Введение
Антитела, или иммуноглобулины (Ig), - это растворимые гликопротеины крови и тканевой жидкости, играющие центральную роль в системе гуморального иммунитета у позвоночных. Антитела синтезируются В-лимфоцитами в ответ на чужеродные биологические и химические вещества (антигены) самой разнообразной структуры с целью их нейтрализации. Благодаря высокой специфичности и высокой аффинности связывания с определенным антигеном, а также возможности возникновения антител к практически неограниченному репертуару антигенов антитела и их производные являются одними из наиболее важных реагентов для использования в фундаментальных, прикладных и медицинских исследованиях.
Классические антитела [1, 2] представляют собой крупные мультимерные белки (IgG ~150 кДа), объединяющие две идентичные тяжелые Н-цепи, которые, в свою очередь, состоят из вариабельного VH, трех константных CH1, СН2, СН3 доменов и шарнирного участка между СН1 и СН2 доменами, и две идентичные легкие L цепи, состоящие из вариабельного, VL, и константного, CL, доменов. Четырехцепочечная молекула объединена посредством нековалентных и ковалентных (дисульфидных) связей между цепями. При помощи протеазы папаина антитела можно расщепить на два фрагмента: Fab (Fragment antigen binding, антиген связывающий фрагмент) и Fc (Fragment crystallizable, фрагмент, способный к кристаллизации). Соответственно, одна область молекулы антител (Fab) определяет ее антигенную специфичность, а другая (Fc) осуществляет эффекторные функции, которые направлены на элиминацию антигена [3, 4]. Домены СН1 и СН2Н-цепи разделены шарнирной областью ("hinge region"), от которой зависит подвижность Fab-фрагмента и взаимодействие молекулы IgG с эффекторными рецепторами иммуноглобулинов, расположенными на клетках. СН2 домен содержит участки связывания как Fcϒ рецепторов, опосредующих клеточную активацию (ADCC и ADCP), так и молекул системы комплемента (CDC). Кроме того, в этом домене присутствует сайт, являющийся местом присоединения углеводов для всех изотипов иммуноглобулинов. СН3-домен в значительной степени определяет стабильность димера IgG, а также взаимодействует с FcRn рецептором на поверхности клеток, определяя фармакокинетические свойства антител и их распределение и метаболизм внутри организма. Комбинация антигенсвязывающих участков (CDR-Complementarity Determining Regions) вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL) формирует участок связывания антигена, в то время как каркасные регионы вариабельных доменов (FR - Framework Regions) и константные домены антител непосредственно не участвуют в распознавании антигена. Минимизированной производной антиген-связывающего фрагмента классических антител является одноцепочечная конструкция, в которой вариабельные домены тяжелой и легкой цепей соединены линкерной последовательностью (scFv).
Важным открытием явилось обнаружение в крови представителей семейства Camelidae (верблюды, ламы, викуньи) в значительном количестве особых неканонических антител с упрощенной структурой [5]. Такие антитела ("heavy chain antibody", HCAb) состоят из димера только одной укороченной тяжелой цепи (без СН1-домена), а легкая цепь при этом отсутствует. Антиген-узнающий участок HCAb формируется лишь одним вариабельным доменом тяжелой цепи (VHH), который непосредственно связан через шарнирную область с Fc-доменом. Часто вместо VHH используют термин "однодоменное антитело", "nanobody" (нанотело), "мини-антитело" или "наноантитело". Как оказалось, такая монодоменная структура в изолированном виде, кроме малых размеров (12-15 кДа), обладает рядом преимуществ перед классическими IgG антителами, а именно агрегационной, химической и термостабильностью. Антитела VHH можно эффективно клонировать и экспрессировать в бактериях и дрожжах. Обладая такими свойствами, они получили технологическое развитие как в терапевтическом направлении, развиваемом компанией "Ablynx", так и области лабораторной и промышленной хроматографии с линейкой носителей CaptureSelect.
Неканонические антитела (HCAb), состоящие из димера только тяжелой цепи иммуноглобулина, впервые обнаружены при электрофоретическом анализе иммуноглобулинов в сыворотке крови различных представителей семейства верблюдовых [5]. Относительная доля HCAb варьирует от примерно 15-25% (всех IgG) у лам и викуний до примерно 60-80% у верблюдов [6].
Как предполагают, неканонические антитела (HCAb), по крайней мере, в случае верблюдовых, представляют собой результат относительно недавней эволюции генов канонических антител. Два константных домена тяжелых цепей, СН2 и СН3, в случае HCAb и классических антител высококонсервативны. В составе антител HCAb домен, соответствующий первому константному СН1-домену классических антител, отсутствует. Геном одногорбого верблюда (вид Dromedary) содержит кластер из примерно пятидесяти VH- и сорока VHH-генеративных генов, за которыми располагаются множественные гены D-сегментов, J-сегментов и гены константных участков (Cμ, Сγ, Сε, Сα). Очевидно, что некоторые из Сγ-генов предназначены для формирования HCAb (мутация приводит к потере СН1-домена), в то время как остальные - для формирования классических антител (с сохраняемым СН1-доменом). Одни и те же гены сегментов D и J могут случайным образом соединяться как с одним из VH-, так и с одним из VHH-генов. Это указывает на то, что гены VH и VHH находятся в одном и том же локусе [7-10].
Организация вариабельных доменов неканонических антител (VHH) и вариабельных доменов (VH) классических антител весьма сходна, т.к. у человека VH-домены подкласса IgG3 имеют особо выраженную гомологию с VH и VHH верблюдовых. В обоих случаях V-домены состоят из четырех консервативных каркасных участков FR, которые окружают три гипервариабельных участка, определяющих комплементарность, CDR. Также в обоих случаях формируется типичная для V-домена иммуноглобулина пространственная структура из двух β-слоев, один из которых состоит из четырех аминокислотных цепочек и второй - из пяти [11, 12]. В этой структуре все три гипервариабельных участка кластеризуются с одной стороны V-домена, где они участвуют в распознавании антигена и располагаются в петлях, соединяющих β-структуры. Однако имеются и важные отличия, связанные с функционированием VHH в формате одного домена. Так, гипервариабельные участки CDR1 и CDR3 VHH заметно увеличены. Часто в гипервариабельных участках VHH обнаруживаются цистеиновые остатки сразу в двух участках (чаще всего - в CDR1 и CDR3, реже - в CDR2 и CDR3). При исследовании кристаллических структур VHH показано, что эти цистеиновые остатки формируют дисульфидные связи, и это дополнительно стабилизирует структуру петель данного антитела [12]. Наиболее явный и воспроизводимый отличительный признак VHH - четыре замены гидрофобных аминокислотных остатков на гидрофильные во втором каркасном участке (Val37Phe, Gly44Glu, Leu45Arg, Trp47Gly, согласно нумерации Kabat и соавт. [13]). Этот каркасный участок у VH-домена высококонсервативен, обогащен гидрофобными аминокислотными остатками и особо важен при образовании связи с вариабельным доменом VL легкой цепи. В этом аспекте VHH-домен сильно отличается: указанные замены гидрофобных аминокислот на гидрофильные делают невозможной ассоциацию VHH и VL. Эти замены также объясняют обычно высокую растворимость VHH (наноантитела) при его получении в виде рекомбинантного белка [14].
Репертуары возможных паратопов (антиген-связывающих структур антитела) HCAb и классических антител, по-видимому, могут заметно отличаться. Так как эти два типа антител сосуществуют в одном организме, то можно предполагать, что они не конкурируют, а взаимно дополняют друг друга. Действительно, не раз отмечалось, что оба типа антител могут возникать параллельно, взаимоисключающе или в разных соотношениях, по отношению к разным эпитопам антигенного материала при иммунизации одного и того же животного. Несмотря на предполагаемое меньшее разнообразие возможных паратопов у однодоменных антител по сравнению с классическими двухдоменными антителами работы многих авторов убедительно продемонстрировали, что HCAb могут быть получены против самых разнообразных эпитопов весьма широкого спектра антигенов [15]. Очевидно, этому способствуют заметно увеличенные гипервариабельные участки CDR1 и CDR3. Следует также отметить удивительно большое (в сравнении с V-доменами классических антител) число соматических гипермутаций в VHH, накапливающихся, по-видимому, в процессе аффинного созревания антител в ходе иммунизации [16]. Рентгеноструктурный анализ показал, что антиген-связывающие петлевые участки VHH способны образовывать необычные для классических V-доменов структуры [12, 16]. Если в случае VH- и VL-доменов классических антител все шесть гипервариабельных участков вносят более или менее одинаковый вклад в связывание антигена, то в случае VHH обычно для формирования паратопа более важен CDR3-участок. Показано, что CDR3-участок в VHH (но не в VH или VL) может образовывать необычные длинные пальцеобразные выступающие структуры, которые могут углубляться в структуру антигена, в частности распознавать активные центры ферментов [12]. Малыми размерами антиген-связывающего участка (VHH) и его способностью формировать необычные выступающие паратопы объясняется возможность получения HCAb, способных распознавать недоступные для классических антител эпитопы, например, при образовании антител, являющихся эффективными ингибиторами ферментов [17].
При всем своем высоком потенциале уникальной в сравнении с классическими IgG антителами специфичности использование монодоменных VHH для терапевтического применения в ряде случаев ограничено из-за быстрого выведения этой малой молекулы из организма. Существует ряд решений по улучшению фармакокинетики VHH структур, которые включают использование химической конъюгации с ПЭГ (PEG) и ковалентное соединение с полипептидом, опосредующим уменьшенный клиренс из крови, такими как сывороточный альбумин человека (HSA) и Fc-фрагмент классического антитела человека, в виде гибридных белков, период полувыведения в крови которых составляет до трех недель [18, 19, 20]. Также показано успешное применение малых пептидов, присоединяемых с помощью методов генной инженерии к VHH и способных к высокоаффиному нековалентному взаимодействию с этими же компонентами (HSA и IgG) в крови человека [21]. Однако технологичность и иммуногенность этих подходов остается пока неясной и находится только на стадии проверки применимости как в клинических исследованиях так и на более ранних стадиях исследований.
Кроме того, наибольшее ограничение при применении антител в качестве препаратов для лечения различных заболеваний накладывает агрегационная и химическая стабильность, аффинность и иммуногенность. Т.к. большинство моноклональных антител в настоящее время получают на основе мышиных, применение таких антител у пациентов приводит к развитию иммунного ответа на терапию антителами, например аллергических реакций. Такие типы иммунного ответа могут повлечь за собой, в конечном итоге, потерю эффективности при лечении по меньшей мере и, в худшем случае, к возможным тяжелым анафилактическим реакциям. С другой стороны, агрегационно или химически нестабильные терапевтические антитела при хранении препарата ухудшают его терапевтические свойства и могут усиливать иммуногенность при введении в организм пациента.
В связи с вышесказанным, актуальным является создание антител на основе антител VHH, которые имели бы, по сравнению с известными ранее антителами, улучшенные функциональные и терапевтические свойства, в частности повышенную агрегационную, химическую и термостабильность, и улучшенную аффинность, и в то же время отличались бы легкостью и простотой получения, в т.ч. и в промышленно значимых масштабах.
Предшествующий уровень техники
Из предшествующего уровня техники известны различные конструкции антител, содержащие VHH домен в своем составе.
В публикации РСТ/ЕР 2008/066368 описаны антитела, которые содержат отдельные вариабельные домены, связанные с Fc-фрагментом. В качестве вариабельных доменов могут быть использованы «нанотела», при этом Fc- фрагмент получен из антитела типа IgE. Указанные Fc-фрагмент и домен могут соединяться через линкер, расположенный в шарнирной области.
В заявке на патент US 2009/0202979 раскрыты антитела, включающие полностью или часть антител VHH, непосредственно соединенные с константными частями человеческих антител.
Также известны аминокислотные замены, влияющие на физико-химические и биологические свойства антител.
Например, в заявке US 20110028348 описаны вариабельные домены тяжелых цепей антитела, в которых введены аминокислотные замены в позициях 35, 45, 47, 93-100 и 100а для повышения гидрофильности получаемого антитела.
К настоящему времени разработаны методы оптимизации структуры изолированных VHH и VH монодоменов, уменьшающие их иммуногенность и улучшающие агрегационную стабильность.
Так, Vincke и соавторы [22] обнаружили, что замены Glu-49→Gly и Arg-50→Leu в характерных аминокислотах приводят к получению отдельного домена, который является более стабильным, но при этом менее растворимым. Другие замены в каркасном участке FR-2 Phe-42→Val и Gly/Ala-52→Trp критично влияют на аффинность антитела к антигену вследствие переориентации петли Н3, повышая константу его кинетической диссоциации в 6-10 раз (6,85⋅10-3 1/с). При замене Phe-42→Vаl наблюдалось снижение стабильности получаемых антител. Замена Gly-49 и Leu-50 в VH последовательности приводила к снижению стабильности домена, в то время как гуманизация по Glu-49 и Arg-50 в VHH позволяла получить стабильные вариабельные домены.
Из литературы также известно, что при наличии коротких HCDR3 регионов, нивелирующих экранирующий эффект конформации классических VHH, и внесении VH-характерной замены Trp-47 на Gly-47 и замен Tyr-37 на Val-37, Glu-44 на Gly-44 и Arg-45 на Leu-45, изолированные VHH-домены могут восстанавливать способность к связыванию с VL-доменами [24].
Связь между повышением аггрегационной стабильности терапевтических антител классической IgG структуры и снижением их иммуногенности подкреплена результатами работ многих авторов и рассмотрена в обзоре Hermeling et al., 2004 [25]. Однако не было обнаружено антител, имеющих в своем составе производные VHH - доменов, но связанных с вариабельными доменами легких цепей в составе полноразмерных иммуноглобулинов человека класса IgG.
Таким образом, существует необходимость в создании нового формата антител, которые при этом обладали бы повышенной стабильностью и аффинностью, отличались хорошей экспрессией и имели бы низкую иммуногенность.
Кроме того, ранее не описано подходов к созданию подобных молекул с улучшенной агрегационной стабильностью и повышенной аффинностью, и высоким уровнем экспрессии в клеточных культурах млекопитающих и легкостью в получении.
В связи с вышесказанным, данное изобретение впервые описывает формат антител, включающих производные VHH-доменов, способных связываться с вариабельными доменами легких цепей в составе полноразмерных иммуноглобулинов человека класса IgG с образованием сходной с природной и, следовательно, обладающей низкой иммуногенностью, но в то же время улучшенной агрегационной стабильностью и повышенной аффинностью, и имеющих структуру терапевтического моноклонального антитела.
Описание фигур
Фиг. 1. Схема создания и оптимизации агрегационно стабильного антитела на основе VHH.
Фиг. 2. SDS-гель-электрофорез SDS-гель-электрофорез антител VHHIgG1c различными аминокислотными заменами.
Фиг. 3. Схема синтеза комбинаторной библиотеки Fab ламы.
Фиг. 4. Фагмида для клонирования Fab фаговых дисплейных библиотек.
Фиг. 5А-С. Сенсограммы проверки сравнительных кинетических характеристик VHHFab клонов по связыванию с IL-17A.
Фиг. 5А. Последовательности вариабельных доменов VHH ламы в составе Fab, специфичных против IL-17А человека.
Фиг. 5В. Последовательности вариабельных доменов легких цепей ламы в составе 3VHHFab, специфичных против IL-17A человека.
Фиг. 5С. Сенсограммы проверки сравнительных кинетических характеристик VHHFab клонов по связыванию с IL-17A.
Фиг. 6А. Аминокислотные последовательности 3 различных мутантов VHH.
Фиг. 6В. Агрегационная стабильность 3 мутантов VHH.
Фиг. 6С. Свойства аффиности 3 мутантов VHH.
Фиг. 7. Клеточный тест блокирования продукции IL-6 путем ингибирования IL-17A антагонистами VHHIgG1VK4B11, mut1 VHHIgG1 и mut4 VHHIgG1.
Фиг. 8. Выделение химерных вариантов mut4 VHHFab, содержащие легкие цепи человека.
Фиг. 9. Клеточный тест блокирования продукции IL-6 путем ингибирования IL-17A антагонистами mut4 VHHIgG1VK3c8, mut4 VHHIgG1VK1A7 и mut4 VHHIgG1VK4E12.
Фиг. 10. Белковый электрофорез в денатурирующих условиях препаратов, содержащие антитела с мутациями в позициях 44 и 45 m4 VHHc8.
Фиг. 11. Сравнительный клеточный тест блокирования продукции IL-6 путем ингибирования IL-17A антагонистами m4 VHHc8 с мутациями в позициях 44 и 45 в FR2.
