ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам, пригодным для продуцирования гетерологичных белков в клетках нитчатых грибов.
УРОВЕНЬ ТЕХНИКИ
Посттрансляционная модификация эукариотических белков, в частности, терапевтических белков, таких как иммуноглобулины, часто является необходимой для надлежащего сворачивания и функционирования белка. Поскольку стандартные прокариотические экспрессирующие системы лишены надлежащего аппарата, необходимого для таких модификаций, для получения этих терапевтических белков необходимо использовать альтернативные экспрессирующие системы. Даже когда эукариотические белки не имеют посттрансляционных модификаций, прокариотические экспрессирующие системы часто лишены необходимых белков-шаперонов, требуемых для надлежащего сворачивания. Дрожжи и грибы являются привлекательными возможностями для экспрессии белков, поскольку они могут без труда расти в большом масштабе в простых средах, что обеспечивает низкую стоимость продуцирования, и дрожжи и грибы имеют посттрансляционный аппарат и шапероны, которые выполняют сходные функции с теми, что и в клетках млекопитающих. Более того, доступны инструменты для манипуляции относительно простым генетическим набором клеток дрожжей и грибов, а также более сложных эукариотических клеток, таких как клетки млекопитающих или клетки насекомых (De Pourcq et al., Appl Microbiol Biotechnol, 87(5): 1617-31). Несмотря на эти преимущества, многие терапевтические белки все еще продуцируют в клетках млекопитающих, которые продуцируют терапевтические белки с посттрансляционными модификациями, наиболее сходными с нативными белками человека, в то время как посттрансляционные модификации, в природе осуществляемые дрожжами и грибами, часто отличаются от посттрансляционных модификаций, встречающихся в клетках млекопитающих.
Для решения этой проблемы разрабатывают новые штаммы дрожжей и грибов, которые осуществляют посттрансляционные модификации, которые в наибольшей степени сходны с посттрансляционными модификациями, встречающимися в нативных белках человека. Таким образом, возобновился интерес к использованию клеток дрожжей и грибов для экспрессии более сложных белков. Однако, поскольку промышленность была сфокусирована на технологии культивирования клеток млекопитающих в течение длительного времени, экспрессирующие системы в клетках грибов, таких как Trichoderma, не являются также хорошо установившимися, как культура клеток млекопитающих, и, таким образом, имеют недостатки при экспрессии белков млекопитающих.
Таким образом, в данной области остается потребность в улучшенных клетках нитчатых грибов, таких как клетки грибов Trichoderma, которые могут стабильно продуцировать гетерологичные белки, такие как иммуноглобулины, предпочтительно на высоких уровнях экспрессии.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем описании описаны композиции, включающие клетки нитчатых грибов, такие как клетки грибов Trichoderma, имеющие сниженную или не поддающуюся обнаружению активность по меньшей мере трех протеаз, и имеющие рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, который продуцируется на увеличенных уровнях. Кроме того, в настоящем описании описаны способы улучшения стабильности гетерологичного полипептида и способы получения гетерологичных полипептидов, в которых протеазы не обладают сниженной активностью.
Таким образом, один аспект включает клетки нитчатых грибов, имеющие сниженную или не поддающуюся обнаружению активность по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке нитчатых грибов, в которой протеазы не обладают сниженной активностью. В определенных вариантах осуществления, когда клетка представляет собой клетку Aspergillus, общая активность протеаз снижена до 50% или менее относительно общей активности протеаз соответствующей родительской клетки Aspergillus, в которой протеаза не обладает сниженной активностью. В других вариантах осуществления общая активность протеаз клетки нитчатых грибов снижена до 49% или менее, 31% или менее, относительно общей активности протеаз соответствующей родительской клетки нитчатых грибов, в которой протеазы не обладают сниженной активностью.
В определенных вариантах осуществления уровень экспрессии по меньшей мере трех протеаз снижен или их экспрессия устранена. В определенных вариантах осуществления каждый из генов, кодирующих три протеазы, содержит мутацию, которая снижает или устраняет соответствующую активность протеазы. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, три гена, кодирующих протеазы, представляют собой pep1, tsp1 и slp1. В других вариантах осуществления три гена, кодирующих протеазы, представляют собой gap1, slp1 и pep1.
В определенных вариантах осуществления клетки грибов имеют сниженную или не поддающуюся обнаружению активность четырех эндогенных протеаз; каждый из генов, кодирующих четыре протеазы, содержит мутацию, которая снижает или устраняет активность соответствующей протеазы. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, четыре гена, кодирующих протеазы, представляют собой pep1, tsp1, slp1 и gap1.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, три или четыре гена, кодирующих протеазы, выбраны из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, tsp1, slp1, slp2, slp3, slp7, gap1 и gap2. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, три или четыре гена, кодирующих протеазы, выбраны из pep1, pep3, pep4, tsp1, slp1, slp2, gap1 и gap2. В определенных вариантах осуществления три или четыре гена, кодирующих протеазы, выбраны из pep1, pep2, pep3, pep4, pep5, gap1, gap2, slp1, slp2 и tsp1.
В других вариантах осуществления клетки грибов имеют сниженную или не поддающуюся обнаружению активность пяти эндогенных протеаз; каждый из генов, кодирующих пять протеаз, содержат мутацию, которая снижает или устраняет активность соответствующей протеазы. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, пять генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1 и pep4. В других вариантах осуществления пять генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1 и gap2.
В определенных вариантах осуществления клетки грибов имеют сниженную или не поддающуюся обнаружению активность шести эндогенных протеаз; каждый из генов, кодирующих шесть протеаз, содержит мутацию, которая снижает или устраняет активность соответствующей протеазы. В определенных вариантах осуществления клетка имеет шесть генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и шесть генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1, gap2 и pep4.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетки грибов имеют от трех до шести протеаз, имеющих сниженную или не поддающуюся обнаружению активность, где каждая из от трех до шести протеаз выбрана из pep1, pep2, pep3, pep4, pep5, tsp1, slp1, slp2, slp3, gap1 и gap2.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка имеет семь генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и семь генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1, gap2, pep4 и pep3.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка имеет восемь генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и восемь генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1, gap2, pep4, pep3 и pep5.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов имеет дополнительную протеазу, имеющую сниженную активность, ген, кодирующий дополнительную протеазу, содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и дополнительная протеаза выбрана из pep7, pep8, pep11, pep12, tpp1, gap2, slp3, slp5, slp6, slp7 и slp8.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, гетерологичный полипептид представляет собой полипептид млекопитающих. В определенных вариантах осуществления полипептид млекопитающих является гликозилированным.
В определенных вариантах осуществления полипептид млекопитающих выбран из иммуноглобулина, антитела и его антигенсвязывающих фрагментов, фактора роста, интерферона, цитокина и интерлейкина. В определенных вариантах осуществления полипептид млекопитающих представляет собой иммуноглобулин или антитело. В определенных вариантах осуществления полипептид млекопитающих выбран из инсулиноподобного фактора роста 1 (IGF1), гормона роста человека (hGH) и интерферона-альфа 2b (IFNα2b).
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, гетерологичный полипептид представляет собой полипептид не млекопитающих. В определенных вариантах осуществления полипептид не млекопитающих представляет собой аминопептидазу, амилазу, карбогидразу, карбоксипептидазу, каталазу, целлюлазу, хитиназу, кутиназу, дезоксирибонуклеазу, эстеразу, альфа-галактозидазу, бета-галактозидазу, глюкоамилазу, альфа-глюкозидазу, бета-глюкозидазу, инвертазу, лакказу, липазу, мутаназу, оксидазу, пектинолитический фермент, пероксидазу, фосфолипазу, фитазу, полифенолоксидазу, протеолитический фермент, рибонуклеазу, трансглутаминазу или ксиланазу.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов дополнительно обладает сниженной или не поддающейся обнаружению активностью ALG3, фермента маннозилтрансферазы. В определенных вариантах осуществления ген, кодирующий ALG3, содержит мутацию, которая снижает или устраняет соответствующую активность. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов дополнительно содержит полинуклеотид, кодирующий α-1,2-маннозидазу.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов имеет мутацию, которая снижает экспрессию протеазы, для которой желательно наличие сниженной активности. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация представляет собой делецию в гене, кодирующем протеазу. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация представляет собой делецию части гена, кодирующего каталитический домен протеазы. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов имеет точковую мутацию в части гена, кодирующей каталитический домен протеазы.
В других вариантах осуществления снижение или устранение протеазной активности одной или нескольких протеаз происходит благодаря конструкциям РНК-i, специфичным к i) одной протеазе или ii) двум или более протеазам, выбранным из группы, состоящей из протезы pep-типа, трипсин-подобной сериновой протеазы, протеазы gap-типа, седолизиновой протеазы и протеазы slp-типа. В определенных вариантах осуществления конструкции РНК-i являются специфичными к slp2, slp3, slp5 и/или slp6.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов дополнительно содержит каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II. В определенных вариантах осуществления каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II кодируются полинуклеотидом. В определенных вариантах осуществления каталитический домен N-ацетилглюкозаминилтрансферазы I кодируется первым полинуклеотидом, и каталитический домен N-ацетилглюкозаминилтрансферазы II кодируется вторым полинуклеотидом. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов дополнительно содержит полинуклеотид, кодирующий маннозидазу II и/или галактозилтрансферазу. В определенных вариантах осуществления клетка грибов содержит ферменты, выбранные из группы, состоящей из α-1,2-маннозидазы, N-ацетилглюкозаминилтрансферазы I, N-ацетилглюкозаминилтрансферазы II, маннозидазы II и/или галактозилтрансферазы, причем указанные ферменты дополнительно содержат нацеливающий пептид, например, гетерологичный нацеливающий пептид для надлежащей локализации соответствующего фермента. В определенных вариантах осуществления нацеливающий пептид выбран из SEQ ID NO: 589-594.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов представляет собой клетку грибов Trichoderma, клетку грибов Myceliophthora, клетку грибов Aspergillus, клетку грибов Neurospora, клетку грибов Fusarium или Penicillium, или клетку грибов Chrysosporium. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов представляет собой Trichoderma reesei.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов является диким типом по протеазе pep4.
Другой аспект включает способы повышения стабильности гетерологичного полипептида посредством: a) предоставления клетки нитчатых грибов согласно любому из предшествующих вариантов осуществления; и b) культивирования клетки так, чтобы экспрессировался гетерологичный полипептид, где гетерологичный полипептид обладает повышенной стабильностью по сравнению с гетерологичным полипептидом, полученным в соответствующей родительской клетке нитчатых грибов, в которой протеазы не обладают сниженной активностью, например, поскольку не содержат мутаций генов, кодирующих протеазы. Другой аспект включает способы получения гетерологичного полипептида посредством: a) предоставления клетки нитчатых грибов согласно любому из предшествующих вариантов осуществления; b) культивирования клетки-хозяина так, чтобы экспрессировался гетерологичный полипептид; и c) очистки гетерологичного полипептида. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка нитчатых грибов дополнительно содержит белок-носитель. В определенных вариантах осуществления белок-носитель представляет собой CBH1. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, культивирование проводят в среде, содержащей ингибитор протеаз. В определенных вариантах осуществления культивирование проводят в среде, имеющей один или два ингибитора протеаз, выбранных из SBT1 и химостатина. В определенных вариантах осуществления гетерологичный полипептид, полученный родуцируемый в соответствии со способом, представляет собой гликозилированный полипептид млекопитающих, и по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида состоит из Man3GlcNAc2 N-гликана. В других вариантах осуществления гетерологичный полипептид, полученный в соответствии со способом, представляет собой гликозилированный полипептид млекопитающих, и по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида состоит из комплексного N-гликана. В определенных вариантах осуществления гетерологичный полипептид, полученный в соответствии со способом, представляет собой гликозилированный полипептид млекопитающих, и по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида состоит из гибридного N- гликана. В определенных вариантах осуществления гетерологичный полипептид, полученный в соответствии со способом, представляет собой гликозилированный полипептид млекопитающих, и по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида состоит из N-гликана G1 или G2. Другой аспект включает гетерологичные полипептиды, получаемые способами, как описано выше.
Другой аспект включает клетки грибов Trichoderma, имеющие сниженную или не поддающуюся обнаружению активность по меньшей мере трех протеаз, выбранных из pep1, pep2, pep3, pep4, pep5, tsp1, slp1, slp2, gap1 и gap2, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке грибов Trichoderma.
В определенных вариантах осуществления уровень экспрессии по меньшей мере трех протеаз снижен или их экспрессия устранена в клетке грибов Trichoderma. В определенных вариантах осуществления каждый из генов, кодирующих по меньшей мере три протеазы, содержит мутацию, которая снижает или устраняет активность соответствующей протеазы в клетке грибов Trichoderma. В определенных вариантах осуществления клетка грибов Trichoderma включает три гена, кодирующих протеазы, с мутацией, которая снижает или устраняет протеазную активность, которые выбраны из gap1, slp1 и pep1. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, полипептид млекопитающих в клетке грибов Trichoderma представляет собой антитело или его антигенсвязывающие фрагменты, или иммуноглобулин, и по меньшей мере три протеазы выбраны из pep1, pep3, pep4, tsp1, slp1, slp2, gap1 и gap2. В определенных вариантах осуществления клетка грибов Trichoderma содержит четыре гена, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и четыре гена, кодирующих протеазы с такой мутацией, представляют собой pep1, tsp1, slp1 и gap1. В определенных вариантах осуществления клетка грибов Trichoderma имеет пять генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и пять генов, кодирующих протеазы с такой мутацией, представляют собой pep1, tsp1, slp1, gap1 и pep4. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, полипептид млекопитающих в клетке грибов Trichoderma представляет собой фактор роста, интерферон, цитокин или интерлейкин, и три протеазы со сниженной активностью выбраны из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, gap1, gap2, slp1, slp2, slp7 и tsp1. В определенных вариантах осуществления клетка грибов Trichoderma имеет пять генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и пять генов, кодирующих протеазы с такой мутацией, представляют собой pep1, tsp1, slp1, gap1 и gap2. В определенных вариантах осуществления клетка грибов Trichoderma имеет шесть генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и шесть генов, кодирующих протеазы с такой мутацией, представляют собой pep1, tsp1, slp1, gap1, gap2 и pep4. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma имеет семь генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает или устраняет активность соответствующей протеазы, и семь генов, кодирующих протеазы, представляют собой pep1, tsp1, slp1, gap1, gap2, pep4 и pep3. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma имеет восемь генов, кодирующих протеазы, каждый из которых содержит мутацию, которая снижает активность соответствующей протеазы, и восемь генов, кодирующих протеазы с такой мутацией, представляют собой pep1, tsp1, slp1, gap1, gap2, pep4, pep3 и pep5.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma дополнительно обладает сниженной или не поддающейся обнаружению активностью одной или нескольких дополнительных протеаз. В определенных вариантах осуществления уровень экспрессии одной или нескольких дополнительных протеаз в клетке грибов Trichoderma снижен или их экспрессия устранена. В определенных вариантах осуществления каждый из генов, кодирующих одну или несколько дополнительных протеаз в клетке грибов Trichoderma, имеет мутацию, которая снижает или устраняет активность соответствующей протеазы. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, один или несколько дополнительных генов, кодирующих протеазы, выбраны из pep7, pep8, pep11, pep12, tpp1, gap2, slp3, slp5, slp6, slp7 и slp8.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma дополнительно обладает сниженной или не поддающейся обнаружению активностью ALG3. В определенных вариантах осуществления ген, кодирующий ALG3 в клетке грибов Trichoderma, содержит мутацию, которая снижает или устраняет соответствующую активность. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma дополнительно содержит полинуклеотид, кодирующий α-1,2-маннозидазу. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация снижает или устраняет экспрессию гена в клетке грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация представляет собой делецию гена в клетке грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация представляет собой делецию части гена, кодирующей каталитический домен протеазы в клетке грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, мутация представляет собой точковую мутацию в части гена, кодирующей каталитический домен протеазы в клетке грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma дополнительно содержит каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II. В определенных вариантах осуществления каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II кодируются полинуклеотидом клетки грибов Trichoderma. В определенных вариантах осуществления каталитический домен N-ацетилглюкозаминилтрансферазы I кодируется первым полинуклеотидом и каталитический домен N-ацетилглюкозаминилтрансфераза II кодируется вторым полинуклеотидом клетки грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка грибов Trichoderma дополнительно содержит полинуклеотид, кодирующий маннозидазу II. В определенных вариантах осуществления каждая из протеаз обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO: 1, 17, 37, 58, 66, 82, 98, 118, 129, 166 и 182. В определенных вариантах осуществления общая протеазная активность в клетке грибов Trichoderma снижена до 49% или менее, 31% или менее от общей протеазной активности соответствующей родительской клетки Trichoderma, в которой протеазы не обладают сниженной активностью. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке грибов Trichoderma. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, полипептид млекопитающих продуцируется в полноразмерной версии на уровне, который превышает уровень продукции полипептида полноразмерной версии в соответствующей родительской клетке грибов Trichoderma.
Другой аспект включает способы повышения стабильности гетерологичного полипептида посредством: a) предоставления клетки грибов Trichoderma согласно любому из предшествующих вариантов осуществления; и b) культивирования клетки так, чтобы экспрессировался гетерологичный полипептид, где гетерологичный полипептид обладает повышенной стабильностью по сравнению с клеткой-хозяином, не содержащей мутации генов, кодирующих протеазы. Другой аспект включает способы получения гетерологичного полипептида путем: a) предоставления клетки грибов Trichoderma согласно любому из предшествующих вариантов осуществления; b) культивирования клетки-хозяина так, чтобы экспрессировался гетерологичный полипептид; и c) очистки гетерологичного полипептида. В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка нитчатых грибов дополнительно содержит белок-носитель. В определенных вариантах осуществления белок-носитель представляет собой CBH1.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 представлен гель PAGE, на котором показаны фракции, элюированные при очистке на аффинной колонке аспарагиновых протеаз.
На фиг.2 представлен гель PAGE, на которым показаны результаты инкубации IgG с аспарагиновыми протеазами.
На фиг.3 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штаммов с делецией одной протеазы M181 и M195. На фиг.3A представлен ожидаемый сигнал pep1 ORF: >8 т.п.н. из родительского M127, у трансформантов сигнал отсутствует. На фиг.3B представлен ожидаемый сигнал 5'-фланкирующей области pep1: >8 т.п.н. из родительского M127, 4 т.п.н. из трансформантов. На фиг.3C представлен ожидаемый сигнал 3'-фланкирующей области pep1: >8 т.п.н. из родительского M127, 4,2 т.п.н. из трансформантов.
На фиг.4 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение антитела ритуксимаба в штамме с делецией pep1 M182. На фиг.4A представлен ожидаемый сигнал ORF pep1: >8 т.п.н. из родительского M169, у трансформантов сигнал отсутствует. На фиг.4B представлен ожидаемый сигнал bar: 1,0+1,7 т.п.н. из трансформантов, 3,1 т.п.н. из pTTv41, у M169 сигнал отсутствует. На фиг.4C представлен ожидаемый сигнал bar: 1,8+2,8 т.п.н. из трансформантов, 3,1 т.п.н. из pTTv41, у M169 сигнал отсутствует.
На фиг.5 представлен белковый гель, на котором показаны пиковые фракции при очистке аспарагиновых протеаз из штамма, содержащего pep1, и штамма Δpep1.
На фиг.6A-B представлен иммуноблот, иллюстрирующий, что делеция протеазы pep2 из продуцирующего ритуксимаб штамма М169 увеличивала продукцию (A) легкой и (B) тяжелой цепи в трансформанте 206A (штамм M455). Полосы, соответствующие фрагменту легкой цепи на уровне 18 кДа и фрагменту тяжелой цепи на уровне 38 кДа, были более интенсивными у штамма M455 по сравнению с родительским штаммом M169.
На фиг.7 графически изображена протеазная активность супернатанта из продуцирующего ритуксимаб штамма M169 и трансформантов с делецией протеаз pep2 98A, 116A, 198A, 201A и 206A (M455). Трансформанты 116A, 198A и 206A демонстрируют сниженную протеазную активность в отношении казеина по сравнению с их родительским штаммом М169.
На фиг.8 представлен иммуноблот, демонстрирующий эффекты протеазной активности PEP3 и PEP7 на тяжелую цепь MAB01 и нативный IGF-1. На фиг.8A представлены эффекты протеазной активности на MAB01 при pH 5,5. На фиг.8B представлены эффекты протеазной активности на MAB01 при pH 4,5. На фиг.8C представлены эффекты протеазной активности на нативный IGF-1 при pH 4,5.
На фиг.9 представлен гель PAGE, демонстрирующий протеазу, содержащую фракции, очищенные на аффинной колонке с пептидом SIP.
На фиг.10 представлен иммуноблот, демонстрирующий активность протеазы SIP в отношении тяжелой цепи MAB01.
На фиг.11 графически изображена протеазная активность в отношении казеина с ингибиторами и без них.
На фиг.12 представлен иммуноблот, демонстрирующий уровни продукции тяжелой и легкой цепей MAB01 после удаления каждой из протеаз slp1, slp2, slp3 и gap1. На фиг.12A представлена продукция тяжелой цепи MAB01. На фиг.12B представлена продукция легкой цепи MAB01.
На фиг.13 графически представлена кратность увеличения продукции тяжелой и легкой цепей MAB01 после делеции каждой из протеаз slp1, slp2, slp3 и gap1. Каждый столбик соответствует среднему значению для нескольких клонов, представленных на фиг.12.
На фиг.14 представлен иммуноблот, демонстрирующий уровни продукции MAB01 из штамма M244 с делецией gap2. На фиг.14A представлена продукция тяжелой цепи MAB01 (HC). На фиг.14B представлена продукция легкой цепи MAB01 (LC).
На фиг.15 представлен иммуноблот, демонстрирующий уровни антитела MAB01 после инкубации с супернатантом Pichia, содержащим протеазу GAP2.
На фиг.16 представлен иммуноблот, демонстрирующий уровень деградации IgG1 человека протеазой.
На фиг.17 представлены результаты зимографии антитела MAB02 после аффинной очистки с помощью колонки с аминобензамидином (очищенные фракции) и в образцах супернатантов (супернатант).
На фиг.18 представлено получение штамма M219 с делецией двух протеаз Δpep1Δtsp1. На фиг.18A представлен ожидаемый сигнал ORF tsp1: 6,4 т.п.н. из родительского M196. На фиг.18B представлен ожидаемый сигнал 5'-фланкирующей области tsp1: 3,9 т.п.н. из трансформантов, >8 т.п.н. из M196, 3,9 т.п.н. из pTTv72. На фиг.18C представлен ожидаемый сигнал 3'-фланкирующей области tsp1: 2,8 т.п.н. из трансформантов, >8 т.п.н. из M196, 3,9 т.п.н. из pTTv72.
На фиг.19 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штамма М194 с двойной делецией Δpep1Δtsp2. На фиг.19A представлен ожидаемый сигнал ORF tsp1: т.п.н. из родительского M181. На фиг.19B представлен ожидаемый сигнал bar: 1,4+2,5 т.п.н. из трансформантов, 2,9 т.п.н. pTTv42, у М181 сигнал отсутствует. На фиг.19C представлен ожидаемый сигнал bar: 1,9+3,2 т.п.н. из трансформантов, 2,9 т.п.н. из pTTv42, у М181 сигнал отсутствует.
На фиг.20 графически представлены нормализованные данные о протеазной активности из культуральных супернатантов каждого из супернатантов с делецией протеазы и родительского штамма M124. Протеазную активность измеряли при pH 5,5 в первых 5 штаммах и при pH 4,5 в последних трех штаммах с делецией. Протеазная активность представлена в отношении зеленого флуоресцентного казеина. Штамм с делецией шести протеаз имеет только 6% от протеазной активности родительского штамма дикого типа, и протеазная активность штамма с делецией 7 протеаз была приблизительно на 40% меньшей, чем активность штамма с делецией 6 протеаз.
На фиг.21A представлены результаты зимографии MAB02 с очищенными с помощью аминобензамидина фракциями из супернатантов ферментации. На фиг.21B представлен гель SDS PAGE (7%) с очищенными с помощью аминобензамидина фракциями из супернатантов ферментации.
На фиг.22 представлены результаты зимографии MAB02 с подвергнутыми аффинной очистке с помощью SBTI фракциями, содержащими протеазы. Основная протеолитическая активность, где протеаза деградировала антитело MAB02, выглядит белой. В геле для зимографии разделяли концентрированную фракцию 3 (cf3) и неконцентрированные фракции 1-4 (f1-f4).
На фиг.23 представлен гель SDS PAGE, демонстрирующий подвергнутые аффинной очистке с помощью SBTI фракции, содержащие протеазы. На геле представлены концентрированные фракции cf3 и cf4.
На фиг.24 представлен иммуноблот, демонстрирующий уровень деградации тяжелой цепи ритуксимаба очищенными с помощью SBTI протеазами.
На фиг.25 представлен иммуноблот, демонстрирующий уровень деградации антитела при инкубации в течение ночи с содержащими субтилизин супернатантами Pichia. На фиг.25A представлена деградация протеазой тяжелой цепи ритуксимаба. На фиг.25B представлена деградация протеазой тяжелой цепи MAB01.
На фиг.26 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штамма M277 с делецией трех протеаз. На фиг.26A представлен ожидаемый сигнал ORF slp1: 6,5 т.п.н. только из родительских штаммов (M219, M228). На фиг.26B представлен ожидаемый сигнал 5'-фланкирующей области slp1: 6,5 т.п.н. из родительских штаммов, 3,3 т.п.н. из трансформантов, 4,4 т.п.н. из контрольной плазмиды pTTv126. На фиг.26C представлен ожидаемый сигнал 3'-фланкирующей области slp1: 6,5 т.п.н. из родительских штаммов, 2,3 т.п.н. из трансформантов, 4,4 т.п.н. из контрольной плазмиды pTTv126.
На фиг.27 представлена зимография MAB02, демонстрирующая активность супернатантов штаммов с делецией протеаз. Белые области на окрашенном геле указывают на область активности протеазы.
На фиг.28 графически представлена общая протеазная активность культуральных супернатантов с делецией протеазы по сравнению с активностью М124 дикого типа.
На фиг.29 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штамма M307 с делецией четырех протеаз. На фиг.29A представлен ожидаемый сигнал ORF gap1: 4 т.п.н. только из родительского штамма (M277 2A=M306). На фиг.29B представлен ожидаемый сигнал 5'-фланкирующей области gap1: 5,5 т.п.н. из родительского штамма, 3,4 т.п.н. из трансформантов, 4,1 т.п.н. из контрольной плазмиды pTTv117. На фиг.29C представлен ожидаемый сигнал 3'-фланкирующей области gap1: 5,5 т.п.н. из родительского штамма, 3,1 т.п.н. из трансформантов, 4,1 т.п.н. из контрольной плазмиды pTTv117.
На фиг.30 графически представлена общая протеазная активность в штаммах с тремя и четырьмя делециями по сравнению со штаммом М124 дикого типа.
На фиг.31 графически представлена протеазная активность с течением времени для штамма M304 с тремя делециями и штамма M371 с четырьмя делециями.
На фиг.32 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штамма M369 с делецией пяти протеаз. На фиг.32A представлен ожидаемый сигнал ORF gap2: 4,9 т.п.н. из родительского штамма (M307). На фиг.32B представлен ожидаемый сигнал 5'-фланкирующей области gap2: 4,9 т.п.н. из родительского штамма, 2,3 т.п.н. из трансформанта, 2,3 т.п.н. из контрольной плазмиды pTTv145. На фиг.32C представлен ожидаемый сигнал 3'-фланкирующей области gap2: 4,9 т.п.н. из родительского штамма, 3,8 т.п.н. из трансформантов, 3,8 т.п.н. из контрольной плазмиды pTTv145. На фиг.32D представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение pyr4- из штамма с делецией пяти протеаз M369, конечного штамма M381 (клон 14). Ожидаемый сигнал на 5'-фланкирующей области представляет собой: 1,5 т.п.н. из всех штаммов, 4,1 т.п.н. из контрольной плазмиды pTTv145. На фиг.32E представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение pyr4- из штамма с делецией пяти протеаз M369 с получением штамма M381 (клон 14). Ожидаемый сигнал 3'-фланкирующей области представляет собой: 3,6 т.п.н. из M307, 2,7 т.п.н. из M369 + клоны с выпетливанием, 3,8 т.п.н. из контрольной плазмиды pTTv145.
На фиг.33 графически представлена протеазная активность супернатантов, полученных после культивирования в течение 5 суток во вращающихся флаконах, проводимого со штаммом M307 с делецией 4 протеаз, штаммом M369 с делецией 5 протеаз и трансформантами с делецией 6 протеаз 10B, 44B, 97A, 97B и 120A. Флуоресцентный казеин инкубировали с разбавленными супернатантами в цитратном буфере при pH 4,5 для обнаружения протеазной активности.
На фиг.34 представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение штаммов M396 и M400 с делецией 6 протеаз. На фиг.34A представлен ожидаемый сигнал ORF pep4: 6,3 т.п.н. из M307 и M369. На фиг.34B представлен ожидаемый сигнал ORF pep4: 6,3 т.п.н. из M307 и M369, у трансформантов сигналы отсутствуют. На фиг.34C представлен ожидаемый сигнал 5'-фланкирующей области pep4: 6,3 т.п.н. из M307 и M369, 4,8 т.п.н. из трансформантов, 4,0 т.п.н. из pTTv181. На фиг.34D представлен ожидаемый сигнал для 3'-фланкирующей области pep4: 6,3 т.п.н. из M307 и M369, 2,1 т.п.н. из трансформантов, 4,0 т.п.н. из pTTv181. На фиг.34E представлен анализ с использованием саузерн-блоттинга, демонстрирующий получение pyr4- из штаммов M396 с делецией 6 протеаз. Ожидаемый сигнал 3'-фланкирующей области pep4 представляет собой: 6,3 т.п.н. из M307 и M369, 2,1 т.п.н. из повторно очищенных трансформантов, 4,9 т.п.н. из клонов с выпетливанием.
На фиг.35 представлен иммуноблот, демонстрирующий количество фрагментов тяжелой цепи ритуксимаба, продуцированных in vitro с помощью протеаз супернатанта.
На фиг.36 представлен иммуноблот, демонстрирующий деградацию тяжелой цепи и легкой цепи образцами супернатанта из обработанных SBTI культур и необработанных контролей. На фиг.36A представлена деградация тяжелой цепи. На фиг.36B представлена деградация легкой цепи.
На фиг.37 представлен иммуноблот, демонстрирующий уровень деградации тяжелой цепи и легкой цепи образцами супернатанта из культур, обработанных химостатином и пепстатином A, или из необработанных контрольных культур. На фиг.37A представлена деградация легкой цепи. На фиг.37B представлена деградация тяжелой цепи.
На фиг.38 представлен процесс очистки антител из культуральных супернатантов T. reesei.
На фиг.39 представлен иммуноблот, демонстрирующий увеличенную стабильность тяжелой цепи (HC) и легкой цепи (LC) антитела из клеток T. reesei, имеющих делецию протеазы pep1. Три модельных антитела исследовали в супернатанте из больших вращающихся флаконов (Δpep1 и М124) и супернатанте ферментации (pH 5,5; 28°C; 20 г/л экстракта пивной дробины, 60 г/л лактозы).
На фиг.40 представлен иммуноблот, демонстрирующий увеличенную продукцию тяжелой цепи ритуксимаба (Rx) из клеток T. reesei, содержащих делецию протеазы tsp1. Трансформанты 12-2A и 12-16A отчетливо демонстрируют больше тяжелых цепей по сравнению с родительским штаммом.
На фиг.41 представлен иммуноблот, демонстрирующий сниженную деградацию тяжелой цепи MAB01 после инкубации в течение ночи с супернатантом штамма M277 с делецией трех протеаз. После инкубации в течение ночи в культуральном супернатанте после культивирования в течение 5 суток, было обнаружено в 2,5 раза больше тяжелых цепей в супернатанте штамма с делецией трех протеаз по сравнению с супернатантом контрольного штамма М124, который не имел делеций протеаз. При инкубации в культуральном супернатанте после культивирования в течение 7 суток, было обнаружено в 4 раза больше тяжелой цепи в супернатанте штамма с делецией трех протеаз по сравнению с супернатантом контрольного штамма М124.
На фиг.43 представлено исследование стабильности тяжелой цепи антитела MAB01 в супернатантах штамма с делецией 6 протеаз. Антитело MAB01 присутствовало в неразбавленном супернатанте в концентрации 0,05 мкг/мкл. 10 мкл каждого образца наносили на 4-20% гель SDS PAGE. Тяжелая цепь была стабильной после инкубации в течение 20 часов при 37°C в супернатанте из штамма с делецией 6 протеаз при pH 4,2. Обнаружение тяжелой цепи проводили с помощью конъюгированного с AP антитела против тяжелой цепи IgG (Sigma #A3188), разбавленного 1:30000 в TBST. Полноразмерная тяжелая цепь мигрировала до уровня 50 кДа на геле.
На фиг.44 представлены образцы гормона роста человека после культивирования в течение 4 суток в культурах в 24-лунках с ингибиторами и без них, и с добавками. Наносили 12 мкл каждого супернатанта. Первичное антитело Acris, номер по каталогу #AM00401PU-N, антитело мыши против hGH (разбавленное до 2 мкг/мл в TBST), и конъюгированное с AP вторичное антитело козы против IgG мыши BioRad (#170-6520), разбавленное 1:10000. Стандарт hGH (200 нг), номер по каталогу Aceam #ab51232. Полноразмерный белок hGH мигрирует до уровня 22 кДа.
На фиг.45 представлена филогенетика аспарагиновых протеаз T. reesei, Myceliophthora thermophila, Neurospora crassa, Penicillium chrysogenum, Aspergillus oryzae, A. nidulans и A. niger. Выравнивание проводили с помощью Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) и дерево строили с помощью BLOSUM62 с использованием среднего расстояния.
На фиг.46 представлена филогенетика субтилизиновых протеаз T. reesei, Myceliophthora thermophila, Neurospora crassa, Penicillium chrysogenum, Aspergillus oryzae, A. nidulans и A. niger. Выравнивание проводили с помощью Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) и дерево строили с помощью BLOSUM62 с использованием среднего расстояния. "Pyr" означает пиролизин, "prKsf3" означает протеазу K, подсемейство 3; prtA, prtK, prJ, prtF и prtBCI означают подсемейства, описанные в Bryant et al. (2009) BMC Evolutionary Biology 9: 168, doi: 10.1186/1471-2148-9-168, фиг.5 и дополнительный документ № 8.
На фиг.47 представлена филогенетика глутаминовых протеаз T. reesei, Myceliophthora thermophila, Neurospora crassa, Penicillium chrysogenum, Aspergillus oryzae, A. nidulans и A. niger. Выравнивание проводили с помощью Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) и дерево строили с помощью BLOSUM62 с использованием среднего расстояния.
На фиг.48 представлена филогенетика седолизиновых протеаз T. reesei, Myceliophthora thermophila, Neurospora crassa, Penicillium chrysogenum, Aspergillus oryzae, A. nidulans и A. niger. Выравнивание проводили с помощью Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) и дерево строили с помощью BLOSUM62 с использованием среднего расстояния. Поскольку slp7 сходен с седолизиновыми протеазами, он включен в дерево. Последовательности Aspergillus fumigatus включены для облегчения определения взаимосвязи между седолизинами. Сокращения sedA/B/C/D/E перед каждой протеазой основаны на Reichard et al. (2006) APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Vol. 72, p. 1739-1748, фиг.4, из которого взят поиск BLAST соответствующей протеазы с A. fumigatus sedolisin.
Фиг.49: A: Схематическое изображение экспрессирующих плазмид pTTv67 и pTTv99. Тяжелая цепь MAB01 содержится в векторе pTTv67 и легкая цепь содержится в векторе pTTv99. B: Схематическое изображение экспрессирующего вектора pTTv223. Тяжелая и легкая цепи MAB01 содержатся в векторе pTTv223.
Фиг.50: A: Анализ с использованием вестерн-блоттинга продукции легкой и тяжелой цепей MAB01 при периодической ферментации при pH 5,2 продуцирующего MAB01 штамма M507. Использованные антитела представляли собой Sigma A3188 против тяжелой цепи (левый блот) и Sigma A3813 против легкой цепи (правый блот), оба в разведениях 1:10000. Коды образцов обозначают время ферментации в сутках. 0,1 мкл супернатанта наносили на каждую дорожку в обоих блотах. B: Анализ с использованием вестерн-блоттинга продукции легкой и тяжелой цепей MAB01 при периодической ферментации продуцирующего MAB01 штамма M507 при pH 5,5. Использованные антитела представляли собой Sigma A3188 против тяжелой цепи (блот слева) и Sigma A3813 против легкой цепи (блот справа), оба в разведениях 1:10000. Коды образцов обозначают время ферментации в сутках. 0,1 мкл супернатанта наносили на каждую дорожку в обоих блотах.
Фиг.51. Анализ с использованием вестерн-блоттинга продукции легкой и тяжелой цепей MAB01 в штаммах M304 при периодической ферментации bio00503b при pH 5,5. Использованные антитела представляли собой Sigma A3188 против тяжелой цепи и Sigma A3813 против легкой цепи. На 8 сутки ферментации M304 в качестве контроля включали bio00477b. Коды образцов обозначают время ферментации в сутках. На оба блота наносили 0,1 мкл супернатанта. Верхний иммуноблот соответствует тяжелой цепи, и нижний иммуноблот соответствует легкой цепи.
Фиг.52. Экспрессирующий вектор для РНК-i pTTv204.
Фиг.53: Иммуноблот для обнаружения продукции тяжелой цепи MAB01 в штаммах, экспрессирующих РНК-i, которая осуществляет нокдаун экспрессии slp2.
На фиг.54A представлено количественное определение уровня экспрессии IFN-α 2b из образца после ферментации M401 в течение 3 суток. 1 мкл/2 мкл/4 мкл супернатанта наносили на 4-20% гель SDS PAGE. Иммуноблоттинг проводили с антителом против IFN-α 2b Aceam (#ab9386), разбавленным до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело козы против IgG мыши от Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Белковые стандарты наносили на гель, соответствующий 50 нг, 100 нг и 200 нг полноразмерного IFN-α 2b. Денситометрическое количественное определение проводили с помощью программного обеспечения Totallab Quant TL100. Для количественного определения наиболее репрезентативным был образец 2 мкл. Полноразмерный контроль IFN-α 2b (100 нг) мигрирует до уровня 19,3 кДа, и связанный с носителем IFN-α 2b мигрирует до уровня 70 кДа.
На фиг.54B представлен анализ с использованием иммуноблоттинга для образцов культур M577 и M652 после ферментации в течение 3-6 суток. 0,2 мкл супернатанта для выращивания наносили на 4-20% гель SDS PAGE. Иммуноблоттинг проводили с антителом против IFN-α 2b Aceam (#ab9386), разбавленным до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело козы против IgG мыши от Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Полноразмерный контроль IFN-α 2b (100 нг) мигрирует до уровня 19,3 кДа, и связанный с носителем IFN-α 2b мигрирует до уровня 70 кДа.
На фиг.55 представлено количественное определение уровня экспрессии IFN-α 2b из образцов на 4 сутки (ферментация M577) и на 3 сутки (ферментация M652). 0,05 мкл и 0,1 мкл супернатанта каждого образца наносили на 4-20% гель SDS PAGE. Иммуноблоттинг проводили с антителом против IFN-α 2b Aceam (#ab9386), разбавленным до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело козы против IgG мыши от Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Белковые стандарты наносили на гель, соответствующий 50 нг, 100 нг и 200 нг полноразмерного IFN-α 2b. Денситометрическое количественное определение проводили с помощью программного обеспечения Totallab Quant TL100. Для количественного определения наиболее репрезентативным был образец 0,1 мкл. Полноразмерный контроль IFN-α 2b (100 нг) мигрирует до уровня 19,3 кДа, и связанный с носителем IFN-α 2b мигрирует до уровня 70 кДа.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение относится к усовершенствованным способам получения рекомбинантных гетерологичных полипептидов в клетках нитчатых грибов, которые обладают сниженной или не обладают активностью по меньшей мере трех протеаз. Настоящее изобретение основано частично на неожиданном открытии, что снижение активности конкретной комбинации эндогенных протеазы в клетках нитчатых грибов увеличивает экспрессию и стабильность различных рекомбинантно экспрессируемых гетерологичных белков, таких как иммуноглобулины и факторы роста. В то время как в других исследованиях были созданы клетки грибов Trichoderma с одной или несколькими инактивированными протеазами, они не предоставили руководства в отношении того, какие протеазы являются наиболее подходящими для увеличения экспрессии и стабильности определенных типов белков, таких как белки млекопитающих. Например, в WO2011/075677 описаны определенные протеазы, которые можно подвергнуть нокауту в Trichoderma, и даже описаны клетки грибов Trichoderma с дефицитом нескольких протеаз. Однако в WO2011/075677 не предоставлено никакого руководства в отношении того, какие из протеаз обладают неблагоприятным влиянием на экспрессию и стабильность белков млекопитающих, таких как иммуноглобулины или факторы роста, поскольку в указанном документе не описаны примеры экспрессии каких-либо белков млекопитающих. Более того, в WO2011/075677 описана только гетерологичная экспрессия одного белка грибов в каждом из трех различных штаммов грибов с дефицитом одной протеазы. Таким образом, специалист в данной области, вероятно, может считать WO 2011/075677 указанием на то, что инактивация каждой единичной протеазы может быть достаточной для продукции гетерологичного белка. Yoon et al. (2009, Appl. Microbiol Biotechnol 82: 691-701, 2010: Appl. Microbiol Biotechnol DOI 10.1007/s00253-010-2937-0) сообщают о конструировании штаммов с разрушением пяти и десяти генов протеаз для продукции гетерологичного белка в Aspergillus oryzae. 10 клеток с разрушением протеаз увеличивают выход продукции химозина только в 3,8 раза, несмотря на высокое количество разрушенных генов протеаз. Van den Hombergh et al. описали штаммы Aspergillus niger с тремя разрушенными генами протеаз. Хотя данные демонстрируют снижение протеазной активности, отсутствует пример какой-либо продукции белков млекопитающих, описанной в настоящем описании.
Заявители неожиданно показали, что несколько протеаз важны для снижения общей протеазной активности, увеличения продукции гетерологичных белков и стабилизации гетерологичных белков после экспрессии в клетках нитчатых грибов, таких как клетки грибов Trichoderma. В частности, авторы изобретения идентифицировали протеазы, которые в действительности экспрессировались в клетках грибов Trichoderma (в противоположность только кодированию их в геноме) посредством очистки этих протеаз и определения того, какие из них обладают активностью, наиболее существенной для деградации гетерологичных белков, таких как белки млекопитающих. Кроме того, авторы изобретения подтвердили, что делеция генов, ответственных за активность конкретных протеаз, обеспечивала существенное снижение общей активности протеаз, которое коррелирует с увеличением стабилизации белка с точки зрения количества и качества белков, продуцируемых в клетках нитчатых грибов, содержащих такие делеции, и обеспечивала увеличение продукции полноразмерных гетерологичных белков в клетках. Также было обнаружено, что клетки грибов Trichoderma, модифицированные способами инженерии для снижения активности по меньшей мере трех генов протеаз, обеспечивали неожиданное синергическое увеличение продукции полноразмерных белков млекопитающих, таких как антитела, терапевтический белок или варианты антител, такие как Fab или однодоменные антитела. Иными словами, количество продуцированного полноразмерного белка млекопитающих превышало сумму количеств, продуцированных в клетках грибов Trichoderma, имеющих делецию только одного или двух генов протеаз. Таким образом, в противоположность WO2011/075677, авторы изобретения показали, что продукцию интактных гетерологичных белков в клетках нитчатых грибов, таких как клетки грибов Trichoderma, можно обеспечивать путем снижения или устранения активности по меньшей мере трех протеаз в клетках.
Таким образом, определенные аспекты настоящего изобретения обеспечивают клетки нитчатых грибов, которые продуцируют увеличенные уровни гетерологичного белка вследствие наличия сниженной или отсутствия активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке нитчатых грибов, в которой протеазы не обладают сниженной активностью. Иными словами, желаемое увеличение уровня продукции гетерологичных белков определяют путем сравнения уровня продукции гетерологичного белка в клетке нитчатых грибов, имеющей сниженную активность по меньшей трех протеаз, с уровнем продукции гетерологичного белка в клетке нитчатых грибов, которая не обладает такой сниженной активностью, но в остальном идентична клетке, проявляющей увеличенный уровень.
Другие аспекты настоящего изобретения относятся к способам повышения стабильности гетерологичного полипептида посредством: a) предоставления клетки нитчатых грибов по настоящему изобретению, имеющей сниженную или отсутствие активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид; и b) культивирования клетки так, чтобы экспрессировался гетерологичный полипептид, где гетерологичный полипептид обладает повышенной стабильностью по сравнению с клеткой-хозяином, не содержащей мутации генов, кодирующих протеазы.
Другие аспекты настоящего изобретения относятся к способам получения гетерологичного полипептида путем: a) предоставления клетки нитчатых грибов по настоящему изобретению, имеющей сниженную или отсутствие активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид; b) культивирования клетки-хозяина так, чтобы экспрессировался гетерологичный полипептид; и c) очистки гетерологичного полипептида.
Определенные аспекты настоящего изобретения также относятся к клеткам грибов Trichoderma, которые продуцируют увеличенные уровни полипептида млекопитающих вследствие наличия сниженной или отсутствия активности по меньшей мере трех протеаз, выбранных из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, tsp1, slp1, slp2, gap1 и gap2, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке грибов Trichoderma, в которой протеазы не обладают сниженной активностью. Иными словами, желаемое увеличение уровня продукции гетерологичного белка определяют путем сравнения уровня продукции гетерологичного белка в клетке грибов Trichoderma, обладающей сниженной активностью по меньшей мере трех протеаз, относительно продукции гетерологичного белка в клетке грибов Trichoderma, которая не обладает такой сниженной активностью, но в остальном идентична клетке, проявляющей увеличенный уровень.
Другие аспекты настоящего изобретения относятся к способам повышения стабильности полипептида млекопитающих путем: a) предоставления клетки грибов Trichoderma по настоящему изобретению, имеющей сниженную активность по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих; и b) культивирования клетки так, чтобы экспрессировался полипептид млекопитающих, где полипептид млекопитающих обладает повышенной стабильностью по сравнению с клеткой-хозяином, не содержащей мутаций в генах, кодирующих протеазы.
Следующие аспекты настоящего изобретения относятся к способам получения полипептида млекопитающих путем: a) предоставления клетки грибов Trichoderma по настоящему изобретению, имеющей сниженную активность по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих; b) культивирования клетки-хозяина так, чтобы экспрессировался полипептид млекопитающих; и c) очистки полипептида млекопитающих.
Определения
Как используют в рамках изобретения, "иммуноглобулин" относится к мультимерному белку, содержащему тяжелую цепь и легкую цепь, ковалентно связанные вместе, и способному к специфическому связыванию с антигеном. Молекулы иммуноглобулинов представляют собой большое семейство молекул, которые включают несколько типов молекул, таких как IgM, IgD, IgG, IgA и IgE.
Как используют в рамках изобретения, "антитело" относится к интактным молекулам иммуноглобулинов, а также к их фрагментам, которые способны связывать антиген. Они включают гибридные (химерные) молекулы антител (см., например, Winter et al. Nature 349: 293-99225, 1991; и патент США № 4816567, 226); фрагменты F(ab')2 и F(ab) и Fv-молекулы; нековалентные гетеродимеры [227, 228]; одноцепочечные молекулы Fv (scFv) (см., например, Huston et al. Proc. Natl. Acad. Sci. U.S.A. 85: 5897-83, 1988); димерные и тримерные конструкции фрагментов антител; миниантитела (см., например, Pack et al. Biochem 31, 1579-84, 1992; и Cumber et al. J. Immunology 149B, 120-26, 1992); молекулы гуманизированных антител (см., например, Riechmann et al. Nature 332, 323-27, 1988; Verhoeyan et al. Science, 239, 1534-36, 1988; и GB 2276169); и любые функциональные фрагменты, полученные из таких молекул, а также антитела, полученные необщепринятыми способами, такими как фаговый дисплей. Предпочтительно, антитела представляют собой моноклональные антитела. Способы получения моноклональных антитела хорошо известны в данной области.
Как используют в рамках изобретения, "пептид" и "полипептид" представляют собой аминокислотные последовательности, включающие множество последовательно расположенных полимеризованных аминокислотных остатков. Для целей настоящего изобретения, как правило, пептиды представляют собой молекулы, включающие вплоть до 50 аминокислотных остатков, и полипептиды включают более 50 аминокислотных остатков. Пептид или полипептид могут включать модифицированные аминокислотные остатки, встречающиеся в природе аминокислотные остатки, не кодируемые кодоном, и не встречающиеся в природе аминокислотные остатки. Как используют в рамках изобретения, "белок" может относиться к пептиду или полипептиду любого размера.
Протеазы по изобретению
Изобретение, описанное в настоящем описании, относится к клеткам нитчатых грибов, таким как клетки грибов Trichoderma, которые продуцируют увеличенные уровни гетерологичного полипептида, такого как полипептид млекопитающих, вследствие наличия сниженной или не поддающейся обнаружению активности по меньшей мере трех протеаз, встречающихся в клетках. Такие протеазы, встречающиеся в клетках нитчатых грибов, которые экспрессируют гетерологичный полипептид, обычно катализируют значительную деградацию экспрессируемых рекомбинантных полипептидов. Таким образом, вследствие снижения или устранения активности протеаз в клетках нитчатых грибов, которые экспрессируют гетерологичный полипептид, стабильность экспрессируемого полипептида увеличивается, что приводит к увеличенному уровню продукции полипептида, и, в некоторых случаях, к улучшенному качеству продуцируемого полипептида (например, полноразмерный вместо деградированного).
Протеазы включают, но не ограничиваются ими, аспарагиновые протеазы, трипсин-подобные сериновые протеазы, субтилизиновые протеазы, глутаминовые протеазы и седолизиновые протеазы. Такие протеазы можно идентифицировать и выделять из клеток нитчатых грибов и исследовать для определения того, влияет ли снижение их активности на продукцию рекомбинантного полипептида из клетки нитчатых грибов. Способы идентификации и выделения протеаз хорошо известны в данной области и включают, но не ограничиваются ими, аффинную хроматографию, зимографию и гель-электрофорез. Затем идентифицированную протеазу можно исследовать путем делеции гена, кодирующего идентифицированную протеазу, из клетки нитчатых грибов, которая экспрессирует рекомбинантный полипептид, такой как гетерологичный полипептид или полипептид млекопитающих, и определения того, приводит ли делеция к снижению общей протеазной активности в клетке, например, до уровня 49% или менее, или 31% или менее, от общей протеазной активности соответствующей родительской клетки нитчатых грибов; и к увеличению уровня продукции экспрессируемого рекомбинантного полипептида, например, который в два раза превышает уровень продукции в соответствующей родительской клетке нитчатых грибов. Способы делеции генов, измерения общей протеазной активности и измерения уровней продуцированного белка хорошо известны в данной области и включают способы, описанные в настоящем описании. "Соответствующая родительская клетка нитчатых грибов" относится к соответствующей клетке, в которой протеазы не обладают сниженной или устраненной активностью.
Аспарагиновые протеазы
Аспарагиновые протеазы представляют собой ферменты, которые используют остаток аспарагиновой кислоты для гидролиза пептидных связей в полипептидах и белках. Как правило, аспарагиновые протеазы содержат два высококонсервативных остатка аспарагиновой кислоты в их активном центре, которые обладают оптимальной активностью в диапазоне кислых значений pH. Аспарагиновые протеазы из эукариотических организмов, таких как грибы Trichoderma, включают пепсины, катепсины и ренины. Такие аспарагиновые протеазы обладают двухдоменной структурой, которая, как полагают, является следствием удвоения гена-предшественника. В соответствии с таким событием удвоения, в целом, укладка каждого из доменов является сходной, хотя последовательности двух доменов начали дивергировать. Каждый домен вносит вклад в один из каталитических остатков аспарагиновой кислоты. Активный центр находится в щели, образованной двумя доменами аспарагиновых протеаз. Эукариотические аспарагиновые протеазы, кроме того, включают дополнительные консервативные дисульфидные мостики, которые могут упрощать идентификацию полипептидов в качестве протеаз аспарагиновой кислоты.
В клетках грибов Trichoderma было идентифицировано девять аспарагиновых протеаз: pep1 (tre74156); pep2 (tre53961); pep3 (tre121133); pep4 (tre77579), pep5 (tre81004) и pep7 (tre58669), pep8 (tre122076), pep11 (tre121306) и pep12 (tre119876).
Pep1
Примеры пригодных протеаз pep1 включают, но не ограничиваются ими, pep1 Trichoderma reesei (SEQ ID NO: 1), gi|11558498 Hypocrea lixii (SEQ ID NO: 2), Trichoderma asperellum gi|47027997 (SEQ ID NO: 3), jgi|Triat2|297887 Trichoderma atroviride (SEQ ID NO: 4), jgi|TriviGv29_8_2|81777 Trichoderma virens (SEQ ID NO: 5), jgi|Trire2|afm:Afu5g13300 Aspergillus fumigatus (SEQ ID NO: 6), gi|94730408 Aspergillus oryzae (SEQ ID NO: 7), gi|322712783 Metarhizium anisopliae (SEQ ID NO: 8), gi|46126795 Gibberella zeae (SEQ ID NO: 9), Fusarium venenatum gi|18448713 (SEQ ID NO: 10), gi|342879173 Fusarium oxysporum (SEQ ID NO: 11), gi|320591399 Grosmannia clavigera (SEQ ID NO: 12), gi|302422750 Verticillium alboatrum (SEQ ID NO: 13), gi|116182964 Chaetomium globosum (SEQ ID NO: 14), gi|85110723 Neurospora crassa (SEQ ID NO: 15), gi|336463990 Neurospora tetrasperma (SEQ ID NO: 16), gi367030924 Myceliophthora thermophila (SEQ ID NO: 491), gi255953325 Penicillium chrysogenum (SEQ ID NO: 492), gi350639535 Aspergillus niger (SEQ ID NO: 493), gi67541436 Aspergillus nidulans (SEQ ID NO: 494) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep1, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 1-16, SEQ ID NO: 491-494. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 1-16, SEQ ID NO: 491-494.
В некоторых вариантах осуществления pep1 представляет собой pep1 T. reesei. Аминокислотная последовательность, кодируемая pep1 T. reesei, указана в SEQ ID NO: 1. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 1. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 1.
Pep2
Примеры пригодных протеаз pep2 включают, но не ограничиваются ими, pep2 Trichoderma reesei (SEQ ID NO: 182), jgi|Triat2|142040 T. atroviride (SEQ ID NO: 183), jgi|TriviGv29_8_2|53481 T. virens (SEQ ID NO: 184), CM01 gi|346326575 Cordyceps militaris (SEQ ID NO: 185), gi85111370 Neurospora crassa (SEQ ID NO: 495) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep2, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 182-185, SEQ ID NO: 495. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 182-185, SEQ ID NO:495.
В некоторых вариантах осуществления pep2 представляет собой pep2 T. reesei. Аминокислотная последовательность, кодируемая pep2 T. Reesei, указана в SEQ ID NO: 182. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 182. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 182.
Pep3
Примеры пригодных протеаз pep3 включают, но не ограничиваются ими, pep3 Trichoderma reesei (SEQ ID NO: 17), T. atroviride jgi|Triat2 (SEQ ID NO: 18), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 19), gi|145583125 Hypocrea lixii (SEQ ID NO: 20), gi|51860175 Trichoderma asperellum (SEQ ID NO: 21), gi|317025164 Aspergillus niger (SEQ ID NO: 22), gi|159122534 Aspergillus fumigatus (SEQ ID NO: 23), gi|134054572 Aspergillus niger (SEQ ID NO: 24), gi|346318620 Cordyceps militaris (SEQ ID NO: 25), gi|310800156 Glomerella graminicola (SEQ ID NO: 26), gi|342871221 Fusarium oxysporum (SEQ ID NO: 27), gi|320591121 Grosmannia clavigera (SEQ ID NO: 28), gi|12002205 Botryotinia fuckeliana (SEQ ID NO: 29), gi|346997107 Thielavia terrestris (SEQ ID NO: 30), gi|156055954 Sclerotinia sclerotiorum (SEQ ID NO: 31), Chaetomium globosum gi|116197829 (SEQ ID NO: 32), gi|336472132 Neurospora tetrasperma (SEQ ID NO: 33), gi|85102020 Neurospora crassa (SEQ ID NO: 34), gi|119467426 Neosartorya fischeri (SEQ ID NO: 35), gi|212534792 Penicillium marneffei (SEQ ID NO: 36), gi367025909 M. thermophila (SEQ ID NO: 496), gi255947264 P. chrysogenum (SEQ ID NO: 497), 391870123 A. oryzae (SEQ ID NO: 498) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep3, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 17-36, SEQ ID NO: 496-498. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 17-36, SEQ ID NO:496-498.
В некоторых вариантах осуществления pep3 представляет собой pep3 T. reesei. Аминокислотная последовательность, кодируемая pep3 T. reesei, указана в SEQ ID NO: 17. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 17. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 17.
Pep4
Примеры пригодных протеаз pep4 включают, но не ограничиваются ими, pep4 Trichoderma reesei (SEQ ID NO: 37), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 38), jgi|Triat2 T. atroviride (SEQ ID NO: 39), gi|193735605 Trichoderma aureoviride (SEQ ID NO: 40), gi|145232965 Aspergillus niger (SEQ ID NO: 41), gi|70999520 Aspergillus fumigatus (SEQ ID NO: 42), gi|121705756 Aspergillus clavatus (SEQ ID NO: 43), gi|302899226 Nectria haematococca (SEQ ID NO: 44), gi|310796316 Glomerella graminicola (SEQ ID NO: 45), gi|346322842 Cordyceps militaris (SEQ ID NO: 46), gi|46138535 Gibberella zeae (SEQ ID NO: 47), gi|322708430 Metarhizium anisopliae (SEQ ID NO: 48), gi|342882947 Fusarium oxysporum (SEQ ID NO: 49), gi|322700747 Metarhizium acridum (SEQ ID NO: 50), gi|346973691 Verticillium dahlia (SEQ ID NO: 51), gi|154309857 Botryotinia fuckeliana (SEQ ID NO: 52), gi|116203505 Chaetomium globosum (SEQ ID NO: 53), gi|347001590 Thielavia terrestris (SEQ ID NO: 54), gi|39973863 Magnaporthe oryzae (SEQ ID NO: 55), gi|296417651 Tuber melanosporum (SEQ ID NO: 56), gi|85094599 Neurospora crassa (SEQ ID NO: 57), gi367031892 gi255947264 M. thermophila (SEQ ID NO: 499), gi255936729 gi255947264 P. chrysogenum (SEQ ID NO: 500), gi169770745 gi255947264 A. oryzae (SEQ ID NO: 501), gi67524891 gi255947264 A. nidulans (SEQ ID NO: 502) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep4, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 37-57, SEQ ID NO: 499-502. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 37-57, SEQ ID NO: 499-502.
В некоторых вариантах осуществления pep4 представляет собой pep4 T. reesei. Аминокислотная последовательность, кодируемая pep4 T. reesei, указана в SEQ ID NO: 37. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 37. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 37.
Pep5
Примеры пригодных генов pep5 включают, но не ограничиваются ими, pep5 Trichoderma reesei (SEQ ID NO: 58), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 59), jgi|Triat2|277859 T. atroviride (SEQ ID NO: 60), gi|322695806 Metarhizium acridum (SEQ ID NO: 61), gi|156071418 Fusarium oxysporum (SEQ ID NO: 62), gi|346324830 Cordyceps militaris (SEQ ID NO: 63), gi|46124247 Gibberella zeae (SEQ ID NO: 64), gi|346978752 Verticillium dahliae (SEQ ID NO: 65), gi367019798 M. thermophila (SEQ ID NO: 503) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep5, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 58-65, SEQ ID NO: 503. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 58-65, SEQ ID NO: 503.
В некоторых вариантах осуществления pep5 представляет собой pep5 T. reesei. Аминокислотная последовательность, кодируемая pep5 T. reesei, указана в SEQ ID NO: 58. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 58. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 58.
Pep7
Примеры пригодных генов pep7 включают, но не ограничиваются ими, pep7 Trichoderma reesei (SEQ ID NO: 186), jgi|Triat2 Trichoderma atroviride (SEQ ID NO: 187), jgi|TriviGv29_8_2 Trichoderma virens (SEQ ID NO: 188), gi|310800487 Glomerella graminicola (SEQ ID NO: 189), gi|322700577 Metarhizium acridum (SEQ ID NO: 190), gi|347003264 Thielavia terrestris (SEQ ID NO: 191), gi|171680938 Podospora anserine (SEQ ID NO: 192), gi|340905460 Chaetomium thermophilum (SEQ ID NO: 193), gi|346975960 Verticillium dahliae (SEQ ID NO: 194), gi|347009870, gi367026634 Myceliophthora thermophila (SEQ ID NO: 195), gi|85090078 Neurospora crassa (SEQ ID NO: 196), gi|39948622 Magnaporthe oryzae (SEQ ID NO: 197), gi|116191517 Chaetomium globosum (SEQ ID NO: 198), gi|39970765 Magnaporthe oryzae (SEQ ID NO: 199), gi67522232 A. nidulans (SEQ ID NO: 504), gi350630464 A. niger (SEQ ID NO: 505), gi317138074 A. oryzae (SEQ ID NO: 506) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep7, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 186-199, SEQ ID NO: 504-506. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 186-199, SEQ ID NO: 504-506.
В некоторых вариантах осуществления pep7 представляет собой pep7 T. reesei. Аминокислотная последовательность, кодируемая pep7 T. reesei, указана в SEQ ID NO: 186. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например. 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 186. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 186.
Pep8
Примеры пригодных генов pep8 включают, но не ограничиваются ими, pep8 EGR48424 Trichoderma reesei (SEQ ID NO: 507), EHK19238 Trichoderma virens (SEQ ID NO: 508), EHK40047 Trichoderma atroviride (SEQ ID NO: 509), EGO53367 Neurospora tetrasperma (SEQ ID NO: 510), XP_003658897 Myceliophthora thermophila (SEQ ID NO: 511), XP_965343 Neurospora crassa (SEQ ID NO: 512), EFZ03501 Metarhizium anisopliae (SEQ ID NO: 513), XP_003656869 Thielavia terrestris (SEQ ID NO: 514), EGU79769 Fusarium oxysporum (SEQ ID NO: 515) и XP_381566 Gibberella zeae (SEQ ID NO: 516), XP_3714540.1 Magnaporthe oryzae (SEQ ID NO: 517), XP_002557331 P. chrysogenum (SEQ ID NO: 518), XP_001822899.1 A. oryzae (SEQ ID NO: 519), XP_664091.1 A. nidulans (SEQ ID NO: 520), EHA24387.1 A. niger (SEQ ID NO: 521) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep8, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 507-521. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 507-521.
В некоторых вариантах осуществления pep8 представляет собой pep8 T. reesei. Аминокислотная последовательность, кодируемая pep8 T. reesei, указана в SEQ ID NO: 507. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 507. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 507.
Pep11
Примеры пригодных генов pep11 включают, но не ограничиваются ими, pep11 EGR49498 Trichoderma reesei (SEQ ID NO: 522), EHK26120 Trichoderma virens (SEQ ID NO: 523), EHK41756 Trichoderma atroviride (SEQ ID NO: 524), EKJ74550 Fusarium pseudograminearum (SEQ ID NO: 525), EFY91821 Metarhizium acridum (SEQ ID NO: 526) и XP_384151 Gibberella zeae (SEQ ID NO: 527), XP_003667387.1 M. thermophila (SEQ ID NO: 528), XP_960328.1 N. crassa (SEQ ID NO: 529) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep11, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 522-529. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 522-529.
В некоторых вариантах осуществления pep11 представляет собой pep8 T. reesei. Аминокислотная последовательность, кодируемая pep11 T. reesei, указана в SEQ ID NO: 522. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 522. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 522.
Pep12
Примеры пригодных генов pep12 включают, но не ограничиваются ими, pep12 EGR52517 Trichoderma reesei (SEQ ID NO: 530), pep12 EHK18859 Trichoderma virens (SEQ ID NO: 531), pep12 EHK45753 Trichoderma atroviride (SEQ ID NO: 532), pep12 EKJ73392 Fusarium pseudograminearum (SEQ ID NO: 533), pep12 XP_388759 Gibberella zeae (SEQ ID NO: 534) и pep12 EFY95489 Metarhizium anisopliae (SEQ ID NO: 535), XP_964574.1 N. crassa (SEQ ID NO: 536), XP_003659978.1 M. thermophila (SEQ ID NO: 537) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза pep12, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 530-537. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 530-537.
В некоторых вариантах осуществления pep8 представляет собой pep12 T. reesei. Аминокислотная последовательность, кодируемая pep12 T. reesei, указана в SEQ ID NO: 530. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 530. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 530.
Трипсин-подобные сериновые протеазы
Трипсин-подобные сериновые протеазы представляют собой ферменты с субстратной специфичностью, сходной с субстратной специфичностью трипсина. Трипсин-подобные сериновые протеазы используют остаток серина для гидролиза пептидных связей в полипептидах и белках. Как правило, трипсин-подобные сериновые протеазы расщепляют пептидные связи после положительно заряженного аминокислотного остатка. Трипсин-подобные сериновые протеазы из эукариотических организмов, таких как грибы Trichoderma, включают трипсин 1, трипсин 2 и мезотрипсин. Такие трипсин-подобные сериновые протеазы, как правило, содержат каталитическую триаду из трех аминокислотных остатков (таких как гистидин, аспартат и серин), которая образует переключатель заряда, который служит для того, чтобы делать серин активного центра нуклеофильным. Эукариотические трипсин-подобные сериновые протеазы, кроме того, включают "оксианионную выемку", образованную атомами водорода амида основной цепи глицина и серина, которая может способствовать идентификации полипептидов в качестве трипсин-подобных сериновых протеаз.
В клетках грибов Trichoderma идентифицирована одна трипсин-подобная сериновая протеаза: tsp1 (tre73897). Как рассмотрено ниже, было продемонстрировано, что tsp1 имеет значительное влияние на экспрессию рекомбинантных полипептидов, таких как иммуноглобулины.
Как рассмотрено ниже в примере 3, сериновые протеазы очищали из Trichoderma, и показано, что они обладают несколькими типами протеазной активности, которая разрушает белки млекопитающих. Среди этих активностей, tsp1 был идентифицирован в качестве трипсин-подобной сериновой протеазы. Затем ген протеазы tsp1 удаляли из клеток грибов Trichoderma, и было продемонстрировано, что удаление tsp1 обеспечивало значительное снижение общей протеазной активности, что приводило к повышению стабилизации белков млекопитающих, продуцированных клетками.
Примеры пригодных протеаз tsp1 включают, но не ограничиваются ими, tsp1 Trichoderma reesei (SEQ ID NO: 66), jgi|Triat2|298187 Trichoderma atroviride (SEQ ID NO: 67), jgi|TriviGv29_8_2 (SEQ ID NO: 68), gi|145583579 Hypocrea lixii (SEQ ID NO: 69), gi|63025000 Hypocrea lixii (SEQ ID NO: 70), gi|156052735 Sclerotinia sclerotiorum (SEQ ID NO: 71), gi|154314937 Botryotinia fuckeliana (SEQ ID NO: 72), gi|169605891 Phaeosphaeria nodorum (SEQ ID NO: 73), Leptosphaeria maculans gi|312219044 (SEQ ID NO: 74), gi|37992773 Verticillium dahliae (SEQ ID NO: 75), gi|l072114 Cochliobolus carbonum (SEQ ID NO: 76), gi|322695345 Metarhizium acridum (SEQ ID NO: 77), gi|4768909 (SEQ ID NO: 78), gi|464963 (SEQ ID NO: 79) Metarhizium anisopliae, gi|46139299 Gibberella zeae (SEQ ID NO: 80), Metarhizium anisopliae (SEQ ID NO: 81), gi67523821 A. nidulans (SEQ ID NO: 538) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза tsp1, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 66-81, SEQ ID NO: 538. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 66-81, SEQ ID NO: 538.
В некоторых вариантах осуществления tsp1 представляет собой tsp1 T. reesei. Аминокислотная последовательность, кодируемая tsp1 T. reesei, указана в SEQ ID NO: 66. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 66. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 66.
Субтилизиновые протеазы
Субтилизиновые протеазы представляют собой ферменты с субстратной специфичностью, сходной с субстратной специфичностью субтилизина. Субтилизиновые протеазы используют остаток серина для гидролиза пептидных связей в полипептидах и белках. Как правило, субтилизиновые протеазы представляют собой сериновые протеазы, которые содержат каталитическую триаду из трех аминокислот: аспартата, гистидина и серина. Расположение этих каталитических остатков является общим с прототипным субтилизином из Bacillus licheniformis. Субтилизиновые протеазы из эукариотических организмов, таких как грибы Trichoderma, включают фурин, MBTPS1 и TPP2. Эукариотические трипсин-подобные сериновые протеазы дополнительно включают остаток аспарагиновой кислоты в оксианионной выемке. Субтилизиновая протеаза slp7 сходна с седолизиновой протеазой tpp1.
В клетках грибов Trichoderma идентифицировано семь субтилизиновых протеаз: slp1 (tre51365); slp1 (tre123244); slp3 (tre123234); slp5 (tre64719), slp6 (tre121495), slp7 (tre123865) и slp8 (tre58698).
Slp1
Примеры пригодных протеаз slp1 включают, но не ограничиваются ими, slp1 Trichoderma reesei (SEQ ID NO: 82), jgi|Triat2 Trichoderma atroviride (SEQ ID NO: 83), jgi|Triat2 Trichoderma atroviride (SEQ ID NO: 84), jgi|TriviGv29_8_2 Trichoderma virens (SEQ ID NO: 85), gi|145583581 Hypocrea lixii (SEQ ID NO: 86), gi|322694632 Metarhizium acridum (SEQ ID NO: 87), gi|342877080 Fusarium oxysporum (SEQ ID NO: 88), gi|46139915 Gibberella zeae (SEQ ID NO: 89), gi|170674476 Epichloe festucae (SEQ ID NO: 90), gi|302893164 Nectria haematococca (SEQ ID NO: 91), gi|336266150 Sordaria macrospore (SEQ ID NO: 92), gi|310797947 Glomerella graminicola (SEQ ID NO: 93), gi|336469805 Neurospora tetrasperma (SEQ ID NO: 94), gi|85086707 Neurospora crassa (SEQ ID NO: 95), gi|145608997 Magnaporthe oryzae (SEQ ID NO: 96), gi|116208730 Chaetomium globosum (SEQ ID NO: 97), gi367029081 M. thermophila (SEQ ID NO: 539) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза slp1, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 82-97, SEQ ID NO: 539. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 82-97, SEQ ID NO: 539.
В некоторых вариантах осуществления slp1 представляет собой slp1 T. reesei. Аминокислотная последовательность, кодируемая slp1 T. reesei, указана в SEQ ID NO: 82. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 82. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 82.
Slp2
Примеры пригодных протеаз slp2 включают, но не ограничиваются ими, slp2 Trichoderma reesei (SEQ ID NO: 98), jgi|Triat2 T. atroviride (SEQ ID NO: 99), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 100), gi|115111226 Hypocrea lixii (SEQ ID NO: 101), gi|70997972 Aspergillus fumigatus (SEQ ID NO: 102), gi|302915240 Nectria haematococca (SEQ ID NO: 103), gi|46105128 Gibberella zeae (SEQ ID NO: 104), gi|68165000 Isaria farinose (SEQ ID NO: 105), gi|310797854 Glomerella graminicola (SEQ ID NO: 106), gi|170674491 Epichloe festucae (SEQ ID NO: 107), gi|322697754 Metarhizium acridum (SEQ ID NO: 108), gi|147225254 Acremonium sp. F11177 (SEQ ID NO: 109), gi|15808807 Ophiostoma piliferum (SEQ ID NO: 110), gi|336463649 Neurospora tetrasperma (SEQ ID NO: 111), gi|340992600 Chaetomium thermophilum (SEQ ID NO: 112), gi|254351265 Metarhizium flavoviride (SEQ ID NO: 113), gi|171680111 Podospora anserine (SEQ ID NO: 114), gi|39943180 Magnaporthe oryzae (SEQ ID NO: 115), gi|156058540 Sclerotinia sclerotiorum (SEQ ID NO: 116), gi|242790441 Talaromyces stipitatus (SEQ ID NO: 117), gi367021472 M. thermophila (SEQ ID NO: 540), gi145237646 A. niger (SEQ ID NO: 541), gi169780712 A. oryzae (SEQ ID NO: 542), gi255955889 P. chrysogenum (SEQ ID NO: 543), gi259489544 A. nidulans (SEQ ID NO: 544), gi85084841 N. crassa (SEQ ID NO: 545) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза slp2, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 98-117, SEQ ID NO: 540-545. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 98-117, SEQ ID NO: 540-545.
В некоторых вариантах осуществления slp2 представляет собой slp2 T. reesei. Аминокислотная последовательность, кодируемая slp2 T. reesei, указана в SEQ ID NO: 98. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 98. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 98.
Slp3
Примеры пригодных протеаз slp3 включают, но не ограничиваются ими, slp2 Trichoderma reesei (SEQ ID NO: 166), jgi|Triat2 T. atroviride (SEQ ID NO: 167), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 168), gi|124295071 Hypocrea koningii (SEQ ID NO: 169), gi|130750164 Purpureocillium lilacinum (SEQ ID NO: 170), gi|16215677 Metarhizium anisopliae (SEQ ID NO: 171), gi|90655148 Hirsutella rhossiliensis (SEQ ID NO: 172), gi|18542429 Tolypocladium inflation (SEQ ID NO: 173), gi|19171215 Metacordyceps chlamydosporia (SEQ ID NO: 174), gi|346321368 Cordyceps militaris (SEQ ID NO: 175), gi|628051 Fusarium sp. (SEQ ID NO: 176), gi|336471881 Neurospora tetrasperma (SEQ ID NO: 177), gi|116197403 Chaetomium globosum (SEQ ID NO: 178), gi|85084841 Neurospora crassa (SEQ ID NO: 179), gi|56201265 Fusarium oxysporum (SEQ ID NO: 180), gi|46114268 Gibberella zeae (SEQ ID NO: 181), gi367026259 M. thermophila (SEQ ID NO: 546), gi67538776 A. nidulans (SEQ ID NO: 547), gi169771349 A. oryzae (SEQ ID NO: 222), gi470729 A. niger (SEQ ID NO: 223) и их гомологи.
Таким образом, в определенных вариантах осуществления протеаза по настоящему изобретению, как правило, протеаза slp3, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 166-181, SEQ ID NO: 546-547, SEQ ID NO: 222-223. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 166-181, SEQ ID NO: 546-547, SEQ ID NO: 222-223.
В некоторых вариантах осуществления slp3 представляет собой slp3 T. reesei. Аминокислотная последовательность, кодируемая slp3 T. reesei, указана в SEQ ID NO: 166. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 166. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 166.
Slp5
Примеры пригодных протеаз slp5 включают, но не ограничиваются ими, slp5 Trichoderma reesei (SEQ ID NO: 200), jgi|Triat2 T. atroviride (SEQ ID NO: 201), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 202), gi|118161442 Hypocrea lixii (SEQ ID NO: 203), gi|342883549 Fusarium oxysporum (SEQ ID NO: 204), gi|46135733 Gibberella zeae (SEQ ID NO: 205), gi|310796396 Glomerella graminicola (SEQ ID NO: 206), gi|302927954 Nectria haematococca (SEQ ID NO: 207), gi|346319783 Cordyceps militaris (SEQ ID NO: 208), gi|85094084 Neurospora crassa (SEQ ID NO: 209), gi|336467281 Neurospora tetrasperma (SEQ ID NO: 210), gi|346971706 Verticillium dahliae (SEQ ID NO: 211), gi|347001418 Thielavia terrestris (SEQ ID NO: 212), gi|145605493 Magnaporthe oryzae (SEQ ID NO: 213), gi367032200 M. thermophila (SEQ ID NO: 548), gi62816282 P. chrysogenum (SEQ ID NO: 549) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза slp5, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 200-213, SEQ ID NO: 548-549. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 200-213, SEQ ID NO: 548-549.
В некоторых вариантах осуществления slp5 представляет собой slp5 T. reesei. Аминокислотная последовательность, кодируемая slp5 T. reesei, указана в SEQ ID NO: 200. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 200. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 200.
Slp6
Примеры пригодных протеаз slp6 включают, но не ограничиваются ими, slp6 Trichoderma reesei (SEQ ID NO: 214), jgi|Triat2 T. atroviride (SEQ ID NO: 215), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 216), gi|29421423 Hypocrea virens (SEQ ID NO: 217), gi|145583127 Hypocrea lixii (SEQ ID NO: 218), gi|30144643 Trichoderma hamatum (SEQ ID NO: 219), gi|2295 Aspergillus fumigatus (SEQ ID NO: 220), gi|115391147 Aspergillus terreus (SEQ ID NO: 221), gi|169771349 Aspergillus oryzae (SEQ ID NO: 222), gi|470729 Aspergillus niger (SEQ ID NO: 223), gi|310794714 Glomerella graminicola (SEQ ID NO: 224), gi|46114946 Gibberella zeae (SEQ ID NO: 225), gi|342873942 Fusarium oxysporum (SEQ ID NO: 226), gi|302884541 Nectria haematococca (SEQ ID NO: 227), gi|119500190 Neosartorya fischeri (SEQ ID NO: 228), gi|302413161 Verticillium alboatrum (SEQ ID NO: 229), gi|310790144 Glomerella graminicola (SEQ ID NO: 230), gi85090020 N. crassa (SEQ ID NO: 550) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза slp6, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 214-230, SEQ ID NO: 550. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 214-230, SEQ ID NO: 550.
В некоторых вариантах осуществления slp6 представляет собой slp6 T. reesei. Аминокислотная последовательность, кодируемая slp6 T. reesei, указана в SEQ ID NO: 214. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 214. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 214.
Slp7
Примеры пригодных протеаз slp7 включают, но не ограничиваются ими, slp7 Trichoderma reesei (SEQ ID NO: 231), jgi|Triat2 T. atroviride (SEQ ID NO: 232), jgi|TriviGv29_8_2 T. virens (SEQ ID NO: 233), gi|322710320 Metarhizium anisopliae (SEQ ID NO: 234), gi|302915000 Nectria haematococca (SEQ ID NO: 235), gi|347009020, gi367024935 Myceliophthora thermophila (SEQ ID NO: 236), gi|46137655 Gibberella zeae (SEQ ID NO: 237), gi|346996549 Thielavia terrestris (SEQ ID NO: 238), gi|145610733 Magnaporthe oryzae (SEQ ID NO: 239), gi67541991 A. nidulans (SEQ ID NO: 551), gi255933786 P. chrysogenum (SEQ ID NO: 552), gi317036543 A. niger (SEQ ID NO: 553), gi169782882 A. oryzae (SEQ ID NO: 554), gi85109979 N. crassa (SEQ ID NO: 555) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза slp7, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 231-239, SEQ ID NO: 551-555. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 231-239, SEQ ID NO: 551-555.
В некоторых вариантах осуществления slp7 представляет собой slp7 T. reesei. Аминокислотная последовательность, кодируемая slp7 T. reesei, указана в SEQ ID NO: 231. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 231. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 231.
Slp8
Примеры пригодных протеаз slp8 включают, но не ограничиваются ими, slp8 Trichoderma reesei (SEQ ID NO: 240), jgi|Triat2|198568 T. atroviride (SEQ ID NO: 241), jgi|TriviGv29_8_2|33902 T. virens (SEQ ID NO: 242) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 240-242. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 240-242.
В некоторых вариантах осуществления slp8 представляет собой slp8 T. reesei. Аминокислотная последовательность, кодируемая slp8 T. reesei, указана в SEQ ID NO: 240. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 240. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 240.
Глутаминовые протеазы
Глутаминовые протеазы представляют собой ферменты, которые гидролизуют пептидные связи в полипептидах и белках. Глутаминовые протеазы не чувствительны к пепстатину A, и, таким образом, их иногда называют нечувствительными к пепстатину кислыми протеазами. Между тем как глутаминовые протеазы ранее объединяли в группу с аспарагиновыми протеазами и часто их вместе называют кислыми протеазами, в настоящее время обнаружено, что глутаминовые протеазы имеют значительно отличающиеся остатки активного центра от аспарагиновых протеаз.
В клетках грибов Trichoderma идентифицировано две глутаминовых протеазы: gap1 (tre69555) и gap2 (tre106661).
Gap1
Примеры пригодных протеаз gap1 включают, но не ограничиваются ими, gap1 Trichoderma reesei (SEQ ID NO: 118), jgi|Triat2|40863 T. atroviride (SEQ ID NO: 119), jgi|TriviGv29_8_2|192684 T. virens (SEQ ID NO: 120), gi|238499183 Aspergillus flavus (SEQ ID NO: 121), gi|145251555 Aspergillus niger (SEQ ID NO: 122), gi|1l5491521 (SEQ ID NO: 123), gi|37154543 (SEQ ID NO: 124), gi|48425531 (SEQ ID NO: 125), gi|351873 (SEQ ID NO: 126) Aspergillus terreus, gi|346997245 Thielavia terrestris (SEQ ID NO: 127), gi|255940586 Penicillium chrysogenum (SEQ ID NO: 128), gi367026504 M. thermophila (SEQ ID NO: 574), gi317150886 A. oryzae (SEQ ID NO: 575), gi85097968 N. crassa (SEQ ID NO: 576), gi131056 A. niger (SEQ ID NO: 577), gi255930123 P. chrysogenum (SEQ ID NO: 578), gi145236956 A. niger (SEQ ID NO: 579), gi169772955 A. oryzae (SEQ ID NO: 580), gi145249222 A. niger (SEQ ID NO: 581), gi67525839 A. nidulans (SEQ ID NO: 582), gi169785367 A. oryzae (SEQ ID NO: 583), gi255955319 P. chrysogenum (SEQ ID NO: 584), gi367019352 M. thermophila (SEQ ID NO: 585), gi391863974 A. oryzae (SEQ ID NO: 586), gi367024513 M. thermophila (SEQ ID NO: 587) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза gap1, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 118-128, SEQ ID NO: 574-587. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 118-128, SEQ ID NO: 574-587.
В некоторых вариантах осуществления gap1 представляет собой gap1 T. reesei. Аминокислотная последовательность, кодируемая gap1 T. reesei, указана в SEQ ID NO: 118. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 118. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 118.
Gap2
Примеры пригодных протеаз gap2 включают, но не ограничиваются ими, gap2 Trichoderma reesei (SEQ ID NO: 129), jgi|Triat2|298116 T. atroviride (SEQ ID NO: 130), jgi|TriviGv29_8_2|30331 (SEQ ID NO: 131), jgi|TriviGv29_8_2|225131 (SEQ ID NO: 132) T. virens, gi|238499183 Aspergillus flavus (SEQ ID NO: 133), gi|145251555 Aspergillus niger (SEQ ID NO: 134), gi|67901056 Aspergillus nidulans (SEQ ID NO: 135), gi|121711990 Aspergillus clavatus (SEQ ID NO: 136), gi|70986250 Aspergillus fumigatus (SEQ ID NO: 137), gi|212534108 Penicillium marneffei (SEQ ID NO: 138), gi|242789335 Talaromyces stipitatus (SEQ ID NO: 139), gi|320591529 Grosmannia clavigera (SEQ ID NO: 140), gi|119474281 Neosartorya fischeri (SEQ ID NO: 141), gi|212527274 Penicillium marneffei (SEQ ID NO: 142), gi|255940586 (SEQ ID NO: 143), gi|131056 Penicillium chrysogenum (SEQ ID NO: 144), gi367030275 M. thermophila (SEQ ID NO: 588) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза gap2, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 129-144, SEQ ID NO: 588. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 129-144, SEQ ID NO: 588.
В некоторых вариантах осуществления gap2 представляет собой gap2 T. reesei. Аминокислотная последовательность, кодируемая gap2 T. reesei, указана в SEQ ID NO: 129. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 129. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 129.
Седолизиновые протеазы
Седолизиновые протеазы представляют собой ферменты, которые используют остаток серина для гидролиза пептидных связей в полипептидах и белках. Седолизиновые протеазы, как правило, содержат уникальную каталитическую триаду из серина, глутамата и аспартата. Седолизиновые протеазы также содержат остаток аспартата в оксианионной выемке. Седолизиновые протеазы из эукариотических организмов, таких как грибы Trichoderma, включают трипептидилпептидазу.
Примеры пригодных протеаз tpp1 включают, но не ограничиваются ими, tpp1 Trichoderma reesei (SEQ ID NO: 145), jgi|Triat2|188756 T. atroviride (SEQ ID NO: 146), jgi|TriviGv29_8_2|217176 T. virens (SEQ ID NO: 147), gi|70993168 Aspergillus fumigatus (SEQ ID NO: 148), gi|169776800 Aspergillus oryzae (SEQ ID NO: 149), gi|145236399 Aspergillus niger (SEQ ID NO: 150), gi|121708799 Aspergillus clavatus (SEQ ID NO: 151), gi|145239871 Aspergillus niger (SEQ ID NO: 152), gi|121714541 Aspergillus clavatus (SEQ ID NO: 153), gi|115387645 Aspergillus terreus (SEQ ID NO: 154), gi|70982015 Aspergillus fumigatus (SEQ ID NO: 155), gi|156045898 Sclerotinia sclerotiorum (SEQ ID NO: 156), gi|154321758 Botryotinia juckeliana (SEQ ID NO: 157), gi|119499774 Neosartorya fischeri (SEQ ID NO: 158), gi|242798348 Talaromyces stipitatus (SEQ ID NO: 159), gi|212541546 Penicillium marneffei (SEQ ID NO: 160), gi|46114460 Gibberella zeae (SEQ ID NO: 161), gi|342890694 Fusarium oxysporum (SEQ ID NO: 162), gi|320592937 Grosmannia clavigera (SEQ ID NO: 163), gi|302406186 Verticillium alboatrum (SEQ ID NO: 164), gi|346971444 Verticillium dahliae (SEQ ID NO: 165), CAE51075.1 A. fumigatus (SEQ ID NO: 556), XP_001820835.1 A. oryzae (SEQ ID NO: 557), XP_002564029.1 P. chrysogenum (SEQ ID NO: 558), XP_664805.1 A. nidulans (SEQ ID NO: 559), XP_002565814.1 P. chrysogenum (SEQ ID NO: 560), XP_003663689.1 M. thermophila (SEQ ID NO: 561), XP_958412.1 N. crassa (SEQ ID NO: 562), XP_001394118.1 A. niger (SEQ ID NO: 563), CAE17674.1 A. fumigatus (SEQ ID NO: 564), XP_001400873.1 A. niger (SEQ ID NO: 565), CAE46473.1 A. fumigatus (SEQ ID NO: 566), XP_002373530.1 A. oryzae (SEQ ID NO: 567), XP_660624.1 A. nidulans (SEQ ID NO: 568), XP_002562943.1 P. chrysogenum (SEQ ID NO: 569), CAE17675.1 A. fumigatus (SEQ ID NO: 570), EAL86850.2 A. fumigatus (SEQ ID NO: 571), XP_961957.1 N. crassa (SEQ ID NO: 572), BAB97387.1 A. oryzae (SEQ ID NO: 573) и их гомологи.
Таким образом, в определенных вариантах осуществления, протеаза по настоящему изобретению, как правило, протеаза tpp1, имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с аминокислотной последовательностью, выбранной из SEQ ID NO: 145-165, SEQ ID NO: 556-573. В некоторых вариантах осуществления протеаза обладает 100% идентичностью с аминокислотной последовательностью, выбранной из SEQ ID NO: 145-165, SEQ ID NO: 556-573.
В некоторых вариантах осуществления tpp1 представляет собой tpp1 T. reesei. Аминокислотная последовательность, кодируемая tpp1 T. reesei, указана в SEQ ID NO: 145. В других вариантах осуществления протеаза по настоящему изобретению имеет аминокислотную последовательность, обладающую 50% или более идентичностью (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или более) с SEQ ID NO: 145. В следующих вариантах осуществления протеаза обладает 100% идентичностью с SEQ ID NO: 145.
Гомологичные протеазы
Другие варианты осуществления настоящего изобретения относятся к снижению активности протеаз, которые являются гомологичными протеазам по настоящему изобретению. "Гомология", как используют в рамках изобретения, относится к сходству последовательностей между эталонной последовательностью и по меньшей мере фрагментом второй последовательности. Гомологи можно идентифицировать любым способом, известным в данной области, предпочтительно, с использованием инструмента BLAST для сравнения эталонной последовательности с одной второй последовательностью или фрагментом последовательности или с базой данных последовательностей. Как описано ниже, BLAST сравнивает последовательности на основе процентной идентичности и сходства.
Термины "идентичный" или процентная "идентичность", в контексте двух или более последовательностей нуклеиновых кислот или аминокислотных последовательностей, относится к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми. Две последовательности являются "по существу идентичными", если две последовательности обладают указанным процентом аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. 29% идентичностью, необязательно 30%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 99% или 100% идентичностью на протяжении указанной области, или, когда не указано, на протяжении всей последовательности), при сравнении и выравнивании для максимального соответствия на протяжении окна сравнения или намеченной области при определении с использованием одного из представленных ниже алгоритмов сравнения последовательностей или путем выравнивания вручную и визуального исследования. Необязательно, идентичность существует на протяжении области, которая имеет длину по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот), или более предпочтительно на протяжении области, которая имеет длину от 100 до 500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
Для сравнения последовательностей, как правило, одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают анализируемые последовательности. При использовании алгоритма сравнения последовательностей анализируемые и эталонные последовательности вводят в компьютер, обозначают координаты подпоследовательностей, если необходимо, и указывают параметры программы алгоритма для последовательностей. Можно использовать параметры по умолчанию или можно назначать альтернативные параметры. Затем на основании параметров программы алгоритм сравнения последовательностей вычисляет процентную идентичность последовательностей для анализируемой последовательности относительно эталонной последовательности. При сравнении двух последовательностей в отношении идентичности, не является необходимым, чтобы последовательности были непрерывными, однако любой пропуск будет обеспечивать штраф, который может снизить общую процентную идентичностью. Для blastn параметрами по умолчанию являются штраф за внесение пропуска=5 и штраф за продолжение пропуска=2. Для blastp параметрами по умолчанию являются штраф за внесение пропуска=11 и штраф за продолжение пропуска=1.
"Окно сравнения", как используют в рамках изобретения, относится к сегменту любого из числа смежных положений, включая, но, не ограничиваясь этим, от 20 до 600, как правило, от приблизительно 50 до приблизительно 200, более конкретно от приблизительно 100 до приблизительно 150, в которых последовательность может быть сравнена с эталонной последовательностью с таким же числом смежных положений после того, как две последовательности оптимально выровнены. Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для сравнения может быть проведено, например, посредством алгоритма локальной гомологии Smith и Waterman (1981), посредством алгоритма выравнивания по гомологии Needleman и Wunsch (1970) J. Mol. Biol. 48: 443, посредством поиска сходства способом Pearson и Lipman, (1988) Proc. Natl. Acad. Sci. USA 85(8): 2444-2448, с помощью компьютеризованного использования этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), или посредством выравнивания вручную и визуального исследования [см., например, Brent et al., (2003) Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (Ringbou Ed)].
Двумя примерами алгоритмов, которые пригодны для определения процентной идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., (1977) Nucleic Acids Res. 25(17): -3402; и Altschul et al., (1990) J. Mol Biol 215(3)-403-410, соответственно. Программное обеспечение для осуществления анализов BLAST является общедоступным через National Center for Biotechnology Information. Этот алгоритм включает первичную идентификацию пар последовательностей с максимальным сходством (HSP) посредством идентификации коротких слов с длиной W в последовательности запроса, которые либо совпадают, либо удовлетворяют некоторому оцениваемому положительно порогу значения Т при выравнивании со словом такой же длины в последовательности из базы данных. Т определяется как порог значений для соседних слов (Altschul et al., выше). Эти изначальные совпадения соседних слов выступают в роли предшественников для начала поиска при обнаружении более длинных HSP, содержащих их. Поиск совпадений слов распространяется в обоих направлениях вдоль каждой последовательности до тех пор, пока суммарное значение для выравнивания может повышаться. Суммарные значения вычисляются с использованием для нуклеотидных последовательностей параметров M (значение компенсации за пару совпавших остатков; всегда >0) и N (значение штрафа для несовпадающих остатков; всегда <0). Для аминокислотных последовательностей для вычисления суммарного значения используется оценочная матрица. Продолжение поиска совпадений слов в каждом направлении останавливают, если суммарное значение для выравнивания снижается по величине X ниже максимально достижимого значения; суммарное значение снижается до нуля или ниже при накоплении одного или нескольких выровненных остатков с отрицательным значением, или если достигнут конец любой последовательности. Параметры алгоритма BLAST W, Т и X определяют чувствительность и скорость выравнивания. Программа BLASTN (для нуклеотидных последовательностей) использует в качестве параметров по умолчанию длину слова (W) 11, ожидаемое значение (Е) 10, М=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей программа BLASTP использует в качестве параметров по умолчанию длину слова 3 и ожидаемые значения (Е) 10 и оценочную матрицу BLOSUM62 [см. Henikoff and Henikoff, (1992) Proc Natl Acad Sci USA 89(22): 10915-10919], фрагменты для выравнивания (В) 50, ожидаемое значение (Е) 10, М=5, N=-4 и сравнение обеих цепей.
Алгоритм BLAST также осуществляет статистический анализ сходства между двумя последовательностями (см., например, Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90(12): 5873-5877). Одной мерой сходства, обеспечиваемой алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая обеспечивает указание вероятности, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями может произойти случайным образом. Например, нуклеиновая кислота рассматривается как сходная с контрольной последовательностью, если наименьшая суммарная вероятность при сравнении анализируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее чем приблизительно 0,2, более предпочтительно менее чем приблизительно 0,01 и наиболее предпочтительно менее чем приблизительно 0,001.
Помимо процента идентичности последовательности, указанного выше, другим показателем того, что две последовательности нуклеиновых кислот или два полипептида по существу идентичны, является то, что полипептид, кодируемый первой нуклеиновой кислотой, является иммунологически перекрестно реактивным с антителами, индуцированными против полипептида, кодируемого второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид, как правило, является по существу идентичным второму полипептиду, например, где два пептида отличаются только консервативными заменами. Другим показателем того, что две последовательности нуклеиновых кислот являются по существу идентичными, является то, что эти две молекулы или комплементарные им последовательности гибридизуются друг с другом в жестких условиях, как описано ниже. Другим показателем того, что две последовательности нуклеиновых кислот являются по существу идентичными, является то, что для амплификации их последовательностей можно использовать одни и те же праймеры.
Как описано в настоящем описании, протеазы по настоящему изобретению также могут включать протеазы, которые представляют собой консервативно модифицированные варианты протеаз, кодируемых генами протеаз, описанными выше. "Консервативно модифицированные варианты", как используют в рамках изобретения, включают индивидуальные замены, делеции или вставки в кодируемую аминокислотную последовательность, которые приводят к замене аминокислоты химически сходной аминокислотой. Таблицы консервативных замен, обеспечивающие функционально сходные аминокислоты, хорошо известны в данной области. Такие консервативно модифицированные замены являются дополнением и не исключают полиморфных вариантов, межвидовых гомологов и аллелей по изобретению. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг для друга: 1) аланин (A), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (T); и 8) цистеин (C), метионин (M) (см., например, Creighton, Proteins (1984)).
На фиг.45-48 представлены филогенетические деревья для аспарагиновых, субтилизиновых, глутаминовых и седолизиновых протеаз выбранных нитчатых грибов.
Способы снижения активности протеаз по изобретению
Следующие аспекты настоящего изобретения относятся к снижению активности протеаз, встречающихся в клетках нитчатых грибов, которые экспрессируют гетерологичный полипептид, такой как полипептид млекопитающих.
Активность протеаз, встречающихся в клетках нитчатых грибов, можно снижать любым способом, известным специалистам в данной области.
В некоторых вариантах осуществления снижения активности протеаз достигают путем снижения экспрессии протеазы, например, посредством модификации промотора или РНК-i.
В других вариантах осуществления снижения активности протеаз достигают путем модификации гена, кодирующего протеазу. Примеры таких модификаций включают, но не ограничиваются ими, мутацию с нокаутом, мутацию с укорочением, точковую мутацию, миссенс-мутацию, мутацию с заменой, мутацию со сдвигом рамки считывания, мутацию со вставкой, мутацию с дупликацией, мутацию с амплификацией, мутацию с транслокацией или мутацию с инверсией, и они приводят к снижению активности соответствующей протеазы. Способы внесения по меньшей мере одной мутации в представляющий интерес ген, кодирующий протеазу, хорошо известны в данной области и включают, но не ограничиваются ими, случайный мутагенез и скрининг, сайт-направленный мутагенез, ПЦР-мутагенез, инсерционный мутагенез, химический мутагенез и облучение.
В определенных вариантах осуществления модифицируют часть гена, кодирующего протеазу, такую как область, кодирующая каталитический домен, кодирующую область или последовательность контроля, требуемая для экспрессии кодирующей области. Такая последовательность контроля гена может представлять собой последовательность промотора или ее функциональную часть, т.е. часть, которая является достаточной для влияния на экспрессию гена. Например, последовательность промотора можно инактивировать, что приводит к отсутствию экспрессии, или нативную последовательность промотора можно заменять более слабым промотором для снижения экспрессии кодирующей последовательности. Другие последовательности контроля для возможной модификации включают, но не ограничиваются ими, лидерную последовательность, последовательность пропептида, сигнальную последовательность, терминатор транскрипции и активатор транскрипции.
Гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать с использованием технологий делеции генов для устранения или снижения экспрессии гена. Способы делеции генов обеспечивают частичное или полное удаление гена, тем самым, устраняя их экспрессию. В таких способах делецию гена можно осуществлять посредством гомологичной рекомбинации с использованием плазмиды, сконструированной так, чтобы она содержала смежные 5'- и 3'-области, фланкирующие ген.
Гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать путем внесения, замены и/или удаления одного или нескольких нуклеотидов в гене или его последовательности контроля, требуемой для транскрипции или трансляции гена. Например, нуклеотиды можно встраивать или удалять для внесения стоп-кодона, для удаления инициирующего кодона или для сдвига рамки считывания в открытой рамке считывания. Такую модификацию можно проводить способами, известными в данной области, включая, но, не ограничиваясь ими, сайт-направленный мутагенез и обеспечиваемый peR мутагенез (см., например, Botstein and Shortie, 1985, Science 229: 4719; Lo et al., 1985, Proceedings of the National Academy of Sciences USA 81: 2285; Higuchi et al., 1988, Nucleic Acids Research 16: 7351; Shimada, 1996, Meth. Mol. Bioi. 57: 157; Ho et al., 1989, Gene 77: 61; Horton et al., 1989, Gene 77: 61; и Sarkar and Sommer, 1990, BioTechniques 8: 404).
Кроме того, гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, можно модифицировать способом разрушения генов посредством встраивания в ген разрушающей конструкции нуклеиновой кислоты, содержащей фрагмент нуклеиновой кислоты, гомологичный гену, который обеспечит дупликацию области гомологии и заключит конструкцию ДНК между дуплицированными областями. Такое разрушение гена может устранять экспрессию гена, если встроенная конструкция отделяет промотор гена от кодирующей области или прерывает кодирующую последовательность, так чтобы образовывался нефункциональный продукт гена. Разрушающая конструкция может представлять собой просто ген селективного маркера, сопровождаемый 5'- и 3'-областями, гомологичными гену. Селективный маркер обеспечивает идентификацию трансформантов, содержащих разрушенный ген.
Гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать способом конверсии генов (см., например, Iglesias and Trautner, 1983, Molecular General Genetics 189: 573-76). Например, при конверсии генов нуклеотидную последовательность, соответствующую гену, подвергают мутагенезу in vitro с получением дефектной нуклеотидной последовательности, которую затем трансформируют в штамм Trichoderma с получением дефектного гена. Посредством гомологичной рекомбинации дефектная нуклеотидная последовательность заменяет эндогенный ген. Может быть желательным, чтобы дефектная нуклеотидная последовательность также содержала маркер для селекции трансформантов, содержащих дефектный ген.
Гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать с помощью общепринятых антисмысловых способов с использованием нуклеотидной последовательности, комплементарной нуклеотидной последовательности гена (см., например, Parish and Stoker, 1997, FEMS Microbiology Letters 154: 151-157). В частности, экспрессию гена клетками нитчатых грибов можно снижать или инактивировать путем введения в клетки нуклеотидной последовательности, комплементарной нуклеотидной последовательности гена, которая может транскрибироваться в штамме и способна гибридизоваться с мРНК, продуцируемой в клетках. Таким образом, в условиях, позволяющих комплементарной антисмысловой нуклеотидной последовательности гибридизоваться с мРНК, количество транслируемого белка снижается или транслируемый белок устраняется.
Кроме того, гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать общепринятыми способами РНК-интерференции (РНК-i) (см., например, WO 2005/056772 и WO 2008/080017).
Гены, кодирующие протеазы по настоящему изобретению, которые присутствуют в клетках нитчатых грибов, которые экспрессируют рекомбинантный полипептид, также можно модифицировать случайным или специфическим мутагенезом с использованием способов, хорошо известных в данной области, включая, но не ограничиваясь ими, химический мутагенез (см., например, Hopwood, The Isolation of Mutants in Methods in Microbiology (J.R. Norris and D.W. Ribbons, eds.) pp. 363-433, Academic Press, New York, 25 1970). Модификацию гена можно проводить, подвергая клетки нитчатых грибов мутагенезу и скринингу мутантных клеток, в которых экспрессия гена снижена или инактивирована. Мутагенез, который может быть специфическим или случайным, можно проводить, например, используя пригодный физический или химический агент мутагенеза, используя подходящий олигонуклеотид, подвергая последовательность ДНК обеспечиваемому peR мутагенезу или с помощью любой их комбинации. Примеры физических и химических агентов мутагенеза включают, но не ограничиваются ими, облучение ультрафиолетовым (УФ) излучением, гидроксиламин, N-метил-N'-нитро-N-нитрозогуанидин (MNNG), N-метил-N'-нитрозогуанидин (NTG), O-метилгидроксиламин, азотистую кислоту, этилметансульфонат (EMS), бисульфит натрия, муравьиную кислоту и нуклеотидные аналоги. Когда такие агенты используют, мутагенез, как правило, проводят путем инкубации клеток Trichoderma, подлежащих мутагенезу, в присутствии выбранного агента мутагенеза в подходящих условиях, а затем выбора мутантов, проявляющих сниженную экспрессию или не проявляющих экспрессию гена.
В определенных вариантах осуществления по меньшей мере одна мутация или модификация в представляющем интерес гене, кодирующем протеазу, по настоящему изобретению приводит к модифицированной протеазе, которая не обладает поддающейся обнаружению протеазной активностью. В других вариантах осуществления по меньшей мере одна модификация в гене, кодирующем протеазу, по настоящему изобретению приводит к модифицированной протеазе, которая обладает по меньшей мере на 25% меньшей, по меньшей мере на 50% меньшей, по меньшей мере на 75% меньшей, по меньшей мере на 90% меньшей, по меньшей мере на 95% меньшей, по меньшей мере на 100% меньшей, по меньшей мере на 200% меньшей, по меньшей мере на 300% меньшей, по меньшей мере на 400% меньшей, по меньшей мере на 500% меньшей, по меньшей мере на 600% меньшей, по меньшей мере на 700% меньшей, по меньшей мере 800% меньшей, по меньшей мере 900% меньшей, по меньшей мере на 1000% или на более высокий процент меньшей протеазной активностью по сравнению с соответствующей немодифицированной протеазой.
В определенных вариантах осуществления, например, в клетке Trichoderma, по меньшей мере одна мутация или модификация в гене, кодирующем протеазу, по настоящему изобретению приводит к снижению общей активности протеазы до 49% или менее, как правило, в случае мутации по меньшей мере в 2 различных генах протеаз, или до 31% или менее, как правило, в случае мутации по меньшей мере в 3 различных генах протеаз, или до 13% или менее, как правило, в случае мутации по меньшей мере в 4 различных генах протеаз, или до 10% или менее, как правило, в случае мутации по меньшей мере в 5 различных генах протеаз, или до 6,3% или менее, как правило, в случае мутации по меньшей мере в 6 различных генах протеаз, или до 5,5% или менее, как правило, в случае мутации по меньшей мере в 7 различных генах протеаз, относительно общей протеазной активности соответствующей исходной клетки Trichoderma.
Гетерологичные полипептиды по изобретению
Изобретение, описанное в настоящем описании, кроме того, относится к увеличению продукции гетерологичных полипептидов в клетках нитчатых грибов, которые экспрессируют такие гетерологичные полипептиды, путем снижения активности протеаз, встречающихся в клетках.
Как используют в рамках изобретения, "гетерологичный полипептид" относится к полипептиду, который не встречается в природе (т.е. не эндогенный) в клетке нитчатых грибов по настоящему изобретению, или который экспрессируется на повышенном уровне в клетке нитчатых грибов по сравнению с эндогенной версией полипептида. В определенных вариантах осуществления гетерологичный полипептид представляет собой полипептид млекопитающих. В других вариантах осуществления гетерологичный полипептид представляет собой полипептид не млекопитающих.
Полипептиды млекопитающих
Полипептиды млекопитающих по настоящему изобретению могут представлять собой любой полипептид млекопитающих, обладающий представляющей интерес биологической активностью. Как используют в рамках изобретения, "полипептид млекопитающих" представляет собой полипептид, который естественным образом экспрессируется у млекопитающего, полипептид, который происходит из полипептида, который естественным образом экспрессируется у млекопитающего, или их фрагмент. Полипептид млекопитающих также включает пептиды и олигопептиды, которые сохраняют биологическую активность. Полипептиды млекопитающих по настоящему изобретению также могут включать два или более полипептидов, которые скомбинированы с образованием кодируемого продукта. Полипептиды млекопитающих по настоящему изобретению, кроме того, могут включать слитые полипептиды, которые содержат комбинацию частичных или полных аминокислотных последовательностей, полученных по меньшей мере из двух различных полипептидов. Полипептиды млекопитающих также могут включать встречающиеся в природе аллельные и модифицированные способами инженерии версии любого из описанных полипептидов млекопитающих и гибридных полипептидов млекопитающих.
Полипептид млекопитающих может представлять собой естественным образом гликозилированный полипептид или естественным образом негликозилированный полипептид.
Примеры пригодных полипептидов млекопитающих включают, но не ограничиваются ими, иммуноглобулины, антитела, антигены, противомикробные пептиды, ферменты, факторы роста, гормоны, интерфероны, цитокины, интерлейкины, иммуномодуляторы, нейротрансмиттеры, рецепторы, репортерные белки, структурные белки и факторы транскрипции.
Конкретные примеры пригодных полипептидов млекопитающих включают, но не ограничиваются ими, иммуноглобулины, тяжелые цепи иммуноглобулинов, легкие цепи иммуноглобулинов, моноклональные антитела, гибридные антитела, фрагменты антител F(ab')2, фрагменты антител F(ab), Fv-молекулы, одноцепочечные Fv-антитела, димерные фрагменты антител, тримерные фрагменты антител, функциональные фрагменты антител, иммуноадгезины, инсулиноподобный фактор роста 1, гормон роста, инсулин, интерферон-альфа 2b, фибробластный фактор роста 21, сывороточный альбумин человека, антитела животных семейства верблюжьих и/или фрагменты антител, однодоменные антитела, мультимерные однодоменные антитела и эритропоэтин.
Другие примеры пригодных белков млекопитающих включают, но не ограничиваются ими, оксидоредуктазу, трансферазу, гидролазу, лиазу, изомеразу, лигазу, аминопептидазу, амилазу, карбогидразу, карбоксипептидазу, каталазу, гликозилтрансферазу, дезоксирибонуклеазу, эстеразу, галактозидазу, бета-галактозидазу, глюкозидазу, глюкуронидазу, глюкуроноилэстеразу, галопероксидазу, инвертазу, липазу, оксидазу, фосфолипазу, протеолитический фермент, рибонуклеазу, урокиназу, альбумин, коллаген, тропоэластин и эластин.
Полипептиды не млекопитающих
Полипептиды не млекопитающих по настоящему изобретению могут представлять собой любой полипептид не млекопитающих, обладающий представляющей интерес биологической активностью. Как используют в рамках изобретения, "полипептид не млекопитающих" представляет собой полипептид, который естественным образом экспрессируется в организме, не являющемся млекопитающим, таком как клетка грибов, полипептид, который происходит из полипептида, который естественным образом экспрессируется в организме, не являющемся млекопитающим, или их фрагмент. Полипептид не млекопитающих также включает пептиды и олигопептиды, которые сохраняют биологическую активность. Полипептиды не млекопитающих по настоящему изобретению также могут включать два или более полипептидов, которые скомбинированы с образованием кодируемого продукта. Полипептиды не млекопитающих по настоящему изобретению, кроме того, могут включать слитые полипептиды, которые содержат комбинацию частичных или полных аминокислотных последовательностей, полученных из по меньшей мере двух различных полипептидов. Полипептиды не млекопитающих также могут включать встречающиеся в природе аллельные и полученные способами инженерии варианты любого из описанных полипептидов не млекопитающих и гибридные полипептиды не млекопитающих.
Примеры пригодных полипептидов не млекопитающих включают, но не ограничиваются ими, аминопептидазы, амилазы, карбогидразы, карбоксипептидазы, каталазы, целлюлазы, хитиназы, кутиназы, дезоксирибонуклеазы, эстеразы, альфа-галактозидазы, бета-галактозидазы, глюкоамилазы, альфа-глюкозидазы, бета-глюкозидазы, инвертазы, лакказы, липазы, мутаназы, оксидазы, пектинолитические ферменты, пероксидазы, фосфолипазы, фитазы, полифенолоксидазы, протеолитические ферменты, рибонуклеазы, трансглутаминазы и ксиланазы.
Получение гетерологичных полипептидов
Представляющий интерес гетерологичный полипептид продуцируется клетками нитчатых грибов по настоящему изобретению, содержащими по меньшей мере три протеазы, имеющие сниженную активность, посредством культивирования клеток в питательной среде для продуцирования гетерологичного полипептида с использованием способов, известных в данной области. Например, клетки можно культивировать посредством культивирования во вращающихся флаконах, мелкомасштабной или крупномасштабной ферментации (включая непрерывную, периодическую, периодическую ферментацию с подпиткой или ферментацию в твердом состоянии) в лабораторных или промышленных ферментерах, проводимую в подходящей среде и в условиях, позволяющих экспрессию и/или выделение полипептида. Культивирование происходит в пригодной питательной среде, содержащей источники углерода и азота и неорганические соли, с использованием методик, известных в данной области. Подходящие среды доступны от коммерческих поставщиков или их можно получать в соответствии с опубликованными композициями (например, в каталогах American Type Culture Collection). Секретируемый полипептид можно выделять прямо из среды. Если полипептид не секретируется, его можно получать из клеточных лизатов.
Представляющий интерес гетерологичный полипептид, продуцируемый клеткой нитчатых грибов по настоящему изобретению, содержащий по меньшей мере три протеазы, имеющие сниженную активность, можно выявлять с использованием способов, известных в данной области, которые являются специфичными к гетерологичному полипептиду. Эти способы выявления могут включать, но не ограничиваются ими, применение специфических антител, высокоэффективную жидкостную хроматографию, капиллярную хроматографию, получение продукта фермента, исчезновение субстрата фермента и SDS-PAGE. Например, для определения активности фермента можно использовать ферментный анализ. Методики определения активности фермента известны в данной области для многих ферментов (см., например, O. Schomburg and M. Salzmann (eds.), Enzyme Handbook, Springer-Verlag, New York, 1990).
Полученный гетерологичный полипептид можно выделять способами, известными в данной области. Например, представляющий интерес гетерологичный полипептид можно выделять из культуральной среды по общепринятым методикам, включающим, но не ограничивающимся ими, центрифугирование, фильтрацию, экстракцию, распылительную сушку, упаривание и преципитацию. Затем выделенный гетерологичный полипептид можно далее очищать с помощью различных методик, известных в данной области, включая, но не ограничиваясь ими, хроматографию (например, ионообменную, аффинную, гидрофобную, хроматофокусирующую и эксклюзионную), методики электрофореза (например, препаративное изоэлектрическое фокусирование (IEF), дифференциальная растворимость (например, преципитация с сульфатом аммония) или экстракции (см., например, Protein Purification, J.-C. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989).
Получение полинуклеотидов, кодирующих гетерологичные полипептиды
Последовательности гетерологичных полинуклеотидов по настоящему изобретению получают любым пригодным способом, известным в данной области, включая, но не ограничиваясь ими, прямой химический синтез или клонирование. Для прямого химического синтеза образование полимера нуклеиновых кислот, как правило, вовлекает последовательное присоединение 3'-блокированных и 5'-блокированных мономеров нуклеотидов к концевой 5'-гидроксильной группе растущей нуклеотидной цепи, где каждое присоединение осуществляется посредством нуклеофильной атаки концевой 5'-гидроксильной группы растущей цепи на 3'-положение присоединяемого мономера, который, как правило, представляет собой производное фосфора, такое как фосфотриэфир, фосфорамидит и т.п. Такая технология известна средним специалистам в данной области и описана в соответствующих справочниках и литературе [например, в Matteucci et al., (1980) Tetrahedron Lett 21: 719-722; патенты США №№ 4500707; 5436327 и 5700637]. Кроме того, желаемые последовательности можно выделять из природных источников путем расщепления ДНК с использованием соответствующих ферментов рестрикции, разделения фрагментов с использованием гель-электрофореза, и, после этого, выделения желаемой последовательности нуклеиновой кислоты из геля способами, известными средним специалистам в данной области, такими как использование полимеразной цепной реакции (ПЦР; например, патент США № 4683195).
Каждый гетерологичный полинуклеотид по настоящему изобретению может быть включен в экспрессирующий вектор. "Экспрессирующий вектор" или "вектор" относится к соединению и/или композиции, которые трансдуцируют, трансформируют или инфицируют клетку-хозяина, тем самым, обеспечивая экспрессию клеткой нуклеиновых кислот и/или белков, отличных от нуклеиновых кислот и/или белков, нативных для клетки, или способом, не являющимся нативным для клетки. "Экспрессирующий вектор" содержит последовательность нуклеиновых кислот (обычно РНК или ДНК), подлежащую экспрессии клеткой-хозяином. Необязательно, экспрессирующий вектор также включает материалы, способствующие достижению вхождения нуклеиновой кислоты в клетку-хозяина, такие как вирус, липосома, белковая оболочка и т.п. Экспрессирующие векторы, предусматриваемые для применения в рамках настоящего изобретения, включают экспрессирующие векторы, в которые может быть встроена последовательность нуклеиновой кислоты, вместе с любыми предпочтительными или требуемыми функциональными элементами. Кроме того, экспрессирующий вектор должен представлять собой вектор, который может быть перенесен в клетку-хозяина и может реплицироваться в ней. Предпочтительными экспрессирующими векторами являются плазмиды, в частности, плазмиды с участками рестрикции, которые подтверждены документально и которые содержат функциональные элементы, предпочтительные или требуемые для транскрипции последовательности нуклеиновой кислоты. Такие плазмиды, а также другие экспрессирующие векторы, хорошо известны в данной области.
Включение индивидуальных полинуклеотидов можно осуществлять известными способами, которые включают, например, использование ферментов рестрикции (таких как BamHI, EcoRI, HhaI, XhoI, XmaI и т.д.) для расщепления конкретных участков в экспрессирующим векторе, например, плазмиде. Фермент рестрикции образует одноцепочечные концы, которые могут отжигаться с полинуклеотидом, имеющим или синтезированным так, чтобы он имел конец с последовательностью, комплементарной концам расщепленного экспрессирующего вектора. Отжиг проводят с использованием соответствующего фермента, например, ДНК-лигазы. Как будет понятно специалистам в данной области, как экспрессирующий вектор, так и желаемый полинуклеотид часто расщепляются тем же ферментом рестрикции, что, тем самым, обеспечивает то, что концы экспрессирующего вектора и концы полинуклеотида комплементарны друг другу. Кроме того, можно использовать ДНК-линкеры для облегчения связывания последовательностей нуклеиновых кислот в экспрессирующий вектор.
Также можно комбинировать ряд индивидуальных полинуклеотидов с использованием способов, которые известны в данной области (например, патент США № 4683195).
Например, каждый из желаемых полинуклеотидов можно первоначально получать в отдельной ПЦР. После этого конструируют специфические праймеры так, чтобы концы продуктов ПЦР содержали комплементарные последовательности. Когда продукты ПЦР смешивают, денатурируют и подвергают повторному отжигу, цепи, имеющие совпадающие последовательности на их 3'-концах, перекрываются и могут действовать в качестве праймеров друг для друга. Удлинение этого перекрывания ДНК-полимеразой продуцирует молекулу, в которой исходные последовательности "сплайсированы" вместе. Таким образом, ряд индивидуальных полинуклеотидов можно "сплайсировать" вместе, а затем одновременно трансдуцировать в клетку-хозяина. Таким образом, осуществляется экспрессия каждого из множества полинуклеотидов.
Затем индивидуальные полинуклеотиды или "сплайсированные" полинуклеотиды встраивают в экспрессирующий вектор. Настоящее изобретение не ограничивается способом, посредством которого полинуклеотид встраивают в экспрессирующий вектор. Специалисту в данной области будут знакомы необходимые стадии для включения полинуклеотида в экспрессирующий вектор. Типичный экспрессирующий вектор содержит желаемый полинуклеотид, которому предшествуют одна или несколько регуляторных областей, вместе с участком связывания рибосомы, например, нуклеотидной последовательностью, которая имеет длину 3-9 нуклеотидов и расположена на 3-11 нуклеотидов выше кодона инициации в E. coli. См. Shine and Dalgarno (1975) Nature 254(5495): 34-38 и Steitz (1979) Biological Regulation and Development (ed. Goldberger, R. R), 1: 349-399 (Plenum, New York).
Термин "функционально связанный", как используют в рамках изобретения, относится к конфигурации, в которой последовательность контроля помещена в соответствующее положение относительно кодирующей последовательности в последовательности ДНК или полинуклеотиде, так чтобы последовательность контроля направляла экспрессию полипептида.
Регуляторные области включают, например, области, которые содержат промотор и оператор. Промотор функционально связан с желаемым полинуклеотидом, тем самым, инициируя транскрипцию полинуклеотида через фермент РНК-полимеразу. Оператор представляет собой последовательность нуклеиновых кислот, соседних с промотором, которая содержит белок-связывающий домен, с которым может связываться белок-репрессор. В отсутствие белка-репрессора, транскрипция инициируется через промотор. Когда он присутствует, белок-репрессор, специфичный к белок-связывающему домену оператора, связывается с оператором, тем самым, ингибируя транскрипцию. Таким образом, осуществляют контроль транскрипции, исходя из конкретных используемых регуляторных областей и присутствия или отсутствия соответствующего белка-репрессора Примеры включают лактозные промоторы (белок-репрессор Lad меняет конформацию при контактировании с лактозой, тем самым, препятствуя связыванию белка-репрессора Lad с оператором) и триптофановые промоторы (когда он находится в комплексе с триптофаном, белок-репрессор TrpR имеет конформацию, которая связывает оператор; в отсутствие триптофана белок-репрессор TrpR имеет конформацию, которая не связывается с оператором). Другим примером является промотор tac (см. de Boer et al., (1983) Proc Natl Acad Sci USA 80(1): 21-25). Как будет понятно специалистам в данной области, эти и другие экспрессирующие векторы можно использовать в рамках настоящего изобретения, и настоящее изобретение не ограничивается в этом отношении.
Хотя для включения желаемых последовательностей можно использовать любой подходящий экспрессирующий вектор, легкодоступные экспрессирующие векторы включают, но неограничиваются ими: плазмиды, такие как pSClO1, pBR322, pBBR1MCS-3, pUR, pEX, pMR100, pCR4, pBAD24, pUC19, pRS426; и бактериофаги, такие как фаг М13 и фаг λ. Безусловно, такие экспрессирующие векторы могут быть пригодными только для конкретных клеток-хозяев. Однако специалист в данной области может без труда определить посредством стандартного экспериментирования, пригоден ли конкретный экспрессирующий вектор для какой-либо данной клетки-хозяина. Например, экспрессирующий вектор можно вводить в клетку-хозяина, которую затем можно подвергать мониторингу в отношении жизнеспособности и экспрессии последовательностей, содержащихся в векторе. Кроме того, может быть приведена отсылка на соответствующие справочники и литературу, где описаны экспрессирующие векторы и их пригодность для какой-либо конкретной клетки-хозяина.
Пригодные экспрессирующие векторы для целей настоящего изобретения, включая экспрессию желаемого гетерологичного полипептида, фермента и одного или нескольких каталитических доменов, описанных в настоящем описании, включают экспрессирующие векторы, содержащие полинуклеотид, кодирующий желаемый гетерологичный полипептид, фермент или каталитический домен(ы), функционально связанные с конститутивным или индуцибельным промотором. Примеры особенно пригодных промоторов для функционального связывания с такими полинуклеотидами, включают промоторы из следующих генов: gpdA, cbh1, амилаза TAKA Aspergillus oryzae, аспарагиновая протеаза Rhizomucor miehei, нейтральная альфа-амилаза Aspergillus niger, устойчивая к кислотам альфа-амилаза Aspergillus niger, глюкоамилаза Aspergillus niger (glaA), glaA Aspergillus awamori, липаза Rhizomucor miehei, щелочная протеаза Aspergillus oryzae, триозофосфатизомераза Aspergillus oryzae, ацетамидаза Aspergillus nidulans, ацетамидаза Aspergillus oryzae, трипсин-подобная протеаза Fusarium oxysporum, эндо-α-L-арабиназа (abnA) грибов, α-L-арабинофуранoзидаза A (abfA) грибов, α-L-арабинофуранозидаза B (abfB) грибов, ксиланаза (xlnA) грибов, фитаза грибов, ATP-синтетаза грибов, субъединица 9 (oliC) грибов, триозофосфатизомераза (tpi) грибов, алкогольдегидрогеназа (adhA) грибов, α-амилаза (amy) грибов, амилоглюкозидаза (glaA) грибов, ацетамидаза (amdS) грибов, глицеральдегид-3-фосфатдегидрогеназа (gpd) грибов, алкогольдегидрогеназа дрожжей, лактаза дрожжей, 3-фосфоглицераткиназа дрожжей, триозофосфатизомераза дрожжей, бактериальная α-амилаза, бактериальный Spo2 и SSO. Примеры таких пригодных экспрессирующих векторов и промоторов также описаны в PCT/EP2011/070956, полное содержание которой включено в настоящее описание в качестве ссылки.
Фармацевтические композиции, содержащие гетерологичные полипептиды, продуцируемые клетками нитчатых грибов по изобретению
В другом аспекте настоящее изобретение относится к композиции, например, фармацевтической композиции, содержащий один или несколько представляющих интерес гетерологичных полипептидов, таких как полипептиды млекопитающих, продуцируемые клетками нитчатых грибов по настоящему изобретению, имеющие сниженную активность по меньшей мере трех протеаз и, кроме того, содержащие рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, составленный вместе с фармацевтически приемлемым носителем. Фармацевтические композиции по изобретению также можно вводить в комбинированной терапии, т.е. комбинировать с другими средствами. Например, комбинированная терапия может включать представляющий интерес полипептид млекопитающих в комбинации по меньшей мере с одним другим лекарственным средством.
Как используют в рамках изобретения, "фармацевтически приемлемый носитель" включает любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, обеспечивающие изотоничность и замедляющие всасывание средства, и т.п., которые являются физиологически совместимыми. Предпочтительно, носитель является пригодным для внутривенного, внутримышечного, подкожного, парентерального, внутриспинномозгового или эпидермального введения (например, посредством инъекции или инфузии). В зависимости от пути введения, активное соединение, т.е. представляющий интерес полипептид млекопитающих, можно покрывать материалом, защищающим соединение от действия кислот и других природных условий, которые могут инактивировать соединение.
Фармацевтические соединения по настоящему изобретению могут включать одну или несколько фармацевтически приемлемых солей. "Фармацевтически приемлемая соль" относится к соли, которая сохраняет требуемую биологическую активность исходного соединения и не оказывает каких-либо нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J Pharm. Sci. 66: 1-19). Примеры таких солей включают кислотно-аддитивные соли и основно-аддитивные соли. Кислотно-аддитивные соли включают соли, образованные из нетоксичных неорганических кислот, таких как хлористоводородная, азотная, фосфорная, серная, бромистоводородная, йодистоводородная, фосфорная и т.п., а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенилзамещенные алканоевые кислоты, гидроксиалканоевые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Основно-аддитивные соли включают соли, образованные из щелочноземельных металлов, таких как натрий, калий, магний, кальций и т.п., а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и т.п.
Фармацевтическая композиция по изобретению также может включать фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: (1) растворимые в воде антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; (2) растворимые в маслах антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) хелатирующие металлы агенты, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, виннокаменная кислота, фосфорная кислота и т.п.
Примеры пригодных водных и неводных носителей, которые можно использовать в фармацевтических композициях по настоящему изобретению, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.), и их пригодные смеси, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Надлежащую текучесть можно поддерживать, например, с использованием материалов покрытий, таких как лецитин, посредством поддержания требуемого размера частиц в случае дисперсий, и с использованием поверхностно-активных веществ.
Эти композиции также могут содержать адъюванты, такие как консерванты, смачивающие вещества, эмульгаторы и диспергирующие вещества. Предотвращение наличия микроорганизмов можно обеспечивать как методиками стерилизации, так и включением различных антибактериальных и противогрибковых средств, например, парабена, хлорбутанола, фенолсорбиновой кислоты и т.п. Также может быть желательным включение в композиции обеспечивающих изотоничность веществ, таких как сахара, хлорид натрия и т.п. Кроме того, более длительного всасывания инъецируемой фармацевтической формы можно достигнуть путем включения средств, которые замедляют всасывание, таких как моностеарат алюминия и желатин.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для быстрого изготовления стерильных инъецируемых растворов или дисперсий. Применение таких сред и средств для фармацевтически активных веществ известно в данной области. За исключением случаев, когда какая-либо общепринятая среда или средство являются несовместимыми с активным соединением, предусматривается их применение в фармацевтических композициях по настоящему изобретению. Также в композиции могут быть включены дополнительные активные соединения.
Терапевтические композиции, как правило, должны быть стерильными и стабильными в условиях изготовления и хранения. Композиция может быть изготовлена в качестве раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль, и т.п.), и их пригодные смеси. Надлежащую текучесть можно поддерживать, например, с использованием покрытия, такого как лецитин, посредством поддержания требуемого размера частиц в случае дисперсии и с использованием поверхностно-активных веществ. Во многих случаях является предпочтительным включение в композицию обеспечивающих изотоничность средств, например, сахаров, полиспиртов, таких как маннит, сорбит или хлорид натрия. Более длительного всасывания инъецируемых композиций можно достигать посредством включения в композицию средства, которое замедляет всасывание, например, солей моностеарата и желатина.
Стерильные инъецируемые растворы можно получать путем включения активного соединения в требуемом количестве в соответствующий растворитель с одним или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизацией микрофильтрацией. Как правило, дисперсии получают включением активного соединения в стерильный носитель, который содержит основную дисперсионную среду и другие требуемые ингредиенты из перечисленных выше ингредиентов. В случае стерильных порошков для получения стерильных инъецируемых растворов, предпочтительными способами изготовления являются вакуумная сушка и сублимационная сушка (лиофилизация), которые приводят к порошку активного ингредиента с любым дополнительным требуемым ингредиентом из ранее стерилизованного фильтрацией его раствора.
Количество активного ингредиента, которое можно комбинировать с материалом носителя с получением единичной дозированной формы варьирует в зависимости от индивидуума, подвергаемого лечению и конкретного способа введения. Количество активного ингредиента, которое можно комбинировать с материалом носителя для получения одной дозированной формы, как правило, представляет собой количество композиции, которое обеспечивает терапевтический эффект. Как правило, из ста процентов, это количество варьирует от приблизительно 0,01 процента до приблизительно девяноста девяти процентов активного ингредиента, предпочтительно от приблизительно 0,1 процентов до приблизительно 70 процентов, наиболее предпочтительно от приблизительно 1 процента до приблизительно 30 процентов активного ингредиента в комбинации с фармацевтически приемлемым носителем.
Режимы дозирования корректируют для обеспечения оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить однократный болюс, можно вводить несколько разделенных доз в течение времени или дозу можно пропорционально снижать или повышать, если это необходимо для терапевтического случая. Особенно преимущественным является изготовление парентеральных композиций в единичной дозированной форме для простоты введения и единообразия дозировки. Единичная дозированная форма, как используют в рамках настоящего изобретения, относится к физически дискретным единицам, подходящим в качестве однократных дозировок для индивидуумов, подлежащих лечению; причем каждая единица содержит заданное количество активного соединения, вычисленное для достижения требуемого терапевтического носителя, вместе с требуемым фармацевтическим носителем. Характеристики единичных дозированных форм по настоящему изобретению определяются и прямо зависят от (a) уникальных характеристик активного соединения и конкретного терапевтического эффекта, подлежащего достижению, и (b) ограничений, свойственных данной области, в отношении составления активного соединения для лечения чувствительности индивидуумов.
Для введения представляющего интерес полипептида млекопитающих, в частности, где полипептид млекопитающих представляет собой антитело, дозировка варьирует приблизительно от 0,0001 до 100 мг/кг, и более обычно от 0,01 до 5 мг/кг, при пересчете на массу тела. Например, дозировка может составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела, или она может находиться в диапазоне 1-10 мг/кг. Иллюстративный режим лечения охватывает введение один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в месяц, один раз в три 3 месяца или один раз в период от трех до 6 месяцев. Определенные режимы дозирования для антитела могут включать 1 мг/кг массы тела или 3 мг/кг массы тела посредством внутривенного введения, причем антитело вводят с использованием одной из следующих схем дозирования: (i) раз в четыре недели - шесть дозировок, затем раз в три месяца; (ii) раз в три недели; (iii) 3 мг/кг массы тела один раз, а затем 1 мг/кг массы тела раз в три недели.
Альтернативно, представляющий интерес полипептид млекопитающих можно вводить в качестве состава с замедленным высвобождением, в случае которого требуется менее частое введение. Дозировка и частота варьируют в зависимости от времени полужизни антитела у пациента. Как правило, наиболее длительное время полужизни демонстрируют антитела человека, за ними следуют гуманизированные антитела, химерные антитела и не являющиеся человеческими антитела. Дозировка и частота могут варьировать, в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических способах применения вводят относительно низкую дозировку с относительно нечастыми интервалами на протяжении длительного периода времени. Некоторые пациенты продолжают получать лечение до конца жизни. В терапевтических способах применения иногда требуется относительно высокая дозировка с относительно короткими интервалами до тех пор, пока прогрессирование заболевания не снизится или не прекратится, и предпочтительно до тех пор, пока у пациента не будет выявлено частичное или полное смягчение симптомов заболевания. После этого, пациенту можно проводить профилактический режим.
Истинные уровни дозировок активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать, чтобы получить количество активного ингредиента, которое является эффективным для достижения требуемого терапевтического ответа для конкретного пациента, композиции и способа введения, не являясь токсичным для пациента. Выбранный уровень дозировки может зависеть от множества фармакокинетических факторов, включая активность конкретных композиций по настоящему изобретению, или их сложных эфиров, солей или амидов, способ введения, время введения, скорость выведения конкретного используемого соединения, длительность лечения, другие лекарственные средства, соединения и/или материалы, применяемые в сочетании с конкретными используемыми композициями, возраст, пол, массу тела, состояние, общее состояние здоровья и предшествующий медицинский анамнез пациента, подвергаемого лечению, и сходные факторы, хорошо известные в области медицины.
"Терапевтически эффективная дозировка" иммуноглобулина по настоящему изобретению предпочтительно приводит к снижению тяжести симптомов заболевания, увеличению частоты и длительности бессимптомных периодов заболевания, или предупреждению ухудшения или нетрудоспособности вследствие заболевания. Например, для лечения опухолей "терапевтически эффективная дозировка" предпочтительно ингибирует клеточный рост или рост опухоли по меньшей мере приблизительно на 20%, более предпочтительно по меньшей мере приблизительно на 40%, еще более предпочтительно по меньшей мере приблизительно на 60%, и еще более предпочтительно по меньшей мере приблизительно на 80% относительно индивидуумов без лечения. Способность соединения ингибировать рост опухоли можно оценивать в модельной системе на животных, предсказывающей эффективность в опухолях человека. Альтернативно, это свойство композиции можно оценивать путем исследования способности соединения к ингибированию, такое ингибирование можно измерять in vitro в анализах, известных квалифицированному специалисту. Терапевтически эффективное количество терапевтического соединения может снижать размер опухоли, или иным образом смягчать симптомы у индивидуума. Специалист в данной области способен определить такие количества на основании таких факторов, как размеры индивидуума, тяжесть симптомов у индивидуума, и конкретная выбранная композиция или путь введения.
Композицию по настоящему изобретению можно вводить одним или несколькими путями введения с использованием одного или нескольких из множества способов, известных в данной области. Как будет понятно специалисту в данной области, путь и/или способ введения может варьировать в зависимости от требуемых результатов. Определенные пути введения для связывающих частей по изобретению включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный, внутриспинномозговой или другие парентеральные пути введения, например посредством инъекции или инфузии. Выражение "парентеральное введение", как используют в рамках изобретения, означает способ введения, отличный от энтерального и местного введения, обычно посредством инъекции, и включает, но не ограничивается ими, внутривенную, внутримышечную, внутриартериальную, интратекальную, интракапсулярную, внутриглазничную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, подкапсулярную, субарахноидальную, интраспинальную, эпидуральную и внутригрудинную инъекцию и инфузию.
Альтернативно, полипептид млекопитающих по настоящему изобретению можно вводить непарентеральным способом, таким как местный, эпидермальный или мукозальный путь введения, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
Активные соединения можно изготавливать с носителями, которые препятствуют быстрому высвобождению соединения, такими как состав для контролируемого высвобождения, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы для доставки. Можно использовать биологически деградируемые биологически совместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Множество способов получения таких составов запатентовано или общеизвестно специалистам в данной области. (См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978).
Терапевтические композиции можно вводить с помощью медицинских устройств, известных в данной области. Например, в определенном варианте осуществления терапевтическую композицию по настоящему изобретению можно вводить с помощью безыгольного устройства для подкожной инъекции, такого как устройства, описанные в патентах США №№ 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556. Примеры хорошо известных имплантатов и модулей, пригодных для настоящего изобретения, описаны в: патенте США № 4487603, в котором описан имплантируемый насос для микроинфузий для высвобождения лекарственного средства с контролируемой скоростью; патенте США № 4486194, в котором описано терапевтическое устройство для введения лекарственных средств через кожу; патенте США № 4447233, в котором описан насос для инфузии лекарственного средства для доставки лекарственного средства с конкретной скоростью инфузии; патенте США № 4447224, в котором описано имплантируемое устройство для инфузии с вариабельным потоком для непрерывной доставки лекарственного средства; патенте США № 4439196, в котором описана осмотическая система для доставки лекарственного средства, имеющая многокамерные отделения; и патенте США № 4475196, в котором описана осмотическая система для доставки лекарственного средства.
В определенных вариантах осуществления полипептиды млекопитающих по настоящему изобретению применяют для лечения любого заболевания, которое можно лечить терапевтическими антителами.
Клетки нитчатых грибов по изобретению
Изобретение, описанное в настоящем описании, относится к увеличению уровней продукции гетерологичных полипептидов, таких как полипептиды млекопитающих, в клетках нитчатых грибов путем снижения или устранения активности по меньшей мере трех протеаз, встречающихся в клетках, которые экспрессируют гетерологичные полипептиды, и которые катализируют деградацию гетерологичных полипептидов. Снижение или устранение активности протеаз, встречающихся в клетках нитчатых грибов, которые экспрессируют гетерологичные полипептиды, увеличивает стабильность экспрессированных рекомбинантных полипептидов, что приводит к увеличению уровня продукции гетерологичных полипептидов. Активность протеаз, встречающихся в клетках нитчатых грибов, может быть снижена, например, путем модификации генов, кодирующих протеазы.
"Клетки нитчатых грибов" включают клетки из всех нитчатых форм подотдела Eumycota и Oomycota (как описано Hawksworth et al., Ainsworth and Bisby's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK). Клетки нитчатых грибов, главным образом, характеризуются мицелярной стенкой, состоящей из хитина, целлюлозы, глюкана, хитозана, маннана и других комплексных полисахаридов. Вегетативный рост осуществляется путем удлинения гифа, и метаболизм углерода является облигатно аэробным. Напротив, вегетативный рост у дрожжей, таких как Saccharomyces cerevisiae, осуществляется почкованием одноклеточного слоевища, и катаболизм углерода может быть ферментативным.
В рамках настоящего изобретения можно использовать любую клетку нитчатых грибов при условии, что она остается жизнеспособной после трансформации последовательностью нуклеиновых кислот и/или является модифицированной или мутантной для снижения протеазной активности. Предпочтительно, на клетку нитчатых грибов не оказывает отрицательного влияния трансдукция необходимых последовательностей нуклеиновых кислот, последующая экспрессия белков (например, белки млекопитающих) или образовавшиеся промежуточные соединения.
Примеры пригодных клеток нитчатых грибов включают, но не ограничиваются ими, клетки штамма Acremonium, Aspergillus, Fusarium, Humicola, Mucor, Myceliophthora, Neurospora, Penicillium, Scytalidium, Thielavia, Tolypocladium или Trichoderma. В определенных вариантах осуществления клетка нитчатых грибов происходит из штамма Trichoderma sp., Acremonium, Aspergillus, Aureobasidium, Cryptococcus, Chrysosporium, Chrysosporium lucknowense, Filibasidium, Fusarium, Gibberella, Magnaporthe, Mucor, Myceliophthora, Myrothecium, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Piromyces, Schizophyllum, Talaromyces, Thermoascus, Thielavia или Tolypocladium.
Клетки грибов Aspergillus по настоящему изобретению могут включать, но не ограничиваться ими, Aspergillus aculeatus, Aspergillus awamori, Aspergillus clavatus, Aspergillus flavus, Aspergillus joetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae или Aspergillus terreus.
Клетки грибов Neurospora по настоящему изобретению могут включать, но не ограничиваться ими, Neurospora crassa.
В определенных вариантах осуществления клетка нитчатых грибов не является клеткой Aspergillus.
В определенных вариантах осуществления клетка нитчатых грибов выбрана из группы, состоящей из Trichoderma (T. reesei), Neurospora (N. crassa), Penicillium (P. chrysogenum), Aspergillus (A. nidulans, A. niger и A. oryzae), Myceliophthora (M. thermophila) и Chrysosporium (C. lucknowense).
В определенных вариантах осуществления клетка нитчатых грибов представляет собой клетку грибов Trichoderma. Клетки грибов Trichoderma по настоящему изобретению могут происходить из штамма Trichoderma дикого типа или его мутанта. Примеры пригодных клеток грибов Trichoderma включают, но не ограничиваются ими, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei, Trichoderma atroviride, Trichoderma virens, Trichoderma viride; и их альтернативные половые формы (т.е. Hypocrea).
Общие способы разрушения генов и культивирования клеток нитчатых грибов описаны, например, для Penicillium, в Kopke et al. (2010) Application of the Saccharomyces cerevisiae FLP/FRT recombination system in filamentous fungi for marker recycling and construction of knockout strains devoid of heterologous genes. Appl Environ Microbiol. 76(14): 4664-74. doi: 10.1128/AEM.00670-10, для Aspergillus, в Maruyama и Kitamoto (2011), Targeted Gene Disruption in Koji Mold Aspergillus oryzae, в James A. Williams (ed.), Strain Engineering: Methods and Protocols, Methods in Molecular Biology, vol. 765, DOI 10.1007/978-l-61779-197-0_27; для Neurospora, в Collopy et al. (2010) High-throughput construction of gene deletion cassettes for generation of Neurospora crassa knockout strains. Methods Mol Biol. 2010; 638: 33-40. doi: 10.1007/978-1-60761-611-5_3; и для Myceliophthora или Chrysosporium, PCT/NL2010/000045 и PCT/EP98/06496.
Компоненты клетки нитчатых грибов
Определенные аспекты настоящего изобретения относятся к клеткам нитчатых грибов, имеющим сниженную или не поддающуюся обнаружению активность по меньшей мере трех протеаз и имеющим рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, который продуцируется на увеличенных уровнях, например на уровнях, увеличенных по меньшей мере в два раза. Другие аспекты настоящего изобретения относятся к клеткам грибов Trichoderma, которые обладают сниженной или не поддающейся обнаружению протеазной активностью по меньшей мере трех протеаз, выбранных из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, tsp1, slp1, slp2, slp7, gap1 и gap2, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих, продуцируемый на уровне, который по меньшей мере в 2 раза превышает уровень продукции полипептида в соответствующей родительской клетке грибов Trichoderma. В определенных вариантах осуществления клетки нитчатых грибов или клетки грибов Trichoderma имеют сниженную активность или не имеют активности по меньшей мере четырех, по меньшей мере пяти, по меньшей мере шести, по меньшей мере семи, по меньшей мере восьми, по меньшей мере девяти, по меньшей мере десяти, по меньшей мере одиннадцати, по меньшей мере двенадцати или более протеаз.
Сниженная экспрессия протеаз
Сниженная активность по меньшей мере трех протеаз в клетках нитчатых грибов или клетках грибов Trichoderma по настоящему изобретению может быть результатом сниженной или устраненной экспрессии протеаз. В некоторых вариантах осуществления сниженная или устраненная экспрессия по меньшей мере трех протеаз является результатом модификации каталитического домена, кодирующей области или последовательности контроля, требуемой для экспрессии кодирующей области генов, кодирующих каждую из протеаз. В других вариантах осуществления сниженная или устраненная экспрессия протеаз является результатом внесения, замены и/или удаления одного или нескольких нуклеотидов в генах, или их последовательности контроля, требуемой для транскрипции или трансляции генов, кодирующих каждую из протеаз.
В следующих вариантах осуществления сниженная или устраненная экспрессия протеаз является результатом встраивания в гены, кодирующие каждую из протеаз, разрушающих конструкций нуклеиновых кислот, каждая из которых содержит фрагмент нуклеиновой кислоты, гомологичный каждому из генов, что приводит к дупликации области гомологии и встраиванию конструкции ДНК между дуплицированными областями. В других вариантах осуществления сниженная или устраненная экспрессия протеаз является результатом конверсии генов, кодирующих каждую из протеаз. В других вариантах осуществления сниженная или устраненная экспрессия протеаз является результатом антисмысловых полинуклеотидов или конструкций РНК-i, которые специфичны к каждому из генов, кодирующих каждую из протеаз. В одном варианте осуществления конструкция РНК-i является специфичной к гену, кодирующему аспарагиновую протеазу, такую как протеаза pep-типа, трипсин-подобные сериновые протеазы, такие как tsp1, глутаминовую протеазу, такую как протеаза gap-типа, субтилизиновую протеазу, такую как протеаза slp-типа, или седолизиновую протеазу, такой как протеаза tpp1 или slp7. В одном варианте осуществления конструкция РНК-i является специфичной к гену, кодирующему протеазу slp-типа. В одном варианте осуществления конструкция РНК-i является специфичной к гену, кодирующему slp2, slp3, slp5 или slp6. В одном варианте осуществления конструкция РНК-i является специфичной к двум или более протеазам. В одном варианте осуществления две или более протеазы представляют собой любую из протеаз pep-типа, любую из трипсин-подобных сериновых протеаз, любую из протеаз slp-типа, любую из протеаз gap-типа и/или любую из седолизиновых протеаз. В одном варианте осуществления две или более протеазы представляют собой slp2, slp3, slp5 и/или slp6. В одном варианте осуществления конструкция РНК-i содержит любую из последовательностей нуклеиновых кислот, представленных в таблице 22.2.
В некоторых вариантах осуществления каждый из генов, кодирующих протеазы, содержит мутацию, которая снижает или устраняет активность соответствующей протеазы. В других вариантах осуществления мутация снижает или устраняет экспрессию каждой из протеаз. В следующих вариантах осуществления мутация представляет собой мутацию с нокаутом, мутацию с укорочением, точковую мутацию, миссенс-мутацию, мутацию с заменой, мутацию со сдвигом рамки считывания, мутацию с инсерцией, мутацию с дупликацией, мутацию с амплификацией, мутацию с транслокацией, мутацию с инверсией, которая снижает или устраняет активность соответствующей протеазы.
В некоторых вариантах осуществления мутация представляет собой делецию гена, кодирующего протеазу. В других вариантах осуществления мутация представляет собой делецию части кодирующего протеазу гена, кодирующей каталитический домен протеазы. В других вариантах осуществления мутация представляет собой точковую мутацию в части кодирующего протеазу гену, кодирующей каталитический домен протеазы.
Комбинации генов протеаз
Клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению могут содержать по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять или более аспарагиновых протеаз, трипсин-подобных сериновых протеаз, субтилизиновых протеаз и/или глутаминовых протеаз. В определенных вариантах осуществления протеазы кодируются генами протеаз pep-типа, генами протеаз gap-типа или генами протеаз slp-типа. В некоторых вариантах осуществления гены протеаз pep-типа выбраны из pep1, pep2, pep3, pep4, pep5, pep8, pep11 и pep12. В других вариантах осуществления гены протеаз gap-типа выбраны из gap1 и gap2. В следующих вариантах осуществления гены протеаз slp-типа выбраны из slp1, slp2, slp3 и slp7; или выбраны из slp1, slp2, slp3, slp5, slp6, slp7 и slp8. В определенных предпочтительных вариантах осуществления ген протеазы slp-типа представляет собой slp1.
В других вариантах осуществления протеазы кодируются генами, выбранными из pep1, pep2, pep3, pep4, pep5, pep7, pep8, pep11, pep12, tsp1, slp1, slp2, slp3, slp5, slp6, slp7, slp8, gap1, gap2 и tpp1. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии по меньшей мере трех или по меньшей мере четырех генов, кодирующих протеазы, выбранные из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, tsp1, slp1, slp2, slp3, slp7, gap1 и gap2. В определенных вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии по меньшей мере трех генов, кодирующих протеазы, выбранных из pep1, tsp1 и slp1. В других вариантах осуществления клетка нитчатых грибов или клетка Trichoderma имеет сниженные уровни экспрессии или не имеет экспрессии по меньшей мере трех генов, кодирующих протеазы, выбранных из gap1, slp1 и pep1. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1 и gap1. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1 и pep4. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4 и slp1. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1 и slp3. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3 и pep3. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3 и pep2. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3, pep2 и pep5. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3, pep2, pep5 и tsp1. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3, pep2, pep5, tsp1 и slp7. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3, pep2, pep5, tsp1, slp7 и slp8. В некоторых вариантах осуществления клетка нитчатых грибов, например, клетка Trichoderma, имеет сниженные уровни экспрессии или не имеет экспрессии генов, кодирующих протеазы slp2, pep1, gap1, pep4, slp1, slp3, pep3, pep2, pep5, tsp1, slp7, slp8 и gap2.
В определенных вариантах осуществления клетка нитчатых грибов имеет по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять или более протеаз со сниженной протеазной активностью, где каждая из соответствующих протеаз с активностью дикого типа имеет аминокислотную последовательность, которая по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотным последовательностям SEQ ID NO: 1-16; 17-36; 37-57; 58-65; 66-81; 82-97; 98-117; 118-128; 129-144; 166-181; 182-185; или SEQ ID NO: 491-588. В вариантах осуществления, где клетка нитчатых грибов представляет собой клетку грибов Trichoderma со сниженной протеазной активностью в одной или нескольких протеазах, где каждая из соответствующих протеаз с активностью дикого типа имеет аминокислотную последовательность, которая по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотным последовательностям SEQ ID NO: 1, 17, 37, 58, 66, 82, 98, 118, 129, 166 или 182; или SEQ ID NO: 507, SEQ ID NO: 522 или SEQ ID NO: 530.
Гетерологичные полипептиды
Клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению содержат рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид. В определенных вариантах осуществления гетерологичный полипептид представляет собой полипептид млекопитающих. В других вариантах осуществления гетерологичный полипептид представляет собой полипептид не млекопитающих.
В вариантах осуществления, где клетка нитчатых грибов содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих, полипептид млекопитающих может представлять собой негликозилированный полипептид млекопитающих, гликозилированный полипептид млекопитающих или их комбинации, включая, но не ограничиваясь ими, иммуноглобулин, антитело, фактор роста и интерферон. В некоторых вариантах осуществления полипептид млекопитающих представляет собой иммуноглобулин или антитело. В вариантах осуществления, где клетка нитчатых грибов содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, клетка нитчатых грибов, например, клетка грибов Trichoderma, может иметь сниженную экспрессию или может не иметь экспрессии по меньшей мере трех или по меньшей мере четырех генов, кодирующих протеазы, выбранных из pep1, pep3, pep4, pep8, pep11, pep12, tsp1, slp1, slp2, slp7, gap1 и gap2. В определенных предпочтительных вариантах осуществления клетка, например, клетка грибов Trichoderma, содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию или не имеет экспрессии генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1, gap1, pep4, pep3, pep2, pep5 и gap2. В определенных предпочтительных вариантах осуществления клетка, например, клетка грибов Trichoderma, содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию или не имеет экспрессии генов, кодирующих протеазы pep1, tsp1, slp1 и gap1. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, tsp1, slp1, gap1 и pep4. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2 и slp3. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3 и tsp1. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1 и pep1. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1 и gap1. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1, gap1 и pep4. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1, gap1, pep4 и pep3. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1, gap1, pep4, pep3 и pep2. В других вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий иммуноглобулин или антитело, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, slp3, tsp1, pep1, gap1, pep4, pep3, pep2 и pep5.
В других вариантах осуществления клетка нитчатых грибов содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин или интерлейкин. В вариантах осуществления, где клетка нитчатых грибов, например, клетка грибов Trichoderma, содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, клетка нитчатых грибов может иметь сниженную экспрессию или не иметь экспрессии по меньшей мере трех или по меньшей мере четырех генов, кодирующих протеазы, выбранные из pep1, pep2, pep3, pep4, pep5, pep8, gap1, gap2, slp1, slp2, slp7 и tsp1. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, tsp1, slp1, gap1 и gap2. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы slp1, slp2, pep1, gap1, pep4, slp7, pep2, pep3, pep5, tsp1 и gap2. В других вариантах осуществления клетка, например, клетка грибов Trichoderma, содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, tsp1, slp1, gap1, gap2 и pep4. В следующем варианте осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, и имеет сниженную экспрессию генов протеаз pep-типа, выбранных из pep1, pep2, pep3, pep4 и pep5. В определенных предпочтительных вариантах осуществления фактор роста представляет собой IGF-1, или интерферон представляет собой интерферон-α 2b. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1 и pep4. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4 и slp7. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7 и slp2. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7, slp2 и pep2. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7, slp2, pep2 и pep3. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7, slp2, pep2, pep3 и pep5. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7, slp2, pep2, pep3, pep5 и slp1. В определенных вариантах осуществления клетка содержит рекомбинантный полинуклеотид, кодирующий фактор роста, интерферон, цитокин, сывороточный альбумин человека или интерлейкин, и имеет сниженную экспрессию генов, кодирующих протеазы pep1, gap1, pep4, slp7, slp2, pep2, pep3, pep5, slp1 и tsp1.
В определенных вариантах осуществления полипептид млекопитающих продуцируется на уровне, который по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз, по меньшей мере в 20 раз, по меньшей мере в 25 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз, по меньшей мере в 70 раз, по меньшей мере в 75 раз, по меньшей мере в 80 раз, по меньшей мере в 90 раз, по меньшей мере в 100 раз или более превышает уровень продукции полипептида в соответствующей родительской клетке нитчатых грибов без сниженной протеазной активности. В других вариантах осуществления полипептид млекопитающих продуцируется в качестве полноразмерной версии на уровне, превышающем уровень полноразмерной версии полипептида в соответствующей исходной клетке нитчатых грибов.
В вариантах осуществления, где клетка нитчатых грибов содержит рекомбинантный полинуклеотид, кодирующий полипептид не млекопитающих, полипептид не млекопитающих может представлять собой аминопептидазу, амилазу, карбогидразу, карбоксипептидазу, каталазу, целлюлазу, хитиназу, кутиназу, дезоксирибонуклеазу, эстеразу, альфа-галактозидазу, бета-галактозидазу, глюкоамилазу, альфа-глюкозидазу, бета-глюкозидазу, инвертазу, лакказу, липазу, мутаназу, оксидазу, пектинолитический фермент, пероксидазу, фосфолипазу, фитазу, полифенолоксидазу, протеолитический фермент, рибонуклеазу, трансглутаминазу или ксиланазу. В вариантах осуществления, где клетка нитчатых грибов содержит рекомбинантный полинуклеотид, кодирующий полипептид не млекопитающих, клетка нитчатых грибов может иметь сниженную или не поддающуюся обнаружению экспрессию по меньшей мере трех, по меньшей мере четырех, по меньшей мере пяти или по меньшей мере шести генов, кодирующих протеазы, выбранные из pep1, pep2, pep3, pep4, pep5, pep8, pep11, pep12, tsp1, slp1, slp2, slp3, gap1 и gap2. В определенных вариантах осуществления полипептид не млекопитающих продуцируется на уровне, который по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз, по меньшей мере в 20 раз, по меньшей мере в 25 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз, по меньшей мере в 70 раз, по меньшей мере в 75 раз, по меньшей мере в 80 раз, по меньшей мере в 90 раз, по меньшей мере в 100 раз или более превышает уровень продукции полипептида в соответствующей родительской клетке нитчатых грибов. В других вариантах осуществления полипептид не млекопитающих продуцируется в качестве полноразмерной версии на уровне, превышающем уровень продукции полноразмерной версии полипептида в соответствующей родительской клетке нитчатых грибов.
Сниженная активность дополнительных протеаз
В некоторых вариантах осуществления клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению также имеют сниженную активность одной или нескольких дополнительных протеаз. В определенных вариантах осуществления уровень экспрессии одной или нескольких дополнительных протеаз является сниженным. В определенных предпочтительных вариантах осуществления каждый из генов, кодирующих одну или несколько дополнительных протеаз, содержит мутацию, которая снижает активность соответствующей протеазы. Один или несколько дополнительных генов, кодирующих протеазы, может представлять собой pep7, tpp1, gap2, slp3, slp5, slp6, slp7 или slp8.
В определенных вариантах осуществления, когда клетки нитчатых грибов представляют собой клетку Aspergillus, общая протеазная активность снижена до 50% или менее от общей протеазной активности в соответствующей родительской клетке Aspergillus, в которой протеазы не обладают сниженной активностью.
В определенных вариантах осуществления общая протеазная активность снижена в клетке по настоящему изобретению, например, клетке Trichoderma, до 49% или менее, 31% или менее, 13% или менее, 10% или менее, 6,3% или менее или 5,5% или менее от общей протеазной активности в соответствующей родительской клетке нитчатых грибов, в которой протеазы не обладают сниженной активностью.
Дополнительные рекомбинантные модификации
В определенных вариантах осуществления клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению также имеют сниженную активность долихил-P-Man:Man(5)GlcNAc(2)-PP-долихилманнозилтрансферазы. Долихил-P-Man:Man(5)GlcNAc(2)-PP-долихилманнозилтрансфераза (EC 2.4.1.130) переносит альфа-D-маннозильный остаток из долихил-фосфат-D-маннозы на связанный с мембранным липидом олигосахарид. Главным образом, фермент долихил-P-Man:Man(5)GlcNAc(2)-PP-долихилманнозилтрансфераза кодируется геном alg3. Таким образом, в определенных вариантах осуществления клетка нитчатых грибов имеет сниженную активность ALG3, которая представляет собой активность, кодируемую геном alg3. В некоторых вариантах осуществления ген alg3 содержит мутацию, которая снижает соответствующую активность ALG3. В определенных вариантах осуществления ген alg3 удален из клетки нитчатых грибов.
В других вариантах осуществления клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению, кроме того, содержат полинуклеотид, кодирующий α-1,2-маннозидазу. Полинуклеотид, кодирующий α-1,2-маннозидазу, может быть эндогенным в клетке-хозяине, или он может быть гетерологичным для клетки-хозяина. Эти полинуклеотиды являются особенно пригодными для клетки нитчатых грибов, экспрессирующих гликаны с высоким содержанием маннозы, перенесенные из аппарата Гольджи в ER без эффективного расщепления экзо-α-2-маннозидазой. α-1,2-маннозидаза может представлять собой фермент маннозидазу типа I, принадлежащий семейству гликозидгидролаз 47 (cazy.org/GH47_all.html). В определенных вариантах осуществления α-1,2-маннозидаза представляет собой фермент, приведенный на cazy.org/GH47_characterized.html. В частности, α-1,2-маннозидаза может представлять собой фермент ER-типа, который расщепляет гликопротеины, такой как ферменты подсемейства ферментов ER α-маннозидазы I EC 3.2.1.113. Примеры таких ферментов включают α-2-маннозидазу 1B человека (AAC26169), комбинацию маннозидаз ER млекопитающих, или фермент нитчатых грибов, такой как α-1,2-маннозидаза (MDS1) (T. reesei AAF34579; Maras M. et al. J Biotech. 77, 2000, 255). Для экспрессии в ER/Гольджи каталитический домен маннозидазы, как правило, является слитым с нацеливающим пептидом, таким как HDEL, KDEL или часть белка ER или раннего белка аппарата Гольджи, или экспрессируется с эндогенными нацеливающими на ER структурами фермента маннозидазы I животных или растений, см., например, Callewaert et al. 2001 Use of HDEL-tagged Trichoderma reesei mannosyl oligosaccharide 1,2-a-D-mannosidase for N-glycan engineering in Pichia pastoris. FEBS Lett 503: 173-178.
В следующих вариантах осуществления клетки нитчатых грибов или клетки грибов Trichoderma по настоящему изобретению также содержат каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II. Такие каталитические домены пригодны для экспрессии комплексных N-гликанов в клетках не млекопитающих. N-ацетилглюкозаминилтрансфераза I (GlcNAc-TI; GnTI; EC 2.4.1.101) катализирует реакцию UDP-N-ацетил-D-глюкозамин+3-(альфа-D-маннозил)-бета-D-маннозил-R <=> UDP+3-(2-(N-ацетил-бета-D-глюкозаминил)-альфа-D-маннозил)-бета-D-маннозил-R, где R соответствует остальной части N-связанного олигосахарида в гликановом акцепторе. Каталитический домен N-ацетилглюкозаминилтрансферазы I представляет собой любую часть фермента N-ацетилглюкозаминилтрансферазы I, которая способна катилизировать эту реакцию. N-ацетилглюкозаминилтрансфераза II (GlcNAc-TII; GnTII; EC 2.4.1.143) катализирует реакцию UDP-N-ацетил-D-глюкозамин+6-(альфа-D-маннозил)-бета-D-маннозил-R <=> UDP+6-(2-(N-ацетил-бета-D-глюкозаминил)-альфа-D-маннозил)-бета-D-маннозил-R, где R соответствует остальной части N-связанного олигосахарида в гликановом акцепторе. Каталитический домен N-ацетилглюкозаминилтрансферазы II представляет собой любую часть фермента N-ацетилглюкозаминилтрансферазы II, которая способна катализировать эту реакцию. Примеры пригодных каталитических доменов N-ацетилглюкозаминилтрансферазы I и каталитических доменов N-ацетилглюкозаминилтрансферазы II могут быть найдены в международной патентной заявке № PCT/EP2011/070956. Каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II могут кодироваться одним полинуклеотидом. В определенных вариантах осуществления один полинуклеотид кодирует слитый белок, содержащий каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II. Альтернативно, каталитический домен N-ацетилглюкозаминилтрансферазы I может кодироваться первым полинуклеотидом и каталитический домен N-ацетилглюкозаминилтрансферазы II может кодироваться вторым полинуклеотидом.
В вариантах осуществления, где клетка нитчатых грибов или клетка грибов Trichoderma содержит каталитический домен N-ацетилглюкозаминилтрансферазы I и каталитический домен N-ацетилглюкозаминилтрансферазы II, клетка также может содержать полинуклеотид, кодирующий маннозидазу II. Ферменты маннозидазы II способны расщеплять структуры Man5 в GlcNAcMan5 с образованием GlcNAcMan3, и в случае комбинирования с действием каталитического домена GnTII - с образованием G0; и, кроме того, с действием каталитического домена галактозилтрансферазы - с образованием G1 и G2. В определенных вариантах осуществления ферменты маннозидазы типа II принадлежат семейству гликозидгидролаз 38 (cazy.org/GH38_all.html). Примеры таких ферментов включают фермент человека AAC50302, фермент D. melanogaster (Van den Elsen J.M. et al. (2001) EMBO J. 20: 3008-3017), ферменты с 3D-структурой, соответствующей ссылке 1HTY в PDB, и другие упоминаемые в PDB ферменты с каталитическим доменом. Для экспрессии в ER/Гольджи каталитический домен маннозидазы, как правило, подвергают слиянию с N-концевым нацеливающим пептидом, например с использованием нацеливающих пептидов, приведенных в международной патентной заявке № PCT/EP 2011/070956 или SEQ ID NO: 589-594. После трансформации каталитическим доменом маннозидазы типа II, отбирают штамм, эффективно продуцирующий GlcNAc2Man3, GlcNAclMan3 или G0.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка нитчатых грибов дополнительно содержит полинуклеотид, кодирующий переносчик UDP-GlcNAc.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, клетка нитчатых грибов дополнительно содержит полинуклеотид, кодирующий β-1,4-галактозилтрансферазу. Главным образом, β-1,4-галактозилтрансферазы принадлежат семейству гликозилтрансфераз CAZy 7 (cazy.org/GT7_all.html). Примеры пригодных ферментов β4GalT включают β4GalT1, например, фермент жвачных животных Bos taurus AAA30534.1 (Shaper N.L. et al. Proc. Natl. Acad. Sci. U.S.A. 83 (6), 1573-1577 (1986)), фермент человека (Guo S. et al. Glycobiology 2001, 11: 813-20) и фермент Mus musculus AAA37297 (Shaper, N.L. et al. 1998 J. Biol. Chem. 263 (21), 10420-10428). В определенных вариантах осуществления изобретения, где клетка нитчатых грибов содержит полинуклеотид, кодирующий галактозилтрансферазу, клетка нитчатых грибов также содержит полинуклеотид, кодирующий UDP-Gal 4-эпимеразу и/или переносчик UDP-Gal. В определенных вариантах осуществления изобретения, где клетка нитчатых грибов содержит полинуклеотид, кодирующий галактозилтрансферазу, в качестве источника углерода вместо глюкозы при культивировании клетки-хозяина можно использовать лактозу. Культуральная среда может иметь pH от 4,5 до 7,0 или от 5,0 до 6,5. В определенных вариантах осуществления изобретения, где клетка нитчатых грибов содержит полинуклеотид, кодирующий галактозилтрансферазу, и, необязательно, полинуклеотид, кодирующий UDP-Gal 4-эпимеразу и/или переносчик UDP-Gal, в среду для культивирования клеток можно добавлять двухвалентный катион, такой как Mn2+, Ca2+ или Mg2+.
В определенных вариантах осуществления, которые могут быть комбинированы с предшествующими вариантами осуществления, уровень активности альфа-1,6-маннозилтрансферазы в клетке-хозяине снижен по сравнению с уровнем активности в клетке-хозяине дикого типа. В определенных вариантах осуществления клетка нитчатых грибов имеет сниженный уровень экспрессии гена och1 по сравнению с уровнем экспрессии в клетке нитчатых грибов дикого типа.
Другой аспект включает способы продуцирования Man3GlcNAc2 N-гликана [т.е. Manα3(Manα6)Manβ4GlcNAcβ4GlcNAc] в клетке нитчатых грибов, включающие стадии предоставления клетки нитчатых грибов с рекомбинантным полинуклеотидом, кодирующим гетерологичный полипептид, и сниженным уровнем активности маннозилтрансферазы alg3 по сравнению с уровнем активности в клетке нитчатых грибов дикого типа, и культивирования клетки нитчатых грибов с получением Man3GlcNAc2 гликана, где Man3GlcNAc2 гликан составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от нейтральных N-гликанов, секретируемых клеткой нитчатых грибов. В определенном варианте осуществления Man3GlcNAc2 N-гликан соответствует по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от общих N-гликанов гетерологичного полипептида.
Другой аспект включает способы продуцирования комплексного N-гликана (т.е. N-гликан, содержащий концевую структуру GlcNAc2Man3), например, GlcNAc2Man3GlcNAc2 {т.е. G0, т.е. GlcNAcβ2Manα3(GlcNAcβ2Manα6)Manβ4GlcNAcβ4GlcNAc} гликана, в клетках нитчатых грибов, включающие стадии предоставления клетки нитчатых грибов с рекомбинантным полинуклеотидом, кодирующим гетерологичный полипептид, сниженным уровнем активности маннозилтрансферазы alg3 по сравнению с уровнем активности в клетке нитчатых грибов дикого типа, и содержащие дополнительный полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы I, и полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы II, и культивирования клетки нитчатых грибов для продуцирования комплексного N-гликана, например, GlcNAc2Man3GlcNAc2 гликана, где GlcNAc2Man3GlcNAc2 гликан составляет по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от нейтральных N-гликанов, секретируемых клеткой нитчатых грибов. В определенных вариантах осуществления комплексный N-гликан, например, GlcNAc2Man3GlcNAc2 гликан, соответствует по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) общих N-гликанов полипептида. В определенных вариантах осуществления указанные комплексные N-гликаны представляют собой GlcNAcMan3 и/или GlcNAc2Man3.
Другой аспект включает способы продуцирования N-гликана G1 или G2 или их смеси, например GalGlcNAc2Man3GlcNAc2 {т.е. G1, т.е. Galβ4GlcNAcβ2Manα3(GlcNAcβ2Manα6)Manβ4GlcNAcβ4GlcNAc} или GlcNAcβ2Manα3(Galβ4GlcNAcβ2Manα6)Manβ4GlcNAcβ4GlcNAc} и/или Gal2GlcNAc2Man3GlcNAc2 {т.е. G2, т.е. Galβ4GlcNAcβ2Manα3(Galβ4GlcNAcβ2Manα6)Manβ4GlcNAcβ4GlcNAc} гликана в клетке нитчатых грибов, включающие стадии предоставления клетки нитчатых грибов с рекомбинантным полинуклеотидом, кодирующим гетерологичный полипептид и сниженный уровень активности маннозилтрансферазы alg3 по сравнению с уровнем активности в клетке нитчатых грибов дикого типа, и содержащей дополнительный полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы I, полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы II, и полинуклеотид, кодирующий каталитический домен GalT, и культивирования клетки нитчатых грибов с получением N-гликана G1 или G2 или их смеси, где гликан G1 составляет по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от нейтральных N-гликанов, секретируемых клетками нитчатых грибов, или где гликан G2 составляет по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) нейтральных N-гликанов, секретируемых клеткой нитчатых грибов. В определенном варианте осуществления гликан G1 составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида. В определенном варианте осуществления гликан G2 составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) от всех N-гликанов полипептида.
В определенных вариантах осуществления способ продуцирования комплексного N-гликана приводит к образованию смеси различных гликанов. Комплексный N-гликан или Man3GlcNAc2 могут составлять по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%) или по меньшей мере 90% или более такой смеси гликанов. В определенных вариантах осуществления по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% или более N-гликанов полипептида состоят из такой смеси гликанов. В определенных вариантах осуществления способ продуцирования комплекса и N-гликана G1 и/или G2 обеспечивает образование смеси различных гликанов. Комплексный N-гликан, Man3GlcNAc2, G1 и/или G2 может составлять по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% или более такой смеси гликанов. В определенных вариантах осуществления по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% или более N-гликанов полипептида состоят из такой смеси гликанов.
В определенных вариантах осуществления являются желательными способы продуцирования гибридного N-гликана. Как используют в рамках изобретения, термин "гибридный" означает гликан, содержащий как незамещенные концевые остатки маннозы (которые присутствуют в гликанах с высоким содержанием маннозы), так и замещенные остатки маннозы с N-ацетилглюкозаминовой связью, например, GlcNAcβ2Manα3[Manα3(Manα6)Manα6]Manβ4GlcNAcβ4GlcNAc. В таких вариантах осуществления клетку нитчатых грибов, экспрессирующую Man5 {т.е. Manα3[Manα3(Manα6)Manα6]Manβ4GlcNAcβ4GlcNAc}, такую как штамм T. reesei, трансформируют рекомбинантным полинуклеотидом, кодирующим гетерологичный полипептид, и полинуклеотидом, кодирующим каталитический домен N-ацетилглюкозаминилтрансферазы I, и клетку нитчатых грибов культивируют для продуцирования гибридного N-гликана, где гибридный N-гликан составляет по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) нейтральных N-гликанов, секретируемых клеткой нитчатых грибов. В определенном варианте осуществления по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100% (моль.%) N-гликанов полипептида состоит из гибридного N-гликана.
Комплексный гибридный Man3GlcNAc2 N-гликан G1 и G2 может быть связан с молекулой, выбранной из аминокислоты, пептида и полипептида. В определенных вариантах осуществления комплексный гибридный Man3GlcNAc2 N-гликан G1 и G2 связан с гетерологичным полипептидом. В определенных вариантах осуществления гетерологичный полипептид представляет собой гликозилированный белок. В определенном варианте осуществления гликозилированный полипептид представляет собой полипептид млекопитающих. В определенных вариантах осуществления полипептид млекопитающих представляет собой антитело или его антигенсвязывающий фрагмент.
В определенных вариантах осуществления гликозилтрансферазы, например, GnTI, GnTII или GalT или гликозилгидролазы, например, α-1,2-маннозидаза или маннозидаза II, включают нацеливающий пептид, связанный с каталитическими доменами. Термин "связанный", как используют в рамках изобретения, означает, что два полимера аминокислотных остатков в случае полипептида или два полимера нуклеотидов в случае полинуклеотида либо прямо связаны рядом друг с другом, либо находятся одном и том же полипептиде или полинуклеотиде, но разделены встроенными между ними аминокислотными остатками или нуклеотидами. "Нацеливающий пептид", как используют в рамках изобретения, относится к любому количеству последовательно расположенных аминокислотных остатков рекомбинантного белка, которые способны обеспечивать локализацию рекомбинантного белка в эндоплазматической сети (ER) или аппарате Гольджи (Гольджи) в клетке нитчатых грибов. Нацеливающий пептид может быть N-концевым или C-концевым для каталитических доменов. В определенных вариантах осуществления нацеливающий пептид является N-концевым для каталитических доменов. В определенных вариантах осуществления нацеливающий пептид обеспечивает прямое связывание с мембраной ER или аппарата Гольджи. Компоненты нацеливающего пептида могут происходить из любого фермента, который обычно располагается в ER или аппарате Гольджи. Такие ферменты включают маннозидазы, маннозилтрансферазы, гликозилтрансферазы, белки Гольджи 2 типа, и ферменты MNN2, MNN4, MNN6, MNN9, MNN10, MNS1, KRE2, VAN1 и OCH1. Пригодные нацеливающие пептиды описаны в международной патентной заявке № PCT/EP2011/070956. В одном варианте осуществления нацеливающий пептид GnTI или GnTII представляет собой фермент GnTII человека. В других вариантах осуществления нацеливающий пептид происходит из Kre2, Kre2-подобного, Och1, Anp1 и Van1 Trichoderma. В одном варианте осуществления нацеливающий пептид выбран из группы SEQ ID NO: 589-594.
Применения клеток нитчатых грибов по изобретению
Изобретение, описанное в настоящем описании, кроме того, относится к способам использования любой из клеток нитчатых грибов по настоящему изобретению, таких как клетки грибов Trichoderma, которые имеют сниженную или не имеют протеазной активности по меньшей мере трех протеаз и которые содержат рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, такой как полипептид млекопитающих, который продуцируется на увеличенных уровнях, для повышения стабильности гетерологичного полипептида и для получения гетерологичного полипептида. Способы измерения стабильности белка и получения гетерологичного полипептида хорошо известны и включают, но не ограничиваются ими, все способы и технологии, описанные в настоящем описании.
Таким образом, определенные варианты осуществления настоящего изобретения относятся к способам повышения стабильности гетерологичного полипептида посредством: a) предоставления клетки нитчатых грибов по настоящему изобретению, имеющей сниженную или не имеющей активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид; и b) культивирования клетки так, чтобы экспрессировался гетерологичный полипептид, где гетерологичный полипептид обладает повышенной стабильностью по сравнению с клеткой-хозяином, не содержащей мутации генов, кодирующих протеазы. Другие варианты осуществления настоящего изобретения относятся к способам повышения стабильности полипептида млекопитающих посредством: a) предоставления клетки грибов Trichoderma по настоящему изобретению, имеющей сниженную или не имеющей активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих; и b) культивирования клетки так, чтобы экспрессировался полипептид млекопитающих, где полипептид млекопитающих обладает повышенной стабильностью по сравнению с клеткой-хозяином, не содержащей мутации в генах, кодирующих протеазы. Клетка нитчатых грибов или клетка грибов Trichoderma может представлять собой любую клетку, описанную в разделе под названием "Клетки нитчатых грибов по изобретению". Способы измерения стабильности полипептида и культивирования клеток нитчатых грибов и клеток грибов Trichoderma хорошо известны в данной области, и включают, но не ограничиваются ими, все способы и технологии, описанные в настоящем описании.
В определенных вариантах осуществления стабильность гетерологичного полипептида или полипептида млекопитающих увеличена по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 4 раза, по меньшей мере в 5 раз, по меньшей мере в 6 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз, по меньшей мере в 9 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз, по меньшей мере в 20 раз, по меньшей мере в 25 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз, по меньшей мере в 70 раз, по меньшей мере в 75 раз, по меньшей мере в 80 раз, по меньшей мере в 90 раз, по меньшей мере в 100 раз или более раз превышает стабильность гетерологичного полипептида или полипептида млекопитающих, экспрессируемых в соответствующей исходной клетке нитчатых грибов или клетке грибов Trichoderma.
Другие варианты осуществления настоящего изобретения относятся к способам получения гетерологичного полипептида посредством: a) предоставления клетки нитчатых грибов по настоящему изобретению, имеющей сниженную или не имеющей активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид; b) культивирования клетки-хозяина так, чтобы экспрессировался гетерологичный полипептид; и c) очистки гетерологичного полипептида. Следующие варианты осуществления настоящего изобретения относятся к способам получения полипептида млекопитающих посредством: a) предоставления клетки грибов Trichoderma по настоящему изобретению, имеющей сниженную или не имеющей активности по меньшей мере трех протеаз, где клетка дополнительно содержит рекомбинантный полинуклеотид, кодирующий полипептид млекопитающих; b) культивирования клетки-хозяина так, чтобы экспрессировался полипептид млекопитающих; и c) очистки полипептида млекопитающих. Клетка нитчатых грибов или клетка грибов Trichoderma может представлять собой любую клетку, описанную в разделе под названием "Клетки нитчатых грибов по изобретению". Способы культивирования клеток нитчатых грибов и клеток грибов Trichoderma и очистки полипептидов хорошо известны в данной области, и включают, но не ограничиваются ими, все способы и технологии, описанные в настоящем описании.
В определенных вариантах осуществления клетку нитчатых грибов или клетку грибов Trichoderma культивируют в диапазоне pH, выбранном из pH от 3,5 до 7; от pH 3,5 до 6,5; от pH 4 до 6; от pH 4,3 до 5,7; от pH 4,4 до 5,6; и от pH 4,5 до 5,5. В определенных вариантах осуществления для продуцирования антитела клетку нитчатых грибов или клетку грибов Trichoderma культивируют в диапазоне pH, выбранном из от 4,7 до 6,5; pH от 4,8 до 6,0; pH от 4,9 до 5,9; и pH от 5,0 до 5,8.
В некоторых вариантах осуществления гетерологичный полипептид представляет собой полипептид млекопитающих. В других вариантах осуществления гетерологичный полипептид представляет собой полипептид не млекопитающих.
В определенных вариантах осуществления полипептид млекопитающих выбран из иммуноглобулина, тяжелой цепи иммуноглобулина, легкой цепи иммуноглобулина, моноклонального антитела, гибридного антитела, фрагмента антитела F(ab')2, фрагмента антитела F(ab), молекулы Fv, одноцепочечного Fv-антитела, димерного фрагмента антитела, тримерного фрагмента антитела, функционального фрагмента антитела, однодоменного антитела, мультимерных однодоменных антител, иммуноадгезина, инсулиноподобного фактора роста 1, гормона роста, инсулина и эритропоэтина. В других вариантах осуществления белок млекопитающих представляет собой иммуноглобулин или инсулиноподобный фактор роста 1. В других вариантах осуществления белок млекопитающих представляет собой антитело. В следующих вариантах осуществления выход полипептида млекопитающих составляет по меньшей мере 0,5, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, или по меньшей мере 5 грамм на литр. В определенных вариантах осуществления полипептид млекопитающих представляет собой антитело, необязательно IgG1, IgG2, IgG3 или IgG4. В следующих вариантах осуществления выход антитела составляет по меньшей мере 0,5, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, или по меньшей мере 5 граммов на литр. В других вариантах осуществления полипептид млекопитающих представляет собой фактор роста или цитокин. В следующих вариантах осуществления выход фактора роста или цитокина составляет по меньшей мере 0,1, по меньшей мере 0,2, по меньшей мере 0,3, по меньшей мере 0,4, по меньшей мере 0,5, по меньшей мере 1, по меньшей мере 1,5, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 граммов на литр. В следующих вариантах осуществления полипептид млекопитающих представляет собой антитело, и антитело содержит по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% C-конца и N-конца природного антитела без дополнительных аминокислотных остатков. В других вариантах осуществления полипептид млекопитающих представляет собой антитело, и антитело содержит по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% C-конца и N-конца природного антитела, которое не лишено никаких C-концевых или N-концевых аминокислотных остатков.
В определенных вариантах осуществления, в которых полипептид млекопитающих очищен из клеточной культуры, культура, содержащая полипептид млекопитающих, содержит полипептидные фрагменты, которые составляют процент по массе, который составляет менее 50%, менее 40%, менее 30%, менее 20% или менее 10% от массы продуцированных полипептидов. В определенных предпочтительных вариантах осуществления полипептид млекопитающих представляет собой антитело, и полипептидные фрагменты представляют собой фрагменты тяжелой цепи и/или фрагменты легкой цепи. В других вариантах осуществления, в которых полипептид млекопитающих представляет собой антитело и антитело очищают из клеточной культуры, культура, содержащая антитело, содержит свободные тяжелые цепи и/или свободные легкие цепи, которые составляют процент по массе, который составляет менее 50%, менее 40%, менее 30%, менее 20% или менее 10% от массы продуцированного антитела. Способы определения процента по массе полипептидных фрагментов хорошо известны в данной области и включают измерение интенсивности сигнала на SDS-геле.
В следующих вариантах осуществления полипептид не млекопитающих выбран из аминопептидазы, амилазы, карбогидразы, карбоксипептидазы, каталазы, целлюлазы, хитиназы, кутиназы, дезоксирибонуклеазы, эстеразы, альфа-галактозидазы, бета-галактозидазы, глюкоамилазы, альфа-глюкозидазы, бета- глюкозидазы, инвертазы, лакказы, липазы, мутаназы, оксидазы, пектинолитического фермента, пероксидазы, фосфолипазы, фитазы, полифенолоксидазы, протеолитического фермента, рибонуклеазы, трансглутаминазы и ксиланазы.
В определенных вариантах осуществления любого из описанных способов, способ включает дополнительную стадию предоставления одного или нескольких, двух или нескольких, трех или нескольких, четырех или нескольких, или пяти или нескольких ингибиторов протеаз. В определенных вариантах осуществления ингибиторы протеаз представляют собой пептиды, которые коэкспрессируются с полипептидом млекопитающих. В других вариантах осуществления ингибиторы ингибируют по меньшей мере две, по меньшей мере три или по меньшей мере четыре протеазы из семейства протеаз, выбранного из аспарагиновых протеаз, трипсин-подобных сериновых протеаз, субтилизиновых протеаз и глутаминовых протеаз.
В определенных вариантах осуществления любого из описанных способов, клетка нитчатых грибов или клетка грибов Trichoderma также содержит белок-носитель. Как используют в рамках изобретения, "белок-носитель" представляет собой часть белка, который является эндогенным и секретируется на высоком уровне клеткой нитчатых грибов или клеткой грибов Trichoderma. Пригодные белки-носители включают, но не ограничиваются ими, белки-носители маннаназы I (Man5A или MANI) T. reesei, целлобиогидролазы II (Cel6A или CBHII) T. reesei (см., например, Paloheimo et al. Appl. Environ. Microbiol. 2003 December; 69(12): 7073-7082) или целлобиогидролазы I (CBHI) T. reesei. В некоторых вариантах осуществления белок-носитель представляет собой CBH1. В других вариантах осуществления белок-носитель представляет собой укороченный белок CBH1 T. reesei, который включает центральную область CBH1 и часть линкерной области CBH1. В некоторых вариантах осуществления носитель, такой как целлобиогидролаза или ее фрагмент, является слитым с легкой цепью антитела и/или тяжелой цепью антитела. В некоторых вариантах осуществления носитель, такой как целлобиогидролаза или ее фрагмент, является слитым с инсулиноподобным фактором роста 1, гормоном роста, инсулином, интерфероном-альфа 2b, фибробластным фактором роста 21 или сывороточным альбумином человека. В некоторых вариантах осуществления слитый полипептид носитель-антитело содержит участок расщепления Kex2. В определенных вариантах осуществления Kex2 или другой отщепляющий носитель фермент является эндогенным для клетки нитчатых грибов. В определенных вариантах осуществления отщепляющая носитель протеаза является гетерологичной для клетки нитчатых грибов, например, представляет собой другой белок Kex2, происходящий из дрожжей, или протеазу TEV. В определенных вариантах осуществления отщепляющий носитель фермент сверхэкспрессируется.
Следует понимать, что, хотя изобретение описано применительно к определенным конкретным вариантам его осуществления, представленное выше описание предназначено для иллюстрации, но не для ограничения объема изобретения. Другие аспекты, преимущества и модификации в пределах объема изобретения будут очевидны специалистам в области, к которой относится изобретение.
После описания изобретения следующие примеры предоставлены для иллюстрации настоящего изобретения в качестве иллюстрации, но не для ограничения.
ПРИМЕРЫ
ПРИМЕР 1- ИДЕНТИФИКАЦИЯ АСПАРАГИНОВЫХ ПРОТЕАЗ В TRICHODERMA REESEI
В этом примере демонстрируется способность аспарагиновых протеаз из культуральных супернатантов Trichoderma reesei (T. reesei) деградировать тяжелые цепи и легкие цепи антитела.
Очистка аспарагиновой протеазы
Было обнаружено, что протеазную активность в супернатантах T. reesei можно ингибировать с помощью ингибитора аспарагиновых протеаз пепстатин A. Таким образом, пепстатин A (Sigma #P2032) связывали с агарозными гранулами через диаминодипропиламиновый линкер и использовали в качестве аффинной смолы для очистки. Супернатант, полученный периодической ферментацией T. reesei с подпиткой (15 мл), использовали для связывания партиями протеаз со смолой в 35 мл буфера, содержавшего 50 мМ ацетат натрия, 0,2 M NaCl, pH 3,0. Колонку промывали тем же буфером для связывания, и связанный белок удаляли с помощью буфера для элюирования (50 мМ Tris-HCL, 1 M NaCl, pH 8,5). Фракции объемом 0,5 мл собирали. Было очищено всего 42 мкг протеазы. Фракции пиков содержали 0,04 мкг/мкл белка. 30 мкл каждой фракции смешивали с 6 мкл буфера для образца Laemmli, содержавшего β-меркаптоэтанол. Образцы нагревали при 95°C в течение 5 минут, а затем наносили на 4-15% гель PAGE (предварительно залитый гель BioRad mini-protean TGX) вместе с широкодиапазонным предварительно окрашенным маркером молекулярной массы (BioRad). Гель разделяли в подвижном буфере для SDS PAGE в течение 30 минут при 100 В, а затем окрашивали синим красителем GelCode (Thermo Scientific).
Двойную полосу размером 42 кДа очищали на аффинной колонке с пепстатином A (фиг.1) и вырезали из геля SDS PAGE и подвергали расщеплению трипсином в геле с помощью модифицированного трипсина категории, пригодной для секвенирования (Promega #V5111). Затем полученные пептиды экстрагировали из геля и очищали с помощью C18 ZipTip (Millipore #ZTC18M096). Очищенные пептиды анализировали посредством LC-MS/MS на ESI гибридном квадрупольном TOF QSTAR Pulsar (AB Sciex).
Этот анализ обеспечил идентификацию 4 аспарагиновых протеаз, которые имеют высокосходную молекулярную массу. Идентифицированные протеазы включали: pep1 (Tre74156; 42,7 кДа, 42% охват последовательности), pep2 (Tre53961; 42,4 кДа, 15% охват последовательности), pep3 (Tre121133; 49 кДа, 6% охват последовательности) и pep5 (Tre81004; 45 кДа, 9% охват последовательности). Эти аспарагиновые протеазы разделяются до уровня сходной молекулярной массы в геле PAGE. Их сходство аминокислотных последовательностей составляет 51%-64%.
Затем белок (0,8 мкг) из фракции пика (F3) инкубировали с IgG (50 мкг/мл) в натрий-цитратном буфере (50 мМ, pH 5,5) при 37°C в течение 20 часов (фиг.2). Белок инкубировали либо в присутствии, либо в отсутствие 10 мкМ пепстатина A. Смесь антител комбинировали с буфером для образца Laemmli и нагревали при 95°C в течение 5 минут. Затем эти образцы наносили на 4-15% гель для PAGE (предварительно залитый гель BioRad mini-protean TGX) вместе с широкодиапазонным предварительно окрашенным маркером молекулярной массы (BioRad). Гель разделяли в подвижном буфере для SDS PAGE в течение 30 минут при 100 В. IgG не восстанавливали перед разделением на геле. Полноразмерный IgG мигрирует до уровня немного выше маркера массой 200 кДа. Как можно видеть в невосстанавливающем геле на фиг.2, аспарагиновые протеазы были способны обеспечивать мягкую деградацию IgG. Более того, деградация IgG ингибировалась пепстатином A. Активность аспарагиновой протеазы была более ограниченной при pH 5,5, чем при кислых значениях pH, где она имела максимальную активность.
Анализ делеции pep1
Затем аспарагиновую протеазу pep1 исследовали для определения ее содержания в T. reesei. Это осуществляли путем очистки аспарагиновых протеаз из образцов супернатанта, происходящих из штамма с делецией pep1 М182. Штамм с делецией pep1 М182 также продуцирует антитело ритуксимаб.
Штамм с делецией pep1 M181, полученный из основного штамма M124, выращивали в больших культурах с вращающимися флаконами вместе с контрольными флаконами М124. Культуры выращивали в 300 мл TrMM с 4 г/л лактозы, 2 г/л экстракта пивной дробины и 100 мМ PIPPS, pH 5,5. Три различных модельных антитела инкубировали (конечная концентрация 0,05 мкг/мкл) в культуральных супернатантах во вращающихся флаконах (разбавленных до 2 мг/мл в натрий-цитратном буфере pH 5,5) штамма с делецией pep1 и его исходного штамма М124, и в качестве сравнения в культуральном супернатанте после ферментации исходного штамма. Образцы супернатанта (30 мкл) после культивирования в течение 5 суток, содержащие антитело, наносили на 4-15% гель для SDS PAGE и переносили на нитроцеллюлозу для иммуноблоттинга с конъюгированным с AP антителом против тяжелой цепи (Sigma #A3188) или конъюгатом с AP антитела против легкой цепи (Sigma #A3813), разбавленным 1:30000 в TBST. Во время инкубации с антителом в течение ночи в течение 18 часов, супернатант Δpep1 деградировал меньше белка тяжелой цепи по сравнению с контрольным штаммом М124 или супернатантом ферментации (pH 5,5; 28°C; 20 г/л экстракта пивной дробины, 60 г/л лактозы) (фиг.39). Тяжелая цепь была более подвержена деградации по сравнению с легкой цепью. Наибольший эффект стабилизации был очевиден для тяжелых цепей ритуксимаба и MAB01. В тяжелой цепи можно видеть два различных продукта деградации ~48 кДа и ~38 кДа (фиг.39). Происходило только небольшое увеличение стабильности белка легкой цепи в супернатанте Δpep1 по сравнению с контролями (фиг.39).
Получение плазмиды для делеции pep1
Первую конструкцию для делеции pep1 (TreID74156) конструировали для обеспечения удаления селективного маркера из генома Trichoderma reesei после успешного встраивания и, тем самым, рециклирования селективного маркера для последующих делеций генов протеаз. В этом подходе рециклирование маркера, т.е. удаление гена pyr4 из конструкции для делеции, напоминает так называемые бластерные кассеты, разработанные для дрожжей (Hartl, L. and Seiboth, B., 2005, Curr Genet 48: 204-211; и Alani, E. et al., 1987, Genetics 116: 541-545). Сходные бластерные кассеты также были разработаны для нитчатых грибов, включая Hypocrea jecorina (анаморф: T. reesei) (Hartl, L. and Seiboth, B., 2005, Curr Genet 48: 204-211).
Номер TreID относится к идентификационному номеру конкретного гена протеазы из базы данных геномов Joint Genome Institute Trichoderma reesei v2.0. Праймеры для конструирования плазмид для делеции конструировали либо "на глаз", либо с использованием программного обеспечения Primer3 (Primer3 website, Rozen and Skaletsky (2000) Bioinformatics Methods and Protocols: Methods in Molecular Biology. Humana Press, Totowa, NJ, pp 365-386).
Принцип бластерной кассеты с использованием pyr4 в качестве маркерного гена является следующим: pyr4, кодирующий декарбоксилазу оротидин-5'-монофосфата (OMP) T. reesei (Smith, J.L., et al., 1991, Current Genetics 19: 27-33), требуется для синтеза уридина. Штаммы с дефицитом активности декарбоксилазы OMP неспособны расти на минимальной среде без дополнения уридином (т.е. представляют собой ауксотрофы по уридину). Использование 5-фтороротовой кислоты (5-FOA) для получения мутантных штаммов, лишенных активности декарбоксилазы OMP (штаммы pyr4-), основано на преобразовании 5-FOA в токсическое промежуточное соединение 5-фтор-UMP посредством декарбоксилазы OMP. Таким образом, клетки, которые имеют мутантный ген pyr4, являются устойчивыми к 5-FOA, но, кроме того, также являются ауксотрофными по уридину. Устойчивость к 5-FOA в принципе также может быть результатом мутации в другом гене (pyr2, оротатфосфорибозилтрансфераза), и, таким образом, спонтанные мутанты, полученные при этой селекции, необходимо проверять в отношении генотипа pyr4- путем комплементации мутанта геном pyr4. После мутации ген pyr4 можно использовать в качестве ауксотрофного селективного маркера в T. reesei. В бластерной кассете после pyr4 следует прямой повтор размером 308 п.н. 5'-нетранслируемой области pyr4 (5'-UTR) и он окружен 5'- и 3'-фланкирующими областями гена, подлежащего делеции. Селекцию на встраивание кассеты для делеции осуществляют через функцию pyr4. Затем осуществляют удаление маркера pyr4 в присутствии 5-FOA посредством рекомбинации между двумя гомологичными областями (прямой повтор 5'UTR), что приводит к выпетливанию селективного маркера и возможности использования той же бластерной касеты (выпетливание pyr4) в последовательных раундах делеций генов. После выпетливания в локусе остается только последовательность 5'-UTR размером 308 п.н.
Таким образом, селективный маркер pyr4 и 5'-фрагмент с прямым повтором 308 п.н. 5'-UTR pyr4) получали способом ПЦР с использованием плазмиды pARO502 (содержащей геномную копию pyr4 T. reesei) в качестве матрицы. Амплификацию способом ПЦР проводили с помощью полимеразы Phusion и либо буфера HF, либо буфера GC, или с помощью полимеразы Dynazyme EXT. Условия реакции варьировали в зависимости от амплифицируемого фрагмента. Оба фрагмента содержали перекрывающиеся последовательности размером 40 п.н., необходимые для клонирования плазмиды с кассетой с выпетливанием с использованием гомологичной рекомбинации в дрожжах (см. ниже). Для обеспечения возможных дополнительных стадий клонирования, между маркером pyr4 и 5'-прямым повтором помещали участок расщепления AscI и участки NotI помещали так, чтобы они окружали полную бластерную кассету.
1066 п.н. 5'-фланкирующей области и 1037 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep1. Фрагменты получали способом ПЦР. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. ДНК-матрица, используемая при амплификации фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a (ATCC13631).
Для системы гомологичной рекомбинации дрожжей, использованной при клонировании, в соответствующие ПЦР-праймеры помещали перекрывающиеся последовательности для вектора и селективного маркера. Для обеспечения переключения маркеров в конструкции, участки рестрикции NotI встраивали между фланкирующими областями и селективным маркером. Участки рестрикции PmeI помещали между вектором и фланкирующими областями для удаления последовательности вектора перед трансформацией в T. reesei. Остов вектора pRS426 расщепляли ферментами рестрикции (EcoRI и XhoI). Затем фрагменты рестрикции разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов.
Для конструирования плазмиды с делецией, остов вектора и соответствующий маркер, и фрагменты фланкирующих областей трансформировали в Saccharomyces cerevisiae (штамм H3488/FY834). Протокол трансформации дрожжей был основан на способе гомологичной рекомбинации дрожжей, описанном в учебном материале по нокауту в Neurospora Colot and Collopy, (web-сайт с протоколами для генома Neurospora Dartmouth) и лабораторном протоколе Gietz (University of Manitoba, web-сайт лаборатории Gietz). Плазмидную ДНК из трансформантов дрожжей спасали посредством трансформации в Escherichia coli. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга правильной рекомбинации с использованием стандартных лабораторных способов. Несколько клонов с правильными размерами вставок секвенировали и хранили.
В первой плазмиде с делецией pep1 (плазмида pTTv41, таблица 1.1) использовался другой селективный маркер, bar, синтетическая конструкция, содержащая фосфинотрицин-N-ацетилтрансферазу Streptomyces ssp (GenBank ID: AF013602.1, Sweigard et al., 1997, Fungal Genet Newsl 44: 52-53). Фланкирующую область и маркерные фрагменты получали способом ПЦР и собирали в плазмиду с использованием способа рекомбинации дрожжей, описанного выше. Для клонирования второй плазмиды с делецией pep1 (pTTv71, таблица 1.1) маркер bar удаляли из плазмиды с делецией pTTv41 посредством расщепления NotI и заменяли на бластерную кассету pyr4, описанную выше, с использованием системы гомологичной рекомбинации дрожжей. Эти плазмиды с делецией для pep1 (pTTv41 и pTTv71) обеспечивают делецию 1874 п.н. в локусе pep1 и охватывают полную кодирующую последовательность PEP1.

