ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] В настоящей заявке испрашивается приоритет по предварительной заявке США № 62/788 439, поданной 4 января 2019 г., описание которой в полном объеме включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит перечень последовательностей, который был подан в электронной форме в формате ASCII и в полном объеме включен в данный документ посредством ссылки. Указанная ASCII-копия, созданная 3 декабря 2019 г., называется 8325018840SL.txt. и имеет размер 10 636 байт.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение относится к области профилактики и/или лечения болезни Фабри с помощью генной терапии.
УРОВЕНЬ ТЕХНИКИ
[0004] Ген α-галактозидазы A (GLA) кодирует фермент лизосомальной гидролазы, α-галактозидазу A (α-Gal A). α-Галактозидаза представляет собой фермент, который катализирует гидролиз концевых α-галактозильных групп олигосахаридов и полисахаридов.
[0005] Болезнь Фабри представляет собой Х-связанную лизосомную болезнь накопления, вызванную мутациями в гене GLA. Недостаток активности α-Gal A приводит к постепенному систематическому накоплению ее первичного субстрата, глоботриаозилцерамида (Gb3) и ее деацетилированной растворимой формы, глоботриаозилсфингозина (лизо-Gb3). Долгосрочное накопление этих субстратов приводит к заболеванию почек, нарушениям со стороны кожи, заболеваниям сердца, дистрофии роговицы (например, помутнение роговицы и хрусталика) и/или цереброваскулярным заболеваниям с сокращением продолжительности жизни. В зависимости от мутации и остаточного уровня фермента α-Gal A заболевание проявляется как классическая болезнь Фабри с ранним дебютом в детстве/подростковом возрасте или как аттенуированная (взрослая) форма в более позднем возрасте. Классическая болезнь Фабри возникает, когда остаточная активность ферментов составляет <5% (Arends et al., 2017), и обычно встречается у мужчин. Ранние симптомы могут включать периодическую акропарестезию, ангиокератомы, помутнение роговицы и хрусталика, прогрессирующую почечную недостаточность, заболевания сердца и цереброваскулярные нарушения. Аттенуированная или взрослая форма болезни Фабри обычно затрагивает только одну систему органов, обычно сердечную или почечную.
[0006] Как для классической, так и для взрослой формы, существующим стандартом лечения является заместительная ферментная терапия (enzyme replacement therapy, ERT) с применением рекомбинантного α-Gal A, ФАБРАЗИМ (FABRAZYME®) (агалзидаза бета или эквивалент) или шапероновая терапия, которая доступна только для пациентов, чьи мутации поддаются такому лечению. Инфузия рекомбинантного α-Gal A в кровоток обеспечивает перенос его во вторичные ткани посредством поглощения, опосредованного маннозо-6-фосфатным рецептором (перекрестная коррекция). Однако короткий период полужизни рекомбинантного α-Gal A, применяемого в ERT (приблизительно 1 час в плазме) (Clarke et al. 2007), требует продолжительности инфузий, что сопряжено с риском связанных с инфузией реакций у значительной части пациентов (Clarke et al. 2007), некоторые из которых имеют тяжелую форму заболевания. Кроме того, значительный процент пациентов в конечном итоге вырабатывает антитела к рекомбинантному ферменту, которые могут влиять на активность фермента ERT, который, следовательно, может не выводить весь субстрат из таких органов, как почки (Linthorst et al. 2004).
[0007] Сейчас разрабатываются рекомбинантные продукты α-Gal A с более длительным периодом полужизни, которые можно вводить реже. Однако ожидается, что они по-прежнему потребуют длительного введения с сопутствующим риском связанных с инфузией реакций и/или неактивности из-за нейтрализующих антител, и что уровни α-Gal A все еще будут значительно колебаться с течением времени.
[0008] Таким образом, существует потребность в альтернативных методах лечения, направленных на решение неудовлетворенных потребностей при болезни Фабри.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0009] В данном документе описан способ экспрессии в клетке по меньшей мере одного белка α-галактозидазы A (α-Gal A). В некоторых вариантах осуществления указанный способ включает введение экспрессионной конструкции, содержащей мутированную последовательность WPRE, необязательно мутированную последовательность WRPE с мутацией mut6 и трансген GLA, кодирующий по меньшей мере один белок α-Gal A, в клетку таким образом, чтобы белок α-Gal A экспрессировался в клетке.
[0010] В некоторых вариантах осуществления экспрессионная конструкция содержит последовательность GLA дикого типа или кодон-оптимизированную последовательность GLA.
[0011] В некоторых вариантах осуществления экспрессионная конструкция включает один или большее количество из следующих элементов: энхансер, промотор, интрон, последовательность, кодирующую сигнальный пептид, и/или сигнал полиаденилирования, при этом мутированная последовательность WPRE, необязательно мутированная последовательность WRPE с мутацией mut6, и трансген GLA, кодирующий по меньшей мере один белок α-Gal A, расположен между сигнальным пептидом и последовательностью, кодирующей сигнал полиаденилирования.
[0012] В некоторых вариантах осуществления экспрессионная конструкция содержит последовательность SEQ ID №: 9.
[0013] В некоторых вариантах осуществления клетка находится в организме субъекта с болезнью Фабри.
[0014] В некоторых вариантах осуществления клетка находится в организме субъекта мужского пола.
[0015] В некоторых вариантах осуществления экспрессионную конструкцию вводят в фармацевтически приемлемом носителе.
[0016] В некоторых вариантах осуществления фармацевтически приемлемый носитель включает фосфатно-солевой буфер, содержащий CaCl2, MgCl2, NaCl, сахарозу и коллифор (полоксамер) P 188.
[0017] В некоторых вариантах осуществления последовательность экспрессионной конструкции включает последовательность, приведенную в Таблице 1, и при этом указанная экспрессионная конструкция доставляется в клетку вирусным вектором AAV.
[0018] В некоторых вариантах осуществления серотип указанного вирусного вектора AAV представляет собой AAV2/6.
[0019] В некоторых вариантах осуществления экспрессионную конструкцию вводят субъекту в дозе от около 5,0E+12 до 1,0E+14 векторных геномов на килограмм (вг/кг).
[0020] В некоторых вариантах осуществления экспрессионную конструкцию вводят в печень субъекта. В других вариантах осуществления экспрессионный вектор вводят субъекту посредством внутривенной инфузии. В других вариантах осуществления субъекту вводят только одну дозу экспрессионной конструкции.
[0021] В некоторых вариантах осуществления субъекту вводят иммунодепрессант до и/или во время введения экспрессионной конструкции. В некоторых вариантах осуществления указанный иммунодепрессант включает преднизон.
[0022] В некоторых вариантах осуществления экспрессия по меньшей мере одного белка α-галактозидазы A (α-Gal A) сохраняется в течение по меньшей мере 3 месяцев, по меньшей мере 9 месяцев или по меньшей мере 12 месяцев.
[0023] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере от около 2 до около 9 раз по сравнению с субъектами, не получавшими лечение.
[0024] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере на 80% по сравнению с субъектами, не получавшими лечение.
[0025] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта в одном или более из следующего: плазма, печень, сердце, почки или селезенка.
[0026] В некоторых вариантах осуществления экспрессионная конструкция, полученная в клеточной системе HEK293, обеспечивает уровни GLA у субъекта примерно в 21 раз выше по сравнению с уровнями GLA у субъектов, которым вводили экспрессионную конструкцию, полученную в клеточной системе Sf9.
[0027] В некоторых вариантах осуществления активность белка α-Gal A у субъекта примерно в 100-1500 раз выше, чем у физиологического нормального/дикого типа.
[0028] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый трансгеном, активен в почках, печени и сердце субъекта.
[0029] В некоторых вариантах осуществления трансген GLA поддерживается вне хромосом и не интегрируется в геном клетки.
[0030] В некоторых вариантах осуществления одну или большее количество нуклеаз, которые расщепляют эндогенный ген альбумина в клетке печени субъекта, вводят таким образом, что трансген интегрируется в ген альбумина и экспрессируется из него.
[0031] Предлагаются генетически модифицированные клетки, содержащие экзогенный трансген GLA, полученные посредством описанных в данном документе способов. В некоторых вариантах осуществления указанная клетка представляет собой стволовую клетку или клетку-предшественницу. В некоторых вариантах осуществления указанная клетка представляет собой клетку печени или мышечную клетку. В некоторых вариантах осуществления трансген GLA поддерживается вне хромосом и не интегрируется в геном клетки. В некоторых вариантах осуществления трансген GLA интегрируется в геном клетки.
[0032] Также предлагается способ предотвращения, ингибирования или лечения болезни Фабри или одного или большего количества симптомов, ассоциированных с болезнью Фабри. Указанный способ может включать введение экспрессионной конструкции нуждающемуся в этом субъекту, при этом экспрессионная конструкция содержит мутированную последовательность WPRE, необязательно mut6 мутированную последовательность WRPE с мутацией mut6 и трансген GLA, кодирующий по меньшей мере один белок α-Gal A.
[0033] В некоторых вариантах осуществления указанные симптомы включают один или большее количество из следующих элементов: уровни Gb3 выше нормального уровня, уровни лизо-Gb3 выше нормального уровня, заболевание почек, заболевание сердца, акропарестезию, ангиокератомы, боль в желудочно-кишечном тракте, помутнение роговицы и хрусталика или цереброваскулярное заболевание. Как описано в данном документе, исходный уровень может означать любое начальное измерение, то есть измерение, проведенное перед назначением конкретного лечения. В некоторых вариантах осуществления субъект является субъектом мужского пола и имеет активность фермента α-Gal A менее чем около 5%. В некоторых вариантах осуществления экспрессионная конструкция содержит последовательность GLA дикого типа или кодон-оптимизированную последовательность GLA. В некоторых вариантах осуществления экспрессионная конструкция дополнительно содержит один или большее количество из следующих элементов: энхансер, промотор, интрон, последовательность, кодирующую сигнальный пептид, и/или сигнал полиаденилирования, при этом мутированная последовательность WPRE, необязательно мутированная последовательность WRPE с мутацией mut6, и трансген GLA, кодирующий по меньшей мере один белок α-Gal A, расположен между сигнальным пептидом и последовательностью, кодирующей сигнал полиаденилирования. В некоторых вариантах осуществления экспрессионную конструкцию вводят в фармацевтически приемлемом носителе. В некоторых вариантах осуществления фармацевтически приемлемый носитель содержит фосфатно-солевой буфер, содержащий CaCl2, Mg Cl2, NaCl, сахарозу и коллифор (полоксамер) P 188.
[0034] В других вариантах осуществления последовательность экспрессионной конструкции содержит последовательность, приведенную в Таблице 1, и при этом указанная экспрессионная конструкция доставляется в клетку субъекта посредством вирусного вектора AAV. В некоторых вариантах осуществления серотип указанного вирусного вектора AAV представляет собой AAV2/6.
[0035] В некоторых вариантах осуществления экспрессионную конструкцию вводят субъекту в дозе от около 5,0E+12 до 1,0E+14 векторных геномов на килограмм (вг/кг). В некоторых вариантах осуществления экспрессионную конструкцию вводят в печень субъекта. В некоторых вариантах осуществления экспрессионный вектор вводят субъекту посредством внутривенной инфузии. В некоторых вариантах осуществления субъекту вводят только одну дозу экспрессионной конструкции.
[0036] В некоторых вариантах осуществления субъектам вводят иммунодепрессант до и/или во время введения экспрессионной конструкции. В некоторых вариантах осуществления указанный иммунодепрессант включает преднизон. В некоторых вариантах осуществления экспрессия по меньшей мере одного белка α-галактозидазы A (α-Gal A) сохраняется в течение по меньшей мере 3 месяцев, по меньшей мере 9 месяцев или по меньшей мере 12 месяцев.
[0037] В других вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере от около 3 до около 9 раз по сравнению с субъектами, не получавшими лечение.
[0038] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере на 80% по сравнению с субъектами, не получавшими лечение.
[0039] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта в одном или более из следующего: плазма, печень, сердце, почки или селезенка.
[0040] В некоторых вариантах осуществления экспрессионная конструкция изготавливают в клеточной системе HEK293, и при этом уровни GLA у субъекта в 21 раз выше по сравнению с уровнями GLA у субъектов, которым вводили экспрессионную конструкцию, изготовленную в клеточной системе Sf9.
[0041] В некоторых вариантах осуществления активность белка α-Gal A у субъекта примерно в 100-1500 раз выше, чем у нормального/дикого типа.
[0042] В некоторых вариантах осуществления белок α-Gal A, экспрессируемый трансгеном, активен в почках, печени и сердце субъекта.
[0043] В некоторых вариантах осуществления трансген GLA поддерживается вне хромосом и не интегрируется в геном клетки субъекта.
[0044] В некоторых вариантах осуществления указанные способы включают введение одной или большего количества нуклеаз, которые расщепляют эндогенный ген альбумина в клетке печени субъекта, таким образом, что трансген интегрируется в ген альбумина и экспрессируется из него.
[0045] В данном документе описаны композиции, содержащие экспрессионную конструкцию, при этом указанная экспрессионная конструкция содержит мутированную последовательность WPRE, необязательно mut6 мутированную последовательность WRPE, и трансген GLA, кодирующий по меньшей мере один белок α-Gal A, для лечения болезни Фабри.
[0046] В некоторых вариантах осуществления указанная композиция включает фармацевтически приемлемый носитель. Композиция по п. 56, причем фармацевтически приемлемый носитель содержит CaCl2, Mg Cl2, NaCl, сахарозу и коллифор (полоксамер) P 188.
[0047] В некоторых вариантах осуществления указанная композиция включает последовательность GLA дикого типа или кодон-оптимизированную последовательность GLA.
[0048] В некоторых вариантах осуществления указанная композиция включает один или большее количество из следующих элементов: энхансер, промотор, интрон, последовательность, кодирующую сигнальный пептид, и/или сигнал полиаденилирования, при этом мутированная последовательность WPRE, необязательно мутированная последовательность WRPE с мутацией mut6, и трансген GLA, кодирующий по меньшей мере один белок α-Gal A, расположен между сигнальным пептидом и последовательностью, кодирующей сигнал полиаденилирования.
[0049] В некоторых вариантах осуществления указанная композиция включает последовательность, приведенную в Таблице 1, и при этом указанная экспрессионная конструкция доставляется в клетку посредством вирусного вектора AAV. В некоторых вариантах осуществления указанная композиция включает вирусный вектор AAV, имеющий серотип AAV2/6.
[0050] В некоторых вариантах осуществления указанная композиция включает экспрессионную конструкцию, которая содержит от около 5,0E+12 до 1,0E+14 векторных геномов на килограмм веса субъекта (вг/кг).
[0051] В некоторых вариантах осуществления указанная композиция включает экспрессионную конструкцию, которая содержит последовательность SEQ ID №: 9.
[0052] Также предлагается способ продуцирования белка α-Gal A для лечения болезни Фабри, включающий экспрессию белка α-Gal A в выделенной клетке в соответствии со способом по любому из пп. 1-4 и выделение белка α-Gal A, продуцируемого клеткой.
[0053] Представлен вектор доставки, содержащий мутированную последовательность WRPE, необязательно последовательность mut6 WPRE и трансген GLA для применения в описанных в данном документе способах.
[0054] В некоторых вариантах осуществления указанный вектор доставки представляет собой вирусный вектор или липидную наночастицу (lipid nanoparticle, LNP). В некоторых вариантах осуществления указанный вирусный вектор содержит AAV2/6, при этом вирусный вектор доставляет экспрессионную конструкцию по меньшей мере в 50%, по меньшей мере в 60%, по меньшей мере в 70% или по меньшей мере в 80% клеток.
[0055] В данном документе также представлено применение экспрессионной конструкции, вектора AAV и/или генетически модифицированной клетки по любому из предшествующих пунктов для лечения болезни Фабри. В некоторых вариантах осуществления энхансер содержит SEQ ID №: 2, промотор содержит SEQ ID №: 3, интрон содержит SEQ ID №: 4, трансген GLA содержит SEQ ID №: 5, мутированная последовательность WPRE содержит SEQ ID №: 6, а сигнал полиаденилирования содержит SEQ ID № 7.
[0056] В некоторых вариантах осуществления указанная композиция включает энхансер SEQ ID №: 2, промотор SEQ ID №: 3, интрон SEQ ID №: 4, трансген GLA SEQ ID №: 5, мутированная последовательность WPRE SEQ ID №: 6, и сигнал полиаденилирования SEQ ID № 7.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0057] Фиг. 1A демонстрирует схематическое изображение конструкции, кодирующей ген GLA, обозначенной как вариант №4. Вариант №4 конструкции включает энхансер (например, APOE); промотор (например, hAAT); интронную последовательность (например, HBB-IGG); сигнальный пептид (например, GLA); GLA-кодирующую последовательность (например, “GLAco”); и сигнал полиаденилирования (например, bGH).
[0058] Фиг. 1B демонстрирует схематическое изображение конструкции, кодирующей ген GLA, обозначенной как вариант № 21, который включает мутированный посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE) (также известный как «mut 6» или «WPREmut6 v1»). Вариант № 21 конструкции также включает энхансер (например, APOE); промотор (например, hAAT); интронную последовательность (например, HBB-IGG); сигнальный пептид (например, GLA); GLA-кодирующую последовательность (например, “GLAco”); и сигнал полиаденилирования (например, bGH).
[0059] Фиг. 2 демонстрирует график, показывающий активность GLA в плазме у отдельных мышей с нокаутом GLA (GLAKO) в указанных Группах от 2 по 4, получавших вариант №4 конструкции, или контрольных животных, как продемонстрировано, в течение 85 дней. Группа 1 получала буфер для состава («Состав»). Группа 2 получала конструкции в дозе 2,0E+12 вг/кг, Группа 3 получала конструкции в дозе 5,0E+12 вг/кг, а Группа 4 получала конструкции в дозе 5,0E+13 вг/кг.
[0060] Фиг. 3 представляет собой график, демонстрирующий активность GLA в плазме у мышей GLAKO в указанных Группах от 2 по 4, получавших экспрессионные конструкции (экспрессионная конструкция варианта №4) или контрольных животных в течение 85 дней. Группе 1 вводили буфер для состава («Состав»). Группа 2 получала конструкции в дозе 2,0E+12 вг/кг, Группа 3 получала конструкции в дозе 5,0E+12 вг/кг, а Группа 4 получала конструкции в дозе 5,0E+13 вг/кг.
[0061] Фиг. 4A представляет собой график, демонстрирующий активность α-Gal A в лизатах печени указанных групп животных, получавших экспрессионные конструкции, вариант №4, или контрольных животных. Группе 2 вводили 2,0E+12 вг/кг, Группе 3 вводили 5,0E+12 вг/кг, а Группе 4 вводили 5,0E+13 вг/кг.
[0062] Фиг. 4В представляет собой график, демонстрирующий активность α-Gal A в лизатах почек указанных групп животных, получавших экспрессионные конструкции (экспрессионная конструкция, вариант №4), или контрольных животных. Группа 2 получала конструкции в дозе 2,0E+12 вг/кг, Группа 3 получала конструкции в дозе 5,0E+12 вг/кг, а Группа 4 получала конструкции в дозе 5,0E+13 вг/кг.
[0063] Фиг. 4С представляет собой график, демонстрирующий активность α-Gal A в лизатах сердца указанных групп животных, получавших экспрессионные конструкции (экспрессионная конструкция, вариант №4), или контрольных животных. Группе 2 вводили 2,0E+12 вг/кг, Группе 3 вводили 5,0E+12 вг/кг, а Группе 4 вводили 5,0E+13 вг/кг.
[0064] Фиг. 5A представляет собой график, демонстрирующий концентрации субстрата Lyso-Gb3 в плазме, селезенке, печени, сердце и почках в указанных группах мышей GLAKO, получавших экспрессионные конструкции (экспрессионная конструкция, вариант №4), или контрольных животных на день 91 после лечения. Для каждой ткани столбцы слева направо демонстрируют животных Группы 1, которые получали буфер для состава, животных Группы 2, которые получали конструкции в дозе 2,0E+12 вг/кг (10 животных); животных Группы 3, которые получали конструкции в дозе 5,0E+12 вг/кг (9 животных); и животных Группы 4, которые получали конструкции в дозе 5,0E+13 вг/кг (20 животных). Как продемонстрировано, концентрации субстрата лизо-Gb3 определяются ниже в Группах от 2 по 4 по сравнению с контрольной Группой 1 во всех тестируемых тканях. Пунктирной линией также показан нижний предел количественного определения (lower limit of quantification, LLOQ).
[0065] Фиг. 5В представляет собой график, демонстрирующий уровни Gb3 в плазме, селезенке, печени, сердце и почках в указанных Группах животных, получавших экспрессионные конструкции (вариант № 4), или контрольных животных. Для каждой ткани столбцы слева направо демонстрируют животных Группы 1, которые получали буфер для состава, животных Группы 2, которые получали конструкции в дозе 2,0E+12 вг/кг (10 животных); животных Группы 3, которые получали конструкции в дозе 5,0E+12 вг/кг (9 животных); и животных Группы 4, которые получали конструкции в дозе 5,0E+13 вг/кг (20 животных). Как продемонстрировано, концентрации субстрата Gb3 определяются ниже в Группах от 2 по 4 по сравнению с контрольной Группой 1 во всех тестируемых тканях. Пунктирной линией также показан нижний предел количественного определения (lower limit of quantification, LLOQ).
[0066] Фиг. 6A представляет собой график, демонстрирующий процент субстрата Gb3 и лизо-Gb3, остающийся в плазме, селезенке, печени, сердце и почках в указанной Группе животных, получавших экспрессионную конструкцию варианта №4, или контрольных животных.
[0067] Фиг. 6B представляет собой график, демонстрирующий процент субстрата Gb3 и лизо-Gb3, остающийся в плазме, селезенке, печени, сердце и почках в указанной Группе животных, получавших экспрессионную конструкцию варианта №4, или контрольных животных.
[0068] Фиг. 7A представляет собой график, демонстрирующий активность α-Gal A in vitro в супернатанте клеток HepG2 человека, обработанных либо кДНК вариантом № 4 конструкции, либо кДНК вариантом № 21 конструкции (как продемонстрировано на Фиг. 1A и 1B). Активность трансгена увеличивалась по меньшей мере примерно в 9 раз в клетках, обработанных 300000 AAV вг/клетку с применением экспрессионной конструкции, содержащей последовательность WPRE (вариант № 21 конструкции, как продемонстрировано на Фиг. 1B), по сравнению с активностью при применении варианта № 4. конструкции в качестве экспрессионной конструкции. Активность трансгена увеличивалась по меньшей мере примерно в 7 раз в клетках, обработанных 100 000 AAV вг/клетку с применением экспрессионной конструкции, содержащей последовательность WPRE (вариант № 21, как продемонстрировано на Фиг. 1B), по сравнению с активностью при применении варианта № 4 конструкции в качестве экспрессионной конструкции
[0069] Фиг. 7В представляет собой график, демонстрирующий активность α-Gal A in vitro в супернатанте индуцированных плюрипотентных клеток гепатоцитов («гепатоциты iCell»), обработанных либо кДНК вариантом № 4 конструкции, либо кДНК вариантом № 21 конструкции (как продемонстрировано на Фиг. 1A и 1B). Активность трансгена увеличивалась по меньшей мере примерно в 4 раза в клетках, обработанных 30 000 AAV вг/клетку с применением экспрессионной конструкции, содержащей последовательность WPRE (вариант № 21, как продемонстрировано на Фиг. 1B), по сравнению с активностью при применении варианта № 4 в качестве экспрессионной конструкции. Активность трансгена увеличивалась по меньшей мере примерно в 3 раза в клетках, обработанных 100 000 AAV вг/клетку с применением экспрессионной конструкции, содержащей последовательность WPRE (вариант № 21, как продемонстрировано на Фиг. 1B), по сравнению с активностью при применении варианта № 4 в качестве экспрессионной конструкции.
[0070] Фиг. 8 представляет собой график, демонстрирующий повышение активности GLA A с увеличением дозы конструкции в плазме мышей дикого типа, пролеченных вариантом № 21 конструкций в дозе 2,0E+12 вг/кг или 5E+11 вг/кг, или вариантом № 4 конструкций в дозе 2,0E+12 вг/кг или 5E+11 вг/кг или буфером для состава.
[0071] Фиг. 9 представляет собой график, демонстрирующий плазменную активность α-Gal A у мышей C57BL/6 в течение 29 дней после обработки вариантом №21 конструкций в дозе 5,0E+13 вг/кг, вариантом №21 конструкций в дозе 5,0E+12 вг/кг, вариантом №4 конструкций в дозе 5,0Е+13 вг/кг, вариантом №4 конструкций в дозе 5,0Е+12 вг/кг или буфером для состава. Как продемонстрировано, вариант №21 конструкций может продуцировать уровни активности α-Gal A в плазме у мышей C57BL/6, превышающие более чем в 1500 раз физиологически нормальные уровни.
[0072] Фиг. 10 представляет собой изображения гибридизации ДНК in situ, окрашенные для определения геномов вектора AAV в образце печени мыши GLAKO, которой вводили вариант №4 конструкций в дозе 5,0Е+13 вг/кг. Нацеливались на некодирующие последовательности. В этом образце 57,5% клеток печени положительно окрашивались относительно геномов вектора AAV через 90 дней после обработки.
[0073] Фиг. 11 представляет собой изображения гибридизации ДНК in situ, окрашенные для определения геномов вектора AAV в образце печени примата дикого типа, не относящихся к человеку (NPH), которому вводили вариант №4 конструкций в дозе 6,0Е+13 вг/кг. Нацеливались на некодирующие последовательности. В этом образце 57,5% клеток печени положительно окрашивались относительно геномов вектора AAV через 60 дней после обработки.
[0074] Фиг. 12A представляет собой график, демонстрирующий процент гепатоцитов, содержащих hGLA кДНК, у мышей GLAKO, которые получали вариант №4 конструкции в дозах 2E+12 вг/кг, 5E+12 вг/кг, 5E+13 вг/кг или буфер для состава в качестве контроля.
[0075] Фиг. 12В представляет собой график, демонстрирующий процент гепатоцитов, содержащих hGLA кДНК, у яванских макаков NHP, которые получали вариант №4 конструкции в дозах 6E+12 вг/кг, 1E+13 вг/кг, 3E+13 вг/кг, 6E+13 вг/кг или буфер для состава в качестве контроля.
[0076] Фиг. 12С представляет собой график, демонстрирующий процент клеток печени , содержащих hGLA кДНК, у отдельных мышей GLAKO, которые получали вариант №4 конструкции в дозах 2E+12 вг/кг, 5E+12 вг/кг, 5E+13 вг/кг или буфер для состава (“0”) в качестве контроля.
[0077] Фиг. 12D представляет собой график, демонстрирующий процент гепатоцитов, содержащих hGLA кДНК, у отдельных яванских макаков NHP, которые получали вариант №4 конструкции в дозах 6E+12 вг/кг, 1E+13 вг/кг, 3E+13 вг/кг, 6E+13 вг/кг или буфер для состава (“0”) в качестве контроля.
[0078] Фиг. 13A и Фиг. 13B представляют собой графики, демонстрирующие активность hGLA в плазме NHP в зависимости от концентрации белка для отдельных животных, получавших вариант №4 конструкций в дозе 6,0E+12 вг/кг или буфер для состава.
[0079] Фиг. 13С и Фиг. 13D представляют собой графики, демонстрирующие активность hGLA в плазме NHP в зависимости от концентрации белка для отдельных животных, получавших вариант №4 конструкций в дозе 1,0E+13 вг/кг или 3,0E+13 вг/кг.
[0080] Фиг. 13Е и Фиг. 13F представляют собой графики, демонстрирующие активность hGLA в плазме NHP в зависимости от концентрации белка для отдельных животных, получавших вариант №4 конструкций в дозе 6,0E+13 вг/кг или 6,0E+13 вг/кг без иммунодепрессантов.
[0081] Фиг. 14 представляет собой вестерн-блот-анализ hGLA и соответствующих уровней мРНК в образцах печени NHP от отдельных животных на день 60 после введения варианта №4 конструкций в дозах 6,0E+12 вг/кг, 1,0E+13 вг/кг, 3,0E+13 вг/кг, 6,0E+13 вг/кг, 6,0E+13 вг/кг без иммунодепрессантов или буфера для состава. Как продемонстрировано, уровни белка hGLA повышаются с увеличением дозы конструкции, при этом уровни белка коррелируют с уровнями мРНК в большинстве образцов.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0082] В данном документе описаны способы и композиции для лечения или предотвращения болезни Фабри. В описании представлены способы и композиции для введения трансгена GLA, кодирующего белок, который отсутствует или недостаточно экспрессируется у субъекта с болезнью Фабри, так что ген экспрессируется в печени и экспрессируется терапевтический (замещающий) белок. В данном документе также описано изменение клетки (например, предшественника или зрелой эритроцитарной клетки (RBC), iPSC или клетки печени) таким образом, что оно продуцирует высокие уровни терапевтического средства, и введение популяции этих измененных клеток пациенту обеспечит поступление необходимого белка. Трансген может кодировать желаемый белок или структурную РНК, которая будет терапевтически полезна пациенту, нуждающемуся в этом.
[0083] Генная терапия аденоассоциированными вирусными (AAV) векторами продемонстрировала большие перспективы как в доклинических, так и в клинических испытаниях для эффективной доставки терапевтических трансгенов в печень, с сообщениями о стабильных уровнях экспрессии трансгена до шести лет для гемофилии B (Lheriteau E, Davidoff E, Nathwani AC. Haemophilia gene therapy: Progress and challenges. Blood Rev. 2015 Sep;29(5):321-8).
[0084] Одна область, которая является особенно многообещающей, - это возможность добавлять трансген к клетке, чтобы эта клетка экспрессировала продукт, который ранее не продуцировался в этой клетке или продуцировался неоптимально. Примеры применения этой технологии включают вставку гена, кодирующего терапевтический белок, вставку кодирующей последовательности, кодирующей белок, который каким-то образом отсутствует в клетке или у индивидуума, и вставку последовательности, которая кодирует структурную нуклеиновую кислоту, такую как микроРНК.
[0085] Трансгены можно вводить и сохранять в клетках различными способами. Следуя подходу «кДНК», трансген вводят в клетку таким образом, чтобы трансген сохранялся вне хромосомы, а не посредством интеграции в хроматин клетки. Трансген может сохраняться в кольцевом векторе (например, плазмиде или неинтегрирующем вирусном векторе, таком как AAV или лентивирус), при этом указанный вектор может включать транскрипционные регуляторные последовательности, такие как промоторы, энхансеры, сигнальные последовательности полиА, интроны и сплайсинг-сигналы (патент США № 10143760).
[0086] Трансгены могут быть доставлены в клетку различными способами, например, трансген интегрируется в собственный геном клетки и сохраняется там. В последние годы была разработана стратегия интеграции трансгена, которая использует расщепление сайт-специфическими нуклеазами для направленной вставки в выбранный геномный локус (см. например, совместный патент США № 7888121). Нуклеазы, такие как нуклеазы цинковые пальцы (zinc finger nucleases, ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (transcription activator-like effector nuclease, TALEN), или нуклеазные системы, такие как система CRISPR/Cas, направляемая РНК (с использованием сконструированной направляющей РНК), специфичны для целевых генов и могут применяться таким образом, что трансгенная конструкция вставляется либо посредством гомологически направленной репарации (homology directed repair, HDR), либо путем захвата концов во время процессов, управляемых негомологичным концевым соединением (non-homologous end joining, NHEJ). См., например, патенты США №№ 9877988; 9816074; 9616090; 9873894; 9597357; 9567573; 9458205; 9447434; 9394545; 9255250; 9222105; 9206404; 9200266; 9045763; 9005973; 9150847; 8956828; 8945868; 8895264; 8771985; 8703489; 8586526; 8106255; 6534261; 6599692; 6503717; 6689558; 7067317; 7262054; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; патентные публикации США 20030232410 и 20050064474, описания которых в полном объеме включены в данный документ посредством ссылки.
[0087] Трансгены могут быть интегрированы в высокоэкспрессируемый локус «безопасного убежища», такой как ген альбумина (см. патент США № 9394545). Этот подход получил название платформы замещения белка in vivo или IVPRP. Следуя этому подходу, трансген вводят в ген безопасного убежища (например, альбумина) посредством нуклеазо-опосредованной направленной вставки, при этом экспрессия трансгена управляется промотором альбумина. Трансген сконструирован так, чтобы он содержал сигнальную последовательность, способствующую секреции/экскреции белка, кодируемого трансгеном.
[0088] Локусы «безопасного убежища» включают такие локусы, как гены AAVS1, HPRT, альбумина и CCR5 в клетках человека и Rosa26 в клетках мыши. См., например, патенты США №№ 9877988; 9567573; 9447434; 9394545; 9222105; 9206404; 9150847; 8895264; 8771985; 8106255; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; и 8586526; патентные публикации США 20030232410 и 20060063231. Интеграция, опосредованная нуклеазами, дает возможность улучшить экспрессию трансгена, повысить безопасность и долговечность экспрессии по сравнению с классическими подходами к интеграции, основанными на случайной интеграции трансгена, поскольку она позволяет точно позиционировать трансген с минимальным риском сайленсинга генов или активации близлежащих онкогенов. Опосредованная нуклеазой вставка трансгена генов, кодирующих терапевтические белки Фабри, описана в публикации США № 20180117181.
[0089] Хотя доставка трансгена в целевую клетку является одним из препятствий, которое необходимо преодолеть, чтобы полностью задействовать эту технологию, необходимо решить еще одну проблему - гарантировать, что после того, как трансген будет вставлен в клетку и будет экспрессирован, закодированный таким образом генный продукт достигнет необходимого места в организме и будет продуцироваться в локальных концентрациях, достаточных для эффективности. При заболеваниях, характеризующихся отсутствием белка или присутствием аберрантного нефункционального белка, доставка кодируемого трансгеном белка дикого типа может быть чрезвычайно полезной.
