ОБЛАСТЬ И УРОВЕНЬ ТЕХНИКИ
Некоторые варианты настоящего изобретения относятся к новым мультимерным белковым структурам, включая, но не ограничиваясь, мультимерными белковыми структурами α-галактозидазы, и их применению для лечения болезни Фабри.
Лизосомальный фермент α-галактозидаза-А (α-GAL или α-Gal, EC 3.2.1.22) катализирует отщепление галактозы от олигосахаридов, гликопротеинов и гликолипидов при катаболизме макромолекул. Недостаток лизосомальных ферментов приводит к накоплению их субстратов в тканях, данные состояния известны как лизосомальные болезни накопления. У людей отсутствие функциональной α-галактозидазы приводит к накоплению в тканях гликолипидов, содержащих концевые остатки α-галактозы (в первую очередь глоботриаозилцерамида, который также называют «церамид-тригексозид», «CTH» или «Gb3»), что приводит к болезни Фабри. Болезнь Фабри является сцепленным с Х-хромосомой рецессивным заболеванием, впервые описанным в 1898 году, характеризующимся хроническими болями, помутнением зрения, поражением печени и почек, поражением кожи, повреждением сосудов и/или сердечной недостаточностью. Рекомбинантная человеческая α-галактозидаза-А обладает способностью восполнять ферментативную активность у больных, и заместительная терапия с использованием фермента (ERT) α-GAL была одобрена в США в 2003 году для лечения болезни Фабри. α-GAL стал вторым рекомбинантным белком, одобренным для лечения лизосомальной болезни накопления после β-глюкозидазы, применяемой для лечения болезни Гоше.
Эндогенная и рекомбинантная α-Gal катализируют гидролиз терминально галактозилированных гликолипидов в лизосомах клеток таких органов как печень, почки, селезенка, сердце и т.д. Лизосомы характеризуются низким рН, достигающим 4,5. Лизосомальные ферменты, в том числе α-GAL, следовательно, проявляют максимальную активность при указанных низких уровнях рН.
Считается, что современное ERT лечение болезни Фабри, основанное на полученной в клетках млекопитающих рекомбинантной α-GAL, обладает ограниченной эффективностью. Указанное лечение только замедляет развитие болезни, но не остановливает его и не приводит к действительному и полному излечению. Альтернативно, в некоторых случаях ERT с применением коммерческой рекомбинантной α-Gal должно быть прекращено в связи с развитием иммунного ответа на лечение, а в некоторых случаях лечение не может быть начато вследствие проблем с иммуногенностью.
Рентгеноструктурный анализ показал, что человеческая α-GAL представляет собой гомодимерный гликопротеин, в котором каждый мономер состоит из двух доменов, (β/α)8 домена, содержащего активный центр, и С-концевого домена, содержащего восемь антипараллельных β-тяжей в составе двух листов в β-сэндвиче [Garman & Garboczi, J Mol Biol 2004, 337:319-335]. Два мономера расположены в ориентации голова к хвосту, и их димеризация является нековалентной. Два мономеров взаимодействуют, образуя поверхность взаимодействия, которая простирается на 75 Ǻ в ширину и включает площадь 2200 Ǻ2. В формировании поверхности взаимодействия димера участвуют 30 остатков каждого мономера. Два активных центра димера разделяет приблизительно 50 Ǻ.
Кристаллическая структура α-Gal была разрешена для белка без лигандов, а также для белка, связавшего в качестве лиганда галактозу. Указанные две структуры характеризуются небольшими изменениями при связывании лиганда по сравнению отсутствием связания лиганда. Однако, следует отметить, что использование галактозы вместо природного субстрата, глоботриаозилцерамида (Gb3), где последний характеризуется наличием протяженных липидных цепей, способных взаимодействовать с гидрофобным доменом одного мономера, в то время как концевая галактоза способна взаимодействовать с активным центром второго мономера, может привести к невозможности обнаружения кооперативности активных центров. Альтернативно, биохимические характеристики позволяют предположить наличие такой кооперативности, иллюстрирующей важность гомодимерной четвертичной структуры [Bishop & Desnick, J Biol Chem 1981, 256:1307-1316]. Были изучены кинетические свойства человеческой α-Gal и была показана кооперативность между мономерами гомодимерного фермента, каждый из которых имел взаимодействующий каталитический центр. Поэтому было предложено, что ферментативная активность и стабильность фермента может зависеть от его димеризации.
В WO 2009/024977, также поданной заявителями настоящего изобретения, которая включена в настоящий документ в полном объеме посредством отсылки, раскрыты конъюгаты сахарида и биомолекулы, ковалентно связанных между собой через не гидрофобный линкер, а также медицинские применения с использованием таких конъюгатов.
В международной заявке PCT/IL 2010/000956, также поданной заявителями настоящего изобретения, раскрыта методика, в которой используют α-галактозидазу, которая проявляет лизосомальную активность при более высоком рН, чем рН в лизосомах.
Уровень техники также включает Bendele et al. [Toxicological Sciences 1998, 42:152-157], патенты США №№5,256804, 5,580757 и 5766897, Международную заявку PCT/NL 2007/050684 (опубликована как WO 2008/075957) и Seely & Richey [J Chromatography A 2001, 908:235-241].
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с некоторыми вариантами осуществления настоящего изобретения предусмотрены мультимерные белковые структуры, содержащие, по меньшей мере, два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент, где мультимерная белковая структура обладает характеристиками, выбранными из группы, состоящей из:
(a) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в условия человеческой плазмы на один час, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час;
(b) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в условия человеческой плазмы на один час, по меньшей мере, на 10% меньше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час
(c) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в условия человеческой плазмы на один час;
(d) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в лизосомальные условия на одну неделю;
(e) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю, по меньшей мере, на 10% меньше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в лизосомальные условия на одну неделю;
(f) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день;
(g) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в лизосомальные условия которая, по меньшей мере, на 10% больше, чем активность нативной α-галактозидазы немедленно после помещении нативной α-галактозидазы в лизосомальные условия;
(h) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в водный раствор, имеющий рН 7 и температуру 37°C, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы немедленно после помещении нативной α-галактозидазы в водный раствор, имеющий рН 7 и температуру 37°C и
(i) времени полураспада в кровотоке в физиологической системе, которое, по меньшей мере, на 20% больше, чем время полураспада в кровотоке нативной α-галактозидазы.
В соответствии с аспектами некоторых вариантов осуществления настоящего изобретения предусмотрены мультимерные белковые структуры, содержащие, по меньшей мере, два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент, где связывающий фрагмент отсутствует в нативной α-галактозидазе.
В соответствии с аспектами некоторых вариантов осуществления настоящего изобретения предусмотрена фармацевтическая композиция, включающая мультимерную белковую структуру, описанную в настоящем документе, и фармацевтически приемлемый носитель.
В соответствии с аспектами некоторых вариантов осуществления настоящего изобретения предусмотрен способ лечения болезни Фабри, включающий введение субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества мультимерной белковой структуры, описанной в настоящем документе, что приводит к лечению болезни Фабри.
В соответствии с аспектами некоторых вариантов осуществления настоящего изобретения предусмотрен способ получения мультимерной белковой структуры, описанной в настоящем документе, где способ включает взаимодействие α-галактозидазы со сшивающим агентом, который содержит связывающий фрагмент, описанный в настоящем документе, и, по меньшей мере, две реакционноспособные группы.
Согласно некоторым вариантам осуществления изобретения связывающий фрагмент, описанный в настоящем документе, отсутствует в нативной α-галактозидазе. Согласно некоторым вариантам осуществления изобретения, мультимерная белковая структура имеет характеристики, выбранные из группы, состоящей из:
(a) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в условия человеческой плазмы на один час, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час;
(b) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в условия человеческой плазмы на один час, по меньшей мере, на 10% меньше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час
(c) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в условия человеческой плазмы на один час;
(d) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в лизосомальные условия на одну неделю;
(e) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю, по меньшей мере, на 10% меньше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в лизосомальные условия на одну неделю;
(f) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день;
(g) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в лизосомальные условия которая, по меньшей мере, на 10% больше, чем активность нативной α-галактозидазы немедленно после помещении нативной α-галактозидазы в условия, соответствующие условиям в лизосоме;
(h) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в водный раствор, имеющий рН 7 и температуру 37°C, которая, по меньшей мере, на 10% выше, чем активность нативной α-галактозидазы немедленно после помещении нативной α-галактозидазы в водный раствор, имеющий рН 7 и температуру 37°C и
(i) времени полураспада в кровотоке в физиологической системе, которое больше, чем время полураспада в кровотоке нативной α-галактозидазы. Согласно некоторым вариантам осуществления изобретения α-галактозидазная активность мультимерной белковой структуры, которая остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день, далее остается практически неизменной при при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю. Согласно некоторым вариантам осуществления изобретения большее время полураспада в кровотоке мультимерной белковой структуры по сравнению с временем полураспада в кровотоке нативной α-галактозидазы, по меньшей мере, на 20% больше, чем время полураспада в кровотоке нативной α-галактозидазы. Согласно некоторым вариантам осуществления изобретения большее время полураспада в кровотоке мультимерной белковой структуры по сравнению с временем полураспада в кровотоке нативной α-галактозидазы, по меньшей мере, на 50% больше, чем время полураспада в кровотоке нативной α-галактозидазы. Согласно некоторым вариантам осуществления изобретения мультимерная белковая структура характеризуется α-галактозидазной активностью в органе при введении мультимерной белковой структуры позвоночным, где орган выбран из группы, состоящей из селезенки, сердца и почек.
Согласно некоторым вариантам осуществления изобретения мультимерная белковая структура состоит из двух мономеров α-галактозидазы и белковая структуры представляет собой димерную белковую структуру. Согласно некоторым вариантам осуществления изобретения α-галактозидаза представляет собой человеческую α-галактозидазу. Согласно некоторым вариантам осуществления изобретения α-галактозидаза представляет собой растительную рекомбинантную α-галактозидазу. Согласно некоторым вариантам осуществления изобретения α-галактозидаза имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3.
Согласно некоторым вариантам осуществления изобретения α-галактозидаза представляет собой щелочную α-галактозидазу.
Согласно некоторым вариантам осуществления изобретения α-галактозидаза представляет собой кислую α-галактозидазу.
Согласно некоторым вариантам осуществления изобретения связывающий фрагмент содержит поли(алкиленгликоль).
Согласно некоторым вариантам осуществления изобретения поли(алкиленгликоль) содержит, по меньшей мере, две функциональные группы, где каждая функциональная группа образует ковалентную связь с одним из мономеров α-галактозидазы.
Согласно некоторым вариантам осуществления изобретения, по меньшей мере, две функциональные группы являются концевыми группами поли(алкиленгликоля).
Согласно некоторым вариантам осуществления изобретения, по меньшей мере, один связывающий фрагмент имеет общую формулу:
-X1-(CR1R2-CR3R4-Y)n-X2-
где каждый из X1 и Х2 представляет собой функциональную группу, которая образует ковалентную связь, по меньшей мере, с одним мономером α-галактозидазы;
Y представляет собой О, S или NR5;
n представляет собой целое число от 1 до 200,
каждый из R1, R2, R3, R4 и R5 независимо выбран из группы, состоящей из водорода, алкильной, циклоалкильной, алкенильной, алкинильной, алкоксильной, гидроксильной, оксо, тиольной и тиоалкоксильной групп.
Согласно некоторым вариантам осуществления изобретения, по меньшей мере, одна из функциональных групп образует амидную связь с мономером α-галактозидазы.
Согласно некоторым вариантам осуществления изобретения n представляет собой целое число от 5 до 150.
Согласно некоторым вариантам осуществления изобретения n представляет собой целое число от 40 до 70.
Согласно некоторым вариантам осуществления изобретения фармацевтическая композиция дополнительно содержит галактозу.
Согласно некоторым вариантам осуществления изобретения мультимерная белковая структура предназначена для применения в качестве лекарственного средства.
Согласно некоторым вариантам осуществления изобретения лекарственное средство предназначено для лечения болезни Фабри.
Согласно некоторым вариантам осуществления изобретения мультимерная белковая структура предназначена для применения в лечении болезни Фабри. Согласно некоторым вариантам осуществления изобретения способ получения включает взаимодействие димерной α-галактозидазы со сшивающим агентом. Согласно некоторым вариантам осуществления изобретения реакционноспособные группы включают уходящую группу.
Согласно некоторым вариантам осуществления изобретения реакционноспособная группа реагирует с аминогруппой с образованием амидной связи. Согласно некоторым вариантам осуществления изобретения каждая из реакционноспособных групп способна образовывать ковалентную связь между связывающим фрагментом, и, по меньшей мере, одним мономером α-галактозидазы. Согласно некоторым вариантам осуществления изобретения молярное соотношение сшивающего агента к мономеру α-галактозидазы находится в диапазоне от 5:1 до 500:1.
Согласно некоторым вариантам осуществления изобретения молярное соотношение находится в диапазоне от 75:1 до 300:1.
Если не указано иное, все технические и/или научные термины, используемые здесь, имеют тот же смысл, как это обычно понимается специалистом в области техники, к которой относится изобретение. Примеры методов и/или материалов, которые могут быть использованы при практическом применении или тестировании вариантов осуществления изобретения, описаны ниже, хотя могут быть сипользованы методы и материалы, аналогичные или эквивалентные описанным здесь. В случае возникновения патентного спора описание патента, в том числе определения, будет иметь определяющее значение. Материалы, способы и примеры приведены только в иллюстративных целях и не предназначены для ограничения изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Патент или заявка содержит, по меньшей мере, одну Фигуру, выполненную в цвете. Копии данного патента или патентной заявки с цветной Фигурой (Фигурами) будут предоставлены Патентным ведомством по запросу и после необходимой оплаты. Некоторые варианты осуществления изобретения описаны здесь со ссылкой на прилагаемые Фигуры только в качестве примеров. Что касается непосредственно приводимых здесь Фигур, необходимо подчеркнуть, что показанные данные приведены в качестве примеров в иллюстративных целях для демонстрации вариантов осуществления изобретения. В связи с этим, описание, приведенное к Фигурам, делает очевидными для специалистов в данной области варианты воплощения изобретения.
Фиг.1 представляет собой график, показывающий активность Fabrazyme® α-GAL, Replagal® α-GAL и полученной в растениях рекомбинантной человеческой α-GAL-I в зависимости от времени инкубации в искусственных условиях, соответствующих условиям в лизосоме (цитрат-фосфатный буфер, рН 4,6, 37°C);
Фиг.2 представляет собой график, показывающий активность Fabrazyme® α-GAL, Replagal® α-GAL и полученной в растениях рекомбинантной человеческой α-GAL-I по отношению к галактозе (100 мг/мл) как функцию времени инкубации при моделируемых физиологических условиях (рН 7,4, 37°C);
Фиг.3 представляет собой график, показывающий активность Fabrazyme® α-GAL, Replagal® α-GAL и полученной в растениях рекомбинантной человеческой α-GAL-I в зависимости от времени инкубации в плазме крови человека при 37°C;
Фиг.4 представляет собой график, показывающий активность Fabrazyme® α-GAL, Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I и полученной в растениях рекомбинантной α-GAL-I по отношению к галактозе (100 мг/мл) как функцию времени инкубации при моделируемых условиях в лизосоме (цитрат-фосфатный буфер, рН 4,6, 37°C);
Фиг.5 представляет собой схему, изображающую молекулярную структуру примера сшивающего агента бис-N-гидроксисукцинимид-поли(этиленгликоля) (бис-NHS-PEG);
Фиг.6 представляет собой схему, изображающую димерный белок, который был введен в реакцию с бис-NHS-PEG сшивающим агентом;
Фиг.7 представляет собой сканированное изображение SDS-PAGE геля, показывающее полученную в растениях рекомбинантную α-GAL-I, прореагировавшую с бис-NHS-PEG5 (дорожки 1-3), бис-NHS-PEG8 (дорожки 7-9) и бис-NHS-PEG45 (дорожки 4-6) при молярном соотношении бис-NHS-PEG:α-GAL 50:1 (дорожки 1, 4 и 7), 100:1 (дорожки 2, 5 и 8) и 200:1 (дорожки 3, 6 и 9), а также маркеры молекулярной массы (Mw) и не введенную в реакцию полученную в растениях рекомбинантную α-GAL-I в качестве контроля (Std) (стрелки показывают полосу, соответсвующую димеру α-GAL);
Фиг.8 представляет собой сканированное изображение геля после изоэлектрофокусирования, показывающее полученную в растениях рекомбинантную α-GAL-I, прореагировавшую с бис-NHS-PEG5 (дорожки 1-3), бис-NHS-PEG8 (полосы 7-9) и бис-NHS-PEG45 (дорожки 4-6) при молярном соотношении бис-NHS-PEG:α-GAL 50:1 (дорожки 1, 4 и 7), 100:1 (дорожки 2, 5 и 8) и 200:1 (дорожки 3, 6 и 9), а также рН маркеры (М) и не введенную в реакцию полученную в растениях рекомбинантную α-GAL-I в качестве контроля (Std) (стрелками показаны значения рН для различных полос);
Фиг.9 представляет собой MALDI-TOF масс-спектр полученной в растениях рекомбинантной α-GAL-I, сшитой бис-NHS-PEG45 (ось Х указывает значения m/z, также приведены m/z значения пиков);
Фиг.10 представляет собой MALDI-TOF масс-спектр полученной в растениях рекомбинантной α-GAL-I, сшитой бис-NHS-PEG8 (ось Х указывает значения m/z, также приведены m/z значения пиков);
Фиг.11 представляет собой фотографию, показывающую субстрат α-GAL N-додеканоил-нитробензоксадиазол-церамид тригексозид (Gb3-NBD) и продукт реакции α-GAL лактозил церамид-нитробензоксадиазол (лактозил церамид-NBD), визуализируемые с помощью облучения УФ-светом (365 нм) после высокопроизводительной тонкослойной хроматографии после инкубации субстрата Gb3-NBD с полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45 (левая дорожка), с Replagal® α-GAL (средняя дорожка) и без инкубации с α-GAL (правая дорожка);
Фиг.12А, 12В и 12С представляют собой графики, показывающие активность Fabrazyme® α-GAL, Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой 6HC-NHS-PEG5 (Фиг.12А), бис-NHS-PEG8 (Фиг.12В) и бис-NHS-PEG45 (Фиг.12С) при молярном соотношении бис-NHS-PEG:α-GAL 50:1 («1» на Фиг.12А, «7» на Фиг.12В и «4» на Фиг.12С), 100:1 («2» на Фиг.12А, «8» на Фиг.12В и «5» на Фиг.12С) и 200:1 («3» на Фиг.. 12А, «9» на Фиг.12В и «6» на Фиг.12С) в зависимости от времени инкубации при моделируемых условиях в лизосоме (цитрат-фосфатный буфер, рН 4,6, 37°C);
Фиг.13 представляет собой график, показывающий фармакокинетический профиль Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45, в плазме страдающих болезнью Фабри мышей; остаточная активность каждой α-GAL представлена в процентах от максимальной остаточной активности каждой α-GAL как функция от времени после введения α-Gal;
Фиг.14А и 14В представляют собой график (Фиг.14А), показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG8 (prh-альфа-GAL-I-CL8) или бис-NHS-PEG45 (prh-альфа-GAL-I-CL45) в селезенке страдающих болезнью Фабри мышей через 2 часа после инъекции α-GAL, и фотографию Вестерн-блоттинга (Фиг.14В), показывающего Replagal® α-GAL (дорожки 10-12 и 15), полученную в растениях рекомбинантную человеческую α-GAL-I (дорожки 7-9 и 13) и полученную в растениях рекомбинантную человеческую α-GAL-I, сшитую бис-NHS-PEG8 (дорожки 4-6) или бис-NHS-PEG45 (дорожки 1-3 и 14) в селезенке страдающих болезнью Фабри мышей после введения α-GAL (дорожки 1-12) или в качестве контроля 50 нг α-GAL (дорожки 13-15);
Фиг.15А и 15В представляют собой график (Фиг.15А), показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG8 (prh-альфа-GAL-I-CL8) или бис-NHS-PEG45 (prh-альфа-GAL-I-CL45) в печени страдающих болезнью Фабри мышей через 2 часа после инъекции α-GAL, и фотографию Вестерн-блоттинга (Фиг.15В), показывающего Replagal® α-GAL (дорожки 10-12 и 15), полученную в растениях рекомбинантную человеческую α-GAL-I (дорожки 7-9 и 13) и полученную в растениях рекомбинантную человеческую α-GAL-I, сшитую бис-NHS-PEG8 (дорожки 4-6) или бис-NHS-PEG45 (дорожки 1-3 и 14) в печени страдающих болезнью Фабри мышей после введения α-GAL (дорожки 1-12) или в качестве контроля 50 нг α-GAL (дорожки 13-15);
Фиг.16 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG8 (prh-альфа-GAL-I-) или бис-NHS-PEG45 (prh-альфа-GAL-I-CL45), в сердце страдающих болезнью Фабри мышей через 2 часа после введения α-GAL;
Фиг.17 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG8 (prh-альфа-GAL-I-CL8) или бис-NHS-PEG45 (prh-альфа-GLA-I-GL45), в почках страдающих болезнью Фабри мышей через 2 часа после введения α-GAL;
Фиг.18 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG8 (prh-альфа-GAL-I-CL8) или бис-NHS-PEG45 (prh-альфа-GAL-I-CL45), в селезенке страдающих болезнью Фабри мышей через 2 часа, 24 часа, 3 дня и 7 дней после введения α-GAL (эндогенная α-GAL дикого типа (WT) приведена в качестве контроля);
Фиг.19 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45 (prh-альфа-GAL-I-CL45), в печени страдающих болезнью Фабри мышей через 2 часа, 24 часа, 3 дня и 7 дней после введения α-GAL (эндогенная α-GAL дикого типа (WT) приведена в качестве контроля);
Фиг.20 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45 (prh-альфа-GAL-I-CL45), в сердце страдающих болезнью Фабри мышей через 2 часа, 24 часа, 3 дня и 7 дней после введения α-GAL (эндогенная α-GAL дикого типа (WT) приведена в качестве контроля);
Фиг.21 представляет собой график, показывающий активность Replagal® α-GAL, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-альфа-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45 (prh-альфа-GAL-I-CL45), в почках страдающих болезнью Фабри мышей через 2 часа, 24 часа, 3 дня и 7 дней после введения α-GAL (эндогенная α-GAL дикого типа (WT) приведена в качестве контроля);
Фиг.22 представляет собой фотографию SDS-PAGE геля, показывающего полученную в клетках млекопитающих рекомбинантную человеческую α-GAL Replagal® (левая дорожка) и полученную в клетках млекопитающих рекомбинантную человеческую α-GAL Replagal®, прореагировавшую с бис-NHS-PEG45 (средняя дорожка), а также маркеры молекулярной массы (правая дорожка; молекулярные массы маркеров указаны в кДа);
Фиг.23 представляет собой фотографию геля после изоэлектрофокусирования, показывающего полученную в клетках млекопитающих рекомбинантную человеческую α-GAL Replagal® (левая дорожка) и полученную в клетках млекопитающих рекомбинантную человеческую α-GAL Replagal®, прореагировавшую с бис-NHS-PEG45 (средняя дорожка), а также маркеры рН (правая дорожка);
Фиг.24А и 24В представляют собой MALDI-TOF масс-спектры полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® (Фиг.24А) и полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal®, прореагировавшей с бис-NHS-PEG45 (ось Х указывает значения m/z, также приведены m/z значения (в Да) пиков);
Фиг.25 представляет собой график Михаэлиса-Ментен, показывающий скорость (V) гидролиза п-нитрофенил-α-D-галактопиранозида (pNP-G) полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® (Replagal) и полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal®, сшитой бис-NHS-PEG45 (ReplagalCL45), как функцию от концентрации pNP-G;
Фиг.26А и 26В представляют собой графики, показывающие активность полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® (Replagal) и полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal®, сшитой 6HC-NHS-PEG45 (Replagal-CL45), как функцию от времени инкубации в моделируемых лизосомальных условиях (цитрат-фосфатный буфер, рН 4,6, 37°C) (Фиг.26, А) или в плазме крови человека при 37°C (Фиг.26В);
Фиг.27A-27D представляют собой графики, показывающие активность α-GAL Replagal® (R) и α-GAL Replagal®, сшитой бис-NHS-PEG45, (R-CL45) в селезенке (Фиг.27А), печени (Фиг.27В), сердце (Фиг.27С) и почках (Фиг.27D) страдающих болезнью Фабри мышей через 2 часа после введения α-GAL;
Фиг.28A-28D представляют собой графики, показывающие уровни Gb3 в сердце (Фиг.28А), почках (Фиг.28В), печени (Фиг.28С) и селезенке (Фиг.28D) страдающих болезнью Фабри мышей как функцию от времени после инъекции α-GAL Replagal® (R) или α-GAL Replagal®, сшитой бис-NHS-PEG45 (R-CL45);
Фиг.29А и 29В представляют собой сканированное изображение SDS-PAGE геля, показывающего полученную в растениях рекомбинантную человеческую α-GAL-II (Фиг.29А и 29В, дорожка 2) и полученную в растениях рекомбинантную человеческую α-GAL-II, прореагировавшую с бис-NHS-PEG21 (Фиг.29А, дорожка 3), бис-NHS-PEG45 (Фиг.29А, дорожка 4) или бис-NHS-PEG68 (Фиг.29В, дорожка 3), а также маркеры молекулярной массы (Фиг.29А и 29В, дорожка 1, молекулярные массы маркеров указаны в кДа);
Фиг.30А-30С представляют собой MALDI-TOF масс-спектры полученной в растениях рекомбинантной человеческой α-GAL-II (Фиг.30А) и полученной в растениях рекомбинантной человеческой α-GAL-II, сшитой бис-NHS-PEG21 (Фиг.30А) или бис-NHS-PEG45 (Фиг.30С) (ось абсцисс указывает значения m/z, указаны значения m/z (в Да) пиков);
Фиг.31A-31D представляют собой графики, показывающие активность полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® (Replagal), полученной в растениях рекомбинантной человеческой α-GAL-II (prh-альфа-GAL-II) и полученной в растениях рекомбинантной человеческой α-GAL-II, сшитой бис-NHS-PEG21 (prh-альфа-GAL-II-CL21; Фиг.31А и 31С), бис-NHS-PEG45 (prh-альфа-GAL-II-CL45, Фиг.31A-31D) или бис-NHS-PEG68 (prh-альфа-GAL-II-CL68, Фиг.31В и 31D), в зависимости от времени инкубации в моделируемых условиях в лизосоме (цитрат-фосфатный буфер, рН 4,6, 37°C) (Фиг.31А и 31В) или в плазме крови человека при 37°C (Фиг.31С и 31D) (данные, показанные на Фиг.31С и 31D, получены из разных экспериментов);
Фиг.32А и 32В представляют собой графики, показывающие фармакокинетические профили α-GAL Replagal® (Replagal), полученной в растениях рекомбинантной человеческой α-GAL-II (prh-альфа-GAL-II) и полученной в растениях рекомбинантной человеческой α-GAL-II, сшитой бис-NHS-PEG45 (prh-альфа-GAL-II-CL45), в плазме страдающих болезнью Фабри мышей; концентрация каждой α-GAL представлена как функция от времени после инъекции α-GAL (Фиг.32А и 32В представляют те же данные в разные промежутки времени);
Фиг.33A-33L представляют собой графики, показывающие активность α-GAL Replagal® (Replagal), полученной в растениях рекомбинантной человеческой α-GAL-II (prh-альфа-GAL-II) и полученной в растениях рекомбинантной человеческой α-GAL-II, сшитой бис-NHS-PEG45 (prh-альфа-GAL-II-CL45, Фиг.33A-33L) или бис-NHS-PEG21 (prh-альфа-GAL-II-CL21; Фиг.33E-33L) в сердце (Фиг.33A, 33Е и 33I), почках (Фиг.33B, 33F и 33J), печени (Фиг.33C, 33G и 33K) и селезенке (Фиг.33D, 33H и 33L) страдающих болезнью Фабри мышей через 2 часа (Фиг.33A-33H), 7 дней (Фиг.33A-33D и 33I-33L), 14 дней (Фиг.33A-33D) и 28 дней (Фиг.33A-33D) после инъекции α-GAL;
Фиг.34А-34С представляют собой графики, показывающие кинетические параметры Vмакс (Фиг.34А), КМ (Фиг.34 В) и kкат (Фиг.34С) для полученной в растениях рекомбинантной человеческой α-GAL-II (prh-альфа-GAL-II) и полученной в растениях рекомбинантных человеческой α-GAL-II, сшитой бис-NHS-РЕ045 (prh-альфа-GAL-II-CL45) в зависимости от рН;
Фиг.35 представляет собой сканированное изображение SDS-PAGE геля, показывающего полученную в растениях рекомбинантную человеческую α-GAL-I (prh-α-Gal-I) и полученную в растениях рекомбинантную человеческую α-GAL-I, прореагировавшую с метокси-замещенным NHS-PEG, имеющим молекулярный вес 2 кДа (prh-α-Gal-I-PEG 2000), 5 кДа (prh-α-Gal-I-PEG 5000) и 10 кДа (prh-α-Gal-I-PEG 10000), а также маркеры молекулярной массы (левая дорожка; молекулярные массы маркеров указаны в кДа);
Фиг.36А и 36В представляют собой графики, показывающие активность полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Fabrazyme® (Fabrazyme), полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® (Replagal), полученной в растениях рекомбинантной человеческой α-GAL-I и полученной в растениях рекомбинантной человеческой α-GAL-I, прореагировавшей с метокси-замещенным NHS-PEG, имеющим молекулярный вес 2 кДа (α-Gal-I-PEG 2000), 5 кДа (α-Gal-I-PEG 5000) и 10 кДа (α-Gal-I-PEG 10000), в зависимости от времени инкубации при моделировании лизосомальных условий (цитрат-фосфатный буфер, рН 4,6, 37°C) (Фиг.36А) или в плазме крови человека при 37°C (Фиг.36В);
Фиг.37 представляет собой сканированное изображение SDS-PAGE геля, показывающего полученную в растениях рекомбинантную α-GAL-I, прореагировавшую с бис-NHS-PEG2 (дорожки 1-3), бис-NHS-PEG4 (дорожки 4-6), бис-NHS-PEG68 (дорожки 7-9), бис-NHS-PEG150 (дорожки 10-12) и бис-NHS-PEG45 (CL45) при молярном соотношении бис-NHS-PEG:α-GAL 50:1 (дорожки 1, 4, 7 и 10), 100:1 (дорожки 2, 5, 8 и 11) и 200:1 (дорожки 3, 6, 9 и 12), а также маркеры молекулярной массы (MW);
Фиг.38 представляет собой сканированное изображение SDS-PAGE геля, показывающего полученную в растениях рекомбинантную α-GAL-I, прореагировавшую с бис-COOH-PEG12 (дорожки 1-3), бис-COOH-PEG28 (дорожки 4-6), бис-COOH-PEG45 (дорожки 7-9) и бис-NHS-PEG45 (CL45) при молярном соотношении бис-NHS-PEG:α-GAL 50:1 (дорожки 1, 4 и 7), 100:1 (дорожки 2, 5 и 8) и 200:1 (дорожки 3, 6 и 9), а также маркеры молекулярной массы (MW) и несшитую полученную в растениях рекомбинантную α-GAL-I в качестве контроля (con);
Фиг.39 представляет собой график, показывающий активность α-GAL Repla gal®, полученной в растениях рекомбинантной человеческой α-GAL-I (prh-α-GAL-I) и полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45 (prh-α-GAL-I-CL45), бис-NHS-PEG4 (prh-α-GAL-I-CL4), бис-NHS-PEG2 (prh-α-GAL-I-CL2), бис-COOH-PEG45 (prh-α-GAL-I-CLA45), бис-COOH-PEG28 (prh-α-GAL-I-CLA28) или бис-COOH-PEG12 (prh-α-GAL-I-CLA12) как функцию времени инкубации при моделировании лизосомальных условий (цитрат-фосфатный буфер, рН 4,6, 37°C);
Фиг.40А и 40В представляют собой графики, показывающие активность полученной в растениях рекомбинантной человеческой α-GAL-II, сшитой бис-NHS-PEG45, в зависимости от времени инкубации при моделировании лизосомальных условий (цитрат-фосфатный буфер, рН 4,6, 37°C) (Фиг.40А) или в плазме крови человека при 37°C (Фиг.40B) (Фиг.40В показывает активность Replagal®, полученной в клетках млекопитающих рекомбинантной α-GAL и не сшитой полученной в растениях рекомбинантной человеческой α-GAL-II для сравнения);
Фиг.41 представляет собой сканированное изображение SDS-PAGE геля, показывающего полученную в растениях рекомбинантную α-GAL-II из 3 различных партий (дорожки 1-3) и полученную в растениях рекомбинантную α-GAL-II, прореагировавшую с бис-NHS-PEG45, из 5 различных партий (дорожки 4-8), а также маркеры молекулярной массы (MW);
Фиг.42 представляет собой сканированное изображение геля после изоэлектрофокусирования, показывающего полученную в растениях рекомбинантную α-GAL-II из 3 различных партий (дорожки 1-3) и полученную в растениях рекомбинантную α-GAL-II, прореагировавшую с бис-NHS-PEG45, из 5 различных партий (дорожки 4-8), а также рН маркеры (М);
Фиг.43A-43F представляют собой MALDI-TOF масс-спектры полученной в растениях рекомбинантной человеческой α-GAL-II (Фиг.43А) и полученной в растениях человеческой α-GAL-II, сшитой бис-NHS-PEG45, из 5 различных партий (Фиг.43B-43F, соответственно) (ось абсцисс указывает значения m/z, указаны значения m/z (в Да) пиков), и
Фиг.44 представляет собой график, показывающий скорость катализа (V) α-GAL, проявляемой полученной в растениях человеческой α-GAL-II, сшитой бис-NHS-PEG45, из 5 различных партий, в зависимости от концентрации субстрата (п-нитрофенил-α-D-галактопиранозида).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение, в некоторых его вариантах, относится к новым мультимерным белковым структурам, включая, но не ограничиваясь мультимерными белковым структурам α-галактозидазы, и их применению для лечения болезни Фабри. До подробного описания, по меньшей мере, одного из вариантов изобретения необходимо подчеркнуть, что изобретение не ограничивается в его применении деталями, изложенными в следующем описании или проиллюстрированными в Примерах ниже. Изобретение может иметь другие варианты или может быть практически воплощено или осуществлено различными способами.
Недостатки лизосомальных белков (например, дефекты в лизосомальных белках или отсутствие лизосомных белков) могут причинять значительный вред здоровью субъекта (лизосомальные болезни накопления). Заместительная терапия с использованием ферментов (ERT), при которой недостающий белок вводят пациенту, была использована как попытка лечения лизосомальных болезней накопления. Однако, введение недостающего белка не обязательно приводит к значительному и/или стойкому повышению активности белка in vivo.
Болезнь Фабри является примером сцепленной с Х-хромосомой рецессивной (наследуемой) лизосомальной болезни накопления, которая может вызывать широкий спектр системных симптомов. Дефицит лизосомного фермента α-галактозидазы А, возникший в результате мутации, вызывает накопление гликолипида, известного как глоботриаозилцерамид (также известный как Gb3 или церамид тригексозид) в кровеносных сосудах, других тканях и органах. Такое накопление приводит к нарушению их нормального функционирования. Для функционального восполнения недостаточности α-галактозидазы доступны две заместительные терапии с использованием ферментов (ERT). Агалзидаза альфа (Replagal®, Shire) и агалзидаза бета (Fabrazyme®, Genzyme) являются рекомбинантными формами человеческого фермента α-галактозидазы А. Указанные ферменты сложно получать, что обусловливает их высокую стоимость. Недавно загрязнение препаратов на предприятии Genzyme′s Allston, MA, вызвало нехватку агалзидазы бета во всем мире, и вводимые пациентам дозы были уменьшены на треть от рекомендованной дозы.
Как показано здесь, α-галактозидаза проявляет свою максимальную активность при низких значениях рН, характерных для лизосом, в то время как ее активность при более высоких уровнях рН снижена. Так, например, α-галактозидаза, используемая для ERT, будет слабо гидролизовать терминально галактозилированные гликолипиды в сыворотке пациентов, страдающих болезнью Фабри.
Альтернативно, как показано в настоящем документе ниже, даже в лизосомальных условиях активность α-галактозидазы постепенно снижается, хотя и медленнее, чем при более высоких уровнях рН.
Руководствуясь необходимостью решения проблемы снижения активности α-галактозидазы, авторы настоящего изобретения искали стабилизированные формы α-галактозидазы (α-GAL). В частности, авторы настоящего изобретения обнаружили, что стабилизированная форма α-галактозидазы будет проявлять активность в течение более длительного времени, в том числе будет проявлять активность в течение более длительного времени в сыворотке крови. Авторы настоящего изобретения, таким образом, разработали и успешно получили и применили стабилизированную форму нативной α-галактозидазы и, показали, что такая стабилизированная форма обладает улучшенными характеристиками с точки зрения повышения активности и/или длительности проявления активности в условиях в лизосоме и в сыворотке, что обусловливает повышенную активность белка in vivo.
Авторы изобретения продемонстрировали формирование стабилизированной формы α-галактозидазы, которая обладает улучшенными характеристиками, посредством сшивания нативной α-галактозидазы, через формирование новой ковалентной связи между мономерами α-галактозидазы.
Фиг.1 и 4 показывают снижение ферментативной активности в лизосомальных условиях для полученной в растениях рекомбинантной человеческой α-GAL I (prh-α-GAL I), α-GAL Fabrazyme® и α-GAL Replagal®. Фиг.2 и 3 показывают снижение ферментативной активности в искусственных физиологических условиях или в плазме крови человека для тех же форм α-GAL. Фиг.2 и 4 показывают, что галактоза снижает скорость уменьшения активности α-GAL.
На Фиг.5 показаны примеры сшивающих агентов на основе PEG (полиэтиленгликоля) в соответствии с дополнительными вариантами осуществления изобретения. На Фиг.6 показан сшитый димер α-GAL в соответствии с дополнительным вариантом осуществления изобретения.
Фиг.7-10 и 37 показывают, что prh-α-GAL-I прореагировала с примерами сшивающих агентов, содержащих фрагменты N-гидроксисукцинимида. Фиг.38 показывает, что prh-α-GAL-I прореагировала с примерами сшивающих агентов, содержащих карбоксильные группы, после in situ активации N-гидроксисукцинимидом. Фиг.7, 37 и 38 показывают, что реакция со сшивающим агентом приводит к получению α-GAL в основном в форме димера, а не в мономерной форме, как показано в денатурирующих условиях, что свидетельствует о том, что четвертичная структура α-GAL поддерживается ковалентным сшиванием. Фиг.11 показывает, что сшитая α-GAL сохраняет свою ферментативную активность.