Получение штаммов с делецией pep1 М181 и М195
Для обеспечения рециклирования селективного маркера и обеспечения быстрой делеции последующих генов протеаз, pep1 удаляли из M127 (мутант pyr4- основного штамма M124) с использованием бластерной кассеты pyr4, описанной выше. Для удаления последовательности вектора плазмиду pTTv71 (Δpep1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции pep1 использовали для трансформации штамма M127. Получение протопластов и трансформацию для селекции pyr4 осуществляли по существу в соответствии со способами в Penttila et al. (1987, Gene 61: 155-164) и Gruber et al (1990, Curr. Genet. 18: 71-76).
200 клонов отбирали в качестве селективных штриховых культур. 24 трансформанта, растущих быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 1.2, в отношении правильного встраивания с использованием стандартных лабораторных способов. Семь предполагаемых штаммов с разрушением очищали до клонов единичных клеток. Делецию pep1 подтверждали с помощью саузерн-анализов этих клонов (фиг. A) с использованием стандартных лабораторных способов. ДНК для саузерн-анализов экстрагировали с помощью набора Easy-DNA kit для выделения геномной ДНК (Invitrogen). Саузерн-анализы проводили по существу в соответствии с протоколом для гомологичной гибридизации в Sambrook et al. (1989, Molecular Cloning: A laboratory manual. 2nd Ed., Cold Spring Harbor Laboratory Press) с использованием наборов для радиоактивного мечения (32P) HexaLabel Plus или DecaLabel Plus (Fermentas). Схемы расщепления в соответствии с саузерн-анализом разрабатывали с использованием программного обеспечения либо Sci Ed Central для Windows 95 (Clone Manager 5 для Windows 95), либо Geneious Pro 5.3.6, (web-сайт Geneious). Саузерн-анализы также подтвердили, что четыре из клонов представляли собой клоны с одной вставкой (фиг.3B и 3C). Для трех клонов было показано множественное или неточное встраивание кассеты для делеции, и они были исключены из исследования. Два чистых клона были обозначены как штаммы номер М181 (9-20A-1) и М195 (9-35A-1).
Получение продуцирующего ритуксимаб штамма с делецией pep1 М182
Для удаления последовательности вектора плазмиду pTTv41 (Δpep1-bar) расщепляли посредством PmeI, и соответствующий фрагмент очищали из геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции pep1 использовали для трансформации штамма M169 (экспрессирующего оптимизированное антитело ритуксимаб). Получение протопластов и трансформацию проводили в соответствии со способами, описанными в Penttila et al. (1987) и Avalos et al. (1989).
Приблизительно 100 клонов было отобрано в качестве селективных штриховых культур. 24 трансформанта, растущих быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 1.2) в отношении правильного встраивания с использованием стандартных лабораторных способов. Восемь предполагаемых штаммов с разрушением очищали до клонов единичных клеток. Делецию pep1 подтверждали саузерн-анализом пяти клонов (фиг.4A) с использованием стандартных лабораторных способов, описанных выше для M181 и M195. Саузерн-анализы также подтвердили, что четыре из клонов представляли собой клоны с единичной вставкой (фиг.4B и 4C). Для одного клона было показано множественное или неточное встраивание кассеты для делеции, и он был исключен из исследования. Один чистый клон (11-1A) бы обозначен как штамм номер М182.

