Изобретение относится к медицине, микробиологической промышленности, генно-инженерной биотехнологии и может быть использовано для получения в клетках E.coli рекомбинантного белка аполипопротеина Α-I человека. Получаемый рекомбинантный аполипопротеин Α-I может быть использован для замены зрелого белка аполипопротеина Α-I, выделяемого из плазмы крови человека и используемого в качестве компонента или носителя различных существующих и/или разрабатываемых лекарственных средств, а также для получения поликлональных антисывороток и антител на аполипопротеин A-I человека.
Аполипопротеин Α-I (апоА-I) является белком, входящим в состав липопротеинов высокой плотности плазмы крови и принимающим участие в обратном транспорте холестерина. Известно несколько изоформ апоА-I человека, отличающихся точечными мутациями. Самая известная форма получила название апоА-I Милано (aпoAI-М) (1), данная форма отличается от апоА-I лишь одной заменой аминокислоты в 173 позиции - аланина на цистеин (2). Обнаруженную форму aпoAI-М связывают с низким уровнем содержания холестерина в организме и с пониженным риском развития атеросклеротических заболеваний. В связи с этим aпoAI-М вызвал особенный интерес у исследователей.
Основными органами, синтезирующими апоА-I, являются печень и тонкая кишка (3). В этих органах синтезируется предшественник апоА-I - препроапоА-I. ПрепроапоА-I расщепляется внутри клетки, и белок проапоА-I секретируется в плазму. Белок проапо A-I является вариантом белка апоА-I, содержащим с N-конца шесть дополнительных аминокислот (Arg - His - Phe - Trp - Gln - Gln). Далее белок проапоА-I расщепляется специфическим ферментом апоА-I пропептидазой с образованием зрелого белка апоА-I (конечная форма белка) (4). Зрелый белок апоА-I представляет собой полипептид с молекулярной массой 28,1 кДа, состоящий из одной цепи с известной последовательностью, включающей 243 аминокислоты (например, последовательность, зарегистрированная под номером gi4557320 в базе данных National Center for Biotechnology Information (5)).
Известно, что липопротеины высокой плотности и их белковый компонент апоА-I могут образовывать стабильные комплексы и транспортировать, в том числе и через цитоплазматическую мембрану, такие биологически активные вещества, как ксенобиотики, жирорастворимые витамины, стероидные соединения, тиреоидные гормоны, лекарственные препараты цитостатического действия (6-10). Однако транспортной ролью функции апоА-I не ограничиваются. Показано, что апоА-I, подобно транскрипционным факторам, может выступать в роли соактиватора процессов репликации, связываясь с ДНК влиять на экспрессию генов посредствам изменения вторичной структуры ДНК в сайтах связывания (11, 12). Кроме того, белок апоА-I находит широкое применение при конструировании искусственных липосом, выполняя в них роль маркера для захвата клетками-мишенями и рецептор-опосредованного переноса в клетки таких наноконструкций (13-16). В связи с этим, получение белка апоА-I в препаративных количествах, а затем и в промышленных масштабах с целью его дальнейшего практического использования имеет большое значение.
Получение природного апоА-I человека на основе выделения апоА-I-содержащих липопротеинов сыворотки крови с помощью изоплотностного ультрацентрифугирования в растворе KBr (17), с последующим делипидированием липопротеинов высокой плотности органическими растворителями и разделением смеси белков гель-фильтрацией не обеспечивает высокого выхода апоА-I, поэтому себестоимость белка, выделенного таким способом, является высокой. Кроме того, использование плазмы крови человека в качестве источника апоА-I является неприемлемым по этическим соображениям.
Известны системы для получения рекомбинантного белка апоА-I на основе технологий рекомбинантных ДНК в клетках бактерий (18-20), насекомых (21, 22), дрожжей (23) и млекопитающих (клетки яичников китайского хомячка) (24, 25). Общим недостатком упомянутых систем является низкий выход рекомбинантного человеческого аполипопротеина A-I.
Известна рекомбинантная ДНК, содержащая фрагмент гена проаполипопротеина Α-I человека (проапоА-I), полученного в результате объединения последовательности синтетической ДНК, кодирующей аминокислоты от -6 до +14 белка проапоА-I, и фрагмента природной ДНК, кодирующей аминокислоты от +15 до +243 белка проапоА-I, характеризующегося заданной нуклеотидной последовательностью, в которой осуществлена синонимическая замена кодонов в последовательности, кодирующей аминокислоты от -6 до +14 белка проапоА-I человека. С помощью заявленной рекомбинантной ДНК в клетках бактерий (E.coli), дрожжей (Saceharomyces cerevisiae) или насекомых (Spodoptera frugiperda Sf9) получают белок проапоА-I человека, который используют в неизменном виде либо после превращения его в зрелый белок после обработки протеолитическим ферментом апоА-I пропептидазой (26). Замена кодонов, по мнению авторов, приводит к снижению или предотвращению образования булавочных структур соответствующей мРНК, что, в свою очередь, способствует увеличению трансляции такой мРНК и, в конечном счете, увеличению синтеза белка в клетках-продуцентах. Вместе с тем, заявленная рекомбинантная ДНК является универсальной для различных типов клеток продуцентов - бактерии, дрожжи и клетки насекомых, и ее кодоны не оптимизированы для какого-то одного типа клеток продуцентов. По этой причине данная генетическая конструкция рекомбинантной ДНК не позволяет существенно повысить выход рекомбинантного белка проапоА-I из клеток продуцентов. Другим недостатком известной рекомбинантной ДНК является то, что ее использование предопределяет применение дорогостоящего индуктора изопропилтиогалактозида (ИПТГ) при наработке биомассы белка апоА-I, использование дорогостоящего оборудования (высокоскоростная ультрацентрифуга), ртуть содержащего реагента и фермента апоА-I пропептидазы при очистке белка.
Известна рекомбинантная ДНК, содержащая нуклеотидную последовательность, кодирующую мономер или димер белка aпoAI-Милано (апоА-I) или ген aпoAI-М, слитый с бактериальной сигнальной последовательностью отрA, обеспечивающая синтез в клетках E.coli шт. RV308 рекомбинантного белка димера aпoAI-М - aпoAI-M, который затем расщепляют на отдельные белки-мономеры (27). Недостатком известной рекомбинантной ДНК является то, что с ее помощью получают крайне низкий выход белка (1,9 мг с литра культуры клеток), обусловленный необходимостью применения трудоемких многоступенчатых способов его выделения и очистки. Кроме того, получаемый aпoAI-M дополнительно содержит аминокислотный фрагмент отрА (21 а.о.), который может изменять природные свойства aпoAI-М.
Известна рекомбинантная ДНК, содержащая нуклеотидную последовательность, кодирующую белок апоА-I, обеспечивающая синтез апоА-I человека в клетках метилотрофных дрожжей Pichia pastoris (28). При сопоставимом выходе белка апоА-I в клетках-продуцентах Pichia pastoris с клетками E.coli, недостатками рекомбинантной ДНК, структура которой оптимизирована для экспрессии в Pichia pastoris, является то, что в качестве питательного компонента или индуктора используют ядовитое вещество метанол (0,5-5,0%), что ограничивает соответствующими условиями техники безопасности возможность работы с клетками Pichia pastoris. Кроме того, наработка биомассы - клеток Pichia pastoris, происходит в течение нескольких дней (2-6 суток), что в несколько раз дольше, чем использование продуцентов на основе клеток E.coli, и, следовательно, увеличивает затраты на выращивание клеток, что увеличивает себестоимость конечного белка, кодируемого известной рекомбинантной ДНК.