Фиг.12А-12С и 39 показывают, что сшитая prh-α-GAL-I обладает более длительной активностью, чем не-сшитая α-GAL в условиях, моделирующих условия в лизосоме. Повышение устойчивости сильнее выражено для PEG28 и PEG45 линкеров, чем для более коротких PEG линкеров. Фиг.13 показывает, что сшитая prh-α-GAL-I обладает более длительной активностью, чем не сшитая α-GAL в плазме in vivo. Фиг.14А-21, показывают, что сшитая prh-α-GAL-I проявляет повышенную активность in vivo в селезенке, печени, сердце и почках. Повышение активности α-GAL сильнее при применении PEG45 линкера, чем при применении более коротких PEG линкеров. Фиг.15А, 15В и 19 показывают, что хотя сшитая PRH-α-GAL-I проявляет повышенную активность in vivo, повышенная активность не столь отчетливо сосредоточена в печени, как активность α-GAL Replagal®.
Приведенные выше результаты показывают, что сшивание полученной в растениях рекомбинантной человеческой α-GAL-I приводит к получению димера с улучшенной стабильностью, что позволяет более эффективно повышать активность α-GAL при введении in vivo.
Альтернативно, Фиг.22-28D показывают, что сшивание полученной в клетках млекопитающих рекомбинантной человеческой α-GAL приводит к получению ковалентно связанного димера (Фиг.22-24В), который обладает нормальной ферментативной активностью (Фиг.25), а также более длительной активностью в лизосомальных условиях и в пламе (Фиг.26А-26В), а также повышенной активностью in vivo в селезенке, печени, сердце и почках (Фиг.27A-28D).
Альтернативно, Фиг.29A-33L показывают, что сшивание полученной в растениях рекомбинантной человеческой α-GAL II приводит к ковалентно связанному димеру (Фиг.29-30), который обладает более длительной активностью в лизосомальных условиях и в пламе (Фиг.31А-31Б), а также повышенной активностью in vivo в плазме крови и в селезенке, печени, сердце и почках (Фиг.32A-33L). Как показано на Фиг.33A-33L, сшивание с помощью PEG45 линкера было особенно эффективным с точки зрения повышения активности in vivo.
Эти результаты показывают, что положительные эффекты сшивания применимы к различным белкам α-GAL.
Фиг.34А-34С, показывают, что сшивание α-GAL улучшает параметры ферментативного катализа α-GAL, расширяет диапазон рН для активности α-GAL и позволяет α-GAL проявлять активность при рН около 7 или более. Фиг.35-36В показывают, что пегилирование без сшивания не оказывает существенного влияния на активность α-GAL, указывая, что благоприятный эффект сшивания обусловлен непосредственно сшиванием и не является эффектом пегилирования.
Фиг.40-44 показывают, что сшивание α-GAL в соответствии с вариантами осуществления изобретения позволяет добиться хорошей воспроизводимости стабильности (Фиг.40А-40В), степени ковалентного сшивания (Фиг.41-43F) и ферментативных свойств (Фиг.44) сшитой α-GAL. Результаты, представленные здесь, показывают, что ковалентно сшитая мультимерная белковая структура α-галактозидазы характеризуется более высокой стабильностью и повышенной активностью при соответствующих физиологических условиях по сравнению с нативной формой α-галактозидазы.
Таким образом, ковалентно сшитая мультимерная белковая структура может проявлять более высокую активность по сравнению с активностью нативной формы α-галактозидазы, так как активность нативной формы быстрее снижается во времени, чем активность сшитой мультимерной белковой структуры, которая стабилизируется ковалентным сшиванием.
Ковалентно сшитая мультимерная белковая структура может проявлять более высокую активность по сравнению с активностью нативной формы α-галактозидазы также из-за высокой начальной активности (например, в силу увеличения различных параметров активности), т.е. независимо от любого снижения активности с течением времени.
Таким образом, в соответствии с аспектами некоторых вариантов осуществления настоящего изобретения предусмотрена мультимерная белковая структура, содержащая, по меньшей мере, два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент. Согласно некоторым вариантам мультимерная белковая структура обладает более высокой стабильностью по сравнению с нативной α-галактозидазой и/или более высокой начальной активностью по сравнению с нативной α-галактозидазой, как подробно описано ниже.
В настоящем документе термин «мономер», употребляемый по отношению к α-галактозидазе, относится к отдельному полипептиду α-галактозидазы. Полипептид может включать непептидные заместители (например, один или несколько сахаридных фрагментов).
В настоящем документе термин «нативный», употребляемый по отношению к α-галактозидазе, включает в себя белки, содержащие последовательность аминокислот, по существу идентичную (то есть по меньшей мере, на 95% гомологичную, необязательно, по меньшей мере, на 99% гомологичную, и, необязательно, на 100% гомологичную) аминокислотной последовательности природного белка α-галактозидазы. Природная α-галактозидаза может представлять собой белок, выделенный из природного источника, или рекомбинантный белок (например, полученный из клеток млекопитающих, растительных клеток, клеток дрожжей, бактериальных клеток, клеток насекомых).
Термин «нативный», употребляемый по отношению к четвертичной структуре α-галактозидазы (например, димеру α-галактозидазы), включает четвертичную структуру, существенно идентичную структуре природного белка.
В настоящем документе фраза «природный белок» относится к белку в той форме, которая встречается в природе (например, в организме), включая аминокислотную последовательность белка, а также четвертичную структуру белка, если белок существует в мультимерной форме.
Пост-трансляционные модификации (например, гликозилирование) природного белка α-галактозидазы (например, в организме, который экспрессирует природный белок α-галактозидазу) могут присутствовать, отсутствовать или быть изменены в нативной форме α-галактозидазы, упомянутой в настоящем документе. Нативная форме α-галактозидазы (например, рекомбинантная α-галактозидаза) может, необязательно, содержать пост-трансляционные модификации, которые отличаются от пост-трансляционных модификаций природной α-галактозидазы, при условии, что нативная форма α-галактозидазы сохраняет по существу аналогичную аминокислотную последовательность и структуру по сравнению с природной α-галактозидазой, как описано выше.
При этом нативная форма белка может относиться к мономерной структуре (например, мономеру α-галактозидазы) и/или мультимерной структуре (например, димеру α-галактозидазы). Например, димерный белок может быть описан как нативная форма α-галактозидазы, и мономерный полипептид в димерном белке может быть описан как нативная форма мономера α-галактозидазы. Необязательно, мультимерная белковая структура, описанная здесь, представляет собой димерную структуру, как и нативная форма α-галактозидазы. Альтернативно, мультимерная белковая структура содержит более двух мономеров α-галактозидазы. Например, мультимерная белковая структура может представлять собой тетрамер, гексамер или октамер, состоящий из мономеров α-галактозидазы. Мультимерная белковая структура, описанная здесь, включает ковалентные связи, которые связывают мономеры α-галактозидазы и которые отсутствуют в нативной форме α-галактозидазы.
Необязательно, связывающий фрагмент, который связывает мономеры α-галактозидазы, представляет собой группу, которая отсутствует в нативной форме α-галактозидазы (например, представляет собой синтетический связывающий фрагмент).
Так, например, связывающий фрагмент, необязательно, представляет собой фрагмент, который ковалентно связан с боковой цепью, N-концом или С-концом, или группой, возникшей в связи с пост-трансляционной модификацией (например, сахаридным фрагментом) одного мономера α-галактозидазы, а также боковой цепью аминокислоты, N-концом или С-концом, или группой, возникшей в связи с посттрансляционной модификацией (например, сахаридным фрагментом) другого мономера α-галактозидазы. Примеры таких связывающих фрагментов подробно описаны в настоящем документе ниже.
Альтернативно, связывающий фрагмент является частью мономеров α-галактозидазы, с которыми он связан (например, частью боковой цепи, N-конца или С-конца, или группы, возникшей в связи с пост-трансляционной модификацией (например, сахаридного фрагмента) одного мономера, а также частью боковой цепи, N-конца или С-конца. или группы, возникшей в связи с пост-трансляционной модификацией (например, сахаридного фрагмента) другого мономера α-галактозидазы).
Так, например, связывающий фрагмент может представлять собой ковалентную связь (например, амидную связь) между функциональной группой боковой цепи, N-концом, С-концом или функциональной группой фрагмента, возникшего в связи с пост-трансляционной модификацией мономера (например, аминогруппой), и соответсвующей функциональной группой боковой цепи, N-концом, С-концом или функциональной группой фрагмента, возникшего в связи с пост-трансляционной модификацией мономера (например, карбоксильной группой), причем такая ковалентная связь отсутствует в нативной форме α-галактозидазы. Также предусмотрены другие ковалентные связи, такие как, например, сложноэфирная связь (между гидроксильной группой и карбоксильной группой); сложная тиоэфирная связь; эфирная связь (между двумя гидроксильными группами); простая тиоэфирная связь; ангидридная связь (между двумя карбоксильными группами); тиоамидная связь; карбаматная или тиокарбаматная связь.
Необязательно, связывающий фрагмент лишен дисульфидных связей. Однако, связывающие фрагменты, которые включают в себя дисульфидные связи, которые не образуют связь между мономерами (то есть, например, расщепление дисульфидных связей не расщепляет связь между мономерами), находятся в рамках этого варианта осуществления изобретения. Потенциальное преимущество связывающего фрагмента, лишенного дисульфидных связей, состоит в том, что он не поддается расщеплению в мягких восстановительных условиях, как это имеет место для дисульфидных связей. Необязательно, связывающий фрагмент представляет собой непептидный фрагмент (например, связывающий фрагмент состоит не из амидной связи, аминокислоты, дипептида, трипептида, олигопептида или полипептида). Альтернативно, связывающий фрагмент может представлять собой или может включать пептидный фрагмент (например, аминокислоту, дипептид, трипептид, олигопептид или полипептид).
Необязательно, связывающий фрагмент не представляет собой простое линейное продолжение любого из мономеров α-галактозидазы, присоединенное к нему (например, N-конец и С-конец пептидного фрагмента не прикреплены непосредственно к С-концу или N-концу любого из мономеров α-галактозидазы). Альтернативно, связывающий фрагмент получен прямым ковалентным присоединением N-конца мономера α-галактозидазы к С-концу другого мономера α-галактозидазы для получения полипептида слияния. Такой полипептид не будет являться нативной формой α-галактозидазы, хотя он может состоять из двух мономеров α-галактозидазы, по существу, в их нативной форме. Однако, ковалентное связывание мономеров α-галактозидазы, описанное здесь, предпочтительно имеет место в форме, отличной от прямого связывания N-конца с C-концом.
Связывающий фрагмент также упоминается здесь как поперечно связывающий фрагмент. Сшивание мономеров α-галактозидазы связывающим фрагментом также обозначаю в настоящем документе как «поперечное сшивание».
Связывающий фрагмент может представлять собой ковалентную связь, атом или химическую группу (например, C(=O)-O-группу, -O-, -S-, NR-, -N=N-, -NH-C(=O)-NH-, и т.п.), или соединяющий фрагмент (состоящий из цепи химических групп).
Соединяющий фрагмент может представлять собой, например, полимерную или олигомерную группу.
Соединяющий фрагмент представляет собой многофункциональную группу (например, бирадикальную, трирадикальную и т.д), которая присоединяется к
боковой цепи, фрагментам, возникшим в связи с пост-трансляционной модификацией (например, сахаридным фрагментам) и/или концам (например, N-концу, C-концу) двух или более мономеров.
Как проиллюстрировано здесь в разделе Примеры, относительно короткие связывающие фрагменты (например, PEG2, PEG4, PEG5) могут быть менее эффективны, чем более длинные связывающие фрагменты (например, PEG28, PEG45) при сшивании различных мономеров α-галактозидазы.
Таким образом, согласно некоторым вариантам, связывающий фрагмент не является ковалентной связью, атомом или химической группой, а представляет собой протяженный соединяющий фрагмент.
Таким образом, согласно некоторым вариантам, связывающий фрагмент включает, по меньшей мере, 10 атомов в длину, необязательно, по меньшей мере, 20 атомов в длину, необязательно, по меньшей мере, 30 атомов в длину, необязательно, по меньшей мере, 50 атомов в длину, необязательно, по меньшей мере, 100 атомов в длину, и необязательно, по меньшей мере, 200 атомов в длину.
В настоящем документе длина связывающего фрагмента (при выражении в виде числа атомов) относится к длине скелета связывающего фрагмента, т.е. числу атомов, образующих линейную цепочку между остатками каждого из двух мономеров, связанных через связывающий фрагмент.
Необязательно, связывающий фрагмент может быть меньше определенного размера, чтобы избежать появления избыточной части связывающего фрагмента в полученном сшитом белке, которая может влиять на функции белка.
Следовательно, в соответствии с некоторыми вариантами, каждый связывающий фрагмент характеризуется молекулярной массой менее 20 кДа, необязательно, менее 10 кДа, необязательно, менее 5 кДа и, необязательно, менее 3 кДа.
Для облегчения сшивания связывающий фрагмент, необязательно, является существенно гибким, где связи в скелете связывающего фрагмента в основном свободны для вращения, например, представляют собой одинарные связи, которые не сопряжены с двойной связью (например, в отличие от амидной связи) и в которых вращение пространственно не затруднено. По меньшей мере, 70%, необязательно, по меньшей мере, 80% и необязательно, по меньшей мере, 90% (например, 100%) связей в скелете связывающего фрагмента являются свободными для вращения.
В некоторых вариантах связывающий фрагмент содержит цепь поли(алкиленгликоля). Термин «поли(алкиленгликоль)» в настоящем документе включает в себя семейство полиэфирных полимеров, которые имеют следующую общую формулу: -O-[(CH2)m-O-]n-, где m представляет собой число метиленовых групп, присутствующих в каждой единице алкиленгликоля, а n представляет число повторяющихся единиц, и, следовательно, определяет размер или длину полимера. Например, при m=2, полимер называют полиэтиленгликоль и при m=3 полимер называют полипропиленгликоль.
В некоторых вариантах m представляет собой целое число, большее 1 (например, m=2, 3, 4, и т.д).
Необязательно, m может варьировать для различных единиц цепи поли(алкиленгликоля). Например, цепь поли(алкиленгликоля) может включать единицы этиленгликоля (m=2) и пропиленгликоля (m=3), связанные между собой. Поли(алкиленгликоль), необязательно, содержит, по меньшей мере, две функциональные группы (например, как описано в настоящем документе), где каждая функциональная группа образует ковалентную связь с одним из мономеров α-галактозидазы. Функциональные группы, необязательно, представляют собой концевые группы поли(алкиленгликоля), так что вся цепь поли(алкиленгликоля) лежит между двумя функциональными группами. Термин «поли(алкиленгликоль)» включает в себя также его аналоги, в которых атом кислорода заменен на другой гетероатом, такой как, например, S, -NH- и тому подобное. Этот термин также охватывает производные указанных выше вариантов, в которых одна или несколько метиленовых групп, составляющих полимер, замещены. Примеры заместителей метиленовых групп включают, но не ограничиваются алкильной, циклоалкильной, алкенильной, алкинильной, алкоксильной, гидроксильной, оксо, тиольной, тиоалкоксильной и подобными группами.
Фраза «единица алкиленгликоля» в настоящем документе включает в себя -(CH2)m-O-группу или ее аналог, как описано выше, которая является основной цепи поли(алкиленгликоля), где (CH2)m (или ее аналог) связан с гетероатомом, принадлежащим другой единице алкиленгликоля или фрагменту мономера α-галактозидазы (в случае концевой единицы), и где O (или аналогичный ему гетероатом) связан с (CH2)m (или ее аналогом) другой единицы алкиленгликоля или функциональной группой, которая образует связь с мономером α-галактозидазы.
Единица алкиленгликоля может являться разветвленной, так что она связан с 3 или более соседними единицами алкиленгликоля, где каждая из 3 или более соседних единиц алкиленгликоля являются частью цепи поли(алкиленгликоля). Такая разветвленная единица алкиленгликоля связана через гетероатом с одной соседней единицей алкиленгликоля, и каждый гетероатом оставшихся соседних единиц алкиленгликоля связан с атомом углерода разветвленной единицы алкиленгликоля. Альтернативно, гетероатом (например, атом азота) может быть связан более чем с одним атомом углерода единицы алкиленгликоля, частью которой он является, образуя разветвленные единицы алкиленгликоля (например, [(-CH2)m]2N- и т.п).
В примерах вариантов изобретения, по меньшей мере, 50% единиц алкиленгликоля идентичны, например, они включают тот же гетероатом и характеризуются одинаковым значением m. По меньшей мере, 70%, необязательно, по меньшей мере, 90%, и, необязательно, 100% единиц алкиленгликоля являются идентичными. В примерах вариантов изобретения гетероатомы, связанные с одинаковыми единицами алкиленгликоля, представляют собой атомы кислорода. В дополнительных примерах вариантов для идентичных единиц m представляет собой 2.
В одном из вариантов линкер представляет собой линкер с одной неразветвленной цепью и предпочтительно представляет собой полиэтиленгликоль (PEG). В настоящем документе термин «поли(этиленгликоль)» описывает поли(алкиленгликоль), как описано выше, где, по меньшей мере, 50%, по меньшей мере, 70%, по меньшей мере, 90%, предпочтительно 100% единиц алкиленгликоля представляют собой -CH2CH2-O-. Альтернативно, фраза «единица этиленгликоля» определяется здесь как единица -CH2CH2O-.
В соответствии с дополнительными вариантами связывающий фрагмент содержит поли(этиленгликоль) или его аналог, имеющий общую формулу:
-X1-(CR1R2-CR3R4-Y)n-X2-
где каждый из X1 и X2 представляет собой функциональную группу, которая образует ковалентную связь, по меньшей мере, с одним мономером α-галактозидазы;
Y представляет собой O, S или NR5 (необязательно O);
n представляет собой целое число от 1 до 200 (необязательно от 5 до 150 и, необязательно, от 40 до 70), хотя большие значения n также предусмотрены,
каждый из R1, R2, R3, R4 и R5 независимо выбран из группы, состоящей из водорода, алкильной, циклоалкильной, алкенильной, алкинильной, алкоксильной, гидроксильной, оксо, тиольной и тиоалкоксильной групп.
В некоторых вариантах n представляет собой, по меньшей мере, 5, необязательно, по меньшей мере, 8, необязательно, по меньшей мере, 15, необязательно, по меньшей мере, 25 и необязательно, по меньшей мере, 40.
В некоторых вариантах n составляет не более 200, необязательно, не более 150, и, необязательно, не более 70.
Поли(этиленгликоль) или его аналог, необязательно, может содержать сополимер, например, где не все CR1R2-CR3R4-Y единицы в приведенной выше формуле идентичны друг другу.
В некоторых вариантах, по меньшей мере, 50% CR1R2-CR3R4-Y единиц идентичны.
По меньшей мере, 70%, необязательно, по меньшей мере, 90%, и, необязательно, 100% CR1R2-CR3R4-Y единиц идентичны.
Необязательно, связывающий фрагмент разветвлен, например, в одной или нескольких из CR1R2-CR3R4-Y единиц в приведенной выше формуле и, по меньшей мере, один из R1, R2, R3, R4 и R5 представляет собой -(CR1R2-CR3R4-Y)p-X3-, где R1-R4 и Y указаны выше, а p является целым числом, как определено здесь для n (например, составляет от 1 до 200), а Х3 определяется здесь, как это указано для X1 и Х2.
Функциональные группы могут, необязательно, образовывать связь, включая, но не ограничиваясь амидной связью, сложноэфирной связью и/или эфирной связью.
Например, функциональная группа может, необязательно, содержать карбонильную группу, которая образует амидную связь с атомом азота полипептида (например, в остатке лизина или на N-конце), или эфирную связь с атомом кислорода полипептида (например, в остатке серина, треонина или тирозина). Альтернативно или дополнительно, функциональная группа может, необязательно, содержать гетероатом (например, N, S, О), который образует амидную связь, эфирную связь или сложную тиоэфирную связь с карбонильной группой полипептида (например, карбонильной группой остатка глутамата или аспартата или С-конца). Альтернативно или дополнительно, функциональная группа может включать алкильную или арильную группу, присоединенную к полипептиду (например, к гетероатому в полипептиде).
Альтернативно или дополнительно, функциональная группа может, необязательно, содержать атом азота, который образует аминную связь с алкильной группой мономера α-галактозидазы, или мономер α-галактозидазы может, необязательно, содержать атом азота, который образует аминную связь с алкильной группой в функциональной группе. Такая аминная связь может быть образована путем восстановительного аминирования (например, как описано ниже). В некоторых вариантах, по меньшей мере, одна из функциональных групп образует амидную связь с полипептидом (например, через остаток лизина в нем). Функциональные группы могут быть идентичны друг другу или могут быть различными.
В некоторых вариантах, по меньшей мере, одна из функциональных групп присоединена к одной функциональной группе полипептида (например, аминогруппе остатка лизина или N-конца), и, по меньшей мере, одна из функциональных групп присоединена к другой функциональной группе полипептида (например, тиоловой группе остатка цистеина).
В соответствии с дополнительным вариантом мультимерные белковые структуры, описанные в настоящем документе, проявляют высокую стабильность в условиях человеческой плазмы и/или в лизосомальных условиях. В настоящем документе фраза «условия человеческой плазмы» относится к плазме крови человека в качестве среды при температуре 37°C. В настоящем документе фраза «лизосомальные условия» относится к водному раствору с рН 4,6 в качестве среды (например, цитрат-фосфатному буферу, описанному в настоящем документе) при температуре 37°C.
Повышенная стабильность в лизосомальных условиях желательна, поскольку лизосома является мишенью для заместительной терапии α-галактозидазой, так как лизосомы в норме являются местом локализации α-галактозидазы в организме, и условия в лизосоме (например, кислый рН) представляют собой оптимальные условия для активности α-галактозидазы.
Не желая быть связанными какой-либо конкретной теорией, авторы настоящего изобретения полагают, что повышенная стабильность условиях, схожих с условиями в сыворотке крови (например, условиями человеческой плазмы, как описано в настоящем документе) также является желательной, поскольку стабильная α-галактозидаза в крови может проявлять активность по отношению к метаболитам (например, Gb3), присутствующим в крови из-за оттока из клеток. Активная в сыворотке мультимерная белковая структура может, необязательно, быть эффективной в устранении и предотвращении отложения гликосфинголипидов на стенках кровеносных сосудов, которое способствует воспалению [Bodary et al., TCM 17(4):129-133]. Например, при болезни Фабри основные патологические эффекты наступают вследствие накопления Gb3 в эндотелии сосудов, что приводит к окклюзии мелких сосудов, ишемии и инфаркту этих сосудов и ишемии и инфаркту почек, сердца и головного мозга [Desnick et al., 2003, Annals of Internal Medicine, 138(4):338-346]. Альтернативно, повышенная стабильность в сыворотке крови может свести на нет необходимость доставки в лизосомы. Таким образом, ERT может стать гораздо более доступной, так как могут быть использованы надежные экономически эффективные системы хозяев, например, растения.
В соответствии с дополнительными вариантами, высокая стабильность мультимерной белковой структуры в условиях человеческой плазмы такова, что мультимерная белковая структура проявляет α-галактозидазную активность при помещении в условия человеческой плазмы на один час, которая, по меньшей мере, на 10% выше, необязательно, на 20% выше, необязательно, на 50% выше, и, необязательно, на 100% выше, чем α-галактозидазная активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час.
Альтернативно или дополнительно, высокая стабильность мультимерной белков ой структуры в условиях человеческой плазмы такова, что α-галактозидазная активность мультимерной белковой структуры снижается более медленно в условиях человеческой плазмы, чем соответствующая активность нативной α-галактозидазы. Необязательно, мультимерная белковая структура проявляет активность, которая уменьшается при помещении белковой структуры в условия человеческой плазмы на один час на величину, которая, по меньшей мере, на 10% меньше, необязательно, на 20% меньше, необязательно, на 50% меньше и, необязательно, на 80% меньше, чем величина, на которую уменьшается соответствующая активность нативной α-галактозидазы при помещении нативной α-галактозидазы в условия человеческой плазмы на один час.
Следует понимать, что в настоящем документе уменьшение на величину, которая «на 10% меньше», чем уменьшение на 50% относится к уменьшению на 45% (45 на 10% меньше, чем 50), а не к уменьшению на 40% (50% -10%).
Альтернативно или дополнительно, высокая стабильность мультимерной белковой структуры в условиях человеческой плазмы такова, что α-галактозидазная активность мультимерной белковой структуры остается практически неизменной при помещении мультимерной белковой структуры в условия человеческой плазмы на один час и, необязательно, на 2, 4 или даже 6 часов.
В настоящем документе фраза «по существу неизменная» относится к уровню (например, активности), который остается в диапазоне от 50% до 150% от исходного уровня, и, необязательно, уровню, который остается в пределах, по меньшей мере, 60%, необязательно, по меньшей мере, 70%, необязательно, по меньшей мере, 80% и необязательно, по меньшей мере, 90% от исходного уровня.
Необязательно, высокая стабильность мультимерной белковой структуры в лизосомальных условия такова, что мультимерная белковая структура проявляет при помещении в лизосомальные условия в течение определенного периода времени (например, на один день, два дня, 3 дня, неделю) α-галактозидазную активность, которая, по меньшей мере, на 10% выше, необязательно, на 20% выше, необязательно, на 50% выше, и, необязательно, на 100% выше, чем активность нативной α-галактозидазы при помещении нативной α-галактозидазы в лизосомальные условия на тот же заданный период времени.
Альтернативно или дополнительно, высокая стабильность мультимерной белков ой структуры в лизосомальных условия такова, что α-галактозидазная активность мультимерной белковой структуры снижается в лизосомальных условиях медленнее, чем соответствующая активность нативной α-галактозидазы. Необязательно, мультимерная белковая структура проявляет активность, которая уменьшается при помещении белковой структуры в лизосомальные условия на определенный период времени (например, один день, 2 дня, 3 дня, неделю), на величину, которая, по меньшей мере, на 10% меньше, необязательно, на 20% меньше, необязательно, на 50% меньше и, необязательно, на 80% меньше, чем величина, на которую соответствующая активность нативной α-галактозидазы уменьшается при помещении нативной α-галактозидазы в лизосомальные условия на тот же период времени.
Альтернативно или дополнительно, высокая стабильность мультимерной белковой структуры в лизосомальных условия такова, что α-галактозидазная активность мультимерной белковой структуры остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день, на 2 дня, на 3 дня, на одну неделю, на две недели и/или на один месяц.
Как показано в разделе Примеры в настоящем документе, в дополнение к проявлению большей стабильности с течением времени мультимерная белковая структура может характеризоваться параметрами α-галактозидазной активности, которые отличаются от параметров активности нативной α-галактозидазы.
Таким образом, в соответствии с дополнительным вариантами мультимерная белковая структура независимо от снижения активности с течением времени характеризуется наличием α-галактозидазной активности, которая является более высокой, чем α-галактозидазная активность нативной формы белка. Необязательно, активность на 10% выше и, необязательно, на 20% выше, чем соответствующая активность нативной формы.
Для того чтобы охарактеризовать такую активность, активность предпочтительно измерять сразу (например, в течение 1 часа, в течение 15 минут) при помещении нативной α-галактозидазы или мультимерной белковой структуры в условия (например, как описано в настоящем документе), в которых активность существенно снижается, так что измеряемая активность будет отражать активность как таковую, а не степень стабильности.
Необязательно, мультимерная белковая структура характеризуются наличием α-галактозидазной активности в лизосомальных условиях, которая является более высокой, чем соответствующая активность нативной α-галактозидазы.
Альтернативно или дополнительно, мультимерная белковая структура характеризуются наличием α-галактозидазной активности в моделируемых физиологических условиях при нейтральном рН, которая является более высокой, чем соответствующая активность нативной α-галактозидазы. Моделируемые физиологические условия включают водный раствор (например, фосфатный буферный солевой раствор) при температуре 37°C. рН, необязательно, составляет 7.
Альтернативно, рН составляет 7,4.
Активность α-галактозидазы, описанная в данном документе, представляет собой биологическую активность, характерную для α-галактозидазы (например, каталитическую активность, характерную для α-галактозидазы, такую как гидролиз α-концевого галактозильного фрагмента субстрата). В некоторых вариантах каталитическая активность α-галактозидазы характеризуется скоростью катализа при насыщении субстратом (т.е. значением Vmax).
Альтернативно, активность α-галактозидазы представляет собой терапевтическую активность (например, ферментативную активность, имеющую терапевтический эффект), такую как терапевтическая активность в контексте болезни Фабри. Необязательно, терапевтическая активность определяется на экспериментальных животных (например, страдающих болезнью Фабри мышах), и, необязательно, на человеческих пациентах, страдающих болезнью Фабри. Методы определения активности α-галактозидазы известны специалистам в данной области. Как правило, α-галактозидазу (т.е. нативную форму или мультимерную белковую структуру по настоящему документу) вводят в контакт с соединением, которое является известным в уровне техники субстратом α-галактозидазы, после чего количественно определяют величину активности. Соединения, которые позволяют быстро обнаруживать α-галактозидазную активность, известны в данной области и являются коммерчески доступными.
В некоторых вариантах α-галактозидазную активность определяют путем анализа гидролиза 4-метилумбеллиферил-α-D-галактопиранозида (например, как описано в разделе Примеры в настоящем документе).
В некоторых вариантах α-галактозидазную активность определяют путем анализа гидролиза п-нитрофенил-α-D-галактопиранозида (например, как описано в разделе Примеры в настоящем документе).
При сравнении активности мультимерной белковой структуры, описанной в настоящем документе, с активностью нативной α-галактозидазы, нативная α-галактозидаза предпочтительно содержит мономеры α-галактозидазы, существенно идентичные (например, по аминокислотной последовательности и гликозилированию) мономерам α-галактозидазы мультимерной структуры. Согласно некоторым вариантам, мультимерная белковая структура характеризуется временем полураспада в кровотоке в физиологической системе (например, крови, сыворотке и/или плазме человека или лабораторных животных), которое является более длительным (например, по меньшей мере, на 20% более длительным, по меньшей мере, на 50% более длительным, по меньшей мере, на 100% более длительным, по меньшей мере, на 400% более длительным, по меньшей мере, на 900% более длительным) по сравнению со временем полураспада в кровотоке нативной α-галактозидазы.
Повышение времени полураспада в кровотоке, необязательно, может быть связано с более высокой стабильностью in vitro (например, как описано в настоящем документе), более высокой стабильностью w vivo (например, устойчивостью к метаболизму) и/или с другими факторами (например, снижением почечного клиренса).
Время полураспада в кровотоке может быть определено путем взятия проб (например, образцов крови, образцов тканей) из физиологической системы (например, у людей, лабораторных животных) через различные интервалы времени и определения уровня α-галактозидазы в образце с использованием известных в уровне техники методов.
Необязательно, время полураспада рассчитывается как конечный период полураспада (например, как описано в разделе Примеры), где время полураспада представляет собой время, необходимое для уменьшения концентрации (например, концентрации в крови) на 50% после достижения псевдо-равновесного распределения. Конечное время полураспада может быть рассчитано из концевой линейной части графика, показывающего зависимость логарифма концентрации от времени с помощью линейной регрессии зависимости логарифма концентрации от времени (см., например, Toutain & Bousquet-Melou [J Vet Pharmacol Ther 2004, 27:427-39]). Таким образом, конечный период полураспада является мерой снижения концентрации лекарственного средства в плазме в результате элиминирования лекарственного средства, а не уменьшения концентрации вследствие других причин, и конечный период полураспада не обязательно представляет собой время, необходимое для уменьшения количества введенного препарата на половину.
Определение уровня α-галактозидазы (например, мультимерной белковой структуры или нативной α-галактозидазы) может включать в себя обнаружение физического присутствия α-галактозидазы (например, с помощью антител против α-галактозидазы) и/или определение уровня α-галактозидазной активности (например, как описано в настоящем документе).
Согласно некоторым вариантам, мультимерная белковая структура характеризуется проявлением α-галактозидазной активности в органе (например, селезенке, сердце, почках, головном мозге, печени) после введения (например, внутривенного введения) белковой структуры позвоночному животному (например, человеку, мыши), например, позвоночному животному с дефицитом α-галактозидазы (например, человеку, страдающему болезнью Фабри, мыши, страдающей болезнью Фабри). Необязательно, α-галактозидазная активность в органе выше, чем α-галактозидазная активность нативной α-галактозидазы в органе при введении ее эквивалентного количества позвоночному животному.
Активность в органе может являться функцией поглощения α-галактозидазы и/или сохранения α-галактозидазной активности после поглощения. Необязательно, α-галактозидазную активность в органе определяют через 2 часа после введения, и, необязательно, через 24 часа, необязательно, через 3 дня, необязательно, через 7 дней, и, необязательно, через 14 дней после введения. Повышение активности α-галактозидазы в печени в некоторых случаях может быть связано с понижением активности в других частях организма, и, следовательно, с уменьшением биологического эффекта α-галактозидазы. Таким образом, в некоторых вариантах мультимерная белковая структура характеризуется увеличенной α-галактозидазной активностью в органе, отличном от печени. Примеры органов включают селезенку, сердце и почки. В некоторых вариантах мультимерная белковая структура характеризуется увеличенной α-галактозидазной активностью в органе после введения (как описано выше), которая, по меньшей мере, на 20% выше, необязательно, по меньшей мере, на 50% выше, необязательно, по меньшей мере, на 100% выше, и необязательно, по меньшей мере, на 300% выше, чем активность нативной α-галактозидазы после введения эквивалентного ее количества. Как отмечалось выше, авторы настоящего изобретения разработали и успешно получили и применили стабилизированную форму α-галактозидазы за счет использования мультимерной структуры сшитых мономеров α-галактозидазы.
Необязательно, α-галактозидаза является человеческой α-галактозидазой (например, рекомбинантной человеческой α-галактозидазой), например, для обеспечения оптимальной биосовместимости для введения человеку в качестве субъекта. Человеческая α-галактозидаза является коммерчески доступной, например, в виде препаратов Replagal® (агалзидаза альфа, Shire) и Fabrazyme® (агалзидаза бета, Genzyme). В настоящем документе «человеческая α-галактозидаза» относится к α-галактозидазе, содержащей аминокислотную последовательность, по существу идентичную (например, как описано выше) аминокислотной последовательности белка α-галактозидазы, который является природным белком организма человека.
В некоторых вариантах α-галактозидаза представляет собой полученную в растениях рекомбинантную α-галактозидазу. Примеры α-галактозидазы включают полученную в растениях рекомбинантную человеческую α-галактозидазу.
Примеры α-GAL включают, но не ограничиваются α-GAL с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3. Необязательно, α-GAL имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2 и SEQ ID NO: 3.
В настоящем документе «α-галактозидаза» относится к любому белку, который обладает ферментативной активностью (например, гидролитической активностью) в отношении остатков галактозы в Gb3 (например, α-галактозидаза А). Необязательно, «α-галактозидаза» относится к ЕС 3.2.1.22.
α-Галактозидаза согласно вариантам осуществления изобретения может быть очищена (например, из растений или тканей животных) или получена с помощью технологии рекомбинантной ДНК.
Как описано в настоящем документе, активность α-галактозидазы в сыворотке может быть желательна, например, для снижения уровня Gb3 в сыворотке крови.
Таким образом, в некоторых вариантах α-галактозидаза представляет собой щелочную α-галактозидазу.
В настоящем документе фраза «щелочная α-галактозидаза» относится к α-GAL, характеризующейся способностью к гидролизу концевых групп α-галактозы на содержащих галактозу олигосахаридах при значениях рН от нейтральных до основных (например, рН 7-7,5), в частности, при нормальном сывороточном pH (например, при рН около 7,35-7,45).
Следует иметь в виду, что щелочная α-GAL по некоторым вариантам осуществления изобретения может быть активна при значениях рН от нейтральных до основных, но все еще может проявлять активность в условиях кислого рН (т.е. при рН около 4,6). В конкретном варианте фермент является активным при значениях рН от кислых до основных (например, при рН около 4,2-7,5).
В еще одном конкретном варианте фермент является активным при рН около 6,5-7,5. Конкретные примеры щелочных α-галактозидаз, которые могут быть использованы по настоящему документу описаны в заявке США 20070036883, WO 03/097791 и PCT/IL 2010/000956, каждая из которых включена в настоящее описание посредством отсылки в полном объеме.
Таким образом, щелочная α-галактозидаза может являться членом семейства растительных α-галактозидаз, выбранных из группы, состоящей из α-галактозидаз семейств тыквенных, губоцветных, перечных, пасленовых, бобовых, крестоцветных и злаковых.
В соответствии с конкретным вариантом, щелочная α-галактозидаза представляет собой α-галактозидазу дыни.
P.-R.Gaudreault и J.A.Webb в ряде публикаций описали (таких как "Alkaline alpha-galactosidase in leaves of Cucurbita pepo". Plant Sci. Lett. 24, 281-288, 1982, "Partial purification and properties of an alkaline alpha-galactosidase from mature leaves of Cucurbita pepo", Plant Physiol., 71, 662-668, 1983, и "Alkaline alpha-galactosidase activity and galactose metabolism in the family Cucurbitaceae", Plant Science, 45, 71-75, 1986) новую α-галактозидазу, очищенную из молодых листьев Cucurbita pepo, которая имеет оптимальную активность при щелочных условиях (рН 7,5). В дополнение к наличию щелочной α-галактозидазы, они также сообщили о трех кислых формах фермента, причем для кислой и щелочной форм были установлены различные субстратные предпочтения.
Наличие α-галактозидазной активности в щелочной среде наблюдается и в других тканях тыквенных, таких как плодоножки огурцов, молодых плодах столовой тыквы и молодых плодах дыни ("Melons: Biochemical and Physiological Control of Sugar Accumulation", Encyclopedia of Agricultural Science, vol.3, pp.25-37, Arntzen, C.J., et al., Academic Press, New York, 1994).
Bachmann и соавторы сообщили ("Metabolism of the raffinose family oligosaccharides in leaves of Ajuga reptens L.", Plant Physiology 105:1335-1345, 1994), что растение Ajuga reptens (живучка ползучая) из неродственного семейства губоцветных, способное использовать и накапливать стахиозу, также содержит щелочную α- галактозидазу. Этот фермент был частично охарактеризован, и было установлено его высокое сродство к стахиозе. Также листья растения Peperomia camptotricha L., из семейства перечных демонстрируют α-галактозидазную активность в щелочной среде, что позволяет предположить, что они также содержат фермент щелочную α-галактозидазу (Madore, M., "Catabolism of raffinose family oligosaccharides by vegetative sink tissues", Carbon Partitioning and Source-Sink Interactions in Plants, Madore, M., Lucas, W.J. pp.204-214, 1995, American Society of Plant Physiologists, Maryland). Также Gao и Schaffer (Plant Physiol. 1999; 119:979-88, которая включена в настоящий документ посредством отсылки в полном объеме) сообщили о наличии α-галактозидазной активности с щелочными оптимумом рН в неочищенных экстрактах тканей из различных видов, включая членов семейства тыквенные и рода колеус семейства Губоцветных. Конкретные примеры последовательностей растительных щелочных α-галактозидаз представлены в SEQ ID NO: 4, 5 и 13 (Cucumis melo), 6 (Т. tetragonioides), 7 и 12 (Cucumis sativus), 8 и 9 (Zea mays), 10 (Oruza sativa), 11 (Pisum sativum) и 14 (Coffea arabica).