Фиг. 12. Диаграмма стабильности и функциональных свойств для различных мутаций 44 и 45 позиций в регионе FR2 m4 VHHc8, определяющих взаимодействия вариабельного домена тяжелой и легкой цепей.
Фиг. 13А. Определение кинетических параметров взаимодействия BCD 109 к IL-17A из различных видов организмов (на приборе Octet RED 96).
Фиг. 13В. Результаты по исследованию кинетических параметров взаимодействия BCD 109 к IL-17A из различных видов организмов (на приборе Octet RED 96).
Фиг. 14. Хроматограмма антитела BCD 109, полученная до и после проведения термостресса.
Определения
"Моноклональное антитело", как используется в данной заявке, относится к антителу ламы, химерному антителу, гуманизированному антителу или полностью человеческому антителу, если в данной заявке не указано иное. Моноклональные антитела по изобретению могут быть получены с использованием, например, рекомбинантных технологий, технологий фагового дисплея, синтетических технологий или комбинаций таких технологий или других технологий, хорошо известных из уровня техники.
"Моноклональное антитело" относится к антителу, полученному из единой копии или клона, включая, например, любой эукариотический, прокариотический или фаговый клон, а не к способу его получения. "Моноклональное антитело" может быть интактным антителом (содержащим полный или полноразмерный Fc-участок), по существу интактным антителом, частью или фрагментом антитела, содержащими антиген связующую часть, например Fab-фрагмент, Fab'-фрагмент или F(ab')2-фрагмент ламы или химерного, гуманизированного или человеческого антитела. "Fab''-фрагмент содержит вариабельный и константный домен легкой цепи и вариабельный домен и первый константный домен (СН1) тяжелой цепи. "F(ab')2"- фрагменты антитела содержат пару Fab-фрагментов, которые в основном ковалентно связаны возле их С-концов шарнирными цистеинами между ними. Другие химические связывания фрагментов антител также хорошо известны из уровня техники.
Кроме того, "моноклональное антитело", как используется в данной заявке, может быть одноцепочечным Fv-фрагментом, который может быть получен путем связывания ДНК, кодирующей VHH и VL, с линкерной последовательностью. До тех пор пока белок сохраняет способность специфического или предпочтительного связывания своей мишени (например, эпитопа или антигена), он относится к термину "антитело". Антитела могут быть гликозилированными, или не быть таковыми, и входят в рамки изобретения.
Термин "производное" или «вариант» антитела в контексте данной заявки относится к молекуле, аминокислотная последовательность которой отличается от аминокислотной последовательности "родительского" антитела путем добавления, делеции и/или замещения одного или более аминокислотных остатков в последовательности родительского антитела. В предпочтительном из вариантов осуществления вариантное антитело содержит по крайней мере одну аминокислотную (например, от одной до приблизительно десяти и предпочтительно 2, 3, 4, 5, 6, 7 или 8 замен) замену в FR- или CDR-участках родительского антитела. Идентичность или гомологичность относительно последовательности вариантного антитела определена в данной заявке как процент аминокислотных остатков в последовательности вариантного антитела, идентичный остаткам родительского антитела, после выравнивания последовательностей и введения разрывов, при необходимости, для достижения максимального процента идентичности последовательности.
Производное (от родительского) антитело сохраняет способность связывать антиген или предпочтительно эпитоп, с которым связывается родительское антитело или предпочтительно имеет по крайней мере одно свойство или биологическую активность, которая превосходит аналогичные свойства родительского антитела. Например, антитело предпочтительно обладает большей агрегационной стабильностью или сильной аффинностью связывания, улучшенными фармакокинетическими свойствами или повышенной способностью ингибировать биологическую активность антигена, чем родительское антитело.
Термин "VHH производное" в контексте данной заявки относится к производным VHH антител, чья аминокислотная последовательность отличается от аминокислотной последовательности "родительского" VHH антитела путем замещения одного или более аминокислотных остатков в последовательности родительского антитела. В предпочтительном из вариантов осуществления производное VHH антитело содержит по крайней мере одну аминокислотную (например, от одной до приблизительно двадцати и предпочтительно 2, 3, 4, 5, 6, 7 или 8) замену в FR- или CDR-участках родительского антитела.
Производное антитело сохраняет способность связывать антиген или предпочтительно эпитоп, с которым связывается родительское антитело или предпочтительно имеет по крайней мере одно свойство или биологическую активность, которая превосходит аналогичные свойства родительского антитела. Например, антитело предпочтительно обладает большей агрегационной стабильностью или сильной аффинностью связывания, улучшенными фармакокинетическими свойствами или повышенной способностью ингибировать биологическую активность антигена, чем родительское антитело.
"Родительское VHH антитело", или "исходное VHH антитело", или "дикое VHH антитело" в контексте данной заявки относится к VHH антителу, выделенному из иммунизированного или неиммунизированного животного семейства верблюдовых и кодированному аминокислотной последовательностью, которая используется для получения VHH производного. Родительское антитело может иметь каркасную последовательность происхождения из семейства верблюдовых в части вариабельного домена VHH, но предпочтительно каркасная последовательность вариабельного домена легкой цепи имеет полностью или по существу человеческое происхождение.
"Родительское", или "исходное", или "дикое" антитело в контексте данной заявки представляет собой антитело, кодированное аминокислотной последовательностью, которая используется для получения варианта. Родительское антитело может иметь каркасную последовательность происхождения из семейства верблюдовых в части вариабельного домена VHH, но предпочтительно каркасная последовательность вариабельного домена легкой цепи имеет полностью или по существу человеческое происхождение.
Термин "специфически связывает", как используется в данной заявке, относится к той ситуации, при которой один участник пары специфического связывания не связывает в значительной степени молекулы, отличные от его партнера (партнеров) по специфическому связыванию. Термин также применим, когда, например, антигенсвязывающий домен антитела по изобретению является специфическим для конкретного эпитопа, который переносится рядом антигенов, в таком случае специфическое антитело, имеющее антигенсвязывающий домен, будет способно к специфическому связыванию различных антигенов, несущих эпитоп. Соответственно, моноклональное антитело по изобретению специфически связывает IL-17 человека (т.е. IL-17A), в то время как оно специфически не связывает человеческие IL-17B, IL-17C, IL-17D, IL-17E. Более того, моноклональное антитело по изобретению специфически связывает IL-17 человека и IL-17 макаки-крабоеда, но специфически не связывает IL-17 крысы или IL-17 мыши.
Термин "предпочтительно связывает", как используется в данной заявке, относится к ситуации, в которой антитело связывает специфический антиген по крайней мере приблизительно на 20% больше, предпочтительно по крайней мере приблизительно на 50% и в 2, 20, 50 или 100 раз больше, чем оно связывает иной антиген в соответствии с измерениями, проведенными по методикам, известным из уровня техники, например, конкурентного анализа ELISA или KD измерений при помощи анализов Octet прибора. Антитело может предпочтительно связывать один эпитоп в пределах антигена, а не другой эпитоп в пределах того же самого антигена. Соответственно, антитело по изобретению предпочтительно связывает IL-17 человека, но не IL-17 кролика.
Термин "эпитоп" относится к той части молекулы, которая способна распознаваться и связываться с антителом в одном или более антигенсвязывающих участках антитела. Эпитопы часто состоят из химически активных поверхностных групп молекул, таких как аминокислоты или сахарные боковые цепи, и обладают определенными трехмерными структурными характеристиками. Под "ингибирующим эпитопом" и/или "нейтрализующим эпитопом" подразумевается эпитоп, который в контексте интактной антигенной молекулы и при связывании антителом, специфическим к эпитопу, приводит к утрате или к уменьшению биологической активности молекулы или организма, который содержит молекулу, in vivo или in vitro.
Термин "эпитоп", как используется в данной заявке, кроме того, относится к части полипептида, которая обладает антигенной и/или иммуногенной активностью у животного, предпочтительно млекопитающего, например мыши или человека. Термин "антигенный эпитоп", как используется в данной заявке, является частью полипептида, с которой может специфически связываться антитело, определенная любым способом, хорошо известным из уровня техники, например при помощи традиционного иммунного анализа. Антигенные эпитопы не обязательно должны быть иммуногенными, но могут также быть имуногенными. "Иммуногенный эпитоп", как используется в данной заявке, определяется как часть полипептида, который вызывает отклик антитела у животного, как устанавливается любым способом, известным из уровня техники. "Нелинейный эпитоп" или "конформационный эпитоп" содержат несмежные полипептиды (или аминокислоты) в пределах антигенного протеина, с которым антитело, специфическое к эпитопу, связывается.
Выражения "функциональная активность" или "функциональная характеристика" или термины "биологическая активность" или "биоактивность" по отношению к антителу по изобретению используются в данной заявке как взаимозаменяемые и включают, но не ограничиваются приведенными, эпитоп/антигенную аффинность и специфичность, способность нейтрализовать или быть антагонистом активности IL-17 in vivo или in vitro, IC50, стабильность антитела и иммуногенные свойства антитела in vivo. Остальные идентифицируемые из уровня техники биологические свойства или характеристики антитела включают, например, перекрестную реактивность (т.е. с нечеловеческими гомологами белка-мишени или с остальными белками или даже клетками, в общем) и способность сохранять высокие уровни экспрессии протеина в клетках млекопитающих. Вышеуказанные свойства или характеристики могут измеряться или оцениваться с использованием методик, включая, но не ограничиваясь приведенными, анализ ELISA, конкурентный анализ ELISA, Octet анализа, анализы нейтрализации in vitro или in vivo без ограничений, рецепторного связывания, продуцирования и/или секреции цитокина или фактора роста, сигнальную трансдукцию и иммуногистохимию срезов тканей, полученных из различных источников, включая человека, примата или любой другой источник.
Популяция "моноклональных антител" относится к гомогенной или по существу гомогенной популяции антител (т.е. по крайней мере приблизительно 85, 90, 91, 92, 93, 94, 95, 96%, более предпочтительно по крайней мере приблизительно 97 или 98% или еще более предпочтительно по крайней мере 99% антител в популяции будут конкурировать в анализе ELISA за тот же антиген или эпитоп, или более предпочтительно антитела являются идентичными в аминокислотной последовательности).
Полноразмерное антитело, существующее в природе, представляет собой молекулу иммуноглобулина, которая состоит из четырех пептидных цепей, две тяжелые (Н) цепи (приблизительно 50-70 кДа при полной длине) и две легкие (L) цепи (приблизительно 25 кДа при полной длине), взаимосвязанных дисульфидными мостиками. Аминоконцевая часть каждой цепи включает вариабельный домен из приблизительно 100-110 или более аминокислот, которые отвечают за связывание антигена. Карбоксиконцевая часть каждой цепи определяет константный участок, главным образом, отвечающий за функцию эффектора. Легкие цепи классифицируют как каппа или лямбда, и они характеризуются конкретным константным участком. Каждая легкая цепь состоит из вариабельного участка N-концевой легкой цепи (в данной заявке "VL" или "VK") и константного участка легкой цепи, состоящего из одного домена, CL или СК. Тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон, и они определяют изотип антитела, такой как IgG, IgM, IgE IgA и IgD, соответственно, и несколько из них могут быть дополнительно разделены на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1. Каждый тип тяжелой цепи характеризуется конкретным константным участком. Каждая тяжелая цепь состоит из вариабельного участка N-концевой тяжелой цепи (в данной заявке "VH") и константного участка тяжелой цепи. Константный участок тяжелой цепи состоит из трех доменов (CH1, СН2 и СН3) для IgG, IgD и IgA и 4 доменов (CH1, СН2, СН3 и СН4) для IgM и IgE. Вариабельные домены VH, VHH и VL могут быть дополнительно разделены на участки гипервариабельности, названные гипервариабельными участками (CDR), чередующиеся с более консервативными участками, названными каркасными участками (FR). Каждый вариабельный домен состоит из трех CDR и четырех FR, расположенных в следующем порядке от N-конца к С-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
В данной заявке 3 CDR тяжелых цепей обозначены как "HCDR1, HCDR2 и HCDR3", а 3 CDR легких цепей обозначены как "LCDR1, LCDR2 и LCDR3". CDR содержат большинство остатков, которые образуют специфические взаимодействия с антигеном. Нумерацию и позиционирование CDR-аминокислотных остатков осуществляют по Kabat номенклатуре.
Термин «антиген» относится к мишени антигена, против которой антитело может быть иммунореактивным, используется в контексте данной заявки в своем традиционном понятии, как его используют специалисты в данной области техники, и включает, среди прочих, полипептиды, пептиды, полисахариды, гликопротеины, полинуклеотиды (например, ДНК), или химические антигены, рецепторы, интерлейкины. Интерлейкины могут включать в себя интерлейкины разных групп, в т.ч. интерлейкин 1 (альфа и бета), интерлейкин 2, интерлейкин 3, интерлейкин 4, интерлейкин 5, интерлейкин 6, интерлейкин 7, интерлейкин 8, интерлейкин 9, интерлейкин 10, интерлейкин И, интерлейкин 17, интерлейкин 18, интерлейкин 33.
Термин «антиген» также может использоваться для описания материала, используемого для иммунизации животных (например, лам) при продуцировании антител согласно изобретению. В этом контексте «антиген» может иметь более широкое значение и может охватывать очищенные формы антигена и также неочищенные или не полностью выделенные или очищенные препараты антигена, такие как, например, клетки, лизаты клеток или супернатанты, фракции клеток, например клеточные мембраны и т.д. с добавлением гаптенов, конъюгированных с соответствующим носителем белка. Антиген, используемый в процессе иммунизации, не обязательно означает антиген, структурно идентичный мишени антигена, с которым в результате антитело согласно изобретению может связываться. Как правило, антиген, используемый для иммунизации, может представлять собой усеченный вариант мишени антигена, например фрагмент, содержащий иммуногенный эпитоп. Более подробные характеристики антигенов, используемых для иммунизации, описаны в литературе и могут быть хорошо известны специалисту в данной области техники.
Вариабельные участки каждой из пар легкая/тяжелая цепь образуют антигенсвязывающие сайты антитела. Таким образом, интактное IgG антитело имеет два сайта связывания. За исключением бифункциональных или биспецифических антител два сайта связывания являются одинаковыми. Как используется в данной заявке, "антигенсвязывающая часть", или "антигенсвязывающий участок", или "антигенсвязывающий домен" относятся, взаимозаменяемо, к такой части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и обуславаливающие специфичность и аффинность антитела по отношению к антигену. Такая часть антитела включает "каркасные" аминокислотные остатки, необходимые для поддержания надлежащей конформации антигенсвязывающих остатков.
Предпочтительно CDR антигенсвязывающего участка VHH или весь антигенсвязывающий участок антител по изобретению полностью происходит от семейства верблюдовых или по существу происходит от семейства верблюдовых, содержит определенные аминокислотные остатки, измененные, например замещенные разными аминокислотными остатками (см., например, табл. 6) с тем, чтобы оптимизировать конкретные свойства антитела, например KD, koff, IC50. Предпочтительно каркасные участки антитела по изобретению имеют происхождение от семейства верблюдовых или человека или по существу человеческое происхождение (по крайней мере на 80, 85, 90, 95, 96, 97, 98 или 99% человеческое происхождение) и соответствуют номенклатуре Kabat.
«Фрагмент антитела» может представлять собой фрагмент антитела или фрагмент антитела, имеющий активность полноразмерного антитела. Указанный фрагмент антитела может представлять собой F(ab')2, F(ab)2, Fab', Fab Fv и scFv.
"Интерлейкин 17", или также "IL-17" или "IL-17 А", является гомодимерным белком с массой 20-30 кДа. Ген IL-17A человека кодирует 155-аминокислотный протеин, который имеет 19-аминокислотную сигнальную последовательность и 136-аминокислотный зрелый сегмент. Аминокислотная последовательность IL-17A человека на 80%, 63% и 58% идентична аминокислотным последовательностям кролика, мыши и крысы, соответственно. Аминокислотная последовательность IL-17A человека на 97% идентична последовательности интерлейкина-17А макаки-крабоеда.
Термин "антитело" при употреблении в связи с анти-IL-17 моноклональным антителом по изобретению (либо упрощенно, "моноклональное антитело по изобретению"), как используется в данной заявке, относится к моноклональному антителу.
Термин "ингибировать" или "нейтрализовать", как используется в данной заявке, по отношению к функциональной активности антитела по изобретению, означает способность в значительной степени препятствовать, предотвращать, ограничивать, замедлять, прекращать, уменьшать или обращать, например, развитие или тяжесть того, что ингибируют, включая, но не ограничиваясь вышеприведенными, биологическую активность (например, активность IL-17) или свойство, заболевание или состояние. Ингибирование или нейтрализация активности IL-17 в результате связывания антитела по изобретению с IL-17 составляет предпочтительно по крайней мере приблизительно 20, 30, 40, 50, 60, 70, 80, 90, 95% или выше.