Штамм М182 выращивали в минимальной среде для Trichoderma (TrMM), дополненной 20 г/л экстракта пивной дробины, 60 г/л лактозы и 8,1 г/л казаминокислот, при pH 5,5 и 28°C. Семь микрограммов аспарагиновой протеазы было выделено из 15 мл супернатанта. Когда очищенные фракции разделяли на 4-15% геле SDS PAGE (предварительно залитый гель BioRad mini-protean TGX), полоса на уровне 42 кДа, которую ранее наблюдали для исходного штамма, исчезла (фиг.5). Можно было видеть только слабую полосу в области 40 кДа. Полоса размером 40 кДа может соответствовать второстепенным аспарагиновым протеазам. Вторую очистку проводили из культурального супернатанта, где присутствовал pep1. Штамм М169 продуцировал ритуксимаб и не содержал делеции протеазы pep1. Штамм выращивали в минимальной среде для Trichoderma, дополненной 20 г/л экстракта пивной дробины, 60 г/л лактозы и 8,1 г/л казаминокислот, при pH 5,5 и 28°C. 17 мкг аспарагиновой протеазы было очищено из 15 мл супернатанта, и она показала полосу размером 42 кДа на геле SDS PAGE (фиг.5). В соответствии с этим анализом, приблизительно 10 мкг протеазы pep1 продуцируется на 15 мл культурального супернатанта. Это составляет приблизительно 60% от всех аспарагиновых протеаз и только приблизительно 0,04% от общего содержания белка в супернатанте. Эти данные демонстрируют, что pep1 является наиболее распространенной аспарагиновой протеазой в T. reesei.
Анализы других аспарагиновых протеаз
Делеция pep2 продемонстрировала только небольшое улучшение продуцирования тяжелой цепи антитела и снижение общей протеазной активности (фиг.6 и 7).
Таким образом, pep3 и pep5 были следующими важными протеазами для делеции, в частности, в штамме с тройной делецией pep1/tsp1/slp1, поскольку они все еще вносят вклад во вплоть до половины оставшейся протеазной активности в супернатанте штамма с тройной делецией.
Получение плазмиды для делеции pep2
Плазмиду pTTv96 для делеции аспарагиновой протеазы pep2 (TreID0053961) конструировали, по существу как описано для плазмиды pTTv41 для делеции выше. 920 п.н. 5'-фланкирующей области и 1081 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep2. Фрагменты фланкирующих областей получали способом ПЦР с использованием праймеров, приведенных в таблице 1.3. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. ДНК-матрица, использованная для ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Кассету bar получали из pTTv41 расщеплением посредством NotI. Остов вектора представлял собой расщепленный посредством EcoRI/XhoI pRS426, как и в случае pTTv41 выше. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного для pTTv41 выше. Эта плазмида для делеции pep2 (pTTv96) обеспечивает делецию 1437 п.н. в локусе pep2 и охватывает полную кодирующую последовательность PEP2.