В некоторых вариантах α-галактозидаза представляет собой кислую α-галактозидазу. В настоящем документе «кислая α-галактозидаза» относится к α-галактозидазе, характеризующейся способностью к гидролизу концевых групп α-галактозы на содержащих галактозу олигосахаридах при кислых значениях рН (например, при рН 4,2-5), которые присутствуют в лизосоме.
α-Галактозидаза согласно вариантам осуществления изобретения может представлять собой α-галактозидазу человеческого, животного или растительного происхождения при условии отсутствия чрезмерной неблагоприятной иммунологической реакции при введении in vivo (например, при введении растительной α-галактозидазы человеку). Для снижения иммунологической реакции, препарат не человеческой α-галактозидазы (например, растительной α-галактозидазы) может быть введен совместно с человеческой α-галактозидазой (например, кислой человеческой α-галактозидазой). Необязательно, мультимерная белковая структура дополнительно содержит, по меньшей мере, один манноза-6-фосфатный (М6Р) фрагмент. Фрагмент М6Р (или фрагменты) может быть связан с одним или несколькими мономерами α-галактозидазы мультимерной белковой структуры (например, через линкер). Методы и реагенты для введения содержащих М6Р фрагментов в биомолекулы (например, полипептиды) описаны в WO 2009/024977. Как показано в разделе Примеры в настоящем документе, мультимерная белковая структура по настоящему документу может быть легко получена путем взаимодействия α-галактозидазы со сшивающим агентом. Таким образом, в соответствии с другим аспектом вариантов изобретения, предусмотрен способ получения мультимерной белковой структуры, описанной в настоящем документе. Способ включает реакцию α-галактозидазы, приводящую к введению, по меньшей мере, одного связывающего фрагмента, который ковалентно связывает, по меньшей мере, два мономера α-галактозидазы. Необязательно, связывающий фрагмент представляет собой связь (например, амидную связь, дисульфидную связь), которая связывает один мономер α-галактозидазы с другим мономером α-галактозидазы. Необязательно, связь вводят с помощью подходящих условий и/или реагентов. Например, реагенты, которые пригодны для формирования амидной связи из карбоксильной группы и аминогруппы, известны в данной области.
Необязательно, связывающий фрагмент представляет собой фрагмент, который не является производным части α-галактозидазы. Например, связывающий фрагмент может представлять собой олигомер, полимер, остаток небольшой молекулы (например, аминокислоты).
В некоторых вариантах связывающий фрагмент вводят путем взаимодействия α-галактозидазы со сшивающим агентом, который содержит связывающий фрагмент (например, как описано в настоящем документе) и, по меньшей мере, две реакционноспособные группы.
Необязательно, α-галактозидаза взаимодействует с реагентом в условиях, в которых нативная α-галактозидаза присутствует в димерной форме. В некоторых вариантах, сшивающий агент взаимодействует с α-галактозидазой при молярном соотношении в диапазоне от 5:1 до 500:1 (сшивающий агент: мономер α-галактозидазы), необязательно, в диапазоне от 50:1 до 400:1, и, необязательно, в диапазоне от 75:1 до 300:1 (например, около 100:1, около 200:1). Указанный способ, необязательно, дополнительно включает очистку сшитого белка, например, удаление избытка сшивающего агента. Могут быть использованы обычные методы очистки, например, диализ и/или ультрафильтрация с использованием мембран с соответствующими величинами исключения по молекулярной массе и/или дополнительные хроматографические стадии, включая хроматографию по размеру, ионообменную хроматографию, аффинную хроматографию, гидрофобную хроматографию и тому подобное.
Реакционноспособную группу выбирают таким образом, чтобы она подходила для прохождения химической реакции, которая приводит к формированию связи с соответствующей функциональной группой мономера α-галактозидазы. Необязательно, каждая реакционноспособная группа способна образовывать ковалентную связь между связывающим фрагментом, описанным здесь, и, по меньшей мере, одним полипептидом (например, так, чтобы формировать функциональную группу, связанную с полипептидом, как описано в настоящем документе).
Реакционноспособные группы сшивающего агента могут быть идентичны друг другу или могут быть различными.
В настоящем документе фраза «реакционноспособная группа» описывает химическую группу, которая способна вступать в химическую реакцию, которая обычно приводит к формированию связи. Связи в соответствии с вариантами настоящего изобретения предпочтительно представляют собой ковалентные связи (например, для каждой из реакционноспособных групп). Химические реакции, которые приводят к формированию связей, включают, например, реакции нуклеофильного и электрофильного замещения, реакции нуклеофильного и электрофильного присоединения, алкилирования, реакции присоединения-элиминирования, реакции циклоприсоединения, перегруппировки и другие известные органические реакции, которые включают участие функциональных групп, а также их комбинации.
Реакционноспособная группа может, необязательно, включать неактивную часть (например, алкильную), которая может служить, например, для прикрепления реакционноспособной части реакционноспособной группы к связывающему фрагменту (например, поли(алкиленгликолю) или его аналогу), описанному в настоящем документе.
Реакционноспособную группу предпочтительно выбирают таким образом, чтобы было возможно провести ее конъюгирование с α-галактозидазой. Примеры реакционноспособных групп включают, но не ограничиваются карбоксильной (например, -CO2H), тиольной (-SH), аминной (-NH2), галогенной, азидной (-N3), изоцианатной (-NCO), изотиоцианатной (-N=C=S), гидроксильной (-ОН), карбонильной (например, альдегидной), малеимидной, сульфатной, фосфатной, сульфонильной (например, мезильной, тозильной) и др., а также активированными группами, такими как N-гидроксисукцинимидная (NHS) (например, NHS эфиры), сульфо-N-гидроксисукцинимидная, ангидридная, ацилгалогенидная (-C(O)-галоген) и др.
В некоторых вариантах реакционноспособная группа включает в себя уходящую группу, такую как уходящая группа, подверженная нуклеофильному замещению (например, галогенная, сульфатная, фосфатная, карбоксильная, N-гидроксисукцинимидная группа).
Необязательно, реакционноспособная группа может присутствовать в своей активированной форме.
В настоящем документе, фраза «активированная форма» описывает производное химической группы (например, реакционноспособной группы), которое является более реакционноспособным, чем исходная химическая группа, и которое, таким образом, легко способно вступать в химическую реакцию, которая приводит к формированию связи. Активированная форма может содержать особенно подходящую уходящую группу, тем самым способствуя реакции замещения. Например, группа -C(=O)-NHS (N-гидроксисукцинимидиловый эфир, или -C(=O)-O-сукцинимид) является известной активированной формой -С(=O)ОН, так как NHS (N-гидроксисукцинимид) может взаимодействовать с -C(=O)ОН с образованием -С(=O)-NHS, который легко вступает в реакцию с образованием продуктов, характерных для реакций с участием -С(=O)ОН группы, таких как амиды и сложные эфиры.
Реакционноспособная группа может быть присоединена к остальной части связывающего фрагмента (например, поли(алкиленгликолю) или его аналогу) с помощью различных групп, атомов или связей. Они могут включать эфирную связь [например, -O-алкил-], сложноэфирную связь [например, -O-C(=O)-алкил-], карбаматную связь [например, O-C(=O)-NH-алкил-], и т.д. Таким образом, могут быть использованы различные концевые группы.
Следующие не ограничивающие примеры различных групп могут представлять собой реакционноспособные группы, как описано в настоящем документе: -CH2CO2H, -CH2CH2CO2H, -CH2CH2SH, -CH2CH2NH2, -CH2CH2N3, -CH2CH2NCO, -CH2-C(O)-NHS, -CH2CH2-C(=O)-NHS, -C(=O)-CH2-С(=O)-NHS, -CH2CH2-NHC(=O)CH2CH2-малеимид и т.д.
Количество метиленовых групп в каждой из вышеуказанных реакционноспособных групп указано исключительно в качестве примера, и может быть изменено. Реакционноспособная группа также может включать гетероатом на конце цепи поли(алкиленгликоля) (например, -ОН).
В примерах вариантов осуществления настоящего изобретения реакционноспособная группа включает карбоксильную группу (например, активированную карбоксильную группу, такую как ее N-гидроксисукцинимидиловый эфир).
Необязательно, реакционноспособная группа реагирует с аминогруппой α-галактозидазы (например, аминогруппой остатка лизина и/или N-конца) с образованием амидной связи.
В некоторых вариантах, реакция реакционноспособной группы включает восстановительное аминирование, в котором аминогруппа взаимодействует с альдегидной группой с образованием имина, затем имин восстанавливают (например, путем добавления восстановителя, такого как цианоборгидрид натрия) с образованием аминной связи. Реакционноспособная группа может представлять собой аминогруппу, которая реагирует с альдегидной группой α-галактозидазы (например, альдегидной группой на сахаридном фрагменте, присоединенном к полипептиду белка), или реакционноспособная группа может представлять собой альдегидную группу, которая реагирует с аминогруппой α-галактозидазы (например, аминогруппой остатка лизина). Необязательно, до реакции реакционноспособной группы с α-галактозидазой сахаридный фрагмент α-галактозидазы окисляют воздействием окислителя с образованием альдегидной группы. Например, реакция сахарида с периодатом натрия может быть использована для получения пары альдегидных групп в сахаридном фрагменте.
В некоторых вариантах, по меньшей мере, одна из реакционноспособных групп выбрана таким образом, что она способна реагировать с одной функциональной группой мономера α-галактозидазы (например, аминогруппой остатка лизина или N-конца), и, по меньшей мере, одна из реакционноспособных групп выбрана таким образом, чтобы реагировать с другой функциональной группой мономера α-галактозидазы (например, тиоловой группой остатка цистеина).
Необязательно, один или несколько полипептидов, описанных здесь, вступают в реакцию с реагентом для гликозилирования для введения одного или нескольких фрагментов М6Р, чтобы получить содержащую М6Р мультимерную белковую структуру (например, как описано в настоящем документе). Подходящие содержащие М6Р реагенты для гликозилирования и их применение описаны, например, в WO 2009/024977.
В настоящем документе термин «амин» и «амино» относится к -NR′R″ группе, где R′ и R″ выбраны из группы, состоящей из водорода, алкильной, циклоалкильной, гетероалициклической (присоединенной через атом углерода кольца), арильной и гетероарильной (присоединенной через атом углерода кольца) групп. R′ и R″ присоединены через атом углерода. Необязательно, R′ и R″ выбраны из группы, состоящей из водорода и алкильной группы, содержащей от 1 до 4 атомов углерода. Необязательно, R′ и R″ представляют собой водород.
В настоящем документе термин «алкил» относится к насыщенному алифатическому углеводороду, включая группы с неразветвленными и разветвленными цепями. Предпочтительно, алкильная группа включает от 1 до 20 атомов углерода. В настоящем документе при указании числового диапазона, например, «1-20», подразумевается, что группа, в данном случае алкильная группа, может содержать 1 атом углерода, 2 атома углерода, 3 атома углерода, и т.д. вплоть до 20 атомов углерода включительно. Более предпочтительно, алкил представляет собой алкильную группу среднего размера, включающую от 1 до 10 атомов углерода. Наиболее предпочтительно, если не указано иное, алкил представляет собой низший алкил, включающий от 1 до 4 атомов углерода. Алкильная группа может быть замещенной или незамещенной. При наличии заместителей замещающая группа может представлять собой, например, циклоалкильную, алкенильную, алкинильную, арильную, гетероарильную, гетероалициклическую, галогенную, гидроксильную, алкоксильную, арилоксильную, тиогидроксильную, тиоалкоксильную, тиоарилоксильную, сульфинильную, сульфонильную, циано, нитро, азидную, фосфонильную, фосфинильную, оксо, карбонильную, тиокарбонильную, карбаматную, тиокарбаматную, O-карбамильную, N-карбамильную, O-тиокарбамильную, N-тиокарбамильную, C-амидную, N-амидную, C-карбокси, O-карбоксильную, сульфонамидную и аминную группу, как указанные термины определены в настоящем документе.
«Циклоалкильная» группа относится к моноциклической или конденсированной кольцевой группе (т.е. циклам, которые имеют общую пару атомов углерода), содержащей в кольце только атомы углерода, в которой одно или более колец не имеют полностью сопряженной пи-электронной системы. Не ограничивающие примеры циклоалкильных групп включают циклопропан, циклобутан, циклопентан, циклопентен, циклогексан, циклогексадиен, циклогептан, циклогептатриен и адамантан. Циклоалкильная группа может быть замещенной или незамещенной. При наличии заместителей замещающая группа может представлять собой, например, алкильную, алкенильную, алкинильную, арильную, гетероарильную, гетероалициклическую, галогенную, гидроксильную, алкоксильную, арилоксильную, тиогидроксильную, тиоалкоксильную, тиоарилоксильную, сульфинильную, сульфонильную, циано, нитро, азидную, фосфонильную, фосфинильную, оксо, карбонильную, тиокарбонильную, карбаматную, тиокарбаматную, O-карбамильную, N-карбамильную, O-тиокарбамильную, N-тиокарбамильную, C-амидную, N-амидную, C-карбокси, O-карбоксильную, сульфонамидную и аминную группу, как указанные термины определены в настоящем документе.
«Алкенильная» группа относится к алкильной группы, которая состоит, по меньшей мере, из двух атомов углерода и, по меньшей мере, одной углерод-углеродной двойной связи.
«Алкинильная» группа относится к алкильной группе, которая состоит, по меньшей мере, из двух атомов углерода и, по меньшей мере, одной углерод-углеродной тройной связи.
«Арильная» группа относится к моноциклической или конденсированной полициклической группе (т.е. циклам, которые имеют общую пару атомов углерода), имеющей полностью сопряженную пиэлектронную систему. Не ограничивающие примеры арильных групп включают фенил, нафталенил и антраценил. Арильная группа может быть замещенной или незамещенной. При наличии заместителей замещающая группа может представлять собой, например, алкильную, алкенильную, алкинильную, циклоалкильную, арильную, гетероарильную, гетероалициклическую, галогенную, гидроксильную, алкоксильную, арилоксильную, тиогидроксильную, тиоалкоксильную, тиоарилоксильную, сульфинильную, сульфонильную, циано, нитро, азидную, фосфонильную, фосфинильную, оксо, карбонильную, тиокарбонильную, карбаматную, тиокарбаматную, O-карбамильную, N-карбамильную, O-тиокарбамильную, N-тиокарбамильную, C-амидную, N-амидную, C-карбокси, O-карбоксильную, сульфонамидную и аминную группу, как указанные термины определены в настоящем документе.
«Гетероарильная» группа относится к моноциклической или конденсированной полициклической группе (т.е. циклам, которые имеют общую пару атомов углерода), имеющей в кольце (кольцах) один или нескольких атомов, таких, как, например, атом азота, кислорода и серы, и, кроме того, имеющей полностью сопряженную пиэлектронную систему. Не ограничивающие примеры гетероарильных групп включают пиррол, фуран, тиофен, имидазол, оксазол, тиазол, пиразол, пиридин, пиримидин, хинолин, изохинолин и пурин. Гетероарильная группа может быть замещенной или незамещенной. При наличии заместителей замещающая группа может представлять собой, например, алкильную, алкенильную, алкинильную, циклоалкильную, арильную, гетероарильную, гетероалициклическую, галогенную, гидроксильную, алкоксильную, арилоксильную, тиогидроксильную, тиоалкоксильную, тиоарилоксильную, сульфинильную, сульфонильную, циано, нитро, азидную, фосфонильную, фосфинильную, оксо, карбонильную, тиокарбонильную, карбаматную, тиокарбаматную, O-карбамильную, N-карбамильную, O-тиокарбамильную, N-тиокарбамильную, C-амидную, N-амидную, C-карбоксильную, O-карбоксильную, сульфонамидную и аминную группу, как указанные термины определены в настоящем документе.
«Гетероалициклическая» группа относится к моноциклической или конденсированной циклической группе, имеющей в кольце (кольцах) один или несколько атомов, таких как азот, кислород и сера. Кольца также могут иметь одну или несколько двойных связей. Однако, кольца не имеют полностью сопряженной пиэлектронной системы. Гетероалициклическая группа может может быть замещенной или незамещенной. При наличии заместителей замещающая группа может представлять собой, например, алкильную, алкенильную, алкинильную, циклоалкильную, арильную, гетероарильную, гетероалициклическую, галогенную, гидроксильную, алкоксильную, арилоксильную, тиогидроксильную, тиоалкоксильную, тиоарилоксильную, сульфинильную, сульфонильную, циано, нитро, азидную, фосфонильную, фосфинильную, оксо, карбонильную, тиокарбонильную, карбаматную, тиокарбаматную, O-карбамильную, N-карбамильную, O-тиокарбамильную, N-тиокарбамильную, C-амидную, N-амидную, C-карбоксильную, O-карбоксильную, сульфонамидную и аминную группу, как указанные термины определены в настоящем документе.
«Гидроксильная» группа относится к -OH группе.
«Азидная» группа относится к -N=N+=N- группе.
«Алкоксильная» группа относится к -O-алкильной группе и -O-циклоалкильной группе, как определено здесь.
«Арилоксильная» группа относится к -O-арильной группе и -O-гетероарильной группе, как определено здесь.
«Эфирная» группа относится к алкоксильной и арилоксильной группам, где группа связана с алкильной, алкенильной, алкинильной, циклоалкильной, арильной, гетероарильной или гетероалициклической группой.
Эфирная связь описывает -O- связь.
«Тиогидроксильная» или «тиольная» группа относится к -SH группе.
«Тиоалкоксильная» группа относится к -S-алкильной группе и -S-циклоалкильной группе, как определено здесь.
«Тиоарилоксильная» группа относится к -S-арильной группе и -S-гетероарильной группе, как определено здесь.
«Тиоэфирная» группа относится к тиоалкоксильной группе и тиоарилоксильной группе, где группа связана с алкильной, алкенильной, алкинильной, циклоалкильной, арильной, гетероарильной или гетероалициклической группой. Тиоэфирная связь описывает -S- связь.
«Дисульфидная» группа относится -S-тиоалкоксильной группе и -S-тиоарилоксильной группе.
Дисульфидная связи описывает -S-S- связь.
«Карбонильная» группа относится к -C(=O)-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Тиокарбонильная» группа относится к -C(=S)-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Карбоксильная» группа относится к «C-карбоксильной» и «O-карбоксильной» группам.
«C-карбоксильная» группа относится к -C(=O)-O-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«O-карбоксильная» группа относится к R′C(=O)-O-группе, где R′ определяется, как описано в настоящем документе выше.
«Оксо» группа относится к группе =O.
«Карбоксилат» или «карбоксильная» группа относится к C-карбоксильной группе и O-карбоксильной группе, как определено здесь.
«Карбоновая кислота» относится к C-карбоксильной группе, в которой R′ представляет собой водород.
«Тиокарбоксильная» группа или «тиокарбоксилат» относится -C(=S)-O-R′ группе и -O-C(=S)R′ группе.
«Сложный эфир» относится к C-карбоксильной группе, в которой R′ не представляет собой водород.
Эфирная связь относится к -O-C(=O)- связи.
Тиоэфирная связь относится к -O-C(=S)- связи или -S-C(-O) связи.
«Галогенная» группа относится к фтору, хлору, брому или йоду.
«Сульфинильная» группа относится к -S(=O)-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Сульфонильная» группа относится к -S(=O)2-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Сульфонатная» группа относится к -S(=O)2-O-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Сульфатная» группа относится к -O-S(=O)2-O-R′ группе, где R′ определяется, как описано в настоящем документе выше.
«Сульфонамидная» или «сульфонамидо» группа включает в себя S-сульфонамидо и N-сульфонамидо группы, как определено здесь.
«S-сульфонамидо» группа относится к -S(=O)2-NR′R″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«N-сульфонамидо» группа относится к R′S (=O)2-NR″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«O-карбамильная» группа относится к -OC(=O)-NR′R″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«N-карбамильная» группа относится к R′OC(=O)-NR″- группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«Карбамильная» или «карбаматная» группа включает в себя O-карбамильную и N-карбамильную группу.
Карбаматная связь описывает -OC(=O)-NR′- связь, где R′ определяется, как описано в настоящем документе выше.
«O-тиокарбамильная» группа относится к -OC(=S)-NR′R″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«N-тиокарбамильная» группа относится к R′OC(=S)NR″- группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«Тиокарбамильная» или «тиокарбаматная» группа включает в себя O-тиокарбамильную и N-тиокарбамильную группы.
Тиокарбаматная связь описывает -OC(=S)-NR′-связь, где R′ определяется, как описано в настоящем документе выше.
«C-амидо» группа относится к -C(=O)-NR′R″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«N-амидо» группа относится к R′C(=O)-NR″ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«Амидная» группа включает в себя C-амидо и N-амидо группы.
Амидная связь описывает -NR′-C(=O)- связь, где R′ определяется, как описано в настоящем документе выше.
Аминная связь описывает связь между атомом азота в аминогруппе (как определено здесь) и R′ группой аминогруппы.
Тиоамидная связь описывает -NR′-C(=S)- связь, где R′ определяется, как описано в настоящем документе выше.
«Мочевинная» группа относится к -N(R′)-C(=O)-NR″R″′ группе, где каждый из R′ и R″ определяется, как описано в настоящем документе выше, а R″′ определяется, как определено для R′ и R″ в настоящем документе.
«Нитро» группа относится к NO2-группе.
«Циано» группа относится к -C=N группе
Термин «фосфонил» или «фосфонат» описывает -P(=O)(OR′)(OR″) группу, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
Термин «фосфат» описывает -OP(=O)(OR′)(OR″) группу, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
«Фосфорная кислота» является фосфатной группой, в которой каждый из R представляет собой водород.
Термин «фосфинил» описывает -PR′R″ группу, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
Термин «тиомочевина» описывает -N(R′)-C(=S)-NR″- группу, где каждый из R′ и R″ определяется, как описано в настоящем документе выше.
Как описано в настоящем документе, мультимерная белковая структура по настоящему документу может характеризоваться повышенной стабильностью и более высокой и/или более длительной α-галактозидазной активностью в терапевтически важных средах in vivo. Такая мультимерная белковая структура, таким образом, очень полезна для различных медицинских применений, для которых желательна α-галактозидазная активность, включая применение в терапевтических и исследовательских целях.
Таким образом, в соответствии с некоторыми вариантами мультимерная белковая структура, описанная в настоящем документе, предназначена для применения в качестве лекарственного средства, например, лекарственного средства для лечения болезни Фабри.
В соответствии с другим аспектом воплощения изобретения предусмотрен способ лечения болезни Фабри, включающий введение субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества мультимерной белковой структуры, описанной в настоящем документе.
Согласно другому аспекту вариантов осуществления изобретения предусмотрена фармацевтическая композиция, которая содержит мультимерную белковую структуру, как описано выше, и фармацевтически приемлемый носитель. В настоящем документе «фармацевтическая композиция» относится к препарату одной или нескольких мультимерных белковых структур, описанных здесь, с другими химическими компонентами, такими как фармацевтически приемлемые и подходящие носители и наполнители. Целью создания фармацевтической композиции является облегчения введения соединения в организм.
В настоящем документе термин «фармацевтически приемлемый носитель» относится к носителю или разбавителю, который не вызывает значительного негативного ответа организма и не нарушает биологическую активность и свойства вводимого соединения. Не ограничивающие примеры носителей включают пропиленгликоль, физиологический раствор, эмульсии и смеси органических растворителей с водой, а также твердые (например, сухие) и газообразные носители. В настоящем документе термин «наполнитель» относится к инертному веществу, добавляемому к фармацевтической композиции для дальнейшего облегчения введения соединения. Не ограничивающие примеры наполнителей включают карбонат кальция, фосфат кальция, различные сахара и типы крахмала, производные целлюлозы, желатин, растительные масла и полиэтиленгликоли. Фармацевтическая композиция, необязательно, содержит дополнительный компонент, который дополнительно стабилизирует мультимерную белковую структуру α-галактозидазы. Необязательно дополнительным ингредиентом является галактоза. Альтернативно, вместо галактозы может быть использовано производное галактозы (например, содержащий галактозу гликозид). Необязательно, может быть использовано невосстанавливающее производное галактозы. Методики изготовления композиций и введения лекарственных средств можно найти в справочнике "Remington's Pharmaceutical Sciences" Mack Publishing Co., Easton, PA, последнее издание которое включено в настоящий документ посредством отсылки. Фармацевтические композиции по настоящему изобретению могут быть получены известными в данной области способами, например, путем обычного смешивания, растворения, гранулирования, изготовления драже, растирания в порошок, эмульгирования, инкапсулирования, захвата или лиофилизации.
Фармацевтические композиции для применения в соответствии с настоящим изобретением, таким образом, могут быть приготовлены обычными способами с использованием одного или нескольких фармацевтически приемлемых носителей, включая наполнители и вспомогательные вещества, которые облегчают превращение мультимерной белковой структуры в препарат, который может быть применен в фармацевтических целях. Выбор подходящей композиции зависит от выбранного способа введения.
Для инъекции или инфузии мультимерная белковая структура согласно вариантам осуществления изобретения может быть приготовлена в виде водного раствора, предпочтительно в виде раствора в физиологически совместимом буфере, таком как раствор Хенкса, раствор Рингера или физиологический буферный раствор, содержащем или не содержащем органические растворители, такие как пропиленгликоль, полиэтиленгликоль.
Для трансмукозального введения в композиции используют смачивающие агенты. Такие смачивающие агенты, как правило, известны в данной области.
Для перорального введения легко может быть приготовлена композиция мультимерной белковой структуры по изобретению путем объединения мультимерной белковой структуры с фармацевтически приемлемыми носителями, хорошо известными в данной области. Такие носители позволяют готовить композицию мультимерной белковой структуры, описанной здесь, в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, растворов, суспензий и т.п.для перорального введения пациентам. Фармакологические препараты для перорального применения могут быть изготовлены с использованием твердого наполнителя, необязательно, с измельчением полученной смеси и обработкой смеси гранул для получения таблеток или драже после необязательного добавления подходящих вспомогательных веществ. Подходящими наполнителями являются, в частности, наполняющие агенты, такие как сахара, включая лактозу, сахарозу, маннит или сорбит; препараты целлюлозы, такие как, например, кукурузный крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал, желатин, трагакант, метилцеллюлоза, гидроксипропилметилцеллюлоза, натрия карбоксиметилцеллюлоза и/или физиологически приемлемые полимеры, такие как поливинилпирролидон (ПВП). При желании, могут быть добавлены разрыхлители, такие как поперечно сшитый поливинилпирролидон, агар или альгиновая кислота или ее соль, такая как альгинат натрия.
Ядра драже предусмотрены в подходящем покрытии. Для этой цели могут быть использованы концентрированные растворы сахара, которые могут, необязательно, содержать гуммиарабик, тальк, поливинилпирролидон, гель карбопола, полиэтиленгликоль, диоксид титана, растворы глазури и подходящие органические растворители или смеси растворителей. Для идентификации или характеристики различных комбинаций доз активных мультимерных белковых структур к покрытию таблеток или драже могут быть добавлены красители или пигменты.
Фармацевтические композиции, которые можно применять перорально, включают твердые капсулы из двух частей из желатина, а также мягкие герметичные капсулы из желатина и пластификатора, такого как глицерин или сорбит. Твердые капсулы из двух частей могут содержать активные ингредиенты в смеси с наполнителем, таким как лактоза, связующим, таким как крахмал, смазывающим агентом, таким как тальк или стеарат магния, и, необязательно, стабилизатором. В мягких капсулах мультимерная белковая структура может быть растворена или суспендирована в подходящих жидкостях, таких как жирные масла, жидкий парафин или жидкие полиэтиленгликоли. Также могут быть добавлены стабилизаторы. Все композиции для перорального введения должны быть предусмотрены в дозировках, подходящих для выбранного способа введения.
Для буккального введения композиции могут иметь форму таблеток или пастилок, приготовленных обычным способом.
Для введения путем ингаляции мультимерную белковую структуру для применения в соответствии с вариантами осуществления настоящего изобретения обычно применяют в форме аэрозольного спрея (который обычно включает в себя порошкообразный, сжиженный и/или газообразный носитель) из упаковки под давлением или распылителя с использованием подходящего пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана или диоксида углерода. В случае аэрозоля под давлением доза может быть определена с помощью клапана для доставки отмеренного количества. Для применения в ингаляторе или инсуффляторе могут быть предусмотрены капсулы и картриджи, например, изготовленные из желатина, содержащие порошковую смесь мультимерной белковой структуры и подходящей порошковой основы, такой как, например, лактоза или крахмал.
Мультимерная белковая структура, описанная в настоящем документе, может быть введена в композицию для парентерального введения, например, путем болюсной инъекции или инфузии. Препараты для инъекций или инфузий могут быть предусмотрены в стандартной лекарственной форме, например, в ампулах или в многодозовых контейнерах, необязательно, с добавлением консерванта. Композиции могут представлять собой суспензии, растворы или эмульсии в масляных или водных носителях и могут содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты.
Фармацевтические композиции для парентерального введения включают водные растворы препарата мультимерной белковой структуры в водорастворимой форме. Альтернативно, суспензии мультимерной белковой структуры могут быть получены в виде соответствующих масляных суспензий и эмульсий для инъекций (например, эмульсий типа вода-в-масле, масло-в-воде или вода-в-масле в масле). Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические эфиры жирных кислот, такие как этилолеат, триглицериды или липосомы. Водные суспензии для инъекций могут содержать вещества, которые повышают вязкость суспензии, такие как натрий карбоксиметилцеллюлоза, сорбит или декстран. Чтобы обеспечить возможность приготовления высококонцентрированных растворов, необязательно, суспензия может также содержать подходящие стабилизаторы или агенты, повышающие растворимость мультимерной белковой структуры.
Альтернативно, мультимерная белковая структура может быть предусмотрена в форме порошка для восстановления перед использованием подходящим носителем, например, стерильной апирогенной водой.
Мультимерная белковая структура согласно вариантам осуществления настоящего изобретения также может быть предусмотрена в виде ректальных композиций, таких как суппозитории или удерживающие клизмы, приготовленных с использованием, например, обычных оснований для суппозиториев, таких как масло какао или другие глицериды.
Фармацевтические композиции, описанные здесь, также могут содержать подходящие твердые или гелевые носители или наполнители. Примеры таких носителей или наполнителей включают, но не ограничиваются карбонатом кальция, фосфатом кальция, различными сахарами, крахмалом, производными целлюлозы, желатином и полимерами, такими как полиэтиленгликоль.
Фармацевтические композиции, пригодные для применения в контексте настоящего изобретения, включают композиции, в которых активные ингредиенты содержатся в количестве, достаточном для достижения поставленной цели. Более конкретно, терапевтически эффективное количество означает количество мультимерной белковой структуры, эффективное для предотвращения, смягчения или облегчения симптомов болезни или продления жизни субъекта, подлежащего лечению.
Для любых мультимерных белковых структур, используемых в методах по изобретению, терапевтически эффективное количество или доза могут быть первоначально оценены с помощью анализов активности на животных. Например, доза может быть подобрана с помощью модели на животных для достижения диапазона концентраций в кровотоке, который включает IC50, определенную с помощью анализов активности (например, концентрацию тестируемой белковой структуры, которая обеспечивает полумаксимальное увеличение биологической активности мультимерной белковой структуры). Такая информация может быть использована для более точного определения подходящих для человека доз.
Как показано в разделе Примеры ниже, терапевтически эффективное количество мультимерной белковой структуры согласно вариантам осуществления настоящего изобретения может составлять от около 1 мкг/кг массы тела до около 500 мг/кг массы тела.
Токсичность и терапевтическая эффективность мультимерной белковой структуры, описанной здесь, могут быть определены с помощью стандартных фармацевтических тестов на экспериментальных животных, например, через определение EC50, IC50 и LD50 (летальная доза, вызывающая смерть у 50% подопытных животных) для исследуемой белковой структуры. Данные, полученные из анализов активности и исследований на животных, могут быть использованы для подбора диапазона дозировки для применения на людях.
Дозировка может варьировать в зависимости от лекарственной формы и способа введения. Точная композиция, способ введения и дозировка могут быть выбраны лечащим врачом с учетом состояния пациента. (См., например, Fingi et al., 1975, in "The Pharmacological Basis of Therapeutics", Ch.1 p.1).
Дозировка и интервал между введениями могут быть скорректированы индивидуально для обеспечения содержания активного компонента в плазме крови, достаточного для поддержания желаемого эффекта, которое называют минимальной эффективной концентрацией (МЕС). МЕС меняется для каждого препарата, но может быть оценена из данных исследований in vitro, например, из концентрации, необходимой для достижения желаемого уровня активности in vitro. Дозы, необходимые для достижения МЕС, будет зависеть от индивидуальных особенностей и способа введения. Для определения концентрации в плазме могут быть использованы ВЭЖХ анализ или другие виды анализов биологических образцов.
Интервалы между введениями также могут быть определены с помощью МЕС. Препараты следует вводить в соответствии с режимом введения, при котором поддерживается уровень белка в плазме выше МЕС в течение 10-90% времени, предпочтительно в течение 30-90% времени и наиболее предпочтительно в течение 50-90% времени.
В зависимости от тяжести состояния, подлежащего лечению, и ответа на лечение дозировка также может представлять собой однократное введение композиции с замедленным высвобождением, описанной выше, при этом курс лечения может длиться от нескольких дней до нескольких недель или до достижения излечения или облегчения болезненного состояния.
Количество вводимой композиции, очевидно, зависит от субъекта, подвергаемого лечению, тяжести его состояния, способа введения, мнения лечащего врача и т.д.
Композиции по настоящему изобретению могут, необязательно, быть предусмотрены в упаковке или дозирующем устройстве, таком как одобренный FDA (Управление по контролю за продуктами и лекарствами США) набор, который может содержать одну или несколько стандартных лекарственных форм, содержащих активный ингредиент. Упаковка может, например, содержать металлические или пластиковые пленки, включая, но не ограничиваясь блистерной упаковкой или контейнером под давлением (для ингаляций). С упаковкой или дозирующим устройством может быть предусмотрена инструкция для введения. С упаковкой или дозирующим устройством может быть предусмотрено уведомление, связанное с контейнером, в форме, установленной правительственным агентством, регулирующим производство, использование или продажу фармацевтических препаратов, где уведомление отражает одобрение агентством формы композиции для применения на человеке или ветеринарного применения. Такое уведомление, например, может представлять собой маркировку Управления по контролю за продуктами и лекарствами США для отпускаемых по рецепту лекарств или вкладыш для одобренных продуктов. Композиции, содержащие мультимерную белковую структуру согласно вариантам осуществления изобретения, введенную в композицию с совместимым фармацевтическим носителем, могут быть получены, помещены в специальный контейнер и снабжены этикеткой, указывающей их предназначение для лечения указанного состояния или диагноза, как подробно описано здесь. Таким образом, в соответствии с вариантом осуществления настоящего изобретения в зависимости от выбранной мультимерной белковой структуры фармацевтическая композиция, описанная здесь, упакована в упаковочный материал, и к ней прилагается напечатанное указание, содержащееся на упаковочном материале или внутри него, на ее применение для лечения состояния, при котором желательна активность мультимерной белковой структуры, как описано выше. В настоящем документе термин «около» относится к ±10%
Термины «содержит», «содержащий», «включает», «включающий», «имеющий» и однокоренные слова означают понятие «включает, но не ограничивается».
Термин «состоящий из» означает понятие «включает и ограничивается».
Термин «пример» используется здесь для обозначения понятия «служащий в качестве примера, образца или иллюстрации». Любой вариант осуществления, описанный как «пример», не обязательно должен рассматриваться как предпочтительный или выгодный по сравнению с другими вариантами и/или должен исключать наличие признаков из других вариантов.
Термин «необязательно» используется здесь для обозначения понятия «предусмотрен в некоторых вариантах и не предусмотрен в других вариантах». Любой конкретный вариант осуществления изобретения может включать в себя множество «необязательных» признаков, если такие признаки не противоречат друг другу.
В настоящем документе указание понятия в единственном числе также подразумевает его указание во множественном числе, если контекст явно не указывает иное.
Например, термины «соединение» или «по меньшей мере, одно соединение» могут включать в себя множество соединений, включая их смеси.
В данной заявке различные варианты осуществления данного изобретения могут быть представлены с указанием различных диапазонов. Следует иметь в виду, что описание с указанием различных диапазонов применяют для удобства и краткости, такое описание не должно быть истолковано как негибкое ограничение объема изобретения.
Соответственно, при указании диапазона его следует рассматривать как включающего все возможные поддиапазоны, а также отдельные числовые значения в пределах этого диапазона. Например, при указании диапазона, например, от 1 до 6 его следует рассматривать как раскрывающего поддиапазоны, например, от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также как раскрывающего отдельные числовые значения в пределах этого диапазона, например, 1, 2, 3, 4, 5 и 6. Этот подход применяется независимо от ширины диапазона.
Указание численного диапазона в настоящем документе включает любые приведенные численные значения (дробные или целые) в пределах указанного диапазона. Фразы «в диапазоне/диапазон между» первым указанным числом и вторым указанным числом и «в пределах от/в диапазоне от» первого указанного числа «до» второго указанного числа используются здесь взаимозаменяемо и предназначены для включения первого и второго указанных чисел и всех дробных и целых численных значений между ними.
В настоящем документе термин «способ» относится к методам, средствам, методикам и процедурам осуществления поставленной задачи, включая, но не ограничиваясь, известными методами, средствами, методиками и процедурами, либо легко разрабатываемыми из известных методов, средств, методик и процедур специалистами в области химии, фармакологии, биологии, биохимии и медицины.
В настоящем документе термин «лечение» включает остановку, существенное подавление, замедление или обращение прогрессирования состояния, существенное улучшение клинических симптомов состояния или ощущений пациента, или существенное предотвращение появления клинических симптомов или болезненных ощущений пациента.
Подразумевается, что определенные признаки изобретения, которые для ясности описаны в контексте отдельных вариантов изобретения, могут быть предусмотрены также в сочетании в одном варианте. С другой стороны, различные признаки изобретения, которые для краткости описаны в контексте одного варианта, также могут быть предусмотрены отдельно или в любой подходящей подкомбинации или, по возможности, в любом другом описанном варианте изобретения. Некоторые признаки, описанные в контексте различных вариантов, не следует считать существенными признаками данных вариантов, за тем исключением, если вариант не осуществим без данных признаков.
Различные варианты и аспекты настоящего изобретения, как это описано выше и как заявлено в Формуле изобретения, находят экспериментальное подтверждение в следующих Примерах.