Термин "выделенный" или "изолированный" при использовании по отношению к нуклеиновой кислоте или белковому препарату (например, антителу) относится к молекуле нуклеиновой кислоты или белковой молекуле, которые идентифицируют и отделяют по крайней мере от одного контаминантного вещества, с которым она обычно связана в природном источнике. Предпочтительно "выделенное антитело" является антителом, которое по существу не содержит другие антитела, обладающие отличительной антигенной специфичностью (например, фармацевтические композиции согласно изобретению содержат выделенное антитело, которое специфически связывает IL-17 и по существу не содержит антитела, которые специфически связывают антигены, отличные от IL-17).
Термины "Kabat номенклатура" или «номенклатура по Kabat» применяются в данной заявке к системе нумерации аминокислотных остатков, которые являются более вариабельными (т.е. гипервариабельными), чем остальные аминокислотные остатки в вариабельных участках тяжелой и легкой цепи антитела (Kabat et al. Ann. NY Acad. Sci., 190:382-93 (1971); Kabat et al. Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication №91-3242 (1991)).
Полинуклеотид является "функционально связанным", если он имеет функциональные связи с другим полинуклеотидом. Например, промотор или энхансер функционально связаны с кодирующей последовательностью, если они влияют на транскрипцию последовательности. Полипептид "функционально связан" с другим полипептидом, если полинуклеотиды, кодирующие их, связаны функционально, предпочтительно, если они находятся в той же открытой рамке считывания.
Термин «конструкция ДНК» в контексте данной заявки относится к ДНК или ее фрагменту, кодирующей антитело согласно изобретению. Обычно ДНК или ее фрагмент, который кодирует антитело, например описанное в настоящей заявке, функционально (операбельно) связывают в рамке считывания по меньшей мере с одним другим фрагментом ДНК, кодирующим дополнительный полипептид (например, домен или район из рецептора другого цитокина, такого как IL-2-рецептор), и встраивают в подходящий экспрессирующий вектор. Обычно конструкции ДНК конструируют таким образом, что несколько фрагментов ДНК, кодирующих соответствующие сайты антитела, функционально связывают в рамке считывания для получения единой конструкции, кодирующей полностью антитело или его функциональный фрагмент. Например, конструкция ДНК кодировала бы от N-конца до С-конца антитело. Такие антитела могут быть экспрессированы, выделены и исследованы на активность.
Термин "вектор", "плазмида" относится к нуклеиновым кислотам, полученным синтетическим путем и методами генной инженерии и содержат определенный, известный в данной области, набор функциональных элементов последовательности. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены, в то время как остальные векторы могут быть интегрированы в геном клетки-хозяина и реплицироваться вместе с геномом-хозяином. Более того, определенные векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы имеют в данной заявке название "векторы для рекомбинантной экспрессии" (или, упрощенно, "экспрессионные векторы"). Примеры таких векторов хорошо известны из уровня техники.
Как используется в данной заявке, термины "клетка", "клетка-хозяин", "линия клеток" и "клеточная культура" используются как взаимозаменяемые и включают индивидуальную клетку или клеточную культуру, являющиеся реципиентом любого выделенного полинуклеотида по изобретению или любого рекомбинантного вектора (любых рекомбинантных векторов), которые содержат последовательность антитела по изобретению. Клетки-хозяева включают потомство индивидуальной клетки-хозяина, и потомство может не обязательно быть полностью идентичным (по морфологии или полному ДНК комплементу) оригинальной родительской клетке из-за природных, случайных или преднамеренных мутаций и/или изменений. Клетка-хозяин включает клетки, трансформированные, трансдуцированные или инфицированные рекомбинантным вектором, или моноклональное антитело, которое экспрессирует полинуклеотид по изобретению или его легкую или тяжелую цепь. Клетка-хозяин, которая содержит рекомбинантный вектор по изобретению (как стабильно включенный в хромосом-хозяин, так и не включенный), также может называться рекомбинантной клеткой-хозяином". Предпочтительными клетками-хозяевами для использования в изобретении являются СНО клетки (например, АТСС CRL-9096), NS0 клетки, SP2/0 клетки, COS клетки (АТСС, например, CRL-1650, CRL-1651) и HeLa (АТСС CCL-2). Дополнительные клетки-хозяева для использования в изобретении включают растительные клетки, дрожжевые клетки, другие клетки млекопитающих и прокариотические клетки.
Термин «Специфическое связывание» между антителом и мишенью антигена (антигеном) относится к иммунологической специфичности.
Антитело может специфически связываться с мишенью антигена, если оно связывает эпитоп на антигене в большей степени, чем другие эпитопы на антигене. Специфическое связывание не исключает кросс-реактивность с другими антигенами, несущими сходные эпитопы антигенов.
VL домены в антителах согласно изобретению могут быть типа VL лямбда, или VL каппа. Термин «VL домен» относится к обоим изотипам VK каппа и VL лямбда, которые содержат одну или более аминокислотных замен, инсерций или делеций.
Термин «фармацевтическая композиция» относится к композиции и/или составу, содержащему антитело согласно изобретению в терапевтически эффективном количества и вспомогательные вещества (эксцепиенты, разбавители, наполнители, растворители и вспомогательные вещества, например).
Термин «применение» или «способ лечения» относится к возможности применения антитела согласно изобретению или фармацевтической композиции, его содержащей, для лечения, облегчения течения заболеваний, для ускорения ремиссии, снижения частоты рецидивов вследствие заболеваний или нарушений, опосредуемых рецепторами, с которыми может связываться антитело согласно изобретению.
Описание изобретения
Согласно настоящему изобретению, предложены гуманизированные моноклональные антитела, предпочтительно типа IgG, обладающие повышенной аффинностью и улучшенной агрегационной стабильностью, в котором вариабельные домены представлены комбинацией производного VHH с вариабельным доменом легкой цепи VL.
В одном из вариантов реализации производное VHH антитела согласно изобретению может содержать аминокислотные замены в позициях 44Х245Х3, где Х2=G, А, V, S, Т; а Х3=А, V, Т, Н; или их комбинации, а 44 и 45 обозначают позиции для аминокислотных замен. Здесь и далее в тексте заявке при указании позиции для аминокислотной замены применяется система нумерации по Kabat (http://www.bioinf.org.uk/abs/).
В другом варианте реализации антитело согласно изобретению имеет повышенную агрегационную стабильность производного VHH в сравнении с исходным IgG антителом, содержащим VHH, выделенным из иммунизированного животного, при этом указанным животным может быть животное семейства Camelidae.
В еще одном варианте реализации изобретения антитело включает производное VHH, которое представляет собой вариабельный домен тяжелой цепи антитела, выделенный из иммунизированного животного семейства Camelidae. При этом производное VHH может иметь дополнительные аминокислотные замены, характерные для человека в любых позициях, исключая указанные следующие аминокислотные замены в позиции 44 и 45:
a) 44Х2, где Х2=G, А, V, S, Т;
b) 45Х3, где Х3=А, V, Т, Н;
или их комбинации.
В еще одном варианте реализации антитело согласно изобретению включает производное VHH, которое может представлять собой вариабельный домен тяжелой цепи, выделенный из неиммунизированного животного группы Camelidae. При этом производное VHH может иметь дополнительные аминокислотные замены, характерные для человека в любых позициях, исключая указанные следующие аминокислотные замены в позиции 44 и 45:
a) 44Х2, где Х2=G, А, V, S, Т;
b) 45Х3, где Х3=А, V, Т, Н;
или их комбинации.
В еще одном варианте реализации антитело согласно изобретению включает вариабельный домен легкой цепи VL, который является производным антитела человека. В дополнительном варианте реализации вариабельный домен легкой цепи VL является гуманизированным фрагментом антитела животного.
В другом варианте реализации антитело согласно изобретению включает производное VHH, которое содержит цистеин в позиции 44 согласно номенклатуре Kabat, а вариабельный домен легкой цепи VL содержит цистеин в позиции 100 согласно номенклатуре Kabat.
В еще одном варианте реализации изобретения антитело представляет собой антитело любого из изотипов IgG1, IgG2, IgG3 или IgG4.
В другом варианте реализации антитело согласно изобретению содержит неприродный модифицированный фрагмент Fc в составе IgG.
В еще одном варианте реализации антитело согласно изобретению имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и хранении при Т=4°С в течение более чем 6 месяцев содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе. В дополнительном варианте реализации антитело имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и при повышении температуры до 37°С в течение более чем 2 недель содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе. В еще одном дополнительном варианте реализации антитело имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и при повышении температуры до 42°С в течение более чем 48 часов содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе. В другом дополнительном варианте реализации антитело имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и повышении температуры до 50°С в течение более чем 6 часов содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
В одном варианте реализации антитело имеет константу диссоциации KD не более 10-9 М. В дополнительном варианте реализации антитело имеет кинетическую константу ассоциации с антигеном kon(1/Mc) не менее 105 1/Мс. В другом дополнительном варианте реализации антитело имеет кинетическую константу диссоциации с антигеном dis(1/c) не более 10-4 1/с.
В одном варианте реализации согласно изобретению предложен фрагмент антитела. При этом фрагмент антитела может представлять собой легкую цепь, тяжелую цепь, вариабельные домены легкой и/или тяжелой цепи, являющиеся частью последовательности антитела, в том числе и биспецифического варианта антитела. В еще одном варианте реализации фрагмент антитела может представлять собой легкую цепь, тяжелую цепь, вариабельные домены легкой и/или тяжелой цепи, являющиеся компонентами Fab-фрагмента. В другом варианте реализации фрагмент антитела может представлять собой легкую цепь, тяжелую цепь, вариабельные домены легкой и/или тяжелой цепи, являющиеся компонентами scFv-фрагмента.
Согласно изобретению также предложены способы получения антитела согласно изобретению. Указанный способ получения антитела может включать стадии, выбранные из следующих: направленный мутагенез, дисплейные методы, методы генной инженерии, биохимии и высокопроизводительных биотехнологических методов, известных в данной области, которые могут также включать методы направленного мутагенеза VHH домена антитела семейства Camelidae в различных позициях.
Кроме того, согласно изобретению, предложена конструкция ДНК, кодирующая антитело согласно изобретению или его фрагмент. Также предложен экспрессионный вектор, содержащий одну или несколько конструкций ДНК согласно изобретению.
Также в другом аспекте изобретения предложена клеточная линия, которая содержит в клетках экспрессионный вектор или конструкцию ДНК, указанные ранее.
Дополнительно, согласно изобретению предложен способ получения гуманизированного моноклонального антитела или его фрагмента, который включает культивирование клеточной линии в культуральной среде в условиях, достаточных для получения указанного антитела или его фрагмента, с последующим выделением и очисткой полученного антитела или его активного фрагмента.
Кроме того, согласно изобретению предложена фармацевтическая композиция, содержащая антитело или его фрагмент антитела, в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами, разбавителями или носителями. Более подробно технологии получения композиций описаны в специальных руководствах по биотехнологии, например [25].
В еще одном варианте реализации изобретения предложено антитело, которое специфически связывается с IL-17A человека, или его фрагмент в составе активной молекулы, содержащий:
a) производное вариабельного домена тяжелой цепи (VHH) антител животных семейства Camelidae, который содержит 3 гипервариабельных участка HCDR1, HCDR2 и HCDR3, где:
HCDR1 включает аминокислотную последовательность, представленную SEQ ID NO: 1:
G-T-F-A-T-X32-X33-X34-Х35 (согласно номенклатуре Kabat), где
Х32 представляет собой аминокислоту, выбранную из группы, включающей S, N, К, R, Е, W, Q, D, А, V и F;
Х33 представляет собой аминокислоту, выбранную из группы, включающей Р или S;
Х34 представляет собой аминокислоту, выбранную из группы, включающей М и I;
Х35 представляет собой аминокислоту, выбранную из группы, включающей G, N, S, A, L, I, R, V и Q;
HCDR2 включает аминокислотную последовательность SEQ ID NO: 2:
X50-I-X52-X52a-S-G-X55-D-R-I-Y-A-D-S-V-K-G, где
Х50 представляет собой аминокислоту, выбранную из группы, включающей A, G и L;
Х52 представляет собой аминокислоту, выбранную из группы, включающей S, D и Е;
Х52а представляет собой аминокислоту, выбранную из группы, включающей Р и А;
Х55 представляет собой аминокислоту, выбранную из группы, включающей G,S,T, L, R, D, Е, К, А и W;
HCDR3 включает аминокислотную последовательность SEQ ID NO: 3:
С-А-Х94-Х95-Х96-X97-F-X99-X100-Х100а-X100b-Х100с-X100d-Х100е-X100f-D-Y-D-S, где
Х94 представляет собой аминокислоту, выбранную из группы, включающей К, S, Т, V, D и G;
Х95 представляет собой аминокислоту, выбранную из группы, включающей R и К;
Х96 представляет собой аминокислоту, выбранную из группы, включающей G, R, Y, Н, D, W и К;
Х97 представляет собой аминокислоту, выбранную из группы, включающей R, А, V, S, L и Н;
Х99 представляет собой аминокислоту, выбранную из группы, включающей D, Е, G, A, R, V, К и Q;
X100 представляет собой аминокислоту, выбранную из группы, включающей G, S и N;
X100a представляет собой аминокислоту, выбранную из группы, включающей G, Т, Р, V, R, N и К;
X100b представляет собой аминокислоту, выбранную из группы, включающей V, S, Т, L, Y, А, Н, G и I;
Х100с представляет собой аминокислоту, выбранную из группы, включающей Y, W и S;
X100d представляет собой аминокислоту, выбранную из группы, включающей R, V, L, Y, A, W, К, G, Q и I;
Х100е представляет собой аминокислоту, выбранную из группы, включающей Т, L, А и S;
X100f представляет собой аминокислоту, выбранную из группы, включающей Т, L, G, Р, N, A, Q, F, I и D;
b) вариабельный домен легкой цепи (VL) антитела человека или вариабельный домен легкой цепи гуманизированного антитела.
В дополнительном варианте реализации антитело или его фрагмент, которое специфически связывается с IL-17А человека, содержит:
a) производное вариабельного домена тяжелой цепи (VHH), который содержит 3 гипервариабельных участка HCDR1, HCDR2 и HCDR3, где:
HCDR1 включает последовательность G-T-F-A-T-S-P-M-G (SEQIDNO:4);
HCDR2 включает последовательность A-I-S-P-S-G-G- D-R-I-Y-A-D-S-V-K-G (SEQ ID NO: 5);
HCDR3 включает последовательность C-A-V-R-R-R-F-D-G-T-S-Y-Y-T-G-D-Y-D-S (SEQ ID NO: 6);
b) вариабельный домен легкой цепи (VL) антитела человека или вариабельный домен легкой цепи гуманизированного антитела.
В еще одном варианте реализации антитело, которое специфически связывается с IL-17A человека, содержит производное вариабельного домена тяжелой цепи (VHH), который содержит аминокислотную последовательность SEQ ID NO: 7; и
вариабельный домен легкой цепи (VL) антитела человека или вариабельный домен легкой цепи гуманизированного антитела.
В дополнительном варианте реализации изобретения антитело, которое специфически связывается с IL-17A человека, или его фрагмент, содержит производное VHH, где указанный вариабельный домен включает аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности SEQ ID NO: 7.
В другом дополнительном варианте реализации антитело, которое специфически связывается с IL-17A человека, содержит производное вариабельного домена тяжелой цепи (VHH), который включает аминокислотную последовательность SEQ ID NO: 7; и вариабельный домен легкой цепи (VL) антитела человека, который включает аминокислотную последовательность SEQ ID NO: 8.
В еще одном дополнительном варианте реализации антитело, которое специфически связывающееся с IL-17A человека, или его фрагмент, содержит производное вариабельного домена тяжелой цепи (VHH) и вариабельный домен легкой цепи (VL) антитела человека, где указанные вариабельные домены включают аминокислотную последовательность по меньшей мере на 90% идентичную SEQ ID NO: 8.
В другом варианте реализации антитело специфически связывается с IL-17A человека, содержит тяжелую цепь, которая содержит который включает аминокислотную последовательность SEQ ID NO: 9 и вариабельный домен легкой цепи (VL) антитела человека, который включает аминокислотную последовательность SEQ ID NO: 10. В еще одном варианте реализации антитело, которое специфически связывается с IL-17A человека, содержащее тяжелую цепь и легкую цепь, где указанные цепи включают аминокислотные последовательности, по меньшей мере на 90% идентичные SED ID NO: 9 и/или SEQ ID NO: 10.