Известна рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-I человека, содержащая ген зрелого белка апоА-I человека, а также лидерную последовательность белка, включающую шесть остатков гистидина, 15 аминокислотных остатков панкреатической рибонуклеазы и шесть аминокислотных остатков пpο-формы белка апоА-I человека (химерная часть) (29). Клетки-продуценты выращиваются на среде LB, индуцируются 500 мкМ ИПТГ. Химерный белок апоА-I человека из клеток-продуцентов выделяют с помощью одностадийной аффинной хроматографии, затем химерную часть отщепляют с помощью протеазы вируса табачной мозаики (TEV-protease) с образованием зрелого белка апоА-I человека. На заключительном этапе выход очищенного зрелого белка апоА-I составляет 26 мг с литра культуры клеток. Недостатками данной конструкции рекомбинантной ДНК является то, что она обусловливает низкий выход на конечной стадии зрелого белка апоА-I и использование больших количеств индуктора (500 мкМ). Очевидно, это связано с включением в рекомбинантную ДНК гена апоА-I «дикого типа», т.е. гена без оптимизации кодонов для экспрессии в клетках E.coli.
Наиболее близкой к заявленному изобретению является рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-I человека, содержащая участок, кодирующий шесть гистидинов (для очистки рекомбинантного белка с помощью одностадийной аффинной хроматографии на никель-хелатном сорбенте) и собственно мутантный ген апоА-I человека, включающий шесть аминокислот известных как пpο-сегмент человеческого белка апоА-I. Мутантный ген апоА-I человека содержит 30 синонимических замен кодонов относительно «дикого типа» и одну замену кодона, кодирующего во второй позиции зрелой формы белка глютаминовую аминокислоту на аспарагиновую кислоту (Glu2Asp) (30) (см. табл. 1). Химерный белок апоА-I человека, кодируемый данной рекомбинантной ДНК, выделяли из клеток продуцентов с помощью разрушения ультразвуком, затем белок из клеточного лизата выделяли хроматографией на никель-хелатной смоле в присутствии 3М гуанидин гидрохлорида с последующим кислотным гидролизом химерного белка апоА-I человека для получения зрелого белка апоА-I человека. Данный зрелый белок апоА-I человека по первичной структуре полностью идентичен природному зрелому белку апоА-I человека, за исключением отсутствия двух первых аминокислот с N-конца белка. Выход химерного белка апоА-I человека из клеток-продуцентов при их выращивании в лабораторных условиях на среде NCZYM и последующей инкубации с добавлением индуктора ИПТГ до 500 мкМ в течение трех часов достигает 100±20 мг с литра культуры клеток. Прототип обладает следующими недостатками, обусловленными структурой рекомбинантной ДНК:
1) недостаточное количество получаемого на конечной стадии химерного белка апоА-I человека из литра культуры клеток-продуцентов при их выращивании в лабораторных условиях в аналогичных с описанными в заявленном изобретении экспрессирующих системах (одинаковый промотор экспрессирующего вектора - промотор бактериофага Т7, одинаковый индуктор и хозяйские клетки E.coli шт. BL21(DE3));
2) большое количество используемого дорогостоящего индуктора ИПТГ, что сказывается на повышенной себестоимости конечного белка.
Задачей настоящего изобретения является создание рекомбинантной ДНК, обеспечивающей более высокий уровень синтеза аполипопротеина Α-I человека в клетках E.coli в составе плазмидного вектора, обеспечение сходства с природным аполипопротеином Α-I по функциональному назначению, снижение себестоимости конечного белка, кодируемого данной рекомбинантной ДНК.
Сходство рекомбинантного белка апоА-I, полученного с использованием заявленной рекомбинантной ДНК, с природным белком апоА-I необходимо для осуществления адресной доставки в клетки млекопитающих биологически активных соединений и получения поликлональных антител к белку апоА-I.
Решение поставленной задачи достигается тем, что рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-I человека, размером 852 п.н., последовательно включает с 5′-конца фрагмент ДНК, кодирующий 6 а.о. гистидина размером 18 п.н. (SEQ ID NO 1), фрагмент ДНК, кодирующий 6 а.о. прο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий N-концевой фрагмент из 15 а.о. бычьей панкреатической РНКазы А размером 45 п.н. (SEQ ID NO 3), фрагмент ДНК, кодирующий 6 а.о. проформы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий сайт узнавания протеазы вируса табачной мозаики размером 21 п.н. (SEQ ID NO 4), мутантный ген зрелого белка апоА-I человека размером 729 п.н. (SEQ ID NO 5).
Раскрытие изобретения
Согласно заявленному изобретению рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-Ι человека, размером 852 п.н., последовательно включает с 5′-конца фрагмент ДНК, кодирующий 6 а.о. гистидина размером 18 п.н. (SEQ ID NO 1), фрагмент ДНК, кодирующий 6 а.о. прο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий N-концевой фрагмент из 15 а.о. бычьей панкреатической РНКазы А размером 45 п. н. (SEQ ID NO 3), фрагмент ДНК, кодирующий 6 а.о. проформы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий сайт узнавания протеазы вируса табачной мозаики размером 21 п. н., (SEQ ID NO 4), имеющей последовательность аминокислот Glu-Asn-Leu-Tyr-Phe-Gln-/-Gly; мутантный ген зрелого белка апоА-I человека размером 729 п.н. (SEQ ID NO 5).
Предложенная рекомбинантная ДНК в составе плазмидного вектора pET36b(+) («Novagen», США) под регуляторной областью промотора бактериофага Т7 позволяет получать в клетках E.coli шт. BL21(DE3) при использовании низких концентраций индуктора (25 мкМ) высокий выход химерного белка апоА-I человека до 270 мг (в среднем 182±32 мг) из литра культуры клеток, или до 50 мг (39,7±6,2 мг) из 1 г влажной биомассы. Таким образом, при использовании рекомбинантной ДНК согласно настоящему изобретению в составе аналогичной прототипу экспрессирующей системы (одинаковый промотор и хозяйские клетки E.coli) выход химерного белка апоА-I человека почти в два раза больше (в 1,8 раз), чем в прототипе, при концентрации индуктора, меньшей в 20 раз, что позволяет снизить себестоимость целевого белка. При этом зрелый белок апоА-I человека, полученный путем гидролиза химерного белка апоА-I человека, кодируемого заявленной рекомбинантной ДНК, обладает сходством с природным аполипопротеином Α-I человека, что важно для использования его в качестве средства адресной доставки лекарственных средств и биологически активных соединений в клетки органов и тканей человека, получения поликлональных антител к белку апоА-I. Данное сходство продемонстрировано в описании осуществления изобретения.
Изобретение иллюстрируется следующими чертежами.
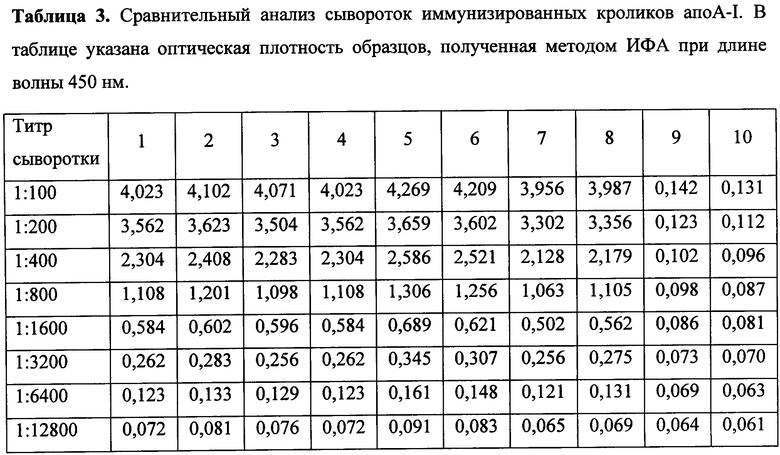
Фиг. 1. Фрагмент электрофореграммы лизатов клеток-продуцентов рекомбинантного химерного белка апоА-I человека в 12% ПААГ, предварительно инкубированных с различной концентрацией индуктора изопропилтиогалактозида (ИПТГ). Дорожки: 1 - очищенный химерный белок апоА-I человека; 2 - лизат не индуцированных клеток; 3-8 - лизаты клеток с добавлением индуктора ИПТГ 0,05, 0,2, 1,0, 5,0, 20 и 100 мкМ соответственно.