ПРИМЕРЫ
Следующие Примеры вместе с приведенным выше описанием иллюстрируют некоторые варианты осуществления изобретения, не ограничивая его.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Бис-N-гидроксисукцинимид-поли(этиленгликоль) (бис-NHS-PEG-) был получен от Iris Biotech GmbH в форме PEG8 и PEG 2000 Далтон (PEG45), PEG5 форма была получена от Pierce, вещества растворяли в диметилсульфоксиде (ДМСО) в концентрации 25 мг/мл;
Лимонная кислота была получена от Sigma;
Кумасси синий G250 был получен от Bio-Rad;
Диметилсульфоксид был получен от Sigma;
D-(+)-галактоза была получена от Sigma;
Человеческая плазма (K3 EDTA) была получена из Bioreclamation Inc.;
4-метилумбеллиферон был получен от Sigma;
4-метилумбеллиферил-α-D-галактопиранозид был получен от Sigma;
N-додеканоил-нитробензоксадиазол-церамид тригексозид (Gb3-NBD) был получен от Matreya;
2-(N-морфолино)этансульфокислота была получена от Merck;
Фосфатный буферный солевой раствор был получен от Sigma;
п-Нитрофенил-α-D-галактопиранозид был получен от Sigma;
Примулин был получен от Sigma; примулин для распыления получали растворением 12,5 мг примулина в 200 мл ацетона и воды (8:2 по объему);
Пиридин был получен от Sigma;
Синапиновая кислота была получена от Sigma;
Карбонат натрия был получен от Sigma;
Фосфат натрия был получен от Sigma;
Таурохолат натрия был получен от Sigma;
Трифторуксусная кислота была получена от Sigma.
Полученная в растениях рекомбинантная человеческая α-GAL-I
Полученная в растениях рекомбинантная человеческая α-GAL (prh-α-GAL), имеющая SEQ ID NO: 1, называемая здесь полученной в растениях рекомбинантной человеческой α-GAL-I (prh-α-GAL-I), была получена, как описано в международной заявке PCT/IL 2008/000576 (опубликована как WO 2008/132743).
Для экспрессии человеческого белка α-GAL-A трансгенный растительный материал получали с использованием растений Nicotiana Benthamiana, инфильтрованных генетической конструкцией, содержащей экспрессионную кассету для α-GAL-A. Процедуру проводили в камере для роста в контролируемых условиях. Затем растительный материал собирали и растворимые белки экстрагировали из растительных клеток. Затем prh-α-GAL-A очищали с помощью процесса очистки, включающего стандартные методы очистки белков с последующим этапом химической модификации для получения сшитого белка. prh-α-GAL-A экстрагировали из растительного материала с помощью гомогенизатора. Растительный дебрис удаляли центрифугированием, белок очищали с использованием стадий осаждения сульфатом аммония и подкисления. Супернатант фильтровали и наносили на гидрофобную колонку, а затем обессоливали и наносили на катионообменную колонку. Элюат после катионообменной колонки концентрировали.
Полученная в растениях рекомбинантная человеческая α-GAL-II
Полученная в растениях рекомбинантная человеческая α-GAL, содержащая смесь α-GAL с последовательностью SEQ ID NO: 2 и α-GAL с последовательностью SEQ ID NO: 3 (без N-концевых аминокислот EF, присутствующих в SEQ ID NO: 1), обозначаемая здесь как prh-α-GAL-II, была получена способом, аналогичным описанному выше для prh-α-GAL-I, но с использованием другой генетической конструкции.
кДНК, кодирующая человеческий белок α-галактозидазу (ЕС 3.2.1.22, GenBank: Х05790), была оптимизирована и синтезирована GENEART AG (Regensburg, Germany). Кодоновый состав последовательности без лидерного пептида (сигнальный пептид, направляющий белок в эндоплазматическую сеть) был адаптирован с учетом встречаемости кодонов в генах Nicotiana tobaccum. При оптимизации из последовательности убирали следующие цис-действующие мотивы: внутренние ТАТА-боксы, хи-сайты и сайты посадки рибосомы, АТ-богатые и GC-богатые участки последовательности, элементы нестабильности РНК (дестабилизирующие мотивы), повторяющиеся последовательности и вторичные структуры РНК, донорные и акцепторные сайты сплайсинга (криптические), точки ветвления. Также убирали районы с очень высоким (>80%) или очень низким (<30%) содержанием GC.
Нуклеотидную последовательность, кодирующую лидерный пептид нативной человеческой α-галактозидазы (сигнальный пептид, направляющий белок в эндоплазматическую сеть) полноразмерного белка человеческой α-галактозидазы (GenBank: X05790) заменяли на нуклеотидную последовательность, кодирующую сигнальный пептид из белка ABPI Arabidopsis, направляющий белок в эндоплазматическую сеть, состоящий из 33 аминокислот (лидерный пептид). Этот сигнальный пептид эффективно обеспечивает направление α-галактозидазы по секреторному пути и отщепляется от полипептида сигнальной пептидазой сразу после перемещения белка в эндоплазматическую сеть. Нуклеотидную последовательность, кодирующую сигнал удержания в эндоплазматической сети SEK.DEL, добавляли на 3′ конец последовательности кДНК, что позволяло экспрессированному белку покидать аппарат Гольджи и эффективно накапливаться в эндоплазматическом ретикулуме.
Интересующий белок экспрессировали с сильного субгеномного вирусного промотора белка оболочки. Указанная система основана на временной амплификации (с помощью инфекции агробактериями) вирусных векторов, доставленных в растения агробактериями. При инфекции агробактериями функциональный в растениях промотор и кДНК, кодирующая вирусный репликон, переносится как Т-ДНК из агробактерии в клетки растений. Т-ДНК транскрибируется in-planta с растительного промотора с получением биологически активной вирусной РНК, которая инициирует саморепликацию.
Для транзиентной экспрессии использовали рекомбинационную систему из 3 векторов, основанную на разработанной ранее системе, как описано [Gleba et al., Vaccine 2005, 23:2042-2048]. В один из векторов вставляли кДНК α-галактозидазы, а два других вектора содержали гены для построения целого вирусного репликона (RdRp и интегразу), создавая таким образом биологически активную вирусную РНК, которая может инициировать саморепликацию.
Растения N. Benthamiana проращивали и растили в коммерческой почвенной смеси (Givaat Ada, IL) с добавлением гранулированного удобрения замедленного высвобождения (Scott Marysville, ОН) в условиях светового режима долгого дня (16 часов света/8 часов темноты) при 24-25°C.
Агробактерии трансформировали векторной системой на основе pICH20866-альфа-GAL с использованием электропорации (2500 V, 5 миллисекунд) [den Dulk-Ra and Hooykaas, Methods Mol Biol 1995, 55:63-72]. Растения инфильтрировали агробактериями, содержащими 3 плазмиды ICON, с помощью вакуумной инфильтрации как стандартного способа, известного в уровне техники. Растения N. benthamiana возрастом 5-6 недель инфильтрировали погружением всех надземных органов растений в бактериальную суспензию с помещением в вакуумную камеру. Примяли вакуум минус (-) 0,8 бар в течение 1 минуты, а затем давление быстро возвращали к атмосферному давлению. Растения возвращали в теплицу еще на 5-7 дней при тех же условиях выращивания.
Образцы листьев Nicotiana benthamiana собирали через 5 дней после инфильтрации и экстрагировали буфером Лэммли для SDS-PAGE или буфером для определения активности (20 мМ лимонная кислота, 30 мМ фосфат натрия, 0,1% бычьего сывороточного альбумина и 0,67% этанола, рН 4,6) для анализа каталитической активности экспрессированного в растениях белка.
Человеческую α-галактозидазу из растительных экстрактов очищали с помощью двухстадийного дифференциального осаждения сульфатом аммония («высаливания»: 1-ая стадия 0,57 М, 2-ая стадияг 2,27 М) с последующей гидрофобной хроматографией (смола Phenyl 650 М) и катионообменной хроматографией.
В связи с различным процессингом лидерной последовательности были получены две последовательности (например, SEQ ID NO: 2 и SEQ ID NO: 3), которые различаются наличием или отсутствием N-концевого глицина.
Определение активности α-GAL с помощью 4-метилумбеллиферил-α-D-галактопиранозида
Активность α-GAL измеряли с помощью 4-метилумбеллиферил-α-D-галактопиранозида как субстрата для гидролиза. Анализ проводили в цитрат-фосфатном буфере (20 мМ лимонная кислота, 30 мМ фосфат натрия, рН 4,6). 10 мкл образца, содержащего тестируемую α-GAL, инкубировали с 40 мкл аналитического буфера, содержащего 5 мМ 4-метилумбеллиферил-α-D-галактопиранозида. Реакционную смесь инкубировали при 37°C в течение 60 минут. 10 мкл реакционной смеси переносили в черный 96-луночный планшет (Greiner), к смеси добавляли 90 мкл раствора для остановки реакции (2 М карбонат натрия), после чего измеряли флуоресценцию при длине волны возбуждения 365 нм и длине волны излучения 450 нм. Интенсивность флуоресценции переводили в концентрацию продукта и затем в активность с помощью калибровочной кривой 4-метилумбеллиферона, продукта реакции.
Определение активности α-GAL с помощью N-додеканоил-нитробензоксадиазол-церамид тригексозида (Gb3-NBD)
Флуоресцентно меченый субстрат N-додеканоил-нитробензоксадиазол-церамид тригексозид (Gb3-NBD) менее липофилен, чем Gb3, что облегчает его использование в ферментативных реакциях in vitro.
10 мкл 0,1 мкг/мкл Gb3-NBD (в воде с 10% этанолом) и 5 мкл 0,2 мг/мл α-GAL добавляли к 85 мкл цитрат-фосфатного буфера при рН 4,6. Конечная концентрация α-GAL составила 10 мкг/мл. Для измерения уровня фона или не каталитической реакции без α-GAL смешивали 90 мкл цитрат-фосфатного буфера с рН 4,6 с 10 мкл 0,1 мкг/мкл Gb3-NBD (в воде с 10% этанола). Реакционную смесь инкубировали в течение 60 минут при 37°C. После инкубации к реакционной смеси добавляли 50 мкл метанола и раствор встряхивали в течение 1 минуты. Затем к смеси добавляли 100 мкл хлороформа и раствор дополнительно перемешивали в течение 1 минуты. Воду и органические растворители удаляли под вакуумом с помощью системы Speed Vac. Остаток растворяли в 80 мкл смеси хлороформ:метанол (1:1). 30 мкл каждого образца наносили на пластины для HPTLC (высокопроизводительная тонкослойная хроматография) Silica Gel 60 (Merck) с использованием системы Linomat V (CAMAG). Образец разделяли на пластинах для HPTLC с использованием раствора хлороформ:метанол: H2O в соотношении 100:42:6 в качестве системы растворителей. Затем пластину высушивали и пятна субстрата и продукта визуализировали с помощью облучения в УФ-свете при длине волны 365 нм.
Определение активности α-GAL с помощью п-нитрофенил-α-D-галактопиранозида (p-NP-G)
Активность α-GAL измеряли с помощью п-нитрофенил-α-D-галактопиранозида как субстрата для гидролиза. Буфер для анализа содержал 20 мМ лимонную кислоту, 30 мМ фосфат натрия, 0,1% BSA (бычий сывороточный альбумин) и 0,67% этанола при рН 4,6. Анализ проводили в 96-луночных планшетах для ИФА (Greiner). 50 мкл образца инкубировали с 150 мкл аналитического буфера, для получения конечной концентрации п-нитрофенил-α-D-галактопиранозида 8 мМ к пробе добавляли 30 мкл субстрата. Реакционную смесь инкубировали при 37°C в течение 90 минут. Через 90 минут в каждую лунку для остановки реакции добавляли 100 мкл 1,98 М раствора карбоната натрия. Количество продукта реакции определяли путем измерения оптической плотности при 405 нм.
Измерение стабильность α-GAL in vitro
Стабильности α-GAL из различных источников определяли при инкубации α-GAL при следующих условиях:
1) моделирование лизосомальных условий: цитрат-фосфатный буфер (20 мМ лимонная кислота, 30 мМ фосфат натрия), pH 4,6, 37°C;
2) моделирование физиологических условий: фосфатный буферный солевой раствор (PBS), pH 7,4, 37°C;
3) плазма крови человека при 37°C.
α-GAL был добавляли в концентрации 1 мкг/мл, определенной по активности α-GAL в растворе, и раствор инкубировали при 37°C. Образцы каждого раствора изымали из эксперимента в заданных временных точках и активность α-GAL измеряли, как описано выше. Значение ферментативной активности сразу после внесения тестируемой α-GAL в каждую среду принимали за 100%, а дальнейшие значения активности в заданных временных точках выражали в процентах от первоначальной активности.
Фармакокинетика α-GAL
Страдающих болезнью Фабри мышей (α-Gal-A -/0) помещали в освещаемое удерживающее устройство из оргстекла, и фермент вводили в хвостовую вену. Образцы крови отбирали в заданных временных точках после инъекции из кончика хвоста или ретро-орбитального синуса, используя гепаринизированные гематокритные пробирки. Плазму разводили в буфере для определения активности с использованием 4-метилумбеллиферил-α-D-галактопиранозида. Анализ с 4-метилумбеллиферил-α-D-галактопиранозидом проводили, как описано выше.
Конечный период полураспада (T1/2) рассчитывали на основе данных по активности в плазме. Полувыведение (период полураспада) представляет собой время, необходимое для снижения концентрации в плазме крови на 50% после достижения псевдоравновесного распределения. Конечный период полураспада рассчитывали из конечной (лог-линейной) части кривой с помощью линейной регрессии зависимости логарифма концентрации от времени [Toutain & Bousquet-Melou, J Vet Pharmacol Ther 2004, 27:427-39].
Биораспределение α-GAL
Страдающим болезнью Фабри мышам (α-Gal-A -/0) вводили внутривенно (в хвостовую вену) α-GAL в дозе 2 мг/кг. Ткани (печень, почки, сердце и селезенку) изымали через 2 часа, 24 часа, 3 дня, 7 дней, 14 дней или 28 дней после инъекции фермента. Уровень α-GAL у нормальных контрольных мышей и у страдающих болезнью Фабри мышей, которым вводили физиологический раствор (без лечения), сравнивали с уровнем α-GAL у страдающих болезнью Фабри мышей, которые получали экзогенный α-GAL. Для определения активности α-GAL в тканях, размороженные образцы тканей помещали в 2 мл полипропиленовые пробирки, содержащие буфер для лизиса (28 мМ лимонная кислота, 44 мМ двухосновный фосфат натрия, 0,5% таурохолат натрия, рН 4,4), как описано Oshima et al. [PNAS 1997, 94:2540-2544]. Пробы гомогенизировали с использованием Tissuelyzer (Retsch ММ400) в течение 10 минут. Дебрис осаждали центрифугированием при 4°C и полученный супернатант анализировали на активность α-GAL с помощью анализа с 4-метилумбеллиферил-α-D-галактопиранозидом, как описано выше. Те же образцы также подвергались анализу методом Вестерн-блоттинг.
Анализ Gb3 in vivo
Эффективность вводимой α-GAL в конечной точке измеряли с помощью анализа уровня Gb3 в тканях животных, позволяющего определить, снижается ли уровень Gb3 под действием активности α-GAL.
Для измерения гидролиза Gb3 из органов-мишеней (например, печени, почек, сердца, селезенки) экстрагировали нейтральные гликосфинголипиды. Образцы ткани по 100 мг гомогенизировали в 1 мл смеси хлороформ:метанол 2:1 (объем/объем) и центрифугировали в течение 20 минут при 13500 оборотах в минуту. Для получения раствора хлороформ:метанол:вода 20:10:2 к 1 мл гомогената добавляли 62 мкл воды. К гомогенату добавляли 10 мкл пиридина до конечной концентрации пиридина 1%. Образец перемешивали в течение 24 часов при 48°C. Растворители и воду удаляется под вакуумом использованием системы SpeedVac. Образец суспендировали в 2,5 мл метанола, к суспензии затем добавляли 250 мкл 1 М КОН в метаноле. Затем образец встряхивали в течение 2 часов при 37°C. Реакцию омыления останавливали добавлением 10 мкл уксусной кислоты. Затем к образцу добавляли 2,5 мл хлороформа с последующим добавлением 2,5 мл холодной воды. Образец энергично встряхивали в течение 5 минут и оставляли на 5 минут для разделения фаз. Верхнюю фазу, состоящую из метанола и воды, отбрасывали, а нижнюю фазу, состоящую из хлороформа и метанола, выпаривали под вакуумом (SpeedVac), а остаток для анализа гликосфинголипидов с помощью HPTLC суспендировали в 300 мкл смеси хлороформ:метанол 1:1 (объем/объем).
Качественный и полуколичественный анализы гликолипидов тканей проводили методом высокоэффективной тонкослойной хроматографии (HPTLC) (CAMAG, Швейцария). HPTLC анализ проводили на стеклянных покрытых силикагелем пластинках для HPTLC silica gel 60 (Merck). Образцы наносили на пластины, используя систему Linomat 5 (CAMAG, Швейцария). Образцы на пластинах разделяли с использованием смеси хлороформ-метанол-вода (60:35:4) в качестве системы растворителей. Нейтральные гликосфинголипиды обнаруживали с помощью распыления реагента примулина. Gb3 идентифицировали с помощью Gb3 из свиных эритроцитов (Matreya) в качестве стандарта и количественно определяли с помощью калибровочной кривой, построенной по полусинтетическому стандарту N-гептадеканоил церамиду тригексозиду (Matreya). Пластины проявляли и окраску соответствующих пятен оценивали количественно с помощью сканера TLC Scanner III (CAMAG, Швейцария) при помощи программного обеспечения winCATS (CAMAG, Швейцария).
SDS-PAGE
SDS-PAGE проводили в восстановительных условиях с использованием системы Bio-Rad Criterion™ и 12% акриламидных гелей. Гель окрашивали кумасси синим G250.
IEF (изоэлектрофокусирование)
IEF проводили с использованием мини-камеры Invitrogen Novex® и коммерческих гелей для IEF, имеющих диапазон рН 3-7 (Invitrogen). Гель окрашивали кумасси синим G250.
Масс-спектрометрия (MALDI-TOF)
MALDI-TOF проводилось с использованием масс-спектрометрической системы Bruker Reflex IV MALDI-TOF (Bruker-Franzen Analytik GmbH, Германия) и насыщенного раствора синапиновая кислота/трифторуксусная кислота (TFA) (0,1% TFA/ацетонитрил (2:1, объем/объем)) в качестве матрицы.
Пример I
Стабильность рекомбинантной α-GAL in vitro
Стабильность рекомбинантной α-GAL in vitro измеряли в различных условиях, как описано выше в разделе Материалы и методы. Были протестированы полученная в растениях рекомбинантная человеческая α-GAL-I, а также коммерческие рекомбинантные человеческие α-GAL Fabrazyme® и Replagal®.
Как показано на Фиг.1, все проверенные типы α-GAL проявляли снижение активности в условиях, имитирующих лизосомальные условия.
Кроме того, как показано на Фиг.2, все проверенные типы α-GAL проявляли снижение активности в условиях, имитирующих физиологические условия. Как показано в настоящем документе ниже, присутствие 100 мг/мл галактозы частично предотвращает потерю активности полученной в растениях рекомбинантной α-GAL-I в таких условиях.
Аналогично, как показано на Фиг.3, все проверенные типы α-GAL проявляли снижение активности при инкубации в плазме крови человека при 37°C.
Как показано на Фиг.4, присутствие 100 мг/мл галактозы частично предотвращает потерю активности полученной в растениях рекомбинантной α-GAL-I в условиях, имитирующих лизосомальные условия.
Эксперименты с хроматографией по размеру (SEC) при лизосомальном и нейтральном уровне рН продемонстрировали изменения в структуре белка (данные не приведены), в то время как анализы методом SDS-PAGE и вестерн блоттинг не показали какой-либо деградации первичной аминокислотной последовательности (данные не приведены).
Эти результаты показывают, что α-GAL теряет активность при лизосомальных условиях и физиологических условиях в связи с изменением белковой структуры α-GAL и что галактоза частично предотвращает эту потерю активности.
Пример II
Перекрестное сшивание полученной в растениях рекомбинаитиой человеческой α-GAL-I агентом бис-N-гидроксисукцинимид-поли(этиленгликолем) (бис-NHS-PEG)
Полученную в растениях рекомбинантную человеческую α-GAL-I (prh-α-GAL-I) сшивали при молярном отношении 50:1, 100:1 и 200:1 к бис-N-гидроксисукцинимид-поли(этиленгликолю) (бис-NHS-PEG) различной молекулярной массы, а именно бис-NHS-PEG5, бис-NHS-PEG8 или бис-NHS-PEG45 (бис-NHS-PEG с PEG массой 2000 Дальтон), структуры которых представлены на Фиг.5.
Бис-NHS-PEG может быть связан с двумя сайтами на белке (например, с остатками лизина), тем самым образуя поперечное сшивание, или с одним сайтом на белке. Эти две формы присоединения изображены на Фиг.6.
100 мкг α-GAL-I в 28,5 мкл 2-(N-морфолино)этансульфокислотного (MES) буфера (25 мМ, рН 6) добавляли к 13,5 мкл фосфатного буфера (100 мМ, рН 8), содержащего 100 мг/мл галактозы.
α-GAL-I сшивали бис-NHS-PEG5 при молярном соотношении белок:реагент 1:50, 1:100 и 1:200, добавляя бис-NHS-PEG5 в 8 мкл ДМСО к раствору α-GAL-I (27,4 мкг раствора α-GAL-I для молярного соотношения 1:50, 54,8 мкг раствора α-GAL-I для молярного соотношения 1:100 и 109,7 мкг раствора α-GAL-I для молярного соотношения 1:200).
α-GAL-I сшивали бис-NHS-PEG45 при молярном соотношении белок:реагент 1:50, 1:100 и 1:200, добавляя бис-NHS-PEG45 в 8 мкл ДМСО к раствору α-GAL-I (103 мкг раствора α-GAL-I для молярного соотношения 1:50, 206 мкг раствора α-GAL-I для молярного соотношения 1:100 и 412 мкг раствора α-GAL-I для молярного соотношения 1:200).
α-GAL-I сшивали бис-NHS-PEG8 при молярном соотношении белок:реагент 1:50, 1:100 и 1:200, добавляя бис-NHS-PEG8 в 11,5 мкл ДМСО к раствору α-GAL-I (37 мкг раствора α-GAL-I для молярного соотношения 1:50, 73 мкг раствора α-GAL-I для молярного соотношения 1:100 и 146 мкг раствора α-GAL-I для молярного соотношения 1: 200).
После добавления бис-NHS-PEG агента к α-GAL-I реакционную смесь перемешивали пипеткой и встряхивали на орбитальном шейкере в течение 2 часов при комнатной температуре.
Во всех реакциях избыток бис-NHS-PEG сшивающего реагента удаляли путем диализа против физиологического раствора (порог пропускания по молекулярной массе 50 кДа).
Выход димера увеличивается с увеличением концентрации белка и концентрации ДМСО, доходя до 30%.
Продукты реакции анализировали с помощью SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), IEF (изоэлектрофокусирования), Вестерн-блоттинга и MALDI-TOF масс-спектрометрии, как описано выше.
Как показано на Фиг.7, стандарт нативной prh-α-GAL-I при гель-электрофорезе был обнаружен в форме мономера (с молекулярной массой 48 кДа), но после реакции prh-α-GAL-I с бис-NHS-PEG prh-α-GAL-I присутствовал в основном в виде димера (с присутствием некоторого количества мономера), что свидетельствует о ковалентном связывании двух мономеров путем сшивания с бис-NHS-PEG.
Как дополнительно показано на Фиг.7, более высокая доля мономерной prh-α-GAL-I наблюдалась при реакции с короткими сшивающими агентами бис-NHS-PEG5 и бис-NHS-PEG8 по сравнению с более длинным сшивающим агентом бис-NHS-PEG45. Бис-NHS-PEG45 дал высокий выход сшитого белка. Эти результаты показывают, что более короткие сшивающие агенты менее эффективны для ковалентного связывания мономеров.
Как дополнительно показано на Фиг.7, для каждого из исследованных сшивающих агентов молекулярная масса мономерной части prh-α-GAL-I увеличилась после реакции с сшивающим агентом. Увеличение молекулярной массы возрастало с увеличением отношения используемого количества сшивающего агента к белку (например, 200:1), а также с увеличением молекулярной массы сшивающего агента (например, бис-NHS-PEG45). Эти результаты показывают, что к мономеру белка, который не был димеризован путем сшивания, был ковалентно присоединен бис-NHS-PEG сшивающий агент, т.е. белок был пегилирован.
Приведенные выше результаты показывают, что использование более высокого молярного избытка сшивающего агента на белок приводит к более высокому уровню модификации α-GAL, включая сшивание с образованием димера и пегилирование белка. Однако, молярное соотношение 100:1 обеспечивает высокий уровень поперечного сшивания, особенно в реакции с использованием реагента бис-NHS-PEG45, так что молярное соотношение 200:1 приводит лишь к незначительному увеличению эффективности сшивания.
Как показано на Фиг.8, реакция prh-α-GAL-I с бис-NHS-PEG уменьшает изоэлектрическую точку (ИЭТ) prh-α-GAL-I, подтверждая тем самым, что бис-NHS-PEG ковалентно связан с prh-α-GAL-I. Присоединение бис-NHS-PEG к prh-α-GAL-I преобразует основные аминогруппы остатков лизина в нейтральные амидные группы, тем самым снижая pI. Снижение pI было более выраженным при использовании большего молярного избытка (например, 200:1) бис-NHS-PEG, что подтверждает приведенные выше результаты, полученные методом SDS-PAGE.
Как дополнительно показано на Фиг.8, pI снижается больше при реакции с бис-NHS-PEG5 и бис-NHS-PEG8 по сравнению с бис-NHS-PEG45.
Этот результат показывает, что реакция с бис-NHS-PEG5 и бис-NHS-PEG8 чаще, чем с бис-NHS-PEG45, приводит к пегилированию, при котором только один конец сшивающего агента присоединен к α-GAL. Сшивающий агент, присоединенный к α-GAL только одним концом, более эффективно снижает pI, потому что такой сшивающий агент в дополнение к преобразованию аминогруппы лизина в амидную группу на присоединяемом конце содержит кислую карбоксильную (-CO2H) группу на несвязанном конце.
Как показано на Фиг.9, реакция prh-α-GAL-I со сшивающим агентом 6HC-NHS-PEG45 увеличивает молекулярный вес димера prh-α-GAL-I с 97 кДа до 113 кДа по данным MALDI-TOF масс-спектрометрии. Увеличение молекулярной массы указывает на добавление приблизительно 8 молекул бис-NHS-PEG45 к димеру prh-α-GAL-I.
Как показано на Фиг.10, реакция prh-α-GAL-I со сшивающим агентом бис-NHS-PEG8 увеличивает молекулярный вес димера prh-α-GAL-I с 97 кДа до 104 кДа по данным MALDI-TOF масс-спектрометрии. Увеличение молекулярной массы указывает на добавление приблизительно 10 молекул бис-NHS-PEG8 к димеру prh-α-GAL-I.
Пример III
Активность сшитой полученной в растениях рекомбинантной человеческой α-GAL-I
Для того, чтобы определить, сохранила ли сшитая полученная в растениях рекомбинантная α-GAL-I (prh-α-GAL-I), описанная в Примере II, ферментативную активность, ферментативную активность сшитой prh-α-GAL-I анализировали с помощью анализа с 4-метилумбеллиферил-α-D-галактопиранозидом, описанного выше.
Как показано в Таблице 1 ниже, prh-α-GAL-I, прореагировавший с бис-NHS-PEG5, бис-NHS-PEG8 или бис-NHS-PEG45 при молярном избытке бис-NHS-PEG реагента 50:1, 100:1 и 200:1 во всех случаях проявлял уровень ферментативной активности, схожий с активносиью нативной prh-α-GAL-I. Как показано в настоящем документе, в некоторых случаях наблюдалось как умеренное уменьшение, так и умеренное увеличение активности, что может быть следствием каких-либо стадий процесса получения. Данные результаты показывают, что сшивание не снижает активность prh-α-GAL-I.
Активность сшитой бис-NHS-PEG45 prh-α-GAL-I также проверяли с помощью анализа с N-додеканоил-NBD-церамид тригексозида, описанного выше, который оценивает активность α-GAL к ее естественному субстрату церамид тригексозиду(Cb3). Полученная клетках млекопитающих рекомбинантная человеческая α-GAL Replagal® анализировали для сравнения.
Как показано на Фиг.11, после инкубации сшитой полученной в растениях рекомбинантной человеческой α-GAL-I с флуоресцентным субстратом почти весь субстрат превращается в продукт, N-додеканоил-нитробензоксадиазол-лактозил церамид, как и после реакции, катализируемой полученной в клетках млекопитающих рекомбинантной α-GAL (Replagal®). Этот анализ с использованием близкого аналога природного субстрата подтверждает, что сшивание не ухудшает ферментативную гидролитическую эффективности prh-α-GAL-I.
Пример IV
Стабильности сшитой полученной в растениях рекомбинантной человеческой α-GAL-I in vitro
Стабильность сшитой полученной в растениях рекомбинантной человеческой α-GAL-I (prh-α-GAL-I), полученный как описано в Примере II, была измерена т vitro в различных условиях, как описано выше в разделе Материалы и методы. Стабильность коммерческих рекомбинантных человеческих α-Gal Fabrazyme® и Replagal® была измерена для сравнения.
Как показано на Фиг.12А-12С, стабильность полученной в растениях рекомбинантной человеческой α-GAL-I в моделируемых лизосомальных условиях была увеличена при сшивании с бис-NHS-PEG5 (Фиг.12А), бис-NHS-PEG8 (Фиг.12В) и бис-NHS-PEG45 (Фиг.12С). Как показано в настоящем документе далее, стабильность сшитой prh-α-GAL-I в течение одной недели превосходила стабильность коммерческих рекомбинантных человеческих α-GAL. После небольшого снижения остаточной активности в течение первых 24 часов сшитая prh-α-GAL-I сохраняла активность даже через 10 дней. Первоначальное снижение активности, наблюдавшееся в течение первых 24 часов, может отражать снижение для части полученной в растениях рекомбинантной человеческой α-GAL-I, которая не вступила в реакцию сшивания.
Как показано на Фиг.12А-12С, сшитая бис-NHS-PEG45 prh-α-GAL-I проявляла самую высокую стабильность в условиях, имитирующих лизосомальные условия. Стабильность полученной в растениях рекомбинантной человеческой α-GAL-I в плазме крови человека при 37°C была также повышена при сшивании бис-NHS-PEG45 (данные не показаны).
Эти результаты показывают, что сшивание α-GAL, как описано в настоящем документе, может повысить эффективность применения α-GAL in vivo за счет увеличения стабильности α-GAL в лизосомах, тем самым позволяя α-GAL действовать в течение более длительного периода времени в лизосомах, и за счет повышения стабильности α-GAL в крови, тем самым увеличивая время полураспада α-GAL в кровотоке.
Пример V
Фармакокинетика и биораспределение сшитой полученной в растениях рекомбинантной человеческой α-GAL-I in vivo
Фармакокинетику и биораспределение растительной рекомбинантной человеческой α-GAL-I (prh-α-GAL-I), сшитой бис-NHS-PEG45 или бис-NHS-PEG8, как описано в Примере II, изучали на страдающих болезнью Фабри мышах, которым вводили 2 мг/кг α-GAL, как описано выше в разделе Материалы и методы. Фармакокинетику и биораспределение не сшитой полученной в растениях рекомбинантной человеческой α-GAL-I и рекомбинантной человеческой α-GAL Replagal® изучали для сравнения. Для фармакокинетического анализа образцы крови были собирали через 1, 3, 5, 10, 20, 30, 40, 60 и 120 минут после инъекции. Для каждого типа α-GAL экспериментальная группа состояла из шести мышей.
Как показано в Таблице 2 ниже, сшивание prh-α-GAL-I бис-NHS-PEG8 и бис-NHS-PEG45 увеличило время полураспада в кровотоке полученной в растениях рекомбинантной человеческой α-GAL-I, где последний реагент оказал более выраженный эффект.
Как показано на Фиг.13 и в Таблице 2, конечное время полураспада prh α-GAL-I, сшитой бис-NHS-PEG45, было значительно больше, чем конечное время полураспада α-GAL Replagal®.
Как показано на Фиг.13, активность полученной в растениях рекомбинантной человеческой α-GAL-I, сшитой бис-NHS-PEG45, на 20 минуте составляла около 40% активности на 1 минуте. Кроме того, сшитая prh-α-GAL-I проявляла активность в плазме даже через 4 часа после инъекции.
Эти результаты показывают, что сшитая prh-α-GAL-I остается активной in vivo в течение относительно длительного времени, что может позволить ферменту достигать дополнительных тканей и органов.
Как показано на Фиг.14А и 14В, уровни полученной в растениях рекомбинантной α-GAL-I, перекрестно сшитой бис-NHS-PEG8 и бис-NHS-PEG45, в селезенке страдающих болезнью Фабри мышей через 2 часа после инъекции были значительно выше, чем уровень не сшитой полученной в растениях рекомбинантной α-GAL-I, а также уровень полученной в клетках млекопитающих рекомбинантной α-GAL Replagal®. Как показано в настоящем документе ниже, уровень prh-α-GAL-I, сшитой бис-NHS-PEG45, был выше по сравнению с уровнем prh-α-GAL-I, сшитой бис-NHS-PEG8. Данные анализа методом Вестерн блоттинг (Фиг.14В) согласуются с результатами анализа биораспределения, полученными путем анализа ферментативной активности α-GAL (Фиг.14А).
Как показано на Фиг.15А и 15В, уровни полученной в растениях рекомбинантной α-GAL-I, перекрестно сшитой бис-NHS-PEG8 и бис-NHS-PEG45, в печени страдающих болезнью Фабри мышей через 2 часа после инъекции были значительно выше, чем уровень не сшитой полученной в растениях рекомбинантной α-GAL-I, но ниже, чем уровень полученной в клетках млекопитающих рекомбинантной α-GAL Replagal®. Как показано в настоящем документе ниже, уровень prh-α-GAL-I, сшитой бис-NHS-PEG45, был несколько выше по сравнению с уровнем prh-α-GAL-I, сшитой бис-NHS-PEG8. Данные анализа методом Вестерн блоттинг (Фиг.15 В) согласуются с результатами анализа биораспределения, полученными путем анализа ферментативной активности α-GAL (Фиг.15А).
Более низкий уровень α-GAL в печени может быть терапевтически выгодным, так как, как правило, около 95% от восполненной ферментативеной активности при заместительной терапии ферментом обнаруживаются в печени, и, следовательно, высокий уровень рекомбинантной α-GAL в печени указывает на более низкий уровень экзогенной α-GAL в органах-мишенях, таких как сердце и почки.
Как показано на Фиг.16, уровни полученной в растениях рекомбинантной α-GAL-I, перекрестно сшитой бис-NHS-PEG8 и бис-NHS-PEG45, в сердце страдающих болезнью Фабри мышей через 2 часа после инъекции были выше, чем уровень не сшитой полученной в растениях рекомбинантной α-GAL-I. Как показано в настоящем документе ниже, уровень prh-α-GAL-I, сшитой бис-NHS-PEG45, был выше по сравнению с уровнем prh-α-GAL-I, сшитой бис-NHS-PEG8, а также по сравнению с уровнем полученной в клетках млекопитающих рекомбинантной α-GAL Replagal®.
Как показано на Фиг.17, уровни полученной в растениях рекомбинантной α-GAL-I, перекрестно сшитой бис-NHS-PEG8 и бис-NHS-PEG45, в почках страдающих болезнью Фабри мышей через 2 часа после инъекции были выше, чем уровень не сшитой полученной в растениях рекомбинантной α-GAL-I. Как показано в настоящем документе ниже, уровень prh-α-GAL-I, сшитой бис-NHS-PEG45, был выше по сравнению с уровнем prh-α-GAL-I, сшитой бис-NHS-РЕС8, а также по сравнению с уровнем полученной в клетках млекопитающих рекомбинантной α-GAL Replagal®.
Аналогично, как показано на Фиг.18-21, уровни полученной в растениях рекомбинантной α-GAL-I, перекрестно сшитой бис-NHS-PEG45, были выше по сравнению с уровнем не сшитой полученной в растениях рекомбинантной α-GAL-I в селезенке (Фиг.18), печени (Фиг.19), сердце (Фиг.20) и почках (Фиг.21) страдающих болезнью Фабри мышей в течение 7 дней после инъекции. Как показано в настоящем документе ниже, уровень prh-α-GAL-I, сшитой бис-NHS-PEG45, был выше по сравнению с уровнем prh-α-GAL-I, сшитой бис-NHS-PEG8, а также по сравнению с уровнем полученной в клетках млекопитающих рекомбинантной α-GAL Replagal® в селезенке, сердце и почках.
Эти результаты показывают, что α-GAL, сшитая бис-NHS-PEG, в частности, бис-NHS-PEG45, характеризуется повышенным поглощением органами, включая почки и сердце, которые являются основными органами-мишенями при лечении болезни Фабри. Эти результаты согласуются с повышением времени полураспада в кровотоке и повышением стабильности сшитой α-GAL.
Пример VI
Перекрестное сшивание полученной в клетках млекопитающих рекомбинантной человеческой α-GAL бис-N-гидроксисукцинимид-поли(этиленгликолем) (бис-NHS-PEG)
Для того, чтобы подтвердить положительные эффекты сшивания, описанные выше, сшивали полученную в клетках млекопитающих рекомбинантную человеческую α-GAL Replagal®, получаемую в линии человеческой фибросаркомы НТ-1080.
333 мкл фосфатного буфера (100 мМ, рН 8) с 100 мг/мл D-(+)-галактозы добавляли к 3,8 мг бис-NHS-PEG45 в 151 мкл раствора в ДМСО (25 мг/мл) и 1,8 мг рекомбинантной человеческой α-GAL Replagal® в 130 мкл цитратного буфера (25 мМ, рН 6). Концентрацию α-GAL Replagal® определяли по анализу активности. Реакционную смесь перемешивали на орбитальном шейкере в течение 2 часов при комнатной температуре. Избыток сшивающего реагента бис-NHS-PEG45 удаляли путем диализа против раствора, используя концентратор Vivaspin 6 с пределом исключения по молекулярной массе 50 кДа. Активность α-GAL сшитой α-GAL Replagal® показала, что концентрация α-GAL составила 3 мг/мл. Продукты реакции анализировали с помощью SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), IEF (изоэлектрофокусирования) и MALDI-TOF масс-спектрометрии, как описано выше. Как показано на Фиг.22, стандарт нативной α-GAL Replagal® при гель-электрофорезе был обнаружен в форме мономера, в то время как после реакции α-GAL Replagal® с бис-NHS-PEG45 α-GAL присутствовал в виде димера, что свидетельствует о ковалентном связывании двух мономеров путем сшивания бис-NHS-PEG. Как показано на Фиг.23, реакуция α-GAL Replagal® с бис-NHS-PEG45 уменьшает изоэлектрическую точку (ИЭТ) α-GAL, тем самым подтверждая, что бис-NHS-PEG ковалентно связан с α-GAL.
Как показано на Фиг.24, реакция α-GAL Replagal® со сшивающим агентом бис-NHS-PEG45 увеличивает молекулярный вес димера α-GAL Replagal® с 103,0 кДа до 121,3 кДа по данным MALDI-TOF масс-спектрометрии. Увеличение молекулярной массы указывает на присоединение к димеру α-GAL приблизительно 9-10 молекул бис-NHS-PEG45, что схоже с результатами, описанными здесь выше для prh-α-GAL-I.