В другом варианте реализации антитело согласно изобретению, которое специфически связывается с IL-17A человека, имеет аффинность связывания с IL-17A человека с KD не более 10-10 М. В еще одном варианте реализации антитело, которое специфически связывается с IL-17A человека, имеет кинетическую константу ассоциации kon(1/Ms) для IL-17A человека не менее 105 1/Ms. В еще одном варианте реализации антитело, которое специфически связывается с IL-17A человека, имеет кинетическую константу диссоциации dis(1/c) для IL-17A человека не более 10-5 1/с. В дополнительном варианте реализации антитело, которое специфически связывается с IL-17A человека и способно ингибировать не менее чем на 50% активность IL-17A человека по любому из параметров, оцениваемых в любых тестах на специфическую активность.
В другом варианте реализации антитело, которое специфически связывается с IL-17A человека, продуцируют в клетках млекопитающих, дрожжевых или бактериальных клетках.
В еще одном варианте реализации антитело, которое специфически связывается с IL-17A человека, дополнительно содержит одну и более аминокислотных замен в Fc-регионе в сравнении с природным вариантом Fc, где указанные замены улучшают физико-химические и фармакокинетические свойства антитела по отношению к антителу с природным Fc-фрагментом и не приводят к потере способности антитела связывать IL-17А.
В другом аспекте изобретения предложена конструкция ДНК, кодирующая антитело, которое специфически связывается с IL-17А человека. Кроме того, предложен экспрессионный вектор, содержащий одну или несколько конструкций ДНК, кодирующих антитело, которое специфически связывается с IL-17A человека. Дополнительно, предложена клетка-хозяин, содержащая вектор для получения антитела, которое специфически связывается с IL-17А человека.
В дополнение, согласно изобретению, предложен способ получения антитела, которое специфически связывается с IL-17A человека, заключающийся в культивировании клетки-хозяина, содержащей конструкцию ДНК, в культуральной среде в условиях, достаточных для получения указанного антитела или его фрагмента, с последующим выделением и очисткой полученного антитела или его активного фрагмента.
Также предложена фармацевтическая композиция, содержащая антитело, которое специфически связывается с IL-17A человека, в комбинации с одним или несколькими фармацевтически приемлемыми эксципиентами, разбавителями или носителями. При этом в качестве дополнительных активных веществ фармацевтическая композиция может содержать дополнительные действующие вещества, выбранные из блокаторов ФНО-α. Данная композиция может быть использована для лечения опосредуемого IL-17A заболевания или нарушения. Опосредуемое IL-17A заболевание или нарушение выбрано из ревматоидного артрита, остеоартрита, ювенильного хронического артрита, септического артрита, артрита Лайма, псориатического артрита, реактивного артрита, спондилоартропатии, системной красной волчанки, болезни Крона, язвенного колита, воспалительного заболевания кишечника, инсулинзависимого сахарного диабета, тиреоидита, астмы, аллергических заболеваний, псориаза, дерматита, склеродермии, реакции "трансплантат против хозяина", отторжения трансплантата органа, острого или хронического иммунного заболевания, связанного с трансплантацией органа, саркоидоза, атеросклероза, диссеминированного внутрисосудистого свертывания, болезни Кавасаки, болезни Грэйвса, нефротического синдрома, синдрома хронической усталости, гранулематоза Вегенера, пурпуры Шенлейна-Геноха, микроскопического васкулита почек, хронического активного гепатита, увеита, септического шока, синдрома токсического шока, септического синдрома, кахексии, инфекционных заболеваний, паразитарных заболеваний, синдрома приобретенного иммунодефицита, острого поперечного миелита, хореи Гентингтона, болезни Паркинсона, болезни Альцгеймера, инсульта, первичного билиарного цирроза, гемолитической анемии, злокачественных опухолей, сердечной недостаточности, инфаркта миокарда, болезни Аддисона, спорадического полигландулярного дефицита типа I и полигландулярного дефицита типа II, синдрома Шмидта, взрослого (острого) респираторного дистресс-синдрома, алопеции, очаговой алопеции, серонегативной артропатии, артропатии, болезни Рейтера, псориатической артропатии, связанной с язвенным колитом артропатии, энтеропатического синовита, связанной с хламидиями, иерсиниями и сальмонеллами артропатии, спондилоартропатии, атероматозного заболевания/артериосклероза, атопической аллергии, аутоиммунного буллезного заболевания, пемфигуса обыкновенного, листовидного пемфигуса, пемфигоида, болезни линейных IgA, аутоиммунной гемолитической анемии, Кумбс-положительной гемолитической анемии, приобретенной перниционзой анемии, ювенильной пернициозной анемии, миалгического энцефалита/синдрома хронической усталости, хронического кожно-слизистого кандидоза, гигантоклеточного артериита, первичного склерозирующего гепатита, криптогенного аутоиммунного гепатита, синдрома приобретенного иммунодефицита, связанных с приобретенным иммунодефицитом заболеваний, гепатита В, гепатита С, вариабельного неклассифицируемого иммунодефицита (вариабельной неклассифицируемой гипогаммаглобулинемии), кардиомиопатии с дилятацией, женского бесплодия, недостаточности яичников, преждевременного угасания функции яичников, фиброзного заболевания легких, криптогенного фиброзного альвеолита, поствоспалительного интерстициального заболевания легких, интерстициального пневмонита, связанного с болезнью соединительной ткани интерстициального заболевания легких, связанного со смешанной болезнью соединительной ткани заболевания легких, связанного с системной склеродермией заболевания легких, связанного с ревматоидным артритом интерстициального заболевания легких, связанного с системной красной волчанкой заболевания легких, связанного с дерматомиозитом/полимиозитом заболевания легких, связанного с болезнью Шегрена, заболевания легких, связанного с анкилозирующим спондилитом заболевания легких, васкулитного диффузного заболевания легких, связанного с гемосидерозом заболевания легких, индуцированного лекарственным средством интерстициального заболевания легких, фиброза, связанного с радиацией фиброза, облитерирующего бронхиолита, хронической эозинофильной пневмонии, заболевания легких с инфильтрацией лимфоцитов, постинфекционного интерстициального заболевания легких, подагрического артрита, аутоиммунного гепатита, аутоиммунного гепатита I типа (классического аутоиммунного или люпоидного гепатита), аутоиммунного гепатита II типа (гепатита, связанного с антителом против LKM), опосредуемой аутоиммунным заболеванием гипогликемии, устойчивости к инсулину типа В с акантокератодермией, гипопаратиреоза, острого иммунного заболевания, связанного с трансплантацией органа, хронического иммунного заболевания, связанного с трансплантацией органа, остеоартроза, первичного склерозирующего холангита, псориаза I типа, псориаза II типа, идиопатической лейкопении, аутоиммунной нейтропении, NOS-болезни почек, гломерулонефрита, микроскопического васкулита почек, болезни Лайма, дискоидной красной волчанки, идиопатического или NOS-мужского бесплодия, аутоиммунитета к сперматозоидам, рассеянного склероза (все подтипы), симпатической офтальмии, легочной гипертензии, вторичной для болезни соединительной ткани, синдрома Гудпасчера, легочного проявления узелкового полиартериита, острой ревматической атаки, ревматоидного спондилита, болезни Стилла, системной склеродермии, синдрома Шенгрена, болезни/артериита Такаясу, аутоиммунной тромбоцитопении, идиопатической тромбоцитопении, аутоиммунного заболевания щитовидной железы, гипертиреоидизма, зобного аутоиммунного гипотиреоза (болезнь Хашимото), атрофического аутоиммунного гипотиреоза, первичной микседемы, факогенного увеита, первичного васкулита, витилиго, острого заболевания печени, хронического заболевания печени, алкогольного цирроза, индуцированного алкоголем повреждения печени, холестаза, идиосинкразического заболевания печени, индуцированного лекарственным средством гепатита, неалкогольного стеатогепатита, аллергии и астмы, стрептококковой инфекции группы В (GBS), психических расстройств (включая депрессию и шизофрению), опосредуемых типом Th2 и типом Th1 заболеваний, острой и хронической боли (различные формы боли), и злокачественных опухолей, таких как рак легкого, молочной железы, желудка, мочевого пузыря, толстого кишечника, поджелудочной железы, яичника, предстательной железы и прямой кишки и гемопоэтические злокачественные опухоли (лейкоз и лимфома), абеталипопротеинемии, акроцианоза, острых и хронических паразитарных и инфекционных процессов, острого лейкоза, острого лимфобластного лейкоза, острого миелоидного лейкоза, острой или хронической бактериальной инфекции, острого панкреатита, острой почечной недостаточности, аденокарцином, эктопической систолы предсердий, СПИД-дементного комплекса, индуцированного алкоголем гепатита, аллергического конъюнктивита, аллергического контактного дерматита, аллергического ринита, отторжения аллотрансплантата, дефицита альфа-1-антитрипсина, бокового амиотрофического склероза, анемии, стенокардии, дегенерации клеток передних рогов спинного мозга, терапию против CD3, антифосфолипидного синдрома, реакций гиперчувствительности против рецепторов, аортальных и периферических аневризм, расслоения аорты, артериальной гипертензии, артериосклероза, артериовенозного свища, атаксии, фибрилляции предсердий (постоянной или пароксизмальной), трепетания предсердий, атриовентрикулярной блокады, В-клеточной лимфомы, отторжения трансплантата кости, отторжения трансплантата костного мозга (ВМТ), блокады пучка Гиса, лимфомы Беркитта, ожогов, аритмий сердца, синдрома оглушения сердца, опухолей сердца, кардиомиопатии, воспалительных ответов на искусственное кровообращение, отторжения трансплантата хряща, дегенерации коры головного мозга, нарушений мозжечка, хаотической или многоочаговой тахикардии предсердий, связанных с химиотерапией нарушений, хронического миелоцитарного лейкоза (CML), хронического алкоголизма, хронических воспалительных патологий, хронического лимфоцитарного лейкоза (ХЛЛ), хронического обструктивного заболевания легких (ХОЗЛ), хронической интоксикации салицилатами, карциномы ободочной и прямой кишки, застойной сердечной недостаточности, конъюнктивита, контактного дерматита, легочного сердца, болезни коронарных артерий, болезни Крейтцфельда-Якоба, культурально-негативного сепсиса, кистозного фиброза, связанных с цитокиновой терапией нарушений, деменции боксеров, демиелинизирующих заболеваний, геморрагической лихорадки денге, дерматита, дерматологических состояний, диабета, сахарного диабета, диабетического атеросклеротического заболевания, диффузного заболевания с тельцами Леви, застойной кардиомиопатии с дилятацией, нарушений базальных ганглиев, синдрома Дауна в среднем возрасте, двигательных нарушений, индуцированных лекарственным средством, которое блокирует дофаминовые рецепторы ЦНС, чувствительности к лекарственным средствам, экземы, энцефаломиелита, эндокардита, эндокринопатии, эпиглоттита, инфекции вирусом Эпштейна-Барр, эритромелалгии, экстрапирамидальных и мозжечковых нарушений, семейного гематофагоцитарного лимфогистиоцитоза, отторжения имплантата эмбрионального тимуса, наследственной атаксии Фридрейха, функциональных нарушений периферических артерий, грибкового сепсиса, газовой гангрены, язвы желудка, гломерулонефрита, отторжения трансплантата любого органа или ткани, грамотрицательного сепсиса, грамположительного сепсиса, гранулем вследствие внутриклеточных организмов, волосатоклеточного лейкоза, болезни Галлервордена-Шпатца, тиреоидита Хашимото, сенной лихорадки, отторжения трансплантата сердца, гемахроматоза, гемодиализа, гемолитического уремического синдрома/тромболитической тромбоцитопенической пурпуры, кровопотери, гепатита (А), аритмий пучка Гиса, ВИЧ-инфекции/ВИЧ-невропатии, болезни Ходжкина, гиперкинетических двигательных нарушений, реакций гиперчувствительности, связанного с гиперчувствительностью пневмонита, гипертензии, гипокинетических двигательных нарушений, обследования гипоталамо-гипофизарно-надпочечниковой системы, идиопатической болезни Аддисона, идиопатическоого фиброза легких, опосредуемой антителами цитотоксичности, астении, младенческой спинальной мышечной атрофии, воспаления аорты, вируса гриппа А, облучения ионизирующей радиацией, иридоциклита/увеита/оптического неврита, повреждения при ишемии-реперфузии, ишемического инсульта, ювенильного ревматоидного артрита, ювенильной спинальной мышечной атрофии, саркомы Капоши, отторжения трансплантата почки, легионеллеза, лейшманиоза, лепры, повреждений кортикоспинальной системы, жирового отека, отторжения трансплантата печени, лимфатического отека, малярии, злокачественной лимфомы, злокачественного гистиоцитоза, злокачественной меланомы, менингита, менингококкемии, метаболических/идиопатических заболеваний, мигрени, митохондриального полисистемного нарушения, смешанной болезни соединительной ткани, моноклональной гаммапатии, множественной миеломы, полисистемной дегенерации (Менцеля, Дежерина-Тома, Шая-Дрейджера и Мачадо-Джозефа), миастении, внутриклеточных Mycobacterium avium, Mycobacterium tuberculosis, миелодиспластического синдрома, инфаркта миокарда, ишемических нарушений миокарда, карциномы носоглотки, хронического заболевания легких новорожденных, нефрита, нефроза, нейродегенеративных заболеваний, нейрогенных мышечных атрофий I, нейтропенической лихорадки, неходжкинских лимфом, окклюзии брюшного отдела аорты и ее ветвей, окклюзионных нарушений артерий, терапии ОКТ3®, орхита/эпидидимита, орхита/возвратных процедур после вазэктомии, органомегалии, остеопороза, отторжения трансплантата поджелудочной железы, карциномы поджелудочной железы, паранеопластического синдрома/гиперкальцемии при злокачественной опухоли, отторжения трансплантата паращитовидной железы, воспалительного заболевания органов таза, круглогодичного ринита, заболевания перикарда, периферического артериосклеротического заболевания, периферических сосудистых нарушений, перитонита, пернициозной анемии, пневмонии Pneumocystiscarinii, пневмонии, синдрома POEMS (полиневропатия, органомегалия, эндокринопатия, моноклональная гаммапатия и синдром кожных изменений), постперфузионного синдрома, синдрома после искусственного кровообращения, посткардиотомного синдрома после инфаркта миокарда, преэклампсии, прогрессирующего супрануклеарного паралича, первичной гипертензии легких, лучевой терапии, феномена и болезни Рейно, болезни Рейно, болезни Рефсума, регулярной тахикардии с узким комплексом QRS, вазоренальной гипертензии, реперфузионного повреждения, рестриктивной кардиомиопатии, сарком, склеродермии, сенильной хореи, сенильной деменции с тельцами Леви, серонегативных артропатий, шока, серповидноклеточной анемии, отторжения аллотрансплантата кожи, синдрома кожных изменений, отторжения трансплантата тонкого кишечника, солидных опухолей, специфических аритмий, спинальной атаксии, спиномозжечковых дегенераций, стрептококкового миозита, структурных повреждений мозжечка, подострого склерозирующего панэнцефалита, обмороков, сифилиса сердечно-сосудистой системы, системной анафилаксии, синдрома системного воспалительного ответа, ювенильного ревматоидного артрита с системным началом, Т-клеточного или FAB ALL, телеангиэктазии, облитерирующего тромбоангита, тромбоцитопении, токсичности, трансплантации, травмы/кровопотери, реакций гиперчувствительности типа III, гиперчувствительности типа IV, нестабильной стенокардии, уремии, уросепсиса, крапивницы, заболеваний клапанов сердца, варикоза вен, васкулита, заболеваний вен, венозного тромбоза, фибрилляции желудочков, вирусных и грибковых инфекций, энцефалита с высоким риском смертельного исхода/асептического менингита, гемофагоцитарного синдрома с высоким риском смертельного исхода, синдрома Вернике-Корсакова, болезни Вилсона, отторжения ксенотрансплантата любого органа или ткани, острого коронарного синдрома, острого идиопатического полиневрита, острой воспалительной демиелинизирующей полирадикулоневропатии, острой ишемии, болезни Стилла взрослых, очаговой алопеции, анафилаксии, синдрома антифосфолипидных антител, апластической анемии, артериосклероза, атопической экземы, атопического дерматита, аутоиммунного дерматита, аутоиммунного нарушения, связанного со стрептококковой инфекцией, аутоиммунной энтеропатии, аутоиммунной потери слуха, аутоиммунного лимфопролиферативного синдрома, аутоиммунного миокардита, аутоиммунного преждевременного угасания функции яичников, блефарита, бронхоэктазов, буллезного пемфигоида, сердечно-сосудистого заболевания, катастрофического антифосфолипидного синдрома, глютеиновой болезни, шейного спондилеза, хронической ишемии, рубцового пемфигоида, клинически изолированного синдрома (cis) с риском рассеянного склероза, конъюнктивита, психиатрического нарушения с началом в детском возрасте, хронического обструктивного заболевания легких (ХОЗЛ), дакриоцистита, дерматомиозита, диабетической ретинопатии, сахарного диабета, грыжи межпозвоночного диска, пролапса межпозвоночного диска, индуцированной лекарственным средством иммунной гемолитической анемии, эндокардита, эндометриоза, эндофтальмита, эписклерита, полиформной эритемы, тяжелой полиформной эритемы, гестационного пемфигоида, синдрома Гийена-Барре, сенной лихорадки, синдрома Хьюза, идиопатической болезни Паркинсона, идиопатической интерстициальной пневмонии, опосредуемой IgE аллергии, иммунной гемолитической анемии, миозита с тельцами включения, инфекционного воспалительного заболевания глаз, воспалительного демиелинизирующего заболевания, воспалительного заболевания сердца, воспалительного заболевания почек, идиопатического пневмосклероза/идиопатического легочного фиброза, ирита, кератита, сухого кератоконъюнктивита, болезни Куссмауля или болезни Куссмауля-Мейера, паралича Ландри, гистиоцитоза клеток Лангерганса, синдрома мраморной кожи, дегенерации желтого пятна, микроскопического полиангиита, болезни Бехтерева, нарушений двигательных нейронов, пемфигоида слизистых оболочек, полиорганной недостаточности, миастении, миелодиспластического синдрома, миокардита, нарушений корешков нервов, невропатии, не-А не-В гепатита, оптического неврита, остеолизиса, рака яичника, олигоартикулярного ювенильного ревматоидного артрита, окклюзионого заболевания периферических артерий, заболевания периферических сосудов, заболевания периферических артерий (PAD), флебита, узелкового полиартериита (или нодозного полиартериита), полихондрита, ревматической полимиалгии, полиоза, полиартикулярного ювенильного ревматоидного артрита, синдрома полиэндокринной недостаточности, полимиозита, ревматической полимиалгии (PMR), синдрома после искусственного кровообращения, первичного паркинсонизма, рака предстательной железы и прямой кишки и гемопоэтических злокачественных опухолей (лейкоз и лимфому), простатита, истинной эритроцитарной аплазии, первичной недостаточности надпочечников, рецидивирующего оптического нейромиелита, рестеноза, ревматической болезни сердца, SAPHO (синовит, акне, пустулез, гиперостоз и остеит), склеродермии, вторичного амилоидоза, шокового легкого, склерита, ишиаса, вторичной недостаточности надпочечников, связанного с кремний органическими соединениями заболевания соединительных тканей, дерматоза Снеддона-Уилкинсона, анкилозирующего спондилита, синдрома Стивенса-Джонсона, синдрома системного воспалительного ответа, височного артериита, токсоплазменного ретинита, токсического эпидермального некролиза, поперечного миелита, TRAPS (связанный с рецептором фактора некроза опухоли периодический синдром), аллергической реакции I типа, диабета типа II, крапивницы, обычной интерстициальной пневмонии (UIP), васкулита, весеннего конъюнктивита, вирусного ретинита, синдрома Фогта-Коянаги-Харада (синдром VKH), влажной дегенерации желтого пятна, заживления ран и связанной с иерсиниями и сальмонеллами артропатии. Фармацевтическая композиция, содержащая антитело, которое специфически связывается с IL-17А человека, может вводиться в терапевтически эффективном количестве для лечения опосредованного IL-17A заболевания или нарушения.
