SEQ ID NO: 204, SEQ ID NO: 205, SEQ ID NO: 206, SEQ ID NO: 207, SEQ ID NO: 208, SEQ ID NO: 209, SEQ ID NO: 210, SEQ ID NO: 211, SEQ ID NO: 223, SEQ ID NO: 230, SEQ ID NO: 231, SEQ ID NO: 232, SEQ ID NO: 233, SEQ ID NO: 234, SEQ ID NO: 234, SEQ ID NO: 236, SEQ ID NO: 237, SEQ ID NO: 238, SEQ ID NO: 239, SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242, SEQ ID NO: 243, SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 245, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 276 и SEQ ID NO: 277, и аминокислотных последовательностей, идентичных на по крайней мере 90%, 95%, 97%, 98% или 99% одной из приведенных последовательностей.
Хотя все возможные спаривания VH-доменов и VL-доменов, выбираемых из групп последовательностей VH- и VL-доменов, перечисленных выше, являются допустимыми и находятся в объеме настоящего изобретения, особенно предпочтительными являются некоторые сочетания VH и VL, которые являются встречающимися в природе сочетаниями в одном Fab, демонстрирующем связывание с высокой аффинностью с CD70 человека. Соответственно, предпочтительными антителами против CD70, или их антигенсвязывающими фрагментами, демонстрирующими связывание с высокой аффинностью с CD70 человека, являются те, которые включают сочетание вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL), причем сочетание выбирают из группы, состоящей из:
(i) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 223, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 241;
(ii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 177, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 189;
(iii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 190;
(iv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 179, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 191;
(v) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 179, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 192;
(vi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 180, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 193;
(vii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 181, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 194;
(viii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 182, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 195;
(ix) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 183, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 196;
(x) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 184, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 197;
(xi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 185, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 198;
(xii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 186, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 199;
(xiii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 187, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 200;
(xiv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 201;
(xv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 202;
(xvi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 203;
(xvii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 204;
(xviii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 180, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 205;
(xix) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 180, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 206;
(xx) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 188, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 207;
(xxi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 188, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 208;
(xxii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 188, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 209;
(xxiii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 188, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 210;
(xxiv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 188, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 211;
(xxv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 274, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 276;
(xxvi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 275, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 277.
В случае каждого из конкретных сочетаний VH/VL, перечисленных выше, является также допустимым и находится в объеме настоящего изобретения сочетание VH-домена, имеющего аминокислотную последовательность, идентичную на по крайней мере 90%, 92%, 95%, 97% или 99% приведенной последовательности VH-домена, с VL-доменом, имеющим аминокислотную последовательность, идентичную на по крайней мере 90%, 92%, 95%, 97% или 99% приведенной последовательности VL-домена.
В предыдущем параграфе, и здесь где-либо в другом месте, структура антител/антигенсвязывающих фрагментов определяется на основе % идентичности последовательности с приведенной эталонной последовательностью (с заданной SEQ ID NO). В этом контексте % идентичности двух аминокислотных последовательностей можно определить посредством сравнения этих двух последовательностей, которые оптимальным образом совмещены, и в случае которых сравнимая аминокислотная последовательность может включать добавления или делеции относительно эталонной последовательности с целью оптимального совмещения этих двух последовательностей. Процент идентичности рассчитывают путем определения числа идентичных положений, в которых аминокислотный остаток идентичен в двух последовательностях, деления этого числа идентичных положений на общее число положений в окне сравнения и умножения полученного результата на 100 для получения процента идентичности этих двух последовательностей. Типично окно сравнения будет соответствовать всей длине сравниваемой последовательности. Например, можно использовать программу BLAST, «BLAST 2 sequences» (Tatusova et al., "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol Lett. 174: 247-250), имеющуюся на сайте http://www.ncbi.nlm.nih.gov/gorf/bl2.html, с использованием параметров, которые заданы по умолчанию (в частности, параметров: «штрафа за начало пропуска»: 5, и «штрафа за расширение пропуска»: 2; при этом выбранной матрицей является, например, матрица «BLOSUM 62», предлагаемая этой программой), при этом процент идентичности двух сравнимых последовательностей рассчитывается непосредственно этой программой.
Самыми предпочтительными антителами против CD70, обеспечиваемыми здесь, которые демонстрируют особенно преимущественное сочетание характеристик, включая с очень высокой аффинностью с CD70 человека, являются те, которые основаны на антигенсвязывающей части происходящего от ламы Fab, обозначенного 27B3 в сопроводительных примерах, плюс приведенные к человеческому зародышевому типу варианты 27B3, включая приведенные к зародышевому типу варианты, идентифицированные в сопроводительных примерах. 27B3 и его приведенные к зародышевому типу варианты, особенно варианты на основе CDR или полных вариабельных доменах варианта 41D12, демонстрируют чрезвычайно преимущественное сочетание характеристик, суммированных, как указано далее: связывания с высокой аффинностью с рекомбинантным CD70 человека, сильного связывания с CD70 на клеточной поверхности, специфического связывания с CD70, экспрессируемым в линиях раковых клеток, особенно линиях клеток, которые экспрессируют CD70 в «низкой копийности», и сильного связывания с раковыми клетками, выделенными из образцов от пациентов (CLL), сильного блокирования взаимодействия CD70/CD27, сильной эффекторной функции - особенно в случае экспрессии в виде химеры с константными областями IgG1 человека и особенно при экспрессии в виде нефукозилированного IgG1, перекрестной реактивности с гомологами CD70 у макака-резус и яванского макака, дающей возможность проведения токсикологических исследований у видов приматов, связывания как со встречающимся в природе (т.е. на клеточной поверхности), так и с подвергнутым тепловой денатурации CD70, частичной интернализации или низких уровней интернализации в некоторые линии раковых клеток. Все из этих характеристик в сочетании делают 41D12, и, конечно, другие варианты 27B3 и другие антитела против CD70, описываемые здесь, которые демонстрируют схожие характеристики, превосходным кандидатом на терапевтическое применение для лечения связанных с CD70 заболеваний, в частности, CD70-экспрессирующих раков и иммунологических нарушений.
27B3 и его варианты характеризуются присутствием последовательности CDR3 вариабельного домена тяжелой цепи, представленной в виде SEQ ID NO: 50 (DAGYSNHVPIFDS).
Соответственно, предпочтительными вариантами антитела против CD70, или его антигенсвязывающего фрагмента, являются те, которые включают вариабельный домен тяжелой цепи, причем CDR3 вариабельного домена тяжелой цепи включает аминокислотную последовательность SEQ ID NO: 50 или ее вариант или состоит из нее или ее варианта, причем вариант последовательности включает одну, две или три аминокислотных замены в приведенной последовательности.
Более предпочтительными вариантами осуществления являются антитела или их антигенсвязывающие фрагменты, которые включают сочетание CDR тяжелой цепи, одинаковое с таковым в 27B3, или приведенных к человеческому зародышевому типу вариантах 27B3. Соответственно, антитело или его антигенсвязывающий фрагмент может включать вариабельный домен тяжелой цепи, причем
CDR3 вариабельного домена тяжелой цепи включает аминокислотную последовательность SEQ ID NO: 50 или ее вариант или состоит из нее или ее варианта;
CDR2 вариабельного домена тяжелой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 27, SEQ ID NO: 249 и их вариантов, или состоит из нее; и
CDR1 вариабельного домена тяжелой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 11, SEQ ID NO: 248 и их вариантов, или состоит из нее, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности.
Любое сочетание HCDR3, HCDR2 и HCDR1 из приведенных групп последовательностей CDR является допустимым и находится в объеме настоящего изобретения, однако некоторые сочетания являются особенно предпочтительными. Соответственно, в предпочтительных вариантах осуществления антитело или его антигенсвязывающий фрагмент может включать вариабельный домен тяжелой цепи, в котором сочетание HCDR3, HCDR2 и HCDR1 выбирают из следующих сочетаний:
(а) CDR3 вариабельного домена тяжелой цепи включает SEQ ID NO: 50 (DAGYSNHVPIFDS) или ее вариант или состоит из нее или ее варианта; CDR2 вариабельного домена тяжелой цепи включает SEQ ID NO: 27 (DINNEGGTTYYADSVKG) или ее вариант или состоит из нее или ее варианта; и CDR1 вариабельного домена тяжелой цепи включает SEQ ID NO: 11 (VYYMN) или ее вариант или состоит из нее или ее варианта, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности;
(b) CDR3 вариабельного домена тяжелой цепи включает SEQ ID NO: 50 (RDAGYSNHVPIFDS) или ее вариант или состоит из нее или ее варианта; CDR2 вариабельного домена тяжелой цепи включает SEQ ID NO: 249 (DINNEGGATYYADSVKG) или ее вариант или состоит из нее или ее варианта; и CDR1 вариабельного домена тяжелой цепи включает SEQ ID NO: 11 (VYYMN) или ее вариант или состоит из нее или ее варианта, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности;
(c) CDR3 вариабельного домена тяжелой цепи включает SEQ ID NO: 50 (DAGYSNHVPIFDS) или ее вариант или состоит из нее или ее варианта; CDR2 вариабельного домена тяжелой цепи включает SEQ ID NO: 27 (DINNEGGTTYYADSVKG) или ее вариант или состоит из нее или ее варианта; и CDR1 вариабельного домена тяжелой цепи включает SEQ ID NO: 248 (GYYMN) или ее вариант или состоит из нее или ее варианта, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности.
В предпочтительных вариантах осуществления антитело или его антигенсвязывающий фрагмент также включает вариабельный домен легкой цепи (VL), спаренный с вариабельным доменом тяжелой цепи. В предпочтительных вариабельных доменах легкой цепи CDR3 вариабельного домена легкой цепи включает SEQ ID NO: 160 или ее вариант или состоит из нее или ее варианта;
CDR2 вариабельного домена легкой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 119, SEQ ID NO: 110 и вариантов приведенных последовательностей, или состоит из нее; и
CDR1 вариабельного домена легкой цепи включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 87, SEQ ID NO: 250, SEQ ID NO: 251, SEQ ID NO: 252, SEQ ID NO: 253 и вариантов приведенных последовательностей, или состоит из нее, и
причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенных последовательностях.
Любое сочетание LCDR3, LCDR2 и LCDR1 из приведенных групп последовательностей CDR легкой цепи является допустимым и находится в объеме настоящего изобретения, однако некоторые сочетания являются особенно предпочтительными. Соответственно, в предпочтительных вариантах осуществления антитело или его антигенсвязывающий фрагмент может включать вариабельный домен легкой цепи, в котором сочетание LCDR3, LCDR2 и LCDR1 выбирают из следующих сочетаний:
(a) CDR3 вариабельного домена легкой цепи включает SEQ ID NO: 160 (ALFISNPSVE) или ее вариант или состоит из нее или ее варианта; CDR2 вариабельного домена легкой цепи включает SEQ ID NO: 119 (NTNTRHS) или ее вариант или состоит из нее или ее варианта; и CDR1 вариабельного домена легкой цепи включает SEQ ID NO: 250 (GLKSGSVTSDNFPT) или ее вариант или состоит из нее или ее варианта, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности;
(b) CDR3 вариабельного домена легкой цепи включает SEQ ID NO: 160 (ALFISNPSVE) или ее вариант или состоит из нее или ее варианта; CDR2 вариабельного домена легкой цепи включает SEQ ID NO: 119 (NTNTRHS) или ее вариант или состоит из нее или ее варианта; и CDR1 вариабельного домена легкой цепи включает SEQ ID NO: 87 (GLKSGSVTSTNFPT) или ее вариант или состоит из нее или ее варианта, причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности.
Другие предпочтительные сочетания CDR легкой цепи для «приведенных к человеческому зародышевому типу» вариантов 27B3 представлены в таблице 15A.
Наиболее предпочтительные варианты антитела против CD70 или его антигенсвязывающего фрагмента включают вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), причем сочетанием 6 CDR, которые образуют центр связывания с CD70 человека, является следующее сочетание:
CDR3 вариабельного домена тяжелой цепи включает SEQ ID NO: 50 (DAGYSNHVPIFDS) или ее вариант или состоит из нее или ее варианта;
CDR2 вариабельного домена тяжелой цепи включает SEQ ID NO: 27 (DINNEGGTTYYADSVKG) или ее вариант или состоит из нее или ее варианта;
CDR1 вариабельного домена тяжелой цепи включает SEQ ID NO: 11 (VYYMN) или ее вариант или состоит из нее или ее варианта,
CDR3 вариабельного домена легкой цепи включает SEQ ID NO: 160 (ALFISNPSVE) или ее вариант или состоит из нее или ее варианта;
CDR2 вариабельного домена легкой цепи включает SEQ ID NO: 119 (NTNTRHS) или ее вариант или состоит из нее или ее варианта;
CDR1 вариабельного домена легкой цепи включает SEQ ID NO: 250 (GLKSGSVTSDNFPT) или ее вариант или состоит из нее или ее варианта,
причем варианты последовательностей включают одну, две или три аминокислотных замены (например, консервативных замены, замены с целью гуманизации или варианты аффинности) в приведенной последовательности.
Другими предпочтительными сочетаниями 6 CDR являются встречающиеся в природе сочетания, которые встречаются в приведенных к человеческому зародышевому типу вариантах 27B3, перечисленных в таблицах 14A (тяжелые цепи) и 15A (легкие цепи).
В вышеприведенных предпочтительных вариантах осуществления, основанных на 27B3 и его приведенных к зародышевому типу вариантах, антитело предпочтительно включает CH1-домен, шарнирную область, CH2-домен и CH3-домен антитела человека, в частности, IgG1, IgG2, IgG3 или IgG4 человека. Наиболее предпочтительным вариантом осуществления является IgG1 человека. Кроме того, предпочтительным является конструирование с максимизацией эффекторной функции IgG1 человека в одной или более из антителозависимой клеточноопосредованной цитотоксичности (ADCC), комплементзависимой цитотоксичности (CDC) или антителозависимого клеточного фагоцитоза (ADCP). Особенно предпочтительным является нефукозилированный IgG1 человека, например, нефукозилированный IgG1, полученный с использованием PotelligentTM технологии BioWa Inc.
Дальнейшие предпочтительные антитела против CD70, демонстрирующие связывание с высокой аффинностью с CD70 человека, основанные на Fab, обозначенном 27B3, и приведенных к человеческому зародышевому типу вариантах 27B3, включают выделенные антитела или их антигенсвязывающие фрагменты, включающие вариабельный домен тяжелой цепи, имеющий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 178, SEQ ID NO: 212, SEQ ID NO: 213, SEQ ID NO: 214, SEQ ID NO: 215, SEQ ID NO: 216, SEQ ID NO: 217, SEQ ID NO: 218, SEQ ID NO: 219, SEQ ID NO: 220, SEQ ID NO: 221, SEQ ID NO: 222, SEQ ID NO: 223, SEQ ID NO: 224, SEQ ID NO: 225, SEQ ID NO: 226, SEQ ID NO: 227, SEQ ID NO: 228, SEQ ID NO: 229, SEQ ID NO: 274 и SEQ ID NO: 275, и аминокислотных последовательностей, идентичных на по крайней мере 90%, 95%, 97%, 98% или 99% одной из приведенных последовательностей, и необязательно включающие вариабельный домен легкой цепи, имеющий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 201, SEQ ID NO: 230, SEQ ID NO: 231, SEQ ID NO: 232, SEQ ID NO: 233, SEQ ID NO: 234, SEQ ID NO: 234, SEQ ID NO: 236, SEQ ID NO: 237, SEQ ID NO: 238, SEQ ID NO: 239, SEQ ID NO: 240, SEQ ID NO: 241, SEQ ID NO: 242, SEQ ID NO: 243, SEQ ID NO: 244, SEQ ID NO: 245, SEQ ID NO: 245, SEQ ID NO: 247, SEQ ID NO: 248, SEQ ID NO: 276 и SEQ ID NO: 277, и аминокислотных последовательностей, идентичных на по крайней мере 90%, 95%, 97%, 98% или 99% одной из приведенных последовательностей.
Хотя все возможные спаривания VH-доменов и VL-доменов, выбираемых из групп последовательностей VH- и VL-доменов, перечисленных выше, являются допустимыми и находятся в объеме настоящего изобретения, особенно предпочтительными являются некоторые сочетания VH и VL, которые являются встречающимися в природе сочетаниями в одном Fab, демонстрирующем связывание с высокой аффинностью с CD70 человека. В случае приведенных к зародышевому типу вариантов 27B3, приведенных в таблицах 14B и 15B, может быть предпочтительным сохранение исходного спаривания VH/VL. Соответственно, предпочтительными антителами против CD70, или их антигенсвязывающими фрагментами, демонстрирующими связывание с высокой аффинностью с CD70, являются те, которые включают сочетание вариабельного домена тяжелой цепи и вариабельного домена легкой цепи, причем сочетание выбирают из группы, состоящей из:
(i) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 223, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 241;
(ii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 178, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 190;
(iii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 212, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 230;
(iv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 213, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 231;
(v) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 214, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 232;
(vi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 215, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 235;
(vii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 216, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 234;
(viii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 217, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 235;
(ix) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 218, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 236;
(x) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 219, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 237;
(xi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 220, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 238;
(xii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 221, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 239;
(xiii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 222, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 240;
(xiv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 224, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 242;
(xv) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 225, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 243;
(xvi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 226, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 244;
(xvii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 227, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 245;
(xviii) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 228, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 246;
(xix) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 229, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 247;
(xx) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 223, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 244;
(xxi) вариабельного домена тяжелой цепи, включающего аминокислотную последовательность SEQ ID NO: 223, и вариабельного домена легкой цепи, включающего аминокислотную последовательность SEQ ID NO: 245.
В случае каждого из конкретных сочетаний VH/VL, перечисленных выше, является также допустимым и находится в объеме настоящего изобретения сочетание VH-домена, имеющего аминокислотную последовательность, идентичную на по крайней мере 90%, 92%, 95%, 97% или 99% приведенной последовательности VH-домена, с VL-доменом, имеющим аминокислотную последовательность, идентичную на по крайней мере 90%, 92%, 95%, 97% или 99% приведенной последовательности VL-домена.
Наиболее предпочтительным вариантом осуществления является антитело против CD70, или его антигенсвязывающий фрагмент, основанное на встречающемся в природе сочетании VH/VL приведенного к человеческому зародышевому типу варианта, обозначенного 41D12. Соответственно, здесь также обеспечивается антитело или его антигенсвязывающий фрагмент, включающее(ий) вариабельный домен тяжелой цепи (VH), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 223, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее, и вариабельный домен легкой цепи (VL), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 241, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее.
Варианты осуществления, в которых аминокислотная последовательность VH-домена идентична менее чем на 100% последовательности, представленной в виде SEQ ID NO: 223, могут, тем не менее, включать CDR тяжелой цепи, которые идентичны HCDR1, HCDR2 и HCDR3 SEQ ID NO: 223 (SEQ ID NO: 11, 27 и 50, соответственно) при проявлении вариации аминокислотной последовательности в каркасных областях. Так же, варианты осуществления, в которых аминокислотная последовательность VL-домена идентична менее чем на 100% последовательности, представленной в виде SEQ ID NO: 241, могут, тем не менее, включать CDR легкой цепи, которые идентичны LCDR1, LCDR2 и LCDR3 SEQ ID NO: 241 (SEQ ID NO: 250, 116 и 160, соответственно) при проявлении вариации аминокислотной последовательности в каркасных областях.
В вышеприведенных предпочтительных вариантах осуществления, основанных на VH- и VL-доменах 41D12, или его вариантов, антитело предпочтительно включает CH1-домен, шарнирную область, CH2-домен и CH3-домен антитела человека, в частности, IgG1, IgG2, IgG3 или IgG4 человека. Наиболее предпочтительным вариантом осуществления является IgG1 человека. Кроме того, предпочтительным является конструирование с максимизацией эффекторной функции IgG1 человека в одной или более из ADCC, CDC или ADCP. Особенно предпочтительным является нефукозилированный IgG1 человека, предпочтительно полученный с использованием экспрессионной системы PotelligentTM.
Другим преимущественным вариантом осуществления является антитело или его антигенсвязывающий фрагмент, включающее(ий) вариабельный домен тяжелой цепи (VH), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 225, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее, и вариабельный домен легкой цепи (VL), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 243, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее.
Другим преимущественным вариантом осуществления является антитело или его антигенсвязывающий фрагмент, включающий вариабельный домен тяжелой цепи (VH), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 226, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее, и вариабельный домен легкой цепи (VL), включающий аминокислотную последовательность, выбираемую из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO: 244, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее.
Свойства/характеристики антител против CD70
В вышеотмеченных аспектах и вариантах осуществления, каждое из антител против CD70, или их антигенсвязывающих фрагментов, может демонстрировать одну или более, или любое сочетание, следующих характеристик или свойств:
Антитело или антигенсвязывающий фрагмент может связываться с CD70 человека с высокой аффинностью, демонстрируя, при исследовании в виде Fab-фрагмента, скорость диссоциации от CD70 человека, составляющую 7×10-4 сек-1 или менее, предпочтительно 5×10-4 сек-1 или менее, а обычно находящуюся в диапазоне от 0,4×10-4 сек-1 до 4,8×10-4 сек-1.
Антитело или антигенсвязывающий фрагмент может связываться с CD70 человека с высокой аффинностью и ингибировать взаимодействие между CD70 и CD27. Альтернативно, антитело или антигенсвязывающий фрагмент может связываться с CD70 человека, но не ингибировать взаимодействие между CD70 и CD27.
Антитело или антигенсвязывающий фрагмент может связываться с высокой аффинностью с CD70 человека на поверхности клеток, экспрессирующих CD70.
Антитело или антигенсвязывающий фрагмент может связываться с CD70 человека на поверхности клеток, экспрессирующих CD70, и подвергаться медленной или лишь частичной интернализации. Ключевым аспектом настоящего изобретения являются данные наблюдений, что антитела против CD70, в действительности, очень плохо интернализуются в широкий ряд линий CD70-экспрессирующих клеток, включая множество линий CD70-экспрессирующих раковых клеток. Эти данные наблюдений прямо противоположны предшествующим опубликованным сообщениям о том, что антитела против CD70 быстро интернализуются после связывания с линиями клеток почечноклеточной карциномы (смотрите Adam et al., British Journal of Cancer (2006) 95: 298-306; и WO 2007/038637), и непосредственно влекут за собой терапевтическое применение антител. Данные наблюдений, что антитела против CD70 очень плохо интернализуются после связывания с раковыми клетками, служат сильным подтверждением вывода, что терапевтические стратегии лечения множества CD70-экспрессирующих раков и, конечно, связанных с CD70 иммунологических заболеваний должны основываться на связывании с очень высокой аффинностью с CD70, вместе с эффекторной функцией антител, в частности, любой одной или более из ADCC, CDC или ADCP, а не на использовании иммуноконъюгатов, в которых антитело против CD70 связано с терапевтическим средством, например, составляющей в виде цитотоксического или цитостатического агента.
Антитело или антигенсвязывающий фрагмент может связываться в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO: 342) в CD70 человека.
Антитело или антигенсвязывающий фрагмент может проявлять перекрестную реактивность с CD70 обезьяньего происхождения, в частности, гомологами CD70 у макака-резус (Macaca mulatta) и яванского макака (Macaca cynomolgus).
Антитело или антигенсвязывающий фрагмент может связываться как со встречающимся в природе CD70 человека (например, CD70, представленным на поверхности клетки, такой как линия клеток или CD70-экспрессирующая клетка, изолированная от являющегося человеком пациента), так и с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
Антитело или антигенсвязывающий фрагмент может дать очень высокие выходы продукции (>4 г/л) в системах для рекомбинантной экспрессии антител, такой как, например, линия клеток CHK1SV (составляющая собственность BioWa/Lonza), по сравнению с составляющим 1-2 г/л первоначальным средним значением для терапевтических продуктов в виде антител, что приводит к значительным снижениям затрат на производство.
Антитело или антигенсвязывающий фрагмент может быть в высокой степени стабильным в условиях хранения при 37°C и в циклах замораживания-оттаивания, что также является основным фактором снижения затрат.
Антитело может проявлять одну или более эффекторных функций, выбираемых из антителозависимой клеточноопосредованной цитотоксичности (ADCC), комплементзависимой цитотоксичности (CDC) и антителозависимого клеточноопосредованного фагоцитоза (ADCP), по отношению к клеткам, на клеточной поверхности которых представлен белок CD70 человека.
Антитело может проявлять ADCC по отношению к CD70-экспрессирующим клеткам, например, раковым клеткам или другим злокачественным клеткам, или иммуноцитам.
Антитело может проявлять увеличенную функцию, относящуюся к ADCC, по сравнению с эталонным антителом, являющимся эквивалентным антителом, включающим природный Fc-домен человека. В неограничивающем варианте осуществления относящаяся к ADCC функция может быть увеличенной в по крайней мере 10x по сравнению с эталонным антителом, включающим природный Fc-домен человека. В этом контексте «эквивалентное» может, как предполагают, означать, что антитело с увеличенной функцией, относящейся к ADCC, проявляет по существу идентичную специфичность связывания с антигенами и/или разделяет идентичную аминокислотную последовательность с эталонным антителом, за исключением любых модификаций (относительно природного Fc человека), осуществленных с целью увеличения ADCC.
Антитело или антигенсвязывающий фрагмент может ингибировать рост опухолей in vivo в модели с использованием ксенотрансплантата опухоли, в отсутствие конъюгации с цитотоксическим или цитостатическим агентом. В неограничивающем варианте осуществления относящаяся к ингибированию роста опухолей функция может быть увеличенной в по крайней мере 10 раз по сравнению с эталонным антителом SGN70.
Антитело или антигенсвязывающий фрагмент может индуцировать апоптоз CD70-экспрессирующих клеток.
Антитело может содержать шарнирную область, CH2-домен и CH3-домен IgG человека, наиболее предпочтительно IgG1, IgG2, IgG3 или IgG4 человека.
Антитело может включать модификации в Fc-области, такие как модификации, которые увеличивают эффекторную функцию антитела, как объяснено здесь в другом месте. В частности, антитело может быть нефукозилированным IgG.
В дальнейших аспектах настоящим изобретением обеспечиваются полинуклеотидные молекулы, которые кодируют перечисленные выше антитела против CD70 и их антигенсвязывающие фрагменты, вдобавок к экспрессионным векторам, включающим полинуклеотиды, клеткам-хозяевам, содержащим векторы, и способам рекомбинантной экспрессии/продукции антител против CD70.
Тем не менее, в дальнейшем аспекте настоящим изобретением обеспечивается фармацевтическая композиция, включающая одно из антител против CD70, описанных выше, и фармацевтически приемлемый носитель или наполнитель.
Тем не менее, в дальнейшем аспекте настоящее изобретение имеет отношение к способам терапевтического лечения, используя вышеперечисленные антитела против CD70, в частности, лечения рака.
Краткое описание чертежей
Настоящее изобретение будет еще понятнее с учетом следующих экспериментальных примеров и сопроводительных фигур, в случае которых:
На фиг. 1 представлен иммунный ответ, проверенный в ELISA с использованием рекомбинантного CD70, у лам, иммунизированных клетками 786-О (верх) и клетками Raji (низ).
На фиг. 2 показано определяемое с помощью ELISA ингибирование связывания CD27 с CD70 с помощью происходящих от ламы Fab против CD70 и эталонных Fab против CD70.
Фиг. 3 является графическим представлением сигнала в случае происходящих от ламы Fab, проверенных в ELISA связывания (черные прямоугольники) или в ELISA ингибирования (белые прямоугольники).
На фиг. 4 показано определяемое с помощью ELISA ингибирование связывания CD27 с CD70 человека (A&B) или CD70 макака-резус (C) с помощью химерных лама-человек мАт против CD70 и эталонных мАт против CD70.
На фиг. 5 показано связывание химерных лама-человек мАт против CD70 с клетками 786-О (A) или клетками MHH-PREB-1 (B) с помощью анализа с использованием FACS (клеточного сортера с возбуждением флуоресценции).
На фиг. 6 показано ингибирование с помощью специфических для CD70 химерных лама-человек мАт в анализе эффективности с использованием сокультуры на основе клеток Raji.
На фиг. 7 представлены результаты анализа ADCC в отношении клеток 786-О на основе высвобождения Cr51.
На фиг. 8 представлены результаты анализа CDC в отношении клеток U266 в присутствии 9% сыворотки человека.
На фиг. 9 показана эффективность химерных лама-человек мАт против CD70 в анализе ADCP на клетках 786-О.
На фиг. 10 показана интернализация антител, оцениваемая в виде MFI (средней интенсивности флуоресценции) «Вне» для различных химерных лама-человек мАт против CD70 в зависимости от времени на клетках 786-О в двух независимых экспериментах.
На фиг. 11 показано выживание мышей в модели с использованием ксенотрансплантата клеток Raji после лечения химерным лама-человек мАт против CD70 41D12, подобранным по изотипу контрольным антителом и контрольным антителом с недействующим Fc.
На фиг. 12 представлены совмещения аминокислотных последовательностей VH и VL клона 27B3 с аминокислотными последовательностями генных сегментов зародышевой линии человека VH3-38 и VL8-61, соответственно.
На фиг. 13 представлен анализ с помощью гель-фильтрации образцов приведенных к зародышевому типу вариантов мАт 27B3, отобранных после инкубации в течение 5 недель при 37°C.
На фиг. 14 показана активность связывания с CD70, определяемая с использованием Biacore, образцов приведенных к зародышевому типу мАт против CD70, отобранных в различные моменты времени после инкубации при различных температурах.
На фиг. 15 показана аффинность мАт против CD70 к линиям CD70-экспрессирующих раковых клеток.
На фиг. 16 показана аффинность мАт против CD70 к клеткам от пациентов с CLL, экспрессирующим CD70.
На фиг. 17 представлен лизис клеток SU-DHL-6, с которыми связались мАт против CD70.
На фиг. 18 показано определяемое с помощью ELISA ингибирование связывания CD27 с CD70 с помощью мАт против CD70.
На фиг. 19 представлено совмещение последовательностей CD70 различных видов.
На фиг. 20 показана аффинность мАт против CD70 к клеткам U266 человека, клеткам LCL8864 макака-резус и клеткам HSC-F яванского макака.
На фиг. 21 показано определяемое с помощью ELISA ингибирование связывания CD27 с CD70 человека, макака-резус и яванского макака с помощью мАт против CD70.
На фиг. 22 показано определяемое с помощью ELISA связывание мАт против CD70 с денатурированным рекомбинантным CD70.
На фиг. 23 представлены последовательности химерных CD70, использованные для картирования эпитопа.
На фиг. 24 показана интернализация мАт против CD70 в зависимости от времени в клетки 786-О.
Определения
«Антитело» или «Иммуноглобулин» - Как здесь используется, термин «иммуноглобулин» включает полипептид, содержащий комбинацию двух тяжелых и двух легких цепей, независимо от того, обладает ли он какой-либо релевантной специфической иммунореактивностью. «Антитела» относятся к таким сборкам, которые обладают значительной известной специфической иммунореактивностью с представляющим интерес антигеном (например, CD70 человека). Как здесь используется, термин «антитела против CD70» относится к антителам, которые проявляют иммунологическую специфичность по отношению к белку CD70 человека. Как объясняется здесь в другом месте, «специфичность» по отношению к CD70 человека не исключает перекрестную реакцию с видовыми гомологами CD70. Антитела и иммуноглобулины включают легкие и тяжелые цепи, с ковалентной связью между их цепями или без нее. Базовые структуры иммуноглобулинов в системах позвоночных относительно хорошо изучены.
Общий термин «иммуноглобулин» включает пять отдельных классов антител, которые можно различить биохимически. Все пять классов антител находятся в объеме настоящего изобретения, следующее обсуждение будет большей частью направлено на класс IgG молекул иммуноглобулинов. Что касается IgG, иммуноглобулины включают две идентичные легкие полипептидные цепи с молекулярной массой, составляющей приблизительно 23000 Дальтон, и две идентичные тяжелые цепи с молекулярной массой, составляющей 53000-70000. Четыре цепи соединены с помощью дисульфидных связей в конфигурации «Y», причем легкие цепи «заключают в скобки» тяжелые цепи, начинающиеся в сужении «Y» и продолжающиеся по всей вариабельной области.
Легкие цепи антитела относят либо к каппа, либо к лямбда (κ, λ). Каждый класс тяжелых цепей может быть связан с легкой цепью либо каппа, либо лямбда. Как правило, легкие и тяжелые цепи ковалентно связываются друг с другом, и «хвостовые» части двух тяжелых цепей связываются друг с другом с помощью ковалентных дисульфидных связей или нековалентных связей при создании иммуноглобулинов с помощью или гибридом, B-клеток, или генетически сконструированных клеток-хозяев. В тяжелой цепи аминокислотные последовательности направляются от N-конца в раздвоенных концах конфигурации Y к C-концу внизу каждой цепи. Квалифицированные в данной области техники специалисты примут во внимание, что тяжелые цепи относятся к гамма, мю, альфа, дельта или эпсилон (γ, µ, α, δ, ε) с некоторым количество подклассов среди них (например, γ1-γ4). Именно природа этой цепи определяет «класс» антитела как IgG, IgM, IgA, IgD или IgE, соответственно. Подклассы (изотипы) иммуноглобулинов, например, IgG1, IgG2, IgG3, IgG4, IgA1 и т.д., хорошо охарактеризованы и, как известно, придают функциональную специализацию. Модифицированные варианты каждого из этих классов и изотипов являются легко распознаваемыми квалифицированным специалистом ввиду настоящего описания и, соответственно, находятся в объеме настоящего изобретения.
Как указано выше, вариабельная область антитела позволяет антителу селективно распознавать эпитопы антигенов и специфически связываться с ними. Т.е. VL-домен и VH-домен антитела объединяются с образованием вариабельной области, которая определяет пространственный антигенсвязывающий центр. Эта четвертичная структура антитела образует антигенсвязывающий центр, присутствующий на конце каждого плеча Y. Конкретнее, антигенсвязывающий центр определяется тремя определяющими комплементарность участками (CDR) в каждой из VH- и VL-областей.
«Белок CD70» или «антиген CD70» - Как здесь используются, термины «белок CD70» или «антиген CD70» или «CD70» используются взаимозаменяемо и относятся к члену семейства лигандов TNF, который является лигандом для TNFRSF27/CD27. Термины «белок CD70 человека» или «антиген CD70 человека» или «CD70 человека» используются взаимозаменяемо для ссылки конкретно на гомолог у человека, в том числе природный белок CD70 человека, экспрессируемый обычно в организме человека и/или представленный на поверхности культивируемых линий клеток человека, а также рекомбинантные формы и его фрагменты. Конкретные примеры CD70 человека включают полипептид, имеющий аминокислотную последовательность, представленную под № доступа к эталонным последовательностям NCBI-NP_001243, или его экстраклеточный домен.
«Связывающий центр» - Как здесь используется, термин «связывающий центр» включает район полипептида, который ответствен за селективное связывание с представляющим интерес антигеном-мишенью (например, CD70 человека). Связывающие домены включают по крайней мере один связывающий центр. Приводимые в качестве примера связывающие домены включают вариабельный домен антитела. Молекулы антител по настоящему изобретению могут включать один связывающий центр или множество (например, два, три или четыре) связывающих центров.