Пример VII
Активность сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL
Для того, чтобы определить, повлияло ли сшивание полученной в клетках млекопитающих рекомбинантной α-GAL, описанной в Примере VI, на ее ферментативную активность, ферментативную активность сшитой α-GAL анализировали с помощью анализа с 4-нитрофенил-α-D-галактопиранозидом (pNP-G), описанного выше.
Как показано на Фиг.25 и в Таблице 3, полученная в клетках млекопитающих рекомбинантная человеческая α-GAL, сшитая бис-NHS-PEG45, проявляли параметры ферментативной активности, которые очень схожи с параметрами активности нативной пол ученной в клетках млекопитающих рекомбинантной человеческой α-GAL. Эти результаты показывают, что сшивание существенно не влияет на активность или каталитический механизм полученной в клетках млекопитающих рекомбинантной человеческой α-GAL.
Пример VIII
Стабильность сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL in vitro
Стабильность сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal® in vitro, полученной, как описано в Примере VI, измеряли в различных условиях, как описано выше в разделе Материалы и методы. Чтобы оценить эффект сшивания для сравнения измеряли стабильность не сшитой α-GAL Replagal®.
Как показано на Фиг.26А и 26В, стабильность полученной в клетках млекопитающих рекомбинантной человеческой α-GAL при моделируемых лизосомальных условиях (Фиг.26А) и в плазме крови человека (Фиг.26В) была значительно повышена при сшивании бис-NHS-PEG45. Сшитая полученная в клетках млекопитающих рекомбинантная человеческая α-GAL проявляла более высокую стабильность в условиях, имитирующих лизосомальные условия, чем в плазме. Эти результаты показывают, что сшивание α-GAL, как описано в настоящем документе, может стабилизировать рекомбинантную α-GAL, полученную из множества источников и систем экспрессии.
Пример IX
Фармакокинетика и биораспределение сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL in vivo Фармакокинетику и биораспределение сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL, описанной в Примере VI, исследовали путем измерения активности α-GAL в селезенке, печени, сердце и почках страдающих болезнью Фабри мышей через 2 часа, 7, 14 и 28 дней после инъекции, а также измерения уровней Gb3 в этих органах, как описано выше в разделе Материалы и методы. Для сравнения исследовали биораспределение не сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal®. Как показано на Фиг.27A-27D, сшитая полученная в клетках млекопитающих рекомбинантная α-GAL снижает уровень Gb3 в селезенке (Фиг.27А), почках (Фиг.27Б), печени (Фиг.27С) и почках (Фиг.27D) страдающих болезнью Фабри мышей значительно сильнее, чем не сшитая полученная в клетках млекопитающих рекомбинантная α-GAL.
Как показано на Фиг.28A-28D, сшитая полученная в клетках млекопитающих рекомбинантная α-GAL снижает уровень Gb3 в сердце (Фиг.28А), почках (Фиг.28Б), печени (Фиг.28С) и селезенке (Фиг.28D) страдающих болезнью Фабри мышей в течение 28 дней после инъекции. Сшитая полученная в клетках млекопитающих рекомбинантная α-GAL снижает уровень Gb3 в большей степени, чем не сшитая рекомбинантная α-GAL в почках (Фиг.28В) и селезенке (Фиг.28D) страдающих болезнью Фабри мышей, и снижает уровень Gb3 приблизительно в такой же степени, как и не сшитая по лученная в клетках млекопитающих рекомбинантная α-GAL в сердце (Фиг.28А) и печени (Фиг.28С).
Эти результаты показывают, что сшивание бис-NHS-PEG приводит к значительному увеличению поглощения рекомбинантной α-GAL из различных источников и систем экспрессия органами, включая почки и сердце, которые являются основными органами-мишенями при лечении болезни Фабри. Эти результаты также показывают, что сшивание бис-NHS-PEG приводит к более существенному снижению уровня Gb3 в органах.
Пример Х
Перекрестное сшивание полученной в растениях рекомбинантной человеческой α-GAL-II бис-N-гидроксисукцинимид-поли(этиленгликолем)(бис-NHS-PEG)
Полученную в растениях рекомбинантную человеческую α-GAL-II (prh-α-GAL-II), в которой отсутствуют аминокислоты EF, присутствующие на N-конце prh-α-GAL-I, сшивали бис-NHS-PEG45, бис-NHS-PEG21 или бис-NHS-PEG68 в молярном соотношении бис-NHS-PEG к α-GAL 200:1 согласно протоколу, описанному в Примере II.
prh-α-GAL-II сохранила свою биологическую активность после сшивания бис-NHS-PEG (данные не показаны).
Продукты реакции анализировали с помощью SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), IEF (изоэлектрофокусирования) и MALDI-TOF масс-спектрометрии, как описано выше. Как показано на Фиг.29А-29В, стандарт нативной prh-α-GAL-II при гель-электрофорезе был обнаружен в форме мономера, в то время как после реакции prh-α-GAL-II с бис-NHS-PEG45 или бис-NHS-PEG21 (Фиг.29А), или бис-NHS-PEG68 (Фиг.29В), prh-α-GAL-II присутствовал в основном в виде димера (с присутствием некоторого количества мономера), что свидетельствует о ковалентном связании двух мономеров за счет сшивания сшивающим агентом бис-NHS-PEG. Как показано далее на Фиг.29А-29В, для каждгой из проверенных сшивающих агентов молекулярная масса мономерной prh-α-GAL-II увеличивалась после реакции с сшивающим агентом. Увеличение молекулярной массы было большим для бис-NHS-PEG45 по сравнению с бис-NHS-PEG21 (Фиг.29А), и было наибольшим для бис-NHS-PEG68 (сравни Фиг.29А с Фиг.29В). Эти результаты показывают, что мономеры, которые не были димеризованы путем сшивания, были ковалентно присоединены к сшивающему агенту бис-NHS-PEG, т.е. белок был пегилирован. Как показано на Фиг.30А-30С, реакция prh-α-GAL-II со сшивающим агентом бис-NHS-PEG21 увеличивает молекулярный вес димера prh-α-GAL-II с 95 кДа (Фиг.30А) до 109 кДа (Фиг.30В), в то время как реакция prh-α-GAL-II со сшивающим агентом бис-NHS-PEG45 увеличивает молекулярный вес димера prh-α-GAL-II до 114 кДа (Фиг.30С) по данным MALDI-TOF масс-спектрометрии. Увеличение молекулярного веса указывает на добавление приблизительно 13 молекул бис-NHS-PEG21 или около 9 молекул бис-NHS-PEG45 к димеру prh-α-GAL-II.
Пример XI
Стабильность сшитой полученной в растениях рекомбинантной человеческой α-GAL-II in vitro
Стабильность сшитой полученной в растениях рекомбинантной человеческой α-GAL-II (prh-α-GAL-II), полученной, как описано в Примере X, измеряли в различных условиях in vitro, как описано выше в разделе Материалы и методы. Для сравнения измеряли стабильность коммерческой рекомбинантной человеческой α-GAL Replagal®.
Как показано на Фиг.31A-31D, стабильность полученной в растениях рекомбинантной человеческой α-GAL-II была повышена в моделируемых лизосомальных условиях (Фиг.31А и 31В) и в плазме крови человека (Фиг.31С и 31D) за счет сшивания бис-NHS-PEG8 (Фиг.31В и 31D), бис-NHS-PEG45 (Фиг.31A-31D) или бис-NHS-PEG21 (Фиг.31А и 31С). Различные сшивающие агенты повышают стабильность prh-α-GAL-II в сопоставимой степени. Как показано в настоящем документе, стабильность сшитой prh-α-GAL-II была больше, чем стабильность рекомбинантной человеческой α-GAL Replagal®. Сшитая prh-α-GAL-II проявляла более высокую стабильность в условиях, имитирующих лизосомальные условия, а также в условиях плазмы.
Как показано на Фиг.31A-31D, не сшитая prh-α-GAL-II значительно более стабильна, чем не сшитая prh-α-GAL-I (см. Фиг.1 и 3 для сравнения) в моделируемых лизосомальных условиях (Фиг.1 и 31А-31В) и в плазме крови человека (Фиг.3 и 31С-31D), хотя prh-α-GAL-II по-прежнему характеризуется некоторой нестабильностью. Эти результаты показывают, что сшивание α-GAL, как описано в настоящем документе, может стабилизировать различные типы α-Gal.
Пример XII
Фармакокинетика и биораспределение сшитой полученной в растениях рекомбинантной человеческой α-GAL-II in vivo
Фармакокинетика и биораспределение сшитой PEG45 и сшитой PEG21 полученной в растениях рекомбинантной человеческой α-GAL-II (prh-α-GAL-II), описанной в Примере X, определяли с помощью измерения активности α-GAL в плазме крови и органах, как описано выше в разделе Мате риалы и методы. Для сравнения были исследованы Фармакокинетика и биораспределение не сшитой полученной в клетках млекопитающих рекомбинантной человеческой α-GAL Replagal®.
Для фармакокинетического анализа образцы крови собирали через 1, 5, 10, 30, 60, 120, 240, 480 и 1440 минут после инъекции α-GAL страдающим болезнью Фабри мышам в дозе 1 мг/кг.
Биораспределение α-GAL изучали, изымая печень, почки, сердце и селезенку у страдающих болезнью Фабри мышей через 2 часа, 7 дней, 14 дней и 28 дней после инъекции 2 мг/кг α-GAL.
Как показано на Фиг.32А и 32В и в Таблице 4, сшивание prh-α-GAL-II бис-NHS-PEG45 значительно увеличивает время полураспада prh-α-GAL-II в кровотоке, давая время полураспада в кровотоке, которое значительно превосходит время полураспада в кровотоке полученной в клетках млекопитающих рекомбинантной α-GAL или не сшитой prh-α-GAL-II.
