Изобретение относится к области биохимии, а именно к биологически активным пептидам, обладающим антимикробным действием, которые могут найти применение в медицине, ветеринарии и пищевой промышленности.
Одной из актуальных задач современной медицины является разработка лекарственных средств для борьбы с возбудителями инфекционных заболеваний, обладающими устойчивостью к существующим антибиотикам. В пищевой биотехнологии создание новых консервантов, подавляющих рост микроорганизмов, позволяет увеличить сроки хранения продуктов, снижает стоимость технологического процесса и увеличивает производительность путем сокращения времени и снижения температуры тепловой обработки.
На протяжении последних трех десятилетий XX века арсенал доступных антибиотиков обогащался лишь за счет новых представителей уже известных классов этих веществ. В 2000-е годы после длительного перерыва на рынке начали появляться первые антибиотики с принципиально новой структурой: оксазолидины (линезолид), циклические липопептиды (даптомицин), глицилциклины (тайгециклин) [Song J.H. What′s new on the antimicrobial horizon? // Int. J. Antimicrob. Agents. - 2008. - 32(4). - S207-13].
Одним из перспективных классов соединений для создания антибиотиков, активных в отношении резистентных микроорганизмов, являются антимикробные пептиды [Bommarius В., Kalman D. Antimicrobial and host defense peptides for therapeutic use against multidrug-resistant pathogens: new hope on the horizon // IDrugs. - 2009. - 12(6). - 376-80]. Антимикробные пептиды прокариотического происхождения получили название бактериоцинов. На основании наиболее общих структурных признаков бактериоцины грам-положительных бактерий подразделяют на четыре класса: I) лантибиотики (низин, эпидермин); II) термостабильные пептиды с молекулярной массой менее 10 кДа (сакацины, плантарицины); III) термолабильные белки с молекулярной массой менее 30 кДа, близкие к колицинам (гельветицин, миллерицин); IV) сложные бактериоцины, включающие углеводную или липидную часть [Asaduzzaman SM, Sonomoto К. Lantibiotics: diverse activities and unique modes of action. // J. Biosci. Bioeng. - 2009. - 107(5). -P.475-87]. Лантибиотики - синтезируемые на рибосомах и содержащие существенные пострансляционные модификации пептиды, отличительной особенностью строения которых является наличие остатков лантионина, метиллантионина и дегидратированных аминокислот. Тиоэфирные связи, образующиеся в результате взаимодействия остатков цистеина с остатками 2,3-дидегидроаланина и 2,3-дидегидроаминомасляной кислоты, играют ключевую роль в проявлении антибактериальных свойств [McAuliffe О., Ross R.P., Hill С.Lantibiotics: structure, biosynthesis and mode of action// FEMS Microbiol. Rev. - 2001. - 25(3). - P.285-308].
Известен лантибиотик лактицин 3147 LtnA1, выделенный из бактерии Lactococcus lactis, имеющий молекулярную массу 3322 Да и обладающий активностью в отношении грамположительных бактерий. Молекула лактицина 3147 LtnA1 содержит 30 аминокислотных остатков, в том числе остатки 2,3-дидегидроаминомасляной кислоты и 2-аминомасляной кислоты. Структура молекулы стабилизирована четырьмя тиоэфирными связями [Martin NI, Sprules Т, Carpenter MR, Cotter PD, Hill C, Ross RP, Vederas JC. Structural characterization of lacticin 3147, a two-peptide lantibiotic with synergistic activity. Biochemistry. - 2004. - 43(11). - P.3049-56].
Известен наиболее близкий к заявленному выделенный из бактерии Bacillus licheniformis ATCC 14580 пептид лихеницидин Bliα с молекулярной массой 3250,5 Да, активный в отношении грамположительных бактерий [Begley M., Cotter P.D., Hill С., Ross R.P. Rational genome mining for LanM proteins leads to the identification of a novel two peptide lantibiotic, lichenicidin // Appl. Environ. Microbiol. - 2009]. Модель молекулы лихеницидина Bliα была построена с использованием стратегии in silico на основании нуклеотидной последовательности, кодирующей предшественник пептида, и данных по строению известных лантибиотиков. Вероятная структура Bliα включает 32 аминокислотных остатка, в том числе остатки 2-аминомасляной кислоты, 2,3-дидегидроаланина, 2,3-дидегидроаминомасляной кислоты, и стабилизирована четырьмя тиоэфирными связями.