[0090] Лизосомные болезни накопления (Lysosomal storage diseases, LSD) представляют собой группу редких метаболических моногенных заболеваний, характеризующихся отсутствием функциональных индивидуальных лизосомных белков, обычно участвующих в расщеплении ненужных липидов, гликопротеинов и мукополисахаридов. Эти заболевания характеризуются накоплением этих соединений в клетке, поскольку она не может переработать их для повторного использования из-за неправильного функционирования определенного фермента. Наиболее распространенными примерами являются болезнь Гоше (дефицит глюкоцереброзидазы - название гена: GBA), болезнь Фабри (дефицит α-галактозидазы A - GLA), синдром Хантера (дефицит идуронат-2-сульфатазы - IDS), Hurler (дефицит альфа-L идуронидазы - IDUA), болезнь Помпе (альфа-глюкозидазы (GAA)) и болезни Ниманна-Пика (дефицит сфингомиелин фосфодиэстеразы 1 - SMPD1). В совокупности, LSD встречаются в популяции примерно в 1 из 7000 новорожденных. См. также, патенты США №№ 9877988 и 9956247 и публикацию США № 20160060656.
[0091] Например, болезнь Фабри представляет собой X-связанное нарушение метаболизма гликосфинголипидов, вызванное дефицитом фермента α-галактозидазы A (α-GalA). Это заболевание ассоциируется с прогрессирующим отложением гликосфинголипидов, включая глоботриаозилцерамид (также известный как GL-3 и Gb3) и глоботриаозилсфингозин (лизо-Gb3), галабиоазилцерамид и вещества группы B. Симптомы болезни разнообразны и могут включать жжение, покалывание (акропарестезия) или эпизоды сильной боли, которые называются «кризами Фабри», которые могут длиться от нескольких минут до нескольких дней. Другие симптомы включают нарушение потоотделения, плохую переносимость физических нагрузок, красновато-пурпурную сыпь, называемую ангиокератомой, аномалии глаз, нарушения желудочно-кишечного тракта, проблемы с сердцем, такие как увеличенное сердце и сердечный приступ, проблемы с почками, которые могут привести к почечной недостаточности, а также проблемы ЦНС, при этом в целом ожидаемая продолжительность жизни пациентов с Фабри значительно сокращается.
[0092] Современное лечение болезни Фабри может включать заместительную ферментную терапию (ERT) с применением двух разных препаратов α-GalA, агалсидазы бета или агалсидазы альфа человека, что требует дорогостоящих и длительных инфузий (обычно около 0,2-1 мг/кг) пациенту каждые две недели. Такое лечение предназначено только для купирования симптомов и не является излечивающим. Соответственно, пациенту необходимо вводить повторные дозы этих белков до конца его жизни, и у него потенциально могут развиться нейтрализующие антитела к введенному белку.
[0093] Кроме того, с ERT ассоциируются нежелательные реакции, включая иммунные реакции, такие как развитие антител анти-α-GalA у субъектов, получавших препараты α-GalA. Фактически, у 50% мужчин, получавших агалсидазу альфа, и у 88% мужчин, получавших агалсидазу бета, формировались антитела против α-GalA. Важно отметить, что значительная часть этих антител является нейтрализующими антителами и, следовательно, снижает терапевтический эффект лечения (Meghdari et al. (2015) PLoS One 10(2):e0118341. Doi:10.1371/journal.pone.0118341). Кроме того, не у всех пациентов ERT останавливает прогрессирование заболевания.
[0094] Таким образом, способы и композиции можно применять для экспрессии из трансгена одного или большего количества терапевтически полезных белков α-GalA из конструкции кДНК, доставленной, например, вирусным вектором или вставленной в любой локус (например, высокоэкспрессируемый локус альбумина) для замены фермента, который является дефектным и/или отсутствует при болезни Фабри. Кроме того, в данном описании представлены способы и композиции для лечения (включая облегчение одного или большего количества симптомов) болезни Фабри путем встраивания трансгенных последовательностей в высокоэкспрессируемые локусы в клетках, таких как клетки печени. В описание включены способы и композиции для доставки трансгена, кодирующего α-GalA, посредством вирусного вектора в печень нуждающегося в этом субъекта, при этом вирус может быть введен путем инъекции в систему периферической вены или путем прямой инъекции в кровеносный сосуд, направляющийся в печень (например, воротная вена). Способы и композиции можно применять для индукции встраивания трансгена в локус безопасного убежища (например, альбумина) или можно применять для обеспечения внехромосомного сохранения конструкции вирусной кДНК в клетке печени. В любом случае трансген высоко экспрессируется и обеспечивает терапевтический эффект нуждающемуся в этом пациенту с болезнью Фабри.
[0095] Кроме того, трансген можно ввести в клетки, полученные от пациента, например, полученные от пациента индуцированные плюрипотентные стволовые клетки (induced pluripotent stem cells, iPSC) или другие типы стволовых клеток (эмбриональные или гематопоэтические) для применения в возможной имплантации. Особенно полезным является введение терапевтического трансгена в гемопоэтическую стволовую клетку для имплантации пациенту, нуждающемуся в этом. По мере того, как стволовые клетки дифференцируются в зрелые клетки, они будут содержать высокие уровни терапевтического белка для доставки в ткани.
Общие сведения
[0096] При практической реализации способов, а также приготовлении и применении композиций, описанных в данном документе, используют, если не указано иное, обычные методы в молекулярной биологии, биохимии, структуре и анализе хроматина, вычислительной химии, культуре клеток, рекомбинантной ДНК и связанных областях, которые находятся в пределах уровня знаний специалиста обычной квалификации в данной области техники. Эти методы в полное мере писаны в литературе. См., например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 и Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, “Chromatin” (P.M. Wassarman and A. P. Wolffe, eds.), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, “Chromatin Protocols” (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
[0097] Термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» применяются взаимозаменяемо и относятся к дезоксирибонуклеотидному или рибонуклеотидному полимеру в линейной или кольцевой конформации, а также в одноцепочечной или двухцепочечной форме. Для целей настоящего изобретения эти термины не должны толковаться как ограничивающие длину полимера. Указанные термины могут охватывать известные аналоги природных нуклеотидов, а также нуклеотиды, модифицированные по основанию, сахару и/или фосфатному фрагменту (например, фосфортиоатным каркасам). В общем, аналог конкретного нуклеотида имеет такую же специфичность спаривания оснований; то есть аналог A будет иметь пару оснований с T.
[0098] Термины «полипептид», «пептид» и «белок» применяются в данном документе взаимозаменяемо для обозначения полимера состоящего из аминокислотных остатков. Этот термин также применяется к полимерам аминокислот, в которых одна или большее количество аминокислот являются химическими аналогами или модифицированными производными соответствующих встречающихся в природе аминокислот.
[0099] «Связывание» относится к специфичному для последовательности нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты связывающего взаимодействия должны быть специфичными для последовательности (например, контакты с фосфатными остатками в основном каркасе ДНК), если взаимодействие в целом является специфичным для последовательности. Такие взаимодействия обычно характеризуются константой диссоциации (Kd) of 10-6 M-1 или ниже. «Аффинность» относится к силе связывания: повышенная аффинность связывания коррелирует с более низким значением Kd.
[0100] «Связывающий домен» представляет собой молекулу, которая способна нековалентно связываться с другой молекулой. Связывающая молекула может связываться, например, с молекулой ДНК (ДНК-связывающий белок, такой как белок цинковые пальцы или белок TAL-эффекторного домена или одиночная направляющая РНК), молекулой РНК (РНК-связывающий белок) и/или белковой молекулой (белок-связывающий белок). В случае молекулы, связывающей белок, она может связываться сама с собой (с образованием гомодимеров, гомотримеров и т.д.) и/или может связываться с одной или большим количеством молекул другого белка или белков. Связывающая молекула может обладать более чем одним типом связывающей активности. Например, белки «цинковые пальцы» обладают ДНК-связывающей, РНК-связывающей и белок-связывающей активностью. Таким образом, ДНК-связывающие молекулы, включая ДНК-связывающие компоненты искусственных нуклеаз и факторов транскрипции, включают, помимо прочего, ZFP, TALE и sgРНК.
[0101] «ДНК-связывающий белок цинковые пальцы» (или связывающий домен) представляет собой белок или домен в более крупном белке, который связывает ДНК специфическим для последовательности образом через один или большее количество цинковых пальцев, которые представляют собой участки аминокислотной последовательности в связывающем домене, структура которого стабилизируется за счет координации иона цинка. Термин «белок цинковые пальцы», связывающий ДНК, часто сокращенно называют «белок цинковые пальцы» или ZFP. Искусственные нуклеазы и факторы транскрипции могут включать ДНК-связывающий домен ZFP и функциональный домен (домен нуклеазы для ZFN или домен регуляции транскрипции для ZFP-TF). Термин «нуклеаза цинковые пальцы» включает один ZFN, а также пару ZFN, которые димеризуются для расщепления целевого гена.
[0102] «ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или большее количество повторяющихся доменов/единиц TALE. Повторяющиеся домены участвуют в связывании TALE с его родственной целевой последовательностью ДНК. Одна «повторяющаяся единица» (также называемая «повтором») обычно имеет длину 33-35 аминокислот и демонстрирует по меньшей мере некоторую гомологию последовательности с другими повторяющимися последовательностями TALE в пределах встречающегося в природе белка TALE. См., например патент США № 8586526. Искусственные нуклеазы и факторы транскрипции могут включать ДНК-связывающий домен TALE и функциональный домен (домен нуклеазы для TALEN или домен регуляции транскрипции для TALEN-TF). Термин «TALEN» включает один TALEN, а также пару TALEN, которые димеризуются для расщепления целевого гена.
[0103] «Цинковый палец» и TALE-связывающие домены могут быть «сконструированы» для связывания с заранее определенной нуклеотидной последовательностью, например, путем конструирования (изменения одной или большего количества аминокислот) области спирали распознавания встречающегося в природе белка цинкового пальца или TALE. Следовательно, сконструированные ДНК-связывающие белки (цинковые пальцы или TALE) представляют собой белки, не встречающиеся в природе. Неограничивающими примерами способов конструирования ДНК-связывающих белков являются разработка и отбор. Разработанный ДНК-связывающий белок представляет собой белок, не встречающийся в природе, конструкция/состав которого основывается главным образом на рациональных критериях. Рациональные критерии для разработки включают применение правил замены и компьютеризированных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих дизайнах ZFP и/или TALE и данных связывания. См., например, патенты США №№ 8568526; 6140081; 6453242 и 6534261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0104] «Отобранный» белок цинковые пальцы или TALE представляет собой белок, не встречающийся в природе, продукция которого в основном является результатом эмпирического процесса, такого как фаговый дисплей, ловушка взаимодействия или гибридный отбор. См. например, патенты США №№ 8586526; 5789538; US 5925523; US 6007988; US 6013453; US 6 200 759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084.
[0105] «Рекомбинация» относится к процессу обмена генетической информацией между двумя полинуклеотидами. Для целей этого изобретения «гомологичная рекомбинация (homologous recombination, HR)» относится к специализированной форме такого обмена, который имеет место, например, во время репарации двухцепочечных разрывов в клетках с помощью механизмов гомологически-направленной репарации. Этот процесс, требующий гомологии нуклеотидной последовательности, использует «донорную» молекулу для репарации матрицы «целевой» молекулы (то есть той, которая испытала двухцепочечный разрыв) и под разными наименованиями известен как «некроссоверная конверсия гена» или «конверсия гена короткого тракта», поскольку приводит к передаче генетической информации от донора к цели. Не ограничиваясь какой-либо конкретной теорией, считают, что такой перенос может включать в себя коррекцию несоответствия гетеродуплексной ДНК, которая образуется между поврежденной мишенью и донором, и/или «зависимый от синтеза отжиг цепи», в котором донор используется для повторного синтеза генетической информации, которая станет частью цели, и/или связанных процессов. Такая специализированная HR часто приводит к изменению последовательности целевой молекулы, так что часть или вся последовательность донорного полинуклеотида включается в целевой полинуклеотид.
[0106] В способах по настоящему изобретению одна или большее количество целевых нуклеаз-мишеней, как описано в данном документе, создают двухцепочечный разрыв в целевой последовательности (например, клеточном хроматине) в заранее определенном сайте и «донорный» полинуклеотид, имеющий гомологию с нуклеотидной последовательностью в области разрыва, может быть введен в клетку. Было продемонстрировано, что наличие двухцепочечного разрыва облегчает интеграцию донорной последовательности. Донорная последовательность может быть физически интегрирована или, в альтернативном варианте, донорный полинуклеотид применяют в качестве матрицы для репарации разрыва посредством гомологичной рекомбинации, что приводит к введению всей или части нуклеотидной последовательности, как у донора, в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может быть изменена и, в некоторых вариантах осуществления, может быть преобразована в последовательность, присутствующую в донорном полинуклеотиде. Таким образом, термины «заменить» или «замена» можно понимать как замену одной нуклеотидной последовательности другой (то есть, замену последовательности в информационном смысле), что не обязательно требует физической или химической замены одного полинуклеотида другим.
[0107] В любом из описанных в данном документе способов можно применять дополнительные пары белков «цинковые пальцы» или TALEN для дополнительного двухцепочечного расщепления дополнительных целевых сайтов внутри клетки.
[0108] В определенных вариантах осуществления способов целевой рекомбинации и/или замены и/или изменения последовательности в области клеточного хроматина, представляющей интерес, хромосомная последовательность изменяется путем гомологичной рекомбинации с экзогенной «донорной» нуклеотидной последовательностью. Такая гомологичная рекомбинация стимулируется наличием двухцепочечного разрыва в клеточном хроматине, если присутствуют последовательности, гомологичные области разрыва.
[0109] В любом из описанных в данном документе способов первая нуклеотидная последовательность («донорная последовательность») может содержать последовательности, которые гомологичны, но не идентичны геномным последовательностям в области, представляющей интерес, тем самым стимулируя гомологичную рекомбинацию для вставки неидентичной последовательности в область, представляющую интерес. Таким образом, в определенных вариантах осуществления части донорной последовательности, которые гомологичны последовательностям в области, представляющей интерес, демонстрируют от около 80 до 99% (или любое целое число между ними) идентичности последовательности с геномной последовательностью, которая заменяется. В других вариантах осуществления гомология между донорной и геномной последовательностями выше 99%, например, если отличается только 1 нуклеотид, как между донорной и геномной последовательностями из более чем 100 смежных пар оснований. В определенных случаях негомологичная часть донорной последовательности может содержать последовательности, не присутствующие в области, представляющей интерес, так что новые последовательности вводятся в область, представляющую интерес. В этих случаях негомологичная последовательность обычно фланкируется последовательностями из 50-1000 пар оснований (или любым целым значением между ними) или любым количеством пар оснований, превышающим 1000, которые гомологичны или идентичны последовательностям в области, представляющей интерес. В других вариантах осуществления донорная последовательность негомологична первой последовательности и вставлена в геном с помощью механизмов негомологичной рекомбинации.
[0110] Любой из описанных в данном документе способов можно применять для частичной или полной инактивации одной или большего количества целевых последовательностей в клетке путем направленной интеграции донорной последовательности, которая нарушает экспрессию гена(ов), представляющего(их) интерес. Также представлены клеточные линии с частично или полностью инактивированными генами.
[0111] Кроме того, описанные в данном документе способы целевой интеграции также можно применять для интеграции одной или большего количества экзогенных последовательностей. Экзогенная последовательность нуклеиновой кислоты может содержать, например, один или большее количество генов или молекул кДНК, или любой тип кодирующей или некодирующей последовательности, а также один или большее количество контрольных элементов (например, промоторов). Кроме того, экзогенная последовательность нуклеиновой кислоты может продуцировать одну или большее количество молекул РНК (например, малые шпильчатые РНК (shРНК), ингибирующие РНК (РНКi), микроРНК (miРНК) и т.д.).
[0112] «Расщепление» относится к разрыву ковалентного каркаса молекулы ДНК. Расщепление можно инициировать множеством способов, включая, помимо прочего, ферментативный или химический гидролиз фосфодиэфирной связи. Возможно как одноцепочечное расщепление, так и двухцепочечное расщепление, при этом двухцепочечное расщепление может происходить в результате двух различных событий одноцепочечного расщепления. Расщепление ДНК может привести к образованию либо тупых концов, либо смещенных концов. В определенных вариантах осуществления гибридные полипептиды применяют для целенаправленного расщепления двухцепочечной ДНК.
[0113] «Полудомен расщепления» представляет собой полипептидную последовательность, которая вместе со вторым полипептидом (идентичным или различным) образует комплекс, обладающий активностью расщепления (предпочтительно активностью двухцепочечного расщепления). Термины «первый и второй полудомены расщепления»; «+ и - полудомены расщепления» и «правый и левый полудомены расщепления» применяются взаимозаменяемо для обозначения пар полудоменов расщепления, которые димеризуются.
[0114] «Сконструированный полудомен расщепления» представляет собой полудомен расщепления, который был модифицирован таким образом, чтобы образовывать облигатные гетеродимеры с другим полудоменом расщепления (например, другим сконструированным полудоменом расщепления). См. патенты США №№ 7888121; 7914796; 8034598 и 8823618, которые в полном объеме включены в данный документ посредством ссылки.
[0115] Термин «последовательность» относится к нуклеотидной последовательности любой длины, которая может быть ДНК или РНК; может быть линейной, кольцевой или разветвленной и при этом может быть одноцепочечной или двухцепочечной. Термин «донорная последовательность» относится к нуклеотидной последовательности, которая вставляется в геном. Донорная последовательность может иметь любую длину, например, от 2 до 10000 нуклеотидов (или любое целое число между ними или выше), предпочтительно от около 100 до 1000 нуклеотидов в длину (или любое целое число между ними), более предпочтительно от около 200 до 500 нуклеотидов в длину.
[0116] «Ассоциированный с заболеванием ген» представляет собой ген, который каким-либо образом дефектен при моногенном заболевании. Неограничивающие примеры моногенных заболеваний включают тяжелый комбинированный иммунодефицит, муковисцидоз, гемофилии, лизосомные болезни накопления (например, болезнь Гоше, Херлера, Хантера, Фабри, Ниманна-Пика, Тея-Сакса и др.), серповидно-клеточную анемию и талассемию.
[0117] «Хроматин» представляет собой структуру нуклеопротеина, составляющую клеточный геном. Клеточный хроматин включает нуклеиновую кислоту, в первую очередь ДНК, и белок, включая гистоны и негистоновые хромосомные белки. Большая часть хроматина эукариотических клеток существует в форме нуклеосом, при этом ядро нуклеосомы содержит приблизительно 150 пар оснований ДНК, связанных с октамером, содержащим по два гистона Н2А, Н2В, Н3 и Н4; и линкерная ДНК (переменной длины в зависимости от организма) простирается между ядрами нуклеосом. Молекула гистона H1 обычно ассоциирована с линкерной ДНК. Для целей настоящего изобретения термин «хроматин» предназначен для охвата всех типов клеточных нуклеопротеидов, как прокариотических, так и эукариотических. Клеточный хроматин включает как хромосомный, так и эписомальный хроматин.
[0118] «Хромосома» представляет собой комплекс хроматина, включающий весь геном клетки или его часть. Геном клетки часто характеризуется ее кариотипом, который представляет собой совокупность всех хромосом, составляющих геном клетки. Геном клетки может включать одну или большее количество хромосом.
[0119] «Эписома» представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеидный комплекс или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписомов включают плазмиды и определенные вирусные геномы.
[0120] «Целевой сайт» или «целевая последовательность» представляет собой последовательность нуклеиновой кислоты, которая определяет часть нуклеиновой кислоты, с которой связывающая молекула будет связываться, при условии, что существуют достаточные условия для связывания.
[0121] «Экзогенная» молекула представляет собой молекулу, которая обычно не присутствует в клетке, но может быть введена в клетку с помощью одного или большего количества из следующих способов: генетический, биохимический или других. «Нормальное присутствие в клетке» определяется с учетом конкретного этапа развития и условий окружающей среды клетки. Так, например, молекула, которая присутствует только во время эмбрионального развития мышцы, является экзогенной молекулой по отношению к взрослой мышечной клетке. Точно так же молекула, индуцированная тепловым шоком, является экзогенной молекулой по отношению к клетке, не подвергшейся тепловому шоку. Экзогенная молекула может включать, например, функционирующую версию неисправно действующей эндогенной молекулы или неисправно действующую версию нормально функционирующей эндогенной молекулы.
[0122] Экзогенная молекула может быть, среди прочего, малой молекулой, такой как полученная в результате комбинаторного химического процесса, или макромолекулой, такой как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любым модифицированным производным вышеуказанной молекулы или любым комплексом, содержащим одну или большее количество из вышеуказанных молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одноцепочечными или двухцепочечными; могут быть линейным, разветвленным или круглым; и могут быть любой длины. Нуклеиновые кислоты включают те, которые способны образовывать дуплексы, а также нуклеиновые кислоты, образующие триплекс. См., например, патенты США №№ 5176996 и 5422251. Белки включают, помимо прочего, ДНК-связывающие белки, факторы транскрипции, факторы ремоделирования хроматина, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликасы.
[0123] Экзогенная молекула может быть молекулы того же типа, что и эндогенная молекула, например, экзогенный белок или нуклеиновая кислота. Например, экзогенная нуклеиновая кислота может содержать инфицированный вирусный геном, плазмиду или эписому, введенные в клетку, или хромосому, которая обычно не присутствует в клетке. Способы введения экзогенных молекул в клетки известны специалистам в данной области и включают, помимо прочего, липид-опосредованный перенос (то есть липосомы, включая нейтральные и катионные липиды), электропорацию, прямую инъекцию, слияние клеток, бомбардировку частицами, соосаждение фосфатом кальция, перенос, опосредованный DEAE-декстраном, и перенос, опосредованный вирусным вектором. Экзогенная молекула также может быть молекулой того же типа, что и эндогенная молекула, но происходить от другого вида, чем клетка. Например, последовательность нуклеиновой кислоты человека может быть введена в клеточную линию, первоначально полученную от мыши или хомяка.
[0124] Напротив, «эндогенная» молекула представляет собой молекулу, которая обычно присутствует в конкретной клетке на определенном этапе развития в определенных условиях окружающей среды. Например, эндогенная нуклеиновая кислота может включать хромосому, геном митохондрии, хлоропласта или другой органеллы или встречающуюся в природе эписомальную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы и ферменты транскрипции.
[0125] Слитая молекула представляет собой молекулу, в которой две или большее количество субъединичных молекул связаны, предпочтительно ковалентно. Субъединичные молекулы могут быть одного химического типа молекулы или могут быть разными химическими типами молекул. Примеры первого типа слитой молекулы включают, помимо прочего, слитые белки (например, слияние ZFP или TALE ДНК-связывающего домена и одного или большего количества доменов активации) и слитых нуклеиновых кислот (например, нуклеиновая кислота, кодирующая слитый белок, описанный выше). Примеры второго типа слитой молекулы включают, помимо прочего, слияние триплекс-образующей нуклеиновой кислоты и полипептида, а также слияние белка, связывающегося с малой бороздой, и нуклеиновой кислоты.
[0126] Экспрессия слитого белка в клетке может быть результатом доставки слитого белка в клетку или оставки полинуклеотида, кодирующего слитый белок, в клетку, при этом полинуклеотид транскрибируется, а транскрипт транслируется, с образованием слитого белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептида также могут быть вовлечены в экспрессию белка в клетке. Способы доставки полинуклеотидов и полипептидов в клетки представлены в другом месте в этом описании.
[0127] «Ген» для целей настоящего изобретения включает область ДНК, кодирующую продукт гена (см. ниже), а также все области ДНК, которые регулируют продуцирование продукта гена, независимо от того, являются ли такие регуляторные последовательности смежными с кодирующими и/или транскрибируемыми последовательностями. Соответственно, ген включает, но не обязательно ограничивается этим, промоторные последовательности, терминаторы, регуляторные последовательности трансляции, такие как сайты связывания рибосом и внутренние сайты посадки рибосомы, энхансеры, сайленсеры, инсуляторы, граничные элементы, источники репликации, сайты прикрепления матрикса и области локус-контроля.
[0128] «Экспрессия гена» относится к преобразованию информации, содержащейся в гене, в продукт гена. Генный продукт может быть продуктом прямой транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловая РНК, рибозим, структурная РНК или любой другой тип РНК) или белком, полученным путем трансляции мРНК. Генные продукты также включают РНК, которые модифицируются такими процессами, как кэппирование, полиаденилирование, метилирование и редактирование, а также белки, модифицированные, например, метилированием, ацетилированием, фосфорилированием, убиквитинированием, ADP-рибозилированием, миристилированием и гликозилированием.
[0129] «Ген GLA» кодирует α-галактозидазу, фермент, расщепляющий глоботриаозилцерамид. Генетическая мутация в гене GLA приводит к нарушению функции фермента α-галактозидазы. Ген GLA расположен в Xq22.1, которое представляет собой длинное (q) плечо X-хромосомы в положении 22.1. Ген GLA может также называться AGAL_HUMAN, агалсидаза альфа, альфа-D-галактозидаза A, альфа-D-галактозидаза, галактозидаза, альфа-галактозидаза, альфа-галактозидаза A, церамидетрихексозидаза, GALA, галактозидаза, альфа или мелибиаза.
[0130] «Модуляция» экспрессии гена относится к изменению активности гена. Модуляция экспрессии может включать, помимо прочего, активацию гена, оптимизацию гена и репрессию гена. Редактирование генома (например, расщепление, изменение, инактивация, случайная мутация) можно применять для модуляции экспрессии. Инактивация гена относится к любому снижению экспрессии гена по сравнению с клеткой, которая не включает систему ZFP, TALE или CRISPR/Cas, как описано в данном документе. Таким образом, инактивация гена может быть частичной или полной.
[0131] «Представляющая интерес область» представляет собой любую область клеточного хроматина, такую как, например, ген или некодирующая последовательность внутри гена или рядом с ним, в которой желательно связывать экзогенную молекулу. Связывание может применяться для целевого расщепления ДНК и/или целевой рекомбинации. Область, представляющая интерес, может находиться, например, в хромосоме, эписоме, органеллярном геноме (например, митохондриальном, хлоропластном) или в геноме инфицированного вируса. Область, представляющая интерес, может находиться внутри кодирующей области гена, внутри транскрибируемых некодирующих областей, таких как, например, лидерные последовательности, концевые последовательности или интроны, или внутри нетранскрибируемых областей, расположенных выше или ниже кодирующей области. Область, представляющая интерес, может быть такой малой по длине, как одна пара нуклеотидов, или до 2000 пар нуклеотидов в длину, или любое целое значение пар нуклеотидов.
[0132] «Эукариотические» клетки включают, помимо прочего, грибковые клетки (такие как дрожжи), клетки растений, клетки животных, клетки млекопитающих и клетки человека (например, клетки печени, мышечные клетки, эритроциты (RBC), Т-клетки и т. д.), включая стволовые клетки (плюрипотентные и мультипотентные).
[0133] «Красные кровяные клетки» (Red Blood Cell, RBC) или эритроциты представляют собой терминально дифференцированные клетки, полученные из гемопоэтических стволовых клеток. В них отсутствуют нуклеаза и большинство клеточных органелл. RBC содержат гемоглобин, переносящий кислород из легких в периферические ткани. Фактически, 33% отдельного взятого эритроцитов - это гемоглобин. Эритроциты также переносят CO2, продуцируемый клетками во время метаболизма, из тканей и обратно в легкие для высвобождения во время выдоха. RBC образуются в костном мозге в ответ на гипоксию крови, которая опосредуется высвобождением эритропоэтина (EPO) почками. EPO вызывает увеличение количества проэритробластов и сокращает время, необходимое для полного созревания эритроцитов. Примерно через 120 дней, поскольку эритроциты не содержат ядра или каких-либо других регенеративных способностей, клетки удаляются из кровотока либо за счет фагоцитарной активности макрофагов в печени, селезенке и лимфатических узлах (~ 90%), либо за счет гемолиза в плазме (~ 10%). После поглощения макрофагами, химические компоненты эритроцитов разрушаются в вакуолях макрофагов из-за действия лизосомальных ферментов. RBC, in vitro или in vivo, могут происходить из генетически модифицированных стволовых клеток или клеток-предшественников RBC, как описано в данном документе.
[0134] «Секреторные ткани» представляют собой те ткани животного, которые секретируют продукты из отдельной клетки в просвет определенного типа, который обычно происходит из эпителия. Примеры секреторных тканей, расположенных в желудочно-кишечном тракте, включают клетки, выстилающие кишечник, поджелудочную железу и желчный пузырь. Другие секреторные ткани включают печень, ткани, ассоциированные с глазом и слизистыми оболочками, например, слюнные железы, молочные железы, предстательная железа, гипофиз и другие органы эндокринной системы. Кроме того, секреторные ткани включают отдельные клетки тканевого типа, способные к секреции.
[0135] Термины «оперативная связь» и «оперативно связанный» (или «оперативно связанный») применяются взаимозаменяемо со ссылкой на сопоставление двух или большего количества компонентов (таких как элементы последовательности), в которых компоненты расположены так, что оба компонента функционируют нормально и допускают возможность того, что по меньшей мере один из компонентов может выполнять функцию, выполняемую по меньшей мере одним из других компонентов. В качестве иллюстрации, транскрипционная регуляторная последовательность, такая как промотор, оперативно связана с кодирующей последовательностью, если транскрипционная регуляторная последовательность контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или большего количества факторов регуляции транскрипции. Последовательность, регулирующая транскрипцию, обычно оперативно связана в cis с кодирующей последовательностью, но не обязательно должна непосредственно примыкать к ней. Например, энхансер представляет собой последовательность, регулирующую транскрипцию, которая оперативно связана с кодирующей последовательностью, даже если они не являются смежными.
[0136] Что касается слитых полипептидов, термин «оперативно связанный» может относиться к тому факту, что каждый из компонентов выполняет ту же функцию при связывании с другим компонентом, как если бы он не был таким образом связан. Например, в отношении слитого полипептида, в котором ДНК-связывающий домен ZFP, TALE или Cas слит с доменом активации, ДНК-связывающий домен ZFP или TALE и домен активации находятся в оперативной связи, если в слитом полипептиде часть ДНК-связывающего домена ZFP или TALE способна связывать свой целевой сайт и/или свой сайт связывания, в то время как домен активации способен повышать экспрессию гена. Когда слитый полипептид, в котором ДНК-связывающий домен ZFP или TALE слит с доменом расщепления, ДНК-связывающий домен ZFP или TALE и домен расщепления находятся в оперативной связи, если в слитом полипептиде часть ДНК-связывающего домена ZFP или TALE способна связываться со своим целевым сайтом и/или своим связывающим сайтом, в то время как домен расщепления способен расщеплять ДНК вблизи целевого сайта.
[0137] «Функциональный фрагмент» белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, последовательность которых не идентична полноразмерному белку, полипептиду или нуклеиновой кислоте, но сохраняет ту же функцию, что и полноразмерный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может иметь больше, меньше или такое же количество остатков, что и соответствующая нативная молекула, и/или может содержать одну или большее количество аминокислотных или нуклеотидных замен. Способы определения функции нуклеиновой кислоты (например, кодирующей функции, способности гибридизоваться с другой нуклеиновой кислотой) хорошо известны в данной области техники. Точно так же хорошо известны способы определения функции белков. Например, ДНК-связывающая функция полипептида может быть определена, например, с помощью анализов степени связывания с фильтром, изменения электрофоретической подвижности или иммунопреципитации. Расщепление ДНК можно оценить с помощью гель-электрофореза. См. Ausubel et al., выше. Способность белка взаимодействовать с другим белком может быть определена, например, с помощью коиммунопреципитации, двухгибридных анализов или комплементации, как генетической, так и биохимической. См., например, Fields et al. (1989) Nature 340:245-246; Патент США № 5585245 и публикацию международного патента WO 98/44350.
[0138] «Вектор» способен переносить последовательности генов в целевые клетки. Как правило, «векторная конструкция», «экспрессионный вектор», «вектор для переноса гена» и «экспрессионная конструкция» означают любую конструкцию нуклеиновой кислоты, способную управлять экспрессией гена, представляющего интерес, и которая может переносить генные последовательности в целевые клетки. Таким образом, этот термин включает клонирующие и экспрессионные носители, а также интегрирующиеся векторы.
[0139] «Репортерный ген» или «репортерная последовательность» относится к любой последовательности, которая продуцирует белковый продукт, который легко измеряется, предпочтительно, хотя и не обязательно, в обычном анализе. Пригодные репортерные гены включают, помимо прочего, последовательности, кодирующие белки, которые опосредуют устойчивость к антибиотикам (например, устойчивость к ампициллину, устойчивость к неомицину, устойчивость к G418, устойчивость к пуромицину), последовательности, кодирующие окрашенные, флуоресцентные или люминесцентные белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люцифераза), а также белки, которые опосредуют усиленный рост клеток и/или амплификацию гена (например, дигидрофолатредуктаза). Теги эпитопа включают, например, одну или большее количество копий FLAG, His, myc, Tap, HA или любую детектируемую аминокислотную последовательность. «Экспрессионные теги» включают последовательности, которые кодируют репортеры, которые могут быть оперативно связаны с желаемой последовательностью гена, чтобы контролировать экспрессию гена, представляющего интерес.
[0140] Термины «субъект» и «пациент» применяются взаимозаменяемо и относятся к млекопитающим, таким как пациенты-люди и приматы, не являющиеся людьми, а также к экспериментальным животным, таким как кролики, собаки, кошки, крысы, мыши и другие животные. Соответственно, в данном контексте термин «субъект» или «пациент» означает любого пациента-млекопитающего или субъекта, которому могут быть введены измененные клетки, описанные в данном документе, и/или белки, продуцируемые измененными клетками, описанными в данном документе. Субъекты по настоящему изобретению включают тех субъектов, у которых есть LSD.
[0141] В данном документе описаны способы и композиции для лечения и/или предотвращения болезни Фабри. В данном документе описаны способы вставки последовательности трансгена в пригодную целевую клетку (например, клетку от субъекта с болезнью Фабри), при этом трансген кодирует по меньшей мере один белок (например, по меньшей мере один белок α-GalA), который лечит указанное заболевание. Способы могут быть in vivo (доставка последовательности трансгена в клетку живого субъекта) или ex vivo (доставка модифицированных клеток живому субъекту). В данном документе также описаны способы трансфекции и/или трансдукции пригодной целевой клетки с помощью системы экспрессии, при которой трансген, кодирующий α-GalA, экспрессирует белок, который лечит указанное заболевание (например, облегчает один или большее количество симптомов, ассоциированных с заболеванием). Белок α-GalA может выделяться (секретироваться) из целевой клетки, так что он может воздействовать или поглощаться другими клетками, которые не содержат трансген (перекрестная коррекция). В настоящем изобретении также предлагаются способы продуцирования клетки (например, зрелой или недифференцированной клетки), которая продуцирует высокие уровни α-GalA, при этом введение популяции этих измененных клеток пациенту будет обеспечивать поступление этого белка, необходимого для лечения заболевания или патологического состояния. Кроме того, предлагаются способы продуцирования клетки (например, зрелой или недифференцированной клетки), которая продуцирует высокоактивную (терапевтическую) форму α-GalA, при этом введение или создание популяции этих измененных клеток в организм пациента будет обеспечивать необходимую активность белка для лечения (например, уменьшения или устранения одного или большего количества симптомов) болезни Фабри. Высокоактивная форма α-GalA, полученная, как описано в данном документе, также может быть выделена из клеток, как описано в данном документе, и введена пациенту, нуждающемуся в этом, с помощью стандартных процедур замещения ферментов, известных специалисту в данной области техники.