Также предложен способ лечения опосредованного IL-17А заболевания или нарушения с помощью антитела, которое специфически связывается с IL-17A человека. Способ лечения может дополнительно включать введение блокаторов ФНО-α.
Следующие примеры иллюстрируют настоящее изобретение, но не предназначены для ограничения настоящего изобретения в какой-либо мере.
В описание настоящей заявке включены все источники информации посредством ссылок.
Примеры
Пример 1
Схема получения антитела на основе VHH
На Фиг. 1 представлена принципиальная схема создания и оптимизации антитела на основе VHH.
Пример 2
Продукция рекомбинантных антигенов и антител в суспензионной культуре клеток млекопитающих
Антитела и антигены продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия СНО-К1), согласно опубликованным протоколам [26; 27]. Использовали клетки, конститутивно экспрессирующие ген белка EBNA1 (Epstein-Barrvirus nuclear antigen 1). Суспензионное культивирование осуществляли в колбах на орбитальном шейкере, с использованием бессывороточных сред производства компании Life Technologies Corporation, согласно инструкциям производителя. Для транзиентной экспрессии клетки в концентрации 2⋅106/мл трансфецировали с помощью линейного полиэтиленимина (ПЭИ «МАХ», компания «Polysciences»). Соотношение ДНК/ПЭИ составляло 1:3-1:10. Через 5-7 дней после трансфекции культуральную среду центрифугировали при 2000g в течение 20 минут и фильтровали через фильтр с размером пор 0,22 мкм. Целевые белки выделяли из культуральной жидкости с помощью аффинной хроматографии.
Рекомбинантный белок IL-17A, содержащий шесть аминокислот His на С-конце белка, выделяли и очищали из культуральной жидкости, используя смолу Profmity IMAC Ni-charged (компании Bio-Rad). До очистки в культуральную жидкость добавляли NiCl2 до концентрации 1 мМ. Далее добавляли в культуральную жидкость 5 мл смолы Profinity IMAC Ni-charged и перемешивали на качалке 1 час при комнатной температуре. Переносили сорбент на колонку Thermo scientific Polypropylene columns объемом 5 мл, промывали 5 колоночными объемами ФСБ, чтобы вымыть неспецифически связывающие компоненты. Связанный антиген элюировали, используя 0,3 М имидазол, рН8, 150 мМ NaCl. Далее белок переводили в ФСБ (рН 7,4) с помощью диализа, используя технологию SnakeSkin Dialysis Tubing, фильтровали (0,22 мкм), переносили в пробирки и хранили при -70°С. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (Фиг. 2).
Исследуемые и контрольное антитела IgGl очищали на колонке HiTrapr ProteinAFF объемом 1 мл (GE Healthcare) согласно методике, описанной выше для антигена IL-17A-Fc. Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза (Фиг. 2).
Пример 3
Иммунизация ламы IL-17A человека и создание Fab библиотеки фаговых антител ламы
Животное Lama Glama последовательно иммунизировали 5 раз путем подкожного введения антигенного материала, смешанного с равным объемом полного (при первой инъекции) или неполного (при остальных инъекциях) адъюванта Фрейнда. В качестве антигена использовали смесь рекомбинантных белков (по 0,2 мг каждого из белков на 1 инъекцию), одним из которых был человеческий IL-17A (набор от R&D Systems). Вторую инъекцию (стадию иммунизации) проводили через 3 недели после первой, затем с интервалом в две недели проводили еще три иммунизации. Взятие крови (50 мл) проводили через 5 дней после каждой инъекции, начиная с третьей.
Отобранную кровь ламы после иммунизации разводили в 2 раза раствором ФСБ, содержащим 1 мМ ЭДТА. 35 мл разбавленного раствора крови наслаивали на среду Histopaque® -1077 (от Sigma) с плотностью 1,077 г/мл объемом 15 мл и проводили центрифугирование в течение 20 мин при 800g. Мононуклеарные клетки (лимфоциты и моноциты) отбирали из интерфазной зоны плазма/среда Histopaque, после чего промывали раствором ФСБ, содержащим 1 мМ ЭДТА.
Суммарную РНК мононуклеарных клеток ламы выделяли с помощью набора RNeasy Mini Kit согласно предлагаемому протоколу (QIAGEN). Концентрацию РНК определяли с помощью Nanovue (GE Healthcare) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле.
Реакцию обратной транскрипции проводили с использованием набора MMLV RT kit (от Evrogen) согласно рекомендуемому протоколу с использованием обратной транскриптазы MMuLV и рандом-гексамерных олигонуклеотидов в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции для получения генов вариабельных доменов, фланкированных сайтами рестрикции, с использованием набора олигонуклеотидов и протоколам авторов [27; 28; 29]. Далее соединение генов вариабельных доменов легких и тяжелых цепей в один фрагмент проводили путем последовательных реакций рестрикции, лигирования и амплификации, согласно схеме, приведенной на Фиг. 3. Гены тяжелых цепей соединяли с генами каппа и отдельно с генами лямбда легких цепей. При этом расчетное количество молекул матрицы во всех реакциях составляло не менее 1011. Полученный ДНК препарат VL-CK-VH обрабатывали рестриктазами NheI/Eco91I и лидировали в оригинальную фагмиду рН5. Строение полученной фагмиды показано на Фиг. 4. Продукты лигирования трансформировали в электрокомпетентные клетки штамма SS320, полученные согласно протоколам [30]. Репертуар производной каппа Fab библиотеки составил 5,1*108 и лямбда Fab библиотеки - 3,7*108, соответственно. Препарат фага нативных библиотек был приготовлен согласно процедуре, описанной ранее [31].
Пример 4
Селекция Fab-библиотек фаговых антител
Специфичные анти-IL-17А фаговые Fab-антитела выделяли из фаговой Fab-дисплейной библиотеки на рекомбинантном IL-17 А человека (R&DSystems), осуществляя серии циклов селекции, как описано ранее [27; 31; 32]. Для осуществления селекции методом пэннинга IL-17A человека в 50 мМ карбонатном буфере (рН 9,5) адсорбировали в течение ночи при 4°С на поверхности сорбирующих пробирок HighSorb (Nunc). Пробирки промывали ФСБ (рН 7,4), затем блокировали раствором ФСБ (рН 7,4) - обезжиренное молоко (0,5% мас./об.) в течение 1 ч. Далее 2-4 мл раствора фагов в ФСБ (рН 7,4) - обезжиренное молоко (0,5% мас./об.), в концентрации 1012 фаговых частиц на 1 мл добавляли в пробирку с антигеном и инкубировали в течение 1 ч при перемешивании. Несвязавшиеся фаги удаляли в ходе серий циклов промывки с использованием раствора ФСБ (рН 7,4)-Твин 20 (0,1% об./об.). Количество отмывок увеличивали от первого раунда к третьему раунду 20-30-40 раз, соответственно. Оставшиеся связанными фаговые частицы элюировали 100 мМ раствором Gly-HCl (рН 2,5) в течение 15 мин при перемешивании, затем нейтрализовали 1 MTRIS-HCl (рН7.6). Бактерии штамма Е. coli TG1 инфицировали полученными фагами, затем фаги выделяли и использовали в следующем цикле селекции.
После второго и третьего раунда селекции на IL-17A проведенный ИФА анализ препарата поликлонального фага показал значительное обогащение. Полученный пул клонов, обогащенных Fab человека, специфичных против IL-17A, был реклонирован в экспрессионную плазмидур LL, содержащую myctag и His6 tag на С-конце гена СН1 тяжелой цепи.
Пример 5
Анализ специфического связывания Fab с IL-17A человека
ИФА (ELISA) использовали для измерения связывания исследуемых Fab-фрагментов с IL-17A человека. В качестве позитивного контроля использовали Fab антитела AIN457 с опубликованной последовательностью (от Novartis). Для анализа специфического связывания лунки планшетов ELISA (от Nunc ImmunoMaxisorp) покрывали 50 мкл (0,5 мкг/мл в 1X покрывающем карбонатном буфере) IL-17A, герметично закрывали и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартному ИФА протоколу с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем GenetixQ-pix2xt (от Molecular Device) и Тесаn Freedom EVO 200 (от Тесаn). Для блокирования неспецифического связывания добавляли блокирующий буфер ВВ (200 мкл 0,5% нежирного молока в ФСБ). Планшеты инкубировали на шейкере в течение часа при комнатной температуре. После отмывок ФСБ-Твином добавляли по 50 мкл на лунку тестируемого клеточного супернатанта, содержащего исследуемый Fab, смешанного с равным объемом блокирующего буфера. Планшеты снова инкубировали, встряхивая, один час при комнатной температуре, после чего каждую лунку планшетов пять раз промывали буфером ФСБ-Твин. После промывания добавляли (50 мкл/лунку) античеловеческого Fab HRP-конъюгированного вторичного антитела (от Pierce-ThermoScientific) в соотношении 1:5000 в ФСБ-Твин. Планшеты встряхивали на ротационном шейкере (50 мин, комнатная температура) и промывали пять раз буфером ФСБ-Твин, как описано выше. Колориметрический сигнал развивали добавлением ТМВ (100 мкл/лунку) до насыщения (в среднем 3-5 мин), затем дальнейшее развитие останавливали добавлением стопового раствора (100 мкл/лунку, 10% серная кислота). Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала. Клоны, у которых цветовой сигнал превышал фоновый более чем в 5 раз, были проверены в конкурентном ИФА анализе для выявления антагонистических Fab, блокирующих взаимодействие IL-17A лиганда и рецептора.
Пример 6
Конкурентный ИФА анализ блокирования взаимодействия IL17A лиганда и IL17R рецептора
Конкурентный ИФА (ELISA) был использован для проверки антагонистической способности отобранных ранее специфичных Fab против IL-17A человека. В качестве позитивного контроля антагониста использовали Fab антитела AIN457c опубликованной последовательностью (от Novartis). Рецептор IL-17RA-Fc (от R&D Systems) иммобилизовали в лунки планшетов ELISA (от Nunc Immuno Maxisorp) по 50 мкл с концентрацией 1 мкг/мл в IX покрывающем карбонатном буфере и инкубировали в течение ночи при 4°С. Все последующие этапы проводили по стандартным ИФА протоколам с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем GenetixQ-pix2xt (от Molecular Device) и Тесаn Freedom EVO 200 (от Tecan). Для блокирования неспецифического связывания добавляли блокирующий буфер ВВ (200 мкл 0,5% нежирного молока в ФСБ). Планшеты инкубировали на шейкере в течение часа при комнатной температуре.
Параллельно 50 мкл тестируемого клеточного супернатанта, содержащего исследуемый Fab, смешивали в несвязывающем 96-луночном планшете с 50 мкл IL-17A-His6-Flag с концентрацией 0,4 мкг/мл в 1% молоке на ФСБ-Твин. Инкубировали 1 час при 37°С и перемешивании на шейкере при 500 об/мин.
После отмывок от блокирующего буфера в планшет, содержащий IL-17RA-Fc рецептор, переносили описанную выше реакционную смесь Fab и IL-17A-His6-Flag лиганда в расчете 90 мкл на лунку. Планшеты снова инкубировали, встряхивая, 45 мин при комнатной температуре, после чего каждую лунку планшетов пять раз промывали буфером ФСБ-Твин. Добавляли 50 мкл/лунку анти-FLAG мышиного М2 антитела (от Sigma) при концентрации 1 мкг/мл. Инкубировали 45 мин при комнатной температуре, после чего каждую лунку планшетов 5 раз промывали буфером ФСБ-Твин. Добавляли 50 мкл/лунку антимышиного-IgG HRP-конъюгированного вторичного антитела (от Pierce-TheimoScientific) в разведении 1:5000 в ФСБ-Твин. Планшеты встряхивали на ротационном шейкере 45 мин при комнатной температуре и промывали 5 раз буфером ФСБ-Твин, как описано выше. Колориметрический сигнал развивали добавлением ТМВ (100 мкл/лунку) до насыщения (в среднем 3-5 мин), затем дальнейшее развитие останавливали добавлением стопового раствора (100 мкл/лунку, 10% серная кислота). Цветовой сигнал измеряли при длине волны 450 нм, используя подходящий планшет-ридер Tecan-Sunrise (от Tecan). Степень связывания антитела была пропорциональна продуцированию цветового сигнала.
Клоны, показавшие блокирование на уровне контрольного Fab антитела AIN457, были отмечены как позитивные и использовались в дальнейших анализах. Гены вариабельных доменов позитивных клонов были секвенированы, согласно стандартным протоколам на аппарате Applied Biosystems 3130 Genetic Analyzer (Applied Biosystems) и проанализированы. Клоны, содержащие в составе 3 VHHFab вариабельный домен, были взяты в дальнейшие исследования (Фиг. 5В). Также обнаружили, что клон 3VHHFab представлен в комбинации с двадцатью тремя различными по последовательности доменами легких цепей, из которых три приведены на рисунке (Фиг. 5В), что может свидетельствовать о его относительной структурной толерантности и о том, что именно VHH домен, а не легкая цепь вносит вклад во взаимодействие с IL-17A.