«Происходящий от (из)» - Как здесь используется, термин «происходящий из» обозначенного белка (например, антитела против CD70 или его антигенсвязывающего фрагмента) относится к происхождению полипептида. В одном варианте осуществления полипептидной или аминокислотной последовательностью, которая происходит из конкретного исходного полипептида, является последовательность CDR или родственная ей последовательность. В одном варианте осуществления аминокислотная последовательность, которая происходит из конкретного исходного полипептида, не является непрерывной. Например, в одном варианте осуществления один, два, три, четыре, пять или шесть CDR происходят из исходного антитела. В одном варианте осуществления полипептидная или аминокислотная последовательностью, которая происходит из конкретной исходной полипептидной или аминокислотной последовательности, имеет аминокислотную последовательность, которая по существу идентична исходной последовательности, или ее части, состоящей из по крайней мере 3-5 аминокислот, по крайней мере 5-10 аминокислот, по крайней мере 10-20 аминокислот, по крайней мере 20-30 аминокислот или по крайней мере 30-50 аминокислот, или которую специалист со средним уровнем компетентности в данной области техники может идентифицировать иным способом как последовательность, происходящую из исходной последовательности. В одном варианте осуществления одну или более последовательностей CDR, происходящих из исходного антитела, изменяют для получения вариантов последовательностей CDR, например, вариантов аффинности, причем варианты последовательностей CDR сохраняют CD70-связывающую активность.
«Происходящий от верблюдовых» - В некоторых предпочтительных вариантах осуществления молекулы антител против CD70 по настоящему изобретению включают аминокислотные последовательности каркасных областей и/или аминокислотные последовательности CDR, происходящие из обычного антитела животного семейства верблюдовых, индуцированного путем активной иммунизации животного семейства верблюдовых антигеном CD70. Однако антитела против CD70, включающие происходящие от верблюдовых аминокислотные последовательности, можно сконструировать для включения последовательностей каркасных областей и/или константных областей, происходящих из аминокислотной последовательности человека (т.е. антитела человека) или других видов млекопитающих, не являющихся животными семейства верблюдовых. Например, каркасная область, часть тяжелой цепи и/или часть шарнирной области человека или не являющегося человеком примата могут быть включены в рассматриваемые антитела против CD70. В одном варианте осуществления одна или более аминокислот не верблюдовых может присутствовать в каркасной области «происходящего от верблюдовых» антитела против CD70, например, аминокислотная последовательность каркасной области верблюдовых может включать одну или более мутаций аминокислот, в результате которых присутствует соответствующий человеку или не являющемуся человеком примату аминокислотный остаток. Кроме того, происходящие от верблюдовых VH- и VL-домены, или их гуманизированные варианты, можно связать с константными доменами антител человека для создания химерной молекулы, как подробно описано здесь в другом месте.
«Консервативная аминокислотная замена» - «Консервативной аминокислотной заменой» является замена, в случае которой аминокислотный остаток заменяют аминокислотным остатком, имеющим схожую боковую цепь. В данной области техники были установлены семейства аминокислотных остатков, имеющих схожие боковые цепи, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глютаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глютамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, заменимый аминокислотный остаток в полипептиде иммуноглобулина может быть заменен другим аминокислотным остатком из того же семейства боковых цепей. В другом варианте осуществления ряд аминокислот может быть заменен структурно схожим рядом, который отличается порядком и/или составом членов семейства боковых цепей.
«Часть тяжелой цепи» - Как здесь используется, термин «часть тяжелой цепи» включает аминокислотные последовательности, происходящие из константных доменов тяжелой цепи иммуноглобулина. Полипептид, включающий часть тяжелой цепи, включает по крайней мере один из CH1-домена, шарнирного домена (например, верхней, срединной и/или нижней части шарнирной области), CH2-домена, CH3-домена или их варианта или фрагмента. В одном варианте осуществления антитело или антигенсвязывающий фрагмент по настоящему изобретению может включать Fc-часть тяжелой цепи иммуноглобулина (например, часть шарнирной области, CH2-домен и CH3-домен). В другом варианте осуществления в антителе или антигенсвязывающем фрагменте по настоящему изобретению может отсутствовать по крайней мере часть константного домена (например, весь CH2-домен или его часть). В некоторых вариантах осуществления по крайней мере один, а предпочтительно все из константных доменов происходят из тяжелой цепи иммуноглобулина человека. Например, в одном предпочтительном варианте осуществления часть тяжелой цепи включает полностью человеческий шарнирный домен. В других предпочтительных вариантах осуществления часть тяжелой цепи включает полностью человеческую Fc-часть (например, последовательности шарнирной области, CH2- и CH3-доменов из иммуноглобулина человека).
В некоторых вариантах осуществления являющиеся составными частями части тяжелой цепи константные домены происходят из различных молекул иммуноглобулинов. Например, часть тяжелой цепи полипептида может включать CH2-домен, происходящий из молекулы IgG1, и шарнирную область, происходящую из молекулы IgG3 или IgG4. В других вариантах осуществления константные домены являются химерными доменами, включающими части из различных молекул иммуноглобулинов. Например, шарнирная область может включать первую часть из молекулы IgG1, а вторую часть из молекулы IgG3 или IgG4. Как изложено выше, специалисту со средним уровнем компетентности в данной области техники будет понятно, что константные домены части тяжелой цепи можно модифицировать из условия, чтобы они отличались по аминокислотной последовательности от встречающейся в природе молекулы иммуноглобулина (дикого типа). Т.е. описываемые здесь полипептиды по настоящему изобретению могут включать изменения или модификации в одном или более константных доменах тяжелой цепи (CH1, шарнирной области, CH2 или CH3) и/или константном домене легкой цепи (CL). Приводимые в качестве примера модификации включают добавления, делеции или замены одной или более аминокислот в одном или более доменах.
«Химерный» - «Химерный» белок включает первую аминокислотную последовательность, связанную со второй аминокислотной последовательностью, с которой она не связана в природных условиях. Аминокислотные последовательности могут обычно существовать в отдельных белках, которые сводят в слитый полипептид, или они могут обычно существовать в одном и том же белке, но размещены по-новому в слитом полипептиде. Химерный белок можно создать, например, посредством химического синтеза или посредством создания полинуклеотида, в котором пептидные районы кодируются в желаемой взаимосвязи, и трансляции с него. Приводимые в качестве примера химерные антитела против CD70 включают слитые белки, включающие происходящие от верблюдовых VH- и VL-домены, или их гуманизированные варианты, слитые с константными доменами антитела человека, например, IgG1, IgG2, IgG3 или IgG4 человека.
«Вариабельная область» или «вариабельный домен» - Термины «вариабельная область» и «вариабельный домен» используются здесь взаимозаменяемо и, как предполагается, имеют эквивалентное значение. Термин «вариабельный» относится к тому факту, что некоторые части вариабельных доменов VH и VL в значительной степени отличаются по последовательности в антителах и используются при связывании и определении специфичности каждого конкретного антитела в отношении антигена - его мишени. Однако вариабельность распределена неравномерно по вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых «гипервариабельными петлевыми участками», в каждом из VL-домена и VH-домена, которые образуют часть антигенсвязывающего центра. Первый, второй и третий гипервариабельные петлевые участки домена легкой цепи Vлямбда называют здесь L1(λ), L2(λ) и L3(λ), и их можно определить как участки, включающие остатки 24-33 (L1(λ), состоящий из 9, 10 или 11 аминокислотных остатков), 49-53 (L2(λ), состоящий из 3 остатков) и 90-96 (L3(λ), состоящий из 5 остатков) в VL-домене (Morea et al., Methods 20: 267-279 (2000)). Первый, второй и третий гипервариабельные петлевые участки домена легкой цепи Vкаппа называют здесь L1(κ), L2(κ) и L3(κ), и их можно определить как участки, включающие остатки 25-33 (L1(κ), состоящий из 6, 7, 8, 11, 12 или 13 остатков), 49-53 (L2(κ), состоящий из 3 остатков) и 90-97 (L3(κ), состоящий из 6 остатков) в VL-домене (Morea et al., Methods 20: 267-279 (2000)). Первый, второй и третий гипервариабельные петлевые участки VH-домена называют здесь H1, H2 и H3, и их можно определить как участки, включающие остатки 25-33 (H1, состоящий из 7, 8 или 9 остатков), 52-56 (H2, состоящий из 3 или 4 остатков) и 91-105 (H3, высоковариабельный по длине) в VH-домене (Morea et al., Methods 20: 267-279 (2000)).
Кроме особо оговоренных случаев, термины L1, L2 и L3 относятся, соответственно, к первому, второму и третьему гипервариабельным петлевым участкам VL-домена и охватывают гипервариабельные петлевые участки, получаемые из обоих изотипов Vкарра и Vлямбда. Термины H1, H2 и H3 относятся, соответственно, к первому, второму и третьему гипервариабельным петлевым участкам VH-домена и охватывают гипервариабельные петлевые участки, получаемые из любого из известных изотипов тяжелых цепей, включая γ, ε, δ, α или µ.
Каждый из гипервариабельных петлевых участков L1, L2, L3, H1, H2 и H3 может включать часть «определяющего комплементарность участка» или «CDR», определяемого ниже. Термины «гипервариабельный петлевой участок» и «определяющий комплементарность участок» не являются, строго говоря, синоминами, поскольку гипервариабельные петлевые участки (HV) определяют на основе структуры, тогда как определяющие комплементарность участки (CDR) определяют на основе вариабельности последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD., 1983), и границы HV и CDR могут различаться в некоторых VH- и VL-доменах.
CDR VL- и VH-доменов можно обычно определить как участки, включающие следующие аминокислоты: остатки 24-34 (CDRL1), 50-56 (CDRL2) и 89-97 (CDRL3) в вариабельном домене легкой цепи и остатки 31-35 или 31-35b (CDRH1), 50-65 (CDRH2) и 95-102 (CDRH3) в вариабельном домене тяжелой цепи (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Таким образом, HV могут содержаться в соответствующих CDR, и ссылки здесь на «гипервариабельные петлевые участки» VH- и VL-доменов должны интерпретироваться как ссылки, также охватывающие соответствующие CDR, и наоборот, кроме особо оговоренных случаев.
Более высококонсервативные части вариабельных доменов называются каркасной областью (FR), определяемой ниже. Каждый из вариабельных доменов встречающихся в природе тяжелых и легких цепей включает четыре FR (FR1, FR2, FR3 и FR4, соответственно), в значительной степени принимающих конформацию в виде β-листа, которые соединены с помощью трех гипервариабельных петлевых участков. Гипервариабельные петлевые участки в каждой цепи удерживаются вместе в непосредственной близости с помощью FR и, вместе с гипервариабельными петлевыми участками другой цепи, участвуют в образовании антигенсвязывающего центра антител. Структурный анализ антител выявил взаимосвязь между последовательностью и конфигурацией связывающего центра, образуемого определяющими комплементарность участками (Chothia et al., J. Mol. Biol. 227: 799-817 (1992)); Tramontano et al., J. Mol. Biol, 215: 175-182 (1990)). Несмотря на высокую вариабельность их последовательностей, пять из шести петлевых участков принимают лишь небольшой набор конформаций основной цепи, называемых «каноническими структурами». Эти конформации, прежде всего, определяются длиной петлевых участков и, во-вторых, присутствием ключевых остатков в определенных положениях в петлевых участках и в каркасных областях, которые определяют конформацию благодаря их укладке, образованию водородных связей и способности принимать необычные конформации основной цепи.
«CDR» - Как здесь используется, термин «CDR» или «определяющий комплементарность участок» означает не являющиеся непрерывными антигенсвязывающие центры, обнаруживаемые в вариабельной области полипептидов как тяжелой, так и легкой цепей. Эти особые участки были описаны Kabat и др. в J. Biol. Chem. 252, 6609-6616 (1977) и Kabat и др. в Sequences of protein of immunological interest. (1991), и Chothia и др. в J. Mol. Biol. 196: 901-917 (1987), и MacCallum и др. в J. Mol. Biol. 262: 732-745 (1996), в которых определения включают перекрытие или подмножества аминокислотных остатков после сравнения друг с другом. Аминокислотные остатки, которые охватывают CDR в соответствии с определениями в каждом из вышеприведенных ссылочных документов, выложены для сравнения. Предпочтительно «CDR» является CDR, определенный Kabat на основе сравнения последовательностей.
Определения CDR
2Нумерация остатков в соответствии с номенклатурой Chothia и др., выше.
3Нумерация остатков в соответствии с номенклатурой MacCallum и др., выше.
«Каркасная область» - Термин «каркасная область» или «FR-область», используемый здесь, включает аминокислотные остатки, которые являются частью вариабельной области, но не являются частью CDR (например, используя определение CDR по Kabat). Следовательно, каркасная область вариабельной области имеет длину, составляющую приблизительно 100-120 аминокислот, но включает лишь аминокислоты вне CDR. Ради конкретного примера вариабельного домена тяжелой цепи и CDR, определяемых согласно Kabat и др., каркасная область 1 соответствует домену вариабельной области, охватывающему аминокислоты 1-30; каркасная область 2 соответствует домену вариабельной области, охватывающему аминокислоты 36-49; каркасная область 3 соответствует домену вариабельной области, охватывающему аминокислоты 66-94, и каркасная область 4 соответствует домену вариабельной области, охватывающему аминокислоты с 103 до конца вариабельной области. Каркасные области в случае легкой цепи также разделены каждым из CDR вариабельной области легкой цепи. Так же, используя определение CDR по Chothia и др. или McCallum и др., границы каркасных областей отделяют концы соответствующих CDR, определенные выше. В предпочтительных вариантах осуществления CDR являются CDR, определяемые согласно Kabat.
Во встречающихся в природе антителах шесть CDR, присутствующих в каждом мономерном антителе, являются короткими, не являющимися непрерывными последовательностями аминокислот, которые особым образом позиционируются для образования антигенсвязывающего центра по мере принятия антителом своей трехмерной конфигурации в водной среде. Оставшаяся часть вариабельных доменов тяжелой и легкой цепей демонстрирует меньшую межмолекулярную вариабельность по аминокислотной последовательности и называется каркасными областями. Каркасные области в значительной степени принимают конформацию в виде β-листа, а CDR образуют петли, которые соединяют, и в некоторых случаях образуют часть, структуру(ы) в виде β-листа. Таким образом, эти каркасные области функционируют для образования каркаса, который обеспечивает позиционирование шести CDR в правильной ориентации с помощью нековалентных взаимодействий между цепями. Антигенсвязывающий центр, образованный позиционированными CDR, определяет поверхностную комплементарность эпитопу иммунореактивного антигена. Эта комплементарная поверхность стимулирует нековалентное связывание антитела с эпитопом иммунореактивного антигена. Специалист со средним уровнем компетентности в данной области техники может без труда определить местоположение CDR.
«Шарнирная область» - Как здесь используется, термин «шарнирная область» включает часть молекулы тяжелой цепи, которая соединяет CH1-домен с CH2-доменом. Эта шарнирная область включает приблизительно 25 остатков и является гибкой, позволяя тем самым двум N-концевым антигенсвязывающим центрам независимо изменять положение. Шарнирные области можно подразделить на три отличных домена: верхний, срединный и нижний шарнирные домены (Roux et al. J. Immunol. 1998 161: 4083). Антитела против CD70, включающие «полностью человеческую» шарнирную область, могут содержать одну из последовательностей шарнирной области, представленных в таблице 2 ниже.
Последовательности шарнирной области человека
(SEQ ID NO:320)
(SEQ ID NO:321)
(SEQ ID NO:322)
(SEQ ID NO:323)
(SEQ ID NO:324)
(SEQ ID NO:325)
(SEQ ID NO:326)
(SEQ ID NO:327)
(SEQ ID NO:328)
(SEQ ID NO:329)
(SEQ ID NO:330)
(SEQ ID NO:331)
«CH2-домен» - Как здесь используется, термин «CH2-домен» включает часть молекулы тяжелой цепи, которая простирается, например, от приблизительно остатка 244 до остатка 360 антитела, используя общепринятые схемы нумерации (остатки 244-360 в соответствии с системой нумерации по Kabat и остатки 231-340 в соответствии с EU системой нумерации, Kabat EA et al. Sequences of Proteins of Immunological Interest. Bethesda, US Department of Health and Human Services, NIH. 1991). CH2-домен является уникальным в том отношении, что он не является близко спаренным с другим доменом. Точнее, две N-связанные разветвленные углеводные цепи помещены между двумя CH2-доменами интактной природной молекулы IgG. Также убедительно подтверждено документально, что CH3-домен простирается от CH2-домена до C-конца молекулы IgG и включает приблизительно 108 остатков.
«Фрагмент» - Термин «фрагмент» относится к части антитела или цепи антитела, включающей меньшее количество аминокислотных остатков, чем интактное или полное антитело или цепь антитела. Термин «антигенсвязывающий фрагмент» относится к полипептидному фрагменту иммуноглобулина или антитела, который связывается с антигеном или конкурирует с интактным антителом (т.е. с интактным антителом, от которого он произошел) за связывания с антигеном (т.е. специфическое связывание с CD70 человека). Как здесь используется, термин «фрагмент» молекулы антитела включает антигенсвязывающие фрагменты антител, например, вариабельный домен легкой цепи антитела (VL), вариабельный домен тяжелой цепи антитела (VH), одноцепочечное антитело (scFv), F(ab′)2-фрагмент, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент и однодоменный фрагмент антитела (DAb). Фрагменты можно получить, например, посредством химической или ферментативной обработки интактного или полного антитела или цепи антитела или рекомбинантным способом.
«Валентность» - Как здесь используется, термин «валентность» относится к числу потенциальных мишень-связывающих центров в полипептиде. Каждый мишень-связывающий центр специфически связывает одну молекулу-мишень или специфический участок молекулы-мишени. Когда полипептид включает более одного мишень-связывающего центра, каждый мишень-связывающий центр может специфически связываться с одинаковыми или различными молекулами (например, может связываться с различными лигандами или различными антигенами, или различными эпитопами одного и того же антигена). Рассматриваемые связывающие молекулы имеют по крайней мере один связывающий центр, специфический для молекулы CD70 человека.
«Специфичность» - Термин «специфичность» относится к способности связываться с (например, иммунореагировать с) конкретной мишенью, например, CD70. Полипептид может быть моноспецифическим и содержать один или более связывающих центров, которые специфически связываются с мишенью, или полипептид может быть полиспецифическим и содержать два или более связывающих центров, которые специфически связываются с одинаковыми или различными мишенями. В одном варианте осуществления антитело по настоящему изобретению является специфическим в отношении более одной мишени. Например, в одном варианте осуществления полиспецифическая связывающая молекула по настоящему изобретению связывается с CD70 и второй молекулой, представленной на опухолевой клетке. Типичные антитела, которые включают антигенсвязывающие центры, которые связываются с антигенами, представленными на опухолевых клетках, известны в данной области техники, и один или более CDR таких антител могут быть включены в антитело по настоящему изобретению.
«Синтетические» - Как здесь используется, термин «синтетические», применительно к полипептидам, включает полипептиды, которые включают аминокислотную последовательность, не встречающуюся в природе. Например, не встречающиеся в природе полипептиды, которые являются модифицированными формами встречающихся в природе полипептидов (например, включающие мутацию, такую как добавление, замена или делеция) или которые включают первую аминокислотную последовательность (которая может быть или может не быть встречающейся в природе), которая связана в линейном порядке аминокислот со второй аминокислотной последовательностью (которая может быть или может не быть встречающейся в природе), с которой она не связана в природе.
«Сконструированные» - Как здесь используется, термин «сконструированные» включает искусственное манипулирование молекулами нуклеиновых кислот или полипептидов (например, с помощью рекомбинантных методов, in vitro синтеза пептидов, с помощью ферментативного или химического соединения пептидов или кое-какой комбинации этих методов). Предпочтительно, антитела по настоящему изобретению являются сконструированными, в том числе, например, гуманизированными и/или химерными антителами, и антителами, которые были сконструированы для улучшения одной или более характеристик, таких как связывание с антигеном, стабильность/полупериод существования или эффекторная функция.
«Модифицированное антитело» - Как здесь используется, термин «модифицированное антитело» включает синтетические формы антител, которые измены из условия, чтобы они были не встречающимися в природе, например, антитела, которые включают по крайней мере части двух тяжелых цепей, но не две полные тяжелые цепи (такие как антитела с делецией домена(ов) или минитела); полиспецифические формы антител (например, биспецифические, триспецифические и т.д.), измененные для связывания с двумя или более различными антигенами или с различными эпитопами одного антигена); молекулы тяжелых цепей, соединенные в молекулы scFv, и т.п. Молекулы scFv известны в данной области техники и описываются, например, в патенте США с № 5892019. Кроме того, термин «модифицированное антитело» включает поливалентные формы антител (например, трехвалентные, четырехвалентные и т.д. антитела, которые связываются с тремя или более копиями одного и того же антигена). В другом варианте осуществления модифицированным антителом по настоящему изобретению является слитый белок, включающий по крайней мере часть одной тяжелой цепи, в которой отсутствует CH2-домен, и включающий связывающий домен полипептида, включающего связывающую часть одного члена пары рецептор-лиганд.
Термин «модифицированное антитело» может также здесь использоваться для ссылки на варианты аминокислотных последовательностей антитела против CD70. Специалисту со средним уровнем компетентности в данной области техники будет понятно, что антитело против CD70 можно модифицировать с созданием варианта антитела против CD70, которое отличается по аминокислотной последовательности от антитела против CD70, на основе которого оно было получено. Например, можно осуществить замены нуклеотидов или аминокислот, приводящие к консервативным заменам в остатках заменимых аминокислот (например, в остатках CDR и/или каркасных областей). Аминокислотные замены могут включать замену одной или более аминокислот встречающейся в природе или неприродной аминокислотой.
«Замены с целью гуманизации» - Как здесь используется, термин «замены с целью гуманизации» относятся к аминокислотным заменам, в случае которых аминокислотный остаток, присутствующий в конкретном положении в VH- или VL-домене антитела против CD70 (например, происходящего от верблюдовых антитела против CD70), заменяют аминокислотным остатком, который встречается в эквивалентном положении в эталонном VH- или VL-домене человека. Эталонным VH- или VL-доменом человека может быть VH- или VL-домен, кодируемый геном человека зародышевой линии. Замены с целью гуманизации могут быть осуществлены в каркасных областях и/или CDR антитела против CD70, определенного здесь.
«Гуманизированные варианты» - Как здесь используется, термин «гуманизированный вариант» относится к варианту антитела, который содержит одну или более «замен с целью гуманизации» по сравнению с эталонным антителом против CD70, причем часть эталонного антитела (например, VH-домен и/или VL-домен или их части, содержащие по крайней мере один CDR) имеет аминокислотную последовательность, происходящую от не являющегося человеком вида, а «замены с целью гуманизации» осуществляются в аминокислотной последовательности, происходящей от не являющегося человеком вида.
«Приведенные к зародышевому типу варианты» - Термин «приведенный к зародышевому типу вариант» используется здесь для ссылки, в частности, на «гуманизированные варианты», в которых «замены с целью гуманизации» приводят к замене одного или более аминокислотных остатков, присутствующих в конкретном положении(ях) в VH- или VL-домене антитела против CD70 (например, происходящего от верблюдовых антитела против CD70), аминокислотным остатком, который встречается в эквивалентном положении в эталонном VH- или VL-домене человека, кодируемом геном человека зародышевой линии. Как правило, в случае любого конкретного «приведенного к зародышевому типу варианта» аминокислотные остатки, введенные путем замены в приводимый к зародышевому типу вариант, для замены взяты исключительно, или преимущественно, из одного VH- или VL-домена, кодируемого геном человека зародышевой линии. Термины «гуманизированный вариант» или «приведенный к зародышевому типу вариант» часто используются здесь взаимозаменяемо. Введение одной или более «замен с целью гуманизации» в происходящий от верблюдовых (например, происходящий от ламы) VH- или VL-домен приводит к созданию «гуманизированного варианта» происходящего от верблюдовых (ламы) VH- или VL-домена. Если введенные в него путем замены аминокислотные остатки происходят преимущественно или исключительно из последовательности одного VH- или VL-домена, кодируемой геном человека зародышевой линии, то результатом может быть «приведенный к человеческому зародышевому типу вариант» происходящего от верблюдовых (ламы) VH- или VL-домена.
«Варианты аффинности» - Как здесь используется, термин «вариант аффинности» относится к варианту антитела, который демонстрирует одну или более замену в аминокислотной последовательности по сравнению с эталонным антителом против CD70, причем вариант аффинности проявляет измененную аффинность к белку CD70 человека по сравнению с эталонным антителом. Типично, варианты аффинности будут проявлять измененную аффинность к CD70 человека, по сравнению с эталонным антителом против CD70. Предпочтительно вариант аффинности будет проявлять увеличенную аффинность к CD70 человека, по сравнению с эталонным антителом против CD70. Увеличение может выражаться в виде меньшего значения KD, для CD70 человека, или меньшей скорости диссоциации от CD70 человека, и изменении характера перекрестной реактивности с гомологами CD70 у не являющихся людьми видов. Варианты аффинности типично демонстрируют одну или более замен в аминокислотной последовательности CDR, по сравнению с эталонным антителом против CD70. Такие замены могут приводить к замене исходной аминокислоты, присутствующей в конкретном положении в CDR, отличным аминокислотным остатком, который может быть встречающимся в природе аминокислотным остатком или не встречающимся в природе аминокислотным остатком. Аминокислотные замены могут быть консервативными или неконсервативными.
«Высокая степень гомологии с последовательностями человека» - Антитело, включающее вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), будет считаться имеющим высокую степень гомологии с последовательностями человека, если VH-домены и VL-домены, вместе взятые, демонстрируют составляющую по крайней мере 90% идентичность аминокислотных последовательностей с ближе всего соответствующими последовательностями VH и VL человека зародышевого типа. Антитела, обладающие высокой степенью гомологии с последовательностями человека, могут включать антитела, включающие VH- и VL-домены природных нечеловеческих антител, которые демонстрируют достаточно высокий % идентичности последовательностей с последовательностями человека зародышевого типа, в том числе, например, антитела, включающие VH- и VL-домены обычных антител верблюдовых, а также сконструированные, особенно гуманизированные или приведенные к зародышевому типу, варианты таких антител, а также «полностью человеческие» антитела.
В одном варианте осуществления VH-домен антитела с высокой степенью гомологии с последовательностями человека может демонстрировать составляющую 80% или выше идентичность аминокислотной последовательности или гомологию последовательности с одним или более VH-доменов человека при сравнении каркасных областей FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотных последовательностей или гомология последовательностей между VH-доменом полипептида по настоящему изобретению и ближе всего соответствующей последовательностью VH-домена человека зародышевого типа может составлять 85% или выше, 90% или выше, 95% или выше, 97% или выше, или вплоть до 99% или даже 100%.
В одном варианте осуществления VH-домен антитела с высокой степенью гомологии с последовательностями человека может содержать одно или более (например, 1-10) несоответствий в аминокислотной последовательности при сравнении каркасных областей FR1, FR2, FR3 и FR4, по сравнению с ближе всего соответствующей последовательностью VH человека.
В другом варианте осуществления VL-домен антитела с высокой степенью гомологии с последовательностями человека может демонстрировать составляющую 80% или выше идентичность последовательности или гомологию последовательности с одним или более VL-доменов человека при сравнении каркасных областей FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотных последовательностей или гомология последовательностей между VL-доменом полипептида по настоящему изобретению и ближе всего соответствующей последовательностью VL-домена человека зародышевого типа может составлять 85% или выше, 90% или выше, 95% или выше, 97% или выше, или вплоть до 99% или даже 100%.
В одном варианте осуществления VL-домен антитела с высокой степенью гомологии с последовательностями человека может содержать одно или более (например, 1-10) несоответствий в аминокислотной последовательности при сравнении каркасных областей FR1, FR2, FR3 и FR4, по сравнению с ближе всего соответствующей последовательностью VL человека.
До расчета процента идентичности по последовательности между антителом с высокой степенью гомологии с последовательностями человека и VH и VL человека зародышевого типа можно определить канонические складчатые структуры, что позволяет идентифицировать семейство сегментов зародышевой линии человека с идентичным сочетанием канонических складчатых структур для H1 и H2 или L1 и L2 (и L3). Впоследствии член семейства сегментов зародышевой линии человека, который обладает наибольшей степенью гомологии последовательности с вариабельной областью представляющего интерес антитела, выбирают для подсчета гомологии последовательностей. Определение канонических классов гипервариабельных петлевых участков L1, L2, L3, H1 и H2 согласно Chothia можно выполнить с помощью инструментальных программных средств биоинформатики, общедоступных на веб-странице www.bioinf.org.uk/abs/chothia.html.page. Выводные данные программы указывают на требования к ключевым остаткам в файле данных. В этих файлах данных положения ключевых остатков представлены вместе с допустимыми аминокислотами в каждом положении. Последовательность вариабельной области представляющего интерес антитела предоставляется в качестве вводных данных и сначала совмещается с консенсусной последовательностью антитела для установления схемы нумерации согласно Kabat. При анализе канонических складчатых структур используется ряд образцов ключевых остатков, получаемый с помощью автоматизированного способа, разработанного Martin и Thornton (Martin et al., J. Mol. Biol. 263: 800-815 (1996)).
С использованием конкретного известного V-сегмента зародышевой линии человека, в котором используется то же сочетание канонических складчатых структур для H1 и H2 или L1 и L2 (и L3), можно определить в наибольшей степени соответствующий член семейства по показателю гомологии последовательности. С помощью инструментальных программных средств биоинформатики можно определить процент идентичности по последовательности между аминокислотными последовательностями каркасных областей VH- и VL-доменов представляющего интерес антитела и соответствующими последовательностями, кодируемыми генами человека зародышевой линии, но можно также использовать совмещение последовательностей фактически вручную. Последовательности иммуноглобулинов человека можно идентифицировать на основе нескольких баз данных, относящихся к белкам, таких как VBase (http://vbase.mrc-cpe.cam.ac.uk/) или база данных Pluckthun/Honegger (http://www.bioc.unizh.ch/antibody/Sequences/Germlines). Для сравнения последовательностей человека с V-сегментами для VH- и VL-доменов в представляющем интерес антителе можно использовать алгоритм для совмещения последовательностей, например, доступный через веб-страницы вроде www.expasy.ch/tools/#align, но также можно выполнить совмещение вручную с ограниченным рядом последовательностей. Последовательности легкой и тяжелой цепей человека зародышевого типа из семейства с одинаковыми сочетаниями канонических складчатых структур и с наибольшей степенью гомологии с каркасными областями 1, 2 и 3 каждой цепи выбирают и сравнивают с представляющей интерес вариабельной областью; также проверяют FR4 на фоне JH- и JK- или JL-сегментов человека зародышевой линии.
Обратите внимание, что при расчете общего процента гомологии последовательностей остатки FR1, FR2 и FR3 оцениваются, используя ближе всего соответствующую последовательность из семейства сегментов человека зародышевой линии с идентичным сочетанием канонических складчатых структур. Подсчитываются лишь остатки, отличные от ближе всего соответствующей последовательности или других членов того же семейства с тем же сочетанием канонических складчатых структур (NB - исключая любые кодируемые праймером различия). Однако с целью гуманизации остатки в каркасных областях, идентичные членам других семейств сегментов человека зародышевой линии, которые не имеют то же сочетание канонических складчатых структур, могут считаться «человеческими», несмотря на тот факт, что они оцениваются как «негативные» в соответствии со строгими условиями, описанными выше. Это допущение основано на подходе «смешивания и соответствия» для гуманизации, в котором каждую из FR1, FR2, FR3 и FR4 по отдельности сравнивают с ближе всего соответствующей ей последовательностью человека зародышевого типа, и поэтому гуманизированная молекула содержит комбинацию различных FR, как это делали Qu и его коллеги (Qu et al., Clin. Cancer Res. 5: 3095-3100 (1999)) и Ono и его коллеги (Ono et al., Mol. Immunol. 36: 387-395 (1999)). Границы отдельных каркасных областей можно определить, используя схему нумерации IMGT, которая является модификацией схемы нумерации Chothia (Lefranc et al., NAR 27: 209-212 (1999); imgt.cines.fr).
Антитела с высокой степенью гомологии с последовательностями человека могут включать гипервариабельные петлевые участки или CDR, имеющие человеческие или подобные человеческим канонические складчатые структуры, как подробно обсуждается ниже. В одном варианте осуществления по крайней мере один гипервариабельный петлевой участок или CDR или в VH-домене, или в VL-домене антитела с высокой степенью гомологии с последовательностями человека может быть получен из VH- или VL-домена нечеловеческого антитела, например, обычного антитела вида семейства Camelidae, и все еще демонстрировать предсказанную или фактическую каноническую складчатую структура, которая по существу идентична канонической складчатой структуре, которая встречается в антителах человека.
В данной области техники общепризнано, что, хотя первичные аминокислотные последовательности гипервариабельных петлевых участков, присутствующих как в VH-домене, так и в VL-домене, кодируемых генами человека зародышевой линии, являются, по определению, в высокой степени вариабельными, все гипервариабельные петлевые участки, за исключением CDR H3 VH-домена, принимают лишь небольшое число отличных структурных конформаций, называемых каноническими складчатыми структурами (Chothia et al., J. Mol. Biol. 196: 901-917 (1987); Tramontano et al. Proteins 6: 382-394 (1989)), которые зависят как от длины гипервариабельного петлевого участка, так и от присутствия так называемых канонических аминокислотных остатков (Chothia et al., J. Mol. Biol. 196: 901-917 (1987)). Фактические канонические структуры гипервариабельных петлевых участков в интактных VH- или VL-доменах можно определить с помощью структурного анализа (например, рентгеновской кристаллографии), но возможно также предсказать каноническую структуру на основе ключевых аминокислотных остатков, которые характерны для конкретной структуры (обсуждаемой, кроме того, ниже). По существу, специфический набор остатков, который определяет каждую каноническую структуру, ставит «подпись», которая дает возможность узнать каноническую структуру в гипервариабельных петлевых участках VH- или VL-домена с неизвестной структурой; следовательно, канонические структуры можно предсказать на основе лишь первичной аминокислотной последовательности.
Предсказанные канонические складчатые структуры для гипервариабельных петлевых участков любой конкретной последовательности VH или VL в антителе с высокой степенью гомологии с последовательностями человека можно проанализировать, используя алгоритмы, которые являются общедоступными на www.bioinf.org.uk/abs/chothia.html, www.biochem.ucl.ac.uk/~martin/antibodies.html и www.bioc.unizh.ch/antibody/Sequences/Germlines/Vbase_hVk.html. Эти инструментальные программные средства позволяют совместить запрашиваемые последовательности VH или VL с последовательностями VH- или VL-доменов человека с известной канонической структурой, и предсказать каноническую структуру для гипервариабельных петлевых участков запрашиваемой последовательности.
В случае VH-домена петлевые участки H1 и H2 могут оцениваться как участки, имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, которая, как известно, встречается в антителах человека, если удовлетворяется по крайней мере первый, а предпочтительно оба, из следующих критериев:
1. Определяемая по числу остатков длина, одинаковая с таковой в ближе всего соответствующем классе канонических структур человека.
2. Составляющая по крайней мере 33% идентичность, предпочтительно составляющая по крайней мере 50% идентичность с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур H1 и H2 человека.
(Обратите внимание, что для целей вышеупомянутого анализа петлевые участки H1 и H2 рассматривают по отдельности, и каждый из них сравнивают с ближе всего соответствующим классом канонических структур человека.)
