ПРИОРИТЕТ
[01] Настоящая заявка заявляет приоритет предварительной заявки США с серийным № 61/377030, поданной 25 августа 2010, которая включена ссылкой в ее полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[02] Настоящие штаммы и способы относятся к генетическим мутациям у мицелиальных грибов, которые дают начало вариантам, имеющим измененные характеристики роста. Такие варианты хорошо подходят для роста в погруженных культурах, например, для крупномасштабного производства ферментов и других белков или метаболитов для коммерческих применений.
ССЫЛКИ
[03] Следующие ссылки и дополнительная ссылка, цитируемая в данном документе, таким образом, включены ссылкой:
Caracuel, Z. et al. (2005) Molecular Plant-Microbe Interactions 18:1140-47.
Hughes, H. and Stephens, D.J. (2008) Cell Biol. 129:129-51.
Karhinen, L. et al. (2005) Traffic 6:562-74.
Mouyna, I. et al. (2005) Molecular Microbiology 56:1675-88.
Passolunghi, S. et al. (2010) Microbial Cell Factories 9:7-17.
Peng, R. et al. (2000) J. Biol. Chem. 275:11521-28.
Roberg, K.J. et al. (1999) J. Cell. Biol. 145:659-72.
Shimoni, Y. et al. (2000) J. Cell. Biol. 151:973-84.
Turchini, A. et al. (2000) J. Becteriol. 182:1167-71.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[04] Мицелиальные грибы способны экспрессировать до высоких уровней нативные и гетерологичные белки, что делает их хорошо подходящими для крупномасштабного производства ферментов и других белков для промышленных применений. Мицелиальные грибы обычно выращивают в погруженных культурах мицелия в биореакторах, которые адаптированы для введения и распределения кислорода и питательных веществ в культуральной среде (т.е. бульоне). Морфологические характеристики мицелия воздействуют на реологические свойства бульона, тем самым воздействуя на производительность биореактора.
[05] Как правило, чем выше вязкость бульона, тем меньше однородность распределения кислорода и питательных веществ, и тем больше энергии необходимо для перемешивания культуры. В некоторых случаях вязкость бульона становится достаточно высокой, чтобы существенно препятствовать растворению кислорода и питательных веществ, тем самым неблагоприятно воздействуя на рост грибов. Дополнительно, мощность, необходимая для перемешивания и аэрирования вязкого бульона, может существенно увеличить себестоимость производства, и привести к более высоким капиталовложениям в отношении двигателей и источников мощности.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[06] Описаны штаммы и способы, относящиеся к мицелиальным грибам, имеющим генетические изменения, которые приводят к фенотипам измененной вязкости.
[07] В одном аспекте обеспечивается вариантный штамм мицелиального гриба, полученный из родительского штамма, вариантный штамм, включающий генетическое изменение, которое побуждает клетки вариантного штамма продуцировать измененное количество функционального белка Sfb3 по сравнению с клетками родительского штамма, где клетки вариантного штамма продуцируют, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует измененного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает измененное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[08] В некоторых вариантах осуществления измененное количество функционального белка Sfb3 представляет собой уменьшенное количество, и вариантный штамм продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[09] В некоторых вариантах осуществления генетическое изменение включает разрушение гена sfb3, присутствующего в родительском штамме. В некоторых вариантах осуществления разрушение гена sfb3 является результатом делеции всего гена sfb3 или части гена sfb3. В некоторых вариантах осуществления разрушение гена sfb3 является результатом делеции части геномной ДНК, включающей ген sfb3. В некоторых вариантах осуществления разрушение гена sfb3 является результатом мутагенеза гена sfb3.
[10] В некоторых вариантах осуществления разрушение гена sfb3 выполняют, используя сайт-специфическую рекомбинацию. В некоторых вариантах осуществления разрушение гена sfb3 выполняют в комбинации с введением селектируемого маркера в генетический локус гена sfb3. В некоторых вариантах осуществления разрушение гена sfb3 представляет собой первичную генетическую детерминанту для придания фенотипа уменьшенной вязкости вариантному штамму.
[11] В некоторых вариантах осуществления вариантный штамм не продуцирует функциональный белок Sfb3. В некоторых вариантах осуществления вариантный штамм не продуцирует белок Sfb3.
[12] В некоторых вариантах осуществления вариантный штамм дополнительно включает ген, кодирующий белок, представляющий интерес.
[13] В некоторых вариантах осуществления вариантный штамм продуцирует, по сути, такое же количество белка на единицу количества биомассы, как родительский штамм. В некоторых вариантах осуществления вариантный штамм продуцирует, по сути, такое же количество белка, представляющего интерес, на единицу количества биомассы, как родительский штамм.
[14] В некоторых вариантах осуществления белок Sfb3 включает аминокислотную последовательность IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток).
[15] В некоторых вариантах осуществления мицелиальный гриб представляет виды Pezizomycotina. В некоторых вариантах осуществления мицелиальный гриб представляет собой Trichoderma reesei.
[16] В другом аспекте обеспечивается способ получения вариантного штамма клеток мицелиального гриба, включающий: введение генетического изменения в родительский штамм клетки мицелиального гриба, такое генетическое изменение изменяет продукцию функционального белка Sfb3 по сравнению с клетками родительского штамма, тем самым давая вариантную клетку мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует измененного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает измененное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[17] В некоторых вариантах осуществления генетическое изменение снижает или предотвращает продукцию функционального белка Sfb3, тем самым давая вариантную клетку мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[18] В некоторых вариантах осуществления генетическое изменение включает разрушение гена sfb3 в родительской клетке мицелиального гриба, используя генетическую манипуляцию.
[19] В некоторых вариантах осуществления генетическое изменение включает делецию гена sfb3 в родительской клетке мицелиального гриба, используя генетическую манипуляцию.
[20] В некоторых вариантах осуществления генетическое изменение выполняют, используя сайт-специфическую генетическую рекомбинацию. В некоторых вариантах осуществления разрушение гена sfb3 выполняют в комбинации с введением селектируемого маркера в генетический локус гена sfb3.
[21] В некоторых вариантах осуществления вариантный штамм продуцирует, по сути, такое же количество белка на единицу количества биомассы, как родительский штамм. В некоторых вариантах осуществления вариантный штамм продуцирует, по сути, такое же количество белка, представляющего интерес, на единицу количества биомассы, как родительский штамм.
[22] В некоторых вариантах осуществления белок Sfb3 включает аминокислотную последовательность IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток).
[23] В некоторых вариантах осуществления мицелиальный гриб представляет собой виды Pezizomycotina. В некоторых вариантах осуществления мицелиальный гриб представляет собой Trichoderma reesei.
[24] В некоторых вариантах осуществления родительский штамм дополнительно включает ген, кодирующий белок, представляющий интерес. В некоторых вариантах осуществления ген, кодирующий белок, представляющий интерес, присутствует в родительском штамме перед введением генетического изменения, которое снижает или предотвращает продукцию функционального белка Sfb3.
[25] В другом аспекте обеспечивается белок, представляющий интерес, полученный с помощью описанного выше вариантного штамма.
[26] В другом аспекте обеспечивается вариантный штамм мицелиального гриба, полученный с помощью описанного выше способа.
[27] В другом аспекте обеспечивается вариантный штамм мицелиального гриба, полученный из родительского штамма, вариантный штамм, включающий: (a) генетическое изменение, которое приводит к (i) необходимости в уменьшенном перемешивании в погруженной культуре для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддержанию повышенного содержания растворенного кислорода в погруженной культуре при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма, и (b) ген, кодирующий белок, представляющий интерес, где ген, кодирующий белок, представляющий интерес, присутствует в вариантном штамме перед генетическим изменением в (a).
[28] В некоторых вариантах осуществления генетическое изменение включает разрушение гена sfb3, присутствующего в родительском штамме. В некоторых вариантах осуществления разрушение гена sfb3 выполняют в комбинации с введением селектируемого маркера в генетический локус sfb3 гена.
[29] В другом аспекте обеспечивается способ для скрининга вариантных клеток мицелиального гриба в отношении фенотипа измененной вязкости, включающий: (a) мутагенез клеток родительского штамма мицелиальных грибов для получения вариантных клеток; (b) скрининг вариантных клеток в отношении измененной чувствительности к флуорохромному красителю; и (c) выбор вариантных клеток, которые имеют измененную чувствительность к флуорохромному красителю; где измененная чувствительность к флуорохромному красителю коррелирует со способностью вариантных клеток мицелиальных грибов продуцировать, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует измененного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает измененное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[30] В некоторых вариантах осуществления измененная чувствительность является увеличенной чувствительностью, и вариантная клетка мицелиального гриба продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний, по сравнению с клетками родительского штамма. В некоторых вариантах осуществления флуорохромный краситель представляет собой калькофлуор белый.
[31] В некоторых вариантах осуществления мутагенез клеток выполняют с помощью генетической рекомбинации. В некоторых вариантах осуществления мутагенез клеток выполняют в комбинации с введением селектируемого маркера в генетический локус гена sfb3.
[32] В другом аспекте обеспечивается способ определения полипептида Sfb3 в видах мицелиального гриба Pezizomycotina, включающий следующее: (a) получение аминокислотной последовательности из видов мицелиального гриба Pezizomycotina; и (b) скрининг аминокислотной последовательности в отношении присутствия смежной аминокислотной последовательности IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток); (c) где присутствие SEQ ID NO: 9 в аминокислотной последовательности из видов мицелиального гриба Pezizomycotina показывает, что аминокислотная последовательность из видов мицелиального гриба Pezizomycotina представляет собой полипептид sfb3.
[33] В другом аспекте обеспечивается выделенный полипептид sfb3, определенный с помощью описанного выше способа.
[34] В еще дополнительном аспекте обеспечивается способ получения белка, представляющего интерес, в клетках мицелиального гриба, включающий введение в родительские клетки мицелиального гриба гена, кодирующего белок, представляющий интерес, и генетического изменения, которое снижает количество или активность белка Sfb3 в клетках, тем самым давая вариантную клетку мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, включающий белок, представляющий интерес, который (i) требует измененного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с клетками родительского штамма, и/или (ii) поддерживает измененное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма, и где белок, представляющий интерес получают, по сути, на таком же уровне в вариантных клетках по сравнению с родительскими клетками.
[35] В некоторых вариантах осуществления белок, представляющий интерес, представляет собой более одного белка (или один, или несколько белков), представляющего интерес, и каждый из более одного белка, представляющего интерес, получают, по сути, на таких же относительных уровнях в вариантных клетках по сравнению с родительскими клетками. В конкретном варианте осуществления каждый из более одного белка, представляющего интерес, выбирают из целлюлаз и гемицеллюлаз.
[36] В родственном аспекте обеспечивается белок, представляющий интерес, полученный таким способом. В еще одном родственном аспекте обеспечивается композиция, включающая более одного белка, представляющего интерес, полученного таким способом. В некоторых вариантах осуществления композиция представляет собой композицию общей целлюлазы.
[37] Эти и другие аспекты и варианты осуществления настоящих штаммов и способов будут очевидными из описания, включающего приложенные фигуры.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[38] Фигура 1 представляет собой карту плазмиды pCR-BluntII-hph-loxP#4.
[39] Фигура 2 представляет собой карту плазмиды pCR-Blunt II-TOPO 889092.
[40] Фигура 3, секции A-D представляют собой изображения культуральных планшетов, показывающие морфологию колонии штаммов T. reesei MorphΔsfb3 и 29-9Δsfb3 на среде, содержащей конго красный. (A) Субпопуляция кандидатов MorphΔsfb3 и (B) соответствующие контроли. (C) Субпопуляция кандидатов 29-9Δsfb3 и (D) соответствующие контроли.
[41] Фигура 4 представляет собой карту плазмиды pTrex-Tel-pyrG13/pDONR221/0927853cre, используемой, чтобы транзиторно экспрессировать ген cre в штамме Morph Δsfb3.
[42] Фигура 5 представляет собой изображение, показывающее потерю устойчивости к гигромицину Б и способность расти на ацетамиде у кандидатов после транзиторной экспрессии плазмиды pTrex-Tel-pyrG13/pDONR221/0927853cre. Верхний ряд (A-C): контрольные штаммы Morph и Morph Δsfb3 на указанных средах. Нижний ряд (D-F): кандидаты после транзиторной экспрессии плазмиды на указанных средах.
[43] Фигура 6 представляет собой карту плазмиды pNSP23.
[44] Фигура 7 представляет собой карту одной из четырех плазмид, используемых для комплементации в 70H2, при этом эта плазмида содержит ген sfb3 дикого типа с нативным промотором и терминатором.
[45] Фигура 8 представляет собой изображение, показывающее рост трансформантов 70H2 и 29-9 на PDA с гигромицином Б через четыре дня роста при 28°C (первый перенос из трансформационных планшетов). (A-C) 70H2+sfb3 дикого типа из 29-9. (D-F) 70H2+sfb3 дикого типа с нативным промотором и терминатором из 29-9. (G) 29-9+только вектор. (H-J) 70H2+sfb3 из 70H2. (K-M) 70H2+sfb3 с нативным промотором и терминатором из 70H2. (N) 70H2+только вектор.
[46] Фигура 9 представляет собой карту плазмиды pCR-BluntII-TOPO 949096.
[47] Фигура 10 представляет собой изображение, показывающее (A) рост трансформантов из 70H2, трансформированных pCR-BluntII-TOPO 949096, содержащей ген sfb3 дикого типа, полученный из штамма 29-9. Штаммы инкубировали на VMH (минимальной среде Фогеля, содержащей гигромицин Б) при 28°C. Два кандидата имеют фенотип дикого типа (W), два имеют фенотип 70H2 (H) и два имеют промежуточный фенотип (I). (B) Сравнение 70H2sfb3#24, 70H2 и 29-9 на VM (минимальной среде Фогеля) через 4 дня роста при 28°C с последующими 3 днями роста при комнатной температуре.
[48] Фигура 11 показывает морфологии колоний H3A и H3A Δsfb3 #1009 на указанных средах через 4 дня роста при 28°C.
[49] Фигура 12 показывает изображения гиф H3A и H3A Δsfb3 #1009 в жидкой культуре через указанные периоды времени при 28°C.
[50] Фигура 13 показывает выравнивание аминокислотных последовательностей белков Sfb3 из S. cerevisiae (SEQ ID NO: 1) и T. reesei (SEQ ID NO: 2).
[51] Фигура 14 показывает выравнивание аминокислотных последовательностей белков Sfb3 из S. cerevisiae (SEQ ID NO: 1), T. reesei (SEQ ID NO: 2) и A. oryzae (SEQ ID NO: 3).
ПОДРОБНОЕ ОПИСАНИЕ
I. Обзор
[52] Настоящие штаммы и способы относятся к вариантным клеткам мицелиального гриба, имеющим генетические модификации, которые воздействуют на их морфологию и характеристики роста. Когда вариантные клетки выращивают в погруженной культуре, они продуцируют клеточный бульон, который имеет несхожие реологические свойства по сравнению с клеточным бульоном, включающим клетки родительского штамма. Некоторые из этих вариантных штаммов хорошо подходят для крупномасштабного производства ферментов и других коммерчески важных белков.