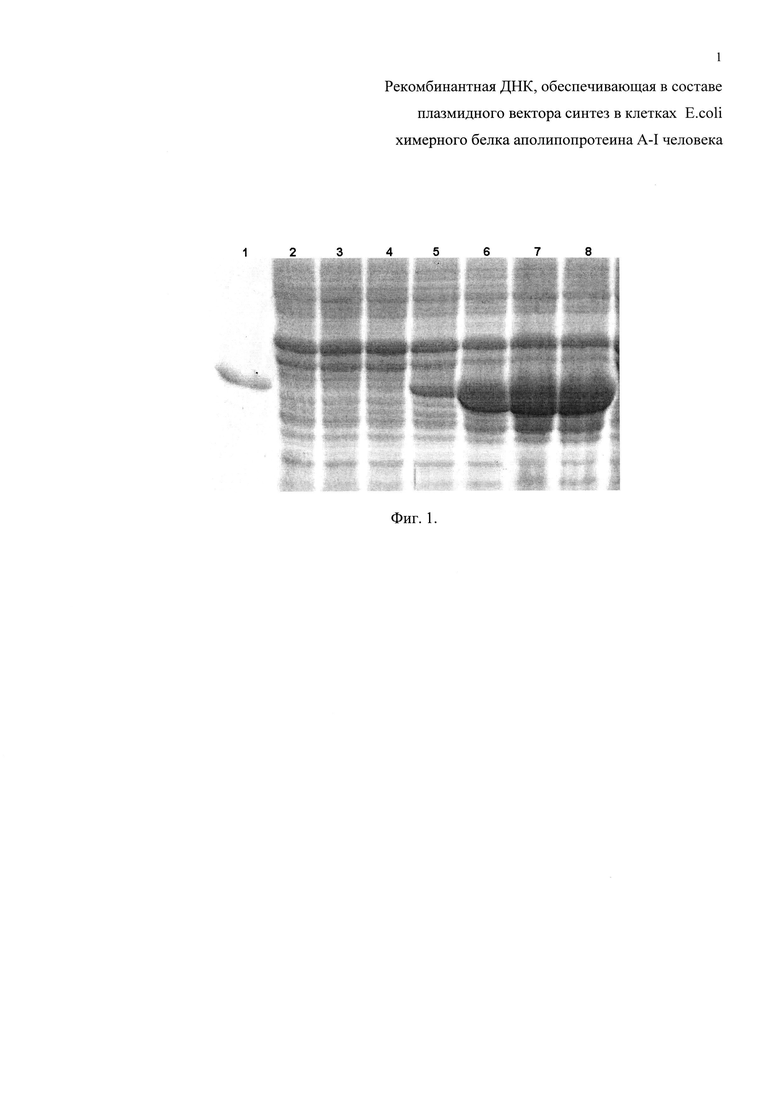
Фиг. 2. Электрофореграмма лизатов клеток-продуцентов химерного белка апоА-I человека в 12% ПААГ, выращенных на средах LB и NCZYM и инкубированных с различной концентрацией ИПТГ. Дорожки: 1 - очищенный химерный белок апоА-I; 2 - лизат не индуцированных клеток-продуцентов, выращенных в среде LB; 3-5 - лизат клеток продуцентов, выращенных в среде LB с добавлением ИПТГ 20, 100 и 500 мкМ соответственно; 6 - лизат не индуцированных клеток-продуцентов, выращенных в среде NZCYM; 7-9 - лизат клеток-продуцентов, выращенных в среде NCZYM с добавлением ИПТГ 20, 100 и 500 мкМ соответственно.
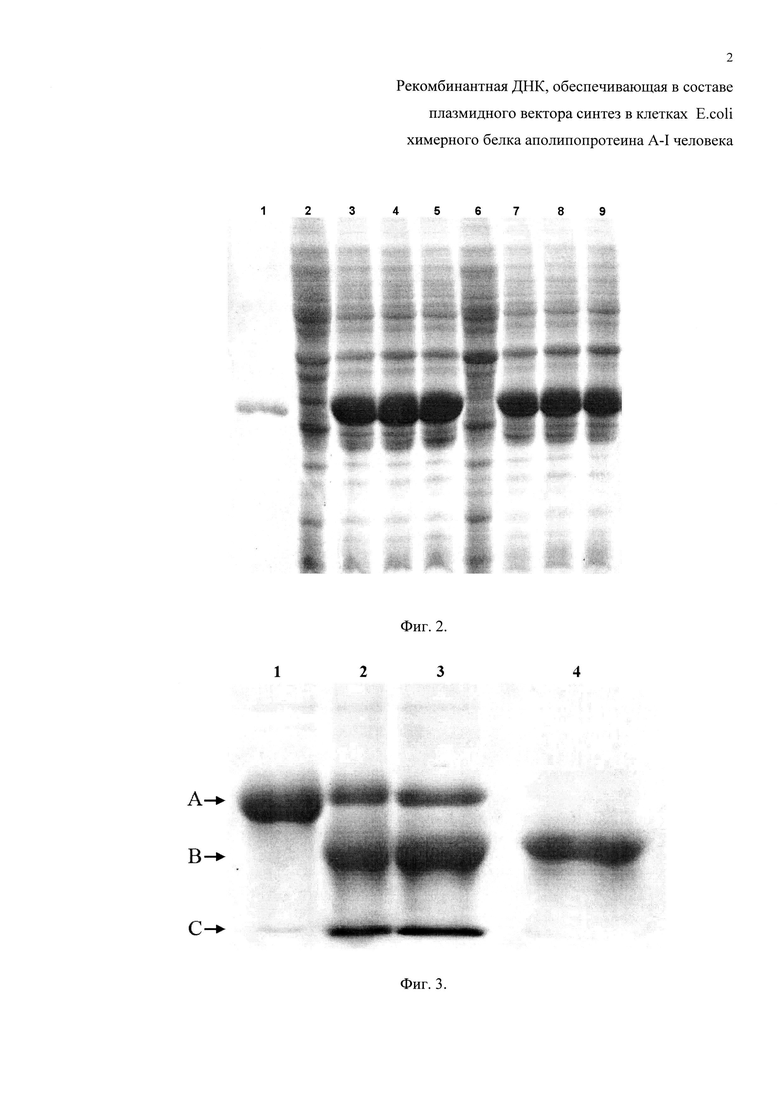
Фиг. 3. Фрагмент электрофореграммы анализа образцов гидролизованного химерного белка апоА-I в 12% ПААГ. Дорожки: 1 - очищенный химерный белок апоА-I человека; 2 - продукты кислотного гидролиза химерного белка апоА-I человека (по примеру 4.1); 3 - продукты ферментативного гидролиза химерного белка апоА-I человека (по примеру 4.2); 4 - природный апоА-I человека. Подписи стрелок: А - химерный белок апоА-I человека (~33,4 кДа); В - зрелый белок апоА-I человека (~28,0 кДа); С - отщепленная «лидерная последовательность» (~5,4 кДа).