Вышеупомянутый анализ основывается на предсказании канонической структуры петлевых участков H1 и H2 представляющего интерес антитела. Если фактические структуры петлевых участков H1 и H2 в представляющем интерес антителе известны, например, на основе рентгеновской кристаллографии, то петлевые участки H1 и H2 в представляющем интерес антителе можно также оценивать как участки, имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, которая, как известно, встречается в антителах человека, если длина петлевого участка отличается от таковой в ближе всего соответствующем классе канонических структур человека (типично на ±1 или ±2 аминокислот), но фактическая структура петлевых участков H1 и H2 в представляющем интерес антителе соответствует канонической складчатой структуре человека.
Ключевые аминокислотные остатки, обнаруживаемые в классах канонических структур человека, для первого и второго гипервариабельных петлевых участков VH-доменов человека (H1 и H2) описаны Chothia и др. в J. Mol. Biol. 227: 799-817 (1992), содержание которого включено сюда в его полном объеме посредством ссылки. В частности, в таблице 3 на стр. 802 Chothia et al., которая, в частности, включена сюда посредством ссылки, перечислены предпочтительные аминокислотные остатки в ключевых местах канонических структур H1, обнаруживаемых в зародышевой линии развития иммуноглобулинов человека, тогда как в таблице 4 на стр. 803, которая также, в частности, включена сюда посредством ссылки, перечислены предпочтительные аминокислотные остатки в ключевых местах канонических структур CDR H2, обнаруживаемых в зародышевой линии развития иммуноглобулинов человека.
В одном варианте осуществления как H1, так и H2 в VH-домене антитела с высокой степенью гомологии с последовательностями человека демонстрируют предсказанную или фактическую каноническую складчатую структуру, которая идентична канонической складчатой структуре, которая встречается в антителах человека.
Антитела с высокой степенью гомологии с последовательностями человека могут включать VH-домен, в котором гипервариабельные петлевые участки Н1 и Н2 образуют сочетание канонических складчатых структур, которое идентично сочетанию канонических складчатых структур, которое, как известно, встречается в по крайней мере одном VH-домене человека зародышевого типа. Было установлено, что лишь некоторые сочетания канонических складчатых структур в H1 и H2 фактически встречается в VH-доменах, кодируемых генами человека зародышевой линии. В одном варианте осуществления H1 и H2 в VH-домене антитела с высокой степенью гомологии с последовательностями человека можно получить из VH-домена не являющегося человеком вида, например, вида семейства Camelidae, и все еще образовывать сочетание предсказанных или фактических канонических складчатых структур, которое идентично сочетанию канонических складчатых структур, которое, как известно, встречается в VH-домене человека зародышевого типа или соматически мутированном VH-домене. В неограничивающих вариантах осуществления H1 и H2 в VH-домене антитела с высокой степенью гомологии с последовательностями человека можно получить из VH-домена не являющегося человеком вида, например, вида семейства Camelidae, и образовывать одно из следующих сочетаний канонических складчатых структур: 1-1, 1-2, 1-3, 1-6, 1-4, 2-1, 3-1 и 3-5.
Антитело с высокой степенью гомологии с последовательностями человека может содержать VH-домен, который демонстрирует высокую степень и идентичности последовательности, и гомологии последовательности с VH человека, и который содержит гипервариабельные петлевые участки, проявляющие структурную гомологию с VH человека.
Может быть преимущественным введение в канонические складчатые структуры, присутствующие в H1 и H2 в VH-домене антитела с высокой степенью гомологии с последовательностями человека и в их сочетание «поправки» на последовательность VH человека зародышевого типа, которая представляет собой последовательность, ближе всего соответствующую VH-домену антитела с высокой степенью гомологии с последовательностями человека по показателю общей идентичности первичной аминокислотной последовательности. В качестве примера, если ближе всего соответствующая последовательность является последовательностью, ближе всего соответствующей VH3-домену человека зародышевого типа, то может быть преимущественным создание сочетания канонических складчатых структур H1 и H2, которое также встречается в природе в VH3-домене человека. Это может быть особенно важным в случае антител с высокой степенью гомологии с последовательностями человека, которые происходят от не являющихся человеком видов, например, антител, содержащих VH- и VL-домены, которые происходят из обычных антител верблюдовых, особенно антител, содержащих гуманизированные VH- и VL-домены верблюдовых.
Таким образом, в одном варианте осуществления VH-домен антитела против CD70 с высокой степенью гомологии с последовательностями человека может демонстрировать составляющую 80% или выше, 85% или выше, 90% или выше, 95% или выше, 97% или выше или вплоть до 99% или даже 100% идентичность последовательности или гомологию последовательности с VH-доменом при сравнении каркасных областей FR1, FR2, FR3 и FR4, и, кроме того, H1 и H2 в этом же антителе получены из нечеловеческого VH-домена (например, происходящего от вида семейства Camelidae), но образуют сочетание предсказанных или фактических канонических складчатых структур, которое является одинаковым с сочетанием канонических складчатых структур, которое, как известно, встречается в природе в том же VH-домене человека.
В других вариантах осуществления L1 и L2 в VL-домене антитела с высокой степенью гомологии с последовательностями человека можно получить из VL-домена не являющегося человеком вида (например, происходящего от верблюдовых VL-домена), и каждый демонстрирует предсказанную или фактическую каноническую складчатую структуру, которая по существу идентична канонической складчатой структуре, которая встречается в антителах человека.
Как и в случае VH-доменов, гипервариабельные петлевые участки VL-доменов обоих типов Vлямбда и Vкаппа могут принимать ограниченный ряд конформаций или канонических структур, определяемых частично длиной, а также присутствием ключевых аминокислотных остатков в определенных положениях канонических структур.
В представляющем интерес антителе, обладающем высокой степенью гомологии с последовательностью человека, петлевые участки L1, L2 и L3, полученные из VL-домена не являющегося человеком вида, например, вида семейства Camelidae, могут оцениваться как участки, имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, которая, как известно, встречается в антителах человека, если удовлетворяется по крайней мере первый, а предпочтительно оба, из следующих критериев:
1. Определяемая по числу остатков длина, одинаковая с таковой в ближе всего соответствующем классе канонических структур человека.
2. Составляющая по крайней мере 33% идентичность, предпочтительно составляющая по крайней мере 50% идентичность с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур L1 или L2 человека, из набора для или Vлямбда, или Vкаппа.
(Обратите внимание, что для целей вышеупомянутого анализа петлевые участки L1 и L2 рассматривают по отдельности, и каждый из них сравнивают с ближе всего соответствующим классом канонических структур человека.)
Вышеупомянутый анализ основывается на предсказании канонической структуры петлевых участков L1, L2 и L3 в VL-домене представляющего интерес антитела. Если фактическая структура петлевых участков L1, L2 и L3 известна, например, на основе рентгеновской кристаллографии, то петлевые участки L1, L2 и L3, происходящие из представляющего интерес антитела, можно также оценивать как участки, имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, которая, как известно, встречается в антителах человека, если длина петлевого участка отличается от таковой в ближе всего соответствующем классе канонических структур человека (типично на ±1 или ±2 аминокислот), но фактическая структура петлевых участков Camelidae соответствует канонической складчатой структуре человека.
Ключевые аминокислотные остатки, обнаруживаемые в классах канонических структур человека, для CDR доменов Vлямбда и Vкаппа человека описаны в Morea et al. Methods, 20: 267-279 (2000) и Martin et al., J. Mol. Biol., 263: 800-815 (1996). Набор структур домена Vкаппа человека также описан в Tomlinson et al. EMBO J. 14: 4628-4638 (1995), а набор структур домена Vлямбда - в Williams et al. J. Mol. Biol., 264: 220-232 (1996). Содержание всех этих документов должно быть включено сюда посредством ссылки.
L1 и L2 в VL-домене антитела с высокой степенью гомологии с последовательностями человека могут образовывать сочетание предсказанных или фактических канонических складчатых структур, которое идентично сочетанию канонических складчатых структур, которое, как известно, встречается в VL-домене человека зародышевого типа. В неограничивающих вариантах осуществления L1 и L2 в домене Vлямбда антитела с высокой степенью гомологии с последовательностями человека (например, антитела, содержащего происходящий от верблюдовых VL-домен или его гуманизированный вариант) могут образовывать одно из следующих сочетаний канонических складчатых структур: 11-7, 13-7(A,B,C), 14-7(A,B), 12-11, 14-11 и 12-12 (как описано в Williams et al. J. Mol. Biol. 264: 220-232 (1996) и как представлено на http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVL.html). В неограничивающих вариантах осуществления L1 и L2 в домене Vкаппа могут образовывать одно из следующих сочетаний канонических складчатых структур: 2-1, 3-1, 4-1 и 6-1 (как описано в Tomlinson et al. EMBO J. 14: 4628-4638 (1995) и как представлено на http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVK.html).
В дальнейшем варианте осуществления все три из L1, L2 и L3 в VL-домене антитела с высокой степенью гомологии с последовательностями человека могут демонстрировать по существу человеческую структуру. Предпочтительно, когда VL-домен антитела с высокой степенью гомологии с последовательностями человека демонстрирует высокую степень и идентичности последовательности, и гомологии последовательности с VL человека, а также когда гипервариабельные петлевые участки в VL-домене проявляют структурную гомологию с VL человека.
В одном варианте осуществления VL-домен антитела против CD70 с высокой степенью гомологии с последовательностями человека может демонстрировать составляющую 80% или выше, 85% или выше, 90% или выше, 95% или выше, 97% или выше или вплоть до 99% или даже 100% идентичность последовательности с VL-доменом при сравнении каркасных областей FR1, FR2, FR3 и FR4, и, кроме того, гипервариабельный петлевой участок L1 и гипервариабельный петлевой участок L2 могут образовывать сочетание предсказанных или фактических канонических складчатых структур, которое является одинаковым с сочетанием канонических складчатых структур, которое, как известно, встречается в природе в том же VL-домене человека.
Конечно, предусматривается, что VH-домены, демонстрирующие высокую степень идентичности последовательностей/гомологии последовательностей с VH человека, а также структурную гомологию с гипервариабельными петлевыми участками VH человека, будут объединены с VL-доменами, демонстрирующими высокую степень идентичности последовательностей/гомологии последовательностей с VL человека, а также структурную гомологию с гипервариабельными петлевыми участками VL человека, для обеспечения антител с высокой степенью гомологии с последовательностями человека, содержащих спаривания VH/VL (например, спаривания происходящих от верблюдовых VH/VL) с максимальной гомологией по последовательности и структуре с кодируемыми генами человеком спариваниями VH/VL.
Как резюмировано выше, настоящее изобретение относится, по крайней мере частично, к антителам, и их антигенсвязывающим фрагментам, которые связываются с CD70 человека с высокой аффинностью. Свойства и характеристики антител против CD70, и фрагментов антител, в соответствии с настоящим изобретением теперь будут описаны с дополнительными подробностями.
Связывание с CD70 и аффинность к нему
В некоторых аспектах обеспечиваемые здесь антитела, и их антигенсвязывающие фрагменты, связываются с CD70 человека с высокой аффинностью. Антитела, или их антигенсвязывающие фрагменты, которые связываются с CD70 человека с высокой аффинностью, могут демонстрировать аффинность (KD) к CD70 человека, а точнее к экстраклеточному домену CD70 человека, составляющую приблизительно 10 нМ или меньше, или 1 нМ или меньше, или 0,1 нМ или меньше, или 10 пМ или меньше.
Антитело против CD70 (или антигенсвязывающий фрагмент) может включать VH- и VL-домены, которые, при их представлении в форме Fab, демонстрируют скорость диссоциации в случае связывания с CD70 человека, составляющую менее 7×10-4 сек-1, предпочтительно менее 5×10-4 сек-1, или менее 2×10-4 сек-1, менее 1×10-4 сек-1. Типично скорость диссоциации будет находиться в диапазоне от 0,4×10-4 сек-1 до 4,8×10-4 сек-1. Аффинность (KD) и скорость диссоциации (Koff) можно определить, используя стандартные методы, хорошо известные квалифицированным в данной области техники специалистам, такие как, например, поверхностный плазмонный резонанс (BIAcore), как описано в сопроводительных примерах. Вкратце, 50 мкл исследуемого образца добавляют к 200 мкл PBS + 0,02% Tween, и разводят 1/400, как описано ниже: 5 мкл образца (1 мг/мл) + 195 мкл HBS-EP+ (буфера для Biacore), с дальнейшим разведением: 10 мкл + 90 мкл HVS-EP+ = 2,5 мкг/мл. Анализ с использованием Biacore выполняют при комнатной температуре, используя покрытый CD70 с высокой плотностью чип CM5 (4000 RU (резонансных единиц)), используя предоставляемый производителем протокол.
В связи с этим следует отметить, что хотя скорость диссоциации определяют для сочетаний VH и VL в форме Fab, это не означает, что VH- и VL-домены вносят равный вклад в связывание с CD70. В случае многих сочетаний VH/VL именно VH-домен вносит основной вклад в связывание с CD70, при этом VL-домен вносит вклад в растворимость и/или стабильность спаривания VH/VL.
Связывание антител против CD70, описываемых здесь, или их Fab-фрагментов, с рекомбинантным CD70 человека можно также оценить с помощью ELISA.
Описываемые здесь антитела против CD70 могут, кроме того, демонстрировать связывание с CD70, представленного на поверхности интактных клеток, например, клеток карциномы почки 786-О и других линий раковых клеток. Связывание антител против CD70 с CD70-экпрессирующими клетками можно оценить с помощью проточной цитометрии. Представленные в сопроводительных примерах результаты показывают, что предпочтительные антитела против CD70, включая вариант 41D12 (при экспрессии в виде нефукозилированного IgG1 ARGX-110), демонстрируют очень сильное связывание с CD70 на клеточной поверхности. Действительно, в случае множества линий клеток ARGX-110 демонстрирует более сильное связывание, чем антитело SGN70 известного уровня техники в качестве препарата сравнения.
В частности, достойно внимания, что ARGX-110 демонстрирует сильное связывание с линиями клеток, которые экспрессируют CD70 в низкой копийности, включая, среди прочих, линии клеток Raji, SU-DHL-6, MHHPREB 1, Mino, Mecl, JVM-2, HH и EBC-1, а также раковыми клетками, изолированными от пациентов (например, пациентов с CLL), в обоих случаях связывание ARGX-110 заметно сильнее связывания SGN70. Увеличенное связывание с линиями клеток с низкой копийностью CD70 и материалами от пациентов с CLL может в значительной степени служить отражением очень высокой аффинности ARGX-110 к рекомбинантному CD70. Эти характеристики связывания поддерживают особую полезность ARGX-110 (на основе 41D12) и других описываемых здесь антител против CD70 со схожими свойствами при лечении рака, особенно при использовании в виде IgG, проявляющего сильную эффекторную функцию, (например, нефукозилированного IgG1), а не в виде иммуноконъюгата, связанного с составляющей в виде цитотоксического или цитостатического агента. Значение связывания с высокой аффинностью (т.е. более высокой аффинностью, чем та, которая может быть достигнута при использовании антител против CD70 известного уровня техники) с рекомбинантным CD70 и CD70 на клеточной поверхности по отношению к клинической полезности антител показано в сопроводительных примерах с помощью экспериментов, в которых в образцы PBMC внедряют линии раковых клеток, а затем обрабатывают антителами против CD70. В этой системе обработка ARGX-110 приводила к значительно большему лизису раковых клеток-мишеней, чем обработка антителами против CD70 MDX1411 и SGN70 в качестве препарата сравнения.
Описываемые здесь антитела против CD70 демонстрируют высокую аффинность к CD70 человека, а точнее к экстраклеточному домену CD70 человека, но не исключается перекрестная реактивность с гомологами CD70 у не являющихся людьми видов. Действительно, преимущественным свойством многих из описываемых здесь антител против CD70, включая 27B3 и его приведенные к зародышевому типу варианты, особенно 41D12, является то, что помимо связывания с высокой аффинностью с CD70 человека они также демонстрируют перекрестную реактивность с гомологами CD70 у обезьян, особенно гомологами CD70 у макака-резус и яванского макака.
В связи с этим можно считать, что антитело против CD70 демонстрирует перекрестную реактивность с гомологами CD70 у обезьян, например, гомологами CD70 у макака-резус и яванского макака, если разница в IC50 в случае CD70 человека в сравнении с CD70 у обезьян (макака-резус или яванского макака) в ELISA ингибирования взаимодействия CD70-CD27, таком как тот, который описан в сопроводительном примере 20.2, является менее чем 5-кратной, предпочтительно менее чем 3-кратной или менее чем 2-кратной. Аффинность, а следовательно, эффективность блокирования, к CD70 человека и обезьян должна быть вполне сравнимой.
Взаимодействие между CD70 и CD27
В некоторых аспектах обеспечиваемые здесь антитела против CD70, или их антигенсвязывающие фрагменты, могут связываться с CD70 человека с высокой аффинностью и блокировать взаимодействие между CD70 и CD27.
Способность антител против CD70, или их антигенсвязывающих фрагментов, к блокированию связывания CD70 с CD27 можно определить с помощью ELISA, используя или захваченный рекомбинантный CD70 (Flag-TNC-CD70), или непосредственно использованный для покрытия CD70 и рекомбинантный CD27-Fc. Описываемые здесь антитела против CD70 могут ингибировать взаимодействие между CD70 и CD27 с IC50, составляющей 300 нг/мл или менее, или 200 нг/мл или менее, или 110 нг/мл или менее, или 70 нг/мл или менее, или 50 нг/мл или менее.
Способность антител против CD70, или их антигенсвязывающих фрагментов, к блокированию связывания CD70 с CD27 можно также определить в анализе на основе сокультивирования клеток Raji (B-клеточной лимфомы человека) и клеток HT1080-CD27 (линии эпителиальных клеток человека, трансфицированных CD27), как описано в сопроводительных примерах. Описываемые антитела против CD70, которые ингибируют взаимодействие между CD70 и CD27, могут демонстрировать IC50 в этом анализе на основе сокультивирования, составляющую 500 нг/мл или менее, или 300 нг/мл или менее, или 100 нг/мл или менее, или 50 нг/ил или менее, или 30 нг/мл или менее.
Предпочтительные антитела против CD70 на основе 27B3 и его приведенных к зародышевому типу вариантов, включая 41D12 (ARGX-110), демонстрируют сильное блокирование взаимодействия CD70/CD27 в системе ELISA. Действительно, ARGX-110 является значительно более сильным в блокировании взаимодействий CD70/CD27, чем антитела SGN70 и MDX1411 в качестве препаратов для сравнения. Взаимодействие между CD70 и CD27 может вносить вклад в выживание опухолевых клеток, их пролиферацию и/или подавление иммунной системы в опухолевом микроокружении. Соответственно, сильное ингибирование взаимодействия CD70/CD27, помимо связывания с CD70 с высокой аффинностью, может способствовать получению улучшенного клинического результата при лечении некоторых CD70-экспрессирующих раков или иммунологических нарушений, например, аутоиммунных заболеваний и раков, которые коэкспрессируют CD70 и CD27.
В других вариантах осуществления антитела против CD70, или их антигенсвязывающие фрагменты, могут связываться с CD70 человека, но не ингибируют взаимодействие между CD70 и CD27. Антитела против CD70, которые связываются с CD70, но не ингибируют взаимодействие CD70 и CD27, можно подвергнуть оценке на основе ELISA, используя или захваченный рекомбинантный CD70 (Flag-TNC-CD70), или непосредственно использованный для покрытия CD70 и рекомбинантный CD27-Fc. Как продемонстрировано в сопроводительных примерах, был идентифицирован ряд Fab, которые связываются с CD70, но не ингибируют взаимодействие CD70/CD27 в значительной степени. В частности, клон Fab, идентифицированный здесь как 59D10, демонстрирует сильное связывание с рекомбинантным CD70, как определено с помощью Biacore, но не блокирует взаимодействие CD70/CD27 в значительной степени, при определении с помощью ELISA. Описываемые здесь антитела против CD70, которые связываются с CD70, но не ингибируют взаимодействие между CD70 и CD27, могут, тем не менее, обладать эффекторными функциями интактных антител, т.е. одной или более из ADCC, CDC, ADCP или ADC, и ингибировать рост опухолевых клеток in vivo. Антитела против CD70, которые связываются с CD70 с высокой аффинностью, но являются не блокирующими, могут преимущественно использоваться в качестве антител с одним плечом, или ПЭГилированных продуктов Fab, или в любой другой форме антитела, которая обеспечивает строго одновалентное взаимодействие с клеткой-мишенью, экспрессирующей CD70.
Эпитопы CD70
В некоторых аспектах описываемые здесь антитела против CD70 связываются с эпитопами в экстраклеточном домене CD70 человека.
Термин «эпитоп» относится к части(ям) антигена (например, CD70 человека), которая контактирует с антителом. Эпитопы могут быть линейными, т.е. влекущими за собой связывание с одной последовательностью аминокислот, или конформационными, т.е. влекущими за собой связывание с двумя или более последовательностями аминокислот в различных областях антигена, которые могут не быть непременно непрерывными.
Обеспечиваемые здесь антитела против CD70 могут связываться с различными (перекрывающимися или не перекрывающимися) эпитопами в экстраклеточном домене белка CD70 человека. Например, Fab, обозначенные 1C2 и 9E1 в сопроводительных примерах, несомненно, связываются с различными, не перекрывающимися эпитопами CD70 человека.
Как отмечено здесь в другом месте, предпочтительное антитело против CD70 41D12 (ARGX-110) демонстрирует особенно полезное сочетание характеристик связывания, которое не демонстрирует какое-либо известное из предшествующего уровня техники антитело против CD70, а именно:
(a) связывание в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO: 342) в CD70 человека;
(b) перекрестную реактивность с гомологами CD70 у макака-резус (Macaca mulatta) и яванского макака (Macaca cynomolgus);
(c) связывание со встречающимся в природе CD70 человека и с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
В случае любого конкретного антитела против CD70 способность к связыванию с CD70 человека в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO: 342) специалист со средним уровнем компетентности в данной области техники может без труда определить, используя, например, анализ связывания с химерным мышь-человек CD70, описанный в сопроводительном примере 20.4.
Перекрестную реактивность с гомологами CD70 у обезьян специалист со средним уровнем компетентности в данной области техники может без труда определить с помощью, например, анализа с использованием FACS, поверхностного плазмонного резонанса (BiacoreTM), или используя ELISA ингибирования взаимодействия CD70-CD27. В связи с этим можно считать, что антитело против CD70 демонстрирует перекрестную реактивность с гомологом CD70 у обезьяны, например, гомологами CD70 у макака-резус и яванского макака, если разница в IC50 в случае CD70 человека в сравнении с CD70 у обезьян (макака-резус или яванского макака) в ELISA ингибирования взаимодействия CD70-CD27, таком как тот, который описан в сопроводительном примере 20.2, является менее чем 5-кратной, предпочтительно менее чем 3-кратной или менее чем 2-кратной.
Описываемые здесь антитела против CD70 могут также демонстрировать способность к связыванию как со встречающимся в природе CD70 человека, так и с подвергнутым тепловой денатурации рекомбинантным CD70 человека. В связи с этим на связывание со встречающимся в природе CD70 человека указывает связывание с CD70, представленным на поверхности CD70-экспрессирующей клетки, такой как любая из линий CD70+ клеток, описанных в сопроводительных примерах, или даже природными CD70-экспрессирующими клетками, выделенными из материала, полученного от пациента, (например, клетками, изолированными от пациентов с CLL, как в сопроводительных примерах), активированными T-клетками и т.д. Таким образом, специалист со средним уровнем компетентности в данной области техники может без труда проверить связывание со встречающимся в природе CD70 человека.
Связывание с подвергнутым тепловой денатурации рекомбинантным CD70 человека можно проверить, как описано в сопроводительном примере 20.3. Предпочтительно, чтобы антитело против CD70 демонстрировало ОП при 620 нм, составляющую 0,6 или выше, в этой анализе, для демонстрации значимого связывания с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
Частичная или медленная интернализация
Описываемые здесь антитела против CD70, включая предпочтительное антитело против CD70 41D12 (ARGX-110), демонстрируют частичную интернализацию в линии клеток карциномы почки, что означает, что при исследовании интернализации антител в клетки карциномы почки 786-О значительная часть связавшегося антитела, т.е. по крайней мере 30-40%, остается снаружи после инкубации в течение 6 часов и даже 24 часов при 37°C. Кроме того, антитела против CD70 могут также демонстрировать медленную скорость интернализации. В связи с этим считают, что антитело против CD70 демонстрирует медленную скорость интернализации, если оно подвергается интернализации с меньшей скоростью, чем эталонное антитело против CD70 9D1 (VH SEQ ID NO: 178; VL SEQ ID NO: 190).
Как показано в сопроводительных примерах, различные линии раковых клеток демонстрируют значимые различия в степени интернализации антител против CD70. Особенно важно, что большинство линий раковых клеток демонстрируют составляющую менее 30%, и даже мене 10%, интернализацию связавшегося антитела против CD70, даже после инкубации в течение 6 часов с ARGX-110. Эти результаты несомненно служат подтверждением полезности антител против CD70 с сильной эффекторной функцией (включая варианты, сконструированные для увеличения эффекторной функции) при лечении CD70-экспрессирующих раков. В научной литературе ранее было сообщение о том, что CD70 является «интернализующейся» мишенью; поэтому была предположена разработка иммуноконъюгатов антитело против CD70-лекарственное средство для лечения как CD70-экспрессирующих раков, так и иммунологических заболеваний. Поэтому представленные здесь результаты, которые неопровержимо показывают, что интернализация связавшихся с клеточной поверхностью антител против CD70 является редким событием, и что в большинство линий CD70-экспрессирующих клеток связавшееся антитело против CD70 не подвергается в значительной степени интернализации, являются очень неожиданными.
Происходящие от верблюдовых антитела против CD70
Тем не менее, в других аспектах описываемые здесь антитела или их антигенсвязывающие фрагменты могут включать по крайней мере один гипервариабельный петлевой участок или определяющий комплементарность участок, полученный из VH-домена или VL-домена вида семейства Camelidae. В частности, антитело или антигенсвязывающий фрагмент может включать VH- и/или VL-домены, или их CDR, антител, полученных в результате активной иммунизации беспородных животных семейства верблюдовых, например, лам, антигеном CD70 человека.
Под «гипервариабельным петлевым участком или определяющим комплементарность участком, полученным из VH-домена или VL-домена вида семейства Camelidae», подразумевается то, что гипервариабельный петлевой участок (HV) или CDR имеет аминокислотную последовательность, которая идентична, или по существу идентична, аминокислотной последовательности гипервариабельного петлевого участка или CDR, который кодируется геном иммуноглобулина Camelidae. В этом контексте «ген иммуноглобулина» включает гены зародышевой линии, гены иммуноглобулинов, которые были подвергнуты реаранжировке, а также соматически мутированные гены. Таким образом, аминокислотная последовательность HV или CDR, полученного из VH- или VL-домена вида семейства Camelidae, может быть идентична аминокислотной последовательности HV или CDR, присутствующем в зрелом обычном антителе Camelidae. Термин «полученный из» в этом контексте означает структурную взаимосвязь, в том смысле, что HV или CDR антитела против CD70 включают аминокислотную последовательность (или ее легкие варианты), которая первоначально кодировалась геном иммуноглобулина Camelidae. Однако это не означает непременно особую взаимосвязь на основе процесса продукции, используемого для приготовления антитела против CD70.
Происходящие от верблюдовых антитела против CD70 могут происходить от любого вида верблюдовых, включая, среди прочих, ламу, одногорбого верблюда, альпаку, вигунью, гуанако или верблюда.
Антитела против CD70, включающие происходящие от верблюдовых VH- и VL-домены, или их CDR, обычно являются рекомбинантно экспрессированными полипептидами, и могут быть химерными полипептидами. Термин «химерный полипептид» относится к искусственному (не встречающемуся в природе) полипептиду, который создают путем соединения двух или более пептидных фрагментов, которые в иных случаях не встречаются рядом. В это определение включены «межвидовые» химерные полипептиды, созданные путем соединения пептидных фрагментов, кодируемых генами двух или более видов, например, животного семейства верблюдовых и человека.
Происходящие от верблюдовых CDR могут включать одну из последовательностей CDR, представленных в виде SEQ ID NO: 49-59, 262 и 263 (CDR3 тяжелой цепи) или SEQ ID NO: 26-37, 249, 258 или 259 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20, 248, 256 или 257 (CDR1 тяжелой цепи), или одну из последовательностей CDR, представленных в виде SEQ ID NO: 148-168, 271 или 273 (CDR3 легкой цепи), или SEQ ID NO: 109-128 или 270 (CDR2 легкой цепи), или SEQ ID NO: 77-95 или 250-253, 267 и 268 (CDR1 легкой цепи).
В одном варианте осуществления весь VH-домен и/или весь VL-домен может быть получен от вида семейства Camelidae. В конкретных вариантах осуществления происходящий от верблюдовых VH-домен может включать аминокислотную последовательность, представленную в виде SEQ ID NO: 177-188, 212-223, 274 или 275, тогда как происходящий от верблюдовых VL-домен может включать аминокислотную последовательность, представленную в виде SEQ ID NO: 189-211, 230-245, 276 или 277 (VL). Происходящий от верблюдовых VH-домен и/или происходящий от верблюдовых VL-домен можно затем представить для создания белка, в ходе которого в аминокислотную последовательность верблюдовых вводят одну или более замен, вставок или делеций аминокислот. Эти создаваемые изменения предпочтительно включают аминокислотные замены относительно последовательности верблюдовых. Такие изменения включают «гуманизацию» или «приведение к зародышевому типу», в ходе которой один или более аминокислотных остатков в кодируемом геном верблюдовых VH- или VL-домене заменяют эквивалентными остатками из гомологичного, кодируемого геном человека VH- или VL-домена.
Выделенные VH- и VL-домены верблюдовых, полученные посредством активной иммунизации животного семейства верблюдовых (например, ламы) антигеном CD70 человека, могут использовать в качестве основы для конструирования антигенсвязывающих полипептидов в соответствии с настоящим изобретением. Начиная с интактных VH- и VL-доменов верблюдовых, можно осуществить одну или более замен, вставок или делеций аминокислот, которые отличаются от исходной последовательности верблюдовых. В некоторых вариантах осуществления такие замены, вставки или делеции могут присутствовать в каркасных областях VH-домена и/или VL-домена. Целью таких изменений первичной аминокислотной последовательности может быть уменьшение предположительно невыгодных свойств (например, иммуногенности у являющегося человеком хозяина (так называемая гуманизация)), мест потенциальной гетерогенности продуктов и/или нестабильности (гликозилирования, дезамидирования, изомеризации и т.д.) или усиление некоего другого выгодного свойства молекулы (например, растворимости, стабильности, биодоступности и т.д.). В других вариантах осуществления изменения первичной аминокислотной последовательности можно осуществить в одном или более из гипервариабельных петлевых участков (или CDR) VH- и/или VL-доменов антител Camelidae, полученных посредством активной иммунизации. Такие изменения могут быть введены для увеличения аффинности к антигену и/или специфичности связывания с антигеном, или для уменьшения предположительно невыгодных свойств, например, иммуногенности у являющегося человеком хозяина (так называемая гуманизация), мест потенциальной гетерогенности продуктов и/или нестабильности, гликозилирования, дезамидирования, изомеризации и т.д., или для усиления некоего другого выгодного свойства молекулы, например, растворимости, стабильности, биодоступности и т.д.
Таким образом, в одном варианте осуществления настоящим изобретением обеспечивается вариант антитела против CD70, который содержит по крайней мере одну аминокислотную замену в по крайней мере одной каркасной области или CDR-участке или в VH-домене, или в VL-домене по сравнению с происходящим от верблюдовых VH- или VL-доменом, примеры которого включают, но без ограничения, VH-домены верблюдовых, включающие аминокислотные последовательности, представленные в виде SEQ ID NO: 177-188, 212-223, 274 или 275, и VL-домены верблюдовых, включающие аминокислотные последовательности, представленные в виде SEQ ID NO: 189-211, 230-245, 276 или 277.
В других вариантах осуществления обеспечиваются «химерные» молекулы антител, включающие происходящие от верблюдовых VH- и VL-домены (или их сконструированные варианты) и один или более константных доменов антитела не верблюдовых, например, кодируемых генами человека константных доменов (или их сконструированных вариантов). В таких вариантах осуществления предпочтительно, когда и VH-домен, и VL-домен получены от одного и того же вида семейства верблюдовых, например, и VH, и VL могут происходить от Lama glama, или и VH, и VL могут происходить от Lama pacos (до внесения создаваемой вариации аминокислотной последовательности). В таких вариантах осуществления и VH-домен, и VL-домен могут происходить от одного животного, в частности, одного животного, которое было подвергнуто активной иммунизации антигеном CD70 человека.
В качестве альтернативы создаваемым изменениям первичной аминокислотной последовательности VH- и/или VL-доменов Camelidae, отдельные, происходящие от верблюдовых гипервариабельные петлевые участки или CDR, или их комбинации, могут быть выделены из VH/VL-доменов верблюдовых и перенесены в альтернативную (т.е. не Camelidae) каркасную область, например, каркасную область VH/VL человека, с помощью пересадки CDR. В частности, неограничивающие варианты происходящих от верблюдовых CDR могут быть выбраны из CDR, имеющих аминокислотные последовательности, представленные в виде SEQ ID NO: 49-59 (CDR3 тяжелой цепи), или SEQ ID NO: 26-37 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20 (CDR1 тяжелой цепи), или одну из последовательностей CDR, представленных в виде SEQ ID NO: 148-168 (CDR3 легкой цепи), или SEQ ID NO: 109-128 (CDR2 легкой цепи), или SEQ ID NO: 77-95 (CDR1 легкой цепи).
Антитела против CD70, включающие происходящие от верблюдовых VH- и VL-домены, или их CDR, могут быть в различных вариантах осуществления, в которых присутствует как VH-домен, так и VL-домен. Термин «антитело» здесь используется в самом широком значении и охватывает, но без ограничения, моноклональные антитела (в том числе полноразмерные моноклональные антитела), поликлональные антитела, полиспецифические антитела (например, биспецифические антитела) при условии, что они проявляют соответствующую иммунологическую специфичность по отношению к белку CD70 человека. Термин «моноклональное антитело», как здесь используется, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными за исключением возможных, встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифическими, направленными против одной области детерминанты. Кроме того, в отличие от обычных препаратов (поликлональных) антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов) антигена, каждое моноклональное антитело направлено против одной детерминанты или эпитопа антигена.
«Фрагменты антитела» включают часть полноразмерного антитела, как правило, его антигенсвязывающий или вариабельный домен. Примеры фрагментов антител включают Fab, Fab′, F(ab′)2, биспецифические Fab′- и Fv-фрагменты, диатела, линейные антитела, одноцепочечные молекулы антител, одноцепочечный вариабельный фрагмент (scFv) и полиспецифические антитела, образованные из фрагментов антител (смотрите Holliger and Hudson, Nature Biotechnol. 23: 1126-1136 (2005), содержание которого включено сюда посредством ссылки).