Изобретение решает задачу расширения ассортимента антимикробных пептидов.
Поставленная задача решается за счет пептида LanA1, обладающего антимикробной активностью, общей формулы:

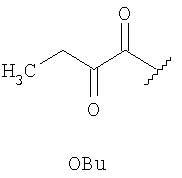
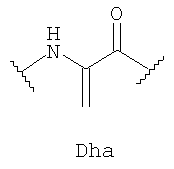
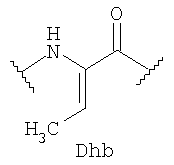
где OBu - остаток 2-оксомасляной кислоты, Abu - остаток 2-аминомасляной кислоты, Dha - остаток 2,3-дидегидроаланина, Dhb - остаток 2,3-дидегидроаминомасляной кислоты.
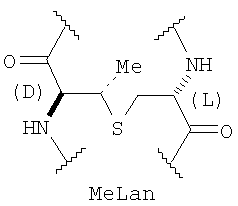
Структура пептида включает N-концевой остаток 2-оксомасляной кислоты и 31 аминокислотный остаток, соединенные в последовательности амидными связями. В состав аминокислотной последовательности наряду с остатками генетически кодируемых аминокислот входят остатки неканонических аминокислот Abu, Dha и Dhb. Кроме того, структуру пептида стабилизируют четыре тиоэфирные связи: остатки Abu3 и Ala7, Abu22 и Ala27, Abu24 и Ala31 соединены попарно тиоэфирными связями с образованием трех остатков 3-метиллантионина (MeLan), а остатки Ala11 и Ala21 соединены тиоэфирной связью с образованием остатка лантионина (Lan). Таким образом, заявляемый пептид относится к классу лантибиотиков.

Наличие указанных неканонических аминокислотных остатков и тиоэфирных связей, а также N-концевого остатка 2-оксомасляной кислоты делает невозможным определение первичной структуры пептида традиционными методами - ступенчатой деградацией аминокислотной цепи по Эдману и масс-спектрометрическим анализом. Полная первичная структура не может быть выведена только на основании данных по нуклеотидной последовательности, кодирующей пептид и его предшественник, в том числе с привлечением информации по строению других представителей класса лантибиотиков и имеющихся в настоящее время сведений о специфичности модифицирующих ферментов. Полная первичная структура заявляемого пептида может быть достоверно определена путем сопоставления результатов секвенирования нуклеотидной последовательности, кодирующей предшественник пептида, и данных ЯМР-спектроскопии.
Молекулярная масса пептида LanA1 составляет 3249,5 Да.
Заявляемый пептид LanA1 обладает рядом существенных отличий от своего ближайшего аналога - лихеницидина Bliα. Так, в структуре лихеницидина Bliα в качестве N-концевого остатка присутствует аланин, а заявляемый пептид содержит в этом положении остаток 2-оксомасляной кислоты. Кроме того, в лихеницидине Bliα остатки Ala1 и Ala7 соединены тиоэфирной связью с образованием лантионина (Lan), а в заявляемом пептиде такая связь соединяет остатки Abu3 и Ala7 с образованием 3-метиллантионина (MeLan). Преимуществом заявляемого пептида LanA1 является возможность его использования в отношении чувствительных штаммов микроорганизмов, обладающих устойчивостью к пептиду-аналогу. Преимуществом заявляемого пептида LanA1 является также возможность его получения из культуральной жидкости штамма-продуцента Bacillus licheniformis VK21, а не из клеточного осадка, что способно упростить очистку пептида.