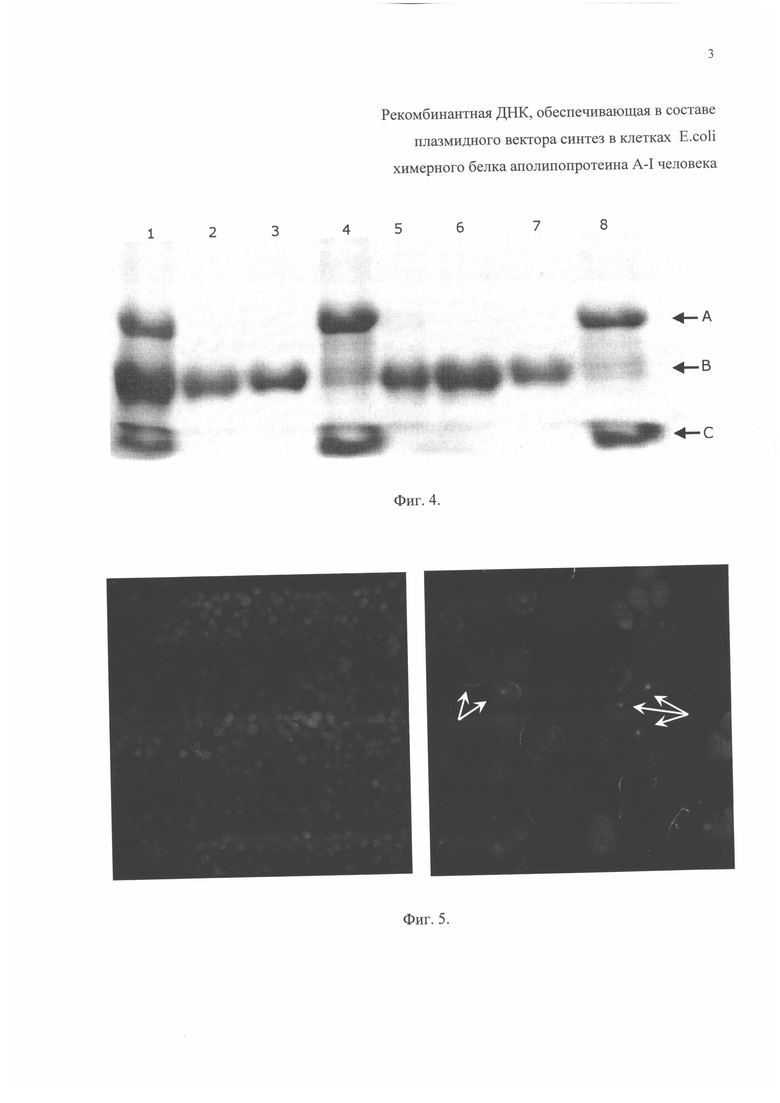
Фиг. 4. Фрагмент электрофореграммы анализа в 12% ПААГ хроматографических фракций очистки зрелого белка апоА-I человека из смеси продуктов гидролиза химерного белка апоА-I человека. Дорожки: 1 - продукты гидролиза химерного белка апоА-I человека; 2 и 3 - зрелый белок апоА-I человека, очищенный от продуктов кислотного гидролиза химерного белка апоА-I человека (первый хроматографический пик); 5, 6 и 7 - зрелый белок апоА-I человека, очищенный от продуктов ферментативного гидролиза химерного белка апоА-I человека (первый хроматогрофический пик); 4 и 8 - химерный белок апоА-I человека ~33,4 кДа и отщепленный белок («лидерная последовательность») ~5,4 кДа (второй хроматографический пик). Подписи стрелок: А - химерный белок апоА-I человека (~33,4 кДа); В - зрелый белок апоА-I человека (~28,0 кДа); С - отщепленная «лидерная последовательность» (~5,4 кДа).
Фиг. 5. Клетки гепатоцитов, инкубированных в течение 3 часов с конъюгатом зрелого белка апоА-I с флуоресцентной меткой ФИТЦ (50 мкг на лунку). Флуоресцентная микроскопия, х400.
Таблица 1. Сравнение кодонов в гене зрелого белка апоА-I человека («дикий тип», gi4557320), мутантном гене, представленном в прототипе и мутантном гене настоящего изобретения.
Примечание. В таблице указаны только те кодоны, которые были изменены в заявленной рекомбинантной ДНК относительно гена зрелого белка апоА-I человека («дикий тип») либо прототипа; в остальных случаях и в прототипе и в гене настоящего изобретения кодоны оставались одинаковыми со структурой гена «дикого типа». Номер указывает на позицию аминокислоты в зрелой форме белка. Частота встречаемости кодонов приведена на 1000 кодонов для генома E.coli.
Таблица 2. Состав питательной среды LB.
Таблица 3. Сравнительный анализ сывороток кроликов, иммунизированных белком апоА-I человека. В таблице указана оптическая плотность образцов, полученная методом ИФА при длине волны 450 нм.
Столбцы: 1 и 2 - результаты анализа взаимодействия природного апоА-I человека с сывороткой кролика, иммунизированного зрелым белком апоА-I, полученным с использованием рекомбинантной ДНК настоящего изобретения; 3 и 4 - результаты анализа взаимодействия зрелого белка апоА-I человека, полученного с использованием рекомбинантной ДНК настоящего изобретения, с сывороткой кролика, иммунизированного природным белком апоА-I человека;
5 и 6 - результаты анализа взаимодействия природного белка апоА-I человека с сывороткой кролика, иммунизированного этим же белком; 7 и 8 - результаты анализа взаимодействия зрелого белка апоА-I человека, полученного с использованием рекомбинантной ДНК настоящего изобретения, с сывороткой кролика, иммунизированного этим же белком.
Осуществление изобретения
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Получение рекомбинантной ДНК, обеспечивающей в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина A-I человека.
Рекомбинантную ДНК настоящего изобретения синтезируют из нуклеотидов по принципу “de novo” в составе плазмиды pBluescript. При синтезе, в структуру гена зрелой формы белка апоА-I, относительно «дикого типа» вносят 33 синонимических замены кодонов и одну замену кодона, кодирующего во второй позиции зрелого белка глютаминовую аминокислоту на аспарагиновую (Glu2Asp) (таблица 1). Полученную плазмиду pBluescript, содержащую в своей структуре рекомбинантную ДНК настоящего изобретения (pBS-aпoA), используют для амплификации рекомбинантной ДНК в необходимых количествах с помощью известных микробиологических методов. Для этого плазмидой pBS-aпoA трансформируют клетки E.coli, выращивают клетки на среде LB (состав указан в таблице 2) в необходимом объеме и собирают в виде осадка. Далее из биомассы выделяют плазмиду pBS-aпoA и вырезают из нее рекомбинантную ДНК настоящего изобретения с помощью эндонуклеаз рестрикции NdeI и XhoI. Рекомбинантную ДНК настоящего изобретения отделяют от фрагментов плазмиды pBluescript с помощью электрофореза в агарозном геле с последующим извлечением рекомбинантной ДНК из геля известными методами. Полученная таким образом рекомбинантная ДНК может быть встроена в любую плазмиду по сайтам рестрикции NdeI и XhoI, в том числе и в вектор, предназначенный для экспрессии генов.
Пример 2. Клонирование рекомбинантной ДНК настоящего изобретения в составе экспрессирующего вектора в клетках E.coli - получение продуцента химерного белка апоА-I.