В неограничивающих вариантах осуществления антитела против CD70, включающие происходящие от верблюдовых VH- и VL-домены, или их CDR, могут включать CH1-домены и/или CL-домены, аминокислотная последовательность которых является полностью или по существу человеческой. Если антигенсвязывающий полипептид по настоящему изобретению является антителом, предназначенным для терапевтического применения для людей, типично вся константная область антитела, или по крайней мере его часть, имеет полностью или по существу человеческую аминокислотную последовательность. Следовательно, один или более или любое сочетание из CH1-домена, шарнирной области, CH2-домена, CH3-домена и CL-домена (и CH4-домена в случае его присутствия) может быть полностью или по существу человеческим, что касается его аминокислотной последовательности.
Преимущественно, все из CH1-домена, шарнирной области, CH2-домена, CH3-домена и CL-домена (и CH4-домена в случае его присутствия) могут иметь полностью или по существу человеческую аминокислотную последовательность. В связи с константной областью гуманизированного или химерного антитела, или его фрагмента, термин «по существу человеческая» относится к составляющей по крайней мере 90%, или по крайней мере 92%, или по крайней мере 95%, или по крайней мере 97%, или по крайней мере 99% идентичности аминокислотной последовательности с константной областью человека. Термин «человеческая аминокислотная последовательность» в этой связи относится к аминокислотной последовательности, которая кодируется геном иммуноглобулина человека, который включает гены зародышевой линии, подвергнутые реаранжировке и соматически мутированные гены. В настоящем изобретении также предусматриваются полипептиды, включающие константные домены с «человеческой» последовательностью, которые были изменены, в результате одного или более добавлений, делеций или замен аминокислот, относительно человеческой последовательности, исключая те варианты осуществления, в случае которых определенно требуется присутствие «полностью человеческой» шарнирной области.
Присутствие «полностью человеческой» шарнирной области в антителах против CD70 по настоящему изобретению может быть полезным, и чтобы минимизировать иммуногенность, и чтобы оптимизировать стабильность антитела.
Как обсуждается здесь в другом месте, предусматривается, что одна или более замен, вставок или делеций аминокислот могут быть осуществлены в константной области тяжелой и/или легкой цепи, в частности, в Fc-области. Аминокислотные замены могут приводить к замене замещаемой аминокислоты отличной, встречающейся в природе аминокислотой, или неприродной или модифицированной аминокислотой. Также допускаются другие структурные модификации, такие как, например, изменения характера гликозилирования (например, в результате добавления или делеции сайтов N- или O-гликозилирования). В зависимости от предполагаемого использования антитела, может быть желательным модифицирование антитела по настоящему изобретению, что касается его характеристик связывания с Fc-рецепторами, например, для модулирования эффекторной функции. Например, в Fc-область может быть введен остаток(и) цистеина, делающий тем самым возможным образование дисульфидной связи между цепями в этой области. Созданное таким образом гомодимерное антитело может обладать улучшенной способностью к интернализации и/или увеличенной способностью к комплемент-опосредованному уничтожению клеток и увеличенной антителозависимой клеточноопосредованной цитотоксичностью (ADCC). Смотрите Caron et al., J. Exp. Med. 176: 1191-1195 (1992) и Shopes, B. J. Immunol. 148: 2918-2922 (1992). Альтернативно, можно сконструировать антитело против CD70, которое имеет две Fc-области и может в силу этого обладать увеличенными способностями к комплемент-опосредованному лизису и ADCC. Смотрите Stevenson et al., Anti-Cancer Drug Design 3: 219-230 (1989). В настоящем изобретении также предусматриваются иммуноконъюгаты, включающие описываемое здесь антитело, конъюгированное с цитотоксическим агентом, таким как химиотерапевтическое средство, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения, или их фрагменты), или радиоактивным изотопом (т.е. радиоконъюгат). Могут быть также сконструированы Fc-области для удлинения полупериода существования, как описано в Chan and Carter, Nature Reviews: Immunology, Vol.10, pp301-316, 2010, который включен сюда посредством ссылки.
Тем не менее, в другом варианте осуществления Fc-область модифицируют для увеличения способности антитела к опосредованию антителозависимой клеточной цитотоксичности (ADCC) и/или для увеличения аффинности антитела к Fcγ-рецептору посредством изменения одной или более аминокислот.
Тем не менее, в другом варианте осуществления изменяют гликозилирование антитела. Например, можно создать негликозилированное антитело (т.е. антитело без гликозилирования). Гликозилирование меняют, например, для увеличения аффинности антитела к являющемуся мишенью антигену CD70. Такие углеводные модификации можно осуществить посредством, например, изменения одного или более сайтов гликозилирования в последовательности антитела. Например, может быть осуществлена одна или более замен аминокислот, которые приводят к удалению одного или более сайтов гликозилирования в каркасных областях вариабельных областей для исключения в силу этого гликозилирования в этом сайте. Такое отсутствие гликолизирования может увеличить аффинность антитела к антигену.
Также предусматриваются варианты антител против CD70, имеющие измененный тип гликозилирования, такие как гипофукозилированное антитело, имеющее уменьшенное число фукозильных остатков, или нефукозилированное антитело (описанное в Natsume et al., Drug Design Development and Therapy, Vol.3, pp7-16, 2009), или антитело, имеющее увеличенное число GlcNac-биссекторных структур. Было установлено, что такие измененные характеры гликозилирования увеличивают относящуюся к ADCC активность антител, приводя обычно к 10-кратному увеличению ADCC по сравнению с такой эквивалентного антитела, включающего «природный» человеческий Fc-домен. Такие углеводные модификации можно осуществить посредством, например, экспрессии антитела в клетке-хозяине с измененным ферментативным механизмом гликозилирования (как описано в Yamane-Ohnuki and Satoh, mAbs 1:3, 230-236, 2009). Примерами нефукозилированных антител с увеличенной функцией, относящейся к ADCC, являются те, которые продуцированы, используя PotelligentTM технологию BioWa Inc.
Настоящее изобретение может, в некоторых вариантах осуществления, включать химерные Camelidae/человек антитела и, в частности, химерные антитела, в которых VH- и VL-домены являются последовательностью целиком от верблюдовых (например, ламы или альпака), а оставшаяся часть антитела является полностью человеческой последовательностью. Антитела против CD70 могут включать антитела, включающие «гуманизированные» или «приведенные к зародышевому типу» варианты происходящих от верблюдовых VH- и VL-доменов, или их CDR, и химерные животное семейства верблюдовых/человек антитела, в которых VH- и VL-домены содержат одну или более аминокислотных замен в каркасных областях по сравнению с VH- и VL-доменами антител верблюдовых, полученных в результате активной иммунизации животного семейства верблюдовых антигеном CD70 человека. Такая «гуманизация» увеличивает % идентичности последовательности с VH- и VL-доменами человека зародышевого типа в результате замены несоответствующих аминокислотных остатков в исходном VH- или VL-домене Camelidae эквивалентным остатком, обнаруживаемым в кодируемом геном человека зародышевой линии VH- или VL-домене.
Антитела против CD70 могут также быть антителами с пересаженными CDR, в которых CDR (или гипервариабельные петлевые участки), происходящие из антитела верблюдовых, например, антитела против CD70 животного семейства верблюдовых, индуцированного в результате активной иммунизации белком CD70 человека, или иначе кодируемого геном верблюдовых, пересажены в каркасную область VH и VL человека, при этом оставшаяся часть антитела также полностью человеческого происхождения. Такие антитела против CD70 с пересаженными CDR могут содержать CDR, имеющие аминокислотные последовательности, представленные в виде SEQ ID NO: 49-59, 262 или 263 (CDR3 тяжелой цепи), или SEQ ID NO: 26-37, 249, 258 или 259 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20, 248, 256 или 257 (CDR1 тяжелой цепи), или одну из последовательностей CDR, представленных в виде SEQ ID NO: 148-168, 271 или 273 (CDR3 легкой цепи), или SEQ ID NO: 109-128 или 270 (CDR2 легкой цепи), или SEQ ID NO: 77-95 или 250-253, 267 и 268 (CDR1 легкой цепи).
Описанные выше гуманизированные, химерные антитела против CD70 и антитела против CD70 с пересаженными CDR, в частности, антитела, включающие гипервариабельные петлевые участки или CDR антител, полученных в результате активной иммунизации животного семейства верблюдовых антигеном CD70 человека, можно без труда продуцировать, используя обычные методы манипулирования рекомбинантными ДНК и экспрессии с них, используя прокариотические и эукариотические клетки-хозяева, сконструированные для продукции представляющего интерес полипептида и включающие, но без ограничения, бактериальные клетки, дрожжевые клетки, клетки млекопитающих, клетки насекомых, клетки растений, некоторых из которых описаны здесь и проиллюстрированы в сопроводительных примерах.
Происходящие от верблюдовых антитела против CD70 включают варианты, в которых гипервариабельный петлевой участок (участки) или CDR(и) VH-домена и/или VL-домена получены из обычного антитела животного семейства верблюдовых, индуцированного против CD70 человека, но в которых по крайней мере один из указанных (происходящих от верблюдовых) гипервариабельных петлевых участков или CDR был сконструирован так, что он включает одну или более замен, добавлений или делеций аминокислот относительно кодируемой геном верблюдовых последовательности. Такие изменения включают «гуманизацию» гипервариабельных петлевых участков/CDR. Происходящие от верблюдовых HV/CDR, сконструированные таким способом, могут, тем не менее, демонстрировать аминокислотную последовательность, которая «в значительной степени идентична» аминокислотной последовательности кодируемого геном верблюдовых HV/CDR. В этом контексте «идентичность в значительной степени» может допускать не более одной или более двух несоответствий по аминокислотной последовательности с кодируемым геном верблюдовых HV/CDR. Конкретные варианты антитела против CD70 могут содержать гуманизированные варианты последовательностей CDR, представленных в виде SEQ ID NO: 49-59, 262 или 263 (CDR3 тяжелой цепи), или SEQ ID NO: 26-37, 249, 258 или 259 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20, 248, 256 или 257 (CDR1 тяжелой цепи), или одной из последовательностей CDR, представленных в виде SEQ ID NO: 148-168, 271 или 273 (CDR3 легкой цепи), или SEQ ID NO: 109-128 или 270 (CDR2 легкой цепи), или SEQ ID NO: 77-95 или 250-253, 267 и 268 (CDR1 легкой цепи).
Происходящие от верблюдовых антитела против CD70, обеспечиваемые здесь, могут быть любого изотипа. Антитела, предназначенные для терапевтического применения для людей, будут типично типа IgA, IgD, IgE, IgG, IgM, часто типа IgG, в последнем случае они могут относиться к любому из четырех подклассов IgG1, IgG2a и b, IgG3 или IgG4. В пределах каждого из этих подклассов допускается осуществление одной или более замен, вставок или делеций аминокислот внутри Fc-части или осуществление других структурных модификаций, например, для усиления или ослабления Fc-зависимых функциональностей.
Гуманизация происходящих от верблюдовых VH- и VL-доменов
Обычные антитела верблюдовых дают выгодную исходную точку для приготовления антител, полезных в качестве терапевтических средств для человека, вследствие следующих факторов, обсуждавшихся в US 12/497239, который включен сюда посредством ссылки:
1) высокий % гомологии по последовательности между VH- и VL-доменами верблюдовых и их аналогами у человека;
2) высокая степень структурной гомологии между CDR VH- и VL-доменов верблюдовых и их аналогами у человека (т.е. подобных человеческим канонических складчатым структур и подобных человеческим сочетаний канонических складчатым структур).
Основа в виде антител верблюдовых (например, ламы) дает также значительное преимущество по показателю функционального разнообразия антител против CD70, которые можно получить.
Полезность антител против CD70, включающих VH- и/или VL-домены верблюдовых, для лечения людей может быть, кроме того, повышена с помощью «гуманизации» природных VH- и VL-доменов верблюдовых, например, для придания им меньшей иммуногенности у являющегося человеком хозяина. Общей целью гуманизации является получение молекулы, в которой VH- и VL-домены проявляют минимальную иммуногенность после введения являющемуся человеком субъекту, при сохранении специфичности и аффинности антигенсвязывающего центра, образуемого родительскими VH- и VL-доменами.
Один подход к гуманизации, так называемое «приведение к зародышевому типу», включает осуществление изменений в аминокислотной последовательности VH- или VL-домена верблюдовых для ее приведения ближе к последовательности VH- или VL-домена зародышевого типа.
Определение гомологии между VH (или VL)-доменом верблюдовых и VH (или VL)-доменами человека является важнейшей стадией в процессе гуманизации, и для выбора аминокислотных остатков верблюдовых, которые должны быть изменены (в конкретном VH- или VL-домене) и для выбора соответствующего аминокислотного остатка(ов) для замены.
Подход к гуманизации обычных антител верблюдовых был разработан на основе совмещения широкого ряда новых последовательностей VH (и VL)-доменов верблюдов, типично соматически мутированных VH (или VL)-доменов, которые, как известно, связываются с антигеном-мишенью, с последовательностями VH (или VL) человека зародышевого типа, консенсусными последовательностями VH (и VL) человека, а также информации о последовательностях зародышевого типа, имеющейся для Llama pacos. Следующие места обрисовывают в общих чертах принципы, которые можно применять для (i) выбора аминокислотных остатков верблюдовых для замены в происходящем от верблюдовых VH- или VL-домене или его CDR, и (ii) выбора аминокислотных остатков человека для подстановки при гуманизации любого конкретного VH (или VL)-домена верблюдовых. Этот подход можно использовать для приготовления гуманизированных вариантов происходящих от верблюдовых CDR, имеющих аминокислотные последовательности, представленные в виде SEQ ID NO: 49-59, 262 или 263 (CDR3 тяжелой цепи), или SEQ ID NO: 26-37, 249, 258 или 259 (CDR2 тяжелой цепи), или SEQ ID NO: 10-20, 248, 256 or 257 (CDR1 тяжелой цепи), или одну из последовательностей CDR, представленных в виде SEQ ID NO: 148-168, 271 или 273 (CDR3 легкой цепи), или SEQ ID NO: 109-128 или 270 (CDR2 легкой цепи), или SEQ ID NO: 77-95 или 250-253, 267 или 268 (CDR1 легкой цепи), а также для гуманизации происходящих от верблюдовых VH-доменов, имеющих последовательности, представленные в виде SEQ ID NO: 177-188, 212-223, 274 или 275, и происходящих от верблюдовых VL-доменов, имеющих последовательности, представленные в виде SEQ ID NO: 189-211, 230-245, 276 или 277.
Стадия 1. Выбрать семейство сегментов человека (зародышевой линии) и член этого семейства, который демонстрирует наибольшую степень гомологии/идентичности со зрелой последовательностью верблюдовых, подвергаемой гуманизации. Общий способ идентификации ближе всего соответствующего сегмента зародышевой линии человека для любого конкретного VH (или VL)-домена верблюдовых изложен ниже.
Стадия 2. Выбрать конкретный член семейства сегментов зародышевой линии человека, к которому будут осуществлять приведение. Предпочтительно он является членом семейства сегментов зародышевой линии с наибольшей гомологией или другим членом этого же семейства сегментов зародышевой линии.
Стадия 3. Определить предпочтительные положения, рассматриваемые для приведения к зародышевому типу, на основе таблицы использования аминокислот для последовательности верблюдовых зародышевого типа, которая ближе всего к таковой выбранного сегмента зародышевой линии человека.
Стадия 4. Попытаться изменить аминокислоты в последовательности верблюдовых зародышевого типа, которые отличаются от таковых в ближе всего соответствующей последовательности сегмента зародышевой линии человека; приведение к зародышевому типу остатков FR является предпочтительным по сравнению с остатками CDR.
a. Предпочтительными являются положения, которые отличаются от таковых в последовательности выбранного сегмента зародышевой линии человека, к которой осуществляют приведение, в которых аминокислота, обнаруживаемая в последовательности верблюдовых, не соответствует таковой в последовательности выбранного сегмента зародышевой линии и не обнаруживается в других сегментах зародышевой линии того же субкласса (и для кодируемых V, и для кодируемых J аминокислот FR).
b. Положения, отличающиеся от таковых в выбранном члене семейства сегментов зародышевой линии человека, но которые используются у других членов этого же семейства сегментов зародышевой линии, могут также рассматриваться в процессе приведения к зародышевому типу.
c. Могут также рассматриваться дополнительные несоответствия (например, вследствие дополнительных соматических мутаций) по отношению к выбранному члену семейства сегментов зародышевой линии человека.
Следующий подход может использоваться для определения ближе всего соответствующего сегмента зародышевой линии человека для конкретного VH (или VL)-домена верблюдовых.
До расчета процента идентичности по последовательности между VH и VL Camelidae и VH и VL человека зародышевого типа можно определить сначала канонические складчатые структуры, что позволяет идентифицировать семейство сегментов зародышевой линии человека с идентичным сочетанием канонических складчатых структур для H1 и H2 или L1 и L2 (и L3). Впоследствии член семейства сегментов зародышевой линии человека, который обладает наибольшей степенью гомологии последовательности с представляющей интерес вариабельной областью Camelidae, можно выбрать для подсчета гомологии последовательностей. Определение канонических классов гипервариабельных петлевых участков L1, L2, L3, H1 и H2 согласно Chothia можно выполнить с помощью инструментальных программных средств биоинформатики, общедоступных на веб-странице www.bioinf.org.uk/abs/chothia.html.page. Выводные данные программы указывают на требования к ключевым остаткам в файле данных. В этих файлах данных положения ключевых остатков представлены вместе с допустимыми аминокислотами в каждом положении. Последовательность вариабельной области антитела предоставляется в качестве вводных данных и сначала совмещается с консенсусной последовательностью антитела для установления схемы нумерации согласно Kabat. При анализе канонических складчатых структур используется ряд образцов ключевых остатков, получаемый с помощью автоматизированного способа, разработанного Martin и Thornton (Martin et al., J. Mol. Biol. 263: 800-815 (1996)). Границы отдельных каркасных областей можно определить, используя схему нумерации IMGT, которая является модификацией схемы нумерации Chothia (Lefranc et al., NAR 27: 209-212 (1999); imgt.cines.fr).
С использованием конкретного известного V-сегмента зародышевой линии человека, в котором используется то же сочетание канонических складчатых структур для H1 и H2 или L1 и L2 (и L3), можно определить в наибольшей степени соответствующий член семейства по показателю гомологии последовательности. С помощью инструментальных программных средств биоинформатики можно определить процент идентичности по последовательности между аминокислотными последовательностями каркасных областей VH- и VL-доменов Camelidae и соответствующими последовательностями, кодируемыми генами человека зародышевой линии, но можно также использовать совмещение последовательностей фактически вручную. Последовательности иммуноглобулинов человека можно идентифицировать на основе нескольких баз данных, относящихся к белкам, таких как VBase (http://vbase.mrc-cpe.cam.ac.uk/) или база данных Pluckthun/Honegger (http://www.bioc.unizh.ch/antibody/Sequences/Germlines). Для сравнения последовательностей человека с V-сегментами для VH- или VL-доменов Camelidae можно использовать алгоритм для совмещения последовательностей, например, доступный через веб-страницы вроде www.expasy.ch/tools/#align, но также можно выполнить совмещение вручную с ограниченным рядом последовательностей. Последовательности легкой и тяжелой цепей человека зародышевого типа из семейств с одинаковыми сочетаниями канонических складчатых структур и с наибольшей степенью гомологии с каркасными областями 1, 2 и 3 каждой цепи выбирают и сравнивают с представляющей интерес вариабельной областью Camelidae; также проверяют FR4 на фоне JH- и JK- или JL-сегментов зародышевой линии человека.
Обратите внимание, что при расчете общего процента гомологии последовательностей остатки FR1, FR2 и FR3 оцениваются, используя ближе всего соответствующую последовательность из семейства сегментов зародышевой линии человека с идентичным сочетанием канонических складчатых структур. Подсчитываются лишь остатки, отличные от ближе всего соответствующей последовательности или других членов того же семейства с тем же сочетанием канонических складчатых структур (NB - исключая любые кодируемые праймером различия). Однако с целью гуманизации остатки в каркасных областях, идентичные членам других семейств сегментов зародышевой линии человека, которые не имеет то же сочетание канонических складчатых структур, могут считаться «человеческими», несмотря на тот факт, что они оцениваются как «негативные» в соответствии со строгими условиями, описанными выше. Это допущение основано на подходе «смешивания и соответствия» для гуманизации, в котором каждую из FR1, FR2, FR3 и FR4 по отдельности сравнивают с ближе всего соответствующей ей последовательностью человека зародышевого типа, и поэтому гуманизированная молекула содержит комбинацию различных FR, как это делали Qu и его коллеги (Qu et la., Clin. Cancer Res. 5: 3095-3100 (1999)) и Ono и его коллеги (Ono et al., Mol. Immunol. 36: 387-395 (1999)).
Перекрестно-конкурирующие антитела
Моноклональными антителами или их антигенсвязывающими фрагментами, которые «перекрестно конкурируют» с молекулами, описываемыми здесь, являются те, которые связываются с CD70 человека в центре(ах), который является идентичным, или перекрывающимся с, центру(ом), с которым связываются антитела против CD70 по настоящему изобретению. Конкурирующие моноклональные антитела или их антигенсвязывающие фрагменты можно идентифицировать, например, посредством анализа конкуренции антител. Например, образец очищенного или частично очищенного CD70 человека можно связать с твердой подложкой. Затем добавляют соединение - антитело или его антигенсвязывающий фрагмент по настоящему изобретению и моноклональное антитело или его антигенсвязывающий фрагмент, который, как полагают, способен конкурировать с таким соединением - антителом по настоящему изобретению. Одна из двух молекул является помеченной. Если меченое соединение и немеченое соединение связываются с отдельными и различными центрами CD70, меченое соединение будет связываться в одинаковой степени, независимо от того, присутствует ли предположительно конкурирующее соединение. Однако, если центры взаимодействия являются идентичными или перекрывающимися, немеченое соединение будет конкурировать, и количество связавшегося с антигеном меченого соединения будет уменьшаться. Если немеченое соединение присутствует в избыточном количестве, очень незначительное количество, если это имеет место, меченого соединения будет связываться. Для целей настоящего изобретения конкурирующими моноклональными антителами или их антигенсвязывающими фрагментами являются те, которые уменьшают связывание с CD70 соединений - антител по настоящему изобретению на приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% или приблизительно 99%. Детали процедур выполнения таких анализов конкуренции хорошо известны в данной области техники и можно найти, например, в Harlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, pages 567-569, ISBN 0-87969-314-2. Такие анализы можно сделать количественными посредством использования очищенных антител. Стандартную кривую строят посредством титрования одного антитела с его же помощью, т.е. одно и то же антитело используют и для мечения, и в качестве конкурента. С помощью титрования определяют способность немеченого конкурирующего моноклонального антитела или его антигенсвязывающего фрагмента к ингибированию связывания меченой молекулы с планшетом. Результаты представляют, строя графики, и сравнивают концентрации, необходимые для достижения желаемой степени ингибирования связывания.
Предпочтительными вариантами осуществления являются антитела, которые перекрестно конкурируют за связывание с CD70 человека с антителами, включающими происходящий от ламы Fab 27B3, и его приведенные к зародышевому типу варианты, включая, в частности, 41D12 (ARGX-110), и которые демонстрируют то же сочетание характеристик связывания, а именно:
a) связывание в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO: 342) в CD70 человека;
(b) перекрестную реактивность с гомологами CD70 у макака-резус (Macaca mulatta) и яванского макака (Macaca cynomolgus);
(c) связывание со встречающимся в природе CD70 человека и с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
Кодирующие антитела против CD70 полинуклеотиды
Настоящим изобретением также обеспечиваются молекулы полинуклеотидов, кодирующие антитела против CD70 по настоящему изобретению, также векторы экспрессии, содержащие нуклеотидные последовательности, которые кодируют антитела против CD70 по настоящему изобретению, функционально связанные с регуляторными последовательностями, которые допускают экспрессию антигенсвязывающего полипептида в клетке-хозяине или бесклеточной экспрессионной системе, и клетка-хозяин или бесклеточная экспрессионная система, содержащая этот вектор экспрессии.
Молекулы полинуклеотидов, кодирующие антитела против CD70 по настоящему изобретению, включают, например, рекомбинантные молекулы ДНК. Термины «нуклеиновая кислота», «полинуклеотид» или «молекула полинуклеотида» используются здесь взаимозаменяемо и относятся к любой молекуле ДНК или РНК, либо одноцепочечной, либо двухцепочечной, и, в случае одноцепочечной молекулы, молекуле с комплементарной ей последовательностью. При обсуждении молекул нуклеиновых кислот последовательность или структура конкретной молекулы нуклеиновой кислоты может описываться здесь в соответствии с общепринятым правилом предоставления последовательности в направлении от 5′ к 3′. В некоторых вариантах осуществления настоящего изобретения нуклеиновые кислоты или полинуклеотиды являются «выделенными». Этот термин, в случае применения к молекуле нуклеиновой кислоты, относится к молекуле нуклеиновой кислоты, которая отделена от последовательностей, с которыми она непосредственно соседствует во встречающемся в природе геноме организма, в котором она порождена. Например, «выделенная нуклеиновая кислота» может включать молекулу ДНК, встроенную в вектор, такой как плазмидный или вирусный вектор, или интегрированную в геномную ДНК прокариотической или эукариотической клетки или не являющегося человеком организма-хозяина. В случае применения к РНК, «термин «выделенный полинуклеотид» относится в основном к молекуле РНК, кодируемой выделенной молекулой РНК, определенной выше. Альтернативно, термин может относиться к молекуле РНК, которая была очищена/отделена от других нуклеиновых кислот, с которыми она была бы связана в ее природном состоянии (т.е. в клетке или тканях). Выделенный полинуклеотид (либо ДНК, либо РНК) может, кроме того, представлять собой молекулу, продуцированную непосредственно биологическим способом или способом синтеза и отделенную от других компонентов, присутствующих во время ее продукции.
В одном варианте осуществления настоящим изобретением обеспечиваются нуклеотидные последовательности, которые кодируют VH-домен и VL-домен приведенного к зародышевому типу, происходящего от ламы Fab, обозначенного здесь 41D12, причем VH-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 223, а VL-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 241. В предпочтительном варианте осуществления нуклеотидная последовательность, кодирующая VH-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 223, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 344, а нуклеотидная последовательность, кодирующая VL-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 241, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 345.
В одном варианте осуществления настоящим изобретением обеспечиваются нуклеотидные последовательности, которые кодируют VH-домен и VL-домен приведенного к зародышевому типу, происходящего от ламы Fab, обозначенного здесь 57В6, причем VH-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 274, а VL-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 276. В предпочтительном варианте осуществления нуклеотидная последовательность, кодирующая VH-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 274, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 346, а нуклеотидная последовательность, кодирующая VL-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 276, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 347.
В одном варианте осуществления настоящим изобретением обеспечиваются нуклеотидные последовательности, которые кодируют VH-домен и VL-домен приведенного к зародышевому типу, происходящего от ламы Fab, обозначенного здесь 59D10, причем VH-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 275, а VL-домен имеет аминокислотную последовательность, представленную в виде SEQ ID NO: 277. В предпочтительном варианте осуществления нуклеотидная последовательность, кодирующая VH-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 275, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 348, а нуклеотидная последовательность, кодирующая VL-домен, имеющий аминокислотную последовательность, представленную в виде SEQ ID NO: 277, включает нуклеотидную последовательность, представленную в виде SEQ ID NO: 349.
Для рекомбинантной продукции антитела против CD70 в соответствии с настоящим изобретением рекомбинантный полинуклеотид, кодирующий его, можно приготовить (используя стандартные методы молекулярной биологии) и встроить в реплицируемый вектор для экспрессии в выбранной клетке-хозяине, или бесклеточной экспрессионной системе. Подходящими клетками-хозяевами могут быть прокариот, дрожжи или клетки высших эукариот, особенно клетки млекопитающих. Примерами применимых линий клеток млекопитающих-хозяев являются линия CV1 клеток почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36: 59 (1977)); клетки почки детеныша хомячка (BHK, ATCC CCL 10); клетки/-DHFR яичника китайского хомячка (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23: 243-251 (1980)); клетки миеломы мыши SP2/0-AG14 (ATCC CRL 1581; ATCC CRL 8287) или NS0 (№ в коллекциях культур HPA-85110503); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы линии buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и линия клеток гепатомы человека (Hep G2), а также DSM линия клеток PERC-6. Векторы экспрессии, подходящие для использования в каждой из этих клеток-хозяев, также, как правило, хорошо известны в данной области техники.
Следует отметить, что термин «клетка-хозяин», как правило, относится к культивируемой линии клеток. Люди в целом, в которых введен вектор экспрессии, кодирующий антигенсвязывающий полипептид в соответствии с настоящим изобретением, однозначно исключены из определения «клетки-хозяина».
Продукция антител
В важном аспекте настоящим изобретением также обеспечивается способ продукции антитела против CD70 по настоящему изобретению, который включает культивирование клетки-хозяина (или бесклеточной экспрессионной системы), содержащей полинуклеотид (например, вектор экспрессии), кодирующий антитело против CD70, в условиях, которые допускают экспрессию антитела против CD70, и выделение экспрессированного антитела против CD70. Этот процесс рекомбинантной экспрессии может использоваться для высокомасштабной продукции антител против CD70 в соответствии с настоящим изобретением, в том числе моноклональных антител, предназначенных для терапевтического применения для людей. Векторы, линии клеток и способы продукции, подходящие для высокомасштабного производства рекомбинантных антител, подходящих для in vivo терапевтического применения, имеются, как правило, в наличии в данной области техники и будут хорошо известны квалифицированному специалисту.
Как отмечено в другом месте, обеспечиваемые здесь антитела против CD70, включая, в частности, антитела на основе Fab 27B3 и его приведенных к зародышевому типу вариантах, таких как 41D12 (ARGX-110), демонстрируют характеристики, которые особенно выгодны для высокомасштабного промышленного производства. В частности, очень высокий уровень продукции (>4 г/л), достигаемый в системе рекомбинантной экспрессии в промышленном масштабе, будет значительно снижать затраты производства.
Предпочтительные варианты осуществления также демонстрируют термостабильность при проверке при 37°C, а также на протяжении нескольких циклов замораживания-оттаивания, что снова является очень выгодным для промышленного производства и хранения продукта для клинического применения. Как ни удивительно, некоторые из приведенных к человеческому зародышевому типу вариантов Fab 27B3, в том числе 41D12 и 40F1, демонстрируют увеличенную термостабильность по сравнению с самим 27B3.
Соответственно, в дальнейшем аспекте настоящим изобретением обеспечивается выделенное антитело, или его антигенсвязывающий фрагмент, которое связывается с CD70 человека, при этом указанное антитело или фрагмент включает вариабельный домен тяжелой цепи, соответствующий аминокислотной последовательности, представленной в SEQ ID NO: 178, при условии, что по крайней мере одна аминокислота в положении согласно Kabat, выбираемом из группы, состоящей из H6, H18, H24, H31, H56, H74, H74, H77, H79, H83, H84, H89, H93, H94, H108, H110 и H112, заменена другой аминокислотой.
Это антитело, или его антигенсвязывающий фрагмент, может демонстрировать большую термостабильность, чем антитело, или его антигенсвязывающий фрагмент, включающее вариабельный домен тяжелой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 178.
В дальнейшем варианте осуществления обеспечивается антитело против CD70 или его антигенсвязывающий фрагмент, которое(ый) включает вариабельный домен легкой цепи, соответствующий аминокислотной последовательности, представленной в SEQ ID NO: 201, при условии, что по крайней мере одна аминокислота в положении согласно Kabat, выбираемом из группы, состоящей из L2, L11, L12, L25, L26, L30, L46, L53, L60, L61, L67, L68, L76, L80, L81, L85 и L87, заменена другой аминокислотой.
Это антитело, или его антигенсвязывающий фрагмент, может демонстрировать большую термостабильность, чем антитело, или его антигенсвязывающий фрагмент, включающее вариабельный домен тяжелой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 201.
В конкретном варианте осуществления антитело против CD70, или его антигенсвязывающий фрагмент, может включать:
a) вариабельный домен тяжелой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 178, включающей одну или более аминокислотных замен в положениях согласно Kabat, выбираемых из группы, состоящей из H6, H18, H24, H31, H56, H74, H74, H77, H79, H83, H84, H89, H93, H94, H108, H110 и H112; и
b) вариабельный домен легкой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 201, включающей одну или более аминокислотных замен в положениях согласно Kabat, выбираемых из группы, состоящей из L2, L11, L12, L25, L26, L30, L46, L53, L60, L61, L67, L68, L76, L80, L81, L85 и L87.
Это антитело, или его антигенсвязывающий фрагмент, по пункту 24 может демонстрировать большую термостабильность, чем антитело, или его антигенсвязывающий фрагмент, включающее вариабельный домен тяжелой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 178, и вариабельный домен легкой цепи с аминокислотной последовательностью, представленной в SEQ ID NO: 201.
Терапевтическая ценность антител против CD70
Обеспечиваемые здесь антитела против CD70, или их антигенсвязывающие фрагменты, могут использоваться для ингибирования роста злокачественных опухолевых клеток in vivo и, следовательно, применимы при лечении CD70-экспрессирующих раков.
Соответственно, дальнейшие аспекты настоящего изобретения относятся к способам ингибирования роста опухолевых клеток у являющегося человеком пациента, а также способам лечения или предупреждения рака, которые включают введение нуждающемуся в этом пациенту терапевтически эффективного количества антитела против CD7 или его антигенсвязывающего фрагмента, которые здесь описываются.
В другом аспекте настоящего изобретения обеспечивается антитело против CD70 или антигенсвязывающий фрагмент, которые здесь описываются, для применения для ингибирования роста CD70-экспрессирующих опухолевых клеток у являющегося человеком пациента.
Тем не менее, в дальнейшем аспекте настоящего изобретения обеспечивается антитело против CD70 или антигенсвязывающий фрагмент, которые здесь описываются, для применения для лечения или предупреждения рака у являющегося человеком пациента.
Предпочтительные раки, рост которых можно ингибировать, используя описываемые здесь антитела против CD70, включают рак почки (например, почечноклеточную карциному), рак молочной железы, опухоли головного мозга, хронические или острые лейкозы, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфолейкоз, лимфомы (например, лимфому Ходжкина и неходжкинскую лимфому, лимфоцитарную лимфому, первичную лимфому ЦНС, T-клеточную лимфому), карциномы носоглотки, меланому {например, метастатическую злокачественную меланому), рак предстательной железы, рак ободочной кишки, рак легкого, рак кости, рак поджелудочной железы, рак кожи, рак головы или шеи, злокачественную меланому кожи или в области сосудистой оболочки глаза, рак матки, рак яичника, рак прямой кишки, рак анальной области, рак желудка, рак яичек, рак матки, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, карциному влагалища, карциному вульвы, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркому мягкой ткани, рак мочеиспускательного канала, рак пениса, солидные опухоли у детей, рак мочевого пузыря, рак почки или мочеточника, карциному почечной лоханки, неоплазму центральной нервной системы (ЦНС), опухолевый ангиогенез, опухоль оси позвоночника, глиому ствола головного мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, мезотелиому.