II. Определения
[53] Перед описанием настоящих штаммов и способов подробно, следующие выражения определяют для ясности. Выражения, которые не были определены, должны соответствовать их общепринятым значениям, как используется в соответствующей области техники.
[54] Как используется в данном документе, «Trichoderma reesei» относится к мицелиальному грибу типа Ascomycota, подтипа Pezizomycotina. Этот организм был ранее классифицирован как Trichoderma longibrachiatum, а также как Hypocrea jecorina.
[55] Как используется в данном документе, фраза «вариантный штамм клеток мицелиального гриба» или сходные фразы, относятся к штаммам клеток мицелиального гриба, которые получены (т.е. получены из или могут быть получены из) из родительского (или эталонного) штамма, принадлежащего Pezizomycotina, например, с помощью генетической манипуляции.
[56] Как используется в данном документе, выражение «белок, представляющий интерес» относится к полипептиду, который необходимо экспрессировать в мицелиальном грибе, необязательно на высоких уровнях и для цели промышленного внедрения. Такой белок может быть ферментом, субстрат-связывающим белком, поверхностно-активным белком, структурным белком или подобным.
[57] Как используется в данном документе, фраза «по сути, свободный от активности» или подобные фразы, означает, что определенная активность является, либо не обнаруживаемой в смеси, либо присутствующей в количестве, которое не будет препятствовать предназначенной цели смеси.
[58] Как используется в данном документе, выражения «полипептид» и «белок» используют взаимозаменяемо для обозначения полимеров любой длины, включающих аминокислотные остатки, связанные пептидными связями. В данном документе используют общепринятые однобуквенные или трехбуквенные коды для аминокислотных остатков. Полимер может быть линейным или разветвленным, он может включать модифицированные аминокислоты, и он может быть прерван не аминокислотами. Выражения также включают аминокислотный полимер, который был модифицирован естественным образом или путем вмешательства; например, формированием дисульфидной связи, гликозилированием, липидизацией, ацетилированием, фосфорилированием или любой другой манипуляцией или модификацией, такой как конъюгация с метящим компонентом. Также в определение включены, например, полипептиды, содержащие один или более аналогов аминокислоты (включая, например, ненатуральные аминокислоты и т.д.), а также другие модификации, известные из уровня техники.
[59] Как используется в данном документе, функционально и/или структурно подобные белки рассматриваются как «родственные белки». Такие белки могут быть получены из организмов различных родов и/или видов, или даже различных классов организмов (например, бактерии и гриба). Родственные белки также включают гомологи, определенные с помощью анализа первичной последовательности, определенные с помощью анализа вторичной или третичной структуры или определенные с помощью иммунологической перекрестной реактивности.
[60] Как используется в данном документе, выражение «производный полипептид/белок» относится к белку, который получен из или может быть получен из белка путем добавления одной или нескольких аминокислот к любому из двух или к обоим N- и C-терминальному(ым) концу(ам), замещения одной или нескольких аминокислот на одном или нескольких различных сайтах в аминокислотной последовательности, делеции одной или нескольких аминокислот на любом из двух или обоих концах белка или на одном или нескольких сайтах в аминокислотной последовательности, и/или вставки одной или нескольких аминокислот на одном или нескольких сайтах в аминокислотной последовательности. Получение белковой производной может быть достигнуто путем модификации последовательности ДНК, которая кодирует нативный белок, трансформации этой последовательности ДНК в подходящем хозяине и экспрессии модифицированной последовательности ДНК для образования производного белка.
[61] Родственные (и производные) белки включают «вариантные белки». Вариантные белки отличаются от эталонного/родительского белка (например, белка дикого типа) замещением, делецией и/или вставкой на небольшое число аминокислотных остатков. Число отличающихся аминокислотных остатков между вариантным и родительским белком может быть одним или больше, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50 или более аминокислотных остатков. Вариантные белки могут разделять по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 91%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 93%, по меньшей мере приблизительно 94%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, или даже по меньшей мере приблизительно 99%, или более идентичности аминокислотной последовательности с эталонным белком. Вариантный белок может также отличаться от эталонного белка в выбранных мотивах, доменах, эпитопах, консервативных участках и подобном.
[62] Как используется в данном документе, выражение «аналогичная последовательность» относится к последовательности в белке, который обеспечивает такую же функцию, третичную структуру и/или консервативные остатки, как и белок, представляющий интерес (т.е. типично исходный белок, представляющий интерес). Например, в эпитопных участках, которые содержат α-спираль или β-листовую структуру, замещающие аминокислоты в аналогичной последовательности предпочтительно поддерживают такую же специфическую структуру. Выражение также относится к нуклеотидным последовательностям, а также аминокислотным последовательностям. В некоторых вариантах осуществления аналогичные последовательности разрабатывают так, чтобы замещающие аминокислоты приводили в результате к вариантному ферменту, показывающему подобную или улучшенную функцию. В некоторых вариантах осуществления третичная структура и/или консервативные остатки аминокислот в белке, представляющем интерес, расположены на или вблизи сегмента или фрагмента, представляющего интерес. Таким образом, при условии, что сегмент или фрагмент, представляющий интерес содержит, например, α-спираль или β-листовую структуру, то замещающие аминокислоты предпочтительно поддерживают эту специфическую структуру.
[63] Как используется в данном документе, выражение «гомологичный белок» относится к белку, который имеет активность и/или структуру, подобную эталонному белку. Предполагается, что гомологи не обязательно являются эволюционно родственными. Таким образом, предполагается, что выражение включает одинаковый(е), подобный(е) или соответствующий(е) фермент(ы) (т.е. в отношении структуры и функции), полученный(е) из различных организмов. В некоторых вариантах осуществления необходимо определить гомолог, который имеет четвертичную, третичную и/или первичную структуру, подобную эталонному белку. В некоторых вариантах осуществления гомологичные белки вызывают иммунологический(е) ответ(ы), подобный(е) эталонному белку. В некоторых вариантах осуществления гомологичные белки конструируют, чтобы получать ферменты с необходимой(ыми) активностью(ями).
[64] Степень гомологии между последовательностями может быть определена с использованием любого приемлемого способа, известного из уровня техники (см., например, Smith and Waterman (1981) Adv. Appl. Math. 2:482; Needleman and Wunsch (1970) J. Mol. Biol., 48:443; Pearson and Lipman (1988) Proc. Natl. Acad. Sci. USA 85:2444; программы, такие как GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package (Genetics Computer Group, Мэдисон, Висконсин); и Devereux et al. (1984) Nucleic Acids Res. 12:387-95).
[65] Например, PILEUP представляет собой пригодную программу для определения уровней гомологии последовательности. PILEUP создает выравнивание множественных последовательностей из группы родственных последовательностей, используя прогрессивные, парные выравнивания. Она может также построить дерево, показывающее взаимоотношения образования кластеров, используемые для создания выравнивания. PILEUP использует упрощение способа прогрессивного выравнивания Фенга и Дулитла (Feng and Doolittle (1987) J. Mol. Evol. 35:351-60). Способ подобен таковому, описанному Хиггинсом и Шарпом ((1989) CABIOS 5:151-53). Пригодные параметры PILEUP, включая вес гэпа по умолчанию 3,00, вес длины гэпа по умолчанию 0,10 и взвешенные концевые гэпы. Другой пример пригодного алгоритма представляет собой алгоритм BLAST, описанный Altschul et al. ((1990) J. Mol. Biol. 215:403-10) и Karlin et al. ((1993) Proc. Natl. Acad. Sci. USA 90:5873-87). Одна особенно пригодная программа BLAST представляет собой программу WU-BLAST-2 (см., например, Altschul et al. (1996) Meth. Enzymol. 266:460-80). Параметры “W,” “T,” и “X” определяют чувствительность и скорость выравнивания. Программа BLAST использует в качестве параметров по умолчанию слово-длина (W) 11, выравнивания (B) 50, ожидание (E) 10, M′5, N′-4 и сравнение обоих цепей матрицы замен BLOSUM62 (см., например, Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915).
[66] Как используется в данном документе, фразы «по сути, подобные» и «по сути, идентичные» в контексте по меньшей мере двух нуклеиновых кислот или полипептидов, как правило означает, что полинуклеотид или полипептид включает последовательность, которая имеет по меньшей мере приблизительно 70% идентичности, по меньшей мере приблизительно 75% идентичности, по меньшей мере приблизительно 80% идентичности, по меньшей мере приблизительно 85% идентичности, по меньшей мере приблизительно 90% идентичности, по меньшей мере приблизительно 91% идентичности, по меньшей мере приблизительно 92% идентичности, по меньшей мере приблизительно 93% идентичности, по меньшей мере приблизительно 94% идентичности, по меньшей мере приблизительно 95% идентичности, по меньшей мере приблизительно 96% идентичности, по меньшей мере приблизительно 97% идентичности, по меньшей мере приблизительно 98% идентичности, или даже по меньшей мере приблизительно 99% идентичности, или более, по сравнению с эталонной последовательностью (т.е. дикого типа). Идентичность последовательности может быть определена с использованием известных программ, таких как BLAST, ALIGN и CLUSTAL, используя стандартные параметры. (см., например, Altschul, et al. (1990) J. Mol. Biol. 215:403-410; Henikoff et al. (1989) Proc. Natl. Acad. Sci. USA 89:10915; Karin et al. (1993) Proc. Natl. Acad. Sci USA 90:5873; и Higgins et al. (1988) Gene 73:237-244). Программное обеспечение для проведения анализов BLAST является общедоступным через Национальный центр биотехнологической информации. Также базы данных могут быть выбраны, используя FASTA (Pearson et al. (1988) Proc. Natl. Acad. Sci. USA 85:2444-48). Одним показателем того, что два полипептида являются, по сути, идентичными, является то, что первый полипептид является иммунологически перекрестно реактивным со вторым полипептидом. Обычно, полипептиды, которые отличаются на консервативные аминокислотные замещения, являются иммунологически перекрестно реактивными. Таким образом, полипептид является, по сути, идентичным второму полипептиду, например, когда два пептида отличаются только на консервативное замещение. Другим показателем того, что две последовательности нуклеиновых кислот являются, по сути, идентичными, является то, что две молекулы гибридизуются друг с другом при строгих условиях (например, в пределах диапазона среды до высокой строгости).
[67] Как используется в данном документе, выражение «ген» является синонимом выражению «аллель» по отношению к нуклеиновой кислоте, которая кодирует и направляет экспрессию белка или РНК. Вегетативные формы мицелиальных грибов являются, как правило, гаплоидными, следовательно, одна копия определенного гена (т.е. один аллель) является достаточной для придания определенного фенотипа.
[68] Как используется в данном документе, гены, белки или штамм «дикого типа» и «нативные» гены, белки или штаммы, являются таковыми, обнаруживаемыми в природе.
[69] Как используется в данном документе, «делеция гена» относится к его удалению из генома клетки хозяина. Если ген включает контрольные элементы (например, энхансерные элементы), которые не расположены сразу же смежно с кодирующей последовательностью гена, то делеция гена относится к делеции кодирующей последовательности, и необязательно смежным последовательностям промотора и/или терминатора.
[70] Как используется в данном документе, «разрушение гена» относится в широком смысле к любой генетической или химической манипуляции, которая, по сути, не допускает продуцирование функционального генного продукта клеткой, например, белка клеткой-хозяином. Иллюстративные способы разрушения включают полную или частичную делецию любой части гена, включая кодирующую полипептид последовательность, промотор, энхансер или другой регуляторный элемент или мутагенез того же, где мутагенез включает замещения, вставки, делеции, инверсии и их комбинации и вариации любых из этих мутаций, которые, по сути, препятствуют продукции функционального генного продукта.
[71] Как используется в данном документе, «генетическая манипуляция» относится к изменению предварительно выбранной целевой последовательности нуклеиновой кислоты, например, используя макромолекулы (т.е. ферменты и/или нуклеиновые кислоты), которые действуют на предварительно выбранную последовательность нуклеиновой кислоты. В этом смысле генетическая манипуляция отличается от химической манипуляции, в которой малые молекулы используют для случайно воздействующих изменений на последовательность нуклеиновой кислоты, которая не является предварительно выбранной.
[72] Как используется в данном документе, «генетическое изменение» представляет собой изменение в ДНК клетки, которое является результатом генетической манипуляции и отличается от изменения в ДНК клетки, которое является результатом химической манипуляции.
[73] Как используется в данном документе, «аэробная ферментация» относится к росту в присутствии кислорода.
[74] Как используется в данном документе, выражение «клеточный бульон» относится в совокупности к среде и клеткам в жидкой/погруженной культуре.
[75] Как используется в данном документе, выражение «клеточная масса» относится к клеточному компоненту (включая интактные и лизированные клетки), присутствующему в жидкой/погруженной культуре. Клеточная масса может быть выражена в сухом или влажном весе.
[76] Как используется в данном документе, выражение «реология» относится к разделу физики, который касается деформации и потоку вещества.
[77] Как используется в данном документе, «вязкость» представляет собой меру устойчивости жидкости к деформации путем механического усилия, такого как напряжение при сдвиге или напряжение при растяжении. В данном контексте вязкость относится к устойчивости клеточного бульона, включающего клетки мицелиальных грибов к механическому усилию, например, как обеспечивается ротором/рабочим колесом. Так как вязкость клеточного бульона может быть сложно измерить напрямую, могут быть использованы непрямые измерения вязкости, такие как содержание растворенного кислорода культурального бульона при предварительно выбранном количестве перемешиваний, количество перемешиваний, необходимых для поддержания предварительно выбранного содержания растворенного кислорода, количество мощности, необходимой для перемешивания клеточного бульона для поддержания предварительно выбранного содержания растворенного кислорода или даже морфология колонии на твердой среде.
[78] Как используется в данном документе, вариантный штамм клеток мицелиальных грибов «с измененной вязкостью» представляет собой вариантный штамм, который продуцирует клеточный бульон, который имеет уменьшенную или увеличенную вязкость (т.е. уменьшенную или увеличенную устойчивость к напряжению при сдвиге или при растяжении) по сравнению с эквивалентным клеточным бульоном, полученным с помощью родительского штамма. Как правило, эквивалентные клеточные бульоны имеют сопоставимые клеточные массы. Предпочтительно, разница между вариантным штаммом, с измененной вязкостью и родительским штаммом, учитывая любое прямое или непрямое измерение вязкости, составляет по меньшей мере 10%, по меньшей мере 15%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, или даже по меньшей мере 50%, или более. Способы сравнения вязкости бульона клеток мицелиальных грибов описаны в данном документе. Как правило, сопоставимые (или эквивалентные) клеточные бульоны имеют сопоставимые клеточные массы.