[0142] В данном документе описаны способы и композиции для экспрессии по меньшей мере одного белка α-галактозидазы A (α-Gal A). Композиции и способы могут применяться in vitro, in vivo или ex vivo, и включают введение трансгена GLA (например, кДНК с последовательностями GLA дикого типа или кодон-оптимизированными последовательностями GLA), кодирующего по меньшей мере один белок α-Gal A, в клетку таким образом, чтобы белок α-Gal A экспрессировался в клетке. В определенных вариантах осуществления клетка находится в организме субъекта с болезнью Фабри. В любом из описанных в данном документе способов трансген можно вводить в печень субъекта. Необязательно, указанные способы дополнительно включают введение одной или большего количества нуклеаз, которые расщепляют эндогенный ген альбумина в клетке печени субъекта, таким образом, что трансген интегрируется в ген альбумина и экспрессируется из него. В любом из описанных в данном документе способов белок α-Gal A, экспрессируемый из трансгена, может снизить количество гликосфинголипидов у субъекта по меньшей мере примерно в 2 раза по сравнению с нелеченными субъектами или субъектами, которые получали буфер для состава или другой носитель. Трансген GLA может дополнительно содержать дополнительные элементы, включая, например, сигнальный пептид и/или один или большее количество контрольных элементов. В определенных вариантах осуществления трансген GLA (например, конструкция кДНК) дополнительно включает последовательность WPRE дикого типа или сконструированную последовательность WPRE, например, мутированную последовательность WPRE, содержащую мутации WPRE mut6, описанные в публикации Zanta-Boussif et al. (2009) Gene Therapy 16:605-619 и в патенте США № 10179918. В некоторых вариантах осуществления мутации mut6 выполнены в элементе J04514 WPRE, тогда как в других вариантах они выполнены в элементе J02442.1 WPRE (Ong et al. (2017) doi.org/10.1101/126904). В некоторых вариантах осуществления экспрессионная конструкция GLA содержит конструкцию, продемонстрированную на Фиг. 1В (вариант №21). Экспрессионные конструкции, содержащие WPRE, как описано в данном документе, приводят к улучшенной экспрессии и активности трансгена по сравнению с экспрессионными конструкциями, не включающими последовательности WPRE (например, 1-кратное, 2-кратное, 3-кратное, 4-кратное, 5-кратное, 6 -кратное, 7-кратное, 8-кратное, 9-кратное, 10-кратное или более увеличение экспрессии или активности). В определенных вариантах осуществления экспрессионная конструкция представляет собой конструкцию, приведенную в Таблице 1.
[0143] В одном аспекте данного изобретения описан способ экспрессии трансгена, кодирующего один или большее количество корректирующих трансгенов GLA, в клетке субъекта. Трансген может быть вставлен в геном пригодной целевой клетки (например, клетки крови, клетки печени, клетки мозга, стволовой клетки, клетки-предшественника и т. д.), так что продукт α-GalA, кодируемый этим корректирующим трансгеном, стабильно интегрируется в геном клетки (этот подход также называется «IVPRP») или, альтернативно, трансген может сохраняться в клетке вне хромосомы (этот подход также называется «кДНК»). В одном варианте осуществления корректирующий трансген GLA вводят (стабильно или вне хромосомно) в клетки клеточной линии для продуцирования in vitro замещающего белка, при этом белок (необязательно очищенный и/или выделенный) затем может быть введен субъекту для лечения субъекта с болезнью Фабри (например, путем ослабления и/или устранения одного или большего количества симптомов, ассоциированных с болезнью Фабри). В определенных вариантах осуществления продукт α-GalA, кодируемый этим корректирующим трансгеном, повышает активность α-GalA в ткани субъекта на любое количество по сравнению с нелеченными субъектами, например, примерно в 2-2000 раз (или любое значение между ними), включая, помимо прочего, в 2-100 раз (или любое значение между ними, включая 10-, 20-, 30-, 40-, 50-, 60-, 70-, 80-, 90-, 100- раз), в 100-500 раз (или любое значение между ними), в 500-1000 раз (или любое значение между ними) или в 1000-2000 раз или более.
[0144] В другом аспекте в настоящем изобретении описаны способы ex vivo или in vivo лечения субъекта с болезнью Фабри (например, путем ослабления и/или устранения одного или большего количества симптомов, ассоциированных с болезнью Фабри), способы, включающие вставку трансгена GLA в клетку, как описано в данном документе (подходы кДНК и/или IVPRP), так что белок продуцируется у субъекта с болезнью Фабри. В определенных вариантах осуществления трансген GLA является частью конструкции, как продемонстрировано в Таблице 1. В определенных вариантах осуществления выделенные клетки, содержащие трансген GLA, можно применять для лечения пациента, нуждающегося в этом, например, путем введения клеток субъекту с болезнью Фабри. В других вариантах осуществления корректирующий трансген GLA вставляют в целевую ткань в организме, так что замещающий белок продуцируется in vivo. В некоторых вариантах осуществления корректирующий трансген вставляют в геном клеток в целевую ткань, тогда как в других предпочтительных вариантах осуществления корректирующий трансген вставляют в клетки целевой ткани, где он сохраняется в клетках вне хромосом. В любом из описанных в данном документе способов экспрессированный белок α-GalA может выводиться из клетки для воздействия на вторичные мишени или поглощаться ими, в том числе другими клетками в других тканях (например, путем экспорта в кровь), в которых отсутствует трансген GLA (перекрестная коррекция). В некоторых случаях первичной и/или вторичной целевой тканью является печень. В некоторых случаях первичной и/или вторичной целевой тканью является головной мозг. В некоторых случаях первичной и/или вторичной целевой тканью является кровь (например, сосудистая сеть). В некоторых случаях первичной и/или вторичной целевой тканью является скелетная мышца.
[0145] В определенных вариантах осуществления способы и композиции, описанные в данном документе, применяются для уменьшения количества гликосфинголипидов, включая глоботриаозилцерамид (также известный как GL-3 и Gb3) и глоботриаозилсфингозин (лизо-Gb3), галабиоазилцерамид, депонированный в тканях субъекта, страдающего болезнью Фабри. В определенных вариантах осуществления продукт α-GalA, кодируемый этим корректирующим трансгеном, снижает уровень гликосфинголипидов в ткани субъекта на любое количество по сравнению с нелеченными субъектами, например, примерно в 2-100 раз (или любое другое значение между ними), включая, помимо прочего, в 2-100 раз (или любое значение между ними, включая 10-, 20-, 30-, 40-, 50-, 60-, 70-, 80-, 90-, 100- раз). В определенных вариантах осуществления продукт α-GalA, кодируемый этим корректирующим трансгеном, снижает гликосфинголипиды в ткани субъекта на любое количество по сравнению с нелеченными субъектами, например, по меньшей мере, на около 10%, 20%, 30%, 40%, 50. %, 60%, 70%, 80%, 90% или около 100%.
[0146] В любом из описанных в данном документе способов корректирующий трансген GLA включает последовательность дикого типа функционирующего гена GLA, в то время как в других вариантах осуществления последовательность корректирующего трансгена GLA изменена некоторым образом для придания повышенной биологической активности (например, оптимизированные кодоны для увеличения биологической активности и/или изменения транскрипционных и трансляционных регуляторных последовательностей для улучшения экспрессии генов). В некоторых вариантах осуществления ген GLA модифицируют для улучшения характеристик экспрессии. Такие модификации могут включать, помимо прочего, вставку сайта начала трансляции (например, метионина), добавление оптимизированной последовательности Kozak, вставку сигнального пептида и/или оптимизацию кодонов. В некоторых вариантах осуществления сигнальный пептид может быть выбран из сигнального пептида альбумина, сигнального пептида F.IX, сигнального пептида IDS и/или сигнального пептида α-GalA.
[0147] В определенных аспектах доноры являются донорами кДНК. Доноры кДНК обычно включают энхансерную последовательность, промоторную последовательность, интронную последовательность, сигнальный пептид, кодирующую последовательность GLA, сигнал полиаденилирования и, необязательно, последовательность WPRE дикого типа или мутированную последовательность. Неограничивающие типовые доноры кДНК схематично продемонстрированы на Фиг. 1A и Фиг. 1B.
[0148] В конструкциях кДНК можно применять любой промотор, энхансер, интрон, сигнальный пептид, последовательность, кодирующую GLA или полиА, и необязательную последовательность WPRE. В некоторых вариантах осуществления энхансер и/или промотор являются специфичными для печени, например, состоят из энхансера ApoE человека и промотора анти-α1-антитрипсина человека (hAAT) (Miao CH et al. (2000) Mol. Ther. 1(6): 522-532 (200)). В некоторых вариантах осуществления специфический для печени промотор содержит одну или большее количество последовательностей энхансера ApoE (например, 1, 2, 3 и/или 4; см. публикацию Okuyama et al. (1996) Hum Gen Ther 7(5):637-45). В некоторых вариантах осуществления промотор связан с интроном. В некоторых вариантах осуществления интрон представляет собой химерный интрон HBB-IGG, содержащий 5’ донорный сайт из первого интрона гена β-глобина человека и ответвление, а также 3’ акцепторный сайт из интрона вариабельной области тяжелой цепи гена иммуноглобулина. В некоторых вариантах осуществления промотор ApoE/hAAT является специфически и высокоактивным в гепатоцитах, предполагаемой целевой ткани, но неактивен в типах клеток и тканей, не относящихся к печени; а также снижает или предотвращает экспрессию и активность в тканях, не являющихся целевыми тканями. В определенных вариантах осуществления сигнальный пептид содержит сигнальный пептид GLA, а сигнал полиаденилирования содержит последовательность полиА SPA51 или bGH. Необязательная последовательность WPRE может быть любой последовательностью WPRE дикого типа или мутированной последовательностью. См., например патент США № 10179918. В определенных вариантах осуществления последовательность WPRE содержит мутированную WPRE, например, последовательность mut6 WPRE.
[0149] Описанные в данном документе экспрессионные векторы кДНК могут быть доставлены с помощью любого пригодного вектора, включая вирусные векторы, такие как AAV любого серотипа (например, AAV2, AAV6 или AAV2/6).
[0150] В определенных вариантах осуществления экспрессионная последовательность (то есть экспрессионный вектор или экспрессионная конструкция) содержит элементы и последовательность варианта № 21, изображенного на Фиг. 1B, и как продемонстрировано ниже в Таблице 1.
Таблица 1. Элементы варианта № 21 кДНК и полная последовательность
GGCGACCTTTGGTCGCCCGGCCTCAGTGAGCGAGCGAGCGCGCAGAGAGG
GAGTGGCCAACTCCATCACTAGGGGTTCCT (SEQ ID №:1)
CCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCC ACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTG CAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAG CTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCC ACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGT
GGTTTAGGTAGTGTGAGAGGG (SEQ ID №:2)
CCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGT (SEQ ID №:3)
TTACTGACATCCACTTTGCCTTTCTCTCCACAG (SEQ ID №:4)
CTTCCTCGCTCTCGTTTCTTGGGACATCCCTGGCGCTAGGGCA GTTTATGTGCAATCTGGATTGCCAGGAGGAGCCGGACTCATGCATCTCGG AGAAGCTGTTCATGGAGATGGCGGAACTTATGGTATCGGAGGGATGGAAG GATGCCGGGTATGAGTATCTCTGTATCGACGATTGTTGGATGGCTCCCCA GAGAGACTCCGAGGGACGACTCCAAGCGGACCCCCAGCGCTTTCCACATG GCATTCGACAGCTCGCCAATTACGTGCACTCGAAGGGGTTGAAGTTGGGA ATCTACGCAGATGTGGGCAACAAAACGTGTGCGGGGTTCCCGGGGTCGTT
TGGATACTACGATATTGATGCGCAGACGTTTGCTGACTGGGGTGTCGATC TTTTGAAATTTGATGGCTGTTACTGTGATTCGTTGGAAAACCTGGCGGAT GGATACAAGCATATGTCACTCGCCTTGAACCGGACAGGTCGCTCAATCGT ATACAGCTGCGAATGGCCCCTCTATATGTGGCCCTTCCAAAAGCCCAATT ACACAGAGATTCGGCAGTATTGCAATCACTGGAGGAACTTTGCCGATATT GACGACAGCTGGAAATCCATCAAGTCCATTCTCGATTGGACGAGCTTCAA CCAGGAGCGCATCGTGGACGTGGCAGGACCCGGAGGTTGGAACGATCCGG ACATGCTCGTAATTGGGAATTTCGGGCTTAGCTGGAATCAGCAAGTCACC CAAATGGCGCTGTGGGCCATCATGGCAGCTCCTCTCTTTATGTCGAATGA TCTGCGGCATATCTCGCCCCAGGCAAAGGCTCTTTTGCAAGACAAGGACG TCATCGCAATCAATCAGGACCCATTGGGGAAACAGGGATATCAACTTCGC CAGGGTGACAATTTCGAAGTATGGGAGAGGCCGCTTAGCGGGCTGGCGTG GGCGGTCGCGATGATTAACCGGCAGGAAATCGGAGGGCCTCGCTCGTATA CCATCGCAGTGGCCTCACTGGGCAAAGGAGTGGCGTGCAATCCGGCCTGC TTCATCACCCAGTTGTTGCCCGTCAAAAGAAAGCTGGGTTTCTACGAGTG GACATCCAGACTTAGATCACACATTAACCCTACTGGTACGGTGTTGCTCC
AGCTCGAAAACACAATGCAGATGTCGTTGAAAGACCTGCTGTAA (SEQ ID №:5)
J04514
CTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG (SEQ ID №:6)
GCTGGGGATGCGGTGGGCTCTATGG (SEQ ID №:7)
GCGCGCAG (SEQ ID №:8)
CTGCGCGCTC GCTCGCTCAC TGAGGCCGCC CGGGCAAAGC CCGGGCGTCG 50
GGCGACCTTT GGTCGCCCGG CCTCAGTGAG CGAGCGAGCG CGCAGAGAGG 100
GAGTGGCCAA CTCCATCACT AGGGGTTCCT GCGGCCTAGT AGGCTCAGAG 150
GCACACAGGA GTTTCTGGGC TCACCCTGCC CCCTTCCAAC CCCTCAGTTC 200
CCATCCTCCA GCAGCTGTTT GTGTGCTGCC TCTGAAGTCC ACACTGAACA 250
AACTTCAGCC TACTCATGTC CCTAAAATGG GCAAACATTG CAAGCAGCAA 300
ACAGCAAACA CACAGCCCTC CCTGCCTGCT GACCTTGGAG CTGGGGCAGA 350
GGTCAGAGAC CTCTCTGGGC CCATGCCACC TCCAACATCC ACTCGACCCC 400
TTGGAATTTC GGTGGAGAGG AGCAGAGGTT GTCCTGGCGT GGTTTAGGTA 450
GTGTGAGAGG GGTACCCGGG GATCTTGCTA CCAGTGGAAC AGCCACTAAG 500
GATTCTGCAG TGAGAGCAGA GGGCCAGCTA AGTGGTACTC TCCCAGAGAC 550
TGTCTGACTC ACGCCACCCC CTCCACCTTG GACACAGGAC GCTGTGGTTT 600
CTGAGCCAGG TACAATGACT CCTTTCGGTA AGTGCAGTGG AAGCTGTACA 650
CTGCCCAGGC AAAGCGTCCG GGCAGCGTAG GCGGGCGACT CAGATCCCAG 700
CCAGTGGACT TAGCCCCTGT TTGCTCCTCC GATAACTGGG GTGACCTTGG 750
TTAATATTCA CCAGCAGCCT CCCCCGTTGC CCCTCTGGAT CCACTGCTTA 800
AATACGGACG AGGACAGGGC CCTGTCTCCT CAGCTTCAGG CACCACCACT 850
GACCTGGGAC AGTCAGGTAA GTATCAAGGT TACAAGACAG GTTTAAGGAG 900
ACCAATAGAA ACTGGGCTTG TCGAGACAGA GAAGACTCTT GCGTTTCTGA 950
TAGGCACCTA TTGGTCTTAC TGACATCCAC TTTGCCTTTC TCTCCACAGG 1000
CAATTGATCC CCCTGATCTG CGGCCTCGAC GGTATCGATA AGCTTGCCAC 1050
CATGCAACTT AGGAACCCCG AACTTCATCT TGGCTGCGCC CTGGCCCTCC 1100
GCTTCCTCGC TCTCGTTTCT TGGGACATCC CTGGCGCTAG GGCACTCGAC 1150
AACGGCCTCG CGCGGACTCC TACGATGGGA TGGTTGCACT GGGAAAGGTT 1200
TATGTGCAAT CTGGATTGCC AGGAGGAGCC GGACTCATGC ATCTCGGAGA 1250
AGCTGTTCAT GGAGATGGCG GAACTTATGG TATCGGAGGG ATGGAAGGAT 1300
GCCGGGTATG AGTATCTCTG TATCGACGAT TGTTGGATGG CTCCCCAGAG 1350
AGACTCCGAG GGACGACTCC AAGCGGACCC CCAGCGCTTT CCACATGGCA 1400
TTCGACAGCT CGCCAATTAC GTGCACTCGA AGGGGTTGAA GTTGGGAATC 1450
TACGCAGATG TGGGCAACAA AACGTGTGCG GGGTTCCCGG GGTCGTTTGG 1500
ATACTACGAT ATTGATGCGC AGACGTTTGC TGACTGGGGT GTCGATCTTT 1550
TGAAATTTGA TGGCTGTTAC TGTGATTCGT TGGAAAACCT GGCGGATGGA 1600
TACAAGCATA TGTCACTCGC CTTGAACCGG ACAGGTCGCT CAATCGTATA 1650
CAGCTGCGAA TGGCCCCTCT ATATGTGGCC CTTCCAAAAG CCCAATTACA 1700
CAGAGATTCG GCAGTATTGC AATCACTGGA GGAACTTTGC CGATATTGAC 1750
GACAGCTGGA AATCCATCAA GTCCATTCTC GATTGGACGA GCTTCAACCA 1800
GGAGCGCATC GTGGACGTGG CAGGACCCGG AGGTTGGAAC GATCCGGACA 1850
TGCTCGTAAT TGGGAATTTC GGGCTTAGCT GGAATCAGCA AGTCACCCAA 1900
ATGGCGCTGT GGGCCATCAT GGCAGCTCCT CTCTTTATGT CGAATGATCT 1950
GCGGCATATC TCGCCCCAGG CAAAGGCTCT TTTGCAAGAC AAGGACGTCA 2000
TCGCAATCAA TCAGGACCCA TTGGGGAAAC AGGGATATCA ACTTCGCCAG 2050
GGTGACAATT TCGAAGTATG GGAGAGGCCG CTTAGCGGGC TGGCGTGGGC 2100
GGTCGCGATG ATTAACCGGC AGGAAATCGG AGGGCCTCGC TCGTATACCA 2150
TCGCAGTGGC CTCACTGGGC AAAGGAGTGG CGTGCAATCC GGCCTGCTTC 2200
ATCACCCAGT TGTTGCCCGT CAAAAGAAAG CTGGGTTTCT ACGAGTGGAC 2250
ATCCAGACTT AGATCACACA TTAACCCTAC TGGTACGGTG TTGCTCCAGC 2300
TCGAAAACAC AATGCAGATG TCGTTGAAAG ACCTGCTGTA ATCTAGAGGA 2350
TCTCGAGAGA TCTAATCAAC CTCTGGATTA CAAAATTTGT GAAAGATTGA 2400
CTGGTATTCT TAACTATGTT GCTCCTTTTA CGCTATGTGG ATACGCTGCT 2450
TTAATGCCTT TGTATCATGC TATTGCTTCC CGTATGGCTT TCATTTTCTC 2500
CTCCTTGTAT AAATCCTGGT TGCTGTCTCT TTATGAGGAG TTGTGGCCCG 2550
TTGTCAGGCA ACGTGGCGTG GTGTGCACTG TGTTTGCTGA CGCAACCCCC 2600
ACTGGTTGGG GCATTGCCAC CACCTGTCAG CTCCTTTCCG GGACTTTCGC 2650
TTTCCCCCTC CCTATTGCCA CGGCGGAACT CATCGCCGCC TGCCTTGCCC 2700
GCTGCTGGAC AGGGGCTCGG CTGTTGGGCA CTGACAATTC CGTGGTGTTG 2750
TCGGGGAAAT CATCGTCCTT TCCTTGGCTG CTCGCCTGTG TTGCCACCTG 2800
GATTCTGCGC GGGACGTCCT TCTGCTACGT CCCTTCGGCC CTCAATCCAG 2850
CGGACCTTCC TTCCCGCGGC CTGCTGCCGG CTCTGCGGCC TCTTCCGCGT 2900
CTTCGCCTTC GCCCTCAGAC GAGTCGGATC TCCCTTTGGG CCGCCTCCCC 2950
GCCTGGGATC TCTGTGCCTT CTAGTTGCCA GCCATCTGTT GTTTGCCCCT 3000
CCCCCGTGCC TTCCTTGACC CTGGAAGGTG CCACTCCCAC TGTCCTTTCC 3050
TAATAAAATG AGGAAATTGC ATCGCATTGT CTGAGTAGGT GTCATTCTAT 4000
TCTGGGGGGT GGGGTGGGGC AGGACAGCAA GGGGGAGGATTGGGAAGACA4050
ATAGCAGGCA TGCTGGGGAT GCGGTGGGCT CTATGGACCG GTCTCGAGAT 4100
CCACTAGGGC CGCAGGAACC CCTAGTGATG GAGTTGGCCA CTCCCTCTCT 4150
GCGCGCTCGC TCGCTCACTG AGGCCGCCCG GGCTTTGCCC GGGCGGCCTC 4200
AGTGAGCGAG CGAGCGCGCA G (SEQ ID №:9) 4221
[0151] Экспрессионная конструкция из Таблицы 1, которая содержит последовательность WPRE, может быть легко получена в клиническом масштабе, и было показано, что она демонстрирует улучшенную активность GLA, например, по меньшей мере примерно в 3 раза, примерно в 4 раза, примерно в 5 раз, примерно в 6 раз , примерно в 7 раз, примерно в 8 раз, примерно в 9 раз , примерно в 10 раз, примерно в 11 раз, примерно в 12 раз, примерно в 13 раз, примерно в 14 раз, примерно в 15 раз, примерно в 16 раз, примерно в 17 раз, примерно в 18 раз, примерно в 19 раз, примерно в 20 раз по сравнению с экспрессионными конструкциями, которые не включают последовательность WPRE.
[0152] В другом аспекте настоящего изобретения описан экспрессионный вектор нуклеазы (например, ZFN, пара ZFN, TALEN, пара TALEN и/или система CRISPR/Cas), содержащий полинуклеотид, кодирующий одну или большее количество нуклеаз, как описано в данном документе, оперативно связанный с промотором. В одном варианте осуществления указанный экспрессионный вектор представляет собой вирусный вектор. В дополнительном аспекте настоящего изобретения описан экспрессионный вектор GLA, содержащий полинуклеотид, кодирующий α-GalA, как описано в данном документе, оперативно связанный с промотором. В одном варианте осуществления указанный экспрессионный вектор представляет собой вирусный вектор
[0153] В другом аспекте настоящего изобретения описана клетка-хозяин, содержащая одну или большее количество экспрессионных векторов нуклеаз (например, ZFN, пара ZFN, TALEN, пара TALEN и/или система CRISPR/Cas) и/или экспрессионных векторов α-GalA, как описано в данном документе. Клетка-хозяин может быть стабильно трансформирована, или временно трансфицирована, или может быть их комбинацией с одним или большим количеством экспрессионных векторов нуклеазы. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку печени.
[0154] В других вариантах осуществления предлагаются способы замены геномной последовательности в любом целевом гене терапевтическим трансгеном GLA, как описано в данном документе, например, с применением нуклеазы (например, ZFN, пара ZFN, TALEN, пара TALEN и/или система CRISPR/Cas). (или один или большее количество векторов, кодирующих указанную нуклеазу), как описано в данном документе, и «донорную» последовательность или трансген GLA, который вставляют в ген после целенаправленного расщепления нуклеазой. Последовательность GLA может присутствовать в векторе, несущем нуклеазу (или ее компонент), присутствовать в отдельном векторе (например, в векторе Ad, AAV или LV или мРНК) или, альтернативно, может быть введена в клетку с применением другого механизма доставки нуклеиновой кислоты. Такая вставка донорной нуклеотидной последовательности в целевой локус (например, высокоэкспрессируемый ген, ассоциированный с заболеванием ген, другой ген безопасного убежища и т.д.) приводит к экспрессии трансгена GLA под контролем эндогенных генетических контрольных элементов целевого локуса (например, альбумин, глобин и др.). В некоторых аспектах вставка трансгена GLA, например, в целевой ген (например, альбумина), приводит к экспрессии интактной последовательности белка α-GalA и не содержит каких-либо аминокислот, кодируемых целевым геном (например, альбумина). В других аспектах экспрессированный экзогенный белок α-GalA представляет собой слитый белок и содержит аминокислоты, кодируемые трансгеном GLA и эндогенным локусом, в который вставлен трансген GLA (например, из эндогенного целевого локуса или, альтернативно, из последовательностей трансгена, кодирующего последовательности целевого локуса). Мишенью может быть любой ген, например, ген безопасного убежища, такой как ген альбумина, ген AAVS1, ген HPRT; ген CCR5; или высокоэкспрессируемый ген, такой как ген глобина в клетке-предшественнике эритроцитов (например, бета-глобин или гамма-глобин). В некоторых случаях эндогенные последовательности будут присутствовать на амино (N)-концевой части экзогенного белка α-GalA, в то время как другие эндогенные последовательности будут присутствовать на карбокси (C)-концевой части экзогенного белка α-GalA. В других случаях эндогенные последовательности будут присутствовать как на N-, так и на C-концевых частях экзогенного белка α-GalA. В некоторых вариантах осуществления эндогенные последовательности кодируют сигнальный пептид секреции, который удаляется в процессе секреции белка α-GalA из клетки. Эндогенные последовательности могут включать полноразмерные эндогенные последовательности дикого типа или мутантные эндогенные последовательности или, альтернативно, могут включать частичные эндогенные аминокислотные последовательности. В некоторых вариантах осуществления слияние эндогенный ген-трансген локализовано в эндогенном локусе внутри клетки, тогда как в других вариантах осуществления слияние эндогенная последовательность - кодирующая трансген последовательность вставлено в другой локус в геноме (например, слияние GLA - последовательность трансгена вставлено в альбумин, локус HPRT или CCR5). В некоторых вариантах осуществления трансген GLA экспрессируется таким образом, что терапевтический белковый продукт α-GalA сохраняется в клетке (например, в клетке-предшественнике или зрелой клетке). В других вариантах осуществления GLA трансген слит с внеклеточным доменом мембранного белка таким образом, что при экспрессии слияние трансген - α-GalA приведет к поверхностной локализации терапевтического белка. В некоторых аспектах отредактированные клетки дополнительно содержат трансмембранный белок для доставки клеток к конкретному типу ткани. В одном аспекте трансмембранный белок содержит антитело, в то время как в других аспектах трансмембранный белок содержит рецептор. В определенных вариантах осуществления клетка является предшественником (например, CD34+или гемопоэтической стволовой клеткой) или зрелым RBC (эритроцитом) (происходящим от генетически модифицированной клетки, продуцирующей GAL, как описано в данном документе). В некоторых аспектах терапевтический белковый продукт α-GalA, кодируемый трансгеном, экспортируется из клетки, чтобы воздействовать на клетки, лишенные трансгена, или поглощаться ими. В определенных вариантах осуществления клетка представляет собой клетку печени, которая высвобождает терапевтический белок α-GalA в кровоток для воздействия на дистальные ткани (например, почки, селезенку, сердце, головной мозг, кожу и т. д.).
[0155] В одном варианте осуществления GLA трансген экспрессируется с промотора альбумина после встраивания в локус альбумина. Биологическое вещество, кодируемое GLA трансгеном, затем может быть высвобождено в кровоток, если трансген вставлен в гепатоцит in vivo. В некоторых аспектах GLA трансген доставляется в печень in vivo в вирусном векторе посредством внутривенного введения. В некоторых вариантах осуществления донорный GLA трансген содержит консенсусную последовательность Kozak перед кодирующей последовательностью α-GalA (Kozak (1987) Nucl Acid Res 15(20):8125-48), так что в экспрессируемом продукте отсутствует сигнальный пептид альбумина. В некоторых вариантах осуществления донорный α-GalA трансген содержит альтернативный сигнальный пептид, такой как пептид из генов альбумина, IDS или F9, вместо нативной сигнальной последовательности GLA.
[0156] В еще одном дополнительном аспекте настоящего изобретения предлагается способ сайт-специфической интеграции последовательности нуклеиновой кислоты в эндогенный локус (например, ассоциированный с заболеванием, высоко экспрессируемый, такой как локус альбумина в клетке печени или локус глобина в RBC клетках-предшественниках хромосомы, например, в хромосоме не относящегося к человеку эмбриона. В некоторых вариантах осуществления указанный способ включает: (а) инъекцию эмбриону, не относящемуся к человеку, (i) по меньшей мере одного ДНК-вектора, при этом ДНК-вектор содержит вышерасположенную последовательность и нижерасположенную последовательность, фланкирующую последовательность нуклеиновой кислоты, кодирующую α-GalA, которая должна быть интегрирована, и (ii) по меньшей мере одной молекулы полинуклеотида, кодирующей по меньшей мере одну нуклеазу (цинковый палец, пара ZFN, нуклеаза TALE, пара TALEN или система CRISPR/Cas), которая распознает сайт интеграции в целевом локусе, и (b) культивирование эмбриона для обеспечения экспрессии нуклеазы (ZFN, TALEN, и/или система CRISPR/Cas, при этом двухцепочечный разрыв, введенный нуклеазой в сайт интеграции, репарируется посредством гомологичной рекомбинации с ДНК-вектором, чтобы интегрировать последовательность нуклеиновой кислоты в хромосому. В некоторых вариантах осуществления полинуклеотид, кодирующий нуклеазу, представляет собой РНК.
Нуклеазы
[0157] Любую нуклеазу можно применять на практике аспектов способов, описанных в данном документе, включая, помимо прочего, по меньшей мере один из следующих элементов: ZFN, TALEN, хоуминг-эндонуклеазы и системы, содержащие направляющие РНК CRISPR/Cas и/или Ttago, которые пригодны для расщепления донорной молекулы in vivo, несущей трансген, и нуклеазы для расщепления генома клетки, так что трансген целенаправленно интегрируется в геном. Таким образом, в данном документе описаны композиции, содержащие одну или большее количество нуклеаз, которые расщепляют выбранный ген, расщепление которого приводит к геномной модификации гена (например, вставки и/или делеции в расщепленный ген). В определенных вариантах осуществления одна или большее количество нуклеаз являются такими, которые встречаются в природе. В других вариантах осуществления одна или большее количество нуклеаз являются такими, которые не встречаются в природе, то есть сконструированы в ДНК-связывающей молекуле (также называемой ДНК-связывающим доменом) и/или в домене расщепления. Например, ДНК-связывающий домен природной нуклеазы может быть изменен для связывания с выбранным целевым сайтом (например, ZFP, TALE и/или sgРНК CRISPR/Cas, которые сконструированы для связывания с выбранным целевым сайтом). В других вариантах осуществления нуклеаза содержит гетерологичные ДНК-связывающие домены и домены расщепления (например, нуклеазы цинковые пальцы; ДНК-связывающие белки с эффекторным доменом TAL; ДНК-связывающие домены мегануклеазы с гетерологичными доменами расщепления). В других вариантах осуществления нуклеаза содержит систему, такую как CRISPR/Cas системы Ttago.
ДНК-связывающие домены
[0158] В некоторых вариантах осуществления, в композиции и способах, описанных в настоящем документе, применяется ДНК-связывающий домен мегануклеазы (хоуминг-эндонуклеаза) для связывания с донорной молекулой и/или связывания с представляющей интерес областью в геноме клетки. Встречающиеся в природе мегануклеазы распознают сайты расщепления из 15-40 пар оснований и обычно группируются в четыре семейства: семейство LAGLIDADG («LAGLIDADG», описанное как SEQ ID №: 10), семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Примеры самонаводящихся эндонуклеаз включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Их последовательности распознавания известны. См. также патент США № 5420032; патент США № 6833252;; Belfort et al. (1997) Nucleic AcidsRes.25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet.12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог биолабораторий Новой Англии. Кроме того, специфичность хоуминг-эндонуклеаз и мегануклеаз относительно связывания ДНК может быть сконструирована для связывания неприродных целевых сайтов. См., например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy7:49-66; патент США № 8021867. ДНК-связывающие домены хоуминг-эндонуклеаз и мегануклеаз могут быть изменены в контексте нуклеазы в целом (то есть так, что нуклеаза включает родственный домен расщепления) или могут быть слиты с гетерологичным доменом расщепления.