Описываемые здесь антитела против CD70 могут также использоваться для лечения субъекта с онкогенным нарушением, характеризующимся присутствием опухолевых клеток, экспрессирующих CD70, включая, например, почечноклеточные карциномы (RCC), такие как светлоклеточная RCC, глиобластому, рак молочной железы, опухоли головного мозга, карциномы носоглотки, неходжкинскую лимфому (NHL), острый лимфолейкоз (ALL), хронический лимфолейкоз (CLL), лимфому Беркитта, анапластические крупноклеточные лимфомы (ALCL), множественную миелому, T-клеточные лимфомы кожи, нодулярные лимфомы из мелких клеток с расщепленными ядрами, лимфоцитарные лимфомы, периферические T-клеточные лимфомы, лимфомы Леннерта, иммунобластные лимфомы, T-клеточный лейкоз/лимфомы (ATLL), T-клеточный лейкоз у взрослых (T-ALL), центробластные/центроцитические (cb/cc) фолликулярные лимфомы-раки, диффузные В-крупноклеточные лимфомы, напоминающую ангиоиммунобластическую лимфаденопатию (AILD) T-клеточную лимфому, связанные с ВИЧ-инфекцией лимфомы полостей тела, эмбриональные карциномы, недифференцированные карциномы носоглотки (например, опухоль Шминке), болезнь Кастлемана, саркому Капоши, множественную миелому, макроглобулинемию Вальденстрема, лимфому из клеток ткани и другие B-клеточные лимфомы.
Конкретные варианты осуществления относятся к лечению любого из вышеперечисленных раков антителом, включающим происходящий от ламы Fab 27B3, или один из его приведенных к зародышевому типу вариантов, включая, в частности, 41D12 (ARGX-110). В частности, любой из вышеперечисленных раков можно лечить, используя антитело, включающее Fab-области 41D12 (приведенного к зародышевому типу варианта 27B3), слитые с константными областями IgGl человека. В последнем варианте осуществления можно, кроме того, сконструировать константную область IgG1 человека (например, посредством добавления точечных мутаций) для максимизации эффекторных функций антитела, или антитело 41D12-IgG1 может быть нефукозилированным, снова для увеличения эффекторной функции антитела.
Как отмечено здесь в другом месте, описываемые здесь антитела против CD70, особенно 27B3 и его приведенные к зародышевому типу варианты, демонстрируют особенно сильное связывание с линиями раковых клеток человека, в которых CD70 представлен на клеточной поверхности в низкой копийности, такие как, например, линия клеток B-крупноклеточной лимфомы SU-DHL-6, линия клеток хронического лимфолейкоза JVM-2, линия клеток T-клеточной лимфомы кожи HH, линия клеток карциномы желудка MKN-45, линии клеток карциномы легкого A549 и EBC-1, линии клеток меланомы WM852 и WM793, линия клеток глиобластомы GaMG и линии клеток карциномы яичника OAW-42, SKOv3 и OVCAR3. Действительно, аффинность 41D12 (ARGX-110) к этим линиям раковых клеток с низкой копийностью CD70 значительно выше таковой антител известного уровня техники, таких как SGN70 и MDX1411, в качестве препаратов сравнения.
«Увеличенная» аффинность к линиям раковых клеток с низкой копийностью CD70 имеет прямое отношение к клиническому лечению соответствующих раков, как показано с помощью экспериментов с внедрением клеток в сопроводительных примерах. Когда клетки линии раковых клеток с низкой копийностью CD70 (например, линии клеток диффузной B-крупноклеточной лимфомы SU-DHL-6) внедряют в только что выделенные от здоровых доноров PBMC, клетки линии раковых клеток предпочтительно истощаются с помощью антитела против CD70 ARGX-110 в качестве примера.
Соответственно, дальнейший аспект настоящего изобретения относится к способу лечения или предупреждения рака, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества антитела против CD70 или антигенсвязывающего фрагмента, которые здесь описываются, причем раком является рак, который демонстрирует экспрессию CD70 в низкой копийности.
Настоящим изобретением также обеспечивается антитело против CD70 или антигенсвязывающий фрагмент, которые описываются здесь, для применения для лечения или предупреждения рака у являющегося человеком пациента, причем раком является рак, который демонстрирует экспрессию CD70 в низкой копийности.
В каждом из этих аспектов рак с низкой копийностью CD70 предпочтительно выбирают из группы, состоящей из B-крупноклеточной лимфомы, хронического лимфолейкоза, T-клеточной лимфомы кожи, рака желудка, рака легкого, меланомы, глиобластомы и рака яичника.
Конкретные варианты осуществления относятся к лечению раков с низкой копийностью CD70 антителом, включающим происходящий от ламы Fab 27B3, или один из его приведенных к зародышевому типу вариантов, включая, в частности, 41D12 (ARGX-110). В частности, любой из вышеперечисленных раков с низкой копийностью CD70 можно лечить, используя антитело, включающее Fab-области 41D12 (приведенного к зародышевому типу варианта 27B3), слитые с константными областями IgGl человека.
Описываемые здесь антитела против CD70 также применимы для лечения иммунологических нарушений, характеризующихся экспрессией CD70. Конкретные примеры таких иммунологических нарушений включают нарушения: ревматоидный артрит, аутоиммунные заболевания с демиелинизацией (например, рассеянный склероз, аллергический энцефаломиелит), эндокринную офтальмопатию, увеоретинит, системную красную волчанку, злокачественную миастению, болезнь Грейвса, гломерулонефрит, аутоиммунное гепатологическое нарушение, воспалительное заболевание кишечника (например, болезнь Крона, неспецифический язвенный колит, глютеновую болезнь), анафилаксию, аллергическую реакцию, синдром Гужеро-Шегрена, сахарный диабет типа I, первичный билиарный цирроз, грануломатоз Вегенера, фибромиалгию, полимиозит, дерматомиозит, множественную эндокринную недостаточность, синдром Шмидта, аутоиммунный увеит, болезнь Аддисона, воспаление надпочечника, тиреоидит, тиреоидит Хашимото, аутоиммунное заболевание щитовидной железы, злокачественную анемию, атрофию желудка, хронический гепатит, волчаночный гепатит, атеросклероз, подострую кожную красную волчанку, гипопаратиреоз, синдром Дресслера, аутоиммунную тромбоцитопению, идиопатический тромбоцитопенический акроангиотромбоз, гемолитическую анемию, обыкновенную пузырчатку, пузырчатку, герпетиформный дерматит, очаговую алопецию, пемфигоид, склеродерму, прогрессирующий системный склероз, синдром CREST (кальциноз, виброболезнь, нарушение моторики пищевода, склеродактилия и телеангиэктазия), аутоиммунное бесплодие у мужчин и женщин, анкилозирующий спондилоартрит, неспецифический язвенный колит, смешанное заболевание соединительной ткани, узелковый полиартериит, системный некротизирующий васкулит, атопический дерматит, атопический ринит, синдром Гудпасчера, болезнь Чага, саркоидоз, ревматическую лихорадку, астму, повторяющиеся выкидыши, антифосфолипидный синдром, легкое «фермера», эритему многоформную, посткардиотомический синдром, синдром Кушинга, аутоиммунный хронический активный гепатит, легкое «птицевода», токсический эпидермальный некролиз, синдром Альпорта, альвеолит, аллергический альвеолит, фиброзный альвеолит, интерстициальное заболевание легких, узелковую эритему, пиодермию гангренозную, реакцию после трансфузии, артериит Такаши, ревматическую полимиалгию, височный артериит, шистосомоз, гигантоклеточный артериит, аскаридоз, аспергиллез, синдром Самптера, экзему, лимфоматоидный грануломатоз, болезнь Бехчета, синдром Каплана, болезнь Кавасаки, денге, энцефаломиелит, эндокардит, эндомиокардиальный фиброз, эндофтальмит, стойкую повышающуюся эритему, псориаз, эритробластоз плода, эозинофильный фасциит, синдром Шульмана, синдром Фелти, филяриоз, циклит, хронический циклит, гетерохронный циклит, циклит Фучи, IgA-нефропатию, пурпуру Геноха-Шенлейна, гомологичную болезнь, отторжение трансплантата, кардиомиопатию, синдром Итона-Ламберта, рецидивирующий полихондрит, криоглобулинемию, синдром Эванса и аутоиммунную гонадную недостаточность, нарушения функции B-лимфоцитов (например, системную красную волчанку, синдром Гудпасчера, ревматоидный артрит или сахарный диабет типа I), Th1-лимфоцитов (например, ревматоидный артрит, рассеянный склероз, псориаз, синдром Гужеро-Шегрена, тиреоидит Хашимото, болезнь Грейвса, первичный билиарный цирроз, грануломатоз Вегенера, туберкулез или гомологичную болезнь), или Th2-лимфоцитов (например, атопический дерматит, системную красную волчанку, атопическую астму, риноконъюнктивит, аллергический ринит, синдром Оменна, системный склероз или хроническую гомологичную болезнь), синдром Черджа-Стросса, микроскопический полиангиит и артериит Такаясу.
Конкретные варианты осуществления относятся к лечению любого из вышеперечисленных иммунологических нарушений антителом, включающим происходящий от ламы Fab 27B3, или один из его приведенных к зародышевому типу вариантов, включая, в частности, 41D12 (ARGX-110). В частности, любое из вышеперечисленных иммунологических нарушений можно лечить, используя антитело, включающее Fab-области 41D12 (приведенного к зародышевому типу варианта 27B3), слитые с константными областями IgGl человека. В последнем варианте осуществления можно, кроме того, сконструировать константную область IgG1 человека (например, посредством добавления точечных мутаций) для максимизации эффекторных функций антитела, или антитело 41D12-IgG1 может быть нефукозилированным, снова для увеличения эффекторной функции антитела.
Как здесь используется, термин «лечение» означает замедление, прерывание, остановку, контролирование, улучшение, прекращение, уменьшение тяжести симптома, нарушения, состояния или заболевания, но не обязательно включает общее устранение всех связанных с заболеванием симптомов, состояний или нарушений.
Для терапевтического применения для людей описываемые здесь антитела против CD70 могут вводиться являющемуся человеком субъекту, нуждающемуся в лечении, в «эффективном количестве». Термин «эффективное количество» относится к количеству или дозе антитела против CD70, которое, после введения однократной дозы или многократных доз являющемуся человеком пациенту, обеспечивает терапевтическую эффективность в лечении заболевания. Терапевтически эффективные количества антитела против CD70 могут включать количество в диапазоне от приблизительно 0,1 мг/кг до приблизительно 20 мг/кг на одну дозу. Терапевтически эффективное количество для любого отдельного пациента может определить специалист в области здравоохранения посредством слежения за эффектом антитела против CD70 на биомаркер, такой как CD70 на клеточной поверхности в опухолевых тканях, или симптом, такой как регрессия опухоли и т.д. Количество антитела, вводимое в любой конкретный момент времени, может варьировать, так что на протяжении лечения вводятся оптимальные количества антитела против CD7, либо применяемого отдельно, либо в комбинации с любым другим терапевтическим средством.
Также предусматривается введение описываемых здесь антител против CD70, или фармацевтических композиций, включающих такие антитела, в комбинации с любым другим противораковым лечением, в качестве комбинированной терапии.
Применение антител, которые плохо интернализуются, для истощения CD70-экспрессирующих клеток
Неожиданным обнаружением, описываемым здесь, является то, что антитела против CD70, связавшиеся с CD70, представленным на поверхности линий раковых клеток, плохо интернализуются. Это является неожиданным результатом, поскольку в известном уровне техники CD70 ранее описывался как «интернализующаяся мишень». Хотя между различными типами раковых клеток существует вариация в точной степени интернализации связавшихся антител против CD70, не обнаружен тип раковых клеток, в который «полностью интернализуется» связавшееся антитело против CD70. Например, в линии клеток рака почки, ранее описанные как быстро интернализующие клетки, в действительности, как установлено, интернализуется максимум приблизительно 70% связавшегося антитела ARGX-110. Во многие другие линии раковых клеток интернализуется менее 30%, менее 20% или даже менее 10% связавшегося ARGX-110.
Вариация в степени интернализации между различными линиями раковых клеток в значительной степени зависит от показания-заболевания, при этом схожие результаты отмечены, используя различные антитела против CD70. Однако плохая интернализация в некоторые линии клеток является особенно явной при использовании антитела ARGX-110. Отмечаемая низкая степень интернализации во многие раковые клетки дает обоснование для применения антител против CD70 с сильной эффекторной функцией для истощения (например, уничтожения или ингибирования роста) раковых клеток in vivo. Поскольку связавшиеся антитела против CD70 относительно плохо интернализуются во многие раковые клетки, антитело будет оставаться связанным с клеточной поверхностью и может в силу этого инициировать эффекторные функции (ADCC, CDC, ADCP) при взаимодействиях с иммунной системой являющегося человеком пациента.
Соответственно, дальнейший важный аспект настоящего изобретения относится к способу истощения CD70-экспрессирующих клеток у являющегося человеком пациента, включающий введение указанному пациенту эффективного количества антитела, которое связывается с CD70 человека, причем указанные CD70-экспрессирующие клетки демонстрируют составляющую 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее или 5% или менее интернализацию указанного антитела после периода времени, составляющего 6 часов, и причем указанное антитело проявляет по крайней мере одну эффекторную функцию, выбираемую из группы, состоящей из ADCC, CDC и ADCP.
В этом способе антитело против CD70 (с активными эффекторными функциями) используется для истощения CD70-экспрессирующих клеток. Это истощение CD70-экспрессирующих клеток может лежать в основе терапевтического или профилактического лечения, например, лечения или предупреждения CD70-экспрессирующих раков, или лечения или предупреждения связанных с CD70 иммунологических нарушений. Точный механизм истощения CD70-экспрессирующих клеток может зависеть от типа эффекторных функций, проявляемых антителом против CD70, но будет зависеть от сильного (с высокой аффинностью) связывания антитела против CD70 с CD70 на клеточной поверхности, вместе с плохой интернализацией связавшегося антитела.
В некоторых вариантах осуществления CD70-экспрессирующими клетками являются CD70-экспресссирующие раковые клетки. В частности, CD70-экспрессирующие раковые клетки могут относиться к типу рака, выбираемому из группы, состоящей из лимфомы Беркитта, B-крупноклеточной лимфомы, лимфомы Ходжкина, неходжкинской лимфомы, лимфомы из клеток ткани, хронического лимфолейкоза, карциномы поджелудочной железы, карциномы желудка, глиобластомы и карциномы легкого. Было установлено, что в линии клеток, происходящие из каждого из этих раков, интернализуется 30% или менее связавшегося антитела против CD70 в течение периода времени, составляющегося 6 часов. В дальнейшем варианте осуществления CD70-экспрессирующие раковые клетки могут относиться к типу рака, выбираемому из группы, состоящей из лимфомы Беркитта, B-крупноклеточной лимфомы, лимфомы Ходжкина, неходжкинской лимфомы, лимфомы из клеток ткани, хронического лимфолейкоза, карциномы желудка, глиобластомы и карциномы легкого. Было установлено, что в линии клеток, происходящие из каждого из этих раков, интернализуется 20% или менее связавшегося антитела против CD70 в течение периода времени, составляющегося 6 часов. В дальнейшем варианте осуществления CD70-экспрессирующие раковые клетки могут относиться к типу рака, выбираемому из группы, состоящей из лимфомы Беркитта, B-крупноклеточной лимфомы, лимфомы из клеток ткани, хронического лимфолейкоза, карциномы желудка и карциномы легкого. Было установлено, что в линии клеток, происходящие из каждого из этих раков, интернализуется 10% или менее связавшегося антитела против CD70 в течение периода времени, составляющегося 6 часов.
В других вариантах осуществления CD70-экспрессирующими клетками могут быть CD70-экспрессирующие активированные T-клетки, которые не демонстрируют какую-либо выявляемую интернализацию связавшихся антител против CD70 (в частности, интернализация ARGX-110 не наблюдается в течение периода времени, составляющего 6 часов). Эти данные наблюдений, что связавшиеся антитела против CD70 не интернализуются в CD70+ активированные T-клетки, служат сильной поддержкой для применения антител против CD70 с сильной эффекторной функцией (ADCC, CDC или ADCP), включая, но без ограничения, ARGX-110, при лечении иммунологических заболеваний, опосредуемых CD70+ активированными T-клетками, в том числе таких иммунологических нарушений, как, например, ревматоидный артрит, псориаз, SLE и т.д.
В вышеперечисленных вариантах осуществления может использоваться по существу любое антитело против CD70. В конкретных вариантах осуществления антителом может быть любое антитело против CD70, которое, как было экспериментально установлено, демонстрируют составляющую 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее или 5% или менее интернализацию после периода времени, составляющего 6 часов, в линию CD70-экспрессирующих раковых клеток, выбираемую из группы, состоящей из Raji, SU-DHL-6, MHHPREB1, Mino, Mecl, JVM-2, MKN-45, A549, U87MG, PANC-1, PANC-89, L428, Granta 519, Rec-1 и EBC-1. В действительности, анализ интернализации в линии клеток, описанный в сопроводительных примерах, может использоваться в качестве фильтра, как для определения того, следует ли оказывать на конкретные показания-заболевания воздействие антителами против CD70, обладающими сильной эффекторной функцией, или конъюгатами антитело против CD70-лекарственное средство (обсуждаемыми ниже), так и для идентификации антител против CD70, подходящих для применения при лечении конкретных показаний-заболеваний.
Соответственно, настоящим изобретением также обеспечивается способ истощения CD70-экспрессирующих раковых клеток у являющегося человеком пациента, включающий введение указанному пациенту эффективного количества антитела, которое связывается с CD70 человека, причем указанное антитело демонстрирует составляющую 30% или менее интернализацию после периода времени, составляющего 6 часов, в линию CD70-экспрессирующих раковых клеток, выбираемую из группы, состоящей из Raji, SU-DHL-6, MHHPREB1, Mino, Mec1, JVM-2, MKN-45, A549, U87MG, PANC-1, PANC-89, L428, Granta 519, Rec-1 и EBC-1, и причем указанное антитело проявляет по крайней мере одну эффекторную функцию, выбираемую из группы, состоящей из ADCC, CDC и ADCP. В одном варианте осуществления антителом, используемым для истощения CD70-экспрессирующих раковых клеток у являющегося человеком пациента, может быть антитело, которое демонстрирует составляющую 10% или менее интернализацию после периода времени, составляющего 6 часов, в линию CD70-экспрессирующих раковых клеток, выбираемую из группы, состоящей из Raji, SU-DHL-6, Granta 519, Rec-1, Mec1, JVM-2, MKN-45, A549 и EBC-1, а также проявляет по крайней мере одну эффекторную функцию, выбираемую из группы, состоящей из ADCC, CDC и ADCP.
В конкретных вариантах осуществления этого аспекта настоящего изобретения может использоваться любое из описываемых здесь антител против CD70, которые связываются с CD70 человека с очень высокой аффинностью. В предпочтительных вариантах осуществления может использоваться антитело, включающее происходящий от ламы Fab 27B3, или один из его приведенных к зародышевому типу вариантов, включая, в частности, 41D12 (ARGX-110). В особенно предпочтительных вариантах осуществления используется антитело, включающее Fab-области 41D12 (приведенного к зародышевому типу варианта 27B3), слитые с константными областями IgG1 человека. В последнем варианте осуществления можно, кроме того, сконструировать (например, посредством добавления точечных мутаций) константную область IgG1 человека для максимизации эффекторных функций антитела, или антитело 41D12-IgG1 может быть нефукозилированным, снова для увеличения эффекторной функции антитела.
Поскольку обоснование применения плохо интернализующихся антител против CD70 для истощения CD70-экспрессирующих клеток опирается на эффекторные функции антител, предпочтительно, когда антитела, используемые в этом аспекте настоящего изобретения, не являются иммуноконъюгатами, в которых антитело против CD70 связано с фармацевтическим агентом, цитотоксическим агентом, цитостатическим агентом, составляющей в виде лекарственного средства и т.д. Действительно, особым преимуществом этого аспекта настоящего изобретения является отмена необходимости использовать конъюгаты антитело-лекарственное средство, которая является желательной и с точки зрения безопасности пациента (отменяет введение потенциально токсичных агентов), и с экономической точки зрения (отменяет дорогостоящий синтез конъюгатов антитело-лекарственное средство).
Использование для уничтожения клеток конъюгатов антитело-лекарственное средство со значительной интернализацией антител против CD70
В примерах настоящего изобретения демонстрируется, что существуют значительные различия от одной клетки к другой в степени интернализации антител против CD70, и что эти различия в значительной степени зависят от типа клетки, а не от природы антитела против CD70 (хотя различия особенно заметны при использовании антитела ARGX-110). В случае тех типов клеток, которые демонстрируют значительную степень интернализации связавшегося антитела против CD70, т.е. составляющую 30% или более, предпочтительно 40% или более, более предпочтительно 50% или более, или даже 60% или более интернализацию через 6 часов, альтернативная стратегия лечения может основываться на использовании конъюгатов антитело против CD70-лекарственное средство.
Соответственно, дальнейший важный аспект настоящего изобретения относится к способу истощения CD70-экспрессирующих клеток у являющегося человеком пациента, включающему введение указанному пациенту эффективного количества конъюгата антитело-лекарственное средство, включающего антитело, которое связывается с CD70 человека, и составляющую в виде цитотоксического или цитостатического агента, причем указанные CD70-экспрессирующие клетки демонстрируют составляющую 30% или более, 40% или более, 50% или более, или 60% или более интернализацию указанного антитела, которое связывается с CD70 человека, после периода времени, составляющего 6 часов.
В конкретных вариантах осуществления в CD70-экспрессирующие клетки может интернализоваться между 40% и 70%, предпочтительно между 50% и 70% антитела против CD70, которое лежит в основе конъюгата антитело-лекарственное средство, в пределах периода времени, составляющего 6 часов.
В этом способе конъюгат антитело против CD70-лекарственное средство используется для истощения CD70-экспрессирующих клеток. Это истощение CD70-экспрессирующих клеток может лежать в основе терапевтического или профилактического лечения, например, лечения или предупреждения CD70-экспрессирующих раков, или лечения или предупреждения связанных с CD70 иммунологических нарушений. Точный механизм истощения CD70-экспрессирующих клеток может зависеть от природы конъюгата антитело-лекарственное средство, т.е. от того, является ли составляющая в виде «лекарственного средства» цитотоксической или цитостатической, но будет зависеть от сильного (с высокой аффинностью) связывания антитела против CD70 с CD70 на клеточной поверхности, вместе со значительной интернализацией связавшегося антитела.
В некоторых вариантах осуществления CD70-экспрессирующими клетками являются CD70-экспресссирующие раковые клетки. В частности, CD70-экспрессирующие раковые клетки могут относиться к типу рака, выбираемому из группы, состоящей из Т-клеточной лимфомы кожи, множественной миеломы, почечноклеточной карциномы, астроцитомы, меланомы и карциномы яичника. Было установлено, в линии клеток, происходящие из каждого из этих раков, интернализуется более 30% связавшегося антитела против CD70 в течение периода времени, составляющегося 6 часов. В дальнейшем варианте осуществления CD70-экспрессирующие раковые клетки могут относиться к типу рака, выбираемому из группы, состоящей из почечноклеточной карциномы, астроцитомы, меланомы и карциномы яичника. Было установлено, что в линии клеток, происходящие из каждого из этих раков, интернализуется более 50% связавшегося антитела против CD70 в течение периода времени, составляющегося 6 часов.
В конкретных вариантах осуществления этого аспекта настоящего изобретения конъюгат антитело-лекарственное средство может основываться на любом из описываемых здесь антител против CD70, которые связываются с CD70 человека с очень высокой аффинностью. В предпочтительных вариантах осуществления конъюгат антитело-лекарственное средство может основываться на антителе, включающем происходящий от ламы Fab 27B3, или один из его приведенных к зародышевому типу вариантов, включая, в частности, 41D12 (ARGX-110). В данной области техники в целом известны особенности конъюгатов антитело-лекарственное средство, предназначенных для терапевтического применения для людей, в частности, что касается природы составляющей в виде «лекарственного средства», например, цитотоксического или цитостатического агента, и способа соединения составляющей в виде лекарственного средства с составляющей в виде антитела против CD70. Примеры конъюгатов антитело против CD70-лекарственное средство приведены, например, в WO 2004/073656.
Фармацевтические композиции
В объем настоящего изобретения включены фармацевтические композиции, содержащие одно антитело против CD70 по настоящему изобретению, или его антигенсвязывающий фрагмент, или их комбинацию, приготовленные с использованием одного или более фармацевтически приемлемых носителей или наполнителей. Такие композиции могут включать одно антитело против CD70 или их комбинацию (например, два или более различных антител).
Методы приготовления антител в виде препаратов для терапевтического применения для людей хорошо известны в данной области техники и проанализированы, например, в Wang et al., Journal of Pharmaceutical Sciences, Vol.96, pp1-26, 2007.
Включение посредством ссылки
В вышеприведенном описании и на всем протяжении следующих примеров приводятся различные публикации, каждая из которых включена сюда посредством ссылки в ее полном объеме.
Примеры
Настоящее изобретение будет еще понятнее с учетом следующих не ограничивающих экспериментальных примеров.
Пример 1: Иммунизация лам
Иммунизацию лам и получение лимфоцитов периферической крови (PBL), а также последующую экстракцию РНК и амплификацию фрагментов антитела выполняли, как описано De Haard и его коллегами (De Haard H, et al., J. Bact. 187: 4531-4541, 2005). Двух лам иммунизировали CD70-экспрессирующими клетками 786-О (ATCC-CRL-1932) и двух лам - CD70-экспрессирующими клетками Raji (ATCC-CCL-86). Клетки были свежеприготовленными для каждой иммунизации и подтвержденными в отношении экспрессии CD70 с помощью анализа с использованием FACS. Иммунизацию лам осуществляли с использованием приблизительно 107 живых клеток, вводимых внутримышечно в шею, один раз в неделю в течение шести недель. Неполный адъювант Фрейнда инъецировали в мышцы шеи в нескольких сантиметрах от места клеточной иммунизации.
Образцы крови объемом 10 мл отбирали до и после иммунизации для исследования иммунного ответа. Сыворотки от лам проверяли на присутствие антител против рекомбинантного CD70 (Flag-TNC-CD70) с помощью ELISA до иммунизации (в день 0) и после иммунизации (в день 55 и 69). Следует отметить, что в качестве антитела для выявления использовали козье антитело против IgG1/2 ламы (Bethyl, A160-100P), которое не выявляло различия между обычными антителами и антителами в виде тяжелых цепей, что означает, что определяется ответ на CD70 от всех IgG. Результаты представлены на фиг. 1.
Через три-четыре дня после последней иммунизации 400 мл крови отбирали для экстракции тотальной РНК из PBL, используя градиент плотности фиколла-пака для выделения PBL и способ, описанный в Chomczynski P, et al., Anal. Biochem. 162: 156-159, 1987, для приготовления РНК. В среднем достигались выходы РНК, составляющие 450 мкг, из которых 80-мкг аликвоту использовали для неспецифического синтеза кДНК и последующей амплификации генных сегментов VHCH1, VλCλ и VκCκ.
Пример 2: Конструирование библиотек
Конструировали независимые библиотеки VλCλ и VκCκ, используя двухстадийную РНК, в случае которой выполняли 25 циклов с использованием немеченых праймеров, а затем 10 циклов, используя меченый вариант этих праймеров (De Haard H, et al., Biol. Chem. 274, 1999). Параллельно создавали библиотеки VHCH1, используя тот же подход. Размеры отдельных библиотек находились между 107 и 1010 бляшкообразующих единиц. Затем легкую цепь библиотек VλCλ и VκCκ реклонировали по отдельности в VHCH1-экспрессирующий вектор для создания, соответственно, библиотек «Лямбда» и «Каппа» Fab ламы. Альтернативно, расщепленные ампликоны VHCH1 непосредственно клонировали в библиотеки VλCλ и VκCκ, избегая конструирования отдельной библиотеки VHCH1. Поскольку с помощью перетасовки легких цепей, как правило, получают варианты с большей аффинностью, чем с помощью перетасовки тяжелых цепей, было выбрано создание библиотек предварительно клонированных легких цепей и непосредственное клонирование ампликонов тяжелых цепей в эти первичные репертуары.
Размеры конечных библиотек фагового дисплея находился между 5×108 и 2×109 бляшкообразующих единиц. Контроль качества библиотек путем расчета процента клонов, содержащих полноразмерные Fab, обычным образом выполняли, используя ПЦР.
Пример 3: Отборы и скрининг
Два-три цикла отборов фагов осуществляли с использованием или захваченного рекомбинантного CD70 (Flag-TNC-CD70), или непосредственно использованного для покрытия CD70, используя стандартные протоколы. Элюирование связавшегося фага осуществляли с помощью трипсина.
Исходя из отобранного фага, трансфицированного в TG1, наугад отбирали отдельные колонии для выращивания в 96-луночных планшетах. После индукции с помощью IPTG в соответствии со стандартными протоколами готовили периплазматические фракции, содержащие Fab. При всех отборах и для всех четырех лам устанавливали несколько клонов, которые были способны экспрессировать Fab, способные к ингибированию взаимодействия между CD27 и CD70 в соответствии с определением с помощью ELISA ингибирования (фиг. 2 и 3). В этом анализе Flag-TNC-CD70 захватывали с помощью мАт против Flag, и связывание CD27-Fc выявляли через посредство биотинилированного мАт против CD27 и стрептавидина-HRP в отсутствие или в присутствии Fab против CD70 (фиг. 2 и 3) или мАт против CD70 (фиг. 4). На фиг. 2B можно видеть, что 27B3, которое содержит VH, идентичную таковой 9D1, но содержит слегка отличную легкую цепь, как представляется, блокирует связывание CD27 с CD70 более эффективно, что согласуется с меньшей скоростью диссоциации этого Fab, как определено с использованием Biacore (не представленные данные). В таблице 3 представлена эффективность блокирования (представленная в виде IC50) для различных химерных лама-человек мАт, проверенных в ELISA ингибирования с использованием CD70. Fab 27B3 характеризуется IC50, составляющей приблизительно 0,4 нМ, которая даже, как представляется, уменьшается после приведения к формату - химерный лама-человек IgG1, приводящему к IC50, составляющей 0,25 нМ, тогда как мАт 9D1 с другой легкой цепью является в 3 раза менее эффективным (0,73 нМ). Фиг. 3 является совокупностью эксперимента, в котором проверяли Fab ламы против CD70 на блокирующую активность (в ELISA ингибирования, смотрите белые прямоугольники) и на связывание с CD70 (ELISA связывания, черные прямоугольники). Для ELISA связывания Flag-TNC-CD70 захватывают с помощью мАт против Flag, и связывание Fab выявляют, используя мАт против myc, конъюгированное с HRP, которое распознает метку MYC, слитую с C-концом Fd-фрагмента. С целью сравнения параллельно проверяли известные моноклональные антитела против CD70 (SGN70, описанное в US 2010/0129362 A1, клон 1F6; и MDX69A7, описанное в WO2008/074004, фиг. 5A и 5B). Было идентифицировано множество происходящих от ламы Fab-клонов, которые демонстрируют только связывание в ELISA и являются не блокирующими (смотрите фиг. 3 - например, Fab-клоны 45H8, 45C4, 46A2, 46G7, 46E4, 46F12 и 46G12). Представляет интерес клон 59D10, который в виде Fab не обладает блокирующей активностью, но после превращения в химерный IgG был антагонистическим (смотрите таблицу 3). Аминокислотные последовательности VH- и VL-доменов связывающих и блокирующих, происходящих от ламы Fab-клонов представлены здесь в другом месте (таблицы 6-9).
Эффективность блокирования, представленная в виде IC50 (нг/мл), для отдельных SIMPLE антител (мАт) и эталонов сравнения SGN70 и MDX69A7, определенная в ELISA ингибирования с использованием CD70, используя рекомбинантный CD70 человека.
мАт 27B3 и 57B6 также проверяли на их способность к ингибированию взаимодействия между рекомбинантным CD70 макака-резус (FLAG-TNC-резус CD70) и CD27. Значение IC50, определенное с помощью ELISA ингибирования, описанного выше, составляло 150 и 135 нг/мл, соответственно.
Пример 4: Аффинность к CD70 человека и макака-резус
Скорости диссоциации очищенных, происходящих от ламы блокирующих Fab против CD70 определяли, используя Biacore. Рекомбинантный CD70 человека подвергали иммобилизации на чипе CM5 Biacore. Иммобилизацию выполняли в соответствии со способом, предоставляемым Biacore, и посредством использования набора NHS/EDC (Biacore AB): после активации чипа готовили 50 мкг/мл раствор рекомбинантного CD70 в 10 мМ ацетатном буфере с pH=5, и инъецировали 1 мкл этого раствора (50 нг), что приводило к поверхностной плотности, составляющей приблизительно 1000 RU.
Fab в случае мАт против CD70 ARGX-110, MDX2H5 и SGN70 готовили посредством расщепления трипсином (SGN70 и MDX69A7 описаны здесь в другом месте, MDX2H5, также называемое MDX1411, описано в WO2008/074004, фиг. 1A и 1B). Fab, в концентрации, составляющей приблизительно 100-400 мкг/мл, разводили в 6 раз в Hepes-забуференном солевом растворе (0,1 M Hepes, 1,5 M NaCl, 30 мМ EDTA, 0,5% (в объемном отношении) поверхностно-активного вещества P20). Их инъецировали (30 мкл) и пропускали через проточные ячейки со скоростью потока, составляющей 30 мкл/мин. После связывания Fab с CD70 за скоростью диссоциации следили в течение периода времени, составляющего 10 минут. После диссоциации поверхности проточных ячеек регенерировали посредством инъекции 5 мкл 10 мМ NaOH. Иногда требовались многократные инъекции NaOH для регенерации поверхностей в зависимости от аффинности Fab. Расчет скоростей диссоциации выполняли, используя программное обеспечение BIAevaluation. Сначала вычитали сенсограмму для контрольных анализов без покрытия из тех, которые были получены с использованием покрытой проточной ячейки. Затем скорость диссоциации определяли для интервала времени, равного 10 минутам, используя приложение для подгонки кинетики, и рассчитывали значение Kd. Кроме того, также была определена скорость диссоциации CD27 от CD70. Скорости диссоциации суммированы в таблице 4.
Все исследованные происходящие от ламы Fab связываются с видимыми очень низкими скоростями диссоциации (т.е. с высокой аффинностью) с рекомбинантным CD70 человека и с рекомбинантным CD70 макака-резус, основываясь на скоростях диссоциации в Biacore в диапазоне 0,4-4,8×10-4 сек-1. Скорости диссоциации происходящих от ламы Fab от CD70 человека намного ниже таковых полученных из эталонных мАт Fab (7-30×10-4 сек-1). Установлено, что происходящие от ламы Fab с самыми низкими скоростями диссоциации имеют скорости диссоциации, сравнимые с таковой CD27 в случае его взаимодействия с CD70 (0,8×10-4 сек-1). Следует отметить, что нижний предел определения скоростей диссоциации, используя Biacore, составляет приблизительно 0,4×10-4 сек-1. Отсюда происходящие от ламы Fab со скоростями диссоциации, находящими вблизи этого предела, определенными с помощью Biacore, могут, в действительности, демонстрировать даже более высокую аффинность к CD70.
Скорости диссоциации происходящих от ламы Fab и эталонных Fab от CD70 человека или макака-резус
CD70 человека
CD70 макака-резус
Пример 5: Связывание химерных лама-человек мАт против CD70 человека с CD70-позитивными клетками
VH и VL представляющих интерес, происходящих от ламы Fab-клонов сливали с константными областями IgG1 человека и с Cλ человека (все вариабельные области легкой цепи происходят от ламы) и продуцировали в виде двухвалентных химерных моноклональных антител в системе, описанной в заявке на патент WO 2009/145606. Химерные лама-человек мАт очищали на белке A с последующей гель-фильтрацией.