[79] Как используется в данном документе, вариантный штамм клеток мицелиальных грибов «с уменьшенной вязкостью» представляет собой вариантный штамм, который продуцирует клеточный бульон, который имеет уменьшенную вязкость (т.е. уменьшенную устойчивость к напряжению при сдвиге или при растяжении) по сравнению с эквивалентным клеточным бульоном, полученным с помощью родительского штамма. Предпочтительно, различие между вариантным штаммом, с измененной вязкостью и родительским штаммом, учитывая любое прямое или непрямое измерение вязкости, составляет по меньшей мере 10%, по меньшей мере 15%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, или даже по меньшей мере 50%, или более.
[80] Как используется в данном документе, «первичная генетическая детерминанта» относится к гену или его генетической манипуляции, который(ая) необходим(а) и достаточен(чна) для придания определенного фенотипа в отсутствии других генов или их генетических манипуляций.
[81] Как используется в данном документе, «функциональный полипептид/белок» представляет собой белок, который обладает активностью, такой как ферментативная активность, связывающая активность, поверхностно-активное свойство или подобное, и который не был подвергнут мутагенезу, процессингу или модифицирован иным образом, чтобы аннулировать или снизить эту активность. Как определено, функциональные полипептиды могут быть термостабильными или термолабильными.
[82] Как используется в данном документе, «функциональный ген» представляет собой ген, способный быть использованным клеточными компонентами для продукции активного генного продукта, обычно белка. Функциональные гены являются противоположностью разрушенным генам, которые модифицированы так, что они не могут быть использованы клеточными компонентами для продукции активного генного продукта.
[83] Как используется в данном документе, вариантные клетки (или вариантный штамм) «поддерживают или сохраняют высокий уровень экспрессии и/или секреции белка» по сравнению с родительскими клетками (или родительским штаммом), если различие в экспрессии белка между вариантными клетками и родительскими клетками составляет менее приблизительно 20%, менее приблизительно 15%, менее приблизительно 10%, менее приблизительно 7%, менее приблизительно 5% или даже менее приблизительно 3%.
[84] Как используется в данном документе, клетки-хозяины были «модифицированы, чтобы препятствовать продукции Sfb3», если они были генетически или химически изменены, чтобы препятствовать продукции функционального полипептида Sfb3, который проявляет характеристику активности белка Sfb3 дикого типа, в частности, активность, которая стимулирует элонгацию гиф или иным образом увеличивает вязкость мицелиального гриба в жидкой культуре. Такие модификации включают, но без ограничения, делецию гена sfb3, разрушение гена sfb3, модификацию гена sfb3 так, чтобы в кодируемом полипептиде не было вышеупомянутой активности, модификации гена sfb3, чтобы воздействовать на посттрансяционный процессинг или стабильность и их комбинации.
[85] Как используется в данном документе, «белок, представляющий интерес», представляет собой белок, который необходимо получить в погруженной культуре клеток мицелиальных грибов. Как правило, белки, представляющие интерес, являются коммерчески важными для промышленного или фармацевтического применения, что делает их желаемыми для получения в больших количествах. Белки, представляющие интерес, должны отличаться от огромного количества других белков, экспрессируемых клетками мицелиальных грибов, которые, как правило, не представляют интерес в качестве продуктов и, главным образом, рассматриваются фоновыми белковыми примесями.
[86] Как используется в данном документе, вариантные клетки (или вариантный штамм) производят(ит) «по сути, такое же количество» белка на единицу количества биомассы, как родительские клетки (или родительский штамм), если количество белка, продуцируемое вариантными клетками, является уменьшенным не более чем на 20%, уменьшенным не более чем на 15%, уменьшенным не более чем на 10%, уменьшенным даже не более чем на 5% по сравнению с количеством белка, продуцируемым родительскими клетками, где количество белка нормализуют к общему количеству биомассы клеток, исходя из которого измеряют продукцию белка, где биомасса может быть выражена на основе, или влажного (например, клеточный осадок), или сухого веса.
[87] Как используется в данном документе, количество белка, представляющего интерес, экспрессированного вариантными клетками и родительскими клетками является «по сути, подобным», если различие в экспрессии между вариантными клетками и родительскими клетками составляет менее приблизительно 20%, менее приблизительно 15%, менее приблизительно 10%, менее приблизительно 5%, менее приблизительно 4%, менее приблизительно 3%, менее приблизительно 2% или даже менее приблизительно 1%.
[88] Как используется в данном документе, «флуорохромы» являются флуоресцентными красителями. Предпочтительные флуорохромы связываются с целлюлозой и/или хитином в клеточных стенках грибов.
[89] Как используется в данном документе, единственное число включает ссылки на формы множественного числа, если только контекст четко не диктует иное. Все ссылки, цитируемые в данном документе, тем самым включены ссылкой в их полноте. Следующие сокращения/аббревиатуры имеют следующие значения, если только не определено иное:
III. Фенотип уменьшенной вязкости у мицелиальных грибов
[90] Предыдущие попытки разработать штаммы Trichoderma reesei с уменьшенной вязкостью включали химический мутагенез, с последующим скринингом полученных мутантов (иногда в данном документе называемых «штаммы») в отношении чувствительности к калькофлуору белому, флуорохромному красителю, который связывается с целлюлозой и хитином в клеточных стенках грибов. Чувствительность к калькофлуору белому связана с изменениями в морфологии дрожжей, хотя значимость чувствительности к калькофлуору белому у мицелиальных грибов была до настоящего момента неизвестна. Таким образом, родительский штамм Trichoderma reesei Morph TrglaA (29-9) был подвергнут химическому мутагенезу и было обнаружено, что один полученный штамм (т.е. 70H2) проявляет уменьшенную скорость колониального роста на агарозных планшетах, уменьшенную споруляцию, измененную морфологию и уменьшенную вязкость в жидкой среде, при этом поддерживая высокий уровень экспрессии и секреции белка. Сравнительный анализ геномной последовательности выявил мутации во множественных генах в штамме 70H2 по сравнению с родительским штаммом 29-9.
[91] В то время как штамм 70H2 демонстрировал фенотип «уменьшенной вязкости», он не имел полностью установленного генома, и ген или гены, ответственные за фенотип уменьшенной вязкости, были неизвестны. Более того, в то время как 70H2 мог быть использован как штамм хозяина для введения экзогенных генов для высоких уровней экспрессии, он не был приспособлен для введения гена или генов, ответственных за фенотип уменьшенной вязкости в другие штаммы.
IV. Изменения в продукции Sfb3 воздействуют на клеточную вязкость у мицелиального гриба
[92] Сейчас было обнаружено, что изменения в продукции Sfb3 воздействуют на клеточную вязкость у мицелиального гриба. Это открытие имеет значительные последствия для применения мицелиальных грибов для экспрессии коммерчески важных белков.
[93] Ген Sfb3 (также известный как Lst1) был ранее охарактеризован только в почкующихся дрожжах (т.е. Saccharomyces cerevisiae), где он кодирует белок, связанный с белковой оболочкой COPII, которая окружает транспортные везикулы, которые несут белки от эндоплазматического ретикулума к аппарату Гольджи. Sfb3, а также Sfb2, являются гомологами Sec24, все из этих генов связанны с упаковкой специфических транспортных белков в везикулы.
[94] В то время как Sec24 является важным геном в дрожжах, Sfb3 и Sfb2 не являются таковыми, хотя делеция Sfb3 в дрожжах, как известно, воздействует на транспорт транспортного белка плазматической мембраны (Pma1p) и глюканозилтрансферазы (Gas1p), которая вовлечена в синтез клеточной стенки.
[95] Использование BLAST для изучения общедоступной геномной последовательности Trichoderma reesei, используя аминокислотные последовательности Sec24p, Sfb3p или Sfb2p из S. cerevisiae в качестве последовательностей запроса, выявляет, что T. reesei имеет один ген, который является наиболее близко гомологичным дрожжевому Sec24, и один ген, который является наиболее близко гомологичным дрожжевому Sfb3. Не был определен никакой другой гомолог предполагающий, что T. reesei не имеет гена, эквивалентного Sfb2.
[96] Использование BLAST для изучения общедоступных геномных последовательностей видов Pezizomycotina, используя аминокислотную последовательность Sfb3 из T. reesei в качестве запроса, демонстрирует общий паттерн. То есть, каждый гриб имеет четкий гомолог каждого из Sfb3 и Sec24, но дополнительный гомолог, более близко родственный дрожжевому Sfb2, не присутствует в геномах этих мицелиальных аскомицетов.
[97] Гомологи белков Sfb3 обнаруживают в мицелиальных грибах, например, Trichoderma reesei и Aspergillus oryzae, хотя функция этих белков была до сих пор не известна. Аминокислотные последовательности белков Sfb3 из S. cerevisiae (SEQ ID NO: 1), T. reesei (SEQ ID NO: 2), A. oryzae (SEQ ID NO: 3), A. niger (SEQ ID NO: 4), P. funiculosum (SEQ ID NO: 5), P. chrysogenum (SEQ ID NO: 6), N. Crassa (SEQ ID NO: 7) и F oxysporum (SEQ ID NO: 8) показаны ниже, в качестве примеров: SEQ ID NO: 4-8 были получены из общедоступных баз данных грибных геномов, но не имеют номеров доступа.
[98] Аминокислотная последовательность Sfb3 из Saccharomyces cerevisiae (SEQ ID NO: 1):


[99] Аминокислотная последовательность Sfb3 из Trichoderma reesei (SEQ ID NO: 2):

[100] Аминокислотная последовательность RIB40 Sfb3 из Aspergillus oryzae (GI: 83766074; SEQ ID NO: 3):


[101] Аминокислотная последовательность Sfb3 из Aspergillus niger (SEQ ID NO: 4)

[102] Аминокислотная последовательность Sfb3 из Penicillium funiculosum (SEQ ID NO: 5)


[103] Аминокислотная последовательность Sfb3 из Penicillium chrysogenum (SEQ ID NO: 6)

[104] Аминокислотная последовательность Sfb3 из Neurospora crassa (SEQ ID NO: 7)

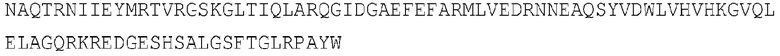
[105] Аминокислотная последовательность Sfb3 из Fusarium oxysporum (SEQ ID NO: 8)

[106] Выравнивание аминокислотных последовательностей белков Sfb3 из S. cerevisiae (SEQ ID NO: 1) и T. reesei (SEQ ID NO: 2) показано на Фигуре 13. Эти последовательности имеют приблизительно 30% идентичности аминокислотной последовательности. Напротив, белки Sfb3 из T. reesei и A. oryzae имеют приблизительно 58% идентичности аминокислотной последовательности. Выравнивание аминокислотных последовательностей белков Sfb3 из S. cerevisiae (SEQ ID NO: 1), T. reesei (SEQ ID NO: 2), и A. oryzae (SEQ ID NO: 3) показано на Фигуре 14.
[107] Выравнивание аминокислотных последовательностей белков Sfb3 из приблизительно 40 видов Pezizomycotina выявило специфическую аминокислотную последовательность, т.е. IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток), которая является близкой к C-концу белков Sfb3, и не обнаруживается в белках Sec24. Эта консенсусная последовательность может быть использована для определения белков Sfb3 в других членах Pezizomycotina.
[108] Отдельные исследования показали, что мутация гена gas1 (или гена gel1 как он известен в Aspergillus fumigatus) воздействует на структуру клеточной стенки гриба и приводит к морфологическим изменениям, а также гиперчувствительности к калькофлуору белому, конго красному и додецилсульфату натрия.
[109] Не ограничиваясь теорией, полагают, что изменение экспрессии Sfb3 и/или активности у мицелиальных грибов препятствует транспорту белков, вовлеченных в синтез клеточной стенки, тем самым изменяя структуру клеточной стенки и давая более компактную клеточную морфологию, характеризующуюся более короткими гифами и более дрожжеподобным внешним видом. Вероятный кандидат для белка, вовлеченного в синтез клеточной стенки, представляет собой Gas1/Gel1.
[110] Вариантные штаммы мицелиальных грибов, которые проявляют фенотип измененной вязкости в жидкой среде, могут быть хорошо подходящими для широкомасштабной продукции коммерчески важных белков. В то время как настоящие штаммы и способы представлены для примера, используя мицелиальный гриб T. reesei, функция белка Sfb3 в Pezizomycotina, как ожидается, является консервативной. Следовательно, настоящие штаммы и способы ни в какой мере не ограничены T. reesei.
V. Штамм мицелиального гриба с измененной продукцией белка Sfb3
[111] В одном аспекте обеспечивается вариантный штамм мицелиального гриба, полученный из родительского штамма, вариантный штамм, включающий генетическое изменение, которое побуждает клетки вариантного штамма продуцировать измененное количество функционального белка Sfb3 по сравнению с клетками родительского штамма. Клетки вариантного штамма впоследствии продуцируют, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который требует измененного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода, или клеточную массу, которая поддерживает измененное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
[112] В некоторых случаях генетическое изменение побуждает клетки вариантного штамма продуцировать уменьшенное количество функционального белка Sfb3 по сравнению с клетками родительского штамма, и полученный клеточный бульон требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода, или поддерживает более высокое содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма. В таких случаях считается, что клеточная масса вариантного штамма проявляет уменьшенную вязкость по сравнению с клеточной массой родительского штамма, что объясняет наблюдения, относящиеся к содержанию растворенного кислорода и перемешиванию.
[113] Снижение в количестве функционального белка Sfb3 может быть результатом разрушения гена sfb3, присутствующего в родительском штамме. Так как разрушение гена sfb3 представляет собой первичную генетическую детерминанту для придания фенотипа уменьшенной вязкости вариантному штамму, то необходимо, чтобы такие вариантные штаммы включали только разрушенный ген sfb3, в то время как все другие гены могут оставаться интактными. В некоторых случаях вариантные штаммы могут необязательно включать дополнительные генетические изменения по сравнению с родительским штаммом, из которого они получены. Такие дополнительные генетические изменения не являются необходимыми для придания снижения вязкости, но могут придавать другое преимущество штамму.
[114] Разрушение гена sfb3 может быть проведено, используя любые подходящие способы, которые, по сути, препятствуют экспрессии функционального генного продукта sfb3, т.е. белка Sfb3. Иллюстративные способы разрушения включают полную или частичную делецию гена sfb3, включая полную или частичную делецию, например, последовательности, кодирующей Sfb3, промотора, терминатора, энхансера или другого регуляторного элемента. Разрушение гена sfb3 может также быть проведено с помощью полной или частичной делеции части хромосомы, которая включает любую часть гена sfb3. Конкретные способы разрушения гена sfb3 включают проведение нуклеотидных замещений или вставок в любую часть гена sfb3, например, последовательность, кодирующую Sfb3, промотор, терминатор, энхансер или другой регуляторный элемент. Предпочтительно, делеции, вставки и/или замещения (совместно названные мутациями) проводят с помощью генетической манипуляции, используя техники молекулярной биологии, специфические к последовательностям, что противоположно химическому мутагенезу, который, как правило, не нацелен на специфические последовательности нуклеиновых кислот.