Получение продуцирующего ритуксимаб штамма с делецией pep2 M455
Для удаления последовательности вектора плазмиду pTTv96 (Δpep2-bar) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 6 мкг кассеты для делеции pep2 использовали для трансформации штамма M169 (экспрессирующего оптимизированное антитело ритуксимаб). Получение протопластов и трансформацию проводили, как описано для М182 выше, с использованием селекции на основе bar.
Более 200 клонов было отобрано в качестве селективных штриховых культур. 29 трансформантов росли хорошо в качестве вторых штриховых культур. Наилучшие 10 трансформантов, растущих быстро в качестве селективных штриховых культур, подвергали скринингу в отношении правильного встраивания способом ПЦР, с использованием праймеров, приведенных в таблице 1.4, с использованием стандартных лабораторных способов. Кассета для делеции встроилась надлежащим образом в 9 из 10 проанализированных клонов. Открытая рамка считывания была удалена в 9 из 10 трансформантов, проанилизированных способом ПЦР. Пять клонов с разрушением очищали до клонов единичных клеток. Один чистый трансформант (206A) был обозначен как штамм номер M455.

Анализ продуцирующего ритуксимаб штамма с делецией pep2
Штамм M455, 4 других трансформанта с делецией pep2 и исходный продуцирующий ритуксимаб штамм М169 выращивали в культурах во вращающихся флаконах в минимальной среде для Trichoderma (TrMM), дополненной 20 г/л экстракта пивной дробины, 40 г/л лактозы, 100 мМ PIPPS и 8,1 казаминокислот, при pH 5,5 и 28°C. Для анализа эффекта на продукцию ритуксимаба 30 мкл супернатанта из образцов после культивирования в течение 5 суток подвергали иммуноблоттингу. Обнаружение тяжелой цепи проводили с помощью конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188), разбавленного 1:10000 в TBST. Обнаружение легкой цепи проводили с помощью конъюгированного с AP антитела против легкой цепи каппа (Sigma #A3813). Небольшое увеличение продукции тяжелой цепи наблюдали для трансформанта 206A (фиг.6). Тяжелая цепь была фрагментирована, однако количество полноразмерной тяжелой цепи и фрагмента массой 38 кДа было немного увеличено относительно родительского штамма. Кроме того, общую протеазную активность измеряли с помощью сукцинилированного казеина (набор для анализа протеаз QuantiCleave, Pierce #23263) в соответствии с протоколом изготовителя. Трансформант 206A/M455 продемонстрировал наибольшее снижение протеазной активности по сравнению с активностью родительского штамма M169 (фиг.7). Общая протеазная активность в супернатанте была снижена на 10% для M455.
Анализ экспрессированных в Pichia аспарагиновых протеаз
Аспарагиновые протеазы T. reesei pep3 (tre121133) и pep7 (tre58669), экспрессируемые в Pichia, также исследовали in vitro, путем измерения деградации антитела MAB01 и IGF-1. Деградацию MAB01 и IGF-1 посредством pep3 и pep7 анализировали с помощью иммуноблоттинга. Аспарагиновые протеазы продуцировались в супернатантах Pichia. Супернатанты Pichia разбавляли до концентрации 1x, а затем смешивали с 50 мМ натрий-цитратным буфером, pH 5,5. MAB01 добавляли в каждую реакционную смесь так, чтобы конечная концентрация составляла 0,05 мкг/мкл. IGF-1 добавляли в каждую реакционную смесь так, чтобы конечная концентрация составляла 0,30 мкг/мкл. Затем проводили взятие образца объемом 10 микролитров каждой реакционной смеси и добавляли их к 3 мкл буфера для образца Laemmli с β-меркаптоэтанолом. Образцы нагревали при 95°C в течение 5 минут, а затем их наносили на 4-15% гель PAGE (предварительно залитый гель BioRad mini-protean TGX) вместе с предварительно окрашенным маркером молекулярной массы all blue precision plus (BioRad). Гель PAGE разделяли в течение 30 минут при 200 В. Затем белки в геле подвергали электропереносу на нитроцеллюлозный фильтр при 100 В в течение 1 часа. Затем содержащий белок нитроцеллюлозный фильтр блокировали 5% сухим молоком в забуференном Tris физиологическом растворе с 0,1% tween (TBST) в течение 1 часа при встряхивании при комнатной температуре. Затем блокированные мембраны обрабатывали антителом. Содержащие MAB01 мембраны обрабатывали конъюгатом антитела против тяжелой цепи IgG с AP (Sigma #A3188), разбавленным 1:30000 в TBST. Образцы IGF-1 анализировали с использованием первичного антитела против IGF-1 (1:2000 в TBST) и конъюгированным с AP вторичным антителом против IgG (1:5000 в TBST). Во всех случаях инкубацию антител проводили в течение 1 часа при комнатной температуре на устройстве для встряхивания. Затем мембраны промывали посредством 3 замен TBST в течение 20 минут, в каждом случае на устройстве для встряхивания. Мембраны проявляли с помощью субстрата щелочной фосфатазы BCIP/NBT (Promega #S3771) в течение вплоть до 5 минут. Как показано на фиг.8, протеаза pep3 имела низкую активность деградации MAB01 при pH 5,5 после инкубации в течение ночи при 37°C, однако активность была более высокой при pH 4,5. Протеаза pep7 имела только минимальную активность деградации антитела при pH 4,5.
Выделение дополнительных аспарагиновых протеаз с использованием пептида SIP
Несколько дополнительных аспарагиновых протеаз выделяли из штамма T. reesei M277 с тройной делецией протеаз (pep1, tsp1, slp1). Штамм M277 не экспрессирует гетерологичных белков. Штамм с делецией M277 получали, как описано в примере 4 ниже. Штамм выращивали в минимальной среде для Trichoderma, дополненной 20 г/л экстракта пивной дробины, 60 г/л лактозы и 9 г/л казаминокислот при pH 5,5 и 28°C. Аспарагиновые протеазы выделяли посредством аффинной очистки с использованием пептида SIP (Ac-Phe-Lys-Phe-(AHPPA)-Leu-Arg-NH2) (Kataoka Y. et al. 2005 FEBS Letters 579, pp. 2991-2994). Пептид SIP конъюгировали с активированной NHS агарозной смолой (Pierce #26196) с использованием протокола, предоставленного изготовителем. Аффинную смолу SIP использовали для очистки протеаз. Затем супернатант (15 мл) после ферментации штамма T. reesei M277 использовали для связывания протеаз партиями со смолой в 35 мл буфера, содержавшего 50 мМ ацетат натрия, 0,2 M NaCl, pH 3,0 (после условий ферментации pH 5,5; 28°C; 9 г/л казаминокислот; 20 г/л экстракта пивной дробины, 60 г/л лактозы). Колонку промывали тем же буфером для связывания, и связанный белок удаляли с помощью буфера для элюирования (50 мМ Tris-HCl, 1 M NaCl, pH 8,5). Затем собирали фракции объемом 0,5 мл.
Затем 30 мкл каждой очищенной фракции разделяли на 4-15% геле SDS PAGE (предварительно залитый гель BioRad mini-protean TGX) и окрашивали в течение ночи с помощью GelCode blue (Thermo Scientific). Этот гель SDS PAGE продемонстрировал преобладающие полосы размером 42 кДа и слабую полосу размером приблизительно 25 кДа (фиг.9). Затем полосы из геля вырезали и подвергали расщеплению трипсином в геле с помощью модифицированного трипсина категории, пригодной для секвенирования (Promega #V5111). Полученные пептиды экстрагировали из геля и очищали с помощью C18 ZipTip (Millipore #ZTC18M096). Очищенные пептиды анализировали с помощью LC-MS/MS на ESI гибридном квадрупольном TOF QSTAR Pulsar, (AB Sciex). Этот анализ показал, что PEP2, PEP3, PEP4 и PEP5 присутствовали в образце вместе с GAP1 и SLP2. Полагают, что слабая полоса на уровне 25 кДа соответствует глутаминовой протеазе GAP1.
Затем очищенные с помощью SIP протеазы исследовали в отношении их способности деградировать тяжелую цепь антитела MAB01. Очищенные протеазы SIP инкубировали в течение ночи с MAB01 в конечной концентрации 0,05 мкг/мкл в натрий-цитратном буфере при 37°C. Образцы инкубировали при pH 4,0 и pH 5,5 и как в присутствии, так и в отсутствие ингибиторного пептида SIP. После этого проводили взятие образцов реакционной смеси. Собранные образцы анализировали иммуноблоттингом с конъюгатом антитела против тяжелой цепи IgG с AP (Sigma A3188), разбавленным 1:30000 в TBST. Результаты иммуноблоттинга показали, что протеазы имели высокую протеазную активность против тяжелой цепи MAB01 при инкубации при pH 4,0 и сниженную активность при pH 5,5 (фиг.10). Кроме того, как аспарагиновая, так и глутаминовая протеазная активность ингибировались путем инкубации с пептидом SIP (фиг.10).
Анализ очищенных с помощью SIP аспарагиновых протеаз
Затем протеазную активность исследовали в отношении казеина, как в присутствии, так и в отсутствие ингибиторов протеаз. Протеазную активность в отношении казеина исследовали с использованием набора для анализа протеаз EnzChek (Molecular probes #E6638, субстрат зеленый флуоресцентный казеин). Рабочий исходный раствор получали разбавлением исходного раствора до 10 мкг/мл в 50 мМ цитрате натрия, pH 5,5. Очищенные фракции протеаз (10 мкл) разбавляли 40 мкл цитрата натрия, pH 5,5. 100 мкл разбавленного субстрата комбинировали с разбавленными фракциями протеаз в 96-луночном планшете для образца. Затем планшет накрывали и держали при 37°C в течение от одного до трех часов. Данные флуоресценции получали через один, два или три часа с помощью флуоресцентного устройства для считывания планшетов Varioskan (Thermo Scientific) с использованием возбуждения при 485 нм и испускания при 530 нм.
В качестве ингибиторов использовали ингибиторный пептид SIP, пепстатин A, пептид LIP, SBTI и химостатин. Ингибиторный пептид SIP ингибировал как аспарагиновую, так и глутаминовую протеазы; пепстатин A ингибировал только аспарагиновые протеазы; пептид LIP ингибировал только глутаминовую протеазу; SBTI был способен ингибировать SLP2 и PEP4, и химостатин ингибировал SLP2. SIP, LIP и пепстатин A использовали в концентрации 60 мкМ, и SBTI использовали в концентрации 200 мкг/мл. Для различения аспарагиновых и глутаминовых протеаз, в качестве ингибитора использовали пепстатин A, поскольку он не ингибирует глутаминовые протеазы.
Когда исследовали расщепление казеина, значительная часть активности протеазы SIP ингибировалась пепстатином A (фиг.11). Результаты исследований деградации казеина указывают на то, что значительная часть активности при pH 5,5 в очищенных фракциях исходит от аспарагиновых протеаз. Пептид LIP, который представляет собой пропептид GAP1, ингибировал протеазную активность значительно меньше по сравнению с ингибитором SIP. SBTI и химостатин были способны ингибировать протеазу SLP2 в очищенном образце.
Эти результаты подтверждают заключение, что существует 4 аспарагиновых протеаз, присутствующих во фракции SIP (PEP2, PEP3, PEP4 и PEP5).
ПРИМЕР 2 - ИДЕНТИФИКАЦИЯ ГЛУТАМИНОВЫХ ПРОТЕАЗ
В этом примере продемонстрирована способность глутаминовых протеаз из культуральных супернатантов Trichoderma reesei (T. reesei) деградировать тяжелые цепи и легкие цепи антитела.
Анализ делеции gap1
Ранее было определено, что в геноме T. reesei существует четыре последовательности глутаминовых протеаз. Наиболее распространенной глутаминовой протеазой является gap1 (tre69555) при определении по профилю транскрипции. Таким образом, протеазу gap1 очищали из супернатанта T. reesei после аффинной хроматографии с пептидом SIP, как описано в примере 1.
Затем делецию gap1 обеспечивали с использованием продуцирующего антитело MAB01 штамма T. reesei M244 (Δpep1).
Получение плазмиды для делеции gap1
Плазмиду pTTv117 для делеции глутаминовой протеазы gap1 (TreID69555) конструировали, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1. 1000 п.н. 5'-фланкирующей области и 1100 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции gap1. Фрагменты фланкирующих областей получали способом ПЦР с использованием праймеров, приведенных в таблице 2.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. ДНК-матрица, использованная для ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получали из pTTv71 посредством расщепления NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции gap1 (pTTv1l7) обеспечивала делецию 1037 п.н. в локусе gap1, и она охватывает полную кодирующую последовательность Gap1.

Получение продуцирующего MAB01 штамма с двойной делецией Δpep1Δgap1 M296
Для получения продуцирующего антитело MAB01 штамма для второй делеции протеазы, штамм с делецией pep1 М181 (пример 1) трансформировали конструкциями легкой и тяжелой цепей MAB01 (pTTv98+pTTv67) с использованием гигромицина и ацетамида при селекции. Этот штамм MAB01 с делецией pep1 был обозначен номером M244. Удаление бластерной кассеты pyr4 из локуса pep1 осуществляли, по существу как описано в примере 3 ниже для M195 (при получении штамма с двойной делецией протеазы M219). Этот штамм pyr4- был обозначен номером M285 и его использовали в качестве исходного штамма для последующей делеции протеазы.
Для удаления последовательности вектора плазмиду pTTv117 (Δgap1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции gap1 использовали для трансформации штамма M285 (продуцирующий антитело MAB01 штамм M244 pyr4-, на основе штамма Δpep1 M181). Получение протопластов и трансформацию проводили с использованием селекции pyr4, по существу как описано для штаммов с делецией pep1 M181 и M195 в примере 1.
Колонии из чашек для трансформации были отобраны в качестве селективных штриховых культур. Клоны, растущие быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 2.2, в отношении правильного встраивания с использованием стандартных лабораторных способов. Предполагаемые клоны с разрушением очищали до клонов единичных клеток.

Анализ продуцирующего MAB01 штамма с двойной делецией Δpep1Δgap1
Штамм с двойной делецией (Δpep1Δgap1) выращивали в культуре во вращающемся флаконе объемом 2 литра, содержавшей 300 мл минимальной среды для Trichoderma, дополненной 40 г/л лактозы, 20 г/л экстракта пивной дробины и 9 г/л казаминокислот, и забуференной до pH 5,5 посредством 100 мМ PIPPS. Затем штамм Δgap1 исследовали в отношении продукции тяжелой цепи и легкой цепи MAB01 (фиг.12). Штамм Δgap1 сравнивали со штаммами, имеющими делеции в каждом из slp1, slp2 и slp3. Штамм Δpep1 M244 использовали в качестве контроля. Образцы представляли собой образцы культур во вращающихся флаконах на 7 сутки. Образцы анализировали иммуноблоттингом с конъюгатом антитела против тяжелой цепи IgG (Sigma #A3188) или против легкой цепи (Sigma #A3812) с AP (фиг.12). Делеция gap1 приводила к увеличению в 2 раза продукции тяжелой цепи и увеличению в 1,6 раза продукции легкой цепи по сравнению с контрольным штаммом M244 (фиг.13).
Анализ делеции gap2
На основе данных о профиле транскрипции штамма Trichoderma reesei М194, вторая из наиболее распространенных протеаз была идентифицирована как gap2 (tre106661). Таким образом, протеазу gap2 также удаляли из штамма M244 (Δpep1) с использованием конструкции для делеции pTTv145.
Получение плазмиды для делеции gap2
Плазмиду pTTv145 для делеции глутаминовой протеазы gap2 (TreID106661) конструировали, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1. 1021 п.н. 5'-фланкирующей области и 1010 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции gap2. В этой плазмиде фрагмент прямого повтора бластерной кассеты pyr4, представляющий собой 5'-UTR pyr4, заменяли на прямой повтор размером 320 п.н. с конца 5'-фланкирующей области gap2 и не добавляли участка AscI между pyr4 и 5'-прямым повтором. Этот тип бластерной кассеты не должен оставлять никакой дополнительной последовательности в локусе удаленного гена после вырезания. Фрагменты получали способом ПЦР с использованием праймеров, приведенных в таблице 2.3. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. ДНК-матрица, использованная для ПЦР фланкирующих областей, представляла собой штамм T. reesei дикого типа QM6a. Маркерный ген pyr4 получали из pHHO5 посредством расщепления NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции gap2 (pTTv145) обеспечивает делецию 944 п.н. в локусе gap2 и охватывает полную кодирующую последовательность GAP2.

Получение продуцирующих MAB01 штаммов M360 с двумя делециями Δpep1Δgap2
Для получения продуцирующего антитело MAB01 штамма для делеции второй протеазы, штамм с делецией pep1 М181 (пример 1) трансформировали конструкциями легкой и тяжелой цепей MAB01 (pTTv98+pTTv67) с использованием гигромицина и ацетамида при селекции. Удаление бластерной кассеты pyr4 из локуса pep1 осуществляли, по существу как описано в примере 3 ниже для M195 (в отношении получения штамма с делецией двух протеаз M219). Этот штамм pyr4- был обозначен номером M285 и его использовали в качестве исходного штамма для последующей делеции протеазы.
Для удаления последовательности вектора плазмиду pTTv145 (Δgap2-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции gap2 использовали для трансформации штамма M285 (продуцирующий антитело MAB01 pyr4- штамм M244, на основе штамма Δpep1 M181). Получение протопластов и трансформацию проводили с использованием селекции pyr4, по существу как описано для штаммов M181 и M195 в примере 1.
Колонии из чашек для трансформации отбирали в качестве селективных штриховых культур. Клоны, растущие быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 2.4, в отношении правильного встраивания с использованием стандартных лабораторных способов. Предполагаемые штаммы с разрушением очищали до клонов единичных клеток.

Анализ продуцирующего MAB01 штамма с двойной делецией Δpep1Δgap2
Получали несколько трансформантов с делецией. Культуральные супернатанты этих трансформантов разделяли на 4-15% геле SDS PAGE, а затем тяжелую цепь антитела MAB01 анализировали иммуноблоттингом с помощью конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188), и обнаружение легкой цепи проводили с помощью конъюгированного с AP антитела против легкой цепи (Sigma #A3812). Результаты иммуноблоттинга демонстрируют, что делеция gap2 обеспечивала увеличение продукции тяжелой и легкой цепей MAB01 в несколько раз (фиг.14).
Анализ экспрессируемого Pichia gap2
Также исследовали супернатанты Pichia, содержащие gap2 Trichoderma reesei in vitro. Содержащий gap2 супернатант и антитело MAB01 разбавляли в натрий-цитратных буферах, доведенных до pH 4,0, 4,5, 5,0 и 5,5, и инкубировали в течение 20 часов при 37°C. Взятие образцов проводили через 0 минут и через 20 часов. Продукцию тяжелой цепи MAB01 анализировали посредством иммуноблоттинга с использованием конъюгата с AP антитела против тяжелой цепи IgG (Sigma #A3188). Результаты иммуноблоттинга демонстрируют, что gap2 имела максимальную протеолитическую активность против тяжелой цепи MAB01 при pH 4,0 (фиг.15). Хотя протеазная активность gap2 была низкой при pH 5,5 (фиг.15), через 4 суток она была способна продемонстрировать значительную активность в отношении тяжелой цепи. Протеаза gap2 продуцировала продукты деградации размером приблизительно 25 кДа, что указывает на то, что она обладает протеолитической активностью в шарнирной области тяжелой цепи.
ПРИМЕР 3 - ИДЕНТИФИКАЦИЯ СЕРИНОВЫХ ПРОТЕАЗ
Этот пример демонстрирует способность сериновых протеаз из Trichoderma reesei (T. reesei) деградировать тяжелые цепи и легкие цепи антител.
Очистка сериновых протеаз
Сериновые протеазы составляют большое семейство протеаз, которые идентифицированы в качестве деградирующих антитела ферментов. Таким образом, сериновые протеазы очищали из супернатанта Trichoderma. Сериновые протеазы сначала подвергали аффинной очистке из культуральных супернатантов ферментации со смолой sepharose 4 fast flow с п-аминобензамидином (GE healthcare # 17-5123-10). 15 мл культурального супернатанта ферментации связывали партиями со смолой в 35 мл буфера для связывания (0,05 M Tris-HCl, 0,5 M NaCl, pH 7,4). После заполнения и промывания колонки тем же буфером для связывания колонку элюировали 0,05 M глицином, pH 3,0. Затем фракции нейтрализовывали 1M Tris HCl, pH 8,8.
Всего 1,7 мг белка было очищено на аффинной колонке. Когда фракции пиков разделяли на 4-15% геле SDS-PAGE, наблюдали несколько основных полос (~110 кДа, 53 кДа, 39 кДа, 29 кДа) и множество полос меньшего размера. Затем смесь белков фракции пика (F4) исследовали в отношении протеазной активности посредством инкубации образца F4 с IgG1 человека в натрий-цитратном буфере (50 мМ, pH 5,5) при 37°C в течение 20 часов. Образцы инкубировали как в присутствии, так и в отсутствие ингибитора сериновых протеаз PMSF (5 мМ). Затем инкубированные образцы анализировали иммуноблоттингом с конъюгированным с AP антителом против тяжелой цепи IgG (Sigma #A3188) и конъюгированным с AP антителом против легкой цепи IgG (Sigma #A3812), разбавленным 1:30000 в TBST. Результаты иммуноблоттинга показали, что очищенная фракция белков F4 полностью деградировала IgG (фиг.16). Кроме того, обработка PMSF была способна ингибировать значительную часть деградации, что указывает на то, что протеазная активность во фракции F4, которая ответственна за деградацию IgG, в основном представляла собой активность сериновых протеаз.
Для идентификации того, какие белки в очищенных фракциях проявляли протеазную активность, фракции пиков разделяли на геле для зимографии SDS PAGE (12%) с IgG (0,5 мг/мл MAB02). Очищенные фракции и неочищенные образцы супернатанта разделяли на геле для зимографии в денатурирующих условиях. После разделения геля белки в геле ренатурировали путем инкубации геля в 1% triton X-100 для удаления SDS. Затем гелю для зимографии позволяли инкубироваться в течение ночи в реакционном буфере (50 мМ цитрат натрия, pH 5,5) так, чтобы протеазы могли деградировать IgG в геле. Затем гель окрашивали GelCode blue для установления степени окрашивания IgG. Активные протеазы образовывали отчетливую полосу без окрашивания (фиг.17).
На геле для зимографии IgG присутствовали две отчетливо видимых полосы на уровне приблизительно 29 кДа и 65 кДа. Однако полоса на уровне 29 кДа была значительно более выраженной, что указывает на то, что она может быть ответственна за основную часть активности сериновых протеаз в образце. Эти полосы были единственными видимыми двумя полосами в образце неочищенного супернатанта, и были более выраженными в очищенных фракциях (фиг.17). Когда образец протеазы предварительно обрабатывали PMSF, известным ингибитором сериновых протеаз, отчетливые белые полосы выглядели серыми и не были видны, что указывает на то, что полосы соответствуют ферментам сериновым протеазам (фиг.17).
Идентификация сериновой протеазы TSP1 массой 29 кДа
Из соответствующего геля SDS PAGE без MAB02, полосу размером 29 кДа вырезали из геля и подвергали расщеплению трипсином в геле с помощью модифицированного трипсина категории, пригодной для секвенирования (Promega #V5111). В очищенных фракциях полосу размером 29 кДа наблюдали в качестве отчетливой полосы белка. Затем эту отчетливую полосу выделяли. Полученные пептиды экстрагировали из геля и очищали с помощью C18 ZipTip (Millipore #ZTC18M096). Очищенные пептиды анализировали с помощью LC-MS/MS на ESI гибридном квадрупольном TOF QSTAR Pulsar (AB Sciex). Результаты анализа массы отчетливо идентифицировали полосу размером 29 кДа как трипсин-подобную сериновую протеазу TSP1 (tre73897, 35% охват последовательности).
Анализ делеции tsp1
Затем ген, кодирующий TSP1 (tsp1), удаляли из продуцирующего антитело ритуксимаб штамма M169 с получением M183 (Δtsp1). Культивирование во вращающихся флаконах проводили с M169 и трансформантами штамма с делецией tsp1 для измерения эффекта на экспрессию ритуксимаба. Культуры выращивали в 300 мл TrMM с 4 г/л лактозы, 2 г/л экстракта пивной дробины и 100 мМ PIPPS, pH 5,5. Образцы супернатанта (30 мкл) после 5 суток наносили на 4-15% гель SDS PAGE и переносили на нитроцеллюлозу для иммуноблоттинга с конъюгированным с AP антителом против тяжелой цепи (Sigma #A3188), разбавленным 1:10000 в TBST. Два трансформанта штамма с делецией tsp1 продемонстрировали отчетливое увеличение экспрессии тяжелой цепи ритуксимаба по сравнению с исходным контрольным штаммом (фиг.40).
Конструкцию для делеции первого гена протеазы pep1 (TreID74156) конструировали, как описано выше в примере 1.
Получение плазмид для делеции tsp1
Плазмиды для делеции щелочной трипсин-подобной сериновой протеазы tsp1 (TreID71322/TreID73897, Dienes et al., 2007, Enz Microb Tech 40: 1087-1094) конструировали, по существу как описано для плазмид для делеции pep1 в примере 1. 953 п.н. 5'-фланкирующей области и 926 п.н. 3'-фланкирующей области выбирали в качестве основы плазмид для делеции tsp1. Что касается pep1, в первой плазмиде для делеции tsp1 (pTTv42) использовали bar в качестве селективного маркера. Фрагменты фланкирующих областей получали способом ПЦР с использованием праймеров, приведенных в таблице 3.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. ДНК-матрица, использованная для ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Маркер bar получали из pTTv41 (пример 1) расщеплением посредством NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1.
Для клонирования второй плазмиды для делеции tsp1 (pTTv72), маркер bar удаляли из плазмиды для делеции pTTv42 расщеплением посредством NotI. Бластерную кассету pyr4 получали из pTTv71 (пример 1) расщеплением посредством NotI, лигировали в расщепленную NotI pTTv42 и трансформировали в E. coli с использованием стандартных лабораторных способов. Несколько трансформантов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга надлежащего лигирования и ориентации бластерной кассеты pyr4 с использованием стандартных способов. Один клон с правильным размером и ориентацией вставки секвенировали и хранили. Эти плазмиды для делеции tsp1 (pTTv42 и pTTv72) обеспечивают делецию 1252 п.н. в локусе tsp1 и охватывают полную кодирующую последовательность TSP1.

Получение штамма M219 с двойной делецией pep1tsp1
Для повторного использования pyr4 в качестве селективного маркера проводили удаление бластерной кассеты pyr4 из штамма с делецией pep1 М195. Споры высевали на чашки с минимальной средой, содержавшей 20 г/л глюкозы, 2 г/л протеозопептона, 1 мл/л Triton X-100, 5 мМ уридин и 1,5 г/л 5-FOA, pH 4,8. Устойчивые к 5-FOA колонии отбирали через 5-7 суток в 0,9% NaCl, суспендировали тщательно посредством встряхивания и фильтровали через заполненный ватой наконечник пипетки. Для очистки клонов до клонов единичных клеток фильтраты вновь высевали на чашки, описанные выше. Очищенные клоны образовывали споры на чашках, содержавших 39 г/л агарозы с картофельной декстрозой. Эти клоны исследовали в отношении ауксотрофии по уридину путем посева спор на чашки с минимальной средой (20 г/л глюкозы, 1 мл/л Triton X-100), где не наблюдали роста, что указывает на то, что выбранные клоны были pyr4-. Затем все клоны исследовали способом ПЦР (с использованием праймеров, приведенных в таблице 3.2) для удаления бластерной кассеты, и было показано, что они являются правильными. Клон (9-35A-1A-a), использованный для получения штамма с делецией двух протеаз (M219), был обозначен как штамм номер M196 (Δpep1, pyr4-).
Для удаления последовательности вектора, плазмиду pTTv72 (Δtsp1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции tsp1 использовали для трансформации M196 (Δpep1, pyr4-). Получение протопластов и трансформацию проводили с использованием селекции с помощью pyr4, по существу как описано для штаммов с делецией pep1 M181 и M195 в примере 1.
Более 100 колоний отобрали и 48 подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 3.2, в отношении правильного встраивания кассеты для делеции, а также в отношении делеции ORF tsp1 с использованием стандартных лабораторных способов. Четыре предполагаемых клона Δtsp1 очищали до клонов единичных клеток. Делецию tsp1 подтверждали саузерн-анализом этих клонов (фиг.18A) с использованием стандартных лабораторных способов, описанных в примере 1 для M181 и M195. Саузерн-анализы также показали, что только четыре трансформанта (два параллельных клона из двух трансформантов, клоны 16-5AA, 16-5BA, 16-11AA, 16-11BA, фиг.18B и 18C) представляли собой клоны с единичной вставкой. Было определено, что другие клоны содержат дополнительные клоны где-либо еще в геноме, и они были исключены из исследования. Для исключения того, что слабый сигнал, наблюдаемый на фиг.18 для ORF tsp1 в трансформантах, может происходить из гена tsp1, делецию ORF tsp1 подтверждали способом ПЦР с использованием праймеров, приведенных в таблице 3.2. Не было получено сигнала для ORF tsp1. Клон (16-5AA), использованный для удаления бластерной кассеты pyr4 (и для получения штамма M277 с тройной делецией) был обозначен как штамм номер M219 (Δpep1Δtsp1).
Получение продуцирующих MAB01 штаммов M252 с двойной делецией Δpep1Δtsp1
Для удаления последовательности вектора плазмиду pTTv42 (Δtsp1-bar) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции tsp1 использовали для трансформации штамма М181 (Δpep1, пример 1). Получение протопластов и трансформацию проводили с использованием селекции с помощью bar, по существу как описано для штамма с делецией pep1 М182 в примере 1.
Колонии, выросшие на чашках для трансформации, отбирали в качестве селективных штриховых культур. Клоны, растущие быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 3.2, в отношении правильного встраивания с использованием стандартных лабораторных способов. Предполагаемые клоны с разрушением очищали до клонов единичных клеток. Делецию tsp1 подтверждали саузерн-анализом этих клонов (фиг.19A) с использованием стандартных лабораторных способов, описанных в примере 1 для M181 и M195. Также подтверждали, что все клоны являются клонами с одной вставкой (фиг.19B и 19C). Один клон с делецией двух протеаз (13-172D) был обозначен номером M194.

Штамм с делецией двух протеаз M194 использовали для получения экспрессирующих антитело MAB01 штаммов M247 и M252 ниже. Конструирование штамма M247 осуществляли путем трансформации M194 конструкциями тяжелой и легкой цепей MAB01 (pTTv101+pTTv102). Штамм M252 конструировали путем трансформации M194 конструкциями тяжелой и легкой цепей MAB01 (pTTv99+pTTv67). Обе трансформации были основаны на селекции с помощью гигромицина и ацетамида.
Анализ продуцирующего MAB01 штамма M252 с двойной делецией Δpep1Δtsp1
Было показано, что продуцирующий антитело MAB01 штамм с двойной делецией (Δpep1Δtsp1) продуцирует 261 мг/л антитела, с 43% полноразмерного антитела, при выращивании в ферментере. Затем протеазную активность штамма исследовали путем выращивания штамма с минимальной среде Trichoderma, дополненной 20 г/л экстракта пивной дробины, 60 г/л лактозы и 9 г/л казаминокислот при pH 5,5 и 22°C. Было определено, что общая протеазная активность в отношении казеина у этого штамма является в 2,0 раза меньшей, чем у штамма M124 дикого типа (фиг.20).
Идентификация сериновой протеазы SLP1 массой 65 кДа
Протеазу, обладающую активностью, массой приблизительно 65 кДа было труднее идентифицировать вследствие ее низкого уровня экспрессии и сходства по размеру с несколькими белками с высоким уровнем экспрессии. Ранее было идентифицировано, что белки с высоким уровнем экспрессии представляют собой CBHI, CBHII, CIP2 и ксиланазу 4. Для лучшего отделения протеазы массой 65 кДа от белков с высокой экспрессией протеазы осуществили усовершенствование. Усовершенствование включало использование более низкопроцентного геля (7%) SDS PAGE для зимографии и стандартных гелей SDS PAGE для разделения образцов в течение более длительного времени так, чтобы маркер молекулярной массы размером 54 кДа был внизу гелей. Кроме того, супернатант ферментации продуцирующего антитело ритуксимаб трансформанта T. reesei также использовали для очистки сериновых протеаз. Продуцирующий антитело ритуксимаб трансформант представляет собой штамм M169, который продуцирует ритуксимаб и лишен делеций протеаз. Штамм выращивали в минимальной среде для Trichoderma, дополненной 20 г/л экстракта пивной дробины и 60 г/л лактозы при pH 5,5 и 28°C. CBHI, продуцируемый в этой культуре, лишен связывающего целлюлозу домена; таким образом он приблизительно на 10 кДа меньше. Однако М169 не демонстрировал отчетливой полосы, соответствующей протеазе массой 65 кДа (фиг.21). Таким образом, основную область вырезали и подвергали расщеплению трипсином в геле с помощью модифицированного трипсина категории, пригодной для секвенирования (Promega #V5111). Полученные пептиды экстрагировали из геля и очищали с помощью C18 ZipTip (Millipore #ZTC18M096). Очищенные пептиды анализировали посредством LC-MS/MS на ESI гибридном квадрупольном TOF QSTAR Pulsar (AB Sciex).
Анализ пептидов показал, что вторым белком с наилучшей оценкой была протеаза tre51365. Белком с наилучшей оценкой была ксиланаза 4, которая была контаминантом в образце. Субтилизиновая протеаза tre51365, в настоящее время называемая SLP1, была обнаружена в 3 независимых образцах после трех отдельных очисток. В образце с наилучшей оценкой было обнаружено 6 пептидов, и они были отсеквенированы посредством LC-MS/MS. Охват последовательности составлял 8%, поскольку нативный ген протеазы кодирует 882 аминокислоты, которые составляют протеазу массой 93 кДа. При зимографии с желатином можно было наблюдать слабую полосу на уровне ~90 кДа с "размазыванием" до 65 кДа, что указывает на то, что протеаза SLP1 сама по себе претерпевает гидролиз, но сохраняет значительную часть ее активности.
Получение плазмиды для делеции slp1
Затем ген, кодирующий SLP1 (slp1), удаляли из штамма, продуцирующего антитело MAB01 M244 (Δpep1).
Плазмиду для делеции субтилизин-подобной протеазы slp1 (TreID51365) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1. 1094 п.н. 5'-фланкирующей области и 1247 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции slp1. Фрагменты получали способом ПЦР с использованием праймеров, приведенных в таблице 3.3. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, использованная для ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получали из pTTv71 (пример 1) расщеплением посредством NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции slp1 (pTTv126) обеспечивает делецию 2951 п.н. в локусе slp1 и охватывает полную кодирующую последовательность SLP1.

Получение продуцирующего MAB01 штамма M298 и M299 с делецией Δpep1Δslp1
Для получения продуцирующего антитело MAB01 штамма для делеции второй протеазы штамм с делецией pep1 М181 (пример 1) трансформировали конструкциями легкой и тяжелой цепей MAB01 (pTTv98+pTTv67) с использованием гигромицина и ацетамида при селекции. Удаление бластерной кассеты pyr4 из локуса pep1 осуществляли, по существу как описано для M195 выше (в отношении получение штамма с делецией двух протеаз M219). Этот штамм pyr4- был обозначен номером M285 и его использовали в качестве исходного штамма для последующей делеции протеазы.
Для удаления последовательности вектора, плазмиду pTTv126 (Δslp1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции slp1 использовали для трансформации M285 (продуцирующий антитело MAB01 штамм pyr4- M244, на основе штамма Δpep1 M181). Получение протопластов и трансформацию проводили с использованием селекции на основе pyr4, по существу как описано для штаммов с делецией pep1 М181 и М195 в примере 1.
Колонии, растущие на чашках для трансформации, отбирали в качестве селективных штриховых культур. Клоны, растущие быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 3.4, в отношении правильного встраивания с использованием стандартных лабораторных способов. Предполагаемые клоны с разрушением очищали до клонов единичных клеток.

Анализ продуцирующего MAB01 штамма с двойной делецией Δpep1Δslp1 M298/M299
Делеция slp1 в штамме M244 продемонстрировала ожидаемое увеличение продукции тяжелой и легкой цепей (фиг.12 и 13). Штамм с делецией slp1 (Δpep1Δslp1) выращивали в культуре во вращающихся флаконах объемом 2 литра, содержавшей 300 мл минимальной среды для Trichoderma, дополненной 40 г/л лактозы, 20 г/л экстракта пивной дробины и 9 г/л казаминокислот, и забуференной до pH 5,5 посредством 100 мМ PIPPS. Как описано в примере 2 выше, культуральные супернатанты разделяли на 4-15% геле PAGE и подвергали иммуноблоттингу для обнаружения тяжелой и легкой цепей MAB01. Тяжелая цепь продуцировалась на уровнях, которые в 2,8 раза превышали уровни продукции исходного штамма M244 (фиг.13). Легкая цепь продуцировалась на уровнях, которые в 1,8 раза превышали уровни продукции исходного штамма M244 (фиг.13).
Идентификация дополнительных сериновых протеаз
Дополнительные деградирующие антитело сериновые протеазы идентифицировали с использованием других аффинных лигандов. Ингибитор трипсина сои (SBTI) эффективно стабилизирует тяжелую и легкую цепи антитела. Таким образом, он способен ингибировать протеазы, которые ответственны за расщепление антитела. Таким образом, для идентификации этих протеаз проводили аффинную очистку с SBTI, связанным с агарозой (Sigma #T0637).
Для идентификации протеаз использовали штамм T. reesei M44. Штамм M44 представляет собой штамм дикого типа без экспрессии гетерологичного белка. Штамм M44 выращивали в минимальной среде для Trichoderma, дополненной 20 г/л экстракта пивной дробины и 60 г/л лактозы, при pH 5,5 и 28°C. Образец супернатанта культуры M44 объемом 20 мл после 217 часов инкубировали с аффинной смолой с SBTI-агароза (1 мл) в 30 мл буфера для связывания (50 мМ Tris, 0,5 M NaCl, pH 7,5) (pH 5,5; 28°C; 20 г/л экстракта пивной дробины, 60 г/л лактозы). Супернатант и буфер для связывания смешивали в 50-мл конической пробирке и встряхивали при комнатной температуре в течение 1 часа. Затем смесь выливали в стеклянную колонку и промывали 200 мл буфера для связывания. Далее использовали 50 мл буфера с высоким содержанием соли (1 M NaCl) для дальнейшего устранения неспецифических взаимодействий. Наконец, колонку вновь промывали 100 мл исходного буфера для связывания/промывания. Затем колонку элюировали 0,8 M бензамидином HCl в 50 мМ Tris, pH 5,0. Фракции собирали объемами 0,5 мл и подвергали анализу белка с использованием реагента BioRad Bradford с бычьим иммуноглобулином в качестве стандарта.
Из всех собранных фракций 190 мкг белка было очищено из аффинной колонки SBTI. Фракцию пика промывали в центрифужном фильтре для ультрафильтрации vivaspin (Sartorius-stedim) с предельным значением молекулярной массы 10 кДа для удаления ингибитора бензамидина и концентрирования фракций. Концентрированные фракции (cf3 и cf4) и неконцентрированные фракции (f1-f4) наносили на гель для зимографии MAB02 (как описано выше) и на обычный гель SDS PAGE для анализа. Результаты зимографии показывают, что существует два заметных типа протеолитической активности (фиг.22). Наиболее преобладающая полоса была видна на уровне приблизительно 40 кДа и более слабая полоса была видна на уровне приблизительно 26 кДа (фиг.22). На геле для зимографии полосы с более темным окрашиванием фланкировали белую полосу активности на зимограмме. При сравнении этого с концентрированными фракциями, нанесенными на гель SDS PAGE, эти двойные полосы можно было видеть на уровне приблизительно 38 кДа (фиг.23). Гель PAGE представлял собой 4-15% градиентный гель, и гель для зимографии был 12%, так что относительные размеры могут немного отличаться. На геле PAGE полосу белка можно было отчетливо наблюдать в области 26 кДа, что соответствует размеру второй более слабой активности на зимограмме.
Для дальнейшего анализа протеолитической активности очищенной протеазы cf3, фракцию исследовали в отношении ее способности деградировать тяжелую цепь антитела ритуксимаба. 5 мкл образца cf3 инкубировали в натрий-цитратном буфере pH 5,5 с 0,05 мкг/мл ритуксимаба. Затем инкубированные образцы анализировали иммуноблоттингом с использованием конъюгированного с AP специфичного к тяжелой цепи IgG человека антитела (Sigma #A3188), разбавленного 1:30000 в TBST. Результаты иммуноблоттинга демонстрируют, что протеазы сразу деградировали тяжелую цепь антитела ритуксимаба. Полноразмерная тяжелая цепь ритуксимаба мигрирует до уровня только немного выше 50 кДа, в то время как первоначальный продукт деградации мигрировал до уровня приблизительно 45 кДа (фиг.24). Кроме того, инкубация в течение ночи обеспечила дополнительный продукт массой 38 кДа (фиг.24).
Протеазы, ответственные за активность на зимограмме, идентифицировали после секвенирования пептида с помощью LC-MS/MS. Содержавшие белок срезы геля вырезали из геля SDS PAGE, представленного на фиг.23, и подвергали расщеплению трипсином в геле с помощью модифицированного трипсина категории, пригодной для секвенирования (Promega #V5111). Затем полученные пептиды экстрагировали из геля и очищали с помощью C18 ZipTip (Millipore #ZTC18M096). Очищенные пептиды анализировали посредством LC-MS/MS на ESI гибридном квадрупольном TOF QSTAR Pulsar (AB Sciex).
Протеазой с наилучшей оценкой соответствия была субтилизин-подобная протеаза slp2 (tre123244). Было выявлено и отсеквенировано два пептида из slp2, охватывающих 6% всей длины последовательности. Полноразмерная протеаза slp2 имеет размер 58 кДа, однако является обычным, что активная протеаза может иметь меньший размер.
Также существовали другие протеазы, встречающиеся в соседних областях. Анализ нижней области на уровне 26 кДа идентифицировал трипсиновую серин-подобную протеазу tsp1 (tre73897). Это соответствовало слабой выявленной активности на зимограмме. Как описано выше, эта протеаза была идентифицирована посредством аффинной очистки с помощью аминобензамидина.
Кроме того, всю подвергнутую аффинной очистке фракцию SBTI расщепляли трипсином в растворе для определения содержания всех протеаз в образце. Другие идентифицированные протеазы включали протеазу slp7 tre123865 (60 кДа); протеазу pep4 tre77579 (42 кДа); и протеазу slp8 tre58698 (41 кДа).
Субтилизиновые протеазы Trichoderma reesei slp5, slp6 и slp7 сверхпродуцировали в супернатантах Pichia для исследования их активности в отношении тяжелых цепей антител ритуксимаба и MAB01 (фиг.25). Имитирующий супернатант для ритуксана сравнивали с супернатантами, содержавшими slp5 и slp6 (фиг.25 A). Имитирующий супернатант для MAB01 сравнивали с супернатантами, содержавшими slp7 (фиг.25B). Ритуксимаб и MAB01 добавляли к содержавшим протеазу супернатантам Pichia и инкубировали в течение ночи при 37°C. Затем проводили взятие образцов и анализировали их иммуноблоттингом с помощью конъюгированного с AP антитела против тяжелой цепи. Этот анализ показал, что протеаза slp6 продемонстрировала высокую активность деградации тяжелой цепи ритуксимаба и слабую активность деградации MAB01, по сравнению с имитирующими контрольными супернатантами (фиг.25).
Получение плазмид для делеции slp2 и slp3
Исходя из представленных выше результатов, каждый из генов протеаз slp2 и slp3 удаляли из продуцирующего антитело MAB01 штамма M244.
Плазмиды для делеции субтилизин-подобных протеаз slp2 (TreID123244) и slp3 (TreID123234) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1. 1000 п.н. 5'- и 1100 п.н. 3'-фланкирующих областей выбирали в качестве основы плазмиды для делеции slp2. Для slp3 выбирали 1000 п.н. 5'- и 1100 п.н. 3'-фланкирующих областей. Фрагменты получали способом ПЦР с использованием праймеров, приведенных в таблице 3.5. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, использованная в ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получали из pTTv71 (пример 1) расщеплением NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиды конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Плазмида для делеции slp2 (pTTv115) обеспечивает делецию 2114 п.н. в локусе slp2 и охватывает полную кодирующую последовательность SLP2. Плазмида для делеции slp3 (pTTv116) обеспечивает делецию 1597 п.н. в локусе slp3 и охватывает полную кодирующую последовательность SLP3.

Получение продуцирующих MAB01 штаммов с делециями Δpep1Δslp2 и Δpep1Δslp3 M292 и M295
Для получения продуцирующего антитело MAB01 штамма для делеции второй протеазы, штамм с делецией pep1 М181 (пример 1) трансформировали конструкциями легкой и тяжелой цепей MAB01 (pTTv98+pTTv67) с использованием гигромицина и ацетамида при селекции. Удаление бластерной кассеты pyr4 из локуса pep1 осуществляли, по существу как описано для M195 выше (в отношении получения штамма с делецией двух протеаз M219). Этот штамм pyr4- обозначили номером M285 и использовали для последующих делеций протеаз.
Для удаления последовательности вектора, плазмиды pTTv115 (Δslp2-pyr4) и pTTv116 (Δslp3-pyr4) расщепляли PmeI, и соответствующие фрагменты очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг любой из кассет для делеции использовали для трансформации M285 (продуцирующий антитело MAB01 штамм pyr4- M244, на основе штамма Δpep1 М181) по отдельности. Получение протопластов и трансформацию проводили с использованием селекции на основе pyr4, по существу как описано для штаммов с делецией pep1 M181 и M195 в примере 1.
Колонии, растущие на чашках для трансформации, отбирали в качестве селективных штриховых культур. Клоны, растущие быстро в качестве селективных штриховых культур, подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 3.6, в отношении правильного встраивания с использованием стандартных лабораторных способов. Предполагаемые клоны с разрушением очищали до клонов единичных клеток. Чистых клонов не получали даже после повторяющихся стадий очистки.
Анализ продуцирующих MAB01 штаммов с двойной делецией Δpep1Δslp2 и Δpep1Δslp3 M292 и M295
Штамм M292 (Δpep1Δslp2) и штамм M295 (Δpep1Δslp3) выращивали вместе с родственными трансформантами в культуре во вращающемся флаконе объемом 2 литра, содержавшей 300 мл минимальной среды для Trichoderma, дополненной 40 г/л лактозы, 20 г/л экстракта пивной дробины и 9 г/л казаминокислот, и забуференной до pH 5,5 посредством 100 мМ PIPPS. Культуральные супернатанты разделяли на 4-15% геле PAGE и проводили анализ с использованием иммуноблоттинга для обнаружения тяжелой цепи и легкой цепей MAB01. Результаты демонстрируют, что обе делеции повышали стабильность MAB01 (фиг.12 и 13). Делеция Δslp2 увеличивала экспрессию тяжелой цепи MAB01 в культуре во вращающемся флаконе приблизительно в 2,4 раза на 7 сутки по сравнению с исходным штаммом M244 (фиг.13). Δslp3 увеличивала экспрессию тяжелой цепи MAB01 в больших вращающихся флаконах приблизительно в 1,5 раза и экспрессию легкой цепи MAB01 приблизительно 1,7 раза по сравнению с родительским штаммом M244 (фиг.13). Более того, по сравнению с Δslp3 и Δgap1, Δslp2 продемонстрировала наиболее высокую кратность увеличения экспрессии тяжелой цепи MAB01 относительно экспрессии тяжелой цепи MAB01 в родительском штамме M244 (фиг.13).
Когда slp2 удаляли из штамма с несколькими делециями M306 (Δpep1Δtsp1Δslp1), делеция slp2 приводила к снижению споруляции и более медленному росту по сравнению с исходным штаммом.
ПРИМЕР 4 - ШТАММЫ TRICHODERMA С НЕСКОЛЬКИМИ ДЕЛЕЦИЯМИ ПРОТЕАЗЫ
В этом примере продемонстрирована увеличенная продукция антител и стабильность штаммов Trichoderma reesei (T. reesei), содержащих несколько делеций генов протеаз, идентифицированных выше в примерах 1-3.
Получение штамма с тройной делецией Δpep1Δtsp1Δslp1
Штамм T. reesei, имеющий тройную делецию Δpep1Δtsp1Δslp1, получали и исследовали в отношении улучшения продуцирования антитела. Штамм также использовали для дальнейших раундов делеций протеаз.
Получение штамма с тремя делециями M277
Для получения не содержащего маркера штамма с делецией трех протеаз, использовали выпетливание маркера pyr4 в штамме M219, по существу как описано выше для выпетливания pyr4 из штамма с делецией одной протеазы Δpep1. Три последовательных стадии селекции с 5-FOA проводили, чтобы убедиться, что выбранные клоны происходили из единичных клеток. Выпетливание pyr4 подтверждали в конечных клонах способом ПЦР (с использованием праймеров, приведенных в таблице 3.1); не наблюдали специфических сигналов в случае праймеров, отжигающихся на подвергнутую выпетливанию часть pyr4. Выпетливание далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. Клон, использованный для получения штамма с тремя делециями, обозначили номером M228 (Δpep1Δtsp1,pyr4-).
Плазмида для делеции pTTv126 для третьего гена протеазы, субтилизин-подобной протеазы slp1 (TreID51365), описана выше (таблица 3.3). Эта плазмида для делеции обеспечивает делецию 2951 п.н. в локусе slp1 и охватывает полную кодирующую последовательность SLP1.
Для удаления последовательности вектора плазмиду pTTv126 (выпетливание Δslp1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции slp1 использовали для трансформации M228 (Δpep1Δtsp1, pyr4-), как описано выше. Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов М181 и M195 с использованием селекции на основе pyr4.
200 клонов было отобрано в качестве первых штриховых культур. 48 из этих штриховых культур подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.1, в отношении правильного встраивания с использованием стандартных лабораторных способов. Пять предполагаемых клонов с разрушением трех протез (Δpep1Δtsp1Δslp1) очищали до клонов единичных клеток. Делецию slp1 подтверждали саузерн-анализом пяти клонов (фиг.26A). Саузерн-анализ проводили, как описано в примере 1. Саузерн-анализы также подтвердили, что три из клонов представляли собой клоны с единичной вставкой (фиг.26B и 26C). Было показано, что два других клона содержат дополнительные копии где-либо в геноме, и они были исключены из исследования. Клон, используемый для удаления бластерной кассеты pyr4 (и для получения штамма с делецией четырех протеаз M307 ниже), был обозначен номером штамма M277 (Δpep1Δtsp1Δslp1).
Получение продуцирующего MAB01 штамма с делецией трех протеаз M304
Для получения продуцирующего антитело MAB01 штамма с делецией третьей протеазы, штамм с делецией двух протеаз pep1/tsp1 M194 (пример 3) трансформировали конструкциями легкой и тяжелой цепей MAB01 (pTTv99+pTTv67) с использованием гигромицина и ацетамида при селекции. Штамм MAB01 с двойной делецией pep1/tsp1 обозначили номером M252. Удаление бластерной кассеты pyr4 из локуса pep1 осуществляли, по существу как описано в примере 3 для M195 (в отношении получения штамма с делецией двух протеаз M219). Этот pyr4- штамм обозначили номером M284 и использовали для родительского штамма для последующей делеции протеазы.
Делецию третьей протеазы в M284 обеспечивали с использованием конструкции pTTv128 для делеции slp1. Эта конструкция содержит нативную сверхэкспрессирующую KEX2 кассету, нацеленную на локус slp1. Трансформацию осуществляли, по существу в соответствии с протоколом, описанным в примере 1, для штаммов М181 и М195 с использованием селекции на основе pyr4. Полученный штамм представляет собой продуцирующий MAB01 штамм с тремя делециями протеаз M304.
Анализ продуцирующего MAB01 штамма с тремя делециями протеаз M304
Было показано, что продуцирующий антитело MAB01 штамм M304 с тремя делециями протеаз (Δpep1Δtsp1Δslp1) продуцирует антитело MAB01 с выходом вплоть до 3,5 г/л в культуре (pH 5,5; 28->22°C; 60 г/л пивной дробины, 30 г/л глюкозы, 60 г/л лактозы + подпитка лактозой), и качеством продукта вплоть до 84% полноразмерного IgG (см. пример 6 ниже). Протеазную активность штамма также исследовали путем выращивания штамма в минимальной среде для Trichoderma, дополненной 60 г/л твердой пивной дробины, 30 г/л глюкозы и 60 г/л лактозы при pH 5,5. Культуру выращивали при 30°C, а затем переключали на 22°C для фазы продуцирования. Периодическое культивирование с подпиткой проводили с подпиткой лактозой. Было определено, что общая протеазная активность в отношении казеина в этом штамме является приблизительно в 3,2 раза меньшей по сравнению со штаммом дикого типа M124 (фиг.20).
Сравнение штаммов с одной, двумя и тремя делециями
Сравнивали относительную протеазную активность культуральных супернатантов штамма с делецией одной протеазы (Δpep1) M181 (см. пример 1), штамма с делецией двух протеаз (Δpep1Δtsp1) M219 (см. пример 3) и штамма с делецией трех протеаз (Δpep1Δtsp1Δslp1) M277. Эти штаммы с делециями сравнивали со штаммом дикого типа М124. Три штамма с делецией протеаз выращивали во вращающихся флаконах объемом 2 литра с 300 мл TrMM, содержавшей 40 г/л лактозы, 20 г/л экстракта пивной дробины и 100 мМ PIPPS при pH 5,5. Взятие образцов проводили на 3, 5, 7 и 10 сутки. Каждый из образцов культурального супернананта M124, M181, M219 и M277 на 7 сутки разбавляли 1:2 в натрий-цитратном буфере (50 мМ, pH 5,5) и 30 мкл наносили на 12% гель для зимографии SDS PAGE, содержавший MAB02. Гель SDS PAGE разделяли при 100 В в течение 45 минут. Затем гель инкубировали в 2,5% Triton X-100 в течение одного часа, а затем промывали несколько раз буфером для реакции (50 мМ цитрат натрия, pH 5,5). Затем гель для зимографии оставляли в течение ночи при встряхивании в буфере для реакции. На следующее утро гель окрашивали реагентом для окрашивания GelCode Blue. Области, где антитело MAB02 было деградировано, выглядели как белые пятна на окрашенном синим цветом геле.
Два типа активности протеазы наблюдали в контрольных образцах М124 и Δpep1 М181 (фиг.27). Наиболее выраженную активность наблюдали на уровне 65-90 кДа, что соответствует slp1. Более слабую активность наблюдали на уровне приблизительно 28 кДа, что соответствует tsp1. Как и ожидалось, штамм Δpep1Δtsp1 M219 не продуцировал полосу зимограммы на уровне 28 кДа. Аналогично, штамм Δpep1Δtsp1Δslp1 M277 не проявлял никакой активности на зимограмме. Размер активной slp1, по-видимому, варьирует, поскольку она была все еще активна после расщепления до 65 кДа, даже несмотря на то, что ее зрелый размер составляет 90 кДа. Варьирование размера можно наблюдать на фиг.27.
Общую протеазную активность в отношении сукцинилированного казеина из супернатантов культур штаммов с делециями M181, M219 и M277 также измеряли в образцах на 3 сутки, 5 сутки и 7 сутки. Сначала супернатанты перед анализом разбавляли до 2 мг/мл общего белка в 50 мМ цитрате натрия, pH 5,5. 50 мкл разбавленного супернатанта наносили на 96-луночный планшет и добавляли 50 мкл сукцинилированного казеина для начала реакции. Для каждого образца использовали контроль фонового уровня с буфером вместо казеина. После добавления казеина протеазной реакции позволяли протекать в течение 1 часа при 37°C. Для развития окраски реакции в каждую лунку добавляли 50 мкл реагента TNBSA, и планшет инкубировали в течение 16 часов при 37°C. Для всего планшета измеряли поглощение при 450 нм. Неспецифический фоновый сигнал вычитают из конкретного показателя активности протеазы. Как показано на фиг.28, образцы супернатантов из трех штаммов с делецией протеаз обладали меньшей протеазной активностью относительно штамма дикого типа М124.
Супернатант культур M277 и М124 (5 и 7 сутки) разбавляли до 6 мг/мл в 50 мМ натрий-цитратном буфере. К этим разбавленным супернатантам добавляли антитело MAB01 до конечной концентрации 0,05 мкг/мкл. Эти реакционные смеси инкубировали при 37°C в течение ночи. Взятие образцов реакции проводили в нулевой момент времени, через 1 час и после инкубации в течение ночи. 20 мкл образцов наносили на 4-15% гель SDS PAGE и разделяли при 200 вольт в течение 40 минут. Гель переносили при 100 вольт в течение 1 часа на нитроцеллюлозу для иммуноблоттинга. Мембрану блокировали 5% молоком в TBST в течение одного часа. Обнаружение тяжелой цепи MAB01 проводили с помощью конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188), разбавленного 1:30000 в TBST. После промывания мембраны посредством TBST, блот подвергали развитию окраски с помощью субстрата AP (Promega). При сравнении образцов, инкубированных в течение ночи, было очевидно, что тяжелая цепь больше деградировалась в супернатанте штамма M124. M124 не имеет делеций протеаз. Штамм M277 с делецией 3 протеаз продуцировал тяжелую цепь MAB01, которая была в значительной степени стабильной. На 5 сутки было в 2,5 раза большей тяжелой цепи в супернатанте M277 после инкубации в течение ночи. В случае супернатанта на 7 сутки, было обнаружено в 4 раза больше тяжелой цепи (фиг.41).
Штамм с четырьмя делециями M307
Штамм M307, имеющий четыре делеции Δpep1Δtsp1Δslp1Δgap1 получали и использовали для дальнейших раундов делеций проетаз.
Получение штамма с делецией четырех протеаз M307
Для получения не содержащего маркера штамма с делецией четырех протеаз использовали удаление бластерной кассеты pyr4 из штамма M277, по существу как описано в примере 3, для удаления бластерной кассеты pyr4 из штамма с делецией одной протеазы M195 (Δpep1). Три последовательных стадии селекции с 5-FOA проводили для обеспечения того, чтобы отобранные клоны происходили из единичных клеток. Подтверждение удаления бластерной кассеты в конечных клонах проводили способом ПЦР с использованием праймеров, приведенных в таблице 4.2, с использованием стандартных лабораторных способов. Не наблюдали специфических сигналов в случае праймеров, отжигающихся на удаленной части pyr4. Удаление далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. На чашках без добавления уридина роста не наблюдали. Клон, использованный для получения штамма с делецией четырех протеаз, был обозначен как штамм номер M306 (Δpep1Δtsp1Δslp1, pyr4-).
Плазмида pTTv117 для делеции четвертого гена протеазы, глутаминовой протеазы gap1 (TreID69555), описана в примере 2 (таблица 2.1). Эта плазмида для делеции обеспечивает делецию 1037 п.н. в локусе gap1 и охватывает полную кодирующую последовательность GAP1.
Для удаления последовательности вектора плазмиду pTTv117 (Δgap1-pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции gap1 использовали для трансформации M306 (Δpep1Δtsp1Δslp1, pyr4-) выше. Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов М181 и М195, с использованием селекции на основе pyr4.
150 клонов было отобрано в качестве первых штриховых культур. 48 из этих штриховых культур подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.2 в отношении правильного встраивания с использованием стандартных лабораторных способов. Восемь предполагаемых клонов с разрушением для четырех протеаз (Δpep1Δtsp1Δslp1Δgap1) очищали до клонов единичных клеток. Делецию gap1 подтверждали саузерн-анализом восьми клонов (фиг.29 A). Саузерн-анализы проводили, как описано в примере 1. Саузерн-анализы также подтвердили, что три из клонов представляли собой клоны с единичной вставкой (фиг.29B и 29C). Было показано, что пять других клонов содержат дополнительные копии где-либо еще в геноме, и они были исключены из исследования. Клон, использованный для удаления бластерной кассеты pyr4 (и для получения штамма с делецией пяти протеаз M369 ниже), был обозначен как штамм номер M307 (Δpep1Δtsp1Δslp1Δgap1).