Пептид LanA1 с заявленной структурой обладает антимикробной активностью в отношении грамположительных бактерий Bacillus subtilis, штаммы L1 и 1621; Bacillus pumilus, штамм 2001; Bacillus globigii, штамм I; Bacillus amyloliquefaciens, штамм I; Bacillus megaterium, штамм VKM41; Mycobacterium smegmatis, штамм 1171; Mycobacterium phlei, штамм 1291; Micrococcus luteus, штамм В1314; Staphylococcus aureus, штамм 209p; Rhodococcus sp., штамм SS1.
Пептид LanA1 может быть получен из природного источника - термофильной грамположительной бактерии Bacillus licheniformis VK21.
Изобретение иллюстрируют графические материалы.
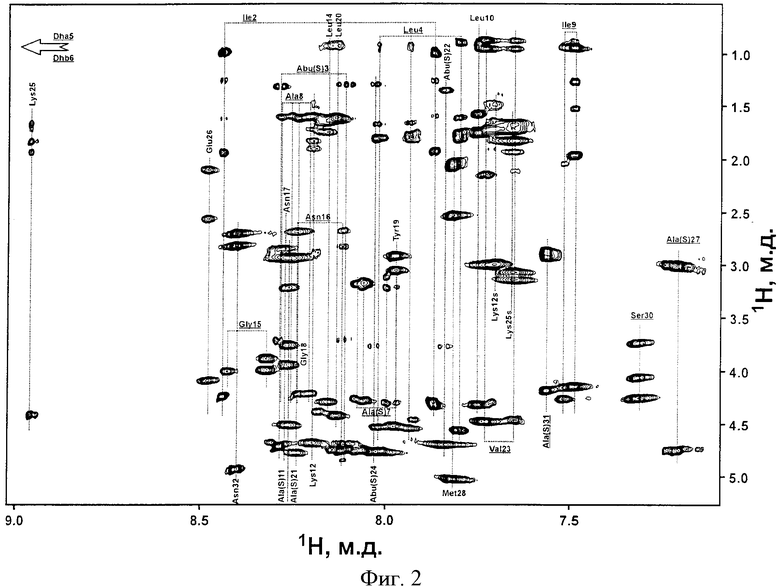
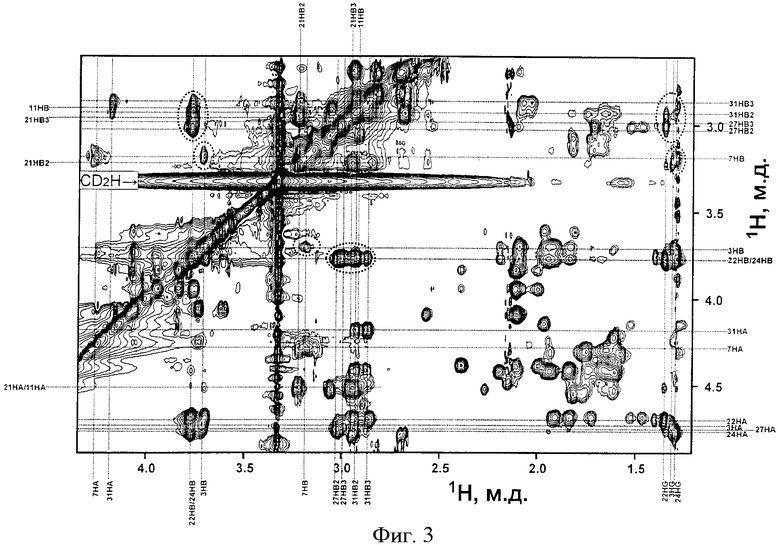
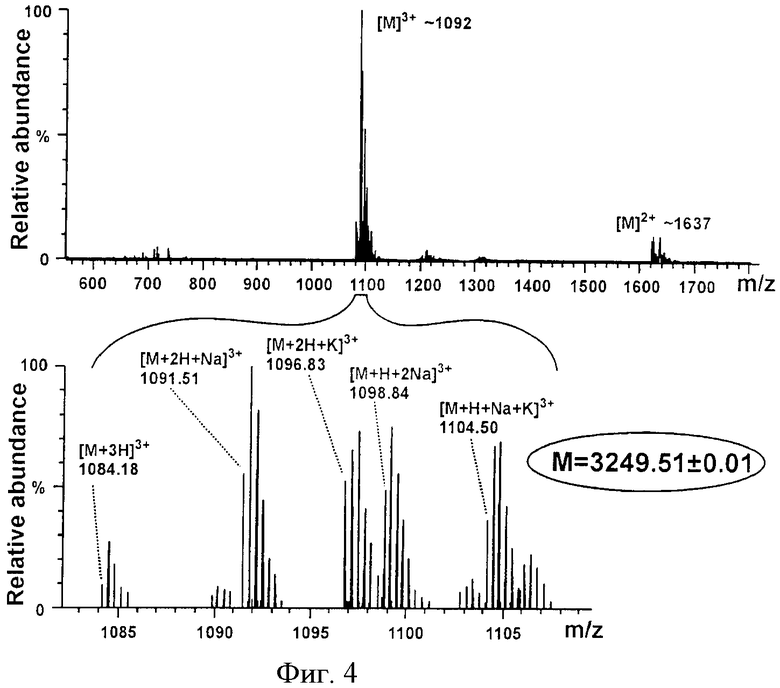
На фиг.1 представлена хроматограмма очистки LanA1 на колонке Xterra MS C18. На фиг.2 представлен фрагмент двумерного ЯМР-спектра TOCSY пептида LanA1 в метаноле (27°С, рН 3,5): вертикальными пунктирными линиями показаны спиновые системы отдельных остатков пептида; для некоторых сигналов пептида наблюдается наличие двух и более спиновых систем, что связано с конформационной гетерогенностью N-концевого фрагмента пептида, а именно cis-trans изомерией пептидной связи Lys12-Pro13 и наличием нескольких конформаций у участка молекулы OBu1-Ile7; сигналы отдельных конформаций пептида соединены горизонтальными пунктирными линиями, а названия соответствующих остатков молекулы подчеркнуты. На фиг.3 представлен фрагмент двумерного ЯМР-спектра NOESY пептида LanA1 в метаноле (27°С, рН 3,5); частоты сигналов остатков Lan и MeLan показаны пунктирными линиями; ЯЭО кросс-пики, позволяющие сделать вывод о расположении тиоэфирных связей в молекуле пептида, выделены пунктирными эллипсами. На фиг.4 представлен масс-спектр высокого разрешения пептида LanA1.
