Настоящее изобретение относится к области микробиологии. Более конкретно, изобретение относится к определению характеристик микроорганизмов из образца с использованием масс-спектрометрии.
Со времен открытия микроорганизмов Пастером, микроорганизмы исследовали микроскопией и биохимическими анализами. Эти общепринятые способы часто являются длительными и трудоемкими, и вскоре стали искать альтернативы для анализа. Таким образом, анализ бактерий масс-спектрометрией был инициирован уже в 1975 г. J. Anhalt и C. Fenselau [1].
После этих предварительных исследований последовали исследования газовой хроматографией в сочетании с масс-спектрометрией (GC-MS) жирных кислот стенки микроорганизмов [2]. Этот способ был распространен под названием FAME, что означает метиловый сложный эфир жирной кислоты. В настоящее время он является эталонным способом для таксономических исследований. Однако его использование остается ограниченным определенными специализированными лабораториями, которые осуществляют обработку образца сапонификацией, гидролизом и преобразованием в производное.
В 1996 году работа M. Claydon et al. [3] и также T. Krishnamurthy и P. Ross [4] продемонстрировала возможность идентификации различных видов бактерий с помощью масс-спектрометра типа MALDI-TOF (времяпролетная лазерная десорбция-ионизация в присутствии матрицы). Анализ сочетает в себе получение спектра масс и интерпретацию его с помощью специализированного программного обеспечения. Он является чрезвычайно простым и его можно проводить за несколько минут. Однако он только совсем недавно начал распространяться среди медицинских испытательных лабораторий [5]. Его клиническое применение в настоящее время ограничено до идентификации видов бактерий и дрожжей. Его не используют ни для типирования, ни для идентификации устойчивости к антимикробным средствам, ни для анализа вирулентности.
Однако определение характеристик микроорганизмов является основополагающим как в клинической области, так и в промышленной области. Таким образом, например, идентификация устойчивости к антимикробным средствам, таким как антибиотики, и детекция факторов вирулентности являются неотъемлемыми элементами для обеспечения оптимального лечения пациентов. Аналогично, типирование является важным для эпидемиологических исследований и для борьбы с нозокомиальными заболеваниями.
Для удовлетворения этих потребностей были предложены другие способы масс-спектрометрии, в частности, тандемная масс-спектрометрия. В качестве примера может быть упомянута работа C. Fenselau et al. по идентификации β-лактамазы с помощью квадрупольного TOF (Q-TOF) [6], работа D. Ding et al. по детекции стафилококкового энтеротоксина C2 (фактор вирулентности SEC2) с тройным квадруполем [7], или также работа R. Everley et al. по типированию Clostridium с помощью Q-TOF [8].
Однако результаты этих исследований не применимы для повседневного клинического применения. Их можно получать с помощью исследовательского оборудования, требующего высококвалифицированных специалистов. Время анализа, которое часто отнимает более одного часа на образец, несовместимо с рабочей нагрузкой микробиологической исследовательской лаборатории. Наконец, данные, полученные различными бригадами, отвечают на конкретный вопрос, но не отвечают одновременно всем клиническим потребностям.
Позднее S. Hofstadler et al. предложили способ, который удовлетворяет всем клиническим потребностям [9]. Они комбинировали амплификацию микробного генома способом ПЦР с детекцией продуктов ПЦР ионизацией электрораспылением-TOF (ESI-TOF). Этот способ в настоящее время полностью автоматизирован [10]. Однако он требует амплификации способом ПЦР с недостатками, присущими молекулярной биологии, а именно, стоимостью зондов, выходом экстракции, и т.д.
В этом контексте, задачей настоящего изобретения является предложение способа определения характеристик микроорганизмов, а именно, идентификации и определения характеристик типирования, устойчивости по меньшей мере к одному противомикробному средству и фактора вирулентности, что позволяет преодолеть недостатки способов уровня техники, а именно, предоставить способ, который является не дорогостоящим, без реагентов, специфичных для каждого вида, в частности, по сравнению со способами молекулярной биологии, что приводит к более короткому времени выполнения, составляющему менее одного часа, и который можно повседневно использовать клинически, без необходимости в высококвалифицированных специалистах. Более того, весь способ определения характеристик микроорганизмов можно преимущественно проводить на одном и том же масс-спектрометре, тем самым упрощая оборудование микробиологической испытательной лаборатории.
В связи с этим, изобретение относится к новому способу определения характеристик по меньшей мере одного микроорганизма из образца, который включает идентификацию указанного по меньшей мере одного микроорганизма и определение характеристик типирования, потенциальной устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности, отличающемуся тем, что определение характеристик типирования, устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности для указанного по меньшей мере одного микроорганизма осуществляют масс-спектрометрией с использованием белков, пептидов и/или метаболитов в качестве маркеров указанных характеристик типирования, устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности.
Таким образом, способ по изобретению является таким, что по меньшей мере для трех из признаков для определения характеристик микроорганизма применим способ масс-спектрометрии с использованием, в качестве маркеров, белков, пептидов или метаболитов, типичных для микроорганизмов, подвергаемых охарактеризации.
Микроорганизмы, которые могут быть охарактеризованы способом по изобретению, все являются патогенными или непатогенными микроорганизмами, встречающимися как в промышленности, так и клинически. Они могут представлять собой бактерии, вирусы, простейших или дрожжи.
Выражение "маркеры характеристик типирования, устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности" означает молекулы, белкового или метаболического происхождения, которые характерны для указанных свойств.
Выражение "типирование микроорганизма" означает дифференциацию нескольких штаммов одного вида. Типирование имеет эпидемиологическое значение; клиницисту известно, происходит ли штамм, выделенный от пациента, из того же источника, что и другие штаммы, которые предположительно являются идентичными и выделены от других пациентов или из окружающей среды. Таким образом, это дает возможность установления очага инфекции в больнице или при пищевом отравлении. В качестве неограничивающих примеров маркеров характеристик типирования в бактериях могут быть упомянуты пептиды, имеющие характерные мутации, такие как продукты транскрипции генов adk, fumC, gyrB, icd, mdh, purA и recA из Escherichia coli, и продукты транскрипции генов arc, aroE, glpF, gmk, pta, tpi и yqiL из Staphylococcus aureus. В качестве неограничивающих маркеров характеристик типирования в простейших могут быть упомянуты продукты гена хитиназы Entamoeba histolytica и E. dispar. В качестве неограничивающих примеров маркеров характеристик типирования в вирусах, могут быть упомянуты продукты гена полимеразы вируса иммунодефицита человека. Наконец, в качестве неограничивающих примеров маркеров характеристик типирования в дрожжах, могут быть упомянуты продукты транскрипции фрагментов генов aat1a, acc1, adp1, mpib, sya1, vps13 и zwf1b Candida albicans.
Выражение "определение устойчивости по меньшей мере к одному антимикробному средству" означает определение чувствительности микроорганизма к уничтожению антимикробным средством. Таким образом, если микроорганизм представляет собой бактерию, то антимикробное средство, против которого он может развить устойчивость, является антибиотиком, если он представляет собой простейшее, то антимикробное средство является антипаразитарным средством, если он представляет собой вирус, то антимикробное средство является антивирусным средством, и если он представляет собой дрожжи, то антимикробное средство является противогрибковым средством. Белки, вовлеченные в механизмы устойчивости, отличаются в зависимости от семейства и вида. В качестве неограничивающих примеров маркеров устойчивости по меньшей мере к одному антибиотику, которые применимы в бактериях, могут быть упомянуты продукты транскрипции гена mecA Staphylococcus aureus, придающие устойчивость к метициллину и позволяющие определения, является ли штамм чувствительным к метициллину (штаммы MRSA) или чувствительными метициллину (штаммы MSSA). Также может быть упомянут белок TEM-2, который позволяет определить, является ли штамм Escherichia coli устойчивым к пенициллинам, но чувствительным к другим классам антибиотиков, таким как цефалоспорины или карбапенемы. Другим маркером является фермент, называемый KPC (по карбапенемазы Klebsiella pneumoniae), который придает устойчивость к карбапенемам. Другим примером маркера устойчивости для Staphylococcus aureus является метаболический профиль, типичный для устойчивости к ванкомицину, как описано E. Alexander et al., в постере "Metabolomics-based approach to antibiotic resistance in Staphylococcus aureus", представленном на конференции ASMS, 2009. В качестве неограничивающего примера маркеров устойчивости по меньшей мере к одному антипаразитарному средству, применимому у простейших, могут быть упомянуты железосодержащая супероксиддисмутаза (Fe-SOD) и пероксиредоксин, увеличенная экспрессия которых придает устойчивость к метронидазолу. В качестве неограничивающего примера маркера устойчивости по меньшей мере к одному противовирусному средству, применимому у вирусов, могут быть упомянуты мутации фермента обратной транскриптазы вируса иммунодефицита человека, придающие чувствительность к нуклеозидным ингибиторам обратной транскриптазы. Наконец, в качестве неограничивающего примера маркеров устойчивости по меньшей мере к одному противогрибковому средству, применимому в дрожжах, может быть упомянута мутация фермента 1,3-b-D-глюкансинтазы Candida albicans, которая обеспечивает сниженную чувствительность к эхинокандинам. В качестве другого примера может быть упомянута устойчивость к азольным противогрибковым средствам в Candida albicans, в частности, устойчивость к флуконазолу. Мишенью флуконазола является фермент ланостериндеметилаза, вовлеченный в синтез эргостерина, главного компонента стенки грибов. Устойчивость к флуконазолу может быть связана с появлением точковых мутаций в гене erg11, кодирующем ланостериндеметилазу.
Следует отметить, что специфические маркеры устойчивости также можно использовать в качестве маркеров типирования, как продемонстрировано заявителем.
Выражение "определение вирулентности микроорганизма" означает оценку патогенного, вредоносного и сильнодействующего характера микроорганизма. В качестве неограничивающих примеров маркера вирулентности в бактериях могут быть упомянуты PVL (лейкоцидин Пантон-Валентайна), цитолитический токсин с двумя синергическими компонентами (LukFet LukS), присутствующий в Staphylococcus aureus, который является одним из наиболее вирулентных токсинов, вызывающих кожные состояния, обширный целлюлит, остеомиелит и некротизирующую пневмонию и вовлечен в вирусные суперинфекции. Другие примеры включают аутолизин и пневмолизин, присутствующие в Streptococcus pneumaniae - виде, ответственном за инфекции дыхательных путей, менингит и бактериемию, и также токсины A и B Clostridium difficile, комменсальной бактерии кишечника, которые вызывают модификацию проницаемости эпителия кишечника (токсин A), или прямо действуют на клетки эпителия (токсин B), или снижают прохождение через кишечник и всасывание в кишечнике с течением времени, вызывая диарею (комбинированное действие токсинов A и B). Также в качестве примера могут быть упомянуты токсины Шига Stx1 и Stx2, присутствующие в Escherichia coli. Эти два цитотоксина считаются важными факторами вирулентности энтерогеморрагических Escherichia coli. Они ответственны за осложнения, такие как язвенный колит или гемолитический-уремический синдром. В качестве неограничивающего примера маркера вирулентности у простейших могут быть упомянуты антиоксиданты (Fe-гидрогеназа 2, пероксиредоксин, супероксиддисмутаза), присутствующие в Entamoeba histolytica - виде, ответственном за дизентерию и абсцессы печени. В качестве неограничивающего примера маркера вирулентности в вирусах может быть упомянут вариант белка Nef в вирусе иммунодефицита человека типа 1, более патогенном типе у человека. Наконец, в качестве неограничивающего примера маркера вирулентности в дрожжах могут быть упомянуты липаза 8 в Candida albicans - виде, ответственном за поверхностный кандидоз, а также за септицемический и диссеминированный кандидоз.
Следует отметить, что также в качестве маркера типирования можно использовать специфические маркеры вирулентности, как продемонстрировано заявителем.
Способ по изобретению можно осуществлять для определения характеристик бактерий, тогда указанное антимикробное средство является антибиотиком, что является вариантом осуществления изобретения. Таким образом, например, в качестве бактерий, которые могут быть охарактеризованы способом по изобретению, могут быть упомянуты:
- Escherichia coli с использованием TEM-2 в качестве маркера типирования, а также токсинов Шига, OmpA в качестве маркеров вирулентности и типирования.
- Enterococcus faecalis и faecium с использованием VanA и VanB для резистентности и типирования, и также ESP (поверхностный энтерококковый белок) для вирулентности и типирования, или еще
- Staphylococcus aureus с использованием белка, известного как иммуноглобулин G-связывающий белок A (также известный как белок A) для типирования, белка PBP2a для устойчивости или типирования, и также белка PVL для вирулентности или также типирования.
В качестве других микроорганизмов, которые можно охарактеризовывать способом по изобретению, могут быть упомянуты:
- Candida albicans с использованием фермента 1,3-b-D-глюкансинтазы или фермента ланостериндеметилазы в качестве маркера устойчивости и типирования, и также липазы 8 в качестве маркера вирулентности и типирования.
Образец, для которого может быть осуществлен способ по изобретению, представляет собой любой образец, способный содержать заданный микроорганизм. Образец может быть биологического происхождения, иными словами животного, растительного происхождения или происхождения из человека. Он может соответствовать образцу биологической жидкости (например, цельная кровь, сыворотка, плазма, моча, цереброспинальная жидкость, органический секрет), образцу ткани или выделенным клеткам. Этот образец можно использовать как есть, при условии, что маркеры для охарактеризации микроорганизмов доступны в тестируемом образце, или также его можно подвергать, перед анализом, приготовлению по типу обогащения, экстракции, концентрирования, очистки и/или культивирования, согласно способам, известным специалистам в данной области.
Образец может быть промышленного происхождения, т.е. согласно неисчерпывающему перечню, образец воздуха, образец воды, образец, взятый с поверхности, объект или изготовленный продукт, или продукт пищевого происхождения. Среди образцов пищевого происхождения могут быть упомянуты, неисчерпывающим образом, образец молочного продукта (йогурт, сыры), мяса, рыбы, яиц, фрукта, овоща, воды или напитка (молоко, фруктовый сок, содовая, и т.д.). Эти образцы пищевого происхождения также могут быть взяты из соусов или приготовленных блюд. Наконец, образец пищи может происходить из корма для животных, такого как, в частности, крупа для животных.
Когда маркеры для определения характеристик микроорганизмов имеют белковое происхождение, перед детекцией масс-спектрометрией, образец, подлежащий анализу, предпочтительно предварительно обрабатывают для получения пептидов из всех белков, присутствующих в образце, так чтобы фрагментировать эти белки в пептиды, например, путем расщепления протеолитическим ферментом (протеазой), или под действием химического реагента. Действительно, белки можно расщеплять путем физико-химической обработки, путем биологической обработки или путем комбинации двух обработок. Среди обработок, которые можно использовать, может быть упомянута обработка гидроксильными радикалами, в частности, H2O2. Обработка гидроксильными радикалами вызывает расщепление пептидных связей, которое происходит случайным образом по любой пептидной связи белка. Концентрация гидроксильных радикалов обуславливает число произведенных расщеплений и, таким образом, длину полученных пептидных фрагментов. Также можно использовать другие виды химической обработки, например, обработку цианоген бромидом (CNBr), которая специфично разрушает пептидные связи на уровне карбоксильной группы метионильных остатков. Также возможно осуществлять частичное расщепление кислотой по аспартильным остаткам путем нагревания при 1000°C раствора белков в трифторуксусной кислоте.