Получение продуцирующего MAB01 штамма с делецией четырех протеаз M371
Для получения продуцирующего антитело MAB01 штамма с делецией четырех протеаз, осуществляли удаление бластерной кассеты pyr4 из локуса slp1 штамма M304, по существу как описано в примере 3 для M195 (в отношении получения штамма с делецией двух протеаз M219). Этот штамм pyr4- был обозначен номером M317 и его использовали в качестве исходного штамма для последующей делеции протеаз.
Делецию четвертой протеазы в M317 осуществляли с использованием конструкции pTTv117 для делеции gap1, как описано выше. Трансформацию осуществляли по существу в соответствии с протоколом, описанным в примере 1, для штаммов M181 и M195 с использованием селекции на основе pyr4. Полученный штамм представляет собой продуцирующий MAB01 штамм с делецией четырех протеаз M371.
Анализ штамма с делецией четырех протеаз
Затем общую протеазную активность культурального супернатанта штамма с четырьмя делециями M307 измеряли и сравнивали с культуральными супернатантами штамма с тремя делециями M277 и штамма дикого типа M124. Каждый штамм выращивали во вращающихся флаконах объемом 2 литра с 300 мл TrMM, содержавшего 40 г/л лактозы, 20 г/л экстракта пивной дробины и 100 мМ PIPPS при pH 5,5. Образцы супернатанта на 7 сутки отбирали для общего анализа протеаз. Общие концентрации белка в супернатантах измеряли с использованием анализа BCA с бычьим иммуноглобулином в качестве стандарта. Супернатанты подвергали серийному разведению 1:2 в натрий-цитратном буфере при pH 5,5. Разбавленные супернатанты добавляли к флуоресцентно меченному субстрату казеину и инкубировали при 37°C. Флуоресценцию измеряли через 1 час с возбуждением при 485 нм и испусканием при 530 нм. Результаты показали, что уровень протеазной активности в штамме с тремя делециями M277 был в 3 раза меньшим, чем у штамма дикого типа М124, и в штамме с четырьмя делециями M307 он был в 8 раз меньшим, чем у штамма дикого типа M124 (фиг.30).
Кроме того, на фиг.20 обобщенно представлена общая протеазная активность в отношении казеина в штамме с одной делецией М188, штамме с двумя делециями M219, штамме с тремя делециями M277 и штамме с четырьмя делециями M307 по сравнению со штаммом дикого типа М124. Единичная делеция pep1 снижала протеазную активность в 1,7 раза, две делеции pep1/tsp1 снижали протеазную активность в 2 раза, три делеции pep1/tsp1/slp1 снижали протеазную активность в 3,2 раза, и четыре делеции pep1/tsp1/slp1/gap1 снижали протеазную активность в 7,8 раз по сравнению со штаммом дикого типа М124 (фиг.20).
Продуцирующий антитело MAB01 штамм M371 содержит четыре делеции Δpep1Δtsp1Δslp1Δgap1. Штамм выращивали в ферментере и сравнивали с продуцирующим MAB01 штаммом с тремя делециями в тех же условиях. Периодическое культивирование проводили для штамма M371, который продуцировал MAB01 и имел делеции протеаз pep1, tsp1, slp1 и gap1 и сверхэкспрессию kex2. Штамм выращивали в минимальной среде для Trichoderma, дополненной 40 г/л твердой пивной дробины, 40 г/л глюкозы и 40 г/л лактозы при pH 5,5. Культуру выращивали при 30°C, а затем переключали на 22°C для фазы продуцирования. Периодическое культивирование проводили с помощью штамма M304, который продуцировал MAB01 и имел делеции протеаз pep1, tsp1 и slp1 и сверхэкспрессию kex2. Штамм выращивали в минимальной среде для Trichoderma, дополненной 40 г/л твердой пивной дробины, 40 г/л глюкозы и 40 г/л лактозы при pH 5,5. Культуру выращивали при 30°C, а затем переключали на 22°C для фазы продуцирования.
Вычисленный выход полноразмерного антитела был на 20% более высоким в штамме с делецией gap1 в образце, полученном на 6 сутки. В тех же условиях штамм с четырьмя делециями продуцировал 1,9 г/л (897 мг/л полноразмерного антитела), и штамм с тремя делециями продуцировал 1,3 г/л (731 мг/л полноразмерного антитела). В супернатантах ферментера измеряли общую протеазную активность в отношении казеина. Образцы супернатанта разбавляли в натрий-цитратном буфере, pH 5,5, так чтобы общая концентрация белка составила 0,15 мг/мл для всех образцов. К этому разбавленному супернатанту добавляли 10 мкг/мл казеина BODIPY для начала анализа протеаз. Образцы каждых суток культивирования сравнивали между двумя различными штаммами. Результаты показывают, что в штамме с делецией gap1 на 5 сутки происходило снижение общей протеазной активности вплоть до 30% (фиг.31). На 6 сутки протеазная активность была на 20% более низкой, что коррелирует с 20% увеличением выхода антитела на эти сутки.
Штамм с пятью делециями
Штамм M369, имеющий пять делеций Δpep1Δtsp1Δslp1Δgap1Δgap2, получали и использовали для дальнейших раундов делеций протеаз.
Получение штамма с делецией пяти протеаз M369
Для получения не содержащего маркера штамма с делецией пяти протеаз использовали удаление бластерной кассеты pyr4 в штамме M307, по существу как описано в примере 3, для удаления бластерной кассеты pyr4 из штамма с делецией одной протеазы M195 (Δpep1). Проводили три последовательных стадии селекции с 5-FOA для обеспечения того, чтобы отобранные клоны происходили из единичных клеток. Конечные клоны подтверждали в отношении удаления бластерной кассеты способом ПЦР с использованием праймеров, приведенных в таблице 4.3, с помощью стандартных лабораторных способов. Не наблюдали специфических сигналов для праймеров, отжигающихся на удаленной части бластерной кассеты pyr4. Удаление далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. На чашках без добавления уридина роста не наблюдали. Клон, использованный для получения штамма с делецией пяти протеаз, был обозначен как штамм номер M321 (Δpep1Δtsp1Δslp1Δgap1, pyr4-).
Плазмида pTTv145 для делеции пятого гена протеазы, глутаминовой протеазы gap2 (TreID106661), описана в примере 2 (таблица 2.3). Эта плазмида для делеции обеспечивает делецию 944 п.н. в локусе gap2 и охватывает полную кодирующую последовательность GAP2.
Для удаления последовательностей вектора, плазмиду pTTv145 (Δgap2-выпетливание pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции gap2 использовали для трансформации M321 (Δpep1Δtsp1Δslp1Δgapl, pyr4-), как описано выше. Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
100 клонов было отобрано в качестве первых штриховых культур. Все 20 выросших штриховых культур подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.3, в отношении правильного встраивания с использованием стандартных лабораторных способов. 10 предполагаемых клонов с разрушением для пяти протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2) очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР. Только один очищенный клон был отрицательным в отношении ORF gap2. Делецию gap2 подтверждали саузерн-анализом клона (фиг.32A). Саузерн-анализы проводили, как описано в примере 1. Саузерн-анализы также показали клоны, которые содержат дополнительную копию кассеты для делеции где-либо в геноме или имеют некоторые внутренние перестройки в этом локусе (фиг.32B и 32C). Поскольку этот клон был единственным полученным клоном с делецией пяти протеаз, его отбирали для дальнейшего применения (фиг.32D и 32E). Клон 14 представлял собой клон, использованный для удаления бластерной кассеты pyr4 и для получения штаммов с делецией 6 протеаз M396 и M400, ниже (фиг.32E). Этот клон был обозначен как штамм номер M369 (Δpep1Δtsp1Δslp1Δgap1Δgap2).

Анализ штамма с делецией пяти протеаз
Протеазную активность штамма M369 измеряли в отношении его родительского штамма M307. Делеция протеазы gap2 приводила к меньшей протеазной активности в отношении казеина на 23% (фиг.33).
Штамм с 6 делециями
Штамм с 6 делециями протеаз, имеющий делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4 получали и использовали для дальнейших раундов делеций протеаз.
Получение плазмид с делецией pep4
Плазмиду pTTv181 для делеции шестого гена протеазы, аспарагиновой протеазы pep4 (TreID77579), конструировали, по существу как описано для плазмиды Δpep1 pTTv71 в примере 1. 959 п.н. 5'-фланкирующей области и 992 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep4. Что касается pep1, первая плазмида для делеции pep4 (pTTv43, таблица 4.4) содержала другой селективный маркер, bar, который был заменен бластерной кассетой pyr4. Бластерную кассету получали из pTTv71 посредством расщепления NotI, лигировали с разрезанной NotI pTTv43, а затем трансформировали в E. coli с использованием стандартных способов. Несколько трансформантов культивировали, выделяли плазмидную ДНК и расщепляли для скрининга правильного лигирования и ориентации бластерной кассеты pyr4 с использованием стандартных лабораторных способов. Один клон с правильным размером вставки и ориентацией секвенировали и хранили (pTTv73, таблица 4.4). Бластерную кассету немного изменяли еще раз: фрагмент прямого повтора, использованный для удаления pyr4, изменяли с 308 п.н. 5'-UTR pyr4 на прямой повтор 300 п.н. с конца 5'-фланкирующей области pep4 (как и в pTTv145, gap2-pyr4). Это осуществляли путем удаления существующей бластерной кассеты pyr4 из pTTv73 расщеплением посредством NotI. Ген pyr4 амплифицировали способом ПЦР с использованием pTTv73 в качестве матрицы с использованием праймеров, приведенных в таблице 4.4. Для системы гомологичной рекомбинации дрожжей, использованной при клонировании, перекрывающиеся последовательности для вектора помещали в соответствующие праймеры для ПЦР. Для обеспечения переключения маркеров в конструкции участки рестрикции NotI вносили по обеим сторонам селективного маркера pyr4 и для дополнительных стадий клонирования AscI вносили между 5'-прямым повтором и 3'-фланкирующей областью pep4. Этот тип бластерной кассеты не должен оставлять никакой дополнительной последовательности в локусе удаленного гена после вырезания. 5'-прямой повтор pep4 размером 300 п.н. амплифицировали способом ПЦР с использованием штамма T. reesei дикого типа QM6a в качестве матрицы. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Несколько клонов, полученных рекомбинацией, культивировали, и плазмидную ДНК выделяли и расщепляли для скрининга надлежащей рекомбинации с использованием стандартных способов. Эти плазмиды для делеции pep4 (pTTv43, pTTv73 и pTTv181, таблица 4.4) обеспечивают делецию 1413 п.н. в локусе pep4 и охватывают полную кодирующую последовательность PEP4.

Получение штаммов с делецией 6 протеаз M396 и M400
Для получения не содержащего маркер штамма с делецией 6 протеаз использовали удаление маркера pyr4 в штамме M369, по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Проводили три последовательных стадии селекции с 5-FOA для обеспечения того, чтобы отобранные клоны происходили из единичных клеток. Конечные клоны подтверждали способом ПЦР с использованием праймеров, приведенных в таблице 4.5, с помощью стандартных лабораторных способов. Сигнал, соответствующий успешному удалению бластерной кассеты, получали для всех клонов. Удаление далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. На чашках без добавления уридина роста не наблюдали. Саузерн-анализ шести предполагаемых клонов pyr4- подтвердил удаление бластерной кассеты для трех клонов. Кроме того, саузерн-анализ показал, что эти три клона утратили дополнительные сигналы, наблюдаемые для фланкирующих областей gap2 в родительском M369. Таким образом, эти клоны должны иметь ожидаемую геномную организацию в локусе gap2. Клон, использованный для получения штамма с делецией 6 протеаз, был обозначен как штамм номер M381 (Δpep1Δtsp1Δslp1Δgap1Δgap2, pyr4-).
Для удаления последовательности вектора плазмиду pTTv181 (Δpep4-выпетливание pyr4) расщепляли посредством PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции pep4 использовали для трансформации M381 (Δpep1Δtsp1Δslp1Δgap1Δgap2, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
Свыше 200 трансформантов было отобрано в качестве первых штриховых культур. 32 растущих штриховых культур подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 4.5) в отношении правильного встраивания. Семь клонов обеспечили ожидаемые сигналы и их очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.5. Делецию pep4 подтверждали саузерн-анализом пяти клонов (фиг.34A и 34B) с использованием стандартных лабораторных способов, описанных в примере 3 для М181 и M195. Саузерн-анализы также показали, что все трансформанты (фиг.34C и 34D) представляли собой клоны с единичной вставкой. Для исключения того, что слабый сигнал, наблюдаемый при скрининге способом ПЦР в отношении ORF pep4 в трансформантах, происходил из гена pep4, три клона очищали далее посредством стадий единичных клеток и повторно анализировали с помощью саузерн-гибридизации и ПЦР. Не было получено сигнала для ORF pep4 ни в одном из анализов, что указывает на чистоту штамма. Клон 25-120A, использованный для удаления бластерной кассеты pyr4 (и для получения штамма с делецией 7 протеаз), был обозначен как штамм номер M396, и клон после повторной очистки 25-120A-a был обозначен как штамм номер M400.

Анализ протеазной активности в штаммах с делецией 4, 5 и 6 протеаз
Трансформанты штамма с делецией четырех протеаз M307, штамма с делецией пяти протеаз M369 и штамма с делецией 6 протеаз культивировали в культурах во вращающихся флаконах. Взятие образцов супернатанта проводили из культур в больших вращающихся флаконах в TrMM с 20 г/л пивной дробины и 40 г/л лактозы, забуференной 100 мМ PIPPS при pH 4,8. Значение pH на 5 сутки составляло ~4,25. Исследованные трансформанты с делецией 6 протеаз не были конечным штаммом, так что существовало варьирование вследствие чистоты спор. Они были некоторыми из лучших трансформантов, однако затем проводили дальнейшую очистку спор. Супернатанты после 5 суток разбавляли 1:3 в 50 мМ натрий-цитратном буфере, pH 4,5. К этому разбавленному супернатанту добавляли казеин FL BODIPY (10 мкг/мл) и инкубировали вместе при 37°C в течение 4 часов. Анализ протеазной активности проводили, как описано в протоколе изготовителя (набор для анализа протеаз enzCheck #E6638, Molecular Probes). Результаты анализа протеазной активности можно видеть на фиг.33.
Происходило небольшое снижение протеазной активности, когда штамм с делецией пяти протеаз M369 выращивали в кислых условиях. Делеция gap2 в штамме обеспечила снижение протеазной активности в отношении казеина на 23%. В штаммах с делецией 6 протеаз аспарагиновая протеаза pep4 была удалена в 5 из исследованных трансформантов. Наилучший трансформант продемонстрировал 35% снижение по сравнению с его родительским штаммом M369.
Получение штамма с 7 делециями
Штамм с делецией 7 протеаз, имеющий делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3, получали и использовали для дальнейших раундов делеций протеаз.
Получение плазмид для делеции pep3
Первую плазиду pTTv188 для делеции седьмого гена протеазы, аспарагиновой протеазы pep3 (TreID121133), конструировали, по существу как описано для плазмиды Δpep1 pTTv41 в примере 1. 1215 п.н. 5'-фланкирующей области и 1082 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep3. Как и для плазмид для делеции gap2 (pTTv145) и pep4 (pTTv181), выше, в этой плазмиде фрагмент прямого повтора представляет собой участок размером 300 п.н. с конца 5'-фланкирующей области pep3. Фрагменты получали способом ПЦР с использованием праймеров, приведенных в таблице 4.6. Что касается pTTv181 (Δpep4-pyr4), выше, для обеспечения переключения маркеров в конструкции участки рестрикции NotI встраивали по обеим сторонам селективного маркера pyr4, и для дополнительных стадий клонирования участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью pep3. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, использованная для ПЦР фланкирующих областей, представляла собой штамм T. reesei дикого типа QM6a. Маркерный ген pyr4 получали из pTTv181 расщеплением посредством NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1.
Вторую плазмиду для делеции аспарагиновой протеазы pep3 (TreID121133) pTTv192 конструировали с использованием плазмиды pTTv188, выше, в качестве остова. Эта вторая плазмида содержит нативную сверхэкспрессирующую KEX2 кассету (TreID123156) и в ней используется ген ацетамидазы (AmdS) из Aspergillus nidulans в качестве селективного маркера. Бластерную кассету pyr4 удаляли из pTTv188 посредством двойного расщепления NotI-AscI. Фрагменты для промотора cDNA1 (матрица: плазмидная ДНК pTHN3), нативного kex2 (матрица: геномная ДНК T. reesei QM6a), терминатора trpC (матрица: плазмидная ДНК pHHO2) и маркера AmdS (матрица: плазмидная ДНК pHHO1) получали способом ПЦР с использованием праймеров, приведенных в таблице 4.6. Как и для pTTv188, выше, для обеспечения переключения маркеров в конструкции участки рестрикции вносили по обеим сторонам селективного маркера AmdS. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1.
Третью плазмиду для делеции аспарагиновой протеазы pep3 (TreID121133) pTTv205 конструировали с использованием плазмиды pTTv192, описанной выше, в качестве остова. Маркер AmdS удаляли из pTTv192 расщеплением посредством NotI. Фрагменты для новой бластерной кассеты pyr4 (расположенные после сверхэкспрессирующей кассеты KEX2) получали способом ПЦР с использованием праймеров, приведенных в таблице 4.6. В этой бластерной кассете прямой повтор представляет собой участок размером 300 п.н. от начала 3'-фланкирующей области pep3 и расположенный перед геном pyr4. Что касается pTTv192, выше, для обеспечения переключения маркера в конструкции, участки рестрикции NotI встраивали по обеим сторонам бластерной кассеты pyr4. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1.
Эти плазмиды для делеции pep3 (pTTv188, pTTv192 и pTTv205, таблица 4.6) обеспечивают делецию 2590 п.н. в локусе pep3 и охватывают полную кодирующую последовательность PEP3.
Получение штаммов с делецией 7 протеаз
Для получения не содержащего маркера штамма с делецией 7 протеаз использовали удаление бластерной кассеты pyr4 из штамма с 6 делециями M396, по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Четыре последовательных стадии селекции с 5-FOA проводили для обеспечения того, чтобы отобранные клоны происходили из единичных клеток.
Конечные клоны подтверждали способом ПЦР с использованием праймеров, приведенных в таблице 4.7, с использованием стандартных лабораторных способов. Получали сигнал, соответствующий успешному удалению бластерной кассеты. Удаление бластерной кассеты далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. На чашках без добавления уридина роста не наблюдали. Саузерн-анализ четырех предполагаемых клонов pyr4- подтвердил удаление бластерной кассеты для всех клонов (фиг.34E). Клон (25-120A-62), использованный для получения штамма с делецией 7 протеаз, был обозначен как штамм номер M402 (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4, pyr4-).
Проводили две параллельных трансформации; одну с конструкцией для делеции в pTTv188 (стандартная делеция pep3) и другую с pTTv205 (включена сверхэкспрессия KEX2). Для удаления последовательности вектора плазмиды pTTv188 и pTTv205 расщепляли PmeI, и соответствующие фрагменты очищали на агарозаном геле с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг каждой из кассет для делеции использовали для трансформации M402 (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195, с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 4.7) для надлежащего встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.7.
Делецию pep3 подтверждали саузерн-анализом выбранных клонов с использованием способов, описанных в примере 1. Выбранные клоны использовали для удаления бластерной кассеты pyr4 и для получения штаммов с делецией 8 протеаз (фиг.34E).
Получение штамма с 8 делециями
Получали штамм с делецией 8 протеаз, имеющий делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5.
Получение плазмиды с делецией pep5
Первую плазмиду для делеции гена восьмой протеазы, аспарагиновой протеазы pep5 (TreID81004), конструировали, по существу как описано для плазмиды Δpep1 pTTv41 в примере 1, однако встраивали дополнительную кассету со вторым селективным маркером (bar, пример 1) после гена pyr4 с получением плазмиды для делеции с бластерной кассетой с двойным селективным маркером. Система двойного маркера обеспечивает a) использование, например, bar (или hph, кассета для устойчивости к гигромицину) в качестве первоначального маркера устойчивости и более быструю селекцию; b) трансформацию штаммов pyr4+ (без необходимости в получении pyr4- перед трансформацией); и c) удаление обоих маркеров из трансформантов с использованием 5-фтороротовой кислоты (подобно удалению стандартной бластерной кассеты pyr4) и одновременное проведение мутагенеза эндогенного pyr4 с получением не содержащего маркера штамма pyr4-.
Вторую плазмиду для делеции аспарагиновой протеазы pep5 (TreID81004) pTTv229 конструировали с использованием плазмиды pTTv202, описанной выше, в качестве остова. Двойной маркер pyr4-bar удаляли из pTTv202 расщеплением посредством NotI. Маркерный ген pyr4 получали из pTTv181 расщеплением посредством NotI. Клонирование плазмиды pTTv229 проводили с помощью стандартного лигирования с использованием ДНК-лигазы T4 при комнатной температуре. Часть смеси для лигирования трансформировали в E. coli с помощью электропорации. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга надлежащего лигирования с использованием стандартных лабораторных способов. Правильное лигирование и ориентацию маркера далее подтверждали секвенированием. Эти плазмиды для делеции pep5 (pTTv202 и pTTv229, таблица 4.8) обеспечивают делецию 1687 п.н. в локусе pep5 и охватывают полную кодирующую последовательность PEP5.
1348 п.н. 5'-фланкирующей области и 1164 п.н. 3'- фланкирующей области выбирали в качестве основы плазмиды для делеции pep5. Участок размером 300 п.н. с конца 5'-фланкирующей области pep5 использовали в качестве фрагмента прямого повтора. Эти фрагменты, а также кассету с вторым селективным маркером bar (пример 1) амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 4.8. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Для обеспечения удаления полной кассеты с двойным маркером, участки рестрикции NotI встраивали на обеих сторонах кассеты с двойным маркером, и между двумя селективными маркерами встраивали участок AsiSI. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью pep5. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Селективный маркер pyr4 получали из pTTv181 (Δpep4-pyr, выше) расщеплением посредством NotI. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции pep5 (pTTv202, таблица 4.8) обеспечивает делецию 1687 п.н. в локусе pep5 и охватывает полную кодирующую последовательность PEP5.

Получение штамма с делецией 8 протеаз
Для получения не содержащего маркера штамма с делецией 8 протеаз использовали удаление маркера pyr4 из штамма с 7 делециями M486 (34-14A-a, pTTv205 в M402), по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Четыре последовательных стадии селекции 5-FOA проводили для обеспечения того, чтобы все выбранные клоны происходили из единичных клеток.
Конечные клоны подтверждали способом ПЦР с использованием праймеров, приведенных в таблице 4.9, с использованием стандартных лабораторных способов. Сигнал, соответствующий успешному удалению бластерной кассеты, получали для большинства клонов. Удаление бластерной кассеты далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. На чашках без добавления уридина роста не наблюдали. Саузерн-анализы предполагаемых клонов pyr4- подтвердили удаление бластерной кассеты.
Для удаления последовательности вектора плазмиду pTTv229 расщепляли посредством PmeI+XbaI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации клона штамма с делецией 8 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов М181 и М195 с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 4.9) в отношении надлежащего встраивания. Клоны, дающие ожидаемые сигналы, очищают до клонов единичных клеток и подвергают повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 4.9. Делецию pep5 подтверждают саузерн-анализами выбранных клонов с использованием способов, описанных в примере 1.

ПРИМЕР 5 - УВЕЛИЧЕННАЯ ПРОДУКЦИЯ АНТИТЕЛА С ПОМОЩЬЮ ИНГИБИТОРОВ ПРОТЕАЗ
В этом примере продемонстрирована способность ингибиторов протеаз увеличивать продукцию полноразмерного антитела в продуцирующих штаммах Trichoderma reesei.
Исходя из информации о том, что тяжелая цепь расщепляется трипсиновыми и химотрипсиновыми протеазами, ингибиторы этих двух классов ферментов исследовали в отношении деградации антитела в экспериментах как in vitro, так и в культуре, с использованием продуцирующего антитела штамма T. reesei. Ингибиторы ингибитора трипсина сои (SBTI) и химостатина исследовали, поскольку ранее в экспериментах in vitro было показано, что они стабилизируют тяжелые цепи антитела.
Обработка ингибитором in vitro
Химостатин и SBTI анализировали in vitro с помощью культурального супернатанта. Супернатант культуры из ферментера с периодической подпиткой разбавляли до 6 мг/мл натрий-цитратным буфером, pH 5,5 (pH 5,5; 28°C; 20 г/л экстракта пивной дробины, 60 г/л лактозы). Периодическое культивирование с подпиткой проводили для штамма T. reesei дикого типа M44, который не обладает экспрессией гетерологичного белка. Штамм выращивали в минимальной среде для Trichoderma, дополненной 20 г/л экстракта пивной дробины и 60 г/л лактозы при pH 5,5 и 28°C.
К этом разбавленному супернатанту добавляли 0,05 мкг/мкл ритуксимаба, 100 мкМ химостатин, 1 мг/мл SBTI, или комбинацию обоих ингибиторов в общем объеме 50 мкл и взятие образцов проводили через 0, 1 и 19 часов для оценки ранней и поздней деградации тяжелой цепи антитела ритуксимаба. Полученные продукты тяжелой цепи анализировали иммуноблоттингом с использованием и конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188), разбавленного 1:30000 в TBST (фиг.35). Первоначальные продукты деградации, полученные из тяжелой цепи, имели массу приблизительно 42 кДа и 38 кДа, что наблюдали на дорожке необработанного контроля через 1 час (фиг.35). Дополнительные фрагменты образовывались через 19 часов, причем два основных продукта оставались. Обработка химостатином ингибировала первоначальное образование фрагмента массой 42 кДа, в то время как обработка SBTI ингибировала образование фрагмента массой 38 кДа (фиг.35). Комбинирование этих двух соединений ингибировало приблизительно 96% первоначальной деградации тяжелой цепи и приблизительно 75% деградации через 19 часов (фиг.35). Эти результаты демонстрируют, что эти два ингибитора были способны эффективно стабилизировать тяжелую цепь антитела ритуксимаба.
Обработка культур T. reesei ингибиторами
Эффективность ингибиторов также исследовали с помощью продуцирующего антитело ритуксимаб штамма, который содержит линкерную последовательность VALEKR и делецию pep1. Этот штамм выращивали в трех экземплярах в небольших флаконах, как в присутствии, так и в отсутствие химостатина, SBTI или пепстатина A. Небольшие вращающиеся флаконы содержали 50 мл TrMM вместе с лактозой (40 г/л), экстрактом пивной дробины (20 г/л) и буфером с 100 мМ PIPPS при pH 5,5. Ингибитор SBTI добавляли к культуре до конечных концентраций либо 100 мкг/мл, либо 500 мкг/мл. Химостатин использовали в концентрации 100 мкМ, и пепстатин A использовали в концентрации 10 мкМ. Каждый из трех ингибиторов добавляли к культурам каждые сутки на 2, 3, 4 и 5 сутки.
За ростом культур наблюдали каждые сутки по pH, начиная с 2 суток до 7 суток. Для культур, выращенных с SBTI, отсутствовали значительные отличия в pH культур по сравнению с необработанными контролями. В забуференных PIPPS культурах значение pH снижалось от первоначального значения pH 5,5 до pH 4,8 после 6 суток. Культуры с химостатином и пепстатином A подвергали мониторингу вплоть до 7 суток. На 7 сутки среднее значение pH для контрольных бутылок составляло 4,6, в то время как среднее значение pH для обработанных химостатином культур составляло 4,9, и среднее значение pH для обработанных пепстатином A культур составляло 5,0. Таким образом, для культур, обработанных химостатином и пепстатином A, происходило небольшое снижение роста.
Образцы культуральных супернатантов (30 мкл) также собирали на 3, 4 и 5 сутки для анализа продукции антител. Анализ проводили с помощью иммуноблоттинга с использованием конъюгированного с AP антитела против тяжелой цепи IgG (Sigma #A3188) и конъюгированного с AP антитела против легкой цепи (Sigma #A3812). Каждое антитело разбавляли 1:30000 в TBST. Полноразмерная тяжелая цепь ритуксимаба имеет массу приблизительно 51 кДа, слитая конструкция легкой цепи ритуксимаба с CHBI имеет массу приблизительно 100 кДа, и свободная легкая цепь ритуксимаба имеет массу приблизительно 28 кДа.
Результаты анализа с использованием иммуноблоттинга образцов культурального супернатанта, обработанных SBTI, представлены на фиг.36. На все сутки, когда проводили взятие образцов, больше полноразмерной тяжелой цепи ритуксимаба и больше продукта деградации массой 38 кДа присутствовало в обработанных SBTI культурах, чем в необработанных культурах. На 5 сутки продукция тяжелой цепи ритуксимаба была в несколько раз более высокой в обработанных SBTI культурах, чем в необработанных культурах (фиг.36A). Таким образом, использование SBTI имело положительный эффект на повышение продукции тяжелой цепи ритуксимаба. Также происходило незначительное увеличение общей продукции легкой цепи ритуксимаба, в частности, в случае связанной с носителем легкой цепи (фиг.36B).
Результаты анализа с использованием иммуноблоттинга образцов культурального супернатанта, обработанных химостатином и пепстатином A, представлены на фиг.37. Обработанные химостатином культуры продемонстрировали сходные результаты с результатами, наблюдаемыми для SBTI (фиг.37). Тяжелая цепь ритуксимаба была стабилизирована на 5 сутки (фиг.37B). По сравнению с необработанными контрольными культурами, химостатин увеличивал количество продуцируемой полноразмерной тяжелой цепи ритуксимаба, хотя все еще наблюдали основной продукт деградации при 38 кДа.
В общем, оказалось, что обработка SBTI была более эффективной в отношении обеспечения более высокой продукции белка, чем обработка химостатином. Однако обработка химостатином обеспечивала более высокое соотношение полноразмерной тяжелой цепи ритуксимаба и фрагмента. Как видно из фиг.37B, третий образец культуры с химостатином продемонстрировал приблизительно 90% полноразмерной тяжелой цепи ритуксимаба по сравнению с 10% фрагмента тяжелой цепи. Таким образом, комбинация обработки SBTI и химостатином может быть в высокой степени полезна для обеспечения более высокого выхода продукции антитела.
ПРИМЕР 6 - ПРОДУЦИРОВАНИЕ АНТИТЕЛА ИЗ T. REESEI
В этом примере количественно определено количество антитела, продуцируемого в штаммах с делецией протеазы T. reesei, описанных выше в примерах 1-4.
Очистка антитела
Культуральный супернатант каждого из штаммов T. reesei с делецией протеаз, приведенных в таблице 6.1, фильтровали через 0,45-мкм шприцевой фильтр и доводили до композиции буфера для связывания путем добавления 1/50 объема 1 M фосфата натрия, pH 7, перед очисткой. Аффинную колонку подсоединяли к AKTA Purifier и очистку проводили в соответствии с инструкциями изготовителя. Использовали следующие условия хроматографии: скорость потока: 1 мл/мин; детекция: 280 нм; инжекционная петля: 5 мл; буфер A: 20 мМ фосфат натрия, pH 7; буфер B: 0,1 M глицин-HCl, pH 2,7. Изократический прогон с помощью буфера A проводили до начала элюирования, которое осуществляли с помощью 5 мл буфера B. Колонку уравновешивали по меньшей мере 5 мл буфера A перед каждым анализом. 1 мл культурального супернатанта инжектировали для количественных прогонов. Фракции объемом 0,5 мл собирали в пробирки, содержавшие 40 мкл 0,5 M Tris, pH 9, на стадии элюирования. Антитело элюировалось резко двумя фракциями, которые объединяли в один образец объемом 1 мл. Из образцов с наиболее высокими площадями пиков в каждой серии образцов (ферментаций) проводили прогон с объемом инъекции 5-10 мл с получением более концентрированного образца для анализа гель-фильтрации.
Для количественного определения приготавливали серию разведений стандарта антитела и прогоняли в объеме 1 мл в колонке HiTrap с белком G, аналогично проанализированным образцам. Стандартную кривую (10-500 мкг) строили на основе площадей пиков, измеренных при 280 нм, и использовали для количественного определения антител MAB01 или ритуксимаба, выделенных из культуральных супернатантов. Качество очищенных образцов проверяли с помощью SDS-PAGE.
Профили гель-фильтрации образцов, очищенных с белком G
250 мкл образца каждого очищенного антитела MAB01 и ритуксимаба прогоняли в Tris-буферном физиологическом растворе (25 мМ Tris, 140 мМ NaCl и 3 мМ KCl, pH7,4) в колонке для гель-фильтрации Superdex 200 10/300 GL (Amersham Biosciences), подсоединенной к системе ВЭЖХ AKTA Purifier HPLC. Скорость потока составляла 0,75 мл/мин, и поглощение измеряли при 280 нм. Фракции (0,75 мл) собирали в ходе всего прогона. Фракции, демонстрировавшие только один пик, концентрировали и охарактеризовывали на стандартном геле для SDS-PAGE (фиг.38). Процент каждого элюированного пика вычисляли путем деления площади пика на общую площадь образца, измеренную при 280 нм.
Процесс очистки антитела представлен на фиг.38.
Количественное определение антитела
Количество антител, продуцированных штаммами T. reesei с делецией протеаз, обобщенно представлено в таблице 6.1.
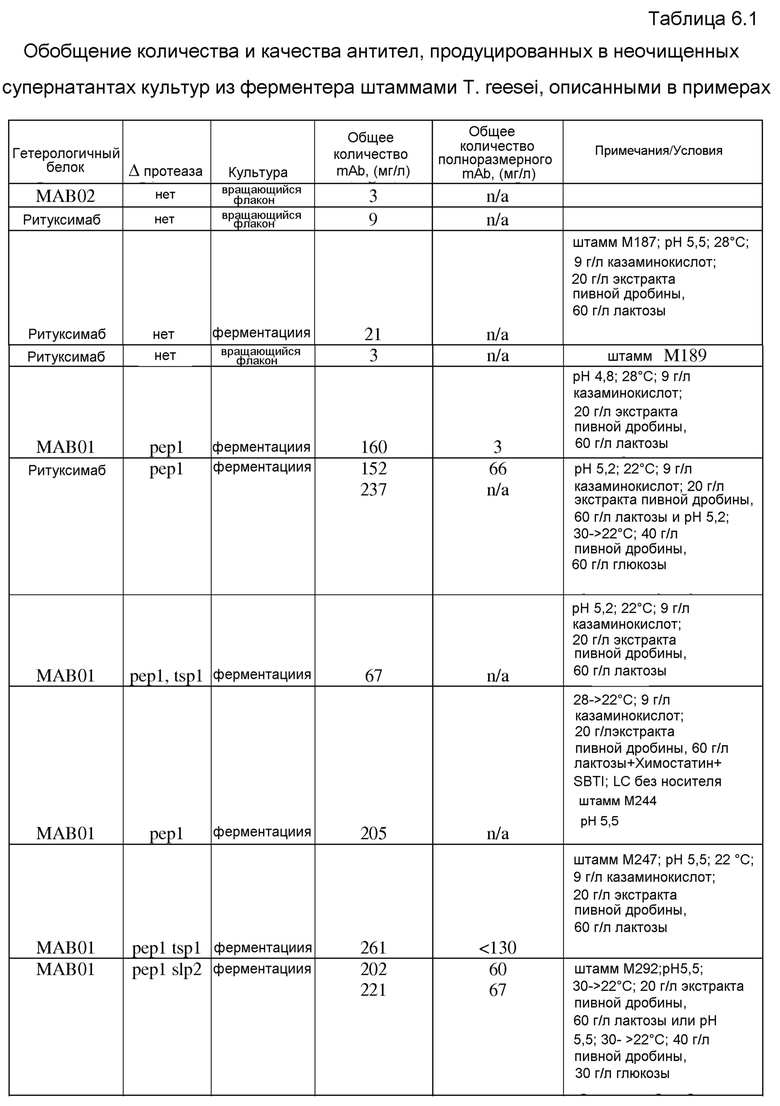
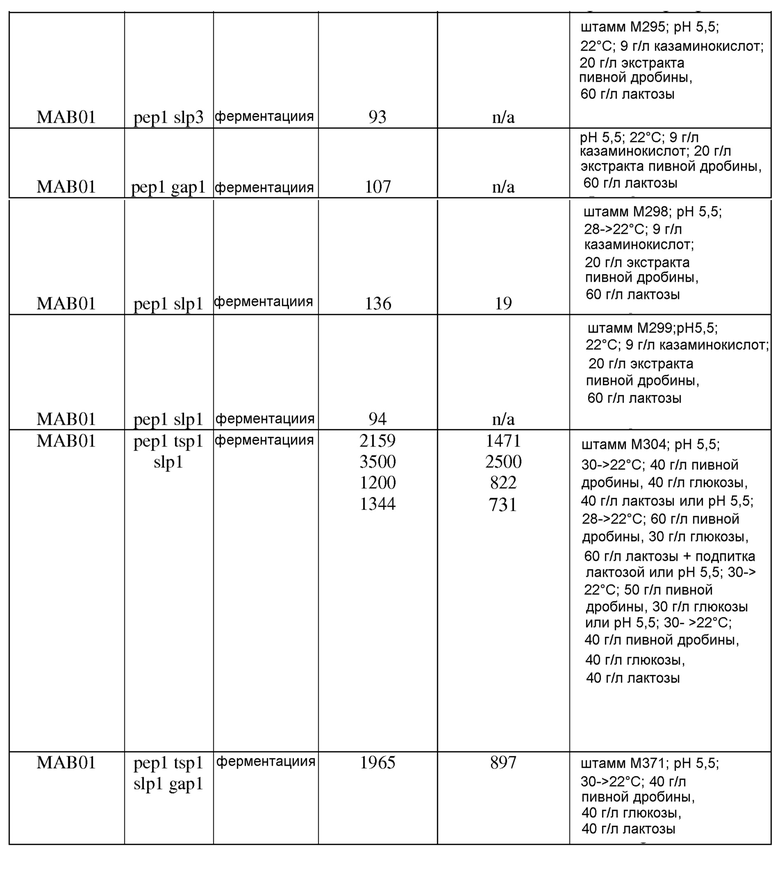
В таблице 6.1 общее количество антитела (mAb) представляет собой количество белка, который очищали из культурального супернатанта. После очистки белка антитело прогоняли в эксклюзионной хроматографии для измерения количества полноразмерного собранного антитела. Затем это количество было обозначено как "полное mAb".
Как показано в таблице 6.1, штамм M304 с тремя делециями (Δpep1Δtsp1Δslp1) обеспечивал выход антитела 3500 мг/л тотального IgG, причем 2500 мг/л были надлежащим образом собраны в полноразмерное антитело MAB01. Это соответствует 71% полноразмерного антитела. Увеличение процента полноразмерного антитела было результатом делеции slp1. В противоположность штамму M304, штамм M247 с двумя делециями (Δpep1Δtsp1) обеспечивал выход продукции полноразмерного антитела 43% (pH 5,5; 22°C; 9 г/л казаминокислот; 20 г/л экстракта пивной дробины, 60 г/л лактозы). Таким образом, можно прямо видеть, что добавление делеции Δslp1 значительно увеличивает качество продукта (на 25%).
Улучшение качества антитела в продуцирующем MAB01 штамме M507
Получали два продуцирующих MAB01 штамма M304 в генетическом фоне с делецией 3 протеаз и M507 в генетическом фоне с делецией 7 протеаз. Штамм M304 конструировали с помощью отдельных кассет для тяжелой и легкой цепей (фиг.49). Тяжелую цепь встраивали в локус cbh1 и легкую цепь встраивали в локус egl1. Штамм M507 получали путем встраивания тандемной кассеты, содержащей как тяжелую, так и легкую цепь, в локус CBHI (фиг.49). Тандемный вектор MAB01 ориентирует тяжелую и легкую цепи в противоположных направлениях. Для легкой цепи использовали участок расщепления NVISKR и для тяжелой цепи использовали участок расщепления DGETVVKR. В штамме M304 было удалено 3 протеазы: pep1, tsp1 и slp1. В M507 было удалено 7 протеаз: pep1, tsp1, slp1, gap1, gap2, pep4, pep3. Оба штамма сверхэкспрессировали протеазу kex2.
Продуцирующий MAB01 двунаправленный тандемный вектор pTTv223 трансформировали в штамм с делецией семи протеаз M486 со сверхэкспрессией kex2 с использованием стандартной трансформации протопластов. Трансформанты отбирали на чашках с ацетамидом-тритоном, и первые штриховые культуры подвергали скринингу способом ПЦР в отношении 5'- и 3'-встраивания маркера AmdS в локус cbh1. Двойные положительные трансформанты очищали посредством культивирования отдельных спор, и исходные растворы спор получали на чашках PD, дополненных ампициллином.
Штаммы M304 и M507 культивировали в ферментерах с 30 г/л глюкозы, 60 г/л пивной дробины, 60 г/л лактозы с подачей лактозы при 28°C и позднее культуру переключали на 22°C. Штамм M507 культивировали при pH 5,2 (культивирование bio00541b) и pH 5,5 (культивирование bio00543b). Штамм M304 выращивали при pH 5,5 (культивирование bio00503b). Образец для ферментации bio00477b M304 включали в качестве контроля в иммуноблот bio00503b. Культивирование bio00477b проводили с помощью той же среды и в условиях, как описано для bio00503b.
Штамм M507 культивировали как при pH 5,2, так и при pH 5,5 для исследования эффекта pH на продукцию антитела. Образцы супернатанта после ферментации при pH 5,2 анализировали с использованием вестерн-блоттинга, и они показаны на фиг.50A, и образцы после ферментации при pH 5,5 представлены на фиг.50B. Продуцированное антитело выглядело довольно сходным при обоих культивированиях M507. В условиях при pH 5,2 присутствовало несколько больше легкой цепи. При обоих культивированиях присутствовали фрагменты тяжелой цепи. Штамм M304 культивировали при pH 5,5 и результаты можно видеть на фиг.51. Количество продуцированной полноразмерной тяжелой цепи падает после 7 суток для штамма M304. Количество тяжелой цепи снижается после 9 суток для штамма M507. Другим отличием между штаммами было количество продуцированной легкой цепи. M304 продуцировал больше.
Концентрации очищенного с помощью белка G иммуноглобулина после прогонов трех ферментационных культур можно видеть в таблице 6.2. Наиболее высокая общая концентрация антитела для штамма M304 составила 3,1 г/л на 9 сутки. Наиболее высокие концентрации для штамма M507 на 10 сутки составили 3,0 г/л при pH 5,2 и 2,8 г/л при pH 5,5. После эксклюзионной хроматографии для каждого образца вычисляли количество полноразмерного антитела (таблица 6.3). Наиболее высокое количество полноразмерного антитела составляло 2,0 г/л для ферментаций M507 на 8 сутки (pH 5,2) и на 9 сутки (pH 5,5). M304 продуцировал сходный уровень полноразмерного антитела 2,0 г/л на 8 сутки.
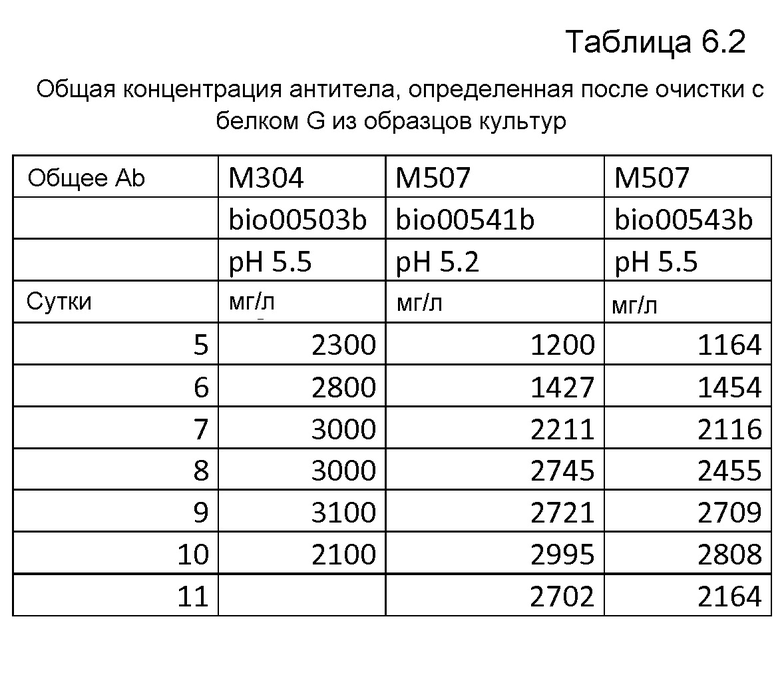
Различие между штаммами M304 и M507 становится очевидным с учетом процента продуцированного полноразмерного антитела в ходе культивирования. Процент полноразмерного антитела был более высоким в случае M507 по сравнению со штаммом M304. Штамм M507, выращенный при pH 5,5, продуцировал антитело наиболее высокого качества, где 78% антитела являлось полноразмерным на 7 сутки. M304 достигало 68% на 6 сутки, а затем качество продукта снижалось по сравнению с M507. Продукт M507 был полноразмерным на 73% до 9 суток.

Измерение протеазной активности
Протеазную активность в супернатанте сравнивали между штаммами M304 и M507, выращенными в тех же условиях. Культивирование Triab62 и Triab67 проводили при pH 5,5 в 30 г/л глюкозы, 60 г/л лактозы, 20 г/л цельной пивной дробины, 20 г/л экстракта пивной дробины с подпиткой лактозой, при 28°C и позднее переключали на 22°C в культуре.
Концентрации белка определяли во всех образцах супернатантов на 2-7 сутки. Все супернатанты разбавляли в натрий-цитратном буфере, pH 5,5, так чтобы все образцы имели общую концентрацию белка 0,625 мг/мл. 100 мкл всех разбавленных супернатантов добавляли в черный 96-луночный планшет с использованием 3 повторяющихся лунок на образец. 100 мкл разбавленного исходного раствора казеина FL (10 мкг/мл), изготовленного в натрий-цитратном буфере, pH 5,5, добавляли в каждую содержащую супернатант лунку. Планшеты инкубировали покрытыми в пластиковом мешке при 37°C. Флуоресценцию из лунок измеряли через 2, 3 и 4 часа. Считывание данных проводили на флуоресцентном устройстве для считывания планшетов Varioskan с использованием возбуждения при 485 нм и испускания при 530 нм.
Протеазная активность в супернатанте из штамма с делецией 7 протеаз M507 была в 2-2,5 раз более низкой, чем из M304 (делеции 3 протеаз), см. таблицу 6.5. Удаленные кислые протеазы (gap1, gap2, pep4, pep3) вносят вклад в это улучшение. Общая протеазная активность в штамме с 7 делециями была значительно более низкой в случае субстрата казеина. Эти данные в основном коррелируют с результатами, наблюдаемыми в случае процента полноразмерного антитела. Более низкая протеазная активность приводит к более высокому качеству антитела.
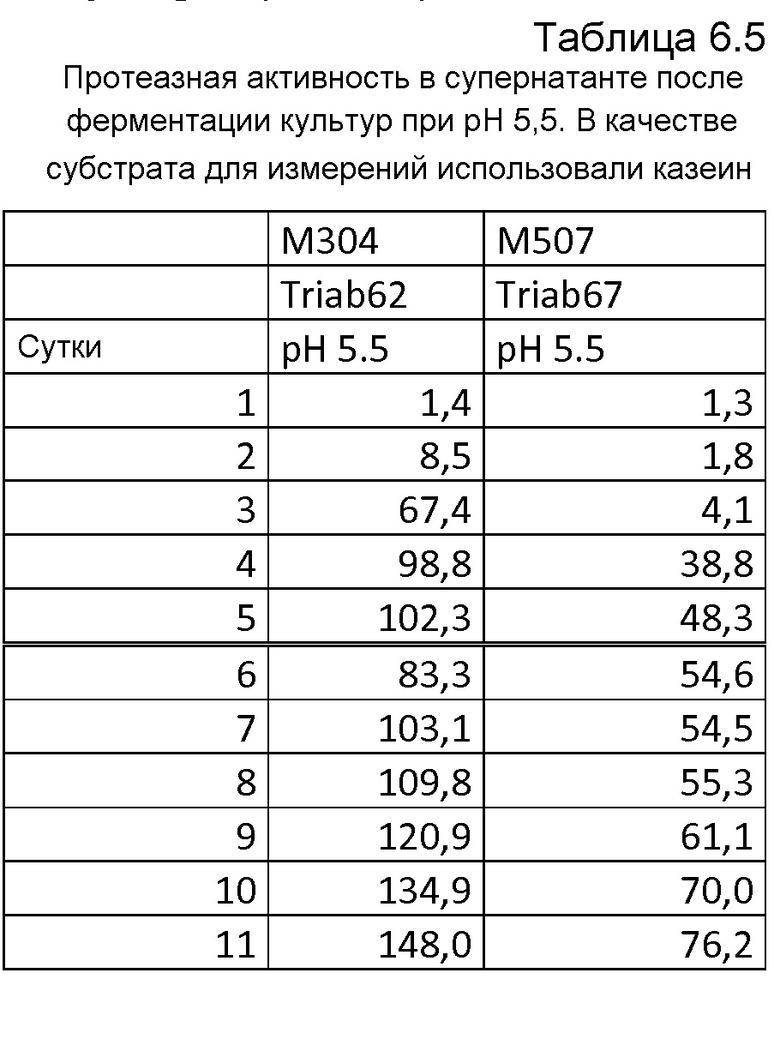
ПРИМЕР 7 - ПОЛУЧЕНИЕ НЕ ЯВЛЯЮЩИХСЯ АНТИТЕЛОМ БЕЛКОВ
Устойчивость к протеазам модельных белков IGF1, hGH и IFNa2b анализировали путем добавления их в супернатант из штамма с делецией 6 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4) M400. Супернатант собирали из большой культуры во вращающихся флаконах CAH15. Неразбавленный супернатант после культивирования во вращающихся флаконах CAH15 инкубировали с очищенными модельными белками с пепстатином A и без него (50 мкМ) и ингибиторами SBTI (0,2 мг/мл) в течение 20 часов при 37°C. Значение pH культурального супернатанта на 5 сутки составляло приблизительно 4,2. Образец реакционной смеси, содержащий 0,05 мкг/мкл модельного белка, отбирали через 20 часов. В качестве контроля в виде буфера использовали 50 мМ цитрат натрия, pH 4,0, с добавлением модельных белков (0,05 мкг/мкл).
Из каждой реакционной смеси 10 мкл наносили на 18% гель SDS PAGE и разделяли в течение 30 минут при 200 В. Затем белки в геле переносили на нитроцеллюлозу для иммуноблоттинга. Нитроцеллюлозную мембрану блокировали в течение 1 часа при комнатной температуре 5% молоком в буфере TBST. Отдельные зонды обрабатывали специфическим первичным антителом для обнаружения соответствующего модельного белка в течение 1 часа при комнатной температуре на устройстве для встряхивания. Использовали антитело мыши против IGF1 (R&D systems #mab291) в количестве 2 мкг/мл, разбавленное в TBST. Использовали антитело мыши против rhGH (Abeam #ab51232) в количестве 2 мкг/мл, разбавленное в TBST. Использовали антитело мыши против IFNa2b (Abeam #ab9386) в количестве 1 мкг/мл, разбавленное в TBST. После кратковременного промывания мембран блотов посредством TBST, добавляли вторичное антитело в течение 1 часа при встряхивании при комнатной температуре. Вторичное конъюгированное с AP антитело козы против антител мыши (Biorad #170-6520) разбавляли 1:10000 в TBST.
При инкубации в течение ночи в супернатанте, полноразмерные белки исследовали в отношении hGH, IFNa2b и IGF1, хотя большинство из них оказалось деградированными (фиг.42). Присутствовал преобладающий продукт деградации для гормона роста и IFNa2b размером приблизительно 15 кДа. Однако эти 3 модельных белка были значительно стабилизированными после обработки супернатанта ингибитором аспарагиновых протеаз пепстатином A. Этот ингибитор блокировал ключевые протеазы, ответственные за основную часть активности протеазы. SBTI обеспечивал только небольшую пользу в отношении стабильности продукта. Хотя оптимум pH для SBTI является более высоким, чем использовали в эксперименте (pH 4,2 против оптимального значения pH 8,0), связывание этих ингибиторов с их протеазами-мишенями, следовательно, может не быть наиболее эффективным.
Пепстатин A эффективно ингибирует аспарагиновые протеазы. Из исследований по аффинной очистке с пепстатином A известно, что остальные аспарагиновые протеазы в супернатанте представляют собой pep2, pep3 и pep5. Таким образом, если остальные 2 или 3 аспарагиновые протеазы удалить, супернатант будет практически свободным от активности аспарагиновых протеаз. Для продуцирования этих модельных белков аспарагиновые протеазы pep2, pep3 и pep5 можно считать основными протеазами.
Тот же эксперимент по добавлению, как и с MAB01, проводили для исследования его стабильности в супернатанте с делецией 6 протеаз с ингибиторами и без них (фиг.43). Взятие образцов проводили, как описано выше, и их подвергали иммуноблоттингу с конъюгированным с AP антителом против тяжелой цепи (Sigma #A3188). После инкубации в течение 20 часов не было значительной деградации тяжелой цепи. Не было очевидной пользы применения этих ингибиторов. Антитело было стабильным в этом супернатанте при pH 4,2. Продукция MAB01 в более кислых условиях, таких как pH 4,5, вероятно может увеличить выход продукции или по меньшей мере снизить уровень расщепления тяжелой цепи, которое могло произойти.
Для оценки того, какие ингибиторы наилучшим образом стабилизируют продукцию hGH, осуществляли культивирование этих штаммов в 24 ячейках. Штамм M369 с гормоном роста человека (Δpep1Δtsp1Δslp1Δgap1Δgap2) выращивали вместе с отдельными компонентами или с комбинацией следующих: трипсин и субтилизин ингибитор SBTI, ингибирующий кислые протеазы пептид SIP, ингибирующий кислые протеазы пептид LIP, свободный ингибирующий аспарагиновую протеазу пептид пепстатин A, пепстатин A, иммобилизованный на агарозных гранулах, ингибитор трипсина и субтилизина BBI из лимской фасоли, ингибитор субтилизина химостатин и BSA. В качестве контрольных лунок были выбраны три независимые лунки, где не добавляли ингибиторов или добавок. Эти два штамма выращивали в 3 мл TrMM с цитратом диаммония без сульфата аммония, 100 мМ PIPPS, 20 г/л экстракта пивной дробины, 40 г/л лактозы, с доведением до pH 4,5. Планшеты с 24 ячейками встряхивали при 800 об/мин, 85% влажности и 28°C. Культуры выращивали в течение 6 суток и накрывали воздухопроницаемой мембраной.
Ингибиторы добавляли сначала на 1 сутки, а затем каждые сутки, начиная на 3 сутки. Образцы объемом 100 мкл извлекали из лунок с культурой, начиная на 3 сутки. Мицелий осаждали центрифугированием в течение 5 минут при 13k и супернатант собирали. Из культурального супернатанта 12 мкл наносили на 4-20% гель SDS PAGE и проводили иммуноблоттинг на нитроцеллюлозе с антителом мыши против hGH (2 мкг/мл) и конъюгированным с AP антителом козы против IgG мыши, разбавленным 1:10000 в TBST.
На 4 сутки гормон роста человека все еще можно было наблюдать в культуральных супернатантах на всех 3 контрольных дорожках (фиг.44). Две из контрольных дорожек продемонстрировали слабую полосу, и одна контрольная дорожка продемонстрировала светлую полосу. Сразу исследовали эффект ингибиторов и добавок. Ингибитор/добавки, которые имели большой эффект, выделены красным цветом, а ингибитор/добавки с наилучшим эффектом обозначены звездочкой. Пепстатин A имел отрицательный эффект на продуцирование гормона роста. При использовании в концентрации 5 или 20 мкМ, продукция hGH, по-видимому, отсутствовала. По-видимому, он имеет некоторый токсический эффект на продуцирование. Только когда пепстатин иммобилизовывали на агарозных гранулах, этот эффект устранялся. Одной из наилучших обработок были гранулы с пепстатином A вместе с 0,2 мг/мл SBTI (см. третья звездочка на блоте на фиг.44). В случае только SBTI (0,2 мг/мл) продукция была улучшенной, однако присутствовала значительная полоса деградации на уровне 18 кДа, которая, по-видимому, образовывалась под действием аспарагиновых протеаз. Как и ожидалось, уровень этого продукта деградации снижался при добавлении гранул с пепстатином A или пептида-ингибитора SIP. Пептид SIP был полезен даже при использовании отдельно в концентрации 20 или 50 мкМ. Происходило заметное увеличение количества hGH, когда использовали пептид SIP, однако наибольшее увеличение происходило при использовании в комбинации с SBTI или BSA. Когда использовали химостатин 5 мкМ и 25 мкМ, это также увеличивало количество наблюдаемого полноразмерного продукта. Дополнение культуры BSA (0,25%) отдельно способствует продуцированию, однако оно не препятствовало образованию большого количества продукта деградации.
При оценке уровней экспрессии относительно 200 нг контрольного образца, в контрольных лунках продуцировалось 3-6 мг/л hGH, обработка BSA (0,25%)/SIP (50 мкМ)/SBTI (0,2 мг/мл) обеспечивала продуцирование 24,5 мг/л, обработка SIP (50 мкМ)/SBTI (0,2 мг/мл) обеспечивала продуцирование 26,6 мг/л, и добавление гранул с пепстатином A/SBTI (0,2 мг/мл) обеспечивало 24,5 мг/л hGH. Таким образом, использование комбинации ингибиторов и добавок было наиболее эффективным, увеличивая уровни продукции по меньшей мере в 4 раза. Критическим параметром было включение в смесь ингибитора аспарагиновой протеазы.
ПРИМЕР 8 - ПОЛУЧЕНИЕ T. REESEI С ДЕФИЦИТОМ PEP7
Плазмиду для делеции аспарагиновой протеазы pep7 (TreID58669) конструируют, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1. 1062 п.н. 5'-фланкирующих областей и 1121 п.н. 3'-фланкирующей области выбирают в качестве основы плазмиды для делеции pep7. Фрагменты продуцируют способом ПЦР с использованием праймеров, приведенных в таблице 8.1. Продукты разделяют агарозным гель-электрофорезом, и соответствующие фрагменты выделяют из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, подлежащая применению в ПЦР фланкирующих областей, происходит из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получают из pTTv71 (пример 1) посредством расщепления NotI. Остов вектора представляет собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструируют с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции pep7 обеспечивает делецию в локусе pep7 и охватывает полную кодирующую последовательность PEP7.