Изобретение иллюстрируют примеры.
Пример 1.
Культивирование клеток штамма Bacillus licheniformis VK21
Культуру выращивают на LB-агаре при 45°С в течение суток, после чего смывают 10 мл ростовой среды C2Mn (казаминовые кислоты - 0,15%; KCl - 0,02%; K2HPO4 - 0,3%; KH2PO4 - 0,1%; MgSO4×7H2O - 0,02%; MnSO4×5H2O - 0,001%). Аликвоты клеточной суспензии объемом 1,5 мл вносят в колбы на 750 мл, содержащие 150 мл среды C2Mn, и инкубируют на роторной качалке при температуре 45°С и скорости вращения 200 об/мин в течение 6 ч до достижения культурой оптической плотности OD620 1,5 (выход культуры на стационарную фазу роста).
Пример 2.
Выделение антимикробного пептида LanA1 из культуральной жидкости термофильной грамположительной бактерии Bacillus licheniformis VK21
По окончании культивирования клетки термофильной бактерии Bacillus licheniformis VK21 отделяют центрифугированием при 8000 g в течение 50 мин. Супернатант концентрируют в 20 раз на роторно-вакуумном испарителе. К полученному раствору добавляют два объема ацетона и удаляют осадок центрифугированием при 8000 g в течение 40 мин. Полученный супернатант упаривают на роторно-вакуумном испарителе досуха и перерастворяют в 50 мл воды. Далее проводят однократную экстракцию равным объемом н-бутанола, после чего водную фазу удаляют с помощью делительной воронки, а полученный органический экстракт упаривают досуха.