Обработка белков ферментативным расщеплением, тем не менее, является предпочтительной по сравнению с физико-химической обработкой, поскольку она в большей степени сохраняет структуру белков, и ее легче контролировать. Термин "ферментативное расщепление" означает единичное или комбинированное действие одного или нескольких ферментов в подходящих условиях реакции. Ферменты, которые осуществляют протеолиз, называемые протеазами, расщепляют белки в конкретных участках. Каждая протеаза, как правило, распознает последовательность аминокислот, по которой она всегда осуществляет одинаковое расщепление. Определенные протеазы распознают единичную аминокислоту или последовательность из двух аминокислот, между которыми они проводят расщепление, другие протеазы распознают только более длинные последовательности. Эти протеазы могут представлять собой эндопротеазы или экзопротеазы. Среди известных протеаз могут быть упомянуты, как описано в WO 2005/098071, следующие:
- специфические ферменты, такие как трипсин, который расщепляет пептидную связь на уровне карбоновой группы остатков Arg и Lys, эндолизин, который расщепляет пептидную связь по группе -CO лизина, химотрипсин, который гидролизует пептидную связь на уровне карбоновой группы ароматических остатков (Phe, Tyr и Trp), пепсин, который расщепляет на уровне группы NH2 ароматических остатков (Phe, Tyr и Trp), протеаза V8 штамма V8 Staphylococcus aureus, которая расщепляет пептидную связь на уровне карбоновой группы остатка Glu;
- неспецифические ферменты, такие как термолизин, происходящий из бактерии Bacillus thermoproteolyticus, который гидрлизует пептидную связь группы NH2 гидрофобных аминокислот (Xaa-Leu, Xaa-Ile, Xaa-Phe), субтилизин и проназа, которые являются бактериальными протеазами, которые гидролизуют практически все связи и могут конвертировать белки в олигопептиды в контролируемых условиях реакции (концентрация фермента и время реакции).
Несколько протеаз можно использовать одновременно, если их способы действия являются совместимыми, или их можно использовать последовательно. В контексте изобретения, расщепление образца предпочтительно проводят под действием фермента протеазы, например, трипсина.
Получение пептидов с использованием химического реагента или протеазы можно осуществлять путем простой реакции в растворе. Также его можно проводить с помощью микроволновой печи [11] или под давлением [12], или альтернативно с помощью ультразвукового устройства [13]. В последних трех случаях протокол является значительно более быстрым.
Среди пептидов, полученных таким образом, пептиды, специфичные для белка, называют протеотипическими пептидами. Именно их анализируют масс-спектрометрией.
Согласно одному варианту осуществления изобретения, маркеры для определения характеристик представляют собой белки микроорганизма, подлежащего охарактеризации. В частности, указанные белки расщепляют на пептиды, предпочтительно ферментом, более предпочтительно трипсином.
Аналогично, образец, содержащий маркеры для охарактеризации белка, также можно предварительно обрабатывать для очистки. Когда маркеры имеют белковое происхождение, эту предварительную обработку для очистки можно осуществлять до или после стадии получения пептидов, как описано выше.
Предварительная обработка для очистки образца широко известна специалистам в данной области и, в частности, ее можно осуществлять центрифугированием, фильтрацией, электрофорезом или способами хроматографии. Эти способы разделения можно использовать отдельно или в комбинации друг с другом для осуществления многомерного разделения. Например, многомерную хроматографию можно использовать путем комбинирования разделения ионообменной хроматографией с обращенно-фазовой хроматографией, как описано T. Fortin et al. [14] или H. Keshishian et al. [15]. В этих публикациях хроматографическая среда может находиться в колонке или кассете (твердофазная экстракция).
Электрофоретическая или хроматографическая фракция (или время удержания в одномерной или многомерной хроматографии) протеотипических пептидов является характерной для каждого пептида и осуществление этих способов, таким образом, дает возможность выбрать протеотипический пептид(ы), подлежащий оценке. Такое фракционирование полученных пептидов позволяет увеличить специфичность последующего анализа масс-спектрометрией.
Альтернативой электрофорезу или способам хроматографии, для фракционирования пептидов, является специфическая очистка N-гликопептидов ([16] и патентная заявка WO 2008/066629). Тем не менее, такая очистка позволяет только количественное определение пептидов, подвергнутых посттрансляционной модификации по типу N-гликозилирования. Однако не все белки являются гликозилированными, что, таким образом, ограничивает их применение.
Масс-спектрометрия, используемая в способе по изобретению, широко известна специалистам в данной области в качестве эффективного инструмента для анализа и детекции различных типов молекул. Как правило, любой тип молекулы может быть ионизирован и подвергнут детекции в качестве функции ее молекулярной массы с использованием масс-спектрометра. В зависимости от природы подлежащей детекции молекулы белкового или метаболического происхождения могут быть более пригодны определенные способы масс-спектрометрии. Тем не менее, какой бы способ масс-спектрометрии ни использовали для детекции, она включает стадию ионизации молекулы-мишени с получением ионов, называемых молекулярными ионами, в настоящем случае стадию ионизации маркеров для определения характеристик, и стадию разделения полученных молекулярных ионов в качестве функции их массы.
Таким образом, все масс-спектрометры включают:
i) источник ионизации, предназначенный для ионизации маркеров, присутствующих в образце, подлежащем анализу, т.е. для придания этим маркерам положительного или отрицательного заряда;
ii) масс-анализатор, предназначенный для разделения ионизированных маркеров, или молекулярных ионов, согласно их отношению массы к заряду (m/z);
iii) детектор, предназначенный для измерения сигнала, генерируемого либо прямо молекулярными ионами, либо ионами, образованными из молекулярных ионов, как подробно описано далее в настоящем описании.
Стадию ионизации, необходимую для проведения масс-спектрометрии, можно проводить с помощью любого способа, известного специалистам в данной области. Источник ионизации дает возможность приведения молекул, подлежащих анализу, в ионизированное и газообразное состояние. Источник ионизации можно использовать либо в положительном режиме для исследования положительных ионов, либо в отрицательном режиме для исследования отрицательных ионов. Существует несколько типов источников и их можно использовать в зависимости от желаемого результата и анализируемых молекул. В частности, могут быть упомянуты:
- ионизация электронами (EI), химическая ионизация (CI) и химическая ионизация с десорбцией (DCI),
- бомбардировка ускоренными атомами (FAB), бомбардировка метастабильными атомами (MAB) или бомбардировка ионами (SIMS, LSIMS),
- индуктивно сопряженная плазма (ICP),
- химическая ионизация при атмосферном давлении (APCI) и фотоионизация при атмосферном давлении (APPI),
- электрораспылительная ионизация (ESI),
- лазерная десорбция-ионизация в присутствии матрицы (MALDI), усиленная поверхностью лазерная десорбция-ионизация (SELDI) или десобрация/ионизация на кремнии (DIOS),
- ионизация-десорбация путем взаимодействия с метастабильными частицами (DART).
В частности, ионизацию можно осуществлять следующим образом: образец, содержащий молекулы-мишени, подают в источник ионизации, где молекулы подвергаются ионизации в газообразном состоянии и, таким образом, конвертируются в молекулярные ионы, которые соответствуют исходным молекулам. Источник ионизации электрораспылительного типа (ESI, электрораспылительная ионизация) позволяет ионизировать молекулу, одновременно вызывая ее переход из жидкого состояния в газообразное состояние. Затем полученные молекулярные ионы соответствуют молекулам, присутствующим в жидком состоянии, с, в положительном режиме, одним, двумя или даже тремя дополнительными протонами или более, и, таким образом, несут один, два или даже три заряда или более. Например, когда молекула-мишень представляет собой белок, ионизация протеотипических пептидов, полученных после фракционирования белка-мишени, посредством источника электрораспылительного типа, действующего в положительном режиме, приводит к полипептидным ионам в газообразном состоянии, с одним, двумя или даже тремя дополнительными протонами или более и которые, таким образом, несут один, два или даже три заряда или более и позволяют изменение с жидкого состояния на газообразное состояние [17]. Этот тип источника особенно пригоден, когда молекулы-мишени или полученные протеотипические пептиды предварительно разделяют обращенно-фазовой жидкостной хроматографией. Тем не менее, выход ионизации молекул, присутствующих в образце, может варьировать, в зависимости от концентрации и природы различных присутствующих частиц. Это явление приводит к эффекту матрицы, известному специалистам в данной области.
Источник ионизации MALDI позволяет ионизировать молекулы из образца в твердом состоянии.
Масс-анализатор, в котором осуществляют стадию разделения ионизированных маркеров в качестве функции их отношения масса/заряд (m/z), представляет собой любой масс-анализатор, известный специалистам в данной области. Могут быть упомянуты анализаторы низкого разрешения, квадрупольного (Q) типа, типа 3D ионной ловушки (IT) или типа линейной ионной ловушки (LIT), также называемые анализаторами с ионной ловушкой и анализаторами высокого разрешения, для измерения точной массы анализируемых соединений, и в которых используются, в частности, магнитный сектор, сопряженный с электрическим сектором - времяпролетные (TOF).
Разделение молекулярных ионов согласно их отношению m/z может быть осуществлено за один раз (однократная масс-спектрометрия или MS), или также можно проводить несколько последовательных разделений посредством MS. Когда осуществляют два последовательных разделения MS, анализ называется MS/MS или MS2. Когда осуществляют три последовательных разделения MS, анализ называется MS/MS/MS или MS3, и, в более общем смысле, когда проводят n последовательных разделений MS, анализ называется MSn.
Среди способов, осуществляющих несколько последовательных разделений, особенно пригодны для разделения MS2 режим SRM (мониторинг отдельных реакций) в случае детекции или анализа одной молекулы-мишени, или режим MRM (мониторинг множественных реакций) в случае детекции или анализа нескольких молекул-мишеней. Аналогично, режим MRM3 особенно пригоден для разделения MS/MS/MS. В этом случае используют термин "направленная масс-спектрометрия".
В случае детекции в однократном режиме MS, полученное отношение масса/заряд коррелирует с молекулой-мишенью, подлежащей детекции.
В случае детекции в режиме MS/MS, в основном добавляют две стадии по сравнению с анализом MS, которые представляют собой:
i) фрагментацию молекулярных ионов, называемых ионами-предшественниками, с получением ионов, называемых фрагментарными ионами 1 поколения, и
ii) разделение ионов, называемых фрагментарными ионами 1 поколения, согласно их массе (m/z)2, причем отношение (m/z)1 соответствует отношению (m/z) ионов-предшественников.
Таким образом, получают отношение масса/заряд фрагментарных ионов 1 поколения, которое коррелирует с молекулой-мишенью, подлежащей детекции. Термин "фрагментарный ион первого поколения" означает ион, образующийся из иона-предшественника с последующей стадией фрагментации и отношением массы к заряду m/z, отличающимся от иона-предшественника.
Пары (m/z)1 и (m/z)2 обозначают переходы и являются характерными для характеристических ионов, подлежащих детекции.
Выбор характеристических ионов, подлежащих детекции для коррелирования с молекулой-мишенью, осуществляют специалисты в данной области в соответствии со стандартными способами. Их выбор преимущественно приводит к анализам, которые являются настолько чувствительными, насколько возможно, насколько специфичными, насколько возможно, и настолько надежными, насколько возможно, с точки зрения воспроизводимости и достоверности. В способах, разработанных для отбора протетипических пептидов (m/z)1, и фрагмента первого поколения (m/z)2, выбор в основном основан на интенсивности ответа. Для дальнейших деталей см. V. Fusaro et al. [18]. Для предсказания всех возможных пар переходов специалисты в данной области могут использовать коммерческое программное обеспечение, такое как программное обеспечение MIDAS и программное обеспечение MRM Pilot от Applied Biosystems или MRMaid [19]. Также можно использовать базу данных, называемую PeptideAtlas, сконструированную F. Desiere et al. [20] для объединения всех MRM-переходов пептидов, описанных научным сообществом. Эта база данных PeptideAtlas свободно доступна через Интернет. Для небелковых молекул также можно использовать базы данных, например, такие как база данных, доступная через программное обеспечение Cliquid от компании Applied Biosystems (США).
Альтернативный подход для выбора протеотипических пептидов, (m/z)1 и (m/z)2, состоит в использовании секторов фрагментации MS/MS, полученных в других исследованиях. Эти исследования могут представлять собой, например, фазы разработки и идентификации биомаркеров посредством протеомного анализа. Этот подход был предложен Thermo Scientific в ходе встречи пользователей [19]. Это дает возможность получить список предполагаемых переходов из пептидов, идентифицированных экспериментально с помощью программного обеспечения SIEVE (Thermo Scientific). Определенные критерии подробно описаны J. Mead et al. [19] для выбора ионов (m/z)1 и (m/z)2 и подробно описаны далее:
- Следует избегать пептидов с внутренними участками расщепления, т.е. с внутренним лизином или аргинином, если за лизином и аргинином не следует пролин.
- Следует избегать пептидов с аспарагином или глутамином, поскольку они могут подвергаться дезаминированию.
- Следует избегать пептидов с N-концевым глутамином или глутаминовой кислотой, поскольку они могут самопроизвольно образовывать цикл.
- Следует избегать пептидов с метионином, поскольку они могут окислиться.
- Следует избегать пептидов с цистеином, поскольку они могут необратимо модифицироваться в ходе возможной стадии денатурации, восстановления и блокирования тиольных функциональных групп.
- Пептиды с пролином можно считать благоприятными, поскольку они, как правило, образуют интенсивные фрагменты в MS/MS с одним в высокой степени преобладающим пиком. Однако один в высокой степени преобладающий фрагмент не дает возможности подтвердить тип перехода в комплексной смеси. Действительно, только одновременное присутствие нескольких характеристических фрагментов дает возможность подтвердить, что действительно выявлен желаемый ион-предшественник.
- Следует избегать пептидов, имеющих пролин рядом с C-концом (положение n-1) или во втором положении относительно C-конца (положение n-2), поскольку в этом случае размер фрагментарного пептида первого поколения обычно считается слишком малым, чтобы быть достаточно специфическим.
- Выбор фрагментов, имеющих массу, превышающую массу предшественника, является предпочтительным для обеспечения специфичности. Для этого необходимо выбрать ион-предшественник с двойным зарядом и выбрать наиболее интенсивный фрагментарный ион первого поколения, имеющий массу, большую чему предшественника, т.е. фрагментарный ион первого поколения с одним зарядом.
Фрагментацию выбранных ионов-предшественников проводят в ячейке для фрагментации, такой как модели тройного квадрупольного типа [21], или типа ионной ловушки [22] или времяпролетного (TOF) типа [23], что также позволяет разделить ионы. Фрагментацию(и) обычно проводят путем столкновения с инертным газом, таким как аргон или азот, в электрическом поле, путем фотовозбуждения или фотодиссоциации с использованием интенсивного источника света, столкновения с электронами или радикальными частицами, путем применения разности потенциалов, например, во времяпролетной трубе, или с помощью любого другого способа активации. Характеристики электрического поля обуславливают интенсивность и природу фрагментации. Таким образом, электрическое поле, применяемое в присутствии инертного газа, например, в квадруполе, обуславливает энергию столкновения, передаваемую ионам. Эта энергия оптимизируется специалистами в данной области для увеличения чувствительности перехода, подлежащего анализу. В качестве примера можно варьировать энергию столкновения между 5 и 180 e -В в q2 в масс-спектрометре AB SCIEX QTRAP® 5500 от компании Applied Biosystems (Foster City, США). Аналогично специалисты в данной области могут оптимизировать длительность стадии столкновения и энергию возбуждения, например, в ионной ловушке, для обеспечения более чувствительного анализа. В качестве примера, можно варьировать эту длительность, называемую временем возбуждения, от 0,010 до 50 мс и энергию возбуждения от 0 до 1 (произвольная единица) в Q3 в масс-спектрометре AB SCIEX QTRAP® 5500 от компании Applied Biosystems.
Наконец, детекцию выбранных характеристических ионов проводят общепринятыми способами, в частности с помощью детектора и системы обработки. Детектор собирает ионы и генерирует электрический сигнал, интенсивность которого зависит от количества собранных ионов. Затем полученный сигнал амплифицируют так, чтобы он мог быть обработан компьютером. Компьютерная система для обработки данных позволяет конвертировать информацию, полученную детектором, в спектр масс.