[115] Мутации в гене sfb3 могут снижать эффективность промотора sfb3, снижать эффективность энхансера sfb3, препятствовать сплайсингу или редактированию мРНК sfb3, препятствовать трансляции мРНК sfb3, вводить стоп-кодон в последовательность, кодирующую Sfb3, чтобы препятствовать трансляции полноразмерного белка Sfb3, изменять кодирующую последовательность белка Sfb3, чтобы давать менее активный или неактивный белок или снижать взаимодействие Sfb3 с другими компонентами клеточной стенки, изменять кодирующую последовательность белка Sfb3, чтобы давать менее стабильный белок или нацеливаться на белок для разрушения, вызывать неправильное сворачивание белка Sfb3 или неправильную его модификацию (например, с помощью гликозилирования) или препятствовать клеточной миграции белка Sfb3.
[116] Как правило, целью этих и других генетических манипуляций является снижение или препятствование экспрессии функционального белка Sfb3, или снижение или препятствование нормальной биологической активности белка Sfb3, тем самым давая морфологическое изменение, которое приводит к фенотипу уменьшенной вязкости.
[117] В других случаях генетическое изменение увеличивает или восстанавливает экспрессию функционального белка Sfb3, или увеличивает нормальную биологическую активность белка Sfb3, тем самым давая морфологическое изменение, которое приводит к фенотипу увеличенной или восстановленной вязкости. Иллюстративные генетические изменения, которые увеличивают или восстанавливают функцию Sfb3, являются такими, которые вводят дополнительные копии гена sfb3 в клетку, увеличивают эффективность промотора sfb3, энхансера или другого контрольного элемента, увеличивают трансляцию мРНК, кодирующей белок Sfb3, увеличивают стабильность мРНК, кодирующей белок Sfb3, вводят изменения в ген sfb3, которые увеличивают активность или стабильность белка Sfb3, вводят изменения в ген sfb3, которые модулируют взаимодействие с другими белками или компонентами клеточной стенки и подобное. Другие генетические изменения, которые увеличивают или восстанавливают функцию Sfb3, являются такими, которые реверсируют эффект генетических изменений, которые снижают или препятствуют экспрессии функционального белка Sfb3.
[118] Клетки мицелиального гриба для манипуляции и применения, как описано, являются, как правило, из типа Ascomycota, подтипа Pezizomycotina, в частности, грибы, которые имеют состояние вегетативных гиф и включают гомолог гена sfb3. Такие организмы включают клетки мицелиального гриба, используемые для получения коммерчески важных промышленных и фармацевтических белков, включая, но без ограничения, виды Trichoderma, виды Aspergillus, виды Fusarium, виды Scedosporium, виды Penicillium, виды Chrysosporium, виды Cephalosporium, виды Talaromyces, виды Geosmithia, виды Myceliophthora и виды Neurospora. Конкретные организмы включают, но без ограничения, Trichoderma reesei (ранее классифицированный как Trichoderma longibrachiatum и Hypocrea jecorina), Aspergillus niger, Aspergillus fumigatus, Aspergillus itaconicus, Aspergillus oryzae, Aspergillus nidulans, Aspergillus terreus, Aspergillus sojae, Aspergillus japonicus, Scedosporium prolificans, Neurospora crassa, Penicillium funiculosum, Penicillium chrysogenum, Talaromyces (Geosmithia) emersonii, Fusarium venenatum, Myceliophthora thermophila и Chrysosporium lucknowense.
[119] В некоторых вариантах осуществления, например, где мицелиальный гриб представляет T. reesei, ген sfb3 кодирует белок, имеющий аминокислотную последовательность SEQ ID NO: 2. В других вариантах осуществления ген sfb3 кодирует белок, имеющий определенную степень идентичности полной аминокислотной последовательности аминокислотной последовательности SEQ ID NO: 2, например, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 91%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 93%, по меньшей мере приблизительно 94%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98% или даже по меньшей мере приблизительно 99% идентичности.
[120] В некоторых вариантах осуществления, например, где мицелиальный гриб представляет собой A. oryzae, ген sfb3 кодирует белок, имеющий аминокислотную последовательность SEQ ID NO: 3. В других вариантах осуществления ген sfb3 кодирует белок, имеющий определенную степень идентичности полной аминокислотной последовательности аминокислотной последовательности SEQ ID NO: 3, например, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 91%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 93%, по меньшей мере приблизительно 94%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98% или даже по меньшей мере приблизительно 99% идентичность.
[121] В некоторых вариантах осуществления, например, где мицелиальный гриб представляет собой T. reesei, ген sfb3 имеет нуклеотидную последовательность SEQ ID NO: 10, показанную ниже. В других вариантах осуществления ген sfb3 имеет нуклеотидную последовательность с определенной степенью идентичности полной нуклеотидной последовательности нуклеотидной последовательности SEQ ID NO: 10, например, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 91%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 93%, по меньшей мере приблизительно 94%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98% или даже по меньшей мере приблизительно 99% идентичность.
[122] Последовательность ДНК sfb3 из Trichoderma reesei (SEQ ID NO: 10; 2 интроны подчеркнуты):
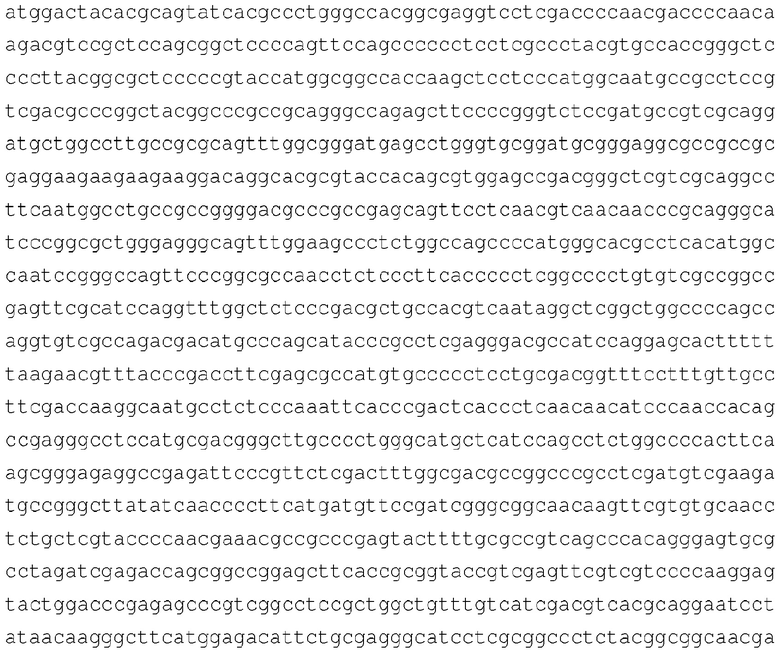


VI. Способ изменения фенотипа вязкости клеток мицелиальных грибов
[123] В другом аспекте обеспечивается способ изменения морфологии клеток мицелиальных грибов. Вариантные клетки мицелиальных грибов проявляют измененную морфологию роста на твердой среде и продуцируют клеточные массы, имеющие различные вязкости, когда выращиваются в погруженной культуре.
[124] В некоторых случаях способ включает разрушение гена sfb3 в родительском штамме, используя соответствующие генетические или химические способы, где в ходе аэробной ферментации вариантный штамм продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода или поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма. Такие способы могут быть использованы для разрушения гена sfb3 любым способом, описанным выше и где-либо еще. Предпочтительно, разрушение гена sfb3 выполняют с помощью генетической манипуляции, используя последовательность-специфические техники молекулярной биологии, в противоположность химическому мутагенезу, который, как правило, не нацелен на специфические последовательности нуклеиновых кислот.
[125] В некоторых вариантах осуществления родительский штамм, в который вводят фенотип уменьшенной вязкости, уже включает ген, представляющий интерес, предназначенный, чтобы экспрессироваться на высоких уровнях. Таким образом, настоящие способы избегают необходимости вводить ген, представляющий интерес, в предварительно существующий штамм с уменьшенной вязкостью для продукции. Таким образом, настоящие способы могут быть использованы для получения вариантного штамма клеток мицелиальных грибов с уменьшенной вязкостью из родительского штамма, уже включающего ген, представляющий интерес.
[126] В другом аспекте также обеспечивается способ для скрининга клеток мицелиальных грибов в отношении фенотипа измененной вязкости. Способ включает скрининг секции клеток мицелиальных грибов (например, подвергнутые мутагенезу клетки или полевые изоляты) в отношении измененной чувствительности к флуорохромному красителю, где измененная чувствительность к флуорохромному красителю показывает, что вариантные клетки продуцируют, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который требует больше или меньше перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода, и/или поддерживает повышенное или пониженное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма. Таким образом, чувствительность к флуорохромному красителю может быть использована для определения вариантных клеток мицелиальных грибов, которые имеют фенотип измененной вязкости. В некоторых случаях, способ включает скрининг секции клеток мицелиальных грибов (например, подвергнутые мутагенезу клетки или полевые изоляты) в отношении увеличенной чувствительности к флуорохромному красителю, где увеличенная чувствительность к флуорохромному красителю показывает, что вариантные клетки продуцируют, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода, и/или поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма. Таким образом, чувствительность к флуорохромному красителю может быть использована для определения вариантных клеток мицелиальных грибов, которые имеют фенотип уменьшенной вязкости.
[127] Иллюстративные флуорохромы связываются с целлюлозой и/или хитином в клеточных стенках мицелиальных грибов и включают, но без ограничения, калькофлуор белый (CAS № 4193-55-9), конго красный (CAS № 573-58-0), солофенил флавин (CAS № 61725-08-4), понтамин прочный алый (CAS № 79770-29-9) и примулин (CAS № 30113-37-2).
[128] Конкретная генетическая техника, используемая для разрушения гена sfb3 в родительском штамме мицелиального гриба, как правило, не является критической для способа, при условии, что техника нацелена на ген sfb3 последовательность-специфическим образом. Иллюстративными способами являются сайт-специфическая рекомбинация, нацеленная вставка гена, применение подвижных элементов, трансдукция с помощью вирусов и применение РНК-опосредованного генного сайленсинга (Raponi M. and Arndt, G.M. (2003) Nucleic Acids Research 31:4481-89l Nakayashiki H. and Nguyen, Q.B. (2008) Current Opinion in Microbiology 11:494-502; Kuck, U. and Hoff, B. (2010) Applied and Environmental Biotechnology 86:51-62).
[129] При необходимости, разрушение гена sfb3 может быть осуществлено с помощью одновременной или последовательной вставки, например, селектируемого маркера, флуоресцентного или другого различимого маркера, сайта клонирования или кассеты клонирования, «отпечатков пальцев» последовательности, чтобы обеспечить последовательное определение штамма или другой генетической модификации, чтобы добавить отличительную особенность или функциональность штамму. В некоторых случаях, может быть необходимым ввести ген, представляющий интерес, предназначенный для высокого уровня экспрессии, в штамм с уменьшенной вязкостью в сайт разрушения гена sfb3. В таких случаях введение фенотипа уменьшенной вязкости и введение гена, представляющего интерес, может быть проведено одновременно.
VII. Применимость
[130] Применение штаммов мицелиальных грибов с уменьшенной вязкостью известно для улучшения распределения кислорода и питательных веществ в погруженной культуре, снижения количества мощности, необходимой для перемешивания погруженной культуры, и увеличения клеточной массы, присутствующей в культуре, что приводит к увеличенной продукции белка. Тем не менее, настоящие вариантные штаммы мицелиального гриба предлагают значительные преимущества сверх описанных выше штаммов с уменьшенной вязкостью.
[131] Во-первых, настоящие штаммы могут иметь полностью определенный геном, что делает их хорошо подходящими для последующей генетической манипуляции, комплементации, скрещивания и подобного. Во-вторых, настоящие штаммы не являются неблагоприятно воздействующими на продукцию секретируемого белка. В-третьих, штаммы с уменьшенной вязкостью могут быть получены, по сути, из любого родительского штамма, включая родительские штаммы, которые уже продуцируют белок, предназначенный для высокого уровня экспрессии (т.е. белок, представляющий интерес), уже кодируют селектируемый маркер или уже включают другие признаки, которые являются желательными для продуцирующего хозяина. Таким образом, настоящий штамм и способы устраняют необходимость в переносе гена, кодирующего белок, представляющий интерес, в ранее существующий продуцирующий штамм с уменьшенной вязкостью.
[132] Настоящие штаммы и способы находят применение в продукции коммерчески важного белка в погруженных культурах мицелиальных грибов. Коммерчески важные белки включают, например, целлюлазы, ксиланазы, пектиназы, лиазы, пектиназы, протеазы, амилазы, пуллуланазы, липазы, эстеразы, пергидролазы, трансферазы, лакказы, каталазы, оксидазы, редуктазы, гидрофобин и другие ферменты и неферментные белки, способные экспрессироваться в мицелиальных грибах. Такие белки могут быть для промышленного или фармацевтического применения.
[133] Эти и другие аспекты и варианты осуществления настоящих штаммов и способов будут очевидными для специалиста в данной области техники с учетом настоящего описания. Следующие примеры предусмотрены, чтобы дополнительно проиллюстрировать, но без ограничения, штаммы и способы.
ПРИМЕРЫ
[134] Для способствования прочтению следующих примеров, общие названия, номера коллекции штаммов Genencor (GICC#) и выбранные признаки исходных штаммов мицелиальных грибов перечислены в Таблице 1. Такая же информация для созданных штаммов мицелиальных грибов перечислена в Таблице 2. Праймеры нуклеиновых кислот, используемые в примерах, перечислены в Таблице 3. Последовательности малыми прописными буквами представляют собой нуклеотиды, добавленные для обеспечения прямой обработки ПЦР-амплифицированного фрагмента. Последовательности курсивом представляют собой сайты распознавания рестрикционными ферментами. Последовательности жирным шрифтом представляют собой сайты loxP. Подчеркнутые последовательности CACC добавляли к соответственным праймерам, чтобы обеспечить включение амплифицированного ДНК-фрагмента в исходный вектор GATEWAY.