Как показано на Фиг.33A-33L, сшивание prh-α-GAL-II бис-NHS-PEG45 увеличивает поглощение prh-α-GAL-II в сердце (Фиг.33А), почках (Фиг.33Б), печени (Фиг.33С) и селезенке (Фиг.33D) страдающих болезнью Фабри мышей, хотя увеличение поглощения печенью проявляется в меньшей степени. Как показано на Фиг.33E-33L, сшивание prh-α-GAL-II бис-NHS-PEG21 также увеличивает поглощение prh-α-GAL-II в сердце (Фиг.33Е и 33I), почках (Фиг.33F и 33J), печени (Фиг.33G и 33К) и селезенке (Фиг.33H и 33L) страдающих болезнью Фабри мышей, хотя такое увеличение не всегда было очевидно в первой точке после 2 часов.
Как показано в настоящем документе, уровень сшитой prh-α-GAL-II был выше уровня полученной в клетках млекопитающих рекомбинантной α-GAL в сердце (Фиг.33А, 33Е и 33I), почках (Фиг.33В, 33F и 33J) и селезенке (Фиг.33D, 33Н и 33L) страдающих болезнью Фабри мышей, но был ниже по сравнению с уровнем полученной в клетках млекопитающих рекомбинантной α-GAL в печени (Фиг.33С, 33G и 33K).
Эти результаты показывают, что сшитая prh-α-GAL-II характеризуется значительно повышенной активностью α-GAL в плазме и в различных органах, особенно в органах, отличных от печени.
Пример XIII
Влияние рН на активность полученной в растениях рекомбинантной человеческой α-GAL рН среды оказывает существенное влияние на стабильность и кинетику лизосомальных ферментов, таких α-GAL. рН может влиять на связывание субстрата с ферментом. рН может также повлиять на протонирования и депротонирования каталитических групп, таких как карбоксильные или аминогруппы, которые входят в состав активного центра фермента, и таким образом повлиять на кинетическое поведение фермента. Стабильность третичной или четвертичной структуры ферментов также зависит от рН, что влияет на скорость ферментативной реакции, особенно при очень кислых или щелочных значениях рН. Для изучения рН-зависимости активности α-GAL и эффекта сшивания на рН-зависимость активность сшитой PEG45 и не сшитой полученной в растениях рекомбинантной человеческой α-GAL-II определяли при различных значениях рН с использованием субстрата pNP-G. Измерения проводили в растворх 20 мМ цитрата и 30 мМ фосфата натрия.
Кинетические параметры, характеризующие активность α-GAL при различных значениях рН, приведены ниже в Таблице 5 и на Фиг.34А-34С. Как показано на Фиг.34А-34С, сшивание α-GAL-II увеличивает Vmax (Фиг.34А) и kкат (Фиг.34С) и не оказывает существенного влияния на KM (Фиг.34В).
Повышение параметров Vmax и kкат указывает на увеличение каталитической активности. Это увеличение особенно важно при значениях рН, по меньшей мере, около 7, где каталитическая активность не сшитой α-GAL-II является незначительной. Км представляет собой кинетический параметр, связанной с аффинностью фермент/субстрат. Отсутствие значимого эвлияния сшивания на значение Км указывает, что сшивание не оказывает существенного влияния на аффинность α-GAL к pNP-G субстрату.
Пример XIV
Влияние пегилирования на стабильность α-GAL
Для того, чтобы определить, обусловлен ли стабилизирующий эффект PEG сшивающих агентов свойствами PEG или воздействием сшивания, был исследован эффект пегилирования как такового на стабильность α-GAL.
Полученную в растениях рекомбинантную человеческую α-GAL-I вводили в реакцию с активированным N-гидроксисукцинимидом (NHS) метоксизамещенным ПЭГ различной молекулярной массы (2, 5 и 10 кДа). Такие реагенты PEG имеют одну группу NHS, и, следовательно, пегилируют белок без образования сшивания.
Продукты реакции анализировали с помощью SDS-PAGE.
Как показано на Фиг.35, метоксизамещенные пегилирующие агенты пегилировали α-GAL (проявляется как увеличение молекулярной массы α-GAL), но не приводили к образованию димеров α-GAL, указывая, что α-GAL не была сшита.
Как показано на Фиг.36А и 36В, пегилирование полученной в растениях рекомбинантной человеческой α-GAL-I без образования сшивания существенно не повышало стабильность полученной в растениях рекомбинантной α-GAL в моделируемых лизосомальных условиях (Фиг.36А) или в плазме крови человека (Фиг.36В).
Эти результаты показывают, что стабилизирующий эффект сшивания, описанный выше, не является результатом пегилирование как такового.
Пример XV Влияние длины цепи ПЭГ на активность сшитой α-GAL
Для того, чтобы оценить влияние длины цепи ПЭГ сшивающего агента на активность α-GAL, полученную в растениях рекомбинантную человеческую α-GAL-I перекрестие сшивали бис-NHS-PEG2, бис-NHS-PEG4, бис-NHS-PEG68 и бис-NHS-PEG150 агентами, по существу, как описано в Примере II (для PEG68 и PEG150 длина цепи является приблизительной). A-GAL-I сшивали при молярном соотношении бис-NHS-PEG:α-GAL 50:1, 100:1 и 200:1. Продукты реакции анализировали с помощью SDS-PAGE, как описано выше. Для сравнения также анализировали α-GAL-I, сшитую бис-NHS-PEG45, как описано в Примере II.
Как показано на Фиг.37, SDS-PAGE анализ показал, что все бис-NHS-PEG агенты сшивали α-GAL, что приводило к ковалентно сшитому димеру, и что сшивание было более эффективным при использовании молярного соотношения 200:1. Ферментативную активность сшитой α-GAL-I определяли, как описано в Примере III. Результаты представлены в Таблице 6.
Как показано в Таблице 6, сшивание PEG2 и PEG4 умеренно снижает активность α-GAL (приблизительно на 30-50%), в то время перекрестное сшивание ПЭГ с длиной цепью не оказывает существенного влияния на активность α-GAL. Эти результаты показывают, что перекрестное сшивание цепями PEG, более длинными, через PEG4, предпочтительно с точки зрения сохранения активности сшитой α-GAL.
Пример XVI Перекрестное сшивание α-GAL с использованием бис-COOH-PEG агентов
В качестве альтернативы описанному выше сшиванию α-GAL с использованием заранее подготовленных (например, коммерчески доступных) бис-NHS-PEG агентов, α-GAL была сшита бис-COOH-PEG агентами путем активации карбоксильной (то есть COOH) группы in situ непосредственно перед проведением реакции сшивания.
Бис-COOH-PEG12, бис-COOH-PEG28 и бис-COOH-PEG45 активировали взаимодействием с 1,1 молярными эквивалентами из расчета на карбоксильные группы (т.е., 2,2 молярными эквивалентами на бис-COOH-PEG) NHS (N-гидроксисукцинимида) и EDC (1-этил-3-(3-диметиламинопропил)карбодиимида). Затем реакционную смесь встряхивали в ДМСО в течение 30 минут при комнатной температуре. Активированный бис-COOH-PEG, который являлся по существу бис-NHS-PEG, затем вводили в реакцию с полученной в растениях рекомбинантной человеческой α-GAL-I при молярном соотношении 50:1, 100:1 и 200:1, как описано в Примере II. Продукты реакции анализировали с помощью SDS-PAGE, как описано выше. α-GAL-I, сшитую бис-NHS-PEG45, как описано в Примере II, также анализировали для сравнения.
Как показано на Фиг.38, SDS-PAGE анализ показал, что все бис-COOH-PEG агенты в некоторой степени сшивали α-GAL, но это сшивание было более эффективным при использовании молярного соотношения 200:1.
Ферментативную активность сшитой α-GAL-I определяли, как описано в Примере III.
Результаты приведены в Таблице 7.
Как показано в Таблице 7, сшивание каждым из исследованных бис-COOH-PEG агентов приводило к α-GAL с активностью, близкой к ожидаемой.
Эти результаты показывают, что сшивание бис-COOH-PEG агентами не снижает активность α-GAL по сравнению с сшиванием бис-NHS-PEG агентами.
Эти результаты также подтверждают описанные выше результаты, показывающие что перекрестное сшивание цепями PEG, более длинными, чем PEG4, существенно не снижает активность сшитой α-GAL.
Пример XVII
Влияние длины и типа сшивающего агента на стабильность сшитой полученной в растениях рекомбинантной человеческой α-GAL-I in vitro
Для дальнейшего исследования влияния длины цепи на стабильность сшитой α-GAL, а также для сравнения стабильности α-GAL, сшитой бис-COOH-PEG агентами (например, как описано в Примере XVI) со стабильностью α-GAL, сшитой бис-NHS-PEG агентами, стабильность полученной в растениях рекомбинантной человеческой α-GAL-I (prh-α-GAL-I), сшитой бис-NHS-PEG2, бис-NHS-PEG4, бис-COOH-PEG12, бис-СООН28 и бис-COOH-PEG45, полученный как описано в примерах XV и XVI, была измерена в различных условиях in vitro, как описано выше в разделе Материалы и методы, по сравнению со стабильностью prh-α-GAL-I, сшитой бис-NHS-PEG45, как описано в Примере II. Для сравнения была измерена стабильность коммерческой рекомбинантной человеческой α-GAL Replagal® и не сшитой prh-GAL-I.
Как показано на Фиг.39, стабильность полученной в растениях рекомбинантной человеческой α-GAL-I в искусственных условиях в лизосоме была увеличена при сшивании каждым из агентов бис-NHS-PEG и бис-COOH-PEG. Как показано в настоящем документе, стабильность сшитой prh-α-GAL-I коррелирует с длиной сшивающих цепей ПЭГ, где бис-NHS-PEG45 и бис-COOH-PEG45 обеспечивают наибольшую стабильность, и бис-NHS-PEG2 обеспечивает наименьшую стабильность. Однако, сшивание бис-COOH-PEG45 приводило лишь к незначительно большей стабильности, чем сшивание бис-COOH-PEG28, что позволяет предположить, что стабильность не зависит от длины цепи ПЭГ после определенной длины цепи.
Как показано на Фиг.39, сшивание бис-NHS-PEG45 приводило к несколько большей стабильности, чем сшивание бис-COOH-PEG45. Указанный эффект может быть результатом неполной активации бис-COOH-PEG агента. Однако разница в стабильности была незначительной.