[0159] В других вариантах осуществления ДНК-связывающий домен одной или большего количества нуклеаз, применяемых в описанных в данном документе способах и композициях, включает природный или сконструированный (не встречающийся в природе) эффекторный ДНК-связывающий домен TAL. См., например, патент США № 8586526, в полном объеме включенный в настоящий документ посредством ссылки. Патогенные для растений бактерии рода Xanthomonas, как известно, вызывают многие заболевания у важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от системы консервативной секреции типа III (T3S), которая вводит более 25 различных эффекторных белков в растительную клетку. Среди этих введенных белков есть эффекторы, подобные активаторам транскрипции (TAL), которые имитируют активаторы транскрипции растений и манипулируют транскриптомом растений (см. публикацию Kay et al. (2007) Science 318:648-651). Эти белки содержат домен связывания ДНК и домен активации транскрипции. Одним из наиболее хорошо охарактеризованных TAL-эффекторов является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (см. публикацию Bonas et al. (1989) Mol Gen Genet 218: 127-136 и WO2010079430). TAL-эффекторы содержат централизованный домен тандемных повторов, каждый из которых содержит приблизительно 34 аминокислоты, которые являются ключевыми для специфичности связывания ДНК этих белков. Кроме того, они содержат последовательность ядерной локализации и кислотный домен активации транскрипции (обзор см. в публикации Schornack S, et al (2006) J Plant Physiol 163(3): 256-272). Кроме того, в фитопатогенных бактериях Ralstonia solanacearum были обнаружены два гена, обозначенные brg11 и hpx17, которые гомологичны семейству AvrBs3 Xanthomonas в биоваре 1 штамма GMI1000 и в биоваре 4 штамма RS1000 R. solanacearum (см. публикацию Heuer et al (2007) Appl and Envir Micro 73(13): 4379-4384). Эти гены на 98,9% идентичны по нуклеотидной последовательности друг другу, но отличаются делецией 1,575 п.н. в повторяющемся домене hpx17. Однако оба генных продукта имеют менее 40% идентичности последовательности с белками семейства AvrBs3 Xanthomonas. См., например, патент США № 8586526, в полном объеме включенный в настоящий документ посредством ссылки.
[0160] Специфичность этих эффекторов TAL зависит от последовательностей, обнаруженных в тандемных повторах. Повторяющаяся последовательность составляет приблизительно 102 п.н., при этом повторы обычно на 91-100% гомологичны друг другу (Bonas et al, ibid). Полиморфизм повторов обычно находится в положениях 12 и 13, и, по-видимому, существует взаимно однозначное соответствие между идентичностью гипервариабельных двойных остатков (RVD) в положениях 12 и 13 с идентичностью смежных нуклеотидов в целевой последовательности TAL-эффектора (см. публикации Moscou and Bogdanove, (2009) Science 326:1501 и Boch et al (2009) Science 326:1509-1512). Экспериментально естественный код распознавания ДНК этих TAL-эффекторов был определен таким образом, что последовательность HD в положениях 12 и 13 приводит к связыванию с цитозином (C), NG связывается с T, NI с A, C, G или T, NN связывается с A или G, а ING связывается с T. Эти ДНК-связывающие повторы были собраны в белки с новыми комбинациями и количеством повторов, чтобы создать искусственные факторы транскрипции, которые могут взаимодействовать с новыми последовательностями и активировать экспрессию неэндогенного репортерного гена в растительных клетках (Boch et al, ibid). Сконструированные белки TAL были связаны с половинным доменом расщепления FokI для получения слияния нуклеаза - домен TAL-эффектора (TALEN), проявляющего активность в анализе с дрожжевым геном-репортером (мишень на основе плазмиды). См., например, патент США № 8586526; Christian et al ((2010) Genetics epub 10.1534/genetics.110.120717).
[0161] В некоторых вариантах осуществления ДНК-связывающий домен одной или большего количества нуклеаз, применяемых для расщепления in vivo и/или целевого расщепления генома клетки, включает белок цинковые пальцы. Предпочтительно белок цинковые пальцы не является встречающимся в природе в том смысле, что он сконструирован так, чтобы связываться с выбранным целевым сайтом. См., например, Beerli et al. (2002) Nature Biotechnol.20:135-141; Pabo et al. (2001) Ann. Rev. Biochem.70:313-340; Isalan et al. (2001) Nature Biotechnol.19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol.12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol.10:411-416; патенты США №№ 6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273; 7888121; 7972854; и патентную публикацию США № 20050267061, все из которых в полном объеме включены в данный документ посредством ссылки.
[0162] Сконструированный связывающий домен цинковые пальцы может иметь новую специфичность связывания по сравнению со встречающимся в природе белком цинковые пальцы. К генно-инженерным методам относятся, помимо прочего, рациональный дизайн и различные типы отбора. Рациональный дизайн включает, например, применение баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности «цинковых пальцев», в которых каждая триплетная или квадруплетная нуклеотидная последовательность ассоциирована с одной или большим количеством аминокислотных последовательностей цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, совместные патенты США 6453242 и 6534261, которые в полном объеме включены в данный документ посредством ссылки.
[0163] Примеры способов отбора, включая фаговый дисплей и двухгибридные системы, описаны в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759; и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; и WO 01/88197. Кроме того, повышение специфичности связывания для связывающих доменов цинковых пальцев описано, например, в совместной заявке WO 02/077227.
[0164] Кроме того, как описано в этих и других ссылках, домены «цинковые пальцы» и/или белки «цинковые пальцы» с множеством «пальцев» могут быть связаны вместе с применением любых пригодных линкерных последовательностей, включая, например, линкеры из 5 или большего количества аминокислот в длину. См. также патенты США №№ 8772453; 6479626; 6903185; и 7153 949 для иллюстративных линкерных последовательностей. Описанные в данном документе белки могут включать любую комбинацию пригодных линкеров между отдельными цинковыми пальцами белка.
[0165] Выбор целевых сайтов; ZFP и способы разработки и конструирования слитых белков (и полинуклеотидов, кодирующих их) известны специалистам в данной области техники и подробно описаны в патентах США №№ 6140081; 5789538; 6453242; 6534261; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0166] Кроме того, как описано в этих и других ссылках, домены «цинковые пальцы» и/или белки «цинковые пальцы» с множеством «пальцев» могут быть связаны вместе с применением любых пригодных линкерных последовательностей, включая, например, линкеры из 5 или большего количества аминокислот в длину. См. также патенты США №№ 6479626; 6903185; и 7153949 для иллюстративных линкерных последовательностей длиной 6 или более аминокислот. Описанные в данном документе белки могут включать любую комбинацию пригодных линкеров между отдельными цинковыми пальцами белка.
[0167] В определенных вариантах осуществления ДНК-связывающий домен является частью системы нуклеаз CRISPR/Cas, включая, например, одиночную направляющую РНК (sgРНК). См., например, патенты США №№ 8697359 и 9873894. Локус CRISPR (сгруппированные с регулярными интервалами короткие палиндромные повторы), который кодирует компоненты РНК системы, и локус Cas (ассоциированный с CRISPR), кодирующий белки (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575; Makarova et al., 2002. Nucleic Acids Res. 30: 482-496; Makarova et al., 2006. Biol. Direct 1: 7; Haft et al., 2005. PLoS Comput. Biol. 1: e60), составляют последовательности генов системы нуклеаз CRISPR/Cas. Локусы CRISPR в микробных организмах-хозяевах содержат комбинацию CRISPR-ассоциированных (Cas) генов, а также некодирующих элементов РНК, способных программировать специфичность CRISPR-опосредованного расщепления нуклеиновой кислоты.
[0168] CRISPR типа II является одной из наиболее хорошо охарактеризованных систем и выполняет целевой двухцепочечный разрыв ДНК в четыре последовательных этапа. Во-первых, две некодирующие РНК, матрица пре-crРНК и tracrРНК, транскрибируются из локуса CRISPR. Во-вторых, tracrРНК гибридизуется с повторяющимися областями pre-crРНК и опосредует процессинг pre-crРНК в зрелые crРНК, содержащие индивидуальные спейсерные последовательности. В-третьих, комплекс зрелая crРНК : tracrРНК направляет Cas9 к целевой ДНК посредством спаривания оснований по Уотсону-Крику между спейсером на crРНК и протоспейсером на целевой ДНК рядом с соседним мотивом протоспейсера (PAM), что является дополнительным требованием для распознавания цели. Наконец, Cas9 опосредует расщепление целевой ДНК с образованием двухцепочечного разрыва внутри протоспейсера. Деятельность системы CRISPR/Cas состоит из трех этапов: (i) вставка последовательностей чужеродной ДНК в матрицу CRISPR для предотвращения будущих атак в процессе, называемом «адаптацией», (ii) экспрессия соответствующих белков, а также экспрессия и обработка матрицы с последующей (iii) РНК-опосредованной интерференцией с чужеродной нуклеиновой кислотой. Таким образом, в бактериальной клетке некоторые из так называемых белков «Cas» участвуют в естественной функции системы CRISPR/Cas и выполняют такие функции, как вставка чужеродной ДНК и т.д.
[0169] В определенных вариантах осуществления белок Cas может быть «функциональным производным» встречающегося в природе белка Cas. «Функциональное производное» полипептида с нативной последовательностью представляет собой соединение, обладающее качественным биологическим свойством, общим с полипептидом с нативной последовательностью. «Функциональные производные» включают, помимо прочего, фрагменты нативной последовательности и производные полипептида с нативной последовательностью и его фрагментов при условии, что они обладают биологической активностью, общей с соответствующим полипептидом с нативной последовательностью. Рассматриваемая в данном документе биологическая активность представляет собой способность функционального производного гидролизовать субстрат ДНК на фрагменты. Термин «производное» охватывает как варианты аминокислотной последовательности полипептида, так и их ковалентные модификации и слияния. Пригодные производные полипептида Cas или его фрагмента включают, помимо прочего, мутанты, слияния, ковалентные модификации белка Cas или его фрагмента. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или его фрагмент, может быть получен из клетки или синтезирован химически или с помощью комбинации этих двух процедур. Клетка может быть клеткой, которая естественным образом продуцирует белок Cas, или клеткой, которая естественным образом продуцирует белок Cas и генетически сконструирована для продукции эндогенного белка Cas с более высоким уровнем экспрессии или для продукции белка Cas из экзогенно введенной нуклеиновой кислоты, при этом указанная кислота кодирует Cas, который является таким же или отличным от эндогенного Cas. В некоторых случаях клетка естественным образом не продуцирует белок Cas, а генетически сконструирована для продукции белка Cas. Дополнительные неограничивающие примеры нуклеаз, направляемых РНК, которые можно применять в дополнение к и/или вместо белков Cas, включают белки CRISPR класса 2, такие как Cpf1. См., например, Zetsche et al. (2015) Cell 163:1-13.
[0170] Система CRISPR-Cpf1, идентифицированная у Francisella spp, представляет собой систему CRISPR-Cas класса 2, которая опосредует надежную интерференцию ДНК в клетки человека. Хотя функционально консервативные, Cpf1 и Cas9 различаются по многим аспектам, в том числе по направляющим РНК и субстратной специфичности (см. публикацию Fagerlund et al., (2015) Genom Bio 16:251). Основное различие между белками Cas9 и Cpf1 состоит в том, что Cpf1 не использует tracrРНК и, следовательно, требует только crРНК. FnCpf1 crРНК имеют длину 42-44 нуклеотида (19-нуклеотидный повтор и 23-25-нуклеотидный спейсер) и содержат один элемент петля-на-стебле, который выдерживает изменения последовательности, сохраняющие вторичную структуру. Кроме того, Cpf1 crРНК являются значительно короче, чем sgРНК, сконструированные из ~ 100 нуклеотидов и необходимые для Cas9, а потребности PAM для FnCpfl - это 5′-TTN-3′ и 5′-CTA-3′ на смещенной цепи. Хотя и Cas9, и Cpf1 образуют двухцепочечные разрывы в целевой ДНК, Cas9 использует свои RuvC- и HNH-подобные домены для создания разрезов с тупым концом в затравочной последовательности направляющей РНК, тогда как Cpf1 использует RuvC-подобный домен для создания ступенчатых разрезы вне затравки. Поскольку Cpf1 осуществляет ступенчатые разрезы от критической затравочной области, NHEJ не нарушает целевой сайт, таким образом гарантируя, что Cpf1 может продолжать разрезать тот же сайт до тех пор, пока не произойдет желаемое событие рекомбинации HDR. Таким образом, в описанных в данном документе способах и композициях подразумевается, что термин «Cas» включает белки как Cas9, так и Cfp1. Таким образом, в данном контексте термин «система CRISPR/ as» относится как к системам CRISPR/Cas, так и/или CRISPR/Cfp1, включая системы как нуклеаз, так и/или факторов транскрипции.
[0171] В некоторых вариантах осуществления ДНК-связывающий домен является частью системы TtAgo (см. публикацию Swarts et al., ibid; Sheng et al., ibid). У эукариот сайлесинг гена опосредуется белками семейства Argonaute (Ago). В этой парадигме Ago связан с малыми (19-31 н.) РНК. Этот сайленсинговый комплекс белок-РНК распознает целевые РНК через спаривание оснований по Уотсона-Крику между малой РНК и мишенью и эндонуклеолитически расщепляет целевую РНК (Vogel (2014) Science 344:972-973). Напротив, прокариотические белки Ago связываются с малыми одноцепочечными фрагментами ДНК и, вероятно, функционируют для обнаружения и удаления чужеродной (часто вирусной) ДНК (Yuan et al., (2005) Mol. Cell 19, 405; Olovnikov, et al. (2013) Mol. Cell 51, 594; Swarts et al., ibid). Типовые прокариотические белки Ago включают протеины из Aquifex aeolicus, Rhodobacter sphaeroides и Thermus thermophilus.
[0172] Одним из наиболее хорошо охарактеризованных прокариотических белков Ago является белок из T. thermophilus (TtAgo; Swarts et al. ibid). TtAgo ассоциируется с одноцепочечными фрагментами ДНК длиной 15 или 13-25 нуклеотидов с 5’-фосфатными группами. Эта «направляющая ДНК», связанная посредством TtAgo, служит для направления комплекса белок-ДНК для связывания комплементарной последовательности ДНК по Уотсону-Крика в третьесторонней молекуле ДНК. После того как информация о последовательности в этих направляющих ДНК позволила идентифицировать целевую ДНК, комплекс TtAgo - направляющая ДНК расщепляет целевую ДНК. Такой механизм также поддерживается структурой комплекса TtAgo - направляющая ДНК, связанного с его целевой ДНК (G. Sheng et al., ibid). Ago из Rhodobacter sphaeroides (RsAgo) обладает аналогичными свойствами (Olivnikov et al. ibid).
[0173] Экзогенные направляющие ДНК с произвольной последовательностью ДНК могут быть загружены в белок TtAgo (Swarts et al. ibid.). Поскольку специфичность расщепления TtAgo направляется направляющей ДНК, комплекс TtAgo-ДНК, образованный с экзогенной, определяемой исследователем направляющей ДНК, будет направлять расщепление целевой ДНК посредством TtAgo на дополнительную целевую ДНК, указанную исследователем. Таким образом можно создать целевой двухцепочечный разрыв ДНК. Применение системы TtAgo - направляющая ДНК (или систем ортологичный Ago - направляющая ДНК от других организмов) позволяет целенаправленно расщеплять геномную ДНК внутри клеток. Такое расщепление может быть одноцепочечным или двухцепочечным. Для расщепления геномной ДНК млекопитающих было бы предпочтительно применять версию кодона TtAgo, оптимизированную для экспрессии в клетках млекопитающих. Кроме того, может быть предпочтительнее обрабатывать клетки комплексом TtAgo - ДНК, образованным in vitro, при этом белок TtAgo слит с проникающим в клетки пептидом. Кроме того, может быть предпочтительнее применять версию белка TtAgo, которая была изменена посредством мутагенеза, чтобы иметь улучшенную активность при 37 градусах Цельсия. TtAgo-РНК-опосредованное расщепление ДНК можно применять для воздействия на множество процессов, включая нокаут гена, целевое добавление гена, коррекцию гена, целенаправленная делеция гена с помощью стандартных в данной области техники методов для использования разрывов ДНК.
[0174] Таким образом, нуклеаза включает ДНК-связывающий домен, который специфически связывается с целевым сайтом в любом гене, в который желательно вставить донор (трансген).
Домены расщепления
[0175] Любой пригодный домен расщепления можно оперативно связать с ДНК-связывающим доменом с образованием нуклеазы. Например, ДНК-связывающие домены ZFP были слиты с нуклеазными доменами для создания ZFN - функционального объекта, способного распознавать предполагаемую нуклеиновую кислоту-мишень через свой сконструированный (ZFP) ДНК-связывающий домен и вызывать разрезание ДНК рядом с сайтом связывания ZFP посредством нуклеазной активности. См., например, публикацию Kim et al. (1996) Proc Natl Acad Sci USA 93(3):1156-1160. Термин «ZFN» включает пару ZFN, которые димеризуются для расщепления целевого гена. Совсем недавно ZFN стали применять для модификации генома у множества организмов. См., например, патенты США №№ 7888121; 8409861; 8106255 и 9447434. Аналогичным образом TALE ДНК-связывающие домены были слиты с нуклеазными доменами для создания TALEN. См., например, патент США № 8586526. Также описаны системы нуклеаз CRISPR/Cas, содержащие одиночные направляющие РНК (sgРНК), которые связываются с ДНК и связываются с доменами расщепления (например, доменами Cas), чтобы вызвать целевое расщепление. См. также патенты США №№ 8697359 и 8932814, а также патент США № 9873894.
[0176] Как отмечалось выше, домен расщепления может быть гетерологичным ДНК-связывающему домену, например ДНК-связывающему домену цинковых пальцев и домену расщепления из нуклеазы или TALEN ДНК-связывающему домену и домену расщепления из нуклеазы; sgРНК ДНК-связывающему домену и домену расщепления из нуклеазы (CRISPR/Cas); и/или ДНК-связывающему домену мегануклеазы и домену расщепления из другой нуклеазы. Гетерологичные домены расщепления можно получить из любой эндонуклеазы или экзонуклеазы. Примеры эндонуклеаз, из которых может быть получен домен расщепления, включают, помимо прочего, рестрикционные эндонуклеазы и хоуминг-эндонуклеазы. См., например, 2002-2003 Catalogue, New England Biolabs, Beverly, MA; и публикацию Belfort et al. (1997) Nucleic Acids Res.25:3379-3388. Известны дополнительные ферменты, расщепляющие ДНК (например, нуклеаза S1; нуклеаза маша; панкреатическая ДНКаза I; микрококковая нуклеаза; дрожжевая эндонуклеаза HO; см. также публикацию Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). Один или большее количество из этих ферментов (или их функциональных фрагментов) можно применять в качестве источника доменов расщепления и полудоменов расщепления.
[0177] Точно так же полудомен расщепления может быть получен из любой нуклеазы или ее части, как указано выше, которая требует димеризации для активности расщепления. Обычно для расщепления требуются два слитых белка, если слитые белки содержат полудомены расщепления. В альтернативном варианте можно применять один белок, содержащий два полудомена расщепления. Два полудомена расщепления могут происходить из одной и той же эндонуклеазы (или ее функциональных фрагментов), или каждый полудомен расщепления может происходить из другой эндонуклеазы (или ее функциональных фрагментов). Кроме того, целевые сайты для двух слитых белков предпочтительно расположены относительно друг друга так, что связывание двух слитых белков с их соответствующими целевыми сайтами помещает полудомены расщепления в пространственную ориентацию друг к другу, что позволяет полудоменам расщепления образовывать функциональный домен расщепления, например, путем димеризации. Таким образом, в определенных вариантах осуществления ближние края целевых сайтов разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако между двумя целевыми сайтами может находиться любое целое число нуклеотидов или пар нуклеотидов (например, от 2 до 50 пар нуклеотидов или более). Обычно сайт расщепления находится между целевыми сайтами.
[0178] Рестрикционные эндонуклеазы (рестрикционные ферменты) присутствуют у многих видов и способны к последовательному связыванию с ДНК (в сайте распознавания) и расщеплению ДНК в сайте связывания или рядом с ним. Определенные рестрикционные ферменты (например, тип IIS) расщепляют ДНК в сайтах, удаленных от сайта распознавания, и имеют разделяемые домены связывания и расщепления. Например, фермент типа IIS Fok I катализирует двухцепочечное расщепление ДНК на 9 нуклеотидах от его сайта распознавания на одной цепи и 13 нуклеотидах от его сайта распознавания на другой. См., например, патенты США №№ 5356802; 5436150 и 5487994; а также публикации Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31,978-31,982. Таким образом, в одном варианте осуществления слитые белки содержат домен расщепления (или полудомен расщепления) по меньшей мере из одного рестрикционного фермента типа IIS и один или большее количество связывающих доменов цинковых пальцев, которые могут быть или могут не быть сконструированы.
[0179] Примером рестрикционного фермента типа IIS, домен расщепления которого отделяется от связывающего домена, является Fok I. Этот конкретный фермент активен как димер. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10,570-10,575. Соответственно, для целей настоящего изобретения часть фермента Fok I, применяемая в описанных слитых белках, считается полудоменом расщепления. Таким образом, для целенаправленного двухцепочечного расщепления и/или целевой замены клеточных последовательностей с применением слияний цинковые пальцы - Fok I, два слитых белка, каждый из которых содержит полудомен расщепления FokI, можно применять для восстановления каталитически активного домена расщепления. В альтернативном варианте, можно также применять одну молекулу полипептида, содержащую домен связывания цинковые пальцы и два полудомена расщепления Fok I. Параметры для целевого расщепления и целевого изменения последовательности с применением слияний цинковые пальцы - Fok I описаны в другом месте в этом документе.
[0180] Домен расщепления или полудомен расщепления может представлять собой любую часть белка, которая сохраняет активность расщепления или способность к мультимеризации (например, димеризации) с образованием функционального домена расщепления.
[0181] Типовые рестрикционные ферменты типа IIS описаны в патенте США 7888121, включенном в данный документ в полном объеме. Дополнительные рестрикционные ферменты также содержат разделяемые домены связывания и расщепления, и они рассматриваются в данном описании. См., например, публикацию Roberts et al. (2003) Nucleic Acids Res.31:418-420.
[0182] В определенных вариантах осуществления домен расщепления содержит один или большее количество сконструированных полудоменов расщепления (также называемых мутантами домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в патентах США №№ 8772453; 8,623,618; 8409861; 8034598; 7914796; и 7888121, описания которых в полном объеме включены в настоящий документ посредством ссылки. Все аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 FokI являются мишенями для влияния на димеризацию полудоменов расщепления FokI.
[0183] Типовые сконструированные полудомены расщепления FokI, которые образуют облигатные гетеродимеры, включают пару, в которых первый полудомен расщепления включает мутации на аминокислотных остатках в положениях 490 и 538 FokI, а второй полудомен расщепления включает мутации на аминокислотных остатках 486 и 499.
[0184] Таким образом, в одном варианте осуществления мутация в положении 490 заменяет Glu (E) на Lys (K); мутация в положении 538 заменяет Iso (I) на Lys (K); мутация в положении 486 заменяет Gln (Q) на Glu (E); и мутация в положении 499 заменяет Iso (I) на Lys (K). В частности, сконструированные полудомены расщепления, описанные в данном документе, были получены путем мутации в положениях 490 (E → K) и 538 (I → K) в одном полудомене расщепления с получением сконструированного полудомена расщепления, обозначенного «E490K:I538K» («K K ») и путем мутации в положениях 486 (Q → E) и 499 (I → L) в другом полудомене расщепления с получением сконструированного полудомена расщепления, обозначенного как «Q486E:I499L», («EL»). Сконструированные полудомены расщепления, описанные в данном документе, представляют собой облигатные гетеродимерные мутанты, в которых аберрантное расщепление минимизировано или устранено. Патенты США №№ 7914796 и 8034598, содержание которых в полном объеме включено в данный документ посредством ссылки. В определенных вариантах осуществления сконструированный полудомен расщепления содержит мутации в положениях 486, 499 и 496 (пронумерованных относительно FokI дикого типа), например, мутации, которые заменяют остаток Gln (Q) дикого типа в положении 486 на Glu (E ), остаток Iso (I) дикого типа в положении 499 на остаток Leu (L) и остаток Asn (N) дикого типа в положении 496 на остаток Asp (D) или Glu (E) (также называемые как домены «ELD» и «ELE» соответственно). В других вариантах осуществления сконструированный полудомен расщепления содержит мутации в положениях 490, 538 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на Lys (K), остаток Iso (I) дикого типа в положении 538 на остаток Lys (K) и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или Arg (R) (также называемые как домены «KKK» и «KKR» соответственно). В других вариантах осуществления сконструированный полудомен расщепления содержит мутации в положениях 490 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K) и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или Arg (R) (также называемые как домены «KIK» и «KIR» соответственно). См., например, патент США № 8772453. В других вариантах осуществления сконструированный полудомен расщепления содержит мутации “Sharkey” (см. публикацию Guo et al., (2010) J. Mol. Biol. 400(1):96-107).
[0185] Сконструированные полудомены расщепления, описанные в данном документе, могут быть получены с помощью любого пригодного способа, например, посредством сайт-направленного мутагенеза полудоменов расщепления дикого типа (Fok I), как описано в патентах США №№ 8623618; 8409861; 8034598; 7914796; и 7888121.
[0186] Способы и композиции также применяются для увеличения специфичности пары нуклеаз в отношении намеченной мишени по сравнению с другими сайтами непреднамеренного расщепления, известными как нецелевые сайты (см. публикации патентов США № 20170218349 и 20180087072). Таким образом, описанные в данном документе нуклеазы могут содержать мутации в одной или большим количестве каркасных областей их ДНК-связывающих доменов и/или одну или большее количество мутаций в их доменах расщепления нуклеазами. Эти нуклеазы могут включать мутации аминокислоты в ДНК-связывающем домене ZFP («каркас ZFP»), который может неспецифично взаимодействовать с фосфатами в каркасе ДНК, но они не содержат изменений в спиралях распознавания ДНК. Таким образом, ZFP может включать мутации катионных аминокислотных остатков в каркасе ZFP, которые не требуются для специфичности нуклеотидной мишени. В некоторых вариантах осуществления эти мутации в каркасе ZFP содержат мутацию катионного аминокислотного остатка с получением нейтрального или анионного аминокислотного остатка. В некоторых вариантах осуществления эти мутации в каркасе ZFP содержат мутацию полярного аминокислотного остатка с получением нейтрального или неполярного аминокислотного остатка. В предпочтительных вариантах осуществления мутации производят в положении (-5), (-9) и/или положении (-14) относительно спирали связывания ДНК. В некоторых вариантах осуществления цинковый палец может содержать одну или большее количество мутаций в (-5), (-9) и/или (-14). В дополнительных вариантах осуществления один или большее количество «цинковых пальцев» в белке с множеством «цинковых пальцев» могут содержать мутации в (-5), (-9) и/или (-14). В некоторых вариантах осуществления аминокислоты в (-5), (-9) и/или (-14) (например, аргинин (R) или лизин (K)) мутированы в аланин (A), лейцин (L), Ser (S), Asp (N), Glu (E), Tyr (Y) и/или глутамин (Q).
[0187] В определенных вариантах осуществления сконструированные полудомены расщепления происходят из домена нуклеазы FokI и содержат мутацию в одном или более аминокислотных остатков 416, 422, 447, 448 и/или 525, пронумерованных относительно полноразмерного домена дикого типа FokI. В некоторых вариантах осуществления мутации в аминокислотных остатках 416, 422, 447, 448 и/или 525 вводятся в FokI«ELD», «ELE», «KKK», «KKR», «KK», «EL», «KIK», «KIR» и/или Sharkey, как описано выше.
[0188] Кроме того, в данном документе описаны способы повышения специфичности активности расщепления посредством независимого титрования сконструированных партнеров полудомена расщепления нуклеазного комплекса. В некоторых вариантах осуществления соотношение двух партнеров (полудоменов расщепления) определяется как 1:2, 1:3, 1:4, 1:5, 1:6, 1:8, 1:9, 1:10 или 1:20, или любое значение между ними. В других вариантах осуществления соотношение двух партнеров составляет больше 1:30. В других вариантах осуществления два партнера имеют соотношение, которое выбрано отличающимся от 1:1. При применении отдельно или в комбинации способы и композиции, описанные в настоящем документе, обеспечивают необычное и неожиданное повышение специфичности нацеливания за счет снижения активности нецелевого расщепления. Нуклеазы, применяемые в этих вариантах осуществления, могут включать ZFN, пару ZFN, TALEN, пару TALEN, CRISPR/Cas, CRISPR/dCas и TtAgo или любую их комбинацию.
[0189] В альтернативном варианте, нуклеазы могут быть собраны in vivo в целевом сайте нуклеиновой кислоты с применением так называемой технологии «расколотого фермента» (см, например, публикацию патента США № 20090068164). Компоненты таких расколотых ферментов могут быть экспрессированы либо в отдельных экспрессионных конструкциях, либо могут быть связаны в одной открытой рамке считывания, где отдельные компоненты разделены, например, с помощью саморасщепляющегося пептида 2А или последовательности IRES. Компоненты могут представлять собой отдельные связывающие домены «цинковые пальцы» или домены связывающего домена мегануклеазы с нуклеиновой кислотой.
[0190] Перед применением нуклеазы можно сканировать на активность, например, в хромосомной системе на основе дрожжей, как описано в патенте США № 8563314. Экспрессия нуклеазы может находиться под контролем конститутивного промотора или индуцибельного промотора, например, промотора галактокиназы, который активируется (де-репрессируется) в присутствии рафинозы и/или галактозы и репрессируется в присутствии глюкозы.
[0191] Система CRISPR/Cas, связанная с Cas9, включает два некодирующих компонента РНК: tracrРНК и матрицу пре-crРНК, содержащую направляющие последовательности нуклеаз (спейсеры), разделенные идентичными прямыми повторами (DR). Чтобы применять систему CRISPR/Cas для выполнения геномной инженерии, должны присутствовать обе функции этих РНК (см. публикацию Cong et al, (2013) Sciencexpress 1/10.1126/science 1231143). В некоторых вариантах осуществления tracrРНК и pre-crРНК поставляются через отдельные экспрессионные конструкции или как отдельные РНК. В других вариантах осуществления конструируют химерную РНК, при этом сконструированная зрелая crРНК (придающая целевую специфичность) сливают с tracrРНК (обеспечивая взаимодействие с Cas9) для создания химерного гибрида cr-РНК-tracrРНК (также называемого одиночной направляющей РНК) (см. публикацию Jinek ibid and Cong, ibid).
Целевые сайты
[0192] Как подробно описано выше, домены ДНК могут быть сконструированы для связывания с любой выбранной последовательностью в локусе, например с альбумином или другим геном безопасного убежища. Сконструированный ДНК-связывающий домен может иметь новую специфичность связывания по сравнению со встречающимся в природе ДНК-связывающим доменом. К генно-инженерным методам относятся, помимо прочего, рациональный дизайн и различные типы отбора. Рациональный дизайн включает, например, применение баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности (например, цинковые пальцы), в которых каждая триплетная или квадруплетная нуклеотидная последовательность ассоциирована с одной или большим количеством аминокислотных последовательностей ДНК-связывающего домена, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, совместные патенты США 6453242 и 6534261, которые в полном объеме включены в данный документ посредством ссылки. Также можно выполнить рациональное проектирование TAL-эффекторных доменов. См., например, патент США № 8586526.
[0193] Примеры способов отбора, применимые к ДНК-связывающим доменам, включая фаговый дисплей и двугибридные системы, описаны в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759; и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; и WO 01/88197 и GB 2338237.
[0194] Выбор целевых сайтов; нуклеазы и способы разработки и конструирования слитых белков (и полинуклеотидов, кодирующих их) известны специалистам в данной области техники и подробно описаны в патентах США №№ 7888121 и 8409891, которые в полном объеме включены в данное описание посредством ссылки.
[0195] Кроме того, как описано в этих и других ссылках, ДНК-связывающие домены (например, белки «цинковые пальцы» с множеством «пальцев») могут быть связаны вместе с применением любых пригодных линкерных последовательностей, включая, например, линкеры из 5 или большего количества аминокислот в длину. См., например, патенты США №№ 9567609; 6479626; 6903185; и 7153949 для иллюстративных линкерных последовательностей длиной 6 или более аминокислот. Описанные в данном документе белки могут включать любую комбинацию пригодных линкеров между отдельными ДНК-связывающими доменами белка.
Доноры
[0196] Как отмечалось выше, в настоящем изобретении представлены способы и композиции для введения экзогенной последовательности (также называемой «донорной конструкцией», «донорной последовательностью» или «донором») субъекту, например, для коррекции мутантного гена или для повышения экспрессии гена, кодирующего белок, отсутствующий или дефицитный при болезни Фабри (например, α-GalA).
[0197] Совершенно очевидно, что донорная последовательность обычно является не идентичной геномной последовательности, в которую она помещена. Донорная последовательность может содержать негомологичную последовательность, фланкированную двумя областями гомологии («плечами гомологии»), чтобы обеспечить эффективную HDR в месте, представляющем интерес. Кроме того, донорные последовательности могут включать векторную молекулу, содержащую последовательности, которые не гомологичны области, представляющей интерес, в клеточном хроматине. Донорная молекула может содержать несколько прерывистых областей гомологии с клеточным хроматином. Например, для целенаправленной вставки последовательностей, обычно не присутствующих в области, представляющей интерес, указанные последовательности могут присутствовать в молекуле донорной нуклеиновой кислоты и фланкированы областями, гомологичными последовательности в области, представляющей интерес.
[0198] В данном документе описаны способы целенаправленной вставки трансгена, кодирующего белок α-GalA, для вставки в выбранное место. Трансген GLA может кодировать полноразмерный белок α-GalA или может кодировать усеченный белок α-GalA. Полинуклеотиды для вставки также могут называться «экзогенными» полинуклеотидами, «донорными» полинуклеотидами или молекулами или «трансгенами». Неограничивающие типовые донорные конструкции GLA продемонстрированы на Фиг. 1А и 1B.
[0199] Донорный полинуклеотид может быть ДНК или РНК, одноцепочечным и/или двухцепочечным и может быть введен в клетку в линейной или кольцевой форме. См., например, патенты США №№ 8703489 и 9255259. Донорная последовательность (последовательности) также может содержаться в ДНК MC, которая может быть введена в клетку в кольцевой или линейной форме. См., например, публикацию патента США № 20140335063. При введении в линейной форме, концы донорной последовательности можно защитить (например, от экзонуклеолитического расщепления) способами, известными специалистам в данной области техники. Например, один или большее количество дидезоксинуклеотидных остатков добавляют к 3’-концу линейной молекулы и/или самокомплементарные олигонуклеотиды лигируют с одним или обоими концами. См., например, Chang et al. (1987) Proc. Natl. Acad. Sci. USA 84:4959-4963; Nehls et al. (1996) Science 272:886-889. Дополнительные способы защиты экзогенных полинуклеотидов от расщепления включают, помимо прочего, добавление концевых аминогрупп и применение модифицированных межнуклеотидных связей, таких как, например, тиофосфаты, фосфорамидаты и остатки О-метилрибозы или дезоксирибозы.