Клетки 786-О инкубировали с рядом разведений 1/5 химерных лама-человек ингибирующих мАт против CD70 (20 мкг/мл-0,25 нг/мл), и связывание мАт с клетками выявляли, используя антитело против Fc человека, конъюгированное с FITC (разведенный 1/500 конъюгат AF006 от поставщика Binding Site). Флуоресценцию 10000 клеток/условие измеряли, используя проточный цитометр, и строили графики медианы флуоресценции. Результаты представлены на фиг. 5. Большинство мАт связываются с высокой аффинностью с клетками 786-О. SIMPLE антитело 1C2, которое в виде Fab, как установлено, характеризуется наименьшей скоростью диссоциации и наибольшей эффективностью блокирования, было не способно к распознанию представленного на клеточной поверхности CD70 (не представленные данные) и поэтому не исследовалось более. SIMPLE антитело 27B3 обладало наибольшей аффинностью к связанному с клеткой рецептору с EC50, составляющей 0,24 нМ.
В дальнейшем эксперименте происходящие из NHL клетки MHH-PREB-1 (105 клеток) инкубировали с градиентом концентрации химерных лама-человек ингибирующих мАт против CD70 (20 мкг/мл-0,25 нг/мл), и связывание мАт с клетками выявляли, используя антитело против IgG человека, конъюгированное с FITC. Результаты представлены на фиг. 5B. Значения EC50 для 57B6 и 59D10 составляли 83 и 74 нг/мл, соответственно.
Пример 6: Ингибирование в экспериментах с использования сокультивирования с помощью химерных лама-человек мАт против CD70
Для определения эффективности блокирования в анализе на основе клеток использовали систему, описанную Wyzgol и его коллегами (J. Immunol. (2009) 183: 1851-1861). Линия клеток фибросаркомы HT1080, трансфицированная CD27 человека, секретирует IL-8 после лигирования с тримерным рекомбинантным CD70, и блокирование этого взаимодействия с помощью нейтрализующего мАт можно определить по уменьшению уровней цитокина. Использовали модифицированный вариант анализа, в котором линия клеток Raji В-клеточной лимфомы служила в качестве источника CD70, хотя молекула представлена на клеточной поверхности и, следовательно, является более соответствующей, чем растворимый CD70.
Клетки Raji подвергали культивированию и проверяли на экспрессию CD70 с помощью анализа с использованием FACS. Затем клетки смешивали в 96-луночном планшете для культивирования тканей (50000 клеток/лунку) с мАт против CD70 и инкубировали при комнатной температуре в течение 1 часа. Клетки HT1080-CD27 добавляли (10000 клеток/лунку) и тщательно перемешивали. Сокультуры переносили в термостат и выращивали в течение ночи при 37°C. Супернатанты собирали и анализировали на содержание в них IL-8 с помощью ELISA. Результаты для SIMPLE антител представлены на фиг. 6 и сравнены с эталонными мАт против CD70. Значения IC50 для химерных лама-человек мАт против CD70 находятся между 3 и 517 нг/мл по сравнению с 42 нг/мл для SGN70 и 143 нг/мл для MDX69A7 (смотрите таблицу 5). SIMPLE антитела 27B3 (IC50=33 пМ) и 9G2 (IC50=20 пМ) являются в 10-20 раз более эффективными, чем эталон SGN70 (IC50=370 пМ), который, с другой стороны, является в 3 раза эффективнее эталона MDX69A7 (1 нМ). Поэтому можно сделать вывод, что химерные лама-человек мАт являются очень эффективными в блокировании взаимодействия между CD27 и CD70 и превосходят эталонные антитела в этом в высокой степени релевантном биоанализе.
Нейтрализующая активность, представленная в виде значения IC50 (нг/мл), химерных лама-человек мАт против CD70 и эталонных мАт, проверенных в анализе с использованием сокультуры на основе клеток Raji
Пример 7: Относящаяся к антителозависимой клеточноопосредованной цитотоксичности активность
Антителозависимую клеточноопосредованную цитотоксичность (ADCC) определяли, используя стандартный анализ высвобождения Cr51, который был описан McEarchern и др. (Blood (2007) 109: 1185-1192). Мононуклеарные клетки периферической крови человека (PBMC) очищали от гепаринизированной цельной крови с помощью стандартного разделения с использованием фиколла и использовали в качестве источника NK-клеток (клеток - природных киллеров) (т.е. эффекторных клеток). Использовали кровь нескольких отдельных доноров. Клетки суспендировали в концентрации 2×106/мл в среде, содержащей 200 Е/мл IL-2 человека (для стимуляции NK-клеток), и инкубировали в течение ночи при 37°C. На следующий день прикрепленные и неприкрепленные клетки собирали и промывали один раз в среде для культивирования. Клетки-мишени представляли собой клетки 786-О (RCC) и были помечены Cr51. Отношение мишень:эффекторная клетка составляло 1:20, и инкубацию выполняли в течение 2 часов с антителом, присутствующим в различных концентрациях, в среде для культивирования. Химерные лама-человек мАт, а также эталонные антитела проверяли в отношении относящейся к ADCC активности на клетках 786-О в различных концентрациях.
Процент лизиса определяли с помощью уравнения: % лизиса = (число импульсов в минуту в образце - число импульсов в минуту в случае самопроизвольного высвобождения)/(число импульсов в минуту в случае максимального высвобождения - число импульсов в минуту в случае самопроизвольного высвобождения)×100.
Результаты показывают, что все из химерных лама-человек мАт индуцируют лизис клеток-мишеней через относящуюся к ADCC активность (фиг. 7). SIMPLE мАт 27B3 обладает активностью, составляющей приблизительно 1 мкг/ил, и, следовательно, является в 10 раз более сильным, чем вариант аффинности 9D1, который характеризуется IC50, сравнимой с таковой эталона SGN70, который, с другой стороны, является в 2,5 раз более сильным в ADCC, чем эталон MDX69A7. В SIMPLE антителе 27B3 наибольшая эффективность блокирования в классе рецептор-лиганд сочетается с лучшей активностью ADCC, и, следовательно, оно имеет подходящий терапевтический профиль.
Пример 8: Относящаяся к комплементзависимой цитотоксичности активность специфических для CD70 мАт
Относящиеся к комплементзависимой цитотоксичности (CDC) характеристики антител определяли в стандартном анализе, описанном McEarchern и др. (Blood (2007) 109: 1185-1192). В первом контрольном эксперименте клетки U266 (MM) и MHH-PREB1 (NHL) были проверены на присутствие антигена CD70 с помощью анализа с использованием FACS. В следующем эксперименте химерное лама-человек мАт против CD70 проверяли в отношении относящейся к CDC активности на обеих линиях клеток в 3 различных концентрациях и в присутствии 3, 6 или 9% сыворотки человека (в качестве источника комплемента человека). Был сделан вывод, что клетки U266 дают наилучшее отношение сигнал/шум в присутствии 9% сыворотки.
В следующем эксперименте клетки U266 смешивали с химерными лама-человек мАт (фиг. 8, верхняя панель) или эталонными мАт против CD70 (фиг. 8, нижняя панель) в концентрации 0,001-20 мкг/мл в отсутствие или в присутствии 9% сыворотки человека и инкубировали в течение 2 ч при 37ºC. Клетки осаждали, и добавляли йодид пропидия для определения количества мертвых клеток. Лизис клеток определяли в FACS. Относящаяся к CDC активность была подтверждена в случае всех химерных лама-человек мАт на клетках U266 в присутствии 9% сыворотки человека. SIMPLE антитело 27B3 характеризуется IC50, схожей с таковой эталона SGN70, в то время как MDX69A7 снова обладает в 10 раз меньшей активностью.
Пример 9: Относящаяся к антителозависимому клеточному фагоцитозу активность
Для анализа относящихся к антителозависимому клеточному фагоцитозу (ADCP) характеристик SIMPLE мАт осуществляли анализ, описанный McEarchern и др. (Blood (2007) 109: 1185-1192). В этом анализе мононуклеарные клетки периферической крови человека (PBMC) очищали от гепаринизированной цельной крови с помощью стандартного разделения с использованием фиколла. Использовали кровь нескольких отдельных доноров. Клетки инкубировали в RPMI, содержащей 10% FCS, в матрасе для культивирования тканей в течение ночи при 37°C. Не прикрепленные клетки удаляли, а прикрепленные клетки культивировали в течение 15 дней в среде 75mlX-VIVO 15, содержащей 500 Е/мл rhGM-CSF (7,5 мкг/75 мл). Через 15 дней собирали прикрепленные клетки (происходящие от моноцитов макрофаги (MDM).
8,0×104 клеток-мишеней (клеток 786-О), нагруженных красным красителем PKH26, инкубировали с мАт в течение 30 минут в 96-луночном планшете с V-образным дном. Затем добавляли 2,0×104 клеток MDM (отношение мишень:эффекторная клетка (786-О:MDM)=4:1), и после инкубации в течение 1 часа при 37°C клетки подвергали центрифугированию, ресуспендировали в 100 мкл PBS с 1% BSA и переносили в 96-луночный планшет с V-образным дном. Моноклональное антитело против CD11b (конъюгированное с FITC) для окрашивания макрофагов добавляли на 30 мин при сохранении планшетов на льду, клетки затем промывали дважды PBS и фиксировали с использованием 1% параформальдегида (в PBS). Образцы анализировали с помощью FACS, в котором фагоцитоз оценивали, исходя из двояко окрашенных клеток. Результаты представлены на фиг. 9, и они показывают, что все исследованные химерные лама-человек мАт являются эффективными в индукции ADCP. SIMPLE антитело 27B3 индуцирует фагоцитоз вплоть до 50% меченых опухолевых клеток макрофагами в концентрациях, составляющих 1 мкг/мл, с IC50, составляющей приблизительно 100 нг/мл (0,4 нМ).
Пример 10: Интернализация мАт против CD70 в линию опухолевых клеток 786-О
Было установлено, что связывание антител против CD70 с линией клеток, происходящих из карциномы почки, 786-О приводит к быстрой (в пределах 1 часа) интернализации комплекса антитело-рецептор (Adam et al., Br J Cancer (2006) 95: 298-306). Для исследования SIMPLE антител и эталонных мАт, направленных против CD70, в отношении интернализации клетки 786-O культивировали в 96-луночном титрационном микропланшете и инкубировали в течение ночи при 37°C. 2,5 мкг/мл мАт добавляли и инкубировали с клетками в течение 0-24 часов при 37°C. Планшеты промывали 3 раза по 5 мин буфером для удаления (150 мМ NaCl, 100 мМ глицин, pH=2,5, при этом закодированный «В» представляет количество мАт, подвергнутого интернализации через посредство CD70), или PBS (при этом закодированный «Вне» представляет количество мАт, связавшегося с рецептором на внешней части клетки). Впоследствии клетки фиксировали с использованием 4% параформальдегида в течение 30 мин при комнатной температуре, промывали PBS и инкубировали в течение 5 мин с 0,2% Triton-X-100 («В») или PBS («Вне»). Затем клетки дважды промывали и инкубировали при комнатной температуре в течение 10 мин с 100 мМ глицином с последующей инкубацией в течение 30 мин с PBS+1% BSA. Наконец, клетки окрашивали козьим антителом против Fc человека (Jackson immunoresearch 109-005-098) и антикозьим Ат IRDYE800 (Li-cor 926-32214) до анализа на инфракрасном сканере Li-Cor Odyssey. Антитело, остающееся на внешней части клетки, т.е. не интернализованное, оценено в виде средней MFI «Вне» на фиг. 10. Результаты показывают, что некоторые мАт остаются дольше на поверхности клеток, чем другие мАт (больший сигнал). Однако ни одно из мАт не интернализуется полностью: в начале (между 0 и 2 часами) интернализация мАт идет очень быстро, но затем, по-видимому, достигается стационарная фаза, в которой 30-40% мАт остаются на внешней части клетки, даже после инкубации в течение 24 часов при 37°C. SIMPLE антитело 27B3 интернализуется настолько же быстро, как эталонные антитела SGN70 и MDX69A7. Интересно отметить, что SIMPLE антитела 9E1 и 19G10, но также и другие, остаются на высоких уровнях на внешней части клеток, даже после инкубации в течение 24 часов (нижняя панель фиг. 10). Для эффективного вызова эффектов, относящихся к ADCC, CDC и ADCP, важно, чтобы высокие уровни специфических для CD70 антител сохранялись на внешней части целевых раковых клеток для рекрутмента эффекторных иммуноцитов, которые ответственны за уничтожение клеток. С другой стороны, быстро интернализующиеся антитела перспективны в виде конъюгатов антитело-лекарственное средство (ADC).
Пример 11: In vivo эффективность химерных лама-человек мАт в модели с использованием ксенотрансплантата опухоли
Для порождения диссеминированного заболевания 106 клеток Raji в 0,2 мл PBS инъецировали в латеральную хвостовую вену мышей с тяжелой комбинированной иммунной недостаточностью (SCID) C.B.-17 (Harlan, Indianapolis, IN). После инъекции всех мышей объединяли, а затем размещали по случайному закону в различные группы лечения, с 9 мышами в каждой группе. Антитела вводили в 0,5 мл в день 1, 4, 8, 11, 15 после имплантации клеток с помощью внутрибрюшинного введения. За мышами наблюдали по крайней мере дважды в неделю, и их умерщвляли, когда они демонстрировали признаки заболевания, включая составляющее 15%-20% уменьшение веса, согнутую позу, вялость, краниальную опухоль или дегидратацию. Мыши получали лечение мАт 41D12 в зависимости доза-эффект. Кроме того, вариант с недействующим Fc 41D12 исследовали в дозе 10 мг/кг. Вариант недействующим Fc был описан ранее в литературе и не обладает относящейся к ADCC и CDC активностью и намного более низкой активностью, относящейся к ADCP (McEarchern et al., Clin Cancer Res (2008) 14(23): 7763-7772). Результаты представлены в виде кривых выживания на фиг. 11. Эти данные показывают, что медиана времени выживания в случае контрольных мышей равна 27 дням. Медиана времени выживания увеличивалась до по крайней мере 67 дней в случае введения доз, равных или превышающих 0,1 мг/кг 41D12. Доза, составляющая 0,01 мг/кг, является все еще эффективной. Вариант с недействующим Fc 41D12 не обладает эффективностью in vivo, что показывает важность ADCC, CDC и ADCP в этой модели.
Пример 12: Экспрессия нефукозилированных мАт
Было установлено, что антитела с уменьшенным числом фукозильных остатков увеличивают ADCC в 10-1000 раз (Iida et al., Clin Cancer Res. (2006) 12(9): 2879-2887). В линии клеток CHO Ms704-PF и MS705, у которых отсутствует ген фукозилтрансферазы (FUT8, BioWa, Inc.), вводили посредством электропорации вектор для экспрессии в эукариотических клетках, кодирующий тяжелую и легкую цепи химерных лама-человек мАт против CD70. Устойчивые к лекарственному средству клоны отбирали по росту в среде для СНО Excell 325-PF. Клоны скринировали на предмет продукции IgG с помощью стандартного ELISA связывания с CD70.
Последовательность специфического для CD70,
происходящего от ламы VH
Последовательность специфического для CD70, происходящего от ламы VL (Q, выделенная жирным шрифтом, кодируется терминирующим кодоном)
Полноразмерный, происходящий от ламы VH
Полноразмерный, происходящий от ламы VL (Q, выделенная жирным шрифтом, кодируется терминирующим кодоном)
Пример 13: Приведение к зародышевому типу мАт против CD70 27B3
Генные сегменты зародышевой линии человека с одинаковой канонической складчатой структурой и наибольшей степенью идентичности аминокислотной последовательности с VH- и VL-областями мАт 27B3 были идентифицированы в результате сравнения с известными последовательностями генов зародышевой линии человека. Ближе всего соответствующими мАт 27В3 VH- и VL-областями человека были VH3-48 человека (с составляющей 89,7% идентичностью по FR) и VL8-61 человека (с составляющей 86,1% идентичностью по FR), соответственно. Ближе всего соответствующими последовательностями генных сегментов зародышевой линии человека JH и JL были JH5 и JL7, соответственно. Совмещение последовательностей VH- и VL-областей мАт 27B3 с ближе всего соответствующими последовательностями VH и VL зародышевого типа человека представлено на фиг. 12. В результате сравнения V-областей SIMPLE антитела 27B3 и последовательностей зародышевой линии человека было идентифицировано 12 возможных мутацией с целью гуманизации в VH-области и 13 в VL-области (2 в CDR1 и 1 в CDR2). Обзор мутаций в целью гуманизации в последовательностях VH и VL 27B3, вместе с необходимыми для охвата вносимого разнообразия размерами библиотек, представлен в таблице 10 и 11.
Намеченные мутации в аминокислотной последовательности VH 27В3. ¶ обозначает размер библиотеки, необходимый для охвата всех возможных мутантов
Намеченные мутации в аминокислотной последовательности VL 27В3. ¶ обозначает размер библиотеки, необходимый для охвата всех возможных мутантов. * обозначает мутации в участках CDR1 или CDR2
Приведенные к зародышевому типу библиотеки VH_27B3 и VL_27B3, охватывающие все комбинации мутаций с целью гуманизации, были созданы с помощью компоновки на основе ПЦР (смотрите, например, Stemmer et al., Gene (1995) 164: 49-53). Перекрывающиеся олигонуклеотиды со специфическими мутациями в определенных положениях были скомпонованы с помощью ПЦР. Библиотека содержала аминокислоты и человека, и ламы в каждом мутированном положении для предотвращения полной утраты связывания в том случае, если остаток ламы дикого типа является важным для связывания с высокой аффинностью (смотрите, например, Baca et al., J. Biol. Chem. (1997) 272: 10678-10684; Tsurushita et al., J. Immunol. Methods (2004) 295: 9-19). Библиотека VH содержала приблизительно 1×1010 клонов, а библиотека VL - приблизительно 8×109 клонов. Библиотеки VH и VL объединяли и реформатировали в одну библиотеку Fab размером, составляющим 1×1010 клонов (91% со вставкой полноразмерного Fab), подходящим для скрининга фагового дисплея. Разнообразие, необходимое для охвата всех возможных мутации, отмеченных в таблицах 10 и 11, превышало таковое в первичных библиотеках тяжелой цепи и легкой цепи, но также объединенная библиотека Fab была достаточно большой для охвата всех возможных перестановок (1,89×107).
Пример 14: Отбор приведенных к зародышевому типу Fab 27B3 с высокой степенью идентичности по FR с последовательностями человека
Фаговый дисплей использовали для отбора приведенных к зародышевому типу Fab 27B3 как с высокой аффинностью к CD70, так и с высокой степенью идентичности по FR с последовательностями человека. А именно, Flag-TNC-CD70 биотинилировали и инкубировали в различных концентрациях с различными количествами экспрессирующего Fab фага. Комплексы фага с Flag-TNC-CD70 затем улавливали на покрытый стрептавидином титрационный микропланшет и промывали небиотинилированным Flag-TNC-CD70 при 37ºC. Фаги элюировали с помощью трипсина и использовали для инфицирования клеток TG1. Выполняли четыре цикла отбора, при этом в каждом цикле увеличивали жесткость отмывки. Детали условий, использованных для каждого цикла отбора, представлены в таблице 12.
Условия отбора приведенных к зародышевому типу Fab 27В3
Фаги, элюированные в циклах 3, 4 и 5 отбора, трансфицировали в TG1 и высевали на чашки в виде клонов; отдельные клоны отбирали наугад для дальнейшего анализа. Периплазматические фракции, содержащие Fab, готовили, исходя из индуцированных с помощью IPTG мелкомасштабных культур, скорость диссоциации каждого Fab определяли, используя покрытый CD70 чип CM5 в анализе связывания с использованием Biacore. Клоны с Fab, характеризующимися скоростями диссоциации, схожими с таковыми 27B3, секвенировали. Скорости диссоциации для клонов с превышающей 94% общей идентичностью с последовательностями FR человека представлены в таблице 13, вместе с процентом идентичности с FR человека индивидуальных клонов (VH, VL и в целом).
Полные аминокислотные последовательности секвенированных Fab-клонов представлены в таблицах 14 и 15, а варианты аминокислотных последовательностей CDR 27B3 представлены в таблице 16. Проверка этих последовательностей показывает, что каркасная область 1 легкой цепи часто содержит дополнительные (нечеловеческие) мутации в районе праймера в аминокислоте 2 (A вместо T). Были созданы варианты клонов 35G3, 40F1 и 39C3 (клоны 53C1, 53B1 и 53E1, соответственно), которые имеют соответствующую человеку аминокислоту в положении 2. Кроме того, клон 41D12 был далее приведен к зародышевому типу посредством объединения тяжелой цепи 41D12 с легкой цепью 40F1 (53A2) или с легкой цепью 53B1 (53H1).
Скорость диссоциации приведенных к зародышевому типу клонов Fab 27В3 с превышающей 94% общей идентичностью с последовательностями FR человека
[10-4 сек-1]
Аминокислотные последовательности VH приведенных к зародышевому типу клонов 27B3 с превышающей 94% общей идентичностью с последовательностями человека, при этом идентификаторы последовательностей разбиты согласно FR и CDR. Мутации с целью гуманизации являются затемненными, а вводимые праймером мутации представлены жирным шрифтом.
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:274
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:248
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:275
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:275
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
SEQ ID NO:2
SEQ ID NO:11
SEQ ID NO:22
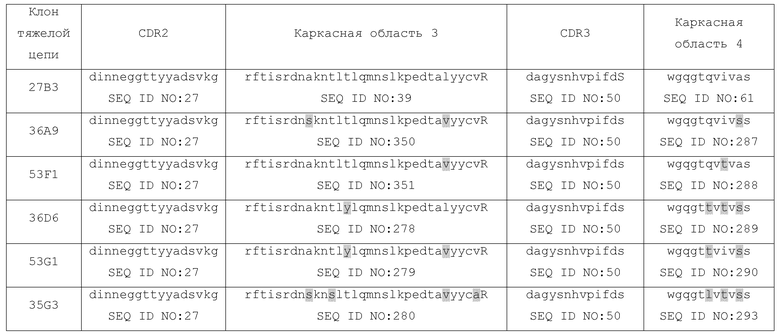


Аминокислотные последовательности VH приведенных к зародышевому типу клонов 27B3 с превышающей 94% общей идентичностью с последовательностями человека, при этом идентификаторы последовательностей для полноразмерной аминокислотной последовательности VH.
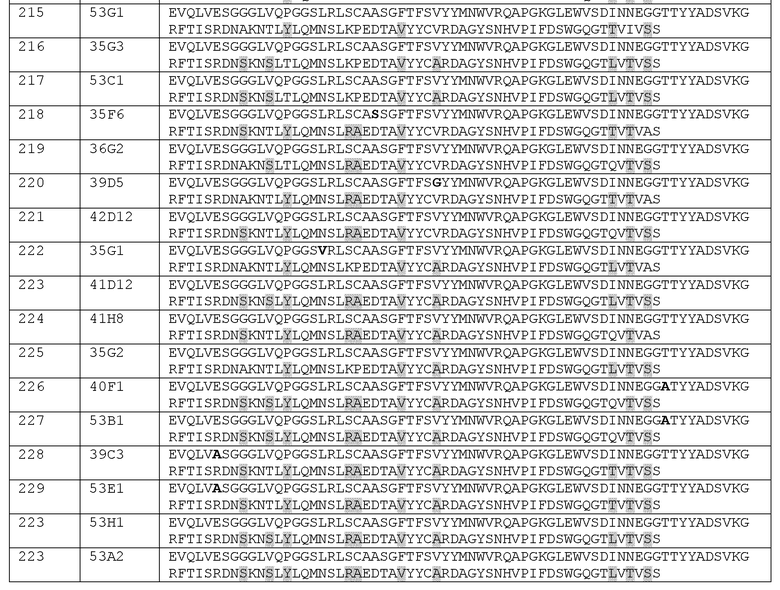
Аминокислотные последовательности VL приведенных к зародышевому типу клонов 27B3 с превышающей 94% общей идентичностью с последовательностями человека, при этом идентификаторы последовательностей разбиты согласно FR и CDR. Мутации с целью гуманизации являются затемненными, а вводимые праймером мутации представлены жирным шрифтом.
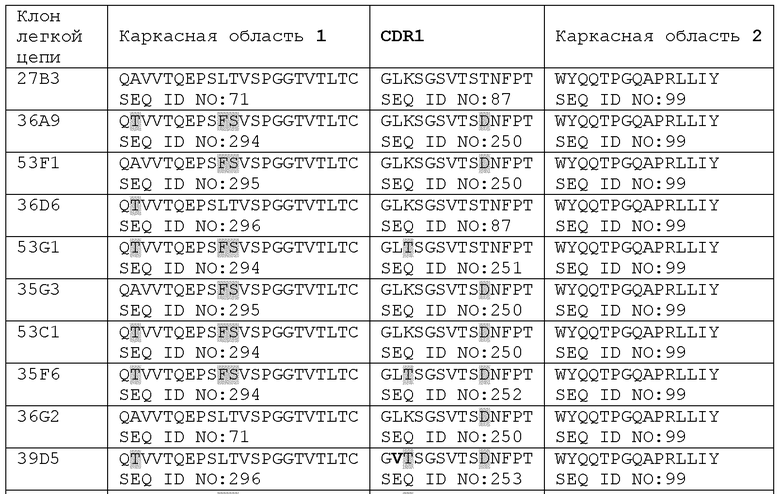


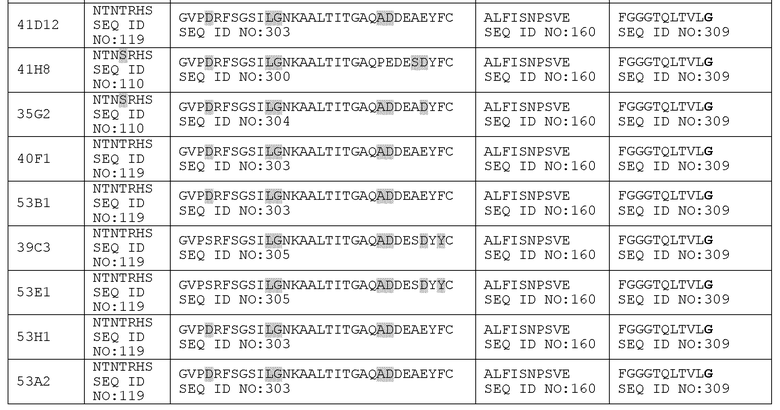
Аминокислотные последовательности VL приведенных к зародышевому типу клонов 27B3 с превышающей 94% общей идентичностью с последовательностями человека, при этом идентификаторы последовательностей для полноразмерной последовательности VL. Мутации с целью гуманизации являются затемненными, а вводимые праймером мутации представлены жирным шрифтом.


Пример 15: Термостабильность приведенных к зародышевому типу вариантов мАт 27B3
Несколько приведенных к зародышевому типу вариантов мАт 27B3 экспрессировали в клетках HEK293 в виде полноразмерных молекул IgG1 человека. После очистки с использованием белка A оценивали термостабильность клонов приведенных к зародышевому типу мАт 27B3. А именно, каждое мАт инкубировали при различных температурах (разделенных составляющими 2 или 5°C интервалами) в течение 1 часа в концентрации, составляющей 100 мкг/мл, в буфере PBS. Температуру мАт затем понижали контролируемым образом (понижение до 25°C в течение 2 часов, с последующей инкубацией в течение ночи при 4°C), а для удаления агрегатов использовали центрифугирование. Количество активного мАт (в виде процента от общей концентрации мАт) в супернатанте определяли с помощью Biacore.
Результаты (представленные в таблице 17) показывают, что большинство клонов имеет температуру плавления (Tm), схожую с таковой для 27B3 дикого типа. Неожиданно было установлено, что клоны 41D12 и 35G2 обладают повышенной термостабильностью по сравнению с 27B3.
Термостабильность приведенных к зародышевому типу
вариантов мАт 27B3
Термостабильность приведенных к зародышевому типу вариантов мАт 27B3: 41D12, 35G2 и 40F1 в дальнейшем исследовали, используя ряд методов. Аликвоты (1 мл) исследуемых мАт в концентрации, составляющей 100 мкг/мл, готовили в PBS Delbucco, содержащем 0,02% Tween. Также готовили аликвоту, состоящую только из буфера (PBS Delbucco, содержащего 0,02% Tween), в качестве отрицательного контроля и аликвоту, состоящую из мАт в PBS Delbucco, содержащем 0,02% Tween, в качестве положительного контроля. В случае каждого препарата мАт аликвоты хранили при 4°C, при комнатной температуре и при 37°C в течение периода времени, составляющего 0-8 недель. Образцы объемом 50 мкл отбирали из аликвот в дни 1, 7, 14, 21, 28, 35, 56 и 92 для анализа. Образец хранили при -20°C и использовали в качестве контрольной точки.
15.1 Анализ образцов с помощью гель-фильтрации
Образцы, отобранные в каждый момент времени, также анализировали с помощью гель-фильтрации, используя колонку Superdex200 10/300 GL. Исследуемый образец объемом 50 мкл добавляли к 200 мкл PBS+0,02% Tween-20. Отбирали образец (125 мкл) для анализа с помощью гель-фильтрации. До анализа образцы центрифугировали для удаления больших агрегатов. Определяли % мономерного пика и площадь под кривой мономерного пика, а также удерживаемый объем. Результаты представлены в таблицах 19, 20 и 21.
Результаты исследования стабильности в PBS - % мономерного пика
Результаты исследования стабильности в PBS - площадь под кривой мономерного пика (mAU*мл)
Результаты исследования стабильности в
PBS - удерживаемый объем (мл)
Результаты анализа с помощью гель-фильтрации образцов 35G2, 40F1 и 41D12, отобранных после инкубации в течение 5 недель при 37°C, представлены на фиг. 13. Два небольших пика видны в случае большего времени удержания (указывающего на присутствие некоторых агрегатов) и меньшего времени удержания (указывающего на присутствие продукта некоторой деградации). Результаты показывают, что большая часть белка является интактной и не агрегирует.
Анализ образцов с помощью BIACORE
Образцы, отобранные в определенные моменты времени, проверяли на активность связывания с CD70, используя Biacore. Образец объемом 50 мкл добавляли к 200 мкл PBS+0,02% Tween-20, и разводили 1/400 следующим образом: 5 мкл образца (1 мг/мл)+195 мкл HBS-EP+ (буфера Biacore), а затем разводили 10 мкл+90 мкл HVS-EP+=2,5 мкг/мл. Анализ с использованием Biacore выполняли, используя покрытый CD70 с высокой плотностью чип CM5 (4000 RU). Результаты для мАт 40F1, 35G2 и 41D12 представлены на фиг. 16. Контрольным образцом является 100% образец.
Результаты показывают, что мАт теряют некоторую активность (аффинность к антигену CD70) со временем после инкубации при 37°C. Для 41D12 графически представлены оба спада и максимальный сигнал (R0) (смотрите фиг. 16C и D).
Затем исследовали стабильность после замораживания-оттаивания приведенных к зародышевому типу мАт 27B3: 41D12, 35G2 и 40F1. По этой причине 0,7-мл аликвоту мАт (в концентрации 5 мг/мл) замораживали в течение по крайней мере 6 часов при -20°C и оттаивали в течение 1 часа при комнатной температуре. Этот цикл повторяли 9x (а значит всего 10 циклов замораживания-оттаивания), и образцы анализировали с помощью гель-фильтрации, как описано выше. Результаты показывают, что мАт являются стабильными после замораживания-оттаивания на протяжении 10 циклов: продукты деградации или агрегаты не обнаруживались при гель-фильтрации.
Результаты исследования стабильности в PBS с помощью гель-фильтрации (слева = значение после 10 циклов замораживания/оттаивания, справа = значение для контрольного образца)
Образцы также проверяли на активность в Biacore, как описано выше. Результаты показывают, что после 10 циклов замораживания-оттаивания мАт все еще обладают 97% своей активности (в случае всех трех проверенных мАт).
Активность в Biacore после 10 циклов
замораживания-оттаивания (%)
Пример 16: Относящаяся к ADCC активность приведенных к зародышевому типу мАт 27B3
Несколько приведенных к зародышевому типу вариантов 27B3 экспрессировали в клетках HEK293 в виде полноразмерных молекул IgG1 человека. После очистки с использованием белка A оценивали относящуюся к ADCC активность приведенных к зародышевому типу мАт 27B3. ADCC определяли, как описано выше в примере 7.
Относительные IC50 для приведенных к зародышевому типу мАт 27B3, относительно IC50 родительского 27B3, приведены в таблице 18 и для относящейся к ADCC активности, и для эффективности блокирования взаимодействия CD70/CD27 в анализе с использованием сокультуры на основе клеток Raji (как описано в примере 6). Цифра <1,0 означает уменьшенную IC50 для приведенного к зародышевому типу варианта относительно родительского 27B3. Исходя из этих данных, можно сделать вывод, что у приведенного к зародышевому типу варианта 41D12 сохранилась относящаяся к ADCC активность, а также блокирующая взаимодействие CD70/CD27 активность вместе с наилучшей Tm.
Относительные IC50 приведенных к зародышевому типу вариантов 27B3 в анализе ADCC на основе 786-О и в биоанализе нейтрализации на основе клеток Raji, относительно IC50 родительского клона мАт
Пример 17
Конструирование и отбор линий клеток, экспрессирующих нефукозилированнное мАт против CD70b, ARGX-110
Создавали вектор с двумя генами, кодирующими аминокислотные последовательности VH и VL приведенного к зародышевому типу клона 27B3-41D12 (аллотипа za). Нуклеотидные последовательности, кодирующие тяжелую цепь (SEQ ID NO: 344) и легкую цепь (SEQ ID NO: 345) 41D12, представлены в таблице 30 ниже.
Клетки CHOK1SV Potelligent® стабильно трансфицировали вектором с двумя генами с помощью электропорации. Выполняли шесть циклов трансфекции. В каждом цикле клетки в результате электропорации добавляли к 200 мл среды с определенным химическим составом без животных компонентов (CDACF) CM63 и высевали в сорок 96-луночных планшетов в объеме 50 мкл на лунку. На следующий день после трансфекции 150 мкл среды CM63, содержащей селективный агент L-метионин сульфоксимин (MSX), добавляли в каждую лунку для обеспечения конечной концентрации MSX, составляющей 50 мкМ. После инкубации в течение приблизительно 3, 4 и 5 недель планшеты скринировали на предмет присутствия колоний, используя отражение клонирования. Любые идентифицированные колонии исследовали дополнительно, используя микроскоп, для определения, произошла ли колония от одной или множества клеток.
Двести сорок пять колоний были идентифицированы и скринированы на предмет продукции антител, используя способ на основе правила октетов. Концентрации антител находились в диапазоне от 0 до 172 мкг/мл. Из 245 подвергнутых скринингу линий клеток 214 были позитивными по продукции антител. Линии клеток упорядочивали на основе продуктивности, и отбирали первые 70 линий клеток. Культуры этих 70 линий клеток вначале наращивали в статической культуре, а впоследствии в суспензионной культуре. Тридцать линий клеток, которые продемонстрировали приемлемый рост, были отобраны и оценены далее при скрининге на предмет продуктивности с использованием периодической культуры во встряхиваемой колбе. Культуры газировали в дни 4, 6, 8 и 10 и собирали в день 12. Концентрацию антитела в образцах собранных супернатантов определяли с помощью HPLC, используя колонку, содержащую белок A. Концентрации антител в образцах 30 линий клеток находились в диапазоне от 109 до 877 мг/л. Результаты оценки с использованием периодической культуры во встряхиваемой колбе использовали для упорядочивания линий клеток, на основе продуктивности. Первые 20 линий клеток были отобраны для дальнейшей оценки.