ПРИМЕР 9- ПОЛУЧЕНИЕ T. REESEI С ДЕФИЦИТОМ SLP5
Плазмиду для делеции аспарагиновой протеазы slp5 (TreID64719) конструируют, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1. 1044 п.н. 5'-фланкирующих областей и 1003 п.н. 3'-фланкирующей области выбирают в качестве основы плазмиды для делеции slp5. Фрагменты продуцируют способом ПЦР с использованием праймеров, приведенных в таблице 9.1. Продукты разделяют с помощью агарозного гель-электрофореза, и соответствующие фрагменты выделяют из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, подлежащая применению в ПЦР фланкирующих областей, происходит из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получают из pTTv71 (пример 1) с помощью расщепления NotI. Остов вектора представляет собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструируют с использованием способа гомологичной рекомбинации дрожжей, описанной в примере 1. Эта плазмида для делеции slp5 обеспечивает делецию в локусе slp5 и охватывает полную кодирующую последовательность SLP5.

ПРИМЕР 10 - ПОЛУЧЕНИЕ T. REESEI С ДЕФИЦИТОМ SLP6
Плазмиду для делеции аспарагиновой протеазы slp6 (TreID121495) конструируют, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1. 1192 п.н. 5'-фланкирующих областей и 1114 п.н. 3'-фланкирующей области выбирают в качестве основы плазмиды для делеции slp6. Фрагменты продуцируют способом ПЦР с использованием праймеров, приведенных в таблице 10.1. Продукты разделяют агарозным гель-электрофорезом, и соответствующие фрагменты выделяют из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Матрица, подлежащая применению в ПЦР фланкирующих областей, происходит из штамма T. reesei дикого типа QM6a. Бластерную кассету pyr4 получают из pTTv71 (пример 1) с помощью расщепления NotI. Остов вектора представляет собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструируют с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции slp6 обеспечивает делецию в локусе slp6 и охватывает полную кодирующую последовательность SLP6.

ПРИМЕР 11- ПОЛУЧЕНИЕ T. REESEI С ДЕФИЦИТОМ SLP7
Получение плазмиды для делеции slp7
Плазмиду pTTv269 для делеции сериновой протеазы slp7 (tre123865) конструировали, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1, за исключением того, что маркер, использованный для селекции, представлял собой pyr4-hgh из pTTv194.
949 п.н. 5'-фланкирующей области и 1025 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции slp7 pTTv269. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 11.1. Матрица, использованная в ПЦР фланкирующих областей, происходила из штамма T. reesei дикого типа QM6a. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Кассету pyr4-hgh получали из pTTv194 (Δpep4-pyr4-hgh) с помощью расщепления NotI. Для обеспечения удаления маркерной кассеты по обеим сторонам кассеты встраивали участки рестрикции NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv269 конструировали с 5'-фланкирующей областью, 3'-фланкирующей областью, маркером pyr4-hgh и остовом вектора с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции slp7 (pTTv269, таблица 11.1) обеспечивает делецию 2019 п.н. в локусе slp7 и охватывает полную кодирующую последовательность SLP7.

ПРИМЕР 12 - ПОЛУЧЕНИЕ T. REESEI С ДЕФИЦИТОМ SLP8
Получение плазмиды для делеции slp8
Плазмиду pTTv330 для делеции субтилизин-подобной протеазы slp8 (tre58698) конструировали, по существу как описано для плазмиды pTTv41 для делеции pep1 в примере 1, за исключением того, что маркер, использованный для селекции, представлял собой двойной маркер pyr4-hph.
975 п.н. 5'-фланкирующей области и 1038 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции slp8. Участок размером 298 п.н. с конца 5'-фланкирующей области slp8 использовали в качестве фрагмента прямого повтора. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 12.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Кассету pyr4-hph получали из pTTv210 (Δsep1-pyr4-hph) расщеплением с помощью NotI. Для обеспечения удаления полной кассеты двойного маркера вносили участки рестрикции NotI по обеим сторонам кассеты двойного маркера. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью slp8. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции slp8 (pTTv330, таблица 12.1) обеспечивает делецию 1433 п.н. в локусе slp8 и охватывает полную кодирующую последовательность SLP8.

ПРИМЕР 13 - ГОМОЛОГИ ПРОТЕАЗ
Идентифицировали гомологи pep1, pep2, pep3, pep4, pep5 и pep7; tsp1; slp1, slp2, slp3, slp5, slp6, slp7 и slp8; gap1 и gap2; и tpp1 T. reesei из других организмов.
Поиск BLAST проводили с использованием Natioinal Center for Biotechnology Information (NCBI) неизбыточной базы данных аминокислотных последовательностей с использованием аминокислотных последовательностей протеаз Trichoderma reesei в качестве последовательностей запроса. Альтернативно проводили поиск FASTA с помощью UniProt Knowledgebase от European Bioinformatics Institute (EBI). Поиск в BLAST Trichoderma virens и Trichoderma atroviride проводили с использованием web-сайта DOE Joint Genome Institute (Trichoderma virens Gv29-8 v2.0 и Trichoderma atroviride v2.0, соответственно). Наиболее совпадающие последовательности после поиска в BLAST выравнивали с использованием инструмента для выравнивания ClustalW2, предоставленного EBI. Также с использованием выравнивания последовательностей строили филогенетиеские деревья.
На фиг.45 представлено филогенетическое дерево аспарагиновых протеаз выбранных нитчатых грибов.
На фиг.46 представлено филогенетическое дерево субтилизиновых протеаз выбранных нитчатых грибов.
На фиг.47 представлено филогенетическое дерево глутаминовых протеаз выбранных нитчатых грибов.
На фиг.48 представлено филогенетическое дерево седолизиновых протеаз выбранных нитчатых грибов.
ПРИМЕР 14 - ПОЛУЧЕНИЕ ШТАММОВ С ДЕЛЕЦИЕЙ 9 ПРОТЕАЗ
Получение штамма с делецией 9 протеаз, имеющего делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2
Получение новых плазмид для делеции pep2
Первую плазмиду pTTv213 для делеции аспарагиновой протеазы pep2 (tre0053961) конструировали, по существу как описано для плазмиды Δpep1 pTTv41 в примере 1, однако дополнительную вторую кассету с селективным маркером, содержащую ген фосфотрансферазы гигромицина (hph), помещали после гена pyr4, получая плазмиду для делеции с бластерной кассетой с двойным селективным маркером. Система двойного маркера обеспечивает a) использование, например, hph в качестве первоначального маркера устойчивости и более быструю селекцию; b) трансформацию штаммов pyr4+ (без необходимости в получении pyr4- перед трансформацией); и c) удаление обоих маркеров из трансформантов с использованием 5-фтороротовой кислоты (подобно удалению стандартной бластерной кассеты pyr4) и одновременное проведение мутагенеза эндогенного pyr4 с получением не содержащего маркера штамма pyr4-. В дополнение к двойному маркеру, первая плазмида для делеции также содержала сверхэкспрессирующую кассету для нативной KEX2 (tre123561; промотор cDNA1, терминатор cbh2).
Вторую плазмиду для делеции аспарагиновой протеазы pep2 (tre0053961) pTTv232 конструировали с использованием плазмиды pTTv213, описанной выше, в качестве остова. Кассету, сверхэкспрессирующую kex2 (pcDNA1-kex2-tcbh2), удаляли из pTTv213 посредством расщепления AscI. Клонирование плазмиды pTTv232 проводили с помощью стандартного лигирования (самолигирование) с использованием ДНК-лигазы T4 при комнатной температуре. Часть смеси для лигирования трансформировали в E. coli способом электропорации. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга правильного лигирования с использованием стандартных лабораторных способов. Правильное лигирование далее подтверждали секвенированием.
Третью плазмиду для делеции аспарагиновой протеазы pep2 (tre0053961) pTTv246 конструировали с использованием плазмиды pTTv232, описанной выше, в качестве остова. Двойной маркер pyr4-hph удаляли из pTTv232 расщеплением посредством NotI. Маркерный ген pyr4 получали из pTTv181 (Δpep4-pyr4, выше) расщеплением посредством NotI. Клонирование плазмиды pTTv246 проводили с помощью стандартного лигирования с использованием ДНК-лигазы T4 при комнатной температуре. Часть смеси для лигирования трансформировали в E. coli с помощью электропорации. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга надлежащего лигирования с использованием стандартных лабораторных способов. Правильное лигирование и ориентацию маркера далее подтверждали секвенированием.
1000 п.н. 5'-фланкирующей области и 1020 п.н. 3'-фланкирующей области выбирали в качестве основы плазмид для делеции pep2. Участок длиной 300 п.н. с конца 5'-фланкирующей области pep2 использовали в качестве фрагмента прямого повтора. Эти фрагменты, а также кассету со вторым селективным маркером (hph), промотор cDNA1, нативный ген kex2 и терминатор cbh2 амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 14.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Селективный маркер pyr4 получали из pTTv181 (Δpep4-pyr4, выше) с помощью расщепления NotI. Для обеспечения удаления полной двойной маркерной кассеты в pTTv213, по обеим сторонам кассеты с двойным маркером встраивали участки рестрикции NotI, и между двумя селективными маркерами встраивали участок SwaI. Участки AscI встраивали по обеим сторонам сверхэкспрессирующей кассеты kex2 (между 5'- прямым повтором и 3'-фланкирующей областью pep2). Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv213 конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эти плазмиды для делеции pep2 (pTTv213, pTTv232 и pTTv246, таблица 14.1) обеспечивают делецию 1580 п.н. в локусе pep2 и охватывают полную кодирующую последовательность PEP2.

Получение штамма с делецией 9 протеаз с pep2 (tre53961); M574
Для получения не содержащего маркера штамма с делецией 9 протеаз использовали удаление маркера pyr4 из штамма M504 с 8 делециями (38-48A, pTTv229 в M496), по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Последовательные стадии селекции с 5-FOA проводили для обеспечения того, чтобы все клоны происходили из единичных клеток.
Конечные клоны проверяли способом ПЦР с использованием праймеров, приведенных в таблице 14.2, с использованием стандартных лабораторных способов. Сигнал, соответствующий успешному удалению бластерной кассеты, был получен для большинства клонов. Удаление бластерной кассеты далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. Не наблюдали роста на чашках без добавления уридина. Полученный штамм, использованный для получения штамма с делецией 9 протеаз, был обозначен как штамм номер M521.
Для удаления последовательности вектора плазмиду pTTv246 (Δpep2-pyr4) расщепляли посредством MssI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации клона M521 с делецией 8 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 14.2) в отношении правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 14.2. Делецию pep2 подтверждали саузерн-анализом выбранных клонов (данные не представлены) с использованием способов, описанных в примере 1. Клон 41-45G обозначили как штамм номер M574.

Получение штамма с делецией 9 протеаз, имеющего делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep12
Получение плазмид для делеции pep12
Первую плазмиду pTTv209 для делеции аспарагиновой протеазы pep12 (tre119876) конструировали, по существу как описано для плазмиды Δpep1 pTTv41 в примере 1, однако дополнительную вторую кассету с селективным маркером, синтетическую конструкцию, содержащую ген фосфинотрицин N-ацетилтрансферазы (bar) Streptomyces ssp., помещали после гена pyr4, получая плазмиду для делеции с бластерной кассетой с двойным селективным маркером. Система двойного маркера обеспечивает a) использование, например, bar в качестве первоначального маркера устойчивости и более быструю селекцию; b) трансформацию штаммов pyr4+ (без необходимости в получении pyr4- перед трансформацией); и c) удаление обоих маркеров из трансформантов с использованием 5-фтороротовой кислоты (подобно удалению стандартной бластерной кассеты pyr4) и одновременное проведение мутагенеза эндогенного pyr4 с получением не содержащего маркера штамма pyr4-.
Вторую плазмиду для делеции аспарагиновой протеазы pep12 (tre119876) pTTv245 конструировали с использованием плазмиды pTTv209, описанной выше, в качестве остова. Двойной маркер pyr4-bar удаляли из pTTv209 расщеплением посредством NotI. Новый маркерный ген pyr4 получали из pTTv181 (Δpep4-pyr4, выше) расщеплением посредством NotI. Клонирование плазмиды pTTv245 проводили с помощью стандартного лигирования с использованием ДНК-лигазы T4 при комнатной температуре. Часть смеси для лигирования трансформировали в E. coli с помощью электропорации. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга правильного лигирования с использованием стандартных лабораторных способов. Правильное лигирование и ориентацию маркера далее подтверждали секвенированием.
1019 п.н. 5'-фланкирующей области и 895 п.н. 3'-фланкирующей области выбирали в качестве основы плазмид для делеции pep12. Участок из 300 п.н. с конца 5'-фланкирующей области pep12 использовали в качестве фрагмента прямого повтора. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 14.3. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Двойной маркер (pyr4-bar) вырезали из pTTv202 (Δpep5-pyr4-bar) с помощью NotI. Для обеспечения удаления полной двойной маркерной кассеты участки рестрикции NotI вносили на обеих сторонах кассеты с двойным маркером. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью pep12. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv209 конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эти плазмиды для делеции pep12 (pTTv209 и pTTv245, таблица 14.3) обеспечивают делецию 2198 п.н. в локусе pep12 и охватывают полную кодирующую последовательность PEP12.

Получение штамма с делецией 9 протеаз с pep12 (tre119876); M575
Для получения не содержащего маркера штамма с делецией 9 протеаз использовали удаление маркера pyr4 из штамма с 8 делециями M504 (38-48A, pTTv229 в M496), по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Последовательные стадии селекции с 5-FOA проводили, чтобы убедиться, что выбранные клоны происходили из единичных клеток.
Конечные клоны проверяли способом ПЦР с использованием праймеров, приведенных в таблице 14.4, с помощью стандартных лабораторных способов. Сигнал, соответствующий успешному удалению бластерной кассеты, получили для большинства клонов. Удаление бластерной кассеты далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. Не наблюдали роста на чашках с добавлением уридина. Полученный штамм, использованный для получения штамма с делецией 9 протеаз, был обозначен как штамм номер M521.
Для удаления последовательности вектора плазмиду pTTv245 (Δpep12-pyr4) расщепляли посредством MssI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации клона штамма с делецией 8 протеаз M521 (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 14.4) в отношении правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 14.4. Делецию pep12 подтверждали саузерн-анализом выбранных клонов (данные не представлены) с использованием способов, описанных в примере 1. Клон 42-45B был обозначен как штамм номер M575.

ПРИМЕР 15 - ПОЛУЧЕНИЕ ШТАММА С ДЕЛЕЦИЕЙ 10 ПРОТЕАЗ
Получение штамма с делецией 10 протеаз, имеющего делеции Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2Δpep11
Получение плазмиды для делеции pep11
Плазмиду pTTv312 для делеции аспарагиновой протеазы pep11 (tre121306) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1.
956 п.н. 5'-фланкирующей области и 943 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep11. Участок из 307 п.н. с конца 5'-фланкирующей области pep11 использовали в качестве фрагмента прямого повтора. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 15.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. pyr4 получали из pTTv181 (Δpep4-pyr4, выше) с помощью расщепления NotI. Для обеспечения удаления кассеты с маркером, участки рестрикции NotI вносили на обеих сторонах кассеты. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью pep11. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции pep11 (pTTv312, таблица 15.1) обеспечивает делецию 2624 п.н. в локусе pep11 и охватывает полную кодирующую последовательность PEP11.

Получение штамма с делецией 10 протеаз с pep11 (tre121306); M658
Для получения не содержащего маркера штамма с делецией 10 протеаз использовали удаление маркера pyr4 из штамма с 9 делециями M574 (41-45G, pTTv246 в M521), по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Последовательные стадии селекции с 5-FOA проводили, чтобы убедиться, что выбранные клоны происходили из единичных клеток.
Конечные клоны проверяли способом ПЦР с использованием праймеров, приведенных в таблице 15.2, с помощью стандартных лабораторных способов. Сигнал, соответствующий успешному удалению бластерной кассеты, получили для большинства клонов. Удаление бластерной кассеты далее подтверждали путем посева клонов на чашки с минимальной средой с 5 мМ уридином или без него. Не наблюдали роста на чашках с добавлением уридина. Полученный штамм, использованный для получения штамма с делецией 10 протеаз, был обозначен как штамм номер M597.
Для удаления последовательности вектора плазмиду pTTv312 (Δpep11-pyr4) расщепляли посредством MssI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации клона штамма с делецией 9 протеаз M597 (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 15.2) в отношении правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 15.2. Делецию pep11 подтверждали саузерн-анализом выбранных клонов (данные не представлены) с использованием способов, описанных в примере 1. Клон 47-62B был обозначен как штамм номер M632. Проводили дополнительную стадию очистки единичных клеток штамма M632 с получением штамма с делецией 10 протеаз M658.

ПРИМЕР 16 - ПОЛУЧЕНИЕ ПЛАЗМИДЫ С ДЕЛЕЦИЕЙ TPP1
Плазмиду pTTv331 для делеции трипептидилпептидазы tpp1 (tre82623) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1, за исключением того, что маркер, используемый для селекции, представлял собой двойной маркер pyr4-hph.
1245 п.н. 5'-фланкирующей области и 1025 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции tpp1. Участок размером 311 п.н. с конца 5'-фланкцирующей области tpp1 использовали в качестве фрагмента прямого повтора. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 16.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Pyr4-hph получали из pTTv210 (Δsep1-pyr4-hph) с помощью расщепления NotI. Для обеспечения удаления полной двойной маркерной кассеты участки рестрикции NotI встраивали по обеим сторонам кассеты с двойным маркером. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью tpp1. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции tpp1 (pTTv331, таблица 16.1) обеспечивает делецию 2152 п.н. в локусе tpp1 и охватывает полную кодирующую последовательность TPP1.

ПРИМЕР 17 - ПОЛУЧЕНИЕ ПЛАЗМИД ДЛЯ ДЕЛЕЦИИ PEP8
Другую плазмиду pTTv319 для делеции аспарагиновой протеазы pep8 (tre122076) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1.
Вторую плазмиду для делеции аспарагиновой протеазы pep8 (tre122076) pTTv328 конструировали с использованием плазмиды pTTv319, описанной выше, в качестве остова. Маркер pyr4 удаляли из pTTv319 с помощью расщепления NotI. Pyr4-hph получали из pTTv210 (Δsep1-pyr4-hph) с помощью расщепления NotI. Клонирование плазмиды pTTv328 проводили с помощью стандартного лигирования с использованием ДНК-лигазы T4 при комнатной температуре. Часть смеси для лигирования трансформировали в E. coli с помощью электропорации. Несколько клонов культивировали, плазмидную ДНК выделяли и расщепляли для скрининга правильного лигирования с использованием стандартных лабораторных способов. Правильное лигирование и ориентацию маркера далее подтверждали секвенированием.
1095 п.н. 5'-фланкирующей области и 988 п.н. 3'-фланкирующей области выбирали в качестве основы плазмид для делеции pep8. Участок размером 324 п.н. с конца 5'-фланкирующей области pep8 использовали в качестве фрагмента прямого повтора. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 17.1. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Селективный маркер pyr4, использованный в pTTv319, получали из pTTv181 (Δpep4-pyr4, выше) с помощью расщепления NotI. Для обеспечения удаления кассеты с маркером pyr4 по обеим сторонам кассеты встраивали участки рестрикции NotI. Участок AscI встраивали между 5'-прямым повтором и 3'-фланкирующей областью pep8. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv319 конструировали с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эти плазмиды для делеции pep8 (pTTv319 и pTTv328, таблица 17.1) обеспечивают делецию 1543 п.н. в локусе pep8 и охватывают полную кодирующую последовательность PEP8.

Третью плазмиду pTTv266 для делеции аспарагиновой протеазы pep8 (tre122076) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1, за исключением того, что маркер, использованный для селекции, представлял собой pyr4-hgh из pTTv194.
1095 п.н. 5'-фланкирующей области и 988 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды pTTv266 для делеции pep8. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 17.2. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Селективный маркер pyr4-hgh, использованный в pTTv266, получали из pTTv194 (Δpep4-pyr-hgh, выше) с помощью расщепления NotI. Для обеспечения удаления кассеты с маркером pyr4-hgh по обеим сторонам кассеты встраивали участки рестрикции NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv266 конструировали с 5'-фланкирующей областью, 3'-фланкирующей областью, маркером pyr4-hgh и остовом вектора с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Плазмиды для делеции pep8 (pTTv266, таблица 17.2) обеспечивают делецию 1543 п.н. в локусе pep8 и охватывают полную кодирующую последовательность PEP8.

ПРИМЕР 18 - ВНЕСЕНИЕ ДЕЛЕЦИЙ ПРОТЕАЗ В ШТАММЫ, ПРОДУЦИРУЮЩИЕ ИНТЕРФЕРОН
Получение штамма с делецией 5 протеаз, продуцирующего IFN-α 2b
Для получения штамма, продуцирующего IFN-α 2b с делецией пяти протеаз, штамм с делецией пяти протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2 M369 трансформировали экспрессирующей кассетой для IFN-α 2b (pTTv173) с использованием гигромицина при селекции. Этот продуцирующий IFN-α 2b штамм с делециями пяти протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2 был обозначен номером M401.
Анализ продуцирующего IFN-α 2b штамма M401 с делецией 5 протеаз
Для исследования уровня экспрессии IFN-α 2b продуцирующий IFN-α 2b штамм M401 с делецией 5 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2) культивировали в условиях культивирования, pH 4,5; 28->22°C; 30 г/л глюкозы, 60 г/л лактозы, 20 г/л цельной пивной дробины и 20 г/л экстракта пивной дробины. Для анализа уровня экспрессии IFN-α 2b образцы культуры, полученные на 3 сутки, подвергали количественному иммуноблоттингу (фиг.54A). Образцы анализировали путем сравнения со стандартной кривой для IFN-α 2b, и денситометрическое количественное определение проводили с помощью программного обеспечения Totallab Quant TL100. Иммуноблоттинг проводили с помощью антитела против IFN-α 2b Aceam (#ab9386), разбавленного до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело от козы против IgG мыши Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Белковые стандарты наносили на гель, соответствующий 50 нг, 100 нг, 200 нг и 400 нг IFN-α 2b. Анализ продемонстрировал, что M401 продуцировал IFN-α 2b с выходом вплоть до 51,9 мг/л и 52% продукта отщеплялось от молекулы-носителя.
Получение продуцирующего IFN-α 2b штамма M577 с делецией 8 протеаз
Для получения штамма, продуцирующего IFN-α 2b с делецией восьми протеаз, штамм с делецией восьми протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5 M504 трансформировали экспрессирующей кассетой для IFN-α 2b (pTTv254) с использованием ацетамида при селекции. Этот продуцирующий IFN-α 2b штамм с делециями восьми протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5 был обозначен номером M577.
Анализ продуцирующего IFN-α 2b штамма M577 с делецией 8 протеаз
Для исследования уровня экспрессии IFN-α 2b, продуцирующий IFN-α 2b штамм M577 с делецией 8 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5) культивировали в условиях культивирования, pH 4,5; 28->22°C; 2% дрожжевой экстракт, 4% целлюлозы, 8% целлобиозы и 4% сорбозы. Для исследования экспрессии IFN-α 2b, образцы М577 после ферментации подвергали иммуноблоттингу (фиг.54B). Для анализа уровня экспрессии IFN-α 2b, образцы культуры, полученные на 4 сутки, подвергали количественному иммуноблоттингу (фиг.55). Образец анализировали путем сравнения со стандартной кривой для IFN-α 2b, и денситометрическое количественное определение проводили с помощью программного обеспечения Totallab Quant TL100. Иммуноблоттинг проводили с помощью антитела против IFN-α 2b Aceam (#ab9386), разбавленного до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело от козы против IgG мыши Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Белковые стандарты наносили на гель, соответствующий 50 нг, 100 нг и 200 нг IFN-α 2b. Анализ продемонстрировал, что M577 продуцировал IFN-α 2b с выходом вплоть до 1780 мг/л, и 66,5% продукта отщеплялось от молекулы-носителя. Штамм M577 с делецией 8 протеаз продуцировал в 34 раза больше IFN-α 2b, чем штамм M401 с делецией 5 протеаз.
Получение продуцирующего IFN-α 2b штамма M652 с делецией 9 протеаз
Для получения штамма, продуцирующего IFN-α 2b с делецией девяти протеаз, штамм с делецией девяти протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2 M574 трансформировали экспрессирующей кассетой для IFN-α 2b (pTTv173) с использованием гигромицина при селекции. Этот продуцирующий IFN-α 2b штамм с делециями девяти протеаз Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2 был обозначен номером M652.
Анализ продуцирующего IFN-α 2b штамма M652 с делецией 9 протеаз
Для исследования уровня экспрессии IFN-α 2b продуцирующий IFN-α 2b штамм M652 с делецией 9 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5Δpep2) культивировали в условиях культивирования, pH 4,5; 28->22°C; 2% дрожжевой экстракт, 4% целлюлозы, 8% целлобиозы и 4% сорбозы. Для исследования экспрессии IFN-α 2b образцы М652 после ферментации подвергали иммуноблоттингу (фиг.54B). Для анализа уровня экспрессии IFN-α 2b, образец культуры, полученный на 3 сутки, подвергали количественному иммуноблоттингу (фиг.55). Образец анализировали путем сравнения со стандартной кривой для IFN-α 2b, и денситометрическое количественное определение проводили с помощью программного обеспечения Totallab Quant TL100. Иммуноблоттинг проводили с помощью антитела против IFN-α 2b Aceam (#ab9386), разбавленного до 1 мкг/мл в TBST. Вторичное антитело представляло собой конъюгированное с AP вторичное антитело от козы против IgG мыши Bio-rad (#170-6520), разбавленное 1:5000 в TBST. Белковые стандарты наносили на гель, соответствующий 50 нг, 100 нг и 200 нг IFN-α 2b. Анализ продемонстрировал, что M652 продуцировал IFN-α 2b с выходом вплоть до 1928 мг/л, и 85% продукта отщеплялось от молекулы-носителя. Штамм M652 с делецией 9 протеаз продуцировал немного больше IFN-α 2b, чем M577 с делецией 8 протеаз, и в 37 раз больше IFN-α 2b, чем штамм M401 с делецией 5 протеаз.
Получение штамма M670 с делецией 9 протеаз с делецией pep8 (tre122076) из продуцирующего интерферон штамма M577
Для удаления кассеты для делеции плазмиду pTTv266 (Δpep8-pyr4-hgh) расщепляли посредством PmeI, и правильный фрагмент очищали с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации штамма с делецией 8 протеаз M577 (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5). Штамм M577 продуцирует интерферон-альфа 2b. Получение протопластов и трансформацию проводили, по существу как описано в примере 1, с использованием селекции с помощью гигромицина.
Трансформанты отбирали и высевали штрихами на чашки для селекции. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 18.1) для правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 18.1. Клон 82-9 был обозначен как штамм номер M670.

Получение плазмиды для делеции pep11
Плазмиду pTTv268 для делеции аспарагиновой протеазы pep11 (tre121306) конструировали, по существу как описано для плазмиды для делеции pep1 pTTv41 в примере 1, за исключением того, что маркер, используемый для селекции, представлял собой pyr4-hgh из pTTv194.
956 п.н. 5'-фланкирующей области и 957 п.н. 3'-фланкирующей области выбирали в качестве основы плазмиды для делеции pep11 pTTv268. Эти фрагменты амплифицировали способом ПЦР с использованием праймеров, приведенных в таблице 18.2. Продукты разделяли агарозным гель-электрофорезом, и соответствующие фрагменты выделяли из геля с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Pyr4-hgh получали из pTTv194 (Δpep4-pyr-hgh) с помощью расщепления NotI. Для обеспечения удаления кассеты с маркером по обеим сторонам кассеты встраивали участки рестрикции NotI. Остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, как и в примере 1. Плазмиду pTTv268 конструировали с 5'-фланкирующей областью, 3'-фланкирующей областью, маркером pyr4-hgh и остовом вектора с использованием способа гомологичной рекомбинации дрожжей, описанного в примере 1. Эта плазмида для делеции pep11 (pTTv268, таблица 18.2) обеспечивает делецию локуса pep11 и охватывает полную кодирующую последовательность PEP11.

Получение штамма M672 с делецией 9 протеаз с pep11 (tre121306), удаленным из продуцирующего интерферон штамма M577
Для удаления кассеты для делеции плазмиду pTTv268 (Δpep11-pyr4-hgh) расщепляли посредством PmeI, и правильный фрагмент очищали с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации штамма M577 с делецией 8 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5). Штамм M577 продуцирует интерферон альфа 2b. Получение протопластов и трансформацию проводили, по существу как описано в примере 1, с использованием селекции с помощью гигромицина.
Трансформанты отбирали и высевали штрихами на чашки для селекции. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 18.3) в отношении правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и повторно подвергали скринингу способом ПЦР с использованием праймеров, приведенных в таблице 18.3. Клон 33-9 был обозначен как штамм номер M672.

Получение штамма M673 с делецией 9 протеаз с slp7 (tre123865), удаленным из продуцирующего интерферон штамма M577
Для удаления кассеты для делеции плазмиду pTTv269 (Δslp7-pyr4-hgh) расщепляли посредством PmeI, и соответствующий фрагмент очищали с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации штамма M577 с делецией 8 протеаз (Δpep1Δtsp1Δslp1Δgap1Δgap2Δpep4Δpep3Δpep5). Штамм M577 продуцирует интерферон-альфа 2b. Получение протопластов и трансформацию проводили, по существу как описано в примере 1, с использованием селекции с помощью гигромицина.
Трансформанты отбирали и наносили штрихами на чашки для селекции. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 18.4) в отношении правильного встраивания. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 18.4. Клон 5-64 был обозначен как штамм номер M673.

ПРИМЕР 19 - ПОЛУЧЕНИЕ ПРОДУЦИРУЮЩИХ G0 ШТАММОВ M627 И M629
Векторы pTTg156 и pTTg173 конструировали встраиванием кассеты с двойным селективным маркером (маркерный ген устойчивости к гигромицину (hph) между промотором pki1 и терминатором cbh2 в дополнение к экспрессирующей кассете pyr4) в промежуточные векторы pTTg145 и pTTg146. Промежуточные векторы конструировали посредством рекомбинантного клонирования дрожжей, кассеты с маркерами добавляли общепринятым клонированием путем расщепления NotI и лигирования.
Стратегия получения фрагментов промежуточных векторов представлена в таблице 19.1 ниже. Праймеры, использованные для получения фрагментов, приведены в таблице 19.2 ниже. После конструирования фрагментов pTTg145 и pTTg146 в соответствии с планом, представленным в таблице 19.1, их подвергали электрофорезу в дрожжи Saccharomyces cerevisiae FY834 для сборки плазмиды посредством гомологичной рекомбинации. Дрожжевые клетки высевали на SC-ura для культивирования в течение 2-3 суток при +30°C. Затем колонии объединяли из планшетов и объединенные плазмиды очищали способом экстракции с фенолом/хлороформом. Объединенные плазмиды трансформировали в компетентные клетки E. coli электропорацией. Подвергнутые электропорации клетки высевали на чашки для селекции с LB+amp, культивировали при +37°C в течение ночи, и колонии подвергали скринингу способом ПЦР. Затем положительные колонии высевали штрихами на свежие чашки в качестве чистых культур, единичные колонии культивировали в жидкой среде с LB+amp, и потенциальные промежуточные векторы pTTg145 и pTTg146 очищали в соответствии со стандартными протоколами. Плазмиды анализировали рестрикционным анализом и последовательности подтверждали секвенированием.
Затем кассету с маркером встраивали в векторы общепринятым расщеплением NotI из pTTg163 и лигированием в линеаризованные с помощью NotI промежуточные векторы pTTg145 и pTTg146.
Стратегия конструирования промежуточных векторов pTTg145-pTTg149. pTTv141 и pTTv11 описаны в международной патентной заявке № PCT/EP2011/070956. Плазмиды pTTg124 и pTTv225 использовали для конструирования фрагментов, указанных в таблице ниже
с помощью SacII/NdeI
с матрицы pTTv11 GP340, GP336

Экспрессирующий MAB01 штамм Trichoderma reesei M507 с делецией 7 протеаз трансформировали PmeI-фрагментами pTTg156 и pTTg173, нацеленными на локус alg3. Различное количество трансформантов (100-170, в зависимости от конструкции) отбирали на чашки для селекции. Исходя из скрининга способом ПЦР с набором Phire Plant Direct PCR kit (Finnzymes F-130), клоны с положительными результатами в отношении 5'- и 3'-встраивания отбирали для посева отдельных спор и повторного скрининга в отношении встраивания и делеции alg3 (5 клонов после трансформации pTTg156, 3 клона после трансформации pTTg173). Праймеры, использованные для скрининга, приведены в таблице 19.3 ниже.
Подвергнутые скринингу способом ПЦР штаммы в конечном итоге подвергали культивированию во вращающихся флаконах и анализу гликанов. Конечные штаммы были обозначены как M629 (трансформант pTTg173) и M627 (трансформант pTTg156).
Праймеры, использованные для скрининга встраивания alg3 в pTTg156 и pTTg173 M507
5'-стороны, прямой, вне фланкирующей области
5'-стороны, обратный, из
pGPDA
3'-стороны, обратный, вне фланкирующей области
(продукт размером 1492 п.н.)
(продукт размером 1492 п.н.)
Ферментация с WSG для штаммов M627 и M629 и анализ гликанов
Штаммы T. reesei M627 и M629 ферментировали в 4% WSG, 2% глюкозе, 4% целлобиозе, 6% лактозе, pH 5,5, и взятие образцов проводили на 3-6 сутки. Титры антител представлены в таблице 19.4.
Для анализа N-гликанов MAB01 очищали из культуральных супернатантов с использованием 96-луночного фильтрующего планшета с белком G HP MultiTrap (GE Healthcare) в соответствии с инструкциями изготовителя. Концентрации антител определяли посредством УФ-поглощения против стандартной кривой для MAB01.
N-гликаны высвобождали из преципитированного с EtOH и денатурированного с помощью SDS антитела с использованием PNGase F (ProZyme Inc.) в 20 мМ натрий-фосфатном буфере, pH 7,3, в реакции в течение ночи при +37°C. Высвободившиеся N-гликаны очищали с помощью Hypersep C-18 и Hypersep Hypercarb (Thermo Scientific) и анализировали с помощью MALDI-TOF MS. Результаты представлены в таблице 19.6. В штаммах M627 и M629 уровни G0 варьировали от 24,3% до 41,7%, в штамме M507 G0 не было обнаружено.
Культивирование с WSG во вращающихся флаконах штаммов M627 и M629 и анализ гликанов
Штаммы T. reesei M627 и M629 культивировали во вращающихся флаконах в TrMM, 4% лактозе, 2% SGE, 100 мМ PIPPS, pH 5,5, при +28°C. Взятие образцов проводили на 5 сутки.
Для анализа N-гликанов MAB01 очищали из культуральных супернатантов с использованием 96-луночных фильтрующих планшетов с белком G HP MultiTrap (GE Healthcare) в соответствии с инструкциями изготовителя. Концентрации антитела в элюентах белка G определяли по УФ-поглощению против стандартной кривой для MAB01 (таблица 19.5). Титры в культуральной среде не измеряли.
Высвобождение N-гликанов проводили, как описано выше, результаты представлены в таблице 19.7. Уровни G0 составляли 21,1 и 56,9% для M627 и M629, соответственно.
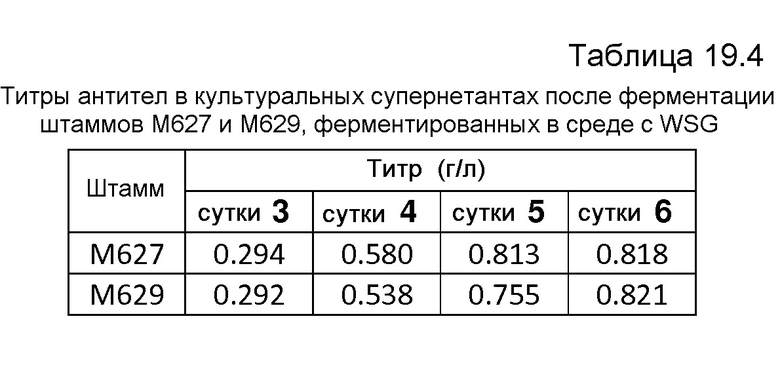

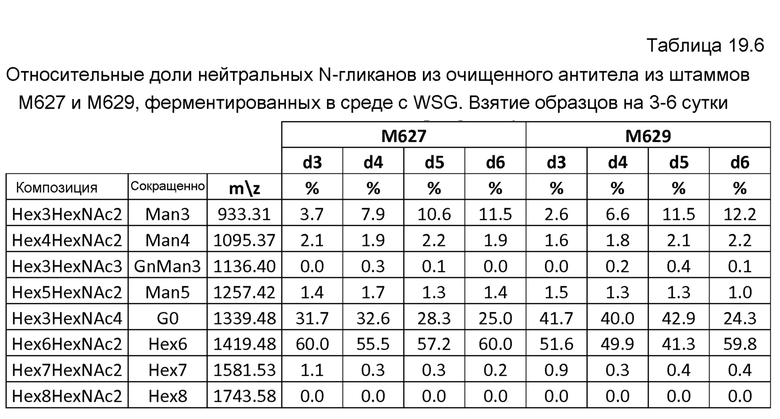
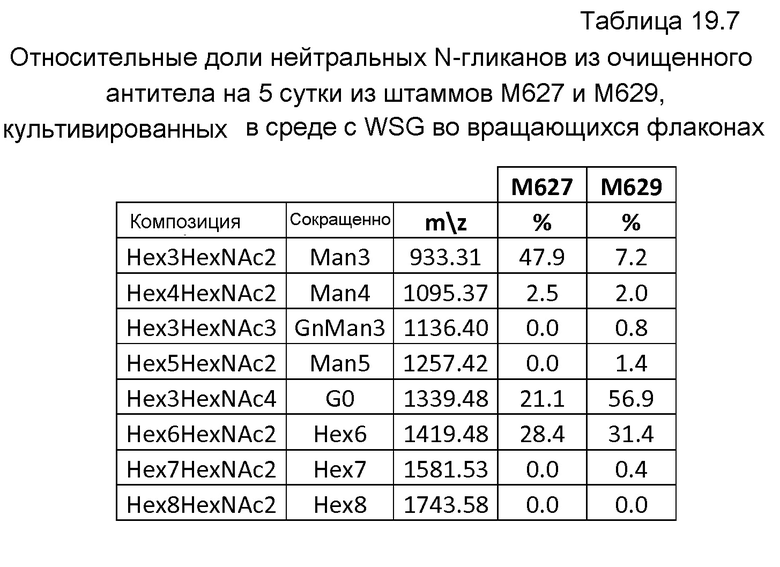
ПРИМЕР 20 - ПОЛУЧЕНИЕ ПРОДУЦИРУЮЩИХ GlcNAcMan5 ШТАММОВ С РАЗЛИЧНЫМИ ПРОМОТОРАМИ
Векторы для GnTI с различными промоторами описаны в таблице 20.1. Векторы были нацелены в локус egl2 T. reesei.
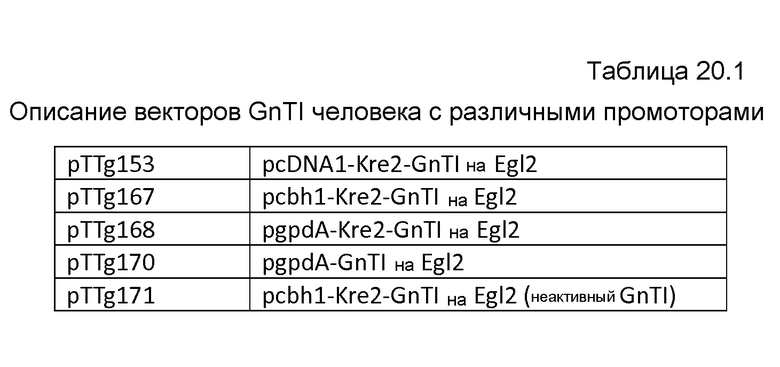
Материалы и способы
Стратегия получения фрагментов промежуточных векторов, представленных в таблице 20.1, представлена в таблице 20.2 ниже, и праймеры, использованные для получения фрагментов, приведены в таблице 20.3. Фрагменты амплифицировали способом ПЦР и продукты очищали из агарозного геля. Расщепленный вектор pTTg152 очищали из геля. Все амплификации способом ПЦР проводили с помощью высокоточной полимеразы Phusion (Finnzymes). Фрагменты для pTTg153-pTTgl71 вводили способом электропорации в дрожжи Saccharomyces cerevisiae FY834 для сборки плазмиды способом гомологичной рекомбинации. Дрожжевые клетки высевали на SC-ura для культивирования в течение 2-3 суток при +30°C. Затем колонии объединяли из планшетов и объединенные плазмиды очищали общепринятым способом экстракции с фенолом/хлороформом. Объединенные плазмиды трансформировали в компетентные клетки E. coli электропорацией. Подвергнутые электропорации клетки высевали на чашки для селекции с LB+amp, культивировали при +37°C в течение ночи, и колонии подвергали скринингу способом ПЦР. Затем положительные колонии высевали штрихами на свежие чашки в качестве чистых культур, единичные колонии культивировали в жидкой среде с LB+amp, и потенциальные векторы pTTg153-pTTg171 очищали в соответствии со стандартными протоколами. Плазмиды анализировали рестрикционным анализом и последовательности подтверждали секвенированием.
Стратегия конструирования векторов. Векторы pTTv225, pTTv77, pTTg91 использовали для конструирования фрагментов, указанных ниже. Векторы pTTv110, pTTv141, pTTv11 и pTTv13 описаны в международной патентной заявке № PCT/EP2011/070956.
>последовательность фрагмента 3'-фланкирующей области egl2
>Амплификация фрагмента промотор cDNA1 – сигнал для наце-ливания Kre2
Амплификация фрагмента неактивного GntI

Экспрессирующий MAB01 штамм Trichoderma reesei M507 с делецией 7 протеаз трансформировали PmeI-фрагментами векторов pTTg153-pTTg171, нацеленных на локус egl2. Различные количества трансформантов отбирали на чашки для селекции. На основе скрининга способом ПЦР с набором Phire Plant Direct PCR kit (Finnzymes F-130), клоны с положительными результатами в отношении 5'- и 3'-встраивания отбирали для посева единичных спор и повторного скрининга в отношении встраивания и делеции egl2. Подвергнутые скринингу способом ПЦР штамм в итоге подвергали культивированию во вращающихся флаконах и анализу гликанов.
Культивирование во вращающихся флаконах штамма M507, трансформированного GnTI и промоторными конструкциями
Штамм M507, трансформированный векторами, представленными в таблицах 20.1 и 20.2, культивировали во вращающихся флаконах в TrMM, 4% лактозе, 2% SGE, 100 мМ PIPPS, pH 5,5, при +28°C, и взятие образцов проводили на 5 сутки. Неактивную конструкцию GnTI исследовали для определения возможных эффектов гликанов GlcNAcMan5 на рост T. reesei.
Для анализа N-гликанов MAB01 очищали, определяли концентрации, и N-гликаны анализировали с помощью MALDI-TOF MS, как описано выше. Анализ N-гликанов MAB01 показал, что уровни GnМan5 находились в диапазоне от 8 до 79,2% от общего содержания гликанов (таблицы 20.4 и 20.5A и B). Неактивный GnTI обеспечивал гликозилирование дикого типа, как и ожидалось.

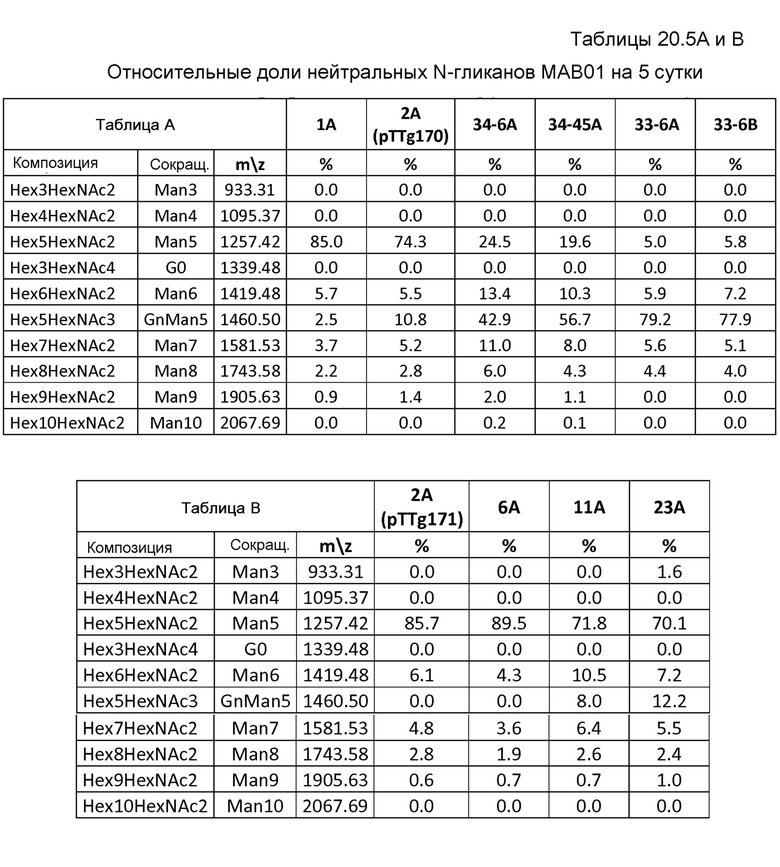
Культуры в ферментере штамма M507, трансформированного конструкциями GnTI
Штаммы M702, M704, M706, M710, M712, M716 и M507 T. reesei ферментировали в 4% WSG, 2% Glc, 4% целлобиозе, 6% лактозе, pH 5,5, и взятие образцов проводили на 3-6 сутки. Титры антител представлены в таблице 20.6. N-гликаны отщепляли и анализировали, как описано выше, с использованием PNGase F.
Анализ N-гликанов MAB01 продемонстрировал, что уровни GnМan5 находились в диапазоне от 1,8 до 68,5% от общего уровня гликанов (таблицы 20.7A, B и C). Неактивный GnTI обеспечивал гликозилирование дикого типа, как и ожидалось, как и контрольный штамм M507.

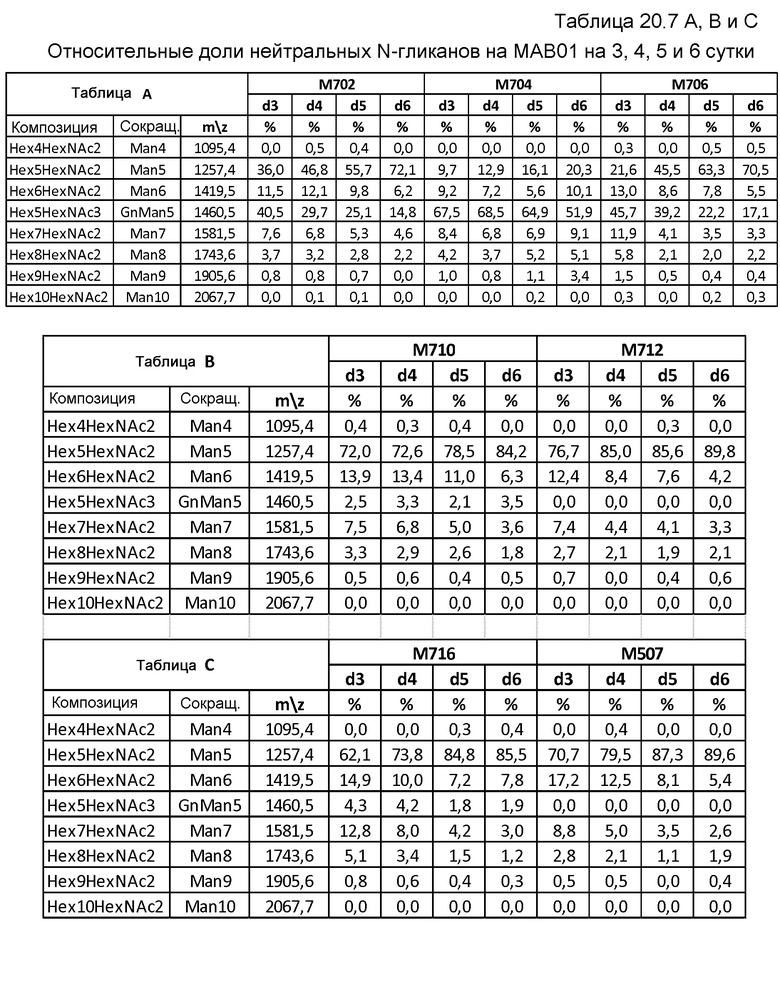
ПРИМЕР 21 - ПОЛУЧЕНИЕ ПРОДУЦИРУЮЩИХ GlcNAcMan5 ШТАММОВ С РАЗЛИЧНЫМИ НАЦЕЛИВАЮЩИМИ ПЕПТИДАМИ
Получение плазмид
Все из плазмид, использованных для получения штаммов GNT1 с различными нацеливающими на аппарат Гольджи пептидами (pTTv274, pTTv275, pTTv276, pTTv278, pTTv279, pTTv280), были основаны на общей родительской плазмиде pTTv265, которая содержит GNT1 человека (P26572) с N-концевым укорочением из 38 аминокислот. Происходящие из pTTv265 плазмиды обобщенно представлены в таблице 21.1.
Плазмиды, происходящие из нацеливающих на аппарат Гольджи плазмид pTTv274, pTTv275, pTTv276, pTTv278, pTTv279, pTTv280. Нацеливающие на аппарат Гольджи пептиды к pTTv265 с получением указанных выше плазмид
Пошаговое описание получения плазмид
Плазмида pTTv77 содержит 5'- и 3'-фланкирующие области egl2 (tre120312) для направленного встраивания в геном T. reesei и промотор cbh1 для экспрессии генов. Встраивание плазмиды pTTv77 в геном обеспечивает делецию 2456 п.н. в локусе egl2. Области 1020 п.н. и 1024 п.н. из локуса egl2 амплифицировали для 5'- и 3'-фланкирующих областей. 2176 п.н. из локуса cbh1 амплифицировали для промоторного фрагмента. Матрица, использованная в реакции ПЦР, представляла собой геномную ДНК T. reesei. Праймеры, использованные в реакциях ПЦР, представлены в таблице 21.2. Бластерная кассета для селективного маркера pyr4 представляла собой фрагмент NotI из pTTv71, описанного выше, и остов вектора представлял собой расщепленный EcoRI/XhoI pRS426, описанный выше. Все фрагменты очищали с использованием стандартных лабораторных способов, и плазмиду клонировали способом рекомбинации дрожжей, как описано в примерах. После спасения плазмиды в E. coli в нескольких клонах подтверждали правильную рекомбинацию. Сохраненный клон подтверждали секвенированием.

Плазмида pTTv256 основана на плазмиде pTTv77. В плазмиде pTTv256 промотор cbh1 был заменен на gpdA. Для клонирования pTTv256 плазмиду pTTv77 расщепляли посредством FseI/PacI для высвобождения промотора cbh1. Новый промотор, gpdA A. nidulans, высвобождался из плазмиды посредством расщепления FseI/PacI. Очистку и клонирование фрагмента проводили с использованием стандартных лабораторных способов. Несколько клонов подтверждали в отношении правильного лигирования. Сохраненный клон подтверждали секвенированием.
Плазмида pTTv264 основана на плазмиде pTTv256. В pTTv264 селективный маркер заменяли с бластерной кассеты pyr4 на селективный маркер hygR. Для клонирования pTTv264 плазмиду pTTv256 расщепляли посредством NotI для высвобождения бластерной кассеты pyr4. Маркер hygR амплифицировали способом ПЦР с использованием модифицированной pRLMex30 в качестве матрицы с праймерами, представленными в таблице 21.3. Все фрагменты очищали с использованием стандартных лабораторных способов, и плазмиду клонировали способом рекомбинации дрожжей, описанным в примерах. После спасения плазмиды в E. coli несколько клонов подтверждали в отношении правильной рекомбинации, и сохраненный клон подтверждали секвенированием.

Плазмида pTTv265 основана на плазмиде pTTv264, описанной выше. В pTTv265 GNT1 человека с N-концевым укорочением 114 нуклеиновых кислот (38 аминокислот) добавляли под контроль промотора gpdA. Для клонирования pTTv265 плазмиду pTTv264 линеаризовывали с помощью PacI. GNT1 человека амплифицировали способом ПЦР из синтетической плазмиды pTTv11, содержащей полноразмерный ген GNT1 человека (P26572, pTTv11 также описан в международной патентной заявке № PCT/EP2011/070956). Праймеры, использованные для амплификации, представлены в таблице 21.4. Все фрагменты очищали с использованием стандартных лабораторных способов. Плазмиду клонировали способом рекомбинации дрожжей, описанным выше. После спасения плазмиды в E. coli в нескольких клонах подтверждали правильную рекомбинацию, и сохраненный клон подтверждали секвенированием.
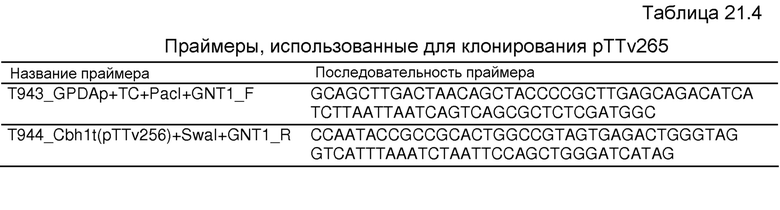
Аминокислотная последовательность GnTI человека с N-концевым укорочением на 38 аминокислот в плазмиде pTTv11 для конструирования pTTv265.
SVSALDGDPASLTREVIRLAQDAEVELERQRGLLQQIGDALSSQRGRVPTAAP PAQPRVPVTPAPAVIPILVIACDRSTVRRCLDKLLHYRPSAELFPIIVSQDCGHEETAQAIA SYGSAVTHIRQPDLSSIAVPPDHRKFQGYYKIARHYRWALGQVFRQFRFPAAVVVEDDL EVAPDFFEYFRATYPLLKADPSLWCVSAWNDNGKEQMVDASRPELLYRTDFFPGLGWL LLAELWAELEPKWPKAFWDDWMRRPEQRQGRACIRPEISRTMTFGRKGVSHGQFFDQH LKFIKLNQQFVHFTQLDLSYLQREAYDRDFLARVYGAPQLQVEKVRTNDRKELGEVRV QYTGRDSFKAFAKALGVMDDLKSGVPRAGYRGIVTFQFRGRRVHLAPPPTWEGYDPSWN
Все из плазмид pTTv274, pTTv275, pTTv276, pTTv278, pTTv279 и pTTv280 были основаны на плазмиде pTTv265, описанной выше. В этих плазмидах различные нацеливающие на аппарат Гольджи пептиды добавляли к предшествующему гену GNT1 человека с N-концевым укорочением. Для клонирования этих плазмид pTTv265 линеаризовывали с помощью PacI. Различные нацеливающие на аппарат Гольджи пептиды амплифицировали способом ПЦР с использованием праймеров, представленных в таблице 21.5A. ДНК-матрица для GNT2 (pTTv274) представляла собой синтетическую плазмиду, содержащую ген GNT2 человека с оптимизацией кодонов из международной патентной заявки № PCT/EP2011/070956. Матрицей для других нацеливающих на аппарат Гольджи пептидов (pTTv276, pTTv278, pTTv279, pTTv280) была геномная ДНК T. reesei. KRE2 (pTTv275) продуцировали способом ПЦР с использованием отжигающихся праймеров, представленных в таблице 21.5A. Все фрагменты очищали с использованием стандартных лабораторных способов, и плазмиды клонировали способом рекомбинации дрожжей, как описано в примерах. После спасения плазмиды в E. coli в нескольких клонах после каждого клонирования подтверждали правильную рекомбинацию, и сохраненные клоны подтверждали секвенированием.