[0200] Полинуклеотид может быть введен в клетку как часть молекулы вирусного или невирусного вектора, имеющей дополнительные последовательности, такие как, например, точки начала репликации, промоторы и гены, кодирующие резистентность к антибиотикам. Более того, донорные полинуклеотиды могут быть введены в виде оголенной нуклеиновой кислоты, в виде комплекса нуклеиновой кислоты с агентом, таким как липосома или полоксамер, или могут быть доставлены вирусами (например, аденовирусом, AAV, вирусом герпеса, ретровирусом, лентивирусом и лентивирусом, дефектным по интегразе (IDLV)).
[0201] Донор может быть вставлен так, чтобы его экспрессия управлялась эндогенным промотором в сайте интеграции, а именно промотором, который управляет экспрессией эндогенного гена, в который вставлен донор (например, высокоэкспрессируемый альбумин, AAVS1, HPRT и т. д.). Однако будет очевидно, что донор может включать промотор и/или энхансер, например, конститутивный промотор или индуцибельный или тканеспецифический промотор. В некоторых вариантах осуществления донор сохраняется в клетке в экспрессионной плазмиде, так что ген экспрессируется вне хромосом.
[0202] Донорная молекула может быть вставлена в эндогенный ген таким образом, чтобы экспрессировался весь эндогенный ген, часть или ни одно из них. Например, описанный в данном документе трансген может быть вставлен в альбумин или другой локус, так что некоторые (N-концевой и/или C-концевой трансген, кодирующий лизосомальный фермент) или ни одна из эндогенных последовательностей альбумина не экспрессируются, например, в виде слияния с трансгеном, кодирующим белок (белки) α-GalA. В других вариантах осуществления трансген (например, с дополнительными кодирующими последовательностями, такими как альбумин или без них) интегрируют в любой эндогенный локус, например, в ген безопасного убежища.
[0203] Когда эндогенные последовательности (эндогенный или часть трансгена) экспрессируются с трансгеном, эндогенные последовательности (например, альбумин и т.д.) могут быть полноразмерными последовательностями (дикого типа или мутантными) или частичными последовательностями. Предпочтительно эндогенные последовательности являются функциональными. Неограничивающие примеры функции этих полноразмерных или частичных последовательностей (например, альбумина) включают увеличение периода полужизни в сыворотке полипептида, экспрессируемого трансгеном (например, терапевтического гена) и/или действующего в качестве носителя.
[0204] Кроме того, хотя это не требуется для экспрессии, экзогенные последовательности могут также включать транскрипционные или трансляционные регуляторные последовательности, например, промоторы, энхансеры, инсуляторы, внутренние сайты посадки рибосомы, последовательности, кодирующие пептиды 2А, и/или сигналы полиаденилирования.
[0205] Экзогенные последовательности, связанные с трансгеном, также могут включать сигнальные пептиды, способствующие процессингу и/или секреции кодируемого белка. Неограничивающие примеры этих сигнальных пептидов включают пептиды из альбумина, IDS и фактора IX.
[0206] В некоторых вариантах осуществления экзогенная последовательность (донор) содержит слияние белка, представляющего интерес, и, в качестве его партнера по слиянию, внеклеточный домен мембранного белка, в результате чего слитый белок располагается на поверхности клетки. Это позволяет белку, кодируемому трансгеном, потенциально действовать в сыворотке. В случае болезни Фабри фермент α-GalA, кодируемый трансгеном, воздействует на продукты метаболизма, которые накапливаются в сыворотке крови из своего местоположения на поверхности клетки (например, RBC). Кроме того, если RBC поглощается макрофагами селезенки, что является нормальным процессом расщепления, лизосома, образующаяся, когда макрофаг поглощает клетку, подвергнет связанный с мембраной слитый белок воздействию высоких концентраций продуктов метаболизма в лизосоме при рН, более благоприятном для этого фермента. Неограничивающие примеры потенциальных партнеров по слиянию продемонстрированы ниже в Таблице 2.
Таблица 2. Примеры потенциальных партнеров по слиянию
[0207] В некоторых случаях экспрессионная конструкция может содержать модифицированный эндогенный ген GLA. Например, оптимизация кодонов может быть выполнена на эндогенном гене. Кроме того, хотя ответ антител на заместительную ферментную терапию варьируется в зависимости от конкретного терапевтического фермента, о котором идет речь, и от конкретного пациента, значительный иммунный ответ наблюдался у многих пациентов с болезнью Фабри, получающих заместительную ферментную терапию с применением α-GalA дикого типа. Считается, что трансген обеспечивает получение терапевтического белка, когда он увеличивает количество белка (и/или его активность) по сравнению с субъектами без трансгена. Кроме того, значение этих антител для эффективности лечения также варьируется (см. публикацию Katherine Ponder, (2008) J Clin Invest 118(8):2686). Таким образом, способы и композиции, описанные в данном документе, могут включать создание экспрессионных конструкций с модифицированными последовательностями по сравнению с GLA дикого типа, включая, помимо прочего, модификации, которые вызывают функционально молчащие аминокислотные изменения в сайтах, которые, как известно, являются примирующими эпитопами для эндогенных иммунных ответов, и/или укорочения, так что полипептид, продуцируемый такой последовательностью, является менее иммуногенным.
[0208] Пациенты с болезнью Фабри часто имеют неврологические нарушения из-за отсутствия в головном мозге фермента α-GalA. К сожалению, часто бывает трудно доставить терапевтические препараты в головной мозг через кровь из-за непроницаемости гематоэнцефалического барьера. Таким образом, способы и композиции можно применять в сочетании со способами повышения доставки терапевтического средства в головной мозг, включая, помимо прочего, способы, которые вызывают временное открытие плотных контактов между клетками капилляров головного мозга, например, временное осмотическое нарушение за счет внутрикаротидного введения гипертонического раствора маннита, применение сфокусированного ультразвука и введение аналога брадикинина (Matsukado et al. (1996) Neurosurgery 39:125). В альтернативном варианте, терапевтические средства могут быть разработаны для использования рецепторов или транспортных механизмов для специфической транспортировки в головной мозг. Примеры конкретных рецепторов, которые можно использовать, включают рецептор трансферрина, рецептор инсулина или белки 1 и 2, связанные с рецептором липопротеинов низкой плотности (LRP-1 и LRP-2). Известно, что LRP взаимодействует с рядом секретируемых белков, таких как apoE, tPA, PAI-1 и т. д., и поэтому слияние последовательности распознавания одного из этих белков для LRP может облегчить транспорт фермента в головной мозг после экспрессии в печени терапевтического белка и секреции его в кровоток (см. публикацию Gabathuler, (2010) ibid).
[0209]
Клетки
[0210] В настоящем изобретении также предлагаются генетически модифицированные клетки (например, стволовые клетки, клетки-предшественники, клетки печени, мышечные клетки и т.д.), содержащие экзогенный трансген GLA (интегрированный или внехромосомный), включая клетки, полученные посредством описанных в данном документе способов Эти клетки можно применять для доставки белка α-Gal A субъекту с болезнью Фабри, например, путем введения клетки (клеток) нуждающемуся в этом субъекту или, в альтернативном варианте, путем выделения белка α-Gal A, продуцируемого клеткой, и введения белка нуждающемуся в этом субъекту (ферментативная терапия). В альтернативном варианте, клетки могут быть получены in vivo у субъекта путем введения экспрессионных конструкций, как описано в данном документе. Таким образом, в настоящем изобретении предлагаются выделенные и генетически модифицированные клетки in vivo. Также предлагаются векторы (например, вирусные векторы, такие как AAV или Ad, или липидные наночастицы), содержащие трансген GLA, для применения в любом из способов, описанных в данном документе, в том числе для применения при лечении болезни Фабри.
[0211] В любом из описанных в данном документе способов трансген GLA может быть вставлен в геном целевой клетки с помощью нуклеазы. Неограничивающие примеры пригодных нуклеаз включают нуклеазы цинковые пальцы (ZFN), TALEN (активаторы транскрипции, подобные белковым нуклеазам) и/или системы нуклеаз CRISPR/Cas, которые включают ДНК-связывающую молекулу, которая связывается с целевым сайтом в области, представляющей интерес (например, ген, ассоциированный с заболеванием, ген с высокой степенью экспрессии, ген альбумина или другой ген безопасного убежища), в геноме клетки и одном или более доменов нуклеазы (например, домен расщепления и/или полудомен расщепления). Домены расщепления и полудомены расщепления могут быть получены, например, из различных рестрикционных эндонуклеаз, белков Cas и/или хоуминг-эндонуклеаз. В определенных вариантах осуществления домен цинковые пальцы распознает целевой сайт в гене альбумина или гене глобина в клетках-предшественниках эритроцитов (RBC). См., например, патент США № 9877988, в полном объеме включенный в настоящий документ посредством ссылки. В других вариантах осуществления нуклеаза (например, ZFN, TALEN и/или система CRISPR/Cas) связывается и/или расщепляет ген безопасного убежища, например, ген CCR5, ген PPP1R12C (также известный как AAVS1), ген альбумина, HPRT или ген Rosa. См., например, патенты США №№ 9877988; 9567573; 9447434; 9394545; 9222105; 9206404; 9150847; 8895264; 8771985; 8106255; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; и 8586526; патентные публикации США 20030232410 и 20060063231. Нуклеазы (или их компоненты) могут быть представлены в виде полинуклеотида, кодирующего одну или большее количество нуклеаз (например, ZFN, TALEN и/или систему CRISPR/Cas), описанных в данном документе. Полинуклеотид может представлять собой, например, мРНК. В некоторых аспектах мРНК может быть химически модифицирована (см., например, Kormann et al., (2011) Nature Biotechnology 29(2):154-157). В других аспектах мРНК может содержать кэп ARCA (см. патенты США 7074596 и 8153773). В дополнительных вариантах осуществления мРНК может содержать смесь немодифицированных и модифицированных нуклеотидов (см. публикацию патента США 20120195936). В других вариантах осуществления мРНК может содержать элемент WPRE (см. патент США № 10179918).
[0212] В другом аспекте настоящего изобретения предлагаются генетически модифицированные клетки (например, стволовые клетки, клетки-предшественники, клетки печени, мышечные клетки и т.д.) с желаемым трансгеном GLA (необязательно интегрированным с помощью нуклеазы). В некоторых аспектах отредактированные стволовые клетки или клетки-предшественники затем размножают и могут индуцировать для дифференцировки в зрелые отредактированные клетки ex vivo, и затем переносят в организм пациента. Таким образом, можно применять клетки, происходящие от генетически отредактированных (модифицированных) стволовых клеток или клеток-предшественников, продуцирующих GLA, как описано в данном документе. В других аспектах отредактированные предшественники (например, стволовые клетки CD34 +) вводят в трансплантат костного мозга, который после успешной имплантации пролиферирует, продуцируя отредактированные клетки, которые затем дифференцируются и созревают in vivo и содержат биологические вещества, экспрессируемые трансгеном GLA. В некоторых вариантах осуществления отредактированные стволовые клетки CD34+ вводят пациенту внутривенно, так что отредактированные клетки мигрируют в костный мозг, дифференцируются и созревают, продуцируя белок α-Gal A. В других аспектах отредактированные стволовые клетки представляют собой мышечные стволовые клетки, которые затем вводят в мышечную ткань. В некоторых аспектах сконструированная нуклеаза представляет собой нуклеазу цинковые пальцы (ZFN) (термин «ZFN» включает пару ZFN), а в других - нуклеаза представляет собой нуклеазу TALE (TALEN) (термин «TALEN» включает пару TALEN), а в других аспектах применяют систему CRISPR/Cas. Нуклеазы могут быть сконструированы так, чтобы иметь специфичность в отношении локуса безопасного убежища, гена, ассоциированного с заболеванием, или гена, который высоко экспрессируется в клетках. Только в качестве неограничивающего примера, локус безопасного убежища может быть сайтом AAVS1, геном CCR5, геном альбумина или геном HPRT, тогда как ген, ассоциированный с заболеванием, может быть геном GLA, кодирующим α-галактозидазу А.
[0213] Трансген GLA может быть полноразмерным или модифицированным и может экспрессироваться вне хромосом или может быть целенаправленно интегрирован в геном клетки с помощью одной или большего количества нуклеаз. В отличие от случайной интеграции, целенаправленная интеграция, опосредованная нуклеазами, гарантирует, что трансген будет интегрирован в указанный ген. Трансген может быть интегрирован в любое место целевого гена. В определенных вариантах осуществления трансген интегрируют в сайт связывания и/или расщепления нуклеазы или рядом с ним, например, в пределах 1-300 (или любого количества пар оснований между ними) пар оснований выше или ниже сайта расщепления и/или сайта связывания, более предпочтительно в пределах 1-100 пар оснований (или любого количества пар оснований между ними) с любой стороны сайта расщепления и/или связывания, даже более предпочтительно в пределах от 1 до 50 пар оснований (или любого количества пар оснований между ними) с любой стороны сайта расщепления и/или связывания. В определенных вариантах осуществления интегрированная последовательность не включает никаких векторных последовательностей (например, последовательностей вирусного вектора).
[0214] Любой тип клеток может быть генетически модифицирован, как описано в данном документе, для включения трансгена, включая, помимо прочего, клетки или клеточные линии. Другие неограничивающие примеры генетически модифицированных клеток, как описано в данном документе, включают Т-клетки (например, CD4 +, CD3 +, CD8+и т. д.); дендритные клетки; В-клетки; аутологичные клетки (например, полученные от пациента), мышечные клетки, клетки головного мозга и т.п. В определенных вариантах осуществления клетки являются клетками печени и модифицированы in vivo. В определенных вариантах осуществления клетки представляют собой стволовые клетки, включая гетерологичные плюрипотентные, тотипотентные или мультипотентные стволовые клетки (например, клетки CD34 +, индуцированные плюрипотентные стволовые клетки (iPSC), эмбриональные стволовые клетки и т.п.). В определенных вариантах осуществления клетки, как описано в данном документе, представляют собой стволовые клетки, полученные от пациента.
[0215] Клетки, как описано в данном документе, являются пригодными для лечения и/или предотвращения болезни Фабри у субъекта с этим нарушением, например, с помощью терапии in vivo. Также предлагаются методы лечения ex vivo, например, когда клетки, модифицированные нуклеазой, могут быть размножены и затем повторно введены пациенту с помощью стандартных методик. См., например, публикацию Tebas et al. (2014) New Eng J Med 370(10):901. В случае стволовых клеток, после инфузии субъекту также происходит дифференцировка этих предшественников in vivo в клетки, экспрессирующие функциональный белок (от встроенного донора).
[0216] Также предлагаются фармацевтические композиции, содержащие описанные в данном документе клетки. Кроме того, перед введением пациенту клетки могут быть криоконсервированы.
Доставка
[0217] Описанные в данном документе экспрессионные конструкции кДНК, нуклеазы, полинуклеотиды, кодирующие эти нуклеазы, донорные полинуклеотиды и/или композиции (например, клетки, белки, полинуклеотиды и т.д.), могут быть доставлены in vivo или ex vivo с помощью любых пригодных для этого способов.
[0218] Способы доставки нуклеаз, как описано в данном документе, описаны, например, в патентах США №№ 6453242; 6503717; 6534261; 6599692; 6607882; 6689558; 6824978; 6933113; 6979539; 7013219; и 7163824, описания которых в полном объеме включены в данный документ посредством ссылки.
[0219] Экспрессионные конструкции и/или нуклеазы, описанные в данном документе, также могут быть доставлены с помощью векторов, содержащих последовательности, кодирующие один или большее количество белков цинковые пальцы, TALEN и/или Cas. Можно применять любые векторные системы, включая, помимо прочего, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксвирусные векторы; герпесвирусные векторы и аденоассоциированные вирусные векторы и т.д. См. также патенты США №№ 6534261; 6607882; 6824978; 6933113; 6979539; 7013219; и 7163824, которые в полном объеме включены в данный документ посредством ссылки. Кроме того, будет очевидно, что любой из этих векторов может содержать одну или большее количество последовательностей, необходимых для лечения. Таким образом, когда в клетку вводят одну или большее количество нуклеаз и донорную конструкцию, нуклеазы и/или донорный полинуклеотид могут переноситься в одном и том же векторе или в разных векторах. При применении множества векторов каждый вектор может содержать последовательность, кодирующую одну или большее количество нуклеаз и/или донорных конструкций.
[0220] Для введения экспрессионных конструкций кДНК или нуклеиновых кислот, кодирующих нуклеазы и/или экспрессионные конструкции, в клетки и целевые ткани можно применять обычные способы переноса генов на вирусной и невирусной основе (например, клетки млекопитающих). Системы доставки с невирусными векторами включают плазмиды ДНК, оголенную нуклеиновую кислоту и нуклеиновую кислоту в комплексе с носителем для доставки, таким как липосома или полоксамер. Системы доставки с вирусными векторами включают ДНК и РНК вирусов, которые после доставки в клетку имеют либо эписомальные, либо интегрированные геномы. Для обзора процедур генной терапии см. публикации Anderson, Science 256:808-813 (1992); Nabel & Felgner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., в Current Topics in Microbiology and Immunology Doerfler and Böhm (eds.) (1995); и Yu et al., Gene Therapy 1:13-26 (1994).
[0221] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистику, виросомы, липосомы, иммунолипосомы, поликатион или конъюгаты липид:нуклеиновая кислота, оголенная ДНК, искусственные вирионы и усиленный агентом захват ДНК. Сонопорацию с помощью, например, системы Sonitron 2000 (Rich-Mar) также можно применять для доставки нуклеиновых кислот.
[0222] Дополнительные примеры систем доставки нуклеиновых кислот включают системы, предоставляемые Amaxa Biosystems (Кельн, Германия), Maxcyte, Inc. (Роквилл, Мэриленд), BTX Molecular Delivery Systems (Холлистон, Массачусетс) и Copernicus Therapeutics Inc, (см., например, US6008336). Липофекция описана, например, в патентах США №№ 5049386; 4946787; и 4897355), а реагенты для липофекции доступны в коммерческой продаже (например, Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые являются пригодными для эффективной липофекции полинуклеотидов с распознаванием рецепторов, включают липиды из Felgner, WO 91/17424, WO 91/16024.
[0223] Получение комплексов липид:нуклеиновая кислота, включая целевые липосомы, такие как иммунолипидные комплексы, хорошо известно специалисту в данной области техники (см., например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0224] Композиции кДНК и/или нуклеаз, описанные в данном документе, также могут быть доставлены с помощью наночастиц, например липидных наночастиц (LNP). См., например, Lee et al. (2016) Am J Cancer Res 6(5):1118-1134; патент США № 10166298; и публикацию США № 20180185516.
[0225] Дополнительные способы доставки включают применение упаковки доставляемых нуклеиновых кислот в средства доставки EnGeneIC (EDV). Эти EDV специфически доставляются к целевым тканям с помощью биспецифических антител, в которых одно плечо антитела имеет специфичность к целевой ткани, а другое - к EDV. Антитело переносит EDV на поверхность целевой клетки, а затем EDV поступает в клетку посредством эндоцитоза. Попав в клетку, содержимое освобождается (см. публикацию MacDiarmid et al. (2009) Nature Biotechnology 27(7):643).
[0226] Применение систем на основе вирусной РНК или ДНК для доставки нуклеиновых кислот, кодирующих сконструированные ZFP, позволяет использовать преимущества высокотехнологичных процессов для нацеливания вируса на определенные клетки в организме и доставки вирусной полезной нагрузки в ядро. Вирусные векторы можно вводить непосредственно субъектам (in vivo) или их можно применять для обработки клеток in vitro, при этом модифицированные клетки вводят субъектам (ex vivo). Обычные вирусные системы для доставки ZFP включают, помимо прочего, ретровирусные, лентивирусные, аденовирусные, аденоассоциированные векторы, векторы вируса осповакцины и простого герпеса для переноса генов. Интеграция в геном хозяина возможна с помощью способов переноса генов ретровируса, лентивируса и аденоассоциированного вируса, что часто приводит к долговременной экспрессии встроенного трансгена. Кроме того, высокая эффективность трансдукции наблюдалась во многих различных типах клеток и целевых тканях.
[0227] Тропизм ретровируса можно изменить путем включения белков чужой оболочки, увеличивая потенциальную целевую популяцию целевых клеток. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны трансдуктировать или инфицировать неделящиеся клетки и обычно продуцируют высокие вирусные титры. Выбор системы переноса ретровирусного гена зависит от целевой ткани. Ретровирусные векторы состоят из цис-действующих длинных концевых повторов с упаковочной способностью до 6-10 т.п.н. чужеродной последовательности. Минимальных цис-действующих LTR достаточно для репликации и упаковки векторов, которые затем применяются для интеграции терапевтического гена в целевую клетку для обеспечения постоянной экспрессии трансгена. Широко применяемые ретровирусные векторы включают векторы на основе вируса лейкемии мышей (MuLV), вируса лейкемии обезьян гиббонов (GaLV), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита человека (ВИЧ) и их комбинаций (см., например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991)).
[0228] В применениях, в которых предпочтительна временная экспрессия, можно использовать системы на основе аденовирусов. Векторы на основе аденовирусов обладают очень высокой эффективностью трансдукции во многих типах клеток и не требуют деления клеток. С такими векторами были получены высокий титр и высокие уровни экспрессии. Этот вектор можно производить в больших количествах в относительно простой системе. Векторы аденоассоциированного вируса («AAV») также используются для трансдукции клеток нуклеиновыми кислотами-мишенями, например, при продукции нуклеиновых кислот и пептидов in vitro, а также для процедур генной терапии in vivo и ex vivo (см., например, West et al., Virology 160:38-47 (1987); патент США № 4797368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Конструирование рекомбинантных векторов AAV описано в ряде публикаций, в том числе в патентах США № 5173414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS 81:6466-6470 (1984); и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0229] В настоящее время доступны по меньшей мере шесть подходов с вирусными векторами для переноса генов в клинических исследованиях, которые используют подходы, которые включают комплементацию дефектных векторов генами, вставленными в линии вспомогательных клеток, для создания трансдуцирующего агента.
[0230] pLASN и MFG-S являются примерами ретровирусных векторов, которые применяли в клинических исследованиях (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS 94:22 12133-12138 (1997)). PA317/pLASN был первым терапевтическим вектором, использованным в клиническом исследовании генной терапии (Blaese et al., Science 270:475-480 (1995)). Эффективность трансдукции 50% или выше наблюдалась для упакованных векторов MFG-S. (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0231] Рекомбинантные аденоассоциированные вирусные векторы (rAAV) представляют собой многообещающие альтернативные системы доставки генов на основе дефектного и непатогенного парвовирусного аденоассоциированного вируса 2 типа. Все векторы получены из плазмиды, которая сохраняет только инвертированные концевые повторы AAV длиной 145 п.о., фланкирующие экспрессионную кассету трансгена. Эффективный перенос генов и стабильная доставка трансгена за счет интеграции в геномы трансдуцированной клетки являются ключевыми характеристиками этой векторной системы. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). Также можно применять другие серотипы AAV, включая, в качестве неограничивающего примера, AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV 8.2, AAV9 и AAV rh10 и псевдотипы AAV, такие как AAV2/8, AAV2/5 и AAV2/6.
[0232] AAV можно изготавливать в клиническом масштабе с помощью ряда различных процессов. Примеры систем, которые можно применять, включают (1) трансфекцию плазмидной ДНК в клетки млекопитающих, (2) Ad-инфицирование стабильных клеточных линий млекопитающих, (3) инфицирование клеток млекопитающих рекомбинантными вирусами простого герпеса (rHSV) и (4) инфицирование клеток насекомых (клетки Sf9) рекомбинантными бакуловирусами (см. публикацию Penaud-Budloo et al. (2018) Mol Ther Methods Clin Dev. 8: 166-180 для обзора).
[0233] Рекомбинантные аденовирусные векторы (Ad) с дефицитом репликации могут быть получены с высоким титром и могут легко инфицировать ряд различных типов клеток. Большинство аденовирусных векторов сконструированы таким образом, что трансген заменяет гены Ad E1a, E1b и/или E3; впоследствии дефектный по репликации вектор размножается в клетках 293 человека, которые обеспечивают функцию удаленного гена в транс. Ad-векторы могут преобразовывать различные типы тканей in vivo, включая неделящиеся дифференцированные клетки, такие как клетки печени, почек и мышц. Обычные Ad-векторы обладают большой транспортирующей способностью. Пример применения Ad-вектора в клиническом исследовании включал полинуклеотидную терапию для противоопухолевой иммунизации путем внутримышечной инъекции (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры применения аденовирусных векторов для переноса генов в клинических исследованиях включают: Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0234] Упаковочные клетки применяются для образования вирусных частиц, способных инфицировать клетку-хозяина. Такие клетки включают клетки 293, которые упаковывают аденовирус, и клетки ψ2 или клетки PA317, которые упаковывают ретровирус. Вирусные векторы, применяемые в генной терапии, обычно генерируются линией клеток-продуцентов, которые упаковывают вектор нуклеиновой кислоты в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в организм хозяина (если это применимо), другие вирусные последовательности заменяются экспрессионной кассетой, кодирующей белок, который должен быть экспрессирован. Недостающие вирусные функции поставляются в транс с помощью упаковочной клеточной линии. Например, векторы AAV, применяемые в генной терапии, обычно содержат только последовательности инвертированных концевых повторов (ITR) из генома AAV, которые необходимы для упаковки и интеграции в геном хозяина. Вирусная ДНК упакована в клеточную линию, которая содержит вспомогательную плазмиду, кодирующую другие гены AAV, а именно rep и cap, но не содержащую последовательностей ITR. Клеточная линия также инфицируют аденовирусом в качестве помощника (хелпера). Хелперный вирус способствует репликации вектора AAV и экспрессии генов AAV из хелперной плазмиды. Хелперная плазмида не упакована в значительных количествах из-за отсутствия последовательностей ITR. Контаминацию деновирусом можно снизить, например, с помощью тепловой обработки, к которой аденовирус более чувствителен, чем AAV.
[0235] Во многих применениях генной терапии желательно, чтобы вектор генной терапии доставлялся с высокой степенью специфичности к конкретному типу ткани. Соответственно, вирусный вектор можно модифицировать так, чтобы он обладал специфичностью для данного типа клеток, путем экспрессии лиганда в виде белка слияния с белком вирусной оболочки на внешней поверхности вируса. Лиганд выбирают так, чтобы он имел аффинность к рецептору, о котором известно, что он присутствует в типе клеток, представляющем интерес. Например, Han et al., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995), сообщил, что вирус лейкемии мышей Молони может быть модифицирован для экспрессии человеческого херегулина, слитого с gp70, при этом рекомбинантный вирус инфицирует определенные клетки рака молочной железы человека, экспрессирующие рецептор эпидермального фактора роста человека. Этот принцип может быть распространен на другие пары вирус-целевая клетка, в которых целевая клетка экспрессирует рецептор, а вирус экспрессирует слитый белок, содержащий лиганд для рецептора клеточной поверхности. Например, нитчатый фаг может быть сконструирован для отображения фрагментов антител (например, FAB или Fv), обладающих специфической аффинностью связывания практически с любым выбранным клеточным рецептором. Хотя приведенное выше описание применимо в первую очередь к вирусным векторам, те же принципы могут быть применены к невирусным векторам. Такие векторы могут быть сконструированы так, чтобы они содержали специфические последовательности захвата, которые способствуют захвату конкретными целевыми клетками.
[0236] Векторы для генной терапии можно доставлять in vivo путем введения отдельному пациенту, обычно путем системного введения (например, внутривенная, внутрибрюшинная, внутримышечная, подкожная или внутричерепная инфузия) или путем местного нанесения как описано ниже. В альтернативном варианте векторы могут быть доставлены в клетки ex vivo, такие как клетки, эксплантированные от отдельного пациента (например, лимфоциты, клетки аспиратов костного мозга, клетки биоптатов ткани) или универсальные донорные гемопоэтические стволовые клетки, с последующей реимплантацией клеток пациенту, обычно после отбора клеток, в которые включен вектор.
[0237] Векторы (например, ретровирусы, аденовирусы, липосомы и т.д.), содержащие нуклеазы и/или донорные конструкции (экспрессионные конструкции), также можно вводить непосредственно в организм для трансдукции клеток in vivo. В альтернативном варианте можно вводить оголенную ДНК. Введение осуществляют любым из способов, обычно применяемых для приведения молекулы в окончательный контакт с кровью или клетками ткани, включая, помимо прочего, инъекцию, инфузию, местное нанесение и электропорацию. Пригодные способы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области техники, и, хотя для введения конкретной композиции можно применять более одного пути, конкретный путь часто может обеспечить более немедленную и более эффективную реакцию, чем другой путь.
[0238] Векторы, пригодные для введения полинуклеотидов, описанных в данном документе, включают неинтегрирующиеся лентивирусные векторы (non-integrating lentivirus vectors, IDLV). См., например, публикации Ory et al. (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull et al. (1998) J. Virol. 72:8463-8471; Zuffery et al. (1998) J. Virol. 72:9873-9880; Follenzi et al. (2000) Nature Genetics 25:217-222.
[0239] Фармацевтически приемлемые носители частично определяются конкретной вводимой композицией, а также конкретным способом, применяемым для введения композиции. Соответственно, доступно большое количество пригодных составов фармацевтических композиций, как описано ниже (см., например, Remington’s Pharmaceutical Sciences, 17th ed., 1989).
[0240] Очевидно, что последовательности, кодирующие нуклеазу, и донорные конструкции могут быть доставлены с помощью одинаковых или разных систем. Например, донорный полинуклеотид может переноситься плазмидой, в то время как одна или большее количество нуклеаз могут переноситься вектором AAV. Кроме того, разные векторы можно вводить одним и тем же или разными путями (внутримышечная инъекция, инъекция в хвостовую вену, другая внутривенная инъекция, внутрибрюшинное введение и/или внутримышечная инъекция. Векторы могут быть доставлены одновременно или в любом последовательном порядке.
[0241] Составы для введения как ex vivo , так и in vivo включают суспензии в жидкости или эмульгированные жидкости. Активные ингредиенты часто смешивают с вспомогательными веществами, которые фармацевтически приемлемы и совместимы с активным ингредиентом. Пригодные вспомогательные вещества включают, например, воду, физиологический солевой раствор, декстрозу, глицерин, этанол и т.п. и их комбинации. Кроме того, композиция может содержать незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, pH буферные агенты, стабилизирующие агенты или другие реагенты, которые повышают эффективность фармацевтической композиции.
Применения
[0242] В описанных в настоящем документе способах предлагается лечение и/или предотвращение болезни Фабри (например, лизосомной болезни накопления). Лечение может включать введение корригирующего ассоциированного с заболеванием трансгена GLA в локус безопасного убежища (например, альбумин) в клетке для экспрессии необходимого фермента и высвобождения его в кровоток. Корригирующий трансген, кодирующий α-GalA, может кодировать белок дикого типа или модифицированный белок; и/или может содержать кодон-оптимизированный трансген GLA; и/или трансген, в котором эпитопы могут быть удалены без функционального изменения белка. В некоторых случаях указанные способы включают введение эписомы, экспрессирующей трансген, кодирующий α-GalA, в клетку для экспрессии необходимого фермента и высвобождения его в кровоток. Особенно полезным является введение в секреторную клетку, такую как клетка печени, для высвобождения продукта в кровоток. Способы и композиции также могут применяться в любых обстоятельствах, когда желательно обеспечить трансген GLA, кодирующий одно или большее количество терапевтических средств, в гемопоэтических стволовых клетках таким образом, чтобы зрелые клетки (например, RBC), полученные из (происходящие от) этих клеток, содержали терапевтический белок α-GalA. Эти стволовые клетки могут быть дифференцированы in vitro или in vivo и могут быть получены из универсального донорного типа клетки, который можно применять для всех пациентов. Кроме того, клетки могут содержать трансмембранный белок для перемещения клеток в организме. Лечение также может включать применение клеток пациента, содержащих терапевтический трансген, при этом и указанные клетки развиваются ex vivo и затем вводятся обратно пациенту. Например, HSC, содержащие пригодный трансген, кодирующий α-GalA, можно ввести пациенту с помощью трансплантата костного мозга. В альтернативном варианте, стволовые клетки, такие как мышечные стволовые клетки или iPSC, которые были отредактированы с помощью трансгена, кодирующего α-GalA, также могут быть введены в мышечную ткань.
[0243] Таким образом, эта технология может быть применена при патологическом состоянии, когда у пациента наблюдается дефицит некоторого белка из-за проблем (например, проблемы на уровне экспрессии или проблемы с белком, экспрессированным на субуровне или являющимся нефункционирующим). Особенно полезной является экспрессия трансгенов для коррекции или восстановления функциональности у субъектов с болезнью Фабри.
[0244] В качестве неограничивающих примеров реализованы различные способы получения функционального белка α-Gal A для замены дефектного или отсутствующего белка α-Gal A, которые применяются для лечения болезни Фабри. Доноры нуклеиновых кислот, кодирующие белки, могут быть вставлены в локус безопасного убежища (например, альбумин или HPRT) и экспрессированы либо с применением экзогенного промотора, либо с применением промотора, присутствующего в безопасном убежище. Особенно полезным является введение трансгена GLA в локус альбумина в клетке печени, при этом указанный трансген GLA дополнительно содержит последовательности, кодирующие сигнальный пептид, который опосредует секрецию экспрессированного белка α-Gal A из клетки печени в кровоток. В альтернативном варианте можно использовать доноров для коррекции дефектного гена in situ. Желаемый трансген, кодирующий α-GalA, может быть вставлен в стволовую клетку CD34+ и возвращен пациенту во время трансплантации костного мозга. Наконец, донор нуклеиновой кислоты может быть вставлен в стволовую клетку CD34+ в локусе бета-глобина, так что зрелый эритроцит, полученный из этой клетки, имеет высокую концентрацию биологического вещества, кодируемого донором нуклеиновой кислоты. Затем RBC, содержащие биологическое вещество, могут быть нацелены на нужную ткань через трансмембранные белки (например, рецептор или антитело). Кроме того, RBC могут быть сенсибилизированы ex vivo посредством электросенсибилизации, чтобы сделать их более восприимчивыми к разрушению после воздействия источника энергии (см. WO2002007752).