Относящиеся к росту и продуктивности данные для этих 20 отобранных линий клеток, экспрессирующих антитело ARGX-110, получали в подпитываемых культурах во встряхиваемых колбах, используя среду CDACF, и результаты представлены в таблице 24. Концентрации антител при сборе находились в диапазоне от 857 до 3922 мг/л, как определено на колонке с белком А для HPLC. Линия клеток F13 была выбрана для инокуляции в биореактор с одноразовым мешком на 10 л для клеток. Была достигнута составляющая 4327 мг/л концентрация антитела при сборе. В случае этой линии клеток была достигнута составляющая 2711 мг/л концентрация антитела при сборе в подпитываемой культуре во встряхиваемом колбе, и она имела вторую наибольшую скорость специфической продукции (2,22 пг/клетку/ч) из всех 6 оцененных линий клеток (B1, D4, D5, Fl, F13 и F18). Эта линия клеток также продемонстрировала приемлемые характеристики роста.
Сводка данных, относящихся к росту и продуктивности 20 отобранных линий клеток, экспрессирующих антитело ARGX-110, выращиваемых в подпитываемых культурах в CDACF во встряхиваемых колбах.
(2) Определенная с помощью HPLC на колонке с белком А.
(3) Рассчитанная с помощью линейно-регрессионного анализа концентрации антитела по отношению к интегралу во времени концентрации жизнеспособных клеток.
Пример 18: Аффинность мАт против CD70
18.1 Аффинность к линиям раковых клеток, экспрессирующих CD70
Несколько линий раковых клеток были проверены с помощью анализа с использованием FACS в отношении связывания ARGX-110 и SGN70 (описанного в US2010/0129362) в насыщающих концентрациях мАт (по крайней мере 2 мкг/мл). Это выполняли или при 4°C (при инкубации клеток с мАт в течение 1 часа), или при 37°C (при инкубации клеток с мАт в течение 15 минут). Связывание выявляли, используя антитело против Fc IgG1 человека, конъюгированное с FITC (AF006, Binding Site), или антитело против Fc IgG1 человека, конъюгированное с PE (eBioscience, 12-4998). Флуоресценцию измеряли, используя проточный цитометр. Результаты сведены в таблицу 25. Третий столбец таблицы показывает, связывается ли ARGX-110 с низкой (+), средней (++) или высокой (+++) аффинностью с различными исследуемыми линиями клеток.
В правом столбце полученный в FACS сигнал (MFI) для ARGX-110 разделен на полученный в FACS сигнал (MFI) для SGN70 в случае эксперимента, проведенного при 37°C. Эти результаты показывают, что ARGX-110 связывается с большей аффинностью с клетками, чем SGN70, особенно в случае линий клеток с низкой копийностью CD70, в случае которых можно ожидать, что связывание с высокой аффинностью антитела можно легче уловить.
Связывание мАт против CD70 ARGX-110 и SGN70 с линиями раковых клеток
В дальнейшем эксперименте определяли видимую аффинность мАт против CD70, ARGX-110, SGN70 и MDX1411 [смотрите выше], во всем диапазоне концентраций, к различным линиям клеток. Клетки инкубировали в течение 1 часа при 4°C с рядом разведений связывающихся с CD70 мАт, и связывание выявляли, используя антитело против Fc IgG1 человека, конъюгированное с FITC, или антитело против Fc IgG1 человека, конъюгированное с PE. Флуоресценцию измеряли, используя проточный цитометр, и графически представляли медиану флуоресценции. Результаты представлены на фиг. 15.
Результаты показывают, что аффинность трех различных мАт к CD70 на клетках является сравнимой, но в случае некоторых линий клеток, SU-DHL-6, A549 и MKN45, связывание ARGX-110 лучше такового MDX1411 и SGN70. В случае всех мАт EC50 для связывания с SU-DHL-6, A549 и MKN45 намного больше EC50 для других линий клеток вроде U266 и многих других, что, вероятно, означает, что мАт связывается только одним плечом (Fab), дольше не создавая возможность для проявления авидности и приводя в силу этого к более низким аффинностям. Действительно, при проверке в Biacore аффинность Fab ARGX-110 в сравнение с SGN70 и MDX1411 намного выше (смотрите ниже).
18.2 Аффинность к клеткам от пациентов, экспрессирующим CD70
Первичные клетки, полученные от пациентов с хроническим лимфолейкозом (CLL) (2 пациента, подверженных высокому риску, 1 пациент, подверженный низкому риску), засевали в количестве 250000 клеток/лунку в круглодонные 96-луночные планшеты в RPMI 1640+10% FBS, и добавляли мАт в концентрации, составляющей 5 мкг/мл, в RPMI+10% FBS. После инкубации в течение вплоть до 5 часов при 37°C клетки дважды промывали охлажденным на льду PBS. Меченное Alexa Fluor 647 козье антитело против IgG человека (Invitrogen каталожный # 21445) разводили 1/500 в PBS/1% BSA и инкубировали в течение 20 минут при 4°C. Планшет дважды промывали охлажденным на льду PBS, и 4% параформальдегид добавляли на 15 минут при комнатной температуре для фиксации клеток. Сигналы измеряли с использованием FACS. Результаты представлены на фиг. 16. Результаты показывают, что ARGX-110 хорошо связываются с клетками от пациентов с CLL, тогда как SGN70 почти не дало сигналов, обусловленных связыванием.
18.3 Аффинность Fab против CD70 к рекомбинантному CD70 в Biacore
Рекомбинантный CD70 человека подвергали иммобилизации на чипе CM5 Biacore. Иммобилизацию выполняли в соответствии со способом, предоставляемым Biacore, и посредством использования набора NHS/EDC (Biacore AB): после активации чипа готовили 50 мкг/мл раствор рекомбинантного CD70 в 10 мМ ацетатном буфере с pH=5, и инъецировали 1 мкл этого раствора (50 нг), что приводило к поверхностной плотности, составляющей приблизительно 1000 RU.
Fab в случае мАт против CD70 ARGX-110, MDX1411 и SGN70 готовили посредством расщепления трипсином. Эти Fab, в концентрации, составляющей приблизительно 100-400 мкг/мл, разводили в 6 раз в Hepes-забуференном солевом растворе (0,1 M Hepes, 1,5 M NaCl, 30 мМ EDTA, 0,5% (в объемном отношении) поверхностно-активного вещества P20). Их инъецировали (30 мкл) и пропускали через проточные ячейки со скоростью потока, составляющей 30 мкл/мин. После связывания Fab с CD70 за скоростью диссоциации следили в течение периода времени, составляющего 10 минут. После диссоциации поверхности проточных ячеек регенерировали посредством инъекции 5 мкл 10 мМ NaOH. Иногда требовались многократные инъекции NaOH для регенерации поверхностей в зависимости от аффинности Fab. Расчет скоростей диссоциации выполняли, используя программное обеспечение BIAevaluation. Сначала вычитали сенсограмму для контрольных анализов без покрытия из тех, которые были получены с использованием покрытой проточной ячейки. Затем скорость диссоциации определяли для интервала времени, равного 10 минутам, используя приложение для подгонки кинетики, и рассчитывали значение Kd. Скорости диссоциации сведены в таблицу 26.
Скорость диссоциации Fab в случае связывания с CD70 человека
18.4 Эксперименты по внедрению для оценки лизиса клеток SU-DHL-6, с которыми связались мАт против CD70
Клетки SU-DHL-6 (линии клеток диффузной B-крупноклеточной лимфомы) пипетировали в лунки в количестве 50000 клеток/лунку, которые затем внедряли в 300000 только что выделенных от здоровых доноров PBMC/лунку, и добавляли мАт в различных концентрациях. Образцы инкубировали в течение 2 дней и анализировали с помощью FACS на предмет истощения линии клеток (определяемого с помощью АРС против CD19 человека (eBioscience 17-0199-42)). Наибольшая эффективность в уничтожении клеток была получена с ARGX-110, которая также дает наибольший % лизиса. Результаты представлены на фиг. 17.
Пример 19: Блокирующая взаимодействие CD27-CD70 активность мАт против CD70
Взаимодействие между CD70 и CD27 может вносить вклад в выживание опухолевых клеток, их пролиферацию и/или подавление иммунной системы в опухолевом микроокружении. По этой причине оценивали способность мАт против CD70 к блокированию взаимодействия между CD70 и CD27 с помощью ELISA.
В этом анализе титрационный микропланшет (Nunc Maxisorb) сначала покрывали 100 мкл моноклонального антитела против FLAG M2 (Sigma Aldrich, F3165) в 1250-кратном разведении в PBS (для достижения конечной концентрации, составляющей приблизительно 3,5 мкг/мл) в течение ночи при 4°C. Планшет промывали один раз PBS-Tween и инкубировали при комнатной температуре в течение 2 часов с 300 мкл PBS-1% казеин. Планшет промывали еще три раза PBS-Tween. 100 мкл 5 нг/мл (80 пМ) Flag-TNC-CD70 (The Journal of Immunology, 2009, 183: 1851-1861), разведенного в PBS-0,1% казеин, добавляли в планшет и инкубировали при комнатной температуре в течение 1 часа при встряхивании. Планшет промывали пять раз PBS-Tween. В планшет добавляли исследуемое мАт против CD70; различные концентрации успешно получали посредством разведения маточного раствора антитела в PBS-0,1% казеин. Сразу же после этого добавляли 50 мкл 1 мкг/мл (конечная концентрация =6,5 нМ) рекомбинантной химеры CD27 человека-Fc (R&D systems 382-CD, М.м.=46,5 кДа), и осуществляли инкубацию при комнатной температуре в течение 1 часа при встряхивании. Планшет промывали пять раз PBS-Tween. 100 мкл биотинилированного антитела против CD27 (eBioscience 13-0271), разведенного в 500 раз в PBS-0,1% казеин, добавляли, и планшет инкубировали при комнатной температуре в течение еще одного часа, при встряхивании. Планшет промывали пять раз PBS-Tween. В планшет добавляли 100 мкл стрептавидина-HRP (Jackson Immunoresearch 016-030-084), разведенного в 5000 раз в PBS-0,1% казеин, и планшет инкубировали при комнатной температуре в течение еще одного часа, при встряхивании. Планшет промывали пять раз PBS-Tween. В планшет добавляли 100 мкл TMB, и измеряли ОП при 620 нм.
Используя этот анализ, блокирующая эффективность ARGX-110 была сравнена с эффективностью SGN70 и MDX1411. Как показано на фиг. 18, ARGX-110 является намного более эффективным (в приблизительно 100 раз) в блокировании взаимодействия между CD70 и CD27, чем два эталонных мАт (IC50 для ARGX-110=67 нг/мл, для MDX1411=5500 нг/мл и для SGN70=4972 нг/мл).
Пример 20: Относящиеся к связыванию характеристики мАт против CD70
20.1 Перекрестная реактивность с CD70 не являющегося человеком вида
Определение перекрестной реактивности мАт против CD70 с CD70 животного полезно с целью определения того, модели на каких животных могут использоваться для in vivo исследований обоснованности концепции, и какие виды в наибольшей степени подходят для токсикологических исследований. CD70 макака-резус идентичен на 94% CD70 человека, а CD70 яванского макака идентичен на 95% CD70 человека. Совмещение последовательностей различных видов представлено на фиг. 19. Последовательности CD70 мыши и крысы также включены для подчеркивания различий по сравнению с последовательностью человека.
мАт против CD70 ARGX-110, SGN70 и MDX1411 проверяли в отношении связывания с U266 (линией клеток множественной миеломы человека), клетками LCL8664 (линией клеток B-клеточной лимфомы макака-резус, ATCC-CRL-1805) и с клетками HSC-F (линией Т-клеток яванского макака, происходящих из эмбриональной селезенки лимфоцитов, Japanese health science foundation) в ряде разведений, начиная с концентрации, составляющей 10-20 мкг/мл.
Выявление осуществляли, используя козье антитело против IgG1 человека, конъюгированное с FITC. Образцы анализировали с использованием FACS. Результаты представлены на фиг. 20. Копийность CD70 в обеих линиях клеток обезьян являются очень низкой (слабые сигналы в FACS). Однако ARGX-110 обладает высокой аффинностью к CD70 макака-резус и яванского макака по сравнению с аффинностью SGN70 и MDX1411 к CD70 этих видов.
20.2 Блокирующая активность мАт против CD70
Способность мАт против CD70 (ARGX-110, SGN70 и MDX1411) к блокированию взаимодействия между CD70 и CD27 проверяли, используя ELISA блокирования, описанный в примере 19 выше. Анализ адаптировали так, чтобы измерить блокирующую активность каждого антитела в случае использования Flag-TNC-CD70 человека, макака-резус и яванского макака (Wyzgol, et al., J. Immunol., 2009, 183: 1851-1861).
В ELISA использовали точно такие же условия в случае использования CD70 различных видов, чтобы можно было непосредственно сравнить значения IC50 между различными видами. Результаты представлены на фиг. 21 и совместимы с результатами, отмеченными, используя клетки человека, макака-резус и яванского макака. ARGX-110 обладает неизмененной активностью, и, следовательно, неизмененной аффинностью к CD70 обезьян по сравнению с CD70 человека, тогда как SGN70 и особенно MDX1411 обладает меньшей аффинностью к CD70 обезьян, чем к CD70 человека. Значения IC50 сведены в таблицу 27.
IC50 для блокирования взаимодействия CD27-CD70 в ELISA в случае CD70 от различных видов
20.3 Связывание с подвергнутым денатурации CD70
Связывание мАт против CD70 с денатурированным рекомбинантным CD70 определяли с помощью ELISA. Рекомбинантный CD70 подвергали денатурации нагреванием в течение 5 минут при 95°C, с последующим немедленным охлаждением на льду в течение 5 минут. Титрационный микропланшет покрывали 5 мкг/мл CD70, подвергнутого или не подвергнутого денатурации. Планшет блокировали, и вносили исследуемые мАт против CD70 в концентрации 10 мкг/мл. После промывки связывание мАт выявляли, используя антитело против IgG человека, конъюгированное с HRP. В качестве положительного контроля для покрытия, вместо использования мАт против CD70, использовали мАт мышиного происхождения против Flag, и выявление осуществляли с помощью антимышиного антитела, конъюгированного с HRP. TMB использовали в качестве субстрата, и измеряли ОП при 620 нм. Результаты этого ELISA представлены на фиг. 22. Результаты показывают, что ARGX-110 связывается с эпитопом, отличным от такового для SGN70, MDX1411 (MDX2H5) и MDX69A7, поскольку ARGX-110 все еще связывается с денатурированным CD70, тогда как другие мАт нет. Среди других мАт некоторые (fe 5F4, 9G2, 57B6) также связываются с денатурированным CD70.
20.4 Картирование эпитопа, используя химеры мышь-человек
Рекомбинантные мышь-человек белки в виде экстраклеточного домена (ECD) CD70 конструировали посредством замены доменов CD70 человека и мыши для картирования домена, распознаваемого мАт. Конструирование осуществляли, используя стандартные методологии для создания рекомбинантных ДНК и ПЦР. Химерные мышь-человек варианты клонировали в вектор для экспрессии в эукариотических клетках с Flag-меткой для улавливания в ELISA и TNC для тримеризации белка (Flag-TNC-CD70). Белки экспрессировались в виде растворимых белков в клетках HEK293. Последовательность различных химер представлена на фиг. 23.
Титрационный микропланшет Maxisorb (Nunc) покрывали 3,5 мкг/мл мАт мыши M2 против Flag (Sigma Aldrich, F3165), и улавливали различные химеры. Использовали ряд разведений исследуемых мАт против CD70, начиная с концентрации, составляющей 10 мкг/мл, и осуществляя 3-кратные разведения. Спустя 2 часа связывание выявляли, используя антитело против Fc человека, конъюгированное с HRP, в 16000-кратном разведение. Окрашивание выполняли с использованием ABTS, и измеряли ОП при 405 нм. В качестве положительного контроля использовали CD27-Fc, который способен связываться с CD70 и человека, и мыши с достаточной аффинностью, а также с химерными человек-мышь вариантами CD70. Значения EC50 для связывания сведены в таблицу 28.
Связывание ARGX-110 с химерными человек-мышь вариантами CD70, представленными на фиг. 23
Эти результаты показывают, что ARGX-110 может связываться с Flag-TNC-CD70 человека и химерами 1, 2.1, 2.2, 2.3 и 2.4, но не связывается с химерами 2, 3 и 4 и Flag-TNC-CD70 мыши. Это означает, что эпитоп для ARGX-110 находится в пределах следующей аминокислотной последовательности: HIQVTLAICSS (SEQ ID NO: 342).
Пример 21: Интернализация CD70 в различные линии опухолевых клеток и первичные опухолевые клетки
Было установлено, что связывание антител против CD70 с линиями клеток, происходящих из карциномы почки, 786-О и A498 приводит к быстрой (в пределах 1 часа) интернализации комплекса антитело-рецептор (Adam et al., Br J Cancer (2006) 95: 298-306). Для исследования мАт ARGX-110, SGN70 и MDX1411 в отношении интернализации клетки 786-O культивировали в 96-луночном титрационном микропланшете и инкубировали в течение ночи при 37°C. 2,5 мкг/мл мАт добавляли и инкубировали с клетками в течение 0-24 часов при 37°C. Планшеты промывали 3 раза по 5 мин буфером для удаления (150 мМ NaCl, 100 мМ глицин, pH=2,5; при этом закодированный «В» представляет количество мАт, подвергнутого интернализации через посредство CD70), или PBS (при этом закодированный «Вне» представляет количество мАт, связавшегося с рецептором на внешней части клетки). Впоследствии клетки фиксировали с использованием 4% параформальдегида в течение 30 мин при комнатной температуре, промывали PBS и инкубировали в течение 5 мин с 0,2% Triton-X-100 («В») или PBS («Вне»). Затем клетки дважды промывали и инкубировали при комнатной температуре в течение 10 мин с 100 мМ глицином с последующей инкубацией в течение 30 мин с PBS+1% BSA. Наконец, клетки окрашивали козьим антителом против Fc человека (Jackson immunoresearch 109-005-098) и антикозьим Ат IRDYE800 (Li-cor 926-32214) до анализа на инфракрасном сканере Li-Cor Odyssey. % мАт «Вне» в зависимости от времени представлен на фиг. 24.