Сухой экстракт перерастворяют в 50 мл буфера A1 (30 мМ ацетат аммония, рН 5,6, 30% ацетонитрил) и наносят на колонку с носителем Silasorb C8, уравновешенную буфером A1. После тщательного промывания колонки 100 мл буфера A1 связавшиеся с носителем вещества элюируют 50 мл буфера B1 (30 мМ ацетат аммония, рН 5,6, 80% ацетонитрил) при скорости потока 2 мл/мин.
Полученный элюат разбавляют 30 мМ ацетатом аммония, рН 5,6, снижая концентрацию ацетонитрила до 30%, и наносят на колонку с носителем Diasorb-130C8T. После промывания колонки буфером A1 до выхода на нулевую линию проводят разделение в линейном градиенте буфера B1 от 0 до 100% в течение 95 мин (1,05% мин-1) при скорости потока 2 мл/мин. Детектирование ведут при длине волны 214 нм.
Активную фракцию, собранную в период с 50 по 55 мин, концентрируют и подвергают дальнейшему фракционированию с помощью обращенно-фазовой высокоэффективной жидкостной хроматографии на колонке Xterra MS C18, уравновешенной буфером А2 (5% ацетонитрил, 60% метанол), в линейном градиенте буфера В2 (35% ацетонитрил, 60% метанол) от 0 до 100% в течение 40 мин (2,5% мин-1) при скорости потока 0,5 мл/мин. Детектирование ведут при длине волны 214 нм.
Содержание пептидов во фракциях оценивают, используя MALDI-времяпролетный масс-спектрометрический анализ на приборе Reflex III (Bruker Daltonics). В качестве матрицы используют 0,15М 2,5-дигидроксибензойную кислоту в смеси, содержащей 25% метанол и 0,1% трифторуксусную кислоту. Образец облучают УФ-лазером с длиной волны 337 нм.
В результате хроматографического разделения на колонке Xterra MS C18 в активной фракции выявляют основной пик, выходящий с колонки при концентрации ацетонитрила 17% (фиг.1).
Пример 3.
Определение нуклеотидной последовательности, кодирующей предшественник пептида LanA1
Геномную ДНК Bacillus licheniformis VK21 выделяют из 20 мл бактериальной культуры, которую получают по методике, описанной в примере 1. Культуральную жидкость центрифугируют при 3000 g в течение 5 мин. Клеточный осадок промывают 0,85% раствором хлорида натрия, затем проводят повторное центрифугирование. Осадок ресуспендируют 1 мл TEN-буфера [0,15М NaCl, 0,1 М Tris-HCl (pH 8,0), 0,1 М EDTA], в котором предварительно растворяют лизоцим в концентрации 4 мг/мл. Полученную суспензию инкубируют в течение 45 мин при 37°С, после чего добавляют 0,25 мл 8,5% додецилсульфата натрия и выдерживают 30 мин при 75°С. К полученному лизату добавляют 375 мкл 5 М ацетата калия (pH 5,2), инкубируют смесь в течение 20 мин при 4°С и затем центрифугируют при 10000 g в течение 5 мин. Образовавшийся супернатант сливают, осадок растворяют в 100 мкл буфера ТЕ [10 mМ Tris-HCl (pH 8,0), 1 mМ EDTA]. Далее проводят очистку полученной геномной ДНК смесью фенол-хлороформ (1:1), осаждают ДНК спиртом и перерастворяют в 200 мкл воды.