[0245] В некоторых применениях эндогенный ген может быть отключен с помощью способов и композиций, описанных в данном документе. Примеры этого аспекта включают отключение регулятора аберрантного гена или аберрантного гена, ассоциированного с заболеванием. В некоторых применениях аберрантный эндогенный ген может быть заменен, функционально или in situ, версией гена дикого типа. Вставленный ген также может быть изменен для улучшения экспрессии терапевтического белка α-GalA или для снижения его иммуногенности. В некоторых применениях вставленный трансген, кодирующий α-GalA, представляет собой слитый белок для увеличения его транспорта в выбранную ткань, такую как головной мозг.
[0246] Следует понимать, что пригодные доноры GLA не ограничиваются донорами, приведенными ниже, но включают любой трансген GLA.
[0247] В описании также представлены способы и композиции для продуцирования клетки (например, RBC), несущей терапевтический белок α-GalA, для лечения болезни Фабри, которые можно универсально применять для всех пациентов в качестве аллогенного продукта. Это позволяет разработать единый продукт, например, для лечения пациентов с болезнью Фабри. Эти носители могут содержать трансмембранные белки, способствующие перемещению клетки. В одном аспекте трансмембранный белок содержит антитело, в то время как в других аспектах трансмембранный белок содержит рецептор.
[0248] В некоторых вариантах осуществления донор трансгена GLA трансфицируют или трансдуцируют в клетку для эписомального или внехромосомного сохранения трансгена. В некоторых аспектах донор трансгена GLA сохраняется в векторе, содержащем регуляторные домены для регулирования экспрессии донора трансгена. В некоторых случаях регуляторные домены для регуляции экспрессии трансгена представляют собой домены, эндогенные по отношению к экспрессируемому трансгену, тогда как в других случаях регуляторные домены являются гетерологичными трансгену. В некоторых вариантах осуществления трансген GLA сохраняется в вирусном векторе, в то время как в других он сохраняется в плазмиде или мини-кольце. В некоторых вариантах осуществления вирусный вектор представляет собой AAV, Ad или LV. В дополнительных аспектах вектор, содержащий донор трансгена, доставляется в пригодную целевую клетку in vivo, так что терапевтический белок α-GalA, кодируемый донором трансгена, высвобождается в кровоток, когда вектор донора трансгена доставляется в гепатоцит.
[0249] В другом варианте осуществления предлагаются клетки-предшественники (мышечные стволовые клетки, клетки-предшественники или клетки гемопоэтических стволовых клеток CD34+ (HSPC)), в которые был вставлен трансген GLA, так что зрелые клетки, полученные из этих предшественников, содержат высокие уровни продукта α-GalA, кодируемого трансгеном. В некоторых вариантах осуществления эти предшественники представляют собой индуцированные плюрипотентные стволовые клетки (iPSC).
[0250] В некоторых вариантах осуществления указанные способы можно применять in vivo в системах трансгенных животных. В некоторых аспектах трансгенное животное может быть использовано при разработке модели, в которой трансген кодирует белок α-GalA человека. В некоторых случаях трансгенное животное может быть нокаутировано в соответствующем эндогенном локусе, что позволяет создать систему in vivo, в которой белок человека можно изучать изолированно. Такие трансгенные модели можно применять в целях скрининга для идентификации малых молекул, или больших биомолекул, или других объектов, которые могут взаимодействовать с белком человека, представляющим интерес, или модифицировать его. В некоторых аспектах трансген GLA интегрируют в выбранный локус (например, высокоэкспрессируемый или безопасное убежище) в стволовую клетку (например, эмбриональную стволовую клетку, индуцированную плюрипотентную стволовую клетку, печеночную стволовую клетку, нервную стволовую клетку и т. д.) или эмбрион животного, не являющегося человеком, полученный любым из способов, описанных в данном документе, и тех, которые являются стандартными в данной области техники, а затем эмбрион имплантируют так, что рождается живое животное. Затем животное доводят до половой зрелости и дают возможность произвести потомство, в котором по меньшей мере часть потомства будет содержать интегрированный трансген GLA.
[0251] В любом из предыдущих вариантов осуществления способы и соединения можно комбинировать с другими терапевтическими агентами для лечения субъектов с болезнью Фабри. В некоторых вариантах осуществления способы и композиции включают применение молекулярного шаперона (Hartl et al (2011) Nature 465: 324-332) для обеспечения правильного фолдинга белка Фабри. В некоторых аспектах шаперон может быть выбран из хорошо известных шаперонных белков, таких как AT1001 (Benjamin et al (2012) Mol Ther 20(4):717-726), AT2220 (Khanna et al (2014) PLoS ONE 9(7): e102092, doi:10.1371), и Migalastat (Benjamin et al (2016) Genet Med doi: 10.1038/gim.2016.122). В некоторых аспектах способы и композиции применяют в комбинации со способами и композициями для обеспечения проникновения через гематоэнцефалический барьер. В других аспектах способы и композиции применяют в комбинации с соединениями, о которых известно, что они подавляют иммунный ответ субъекта.
[0252] Также предлагается набор, содержащий систему нуклеазы и/или донор GLA, как описано в данном документе. Указанный набор может содержать нуклеиновые кислоты, кодирующие одну или большее количество нуклеаз (ZFN, пары ZFN, TALEN, пары TALEN и/или систему CRISPR/Cas) (например, Молекулы РНК или системы ZFN, TALEN и/или CRISPR/Cas, кодирующие гены, содержащиеся в пригодном экспрессионном векторе), донорные молекулы, экспрессионные векторы, кодирующие линии клеток-хозяев, пригодные для однонаправленной РНК, инструкции по выполнению описанных в данном документе способов и т.п.
[0253] Эти и другие аспекты будут очевидны специалисту в данной области техники в свете описания в целом.
[0254] Если в данном документе не определено иное, научные и технические термины, применяемые в связи с настоящим описанием, имеют значения, которые обычно понимаются специалистами в данной области техники. Типовые способы и материалы описаны ниже, хотя способы и материалы, аналогичные или эквивалентные описанным в данном документе, также могут применяться на практике или при тестировании настоящего изобретения. В случае конфликта, данное описание, включая определения, является превалирующим. Обычно номенклатура, применяемая в связи с кардиологией, медициной, медицинской и фармацевтической химией и клеточной биологией, и описанные в данном документе способы, хорошо известны и широко применяются в данной области техники. Ферментативные реакции и способы очистки проводят в соответствии со спецификациями производителя, как это обычно осуществляется в данной области техники или как описано в данном документе. Кроме того, если иное не требуется по контексту, термины в единственном числе включают множественное число, а термины во множественном числе включают единственное число. В данном описании и вариантах осуществления слова «иметь» и «содержать» или варианты, такие как «имеет», «имеющий», «содержит» или «содержащий», будут пониматься как подразумевающие включение указанного целого числа или группы целых чисел, но не исключая при этом любого другого целого числа или группы целых чисел. Все публикации и другие ссылки, упомянутые в данном документе, полностью включены в данный документ посредством ссылки. Хотя в данном документе цитируется ряд документов, эта ссылка не является признанием того, что какой-либо из этих документов является частью общеизвестных знаний в данной области техники. В данном контексте термин «приблизительно» или «около» применительно к одному или большему количеству значений, представляющих интерес, относится к значению, которое аналогично установленному эталонному значению. В определенных вариантах осуществления указанный термин относится к диапазону значений, которые находятся в пределах 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1% или менее в любом направлении (больше или меньше) заявленного эталонного значения, если иное не указано или иное не очевидно из контекста.
[0255] Следующие примеры представлены для лучшего понимания этого изобретения. Эти примеры приведены только в целях иллюстрации и не должны рассматриваться как ограничивающие объем изобретения каким-либо образом.
Пример 1
Высокая активность α-Gal A в плазме у мышей GLAKO, получавших вариант №4 экспрессионной конструкции, сохранялась в течение 3 месяцев
[0256] Образцы варианта №4 экспрессионной конструкции, как продемонстрировано на Фиг. 1A, вводили самцам мышей GLAKO для оценки фармакодинамической активности и биораспределения после однократного внутривенного введения.
[0257] Возраст самцов мышей GLAKO на момент начала исследования составлял 8-12 недель. Животные (n=10-20 самцов/на группу) получали буфер для состава, содержащий фосфатно-солевой буферный раствор (PBS), содержащий CaCl2, MgCl2, NaCl, сахарозу и коллифор (полоксамер) P 188 (контрольные мыши) или один из трех уровней дозы варианта № 4 (2,0E+12, 5,0E+12 или 5,0E+13 вг/кг, соответственно; n=10/на группу) в виде однократного внутривенного введения 200 мкл в хвостовую вену в День 1. За мышами наблюдали в течение 3 месяцев. Результаты фармакокинетических оценок (активность α-Gal A в плазме) представлены на Фиг. 2 для отдельных мышей, а средние значения по группе (среднее+СО) представлены на Фиг. 3. Как продемонстрировано, активность α-Gal A в плазме измеряется дозой AAV/конструкции. Кроме того, активность α-Gal A в плазме более чем в 300 раз превышала физиологическую нормальную активность или активность α-Gal A у субъекта дикого типа (немутантного). (* На Фиг. 3 указывает, что один выброс был удален из-за превышения производительности).
[0258] Однократное введение возрастающих количеств AAV hGLA кДНК без WPRE (вариант № 4) осуществляли с помощью процесса изготовления в клиническом масштабе, что приводило к супрафизиологической экспрессии α-GalA в плазме (более чем в 300 раз больше WT) ко Дню 15 исследования, при этом AAV hGLA кДНК хорошо переносился и оставался стабильным в течение 3 месяцев после инъекции. Дозозависимое увеличение активности α-GalA было достигнуто в печени, сердце и почках с соответствующим снижением Gb3/лизо-Gb3.
[0259] Продуцируемая печенью α-Gal A секретировалась в кровоток и поглощалась вторичной тканью. Фиг. 4А демонстрирует активность тканевой α-Gal A в лизатах печени. Фиг. 4В демонстрирует активность тканевой α-Gal A в лизатах почек. Фиг. 4С демонстрирует активность тканевой α-Gal A в лизатах сердца.
Пример 2
Высокий уровень активности α-Gal A приводит к соответствующему снижению
субстратов Фабри
[0260] Вариант №4 состава конструкции вводили внутривенно мышам GLAKO в дозах 0 мкг/кг, 2,0E+12 вг/кг, 5,0E+12 вг/кг или 5,0E+13 вг/кг для оценки уровня субстратов Фабри в плазме и ткани мыши. Ткани собирали при некропсии в День 91 после введения дозы и анализировали на уровни α-Gal A субстрата Gb3 (изоформы C22:0 и C24:0) и его деацилированной формы лизо-Gb3 с использованием ЖХ-МС. Вкратце, ткани взвешивали и механически разрушали в жидкости для разрушения тканей (5% MeOH, 95% воды и 0,1% уксусной кислоты) в соотношении 5 мл жидкости на мг ткани. Затем 10 мкл суспензии плазмы или ткани добавляли к 90 мкл растворителя для осаждения (MeOH с внутренним стандартом N-трикозаноилцерамида тригексозида (C23:0, Matreya), добавленным в раствор) в силиконизированной пробирке, встряхивали и помещали на качающийся планшет при комнатной температуре на 30 минут. Затем образцы центрифугировали и 10 мкл образца добавляли к 90 мкл одиночной холостой матрицы (DMSO/MeOH 1:1+0,1% FA) в стеклянном флаконе для ЖХ-МС. Образцы анализировали на длину цепи Gb3 24:0, преобладающие виды Gb3, присутствующие у мышей GLAKO, и измеряли по стандартной кривой, составленной для церамида тригексозида (Gb3, Matreya).
[0261] Глоботриаозилсфингозин (лизо-Gb3) измеряли аналогичным образом с применением глюкозилсфингозина (Matreya) в качестве внутреннего стандарта и тригексозида лизо-церамида (лизо-Gb3, Matreya) для построения стандартной кривой. Данные представляют собой среднее значение+СО от 9 до 20 животных/на группу, как указано в обозначениях. Субстрат Фабри глоботриаозилцерамид (Gb3) измеряли в отобранной плазме и тканях мышей с помощью масс-спектрометрии.
[0262] Постоянная продукция α-Gal A должна способствовать уменьшению и потенциальному клиренсу субстратов болезни Фабри, Gb3 и лизо-Gb3. Дозозависимое снижение уровней субстрата Фабри Gb3 и лизо-Gb3 было обнаружено в плазме, печени, сердце, почках и селезенке, как продемонстрировано на Фиг. 5A и Фиг. 5В. В большинстве образцов животных в группе с высокой дозой, уровни Gb3 в тканях были снижены на 80% или более по сравнению с образцами животных в контрольной группе, получающей состава, как продемонстрировано на фиг. 5В.
[0263] При высоком уровне дозы, уровни Gb3 в сердце и почках снижались до около 10% у нелеченных животных, как продемонстрировано на Фиг. 6А У леченных субъектов уровни Gb3 были ниже нижнего уровня количественного определения, как продемонстрировано на Фиг. 6В.
Пример 3
Вариант №21 экспрессионного вектора продуцирует активность α-Gal A в плазме in vitro и in vivo
[0264] Уровни и активность секретируемой α-Gal A человека оценивали в различных первичных клетках и линиях мышей, яванских макак и человека после трансдукции варианта №4 или варианта№21 экспрессионных векторов. Вариант №4 или вариант №21 экспрессионных векторов продуцировали в 1) клетках HEK293 или 2) клеточной линии Sf9 насекомых.
[0265] Клетки HepG2 и гепатоциты, полученные из iPSC (гепатоциты iCell), трансдуцировали с помощью стандартных методик и, как описано в публикации США № 20180117181. Вкратце, клетки высевали с различной плотностью на лунку и трансдуцировали с множественностью заражения (MOI) в диапазоне от 100000 до 600000 вг/на клетку варианта № 21 экспрессионной конструкции или варианта № 4 экспрессионной конструкции. Образцы супернатанта собирали со Дня 3 по День 7, и ферментативную активность α-Gal A оценивали с помощью флуорометрического анализа активности α-Gal A и в осадках клеток, собранных в конце исследования (День 6 или 7).
[0266] Подход кДНК может включать применение экспрессионной конструкции с доставкой AAV, содержащей энхансер APOE, связанный с промотором hAAT (Okuyama et al (1996) Hum Gene Ther 7(5):637-45), интрон HBB-IGG (химерный интрон, состоящий из 5'-донорного сайта из первого интрона гена бета-глобина человека и ответвления, а также 3'-акцепторного сайта из интрона вариабельной области тяжелой цепи гена иммуноглобулина), сигнальный пептид, кодирующую последовательность (при этом кодирующая последовательность необязательно является кодон-оптимизированной) и сигнальная последовательность поли A бычьего гормона роста (например, bGH или SPA51).
[0267] Клетки HepG2/C3A (также называемые клетками HepG2) (ATCC CRL 10741) поддерживали в минимальной необходимой среде (MEM) с солями Эрла и L-глутамином (Corning) с 10% фетальной бычьей сывороткой (FBS) (Life Technologies) и 1X Пенициллин Стрептомицин Глютамин (Life Technologies) и инкубировали при 37°C и 5% CO2. Пассаж клеток осуществляли каждые 3-4 дня.
[0268] Для трансдукции клетки промывали и трипсинизировали 0,25% трипсином/2,21 мМ ЭДТК (Corning) и повторно суспендировали в питательной среде. Небольшую аликвоту смешивали в соотношении 1:1 с 0,4% (масс/об.) раствором трипанового синего в фосфатно-солевом буфере (PBS; Corning) и подсчитывали на автоматическом счетчике клеток TC20 (Bio Rad). Клетки ресуспендировали при плотности 2e5 на мл в среде для выращивания и высевали в 24-луночный планшет (Corning) при 1e5 в 0,5 мл среды на лунку. Рекомбинантные частицы AAV2/6 смешивали при соответствующей множественности инфицирования (MOI) с питательной средой и добавляли к клеткам. Значение MOI для конструкций GLA кДНК составляло 3e4, 1e5, 3e5 или 1e6 вг/на клетку.
[0269] После трансдукции клетки оставляли в культуре на 6-10 дней. Супернатант собирали в Дни 3, 5, 7 и 10 (если это применимо) и заменяли свежей средой. После последнего этапа сбора супернатанта клетки трипсинизировали и ресуспендировали, как описано выше, затем центрифугировали для создания осадка клеток, промывали PBS и хранили при -80°C.
[0270] Активность α-GalA оценивали с помощью флуорометрического анализа с применением синтетического субстрата 4-метилумбеллиферил-α-D-галактопиранозида (4MU-α-Gal, Sigma).
[0271] Вкратце, 10 микролитров супернатанта клеточной культуры HepG2 смешивали с 40 мкл 5 мМ 4MU-α-Gal, растворенного в фосфатном буфере (0,1 М цитрат/0,2 М фосфатный буфер, pH 4,6, 1% Triton X-100). Реакции инкубировали при 37°C и останавливали добавлением 100 мкл 0,5 М глицинового буфера, pH 10,3. Высвобождение 4-метилумбеллиферона (4 MU) определяли путем измерения флуоресценции (Ex365/Em450) с помощью ридера флуоресценции SpectraMax Gemini XS (Molecular Devices, Саннивейл, штат Калифорния).
[0272] Стандартная кривая строилась с применением серийных 2-кратных разведений 4 MU. Полученные данные подгоняли под логарифмическую кривую; концентрация 4 MU в тестируемых образцах рассчитывали с помощью этой кривой наилучшего соответствия. Ферментативную активность выражали в нмоль 4 MU, высвобождаемых за час времени инкубации анализа, на мл супернатанта клеточной культуры (нмоль/час/мл).
[0273] Обращаясь к Фиг. 7A и Фиг. 7B, вариант №21 экспрессионной конструкции имеет улучшенную активность α-Gal A по сравнению с вариантом №4 экспрессионного вектора AAV GLA in vitro. В клетках HepG2 активность α-Gal A в супернатанте увеличивалась примерно в 4-9 раз, как продемонстрировано на Фиг. 7А. В гепатоцитах человека, полученных из iPSC активность α-Gal A в супернатанте увеличивалась примерно в 3-5 раз, как продемонстрировано на Фиг. 7B.
[0274] Эпизомальные векторы AAV (серотип 2/6), кодирующие GLA кДНК человека (hGLA), управляемую печеночно-специфическим промотором, у которого отсутствует (вариант № 4) или имеется мутированная последовательность WPRE (вариант № 21), вводили животным в различных дозах. На Фиг. 8 продемонстрировано повышение активности GLA A с увеличением дозы конструкции в плазме мышей дикого типа, пролеченных вариантом № 21 конструкций в дозе 2,0E+12 вг/кг или 5E+11 вг/кг, или вариантом № 4 конструкций в дозе 2,0E+12 вг/кг или 5E+11 вг/кг или буфером для состава. Полученные результаты указывают на улучшение плазменной активности у мышей дикого типа примерно в 7-9 раз за 28 дней.
[0275] Вариант № 4 экспрессионной конструкции сравнивали с конструкцией кДНК, включающей последовательность WPRE (вариант № 21), в одномесячном исследовании с применением двух разных доз AAV (AAV несущий кДНК донор) у мышей C57BL/6 дикого типа. В таблице 1 выше приведена полная последовательность применяемой конструкции. Конструкция, содержащая кДНК с последовательностью WPRE, продуцировала в среднем в 7 раз более высокие уровни активности α-GalA в плазме на 28 день исследования, по сравнению с мышами, которым вводили ту же дозу исходной (не содержащей WPRE) кДНК.
[0276] При применении варианта № 21 (конструкция, содержащая WPRE) наблюдалось 4-9-кратное увеличение активности GLA в супернатанте обработанных клеток HepG2 по сравнению с вариантом № 4 (без WPRE) и при применении варианта № 21 (конструкция, содержащая WPRE) наблюдали 3-5-кратное увеличение активности GLA в супернатанте гепатоцитов, полученных из индуцированных плюрипотентных клеток (iCells), по сравнению с вариантом № 4 (без WPRE). Кроме того, у мышей, получавших вариант № 21 (конструкция, содержащая WPRE), наблюдали 7-9-кратное увеличение активности GLA в плазме по сравнению с вариантом № 4 (без WRPE).
[0277] Высокие уровни активности α-GalA, наблюдаемые в этих исследованиях, наряду с сопутствующим заметным снижением накопленного Gb3/лизо-Gb3 в ключевых тканях модели мыши GLAKO, демонстрируют, что AAV-опосредованное нацеливание на гепатоциты приводит к терапевтическим уровням α-GalA человека у субъектов, в том числе с помощью производственных процессов в клиническом масштабе, которые обеспечивают быстрое и эффективное продуцирование терапевтических векторов.
[0278] Терапевтические уровни белка α-Gal A для лечения болезни Фабри генерируют in vivo с помощью подхода кДНК, включая последующее продуцирование экспрессионного вектора в клиническом масштабе.
[0279] Результаты, представленные на Фиг. 9 и в Таблице 3 ниже демонстрируют, что вариант № 21 экспрессионной конструкции продуцирует активность α-Gal A в плазме до 1500 раз выше физиологической нормы (wt) in vivo. Вариант № 4 экспрессионной конструкции вводили путем инъекции в хвостовую вену мышам C57BL/6 в дозе 5,0E+12 и 5,0E+13 вг/кг. Вариант № 21 экспрессионной конструкции вводили путем инъекции в хвостовую вену мышам C57BL/6 в дозе 5,0E+12, 5,0E+13 и 5,0E+14 (не продемонстрировано). Образцы плазмы собирали за одну неделю до введения дозы и в Дни 8, 15, 22 и 29, а затем оценивали ферментативную активность α-Gal A с помощью флуорометрического анализа. Точки данных представляют собой средний ответ +/- СО на дозу. Нижний предел количественного определения (LLOQ) составляет 2,5 нмоль/час/мл.
Таблица 3. Активность α-Gal A в плазме на день 29
[0280] Фиг. 9 демонстрирует плазменную активность α-Gal A у мышей C57BL/6 в течение 29 дней после обработки вариантом №21 конструкций в дозе 5,0E+13 вг/кг, вариантом №21 конструкций в дозе 5,0E+12 вг/кг, вариантом №4 конструкций в дозе 5,0Е+13 вг/кг, вариантом №4 конструкций в дозе 5,0Е+12 вг/кг или буфером для состава.
[0281] В соответствии с данными in vitro, уровни GLA в плазме и печени были выше у животных, которым вводили Вариант № 4 экспрессионной конструкции, изготовленной в клетке HEK293, по сравнению с клеточной системой Sf9 (до 21 раз выше).
Пример 4
Применение варианта №4 экспрессионного вектора приводила к высоким уровням трансдукции гепатоцитов у мышей GLAKO и приматов, не относящихся к человеку
[0282] Чтобы оценить уровни копий экспрессионной конструкции в гепатоцитах после внутривенного введения варианта # 4 экспрессионной конструкции, фиксированные формалином и залитые парафином (FFPE) образцы печени от подгруппы животных оценивали с помощью гибридизации in situ (ISH) BASESCOPE™ . После окрашивания ISH, проводили количественный анализ изображения с помощью программного обеспечения HALO™. Нацеливались на некодирующие последовательности. Зонд гена домашнего хозяйства, PPIB (циклофилин B), применяли в качестве маркера положительного контроля для контроля качества образца и для оценки качества РНК в образцах ткани. Бактериальный ген DapB применяли в качестве отрицательного контроля. Для всех образцов были получены полуколичественные баллы (шкала от 0 до 4) для оценки качества образцов и определения, прошел/не прошел образец контроль качества. Балл PPIB (циклофилин B; контроль гена домашнего хозяйства) составлял преимущественно 3 для образцов, что указывает на хорошее качество РНК. Баллы DapB (контроль бактериального гена) составляли в основном 0, что указывает на отсутствие или незначительный неспецифический фон. Сигнал специфического окрашивания ДНК определяется как темные (красные) пунктирные точки в ядре клетки. Образцы контрастировали гематоксилином Гилла, показанным светло-серым (синим цветом).
[0283] Репрезентативные изображения ISH печени мышей GLAKO, которым вводили 5,0+13 вг/кг варианта №4 экспрессионного вектора, при различных увеличениях представлены на Фиг. 10. Репрезентативные изображения ISH окрашивания печени NHP, которым вводили 6,0+13 вг/кг варианта №4 экспрессионного вектора, при различных увеличениях представлены на Фиг. 11. Сигнал специфического окрашивания ДНК определяется как темно серые пунктирные точки в ядре клетки. Образцы контрастировали гематоксилином Гилла, показанным светло-серым. Как продемонстрировано, 57,5% клеток гепатоцитов мыши были положительными в отношении экспрессионного вектора, с 2,34 точками/на клетку и H-баллом 126,82 в репрезентативном образце на Фиг. 10. Впечатляет, что 72,9% клеток гепатоцитов NHP были положительными в отношении экспрессионной конструкции, с 3,20 точками/на клетку и H-баллом 175,39 в репрезентативном образце на Фиг. 11.
[0284] В целом, зависимость доза-ответ в клетках печени мыши наблюдалась для всех оцениваемых параметров, включая% положительных клеток, среднее количество точек на клетку и H-балл. Для определения H-балла, клетки разделяли на 5 сегментов в зависимости от количества точек на клетку, а затем рассчитывали путем суммирования процента клеток в каждой ячейке в соответствии со взвешенной формулой.
[0285] Процент гепатоцитов мышей GLAKO, содержащих hGLA кДНК, у мышей GLAKO, которые получали вариант №4 конструкции в дозах 2E+12 вг/кг, 5E+12 вг/кг, 5E+13 вг/кг или буфер для состава в качестве контроля, продемонстрирован на Фиг. 12А. Процент гепатоцитов, содержащих hGLA кДНК, у отдельных субъектов представлен на Фиг. 12C. Подобным образом, Фиг. 12В представляет собой график, демонстрирующий процент гепатоцитов, содержащих hGLA кДНК, у яванских макаков NHP, которые получали вариант №4 конструкции в дозах 6E+12 вг/кг, 1E+13 вг/кг, 3E+13 вг/кг, 6E+13 вг/кг или буфер для состава в качестве контроля. Фиг. 12D демонстрирует процент гепатоцитов, содержащих hGLA кДНК, у отдельных субъектов NHP.
[0286] Исследования гибридизации in situ, измеряющие уровни hGLA ДНК конструкции в печени, продемонстрировали зависимость доза-ответ в гепатоцитах мыши и NHP и подтвердили перенос ДНК в ядра. Высокая доза (5,0E+13 вг/кг) для мышей обеспечила диапазон от 28% до 58% положительно окрашенных клеток, а высокая доза (6,0E+13 вг/кг; с иммуносупрессией) в исследовании NHP обеспечила диапазон от 61% до 73% положительно окрашенных клеток. У другого NHP (без иммуносупрессии) получили 49% положительно окрашенных клеток.
Пример 5
Белок α-Gal A и активность фермента в яванских макак NHP после однократного внутривенного введения варианта №4 экспрессионной конструкции
[0287] Вариант №4 экспрессионной конструкции оценивали в NHP относительно фармакологии и токсикологии. Однократную внутривенную дозу варианта №4 экспрессионной конструкции вводили самцам яванских макак в количестве 0 (n=2), 6,0E+12, 1,0E+13, 3,0E+13 или 6,0E+13 вг/кг (n=3 /на группу). С целью смягчения возможного иммунного ответа на экспрессионный вектор и/или α-Gal A человека, животные получали ритуксимаб (10 мг/кг; в/в) перед введением экспрессионной конструкции и метилпреднизолон (10 мг/кг; внутримышечно) ежедневно на протяжении всего исследования. Дополнительная группа получала вариант №4 экспрессионной конструкции в наивысшей дозе (6,0Е+13 вг/кг), но без введения иммунодепрессанта. Применяемый вариант №4 экспрессионной конструкции был изготовлен в процессе клинического производства GMP с применением платформы бакуловирус/клетки Sf9.
[0288] Кровь собирали перед введением дозы (5 моментов времени) и в Дни 7, 14, 21, 28, 35, 42, 49 и 56 и обрабатывали для получения плазмы. В этих образцах плазмы оценивали уровни белка α-Gal A человека и активность α-Gal A. При некропсии на День 56 собирали 4 сегмента печени (по 2 сегмента левой и правой боковых долей) и 2 сегмента селезенки для оценки активности α-Gal A. Результаты представлены на Фиг. 13A - 13F.
[0289] Уровни циркулирующего белка α-Gal A и активность α-Gal A в плазме обычно определяли ко Дню 7, при этом уровни и активность белка достигали пика между Днями 7 и 21, и при этом не было четкого ответа на дозу. Животные, которым вводили вариант №4 экспрессионной конструкции без иммуносупрессанта, обычно имели более низкие уровни и активность белка α-Gal A, чем животные, которым вводили вариант №4 экспрессионной конструкции с иммуносупрессантом. Это отсутствие сильного ответа на дозу и клиренса активности α-Gal A и уровней белка согласуется с возникающим иммунным ответом против α-Gal A человека (белка человека, вводимого животному), что подтверждается присутствием антител к α-Gal A человека. Несмотря на пониженные уровни α-Gal A человека, некоторые животные поддерживали высокие уровни α-Gal A человека (активности и белка). У одного животного, получавшего высокую дозу (6.0E+13 IS), уровни 193 нмоль/час/мл определялись на День 56, в то время как уровни у животных, получавших носитель, не обнаруживались (<10 нмоль/час/мл). Временный характер этого ответа у некоторых животных, вероятно, был связан с ожидаемым иммунным ответом на человеческий фермент α-Gal A (белок человека, вводимый животному).
[0290] Кроме того, в исследовании NHP образцы для анализа шеддинга векторов оценивали методом кПЦР. Низкие уровни вектора hGLA измеряли в слюне, моче и кале некоторых животных, получавших вариант №4, до Дня 4 (моча) или до Дня 14 (слюна, кал). На День 60 в этих биологических жидкостях не обнаруживались уровни вектора hGLA.
Пример 6
hGLA и соответствующие уровни мРНК в печени NHP
[0291] Вестерн-блот-анализ hGLA и соответствующих уровней мРНК в образцах печени NHP от отдельных животных выполняли на день 60 после введения варианта №4 конструкций в дозах 6,0E+12 вг/кг, 1,0E+13 вг/кг, 3,0E+13 вг/кг, 6,0E+13 вг/кг, 6,0E+13 вг/кг без иммунодепрессантов или буфера для состава. Как продемонстрировано на Фиг. 14, уровни белка hGLA повышаются с увеличением дозы конструкции, при этом уровни белка коррелируют с уровнями мРНК в большинстве образцов.
Пример 7
Оценка безопасности, переносимости и фармакодинамики варианта №21 экспрессионной конструкции у людей
[0292] Будет проведено исследование для оценки безопасности и переносимости варианта №21 экспрессионной конструкции у людей Кроме того, будут измеряться фармакодинамика α-Gal A и присутствие его субстратов в плазме, моче и тканях в динамике. Влияние варианта №21 экспрессионной конструкции на введение ERT для субъектов на ERT, функцию почек, иммунный ответ и шеддинг вирусной векторной ДНК также можно оценить в динамике.
[0293] В целом вариант №21 экспрессионной конструкции и вариант №4 экспрессионной конструкции хорошо переносились в мышиной модели болезни Фабри (GLAKO), мышами дикого типа (C56BL/6) и NHP яванскими макаками. У мышей GLAKO не было никаких нежелательных признаков, связанных с однократным внутривенным введением варианта №4 экспрессионной конструкции, в дозе до 5,0Е+13 вг/кг, наивысшего испытанного уровня дозы. У мышей C57BL/6 предварительный анализ продемонстрировал, что вариант №21 экспрессионной конструкции хорошо переносился вплоть до самой высокой испытанной дозы, 1,5E+14 вг/кг. Для NHP, данные, связанные с вариантом №21 экспрессионной конструкции, были ограничены животными, которые не получали иммуносупрессивную терапию (6,0Е+13 вг/кг). Эти данные заключались в увеличении лимфоидной клеточности в лимфоидной ткани и селезенке и, вероятно, соответствовали иммунному ответу, связанному с введением hGLA и/или rAAV2/6. В этих исследованиях уровень дозы, не приводящий к развитию наблюдаемого нежелательного эффекта (no-observed-adverse-effect level , NOAEL), составлял 6,0E+13 вг/кг, с иммуносупрессивной схемой или без нее, самый высокий протестированный уровень дозы.
[0294] В исследовании применяется конструкция рекомбинантного вектора (например, rAAV2/6), кодирующая кДНК для α-Gal A человека. Векторная конструкция кодирует специфический для печени промотор, а rAAV2/6 проявляет тропизм к печени, что обеспечивает возможность долгосрочного и стабильного продуцирования печенью α-Gal A у субъектов с болезнью Фабри после введения однократной дозы. Можно применять различные серотипы AAV, включая AAV2, 5, 6 и 8. Серотип rAAV2/6 был выбран для применения в этом и в описанных выше примерах на основании предыдущих данных NHP, демонстрирующих, что AAV2/6 был в первую очередь гепатотропным с аналогичным биораспределением для AAV2/8 и векторы AAV2/6 и AAV2/8 обеспечивали аналогичные уровни экспрессии циркулирующего трансгена FIX. Предварительные данные о клинической безопасности были собраны у 13 субъектов, которым вводили исследуемые препараты в 3 исследованиях, и позволяют предположить, что инфузии с этим серотипом AAV2/6 хорошо переносятся (данные не показаны).
[0295] Исследования на мышиной модели болезни Фабри с внутривенным введением rAAV2/6, кодирующего hGLA кДНК, демонстрируют получение терапевтических уровней (более чем в 300 раз превышающих дикий тип) α-Gal A. Одноразовое применение экспрессионного вектора сводит к минимуму частоту реакций, связанных с инфузией. Продуцирование терапевтических уровней α-Gal A у людей может способствовать снижению и потенциальному клиренсу субстратов болезни Фабри Gb3 и лизо-Gb3 и может снизить риск развития антител к ферменту, продуцируемому из-за постоянного продукции фермента, а не пика и впадины, наблюдаемых при ERT. Вариант №21 экспрессионной конструкции была разработана для обеспечения стабильной, долгосрочной продукции α-Gal A на терапевтических уровнях у субъектов с болезнью Фабри. Постоянное продукция α-Gal A у людей может способствовать уменьшению и клиренсу субстратов Gb3 и лизо-Gb3 болезни Фабри.