Принципом режима SRM или также режима MRM является определенный выбор иона-предшественника для фрагментации, а затем определенный выбор одного из фрагментарных ионов. Для таких применений обычно используют устройства тройного квадрупольного типа или гибриды тройной квадруполь-ионная ловушка.
В случае тройного квадрупольного устройства (Q1q2Q3), используемого в режиме MS2, для цели анализа или детекции белка-мишени, первый квадруполь (Q1) позволяет фильтровать молекулярные ионы, соответствующие протеотипическим пептидам, характерным для белка, подлежащего анализу и полученным в ходе предшествующей стадии расщепления согласно их отношению массы к заряду (m/z). Только пептиды, имеющие искомое отношение масса/заряд протеотипического пептида, отношение, называемое (m/z)1, передаются на второй квадруполь (q2) и играют роль ионов-предшественников для последующей фрагментации. Анализатор q2 позволяет фрагментировать пептиды с отношением масса/заряд (m/z)1 на фрагментарные ионы первого поколения. Фрагментацию обычно обеспечивают путем столкновения пептидов-предшественников посредством инертного газа, например, азота или аргона, в q2. Фрагментарные ионы первого поколения передаются в третий квадруполь (Q3), который фильтрует фрагментарные ионы первого поколения согласно конкретному отношению массы к заряду, которое называют (m/z)2. Только фрагментарные ионы первого поколения, имеющие искомое отношение масса/заряд характеристического фрагмента протеотипического пептида (m/z)2 переходят в детектор для детекции или даже количественного определения.
Этот режим работы имеет двойную селективность, в отношении отбора, с одной стороны, иона-предшественника и отбора, с другой стороны, фрагментарного иона первого поколения. Таким образом, масс-спектрометрия в режиме SRM или MRM является преимущественной для количественного определения.
Когда масс-спектрометрия, проводимая в способе по изобретению, представляет собой масс-спектрометрию (MS2, MS3, MS4 или MS5), несколько масс-анализаторов могут быть сопряжены друг с другом. Например, первый анализатор разделяет ионы, ячейка для столкновения позволяет фрагментировать ионы, и второй анализатор разделяет фрагментарные ионы. Некоторые анализаторы, такие как ионные ловушки или FT-ICR, представляют собой несколько анализаторов в одном и позволяют фрагментировать ионы и анализировать фрагменты непосредственно.
Согласно предпочтительным вариантам осуществления изобретения способ по изобретению включает одну или несколько из следующих характеристик:
- масс-спектрометрия, проведенная для характеристик типирования, потенциальной устойчивости по меньшей мере к одному антимикробному средству, и фактора вирулентности, представляет собой спектрометрию типа MS/MS, которая имеет преимущество образования фрагмента, специфического для молекулы, подлежащей детекции или количественному определению, и, таким образом, обеспечения способа анализа с большей специфичностью;
- спектрометрия MS/MS представляет собой MRM, которая имеет преимущество использования времени цикла анализа в масс-спектрометре, составляющего несколько десятых миллисекунд, что позволяет проводить детекцию или количественное определение с большей чувствительностью и мультиплексным образом большого количества различных молекул;
- определение характеристик типирования, устойчивости к антимикробному средству и фактора вирулентности проводят в одном и том же устройстве для масс-спектрометрии, предпочтительно одновременно, что имеет преимущество снижения времени анализа и стоимости оборудования; этот также упрощает переработку и выдачу результатов.
В дополнение к определению характеристик типирования, устойчивости к антимикробному средству и фактора вирулентности, целесообразно идентифицировать микроорганизм(ы), присутствующий в образце, подлежащем тестированию.
Способы идентификации микроорганизмов широко известны специалистам в данной области, как описано, например, P. R. Murray el al., в Manual of Clinical Microbiology, 2007, 9 издание, и в частности в томе I, разделе III, главах 15 и 16 для бактерий и дрожжей, в томе II, разделе VI, главе 82 для вирусов и томе II, разделе X, главе 135 для простейших. В качестве примера общепринятых способов идентификации, может быть упомянуто определение биологического профиля, с использованием, например, идентификационных карт Vitek 2 (bioMerieux), или альтернативно с использованием способов молекулярной биологии с критериями идентификации на основе исследования присутствия определенных генов, и на основе исследования их последовательности.
Идентификацию можно проводить прямо из образца, в котором проводят идентификацию, или микроорганизмы, содержащиеся в образце, можно культивировать способами, хорошо известными специалистам в данной области, с помощью культуральной среды и оптимальных условий культивирования, адаптированных в соответствии с видом выявляемых микроорганизмов, как описано P.R. Murray et al., in Manual of Clinical Microbiology, 2007, 9 издание, том I, раздел III, глава 14, в частности том I, раздел IV, глава 21 для бактерий, том II, раздел VI, глава 81 для вирусов, том II, раздел VIII, глава 117 для дрожжей, и том II, раздел X, глава 134 для простейших.
Таким образом, как правило, в случае идентификации с помощью биохимического способа бактерии в образце, сначала необходимо получить ее в чистой культуре, например, после инокуляции на агар. Способы молекулярной биологии (ПЦР) в определенных случаях можно применять непосредственно к образцу, подлежащему анализу.
Вместо культивирования микроорганизмов, их можно концентрировать путем улавливания непосредственно из образца с помощью активных поверхностей. Такой способ описан W.-J. Chen et al. [11], которые улавливали различные виды бактерий с помощью магнитных гранул с Fe3O4/TiO2-активированной поверхностью. Также возможно улавливание другими способами, такими как улавливание с помощью лектинов [24], или посредством антител [25], или с помощью ванкомицина [26]. Улавливание позволяет концентрировать микроорганизмы и, таким образом, уменьшать или даже устранять стадию культивирования. Это приводит к существенной экономии времени.
Идентификацию также можно проводить посредством масс-спектрометрии, согласно способам, описанным ранее, предпочтительно посредством MS, MS/MS или же MS с последующей спектрометрией типа MS/MS, которая составляет вариант осуществления изобретения. В этом случае также образец можно подвергать предварительной стадии культивирования, такой как инокуляция на агар.
Использование способа идентификации посредством MS является преимущественным, поскольку его можно проводить в течение нескольких минут и он требует масс-спектрометра с одним анализатором, т.е. устройства, которое является менее сложным, чем тандемный масс-спектрометр, используемый при MS/MS.
Использование способа идентификации посредством MS с последующей спектрометрией типа MS/MS также является преимущественным. Оно позволяет быть уверенным в типе наблюдаемых в MS ионов, что увеличивает специфичность анализа.
Использование способа идентификации посредством MS/MS типа MRM имеет преимущество большей чувствительности и большей простоты, чем общепринятые подходы MS, а затем MS/MS. Этот способ не требует ни эффективного программного обеспечения для обработки информации между получением MS-спектра и MS/MS-спектра, ни какого-либо изменения регуляции параметров устройства для связывания спектров MS и MS/MS.
Способ идентификации посредством MS можно проводить с электрораспылительным источником на неочищенном образце, как описано S. Vaidyanathan et al. [27] или же R. Everley et al. [8] после хроматографического разделения. Затем различные диапазоны m/z позволяют идентифицировать микроорганизмы. S. Vaidyanathan et al. использовали окно между 200 и 2000 Th, и R. Everley et al. окно между 620 и 2450 Th. Спектры масс также могут быть подвергнуты обратной свертке для доступа к массе белков независимо от их заряженного состояния. Таким образом R. Everley et al. использовали массы приблизительно от 5000 до 50000 Да. Альтернативно способ идентификации посредством MS также можно проводить посредством MALDI-TOF, как описано Claydon et al. [3] и T. Krishnamurthy и P. Ross [4]. Анализ сочетает в себе получение спектра масс и интерпретацию с помощью специализированного программного обеспечения. Он чрезвычайно прост и его можно проводить за несколько минут. Этот способ идентификации в настоящее время распространяется в медицинских испытательных лабораториях [28].
Идентификация бактерий посредством MS, а затем MS/MS, через их белки, присутствующие образце, широко использовалась многими бригадами. В качестве примера, могут быть упомянуты недавние исследования N. Manes et al. [29], которые исследовали пептидом Salmonella enterica, или исследования R. Nandakumar et al. [30] или L. Hernychova et al. [31], которые исследовали протеом бактерий после расщепления белков трипсином. Общепринятый подход состоит в i) получении MS-спектра, ii) последовательном выборе каждого иона-предшественника, наблюдаемого на MS-спектре с интенсивным сигналом, iii) последовательной фрагментации каждого иона-предшественника и получения его MS/MS-спектра, iv) поиска в базе данных белков, таких как SWISS-PROT или NCBI, с помощью программного обеспечения, такого как Mascot (Matrix Science, London, Великобритания) или SEQUEST (Thermo Scientific, Waltham, США), для идентификации пептида, имеющего высокую вероятность соответствия наблюдаемому спектру MS/MS. Этот способ может привести к идентификации микроорганизма, если идентифицирован белок или пептид, характерный для данного вида.
Согласно другому варианту осуществления идентификацию указанного по меньшей мере одного микроорганизма проводят с помощью общепринятого способа идентификации, и способ по изобретению включает дополнительную стадию подтверждения идентификации указанного по меньшей мере одного микроорганизма, которую проводят с помощью масс-спектрометрии, согласно способам, ранее описанным для идентификации микроорганизмов.
Согласно одному конкретному варианту осуществления масс-спектрометрия стадии подтверждения представляет собой масс-спектрометрию типа MS/MS, предпочтительно MRM.
Одно из преимуществ использования масс-спектрометрии состоит в том, что она, в частности, пригодна для количественного определения молекул, в настоящем случае маркеров характеристик типирования и устойчивости по меньшей мере к одному антимикробному средству. Для этого используют выявляемую силу тока, которая пропорциональна количеству молекулы-мишени. Измеренная таким образом сила тока может служить в качестве количественной меры для определения количества присутствующей молекулы-мишени, которое характеризуется его выражением в единицах международной системы (СИ) типа моль/м3 или кг/м3, или множествами или подмножествами этих единиц, или обычными производными единиц СИ, включая их множества или подмножества. В качестве неограничивающего примера, единицы, такие как нг/мл или фмоль/л являются единицами, характеризующими количественную меру.
Тем не менее, необходима калибровка для возможности коррелирования площади измеренного пика, соответствующего интенсивности тока, индуцированного детектированными ионами, с количеством молекулы-мишени, подлежащей анализу. Для этого в контексте изобретения можно осуществлять калибровку, обычно используемую в масс-спектрометрии. Анализы MRM обычно калибруют с использованием внешних стандартов или, предпочтительно, с использованием внутренних стандартов, как описано T. Fortin et al. [14]. Когда молекула-мишень представляет собой протеотипический пептид, что позволяет анализировать представляющий интерес белок, корреляцию между количественной мерой и количеством протеотипического белка-мишени, и, следовательно, представляющего интерес белка, получают путем калибровки измеренного сигнала относительно стандартного сигнала, для которого количество, подлежащее анализу, известно. Калибровку можно осуществлять с помощью калибровочной кривой, например, полученной путем последовательных инъекций стандартного протеотипического пептида при различных концентрациях (внешняя калибровка) или, предпочтительно, путем внутренней калибровки с использованием тяжелого пептида в качестве внутреннего стандарта, например, согласно способам AQUA, QconCAT или PSAQ, подробно описанным ниже. Термин "тяжелый пептид" означает пептид, соответствующий протеотипическому пептиду, но в котором один или несколько атомов углерода-12 (12C) заменен(ы) углеродом-13 (13C), и/или один или несколько атомов азота-14 (14N) заменен(ы) азотом-15 (15N).
Использование тяжелых пептидов в качестве внутренних стандартов (AQUA) также предложено в патентной заявке US 2004/0229283. Принцип состоит в искусственном синтезе протеотипических пептидов с аминокислотами, содержащими изотопы, которые тяжелее обычных природных изотопов. Такие аминокислоты получают, например, путем замены некоторых из атомов углерода-12 (12C) углеродом-13 (13C), или замены некоторых из атомов азота-14 (14N) азотом-15 (15N). Синтезированный таким образом искусственный пептид (AQUA) имеет точно такие же физико-химические свойства, как и природный пептид (за исключением более высокой массы). Его обычно добавляют, при данной концентрации, к образцу перед анализом масс-спектрометрией, например, между обработкой, приводящей к расщеплению белков представляющего интерес образца, и фракционированием полученных пептидов после стадии обработки. В результате пептид AQUA совместно очищают с природным пептидом, подлежащим анализу, в процессе фракционирования пептидов. Таким образом, для анализа два пептида инъецируют в масс-спектрометр одновременно. Затем они могут быть подвергнуты одинаковому выходу ионизации в источнике. Сравнение площадей пика для природных пептидов и пептидов AQUA, концентрация которых известна, позволяет вычислить концентрацию природного пептида, и, таким образом, далее концентрацию белка, подвергаемого анализу. Вариант способа AQUA был предложен J.-M. Pratt et al. [32] под названием QconCat. Этот вариант также описан в патентной заявке WO 2006/128492. Он состоит в связывании различных пептидов AQUA и получении искусственного полипептида в форме тяжелого рекомбинантного белка. Рекомбинантный белок синтезируют с аминокислотами, содержащими тяжелые изотопы. Таким образом, можно получить стандарт для калибровки, одновременно анализирующий несколько белков, с более низкой стоимостью. Сначала добавляют стандарт QconCAT перед обработкой, приводящей к расщеплению белков, и перед стадиями фракционирования, денатурации и восстановления белка, а затем блокирования тиольных функциональных групп белков, если они присутствуют. Таким образом, стандарт QconCAT подвергается тому же циклу обработки, который приводит к расщеплению белков, что и природный белок, тем самым позволяя учитывать выход после стадии обработки, приводящей к расщеплению белков. Это является следствием того, что обработка, в частности путем расщепления, природного белка может быть не полной. В этом случае, использование стандарта AQUA приведет к недооцененному количеству природного белка. Таким образом, для абсолютного анализа, может быть важным учет выхода обработки, приводящей к расщеплению белков. Однако V. Brun et al. [33] показали, что иногда стандарты QconCAT не воспроизводят точно выход обработки, в частности путем расщепления, природного белка, несомненно, вследствие отличающейся трехмерной конформации белка QconCAT.
Таким образом, V. Brun et al. [33] предложили использовать способ, называемый PSAQ, который описан в патентной заявке WO 2008/145763. В этом случае, внутренним стандартом является рекомбинантный белок, который имеет ту же последовательность, что и природный белок, но синтезирован с тяжелыми аминокислотами. Синтез осуществляют ex vivo с тяжелыми аминокислотами. Этот стандарт имеет точно такие же физико-химические свойства, что и природный белок (за исключением более высокой массы). Его добавляют вначале, до стадии фракционирования, когда указанная стадия присутствует. Таким образом, его очищают совместно с нативным белком в процессе стадии фракционирования белков. Он обладает тем же выходом после обработки, в частности, путем расщепления, что и нативный белок. Тяжелый пептид, полученный после расщепления, также совместно очищают с природным пептидом, если проводят стадию фракционирования пептида. Таким образом, два пептида инжектируют одновременно в масс-спектрометр для количественного анализа. Таким образом, они подвергаются одинаковым выходам ионизации в источнике. Сравнение площадей пиков природных пептидов и эталонных пептидов в способе PSAQ позволяет вычислить концентрацию белка, подлежащего анализу, учитывая все стадии этого способа анализа.
Все эти способы, а именно AQUA, QconCAT или PSAQ или любой другой способ калибровки, используемые в анализах с помощью масс-спектрометрии и, в частности, в анализах MRM или MS, могут быть осуществлены для проведения калибровки, в контексте изобретения.
Согласно одному предпочтительному варианту осуществления способ по изобретению позволяет определение характеристик Staphylococcus aureus.
В частности, для определения характеристик Staphylococcus aureus используется по меньшей мере один из следующих пептидов:
1. Для типирования:
по меньшей мере один пептид, принадлежащий белку A, имеющему следующую последовательность SEQ ID NO:1:


причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:2, 3, 4, 5, 6, 7 и 8, как определено в настоящем описании далее:
2. Для потенциальной устойчивости по меньшей мере к одному антибиотику
по меньшей мере один пептид, принадлежащий белку PBP2a, имеющему следующую последовательность SEQ ID NO:9:


причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:10-17, как определено в настоящем описании далее:
3. Для вирулентности:
по меньшей мере один пептид, принадлежащий белку PVL, субъединицам LukS и LukF, имеющим следующие последовательности SEQ ID NO:18 и 22, соответственно:


причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательности SEQ ID NO:19, 20, 21, 23 и 24, как определено в настоящем описании далее:
4. Для идентификации:
по меньшей мере один пептид, принадлежащий 50S рибосомальному белку L30 (RL30), 50S рибосомальному белку L331 (RL331), стафилококковому секреторному антигену ssaA2 (SSAA2), белку UPF0337 SA0772 (Y772), бифункциональному аутолизину (ATL), фактору элонгации Tu (EFTU), возможной трансгликолазе isaA (ISAA) и белку UPF0457 SA1975.1 (Y197A), имеющим следующие последовательности SEQ ID NO:25, 28, 31, 33, 36, 39, 41 и 43, соответственно:




причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательности SEQ ID NO:26, 27, 29, 30, 32, 34, 35, 37, 38, 40, 42, 44 и 45, как определено в настоящем описании далее:
Следует отметить, как указано выше, что для типирования в способе по изобретению также может использоваться по меньшей мере один пептид, имеющий последовательность SEQ ID NO:10, 11, 12, 13, 14, 15, 16, 17, 19, 20, 21, 23, 24, 26, 27, 29, 30, 32, 34, 35, 37, 38, 40, 42, 44 и 45, которые применимы для определения возможной устойчивости по меньшей мере к одному антибиотику, или вирулентности, как указано выше.
Безусловно, термин "по меньшей мере один пептид" означает по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или более пептидов, характерных для маркера, который намереваются выявить. Предпочтительно, используют по меньшей мере два или даже по меньшей мере три, или даже по меньшей мере четыре, пептида на характеристику.
Согласно другому предпочтительному варианту осуществления способ по изобретению позволяет определение характеристик Escherichia coli.
В частности, для определения характеристик Escherichia coli используется по меньшей мере один из следующих пептидов:
1. Для типирования:
по меньшей мере один пептид, принадлежащий белку аспартат-аммиак-лиазы (ASPA), альфа-субъединице ATP-синтазы (ATPA), шаперонину массой 10 кДа (CH10), шаперонину массой 60 кДа (CH60), ДНК-связывающему белку HU-бета (DBHB), глутаматдекарбоксилазе (DCEB), флавопротеиновой субъединице сукцинатдегидрогеназы (DHSA), белку защиты ДНК во время голодания (DPS), ДНК-связывающему белку H-NS (HNS), малатдегидрогеназе (MDH), фосфоглицераткиназе (PGK), фосфорибозиламиноимидазол-сукцинокарбоксамидсинтазе (PUR7), 50S рибосомальному белку L4 (RL4), 30S рибосомальному белку S1 (RS1), белку UPF0076 yjgF (YJGF), имеющим следующие последовательности SEQ ID NO:138-152, соответственно:


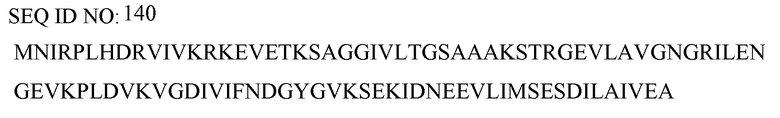

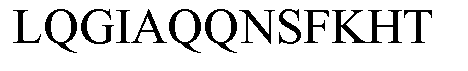







причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:67-84, как определено в настоящем описании далее:
2. Для потенциальной устойчивости по меньшей мере к одному антибиотику
по меньшей мере один пептид, принадлежащий белку бета-лактамазы TEM-2 (TEM-2), имеющему следующую последовательность SEQ ID NO:126:

причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:62-66, как определено в настоящем описании ниже:
3. Для вирулентности:
по меньшей мере один пептид, принадлежащий белку субъединицы A токсина 1 Шига (STX1A), белку субъединицы A токсина 2 Шига (STX2A) или обоим из них, имеющим последовательности SEQ ID NO:153 и 154, соответственно:

причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательности SEQ ID NO:85, 86 и 87, как определено в настоящем описании далее:
4. Для идентификации:
по меньшей мере один пептид, принадлежащий аконитатгидратазе 2 (ACON2), L-аспарагиназе 2 (ASPG2), 3-оксоацил-[ацил-белок-переносчик]-синтазе 1 (FABB), глутамин-связывающему периплазматическому (GLNH), молибдат-связывающему периплазматическому белку (MODA), компоненту ацетилтрансферазы остатка дигидролипоиллизина в комплексе пируватдегидрогеназы (ODP2), белку наружной мембраны C (OMPC), формиатацетилтрансферазе 1 (PFLB), [ADP-образующей] субъединице альфа сукцинил-CoA-лигазы (SUCD), транскетолазе 1 (TKT1), белку UPF0381 yfcZ (YFCZ), неохарактеризованному белку ygaU (YGAU), имеющим следующие последовательности SEQ ID NO:127-135, 176, 136 и 137, соответственно:







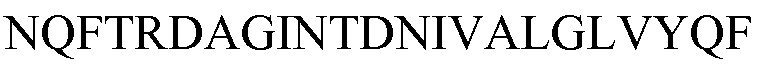





причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:46-61, как определено в настоящем описании ниже:
Следует отметить, как указано ранее, для типирования в способе по изобретению также может использоваться по меньшей мере один пептид, имеющий последовательность SEQ ID NO:46-66 и 85-87, которые применимы для определения потенциальной устойчивости по меньшей мере к одному антибиотику, для идентификации или для вирулентности, как указано выше.
Согласно другому предпочтительному варианту осуществления, способ по изобретению позволяет определение характеристик Candida albicans.
В частности, для определения характеристик Candida albicans используется по меньшей мере один из следующих пептидов:
1. Для типирования:
по меньшей мере один пептид, принадлежащий белку алкогольдегидрогеназы 1 (ADH1), белку фруктоза-бифосфатальдолазы (ALF), белку ланостерин-14-альфа-деметилазе (CP51), белку F-box COS111 (CS111), фактору элонгации 1-бета (EF1B), енолазе 1 (ENO1), глицеральдегид-3-фосфат-дегидрогеназе (G3P), белку теплового шока SSA1 (HSP71), белку теплового шока SSB1 (HSP75), пируваткиназе (KPYK), липазе 8 (LIP8), фактору связывания полибелка 1 (MBF1), фактору ядерного транспорта 2 (NTF2), фосфоглицераткиназе (PGK), пептидил-пролил-цис-транс-изомеразе (PPIA), 60S рибосомальному белку L13 (RL13), 60S рибосомальному белку L28 (RL28), 60S рибосомальному белку L36 (RL36), 40S рибосомальному белку S22 (RS22), триозофосфатизомеразе (TPIS), имеющим следующие последовательности SEQ ID NO:155-175, соответственно:




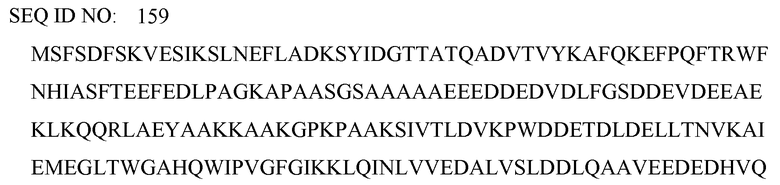












причем указанные пептиды выбраны из пептидов, имеющих последовательность SEQ ID NO:88-125, как определено в настоящем документе далее:
2. Для потенциальной устойчивости по меньшей мере к одному антибиотику
по меньшей мере один пептид, принадлежащий белку ланостерин-14-альфа-деметилазы (CP51), имеющему последовательность SEQ ID NO:157, причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательности SEQ ID NO:117-122 и 177, как определено выше.
3. Для вирулентности:
по меньшей мере один пептид, принадлежащий белку липазы 8 (LIP8), имеющему последовательность SEQ ID NO:163, причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:123-125, как определено выше.
4. Для идентификации:
по меньшей мере один пептид, принадлежащий белку алкогольдегидрогеназы 1 (ADH1), белку фруктоза-бифосфатальдолазы (ALF), белку F-box COS111 (CS111), фактору элонгации 1-бета (EF1B), енолазе 1 (ENO1), глицеральдегид-3-фосфат-дегидрогеназе (G3P), белку теплового шока SSA1 (HSP71), белку теплового шока SSB1 (HSP75), пируваткиназе (KPYK), фактору связывания полибелка 1 (MBF1), фактору ядерного транспорта 2 (NTF2), фосфоглицераткиназе (PGK), пептидил-пролил-цис-транс-изомеразе (PPIA), 60S рибосомальному белку L13 (RL13), 60S рибосомальному белку L28 (RL28), 60S рибосомальному белку L36 (RL36), 40S рибосомальному белку S22 (RS22), триозофосфатизомеразе (TPIS), имеющим последовательности SEQ ID NO:155, 156, 158-164 и 166-175, причем указанные пептиды выбраны, предпочтительно, из пептидов, имеющих последовательность SEQ ID NO:88-116, как определено выше.
Пептиды, которые применимы для целей изобретения, имеющие последовательность SEQ ID NO:2, 3, 4, 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 19, 20, 21, 23, 24, 26, 27, 29, 30, 32, 34, 35, 37, 38, 40, 42, 44-125 и 177, являются новыми и составляют другой объект изобретения.
Способ по изобретению и его преимущества представлены в остальной части настоящего описания, касающегося различных неограничивающих примеров осуществления способа по изобретению.
Пример 1
Идентификация микроорганизмов из образца посредством биохимического профиля
1. Культивирование образца на культуральной среде
Оптимальные культуральные среды и оптимальные условия культивирования отличаются, в зависимости от вида микроорганизма. По умолчанию, образец инокулируют в различные среды:
- Агар Columbia с кровью овцы (bioMerieux, № 43041) в течение от 18 до 24 ч при 35°C, в анаэробных условиях или без них;
- Агар TSA (bioMerieux, № 43011) в течение от 18 до 24 ч при 37°C.
2. Идентификация микроорганизмов
Идентификацию осуществляют следующим образом:
1. Выбор отдельных колоний.
2. В асептических условиях перенести 3,0 мл водного солевого физиологического раствора (содержащего 0,45-0,50% NaCl, при pH 4,5-7,0) в прозрачную пластмассовую (полистирол) тестовую пробирку.
3. С использованием стерильной ватной палочки или стерильного тампона, перенести достаточное количество идентичных колоний в пробирку с солевым раствором, приготовленным на стадии 2 и довести бактериальную суспензию до 0,50-0,63 величины McFarland с помощью калиброванного VITEK 2 DENSICHEK.
4. Расположить пробирку с бактериальной суспензией и идентификационную карту VITEK 2 на кассете VITEK 2.
5. Поместить кассету в устройство VITEK 2.
6. Действия по заполнению, герметизации, инкубации и считыванию являются автоматическими.
7. Получение биохимического профиля.
8. Идентификация с помощью системы VITEK 2, проводимая путем сравнения с биохимическими профилями известных штаммов.
Пример 2
Идентификация микроорганизмов из образца с помощью MALDI-TOF
Идентификацию проводят следующим образом:
1. Перенесение с использованием 1-мл петли части колонии микроорганизмов, полученной согласно примеру 1, и гомогенное нанесение на планшет для масс-спектрометрии с помощью MALDI-TOF.
2. Покрытие нанесенного образца 1 мкл матрицы. Используемая матрица представляет собой насыщенный раствор HCCA (альфа-циано-4-гидроксикоричная кислота) в органическом растворителе (50% ацетонитрил и 2,5% трифторуксусная кислота).
3. Высушивание при температуре окружающей среды.
4. Установка планшета в масс-спектрометр.
5. Получение спектра масс.
6. Сравнение полученного спектра со спектрами, содержащимися в базе знаний.
7. Идентификация микроорганизма путем сравнения полученных пиков с пиками из базы знаний.
Пример 3
Идентификация микроорганизмов из образца с помощью ESI-MS
Идентификацию осуществляют следующим образом:
1. Взятие образца колонии микроорганизма, полученной согласно примеру 1, и суспендирование в 100 мкл деминерализованной воды.
2. Центрифугирование при 3000×g в течение 5 минут.
3. Удаление супернатанта.
4. Ресуспендирование в 100 мкл дименирализованной воды.
5. Центрифугирование при 3000×g в течение 5 минут.
6. Удаление супернатанта.
7. Ресуспендирование в 100 мкл смеси ацетонитрила, деминерализованной воды и муравьиной кислоты (50/50/0,1%).
8. Фильтрация с помощью фильтра, имеющего размер пор 0,45 мкм.
9. Инжекция в масс-спектрометр в режиме однократной MS.
10. Получение спектра масс.
11. Сравнение полученного спектра со спектрами, содержащимися в базе знаний.
12. Идентификация микроорганизма, исходя из эталонных спектров.
Пример 4
Получение расщепленных белков из микроорганизмов
Обычно, осуществляют следующий протокол из 11 стадий:
1. Взятие образца колонии микроорганизмов, полученной согласно примеру 1, и суспендирование в 10-100 мкл раствора 6M гуанидина гидрохлорида, 50 мМ Tris-HCl, pH=8,0.
2. Добавление дитиотреитола (DTT) для получения конечной концентрации 5 мМ.
3. Восстановление в течение 20 минут при 95°C на водяной бане.
4. Охлаждение пробирок до температуры окружающей среды.
5. Добавление йодацетамида до конечной концентрации 12,5 мМ.
6. Алкилирование в течение 40 минут при температуре окружающей среды и в темноте.
7. Разбавить в 6 раз 50 мМ раствором NH4HCO3, pH=8,0, для получения конечной концентрации гуанидина гидрохлорида 1 M.
8. Добавление 1 мкг трипсина.
9. Расщепление при 37°C в течение 6 часов в течение ночи.
10. Добавление муравьиной кислоты до pH менее 4 для остановки реакции.
11. Ультрацентрифугирование при 100000 g в течение 30 минут.
Пример 5
Определение характеристик образцов S. aureus :
После установления одного или нескольких видов в образцах согласно любому из способов, описанных в примерах 1-3, анализируют виды, приведенные ниже.
Тринадцать штаммов S. aureus анализируют для подтверждения их идентификации и для установления их характеристик:
Тот же способ анализа применяют для видов, которые не относятся к виду S. aureus, чтобы они служили в качестве отрицательного контроля:
Каждый образец обрабатывают согласно примеру 4, затем расщепленные белки объемом 5 мкл инъецируют и анализируют согласно следующим условиям:
- Хроматографическая система Agilent 1100 series от компании Agilent Technologies (Massy, Франция).
- Колонка Waters Symmetry C18, внутренний диаметр 2,1 мм, длина 100 мм, размер частиц 3,5 мкм.
- Растворитель A: H2O + 0,1% муравьиная кислота.
- Растворитель B: ACN + 0,1% муравьиная кислота.
- градиент ВЭЖХ, определенный в таблице 1 ниже:
- Элюат на выходе из хроматографической колонки прямо инъецируют в источник ионизации масс-спектрометра QTRAP® 5500 от компании Applied Biosystems (Foster City, США).
- Пептиды, полученные путем расщепления белков микроорганизма, анализируют масс-спектрометрией в режиме MRM. Проводят детекцию только пептидов, указанных в таблице 2. Для этого проводят детекцию фрагмента(ов) первого поколения, указанного в таблице 2. Применение, для которого каждый переход, т.е. каждый пептид, ассоциированный с его фрагментом первого поколения, позволяет получить ответ, указано в столбце "клинический интерес" таблице 2 с помощью букв I, T, R и V. I обозначает подтверждение идентификации микроорганизма, T обозначает типирование, R обозначает устойчивость по меньшей мере к одному антибиотику и V обозначает детекцию факторов вирулентности.
Состояние заряда пептида-предшественника, его время удержания и переходы, т.е. отношения (m/z)1 в Q1 и (m/z)2 в Q3, указаны в таблице 3. Энергия столкновения, используемая для фрагментации иона-предшественника, также указана в таблице 3.
- Другие использованные параметры устройства являются следующими:
Измеряли площади, полученные для каждого из исследованных переходов и для каждого из исследованных микроорганизмов. Все переходы с площадью, превышающей или равной 1000 (произвольные единицы), считаются положительными и обозначены как "1" в таблицах 4A и 4B. Все переходы с площадью менее 1000 считаются отрицательными и обозначены как 0 в таблицах 4A и 4B. Когда не наблюдали пика сигнала, переход обозначали как отрицательный.
Далее, число положительных переходов суммировано для применений I, R и V и представлено в таблице 5.