Пример 7
Сравнительный скрининг анти-IL-17А VHHFab кандидатов по кинетической константе диссоциации koff (kdis)
Сравнительный скрининг koff для анти-IL-17А Fab кандидатов проводили с использованием прибора Pall Forte Bio Octet Red 96. Анти FABCH1 биосенсоры в течение 30 минут регидратировались в рабочем буфере, содержащем 10 мМ ФСБ, рН 7.2-7.4, 0.1% Твин-20, 0,1% БСА. В исследуемые образцы супернатантов Е. coli добавляли 10х-рабочий буфер до конечной концентрации 1х. Затем анти FABCH1 биосенсоры погружали в супернатанты Е. coli, содержащие Fab-фрагменты кандидатов антител, на 12 часов при 4°С. Сенсоры с иммобилизованными на поверхности Fab-фрагментами переносили в лунки с рабочим буфером, где прописывалась базовая линия (60 с.). Далее сенсоры переносили в лунки с раствором аналита (IL-17A, 30 мкг/мл) для ассоциации комплекса антиген-антитело (300 с.). Затем сенсоры возвращали в лунки, содержащие рабочий буфер, для последующей стадии диссоциации (300 с.). После каждого эксперимента использованные сенсоры регенерировали путем трехкратного помещения их в буфер для регенерации (Gly-HCl, рН 1,7) после чего они могли быть использованы в следующем эксперименте. Анализ полученных кривых проводили с помощью программного обеспечения Octet Data Analysis (версия 7.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1.
Результаты koff-скрининга анти-IL-17А Fab кандидатов представлены на Фиг. 5С и таблице 1. Продемонстрировано специфическое высокоаффиное связывание всех уникальных VHHFab с IL-17A человека, при этом 3 VHHFab демонстрировали очень быструю kon и очень медленную kdis(1/c), выходящую за пределы чувствительности прибора.

Таким образом, на основании данных анализов кандидат 3VHHVK4B11 был отобран для дальнейшего исследования.
Пример 8
Создание VHHIgG1 антител, с мутациями в участке FR1 и FR2 вариабельного домена VHH
Гены вариабельного домена легкой и тяжелой VHH цепей кандидата 3VHHVK4B11 были клонированы в плазмиды рЕЕ-НсирЕЕ-Lc для совместной транзиентной экспрессии в клетках CHO-EBNA, как описано в Примере 3. Далее, методом олигонуклеотид-направленного мутагенеза вводили замены в позиции 44 и 45 (по номенклатуре Kabat) с помощью PfuUltraHS polymerase (Stratagene) согласно протоколу [Q5® Site-Directed Mutagenesis Kit (от NEB)] и методике [34], используя в качестве матрицы плазмиду pEE-3VHH. Продукты ПЦР фракционировали на легкоплавкой агарозе и очищали на колонках. После реакции лигирования ДНК трансформировали в E. coli. После отбора корректных по последовательности мутантных клонов плазмиды с мутациями в 3VHH котрансфецировали с pEE-LcVK4B11(см. Фиг. 6А, таблицу 2).

Пример 9
Сравнительный анализ кинетических характеристик VHHFab антител, с мутациями в участке FR1 и FR2 вариабельного домена VHH
Сравнительный скрининг koff для анти-IL-17А VHHFab кандидатов скрининг производился с использованием прибора Pall Forte Bio Octet Red 96 по стандартному протоколу (см. Пример 8). В результате было обнаружено заметное снижение величин koff для mut1, mut2 и mut4 и менее значительное снижение для mut3 в сравнении с диким 3VHHFab (см. Фиг. 6В).
Пример 10
Сравнительный анализ термоагрегационных характеристик 3VHHIgG1VK4B11 полноразмерных антител, с мутациями в участке FR1 и FR2 вариабельного домена VHH
Сравнительный анализ агрегационных характеристик для анти-IL-17А VHHIgG1 кандидатов проводили по следующей методике. Препарат антитела VHHIgG1 с концентрацией около 10 мг/мл в ФСБ буфере прогревали в течение 6 часов при температуре 50°С. Определение агрегации после термостресса проводили методом гель-фильтрационной высокоэффективной хроматографии. Хроматографию проводили на системе ВЭЖХ (Agilent) 1100 на колонке Tosoh TSK-Gel G3000SWXL, 7.8 мм × 30 см, кат. №08541 с предколонкой Tosoh TSKgel Guard SWXL, 6.0 мм × 4.0 см, с диаметром частиц 7 мкм, кат. №08543. Элюирование проводили в изократическом режиме подвижной фазой: 50 мМ NaФб, 0,3 М NaCl, рН 7.0 на скорости потока 0,5 мл/мин. Детектирование проводили на длинах волн 214 и 280 нм. Образцы антител разводили буфером ФСБ, рН 7.5, до концентрации ~1 мг/мл. Объем вводимой пробы составлял 10 мкл. Предварительно была хроматографирована калибровочная смесь Gel filtration standard (от Bio-Rad), кат. №151-1901.
Представленные на Фиг. 6С хроматограммы и сводная таблица 3 демонстрируют, что все мутанты в различной степени агрегируют в условиях термостресса, причем минимально стабилен дикий вариант, а максимальной стабильностью обладает mut1 с заменами E44G+R45L, характерными для классической структуры VH.
Кроме того, для изучения сравнительной термостабильности полученных препаратов также применили методику Thermofluor (также известную как Thermal shift assay), которая отражает температуру плавления белка, фиксируя изменение флюоресценции специального красителя SYPROOrange с гидрофобными участками денатурированного белка [35]. Используя прибор StepOneReal-TimePCRSystem (Applied Biosystems) и рекомендуемый протокол, было проведено исследование препаратов мутантов. Полученные результаты исследований приведены в таблице 3. Они в значительной степени коррелируют с результатами термостресс-теста, подтверждая повышенную стабильность mut1 и mut4 в сравнении с исходным вариантом 3VHHIgG1VK4B11.

Пример 11
Клеточный тест блокирования мутантов анти-IL17А 3VHHIgG1VK4B11 функции IL-17A индуцировать продукцию IL-6
Способность IL-17 индуцировать производство IL-6 человеческими клетками линии НТ1080 (ATCC:CCL-121) использовали для анализа нейтрализующей активности VHHIgG1 кандидатов mut1 и mut4 в отношении человеческого рекомбинантного IL-17. Клетки выращивали в питательной среде ДМЕМ с добавлением 10% инактивированной эмбриональной сыворотки, гентамицином и глутамином. Клетки в количестве 5⋅104 на лунку высевали в 96-луночные плоскодонные планшеты для культуры клеток. Клетки оставляли в течение 5 часов для прикрепления. Смесь рекомбинантного 40 нг/мл IL-17 и 20 нг/мл ФНО-α инкубировали с разведениями антитела VHHIgG1 в течение часа при 37°С. Затем смесь цитокина и антитела добавляли к клеткам и оставляли на ночь. Производство IL-6 в культуре клеток НТ1080 было пропорционально добавленному количеству IL-17. Количество высвобожденного IL-6 в клеточных супернатантах определяли методом ELISA c помощью системы «DuoSet ELISA development system human IL6» (от RD System, Cat. № DY206). Результаты анализа антагонистических свойств кандидатов VHHIgG1 приведены на Фиг. 7 в сравнении с антителом AIN457 (анти-IL-17А антитело от Novartis). mutl показал величину IC50, что было почти в 30 раз выше, чем в исходном варианте, тогда как вариант mut4 сохранил практически полностью свои ингибирующие свойства. Величина IC50 для mut4 кандидата составила 30±10 пМ. По результатам этого исследования данный кандидат по совокупности физико-химических и биологических характеристик был выбран для дальнейшей работы по оптимизации свойств.
Пример 12
Конструирование mut4 VHHFab антител, содержащих легкие цепи человека
Суммарную РНК В-лимфоцитов из крови 550 человеческих доноров выделяли с помощью набора RNeasy Mini Kit согласно предлагаемому протоколу (от QIAGEN). Концентрацию РНК определяли с помощью набора Nanovue (от GE Healthcare) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле.
Реакцию обратной транскрипции проводили с использованием набора MMLV RT kit (от Evrogen) согласно рекомендуемому протоколу с использованием обратной транскриптазы MMuLV и рандом-гексамерных олигонуклеотидов в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции для получения генов вариабельных доменов, фланкированных сайтами рестрикции, с использованием набора олигонуклеотидов и протоколам авторов [27]. Создание химерных Fab, специфичных против IL-17A, проводили согласно технологии, описанной в WO 93/06213, на основе оригинальной фагмиды рН5, описанной выше. Для этого соединение генов вариабельных доменов легких цепей человека с геном вариабельного домена mut4 VHH в один фрагмент проводили путем последовательных реакций рестрикции, лигирования и амплификации, согласно схеме, приведенной на Фиг. 8. При этом расчетное количество молекул матрицы генов вариабельных доменов человека во всех реакциях составляло не менее 1012. Полученный ДНК препарат VL-CK-VH обрабатывали рестриктазами NheI/Eco91I и лигировали в оригинальную фагмиду рН5. Продукты лигирования трансформировали в электрокомпетентные клетки штамма SS320, полученные согласно протоколам [30]. Репертуар химерной mut4 VHH Fab-библиотеки на основе смешанных каппа и лямбда цепей человека составил 2,8*109 трансформантов. Препараты фага химерных Fab-библиотек были приготовлены согласно процедуре, описанной ранее [27].
Селекции полученных фаговых химерных Fab-библиотек проводили в условиях, аналогичных описанным выше (см. Пример 5), за исключением дополнительной инкубации связанных с IL-17A фаговых антител в присутствии 20 мкг/мл растворенного IL17A при 37°С в течение 12 часов.
После второго раунда селекции на IL-17A было замечено значительное обогащение библиотеки. Полученные в результате этого пулы клонов обогащенных химерных mut4 VHH Fab-библиотек были использованы в скрининге на IL-17A согласно стандартному протоколу (см. Пример 6). Результирующие позитивные клоны были секвенированы. На Фиг. 8В приведены четыре последовательности вариабельных домена легких цепей VK1A7, VK3c18, VK3cl18 и VK4clE12 человека из высокоаффиных mut4 VHHFab. Последовательности принадлежат к различным каппа VK1, VK3 и VK4 гермлайн человека с минимальным количеством соматических мутаций. Следует добавить, что последовательность VK4clE12 человека в значительной степени гомологична ранее отобранной легкой цепи VK4B11 ламы с характеристическими отличиями.
Пример 13
Сравнительный анализ термоагрегационных характеристик mut4 VHHIgG1 полноразмерных антител, с различными вариантами легких цепей
Гены вариабельного домена тяжелых VHH цепей кандидата mut4 VHH были клонированы в плазмиды рЕЕ-Нс, а гены вариабельных доменов легких цепей VK1A7, VK3cl8,VK3cl18, VK3A4, VK4E12 человека домена легкой цепи - в pEE-Lc для совместной транзиентной экспрессии в клетках СНО-EBNA, как описано в Примере 3. Далее полученные антитела подвергали термострессу и анализировали агрегационный профиль согласно Примеру 9. Результаты приведены в сводной таблице 4.
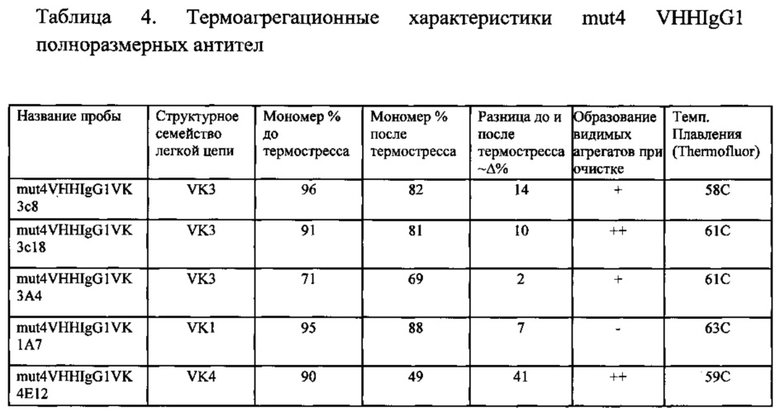
Кроме того, для изучения сравнительной термостабильности полученных препаратов применили методику Thermofluor, аналогично описанной в Примере 11. На основании полученных данных можно сделать вывод, что были отобраны более стабильные пары mut4 VHH c легкими цепями человека, чем исходный вариант с легкой цепью ламы VK4B11. Таким образом, лучшие характеристики сравнительной агрегационной стабильности продемонстрировали комбинации mut4 VHHIgG1VK1A7 и mut4 VHHIgG1 VK3cl8.
Пример 14
Клеточный тест блокирования функции IL-17A индуцировать продукцию IL-6 с помощью mut4 VHHIgG1VK3c8 и mut4 VHHIgG1VK1A7
Способность IL-17 индуцировать производство IL-6 человеческими клетками линии НТ1080 (АТСС:CCL-121) использовали для анализа нейтрализующей активности VHHIgG1 кандидатов mut4 VHHIgG1VK3c8 и mut4 VHHIgG1VK1A7 в отношении человеческого рекомбинантного IL-17A, как показано в Примере 12. На Фиг. 9 приведены результаты такого исследования на блокирование. Следует отметить, что у варианта mut4 VHHIgG1VK1A7, отличающегося максимальной стабильностью, наблюдали значительное падение величины IC50, тогда как вариант mut4 VHHIgG1VK3c8, отличающийся средней стабильностью, почти в несколько раз превысил величину ингибирования более стабильного варианта mut4 VHHIgG1VK1A7.
Пример 15
Создание mut4 VHHIgG1 полноразмерных антител с различными мутациями в позициях 44 и 45 FR2 вариабельного домена VHH и сравнительный анализ их агрегационных и функциональных характеристик
С целью дальнейшего повышения агрегационной стабильности методом олигонуклеотид-направленного мутагенеза были проведены замены в позициях 44 и 45 FR2 региона VHH для кандидата mut4 VHHIgG1VK3c8 (для упрощения здесь и далее - m4VHHc8). В исследование не были включены варианты с ароматическими, алифатическими и позитивными аминокислотами в позиции 44 как потенциально иммуногенные и структурно запрещенные. Мутанты были транзиентно наработаны, как описано в Примере 3 (Фиг. 8). Хотя количество препарата для различных мутантов варьировало от одного эксперимента к другому, следует отметить, что выходы внутри одного эксперимента для различных замен являлись соразмерными. Полученные варианты мутантов подвергали термострессу. В таблице 5 приведены результаты термостресс-теста для различных мутантов. Как видно по результатам, наименее стабильными мутантами являются 44G45G и 44G45D, которые имеют сниженную стабильность по сравнению с исходной комбинацией 44G45R в составе m4VHHc8. Такие варианты, как 44G45N, 44V45T, 44D45T, 44Т45Т, 44D45V, сохранили в значительной степени свою стабильность. В случае комбинаций остальных мутаций наблюдали добавочный эффект в увеличении агрегационной стабильности m4VHHc8, причем наиболее стабильными оказались варианты с ароматическими и алифатическими группами в позиции 45. Однако варианты с малыми гидрофильными и гидрофобными аминокислотами в обоих позициях, такие как 44G45V, 44G45T, 44V45V, 44A45V, 44T45V, 44А45Т, 44S45Т, также продемонстрировали значительную стабильность.