Оценки исследования
[0296] Оценка может включать в себя случаи нежелательных явлений, возникающих в связи с лечением (treatment-emergent adverse events, TEAE), рутинные гематологические, биохимические исследования и функцию печени, показатели жизненно важных функций, ЭКГ и ЭхоКГ, серийное тестирование альфа-фетопротеина (AFP) и МРТ печени (или эквивалентную визуализацию) для мониторинга состояния любого новообразования в печени. Кроме того, в определенные моменты времени в течение 1 года можно измерить изменение от исходного уровня следующих показателей: активность α-Gal A в плазме; уровни Gb3 в плазме; уровни лизо-Gb3 в плазме; частота инфузии FABRAZYME® (или эквивалентной ERT); расчетная скорость клубочковой фильтрации (рСКФ), рассчитанную по уровням креатинина в крови; массу левого желудочка, измеренную с помощью магнитно-резонансной томографии сердца (МРТ), отношение общего белка и альбумина к креатинину в моче; уровни α-Gal A и Gb3, измеренные в ткани; уровни субстрата, измеренные в тканях и моче; биомаркеры почечной функции в моче; нейропатическая боль, измеренную с помощью Краткого опросника оценки боли (Brief Pain Inventory, BPI), частота применения обезболивающих средств; симптомы со стороны желудочно-кишечного тракта (ЖКТ), измеренные по шкале оценки симптомов ЖКТ; индекс степени тяжести Майнца (Mainz Severity Score Index, MSSI); качество жизни (quality of life, QOL), сообщаемое пациентом и измеренное с помощью опросника SF-36; иммунный ответ на rAAV2/6 и α-Gal A; и клиренс вектора rAAV, который можно измерить по уровню генома вектора в крови, плазме, слюне, моче, стуле и сперме.
Критерии включения и исключения субъектов
[0297] Субъектами в исследовании могут быть мужчины в возрасте ≥18 лет с классической болезнью Фабри. Следует набирать субъектов мужского пола с классической болезнью Фабри, чтобы убедиться, что любой остаточный уровень фермента не влияет на измерение уровней ферментов, продуцируемых трансгеном кДНК.
[0298] Более конкретно, критерии включения субъектов могут включать: (1) субъекты с документально подтвержденным диагнозом классической болезни Фабри, определяемым по активности α-Gal A < 5% в плазме или лейкоцитах и по одной или большим количеством из следующих симптоматических характеристик классической болезни Фабри: i) воронковидная кератопатия (cornea verticillata); ii) акропарестезия, iii) ангидроз, iv) ангиокератома (если имеется документально подтвержденная кластерная периумбиликальная ангиокератома, одного этого симптома достаточно, поскольку он является патогномоничным признаком классической болезни Фабри); (2) субъект, получающий ERT (схема 14 дней [±1 день]); или субъект получающий ERT, чья активность -Gal A составляет >5%; или является ERT-наивным; или является псевдо-наивным ERT и не получал лечение ERT в течение последних 6 месяцев до получения согласия; (3) для субъектов, получающих ERT, ERT должна вводиться в стабильной дозе (что определяется тем, что пациент не пропустил более 3 доз ERT в течение 6 месяцев до получения согласия) и по схеме (14 дней ± 1 день в течение по меньшей мере 3 месяцев до начала участия в исследовании); (4) субъект с мутацией, которая указывает на классическую болезнь Фабри (то есть внесена в базу данных, например www.dbfgp.org); (5) субъект, который имеет активность α-Gal A, определяемую ниже нижнего предела нормального диапазона анализа; (6) субъекты мужского пола в возрасте ≥18 лет; (7) половозрелые субъекты должны согласиться использовать презерватив и воздерживаться от донорства спермы с момента введения экспрессионной конструкции до тех пор, пока минимум 3 последовательных образца спермы не станут отрицательными на AAV после введения исследуемого препарата и как минимум через 90 дней после введения исследуемого лечения; а также (8) подписанное письменное информированное согласие субъекта.
[0299] Для субъектов, у которых нет задокументированного диагностического уровня активности α-Gal A, следует взять образец крови для измерения уровней активности α-Gal A (в плазме и/или лейкоцитах). Для тех субъектов, которые находятся на ERT, этот анализ крови должен быть взят по меньшей мере через 13 дней после их последней инфузии ERT (впадина). i. Если уровень активности α-Gal A у субъекта составляет >5% и субъект принимает ERT, этот уровень активности фермента может быть связан с остаточной активностью α-Gal A после последней инфузии ERT. В этом случае диагноз классической болезни Фабри может быть подтвержден при соблюдении следующих трех критериев:
[0300] а. две или большее количество из следующих документально подтвержденных симптоматических характеристик классической болезни Фабри: воронковидная кератопатия, акропарестезия, ангидроз, ангиокератома. Если имеется документально подтвержденная кластерная периумбиликальная ангиокератома, одного этого симптома достаточно, поскольку он является патогномоничным признаком классической болезни Фабри;
[0301] b. мутация, которая указывает на классическую болезнь Фабри (то есть внесена в базу данных, например www.dbfgp.org); а также
[0302] c. активность α-Gal A определяется ниже нижнего предела нормального диапазона анализа.
[0303] Секвенирование гена болезни Фабри может быть выполнено при скрининге для подтверждения того, что субъекты имеют мутацию в гене GLA. Анализ можно проводить на образцах крови или слюны. Если это возможно, можно применять результаты секвенирования генов, полученные до исследования.
[0304] Тестирование на ВИЧ, ВГА, ВГВ, ВГС и ТБ может проводиться во время скрининга. Субъекты с диагнозом ВИЧ-инфекции или признаками активной инфекции ВГА, ВГВ, ВГС или ТБ могут не иметь возможности участия в этом исследовании.
[0305] Уровень нейтрализующих антител к AAV6 можно измерить при скрининге для оценки ранее существовавшего иммунного ответа субъекта на AAV6. Субъекты с повышенным уровнем ранее существовавших нейтрализующих антител к AAV6 могут не иметь возможности участия в этом исследовании. Если введение дозы не было завершено в течение 3 месяцев после скрининга, следует повторить анализ нейтрализации AAV6 в сыворотке.
[0306] Если возможно, можно применять диагностические результаты уровня активности α-Gal A в плазме или лейкоцитах, полученные до исследования. Для субъектов, у которых нет задокументированного диагностического уровня активности α-Gal A, следует взять образец крови для измерения уровней активности α-Gal A (в плазме и/или лейкоцитах). Для тех субъектов, которые находятся на ERT, этот анализ крови должен быть взят по меньшей мере через 13 дней после их последней инфузии ERT.
[0307] Рентгенологическое исследование грудной клетки (также известное как рентгенограмма грудной клетки) может быть осуществлено оценки общего состояния здоровья и соответствия критериям исследования. Если по медицинским показаниям не указано иное, рентгеновский снимок грудной клетки, выполненный в пределах 6 месяцев до включения в исследование, может быть использован для определения возможности участия субъекта в этом исследовании. Физикальному осмотру должен подвергаться каждый субъект, при этом указанный физикальный осмотр должен включать как минимум: общее визуальное состояние, осмотр головы, глаз, ушей, носа и горла (ЛОР-органы); а также физикальное обследование сердечно-сосудистой, кожной, респираторной, желудочно-кишечной, опорно-двигательной и неврологической систем.
[0308] Критерии исключения для субъектов могут включать субъектов, которые: (1) заведомо не отвечают на ERT по мнению исследователя центра и Медицинского монитора (например, не зарегистрировано снижение уровня субстрата на ERT); (2) получают текущее лечение мигаластатом (Galafold™) или предшествующее лечение в течение 3 месяцев с момента подписания информированного согласия, (3) имеют положительный нейтрализующий ответ антител на AAV (например, AAV6), (4) имеют интеркуррентное заболевание, которое, по мнению исследователя на сайте или медицинского монитора, предположительно ухудшит оценку безопасности или эффективности в течение периода наблюдения в исследовании; (5) имеют рСКФ ≤ 60 мл/мин/1,73 м2; (6) имеют класс III или выше по классификации Нью-Йоркской кардиологической ассоциации; (7) имеют активную инфекцию вируса гепатита А (ВГА), вируса гепатита В (ВГВ), вируса гепатита С (ВГС) (отрицательная ДНК ВГС) или вируса иммунодефицита человека (ВИЧ), по данным количественной полимеразной цепной реакции (кПЦР) или активную инфекцию туберкулеза (ТБ); (8) имеют в анамнезе заболевания печени, такие как вторичный стеатоз, неалкогольный стеатогепатит (НАСГ) и цирроз, холангит, заболевание желчевыводящих путей в пределах 6 месяцев до подписания информированного согласия; кроме синдрома Жильбера; аномально циркулирующего AFP; (9) для субъектов, получающих ERT, которые имеют недавнюю или продолжающуюся гиперчувствительность к лечению ERT, в пределах 6 месяцев до подписания информированного согласия, что проявляется значительной инфузионной реакцией на ERT по мнению исследователя и медицинского монитора; (10); маркеры воспаления печени или явных или скрытых причин дисфункции печени, что подтверждается одним или большим количеством из следующих признаков: (i) альбумин ≤3,5 г/дл; (ii) общий билирубин > верхнего предела нормы (upper limit of normal, ULN) и прямой билирубин ≥0,5 мг/дл; (iii) щелочная фосфатаза (ALP) > 2,0 x ULN; (iv) аланинаминотрансфераза (ALT) > 1,5 x ULN; (11) субъект в настоящее время или в анамнезе принимал системные (внутривенные или пероральные) иммуномодуляторы или стероиды в течение последних 6 месяцев (разрешено местное лечение, например, астмы или экземы) (периодическое применение системных стероидов может быть разрешено после обсуждения с медицинским монитором); (12) имеют противопоказания к применению кортикостероидов для подавления иммунитета; (13) имеют в анамнезе злокачественные новообразования, кроме немеланомного рака кожи; (14) употребляли алкоголь или наркотики в анамнезе ; (15) участвовали в предшествующем интервенционном исследовании лекарственных препаратов или медицинских изделий в течение последних 3 месяцев до подписания информационного согласия (за исключением имплантируемых петлевых регистраторов, как в исследовании RaILRoAD); (16) получали ранее лечение продуктом генной терапии; (17) известная гиперчувствительность к компонентам состава ST-920; (18) любая другая причина, которая, по мнению исследователя центра или медицинского монитора, сделала бы субъекта непригодным для участия в исследовании.
Сопутствующая лекарственная терапия
[0309] Разрешены все лекарственные средства, кроме потенциально гепатотоксичных. Гепатотоксические агенты, такие как диклофенак, амиодарон, хлорпромазин, флуконазол, изониазид, рифампицин, вальпроевая кислота, высокие дозы ацетаминофена (4-8 г/день) и т.д., а также гепатотоксические травяные добавки, такие как крестовник/кроталария, дубровник в чае, чапаррель, цзинь бу хуан, ма-хуан (китайские травы) и др. не следует принимать в период исследования. Для субъектов, получающих ERT, ERT следует вводить в стабильной дозе (определяется как отсутствие пропуска более 3 доз ERT в течение последних 6 месяцев до подписания информированного согласия) и по схеме (14 дней ± 1 день в течение по меньшей мере 3 месяцев до зачисление в исследование). Субъекты должны продолжать получать ERT в стабильной дозе и по схеме (14 дней ± 1 день) во время исследования в соответствии со стандартом лечения, если им ERT не будет отменена.
Когорты доз
[0310] Начальная доза будет составлять 5,0E+12 вг/кг, и любое повышение дозы до следующего уровня будет производиться после анализа данных предыдущей когорты и/или других клинических исследований, в которых применяется терапия на основе rAAV2/6 in vivo, и на основе рекомендации Комитета по мониторингу безопасности (Safety Monitoring Committee, SMC), который может включать внешних экспертов в предметной области, медицинских мониторов исследований и исследователей в исследовательских центрах. В данном контексте представители SMC должны обладать соответствующими медицинскими и научными знаниями и обеспечивать надзор за безопасностью исследования. Кроме того, в зависимости от наблюдаемых уровней активности ферментов и профиля безопасности субъектов, которым вводили дозу, SMC может рекомендовать повышение дозы до промежуточного уровня дозы 3,0E+13 вг/кг, что в 3 раза превышает дозу в когорте 2 вместо 5-кратного увеличения дозы 5,0E+13 вг/кг в когорте 3. Также можно рассмотреть дозу около 1,0E+14 вг/кг. Три когроты доз приведены в Таблице 4.
Таблица 4. Когорты доз
(векторные геномы
[вг]/кг)
*rAAV=рекомбинантный аденоассоциированный вирус.
[0311] Будут включены субъекты в возрасте ≥18 лет, соответствующие всем критериям включения/исключения. По меньшей мере, два субъекта будут распределены в каждую из трех дозовых когорт с возможным расширением любой когорты с дополнительными 4 взрослыми субъектами, всего до 18 субъектов, после рассмотрения SMC. Экспрессионный вектор можно вводить посредством внутривенной инфузии. В пределах каждой когорты лечение будет поэтапным, так что каждому последующему субъекту нельзя будет вводить дозу по меньшей мере до около 2 недели после того, как предыдущему субъекту было осуществлено введение дозы. Повышение дозы до следующего уровня может произойти не ранее чем через около 4 недель после того, как последний субъект в предыдущей когорте получит дозу, а данные о безопасности всей предыдущей когорты будут рассмотрены SMC.
[0312] Субъекты, которые получали ERT до включения в исследование, должны продолжать получать ERT во время исследования и придерживаться своей текущей дозы и схемы (14 дней ± 1 день) в соответствии со стандартом лечения, если им не будет отменена ERT. Для субъектов, получающих ERT, тестирование исходных уровней ферментов и субстратов будет скоординировано таким образом, чтобы образцы можно было брать в 2 отдельных случаях утром при самом минимальном уровне, что определяется как 14 дней (+/- 1 день) после предыдущей инфузии ERT. Дополнительный момент времени будет принят ранее во время периода скрининга, следовательно, будет 3 момента времени для оценки остаточных уровней α-Gal A при самых минимальных значениях до введения генной терапии. Эти 3 образца следует брать при самых минимальных значениях и предпочтительно в одно и то же время в течение дня (например, утром), чтобы свести к минимуму неспецифические факторы, потенциально влияющие на уровни ферментов.
[0313] Чтобы свести к минимуму потенциальный иммунный ответ на капсидный белок rAAV, избежать потери экспрессии трансгена в случае поражения печени и сохранить функцию печени, можно профилактически вводить преднизон или эквивалентный кортикостероид, начиная примерно за 2 дня до инфузии экспрессионного вектора, и его дозу можно постепенно снижать. в течение около 20 недель.
[0314] Экспрессионный вектор может быть введен с помощью шприцевого насоса или инфузионного насоса для внутривенного вливания (см. Study Pharmacy Manual). Общие объемы будут зависеть от когорты, в которую определен пациент, и массы тела (кг) на исходном уровне. Экспрессионный вектор можно вводить через внутривенный катетер с контролируемой скоростью и мониторингом жизненно важных функций субъекта (температура, частота сердечных сокращений, частота дыхания и артериальное давление), пока субъект находится в больнице или учреждении неотложной помощи, где субъект может оставаться для наблюдения по меньшей мере на 24 часа после завершения инфузии экспрессионного вектора. Субъект может быть выписан, когда все показатели жизненно важных функций стабилизируются, и любые нежелательные явления (adverse events, AE) будут купированы или состояние субъекта будет расценено как стабилизированное в соответствии с заключением исследователя.
[0315] После инфузии экспрессионного вектора, визиты в рамках исследования можно проводить в День 8; на Неделях 2, 4, 6, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48 и 52. На визитах в рамках исследования на Неделе 28, 32, 40, 44 и 48 проводятся оценки, которые не требуют физического присутствия субъекта в клиническом центре, и поэтому могут осуществляться удаленно. Оценки относительно AE и приема сопутствующих лекарственных средств можно проводить удаленно по телефону.
[0316] Функциональные тесты печени (АСТ, АЛТ, ГГТ, общий и прямой билирубин, ЩФ, ЛДГ, альбумин и уровни общего белка) можно проводить для мониторинга AAV-опосредованной иммуногенности два раза в неделю в течение примерно первых 20 недель после инфузии экспрессионного вектора, пока субъект принимает преднизон или аналогичный кортикостероид, и можно осуществлять удаленно. Образцы крови для функциональных тестов печени могут быть взяты с интервалом 2-4 дня, когда это возможно, за исключением первой недели, когда они могут быть взяты во время Визитов в День 2 и День 8. Затем функциональные тесты печени можно проводить еженедельно в течение четырех недель после прекращения иммуносупрессии (Недели 21-24), а затем ежемесячно, чтобы они совпадали визитами в рамках исследования (Недели 28-52).
[0317] Если, несмотря на предварительное лечение преднизоном или эквивалентным кортикостероидом, есть признаки повышения уровня АЛТ, прием дозы преднизона или эквивалентного кортикостероида будет продолжена (преднизон 1 мг/кг [максимум 60 мг] или эквивалента; перорально или внутривенно и/или будет увеличена в каждом конкретном случае, при этом уровень ферментов печени можно оценивать два раза в неделю до нормализации указанного уровня ферментов печени, а затем в соответствии с протоколом.
[0318] Для первых 2 субъектов в каждой когорте лечение может быть поэтапным, так что каждому последующему субъекту не будут проводить инфузию до тех пор, пока не истечет период наблюдения за предыдущим субъектом, составляющий по меньшей мере 2 недели. Повышение дозы до следующего уровня может произойти не ранее чем через по меньшей мере 4 недели после того, как 2 субъекта из предыдущей когорты получат дозу, а данные о безопасности этих 2 субъектов из предыдущей когорты будут рассмотрены SMC.
[0319] Введение и повышение дозы можно приостановить, если соблюдать правила приостановки.
[0320] Применение экспрессионного вектора может устранить необходимость в ERT за счет вектора rAAV, кодирующего кДНК для α-Gal A человека, что приводит к долговременной печеночно-специфической экспрессии α-Gal A у субъектов с болезнью Фабри. Субъекты, которым ERT была отменена, будут тщательно контролироваться на предмет любых AE, показателей жизненно важных функций, любых изменений лабораторных оценок безопасности и уровней α-Gal A и субстратов по сравнению с исходным уровнем. Возможность отмены ERT следует рассматривать по прошествии четырех недель, чтобы дать достаточно времени для трансдукции целевых клеток печени. Субъекты, которым ERT была отменена, будут тщательно контролироваться на предмет любых клинических симптомов, включая усталость и невропатическую боль, любых AE, показателей жизненно важных функций, любых изменений в лабораторных оценках безопасности, включая функциональные тесты печени и уровни α-Gal A и субстратов (Gb3 и лизо-Gb3) по сравнению с исходным уровнем. Отмена ERT может быть произведена Исследователем центра после консультации со Спонсором и должна быть рассмотрена для субъектов, которые желают отменить прием ERT и соответствуют следующим критериям:
(1) у пациентов прошло ≥4 недель после введения ST-920;
(2) пациенты являются стабильными с медицинской точки зрения и могут переносить временное прекращение приема ERT по мнению исследователя центра;
(3) пациенты соглашаются на усиленный мониторинг безопасности и дополнительные лабораторные исследования до визита последующего наблюдения при отмене ERT;
(4) нет необходимости повторно начинать прием ERT после визита последующего наблюдения при отмене ERT. Тем не менее ERT может быть возобновлена в любое время в зависимости от клинических обстоятельств или по решению исследователя центра.
[0321] Отмена ERT может быть осуществлена повторно, если ранее она была безуспешной, при условии, что это будет сделано по меньшей мере через 12 недель после предыдущей попытки, если субъект желает этого, и может выполнена по усмотрению Исследователя центра и после консультации со Спонсором.
[0322] Продолжительность участия в исследовании может составлять до 76 недель для каждого субъекта, разделенных на до 8 недель для периода скрининга, до 12 недель для периода исходного уровня и 52 недели для периода последующего наблюдения после введения дозы. Набор планируется от 9 до 12 месяцев. Субъектов следует поощрять к участию в дополнительном отдельном долгосрочном исследовании последующего наблюдения на срок до 4 лет.
[0323] Набор в исследовании должен быть приостановлен, если выполняется любой из следующих критериев, и SMC может собраться для вынесения рекомендаций относительно правильного алгоритма действий: (1) любое нежелательное явление 3 степени или выше с по меньшей мере обоснованной возможностью причинной связи с составом экспрессионного вектора; (2) серьезное нежелательное явление (SAE) с по меньшей мере обоснованной возможностью причинной связи с составом экспрессионного вектора; (3) смерть субъекта-человека; (4) развитие злокачественного новообразования.
[0324] AE, возникшие в связи с лечением, можно обобщить в целом и по дозовой когорте. Для каждого субъекта в обобщении можно использовать максимальную тяжесть каждого AE по степени тяжести. Кроме того, можно обобщить все SAE и AE, связанные с исследуемым лечением. Для других оценок безопасности, данные могут быть обобщены для каждого момента времени. Отклонение от исходных значений можно рассчитать для непрерывных параметров и обобщить по моменту времени. Для выбранных параметров также можно построить таблицы сдвигов.
[0325] Для того, чтобы оценить, продуцируется ли α-Gal A и является ли она активной, следует измерить активность α-Gal A в плазме. Измерения уровня α-Gal A можно проводить в плазме, сыворотке, цельной крови, сухих пятнах крови, лейкоцитах или других компонентах крови. Для субъектов, получающих ERT, образцы следует получать через 14 дней (± 1 день) после предыдущего введения ERT. Дополнительные образцы также могут быть получены в ходе исследования, чтобы улучшить понимание фармакокинетики фермента и гарантировать, что образцы, полученные до ERT, имеют самые низкие показатели.
[0326] Gb3 представляет собой тип гликосфинголипида, который накапливается в кровеносных сосудах, тканях и органах при болезни Фабри из-за дефицита α-Gal A. Уровни Gb3 в плазме, моче и других тканях могут быть измерены в ходе этого исследования для оценки воздействия лечения и уровней α-Gal A. Для субъектов, получающих ERT, образцы следует получать через 14 дней (± 1 день) после предыдущего введения ERT.
[0327] Лизо-Gb3 представляет собой растворимую форму субстрата Gb3. Уровни лизо-Gb3 в плазме, моче и других тканях можно измерять на протяжении всего этого исследования, чтобы оценить влияние лечения и уровни α-Gal A. Для субъектов, получающих ERT, образцы следует получать через 14 дней (± 1 день) после предыдущего введения ERT.
[0328] В каждый момент времени отбора образцов, фактическое значение и отклонение от исходного показателя для уровней α-Gal A и Gb3 и лизо-Gb3 можно обобщить с помощью описательной статистики и нанести на график в динамике относительно дозовой когорты. Для субъектов, которым отменяли ERT, изменения частоты и дозы инфузий ERT до и после отмены ERT можно оценить и обобщить с учетом общей дозы и количества инфузий в годовом исчислении. Также может быть проанализирована продолжительность периода отмены ERT. Клиренс AAV, измеренный с помощью векторных геномов в различных образцах (плазма, слюна, моча, стул и сперма), может быть нанесен на график в динамике относительно дозовой когорты.
[0329] Как продемонстрировано на Фиг. 1A, вектор rAAV содержит вариант №21 экспрессионной кассеты hGLA (3321 п.н.), которая включает печеночно-специфические регуляторные элементы, которые управляют экспрессией трансгена hGLA. Трансген hGLA находится под контролем энхансера и области, контролирующей печень, из гена аполипопротеина E (ApoE) человека и промотора α-1-антитрипсина человека (hAAT). Энхансер ApoE и промотор hAAT являются специфическими и высокоактивными в печени, предполагаемой целевой ткани, но неактивны в типах клеток и тканей, не относящихся к печени, и таким образом предотвращают экспрессию и активность hGLA в тканях, не являющихся мишенями. Модифицированный химерный интрон (трансген HBB-IghGLA содержит кодон-оптимизированный фермент hGLA α-Gal A.
[0330] Вариант № 21 содержит мутированную форму посттранскрипционного регуляторного элемента вируса гепатита сурка (WHV) (WPREmut6). WPREmut6 представляет собой последовательность ДНК длиной 592 п.н., содержащую промоторную область белка WHV X, за которой следует усеченная форма самого белка X с точечными мутациями в предполагаемой области промотора и стартовый кодон открытой рамки считывания белка X для предотвращения экспрессии белка X (mut6). Последовательность поли A является производным сигнала полиаденилирования бычьего гормона роста. Добавление элемента WPREmut6 приводило к увеличению продукции белка α-Gal A. Действительно, была отмечена более высокая эффективность варианта №21 экспрессионной конструкции по сравнению с вариантом №4 экспрессионной конструкции (в которой отсутствует элемент WPREmut6).
[0331] Вариант №21 экспрессионной конструкции может быть составлен с концентрацией приблизительно 1,0E+13 вг/мл в фосфатно-солевом буфере (PBS), содержащем CaCl2, MgCl2, NaCl, сахарозу и коллифор (Poloxamer) P 188, разлитого во флаконы по 2 мл, 5 мл или 10 мл и т.д. и хранящегося при температуре ≤-65°C. Флаконы имеют алюминиевое уплотнение с откидной торцевой крышкой.
[0332] Вектор rAAV экспрессионной конструкции может быть упакован с капсидным серотипом AAV2/6 с применением экспрессионной системы клетки насекомых Sf9/рекомбинантный бакуловирус (Sf9/rBV). В альтернативном варианте, вектор rAAV экспрессионной конструкции может быть упакован с капсидным серотипом AAV2/6 с применением экспрессионной системы млекопитающих, например, HEK293.
[0333] Исследования на мышах с моделями болезни Фабри, на мышах дикого типа и яванских макаках NHP демонстрируют возможность безопасного продуцирования устойчивых и потенциально эффективных уровней α-Gal A после применения варианта №21 экспрессионного вектора.
[0334] Каких-либо нежелательных эффектов не отмечалось у мышей при уровнях доз до 1,5E+14 вг/кг и у NHP при уровнях доз до 6,0E+13 вг/кг, то есть при наивысших уровнях введенных доз, соответственно. Следовательно, клиническая начальная доза 5,0E+12 вг/кг поддерживается 30-кратной безопасной дозой для мышей, и 12-кратной безопасной дозой для NHP.
[0335] Измеримые уровни α-Gal A ожидаются у людей при дозе 5,0E+12 вг/кг на основании выраженной фармакодинамической реакции, отмеченной у мышей с болезнью Фабри, получавших 2,0E+12 вг/кг.
[0336] Все патенты и публикации, на которые делается ссылка в данном документе, включены в этот документ посредством ссылки в полном объеме.
[0337] Несмотря на то, что настоящее изобретение было довольно подробно описано в качестве иллюстрации и примера в целях ясности понимания, для специалистов в данной области техники будет очевидно, что различные изменения и модификации могут быть осуществлены на практике без отклонения от сущности или объема настоящего изобретения. Соответственно, приведенные выше описания и примеры не следует рассматривать как ограничивающие.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANGAMO THERAPEUTICS, INC.