Получение штаммов с различными нацеливающими на аппарат Гольджи пептидами для GNT1
Фрагменты для трансформаций высвобождали из плазмид pTTv274 (GNT2), pTTv275 (KRE2), pTTv276 (KRE2-подобный), pTTv278 (OCH1), pTTv279 (ANP1) и pTTv280 (VAN1) (Таблицы 21.5A и 21.5B) с помощью PmeI. Все фрагменты трансформировали по отдельности в экспрессирующий MAB01 штамм M507, и трансформации протопластов осуществляли, по существу как описано в примерах для селекции с гигромицином.
Хорошо растущие клоны на селективных штриховых культурах подвергали скринингу в отношении 5'- и 3'-встраивания в локус egl2. Положительные по двойному встраиванию клоны дополнительно подвергали скринингу в отношении утраты ORF egl2. Клоны, дающие желаемые результаты, очищали посредством посева единичных спор и способом ПЦР подтверждали, что происходящие из единичных спор клоны являются чистыми штаммами с встраиванием. Полученные штаммы приведены в таблице 21.6 ниже.
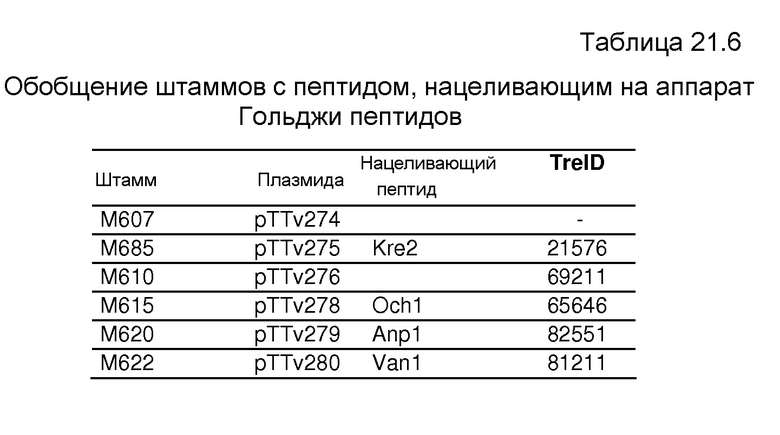
Ферментация штаммов M507, M607, M610, M615, M620, M622 и M685 и анализ гликанов
Штаммы M507, M607, M610, M615, M620, M622 и M685 T. reesei ферментировали в 4% цельной пивной дробине, 2% глюкозе, 4% целлобиозе, 6% лактозе, pH 5,5, и взятие образцов проводили на 3-6 сутки. Титры антител представлены в таблице 21.7. N-гликаны отщепляли и анализировали, как описано выше, с использованием PNGase F.
Анализ MAB01 с N-гликанами показал, что уровни GnМan5 находились в диапазоне от 4 до 66% от общего содержания гликанов (таблица 21.8A, B и C). Контрольный штамм M507 продемонстрировал гликозилирование дикого типа, как и ожидалось.
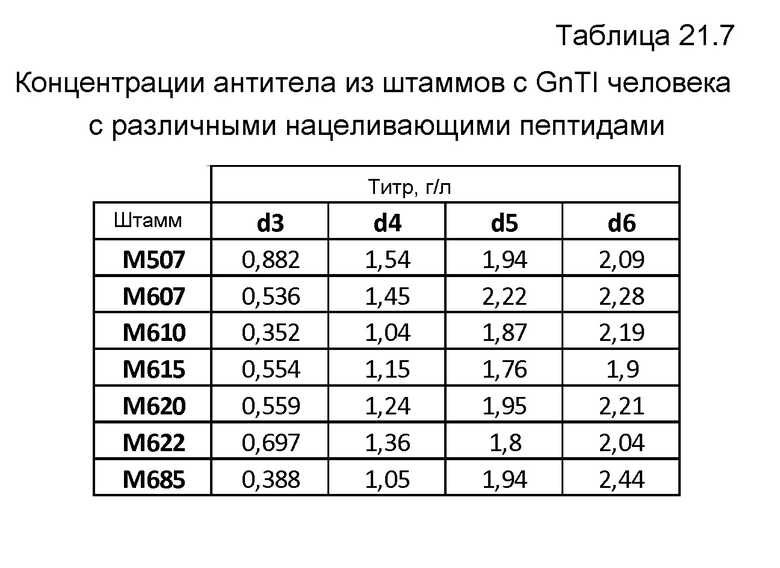
ПРИМЕР 22 - ПОДАВЛЕНИЕ ГЕНОВ SLP С ПОМОЩЬЮ РНК-I И ДЕЛЕЦИИ SLP2
Конструировали три подавляющих конструкции для нокдауна экспрессии slp2 (tre123244). Эти конструкции РНК-i содержат промотор gpdA, направленную вставку в локус протеазы pep2 (tre53961) и маркер для выпетливания pyr4 с прямым 3'-повтором pep2. Две коротких последовательности-мишени размером 19 п.н. и большую последовательность размером 448 п.н. встраивали в этот вектор с получением pTTv217, pTTv218 и pTTv263, соответственно. Эти векторы конструировали для нокдауна экспрессии slp2 и для снижения его протеазной активности. Векторы для РНК-i трансформировали в pyr4- версию продуцирующего MAB01 штамма M507. Вектор pTTv204 представлен на фиг.52.
Экспрессирующий вектор для РНК-i pTTv204 линеаризовывали с помощью фермента рестрикции AsiSI. Праймеры T846 и T847 отжигали вместе и встраивали посредством рекомбинации дрожжей в вектор pTTv204. Праймеры представлены в таблице 22.1. В полученном векторе pTTv217 содержится последовательность-мишень из 19 п.н. Праймеры T848 и T849 отжигали вместе и встраивали в линеаризованный вектор pTTv204 с получением вектора РНК-i pTTv218. Этот вектор содержит последовательность-мишень из 19 п.н. Праймеры представлены в таблице 22.1. Последовательности-мишени представлены в таблице 22.2.
Вектор pTTv263 получали в виде двух фрагментов и встраивали в вектор pTTv204. Праймеры T965 и T967 использовали для амплификации смыслового фрагмента размером 506 п.н., включающего последовательность интрона размером 58 п.н. в гене slp2. Вектор pTTv204 раскрывали с помощью фермента рестрикции AsiSI и смысловой фрагмент размером 506 п.н. встраивали в вектор посредством рекомбинации дрожжей. Праймеры T1006 и T1007 использовали для амплификации антисмыслового фрагмента размером 448 п.н. Антисмысловой фрагмент расщепляли ферментами рестрикции FseI и AscI. Вектор, включающий смысловой фрагмент, также расщепляли посредством FseI и AscI. Вектор и антисмысловой фрагмент лигировали вместе с получением вектора pTTv263. Праймеры приведены в таблице 22.1. Последовательность-мишень представлена в таблице 22.2.
Векторы для РНК-i pTTv217, pTTv218, pTTv263 расщепляли с помощью PmeI для высвобождения экспрессирующей кассеты. Фрагменты разделяли с помощью агарозного гель-электрофореза, и правильные фрагменты выделяли с помощью набора для экстракции из геля (Qiagen) с использованием стандартных лабораторных способов. Приблизительно 5 мкг экспрессирующей кассеты использовали для трансформации экспрессирующего антитело MAB01 штамма M507 (версия pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195 с использованием селекции на основе pyr4.
Короткую последовательность-мишень в векторе pTTv217 конструировали, чтобы влиять специфически только на slp2. Последовательность-мишень pTTv218 была гомологична slp3, slp5 и slp6. Большая последовательность-мишень размером 448 п.н. в векторе pTTv263 была предназначена влиять на несколько субтилизинов. Последовательности-мишени в этих векторах приведены в таблице 22.2. Полученные штаммы с нокдауном M665, M666 и M667 культивировали в мелкомасштабных культурах.
Несколько трансформантов pTTv217, pTTv218 и pTTv263 выращивали в культурах в 24 ячейках для сравнения продуцирования ими MAB01 относительно контрольного штамма M507. Штаммы выращивали в TrMM с цитратом диаммония без сульфата аммония, 100 мМ PIPPS, 2% экстрактом пивной дробины, 4% лактозой при pH 5,5. Для каждого трансформанта использовали дублированные лунки. Образцы из культур в 24 ячейках, взятые на 6 сутки, использовали для иммуноблоттинга. Супернатант разбавляли натрий-цитратным буфером, pH 5,5, так чтобы 0,5 мкл каждого супернатанта можно было наносить на 4-15% гель Criterion. Смешивали с LSB+BME и нагревали при 95°C в течение 5 минут. Белки переносили на нитроцеллюлозу с помощью устройства для блоттинга Criterion при 100 вольт в течение 30 минут. Нитроцеллюлозную мембрану блокировали 5% молоком в TBST в течение 1 часа. Обнаружение тяжелой цепи проводили с помощью конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188), разбавленного 1:10000 в TBST. После инкубации в течение 1 часа с антителом для обнаружения блот промывали TBST, и мембрану проявляли субстратом AP (Promega).
Результаты можно видеть на фиг.53. Трансформант 217.12G продуцировал немного более высокие количества тяжелой цепи по сравнению с M507 или вторым трансформантом 217.12E. Наиболее заметное улучшение наблюдали для трансформантов pTTv218. Три трансформанта имели значительно более высокие количества по сравнению с контролем. 218.25F заметно выделялся. Результаты для трансформантов pTTv263 в большей степени варьировали. Два трансформанта продуцировали очень мало тяжелой цепи антитела. Трансформант 263.110F, по-видимому, продуцировал в два раза больше тяжелой цепи относительно контроля.
Конструкции, которые были нацелены на несколько протеаз, были более успешными в отношении увеличения экспрессии тяжелой цепи. В общем, трансформанты pTTv218 были неизменно лучшими, чем контроль M507. Отсутствие продуцирования, наблюдаемое в двух из трансформантов pTTv263, указывало на то, что РНК-i действовала чрезмерно. Когда ген slp2 удаляли, рост штамма снижался и, таким образом, также снижалась экспрессия антитела. Трансформанты 263.36A и 263.124C росли очень плохо и экспрессировали очень мало slp2. Это было подтверждено исследованиями во вращающихся флаконах и исследованиями с помощью qPCR.
Показатели массы сухого вещества в культурах во вращающихся флаконах можно видеть в таблице 22.5. Штаммы выращивали в TrMM с цитратом диаммония без сульфата аммония, 100 мМ PIPPS, 2% экстракта пивной дробины, 4% лактозы при pH 5,5. Для каждого трансформанта использовали по два флакона. Рост трансформанта 263.124C был затрудненным. В общем, происходило небольшое уменьшение роста во всех штаммах, которые экспрессировали РНК-i. Этот эффект может быть связан с более низкими уровнями экспрессии slp2.
Для подтверждения того, что экспрессия slp2 действительно снижалась посредством экспрессии РНК-i, исследования qPCR проводили на мицелии из исследования во вращающихся флаконах. РНК очищали из мицелия культуры во вращающихся флаконах, кДНК синтезировали и проводили анализ qPCR. Мониторинг экспрессии slp2, slp3, slp5, slp6 и gpd1 осуществляли с помощью специфичных к гену праймеров. Кратность изменений измеряли относительно контрольных штаммов. Экспрессию нормализовывали относительно gpd1.
Трансформант 263.124C продемонстрировал наибольшее подавление slp2 (таблица 22.6). Высокий уровень РНК-i индуцировал подавление гена slp2 в 36 раз, практически до его выключения. Другие трансформанты продемонстрировали значительно более мягкую активность нокдауна, находящуюся в диапазоне от 1,2 до 2,5 раза. Более мягкий нокдаун является более предпочтительным, поскольку штамм растет лучше и может продуцировать высокие уровни антитела.
В случае двух трансформантов было более заметно, на какие другие субтилизины влияла экспрессия РНК-i. В трансформанте 263.124C было очевидно, что slp3 и slp6 также подвергались нокдауну в 6 и 2,3 раз, соответственно. В случае штамма с более мягким нокдауном 218.25F, как slp2, так и slp3 продемонстрировали экспрессию, сниженную в 1,7 и 1,8 раза.
Получение штамма с 8 делециями M646, включающего делецию Slp2
Штамм M646 с делецией slp2 получали путем трансформации кассеты для делеции pTTv115 в M564 (pyr4- версия M507). Штамм pyr4- M564 получали, по существу как описано в примере 3 для удаления бластерной кассеты pyr4 из штамма M195 (Δpep1). Последовательные стадии селекции 5-FOA проводили для обеспечения того, чтобы все выбранные клоны происходили из единичных клеток.
Кассету для делеции, содержащую фланкирующие области slp2 и маркер pyr4, удаляли из вектора путем расщепления PmeI, и соответствующий фрагмент очищали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen). Приблизительно 5 мкг кассеты для делеции использовали для трансформации продуцирующего MAB01 штамма M564 (Δpep1Δtsp1Δslp1ΔgaplΔgap2Δpep4Δpep3, pyr4-). Получение протопластов и трансформацию проводили, по существу как описано в примере 1 для штаммов M181 и M195, с использованием селекции на основе pyr4.
Трансформанты отбирали в качестве первых штриховых культур. Растущие штриховые культуры подвергали скринингу способом ПЦР (с использованием праймеров, приведенных в таблице 22.3) в отношении правильного встраивания и утраты ORF slp2. Клоны, дающие ожидаемые сигналы, очищали до клонов единичных клеток и подвергали повторному скринингу способом ПЦР с использованием праймеров, приведенных в таблице 22.3. Правильный клон был обозначен как штамм M646.
Ферментация штаммов M507, M665, M666, M667 и M646
Штамм M507 культивировали в ферментере для культивирования серии FTR104 в тех же условиях, что и M665, M666, M667 и M646. M646 представлял собой штамм с делецией slp2. M665, M666 и M667 представляли собой штаммы с подавлением РНК-i. Культивирование FTR104 проводили в минимальной среде для Trichoderma (TrMM) с 20 г/л дрожжевого экстракта, 40 г/л целлюлозы, 80 г/л целлобиозы и 40 г/л сорбозы при pH 5,5. Температуру сдвигали от 28°C до 22°C после 48 часов. Культуры выращивали в течение 6 суток. Минимальная среда Trichoderma содержит 5 г/л сульфата аммония, 5 г/л гидрофосфата калия, 1 мл/л микроэлементов, 4,1 мл 1 M хлорида кальция на л, и 2,4 мл 1 M сульфата магния на л среды.
Общие концентрации антитела определяли на 3-6 сутки. На 6 сутки штамм M667 достигал 3,81 г/л, см. таблицу 22.8. После 5 суток экспрессия антитела снижалась в культурах M507, M665 и M666. На 6 сутки штамм M507 продуцировал 2,2 г/л, M665 достигал 2,7 г/л, и M666 продуцировал 2,8 г/л. Таким образом, штаммы с небольшими последовательностями-мишенями РНК-i продуцировали немного больше антитела, чем M507, что указывает на то, что подавление эффективно в этих штаммах. Штамм M646 с делецией slp2 рос медленнее, чем другие штаммы. Штамм с делецией slp2 продуцировал немного больше 2 г/л на 6 сутки.
Ферментация штаммов M507, M665, M666 и M667
217.12G (M665), 218.25F (M666) и 263.110F (M667) выращивали в ферментерах объемом 1 л с 30 г/л глюкозы, 60 г/л лактозы, 20 г/л WSG, 20 г/л SGE с подпиткой лактозой при pH 5,5, начиная при 28°C, и позднее переключая культуру на 22°C. Экспрессию тяжелой и легкой цепей MAB01 анализировали с использованием иммуноблоттинга из образцов супернатанта, собираемых каждые сутки в процессе культивирования. Супернатанты разбавляли в цитратном буфере, pH 5,5, так, чтобы на лунку можно было наносить 0,1 мкл. Добавляли LSB+BME и их нагревали вместе в течение 5 минут при 95°C. Образцы наносили на 4-15% гель Criterion SDS PAGE. Белки переносили на нитроцеллюлозу с помощью устройства для блоттинга Criterion при 100 вольт в течение 30 минут. Нитроцеллюлозную мембрану блокировали 5% молоком в TBST в течение 1 часа. Обнаружение тяжелой цепи проводили с помощью конъюгированного с AP антитела против тяжелой цепи (Sigma #A3188) и против легкой цепи (Sigma #A3813), разбавленных 1:10000 в TBST. После инкубации в течение 1 часа с антителом для обнаружения блот промывали TBST, и мембрану проявляли субстратом AP (Promega).
Общую экспрессию антитела измеряли после очистки с белком G и величины представлены в таблице 22.4, вместе с результатами для двух контрольных штаммов. Штамм M507 культивировали с ингибитором SBTI и без него в тех же условиях. Уровни экспрессии MAB01 в штамме M667 были более высокими, чем уровни экспрессии, измеренные в родительском штамме M507. На 9 сутки, например, уровень экспрессии был в два раза более высоким для M667. Уровни экспрессии, выявленные для M667, были сходны с уровнями в случае культивирования, проведенного при добавлении SBTI. Штаммы M665 и M666 продуцировали уровни, немного более низкие или сходные с контролем. Существовало очевидное увеличение в 2 раза экспрессии антитела по сравнению со стандартным штаммом M507.
Протеазную активность в культурах, приведенных в таблице 22.4, измеряли для определения того, как РНК-i влияла на общую протеазную активность. Показатели протеазной активности с казеином в качестве субстрата можно видеть в таблице 22.9. Измеряли общие концентрации белка в образцах супернатанта. Общую концентрацию белков для всех образцов нормализовывали в натрий-цитратном буфере, pH 5,5, до 0,625 мг/мл для всех суток культивирования.
100 мкл всех разбавленных супернатантов добавляли в 96-луночный планшет. Приготавливали три повторяющихся лунки на образец. Добавляли 100 мкл разбавленного исходного раствора казеина FL (10 мкг/мл), приготовленного в натрий-цитратном буфере, pH 5,5. Исходный раствор казеина из флакона разбавляли до 1000 мкг/мл в 200 мкл PBS. Для каждого образца включали контроль фона с 100 мкл разбавленного супернатанта и 100 мкл натрий-цитратного буфера, pH 5,5. Инкубированные планшеты, содержавшие супернатанты и субстрат, помещали в пластиковый мешок при 37°C. Флуоресценцию измеряли в планшетах после инкубации в течение 4 часов. Считывание проводили на флуоресцентном устройстве для считывания планшетов с использованием возбуждения при 485 нм и испускания при 530 нм.
Протеазная активность в супернатанте штамма M665 была наиболее низкой из всех. На протяжении культивирования она была практически в два раза ниже M507. Активность вектора со шпилькой в M667 также была низкой, однако она начала снижаться после 5 суток и была наиболее низкой на 10 сутки. Это соответствовало времени, когда продукция антитела для штамма M667 была наиболее высокой, на 10 сутки. В конце культивирования как культуральный супернатант M665, так и культуральный супернатант M667 имели половину протеазной активности по сравнению с контролем M507. Когда культуру M507 дополняли ингибитором протеаз SBTI, протеазная активность также падала с 6 суток до 8 суток и оставалась более низкой, чем у штамма M507. Низкая протеазная активность в конце культивирования объясняет, почему штамм M667 продуцировал в два раза больше антитела по сравнению со штаммом M507.




 .
.

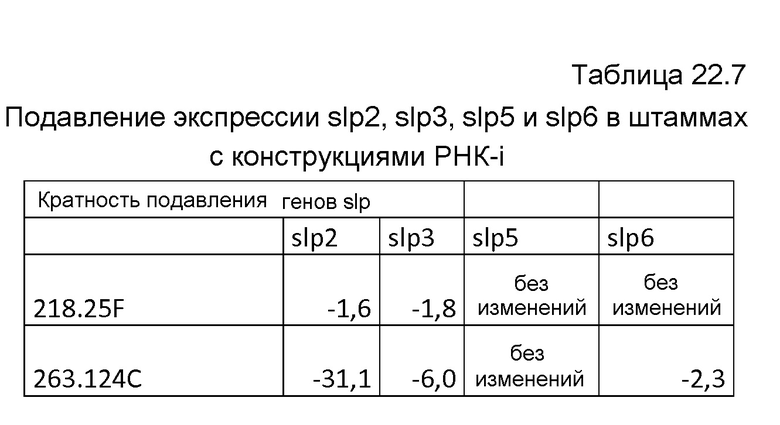
ПРИМЕР 23 - ПОЛУЧЕНИЕ ЭКСПРЕССИРУЮЩИХ ФРАГМЕНТЫ АНТИТЕЛ ШТАММОВ TRICHODERMA REESEI
Получали семь штаммов Trichoderma reesei для экспрессии фрагментов антител (Fab, мультимерные однодоменные антитела (sd-Ab) и scFV) для различных генетических фонов в отношении делеции протеаз, приведенных в таблице 23. Архитектура генетических экспрессирующих кассет, использованных для этой цели, была основана на регуляторных элементах (промотор и терминатор) гена целлобиогидролазы I (cbh1). Каталитический домен белка CBHI модифицировали для удаления последовательностей интронов и использовали в качестве партнера по слиянию для увеличения экспрессии и секреции фрагментов антител. Мотив для распознавания протеазы Kex2 встраивали между партнерами по слиянию для облегчения косекреторного освобождения фрагментов антител от белка-носителя CBHI. Экспрессирующие кассеты были фланкированы гомологичными областями для обеспечения направленного встраивания в локус cbh1 Trichoderma reesei. Всю конструкцию хранили в клонирующем векторе.
Для получения фланкированных экспрессирующих кассет для трансформации соответствующие фрагменты высвобождали из их соответствующих остовов векторов с помощью рестрикционного расщепления PmeI и очищали с использованием набора illustra GFX PCR DNA and Gel Band Purification Kit (GE Healthcare).
Как указано в таблице 23, штаммы T. reesei с делецией протеаз трансформировали очищенными экспрессирующими кассетами с использованием опосредуемой PEG трансформации протопластов. Трансформанты отбирали по устойчивости к гигромицину B или прототрофии ацетамидазы путем посева их на среду, содержавшую гигромицин B в качестве селективного агента или ацетамида в качестве единственного источника азота, соответственно. Каждый из вплоть до 48 трансформантов подвергали скринингу способом ПЦР в отношении гомологичного встраивания экспрессирующей кассеты в локус cbh1 с использованием прямого праймера снаружи фрагмента 5'-фланкирующей области в конструкции и обратного праймера внутри модифицированного каталитического домена CBHI (5'-встраивание) в качестве прямого праймера внутри селективного маркера для гигромицина B или ацетамидазы, соответственно, и обратного праймера вне фрагмента 3'-фланкирующей области (3'-встраивание). После каждой трансформации от пяти до семи независимых трансформантов, для которых скрининг способом ПЦР показал правильное встраивание конструкции в локус cbh1, отбирали для очистки единичных спор с получением одноядерных клонов. Надлежащее встраивание кассеты для разрыва повторно подтверждали способом ПЦР с использованием тех же комбинаций праймеров, которые описаны выше, и отсутствие исходного локуса CBHI подтверждали с использованием комбинации праймеров, нацеленных на открытую рамку считывания cbh1. Правильное встраивание кассеты для разрыва, кроме того, подтверждали для всех клонов с использованием саузерн-гибридизации. Геномную ДНК одноядерных клонов, а также исходного штамма, независимо расщепляли комбинациями двух различных ферментов рестрикции и исследовали в отношении 5'- и 3'-фланкирущих областей гена cbh1 для подтверждения модификации локуса cbh1, как и ожидалось.
Анализ экспрессии и титра фрагментов антител (Fab, однодоменные антитела и scFV)
Экспрессии фрагментов антител способствовали с помощью промотора целлобиогидролазы I. Штаммы выращивали периодической ферментацией в течение 7 суток в среде, содержащей 2% дрожжевой экстракт, 4% целлюлозу, 8% целлобиозу, 4% сорбозу, 5 г/л KH2PO4 и 5 г/л (NH4)2SO4. pH культуры контролировали на уровне pH 5,5 (корректируемом с помощью NH4OH) и температуру постоянно поддерживали на уровне 28°C. Ферментацию проводили в 4 параллельных стеклянных реакторах на 2 л (DASGIP) с объемом культуры 1 л. В ходе ферментации проводили взятие образцов культурального супернатанта и их хранили при -20°C. Образцы собирали каждые сутки в течение всего культивирования, и уровни продукции анализировали аффинной жидкостной хроматографией для всех молекул. Для каждого фрагмента антитела максимальный титр, ID штамма и генетический фон по делеции протеаз указаны в таблице 23.
Определение титра
Mab и sdAb
Концентрации Mab и sdAb количественно определяли с помощью ВЭЖХ - хроматографии с белком A, которая основана на аффинной хроматографии с УФ-детекцией. Fc-домен иммуноглобулинов человека G-класса (подтип-класс: IgG1, IgG2, IgG4, за исключением IgG3) специфично связывается с белком A, который ковалентно связан со стационарной фазой. Аффинность связывания белка A с Fc-доменом зависит от pH. После связывания при pH 7,5 моноклональное антитело элюировали в кислых условиях при pH 2,0 и его обнаружение проводили при 280 нм.
Fab
Концентрации Fab количественно определяли с помощью ВЭЖХ - хроматографиии против лямбда-цепи, которая основана на аффинной хроматографии с УФ-детекцией. Лямбда-цепь Fab-фрагментов человека связывается специфично с происходящим из верблюдов лигандом лямбда-цепи, который ковалентно связан со стационарной фазой. После связывания при pH 7,5, моноклональное антитело элюировали в кислых условиях при pH 1,4 и его обнаружение проводили при 280 нм.
scFV2
Концентрации scFV2 количественно определяли с помощью очистки HiTrap с белком L с использованием системы AktaTM avant и последующей УФ-детекции. Часть легкой цепи каппа scFV2 специфично связывается с лигандом в виде белка L, который ковалентно связан со стационарной фазой. После связывания при pH 7,2, scFV2 элюировали в кислых условиях с помощью 0,1 мМ HCl и его обнаруживали при 280 нм.
scFV1-His
Концентрации scFV1-His количественно определяли с помощью очистки HisTrap HP с использованием системы AktaTM avant и последующей UV-детекции. Гистидиновая метка белка специфично связывается с Ni-сефарозой. После связывания белок элюировали с использованием 500 мМ имидазола и его подвергали обнаружению при 280 нм.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВАЯ ПРОТЕАЗА ГРИБОВ И ЕЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2566549C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ФЕРМЕНТ ПРОТЕАЗА И ЕГО ПРИМЕНЕНИЯ | 2012 |
|
RU2611043C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |
| МИЦЕЛИАЛЬНЫЕ ГРИБЫ, ИМЕЮЩИЕ ФЕНОТИП ИЗМЕНЕННОЙ ВЯЗКОСТИ | 2011 |
|
RU2608639C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
| МОЮЩАЯ КОМПОЗИЦИЯ | 2015 |
|
RU2737535C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |





















































































































































































































































































































































































































































































































































































































































































































































































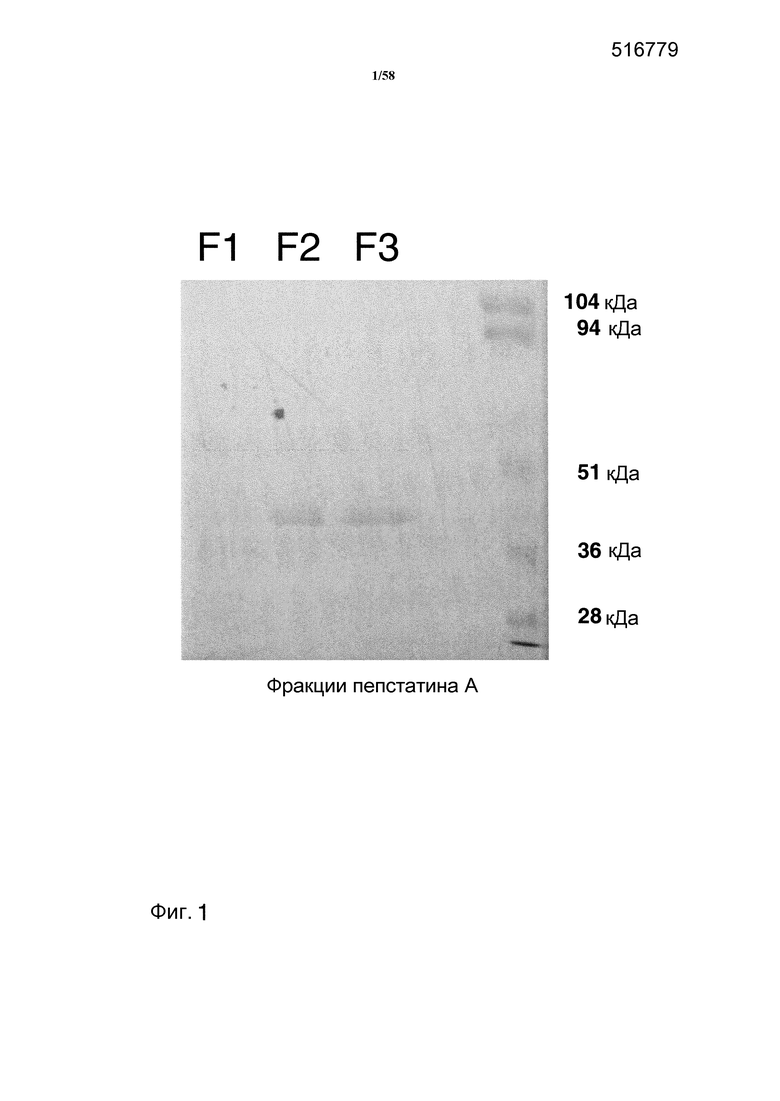


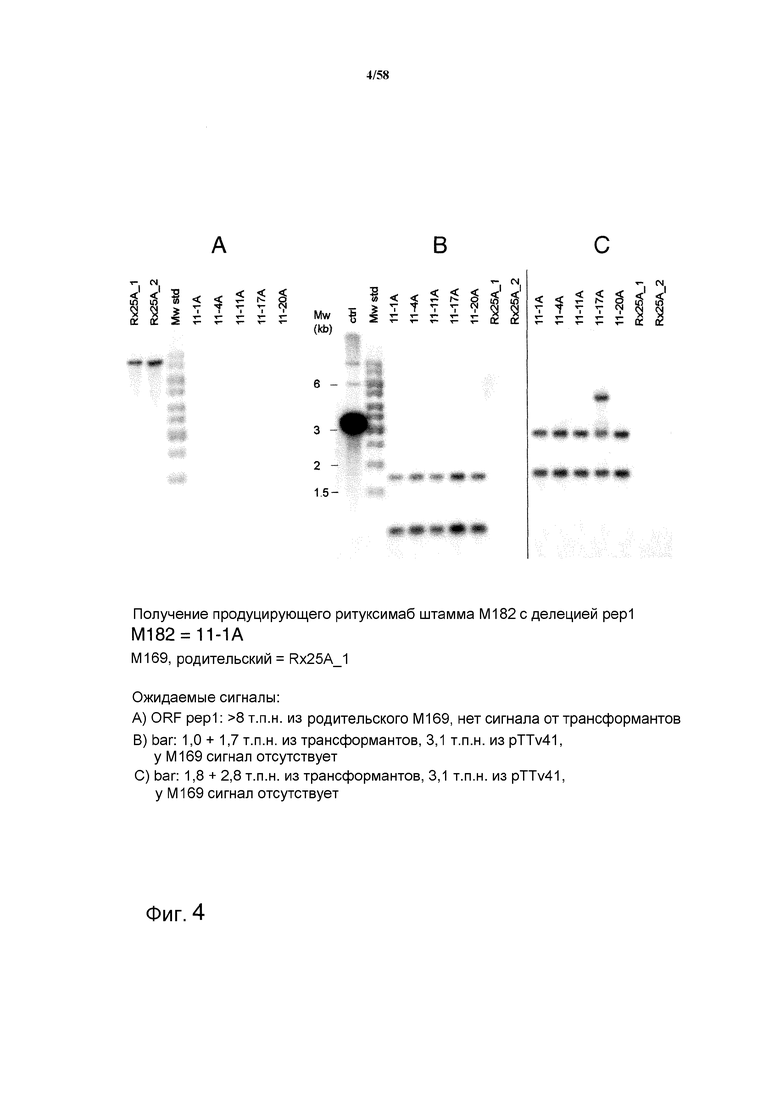
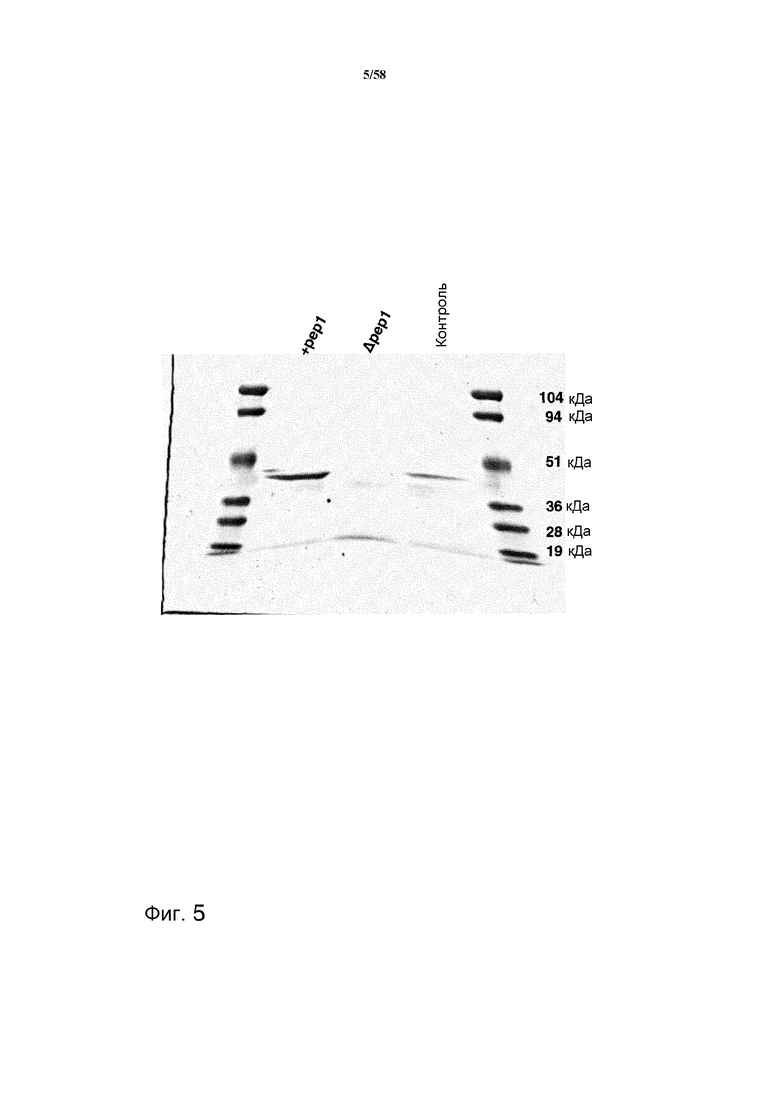


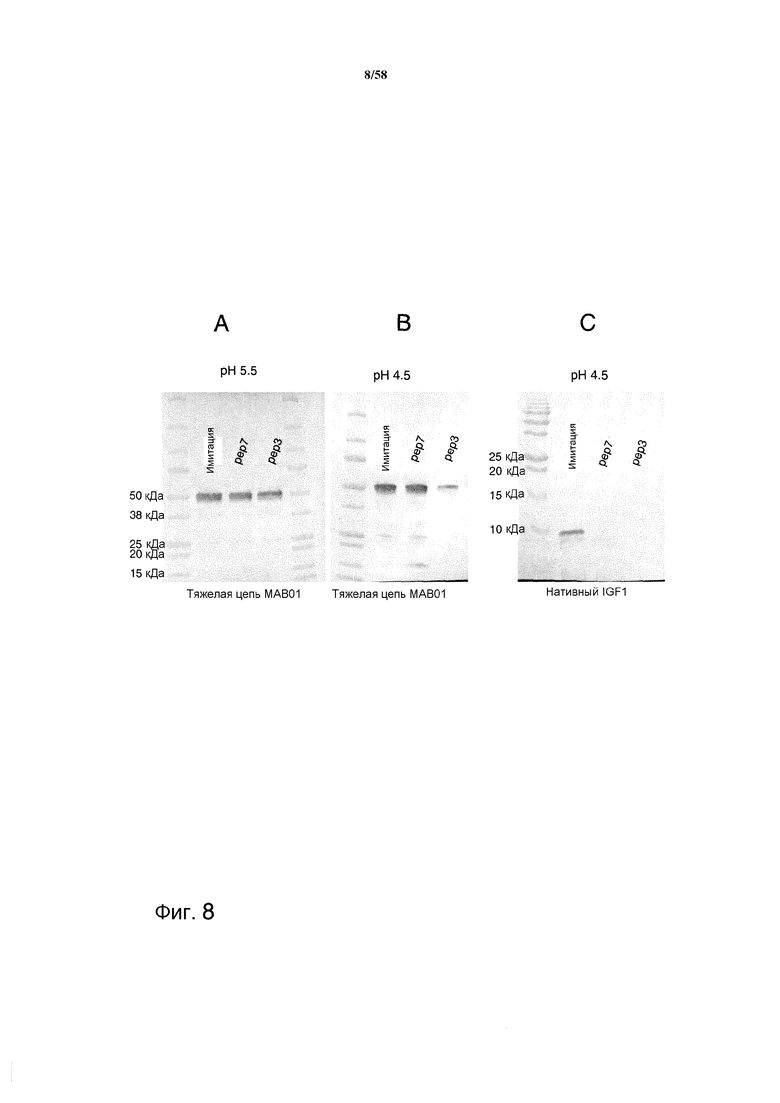
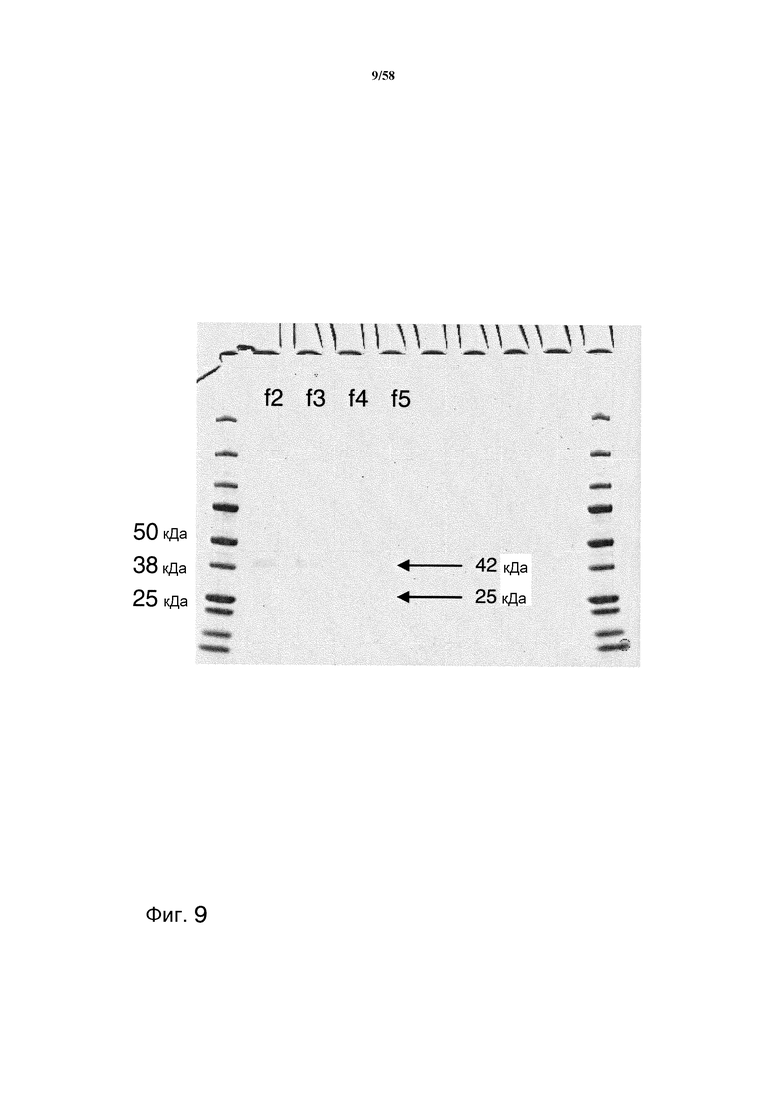

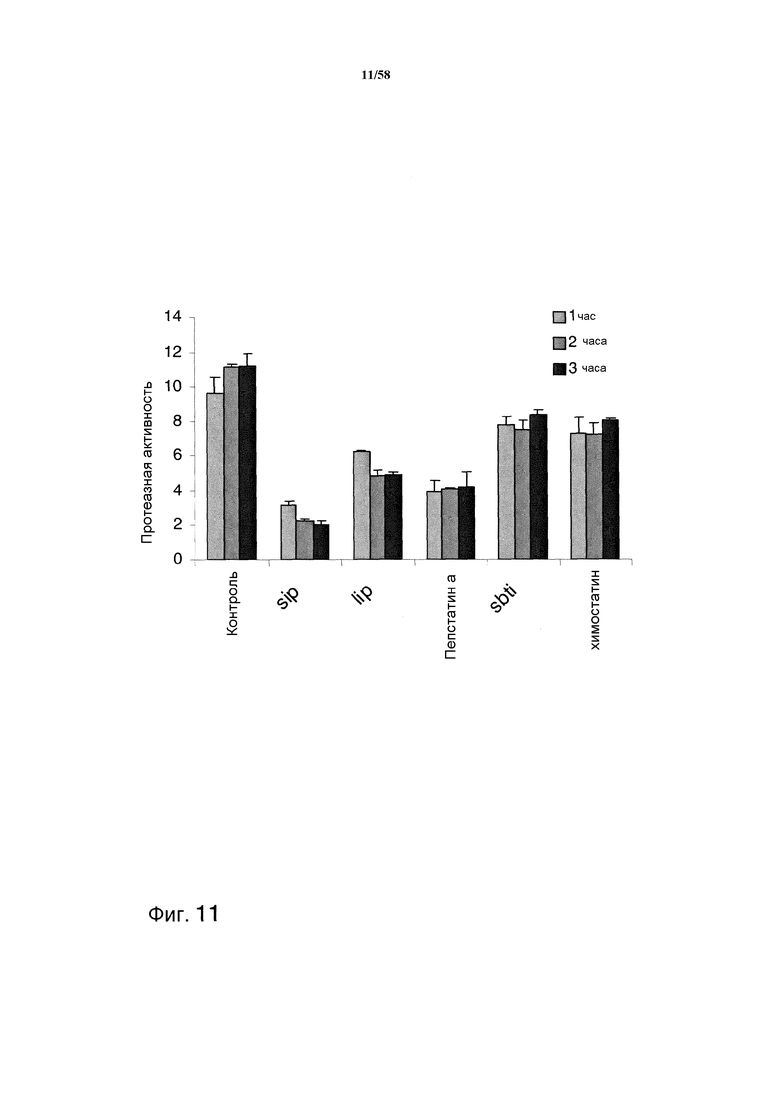
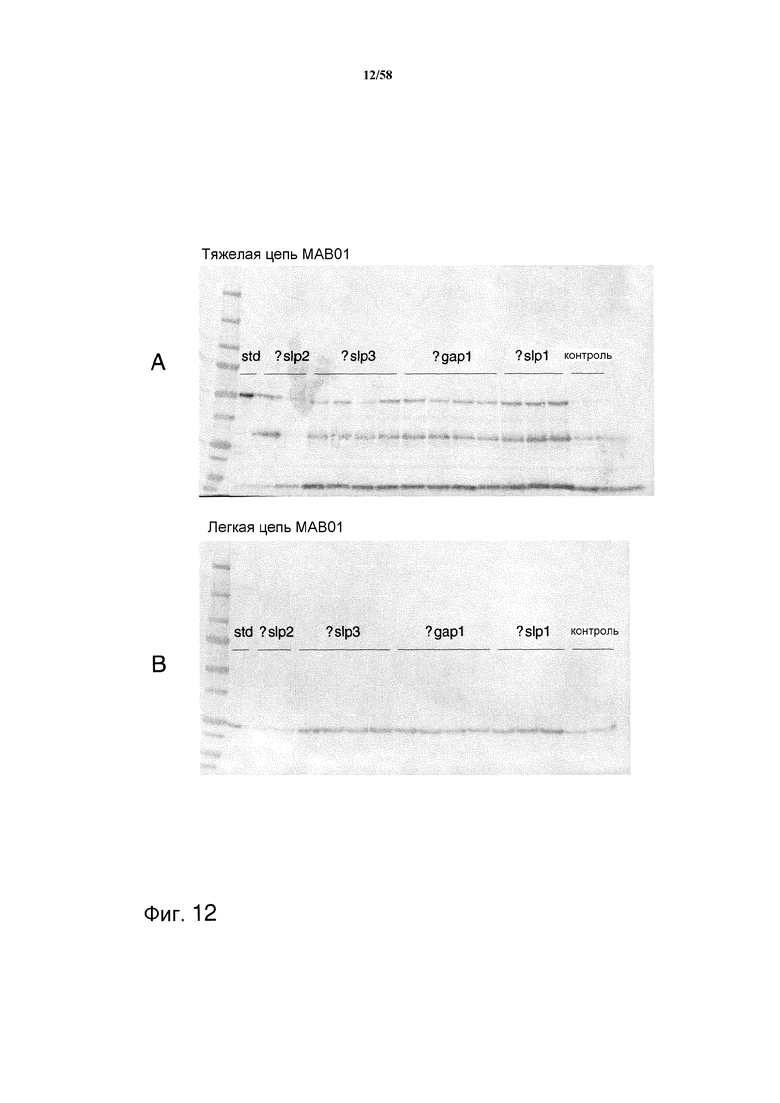
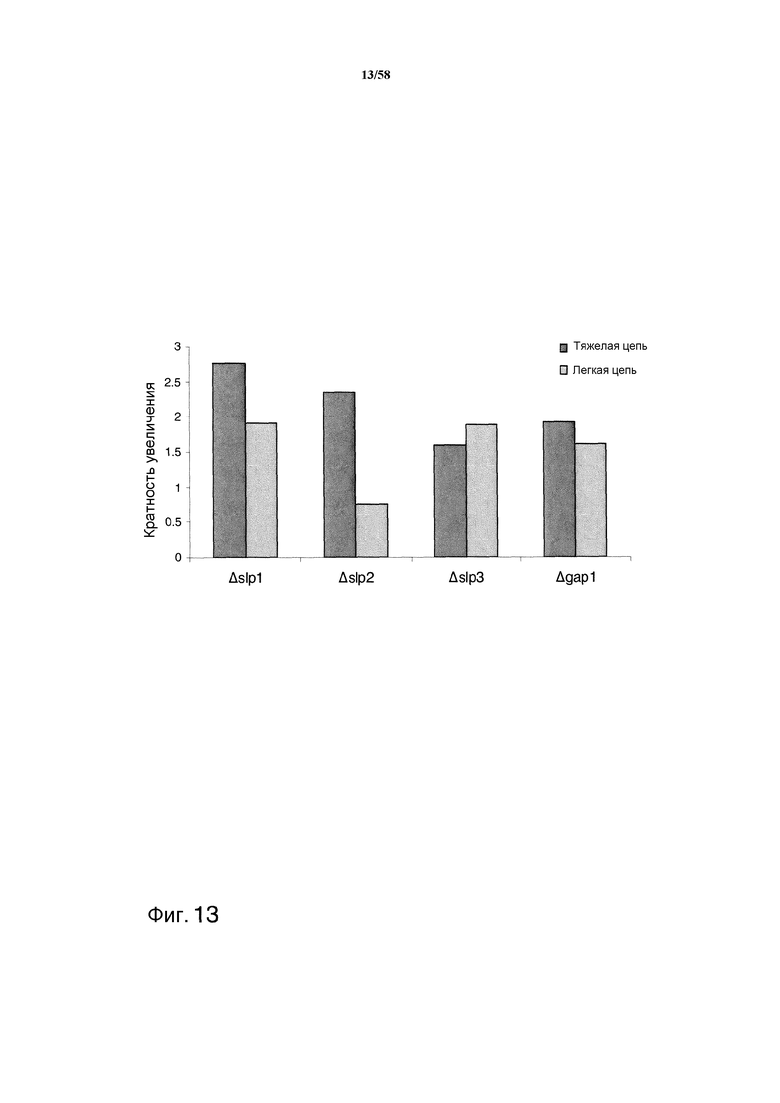
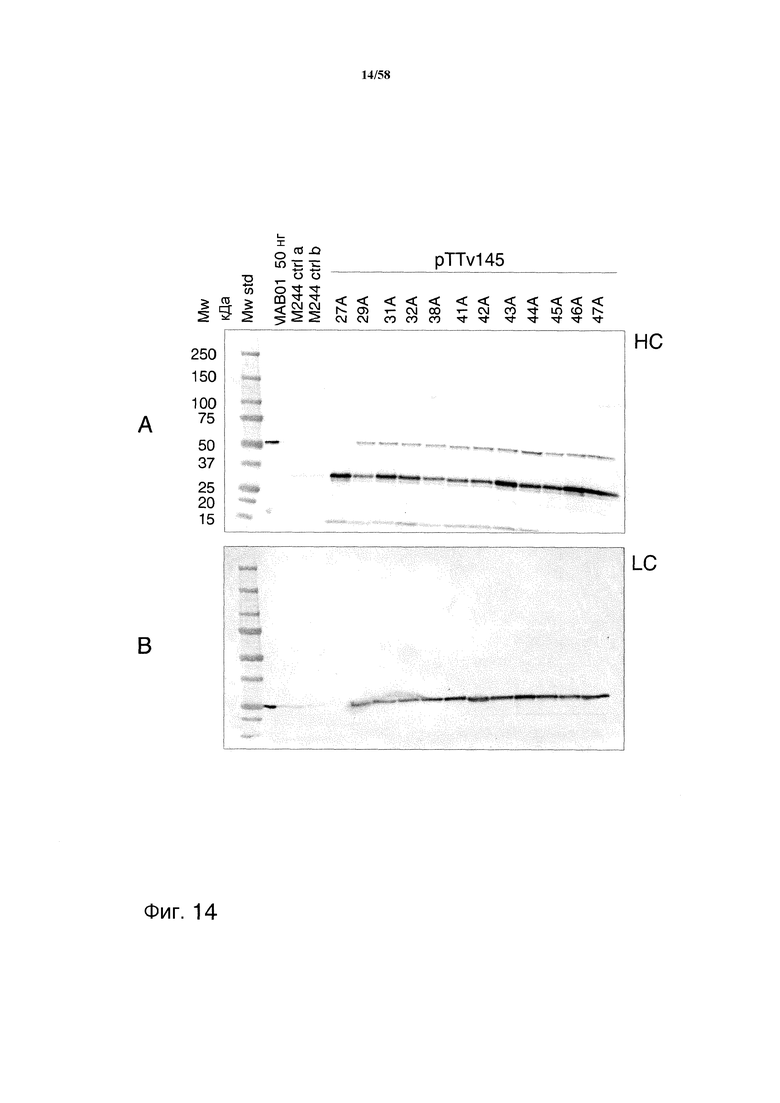
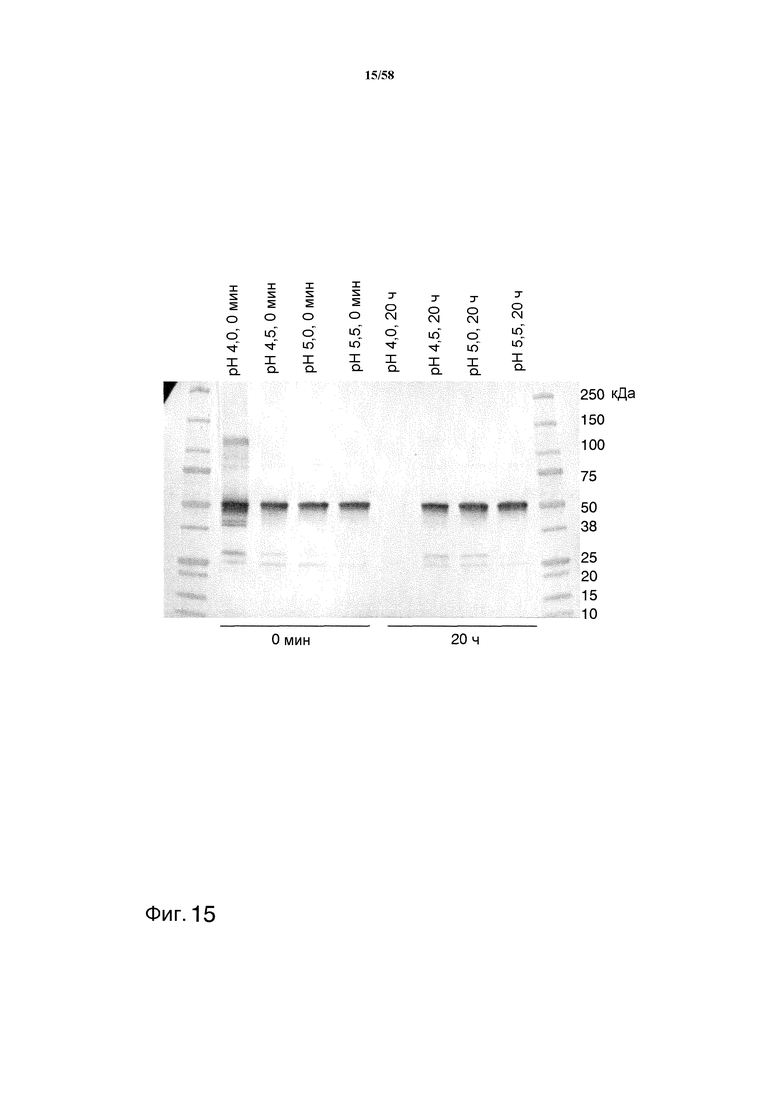

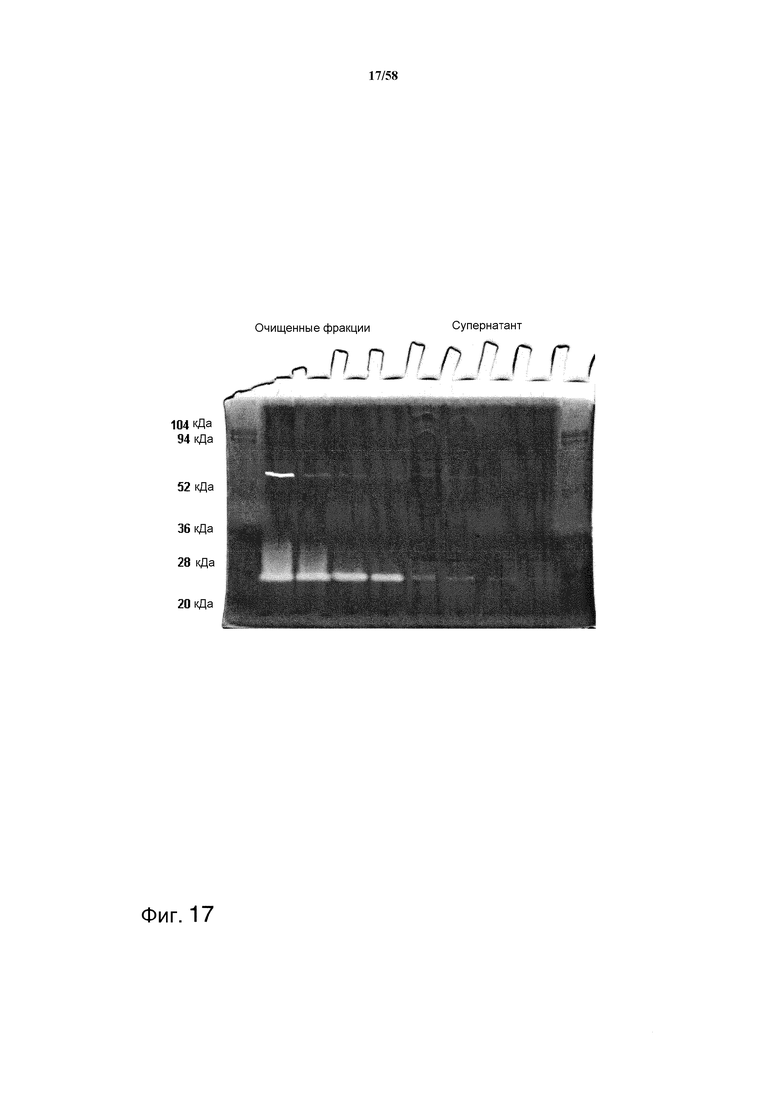
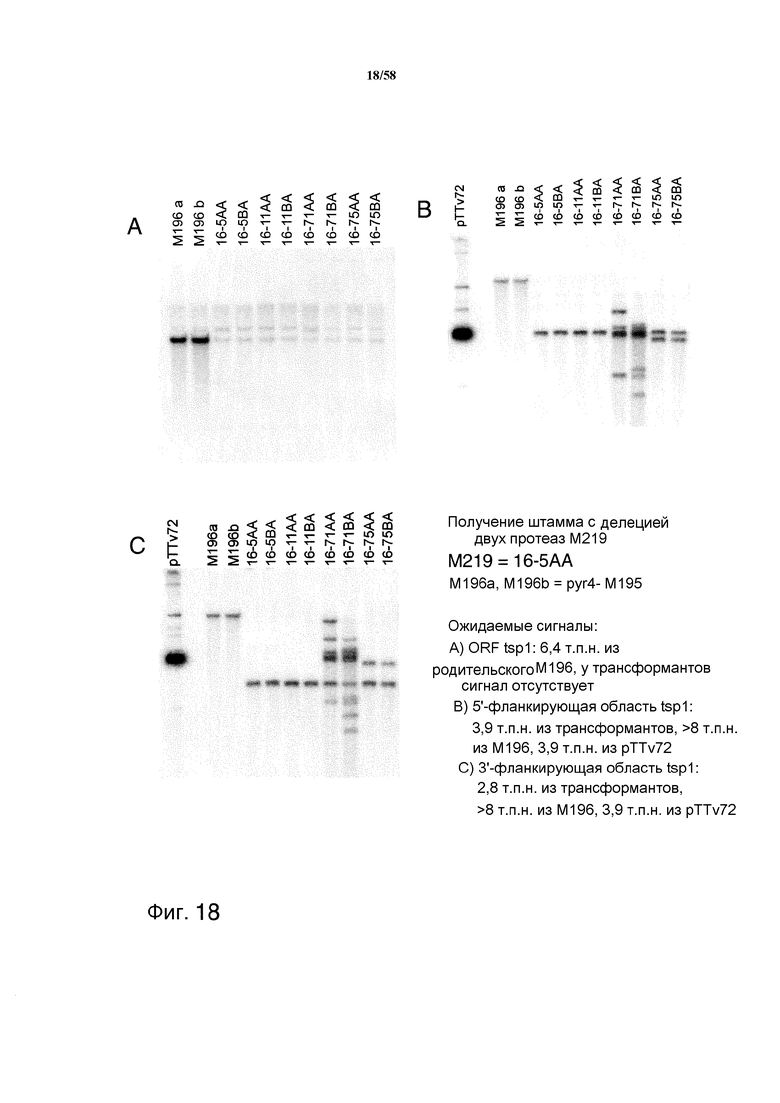

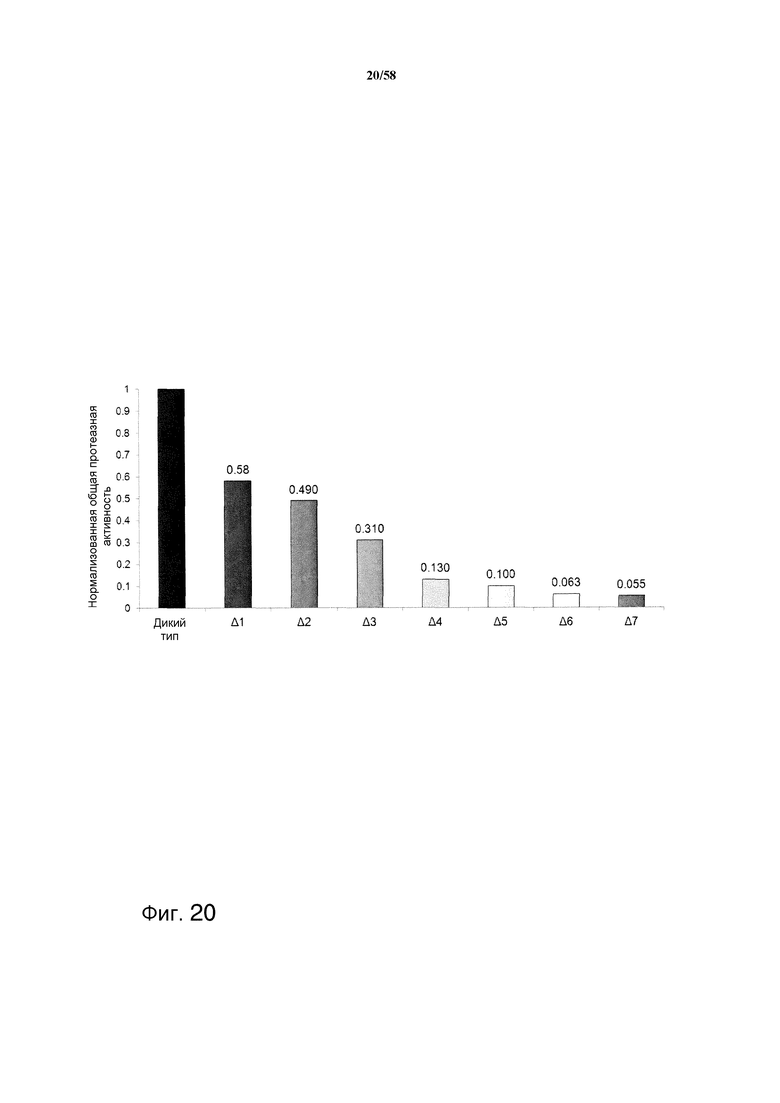
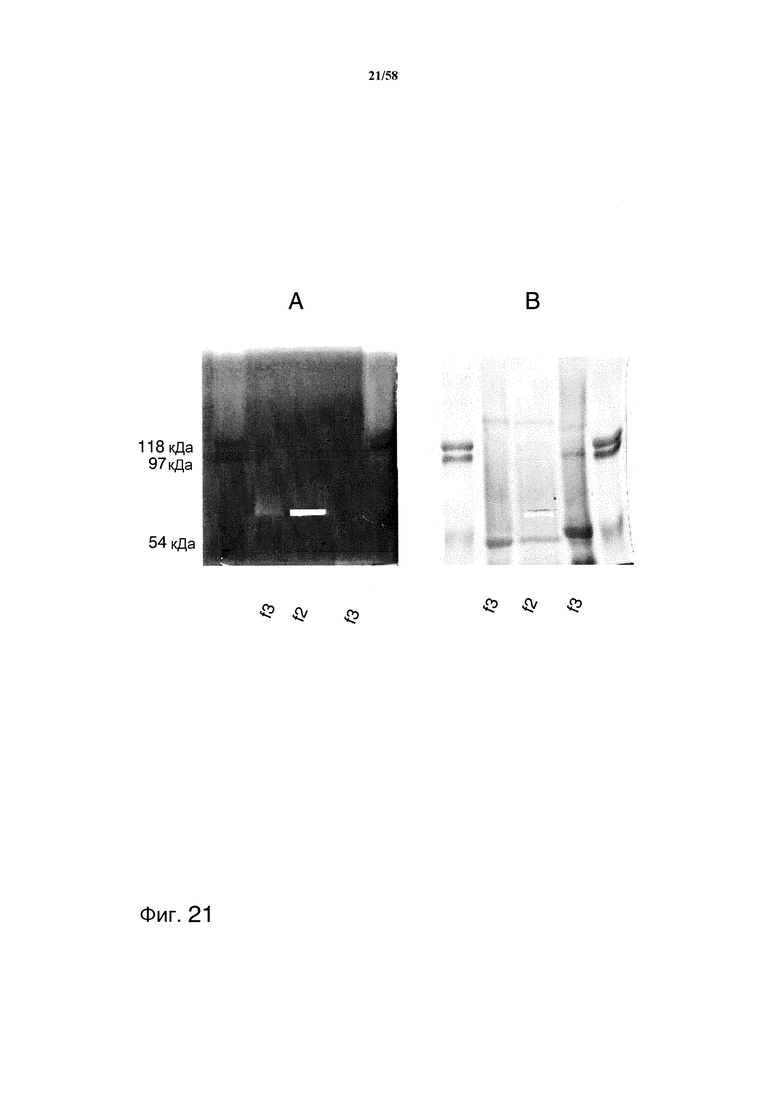
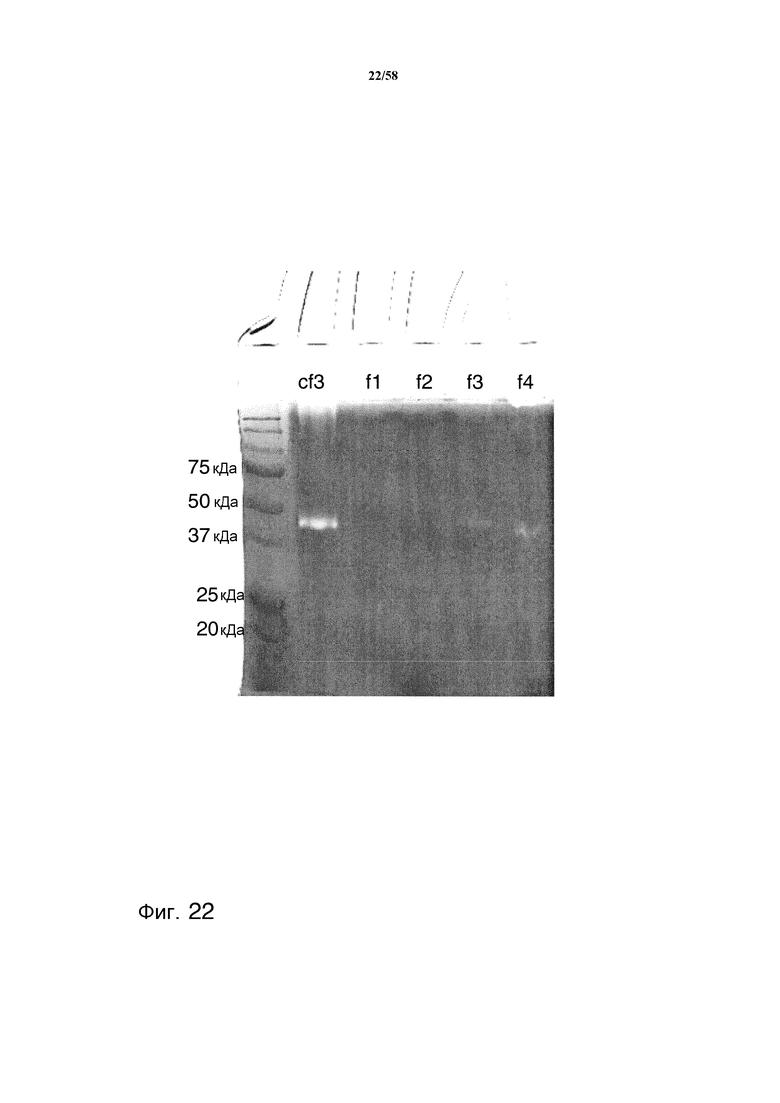
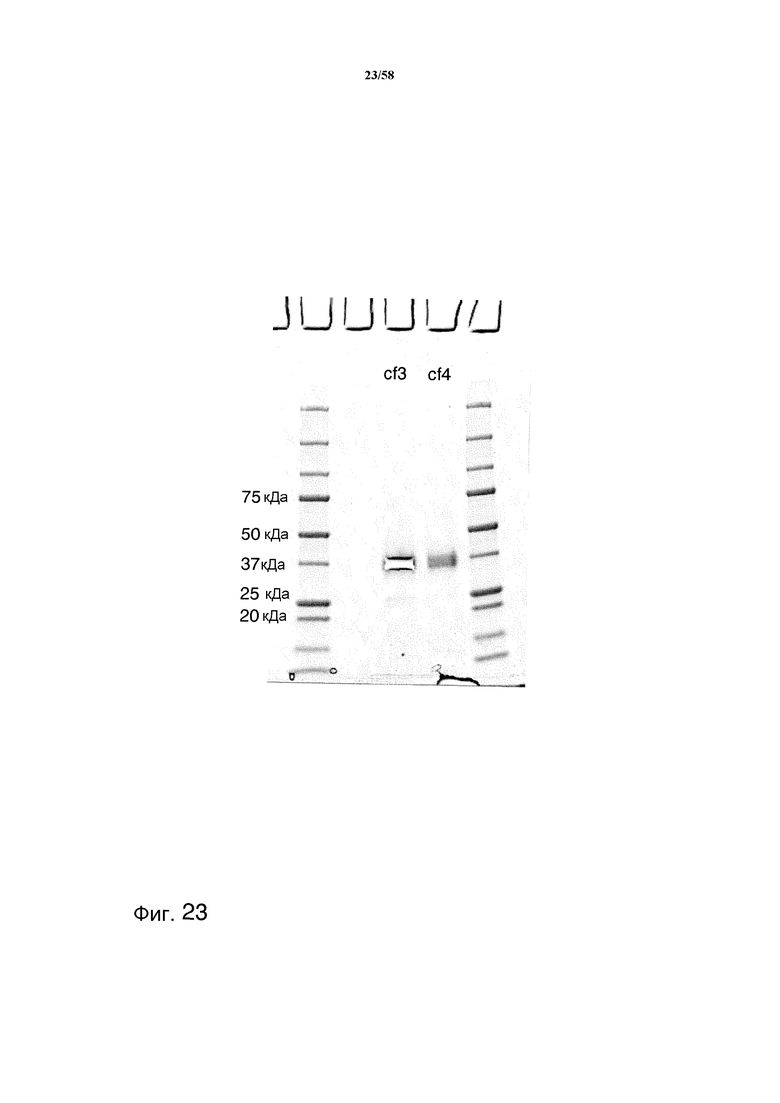
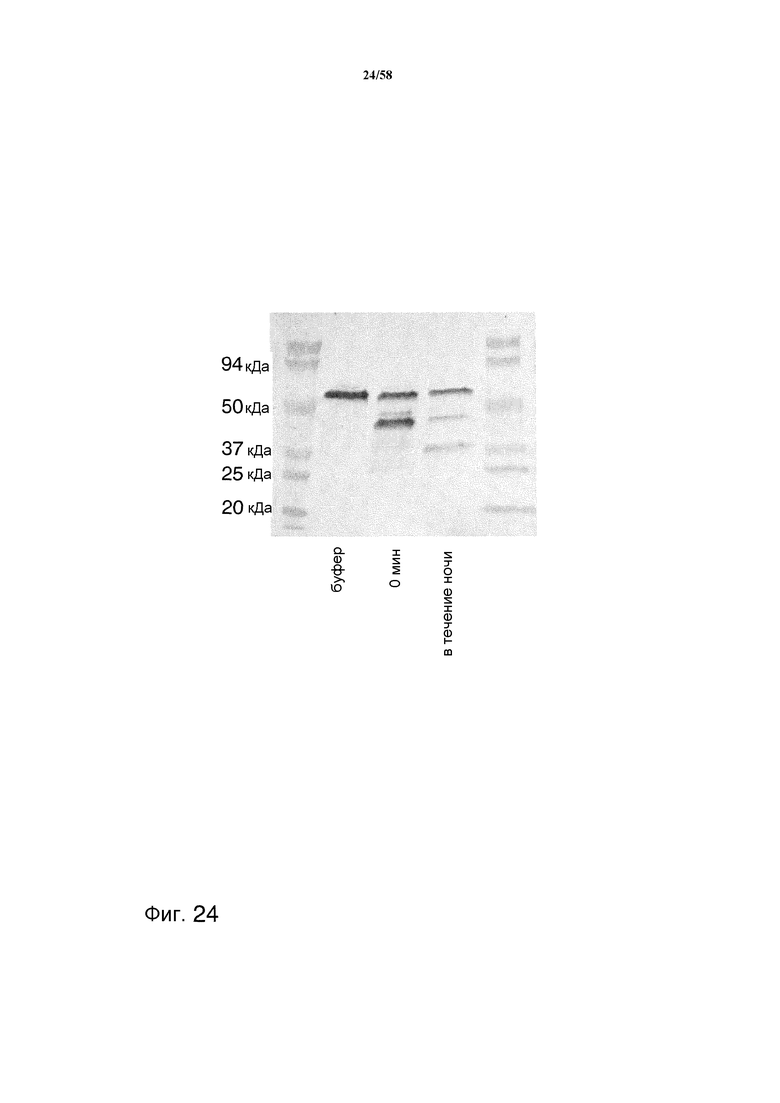
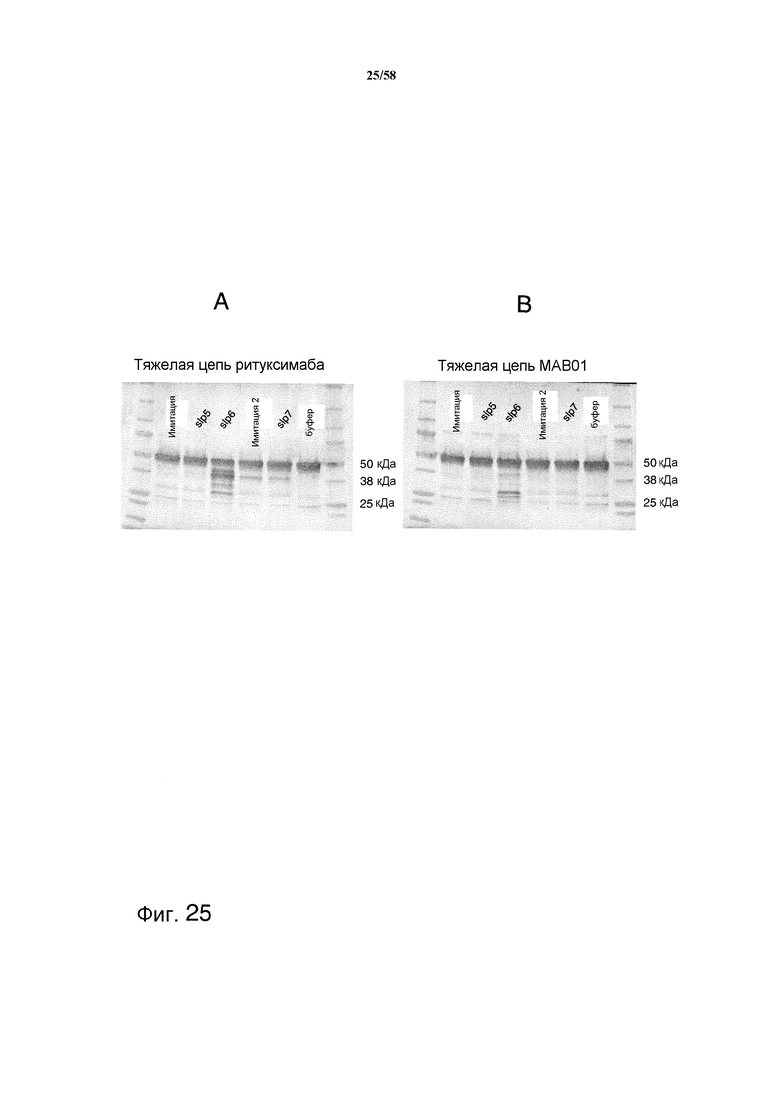