Предварительно существующие штаммы Trichoderma reesei, используемые в примерах
Штаммы Trichoderma reesei, созданные в этом исследовании
Последовательности праймеров, используемых в примерах
[135] Среды и другие маточные растворы, используемые в примерах, описаны ниже:
[136] МИНИМАЛЬНАЯ СРЕДА ФОГЕЛЯ (VM) 1 л
[137] 50x РАСТВОР ФОГЕЛЯ
[138] РАСТВОР МИКРОЭЛЕМЕНТОВ ФОГЕЛЯ
[139] РАСТВОР БИОТИНА ФОГЕЛЯ 1 л
[140] VM СРЕДА С ГИГРОМИЦИНОМ (VMH) 1 л
[141] VM СРЕДА С СОРБИТОЛОМ И ГИГРОМИЦИНОМ (VMSH) 1 л
[142] КАРТОФЕЛЬНЫЙ АГАР С ДЕКСТРОЗОЙ (PDA) 1 л
[143] СРЕДА С КОНГО КРАСНЫМ (CR) 1 л
[144] 1% МАТОЧНЫЙ РАСТВОР КОНГО КРАСНОГО 25 мл
[145] СРЕДА С YEG 1 л
[146] МИНИМАЛЬНАЯ СРЕДА С ГЛИЦИНОМ И T. reesei 1 л
[147] МИКРОЭЛЕМЕНТЫ T. reesei 400x 1 л
Пример 1. Делеция sfb3 из штаммов Morph и 29-9 T. reesei для получения фенотипа уменьшенной вязкости
1.1. Создание кассеты делеции Δsfb3
[148] Последовательности ДНК, фланкирующие 5΄- конец гена sfb3 из T. reesei, амплифицировали с праймерной парой AVG88/AVG89. Амплификация фрагмента с этой праймерной парой вводила сайт SalI на 5΄- конец фрагмента и сайт BglII на 3΄- конец фрагмента. Кассету устойчивости к гигромицину Б, фланкированную параллельными сайтами loxP, амплифицировали из плазмиды pCR-Blunt II-hph- (Фигура 1) с праймерной парой AVG90/AVG91, вводя сайт BglII на 5΄- конец фрагмента и сайт NotI на 3’- конец фрагмента. Последовательность ДНК, фланкирующую 3΄- конец sfb3, амплифицировали с праймерной парой AVG92/AVG93, вводя сайт NotI на 5΄- конец и сайт XbaI на 3΄- конец фрагмента.
[149] Три вышеупомянутые фрагмента последовательно лигировали в вектор pCR®-Blunt II-TOPO® (Invitrogen Corp., Карлсбад, Калифорния, США), и полученную плазмиду называли pCR-Blunt II-TOPO 889092 (Фигура 2). Последовательности ДНК на 5΄- и 3΄-флангах гена sfb3 амплифицировали, используя геномную ДНК Morph TrglaA (29-9) в качестве матрицы. Кассета делеции ∆sfb3, содержащая 5΄-фланг sfb3, кассету устойчивости к гигромицину Б, окруженную сайтами loxP на каждом конце, и 3΄-фланг sfb3, амплифицировали из плазмиды pCRBluntII-TOPO 889092 с праймерной парой AVG104/AVG105. Множественные ПЦР-реакции объединяли, очищали, используя набор для очистки ПЦР, и осаждали этанолом для концентрирования амплифицированного фрагмента ДНК. Полученную таким образом ДНК использовали на последующих этапах.
1.2. Создание штаммов 29-9 Δsfb3 и Morph Δsfb3, в которых отсутствует ген sfb3
[150] Штаммы Morph и 29-9 трансформировали кассетой делеции Δsfb3 с помощью PEG-опосредованной трансформации, и помещали на планшеты с минимальной средой Фогеля, содержащей гигромицин Б и сорбитол. Трансформация Trichoderma описана, например, в патенте США № 5246853. Кандидатов (684 для 29-9+Δsfb3 и 348 для Morph+Δsfb3) переносили на минимальную среду Фогеля, содержащую гигромицин Б для отбора кандидатов в отношении устойчивых к гигромицину Б. Устойчивые к гигромицину B трансформанты переносили на минимальную среду Фогеля или PDA, содержащую конго красный, для оценки чувствительности к конго красному. Анализ ПЦР выявил один чувствительный к конго красному кандидат из каждой трансформации, в котором кассета делеции Δsfb3 интегрирована на локусе sfb3 с помощью гомологичной рекомбинации (Фигура 3). Гомологичная интеграция кассеты делеции Δsfb3 на локусе sfb3, как в 29-9 Δsfb3, так и Morph Δsfb3 была верифицирована с помощью амплификации фрагментов ДНК предполагаемого размера, используя праймерные пары AVG108/AVG109, AVG110/AVG111 и AVG108/AVG111. Праймерная пара AVG108/AVG109 амплифицирует фрагмент ДНК, который начинается за пределами 5΄-конца участка AVG88/AVG89 кассеты делеции и заканчивается в кассете устойчивости к гигромицину Б. Праймерная пара AVG110/AVG111 амплифицирует фрагмент ДНК, который начинается в кассете устойчивости к гигромицину Б и заканчивается за пределами 3΄-конца участка AVG92/AVG93 кассеты делеции. Праймерная пара AVG108/AVG110 амплифицирует целую кассету делеции, интегрированную в локус sfb3. Созданные штаммы с подтвержденной гомологичной интеграцией кассеты делеции называли 29-9 Δsfb3 и Morph Δsfb3, соответственно.
1.3. Рост штаммов 29-9, 70H2 и 29-9 Δsfb3 в погруженной культуре
[151] Штаммы 29-9, 70H2 и 29-9 Δsfb3 выращивали в идентичных условиях в погруженной (жидкой) культуре, и сравнивали их фенотипы роста.
[152] Кратко, споры каждого штамма добавляли отдельно к 500 мл среды в 3-литровую колбу, как с боковой, так и нижней перегородками. Среду, содержащую 5 г/л (NH4)2SO4, 4,5 г/л KH2PO4, 1 г/л MgSO4∙7H2O и 14,4 г/л лимонной кислоты, доводили до pH 5,5 с помощью 5% NaOH. После автоклавирования в течение 30 минут, стерильную 60% глюкозу добавляли до конечной концентрации 27,5 г/л, наряду с 2,5 мл/л раствора микроэлементов, содержащим 175 г/л лимонной кислоты, 200 г/л FeSO4∙7H2O, 16 г/л ZnSO4∙7H2O, 3,2 г/л CuSO4∙5H2O, 1,4 г/л MnSO4∙H2O и 0,8 г/л H3BO3. Культуру выращивали в течение 48 часов при 34°C во встряхивающем инкубаторе.
[153] Через 48 часов содержание каждой колбы добавляли отдельно к 15-литровым ферментерам, содержащим 9,5 л среды, содержащей 4,7 г/л KH2PO4, 1,0 г/л MgSO4∙7H2O, 4,3 г/л (NH4)2SO4 и 2,5 мл/л такого же раствора микроэлементов. Эти компоненты стерилизовали вместе нагреванием при 121°C в течение 30 минут. Раствор 60% глюкозы и 0,48% CaCl2∙2H2O отдельно автоклавировали, охлаждали и добавляли к ферментеру до конечной концентрации 75 г/л глюкозы и 0,6 г/л CaCl2∙2H2O. Среду доводили до pH 3,5 с помощью 28% NH3 и температуру поддерживали при 34°C в течение всего периода роста.
[154] Датчик растворенного кислорода (DO) калибровали до 100%, когда не было добавочного давления в свободном пространстве над жидкостью (т.е. 0 бар избыточного давления, 1 бар абсолютного давления). Давление в свободном пространстве над жидкостью затем устанавливали на 0,7 бар (избыточного давления), после чего датчик кислорода давал показание 170% перед добавлением посевной культуры. Ферментер содержал двух-, четырехлопастные турбины, которые обеспечивали перемешивание посредством двигателя с изменяющейся скоростью, который изначальной был установлен на 500 об./мин.
[155] Так как культуры росли, уровни DO падали, по меньшей мере частично, как следствие увеличенной вязкости бульона из-за пролиферации гиф мицелиальных грибов. Когда DO падал ниже 40%, то скорость перемешивания увеличивали для поддержания растворенного кислорода на уровне 40%. Если DO не падал ниже 40%, то не было необходимости увеличивать скорость перемешивания в ходе ферментации, и начальная скорость перемешивания была выше, чем необходимо. Когда глюкоза полностью потреблялась, количество биомассы, полученной в каждом ферментере измеряли, и обнаруживали, что оно, по сути, одинаковое для всех трех штаммов.
[156] Уровень DO в каждом ферментере на установленном уровне перемешиваний и количество перемешиваний, необходимое для поддержания данного уровня DO являются непрямыми измерениями вязкости различных бульонов, вследствие различных фенотипов роста штаммов. Хотя было бы оптимальным изменять только одну переменную (т.е. DO или перемешивание) и измерять другие, необходимо препятствовать падению DO ниже 40% для обеспечения продукции достаточной биомассы в каждом ферментере, тем самым позволяя провести более полноценное сравнение между ростом различных штаммов.
[157] Как правило, где необходимо увеличить скорость перемешивания для поддержания целевого уровня DO, количество перемешиваний может оцениваться с помощью количества мощности, подаваемой к двигателю, управляющего движением турбины ферментера, что обеспечивает показатель, который коррелирует с вязкостью бульона.
[158] В частности, дополнительная мощность, необходимая для перемешивания суспендированной культуры, является пропорциональной скорости перемешивания, повышенной до 3-ей мощности. Таблица 4 показывает самую высокую скорость перемешивания, необходимую для поддержания растворенного кислорода на уровне 40% в конце фазы роста.
Скорость перемешивания, необходимая для поддержания DO на уровне 40% в конце фазы роста
[159] При этих условиях роста исходному штамму, 29-9, необходимо в 2,6 раза больше мощности, чем штаммам, или 70H2, или 29-9 Δsfb3 для того, чтобы поддержать DO на уровне 40% и давать количество биомассы. Штаммы 70H2 и 29-9 Δsfb3 имели подобные свойства вязкости, и полученные подобные уровни белка, представляющего интерес, (TrGA) в суспендированной культуре, демонстрируя, что фенотип роста с уменьшенной вязкостью может быть обеспечен мицелиальному грибу путем разрушения гена sfb3.
1.4. Устранение loxP-фланкированной кассеты устойчивости к гигромицину Б из Morph Δsfb3
[160] loxP-фланкированную кассету устойчивости к гигромицину Б устраняли из штамма Morph Δsfb3 путем транзиторной экспрессии гена cre, который вызывал рекомбинацию между loxP сайтами. Штамм Morph Δsfb3 трансформировали с cre-содержащей теломерной плазмидой pTrex-Tel-pyrG13/pDONR221/0927853cre (Фигура 4), чтобы индуцировать рекомбинацию между сайтами loxP и устранить кассету устойчивости к гигромицину Б. Этот вектор содержит маркерную кассету amdS, обеспечивающую рост на среде, содержащей ацетамид. Трансформанты переносили один раз на среду, содержащую ацетамид, с последующими тремя переносами на среду PDA, чтобы обеспечить потерю cre-содержащей теломерной плазмиды. Трансформанты затем переносили параллельно на среду Фогеля, содержащую гигромицин Б, для оценки того, была ли утрачена кассета гигромицина Б, и среду, содержащую ацетамид, для оценки того, была ли утрачена cre-содержащая плазмида (Фигура 5; PDA=картофельный агар с декстрозой, VMH = минимальная среда Фогеля с гигромицином Б, amdS = минимальная среда с ацетамидом, hph-=чувствительный к гигромицину Б, cre- = чувствительный к присутствию ацетамида в среде). 91,8% трансформантов утратили кассету гигромицина Б и 92,5% трансформантов утратили cre-содержащую плазмиду. Это показывает, что возможно устранить кассету гигромицина Б из интегрированной кассеты делеции Δsfb3.
1.5. Экспрессия гена, представляющего интерес, не нарушается путем разрушения гена sfb3
[161] Кассета экспрессии, кодирующая фермент глюкоамилазу, служила в качестве иллюстративного гена, представляющего интерес, и обеспечивала удобный способ измерения количества белка, секретируемого вариантными штаммами мицелиального гриба. Кассету экспрессии глюкоамилазы (т.е. кассету экспрессии TrglaA), содержащую промотор cbhI, ген глюкоамилазы TrglaA из T. reesei и терминатор cbhI, слитую с маркерной кассетой amdS, ПЦР-амплифицировали с плазмидой pNSP23 (Фигура 6), используя праймерную пару SK745/SK746. Множественные ПЦР-реакции объединяли, очищали, используя набор для очистки ПЦР, и осаждали этанолом для концентрирования амплифицированного фрагмента ДНК. Штамм Morph Δsfb3 трансформировали с кассетой экспрессии TrglaA с помощью PEG-опосредованной трансформации и затем помещали на планшеты с минимальной средой, содержащей ацетамид с сорбитолом. Стабильных кандидатов выбирали на минимальной среде, содержащей ацетамид, и переносили на индукционную среду в формате микротитровального планшета. Штаммы выращивали в микротитровальных планшетах и активность глюкоамилазы в супернатанте исследовали, используя PNPG в качестве субстрата.
[162] Лучшие кандидаты Morph Δsfb3+TrglaA имели глюкоамилазную активность, которая была выше, чем у штамма 29-9 (который также включает кассету экспрессии TrglaA). Глюкоамилазную активность супернатанта основного кандидата верифицировали после роста во встряхиваемых колбах, и подтверждали результаты, полученные в микротитровальных планшетах. Результаты показывают, что делеция sfb3 не нарушает экспрессию или секрецию белка, представляющего интерес.
Пример 2. Комплементация штамма 70H2 с геном sfb3 дикого типа
2.1. Создание конструктов, содержащих ген sfb3
[163] Создавали четыре конструкта, содержащих ген sfb3. Праймерную пару AVG82/AVG83 использовали для амплификации гена sfb3 дикого типа с его нативным промотором и терминатором из геномной ДНК из 29-9 и мутированного гена sfb3 с его нативным промотором и терминатором из геномной ДНК из 70H2. Праймерную пару AVG84/AVG85 использовали для амплификации гена sfb3 дикого типа от старт-кодона до стоп-кодона, используя геномную ДНК из 29-9 и мутированного гена sfb3 от старт-кодона до стоп-кодона, используя геномную ДНК из 70H2. Результатом были четыре ПЦР-амплифицированных фрагмента, содержащих ген sfb3 дикого типа или мутированный ген sfb3, с или без нативного промотора sfb3 и нативного терминатора sfb3. Каждый из этих четырех фрагментов независимо клонировали в pENTR/D-TOPO вектор (Invitrogen, Карлсбад, Калифорния, США).
[164] На следующем этапе четыре pENTR/D-TOPO конструкта переносили на вектор доставки pTrex2g/HygB, используя реакцию LR клоназы. Каждый из четырех конструктов амплифицировали в E. coli, и полученные минипрепараты для каждого конструкта концентрировали в вакууме. ДНК, полученную этим способом, использовали в последующем этапе. Иллюстративный конструкт доставки показан на Фигуре 7.
2.2. Комплементация 70H2 фенотипа с геном sfb3
[165] Каждый из четырех конструктов вектора доставки отдельно трансформировали в 70H2 с помощью PEG-опосредованной трансформации, с последующим помещением на планшеты с минимальной средой Фогеля, с гигромицином Б. Тридцать кандидатов из каждой из четырех трансформаций переносили на PDA с гигромицином Б, и их фенотип сравнивали с 70H2 и 29-9, трансформированными вектором pTrex2g/HygB отдельно (в качестве контролей). Все кандидаты, трансформированные генами sfb3 дикого типа, имели фенотип дикого типа, подобный 29-9 на среде PDA с гигромицином Б. Все кандидаты, трансформированные мутированным геном sfb3, полученным из 70H2, сохраняли фенотип 70H2 (Фигура 8). Эти результаты показали, что ген sfb3 дикого типа, полученный из 29-9, но не мутированный ген sfb3, полученный из 70H2, мог придавать фенотип дикого типа штамму 70H2.