<120> Способы и композиции для лечения болезни Фабри
<130> 4341.019PC01
<140> PCT/US2020/012274
<141> 2020-01-03
<150> 62/788,439
<151> 2019-01-04
<160> 10
<170> PatentIn версия 3.5
<210> 1
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 1
ctgcgcgctc gctcgctcac tgaggccgcc cgggcaaagc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct 130
<210> 2
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 2
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg g 321
<210> 3
<211> 393
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 3
gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga gggccagcta 60
agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg gacacaggac 120
gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg aagctgtaca 180
ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact 240
tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct 300
cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct 360
cagcttcagg caccaccact gacctgggac agt 393
<210> 4
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 4
gtaagtatca aggttacaag acaggtttaa ggagaccaat agaaactggg cttgtcgaga 60
cagagaagac tcttgcgttt ctgataggca cctattggtc ttactgacat ccactttgcc 120
tttctctcca cag 133
<210> 5
<211> 1290
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 5
atgcaactta ggaaccccga acttcatctt ggctgcgccc tggccctccg cttcctcgct 60
ctcgtttctt gggacatccc tggcgctagg gcactcgaca acggcctcgc gcggactcct 120
acgatgggat ggttgcactg ggaaaggttt atgtgcaatc tggattgcca ggaggagccg 180
gactcatgca tctcggagaa gctgttcatg gagatggcgg aacttatggt atcggaggga 240
tggaaggatg ccgggtatga gtatctctgt atcgacgatt gttggatggc tccccagaga 300
gactccgagg gacgactcca agcggacccc cagcgctttc cacatggcat tcgacagctc 360
gccaattacg tgcactcgaa ggggttgaag ttgggaatct acgcagatgt gggcaacaaa 420
acgtgtgcgg ggttcccggg gtcgtttgga tactacgata ttgatgcgca gacgtttgct 480
gactggggtg tcgatctttt gaaatttgat ggctgttact gtgattcgtt ggaaaacctg 540
gcggatggat acaagcatat gtcactcgcc ttgaaccgga caggtcgctc aatcgtatac 600
agctgcgaat ggcccctcta tatgtggccc ttccaaaagc ccaattacac agagattcgg 660
cagtattgca atcactggag gaactttgcc gatattgacg acagctggaa atccatcaag 720
tccattctcg attggacgag cttcaaccag gagcgcatcg tggacgtggc aggacccgga 780
ggttggaacg atccggacat gctcgtaatt gggaatttcg ggcttagctg gaatcagcaa 840
gtcacccaaa tggcgctgtg ggccatcatg gcagctcctc tctttatgtc gaatgatctg 900
cggcatatct cgccccaggc aaaggctctt ttgcaagaca aggacgtcat cgcaatcaat 960
caggacccat tggggaaaca gggatatcaa cttcgccagg gtgacaattt cgaagtatgg 1020
gagaggccgc ttagcgggct ggcgtgggcg gtcgcgatga ttaaccggca ggaaatcgga 1080
gggcctcgct cgtataccat cgcagtggcc tcactgggca aaggagtggc gtgcaatccg 1140
gcctgcttca tcacccagtt gttgcccgtc aaaagaaagc tgggtttcta cgagtggaca 1200
tccagactta gatcacacat taaccctact ggtacggtgt tgctccagct cgaaaacaca 1260
atgcagatgt cgttgaaaga cctgctgtaa 1290
<210> 6
<211> 592
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 6
aatcaacctc tggattacaa aatttgtgaa agattgactg gtattcttaa ctatgttgct 60
ccttttacgc tatgtggata cgctgcttta atgcctttgt atcatgctat tgcttcccgt 120
atggctttca ttttctcctc cttgtataaa tcctggttgc tgtctcttta tgaggagttg 180
tggcccgttg tcaggcaacg tggcgtggtg tgcactgtgt ttgctgacgc aacccccact 240
ggttggggca ttgccaccac ctgtcagctc ctttccggga ctttcgcttt ccccctccct 300
attgccacgg cggaactcat cgccgcctgc cttgcccgct gctggacagg ggctcggctg 360
ttgggcactg acaattccgt ggtgttgtcg gggaaatcat cgtcctttcc ttggctgctc 420
gcctgtgttg ccacctggat tctgcgcggg acgtccttct gctacgtccc ttcggccctc 480
aatccagcgg accttccttc ccgcggcctg ctgccggctc tgcggcctct tccgcgtctt 540
cgccttcgcc ctcagacgag tcggatctcc ctttgggccg cctccccgcc tg 592
<210> 7
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 7
ctgtgccttc tagttgccag ccatctgttg tttgcccctc ccccgtgcct tccttgaccc 60
tggaaggtgc cactcccact gtcctttcct aataaaatga ggaaattgca tcgcattgtc 120
tgagtaggtg tcattctatt ctggggggtg gggtggggca ggacagcaag ggggaggatt 180
gggaagacaa tagcaggcat gctggggatg cggtgggctc tatgg 225
<210> 8
<211> 108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 8
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgcccgggc tttgcccggg cggcctcagt gagcgagcga gcgcgcag 108
<210> 9
<211> 3321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 9
ctgcgcgctc gctcgctcac tgaggccgcc cgggcaaagc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct gcggcctagt aggctcagag gcacacagga gtttctgggc tcaccctgcc 180
cccttccaac ccctcagttc ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc 240
acactgaaca aacttcagcc tactcatgtc cctaaaatgg gcaaacattg caagcagcaa 300
acagcaaaca cacagccctc cctgcctgct gaccttggag ctggggcaga ggtcagagac 360
ctctctgggc ccatgccacc tccaacatcc actcgacccc ttggaatttc ggtggagagg 420
agcagaggtt gtcctggcgt ggtttaggta gtgtgagagg ggtacccggg gatcttgcta 480
ccagtggaac agccactaag gattctgcag tgagagcaga gggccagcta agtggtactc 540
tcccagagac tgtctgactc acgccacccc ctccaccttg gacacaggac gctgtggttt 600
ctgagccagg tacaatgact cctttcggta agtgcagtgg aagctgtaca ctgcccaggc 660
aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact tagcccctgt 720
ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct cccccgttgc 780
ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct cagcttcagg 840
caccaccact gacctgggac agtcaggtaa gtatcaaggt tacaagacag gtttaaggag 900
accaatagaa actgggcttg tcgagacaga gaagactctt gcgtttctga taggcaccta 960
ttggtcttac tgacatccac tttgcctttc tctccacagg caattgatcc ccctgatctg 1020
cggcctcgac ggtatcgata agcttgccac catgcaactt aggaaccccg aacttcatct 1080
tggctgcgcc ctggccctcc gcttcctcgc tctcgtttct tgggacatcc ctggcgctag 1140
ggcactcgac aacggcctcg cgcggactcc tacgatggga tggttgcact gggaaaggtt 1200
tatgtgcaat ctggattgcc aggaggagcc ggactcatgc atctcggaga agctgttcat 1260
ggagatggcg gaacttatgg tatcggaggg atggaaggat gccgggtatg agtatctctg 1320
tatcgacgat tgttggatgg ctccccagag agactccgag ggacgactcc aagcggaccc 1380
ccagcgcttt ccacatggca ttcgacagct cgccaattac gtgcactcga aggggttgaa 1440
gttgggaatc tacgcagatg tgggcaacaa aacgtgtgcg gggttcccgg ggtcgtttgg 1500
atactacgat attgatgcgc agacgtttgc tgactggggt gtcgatcttt tgaaatttga 1560
tggctgttac tgtgattcgt tggaaaacct ggcggatgga tacaagcata tgtcactcgc 1620
cttgaaccgg acaggtcgct caatcgtata cagctgcgaa tggcccctct atatgtggcc 1680
cttccaaaag cccaattaca cagagattcg gcagtattgc aatcactgga ggaactttgc 1740
cgatattgac gacagctgga aatccatcaa gtccattctc gattggacga gcttcaacca 1800
ggagcgcatc gtggacgtgg caggacccgg aggttggaac gatccggaca tgctcgtaat 1860
tgggaatttc gggcttagct ggaatcagca agtcacccaa atggcgctgt gggccatcat 1920
ggcagctcct ctctttatgt cgaatgatct gcggcatatc tcgccccagg caaaggctct 1980
tttgcaagac aaggacgtca tcgcaatcaa tcaggaccca ttggggaaac agggatatca 2040
acttcgccag ggtgacaatt tcgaagtatg ggagaggccg cttagcgggc tggcgtgggc 2100
ggtcgcgatg attaaccggc aggaaatcgg agggcctcgc tcgtatacca tcgcagtggc 2160
ctcactgggc aaaggagtgg cgtgcaatcc ggcctgcttc atcacccagt tgttgcccgt 2220
caaaagaaag ctgggtttct acgagtggac atccagactt agatcacaca ttaaccctac 2280
tggtacggtg ttgctccagc tcgaaaacac aatgcagatg tcgttgaaag acctgctgta 2340
atctagagga tctcgagaga tctaatcaac ctctggatta caaaatttgt gaaagattga 2400
ctggtattct taactatgtt gctcctttta cgctatgtgg atacgctgct ttaatgcctt 2460
tgtatcatgc tattgcttcc cgtatggctt tcattttctc ctccttgtat aaatcctggt 2520
tgctgtctct ttatgaggag ttgtggcccg ttgtcaggca acgtggcgtg gtgtgcactg 2580
tgtttgctga cgcaaccccc actggttggg gcattgccac cacctgtcag ctcctttccg 2640
ggactttcgc tttccccctc cctattgcca cggcggaact catcgccgcc tgccttgccc 2700
gctgctggac aggggctcgg ctgttgggca ctgacaattc cgtggtgttg tcggggaaat 2760
catcgtcctt tccttggctg ctcgcctgtg ttgccacctg gattctgcgc gggacgtcct 2820
tctgctacgt cccttcggcc ctcaatccag cggaccttcc ttcccgcggc ctgctgccgg 2880
ctctgcggcc tcttccgcgt cttcgccttc gccctcagac gagtcggatc tccctttggg 2940
ccgcctcccc gcctgggatc tctgtgcctt ctagttgcca gccatctgtt gtttgcccct 3000
cccccgtgcc ttccttgacc ctggaaggtg ccactcccac tgtcctttcc taataaaatg 3060
aggaaattgc atcgcattgt ctgagtaggt gtcattctat tctggggggt ggggtggggc 3120
aggacagcaa gggggaggat tgggaagaca atagcaggca tgctggggat gcggtgggct 3180
ctatggaccg gtctcgagat ccactagggc cgcaggaacc cctagtgatg gagttggcca 3240
ctccctctct gcgcgctcgc tcgctcactg aggccgcccg ggctttgccc gggcggcctc 3300
agtgagcgag cgagcgcgca g 3321
<210> 10
<211> 9
<212> БЕЛОК
<213> Неизвестно
<220>
<223> Описание неизвестного:
последовательность пептидного мотива семейства 'LAGLIDADG'
<400> 10
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| ПЕЧЕНЬ-СПЕЦИФИЧНЫЕ КОНСТРУКЦИИ, ЭКСПРЕССИОННЫЕ КАССЕТЫ ФАКТОРА VIII И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774874C2 |
| РЕДАКТИРОВАНИЕ ГЕНОВ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННОЙ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2811724C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| ДНК-ВЕКТОРЫ С ЗАМКНУТЫМИ КОНЦАМИ, ПОЛУЧАЕМЫЕ ПУТЕМ БЕСКЛЕТОЧНОГО СИНТЕЗА, И СПОСОБ ПОЛУЧЕНИЯ зкДНК-ВЕКТОРОВ | 2019 |
|
RU2820586C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2691102C2 |
| КОНТРОЛИРУЕМАЯ ЭКСПРЕССИЯ ТРАНСГЕНОВ С ИСПОЛЬЗОВАНИЕМ ДНК-ВЕКТОРОВ С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2019 |
|
RU2816871C2 |
| НАПРАВЛЕННАЯ МОДИФИКАЦИЯ МАЛАТДЕГИДРОГЕНАЗЫ | 2013 |
|
RU2658437C2 |
| В-КЛЕТКИ ДЛЯ ДОСТАВКИ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ IN VIVO И ИХ ДОЗЫ | 2018 |
|
RU2812477C2 |
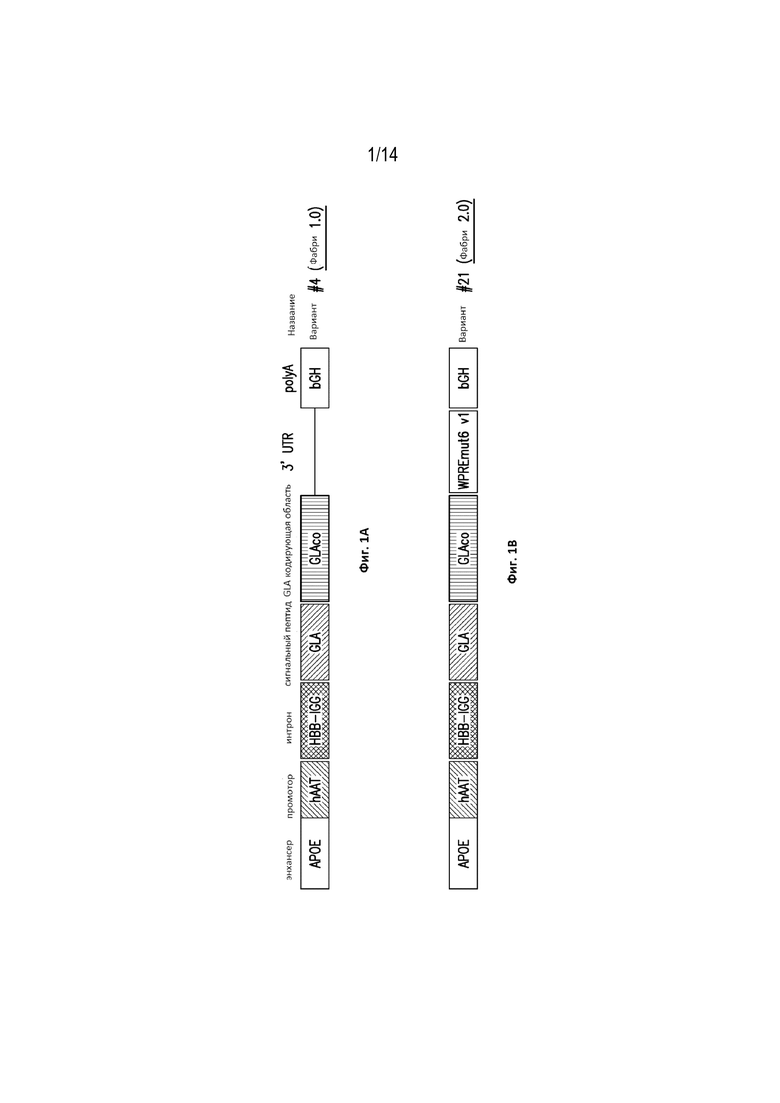

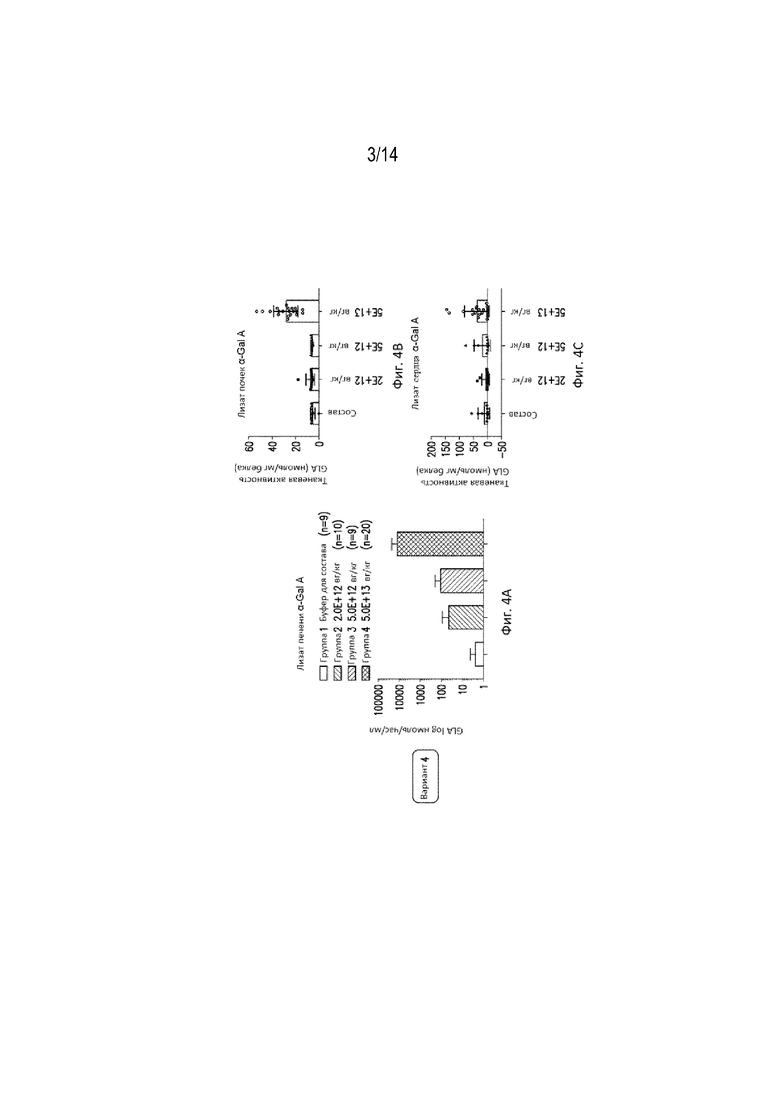

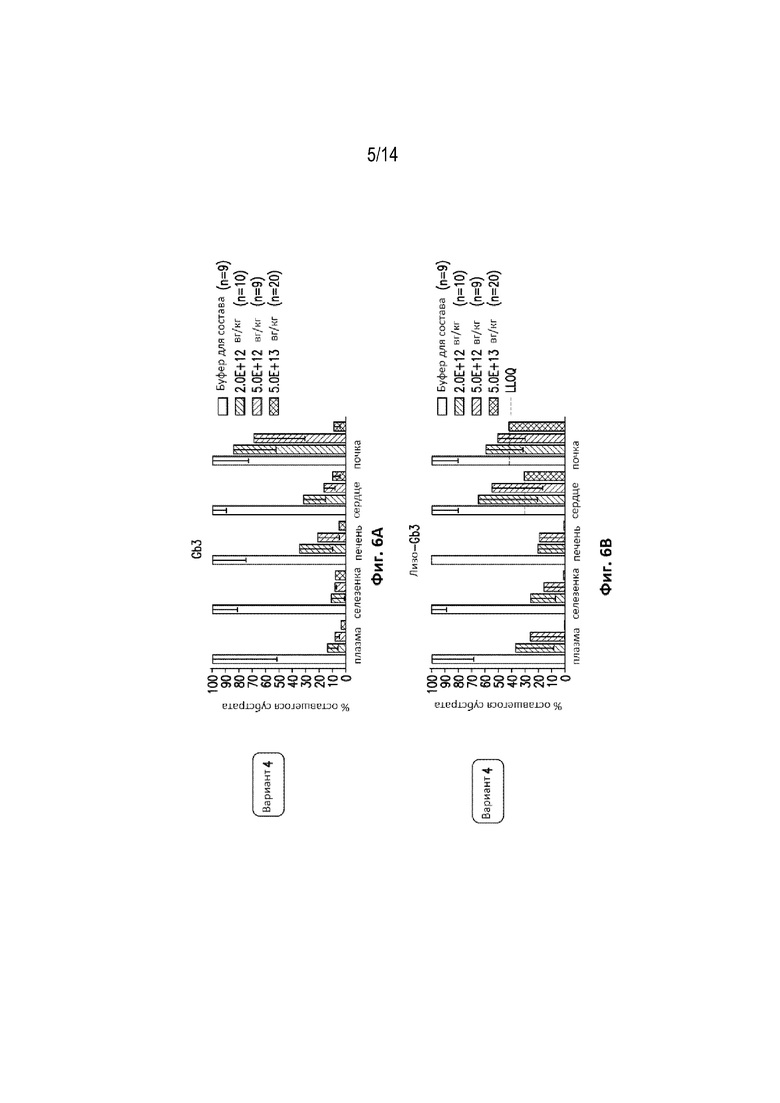

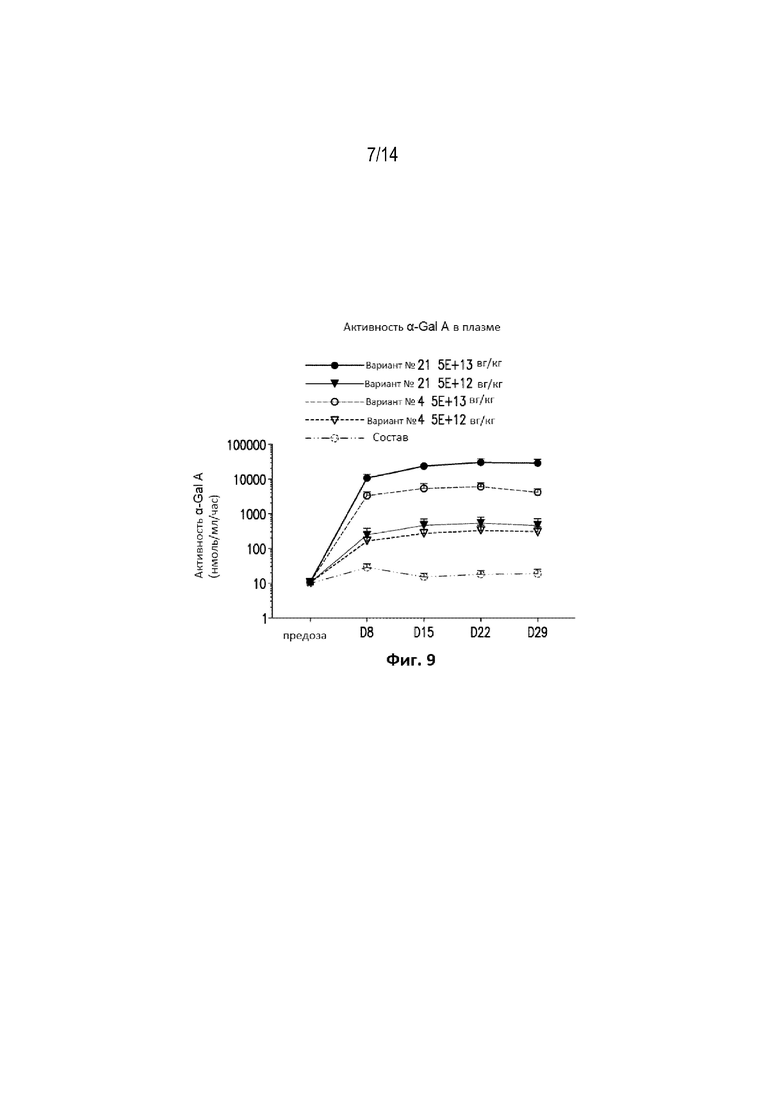


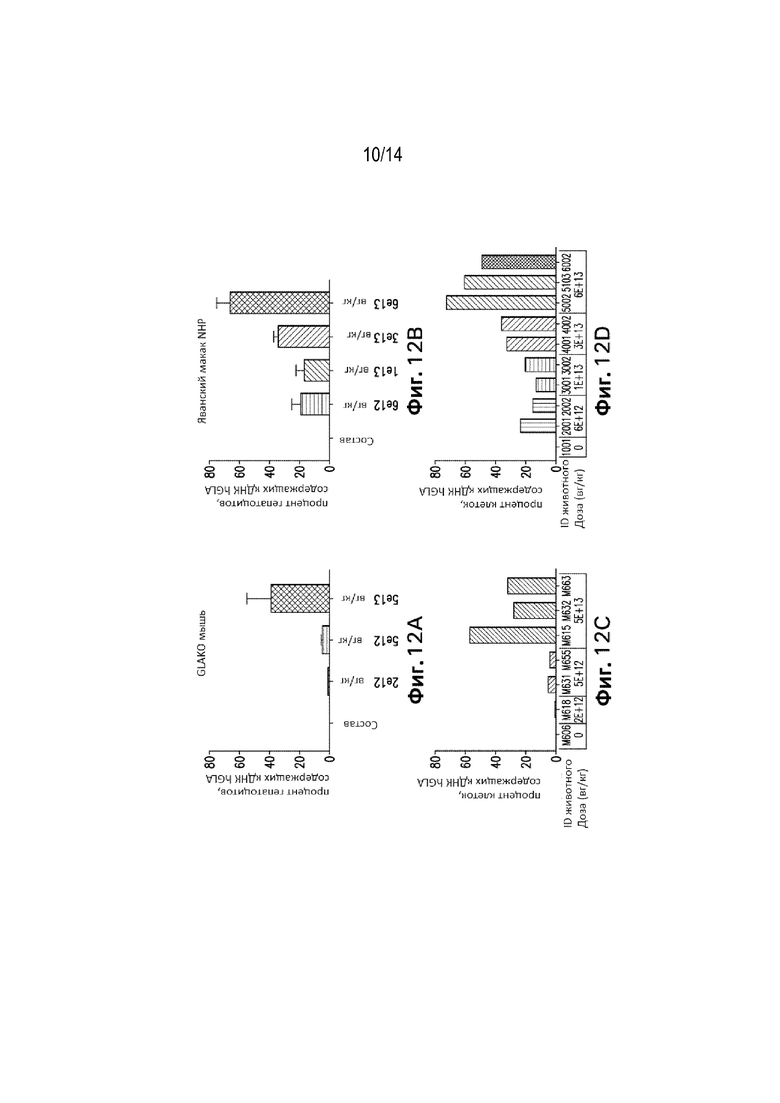
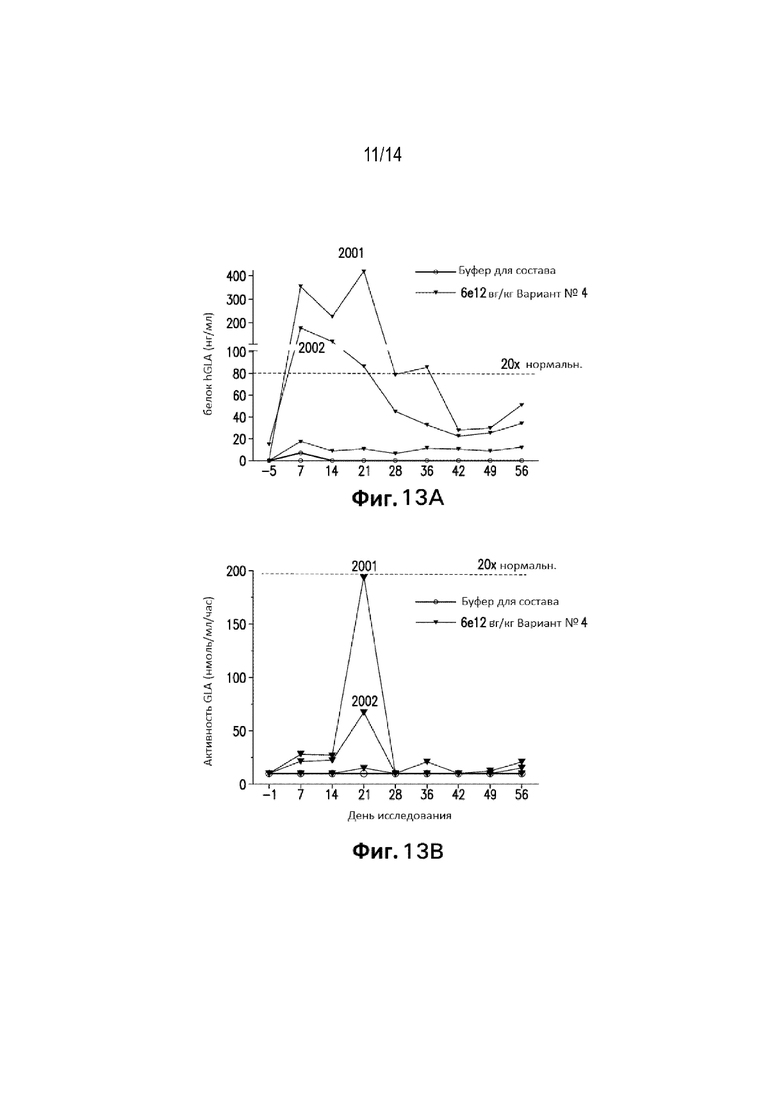
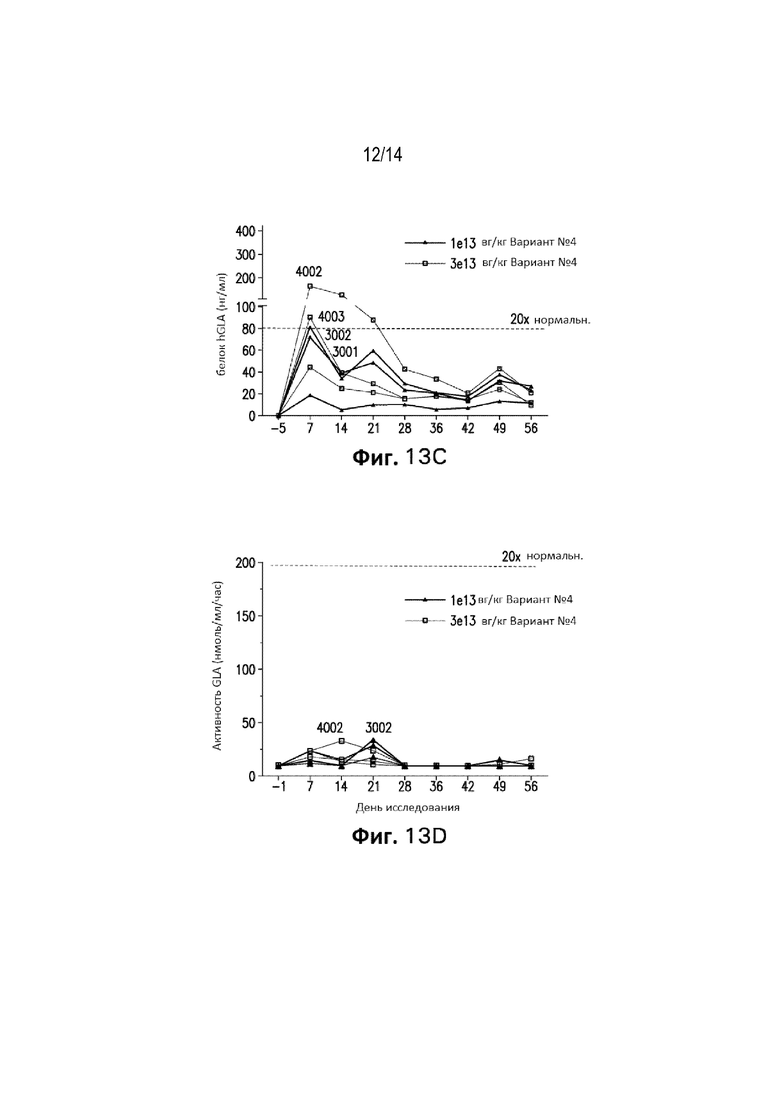

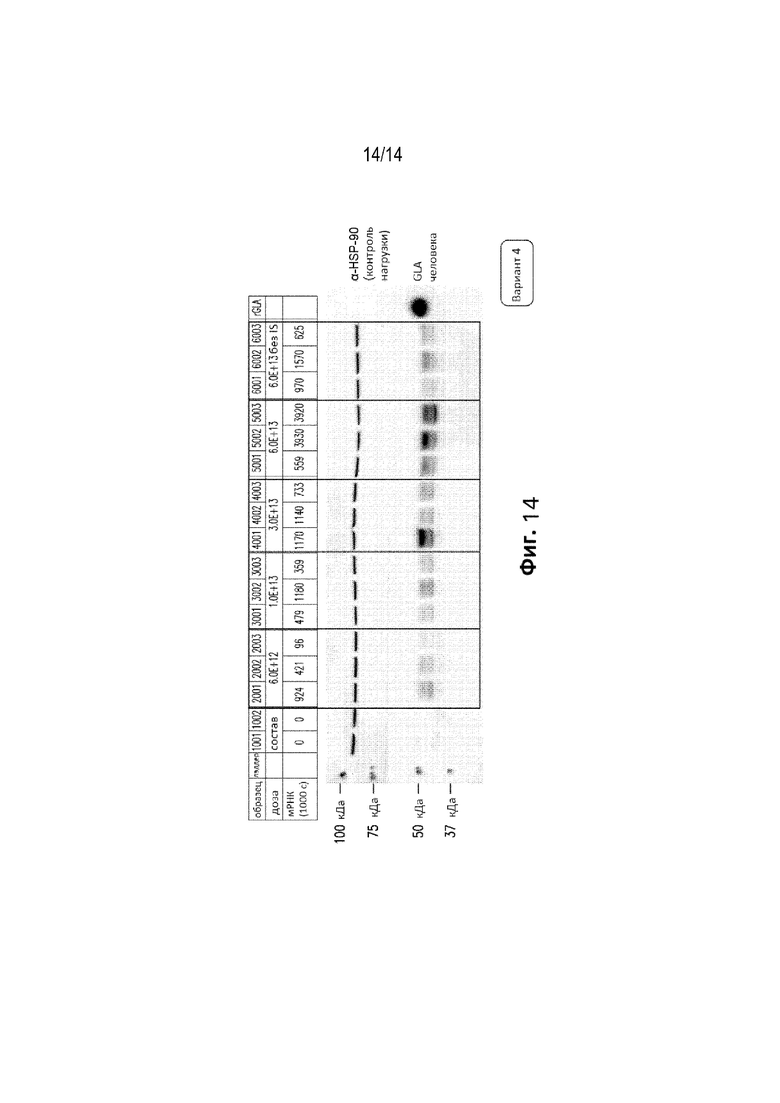
Изобретение относится к биотехнологии. Описана экспрессионная конструкция, содержащая мутированную последовательность посттранскрипционного регуляторного элемента вируса гепатита сурка (WHV)(WRPE) и трансген α-галактозидазы A (α-Gal A), кодирующий по меньшей мере один белок α-Gal A. Также описана экспрессионная конструкция, содержащая энхансер, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 2, промотор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 3, интрон, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4, трансген α-Gal A, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 5, мутированную последовательность WPRE, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 6, и последовательность сигнала poly A, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 7. Представлена композиция для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри, где композиция содержит экспрессионную конструкцию. Описан in vitro способ экспрессии белка α-Gal A в клетке, где способ включает введение экспрессионной конструкции в клетку, вследствие чего белок α-Gal A экспрессируется в клетке. Также описана генетически модифицированная клетка для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри, где клетка содержит экспрессионную конструкцию. Ракрыто применение экспрессионной конструкции, композиции или клетки для изготовления лекарственного средства для применения при профилактике, ингибировании или лечении болезни Фабри или одного или более симптомов, связанных с болезнью Фабри. Раскрыт in vitro способ получения белка α-Gal A для лечения болезни Фабри, где экспрессию белка α-Gal A осуществляют в выделенной клетке, и выделение белка α-Gal A, продуцированного клеткой. Описаны применение композиции, содержащей экспрессионную конструкцию, композиция или композиция, содержащая клетку для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри. Изобретение расширяет арсенал средств для применения при экспрессии белков α-Gal A и для предотвращения, ингибирования или лечения болезни Фабри или одного или более симптомов, ассоциированных с болезнью Фабри. 8 н. и 24 з.п. ф-лы, 14 ил., 4 табл., 7 пр.
1. Экспрессионная конструкция, содержащая мутированную последовательность посттранскрипционного регуляторного элемента вируса гепатита сурка (WHV)(WRPE) и трансген α-галактозидазы A (α-Gal A), кодирующий по меньшей мере один белок α-Gal A.
2. Экспрессионная конструкция по п. 1, где экспрессионная конструкция содержит последовательность α-Gal A дикого типа или кодон-оптимизированную последовательность α-Gal A.
3. Экспрессионная конструкция по п. 1 или 2, где экспрессионная конструкция дополнительно содержит одно или более из следующего: энхансер, промотор, интрон, последовательность, кодирующую сигнальный пептид, и/или последовательность сигнала полиаденилирования (polyA), при этом мутированная последовательность WPRE и трансген α-Gal A, кодирующий по меньшей мере один белок α-Gal A, расположены между последовательностью, кодирующей сигнальный пептид, и последовательностью сигнала polyA.
4. Экспрессионная конструкция по любому из пп. 1-3, где мутированная последовательность WPRE представляет собой мутированную последовательность WRPE с мутацией mut6.
5. Экспрессионная конструкция по любому из пп. 1-4, где экспрессионная конструкция содержит энхансер аполипопротеина E (ApoE), функционально связанный с промотором α-1-антитрипсина (hAAT), интрон гемоглобина бета человека (HBB)-IGG, последовательность, кодирующую сигнальный пептид α-Gal A, мутированную последовательность WPRE и сигнальную последовательность поли A бычьего гормона роста.
6. Экспрессионная конструкция по п. 3, где экспрессионная конструкция содержит одно или более из следующего: энхансер, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 2, промотор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 3, интрон, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4, трансген α-GalA, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 5, мутированную последовательность WPRE, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 6, и/или последовательность сигнала poly A, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 7.
7. Экспрессионная конструкция по любому из пп.1-6, где экспрессионная конструкция содержит нуклеотидную последовательность, представленную SEQ ID NO: 9.
8. Экспрессионная конструкция по любому из пп. 1-7, где экспрессионная конструкция представляет собой экспрессионную конструкцию на основе аденоассоциированного вируса(AAV).
9. Экспрессионная конструкция по п. 8, где серотип экспрессионной конструкции AAV представляет собой AAV2/6.
10. Экспрессионная конструкция, содержащая энхансер, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 2, промотор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 3, интрон, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4, трансген α-GalA, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 5, мутированную последовательность WPRE, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 6, и последовательность сигнала poly A, содержащую нуклеотидную последовательность, представленную SEQ ID NO: 7.
11. Композиция для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри, где композиция содержит экспрессионную конструкцию по любому из пп. 1-10.
12. Композиция по. п. 11, дополнительно содержащая фармацевтически приемлемый носитель.
13. Композиция по п. 12, где фармацевтически приемлемый носитель включает CaCl2, MgCl2, NaCl, сахарозу и коллифор (полоксамер) P 188.
14. In vitro способ экспрессии белка α-Gal A в клетке, где способ включает введение экспрессионной конструкции по любому из пп. 1-10 в клетку, вследствие чего белок α-Gal A экспрессируется в клетке.
15. Генетически модифицированная клетка для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри, где клетка содержит экспрессионную конструкцию по любому из пп. 1-10.
16. Генетически модифицированная клетка по п. 15, где клетка образуется в результате способа по п. 14.
17. Генетически модифицированная клетка по п. 15 или 16, где (a) клетка представляет собой стволовую клетку или клетку-предшественник или (b)клетка представляет собой клетку печени или мышцы.
18. Применение экспрессионной конструкции по любому из пп. 1-10, композиции по любому из пп. 11-13 или клетки по любому из пп. 15-17 для изготовления лекарственного средства для применения при профилактике, ингибировании или лечении болезни Фабри или одного или более симптомов, связанных с болезнью Фабри.
19. Применение по п. 18, где симптомы включают один или более из следующего: уровни Gb3 выше нормального уровня, уровни лизо-Gb3 выше нормального уровня, заболевание почек, заболевание сердца, акропарестезию, ангиокератомы, боль в желудочно-кишечном тракте, помутнение роговицы и хрусталика или цереброваскулярное заболевание.
20. Применение по п. 18 или 19, где экспрессионную конструкцию вводят субъекту посредством внутривенной инфузии.
21. Применение по п. 20, где субъекту вводят только одну дозу экспрессионной конструкции.
22. Применение по п. 20 или 21, где субъекту вводят иммунодепрессант до и/или во время введения экспрессионной конструкции.
23. Применение по п. 22, где иммунодепрессант содержит преднизон.
24. Применение по любому из пп. 20-23, где экспрессия по меньшей мере одного белка α-Gal A сохраняется в течение по меньшей мере 3 месяцев, по меньшей мере 9 месяцев или по меньшей мере 12 месяцев.
25. Применение по любому из пп. 20-24, где белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере в 2 раза.
26. Применение по любому из пп. 20-25, где белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта по меньшей мере на 80%.
27. Применение по любому из пп. 20-26, где белок α-Gal A, экспрессируемый из трансгена, снижает количество гликосфинголипидов у субъекта в одном или более из следующего: плазма, печень, сердце, почки или селезенка.
28. Применение по любому из пп. 20-27, где экспрессионная конструкция, полученная в клеточной системе HEK293, обеспечивает уровни белка α-Gal A у субъекта примерно в 21 раз выше по сравнению с уровнями белка α-Gal A у субъектов, которым вводили экспрессионную конструкцию, полученную в клеточной системе Sf9.
29. Применение по любому из пп. 20-28, где активность белка α-Gal A у субъекта примерно в 100-1500 раз выше, чем у физиологического нормального/дикого типа.
30. Применение по любому из пп. 20-29, где белок α-Gal A, экспрессируемый трансгеном, проявляет активность в почках, печени и/или сердце субъекта.
31. In vitro способ получения белка α-Gal A для лечения болезни Фабри, где способ включает экспрессию белка α-Gal A в выделенной клетке согласно способу по п. 14 и выделение белка α-Gal A, продуцированного клеткой.
32. Применение композиции, содержащей экспрессионную конструкцию по любому из пп. 1-10, композиции по любому из пп. 11-13 или композиции, содержащей клетку по любому из пп. 15-17, для профилактики, ингибирования или лечения болезни Фабри или одного или более симптомов, связанных с болезнью Фабри.
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ МУЛЬТИПОТЕНТНЫХ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ | 2008 |
|
RU2535365C2 |
| US 20030113894 A1, 19.06.2003 | |||
| US 20180117181 A1, 03.05.2018 | |||
| US 20090111106 A1, 30.04.2009. | |||