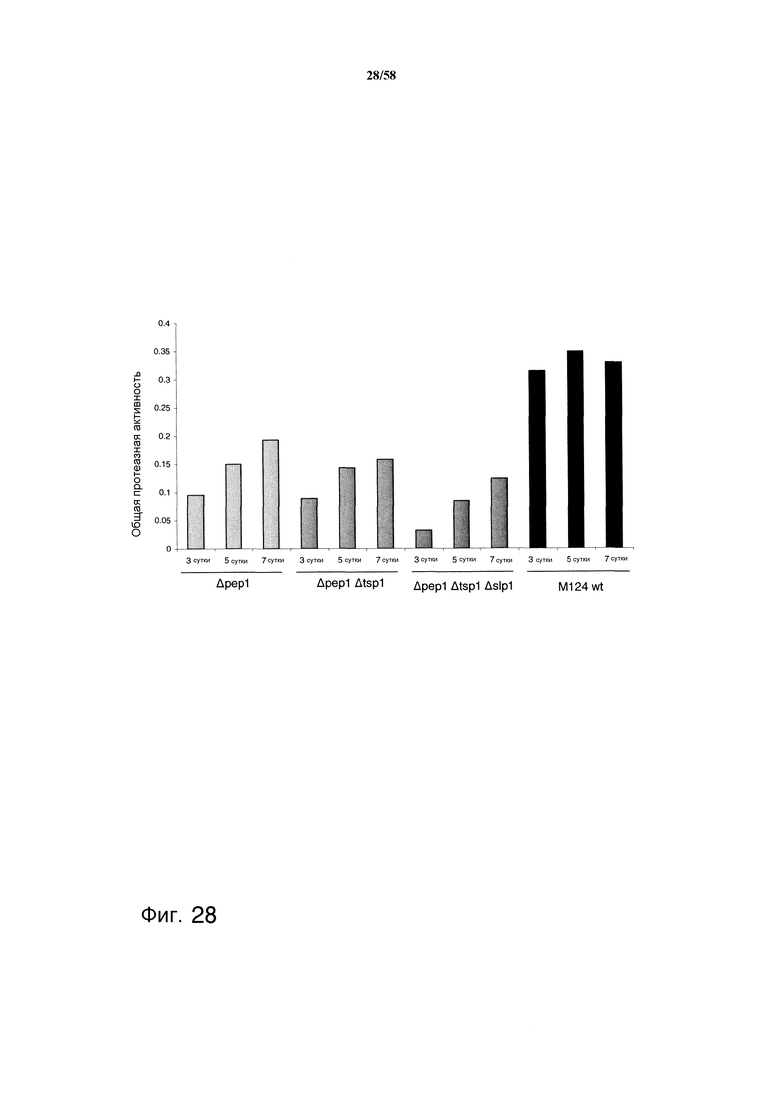

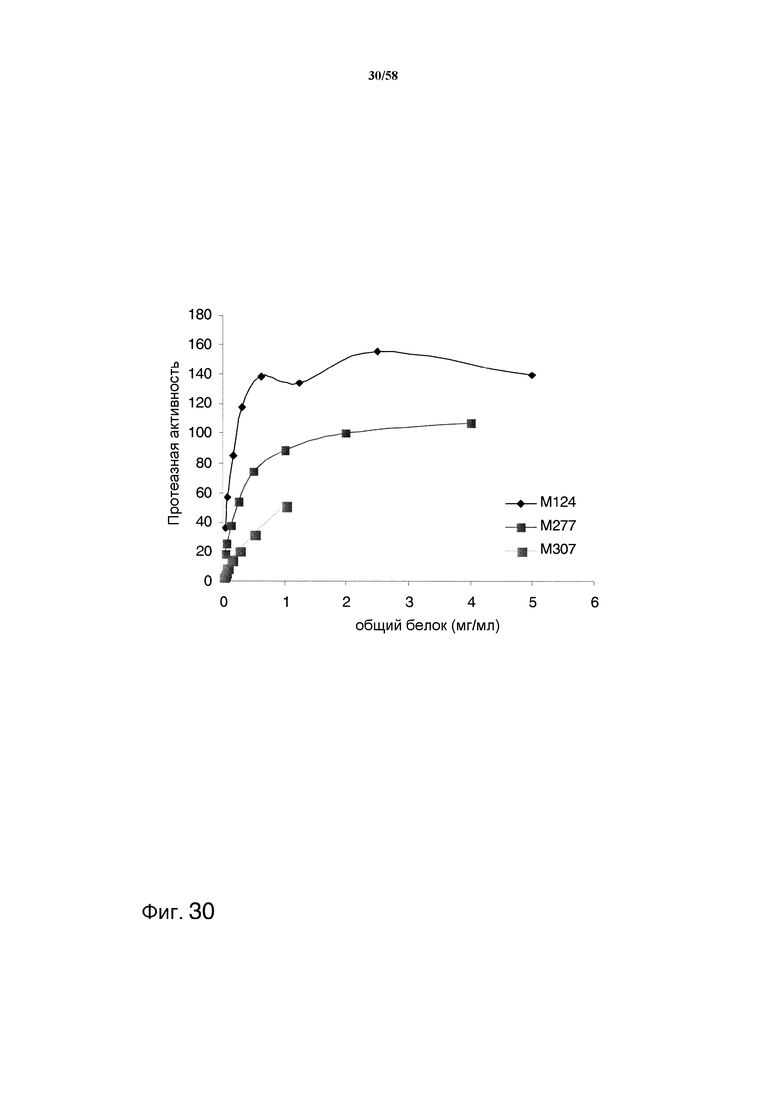

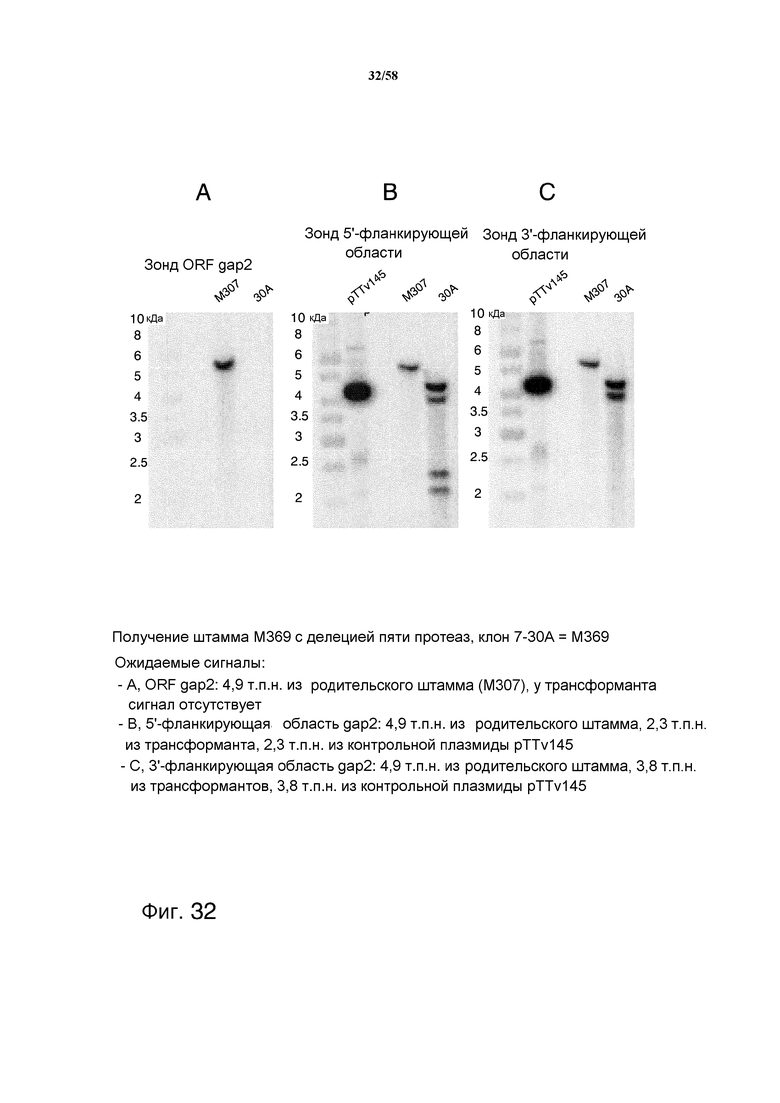


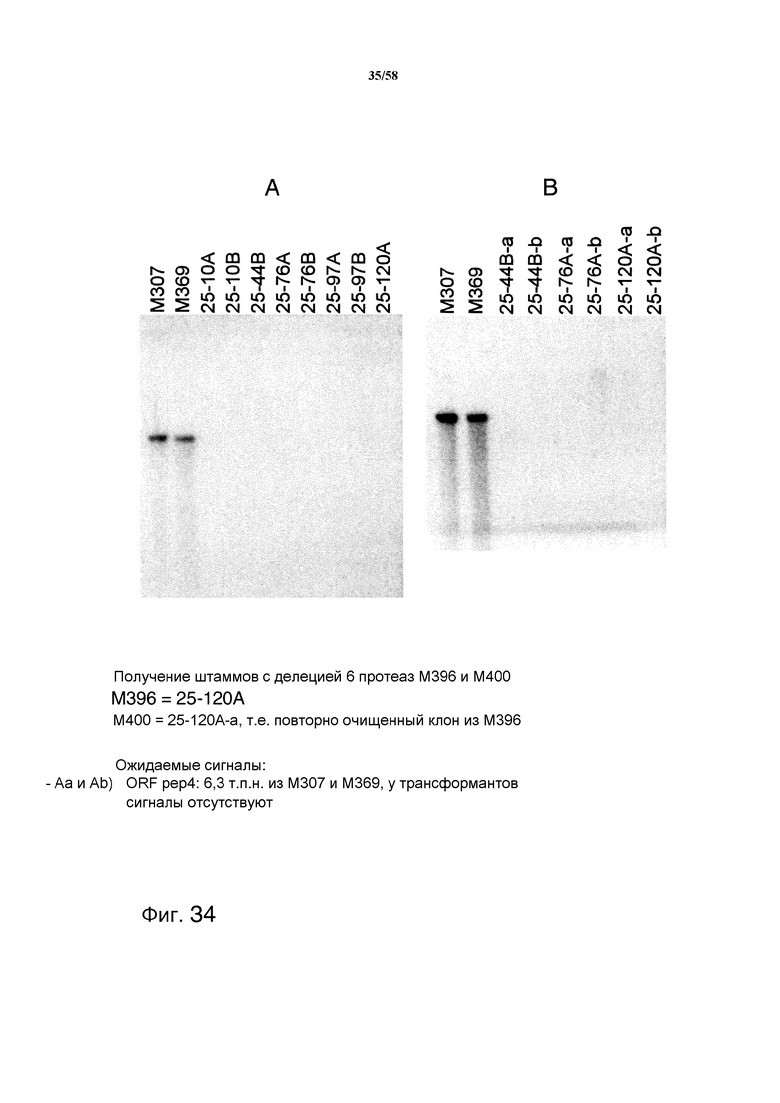
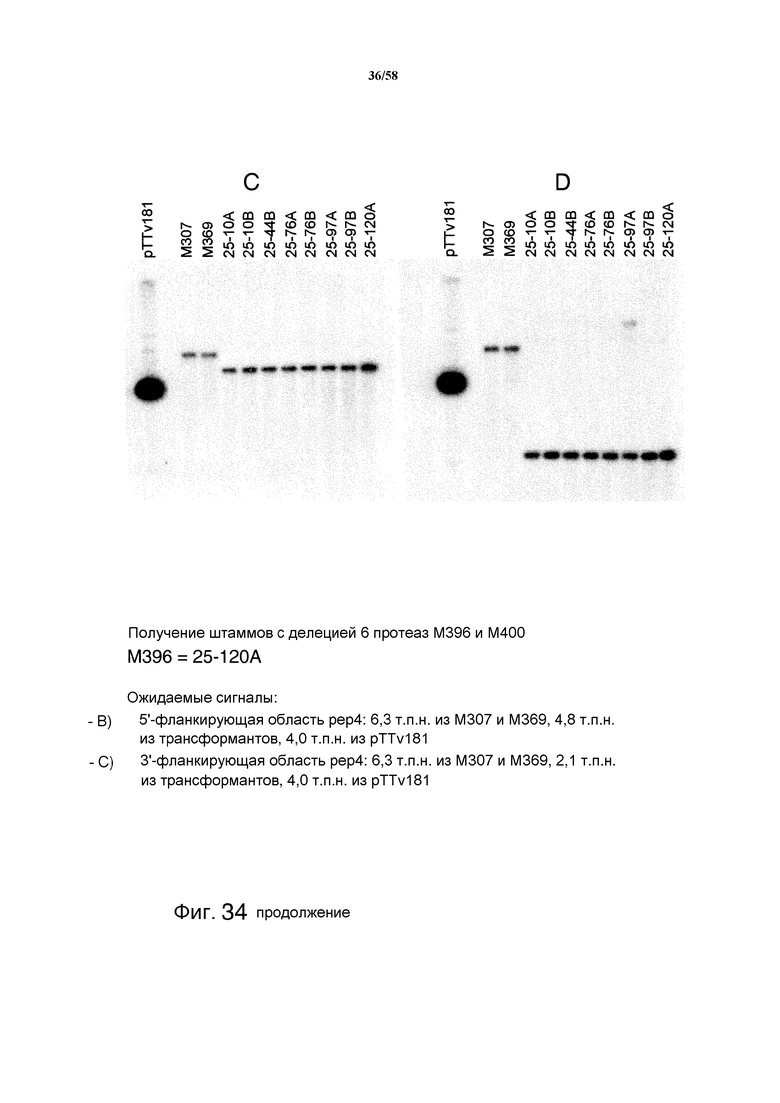


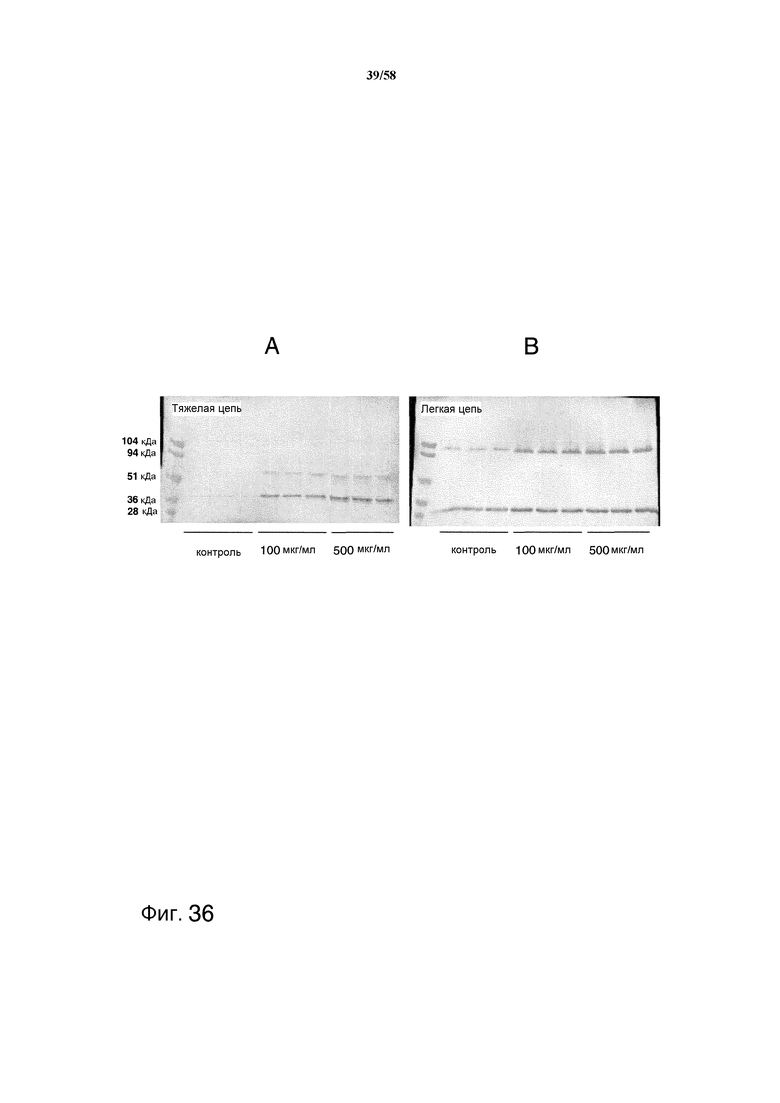
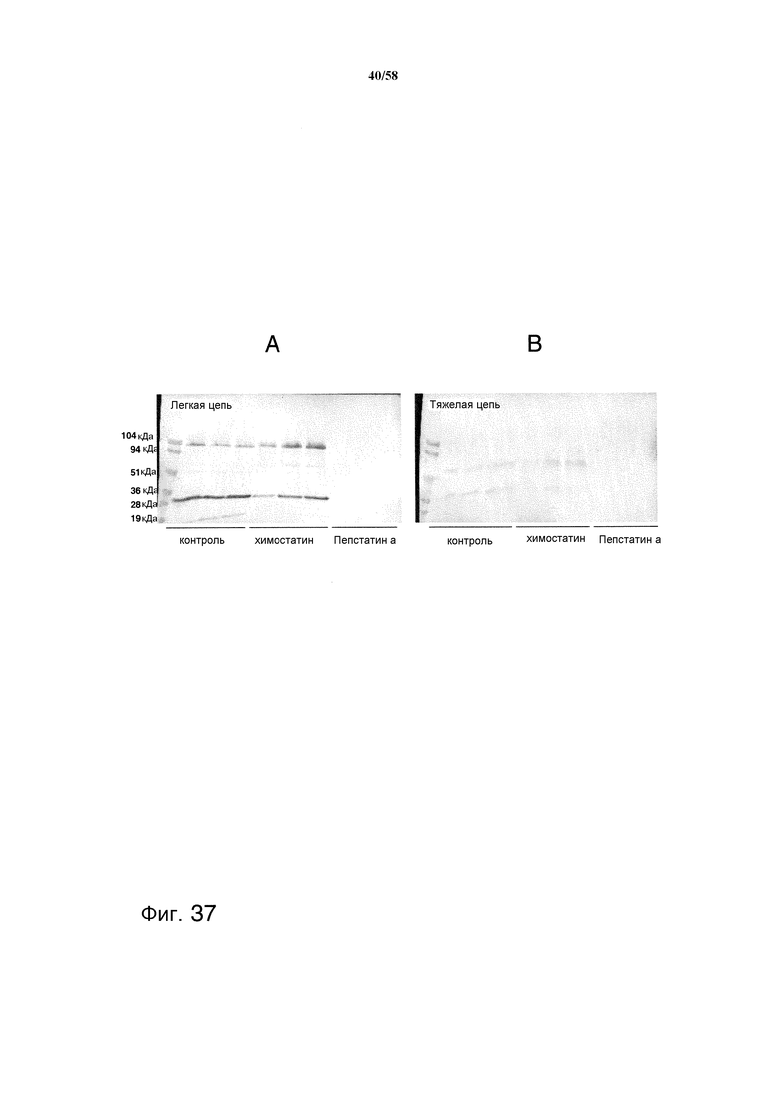
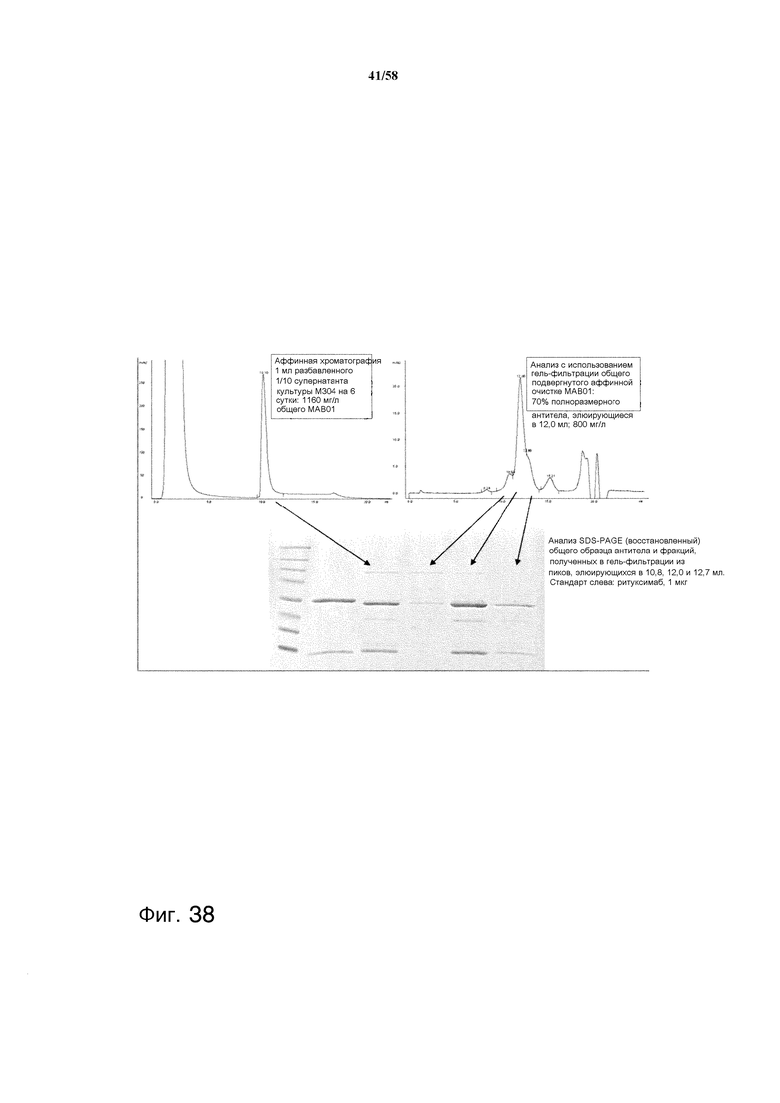
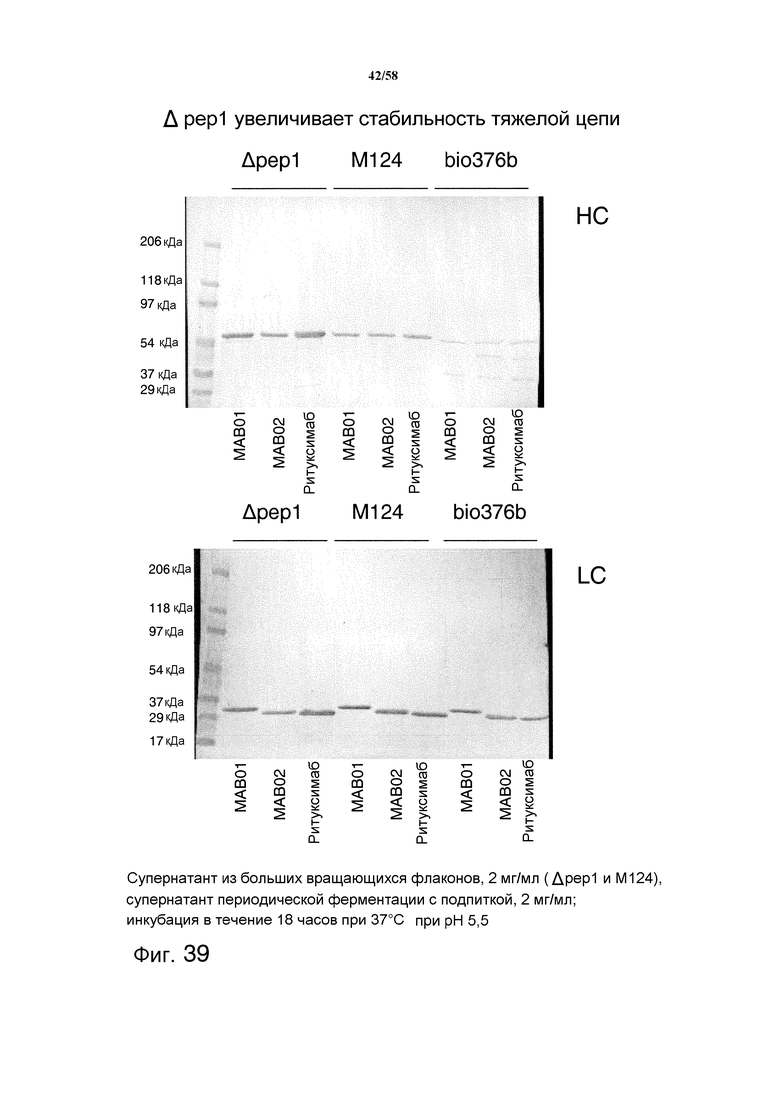
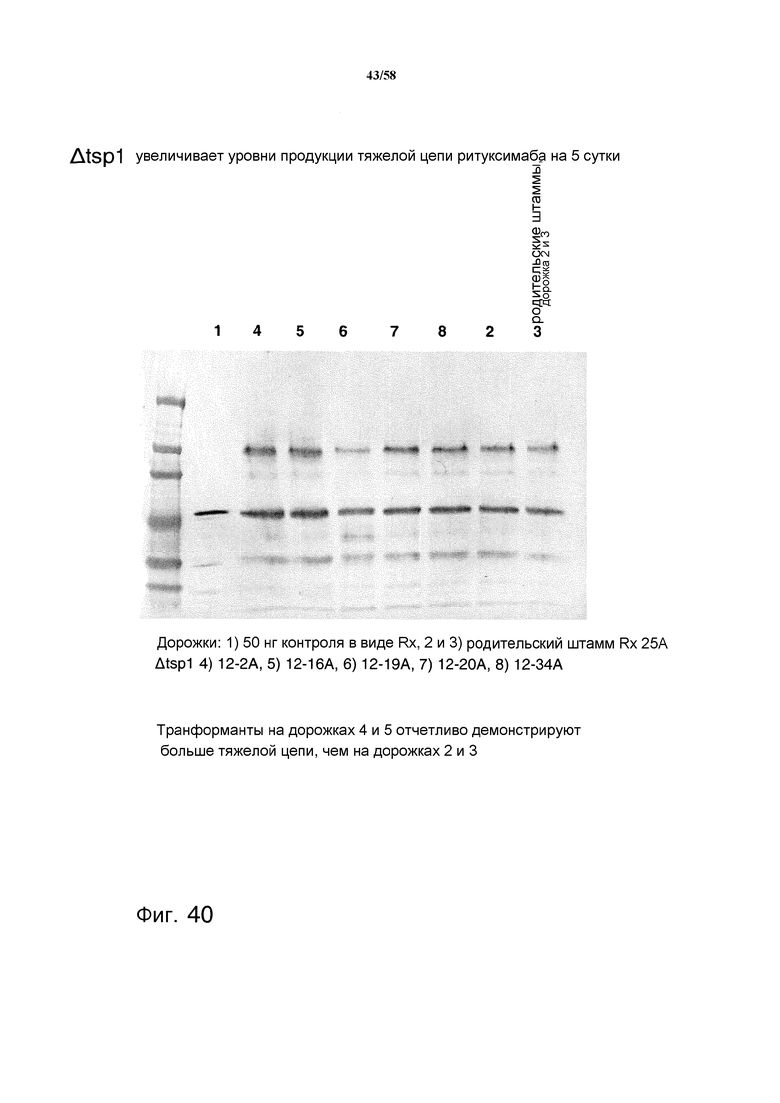
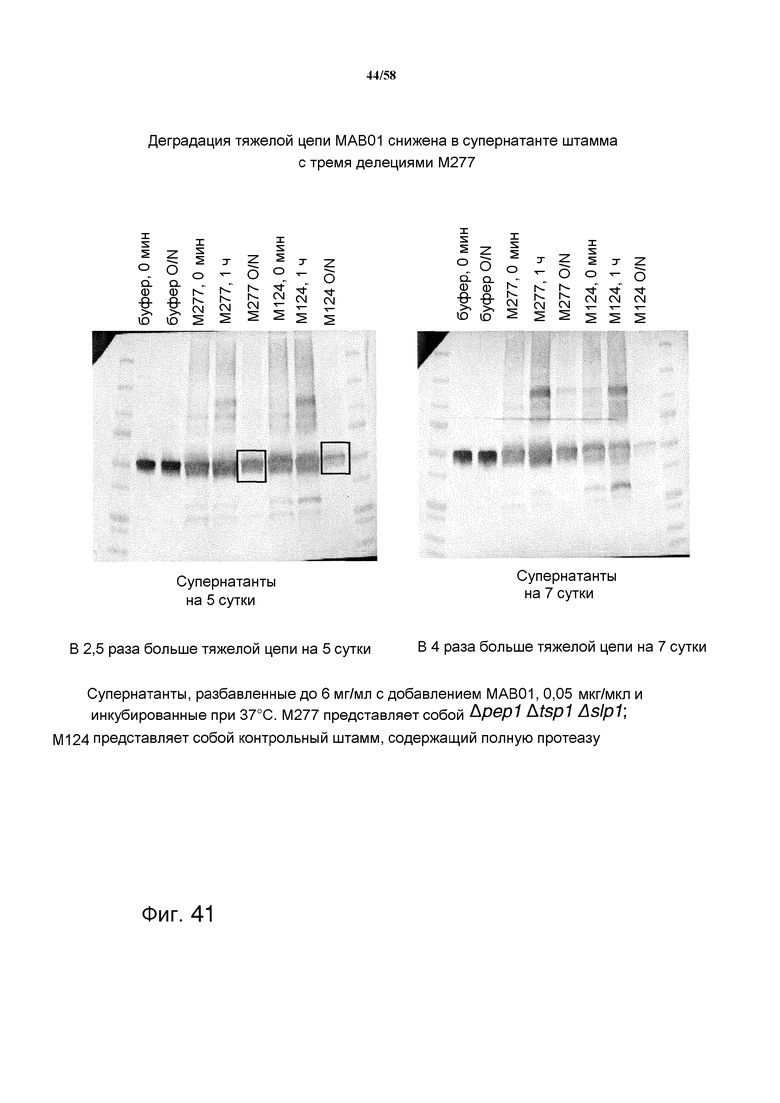
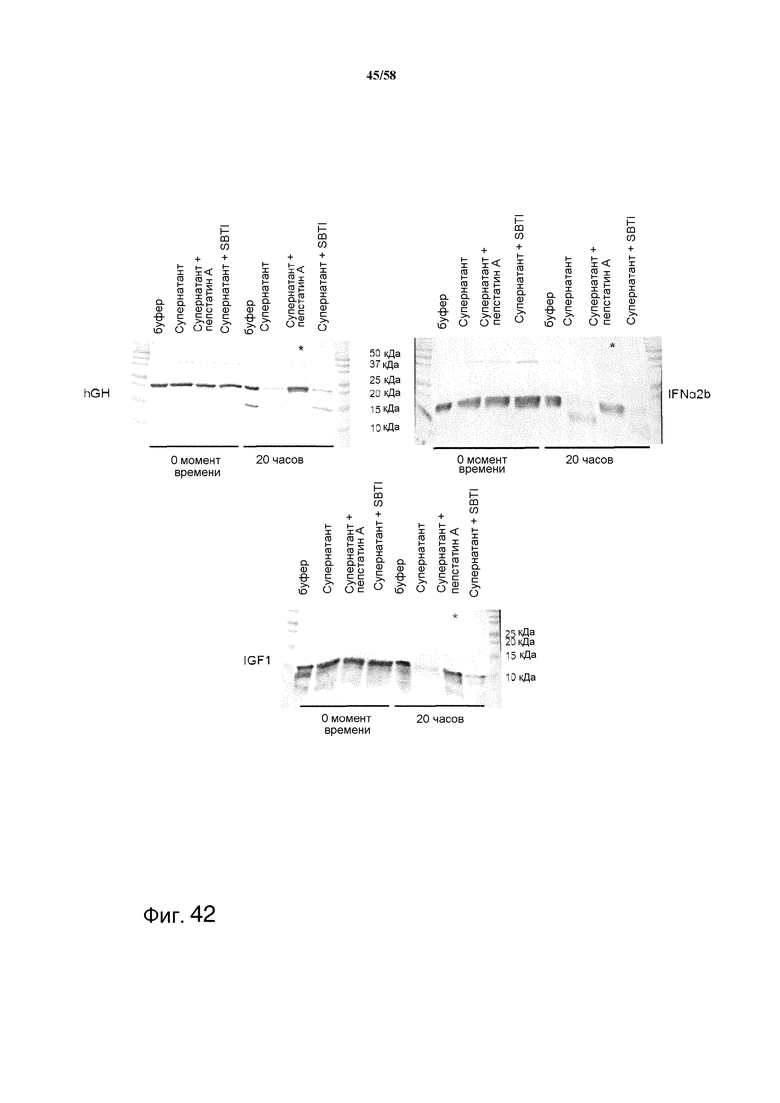






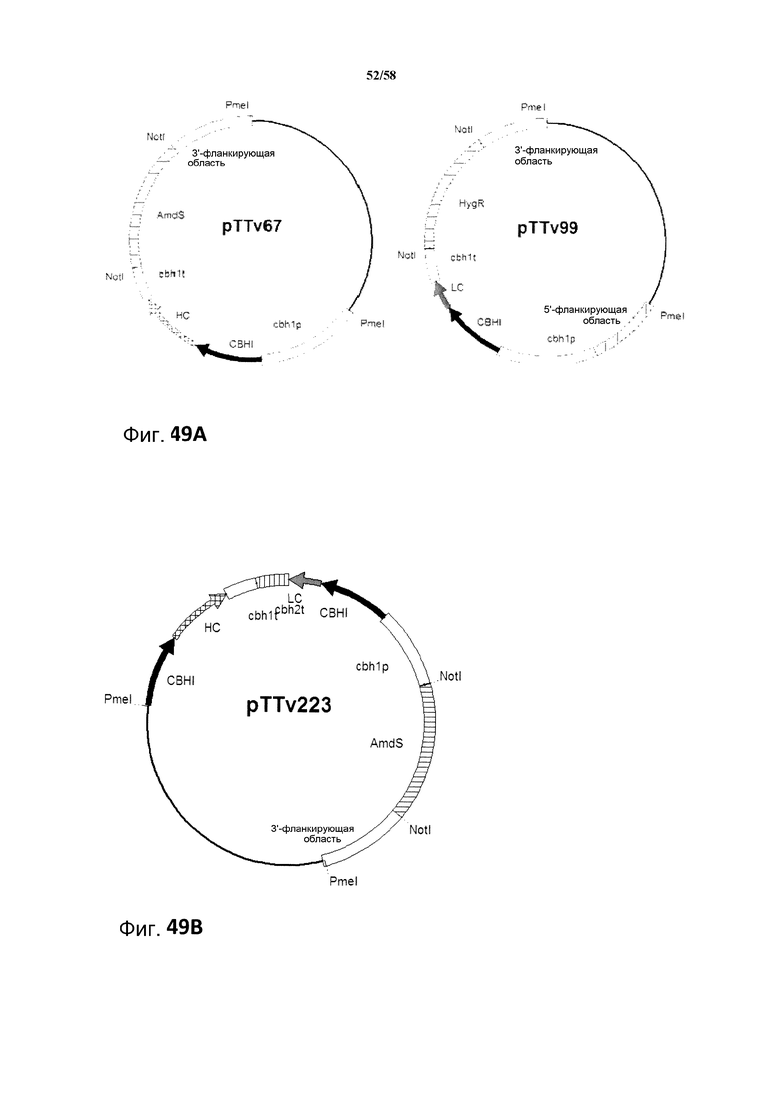
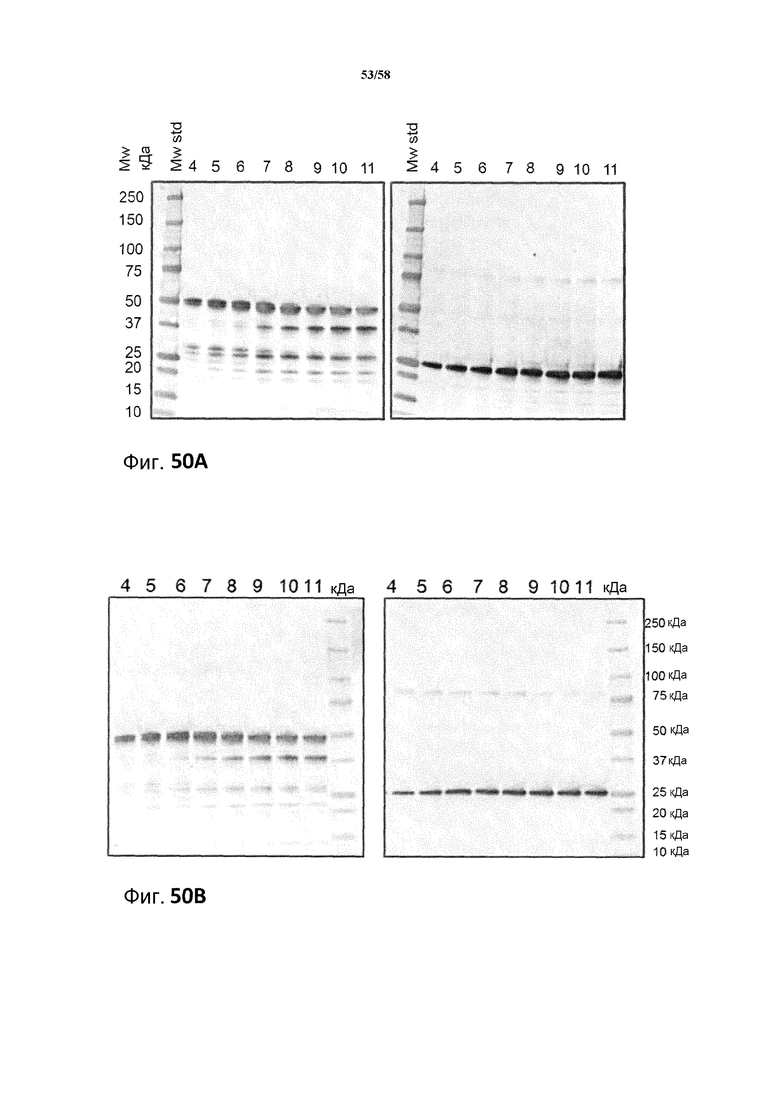
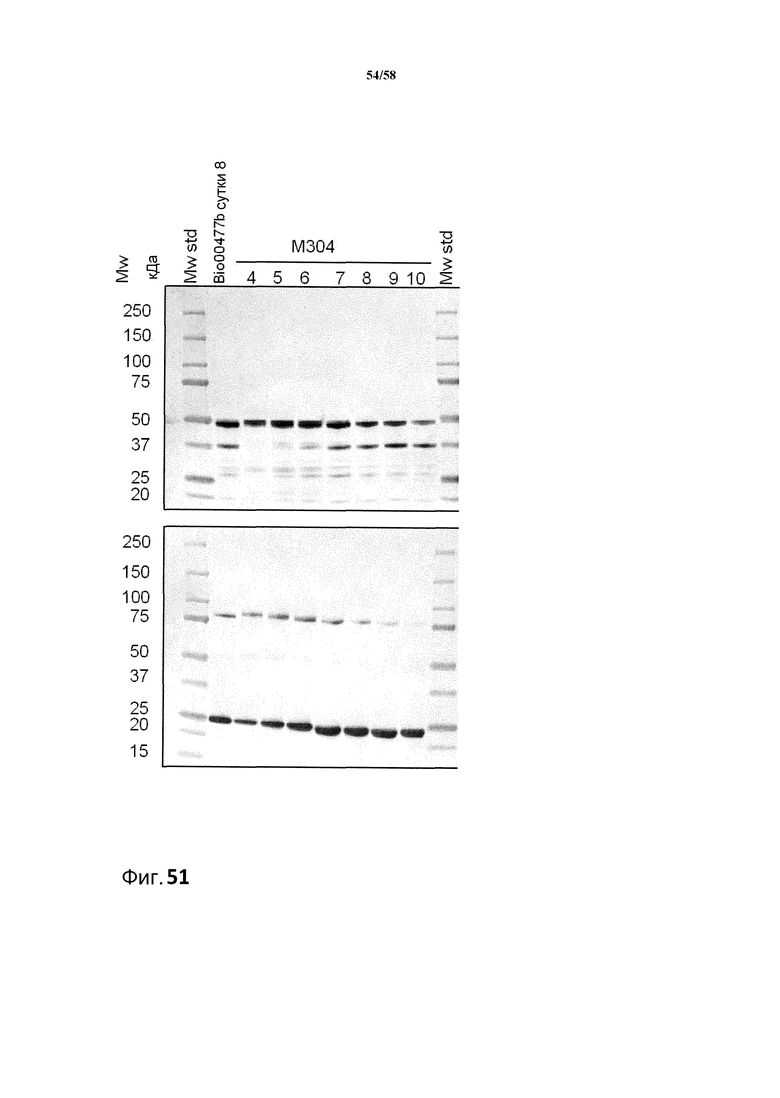
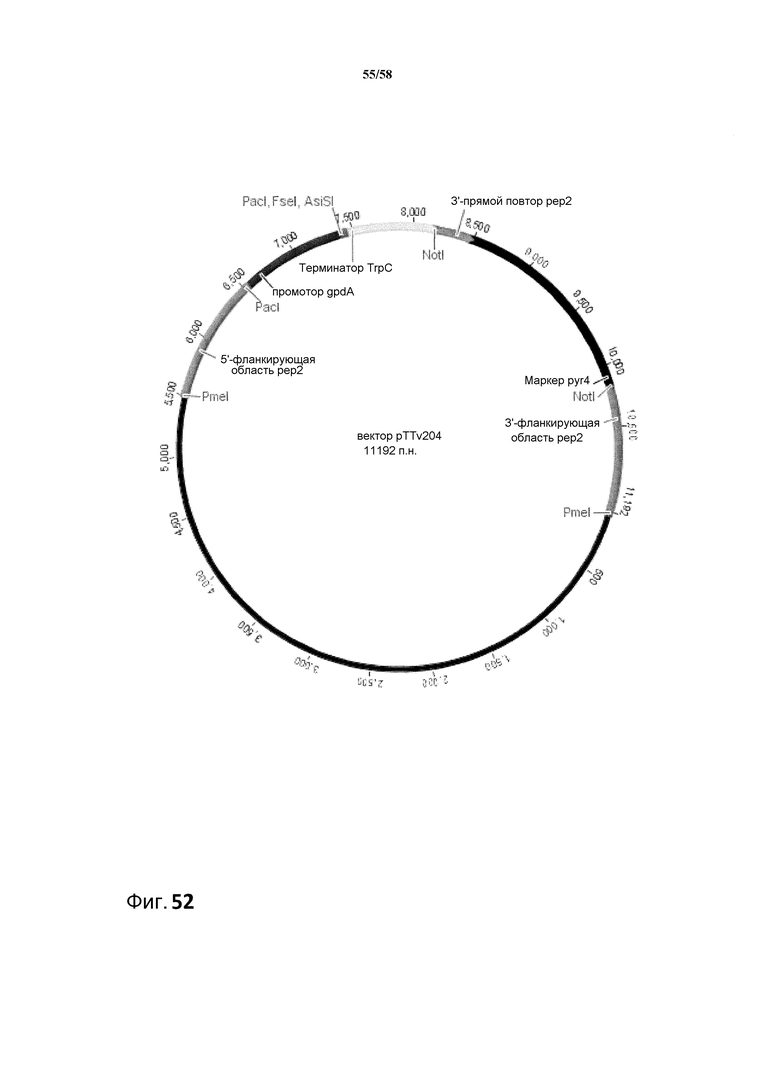
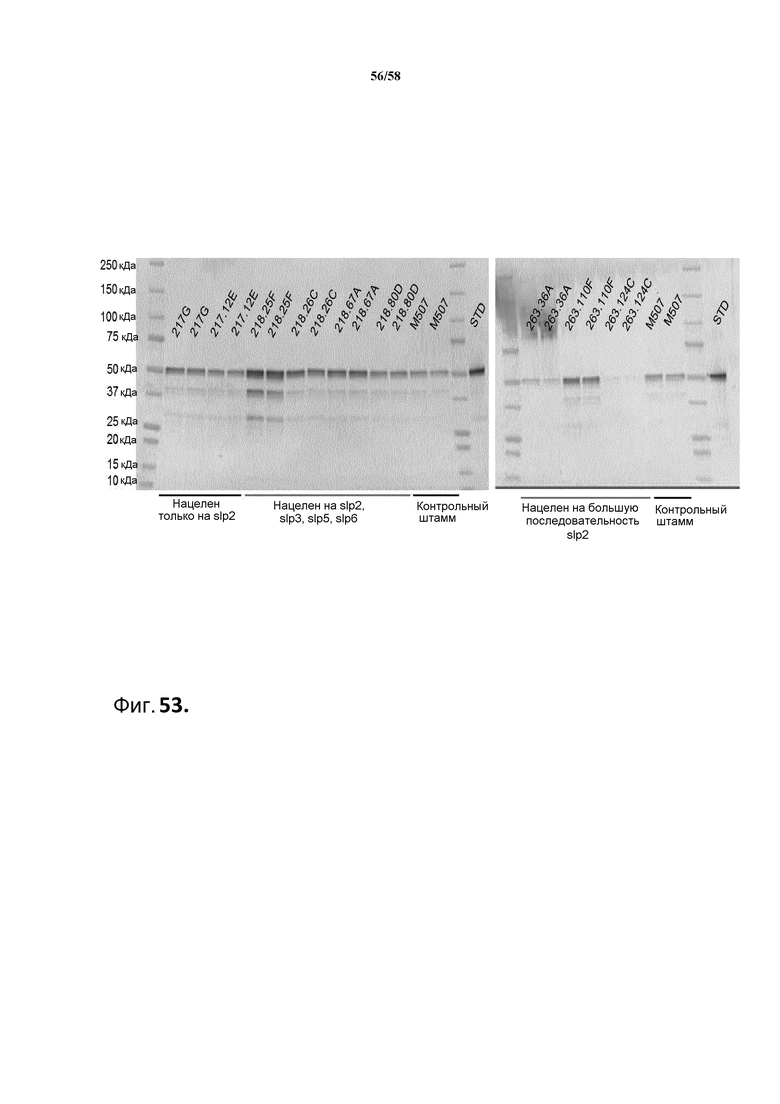
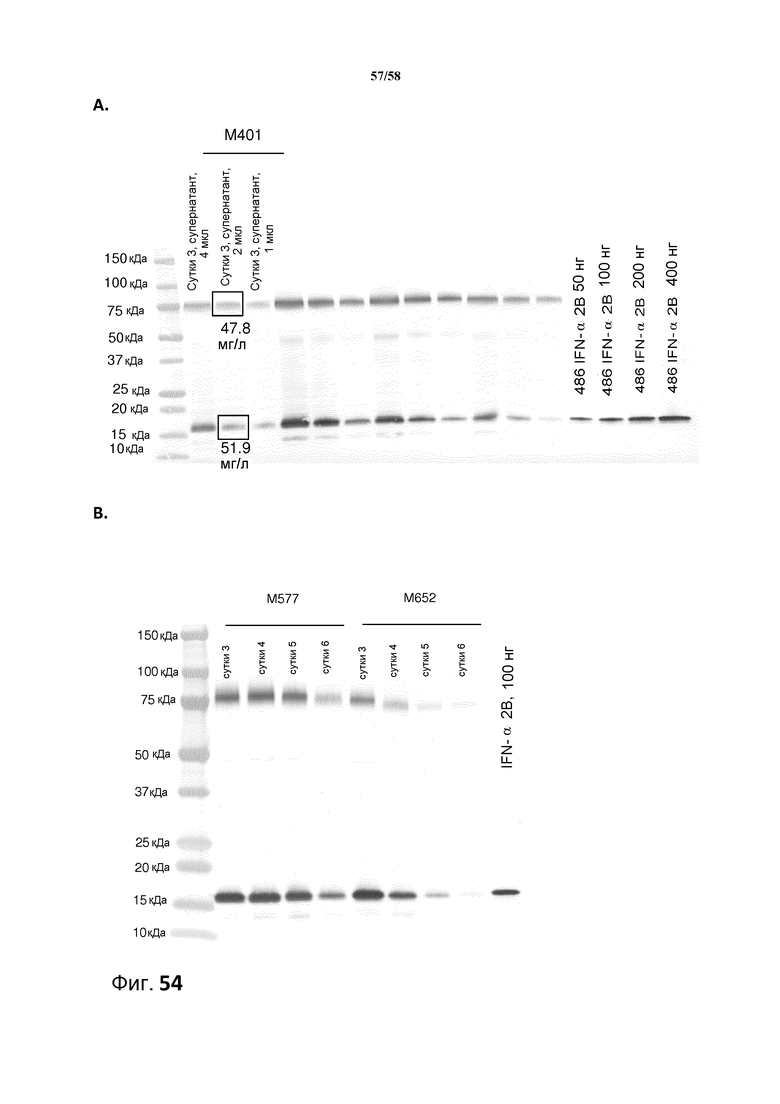

Группа изобретений относится к клетке для получения гетерологичного полипептида, способу повышения стабильности полипептида и способу получения гетерологичного полипептида с использованием указанной клетки. Клетка Trichoderma для получения гетерологичного полипептида содержит по меньшей мере три эндогенных протеазы с устраненной активностью и рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид. Клетка имеет устраненную активность по меньшей мере протеазы gap1 SEQ ID NO: 118 и по меньшей мере двух дополнительных протеаз, выбранных из рер2 SEQ ID NO: 182, рер3 SEQ ID NO: 17, рер4 SEQ ID NO: 37, pep5 SEQ ID NO: 58, pep8 SEQ ID NO: 507, pep11 SEQ ID NO: 522, pep12 SEQ ID NO: 530, tsp1 SEQ ID NO: 66, slp1 SEQ ID NO: 82, slp2 SEQ ID NO: 98, slp3 SEQ ID NO: 166, slp7 SEQ ID NO: 231, pep1 SEQ ID NO: 1 и gap2 SEQ ID NO: 129. При этом полипептид продуцируется на уровне, который по меньшей мере в два раза превышает уровень продукции полипептида в соответствующей родительской клетке Trichoderma, в которой протеазы не имеют устраненную активность. Способ повышения стабильности полипептида включает культивирование вышеуказанной клетки так, чтобы экспрессировался гетерологичный полипептид. При этом гетерологичный полипептид проявляет повышенную стабильность по сравнению с гетерологичным полипептидом, полученным в соответствующей родительской клетке Trichoderma, в которой протеазы не имеют устраненную активность. Способ получения гетерологичного полипептида включает культивирование вышеуказанной клетки так, чтобы экспрессировался гетерологичный полипептид, очистку гетерологичного полипептида. 3 н. и 20 з.п. ф-лы, 55 ил., 81 табл., 23 пр.
1. Клетка Trichoderma для получения гетерологичного полипептида, содержащая по меньшей мере три эндогенных протеазы с устраненной активностью и рекомбинантный полинуклеотид, кодирующий гетерологичный полипептид, где полипептид продуцируется на уровне, который по меньшей мере в два раза превышает уровень продукции полипептида в соответствующей родительской клетке Trichoderma, в которой протеазы не имеют устраненную активность, и где клетка Trichoderma имеет устраненную активность по меньшей мере протеазы gap1 SEQ ID NO: 118 и по меньшей мере двух дополнительных протеаз, выбранных из рер2 SEQ ID NO: 182, рер3 SEQ ID NO: 17, рер4 SEQ ID NO: 37, pep5 SEQ ID NO: 58, pep8 SEQ ID NO: 507, pep11 SEQ ID NO: 522, pep12 SEQ ID NO: 530, tsp1 SEQ ID NO: 66, slp1 SEQ ID NO: 82, slp2 SEQ ID NO: 98, slp3 SEQ ID NO: 166, slp7 SEQ ID NO: 231, pep1 SEQ ID NO: 1 и gap2 SEQ ID NO: 129.
2. Клетка Trichoderma по п.l, где общая протеазная активность снижена до 49% или менее, предпочтительно до 31% или менее, от общей протеазной активности соответствующей родительской клетки Trichoderma, в которой протеазы не имеют устраненную активность.
3. Клетка Trichoderma по п.1, где клетка Trichoderma представляет собой Trichoderma reesei.
4. Клетка Trichoderma по п.1, где клетка Trichoderma не имеет протеазной активности по меньшей мере следующих трех протеаз: gap1, slp1 и pep1.
5. Клетка Trichoderma по п.1, где клетка Trichoderma имеет устраненную протеазную активность по меньшей мере трех или четырех протеаз, выбранных из группы, состоящей из pep1, рер2, рер3, рер4, рер5, рер8, pep11, рер12, tsp1, slp1, slp2, slp3, slp7, gap1 и gap2.
6. Клетка Trichoderma по п.1, где клетка Trichoderma не имеет протеазной активности по меньшей мере в восьми протеазах, где каждый из генов, кодирующих восемь протеаз, был удален для устранения активности соответствующей протеазы, и восемь протеаз представляют собой pep1, tsp1, slp1, gap1, gap2, pep4, рер3 и pep5.
7. Клетка Trichoderma по п.1, где клетка Trichoderma содержит от трех до шести протеаз с устраненной активностью, и каждая из от трех до шести протеаз выбраны из группы, состоящей из pep1, рер2, рер3, рер4, рер5, tsp1, slp1, slp2, slp3, gap1 и gap2.
8. Клетка Trichoderma по п.1, где клетка Trichoderma содержит ген, кодирующий дополнительную протеазу, которая была удалена для устранения активности соответствующей протеазы, и дополнительная протеаза выбрана из группы, состоящей из pep7, рер8, pep11, рер12, tpp1, gap2, slp3, slp5, slp6, slp7 и slp8.
9. Клетка Trichoderma по п.1, где гетерологичный полипептид представляет собой полипептид млекопитающих.
10. Клетка Trichoderma по п.9, где полипептид млекопитающих является гликозилированным.
11. Клетка Trichoderma по п.9, где полипептид млекопитающих выбран из группы, состоящей из антитела и его антигенсвязывающих фрагментов, фактора роста, интерферона, цитокина и интерлейкина.
12. Клетка Trichoderma по п.9, где полипептид млекопитающих выбран из группы, состоящей из инсулиноподобного фактора роста 1 (IGF1), гормона роста человека (hGH) и интерферона-альфа 2b (IFNα2b).
13. Клетка Trichoderma по п.1, где клетка Trichoderma дополнительно содержит ALG3, имеющую устраненную активность.
14. Клетка Trichoderma по п.1, где клетка грибов дополнительно содержит полинуклеотид, кодирующий α-1,2-маннозидазу.
15. Клетка Trichoderma по п.1, дополнительно содержащая каталитический домен N-ацетилглюкозаминилтрансферазы I и, необязательно, каталитический домен N-ацетилглюкозаминилтрансферазы II.
16. Клетка Trichoderma по п.15, дополнительно содержащая первый полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы I, и, необязательно, второй полинуклеотид, кодирующий каталитический домен N-ацетилглюкозаминилтрансферазы II.
17. Клетка Trichoderma по п.1, дополнительно содержащая полинуклеотид, кодирующий маннозидазу II и/или галактозилтрансферазу.
18. Способ повышения стабильности полипептида, включающий
a) наличие клетки Trichoderma по любому из пп. 1-17, и
b) культивирование клетки Trichoderma так, чтобы экспрессировался гетерологичный полипептид,
где гетерологичный полипептид проявляет повышенную стабильность по сравнению с гетерологичным полипептидом, полученным в соответствующей родительской клетке Trichoderma, в которой протеазы не имеют устраненную активность.
19. Способ получения гетерологичного полипептида, включающий
a) наличие клетки Trichoderma по любому из пп. 1-17,
b) культивирование клетки Trichoderma так, чтобы экспрессировался гетерологичный полипептид, и
с) очистку гетерологичного полипептида.
20. Способ по п. 18 или 19, где клетка Trichoderma дополнительно содержит белок-носитель.
21. Способ по п.20, где белок-носитель представляет собой СВН1.
22. Способ по п.19, где культивирование проводят в среде, содержащей ингибитор протеаз.
23. Способ по п.19, где культивирование проводят в среде, содержащей один или два ингибитора протеаз, выбранных из SBT1 и химостатина.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| VAN DEN HOMBERGH J.P.T.W | |||
| et al | |||
| Disruption of three acid proteases in Aspergillus niger effects on proteasespectrum, intracellular proteolysis, and degradation of target proteins // European Journal of Biochemistry, 1997, vol | |||
| Приспособление для картограмм | 1921 |
|
SU247A1 |
| СПОСОБ ПРИГОТОВЛЕНИЯ РУДНО-УГОЛЬНЫХ БРИКЕТОВ | 1922 |
|
SU605A1 |
| SIMS A.H | |||
| et al | |||
| Glutamic protease distribution is limited to filamentous fungi // FEMS Microbiology Letters, 239 (2004), pp.95-101 | |||
| YOON J | |||
| ET AL | |||
| Construction of quintuple protease gene disruptant in Aspergillus oryzae // Applied Microbiology and Biotechnology, 2008, vol | |||
| Машина для разделения сыпучих материалов и размещения их в приемники | 0 |
|
SU82A1 |
| Счетчик оборотов горизонтального вала машины | 1925 |
|
SU691A1 |
| MARTINEZ D | |||
| et al | |||
| Genome sequencing and analysis of the biomass-degrading fungus Ttichoderma reesei (syn | |||
| Hypocrea jecorina) // Nature Biotechnology, 2008, vol.26, no.5, pp.553-560 | |||
| WO 1997012045 A1, 03.04.1997 | |||
| Ударная мельница | 1927 |
|
SU6873A1 |