Плазмидой pBS-aпoA трансформируют клетки E.coli шт. BL21 (DE3) с помощью химической трансформации, например, общеизвестным кальциевым методом (31). Трансформированные клетки выращивают на агаризованной среде LB с добавлением в нее ампициллина 100 мкг/мл. Для работы отбирают несколько клонов. Из одного клона, содержащего плазмиду pBS-aпoA, выращивают ночную культуру в объеме 2 мл в жидкой среде LB при 37°С. На следующее утро ночную культуру добавляют к свежей среде объемом 200 мл, так же содержащей ампициллин в указанном выше количестве, и выращивают при перемешивании до оптической плотности D600=0,6-0,8 о.е. Клетки осаждают центрифугированием 3000 об/мин, 15 минут. Плазмидную ДНК pBS-aпoA выделяют из клеток набором «Plasmid Maxiprep» (ЗАО «Евроген», Россия). Плазмидную ДНК pBS-aпoA гидролизуют эндонуклеазами рестрикции NdeI и XhoI, согласно инструкции фирмы-производителя ферментов (НПО «СибЭнзим», Россия). Вырезанный фрагмент ДНК - рекомбинантную ДНК настоящего изобретения, отделяют от фрагментов плазмиды pBluescript с помощью электрофореза в агарозном геле с последующим извлечением рекомбинантной ДНК из геля набором «Cleanup Standard» (ЗАО «Евроген», Россия). Полученную таким образом рекомбинантную ДНК настоящего изобретения лигируют с плазмидой-вектором pET36b(+) («Novagene», США), предварительно гидролизованной аналогичными ферментами - NdeI и XhoI. Лигирование проводят с помощью фермента Т4 ДНК-лигазы согласно инструкции фирмы производителя (НПО «СибЭнзим», Россия). Лигазной смесью трансформируют клетки E.coli шт. BL21 (DE3) с помощью химической трансформации. Трансформированные клетки выращивают на агаризованной среде LB с добавлением канамицина 30 мкг/мл. Для работы отбирают несколько клонов.
Способность клонов экспрессировать рекомбинантную ДНК настоящего изобретения проверяют электрофоретическим анализом лизата клеток. Из клонов выращивают культуры клеток до оптической плотности D600=0,6-1,0 о.е. и делят пополам. К одной половине добавляют индуктор ИПТГ до 0,5 мМ, вторую половину клеток оставляют как контроль, после чего инкубируют все клетки 4 часа при 37°С. По окончании индукции готовят пробы для анализа в полиакриламидном геле (ПААГ) в денатурирующих условиях по Лемли. Белки окрашивают в растворе Кумаси G250. По наличию банда в электрофореграмме на уровне 33-34 кДа в образце, полученном из клеток, инкубированных с индуктором, судят о способности клеток синтезировать химерный апоА-I человека. Клоны, показавшие положительный результат, отбирают и помещают в музей культур как продуценты химерного белка апоА-I человека, который используют для дальнейших работ. Аналогичным образом клетки-продуценты анализируют на способность синтезировать белок под действием различных концентраций индуктора ИПТГ от 0,1 до 100 мкМ (0,1 мМ), (фиг. 1). При проведении сравнения синтеза белка клетками-продуцентами на различных средах - LB (заявленное решение) и NCZYM (прототип) - различий не выявлено. Пример анализов приведен на фиг. 2.
Пример 3. Получение биомассы клеток-продуцентов для выделения химерного белка апоА-I человека
Из музейной культуры выращивают ночную культуру в среде LB объемом 5 мл при 37°С. На следующий день ночную культуру переносят в 2-х литровую колбу с 500 мл свежей среды LB с добавлением канамицина 30 мкг/мл. Клетки выращивают при перемешивании на ротационном шейкере 200 об/мин, при 37°С до оптической плотности D600=0,8-1,2 о.е. и добавляют индуктор ИПТГ до 25 мкг/мл. Далее клетки инкубируют двумя различными способами:
1) клетки инкубируют 4 часа при 37°С и перемешивании на ротационном шейкере 200 об/мин;
2) перед добавлением индуктора клетки охлаждают до 30°С, после добавления индуктора клетки инкубируют 18 часов при 30°С и перемешивании на ротационном шейкере 200 об/мин.
По окончании инкубации клетки собирают центрифугированием 3000 об/мин, 20 минут. Оптическая плотность клеток в культуре составляет 5,0-7,0 о.е. (D600), выход влажной биомассы («пасты») составляет 2,3±0,5 г (4,6±1,0 г/л). Биомассу замораживают и хранят при -20°С до процедуры выделения химерного белка апоА-I человека.
Пример 4. Выделение и очистка химерного белка апоА-I человека
Выделение и очистку химерного белка апоА-I человека из биомассы (пример 3) проводят с помощью одностадийной аффинной хроматографии лизата клеток на колонке со смолой «Ni-NTA-Superflow» в денатурирующих условиях. За основу метода берут протокол фирмы-производителя смолы - «Quiagen» (США). Биомассу клеток размораживают и добавляют «лизирующий» буфер, 5 мл на 1 г биомассы (6М гуанидинхлорид, 100 мМ NaH2PO4, 10 мМ Трис-HCl, 10 мМ имидазол, рН 8,0), инкубируют суспензию 30 минут при комнатной температуре. Затем суспензию подвергают ультразвуковой обработке (дополнительное разрушение клеток и ДНК). Дебрис осаждают центрифугированием при 18000g в течение 20 мин. Лизат наносят на колонку со смолой, предварительно уравновешенную «лизирующим» буфером. На 1 г биомассы используют 4 мл смолы. Затем колонку промывают «промывочным» буфером (6М мочевина, 100 мМ NaH2PO4, 10 мМ Трис-HCl, 10 мМ имидазол, рН 7,0), пока показание проточного спектрофотометра (А280) не выйдет на постоянный минимальный уровень. После чего колонку промывают «элюирующим» буфером (6М мочевина, 100 мМ NaH2PO4, 10 мМ Трис-HCl, рН 4,5), пик собирают как раствор, содержащий целевой белок. Скорость промывки колонки на всех стадиях составляет 1-2 мл/мин. Собранный раствор белка хранят при 4°С в «элюирующем» буфере до дальнейшего использования.
Выход химерного белка апоА-I человека из одного литра культуры клеток E.coli штамма BL21 (DE3) в среднем составляет 182±32 мг, или 39,7±6,2 мг из 1 г влажной биомассы. Чистота химерного белка апоА-I человека по результатам ПААГ составляет не менее 95%.
Пример 5. Гидролиз химерного белка апоА-I человека, получение зрелого белка апоА-I человека.
Гидролиз химерного белка апоА-I человека проводят двумя различными способами: кислотный гидролиз, ферментативный гидролиз. В случае кислотного гидролиза получают форму 1 зрелого белка апоА-I человека, 241 а.о., в случае ферментативного гидролиза - форму 2 зрелого белка апоА-I человека, 244 а.о.
Пример 5.1. Кислотный гидролиз химерного белка апоА-I человека, получение зрелого белка апоА-I человека формы 1 (241 а.о.).
Химерный белок апоА-I человека, полученный способом, описанным в примере 4, обессоливают диализом в стандартном фосфатно-солевом буфере, дополнительно содержащем 0,1% муравьиной кислоты (на 10 мл раствора белка 1 л буфера, в течение 4 часов при перемешивании). При диализе часть белка выпадает в осадок. После диализа к суспензии белка добавляют 99% муравьиную кислоту до конченой концентрации 45%, при этом белок переходит в растворимую форму. Раствор белка выдерживают при 55°С в течение 4-х часов при периодическом перемешивании, после чего белок снова обессоливают диализом в стандартном фосфатно-солевом буфере (на 10 мл раствора белка 1 л буфера, в течение 4 часов при перемешивании, буфер меняют дважды). Затем к раствору белка, в центрифужных стаканах, добавляют сульфат аммония 40 г на 100 мл раствора. Перемешивают раствор белка до полного растворения сульфата аммония, выдерживают стаканы с раствором при 4°С в течение 60 минут и осаждают белок центрифугированием 30 минут при 15000 об/мин (примерно 27000g). Осадок растворяют в буфере с мочевиной (6М мочевина, 100 мМ NaH2PO4, 10 мМ Трис-HCl, рН 7,0), объем буфера равен ½ от исходного объема химерного белка апоА-I человека. Далее полученный зрелый белок апоА-I человека отделяют от продуктов неполного гидролиза химерного белка апоА-I человека способом, описанным в примере 6.
Пример 5.2. Ферментативный гидролиз химерного белка апоА-I человека, получение зрелого белка апоА-I человека формы 2 (244 а.о.).