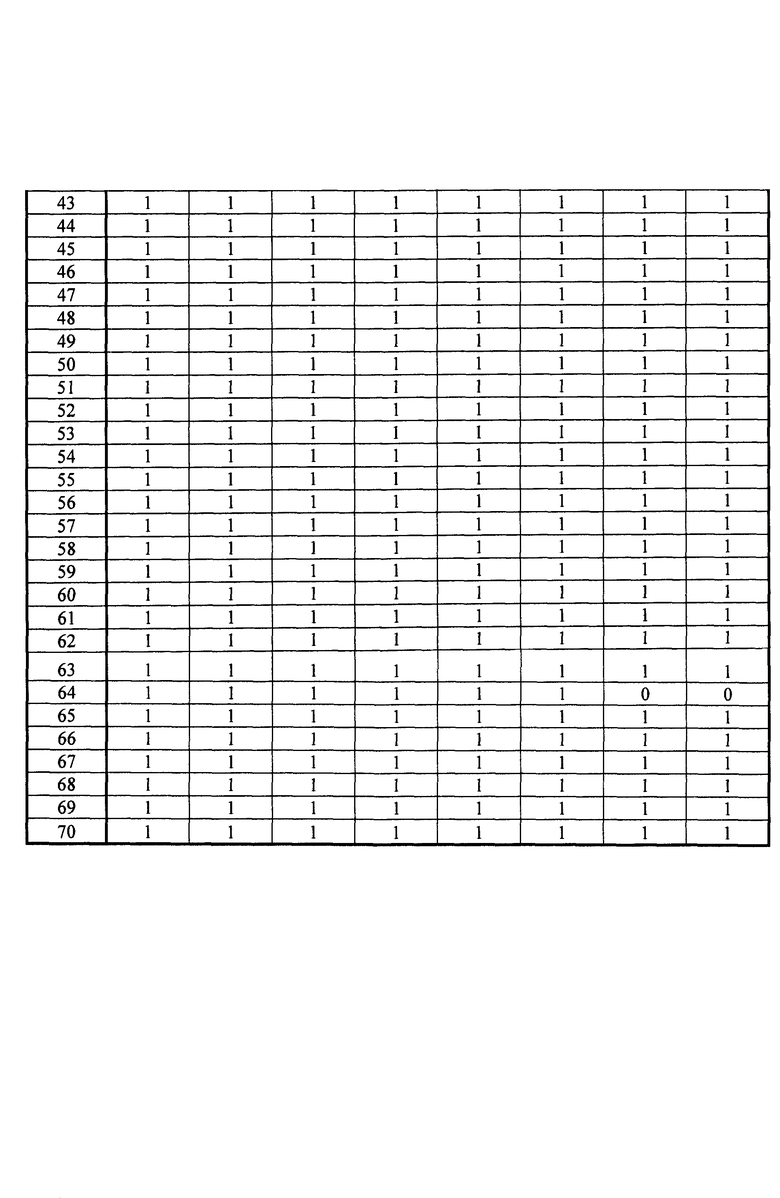
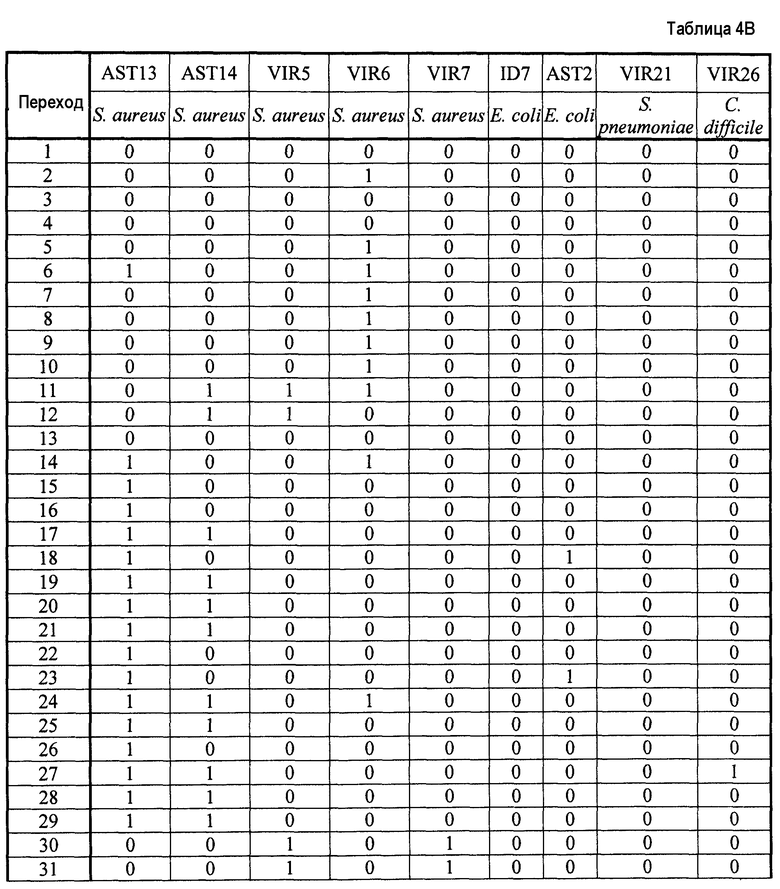
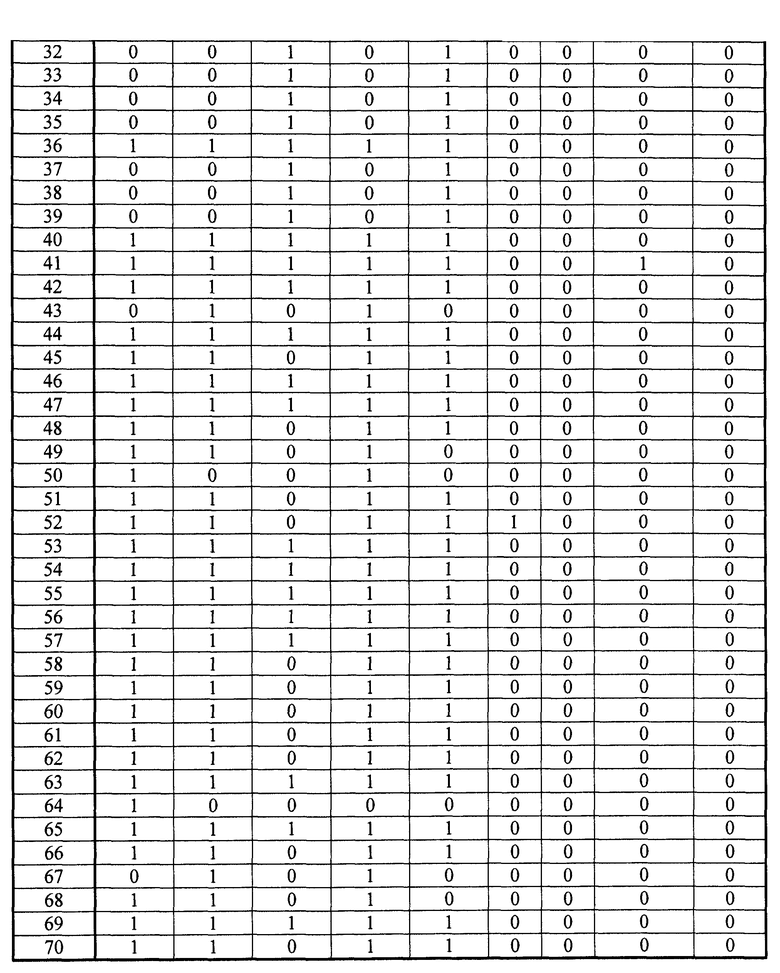
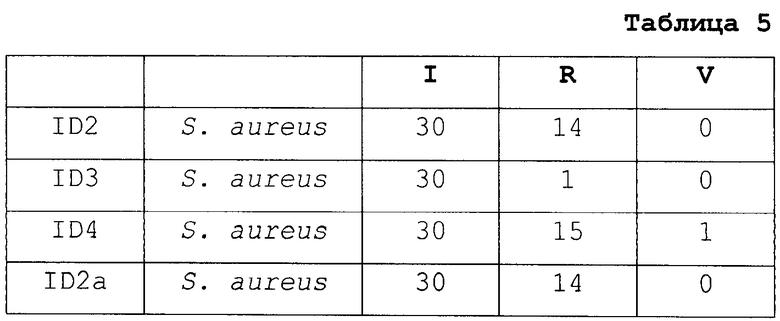

Все образцы S. aureus обладают более чем 12 положительными переходами в категории I. Таким образом, подтверждено, что эти образцы действительно принадлежат виду S. aureus.
С другой стороны, образцы ID7, AST2, VIR21 и VIR26 обладают менее чем двумя положительными переходами в категории I, таким образом, подтверждено, что эти образцы не принадлежат виду S. aureus.
Штаммы ID2, ID4, ID2a, ID4a, AST13 и AST14 S. aureus обладают более чем восемью положительными переходами для категории R, таким образом, они экспрессируют пенициллин-связывающий белок (PBP2a), что является синонимичным механизму устойчивости к пенициллинам группы M (например, метициллину).
С другой стороны, штаммы ID3, ID3a, AST7, AST8, VIR5, VIR6 и VIR7 S. aureus обладают менее чем тремя положительными переходами для категории V, таким образом, они не экспрессируют PBP2a. Таким образом, эти штаммы являются чувствительными к антибиотику, такому как пенициллин группы M.
Штаммы ID7, AST2, VIR21 и VIR26, которые не принадлежат виду S. aureus, также обладают менее чем тремя переходами для категории R, таким образом, они не экспрессируют PBP2a, тем самым подтверждая специфичность способа.
Образцы VIR5 и VIR7 S. aureus обладают более чем девятью положительными переходами в категории V, таким образом, они экспрессируют белок лейкоцидина Пантон-Валентайна (PVL).
С другой стороны, штаммы ID2, ID3, ID4, ID2a, ID3a, ID4a, AST7, AST8, AST13, AST14 и VIR6 S. aureus проявляют менее трех положительных переходов, таким образом, они не экспрессируют PVL. Эти штаммы, таким образом, не обладают вирулентными свойствами, связанными с PVL.
Штаммы ID7, AST2, VIR21 и VIR26, которые не принадлежат виду S. aureus, также обладают менее чем тремя переходами для категории V, таким образом, они не экспрессируют PVL, тем самым подтверждая специфичность способа.
Для типирования переходы категории T каждого штамма сравнивают с переходами других штаммов, считающихся эталонными штаммами. На практике величину 0 приписывают, когда переходы между двумя штаммами классифицируют в одну категорию (положительная или отрицательная) и величину 1 приписывают, когда переходы между двумя штаммами классифицируют в различные категории (положительный переход и отрицательный переход). Величины обобщенно представлены для всех переходов категории T каждой пары штаммов для установления показателя. Показатели представлены в таблице 6:
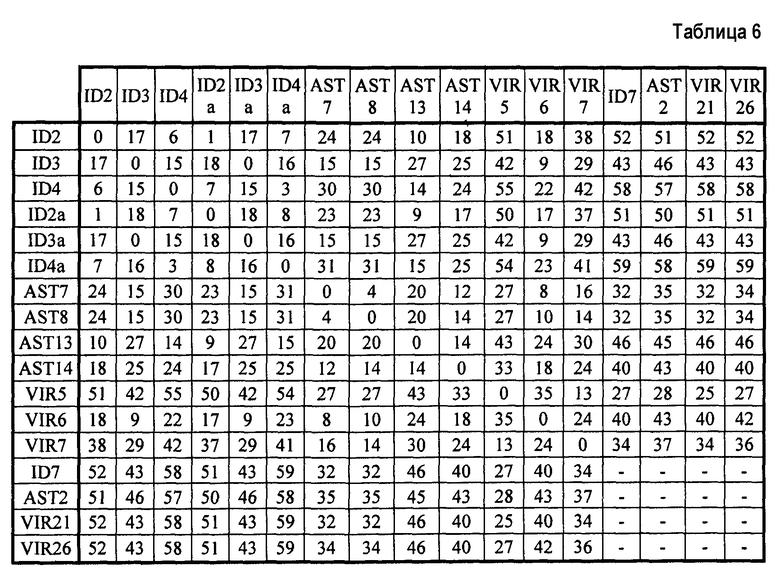
Штаммы, которые имеют показатель, меньший или равный 4 являются штаммами одного типа, штаммы, которые имеют показатель строго выше 4, являются штаммами разного типа.
Таким образом, штаммы ID2 и ID2a, ID3 и ID3a, ID4 и ID4a и AST7 и AST8 являются штаммами одного типа. Все другие штаммы, взятые парами, являются штаммами разного типа. Следует отметить высокие суммы, полученные для штаммов ID7, AST2, VIR21 и VIR26, которые не являются S. aureus, и всех других штаммов, которые являются S. aureus. Эти результаты подтверждают специфичность способа.
Штаммы ID7, AST2, VIR21 и VIR26, безусловно, не являются штаммами одного типа; это было бы абсурдным, они являются штаммами различных видов. Таким образом, эти штаммы нельзя сравнивать друг с другом, и в таблице 6 величина не представлена.
Чрезвычайно преимущественно, показатели выше 25, например, для ID2 и VR7, отражают высокую дивергенцию между штаммами. Показатели от 15 до 25, как для ID2 и AST14, отражают умеренную дивергенцию и показатели от 5 до 15, как для ID2 и ID4a, отражают слабую дивергенцию.
Следовательно, способ, осуществленный таким образом, позволяет не только установить, являются ли два штамма штаммами одного типа, что важно для идентификации общего очага инфекции, но также оценить сходство двух штаммов, что является чрезвычайно важным для эпидемиологических исследований.
Этот пример показывает, что, в высокой степени преимущественно, настоящее изобретение позволяет, за время менее одного часа, которое является очень коротким, подтвердить принадлежность вида, такого как S. aureus, и определить, одновременно в том же анализе, характеристики типирования и потенциальную устойчивость по меньшей мере к одному противомикробному средству и установить существование фактора вирулентности. Эти свойства были установлены с помощью одного и того же устройства, что существенно упрощает анализ и выдачу результатов. Наконец, характеристики бактерий установлены с использованием бактериальных белков, которые отражают существование живых и жизнеспособных микроорганизмов, в отличие от определения характеристик с использованием бактериальной ДНК, которое может быть искажено существованием погибших бактерий.
Пример 6
Протокол расщепления микроорганизмов с помощью стадии обессоливания
Обычно, выполняют следующий протокол из 17 стадий:
Стадии 1-10: те же, что и в примере 4.
11. Объем образца доводят до 1 мл смесью вода/0,1% (об./об.) муравьиная кислота.
12. Уравновешивание колонок Waters Oasis HLB с помощью 1 мл метанола, затем 1 мл смеси H2O/0,1% (об./об.) муравьиная кислота.
13. Нанесение образца, который продвигается под действием силы тяжести.
14. Промывание смесью 1 мл H2O/0,1% (об./об.) муравьиная кислота.
15. Элюирование 1 мл смеси 80% метанола и 20% воды/0,1% (об./об.) муравьиная кислота.
16. Элюат выпаривают с помощью испарителя SpeedVac® SPD2010 (Thermo Electron Corporation, Waltham, Massachusetts, США), в течение двух часов для получения объема приблизительно 100 мкл.
17. Затем элюат отбирают в смесь вода/0,5% (об./об.) раствор муравьиной кислоты, в количестве, достаточном для (QS) 250 мкл.
Пример 7
Определение характеристик образцов E coli :
После установления одного или нескольких видов в образцах согласно одному из способов, описанных в примерах 1-3, анализируют виды, приведенные ниже.
Пятнадцать штаммов E coli анализируют для подтверждения их принадлежности и для определения их характеристик:
Тот же способ анализа применяют для видов, не принадлежащих виду E. coli, чтобы они служили в качестве отрицательного контроля:
- S. aureus
VIR10
- S. pneumoniae
VIR19
- C. difficile
VIR28
Каждый образец обрабатывают согласно примеру 6, затем расщепленные белки объемом 5 мкл инжектируют и анализируют согласно следующим условиям:
- Хроматографическая система Agilent 1100 series от компании Agilent Technologies (Massy, Франция).
- Колонка Waters Symmetry C18, внутренний диаметр 2,1 мм, длина 100 мм, размер частиц 3,5 мкм.
- Растворитель A: H2O + 0,1% муравьиная кислота.
- Растворитель B: ACN + 0,1% муравьиная кислота.
- градиент ВЭЖХ, определенный в таблице 7 ниже:
- Элюат на выходе из хроматографической колонки прямо инъецируют в источник ионизации масс-спектрометра QTRAP® 5500 от компании Applied Biosystems (Foster City, США).
- Пептиды, полученные путем расщепления белков микроорганизма, анализируют масс-спектрометрией в режиме MRM. Проводят детекцию только пептидов, указанных в таблице 8. Для этого проводят детекцию фрагмента(ов) первого поколения, указанного в таблице 8. Применение, для которого каждый переход, т.е. каждый пептид, ассоциированный с его фрагментом первого поколения, позволяет получить ответ, указано в столбце "клинический интерес" таблице 8 с помощью букв I, T, R и V. I обозначает подтверждение идентификации микроорганизма, T обозначает типирование, R обозначает устойчивость по меньшей мере к одному антибиотику и V обозначает детекцию факторов вирулентности.
Состояние заряда пептида-предшественника, его время удержания и переходы, т.е. отношения (m/z)1 в Q1 и (m/z)2 в Q3, указаны в таблице 9. Энергия столкновения, используемая для фрагментации иона-предшественника, также указана в таблице 9.
- Другие использованные параметры устройства являются следующими:
Измеряли площади, полученные для каждого из исследованных переходов и для каждого из исследованных микроорганизмов. Все переходы с площадью, превышающей или равной 2500 (произвольные единицы) считаются положительными и обозначены как "1" в таблицах 10A и 10B. Все переходы с площадью менее 2500 считаются отрицательными и обозначены как 0 в таблицах 10A и 10B. Когда не наблюдали пика сигнала, переход обозначали как отрицательный.
Далее, число положительных переходов суммировано для применений I, R и V и представлено в таблице 11.
В образцы E. coli имеют более 44 положительных переходов в категории I. Таким образом, подтверждено, что все эти образцы принадлежат виду E. coli.
С другой стороны, образцы VIR10, VIR19 и VIR28 имеют менее 3 положительных переходов в категории I, таким образом, подтверждено, что эти образцы не принадлежат виду E. coli.
Штаммы AST2, AST3, AST4 и AST5 E. coli имеют более 14 положительных переходов для категории R, следовательно, они экспрессируют обусловленную плазмидой пенициллиназу TEM-2, что является синонимичным с механизмом устойчивости к пенициллинам.
С другой стороны, штаммы AST1, VIR41, VIR42, VIR43, VIR44, VIR45, ID6, ID7, ID8, ID9 и ID10 E. coli имеют менее 7 положительных переходов для категории R, следовательно, они не экспрессируют обусловленную плазмидой пенициллиназу TEM-2. Таким образом, эти штаммы являются устойчивыми к пенициллинам, в частности аминопенициллинам или пенициллинам A (ампициллин), карбоксипенициллинам или пенициллинам C (тикарциллин) и уреидопенициллину или пенициллину U (пиперациллин).
Штаммы VIR10, VIR19 и VIR28, которые не принадлежат виду E. coli, имеют менее двух переходов для категории К, следовательно, они не экспрессируют TEM-2, что, тем самым, подтверждает специфичность способа.
Образцы VIR41, VIR42, VIR43 и VIR44 E. coli имеют более четырех положительных переходов в категории V, следовательно, они экспрессируют токсины Шига 1 или 2 (STX1 или STX2). Более конкретно, для VIR41, положительными являются переходы 118-125, следовательно, VIR41 одновременно экспрессирует токсин Шига 1 и токсин Шига 2. Для VIR42 и VIR44 положительными являются переходы 121-125, следовательно, они экспрессируют токсин Шига 2. То же самое справедливо для VIR45, который имеет переходы 123 и 124. VIR43, который имеет переходы 118-122, экспрессирует токсин Шига 2. С другой стороны, штаммы AST1, AST2, AST3, AST4, AST5, VIR45, ID6, ID7, ID8, ID9 и ID10 E. coli имеют менее трех положительных переходов, таким образом, они не экспрессируют токсин Шига. Таким образом, эти штаммы не обладают свойствами вирулентности, связанными с токсинами Шига.
Штаммы VIR10, VIR19 и VIR28, которые не принадлежат виду E. coli, также обладают менее чем тремя переходами для категории V, таким образом, они не экспрессируют stx1 или stx2, что, тем самым, подтверждает специфичность способа.
Для типирования переходы категории T каждого штамма сравнивают с переходами других штаммов, считающихся эталонными штаммами. На практике величину 0 приписывают, когда переходы между двумя штаммами классифицируют в одну категорию (положительная или отрицательная) и величину 1 приписывают, когда переходы между двумя штаммами классифицируют в различные категории (положительный переход и отрицательный переход). Величины обобщенно представлены для всех переходов категории T каждой пары штаммов для установления показателя. Показатели представлены в таблице 12:

Штаммы, которые имеют показатель, меньший или равный 4 являются штаммами одного типа, штаммы, которые имеют показатель строго выше 4, являются штаммами разного типа.
Таким образом, штаммы AST2, AST3, AST4 и AST5 являются штаммами одного типа. То же самое является справедливым, соответственно, для штаммов AST1 и ID6 и для штаммов ID7, ID8, ID9 и ID10. Все другие штаммы, взятые парами, являются штаммами разного типа. Следует отметить высокие суммы, полученные для штаммов VIR10, VIR19 и VIR28, которые не являются S. aureus, и всех других штаммов, которые являются S. aureus. Эти результаты подтверждают специфичность способа.
Штаммы VIR10, VIR19 и VIR28 представляют собой штаммы различных видов. Таким образом, эти штаммы нельзя сравнивать друг с другом, и в таблице 12 величина не представлена. Чрезвычайно преимущественно, показатель выше 20, например, между AST4 и VIR41, отражает высокую дивергенцию между штаммами. Показатели от 12 до 20, как между ID7 и VIR42, отражают умеренную дивергенцию и показатели от 4 до 12, как между ID10 и AST1, отражают слабую дивергенцию.
Следовательно, способ, осуществленный таким образом, позволяет не только установить, являются ли два штамма штаммами одного типа, что важно для идентификации общего очага инфекции, но также оценить сходство двух штаммов, что является чрезвычайно важным для эпидемиологических исследований.
Этот пример показывает, что, в высокой степени преимущественно, настоящее изобретение позволяет, за время менее одного часа, которое является очень коротким, подтвердить принадлежность вида, такого как E. coli и определить, одновременно в том же анализе, характеристики типирования и потенциальную устойчивость по меньшей мере к одному противомикробному средству и установить существование фактора вирулентности. Эти свойства были установлены с помощью одного и того же устройства, что существенно упрощает анализ и выдачу результатов. Наконец, характеристики бактерий установлены с использованием бактериальных белков, которые отражают существование живых и жизнеспособных микроорганизмов, в отличие от определения характеристик с использованием бактериальной ДНК, которое может быть искажено существованием погибших бактерий.
Пример 8
Протокол расщепления микроорганизмов в присутствии метанола
Обычно осуществляют следующий протокол, имеющий 17 стадий:
1. Взятие образца колонии микроорганизма, полученной согласно примеру 1, и суспендирование в 400 мкл 50 мМ раствора бикарбоната аммония, pH=8,0.
2. Добавление 600 мкл метанола.
Стадии 3-7: такие же, как стадии 2-6 примера 4.
Стадии 8-17: такие же как стадии 8-17 примера 6.
Пример 9
Определение характеристик образцов C. albicans
После установления одного или нескольких видов в образцах согласно одному из способов, описанных в примерах 1-3, анализируют виды, приведенные ниже.
Семнадцать штаммов C. albicans анализируют для подтверждения их принадлежности и для определения их характеристик:
Тот же способ анализа применяют для видов, не принадлежащих видов C. albicans, чтобы они служили в качестве отрицательного контроля:
- E. coli
VIR43
- S. aureus
VIR5
- E. faecium
AST-VAN8
Каждый образец обрабатывают согласно примеру 8, затем расщепленные белки объемом 5 мкл инжектируют и анализируют согласно тем же условиям, что и в примере 7.
- Пептиды, полученные путем расщепления белков микроорганизма, анализируют масс-спектрометрией в режиме MRM. Проводят детекцию только пептидов, указанных в таблице 13. Для этого проводят детекцию фрагмента(ов) первого поколения, указанного в таблице 13. Применение, для которого каждый переход, т.е. каждый пептид, ассоциированный с его фрагментом первого поколения, позволяет получить ответ, указано в столбце "клинический интерес" таблице 14 с помощью букв I, T, R и V. I обозначает подтверждение идентификации микроорганизма, T обозначает типирование, R обозначает устойчивость по меньшей мере к одному антибиотику и V обозначает детекцию факторов вирулентности.
Состояние заряда пептида-предшественника, его время удержания и переходы, т.е. отношения (m/z)1 в Q1 и (m/z)2 в Q3, указаны в таблице 14. Энергия столкновения, используемая для фрагментации иона-предшественника, также указана в таблице 14.
перехода
Другие использованные параметры устройства, являются такими же, как в примере 7. Измеряли площади, полученные для каждого из исследованных переходов и для каждого из исследованных микроорганизмов. Все переходы с площадью, превышающей или равной 2000 (произвольные единицы) считаются положительными и обозначены как "1" в таблицах 15A и 15B. Все переходы с площадью менее 2000 считаются отрицательными и обозначены как 0 в таблицах 15A и 15B. Когда не наблюдали пика сигнала, переход обозначали как отрицательный.
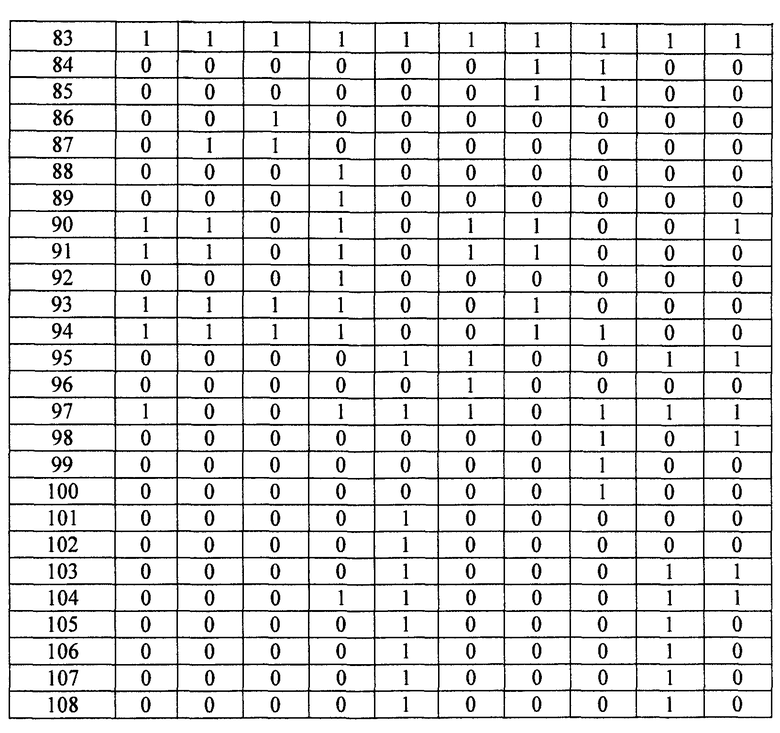
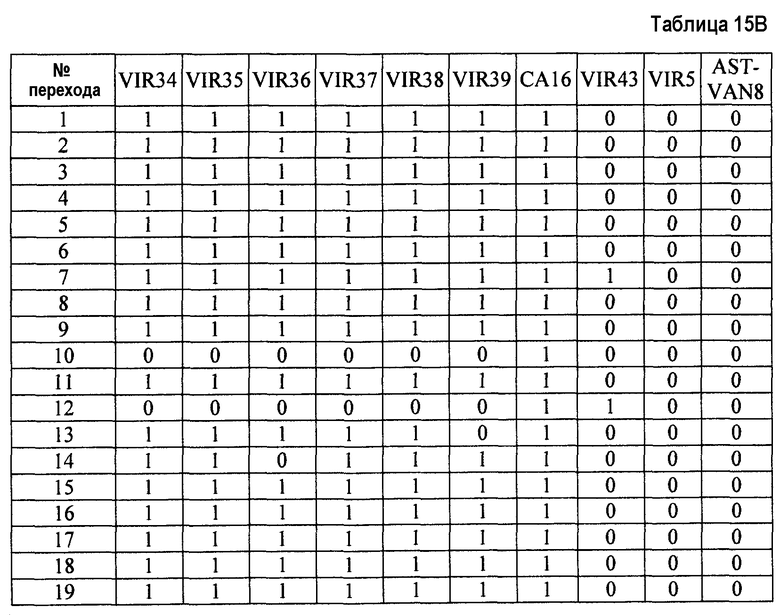
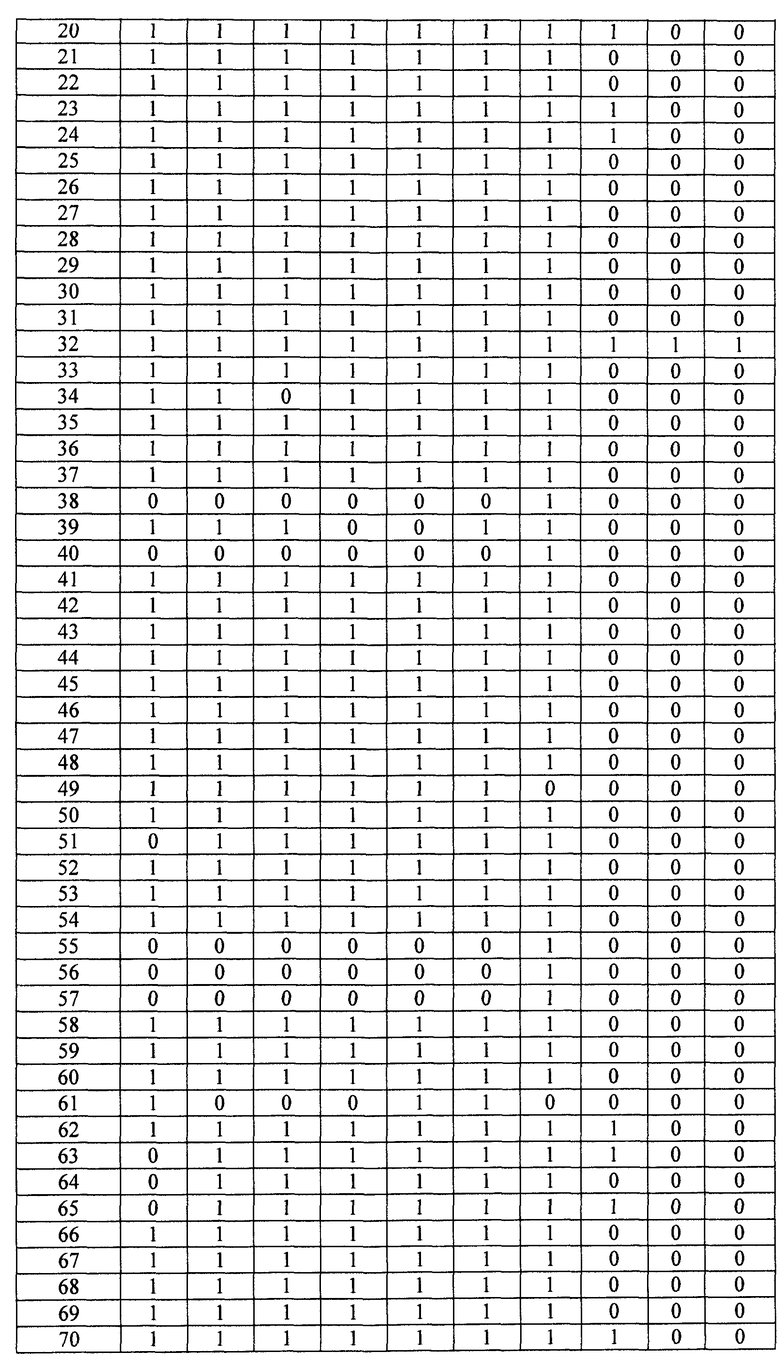
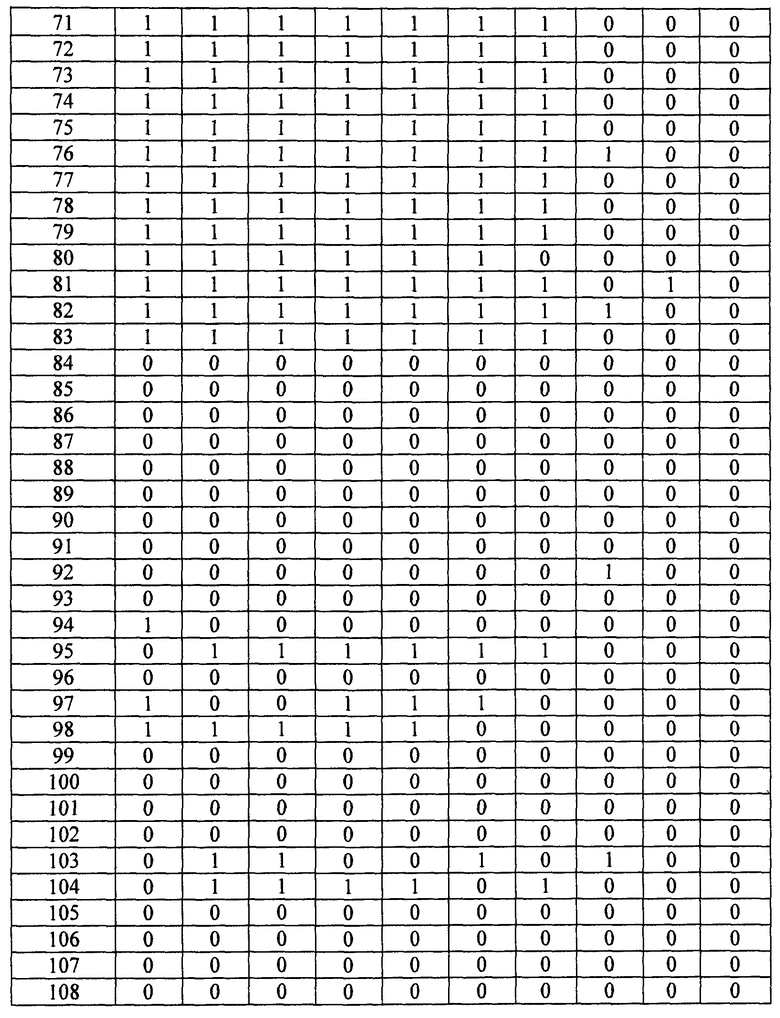
Далее, число положительных переходов суммировано для применений I, R и V и представлено в таблице 16.
Все образцы C. albicans имеют более 60 положительных переходов в категории I. Таким образом, подтверждено, что все эти образцы принадлежат виду C. albicans.
С другой стороны, образцы VIR43, VIR5 и AST-VAN8 имеют менее 13 положительных переходов в категории I, таким образом, подтверждено, что эти образцы не принадлежат виду C. albicans.
Наблюдение переходов 84 и 85 указывает на присутствие мутантного пептида и отражает устойчивость штаммов ATF6 и ATF7.
Наблюдение переходов 86 и 87 указывает на присутствие мутантного пептида и отражает устойчивость штаммов ATF2 и ATF3.
Наблюдение перехода 96 указывает на присутствие мутантного пептида и отражает устойчивость штамма ATF5.
Наблюдение переходов 99 и 100 указывает на присутствие мутантного пептида и отражает устойчивость штамма ATF7.
Наблюдение переходов 88 и 89 указывает на присутствие нативного пептида и отражает чувствительность штамма ATF4.
Наблюдение переходов 93 и 94 указывает на присутствие нативного пептида и отражает чувствительность штаммов ATF1 и ATF4.
Все площади переходов I суммированы с получением суммарного значения SI. Все площади переходов V суммированы с получением суммарного SV. Далее, вычислено отношение SV/SI и умножено на коэффициент умножения MF. Полученные результаты представлены в таблице 17.
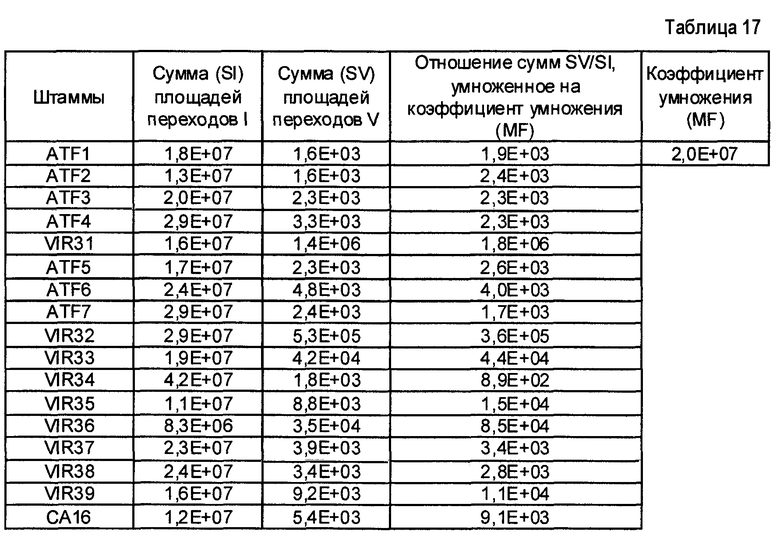
Штаммы VIR31 и VIR32, которые имеют отношение (SV/SI)×MF более 9×104, сверхэкспрессируют липазу 8, следовательно, эти штаммы являются вирулентными. Все другие штаммы имеют отношение менее 9×104, они не сверхэкспрессируют липазу 8 и, таким образом, не являются вирулентными. Интересно, что, поскольку отношение (SV/SI)×MF является более высоким для штамма VIR31, чем для штамма VIR32, штамм VIR31 характеризуется как более вирулентный, чем штамм VIR32.
Для типирования переходы категории T каждого штамма сравнивают с переходами других штаммов, считающихся эталонными штаммами. На практике величину 0 приписывают, когда переходы между двумя штаммами классифицируют в одну категорию (положительная или отрицательная) и величину 1 приписывают, когда переходы между двумя штаммами классифицируют в различные категории (положительный переход и отрицательный переход). Величины обобщенно представлены для всех переходов категории T каждой пары штаммов для установления показателя. Показатели представлены в таблице 18:
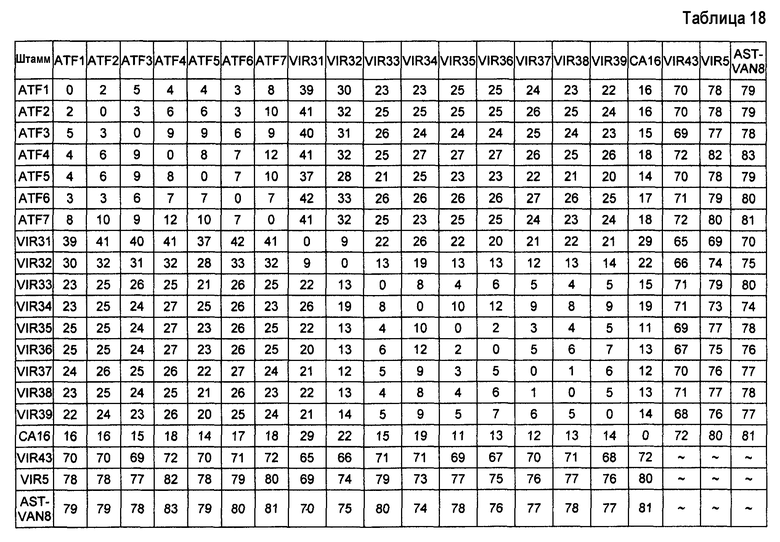
Штаммы, которые имеют показатель, меньший или равный 2 являются штаммами одного типа, штаммы, которые имеют показатель строго выше 2, являются штаммами разного типа.
Таким образом, штаммы ATF1 и ATF2, VIR35 и VIR36, и VIR37 и VIR38 соответственно являются штаммами одного типа. Все другие штаммы, взятые парами, являются штаммами разного типа. Следует отметить высокие суммы, полученные для штаммов VIR43, VIR5 и AST-VAN8, которые не являются S. aureus, и всех других штаммов, которые являются S. aureus. Эти результаты подтверждают специфичность способа.
Штаммы VIR43, VIR5 и AST-VAN8 представляют собой штаммы различных видов. Таким образом, эти штаммы нельзя сравнивать друг с другом и в таблице 18 величина не представлена.
Чрезвычайно преимущественно, показатели более 20, например, от ATF1 до VIR39, отражает высокую дивергенцию между штаммами. Показатели от 14 до 20, как между CA16 и ATF1, отражают умеренную дивергенцию, и показатели от 4 до 14, как между ATF1 и ATF7, отражают слабую дивергенцию.
Следовательно, способ, осуществленный таким образом, позволяет не только установить, являются ли два штамма штаммами одного типа, что важно для идентификации общего очага инфекции, но также оценить сходство двух штаммов, что является чрезвычайно важным для эпидемиологических исследований.
Этот пример показывает, что, в высокой степени преимущественно, настоящее изобретение позволяет, за время менее одного часа, которое является очень коротким, подтвердить принадлежность вида, такого как C. albicans и определить, одновременно в том же анализе, характеристики типирования и потенциальную устойчивость по меньшей мере к одному противомикробному средству и установить существование фактора вирулентности. Настоящее изобретение также позволяет количественный анализ, который является особенно преимущественным, когда характеристики устойчивости по меньшей мере к одному антибиотику или вирулентности связаны с уровнем экспрессии белка или метаболита. Это проиллюстрировано в настоящем описании для количественного анализа липазы 8. Эти свойства были установлены с помощью одного и того же устройства, что существенно упрощает анализ и выдачу результатов. Наконец, установлены характеристики дрожжей с использованием белков дрожжей, которые отражают существование живых и жизнеспособных микроорганизмов, в отличие от определения характеристик с использованием ДНК грибов, которое может быть искажено существованием погибших дрожжей.
Пример 10
Протокол расщепления микроорганизмов в присутствии метанола, пригодный для анализа по меньшей мере одного метаболита
Обычно выполняют следующий протокол, имеющей 19 стадий:
1. Взятие образцов пяти колоний микроорганизмов, полученных согласно примеру 1, и суспендирование в 100 мкл 6 M гидрохлорида гуанидина, 50 мМ растворе Tris-HCl, pH=8,0.
2. Центрифугирование при 15000×g в течение 5 минут.
3. Перенесение осадка в 400 мкл a 50 мМ раствора бикарбоната аммония, pH=8,0.
4. Добавление 600 мкл метанола.
Стадии 5-9: такие же, как стадии 2-6 примера 4.
Стадии 10-19: такие же как стадии 8-17 примера 6.
Пример 11
Определение характеристик образцов C. albicans путем одновременного анализа белков и метаболитов
После установления одного или нескольких видов в образцах согласно одному из способов, описанных в примерах 1-3, анализируют виды, приведенные ниже.
Пять штаммов C. albicans анализируют для подтверждения их принадлежности и для определения их характеристик:
Каждый образец обрабатывают согласно примеру 10, затем расщепленные белки объемом 20 мкл инжектируют и анализируют согласно тем же условиям, что и в примере 7.
Таблицы 13 и 14 идентичны с одним исключением. К способу добавлен номер перехода 109. Он соответствует молекуле эргостерина с клиническим интересом I и R.
Состояние заряда предшественника составляет 1, время удержания составляет 36,3 минут, m/z, отфильтрованное в Q1, составляет 379,4, m/z, отфильтрованное в Q3, составляет 69,2, и энергия столкновения составляет 47.
Параметры масс-спектрометрии идентичны параметрам примера 7, за исключением представленного ниже отличия:
Все переходы 1-108 анализируют аналогично примеру 9. Все площади переходов I суммированы для получения суммы SI. Площадь перехода 109 названа A109. Затем вычислено отношение A109/SI и умножено на коэффициент умножения MF. Полученные результаты приведены в таблице 19.
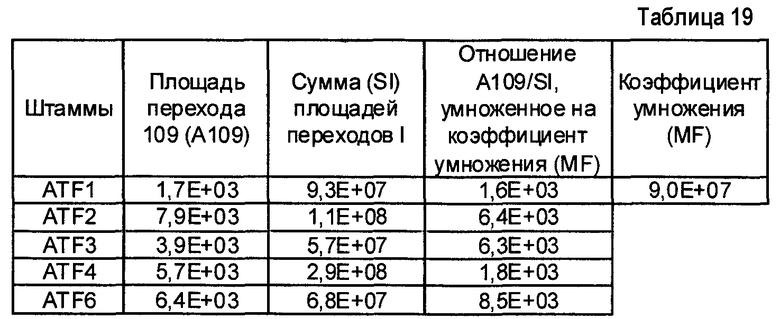
Штаммы ATF2, ATF3 и ATF6, которые имеют отношение (A109/SI)×MF более 2×103, сверхэкспрессирует эргостерин; следовательно, эти штаммы являются устойчивыми. Все другие штаммы имеют отношение менее 2×103, они не экспрессируют эргостерин и, следовательно, не являются устойчивыми.
Этот пример показывает, что, в высокой степени преимущественно, настоящее изобретение позволяет одновременно проанализировать, в одном анализе, за время менее одного часа, которое является очень коротким, и количественно различные соединения, такие как белки, пептиды или метаболиты. Еще более преимущественно, чем анализ белка, количественный анализ метаболита вследствие действия белка характеризует присутствие функционального белка и более широко функциональный путь синтеза. Таким образом, количественное определение эргостерина позволяет в этом случае установить существование механизма устойчивости к флуконазолу, связанного с функциональной и высоко активной ланостериндеметилазы.
Пример 12
Определение характеристик микроорганизмов, присутствующих в образцах мочи
После установления одного или нескольких видов в образцах согласно одному из способов, описанных в примерах 1-3, анализируют пять образцов мочи, контаминированных E. coli (образец мочи 1-5). Также анализировали неконтаминированный шестой образец мочи (образец мочи 6), чтобы он служил в качестве отрицательного контроля.
Осуществляют следующий протокол, имеющий 29 стадии (стадии 5-12 являются необязательными и могут быть пропущены без существенного изменения результатов):
1. Центрифугирование 5 мл контаминированной мочи при 2000×g в течение 30 секунд.
2. Выделение супернатанта.
3. Центрифугирование при 15000×g в течение 5 минут.
4. Удаление супернатанта.
5. Промывание осадка 3 мл дистиллированной воды путем ресуспендирования.
6. Центрифугирование при 15000×g в течение 5 минут.
7. Удаление супернатанта.
8. Контактирование осадка с растворителем в разведении 1/10.
9. Оставление на один час при -20°C.
10. Центрифугирование при 15000×g в течение 5 минут.
11. Удаление супернатанта.
12. Контактирование осадка с растворителем в разведении 1/10.
13. Оставление на один час при -20°C.
14. Центрифугирование при 15000×g в течение 5 минут.
15. Удаление супернатанта.
12. Суспендирование осадка в 10-100 мкл 6 M гуанидина гидрохлорида, 50 растворе мМ Tris-HCl, pH=8,0.
13. Стадии 14-18: такие же, как стадии 2-6 примера 4.
19. Стадии 19-28: такие же, как стадии 8-17 примера 6.
29. Инжектирование 100 мкл подкисленного элюата в хроматографическую систему и масс-спектрометр, согласно протоколу, описанному в примере 7.
Измеряли площади, полученные для каждого из исследованных переходов и для каждого из исследованных микроорганизмов. Поскольку образцы мочи имеют бактериальную нагрузку, которая в значительной степени отличается между образцами, пороговое значение, использованное в примере 7 для утверждения о положительном переходе, нельзя использовать в этом случае. Оно должно быть скорректировано по количеству присутствующих бактерий. В предыдущем примере взятие образцов колонии приводило к сравнимому количеству микроорганизмов. Для этого примера это не так. Таким образом, количество бактерий оценено путем суммирования площадей всех переходов I. Сумма площадей переходов I представлена в таблице 20.