[166] Для подтверждения того, что реверсия фенотипа была вызвана присутствием гена sfb3 дикого типа, а не изменением в хромосомной ДНК, кандидатов переносили четыре раза на среду PDA (неселективные условия), чтобы найти кандидатов, которые утратили вектор, и затем возвращали на селективную среду с гигромицином Б. Для всех кандидатов, которые были нестабильными и утратили плазмиду (на основании потери устойчивости к гигромицину Б), потеря плазмиды коррелировала с повторным возникновением фенотипа 70H2. Эти результаты подтверждают, что sfb3 дикого типа отвечал за восстановление фенотипа дикого типа у 70H2.
2.3. Создание кассеты для замены гена sfb3, содержащей ген sfb3 дикого типа
[167] Последовательность ДНК, содержащую приблизительно 2/3 3′- конца гена sfb3 дикого типа, амплифицировали с праймерной парой AVG94 /AVG95, что ввело сайт SpeI на 5′- конец фрагмента и сайт BglII на 3′- конец фрагмента. Кассету устойчивости к гигромицину Б, фланкированную с помощью сайтов loxP, амплифицировали из плазмиды pCR-Blunt II-hph-loxP#4 (Фигура 1) с праймерной парой AVG90/AVG91, что ввело сайт BglII на 5′- конец фрагмента и сайт NotI на 3′- конец фрагмента. Последовательность ДНК, содержащую 3′- ДНК- последовательность ниже терминаторного участка sfb3, амплифицировали праймерной парой AVG96/AVG97, что ввело сайт NotI на 5′- конец фрагмента и сайт XbaI на 3′- конец фрагмента.
[168] Эти три фрагмента последовательно лигировали в вектор pCR®-Blunt II-TOPO® (Invitrogen Corp., Карлсбад, Калифорния, США), и полученную плазмиду называли pCR-Blunt II-TOPO 949096 (Фигура 9). Две последовательности ДНК, окружающие кассету устойчивости к гигромицину Б, амплифицировали, используя геномную ДНК из 29-9 в качестве матрицы. Кассету для замены гена sfb3, содержащую часть гена sfb3, кассету устойчивости к гигромицину Б, окруженную сайтами loxP на каждом конце, и 3′-последовательность ниже sfb3, амплифицировали из плазмиды pCR-Blunt II-TOPO 889092 с праймерной парой AVG102/AVG103. Множественные ПЦР-реакции объединяли, очищали, используя набор для очистки ПЦР, и осаждали этанолом для концентрирования амплифицированного фрагмента ДНК. ДНК, полученную таким путем, использовали на последующем этапе.
2.4. Комплементация фенотипа 70H2 путем экспрессии гена sfb3 дикого типа из нативного локуса sfb3
[169] Штамм 70H2 трансформировали кассетой для замены гена sfb3 (описанной в разделе 2.3) с помощью PEG-осопредованной трансформации с последующим помещением на планшеты с минимальной средой Фогеля, содержащей гигромицин Б и сорбитол. Пятнадцать кандидатов, которые имели фенотип колонии дикого типа, переносили на минимальную среду Фогеля, содержащую гигромицин Б, чтобы выбрать кандидатов, устойчивых к гигромицину Б с фенотипом колонии дикого типа. Приемлемые кандидаты, устойчивые к гигромицину Б, были выбраны на минимальной среде, содержащей гигромицин Б и оценены в отношении фенотипа колонии дикого типа (Фигура 10).
[170] Гомологичную интеграцию кассеты для замены гена sfb3 в локусе sfb3 70H2 верифицировали путем амплификации фрагментов ДНК предполагаемого размера, используя праймерные пары AVG112/AVG113 и AVG114/AVG115. Праймерная пара AVG112/AVG113 амплифицировала фрагмент ДНК, который начинается за пределами 5′-конца участка AVG94/AVG95 кассеты для замены гена sfb3 и заканчивается в кассете устойчивости к гигромицину Б. Праймерная пара AVG114/AVG115 амплифицировала фрагмент ДНК, который начинается в кассете устойчивости к гигромицину Б и заканчивается за пределами 3′- конца участка AVG96/AVG97 кассеты для замены гена sfb3.
[171] Созданный штамм с подтвержденной гомологичной интеграцией кассеты для замены гена sfb3 называли 70H2+sfb3 дикого типа. Этот штамм имел фенотип, подобный 29-9. Таким образом, замена мутированного гена sfb3 в 70H2 на ген sfb3 дикого типа в нативном локусе sfb3 восстанавливала фенотип дикого типа у 70H2, обеспечивая дополнительное доказательство того, что разрушение гена sfb3 отвечает за фенотип уменьшенной вязкости у 70H2.
Пример 3. Рост штаммов 29-9, 70H2 и 29-9 Δsfb3 в погруженной культуре
[172] Штаммы 29-9, 70H2 и 29-9 Δsfb3 выращивали при идентичных условиях в погруженной (жидкой) культуре, и сравнивали их фенотипы роста. Кратко, споры каждого штамм добавляли отдельно к 500 мл среды в 3-литровую колбу, как с боковой, так и с нижней перегородками. Среду, содержащую 5 г/л (NH4)2SO4, 4,5 г/л KH2PO4, 1 г/л MgSO4∙7H2O и 14,4 г/л лимонной кислоты, доводили до pH 5,5 с помощью 5% NaOH. После автоклавирования в течение 30 минут стерильную 60% глюкозу добавляли до конечной концентрации 27,5 г/л, вместе с 2,5 мл/л раствора микроэлементов, содержащего 175 г/л лимонной кислоты, 200 г/л FeSO4∙7H2O, 16 г/л ZnSO4∙7H2O, 3,2 г/л CuSO4∙5H2O, 1,4 г/л MnSO4∙H2O и 0,8 г/л H3BO3. Культуру выращивали в течение 48 часов при 34°C во встряхивающем инкубаторе.
[173] Через 48 часов содержание каждой колбы добавляли отдельно к 15-литровым ферментерам, содержащим 9,5 л среды, содержащей 4,7 г/л KH2PO4, 1,0 г/л MgSO4∙7H2O, 4,3 г/л (NH4)2SO4 и 2,5 мл/л такого же раствора микроэлементов. Эти компоненты стерилизовали вместе нагреванием при 121°C в течение 30 минут. Раствор 60% глюкозы и 0,48% CaCl2∙2H2O отдельно автоклавировали, охлаждали и добавляли к ферментеру до конечной концентрации 75 г/л глюкозы и 0,6 г/л CaCl2∙2H2O. Среду доводили до pH 3,5 28% NH3 и температуру поддерживали при 34°C в течение всего периода роста.
[174] Датчик растворенного кислорода (DO) калибровали до 100%, когда не было добавочного давления в свободном пространстве над жидкостью (т.е. 0 бар избыточного давления, 1 бар абсолютного давления). Давление в свободном пространстве над жидкостью затем устанавливали на 0,7 бар (избыточного давления), после чего датчик кислорода давал показание 170% перед добавлением посевной культуры. Ферментер содержал двух-, четырехлопастные турбины, которые обеспечивали перемешивание посредством двигателя с изменяющейся скоростью, который изначально был установлен на 500 об./мин.
[175] Так как культуры росли, уровни DO падали, по меньшей мере частично, как следствие увеличенной вязкости бульона из-за пролиферации гиф мицелиальных грибов. Когда DO падал ниже 40%, то скорость перемешивания увеличивали для поддержания растворенного кислорода на уровне 40%. Если DO не падал ниже 40%, то не было необходимости увеличивать скорость перемешивания в ходе ферментации, и начальная скорость перемешивания была выше, чем необходимо. Когда глюкоза полностью потреблялась, количество биомассы, полученной в каждом ферментере, измеряли, и обнаруживали, что оно, по сути, одинаковое для всех трех штаммов.
[176] Уровень DO в каждом ферментере на установленном уровне перемешиваний и количество перемешиваний, необходимое для поддержания установленного уровня DO, являются непрямыми измерениями вязкости различных бульонов, вследствие различных фенотипов роста штаммов. Хотя было бы оптимальным изменять только одну переменную (т.е. DO или перемешивание) и измерять другие, необходимо препятствовать падению DO ниже 40% для обеспечения продукции достаточной биомассы в каждом ферментере, тем самым позволяя провести более полноценное сравнение между ростом различных штаммов.
[177] Как правило, где необходимо увеличить скорость перемешивания для поддержания целевого уровня DO, количество перемешиваний может оцениваться с помощью количества мощности, подаваемой к двигателю, управляющего движением турбины ферментера, что обеспечивает показатель, который коррелирует с вязкостью бульона.
[178] В частности, дополнительная мощность, необходимая для перемешивания суспендированной культуры, является пропорциональной скорости перемешивания, повышенной до 3-ей мощности. Таблица 4 показывает самую высокую скорость перемешивания, необходимую для поддержания растворенного кислорода на уровне 40% в конце фазы роста.
Скорость перемешивания, необходимая для поддержания DO на уровне 40% в конце фазы роста
[179] При этих условиях роста исходному штамму, 29-9, необходимо в 2,6 раза больше мощности, чем штамму, или 70H2, или 29-9 Δsfb3 для того, чтобы поддержать DO на уровне 40% и продуцировать количество биомассы. Важно, что штаммы 70H2 и 29-9 Δsfb3 имели подобные свойства вязкости в суспендированной культуре, демонстрируя, что фенотип роста с уменьшенной вязкостью может быть обеспечен мицелиальному грибу путем разрушения гена sfb3.
Пример 4. Создание штамма Δsfb3 H3A для получения композиции общей целлюлазы
4.1. Создание кассеты делеции ∆sfb3
[180] Последовательности, фланкирующие 5΄- конец гена sfb3 T. reesei, амплифицировали праймерной парой AVG88/AVG89. Амплификация фрагмента с этой праймерной парой вводила сайт SalI на 5΄- конец фрагмента и сайт BglII на 3΄- конец фрагмента. Кассету устойчивости к гигромицину Б, фланкированную параллельными сайтами loxP, амплифицировали из плазмиды pCR-Blunt II-hph-loxP#4 (Фигура 1) с праймерной парой AVG90/AVG91, вводя сайт BglII на 5΄- конец фрагмента и сайт NotI на 3΄- конец фрагмента. Последовательность ДНК, фланкирующую 3΄- конец sfb3, амплифицировали праймерной парой AVG92/AVG93, вводя сайт NotI на 5΄- конец и сайт XbaI на 3΄- конец фрагмента.
[181] Три вышеупомянутых фрагмента последовательно лигировали в вектор pCR®-Blunt II-TOPO® (Invitrogen Corp., Карлсбад, Калифорния, США), и полученную плазмиду называли pCR-Blunt II-TOPO 889092 (Фигура 2). Последовательности ДНК на 5΄- и 3΄-фланги гена sfb3 амплифицировали, используя геномную ДНК Morph TrglaA (29-9) в качестве матрицы. Кассету делеции ∆sfb3, содержащую 5΄-фланг sfb3, кассету устойчивости к гигромицину Б, окруженную сайтами loxP на каждом конце, и 3΄-фланг sfb3, амплифицировали из плазмиды pCRBluntII-TOPO 889092 с праймерной парой AVG104/AVG105. Множественные ПЦР-реакции объединяли, очищали, используя набор для очистки ПЦР, и осаждали этанолом для концентрирования амплифицированного фрагмента ДНК. ДНК, полученную таким образом, использовали на последующих этапах.
4.2. Создание штамма H3A ∆sfb3 #1009, в котором отсутствует ген sfb3
[182] Штамм экспрессии Trichoderma reesei с интегрированным H3A получали согласно описанию PCT/US2010/049849, опубликованной как WO/2011/038019. Штамм H3A трансформировали кассетой делеции ∆sfb3 с помощью PEG (полиэтиленгликоль)-опосредованной трансформации, и поместили на планшеты с минимальной средой Фогеля, содержащей гигромицин Б и сорбитол. PEG-опосредованная трансформация Trichoderma была ранее описана в, например, патенте США № 5246853.
[183] 1020 кандидатов переносили на планшеты с минимальной средой Фогеля, содержащие гигромицин Б для выбора в отношении устойчивости к гигромицину Б. Кандидаты, устойчивые к гигромицину Б, затем переносили на минимальную среду Фогеля или среду PDA, содержащую конго красный для оценки чувствительности к конго красному.
[184] Был проведен анализ ПЦР 43 стабильных кандидатов, показывающий слабую чувствительность к конго красному, и один кандидат, #1009, показал профиль, согласующийся с гомологичной интеграцией кассеты делеции sfb3 в геном H3A, в сочетании с устранением нативного гена sfb3. Гомологичную интеграцию кассеты делеции ∆sfb3 в локусе sfb3 в H3A ∆sfb3 #1009 верифицировали путем амплификации фрагментов ДНК предполагаемого размера, используя праймерные пары AVG108/AVG109; AVG110/AVG111; и AVG108/AVG111. Специфически, праймерная пара AVG108/AVG109 амплифицировала фрагмент ДНК, который начинается за пределами 5΄- конца участка AVG88/AVG89 кассеты делеции и заканчивается в кассете устойчивости к гигромицину Б. Праймерная пара AVG110/AVG111 амплифицировала фрагмент ДНК, который начинается в кассете устойчивости к гигромицину Б и заканчивается за пределами 3΄- конца участка AVG92/AVG93 кассеты делеции. Праймерная пара AVG108/AVG111 амплифицировала целую кассету делеции, интегрированную в локус sfb3. Отсутствие гена sfb3 подтверждали путем отсутствия ПЦР-продукта при использовании праймерной пары AVG160/AVG161, разработанной для амплификации внутреннего фрагмента sfb3.
[185] Когда сравнивали со штаммом H3A хозяина, то штамм H3A ∆sfb3 #1009 имел более ограниченную морфологию колонии на среде, содержащей конго красный, более медленную скорость роста и уменьшенное образование конидий на PDA, и был способен расти на среде, содержащей гигромицин Б (Фигура 11). Также сравнивали жидкие культуры штаммов H3A и H3A ∆sfb3 #1009. Культуры во встряхиваемых колбах обоих штаммов выращивали в 50 мл минимальной среды, содержащей YEG или глицин, при 28°C и 220 об./мин в течение 1 или 6 дней, соответственно. В обеих минимальных средах, содержащих, или YEG, или глицин, штамм H3A ∆sfb3 #1009, как правило, имел укороченные гифы (Фигура 12), подтверждая свойства улучшенной вязкости.