Химерный белок апоА-I человека, полученный способом, описанным в примере 4, разводят фосфатно-солевым буфером с добавлением 6М мочевины в 5-7 раз, затем обессоливают диализом в растворе 50 мМ Трис, 0,5 мМ ЭДТА, рН 8,0 (на 10 мл раствора белка 1 л буфера, в течение 4 часов при перемешивании, буфер меняют дважды). Если после диализа раствор опалесцирует, то его осветляют центрифугированием в течение 10 минут при 15000 об/мин (примерно 27000g). В растворе измеряют оптическую плотность А280, оптическую плотность принимают за концентрацию белка по формуле 1 о.е./мл = 1 мг/мл. К раствору белка добавляют фермент - протеазу вируса табачной мозаики (TEV Protease, Sigma-Aldrich, США, кат. Т4455) в соотношении 1 мг фермента на 100 мг белка и инкубируют раствор при температуре 4°С в течение 18 часов (согласно инструкции фирмы-производителя фермента). Затем раствор переносят в центрифужные стаканы и добавляют сульфат аммония 40 г на 100 мл раствора. Перемешивают раствор белка до полного растворения сульфата аммония, выдерживают стаканы с раствором при 4°С в течение 60 минут и осаждают белок центрифугированием 30 минут при 15000 об/мин (27000g). Осадок растворяют в буфере с мочевиной (6М мочевина, 100 мМ NaH2PO4, 10 мМ Трис-HCl, рН 7,0), объем буфера равен ½ от исходного объема химерного белка апоА-I человека. Далее полученный зрелый белок апоА-I человека отделяют от продуктов неполного гидролиза химерного белка апоА-I человека и фермента способом, описанным в примере 6.
Пример гидролизованного химерного белка апоА-I человека двумя способами представлен на фиг. 2.
Пример 6. Очистка зрелого белка апоА-I человека от продуктов неполного гидролиза химерного белка апоА-I человека.
Выделение и очистку зрелого белка апоА-I из растворов, полученных способами, описанными в примере 5.1 и 5.2, проводят с помощью одностадийной аффинной хроматографии на смоле «Ni-NTA-Superflow» в денатурирующих условиях. За основу метода взят метод, описанный в примере 4. Раствор наносят на колонку со смолой, предварительно уравновешенную «промывочным» буфером. Затем колонку промывают «промывочным» буфером, пока показание проточного спектрофотометра (А280) не выйдет на постоянный минимальный уровень, при этом собирают раствор, соответствующий первому пику, как раствор, содержащий целевой зрелый белок апоА-I. После этого колонку промывают «элюирующим» буфером, пик собирают как раствор, содержащий лидерную последовательность (аминокислотная последовательность, отщепляющаяся от химерного белка апоА-I в процессе гидролиза), фермент TEV-протеазу и часть не гидролизованного химерного белка апоА-I. Промывку колонки на всех стадиях осуществляют со скоростью примерно 1-2 мл/мин. Собранный раствор белка хранят при 4°С в «элюирующем» буфере до дальнейшего использования.
В растворе спектрофотометрическим методом измеряют оптическую плотность А280, раствор белка разбавляют до 1 о.е./мл буфером, содержащим мочевину, и обессоливают диализом в стандартном фосфатно-солевом буфере (на 10 мл раствора белка 1 л буфера, в течение 4 часов при перемешивании, буфер меняют дважды). При необходимости раствор белка хранят при 4°С до дальнейшего использования. Пример анализа очищенного зрелого белка апоА-I человека от продуктов гидролиза представлен на фиг. 4.
Пример 7. Очистка зрелого белка апоА-I от эндотоксинов.
Для очистки зрелого белка апоА-I от эндотоксинов используют известные методы для удаления липополисахаридов из препаратов рекомбинантных белков, полученных из клеток E.coli. Например методы, основанные на использовании полимиксина В и различных поверхностно-активных веществ, таких как тритон Х-114, дезоксихалат натрия и пр. (32). При использовании смола для удаления эндотоксинов «Pierce High Capacity Endotoxin Removal Resin» производства Thermo Fisher Scientific, США на очистку 1 мг белка используют 0,25 мл смолы, процедуру очистки выполняют согласно инструкции фирмы-производителя смолы. На заключительном этапе раствор белка обессоливают диализом в стандартном фосфатно-солевом буфере (на 10 мл раствора белка 1 л буфера, в течение 4 часов при перемешивании) и стерилизуют фильтрованием через насадки с порами 0,22 мкм («Syringe-Diven Filters», Jet Biofilm, Корея). Стерильный чистый белок хранят до экспериментов при 4°С либо замораживают.
Пример 8. Получение кроличьих поликлональных антител к зрелому белку апоА-I человека
Для иммунизации используют молодых кроликов массой 3,0-4,0 кг. Иммунизацию осуществляют двумя белками апоА-I: один вариант получают по методам, описанным в примерах 3, 4, 5.1 (зрелый белок апоА-I форма 1); второй вариант белка выделяют из плазмы крови человека (природный белок) по методике, описанной в работах (33, 34). Перед иммунизацией у животных производят забор крови и получают сыворотку для проведения отрицательной контрольной реакции на антитела. Сыворотку хранят замороженной в течение всей процедуры иммунизации. Иммунизацию проводят по классической схеме: первую иммунизацию проводят с полным адъювантом Фрейнда (1 мл), суммарно используют 200 мкг белка; последующие, вторую, третью и четвертую иммунизации проводят с неполным адъювантом Фрейнда (1 мл) с периодичностью в 10 дней, при этом используют так же по 200 мкг белка. Через неделю после четвертой иммунизации у животных производят забор крови и получают сыворотки, в которых анализируют наличие антител к белку апоА-I.
Для качественного анализа и сравнения в образцах титра антител к апоА-I используют метод твердофазного иммуноферментного анализа. В качестве антигена используют варианты белка апоА-I, применяемые для иммунизации животных, при этом проводят как прямой, так и перекрестный анализ, т.е. к зрелому белку апоА-I (форма 1) применяют антитела, полученные к природному белку и наоборот. Сорбцию антигенов проводят в 50 мМ бикарбонатном буфере рН 8,0 в течение ночи при комнатной температуре. Блокировку планшета осуществляют 0,1% раствором сухого молока. В качестве растворов для инкубации с первыми и вторыми антителами используют стандартный фосфатно-солевой буфер, содержащий 0,1% бычий сывороточный альбумин. Инкубацию с первыми и вторыми антителами проводят при 37°С в течение 30 минут. Промывку планшета осуществляют стандартным фосфатно-солевым буфером, содержащим 0,1% Твин-20. В качестве первых антител используют сыворотки иммунизированных кроликов с различными титрами, в контрольном анализе (отрицательном) используют сыворотки животных до иммунизации. В качестве вторых антител используют антикроличьи поликлональные антитела козы, меченные пероксидазой хрена (Sigma-Aldrich, США, кат. А0545) с рабочим титром 1:10000. Для выявления комплексов антиген-антитело 1-антитело 2 используют раствор 3,3′,5,5′-тетраметилбензидин гидрохлорида. Результаты фиксируют на микропланшетном фотометре для иммуноферментного анализа (Stat Fax 2100, Awareness Technology Inc., США) на одном фильтре (450 нм).
Анализ сывороток, полученных от кроликов, иммунизированных различными вариантами белка апоА-I человека, позволяет выявить специфичный сигнал. Уровень сигнала при одинаковых титрах сыворотки примерно сопоставим и представлен в табл.3. Полученные результаты показывают:
1) специфичность зрелого белка апоА-I человека, получаемого с помощью рекомбинантной ДНК настоящего изобретения, что подтверждает сходство его биологических свойств с природным белком апоА-I человека;
2) возможность использования зрелого белка апоА-I человека, получаемого с помощью настоящего изобретения, для иммунизации животных с целью получения поликлональных антител к человеческому белку апоА-I.