Кроме того, сшивание каждым из агентов бис-NHS-PEG и бис-COOH-PEG повышало стабильность полученной в растениях рекомбинантной человеческой α-GAL-I в плазме крови человека при 37°C (данные не представлены).
Эти результаты являются еще одним свидетельством, что сшивание α-GAL, как описано в настоящем документе, может повысить эффективность α-GAL in vivo за счет увеличения стабильности α-GAL в лизосомах и в крови, и что цепи PEG длиной около 28-45 единиц являются более эффективными для повышения стабильности α-GAL путем сшивания, чем более короткие цепи ПЭГ.
Пример XVIII
Кинетические параметры сшитой полученной в растениях рекомбинантной человеческой α-GAL-II
Чтобы изучить влияние сшивания на кинетические параметры сшитой полученной в растениях рекомбинантной человеческой α-GAL-II, полученной, как описано в Примере X, а также не сшитой полученной в растениях рекомбинантной человеческой α-GAL-II, кинетику их катализа изучали с помощью субстрата pNP-G и анализа кинетики Михаэлиса-Ментен. Измерения проводили в растворе 20 мМ цитрата, 30 мМ фосфата натрия, 0,1% бычьего сывороточного альбумина и 0,67% этанола при рН 4,6. Кинетические параметры рассчитывали с использованием значения содержания белка, основанного на анализе активности.
Как показано в Таблице 8 ниже, сшивание α-GAL-II привело к улучшению кинетических свойств по сравнению с не сшитой α-GAL-II. Константа Михаэлиса (Км) была снижена, что указывает на более высокое сродство фермента к субстрату. Кроме того, отношение kкат/КМ, которое отражает общую эффективность катализа фермента по отношению к субстрату при описанных выше условиях, было увеличено для сшитых форм.
Пример XIX
Воспроизводимость сшивания полученной в растениях рекомбинантной человеческой α-GAL-II
Воспроизводимость сшивания от партии к партии оценивали после приготовления 5 партий растительной рекомбинантной человеческой α-GAL-II (prh-α-GAL-II), сшитой бис-NHS-PEG45 в соотношении 200:1 по способу, аналогичному описанному в Примере II.
В партиях 1, 2, 4 и 5 1 мг prh-α-GAL-II вводили в реакцию с 3,98 мг бис-NHS-PEG. В партии 3 20,5 мг prh-α-GAL-II вводили в реакцию с 80,7 мг бис-NHS-PEG. Ферментативную активность сшитой prh-α-GAL-II определяли, как описано в Примере III. Результаты приведены в Таблице 9 ниже.
Как показано в Таблице 9, измеренная активность была близка к ожидаемой активности для всех 5 партий. В 4 из 5 партий измеренная активность отличалась от ожидаемой активности приблизительно на 10% или меньше.
Эти результаты показывают, что полученная активность сшитой prh-α-GAL-II является относительно предсказуемой и воспроизводимой. Стабильность сшитой prh-α-GAL-II в лизосомальных условиях и в плазме крови человека определяли, как описано выше.
Как показано на Фиг.40А и 40В, стабильность сшитой prh-α-GAL-II характеризовалась хорошей воспроизводимостью как в моделируемых лизосомальных условиях, так и в плазме крови человека.
Сшивание также анализировали методом SDS-PAGE, IEF (изоэлектрофокусирования) и MALDI-TOF масс-спектрометрии, как описано выше. Не сшитую prh-α-GAL-II анализировали для сравнения.
Как показано на Фиг.41, сшитая prh-α-GAL-II из различных партий характеризовалась одинаковой степенью ковалентной димеризации по данным SDS-PAGE анализа.
Как показано на Фиг.42, сшитая prh-α-GAL-II из различных партий характеризовалась одинаковой изоэлектрической точкой при анализе IEF.
Как показано на Фиг.43A-43F, сшитая prh-α-GAL-II из партий 1-5 (Фиг.43B-43F, соответственно) характеризовалась увеличением молекулярной массы димера приблизительно на 20-21 кДа по сравнению с не сшитой prh-α-GAL-II (Фиг.43). Такое уввеличение соответствует присоединению приблизительно 10 молекул PEG к димеру α-GAL. Как дополнительно показано на Фиг.43B-43F, сшитая prh-α-GAL-II из различных партий характеризовалась аналогичными соотношениями мономера и димера.
Эти результаты также показывают хорошую воспроизводимость сшивания α-GAL. Чтобы изучить воспроизводимость получаемой ферментативной активности кинетические параметры сшитой prh-α-GAL-II определяли с помощью субстрата pNP-G и анализа Михаэлиса-Ментен. Измерения проводили в растворе 20 мМ цитрата, 30 мМ фосфата натрия, 0,1% бычьего сывороточного альбумина и 0,67% этанола при рН 4,6. Кинетические параметры рассчитывали с использованием значения содержания белка, основанного на оптической плотности при 280 нм. Как показано на Фиг.44, сшитая prh-α-GAL-II из различных партий проявляла аналогичные профили зависимости скорости катализа от концентрации субстрата. Как показано в Таблице 10 ниже, сшитая prh-α-GAL-II из различных партий характеризовалась хорошей воспроизводимостью величин Vmax и kкат. Величина параметра KM заметнее изменялась между партиями, хотя это может представлять собой артефакт количественной оценки содержания белка. Приведенные выше результаты свидетельствуют о хорошей воспроизводимости ферментативных свойств сшитой α-GAL.
Хотя изобретение было описано в связи с конкретными вариантами осуществления, очевидно, что специалисту в данной области будут очевидны различные альтернативы, модификации и изменения изобретения. Соответственно, настоящее изобретение охватывает все такие альтернативы, модификации и изменения, которые находятся в пределах объема прилагаемой Формулы изобретения.
Все публикации, патенты и заявки, упомянутые в настоящем описании, включены в настоящий документ во всей своей полноте посредством ссылки в той же степени, как если бы для каждой отдельной публикации, патента или заявки было специально и отдельно отмечено о включении ее здесь путем ссылки. Кроме того, цитирование или указание любой ссылки в данной заявке не должно быть истолковано как признание того, что такая ссылка является предшествующим уровнем техники для настоящего изобретения. Используемые заголовки разделов не должны быть истолкованы как во всяком случае ограничивающие.
| название | год | авторы | номер документа |
|---|---|---|---|
| СХЕМА ЛЕЧЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ С ИСПОЛЬЗОВАНИЕМ СТАБИЛИЗИРОВАННОЙ α-ГАЛАКТОЗИДАЗЫ | 2018 |
|
RU2793184C2 |
| ЛЕЧЕНИЕ α-ГАЛАКТОЗИДАЗНОЙ А НЕДОСТАТОЧНОСТИ | 2000 |
|
RU2248213C2 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2009 |
|
RU2535342C2 |
| ПРОИЗВОДСТВО АКТИВНОЙ ВЫСОКОФОСФОРИЛИРОВАННОЙ N-АЦЕТИЛГАЛАКТОЗАМИН-6-СУЛЬФАТАЗЫ ЧЕЛОВЕКА И ЕЕ ПРИМЕНЕНИЕ | 2011 |
|
RU2607376C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2020 |
|
RU2822369C2 |
| ИЗГОТОВЛЕНИЕ АКТИВНЫХ ВЫСОКОФОСФОРИЛИРОВАННЫХ ЛИЗОСОМАЛЬНЫХ ФЕРМЕНТОВ СУЛЬФАТАЗ ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2510820C2 |
| ПРОДУЦИРОВАНИЕ ВЫСОКОМАННОЗНЫХ БЕЛКОВ В РАСТИТЕЛЬНЫХ КУЛЬТУРАХ | 2004 |
|
RU2385928C2 |
| БЕЛКОВЫЙ КОМПЛЕКС, ПОЛУЧЕННЫЙ С ИСПОЛЬЗОВАНИЕМ ФРАГМЕНТА ИММУНОГЛОБУЛИНА, И СПОСОБ ПОЛУЧЕНИЯ ТАКОГО КОМПЛЕКСА | 2004 |
|
RU2356909C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРЕЗЕНТАЦИИ МНОЖЕСТВЕННЫХ АНТИГЕНОВ, ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ И ПРИМЕНЕНИЯ | 2012 |
|
RU2619176C2 |
| СОДЕРЖАЩИЕ ГАЛАКТОЗА-АЛЬФА-1,3-ГАЛАКТОЗУ N-ГЛИКАНЫ В ГЛИКОПРОТЕИНОВЫХ ПРОДУКТАХ, ПОЛУЧЕННЫХ ИЗ КЛЕТОК СНО | 2009 |
|
RU2484142C2 |






































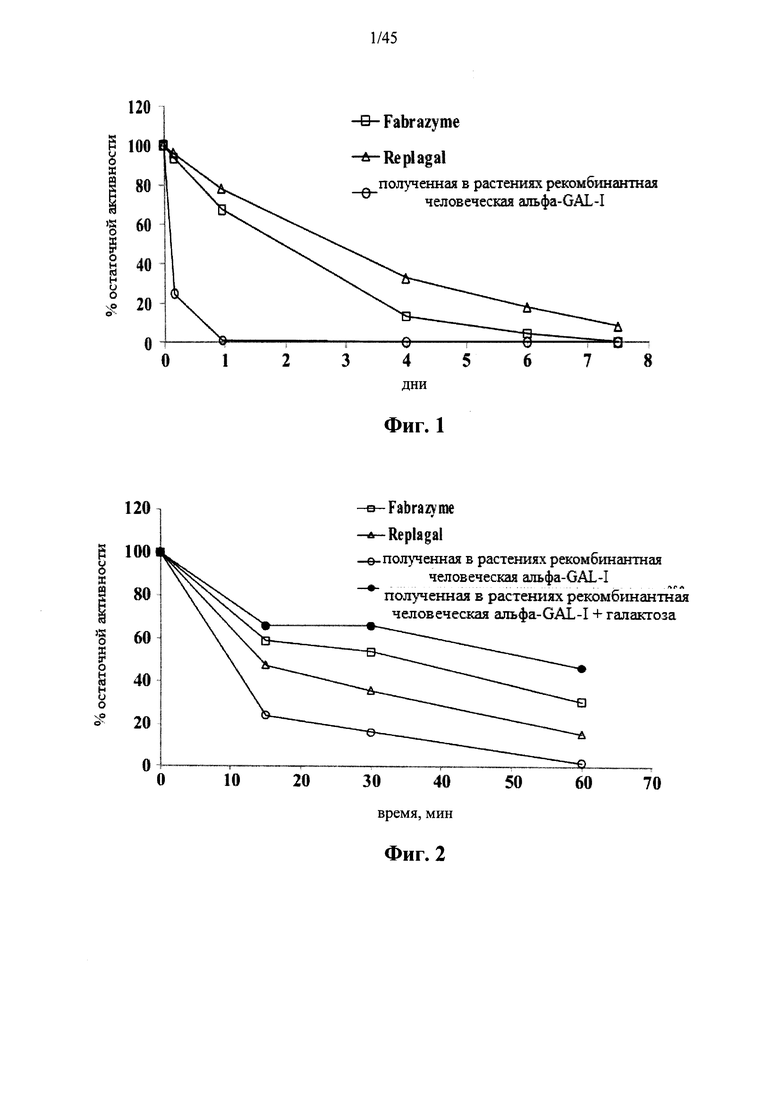
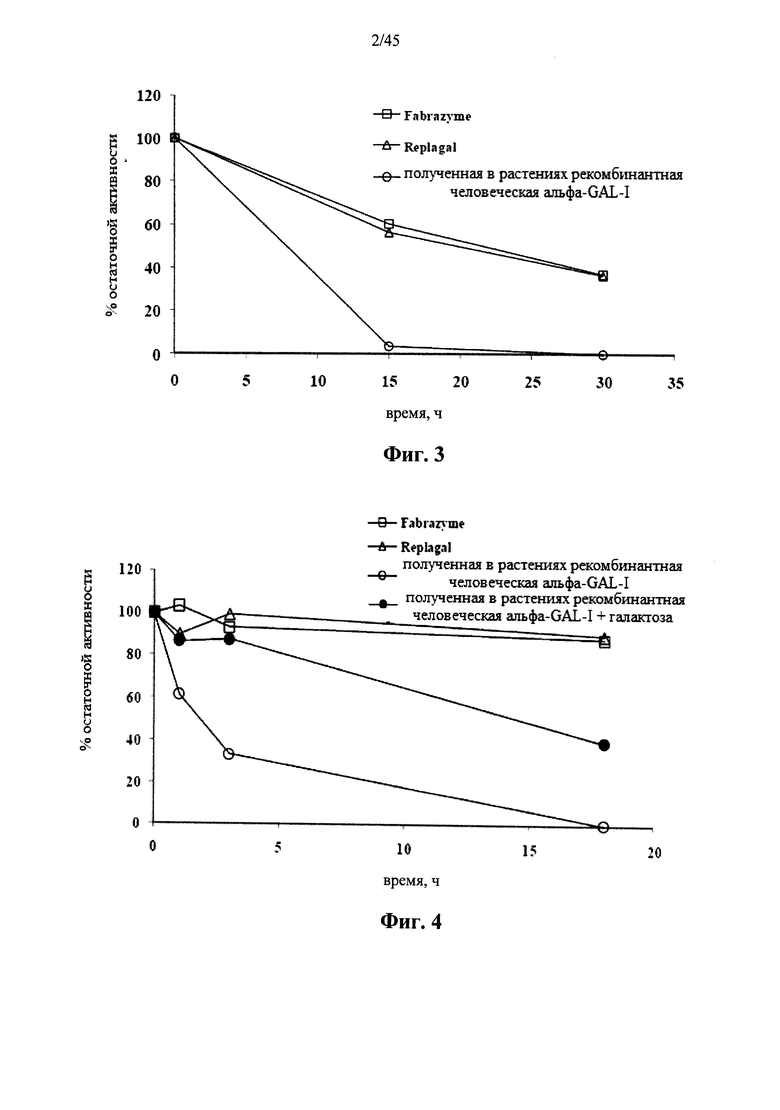
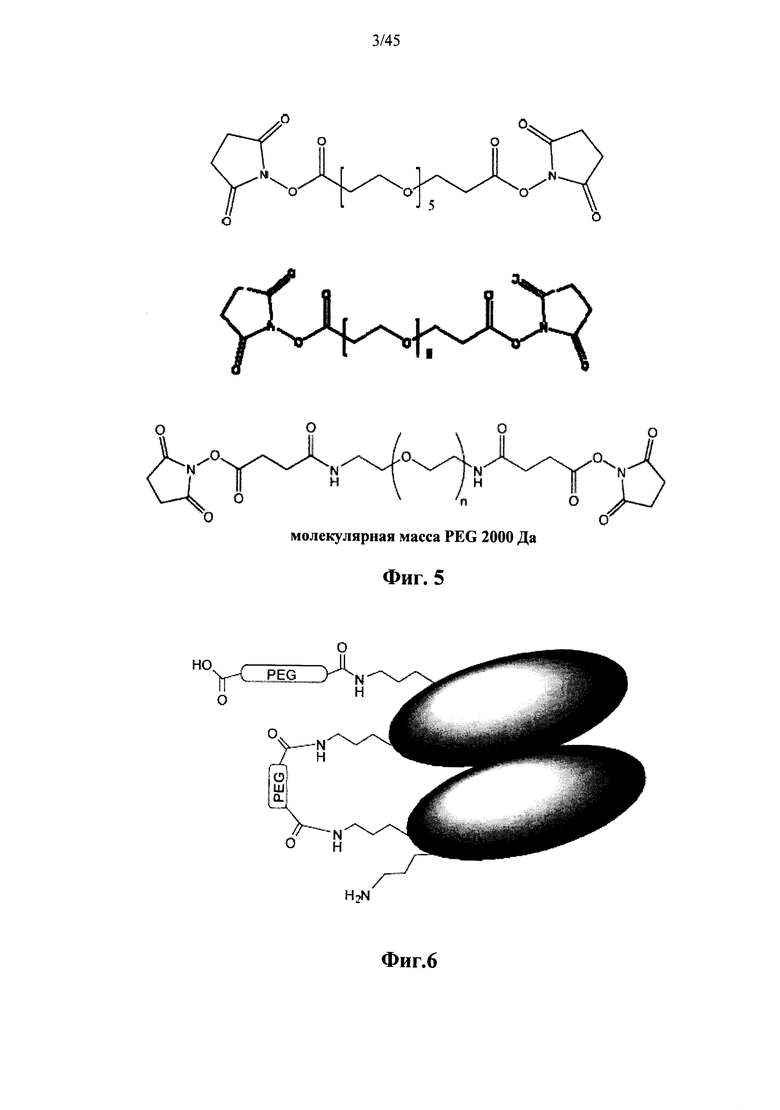
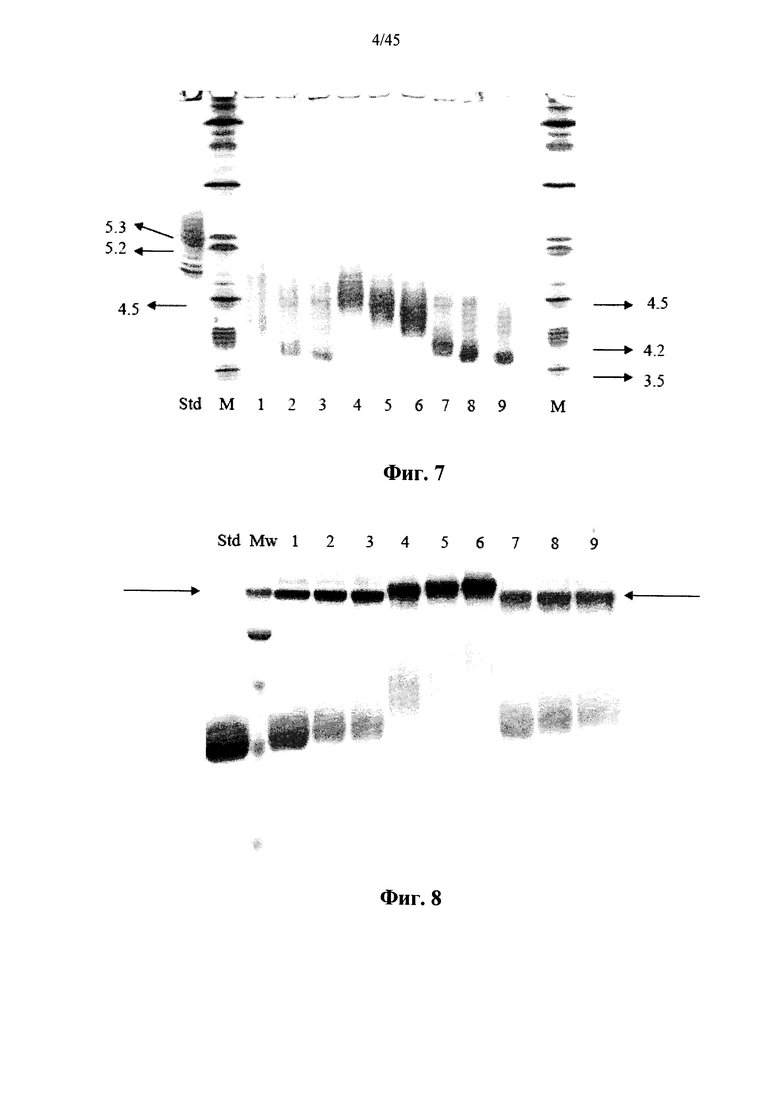
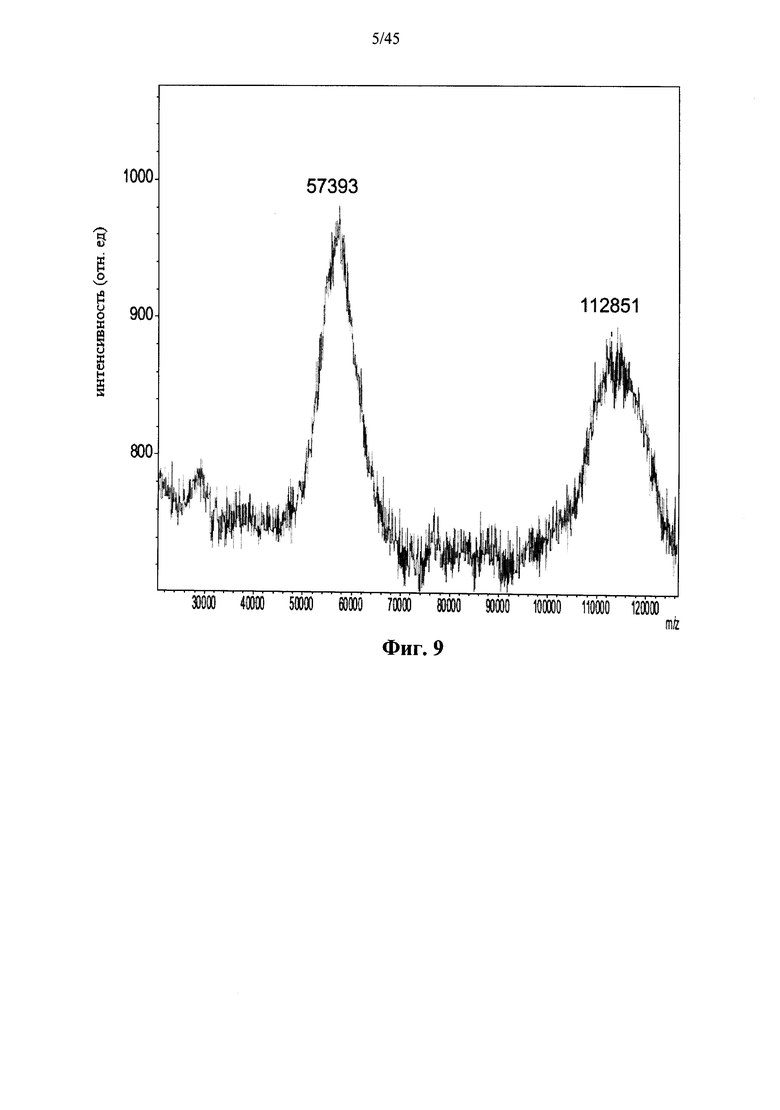
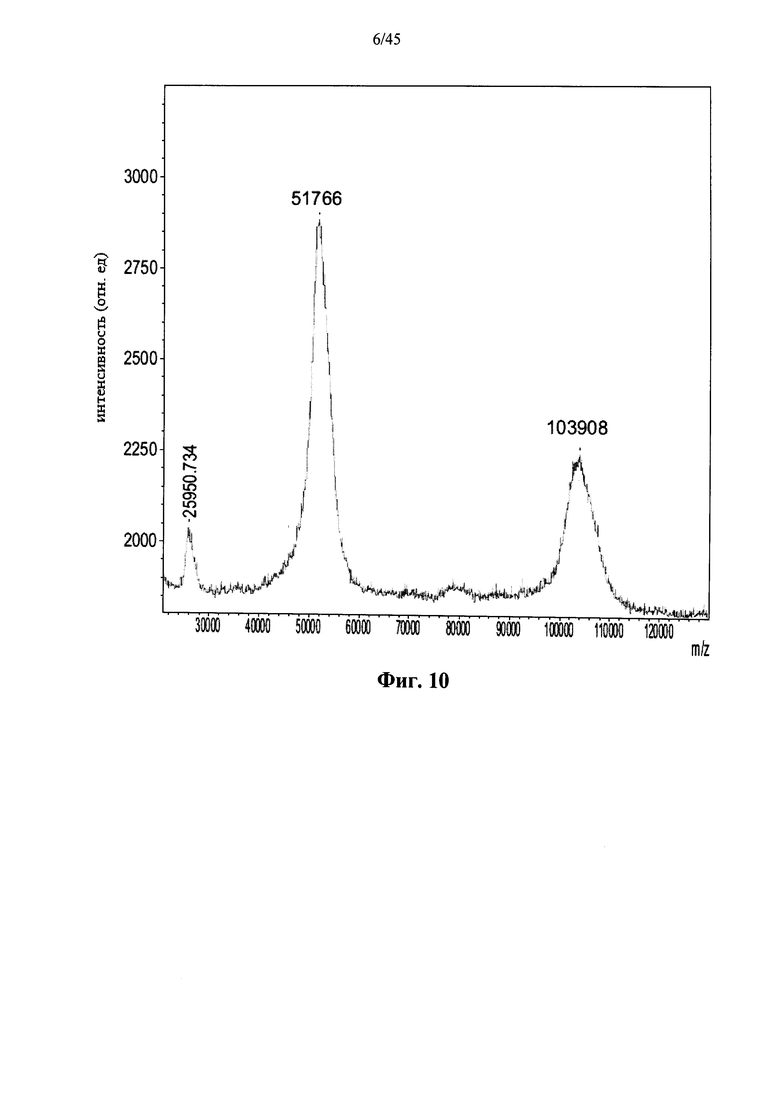

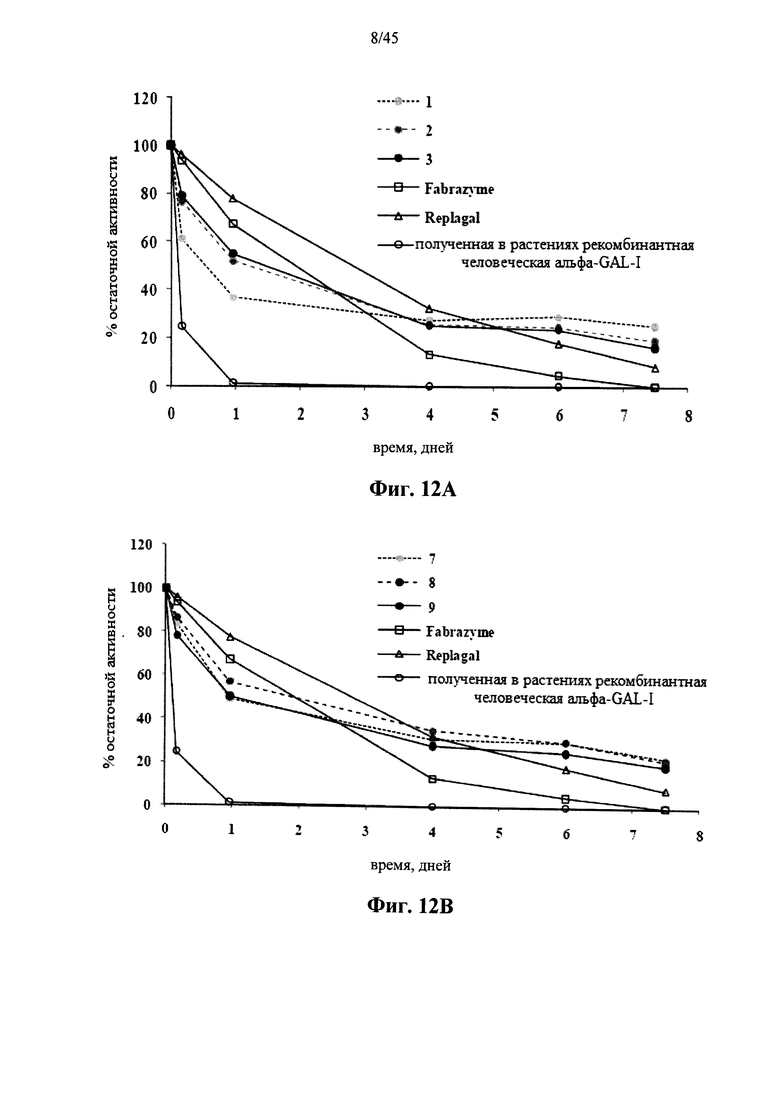
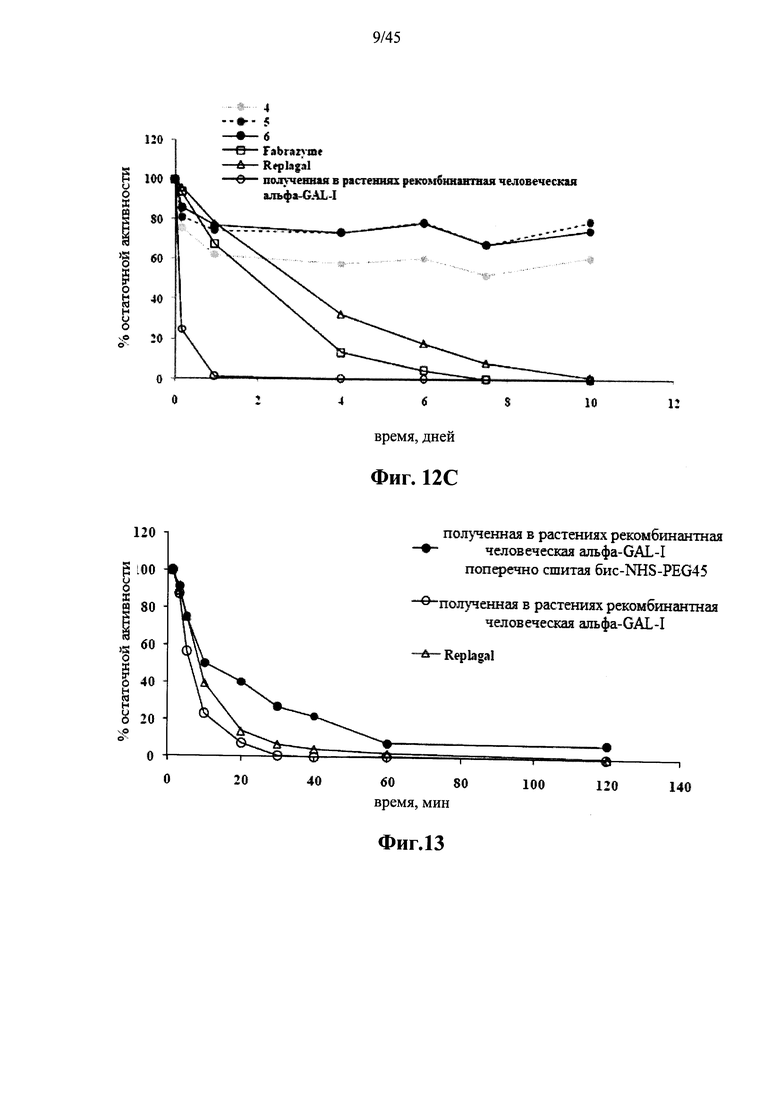
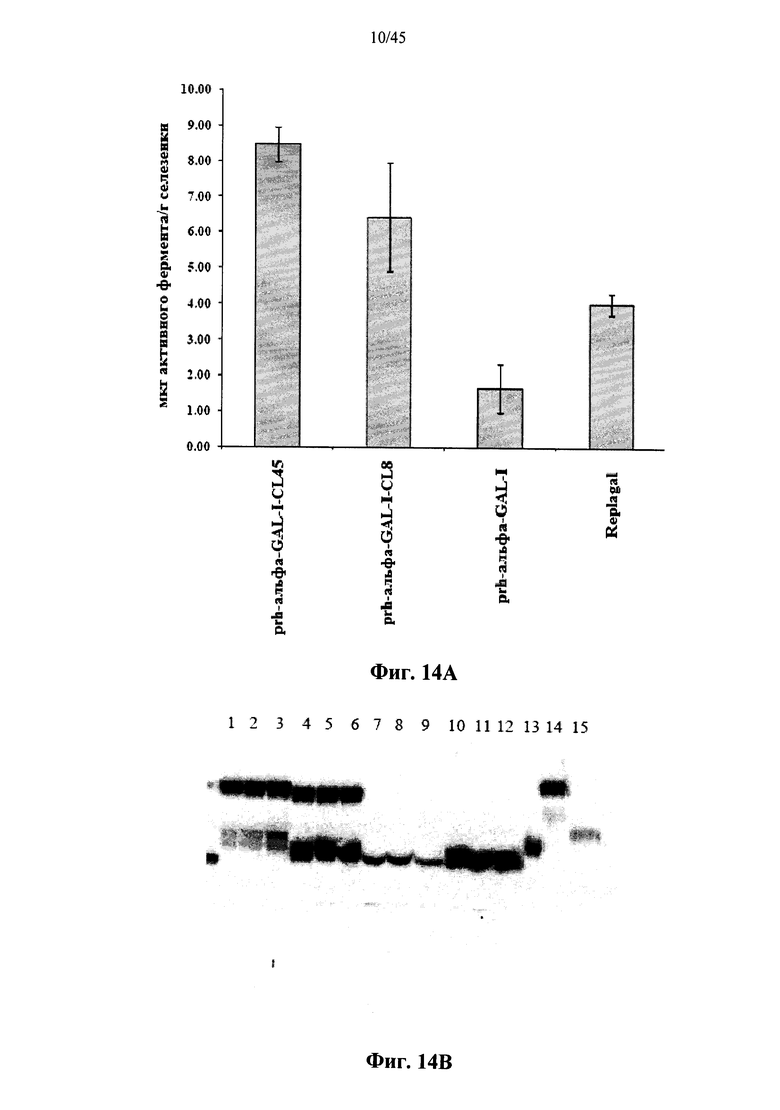
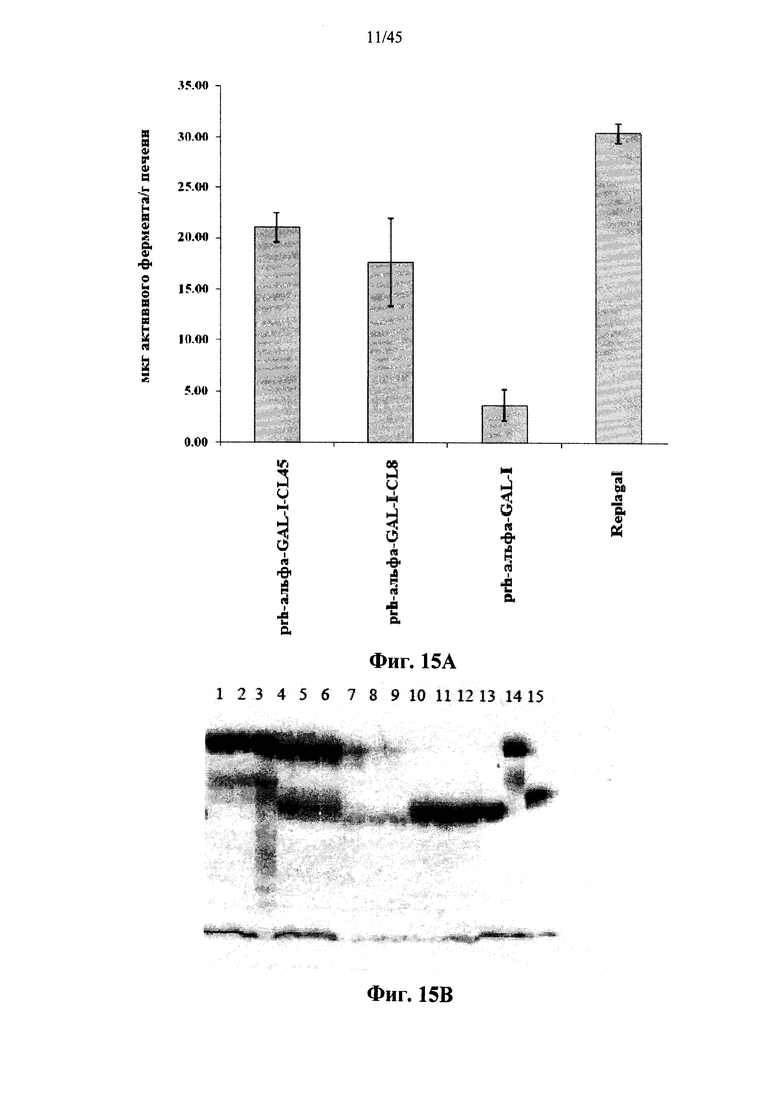
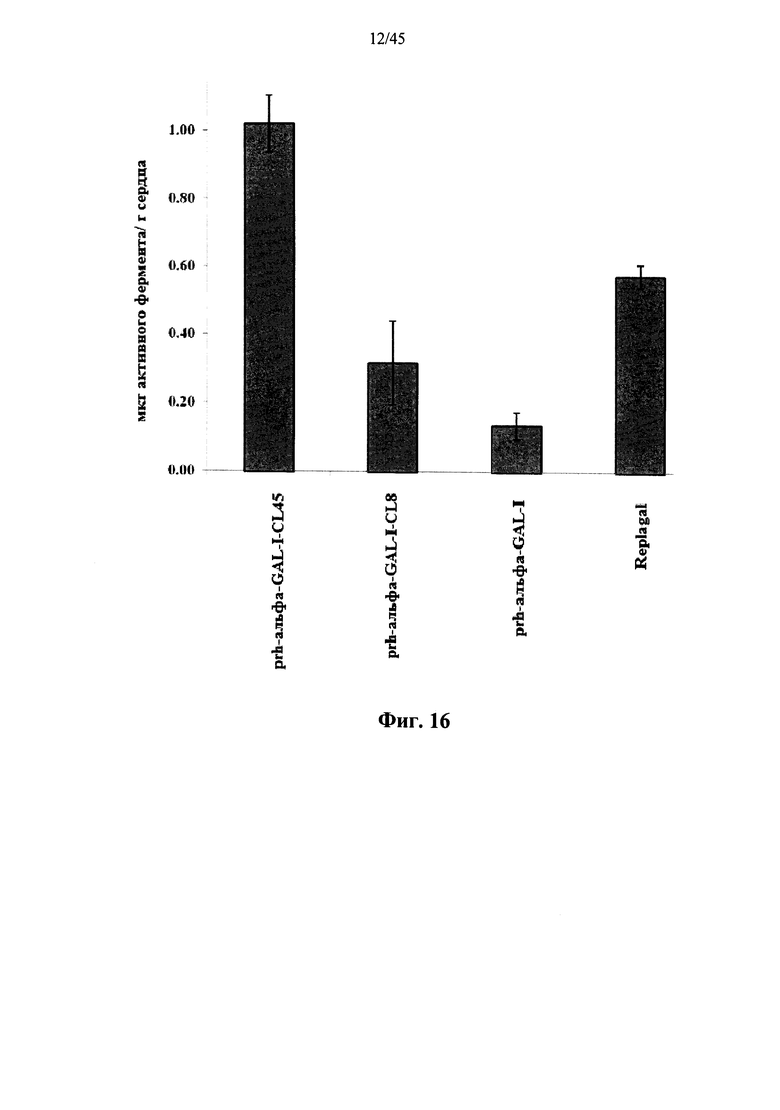
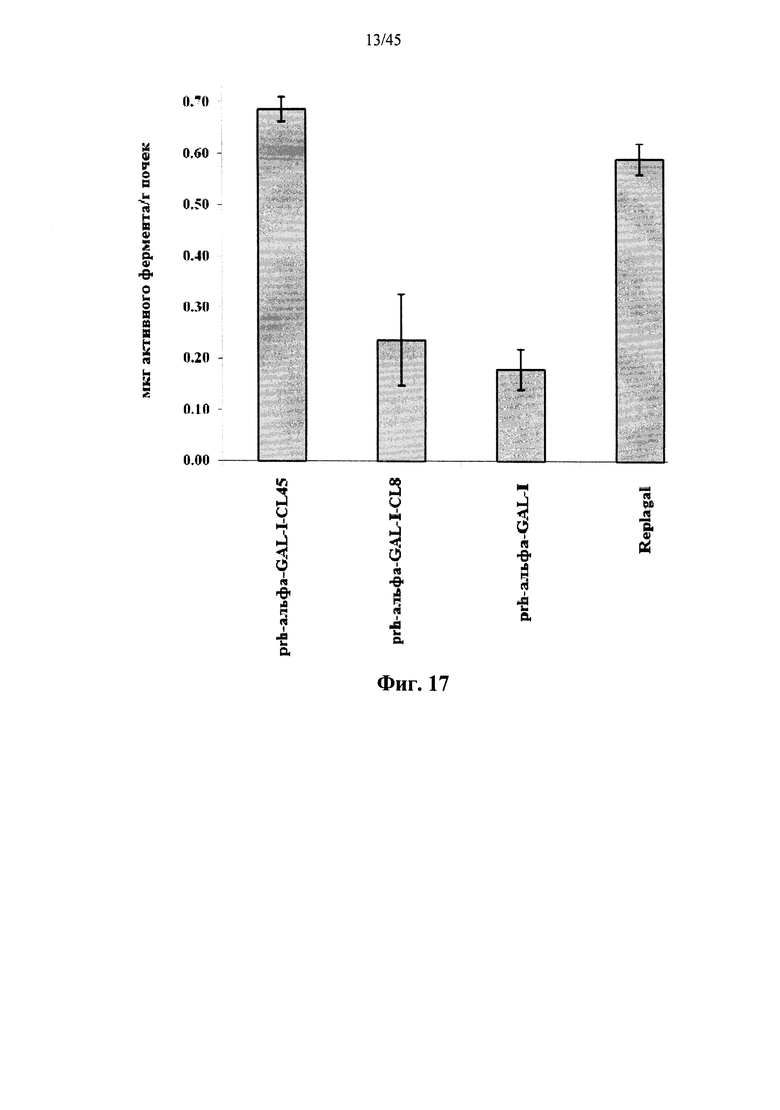
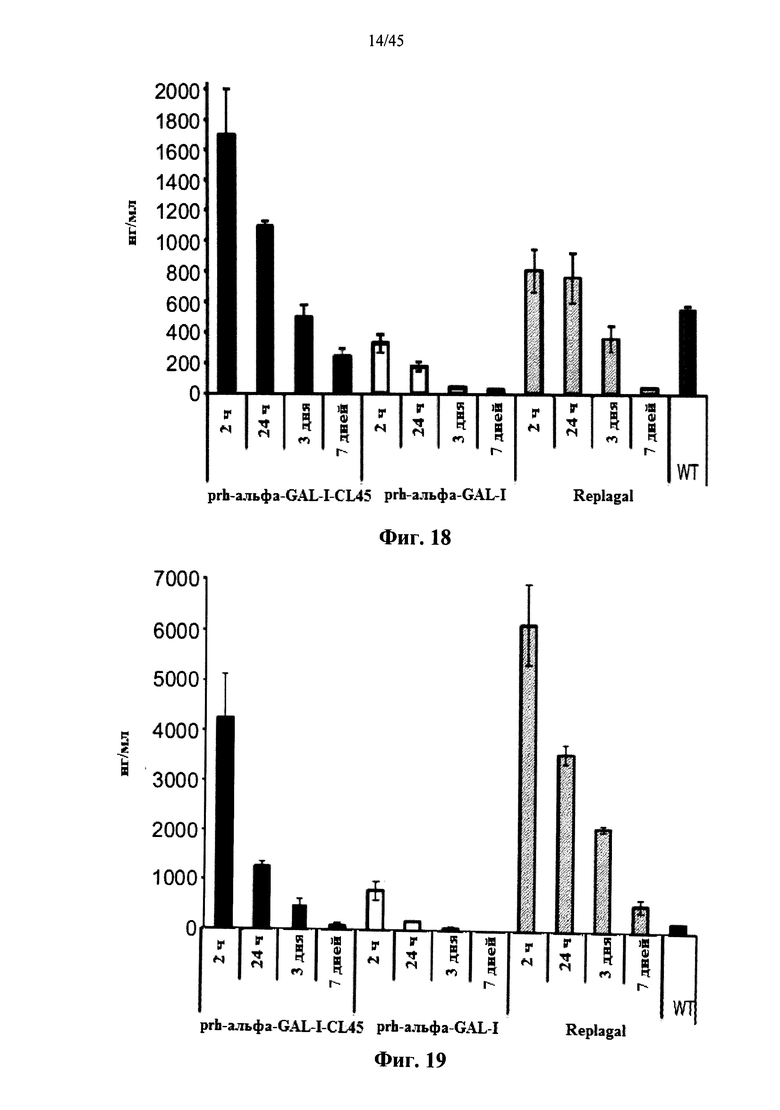
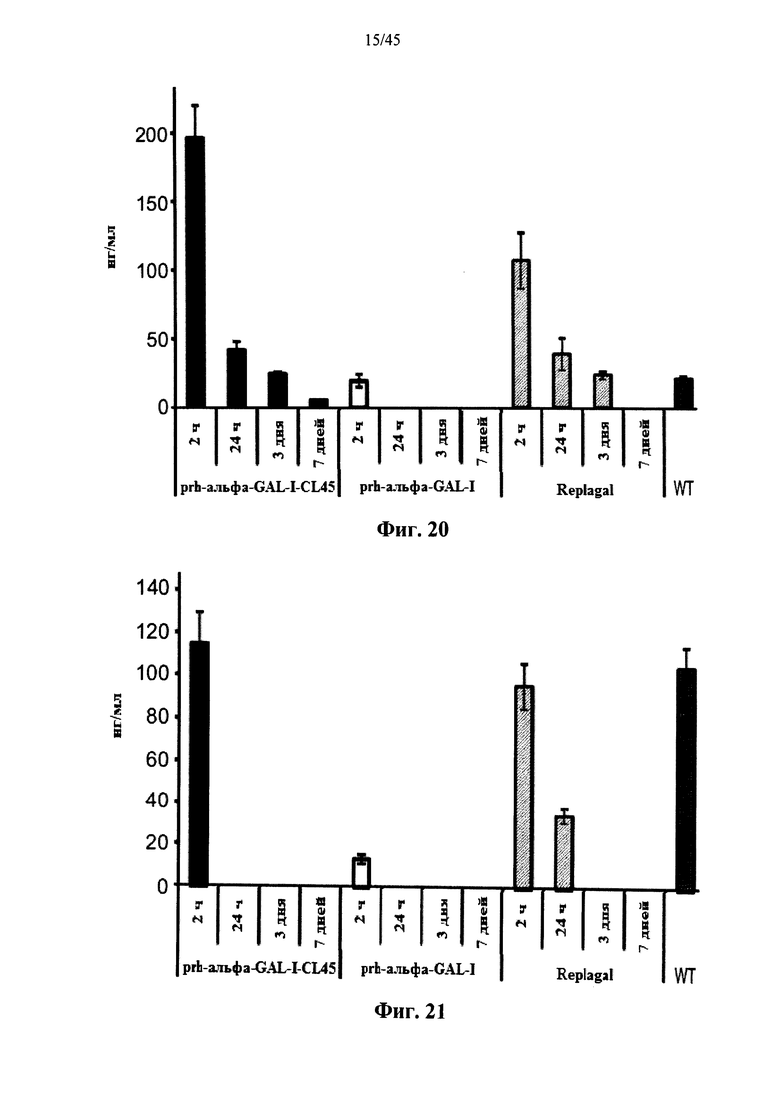
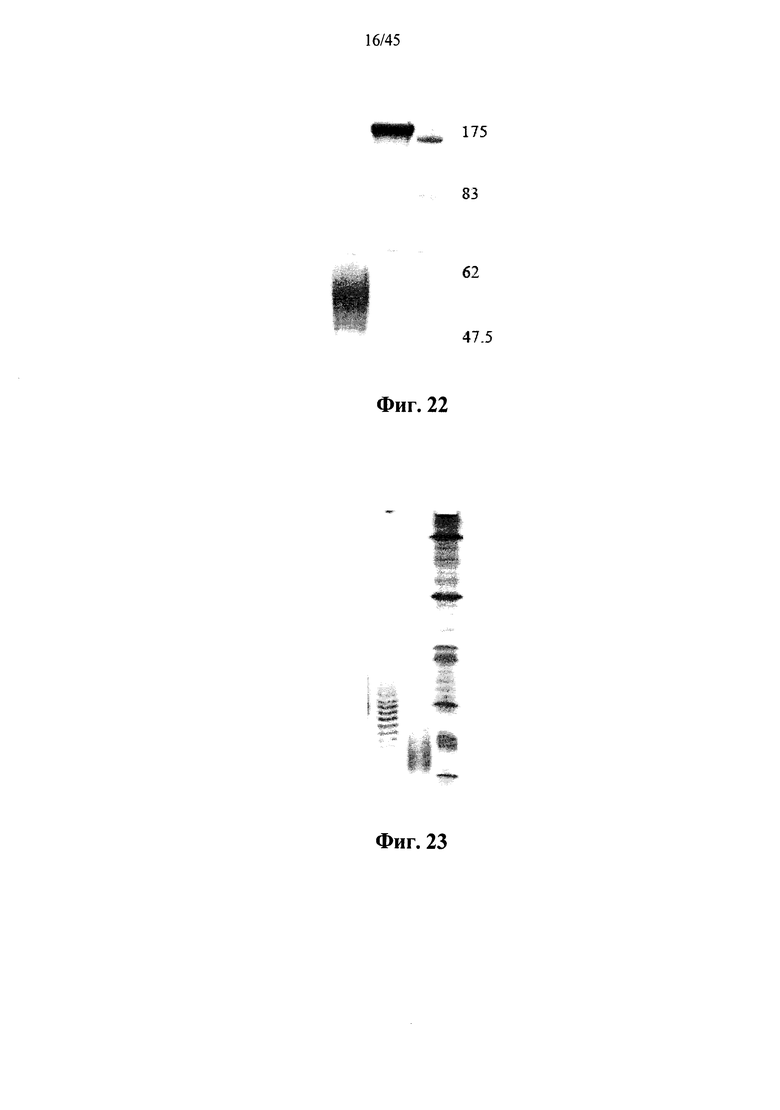
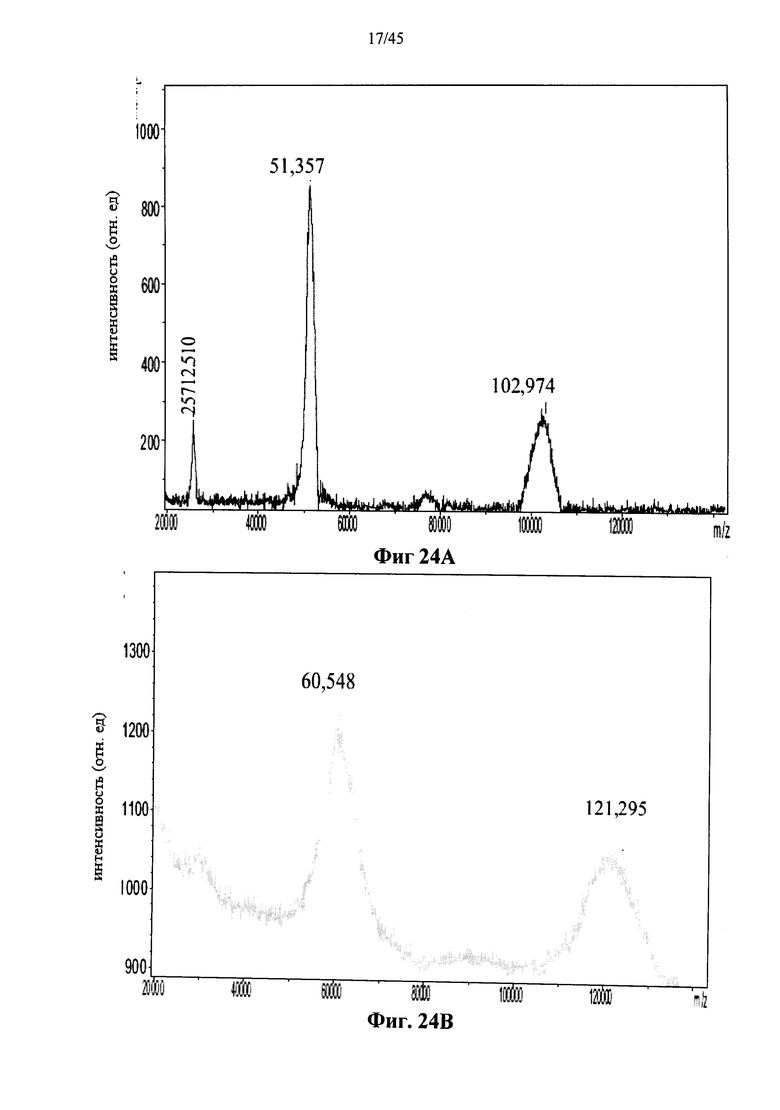
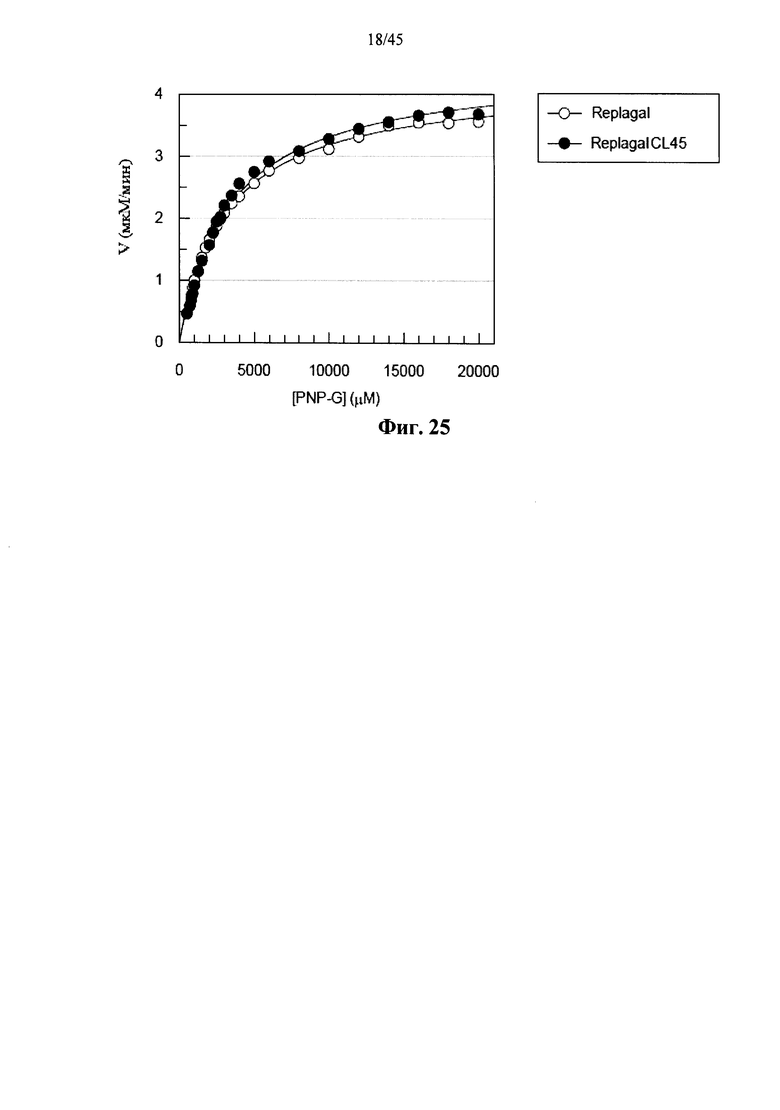
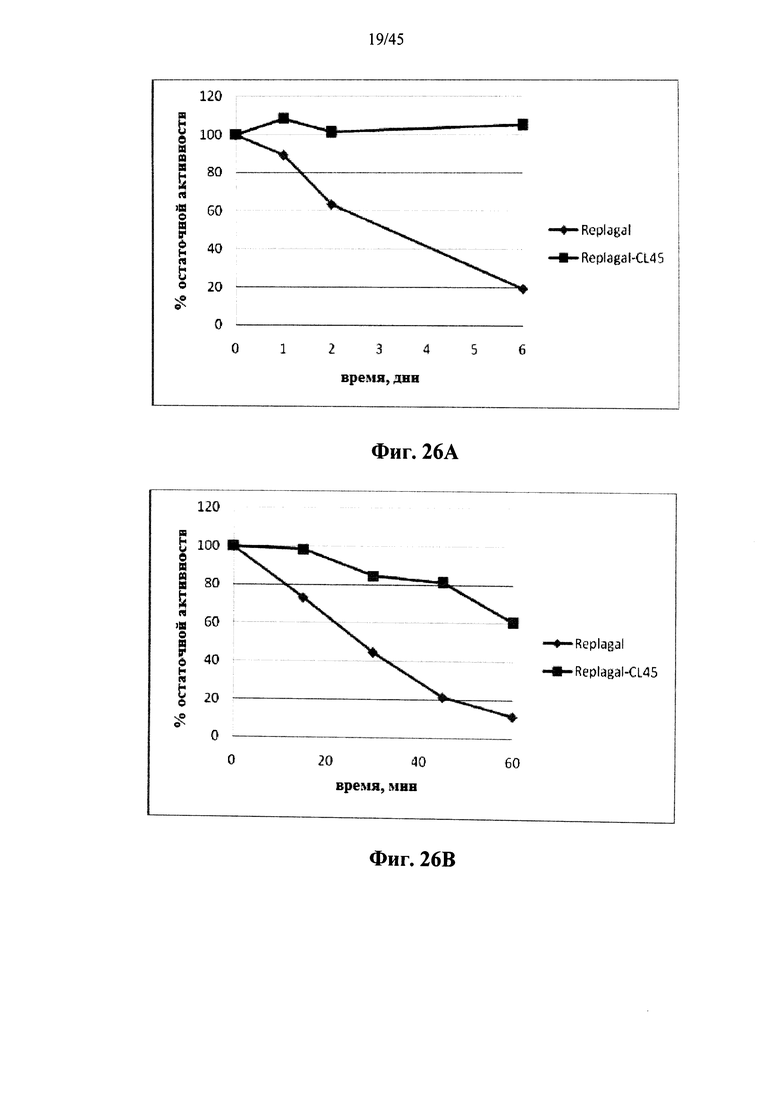
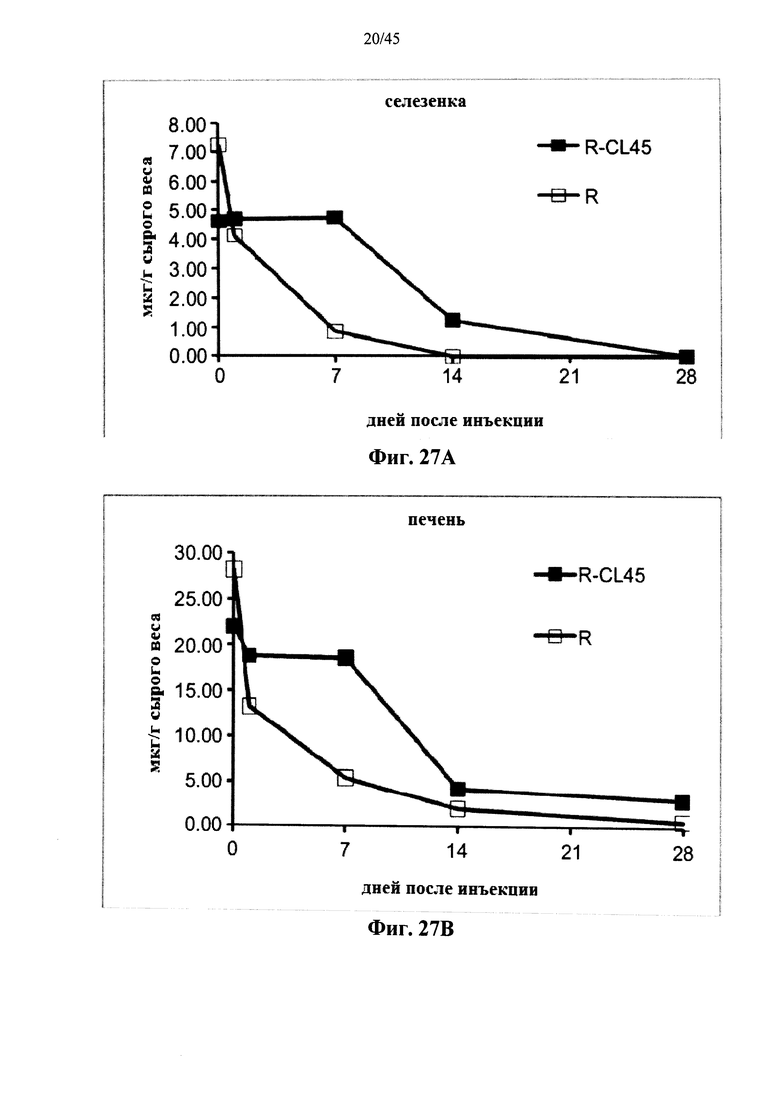
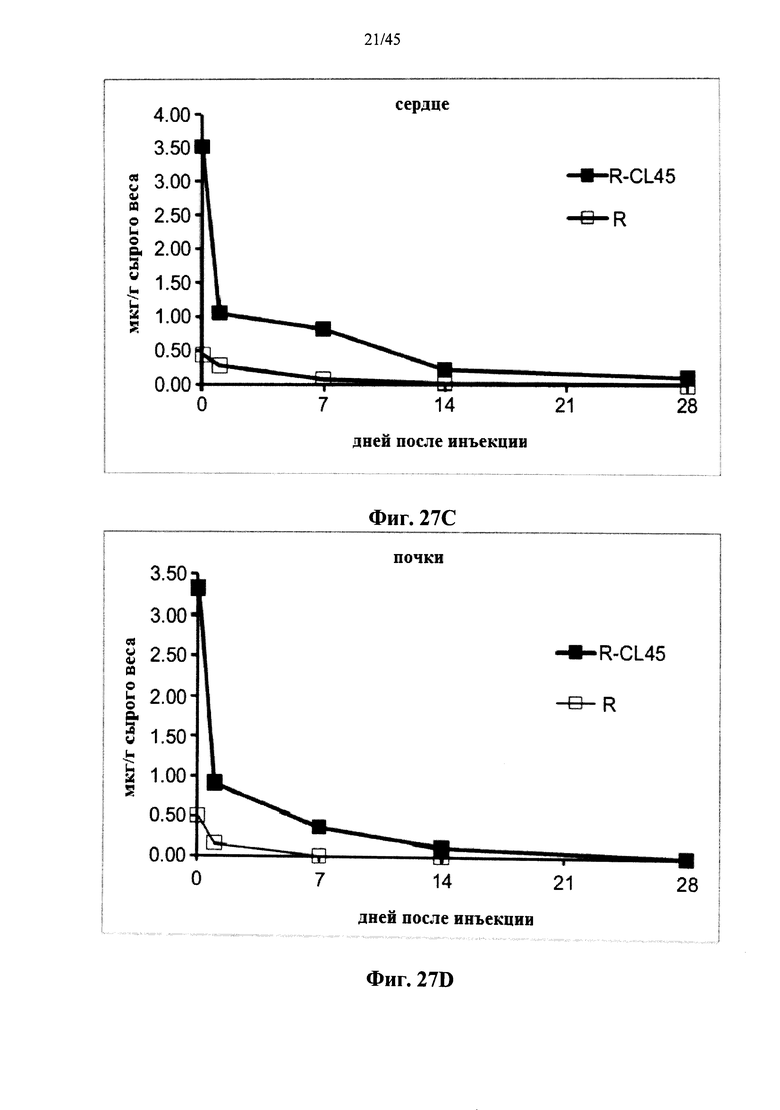
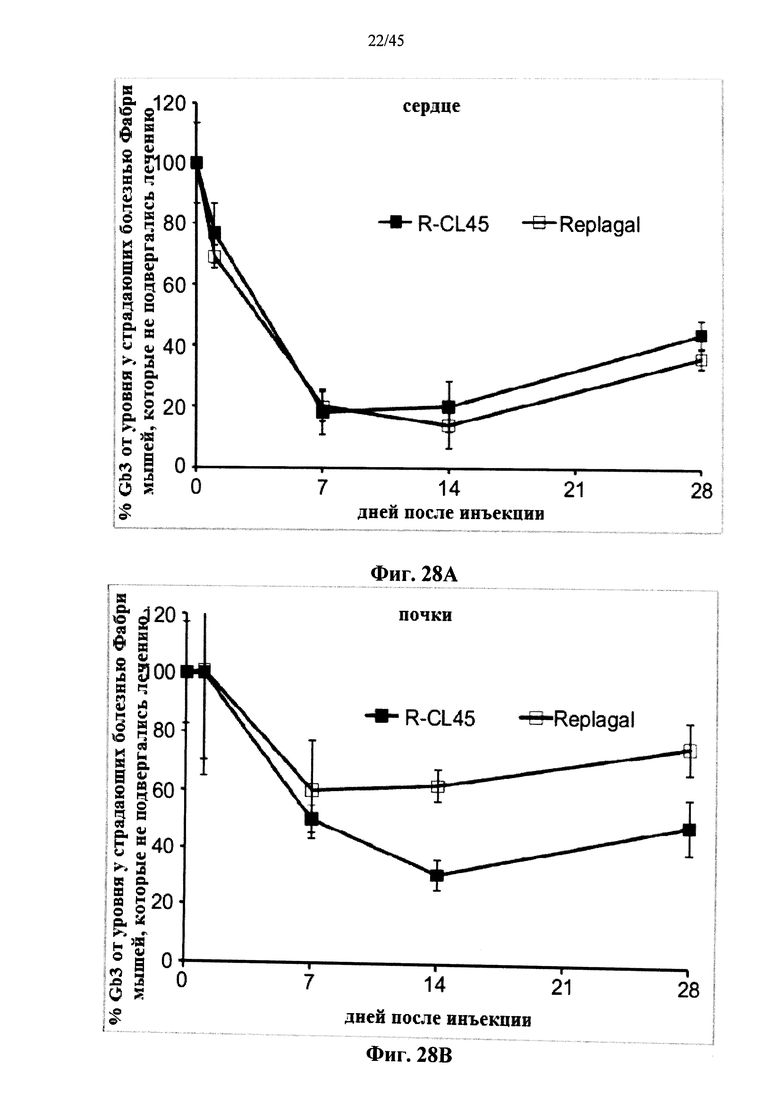
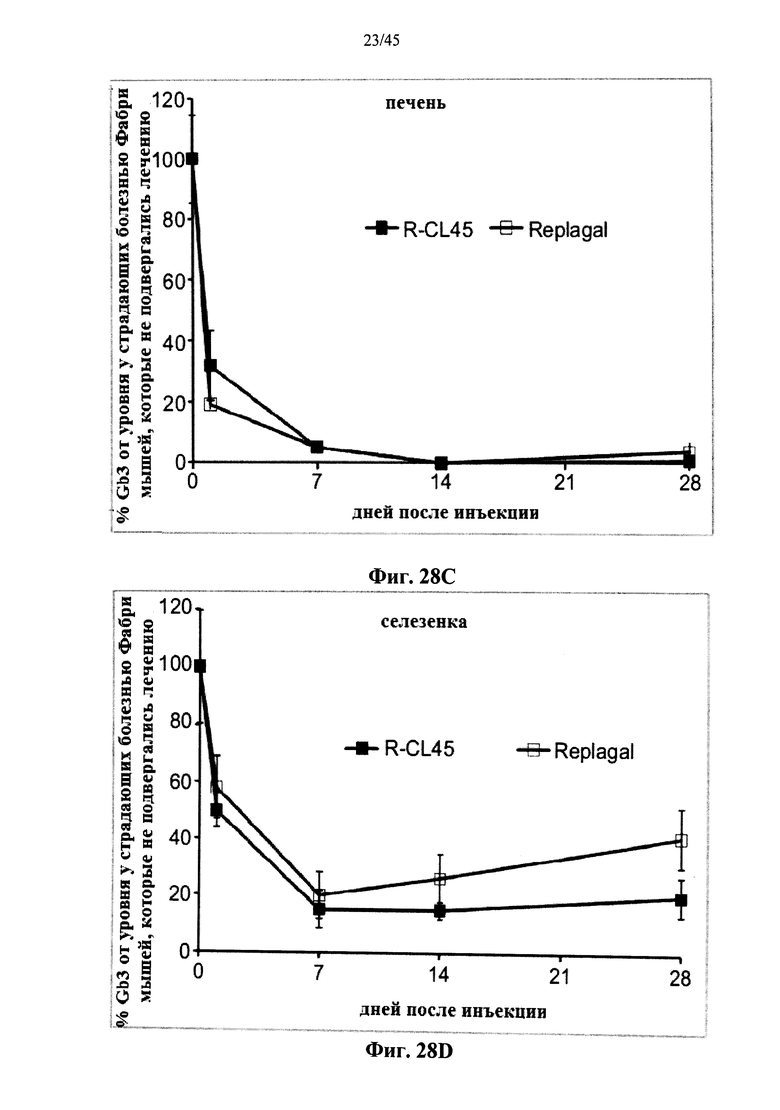
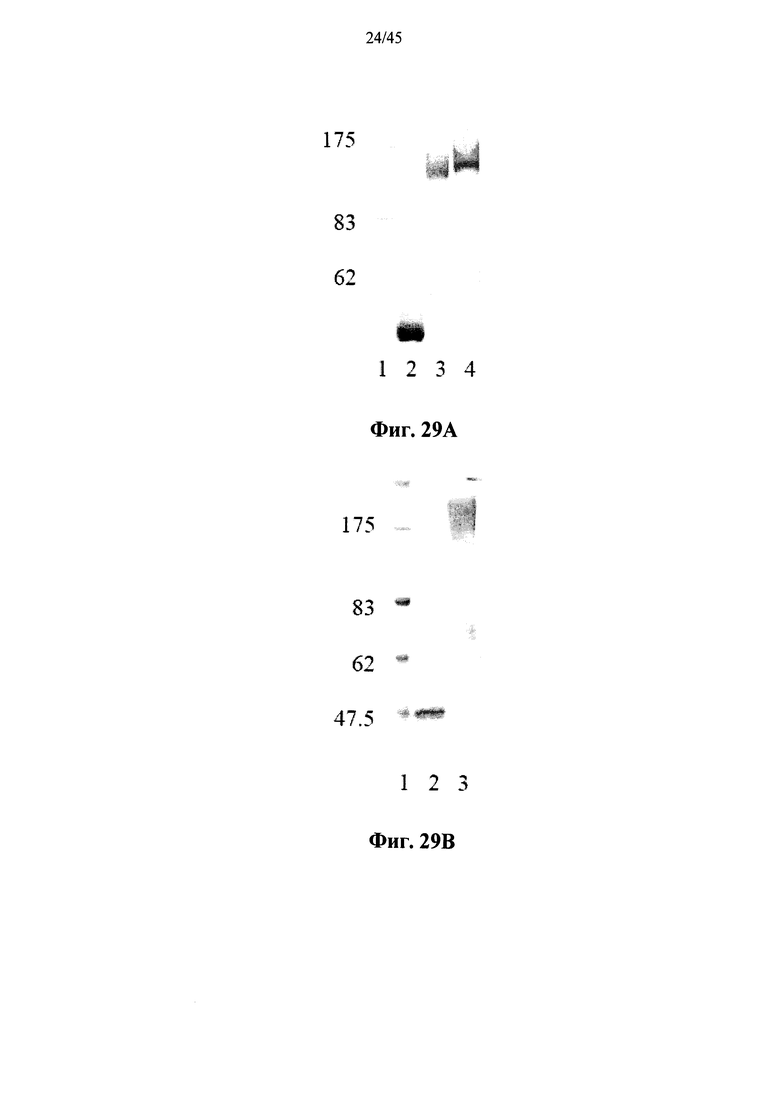
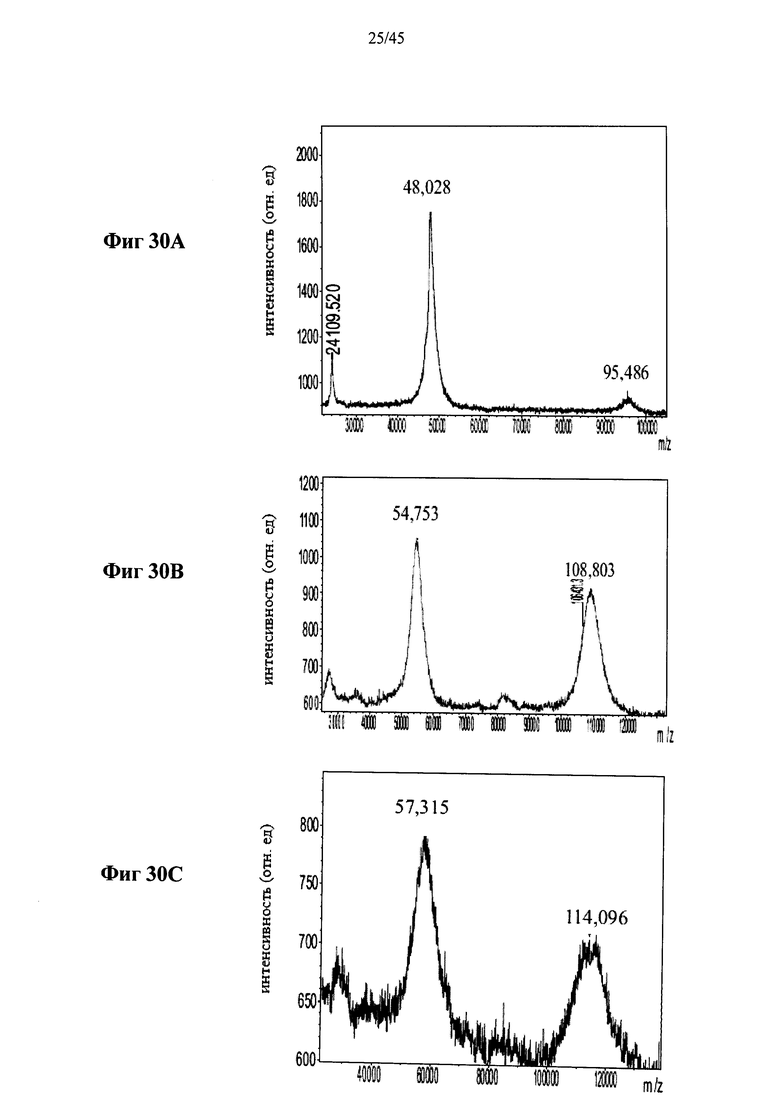
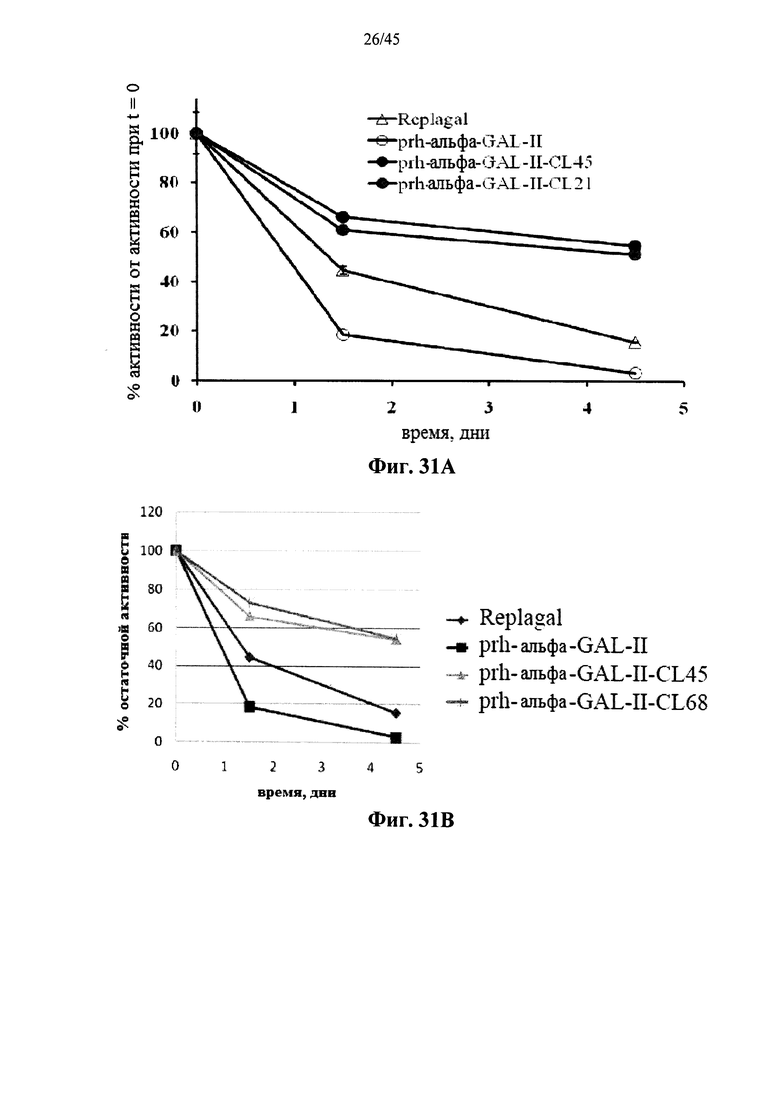
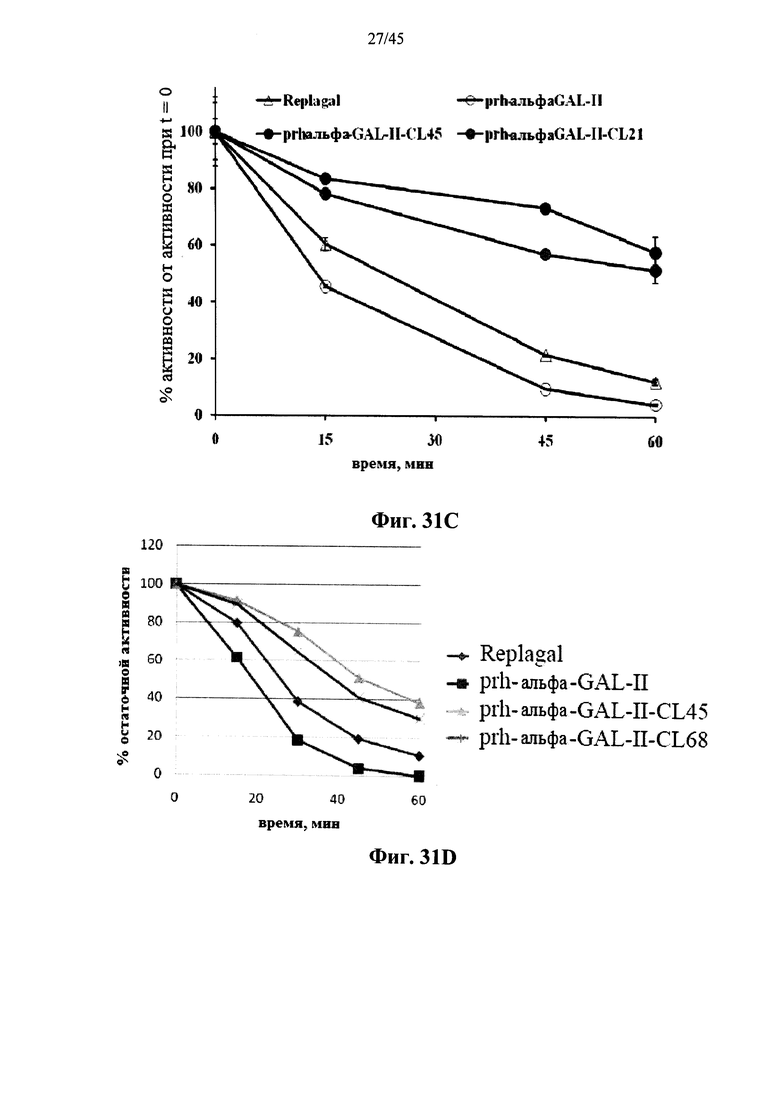
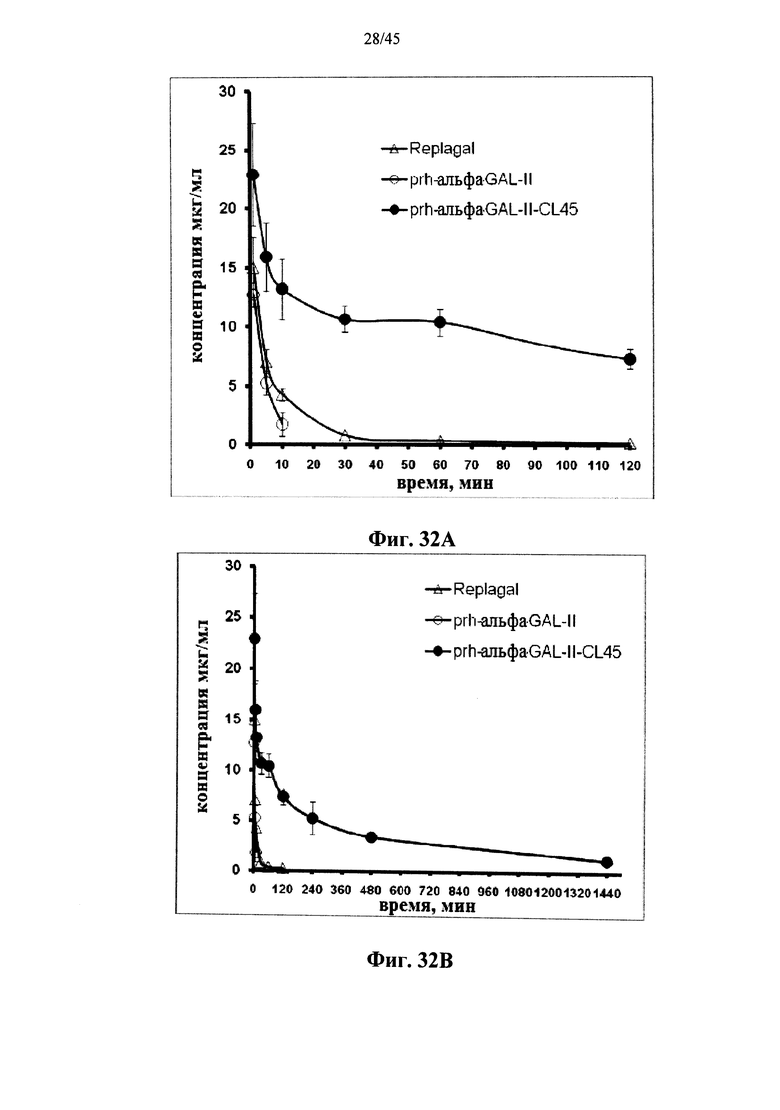
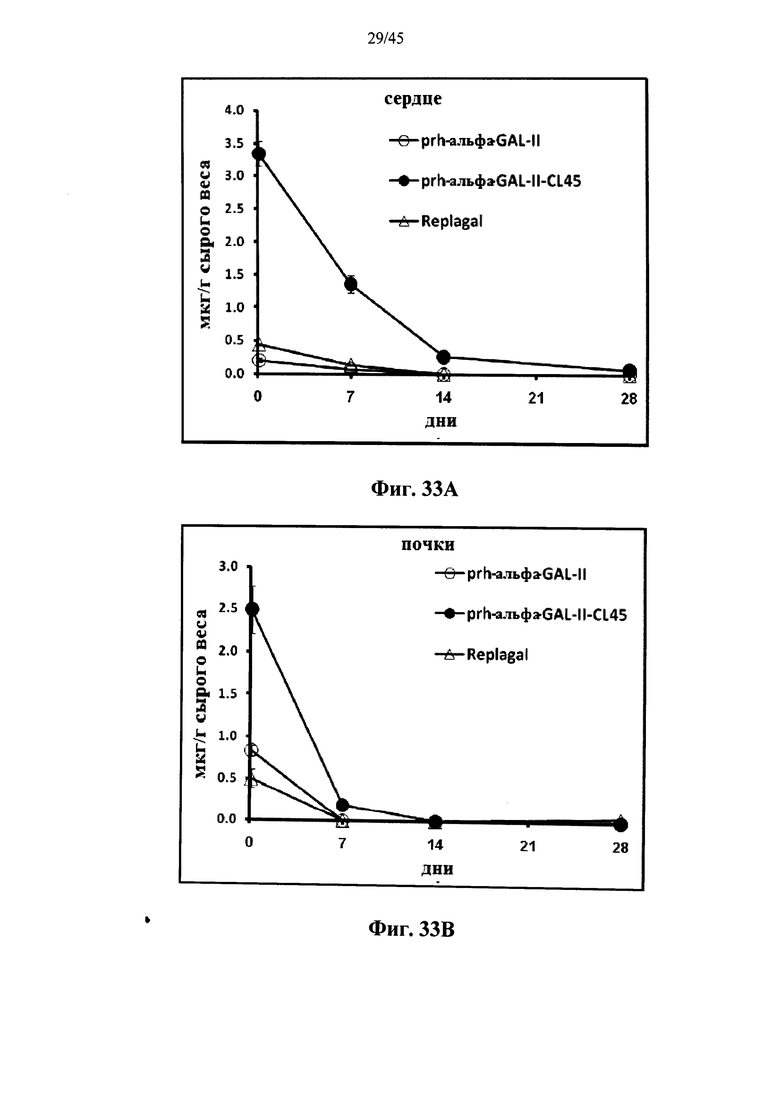
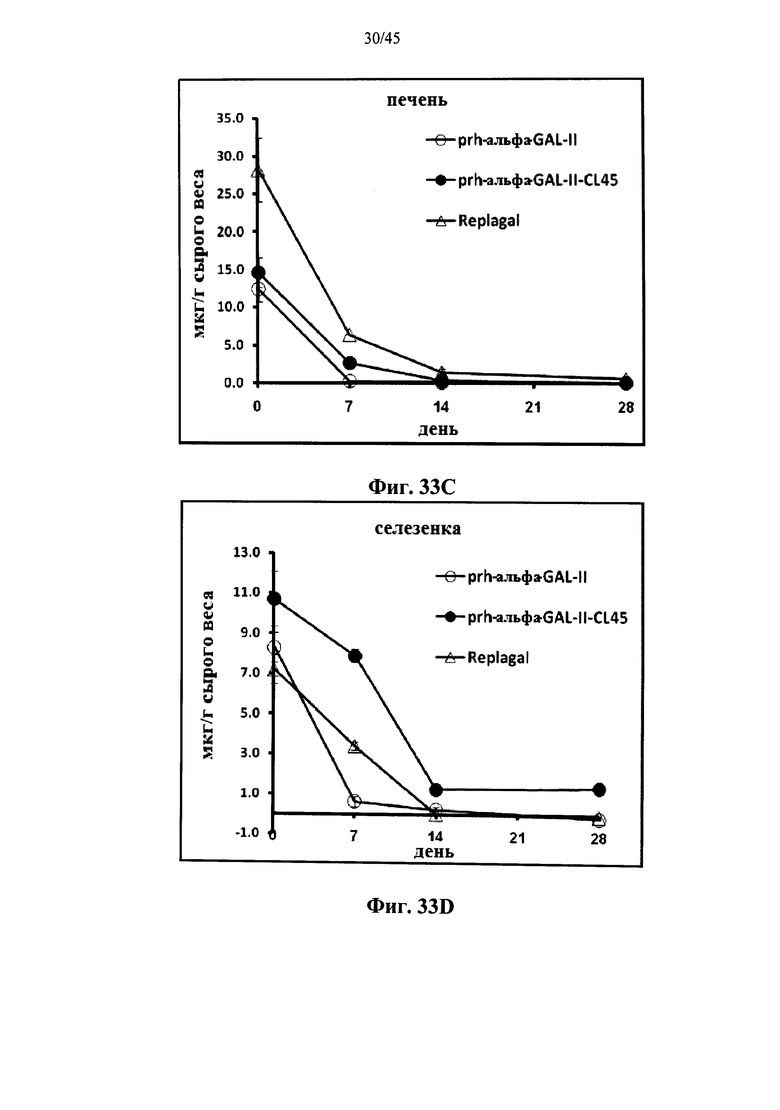
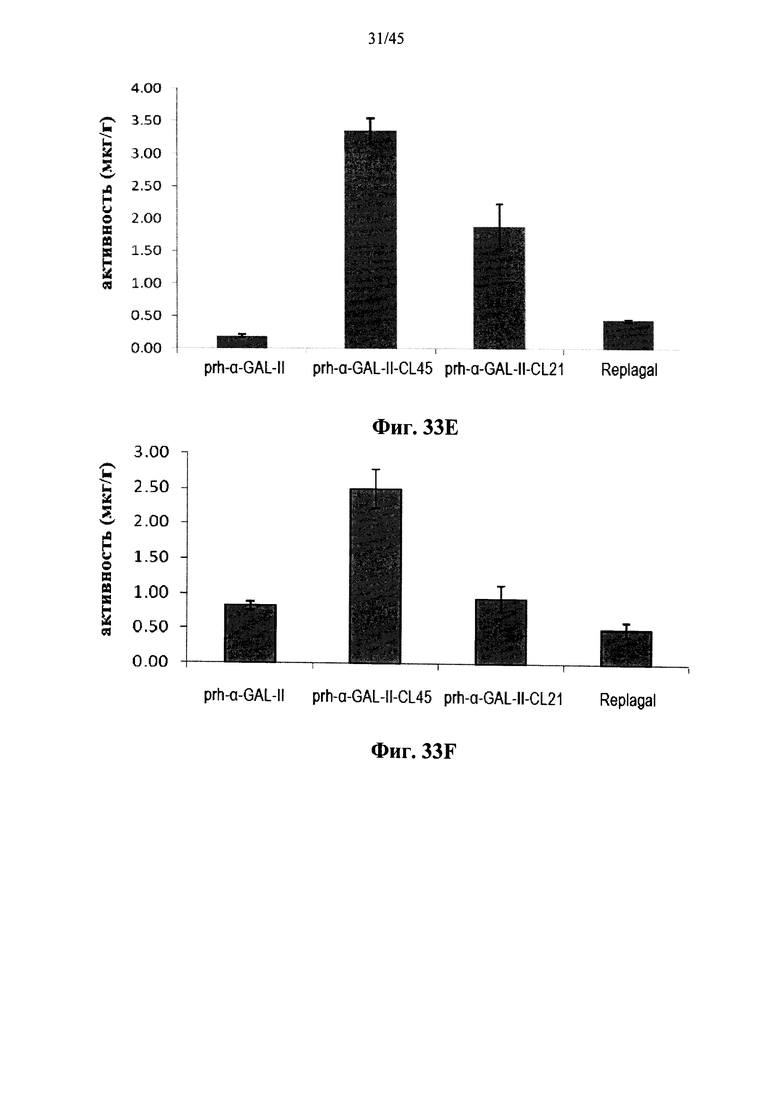
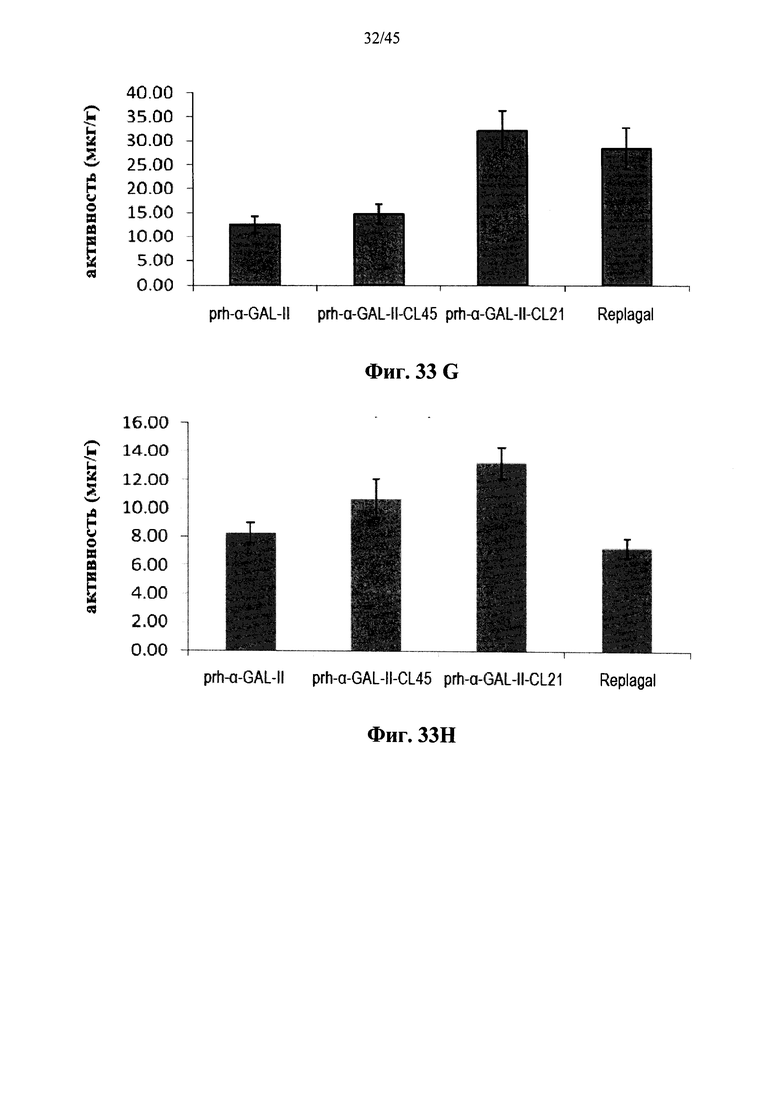
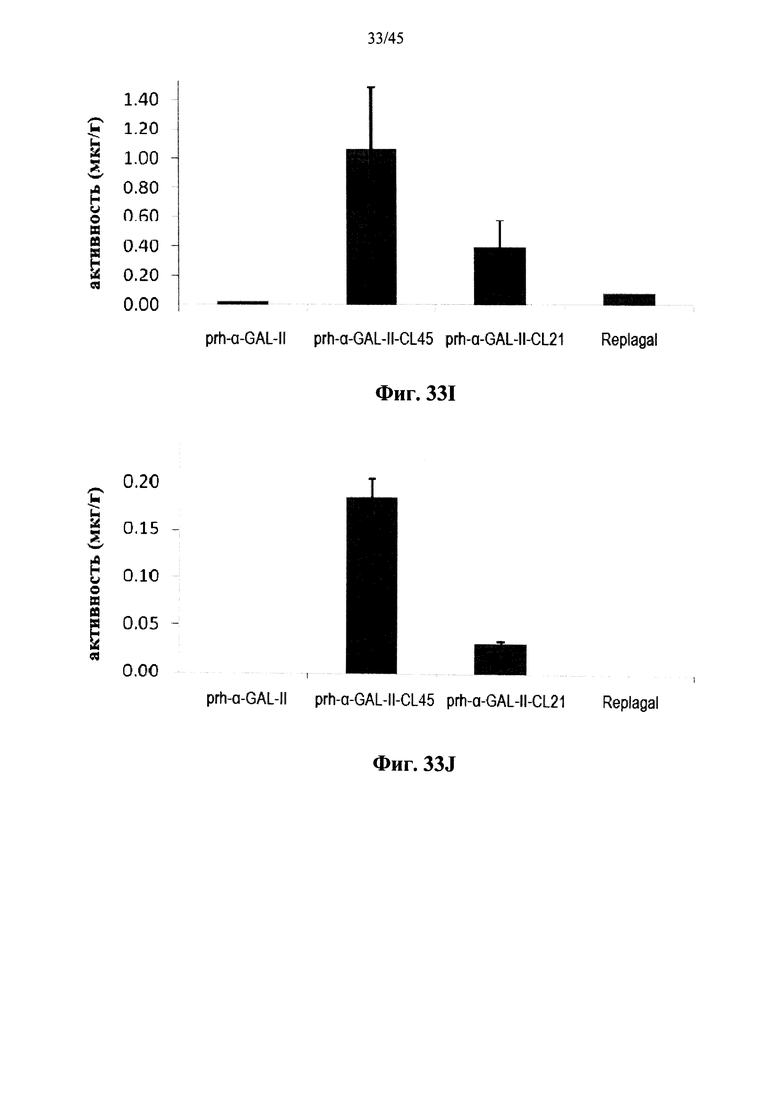
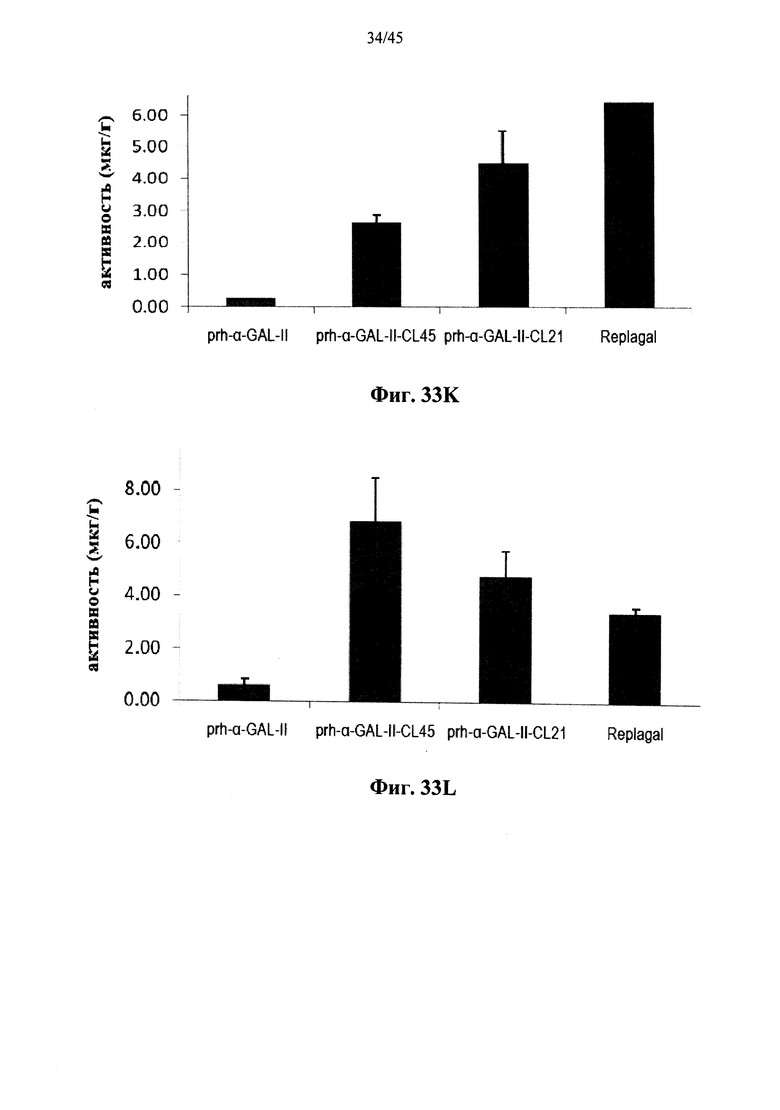
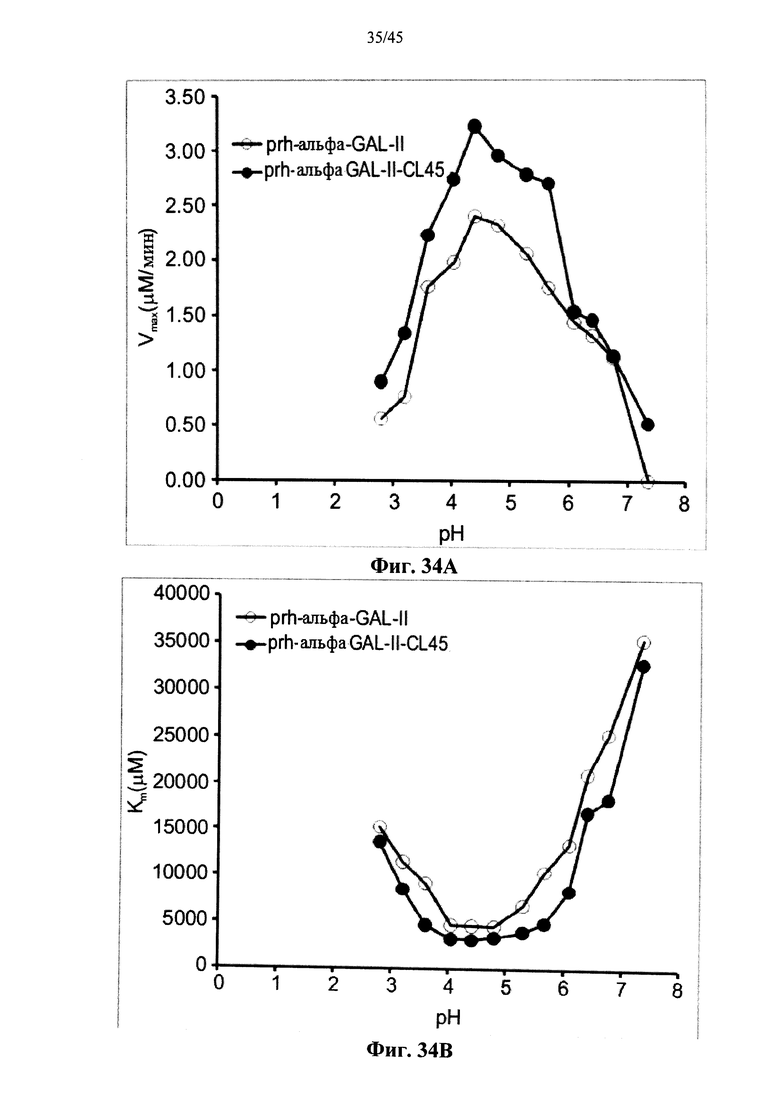
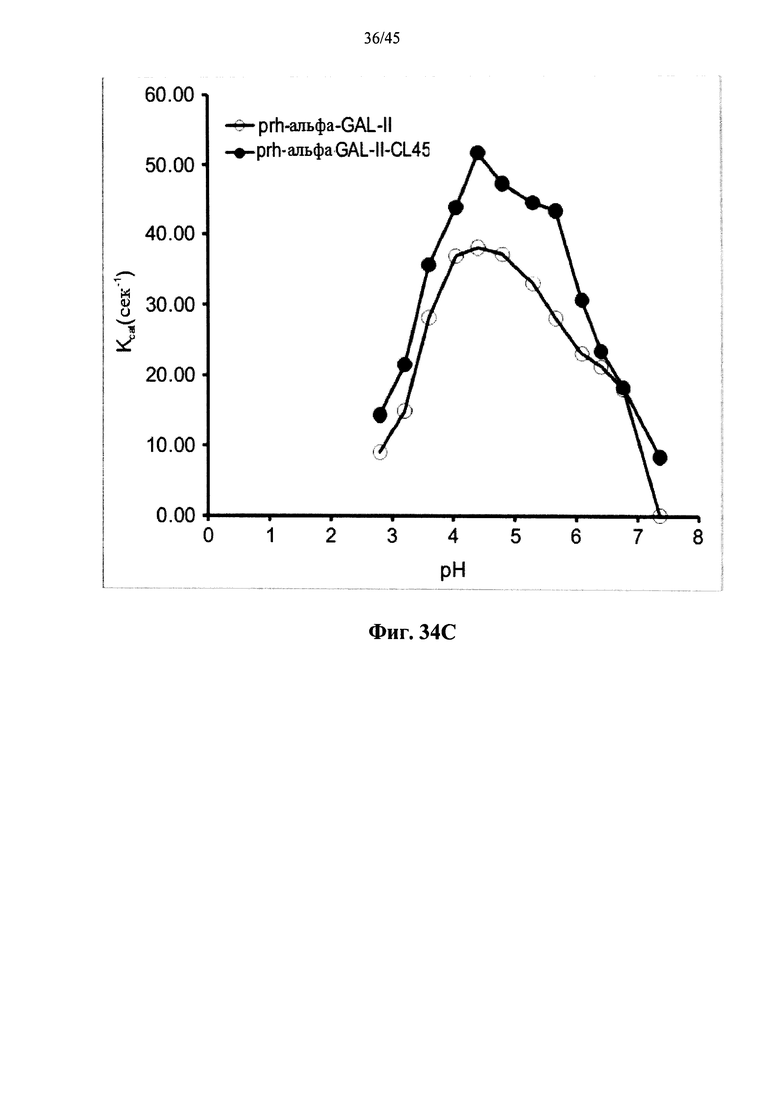
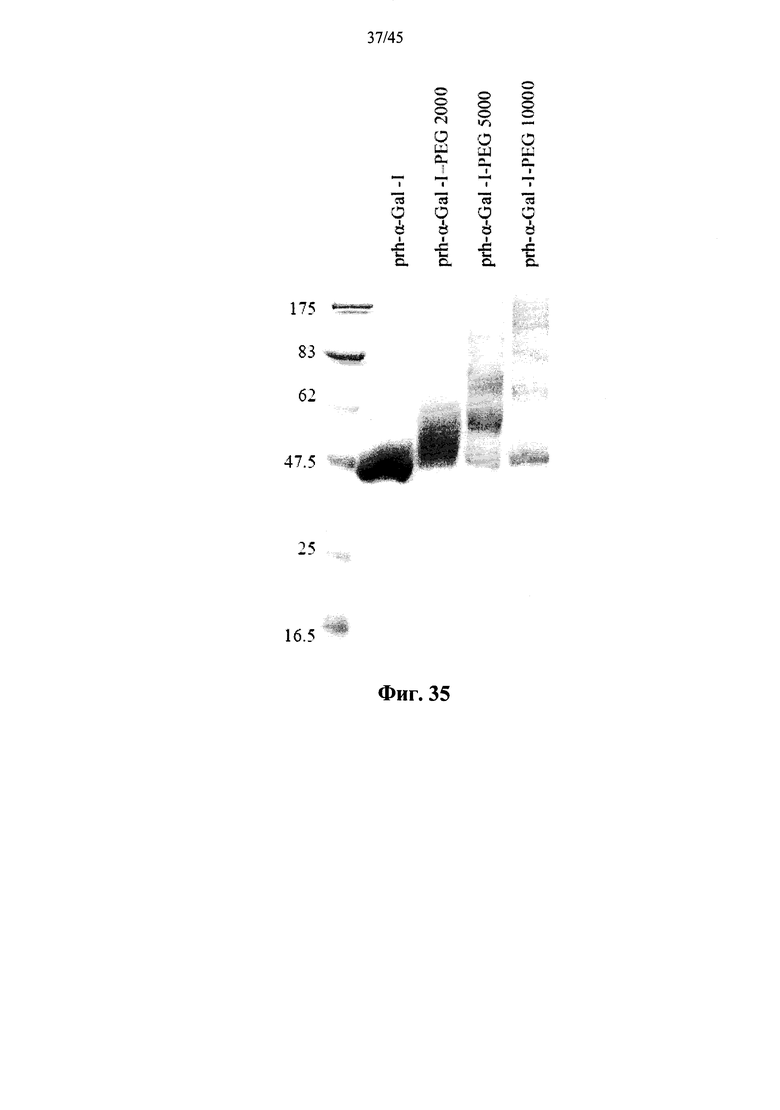
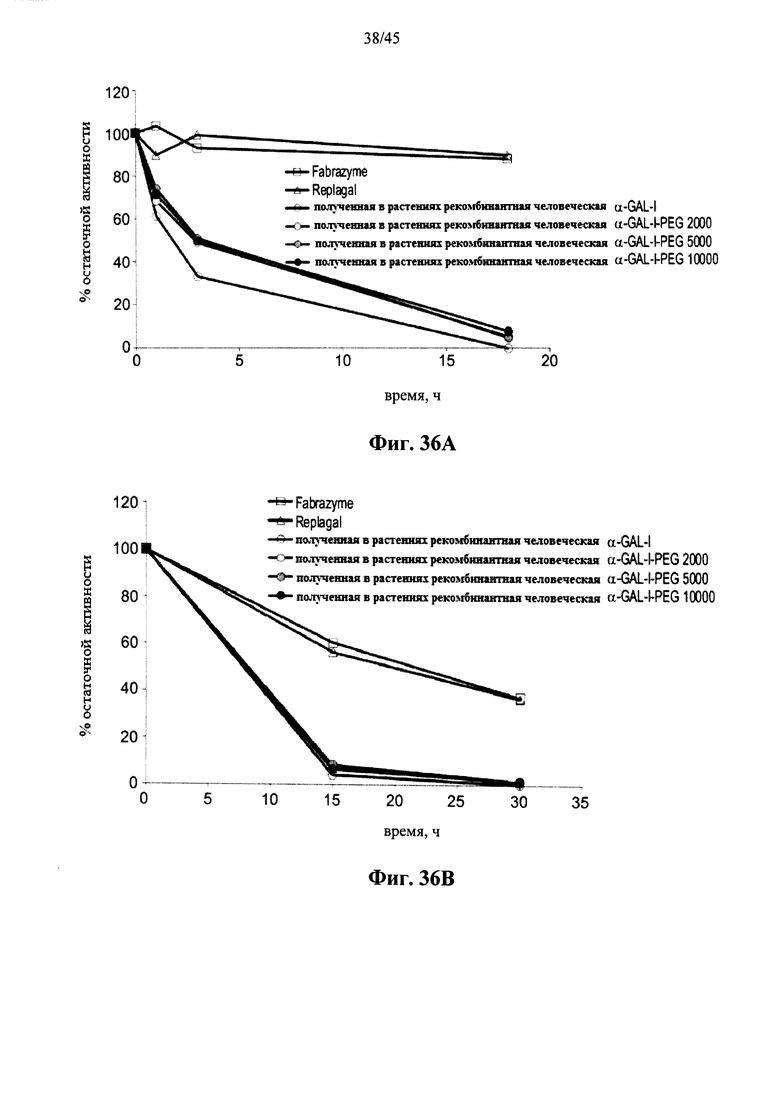
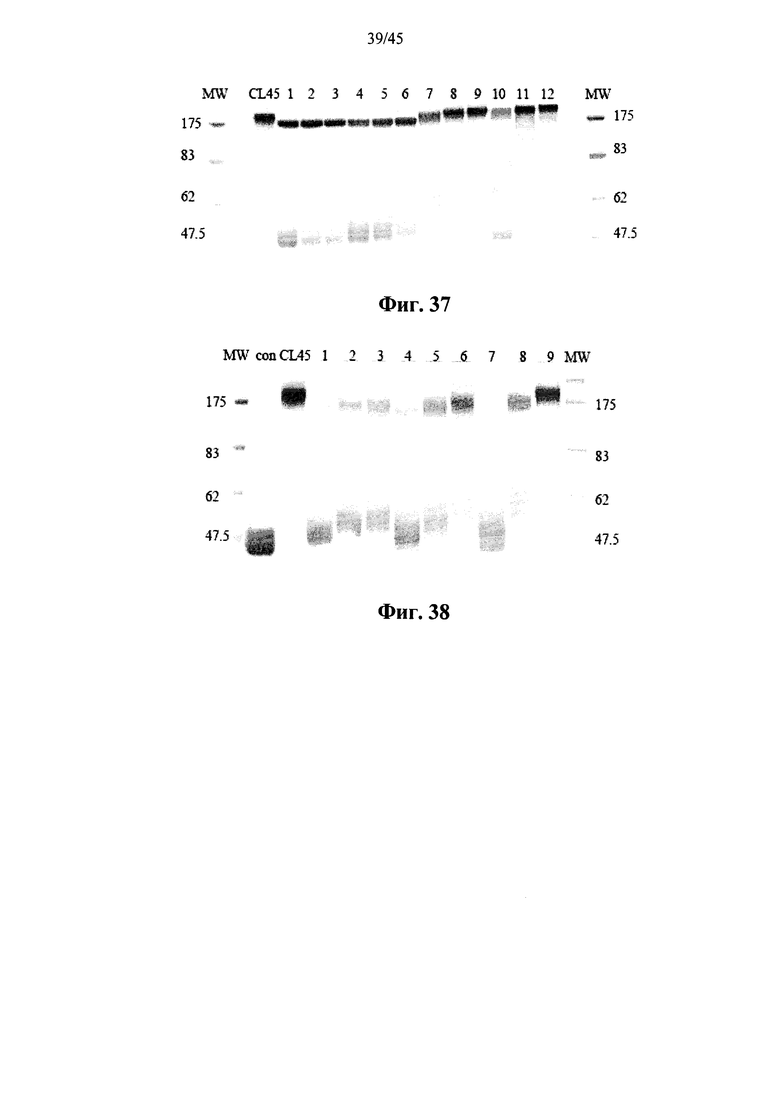
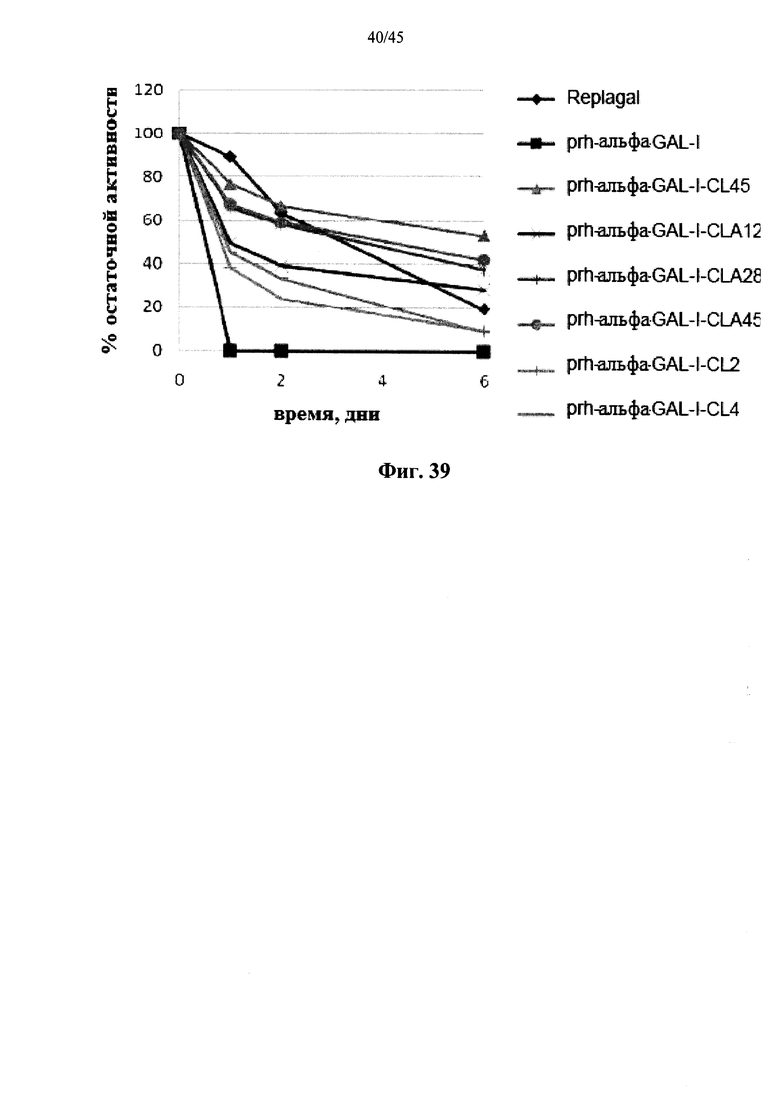
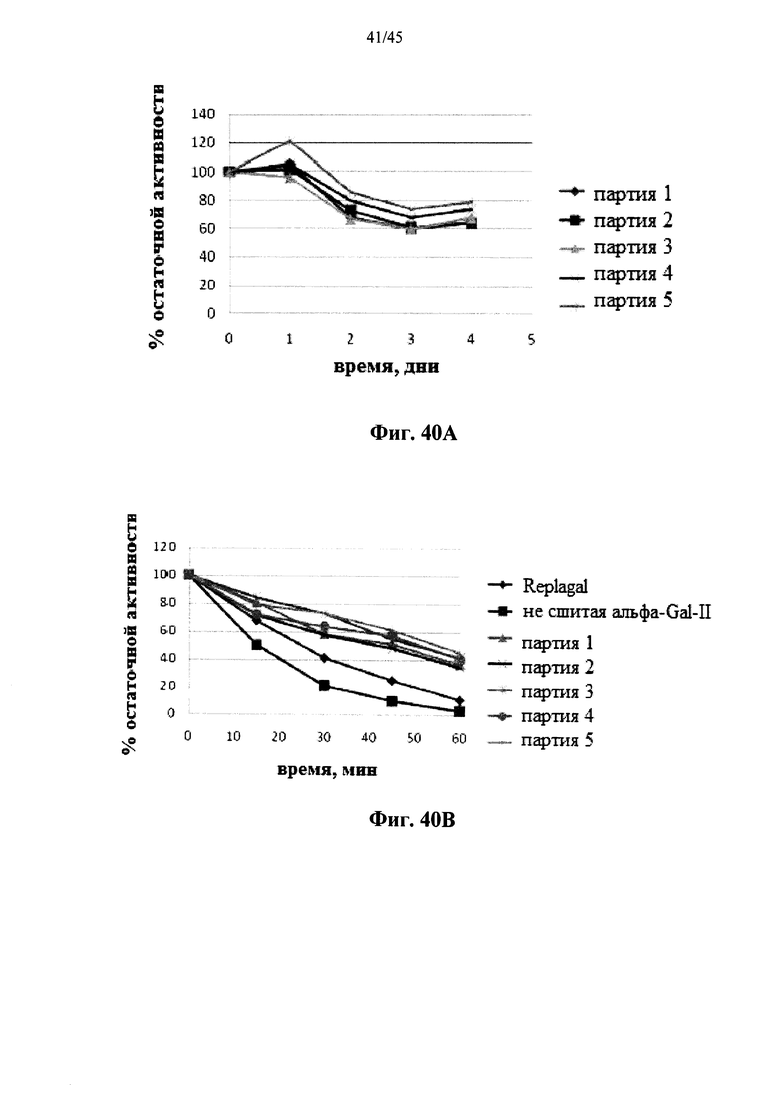
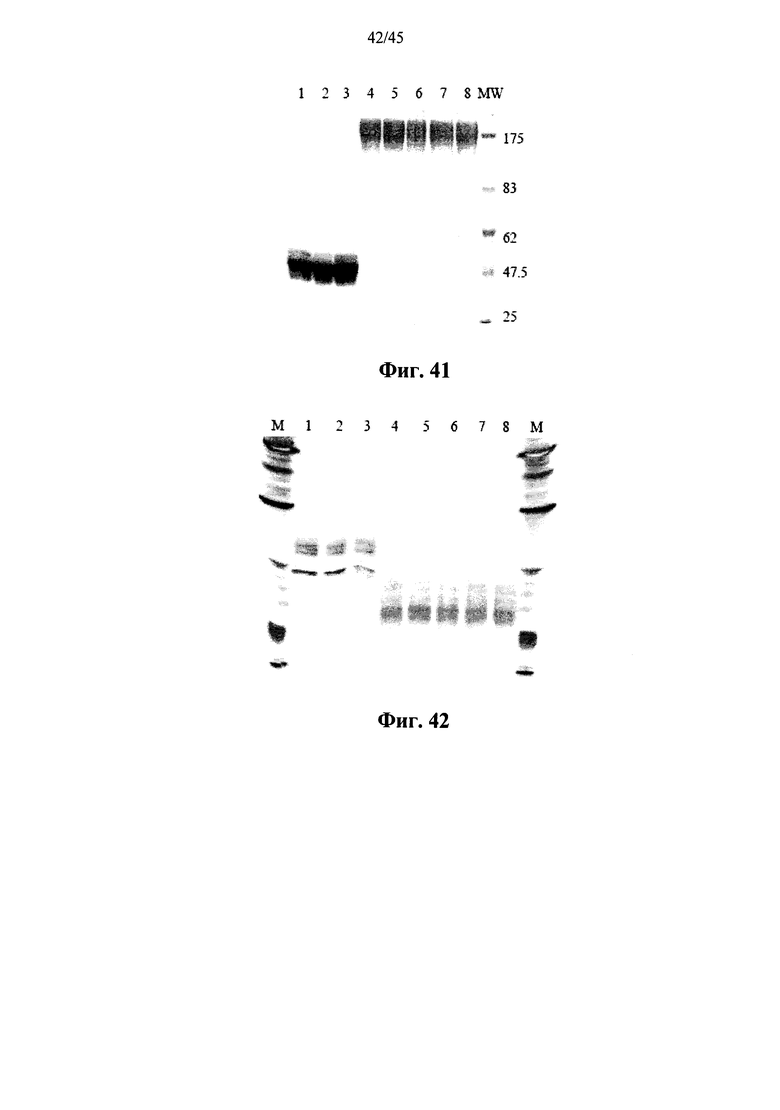
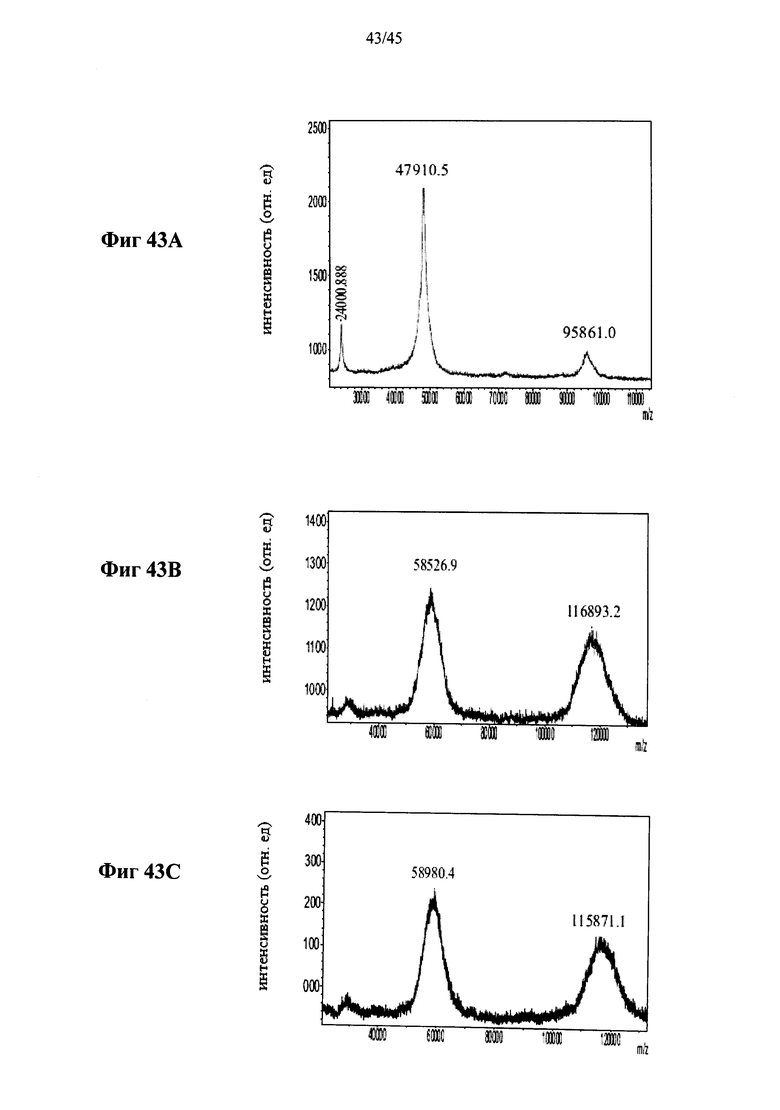
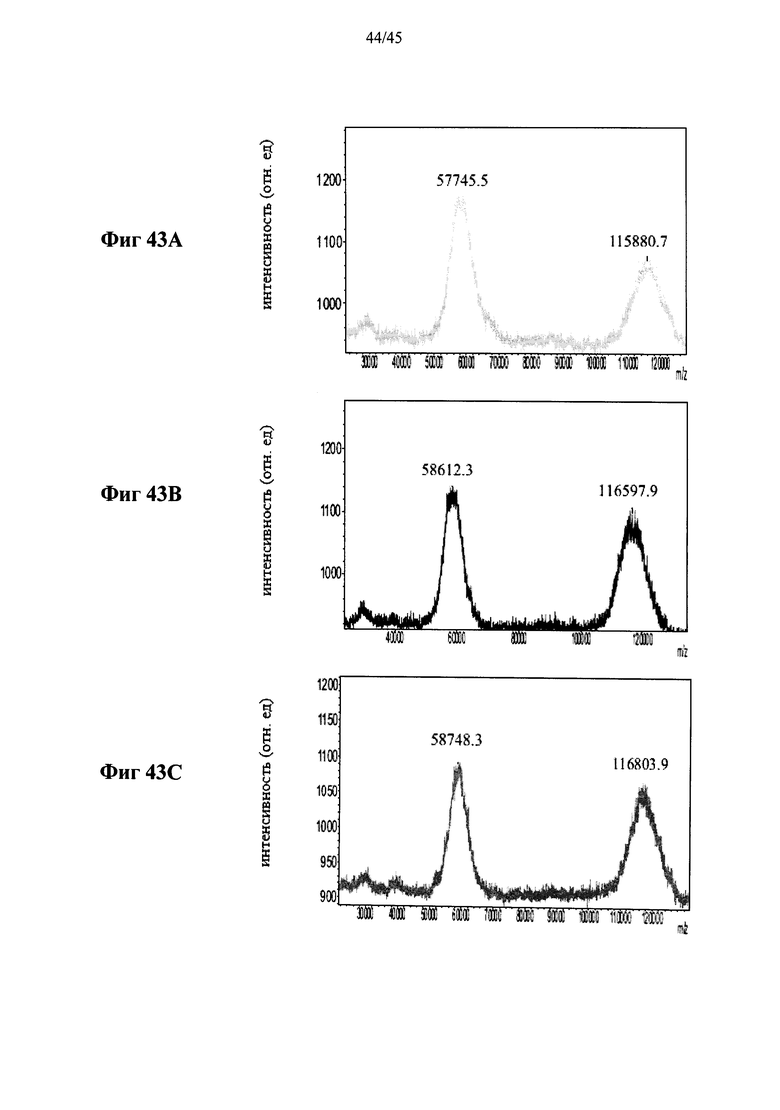
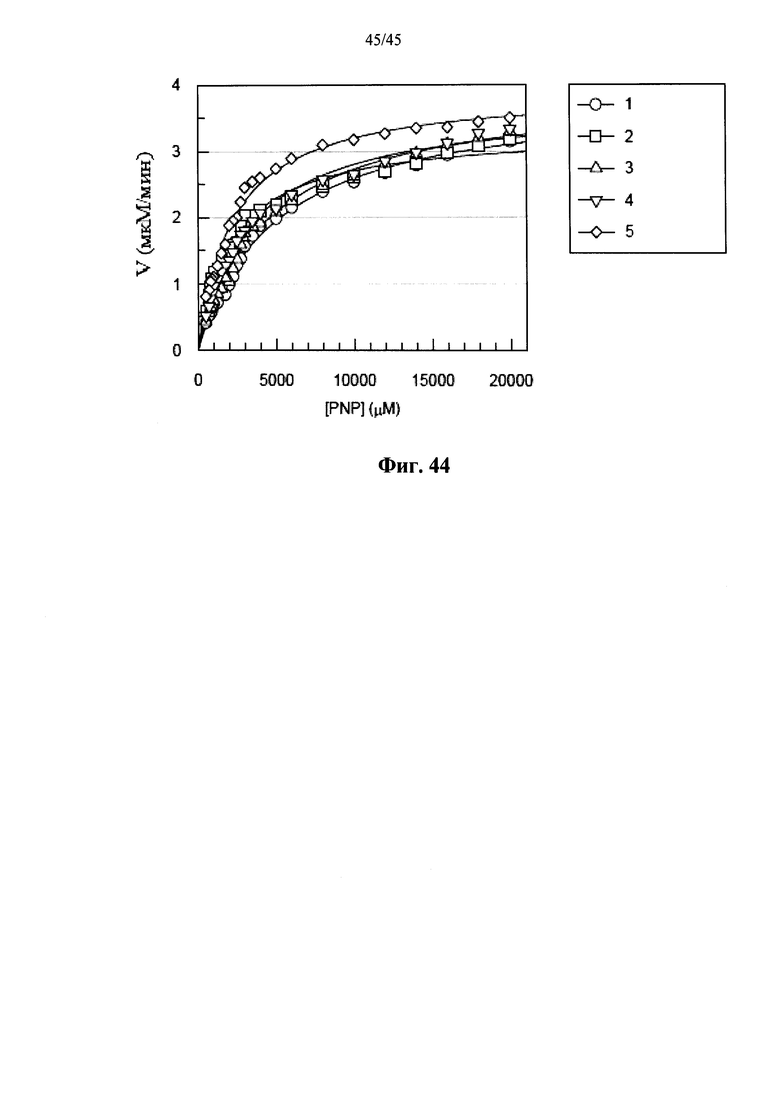