Таким образом, образец 4 обладает бактериальной нагрузкой, которая превышает бактериальную нагрузку образца 3, которая сама по себе имеет более высокую нагрузку, чем в примерах 1 и 5, которые сами по себе имеют более высокую нагрузку, чем пример 2. Образцы 1 и 5 имеют сравнимую бактериальную нагрузку. Образец 6 не содержит E. coli и проявляет нулевую нагрузку, что тем самым, демонстрирует специфичность способа.
Затем площадь каждого перехода стандартизуют делением его площади на площадь всех переходов I. Все переходы, для которых отношение превышает или равно 0,00015 (безразмерное отношение) считаются положительными и обозначены как "1" в таблице 21. Все переходы, для которых отношение является меньшим, чем 0,00015 считаются отрицательными и обозначены как 0 в таблице 21. Когда не наблюдали пика сигнала, переход обозначали как отрицательный.
Далее, число положительных переходов суммировано для применений I, R и V и представлено в таблице 22:
Аналогично тому, как необходимо было модифицировать порог положительности перехода для учета концентрации образцов, количество пептидов, необходимых для определения характеристик штамма, необходимо корректировать по общей концентрации бактерий. Определенные плохо выявляемые пептиды в примере 7 могут быть ниже предела детекции, если количество бактерий составляет менее одной колонии.
Образцы мочи 1-5 обладают более чем 30 положительными переходами в категории I. Таким образом, подтверждено, что все эти образцы контаминированы видами E. coli.
С другой стороны, образце мочи 6 имеет положительный переход в категории I. Таким образом, подтверждено, что он не контаминирован видами E. coli.
Образцы мочи 1, 2, 3 и 5 имеют по меньшей мере пять положительных переходов для категории R, следовательно, они экспрессируют обусловленную плазмидой пенициллиназу TEM-2, что является синонимичным механизму устойчивости к пенициллинам, в частности, аминопенициллинам или пенициллинам A (ампициллин), карбоксипенициллинам или пенициллинам C (тикарциллин) и уреидопенициллину или пенициллину U (пиперациллин).
С другой стороны, образцы мочи 4 и 6 не имеют положительных переходов для категории R, таким образом, они не экспрессируют обусловленную плазмидой пенициллиназу TEM-2. Следовательно, эти штаммы являются устойчивыми к пенициллинам.
Эти результаты устойчивости к ампициллину, тикарциллину и пиперациллину были подтверждены с помощью автоматического устройства VITEK®2, продаваемого заявителем, и карт AST-EXN и AST-N103. Подтверждение на автоматическом устройстве VITEK®2 отняло время, составляющее от 6 до 8 часов, что медленнее чем для настоящего изобретения.
Образцы мочи 1-6 не имеют более одного положительного перехода для категории V. Таким образом, эти образцы мочи не являются зараженными токсинами типа токсина Шига.
Для типирования переходы категории T каждого штамма сравнивают с переходами других штаммов, считающихся эталонными штаммами. На практике величину 0 приписывают, когда переходы между двумя штаммами классифицируют в одну категорию (положительная или отрицательная) и величину 1 приписывают, когда переходы между двумя штаммами классифицируют в различные категории (положительный переход и отрицательный переход). Величины обобщенно представлены для всех переходов категории T каждой пары штаммов для установления показателя. Показатели представлены в таблице 23:
Ни один образец мочи не имеет показатель, меньший или равный 4. Штаммы, инфицирующие образцы мочи 1-5, таким образом, являются штаммами различных типов. Эти образцы мочи были собраны в местной медицинской испытательной лаборатории, где инфекции часто встречаются в различных окружающих условиях. В этих условиях, нечасто наблюдают инфекции мочи, связанные с одним и тем же штаммом.
Этот пример показывает, что, в высокой степени преимущественно, настоящее изобретение позволяет прямо из первичного образца, что является высоко преимущественным, подтвердить принадлежность вида, такого как E. coli, и определить, одновременно в том же анализе, характеристики типирования и потенциальную устойчивость по меньшей мере к одному антибиотику.
Ссылки:
[1] J. Anhalt & C. Fenselau, 1975, Anal. Chem., 47(2):219-225.
[2] A. Fox et al., ed., 1990, Analytical microbiology methods: chromatography and mass spectrometry, Plenum Press, New York, N.Y.
[3] M. Claydon et al., 1996, Nature Biotech. 14:1584-1586.
[4] T. Krishnamurthy & P. Ross, 1996, Rapid Com. Mass Spec., 10:1992-1996.
[5] P. Seng et al. 2009, Clin. Infect. Dis., 49:543-551.
[6] C. Fenselau et al., 2008, Appl. Environ. Microbiol., 904-906.
[7] D. Ding et al. 2009, J. Pharm. Biomed. Anal. 50:79-85.
[8] R. Everley et al., 2009, J. Microbiol. Methods, 77:152-158.
[9] S. Hofstadler et al., 2005, Int. J. Mass Spectrom., 242:23-41.
[10] D. Ecker, 2008, Nat. Rev. Microbiol., 6(7):553-558.
[11] W.-J. Chen et al., 2008, Anal. Chem., 80:9612-9621.
[12] D. Lopez-Ferrer et al., 2008, Anal. Chem., 80:8930-8936.
[13] D. Lopez-Ferrer et al., 2005, J. Proteome res., 4(5):1569-1574.
[14] T. Fortin et al., 2009, Mol. Cell Proteomics, 8(5):1006-1015.
[15] H. Keshishian et al., 2007, Mol. Cell Proteomics, 2212-2229.
[16] J. Stal-Zeng et al., 2007, Mol. Cell Proteomics, 1809-1817.
[17] Gaskell, Electrospray: principles and practise, 1997, J. Mass Spectrom., 32, 677-688).
[18] V. Fusaro et al., 2009, Nature Biotech. 27, 190-198.
[19] J. Mead et al., 15 Nov 2008, Mol. Cell Proteomics, E-pub.
[20] F. Desiere et al., 2006, Nucleic Acids Res., 34(database issue): D655-8.
[21] L. Anderson & C. Hunter, 2006, Mol. Cell Proteomics, 573-588.
[22] B. Han & R. Higgs, 2008, Brief Funct Genomic Proteomic., 7(5):340-54.
[23] K.-Y. Wang et al., 2008, Anal Chem, 80(16) 6159-6167.
[24] J. Bundy & C. Fenselau, 1999, Anal. Chem. 71:1460-1463.
[25] K-C Ho et al., 2004, Anal. Chem. 76:7162-7268.
[26] Y.S. Lin et al., 2005, Anal. Chem., 77:1753-1760.
[27] S. Vaidyanathan et al., 2001, Anal. Chem., 73:4134-4144.
[28] P. Seng et al., 2009, Clin. Infect. Dis., 49:543-551.
[29] Manes N. et al., 2007, Mol. & Cell. Proteomics, 6(4):717-727.
[30] R. Nandakumar et al., 2009, Oral Microbiology Immunology, 24:347-352.
[31] L. Hernychova et al., 2008, Anal. Chem., 80:7097-7104.
[32] J.-M. Pratt et al., 2006, Nat. Protoc., 1:1029-1043.
[33] V. Brun et al., 2007, Mol. Cell Proteomics, 2139-2149.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕПТИД LanA2, ВЫДЕЛЕННЫЙ ИЗ БАКТЕРИИ BACILLUS LICHENIFORMIS VK21, ОБЛАДАЮЩИЙ АНТИМИКРОБНЫМ ДЕЙСТВИЕМ | 2009 |
|
RU2408604C1 |
| ПОСЛЕДОВАТЕЛЬНОСТИ CRISPR БИФИДОБАКТЕРИЙ | 2009 |
|
RU2570562C2 |
| ПЕПТИД LanA1, ВЫДЕЛЕННЫЙ ИЗ БАКТЕРИИ Bacillus licheniformis VK21, ОБЛАДАЮЩИЙ АНТИМИКРОБНЫМ ДЕЙСТВИЕМ | 2009 |
|
RU2408732C1 |
| ИММУНОГЕННЫЕ БЕЛКИ STREPTOCOCCUS | 2008 |
|
RU2518315C2 |
| СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ СТАФИЛОКОККА ЗОЛОТИСТОГО И СВЯЗАННЫХ С НИМИ НАРУШЕНИЙ | 2012 |
|
RU2635462C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ РАЗМНОЖЕНИЯ ВИЧ-1 | 2001 |
|
RU2275379C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2296160C2 |
| ГЕН И БЕЛОК ВИРУЛЕНТНОСТИ И ИХ ИСПОЛЬЗОВАНИЕ | 2000 |
|
RU2240327C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНЫХ АЛКЕНОВ С ПОМОЩЬЮ ФЕРМЕНТАТИВНОГО ДЕКАРБОКСИЛИРОВАНИЯ 3-ГИДРОКСИАЛКАНОЕВЫХ КИСЛОТ | 2009 |
|
RU2571933C2 |

















































































