Поликлональные антитела к зрелому белку апоА-I человека, полученному с использованием заявленной рекомбинантной ДНК, могут быть выделены из сывороток иммунизированных кроликов известными методами, например с помощью сульфат аммонийного осаждения и последующей хроматографии на ДЕАЕ-целлюлозе (35).
Пример 9. Демонстрация проникновения зрелого белка апоА, полученного с использованием заявленной рекомбинантной ДНК, в ядра клеток млекопитающих.
В эксперименте используют зрелый белок апоА-I человека, полученный способами, описанными в примере 5.1 и 5.2. В качестве культуры клеток животных используют монослой нормальных гепатоцитов крыс. Гепатоциты крыс выделяют методом рециркуляторной ферментативной перфузии с использованием 0,03% раствора коллагеназы, согласно методу, использованному в работе Сеглена (36). Выращивание клеток проводят в питательной среде RPMI-1640 с глутатионом (ООО «Биолот», Санкт-Петербург, Россия) и добавлением фетальной сыворотки до 5% и антибиотиков гентамицина (50 мкг/мл) и пенициллина (100 ед/мл). Клетки высевают в 24-х луночный полистироловый планшет, предварительно обработанный раствором коллагена 0,1 мг/мл, из расчета 180000 живых клеток на лунку при объеме среды 1 мл. Клетки инкубируют ночь в СО2-инкубаторе для формирования монослоя в атмосфере, содержащей 5% СО2 и 95% воздуха, при 37°С. На следующие сутки производят замену культуральной среды и инкубацию с конъюгатом зрелого белка апоА-I человека с флуоресцентной меткой ФИТЦ. Указанный конъюгат предварительно получают путем инкубации зрелого белка апоА-I человека с ФИТЦ (флуоресцеин-5-изотиоцианат) в течение ночи в карбонатном буфере рН 9,5, в соотношении 12,5 мкг ФИТЦ к 1 мг белка. Конъюгат отделяют от не прореагировавшего ФИТЦ с помощью гель-фильтрации на Sephadex G25 (1 мг конъюгата на 10 мл смолы, высота колонки 10 см), хроматографию проводят в стандартном фосфатно-солевом буфере. Полученный таким образом конъюгат используют для инкубации с клетками. Визуально конъюгат имеет желто-зеленый оттенок, что свидетельствует о присоединении флуоресцентной метки ФИТЦ к молекулам белка.
Конъюгат инкубируют с клетками в течение различных промежутков времени, от 30 до 180 минут. Концентрацию конъюгата так же варьируют от 5 мкг/мл до 50 мкг/мл. В качестве отрицательного контроля клетки инкубируют с не конъюгированным зрелым белком апоА-I человека. По окончании инкубации клетки фиксируют на покровном стекле и анализируют во флуоресцентном микроскопе. В результате анализа обнаружено зеленое свечение в ядрах некоторых клеток (фиг. 5). Максимальное количество таких клеток наблюдается в образцах, инкубированных 180 минут с концентрацией конъюгата 50 мкг/мл. Наличие выраженных светящихся ядрышек в клетках является прямым доказательством проникновения меченого зрелого белка апоА-I человека в ядра гепатоцитов. На контрольных клетках, инкубированных с не конъюгированным белком апоА-I человека, подобных эффектов не наблюдалось.
Таким образом, результаты показывают, что полученный на основе заявленной рекомбинантной ДНК зрелый белок апоА-I человека проникает в ядра клеток и, соответственно, может выступать переносчиком биологически активных веществ, либо служить маркером для рецептор-опосредованного переноса комплексов, содержащих зрелый белок апоА-I человека, в клетки млекопитающих. Этими результатами подтверждается сходство биологических свойств зрелого белка апоА-I человека, полученного с помощью заявленной рекомбинантной ДНК, с природным белком апоА-I человека.
Список использованных источников
1. Franceschini G., Sirtori C.R., Capurso Α., Weisgraber K.H., Mahley R.W. A-IMilano apoprotein. Decreased high density lipoprotein cholesterol levels with significant lipoprotein modifications and without clinical atherosclerosis in an Italian family // J. Clin. Invest. 1980. V. 66. P. 892-900.
2. Weisgraber K.H., Rail S.C.Jr., Bersot T.P., Mahley R.W., Franceschini G., Sirtori C.R. Apolipoprotein A-I Milano. Detection of normal A-I in affected subjects and evidence for a cysteine for arginine substitution in the variant A-I // J. Miol Chem. 1983. V. 258. P. 2508-2513.
3. Gordon J.I., Sims H.F., Lentz S.R., Edelstein C, Scanu A.M., Strauss A.W., Proteolytic processing of human preproapolipoprotein A-I. A proposed defect in the conversion of pro A-I to A-I in Tangier′s disease // J. Biol. Chem. 1983. V. 258. P. 4037-4044.
4. Saku K., Eckardstein Α., Zhang В., Liu R., Jimi S., Ou J., Ohta T., Assmann G., Arakawa K., In vivo kinetics of human apolipoprotein A-I variants in rabbits // Eur. J. Clin. Invest. 1999. V. 29. P. 196-203.
5. National Center for Biotechnology Information
http://www.ncbi.nlm.nih.gov/nuccore/4557320
6. Суменкова Д.В., Князев Р.А., Поляков Л.М., Панин Л.Е. Влияние липопротеинов и стероидных гормонов на биосинтез белка в клетках асцитной карциномы Эрлиха // Бюлл. СО РАМН. 2010. Т. 30. №2. С. 44-48.
7. Поляков Л.М., Суменкова Д.В., Князев Р.А. Панин Л.Е. Анализ взаимодействия липопротеинов и стероидных гормонов // Биомедицинская химия. 2011. Т. 57. Вып. 3. С. 308-313.
8. Суменкова Д.В., Поляков Л.М., Панин Л.Е. Липопротеины высокой плотности как транспортная форма даунорубицина в клетки гепатомы мышей // Экспериментальная и клиническая фармакология. 2012. Т. 75. №5. С. 32-34.
9. Суменкова Д.В., Поляков Л.М., Панин Л.Е. Влияние комплекса изониазида с аполипопротеином Α-I на активность ферментов лизосом у мышей с моделью туберкулезного воспаления // Экспериментальная и клиническая фармакология. 2012. Т. 75. №11. Р. 28-30.
10. Панин Л.Е., Коваленко Г.Α., Поляков Л.М., Тузиков Ф.В., Тузикова Н.А., Князев Р.А. Особенности взаимодействия аполипопротеина Α-I и его комплексов с тетрагидрокортизолом с прокариотической ДНК // Бюллетень СО РАМН. 2012. Т. 32. №1. С. 38-42.
11. Гимаутдинова О.И., Базалук В.В., Кузнецов П.А., Клейменова Н.И. Влияние структуры ДНК на связывание с комплексом тетрагидрокортизол-aпoA-I; роль образовавшихся аддуктов в транскрипции и копировании // Бюлл. СО РАМН. 2010. Т. 30. №2. С. 23-27.
12. Panin L.E., Gimautdinova O.I., Kuznetsov Р.А., Tuzikov F.V. The interaction mechanism of biologically active complexes dehydroepiandrosterone - or tetrahydrocortisol-apolipoprotein A-I with DNA and their role in enhancement of gene expression and protein biosynthesis in hepatocytes // Current Chemical Biology. 2011. V. 5. N. 1. P. 9-16.
13. Kim S.I., Shin D., Lee H., Ann B.Y., Yoon Y., Kim M. Targeted delivery of siRNA against hepatitis С virus by apolipoprotein A-I-bound cationic liposomes // J Hepatol. 2009. V. 50. P. 479-488.
14. Lee H., Kim S.I., Shin D., Yoon Y., Choi Т.Н., Cheon G.J., Kim M. Hepatic siRNA delivery using recombinant human apolipoprotein A-I in mice // Biochem. Biophys. Res. Commun. 2009. V. 378. P. 192-196.