Группа изобретений относится к области биотехнологии. Представлены варианты мультимерной белковой структуры, которая гидролизует концевой α-галактозильный фрагмент глоботриаозилцерамида и содержит два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент, где мультимерная белковая структура представляет собой димерную белковую структуру. Представлены способы получения указанных вариантов мультимерных белковых структур, включающие взаимодействие α-галактозидазы со сшивающим агентом. Представлена фармецевтическая композиция, включающая указанную мультимерную белковую структуру. Группа изобретений позволяет получать мультимерные белковые структуры с более высокой стабильностью и повышенной активностью in vivo по сравнению с нативной формой α-галактозидазы, которые могут быть использованы при лечении болезни Фабри. 5 н. и 20 з.п. ф-лы, 83 ил., 10 табл., 19 пр.
1. Стабилизированная мультимерная белковая структура, которая гидролизует концевой α-галактозильный фрагмент глоботриаозилцерамида и содержит два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент, где мультимерная белковая структура представляет собой димерную белковую структуру, где указанный связывающий фрагмент имеет общую формулу
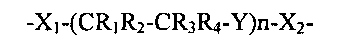
где каждый из X1 и X2 представляет собой функциональную группу, которая образует ковалентную связь с по меньшей мере одним мономером α-галактозидазы;
Y представляет собой О, S или NR5;
n представляет собой целое число от 1 до 200 и
каждый из R1, R2, R3, R4 и R5 независимо выбран из группы, состоящей из водорода, алкильной, циклоалкильной, алкенильной, алкинильной, алкоксильной, гидроксильной, оксо, тиольной и тиоалкоксильной групп.
2. Стабилизированная мультимерная белковая структура, которая гидролизует концевой α-галактозильный фрагмент глоботриаозилцерамида и содержит два мономера α-галактозидазы, ковалентно связанных друг с другом через связывающий фрагмент, белковая структура представляет собой димерную белковую структуру, где связывающий фрагмент отсутствует в нативной α-галактозидазе, содержит поли(алкиленгликоль) и составляет по меньшей мере 20 атомов в длину.
3. Мультимерная белковая структура по п. 2, отличающаяся характеристиками, выбранными из группы, состоящей из:
(а) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в условия человеческой плазмы на один час, которая по меньшей мере на 10% выше, чем активность нативной α-галактозидазы при помещении указанной нативной α-галактозидазы в указанные условия человеческой плазмы на один час;
(b) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в условия человеческой плазмы на один час на процентную величину, которая по меньшей мере на 10% меньше, чем процентная величина, на которую уменьшается активность указанной нативной α-галактозидазы при помещении указанной нативной α-галактозидазы в указанные условия человеческой плазмы на один час;
(c) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в условия человеческой плазмы, на один час;
(d) проявления α-галактозидазной активности при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю, которая по меньшей мере на 10% выше, чем активность нативной α-галактозидазы при помещении указанной нативной α-галактозидазы в указанные лизосомальные условия на одну неделю;
(e) проявления α-галактозидазной активности, которая уменьшается при помещении мультимерной белковой структуры в лизосомальные условия на один день на процентную величину, которая по меньшей мере на 10% меньше, чем процентная величина, на которую уменьшается активность указанной нативной α-галактозидазы при помещении указанной нативной α-галактозидазы в указанные лизосомальные условия на один день;
(f) проявления α-галактозидазной активности, которая остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день;
(g) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в лизосомальные условия, которая по меньшей мере на 10% больше, чем активность нативной α-галактозидазы немедленно после помещения указанной нативной α-галактозидазы в указанные лизосомальные условия;
(h) проявления α-галактозидазной активности немедленно после помещения мультимерной белковой структуры в водный раствор, имеющий рН 7 и температуру 37°C, которая по меньшей мере на 10% выше, чем активность нативной α-галактозидазы немедленно после помещения указанной нативной α-галактозидазы в указанный водный раствор, имеющий рН 7 и температуру 37°C; и
(i) периода полувыведения из крови в физиологической системе, большего, чем период полувыведения из крови указанной нативной α-галактозидазы.
4. Мультимерная белковая структура по п. 3, где указанный период полувыведения из крови мультимерной белковой структуры больше, чем период полувыведения из крови указанной нативной α-галактозидазы и превосходит указанный период полувыведения из крови указанной нативной α-галактозидазы по меньшей мере на 20%.
5. Мультимерная белковая структура по любому из пп. 3 или 4, где указанная α-галактозидазная активность указанной мультимерной белковой структуры, которая остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на один день, также остается практически неизменной при помещении мультимерной белковой структуры в лизосомальные условия на одну неделю.
6. Мультимерная белковая структура по любому из пп. 1-4, характеризующаяся α-галактозидазной активностью в органе при введении указанной мультимерной белковой структуры позвоночному животному, где указанный орган выбран из группы, состоящей из селезенки, сердца и почек.
7. Мультимерная белковая структура по любому из пп. 1-4, где указанная α-галактозидаза является человеческой α-галактозидазой.
8. Мультимерная белковая структура по п. 7, где указанная α-галактозидаза является человеческой α-галактозидазой А.
9. Мультимерная белковая структура по любому из пп. 1-4, 8, где указанная α-галактозидаза является рекомбинантной α-галактозидазой.
10. Мультимерная белковая структура по любому из пп. 1-4, 8, где указанная α-галактозидаза является растительной рекомбинантной α-галактозидазой.
11. Мультимерная белковая структура по п. 7, где указанная человеческая α-галактозидаза выбирается из группы, состоящей из агалзидазы альфа и агалзидазы бета.
12. Мультимерная белковая структура по любому из пп. 1-4, 8, где указанная α-галактозидаза имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3.
13. Мультимерная белковая структура по п. 2, где указанный поли(алкиленгликоль) содержит по меньшей мере две функциональные группы, где каждая функциональная группа образует ковалентную связь с одним из мономеров α-галактозидазы.
14. Мультимерная белковая структура по п. 1, где n представляет собой целое число от 8 до 200.
15. Мультимерная белковая структура по любому из пп. 1 и 13, где по меньшей мере одна из указанных функциональных групп образует амидную связь с мономером α-галактозидазы.
16. Мультимерная белковая структура по п. 1, где n представляет собой по меньшей мере 25.
17. Мультимерная белковая структура по п. 14, где указанная α-галактозидаза имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, причем каждая из функциональных групп образует амидную связь с мономером α-галактозидазы, и n представляет собой целое число от 40 до 70.
18. Мультимерная белковая структура по п. 12, где указанный связывающий фрагмент имеет формулу

причем молекулярная масса полиэтиленгликоля в указанном связывающем фрагменте составляет 2 кДа, а каждая концевая группа указанного связывающего фрагмента образует амидную связь с мономером α-галактозидазы.
19. Мультимерная белковая структура по любому из пп. 1-4, 8, 11, 13, 17 и 18 для применения для лечения болезни Фабри.
20. Способ получения мультимерной белковой структуры по любому из пп. 1, 6-12 и 14-18, включающий взаимодействие α-галактозидазы со сшивающим агентом, который включает указанный связывающий фрагмент, имеющий указанную общую структуру
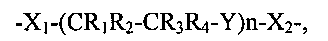
и по меньшей мере две реакционноспособные группы.
21. Способ получения мультимерной белковой структуры по любому из пп. 2-5 и 13, включающий взаимодействие α-галактозидазы со сшивающим агентом, который включает указанный связывающий фрагмент, содержащий поли(алкиленгликоль), составляющий по меньшей мере 20 атомов в длину и отсутствующий в нативной α-галактозидазе, и по меньшей мере две реакционноспособные группы.
22. Способ по п. 20 или 21, где молярное соотношение указанного сшивающего агента к мономерам указанной α-галактозидазы находится в диапазоне от 5:1 до 500:1.
23. Фармацевтическая композиция для лечения болезни Фабри, включающая мультимерную белковую структуру по любому из пп. 1-18 и фармацевтически приемлемый носитель.
24. Фармацевтическая композиция по п. 23, дополнительно содержащая галактозу.
25. Фармацевтическая композиция по любому из пп. 23 и 24, где указанный фармацевтически приемлемый носитель представляет собой водный раствор, содержащий физиологический раствор и физиологически совместимый буфер, и указанная мультимерная белковая структура находится в водорастворимой форме.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| КОНЪЮГАТЫ PEG-УРИКАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 1999 |
|
RU2246318C2 |