Амплификацию участка ДНК, кодирующего предшественник пептида LanA1, проводят с использованием праймеров 1-4 (SEQ ID Nos. 1-4) в два этапа методом «гнездовой» ПЦР. В состав реакционной смеси в обеих реакциях входят 2 мкл 10-кратного буфера для ДНК-полимеразы Taq Advantage 2 (Clontech), 3 мкл раствора эквимолярной смеси dNTP (по 1,25 мкМ), 1 мкл раствора ДНК-матрицы, 0,5 мкл раствора ДНК-полимеразы Taq Advantage 2 (Clontech), по 1 мкл растворов двух ген-специфичных праймеров (10 пмоль/мкл) и 12 мкл воды; суммарный объем смеси составляет 20,5 мкл. В первую реакционную смесь добавляют праймеры 1 (SEQ ID No. 1) и 2 (SEQ ID No. 2) и раствор геномной ДНК В. licheniformis в качестве ДНК-матрицы. Во вторую реакционную смесь добавляют праймеры 3 (SEQ ID No. 3) и 4 (SEQ ID No. 4) и продукты первой реакции в разведении 1:20 в качестве ДНК-матрицы. Температурный режим обеих реакций: 94°С - 2 мин («горячий старт»); далее 30 циклов: 94°С - 30 с, 52°С - 40 с, 72°С - 60 с. Продукты ПЦР разделяют в агарозном геле и визуализируют с помощью бромида этидия. Продукт второй реакции размером 698 п.н. элюируют из агарозы и секвенируют.
Секвенирование ДНК проводят с помощью набора реактивов ABI PRISM BigDye Terminator v.3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе ДНК ABI PRISM 3100-Avant (Applied Biosystem). В результате получают нуклеотидную последовательность SEQ ID No. 5, участок которой кодирует аминокислотную последовательность SEQ ID No. 6 предшественника антимикробного пептида LanA1.
Пример 4.
Определение первичной структуры выделенного пептида LanA1
Для определения структуры выделенного антибиотика применяют метод ЯМР-спектроскопии. Пептидный антибиотик LanA1 растворяют в метаноле (2,0 мг/450 мкл, рН 3,5). ЯМР-спектры пептида получают при температуре 27°С на спектрометре Bruker Avance-700 (Германия) с рабочей частотой 700 МГц. Для измерения ЯМР-спектров используют датчик тройного резонанса (1H, 13C, 15N) с криогенно охлажденной 1Н катушкой (Bruker, Германия).
На основании анализа двумерных 1Н-ЯМР-спектров TOCSY (фиг.2) и NOESY (фиг.3) при помощи стандартной процедуры [Wuthrich К. NMR of Proteins and Nucleic Acids. // New York: John Wiley Sons, 1986] проводят полное отнесение сигналов ядер 1H исследуемого образца. На основании анализа двумерного 13С-ЯМР-спектра HSQC проводят полное отнесение сигналов 13С ядер, атомы которых имеют ковалентно присоединенные протоны.
Сопоставление полученного отнесения с результатами секвенирования нуклеотидной последовательности гена антимикробного пептида LanA1 из штамма VK21 термофильной бактерии Bacillus lichenifbrmis позволяет сделать вывод, что пептидный антибиотик LanA1 имеет следующую первичную структуру:
которая получается путем посттрансляционной модификации пропептида, имеющего последовательность SEQ ID No. 6.
Наличие N-концевой группы OBu1 подтверждают, используя двумерные ЯМР-спектры TOCSY и 13C-HSQC, путем наблюдения характерных сигналов от фрагмента СН3-СН2-(1Н 2,93 и 1,09 мд; 13С 30,1 и 5,8 мд).
Расположение тиоэфирных связей в молекуле пептида LanA1 определяют на основании анализа двумерного ЯМР-спектра NOESY (фиг.3), наблюдая ЯЭО кросс-пики между алифатическими протонами в остатках Lan или MeLan. Наличие ЯЭО кросс-пиков позволяет сделать вывод, что в пептиде LanA1 тиоэфирные связи соединяют попарно остатки Ala11 и Ala21, Abu3 и Ala7, Abu22 и Ala27, Abu24 и Ala31 с образованием одного остатка Lan и трех остатков MeLan.
Пример 5.