4.3. Эффективная продукция композиции общей целлюлазы в H3A ∆sfb3 #1009
[186] Два эксперимента по ферментации провели совместно, один с H3A, а другой с H3A ∆sfb3 #1009, при стандартных условиях ферментации, как описано, например, в PCT/US2010/049849. Скорость перемешивания 500 об./мин использовали в каждом ферментационном чане. Растворенный кислород измеряли в конце фазы роста, используя оптический датчик кислорода Hamilton (Hamilton Company, США, Рено, Невада). Сравнение уровней DO показано в Таблице 5, ниже:
DO после фазы роста
[187] Полученные композиции общей целлюлазы из каждого из экспериментов по ферментации характеризовали, используя способ HPLC, как описано, например, в PCT/US2010/049849. Количества основных целлюлаз/гемицеллюлаз, содержащихся в них, сравнивают рядом в Таблице 6, ниже:
Сравнение целлюлаз/гемицеллюлаз, присутствующих в композиции общей целлюлазы
[188] Обе композиции общей целлюлазы содержали, по сути, одинаковые количества каждого основного компонента, показывая, что экспрессия белка в H3A ∆sfb3 #1009 была подобна таковой у H3A.
[189] Хотя вышеупомянутые композиции и способы были описаны в некоторых деталях для иллюстрации и примеров для целей ясности понимания, специалистам в данной области техники станет очевидно, что могут быть произведены определенные изменения и модификации. Следовательно, описание не должно истолковываться как ограничивающее объем настоящего изобретения, который установлен приложенной формулой изобретения.
[190] Все публикации, патенты и патентные заявки, процитированные в данном документе, настоящим включены ссылкой в их полноте для всех целей и в той же мере, как если бы каждая отдельная публикация, патент или патентная заявка были конкретно и в индивидуальном порядке указаны, чтобы таким образом быть включенными путем ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА НИТЧАТЫХ ГРИБОВ С ДЕФИЦИТОМ ПРОТЕАЗ И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2013 |
|
RU2645252C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| НОВАЯ ЭНДО-(1-4)-β-D-КСИЛАНАЗА ИЗ PENICILLIUM CANESCENS (ВАРИАНТЫ), ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ СЕКРЕТИРУЕМУЮ ЭНДО-(1-4)-β-D-КСИЛАНАЗУ ИЗ PENICILLIUM CANESCENS (ВАРИАНТЫ), И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2009 |
|
RU2412246C2 |
| ГЕНЕТИЧЕСКИЕ ЛОКУСЫ, СВЯЗАННЫЕ С УСТОЙЧИВОСТЬЮ К ПЫЛЬНОЙ ГОЛОВНЕ У МАИСА | 2009 |
|
RU2562864C2 |
| РЕКОМБИНАНТНАЯ ЛАККАЗА ЛИГНИНОЛИТИЧЕСКОГО ГРИБА Trametes sp. И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2008 |
|
RU2394912C2 |
| КЛЕТКА МИЦЕЛИАЛЬНОГО ГРИБА Penicillium canescens - ПРОДУЦЕНТ КСИЛАНАЗЫ И ЛАККАЗЫ, СПОСОБ ПОЛУЧЕНИЯ КОМБИНИРОВАННОГО ФЕРМЕНТНОГО ПРЕПАРАТА КСИЛАНАЗЫ И ЛАККАЗЫ | 2012 |
|
RU2538149C2 |
| МИКРООРГАНИЗМ, СПОСОБНЫЙ ПРОДУЦИРОВАТЬ L- АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2600875C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| ПРИМЕНЕНИЕ АКТИВНОСТИ ЭНДОГЕННОЙ ДНКАЗЫ ДЛЯ ПОНИЖЕНИЯ СОДЕРЖАНИЯ ДНК | 2012 |
|
RU2642290C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ, СОДЕРЖАЩИЕ КЛАСТЕР ГЕНОВ БИОСИНТЕЗА ПИРИПИРОПЕНА И МАРКЕРНЫЙ ГЕН | 2011 |
|
RU2576001C2 |







































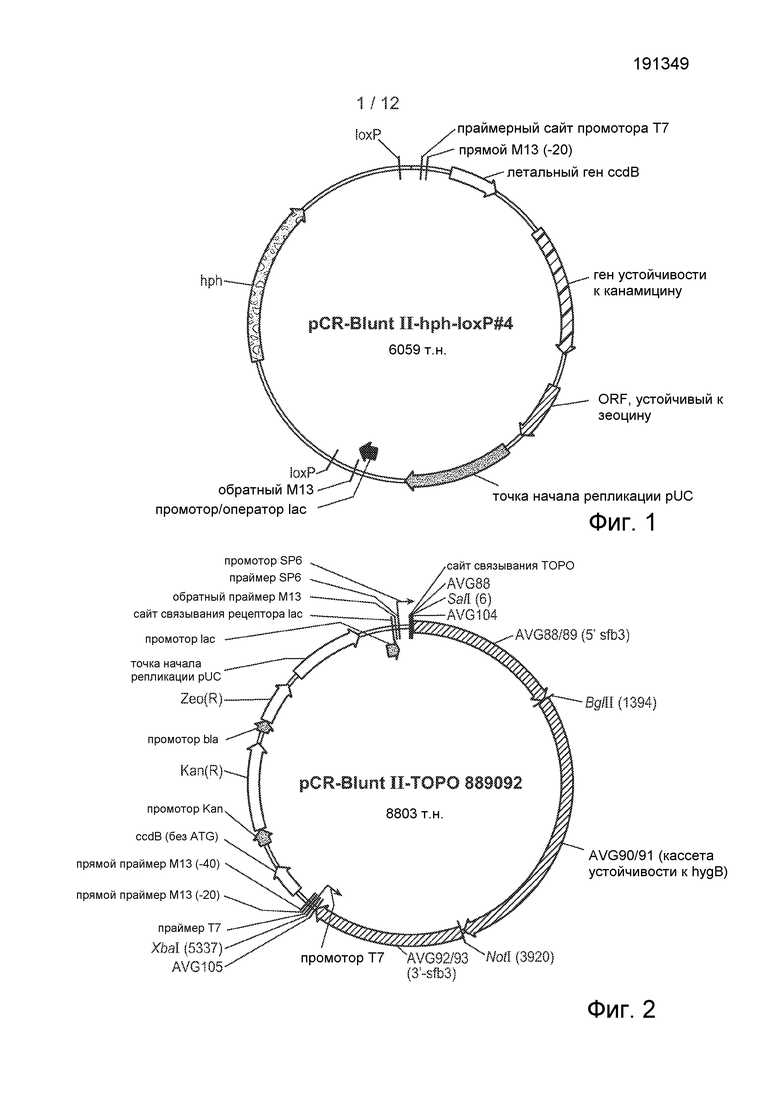
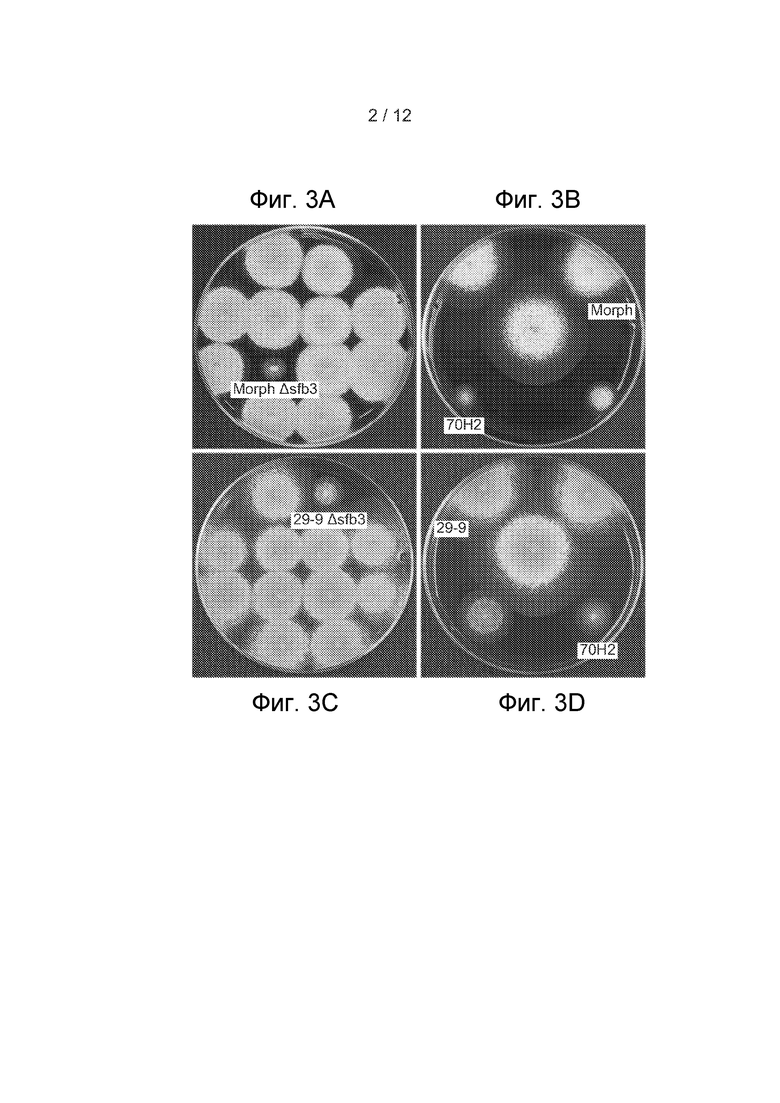
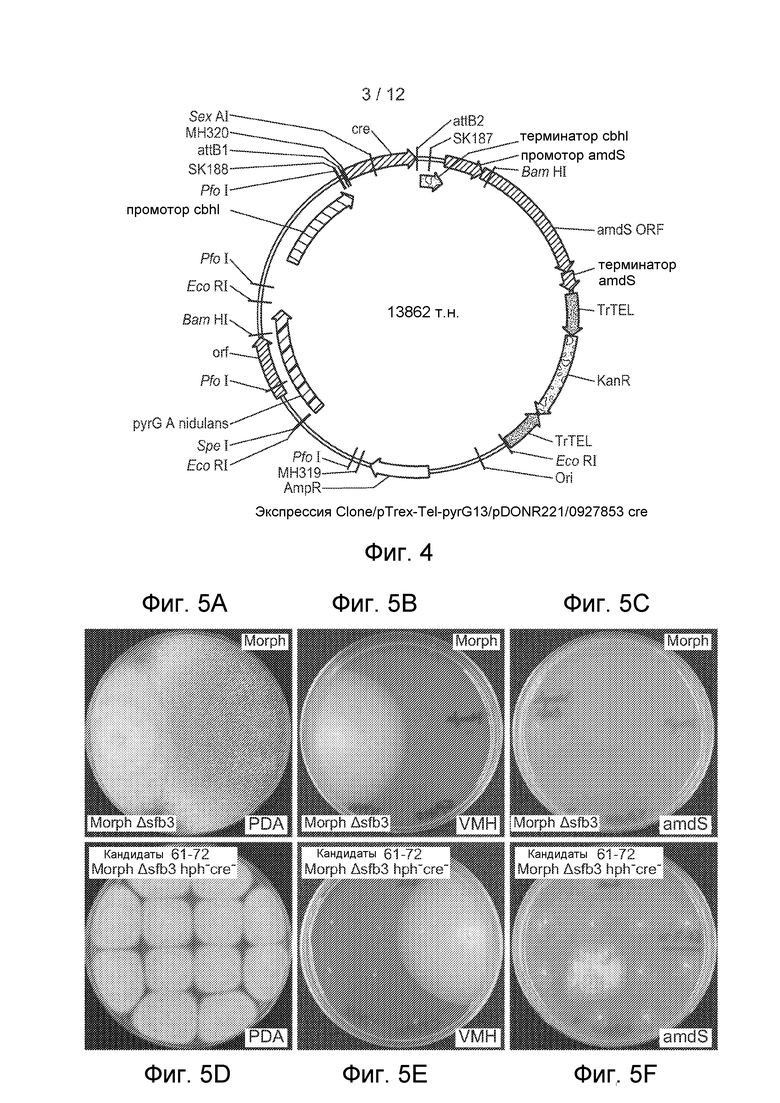
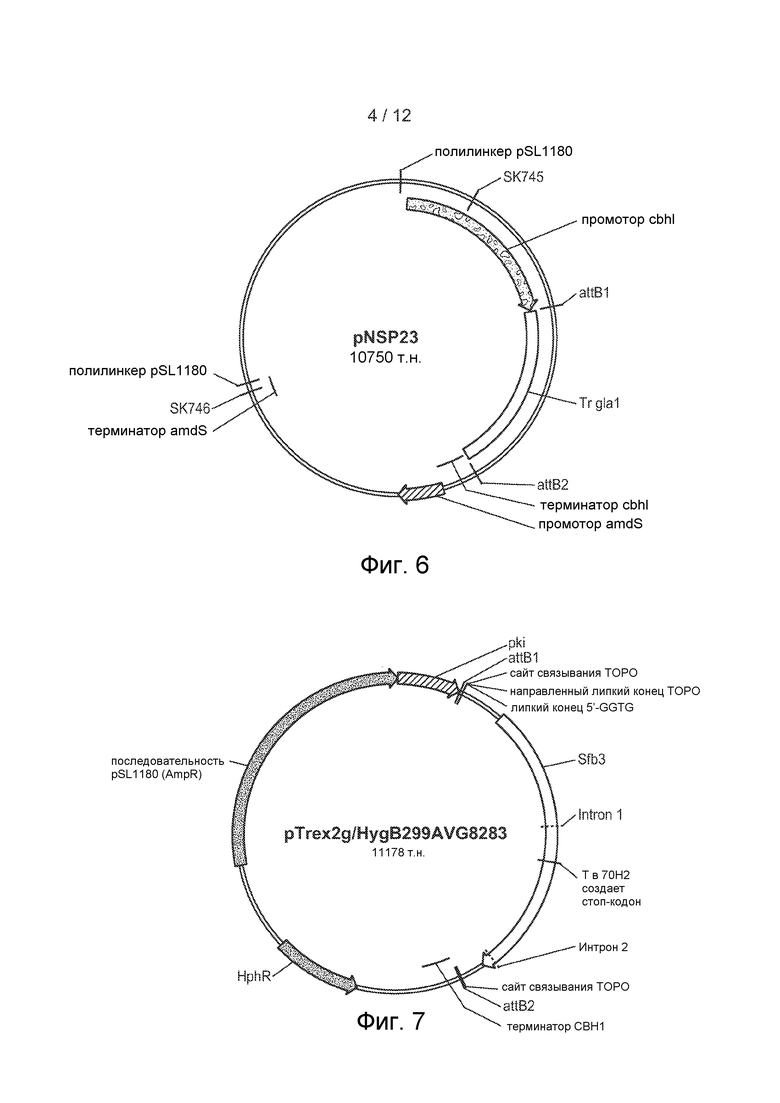
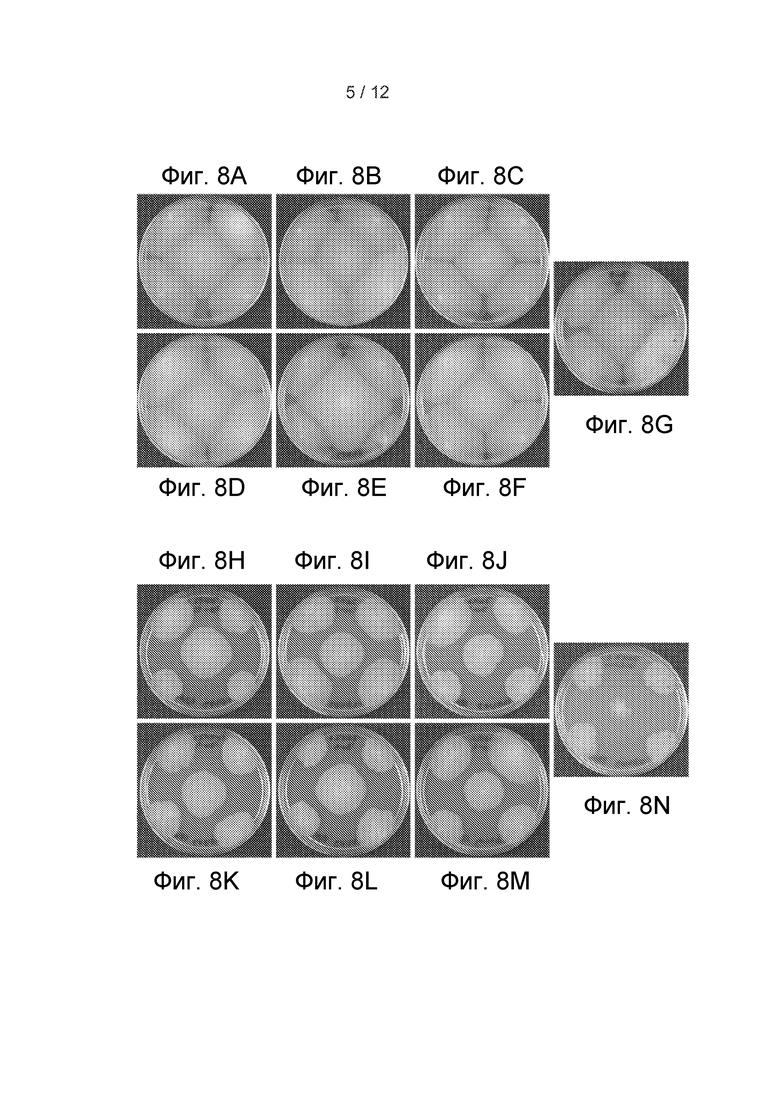
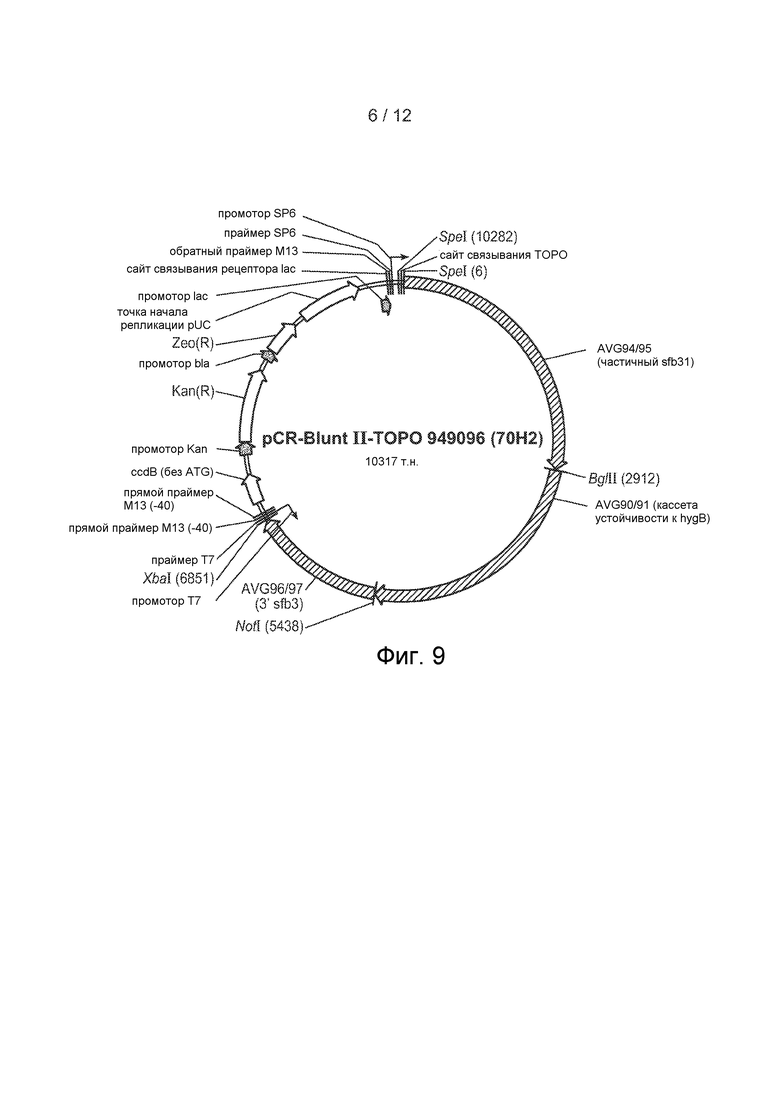
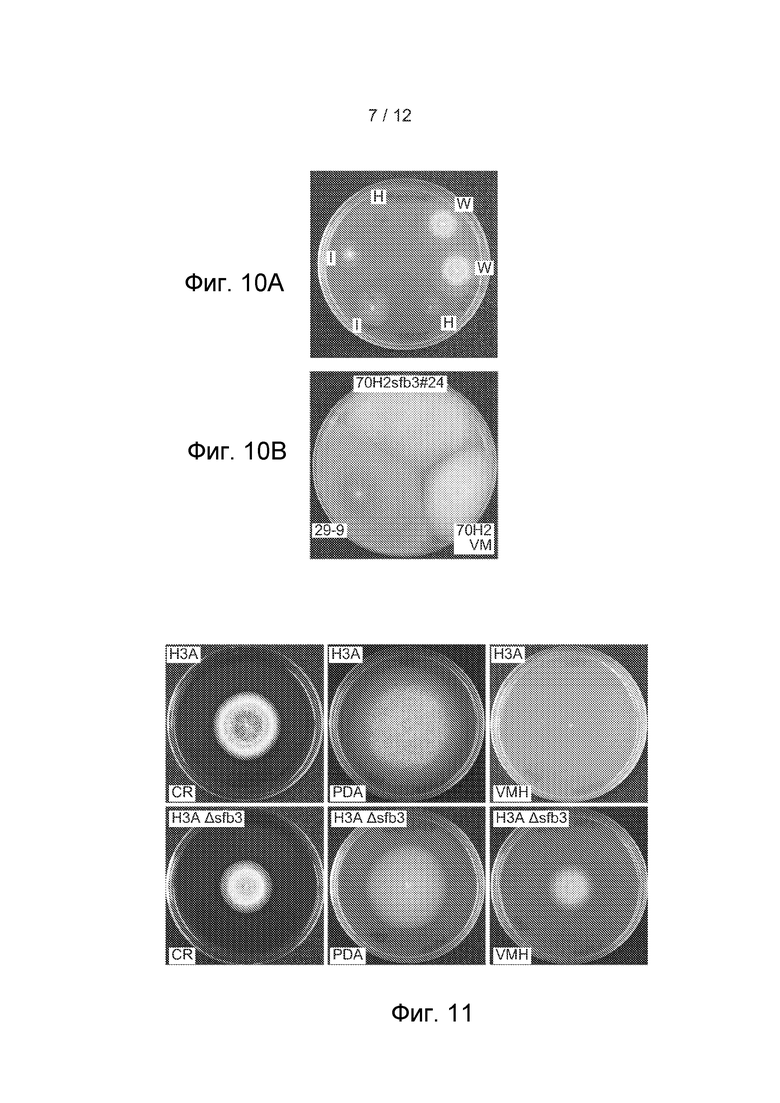





Изобретение относится к биотехнологии и представляет собой клетку-хозяина мицелиального гриба, полученную из родительской клетки, для получения белка, представляющего интерес, где клетка-хозяин включает генетическое изменение в гене Sfb3, которое приводит к продуцированию в клетке уменьшенного количества функционального белка Sfb3 по сравнению с родительскими клетками или его отсутствию, где клетка-хозяин продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует сниженного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с родительскими клетками. Изобретение позволяет уменьшить количество перемешиваний при культивировании клеток, тем самым упростить процесс получения белка, представляющего интерес. 3 н. и 24 з.п. ф-лы, 14 ил., 6 табл., 4 пр.
1. Клетка-хозяин мицелиального гриба, полученная из родительской клетки, для получения белка, представляющего интерес, где клетка-хозяин включает генетическое изменение в гене Sfb3, которое приводит к продуцированию в клетке уменьшенного количества функционального белка Sfb3 по сравнению с родительскими клетками или его отсутствию, где клетка-хозяин продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует сниженного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с родительскими клетками.
2. Клетка-хозяин по п. 1, где количество функционального белка Sfb3 представляет собой уменьшенное количество и клетка-хозяин продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует сниженного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает повышенное содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с клетками родительского штамма.
3. Клетка-хозяин по п. 1 или 2, где генетическое изменение включает разрушение гена sfb3, присутствующего в родительской клетке.