Результаты показывают, что в случае всех мАт интернализация является сравнимой, и что ни одно из мАт не интернализуется полностью. В начале, между 0 и 2 часами, интернализация мАт идет очень быстро, но затем, по-видимому, достигается стационарная фаза, в которой приблизительно 30% мАт остаются на внешней части клетки, даже после инкубации в течение 24 часов при 37°C.
Интернализацию связавшихся с CD70 мАт также определяли, используя другие линии клеток. В этих экспериментах только сигнал от мАт вне клетки измеряли в зависимости от времени, используя анализ с использованием FACS, в соответствии с протоколом, описанным ниже. Было установлено, что этот альтернативный протокол является надежным считыванием по сравнению с описанным выше способом (смотрите фиг. 24, правая панель).
В случае клеток, растущих в суспензионной культуре, клетки центрифугировали, подсчитывали, и осадок ресуспендировали в среде до плотности, составляющей 5×105 клеток/мл, и 100 мкл этой суспензии добавляли в каждую лунку 96-луночного планшета с V-образным дном. В случае прикрепляемых клеток клетки высевали в 96-луночный планшет и выращивали в течение ночи при 37°C, 5% CO2 до полного прикрепления клеток. Клетки пассировали на планшете в плотности, которая делала возможным линейный рост клеток в течение еще 48-ч периода.
Исследуемые мАт разводили и инкубировали с клетками при 37°C, 5% CO2 в течение переменных периодов времени: 24 ч, 8 ч, 6 ч, 4 ч, 2 ч, 1 ч, 30 мин, 15 мин, 5 мин, 0 мин (= отсутствие мАт). После инкубации планшеты промывали на льду холодным буфером для FACS для прекращения реакции интернализации. На этой стадии прикрепленные клетки отсоединяли от планшета, используя раствор для диссоциации клеток (Sigma), и переносили в 96-луночный планшет с V-образным дном. Затем клетки осаждали с помощью центрифугирования при 4°C. Супернатант удаляли посредством плавного перевертывания планшета. Клетки промывали дважды с помощью легкого ресуспендирования клеточного осадка в 100 мкл холодного буфера для FACS. После промывки клеточный осадок ресуспендировали в 100 мкл антитела против IgG человека, конъюгированного с FITC, разведенного 1/500 в буфере для FACS. Планшет инкубировали в течение 30 мин при 4°C, при встряхивании, и клетки промывали еще три раза холодным буфером для FACS. Клетки ресуспендировали в 100 мкл буфера для FACS, и сразу же измеряли флуоресценцию. Строили график зависимости медианы средней интенсивности флуоресценции (MFI) от времени.
Некоторые из экспериментов были повторены два или более раз. Результаты двух экспериментов были очень сопоставимыми. В некоторых экспериментах с использованием клеток U266, SU-DHL-6 и Raji использовали диапазон концентраций (20 нг/мл-5 мкг/мл) для обнаружения, есть ли эффект концентрации мАт на скорость интернализации. Такой эффект не отмечался (не представленные данные).
Результаты для различных исследованных линий клеток представлены в таблице 29 и показывают, что интернализация не является общим явлением, в действительности, она является довольно редким событием. В таблицу 29 сведен процент интернализации через 6 часов для различных линий клеток. Эти результаты показывают, что большая часть ARGX-110 остается связанной с внешней частью клеток, делая клетки более чувствительными к ADCC, CDC и ADCP.
% интернализации мАт против CD70 в различные линии
клеток через 6 часов
Нуклеотидные последовательности, кодирующие отобранные антитела против CD70
SEQ ID NO: 344
AAACTCACCGTGGATAAAAGCAGGTGGCAGCAGGGCAACGTGTTCAGCTGTTCCGTTATGCATGAGGCCCTGCATAACCATTATACCCAGAAGTCTTTGTCCCTCAGTCCAGGAAAGTGA
Козака
лидерная последовательность
Вариабельная область
Константная область лямбда
SEQ ID NO: 345
Козака
лидерная последовательность
Вариабельная область
Константная область лямбда
SEQ ID NO: 346
TCTTGTGCAGCCTCTGGATTCAGCTTCAGTCACTATGCCATGAGCTGGGTCCGCCAGGCT
CCAGGAAAGGGGCTAGAGTGGGTCTCAGGTGATAATACCTACGATGGTGGTACAAGGTAT
CAAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATGGCAAGAACACGCTGTAT
CTGCAAATGAACAGCCTGAAACCTGAGGACACGGCCGTGTATTACTGTGCAAAAGATACT
GGTAGAGGCATTATGGGGGAGTACGGCATGGACTACTGGGGCAAAGGGACCCTGGTCACC
GTCTCCTCA
SEQ ID NO: 347
ACCTGCGGCCTCAAGTCTGGGTCTGTCACTTCCAGTAACTACCCTGCTTGGTACCAGCAG
ACACCAGGCCAGGCTCCCCGATTGCTTATCTACAACACAAACAGCCGTCACTCTGGGGTC
CCCAGTCGCTTCTCCGGATCCATCTCTGGGAACAAAGCCGCCCTCACCATCACGGGGGCC
CAGCCCGAGGACGAGGCCGACTATTACTGTGCTCTGTACATGGGTAGTGGTAGTGCCAAT
GCTATGTTCGGCGGAGGGACCCATCTGACCGTCCTGGGTCA
SEQ ID NO: 348
TCCTGTGCAGCGTCCGAATTGTCCTTCAGTATTTCTGAGATGACCTGGGTCCGCCAGGCT
CCAGGAAAGGGGCTCGAGTGGGTCTCAGGTATTAGTGGTGTAACTGGTGGTAGTAGTACA
AGTTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAACGACAAGAACACG
TTGTATCTACAAATGAACAGCCTGATACCCGAGGACACGGCCGTATATTACTGTGCAACA
ACTAGTGGTACTTACTACTTCATCCCCGAGTATGAGTACTGGGGCCAGGGGACCCAGGTC
ACCGTCTCCTCA
SEQ ID NO: 349
TCCTGTGCAGGAACCAGCAGTGATGTTGGGTATGGATACTATGTCTCCTGGTATCAACAG
TTCCCAGGAATGGCCCCCAAACTCCTGATATATGACGTCAATAAACGGGCCTCAGGGATC
GCTGATCGCTTCTCTGGCTCCAAGGCCGGCAACACTGCCTCCCTGACCATCTCTGGGCTC
CAGTCTGAGGACGAGGCTGATTATTACTGTGCCTCATATAGAAGTAGCGCCAATGCTGTG
TTCGGCGGAGGGACCCATCTGACCGTCCTGGGT
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С CD40 И FAP | 2018 |
|
RU2766234C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
| АНТИТЕЛА К ФАКТОРУ XI И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739946C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| Белок, связанный с гликопротеином человека, композиция, фармацевтическая композиция, вектор экспрессии | 2016 |
|
RU2773819C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 6H8HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2768737C1 |
| АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822496C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 5D3HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2761876C1 |


















































































































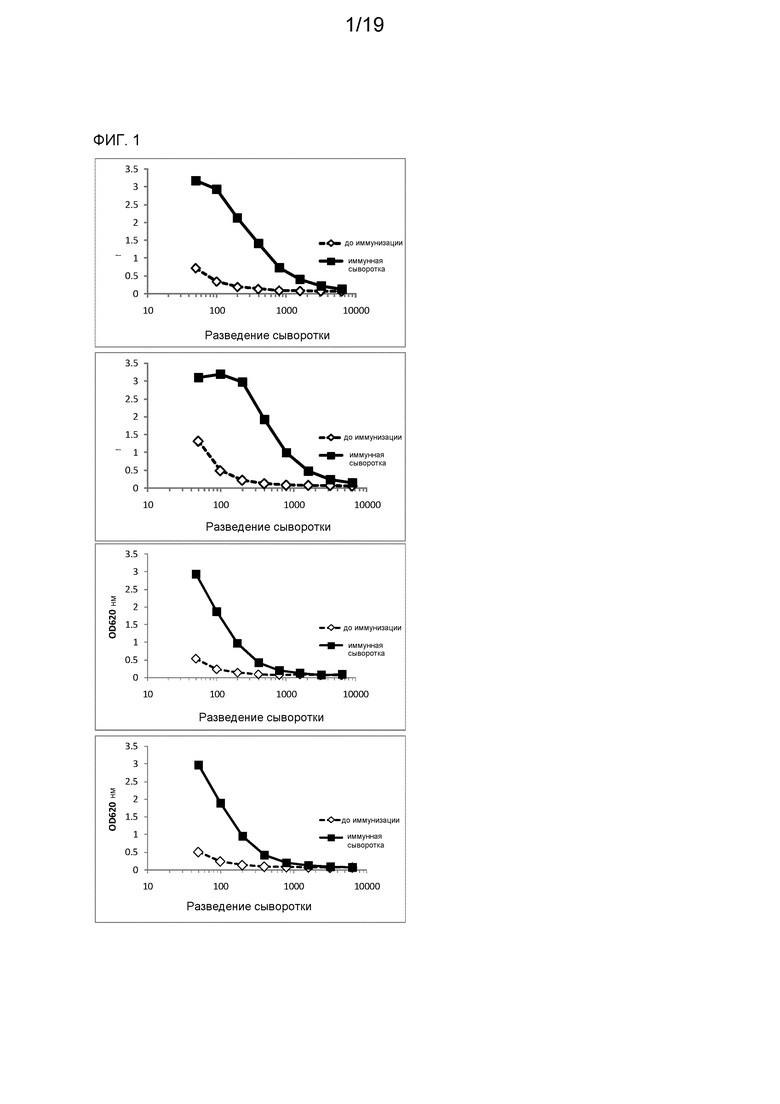
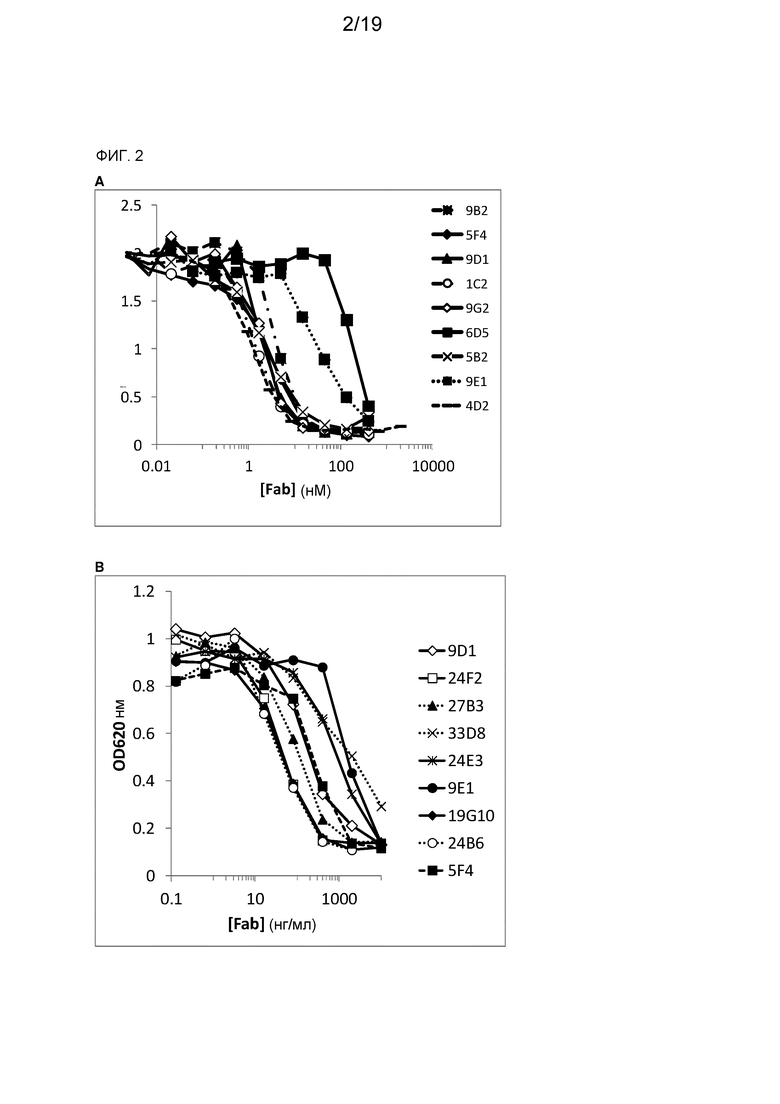
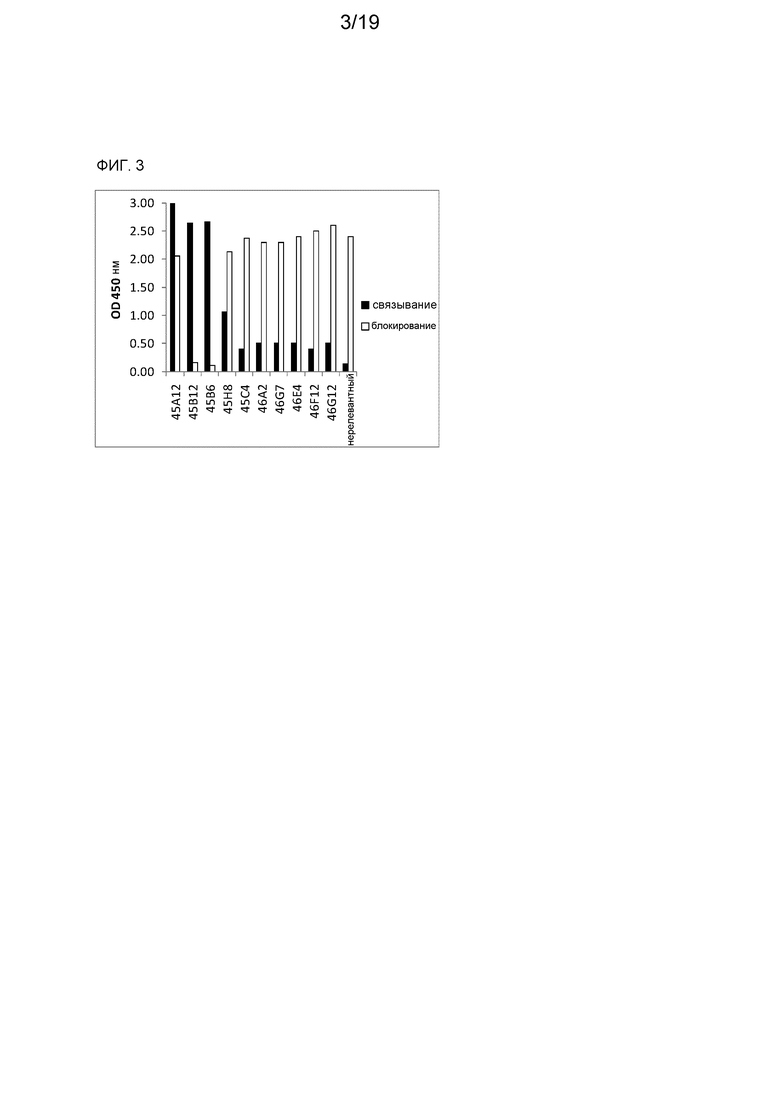


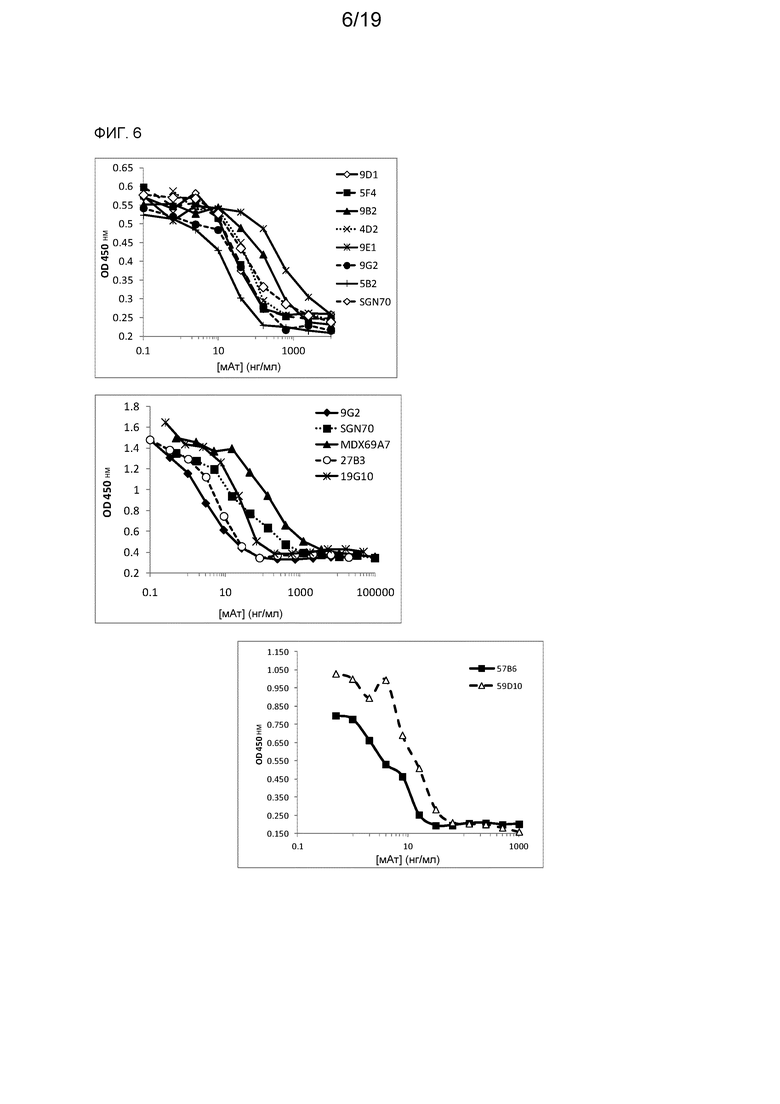
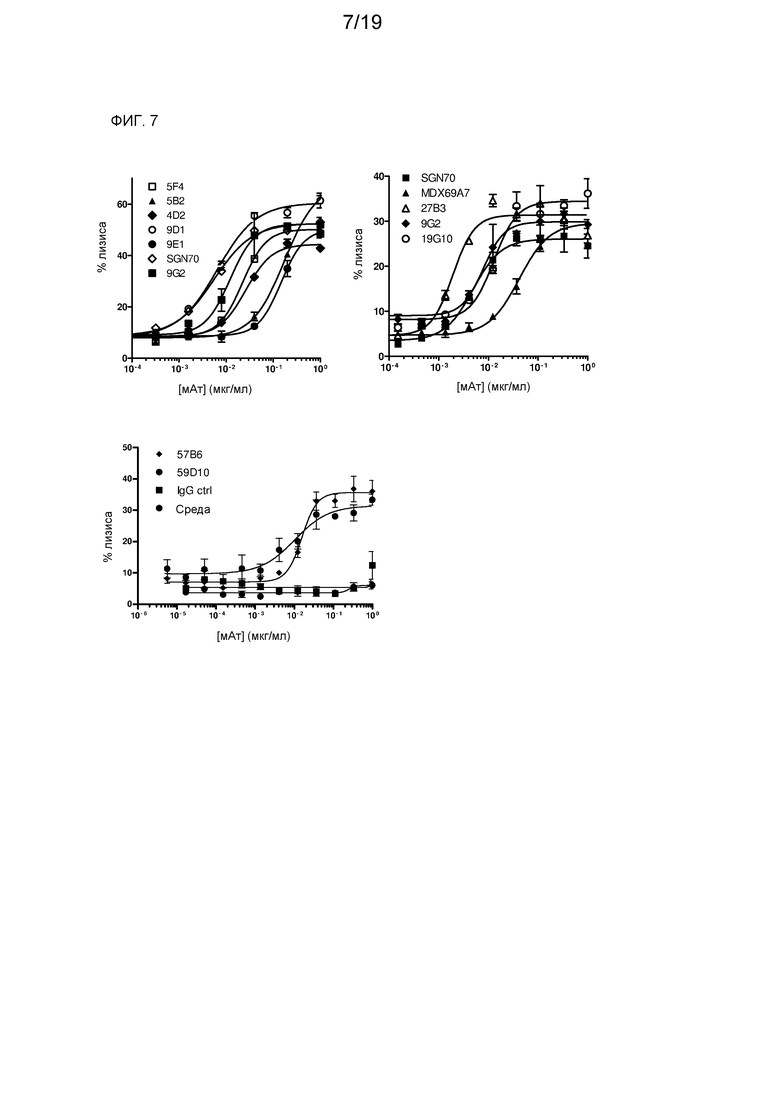
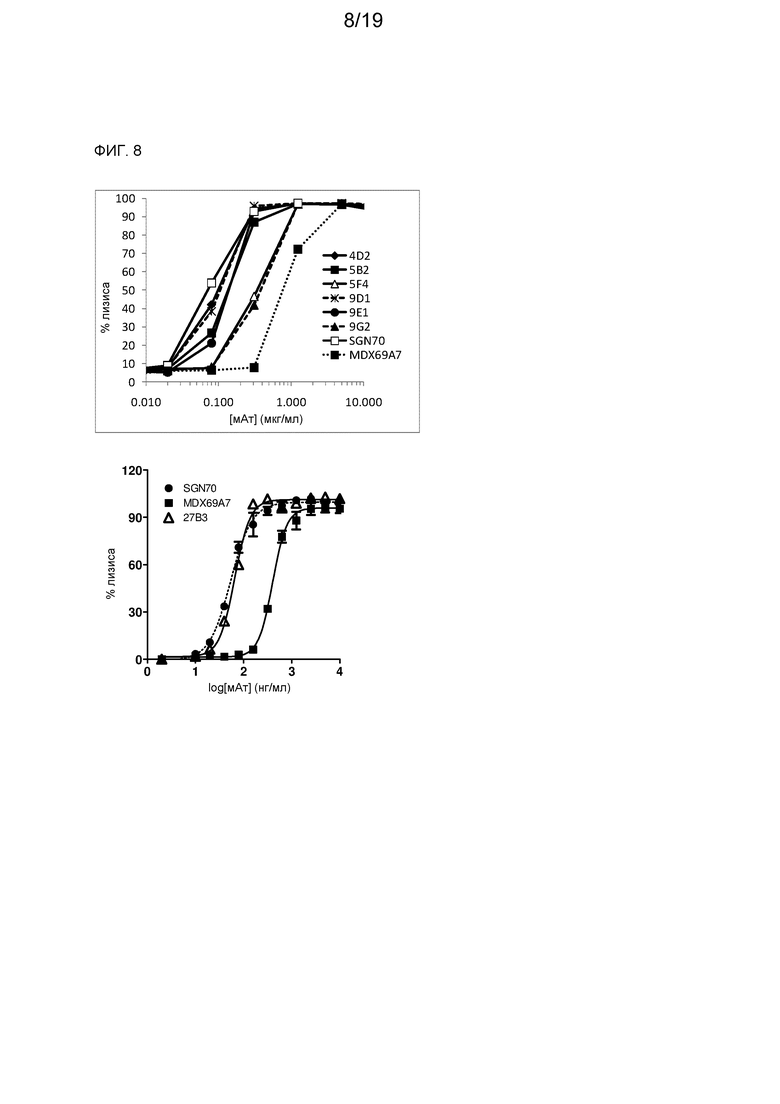
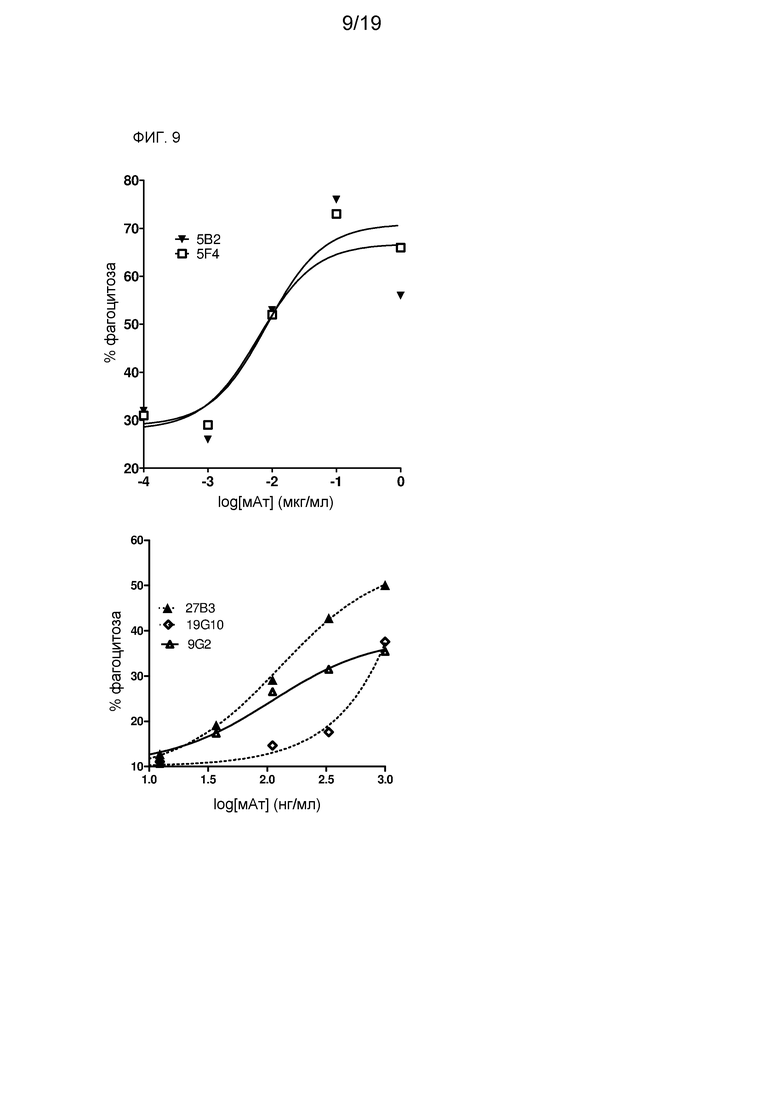
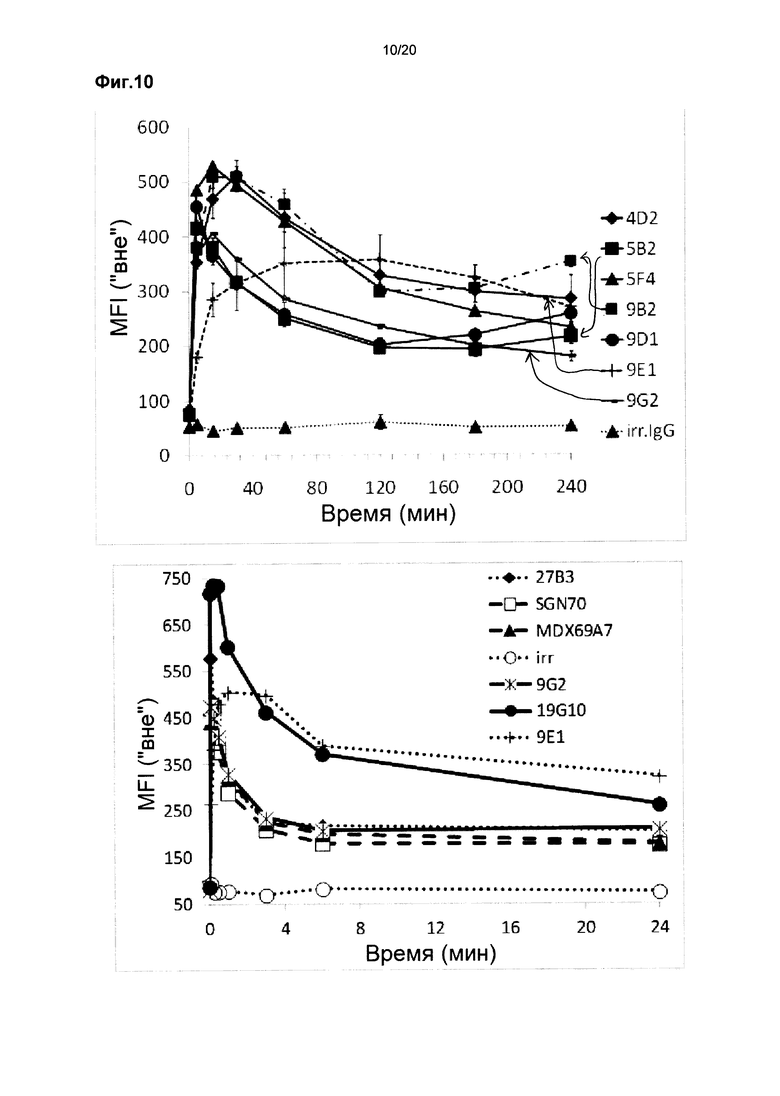
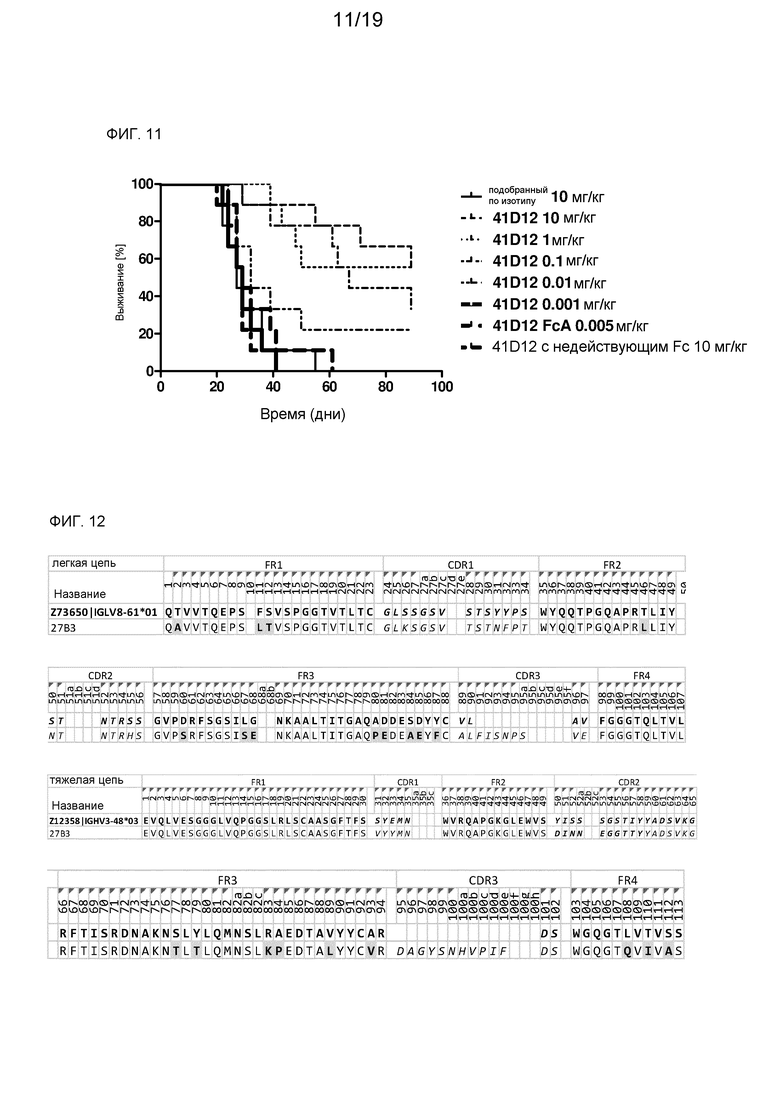

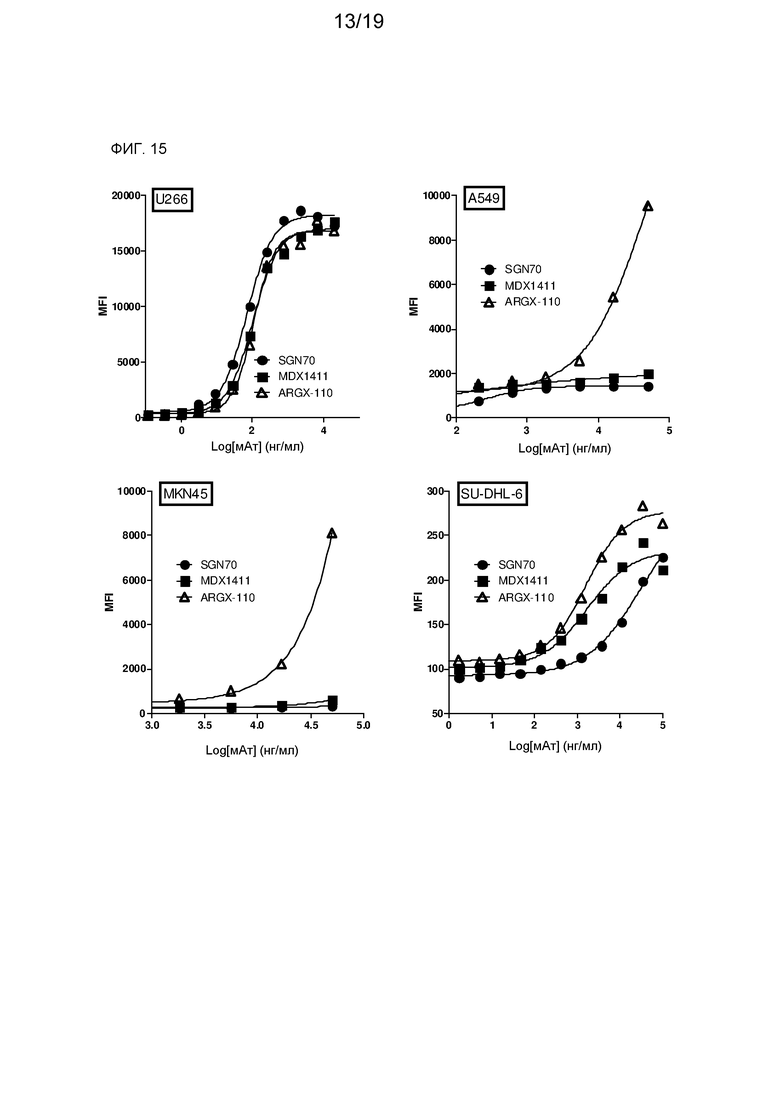
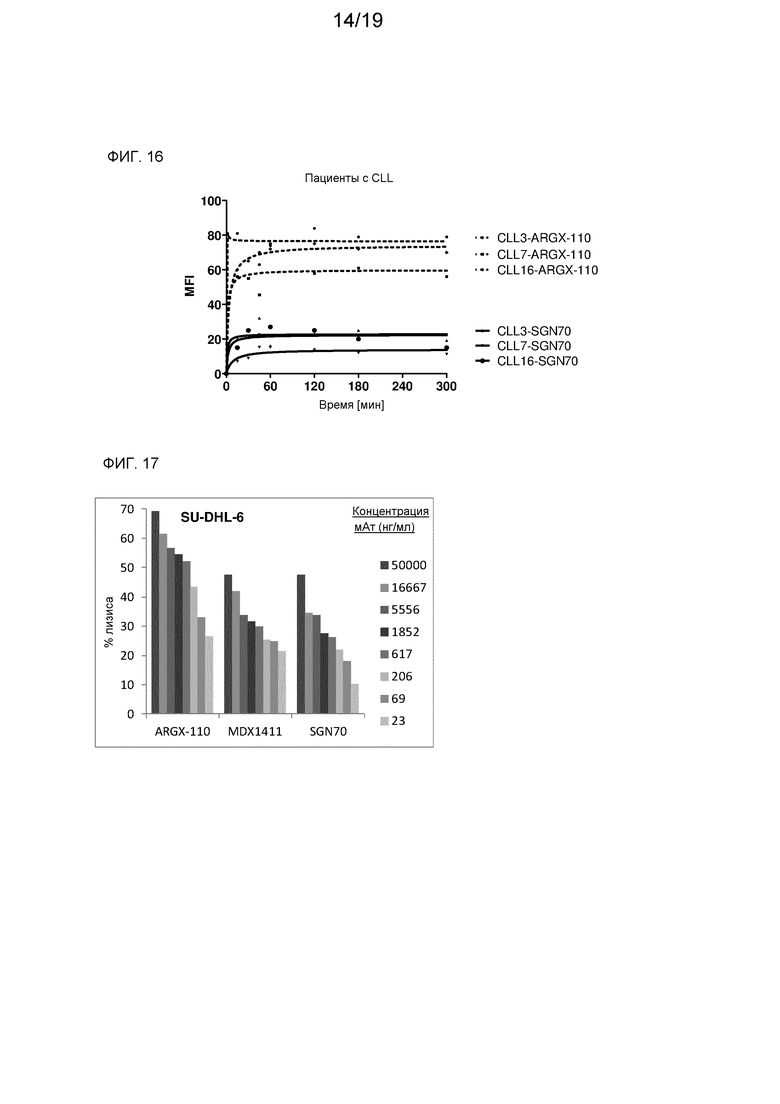
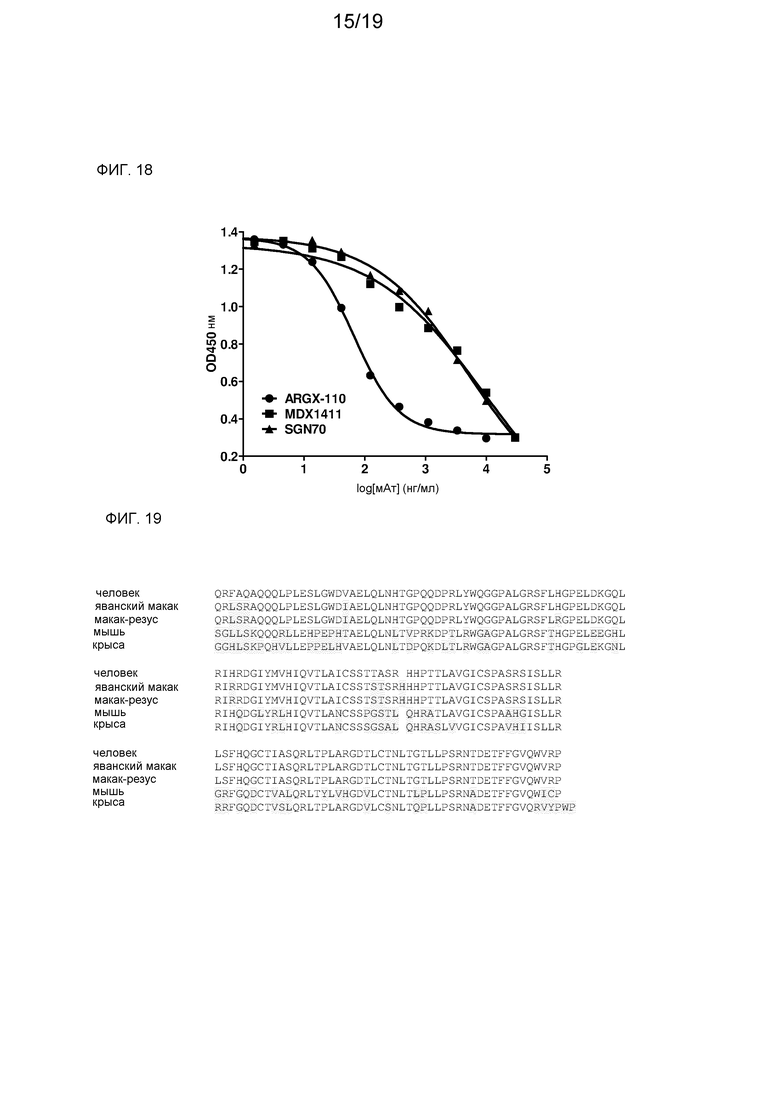

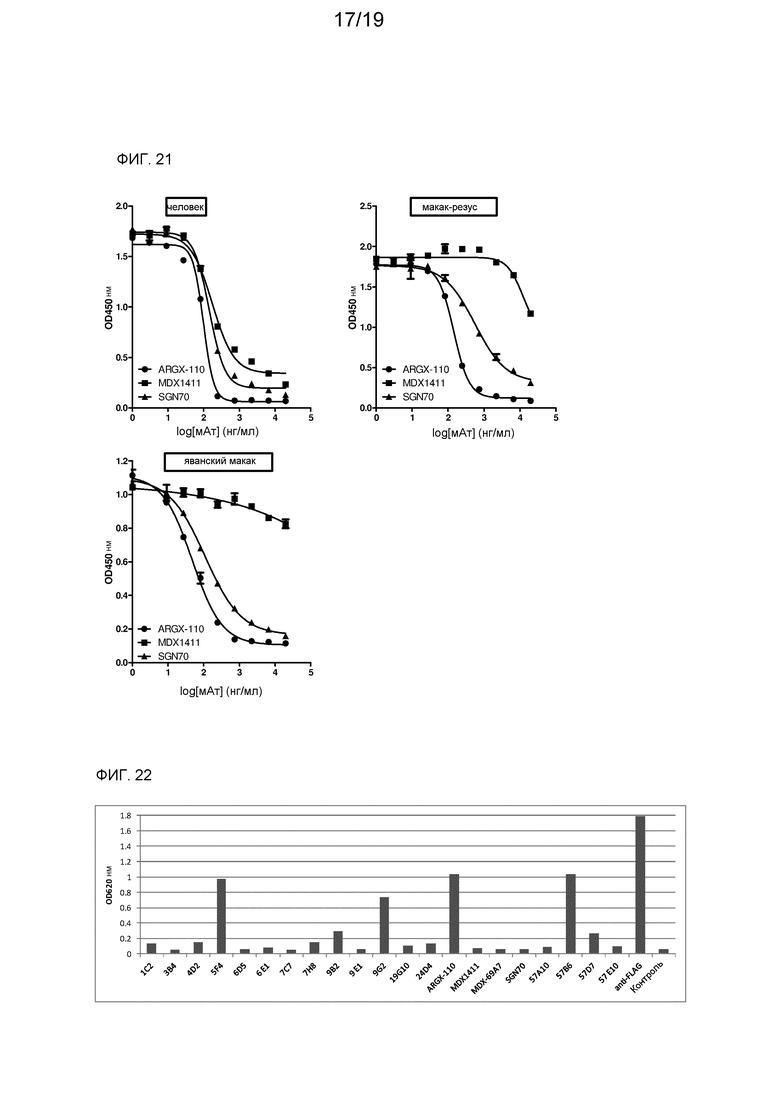


Данное изобретение относится к области иммунологии. Предложены антитело и его антигенсвязывающий фрагмент, которые связываются с CD70 человека, охарактеризованные аминокислотными последовательностями CDR вариабельных доменов тяжелой и легкой цепи. Рассмотрен выделенный полинуклеотид, который кодируют антитело или антигенсвязывающий фрагмент по изобретению, а также вектор экспрессии, клетка-хозяин, бесклеточная экспрессионная система и способ продукции антитела или его антигенсвязывающего фрагмента по изобретению. Также представлена фармацевтическая композиция, способ ингибирования роста опухолевых клеток и способ истощения CD70-экспрессирующих клеток у пациента. Данное изобретение может найти дальнейшее применение в терапии и диагностике заболеваний, в частности рака и аутоиммунных заболеваний, характеризующихся экспрессией CD70. 9 н. и 18 з.п. ф-лы, 24 ил., 29 табл., 21 пр.
1. Антитело или его антигенсвязывающий фрагмент, которое(ый) связывается с CD70 человека, причем указанное антитело или его антигенсвязывающий фрагмент содержит по меньшей мере один вариабельный домен тяжелой цепи (VH) и по меньшей мере один вариабельный домен легкой цепи (VL), содержащие комбинацию CDR3 вариабельного домена тяжелой цепи (HCDR3), CDR2 вариабельного домена тяжелой цепи (HCDR2), CDR1 вариабельного домена тяжелой цепи (HCDR1), CDR3 вариабельного домена легкой цепи (LCDR3), CDR2 вариабельного домена легкой цепи (LCDR2) и CDR1 вариабельного домена легкой цепи (LCDR1), где
CDR3 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:50 (DAGYSNHVPIFDS);
CDR2 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:27 (DINNEGGTTYYADSVKG); и
CDR1 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:11 (VYYMN);
CDR3 вариабельного домена легкой цепи включает или состоит из SEQ ID NO:160;
CDR2 вариабельного домена легкой цепи включает или состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:119 и SEQ ID NO:110; и
CDR1 вариабельного домена легкой цепи включает или состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:87, SEQ ID NO:250, SEQ ID NO:251, SEQ ID NO:252 и SEQ ID NO:253.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, в котором
CDR3 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:50 (DAGYSNHVPIFDS);
CDR2 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:27 (DINNEGGTTYYADSVKG);
CDR1 вариабельного домена тяжелой цепи включает или состоит из SEQ ID NO:11 (VYYMN),
CDR3 вариабельного домена легкой цепи включает или состоит из SEQ ID NO:160 (ALFISNPSVE);
CDR2 вариабельного домена легкой цепи включает или состоит из SEQ ID NO:119 (NTNTRHS);
CDR1 вариабельного домена легкой цепи включает или состоит из SEQ ID NO:250 (GLKSGSVTSDNFPT).
3. Выделенное антитело или его антигенсвязывающий фрагмент по п.2, включающее
вариабельный домен тяжелой цепи (VH), включающий аминокислотную последовательность, выбранную из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO:223, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее, и вариабельный домен легкой цепи (VL), включающий аминокислотную последовательность, выбранную из группы, состоящей из аминокислотной последовательности, представленной в виде SEQ ID NO:241, ее приведенных к зародышевому типу вариантов и вариантов аффинности, и аминокислотных последовательностей, идентичных ей на по крайней мере 90%, 95%, 97%, 98% или 99%, или состоящий из нее.
4. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.1-3, которое демонстрирует следующее сочетание характеристик связывания:
(a) связывание в пределах аминокислотной последовательности HIQVTLAICSS (SEQ ID NO:342) в CD70 человека;
(b) перекрестная реактивность с гомологами CD70 у макака-резус (Масаса mulatta) и яванского макака (Масаса cynomolgus);
(c) связывание со встречающимся в природе CD70 человека и с подвергнутым тепловой денатурации рекомбинантным CD70 человека.
5. Антитело или антигенсвязывающий фрагмент по любому из пп.1-3, которое подвергается частичной интернализации в клетки карциномы почки 786-O.
6. Антитело по любому из пп.1-3, которое является химерным лама-человек антителом, в котором VH- и VL-домены, или один или более их CDR, происходят от ламы, а один или более константных доменов происходят из иммуноглобулина человека.
7. Антитело по п.6, которое включает шарнирную область, CH2-домен и CH3-домен IgG человека, предпочтительно IgG1 человека.
8. Антитело или антигенсвязывающий фрагмент по любому из пп.1-3 или 7, которое проявляет одну или более эффекторных функций, выбираемых из антителозависимой клеточноопосредованной цитотоксичности (ADCC), комплементзависимой цитотоксичности (CDC) и антителозависимого клеточноопосредованного фагоцитоза (ADCP) по отношению к клеткам, экспрессирующим белок CD70 человека на клеточной поверхности.
9. Антитело или антигенсвязывающий фрагмент по п.8, которое проявляет одну или более из антителозависимой клеточноопосредованной цитотоксичности (ADCC), комплементзависимой цитотоксичности (CDC) и антителозависимого клеточноопосредованного фагоцитоза (ADCP) по отношению к CD70-экспрессирующим раковым клеткам.
10. Антитело или антигенсвязывающий фрагмент по п.8, которое проявляет увеличенную функцию, относящуюся к ADCC, по сравнению с эквивалентным антителом, включающим природный Fc-домен человека.
11. Антитело по любому из пп.1-3, 8, 9 или 10, которое является нефукозилированным IgG, предпочтительно нефукозилированным IgG1.
12. Выделенный полинуклеотид, который кодируют антитело или антигенсвязывающий фрагмент по любому из пп.1-11.
13. Вектор экспрессии, включающий полинуклеотид по п.12, функционально связанный с регуляторными последовательностями, которые допускают экспрессию антигенсвязывающего полипептида в клетке-хозяине или бесклеточной экспрессионной системе.
14. Клетка-хозяин, содержащая экспрессионный вектор по п.13, для продукции антитела или его антигенсвязывающего фрагмента по любому из пп.1-11.
15. Бесклеточная экспрессионная система, содержащая экспрессионный вектор по п.13, для продукции антитела или его антигенсвязывающего фрагмента по любому из пп.1-11.
16. Способ продукции рекомбинантного антитела или его антигенсвязывающего фрагмента, который включает культивирование клетки-хозяина по п.14 или бесклеточной экспрессионной системы по п.15 в условиях, которые допускают экспрессию антитела или его антигенсвязывающего фрагмента по любому из пп.1-11, и выделение экспрессированного антитела или антигенсвязывающего фрагмента.
17. Фармацевтическая композиция, включающая терапевтически эффективное количество антитела или антигенсвязывающего фрагмента по любому из пп.1-11 и по крайней мере один фармацевтически приемлемый носитель или наполнитель, для лечения или предупреждения рака, связанного с CD70, у человека.
18. Способ ингибирования роста опухолевых клеток, экспрессирующих CD70, у являющегося человеком пациента, который включают введение нуждающемуся в этом пациенту терапевтически эффективного количества антитела или антигенсвязывающего фрагмента по любому из пп.1-11.
19. Антитело или антигенсвязывающий фрагмент по любому из пп.1-11 для применения для ингибирования роста CD70-экспрессирующих опухолевых клеток у являющегося человеком пациента.
20. Антитело или антигенсвязывающий фрагмент по любому из пп.1-11 для применения для лечения или предупреждения рака у являющегося человеком пациента.
21. Антитело или антигенсвязывающий фрагмент для применения по п.20, в случае которого указанным раком является рак, который демонстрирует экспрессию CD70 в низкой копийности, при этом указанный рак предпочтительно выбирают из группы, состоящей из В-крупноклеточной лимфомы, хронического лимфолейкоза, Т-клеточной лимфомы кожи, рака желудка, рака легкого, меланомы, глиобластомы и рака яичника.
22. Антитело или антигенсвязывающий фрагмент по любому из пп.1-11 для применения для лечения иммунологического нарушения у являющегося человеком пациента.
23. Антитело или антигенсвязывающий фрагмент для применения по п.22, в случае которого иммунологическое нарушение выбирают из группы, состоящей из аутоиммунных заболеваний, системной красной волчанки (SLE), рассеянного склероза, псориаза, ревматоидного артрита, васкулита, грануломатоза Вегенера, синдрома Черджа-Стросса, микроскопического полиангиита, васкулита иммунокомплексного генеза, узелкового полиартериита, болезни Кавасаки, гигантоклеточного артериита, гломерулонефрита, болезни Бехчета и болезни Такаясу.
24. Способ истощения CD70-экспрессирующих клеток у являющегося человеком пациента, включающий введение указанному пациенту эффективного количества антитела по любому из пп.1-11, которое связывается с CD70 человека, где указанные CD70-экспрессирующие клетки демонстрируют составляющую 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее или 5% или менее интернализацию указанного антитела после периода времени, составляющего 6 часов, и где указанное антитело проявляет по крайней мере одну эффекторную функцию, выбранную из группы, состоящей из ADCC, CDC и ADCP, и не является конъюгатом антитело-лекарственное средство.
25. Способ по п.24, в котором CD70-экспрессирующими клетками являются CD70-экспресссирующие раковые клетки.
26. Способ по п.25, в котором CD70-экспрессирующие раковые клетки относятся к типу рака, выбираемому из группы, состоящей из лимфомы Беркитта, В-крупноклеточной лимфомы, лимфомы Ходжкина, неходжкинской лимфомы, лимфомы из клеток ткани, хронического лимфолейкоза, карциномы поджелудочной железы, карциномы желудка, глиобластомы и карциномы легкого.
27. Способ по п.24, в котором CD70-экспрессирующими клетками являются CD70-экспрессирующие активированные Т-клетки.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| MCEARCHERN J.A | |||
| et al | |||
| Деревянный торцевой шкив | 1922 |
|
SU70A1 |
| ADAM P | |||
| J | |||
| et al | |||
| Деревянный торцевой шкив | 1922 |
|
SU70A1 |