Определение молекулярной массы пептида методом масс-спектрометрии
Масс-спектры получают на гибридном масс-спектрометре LTQ FT (Thermo Electron, Германия), который состоит из линейной квадрупольной ионной ловушки и масс-спектрометра ионного циклотронного резонанса с преобразованием Фурье (ИЦР ПФ). В качестве источника ионов используют универсальный источник ионизации Finnigan Ion Max Source (Thermo Electron, Германия) в режиме электрораспыления. Для пептида LanA1 наблюдают набор двузарядных и трехзарядных ионов, соответствующий моноизотопной молекулярной массе пептида 3249,51±0,01 Да (фиг.4). Полученная масса с учетом точности измерения совпадает с расчетной моноизотопной молекулярной массой пептида LanA1 (3249,515 Да). Измеренная масса пептида LanA1 согласуется с расчетной массой пропептида (3374,605 Да) с учетом 7 сайтов дегидратации (-126,074 Да) и одной реакции дезаминирования с присоединением молекулы воды (+0,984 Да). Масс-спектрометрический анализ подтверждает заявляемую структуру пептида LanA1.
Пример 6.
Определение антимикробного действия выделенного пептида LanA1 Антимикробную активность выделенного пептида определяют методом радиальной диффузии пептида в агарозном геле с грамположительными бактериями:
Bacillus subtilis, штаммы L1 и 1621; Bacillus pumilus, штамм 2001; Bacillus globigii, штамм I; Bacillus amyloliquefaciens, штамм I; Bacillus megaterium, штамм VKM41; Mycobacterium smegmatis, штамм 1171; Mycobacterium phlei, штамм 1291; Micrococcus luteus, штамм В1314; Staphylococcus aureus, штамм 209p; Rhodococcus sp., штамм SS1. Тест-культуры выращивают в течение ночи на плотной среде. Выросшие колонии переносят в 3 мл жидкой среды и инкубируют на роторной качалке до достижения культурой оптической плотности OD620 1,2. Аликвоту клеточной суспензии добавляют к 12 мл расплавленной и охлажденной обедненной среды (9 мМ фосфатный буфер, рН 6,5; 0,03% TSB, 1% агарозы) и полученной смесью заливают чашки диаметром 90 мм. Конечная концентрация бактерий составляет 2·105 КОЕ/мл. В слое агарозы высверливают лунки диаметром 2 мм и вносят по 5 мкл раствора образца в 20% метаноле. В качестве контроля используют 20% метанол. Чашки инкубируют, не переворачивая в течение 3 ч, после чего на первый слой агарозы наслаивают 12 мл обогащенной среды (2LB, 1% агарозы). Антимикробное действие пептида фиксируют через 20 ч по наличию зон ингибирования роста тест-культуры (см. таблицу).
Показано, что выделенный пептид LanA1 в количестве 4 мкг/лунку проявляет антимикробную активность и ингибирует рост всех тестируемых грамположительных бактерий. Наиболее чувствительными к выделенному пептиду культурами являются М.luteus и В.megaterium. Наименее выражено ингибирующее действие выделенного пептида в отношении В.globigii, М.smegmatis, М.phlei и Rhodococcus sp.