4. Клетка-хозяин по п. 3, где разрушение гена sfb3 является результатом делеции всего гена sfb3 или части гена sfb3.
5. Клетка-хозяин по п. 3, где разрушение гена sfb3 является результатом делеции части геномной ДНК, включающей ген sfb3.
6. Клетка-хозяин по п. 3, где разрушение гена sfb3 является результатом мутагенеза гена sfb3.
7. Клетка-хозяин по п. 3, где разрушение гена sfb3 выполняют, используя сайт-специфическую рекомбинацию.
8. Клетка-хозяин по п. 3, где разрушение гена sfb3 выполняют в комбинации с введением селектируемого маркера в генетический локус гена sfb3.
9. Клетка-хозяин по п. 3, где разрушение гена sfb3 является первичной генетической детерминантой для придания фенотипа уменьшенной вязкости клетке-хозяину.
10. Клетка-хозяин по п. 1 или 2, где клетка-хозяин не продуцирует функциональный белок Sfb3.
11. Клетка-хозяин по п. 1, где клетка-хозяин не продуцирует белок Sfb3.
12. Клетка-хозяин по п. 1 или 2, где клетка-хозяин включает дополнительный ген, кодирующий белок, представляющий интерес.
13. Клетка-хозяин по п. 1 или 2, где клетка-хозяин продуцирует, по сути, такое же количество белка на единицу количества биомассы, как родительская клетка.
14. Клетка-хозяин по п. 1 или 2, где белок Sfb3 включает аминокислотную последовательность IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток).
15. Клетка-хозяин по п. 1 или 2, где мицелиальный гриб представляет собой виды Pezizomycotina.
16. Клетка-хозяин по п. 1 или 2, где мицелиальный гриб представляет собой Trichoderma reesei.
17. Способ получения клетки-хозяина мицелиального гриба, включающий введение генетического изменения в ген Sfb3 родительской клетки мицелиального гриба, при этом генетическое изменение снижает или предотвращает продукцию функционального белка Sfb3 по сравнению с родительскими клетками, что приводит к получению клетки-хозяина мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует сниженного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с родительскими клетками.
18. Способ по п. 17, где генетическое изменение снижает продукцию функционального белка Sfb3, что приводит к получению клетки-хозяина мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, который (i) требует уменьшенного перемешивания для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с родительскими клетками.
19. Способ по п. 17 или 18, где генетическое изменение включает разрушение гена sfb3 в родительской клетке мицелиального гриба, используя генетическую манипуляцию.
20. Способ по п. 17 или 18, где генетическое изменение включает делецию гена sfb3 в родительской клетке мицелиального гриба, используя генетическую манипуляцию.
21. Способ по п. 17 или 18, где генетическое изменение выполняют, используя сайт-специфическую генетическую рекомбинацию.
22. Способ по п. 17 или 18, где разрушение гена sfb3 выполняют в комбинации с введением селектируемого маркера в генетический локус гена sfb3.
23. Способ по п. 17 или 18, где клетка-хозяин продуцирует, по сути, такое же количество белка на единицу количества биомассы, как родительская клетка.
24. Способ по п. 17 или 18, где белок Sfb3 включает аминокислотную последовательность IQLARQGXDGXEXXXARXLXEDRNXEAXSXVDWL (SEQ ID NO: 9, где X представляет собой любой аминокислотный остаток).
25. Способ по п. 17 или 18, где мицелиальный гриб представляет собой виды Pezizomycotina.
26. Способ по п. 17 или 18, где мицелиальный гриб представляет собой Trichoderma reesei.
27. Способ по п. 17 или 18, где родительская клетка включает дополнительный ген, кодирующий белок, представляющий интерес.
28. Способ по п. 27, где ген, кодирующий белок, представляющий интерес, присутствует в родительской клетке перед введением генетического изменения, которое снижает или предотвращает продукцию функционального белка Sfb3.
29. Способ получения белка, представляющего интерес, в клетке-хозяине мицелиального гриба, включающий введение в родительскую клетку мицелиального гриба гена, кодирующего белок, представляющий интерес, и генетического изменения в ген Sfb3, которое снижает количество или активность белка Sfb3 в клетках, тем самым получая клетку-хозяина мицелиального гриба, которая продуцирует, в ходе аэробной ферментации в погруженной культуре, клеточный бульон, включающий белок, представляющий интерес, который (i) требует сниженного количества перемешиваний для поддержания предварительно выбранного содержания растворенного кислорода по сравнению с родительскими клетками и (ii) поддерживает содержание растворенного кислорода при предварительно выбранном количестве перемешиваний по сравнению с родительскими клетками, и где белок, представляющий интерес, получают, по сути, на том же уровне в клетке-хозяине по сравнению с родительскими клетками.
| US 7504490 B1, 17.05.2009 | |||
| US 20080299619 A1, 04.12.2008 | |||
| ФРАГМЕНТ ДНК МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ ЭНДОГЛЮКАНАЗЫ III И ГРУППА ШТАММОВ PENICILLIUM CANESCENS, СИНТЕЗИРУЮЩИХ ЭНДОГЛЮКАНАЗУ III PENICILLIUM VERRUCULOSUM, СКОНСТРУИРОВАННЫХ МЕТОДАМИ ТРАНСФОРМАЦИИ И ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ НА ОСНОВЕ ЭТОГО ФРАГМЕНТА ДНК | 2001 |
|
RU2238974C2 |