15. Nakayama T., Butler J.S., Sehgal A. et.al. Harnessing a physiologic mechanism for siRNA delivery with mimetic lipoprotein particles // The American Society of Gene & Cell Therapy. 2012. V. 20. P. 1582-1589.
16. Rui M., Tang H., Li Y., Wei X., Xu Y. Recombinant high density lipoprotein nanoparticles for target-specific delivery of siRNA // Pharm. Res. 2013. V. 30. P. 1203-1214.
17. Hatch, F.T. Practical method for plasma lipoprotein analysis / F.T Hatch, R.S. Less // Adv. Lipid Res. 1968. V. 6. P. 2-68.
18. Isacchi Α., Sarmientos P., Lorenzetti R., Soria M., Mature apolipoprotein AI and its precursor proApoAI: influence of the sequence at the 50 end of the gene on the efficiency of expression in Escherichia coli // Gene. 1989. V. 81. P. 129-137.
19. McGuire K.A., Davidson W.S., Jonas A. High yield overexpression and characterization of human recombinant proapolipoprotein A-I // J. Lipid Res. 1996. V. 37. P. 1519-1528.
20. Bergeron J., Frank P.G., Emmanuel F., Latta M., Zhao Y., Sparks D.L., Rassart E., Denefle P., Marcel Y.L. Characterization of human apolipoprotein A-I expressed in Escherichia coli // Biochim. Biophys. Acta. 1997. V. 1344. P. 139-152.
21. Pyle L.E., Barton P., Fujiwara Y., Mitchell Α., Fidge N. Secretion of biologically active human proapolipoprotein A-I in a báculo virus-insect cell system: protection from degradation by protease inhibitors // J. Lipid Res. 1995. V. 36. P. 2355-2361.
22. Sorci-Thomas M.G., Parks J.S., Kearns M.W., Pate G.N., Zhang C., Thomas M.J. High level secretion of wild-type and mutant forms of human proapoA-I using baculovirus-mediated Sf-9 cell expression // J. Lipid Res. 1996. V. 37. P. 673-683.
23. Feng, M.Q., Cai Q.S., Song D.X., Dong J.B., Zhou P. High yield and secretion of recombinant human apolipoprotein AI in Pichia pastoris // Protein Expr. Purif. 2006. V. 46. P. 337-342.
24. Mallory J.B., Kushner P.J., Protter A.A., Cofer C.L., Appleby V.L., Lau K., Schilling J.W., Vigne J.L. Expression and characterization of human apolipoprotein A-I in Chinese hamster ovary cells // J. Biol. Chem. 1987. V. 262. P. 4241-4247.
25. Forte T.M., McCall M.R., Amacher S., Nordhausen R.W., Vigne J.L., Mallory J.B., Physical and chemical characteristics of apolipoprotein A-I-lipid complexes produced by Chinese hamster ovary cells transfected with the human apolipoprotein A-I gene // Biochim. Biophys. Acta. 1990. V. 1047. P. 11-18.
26. Патент на изобретение РФ 2009198 Фрагмент нуклеиновой кислоты, кодирующий человеческий проаполипопротеин А-I. 1994. МПК C12N 15/12.
27. Патент на изобретение РФ №2134696 Димер молекулярного варианта аполипопротеина, способ его получения и фармацевтическая композиция на его основе. 1999, МПК С07К 14/775, А61К 38/16.
28. Заявка на изобретение CN1594582 (A) Process for preparing human apolipoprotein A-I. 2005. МПК C07K 1/14; C12N 1/19; C12N 15/12; C12N 15/63; C12N 15/81; C12P 21/02.
29. Tubb M.R, Smith L.E, Davidson W.S. Purification of recombinant apolipoproteins A-I and A-IV and efficient affinity tag cleavage by tobacco etch virus protease // Journal of Lipid Research. 2009. V. 50. P. 1497-1504.
30. Ryan R.O., Forte T.M., Oda M.N. Optimized bacterial expression of human apolipoprotein A-I // Protein Expression and Purification. 2003. V. 27. P. 98-103.
31. Norgard M.V., KeemK., Monahan J.J. Factors affecting the transformation of Escherichia coli strain chil776 bypBR322 plasmid DNA//Gene. 1978. V. 3. №4. P. 279-292.
32. Magalhaes P.O., Lopes A.M., Mazzola P.G. et. al. Methods of endotoxin removal from biological preparations: a review // J. Pharm. Pharm. Sci. - 2007. - V. 10. - №3. - P. 388-404.
33. Князев P.А. Роль аполипопротеина A-I и аполипопротеина Ε в регуляции биосинтеза белка и нуклеиновых кислот в культуре нормальных и опухолевых гепатоцитов: дисс. канд. биол. наук. Новосибирск, 2007. С. 56-57.
34. Панин Л.Е., Поляков Л.М., Усынин И.Ф., Суменкова Д.В., Князев Р.А. Влияние кортикостероидов в комплексе с аполипопротеином Α-I на биосинтез белка в культуре гепатоцитов // Проблемы эндокринологии. 2009. №3. С. 45-47.
35. Практикум по биохимии: учебное пособие. / Под. ред. Северина С.Е., Соловьевой Г.А. - М.: Изд. МГУ. 1989. С. 308
36. Seglen P. Preparation of isolated rat liver cells // Meth. Cell Biol. 1976. V. 13. P. 29-83.





Изобретение относится к биотехнологии. Предложена рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-I человека, размером 852 п.н. Рекомбинантная ДНК последовательно включает с 5′-конца фрагмент ДНК, кодирующий 6 а.о. гистидина размером 18 п.н. (SEQ ID NO 1), фрагмент ДНК, кодирующий 6 а.о. пpο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий N-концевой фрагмент из 15 а.о. бычьей панкреатической РНКазы А размером 45 п.н. (SEQ ID NO 3), фрагмент ДНК, кодирующий 6 а.о. пpο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий сайт узнавания протеазы вируса табачной мозаики размером 21 п.н. (SEQ ID NO 4), мутантный ген зрелого белка апоА-I человека размером 729 п.н. (SEQ ID NO 5). Выход химерного белка аполипопротеина Α-I человека из 1 л культуры клеток E.coli составляет 182±32 мг, что составляет 39,7±6,2 мг из 1 г биомассы. 5 ил., 3 табл., 9 пр.
Рекомбинантная ДНК, обеспечивающая в составе плазмидного вектора синтез в клетках E.coli химерного белка аполипопротеина Α-I человека, размером 852 п.н., последовательно включающая с 5′-конца фрагмент ДНК, кодирующий 6 а.о. гистидина размером 18 п.н. (SEQ ID NO 1), фрагмент ДНК, кодирующий 6 а.о. пpο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий N-концевой фрагмент из 15 а.о. бычьей панкреатической РНКазы А размером 45 п.н. (SEQ ID NO 3), фрагмент ДНК, кодирующий 6 а.о. пpο-формы белка апоА-I человека размером 18 п.н. (SEQ ID NO 2), фрагмент ДНК, кодирующий сайт узнавания протеазы вируса табачной мозаики размером 21 п.н. (SEQ ID NO 4), мутантный ген зрелого белка апоА-I человека размером 729 п.н. (SEQ ID NO 5).
| RYAN R.O | |||
| ET AL | |||
| Optimized bacterial expression of human apolipoprotein A-I // Protein Expression and Purification, 2003, 27, pp.98-103 | |||
| TUBB M.R | |||
| ET AL | |||
| Purification of recombinant apolipoproteins A-I and A-IV and efficient affinity tag cleavage by tobaccoetch virus protease // Journal of Lipid Research, 2009, vol | |||
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| Плунжерный насос | 1924 |
|
SU1497A1 |
| BURGESS-BROWN | |||