Настоящее изобретение относится к области микробиологии и касается способа определения таких характеристик, как типирование, потенциальная устойчивость по меньшей мере к одному антимикробному средству и фактора вирулентности, по меньшей мере, одного микроорганизма из образца. Представленный способ включает идентификацию указанного микроорганизма и определение характеристик. Определение характеристик осуществляют масс-спектрометрией типа MS/MS в режиме MRM, одновременно, в одном устройстве для масс-спектрометрии, с использованием белков, пептидов и/или метаболитов указанного микроорганизма в качестве маркеров характеристик. Изобретение позволяет быстро и с небольшими затратами определять характеристики микроорганизмов и может быть использовано в повседневной клинической практике. 1 н. и 26 з.п. ф-лы, 23 табл., 12 пр.
1. Способ определения характеристик по меньшей мере одного микроорганизма из образца, который включает
идентификацию указанного по меньшей мере одного микроорганизма и определение характеристик типирования, потенциальной устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности,
отличающийся тем, что
определение характеристик типирования, устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности для указанного по меньшей мере одного микроорганизма осуществляют масс-спектрометрией типа MS/MS в режиме MRM, одновременно, в одном устройстве для масс-спектрометрии, с использованием белков, пептидов и/или метаболитов указанного по меньшей мере одного микроорганизма в качестве маркеров указанных характеристик типирования, устойчивости по меньшей мере к одному антимикробному средству и фактора вирулентности.
2. Способ по п. 1, отличающийся тем, что идентификацию указанного по меньшей мере одного микроорганизма также осуществляют масс-спектрометрией.
3. Способ по п. 2, отличающийся тем, что масс-спектрометрия для идентификации указанного по меньшей мере одного микроорганизма представляет собой спектрометрию типа MS, типа MS/MS или типа MS с последующей спектрометрией типа MS/MS.
4. Способ по п. 1, отличающийся тем, что он включает дополнительную стадию подтверждения идентификации указанного по меньшей мере одного микроорганизма, которую осуществляют с помощью масс-спектрометрии.
5. Способ по п. 4, отличающийся тем, что масс-спектрометрия стадии подтверждения представляет собой масс-спектрометрию типа MS/MS.
6. Способ по п. 5, отличающийся тем, что масс-спектрометрия стадии подтверждения представляет собой MRM.
7. Способ по п. 1, отличающийся тем, что белки указанного микроорганизма расщепляют на пептиды.
8. Способ по п. 7, отличающийся тем, что расщепление осуществляют с помощью фермента.
9. Способ по п. 8, отличающийся тем, что фермент представляет собой трипсин.
10. Способ по п. 1, отличающийся тем, что указанный по меньшей мере один микроорганизм представляет собой бактерию, и антимикробное средство представляет собой антибиотик.
11. Способ по п. 10, отличающийся тем, что бактерия представляет собой Staphylococcus aureus.
12. Способ по п. 11, отличающийся тем, что для определения характеристик типирования используется по меньшей мере один пептид, имеющий последовательность SEQ ID NO: 2, 3, 4, 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 19, 20, 21, 23, 24, 26, 27, 29, 30, 32, 34, 35, 37, 38, 40, 42, 44 или 45.
13. Способ по п. 11 или 12, отличающийся тем, что для определения характеристик потенциальной устойчивости по меньшей мере к одному антибиотику используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 10-17.
14. Способ по п. 11 или 12, отличающийся тем, что для определения вирулентности используется по меньшей мере один пептид, имеющий последовательность SEQ ID NO: 19, 20, 21, 23 или 24.
15. Способ по п. 11 или 12, отличающийся тем, что для идентификации используется по меньшей мере один пептид, имеющий последовательность SEQ ID NO: 26, 27, 29, 30, 32, 34, 35, 37, 38, 40, 42, 44 или 45.
16. Способ по п. 10, отличающийся тем, что бактерия представляет собой Escherichia coli.
17. Способ по п. 16, отличающийся тем, что для определения характеристик типирования используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 46-87.
18. Способ по п. 16 или 17, отличающийся тем, что для определения характеристик потенциальной устойчивости по меньшей мере к одному антибиотику используется по меньшей мере один пептид, имеющий последовательность SEQ ID NO: 85, 86 или 87.
19. Способ по п. 16 или 17, отличающийся тем, что для определения вирулентности используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 62-66.
20. Способ по п. 16 или 17, отличающийся тем, что для идентификации используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 46-61.
21. Способ по п. 1, отличающийся тем, что указанный по меньшей мере один микроорганизм представляет собой дрожжи, и антимикробное средство представляет собой противогрибковое средство.
22. Способ по п. 21, отличающийся тем, что дрожжи представляют собой Candida albicans.
23. Способ по п. 22, отличающийся тем, что для определения свойств типирования используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 88-125 и 177.
24. Способ по п. 22 или 23, отличающийся тем, что для определения свойств потенциальной устойчивости по меньшей мере к одному противогрибковому средству используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 117-122 и 177.
25. Способ по п. 22 или 23, отличающийся тем, что для определения вирулентности используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 123-125.
26. Способ по п. 22 или 23, отличающийся тем, что для идентификации используется по меньшей мере один пептид, имеющий любую из последовательностей SEQ ID NO: 88-116.
27. Способ по п. 1, отличающийся тем, что метаболит представляет собой эргостерин.
| BERNARDO K et al., Identification and discrimination of Staphylococcus aureus strains using matrix-assisted laser desorption/ionization-time of flight mass spectrometry, Proteomics., 2002 Jun, Vol.2, N6, рр.747-53 | |||
| CAMARA JE et al., Discrimination between wild-type and ampicillin-resistant Escherichia coli by matrix-assisted laser |