Изобретение относится к области биотехнологии. Предложен биологически активный пептид общей формулы:
где ОВu - остаток 2-оксомасляной кислоты, Abu - остаток 2-аминомасляной кислоты, Dha - остаток 2,3-дидегидроаланина, Dhb - остаток 2,3-дидегидроаминомасляной кислоты, причем остатки в последовательности соединены амидными связями, остатки Abu3-Ala7, Abu22-Ala27 и Abu24-Ala31 также соединены попарно тиоэфирными связями с образованием трех остатков 3-метиллантионина, остатки Ala11-Ala21 соединены тиоэфирной связью с образованием остатка лантионина. Данный пептид обладает антимикробной активностью в отношении грамположительных бактерий Bacillus subtilis, штаммы L1 и 1621; Bacillus pumilus, штамм 2001; Bacillus globigii, штамм I; Bacillus amyloliquefaciens, штамм I; Bacillus megaterium, штамм VKM41; Mycobacterium smegmatis, штамм 1171; Mycobacterium phlei, штамм 1291; Micrococcus luteus, штамм В 1314; Staphylococcus aureus, штамм 209p; Rhodococcus sp., штамм SS1. 4 ил., 1 табл.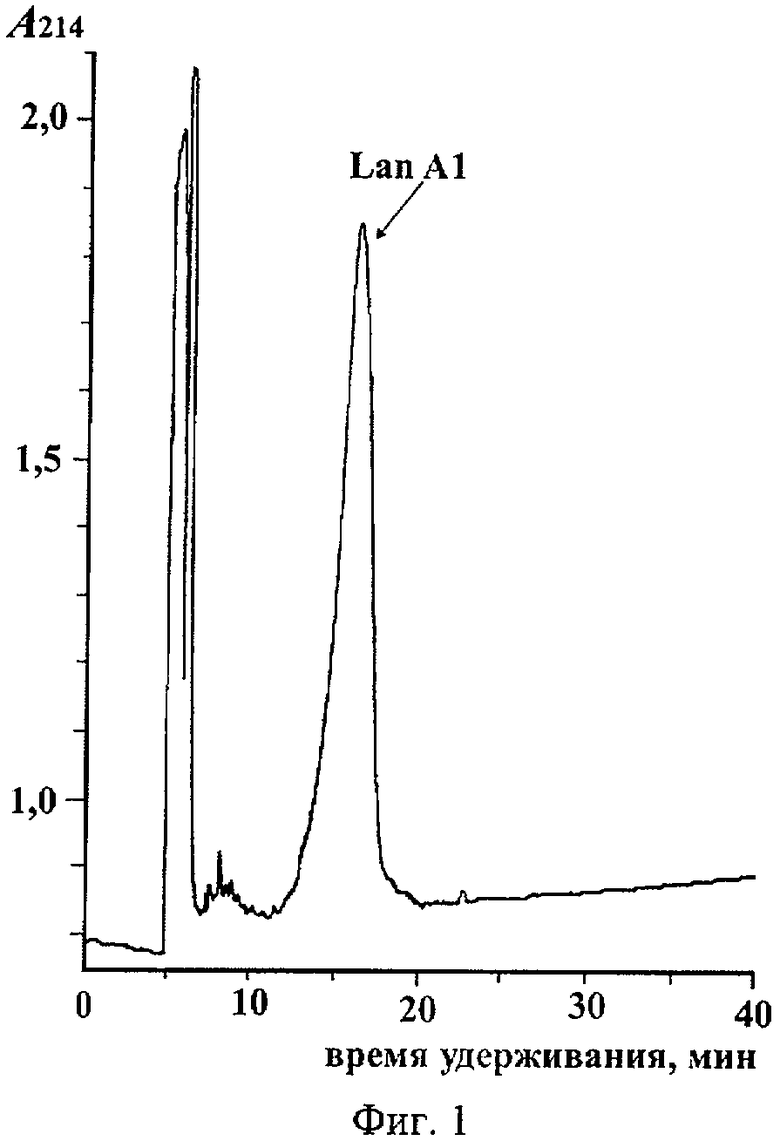
Пептид, обладающий антимикробной активностью, общей формулы:
где ОВu - остаток 2-оксомасляной кислоты, Abu - остаток 2-аминомасляной кислоты, Dha - остаток 2,3-дидегидроаланина, Dhb - остаток 2,3-дидегидроаминомасляной кислоты, причем остатки в последовательности соединены амидными связями, остатки Abu3-Ala7, Abu22-Ala27 и Abu24-Ala31 также соединены попарно тиоэфирными связями с образованием трех остатков 3-метиллантионина, остатки Ala11-Ala21 соединены тиоэфирной связью с образованием остатка лантионина.
| BEGLEY М | |||
| ET.AL | |||
| Identification of a Novel Two-Peptide Lantibiotic, Lichenicidin, following Rational Genome Mining for LanM Proteins | |||
| // Applied and Environmental Microbiology, Vol.75, No | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| MARTIN N.I | |||
| ET | |||
| AL | |||
| Способ обработки нефти | 1913 |
|
SU3147A1 |