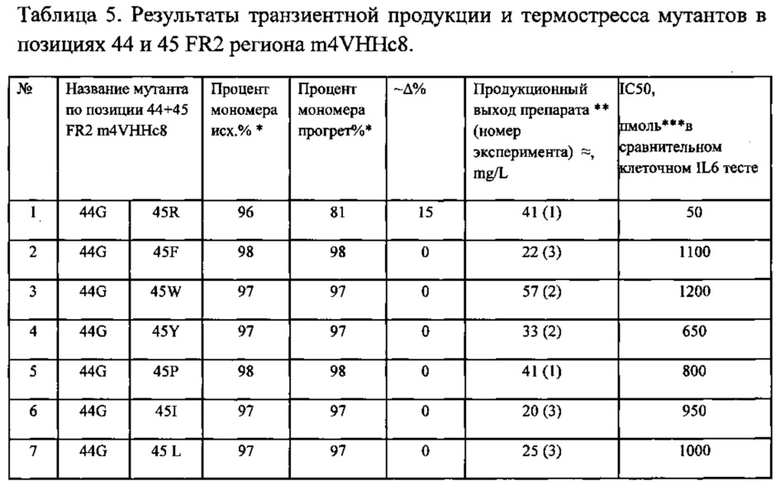
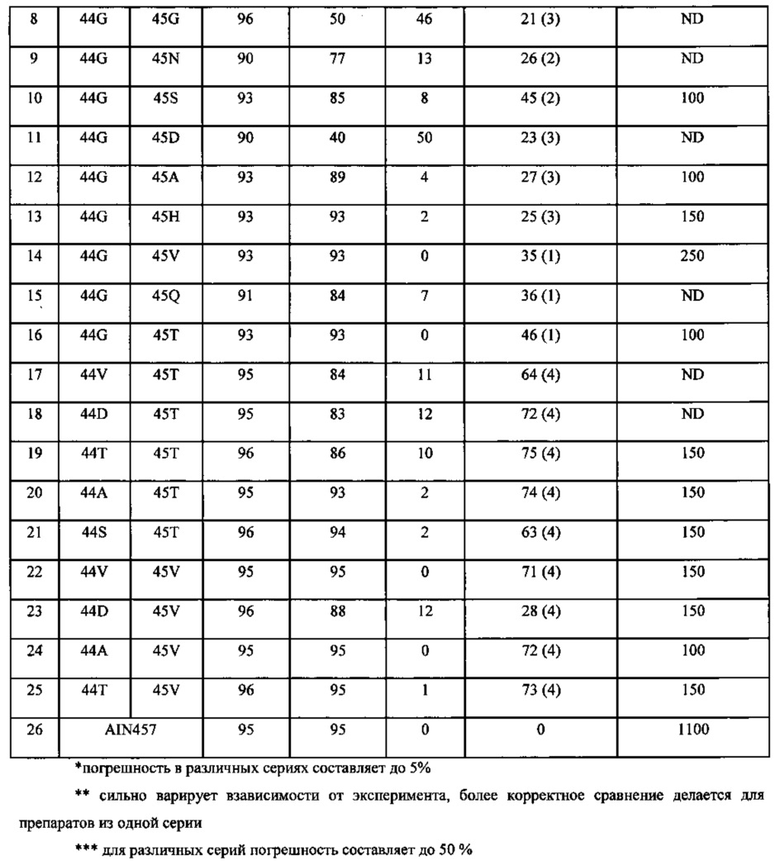
С целью проверки нейтрализующей активности в клеточном тесте аналогично Примеру 12 были проведены эксперименты с различными вариантами мутаций (Фиг. 11). Результаты приведены в таблице 5. Как показано, аминокислоты с малыми размерами в позициях 44 и 45 FR2 региона VHH в значительной степени демонстрировали высокие антагонистические свойства, тогда как большие аминокислоты (ароматические и алифатические) при заменах в позиции 45 приводили к 10-кратному снижению величины IC50. Таким образом, на Фиг. 12 показана диаграмма стабильности и функциональных свойств для мутантов, содержащих различные аминокислотные замены в регионе, определяющие взаимодействие вариабельного домена тяжелой и легкой цепей.
Пример 16
Сканирующий мутагенез позиций в CDRs antiIL17A 3VHH домене
Для введения мутаций в индивидуальные позиции CDR-участков кандидата была использована методика NNK кодоновой рандомизации [26] с помощью Q5® Site-Directed Mutagenesis Kit (от NEB) согласно протоколу, в качестве матрицы используя плазмиду pLL-Fab. Продукты ПЦР фракционировали на легкоплавкой агарозе и очищали на колонках. После реакции лигирования ДНК трансформировали в E.coli экспрессирующий штамм BL21gold (от Stratagene). Полученные индивидуальные клоны наращивали в условиях Fab-экспрессии в 96-луночных планшетах, как описано ранее. Супернатанты, содержащие мутантные Fab, анализировали с использованием методов ИФА, описанных ранее, с использованием высокопроизводительной автоматизированной платформы на базе роботизированных систем Genetix Q-pix2xt и Тесаn Freedom EVO200. Концентрация иммобилизованного IL-17A составляла 0,2 мкг/мл. Окрашивание связанных Fab проводили с разведенным 1:5000 конъюгатом Goat anti-Human IgG (Fab')2 (HRP) (от Pierce) и краской ТМБ+H2O2/ H2SO4 с последующим измерением оптической плотности при длине волны 450 нм.
Результаты сканирующего мутагенеза представлены в Таблице 6. В таблице приведены замены в позициях CDR регионов, которые соответствуют не более чем на 30% уменьшению сигнала связывания мутантных Fabc IL-17A человека в сравнении с "дикой" последовательностью. Таким образом, эти индивидуальные мутанты или любые их комбинации также являются объектом изобретения согласно данной заявке.

Таким образом, проведен скрининг аминокислотных позиций CDR регионов антитела BCD 109, толерантных к заменам на другие аминокислоты. В результате этого обнаружено, что данный набор аминокислотных замен значительно не меняют аффинности антитела к IL-17Ачеловека. Комбинация замен в таких позициях может быть использована для оптимизации различных свойств кандидата.
Пример 17
Создание BCD109 кандидата и определение аффинности к IL-17A из различных видов организмов
Антитело BCD109 было получено из варианта m4VHHc8, содержащего замены 44G45T, описанного в Примере 16, путем внесения гуманизирующих мутаций Q5V, R89V в VHH, не изменяющих свойств стабильности или IC50 (данные не приведены) и трех добавочных мутаций в СН2 домене FcIgG1, 232Y/234T/236E, для улучшения фармакокинетики антитела. Антитело было транзиентно наработано.
Анализ аффинности связывания BCD109 к IL-17A человека, макаки и крысы производился на приборе OctetRed 96 (от ForteBio). BCD109 было неспецифически иммобилизовано на поверхность аминореактивных сенсоров второго поколения (от AR2G) по стандартному протоколу согласно инструкции производителя, для подготовки и иммобилизации AR2G сенсоров. Анализ был произведен при 30°С с использованием ФСБ, содержащим 0,1% Твин-20 и 0,1% БСА в качестве рабочего буфера.
Титрование IL-17A человека, макаки и крысы производили с использованием рабочего буфера от концентрации 126 нМ до 2 нМ с шагом 2.
Кривые связывания с вычетом референсного сигнала анализировали с помощью программного обеспечения Octet Data Analysis (версия 7.0) согласно стандартной процедуре с использованием модели взаимодействия 1:1. Результаты приведены на Фиг. 13А.
Как видно, BCD 109 связывается с рекомбинантным препаратом IL-17A человека и IL-17A макаки с пикомолярной аффиностью (Фиг. 13В). Кроме того, полученный кандидат антитела BCD109 не взаимодействует с IL-17A крысы (данные не приведены).
Пример 18
Определение агрегационной стабильности BCD109 в условиях термостресса
Препарат антитела BCD109 с концентрацией около 10 мг/мл в ФСБ буфере прогревали в течение 6 часов при температуре 50°С, по протоколу, описанном в Примере 9.
Представленные на Фиг. 14 результаты показывают, что антитело BCD109 стабильно в условиях термостресса, при этом образование агрегатов составило менее 5%.
Пример 19
Получение фармацевтической композиции, содержащей антитело согласно изобретению
Антитело BCD109 в концентрации 50 мг/мл растворяли в необходимом количестве воды для инъекций, доводили рН до 5,5 с помощью лимонной кислоты, фильтровали полученный раствор (стерилизующая фильтрация) и запаивали в ампулы.
Полученный препарат был стабилен в течение 6 мес, без образования осадка.
Также антитело BCD109 растворяли в воде, содержащей маннитол в качестве лиопротектора, фильтровали (стерилизующая фильтрация) и подвергали лиофилизации. Полученный порошок упаковывали в стерильные флаконы. Флаконы укупоривали резиновыми пробками и обжимали алюминиевыми колпачками. Полученный препарат антитела можно восстановить из лиофилизата при применения воды для инъекций.
Источники информации
1. Porter R.R. 1973. Structural studies of immunoglobulins. Science. 180, 713-716.
2. Padlan E.A. 1994. Anatomy of the antibody molecule. Mol. Immunol. 31, 169-217.
3. Dwek R.A., Sutton B.J., Perkins S.J., Rademacher T.W. 1984. Structure-function relationships in immunoglobulins. Biochem. Soc. Symp.49, 123-136.
4. Burton D.R. 1985. Immunoglobulin G: functional sites. Mol. Immunol. 22, 161-206.
5. Hamers Casterman C, Atarhouch Т., Muyldermans S., Robinson G., Hamers C, Bajyana Songa E., Bendahman N., Hamers R. 1993. Naturally occurring antibodies devoid of light chains. Nature. 363, 446-448.
6. C.В. Тиллиб "Верблюжьи наноантитела" - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология, 2011, том 45, №1, с. 77-85.
7. Nguyen V.K., Hamers R., Wyns L., Muyldermans S.1999. Loss of splice consensus signal is responsible for the removal of the entire CH1 domain of the functional camel IGG2A heavy-chain antibodies. Mol. Immunol. 36, 515-524.
8. Woolven B.P., Frenken L., van der Logt P., Nicholls P.J. 1999. The structure of the llama heavy chain constant genes reveals a mechanism for heavy-chain antibody formation. Immunogenetics.50, 98-101.
9. Nguyen V.K., Hamers R., Wyns L., Muyldermans S. 2000. Camel heavy-chain antibodies: diverse germline VHH and specific mechanisms enlarge the antigen-binding repertoire. EMBO J. 19, 921-931.
10. De Genst E., Saerens D., Muyldermans S., Conrath K. 2006. Antibody repertoire development in camelids.Develop.Comp. Immunol. 30, 187-198.
11. Muyldermans S., Cambillau C, Wyns L. 2001. Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains. TIBS. 26, 230-235.
12. De Genst E., Silence K., Decanniere K., Loris R., Kinne J., Muyldermans S. 2006. Molecular basis for the preferential cleft recognition by dromedary heavy chain antibodies. Proc. Natl. Acad. Sci. USA. 103, 4586-4591.
13. Kabat E., Wu T.T., Perry H.M., Gottesman K.S., Foeller C. 1991. Sequence of proteins of immunological interest. US Public Health Services. NIH, Bethesda, MD, Publication no. 91-3242.
14. Nguyen V.K., Desmyter A., Muyldermans S. 2001. Functional heavy-chain antibodies in camelidae. Adv. Immunol. 79, 261-296.
15. Muyldermans S., Baral T.N., Retamozzo V.C., et al. 2009. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunopathol. 128 (1-3), 178-183.
16. de Genst E., Silence K., Decanniere K., Loris R., Kinne J., Wyns L., Muyldermans S. 2005. Strong in vivo maturation compensates for structurally restricted H3 loops in antibody repertoires. J. Biol. Chem. 280, 14114-14121.
17. Lauwereys M., Ghahroudi M., Desmyter A., Kinne J., Holzer W., De Genst E., Wyns L., Muyldermans S. 1998. Potent enzyme inhibitors derived from dromedary heavy chain antibodies. EMBO J. 17, 3512-3520.
18. Kontermann RE. 2009 Strategies to extend plasma half-lives of recombinant antibodies. BioDrugs.; 23(2): 93-109.
19. Ken Coppieters, Torsten Dreier, Karen Silence, Hans de Haard, Marc Lauwereys, Peter Casteels, Els Beirnaert, Heidi Jonckheere, 2006 Formatted antitumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis Arthritis & Rheumatology.; 54(6): 1856-66.
20. Gabrielle Richard, Ashley J Meyers, Michael D McLean, Mehdi Arbabi-Ghahroudi, Roger MacKenzie, J Christopher Hall2013. In Vivo Neutralization of α-Cobratoxin with High-Affinity Llama Single-Domain Antibodies (VHHs) and a VHH-Fc Antibody. PLoS One 22; 8(7):e69495.
21. Jan Terje Andersen, Maria Gonzalez-Pajuelo, Stian Foss, Ole J. B. Landsverk, Debora Pinto, Alexander Szyroki, Hans J. de Haard, Michael Saunders, Peter Vanlandschoot Inger Sandlie 2012, Selection of Nanobodies that Target Human Neonatal Fc Receptor. Scientific Reports, 3, 1118.
22. Vincke C1, Loris R, Saerens D, Martinez-Rodriguez S, Muyldermans S, Conrath K. 2009 General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold. J Biol Chem. Jan 30; 284(5): 3273-84.
23. Barthelemy PA, Raab H, Appleton В A, Bond CJ, Wu P, Wiesmann C, Sidhu SS. 2008 Comprehensive analysis of the factors contributing to the stability and solubility of autonomous human VH domains. J Biol Chem. Feb 8; 283(6): 3639-54.
24. Conrath K, Vincke C, Stijlemans B, Schymkowitz J, Decanniere K, Wyns L, Muyldermans S, Loris R. Antigen binding and solubility effects upon the veneering of a camel VHH in framework-2 to mimic a VH. J Mol Biol. 2005 Jul 1; 350(1): 112-25.
25. Hermeling S, Crommelin DJ, Schellekens H, Jiskoot W (2004) Structure-immunogenicity relationships of therapeutic proteins. Pharm Res 21:897-903.
26. Handbook of pharmaceutical manufacturing formulations. Volume 6. Sterile products / Sarfaraz K. Niazi, 2004.
27. J. de Haard H., van Neer N., Anneke Reurs, E. Hufton S., Roovers R., Henderikx P., de Bruïne A., Arends J.-W. and Hoogenboom H. A Large Non-immunized Human Fab Fragment Phage Library That Permits Rapid Isolation and Kinetic Analysis of High Affinity Antibodies. J Biol Chem. 1999 Jun 25 274(26):18218-30.
28. WO 2010/001251.
29. Koch-Nolte F., Reyelt J., Schöβow В., Schwarz N., Scheuplein F., Rothenburg S., Haag F., Alzogaray V., Cauerhff A. and Goldbaum F. Single domain antibodies from llama effectively and specifically block T cell ecto-ADP-ribosyltransferase ART2.2 in vivo. FASEB J., 2007 Nov; 21(13):3490-8.
30. Sidhu S., Lowman H., Cunningham В., Wells J. Phage display for selection of novel binding peptides. J. Methods Enzymol. 2000; 328: 333-63.
31. Marks J., Hoogenboom H., Bonnert Т., McCafferty J., Griffiths A., Winter G. By-passing immunization: Human antibodies from V-gene libraries displayed on phage. J Mol Biol. 1991 Dec 5; 222(3):581-97.
32. Vaughan Т., Williams A., Pritchard K., Osbourn J., Pope A., Earnshaw J., McCafferty J., Hodits R., Wilton J. and Johnson K. Human Antibodies with Sub-nanomolar Affinities Isolated from a Large Non-immunized Phage Display Library. Nat Biotechnol. 1996 Mar; 14 (3): 309-14.
34. Environ Microbiol. 2006 Feb;72(2):l 141-7; MAbs. 2012 May-Jun; 4(3): 341-8.
35. Cummings M., Farnum M., and Nelen M. Universal Screening Methods and Applications of ThermoFluor®. J Biomol Screen 2006 11(7): 854-863.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-IL-17-АНТИТЕЛА, СПОСОБ ИХ ПОЛУЧЕНИЯ И СПОСОБ ПРИМЕНЕНИЯ | 2014 |
|
RU2577228C2 |
| ТРИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IL-17A, IL-17F И ДРУГОЙ ПРОВОСПАЛИТЕЛЬНОЙ МОЛЕКУЛЫ | 2016 |
|
RU2680011C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К IL-5Rα | 2017 |
|
RU2698048C2 |
| БИСПЕЦИФИЧНЫЕ АНТИ-HER2/АНТИ-HER3 АНТИТЕЛА | 2016 |
|
RU2653443C2 |
| ВОДНАЯ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ АНТИ-IL17A АНТИТЕЛА И ЕЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2754760C2 |
| АНТИ-IL-17F АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2605318C2 |
| АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, СПОСОБНЫЙ СВЯЗЫВАТЬСЯ С РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА-6 ЧЕЛОВЕКА | 2016 |
|
RU2656160C2 |
| АНТИ-PD-1-АНТИТЕЛА, СПОСОБ ИХ ПОЛУЧЕНИЯ И СПОСОБ ПРИМЕНЕНИЯ | 2016 |
|
RU2656181C1 |
| МУЛЬТИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО СО СПЕЦИФИЧНОСТЬЮ СВЯЗЫВАНИЯ IL-13 И IL-17 ЧЕЛОВЕКА | 2020 |
|
RU2827946C1 |
| СОДЕРЖАЩИЕ СУПЕРСПИРАЛЬ И/ИЛИ ПРИВЯЗКУ БЕЛКОВЫЕ КОМПЛЕКСЫ И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2573915C2 |











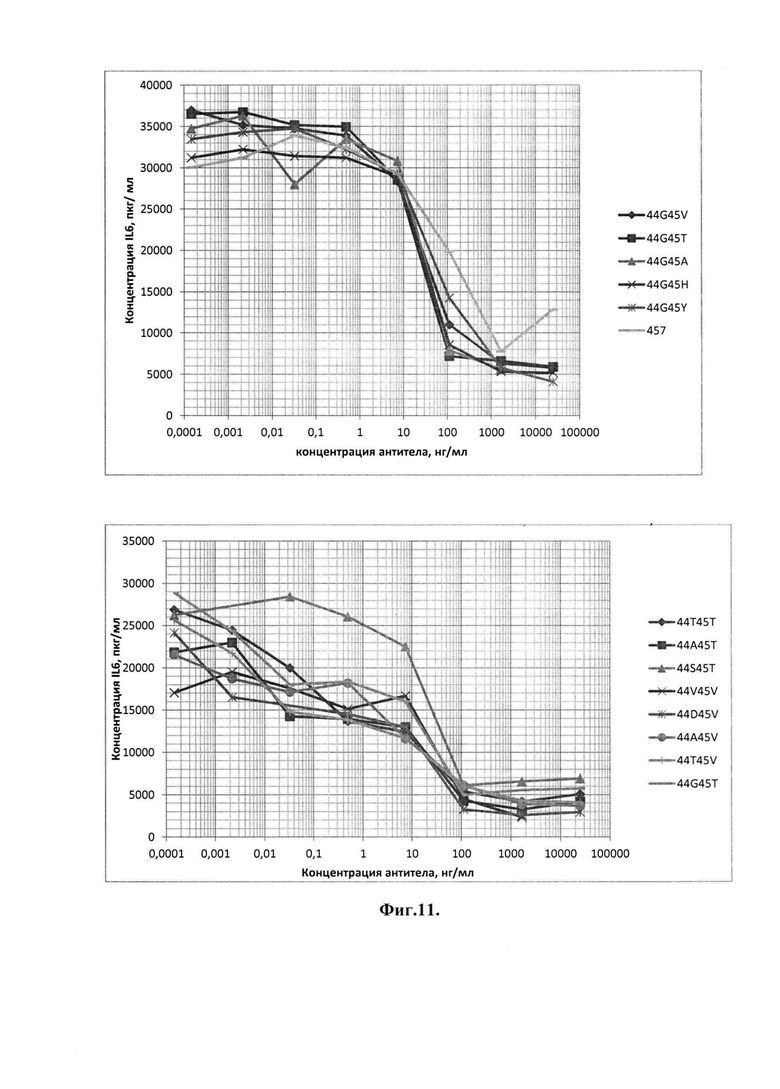




Изобретение относится к области биохимии. Предложено гуманизированное моноклональное антитело типа IgG, связывающееся с интерлейкином 17A человека. Антитело содержит вариабельные домены в виде комбинации производного VHH с вариабельным доменом легкой цепи VL. Указанное производное VHH содержит замены аминокислот в позициях 44 и 45 (по нумерации по Kabat). Также изобретение относится к ДНК, кодирующей указанное антитело, экспрессионному вектору, клеточной линии и способу получения указанного антитела. Кроме того, предложена фармацевтическая композиция для лечения опосредуемого IL-17A заболевания или нарушения, содержащая указанное антитело. Изобретение позволяет получить антитела с повышенной аффинностью и улучшенной агрегационной стабильностью. 7 н. и 18 з.п. ф-лы, 17 ил., 6 табл., 19 пр.
1. Гуманизированное моноклональное антитело типа IgG, обладающее способностью связываться с интерлейкином 17А человека, в котором вариабельные домены представлены комбинацией:
а) производного вариабельного домена тяжелой цепи VHH, отличающегося повышенной агрегационной стабильностью в сравнении с исходным VHH, выделенным из иммунизированного животного семейства Camelidae и в котором гипервариабельные участки HCDR1, HCDR2, HCDR3 отличаются тем, что:
HCDR1 включает аминокислотную последовательность, представленную SEQ ID NO: 1:
G-T-F-A-T-X32-X33-X34-X35, где
Х32 представляет собой аминокислоту, выбранную из группы, включающей S, N, K, R, Е, W, Q, D, А, V и F;
Х33 представляет собой аминокислоту, выбранную из группы, включающей Р или S;
Х34 представляет собой аминокислоту, выбранную из группы, включающей М и I;
Х35 представляет собой аминокислоту, выбранную из группы, включающей G, N, S, A, L, I, R, V и Q;
HCDR2 включает аминокислотную последовательность SEQ ID NO: 2:
X50-I-X52-X52a-S-G-X55-D-R-I-Y-A-D-S-V-K-G, где
Х50 представляет собой аминокислоту, выбранную из группы, включающей A, G и L;
Х52 представляет собой аминокислоту, выбранную из группы, включающей S, D и Е;
Х52а представляет собой аминокислоту, выбранную из группы, включающей Р и А;
Х55 представляет собой аминокислоту, выбранную из группы, включающей G, S, Т, L, R, D, Е, K, А;
HCDR3 включает аминокислотную последовательность SEQ ID NO: 3:
С-А-Х94-Х95-Х96-X97-F-X99-X100-Х100а-X100b-Х100с-X100d-Х100е-X100f-D-Y-D-S, где
Х94 представляет собой аминокислоту, выбранную из группы, включающей K, S, Т, V, D и G;
Х95 представляет собой аминокислоту, выбранную из группы, включающей R и K;
Х96 представляет собой аминокислоту, выбранную из группы, включающей G, R, Y, Н, D, W и K;
Х97 представляет собой аминокислоту, выбранную из группы, включающей R, А, V, S, L и Н;
Х99 представляет собой аминокислоту, выбранную из группы, включающей D, Е, G, A, R, V, K и Q;
X100 представляет собой аминокислоту, выбранную из группы, включающей G, S и N;
X100a представляет собой аминокислоту, выбранную из группы, включающей G, Т, Р, V, R, N и K;
X100b представляет собой аминокислоту, выбранную из группы, включающей V, S, Т, L, А, Н, G и I;
Х100с представляет собой аминокислоту, выбранную из группы, включающей Y, W и S;
X100d представляет собой аминокислоту, выбранную из группы, включающей R, V, L, Y, A, W, K, G, Q и I;
Х100е представляет собой аминокислоту, выбранную из группы, включающей Т, L, А и S;
X100f представляет собой аминокислоту, выбранную из группы, включающей Т, L, G, Р, N, A, Q, F, I и D;
а аминокислотные замены в позициях Х44 Х45 представляют собой соответственно G и A, G и V, G и Т, G и Н, А и V, А и Т, V и V, S и T, Т и V или Т и Т аминокислоту, выбранную из группы, включающей А, V, Т и Н;
b) вариабельного домена легкой цепи VL антитела человека или вариабельного домена легкой цепи гуманизированного антитела.
2. Антитело по п.1, содержащее:
a) производное вариабельного домена тяжелой цепи VHH, который содержит 3 гипервариабельных участка HCDR1, HCDR2 и HCDR3, где:
HCDR1 включает последовательность G-T-F-A-T-S-P-M-G (SEQ ID NO: 4);
HCDR2 включает последовательность A-I-S-P-S-G-G-D-R-I-Y-A-D-S-V-K-G (SEQ ID NO: 5);
HCDR3 включает последовательность C-A-V-R-R-R-F-D-G-T-S-Y-Y-T-G-D-Y-D-S (SEQ ID NO: 6);
b) вариабельный домен легкой цепи VL антитела человека или вариабельный домен легкой цепи гуманизированного антитела.
3. Антитело по п.1, содержащее:
а) производное вариабельного домена тяжелой цепи VHH, который содержит аминокислотную последовательность
QVQLVQSGGGLVQAGGSLRLSCAASGGTFATSPMGWLRQAPGKGTEFVAAISPSGGDRIYADSVKGRFTISRDNAGYFIYLQMNSLKPEDTAVYYCAVRRRFDGTSYYTGDYDSWGQGTLVTVSS (SEQ ID NO: 7);
b) вариабельный домен легкой цепи VL антитела человека или вариабельный домен легкой цепи гуманизированного антитела.
4. Антитело по п.1, содержащее:
a) производное вариабельного домена тяжелой цепи VHH, который включает аминокислотную последовательность SEQ ID NO: 7;
b) вариабельный домен легкой цепи VL антитела человека, который включает аминокислотную последовательность
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYDASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYSYSPVTFGQGTKVEIKR (SEQ ID NO: 8).
5. Антитело по п.1, содержащее:
a) тяжелую цепь, которая содержит аминокислотную последовательность
QVQLVQSGGGLVQAGGSLRLSCAASGGTFATSPMGWLRQAPGKGTEFVAAISPSGGDRIYADSVKGRFTISRDNAGYFIYLQMNSLKPEDTAVYYCAVRRRFDGTSYYTGDYDSWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLYITREPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 9);
b) вариабельный домен легкой цепи VL антитела человека, который включает аминокислотную последовательность
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYDASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYSYSPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 10).
6. Антитело по п.1, отличающееся тем, что указанное антитело является антителом любого из изотипов IgG1, IgG2, IgG3 или IgG4.
7. Антитело по п.6, содержащее неприродный модифицированный фрагмент Fc в составе IgG.
8. Антитело по п.1, отличающееся тем, что имеет такую агрегационную стабильность, что при концентрациях более 10 мг/мл и повышении температуры до 50°С в течение более чем 6 часов содержание агрегатов не увеличивается более чем на 5% от исходного содержания в растворе.
9. Антитело по п.1, отличающееся тем, что имеет константу диссоциации с KD не более 10-9 М.
10. Антитело по п.1, отличающееся тем, что имеет кинетическую константу ассоциации с антигеном kon (1/Ms) не менее 105 1/Мс.
11. Антитело по п.1, отличающееся тем, что имеет кинетическую константу диссоциации с антигеном dis(1/c) не более 10-4 1/с.
12. Антитело по п.1, имеющее аффинность связывания с IL-17A человека с KD не более 10-10 М.
13 Антитело по п.1, отличающееся тем, что кинетическая константа ассоциации kon (1/Мс) для IL-17A человека составляет не менее 105 1/Ms.
14. Антитело по п.1, отличающееся тем, что кинетическая константа диссоциации dis(1/c) для IL-17A человека составляет не более 10-5 1/с.
15. Антитело по п.1, способное ингибировать не менее чем на 50% активность IL-17A человека.
16. Антитело по п.1, отличающееся тем, что его продуцируют в клетках млекопитающих, дрожжевых или бактериальных клетках.
17. Антитело по п.1, отличающееся тем, что дополнительно содержит одну и более аминокислотных замен в Fc-регионе в сравнении с природным вариантом Fc, где указанные замены улучшают физико-химические и фармакокинетические свойства антитела по отношению к антителу с природным Fc-фрагментом и не приводят к потере способности антитела связывать IL-17А.
18. Фрагмент антитела по пп.1-17, характеризующийся тем, что представляет Fab-фрагмент.
19. ДНК, кодирующая антитело по пп.1-17 или его фрагмент по п.18.
20. Экспрессионный вектор для получения антитела по пп.1-17 или его фрагмента по п.18, содержащий одну или несколько ДНК по п.19.
21. Клеточная линия, содержащая в клетках вектор по п.20 или ДНК по п.19 и способная продуцировать антитела по пп.1-17 или их фрагменты по п.18.
22. Способ получения антитела по пп.1-17 или его фрагмента по п.18, включающий культивирование клеточной линии по п.21, в культуральной среде в условиях, достаточных для получения указанного антитела или его фрагмента, с последующим выделением и очисткой полученного антитела или его активного фрагмента.
23. Фармацевтическая композиция, предназначенная для лечения опосредуемого IL-17A заболевания или нарушения, содержащая в терапевтически эффективном количестве не менее 25 мг антитела по пп.1-17 или его фрагмента по п.18, в сочетании с одним или несколькими фармацевтически приемлемыми эксципиентами, разбавителями или носителями.
24. Фармацевтическая композиция по п.23, содержащая дополнительные действующие вещества, выбранные из блокаторов ФНО-α.
25. Фармацевтическая композиция по п.23, где опосредуемое IL-17A заболевание или нарушение выбрано из ревматоидного артрита, остеоартрита, ювенильного хронического артрита, септического артрита, артрита Лайма, псориатического артрита, реактивного артрита, спондилоартропатии, системной красной волчанки, болезни Крона, язвенного колита, воспалительного заболевания кишечника, инсулинзависимого сахарного диабета, тиреоидита, астмы, аллергических заболеваний, псориаза, атопического дерматита, склеродермии, реакции «трансплантат против хозяина», отторжения трансплантата органа, острого или хронического иммунного заболевания, связанного с трансплантацией органа, саркоидоза, болезни Кавасаки, болезни Грэйвса, нефротического синдрома, синдрома хронической усталости, гранулематоза Вегенера, пурпуры Шенлейна-Геноха, микроскопического васкулита почек, хронического активного гепатита, увенита, септического шока, синдрома токсического шока, септического синдрома, кахексии, синдрома приобретенного иммунодефицита, острого поперечного миелита, хореи Гентингтона, болезни Паркинсона, болезни Альцгеймера, инсульта, первичного билиарного цирроза, гемолитической анемии, взрослого (острого) респираторного дистресс-синдрома, алопеции, очаговой алопеции, серонегативной артропатии, артропатии, болезни Рейтера, псориатической артропатии, связанной с язвенным колитом артропатии, атопической аллергии, аутоиммунного буллезного заболевания, пемфигуса обыкновенного, листовидного пемфигуса, пемфигоида, болезни линейных IgA, аутоиммунной гемолитической анемии, Кумбс-положительной гемолитической анемии, приобретенной пернициозной анемии, ювенильной перниционзой анемии, артериита, первичного склерозирующего гепатита, криптогенного аутоиммунного гепатита, фиброзного заболевания легких, криптогенного фиброзного альвеолита, поствоспалительного интерстициального заболевания легких, интерстициального пневмонита, хронической эозинофильной пневмонии, постинфекционного интерстициального заболевания легких, подагрического артрита, аутоиммунного гепатита, аутоиммунного гепатита I типа (классического аутоиммунного или люпоидного гепатита), аутоиммунного гепатита II типа, остеоартроза, первичного склерозирующего холангита, псориаза I типа, псориаза II типа, идиопатической лейкопении, аутоиммунной нейтропении, NOS-болезни почек, гломерулонефрита, микроскопического васкулита почек, дискоидной красной волчанки, идиопатического или NOS-мужского бесплодия, аутоиммунитета к сперматозоидам, рассеянного склероза (все подтипы), симпатической офтальмии, легочной гипертензии, вторичной для болезни соединительной ткани, синдрома Гудпасчера, легочного проявления узелкового полиартериита, острой ревматической атаки, ревматоидного спондилита, болезни Стилла, системной склеродермии, синдрома Шенгрена, болезни/артериита Такаясу, аутоиммунной тромбоцитопении, идиопатической тромбоцитопении, аутоиммунного заболевания щитовидной железы, гипертиреоидизма, зобного аутоиммунного гипотиреоза (болезнь Хашимото), атрофического аутоиммунного гипотиреоза, первичной микседемы, факогенного увеита, первичного васкулита, витилиго, острого заболевания печени, хронического заболевания печени, аллергии и астмы, психических расстройств (включая депрессию и шизофрению), опосредуемых типом Th2 и типом Th1 заболеваний, конъюнктивита, аллергического контактного дерматита, аллергического ринита, дефицита альфа-1-антитрипсина, бокового амиотрофического склероза, анемии, кистозного фиброза, связанных с цитокиновой терапией нарушений, демиелинизирующих заболеваний, дерматита, иридоциклита/увеита/оптического неврита, повреждения при ишемии-реперфузии, ишемического инсульта, ювенильного ревматоидного артрита аутоиммунной энтеропатии, аутоиммунной потери слуха, аутоиммунного лимфопролиферативного синдрома, аутоиммунного миокардита, аутоиммунного преждевременного угасания функции яичников, блефарита.
| АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА К IL-17 | 2005 |
|
RU2426741C2 |
| MUYLDERMANS S | |||
| et al., Recognition of antigens by singledomain antibody fragments: the superfluous luxury of paired domains, TRENDS in Biochemical Sciences, 2001, Vol.26 No.4, pp.230-235 | |||
| CONRATH K | |||
| et al., Antigen Binding and Solubility Effects upon the Veneering of a Camel VHH in Framework-2 to Mimic a VH, J | |||
| Mol | |||
| Biol., 2005, vol.350, pp.112-125; | |||
| РОЙТ А., БРОСТОФФФ ДЖ., МЕЙЛ Д | |||
| Иммунология | |||
| Пер | |||
| с англ | |||
| - М.:Мир, 2000 | |||
| Телефонная трансляция с катодными лампами | 1920 |
|
SU592A1 |

